Область, к которой относится данное изобретение
Настоящее изобретение относится к антителам, например полноразмерным антителам или их антигенсвязывающим частям, пептидам и аптамерам, которые противодействуют активности внеклеточного пробелка конвертаза субтилизин кексин типа 9 (PCSK9), в том числе его взаимодействию с рецептором липопротеинов низкой плотности (LDLR). Конкретнее, данное изобретение относится к композициям, содержащим антагонистические антитела против PCSK9, пептиды и/или аптамеры, и способам применения этих антител, и/или пептидов, и/или аптамеров в качестве лекарственного средства. Антагонистические антитела, пептиды и аптамеры против PCSK9 можно использовать в терапевтических целях для снижения уровней холестерина-LDL в крови и можно использовать для профилактики и/или лечения нарушений метаболизма холестерина и липопротеинов, в том числе семейной гиперхолестеринемии, атерогенной дислипидемии, атеросклероза и, в более общем смысле, сердечно-сосудистого заболевания (CVD).
Предпосылки к созданию изобретения
Миллионы людей в США подвержены риску развития заболеваний сердца и обусловленных ими кардиальных событий. CDV и лежащий в его основе атеросклероз является основной причиной смерти среди всех демографических групп, несмотря на доступность методов лечения, направленных на многочисленные факторы риска. Атеросклероз представляет собой заболевание артерий и является ответственным за ишемическую болезнь сердца, связанную с многочисленными смертельными исходами в промышленно развитых странах. В настоящее время некоторые факторы риска развития ишемической болезни сердца установлены: дислипидемии, артериальная гипертензия, сахарный диабет, курение, недостаток питания, малоподвижный образ жизни и стресс. Самые клинически релевантные и распространенные дислипидемии характеризуются повышением бета-липопротеинов (липопротеины очень низкой плотности (VLDL) и LDL) с гиперхолестеринемией без или с гипертриглицеридемией (Fredrickson et al., 1967, N Engl J Med. 276:34-42, 94-103, 148-156, 215-225, и 273-281). Существует давно ощущаемая значительная неудовлетворенная потребность в отношении лечения CDV с 60-70% сердечно-сосудистых событий, сердечных приступов и инсультов, невзирая на лечение статинами (современный стандарт лечения атеросклероза). Более того, новые стандарты предполагают, что должны быть достигнуты еще более низкие уровни LDL для того, чтобы защитить пациентов с высокой степенью риска от раннего CDV [National Cholesterol Education Program (NCEP), 2004].
PCSK9, также известный как NARC-1, был идентифицирован при некоторых формах семейной гиперхолестеринемии как белок с генетической мутацией. PCSK9 синтезируется в виде зимогена, который в эндоплазматическом ретикулуме подвергается аутокаталитическому процессингу в мотив LVFAQ. Популяционные исследования показали, что некоторые мутации PCSK9 представляют собой мутации «с приобретением функции» и обнаруживаются у индивидуумов с аутосомно-доминантной гиперхолестеринемией, в то время как другие мутации «с потерей функции» (LOF) связаны со сниженным холестерином плазмы крови. Исследования причин заболеваемости и смертности в этой группе отчетливо демонстрируют, что снижение функции PCSK9 значительно снижает риск сердечно-сосудистого заболевания.
В результате важного значения для лечения CDV мутация LOF может повысить чувствительность человека к статинам, обеспечивая эффективность при более низкой дозе (тем самым снижая риски, связанные с безопасностью и устойчивостью) и потенциально достигая более низких уровней холестерина плазмы крови, чем при общепринятых методах лечения.
PCSK9 секретируется в плазму преимущественно гепатоцитами. Генетическая модуляция PCSK9 у мышей подтвердила способность PCSK9 регулировать липиды крови и показала, что его действие направлено на подавление печеночных уровней белка LDLR.
Механизм подавления и место приложения действия, в котором PCSK9 подавляет белок LDLR, не был четко установлен. В случае сверхэкспрессии PCSK9 может действовать и в гепатоцитах, и в виде секретируемого лиганда для LDLR. Существует четкое доказательство, что внеклеточный PCSK9 связывается с клеточной поверхностью LDLR и способствует деградации LDLR во внутриклеточном местоположении. Однако также возможно, что PCSK9 может взаимодействовать с LDLR, когда два белка транслируются внутри эндоплазматического ретикулума (ЭР) и двигаются сквозь эндосомальные компартменты по направлению к клеточной мембране. Maxwell et al., 2005, Curr. Opin. Lipidol. 16:167-172, продемонстрировали, что опосредованный PCSK9 эндоцитоз LDLR и деградация не изменялись под воздействием ингибиторов протеосом и не модулировались различными классами лизосомных и нелизосомных протеаз. Две природные мутации при семейной гиперхолестериемии, S127R и D129G, как сообщалось, были с дефектом аутопроцессинга и секреции, поскольку уровни этих мутантных белков были в значительной степени снижены или не определялись в среде трансфицированных клеток. Еще эти мутанты продемонстрировали увеличенную способность подавлять LDLR, исходя из их идентификации у индивидуумов с высоким уровнем LDL в плазме крови (Homer et al., 2008, Atherosclerosis 196:659-666; Cameron et al., 2006 Human Molecular Genetics 15:1551-1558; Lambert et al., 2006, TRENDS in Endocrinology and Metabolism 17:79-81). Поскольку эти мутанты очевидно не секретируются внеклеточно и все же подавляют LDLR, это убедительно показывает, что внутриклеточное место приложения действия является физиологически важным.
Из информации, доступной в уровне техники и предшествующей настоящему изобретению, остается неясным, будет ли введение антагониста PCSK9 на основе антитела, пептида или аптамера в кровоток для выборочного противодействия внеклеточному PCSK9 эффективным для уменьшения гиперхолестеринемии и связанного инцидента CDV и, если да, то какие свойства антагониста PCSK9 необходимы для такой эффективности in vivo.
Сущность изобретения
Это изобретение относится к антагонистическим антителам, пептидам и аптамерам, которые выборочно взаимодействуют с PCSK9 и ингибируют его функцию. Впервые продемонстрировано, что некоторые антагонисты PCSK9 являются эффективными in vivo для снижения холестерина крови.
В одном варианте осуществления данное изобретение относится к изолированному антагонисту PCSK9, который содержит антитело, пептид или аптамер, который взаимодейтсвует с PCSK9 и при введении пациенту снижает уровень холестерина-LDL в крови пациента. Данный антагонист может представлять собой антитело, например моноклональное антитело или антитело человека, гуманизированное или химерное антитело.
В другом варианте осуществления изобретение относится к изолированному анти-PCSK9 антителу, которое специфически связывается с PCSK9 и которое является полным антагонистом опосредованного PCSK9 действия на уровни LDLR при измерении in vitro посредством анализа подавления LDLR в клетках Huh7, как раскрыто в настоящем описании.
В еще одном варианте осуществления данное изобретение относится к изолированному антителу, которое противодействует внеклеточному взаимодействию PCSK9 с LDLR, как определено по связыванию PCSK9 с LDLR in vitro, и, при введении пациенту, снижает уровень холестерина-LDL в крови пациента. Предпочтительно данное антитело распознает эпитоп на PCSK9 человека, который перекрывается более чем с 75% поверхности PCSK9, который взаимодействует с EGF-подобным доменом LDLR, как описано у Kwon et al., 2008, PNAS, 105:1820-1825.
В еще одном варианте осуществления данное изобретение относится к антителу, которое распознает первый эпитоп на PCSK9, который перекрывается со вторым эпитом, который распознается моноклональным антителом, выбранным из группы, состоящей из 5A10, которое продуцируется гибридомной клеточной линией, депонированной в Американской Коллекции Типовых Культур и имеющей регистрационный номер PTA-8986; 4A5, которое продуцируется гибридомной клеточной линией, депонированной в Американской Коллекции Типовых Культур и имеющей регистрационный номер PTA-8985; 6F6, которое продуцируется гибридомной клеточной линией, депонированной в Американской Коллекции Типовых Культур и имеющей регистрационный номер PTA-8984, и 7D4, которое продуцируется гибридомной клеточной линией, депонированной в Американской Коллекции Типовых Культур и имеющей регистрационный номер PTA-8983.
