Область изобретения
Настоящее изобретение относится к антителам, например, полноразмерным антителам или их антиген-связывающим участкам, которые проявляют рН-зависимое связывание, так что отношение равновесных констант диссоциации (KD) и/или констант скорости диссоциации (koff) при эндосомальном рН/физиологическом рН (например, рН 5,5/рН 7,4 или рН 6,0/рН 7,4) составляет 2 или более.
Предшествующий уровень техники
Моноклональные антитела (mAb) приобрели важные терапевтические возможности для многих заболеваний (Brekke and Sandlie, Nat Rev Drug Discov 2: 52-62, 2003; Maggon, Curr Med Chem 14: 1978-1987, 2007). Большинство mAb, имеющихся в настоящее время на рынке, представляют собой антитела иммуноглобулины G (IgG). Их относительно длительное время полужизни опосредовано связыванием с неонатальным Fc-рецептором (FcRn). Захват IgG в клетку происходит посредством жидкофазного пиноцитоза, и этот IgG затем связывается с FcRn в подкисленном окружении (рН 6,0) эндосомального компартмента (Lobo et al., J Pharm Sci 93: 2645-2668, 2004). Считают, что FcRn-связанный IgG защищен от деградации рециклингом в клеточной поверхности, где нейтральный рН облегчает диссоциацию и высвобождение IgG в кровоток. Несвязанный IgG, напротив, как полагают, переходит в лизосомы и затем распадается (Lencer and Blumberg, Trends Cell Biol 15: 15: 5-9, 2005).
К настоящему времени применяли разные технологии для оптимизации функциональной активности IgG-антитела путем введения конкретных замен с целью снижения дозы и/или частоты приема доз и улучшения эффективности и безопасности (Presta, Curr. Opinion Immunol 20: 460-470, 2008). В общем, оптимизация IgG-антител может быть классифицирована на инженерию Fc константной области для воздействия на связывание антитела с FcRn, FcγR и системой комплемента и инженерию вариабельной области для воздействия на аффинность связывания.
Несколько работ описывают инженерию константной области для увеличения связывания с Fc γ-рецепторами и тем самым усиления эффекторной функции IgG1-антитела (Stavenhagen et al., Cancer Res 67: 8882-8890, 2007; Zaievsky et al., Blood 113: 3735-3743, 2009). Было показано, что замены, такие как S239D/I332E/A330L или F243L/R292P/Y300L/V305I/P396L, в IgG1 усиливают связывание Fc γ-рецептора Illa и проявляют превосходную антителозависимую клеточно-опосредованную цитотоксическую (ADCC) активность in vitro и превосходящую эффективность in vivo по сравнению с IgG1 дикого типа. Следовательно, ожидают, что по сравнению с антителами дикого типа антитела с такими заменами показывают превосходную эффективность в той же самой дозе или сравнимую эффективность в более низкой дозе и/или с меньшей частотой дозирования у человека.
Другой способ снижения дозы и/или частоты дозирования заключается в том, чтобы снизить элиминацию IgG-антитела. Сообщают, что длительное время полужизни IgG-антител зависит от его связывания с FcRn. Таким образом, широко исследуются замены, которые увеличивают аффинность связывания IgG с FcRn при рН 6,0, в то же время сохраняя рН-зависимость взаимодействия, путем инженерии константной области (Ghetie et al., Nature Biotech. 15: 637-640, 1997; Hinton et al., JBC 279: 6213-6216, 2004; Dall'Acqua et al., J Immunol 117: 1129-1138, 2006). Замены, такие как M428L/N434S, ведут к увеличенному времени полужизни и улучшенному фармакодинамическому эффекту в вариантах (Zaievsky et al., Nature Biotech. 28: 157-159, 2010). В некоторых работах сообщали об успешном увеличении времени полужизни путем введения замен, таких как T250Q/M428L или M252Y/S254T/T256E, для увеличения связывания с FcRn при кислом рН. В фармакокинетическом исследовании на приматах, не представляющих собой человека, замена T250Q/M428L в IgG1 показала время полужизни 35 суток, что является значительным увеличением в сравнении с временем полужизни 14 суток для IgG1 дикого типа (Hinton et al., J Immunol 176: 346-356, 2006).
Хотя замены в константной области способны значительно улучшить функции терапевтических IgG-антител, замены в строго консервативной области имеют риск иммуногенности у человека (Presta, supra, 2008; De Groot and Martin, Clin Immunol 131: 189-201, 2009), а замена в последовательности высоко вариабельной области может быть менее иммуногенной. Сообщения относительно вариабельной области включают инженерию остатков гипервариабельных участков (CDR) для улучшения аффинности связывания с антигеном (Rothe et al., Expert Opin Biol Ther 6: 177-187, 2006; Bostrom et al., Methods Mol Biol 525: 353-376, 2009; Thie et al., Methods Mol Biol 525: 309-322, 2009) и инженерию остатков CDR и каркасной области для улучшения стабильности (Wörn and Plückthun, J Mol Biol 305: 989-1010, 2001; Ewert et al., Methods 34: 184-199, 2004) и снижения риска иммуногенности (De Groot and Martin, выше, 2009; Jones et al., Methods Mol Bio 525: 405-423, xiv, 2009). Как сообщалось, улучшенная аффинность к антигену может быть достигнута путем созревания аффинности с использованием фагового или рибосомного дисплея рандомизированной библиотеки. Улучшенная стабильность может быть эффективно получена исходя из рационального дизайна на основе последовательности и структуры. Сниженного риска иммуногенности (деиммунизация) можно добиться разными методологиями гуманизации и удаления Т-клеточных эпитопов, что можно предсказать с использованием технологий in silico или определить анализами in vitro. Кроме того, были сконструированы вариабельные области для снижения pl. Более продолжительное время полужизни наблюдали для этих антител по сравнению с антителами дикого типа, несмотря на сравнимое FcRn-связывание (Igawa et al., PEDS, Advance Access, doi: 10.1093/protein/gzq009, 2010).
Настоящее изобретение относится к инженерии или селекции антител с рН-зависимым связыванием антигена для модификации времени полужизни антитела и/или антигена. Время полужизни антитела IgG2 может быть сокращено, если механизмы антиген-опосредованного клиренса нормально разрушают антитело при связывании с антигеном. Подобным образом, комплекс антиген: антитело может влиять на время полужизни антигена, либо пролонгируя время полужизни посредством защиты антигена от обычных процессов деградации, либо сокращая время полужизни посредством антитело-опосредованной деградации. Настоящее изобретение относится к антителам с более высокой аффинностью в отношении антигена при рН 7,4 по сравнению с эндосомальным рН (то есть, рН 5,5-6,0), так что отношение KD при рН 5,5/ рН 7,4 или при рН 6,0/ рН 7,4 составляет 2 или более.
Изобретение относится к антителу с таким рН-зависимым связыванием со своим антигеном и способам конструирования, получения и применения таких антител. Примеры полезных антител направлены на антигены, такие как пропротеин конвертаза субтилизин/кексин типа 9 (PCSK9), также известный как NARC-1, IgE, Dickkopf-родственный белок 1 (DKK1), Комплемент 5 (С5), склеростин (SOST) и рецептор гранулоцитарно-макрофагеального колониестимулирующего фактора (GMCSF).
PCSK9 был идентифицирован как белок с генетической мутацией при некоторых формах семейной гиперхолестеринемии. PCSK9 синтезируется как зимоген, который подвергается аутокаталитическому процессингу в конкретном мотиве в эндоплазматическом ретикулуме. Исследования популяций показали, что некоторые мутации PCSK9 представляют собой мутации, при которых белковый продукт экспрессии мутантного гена приобретает новые и патологические функции ("gain-of-function"), и обнаруживаются у индивидуумов с аутосомно-доминантной гиперхолестеринемией, в то время как другие мутации с потерей функции ("loss-of-function", LOF) связаны с пониженным уровнем холестерина в плазме крови. Исследования по заболеваемости и смертности в этой группе ясно продемонстрировали, что снижение функции PCSK9 значительно минимизировало риск возникновения сердечнососудистого заболевания.
Краткое изложение сущности изобретения
Настоящее изобретение относится к антителам с рН-зависимым связыванием со своим антигеном, так что аффинность в отношении связывания антигена при физиологическом рН (то есть, рН 7,4) выше чем при эндосомальном рН (то есть, рН 6,0 или 5,5). Другими словами, отношение KD или koff при рН 5,5/ рН 7,4 или при рН 6,0/ рН 7,4 составляет более чем 2, 3, 4, 8, 10, 16, 20, 30, 40 или 100 или более, или находится в интервале между 2, 3, 4, 8, 10, 16, 20, 30, 40 или 100 или более. Такие рН-зависимые антитела преимущественно диссоциируют от антигена в эндосоме. Это может увеличивать время полужизни антитела по сравнению с антителами с эквивалентными значениями KD при рН 7,4, но не проявляющими рН-зависимое связывание, когда антиген является таким, что подвергается антиген-опосредованному клиренсу (например, PCSK9). Антитела с рН-зависимым связыванием могут сокращать общее время полужизни антигена, когда антиген подвергается пониженному клиренсу при связывании с антителом (например, IL6). Антитела с рН-зависимым связыванием могут также пролонгировать это антитело-опосредованное снижение в антигене, который не связан с антителом. Это может быть важным при антагонизации антигена-мишени, обычно присутствующего в высоких уровнях (например, IgE, DKK1, С5 и SOST). Кроме того, такие антитела могут увеличивать время полужизни антигена, когда антиген представляет собой рецептор и этот рецептор имеет повышенный клиренс при связывании с антителом (например, рецептор GMCSF). В любом из воплощений изобретения, описанных ниже, KD и koff могут быть измерены при 25°С или 37°С.
В предпочтительном воплощении антитело с рН-зависимым связыванием, которое специфически связывает антиген с более высокой аффинностью при рН 7,4, чем при рН 6,0, где отношение KD и/или отношение koff при рН 6,0/рН 7,4 и при 25°С составляет более чем 2, 3, 4, 8, 10, 16 или более, или находится в интервале между 2, 3, 4, 8, 10, 16 или более, и где антитело имеет пониженный клиренс из плазмы in vivo при взаимодействии с указанным антигеном по сравнению с антителом, не проявляющим рН-зависимое связывание, которое имеет подобную аффинность в отношении данного антигена при рН 7,4, но имеет сравниваемое отношение KD и/или koff при рН 6,0/рН 7,4 менее чем 2. Предпочтительно, антиген не представляет собой рецептор интерлейкина-6 (IL6R), или предпочтительно, антитело не представляет собой антитело Fv3-m73, Fv4-m73 или H3pl/L73 против IL6R, как раскрыто в WO 2010/106812 или WO 2009/041621.
В другом предпочтительном воплощении антитело с рН-зависимым связыванием, которое специфически связывает антиген с более высокой аффинностью при рН 7,4, чем при рН 6,0, где отношение KD и/или отношение koff при рН 6,0/рН 7,4 и при 25°С составляет более чем 2, 3, 4, 8, 10, 16 или более, или находится в интервале между 2, 3, 4, 8, 10, 16 или более, и где антиген является как мембраносвязанным, так и растворимым in vivo, и где антитело опосредует повышенную локализацию в рецепторе клеточной мембраны по сравнению с антителом, которое имеет подобную аффинность в отношении данного антигена при рН 7,4, но имеет сравниваемое отношение KD и/или koff при рН 6,0/рН 7,4 менее чем 2. Предпочтительно, антиген не представляет собой рецептор интерлейкина-6 (IL6R), или предпочтительно, антитело не представляет собой антитело Fv3-m73, Fv4-m73 или H3pl/L73 против IL6R, как раскрыто в WO 2010/106812 или WO 2009/041621. В другом предпочтительном воплощении антиген представляет собой растворимый рецептор, который является несигнальным рецептором-ловушкой. В дополнительных предпочтительных воплощениях антитело с рН-зависимым связыванием представляет собой конъюгат антитела с лекарственным средством, опосредует антителозависимую клеточно-опосредованную цитотоксичность (ADCC) и/или комплементзависимую цитотоксичность (CDC).
Изобретение включает антитело с рН-зависимым связыванием, которое специфически связывает антиген с более высокой аффинностью при рН 7,4, чем при рН 6,0, где отношение KD и/или отношение koff при рН 6,0/рН 7,4 и при 25°С составляет более чем 2, 3, 4, 8, 10, 16 или более, или находится в интервале между 2, 3, 4, 8, 10, 16 или более, и где снижение in vivo количества не связанного с антителом антигена пролонгировано при взаимодействии с указанным антителом по сравнению с антителом, не проявляющим рН-зависимое связывание, которое имеет подобную аффинность в отношении данного антигена при рН 7,4, но имеет сравниваемое отношение KD и/или koff при рН 6,0/рН 7,4 менее чем 2.
В изобретении предложено антитело с рН-зависимым связыванием, которое специфически связывает антиген с более высокой аффинностью при рН 7,4, чем при рН 6,0, где отношение KD и/или отношение koff при рН 6,0/рН 7,4 и при 25°С составляет более чем 2, 3, 4, 8, 10, 16 или более, или находится в интервале между 2, 3, 4, 8, 10, 16 или более, и где имеется снижение in vivo количества связанного с антителом антигена по сравнению с антителом, не проявляющим рН-зависимое связывание, которое имеет подобную аффинность в отношении данного антигена при рН 7,4, но имеет сравниваемое отношение KD и/или koff при рН 6,0/рН 7,4 менее чем 2. В предпочтительном воплощении антиген представляет собой остеопонтин.
В изобретении также предложено агонистическое антитело с рН-зависимым связыванием, которое специфически связывает антиген с более высокой аффинностью при рН 7,4, чем при рН 6,0, где отношение KD и/или отношение koff при рН 6,0/рН 7,4 и при 25°С составляет более чем 2, 3, 4, 8, 10, 16 или более, или находится в интервале между 2, 3, 4, 8, 10, 16 или более, и где антиген представляет собой рецептор и этот рецептор имеет пониженный клиренс in vivo при взаимодействии с указанным антителом по сравнению с антителом, которое имеет подобную аффинность связывания в отношении данного рецептора при рН 7,4, но имеет сравниваемое отношение KD и/или koff при рН 6,0/рН 7,4 менее чем 2. В предпочтительном воплощении рецептор представляет собой рецептор GMCSF.
В других предпочтительных воплощениях любого из вышеописанных антител отношение KD или отношение koff при рН 6,0/рН 7,4 составляет более чем 20, 30, 40 или 100 или более, или находится в интервале между 20, 30, 40 или 100 или более. В других предпочтительных воплощениях предпочтительные отношения KD или koff при рН 6,0/рН 7,4 находятся в интервале между 2-3, 2-4, 2-8, 2-10, 2-16 или 2-20 или более, или 3-4, 3-8, 3-10, 3-16 или 3-20, или 4-8, 4-10, 4-16 или 4-20 или более, или 8-10, 8-16, 8-20 или более, 10-16, 10-20 или более, или 16-20 или более.
В других предпочтительных воплощениях вышеописанных антител связывание антитела с антигеном при рН 7,4 и при 25°С имеет KD от примерно 0,01 нМ до примерно 100 нМ, или более предпочтительно, от примерно 0,1 нМ до примерно 10 нМ.
В других предпочтительных воплощениях вышеописанных антител связывание антитела с антигеном при рН 7,4 имеет koff от примерно 1×10-4 сек-1 до примерно 1×10-1 сек-1, более предпочтительно от примерно 1×10-3 сек-1 до примерно 1×10-1 сек-1.
В еще одном предпочтительном воплощении вышеописанных антител антиген представляет собой PCSK9. В одном предпочтительном воплощении антитело против PCSK9 не представляет собой антитело H1M300N против PCSK9 (см. US 2010/0166768). В других предпочтительных воплощениях антиген представляет собой IgE, С5 или DKK1 и, в предпочтительных воплощениях, KD находится в интервале от 1,0 нМ до примерно 10 нМ или от 1,0 нМ до примерно 100 нМ.
В изобретении также предложен способ расширения интервала между приемом доз и/или снижения терапевтической дозы для лечения пациента терапевтическим антителом, включающий введение указанному пациенту терапевтически эффективного количества антитела из любых вышеописанных антител по изобретению, где фармакодинамический эффект и/или время полужизни указанного антитела пролонгированы по сравнению с антителом, которое имеет подобную аффинность при рН 7,4, но которое имеет отношение KD и/или отношение koff при рН 6,0/7,4 менее чем 2.
Кроме того, в изобретении предусмотрен способ получения антитела с пролонгированным временем полужизни и/или фармакодинамическим эффектом посредством регулирования аффинности связывания антитела рН-зависимым образом, включающий селекцию по гистидиновым остаткам или другим остаткам гипервариабельного участка (CDR) антитела, которые оптимизируют микроокружение, влияющее на рKа, так что связывание антитела с антигеном имеет отношение KD и/или отношение koff при рН 6,0/рН 7,4, которое составляет более чем 2, 3, 4, 8, 10, 16 или более, или находится в интервале между 2, 3, 4, 8, 10, 16 или более. В данном изобретении также предусмотрены антитела, полученные этим способом, включая антитела с 1, 2, 3, 4, 5 или более гистидиновыми заменами в остатках CDR, которые оптимизируют микроокружение, влияющее на рKа.
В предпочтительном воплощении вышеописанного способа способ дополнительно включает мутагенез антитела до достижения аффинности антитела с KD при рН 7,4 по меньшей мере 100 нМ, как измерено при 25°С. В другом воплощении данного изобретения предложена библиотека антител, богатых гистидиновыми остатками в CDR или другими остатками, которые оптимизируют микроокружение, влияющее на рKа.
В других предпочтительных воплощениях изобретения предложено выделенное антитело, которое специфически связывает PCSK9 и содержит гипервариабельный участок вариабельной области тяжелой цепи 1 (VH CDR1), VH CDR2 и VH CDR3 из аминокислотной последовательности VH, показанной в SEQ ID NO: 4 или 5, или ее варианта, имеющего одну, две, три или более консервативных аминокислотных замен в CDR1, CDR2 и/или CDR3.
В предпочтительном воплощении антитело дополнительно содержит CDR1, CDR2 и CDR3 вариабельной области легкой цепи (VL) из аминокислотной последовательности VL, показанной в SEQ ID NO: 3, или ее варианта, имеющего одну, две, три или более консервативных аминокислотных замен в CDR1, CDR2 и/или CDR3.
В изобретении также предложено выделенное антитело, которое специфически связывает PCSK9 и содержит гипервариабельный участок вариабельной области тяжелой цепи 1 (VH CDR1) с аминокислотной последовательностью, показанной в SEQ ID NO: 6, VH CDR2 с аминокислотной последовательностью, показанной в SEQ ID, NO: 7, и/или VH CDR3 с аминокислотной последовательностью, показанной в SEQ ID NO: 8, или ее вариантом, имеющим одну или более консервативных аминокислотных замен в CDR1, CDR2 и/или CDR3, а также выделенное антитело, которое специфически связывает PCSK9 и содержит VH CDR1 с аминокислотной последовательностью, показанной в SEQ ID NO: 6, VH CDR2 с аминокислотной последовательностью, показанной в SEQ ID NO: 7, и/или VH CDR3 с аминокислотной последовательностью, показанной в SEQ ID NO: 9, или ее вариантом, имеющим одну, две, три или более консервативных аминокислотных замен в CDR1, CDR2 и/или CDR3.
В дополнительном воплощении изобретения предусмотрено выделенное антитело, содержащее гипервариабельный участок вариабельной области легкой цепи 1 (VL CDR1) с аминокислотной последовательностью, показанной в SEQ ID NO: 10, VL CDR2 с аминокислотной последовательностью, показанной в SEQ ID NO: 11, и/или VL CDR3 с аминокислотной последовательностью, показанной в SEQ ID NO: 12, или их вариантами, имеющими одну, две, три или более консервативных аминокислотных замен в CDR1, CDR2 и/или CDR3.
В предпочтительных воплощениях из указанных выше антитело дополнительно содержит VL CDR 1 с аминокислотной последовательностью, показанной в SEQ ID NO: 10, VL CDR2 с аминокислотной последовательностью, показанной в SEQ ID NO: 11, и/или VL CDR3 с аминокислотной последовательностью, показанной в SEQ ID NO: 12, или их вариантами, имеющими одну, две, три или более консервативных аминокислотных замен в CDR1, CDR2 и/или CDR3, предпочтительно, VH область содержит SEQ ID NO: 4 или SEQ ID NO: 5, и VL область содержит SEQ ID NO: 3, или их вариант, имеющий одну, две, три или более консервативных аминокислотных замен в SEQ ID NO: 4, SEQ ID NO: 5 и/или SEQ ID NO: 3.
В другом предпочтительном воплощении антител против PCSK9 по настоящему изобретению антитело имеет одну или более Fc мутаций, предпочтительно, N434S, N434H, M428L-N434H обратимый мутант, M428L-N434A обратимый мутант, T250Q-M428L обратимый мутант и M428L-N434S обратимый мутант.
В другом воплощении изобретения предложено антитело или его антиген-связывающий участок, кодируемые плазмидами, депонированными в Американской коллекции типовых культур (АТСС) и имеющими номер в АТСС No. РТА-10547, или РТА-10548, и/или РТА-10549.
Также в изобретении предусмотрены фармацевтические композиции, содержащие терапевтически эффективное количество любого из вышеописанных антител, клетка-хозяин, которая рекомбинантно продуцирует антитело из любых вышеописанных антител, выделенная нуклеиновая кислота, кодирующая любое из вышеописанных антител, и выделенная нуклеиновая кислота, кодирующая любое из вышеописанных антител.
Также в изобретении предусмотрен способ снижения уровня холестерина в липопротеидах низкой плотности (ЛПНП) в крови субъекта, нуждающегося в этом, включающий введение указанному субъекту терапевтически эффективного количества любого из антител по изобретению, направленных на антиген PCSK9.
Краткое описание графических материалов
На Фиг.1 представлен график, показывающий увеличение концентрации антитела в зависимости от времени как функция отношения KD.
На Фиг.2 представлен график, показывающий уменьшение концентрации свободного лиганда (антигена) от времени как функция отношения KD.
На Фиг.3 представлен график, показывающий влияние изменения koff и kon на концентрацию антитела в зависимости от времени.
На Фиг.4 представлена тепловая карта, показывающая, на сколько суток дольше антитело с рН-зависимым связыванием будет снижать сывороточную концентрацию антигена, как функция KD (R), времени полужизни антигена в сыворотке и концентрации антигена в сыворотке. R эквивалентно отношению KD при эндосомальном рН против физиологического рН.
На Фиг.5 подтверждается прогнозируемость модели рН-зависимого антитела. Эта модель успешно прогнозировала общую концентрацию антитела 5А10 (Фиг.5А). На Фиг.5В представлен график, демонстрирующий эффект 5А10 в отношении LDL от времени.
На Фиг.6 также подтверждается прогнозируемость модели рН-зависимого антитела. Эта модель успешно прогнозировала общую концентрацию антитела 5L1721H23_6L3H3 (6L3H3) (Фиг.6А). На Фиг. 6В представлен график, демонстрирующий эффект 6L3H3 в отношении LDL от времени. Антитело с рН-зависимым связыванием 6L3H3 удлиняло интервал, в котором уровень LDL был снижен, по сравнению с 5А10.
На Фиг.7 показано влияние на общий холестерин от времени при введении разных антител против PCSK9. На Фиг.7А показан дозозависимый эффект 5А10 в отношении общего холестерина. На Фиг. 7В показан дозозависимый эффект рН-зависимого антитела 5L1721H23_6H3. Действие эффекта пролонгировано по сравнению с эффектом 5А10. [mpk означает мг/кг].
На Фиг.8 показан график, демонстрирующий, что антитела с рН-зависимым связыванием 5L1721H23_6H3 и 5L1721H23_6L3H3 имели пониженную деградацию антитела и пролонгированное время полужизни по сравнению с антителами, не проявляющими рН-зависимое связывание. На Фиг.8В показан график, демонстрирующий, что эффект, показанный на Фиг.8А, вызван мишень-опосредованной деградацией. Деградация антитела у мышей с выключенным геном PCSK9 резко повышалась после инъекции PCSK9. Примечания: *один удален из группы 5А10; **концентрации mAb ниже уровня обнаружения заменены значениями уровня обнаружения.
На Фиг.9 показан график, иллюстрирующий влияние рН-чувствительного антагонистического антитела против PCSK9 и не чувствительного к рН антагонистического антитела против PCSK9 на уровни холестерина у обезьян. Хотя никаких резких изменений в уровнях HDL не было обнаружено (Фиг.9А), рН-чувствительные антитела опосредовали более продолжительное снижение уровней LDL по сравнению с независимым от рН антителом L1L3.
На Фиг.10 показан график, демонстрирующий, что антитела против PCSK9 с рН-зависимым связыванием имели пролонгированное время полужизни in vivo по сравнению с независимыми от рН антителами.
На Фиг. 11 представлена тепловая карта, демонстрирующая общее моделирование для рН-зависимого связывания. Такие антитела, направленные против антигенов DKK1, IgE или С5, могут значительно увеличивать количество суток, при которых данный антиген испытывал пониженные антигенные уровни по сравнению с антителом без рН-зависимого связывания.
На Фиг.12 смоделирована динамика изменения концентрации антигена от времени после введения антитела с рН-зависимым связыванием, направленным против антигена IgE.
На Фиг.13 смоделирована динамика изменения концентрации антигена от времени после введения антитела с рН-зависимым связыванием, направленным против антигена DKK1.
На Фиг.14 смоделирована динамика изменения концентрации антигена от времени после введения антитела с рН-зависимым связыванием, направленным против антигена С5.
На Фиг.15 показана модель переноса для антител с рН-зависимым связыванием, используемых для моделирования.
Подробное описание изобретения
Настоящее изобретение относится к антителам с рН-зависимым связыванием со своим антигеном, так что аффинность связывания антигена при физиологическом рН (то есть, рН 7,4) выше чем при эндосомальном рН (то есть, рН 6,0 или 5,5). Другими словами, отношение KD или koff при рН 5,5/рН 7,4 или при рН 6,0/рН 7,4 составляет более чем 2, 3, 4, 8, 10, 16, 20, 30, 40 или 100 или более, или находится в интервале между 2, 3, 4, 8, 10, 16, 20, 30, 40 или 100 или более. Такие рН-зависимые антитела преимущественно диссоциируют от антигена в эндосоме. Это может увеличивать время полужизни антитела в кровотоке по сравнению с антителами с эквивалентными значениями KD при рН 7,4, но не проявляющими рН-зависимое связывание, когда антиген является таким, что подвергается антиген-опосредованному клиренсу (например, PCSK9). Антитела с рН-зависимым связыванием могут уменьшать общее время полужизни антигена, когда антиген подвергается пониженному клиренсу при связывании с антителом (например, IL6). Антитела с рН-зависимым связыванием могут также пролонгировать это снижение в антигене, который не связан с антителом. Это может быть важным при антагонизации антигена-мишени, обычно присутствующего в высоких уровнях (например, IgE, DKK1, С5 и SOST). Кроме того, такие антитела могут увеличивать время полужизни антигена, когда антиген представляет собой рецептор и этот рецептор имеет повышенный клиренс при связывании с антителом (например, рецептор GMCSF).
Если антиген опосредует мишень-опосредованную деградацию, то использование таких антител с рН-зависимым связыванием для достижения диссоциации в эндосоме может увеличивать фармакодинамический эффект антитела, например, когда антиген подвергается мишень-опосредованному клиренсу (например, PCSK9). Антитело с рН-зависимым связыванием диссоциирует от антигена, избегает антиген-опосредованной деградации, может подвергаться рециклингу из клетки посредством FcRn-связывания и будет иметь более продолжительное время полужизни, чем антитело с подобным Ко при рН 7,4, но не проявляющее рН-зависимое связывание.
Использование таких антител с рН-зависимым связыванием также терапевтически полезно, когда растворимый антиген присутствует в высокой концентрации (например, IgE, C5, DKK1 или SOST). При диссоциации от антигена в эндосоме и деградации антигена в лизосоме антитело может подвергаться рециклингу в плазме для связывания дополнительного свободного антигена, может пролонгировать снижение не связанного с антителом антигена и может уменьшать необходмую терапевтическую дозу по сравнению с антителом с подобным KD при рН 7,4, но не проявляющим рН-зависимое связывание.
Кроме того, использование антител с рН-зависимым связыванием может быть полезным, когда антиген присутствует в мембране в связанной, а также свободной форме, например, рецептор, и это желательно для усиления связывания с мембрано-связанной формой. Благодаря диссоциированию от растворимой формы, антитело обладает повышенной возможностью к повторному связыванию с мембранной формой, увеличивая близость антитела к клеточной мембране. При связывании с мембранной формой дивалентным образом эффективная аффинность может быть выше, или эффективная скорость диссоциации может быть медленнее, вследствие эффекта авидности.
