ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
В соответствии с 35 USC § 119 U.S. по настоящей заявке испрашивается приоритет предварительной заявки № 61/364210, поданной 14 июля 2011.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к моноклональным антителам, которые могут использоваться при лечении болезни Альцгеймера. Изобретение также относится к композициям, содержащим моноклональные антитела, и к способам применения композиций в качестве биомаркеров или для диагностики и лечения заболеваний, связанных с бета-амилоидом (Αβ) и диффундирующими лигандами, происходящими из Αβ (ADDL).
УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
Болезнь Альцгеймера (AD) характеризуется прогрессирующей потерей когнитивной функции и накоплением бляшек бета-амилоида (Αβ) в областях, связанных с обучением и запоминанием. Несмотря на то, что одно время считали, что бляшки Αβ играют главную роль в патогенезе AD, растущее число доказательств, позволяет предположить, что диффундирующие лиганды, происходящие из Αβ (ADDL), могут быть ответственны за связанную с заболеванием нейрональную дисфункцию и снижение когнитивных способностей (Walsh and Selkoe, 2004, Protein Pept. Lett., 11:213-228). ADDL представляют собой небольшие, растворимые олигомеры Αβ, которые присутствуют в большом количестве при AD, но не в образцах нормального мозга (McLean et al., 1999, Ann. Neurol., 46: 860-866; Gong et al., 2003, Proc, Natl. Acad. Sci. USA, 100: 10417-10422). Исследования in vitro показали, что ADDL, выделенные из мозга при AD, или синтетические препараты связываются с субпопуляцией кортикальных и гиппокампальных нейронов (Gong et al., 2003; Klein et al., 2004, Neurobiol. Aging, 25: 569-580; Lacor et al., 2004, J. Neurosci., 24: 10191-10200; Shughrue et al., 2010, Neurobiol. Aging, 31:189-202), в то время как обнаруживали незначительное связывание или отсутствие связывания с препаратами фибриллярного, или мономерного Αβ (Lacor et al., 2004; Hepler et al., 2006, Biochemistry, 45: 15157-15167). Кроме того, связывание ADDL с нейронами можно уменьшить с помощью поликлональных (Gong et al., 2003) и моноклональных антител (Lee et al., 2006, J. Biol. Chem., 281:4292-4299; De Felice et al., 2007, Neurobiol. Aging 29: 1334-1347; Shughrue et al., 2010), вызванных против ADDL.
В моделях на грызунах центральное введение ADDL вызывает нарушения у грызуна долговременной потенциации (LTP) и формирования памяти (Walsh et al., 2002, Nature, 416: 535-539; Cleary et al., 2004, Nat. Neurosci., 8: 79-84; Klyubin et al., 2005, Nat. Med., 11: 556-561). Действие олигомеров на LTP ослаблялось, если ADDL вводили совместно с антителом против Αβ или вводили животным, которые были вакцинированы Aβ-пептидом (Rowan et al., 2004, Exp, Gerontol,, 39: 1661-1667). В трансгенной модели AD, такой как трансгенные мыши, которые продуцируют белок-предшественник амилоида (hAPP) человека, наблюдали возрастные когнитивные нарушения при наличии повышенных уровней ADDL (Westerman et al., 2002, J. Neurosci,, 22: 1858-1867; Ashe, 2005, Biochem, Soc. Trans., 33: 591-594; Lee et al., 2006; Lesne et al., 2006, Nature, 440: 352-357). При лечении мышей hAPP антителом против ADDL, наблюдали существенное улучшение когнитивной деятельности без сопутствующего уменьшения количества бляшек Αβ (Lee et al., 2006). Полученные результаты в совокупности позволяют предположить, что ADDL, но не бляшки Αβ, главным образом ответственны за когнитивное нарушение и что применение антител против ADDL может оказаться эффективным в лечении AD. См. также патенты США US 2006/0228349; US 7731962, международный патент WO 2007/050359; патент США US 2007/0218499, международный патент WO 2006/014478; патенты США US 7700099; US 2008/01758835, международный патент WO 2006/055178.
Таким образом, существует потребность в ADDL-селективных терапевтических антителах для профилактики и лечения AD. Настоящее изобретение отвечает указанным потребностям.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к выделенному антителу или его фрагменту, способному к дифференциальному распознаванию многомерной конформации одного или более диффундирующих лигандов (ADDL), происходящих из β-амилоида, для лечения заболеваний, связанных с ADDL, таких как болезнь Альцгеймера (AD). Настоящее изобретение также относится к фармацевтическим композициям, содержащим выделенное антитело по изобретению, либо самостоятельно, либо в сочетании с одним или более терапевтически активными средствами, носителями или разбавителями.
Настоящее изобретение также относится к способам применения выделенного антитела, таким как способы обнаружения ADDL в образце, ингибирования сборки ADDL, выявления терапевтических средств, которые предотвращают связывание ADDL с нейронами и ослабление симптомов заболевания, связанного с ADDL, и в качестве биомаркера, для применения в диагностике заболевания, связанного с ADDL или для обнаружения ADDL в образце.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1 является графическим представлением анализа связывания ELISA панели гуманизированных (h3B3) и аффинно зрелых антител против ADDL (14.2, 7.2, 11.4, 9.2, 13.1, 17.1 и 19.3) и трех антител сравнения (Соед. 1, 2 и 3) с мономерным Αβ, ADDL и фибриллярным Αβ. Фоновый уровень в данном тесте определяли путем удаления захватывающего антитела из анализа ELISA (не mAb). Планки погрешностей представляют стандартную ошибку среднего.
Фигура 2 является графическим представлением анализа связывания ELISA антитела против ADDL 19.3 и антитела 3B3 с ADDL или с мономером Αβ (Αβ1-40), оцениваемым с помощью 11-точечной титрационной кривой.
Фигура 3 является графическим представлением способности антитела против ADDL 19.3 и 3B3 блокировать связывание ADDL с гиппокампальными нейронами в первичной культуре после предварительной инкубации с возрастающей концентрацией антитела. Способность антитела против ADDL 19.3 блокировать связывание ADDL с нейронами уменьшалась после тепловой денатурации антитела. Планки погрешностей представляют стандартную ошибку среднего.
Фигуры 4A-4C представляют собой графические представления анализа связывания ELISA с ADDL антитела против ADDL 19.3 (обозначаемого как WT на фигуре 4A) и двух антител против ADDL, происходящих из 19.3 (фигуры 4B и 4C), после инкубации в течение одного месяца при различных температурах для оценки стабильности антитела. Антитела против ADDL, полученные из 19.3, содержали единичную аминокислотную замену Asn33 в CDR1 легкой цепи на Ser33 (19.3S33) или Thr33 (19.3T33) (SEQ ID NO:55 и 56, соответственно). Замена Asn33 на S33 (фигура 4B) или T33 (фигура 4C) приводила к увеличению стабильности антитела, по сравнению с исходным антителом 19.3.
Фигура 5 является графическим представлением связывания и диссоциации антител против ADDL с иммобилизованным FcRn человека, которые оценивали с помощью BiacoreTM (GE Healthcare, Piscataway, NJ). Уточненная сенсограмма показывает начальное связывание при pH 6,0 и последующую диссоциацию антител при pH 7,3 от 180 секунд. Точку отчета (состояние равновесия) устанавливали на момент времени 5 секунд после окончания связывания при pH 6,0 и значение "% связывания" рассчитывали как
RUравновесие/RUсвязывание(%).
На фигуре 6A показано выравнивание вариабельных областей тяжелой и легкой цепей антитела против ADDL 19.3 с зародышевой линии человека с определяющими комплементарность областями (CDR), указанными жирным шрифтом. Фигура 6B представляет собой трехмерную модель вариабельных областей тяжелой и легкой цепей антитела 19.3, показывающую расположение CDR.
Фигура 7 является графическим представлением фармакокинетического профиля (PK) антител против ADDL 19.3 и 3B3, оцененным у гетерозиготных мышей 276 с FcRn человека (Jackson Laboratory (Bar Harbor, ME) после однократного внутривенного (IV) введения 10 мг/кг. Концентрацию антитела измеряли в различные интервалы времени, чтобы определить время полужизни (t1/2) свободного антитела (19.3: 77±6 часов; 3B3 соответственно: 29±9 часов).
Фигура 8 является графическим представлением PK антитела против ADDL 19.3 (в сыворотке), которую оценивали у шести макак-резус после введения болюсной внутривенной (IV) или подкожной (SC) дозы 5 мг/кг. Время полужизни (t1/2) 254±28 (274±9) часов было установлено после IV введения и 204±49 (219±52) часов после SC дозы.
Фигура 9 является графическим представлением PK антитела против ADDL 19.3, которую оценивали в цереброспинальной жидкости (CSF) приматов (три самца макак-резус) с применением модели подсоединения к мозжечково-мозговой цистерне макак-резус после введения болюсной IV дозы 5 мг/кг. Приблизительно через 48 часов после введения дозы антитело против ADDL 19.3 было представлено в CSF в 0,1% от концентрации в сыворотке.
Фигуры 10A-10D представляют собой изображения способности антитела против ADDL 19.3, в сопоставлении с двумя антителами сравнения (Соед. 1 и Соед. 2), проходить через гематоэнцефалический барьер в модели трансгенных мышей, которые сверхэкспрессируют белок-предшественник амилоида человека (hAPP). Мышам вводили внутривенно (IV) 125I-меченное антитело против ADDL 19.3, или антитело сравнения, и собирали образцы крови, CSF и ткани мозга через два часа после введения дозы. Исходя из распределения радиоактивности, 0,02% антитела против ADDL 19.3 представлено в CSF (Фигура 10A), тогда как 0,19% наблюдали в ткани мозга (Фигура 10B). Аналогичные уровни получали в случае двух антител сравнения. Иммуноцитохимический анализ показал местонахождение антитела против ADDL 19.3 (фигура 10C, стрелки), и концентрирование антитела против ADDL 19.3 было видимым благодаря бляшкам (Фигура 10D). Антитело против ADDL l9.3 способно проникать в мозг и связывать ADDL.
На фигурах 11A-11C отображена способность антитела против ADDL 19.3 блокировать отложение ADDL в растущих бляшках в модели трансгенных мышей, которые сверхэкспрессируют hAPP. Биотинилированные ADDL (bADDL) вводили в гиппокамп 12-месячных мышей в течение четырех недель (одна инъекция в неделю) (фигура 11A) меченные существующие бляшки (только носитель: фигура 11B; антитело 19.3: фигура 11C, кружок). Иммуноцитохимический анализ применяли, чтобы оценить отложения нового вещества (ADDL) (фигуры 11B и 11C).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к антителам или антигенсвязывающему фрагменту, которые связывают диффундирующие лиганды, происходящие из β-амилоида (Aβ) (ADDL), т.е. к антителам против ADDL, и ослабляют связывание ADDL с нейронами. Результаты количественного клеточного анализа показали, что антитела против ADDL преимущественно связывали ADDL, уменьшали связывание ADDL с гиппокампальными нейронами, пересекали гематоэнцефалический барьер, и обладали улучшенным фармакокинетическим (PK) профилем.
В одном из вариантов осуществления настоящее изобретение относится к выделенному антителу или его антигенсвязывающему фрагменту, которое связывает диффундирующие лиганды, происходящие из β-амилоида (ADDL), содержащему:
(a) вариабельную область легкой цепи, содержащую
(i) CDR1 с последовательностью Arg-Ser-Ser-Gln-Ser-Ile-Val-His-Ser-Asn-Gly-Asn-Thr-Tyr-Leu-Glu (SEQ ID NO:1),
(ii) CDR2 с последовательностью Lys-Ala-Ser-Asn-Arg-Phe-Ser (SEQ ID NO:2), и
(iii) CDR3 с последовательностью Phe-Gln-Gly-Ser-Xaa1-Xaa2-Xaa3-Xaa4-Xaa5 (SEQ ID NO:3), в которой Xaa1 представляет собой Arg, Lys или Tyr, Xaa2 представляет собой Val, Ala или Leu, Xaa3 представляет собой Pro, His или Gly, Xaa4 представляет собой Ala, Pro или Val, и Xaa5 представляет собой Ser, Gly или Phe; и
(b) вариабельную область тяжелой цепи, содержащую
(i) CDR1 с последовательностью Gly-Phe-Thr-Phe-Ser-Ser-Phe-Gly-Met-His (SEQ ID NO:4),
(ii) CDR2 с последовательностью Tyr-Ile-Ser-Arg-Gly-Ser-Ser-Thr-Ile-Tyr-Tyr-Ala-Asp-Thr-Val-Lys-Gly (SEQ ID NO:5), и
(iii) CDR3 с последовательностью Gly-Ile-Thr-Thr-Ala-Leu-Asp-Tyr (SEQ ID NO:6).
В одном из вариантов осуществления настоящее изобретение относится к выделенному антителу, или его антигенсвязывающему фрагменту, который связывает диффундирующие лиганды, происходящие из β-амилоида (ADDL), содержащему:
(a) вариабельную область легкой цепи, содержащую
(i) CDR1 с последовательностью Arg-Ser-Ser-Gln-Ser-Ile-Val-His-Ser-Xaa1-Gly-Xaa2-Thr-Tyr-Leu-Glu (SEQ ID NO:53), в которой Xaa1 представляет собой Asn, Ser, Thr, Ala, Asp или Glu, и Xaa2 представляет собой Asn, His, Gln, Ser, Thr, Ala или Asp;
(ii) CDR2 с последовательностью Lys-Ala-Ser-Xaa1-Arg-Phe-Ser (SEQ ID NO:54), в которой Xaa1 представляет собой Asn, Gln, Ser, Thr или Ala и
(iii) CDR3 с последовательностью Phe-Gln-Gly-Ser-Arg-Leu-Gly-Pro-Ser (SEQ ID NO:10); и
(b) вариабельную область тяжелой цепи, содержащую
(i) CDR1 с последовательностью Gly-Phe-Thr-Phe-Ser-Ser-Phe-Gly-Met-His (SEQ ID NO:4),
(ii) CDR2 с последовательностью Tyr-Ile-Ser-Arg-Gly-Ser-Ser-Thr-Ile-Tyr-Tyr-Ala-Asp-Thr-Val-Lys-Gly (SEQ ID NO:5), и
(iii) CDR3 с последовательностью Gly-Ile-Thr-Thr-Ala-Leu-Asp-Tyr (SEQ ID NO:6).
В одном из вариантов осуществления настоящее изобретение относится к выделенному антителу, которое связывает ADDL, а именно к антителу против ADDL, или его антигенсвязывающему фрагменту с вариабельной областью легкой цепи CDR3, которая выбрана из группы, состоящей из 17.1 с последовательностью Phe-Gln-Gly-Ser-Arg-Val-Pro-Ala-Ser (SEQ ID NO:7), 14.2 с последовательностью Phe-Gln-Gly-Ser-Arg-Val-Pro-Pro-Gly (SEQ ID NO:8), 13.1 с последовательностью Phe-Gln-Gly-Ser-Lys-Ala-His-Pro-Ser (SEQ ID NO:9), 19.3 с последовательностью Phe-Gln-Gly-Ser-Arg-Leu-Gly-Pro-Ser (SEQ ID NO:10), 7.2 с последовательностью Phe-Gln-Gly-Ser-Tyr-Ala-Pro-Pro-Gly (SEQ ID NO:11), 9.2 с последовательностью Phe-Gln-Gly-Ser-Arg-Ala-Pro-Pro-Phe (SEQ ID NO:12) и 11.4 с последовательностью Phe-Gln-Gly-Ser-Arg-Val-Pro-Val-Arg (SEQ ID NO:13). В подварианте осуществления изобретения вариабельная область легкой цепи CDR3 представляет собой SEQ ID NO:10.
В другом варианте осуществления настоящего изобретения выделенное антитело против ADDL также содержит вариабельную область легкой цепи SEQ ID NO:15 и вариабельную область тяжелой цепи SEQ ID NO:17.
В другом варианте осуществления настоящего изобретения выделенное антитело против ADDL также содержит константную область тяжелой цепи SEQ ID NO:21.
В одном из вариантов осуществления настоящего изобретения выделенное антитело против ADDL представляет собой моноклональное антитело.
Другой вариант осуществления настоящего изобретения относится к фармацевтической композиции, содержащей выделенное антитело против ADDL, или его антигенсвязывающий фрагмент в смеси с фармацевтически приемлемым носителем.
Другой вариант осуществления настоящего изобретения относится к способу уменьшения связывания ADDL с нейроном, включающему приведение в контакт нейрона с выделенным антителом против ADDL, или его антигенсвязывающим фрагментом так, что связывание диффундирующих лигандов, происходящих из Αβ, с нейроном ослабляется.
Другой вариант осуществления настоящего изобретения относится к способу ингибирования сборки ADDL, включающему приведение в контакт образца, содержащего 1-42 пептида β-амилоида с выделенным антителом против ADDL, или его антигенсвязывающим фрагментом, ингибирующее таким образом сборку ADDL.
Другой вариант осуществления настоящего изобретения относится к способу ингибирования фосфорилирования тау-белка по Ser202/Thr205, включающему приведение в контакт образца, содержащего тау-белок с выделенным антителом против ADDL, или его антигенсвязывающим фрагментом, ингибируя таким образом фосфорилирование тау-белка по Ser202/Thr205.
Другой вариант осуществления настоящего изобретения относится к способу ослабления симптомов заболевания, связанного с ADDL, включающему введение пациенту, который в этом нуждается, эффективного количества фармацевтической композиции, содержащей выделенное антитело против ADDL или его антигенсвязывающий фрагмент.
Другой вариант осуществления настоящего изобретения относится к способу идентификации предполагаемого терапевтического средства, которое уменьшает связывание диффундирующих лигандов, происходящих из β амилоида (ADDL), с нейронами, включающему:
(a) приведение в контакт композиции, содержащей нейрон, с ADDL в присутствии средства;
(b) приведение в контакт композиции с выделенным антителом против ADDL, или с его антигенсвязывающим фрагментом; и
(c) определение количества антитела или антигенсвязывающего фрагмента, связанного в присутствии средства,
где уменьшение количества антитела или антигенсвязывающего фрагмента, связанного в присутствии средства, по сравнению с количеством антитела, связанного в отсутствии средства, показывает, что средство является предполагаемым терапевтическим средством для уменьшения связывания ADDL с нейронами.
Другой вариант осуществления настоящего изобретения относится к способу обнаружения ADDL в образце, включающему приведение в контакт образца с выделенным антителом против ADDL, или с его антигенсвязывающим фрагментом, и определение наличия комплекса, содержащего ADDL и указанное антитело, или антигенсвязывающий фрагмент.
Другой вариант осуществления настоящего изобретения относится к способу диагностики заболевания, связанного с ADDL, включающему приведение в контакт образца с выделенным антителом против ADDL, или его антигенсвязывающим фрагментом, и определение наличия комплекса, содержащего ADDL и указанное выделенное антитело, или его антигенсвязывающий фрагмент, где наличие указанного комплекса является диагностическим для заболевания, связанного с ADDL.
Другой вариант осуществления настоящего изобретения относится к набору для обнаружения ADDL, включающему выделенное антитело против ADDL, или его антигенсвязывающий фрагмент, который связывает ADDL.
Моноклональные антитела, которые селективно распознают многомерные конформации диффундирующих лигандов, происходящих из Αβ (ADDL), известны в данной области (см. патент США №7780963, патент США № 7731962 и патент США № 7811563, которые включены в описание в качестве ссылки в полном объеме), и, как показано, уменьшают связывание ADDL с нейронами в клеточных анализах. Антитела против ADDL способны различать экстракты мозга людей с болезнью Альцгеймера (AD) и контрольные экстракты мозга человека, могут идентифицировать эндогенные олигомеры в срезах мозга AD и на гиппокампальных клетках, и могут нейтрализовать эндогенные и синтетические ADDL в растворе. Антитела против ADDL специфически связываются с одной или более многомерными конформациями ADDL, связывают определенные ADDL, получаемые в результате олигомеризации Αβ42, и в то же время обладают пониженной аффинностью в отношении других пептидов Αβ, включая Αβ1-40.
