Описываемое здесь изобретение было создано при финансовой поддержке Правительства Соединенных Штатов в рамках номера гранта R01 HL62454, предоставленного Национальным Институтом Здравоохранения, и номера гранта DAMRDCW81XWH-04-1-054, предоставленного Министерством Обороны США. Соответственно, Правительство Соединенных Штатов имеет определенные права на это изобретение.
По всей этой заявке для ссылок на различные публикации используются арабские числа в круглых скобках или имя автора и дата публикации в круглых скобках. Полные ссылки на эти публикации можно найти в конце описания настоящего изобретения. Таким образом, описания этих публикаций включены посредством ссылки в эту заявку для более полного описания области техники, к которой относится это изобретение.
Предпосылки создания изобретения
Развитие сосудов
Во время эмбриогенеза у млекопитающих формирование сосудистой системы является ранним и весьма важным процессом. У эмбриона развитие сосудов начинается с плюрипотентного гемангиобласта, возникающего из параксиальной мезодермы и мезодермы боковых пластинок. Гемангиобласт обладает способностью к дифференцировке либо в гемопоэтический предшественник, либо в предшественник эндотелиальных клеток, известный как ангиобласт.
Развитие сосудов начинается с процесса, известного как васкулогенез, посредством чего ангиобласты подвергаются дифференцировке в эндотелиальные клетки и перемещаются вместе с образованием недифференцированного сосудистого сплетения. Эта первоначальная сосудистая сеть состоит из сосудов, которые являются однородными по размеру и целиком состоят из эндотелиальных клеток. Сосудистое сплетение затем подвергается ремоделированию благодаря ангиогенезу.
Ангиогенез включает спраутинг (разрастание) новых сосудов, перемещение этих сосудов в лишенные сосудов области и привлечение дополнительных клеток, перицитов и гладкомышечных клеток (Gale and Yancopoulos, 1999). Гладкомышечные клетки, которые подвергаются дифференцировке и образуют сокращающиеся сосудистые стенки, возникают из множества предшественников, включающих клетки нервного гребешка, мезенхимные клетки и даже эндотелиальные клетки (Owens, 1995). У взрослых ангиогенез вовлечен в фолликулярное развитие, заживление ран и патологические процессы, такие как ангиогенез в опухоли и болезнь сердца.
Семейство Notch и лиганды для Notch
Исследования Drosophila, C. elegans, данио и млекопитающих показали, что путь с участием Notch является эволюционно консервативным механизмом передачи сигнала, который функционирует для модуляции многочисленных выборов судьбы клетки. Для передачи сигнала Notch необходима надлежащая картина (структурирование) клеток, возникающих из всех трех зародышевых листков. В зависимости от клеточного окружения передача сигнала Notch может как ингибировать, так и индуцировать дифференциацию, индуцировать пролиферацию и способствовать выживанию клеток (Artavanis-Tsakonas et al., 1995; Lewis, 1998; Weinmaster, 1997). У Drosophila единственный белок Notch активируется двумя лигандами, Serrate и Delta. У млекопитающих эти семейства расширены до четырех генов Notch (Notch1, Notch2, Notch3 и Notch4) и пяти лигандов, 2 Serrate-подобных белков (Jagged1-2) и 3 Delta (Dl1, 3, 4) (Bettenhausen et al., 1995; Dunwoodie et al., 1997; Gallahan and Callahan, 1997; Lardelli et al., 1994; Lindsell et al., 1995; Shawber et al., 1996a; Shutter et al., 2000a; Uyttendaele et al., 1996; Weinmaster et al., 1992; Weinmaster et al., 1991). Во время эмбриогенеза картины экспрессии рецепторов и лигандов Notch являются динамическими в пространстве и времени. Однако неизвестно, всякие ли лиганды активируют всякие рецепторы.
Сигнализация и функция Notch
Передача сигнала Notch оказывает влияние на множество различных типов выборов судьбы клетки благодаря обеспечению ингибирующих, индукционных или пролиферативных сигналов, зависящих от внешнего окружения (проанализированных в Artavanis-Tsakonas et al., 1995; Greenwald, 1998; Robey, 1997; Vervoort et al., 1997). Эта плейотропная функция означает, что Notch модулирует множество путей передачи сигналов пространственно-временным образом.
В соответствии с регулированием Notch выборов судьбы клетки, как рецепторы, так и лиганды являются белками клеточной поверхности с единичными трансмембранными доменами (фиг.1). Регуляторный внеклеточный домен белков Notch в основном состоит из последовательно расположенных EGF-подобных повторов, которые требуются для связывания лигандов (Artavanis-Tsakonas et al., 1995; Weinmaster, 1998). В С-концевом направлении от EGF-подобных повторов находятся три дополнительных, богатых цистеином повтора, названные LIN12/Notch-повторами (LNR) (Greenwald, 1994). После LNR находится последовательность протеолитического расщепления (RXRR), которую распознает фуриноподобная конвертаза. В случае Notch1 расщепление в этом сайте дает внеклеточный пептид с М.м. 180 килодальтон и внутриклеточный пептид с М.м. 120 килодальтон, которые удерживаются вместе с образованием гетеродимерного рецептора на клеточной поверхности (Blaumueller et al., 1997; Kopan et al., 1996; Logeat et al., 1998).
Внутриклеточный домен Notch (NotchICD, фиг.1) восстанавливает фенотипы Notch, связанные с потерей функции, означая, что эта форма Notch передает сигнал коститутивно (Fortini and Artavanis-Tsakonas, 1993; Lyman and Young, 1993; Rebay et al., 1993; Struhl et al., 1993).
Цитоплазматический домен Notch содержит три идентифицируемых домена: RAM-домен, содержащий анкириновые повторы домен и С-концевой PEST-домен (фиг.1). В результате активации под действием лигандов Notch подвергается двум дополнительным протеолитическим расщеплениям, которые приводят к освобождению цитоплазматического домена (Weinmaster, 1998). Этот пептид Notch перемещается в ядро и взаимодействует с репрессором транскрипции, известным как CSL (CBF, Su (H), Lag-2) и превращает его в активатор транскрипции. Взаимодействие CSL/Notch зависит от присутствия RAM-домена Notch; в то время как для транскрипционной активности требуется также присутствие анкириновых повторов (Hsieh et al., 1996; Hsieh et al., 1997; Roehl et al., 1996; Tamura et al., 1995; Wettstein et al., 1997). И in vivo, и in vitro исследования показывают, что гены HES и Hey являются прямыми мишенями Notch/CSL-зависимой сигнализации (Bailey and Posakony, 1995; Eastman et al., 1997; Henderson et al., 2001; Jarriault et al., 1995; Nakagawa et al., 2000; Wettstein et al., 1997). Продукты генов HES и Hey являются репрессорами транскрипции bHLH, которые связываются с ДНК в N-боксах (Nakagawa et al., 2000; Sasai et al., 1992; Tietze et al., 1992). Было высказано предположение, что Notch также передает сигнал с помощью CSL-независимого пути. В действительности, экспрессия лишь содержащего анкириновые повторы домена необходима и достаточна для некоторых форм передачи сигнала Notch (Lieber et al., 1993; Matsuno et al., 1997; Shawber et al., 1996b).
Наконец, установлено, что PEST-домен вовлечен в белковый обмен с использованием SEL-10/убиквитин-зависимого пути (Greenwald, 1994; Oberg et al., 2001; Rogers et al., 1986; Wu et al., 1998; Wu et al., 2001). Так же как и в рецепторах, внеклеточный домен лигандов Notch также в основном состоит из последовательно расположенных EGF-подобных повторов (фиг.1). Перед этими повторами находится отличный EGF-подобный повтор, известный как DSL (Delta, Serrate, Lag-2), который требуется для связывания лигандов и активации рецепторов (Artavanis-Tsakonas et al., 1995).
Передача сигнала Notch и развитие сосудов
Хотя многие из генов, которые функционируют с индукцией васкулогенеза и ангиогенеза, были идентифицированы, мало известно о том, каким образом определяются выборы судьбы клетки во время развития сосудов. Ряд наблюдений наводит на мысль, что путь передачи сигнала с участием Notch играет роль в определении судьбы клетки и в структурировании сосудистой системы.
Все из Notch1, Notch4, Jagged1 и Dll4 экспрессируются при развитии сосудистой сети, в то время как Notch3 экспрессируется во вспомогательных гладкомышечных клетках (Krebs et al., 2000; Shutter et al., 2000b; Uyttendaele et al., 1996; Villa et al., 2001; Xue et al., 1999). Мыши, у которых отсутствует Jagged1, имеют смертельный исход на эмбриональной стадии и имеют тяжелые пороки развития сосудов (Xue et al., 1999). Нуль-зиготные по Notch1 мыши имеют смертельный исход на эмбриональной стадии и умирают от тяжелых нейрональных дефектов, но также имеют недостатки ангиогенеза (Krebs et al., 2000; Swiatek et al., 1994). Мыши, у которых отсутствует Notch4, рождаются и кажутся нормальными, но эмбрионы, у которых отсутствуют как Notch1, так и Notch4, умирают на E9,5 от значительного кровоизлияния и пороков сосудистого структурирования, что означает, что Notch1 и Notch4 могут быть функционально дублирующими во время развития сосудов (Krebs et al., 2000). Экзогенная экспрессия активированной формы Notch4 в эндотелии также приводила к порокам развития сосудов, схожим с теми, которые наблюдались в случае нуль-зиготных и по Notch1, и по Notch4 мышей, наводя на мысль, что соответствующие уровни передачи сигнала Notch важны для надлежащего развития сосудистой сети на эмбриональной стадии (Uyttendaele et al., 2001).
Вместе взятые, данные, полученные на основе мутантных по Notch/компонентам передачи сигнала Notch мышей, выявляют несколько зависимых от Notch процессов, включающих ремоделирование сосудов, определение эндотелиальных клеток в качестве артериальных/венозных, привлечение гладкомышечных клеток к сосудам и развитие сосудов сердца/оттока от сердца.
Недавние эксперименты показали вовлеченность передачи сигнала Notch в определение эндотелиальных клеток в качестве артериальных/венозных. In situ анализ эмбрионов на E13,5 показал, что экспрессия Notch1, Notch3, Notch4, Dl4, Jagged1 и Jagged2 ограничена артериями и отсутствует в венах (Villa et al., 2001). В соответствии с данными по экспрессии была установлена связь нарушения передачи сигнала Notch у данио с утратой маркера артериальной эндотелиальной клетки эфрина B2; в то время как эктопическая экспрессия активированной формы Notch приводит к утрате маркера венозной эндотелиальной клетки EphB4 в дорсальной аорте (Lawson et al., 2001). Эти данные говорят о том, что передача сигнала Notch может помочь в определении судеб клеток в качестве клеток артерий и вен во время ангиогенеза.
Вместе взятые, данные, полученные на основе мутантных по Notch/компонентам передачи сигнала Notch мышей, выявляют несколько зависимых от Notch процессов, включающих ремоделирование сосудов, определение эндотелиальных клеток в качестве артериальных/венозных, привлечение гладкомышечных клеток к сосудам и развитие сосудов сердца/оттока от сердца.
Также было высказано предположение, что передача сигнала Notch функционирует в сосудистой системе взрослого. У людей имеется связь миссенс-мутаций во внеклеточном домене Notch3 с развитием дегенеративного заболевания сосудов, CADASIL (Caronti et al., 1998; Desmond et al., 1998; Joutel et al., 2000; Joutel et al., 1996). В модели заживления раны увеличение экспрессии Jagged1 наблюдалось в крае раны с регенерацией эндотелия, что говорит о том, что передача сигнала Notch может функционировать во время процессов ангиогенеза у взрослых (Lindner et al., 2001). Вместе взятые, эти данные подтверждают функционирования передачи сигналов Notch в ряде важных стадий во время развития сосудов: при васкулогенезе, структурировании сосудов/ангиогенезе и определении эндотелиальных клеток в качестве артериальных/венозных. Однако молекулярный механизм(ы), посредством которого пути передачи сигнала с участием Notch оказывают влияние на эти различные стадии, все еще не уяснен.
Значение
Shimizu и др. (J. Biol. Chem. 274(46): 32961-32969 (1999)) описывают использование Notch1ECD/Fc, Notch2ECD/Fc и Notch3ECD/Fc в исследованиях связывания. Однако Shimizu и др. не указывают на применение таких белков для ингибирования ангиогенеза.
В патенте США с № 6379925, выданном Kitajewski и др. 30 апреля 2002 года, описывается Notch4 мыши. Однако в нем не описываются основанные на Notch гибридные белки, изложенные в связанной заявке.
Белки Notch играют важную роль в выборах развития, затрагивающих сосудистую сеть, гемопоэтическую систему и нервную систему. Как таковое, понимание их функционирования является ключом к пониманию того, каким образом выборы судьбы клетки и коммитирование контролируются во время развития и в тканях взрослых. На данное число фенотипы сосудов были описаны в нескольких сообщениях о нарушениях генов Notch или лигандов Notch, в которых подчеркивается, что этот путь является основной частью механизма, который определяет развитие сосудов. Была установлена связь аберрантной активности Notch с патологиями у людей, включающими как рак, так и сосудистые нарушения (CADASIL). Исследование Notch при ангиогенезе в опухоли было начато лишь недавно; однако обнаружение авторами настоящего изобретения возможных, находящихся в прямом направлении передачи сигнала мишеней Notch говорит о роли в патологических процессах, связанных с ангиогенезом. Например, была установлена связь VEGFR-3 как с ангиогенезом в опухоли, так и с лимфоангиогенезом в опухоли. С ангиогенезом в опухоли также была связана экспрессия или функционирование нескольких других возможных мишеней Notch, в том числе эфрина B2, Id3, ангиопоэтина 1 и PDGF-B. Понимание роли этих мишеней в функционировании генов Notch будет, несомненно, способствовать дальнейшему исследованию Notch при патологиях у людей.
Краткое изложение сущности изобретения
Этим изобретением предоставляется гибридный белок, включающий сигнальный пептид, EGF-повторы 1-Х внеклеточного домена белка рецептора Notch3 человека, причем X является любым целым числом от 12 до 34, и Fc-часть антитела, связанную с ними.
Этим изобретением предоставляется гибридный белок, включающий сигнальный пептид, EGF-повторы 1-Х внеклеточного домена белка рецептора Notch3 человека, причем X является любым целым числом от 1 до 10, и Fc-часть антитела, связанную с ними.
Этим изобретением предоставляется гибридный белок, включающий сигнальный пептид, по меньшей мере 12 EGF-повторов внеклеточного домена белка рецептора Notch3 человека и Fc-часть антитела, связанную с ними.
Этим изобретением предоставляется гибридный белок, включающий сигнальный пептид, EGF-повторы внеклеточного домена белка рецептора Notch3 человека, причем присутствуют по меньшей мере 12 EGF-повторов, и Fc-часть антитела, связанную с ними.
Этим изобретением предоставляется способ лечения имеющего опухоль субъекта, включающий введение субъекту количества указанного выше гибридного белка, эффективного для лечения субъекта, осуществляя тем самым лечение субъекта, имеющего опухоль.
Этим изобретением предоставляется способ ингибирования ангиогенеза у субъекта, включающий введение субъекту количества указанного выше гибридного белка, эффективного для ингибирования ангиогенеза у субъекта, ингибируя тем самым ангиогенез у субъекта.
Этим изобретением предоставляется способ лечения имеющего рак яичника субъекта, включающий введение субъекту количества указанного выше гибридного белка, эффективного для лечения субъекта, осуществляя тем самым лечение субъекта, имеющего рак яичника.
Этим изобретением предоставляется применение указанного выше гибридного белка для приготовления фармацевтической композиции для лечения имеющего опухоль субъекта.
Этим изобретением предоставляется применение указанного выше гибридного белка для приготовления фармацевтической композиции для ингибирования ангиогенеза у субъекта.
Этим изобретением предоставляется применение указанного выше гибридного белка для приготовления фармацевтической композиции для лечения имеющего рак яичника субъекта.
Этим изобретением предоставляется применение указанного выше гибридного белка для приготовления фармацевтической композиции для лечения субъекта, имеющего нарушение обмена веществ.
Этим изобретением предоставляется способ ингибирования физиологического лимфоангиогенеза или патологического лимфоангиогенеза у субъекта, включающий введение субъекту количества указанного выше гибридного белка, эффективного для ингибирования физиологического лимфоангиогенеза или патологического лимфоангиогенеза у субъекта.
Этим изобретением предоставляется способ ингибирования метастазирования опухоли у субъекта, включающий введение субъекту количества указанного выше гибридного белка, эффективного для ингибирования метастазирования опухоли у субъекта.
Этим изобретением предоставляется способ ингибирования роста вторичной опухоли у субъекта, включающий введение субъекту количества указанного выше гибридного белка, эффективного для ингибирования роста вторичной опухоли у субъекта.
Этим изобретением предоставляется способ ингибирования использования опухолью для своих собственных целей кровеносных сосудов у субъекта, включающий введение субъекту количества указанного выше гибридного белка, эффективного для ингибирования использования опухолью для своих собственных целей кровеносных сосудов у субъекта.
Этим изобретением предоставляется способ лечения рака у субъекта, включающий введение субъекту указанного выше гибридного белка и ингибитора фактора роста сосудистого эндотелия (VEGF) в количестве каждого из них, эффективном для лечения рака у субъекта.
Этим изобретением предоставляется способ лечения рака у субъекта, включающий введение субъекту указанного выше гибридного белка и ингибитора рецептора VEGF в количестве каждого из них, эффективном для лечения рака у субъекта.
Этим изобретением предоставляется способ лечения рака у субъекта, включающий введение субъекту указанного выше гибридного белка и ингибитора тромбоцитарного фактора роста (PDGF) в количестве каждого из них, эффективном для лечения рака у субъекта.
Этим изобретением предоставляется способ лечения рака у субъекта, включающий введение субъекту указанного выше гибридного белка и антагониста рецептора PDGF в количестве каждого из них, эффективном для лечения рака у субъекта.
Этим изобретением предоставляется способ лечения рака у субъекта, включающий введение субъекту указанного выше гибридного белка и ингибитора HER2/neu в количестве каждого из них, эффективном для лечения рака у субъекта.
Этим изобретением предоставляется способ лечения рака молочной железы у субъекта, включающий введение субъекту количества указанного выше гибридного белка, эффективного для лечения рака молочной железы у субъекта.
Этим изобретением предоставляется применение указанного выше гибридного белка для приготовления фармацевтической композиции для лечения имеющего рак молочной железы субъекта.
Краткое описание чертежей
Фиг.1
На этой фигуре демонстрируется схематическая структура Notch и лигандов Notch: Notch1, Notch2, Notch3, Notch4, Jagged-1, Jagged-2, Delta-подобного белка 1, Delta-подобного белка 3, Delta-подобного белка 4.
Фиг.2
На этой фигуре демонстрируется схематическая конструкция основанных на Notch гибридных белков (NotchECD/Fc). Внеклеточный домен (ECD) Notch1, Notch2, Notch3 или Notch4, содержащий EGF-повторы, слит с Fc-частью антитела.
Фиг.3
На этой фигуре демонстрируется анализ сокультивирования для проверки активности основанных на Notch гибридных белков. Notch и реагирующие на Notch транскрипционные факторы-репортеры экспрессируются в «Notch-реагирующей» клетке, HeLa. Лиганды Notch, Jagged-1, Delta-подобный белок 1 или Delta-подобный белок 4, экспрессируются в «лигандпрезентирующей» клетке, 293. Экспрессии добиваются при помощи трансфекции отдельных клеточных популяций, клетки подвергают сокультивированию, а затем анализируют на Notch-зависимую активность репортера.
Фиг.4
На этой фигуре демонстрируется ингибиторная в отношении активации передачи сигнала Notch при взаимодействии между Notch и лигандом Notch активность основанного на Notch гибридного белка. Индукция передачи сигнала Notch была выявлена при сокультивировании экспрессирующих и Notch1, и 3 типа лигандов Notch клеток, и эти индукции были ингибированы при котрансфенции экспрессирующего основанный на Notch гибридный белок вектора в Notch1-экспрессирующие клетки. Следовательно, основанные на Notch гибридные белки могут использоваться в качестве ингибитора Notch на основе ингибирования взаимодействия между Notch и лигандом Notch.
Фиг.5
На этой фигуре демонстрируется экспрессия основанного на Notch1 гибридного белка (Notch1ECD/Fc) в клетках 293. Панель A: экспрессия в клеточном лизате (Lys) или с секрецией в среды (Sup). Панель B: экспрессия перечисленных NECD/Fc в лизатах клеток 293.
Фиг.6
На этой фигуре демонстрируется активация передачи сигнала Notch в HUVEC, инфицированных аденовирусным вектором, кодирующим VEGF-165. Активацию передачи сигнала Notch можно обнаружить посредством использования активности промотора CBF1. Транскрипционная активность промотора CBF1 повышается при связывании Notch-IC с CBF1. Активность промотора CBF1 определяли в HUVEC, которые были инфицированы аденовирусом, кодирующим VEGF-165, при различных множественностях инфицирования (MOI). Индукция промотора CBF1 была однозначно обнаружена в Ad-VEGF-инфицированных HUVEC по сравнению с Ad-LacZ-инфицированными клетками, зависимым от множественности заражения образом. Эти данные показали, что сверхэкспрессия VEGF может активировать передачу сигнала Notch в HUVEC.
Фиг.7
На этой фигуре демонстрируется эффект основанных на Notch гибридных белков на VEGF-индуцируемую активацию передачи сигнала Notch. Коинфицирование аденовирусным вектором, кодирующим основанный на Notch гибридный белок, и аденовирусом, кодирующим VEGF, отчетливо снижало повышение активности промотора CBF1, индуцируемое при инфицировании только Ad-VEGF. В случае инфицирования с составляющей 40 множественностью инфицирования для каждого аденовируса на панели A было обнаружено ингибирование на 60% через 24 ч и ингибирование на 90% через 48 ч после трансфекции гена-репортера. Эта ингибиторная активность Notch-ловушки зависела от множественности инфицирования аденовирусным вектором, кодирующим основанный на Notch гибридный белок.
Фиг.8
На этой фигуре демонстрируется эксперимент, в котором оценивался эффект основанных на Notch гибридных белков на индукцию под действием сверхэкспрессируемого VEGF-165 образования выступов у HUVEC. После подвергания Ad-VEGF-инфицированных HUVEC культивированию на коллагеновом (типа I) геле в течение 8 дней индуцировалось образование выступов в коллагеновый гель. Эта индукция образования выступов под действием сверхэкспрессируемого VEGF была явно ингибирована при коинфицировании аденовирусами, кодирующими основанные на Notch гибридные белки. Сам по себе аденовирус, кодирующий основанный на Notch гибридный белок, оказывал меньший эффект на морфологию.
Фиг.9
На этой фигуре демонстрируется результат подсчета выступов в каждом поле под микроскопом. Инфицирование Ad-VEGF клеток HUVEC увеличило число выступов, зависящее от используемой множественности инфицирования (MOI). Несмотря на то, что в случае аденовирусного вектора для основанного на Notch гибридного белка была использована множественность инфицирования, составляющая половину от таковой в случае Ad-VEGF, Ad-VEGF-индуцируемое образование выступов было явно ингибировано. Эти данные говорят о том, что VEGF индуцировал образование выступов у HUVEC благодаря активации передачи сигнала Notch, и что основанный на Notch гибридный белок мог ингибировать VEGF-индуцируемое образование выступов.
Фиг.10
На этой фигуре демонстрируется аминокислотная последовательность внеклеточного домена белка Notch1 крысы (SEQ ID NO: 1) и последовательность линкера (SEQ ID NO: 2).
Фиг.11
На этой фигуре демонстрируется аминокислотная последовательность внеклеточного домена белка Notch2 крысы (SEQ ID NO: 3) и последовательность линкера (SEQ ID NO: 2).
Фиг.12
На этой фигуре демонстрируется аминокислотная последовательность внеклеточного домена белка Notch3 мыши (SEQ ID NO: 4).
Фиг.13
На этой фигуре демонстрируется аминокислотная последовательность внеклеточного домена белка Notch4 мыши (SEQ ID NO: 5) и последовательность линкера (SEQ ID NO: 2).
Фиг.14A и 14B
На этой фигуре демонстрируется последовательность нуклеиновой кислоты внеклеточного домена гена Notch1 крысы (SEQ ID NO: 6).
Фиг.15A и 15B
На этой фигуре демонстрируется последовательность нуклеиновой кислоты внеклеточного домена гена Notch2 крысы (SEQ ID NO: 7).
Фиг.16A и 16B
На этой фигуре демонстрируется последовательность нуклеиновой кислоты внеклеточного домена гена Notch3 мыши (SEQ ID NO: 8).
Фиг.17A и 17B
На этой фигуре демонстрируется последовательность нуклеиновой кислоты внеклеточного домена гена Notch4 мыши (SEQ ID NO: 9), а также последовательность нуклеиновой кислоты (SEQ ID NO: 10) и аминокислотная последовательность (SEQ ID NO: 2) линкерной последовательности.
Фиг.18A и 18B
На этой фигуре демонстрируется последовательность нуклеиновой кислоты внеклеточного домена гена Notch1 человека (SEQ ID NO: 11).
Фиг.19A и 19B
На этой фигуре демонстрируется последовательность нуклеиновой кислоты внеклеточного домена гена Notch2 человека (SEQ ID NO: 12).
Фиг.20A и 20B
На этой фигуре демонстрируется последовательность нуклеиновой кислоты внеклеточного домена гена Notch3 человека (SEQ ID NO: 13).
Фиг.21A и 21B
На этой фигуре демонстрируется последовательность нуклеиновой кислоты внеклеточного домена гена Notch4 человека (SEQ ID NO: 14).
Фиг.22A-22I
На этих фигурах демонстрируется, что VEGF активирует передачу сигнала Notch с индукцией образования выступов у HUVEC. HUVEC подвергали трансдукции с помощью Ad-VEGF с множественностью инфицирования (MOI) - 40 (фиг.22A, 22H, 22I) или 20 (фиг.22C, 22G). Ad-LacZ был котрансдуцирован в HUVEC с получением одного и того же общего количества аденовируса - с 60 MOI (фиг.22G), 80 MOI (фиг.22A) и 100 MOI (фиг.22H, 22I). На фиг.22А демонстрируется анализ с использованием ОТ-ПЦР экспрессии Notch и лигандов Notch. Числа означают число циклов ПЦР. На фиг.22В демонстрируется эффект трансдуцированного гена VEGF на активность регулируемого CSL промотора гена-репортера. На фиг.22С демонстрируется эффект SU5416 на активность регулируемого CSL промотора гена-репортера, трансактивируемого Ad-VEGF. На фиг.22D демонстрируется конструкция Notch-ловушки (N1ECDFc). На фиг.22E демонстрируется секреция N1ECDFc из HUVEC, трансдуцированных с помощью Ad-N1ECDFc. На фиг.22F демонстрируется действие N1ECDFc, направленное против лиганд-индуцируемой активности регулируемого CSL промотора гена-репортера, в анализе сокультивирования (□: (-); ■: 0,33 нг pHyTC-N1ECDFc; ■: 0,67 нг pHyTC-N1ECDFc). На фиг.22G-I демонстрируется действие N1ECDFc, направленное против Ad-VEGF-трансдуцированных HUVEC. Передачу сигнала Notch активировали с помощью трансдукции Ad-VEGF в HUVEC в отсутствие или при осуществлении котрансдукции Ad-N1ECDFc в указанной дозе. На фиг.22G демонстрируется эффект N1ECDFc на активность регулируемого CSL промотора гена-репортера, трансактивируемого Ad-VEGF. На фиг.22H демонстрируется ингибирование образования выступов у Ad-VEGF-трансдуцированных HUVEC с помощью котрансдукции Ad-N1ECDFc с составляющей 40 множественностью инфицирования. На фиг.22I демонстрируется количественный анализ эффекта N1ECDFc на образование выступов у Ad-VEGF-трансдуцированных HUVEC (□: выступ; ■: число клеток).
Фиг.23A-23J
На этих фигурах демонстрируется, что передача сигнала Notch увеличивает экспрессию Flt1 с индукцией образования выступов у HUVEC. HUVEC подвергали трансдукции с помощью либо Ad-LacZ, либо Ad-N1IC с множественностью инфицирования (MOI), составляющей 40. На фиг.23A-23C демонстрируется эффект ингибиторов рецепторных тирозинкиназ на Notch-индуцируемое образование выступов у HUVEC. Фиг.23A является фотографией образования выступов у Ad-N1IC-трансдуцированных HUVEC, подвергнутых обработке PD166866, ZD1893 в концентрации 1 мкМ и SU5416 в концентрации 0,5 мкМ. На фиг.23B демонстрируется количественный анализ эффекта ингибиторов в концентрации 1 мкМ (□: выступ; ■: количество клеток). На фиг.23C демонстрируется зависимость от дозы эффекта SU5416 (□: выступ; ■: количество клеток). На фиг.23D-E демонстрируется индукция экспрессии Flt-1 в Ad-N1IC-трансдуцированных HUVEC. На фиг.23D демонстрируется анализ с использованием ОТ-ПЦР экспрессии мРНК Flt-1. На фиг.23E демонстрируется анализ с использованием Вестерн-блоттинга экспрессии белка Flt-1. На фиг.23F-G демонстрируется активация Notch-индуцируемого образования выступов у HUVEC при стимуляции PlGF. Ad-N1IC-трансдуцированные HUVEC культивировали на коллагеновом геле с SFM, вместо полной среды, в отсутствие или в присутствии 50 нг/мл PlGF. На фиг.23F демонстрируется PlGF-индуцируемое образование выступов у Ad-N1IC-трансдуцированных HUVEC (острие стрелки: выступы с одной филоподией; стрелка: выступы с множеством филоподий). На фиг.23G демонстрируется количественный анализ эффекта PlGF на образование выступов у Ad-N1IC-трансдуцированных HUVEC (□: выступы с множеством филоподий; ■: все выступы). На фиг.23H-I демонстрируется эффект трансфекции siRNA для Flt-1 на экспрессию Flt1. Ad-N1IC-трансдуцированные HUVEC подвергали трансфекции 200 пмоль либо контрольной siRNA (CT), либо siRNA для Flt-1. На фиг.23H демонстрируется уменьшение экспрессии мРНК Flt-1. На фиг.23I демонстрируется уменьшение экспрессии белка Flt-1. На фиг.23J демонстрируется эффект трансфекции siRNA для Flt-1 на Notch-индуцируемое образование выступов у HUVEC. Ad-N1IC-трансдуцированные HUVEC подвергали трансфекции либо 100, либо 200 пмоль siRNA и культивировали на коллагеновом геле в течение 2 дней.