В другом варианте осуществления данное изобретение относится к антителу PCSK9 человека, где данное антитело распознает эпитоп на PCSK9 человека, содержащей аминокислотные остатки 153-155, 194, 195, 197, 237-239, 367, 369, 374-379 и 381 аминокислотной последовательности PCSK9 последовательности SEQ ID NO: 53. Предпочтительно эпитоп для антитела на PCSK9 человека не содержит один или несколько из аминокислотных остатков 71, 72, 150-152, 187-192, 198-202, 212, 214-217, 220-226, 243, 255-258, 317, 318, 347-351, 372, 373, 380, 382 и 383.
В еще одном варианте осуществления данное изобретение относится к антителу, которое специфически связывается с PCSK9, содержащее первую определяющую комплементарность область (CDR1) VH, с аминокислотной последовательностью, представленной в SEQ ID NO: 8 (SYYMH), VH CDR2 с аминокислотной последовательностью, представленной в SEQ ID NO: 9 (EISPFGGRTNYNEKFKS), и/или VH CDR3 с аминокислотной последовательностью, представленной в SEQ ID NO: 10 (ERPLYASDL), или его вариант, имеющий одну или несколько консервативных аминокислотных замен в указанных последовательностях CDR1, CDR2 и/или CDR3, где данный вариант сохраняет по существу такую же специфичность, как и CDR, определенная указанными последовательностями. Предпочтительно данный вариант содержит приблизительно до десяти аминокислотных замен и более предпочтительно приблизительно до четырех аминокислотных замен.
Данное изобретение дополнительно нацелено на антитело, содержащее VL CDR1 с аминокислотной последовательностью, представленной в SEQ ID NO: 11 (RASQGISSALA), CDR2 с аминокислотной последовательностью, представленной в SEQ ID NO: 12 (SASYRYT), и/или CDR3 с аминокислотной последовательностью, представленной в SEQ ID NO: 13 (QQRYSLWRT), или их вариант, имеющий одну или несколько консервативных аминокислотных замен в указанных последовательностях CDR1, CDR2 и/или CDR3, где данный вариант сохраняет по существу такую же специфичность, как и CDR1, определенная указанными последовательностями. Предпочтительно данный вариант содержит приблизительно до десяти аминокислотных замен и более предпочтительно приблизительно до четырех аминокислотных замен.
В другом варианте осуществления данное изобретение относится к антителу, содержащему специфические последовательности VL CDR1, CDR2 и/или CDR3, или их вариант, имеющий одну или несколько консервативных аминокислотных замен в указанных последовательностях CDR1, CDR2 и/или CDR3, и дополнительно содержащий определяющую комплементарность область CDR1 VH с аминокислотной последовательностью, представленной в SEQ ID NO: 59, 60, или 8, VH CDR2 с аминокислотной последовательностью, представленной в SEQ ID NO: 61 или 9, и/или VH CDR3 с аминокислотной последовательностью, представленной в SEQ ID NO: 10, или их вариант, имеющий одну или несколько консервативных аминокислотных замен в указанных последовательностях CDR1, CDR2 и/или CDR3, где данный вариант сохраняет по существу такую же специфичность связывания, как и CDR1, CDR2 и/или CDR3, определенные указанными последовательностями. Предпочтительно данный вариант содержит приблизительно до двадцати аминокислотных замен и более предпочтительно приблизительно до восьми аминокислотных замен. В другом предпочтительном варианте осуществления данное антитело по данному изобретению имеет вариабельную последовательность тяжелой цепи, содержащую или состоящую из последовательности SEQ ID NO: 54, и вариабельную последовательность легкой цепи, содержащую или состоящую из последовательности SEQ ID NO: 53.
Данное изобретение также обеспечивает гуманизированное антитело, содержащее полипептиды, выбранные из групп, состоящих из SEQ ID NO: 14, SEQ ID NO: 15, или обеих последовательностей SEQ ID NO: 14 и SEQ ID NO: 15, или их варианта, имеющего одну или несколько консервативных аминокислотных замен в указанных последовательностях, где данный вариант сохраняет по существу такую же специфичность связывания, как и антитело, определенное указанной последовательностью (последовательностями). Это также включает антитело с отсутствующим концевым лизином на тяжелой цепи, поскольку он обычно теряется в части антител в процессе производства.
Предпочтительно данный вариант содержит приблизительно до двадцати аминокислотных замен и более предпочтительно приблизительно до восьми аминокислотных замен. Предпочтительно данное антитело дополнительно содержит иммунологически инертную константную область, и/или данное антитело имеет изотип, который отобран из группы, состоящей из IgG2, IgG4, IgG2∆a, IgG4∆b, IgG4∆c, IgG4 S228P, IgG4∆b S228P и IgG4∆c S228P. В другом предпочтительном варианте осуществления данная константная область представляет собой дегликозилированный Fc.
В одном варианте осуществления данное изобретение обеспечивает способ снижения уровня LDL, холестерина-LDL или общего холестерина в крови, сыворотке или плазме пациента, нуждающегося в этом, включающий введение в организм данного пациента терапевтически эффективного количества антагониста по данному изобретению.
В одном варианте осуществления данное изобретение обеспечивает терапевтически эффективное количество антагониста по данному изобретению для использования при снижении уровня LDL, холестерина-LDL или общего холестерина в крови, сыворотке или плазме пациента, нуждающегося в этом. Данное изобретение дополнительно обеспечивает применение терапевтически эффективного количества антагониста по данному изобретению при производстве лекарственного средства для снижения уровня LDL, холестерина-LDL или общего холестерина в крови, сыворотке или плазме пациента, нуждающегося в этом.
В еще одном варианте осуществления данное изобретение обеспечивает способ получения антитела, специфически связывающего PCSK9, который включает: a) обеспечение PCSK9-негативного животного-хозяина; b) иммунизацию указанного PCSK9-негативного животного-хозяина с использованием PCSK9; и c) получение антитела, антителообразующей клетки или нуклеиновой кислоты, кодирующей антитело, от указанного PCSK9-негативного животного-хозяина, и получение антитела из указанной антителообразующей клетки или указанной нуклеиновой кислоты, кодирующей антитело.
Данное изобретение также охватывает способ снижения уровня LDL в крови пациента, нуждающегося в этом, включающий введение в организм данного пациента терапевтически эффективного количества антитела, полученного в соответствии с данным изобретением. Данный пациент далее может быть подвергнут лечению посредством введения статина. В предпочтительном варианте осуществления данным пациентом является человек.
В одном варианте осуществления данное антитело применяется в композиции в виде стерильного водного раствора, имеющего значение рН в пределах от приблизительно 5,0 до приблизительно 6,5, и содержащей антитело от приблизительно 1 мг/мл до приблизительно 200 мг/мл, гистидиновый буфер от приблизительно 1 миллимолярного до приблизительно 100 миллимолярного, полисорбат 80 от приблизительно 0,01 мг/мл до приблизительно 10 мг/мл, трегалозу от приблизительно 100 миллимоль до приблизительно 400 миллимоль и двунатриевый ЭДТА дигидрат от приблизительно 0,01 миллимолярного до приблизительно 1,0 миллимолярного.
В другом варианте осуществления данное изобретение обеспечивает терапевтически эффективное количество антитела, полученного в соответствии с данным изобретением, для использования при снижении уровня LDL в крови пациента, нуждающегося в этом. Данное изобретение дополнительно обеспечивает использование терапевтически эффективного количества данного антитела, полученного в соответствии с данным изобретением, при производстве лекарственного средства для снижения уровня LDL в крови пациента, нуждающегося в этом. Данное терапевтически эффективное количество может сочетаться с терапевтически эффективным количеством статина.
В другом варианте осуществления данное изобретение обеспечивает гибридомную клеточную линию, которая продуцирует PCSK9-специфическое антитело, или его антигенсвязывающий участок, где данная гибридомная клеточная линия выбрана из группы, состоящей из:
4A5, имеющей ATCC регистрационный № PTA-8985;
5A10, имеющей ATCC регистрационный № PTA-8986;
6F6, имеющей ATCC регистрационный № PTA-8984; и
7D4, имеющей ATCC регистрационный № PTA-8983.