Это применимо для использования конъюгатов антитела с лекарственным средством (ADC) при нацеливании на антиген, присутствующий как в мембрано-связанной, так и свободной форме. В FcRn-содержащих клетках эндотелия растворимый антиген будет выводиться при ADC-рециклинге в компартмент плазмы, давая возможность для связывания мембрано-связанного антигена. При использовании антител с рН-зависимым связыванием повышенное связывание с мембрано-связанной формой, дивалентным или моновалентным образом, будет вызывать повышенную интернализацию антитела с мембрано-связанным антигеном и клеточную гибель. При связывании с рецептором дивалентным образом авидность может увеличивать эффективную аффинность или замедлять эффективную скорость диссоциации.
Механизм для ADCC и комплемент-зависимой цитотоксичности (CDC) также может быть задействован при использовании антител с рН-зависимым связыванием. В FcRn-содержащих клетках эндотелия растворимый антиген будет выводиться при ADC-рециклинге в компартмент плазмы, давая возможность для связывания мембрано-связанного антигена. Высвобождение антител из растворимого рецептора будет повышать доступность свободного антитела, которое может затем связываться с мембрано-связанным антигеном и увеличивать клеточную гибель.
Общие методики
При осуществлении настоящего изобретения, если не указано иное, использованы традиционные методики молекулярной биологии (включая рекомбинантные технологии), микробиологии, клеточной биологии, биохимии и иммунологии, которые известны специалисту в данной области техники. Такие методики всесторонне описаны в литературе, например, по молекулярному клонированию: A Laboratory Manual, второе издание (Sambrook et al., 1989) Cold Spring Harbor Press; Oligonucleotide Synthesis (M.J.Gait, ed., 1984); Methods in Molecular Biology, Humana Press; Cell Biology: A Laboratory Notebook (J.E.Cellis, ed., 1998) Academic Press; Animal Cell Culture (R.I. Freshney, ed., 1987); Introduction to Cell and Tissue Culture (J.P.Mather and P.E.Roberts, 1998) Plenum Press; Cell and Tissue Culture: Laboratory Procedures (A.Doyle, J.B.Griffiths, and D.G.Newell, eds., 1993-1998) J.Wiley and Sons; Methods in Enzymology (Academic Press, Inc.); Handbook of Experimental Immunology (D.M.Weir and C.C.Blackwell, eds.); Gene Transfer Vectors for Mammalian Cells (J.M.Miller and M.P.Calos, eds., 1987); Current Protocols in Molecular Biology (F.M.Ausubel et al., eds., 1987); PCR: The Polymerase Chain Reaction, (Mullis et al., eds., 1994); Current Protocols in Immunology (J.E.Coligan et al., eds., 1991); Short Protocols in Molecular Biology (Wiley and Sons, 1999); Immunobiology (C.A.Janeway and P.Travers, 1997); Antibodies (P. Finch, 1997); Antibodies: a practical approach (D.Catty, ed., IRL Press, 1988-1989); Monoclonal antibodies: a practical approach (P.Shepherd and C.Dean, eds., Oxford University Press, 2000); Using antibodies: a laboratory manual (E.Harlow and D.Lane (Cold Spring Harbor Laboratory Press, 1999); The antibodies (M.Zanetti and J.D.Capra, eds., Harwood Academic Publishers, 1995).
Определения
Термин "антитело" означает молекулу иммуноглобулина, способную специфически связываться с мишенью, такой как углевод, полинуклеотид, липид, полипептид и так далее, посредством по меньшей мере одного сайта распознавания антигена, расположенного в вариабельной области молекулы иммуноглобулина. В определение данного термина, как он использован здесь, входят не только интактные поликлональные или моноклональные антитела, но также их любые антиген-связывающие фрагменты (например "антиген-связывающий участок") или одноцепочечные варианты, слитые белки, содержащие антитело, и любая другая модифицированная конфигурация молекулы иммуноглобулина, которая содержит сайт распознавания антигена, в том числе, например, одноцепочечные варианты (scFv) и однодоменные антитела (например, однодоменные антитела человека, семейства верблюдовых или акул), макситела, минитела, интратела, диатела, триатела, тетратела, vNAR и бис-scFv (см., например, Hollinger and Hudson, Nature Biotech 23: 1126-1136, 2005), но не ограничиваясь ими. Антитело включает антитело любого класса, например IgG, IgA или IgM (или их подклассы), и антитело, не относящееся к какому-либо определенному классу. В зависимости от аминокислотной последовательности константного домена тяжелой цепи антитела иммуноглобулины могут быть распределены в разные классы. Существует пять основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, и некоторые из них могут быть дополнительно разделены на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелой цепи, соответствующие разным классам иммуноглобулинов, соответственно называют альфа, дельта, эпсилон, гамма и мю. Структуры субъединиц и трехмерные конфигурации иммуноглобулинов разных классов хорошо известны.
Термин "антиген-связывающий участок" антитела, как он использован здесь, относится к одному или более чем одному фрагменту интактного антитела, который сохраняет способность специфически связывать данный антиген (например, мишень X). Антиген-связывающие функции антитела могут осуществляться фрагментами интактного антитела. Примеры связывающих фрагментов, охватываемых термином "антиген-связывающий участок" антитела, включают Fab, Fab', F(ab')2, Fd-фрагмент, состоящий из VH и СН1 доменов, Fv-фрагмент, состоящий из VL и VH доменов одного плеча антитела, фрагмент однодоменного антитела (dAb) (Ward et al., Nature 341: 544-546, 1989) и выделенный гипервариабельный участок (CDR).
Термин "CDR", как он использован здесь, может быть определен в соответствии с любым из определений Kabat, Chothia, расширенным, АbМ, контактным и/или конформационным определением. Идентичность аминокислотных остатков в конкретном антителе, которые составляют CDR, могут быть определены с использованием способов, хорошо известных в данной области техники. CDR антитела, как они определены здесь, могут быть идентифицированы как гипервариабельные участки, первоначально определенные по Kabat et al. См., например, Kabat et al., 1992, Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, NIH, Washington D.C. Положения CDR также могут быть идентифицированы как петлевые структуры, первоначально описанные Chothia и другими. См., например, Chothia et al., Nature 342: 877-883, 1989. Другие подходы идентификации CDR включают "определение АВМ", которое является компромиссом между Kabat и Chothia и производится с использованием программного обеспечения моделирования АВМ антитела Oxford Molecular's (в настоящее время Accelrys®), или "контактное определение" CDR на основе наблюдаемых антигенных контактов, как изложено в MacCallum et al., J.Mol. Biol. 262: 732-745, 1996. В другом подходе, на который ссылаются здесь как на "конформационное определение" CDR, положения CDR могут быть идентифицированы как остатки, которые делают энтальпические вклады в связывание с антигеном. См., например, Makabe et al., Journal of Biological Chemistry, 283: 1156-1166, 2008. Еще дополнительные определения CDR связывания могут не точно следовать одному из вышеопределенных подходов, но будут тем не менее перекрываться с по меньшей мере частью из CDR Kabat, хотя они могут быть укороченными или удлиненными в свете предсказания или экспериментальных обнаружений, что конкретные остатки или группы остатков или даже целые CDR не влияют значительно на связывание с антигеном. CDR, как он использован здесь, может ссылаться на CDR, определенные по любому из подходов, известных в данной области техники, включая комбинации подходов.
Термин "моноклональное антитело", как он использован здесь, относится к антителу, полученному из популяции по существу гомогенных антител, т.е. индивидуальные антитела, содержащие данную популяцию, являются идентичными, за исключением возможных естественных мутаций, которые могут присутствовать в минорных количествах. Моноклональные антитела являются высокоспецифичными, направленно воздействуя на один антигенный сайт. Кроме того, в противоположность препаратам поликлональных антител, которые как правило включают разные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело направлено против одной детерминанты на антигене. Определение "моноклональное" указывает на природу антитела как полученного из по существу гомогенной популяции антител и не должен трактоваться как требующее продуцирования антитела каким-либо конкретным способом. Например, моноклональные антитела, предназначенные для использования в соответствии с настоящим изобретением, могут быть получены гибридомным методом, первоначально описанным Kohler и Milstein, 1975, Nature 256:495, или могут быть получены методами рекомбинантной ДНК, такими как описано в патенте США 4816567. Моноклональные антитела также могут быть выделены из фаговых библиотек, генерированных с использованием методик, описанных в McCafferty et al., 1990, Nature 348:552-554, например.
Термин "гуманизированное" антитело, как он использован здесь, относится к формам антител, не являющихся человеческими (например, мышиные), которые представляют собой химерные иммуноглобулины, иммуноглобулиновые цепи или их фрагменты (такие как Fv, Fab, Fab', F(ab')2 или другие антиген-связывающие последовательности антител), которые содержат минимальную последовательность, имеющую происхождение из иммуноглобулина, не являющегося человеческим. Предпочтительно, гуманизированные антитела представляют собой человеческие иммуноглобулины (реципиентное антитело), в которых остатки из гипервариабельного участка (CDR) реципиента заменены остатками из CDR видов, не являющихся человеческими (донорное антитело), такими как мышь, крыса или кролик, с желаемой специфичностью, аффинностью и способностью. В некоторых случаях остатки каркасной области (FR) Fv человеческого иммуноглобулина заменены соответствующими остатками, не являющимися человеческими. Кроме того, гуманизированное антитело может содержать остатки, которые не обнаружены ни в реципиентном антителе, ни во вставленных последовательностях CDR или каркасной области, но включены для осуществления дополнительной очистки и оптимизации антитела. В общем, гуманизированное антитело содержит по существу все из по меньшей мере одного и, как правило, двух вариабельных доменов, в которых все или по существу все из CDR участков соответствуют таковым из иммуноглобулина, не являющегося человеческим, и все или по существу все из FR участков являются таковыми из консенсусной последовательности человеческого иммуноглобулина. Гуманизированное антитело оптимально также будет содержать по меньшей мере часть константной области или домена (Fc) иммуноглобулина, как правило ту, которая принадлежит человеческому иммуноглобулину. Предпочтительны антитела, имеющие Fc участки, модифицированные как описано в WO - 99/58572. Другие формы гуманизированных антител имеют один или более чем один CDR (CDR L1, CDR L2, CDR L3, CDR H1, CDR H2 и/или CDR H3), которые изменены относительно исходного антитела, которые также называют один или более чем один CDR, "имеющий происхождение" из одного или более чем одного CDR из исходного антитела.
Термин "человеческое антитело", как он использован здесь, означает антитело, имеющее аминокислотную последовательность, соответствующую последовательности антитела, которое может продуцироваться человеком и/или которое создано с использованием любой из методик получения человеческих антител, известных специалисту в данной области техники, или описанных здесь. Это определение человеческого антитела включает антитела, содержащие по меньшей мере один полипептид человеческой тяжелой цепи или по меньшей мере один полипептид человеческой легкой цепи. Один такой пример представляет собой антитело, содержащее полипептиды мышиной легкой цепи и человеческой тяжелой цепи. Человеческие антитела могут быть получены с использованием различных методик, известных в данной области техники. В одном воплощении человеческое антитело выбрано из фаговой библиотеки, экспрессирующей человеческие антитела (Vaughan et al., 1996, Nature Biotechnology, 14:309-314; Sheets et al., 1998, Proc. Natl. Acad. Sci. (USA) 95:6157-6162; Hoogenboom and Winter, 1991, J. Mol. Biol., 227:381; Marks et al., 1991, J. Mol. Biol., 222:581). Человеческие антитела также могут быть получены путем иммунизации животных, в которых локусы человеческих иммуноглобулинов были трансгенно введены вместо эндогенных локусов, например, мыши, у которых эндогенные иммуноглобулиновые гены были частично или полностью инактивированы. Этот подход описан в патентах США 5545807, 5545806, 5569825, 5625126, 5633425 и 5661016. Альтернативно, человеческое антитело может быть получено путем иммортализации человеческих В-лимфоцитов, которые продуцируют антитело, направленное против антигена-мишени (такие В-лимфоциты могут быть выделены из индивидуума или могут быть иммунизированы in vitro,). См., например, Cole et al. Monoclonal antibodies and Cancer Therapy, Alan R.Liss, p.77, 1985; Boerner et al., 1991, J.Immunol., 147 (1): 86-95; и патент США 5750373.
Термин "вариабельная область" антитела относится к вариабельной области легкой цепи антитела или вариабельной области тяжелой цепи антитела, либо в отдельности, либо в комбинации. Как известно в данной области техники, каждая из вариабельных областей тяжелой и легкой цепи состоит из четырех каркасных областей (FR), соединенных тремя областями, определяющими комплементарность, которые содержат гипервариабельные участки (CDR). Эти CDR в каждой цепи удерживаются вместе в непосредственной близости посредством указанных FR и вместе с CDR из другой цепи вносят вклад в образование антиген-связывающего сайта антител. Существует по меньшей мере две методики определения CDR: (1) подход на основе межвидового разнообразия последовательностей (Kabat et al. Sequences of Proteins of Immunological Interest, (5th ed., 1991, National Institutes of Health, Bethesda MD)); и (2) подход на основе кристаллографических исследований комплексов антиген-антитело (Al-lazikani et al, 1997, J. Molec. Biol. 273: 927-948). CDR, как он использован здесь, может ссылаться на CDR, определенные согласно любому из подходов или комбинации обоих подходов.
Как известно в данной области техники, "константная область" антитела относится к константной области легкой цепи антитела или константной области тяжелой цепи антитела, либо в отдельности, либо в комбинации.
Термин "PCSK9", как он использован здесь, относится к любой форме PCSK9 и его вариантам, которые сохраняют по меньшей мере часть активности PCSK9. Если не указано иное, например конкретной ссылкой на человеческий PCSK9, PCSK9 включает нативную последовательность PCSK9 всех видов млекопитающих, например, человека, собаки, кошки, лошади и коровы. Один из примеров человеческого PCSK9 можно найти как Uniprot Accession Number Q8NBP7 (SEQ ID NO: 16).
Термин "антагонистическое антитело против PCSK9 ", как он использован здесь, относится к антителу, которое способно ингибировать биологическую активность PCSK9 и/или нисходящий(е) путь(и) передачи внутриклеточных сигналов, опосредованный(е) передачей сигнала PCSK9, включая PCSK9-опосредованную регуляцию рецептора липопротеидов низкой плотности (LDLR) по типу обратной связи и PCSK9-опосредованное снижение клиренса липопротеидов низкой плотности (LDL) из крови. рН-Зависимое антагонистическое антитело против PCSK9 охватывает антитела, которые блокируют, антагонизируют, подавляют или снижают (до любой степени, в том числе значительно) биологическую активность PCSK9, включая нисходящие пути передачи внутриклеточных сигналов, опосредованные передачей сигнала PCSK9, например взаимодействие с LDLR и/или индуцирование клеточного ответа на PCSK9. Понятно, что для целей настоящего изобретения термин "антагонистическое антитело против PCSK9" охватывает все идентифицированные выше термины, названия, а также функциональные состояния и характеристики, посредством которых сам PCSK9, биологическая активность PCSK9 (включая его способность опосредовать любой аспект взаимодействия с LDLR, регуляцию LDLR по типу обратной связи и пониженный клиренс LDL из крови, но не ограничиваясь этим), или последствия этой биологической активности по существу аннулируются, снижаются или нейтрализуются в любой значимой степени. В некоторых воплощениях рН-зависимое антагонистическое антитело против PCSK9 связывается с PCSK9 и предотвращает взаимодействие с LDLR. Примеры антагонистических антител против PCSK9 предложены в данном описании.
Термины "полипептид", "олигопептид", "пептид" и "белок" используют в данном описании взаимозаменяемо для ссылки на цепи аминокислот любой длины, предпочтительно относительно короткие (например, 10-100 аминокислот). Цепь может быть линейной или разветвленной и может содержать модифицированные аминокислоты и/или может быть прервана кислотами, не относящимися к аминокислотам. Эти термины также охватывают аминокислотную цепь, которая была модифицирована естественным путем или путем вмешательства; например, образование дисульфидной связи, гликозилирование, липидизация, ацетилирование, фосфорилирование или любая другая манипуляция или модификация, например конъюгирование с агентом мечения. Также в данное определение включены, например, полипептиды, содержащие один или более аналогов аминокислот (включая, например, неприродные аминокислоты и так далее), а также другие модификации, известные в данной области техники. Следует понимать, что полипептиды могут существовать в виде отдельных цепей или ассоциированных цепей.
Как известно в данной области техники, "полинуклеотид" или "нуклеиновая кислота," используемые в данном описании взаимозаменяемо, относятся к цепям нуклеотидов любой длины и включают ДНК и РНК. Нуклеотиды могут представлять собой дезоксирибонуклеотиды, рибонуклеотиды, модифицированные нуклеотиды или основания и/или их аналоги, или любой субстрат, который может быть включен в цепь посредством ДНК- или РНК-полимеразы. Полинуклеотид может содержать модифицированные нуклеотиды, такие как метилированные нуклеотиды и их аналоги. Если присутствует, модификацию нуклеотидной структуре можно придать до или после сборки цепи. Последовательность нуклеотидов может прерываться компонентами, не являющимися нуклеотидами. Полинуклеотид может быть дополнительно модифицирован после полимеризации, например путем конъюгирования с агентом мечения. Другие типы модификаций включают, например, "кэпы", замену одного или более чем одного природного нуклеотида аналогом, внутринуклеотидные модификации, такие как, например, модификации с незаряженными связями (например, метилфосфонаты, фосфотиоэфиры, фосфоамидаты, карбаматы и так далее) и с заряженными связями (например, фосфоротиолаты, фосфордитиоаты и так далее), модификации, содержащие боковые группировки макромолекулы, такие как, например, белки (например, нуклеазы, токсины, антитела, сигнальные пептиды, поли-L-лизин и так далее), модификации с интеркаляторами (например, акридин, псорален и так далее), модификации, содержащие хелаторы (например, металлы, радиоактивные металлы, бор, окислительные металлы и так далее), модификации, содержащие алкилаторы, модификации с модифицированными связями (например, альфа-аномерные нуклеиновые кислоты и так далее), а также немодифицированные формы полинуклеотида(ов). Кроме того, любая из гидроксильных групп, обычно присутствующих в сахарах, может быть заменена, например, фосфонатными группами, фосфатными группами, защищена стандартными защитными группами, или активирована для получения дополнительных связей с дополнительными нуклеотидами, или может быть конъюгирована с твердыми подложками. 5'- и 3'-Концевая группа ОН может быть фосфорилирована или замещена аминами или органическими кэп-группировками из 1-20 атомов углерода. Другие гидроксилы также могут быть дериватизированы до стандартных защитных групп. Полинуклеотиды также могут содержать аналогичные формы рибозных или дезоксирибозных Сахаров, которые в общем известны в данной области техники, включая, например, 2'-O-метил-, 2'-O-аллил-, 2'-фтор- или 2'-азидо-рибозу, карбоциклические аналоги Сахаров, альфа- или бета-аномерные сахара, эпимерные сахара, такие как арабиноза, ксилозы или ликсозы, пиранозные сахара, фуранозные сахара, седогептулозы, ациклические аналоги и нуклеозидные аналоги с удаленными азотистыми основаниями, такие как метилрибозид. Одна или более фосфодиэфирных связей могут быть заменены альтернативными связывающими группами. Эти альтернативные связывающие группы включают воплощения, где фосфат заменен на Р(O)S("тиоат"), P(S)S ("дитиоат"), (O)NR2 ("амидат"), P(O)R, P(O)OR', СО или СН2 ("формацеталь"), в которых каждый R или R' независимо представляет собой Н или замещенный или незамещенный алкил (С1-20), возможно содержащий эфирную (-O-) связь, арил, алкенил, циклоалкил, циклоалкенил или аралдил, но не ограничиваются ими. Не все связи в полинуклеотиде обязательно идентичны. Предшествующее описание применимо ко всем полинуклеотидам, на которые здесь ссылаются, включая РНК и ДНК.
Антитело "специфически связывается" или "предпочтительно связывается" с мишенью, если оно связывается с более высокой аффинностью, авидностью, более быстро и/или с большей продолжительностью, чем оно связывается с другими веществами. Например, антитело, которое специфически или предпочтительно связывается с PCSK9 эпитопом, представляет собой антитело, которое связывает этот эпитоп с большей аффинностью, авидностью, более быстро и/или с большей продолжительностью, чем оно связывается с другими PCSK9 эпитопами или не-РСSК9 эпитопами. Также понятно при прочтении этого определения, что, например, антитело (или группировка или эпитоп), которое специфически или предпочтительно связывается с первой мишенью, может специфически или предпочтительно связываться со второй мишенью или может не связываться с ней. Как таковое, "специфическое связывание" или "предпочтительное связывание" не обязательно требует исключительного связывания (хотя оно может включать его). В общем, но не обязательно, ссылка на связывание означает предпочтительное связывание.
Термин "несигнальный рецептор-ловушка" представляет собой растворимую изоформу рецептора или связывающий белок, который изолирует лиганд от его когнатного рецептора(ов).
Термин "по существу чистый", как он использован здесь, относится к веществу, которое является по меньшей мере на 50% чистым (то есть, не содержит примесей), более предпочтительно по меньшей мере на 90% чистым, более предпочтительно по меньшей мере на 95% чистым, еще более предпочтительно по меньшей мере на 98% чистым и наиболее предпочтительно по меньшей мере на 99% чистым.
Термин "клетка-хозяин" включает индивидуальную клетку или клеточную культуру, которые могут быть или являются реципиентом для вектора(ов) для включения полинуклеотидных вставок. Клетки-хозяева включают потомство от одной клетки-хозяина, и это потомство необязательно может быть полностью идентичным (по морфологии или по геномному ДНК комплементу) относительно исходной родительской клетки в виду естественной, случайной или преднамеренной мутации. Клетка-хозяин включает клетки, трансфицированные in vivo полинуклеотидом(ами) по данному изобретению.
Как известно в уровне техники, термин "Fc-область" используют для определения С-концевой области тяжелой цепи иммуноглобулина. "Fc-область" может представлять собой нативную последовательность Fc-области или вариант Fc-области. Хотя границы Fc-области тяжелой цепи иммуноглобулина могут варьировать, Fc-область тяжелой цепи человеческого IgG обычно определяют как участок от аминокислотного остатка по положению Cys226 или от Рrо230 до его карбоксильного конца. Нумерация остатков в Fc-области соответствует нумерации EU-индекса базы данных Kabat (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md., 1991). Fc-область иммуноглобулина как правило содержит два константных домена: СН2 и СН3.
"Fc-рецептор" и "FcR", как их используют в данной области техники, описывают рецептор, который связывается с Fc-областью антитела. Предпочтительный FcR представляет собой нативную последовательность человеческого FcR. Кроме того, предпочтительным FcR является тот, который связывается с IgG антителом (гамма-рецептор), и включает рецепторы подклассов FcγRI, FcγRII и FcγRIII, в том числе аллельные варианты и, альтернативно, сплайсированные формы этих рецепторов. FcγRII рецепторы включают FcγRIIA ("активирующий рецептор") и FcγRIIB ("ингибирующий рецептор"), которые имеют сходные аминокислотные последовательности, которые отличаются главным образом их цитоплазматическими доменами. Обзор по FcRs приведен в Ravetch and Kinet, 1991, Ann. Rev. Immunol., 9: 457-92; Capel et al., 1994, Immunomethods, 4: 25-34; и de Haas et al., 1995, J. Lab. Clin. Med., 126:330-41. "FcR" также включает неонатальный рецептор FcRn, который отвечает за перенос материнских IgG плоду (Guyer et al., 1976 J. Immunol., 117: 587; и Kirn et al., 1994, J.Immunol., 24: 249).
Термин "конкурировать", как он использован здесь в отношении антитела, означает, что первое антитело или его антиген-связывающий участок связываются с эпитопом таким образом, который является достаточно сходным со связыванием второго антитела или его антиген-связывающего участка, так что результат связывания первого антитела с его когнатным эпитопом обнаружимым образом снижается в присутствии второго антитела по сравнению со связыванием первого антитела в отсутствие второго антитела. Альтернатива, где связывание второго антитела с его эпитопом также обнаружимым образом снижается в присутствии первого антитела, может иметь место, но не является обязательной. То есть, первое антитело может ингибировать связывание второго антитела с его эпитопом без того, чтобы второе антитело ингибировало связывание первого антитела с его соответствующим эпитопом. Однако, когда каждое антитело обнаружимым образом ингибирует связывание другого антитела с его когнатным эпитопом или лигандом, независимо от того, происходит ли это в такой же, в большей или в меньшей степени, такие антитела называют "перекрестно-конкурирующими" друг с другом за связывание их соответствующих эпитопов. И конкурирующие, и перекрестно-конкурирующие антитела охватываются настоящим изобретением. Независимо от механизма, по которому происходит такое конкурирование или перекрестное конкурирование (например, стерическое препятствие, конформационное изменение или связывание с общим эпитопом или его участком), специалисту в данной области будет понятно, основываясь на предложенных здесь учениях, что такие конкурирующие и/или перекрестно-конкурирующие антитела охватываются данным изобретением и могут быть полезными для раскрытых здесь способов.
"Функциональная Fc-область" обладает по меньшей мере одной эффекторной функцией нативной последовательности Fc-области. Репрезентативные "эффекторные функции" включают C1q связывание; CDC; связывание Fc-рецептора; антителозависимую клеточно-опосредованную цитотоксичность; фагоцитоз; регуляцию рецепторов клеточной поверхности (например, В-клеточного рецептора) по типу обратной связи, и так далее. Такие эффекторные функции, как правило, требуют, чтобы Fc-область объединялась со связывающим доменом (например вариабельным доменом антитела) и могла быть оценена с использованием различных анализов, известных в данной области техники, для оценки таких эффекторных функций антитела.
"Нативная последовательность Fc-области" содержит аминокислотную последовательность, идентичную аминокислотной последовательности Fc-области, обнаруживаемой в природе. "Вариант Fc-области" содержит аминокислотную последовательность, которая отличается от нативной последовательности Fc-области в силу по меньшей мере одной аминокислотной модификации, но все еще сохраняет по меньшей мере одну эффекторную функцию нативной последовательности Fc-области. Предпочтительно, вариант Fc-области имеет по меньшей мере одну аминокислотную замену по сравнению с нативной последовательностью Fc-области или с Fc-областью родительского полипептида, например, от примерно одной до примерно десяти аминокислотных замен, предпочтительно от примерно одной до примерно пяти аминокислотных замен в нативной последовательности Fc-области или в Fc-области родительского полипептида. Описанный здесь вариант Fc-области будет предпочтительно иметь по меньшей мере примерно 80% идентичность последовательности с нативной последовательностью Fc-области и/или с Fc-областью родительского полипептида, более предпочтительно по меньшей мере примерно 90% идентичность последовательности с ней, еще более предпочтительно по меньшей мере примерно 95%, по меньшей мере примерно 96%, по меньшей мере примерно 97%, по меньшей мере примерно 98%, по меньшей мере примерно 99% идентичность последовательности с ней.
Под "минимальным ожидаемым уровнем биологического эффекта" (Minimal Anticipated Biological Effect Level (MABEL)) понимают минимальный ожидаемый уровень дозы, приводящий к минимальному биологическому эффекту у людей. Факторы безопасности обычно применяют для расчета для первой дозы у человека исходя из MABEL. Расчет MABEL должен использовать всю релевантную in vitro и in vivo информацию по фармакокинетике и фармакодинамике.
Термины "лечение" и "терапевтически эффективный", как они использованы здесь, представляют подходы для получения полезных или желаемых клинических результатов. Для целей данного изобретения относительно рН-зависимых антагонистических антител против PCSK9, полезные или желаемые клинические результаты включают по меньшей мере одно из следующих: усиление клиренса LDL и снижение заболеваемости или исправление аномальных уровней холестерина и/или липопротеина, приводящих к метаболическим расстройствам и/или расстройствам приема пищи, или в том числе семейной гиперхолестеринемии, атерогенной дислипидемии, атеросклерозу и, в более общем случае, сердечно-сосудистому заболеванию (CVD), но не ограничены ими.
"Снижение заболеваемости" означает любое из снижения тяжести, которое может включать снижение в необходимости (например воздействия) и/или количестве других лекарственных средств и/или терапий, обычно используемых при данном состоянии. Специалисту в данной области техники понятно, что индивидуумы могут отличаться по своему ответу на лечение, и, как таковой, например, "способ снижения заболеваемости" отражает введение рН-зависимого антитела на основе целесообразного ожидания того, что такое введение может вероятно вызывать такое снижение заболеваемости у данного конкретного индивидуума.
"Улучшение" означает сокращение или уменьшение интенсивности одного или более симптомов после получения лечения по сравнению с отсутствием лечения. "Улучшение" также включает укорочение или снижение продолжительности симптома.