Фиг.24A-24E
На этих фигурах демонстрируется, что VEGF регулирует желатиназную активность через передачу сигнала Notch посредством увеличения экспрессии как MMP-9, так и MT1-MMP. На фиг.24A-B демонстрируется зимографический анализ с использованием желатина активности MMP-9 и MMP-2, стимулируемой под действием VEGF, в HUVEC. На фиг.24A демонстрируется эффект N1ECDFc на активность MMP-9. Трансдуцированные HUVEC культивировали на фибриновом геле в указанный день (т.е. D2, D4, D6, D8). Схожие результаты были также получены при использовании коллагенового геля, хотя индукция MMP-9 была более сильной на фибриновом геле, чем на коллагеновом геле (непредставленные данные). На фиг.24B демонстрируется эффект N1ECDFc на активность MMP-2. HUVEC были подвергнуты трансдукции с помощью Ad-N1ECDFc в указанных дозах, и кондиционированные среды собирали от HUVEC, подвергнутых культивированию на коллагеновом геле, в день 4. На фиг.24C-D демонстрируется увеличение экспрессии MMP-9 и MT1-MMP с помощью передачи сигнала Notch. HUVEC были подвергнуты трансдукции с помощью либо Ad-LacZ, либо Ad-N1IC с множественностью инфицирования (MOI), составляющей 40. Числа означают число циклов ПЦР. На фиг.24C демонстрируется анализ с использованием ОТ-ПЦР эффекта передачи сигнала Notch на экспрессию MMP-9 и MMP-2. На фиг.24D демонстрируется индукция экспрессии MT1-MMP и на транскрипционном уровне, и на уровне белка с помощью передачи сигнала Notch. На фиг.24E демонстрируется анализ с использованием ОТ-ПЦР экспрессии MMP-9 и MT1-MMP в Ad-VEGF-HUVEC, котрансдуцированных с помощью Ad-N1ECDFc. HUVEC были подвергнуты трансдукции с помощью Ad-VEGF в отсутствие или при осуществлении котрансдукции Ad-N1ECDFc с множественность инфицирования (MOI) - 40 для каждого вектора. Ad-LacZ котрансдуцировали с получением одного и того же общего количества аденовируса - с 80 MOI.
Фиг.25A-25D
На этих фигурах демонстрируется роль передачи сигнала Notch в VEGF-зависимом in vivo ангиогенезе. На фиг.25A-25D демонстрируется ингибирование VEGF-индуцируемого ангиогенеза с помощью N1ECDFc в исследовании дорсальных альвеолярных мешочков (DAS) у мыши. Представлены репрезентативные фотографии. На фиг.25A демонстрируется ангиогенез в подкожной основе, индуцированный трансфектантом клеток 293, сверхэкспрессирующим VEGF121, (293/VEGF) в сравнение с 293/VEGF, также экспрессирующим Notch-ловушку (основанный на Notch гибридный белок) - N1ECDFc. На фиг.25B демонстрируется количественное определение степени васкуляризации, индуцированной 293/VEGF, в контроле в сравнение с 293, экспрессирующими Notch-ловушку (основанный на Notch гибридный белок) - N1ECDFc. На фиг.25C демонстрируется ангиогенез в подкожной основе, индуцированный Ad-LacZ-инфицированными клетками MDA-MB-231 в сравнение с инфицированными Ad-N1ECDFc (аденовирусом, кодирующим основанный на Notch гибридный белок) клетками MDA-MB-231. Клетки рака молочной железы MDA-MB-231 продуцируют VEGF (непредставленные данные). На фиг.25D демонстрируется количественное определение степени васкуляризации, индуцированной Ad-LacZ-инфицированными клетками MDA-MB-231 в сравнение с инфицированными Ad-N1ECDFc (аденовирусом, кодирующим основанный на Notch гибридный белок) клетками MDA-MB-231.
Фиг.26A и 26B
На этих фигурах демонстрируется пролиферация Ad-VEGF165-трансдуцированых HUVEC. HUVEC подвергали трансдукции с помощью Ad-VEGF165 в указанных дозах. Также осуществляли коинфицирование Ad-LacZ с получением одного и того же общего количества аденовируса - с множественностью инфицирования (MOI), составляющей 40 бляшкообразующих единиц/клетку. HUVEC суспендировали в SFM, дополненной 1% FBS, и затем засевали в количестве 1×104 клеток/лунку в 24-луночные планшеты с 0,4 мл среды. Через 4 дня определяли количества клеток, используя набор CCK-8, и результаты представлены в виде отношения определенных количеств клеток к количеству контрольных клеток, которые были подвергнуты трансдукции с помощью Ad-GFP с множественностью инфицирования (MOI), составляющей 40 бляшкообразующих единиц/клетку. На фиг.26A демонстрируется эффект трансдуцированного гена VEGF на пролиферацию. На фиг.26B демонстрируется ингибиторный эффект SU5416. Ad-VEGF-трансдуцированные HUVEC обрабатывали SU5416 в указанных дозах.
Фиг.27A и 27B
На этих фигурах демонстрируется индукция образования выступов у HUVEC на коллагеновом (типа I) геле. HUVEC подвергали трансдукции с помощью либо Ad-VEGF165, либо AD-N1IC в указанных дозах. Также осуществляли коинфицирование Ad-LacZ с получением одного и того же общего количества аденовируса - с множественностью инфицирования (MOI), составляющей 40 бляшкообразующих единиц/клетку. Трансдуцированные HUVEC культивировали на коллагеновом геле с использованием полной среды. Число выступов определяли под микроскопом в день 7.
Фиг.28A и 28B
На этих фигурах демонстрируется эффект изменения передачи сигнала Notch на пролиферацию клеток. Клетки подвергали трансдукции с помощью указанных аденовирусов. Также осуществляли коинфицирование Ad-GFP с получением одного и того же общего количества аденовируса - с множественностью инфицирования (MOI), составляющей 60 бляшкообразующих единиц/клетку. Через 4 дня определяли количества клеток, используя набор CCK-8, и результаты представлены в виде отношения определенных количеств клеток к количеству контрольных клеток, которые были подвергнуты трансдукции с помощью AD-GFP с множественностью инфицирования (MOI), составляющей 60 бляшкообразующих единиц/клетку. На фиг.28A демонстрируется эффект трансдуцированного N1IC и основанного на Notch гибридного белка на пролиферацию HUVEC. Трансдуцированные HUVEC суспендировали в полной среде и затем засевали в количестве 1×104 клеток/лунку в 24-луночные планшеты с 0,4 мл указанной среды (□: Ad-N1IC; ■: Ad-N1ECDFc). На фиг.28B демонстрируется эффект основанного на Notch гибридного белка на пролиферацию трансфектантов KP1/VEGF. Подвергнутые трансдукции трансфектанты KP1/VEGF суспендировали в среде RPMI1640 и затем засевали в количестве 2×104 клеток/лунку в 24-луночные планшеты с 0,5 мл среды.
Фиг.29
На этой фигуре демонстрируется анализ с использованием ОТ-ПЦР индукции экспрессии PIGF в Ad-N1IC-трансдуцированных HUVEC. HUVEC инфицировали либо Ad-LacZ, либо Ad-N1IC со множественностью инфицирования (MOI), составляющей 40 бляшкообразующих единиц/клетку. Тотальную РНК выделяли из трансдуцированных HUVEC, подвергнутых культивированию на коллагеновом геле в течение 5 дней с использованием полной среды.
Фиг.30A-30C
На этих фигурах демонстрируется ингибирование образования выступов у либо Ad-N1IC-, либо Ad-VEGF-трансдуцированных HUVEC с помощью трансфекции siRNA для Flk-1. На фиг.30А демонстрируется уменьшение экспрессии мРНК и белка Flk-1 в Ad-VEGF-HUVEC с помощью трансфекции 200 пмоль siRNA для Flk-1. Ad-VEGF-HUVEC с множественностью инфицирования (MOI), составляющей 40 бляшкообразующих единиц/клетку, подвергали трансфекции 200 пмоль либо контрольной siRNA (CT), либо siRNA для Flk-1. Через 48 часов после трансфекции выделяли тотальную РНК. Через 48 часов после трансфекции собирали тотальный клеточный лизат от страдающих от недостатка сыворотки при использовании SFM клеток. На фиг.30B и 30C демонстрируется ингибиторный эффект трансфекции siRNA для Flk-1 на либо VEGF-, либо Notch-индуцируемое образование выступов у HUVEC. Либо Ad-N1IC-, либо Ad-VEGF-HUVEC с множественностью инфицирования (MOI), составляющей 40 бляшкообразующих единиц/клетку, подвергали трансфекции 200 пмоль указанной siRNA и культивировали на коллагеновом геле в течение 5 дней. На фиг.30В демонстрируется эффект трансфекции siRNA для Flk-1 на образование выступов у HUVEC (□: Ad-VEGF; ■: Ad-N1IC). На фиг.30C демонстрируется количественный анализ ингибиторного эффекта трансфекции siRNA для Flk-1.
Фиг.31A и 31B
На этих фигурах демонстрируется ингибирование образования выступов у Ad-N1IC-трансдуцированных HUVEC с помощью обработки ингибитором матриксных металлопротеиназ GM6001. Либо Ad-LacZ-, либо Ad-N1IC-HUVEC с множественностью инфицирования (MOI), составляющей 40 бляшкообразующих единиц/клетку, культивировали на коллагеновом геле в течение 5 дней в отсутствие или в присутствии GM6001 в концентрации 50 мкм. На фиг.31A демонстрируется эффект GM6001 на Notch-индуцируемое образование выступов у HUVEC. На фиг.31B демонстрируется количественный анализ ингибиторного эффекта GM6001.
Фиг.32A, 32B и 32C
На этой фигуре демонстрируется полноразмерная нуклеотидная последовательность Notch3 человека (SEQ ID NO: 15), состоящая из последовательности от инициирующего кодона ATG (нуклеотида 1) до стоп-кодона (TGA; нуклеотида 6964). Сигнальный пептид и домены в виде первых 34 EGF-подобных повторов соответствуют нуклеотидам 1-4158 этой последовательности. Нуклеотиды 1-4158 используются для разработки ложных белков Notch3 человека, описываемых здесь. Подчеркнуты нуклеотиды, охватывающие EGF-повторы 1-34.
Фиг.33
На этой фигуре демонстрируется полноразмерная аминокислотная последовательность Notch3 человека (SEQ ID NO: 16), состоящая из последовательности от аминокислоты 1 (M=метионина) до аминокислоты 2555 (K=лизина). Сигнальный пептид и домены в виде первых 34 EGF-подобных повторов соответствуют аминокислотам 1-1386 этой последовательности. Аминокислоты 1-1386 используются для разработки ложных белков Notch3 человека, описываемых в последующих частях. Подчеркнуты аминокислоты, охватывающие EGF-повторы 1-34.
Фиг.34
На этой фигуре демонстрируется схематизация двух ложных белков Notch3 человека, ложного белка h-Notch3(1-34) и ложного белка h-spHC-Notch3(1-34).
Фиг.35
На этой фигуре демонстрируется используемая для создания Fc-метки на ложных белках Notch3 нуклеотидная последовательность Fc-фрагмента человека (SEQ ID NO: 17). 713 нуклеотидов Fc-фрагмента человека слиты с 3'-концом конструкции для ложного белка Notch3, непосредственно после EGF-подобных повторов Notch3. Этот район Fc-фрагмента человека делает возможным обнаружение и очистку ложных белков Notch и служит для стабилизации секретируемых гибридных белков «Notch3 человека-Fc человека».
Фиг.36
На этой фигуре демонстрируется используемая для создания Fc-метки на ложных белках Notch3 аминокислотная последовательность Fc-фрагмента человека (SEQ ID NO: 18). 237 аминокислот Fc-фрагмента человека слиты с C-концом всех конструкций ложных белков Notch3, непосредственно после EGF-подобных повторов Notch3. Этот район Fc-фрагмента человека делает возможным обнаружение и очистку ложных белков Notch и служит для стабилизации секретируемых гибридных белков «Notch3 человека-Fc человека».
Фиг.37
На этой фигуре демонстрируется последовательность слияния человеческий Notch3/Fc для всех конструкций, которые заканчиваются после EGF-повтора 34 Notch 3 человека.
Фиг.38
На этой фигуре демонстрируется анализ сигнальной последовательности сигнального пептида Notch3 человека, который в соответствии с предсказанием охватывает аминокислоты 1-40 Notch3 человека. Это предсказание было сделано, используя программу Signal IP 3.0 Server, предоставленную Technical University of Denmark. В соответствии с предсказанием, относящимся к этим результатам, основной сайт расщепления локализован между аланином 39 (A39) и аланином 40 (A40). Этот сайт расщепления отмечен «/» в аминокислотной последовательности 1-40 Notch3 человека, представленной на этой фигуре.
Фиг.39
На этой фигуре демонстрируется анализ сигнальной последовательности сигнального пептида HC (тяжелой цепи) человека, который в соответствии с предсказанием охватывает аминокислоты 1-22 НС человека. Это предсказание было сделано, используя программу Signal IP 3.0 Server, предоставленную Technical University of Denmark. В соответствии с предсказанием, относящимся к этим результатам, основной сайт расщепления локализован между аланином 21 (A21) и аргинином 22 (R22). Этот сайт расщепления отмечен «/» в аминокислотной последовательности 1-22 HC человека, представленной выше.
Фиг.40A и 40B
На этой фигуре демонстрируется нуклеотидная последовательность ложного белка h-Notch3(1-34) (SEQ ID NO: 31). Подчеркнут предсказанный сигнальный пептид Notch3 человека (нуклеотиды 1-120). EGF-повторы 1-34 Notch3 кодируются нуклеотидами 121-4158. Место слияния, сайт для BglII, находится в положении нуклеотидов 4158-4163. Последовательность Fc-метки подчеркнута и выделена курсивом.
Фиг.41
На этой фигуре демонстрируется аминокислотная последовательность ложного белка h-Notch3(1-34) (SEQ ID NO: 32). Подчеркнут предсказанный сигнальный пептид Notch3 человека (аминокислоты 1-40). EGF-повторы 1-34 Notch3 соответствуют аминокислотам 41-1386. Последовательность Fc-метки подчеркнута и выделена курсивом.
Фиг.42
На этой фигуре демонстрируется аминокислотная последовательность ложного белка h-spHCNotch3(1-34) (SEQ ID NO: 33). Подчеркнут предсказанный сигнальный пептид Нс человека (аминокислоты 1-22). EGF-повторы 1-34 Notch3 соответствуют аминокислотам 22-1386. Последовательность Fc-метки подчеркнута и выделена курсивом.
Фиг.43A и 43B
На этой фигуре демонстрируется нуклеотидная последовательность ложного белка h-spHCNotch3(1-34) (SEQ ID NO: 34). Подчеркнут предсказанный сигнальный пептид HC человека (нуклеотиды 1-66). EGF-повторы Notch3 кодируются нуклеотидами 67-4104. Место слияния, сайт для BglII, находится в положении нуклеотидов 5004-5009. Последовательность Fc-метки подчеркнута и выделена курсивом.
Фиг.44
На этой фигуре демонстрируется экспрессия белков и лигандов Notch в эндотелиальных клетках кровеносных и лимфатических сосудов. Для Notch1-4, Dll1, Dll4 и Jagged1 была проведена ОТ-ПЦР на РНК, выделенной из эндотелиальных клеток кровеносных сосудов (BEC) и эндотелиальных клеток лимфатических сосудов (LEC), очищенных из HMVEC. Notch1, Notch2, Notch4, Dll4 и Jagged1 экспрессировались и в BEC, и в LEC на сходном уровне. Экспрессия Notch3, по-видимому, ограничивается LEC, что наводит на мысль о функционированиях передачи сигнала Notch3 в лимфатическом эндотелии.
Фиг.45
На этой фигуре демонстрируется, что Notch3 коэкспрессируется с маркерами эндотелиальных клеток лимфатических сосудов LYVE-1 и Prox1 у эмбрионов на El3,5. Последовательные срезы толщиной 10 микрон мышиных эмбрионов дня 13,5 подвергали иммуноокрашиванию на каждый из LYVE-1, Prox1 и Notch3. Notch3 экспрессировался в клетках, которые также экспрессировали маркеры эндотелиальных клеток лимфатических сосудов, LYVE-1 и Prox1.
Фиг.46
На этой фигуре демонстрируется, что Prox1 индуцировал экспрессию Notch3 в эндотелиальных клетках кровеносных сосудов. (А) Было проверено, могла ли бы эктопическая экспрессия Prox1 изменить экспрессию белков или лигандов Notch. Через двадцать четыре часа после инфицирования аденовирусным вектором - либо Ad-Prox1, либо Ad-LacZ выделяли тотальную РНК HUVEC, и проводили количественную ОТ-ПЦР для Notch1-4, Dll4 и Jagged1. Prox-1 сильно увеличивал экспрессию Notch3. Значительного влияния на экспрессию Notch1, Notch2, Notch4, Dll4 и Jagged1 не оказывалось. (В) Соединение E (cE), ингибитор пресенлина, который ингибирует передачу сигнала Notch, инкубировали в течение 24 часов с либо Ad-LacZ-, либо Ad-Prox1-инфицированными HUVEC. Выделяли тотальную РНК, и проводили количественную ОТ-ПЦР для определения экспрессии Notch3. Prox1 индуцировал экспрессию Notch3, и эта индукция была ингибирована при добавлении соединения E. Это говорит о том, что индукция Notch3 под действием Prox1 зависит от активации сигнала Notch.
Фиг.47
На этой фигуре демонстрируется, что Prox1 индуцирует гены-мишени Notch в эндотелиальных клетках кровеносных сосудов. HUVEC инфицировали аденовирусами, кодирующими LacZ, Prox1, N1IC или N4/int-3, и через 24 часа после инфицирования выделяли тотальную РНК. Количественную ОТ-ПЦР проводили для генов-мишеней Notch эндотелиальных клеток, VEGFR-3, эфрина B2, Hey1 и Hey2. Подобно активации сигнала Notch1 и Notch4, Prox1 индуцировал все четыре гена (A и B). Об экспрессии Hey1 и Hey2 в лимфатическом эндотелии неизвестно.
Фиг.48
На этой фигуре демонстрируется, что то, Prox1 индуцирует гены-мишени Notch, зависит от передачи сигнала Notch в эндотелиальных клетках кровеносных сосудов. HUVEC инфицировали аденовирусами, кодирующими LacZ, Prox1, N1IC или N4/int-3. Соединение E(cE), ингибитор пресенлина, который ингибирует передачу сигнала Notch, инкубировали в течение 24 часов с либо Ad-LacZ-, либо Ad-Prox1-инфицированными HUVEC, и выделяли тотальную РНК. Проводили количественную ОТ-ПЦР для генов-мишеней Notch эндотелиальных клеток, VEGFR-3, эфрина B2 и Hey2. Prox-1-опосредованная индукцию генов-мишеней Notch, эфрина B2, VEGFR-3 и Hey2, была ингибирована при добавлении соединения Е - ингибитора передачи сигнала Notch. Таким образом, Prox1 регулирует экспрессию эфрина B2, VEGFR-3 и Hey2 через посредство Notch.
Фиг.49
На этой фигуре демонстрируется схема включения N1IC. Активированную форму Notch1 вставляли в локус EF1-альфа, фланкированный двумя сайтами для LoxP. В результате экспрессии Cre-рекомбиназы, кассета neo/tpA утрачивается, и N1IC экспрессируется под контролем убиквитарного промотора EF1-альфа.
Фиг.50
На этой фигуре демонстрируется, что активация Notch в SM22-экспрессирующих гладкомышечных клетках сосудов приводит к смертности у эмбрионов до E10,5. Жизнеспособные SM22Cre/+; EF1αN1IC/+ мыши не зафиксированы в день 21 после рождения (P21) с p-значением, составляющим менее 0,001. В день Е9,5 эмбрионального развития было зафиксировано прогнозируемое число SM22Cre/+; EF1αN1IC/+ эмбрионов, но они серьезно отставали в росте по сравнению с контрольными мышами одного с ними помета (нижняя панель).
Фиг.51
На этой фигуре демонстрируется, что активация Notch в SM22-экспрессирующих гладкомышечных клетках сосудов меняет экспрессию альфа-актина в гладкомышечных клетках. Эмбрионы на E9,5 подвергали в виде тотального препарата иммуноокрашиванию на альфа-актин гладкомышечных клеток. Экспрессия альфа-актина в гладкомышечных клетках была изменена у SM22Cre/+; EF1αN1IC/+ эмбрионов по сравнению с контролями - WT (эмбрионами дикого типа). Таким образом, активация сигнала Notch в гладкомышечных клетках сосудов нарушает развитие сердечно-сосудистой системы у эмбриона.
Подробное описание изобретения
Термины
Как используется в этой заявке, каждый из следующих терминов будет иметь объясненное ниже значение, за исключением тех случаев, когда прямо указано иное.
«Введение» может быть произведено или выполнено, используя любой из способов, известных квалифицированному в данной области техники специалисту. Способы включают, например, введение внутрь пораженных тканей, внутримышечный, подкожный, внутривенный, внутрибрюшинный способы, способ с использованием липосом, через слизистую оболочку, интестинальный, местный, назальный, оральный, анальный, глазной или ушной способы доставки.
Под «присоединенным» следует понимать присоединенный любым способом. В одном варианте осуществления «присоединенный» означает присоединенный с помощью ковалентной связи. В другом варианте осуществления «присоединенный» означает присоединенный нековалентно.
Термины «аминокислота», «аминокислотный остаток» и «остаток» используются здесь взаимозаменяемо для ссылки на аминокислоту, которая включена в состав белка, полипептида или пептида. Аминокислотой может быть, например, встречающаяся в природе аминокислота или аналог встречающейся в природе аминокислоты, который может функционировать образом, подобным таковому встречающейся в природе аминокислоты.
«Антитело» должно включать, без ограничения, (a) молекулу иммуноглобулина, включающую две тяжелые цепи и две легкие цепи и распознающую антиген; (b) молекулу поликлонального или моноклонального антитела; и (c) ее одновалентный или двухвалентный фрагмент. Молекулы иммуноглобулинов могут происходить от любого из общеизвестных классов, включающих, но без ограничения, IgA, секреторный IgA, IgG, IgE и IgM. Подклассы IgG хорошо известны квалифицированным в данной области техники специалистам и включают, но без ограничения, IgG1, IgG2, IgG3 и IgG4 человека. Антитела могут быть как встречающимися в природе, так и не встречающимися в природе. Кроме того, антитела включают химерные антитела, полностью синтетические антитела, одноцепочечные антитела и их фрагменты. Антитела могут быть антителами человека или не человека. Антитела не человека могут быть подвергнуты гуманизации посредством рекомбинантных способов для уменьшения их иммуногенности у людей. Фрагменты антител включают, без ограничения, Fab- и Fc-фрагменты. В одном варианте осуществления «Fc-часть антитела» является полученным посредством расщепления иммуноглобулина папаином кристаллизующимся фрагментом, который состоит из C-концевой половины из двух тяжелых цепей, связанных дисульфидными связями, и известен как «эффекторный район» иммуноглобулина. В другом варианте осуществления «Fc-часть антитела» означает всю, или по существу всю, одну C-концевую половину тяжелой цепи.
«Гуманизированное», по отношению к антителу, означает антитело, в котором некоторые, большая часть или все из аминокислот вне CDR-участка заменены соответствующими аминокислотами, происходящими из молекулы иммуноглобулина человека. Добавления, делеции, вставки, замены или модификации небольшого числа аминокислот допустимы при условии, что они не аннулируют способность антитела к связыванию с конкретным антигеном. Подходящие молекулы иммуноглобулинов человека включают, без ограничения, молекулы IgG1, IgG2, IgG3, IgG4, IgA и IgM. В различных публикациях, например, в патентах США №№ 4816567, 5225539, 5585089 и 5693761 и публикации международной РСТ заявки с № WO 90/07861, приводится описание того, как получить гуманизированные антитела.
Предполагается, что используемый здесь термин «композиция», как например фармацевтическая композиция, охватывает продукт, включающий активный ингредиент(ы) и инертный ингредиент(ы), которые составляют носитель, а также любой продукт, который является следствием, прямым или непрямым, объединения, комплексообразования или агрегации любых двух или более ингредиентов, или диссоциации одного или более ингредиентов, или других типов реакций или взаимодействий одного или более ингредиентов.
Как здесь используется, «эффективное количество» относится к количеству, которое способно лечить субъекта, имеющего опухоль, заболевание или нарушение. Соответственно, эффективное количество будет изменяется в зависимости от подвергаемого лечению субъекта, а также подвергаемого лечению заболевания. Специалист со средним уровнем компетентности в данной области техники может выполнить обычные титрационные эксперименты для определения такого эффективного количества. Эффективное количество соединения будет изменяется в зависимости от субъекта и конкретного используемого пути введения. Исходя из соединения, количество может доставляться непрерывно, например, с помощью непрерывной накачки, или с повременными интервалами (например, в одном или нескольких отдельных случаях). Желаемые временные интервалы между множеством количеств конкретного соединения могут быть определены без чрезмерного экспериментирования квалифицированным в данной области техники специалистом. В одном варианте осуществления эффективное количество находится между приблизительно 1 мкг/кг и 10 мг/кг. В другом варианте осуществления эффективное количество находится между приблизительно 10 мкг/кг и 1 мг/кг. В дальнейшем варианте осуществления эффективное количество составляет 100 мкг/кг.
«Внеклеточный домен», используемый по отношению к белку рецептору Notch, означает весь или часть Notch, который(ая) (i) встречается внеклеточно (т.е. не встречается ни в виде трансмембранной части, ни в виде внутриклеточной части) и (ii) связывается с внеклеточными лигандами, с которыми связывается интактный белок рецептор Notch. Внеклеточный домен Notch может необязательно включать сигнальный пептид. «Внеклеточный домен», «ECD» и «эктодомен» являются синонимами.
«Увеличивающая полупериод существования составляющая» означает составляющую, которая, после функционального присоединения ко второй составляющей, увеличивает полупериод in vivo существования второй составляющей. Увеличивающие полупериод существования составляющие включают, например, Fc-части антител, метки гликозилирования (т.е. гликозилированные полипептиды), полиэтиленгликоль (ПЭГ), полипептиды с присоединенным к ним ПЭГ и липидмодифицирующие полипептиды.
Под «ингибированием» возникновения нарушения или нежелательного биологического процесса» следует понимать либо уменьшение вероятности возникновения нарушения или процесса, либо полное предотвращение возникновения нарушения или процесса. В предпочтительном варианте осуществления ингибирование возникновения нарушения или процесса означает полное предотвращение его возникновения.
«Notch», «белок Notch» и «белок рецептор Notch» являются синонимами. Кроме того, синонимами являются термины «основанный на Notch гибридный белок» и «ложный белок Notch (Notch-ловушка)». Известны и, таким образом, включены посредством ссылки следующие аминокислотные последовательности Notch: Notch1 (№ доступа в Genbank - S18188 (крысы)); Notch2 (№ доступа в Genbank - NP_077334 (крысы)); Notch3 (№ доступа в Genbank - Q61982 (мыши)); и Notch4 (№ доступа в Genbank - T09059 (мыши)). Известны и, таким образом, включены посредством ссылки следующие последовательности нуклеиновых кислот Notch: Notch1 (№ доступа в Genbank - XM_342392 (крысы) и NM_017617 (человека)); Notch2 (№ доступа в Genbank - NM_024358 (крысы), M99437 (человека) и AF308601 (человека)); Notch3 (№ доступа в Genbank - NM_008716 (мыши) и XM_009303 (человека)); и Notch4 (№ доступа в Genbank - NM_010929 (мыши) и NM_004557 (человека)).
Термины «нуклеиновая кислота», «полинуклеотид» и «последовательность нуклеиновой кислоты» используются здесь взаимозаменяемо, и каждый относится к полимеру из дезоксирибонуклеотидов и/или рибонуклеотидов. Дезоксирибонуклеотиды и рибонуклеотиды могут быть встречающимися в природе или их синтетическим аналогами. Под «нуклеиновой кислотой» следует понимать любую нуклеиновую кислоту, в том числе, без ограничения, ДНК, РНК и их гибриды. Основаниями нуклеиновой кислоты, образующими молекулы нуклеиновых кислот, могут быть основания A, C, G, T и U, а также их производные. Производные этих оснований хорошо известны в данной области техники, и их примеры приведены в PCR Systems, Reagents and Consumables (Perkin Elmer Catalogue 1996-1997, Roche Molecular Systems, Inc., Branchburg, New Jersey, USA). Нуклеиновые кислоты включают, без ограничения, антисмысловые молекулы и каталитические молекулы нуклеиновых кислот, такие как рибозимы и ДНКзимы. Нуклеиновые кислоты также включают нуклеиновые кислоты, кодирующие аналоги, фрагменты или производные пептида, которые отличаются от встречающихся в природе форм на предмет идентичности одного или нескольких аминокислотных остатков, (делеционные аналоги, содержащие меньше чем все из точно определенных остатков; содержащие замену(ы) аналоги, в которых один или несколько остатков заменен одной или несколькими остатками; и инсерционные аналоги, в которых один или несколько остатков добавлены к концевой или средней части пептида), которые разделяют некоторые или все из свойств встречающихся в природе форм.
«Функционально присоединенная», по отношению к первой составляющей, присоединенной ко второй составляющей, означает присоединенную таким образом, который позволяет первой составляющей функционировать (например, проявлять способности к связыванию), как если бы она не была присоединена.
Термины «полипептид», «пептид» и «белок» используются здесь взаимозаменяемо, и каждый означает полимер из аминокислотных остатков. Аминокислотные остатки могут быть встречающими в природе или их химическими аналогами. Полипептиды, пептиды и белки могут также включать модификации, такие как гликозилирование, присоединение липида, сульфатирование, гидроксилирование и АДФ-рибозилирование.