Настоящее изобретение относится к антителам против ADDL, в особенности к антителам 17.1, 14.2, 13.1, 19.3, 19.3T33, 19.3S33, 7.2, 9.2 и 11.4, которые, предпочтительно, связывают ADDL и которые были описаны по их специфичности и селективности в отношении ADDL. Важно, что специфичность и селективность указанных антител против ADDL по настоящему изобретению не являлись прогнозируемыми на основе линейного эпитопа Αβ, с которым они связывались, указанная активность не являлась прогнозируемой на основе их способности обнаруживать ADDL при вестерн-блоттинге, или на основе их способности обнаруживать иммуно-окрашенные ADDL, связанные с нейронами. Кроме того, избирательная способность антител против ADDL по настоящему изобретению нейтрализовывать ADDL и блокировать связывание с первичной культурой гиппокампальных нейронов поддерживает предположение, что антитела против ADDL действуют путем связывания с более подходящим, конформационным эпитопом, что предотвращает связывание ADDL с нейронами. В одном из вариантов осуществления настоящего изобретения, антитело против ADDL 19.3, не только блокировало связывание ADDL с первичной культурой нейронов, но также уменьшало ADDL-индуцированные изменения морфологии шипиков гиппокампальных клеток, показатель того, что сопротивление против связывания ADDL с нейронами имеет значительные физиологические последствия, например, выживание нейронов, взаимосвязь нейронов и передача сигнала. Антитело против ADDL 19.3 также обладало улучшенным фармакокинетическим (PK) профилем, по сравнению с ранее известным антителом против ADDL, 3B3, когда его оценивали в моделях in vitro и in vivo. Кроме того, при введении трансгенным мышам, которые сверхэкспрессируют форму белка-предшественника амилоида (hAPP) человека, антитело против ADDL 19.3, как показано, проникает через гематоэнцефалический барьер и концентрируется в мозге. Поскольку ADDL локализуются в мозге и оказывают негативное влияние, поражая нейрональную функцию, специалисту в данной области понятно и известно, что проникновение и концентрирование антитела в мозге было бы полезным при иммунотерапии. В совокупности, указанные данные показывают, что селективные антитела против ADDL, такие как антитело 19.3, могут блокировать связывание ADDL с гиппокампальными нейронами, которые играют критическую роль в процессах обучения и памяти.
Полезность антител против ADDL в лечении AD основана на растущем числе данных, которые позволяют предположить, что ADDL, а не амилоидные бляшки как таковые, играют фундаментальную роль в снижении когнитивных способностей, связанных с данным заболеванием (Walsh and Selkoe, 2004, Protein Pept. Lett.,11: 213-228). ADDL возрастают в мозге при AD, и вызывают снижение показателей в поведенческих и электрофизиологических конечных критериях оценки при центральном введении у грызунов (Walsh, et al., 2002, Nature, 416: 535-539; Cleary, et al., 2004, Nat. Neurosci., 8: 79-84; Klyubin, et al., 2005, Nat. Med., 11: 556-561; Balducci, et al., 2010, Proc. Natl. Acad. Sci. USA, 107: 2295-2300). Дефицит обучения и памяти также наблюдали в модели мышей, экспрессирующих hAPP, с наступлением ухудшения, связанного с возрастающими уровнями ADDL (Westerman, et al., 2002, J. Neurosci., 22: 1858-1867; Ashe, 2005, Biochem. Soc. Trans., 33: 591-594; Lee, et al., 2005, J. Biol. Chem., 281: 4292-4299; Lesne, et al., 2006, Nature, 440: 352-357). Несмотря на то, что клеточные и субклеточные явления, которые опосредуют указанные эффекты на когнитивную деятельность, не изучены полностью, очевидно, что ADDL связываются с синаптическими окончаниями, расположенными на дендритных отростках гиппокампальных нейронов (Lacore, et al., 2004, J. Neurosci., 24: 10191-1022) и изменяют морфологию и количество дендритных шипиков (Lacor et al., 2007, J. Neurosci., 27: 796-807; Shankar, et al., 2007, J. Neurosci., 27: 2866-2875; Shughrue, et al., 2010, Neurobiol. Aging. 31: 189-202). Полученные данные о том, что ADDL связываются с ГАМК-эргическими и глутаматергическими нейронами в гиппокампе (Shughrue, et al., 2010), нейронами критически вовлеченными в процессы обучения и памяти, что приводит к интернализации AMPA-рецепторов (Zhao, et al., 2010, J. Biol. Chem., 285: 7619-7632) также подтверждают предположение, что ADDL прямо или косвенно модулируют указанные нейротрансмиттерные системы (см., например, Venkitaramani, et al., 2007, J. Neurosci., 27: 11832-11837).
В настоящем изобретении панель антител против ADDL, полученных из антитела против ADDL, 3B3 (патент США №7780963 и патент США №7811563, которые включены в описание в качестве ссылки в полном объеме), оценивали по их способности блокировать связывание ADDL в культуре первичных гиппокампальных нейронов. Отобранные моноклональные антитела затем гуманизировали и подвергали аффинному созреванию для дальнейшего описания. Ведущие антитела, отобранные благодаря своей способности связываться с ADDL, затем оценивали в одной концентрации с использованием трехдорожкового теста ELISA для определения связывания антитела с мономерным Αβ, ADDL и фибриллярным Αβ. Как показано на фигуре 1, шесть из семи антител против ADDL со зрелой аффинностью, в особенности антитела 14.2, 7.2, 11.4, 13.1, 17.1 и 19.3 являлись ADDL-предпочтительными, по сравнению с мономерным Αβ и фибриллярным Αβ. Затем использовали одиннадцатиточечную титрационную кривую и ELISA для определения аффинности связывания антител против ADDL с ADDL и мономерным Αβ (Αβ1-40) в широком диапазоне концентраций. Как показано на фигуре 2, антитела против ADDL 3B3 и 19.3 являлись высоко ADDL-селективными. Кроме того, антитела сравнивали в клеточном анализе связывания для определения способности антител блокировать связывание ADDL с нейронами. Как показано на фигуре 3, ADDL, предварительно инкубированные с повышенными концентрациями антител против ADDL 3B3 и 19.3, добавляли в первичную культуру гиппокампальных нейронов, и использовали титрационную кривую для количественного представления способности антитела блокировать связывание ADDL с нейронами. В совокупности указанные результаты показывают, что антитела против ADDL серьезно ослабляют связывание с нейронами в клеточном формате.
Определение аминокислотной последовательности проводили для идентификации потенциальных сайтов деамидирования. Остатки аспарагина и аспарагиновой кислоты, представленные в CDR терапевтических антител, как известно, подвергаются деамидированию с образованием изоаспартата (Valsak and Ionescu, 2008, Curr.Pharm.Biotech., 9:468-481; Aswad et al., 2000, J. Pharm.Biomed.Anal., 21: 1129-1136), образование которого может изменять эффективность связывания антитела и, в свою очередь, уменьшать эффективность антитела при использовании в качестве лекарства. Таким образом, специалисту в данной области следует понимать и учитывать, что присутствие аспарагина и аспарагиновой кислоты в CDR для антитела 19.3 нежелательно. Таким образом, заявители изменяли остаток аспарагина в положении 33 CDR1 легкой цепи для оптимизации стабильности антитела против ADDL 19.3 (Таблица 4B). Создавали производные антитела 19.3 с заменой серином (SEQ ID NO:55), треонином (SEQ ID NO:56) или глутаминовой кислотой (SEQ ID NO:67) аспарагина в положении 33 (SEQ ID NO:1) в CDR1. Также производили замену аспарагиновой кислотой (SEQ ID NO:68) аспарагина в положении 33 в качестве контроля. Указанные изменения будут устранять возможность деамидирования аспарагина в положении 33 в CDR1. Производные 19.3 получали, как описано в Примере 3 и оценивали, как описано в Примере 4, как и производные с заменой серином (SEQ ID NO:55), треонином (SEQ ID NO:56), глутаминовой кислотой (SEQ ID NO:67), и аспарагиновой кислотой (SEQ ID NO:68), для определения стабильности новых конструкций. Как показано на фигурах 4B и 4C, соответственно, два характерных производных, 19.3S33 (SEQ ID NO:55) и 19.3T33 (SEQ ID NO:56), обладали повышенной стабильностью связывания по результатам месячной инкубации при различных температурах. Другие аминокислотные замены в CDR1 легкой цепи для аспарагина в положениях 33 и 35 (SEQ ID NO:53) и в CDR2 легкой цепи для аспарагина в положении 58 (SEQ ID NO:54) представлены в таблицах 4B и 4C для последующей оценки.
Для определения фармакокинетики аффинно зрелых антител против ADDL по настоящему изобретению проводили серии исследований in vitro и in vivo. Показано, что связывание антител с рецептором FcRn при pH 6,0 прогнозируется по периоду полувыведения антитела у человека (Zalevsky, et al., 2010, Nat. Biotech. 28(2): 157-159) и при pH 7,3 (USSN 61/307182). Связывание и диссоциацию антител против ADDL по настоящему изобретению с иммобилизованным FcRn человека оценивали с помощью анализа взаимодействия со свободной меткой, такого как анализ предлагаемый BiacoreTM Life Sciences, BiacoreTM T-100 (GE Healthcare, Piscataway, NJ). Скорректированную сенсограмму используют, чтобы показать начальное связывание при pH 6,0 и затем диссоциацию антител при pH 7,3 от 180 секунд. Точку отчета (состояние равновесия) устанавливали на момент времени 5 секунд после окончания связывания при pH 6,0 и "% связывания" рассчитывали как RUравновесие/RUсвязывание (%). Как показано на фигуре 5, скорость диссоциации в случае гуманизированного 3B3 была значительно ниже, чем у семи антител против ADDL по настоящему изобретению, которые включали антитело 19.3 и три антитела сравнения. Поскольку низкая скорость диссоциации считается индикатором слабой PK in vivo, проводили дополнительное исследование in vivo на FcRn-трансгенных мышах (гетерозиготные по FcRn 276 человека мыши, Jackson Laboratories, Bar Harbor, ME). Когда FcRn-трансгенным мышам вводили 10 мг/кг внутривенно (IV) либо антитело против ADDL 3B3, либо 19.3, то обнаруживали существенное различие в фармакокинетике. Как показано на фигуре 7, время полужизни (t1/2) антитела против ADDL 3B3 являлось относительно небольшим (29±9 часов), что соответствовало прогнозу на основе данных, полученных in vitro с помощью BiacoreTM, тогда как время полужизни антитела против ADDL 19.3 было значительно ниже (77±6 часов). В целом, низкая PK, как видно в случае антитела 3B3, будет препятствовать дальнейшему усовершенствованию антитела при использовании в качестве терапевтического средства, вследствие его низкой биодоступности.
Для подтверждения прогнозируемого времени полужизни антитела против ADDL 19.3 у приматов, проводили исследование фармакокинетики у приматов для определения антитела в группе макак-резус с подсоединением к мозжечково-мозговой цистерне. Животным вводили однократную внутривенную (IV) болюсную или подкожную (SC) инъекцию антитела против ADDL 19.3 (5 мг/кг) и собирали образцы крови после введения антитела. Одновременно собирали образцы CSF из порт-системы, соединенной с мозжечково-мозговой цистерной, в установленные интервалы времени и концентрацию антитела против ADDL 19.3 определяли в сыворотке и в CSF с помощью теста ELISA с применением античеловеческого IgG. При введении животным антитела против ADDL 19.3 путем однократной IV болюсной инъекции, наблюдали t1/2 254±28 часов (фигура 8), в то время как при подкожном введении получали t1/2 204±49 часов. Кроме того, заявители обнаружили, что антитело против ADDL 19.3 способно переходить в CSF приматов, где его концентрация возрастала в течение первых 48 часов и достигала максимума приблизительно 0,1% введенного антитела (фигура 9).
Для определения количества антитела, которое проникает через гематоэнцефалический барьер и поступает в CSF и мозг, антитело против ADDL 19.3 и два антитела сравнения (Соед. 1 и Соед. 2) метили 125I и вводили старым (двенадцатимесячным) мышам, которые сверхэкспрессировали hAPP, моделируя AD на грызунах. Через два часа после IV введения, приблизительно 0,02% антитела 19.3 наблюдали в CSF (фигура 10A), в то время как приблизительно 0,19% антитела 19.3 обнаруживали в мозге (фигура 10B). Аналогичные уровни наблюдали в случае двух антител сравнения (фигура 10A и 10B). При проведении иммуноцитохимического анализа срезов мозга мышей, получавших препарат, и определении местонахождения антитела против ADDL 19.3 (стрелка на фигуре 10C), наблюдали накопление антитела, связанного с депонированием Αβ, в бляшках (фигура 10D). Данный факт продемонстрировал, что антитело против ADDL 19.3 проникало в CSF и накапливалось в мозге. Недавно показано, что экзогенные ADDL откладывались в бляшках при введении мышам, которые сверхэкспрессировали hAPP (Caspar, et al., 2010, Exp. Neurol., 223: 394-400). Таким образом, полученные данные подтверждали, что локализованное антитело против ADDL l9.3, связанное с циркулирующими ADDL, связано с бляшками.
Для дальнейшей оценки эффективности in vivo антител против ADDL, способность антитела 19.3 блокировать депонирование ADDL в растущих бляшках оценивали у hAPP-трансгенных мышей после четырех еженедельных инфузий биотинилированных ADDL (bADDL) в гиппокамп 12-месячных мышей для маркирования существующих бляшек (Фигура 11A). Затем животные получали четыре еженедельные внутривенные инфузии антитела 19.3 (Фигура 11A). Депонирование нового вещества (ADDL) в растущих бляшках оценивали с помощью иммуноцитохимического анализа. Как видно на фигурах 11B и 11C, антитело против ADDL 19.3 существенно снижало депонирование ADDL на периферии существующих бляшек (фигура 11C), по сравнению с мышами, получавшими только носитель (фигура 11B). В совокупности, указанные результаты показали, что антитело против ADDL, в особенности антитело 19.3, способно проходить через гематоэнцефалический барьер, связывать ADDL, и блокировать депонирование нового вещества в растущих бляшках.
Связывание ADDL также может оказывать долговременные эффекты на нейроны. Недавние исследования показали, что связывание ADDL с гиппокампальными нейронами может инициировать сигнальный каскад, который приводит к гиперфосфорилированию тау-белка (De Felice, et al., 2006, Neurobiol. Aging, 29: 394-400). Также показано, что один компонент указанного сигнального каскада, GSK-3β, модулируется связыванием ADDL in vivo и in vitro (Ma, et al., 2006, J. Neurosci. Res., 83: 374-384). Ma, et al., 2006, обнаружили, что пассивная иммунизация мышей hAPP антителом, которое уменьшало количество ADDL, также снижала уровни GSK-3β и фосфорилирование тау-белка в коре головного мозга. Указанный факт служит доказательством связи между Αβ и фосфорилированным тау-белком и позволяет предположить, что связывание ADDL может запускать события, которые приводят к внутриклеточной агрегации тау-белка. Кроме того, полученные данные позволяют предположить, что антитела, которые предотвращают связывание ADDL с нейронами и связанную потерю синаптических шипиков, такие как антитела по настоящему изобретению, могли бы уменьшать когнитивные и/или патологические последствия, связанные с болезнью Альцгеймера и родственные заболевания.
Моноклональные антитела, которые избирательно распознают многомерные конформации диффундирующих лигандов, происходящих из Αβ, т.е. ADDL, теперь созданы. Указанные антитела являлись гуманизированными и, в некоторых вариантах осуществления, аффино зрелыми. Антитела успешно распознают экстракты мозга человека с болезнью Альцгеймера и контрольные экстракты мозга человека, и обнаруживают эндогенные олигомеры в срезах мозга при болезни Альцгеймера и в культивируемых гиппокампальных клетках. Кроме того, антитела по настоящему изобретению нейтрализуют эндогенные и синтетические ADDL в растворе. Так называемые "синтетические" ADDL получают in vitro смешиванием очищенных Αβ1-42 в условиях, которые порождают ADDL. См. патент США № 6218506. Антитела, раскрытые в описании, проявляют высокую степень селективности в отношении ADDL, при минимальном обнаружении мономерного соединения Αβ. Кроме того, указанные антитела избирательно блокируют способность ADDL-содержащих препаратов связывать первичные культуры гиппокампальных нейронов крыс и иммортализованные клеточные линии нейробластомы, и также блокировать сборку ADDL. Данный факт показывает, что указанные антитела обладают различной способностью распознавать многомерную конформацию ADDL, несмотря на сходное распознавание линейных последовательностей и сходные аффинности. Поскольку ADDL, как известно, связываются с субпопуляцией нейронов и нарушают нормальную нейрональную функцию, антитела по настоящему изобретению находят применение в предотвращении связывания ADDL с нейронами и сборке ADDL и, в свою очередь, могут использоваться для лечения ADDL-связанных заболеваний, включая болезнь Альцгеймера.
Таким образом, в одном из вариантов осуществления настоящее изобретение относится к выделенному антителу, которое избирательно распознает одну или более многомерных конформаций ADDL. "Выделенное" антитело по настоящему изобретению относится к антителу, которое по существу свободно от других антител. Однако молекула может включать некоторые дополнительные средства или функциональные группы, которые не оказывают отрицательного воздействия на основные характеристики антитела (например, специфичность связывания, нейтрализующая активность и т.п.).
Антитело, которое способно к специфическому связыванию с одной или более многомерными конформациями ADDL, связывается с определенным ADDL, получаемым в результате олигомеризации Αβ1-42, но не дает перекрестной реакции с другими Αβ пептидами, а именно Αβ1-12, Αβ1-28, Αβ1-40 и Αβ12-28, что определяли с помощью анализов вестерн-блот, как раскрыто в описании, и преимущественно связывает ADDL в растворе. Специфическое связывание между двумя частицами, как правило, относится к значениям аффинности по меньшей мере 106, 107, 108, 109 или 1010 M-1. Значения аффинности выше чем 108 M-1 желательны для достижения специфического связывания.
В конкретных вариантах осуществления, также получают антитело, которое способно к специфическому связыванию с многомерной конформацией одного или более ADDL, т.e. животное иммунизируют многомерными конформациями ADDL. В других вариантах осуществления, антитело, которое способно к специфическому связыванию с многомерной конформацией одного или более ADDL, получают против пептида, образующего полимер с низким значением n-mer, такого как Aβ1-42[Nle35-Dpro37].
Термин "эпитоп" обозначает участок антигена, на который отвечают B- и/или T-клетки или участок молекулы, против которого будет продуцироваться антитело и/или с которым антитело будет связываться. Например, эпитоп может распознаваться антителом, определяющим эпитоп.
Линейный эпитоп представляет собой эпитоп, в котором первичная аминокислотная последовательность содержит распознаваемый эпитоп. Линейный эпитоп обычно включает по меньшей мере 3, и еще чаще, по меньшей мере 5, например, приблизительно 6-10 аминокислот в уникальной последовательности.
Конформационный эпитоп, в отличие от линейного эпитопа, представляет собой эпитоп, в котором первичная последовательность аминокислот, содержащая эпитоп не является единственным определяющим компонентом распознаваемого эпитопа (например, эпитопа, в котором первичная последовательность аминокислот необязательно распознается антителом, определяющим эпитоп). Обычно конформационный эпитоп охватывает большее число аминокислот, чем линейный эпитоп. Что касается распознавания конформационных эпитопов, антитело распознает трехмерную структуру пептида или белка. Например, когда белковая молекула складывается с образованием трехмерной структуры, определенные аминокислоты и/или полипептидная основная цепь, формирующая конформационный эпитоп, сближаются, что дает возможность антителу распознать эпитоп. Способы определения конформации эпитопов включают, но без ограничения, например, рентгеноструктурный анализ, двумерную ядерную магнитно-резонансную спектроскопию и сайт-направленное мечение, и электронную парамагнитную резонансную спектроскопию. См., например, Epitope Mapping Protocols in Methods in Molecular Biology (1996) Vol. 66, Morris (Ed.).
Диффундирующие лиганды, происходящие из β-амилоида или ADDL, относятся к растворимым олигомерам Αβ1-42, которые желательно состоят из агрегатов, состоящих менее чем из восьми или девяти пептидов Αβ1-42 и, как установлено, связаны с болезнью Альцгеймера. Это является отличием от высокомолекулярных промежуточных продуктов агрегации, которые образуют нити мицелл, приводя к образованию фибрилл.