В другом варианте осуществления данное изобретение обеспечивает клеточную линию, которая рекомбинантно продуцирует антитело, которое специфически связывается с PCSK9 и содержит первую определяющую комплементарность область (CDR1) вариабельной области тяжелой цепи (VH), имеющую аминокислотную последовательность, представленную в SEQ ID NO: 8, 59 или 60, VH CDR2, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 9 или 61, и/или VH CDR3, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 10, или их вариант, имеющий одну или несколько консервативных аминокислотных замен в CDR1, CDR2 и/или CDR3, и/или содержит CDR1 вариабельной области легкой цепи (VL) с аминокислотной последовательностью, представленной в SEQ ID NO: 11, VL CDR2 с аминокислотной последовательностью, представленной в SEQ ID NO: 12, и/или VL CDR3 с аминокислотной последовательностью, представленной в SEQ ID NO: 13, или их вариант, имеющий одну или несколько консервативных аминокислотных замен в CDR1, CDR2 и/или CDR3. Предпочтительно данная клеточная линия рекомбинантно продуцирует антитело, содержащее последовательность SEQ ID NO: 53 и/или 54 и более предпочтительно последовательность SEQ ID NO: 14 и/или 15.
Краткое описание чертежей/фигур
На Фигуре 1 представлен эффект анти-PCSK9 антагонистических моноклональных антител 7D4.4, 4A5.G3, 6F6.G10.3 и 5A10.B8 на способность PCSK9 мыши (A) и PCSK9 человека (B) подавлять LDLR в культивируемых клетках Huh7. 6F6.G10.3 представляет собой субклон 6F6, 7D4.4 представляет собой субклон 7D4, 4A5.G3 представляет собой субклон 4A5, и 5A10.B8 представляет собой субклон 5A10.
На Фигуре 2 представлен эффект дозы анти-PCSK9 антагонистических моноклональных антител 6F6.G10.3, 7D4.4, 4A5.G3, 5A10.B8, антитела негативного контроля 42H7 и PBS на блокаду связывания рекомбинантного биотинилированного PCSK9 человека (A) и PCSK9 мыши (B) с иммобилизованным внеклеточным доменом рекомбинантного LDLR in vitro.
На Фигуре 3 представлен эффект дозы анти-PCSK9 антагонистических моноклональных антител 6F6.G10.3, 7D4.4, 4A5.G3 и 5A10.B8 на блокаду связывания рекомбинантного биотинилированного PCSK9 человека (30 нМ) с внеклеточным доменом рекомбинантного LDLR (10 нМ), меченного европием, в растворе с нейтральным значением рН in vitro.
На Фигуре 4 представлено сравнительное связывание анти-PCSK9 антител с эпитопом.
На Фигуре 5 представлены вестерн-блоты связи анти-PCSK9 антител с сывороточным PCSK9 от различных видов.
На Фигуре 6 представлен эффект анти-PCSK9 моноклонального антитела 7D4 на уровни холестерина крови у мышей.
На Фигуре 7 представлен (A) эффект частично антагонистического поликлонального анти-PCSK9 mAb CRN6 на угнетающую регуляцию LDLR и (B) отсутствие эффекта на уровни холестерина у мышей.
На Фигуре 8 изображена динамика эффекта, снижающего уровни холестерина, достигнутого применением анти-PCSK9 антагонистического антитела 7D4 у мышей.
На Фигуре 9 изображен дозозависимый эффект анти-PCSK9 антагонистического mAb 7D4 на снижение общего холестерина, HDL и LDL в сыворотке крови у мышей.
На Фигуре 10 изображен дозозависимый эффект анти-PCSK9 антагонистического антитела 5A10 на снижение уровней холестерина у мышей.
На Фигуре 11 изображен дозозависимый эффект анти-PCSK9 антагонистических антител 4A5 (А) и 6F6 (В) на снижение уровней холестерина у мышей.
На Фигуре 12 показаны вестерн-блоты действия анти-PCSK9 антагонистических антител на уровни LDLR печени.
На Фигуре 13 изображено отсутствие эффекта анти-PCSK9 антагонистического антитела 4A5 на мышиной модели LDLR-/-.
На Фигуре 14 изображен эффект на общий холестерин сыворотки крови многократных введений анти-PCSK9 антагонистических антител у мышей в течение более длительного времени, чем наблюдаемый после введения однократной дозы.
На Фигуре 15 показана динамика эффектов анти-PCSK9 антагонистического антитела 7D4 на липидные показатели в модели на человекообразных обезьянах.
На Фигуре 16 представлены эффект дозы и динамическая реакция анти-PCSK9 антагонистического антитела 7D4 на уровни холестерина сыворотки крови у человекообразных обезьян.
На Фигуре 17 представлено сравнение действия анти-PCSK9 антагонистических антител 4A5, 5A10, 6F6 и 7D4 на уровни холестерина сыворотки крови у человекообразных обезьян.
На Фигуре 18 изображена динамика эффекта анти-PCSK9 антагонистического антитела 7D4 на уровни холестерина плазмы крови у человекообразных обезьян, получавших рацион с 33,4% кКал жира, дополненный 0,1% холестерина.
На Фигуре 19 изображен эффект L1L3 (гуманизированное анти-PCSK9 моноклональное антитело) на угнетающую регуляцию LDLR в клетках Huh7.
На Фигуре 20 изображен эффект дозы гуманизированного антитела L1L3, мышиного предшественника 5A10 и антитела негативного контроля 42H7 на блокаду связывания рекомбинантного биотинилированного PCSK9 человека (A и B) и PCSK9 мыши (C и D) с иммобилизированным внеклеточным доменом рекомбинантного LDLR in vitro при pH 7,5 (A и C) и pH 5,3 (B и D).
На Фигуре 21 представлен эффект на уровни холестерина крови после обработки мышей L1L3 в дозе 10 мг/кг.
На Фигуре 22 изображен эффект введения антитела 5A10 или L1L3 человекообразным обезьянам и измерения изменений HDL сыворотки крови (A) и LDL сыворотки крови (B) в виде функции времени.
На Фигуре 23A изображена кристаллическая структура PCSK9 (светло-серые поверхности), связанного с антителом L1L3 (изображения черного цвета). На Фигуре 23B изображена кристаллическая структура PCSK9 (светло-серые поверхности), связанного с EGF-подобным доменом LDLR (изображения черного цвета) (Kwon et al., PNAS, 105, 1820-1825, 2008). На Фигуре 23C показано изображение площади поверхности PCSK9 с эпитопом для L1L3, отмеченным темно-серым цветом. На Фигуре 23D показано изображение площади поверхности PCSK9 с эпитопом для EGF-подобного домена LDLR, отмеченным темно-серым цветом.
На Фигурах 24A-G представлены замены, осуществленные в CDR антитела 5A10 в процессе созревания и оптимизации аффинности и обеспечения определенных свойств. Также представлено связывание PCSK9, ассоциированное с антителами, имеющими эти замены в CDR. Число после каждой последовательности представляет собой SEQ ID NO, присвоенное для каждой последовательности.
Подробное описание изобретения
Настоящее изобретение относится к антителам, пептидам и аптамерам, которые противодействуют функции внеклеточного PCSK9, в том числе его взаимодействию с LDLR. Конкретнее, данное изобретение относится к способам получения антагонистических антител, пептидов и аптамеров против PCSK9, композиций, содержащих эти антитела, пептиды и/или аптамеры, в качестве лекарственного средства. Антагонистические антитела и пептиды против PCSK9 можно использовать для снижения уровней холестерина-LDL крови и можно использовать для профилактики и/или лечения нарушений метаболизма холестерина и липопротеинов, в том числе семейной гиперхолестеринемии, атерогенной дислипидемии, атеросклероза и, в более общем смысле, CDV.