Как используют здесь, "эффективная дозировка" или "эффективное количество" лекарственного средства, соединения или фармацевтической композиции означают количество, достаточное для получения одного или более полезных или желаемых результатов. Для профилактического применения полезные или желаемые результаты включают устранение или снижение риска, уменьшения тяжести или задержки начала возникновения заболевания, включая биохимические, гистологические и/или поведенческие симптомы заболевания, его осложнения и промежуточные патологические фенотипы, возникающие при развитии заболевания. Для терапевтического применения рН-зависимого антагонистического антитела против PCSK9 полезные или желаемые результаты включают клинические результаты, такие как снижение гиперхолестеринемии или одного или более симптомов дислипидемии, атеросклероза, CVD или коронарной болезни сердца, снижение дозы других лекарственных средств, необходимых для лечения данного заболевания, усиление эффекта другого лекарственного средства и/или задержка прогрессирования заболевания у пациентов. Эффективная дозировка может быть введена за одно или более введений. Для целей данного изобретения эффективная дозировка лекарственного средства, соединения или фармацевтической композиции представляет собой количество, достаточное для осуществления профилактического или терапевтического лечения прямо или опосредованно. Как следует понимать в клиническом контексте, эффективная дозировка лекарственного средства, соединения или фармацевтической композиции может быть достигнута или не обязательно достигнута в сочетании с другими лекарственным средством, соединением или фармацевтической композицией. Так, "эффективная дозировка" может рассматриваться в контексте введения одного или более терапевтических агентов, и индивидуальный агент может рассматриваться как даваемый в эффективном количестве, если в сочетании с одним или более терапевтическими агентами достигается или может быть достигнут желаемый результат.
"Индивидуум" или "субъект" представляет собой млекопитающее, более предпочтительно человека. Млекопитающие также включают сельскохозяйственных животных, спортивных животных, домашних питомцев, приматов, лошадей, собак, кошек, мышей и крыс, но не ограничиваются ими.
"Вектор", как он использован здесь, означает конструкцию, которая способна доставлять и, предпочтительно, экспрессировать один или более генов или последовательностей, представляющих интерес, в клетке-хозяине. Примеры векторов включают вирусные векторы, голые ДНК- или РНК-экспрессионные векторы, плазмидные, космидные или фаговые векторы, ДНК- или РНК-экспрессионные векторы, ассоциированные с катионными конденсирующими агентами, ДНК- или РНК-экспрессионные векторы, инкапсулированные в липосомах, и некоторые эукариотические клетки, такие как клетки-продуценты, но не ограничены ими.
"Экспрессионная контрольная последовательность", как используют здесь, означает последовательность нуклеиновой кислоты, которая направляет транскрипцию нуклеиновой кислоты. Экспрессионная контрольная последовательность может представлять собой промотор, такой как конститутивный или индуцибельный промотор, или энхансер. Экспрессионная контрольная последовательность функциональным образом связана с последовательностью нуклеиновой кислоты, подлежащей транскрипции.
"Фармацевтически приемлемый носитель" или "фармацевтически приемлемый эксципиент", как они использованы здесь, включают любое вещество, которое при объединении с активным ингредиентом обеспечивает сохранение биологической активности этого ингредиента и является инертным для иммунной системы субъекта. Примеры включают любые из стандартных фармацевтических носителей, таких как забуференный фосфатом физиологический раствор, воду, эмульсии, такие как эмульсия масло/вода, и различные типы увлажняющих агентов, но не ограничены ими. Предпочтительные разбавители для аэрозольного или парентерального введения представляют собой забуференный фосфатом физиологический раствор (PBS) или нормальный (0,9%) физиологический раствор. Композиции, содержащие такие носители, изготавливают хорошо известными традиционными способами (см., например, Remington's Pharmaceutical Sciences, 18th edition, A.Gennaro, ed., Mack Publishing Co., Easton, PA, 1990; и Remington, The Science and Practice of Pharmacy, 20th Ed., Mack Publishing, 2000).
Термин "kon", как он использован здесь, относится к константе скорости ассоциации антитела с антигеном. Конкретно, константы скорости (kon и koff) и равновесные константы диссоциации измеряют с использованием Fab фрагментов антитела (то есть, одновалентных) и антигена.
Термин "koff", как он использован здесь, относится к константе скорости диссоциации антитела из комплекса антитело/антиген.
Термин "KD", как он использован здесь, относится к равновесной константе диссоциации взаимодействия антитело-антиген.
Определения констант скорости ассоциации и диссоциации, ka и kd соответственно, для определения отношений KD и koff, проводят с использованием биосенсора на основе поверхностного плазмонного резонанса для характеристики взаимодействия анализируемого вещества/лиганда в условиях, когда анализируемое вещество является моновалентным в отношении связывания лиганда, который иммобилизирован с низкой емкостью на поверхности сенсора посредством реагента захвата. Анализ осуществляют с использованием методологии кинетического титрования, как описано в Karlsson etal., Anal. Biochem 349, 136-147, 2006. Сенсорный чип, реагент захвата и буфер для анализа, используемые для данного анализа, выбирают так, чтобы обеспечить стабильный захват лиганда на сенсорной поверхности, минимизировать неспецифическое связывание анализируемого вещества с поверхностями и получить ответы связывания анализируемого вещества, которые приемлемы для кинетического анализа, в соответствии с рекомендациями Myszka, J. Mol. Recognit 12, 279-284, 1999. Ответы связывания анализируемого вещества по взаимодействию анализируемого вещества/лиганда получают с двойным сравнением и подгоняют под ленгмюровскую модель связывания с массопереносом 1:1 cka, kd и Rmax в качестве универсальных параметров, как описано в Myszka & Morton et al., Biophys. Chem 64, 127-137 (1997). Равновесную константу диссоциации KD выводят из отношения кинетических констант скорости: KD=kd/ka. Такие определения предпочтительно осуществляют при 25°С или 37°С.
А. Способы предупреждения или лечения расстройств
В одном аспекте, касающемся рН-зависимых антагонистических антител против PCSK9, в данном изобретении предложен способ лечения или предупреждения гиперхолестеринемии и/или по меньшей мере одного симптома из дислипидемии, атеросклероза, CVD или сердечно-сосудистого заболевания, у индивидуума, включающий введение указанному индивидууму эффективного количества рН-зависимого антагонистического антитела против PCSK9, которое антагонизирует циркулирование PCSK9.
В дополнительном аспекте в данном изобретении предложено эффективное количество рН-зависимого антагонистического антитела против PCSK9, которое антагонизирует циркулирование PCSK9, для использования в лечении или предупреждении гиперхолестеринемии и/или по меньшей мере одного симптома из дислипидемии, атеросклероза, CVD или сердечнососудистого заболевания, у индивидуума. В изобретении дополнительно предложено применение эффективного количества рН-зависимого антагонистического антитела против PCSK9, которое антагонизирует внеклеточный или циркулирующий PCSK9, в изготовлении лекарственного средства для лечения или предупреждения гиперхолестеринемии и/или по меньшей мере одного симптома из дислипидемии, атеросклероза, CVD или сердечно-сосудистого заболевания, у индивидуума.
Преимущественно, терапевтическое введение антитела приводит в результате к более низким уровням холестерина в крови и/или более низким уровням LDL в крови. Предпочтительно, уровень холестерина в крови и/или уровень LDL в крови по меньшей мере примерно на 10% или 15% ниже, чем до введения. Более предпочтительно, уровень холестерина и/или уровень LDL по меньшей мере примерно на 20% ниже, чем до введения антитела. Еще более предпочтительно, уровень холестерина и/или уровень LDL по меньшей мере примерно на 30% ниже, чем до введения антитела. Преимущественно, уровень холестерина в крови и/или уровень LDL в крови по меньшей мере примерно на 40% ниже, чем до введения антитела. Еще более предпочтительно, уровень холестерина в крови и/или уровень LDL в крови по меньшей мере примерно на 50% ниже, чем до введения антитела. Более предпочтительно, уровень холестерина в крови и/или уровень LDL в крови по меньшей мере на 60% ниже, чем до введения антитела. Наиболее предпочтительно, уровень холестерина в крови и/или уровень LDL в крови по меньшей мере на 70% ниже, чем до введения антитела.
В отношении всех описанных здесь способов, ссылка на рН-зависимые антитела против любого соответствующего антигена также включает композиции, содержащие один или более дополнительных агентов. Эти композиции могут дополнительно содержать подходящие эксципиенты, такие как фармацевтически приемлемые эксципиенты, включая буферы, которые хорошо известны в данной области техники. Настоящее изобретение может быть использовано отдельно или в комбинации с другими общепринятыми способами лечения.
рН-Зависимое антитело может быть введено индивидууму любым подходящим путем. Специалисту в данной области техники понятно, что описанные здесь примеры не предназначены ограничивать объем, а являются только иллюстративными в отношении доступных методик. Соответственно, в некоторых воплощениях рН-зависимое антитело вводят индивидууму в соответствии с известными способами, такими как внутривенное введение, например, в виде болюсной инъекции или посредством непрерывной инфузии в течение периода времени, внутримышечное, внутрибрюшинное, интрацереброспинальное, трансдермальное, подкожное, интраартикулярное, сублингвальное, внутрисуставное введение, введение посредством инсуффляции, интратекальное, пероральное введение, ингаляция или местное введение. Введение может быть системным, например, внутривенное введение, или локализованным. Имеющиеся в продаже небулайзеры для жидких препаратов, включая струйные небулайзеры и ультразвуковые небулайзеры, являются полезными для введения. Жидкие препараты можно распылять непосредственно, а лиофилизированные порошки можно распылять после их разведения. Альтернативно, рН-зависимое антитело можно ввести в состав аэрозоля с использованием фторуглеродов и в дозирующий ингалятор, или ингалировать в виде лиофилизированного и измельченного порошка.
В одном воплощении рН-зависимое антитело вводят посредством методологий сайт-специфической или направленной местной доставки. Примеры методологий сайт-специфической или направленной местной доставки включают различные имплантируемые депо-источники рН-зависимого антитела или катетеры для местной доставки, такие как инфузионные катетеры, постоянные катетеры или игольчатые катетеры, искусственные трансплантаты, адвентициальные повязки, шунты и стенты или другие имплантируемые устройства, сайт-специфические носители, прямая инъекция или прямое нанесение. См., например, публикацию РСТ заявки WO 00/53211 и патент США 5981568.
Различные препараты рН-зависимого антитела могут быть использованы для введения. В некоторых воплощениях рН-зависимое антитело может быть введено в чистом виде. В некоторых воплощениях рН-зависимое антитело и фармацевтически приемлемый эксципиент могут быть введены в состав различных препаратов. Фармацевтически приемлемые эксципиенты известны в данной области техники и представляют собой относительно инертные вещества, которые облетают введение фармакологически эффективного вещества. Например, эксципиент может обеспечить форму или консистенцию или действовать в качестве разбавителя. Подходящие эксципиенты включают стабилизаторы, увлажнители и эмульгаторы, соли для варьирования осмолярности, инкапсулирующие агенты, буферы и усилители проникновения через кожу, но не ограничены ими. Эксципиенты, а также препараты для парентеральной и непарентеральной доставки лекарственного средства, изложены в Remington, The Science and Practice of Pharmacy, 20th Ed., Mack Publishing (2000).
Эти агенты могут быть объединены с фармацевтически приемлемыми носителями, такими как солевой раствор, раствор Рингера, раствор декстрозы и тому подобное. Конкретные режимы дозировки, то есть, доза, время и повторность, будут зависеть от конкретного индивидуума и его истории болезни.
Антитела с рН-зависимым связыванием могут также быть введены посредством ингаляции, как здесь описано. В общем, для введения рН-зависимых антител начальная дозировка кандидата может составлять примерно 2 мг/кг. Для целей настоящего изобретения типичная суточная дозировка может варьировать от примерно 3 мкг/кг до 30 мкг/кг до 300 мкг/кг до 3 мг/кг, до 30 мг/кг, до 100 мг/кг или более, в зависимости от факторов, упомянутых выше. Например, могут быть использованы дозировки примерно 1 мг/кг, примерно 2,5 мг/кг, примерно 5 мг/кг, примерно 10 мг/кг и примерно 25 мг/кг. Для повторных введений в течение нескольких суток или дольше, в зависимости от состояния, лечение продолжают до тех пор, пока не возникнет желаемая степень подавления симптомов, или до тех пор, пока не будут достигнуты достаточные терапевтические уровни, например, до снижения уровней LDL в крови. Примерные режимы дозирования включают введение начальной дозы примерно 2 мг/кг, а затем еженедельной поддерживающей дозы примерно 1 мг/кг антитела или последующей поддерживающей дозы примерно 1 мг/кг через неделю. Однако другие режимы дозирования могут быть полезны, в зависимости от картины фармакокинетического убывания, которую желает достичь практикующий врач. Например, в некоторых воплощениях предусмотрена дозировка от одного до четырех раз в неделю. В других воплощениях предусмотрена дозировка один раз в месяц, или один раз в два месяца, или один раз в три месяца. Успешность этой терапии легко контролировать традиционными способами и анализами. Режим дозировки (включающий используемое антитело) может варьировать в зависимости от времени.
Для целей настоящего изобретения подходящая дозировка рН-зависимого антитела будет зависеть от используемого антитела (или его композиций), типа и тяжести симптомов, подлежащих лечению, независимо от того вводят ли агент для профилактических или терапевтических целей, предыдущей терапии, клинической истории пациента и реакции на агент, уровней антигена в крови пациента, скорости синтеза и клиренса антигена у пациента, скорости клиренса вводимого агента у пациента и усмотрения лечащего врача. Как правило, врач будет вводить рН-зависимое антитело до достижения дозировки, которая дает желаемый результат. Дозу и/или частоту можно варьировать в течение курса лечения. Эмпирические наблюдения, такие как время полужизни, обычно вносят вклад в определение дозировки. Например, антитела, которые совместимы с иммунной системой человека, такие как гуманизированные антитела или полностью человеческие антитела, могут быть использованы для пролонгирования времени полужизни антитела и для предотвращения атаки антитела иммунной системой организма хозяина. Частота введения может быть определена и отрегулирована в течение курса терапии и основана, как правило, но не обязательно, на лечении и/или подавлении и/или уменьшении интенсивности и/или задержки возникновения симптомов, например, гиперхолестеринемии. Альтернативно, препараты с непрерывным высвобождением могут быть полезны. Различные препараты и устройства для достижения непрерывного высвобождения известны в данной области техники.
В одном воплощении дозировки для антагонистического антитела могут быть определены эмпирически у индивидуумов, которым назначают одно или более введений антагонистического антитела. Индивидуумам дают нарастающие дозировки антитела. Для оценки эффективности можно отслеживать индикатор заболевания.
Введение рН-зависимого антитела в соответствии со способом по настоящему изобретению может быть непрерывным или прерывистым, в зависимости, например, от физиологического состояния реципиента, от того является ли цель введения терапевтической или профилактической и других факторов, известных специалистам. Введение рН-зависимого антитела может быть по существу непрерывным в течение - предопределенного периода времени или может происходить в виде ряда разделенных доз.
В некоторых воплощениях присутствует более одного антагонистического антитела. Может присутствовать по меньшей мере одно, по меньшей мере два, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять или более разных антагонистических антител и/или пептидов. В общем, такие антитела или пептиды могут иметь комплементарные активности, которые не влияют неблагоприятным образом друг на друга. рН-Зависимое антитело также может быть использовано в комбинации с другими терапевтическими средствами. рН-Зависимое антитело также может быть использовано в комбинации с другими агентами, которые служат для усиления и/или дополнения эффективности агентов.
Приемлемые носители, эксципиенты или стабилизаторы являются нетоксичными для реципиентов в используемых дозировках и концентрациях и могут содержать буферы, такие как фосфат, цитрат и другие органические кислоты; соли, такие как хлорид натрия; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония, хлорид бензетония; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3-пентанол и м-крезол); низкомолекулярные (менее чем примерно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как этилендиаминтетрауксусная кислота (EDTA); сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы с металлами (например, Zn-белковыё комплексы); и/или неионные поверхностно-активные вещества, такие как TWEEN™, PLURONICS™ или полиэтиленгликоль (PEG).
Липосомы, содержащие рН-зависимое антитело, получают способами, известными в данной области техники, например как описано в Epstein, et al., 1985, Proc. Natl. Acad. Sci. USA 82: 3688; Hwang, et al., 1980, Proc. Natl Acad. Sci. USA 77: 4030; и патентах США 4485045 и 4544545. Липосомы с увеличенным временем циркуляции раскрыты в патенте США 5013556. Особенно полезные липосомы могут быть получены способом выпаривания с обращенной фазой с липидной композицией, содержащей фосфатидилхолин, холестерин и PEG-дериватизированный фосфатидилэтаноламин (PEG-PE). Липосомы продавливают через фильтры с определенным размером пор с получением липосом желаемого диаметра.
Активные ингредиенты также могут быть включены в микрокапсулы, полученные, например, методиками коацервации или полимеризации на границе фаз, например, микрокапсулы из гидроксиметилцеллюлозы или желатина и микрокапсулы из поли-(метилметакрилата), соответственно, в коллоидных системах доставки лекарственного средства (например, липосомы, микросферы альбумина, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсиях. Такие методики раскрыты в Remington, The Science and Practice of Pharmacy, 20th Ed., Mack Publishing (2000).
Могут быть получены препараты с непрерывным высвобождением. Подходящие примеры препаратов с непрерывным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащие антитело, где матрицы находятся в форме изделий определенного размера, например, пленок или микрокапсул. Примеры матриц с непрерывным высвобождением включают полиэфиры, гидрогели (например, поли(2-гидроксиэтил-метакрилат) или поливиниловый спирт)), полилактиды (патент США 3773919), сополимеры L-глутаминовой кислоты и 7-этил-L-глутамата, неразлагаемые сополимеры этиленвинилацетата, разлагаемые сополимеры молочной кислоты и гликолевой кислоты, такие как LUPRON DEPOT™ (инъецируемые микросферы, состоящие из сополимера молочной кислоты и гликолевой кислоты и ацетата лейпролида), изобутират ацетат сахарозы и поли-D-(-)-3-гидроксимасляную кислоту.
Препараты, предназначенные для введения in vivo, должны быть стерильными. Этого легко достичь посредством, например, фильтрования через мембраны для стерильной фильтрации. Терапевтические композиции рН-зависимого антитела, как правило, помещают в контейнер, имеющий стерильное входное отверстие, например, пакет с раствором для внутривенного введения или флакон, имеющий пробку, прокалываемую иглой для подкожной инъекции.
Подходящие эмульсии могут быть приготовлены с использованием имеющихся в продаже жировых эмульсий, таких как Intralipid™, Liposyn™, Infonutrol™, Lipofundin™ и Lipiphysan™. Активный ингредиент может быть растворен в предварительно смешанной эмульсионной композиции, или альтернативно, его можно растворить в масле (например, соевом масле, сафлоровом масле, хлопковом масле, кунжутном масле, кукурузном масле или миндальном масле) и образованную эмульсию затем смешать с фосфолипидом (например, яичными фосфолипидами, соевыми фосфолипидами или соевым лецитином) и водой. Очевидно, что могут быть добавлены другие ингредиенты, например глицерин или глюкоза, для регулирования тоничности эмульсии. Подходящие эмульсии обычно содержат вплоть до 20% масла, например от 5 до 20%. Жировая эмульсия может содержать капли жира размером от 0,1 до 1,0 мкм, в частности от 0,1 до 0,5 мкм, и имеют рН в интервале от 5,5 до 8,0.
Эмульсионные композиции могут представлять собой эмульсии, приготовленные смешиванием рН-зависимого антитела с Intralipid™ или его компонентами (соевым маслом, яичными фосфолипидами, глицерином и водой).
Композиции для ингаляции или инсуффляции включают растворы и суспензии в фармацевтически приемлемых водных или органических растворителях или их смесях и порошки. Жидкие или твердые композиции могут содержать подходящие фармацевтически приемлемые эксципиенты, как изложено выше. В некоторых воплощениях композиции вводят пероральным или назальным респираторным путем для местного или системного эффекта.
Композиции в предпочтительно стерильных фармацевтически приемлемых растворителях можно распылять с помощью газов. Распыляемые растворы можно вдыхать непосредственно из небулайзера, либо небулайзер может быть подсоединен к защитной маске, палатке или аппарату для вентиляции легких с положительным прерывистым давлением. Композиции в виде раствора, суспензии или порошка могут быть введены, предпочтительно перорально или интраназально, из устройств, которые доставляют препарат подходящим образом.
В. рН-Зависимые антитела
Антитела, полезные в настоящем изобретении, могут охватывать моноклональные антитела, политональные антитела, фрагменты антител (например, Fab, Fab', F(ab')2, Fv, Fc и т.д.), химерные антитела, биспецифические антитела, гетероконъюгатные антитела, одноцепочечные (ScFv), их мутанты, слитые белки, содержащие участок антитела (например, однодоменное антитело), человеческие антитела, гуманизированные антитела и любую другую модифицированную конфигурацию молекулы иммуноглобулина, которая содержит сайт распознавания антигена требуемой специфичности, включая гликозилированные варианты антител, варианты аминокислотной последовательности антител и ковалентно модифицированные антитела. Антитела могут быть мышиными, крысиными, человеческими или любого другого происхождения (включая химерные или гуманизированные антитела).
В некоторых воплощениях рН-зависимое антитело представляет собой моноклональное антитело. рН-Зависимое антитело также может быть гуманизированным. В других воплощениях антитело является человеческим.
В некоторых воплощениях антитело содержит модифицированную константную область, такую как константная область, которая является иммунологически инертной, то есть имеющая пониженный потенциал для индуцирования иммунного ответа. В некоторых воплощениях константная область модифицирована, как описано в Eur. J.Immunol., 1999, 29: 2613-2624; публикации РСТ заявки WO 99/58572; и/или заявке на патент UK No.9809951.8. Fc может представлять собой человеческий IgGs или человеческий IgG4. Fc может представлять собой человеческий IgG2, содержащий мутацию А330Р331 на S330S331 (IgG2Δа), где аминокислотные остатки пронумерованы со ссылкой на последовательность IgG2 дикого типа (Eur. J.Immunol., 1999, 29: 2613-2624). В некоторых воплощениях антитело содержит константную область IgG4, содержащую следующие мутации (Armour et al., 2003, Molecular Immunology 40 585-593): E233F234L235 на P233V234A235 (IgG4Δс), где нумерация осуществлена со ссылкой на последовательность lgG4 дикого типа. В еще одном воплощении Fc представляет собой человеческий lgG4 E233F234L235 на P233V234A235 с делецией G236 (IgG4Δb). В другом воплощении Fc представляет собой любой человеческий IgG4 Fc (IgG4, IgG4Δb или IgG4Δс), содержащий петлевую стабилизирующую мутацию S228 на Р228 (Aalberse et al., 2002, Immunology 105, 9-19). В другом воплощении Fc может представлять собой агликозилированный Fc.
В некоторых воплощениях константная область является агликозилированной посредством мутирования остатка присоединения к олигосахариду (такого как Asn297) и/или фланкирования остатков, которые являются частью последовательности распознавания гликозилирования в константной области. В некоторых воплощениях константная область является агликозилированной для N-связанного гликозилирования ферментативно. Константная область может быть агликозилированной для N-связанного гликозилирования ферментативно или путем экспрессии в клетке-хозяине, дефицитной по гликозилированию.
Один путь определения аффинности связывания антитела с антигеном заключается в измерении аффинности связывания монофункциональных Fab фрагментов антитела. Для получения монофункциональных Fab фрагментов антитело (например IgG) можно подвергать расщеплению папаином или экспрессировать рекомбинантно. Аффинность Fab фрагмента антитела может быть определена посредством поверхностного плазменного резонанса (система поверхностного плазменного резонанса (SPR) Biacore3000™, Biacore, INC, Piscataway NJ, оборудованная сенсорными чипами с предварительно иммобилизированным стрептавидином (SA) с использованием подвижного буфера HBS-EP (0,01 М HEPES, рН 7,4, 0,15 NaCl, 3 мМ EDTA, 0,005% об./об. Surfactant P20). Биотинилированный антиген может быть разведен в буфере HBS-EP до концентрации менее чем 0,5 мкг/мл и инъецирован через каналы индивидуального чипа при варьировании времени контакта, для достижения двух интервалов антигенной плотности, либо 50-200 единиц ответа (RU) для подробных кинетических исследований, либо 800-1000 RU для анализов скрининга. Исследования регенерации показали, что 25 мМ NaOH в 25% об./об. этанола эффективно удаляет связанный Fab при сохранении активности антигена на чипе в течение более 200 инъекций. Как правило, серийные разведения (диапазон концентраций 0,1-10х оцененных KD) очищенных образцов Fab инъецировали в течение 1 мин при 100 мкл/мин и обеспечивали время диссоциации вплоть до 2 часов. Концентрации Fab белков определяют путем твердофазного иммуноферментного анализа (ELISA) и/или электрофореза в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE) с использованием известной концентрации Fab (как определено аминокислотным анализом) в качестве стандарта. Кинетические скорости ассоциации (kon) и скорости диссоциации (koff) получают одновременно путем универсальной подгонки данных к ленгмюровской модели связывания 1:1 (Karlsson, R.Roos, H.Fagerstam, L.Petersson, В., 1994. Methods Enzymology 6. 99-110) с использованием программы BIAevaluation. Значения равновесной константы диссоциации (KD) рассчитывают как koff/kon. Этот протокол подходит для использования при определении аффинности связывания антитела с любым антигеном, включая человека или любых других видов млекопитающих (таких как мышь, крыса, примат). Аффинность связывания антитела в общем измеряют при 25°С, но также можно измерять при 37°С.
Антитела могут быть получены любым способом, известным в данной области техники. Путь и режим иммунизации животного-хозяина в общем соответствуют установленным и общепринятым методикам стимулирования и продуцирования антител, как дополнительно здесь описано. Общие методики продуцирования человеческих и мышиных антител известны в данной области техники и/или описаны здесь. Для рН-зависимых антител против PCSK9 предпочтительный в настоящее время способ получения антител включает иммунизацию животных с выключенным геном PCSK9 (PCSK9 -/-), как описано здесь.
Подразумевают, что любого субъекта млекопитающего, включая людей или антитело-продуцирующие клетки из них, можно использовать в качестве основы для продуцирования гибридомных клеточных линий млекопитающих, включая человека. Как правило, животное-хозяин инокулируют внутрибрюшинно, внутримышечно, перорально, подкожно, в подошву и/или внутрикожно некоторым количеством иммуногена, в том числе как описано здесь.
Гибридомы могут быть получены из лимфоцитов и иммортализованных клеток миеломы с использованием общих методик гибридизации соматических клеток согласно Kohler, В. and Milstein, С., 1975, Nature 256: 495-497, или с видоизменениями согласно Buck, D.W., et al., 1982, In Vitro, 18: 377-381. Доступные линии миеломы, включая Х63-Ад8.653 и линии из Salk Institute, Cell Distribution Center, San Diego, Calif., USA, но не ограничиваясь ими, могут быть использованы в гибридизации. Как правило, эта методика включает слияние клеток миеломы и лимфоидных клеток с использованием вещества, способствующего слиянию генов, такого как полиэтиленгликоль, или с помощью электрических средств, хорошо известных специалистам в данной области техники. После слияния клетки отделяют от среды слияния и выращивают в селективной среде роста, такой как среда гипоксантин-аминоптерин-тимидин (HAT), для удаления негибридизованных родительских клеток. Любая из описанных здесь сред, дополненных сывороткой или без нее, может быть использована для культивирования гибридом, которые секретируют моноклональные антитела. В качестве другой альтернативы методикам слияния клеток, иммортализованные В-клетки вируса Эпштейна-Барра (EBV) могут быть использованы для продуцирования моноклональных антител против PCSK9 по изобретению. Гибридомы размножают и субклонируют при желании, и супернатанты анализируют на противоиммуногенную активность традиционными методиками иммуноанализа (например, радиоиммуноанализа, ферментного иммуноанализа или флуоресцентного иммуноанализа).
Гибридомы, которые могут быть использованы в качестве источника антител, охватывают все производные, клетки-предшественники родительских гибридом, которые продуцируют моноклональные антитела, специфичные в отношении PCSK9, или их участок.