Как проиллюстрировано в описании, антитела по настоящему изобретению связывают или распознают по меньшей мере одну многомерную конформацию ADDL. В конкретных вариантах осуществления антитела связываются по меньшей мере с двумя, по меньшей мере с тремя, или по меньшей мере с четырьмя многомерными конформациями ADDL. Многомерные конформации ADDL предназначены для включения димеров, тримеров, тетрамеров, пентамеров, гексамеров, гептамеров, октамеров, наномеров, декамеров и т.п., которые определяют с помощью анализа с применением SDS-PAGE. Поскольку тример, тетрамер и т.п. обозначения могут варьировать в зависимости от применяемого способа количественного анализа (см., например, Bitan, et al., 2005, Amyloid, 12:88-95), определение тримера, тетрамера и т.п., которое используют в описании, соответствует анализу SDS-PAGE. Чтобы проиллюстрировать в описании возможности избирательного связывания антител, обнаружено, что определенные антитела распознают одну многомерную конформацию, например, тетрамеры ADDL (патент США № 7780963, антитела мыши 2D6 и 4E2), в то время как другие антитела распознают несколько многомерных конформаций, например, тримеры и тетрамеры ADDL (патент США № 7780963, антитела мыши 2A10, 2B4, 5F10 и 20C2 и гуманизированное антитело 20C2). По существу антитело по настоящему изобретению обладает олигомерспецифическими характеристиками. В конкретных вариантах осуществления многомерная конформация ADDL ассоциируется со специфической полипептидной структурой, которая дает в результате конформационный эпитоп, который распознается антителом по настоящему изобретению. В других вариантах осуществления антитело по изобретению специфически связывается с многомерной конформацией ADDL, характеризующейся диапазоном величин приблизительно тримера или тетрамера, которые обладают молекулярным весом свыше >50 кДа.
В то время как антитела по настоящему изобретению могут обладать сходными линейными эпитопами, такие линейные эпитопы не отражают полностью характеристики связывания указанных антител, т.е. способность блокировать связывание ADDL с нейронами, предотвращать фосфорилирование тау-белка и ингибировать сборку ADDL, поскольку, как хорошо известно специалисту в данной области, линейный эпитоп может соответствовать только части эпитопа антигена (см., например, Breitling and Dübel, 1999, Recombinant Antibodies, John Wiley & Sons, Inc., NY, pg. 115). Антитела по настоящему изобретению могут отличаться от антител в данной области, поскольку они способны к избирательному распознаванию многомерного ADDL и соответственно к избирательному блокированию связывания ADDL с нейронами, избирательному предотвращению гиперфосфорилирования тау-белка и избирательному ингибированию сборки ADDL.
Антитело, которое используют согласно настоящему изобретению, включает, но без ограничения поликлональные или моноклональные антитела и химерные антитела, антитела человека (например, выделенные из B-клеток), гуманизированные, нейтрализующие, биспецифические или одноцепочечные антитела. В одном из вариантов осуществления антитело по настоящему изобретению является моноклональным антителом. Для получения антител, различных хозяев, включая коз, кроликов, цыплят, крыс, мышей, людей и других можно иммунизировать инъекцией синтетических или природных ADDL. Способы получения антител хорошо известны в данной области. См., например, Kohler and Milstein, 1975, Nature, 256:495-497: Harlow and Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, New York, 1988.
В зависимости от видов хозяев можно использовать различные адъюванты для увеличения иммунологического ответа. Адъюванты, применяемые согласно настоящему изобретению в соответствии с желанием увеличивают природный ответ на ADDL, не вызывая конформационных изменений в иммуногене, который влияет на качественную форму ответа. Особенно подходящие адъюванты включают 3 De-O-ацилированный монофосфорил-липид A (MPLTM; RIBI ImmunoChem Research Inc., Hamilton, MT; см. GB 2220211) и эмульсии масло-в-воде, такие как сквален или арахисовое масло, необязательно в комбинации с иммуностимуляторами, такими как монофосфорил-липид A (см., Stoute, et al., 1997, N. Engl. J. Med., 336:86-91), мурамилпептиды (например, N-ацетилмурамил-L-теонил-D-изоглутамин (thr-MDP), N-ацетилнормурамил-L-аланил-D-изоглутамин (nor-MDP), N-ацетилмурамил-L-аланил-D-изоглутаминил -L-аланин-2-(1'-2'дипальмитоил-sn-глицеро-3-гидроксифосфорилокси)-этиламин (E-PE), N-ацетилглюкозаминил-N-ацетилмурамил-L-Al-D-isoGlu-L-Ala-дипальмитоксипропиламид (DTP-DPP)) или другие компоненты бактериальной клеточной стенки. Конкретные примеры эмульсий масло-в-воде включают MF59 (WO 90/14837), содержащий 5% сквален, 0,5% TWEENTM 80, и 0,5% SPAN 85 (необязательно содержащий различные количества MTP-PE), включенный в субмикронные частицы с помощью микрофлюидайзера, такого как модель микрофлюидайзера 110Y (Microfluidics, Newton, MA); SAF, содержащий 10% сквален, 0,4% TWEENTM 80, 5% PLURONIC®-блокированный полимер L121, и thr-MDP, либо обработанный на микрофлюидайзере с образованием субмикронной эмульсии, либо обработанный на вортексе с образованием эмульсии с частицами большего размера; и адъювантная система RIBITM (RAS) (Ribi ImmunoChem, Hamilton, MT), содержащая 2% сквален, 0,2% TWEENTM 80, и один или более компонентов бактериальной стенки, таких как монофосфориллипид A, димиколат трегалозы (TDM), и каркас клеточной стенки (CWS).
Другой класс адъювантов представляет собой сапониновые адъюванты, такие как STIMULONTM (QS-21, Aquila, Framingham, MA) или частицы, полученные на его основе, такие как ISCOM (комплексы с иммуностимулирующими свойствами) и ISCOMATRIX® (CSL Ltd., Parkville, Australia). Другие подходящие адъюванты включают полный адъювант Фрейнда (CFA), неполный адъювант Фрейнда (IFA), минеральные гели, такие гидроксид алюминия, и поверхностно-активные вещества, такие как лизолецитин, полиолы PLURONIC®, полианионы, пептиды, CpG (WO 98/40100), гемоцианин лимфы улитки, динитрофенол и цитокины, такие как интерлейкины (IL-1, IL-2 и IL-12), колониестимулирующий фактор макрофагов (M-CSF), и фактор некроза опухоли (TNF). Среди адъювантов, применяемых у человека, BCG (bacilli Calmette-Guerin) и Corynebacterium parvum являются особенно предпочтительными.
Антитело для многомерной конформации ADDL получают иммунизацией животного ADDL. Как правило, ADDL можно получить синтетически или путем рекомбинантной экспрессии фрагментов и очистки. Синтетические ADDL можно получить, как раскрыто в описании, или по способам, раскрытым в патентах США № 6218506 и 7811563 или в одновременно рассматриваемых патентных заявках США 2007/0218499, США 2010/0143396, и США 2010/0240868, которые включены в описание в качестве ссылки в полном объеме. Кроме того, ADDL можно сливать с другим белком, таким как гемоцианин лимфы улитки для создания антитела против химерной молекулы. ADDL могут быть конформационно ограничены для создания эпитопа, применяемого, как описано в изобретении, и, кроме того, могут быть соединены с поверхностью, например, прикреплены физически или связаны химически с поверхностью таким образом, чтобы обеспечить получение конформации, которая распознается антителами по настоящему изобретению.
Моноклональные антитела для многомерных конформаций ADDL можно получить с применением любого способа, который обеспечивает продукцию молекул антитела стабильными клеточными линиями в культуре. Они включают, но без ограничения, способ гибридом, способ B-клеточных гибридом человека, и способ EBV-гибридом (Kohler, et al.,1975, Nature 256:495-497; Kozbor, et at, 1985, J. Immunol. Methods 81:31-42; Cote, et al., 1983, Proc.Natl.Acad.Sci. 80:2026-2030; Cole, et al., 1984, Mol. Cell Biol. 62:109-120).
В конкретных вариантах осуществления антитела по настоящему изобретению гуманизируют. Гуманизированные или химерные антитела можно создать с помощью соединения генов антител мыши с генами антител человека для получения молекулы с соответствующей антигенной специфичностью и биологической активностью (см., Morrison, et al., 1984, Proc. Natl. Acad. Sci. 81, 6851-6855; Neuberger, et al., 1984, Nature 312:604-608; Takeda, et al., 1985, Nature 314:452-454; Queen, et al., 1989, Proc. Natl. Acad. Sci. USA 86:10029-10033; WO 90/07861). Например, антитело мыши экспрессируют в виде фрагмента Fv или Fab в фаговом векторе селекции. Ген легкой цепи (и в параллельном эксперименте ген тяжелой цепи) заменяют библиотекой генов антител человека. Затем идентифицируют фаговые антитела, которые по-прежнему связывают антиген. Данный способ, общеизвестный как “перетасовка цепей”, предоставлял гуманизированные антитела, которые должны связываться с тем же эпитопом, что и антитело мыши, из которого они происходят (Jespers, et al., 1994, Biotechnology NY 12:899-903). В качестве альтернативы, перетасовку цепей можно произвести на белковом уровне (см., Figini, et al., 1994, J. Mol. Biol. 239:68-78).
Антитела человека также можно получить с использованием способов фагового дисплея. См., например, международный патент WO 91/17271 и международный патент WO 92/01047. В указанных способах получают библиотеки фага, члены которых демонстрируют различные антитела на своей внешней поверхности. Антитела обычно демонстрируются в виде фрагментов Fv или Fab. Демонстрируемые фагами антитела с желаемой специфичностью отбирают по увеличению аффинности к ADDL. Антитела человека против ADDL также могут быть получены с помощью трансгенных млекопитающих, отличных от человека, содержащих трансгены, кодирующие по меньшей мере сегмент локуса иммуноглобулина человека и инактивированный эндогенный локус иммуноглобулина. См., например, международный патент WO 93/12227 и международный патент WO 91/10741. Антитела человека можно выбрать с помощью экспериментов конкурентного связывания или иным образом, чтобы получить такую же специфичность к эпитопу, как и у определенного антитела мыши. Такие антитела обычно сохраняют полезные функциональные свойства антител мыши. Поликлональные антитела человека также можно получить в форме сыворотки от людей, иммунизированных иммуногенным средством. Необязательно, такие поликлональные антитела могут быть подвергнуты концентрированию с помощью аффинной очистки с применением ADDL в качестве аффинного реагента.
Как иллюстрируется в настоящем описании, гуманизированные антитела также можно получить путем венирования (veneering) или путем изменения поверхности антител мыши. Венирование включает замещение в вариабельных участках тяжелой и легкой цепей только аминокислот, входящих в состав участка, связанного с поверхностью, аминокислотами из гомологичной последовательности антитела человека. Замещение поверхностных аминокислот антитела мыши аминокислотными остатками антитела человека, расположенными в том же положении в гомологичной последовательности человека, как показано, уменьшает иммуногенность антитела мыши, сохраняя, в то время, его способность связывать лиганды. Замена поверхностных остатков аминокислот обычно оказывает незначительный эффект или не оказывает эффекта на внутренние домены, или на междоменные контакты. См., например, патент США № 6797492.
Можно сконструировать антитела человека или гуманизированные антитела, которые содержат константные области IgG, IgD, IgA, IgM или IgE, и любой изотип, включая IgG1, IgG2, IgG3 и IgG4. В конкретных вариантах осуществления антитело по настоящему изобретению представляет собой IgG или IgM, или их комбинацию. В одном конкретном варианте осуществления антитела по настоящему изобретению представляют собой IgG2. Специалистам в данной области понятно, что могут быть использованы другие изоформы. Характерные последовательности указанных изоформ представлены в SEQ ID NO:43-45. Другие варианты осуществления настоящего изобретения включают константную область, образованную селективным включением последовательностей IgG4 человека в стандартную константную область IgG2 человека. Иллюстративный мутантный IgG2 Fc представляет собой IgG2m4, предлагаемый в настоящем описании в виде SEQ ID NO:46. Антитела могут экспрессироваться в виде тетрамеров, содержащих две легких и две тяжелых цепи, в виде отдельных тяжелых цепей и легких цепей, или в виде одноцепочечных антител, в которых вариабельные домены тяжелой и легкой цепей соединены с помощью спейсера. Способы получения одноцепочечных антител хорошо известны в данной области.
Характерные гуманизированные антитела, полученные с помощью пересадки CDR и венирования, раскрыты в патентах США № 7780963, 7731962 и 7811563.
Также рассматривают диатела. Диатело обозначает разработанную конструкцию антитела, полученную путем выделения связывающих доменов (тяжелой и легкой цепей) связывающего антитела, и дополнения связывающей группой, которая соединяет или функционально связывает тяжелую и легкую цепи на одной полипептидной цепи, сохраняя таким образом, связывающую функцию (см., Holliger, et al., 1993, Proc. Natl. Acad. Sci. USA 90:6444; Poljak, 1994. Structure 2:1121-1123). При этом образуется, по существу, радикально укороченное антитело, обладающее только вариабельным доменом, необходимым для связывания антигена. Путем использования линкера, который является настолько коротким, что допускает спаривание двух доменов в одной цепи, указанные домены вынуждают образовывать пару с комплементарными доменами другой цепи и создавать два антигенсвязывающих сайта. Указанные димерные фрагменты антитела, или диатела являются бивалентными и биспецифическими. Специалисту в данной области понятно, что может быть использован любой способ получения диател. Подходящие способы описаны Holliger, et al., 1993, выше; Poljak, 1994, выше; Zhu, et al., 1996, Biotechnology 14:192-196, и патентом США № 6492123, которые включены в описание в качестве ссылки в полном объеме.
Фрагменты выделенного антитела по настоящему изобретению также явным образом включены в настоящее изобретение. Подразумевается, что фрагменты включают Fab-фрагменты, F(ab')2 фрагменты, F(ab') фрагменты, биспецифические scFv-фрагменты, Fv-фрагменты и фрагменты, полученные с помощью экспрессионной библиотеки Fab, а также пептидные аптамеры. Например, фрагменты F(ab')2 получают расщеплением пепсином молекулы антитела по настоящему изобретению, в то время как Fab-фрагменты получают путем восстановления дисульфидных мостиков фрагментов F(ab')2. Альтернативно, можно создать экспрессионные библиотеки Fab для обеспечения быстрой удобной идентификации моноклональных Fab-фрагментов с желаемой специфичностью (см., Huse, et al., 1989, Science, 254: 1275-1281). В конкретных вариантах осуществления фрагменты антитела по настоящему изобретению представляют собой фрагменты нейтрализующих антител, которые сохраняют их участок связывания вариабельной области, т.е. антигенсвязывающий фрагмент. Характерными являются фрагменты F(ab')2, фрагменты F(ab') и Fab-фрагменты. См., в целом, Immunology: Basic Processes, 1985, 2nd edition, J. Bellanti (Ed.) pp. 95-97.
Пептидные аптамеры, которые избирательно распознают многомерные конформации ADDL, можно рационально разработать или провести скрининг в библиотеке аптамеров (например, предоставляемой Aptanomics SA, Lyon, France). В целом, пептидные аптамеры представляют собой синтетические распознающие молекулы, конструкция которых основана на структуре антител. Пептидные аптамеры состоят из вариабельной пептидной петли, присоединенной обоими концами к белковому остову. Указанное двойное структурное ограничение значительно увеличивает аффинность связывания пептидного аптамера до уровней, сравнимых с уровнем антитела (наномолярный диапазон).
Характерные последовательности нуклеиновой кислоты, кодирующие вариабельные области тяжелой и легкой цепей, для применения в получении антитела или фрагментов антитела по настоящему изобретению раскрыты в описании в SEQ ID NO:14 и 16. Специалисту в данной области понятно, что вариабельные области тяжелой цепи, раскрытые в настоящем описании, такие как область, представленная в SEQ ID NO:16, можно использовать в комбинации с любой из вариабельных областей легкой цепи, раскрытых в настоящем описании, для создания антител с модифицированными аффинностями, диссоциацией, эпитопами и т.п.
Антитела или фрагменты антител по настоящему изобретению могут обладать дополнительными функциональными группами, присоединенными к ним. Например, микросфера или микрочастица может быть прикреплена к антителу или к фрагменту антитела, как описано в патенте США № 4493825, раскрытие которого включено в описание в качестве ссылки в полном объеме.
Кроме того, конкретный вариант осуществления включает антитело или фрагменты антитела, которые видоизменяют и проводят отбор на предмет повышенной аффинности к антигену, нейтрализующей активности {т.е. способности блокировать связывание ADDL с нервными клетками или способности блокировать сборку ADDL), или на предмет измененной константы диссоциации. Мутаторные штаммы E. coli (Low, et al., 1996, J. Mol. Biol., 260:359-368), “перетасовка цепей” (Figini, et al., 1994, выше) и мутагенез с помощью ПЦР являются общепризнанными методами изменения молекул нуклеиновой кислоты, кодирующих антитела. В качестве иллюстрации, можно провести отбор на повышенную аффинность путем взаимодействии большого числа фаговых антител с небольшим количеством биотинилированного антигена, так что антитела конкурируют за связывание. В этом случае число молекул антигена должно превышать число фаговых антител, но концентрация антигена должна быть несколько ниже константы диссоциации. Таким образом, преимущественно мутированные фаговые антитела с повышенной аффинностью связываются с биотинилированным антигеном, в то время как большая часть фаговых антител с более слабой аффинностью остается несвязанной. Затем с помощью стрептавидина можно улучшить фаговые антитела из смеси повышенной аффинностью (Schier, et al., 1996, J. Mol. Biol. 255:28-43). Характерные аминокислотные последовательности CDR3 легких цепей с измененной аффинностью раскрыты в описании (см. таблицу 4), причем конкретные варианты осуществления включают аминокислотную последовательность CDR3 легкой цепи SEQ ID NO: 3 и специфические варианты осуществления SEQ ID NO:7-13. Настоящее изобретение также включает альтернативные варианты CDR1 (SEQ ID NO:53) и CDR2 (SEQ ID NO:54) легкой цепи.
Для некоторых терапевтических применений желательно уменьшение диссоциации между антителом и антигеном. Для достижения этого, фаговые антитела связывают с биотинилированным антигеном и добавляют избыток небиотинилированного антигена. По истечении некоторого периода времени можно собрать с помощью стрептавидина преимущественно фаговые антитела с более низкой константой диссоциации (Hawkins, et al., 1992, J. Mol. Biol. 226:889-96).
Различные иммунотесты, включающие тесты, раскрытые в описании, можно использовать для скрининга с целью идентификации антител или их фрагментов, обладающих желаемой специфичностью в отношении многомерной конформации ADDL. Многочисленные протоколы конкурентного связывания (например, ELISA), тесты латекс-агглютинации, радиоиммунные тесты, анализы кинетики (например, анализ BiacoreTM) с применением либо поликлональных, либо моноклональных антител, или их фрагментов, хорошо известны в данной области. Такие иммунотесты обычно включают количественное определение образования комплексов между специфическим антителом и узнаваемым им антигеном. Подходящим является двухстадийный иммунотест на моноклональной основе, с применением моноклональных антител, реактивных в отношении двух не реагирующих перекрестно эпитопов, но также можно использовать тест конкурентного связывания. Такие тесты также можно использовать для обнаружения многомерных конформаций ADDL в образце.
Также могут быть проведены другие тесты с участием антитела или фрагментов антитела для определения биологической активности, например, замещение связывания ADDL с нейронами или в культуре гиппокампальных клеток, или блокировка сборки ADDL для оценки нейтрализующей или фармакологической активности и возможной эффективности в качестве профилактического или терапевтического средства. Такие тесты описаны в настоящем описании и хорошо известны в данной области.