Общие методики
При практическом осуществлении данного изобретения будут задействованы, если не указано иначе, общепринятые методы молекулярной биологии (в том числе рекомбинантные технологии), микробиологии, клеточной биологии, биохимии и иммунологии, которые входят в число навыков в данной области. Такие технологии описаны полностью в источниках литературы, таких как Molecular Cloning: A Laboratory Manual, second edition (Sambrook et al., 1989) Cold Spring Harbor Press; Oligonucleotide Synthesis (M.J. Gait, ed., 1984); Methods in Molecular Biology, Humana Press; Cell Biology: A Laboratory Notebook (J.E. Cellis, ed., 1998) Academic Press; Animal Cell Culture (R.I. Freshney, ed., 1987); Introduction to Cell and Tissue Culture (J.P. Mather and P.E. Roberts, 1998) Plenum Press; Cell and Tissue Culture: Laboratory Procedures (A. Doyle, J. B. Griffiths, and D.G. Newell, eds., 1993-1998) J. Wiley and Sons; Methods in Enzymology (Academic Press, Inc.); Handbook of Experimental Immunology (D.M. Weir and C.C. Blackwell, eds.); Gene Transfer Vectors for Mammalian Cells (J.M. Miller and M.P. Calos, eds., 1987); Current Protocols in Molecular Biology (F.M. Ausubel et al., eds., 1987); PCR: The Polymerase Chain Reaction, (Mullis et al., eds., 1994); Current Protocols in Immunology (J.E. Coligan et al., eds., 1991); Short Protocols in Molecular Biology (Wiley and Sons, 1999); Immunobiology (C.A. Janeway and P. Travers, 1997); Antibodies (P. Finch, 1997); Antibodies: a practical approach (D. Catty., ed., IRL Press, 1988-1989); Monoclonal antibodies: a practical approach (P. Shepherd and C. Dean, eds., Oxford University Press, 2000); Using antibodies: a laboratory manual (E. Harlow and D. Lane (Cold Spring Harbor Laboratory Press, 1999); The Antibodies (M. Zanetti and J.D. Capra, eds., Harwood Academic Publishers, 1995).
Определения
«Антитело» представляет собой молекулу иммуноглобулина, способную специфически связывать мишень, такую как углевод, полинуклеотид, липид, полипептид и тому подобное, с помощью, по меньшей мере, одного участка распознавания антигена, расположенного в вариабельной области данной молекулы иммуноглобулина. Термин охватывает не только интактные поликлональные или моноклональные антитела, но также их фрагменты (такие как Fab, Fab', F(ab')2, Fv), одноцепочечные (ScFv) и доменные антитела и слитые белки, содержащие часть антитела, и любую другую модифицированную конфигурацию молекулы иммуноглобулина, которая содержит участок распознавания антигена. Антитело включает антитело любого класса, например IgG, IgA или IgM (или их подкласс), и данное антитело не должно быть определенного класса. В зависимости от антительной аминокислотной последовательности константного домена его тяжелых цепей, иммуноглобулины могут быть отнесены к различным классам. Существует пять главных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, и некоторые из них можно дополнительно разделить на подклассы (изотипы), например IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелых цепей, которые соответствуют различным классам иммуноглобулинов, называют альфа, дельта, эпсилон, гамма и мю, соответственно. Структуры субъединиц и трехмерные конфигурации различных классов иммуноглобулинов хорошо известны.
Термин «моноклональное антитело» относится к антителу, полученному из популяции по существу гомогенных антител, а именно отдельные антитела, входящие в состав популяции, являются идентичными, за исключением возможных встречающихся в природе мутаций, которые могут быть представлены в незначительных количествах. Моноклональные антитела являются высокоспецифичными, будучи направленными против единственного антигенного участка. Более того, в противоположность препаратам поликлональных антител, которые обычно включают различные антитела, направленные против различных антигенных детерминант (эпитопов), каждое моноклональное антитело направлено против единственной детерминанты на данном антигене. Определение «моноклональное» обозначает характер антитела, как получаемого из, по существу, гомогенной популяции антител, и не должно рассматриваться как требование к получению данного антитела любым определенным способом. Например, моноклональные антитела для использования в соответствии с настоящим изобретением могут быть получены посредством гибридомного способа, впервые описанного у Kohler and Milstein, 1975, Nature 256:495, или могут быть получены посредством методов рекомбинантных ДНК, таких как описанные в патенте США № 4816567. Моноклональные антитела также могут быть изолированы из фаговых библиотек, созданных с использованием технологий, описанных у McCafferty et al., 1990, Nature 348:552-554, например.
Термин «гуманизированное» антитело относится к формам не-человеческих (например, мышиных) антител, которые представляют собой химерные иммуноглобулины, иммуноглобулиновые цепи, или их фрагменты (такие как Fv, Fab, Fab', F(ab')2 или другие антигенсвязывающие подпоследовательности антител), которые содержат минимальную последовательность, полученную из иммуноглобулина, отличного от человеческого. Предпочтительно гуманизированные антитела представляют собой иммуноглобулины человека (антитело-реципиент), в котором остатки из определяющей комплементарность области (CDR) реципиента замещены остатками из CDR видов, не являющихся человеком (антитело-донор), таких как мышь, крыса или кролик, имеющие требуемую специфичность, аффинность и емкость. В некоторых случаях остатки каркасной области Fv (FR) иммуноглобулина человека замещены соответствующими остатками, не происходящими от человека. Более того, гуманизированное антитело может содержать остатки, которые не обнаружены ни в антителе-реципиенте, ни в импортированных CDR или каркасных последовательностях, но включены для дополнительного очищения и оптимизации действия антитела. В большинстве случаев гуманизированное антитело будет содержать по существу целый по меньшей мере один и обычно два вариабельных домена, в которых все или в основном все области CDR соответствуют таковым иммуноглобулина, отличного от человеческого, и все или в основном все области FR представляют собой таковые из консенсусной последовательности иммуноглобулина человека. Оптимально данное гуманизированное антитело также будет содержать, по меньшей мере, часть константной области или домена (Fc) иммуноглобулина, обычно таковой иммуноглобулина человека. Предпочтительными являются антитела, имеющие области Fc, модифицированные как описано в WO 99/58572. Другие формы гуманизированных антител имеют одну или несколько CDR (CDR L1, CDR L2, CDR L3, CDR H1, CDR H2 и/или CDR H3), которые изменены по отношению к исходному антителу, которые также называют один или несколько «полученные из» одной или нескольких CDR из исходного антитела.
Термин «антитело человека» означает антитело, имеющее аминокислотную последовательность, соответствующую таковой антитела, которое вырабатывается у человека и/или которое было получено с использованием любой из технологий для изготовления антител человека, известных специалисту в данной области, или раскрытых здесь. Это определение антитела человека включает антитела, содержащие по меньшей мере один полипептид тяжелой цепи человека или по меньшей мере один полипептид легкой цепи человека. Одним таким примером является антитело, содержащее полипептиды легкой цепи мыши и тяжелой цепи человека. Антитела человека можно получить с помощью различных технологий, известных в уровне техники. В одном варианте осуществления антитело человека отобрано из фаговой библиотеки, где эта фаговая библиотека экспрессирует антитела человека (Vaughan et al., 1996, Nature Biotechnology, 14:309-314; Sheets et al., 1998, Proc. Natl. Acad. Sci. (USA) 95:6157-6162; Hoogenboom and Winter, 1991, J. Mol. Biol., 227:381; Marks et al., 1991, J. Mol. Biol., 222:581). Антитела человека также можно получить в результате иммунизации животных, которым локус иммуноглобулина человека был трансгенно введен в местоположение эндогенного локуса, например мышей, у которых гены эндогенного иммуноглобулина были частично или полностью инактивированы. Такой подход описан в патентах США №№ 5545807; 5545806; 5569825; 5625126; 5633425; и 5661016. Альтернативно антитело человека может быть получено с помощью иммортализованных В лимфоцитов человека, которые вырабатывают антитело, направленное против антигена-мишени (такие В лимфоциты могут быть получены от индивидуума или могут быть иммунизированы in vitro). Смотри, например, Cole et al. Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p. 77, 1985; Boerner et al., 1991, J. Immunol., 147 (1):86-95; и патент США № 5750373.