Гибридомы, которые продуцируют такие антитела, можно выращивать in vitro или in vivo, используя известные методики. Моноклональные антитела могут быть выделены из культуральных сред или жидкостей организма традиционными методиками очистки иммуноглобулинов, такими как осаждение сульфатом аммония, гель-электрофорез, диализ, хроматография и ультрафильтрация, при желании. Если присутствует нежелательная активность, ее можно удалить, например, путем пропускания препарата над адсорбентами, изготовленными из иммуногена, присоединенного к твердой фазе, и элюции или высвобождения желаемых антител из этого иммуногена. Иммунизация животного-хозяина человеческим PCSK9 или фрагментом, содержащим целевую аминокислотную последовательность, конъюгированную с белком, который является иммуногенным у видов, подлежащих иммунизации, например, гемоцианин лимфы улитки, сывороточный альбумин, бычий тироглобулин или ингибитор соевого трипсина, с использованием бифункционального или дериватизирующего агента, например сульфосукцинимидный малеимидобензоиловый эфир (конъюгирование через цистеиновые остатки), N-гидроксисукцинимид (через лизиновые остатки), глутаровый альдегид, ангидрид янтарной кислоты, SOCl2 или R1N=C=NR, где R и R1 представляют собой разные алкильные группы, может привести к популяции антител (например моноклональных антител).
Как только антитела генерированы или отобраны, их можно оптимизировать в отношении рН-зависимого связывания, например как описано здесь в Примере 1.
При желании представляющее интерес рН-зависимое антитело (моноклональное или поликлональное) может быть секвенировано, и полинуклеотидная последовательность затем может быть клонирована в вектор для экспрессии или размножения. Последовательность, кодирующая представляющее интерес антитело, может сохраняться в векторе в клетке-хозяине, и эту клетку-хозяина затем можно размножать и замораживать для использования в будущем. Продуцирование рекомбинантных моноклональных антител в клеточной культуре может быть осуществлено посредством клонирования генов антитела из В-клеток средствами, известными в данной области техники. См., например, Tiller et al., 2008, J.Immunol. Methods 329, 112; патент США 7314622.
В качестве альтернативы, полинуклеотидная последовательность может быть использована для генетической манипуляции для "гуманизации" антитела или для улучшения аффинности или других характеристик антитела. Например, константная область может быть сконструирована с более точным сходством с человеческими константными областями, для того чтобы избежать иммунного ответа, если это антитело используют в клинических испытаниях и лечении людей. Может быть желательно генетически манипулировать последовательностью антитела для получения большей аффинности к антигену и большей эффективности. Специалисту в данной области техники очевидно, что одна или более полинуклеотидных замен могут быть произведены в рН-зависимом антителе с сохранением его антиген-связывающей способности.
Существует четыре общие стадии гуманизации моноклонального антитела. Они представляют собой: (1) определение нуклеотидной и предсказанной аминокислотной последовательности вариабельных доменов легкой и тяжелой цепи исходного антитела; (2) конструирование гуманизированного антитела, то есть, принятие решения, какую каркасную область антитела использовать во время процесса гуманизации; (3) фактическое осуществление процессов/методик гуманизации; и (4) трансфекцию и экспрессию гуманизированного антитела. См., например, патенты США 4816567, 5807715, 5866692, 6331415, 5530101, 5693761, 5693762, 5585089 и 6180370.
Был описан ряд молекул "гуманизированных" антител, содержащих антиген-связывающий сайт, имеющий происхождение из иммуноглобулина, не являющегося человеческим, включая химерные антитела, имеющие V-области грызунов или модифицированные V-области грызунов и их ассоциированные CDR, слитые с человеческими константными доменами. См., например, Winter et al., 1991, Nature 349: 293-299; Lobuglio et al., 1989, Proc. Nat. Acad. Sci. USA 86: 4220-4224; Shaw et al., 1987, J Immunol. 138: 4534-4538; and Brown et al., 1987, Cancer Res. 47: 3577-3583. Другие источники информации описывают CDR грызунов, перенесенные на человеческую каркасную область (FR) перед слиянием с соответствующим человеческим константным доменом антитела. См., например, Riechmann et al.,1988, Nature 332: 323-327; Verhoeyen et al., 1988, Science 239: 1534-1536; и Jones et al., 1986, Nature 321: 522-525. В еще одном источнике информации описаны CDR грызунов, поддерживаемые рекомбинантно сконструированными каркасными областями грызунов. См., например, публикацию европейской заявки № 0519596. Эти "гуманизированные" молекулы конструируют для минимизации нежелательного иммунологического ответа, направленного на молекулы антитела грызунов против человека, который ограничивает продолжительность и эффективность терапевтических применений этих группировок у реципиентов-людей. Например, константная область антитела может быть сконструирована таким образом, чтобы быть иммунологически инертной (например, не запускает лизис комплемента). См., например, публикацию РСТ заявки WO 99/58572; заявку на патент UK No. 9809951.8. Другие способы гуманизации антител, которые также могут быть использованы, раскрыты Daugherty et al., 1991, Nucl. Acids Res. 19: 2471-2476 и в патентах США 6180377, 6054297, 5997867, 5866692, 6210671 и 6350861; а также в публикации РСТ заявки WO 01/27160.
В качестве еще одной альтернативы, полноразмерные человеческие антитела могут быть получены путем использования коммерчески доступных мышей, которые были разработаны для экспрессии специфических человеческих иммуноглобулиновых белков. Трансгенные животные, которые разработаны для продуцирования более желательного или более сильного иммунного ответа, также могут быть использованы для генерирования гуманизированных или человеческих антител. Примеры такой технологии представляют собой Xenomouse™ от Abgenix, Inc. (Fremont, CA), HuMAb-Mouse® и TC Mouse™ от Medarex, Inc. (Princeton, NJ), и Veloclmmune® mouse от Regeneron Pharmaceuticals, Inc. (Tarrytown, NY).
В качестве альтернативы, антитела могут быть получены рекомбинантным путем и экспрессированы с использованием любого способа, известного в данной области техники. В качестве другой альтернативы, антитела могут быть получены рекомбинантным путем с помощью технологии фагового дисплея. См., например, патенты США 5565332, 5580717, 5733743 и 6265150; и Winter et al., 1994, Annu. Rev. Immunol. 12: 433-455. Альтернативно, технология фагового дисплея (McCafferty et al., 1990, Nature 348: 552-553) может быть использована для продуцирования человеческих антител и фрагментов антител in vitro из репертуаров генов вариабельного (V) домена иммуноглобулина от иммунизированных доноров. В соответствии с этой технологией, гены V-домена антитела клонируют в рамке считывания в гене основного или неосновного оболочечного белка нитевидного бактериофага, такого как М13 или fd, и экспонируют в виде функциональных фрагментов антитела на поверхности фаговой частицы. Поскольку нитевидная частица содержит одноцепочечную копию ДНК фагового генома, селекции на основе функциональных свойств антитела также приводят в результате к селекции гена, кодирующего антитело, обладающее этими свойствами. Таким образом, фаг имитирует некоторые свойства В-клетки. Фаговый дисплей может быть осуществлен в ряде форматов; см., например, Johnson, Kevin S. and Chiswell, David J., 1993, Current Opinion in Structural Biology 3: 564-571. Некоторые источники V-генных сегментов могут быть использованы для фагового дисплея. Clackson et al., 1991, Nature 352: 624-628 выделили разнотипный ряд анти-оксазолоновых антител из небольшой произвольной комбинаторной библиотеки V-генов, имеющих происхождение из селезенок иммунизированных мышей. Репертуар V-генов от иммунизированных доноров-людей может быть сконструирован, и антитела к разнотипному ряду антигенов (включая аутоантигены) могут быть выделены по существу согласно методикам, описанным Mark et al., 1991, J.Mol. Biol. 222: 581-597, или Griffith et al., 1993, EMBO J. 12: 725-734. При естественном иммунном ответе гены антитела аккумулируют мутации в высокой степени (соматическая гипермутация). Некоторые из введенных замен будут обеспечивать более высокую афинность, и В-клетки, экспонирующие высокоаффинный поверхностный иммуноглобулин, предпочтительно реплицируются и дифференцируются во время последующей антигенной стимуляции. Этот естественный процесс может быть имитирован посредством технологии, известной как "перетасовка цепей." (Marks et al., 1992, Bio/Technol. 10: 779-783). В этом способе аффинность "первичных" человеческих антител, полученных посредством фагового дисплея, может быть улучшена последовательной заменой генов V-областей тяжелой и легкой цепей репертуарами природных вариантов (репертуаров) генов V-доменов, полученных от иммунизированных доноров. Эта методика обеспечивает продуцирование антител и фрагментов антител с аффинностями в рМ-нМ диапазоне. Стратегия получения очень крупных репертуаров фаговых антител (также известных как "the mother-of-all libraries") была описана Waterhouse et al., 1993, Nucl. Acids Res. 21:2265-2266. Перетасовка генов также может быть использована для получения человеческих антител из антител грызунов, где человеческое антитело имеет аффинности и специфичности, подобные аффинностям и специфичностям исходного антитела грызунов. В соответствии с этим способом, который также называют "импринтинг эпитопа", ген V-домена тяжелой или легкой цепи антител грызунов, полученных технологией фагового дисплея, заменяют репертуаром генов человеческого V-домена, создавая химеры грызун-человек. Селекция по антигену приводит в результате к выделению человеческих вариабельных областей, способных к воссозданию функционального антиген-связывающего сайта, то есть, эпитоп направляет ("отпечатывает") выбор партнера. Когда процесс повторяют с целью замены оставшегося V-домена грызунов, получают человеческое антитело (см. публикацию РСТ заявки WO 93/06213). В отличие от традиционной гуманизации антител грызунов посредством переноса CDR, эта техника обеспечивает полностью человеческие антитела, которые не имеют ни остатков каркасной области, ни остатков CDR, имеющих происхождение от грызунов.
Очевидно, что, хотя вышеописанное обсуждение относится к гуманизированным антителам, общие обсужденные принципы применимы к изготавливаемым по техническим требованиям заказчика антителам для применения, например, у собак, кошек, приматов, лошадей и крупного рогатого скота. Кроме того, очевидно, что один или более аспектов гуманизации описанных здесь антител могут быть объединены, например, перенос CDR, мутация каркасной области и мутация CDR.
Антитела могут быть получены рекомбинантным образом посредством сначала выделения антител и антитело-продуцирующих клеток из животных-хозяев, получения последовательности гена и использования этой последовательности гена для экспрессии антитела рекомбинантным образом в клетках-хозяевах (например, клетках СНО). Другой способ, который может быть использован, заключается в том, чтобы экспрессировать последовательность антитела в растениях (например, табак) или трансгенном молоке. Способы экспрессирования антител рекомбинантным образом в растениях или молоке были описаны. См., например, Peelers, 2001, et al. Vaccine 19: 2756; Lonberg, N. and D.Huszar, 1995, Int. Rev. Immunol 13: 65; and Pollock, et al., 1999, J Immunol Methods 231: 147. Способы получения производных антител, например гуманизированных, одноцепочечных и других, известны в данной области техники.
Иммуноанализы и методики сортировки клеток с проточной цитометрией, такие как сортировка клеток с возбуждением флуоресценции (FACS), также могут быть использованы для выделения антител, которые являются специфичными в отношении желаемого антигена.
Антитела могут быть связаны со многими разными носителями. Носители могут быть активными и/или инертными. Примеры хорошо известных носителей включают полипропилен, полистирол, полиэтилен, декстран, найлон, амилазы, стекло, природные и модифицированные целлюлозы, полиакриламиды, агарозы и магнетит. Природа носителя может быть растворимой или нерастворимой для целей изобретения. Специалисты в данной области техники знают другие подходящие носители для связывания антител или способны их определить, используя стандартные эксперименты. В некоторых воплощениях носитель содержит группировку, которая направлена на миокард.
ДНК, кодирующую моноклональные антитела, легко выделить и секвенировать, используя стандартные методики (например, используя олигонуклеотидные зонды, которые способны специфически связывать гены, кодирующие тяжелые и легкие цепи моноклональных антител). Гибридомные клетки служат в качестве предпочтительного источника такой ДНК. Как только ДНК выделена, ее можно поместить в экспрессионные векторы (такие как экспрессионные векторы, раскрытые в публикации РСТ заявки WO 87/04462), которые затем трансфицируют в клетки-хозяева, такие как клетки Е.coli, клетки COS обезьяны, клетки яичников китайского хомячка (СНО) или клетки миеломы, которые в иных случаях не продуцируют иммуноглобулиновый белок, для достижения синтеза моноклональных антител в рекомбинантных клетках-хозяевах. См., например, публикацию РСТ заявки WO 87/04462. ДНК также может быть модифицирована, например, путем замены кодирующей последовательности константных доменов человеческой тяжелой и легкой цепи вместо гомологических мышиных последовательностей, Morrison et al., 1984, Proc. Nat. Acad. Sci. 81: 6851, или путем ковалентного присоединения к иммуноглобулин-кодирующей последовательности всей или части кодирующей последовательности для неиммуноглобулинового полипептида. Таким же образом получают "химерные" или "гибридные" антитела, которые имеют специфичность связывания описанного здесь моноклонального антитела.
рН-Зависимые антитела и полипептиды, имеющие происхождение из антител, могут быть идентифицированы или охарактеризованы с использованием способов, известных в данной области техники, посредством которых детектируют и/или измеряют снижение, улучшение или нейтрализацию биологической активности. В некоторых воплощениях рН-зависимое антитело или полипептид идентифицируют путем инкубации агента-кандидата с антигеном и мониторинга связывания и/или сопутствующего снижения или нейтрализации биологической активности. Анализ связывания может быть осуществлен с очищенным антигенным полипептидом(ами), или с клетками, экспрессирующими естественным путем, или трансфицированными для экспрессии данного полипептида(ов). В одном воплощении анализ связывания представляет собой конкурентный анализ связывания, где оценивают способность антитела-кандидата конкурировать с известным антагонистом за связывание. Этот анализ может быть осуществлен в различных форматах, включая формат ELISA. В других воплощениях рН-зависимое антитело идентифицируют путем инкубации агента-кандидата с антигеном и мониторинга связывания и сопутствующего ингибирования экспрессии LDLR и/или клиренса холестерина из крови.
После первичной идентификации активность рН-зависимого антитела-кандидата может быть дополнительно подтверждена и уточнена биологическими анализами, которые известны для тестирования целевых биологических активностей. Альтернативно, биологические анализы могут быть использованы для непосредственного скрининга кандидатов. Некоторые из способов идентификации и характеризации антител, пептидов или аптамеров описаны подробно в Примерах.
Антитела с рН-зависимым связыванием могут быть охарактеризованы с использованием способов, хорошо известных в данной области техники. Например, один из способов заключается в идентификации эпитопа, с которым оно связывается, или "картирование эпитопов". - Существует много способов, известных в данной области техники для картирования и характеризации расположения эпитопов в белках, включая выяснение кристаллической структуры комплекса антитело-антиген, конкурентные анализы, анализы экспрессии генных фрагментов и анализы на основе синтетических пептидов, как описано, например, в Главе 11 Hariow and Lane, Using Antibodies, a Laboratory Manual, (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1999). В дополнительном примере картирование эпитопов может быть использовано для определения последовательности, с которой связывается рН-зависимое антитело. Картирование эпитопов доступно от различных источников, например, Pepscan Systems (Edelhertweg 15, 8219 PH Leiystad, The Netherlands). Эпитоп может представлять собой линейный эпитоп, то есть содержится в одной цепи аминокислот, или конформационный эпитоп, образуемый пространственным взаимодействием аминокислот, которые не обязательно могут содержаться в одной цепи. Пептиды варьирующей длины (например, по меньшей мере длиной 4-6 аминокислот) могут быть выделены или синтезированы (например, рекомбинантным путем) и использованы для анализов связывания с антителом. В другом примере эпитоп, с которым связывается рН-зависимое антитело, может быть определен в последовательном скрининге путем использования перекрывающихся пептидов, имеющих происхождение из последовательности антигена, и определения связывания антителом. В соответствии с анализами экспрессии генного фрагмента открытую рамку считывания, кодирующую антиген, фрагментируют случайным образом или посредством специфических генетических конструкций и определяют реактивность экспрессированных фрагментов антигена с антителом, подлежащим тестированию. Генные фрагменты могут, например, быть продуцированы посредством полимеразной цепной реакции (ПЦР) и затем подвергнуты транскрипции и трансляции в белке in vitro в присутствии радиоактивных аминокислот. Связывание антитела с мечеными изотопами фрагментами затем определяют посредством иммунопреципитации и гель-электрофореза. Некоторые эпитопы также могут быть идентифицированы путем использования крупных библиотек случайных пептидных последовательностей, экспонированных на поверхности фаговых частиц (фаговые библиотеки). Альтернативно, определенная библиотека перекрывающихся пептидных фрагментов может быть тестирована на связывание с тестируемым антителом в простых анализах связывания. В качестве дополнительного примера мутагенез антиген-связывающего домена, эксперименты по перестановке доменов и аланин-сканирующий мутагенез могут быть осуществлены для идентификации требуемых остатков, достаточных и/или необходимых для связывания эпитопов. Например, эксперименты по перестановке доменов могут быть осуществлены с использованием мутантного антигена, в котором разные фрагменты полипептида были заменены (переставлены) последовательностями из антигена из других видов или близкородственного, но антигенно отличного белка (такого как другой представитель семейства пропротеин-конвертазы). Путем оценки связывания антитела с этим мутантным антигеном устанавливают важность конкретного антигенного фрагмента для связывания антитела.
Еще один способ, который может быть использован для характеризации рН-зависимого антитела, заключается в использовании конкурентных анализов с другими известными антителами за связывание с тем же антигеном для определения того, связывается ли рН-зависимое антитело с тем же эпитопом, как и другие антитела. Конкурентные анализы хорошо известны специалистам в данной области техники.
Экспрессионный вектор может быть использован для управления экспрессией рН-зависимого антитела. Специалист в данной области техники знаком с введением экспрессионных векторов для достижения экспрессии экзогенного белка in vivo. См., например, патенты США 6436908, 6413942 и 6376471. Введение экспрессионных векторов включает местное или системное введение, включая инъекцию, пероральное введение, генное ружье или введение через катетер и местное введение. В другом воплощении экспрессионный вектор вводят непосредственно в симпатический ствол или ганглий или в коронарную артерию, предсердие, желудочек или перикард.
Направленная доставка терапевтических композиций, содержащих экспрессионный вектор или субгеномные полинуклеотиды, также может быть использована. Технология опосредованной рецептором доставки ДНК описана, например, в Findeis et al., 1993, Trends Biotechnol. 11: 202; Chiou et al., 1994, Gene Therapeutics: Methods And Applications Of Direct Gene Transfer (J.A.Wolff, ed.); Wu et al., 1988, J.Biol. Chem. 263: 621; Wu et al., 1994, J. Biol. Chem. 269: 542; Zenke et al., 1990, Proc. Natl. Acad. Sci. USA 87: 3655; Wu et al., 1991, J. Biol. Chem. 266: 338. Терапевтические композиции, содержащие полинуклеотид, вводят в диапазоне от примерно 100 нг до примерно 200 мг ДНК для местного введения в протоколе генной терапии. Диапазоны концентраций от примерно 500 нг до примерно 50 мг, от примерно 1 мкг до примерно 2 мг, от примерно 5 мкг до примерно 500 мкг и от примерно 20 мкг до примерно 100 мкг ДНК также могут быть использованы в протоколах генной терапии. Терапевтические полинуклеотиды и полипептиды могут быть доставлены с использованием средств для генной доставки. Средства для генной доставки могут быть вирусного и невирусного происхождения (см. в общем Jolly, 1994, Cancer Gene Therapy 1: 51; Kimura, 1994, Human Gene Therapy 5: 845; Connelly, 1995, Human Gene Therapy 1: 185; and Kaplitt, 1994, Nature Genetics 6: 148). Экспрессия таких кодирующих последовательностей может быть индуцирована с использованием эндогенных промоторов млекопитающих или гетерологичных промоторов. Экспрессия кодирующей последовательности может быть конститутивной или регулируемой.
Векторы на основе вирусов для доставки желаемого полинуклеотида и экспрессии в желаемой клетке хорошо известны в данной области техники. Примеры векторов на основе вирусов включают рекомбинантные ретровирусы (см., например, публикации международных заявок WO 90/07936, WO 94/03622, WO 93/25698, WO 93/25234, WO 93/11230, WO 93/10218, WO 91/02805, патенты США 5219740 и 4777127; патент GB 2200651 и патент ЕР 0345242), векторы на основе альфавирусов (например, векторы на основе вируса Синдбис, вируса леса Семлики (АТСС VR-67; ATCC VR-1247), вируса Росс-Ривер (АТСС VR-373; АТСС VR-1246) и вируса венесуэльского энцефалита лошадей (АТСС VR-923; АТСС VR-1250; АТСС VR 1249; АТСС VR-532)), и векторы на основе адено-ассоциированных вирусов (AAV) (см., например, публикации международных заявок WO 94/12649, WO 93/03769, WO 93/19191, WO 94/28938, WO 95/11984 и WO 95/00655), но не ограничены ими. Введение ДНК, связанной с убитым аденовирусом, как описано в Curiel, 1992, Hum. Gene Ther. 3: 147, также может быть использовано.
Также могут быть использованы средства и методы невирусной доставки, включая поликатионные конденсированные ДНК, связанные или не связанные с убитым аденовирусом в отдельности (см., например, Curiel, 1992, Hum. Gene Ther. 3: 147); связанные с лигандом ДНК (см., например, Wu, J., 1989, Biol. Chem. 264:16985); средства для доставки на основе эукариотической клетки (см., например, патент США 5814482; публикации международных заявок WO 95/07994, WO 96/17072, WO 95/30763 и WO 97/42338) и нейтрализация заряда ядра или слияние с клеточными мембранами, но не ограничиваясь ими. Также может быть использована "голая" ДНК. Примеры способов введения "голой" ДНК описаны в публикации международной заявки WO 90/11092 и патенте США 5580859. Липосомы, которые могут действовать в качестве средств генной доставки, описаны в патенте США 5422120; публикациях международных заявок WO 95/13796, WO 94/23697 и WO 91/14445; и в ЕР 0524968. Дополнительные подходы описаны в Philip, 1994, Mol. Cell Biol., 14:2411, и в Woffendin, 1994 Proc. Natl. Acad. Sci. 91:1581.
Данное изобретение охватывает композиции, включая фармацевтические композиции, содержащие описанные здесь антитела или изготовленные способами и имеющие характеристики, описанные здесь. Композиции, как их используют здесь, содержат одно или более рН-зависимых антител и/или один или более полинуклеотидов, содержащих последовательности, кодирующие одно или более этих антител. Эти композиции могут дополнительно содержать подходящие эксципиенты, такие как фармацевтически приемлемые эксципенты, включая буферы, которые хорошо известны в данной области техники.
В изобретении также предложены участки CDR рН-зависимых антител (включая CDR согласно Chothia и Kabat). Определение участков CDR хорошо известно специалисту в данной области техники. Следует понимать, что в некоторых воплощениях CDR могут представлять собой комбинацию CDR согласно Kabat и Chothia (также называемых "объединенные CDR" или "расширенные CDR"). В некоторых воплощениях CDR представляют собой CDR согласно Kabat. В других воплощениях CDR представляют собой CDR согласно Chothia. Другими словами, в воплощениях с более чем одним CDR эти CDR могут быть любыми из CDR согласно Kabat, Chothia, комбинации CDR или их комбинаций.
В изобретении также предложены способы получения любых этих антител или полипептидов. Антитела по данному изобретению могут быть получены методиками, известными в данной области техники. Полипептиды могут быть продуцированы путем протеолитического или другого разложения антител, рекомбинантными способами (а именно, индивидуальные или слитые полипептиды), как описано здесь, или химическим синтезом. Полипептиды антител, особенно более короткие полипептиды вплоть до примерно 50 аминокислот, удобно получать химическим синтезом. Способы химического синтеза хорошо известны в данной области техники и коммерчески доступны. Например, антитело может быть получено с помощью автоматизированного синтезатора полипептидов, использующего твердофазную методику. См. также патенты США 5807715, 4816567 и 6331415.
В качестве другой альтернативы, антитела и пептиды могут быть получены рекомбинантным способом с использованием методик, хорошо известных в данной области техники. В одном воплощении полинуклеотид содержит последовательность, кодирующую вариабельные области тяжелой и/или легкой цепи антитела 4А5, 5А10, 6F6, 7D4 или L1L3. Последовательность, кодирующая интересующее антитело, может сохраняться в векторе в клетке-хозяине, и эта клетка-хозяин затем может быть культивирована и заморожена для использования в будущем. Векторы (включая экспрессионные векторы) и клетки-хозяева дополнительно описаны здесь.
Изобретение также охватывает scFv антител по изобретению. Одноцепочечные фрагменты вариабельной области получают путем связывания вариабельных областей легкой и/или тяжелой цепи, используя короткий связывающий пептид. Bird et al., 1988, Science 242: 423-426. Пример связывающего пептида представляет собой (GGGGS)3 (SEQ ID NO: 22), который образует мостиковую связь приблизительно 3,5 нм между карбоксильной концевой группой одной вариабельной области и аминоконцевой группой другой вариабельной области. Были разработаны и используются линкеры с другими последовательностями. Bird et al., 1988, выше.
Линкеры должны представлять собой короткие гибкие полипептиды и предпочтительно состоять из менее чем примерно 20 аминокислотных остатков. Линкеры, в свою очередь, могут быть модифицированы для дополнительных функций, таких как присоединение лекарств или присоединение к твердым подложкам. Одноцепочечные варианты могут быть получены либо рекомбинантным способом, либо синтетическим путем. Для синтетического продуцирования scFv может быть использован автоматизированный синтезатор. Для рекомбинантного продуцирования scFv подходящая плазмида, содержащая полинуклеотид, который кодирует scFv, может быть введена в подходящую клетку-хозяина, либо эукариотическую, такую как клетки дрожжей, растений, насекомых или млекопитающих, либо прокариотическую, такую как Е.coli. Полинуклеотиды, кодирующие интересующие scFv, могут быть получены стандартными манипуляциями, такими как лигирование полинуклеотидов. Полученные в результате scFv могут быть выделены с использованием стандартных методик очистки белков, известных в данной области техники.
Также охватываются другие формы одноцепочечных антител, такие как диатела. Диатела представляют собой дивалентные биспецифические антитела, в которых VH- и VL-домены экспрессируются на одной полипептидной цепи, но с использованием линкера, который является слишком коротким для того, чтобы обеспечить возможность спаривания между двумя доменами на одной и той же цепи, тем самым заставляя домены спариваться с комплементарными доменами другой цепи и создавая два антиген-связывающих сайта (см., например, Holliger, P., et al., 1993, Proc. Natl. Acad Sci. USA 90: 6444-6448; Poljak, R.J., et al., 1994, Structure 2: 1121-1123).
Например, биспецифические антитела, моноклональные антитела, которые имеют специфичности связывания в отношении по меньшей мере двух разных антигенов, могут быть получены с использованием раскрытых здесь антител. Способы получения биспецифических антител известны в данной области техники (см., например, Suresh et al., 1986, Methods in Enzymology 121:210). Традиционно, рекомбинантное продуцирование биспецифических антител было основано на коэкспрессии двух пар легкая цепь/тяжелая цепь иммуноглобулина, причем две тяжелые цепи имели разные специфичности (Millstein and Cuello, 1983, Nature 305, 537-539).
Согласно одному из подходов получения биспецифических антител, вариабельные домены антитела с желаемыми специфичностями связывания (сайты объединения антитело-антиген) подвергают слиянию с последовательностями константного домена иммуноглобулина. Слияние предпочтительно производят с константным доменом тяжелой цепи иммуноглобулина, содержащим по меньшей мере часть шарнирной, СН2 и СН3 областей. Предпочтительно иметь первую константную область тяжелой цепи (СН1), содержащую сайт, необходимый для связывания легкой цепи, присутствующей в по меньшей мере одном из слияний. ДНК, кодирующие слияния тяжелой цепи иммуноглобулина и при желании легкой цепи иммуноглобулина, встраивают в отдельные экспрессионные векторы и котрансфицируют в подходящий организм-хозяин. Это обеспечивает более высокую гибкость в регулировании взаимных соотношениий трех полипептидных фрагментов в воплощениях, когда неравные отношения трех полипептидных цепей, используемых в конструкте, дают оптимальные выходы. Однако, возможно встраивание кодирующих последовательностей для двух или всех трех полипептидных цепей в один экспрессионный вектор, когда экспрессия по меньшей мере двух полипептидных цепей в равных соотношениях приводит в результате к высоким выходам, или когда соотношения не представляют особой значимости.
В одном подходе биспецифические антитела состоят из гибридной тяжелой цепи иммуноглобулина с первой специфичностью связывания в одном звене и гибридной парой тяжелая цепь/легкая цепь иммуноглобулина (обеспечивающей вторую специфичность связывания) в другом звене. Такая асимметричная структура, с легкой цепью иммуноглобулина только в одной половине биспецифической молекулы, облегчает отделение желаемого биспецифического соединения от нежелательных комбинаций цепей иммуноглобулина. Этот подход описан в публикации международной заявки WO 94/04690.