Антитела и фрагменты антител можно получить и поддерживать в виде гибридом или, альтернативно, можно произвести рекомбинантно в общепринятых системах экспрессии, включая, но без ограничения, E. coli, дрожжи (например, Saccharomyces spp. и Pichia spp.), бакуловирус, клетки млекопитающих (например, миеломные клетки, CHO, COS), растения или трансгенные животные (Breitling and Dübel, 1999, Recombinant Antibodies, John Wiley & Sons, Inc., NY, pp. 119-132). Антитела и фрагменты антител можно выделить любыми подходящими способами, включая, но без ограничения, аффинную хроматографию, молекулы, связывающие иммуноглобулины (например, протеины A, L, G или H), метки, функционально связанные с антителом или фрагментом антитела (например, His-tag, FLAG®-tag, Strep tag, c-myc tag) и т.п. См., Breitling and Dübel, 1999, выше.
Антитело и фрагменты антитела по настоящему изобретению имеют ряд применений, включая диагностику заболеваний, связанных с накоплением ADDL, блокирование или ингибирование связывания ADDL с нервными клетками, блокирование сборки ADDL, профилактическое или терапевтическое лечение заболевания, связанного с ADDL, выявление терапевтических средств, которые предотвращают связывание ADDL с нейронами, и предотвращение гиперфосфорилирования тау-белка в положениях Ser202/Thr205.
Антитело или фрагменты антитела по настоящему изобретению полезны в способе блокирования или ингибирования связывания ADDL с нервными клетками. Указанный способ изобретения осуществляется путем взаимодействия нейрона in vitro или in vivo с антителом или фрагментом антитела по настоящему изобретению, так что связывание ADDL с нейроном блокируется. В конкретных вариантах осуществления антитело или фрагмент антитела по настоящему изобретению обеспечивает по меньшей мере 15%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, или 97% снижение связывания ADDL по сравнению со связыванием ADDL в условиях отсутствия антитела или фрагмент антитела. Степень, в которой антитело может блокировать связывание ADDL с нейроном, можно определить согласно методам, раскрытым в настоящем описании, а именно иммуноцитохимия или тест определения активности щелочной фосфатазы в клетках или любой другой подходящий тест. Антитела, особенно полезные для уменьшения связывания ADDL с нервными клетками, включают характерные антитела против ADDL, представленные в патентах США № 7731962, 7780963 и 7811563, а также их производные и фрагменты.
Антитело и фрагменты антитела по настоящему изобретению также полезны в способе блокирования или ингибирования сборки ADDL. Указанный способ включает приведение в контакт образца, содержащего пептиды β-амилоида 1-42, с антителом или фрагментом антитела по настоящему изобретению, так что ингибируется сборка ADDL. Степень, в которой антитело может блокировать сборку ADDL, можно определить согласно способам, раскрытым в описании, т.е. FRET или флуоресцентная поляризация или любой другой подходящий метод. Антитела, особенно полезные для блокирования сборки ADDL, включают антитела против ADDL, характеризующиеся аминокислотной последовательностью CDR3, изложенной в SEQ ID NO:10, а также ее производными и фрагментами.
Антитела, раскрытые в описании, также могут использоаться в способах профилактики фосфорилирования тау-белка в положениях Ser202/Thr205. Указанный способ включает приведение в контакт образца, содержащего тау-белок, с антителом или фрагментом антитела по настоящему изобретению, так что связывание ADDL с нейронами блокируется, предотвращая тем самым фосфорилирование тау-белка. Степень, в которой антитело может предотвращать фосфорилирование тау-белка в положениях Ser202/Thr205, можно определить согласно методам, раскрытым в описании, или с помощью любого другого подходящего теста.
Блокирование или уменьшение связывания ADDL с нейронами, ингибирование сборки ADDL, и предотвращение фосфорилирования тау-белка в положениях Ser202/Thr205, все выше перечисленное находит применение в способах профилактического или терапевтического лечения заболевания, связанного с накоплением ADDL. Соответственно, настоящее изобретение также включает применение антитела или фрагмента антитела в настоящем описании для профилактики или лечения заболевания, связанного с накоплением ADDL (например, болезнь Альцгеймера или сходные нарушения памяти). Результаты, полученные в данной области, показывают, что повышенные уровни Αβ, но не обязательно агрегированная бляшка, вызывают связанную с болезнью Альцгеймера деменцию и последующие нарушения, связанные с тау-белком. Диффундирующие лиганды, происходящие из Αβ, непосредственно вовлечены в нейротоксичность, связанную с болезнью Альцгеймера. Уровень техники показывает, что ADDL повышаются у трансгенных мышей и пациентов с болезнью Альцгеймера и модулируют функциональную активность, связанную с мнемоническими процессами в животных моделях. Таким образом, удаление указанной формы Αβ могло бы обеспечить ослабление нейротоксичности, связанной с болезнью Альцгеймера. По существу, лечение антителом по настоящему изобретению, которое уменьшает нагрузку ADDL на центральную нервную систему, может оказаться эффективным в лечении болезни Альцгеймера. Пациенты, нуждающиеся в лечении, включают индивидов с повышенным риском заболевания, но без проявляющихся симптомов, а также пациентов, имеющих симптомы в настоящее время. Для болезни Альцгеймера, фактически у каждого человека имеется повышенный риск возникновения болезни Альцгеймера, если он или она живет достаточно долго. Таким образом, антитело или фрагменты антитела по настоящему изобретению можно вводить профилактически общей популяции без необходимости проведения какой-либо оценки риска у подверженного заболеванию пациента. Настоящие способы особенно полезны для индивидов, которые характеризуются известным генетическим риском болезни Альцгеймера. Такие индивиды включают индивидов, имеющих родственников у которых диагностировано заболевание, и индивидов, у которых повышенный риск заболевания определяется путем анализа генетических или биохимических маркеров. Генетические маркеры риска болезни Альцгеймера включают мутации в гене APP, в особенности, мутации в положении 717 и в положениях 670 и 671, обозначаемые как мутации Hardy и Swedish, соответственно. Другие маркеры риска представляют собой мутации в генах пресенилинов, PS1 и PS2, и ApoE4, семейная история болезни Альцгеймера, гиперхолестеринемия или атеросклероз. Индивидов, в настоящее время страдающих от болезни Альцгеймера, можно выявить по характерной деменции, а также по наличию факторов риска, описанных выше. Кроме того, имеется ряд диагностических тестов для выявления индивидов, которые имеют болезнь Альцгеймера. Тесты включают измерение концентраций тау-белка в CSF и Αβ1-42. Индивидов, страдающих от болезни Альцгеймера, также можно выявить с помощью критериев ADRDA или с помощью способа, раскрытого в настоящем описании.
У пациентов, не обнаруживающих симптомов заболевания, лечение можно начинать в любом возрасте (например, 10, 20, 30 лет). Обычно, однако, нет необходимости начинать лечения до достижения пациентом возраста 40, 50, 60 или 70 лет. Лечение обычно включает введение многочисленных доз в течение некоторого периода времени. Мониторинг лечения может осуществляться путем проведения анализа на присутствие ADDL в динамике по времени.
В терапевтических применениях фармацевтическую композицию или лекарство, содержащее антитело или фрагмент антитела по настоящему изобретению, вводят пациенту, потенциально или уже имеющему такое заболевание, связанное с накоплением ADDL, в количестве, достаточном для лечения, или по меньшей мере частичного прекращения симптомов заболевания (биохимических, гистологических и/или поведенческих), включая его осложнения и промежуточные патологические фенотипы во время развития заболевания. Для профилактики фармацевтическую композицию или лекарство, содержащее антитело или фрагмент антитела по настоящему изобретению, вводят пациенту, потенциально имеющему, или в другом случае имеющему повышенный риск заболевания, связанного с накоплением ADDL, в количестве, достаточном для достижения пассивной иммунизации пациента, тем самым устраняя или уменьшая риск, снижая тяжесть, или задерживая начало болезни, включая биохимические, гистологические и/или поведенческие симптомы заболевания, его осложнения и промежуточные патологические фенотипы, представленные во время развития заболевания. В некоторых способах введение средства уменьшает или устраняет миокогнитивное нарушение у пациентов, которые еще не имеют развившейся характерной патологии болезни Альцгеймера. В конкретных вариантах осуществления эффективное количество антитела или фрагмента антитела по настоящему изобретению представляет собой количество, которое обеспечивает по меньшей мере 15%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95% или 97% уменьшение связывания ADDL с нейронами у пациента по сравнению со связыванием ADDL в условиях отсутствия лечения. По существу, уменьшается недостаточность долговременной потенциации/формирования памяти.
Эффективные дозы композиций по настоящему изобретению для лечения описанных выше состояний изменяются в зависимости от многих различных факторов, включая способы введения, физиологическое состояние пациента, является ли пациент человеком или животным, другие вводимые лекарства, и является ли лечение профилактическим или терапевтическим. Обычно пациентом является человек, нечеловекообразные млекопитающие, такие как собаки, или трансгенные млекопитающие также могут получать лечение.
Лечебные дозы, как правило, титруют для оптимизации безопасности и эффективности. Для пассивной иммунизации антителом или фрагментом антитела, подходят диапазоны доз приблизительно 0,0001-100 мг/кг, и более часто 0,01-5 мг/кг веса тела хозяина. Например, дозы могут составлять 1 мг/кг веса тела или 10 мг/кг веса тела, или могут находиться в диапазоне 1-10 мг/кг. В некоторых способах два или более антител по настоящему изобретению с разными специфичностями связывания вводят одновременно, в таком случае доза каждого вводимого антитела находится в указанных пределах. Антитела обычно вводят неоднократно, где интервалы между отдельными дозами могут быть недельными, месячными или годичными. Характерный режим лечения включает подкожные дозы, вводимые один раз в две недели или раз в месяц. Интервалы также могут быть неодинаковыми, в соответствии с количественным определением в крови концентраций антитела к ADDL у пациента. В некоторых способах дозу регулируют для получения концентрации антитела в плазме крови 1-1000 мкг/мл и в некоторых способах 25-300 мкг/мл. Альтернативно, антитело или фрагмент антитела можно ввести в виде состава с замедленным высвобождением, в таком случае требуется менее частое введение препарата. Дозу и частоту введения варьируют в зависимости от периода полувыведения антитела у пациента. В целом, антитела человека и гуманизированные антитела характеризуются более продолжительным периодом полувыведения, чем химерные антитела и нечеловеческие антитела. Как указано выше, доза и частота введения могут варьировать в зависимости от того, является ли лечение профилактическим или терапевтическим. Для профилактики относительно низкую дозу вводят в относительно редкие интервалы в течение длительного периода времени. Некоторые пациенты продолжают принимать лечение в течение остатка своей жизни. Для лечебных целей иногда необходимо применение относительно высоких доз в относительно короткие интервалы времени до тех пор, пока прогрессирование заболевания не уменьшится или не прекратится, и предпочтительно до тех пор, пока у пациента не будет наблюдаться частичное или полное ослабление симптомов заболевания. Соответственно, пациент сможет получать лечение в профилактическом режиме.
Антитело и фрагменты антитела по настоящему изобретению можно вводить в виде компонента фармацевтической композиции или лекарственного средства. Фармацевтические композиции или лекарственные средства, как правило, содержат активное терапевтическое средство и ряд других фармацевтически приемлемых компонентов. См. Remington: The Science and Practice of Pharmacy, Alfonso R. Gennaro, editor, 20th ed. Lippincott Williams & Wilkins: Philadelphia, PA, 2000. Предпочтительная форма зависит от предполагаемого способа введения и терапевтического применения. Фармацевтические композиции могут содержать, в зависимости от желаемого препарата, фармацевтически приемлемые нетоксические носители или разбавители, которые определяют как носители, обычно используемые для создания фармацевтических композиций для введения животным или человеку. Разбавители выбирают таким образом, чтобы они не влияли на биологическую активность комбинации. Примеры таких разбавителей включают дистиллированную воду, физиологический раствор, забуференный фосфатом, растворы Рингера, раствор декстрозы и раствор Ханка.
Фармацевтические композиции также могут содержать большие, медленно метаболизируемые макромолекулы, такие как белки, полисахариды, такие как хитозан, полимолочные кислоты, полигликоливые кислоты и сополимеры (такие как латекс-функционализированная SEPHAROSETM, агароза, целлюлоза и т.п.), полимерные аминокислоты, сополимеры аминокислот и липидные агрегаты (такие как капельки масла или липосомы).
Введение фармацевтической композиции или лекарства по настоящему изобретению можно осуществить разными путями, включая, но без ограничения пероральный, местный, легочный, ректальный, подкожный, внутрикожный, интраназальный, внутричерепной, внутримышечный, внутриглазной или внутриоболочечный путь, или внутрисуставную инъекцию и т.п. Наиболее типичным путем введения является внутривенное введение, за которым следует подкожное, хотя другие пути могут быть столь же эффективны. Внутримышечная инъекция также может быть проведена в мышцы руки или ноги. В некоторых способах средства вводят непосредственно в определенную ткань, где скопились отложения, например, внутричерепная или внутриоболочечная инъекция. В некоторых вариантах осуществления антитело или фрагмент антитела вводят непосредственно внутрь черепа или CSF. В других вариантах осуществления антитело или фрагмент антитела вводят в виде композиции или устройства с замедленным высвобождением, таким как устройство MEDIPADTM.
В случае парентерального введения антитело или фрагменты антитела по настоящему изобретению можно вводить в виде инъекционных доз раствора или суспензии субстанции в физиологически приемлемом растворителе с фармацевтическим носителем, который может представлять собой стерильную жидкость, такую как вода, масла, физиологический раствор, глицерин или этанол. Кроме того, вспомогательные вещества, такие как смачивающие или эмульгирующие средства, сурфактанты, pH-буферные вещества и т.п. могут быть представлены в композиции. Другие компоненты фармацевтических композиций представляют собой компоненты нефтяного, животного, растительного или синтетического происхождения, например, арахисовое масло, соевое масло и минеральное масло. В целом, гликоли, такие как пропиленгликоль или полиэтиленгликоль, являются подходящими жидкими носителями, в особенности для инъекционных растворов. Антитела можно вводить в форме инъекции вещества замедленного всасывания или имплантного препарата, который может быть составлен таким образом, чтобы обеспечить замедленное высвобождение активного ингредиента.
Характерная композиция содержит выделенное антитело или фрагмент антитела по настоящему изобретению, включенный в состав в виде стерильной, прозрачной жидкости в концентрации по меньшей мере 10 мг/мл в изотоническом забуференном физиологическом растворе (10 мМ гистидина, 150 мМ хлорида натрия, 0,01% (масс/об) POLYSORBATE 80, pH 6,0). Характерный препарат антитела расфасован в виде однократной дозы, 0,6 мл стеклянные пузырьки, заполненные 3,3 мл раствора на пузырек. Каждый пузырек закрывают пробкой, покрытой тефлоном, и запечатывают алюминиевым колпачком.
Обычно композиции получают в виде инъекционных форм, либо в виде жидких растворов, либо в виде суспензий; также могут быть получены твердые формы, подходящие для растворения или суспендирования в жидких носителях перед выполнением инъекции. Препарат также может быть эмульгирован или заключен в липосомы, или микрочастицы, такие как полилактид, полигликолид, или сополимер для улучшенной доставки.
В случае суппозиториев, связующие вещества и носители включают, например, полиалкиленгликоли или триглицериды; такие суппозитории могут быть составлены из смесей, содержащих активный ингредиент в диапазоне 0,5%-10%, или более желательно 1%-2%.
Пероральные препараты содержат вспомогательные вещества, такие как маннит, лактоза, крахмал, стеарат магния, сахарин натрия, целлюлоза и карбонат магния с фармацевтической степенью чистоты. Указанные композиции имеют форму растворов, суспензий, таблеток, пилюль, капсул, препаратов с замедленным высвобождением или порошков и содержат 10%-95% активного ингредиента, или чаще 25%-70%.
Местное применение может иметь своим результатом трансдермальную или интрадермальную доставку. Топическое введение можно обеспечить путем совместного введения средства с холерным токсином или нейтрализованными производными или их субъединицами, или другими сходными бактериальными токсинами (см. Glenn, et al. (1998) Nature 391:851). Совместное введение можно осуществить при использовании компонентов в виде смеси или в виде связанных молекул, полученных химическим сшиванием или экспрессией в виде слитого белка.
Альтернативно, трансдермальную доставку можно осуществить методом чрезкожного введения или с применением трансферосом (Paul, et al.s 1995, Eur. J. Immunol. 25:3521-3524; Cevc, et al., 1998, Biochem. Biophys. Acta 1368:201-215).
Антитело или фрагмент антитела по настоящему изобретению необязательно можно вводить в комбинации с другими средствами, которые по меньшей мере частично эффективны в лечении амилоидогенетического заболевания. Например, настоящее антитело можно вводить с существующими паллиативными лекарствами от болезни Альцгеймера, такими как ингибиторы ацетилхолинэстеразы, например, ARICEPTTM, EXELONTM, и REMINYLTM и, с антагонистом NMDA, NAMENDATM. В дополнение к указанным утвержденным лекарственным препаратам, настоящее антитело можно использовать для обеспечения синергического/аддитивного положительного действия в любом из нескольких подходов, разрабатываемых в настоящее время для лечения болезни Альцгеймера, которые включают без ограничения, ингибиторы продукции Αβ и аггрегации.
Антитело или фрагменты антитела по настоящему изобретению также находят применение в идентификации терапевтических средств, которые предотвращают связывание ADDL с нейронами (например, с гиппокампальной клеткой), предотвращая таким образом последующие события, связанные с ADDL. Такой тест осуществляют путем взаимодействия нейрона с ADDL в присутствии средства и с применением антитела или фрагмента антитела по настоящему изобретению для определения связывания ADDL с нейроном в присутствии средства. Как понятно специалисту в данной области, средство, которое блокирует связывание ADDL с нейроном, будет снижать количество ADDL, связанного с нейроном, по сравнению с нейроном, который не взаимодействовал со средством; количество, которое можно обнаружить в иммунотесте с применением антитела или фрагмента антитела по настоящему изобретению. Подходящие иммунотесты для выявления связанных с нейронами ADDL раскрыты в описании.
Средства, скрининг которых можно провести с применением способа, предоставляемого в описании, включают многочисленные химические классы, хотя обычно они представляют собой органические молекулы, предпочтительно, небольшие органические соединения, характеризующиеся молекулярным весом выше 100 и меньше 2500 Дальтон. Средства охватывают функциональные группы, необходимые для структурного взаимодействия с белками, в особенности, для водородного связывания, и обычно включают по меньшей мере аминогруппу, карбонильную, гидроксильную или карбоксильную группу, предпочтительно, по меньшей мере две из указанных функциональных химических групп. Средства часто содержат циклический углерод или гетероциклические структуры и/или ароматические, или полиароматические структуры, замещенные одной или больше указанными выше функциональными группами. Средства также можно найти среди биомолекул, включая пептиды, антитела, сахариды, жирные кислоты, стероиды, пурины, пиримидины, производных, структурных аналогов или их комбинаций. Средства получают из широкого набора источников, включая библиотеки природных и синтетических соединений.
Ряд других реагентов, таких как соли и нейтральные белки можно включить в скрининговые тесты. Также можно использовать реагенты, которые другим путем увеличивают эффективность теста, такие как ингибиторы протеаз, ингибиторы нуклеаз, противомикробные средства и т.п. Смесь компонентов можно добавлять в любом порядке, который обеспечивает необходимое связывание.
Средства, выявленные с помощью анализа скрининга по настоящему изобретению, будут полезны при лечении амилоидогенетических заболеваний и/или таупатий. Кроме того, предполагается, что экспериментальные системы, используемые для иллюстрации указанных концепций, предоставляют средства исследования для оценки, выявления и скрининга новых лекарственных мишеней, связанных с индукцией бета-амилоидом фосфорилирования тау-белка.
Настоящее изобретение также относится к способам обнаружения ADDL и диагностирования заболевания, связанного с накоплением ADDL, с применением антитела или фрагмента антитела. Подразумевается, что заболевание, связанное с накоплением ADDL, включает любое заболевание, при котором накопление ADDL приводит к физиологическому ухудшению долговременной потенциации/формирования памяти. Заболевания указанного типа включают, но без ограничения, болезнь Альцгеймера и сходные нарушения памяти.