Как здесь используется, «фармацевтически приемлемый носитель» означает, что носитель совместим с другими ингредиентами композиции и не является вредным для его реципиента, и охватывает любые из стандартных фармацевтически приемлемых носителей. Такие носители включают, например, 0,01-0,1 M и предпочтительно 0,05 M фосфатный буфер или 0,8% солевой раствор. Кроме того, такие фармацевтически приемлемые носители могут быть водными или неводными растворами, суспензиями и эмульсиями. Примерами неводных растворителей являются пропиленгликоль, полиэтиленгликоль, растительные масла, такие как оливковое масло, и инъецируемые эфиры органических кислот, такие как этилолеат. Водные носители включают воду, спиртовые/водные растворы, эмульсии и суспензии, в том числе солевой раствор и забуференные среды. Носители для парентерального введения включают раствор натрия хлорида, декстрозу в растворе Рингера, декстрозу и натрия хлорид, раствор Рингера с лактатом и нелетучие масла. Носители для внутривенного введения включают жидкость и пищевые добавки, электролитные добавки, такие как те, которые основаны на декстрозе в растворе Рингера, и т.п. Могут также присутствовать консерванты и другие добавки, такие как, например, противомикробные средства, антиоксиданты, хелатообразующие вещества, инертные газы и т.п.
Под «субъектом» следует понимать любой организм, в том числе, без ограничения, млекопитающее, такое как мышь, крыса, собака, морская свинка, африканский хорек, кролик и примат. В предпочтительном варианте осуществления субъектом является человек.
«Лечение» означает либо замедление, остановку или реверсию прогрессирования заболевания или нарушения. Как здесь используется, «лечение» также означает уменьшение интенсивности симптомов, связанных с заболеванием или нарушением. Заболевания включают, но без ограничения, ангиогенез в опухоли, атеросклероз, заживление раны, дегенерацию желтого пятна, ретинопатию недоношенных, преэклампсию, диабетическую ретинопатию, ишемию, удар, сердечно-сосудистое заболевание и псориаз.
Ангиогенез встречается во время процессов заживления ран, менструального цикла у женщин и реструктурирования эндометрия, а также во время эмбрионального развития и роста органов. В патологической ситуации ангиогенез играет важную роль при различных заболеваниях, вроде ревматоидного артрита, псориаза, дегенерации желтого пятна, диабетической ретинопатии, и росте опухоли.
Накопилось большое число in vivo данных, включающих клинические наблюдения, о вовлеченности анормального ангиогенеза в ряд болезненных состояний, включающих ревматоидный артрит, воспаление, рак, псориаз, дегенеративные заболевания глаз и другие.
Единицы, префиксы и символы могут быть указаны в обычно применяемой для них форме СИ. Кроме случаев, оговоренных особо, последовательности нуклеиновых кислот читаются слева направо в направлении от 5' к 3', а аминокислотные последовательности читаются слева направо в направлении от амино-конца к карбоксильному концу. На аминокислоты здесь ссылаются, используя либо общеизвестные трехбуквенные символы, либо однобуквенные символы, рекомендованные IUPAC-IUB Biochemical Nomenclature Commission. Так же на нуклеотиды могут ссылаться, используя общепринятые однобуквенные коды.
Здесь используются следующие сокращения: ECD: внеклеточный домен; IC: внутриклеточный домен; NECD/Fc: основанный на Notch гибридный белок; N1: Notch1; N2: Notch2; N3: Notch3; N4: Notch4; Dll: Delta-подобный белок; EC: эндотелиальные клетки; FGF: фактор роста фибробластов; FGFR: рецептор фактора роста фибробластов; HUVEC: эндотелиальная клетка пупочной вены человека; m.o.i.: множественность инфицирования; VMC: клетки сосудистой стенки; VEGF: фактор роста эндотелиальных клеток сосудов; VEGFR: рецептор фактора роста эндотелиальных клеток сосудов; sp: сигнальный пептид; HC или Hc: тяжелая цепь IgG; PDGF: тромбоцитарный фактор роста; PlGF: плацентарный фактор роста.
Варианты осуществления настоящего изобретения
Этим изобретением предоставляется гибридный белок, включающий сигнальный пептид, EGF-повторы 1-Х внеклеточного домена белка рецептора Notch3 человека, причем X является любым целым числом от 12 до 34, и Fc-часть антитела, связанную с ними.
Этим изобретением предоставляется гибридный белок, включающий сигнальный пептид, EGF-повторы 1-Х внеклеточного домена белка рецептора Notch3 человека, причем X является любым целым числом от 1 до 10, и Fc-часть антитела, связанную с ними.
Этим изобретением предоставляется гибридный белок, включающий сигнальный пептид, по меньшей мере 12 EGF-повторов внеклеточного домена белка рецептора Notch3 человека и Fc-часть антитела, связанную с ними.
Этим изобретением предоставляется гибридный белок, включающий сигнальный пептид, EGF-повторы внеклеточного домена белка рецептора Notch3 человека, причем присутствуют по меньшей мере 12 EGF-повторов, и Fc-часть антитела, связанную с ними.
В одном варианте гибридного белка внеклеточный домен белка рецептора Notch3 включает EGF-подобные повторы 1-34.
В одном варианте гибридного белка Fc-частью антитела является Fc-часть антитела человека.
В одном варианте гибридного белка сигнальным пептидом является сигнальный пептид Notch3 или часть Hc (HC; тяжелой цепи) антитела.
В одном варианте осуществления гибридный белок включает последовательные аминокислоты, последовательность которых определена в SEQ ID NO: 32. В другом варианте осуществления гибридный белок включает последовательные аминокислоты, последовательность которых определена в SEQ ID NO: 33.
В одном варианте осуществления гибридный белок кодируется последовательными нуклеотидами, последовательность которых определена в SEQ ID NO: 31. В другом варианте осуществления гибридный белок кодируется последовательными нуклеотидами, последовательность которых определена в SEQ ID NO: 34.
Этим изобретением предоставляется способ лечения имеющего опухоль субъекта, включающий введение субъекту количества указанного выше гибридного белка, эффективного для лечения субъекта, осуществляя тем самым лечение субъекта, имеющего опухоль.
Этим изобретением предоставляется способ ингибирования ангиогенеза у субъекта, включающий введение субъекту количества указанного выше гибридного белка, эффективного для ингибирования ангиогенеза у субъекта, ингибируя тем самым ангиогенез у субъекта.
Этим изобретением предоставляется способ лечения имеющего рак яичника субъекта, включающий введение субъекту количества указанного выше гибридного белка, эффективного для лечения субъекта, осуществляя тем самым лечение субъекта, имеющего рак яичника.
Этим изобретением предоставляется применение указанного выше гибридного белка для приготовления фармацевтической композиции для лечения субъекта, имеющего сердечнососудистое заболевание. В одном варианте осуществления сердечнососудистым заболеванием является атеросклероз, ишемия или удар.
Этим изобретением предоставляется применение указанного выше гибридного белка для приготовления фармацевтической композиции для лечения имеющего опухоль субъекта.
Этим изобретением предоставляется применение указанного выше гибридного белка для приготовления фармацевтической композиции для ингибирования ангиогенеза у субъекта.
Этим изобретением предоставляется применение указанного выше гибридного белка для приготовления фармацевтической композиции для лечения имеющего рак яичника субъекта.
Этим изобретением предоставляется способ ингибирования физиологического лимфоангиогенеза или патологического лимфоангиогенеза у субъекта, включающий введение субъекту количества указанного выше гибридного белка, эффективного для ингибирования физиологического лимфоангиогенеза или патологического лимфоангиогенеза у субъекта. В одном варианте осуществления патологическим лимфоангиогенезом является лимфоангиогенез в опухоли или метастазирование в лимфатические узлы, которое может зависеть от лимфоангиогенеза в опухоли.
Этим изобретением предоставляется способ ингибирования метастазирования опухоли у субъекта, включающий введение субъекту количества указанного выше гибридного белка, эффективного для ингибирования метастазирования опухоли у субъекта. В одном варианте осуществления метастазирование происходит через кровеносный сосуд, лимфатическую сосудистую сеть или лимфатический узел. Метастазированием опухоли является распространение рака из одного органа в другой не соседний орган.
Этим изобретением предоставляется способ ингибирования роста вторичной опухоли у субъекта, включающий введение субъекту количества указанного выше гибридного белка, эффективного для ингибирования роста вторичной опухоли у субъекта. Ингибированием может также быть ингибирование ангиогенеза в опухоли, связанного со вторичной или метастатической опухолью. В одном варианте осуществления рост вторичной опухоли ингибируется посредством ингибирования ангиогенеза, связанного со вторичной опухолью.
Этим изобретением предоставляется способ ингибирования использования опухолью для своих собственных целей кровеносных сосудов у субъекта, включающий введение субъекту количества указанного выше гибридного белка, эффективного для ингибирования использования опухолью для своих собственных целей кровеносных сосудов у субъекта. Процесс такого использования сосудов является процессом, посредством которого опухолевые клетки связываются с уже существующими сосудами и растут с помощью используемых таким образом сосудов. Этот рост опухолей на используемых таким образом сосудах может происходить в отсутствие антиогенеза в опухоли, предшествовать ему или происходить совместно с ним.
Этим изобретением предоставляется способ лечения рака у субъекта, включающий введение субъекту указанного выше гибридного белка и ингибитора фактора роста сосудистого эндотелия (VEGF) в количестве каждого из них, эффективном для лечения рака у субъекта. В одном варианте осуществления ингибитором VEGF является ингибитор VEGF-A, ингибитор PGIF, ингибитор VEGF-B, ингибитор VEGF-C или ингибитор VEGF-D. Примеры ингибиторов VEGF включают, но без ограничения, бевацизумаб, PTK787, Bay43-9006, SU11248, AG013676, ZD6474, VEGF-ловушку и антитело против VEGFR2. Примеры таких ингибиторов полнее описываются в Ferrara et al., (2004) Nature Reviews Drug Discovery, Vol. 3: 391-400 и Ellis et al. (2008) Nature Reviews Cancer Vol. 8: 579-591, содержание каждого из которых таким образом включено посредством ссылки.
Этим изобретением предоставляется способ лечения рака у субъекта, включающий введение субъекту указанного выше гибридного белка и ингибитора рецептора VEGF в количестве каждого из них, эффективном для лечения рака у субъекта. В одном варианте осуществления ингибитором рецептора VEGF является ингибитор VEGFR-1, ингибитор VEGFR-2, ингибитор VEGFR-3 или ингибитор любой комбинации рецепторов VEGF.
Этим изобретением предоставляется способ лечения рака у субъекта, включающий введение субъекту указанного выше гибридного белка и ингибитора тромбоцитарного фактора роста (PDGF) в количестве каждого из них, эффективном для лечения рака у субъекта. В одном варианте осуществления ингибитором тромбоцитарного фактора роста является ингибитор PDGF-A или ингибитор PDGF-B.
Этим изобретением предоставляется способ лечения рака у субъекта, включающий введение субъекту указанного выше гибридного белка и антагониста рецептора PDGF в количестве каждого из них, эффективном для лечения рака у субъекта. В одном варианте осуществления антагонистом рецептора PDGF является антагонист рецептора-В PDGF.
Этим изобретением предоставляется способ лечения рака у субъекта, включающий введение субъекту указанного выше гибридного белка и ингибитора HER2/neu в количестве каждого из них, эффективном для лечения рака у субъекта.
Этим изобретением предоставляется способ лечения рака молочной железы у субъекта, включающий введение субъекту количества указанного выше гибридного белка, эффективного для лечения рака молочной железы у субъекта.
Этим изобретением предоставляется применение указанного выше гибридного белка для приготовления фармацевтической композиции для лечения имеющего рак молочной железы субъекта.
Этим изобретением также предоставляется первый способ лечения имеющего опухоль субъекта, включающий введение субъекту эффективного количества являющейся объектом настоящего изобретения композиции, включающей внеклеточный домен белка рецептора Notch, функционально присоединенный к увеличивающей полупериод существования составляющей, для осуществления тем самым лечения субъекта.
Этим изобретением также предоставляется второй способ ингибирования ангиогенеза у субъекта, включающий введение субъекту эффективного количества являющейся объектом настоящего изобретения композиции, включающей внеклеточный домен белка рецептора Notch, функционально присоединенный к увеличивающей полупериод существования составляющей, для ингибирования тем самым ангиогенеза у субъекта.
В первом варианте осуществления указанных выше способов белком рецептором Notch является белок рецептор Notch1. В одном варианте осуществления белком рецептором Notch1 является белок рецептор Notch1 человека. В другом варианте осуществления увеличивающей полупериод существования составляющей является Fc-часть антитела. В другом варианте осуществления Fc-частью антитела является Fc-часть антитела человека. В дальнейшем варианте осуществления внеклеточный домен и увеличивающая полупериод существования составляющая находятся внутри одной и той полипептидной цепи.
Во втором варианте осуществления указанных выше способов белком рецептором Notch является белок рецептор Notch2. В одном варианте осуществления белком рецептором Notch2 является белок рецептор Notch2 человека. В другом варианте осуществления увеличивающей полупериод существования составляющей является Fc-часть антитела. В другом варианте осуществления Fc-частью антитела является Fc-часть антитела человека. В дальнейшем варианте осуществления внеклеточный домен и увеличивающая полупериод существования составляющая находятся внутри одной и той полипептидной цепи.
В третьем варианте осуществления указанных выше способов белком рецептором Notch является белок рецептор Notch3. В одном варианте осуществления белком рецептором Notch3 является белок рецептор Notch3 человека. В другом варианте осуществления увеличивающей полупериод существования составляющей является Fc-часть антитела. В другом варианте осуществления Fc-частью антитела является Fc-часть антитела человека. В дальнейшем варианте осуществления внеклеточный домен и увеличивающая полупериод существования составляющая находятся внутри одной и той полипептидной цепи.
В четвертом варианте осуществления указанных выше способов белком рецептором Notch является белок рецептор Notch4. В одном варианте осуществления белком рецептором Notch4 является белок рецептор Notch4 человека. В другом варианте осуществления увеличивающей полупериод существования составляющей является Fc-часть антитела. В другом варианте осуществления Fc-частью антитела является Fc-часть антитела человека. В дальнейшем варианте осуществления внеклеточный домен и увеличивающая полупериод существования составляющая находятся внутри одной и той полипептидной цепи.
В пятом варианте осуществления указанных выше способов субъектом является млекопитающее. В одном варианте осуществления млекопитающим является человек.
В шестом варианте осуществления указанных выше способов ангиогенезом является ангиогенез в опухоли.
В дальнейшем варианте осуществления второго способа субъект имеет опухоль. В другом варианте осуществления субъект поражен патологической сосудистой гиперплазией. В одном варианте осуществления патологической сосудистой гиперплазией является доброкачественная гемангиома. В дальнейшем варианте осуществления субъект поражен пролиферативным заболеванием лимфатических сосудов.
Этим изобретением предоставляется первая являющаяся объектом настоящего изобретения композиция, включающая внеклеточный домен белка рецептора Notch4, функционально присоединенный к увеличивающей полупериод существования составляющей. В одном варианте осуществления внеклеточный домен ковалентно связан с увеличивающей полупериод существования составляющей. В другом варианте осуществления внеклеточный домен и увеличивающая полупериод существования составляющая находятся внутри одной и той полипептидной цепи.
Этим изобретением предоставляется вторая являющаяся объектом настоящего изобретения композиция, включающая внеклеточный домен белка рецептора Notch4, функционально присоединенный к увеличивающей полупериод существования составляющей, и фармацевтически приемлемый носитель.
Этим изобретением, кроме того, предоставляется изделие производства, включающая (i) упаковочный материал, в котором содержится являющаяся объектом настоящего изобретения композиция, включающей внеклеточный домен белка рецептора Notch, функционально присоединенный к увеличивающей полупериод существования составляющей, и (ii) этикетку с указанием того, что композиция предназначена для применения при лечении субъекта, имеющего опухоль или другое нарушение, поддающееся лечению посредством ингибирования ангиогенеза у субъекта.
В первом варианте осуществления указанного выше изделия белком рецептором Notch является белок рецептор Notch1. В одном варианте осуществления белком рецептором Notch1 является белок рецептор Notch1 человека. В другом варианте осуществления увеличивающей полупериод существования составляющей является Fc-часть антитела. В другом варианте осуществления Fc-частью антитела является Fc-часть антитела человека. В дальнейшем варианте осуществления внеклеточный домен и увеличивающая полупериод существования составляющая находятся внутри одной и той полипептидной цепи.
Во втором варианте осуществления указанного выше изделия белком рецептором Notch является белок рецептор Notch2. В одном варианте осуществления белком рецептором Notch2 является белок рецептор Notch2 человека. В другом варианте осуществления увеличивающей полупериод существования составляющей является Fc-часть антитела. В другом варианте осуществления Fc-частью антитела является Fc-часть антитела человека. В дальнейшем варианте осуществления внеклеточный домен и увеличивающая полупериод существования составляющая находятся внутри одной и той полипептидной цепи.
В третьем варианте осуществления указанного выше изделия белком рецептором Notch является белок рецептор Notch3. В одном варианте осуществления белком рецептором Notch3 является белок рецептор Notch3 человека. В другом варианте осуществления увеличивающей полупериод существования составляющей является Fc-часть антитела. В другом варианте осуществления Fc-частью антитела является Fc-часть антитела человека. В дальнейшем варианте осуществления внеклеточный домен и увеличивающая полупериод существования составляющая находятся внутри одной и той полипептидной цепи.
В четвертом варианте осуществления указанного выше изделия белком рецептором Notch является белок рецептор Notch4. В одном варианте осуществления белком рецептором Notch4 является белок рецептор Notch4 человека. В другом варианте осуществления увеличивающей полупериод существования составляющей является Fc-часть антитела. В другом варианте осуществления Fc-частью антитела является Fc-часть антитела человека. В дальнейшем варианте осуществления внеклеточный домен и увеличивающая полупериод существования составляющая находятся внутри одной и той полипептидной цепи.
В другом варианте осуществления указанного выше изделия к композиции примешан фармацевтический носитель. В конечном варианте осуществления субъектом является человек.
Этим изобретением предоставляется способный к репликации вектор, который кодирует полипептид, включающий внеклеточный домен белка рецептора Notch3, функционально присоединенный к увеличивающей полупериод существования составляющей. В одном варианте осуществления увеличивающей полупериод существования составляющей является Fc-часть антитела. В другом варианте осуществления вектор включает, без ограничения, плазмиду, космиду, ретровирус, аденовирус, фаг лямбда или YAC.
Этим изобретением также предоставляется система «хозяин-вектор», которая включает способный к репликации вектор, который кодирует полипептид, включающий внеклеточный домен белка рецептора Notch, функционально присоединенный к увеличивающей полупериод существования составляющей, и подходящую клетку-хозяина. В одном варианте осуществления клеткой-хозяином является эукариотическая клетка. В другом варианте осуществления эукариотической клеткой является клетка CHO. В другом варианте осуществления эукариотической клеткой является клетка HeLa. В дальнейшем варианте осуществления клеткой-хозяином является бактериальная клетка.
Наконец, этим изобретением предоставляется третий способ продуцирования полипептида, включающий выращивание системы «хозяин-вектор», которая включает способный к репликации вектор, который кодирует полипептид, включающий внеклеточный домен белка рецептора Notch, функционально присоединенный к увеличивающей полупериод существования составляющей, и подходящую клетку-хозяина, в условиях, позволяющих продуцировать полипептид, и извлечение продуцированного таким образом полипептида.
Это изобретение проиллюстрировано в следующем части «Детали экспериментов». Эта часть изложена для помощи в понимании настоящего изобретения, но не предназначена для ограничения каким-либо образом настоящего изобретения, изложенного в следующей за ней формуле изобретения, и не должна толковаться как такое ограничение.
ДЕТАЛИ ЭКСПЕРИМЕНТОВ
Первый ряд экспериментов
Основанные на Notch3 человека гибридные белки (ложные белки Notch3)
Сборку ложных белков Notch3 осуществляли, используя последовательности, кодирующие сигнальный пептид, часть внеклеточного домена Notch3, включающую все домены в виде EGF-подобных повторов, и часть белка Fc-фрагмента человека (аминокислоты 1-237). Полноразмерная нуклеотидная последовательность Notch3 человека предоставлена на фиг.32. Полноразмерная аминокислотная последовательность Notch3 человека предоставлена на фиг.33.
Используемым сигнальным пептидом является либо встречающийся в природе сигнальный пептид Notch3, либо сигнальный пептид Hc человека, каждый из которых слит с районом Notch3. Сигнальный пептид делает возможной секрецию ложных белков Notch.
Используемые внеклеточные домены Notch3 предназначены для связывания с лигандами Notch и состоят из всех 34 доменов в виде EGF-подобных повторов белка Notch3 человека или его подмножества.
Fc-метка слита с C-концом конкретного EGF-подобного повтора Notch3 человека и служит для того, чтобы сделать возможной очистку, обнаружение и стабилизацию ложных белков Notch3.
Конструкция в целом ложных белков Notch3 человека, двух разработок, должна кодировать (1) сигнальный пептид для того, чтобы сделает возможной секрецию ложных белков Notch3 во внеклеточные среды эукариотических клеток, используемых для продуцирования белков, (2) часть внеклеточного домена из всех EGF-подобных повторов Notch3 человека для того, чтобы сделать возможным связывание с лигандами Notch, и (3) часть белка Fc-фрагмента для того, чтобы сделать возможным обнаружение.
Следующие две разработки ложных белков Notch3 человека будут описываться и схематизированы на фиг.34.
1) ложный белок h-Notch3(1-34)
2) ложный белок h-spHCNotch3(1-34)
Последовательность Notch3 человека
Полноразмерная нуклеотидная последовательность Notch3 человека, состоящая из последовательности от инициирующего кодона ATG (нуклеотида 1) до стоп-кодона (TGA; нуклеотида 6964), определена на фиг.32. Сигнальный пептид и домены в виде первых 34 EGF-подобных повторов соответствуют нуклеотидам 1-4158 этой последовательности. Нуклеотиды 1-4158 используются для разработки ложных белков Notch3 человека, описываемых в последующих частях. Подчеркнуты нуклеотиды, охватывающие EGF-повторы 1-34.
Полноразмерная аминокислотная последовательность Notch3 человека, состоящая из последовательности от аминокислоты 1 (M=метионина) до аминокислоты 2555 (K=лизина), определена на фиг.33. Сигнальный пептид и домены в виде первых 34 EGF-подобных повторов соответствуют аминокислотам 1-1386 этой последовательности. Аминокислоты 1-1386 используются для разработки ложных белков Notch3 человека, описываемых в последующих частях. Подчеркнуты аминокислоты, охватывающие EGF-повторы 1-34.
Используемая для создания Fc-метки на ложных белках Notch3 последовательность Fc-фрагмента человека
713 нуклеотидов Fc-фрагмента человека, определенных на фиг.35, были слиты с 3'-концом конструкции для ложного белка Notch3, непосредственно после EGF-подобных повторов Notch3. Этот район Fc-фрагмента человека делает возможным обнаружение и очистку ложных белков Notch и служит для стабилизации секретируемых гибридных белков «Notch3 человека-Fc человека».
237 аминокислот Fc-фрагмента человека, продемонстрированных на фиг.36, были слиты с C-концом всех конструкций ложных белков Notch3, непосредственно после EGF-подобных повторов Notch3. Этот район Fc-фрагмента человека делает возможным обнаружение и очистку ложных белков Notch и служит для стабилизации секретируемых гибридных белков «Notch3 человека-Fc человека».
Используемые в ложных белках Notch3 сигнальные пептиды
Две различные последовательности сигнальных пептидов были включены в состав конструкции ложных белков Notch1 человека. Первым сигнальным пептидом является сигнальный пептид Notch3 человека, который в соответствии с предсказанием охватывает аминокислоты 1-40 Notch3 человека. Это предсказание было сделано, используя программу Signal IP 3.0 Server, предоставляемую Technical University of
Denmark. Вторым сигнальным пептидом является сигнальный пептид Hc человека, который в соответствии с предсказанием охватывает аминокислоты 1-22 сигнального пептида тяжелой цепи (НС) IgG человека.
1. Сигнальный пептид Notch3 человека (нуклеотиды 1-20)
MGPGARGRRRRRRPMSPPPPPPPVRALPLLLLLAGPGAA/A (SEQ ID NO: 27)
Аминокислотная последовательность предсказанного сигнального пептида Notch3 человека схематизирована на фиг.37. Относящиеся к предсказанию результаты анализа, в котором используется программа Signal IP 3.0 Server, предоставляемая по сети Technical University of Denmark, представлены на фиг.37. В соответствии с предсказанием, относящимся к этим результатам, основной сайт расщепления локализован между аланином 39 (A39) и аланином 40 (A40). Этот сайт расщепления отмечен «/» в аминокислотной последовательности 1-40 Notch3 человека, представленной выше.
2. Сигнальный пептид HC человека (аминокислоты 1-22)
Аминокислотной последовательностью предсказанного сигнального пептида Hc человека является
MWGWKCLLFWAVLVTATLCTA/R (SEQ ID NO: 29).
Нуклеотидной последовательностью предсказанного сигнального пептида Hc человека является:
Относящиеся к предсказанию результаты анализа, в котором используется программа SignalIP 3.0 Server, предоставляемая по сети Technical University of Denmark, представлены выше. В соответствии с предсказанием, относящимся к этим результатам, основной сайт расщепления локализован между аланином 21 (A21) и аргинином 22 (R22). Этот сайт расщепления отмечен «/» в аминокислотной последовательности 1-22 Нс человека, представленной выше.
Ложный белок h-Notch3 (1-34)
Ложный белок h-Notch3(1-34) обозначает ложный белок Notch3 человека, который включает EGF-подобные повторы 1-34 Notch3.
Аминокислотная последовательность ложного белка h-Notch3(1-34) определена на фиг.41. Подчеркнут предсказанный сигнальный пептид Notch3 человека (аминокислоты 1-40). EGF-повторы 1-34 Notch3 соответствуют аминокислотам 41-1386. Последовательность Fc-метки подчеркнута и выделена курсивом.
Нуклеотидная последовательность ложного белка h-Notch3(1-34) определена на фиг.40. Подчеркнут предсказанный сигнальный пептид Notch3 человека (нуклеотиды 1-120). EGF-повторы 1-34 Notch3 кодируются нуклеотидами 121-4158. Место слияния, сайт для BglII - это нуклеотиды 4158-4163. Последовательность Fc-метки подчеркнута и выделена курсивом.
Ложный белок h-sp Hc Notch3 (1-34)
Ложный белок h-spHcNotch3(1-34) обозначает ложный белок Notch3 человека, который включает EGF-подобные повторы 1-34. Сокращение spHc обозначает, что в этой разработке используется сигнальный пептид Нс человека.
Аминокислотная последовательность ложного белка h-spHcNotch3(1-34) определена на фиг.42. Подчеркнут предсказанный сигнальный пептид Нс человека (аминокислоты 1-22). EGF-повторы 1-34 Notch3 соответствуют аминокислотам 22-1386. Последовательность Fc-метки подчеркнута и выделена курсивом.
Нуклеотидная последовательность ложного белка h-Notch3(1-34) определена на фиг.43. Подчеркнут предсказанный сигнальный пептид Нс человека (нуклеотиды 1-66). EGF-повторы 1-34 Notch3 кодируются нуклеотидами 67-4104. Место слияния, сайт для BglII - это нуклеотиды 4104-4109. Последовательность Fc-метки подчеркнута и выделена курсивом.