Гетероконъюгатные антитела, содержащие два ковалентно связанных антитела, также входят в объем данного изобретения. Такие антитела используют для нацеливания клеток иммуной системы на нежелательные клетки (патент США 4676980) и для лечения HIV-инфекции (публикации международных заявок WO 91/00360 и WO 92/200373; ЕР 03089). Гетероконъюгатные антитела могут быть получены с использованием любых подходящих способов перекрестного сшивания. Подходящие сшивающие агенты и методики хорошо известны в данной области техники и описаны в патенте США 4676980.
Химерные или гибридные антитела также могут быть получены in vitro с использованием известных химических способов синтеза белков, включая те, в которых используют сшивающие агенты. Например, иммунотоксины могут быть сконструированы с использованием реакции дисульфидного обмена или путем образования тиоэфирной связи. Примеры подходящих реагентов для этой цели включают иминотиолат и метил-4-меркаптобутиримидат.
Гуманизированное антитело, содержащее один или более CDR антител 5А10 или 7D4 или один или более CDR, имеющих происхождение из антител 5А10 или 7D4, может быть получено, например, путем использования любых способов, известных в данной области техники. Например, четыре общих стадии могут быть использованы для гуманизации моноклонального антитела. Они являются следующими: (1) определение нуклеотидной и предсказанной аминокислотной последовательности вариабельных доменов легкой и тяжелой цепи исходного антитела; (2) конструирование гуманизированного антитела, то есть, принятие решения, какую каркасную область антитела использовать во время процесса гуманизации; (3) фактическое осуществление процессов/методик гуманизации; и (4) трансфекция и экспрессия гуманизированного антитела. См., например, патенты США 4816567, 5807715, 5866692, 6331415, 5530101, 5693761, 5693762, 5585089 и 6180370.
В рекомбинантных гуманизированных антителах Fc-участок может быть модифицирован, для того чтобы избежать взаимодействия с Fcv-рецептором и иммунной системой и комплементом. Методики получения таких антител описаны в WO 99/58572. Например, константная область может быть сконструирована с более сходными человеческими константными областями, для того чтобы избежать иммунного ответа, если это антитело используют в клинических исследованиях и лечении у людей. См., например, патенты США 5997867 и 5866692.
Гуманизированное антитело, содержащее вариабельные области легкой и тяжелой цепи или один или более CDR антитела или его вариантов либо один или более CDR, имеющих происхождение из этого антитела или его вариантов, может быть получено с использованием любых способов, известных в данной области техники.
Гуманизированные антитела могут быть получены любым способом, известным в данной области техники.
Изобретение охватывает модификации антител и полипептидов по изобретению, включая функционально эквивалентные антитела, которые не оказывают значительного влияния на их свойства, и варианты, которые обладают повышенной или пониженной активностью и/или аффинностью. Например, аминокислотная последовательность может быть мутирована с получением антитела с желаемой аффинностью связывания со своим антигеном. Модификация полипептидов является стандартной практикой в данной области техники, поэтому нет необходимости описывать ее здесь подробно. Модификация полипептидов проиллюстрирована в Примерах. Примеры модифицированных полипептидов включают полипептиды с консервативными заменами аминокислотных остатков, одной или более делениями или вставками аминокислот, которые не изменяют значительно вредно функциональную активность, или которые созревают (усиливают) аффинность полипептида в отношении своего лиганда, или используют химические аналоги.
Вставки аминокислотных последовательностей включают амино- и/или карбокси-концевые слияния, изменяющиеся по длине от одного остатка до полипептидов, содержащих сто или более остатков, а также вставки внутри последовательностей одной или нескольких аминокислотных остатков. Примеры концевых вставок включают антитело с N-концевым метионильным остатком антитела, слитым с эпитопной меткой. Другие варианты вставок в молекулу антитела включают слияние N- или С-конца антитела фермента или полипептида, которые увеличивают время полужизни антитела в кровотоке.
Варианты замен включают удаление по меньшей мере одного аминокислотного остатка в молекуле антитела и вставку другого остатка на его место. Сайты, представляющие наибольший интерес для заместительного мутагенеза, включают гипервариабельные участки, но также подразумеваются изменения в каркасной области (FR). Консервативные замены показаны в Таблице 1 под заголовком "консервативные замены". Если такие замены приводят в результате к изменению биологической активности, тогда более существенные изменения, обозначенные как "примеры замен" или как дополнительно описано ниже со ссылкой на классы аминокислот, могут быть включены и продукты подвергнуты скринингу.
Существенные модификации биологических свойств антитела сопровождаются выбором замен, которые значительно отличаются по их влиянию на сохранение (а) структуры полипептидного каркаса в области замены, например, в виде листовой или спиральной конформации, (б) заряда или гидрофобности молекулы в сайте-мишени или (в) величины боковой цепи. Природные остатки выделяют в группы на основе общих свойств боковой цепи: (1) неполярные: Норлейцин, Met, Ala, Val, Leu, Ile; (2) полярные незаряженные: Cys, Ser, Thr, Asn, Gln; (3) кислые (отрицательно заряженные): Asp, Glu; (4) основные (положительно заряженные): Lys, Arg; (5) остатки, влияющие на ориентацию цепи: Gly, Pro; и (6) ароматические: Trp, Tyr, Phe, His.
Неконсервативные замены осуществляют путем замены представителя одного из этих классов представителем другого класса.
Любой цистеиновый остаток, не вовлеченный в сохранение надлежащей конформации антитела, также может быть замещен, обычно серином, для улучшения окислительной стабильности молекулы и предотвращения аномального перекрестного связывания. Напротив, цистеиновая(ые) связь(и) может(гут) быть добавлена(ы) к антителу для улучшения его стабильности, особенно когда антитело представляет собой фрагмент антитела, такой как Fv фрагмент.
Аминокислотные модификации могут варьировать от изменения или модификации одной или более аминокислот до полной реконструкции области, такой как вариабельная область. Изменения в вариабельной области могут изменять аффинность и/или специфичность связывания. В некоторых воплощениях не более 1-5 консервативных аминокислотных замен сделано в пределах CDR домена. В других воплощениях не более 1-3 консервативных аминокислотных замен сделано в пределах CDR домена. В дополнительных воплощениях CDR домен представляет собой CDR Н3 и/или CDR L3.
Модификации также включают гликозилированные и негликозилированные полипептиды, а также полипептиды с другими посттрансляционными модификациями, такими как, например, гликозилирование с разными сахарами, ацетилирование и фосфорилирование. Антитела гликозилированы в консервативных положениях в их константных областях (Jefferis and Lund, 1997, Chem. Immunol. 65: 111-128; Wright and Morrison, 1997, TibTECH 15: 26-32). Олигосахаридные боковые цепи иммуноглобулинов влияют на функции белков (Boyd et al., 1996, Mol. Immunol. 32: 1311-1318; Wittwe and Howard, 1990, Biochem. 29: 4175-4180) и внутримолекулярное взаимодействие между положениями гликопротеина, которые могут влиять на конформацию и презентированную трехмерную поверхность гликопротеина (Jefferis and Lund, выше; Wyss and Wagner, 1996, Current Opin. Biotech. 7: 409-416). Олигосахариды также могут служить для нацеливания данного гликопротеина на определенные молекулы на основе конкретных структур распознавания. Сообщалось, что гликозилирование антител влияет на ADCC. В частности, сообщалось, что клетки СНО с тетрациклин-регулируемой экспрессией β(1,4)-N-ацетилглюкозаминил-трансферазы III (GnTIII), гликозилтрансферазы, катализирующей образование разрезающей GlcNAc, имели улучшенную активность ADCC (Umana et al., 1999, Nature Biotech. 17: 176-180).
Гликозилирование антител является, как правило, либо N-связанным, либо O-связанным. N-связанное относится к присоединению углеводной группировки к боковой цепи аспарагинового остатка. Трипептидные последовательности аспарагин-Х-серин, аспарагин-Х-треонин и аспарагин-Х-цистеин, где Х представляет собой любую аминокислоту, за исключением пролина, представляют собой последовательности распознавания для ферментативного присоединения углеводной группировки к аспарагиновой боковой цепи. Так, присутствие любой из этих трипептидных последовательностей в полипептиде создает потенциальный сайт гликозилирования. O-связанное гликозилирование относится к присоединению одного из сахаров N-ацетилглюкозамина, галактозы или ксилозы к гидроксиаминокислоте, чаще всего серину или треонину, хотя 5-гидроксипролин или 5-гидроксилизин также могут быть использованы.
Добавление сайтов гликозилирования к антителу обычно сопровождается изменением аминокислотной последовательности, так что она содержит одну или более вышеупомянутых трипептидных последовательностей (для N-связанных сайтов гликозилирования). Изменение также может быть осуществлено путем добавления или замены одного или более сериновых или треониновых остатков к последовательности исходного антитела (для O-связанных сайтов гликозилирования).
Картина гликозилирования антител также может быть изменена без изменения лежащей в основе нуклеотидной последовательности. Гликозилирование в значительной степени зависит от клетки-хозяина, используемой для экспрессии антитела. Поскольку тип клетки, используемый для экспрессии рекомбинантных гликопротеинов, например антител, в качестве потенциальных терапевтических средств, редко представляет собой нативную клетку, можно ожидать вариаций в картине гликозилирования антител (см., например, Hse et al., 1997, J. Biol. Chem. 272: 9062-9070).
Помимо выбора клеток-хозяев, факторы, которые влияют на гликозилирование во время рекомбинантного продуцирования антител, включают режим роста, состав среды, плотность культуры, насыщение кислородом, рН, схемы очистки и тому подобное. Различные способы были предложены для изменения картины гликозилирования, достигаемой в конкретном организме-хозяине, включая введение или сверхэкспрессию определенных ферментов, вовлеченных в продуцирование олигосахаридов (патенты США 5047335, 5510261 и 5278299). Гликозилирование или некоторые типы гликозилирования могут быть удалены из гликопротеина ферментативно, например, с использованием эндогликозидазы Н (Endo H), N-гликозидазы F, эндогликозидазы F1, эндогликозидазы F2, эндогликозидазы F3. В дополнение, рекомбинантная клетка-хозяин может быть генетически сконструирована таким образом, чтобы быть дефектной по процессингу определенных типов полисахаридов. Эти и подобные методики хорошо известны в данной области техники.
Другие способы модификации включают использование методик сочетания, известных в данной области техники, включая ферментативные средства, окислительное замещение и хелатирование, но не ограничиваясь ими. Модификации могут быть использованы, например, для присоединения меток для иммуноанализа. Модифицированные полипептиды получают с использованием установленных процедур в данной области техники и могут быть защищены с использованием стандартных анализов, известных в данной области техники, некоторые из которых описаны ниже и в Примерах.
В некоторых воплощениях изобретения антитело содержит модифицированную константную область, такую как константная область, которая является иммунологически инертной или частично инертной, например, не запускает комплемент-опосредованный лизис, не стимулирует ADCC или не активирует микроглию; или имеет пониженные активности (по сравнению с немодифицированным антителом) в любом одном или более из следующих: запуск комплемент-опосредованного лизиса, стимуляция ADCC или активация микроглии. Разные модификации константной области могут быть использованы для достижения оптимального уровня и/или комбинации эффекторных функций. См., например, Morgan et al., 1995, Immunology 86: 319-324; Lund et al., 1996, J.Immunology 157: 4963-9 157: 4963-4969; Idusogie et al., 2000, J.Immunology 164: 4178-4184; Tao et al., 1989, J. Immunology 143: 2595-2601; and Jefferis et al., 1998, Immunological Reviews 163: 59-76. В некоторых воплощениях константная область модифицирована, как описано в Eur. J.Immunol., 1999, 29: 2613-2624; публикации международной заявки WO 99/58572; и/или заявке на патент UK No. 9809951.8. В других воплощениях антитело содержит человеческую константную область тяжелой цепи IgG2, содержащую следующие мутации: АЗЗОР331 на S330S331 (нумерация аминокислот со ссылкой на последовательность IgG2 дикого типа). Eur. J.Immunol., 1999, 29:2613-2624. В других воплощениях константная область является агликозилированной для N-связанного гликозилирования. В некоторых воплощениях константная область является агликозилированной для N-связанного гликозилирования путем мутирования гликозилированного аминокислотного остатка или фланкирующих остатков, которые являются частью последовательности распознавания N-гликозилирования в константной области. Например, сайт N-гликозилирования N297 может быть мутирован до A, Q, K или Н. См., Tao et al., 1989, J. Immunology 143: 2595-2601; и Jefferis et al., 1998, Immunological Reviews 163: 59-76. В некоторых воплощениях константная область является агликозилированной для N-связанного гликозилирования. Константная область может быть агликозилированной для N-связанного гликозилирования ферментативным путем (например, удаление углевода ферментом пептид-N-гликозидазой (PNGaзой)) или путем экспрессии в клетке-хозяине, дефицитной по гликозилированию.
Другие модификации антител включают антитела, которые были модифицированы, как описано в публикации международной заявки WO 99/58572. Эти антитела содержат, в дополнение к связывающему домену, направленному на молекулу-мишень, эффекторный домен, имеющий аминокислотную последовательность, по существу гомологичную всем или части константного домена тяжелой цепи человеческого иммуноглобулина. Эти антитела способны к связыванию молекулы-мишени, не запуская значительный комплемент-зависимый лизис или клеточно-опосредованную деструкцию мишени. В некоторых воплощениях эффекторный домен способен к специфическому связыванию FcRn и/или FcγRllb. Они, как правило, основаны на химерных доменах, имеющих происхождение из двух или более CН2 доменов тяжелой цепи человеческого иммуноглобулина. Антитела, модифицированные таким образом, особенно подходят для применения в длительной терапии антителами, чтобы избежать воспалительных и других неблагоприятных реакций на стандартную терапию антителами.
Изобретение включает воплощения антител созревшей аффинности. Например, антитела созревшей аффинности могут быть продуцированы способами, известными в данной области техники (Marks et al., 1992, Bio/Technology, 10: 779-783; Barbas et al., 1994, Proc Nat. Acad. Sci, USA 91: 3809-3813; Schier et al., 1995, Gene, 169: 147-155; Yelton et al., 1995, J.Immunol., 155: 1994-2004; Jackson et al., 1995, J.Immunol., 154(7): 3310-9; Hawkins et al., 1992, J. Mol. Biol., 226: 889-896; и публикация международной заявки WO 2004/058184).
Следующие способы могут быть использованы для регулирования аффинности антитела и для характеризации CDR. Один путь характеризации антитела и/или изменения (такого как улучшение) аффинности связывания полипептида, такого как антитело, называют "мутагенез со сканированием библиотеки" (library scanning mutagenesis). В общем, мутагенез со сканированием библиотеки работает следующим образом. Одно или более аминокислотных положений в CDR заменяют двумя или более (например 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20) аминокислотами, используя признанные в данной области способы. В результате генерируются небольшие библиотеки клонов (в некоторых воплощениях по одной для каждого аминокислотного положения, которое анализируют), каждая с двумя или более составляющими (если две или более аминокислоты заменены в каждом положении). В общем, библиотека также включает клон, содержащий нативную (незамененную) аминокислоту. Небольшой ряд клонов, например, примерно 20-80 клонов (в зависимости от множества составляющих библиотеки), из каждой библиотеки подвергают скринингу в отношении аффинности связывания с полипептидом-мишенью (или другой мишенью связывания), и идентифицируют кандидатов с повышенной, такой же или пониженной активностью связывания, или у которых отсутствует связывание. Способы определения аффинности связывания хорошо известны в данной области техники. Аффинность связывания может быть определена с использованием анализа поверхностного плазменного резонанса Biacore, который обнаруживает различия в аффинности примерно в 2 раза или больше. Biacore особенно полезен, когда исходное антитело уже связывается с относительно высокой аффинностью, например KD примерно 10 нМ или ниже. Скринингс использованием анализа поверхностного плазменного резонанса Biacore описан здесь в Примерах.
Аффинность связывания может быть определена с использованием Kinexa Biocensor, анализов сцинтиляционной близости, ELISA, иммуноанализа ORIGEN (IGEN), гашения флуоресценции, переноса флуоресценции и/или дрожжевого дисплея. Аффинность связывания также может быть подвергнута скринингу с использованием подходящего биологического анализа.
В некоторых воплощениях каждое аминокислотное положение в CDR заменено (в некоторых воплощениях по одному за раз) всеми 20 природными аминокислотами с использованием признанных в данной области способов мутагенеза (некоторые из которых описаны здесь). В результате генерируются небольшие библиотеки клонов (в некоторых воплощениях по одной для каждого аминокислотного положения, которое анализируют), каждая с 20 составляющими (если все 20 аминокислот замещены в каждом положении).
В некоторых воплощениях библиотека, подлежащая скринингу, содержит замены по двум или более положениям, которые могут быть в том же CDR или в двух или более CDR. Таким образом, библиотека может содержать замены по двум или более положениям в одном CDR. Библиотека может содержать замены по двум или более положениям в двух или более CDR. Библиотека может содержать замены по 3, 4, 5 или более положениям, причем указанные положения обнаруживают в двух, трех, четырех, пяти или шести CDR. Замены могут быть осуществлены с использованием кодонов с низкой избыточностью. См., например, Таблицу 2 Balintetal., 1993, Gene 137 (1): 109-18).
CDR может представлять собой CDRH3 и/или CDRL3. CDR может представлять собой один или более из CDRL1, CDRL2, CDRL3, CDRH1, CDRH2 и/или CDRH3. CDR может представлять собой CDR no Kabat, CDR no Chothia или расширенный CDR.
Кандидаты с улучшенным связыванием могут быть секвенированы, тем самым идентифицируя мутанта с заменой в CDR, который дает улучшенную аффинность (также называют "улучшенная" замена). Кандидаты, которые связываются, также могут быть секвенированы, тем самым идентифицируя замену в CDR, которая сохраняет связывание.
Может быть осуществлено множество раундов скрининга. Например, кандидаты (каждый из которых содержит аминокислотную замену по одному или более положениям в одном или более CDR) с улучшенным связыванием также полезны для конструирования второй библиотеки, содержащей по меньшей мере исходную и замещенную аминокислоту в каждом улучшенном положении CDR (то есть, аминокислотное положение в этом CDR, при котором мутант с заменой показал улучшенное связывание). Получение, скрининг и селекция этой библиотеки обсуждаются дополнительно ниже.
Мутагенез со сканированием библиотеки также обеспечивает средство для характеризации CDR, а также частоты образования клонов с улучшенным связыванием, таким же связыванием, пониженным связыванием или не проявляющим связывание также дают информацию относительно важности каждого аминокислотного положения для стабильности комплекса антитело-антиген. Например, если положение в CDR сохраняет связывание, когда заменено вплоть до всех 20 аминокислот, тогда это положение идентифицируют как положение, которое вряд ли требуется для связывания антигена. И наоборот, если положение в CDR сохраняет связывание в очень малом проценте замен, тогда это положение идентифицируют как положение, важное для функции CDR. Таким образом, способы мутагенеза со сканированием библиотеки дают информацию относительно положений в CDR, которые могут быть заменены на многие другие аминокислоты (включая все 20 аминокислот), и положений в CDR, которые не могут быть заменены или которые могут быть заменены только на некоторые аминокислоты.
Кандидаты с улучшенной аффинностью могут быть объединены во второй библиотеке, которая включает улучшенную аминокислоту, исходную аминокислоту и может дополнительно включать дополнительные замены по этому же положению, в зависимости от комплексности библиотеки, которая желательна или обеспечивает использование желаемых способов скрининга или селекции. В дополнение, при желании также соседнее аминокислотное положение может быть рандомизировано до по меньшей мере двух или более аминокислот. Рандомизация соседних аминокислот может обеспечить дополнительную конформационную подвижность в мутантном CDR, которая может, в свою очередь, обеспечить или облегчить введение большего количества улучшающих мутаций. Библиотека может также содержать замену по положениям, которые не проявляют улучшенную аффинность в первом раунде скрининга.
Эту вторую библиотеку подвергают скринингу или селекции в отношении представителей библиотеки с улучшенной и/или измененной аффинностью связывания с использованием любого способа, известного в данной области техники, включая скрининг с использованием анализа поверхностного плазменного резонанса Biacore и селекцию с использованием любого способа, известного в данной области техники, включая фаговый дисплей, дрожжевой дисплей и рибосомный дисплей.
Изобретение также охватывает слитые белки, содержащие один или более фрагментов или участков из антител или полипептидов по данному изобретению. В одном воплощении предложен слитый полипептид, который содержит по меньшей мере 10 непрерывных аминокислот вариабельной области легкой цепи, показанной в SEQ ID NO: 3, и/или по меньшей мере 10 аминокислот вариабельной области тяжелой цепи, показанной в SEQ ID NO: 4 или 5. В других воплощениях предложен слитый полипептид, который содержит по меньшей мере примерно 10, по меньшей мере примерно 15, по меньшей мере примерно 20, по меньшей мере примерно 25 или по меньшей мере примерно 30 непрерывных аминокислот вариабельной области легкой цепи и/или по меньшей мере примерно 10, по меньшей мере примерно 15, по меньшей мере примерно 20, по меньшей мере примерно 25 или по меньшей мере примерно 30 непрерывных аминокислот вариабельной области тяжелой цепи. В другом воплощении слитый полипептид содержит один или более CDR. В дополнительных воплощениях слитый полипептид содержит CDR H3 (VH CDR3) и/или CDR L3 (VL CDR3). Для целей данного изобретения слитый белок содержит одно или более антител и другую аминокислотную последовательность, к которой он не присоединен в нативной молекуле, например, гетерологичную последовательность или гомологичную последовательность из другой области. Репрезентативные гетерологичные последовательности включают "метку", такую как метка FLAG или метка 6His, но не ограничены ими. Метки хорошо известны в данной области техники.
Слитый полипептид может быть создан способами, известными в данной области техники, например, синтетическим или рекомбинантным путем. Как правило, слитые белки по данному изобретению получают путем подготовки экспрессирования полинуклеотида, кодирующего их, с использованием рекомбинантных способов, описанных здесь, хотя они также могут быть получены другими средствами, известными в данной области техники, включая, например, химический синтез.
В данном изобретении также предложены композиции, содержащие антитела или полипептиды, конъюгированные (например, связанные) с агентом, облегчающим сцепление с твердой подложкой (например, биотином или авидином). Для простоты дана ссылка в общем на антитела, понимая, что эти способы применимы к любым описанным здесь воплощениям связывания антигенов и/или антагонистов. Конъюгирование в общем ссылается на связывание этих компонентов, как описано здесь. Связывание (которое в общем фиксирует эти компоненты в тесной ассоциации по меньшей мере для введения) может быть достигнуто любым рядом способов. Например, прямое взаимодействие между агентом и антителом возможно, когда каждый обладает заместителем, способным к взаимодействию с другим. Например, нуклеофильная группа, такая как амино или сульфгидрильная группа, на одном может быть способна взаимодействовать с карбонил-содержащей группой, такой как ангидрид или галогенангидрид, или с алкильной группой, содержащей хорошую уходящую группу (например галогенид), на другом.
Антитело или полипептид по данному изобретению может быть связан с метящим агентом, таким как флуоресцентная молекула, радиоактивная молекула или любые другие метки, известные в данной области техники. В данной области техники известны метки, которые в общем обеспечивают (либо непосредственно, либо опосредованно) сигнал.
В изобретении также предложены композиции (включая фармацевтические композиции) и наборы, содержащие, как поясняет данное описание, любое или все из антител и/или полипептидов, описанных здесь.
В изобретении также предложены выделенные полинуклеотиды, кодирующие антитела и пептиды по изобретению, а также векторы и клетки-хозяева, содержащие полинуклеотид.
В другом аспекте изобретения предложены полинуклеотиды, кодирующие любое из антител (включая фрагменты антител) и полипептидов, описанных здесь, таких как антител и полипептидов, имеющих улучшенную эффекторную функцию. Полинуклеотиды могут быть получены и экспрессированы согласно методикам, известным в данной области техники.
В другом аспекте изобретения предложены композиции (такие как фармацевтические композиции), содержащие любой из полинуклеотидов по изобретению. В некоторых воплощениях композиция содержит экспрессионный вектор, содержащий полинуклеотид, кодирующий антитело, как оно описано здесь. В другом воплощении композиция содержит экспрессионный вектор, содержащий полинуклеотид, кодирующий любое из антител или полипептидов, описанных здесь. В дополнительных воплощениях композиция содержит любое или оба из полинуклеотидов, показанных в SEQ ID NO: 23 и SEQ ID NO: 24. Экспрессионные векторы и введение композиций на основе полинуклеотидов дополнительно описаны здесь.
В другом аспекте изобретения предложен способ получения любого из описанных здесь полинуклеотидов.
Полинуклеотиды, комплементарные к любым таким последовательностям, также охватываются настоящим изобретением. Полинуклеотиды могут быть одноцепочечными (кодирующими или антисмысловыми) или двухцепочечными и могут представлять собой ДНК (геномную, кДНК или синтетическую) или РНК молекулы. РНК молекулы включают HnPHK молекулы, которые содержат интроны и соответствуют ДНК молекуле один к одному, и мРНК молекулы, которые не содержат интронов. Дополнительные кодирующие или некодирующие последовательности могут, но не обязательно должны, присутствовать в полинуклеотиде по настоящему изобретению, и полинуклеотид может, но не обязательно должен, быть связан с другими молекулами и/или материалами подложки.
Полинуклеотиды могут содержать нативную последовательность (то есть, эндогенную последовательность, которая кодирует антитело или его участок) или могут содержать вариант такой последовательности. Варианты полинуклеотидов содержат одну или более замен, добавлений, делеций и/или вставок, так что иммунореактивность кодируемого полипептида не нарушается, относительно нативной иммунореактивной молекулы. Влияние на иммунореактивность кодируемого полипептида может в общем быть оценено, как описано здесь. Варианты предпочтительно проявляют по меньшей мере примерно 70% идентичность, более предпочтительно по меньшей мере примерно 80% идентичность, еще более предпочтительно по меньшей мере примерно 90% идентичность и наиболее предпочтительно по меньшей мере примерно 95% идентичность полипептидной последовательности, которая кодирует нативное антитело или его участок.
Две полинуклеотидные или полипептидные последовательности считают "идентичными", если последовательность нуклеотидов или аминокислот в этих двух последовательностях является такой же при выравнивании для максимального соответствия, как описано ниже. Сравнения между двумя последовательностями, как правило, осуществляют путем сравнивания последовательностей над окном сравнения для идентификации и сравнения локальных областей подобия последовательности. "Окно сравнения", как используют здесь, относится к сегменту из по меньшей мере примерно 20 положений подряд, обычно от 30 до примерно 75, или от 40 до примерно 50, в котором последовательность можно сравнивать с эталонной последовательностью из такого же числа положений подряд после того, как эти две последовательности оптимально выровнены.
Оптимальное выравнивание последовательностей для сравнения может быть осуществлено с использованием программы Megalign в программном обеспечении Lasergene suite of bioinformatics (DNASTAR, Inc., Madison, WI), используя параметры ошибок. Эта программа дает некоторые схемы выравнивания, описанные в следующих источниках информации: Dayhoff, M.O., 1978, A model of evolutionary change in proteins,- Matrices for detecting distant relationships. In Dayhoff, M.O. (ed.) Atlas of Protein Sequence and Structure (National Biomedical Research Foundation, Washington DC), Vol.5, Suppl.3, pp.345-358; Hein J., 1990, Unified Approach to Alignment and Phylogenes pp.626-645 Methods in Enzymology vol. 183, (Academic Press, Inc., San Diego, CA); Higgins, D.G. and Sharp, P.M., 1989, CABIOS 5: 151-153; Myers, E.W. and MullerW., 1988, CABIOS 4: 11-17; Robinson, E.D., 1971, Comb. Theor. 11:105; Santou, N., Nes, M., 1987, Mol. Biol. Evol. 4: 406-425; Sneath, P.H.A. and Sokal, R.R., 1973, Numerical Taxonomy the Principles and Practice of Numerical Taxonomy (Freeman Press, San Francisco, CA); Wilbur, W.J. and Lipman, D.J., 1983, Proc. Natl. Acad. Sci. USA 80: 726-730.
Предпочтительно "процент идентичности последовательности" определяют путем сравнения двух оптимально выровненных последовательностей над окном сравнения из по меньшей мере 20 положений, где участок полинуклеотидной или полипептидной последовательности в окне сравнения может содержать добавления или делеции (то есть, пробелы) 20 процентов или менее, обычно от 5 до 15 процентов или от 10 до 12 процентов, по сравнению с эталонными последовательностями (которые не содержат добавлений или делеции) для оптимального выравнивания этих двух последовательностей. Процент рассчитывают путем определения числа положений, при которых идентичные нуклеотидные основания или аминокислотные остатки существуют в обеих последовательностях, с получением числа соответствующих положений, деления этого числа соответствующих положений на общее число положений в эталонной последовательности (то есть размер окна) и умножения результатов на 100 с получением процента идентичности последовательности.