Согласно указанным способам образец, полученный от пациента, приводят в контакт с антителом или фрагментом антитела по настоящему изобретению, и связывание антитела или фрагмента антитела с образцом свидетельствует о наличии ADDL в образце. В контексте настоящего изобретения, подразумевается, что образец обозначает любую жидкость организма или ткань, анализ которой можно осуществить с применением иммунотестов. Подходящие образцы, которые можно анализировать согласно способам по настоящему изобретению включают, но без ограничения, образцы биопсии и образцы жидкости мозга пациента (например, млекопитающего, такого как человек). Для целей in vitro (например, в тестах мониторинга образования олигомеров), образец может представлять собой клеточную линию нервных клеток или образец ткани. Для диагностических целей предполагается, что образец может быть получен от индивида, который предположительно имеет заболевание, связанное с накоплением ADDL, или от индивида с повышенным риском заболевания, связанного с накоплением ADDL, например, индивида с семейным анамнезом, который предрасполагает индивида к заболеванию, связанному с накоплением ADDL.
Обнаружение связывания антитела или фрагмента антитела с ADDL в образце можно осуществить с применением любого стандартного иммунотеста (например, как раскрыто в описании), или альтернативно если фрагмент антитела представляет собой, например, пептидный аптамер, связывание можно обнаружить непосредственно, например, с помощью детектируемого маркерного белка (например, β-галактозидазы, GFP или люциферазы), объединенного с аптамером. Следовательно, наличие или отсутствие комплекса ADDL-антитело связано с наличием или отсутствием, соответственно, ADDL в образце и таким образом с наличием или отсутствием, соответственно, заболевания, связанного с накоплением ADDL. Предполагается, что одно или более антител или фрагментов антитела по настоящему изобретению можно использовать совместно с современными неинвазивными иммунными методами визуализации для существенного увеличения выявления и ранней диагностики заболевания, связанного с накоплением ADDL.
В целях упрощения диагностики, настоящее изобретение также относится к набору, содержащему антитело или фрагмент антитела. Набор включает контейнер, содержащий одно или более антитело, или фрагменты антитела, которые распознают многомерную конформацию ADDL, и инструкции по применению антитела для осуществления связывания с ADDL с образованием комплекса антитело-антиген и обнаружению образования комплекса антитело-антиген, так что присутствие или отсутствие комплекса антитело-антиген находится в корреляции с присутствием или отсутствием ADDL в образце. Примеры контейнеров включают многолуночные планшеты, которые обеспечивают одновременное обнаружение ADDL в многочисленных образцах.
Все ссылки, цитируемые в описании, включены в качестве ссылки в полном объеме.
Ниже следует подробное описание изобретения с помощью неограничивающих примеров.
ПРИМЕРЫ
В описании использовались следующие аббревиатуры: Ab: антитело; Αβ: белок амилоид бета; AD: Болезнь Альцгеймера; ADDL: диффундирующий лиганд, происходящий из β-амилоида (Aβ); Ag: антиген; APP: белок-предшественник амилоида; bADDL: биотинилированные ADDL; CSF: цереброспинальная жидкость; DMSO: диметилсульфоксид; hAPP: белок-предшественник амилоида человека; среда HAT: среда гипоксантин-аминоптерин-тимидин; HFIP: гексафтор-2-пропанол; IV: внутривенный; LB agar: агар с содержанием лизогенного бульона; SC: подкожный; PBS: забуференный фосфатом физиологический раствор; TEA: триэтиламин.
Пример 1
Общие материалы и методы
A. Получение моноклональных антител ADDL
Растворимые олигомеры Αβ, виды которого обозначаются в настоящем описании как "синтетические" ADDL, смешивали в отношении 1:1 с полным адъювантом Фрейнда (первая и вторая вакцинации) или с неполным адъювантом Фрейнда (все последующие вакцинации) и вводили с помощью подкожной (первые две вакцинации), или внутрибрюшинной инъекции трем мышам в общем объеме 1 мл/мышь. Каждая инъекция состояла из очищенных ADDL, эквивалентных 194±25 мкг общего белка. Мышам вводили инъекции приблизительно каждые три недели. После шести инъекций одну мышь умерщвляли и ее селезенку замораживали. Клетки селезенки мыши с самым высоким титром антител в сыворотке затем сливали с клетками миеломы SP2/0 в присутствии полиэтиленгликоля и помещали в шесть 96-луночных планшетов. Клетки культивировали при 37°C с 5% CO2 в течение 10 дней в 200 мкл селективной среды гипоксантин-аминоптерин-тимидин (HAT), которая состоит из обогащенной синтетической среды, такой как среда Дульбекко, модифицированная по способу Исков (IMDM), (Sigma-Aldrich, St. Louis, MO), дополненной 10% фетальной бычьей сыворотки (FBS), 1 мкг/мл HYBRI-MAX® (азасерин-гипоксантин; Sigma-Aldrich, MO), и 30% кондиционированных сред, собранных из клеточной культуры SP2/0. В культуры вносили однократно IMDM (Sigma-Aldrich, St. Louis, MO), дополненную 10% FBS на 10-й день, и культуральные супернатанты убирали на 14-й день для скрининга положительных лунок с помощью ELISA. Положительные культуры затем клонировали методом серийных разведений с вероятностью 0,3 клеток на лунку. Положительные клоны подтверждали с помощью ELISA и затем выращивали. В дальнейшем получали моноклональные антитела и очищали их для применения (QED Bioscience, San Diego, CA).
B. Получение ADDL и bADDL
ADDL получали по ранее описанным способам (Hepler, et al., 2006, Biochemistry, 45: 15157-15167; Shughrue, et al., 2010, Neurobiol. Aging, 31:189-202). Кратко, синтетический пептид Αβ1-42 (American Peptide, Sunnyvale, CA) растворяли в гексафтор-2-пропаноле (HFIP) в концентрации 10 мг/мл, и инкубировали при комнатной температуре (RT) в течение одного часа. Пептидный раствор распределяли по 50 мкл аликвотам в полипропиленовые 1,5 мл микроцентрифужные пробирки. Удаляли HFIP с применением SpeedVac® (Thermo-Fisher Scientific, Waltham, MA), и полученные пептидные пленки хранили в высушенном состоянии при -70°C до использования. Растворяли 0,5 мл высушенной пленки HFIP в 22 мкл безводного диметилсульфоксида (DMSO) с перемешиванием в течение 10 минут на смесителе вортекс. Затем 1 мл холодной среды F12 Ham's без фенола красного (United Biosource, San Francisco, CA) быстро добавляли к смеси DMSO/пептид. Пробирку закрывали, переворачивали, чтобы обеспечить полное перемешивание, и инкубировали в течение ночи при 4°C. На следующее утро образцы центрифугировали в течение десяти минут при 12000xg в микроцентрифуге Beckman (Beckman Coulter, Brea, CA), эксплуатируемой при 2-8°C. Супернатант собирали и фильтровали через ym 50 (отсечение по молекулярной массе 50000 кДа) Centricon® центрифужный фильтр (Millipore, Billerica, MA) для обогащения олигомерных соединений. Биотинилированные ADDL (bADDL) получали с применением тех же способов, но начиная с N-концевого биотинилированного пептида Αβ1-42 (American Peptide, Sunnyvale, CA).
C. Препараты мономеров и фибрилл
Для получения препаратов мономеров пептидную пленку RT Αβ1-40 или Αβ1-42 растворяли в 2 мл 25 мМ боратного буфера (pH 8,5) на мг пептида, делили на аликвоты и замораживали при -70°C до использования. Препараты фибрилл получали добавлением 2 мл 10 мМ соляной кислоты на мг пленки пептида Αβ1-42. Раствор перемешивали на смесителе vortex при самой низкой возможной скорости в течение пяти-десяти минут и полученный препарат хранили при 37°C в течение 18-24 часов перед использованием.
D. Первичные культуры нейронов
Первичные культуры нейронов получали из гиппокампальных и/или кортикальных тканей крыс, приобретенных в BrainBits (Springfield, IL). После диссоциации клетки помещали в концентрации 35000 клеток/лунку в 96-луночные планшеты предварительно покрытые ламинином и поли-D-лизином (Corning Life Sciences, Lowell, MA). Клетки выдерживали при 37°C с 5% CO2 в среде (Neurobasal, дополненная 2% B27, 1% L-глутамин, и 1% pen/strep; Invitrogen, Carlsbad, CA) в течение двух-трех недель и затем использовали в исследованиях связывания.
E. Клеточный анализ связывания ADDL
Для оценки эффекта антител против ADDL на блокирование связывания ADDL, антитела против ADDL смешивали с 500 нМ bADDL, с конечными концентрациями антител в диапазоне 1,8 нМ - 450 нМ. В качестве контроля, денатурированное нагреванием антитело (98°C в течение 30 минут) в аналогичной концентрации смешивали с bADDL. Смеси антитело-bADDL инкубировали в силиконизированных центрифужных микропробирках (Fischer Scientific, Pittsburgh, PA) при 37°C в течение одного часа с постоянным непрерывным вращением при низкой скорости. Смеси затем вносили в первичные культуры гиппокампальных и/или кортикальных клеток и инкубировали при 37°C в течение одного часа. Инкубацию останавливали удалением культуральной среды. Проводили фиксационную и постфиксационную обработку клеток, как описано выше. Затем клетки инкубировали со стрептавидином, конъюгированным со щелочной фосфатазой (AP) при 4°C в течение ночи, отмывали пять раз PBS и проводили реакцию с хемилюминесцентным субстратом Tropix® CDP®-Star (Life TechnologiesTM, Carlsbad, CA) при комнатной температуре в течение 30 минут. Интенсивность связывания bADDL измеряли и записывали с помощью микропланшетного ридера EnVision® (PerkinElmer, Waltham, MA).
F. ELISA
Биотинилированные ADDL (bADDL) или мономерные Αβ1-40 или Αβ1-42 добавляли в покрытый стрептавидином планшет для высокоэффективного анализа (Sigma-Aldrich, St. Louis, MO) с 100 мкл на лунку покрывающего реагента в PBS, по 1 мкМ и инкубировали в течение двух часов при комнатной температуре. Планшеты отмывали PBS с 0,05% Tween (шесть раз) и затем только PBS (три раза) перед блокированием лунок 5% раствором нежирного сухого молока PBS в течение одного часа при комнатной температуре. Затем лунки отмывали и добавляли серийные разведения образцов антитела в планшеты и оставляли для связывания в течение двух часов при комнатной температуре. После инкубации и отмывки связывание антитела определяли с помощью козьего вторичного антитела против IgG-Fc человека, конъюгированного с пероксидазой хрена (HRP) (1:1000; один час при комнатной температуре). Визуализировали HRP-метку с помощью тетраметилбензидина (Virolabs, Chantilly, VA), в качестве субстрата и проводили считывание при 450 нм на микропланшетном ридере.
Пример 2
Селекция антител против ADDL
A. Пэннинг библиотеки гуманизированных антител
Создавали аффинно зрелую библиотеку гуманизированного антитела против ADDL, h3B3, (см. патент США 2006/0228349 и патент США 2008/0175835), в которой часть аминокислотных последовательностей CDR3 легкой цепи подвергали случайному мутагенезу. Для покрытия всей области CDR3 строили две подбиблиотеки. Одна библиотека состояла из родительской вариабельной области тяжелой цепи и измененных аминокислот в левой половине CDR3 легкой цепи, и другая библиотека содержала измененные аминокислоты в правой половине CDR3 легкой цепи. Аналогичную стратегию применяли при случайном мутагенезе CDR тяжелой цепи с тремя подбиблиотеками.
Гуманизированное 3B3 (h3B3) подвергали аффинному созреванию с помощью известных в данной области способов. Вариабельные области h3B3 клонировали в векторе Fab-дисплея (pFab3D). В данном векторе последовательности, кодирующие вариабельные области тяжелой и легкой цепей, помещали в рамку считывания так, чтобы обеспечить их соединение с доменом CH1 константной области и константной области каппа-цепи, соответственно. В Fab3D эпитоп myc и шесть последовательных остатков гистидина находились за последовательностью CH1, которую в свою очередь соединяли с фаговым белком pIII с целью получения дисплея. Проводили случайный мутагенез по всем положениям последовательностей CDR3 тяжелой и легкой цепей с использованием вырожденных последовательностей, используемых в качестве праймеров ПЦР. Чтобы получить соответствующий физический размер, получали подбиблиотеки, каждая из которых фокусируется на 5-6 аминокислотах. ДНК-вектор 3B3 человека (H3B3) использовали в качестве ДНК-матрицы для амплификации как тяжелой, так и легкой цепи с использованием мутантных ПЦР-праймеров (таблица 1). После ПЦР-амплификации синтезированные фрагменты ДНК подвергали электрофорезу в 1,3% агарозном геле, праймеры удаляли и фрагменты вариабельных областей расщепляли ферментами рестрикции: для клонирования вариабельных областей легких цепей использовали участки клонирования BsiWI и Xbal, а для клонирования вариабельных областей тяжелых цепей используют участки XhoI и ApaI.
SEQ ID NO:24
SEQ ID NO:27
Для построения библиотеки аффинной зрелости в векторе фагового дисплея pFab3D, ДНК pFab3D-3B3 расщепляли той же парой ферментов рестрикции, очищали и ПЦР-фрагменты вариабельных областей тяжелых и легких цепей лигировали с помощью лигазы T4 (Invitrogen) в течение ночи при 16°C. Затем продукты лигирования трансфицировали в подходящие для электропорации клетки E. coli TG1 (Stratagene, Agilent Technologies, Santa Clara, CA) и аликвоты бактериальной культуры помещали на планшеты с LB агар-карбенициллином (50 мкг/мл) для определения размера библиотеки. Оставшиеся культуры либо помещали на большой планшет с карбенициллином и инкубировали при 30°C в течение ночи для получения стока библиотеки E. coli, либо инфицировали вспомогательным фагом M13K07 (Invitrogen, Carlsbad, CA, 1011 БОЕ/мл) путем инкубирования при комнатной температуре и при 37°C в течение десяти минут. Затем добавляли среду 2YT с карбенициллином (50 мкг/мл) и инкубировали при 37°C в течение одного часа при взбалтывании. Затем добавляли канамицин (70 мкг/мл) и выращивали культуры в течение ночи при 30°C при перемешивании. Супернатант фаговой культуры титровали и концентрировали осаждением с 20% (об/об) PEG (полиэтиленгликоль)/NaCl, ресуспендировали в PBS, стерилизовали с помощью 0,22 мкм фильтра и делили на аликвоты для пэннинга фаговой библиотеки.
Затем поводили пэннинг фаговой библиотеки, как кратко изложено в таблице 2.
Вхождение фагов из фаговых библиотек Fab-дисплея (100 мкл, приблизительно 1011-12 БОЕ) блокировали с помощью 900 мкл блокирующего раствора (3% раствор нежирного сухого молока в PBS) для уменьшения неспецифического связывания с поверхностью фага. Бусины, покрытые стрептавидином, получали путем накопления 200 мкл суспензии бусин в магнитном сепараторе и удаления супернатантов. Затем бусины суспендировали в 1 мл блокирующего раствора и помещали на роторный смеситель на 30 минут. Для удаления фага, не специфически связывающегося со стрептавидином, блокированную фаговую библиотеку смешивали с блокированными бусинами, покрытыми стрептавидином, и помещали на роторный смеситель на 30 минут. Суспензии фагов, полученных в результате процесса отмены селекции, переносили в новую пробирку и 200 мкл антигена, 10% bADDL добавляли и инкубировали в течение двух часов для связывания антитела и антигена. После инкубации смесь добавляли к блокированным бусинам, покрытым стрептавидином, и инкубировали на роторном смесителе в течение одного часа для иммобилизации комплекса Ab/Ag на бусинах, покрытых стрептавидином. Бусины с иммобилизованными 10% bADDL/фаг комплексами отмывали пять раз PBS/0,05% Tween 20 и затем дважды только PBS. Связанные фаги элюировали из bADDL с помощью 200 мкл 100 мМ TEA (Sigma Aldrich, St. Louis, MO) и инкубировали в течение двадцати минут. Элюированный фаг затем переносили в 50 мл пробирку, нейтрализовали 100 мкл 1M Tris-HCl, pH7,5 и добавляли к 10 мл клеток TG1 E. coli со значением OD 600 нм в диапазоне 0,6-0,8. После инкубации при 37°C с перемешиванием в течение одного часа, аликвоты культуры помещали на планшеты с LB агаром-карбенициллином (50 мкг/мл) для титрования числа фагов на выходе, и оставшиеся бактерии центрифугировали и суспендировали в 500 мкл 2xYT среды (Teknova, Hollister, CA,) высевали на планшеты с агаром для биотеста YT (Teknova, Hollister, CA), содержащие 100 мкг/мл ампициллина и 1% глюкозы. Планшеты для биотеста инкубировали в течение ночи при 30°C.
После каждого раунда пэннинга отдельные колонии выбирали случайным образом для получения фагов в 96-луночных планшетах. Методы получения фагов в 96-луночном планшете аналогичны методам, описанным выше, за исключением того, что не использовали стадию преципитации фагов. Культуральные планшеты, содержащие колонии, которые выращивали в 120 мкл 2xTY среды с 100 мкг/мл ампициллина и 0,1% глюкозы, инкубировали в течение ночи в шейкере HiGro® (Genomic Solutions, Ann Arbor, MI) при 30°C с перемешиванием при 450 об/мин. Фаговые супернатанты (приблизительно 100 мкл) использовали непосредственно для анализа в тесте связывания ADDL в ELISA, описанном выше. Отличие заключается в том, что связывание фага с ADDL выявляли с помощью антитела против M13, конъюгированного с HRP (Amersham Bioscience, GE Healthcare, Waukesha, WI).
Пример 3
Идентификация антител против ADDL
В результате исследования созревания аффинности легких цепей, группа из семи клонов показала высокую активность связывания с ADDL по сравнению с h3B3 в тесте ELISA фаг/Fab (данные не представлены). Семь клонов отбирали для преобразования в IgG и моноклональные антитела получали, и очищали для последующего описания их характеристик.
A. Селекция антитела против ADDL
После проведения пэннинга и скрининга библиотек, как описано в Примере 2, семь главных Fab-клонов (таблицы 3-5) выбирали для преобразования в IgG. В таблице 3 представлено сходство клонов, выбранных из библиотеки созревания аффинности легких цепей, по отношению к родительскому антителу h3B3. В таблице 4A представлено в обобщенном виде число аминокислотных отличий в CDR3 легкой цепи выбранных клонов от CDR3 легкой цепи родительского антитела, h3B3. В таблице 4B представлено в обобщенном виде число аминокислотных отличий в CDR1 легкой цепи выбранных клонов от CDR3 легкой цепи родительского антитела 19.3. В таблице 4C представлено в обобщенном виде число аминокислотных отличий в CDR2 легкой цепи выбранных клонов от CDR3 легкой цепи родительского антитела 19.3. Таблица 5 представляет собой выравнивание участка (положения 21-117) вариабельных областей легкой цепи выбранных клонов и родительского антитела h3B3. Для каждого клона область CDR3 показана жирным шрифтом.