Методы
Конструирование ложных белков Notch3 человека
Для создания вариантов ложных белков Notch3 человека использовали тотальную РНК из либо гладкомышечных клеток аорты человека (AoSMC), либо эндотелиальных клеток пупочной вены человека (HUVEC), которые сверхэкспрессируют Prox1. Обратную транскрипцию тотальной РНК осуществляли с помощью обратной транскриптазы M-MLV и либо рандомизированных гексамерных праймеров, либо специфичных для ложных белков Notch3 праймеров. Затем синтезированную кДНК амплифицировали с использованием специфичных для ложных белков Notch3 прямого (на основе смысловой цепи) и обратного (на основе антисмысловой цепи) праймеров. Конструирование ложного белка Notch3 было осуществлено на основе 4 отдельных ампликонов. 3'-ампликон амплифицировали с использованием обратного праймера, кодирующего рестрикционный сайт для BglII на 5'-конце, для лигирования в сайт для BglII в последовательности Fc-фрагмента для создания химеры «человеческий Notch3/Fc» в одной рамке считывания.
В случае ложных белков Notch3, которые образуют слияние после нуклеотидной последовательности, кодирующей EGF-подобный повтор 34, для создания сайта слияния будет создаваться сайт для BglII, и здесь представлена эта последовательность слияния (Notch3, фиг.37). Это распространяется на разработки ложный белок h-Notch3(1-34) и ложный белок h-spHCNotch3(1-34).
Амплифицированные ПЦР-продукты субклонировали в pBluescript SK II Fc для создания различных химер «человеческий Notch3/Fc». Затем осуществляют челночное перемещение последовательностей ложных белков Notch3/Fc в векторы для экспрессии в клетках млекопитающих (pAd-lox, pCCL или pcDNA3) для экспрессии и очистки ложных белков Notch3 человека.
Второй ряд экспериментов
Материалы и методы
Плазмидные конструкции
Аденовирусные конструкции, кодирующие LacZ, полноразмерный Notch4 или активированную форму Notch4/int3, были описаны ранее (Shawber et al., 2003). кДНК активированной формы Notch1, слитой в одной рамке считывания с метками в виде 6 myc (Kopan et al., 1994), клонировали в аденовирусный экспрессионный вектор pAd-lox. В pAd-lox также клонировали и VEGF165, и N1ECDFc. Аденовирусные штаммы создавали и подвергали титрованию, как описано ранее (Hardy et al., 1997). Ретровирусные экспрессионные векторы pHyTc, кодирующие каждый из LacZ, активированной формы Notch4/int3, J1, Dll1 и Dll4, были описаны ранее (Uyttendaele et al., 2000, Shawber et al., 2003, Das et al., 2004 в печати). Плазмиды, кодирующие внеклеточный домен Notch1 (п.о. 5479-7833, № доступа в Genbank - X57405) и внеклеточный домен Dll4 (п.о. 1-1545, № доступа в Genbank - AF253468, предоставленный Chiron), слитые в одной рамке считывания с myc/His-меткой, были сконструированы в pHyTC.
Используя способы, изложенные в Clin. Exp. Immunol. (1992) 87(1): 105-110, созданы конструкции Notch1ECD, Notch2ECD, Notch3ECD и Notch4ECD в содержащей Fc-фрагмент плазмиде pCMX-sFR1-IgG для создания основанных на Notch гибридных белков, т.е. Notch1ECD/Fc, Notch2ECD/Fc, Notch3ECD/Fc и Notch4ECD/Fc.
Перенос генов с помощью аденовирусных векторов
7,5×105 клеток HUVEC пассажа 3 засевали в покрытые коллагеном типа I 6-луночные планшеты за день до инфицирования аденовирусными векторами. Инфицирование аденовирусным вектором Ad-lacZ, Ad-VEGF165 или Ad-N1ECDFc осуществляли с указанной множественностью инфицирования и инкубировали при 37οC в течение 1 ч с использованием время от времени вихревого движения в отношении планшетов.
Анализы по люциферазному гену-репортеру
Для определения индуцируемой лигандами передачи сигнала Notch выполняли анализы сокультивирования, используя клетки HeLa и клетки Bosc, происходящие от клеток 293. Транзиторные трансфекции выполняли посредством преципитации фосфатом кальция. Клетки HeLa, засеянные за 1-день до трансфекции в 10-см чашки в количестве 1,5×106, трансфецировали 333 нг pBOS Notch1, 333 нг pGA981-6, и 83 нг pLNC lacZ с либо 666 нг pCMV-Fc, либо pHyTC-N1ECDFc (333 нг в случае ×1, 666 нг в случае ×2). Клетки Bosc, засеянные за 1-день до трансфекции в 10-см чашки в количестве 4×106, трансфецировали либо 680 нг pHyTc-Jagged1, pHyTc-Dll1, pHyTc-Dll4, либо pHyTc-x (пустым вектором). Через один день после трансфекции клетки подвергали сокультивированию в трех повторах (HeLa:Bosc, 1:2) в 12-луночных планшетах в течение 24 часов. Клетки собирали, и через 2 дня после трансфекции определяли люциферазную активность, используя набор Enhanced Luciferase assay (BD PharMingen), а β-галактозидазную активность определяли, используя набор Galacto-Light Plus (PE Biosysterns). Все анализы проводили в люминометре с одновременно-раздельной закачкой жидкости Berthold.
Для определения VEGF-индуцируемой передачи сигнала Notch использовали HUVEC, которые были инфицированы аденовирусом. HUVEC, засеянные за 1-день до инфицирования в 6-луночные планшеты в количестве 8,0×105, инфицировали либо Ad-LacZ в качестве контроля, либо Ad-VEGF с указанной множественностью заражения в присутствии или в отсутствие Ad-N1ECD/Fc. Через два дня после инфицирования инфицированные HUVEC пересевали в 24-луночные планшеты в количестве 1,5×105 в трех повторах и культивировали в течение 24 часов, а затем трансфецировали 12,5 нг pRL-SV40 (Promega) и 137,5 нг pGA981-6, используя реагент для трансфекции Effectene (Qiagen). Клетки собирали через или 1, или 2 дня после трансфекции, и люциферазную активность определяли, используя систему анализа по гену-репортеру Dual-Luciferase® (Promega).
Анализ спраутинга
Для получения коллагеновых гелей холодный как лед раствор свиного коллагена типа I (Nitta gelatin, Tokyo, Япония) смешивали со средой 10×RPMI1640 и буфером для нейтрализации в соотношении 8:1:1. 400-мкл аликвоты коллагенового геля затем добавляли в 24-луночные планшеты и позволяли превращаться в гель в течение по меньшей мере 1 часа при 37°C. После инфицирования аденовирусными векторами (выше) HUVEC собирали и засевали в количестве 1,3×105 клеток на лунку в 0,8 мл среды EGM2 сверху коллагенового геля в 24-луночных планшетах. HUVEC становились почти конфлюэнтными через 48 часов после засева. После засева среду меняли каждые 2 дня в течение 1 недели. Наблюдали за спраутингом, и через 8 дней делали фотографии с помощью цифровой камеры Olympus, вмонтированной в микроскоп. Для количественного определения числа врастаний произвольно выбирали 5 полей на каждую лунку, и врастания подсчитывали под микроскопом «вслепую» два исследователя.
Результаты и обсуждение
Гибридные белки NOTCHECD/Fc
Функционирование в качестве антагонистов Notch
Гибридные белки NotchECD/Fc - антагонисты Notch
Авторы настоящего изобретения создали несколько антагонистов Notch (фиг.2). Их стратегией было слияние последовательности, кодирующей EGF-повторы Notch во внеклеточном домене (ECD), с Fc-доменом человека или мыши. В случае такого конструирования создается секретируемый белок без функции передачи сигнала, но который сохраняет лигандсвязывающий домен и, следовательно, должен связываться с лигандами и ингибировать их функционирование. Эти белки упоминаются как «NotchECD/Fc», и были созданы все четыре Notch1-4ECD/Fc. Fc-домен способствует аффинной очистке и обнаружению белка посредством иммуноблоттинга или иммуногистохимии.
Проверка антагонистов Notch
Для определения активации пути с участием Notch на транскрипционном уровне была использована in vitro система сокультивирования (фиг.3) с использованием лигандов, экспрессируемых на одной клетке, и активации рецептора Notch, отмечаемой в другой клетке. Этот анализ сокультивирования был использован для демонстрации того, что Notch1ECD/Fc функционирует с блокированием лиганд-зависимой передачи сигнала Notch (фиг.4). Вектор для экспрессии N1ECD/Fc котрансфецировали в различных соотношениях с полноразмерным геном Notch1 и регулируемым CSL люциферазым геном - репортером в клетки HeLa, с последующим сокультивированием с экспрессирующими лиганды клетками 293. Наблюдалось снижение активации передачи сигнала Notch1 лигандами Notch при экспрессии N1ECD/Fc. Этот эффект демонстрировал зависимость от концентрации; составляющее 2:1 соотношение N1ECD/Fc и Notch1 было эффективнее в ингибировании передачи сигнала, чем соотношение 1:1. Notch1ECD/Fc был способен к блокированию передачи сигнала, опосредуемой Jagged1, Delta-подобным белком 1 или Delta-подобным белком 4.
Экспрессия и очистка антагонистов Notch
С целью очистки белков авторы настоящего изобретения создали линии клеток CHO и HeLa, экспрессирующие NotchECD/FC, используя ретровирусные векторы. Белки N1ECD/Fc секретируются (фиг.5); как установлено в кондиционированных средах, полученных от линий HeLa-NotchECD/Fc и очищенных с использованием агарозы с белком-A(pA). Очищенный на агарозе с pA образец (Sup) и цельноклеточные лизаты (Lys) подвергали анализу с помощью иммуноблоттинга с использованием антитела против α-Fc (фиг.5, панель A), демонстрируя, что N1ECD/Fc секретируется в среды. Аденовирусные векторы для NotchECD/Fc были использованы для инфицирования клеток HeLa, и лизаты этих клеток подвергали анализу с помощью иммуноблоттинга с использованием антител против α-Fc, демонстрируя, что они экспрессируют белки NotchECD/Fc (1, 2, 3, 4) (фиг.5, панель B). В настоящее время проводится очистка N1ECD/Fc из кондиционированных сред от клеток CHO, используя аффинную хроматографию на агарозе с pA.
Определение ингибирования ангиогенеза с использованием основанных на Notch гибридных белков
Активацию передачи сигнала Notch можно обнаружить посредством использования активности промотора CBF1
Функционирование передачи сигнала Notch можно определить посредством измерения транскрипционной активности промотора CBF1, который активируется при связывании Notch-IC с CBF1. Активность промотора CBF1 определяли в HUVEC, которые были инфицированы аденовирусом, кодирующим VEGF-165, при различных множественностях инфицирования (фиг.6). Индукция промотора CBF1 была однозначно обнаружена в Ad-VEGF-инфицированных HUVEC по сравнению с Ad-LacZ-инфицированными клетками зависимым от множественности заражения образом. Эти данные показали, что сверхэкспрессия VEGF может активировать передачу сигнала Notch в HUVEC. Следовательно, VEGF индуцировал сигнализационную активность Notch.
Авторы настоящего изобретения задались вопросом, могут ли основанные на Notch гибридные белки блокировать VEGF-индуцируемую активацию передачи сигнала Notch. Коинфицирование аденовирусом, кодирующим основанный на Notch гибридный белок, с аденовирусом, кодирующим VEGF, отчетливо снижало повышение активности промотора CBF1, индуцируемое при инфицировании только Ad-VEGF (фиг.7). В случае инфицирования с составляющей 40 множественностью инфицирования для каждого аденовируса на фиг.7 (панель A) было обнаружено ингибирование на 60% через 24 ч и ингибирование на 90% через 48 ч после трансфекции гена-репортера, при этом ингибиторная активность ложного белка Notch зависела от множественности инфицирования аденовирусным вектором, кодирующим основанный на Notch гибридный белок.
Основанные на Notch гибридные белки блокируют инициацию спраутинга в ходе ангиогенеза, индуцируемого VEGF
В этом эксперименте был оценен эффект ложного белка Notch на индукцию образования выступов (инициацию спраутинга) под действием сверхэкспрессируемого VEGF-165 в HUVEC. После подвергания Ad-VEGF-инфицированных HUVEC культивированию на коллагеновом (типа I) геле в течение 8 дней индуцировалось образование выступов в коллагеновый гель. Эта индукция образования выступов под действием сверхэкспрессируемого VEGF была явно ингибирована при коинфицировании аденовирусным вектором, кодирующим основанный на Notch гибридный белок (фиг.8). Сам по себе аденовирус, кодирующий основанный на Notch гибридный белок, оказывал меньший эффект на морфологию.
На фиг.9 были подсчитаны выступы в каждом поле, используя микроскоп. Инфицирование Ad-VEGF клеток HUVEC увеличило число выступов, зависящее от используемой множественности инфицирования. Ad-VEGF-индуцируемое образование выступов было явно ингибировано. Эти данные говорят о том, что VEGF индуцировал образование выступов у HUVEC благодаря активации передачи сигнала Notch, и что основанный на Notch гибридный белок мог ингибировать VEGF-индуцируемое образование выступов.
Ссылки, приведенные во втором ряде экспериментов
1. Artavanis-Tsakonas, S., K. Matsuno, and M.E. Fortini. 1995. Notch signaling. Science 268: 225-232.
2. Bailey, A.M., and J.W. Posakony. 1995. Suppressor of hairless directly activates transcription of enhancer of split complex genes in response to Notch receptor activity. Genes & Development 9: 2609-22.
3. Bettenhausen, В., M. Hrabe de Angelis, D. Simon, J.L. Guenet, and A. Gossler. 1995. Transient and restricted expression during mouse embryogenesis of DU, a murine gene closely related to Drosophila Delta. Development 121: 2407-18.
4. Blaumueller, СМ., H. Qi, P. Zagouras, and S. Artavanis-Tsakonas. 1997. Intracellular cleavage of Notch leads to a heterodimeric receptor on the plasma membrane. Cell 90: 281-91.
5. Caronti, В., L. Calandriello, A. Francia, L. Scorretti, M. Manfredi, T. Sansolini, E.M. Pennisi, C. Calderaro, and G. Palladini. 1998. Cerebral autosomal dominant arteriopathy with subcortical infarcts and leucoencephalopathy (CADASIL). Neuropathologieal and in vitro studies of abnormal elastogenesis. Acta Neurol Scand. 98: 259-67.
6. Desmond, D.W., J.T. Moroney, T. Lynch, S. Chan, S.S. Chin, D.C. Shungu, A.B. Naini, and J.P. Mohr. 1998. CADASIL in a North American family: clinical, pathologic, and radiologie findings [see comments]. Neurology 51: 844-9.
7. Dunwoodie, S.L., D. Henrique, S.M. Harrison, and 5 R.S. Beddington. 1997. Mouse Dll3: a novel divergent Delta gene which may complement the function of other Delta homologues during early pattern formation in the mouse embryo. Development 124: 3065-76.
8. Eastman, D.S., R. Slee, E, Skoufos, L. Bangalore, S. Bray, and C. Delidakis. 1997. Synergy between suppressor of Hairless and Notch in regulation of Enhancer of split m gamma and m delta expression. Mol Cell Biol. 17: 5620-5634.
9. Fortini, M.E., and S. Artavanis-Tsakonas. 1993. Notch: neurogenesis is only part of the picture. Cell 75: 1245-7.
10. Gale, N.W., and G.D. Yancopoulos. 1999. Growth factors acting via endothelial cell-specific receptor tyrosine kinases: VEGFs, Angiopoietins, and ephrins in vascular development. Genes and Development 13: 1055-1066.
11. Gallahan, D., and R. Callahan. 1997. The mouse mammary tumor associated gene INT3 is a unique member of the NOTCH gene family (NOTCH4). Oncogene 14: 1883-90.
12. Greenwald, I. 1994. Structure/function studies of lin-12/Notch proteins. Current Opinion in Genetics & Development 4: 556-62.
13. Greenwald, I. 1998. LIN-12/Notch signaling: lessons from worms and flies. Genes Dev. 12: 1751-62.
14. Henderson, A.M., S.J. Wang, A.C. Taylor, M. Aitkenhead, and C.C.W. Hughes. 2001. The basic helix-loop-helix transcription factor HESRl regulates endothelial cell tube formation. J Biol Chem. 276: 6169-6176.
15. Hicks, C, S.H. Johnston, G. diSibio, A. Collazo, T.F. Vogt, and G. Weinmaster. 2 000. Fringe differentially modulates Jaggedl and Deltal signalling through Notchl and Notch2. Nature Cell Biology 2: 515-520.
16. Hsieh, J.J., T. Henkel, P. Salmon, E. Robey, M.G. 20 Peterson, and S.D. Hayward. 1996. Truncated mammalian Notchl activates CBFl/RBPJk-repressed genes by a mechanism resembling that of Epstein-Barr virus EBNA2. Molecular & Cellular Biology 16: 952-9.
17. Hsieh, J.J., D.E. Nofziger, G. Weinmaster, and S.D. Hayward. 1997. Epstein-Barr virus immortalization: Notch2 interacts with CBFl and blocks differentiation. J Virol. 71: 1938-45.
18. Jarriault, S., C. Brou, F. Logeât, E.H. Schroeter, R. Kopan, and A. Israel. 1995. Signaling downstream of activated mammalian Notch. Nature 377: 355-358.
19. Joutel, A., F. Andreux, S. Gaulis, V. Domenga, M. Cecillon, N. Battail, N. Piga, F. Chapon, С. Godfrain, and E. Tournier-Lasserve. 2000. The ectodomain of the Notch3 receptor accumulates within the cerebrovasculature of CADASIL patients [see comments]. J Clin Invest. 105: 597-605.
20. Joutel, A., C. Corpechot, A. Ducros, K. Vahedi, H. Chabriat, P. Mouton, S. Alamowitch, V. Domenga, M. Cecillion, E. Maréchal, J. Maciazek, C. Vayssiere, C. Cruaud, E.A. Cabanis, M. M. Ruchoux, J. Weissenbach, J.F. Bach, M.G. Bousser, and E. Tournier-Lasserve. 1996. Notch3 mutations in 15 CADASIL, a hereditary adult-onset condition causing stroke and dementia. Nature 383: 707-10.
21. Kopan, R., E.H. Schroeter, H. Weintraub, and J.S. Nye. 1996. Signal transduction by activated mNotch: importance of proteolytic processing and its regulation by the extracellular domain. Proc Natl Acad Sei USA 93: 1683-8.
22. Krebs, L.T., Y. Xue, CR. Norton, J.R. Shutter, M. 25 Maguire, J.P. Sundberg, D. Gallahan, V. Closson, J. Kitajewski, R. Callahan, G.H. Smith, K.L. Stark, and T. Gridley. 2000. Notch signaling is essential for vascular morphogenesis in mice. Genes and Development 14: 1343-1352.
23. Lardelli, M., J. Dahlstrand, and U. Lendahl. 1994. The novel Notch homologue mouse Notch3 lacks specific epidermal growth factor-repeats and is expressed in proliferating neuroepithelium. Mechanism of Development 46: 123-136.
24. Lawson, N.D., N. Scheer, V.N. Pham, C. Kim, A.B. Chitnis, J.A. Campos-Ortega, and B.M. Weinstein. 2001. Notch signaling is required for arterial-venous differentiation during embryonic vascular development. Development 128: 3675-3683.
25. Lewis, J. 1998. Notch signaling and the control of cell fate choices in vertebrates. Semin Cell Dev Biol. 9: 583-589.
26. Lieber, T., S. Kidd, E. Alcomo, V. Corbin, and M.W. Young. 1993. Antineurogenic phenotypes induced by truncated Notch proteins indicate a role in signal transduction and may point to a novel function for Notch in nuclei. Genes Dev. 7: 1949-1965.
27. Lindner, v., C. Booth, I. Prudovsky, D. Small, T. Maciag, and L. Liaw. 2001. Members of the Jagged/Notch gene familites are expressed in injured arteries and regulate cell phenotype via alteration in cell matrix and cell-cell interations. Pathology 159: 875-883.
28. Lindseil, C.E., C.J. Shawber, J. Boulter, and G. Weinmaster. 1995. Jagged: A mammalian ligand that activates Notchl. Cell 80: 909-917.
29. Logeât, F., С. Bessia, С. Brou, О. LeBail, S. Jarriault, N.G. Seidah, and A. Israel. 1998. The Notchl receptor is cleaved constitutively by a furin-like convertase. Proc Natl Acad Sei USA 95: 8108-12.
30. Lyman, D., and M.W. Young. 1993. Further evidence for function of the Drosophila Notch protein as a transmembrane receptor. Proc Natl Acad Sei USA 90: 10395-10399.
31. Matsuno, K., M.J. Go, X. Sun, D.S. Eastman, and S. Artavanis-Tsakonas. 1997. Suppressor of Hairless- independent events in Notch signaling imply novel pathway elements. Development 124: 4265-4273.
32. Nakagawa, O., D.G. McFadden, M. Nakagawa, H. Yanagisawa, T. Hu, D. Srivastava, and E.N. Olson. 2000. Members of the HRT family of basic helix-loop- helix proteins act as transcriptional repressors downstream of Notch signaling. Proc Natl Acad Sei USA 97: 13655-13660.
33. Oberg, C, J. Li, A. Pauley, E. Wolf, M. Gurney, and U. Lendahl. 2001. The Notch intracellular domain is ubiquitinated and negatively regulated by the mammalian Sel-10 homolog. J Biol Chem. 276: 35847-35853.
34. Owens, G.K. 1995. Regulation of differentiation of 30 vascular smooth muscle cells. Physiol Rev. 75: 487-527.
35. Rebay, I., R.G. Fehon, and S. Artavanis-Tsakonas. 1993. Specific truncations of Drosophila Notch define dominant activated and dominant negative forms of the receptor. Cell 74: 319-29.
36. Robey, E. 1997. Notch in vertebrates. Curr Opin Genet Dev. 7: 551-7.
37. Roehl, H., M. Bosenberg, R. Blelloch, and J. Kimble. 1996. Roles of the RAM and ANK domains in signaling by the C. elegans GLP-1 receptor. Embo J. 15: 7002-7012.
38. Rogers, S., R. Wells, and M. Rechsteiner. 1986. Amino acid sequences common to rapidly degrade proteins: The PEST hypothesis. Science 234: 364-368.
39. Sasai, Y., R. Kageyama, Y. Tagawa, R. Shigemoto, and S. Nakanishi. 1992. Two mammalian helix-loop-helix factors structurally related to Drosophila hairy and Enhancer of split. Genes & Dev. 6: 2620-2634.
40. Shawber, C, J. Boulter, C.E. Lindsell, and G. Weinmaster. 1996a. Jagged2: a serrate-like gene expressed during rat embryogenesis. Dev Biol. 180: 370-6.
41. Shawber, C, D. Nofziger, J.J. Hsieh, C. Lindsell, 0. Bogler, D. Hayward, and G. Weinmaster. 1996b. Notch signaling inhibits muscle cell differentiation through a CBFl-independent pathway. Development 122: 3765-73.
42. Shimizu, К., S. Chiba, T. Saito, T. Takahashi, К. Kumano, H. Hamada, and H. Hirai. 2002. Integrity of intracellular domain of Notch ligand is indespensable for cleavage required for the release of the Notch2 intracellular domain. Embo J. 21: 294-302.
43. Shutter, J.R., S. Scully, W. Fan, W.G. Richards, J. Kitajewski, G.A. Deblandre, C.R. Kintner, and K.L. Stark. 2000a. Dll4, a novel Notch ligand expressed in arterial endothelium. Genes Dev. 14: 1313-1318.
44. Shutter, J.R., S. Scully, W. Fan, W.G. Richards, J. Kitajewski, G.A. Deblandre, CR. Kitner, and K.L. Stark. 2000b. Dll4, a novel Notch ligand expressed in arterial endothelium. Genes and Development 14: 1313-1318.
45. Struhl, G., K. Fitzgerald, and I. Greenwald. 1993. Intrinsic activity of the Lin-12 and Notch intracellular domains in vivo. Cell 74: 331-45.
46. Swiatek, P.J., C.E. Lindsell, F. Franco del Arno, G. Weinmaster, and T. Gridley. 1994. Notch 1 is essential for postimplantation development in mice. Genes & Development 8: 707-719.
47. Tamura, K., Y. Taniguchi, S. Minoguchi, T. Sakai, T. Tun, T. Furukawa, and T. Honjo. 1995. Physical interaction between a novel domain of the receptor Notch and the transcription factor RBP-J kappa/Su(H). Curr Biol. 5: 1416-1423.
48. Tietze, K., N. Oellers, and E. Knust. 1992. Enhancer of splitD, a dominant mutation of Drosophila, and its use in the study of functional domains of a helix-loop-helix protein. Proc Natl Acad Sei USA 89: 6152-6156.
49. Uyttendaele, H., J. Ho, J. Rossant, and J. Kitajewski. 2001. Vascular patterning defects associated with expression of activated Notch4 in embryonic endothelium. PNAS. 98: 5643-5648.
50. Uyttendaele, H., G. Marazzi, G. Wu, Q. Yan, D. Sassoon, and J. Kitajewski. 1996. Notch4/int-3, a mammary proto-oncogene, is an endothelial cell-specific mammalian Notch gene. Development 122: 2251-9.
51. Vervoort, M., C. Dambly-Chaudiere, and A. Ghysen. 1997. Cell fate determination in Drosophila. Curr Opin Neurobiol. 7: 21-28.
52. Villa, N., L. Walker, C.E. Lindsell, J. Gasson, M.L. Iruela-Arispe, and G. Weinmaster. 2001. Vascular expression of Notch pathway receptors and ligands is restricted to arterial vessels. Mechanisms of Development 108: 161-164.
53. Weinmaster, G. 1997. The Ins and Outs of Notch Signaling. Mol Cel Neurosci. 9: 91-102.
54. Weinmaster, G. 1998. Notch signaling: direct or what? Curr Opin Genet Dev. 8: 436-42.
55. Weinmaster, G., V.J. Roberts, and G. Lemke. 1992. Notch 2: a second mammalian Notch gene. Development 116: 931-941.
56. Weinmaster, G., V.J. Roberts, and G.A. Lemke. 1991. A homolog of Drosophila Notch expressed during mammalian development. Development 113: 199-205.
57. Wettstein, D.A., D.L. Turner, and С Kintner. 1997. The Xenopus homolog of Drosophila Suppressor of Hairless mediates Notch signaling during primary neurogenesis. Development 124: 693-702.
58. Wu, G., E.J. Hubbard, J.K. Kitajewski, and I. Greenwald. 1998. Evidence for functional and physical association between Caenorhabditis elegans SEL-10, a Cdc4p-related protein, and SEL-12 presenilin. Proc Natl Acad Sei USA 95: 15787-91.
59. Wu, G., S.A. Lyapina, I. Das, J. Li, M. Gurney, A. Pauley, I. Chui, R.J. Deshaies, and J. Kitajewski. 2001. SEL-10 is an inhibitor of notch signaling that targets notch for ubiquitin-mediated protein degradation. Mol Cell Biol. 21: 7403-7015.
60. Xue, Y., X. Gao, C.E. Lindsell, C.R. Norton, B. Chang, C. Hicks, M. Gendron-Maguire, E.B. Rand, G. Weinmaster, and T. Gridley. 1999. Embryonic lethality and vascular defects in mice lacking the Notch ligand Jagged1.Hum Mol Genet. 8: 723-30.
Третий ряд экспериментов
VEGF инициирует ангиогенез через активацию передачи сигнала Notch
Пути передачи сигналов с участием как VEGF, так и Notch важны для развития сосудов. Здесь демонстрируется, что VEGF активирует передачу сигнала Notch для инициирования ангиогенеза. VEGF увеличивает экспрессию Delta4 и Notch4, вызывающих активацию передачи сигнала Notch и индуцирующих образование филоподий у культивируемых первичных эндотелиальных клеток. Исследования, в которых используются ингибиторы рецептора VEGF, показывают, что активации передачи сигнала Notch, в свою очередь, усиливает действие VEGF посредством индукции экспрессии VEGFR-1 (Flt-1). Notch опосредует другие элементы деятельности VEGF, включающие индукцию MMP-9 и MT1-MMP. Используя in vivo исследования для моделирования VEGF-индуцируемого образования новых сосудов в коже, авторы настоящего изобретения установили, что секретируемый ингибитор Notch (основанный на Notch гибридный белок) блокирует VEGF-индуцируемое образование новых сосудов и индукцию экспрессии VEGFR-1. Таким образом, передача сигнала Notch необходима для ангиогенеза, регулируемого VEGF, по-видимому, на уровне инициации.
VEGF является основным регулятором развития ангиогенеза, состоящего из множества процессов, таких как деградация экстраклеточного матрикса, образование выступов (образование филоподий), пролиферация, выживание и миграция эндотелиальных клеток. Хотя большая часть стадий могла бы быть объединена с помощью принимающих сигнал от VEGF молекул, неизвестно, каким образом эти стадии координационно регулируются для того, чтобы обеспечить более сложные морфологические события, такие как спраутинг в ходе ангиогенеза. Передача сигнала Notch является эволюционно консервативным механизмом передачи сигнала, который действует для регуляции выборов судьбы клетки (1). В результате связывания лигандом, таким как Jagged и Delta-подобный белок, цитоплазматический домен Notch (NotchIC) высвобождается под действием пресенилина/γ-секретазы, перемещается в ядро, взаимодействует с репрессором транскрипции - CSL (CBF1/Su(H)/lag2) и превращает его в активатор транскрипции (1). О ролях передачи сигнала Notch в развитии сосудов говорили исследования мышей со специализированной мутацией (2). Поскольку активация Notch в эндотелии также нарушает ремоделирование сосудов, надлежащая передача сигнала Notch весьма важна для развития сосудов (3). Хотя релевантность Notch к передаче сигнала от VEGF и предполагается (4-6), все еще неясно, в какой степени передача сигнала Notch играет роль в VEGF-регулируемом ангиогенезе, и принимает ли передача сигнала Notch участие в физиологическом и патологическом ангиогенезе сосудистой сети у взрослых.