Термин «вариабельная область» обозначает вариабельную область легкой цепи антитела или вариабельную область тяжелой цепи антитела, либо по отдельности, либо в сочетании. Как известно в уровне техники, вариабельные области тяжелой цепи и легкой цепи каждая состоит из четырех каркасных областей (FR), соединенных посредством трех определяющих комплементарность областей (CDR), которые содержат гипервариабельные участки. Данные CDR в каждой цепи скреплены в непосредственной близости с помощью данных FR и, с данными CDR из другой цепи, способствуя образованию антигенсвязывающих центров антител. Существуют по меньшей мере два способа CDR: (1) метод, основанный на межвидовой вариативности последовательностей (а именно Kabat et al., Sequences of Proteins of Immunological Interest, (5th ed., 1991, National Institutes of Health, Bethesda MD)); и (2) метод, основанный на кристаллографических исследованиях комплексов антитело-антиген (Al-Iazikani et al, 1997, J. Molec. Biol. 273:927-948). Как используется здесь, CDR можно отнести к CDR, определенным либо с помощью одного метода, либо с помощью комбинации обоих методов.
Как известно в уровне техники, «константная область» антитела относится к константной области легкой цепи антитела, или константной области тяжелой цепи антитела, либо по отдельности, либо в сочетании.
Термин «PCSK9» относится к любой форме PCSK9 и его вариантам, которые сохраняют, по меньшей мере, частичную активность PCSK9. Если не указано иначе, например, посредством определенной ссылки на PCSK9 человека, PCSK9 включает нативные последовательности PCSK9 всех видов млекопитающих, например человека, собачьих, кошачьих, лошадиных и бычьих. Один пример PCSK9 человека обнаружен в банке данных Uniprot с номером доступа Q8NBP7 (SEQ ID NO:16).
Термин «антагонист PCSK9» относится к антителу, пептиду или аптамеру, способному ингибировать биологическую активность PCSK9 и/или подавлять каскад(ы) реакций, опосредованный сигнальным путем PCSK9, в том числе PCSK9-опосредованную угнетающую регуляцию LDLR, и опосредованное PCSK9 понижение клиренса LDL крови. Антагонистическое антитело против PCSK9 охватывает антитела, которые блокируют, противодействуют, супрессируют или уменьшают (до любой степени, в том числе значительной) биологическую активность PCSK9, в том числе нисходящий метаболический путь, опосредованный сигнальным путем PCSK9, например взаимодействие с LDLR и/или выявление клеточного ответа на PCSK9. Для целевого назначения настоящего изобретения будет отчетливо понятно, что термин «антагонистическое антитело против PCSK9» охватывает все ранее определенные термины, названия и функциональные формы и характеристики, в силу чего сам PCSK9, биологическая активность PCSK9 (в том числе, но не ограничиваясь его способностью опосредовать любой аспект его взаимодействия с LDLR, угнетающей регуляцией LDLR и снижением клиренса LDLR крови) или проявления биологической активности, по существу, аннулированы, снижены или нейтрализованы в любой значимой степени. В некоторых вариантах осуществления, антагонистическое антитело против PCSK9 связывает PCSK9 и предотвращает взаимодействие с LDLR. Здесь обеспечены примеры антагонистических антител против PCSK9.
«Полный антагонист» представляет собой антагонист, который, при эффективной концентрации, по существу, полностью блокирует поддающийся измерению эффект PCSK9. Под частичным антагонистом понимают антагонист, который способен частично блокировать поддающийся измерению эффект, но который, даже при наибольшей концентрации, не является полным антагонистом. Под, по существу, полным подразумевают, что поддающийся измерению эффект заблокирован по меньшей мере приблизительно на 80%, предпочтительно по меньшей мере приблизительно на 90%, более предпочтительно по меньшей мере приблизительно на 95% и наиболее предпочтительно по меньшей мере приблизительно на 98% или 99%. Соответствующие «поддающиеся измерению эффекты» описаны здесь и включают угнетающую регуляцию LDLR посредством антагониста PCSK9, как испытано на клетках Huh7 in vitro, снижение уровней общего холестерина в крови (или плазме) in vivo и снижение уровней LDL в крови (или плазме) in vivo.
Термин «клинически значимый» означает, по меньшей мере, снижение на 15% уровней LDL-холестерина в крови у человека или, по меньшей мере, снижение на 15% уровней общего холестерина крови у мышей. Очевидно, что измерения в плазме или сыворотке могут служить в качестве заменителей измерения уровней в крови.
Термин «антагонистический пептид против PCSK9» или «антагонистический аптамер против PCSK9» включает любой общеупотребительный полипептид или аптамер, который блокирует, противодействует, супрессирует или уменьшает (до любой степени, в том числе значительной) биологическую активность PCSK9, в том числе нисходящий метаболический путь, опосредованный сигнальным путем PCSK9, например взаимодействие с LDLR и/или выявление клеточного ответа на PCSK9. Антагонистические пептиды или полипептиды против PCSK9 включают слияния Fc, содержащие LDLR и растворимые части LDLR, или их мутации с более высокой аффинностью к PCSK9.
Термины «полипептид», «олигопептид», «пептид» и «белок» используются здесь взаимозаменяемо для обозначения цепей аминокислот любой длины, предпочтительно относительно коротких (например, 10-100 аминокислот). Данная цепь может быть линейной или разветвленной, может содержать модифицированные аминокислоты, и/или может прерываться не-аминокислотами. Данные термины также включают аминокислотную цепь, которая была модифицирована естественным образом или в результате вмешательства, например образования дисульфидной связи, гликозилирования, липидации, ацетилирования, фосфорилирования, или любой другой манипуляции или модификации, такой как конъюгирование с компонентом-меткой. Также в данное определение включены, например, полипептиды, содержащие один или несколько аналогов аминокислоты (в том числе, например, неестественные аминокислоты и тому подобное), а также другие модификации, известные в уровне техники. Подразумевается, что полипептиды могут встречаться в виде одиночных цепей или в виде объединенных цепей.
Как известно в уровне техники, термины «полинуклеотид» или «нуклеиновая кислота», используемые здесь взаимозаменяемо, относятся к цепям нуклеотидов любой длины и включают ДНК и РНК. Нуклеотиды могут представлять собой дезоксирибонуклеотиды, рибонуклеотиды, модифицированные нуклеотиды или основания, и/или их аналоги, или любые субстраты, которые могут быть встроены в цепь с помощью ДНК- или РНК-полимеразы. Полинуклеотид может содержать модифицированные нуклеотиды, такие как метилированные нуклеотиды и их аналоги. Если присутствует, изменение структуры может быть передано до или после сборки данной цепи. Последовательность нуклеотидов может быть прервана ненуклеотидными компонентами. Полинуклеотид может быть далее изменен после полимеризации, например, с помощью конъюгирования с компонентом-меткой. Модификациями других типов являются, например, «кэпы», замена одного или нескольких природных нуклеотидов их аналогами, межнуклеотидные модификации, такие как, например, модификации путем введения незаряженных соединений (например, метилфосфонатов, фосфотриэфиров, фосфоамидатов, карбаматов и тому подобного) и заряженных соединений (например, фосфортиоатов, фосфордитиоатов и тому подобного), модификации, содержащие боковые группы, такие как, например, белки (например, нуклеазы, токсины, антитела, сигнальные пептиды, поли-L-лизин, и тому подобное), модификации, содержащие интеркалирующие агенты (например, акридин, псорален и тому подобное), модификации, содержащие хелатообразующие агенты (например, металлы, радиоактивные металлы, бор, металлы-окислители и тому подобное), модификации, содержащие алкилирующие агенты, модификации, содержащие модифицированные соединения (например, альфа-аномерные нуклеиновые кислоты и т.п.), а также немодифицированные формы полинуклеотида(ов). Кроме того, любые из гидроксильных групп, изначально присутствующих, могут быть заменены, например, фосфонатными группами, фосфатными группами, защищены стандартными защитными группами, или активированы для получения дополнительных соединений с дополнительными нуклеотидами, или могут быть конъюгированы с твердофазной подложкой. 5' и 3' концевая OH-группа может быть фосфорилирована или заменена на амины или мотивы органических блокирующих групп длиной от 1 до 20 атомов углерода. Другие гидроксильные группы также могут быть дериватизированы стандартными защитными группами. Полинуклеотиды также могут содержать функционально подобные формы сахаров рибоза или дезоксирибоза, которые общеизвестны в уровне техники, в том числе, например, 2'-O-метил-, 2'-O-аллил, 2'-фтор- или 2'-азидо-рибоза, аналоги карбоциклических сахаров, альфа- или бета-аномерные сахара, эпимерные сахара, такие как арабиноза, ксилоза или ликсоза, пиранозные сахара, фуранозные сахара, седогептулозы, ациклические аналоги и аналоги абазических нуклеозидов, такие как метилрибозид. Один или несколько фосфодиэфиров могут быть замещены альтернативными линкерными группами. Эти альтернативные линкерные группы включают, но не ограничиваются ими, варианты осуществления, в которых фосфат заменен на P(O)S («тиоат»), P(S)S («дитиоат»), (O)NR2 («амидат»), P(O)R, P(O)OR', CO или CH2 («формацеталь»), в котором каждый R или R' представляет собой независимо H, или замещенный или незамещенный алкил (1-20 C), необязательно содержащий либо (-O-) соединение, арил, алкенил, циклоалкил, циклоалкенил или аралдил. Не все связи в полинуклеотиде должны быть идентичными. Вышеприведенное описание относится ко всем используемым здесь полинуклеотидам, включая РНК и ДНК.