рованная LC
(SEQ ID NO:1)
(SEQ ID NO:55)
(SEQ ID NO:56)
(SEQ ID NO:57)
(SEQ ID NO:67)
(SEQ ID NO:68)
(SEQ ID NO:59)
(SEQ ID NO:60)
(SEQ ID NO:61)
(SEQ ID NO:62)
14.2 PASISCRSSQSIVHSNGNTYLEWYLQKPGQSPQLLIYKASNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSRVPPGFGQGTKLEIK (SEQ ID NO:34)
13.1 PASISCRSSQSIVHSNGNTYLEWYLQKPGQSPQLLIYKASNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSKAHPSFGQGTKLEIK (SEQ ID NO:35)
19.3 PASISCRSSQSIVHSNGNTYLEWYLQKPGQSPQLLIYKASNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSRLGPSFGQGTKLEIK (SEQ ID NO:36)
7.2 PASISCRSSQSIVHSNGNTYLEWYLQKPGQSPQLLIYKASNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSYAPPGFGQGTKLEIK (SEQ ID NO:37)
9.2 PASISCRSSQSIVHSNGNTYLEWYLQKPGQSPQLLIYKASNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSRAPPFFGQGTKLEIK (SEQ ID NO:38)
11.4 PASISCRSSQSIVHSNGNTYLEWYLQKPGQSPQLLIYKASNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSRVPVRFGQGTKLEIK (SEQ ID NO:39)
h3B3 PASISCRSSQSIVHSNGNTYLEWYLQKPGQSPQLLIYKASNRFSGVPDRFSGSGSGTDFTLKI
B. Преобразование IgG
Модифицированные IgG можно экспрессировать с помощью плазмидных векторов. Векторы экспрессии строили так, чтобы они содержали все необходимые компоненты за исключением вариабельных областей. В основных векторах экспрессия тяжелой и легкой цепей запускалась промотором CMV человека и сигналом полиаденилирования гена гормона роста крупного рогатого скота. В семи клонах, выбранных для преобразования IgG, вариабельная область тяжелой цепи соединена внутри рамки считывания с константной областью тяжелой цепи IgG2 человека (SEQ ID NO:20 и 21) в то время как вариабельная область легкой цепи слита внутри рамки считывания с константной областью легкой цепи каппа (SEQ ID NO:18 и 19). Лидерные последовательности тяжелой (SEQ ID NO:29 и 30) и легкой (SEQ ID NO:31 и 32) цепей, которые опосредуют секрецию антитела в культуральных средах, также слита внутри рамки считывания с вариабельными областями соответствующим образом. Для векторов экспрессии тяжелых цепей, константную область можно выбрать из изотипа другого подкласса, например, IgG1 или IgG2. Находящиеся между лидерной последовательностью и константной областью межгенные последовательности содержат клонирующие последовательности, позволяющие осуществить “бесшовное” слияние в рамке считывания экзогенного вариабельного участка с лидерной последовательностью по 5'-концу и константной областью по 3'-концу с помощью стратегии клонирования “In-Fusion” (Clontech, Mountain View, CA). Наборы In-FusionTM Dry-Down PCR Cloning Kits (Clontech, Mountain View, CA) использовали для ПЦР-амплификации вариабельных областей. Набор dry-down cloning kit содержал все необходимые компоненты для поведения реакции ПЦР. Добавляли ПЦР-праймеры и матрицу ДНК. Экспрессионные векторы содержали oriP из генома вируса EBV. Пару векторов oriP/EBNA1 часто используют для продления присутствия вектора экспрессии внутри трансфицированных клеток и широко применяют для увеличения продолжительности экспрессии (Lindner, et al., 2007, Plasmid 58:1-12), достигая пролонгированной экспрессии в клетках 293EBNA бактериальных последовательностей маркера селекции по канамицину и точки начала репликации в E. coli. После вставки в вектор последовательностей, кодирующих вариабельные участки, IgG непосредственно экспрессировали в клетках млекопитающих. Все вариабельные области тяжелых цепей клонировали в вектор экспрессии IgG1 (pV1 JNSA-BF-HCG1) и вариабельные области легкой цепи клонировали в соответствующий вектор экспрессии цепей каппа или лямбда (pV1 JNSA-GS-FB-LCK).
C. Клонирование антител
Процедура клонирования для векторов экспрессии полученного антитела состояла в следующем. Вариабельные области подвергали ПЦР-амплификации, при которой реакции ПЦР осуществляли в объеме 25 мкл, содержащем высококачественную мастер-смесь ПЦР, объем матрицы 1 мкл и прямой и обратный праймеры: по 1 мкл каждый. Условия ПЦР: 1 цикл при 94°C, 2 минуты; 25 циклов при 94°C, 1,5 минуты; 60°C, 1,5 минуты; 72°C, 1,5 минуты и 72°C, 7 минут; 4°C до удаления. Продукты ПЦР затем расщепляли с помощью DpnI и очищали с помощью набора QIAquick plate kit (Qiagen, Venlo, The Netherlands). 100 нг соответствующих предварительно линеаризированных векторов, кодирующих тяжелую или легкую цепи, отжигали с 10 нг ПЦР-фрагмента, полученного в результате реакции “In-Fusion” (IN-Fusion Dry-Down Cloning Kit, Clontech, Mountain View, CA). Реакционную смесь переносили к XL2 Blue MRF' компетентным клеткам и выращивали в течение ночи на планшетах с агаром, содержащем 50 мкг/мл канамицина. Конструкции легких цепей расщепляли с помощью HindIII+NotI и конструкции тяжелых цепей расщепляли с помощью AspI+HindIII для проверки структуры с помощью рестрикционного анализа. Последовательности ДНК для всех клонов подтверждали секвенированием.
D. Экспрессия антитела в клетках млекопитающих и очистка
Подтвержденные секвенированием конструкции ДНК легких и тяжелых цепей трансфицировали в клетки 293 Freestyle (Invitrogen, Carlsbad, CA). Клетки 293 Freestyle трансфицировали с помощью 293 Transfectin (Invitrogen, Carlsbad, CA). Монослойную культуру клеток EBNA трансфицировали с помощью трансфекционных реагентов на основе PEI. Трансфицированные клетки инкубировали при 37°C/5% CO2 в течение семи дней в среде Opti-MEM, не содержащей сыворотки (Invitrogen, Carlsbad, CA). Среду собирали, осаждали центрифугированием, фильтровали через фильтрационную систему 0,22 мкм (Millipore, Billerica, MA), и затем концентрировали с помощью центрифужного фильтра Centricon® (Millipore, Billerica, MA). Концентрированную среду смешивали 1:1 с буфером для связывания (Pierce, Thermo Fisher Scientific, Rockford, IL), и затем помещали в колонку предварительно уравновешенную протеином A/G (Pierce, Thermo Fisher Scientific, Rockford, IL) или HI trap rProtein A FF от GE Healthcare, Waukesha, WI. Загруженную колонку промывали буфером для связывания и элюировали элюирующим буфером (Pierce, Thermo Fisher Scientific, Rockford, IL). Элюированное антитело немедленно нейтрализовали и диализовали против буфера PBS в течение ночи. Диализованное антитело концентрировали с помощью центрифужного фильтра Amicon (Pierce, Thermo Fisher Scientific, Rockford, IL) и концентрацию белка определяли по значению OD280нм с коэффициентом поглощения 1,34 мг/мл. Очищенное антитело анализировали с применением SDS-PAGE (Invitrogen, Carlsbad, CA), или с помощью набора Protein LabChip (Caliper LifeSciences, Hopkinton, MA). SDS-PAGE проводили при невосстанавливающих условиях.
Мутагенез аспарагина в положении 33 (N33) CDR1 легкой цепи для антитела 19.3 в N33S (SEQ ID NO:55), N33T (SEQ ID NO:56), N33E (SEQ ID NO:67), или N33D (SEQ ID NO:68) осуществляли с помощью сайт-направленного мутагенеза из вектора экспрессии WT pV1 JASN-GS-19.3-LCK с применением набора для сайт-направленного мутагенеза QuikChange II XL (Agilent Technologies, La Jolla, CA). Кодон AAT для N трансформировался в AGT для S в 19.3S33 (SEQ ID NO: 55), ACT для T в 19.3T33 (SEQ ID NO:56), GAA для E в 19.3E33 (SEQ ID NO:67), или GAT для D в 19.3D33 (SEQ ID NO:68), и новые кодоны в указанном положении подтверждали секвенированием ДНК. Для получения полноразмерных антител IgG указанных мутантов, соответствующие плазмиды, содержащие гены легких цепей, соединяли с родственной плазмидой, содержащей ген тяжелой цепи, pV1JNSA-19.3-HCG2, для временной трансфекции в клетки 293 FreeStyle (Invitrogen, Carlsbad, CA). Способы экспрессии и очистки описаны выше в данном примере. Аликвоты очищенных мутантных антител вместе с родительским антителом 19.3 (SEQ ID NO:1) инкубировали в различных концентрациях при 4°C, 25°C или 40°C в течение месяца перед использованием в тестах ELISA, результаты которых представлены на фигурах 4A-4C.
Пример 4
Описание антител против ADDL
Отобранные антитела против ADDL, т.e. антитела, полученные из родительского антитела, h3B3, сначала оценивали в трех-направленном тесте Αβ ELISA для определения связывания антитела с мономерным Αβ, ADDL и фибриллярной формой Αβ. Как показано на фигуре 1, за исключением антитела 9.2, все антитела против ADDL показали преимущественное связывание с ADDL, по сравнению с h3B3, селективными (Соед. 1 и 3: связывали только ADDL), неселективными (Соед. 2: связывало все оцениваемые формы Αβ) препаратами сравнения, и контролем (антитело отсутствует). Антитело 9.2 показало низкое связывание в отношении всех форм Αβ, что позволяет предположить, что на его аффинность связывания пострадала при преобразовании IgG и/или продукции антитела. Строили полную титрационную кривую (фигура 2) для каждого антитела и для антитела h3B3 для определения аффинности связывания в отношении ADDL, в сравнении с аффинностью связывания в отношении мономерного Αβ. Тем не менее, шесть из семи антител со зрелой аффинностью показали предпочтительное связывание в отношении ADDL. Авторы изобретения ранее показали, что некоторые антитела против ADDL, предпочтительно связывающиеся с ADDL, не способны предотвратить связывание ADDL с первичной культурой гиппокампальных нейронов (Shughrue, et al., 2010, Neurobiol. Aging, 31: 189-202, Фигура 1).
Поскольку предпочтительное связывание с ADDL само по себе не может служить точным показателем эффективности, желательно идентифицировать антитела против ADDL, которые также блокируют связывание ADDL с нейронами, которое можно оценить в клеточном тесте связывания, как указано ниже. Антитела, предварительно, инкубировали с ADDL и затем добавляли в первичные культуры гиппокампальных клеток, чтобы оценить блокирование связывания ADDL. Результаты исследования показали, что антитела против ADDL, в особенности антитело 19.3, значительно уменьшали связывание ADDL с нейронами (Фигура 3). Однако заметное снижение активности антител в данном тесте наблюдали, если антитела подвергали термической денатурации (фигура 3).
Определение EC50. В планшеты с высоким связыванием белка (Costar, Corning, Lowell, MA), наносили раствор лиганда-мишени в PBS и оставляли на ночь, при 4°C. Концентрация иммобилизуемого белка в растворе составляла 100 пмоль/лунку в случае Αβ40 (American Peptide, Sunnyvale, CA) и 50 пмоль/лунку в случае ADDL. ADDL получали, как описано в Примере 1B. На следующий день планшеты отмывали пять раз PBS+0,05% Tween-20 (Sigma Aldrich, St. Louis, MO) и блокировали в течение ночи блокирующим буфером с казеином (Thermo Scientific, Waltham, MA) и 0,05% Tween-20. Три характерных антитела, 19.3 (Фиг. A), 19.3S33 (Фиг. 4B), и 19.3T33 (Фиг. 4C), получали как описано в Примере 3, тестировали в количестве от 15 мкг/мл до 0 мкг/мл в 12-точечных тройных сериях разведений. После инкубации в течение 2 часов при комнатной температуре, планшеты отмывали и щелочную фосфатазу, конъюгированную с античеловеческим IgG (ThermoScientific, Waltham, MA) добавляли в 0,08 мкг/мл. После инкубации в течение 45 минут при комнатной температуре, планшеты отмывали и добавляли хемилюминисцентный субстрат Tropix® CDP®-Star (Life TechnologiesTM, Carlsbad, CA). Люминесценцию измеряли через 30 минут на микропланшетном ридере EnVision® (PerkinElmer, Waltham, MA). Построение кривых по точкам выполняли с применением программного обеспечения GraphPad Prism (GraphPad Software, Inc., San Diego, CA).
Пример 5
Связывание FcRn антител против ADDL in vitro
Для описания способности антител против ADDL связываться и диссоциировать с иммобилизованным FcRn человека, семь антител против ADDL оценивали в тесте связывания FcRn с помощью устройства Biacore, системы-заменителя, используемой для оценки PK антитела и расчета конечного периода полувыведения (t1/2) антител у приматов.
Кратко, очищенный белок FcRn человека иммобилизовали на биосенсорный чип устройства Biacore CM5 и PBSP (50 мМ NaPO4, 150 мМ NaCl и 0,05% (об/об) Surfactant 20), pH 7,3 использовали в качестве электропроводного буфера. Разводили mAb PBSP pH 6,0 до концентрации 100 нМ, оставляли для связывания с FcRn в течение 3 мин для достижения состояния равновесия с последующей диссоциацией в электропроводном буфере со значением pH 7,3. Точку получения отчета (состояние равновесия) устанавливали на момент времени 5 секунд после окончания связывания mAb и значение "% связывания" рассчитывали как RUсостояние равновесия/RUсвязывание (%). Заявители обнаружили, что моноклональные антитела (mAb) с одинаковыми Fc-последовательностями, но различными Fab-доменами могут связываться и диссоциировать с FcRn со значительными различиями (данные не представлены). Кроме того, наблюдали очевидную взаимосвязь между диссоциацией при нейтральном значении pH и фармакокинентикой in vivo, в которой mAbs с медленно диссоциирующими фракциями (т.e. с более высоким значением "% связывания") имели тенденцию к проявлению более низких значений t1/2 in vivo. Данную особенность использовали в качестве инструмента скрининга in vitro для определения фармакокинетики антител.
Проводили сравнение семи антител против ADDL вместе с антителом h3B3, двумя антителами, предпочтительно, связывающимися с ADDL (Соед. 1 и 3) и неселективным (Соед. 2: связывает все оцениваемые формы Αβ) антителом сравнения в тесте связывания FcRn. Получали сенсограмму (Фигура 5), на которой показано начальное связывание антитела при pH 6,0 и затем диссоциация антитела при pH 7,3, начиная от 180 секунд. Как показано на фигуре 5, наблюдалось заметное различие между h3B3 и другими оцениваемыми антителами. В то время как антитело h3B3 характеризовалось высоким процентом связывания с FcRn, семь антител против ADDL по настоящему изобретению, а также два антитела сравнения проявляли значительно более низкое связывание.
Пример 6
Описание антитела против ADDL 19.3
Антитело со зрелой аффинностью 19.3 выбирали для дальнейшего описания. Определяли полную последовательность ДНК и расшифрованную аминокислотную последовательность вариабельной области легкой цепи, SEQ ID NO:14 и 15, соответственно. Выравнивание последовательностей вариабельных областей тяжелой (SEQ ID NO:17) и легкой (SEQ ID NO:15) цепей представлено на фигуре 6A, вместе с наиболее близкой последовательностью, полученной из зародышевой линии (SEQ ID NO:47). 3D модель вариабельных областей тяжелой и легкой цепей и расположение шести областей, определяющих комплементарность (CDR), представлена на фигуре 6B.
Проводили анализы BiacoreTM (GE Healthcare, Waukesha, WI) и KinExA (Sapidyne, Boise, ID) для оценки аффинности связывания антитела против ADDL 19.3 с ADDL и определения селективности 19.3 в отношении ADDL в сравнении с мономерным Αβ. Технологии на основе BiacoreTM и KinExA широко используются для измерения аффинности связывания между макромолекулами, такими как антитело и белок-мишень. В технологии поверхностного плазмонного резонанса (SPR), на которой основана работа устройства BiacoreTM, количественные измерения связывающего взаимодействия между одной или более молекулами зависят от иммобилизации молекулы-мишени на поверхности сенсорного чипа. Связывающие партнеры мишени могут быть зарегистрированы, в то время как они проходят через чип. Поверхностный плазмонный резонанс (SPR) выявляет изменения в массе в водном слое в непосредственной близости от поверхности сенсорного чипа путем определения изменений показателя преломления. Когда молекулы в исследуемом растворе связываются с молекулой-мишенью, масса возрастает (ka), когда они диссоциируют, масса уменьшается (kd). Указанный простой принцип лежит в основе сенсограммы-непрерывного мониторинга в реальном масштабе времени ассоциации и диссоциации взаимодействующих молекул. Сенсограмма предоставляет количественную информацию в реальном режиме времени относительно специфичности связывания, активной концентрации молекулы в образце, кинетики и аффинности.
Технология KinExA от Sapidyne Instruments, Boise, Idaho, позволяет измерить константы связывания для описания биомолекулярных событий связывания в жидкой фазе, но не событий связывания между жидкой фазой и твердой фазой. В растворе связывающиеся партнеры достигают состояния равновесия после достаточного времени инкубации. Несвязанные молекулы подсчитываются с помощью титрования, что будет отражать долю молекул, связанных с партнерами. Метод KinExA не требует преобразования изучаемых молекул. В KinExA, реакция, параметры которой нужно измерить, происходит между немодифицированными молекулами в растворе. Поэтому устраняются вопросы, как преобразование меняет "нативные" реакции связывания. Метод KinExA предусматривает широкий диапазон определения констант связывания настолько высоких, как 10-13 M. Программное обеспечение KinExA позволяет проводить анализы данных, которые основаны на точных решениях классических уравнений связывания (kd математика), не на приближениях псевдо-первого порядка. KinExA не требует произвольных преобразований данных или выбора диапазона.
Как показано в таблице 6, антитело 19.3 обладало аффинностью 4,8 нМ к ADDL, по сравнению с 150 нМ аффинностью к мономеру Αβ в тесте BiacoreTM. Селективность антитела 19.3 в отношении ADDL, в тридцать раз превышающая селективность в отношении Αβ-мономера, в значительной степени превосходила селективность исходного антитела, h3B3, в отношении ADDL, которая только в 10 раз превосходила селективность в отношении мономера Αβ.
Аналогичным образом, антитело 19.3 оценивали в тесте определения константы равновесия на основе KinExA. Как показано в таблице 7, антитело 19.3 обладало константой равновесия 2,7 нМ, что соответствует более чем шестикратному предпочтительному связыванию олигомеров ADDL, по сравнению со связыванием мономера Αβ40 в указанном тесте.
Пример 7
Биофизическая характеристика антитела против ADDL 19.3
Биофизическую характеристику для оценки возможности формирования агрегатов антител осуществляли, чтобы показать, что антитела против ADDL стабильны в неблагоприятных условиях и подходят для применения в качестве лекарства. Антитело против ADDL 19.3 концентрировали до >50 мг/мл и вводили в состав ряда препаратов со значением pH в диапазоне 5,0-8,0. Два набора образцов инкубировали при 37°C и 45°C в течение одной недели. Третий набор образцов помещали в -70°C для инициации серии из пяти циклов замораживания/оттаивания. Анализ гель-хроматографии показал, что препараты антитела находились преимущественно (>95%) в мономерном состоянии, с небольшим количеством димеров, что является обычным для препаратов моноклональных антител. Количество димеров и высокомолекулярных олигомеров не возрастало после температурного стресса во всех буферах и не наблюдали фрагментации. Как обобщено в таблице 8, анализ мутности в ближней ультрафиолетовой области спектра также показал отсутствие агрегации. Образцы, подвергнутые замораживанию/оттаиванию, показали увеличение мутности, зависимое от буфера, которое сравнимо с увеличением мутности в препаратах других моноклональных антител. Значение вязкости при 50 мг/мл составляло менее 2 сантипуаз, что свидетельствует о вязкости, приемлемой для инъекций, поскольку значение 20 сантипуаз обычно рассматривают как практическое ограничение для подкожных инъекций. Дифференциальная сканирующая калориметрия также показала подходящую температурную стабильность, причем Fab разворачивался при 72°C и наименее стабильный домен CH2 разворачивался при температуре выше 65°C. В совокупности, антитело 19.3 продемонстрировало очень высокую структурную стабильность с биофизическими свойствами, соответствующими подкожному способу введения.