Рост HUVEC (эндотелиальных клеток пупочной вены человека) зависит от VEGF (фиг.26A и 26B) и связанных с дифференциацией биологических ответов, таких как спраутинг, и может быть оценен на ранней стадии (7). Сначала авторы настоящего изобретения проверили, индуцирует ли VEGF, кодируемый трансдуцированным с помощью аденовируса геном, экспрессию как Notch, так и лигандов Notch в HUVEC, культивируемых с использованием полной среды, содержащей bFGF (фиг.22A), как приведено в публикации (5). Анализ с использованием ОТ-ПЦР показал, что экспрессии мРНК как D14, так и Notch4 была увеличена в подвергнутых с помощью аденовируса трансдукции гена VEGF клетках HUVEC (Ad-VEGF-HUVEC), по сравнению с подвергнутыми с помощью аденовируса трансдукции гена LacZ клетками HUVEC (Ad-LacZ-HUVEC) (фиг.22A). По-видимому, трансдуцированный VEGF не индуцировал экспрессию Jagged1 и Notch1. Трансдуцированный VEGF также активировал передачу сигнала Notch дозозависимым образом при определении активности регулируемого CSL промотора люциферазного гена-репортера (фиг.22B), который был трансактивирован с помощью передачи сигнала Notch (8). По сравнению с пролиферацией передача сигнала Notch активировалась при более высокой дозе Ad-VEGF (фиг.26A). Так как SU5416, являющийся ингибитором киназ VEGFR, уменьшал активность регулируемого CSL промотора люциферазного гена-репортера (фиг.22C), VEGF индуцировал передачу сигнала Notch благодаря активации рецепторной киназы. Поскольку мутанты Notch, в которых отсутствует и трансмембранный, и цитоплазматический домены, функционируют в качестве доминантных отрицательных ингибиторов передачи сигнала Notch (9), авторы настоящего изобретения создали основанные на Notch гибридный белок или ловушку (N1ECDFc) для ингибирования передачи сигнала Notch (фиг.22D). Анализ с использованием Вестерн-блоттинга кондиционированной среды от Ad-N1ECDFc-трансдуцированных HUVEC (Ad-N1ECDFc-HUVEC) показал, что N1ECDFc хорошо экспрессируется и секретируется (фиг.22E). Посредством использования анализа сокультивирования, в котором по сравнению с контрольными клетками Bosc клетки Bosc, экспрессирующие лиганды Notch (или J1, Dl1 или Dl4), активировали передачу сигнала Notch в клетках HeLa, экспрессирующих Notch1, авторы настоящего изобретения определили ингибирование передачи сигнала Notch при трансфекции плазмиды для экспрессии N1ECDFc (фиг.22F). Затем авторы настоящего изобретения проверили, ингибирует ли N1ECDFc активацию передачи сигнала Notch трансдуцированным VEGF в HUVEC (фиг.22G). Котрансдукция Ad-N1ECDFc с Ad-VEGF в HUVEC, несомненно, снижала активность регулируемого CSL промотора люциферазного гена-репортера, индуцируемого под действием VEGF. Gerhardt и др. представили данные, что VEGF контролирует ангиогенез в сетчатке в раннем постнатальном периоде посредством стимулирования удлинения филоподий на концах сосудистых врастаний (10). Во время спраутинга в ходе ангиогенеза возникновение специализированной эндотелиальной клетки, образующей выступы - филоподии, среди неактивных эндотелиальных клеток, вероятно, является одним из ранних событий. Здесь в качестве образования выступов подразумевается возникновение одной эндотелиальной клетки, образующей выступы - филоподии. Образование выступов у первичных эндотелиальных клеток индуцируется при их объемном культивировании на либо фибриновом, либо коллагеновом геле (11). В том случае, когда Ad-VEGF-HUVEC культивировали на коллагеновом геле с использованием полной среды, трансдуцированные HUVEC создавали удлинения филоподий в коллагеновый гель в течение 5 дней (фиг.22H), и число выступов увеличивалось дозозависимым образом (фиг.27A). Активации передачи сигнала Notch с помощью аденовируса, кодирующего активированную форму Notch4 (Ad-Notch4/int3), индуцировала образованием выступов у HUVEC (12), и активация передачи сигнала Notch с помощью аденовируса, кодирующего Notch1, (Ad-N1IC) также индуцировала образованием выступов у HUVEC (фиг.23A и 27B). Поскольку передача сигнала и от VEGF, и от Notch индуцирует образование выступов у HUVEC, авторы настоящего изобретения проверили, ингибирует ли N1ECDFc VEGF-индуцируемое образование выступов у HUVEC (фиг.22H-I). Образование выступов у Ad-VEGF-HUVEC было явно ингибировано при котрансдукции Ad-N1ECDFc. Ни Ad-LacZ-, ни Ad-N1ECDFc-трансдуцированные HUVEC не образовывали выступы (фиг.22H). N1ECDFc ингибировал VEGF-индуцируемое образование выступов у HUVEC без оказания влияния на количество клеток (фиг.22I). Трансдуцированная конструкция для N1ECDFc не приводила к явному изменению пролиферации HUVEC, в то время как пролиферация Ad-N1IC-трансдуцированных HUVEC была ингибирована дозозависимым образом (фиг.28A), в соответствии с ингибиторной в отношении пролиферации эндотелиальных клеток эффективностью передачи сигнала Notch (13).
Для проверки того, находится ли передачи сигнала Notch после VEGF, авторы настоящего изобретения оценили три различных ингибитора рецепторных тирозинкиназ, включающих VEGFR, на N1IC-индуцируемое образование выступов у HUVEC, поскольку эти три фактора роста присутствуют в полной среде (фиг.23A-C). В концентрации, составляющей 1 мкМ, каждое соединение продемонстрировало ингибирование, селективное в отношении каждой киназы (непредставленные данные). Ни PD166866, ни ZD1893 не оказывали влияние на образование выступов у Ad-N1IC-HUVEC, в то время как SU5416 явно ингибировал такое образование (фиг.23A-B). SU5416 селективно ингибировал образование выступов у Ad-N1IC-HUVEC с меньшим снижением жизнеспособности при более низких концентрациях (фиг.23C). Поскольку Taylor и др. представили данные, что Notch уменьшает экспрессию Flk1/KDR/VEGFR2 (14), маловероятно, что Notch действует вместе с Flk1 для стимулирования образования выступов. Поэтому авторы настоящего изобретения проверили, оказывает ли активация передачи сигнала Notch влияние на экспрессию Flt1/VEGFR1 в HUVEC, поскольку SU5416 ингибирует киназную активность и Flt1, и Flk1 (15). Анализ с использованием ОТ-ПЦР показал, что экспрессия мРНК Flt1 была уменьшена в Ad-N1IC-HUVEC, в то время как экспрессия маркера эндотелиальных клеток, мРНК CD31, была соизмеримой с таковой в Ad-LacZ-HUVEC (фиг.23D). Анализ с помощью Вестерн-блоттинга также показал, что экспрессия белка Flt1 была уменьшена в Ad-N1IC-HUVEC (фиг.23E). Поэтому авторы настоящего изобретения проверили, стимулирует ли PlGF, который является селективным лигандом для Flt1, образование выступов у HUVEC, в которых экспрессия Flt1 была увеличена, через активацию передачи сигнала Notch (фиг.23F-G). PlGF увеличивал на 150% число выступов у Ad-N1IC-HUVEC по сравнению с указанным числом в отсутствие PlGF (фиг.23F). Более того, PlGF увеличивал на 250% число выступов у HUVEC, содержащих множество филоподий (фиг.23G). Тогда как уменьшение, используя короткую интерферирующую РНК (siRNA) для Flt1, экспрессии Flt1 ингибировало образование выступов у Ad-N1IC-HUVEC (фиг.23J), трансфекция которых селективно уменьшала экспрессию мРНК Flt1 (фиг.23H) и экспрессию белка Flt1 (фиг.23I). Хотя уменьшение экспрессии Flk1 с помощью siRNA для Flk1 также ингибировало образование выступов у Ad-N1IC-HUVEC (фиг.30B), ингибиторная эффективность siRNA для Flk1 была меньше таковой siRNA для Flt1 (фиг.23J). Эффекты siRNA для Flk1 на образование выступов были более выраженными в случае Ad-VEGF-HUVEC, чем в случае Ad-N1IC-HUVEC (фиг.30B-C). Трансфекция siRNA для Flt1 ингибировала образование выступов и у Ad-N1IC-HUVEC, и у Ad-VEGF-HUVEC в схожей степени (непредставленные данные).
С помощью нескольких исследований установлено, что VEGF регулирует желатиназные активности в эндотелиальных клетках, и твердо доказано, что важность активности желатиназы, вроде MMP-2 и MMP-9, заключается в индукции спраутинга в ходе ангиогенеза (16). Авторы настоящего изобретения проверили, регулирует ли VEGF желатиназную активность через посредство передачи сигнала Notch в HUVEC.
При зимографии с использованием желатина кондиционированная среда от Ad-VEGF-HUVEC продемонстрировала как индукцию и активацию MMP9 с началом обнаружения в день 6 (фиг.24A), так активацию MMP2, которая выявлялась в день 4 (фиг.24B), по сравнению с аналогичной средой от Ad-LacZ-HUVEC. Котрансдукция Ad-N1ECDFc с Ad-VEGF продемонстрировала ингибирование как индукции и активации MMP9 (фиг.24A), так и активации MMP2 (фиг.24B). Анализ с использованием ОТ-ПЦР показал, что экспрессия мРНК MMP9 была увеличена в Ad-N1IC-HUVEC, однако экспрессия мРНК MMP2 была уменьшена в Ad-N1IC-HUVEC (фиг.24C). Поскольку при использовании зимографии с использованием желатина индукция активности MMP2 не была выявлена (фиг.24B), этот результат был правдоподобным результатом. Тогда как экспрессия MT1-MMP, которая способна активировать MMP2 на клеточной поверхности (17), была увеличена как на транскрипционном уровне, так и на уровне белка в Ad-N1IC-HUVEC (фиг.24D). Так как VEGF может регулировать экспрессию как желатиназы, так и MT1-MMP (16), анализ с использованием ОТ-ПЦР показал, что экспрессия и MMP9, и MT1-MMP была увеличена в Ad-VEGF-HUVEC по сравнению с Ad-LacZ-HUVEC, и эта индукция ингибировалась при котрансдукции Ad-N1ECDFc (фиг.24E). Инфицирование только Ad-N1ECDFc не оказывало влияния на экспрессию ни MMP9, ни MT1-MMP в Ad-LacZ-инфицированных HUVEC (непредставленные данные). Необходимость MMP для спраутинга в ходе ангиогенеза была установлена с помощью синтетических ингибиторов MMP (16). GM6001 является один из ингибиторов ММР широкого действия, в том числе MMP2, MMP9 и MT1-MMP (18). GM6001 явно уменьшал образование выступов у Ad-N1IC-HUVEC и на коллагеновом (фиг.31A-B), и на фибриновом геле (непредставленные данные).
При исследовании дорсальных альвеолярных мешочков (DAS) у мыши (19) стабильный трансфектант клеток 293, сверхэкспрессирующий VEGF121, (293/VEGF) значительно индуцировал in vivo ангиогенез (фиг.25A, левая панель). Этот VEGF-индуцируемый ангиогенез был явно ингибирован при коэкспрессии N1ECDFc, по сравнению с 293/VEGF только (фиг.25A). Определяли плотность сосудов, и индекс ангиогенеза представлен на фиг.25B, на которой демонстрируется, что 293/VEGF-индуцируемый ангиогенез ингибируется при коэкспрессии 293/N1ECDFc (фиг.25B).
При исследовании дорсальных альвеолярных мешочков (DAS) у мыши (19) также линия клеток рака молочной железы человека, MDA-MB-231, значительно индуцировала in vivo ангиогенез, по-видимому, через посредство секреции VEGF (фиг.25C, левая панель). Этот VEGF-индуцируемый ангиогенез был явно ингибирован при аденовирусопосредованной экспрессии N1ECDFc, по сравнению с аденовирусом, экспрессирующим LacZ (фиг.25C). Определяли плотность сосудов, и индекс ангиогенеза представлен на фиг.25D, на которой демонстрируется, что MDA-MB-231-индуцируемый ангиогенез ингибируется при экспрессии N1ECDFc.
Flk1 является основным передатчиком положительного сигнала в отношении ангиогенеза благодаря его сильной тирозинкиназной активности у эмбриона, тогда как Flt1, как полагают, является передатчиком отрицательного сигнала в отношении ангиогенеза. Однако у взрослых мышей была установлена положительная роль Flt-1, поскольку in vivo рост LLC, сверхэкспрессирующих P1GF2, был значительно уменьшен у мышей, у которых отсутствовал цитоплазмический киназный домен Flt-1 (20). Notch мог бы функционировать с изменением передачи сигнала VEGF посредством индукции передачи Flt-1 и с модуляцией передачи сигнала Flk-1 либо для индукции удлинения филоподий, либо усиления спраутинга в ходе ангиогенеза, поскольку передача сигнала PlGF/Flt-1 видоизменяла сайт фосфорилирования Flk-1 и усиливала ангиогенез в ишемизированном миокарде (21). Интересно, что передача сигнала Notch также увеличивает экспрессию PlGF (фиг.29). Однако длительная активация передачи сигнала Notch ингибирует возникновение многоклеточных, содержащих просветы врастаний в ходе ангиогенеза, как сообщалось ранее (22). Передача сигнала Notch должна быть выключена после образования выступов/образования филоподий, и, возможно, потребуется транзиторная активация пути с участием Notch. В модели развития рака из бета-клеток поджелудочной железы у трансгенных мышей (мышей Rip1Tag2), в которой ангиогенез в опухоли зависит от VEGF, уровень экспрессии VEGF не увеличен, однако имеет место мобилизация внеклеточного VEGF, хранящегося в матриксе, к рецепторам VEGF. За эту мобилизацию ответственна MMP-9, и прогрессирование опухоли было ингибировано у Rip1Tag23MMP-9-нулевых дважды трансгенных мышей (23). Notch увеличивал экспрессию MMP-9 и, возможно, увеличивает локальный уровень VEGF в месте спраутинга в ходе ангиогенеза. Хотя Notch также увеличивает экспрессию MT1-MMP, внеклеточный MMP-2, возможно, направляется в клеточную мембрану Notch-активированных эндотелиальных клеток. Возможно, Notch определяет место спраутинга в ходе ангиогенеза посредством регуляции желатиназной активности и концентрации VEGF. Так как эндотелиальный MMP-9 регулировался с помощью Flt-1 при специфичном для легких метастазе (20), Flt-1, возможно, принимает непосредственное участие в индукции MMP-9.
Ссылки, приведенные в третьем ряде экспериментов
1. Artavanis-Tsakonas S, Rand MD, Lake RJ. Notch Signaling: Cell Fate Control and Signal Integration in Development. Science 1999; 284(5415): 770-776.
2. Shawber CJ, J. K. Notch function in the vasculature: insights from zebrafish, mouse and man. Bioessays. 2004; 26(3): 225-34.
3. Uyttendaele H, Ho J, Rossant J, J. K. Vascular patterning defects associated with expression of activated Notch4 in embryonic endothelium. Proc Natl Acad Sei USA. 2001; 98(10): 5643-8.
4. Lawson ND, Vogel AM, BM. W. sonic hedgehog and vascular endothelial growth factor act upstream of the Notch pathway during arterial endothelial differentiation. Dev Cell 2002; 3(1): 127-36.
5. Liu ZJ, Shirakawa T, Li Y, Soma A, Oka M, Dotto GP, et al. Regulation of Notchl and Dll4 by vascular endothelial growth factor in arterial endothelial cells: implications for modulating arteriogenesis and angiogenesis. Mol Cell Biol. 2003; 23 (1): 14-25.
6. Gale NW, Dominguez MG, Noguera I, Pan L, Hughes V, Valenzuela DM, et al. Haploinsufficiency of delta like 4 ligand results in embryonic lethality due to major defects in arterial and vascular development. Proc Natl Acad Sei USA. 2004; 101(45): 5949-54.
7. Montesano R, L. О. Phorbol esters induce angiogenesis in vitro from large-vessel endothelial cells. J Cell Physiol. 1987; 130(2): 284-91.
8. Jarriault S, Brou C, Logeât F, Schroeter EH, Kopan R, A. I. Signalling downstream of activated mammalian Notch. Nature. 1995; 377(6547): 355-8.
9. Small D, Kovalenko D, Kacer D, Liaw L, Landriscina M, Di Serio С, et al. Soluble Jagged 1 represses the function of its transmembrane form to induce the formation of the Src-dependent chord-like phenotype. J Biol Chem 2001; 276(34): 32022-30.
10. Gerhardt H, Golding M, Fruttiger M, Ruhrberg С, Lundkvist A, Abramsson A, et al. VEGF guides angiogenic sprouting utilizing endothelial tip cell filopodia. J Cell Biol 2003; 161(6): 1163-77.
11. Koolwijk P, van Erck MG, de Vree WJ, Vermeer MA, Weich HA, Hanemaaijer R, et al. Cooperative effect of TNFalpha, bFGF, and VEGF on the formation of tubular structures of human microvascular endothelial cells in a fibrin matrix. Role of urokinase activity. J Cell Biol 1996; 132(6): 1177-88.
12. Das I, Craig C, Funahashi Y, Jung KM, Kim TW, Byers R, et al. Notch oncoproteins depend on gamma-secretase/presenilin activity for processing and function. J Biol Chem 2004; 279(29): 30771-80.
13. Noseda M, Chang L, McLean G, Grim JE, Clurman BE, Smith LL, et al. Notch activation induces endothelial cell cycle arrest and participates in contact inhibition: role of p21Cipl repression. Mol Cell Biol 2004; 24(20): 8813-22.
14. Taylor KL, Henderson AM, CC. H. Notch activation during endothelial cell network formation in vitro targets the basic HLH transcription factor HESR-1 and downregulates VEGFR-2/KDR expression. Microvasc Res 2002; 64(3): 372-83.
15. Itokawa T, Nokihara H, Nishioka Y, Sone S, Iwamoto Y, Yamada Y, et al. Antiangiogenic effect by SU5416 is partly attributable to inhibition of Flt-1 receptor signaling. Mol Cancer Ther 2002; 1(5): 295-302.
16. Pepper MS. Role of the matrix metalloproteinase and plasminogen activator-plasmin systems in angiogenesis. Arterioscler Thromb Vase Biol 2001; 21(7): 1104-17.
17. Seiki M,- Koshikawa N, I. Y. Role of pericellular proteolysis by membrane-type 1 matrix metalloproteinase in cancer invasion and angiogenesis. Cancer Metastasis Rev 2003; 22(2-3): 129-43.
18. Yamamoto M, Tsujishita H, Hori N, Ohishi Y, Inoue S, Ikeda S, et al. Inhibition of membrane-type 1 matrix metalloproteinase by hydroxamate inhibitors: an examination of the subsite pocket. J Med Chem 1998; 41(8): 1209-17.
19. Funahashi Y, Wakabayashi T, Semba T, Sonoda J, Kitoh K, К. Y. Establishment of a quantitative mouse dorsal air sac model and its application to evaluate a new angiogenesis inhibitor. Oncol Res. 1999; 11(7): 319-29.
20. Hiratsuka S, Nakamura K, Iwai S, Murakami M, Itoh T, Kijima H, et al. MMP9 induction by vascular endothelial growth factor receptor-1 is involved in lung-specific metastasis. Cancer Cell 2002; 2(4): 289-300.
21. Autiero M, Waltenberger J, Communi D, Kranz A, Moons L, Lambrechts D, et al. Role of PlGF in the intra-and intermoleeular cross talk between the VEGF receptors Fltl and Flkl. Nat Med 2003; 9 (7): 936-43.
22. Leong KG, Hu X LL, Noseda M, Larrivee B, Hull C, Hood L, et al. Activated Notch4 inhibitsangiogenesis: role of beta 1-integrin activation. Mol Cell Biol 2002; 22(8): 2830-41.
23. Bergers G, Brekken R, McMahon G, Vu TH, Itoh T, Tamaki K, et al. Matrix metalloproteinase-9 triggers the angiogenic switch during carcinogenesis. Nat Cell Biol 2000; 2 (10): 737-44.
Четвертый ряд экспериментов
Экспрессия белков и лигандов Notch в эндотелиальных клетках кровеносных и лимфатических сосудов
Для Notch1-4, Dll1, Dll4 и Jagged1 была проведена ОТ-ПЦР на РНК, выделенной из эндотелиальных клеток кровеносных сосудов (BEC) и эндотелиальных клеток лимфатических сосудов (LEC), очищенных из HMVEC. Как продемонстрировано на фиг.44, Notch1, Notch2, Notch4, Dll4 и Jagged1 экспрессировались и в BEC, и в LEC на сходном уровне. Экспрессия Notch3, по-видимому, ограничивается LEC, что наводит на мысль о функционированиях передачи сигнала Notch3 в лимфатическом эндотелии.
Notch3 коэкспрессируется с маркерами эндотелиальных клеток лимфатических сосудов LYVE-1 и Prox1 у эмбрионов на El3,5
Последовательные срезы толщиной 10 микрон мышиных эмбрионов дня 13,5 подвергали иммуноокрашиванию на каждый из LYVE-1, Prox1 и Notch3. Как продемонстрировано на фиг.45, Notch3 экспрессировался в клетках, которые также экспрессировали маркеры эндотелиальных клеток лимфатических сосудов, LYVE-1 и Prox1.
Prox1 индуцировал экспрессию Notch3 в эндотелиальных клетках кровеносных сосудов
Было проверено, могла ли бы эктопическая экспрессия Prox1 изменить экспрессию белков или лигандов Notch. Как продемонстрировано на фиг.46, секция A, через двадцать четыре часа после инфицирования аденовирусным вектором - либо Ad-Prox1, либо Ad-LacZ выделяли тотальную РНК HUVEC, и проводили количественную ОТ-ПЦР для Notch1-4, Dll4 и Jagged1. Prox-1 сильно увеличивал экспрессию Notch3. Значительного влияния на экспрессию Notch1, Notch2, Notch4, Dll4 и Jagged1 не оказывалось. Как продемонстрировано на фиг.46, секция B, соединение E (cE), ингибитор пресенлина, который ингибирует передачу сигнала Notch, инкубировали в течение 24 часов с либо Ad-LacZ-, либо Ad-Prox1-инфицированными HUVEC. Выделяли тотальную РНК, и проводили количественную ОТ-ПЦР для определения экспрессии Notch3. Prox1 индуцировал экспрессию Notch3, и эта индукция была ингибирована при добавлении соединения E.
Это говорит о том, что индукция Notch3 под действием Prox1 зависит от активации сигнала Notch.
Prox1 индуцирует гены-мишени Notch в эндотелиальных клетках кровеносных сосудов
HUVEC инфицировали аденовирусами, кодирующими LacZ, Prox1, N1IC или N4/int-3, и через 24 часа после инфицирования выделяли тотальную РНК. Количественную ОТ-ПЦР проводили для генов-мишеней Notch эндотелиальных клеток, VEGFR-3, эфрина B2, Hey1 и Hey2. Подобно активации сигнала Notch1 и Notch4, Prox1 индуцировал все четыре гена (фиг.47, секции A и B). Об экспрессии Hey1 и Hey2 в лимфатическом эндотелии неизвестно.
То, что Prox1 индуцирует гены-мишени Notch, зависит от передачи сигнала Notch в эндотелиальных клетках кровеносных сосудов
HUVEC инфицировали аденовирусами, кодирующими LacZ, Prox1, N1IC или N4/int-3. Соединение E(cE), ингибитор пресенлина, который ингибирует передачу сигнала Notch, инкубировали в течение 24 часов с либо Ad-LacZ-, либо Ad-Prox1-инфицированными HUVEC, и выделяли тотальную РНК. Проводили количественную ОТ-ПЦР для генов-мишеней Notch эндотелиальных клеток, VEGFR-3, эфрина B2 и Hey2. Prox-1-опосредованная индукцию генов-мишеней Notch, эфрина B2, VEGFR-3 и Hey2, была ингибирована при добавлении соединения Е - ингибитора передачи сигнала Notch, как продемонстрировано на фиг.48. Таким образом, Prox1 регулирует экспрессию эфрина B2, VEGFR-3 и Hey2 через посредство Notch.
Пятый ряд экспериментов
Предпосылка
Понимания функционирования Notch для поддержания сосудистого гомеостаза можно добиться на основе нейрососудистого нарушения у человека, церебральной аутосомно-доминантной артериопатии с субкортикальными инфарктами и лейкоэнцефалопатией (CADASIL). Было установлено, что у большинства пациентов имеется корреляция между CADASIL и миссенс-мутацией в Notch3. CADASIL является аутосомно-доминантным нарушением с поздним началом (в среднем возрасте, составляющим 45 лет), характеризующимся мигренью с аурой и повторяющими ударами, которые приводят к психиатрическим симптомам, прогрессирующему снижению когнитивных способностей, деменции и смерти (81). Эти невропатологические симптомы возникают на фоне медленного развития артериопатии, связанной с дезорганизацией и разрушением гладкомышечных клеток сосудов, окружающих церебральные артерии и артериолы. Регрессия гладкомышечных клеток сосудов сопровождается уменьшением толщины сосудистой стенки, утратой внеклеточного матрикса и непрочностью сосудистой стенки (82). Внутри гладкомышечных клеток сосудов существует накопление внеклеточного домена Notch3, а во внеклеточном матриксе - анормальное отложение частиц, называемых гранулярными осмофильными материалами (GOM) (83). При этом нарушении повреждения артерий не ограничиваются головным мозгом и обнаруживаются в артериях кожи и сетчатки (83-85).
Существует корреляция фенотипа CADASIL с экспрессией Notch3 в гладкомышечных клетках сосудов (70, 81). Существует гипотеза, что Notch3 функционирует для поддержания межклеточных взаимодействий или соединения между гладкомышечными клетками сосудов и эндотелиальными клетками артерий. В недавнем исследовании была воссоздана сосудистая патология при CADASIL у трансгенных мышей, которые экспрессируют трансген Notch3, кодирующий связанную с CADASIL мутацию R90C, главным образом, в гладкомышечных клетках сосудов (86). Сосудистая сеть этих мышей продемонстрировала артериопатию - классическую CADASIL, в том числе отложения GOM и накопление Notch3. Однако эти отличительным признакам предшествовало нарушение крепления и адгезии гладкомышечных клеток сосудов к соседним клеткам с последующей деградацией гладкомышечных клеток сосудов. Таким образом, CADASIL является следствием уменьшения контактирования и жизнеспособности гладкомышечных клеток сосудов, а отложение GOM и накопление внеклеточного домена Notch3 являются вторичными результатами этого клеточного ухудшения. В соответствии с ролью Notch3 в выживании клеток экспрессия конститутивно активной формы Notch3 в гладкомышечных клетках аорты крыс вызывала индукцию cFlip, антагониста Fas-зависимого апоптоза (87). Кроме того, эктопическая экспрессия Hey1 в культивируемых гладкомышечных клетках сосудов способствовала выживанию клеток через посредство Akt и, следовательно, ингибировала апоппоз в ответ на истощение сыворотки и лиганд Fas (88). Вместе взятые, эти данные показывают, что Notch3 поддерживает артериальный сосудистый гомеостаз посредством способствования выживанию гладкомышечных клеток сосудов. Являющаяся результатом неплотность артериальной сосудистой стенки могла быть вызвана гибелью гладкомышечных клеток сосудов или неспособностью гладкомышечных клеток сосудов к соединению с соседними с ними эндотелиальными клетками. Нарушение активности Notch3 у мышей может помочь в определении природы этого дефекта.
Специфическая активность связанных с CADASIL мутантных белков Notch3 все еще плохо уяснена. Одно из затруднений в интерпретации функционирования мутантного Notch3 обусловлено противоречивостью in vitro исследований, в которых было установлено, что усеченный цитоплазматический Notch3 может либо ингибировать, либо активировать транскрипционный фактор CSL (89, 90).
Активация передачи сигнала Notch в гладкомышечных клетках сосудов приводит к смертности у эмбрионов
Notch3 экспрессируется и является активным в клетках, которые окружают кровеносные сосуды, гладкомышечных клетках и перицитах. Гладкомышечные клетки важны для функционирования сердечно-сосудистой системы, и они должны быть здоровыми, чтобы предупреждать удар. Перициты могут вносить вклад в рост сосудов в опухоли. Однако Notch1 и Notch4 не должны функционировать в этих типах клеток.
Следовательно, основанные на Notch3 гибридные белки, описываемые здесь, могут быть полезны для предупреждения удара посредством предотвращения анормальной активности Notch3. Кроме того, основанные на Notch3 гибридные белки могут быть полезны для поддержания гладкомышечных клеток сосудов, сдерживания роста или функционирования перицитов в опухоли или для оказания влияния на ангиогенез в сетчатки посредством модулирования функционирования перицитов.
Авторы настоящего изобретения создали трансгенную мышь, которая экспрессирует активированную форму Notch1 (N1IC) под контролем промотора фактора 1-альфа элонгации (EF1α) в тканях, которые экспрессируют Cre-рекомбиназу, названную EFlαN1IC/+ (фиг.49). EF1αN1IC/+ является жизнеспособной и фертильной (фиг.50). Авторы настоящего изобретения экспрессировали N1IC в гладкомышечных клетках сосудов посредством скрещивания EF1αN1IC/+ с линией мышей SM22-Cre (SM22Cre/+). Результирующая SM22Cre/+; EF1αN1IC/+ дважды трансгенная мышь умирала на E9,5 (фиг.50). SM22Cre/+; EF1αN1IC/+ эмбрионы демонстрируют пороки развития миокарда, которые, как полагают авторы настоящего изобретения, ответственны за смертность у эмбрионов. В соответствии с этими пороками развития гладкомышечных клеток миокарда авторы настоящего изобретения наблюдали изменение экспрессии маркера гладкомышечных клеток сосудов, альфа-актина гладкомышечных клеток, у SM22Cre/+; EF1αN1IC/+ трансгенных животных на E9,5 (фиг.51). Эти результаты показывают, что увеличение передачи сигнала Notch в гладкомышечных клетках сосудов нарушает развитие сердечно-сосудистой системы у эмбриона.
Ссылки, приведенные в пятом ряде экспериментов
81. Viitanen M, Kalimo H, CADASIL: Hereditary arteriopathy leading to multiple brain infarcts and dementia. Ann. N. Y. Acad. Sei. 2000; 903: 273-84.
82. Brulin P, Godfraind C, Leteurtre E, Ruchoux MM. Morphometric analysis of ultrastructural vascular changes in CADASIL: analysis of 50 skin biopsy specimens and pathogenic implications. Acta. Neuropathol. 2002; 104: 241-8.
83. Uyama E, Tokunaga M, Suenaga A, Kotorii S, Kamimura K, Takahashi K, Tabira T, Uchino M. Argl33Cys mutation of Notch3 in two unrelated Japanese families with CADASIL. Intern. Med. 2000; 39(9): 732-7.
84. Joutel A, Favrole P, Labauge P, Chabriat H, Lescoat C, Andreux F, Domenga V, Cecillon M, Vahedi K, Ducros A and others. Skin biospy immunostaining with a Notch3 monoclonal antibody for CADASIL diagnosis. Lancet 2001; 358: 2049-51.
85. Smith BW, Henneberry J, Connolly T. Skin biopsy findings in CADASIL. Neurology 2002; 59 (6): 961.
86. Ruchoux MM, Domenga V, Brulin P, Maciazek J, Limol S, Tournier-Lasserve E, Joutel A. Transgenic mice expressing mutant Notch3 develop vascular alterations characteristic of cerebral autosomal dominant arteriopathy with subcortical infarcts and 1euekoencephalopathy. Am. J. Path. 5 2003; 162(1): 329-42.
87. Wang W, Prince CZ, Mou Y, Pollman MJ. Notch3 signaling in vascular smooth muscle cells induces c-FLlP expression via ERK/MAPK activation. J Biol Chem 2002; 277(24): 21723-9.
88. Wang W, Prince CZ, Hu X, Pollman MJ. HRT1 modulates vascular smooth muscle cell proliferation and apoptosis. Biochem Biophys Res Commun 2003; 308(3): 596-601.
89. Beatus P, Lundkvist J, Oberg C, Lendahl U. The 15 Notch3 intracellular domain represses Notchl-mediated activation through Hairy/Enhancer of split (HES) promoters. Development 1999; 126(17): 3925-35.
90. Saxena MT, Schroeter EH, Mumm JS, Kopan R. Murine notch homologs (N1-4) undergo presenilin-dependent proteolysis. J. Biol. Chem. 2001; 276 (43): 40268-73.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИИ ГУМАНИЗИРОВАННЫХ СЛИТЫХ БЕЛКОВ NOTCH И СПОСОБЫ ЛЕЧЕНИЯ | 2008 |
|
RU2532830C2 |
| НУКЛЕОТИДНАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ, КОДИРУЮЩАЯ СЛИТЫЙ БЕЛОК, СОСТОЯЩИЙ ИЗ РАСТВОРИМОГО ВНЕКЛЕТОЧНОГО ФРАГМЕНТА ЧЕЛОВЕЧЕСКОГО Dll4 И КОНСТАНТНОЙ ЧАСТИ ТЯЖЕЛОЙ ЦЕПИ ЧЕЛОВЕЧЕСКОГО IgG4 | 2021 |
|
RU2787060C1 |
| АГОНИСТИЧЕСКИЕ АНТИТЕЛА ПРОТИВ NOTCH3 И ИХ ПРИМЕНЕНИЕ ДЛЯ ЛЕЧЕНИЯ NOTCH3-АССОЦИИРОВАННЫХ ЗАБОЛЕВАНИЙ | 2007 |
|
RU2461569C2 |
| ТЕРАПЕВТИЧЕСКИЕ СПОСОБЫ ЛЕЧЕНИЯ ВАСКУЛЯРНЫХ ЗАБОЛЕВАНИЙ ГЛАЗ АНТАГОНИСТАМИ Dll4 | 2007 |
|
RU2429876C2 |
| СПОСОБЫ ЛЕЧЕНИЯ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ С ИСПОЛЬЗОВАНИЕМ ИНГИБИТОРОВ ПУТИ ПЕРЕДАЧИ СИГНАЛА NOTCH | 2008 |
|
RU2584579C2 |
| АНТИТЕЛА-АНТАГОНИСТЫ ПРОТИВ Notch3 И ИХ ПРИМЕНЕНИЕ ДЛЯ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ СВЯЗАННЫХ С Notch3 ЗАБОЛЕВАНИЙ | 2007 |
|
RU2493167C2 |
| АНТИТЕЛА ПРОТИВ NOTCH1 | 2011 |
|
RU2622083C2 |
| АНТИТЕЛА ПРОТИВ NRR Notch1 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2008 |
|
RU2476443C2 |
| ПЕПТИДЫ НАПРАВЛЕННОГО ДЕЙСТВИЯ НА VEGFR-1/NRP-1 | 2008 |
|
RU2488592C2 |
| АНТИТЕЛА ПРОТИВ NOTCH2 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2009 |
|
RU2580029C2 |























