«Аптамер-антагонист против PCSK9», который содержит нуклеиновую кислоту или белковую последовательность, например, отобран из большого банка случайных последовательностей и специфически связывается с PCSK9. Нуклеиновая кислота данного аптамера представляет собой двухцепочечную ДНК или одноцепочечную РНК. Аптамеры, состоящие из нуклеиновой кислоты, могут включать модифицированные основания или функциональные группы, в том числе, но не ограничиваясь ими, 2'-фторсодержащие нуклеотиды и 2'-O-метил нуклеотиды. Аптамеры могут включать гидрофильные полимеры, например полиэтиленгликоль. Аптамеры могут быть получены способами, известными в уровне техники и выбранными для антагонистического действия против PCSK9 в результате рутинной модификации способов, раскрытых в Примерах.
Антитело, пептид или аптамер «взаимодействуют с» PCSK9 в случае, когда равновесная константа диссоциации равна или меньше 20 нМ, предпочтительно менее чем приблизительно 6 нМ, более предпочтительно менее чем приблизительно 1 нМ, наиболее предпочтительно менее чем приблизительно 0,2 нМ, как измерено с использованием методов, раскрытых в Примере 2.
Эпитоп, который «предпочтительно связывает» или «специфически связывает» (используются здесь взаимозаменяемо) антитело или пептид, представляет собой хорошо изученное в уровне техники понятие, и способы определения таких специфических или предпочтительных связей также хорошо известны в уровне техники. Молекула, как говорят, проявляет «специфическое связывание» или «предпочтительное связывание», если она реагирует или соединяется более часто, более быстро, с более длительной продолжительностью и/или с большей аффинностью с определенной клеткой или субстанцией, чем с альтернативными клетками или субстанциями. Антитело «специфически связывается» или «предпочтительно связывается» с мишенью, если оно связывается с большей аффинностью, авидностью, более легко и/или более длительно, чем с другими субстанциями. Например, антитело, которое специфически или предпочтительно связывается с эпитопом на PCSK9, представляет собой антитело, которое связывается с этим эпитопом с большей аффинностью, авидностью, более легко и/или более продолжительно, чем оно связывается с другими эпитопами на PCSK9 или не-PCSK9 эпитопами. В результате прочтения этого определения также понятно, что, например, антитело (или мотив, или эпитоп), которое специфически или предпочтительно связывается с первой мишенью, может специфически или предпочтительно связываться, или может не связываться, со второй мишенью. По существу, «специфическое связывание» или «предпочтительное связывание» необязательно подразумевает (хотя может включать) эксклюзивное связывание. В большинстве случаев, но необязательно, ссылка на связывание подразумевает предпочтительное связывание.
Термин «по существу чистое» относится к веществу, которое по меньшей мере на 50% чистое (то есть не содержит примесей), более предпочтительно по меньшей мере на 90% чистое, более предпочтительно по меньшей мере на 95% чистое, еще более предпочтительно по меньшей мере на 98% чистое и наиболее предпочтительно по меньшей мере на 99% чистое.
Термин «клетка-хозяин» включает отдельную клетку или клеточную культуру, которая может быть или была реципиентом для вектора (векторов) для встраивания полинуклеотидных вставок. Клетки-хозяева включают потомство единственной клетки-хозяина, и данное потомство может необязательно быть полностью идентичным (по морфологии или по комплементу геномной ДНК) исходной родительской клетке по причине естественной, случайной или преднамеренной мутации. Термин клетка-хозяин включает клетки, трансфицированные in vivo с помощью полинуклеотида (полинуклеотидов) по этому изобретению.
Как известно в уровне техники, термин «Fc-область» используется для обозначения С-концевой области тяжелой цепи иммуноглобулина. «Fc-область» может представлять собой природную последовательность Fc-области или варианта Fc-области. Несмотря на то что границы Fc-области тяжелой цепи иммуноглобулина могут варьироваться, Fc-область тяжелой цепи IgG человека обычно определена участком от аминокислотного остатка в положении Cys226, или от Pro230, до его концевой карбоксильной группы. Нумерация остатков в Fc-области соответствует таковому в индексном указателе EU, как у Кабата (Kabat). Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md., 1991. Fc-область иммуноглобулина обычно содержит два константных домена, CH2 и CH3.
Используемый в уровне техники термин «Fc рецептор» и «FcR» описывает рецептор, который связывается с Fc-областью антитела. Предпочтительный FcR представляет собой природную последовательность FcR человека. Более того, предпочтительным FcR является тот, который связывает антитело IgG (гамма-рецептор) и включает рецепторы подклассов FcγRI, FcγRII и FcγRIII, в том числе аллельные варианты и формы альтернативного сплайсинга этих рецепторов. Рецепторы FcγRII включают FcγRIIA («активирующий рецептор») и FcγRIIB («ингибирующий рецептор»), которые имеют схожие аминокислотные последовательности, отличающиеся, главным образом, своими цитоплазматическими доменами. Обзор FcR представлен у Ravetch and Kinet, 1991, Ann. Rev. Immunol., 9:457-92; Capel et al., 1994, Immunomethods, 4:25-34; и de Haas et al., 1995, J. Lab. Clin. Med., 126:330-41. «FcR» также включает неонатальный рецептор, FcRn, ответственный за перенос материнских IgG плоду (Guyer et al., 1976 J. Immunol., 117:587; и Kim et al., 1994, J. Immunol., 24:249).
Термин «конкурировать», используемый здесь по отношению к антителу, означает, что первое антитело или его антиген-связывающий участок связывается с эпитопом в некотором смысле довольно одинаково со связыванием второго антитела, или его антигенсвязывающего участка, таким образом, результат связывания первого антитела с его сопряженным эпитопом заметно снижен в присутствии второго антитела, по сравнению со связыванием первого антитела в отсутствие второго антитела. Альтернатива, при которой связывание данного второго антитела с его эпитопом также заметно снижено в присутствии данного первого антитела, может иметь место, но необязательно. Таким образом, первое антитело может ингибировать связывание второго антитела с его эпитопом, без ингибирования данным вторым антителом связывания данного первого антитела с его соответствующим эпитопом. Тем не менее, в случае, когда каждое антитело значительно ингибирует связывание другого антитела с его сопряженным эпитопом или лигандом, либо в такой же, большей или меньшей степени, данные антитела называют «перекрестно-конкурирующими» друг с другом за связывание их соответствующего эпитопа (эпитопов). И конкурирующие, и перекрестно-конкурирующие антитела охвачены настоящим изобретением. Вне зависимости от механизма возникновения такой конкуренции или перекрестной конкуренции (например, стерическое несоответствие, конформационное изменение или связывание с общим эпитопом или его частью), специалист в данной области примет во внимание, основываясь на предоставленных здесь сведениях, что такие конкурирующие и/или перекрестно-конкурирующие антитела охвачены и могут быть использованы в способах, раскрытых здесь.