Пример 8
Фармакокинетический анализ 19.3 и Эффективность в модели AD
A. Исследование фармакокинетики у мышей, трансгенных по FcRn человека
Мыши, трансгенные по FcRn человека (гетерозиготные Tg276) (Jackson Laboratories, Bar Harbor, ME) недавно были предложены в качестве полноценной заместительной системы для оценки фармакокинетики моноклональных антител. Для описания фармакокинетики антитела против ADDL 19.3 у мышей, трансгенных по FcRn человека, трое животных получали однократную внутривенную инъекцию антитела 19.3 в количестве 10 мг/кг через хвостовую вену. Затем серии из 10 мкл образцов крови, которые собирали в моменты времени 0, 25, 50, 75, 100, 150, 250 и 350 часов после IV-введения антитела 19.3 или h3B3 и утвержденный иммунотест с применением античеловеческого IgG использовали для определения концентрации антитела в крови. Как показано на фигуре 7, концентрации в крови антитела 19.3 снижалась в бифазной манере с кажущимся значением t1/2 77±6 часов, которое было значительно выше, чем время полужизни родительского антитела, h3B3, составлявшее приблизительно 29±9 часов. Указанные значения периода полувыведения соответствовали различию, рассчитанному с помощью теста связывания FcRn in vitro (фигура 5). Конечный элиминационный период полувыведения определяли с применением некомпартментной модели (WinNonlin®, Pharsight, Sunnyvale, CA) и экспериментальных данных, полученных в моменты времени между 3-м днем и 15-м днем после введения дозы.
B. Фармакокинетическое исследование с участием нечеловекообразных приматов
Для подтверждения расчетного значения t1/2 для антитела 19.3 у приматов, осуществляли фармакокинетическое исследование в отношении антитела против ADDL 19.3 в группе макак-резус с установленной в мозжечково-мозговой цистерне порт-системой. Шести животным (три самца/три самки) вводили однократную внутривенную болюсную или подкожную инъекцию антитела 19.3 (5 мг/кг) и собирали образцы крови после введения антитела. Одновременно образцы CSF забирали из порт-системы, подключенной к мозжечково-мозговой цистерне, в моменты времени 0, 2, 4, 8, 12, 24, 30, 48, 54 и 72 часов, и концентрацию антитела 19.3 в сыворотке и CSF определяли с помощью теста ELISA с античеловеческим IgG. Когда животным вводили однократную IV болюсную инъекцию антитела 19.3, наблюдали t1/2 254±28 часов, в то время как t1/2 204±49 часов наблюдали после подкожного введения (фигура 8). Кроме того, авторы изобретения обнаружили, что антитело 19.3 способно проходить в CSF приматов, где его концентрация возрастала в течение первых 48 часов и достигала максимального значения приблизительно 0,1% введенного количества антитела (Фигура 9).
C. Распределение 125I-меченного антитела против ADDL 19.3 в мозге мышей
В исследовании по определению концентрации антитела, которое проникало в мозг, самцам мышей Tg2576 в возрасте двенадцати месяцев (линия B6; SJL-TgN APPSWE) вводили (через хвостовую вену) 200 мкг 125I-меченного антитела 19.3 (~8 мг/кг), или одно из двух антител сравнения, и образцы крови и CSF собирали через два часа. Остаточную радиоактивность удаляли из сосудов мозга с помощью перфузии сердца PBS перед удалением мозга. Затем образец крови, CSF и мозг целиком помещали в гамма-счетчик для определения количества радиоактивно меченного антитела, представленного в каждом образце. После проведения подсчета, образцы мозга фиксировали в 4% параформальдегиде в течение 48 часов и затем использовали для иммуногистохимического анализа со свободно плавающими срезами. Локализацию антитела 19.3 в мозге мыши определяли с помощью античеловеческого вторичного антитела и стандартного ABC-метода детекции. Указанное значение иммунореактивности затем объединяли с окрашиванием тиофлавином S (краситель для выявления бляшек) для определения совместной локализации антитела и бляшек в мозге мыши.
Как показано на фигурах 10A и 10B, меченное изотопом антитело 19.3 способно проникать через гематоэнцефалический барьер в CSF и мозг мыши. Кроме того, результаты показали, что количество антитела 19.3 повышено в мозге (0,19%), по сравнению с концентрациями, наблюдаемыми в CSF (0,02%). Чтобы определить обусловлена ли указанная концентрация в мозге ассоциацией антитела 19.3 с Αβ, образцы ткани мозга фиксировали и проводили иммуноцитохимическое исследование. Анализ распределения антитела в мозге старых мышей Tg2576, показал, что антитело 19.3 связано с тиофлавин S-положительными амилоидными бляшками в мозге (фигура 10C и 10D). Полученные результаты впервые предоставляли свидетельство, что антитело 19.3 способно проникать в мозг трансгенных мышей и связываться с соединениями Αβ, представляющими интерес.
Пример 9
Модель отложения бляшек
Для дальнейшей оценки способности антитела против ADDL 19.3 уменьшать депонирование ADDL в амилоидных бляшках в мозге, самцов мышей Tg2576 в возрасте двенадцати месяцев (Taconic, NY) канюлировали с каждой стороны раз в неделю и bADDL (50 пмоль/мкл) вводили еженедельно в течение четырех недель в гиппокамп (фигура 11 A). Через неделю после последнего введения bADDL, половине мышей (n=5/введение) вводили (через хвостовую вену) еженедельно, в течение четырех недель PBS, в то время как остальным животным вводили еженедельно 200 мкг антитела против ADDL (приблизительно 8 мг/кг). Всех животных подвергали эвтаназии через неделю после последнего введения и образцы их мозга подвергали иммуноцитометрическому анализу. Для обнаружения bADDL и бляшек, срезы мозга инкубировали с реагентом Streptavidin Alexa Fluor® 594 (Invitrogen, Carlsbad, CA), заключали в гистологические срезы и окрашивали бляшки тиофлавином S. Затем флуоресцентные изображения бляшек фиксировали с помощью устройства для формирования изображений PerkinElmer Rapid Confocal Imager с программным обеспечением UltraVIEW ERS и количественно оценивали различие в росте бляшек. Подробности указанной модели недавно опубликованы (Gaspar et. al., 2010, Exp. Neurol., 223: 394-400). После лечения в течение одного месяца, существенное уменьшение депонирования новых ADDL в существующих бляшках наблюдали у животных, получавших лечение антителом 19.3 (фигура 11C), по сравнению с животными, получавшими только носитель (фигура 11B).