Варианты могут также, или альтернативно, быть по существу гомологичными нативному гену или его участку или комплементу. Такие полинуклеотидные варианты способны к гибридизации в умеренно строгих условиях с природной последовательностью ДНК, кодирующей нативное антитело (или комплементарной последовательностью).
Подходящие "умеренно строгие условия" включают предварительную промывку в растворе 5Х SSC, 0,5% SDS, 1,0 мМ EDTA (рН 8,0); гибридизацию при 50°С-65°С, 5Х SSC, в течение ночи; затем промывку дважды при 65°С в течение 20 минут с каждым из 2Х, 0,5Х и 0,2Х SSC, содержащим 0,1% SDS.
"Очень строгие условия" или "условия высокой строгости", как используют здесь, являются следующими: (1) используют низкую ионную силу и высокую температуру для промывки, например 0,015 М хлорид натрия/0,0015 М цитрат натрия/0,1% додецилсульфат натрия при 50°С; (2) используют во время гибридизации денатурирующий агент, такой как формамид, например, 50% (об./об.) формамид с 0,1% бычьим сывороточным альбумином/0,1% Ficoll/0,1% поливинилпирролидон/буфер 50 мМ фосфата натрия при рН 6,5 с 750 мМ хлоридом натрия, 75 мМ цитратом натрия при 42°С; или (3) используют 50% формамид, 5×SSC (0,75 М NaCl, 0,075 М цитрат натрия), 50 мМ фосфат натрия (рН 6,8), 0,1% пирофосфат натрия, 5 × раствор Денгхардта, ДНК обработанной ультразвуком спермы лосося (50 мкг/мл), 0,1% SDS и 10% декстрансульфат при 42°С, с промывками при 42°С в 0,2×SSC (хлорид натрия/цитрат натрия) и 50% формамид при 55°С, с последующей промывкой высокой строгости, состоящей из 0,1×SSC, содержащего EDTA при 55°С. Специалист в данной области знает, как регулировать температуру, ионную силу и так далее, при необходимости, чтобы приспособиться к факторам, таким как длина зонда и тому подобное.
Специалисту в данной области техники понятно, что в результате вырожденности генетического кода имеется много нуклеотидных последовательностей, которые кодируют полипептид, как описано здесь. Некоторые из этих полинуклеотидов имеют минимальную гомологию нуклеотидной последовательности какого-либо нативного гена. Тем не менее, полинуклеотиды, которые варьируют из-за различий в использовании кодонов, прямо предусмотрены настоящим изобретением. Кроме того, аллели генов, содержащих полинуклеотидные последовательности, предложенные здесь, входят в объем настоящего изобретения. Аллели являются эндогенными генами, которые изменены в результате одной или более мутаций, таких как делеции, добавления и/или замены нуклеотидов. Получающиеся в результате мРНК и белок могут, но не обязательно должны, иметь измененную структуру или функцию. Аллели могут быть идентифицированы с использованием стандартных методик (таких как гибридизация, амплификация и/или сравнение последовательностей из базы данных).
Полинуклеотиды по данному изобретению могут быть получены с использованием химического синтеза, рекомбинантных способов или ПЦР. Способы химического синтеза полинуклеотидов хорошо известны в данной области техники и не требуют подробного описания здесь. Специалист в данной области техники может использовать предложенные здесь последовательности и коммерческий синтезатор ДНК для продуцирования желаемой последовательности ДНК.
Для получения полинуклеотидов с использованием рекомбинантных способов полинуклеотид, содержащий желаемую последовательность, может быть встроен в подходящий вектор, и этот вектор, в свою очередь, может быть введен в подходящую клетку-хозяин для репликации и амплификации, как дополнительно обсуждается здесь. Полинуклеотиды могут быть введены в клетки-хозяева любыми средствами, известными в данной области техники. Клетки трансформируют путем введения экзогенного полинуклеотида прямым захватом, эндоцитозом, трансфекцией, F-скрещиванием или электропорацией. Как только экзогенный полинуклеотид введен, он может сохраняться в клетке в виде неинтегрированного вектора (такого как плазмида) или интегрированного в клеточный геном хозяина. Амплифицированный таким образом полинуклеотид может быть выделен из клетки-хозяина способами, известными в данной области техники. См., например, Sambrook et al., 1989, выше.
Альтернативно, ПЦР делает возможной репродуцирование последовательностей ДНК. Технология ПЦР хорошо известна в данной области техники и описана в патентах США 4683195, 4800159, 4754065 и 4683202, а также в PCR: The Polymerase Chain Reaction, Mullis et al., 1994, eds. (Birkauswer Press, Boston, MA).
РНК может быть получена путем использования выделенной ДНК в подходящем векторе и встраивания его в подходящую клетку-хозяина. Когда клетка реплицируется и ДНК транскрибируется в РНК, эта РНК может быть затем выделена с использованием способов, хорошо известных специалистам в данной области техники, как изложено в Sambrook et al., 1989, выше, например.
Подходящие клонирующие векторы могут быть сконструированы согласно стандартным методикам или могут быть выбраны из огромного количества клонирующих векторов, доступных в данной области техники. В то время как выбранный клонирующий вектор может варьировать в соответствии с клеткой-хозяином, предназначенной для использования, полезные клонирующие векторы обычно имеют способность к саморепликации, могут обладать единственной мишенью для конкретной рестрикционной эндонуклеазы и/или могут нести гены для маркера, который может быть использован в селекции клонов, содержащих этот вектор. Подходящие примеры включают плазмиды и бактериальные вирусы, например, pUC18, pUC19, Bluescript (например, pBS SK+) и его производные, mp18, mp19, pBR322, pMB9, ColE1, pCR1, RP4, фаговые ДНК и шатгл-векгоры, такие как рSА3 и рАТ28. Эти и многие другие клонирующие векторы доступны от коммерческих поставщиков, таких как BioRad, Strategene и Invitrogen.
Экспрессионные векторы в общем представляют собой реплицируемые полинуклеотидные конструкции, которые содержат полинуклеотид по изобретению. Подразумевают, что экспрессионный вектор должен быть реплицируемым в клетках-хозяевах либо в виде эписом, либо в виде интегральной части хромосомной ДНК. Подходящие экспрессионные векторы включают плазмиды, вирусные векторы, включая аденовирусы, аденоассоциированные вирусы, ретровирусы, космиды и векторы экспрессии, раскрытые в публикации международной заявки WO 87/04462, но не ограничены ими. Компоненты вектора могут в общем включать одно или более из следующего: сигнальная последовательность; ориджин репликации; один или более маркерных генов; подходящие элементы регулирования транскрипции (такие как промоторы, энхансеры и терминатор), но не ограничены ими. Для экспрессии (то есть, трансляции) также требуются один или более элементов, регулирующих трансляцию, таких как сайты связывания рибосомы, сайты инициации трансляции и стоп-кодоны.
Векторы, содержащие полинуклеотиды, представляющие интерес, могут быть введены в клетку-хозяина любым из ряда подходящих средств, включая электропорацию, трансфекцию с использованием хлорида кальция, хлорида рубидия, фосфата кальция, DEAE-декстрана или других веществ; бомбардировку микрочастицами; липофекцию и инфекцию (например, где вектор представляет собой инфекционный агент, такой как вирус осповакцины). Выбор векторов для введения или полинуклеотидов часто зависит от характеристик клетки-хозяина.
В данном изобретении также предложены клетки-хозяева, содержащие любой из описанных здесь полинуклеотидов. Любые клетки-хозяева, способные к сверхэкспрессии гетерологичных ДНК, могут быть использованы с целью выделения генов, кодирующих антитело, полипептид или белок, представляющие интерес. Неограничивающие примеры клеток-хозяев включают клетки COS, HeLa, NSO и СНО, но не ограничены ими. См. также публикацию международной заявки WO 87/04462. Подходящие клетки-хозяева, не являющиеся животными, включают прокариоты (такие как Е.coli или В.subtillis) и дрожжи (такие как S.cerevisae, S.pombe или К.lactis). Предпочтительно клетки-хозяева экспрессируют кДНК на уровне примерно в 5 раз выше, более предпочтительно в 10 раз выше, еще более предпочтительно в 20 раз выше по сравнению с соответствующим эндогенным антителом или белком, представляющим интерес, если присутствует, в клетках-хозяевах. Скрининг клеток-хозяев на специфическое связывание с антигеном осуществляют путем иммуноанализа или FACS. Клетка, сверхэкспрессирующая антитело или белок, представляющий интерес, может быть идентифицирована.
С. Композиции
Композиции, используемые в способах по изобретению, содержат эффективное количество рН-зависимого антитела или полипептида-производного рН-зависимого антитела, описанного здесь. Примеры таких композиций, а также способы их изготовления, также описаны здесь в предыдущих разделах и ниже. В одном воплощении композиция содержит одно или более рН-зависимых антител. В других воплощениях рН-зависимое антитело распознает человеческий PCSK9. В других воплощениях рН-зависимое антитело является гуманизированным. В дополнительных воплощениях рН-зависимое антитело содержит константную область, которая не запускает ненужный или нежелательный иммунный ответ, такой как антитело-опосредованный лизис или ADCC. В других воплощениях рН-зависимое антитело содержит один или более CDR антитела (например один, два, три, четыре, пять или, в некоторых воплощениях, все шесть CDR). В некоторых воплощениях рН-зависимое антитело является человеческим.
Композиция, используемая в настоящем изобретении, может дополнительно содержать фармацевтически приемлемые носители, эксципиенты или стабилизаторы (Remington: The Science and Practice of Pharmacy 20th Ed., 2000, Lippincott Williams and Wilkins, Ed. К.Е.Hoover), в форме лиофилизированных препаратов или водных растворов. Приемлемые носители, эксципиенты или стабилизаторы являются нетоксичными для реципиентов в данных дозировках и концентрациях и могут содержать буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония, хлорид бензетония; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3-пентанол и мета-крезол); низкомолекулярные полипептиды (менее чем примерно 10 остатков); белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстраны; хелатирующие агенты, такие как EDTA; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы с металлами (например, комплексы Zn-белок); и/или неионогенные поверхностно-активные вещества, такие как TWEEN™, PLURONICS™ или полиэтиленгликоль (PEG). Фармацевтически приемлемые эксципиенты дополнительно описаны здесь.
В одном воплощении антитело вводят в препарате в виде стерильного водного раствора, имеющего рН в интервале от примерно 5,0 до примерно 6,5 и содержащего от примерно 1 мг/мл до примерно 200 мг/мл антитела, от примерно 1 миллимолярного до примерно 100 миллимолярного гистидинового буфера, от примерно 0,01 мг/мл до примерно 10 мг/мл полисорбата 80, от примерно 100 миллимолярной до примерно 400 миллимолярной трегалозы и от примерно 0,01 миллимолярного до примерно 1,0 миллимолярного дигидрата EDTA динатрия.
рН-Зависимое антитело и его композиции также могут быть использованы в сочетании с другими агентами, которые служат для усиления и/или дополнения эффективности агентов.
D. Наборы
В изобретении также предложены наборы для использования в способах по настоящему изобретению. Наборы по изобретению включают один или более контейнеров, содержащих рН-зависимое антитело (такое как гуманизированное антитело) или описанный здесь пептид, и инструкции для использования в соответствии с любым из описанных здесь способов по изобретению. В общем, эти инструкции содержат описание введения рН-зависимого антитела для вышеописанных терапевтических лечений.
В некоторых воплощениях антитело представляет собой гуманизированное антитело. В других воплощениях антитело является человеческим. В других воплощениях антитело представляет собой моноклональное антитело. Инструкции относительно использования рН-зависимого антитела в целом включают информацию по дозировке, схеме введения и пути введения для предназначенного лечения. Контейнеры могут представлять стандартные единицы дозировки, объемные упаковки (например многодозовые упаковки) или субъединичные дозы. Инструкции, прилагаемые в наборах по изобретению, являются, как правило, письменными инструкциями на маркировке или вкладыше в упаковке (например, листе бумаги, вложенном в набор), но также приемлемы и машиночитаемые инструкции (например инструкции на магнитном носителе или оптическом диске).
Наборы по данному изобретению представлены в подходящих упаковках. Подходящие упаковки включают флаконы, бутыли, сосуды, гибкие упаковки (например, герметично закрытые майларовые или пластиковые пакеты) и им подобные, но не ограничены ими. Также предусмотрены упаковки для использования в комбинации с конкретным устройством, таким как ингалятор, устройством для назального введения (например, распылителем) или устройством для инфузии, таким как минипомпа. Набор может иметь стерильный порт доступа (например, контейнер может представлять собой пакет для внутривенного раствора или флакон с крышкой, прокалываемой иглой для подкожных инъекций). Контейнер также может иметь стерильный порт доступа (например, контейнер может представлять собой пакет для внутривенного раствора или флакон с крышкой, прокалываемой иглой для подкожных инъекций). По меньшей мере один активный агент в композиции представляет собой рН-зависимое антитело. Контейнер (например, предварительно заполненный шприц или аутоинъектор) может дополнительно содержать второй фармацевтически активный агент.
Наборы могут возможно предоставлять дополнительные компоненты, такие как буфер и разъясняющую информацию. Обычно, набор содержит контейнер и маркировку или вкладыш в упаковку на контейнере.
Мутации и модификации
Для экспрессии антител по настоящему изобретению ДНК-фрагменты, кодирующие VH и VL области, могут быть сначала получены с использованием любых описанных выше способов. Различные модификации, например, мутации, делеции и/или вставки также могут быть введены в последовательности ДНК с использованием стандартных способов, известных специалистам в данной области техники. Например, мутагенез может быть осуществлен с использованием стандартных способов, таких как ПЦР-опосредованный мутагенез, при котором мутированные нуклеотиды включают в ПЦР-праймеры, так что ПЦР-продукт содержит желаемые мутации, или сайт-направленный мутагенез.
Один тип замены, например, который может быть сделан, состоит в том, чтобы заменить один или более цистеинов в антителе, которые могут быть химически реактивны, на другой остаток, такой как, без ограничения, аланин или серин. Например, может иметь место замена неканоническим цистеином. Замена может быть осуществлена в CDR или каркасной области вариабельного домена или в константном домене антитела. В некоторых воплощениях цистеин является каноническим.
Антитела также могут быть модифицированы, например, в вариабельных доменах тяжелой и/или легкой цепей, например, для изменения связывающего свойства антитела. Например, мутация может быть осуществлена в одном или более CDR для повышения или снижения KD антитела в отношении антигена, для повышения или снижения koff или для изменения специфичности связывания антитела. Технологии сайт-направленного мутагенеза хорошо известны в данной области техники. См., например, Sambrook et al. and Ausubel et al., выше.
Модификация или мутация также могут быть осуществлены в каркасной области или константном домене для увеличения времени полужизни рН-зависимого антитела. См., например, публикацию международной заявки WO 00/09560. Мутация в каркасной области или константном домене также может быть осуществлена для изменения иммуногенности антитела, для предоставления сайта для ковалентного или нековалентного связывания с другой молекулой или для изменения таких свойств, как фиксация комплемента, FcR-связывание и антителозависимая клеточно-опосредованная цитотоксичность. Согласно изобретению, индивидуальное антитело может иметь мутации в одном или более CDR или каркасных областях вариабельного домена или в константном домене.
В процессе, известном как "гуманизация" (germlining), некоторые аминокислоты в VH и VL последовательностях могут быть мутированы для соответствия тем, которые обнаруживают в природе в VH и VL последовательностях зародышевого типа. В частности, аминокислотные последовательности каркасных областей в VH и VL последовательностях могут быть мутированы для соответствия последовательностям зародышевого типа для снижения риска иммуногенности при введении антитела. Последовательности ДНК зародышевого типа для человеческих VH и VL генов известны в данной области техники (см., например, базу данных по человеческим последовательностям зародышевого типа "Vbase"; см. также Kabat, Е.А., et al. (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publ. No.91-3242; Tomlinson et al., 1992, J. Mol. Biol. 227: 776-798; and Cox et al., 1994, Eur. J.Immunol. 24:827-836.
Другой тип аминокислотных замен, который может быть сделан, состоит в том, чтобы удалить потенциальные протеолитические сайты в антителе.
Такие сайты могут существовать в CDR или каркасной области вариабельного домена или в константном домене антитела. Замены цистеиновых остатков и удаление протеолитических сайтов могут снизить риск гетерогенности в антительном продукте и тем самым увеличить его гомогенность. В еще одном типе аминокислотных замен удаляют пары аспарагин-глицин, которые образуют потенциальные сайты дезамидирования, путем изменения одного или обоих этих остатков. В другом примере С-концевой лизин тяжелой цепи рН-зависимого антитела по изобретению может быть отщеплен. В различных воплощениях тяжелые и легкие цепи антител могут возможно включать сигнальную последовательность.
Как только ДНК-фрагменты, кодирующие VH и VL сегменты по настоящему изобретению, получены, эти ДНК-фрагменты могут быть дополнительно обработаны стандартными рекомбинантными технологиями ДНК, например, для преобразования генов вариабельной области в гены цепи полноразмерного антитела, в гены Fab фрагмента или в ген scFv. При этих манипуляциях VL- или VH-кодирующий ДНК-фрагмент связывают функциональным образом с другим ДНК-фрагментом, кодирующим другой белок, такой как константная область антитела или гибкий линкер. Термин "связан функциональным образом", как используют в данном контексте, означает, что два ДНК-фрагмента соединены таким образом, что аминокислотные последовательности, кодируемые этими двумя ДНК-фрагментами, остаются внутри рамки считывания.
Выделенная ДНК, кодирующая VH область, может быть преобразована в ген полноразмерной тяжелой цепи путем функционального связывания VH-кодирующей ДНК с другой молекулой ДНК, кодирующей константные области тяжелой цепи (СН1, СН2 и СН3). Последовательности генов константной области человеческой тяжелой цепи известны в данной области техники (см., например, Kabat, E.A., et al., 1991, Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, номер публикации NIH 91-3242), и ДНК-фрагменты, охватывающие эти области, могут быть получены стандартной ПЦР-амплификацией. Константная область тяжелой цепи может представлять собой константную область IgG1, IgG2, IgG3, IgG4, IgA, IgE, IgM или IgD, но наиболее предпочтительно предсталяет собой константную область IgG1 или IgG2. Константная область IgG может быть любой из разных аллелей или аллотипов, существование которых известно среди разных индивидуумов, например Gm (1), Gm (2), Gm (3) и Gm (17). Эти аллотипы представляют собой встречающиеся в природе аминокислотные замены в константных областях IgG1. Для гена тяжелой цепи Fab-фрагмента VH-кодирующая ДНК может быть функционально связана с другой молекулой ДНК, кодирующей только константную область тяжелой цепи СН1. Эта константная область тяжелой цепи СН1 может происходить из любого гена тяжелой цепи.
Выделенная ДНК, кодирующая VL область, может быть преобразована в ген полноразмерной легкой цепи (а также ген Fab легкой цепи) путем функционального связывания VL-кодирующей ДНК с другой молекулой ДНК, кодирующей константную область легкой цепи, CL. Последовательности генов константной области человеческой легкой цепи известны в данной области техники (см., например, Kabat, E.A., et al., 1991, Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publ. No.91-3242), и ДНК-фрагменты, охватывающие эти области, могут быть получены стандартной ПЦР-амплификацией. Константная область легкой цепи может представлять собой константную область каппа или лямбда. Константная область каппа может быть любой из разнообразных аллелей, существование которых известно среди разных индивидуумов, например Inv(1), Inv(2) и Inv(3). Константная область лямбда может иметь происхождение из любого из этих трех генов лямбда.
Для создания scFv гена VH- и VL-кодирующие ДНК-фрагменты функционально связывают с другим фрагментом, кодирующим гибкий линкер, например, кодирующий аминокислотную последовательность (Gly4-Ser)3, так что VH и VL последовательности могут быть экспрессированы в виде непрерывного одноцепочечного белка, с VL и VH областями, соединенными гибким линкером (см., например, Bird et al., 1988, Science 242: 423-426; Huston et al., 1988, Proc. Natl. Acad. Sci. USA 85: 5879-5883; McCafferty et al., 1990, Nature 348: 552-554). Одноцепочечное антитело может быть моновалентным, если используют только одну VH и VL, бивалентным, если используют два VH и VL, или поливалентным, если используют более двух VH и VL. Могут быть получены биспецифические или поливалентные антитела, которые специфически связываются с антигеном и с другой молекулой.
В другом воплощении могут быть получены слитое антитело или иммуноадгезин, которые содержат все или часть антитела по изобретению, связанные с другим полипептидом. В другом воплощении только вариабельные домены антитела связаны с полипептидом. В другом воплощении VH домен антитела связан с первым полипептидом, в то время как VL домен антитела связан со вторым полипептидом, который ассоциирован с первым полипептидом таким образом, что VH и VL домены могут взаимодействовать друг с другом с образованием антиген-связывающего сайта. В другом предпочтительном воплощении VH домен отделен от VL домена линкером, так что VH и VL домены могут взаимодействовать друг с другом. Антитело VH-линкер-VL затем присоединяют к полипептиду, представляющему интерес. В дополнение, могут быть созданы слитые антитела, в которых два (или более) одноцепочечных антитела связаны друг с другом. Это является полезным при желании создать дивалентное или поливалентное антитело на одной полипептидной цепи или при желании создать биспецифическое антитело.
В других воплощениях другие модифицированные антитела могут быть получены с использованием рН-зависимого антитела, кодирующего молекулы нуклеиновой кислоты. Например, "каппа-тела" (III et al., 1997, Protein Eng. 10: 949-57), "минитела" (Martin et al., 1994, EMBO J. 13: 5303-9), "диатела" (Holliger et al., 1993, Proc. Natl. Acad. Sci. USA 90: 6444-6448) или "Janusins" (Traunecker et al., 1991, EMBO J. 10: 3655-3659 и Traunecker et al., 1992, Int. J. Cancer (Suppl.) 7: 51-52) могут быть получены с использованием стандартных молекулярно-биологических методик, следуя идеям, изложенным в данном описании.
Биспецифические антитела и антиген-связывающие фрагменты могут быть получены разнообразными способами, включая слияние гибридом или сшивание Fab' фрагментов. См., например, Songsivilai & Lachmann, 1990, Clin. Exp. Immunol. 79: 315-321, Kostelny et al., 1992, J.Immunol. 148: 1547-1553. В дополнение, биспецифические антитела могут быть образованы в виде "диател" или "Janusins." В некоторых воплощениях биспецифическое антитело связывается с двумя разными эпитопами антигена. В некоторых воплощениях модифицированные антитела, описанные выше, получают с использованием одного или более вариабельных доменов или CDR из человеческого антитела, предложенного здесь.
Создание антиген-специфичных антител
ДНК тяжелой и легкой цепей репрезентативных антител по настоящему изобретению, 5L1721H23_6L3 и 5L1721H23_6L3H3, были депонированы в Американской коллекции типовых культур (АТСС) (в соответствии с Будапештским договором) 22 декабря 2009 года, и им были присвоены номера, указанные в Таблице 2. Все ограничения доступности плазмид, депонированных таким образом, для неограниченного круга лиц будут безоговорочно сняты после выдачи патента по настоящему изобретению. Ссылки на тяжелую и легкую цепи антитела 5L1721H23_6L3 означают UC-Н5Н23 и UC-H5L1721-6L3, соответственно. Ссылки на тяжелую и легкую цепи антитела 5L1721H23_6L3H3 означают UC-H5H23-6H3 и UC-H5L1721-6L3, соответственно.
Примеры
Пример 1: Моделирование рН-зависимого связывания антитела с антигеном PCSK9
Использовали компьютерное моделирование для предсказания того, сможет ли антитело с рН-зависимым связыванием антигена влиять на время полужизни антитела, продолжительность снижения концентрации сывороточного PCSK9. Для целей этого моделирования следующие допущения были приняты: доза антитела в крови 1 мкМ; время полужизни антитела (не проявляющего рН-зависимое связывание) 21 сутки; продолжительность моделирования 100 суток; для рН-зависимого антитела Kon=1е5/М/сек, Koff при нейтральном рН=KD*Kon, рН-зависимое связывание моделируют как увеличение Koff в кислых эндосомах; и Koff при кислых рН=R*Koff при нейтральном рН.
Быстроту изменения параметров в модели определяли в контексте модельных параметров с помощью дифференциальных уравнений, описанных ниже. В общем, эта модель принята для работы с уровнями антител от нуля до тех пор, пока не будет достигнуто равновесное состояние; в это время болюс антитела моделируют путем переустановки [mAb} от нуля до уровня, отражающего данную дозу, и далее следуя модели, описанной ниже.
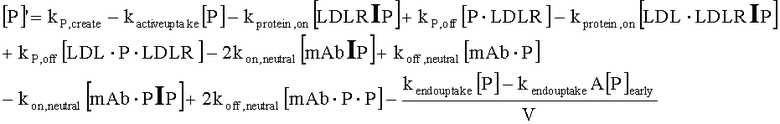
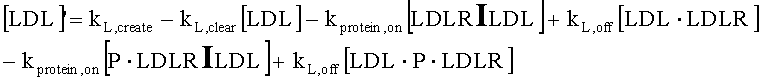

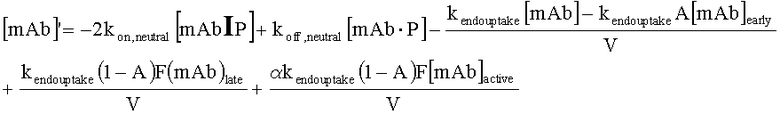
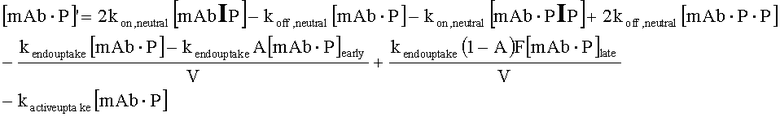
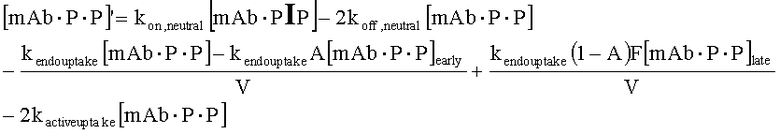
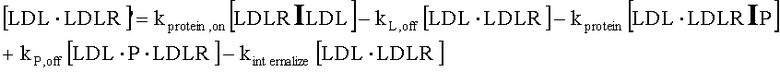
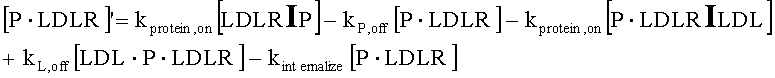
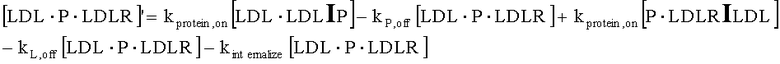
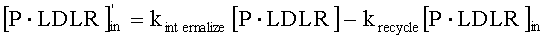
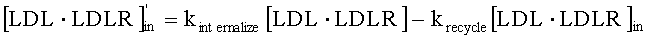
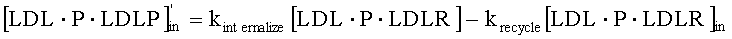
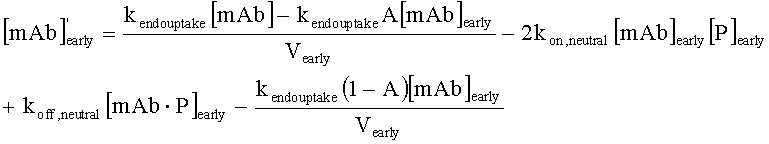
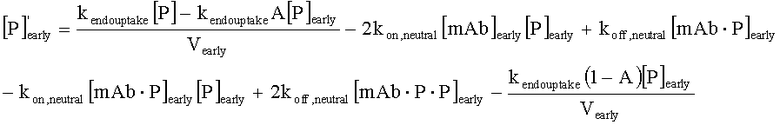
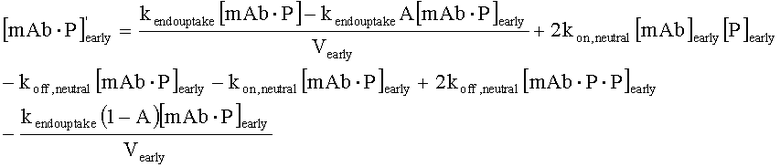
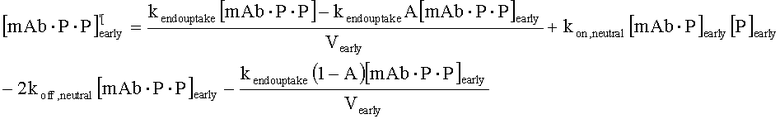
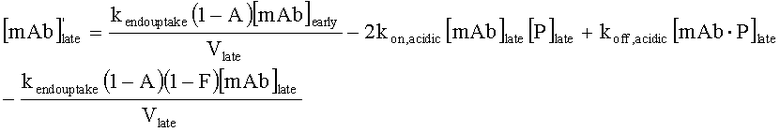
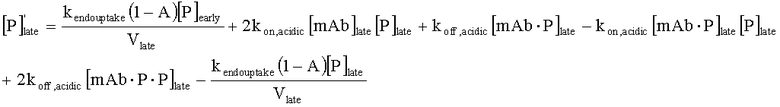
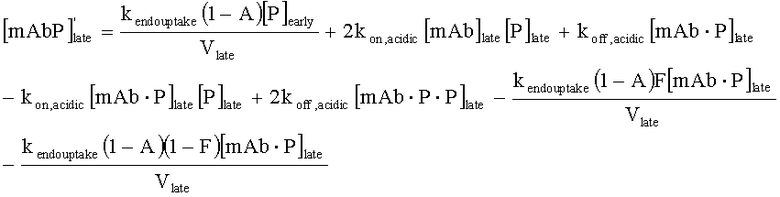
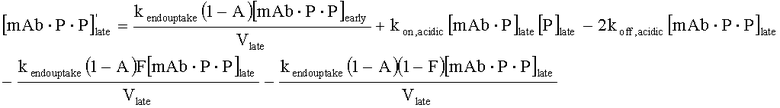
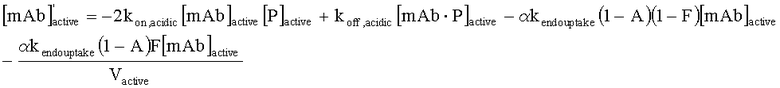
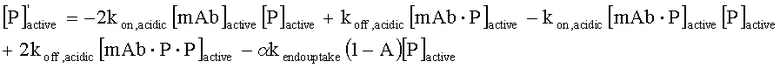
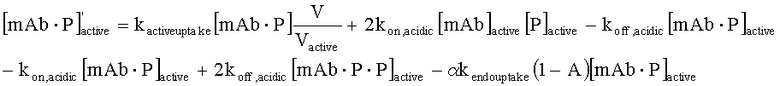
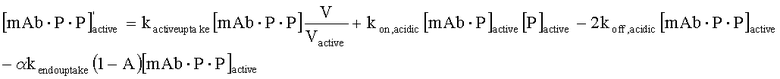
Параметры, используемые в этой модели, описаны ниже в Таблице 3. В некоторых случаях параметры являются производными от других физиологически релевантных или измеряемых величин; они также указаны в таблице ниже.