Под антителом с эпитопом, который «перекрывается» с другим (вторым) эпитом, или с поверхностью на PCSK9, которая взаимодействует с EGF-подобным доменом на LDLR, подразумевают совместное использование области относительно остатков PCSK9, с которыми происходит взаимодействие. Для подсчета процента области перекрывания, например процента области перекрывания эпитопа на PCSK9 для заявленного антитела с поверхностью PCSK9, которая взаимодействует с EGF-подобным доменом на LDLR, площадь поверхности PCSK9 скрыта, когда в комплексе с LDLR расчет произведен на основе остатков. Данная скрытая поверхность также рассчитана для этих остатков в комплексе PCSK9:антитело. Для предотвращения более чем 100% возможного перекрывания площадь поверхности для остатков, которые имеют более обширную скрытую площадь поверхности в комплексе PCSK9:антитело, чем в комплексе LDLR:PCSK9, установлена до значений комплекса LDLR:PCSK9 (100%). Процент площади перекрывания рассчитывается посредством суммирования всех взаимодействующих остатков LDLR:PCSK9 и взвешена по области взаимодействия.
«Функциональная область Fc» обладает, по меньшей мере, одной эффекторной функцией нативной последовательности Fc-области. Примеры «эффекторных функций» включают связывание C1q; комплементзависимую цитотоксичность; связывание рецептора Fc; антителозависимую клеточноопосредованную цитотоксичность; фагоцитоз; угнетающую регуляцию рецепторов клеточной поверхности (например, рецептора В-клетки) и тому подобное. Такие эффекторные функции в большинстве случаев требуют комбинации Fc-области с доменом связывания (например, вариабельный домен антитела) и могут быть оценены с использованием различных исследований, известных в уровне техники для оценки таких эффекторных функций антитела.
«Нативная последовательность Fc-области» содержит аминокислотную последовательность, идентичную аминокислотной последовательности Fc-области, обнаруженной в природе. «Вариант Fc-области» содержит аминокислотную последовательность, которая отличается от нативной последовательности Fc-области по меньшей мере одной аминокислотной модификацией, несмотря на это сохраняет по меньшей мере одну эффекторную функцию нативной последовательности Fc-области. Предпочтительно данный вариант Fc-области имеет по меньшей мере одну аминокислотную замену по сравнению с нативной последовательностью Fc-области или Fc-области родительского полипептида, например, приблизительно от одной и приблизительно до десяти аминокислотных замен и предпочтительно приблизительно от одной и приблизительно до пяти аминокислотных замен в нативной последовательности Fc-области, или в Fc-области родительского полипептида. Вариант Fc-области при этом будет предпочтительно обладать по меньшей мере 80% идентичностью с нативной последовательностью Fc-области и/или с Fc-областью родительского полипептида и наиболее предпочтительно по меньшей мере 90% идентичностью последовательности с этим, более предпочтительно по меньшей мере приблизительно 95%, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98%, по меньшей мере приблизительно 99% идентичностью последовательности с этим.
«Лечение» представляет собой способ получения благоприятных или желательных клинических результатов. В целях настоящего изобретения благоприятные или желательные клинические результаты включают, но не ограничиваются ими, одно или несколько из следующего: увеличение клиренса LDL и уменьшение уровня заболеваемости или уменьшение интенсивности абберантных уровней холестерина и/или липопротеинов в результате нарушения метаболизма и/или нарушений питания, или в том числе семейной гиперхолестеринемии, атерогенной дислипидемии, атеросклероза и, в более общем смысле, сердечно-сосудистого заболевания (CDV).
«Уменьшение уровня заболеваемости» означает уменьшение тяжести, что может включать снижение потребности в, и/или количества (например, воздействия) других лекарственных средств и/или способов лечения, обычно используемых для данного состояния. Как понятно специалисту в данной области, индивидуумы могут различаться по их реакции на лечение, и, по существу, например, «метод уменьшения уровня заболеваемости» отображает применение антагонистического антитела, пептида или аптамера против PCSK9 на основании обоснованного ожидания, что такое применение может, вероятно, быть причиной такого уменьшения заболеваемости у данного конкретного индивидуума.
«Улучшение» означает снижение интенсивности или положительную динамику одного или нескольких симптомов, по сравнению с отсутствием применения антагонистического антитела, пептида или аптамера против PCSK9. «Улучшение» также включает укорочение или снижение продолжительности симптома.
«Эффективная доза» или «эффективное количество» лекарственного средства, соединения или фармацевтической композиции представляет собой количество, достаточное для достижения одного или нескольких благоприятных или желательных результатов. Для профилактического применения благоприятные или желательные результаты включают устранение или снижение риска, уменьшение степени тяжести, или отсрочку манифестации заболевания, включая биохимические, гистологические и/или поведенческие симптомы данного заболевания, его осложнения и промежуточные патологические фенотипы, определяемые во время развития заболевания. Для терапевтического применения благоприятные или желательные результаты включают клинические результаты, такие как уменьшение гиперхолестеринемии или одного или нескольких симптомов дислипидемии, атеросклероза, CDV, или ишемической болезни сердца, снижение дозы других лекарственных средств, необходимых для лечения данного заболевания, повышение эффекта другого лекарственного средства, и/или отсрочка прогрессирования заболевания у пациентов. Эффективная доза может быть введена в организм в одном или нескольких введениях. В целях этого изобретения эффективная доза лекарственного средства, соединения или фармацевтической композиции представляет собой количество, достаточное для осуществления профилактического или терапевтического лечения либо напрямую, либо косвенно. Как понятно в клиническом смысле, эффективная доза лекарственного средства, соединения или фармацевтической композиции может быть, а может и не быть обеспечена в сочетании с другим лекарственным средством, соединением или фармацевтической композицией. Таким образом, «эффективная доза» может быть рассмотрена в контексте применения одного или нескольких терапевтических средств, и единственный терапевтический агент можно рассматривать, как введенный в эффективном количестве, если, в сочетании с одним или несколькими другими средствами, желательный результат достигнут или может быть достигнут.
«Индивидуумом» или «пациентом» является млекопитающее, более предпочтительно человек. Млекопитающие также включают, но не ограничиваются ими, сельскохозяйственных животных, животных-участников спортивных соревнований, домашних питомцев, приматов, лошадей, собак, кошек, мышей и крыс.
Термин «вектор» означает конструкцию, которая способна доставлять, и, предпочтительно, экспрессировать один или несколько представляющих интерес генов или последовательностей в клетке-хозяине. Примеры векторов включают, но не ограничиваются ими, вирусные векторы, векторы, экспрессирующие оголенную ДНК или РНК, плазмиды, космиды или фаговые векторы, векторы, экспрессирующие ДНК или РНК, связанные с катионными конденсирующими агентами, векторы, экспрессирующие ДНК или РНК, инкапсулированные в липосомах, и некоторые эукариотические клетки, такие как клетки-продуценты.
Термин «последовательность, управляющая экспрессией» означает последовательность нуклеиновой кислоты, которая контролирует транскрипцию нуклеиновой кислоты. Последовательностью, управляющей экспрессией, может быть промотор, такой как конститутивный или индуцибельный промотор, или энхансер. Последовательность, контролирующая экспрессию, функционально связана с транскрибируемой последовательностью нуклеиновой кислоты.
Термин «фармацевтически приемлемый носитель» или «фармацевтически приемлемый наполнитель» включает любое вещество, которое, при объединении с активным ингредиентом, позволяет данному ингредиенту сохранить биологическую активность и является нереактивноспособным с иммунной системой пациента. Примеры включают, но не ограничиваются ими, любые стандартные фармацевтические носители, такие как забуференный фосфатом физиологический раствор, воду, эмульсии, такие как эмульсия типа вода в масле, и различные типы увлажняющих средств. Предпочтительными разбавителями для аэрозольного или парентерального введения являются забуференный фосфатом физиологический раствор (PBS) или нормальный физиологический раствор (0,9%). Композиции, содержащие такие носители, разрабатываются с помощью хорошо известных общепринятых методов (смотри, например, Remington's Pharmaceutical Sciences, 18th edition, A. Gennaro, ed., Mack Publishing Co., Easton, PA, 1990; и Remington, The Science and Practice of Pharmacy, 20th Ed., Mack Publishing, 2000).
Термин «kon» относится к константе скорости ассоциации антитела с антигеном. В частности, константы скорости (kon и koff) и равновесные константы диссоциации измеряются с использованием фрагментов антител Fab (а именно одновалентных) и PCSK9.
Термин «koff» относится к константе скорости диссоциации антитела из комплекса антитело/антиген.
Термин «KD» относится к равновесной константе диссоциации взаимодействия антитело-антиген.