| название | год | авторы | номер документа |
|---|---|---|---|
| СРЕДСТВА, ПУТИ ПРИМЕНЕНИЯ И СПОСОБЫ ЛЕЧЕНИЯ СИНУКЛЕОПАТИИ | 2016 |
|
RU2765303C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ | 2011 |
|
RU2603078C2 |
| СПОСОБЫ И РЕАГЕНТЫ ДЛЯ УЛУЧШЕНИЯ ДЕТЕКТИРОВАНИЯ АМИЛОИДНЫХ БЕТА-ПЕПТИДОВ | 2010 |
|
RU2554774C2 |
| ФОСФОСПЕЦИФИЧНЫЕ АНТИТЕЛА, РАСПОЗНАЮЩИЕ ТАУ | 2012 |
|
RU2639537C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА К β-АМИЛОИДНОМУ ПЕПТИДУ | 2008 |
|
RU2475500C2 |
| АНТИТЕЛА, КОТОРЫЕ СВЯЗЫВАЮТСЯ С СОРТИЛИНОМ И ПОДАВЛЯЮТ СВЯЗЫВАНИЕ ПРОГРАНУЛИНА | 2016 |
|
RU2735639C2 |
| АНТИТЕЛА | 2018 |
|
RU2798399C2 |
| АНТИТЕЛО, КОТОРОЕ РАСПОЗНАЕТ ПЕПТИД Т14 АСНЕ | 2016 |
|
RU2729491C2 |
| ПРЕДУПРЕЖДЕНИЕ И ЛЕЧЕНИЕ ЦЕРЕБРАЛЬНОЙ АМИЛОИДНОЙ АНГИОПАТИИ | 2008 |
|
RU2523894C2 |
| АНТИТЕЛА К BCMA, СОДЕРЖАЩИЕ ТОЛЬКО ТЯЖЕЛУЮ ЦЕПЬ | 2018 |
|
RU2816994C2 |























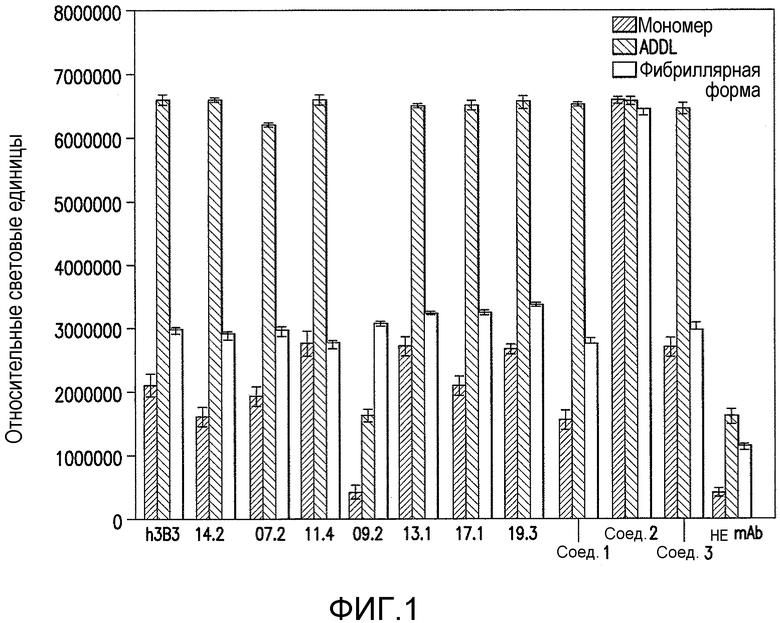
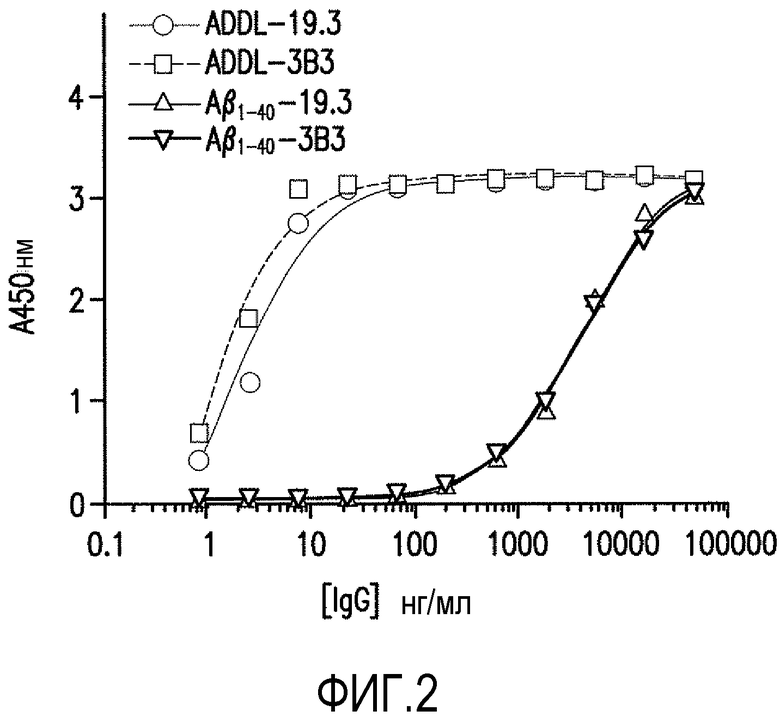
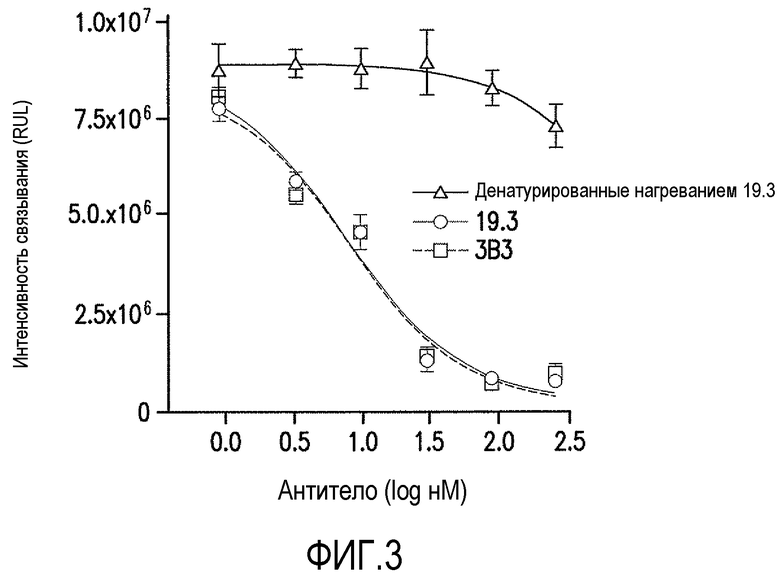
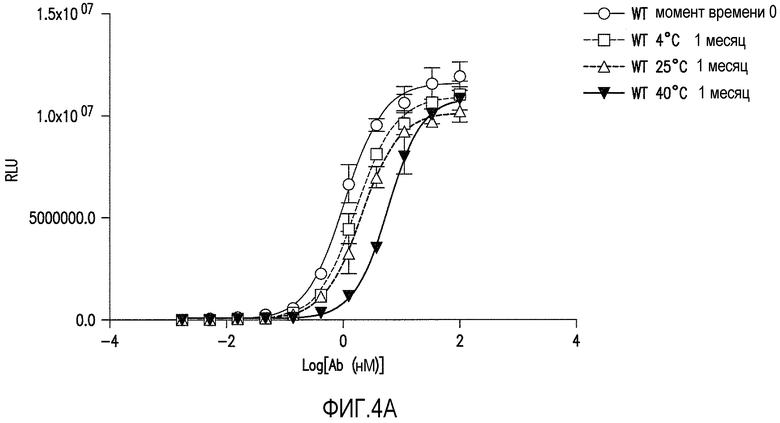
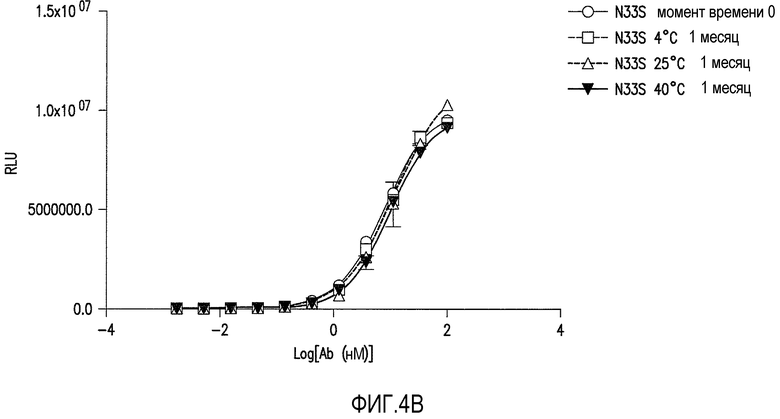
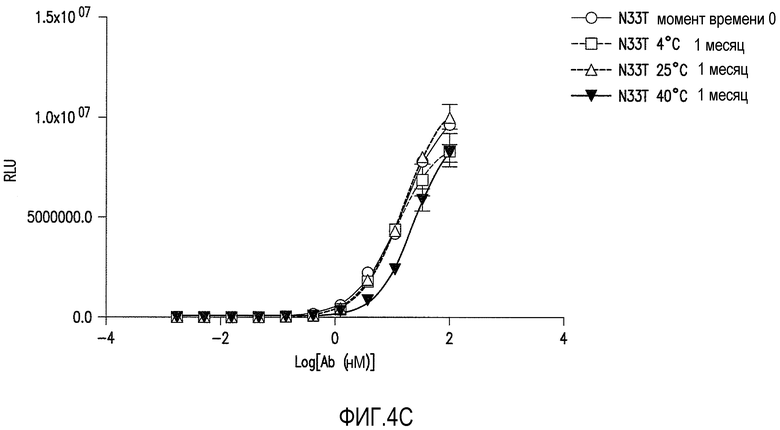
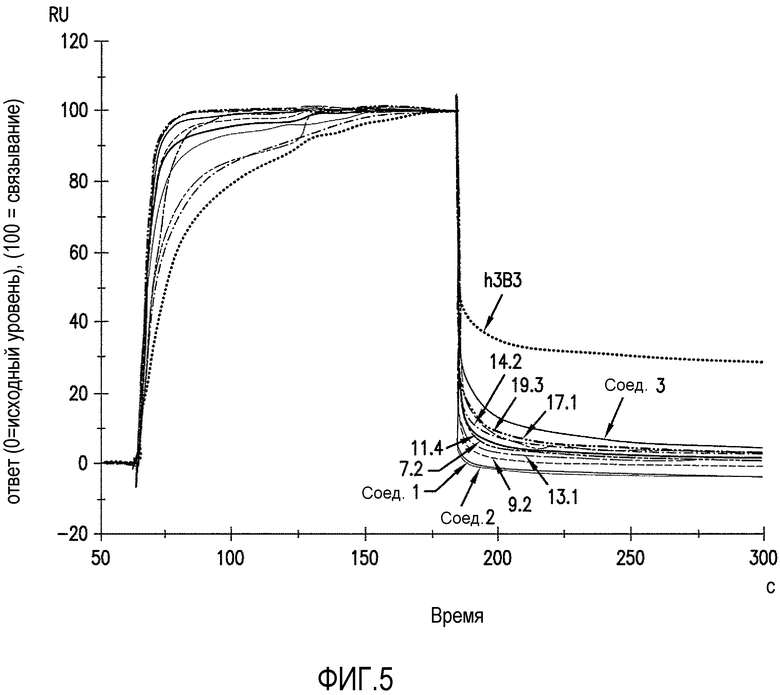

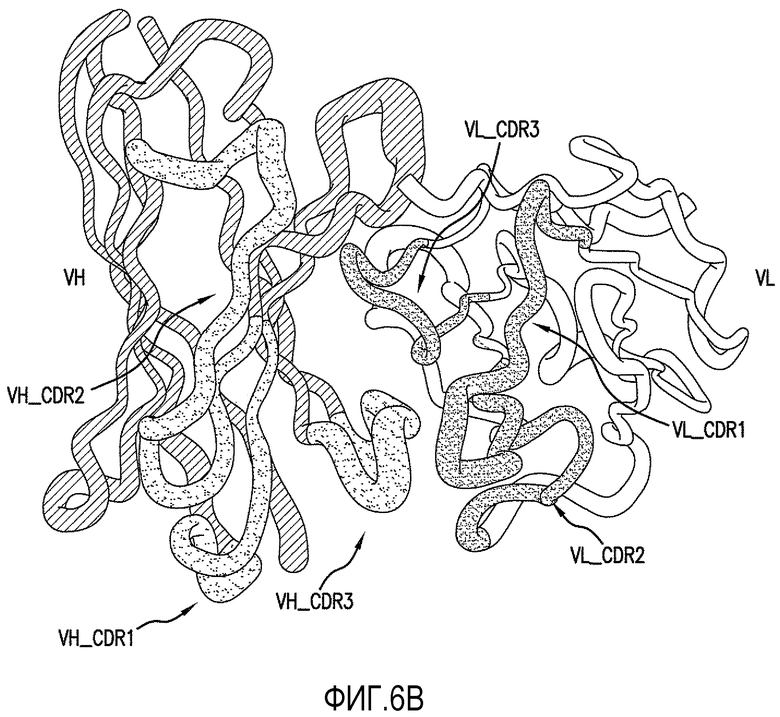
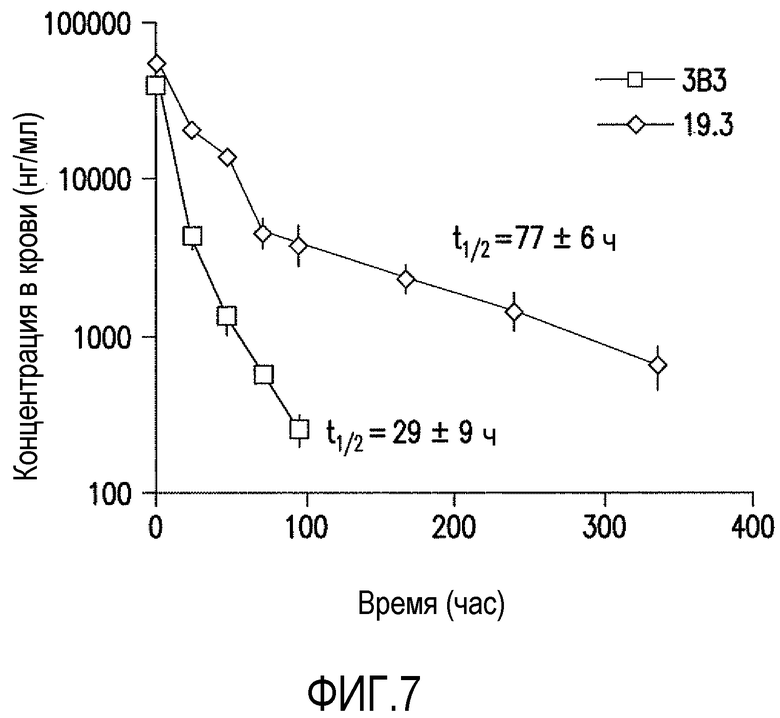
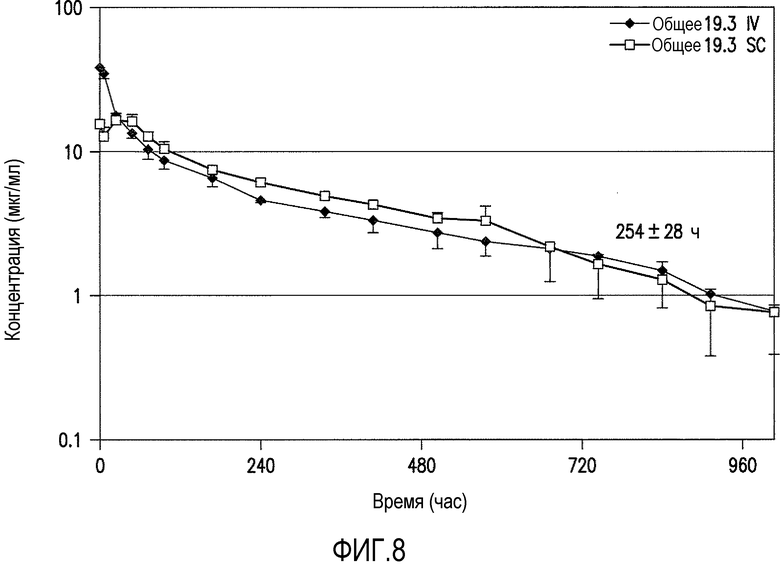
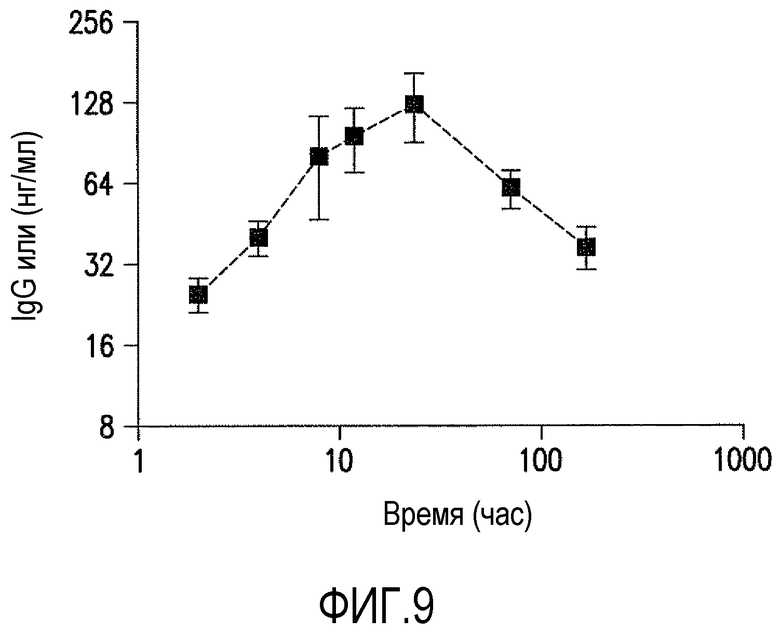
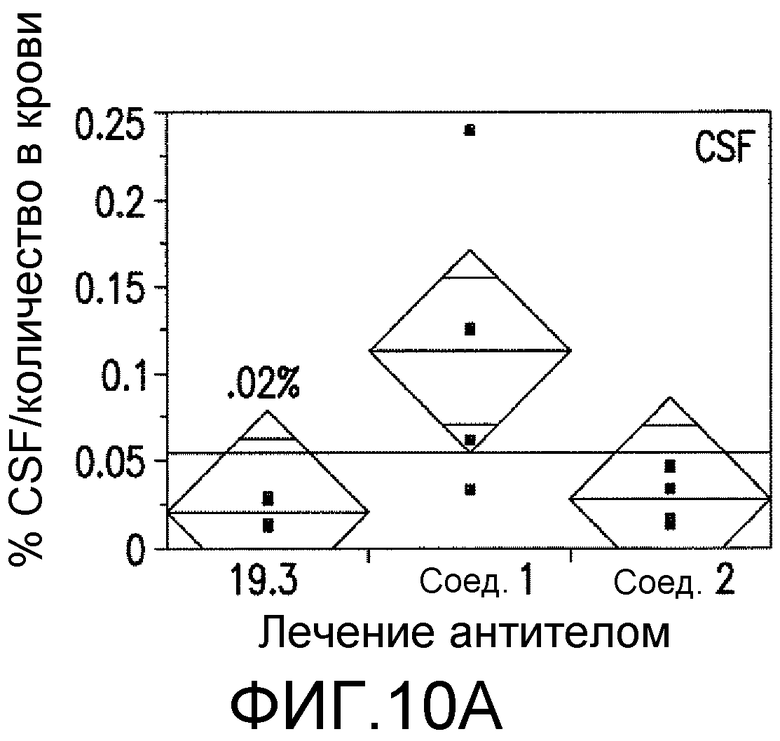
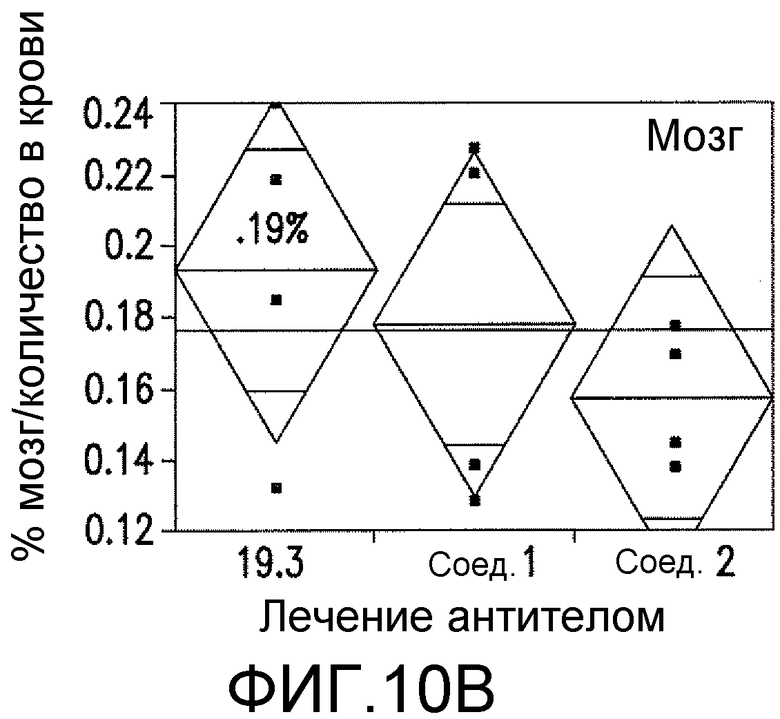
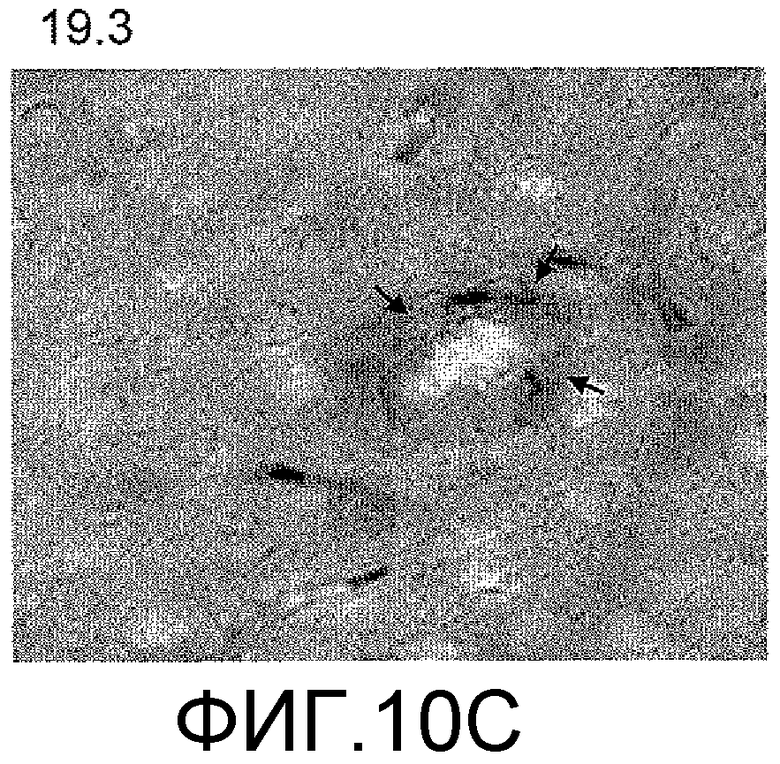
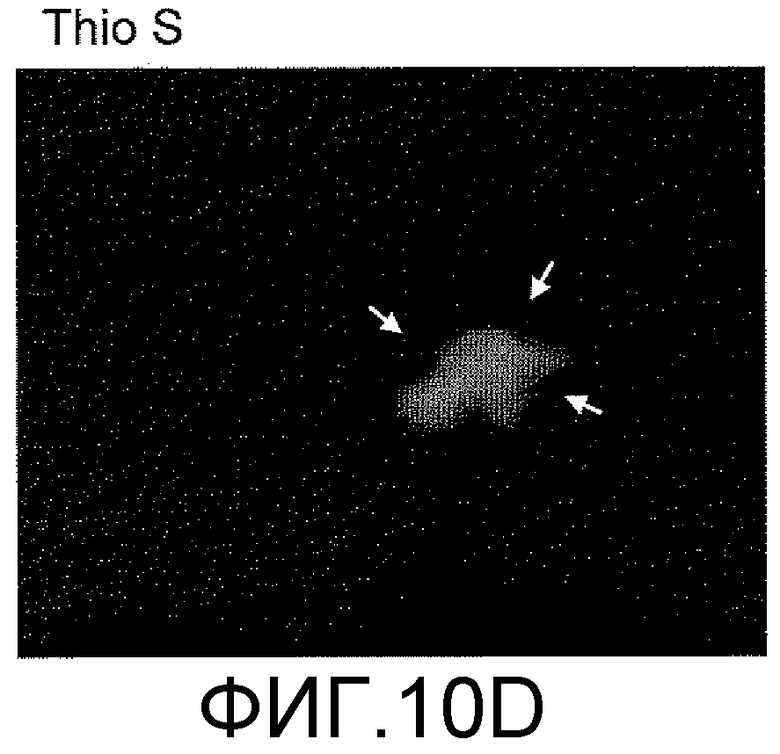
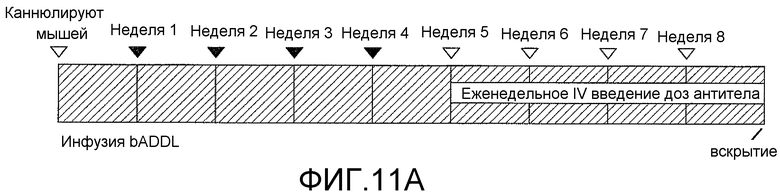
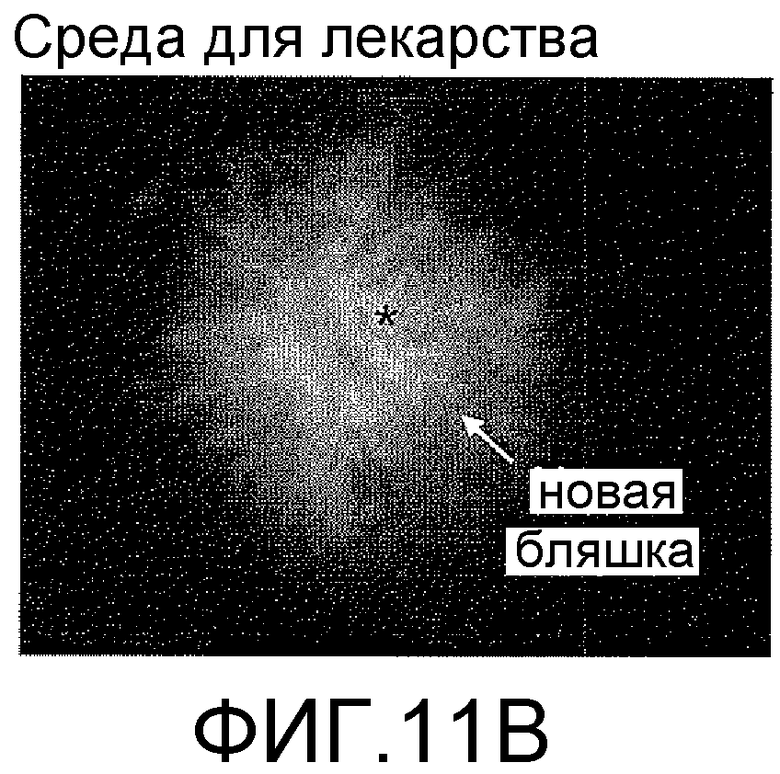
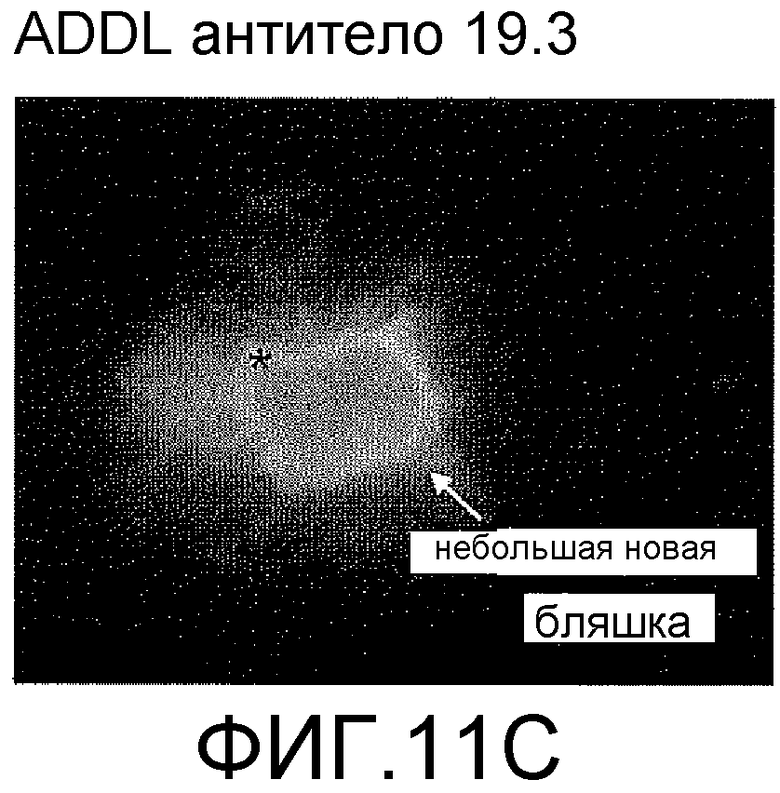
Изобретение относится к биотехнологии, а именно к выделенному антителу или его антигенсвязывающему фрагменту, которые специфически связывают диффундирующие лиганды, происходящие из β-амилоида, а также к его применению. Указанное антитело или его фрагмент содержит вариабельную область легкой цепи, содержащую CDR1 с последовательностью SEQ ID NO:1, CDR2 с последовательностью SEQ ID NO:2, CDR3 с последовательностью SEQ ID NO:10; и вариабельную область тяжелой цепи, содержащую CDR1 с последовательностью SEQ ID NO:4, CDR2 с последовательностью SEQ ID NO:5 и CDR3 с последовательностью SEQ ID NO:6. Антитела являются селективными в отношении ADDL, могут проникать в мозг и являются полезными в способах обнаружения ADDL и диагностики болезни Альцгеймера. Антитела также блокируют связывание ADDL с нейронами, сборку ADDL и фосфорилирование тау-белка. Предложенное изобретение позволяет специфически связывать диффундирующие лиганды, происходящие из β-амилоида. 8 н. и 3 з.п. ф-лы, 19 ил., 8 табл., 9 пр.
1. Выделенное антитело или его антигенсвязывающий фрагмент, которые специфически связывают диффундирующие лиганды, происходящие из β-амилоида, где указанное антитело или фрагмент содержит:
(a) вариабельную область легкой цепи, содержащую
(i) CDR1 с последовательностью SEQ ID NO:1,
(ii) CDR2 с последовательностью SEQ ID NO:2,
(iii) CDR3 с последовательностью SEQ ID NO:10; и
(b) вариабельную область тяжелой цепи, содержащую
(i) CDR1 с последовательностью SEQ ID NO:4,
(ii) CDR2 с последовательностью SEQ ID NO:5 и
(iii) CDR3 с последовательностью SEQ ID NO:6.
2. Выделенное антитело по п. 1, где вариабельная область легкой цепи указанного антитела содержит SEQ ID NO:15 и вариабельная область тяжелой цепи указанного антитела содержит SEQ ID NO:17.
3. Выделенное антитело по п. 1, кроме того, содержащее константную область тяжелой цепи SEQ ID NO:21.
4. Выделенное антитело по п. 1, где антитело представляет собой моноклональное антитело.
5. Фармацевтическая композиция для лечения заболевания, связанного с ADDL, содержащая эффективное количество антитела или антигенсвязывающего фрагмента по п. 1 в смеси с фармацевтически приемлемым носителем.
6. Способ уменьшения связывания диффундирующих лигандов, происходящих из β-амилоида, с нейроном, включающий приведение в контакт нейрона с эффективным количеством антитела или антигенсвязывающего фрагмента по п. 1, так что уменьшается связывание диффундирующих лигандов, происходящих из Аβ, с нейроном.
7. Способ ингибирования сборки диффундирующих лигандов, происходящих из β-амилоида, включающий приведение в контакт образца, содержащего 1-42 пептидов β-амилоида, с эффективным количеством антитела или антигенсвязывающего фрагмента по п. 1, ингибируя тем самым сборку диффундирующих лигандов, происходящих из Aβ.
8. Способ ослабления симптомов заболевания, связанного с диффундирующими лигандами, происходящими из β-амилоида, включающий введение эффективного количества фармацевтической композиции по п. 5.
9. Способ обнаружения диффундирующих лигандов, происходящих из β-амилоида, в образце, включающий приведение в контакт образца с антителом или антигенсвязывающим фрагментом по п. 1 и определение присутствия комплекса, содержащего диффундирующие лиганды, происходящие из β-амилоида, и указанное антитело или антигенсвязывающий фрагмент.
10. Способ диагностики заболевания, связанного с диффундирующими лигандами, происходящими из β-амилоида, включающий приведение в контакт образца с антителом или антигенсвязывающим фрагментом по п. 1 и определение наличия комплекса, содержащего диффундирующие лиганды, происходящие из β-амилоида, и указанное антитело или антигенсвязывающий фрагмент, где комплекс является диагностическим в отношении заболевания, связанного с диффундирующими лигандами, происходящими из β-амилоида.
11. Набор для обнаружения диффундирующих лигандов, происходящих из β-амилоида, содержащий антитело или антигенсвязывающий фрагмент по п. 1 и инструкцию по применению антитела или его антигенсвязывающего фрагмента.
| US2008175835 A1, 24.07.2008.WO 2007050359 A2, 03.05.2007.EA 10469 B1, 29.08.2008.RU 2385161 C2, 27.03.2010.EA 10902 B1, 30.12.2008.EA 11450 B1, 27.02.2009.RU 2353389 C2, 27.04.2009 |