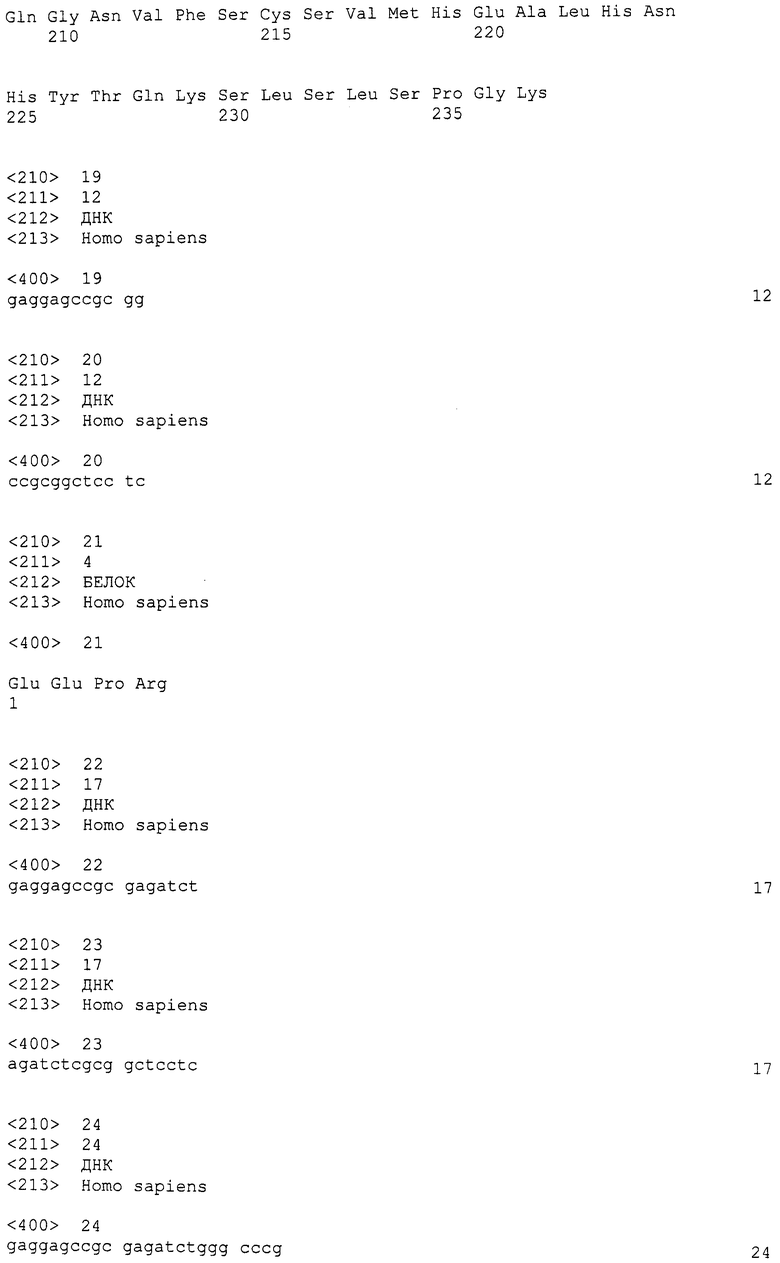




















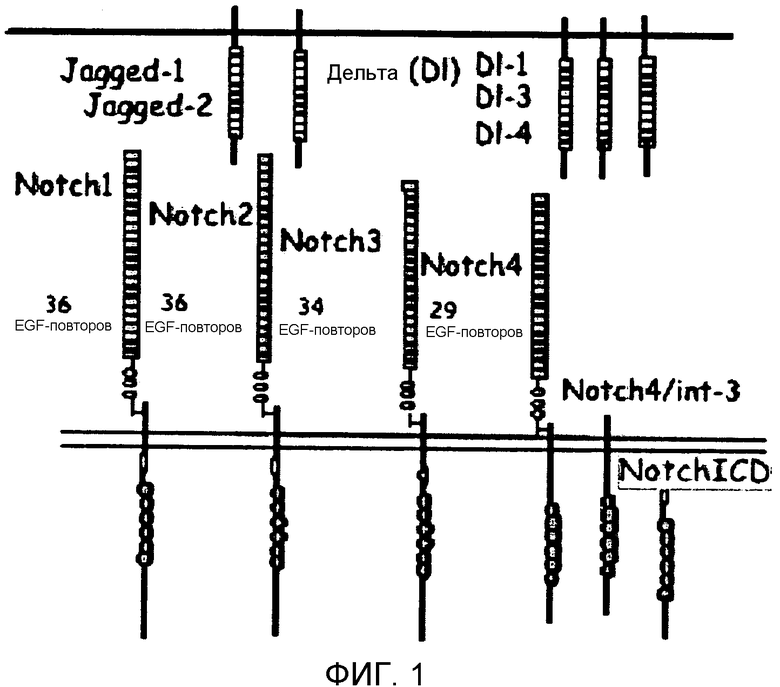
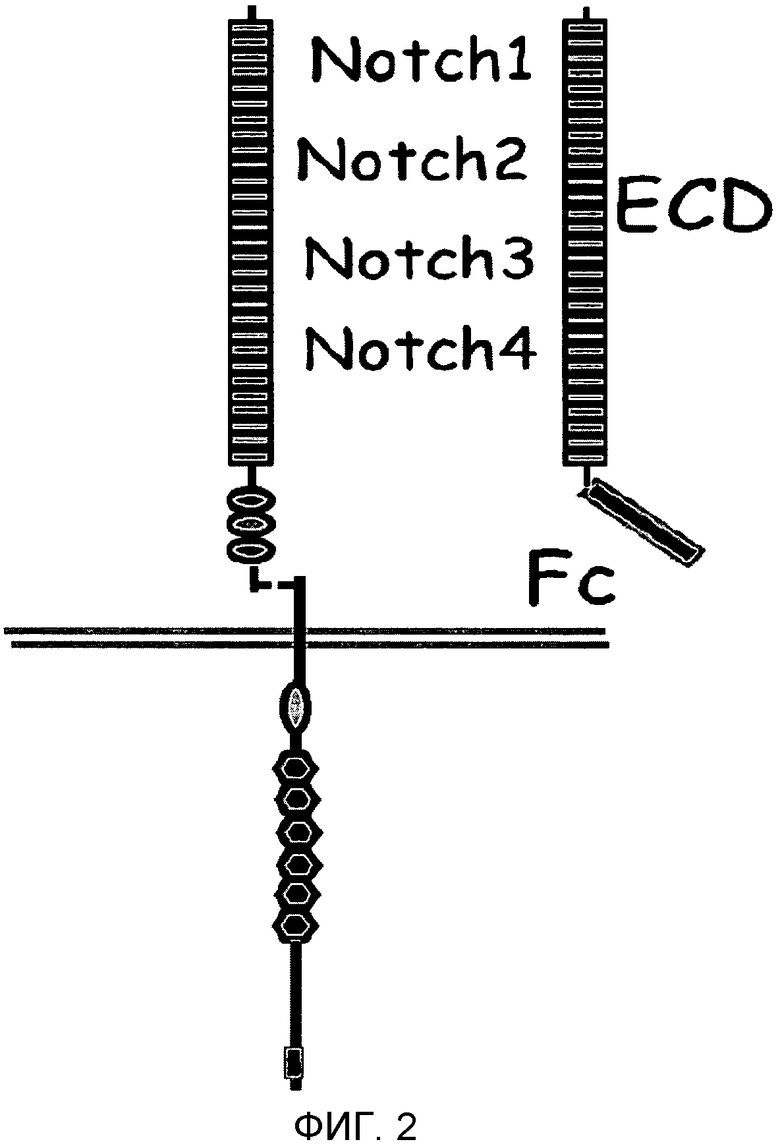
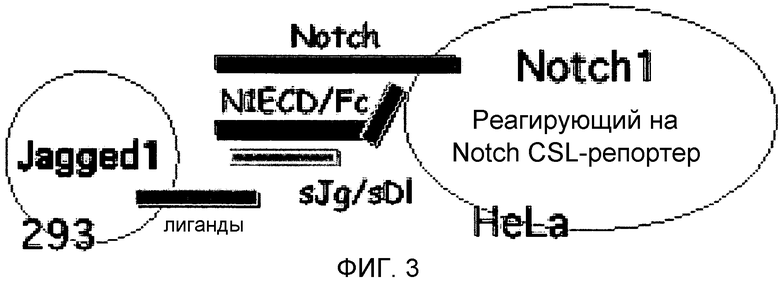
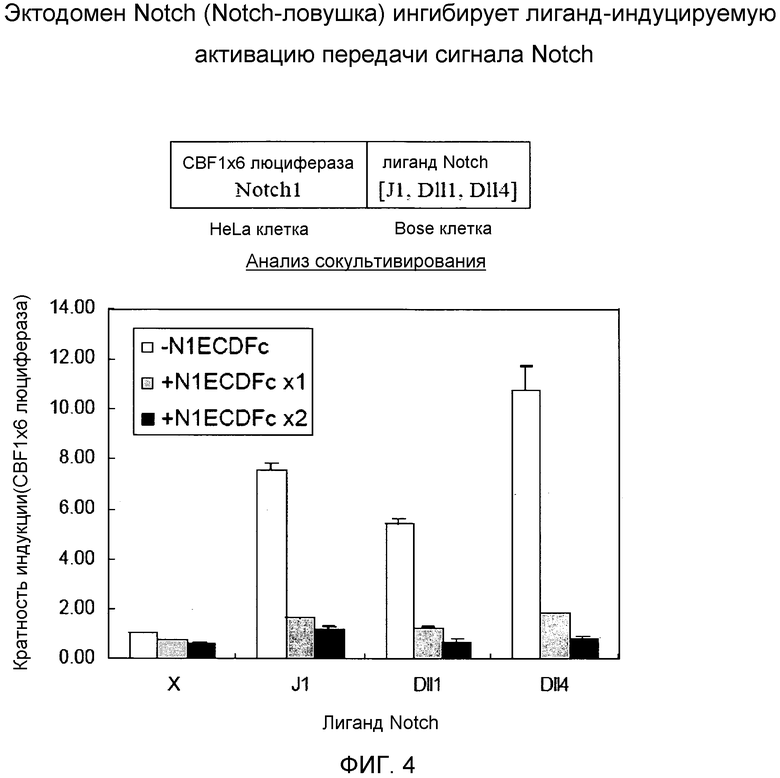
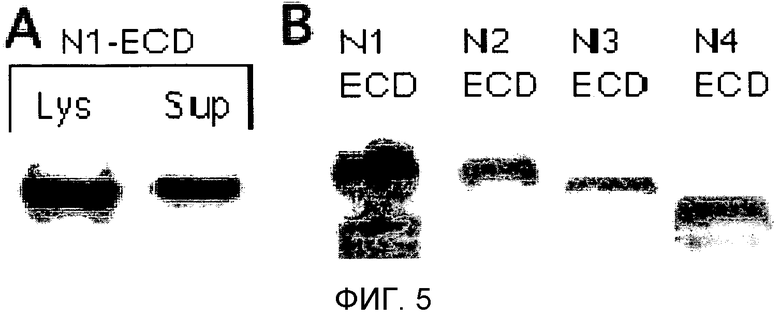
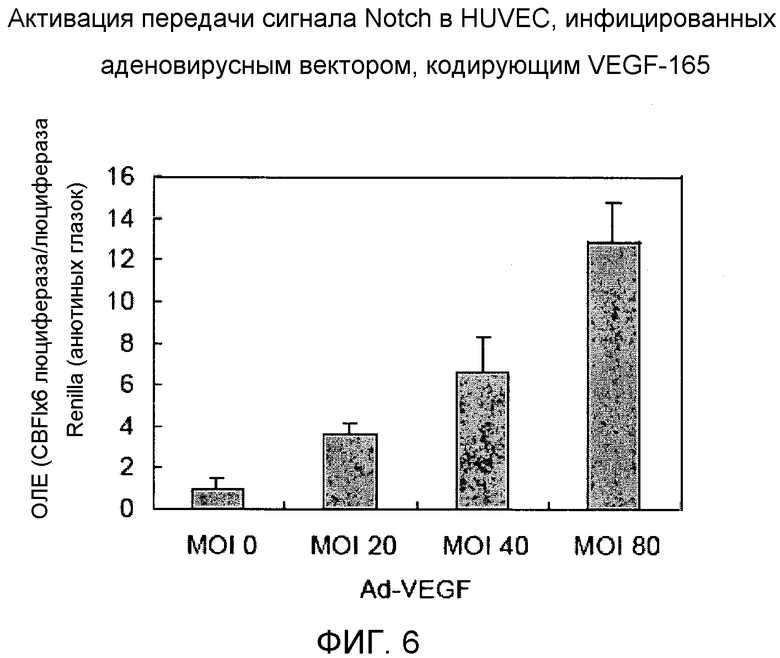
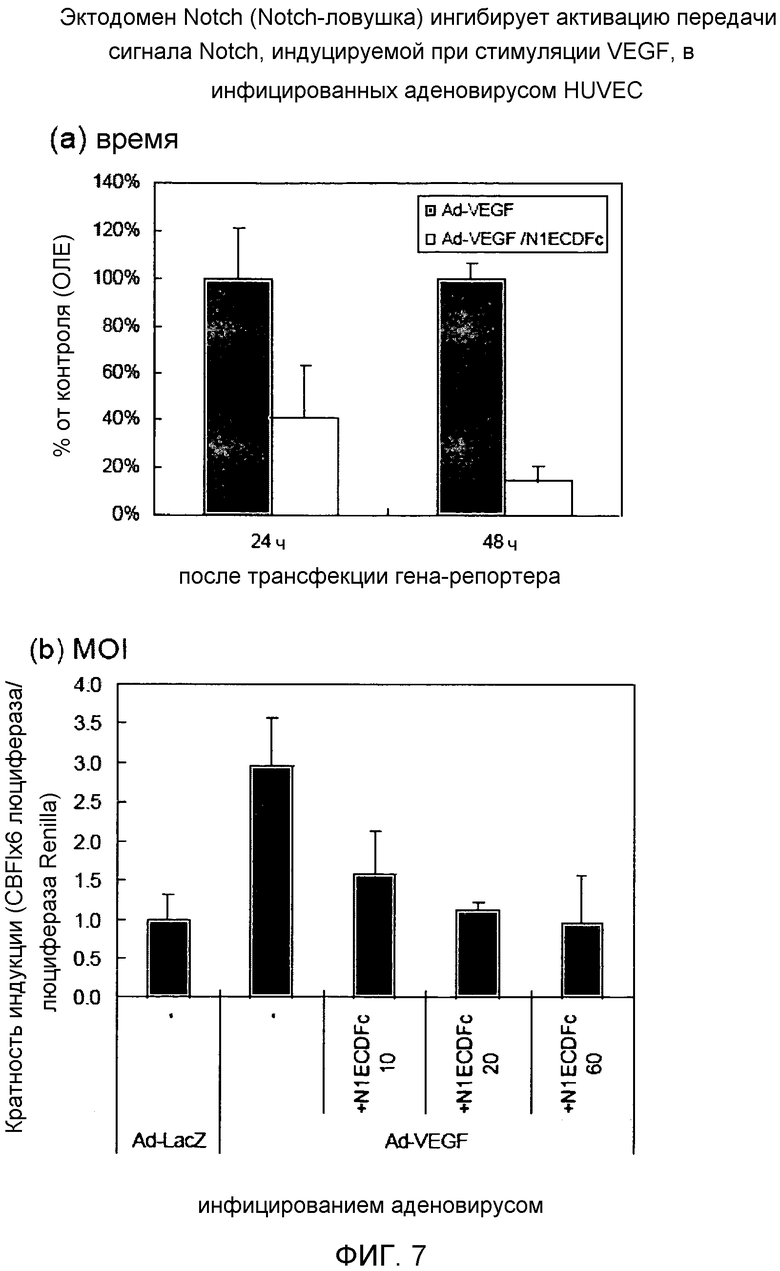

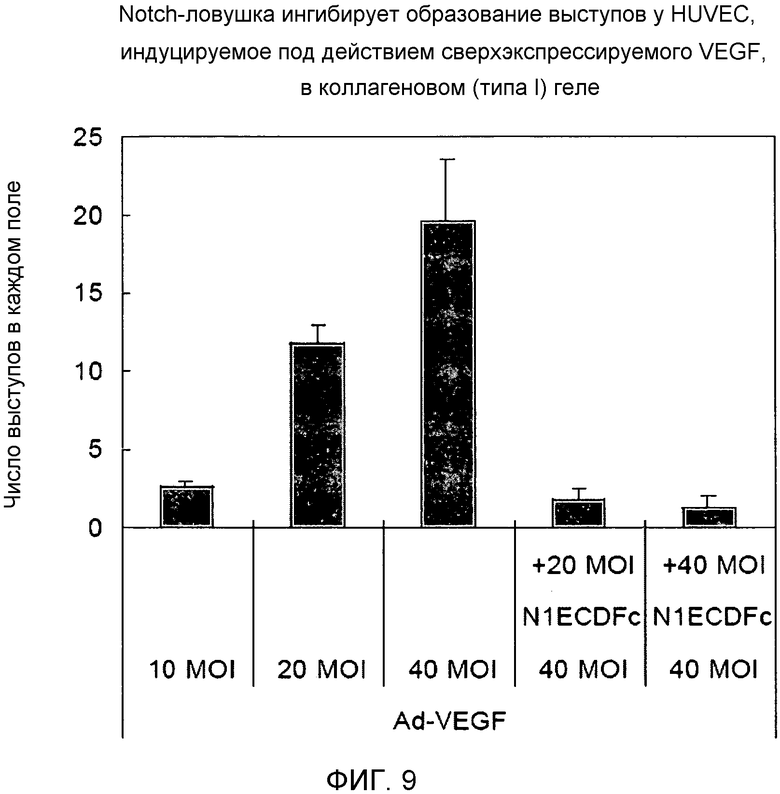













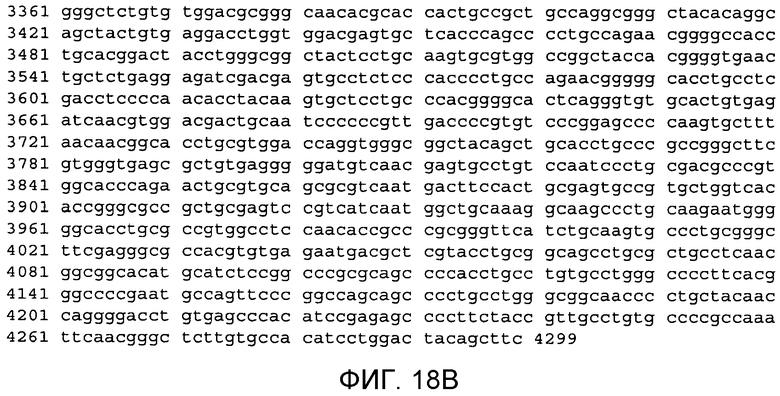





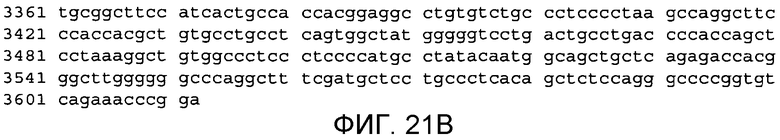

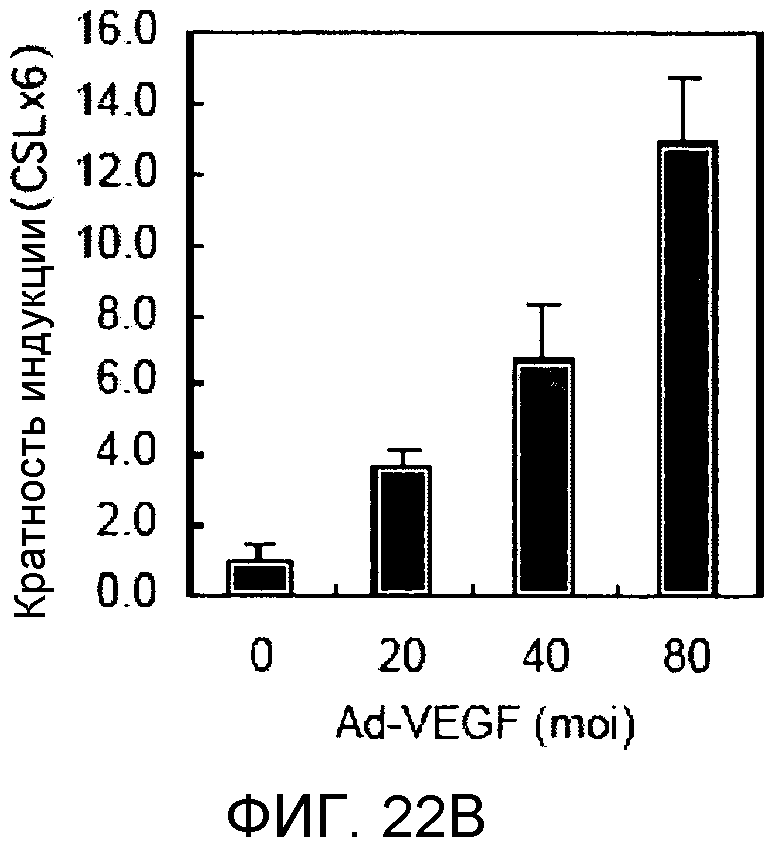
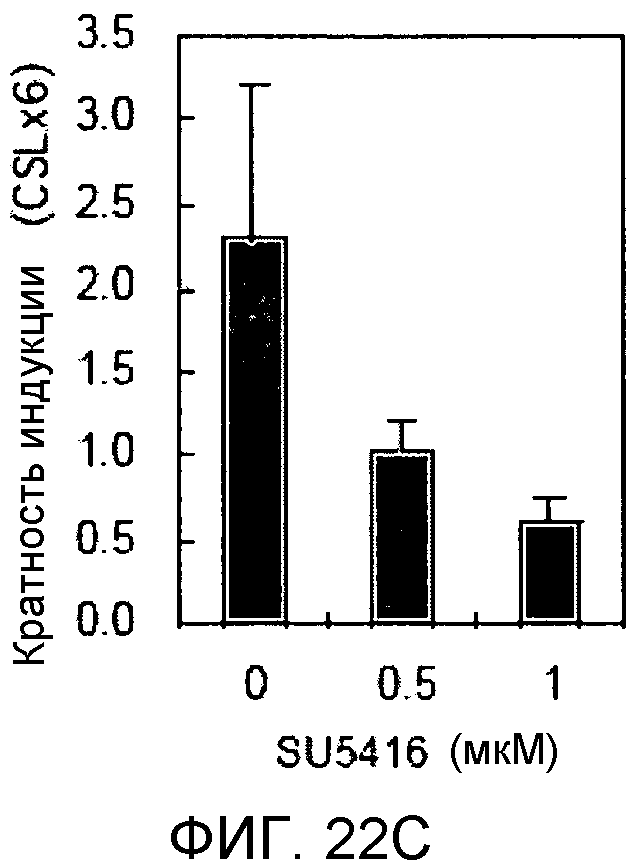
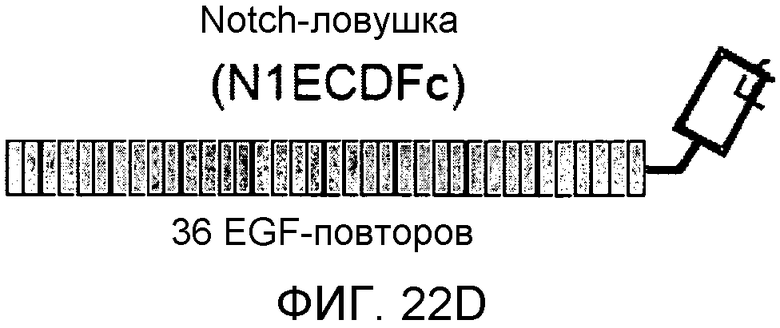

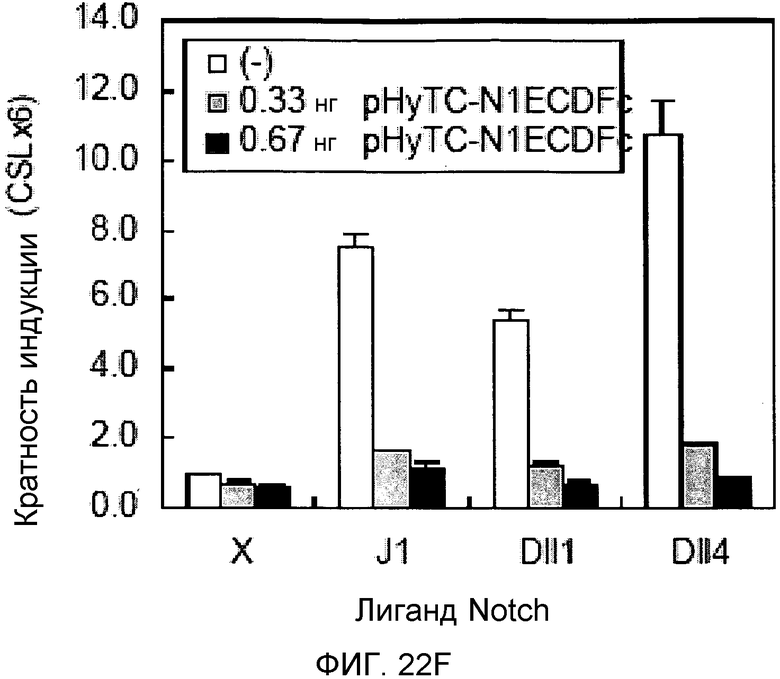
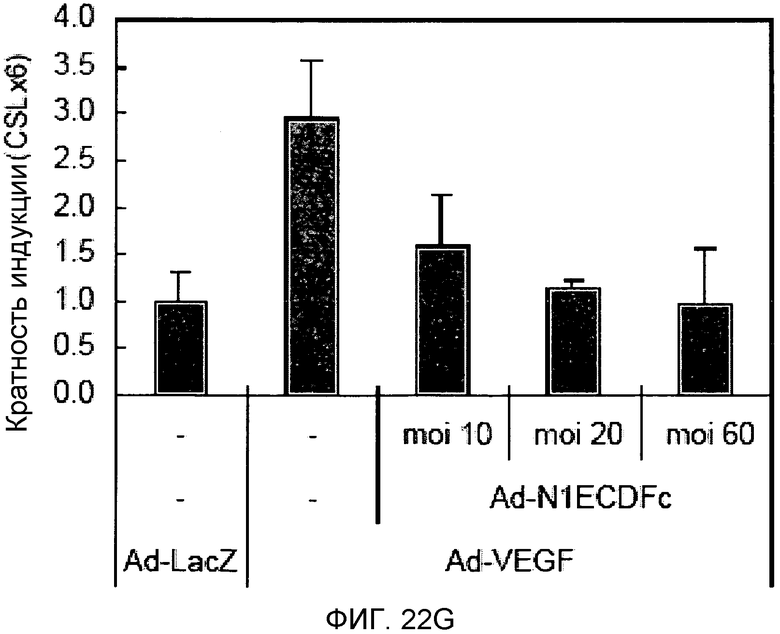
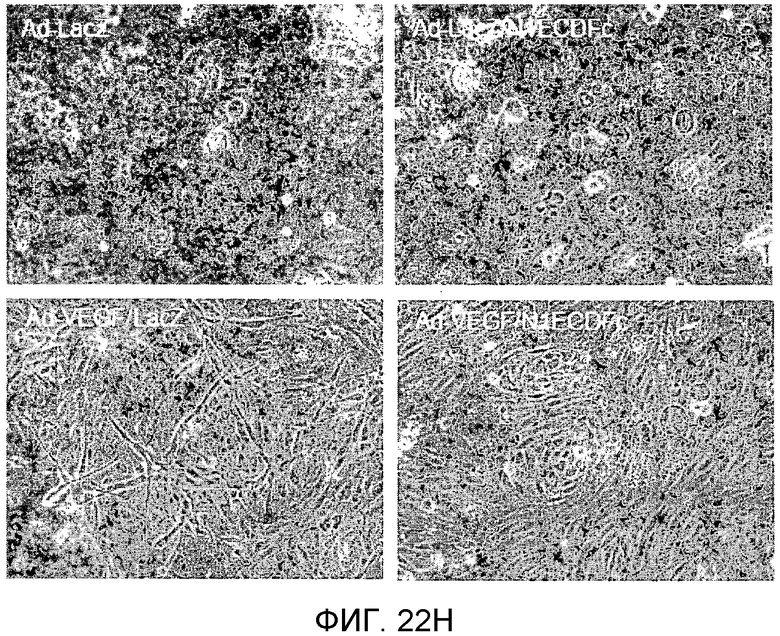
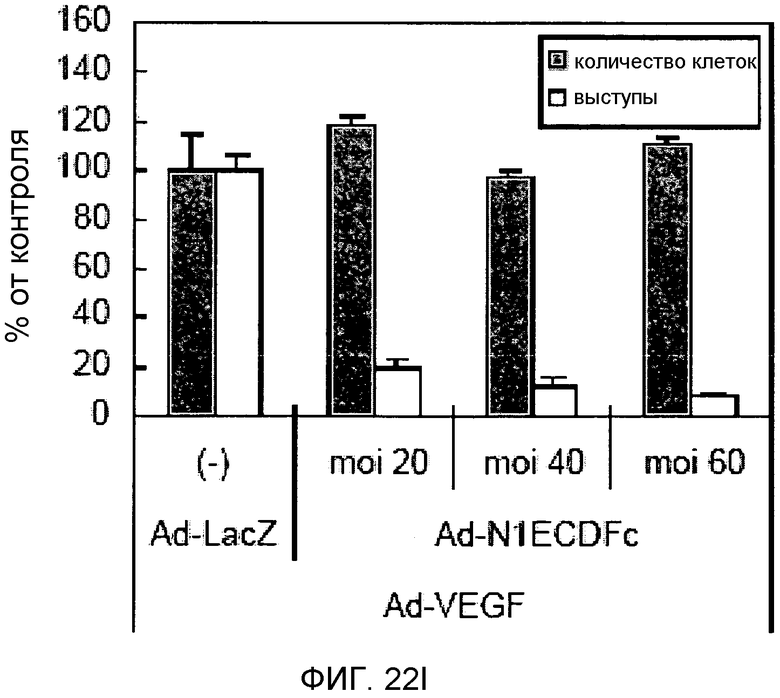
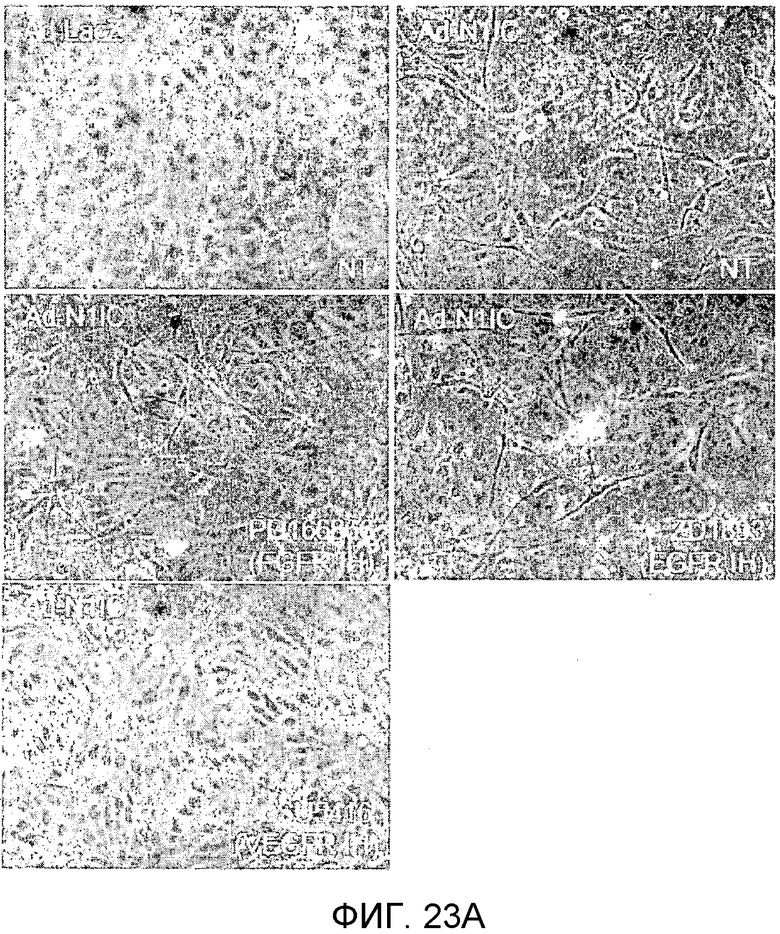