Одна из картин моделирования показана на Фиг.1. Увеличение отношения KD при эндосомальном рН (то есть, рН 5,5 или 6,0)/ физиологическом рН 7,4 от 1 до 2 привело в результате к увеличению концентрации антитела в зависимости от времени, что коррелировало с показателем отношения KD. Когда отношение KD-изменялось, или koff или коп, концентрация свободного от антигена лиганда изменялась (Фиг.2), также как и концентрация антитела (Фиг.3), при корреляции с увеличенным отношением KD (Фиг.2), увеличенным koff (Фиг.3А) и уменьшенным kon (Фиг.3В). Эти результаты не зависели от того, было ли значение эндосомального рН 5,5 или 6,0.
Тепловую карту (Фиг. 4) строили для общей модели антител с рН-зависимым связыванием, эффективно показывая, на сколько суток дольше антитело с рН-зависимым связыванием должно уменьшать сывороточную концентрацию антигена свыше того, которое наблюдалось для антитела, не проявляющего рН-зависимое связывание (нокдаун-эффект). Сутки, при которых наблюдается нокдаун-эффект, определяют интегралом от: 1-(Cserum(t))/(Cserum(t=0)) от суток 0 до суток 100, например, 100% снижение сывороточной концентрации за N суток с последующим возвращением к уровню до обработки будет соответствовать N суткам нокдаун-эффекта. Тепловые карты показывают увеличение суток нокдаун-эффекта для рН-зависимых антител по сравнению с контрольными антителами на основе отношения KD при нейтральном рН и также отношения KD (R). Эффекты наблюдаются для всех моделированных значений KD, в частности от 0,01 до 100 нМ, и более конкретно KD от 0,1 до 10 нМ. Изменения некоторых параметров, например, размер дозы, кинетические параметры антитела, продолжительность модельного эксперимента или другие показатели, приведут к некоторым различиям в показателях, показанных на тепловых картах.
Предсказания моделирующих экспериментов в отношении эффективности антител против PCSK9, с рН-зависимым связыванием или без него, хорошо коррелировали с результатами фактических экспериментов. На Фиг.5А показана общая концентрация антитела в зависимости от времени после введения антитела 5А10, которое не проявляет рН-зависимого связывания и имеет отношение KD 1,1. Соответствующие уровни LDL показаны на Фиг.5В. На Фиг.6А показана общая концентрация антитела в зависимости от времени после введения антитела 5L1721H23_6L3H3, которое проявляет рН-зависимое связывание и имеет отношение KD 14,4. Соответствующие уровни LDL показаны на Фиг.6В. Для этих моделей и экспериментов на Фиг.5 и 6 значение рН было принято 5,5, и экспериментальные значения KD для антител рассчитаны при рН 5,5. Антитело с рН-зависимым связыванием демонстрировало пролонгированную общую концентрацию антитела и пролонгированное ингибирование уровней LDL по сравнению с 5А10. Дополнительное обсуждение общей модели антител с рН-зависимым связыванием показано в Примере 4.
Пример 2: Создание и скрининг антител против PCSK9 с рН-зависимым связыванием PCSK9
Гистидин-сканирующий мутагенез проводили для всех CDR моноклинального антитела h5A10 (также известного как 5А10 hu frame). Это антитело имеет происхождение из мышиного моноклонального антитела 5А10 (также известного как 5А10.В8), но имеет человеческие каркасные области. Последовательность антитела h5A10, использованного в качестве исходной матрицы, показана ниже (CDR выделены жирным шрифтом):
Вариабельная легкая цепь:
DIQMTQSPSSLSASVGDRVTITCKASQDVSTAVAWYQQKPGKAPKLLIYSASYRYT GVPSRFSGSGSGTDFTFTISSLQPEDIATYYCQQRYSTPRTFGQGTKLEIK (SEQ ID NO: 1)
Вариабельная тяжелая цепь:
QVQLVQSGAEVKKPGASVKVSCKASGYTFTSYYMHWVRQAPGQGLEWMGEINPS GGRTNYNEKFKSRVTMTRDTSTSTVYMELSSLRSEDTAVYYCARERPLYAMDYW GQGTTVTVSS (SEQ ID NO: 2)
Антитело h5A10 клонировали в бактериальный экспрессионный вектор, который обеспечивает экспрессию Fabs в периплазме Е. coli. Используя праймеры, каждое положение CDR мутировали до гистидина, получая в результате 58 единичных гистидиновых мутантов антитела h5A10.
Антитела с этими единичными гистидиновыми мутантами экспрессировали в периплазме Е.coli после индукции с помощью изопропилтиогалактозида (IPTG) и очищали с использованием гистидиновой метки на С-конце СН1. Аффинности очищенных Fabs измеряли при рН 5,5 и 7,4 с помощью Biacore. Мутации, влияющие на отношение KD при рН 5,5/рН 7,4, были обнаружены в четырех разных CDR (L1, L2, Н2 и Н3). рН-Зависимое изменение KD варьировало между минимально сниженным и значительно сниженным.
Мутации из трех CDR (L1, L2 и Н2) повышали отношение KD при рН 5,5/рН 7,4, не влияя на аффинность при рН 7,4, и были генерированы антитела, объединяющие CDR, содержащие эти мутации. Для h5A10 и всех мутантов CDR тяжелой цепи CDR1 (Н1) представляет собой GYTFTSYYMH (SEQ ID NO: 6). Для 5А10, единичных, двойных и тройных мутантов и 5L1721H23J3H3 CDR легкой цепи CDR3 (L3) представляет собой QQRYSTPRT (SEQ ID NO: 27). Для других мутантов с созревшей аффинностью и мутантов на основе L1L3 L3 представляет собой QQRYSLWRT (SEQ ID NO: 12). Последовательности CDR антител, содержащие эти разные комбинации мутаций, показаны в Таблице 4 ниже. Числовые идентификаторы последовательностей даны в скобках. 
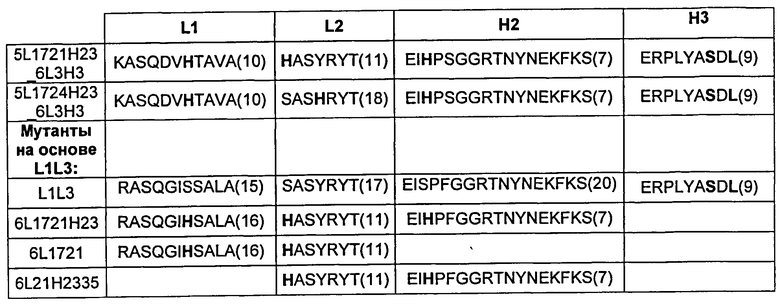
Никаких изменений не было сделано в последовательностях каркасной области для этих антител по сравнению с 5А10.В8. Например, последовательности для вариабельных тяжелых цепей 5L1721H23_6L3 и 5L1721H23_6L3H3 предложены ниже. Одну и ту же вариабельную легкую цепь использовали в каждом из этих антител, и она также дана ниже. CDR выделены жирным шрифтом.

Вариабельная легкая цепь (каппа) 5L1721H23_6L3 и 5L1721H23_6L3H3
gatatccagatgacacagtccccatcctccctgtctgcctctgtgggcgaccgcgtcaccatcacctgcaaggcctctc aggatgtgcatactgctgtagcctggtatcagcagaagccaggcaaagccccaaaactgclgatctaccatgcatcct accgctacactggtgtcccatcacgcttcagtggcagtggctctggtacagatttcaccttcaccattagcagcctgcaa ccagaagatattgccacttattactgccagcaacgttatagtctgtggcgcacgttcggtcaaggcaccaagctggag atcaaa (SEQ ID NO: 23)
Вариабельная тяжелая цепь 5L1721H23_6L3H3
caggtgcagctggtgcagtctggtgctgaggtgaagaagcctggcgcttccgtgaaggtttcctgcaaagcatctggtt acacctttaccagctactatatgcactgggtgcgccaagcccctggtcaaggcctggagtggatgggcgagattcatc ctagcggcggtcgtactaactacaatgagaagttcaagagccgcgtgactatgactcgcgatacctccaccagcact gtctacatggaactgagctctctgcgctctgaggacactgctgtgtattactgtgcccgcgagcgccccctgtatgctag cgacctgtggggccagggtaccacggtcaccgtctcctca (SEQ ID NO: 24)
Эти антитела экспрессировали и очищали для определения их KD при 7,4 и их отношения KD при рН 5,5/рН 7,4, как измерено поверхностным плазменным резонансом с помощью биосенсора Biacore 3000, оборудованного сенсорным чипом исследовательского класса, с использованием подвижного буфера HBS-ЕР (Biacore AB, Uppsala, Sweden - в настоящее время GE Healthcare). Поликлональные анти-Ms IgG кролика подвергали аминосочетанию в насыщающих уровнях на этом чипе, используя стандартный способ с N-гидроксисукцинимидом/ этилдиметиламинопропил-карбодиимидом (NHS/EDC). Этот буфер заменяли на HBS-EP плюс 1 мг/мл BSA плюс 1 мг/мл СМ-декстран. Полноразмерные IgG против PCSK9 разбавляли до примерно 15 мкг/мл и иммобилизировали в течение 1 мин при 5 мкл/мин с получением уровней примерно 500 RU на проточную ячейку, оставляя одну пустой для обеспечения эталонного канала. 3,73-302 нМ hPCSK9 или 2,54-206 нМ mPCSK9 инъецировали в виде 5-членных трехкратных серий в течение 1 мин при 100 мкл/мин. Диссоциацию наблюдали в течение 5 мин. Чип регенерировали после последней инъекции каждого титрования с двумя 30-секундными импульсными введениями 100 мМ фосфорной кислоты. Буферные циклы обеспечивали данные для двойных контролей, которые затем подгоняли в общем к простой модели связывания с использованием программного обеспечения Biaevaluation v.4.1. Аффинности выводили из соотношения кинетических констант скорости (KD=koff/kon). Результаты по аффинности к человеческому PCSK9 показаны в Таблице 5, и результаты по аффинности к мышиному PCSK9 показаны в Таблице 6. Эти данные показывают, что могут быть сконструированы и отобраны антитела, которые имеют более высокую аффинность в отношении человеческого или мышиного PCSK9 при рН 7,4 и более низкую аффинность при рН 5,5.
Аффинность и кинетические параметры для антител h5A10 и 5L1721H23_6L3H3 против PCSK9, а также антитела H1M300N против PCSK9 (см. US 2010/0166768, например в Таблице 7). Все эксперименты осуществляли на биосенсоре Biacore 2000.
Сенсорный чип против человеческого Fc готовили путем активации всех проточных ячеек на сенсорном чипе Biacore CM4 смесью 1:1 (об./об.) 400 мМ EDC и 100 мМ NHS в течение 7 минут со скоростью потока 10 мкл/мин. Реагент против человеческого Fc (козий F(AB')2 фрагмент против человеческого IgG Fc, номер по каталогу Cappel #: 55053) разбавляли до 60 мкг/мл в 10 мМ ацетате натрия с рН 5,0 и вводили во все проточные ячейки в течение 7 минут при 20 мкл/мин. Все проточные ячейки блокировали 100 мМ этилендиамином в 150 мМ боратном буфере с рН 8,5 в течение 7 минут при 10 мкл/мин.
Анализ кинетических параметров проводили с использованием методологии кинетического титрования, как описано в Karlsson et al., Anal. Biochem 349, 136-147 (2006).
То же самое антитело (например, 5L1721H23_6L3H3) иммобилизировали на нисходящих проточных ячейках (проточные ячейки 2, 3 и 4) при 2 мкг/мл со скоростью потока 10 мкл/мин в течение 30 секунд, 60 секунд и 120 секунд для проточных ячеек 2, 3 и 4, соответственно. Проточную ячейку 1 использовали в качестве эталонной поверхности. После иммобилизации антител PCSK9 инъецировали при 30 мкл/мин на все проточные ячейки серией инъекций от низкой до высокой концентрации. Высшая концентрация составляла 200 нМ PCSK9, и фактор разведения был трехкратным. Каждое инъецирование PCSK9 было двухминутным, время диссоциации после инъекции 200 нМ PCSK9 составило 20 минут. Аналогичный ряд инъецирований проводили с подвижным буфером вместо PCSK9 для целей двойного контроля (двойной контроль как описано в Myszka, J. Mol. Recognit 12, 279-284, 1999). После каждого аналитического цикла все проточные ячейки регенерировали тремя 30-секундными инъекциями 75 мМ фосфорной кислоты. Сенсорограммы от проточных ячеек 2 и 3 для данной пары РС8К9/антитело подгоняли в общем под простую ленгмюровскую модель связывания с массопереносом 1:1.
Эксперименты осуществляли при рН 6,0 и 7,4 с использованием простого и подвижного буферов из 10 мМ фосфата натрия, 150 мМ хлорида натрия, 0,05% Tween-20 с рН 6, и 10 мМ фосфата натрия, 150 мМ хлорида натрия, 0,05% Tween-20 с рН 7,4, соответственно.
Пример 3: рН-зависимые PCSK9-связывающие антитела проявляют пролонгированный Фармакодинамический эффект по снижению холестерина
А. рН-зависимое РСSK9-связывающее антагонистическое антитело снижало сывороточный холестерин в течение продолжительного периода времени у мышей.
Для определения того, могут ли рН-зависимые PCSK9-связывающие антагонистические антитела снижать уровни холестерина in vivo в течение продолжительного периода времени по сравнению с антителами, не проявляющими рН-зависимое связывание, исследовали зависимости эффектов от времени чувствительных к кислоте антител 5L1721H23_6H3 и 5L1721H23_6L3H3 и не чувствительных к кислоте антител h5A10 (только в дозе 10 мг/кг) и 5А10.В8 в отношении сывороточного холестерина при инъецировании мышам в дозах 1, 3 или 10 мг/кг. Все четыре антитела имели сходные аффинности связывания, составляющие 5-14 нМ при нейтральном рН (рН 7,4), с мышиным PCSK9. Антитела 5L1721H23_6H3 и 5L1721H23_6L3H3 имели пониженные аффинности 117 нМ и 408 нМ, соответственно, при рН 5,5, в то время как h5A10 и 5А10.В8 имеют KD при рН 5,5, аналогичный значению при 7,4 (5,2 нМ). Самцов линии мышей С57/bl6 возрастом 6-7 недель держали на 12-часовом цикле свет/темнота, брали кровь для сбора приблизительно 70 мкл сыворотки на сутки -7. Антагонистические антитела против PCSK9 и контрольный изотип, соответствующий моноклональному антителу, которое не связывается с какими-либо известными животными белками, инъецировали внутривенно самцам возраста 7 недель линии мышей С57/bl6 и брали образцы сыворотки на сутки 5, 12, 19, 26, 61 и 75 после инъекции. Все образцы сыворотки анализировали на общий холестерин, триглицериды, холестерин HDL, используя оборудование Асе Alera (Alfa Wassermann, West Caldwell, NJ), и уровни холестерина LDL рассчитывали с использованием уравнения Фридвальда. На Фиг.7 показано быстрое и дозозависимое снижение уровней общего холестерина после инъекции антагонистического антитела против PCSK9. Уровни холестерина LDL у мышей были слишком низкими для достоверного измерения и расчета. При дозе 10 мг/кг все четыре антитела снижали HDL-холестерин на 35-40% на сутки 5 и 12, в то время как 5L1721H23_6H3 и 5L1721H23_6L3H3, инъецированные животным, не восстанавливались вплоть до суток 61, h5A10 и 5А10.В8, инъецированные животным, возвращались к исходным уровням на сутки 26 и 33, соответственно.
В. рН-Зависимые РСSK9-связывающие антитела проявляют пролонгированное время полужизни у мышей
Сывороточные концентрации антитела определяли в том же исследовании, что описано в примере b, для определения того, приведут ли в результате рН-чувствительные антитела против PCSK9 к пролонгированию времени полужизни антитела. Нормальные антитела против PCSK9, такие как h5A10 и 5А10.В6, имеют дозозависимое, более короткое время полужизни по сравнению с антителами, которые связываются с другими растворимыми антигенами, из-за PCSK9-опосредованной деградации комплекса антитело/антиген. Как показано на Фиг.8А, рН-зависимое свойство связывания снижало деградацию антитела и пролонгировало время полужизни антител 5L1721H23_6H3 и 5L1721H23_6L3H3 против PCSK9. Для дополнительной иллюстрации того, что пролонгированные РК у 5L1721H23_6H3 и 5L1721H23_6L3H3 являлись результатом пониженного PCSK9-опосредованного клиренса антител, провели аналогичное исследование зависимости эффекта от времени на мышах с выключенным геном PCSK9. Различия в сывороточных концентрациях антитела и скоростях снижения между рН-чувствительными и не чувствительными антителами были незначимы вплоть до дозы 3 мг/кг человеческого PCSK9, инъецированного мышам. После этой инъекции не чувствительное к рН антитело против PCSK9 демонстрировало повышенную деградацию по сравнению с чувствительным к рН антителом и отрицательным контрольным антителом (Фиг.8В). Эти результаты показывают, что наблюдаемая пониженная деградация рН-зависимых PCSK9-связывающих антител против PCSK9 происходит в результате диссоциации антитела от PCSK9 и, таким образом, риска возникновения для этого антитела PCSK9-опосредованной деградации.
С. рН-Чувствительные антагонистические антитела против PCSK9 снижают концентрацию сывороточного холестерина в течение продолжительного времени у обезьян
На Фиг.9В показано влияние рН-чувствительных антагонистических антител 5L1721H23_6H3 и 5L1721H23_6L3H3 против PCSK9 и не чувствительного антитела L1L3 против PCSK9 на уровни сывороточного LDL-холестерина у Cynomolgus в качестве контроля в процентах. Антитела (1,5 мг/кг каждого) вводили на сутки 0 самкам обезьян Cynomolgus посредством внутривенной болюсной инъекции. LDL-холестерин снижался на 50% от исходного уровня в сутки во всех трех группах лечения антителом. В то время как уровни LDL-холестерина возвращались к исходному уровню к 10 суткам после введения не чувствительного к рН антитела, уровни LDL-холестерина оставались подавленными вплоть до 21 суток у обезьян, которых лечили чувствительными к рН антителами. Уровни HDL оставались по существу неизмененными в результате лечения антителом (Фиг.9А). На Фиг.10 продемонстрировано, что время полужизни чувствительных к рН антител против PCSK9 было пролонгированным по сравнению с не чувствительным к PHL1L3.
Пример 4: Общее моделирование для антител с рН-зависимым связыванием антигена
Компьютерное моделирование использовали для предсказания того, может ли антитело с рН-зависимым связыванием со своим родственным антигеном влиять на время полужизни антитела и/или продолжительность снижения количества или сывороточной концентрации антигена. Для целей этого моделирования следующие допущения были сделаны: 1) антитело, 1 мкМ доза антитела в крови, время полужизни антитела 21 сутки; 2) продолжительность модельного эксперимента 100 суток; связывание антитела и рН-зависимое связывание, Kon равен 1е5/М/сек, Koff при нейтральном рН равен KD* Kon; рН-зависимое связывание смоделировано как повышение Koff в кислых эндосомах, Koff при кислом рН равен R* Koff при нейтральном рН.
На Фиг.15 подробно описана модель переноса для антител с рН-зависимым связыванием, использованная для моделирования, и уравнения, определяющие эту модель, являются следующими:

Параметры, используемые в этой модели, описаны ниже в Таблице 8.
Тепловые карты, показанные на Фиг.4, иллюстрируют, на сколько больше антитело с рН-зависимым связыванием влияет на нокдаун-эффект в отношении антигена свыше нокдаун-эффекта, проявляемого антителом без рН-зависимого связывания. Это количество, при котором наблюдается нокдаун-эффект, определяют путем измерения области между кривыми свободного антигена для двух антител в течение 100-суточного моделирования. Это можно рассматривать в качестве того, насколько дольше антитело с рН-зависимым связыванием будет влиять на нокдаун-эффект в отношении антигена свыше нокдаун-эффекта, проявляемого антителом без рН-зависимого связывания, хотя эти две меры определения не имеют точной корреляции. Более светлые области на карте отражают более длительный и/или более сильный нокдаун-эффект в отношении свободного антигена. Тепловая карта на Фиг.11 показывает, как потенциальные антитела с рН-зависимым связыванием антигена будут влиять на нокдаун-эффект в отношении антигена, если антиген представляет собой DKK1, IgE или С5. Для каждого из этих антигенов антитело с рН-зависимым связыванием может удлинять нокдаун-эффект в отношении антигена.
На Фиг.12 показано более подробное моделирование для антитела с рН-зависимым связыванием с IgE. Увеличенная продолжительность нокдаун-эффекта в отношении IgE имеет место у антитела, имеющего KD примерно 1 нМ или больше и отношение KD при рН 6,0/ рН7,4 (R), равное 3 или более.
Аналогичный анализ для антигена DKK1 показан на Фиг.13 и для С5 на Фиг.14. Для DKK1 идеальный интервал KD составляет 1,0-100 нМ со всеми отношениями KD в интервале 3-30. Для С5 идеальный интервал KD также составляет 1,0-100 нМ для интервала отношений KD 3-30.
Пример 5: Создание и скрининг антител против IgE с рН-зависимым связыванием IdE
Осуществляли гистидиновые замены в положениях подчеркнутых остатков в антителе 5.948-H100Y против IgE, которое имеет следующие аминокислотные последовательности вариабельной легкой цепи (VL) и вариабельной тяжелой цепи (VH) (CDR выделены жирным шрифтом):

Антитела тестировали в отношении связывания IgE при 25°С. Для определения аффинностей и кинетических констант все эксперименты проводили на биосенсоре Biacore T200.
Поверхность с иммобилизованным биотином готовили путем иммобилизации NeutrAvidin: все проточные ячейки сенсорного чипа Biacore СМ4 активировали смесью 1:1 (об./об.) 400 мМ EDC и 100 мМ NHS в течение 7 минут со скоростью потока 10 мкл/мин. NeutrAvidin (Pierce, продукт #: 31000) разбавляли до 100 мкг/мл в 10 мМ ацетате натрия с рН 4,5 и вводили во все проточные ячейки в течение 7 минут при 20 мкл/мин. Все проточные ячейки блокировали 100 мМ этилендиамином в 150 мМ боратном буфере с рН 8,5 в течение 7 минут при 10 мкл/мин.
IgE-конструкт (человеческий IgE-Fc (Ch2-Ch3-Ch4), экспрессированный и очищенный собственными средствами) минимально биотинилировали путем инкубации IgE с эквимолярным количеством EZ-Link NHS-LC-LC-биотин (Thermo Scientific, номер по каталогу # 21343) в течение 30 мин при комнатной температуре. Свободный биотин удаляли посредством буферного обмена.
Все эксперименты Biacore осуществляли с использованием двух из четырех проточных ячеек (1 и 2 или 3 и 4) сенсорного чипа в параллельных опытах. Нисходящую проточную ячейку, использовали в качестве аналитической проточной ячейки, в то время как другую проточную ячейку использовали в качестве эталонной поверхности. Таким образом, биотинилированный IgE (B-IgE) иммобилизировали на аналитической проточной ячейке, и каждый очищенный Fab фрагмент вводили над обоими. Сенсорограмму от эталонной проточной ячейки вычитали из таковой от аналитической проточной ячейки.
B-IgE разбавляли в подвижном буфере до конечной концентрации 5 мкг/мл непосредственно после иммобилизации. При рН 6,0 B-IgE вводили при 10 мкл/мин в течение 60 секунд. При рН 7,4 B-IgE вводили в течение 80 секунд для получения подобных уровней иммобилизации. Оценку кинетики проводили с использованием методологии кинетического титрования, как описано в Karlsson et al., Anal Biochem 349: 136-147, 2006.
После иммобилизации B-IgE очищенный Fab фрагмент вводили при 30 мкл/мин на аналитическую и эталонную проточные ячейки в виде серии инъекций от низкой до высокой концентрации. Высшая концентрация Fab находилась в интервале от 30 нМ до 200 нМ, и фактор разведения был трехкратным. Каждое разведение Fab вводили в течение 2 минут, с последующим конечным временем диссоциации 30 минут после последней инъекции. Аналогичный набор инъекций осуществляли с использованием подвижного буфера вместо Fab для целей двойного сравнения (двойное сравнение, как описано в Myszka, J Mol Recognit 12: 279-284, 1999). Сенсорограммы двойного сравнения подгоняли в общем к простой ленгмюровской модели связывания с массопереносом 1:1.
Эксперименты осуществляли при рН 6,0 и 7,4 с использованием буфера для образцов и подвижного буфера из 10 мМ фосфата натрия, 150 мМ хлорида натрия, 0,05% Tween-20, 1 мг/мл BSA, рН 6, и 10 мМ фосфата натрия, 150 мМ хлорида натрия, 0,05% Tween-20, 1 мг/мл BSA, рН 7,4, соответственно.
Как показано ниже в Таблице 9, гистидиновая замена в IgE антителе 5.948-H100Y успешно привела к антителу с более быстрой koff при рН 6,0 и более высокими отношениями KD и koff при рН 6,0/рН 7,4.
Описания всех цитированных здесь источников информации включены в данное описание посредством ссылки.
| название | год | авторы | номер документа |
|---|---|---|---|
| ЛЕЧЕНИЕ ИНТЕРСТИЦИАЛЬНОГО ЦИСТИТА | 2008 |
|
RU2570559C2 |
| АНТАГОНИСТЫ PCSK9 | 2009 |
|
RU2528735C2 |
| АНТАГОНИСТЫ PCSK9 | 2014 |
|
RU2618869C2 |
| СПОСОБЫ ЛЕЧЕНИЯ БОЛИ, ВЫЗВАННОЙ ОСТЕОАРТРИТОМ, ПУТЕМ ВВЕДЕНИЯ АНТАГОНИСТА ФАКТОРА РОСТА НЕРВОВ И СОДЕРЖАЩИХ ЕГО КОМПОЗИЦИЙ | 2006 |
|
RU2429013C2 |
| АНТИТЕЛА ПРОТИВ TNF-α И ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2595379C2 |
| СПОСОБЫ ЛЕЧЕНИЯ ХРОНИЧЕСКОЙ БОЛИ | 2009 |
|
RU2522493C2 |
| АНТИТЕЛА ПРОТИВ VEGF И ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2567639C2 |
| СПОСОБЫ ЛЕЧЕНИЯ ГЛАЗНЫХ БОЛЕЗНЕЙ | 2008 |
|
RU2434639C2 |
| СПОСОБЫ ЛЕЧЕНИЯ ПСОРИАЗА | 2008 |
|
RU2475265C2 |
| МОЛЕКУЛЫ АНТИТЕЛ С УЛУЧШЕННЫМИ СВОЙСТВАМИ | 2009 |
|
RU2430111C1 |
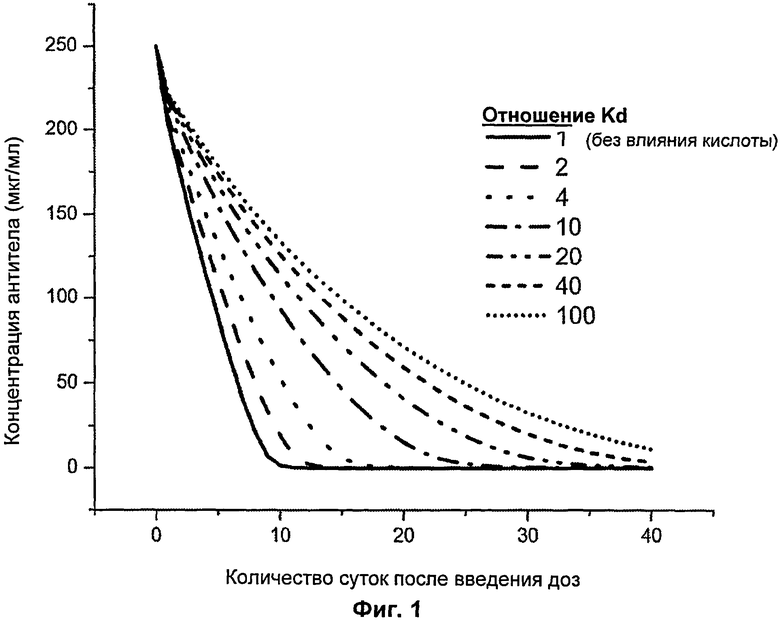
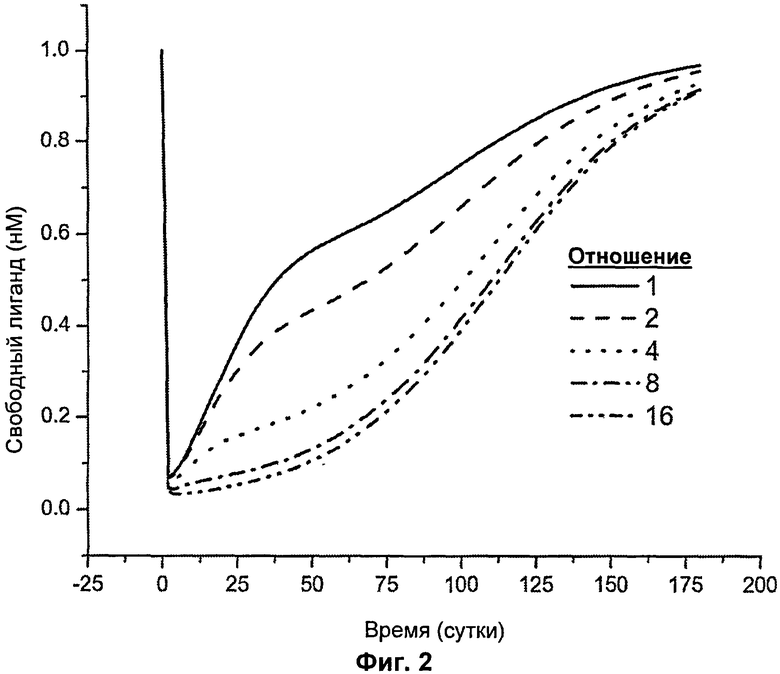
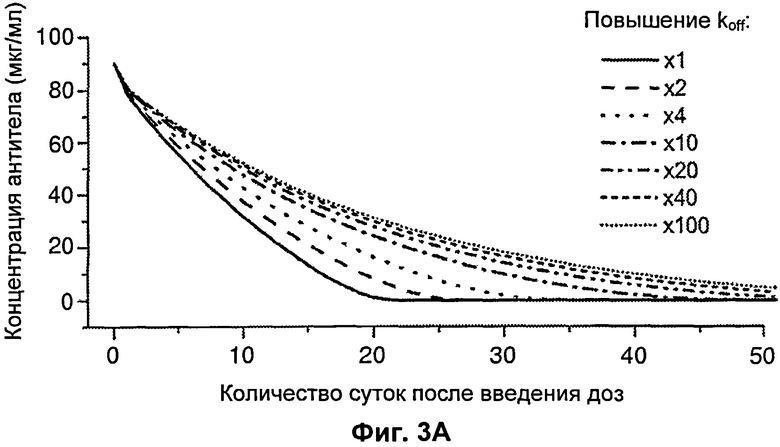
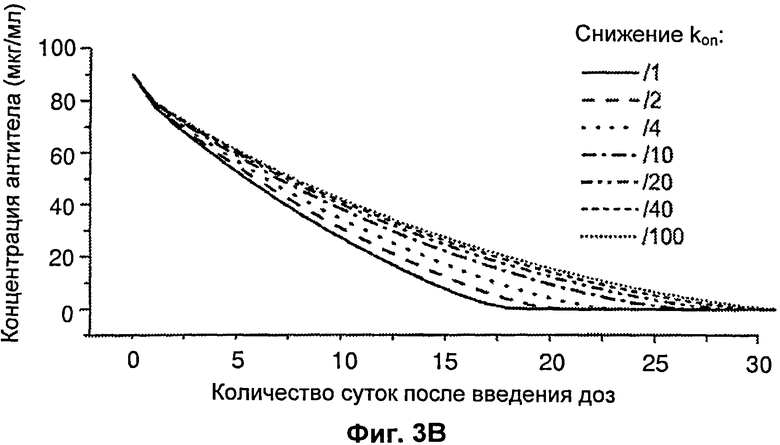
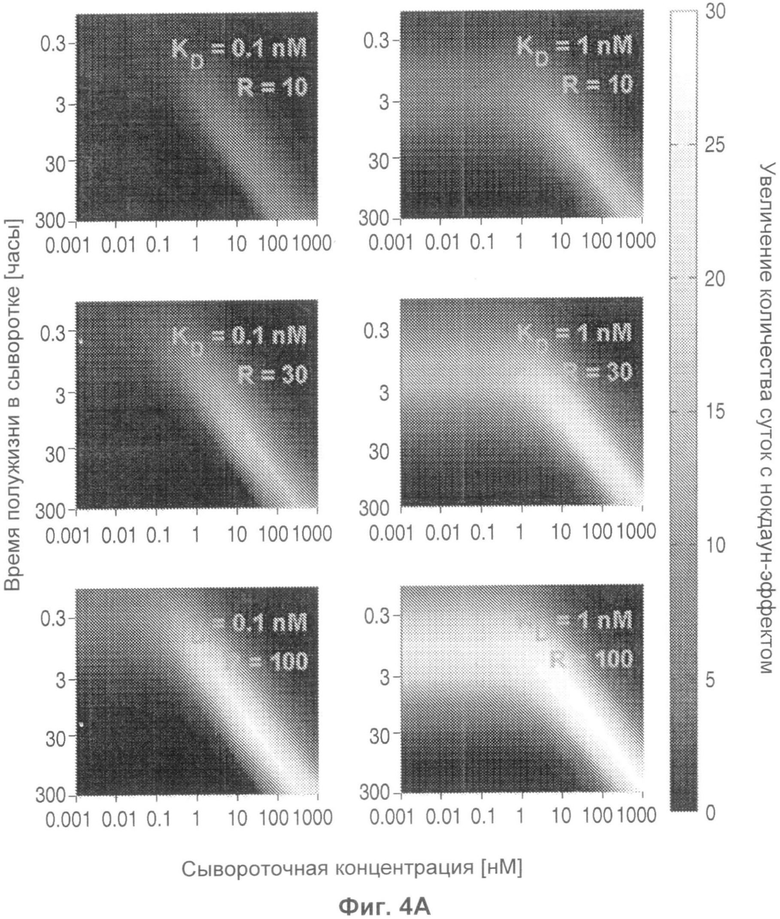
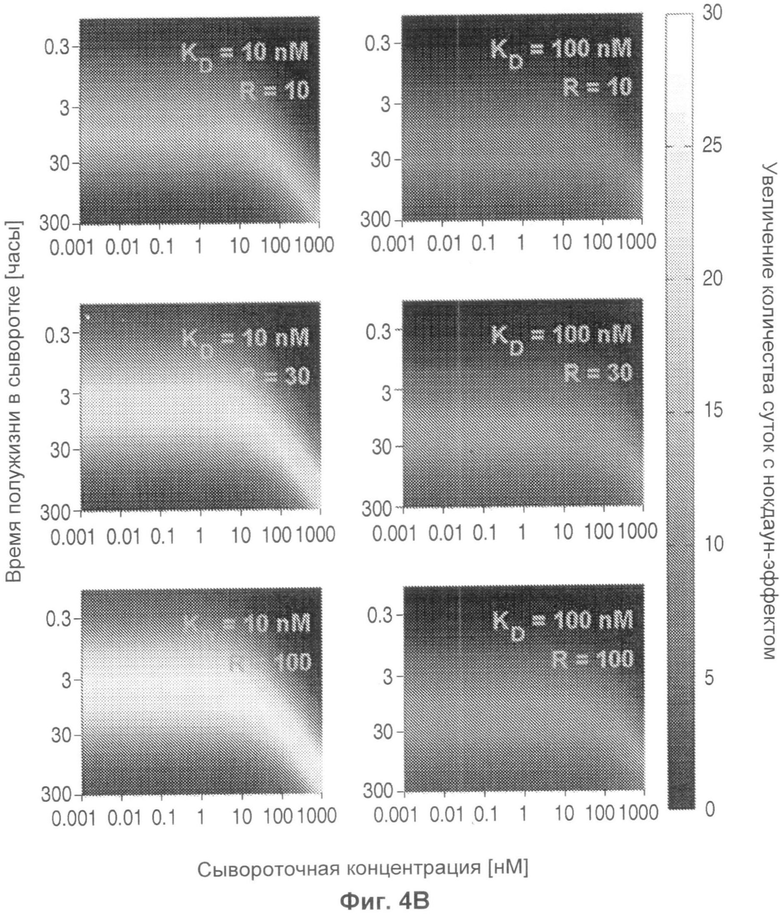
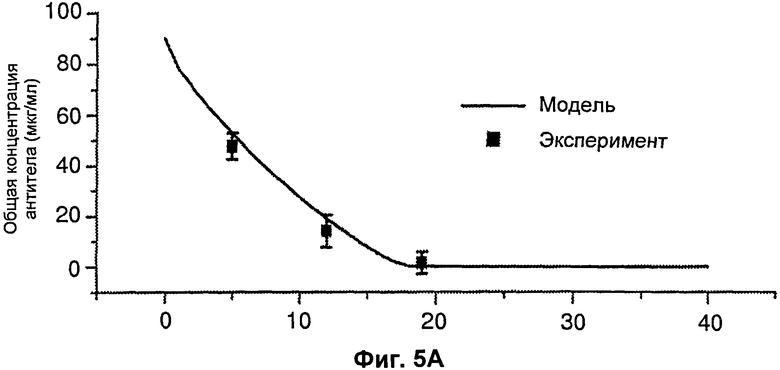
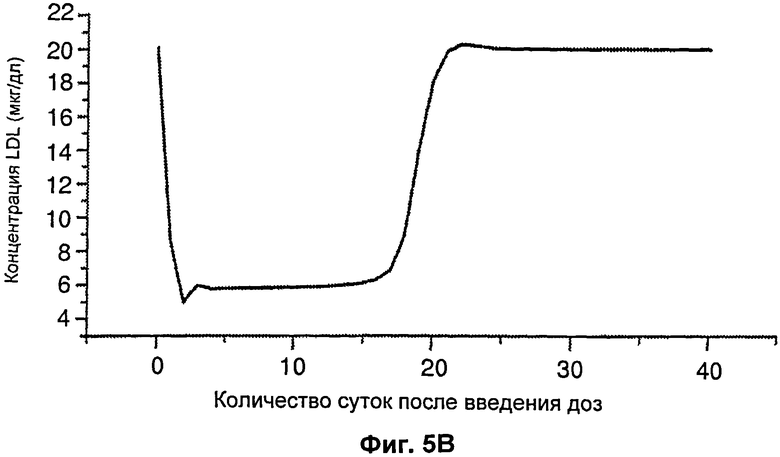
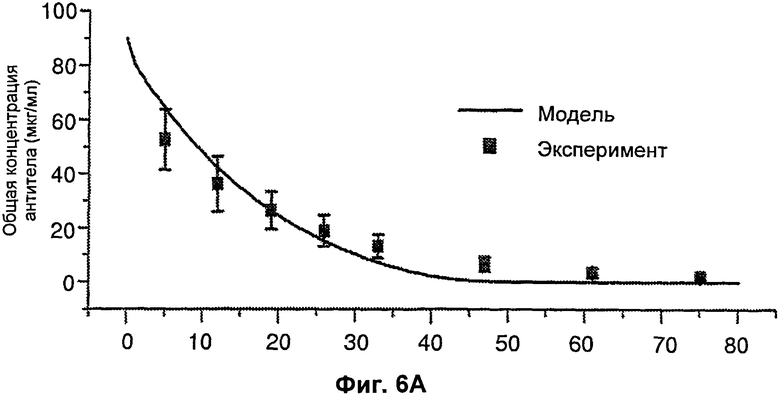
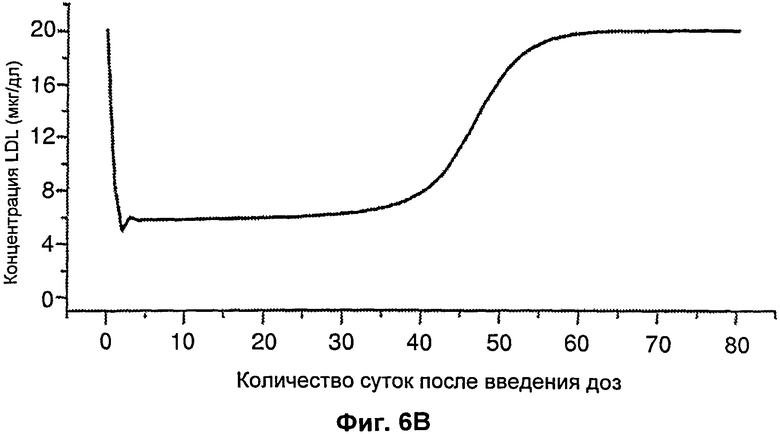
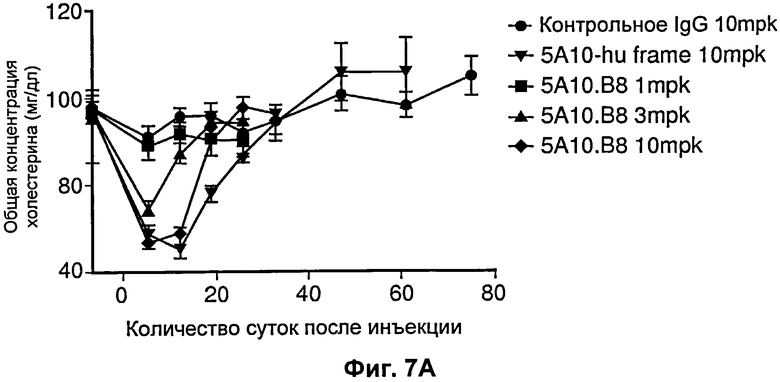
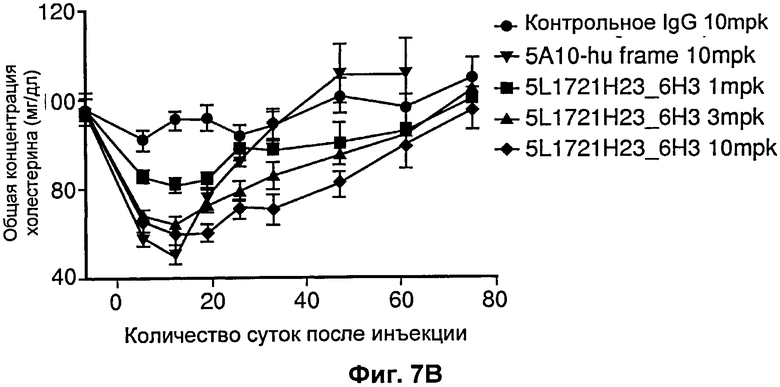

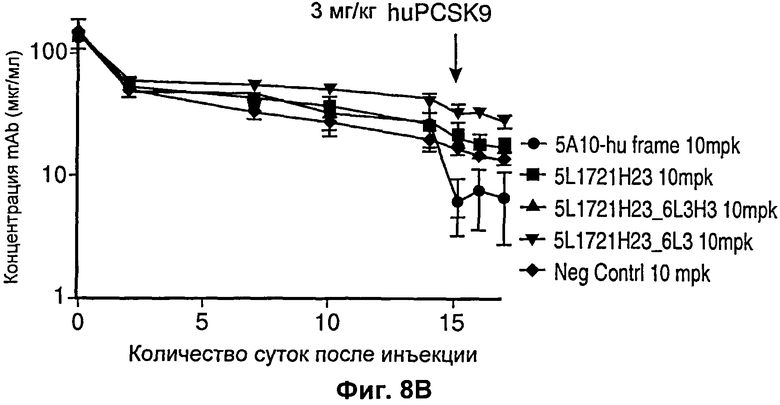
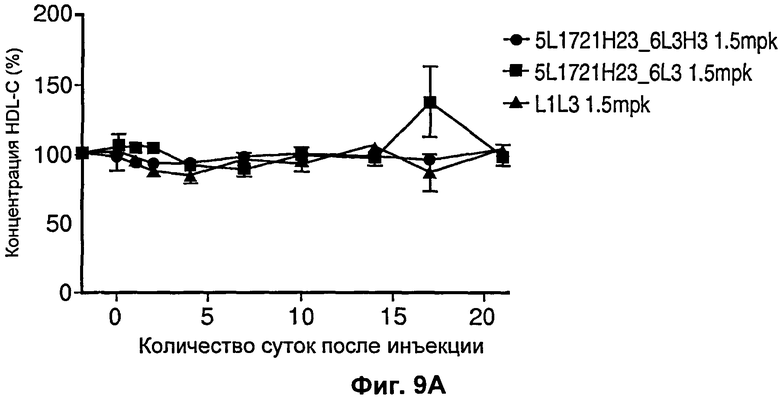
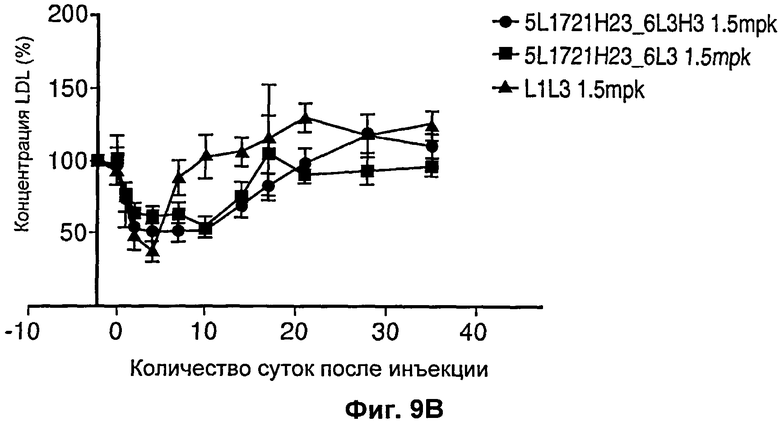
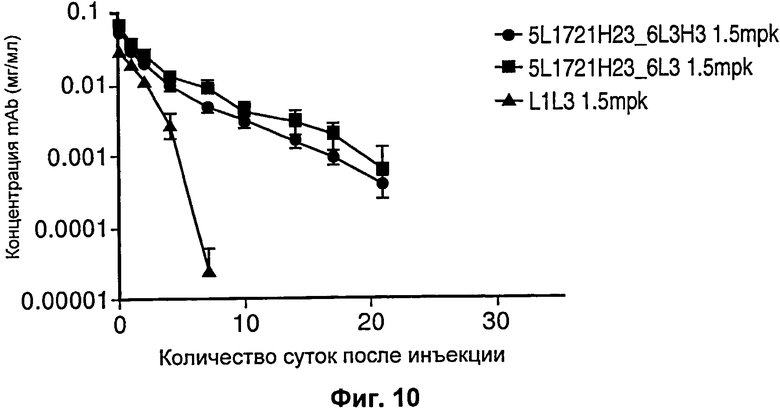

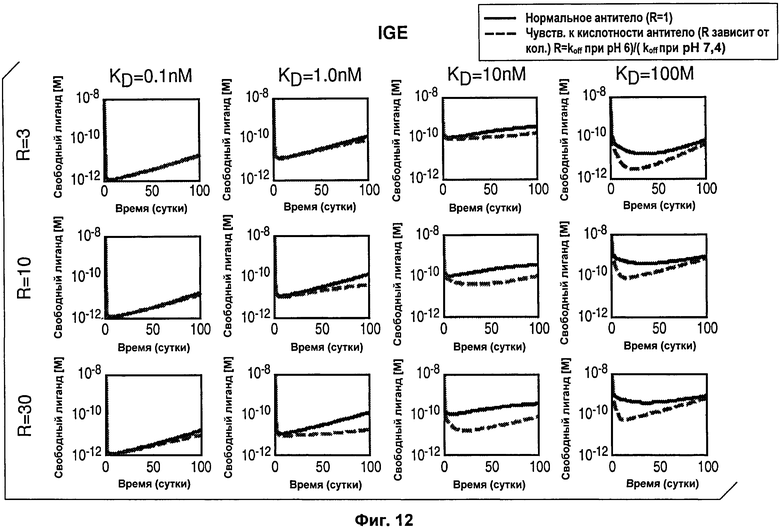
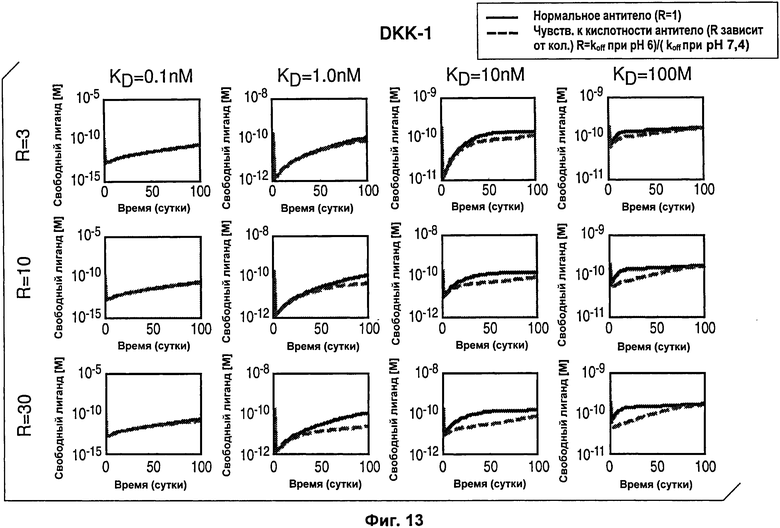

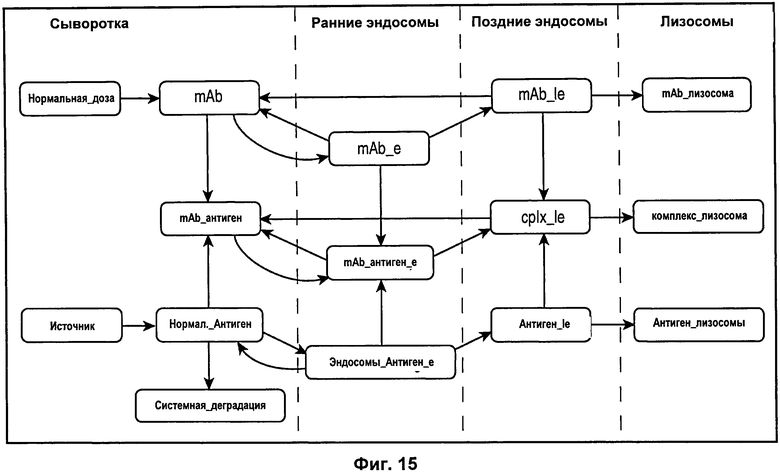
Группа изобретений относится к области медицины и фармакологии и касается выделенного антитела, которое специфически связывает человеческий PCSK9, содержащее: гипервариабельный участок вариабельной области тяжелой цепи 1 (VH CDR1) с аминокислотной последовательностью, показанной в SEQ ID NO: 6; VH CDR2 с аминокислотной последовательностью, показанной в SEQ ID NO: 7; VH CDR3 с аминокислотной последовательностью, показанной в SEQ ID NO: 8 или 9, и дополнительно содержащее: CDR1 вариабельной области легкой цепи (VL) с аминокислотной последовательностью, показанной в SEQ ID NO: 10; VL CDR2 с аминокислотной последовательностью, показанной в SEQ ID NO: 11; и VL CDR3 с аминокислотной последовательностью, показанной в SEQ ID NO: 12, а также антитела, кодируемого плазмидами, депонированными в Американской коллекции типовых культур (АТСС) и имеющими номер в АТСС No. РТА-10547 или РТА-10548 и РТА-10549, где указанное антитело специфически связывает человеческий PCSK9, и способа снижения уровня холестерина липопротеидов низкой плотности в крови субъекта, включающий введения антитела. Группа изобретений обеспечивает пролонгированное действие антител при лечении гиперхолестеринемии. 7 н. и 3 з.п. ф-лы, 5 пр., 9 табл., 15 ил.
1. Выделенное антитело, которое специфически связывает человеческий PCSK9, содержащее: гипервариабельный участок вариабельной области тяжелой цепи 1 (VH CDR1) с аминокислотной последовательностью, показанной в SEQ ID NO: 6; VH CDR2 с аминокислотной последовательностью, показанной в SEQ ID NO: 7; VH CDR3 с аминокислотной последовательностью, показанной в SEQ ID NO: 8 или 9, и дополнительно содержащее: CDR1 вариабельной области легкой цепи (VL) с аминокислотной последовательностью, показанной в SEQ ID NO: 10; VL CDR2 с аминокислотной последовательностью, показанной в SEQ ID NO: 11; и VL CDR3 с аминокислотной последовательностью, показанной в SEQ ID NO: 12.
2. Выделенное антитело по п. 1, где VH CDR3 имеет аминокислотную последовательность SEQ ID NO: 9.
3. Выделенное антитело по п. 1, где VH-область содержит SEQ ID NO: 4 или SEQ ID NO: 5, и VL-область содержит SEQ ID NO: 3.
4. Выделенное антитело по п. 3, где VH-область содержит SEQ ID NO: 5.
5. Антитело, кодируемое плазмидами, депонированными в Американской коллекции типовых культур (АТСС) и имеющими номер в АТСС No. РТА-10547 или РТА-10548 и РТА-10549, где указанное антитело специфически связывает человеческий PCSK9.
6. Фармацевтическая композиция для снижения уровня холестерина липопротеидов низкой плотности (ЛПНП), содержащая терапевтически эффективное количество антитела по любому из пп. 1-5.
7. Клетка-хозяин, которая рекомбинантно продуцирует антитело по любому из пп. 1-5.
8. Выделенная нуклеиновая кислота, кодирующая антитело по любому из пп. 1-5.
9. Выделенная плазмида, депонированная в АТСС и имеющая номер в АТСС No. РТА-10547, РТА-10548 или РТА-10549, где указанные плазмиды РТА-10547 или РТА-10548 и РТА-10549 кодируют антитело, которое специфически связывает человеческий PCSK9.
10. Способ снижения уровня холестерина липопротеидов низкой плотности (ЛПНП) в крови субъекта, нуждающегося в этом, включающий введение указанному субъекту терапевтически эффективного количества антитела по любому из пп. 1-5.
| WO2009125825 A1, 15.10.2009 | |||
| WO2008063382A2, 29.05.2008 | |||
| US20100040610 A1, 18.02.2010 | |||
| WO2008057459A2, 15.05.2008 | |||
| ПОЛИПЕПТИДНЫЕ ВАРИАНТЫ С ИЗМЕНЕННОЙ ЭФФЕКТОРНОЙ ФУНКЦИЕЙ | 2005 |
|
RU2367667C2 |