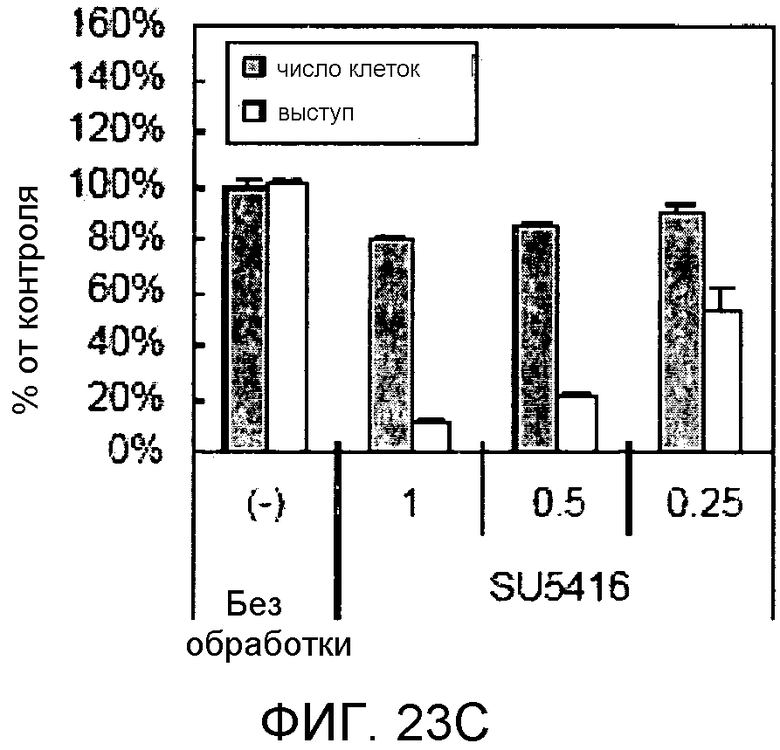
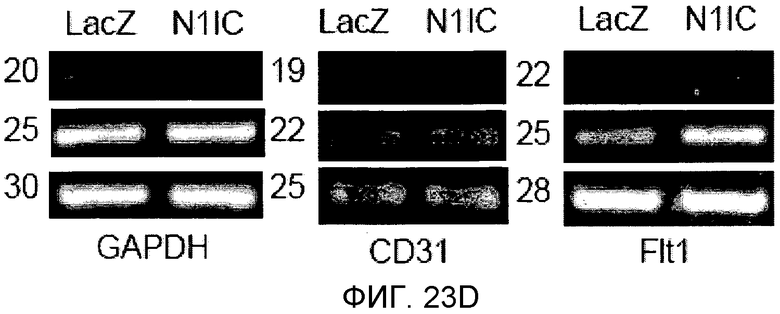

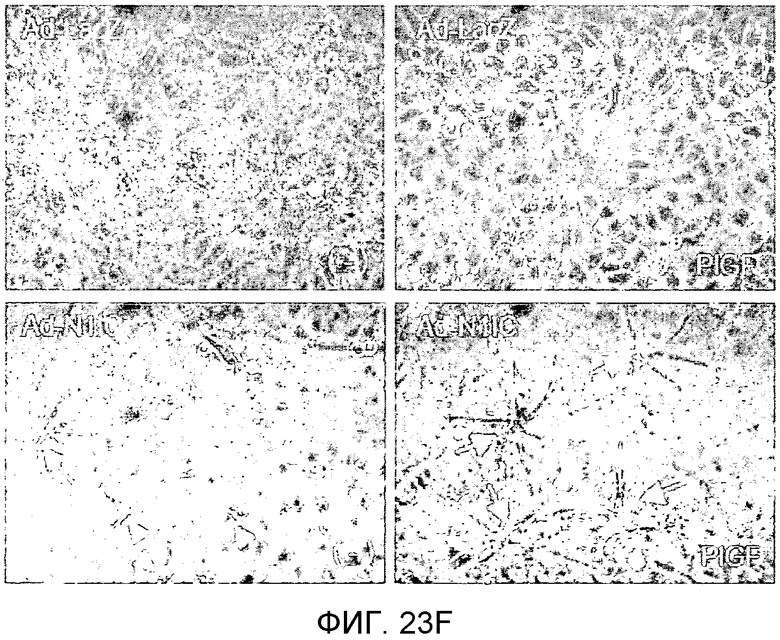
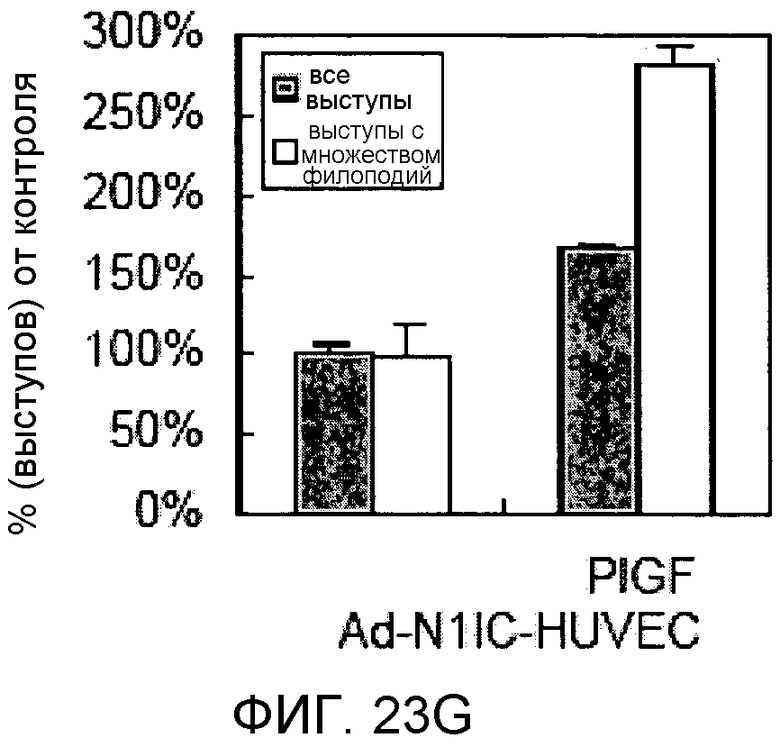
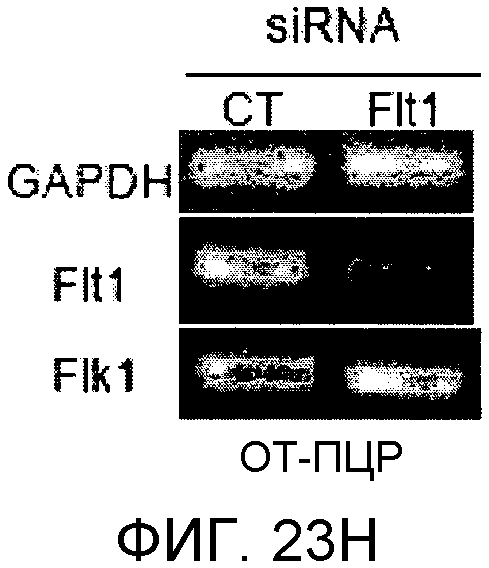
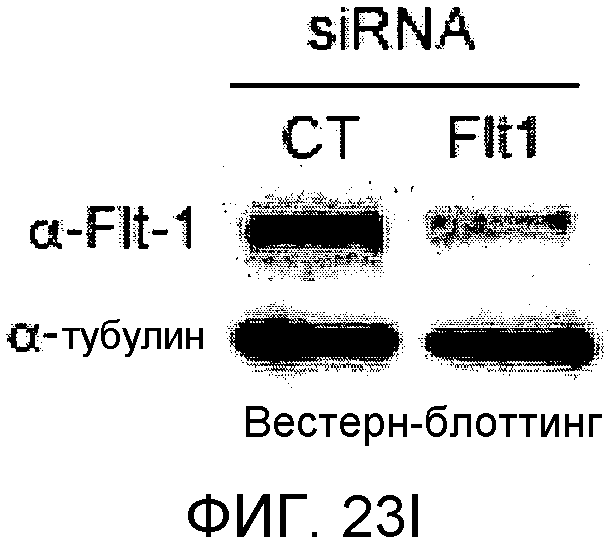
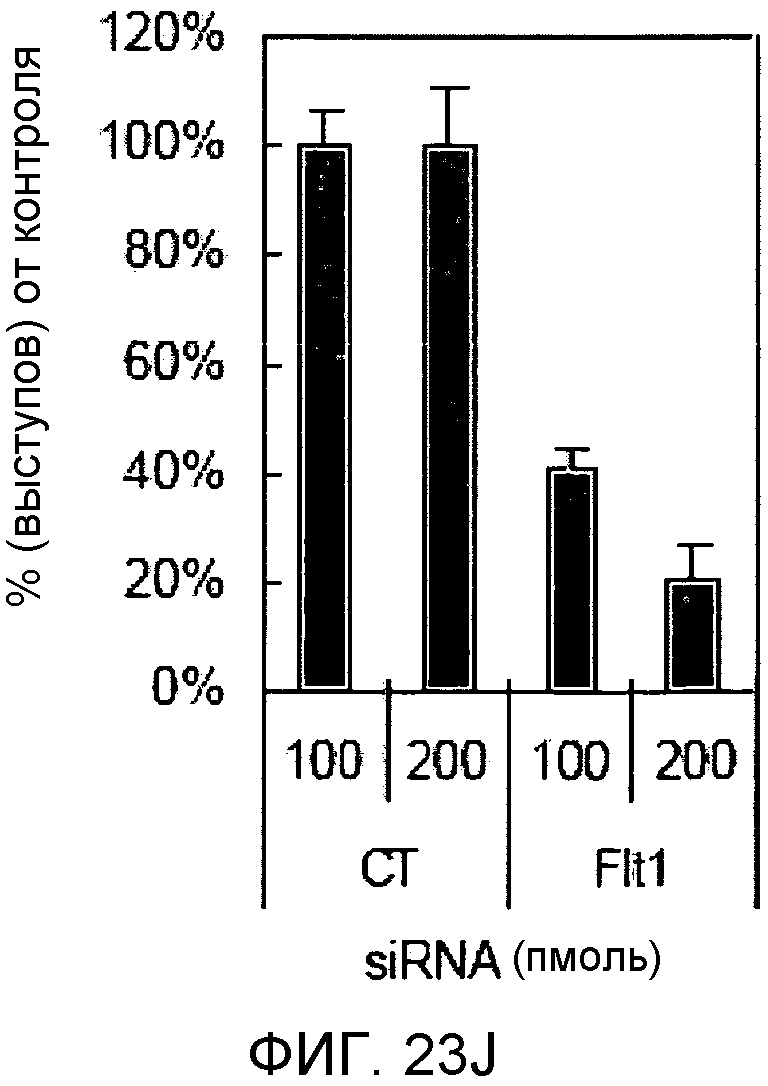

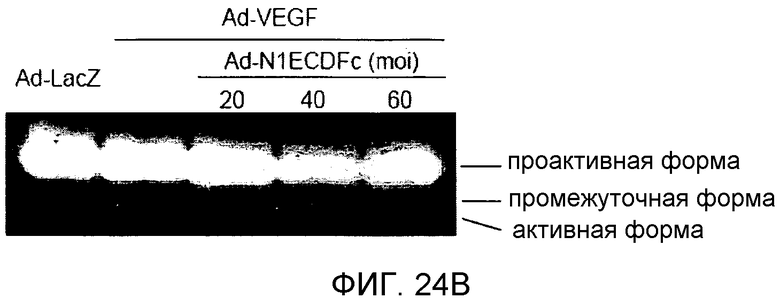
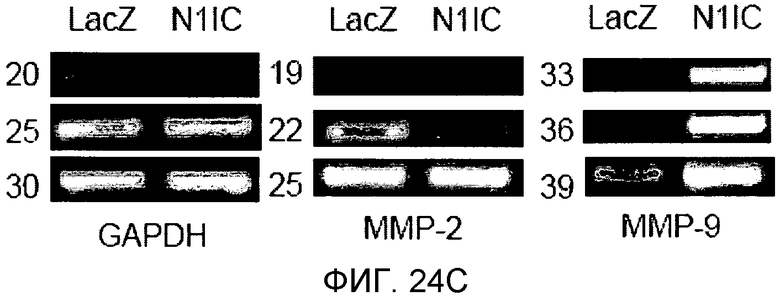
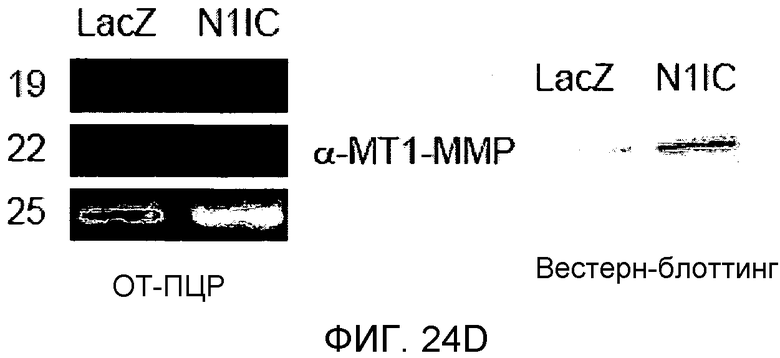

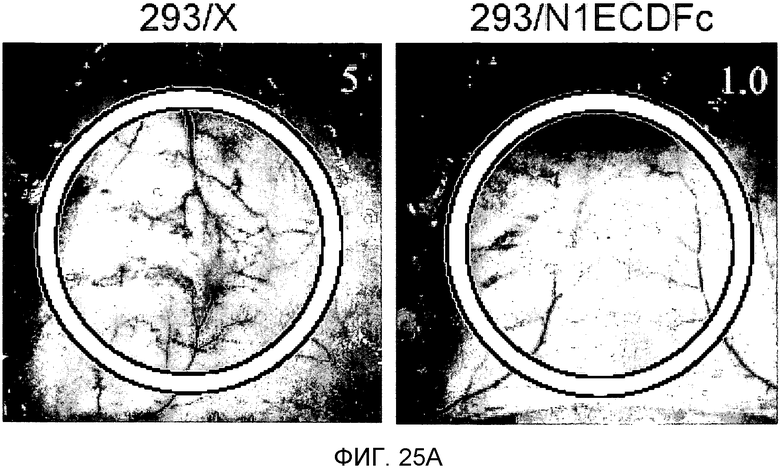
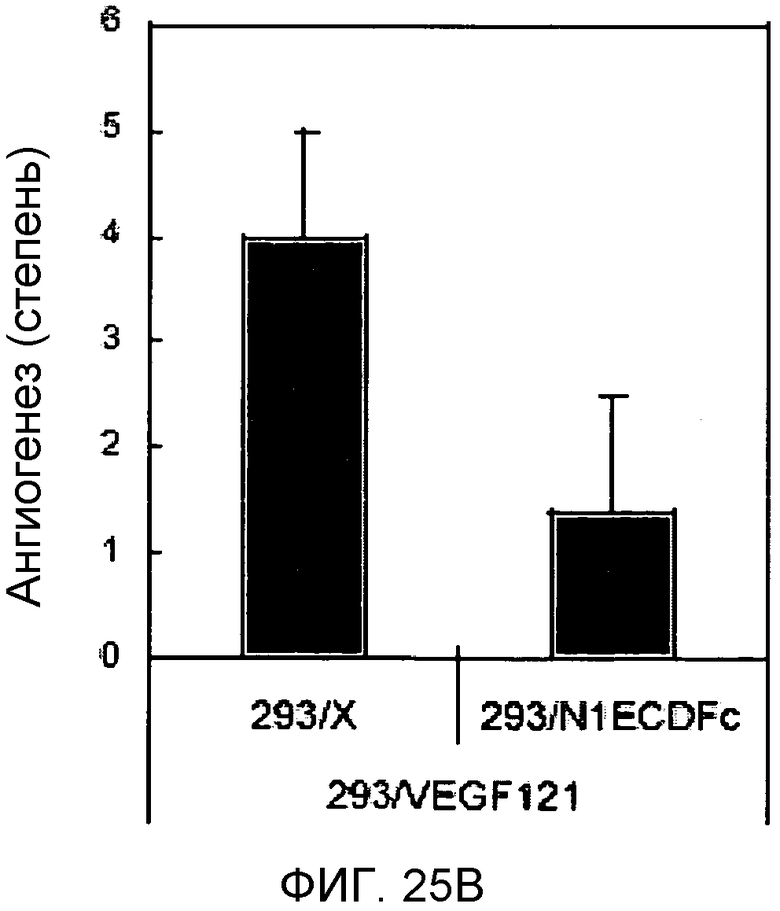
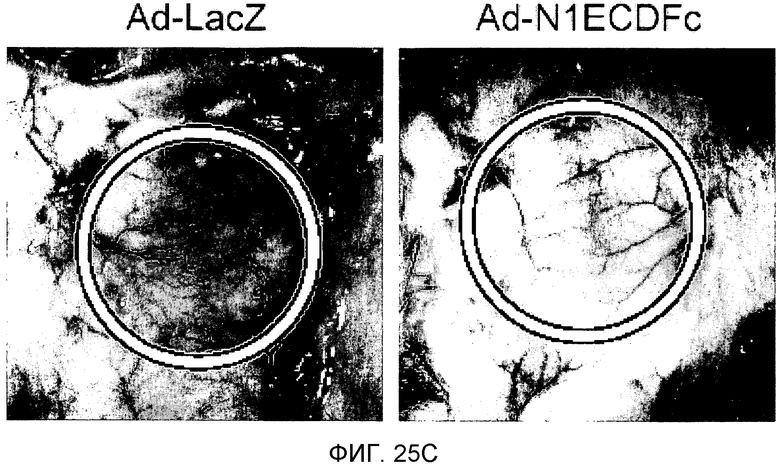

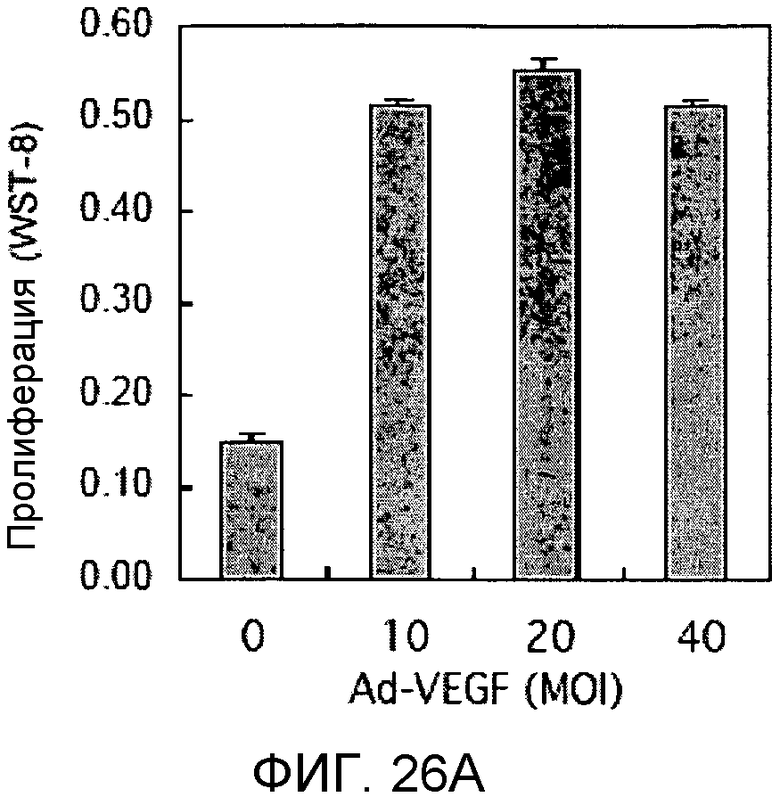

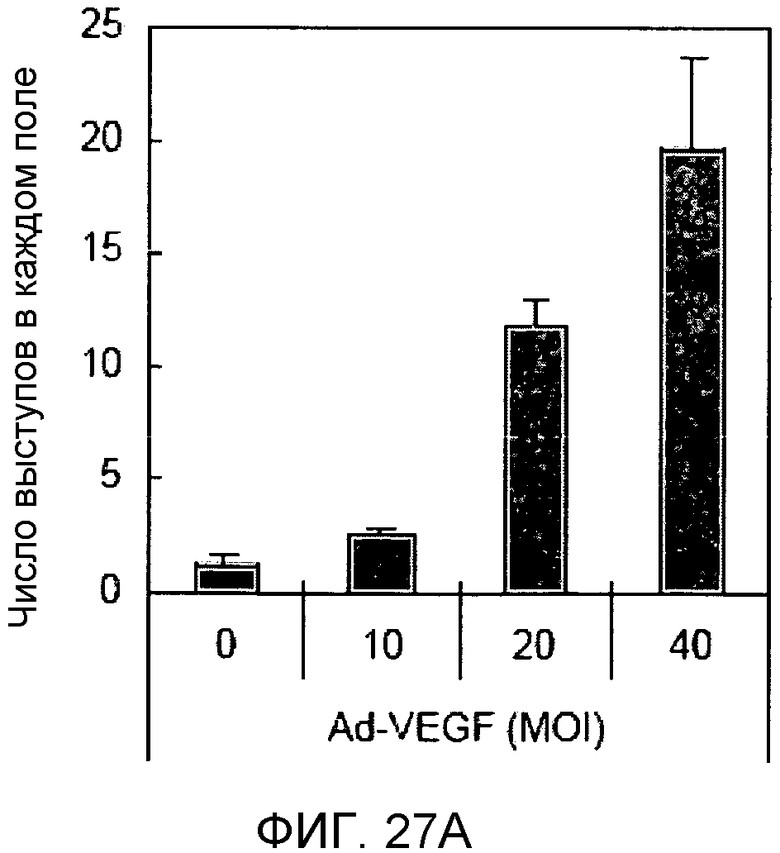
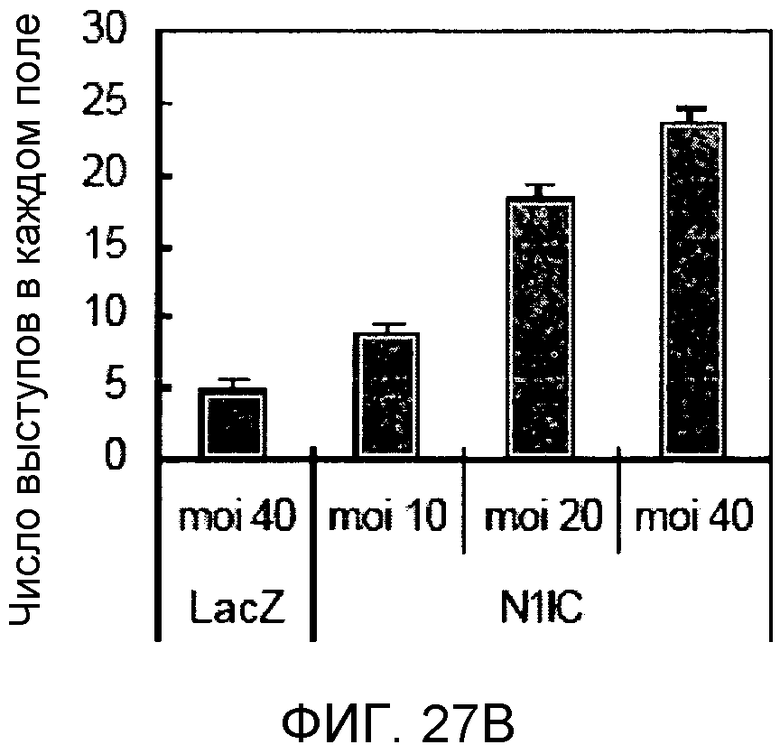
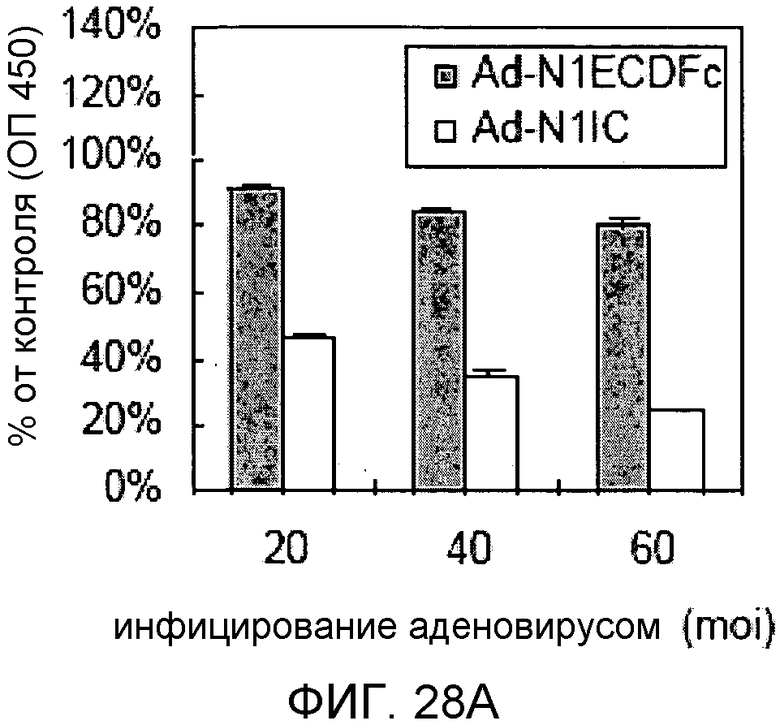
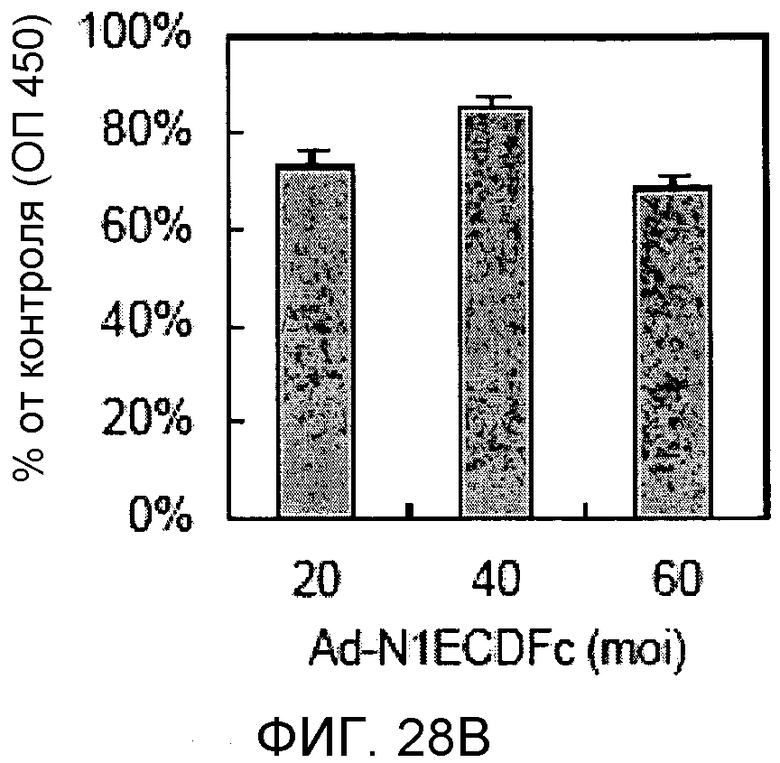

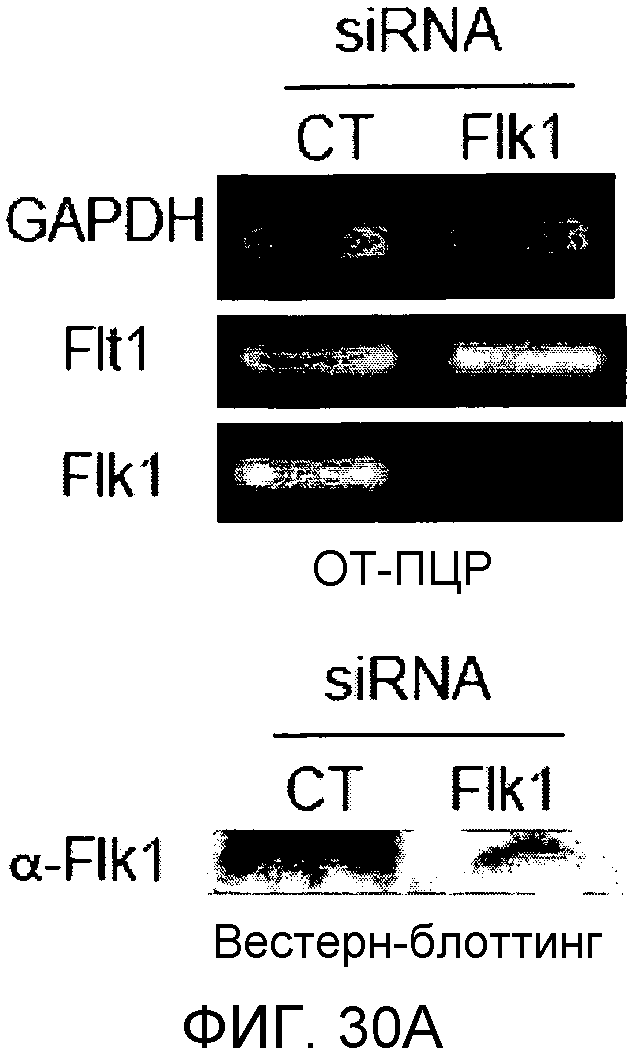
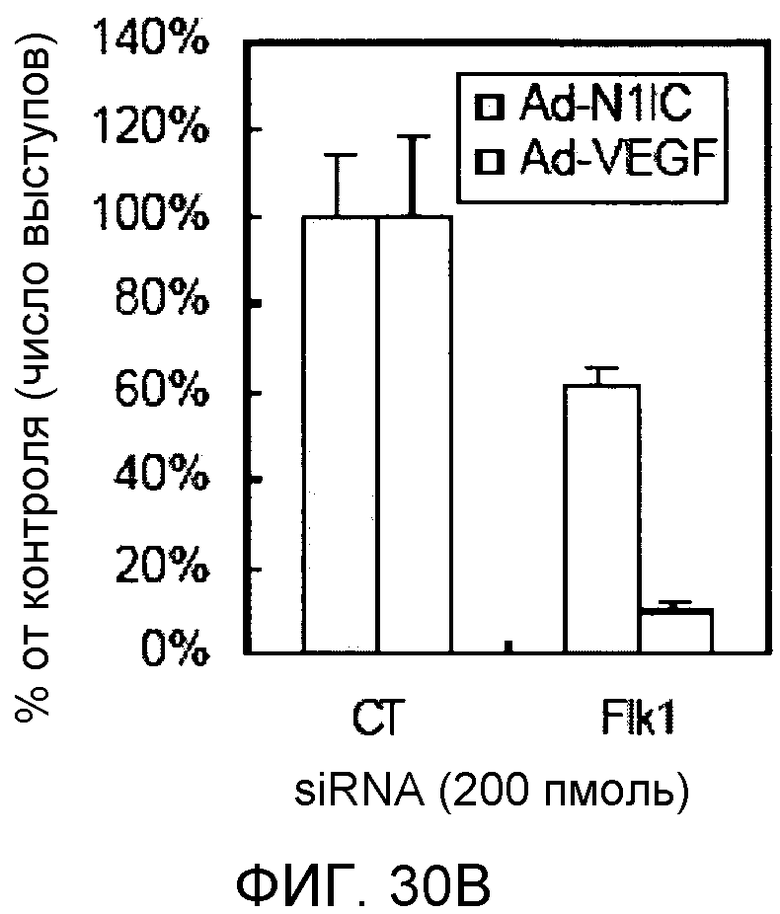
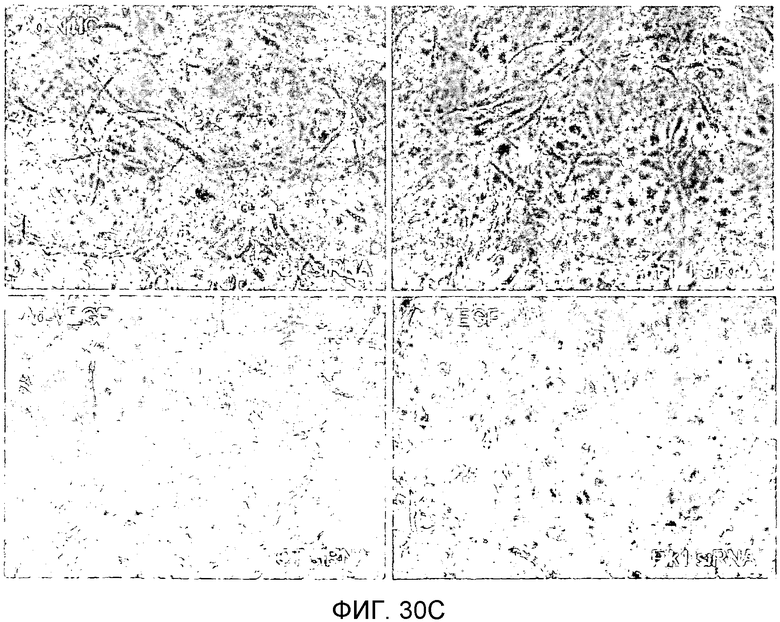




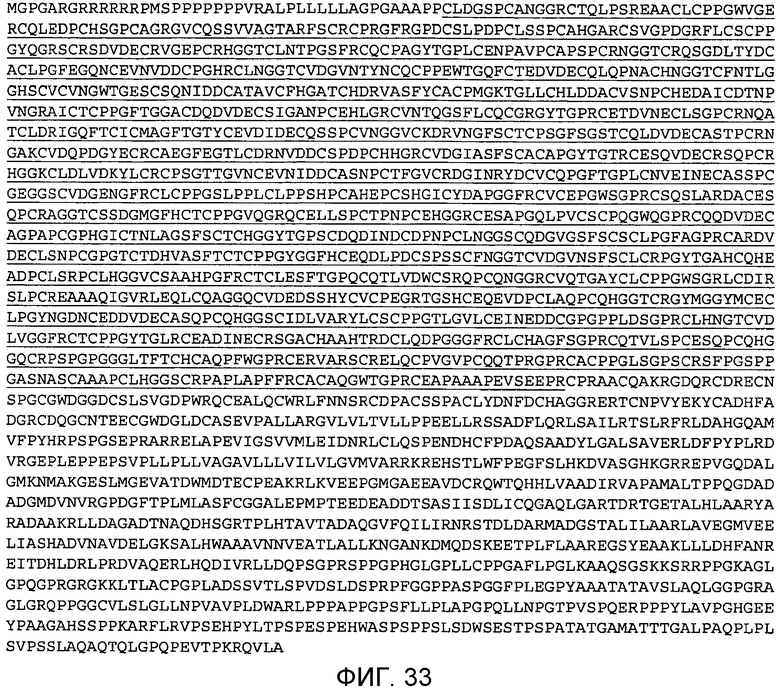
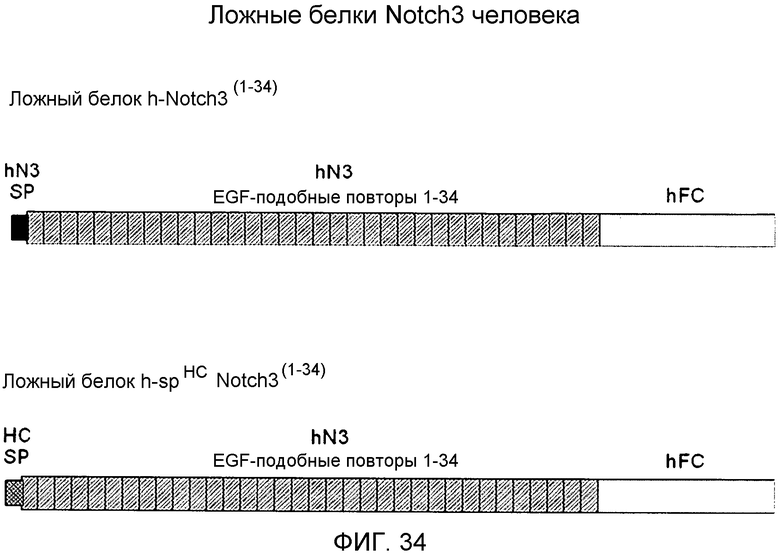



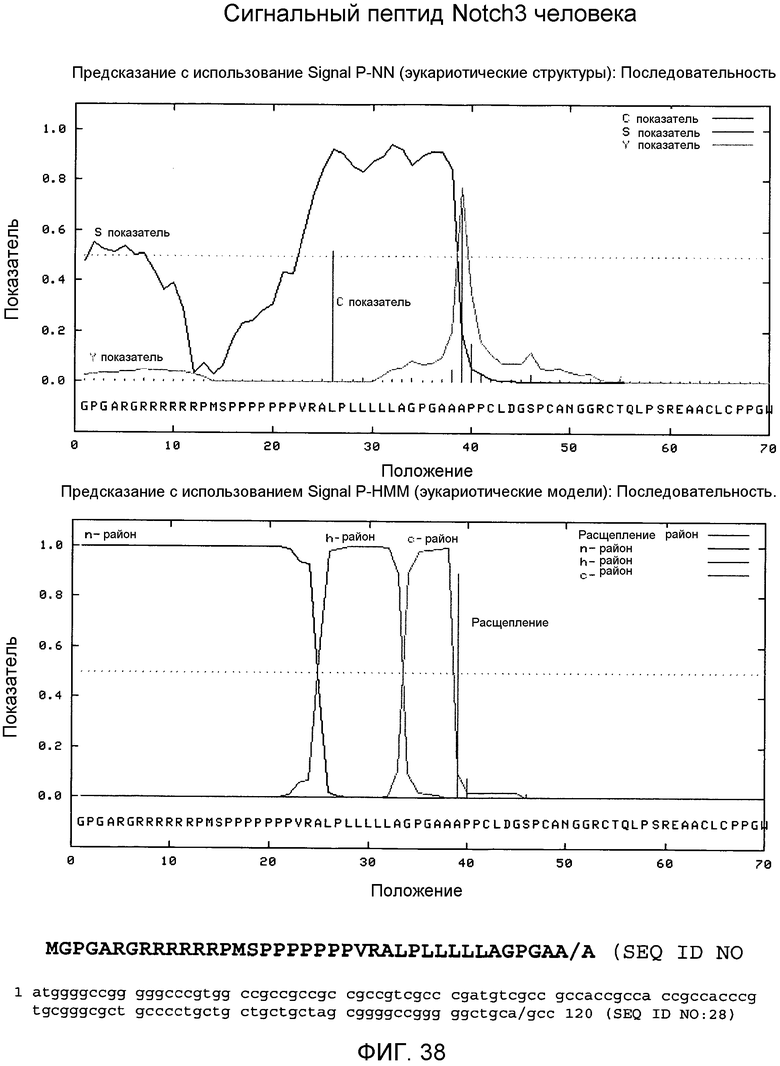
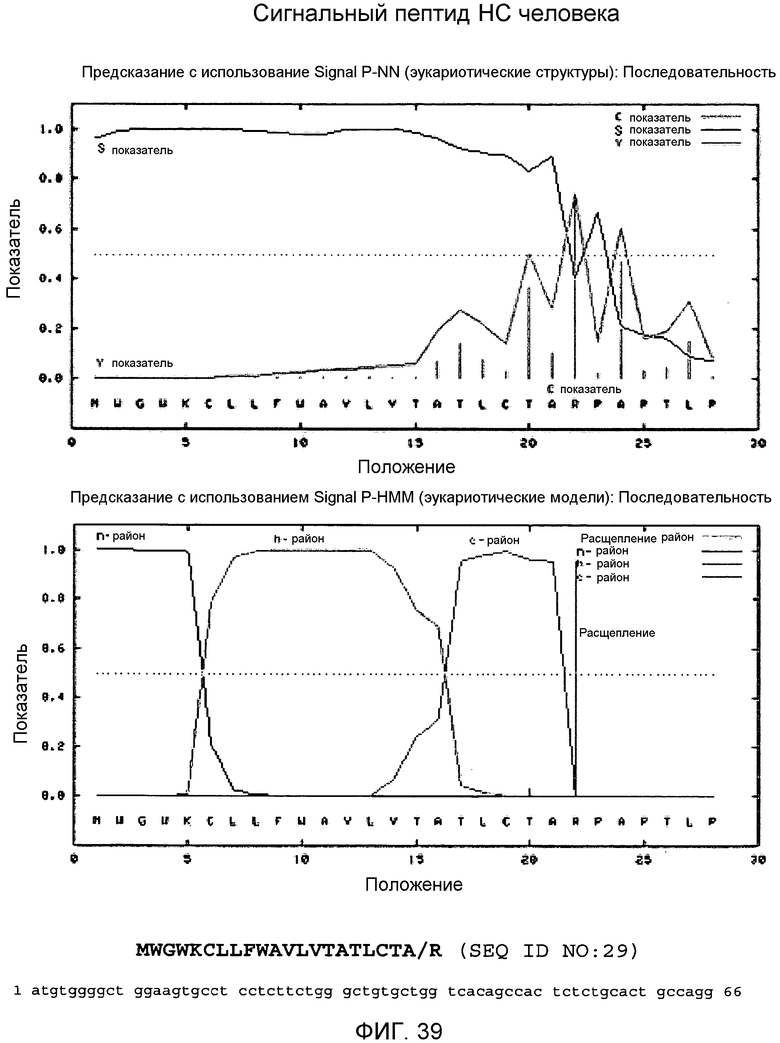


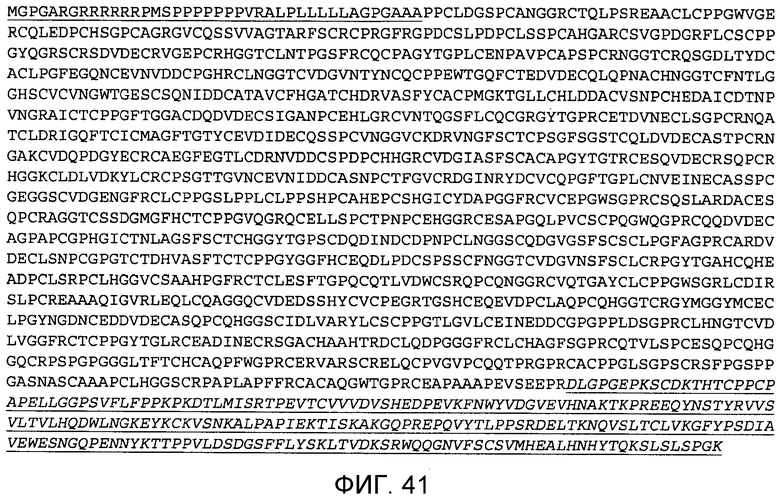



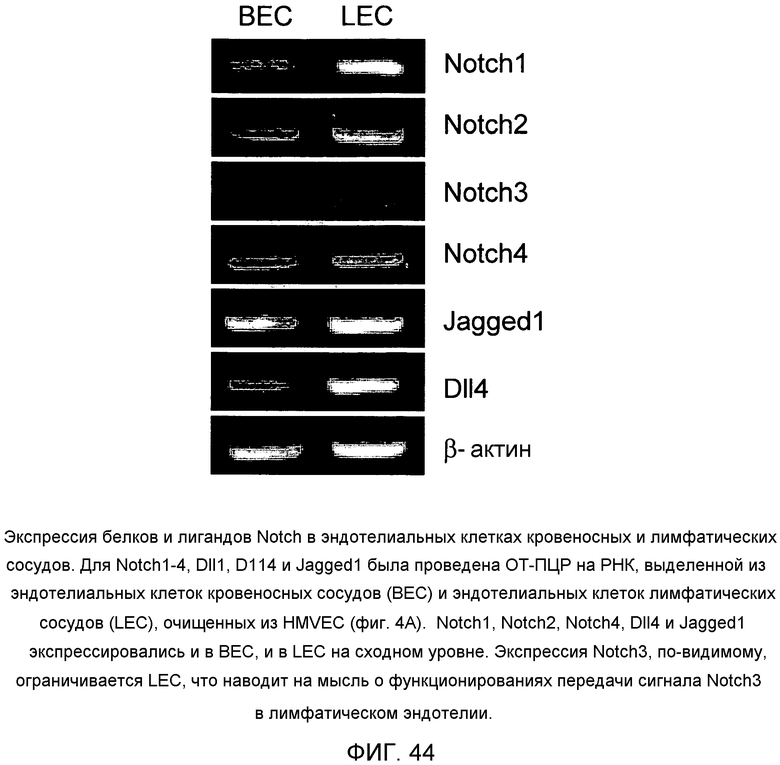
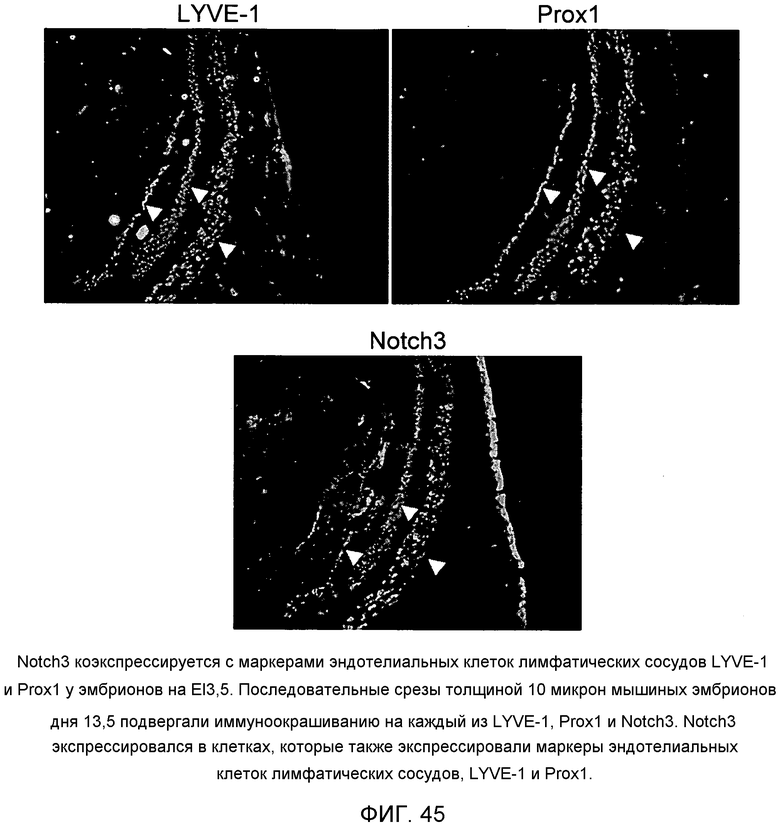
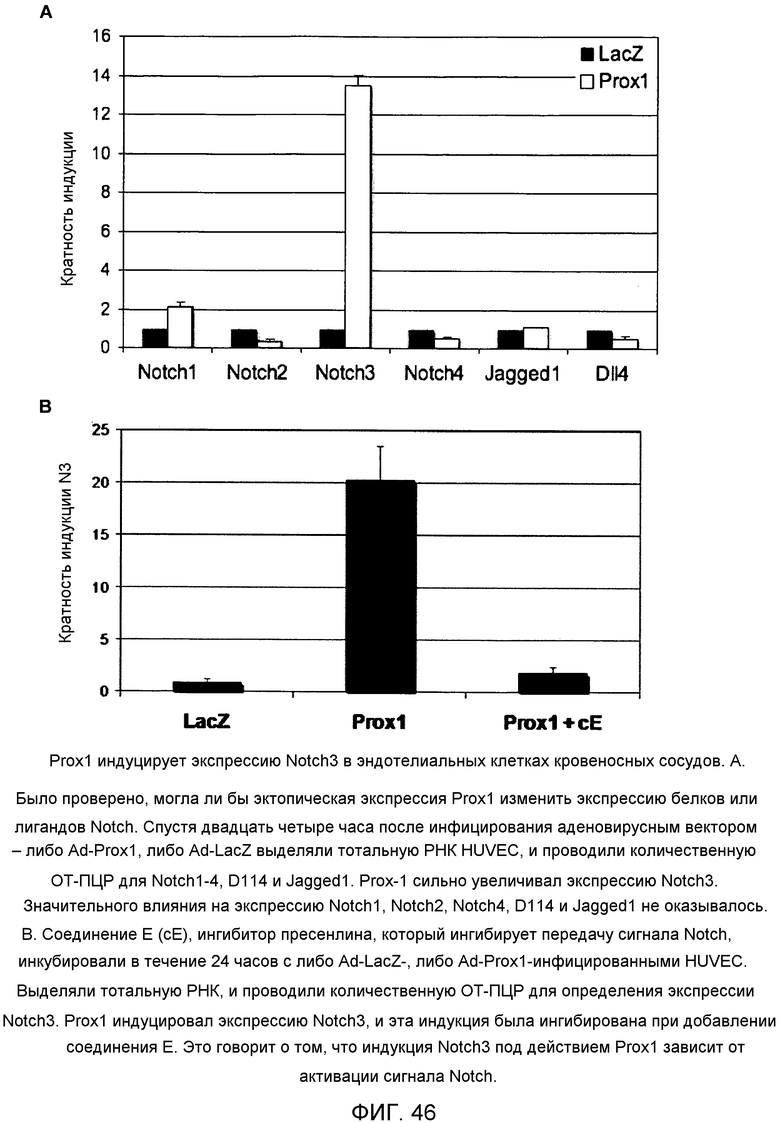
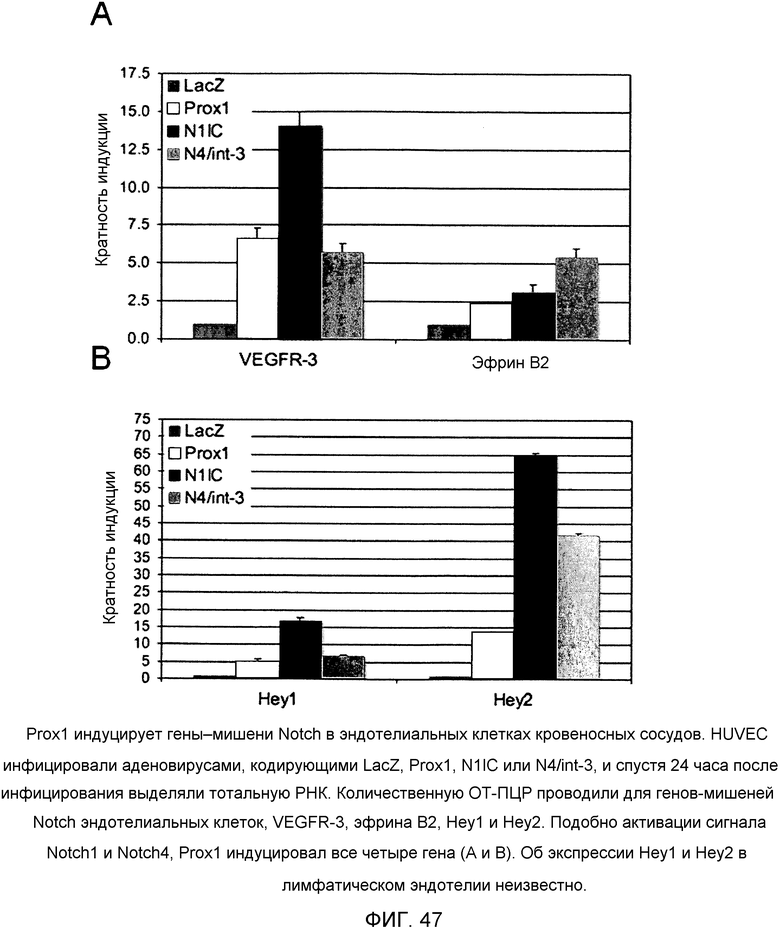
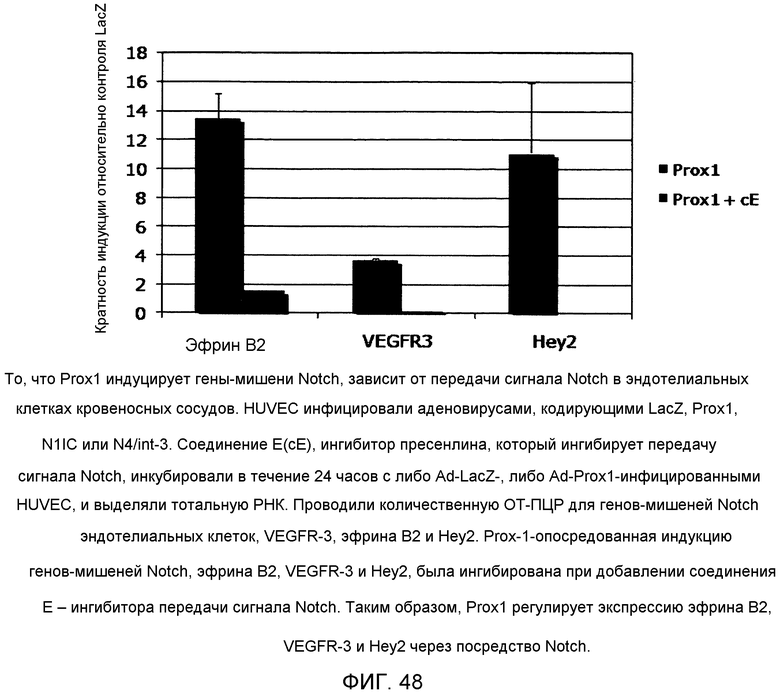
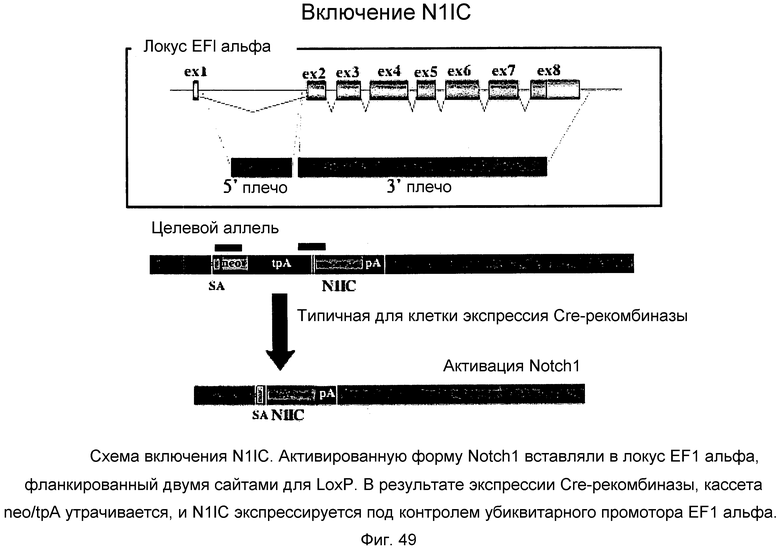

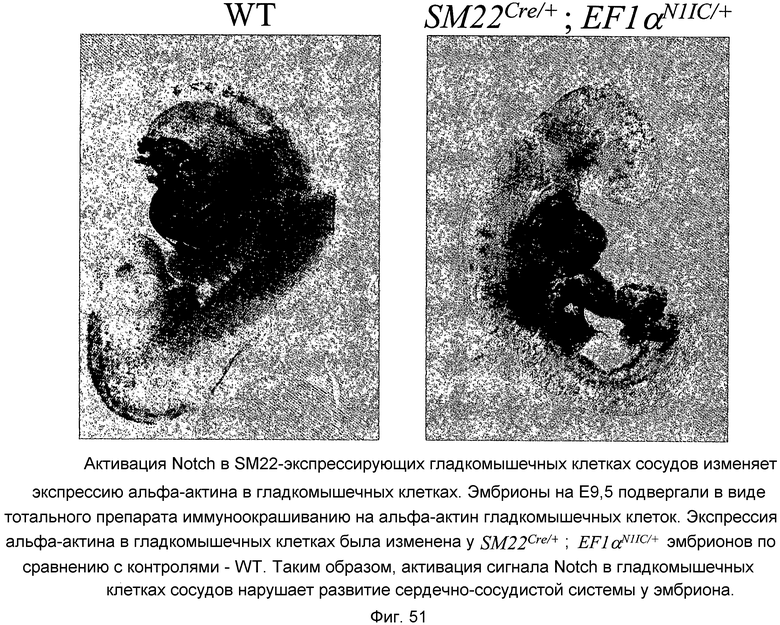
Изобретение относится к области биотехнологии. Описан гибридный белок, включающий сигнальный пептид, EGF-повторы 1-Х внеклеточного домена белка рецептора Notch3 человека, причем X является любым целым числом от 1 до 10 или 12 до 33, и Fc-часть антитела, связанную с ними. Также описаны способ лечения субъекта, имеющего опухоль, способ ингибирования ангиогенеза у субъекта, способ лечения субъекта, имеющего рак яичника, и способ лечения субъекта, имеющего нарушение обмена веществ. Способы включают введение субъекту эффективного для лечения количества указанного выше гибридного белка. Кроме того, раскрывается применение указанного выше гибридного белка для приготовления фармацевтической композиции для лечения имеющего опухоль субъекта, для ингибирования ангиогенеза у субъекта, для лечения имеющего рак яичника субъекта и для лечения имеющего нарушение обмена веществ субъекта. Предложенная группа изобретений может быть использована в медицине. 6 н. и 15 з.п. ф-лы, 93 ил.
1. Гибридный белок антагонист Notch3, последовательность которого (a) идентична последовательности EGF-повторов 1-X внеклеточного домена белка рецептора Notch3 человека, где X является любым целым числом от 1 до 10 или от 12 до 33, за которой следует (b) последовательность, идентичная последовательности Fc-части антитела, причем (b) расположена на карбоксильном конце (а), и где (b) присоединена к (a) либо непосредственно, либо посредством линкера.
2. Гибридный белок антагонист Notch3, последовательность которого (a) идентична последовательности EGF-повторов 1-Х внеклеточного домена белка рецептора Notch3 человека, где X является любым целым числом от 12 до 33, за которой следует (b) последовательность, идентичная последовательности Fc-части антитела, причем (b) расположена на карбоксильном конце (a), и где (b) присоединена к (a) либо непосредственно, либо посредством линкера.
3. Гибридный белок по п. 1 или 2, в котором Fc-частью антитела является Fc-часть антитела человека.
4. Гибридный белок по любому из пп. 1-3, в котором (b) присоединена к (a) посредством линкера.
5. Гибридный белок по любому из пп. 1-3, в котором (b) присоединена непосредственно к (a).
6. Гибридный белок по любому из пп. 1 или 3-5, в котором X является любым целым числом от 12 до 33.
7. Применение гибридного белка по любому из пп. 1-6 для приготовления фармацевтической композиции для лечения субъекта, страдающего нарушением обмена веществ.
8. Применение по п. 7, где нарушением обмена веществ является диабет, ожирение или атеросклероз.
9. Применение по п. 7, где нарушением обмена веществ является ишемия, инсульт или сердечно-сосудистое заболевание.
10. Применение гибридного белка по любому из пп. 1-6 для приготовления фармацевтической композиции для лечения субъекта, страдающего опухолью.
11. Применение по п. 10, где опухоль представляет собой рак яичника.
12. Применение по п. 10, где опухоль представляет собой рак молочной железы.
13. Применение гибридного белка по любому из пп. 1-6 для приготовления фармацевтической композиции для ингибирования ангиогенеза, физиологического лимфангиогенеза, патологического лимфангиогенеза, метастазирования опухоли, роста вторичной опухоли или использования опухолью для своих собственных целей кровеносных сосудов.
14. Применение гибридного белка по любому из пп. 1-6 для приготовления фармацевтической композиции для лечения субъекта, страдающего злокачественным новообразованием, где гибридный белок вводят в дополнение к (i) ингибитору фактора роста сосудистого эндотелия (VEGF), (ii) ингибитору рецептора VEGF, (iii) ингибитору PDGF, (iv) антагонисту рецептора PDGF или (v) ингибитору HER2/neu.
15. Применение по п. 14, где гибридный белок вводят в дополнение к ингибитору VEGF, и ингибитором VEGF является ингибитор VEGF-A, PGIF, VEGF-B, VEGF-C или VEGF-D.
16. Применение по п. 14, где гибридный белок вводят в дополнение к ингибитору рецептора VEGF, и ингибитором рецептора VEGF является ингибитор VEGFR-1 или ингибитор VEGFR-2.
17. Применение по п. 14, где гибридный белок вводят в дополнение к ингибитору PDGF.
18. Применение по п. 14 или 17, где ингибитором PDGF является ингибитор PDGF-A или ингибитор PDGF-B.
19. Применение по п. 14, где гибридный белок вводят в дополнение к антагонисту рецептора PDGF.
20. Применение по п. 14 или 19, где антагонистом рецептора PDGF является антагонист рецептора-B PDGF.
21. Применение по п. 14, где гибридный белок вводят в дополнение к ингибитору HER2/neu.
| Способ очистки нефти и нефтепродуктов от сероводорода | 1988 |
|
SU1616958A1 |