A. Способы профилактики или лечения заболеваний, связанных с гиперхолестеринемией
В одном аспекте данное изобретение относится к способу лечения или профилактики гиперхолестеринемии, и/или, по меньшей мере, одного симптома дислипидемии, атеросклероза, CDV или ишемической болезни сердца у индивидуума, включающий введение в организм данного индивидуума эффективного количества антагонистического антитела или пептида или аптамера против PCSK9, противодействующего циркулирующему PCSK9.
В дополнительном аспекте данное изобретение обеспечивает эффективное количество антагонистического антитела, пептида или аптамера против PCSK9, противодействующего циркулирующему PCSK9, для использования при лечении или профилактике гиперхолестеринемии, и/или по меньшей мере одного симптома дислипидемии, атеросклероза, CDV или ишемической болезни сердца у индивидуума. Данное изобретение дополнительно обеспечивает использование эффективного количества антагонистического антитела, пептида или аптамера против PCSK9, противодействующего внеклеточному или циркулирующему PCSK9, при промышленном производстве лекарственного средства для лечения или профилактики гиперхолестеринемии и/или по меньшей мере одного симптома дислипидемии, атеросклероза, CDV или ишемической болезни серца у индивидуума.
Предпочтительно терапевтическое применение антитела, пептида или аптамера приводит в результате к снижению холестерина крови и/или снижению LDL крови. Предпочтительно холестерин крови и/или LDL крови по меньшей мере приблизительно на 10% или 15% ниже, чем до введения. Более предпочтительно холестерин крови и/или LDL крови по меньшей мере приблизительно на 20% ниже, чем до введения антитела. Даже более предпочтительно холестерин крови и/или LDL крови по меньшей мере приблизительно на 30% ниже, чем до введения антитела. Выгодно, если холестерин крови и/или LDL крови по меньшей мере приблизительно на 40% ниже, чем до введения антитела. Более выгодно, если холестерин крови и/или LDL крови по меньшей мере приблизительно на 50% ниже, чем до введения антитела. Очень предпочтительно, если холестерин крови и/или LDL крови по меньшей мере приблизительно на 60% ниже, чем до введения антитела. Наиболее предпочтительно, если холестерин крови и/или LDL крови по меньшей мере приблизительно на 70% ниже, чем до введения антитела.
По отношению ко всем методам, описанным здесь, ссылка на антагонистическое антитело, пептид или аптамер против PCSK9 также включает композиции, содержащие одно или несколько дополнительных веществ. Эти композиции могут дополнительно содержать подходящие инертные наполнители, такие как фармацевтически приемлемые наполнители, в том числе буферы, которые хорошо известны в уровне техники. Настоящее изобретение можно применять самостоятельно или в сочетании с другими общепринятыми способами лечения.
Антагонистическое антитело, пептид или аптамер против PCSK9 можно вводить в организм индивидуума посредством любого подходящего пути введения. Специалисту в данной области должно быть очевидно, что примеры, описанные здесь, предназначены не для ограничения, а для иллюстрации доступных способов. Соответственно в некоторых вариантах осуществления антагонистическое антитело пептид или аптамер против PCSK9 вводится в организм индивидуума в соответствии с известными способами, такими как внутривенное введение, например, в виде болюса или посредством продолжительной инфузии за какой-то период времени, внутримышечно, внутрибрюшинно, внутрь спинного мозга, чрескожно, подкожно, интраартикулярно, сублингвально, внутрисуставно, посредством инсуффляции, подоболочечно, перорально, посредством ингаляции или местных путей введения. Применение может быть системным, например внутривенное введение, или местным. Эффективными для применения являются коммерчески доступные ингаляторы для жидких композиций, в том числе эжекторные ингаляторы и ультразвуковые ингаляторы. Жидкие композиции могут быть непосредственно введены посредством ингаляции, и лиофилизированный порошок может быть введен распылением после восстановления. Альтернативно антагонистические антитела, пептиды или аптамеры против PCSK9 могут быть аэролизованы с использованием фторуглеродной композиции и дозирующего ингалятора или ингалированы в виде лиофилизированного и измельченного порошка.
В одном варианте осуществления антагонистическое антитело, пептид или аптамер против PCSK9 применяется с помощью сайт-специфических технологий или технологий целевой местной доставки. Примеры сайт-специфических технологий или технологий целевой местной доставки включают различные имплантируемые депо-источники антагонистического антитела, пептида или аптамера против PCSK9, или катетеры для местной доставки, такие как катетеры для инфузии, постоянные катетеры, или игольчатые катетеры, синтетические импланты, адвентициальные манжеты, шунты и стенты, или другие имплантируемые устройства, сайт-специфические носители, прямое впрыскивание или направленное введение. Смотри, например, PCT публикацию № WO 00/53211 и патент США № 5981568.
Различные композиции антагонистического антитела, пептида или аптамера против PCSK9 можно использовать для введения. В некоторых вариантах осуществления может быть введено беспримесное антагонистическое антитело, пептид или аптамер против PCSK9. В некоторых вариантах осуществления антагонистическое антитело, пептид или аптамер против PCSK9 и фармацевтически приемлемый наполнитель могут быть в различных композициях. Фармацевтически приемлемые наполнители известны в уровне техники, и они являются относительно интертными веществами, которые облегчают введение фармацевтически эффективных веществ. Например, наполнитель может придавать форму или консистенцию или действовать в качестве разбавителя. Подходящие наполнители включают, но не ограничиваются ими, стабилизирующие вещества, увлажняющие или эмульгирующие вещества, соли для изменения осмолярности, инкапсулирующие вещества, буферы и вещества, усиливающие проникновение через кожу. Наполнители, а также композиции для парентеральной и непарентеральной доставки лекарственного средства описаны у Remington, The Science and Practice of Pharmacy, 20th Ed., Mack Publishing (2000).
Эти агенты можно скомбинировать с фармацевтически приемлемыми носителями, такими как физиологический раствор, раствор Рингера, раствор декстрозы и тому подобное. Конкретный режим дозирования, то есть доза, момент времени и повторяемость, будет зависеть от конкретного индивидуума и анамнеза заболевания данного индивидуума.
Антитела против PCSK9 также можно вводить посредством ингаляции, как описано здесь. В большинстве случаев для применения антител против PCSK9, первичная возможная доза может составлять приблизительно 2 мг/кг. В контексте настоящего изобретения типичная дневная доза может находиться в диапазоне от приблизительно 3 мкг/кг до приблизительно 30 мкг/кг, до 300 мкг/кг, до 3 мг/кг, до 30 мг/кг, до 100 мг/кг или более, в зависимости от вышеупомянутых факторов. Например, можно применять дозу приблизительно 1 мг/кг, приблизительно 2,5 мг/кг, приблизительно 5 мг/кг, приблизительно 10 мг/кг и приблизительно 25 мг/кг. Для повторных введений на протяжении нескольких дней или более, в зависимости от состояния, лечение продолжается до тех пор, пока не произойдет желаемая супрессия симптомов, или до тех пор, пока не будут достигнуты удовлетворительные терапевтические уровни, например, для уменьшения уровней LDL крови. Примерный режим дозировки включает введение первичной дозы приблизительно 2 мг/кг, с последующей еженедельной поддерживающей дозой антитела против PCSK9 приблизительно 1 мг/кг, или с последующей дозой приблизительно 1 мг/кг раз в две недели. Однако можно применять другие режимы дозировки, в зависимости от характера фармакокинетического распада. Например, в некоторых вариантах осуществления предусмотрено введение дозы от одного до четырех раз в неделю. В других вариантах осуществления предусмотрено введение дозы один раз месяц, или один раз в два месяца, или один раз в три месяца. Прогресс такого лечения без труда отслеживается с использованием общепринятых методов и анализов. Режим дозирования (в том числе используемого антагониста(ов) PCSK9) может варьироваться со временем.
В контексте настоящего изобретения адекватная доза антагонистического антитела, пептида или аптамера против PCSK9 будет зависеть от применяемого антагонистического антитела, пептида или аптамера против PCSK9 (или их комбинаций), типа и тяжести подвергающихся лечению симптомов, применяется ли данное вещество в профилактических или терапевтических целях, предшествующей терапии, клинического анамнеза пациента и реакции на данное вещество, уровней PCSK9 крови у пациента, скорости синтеза и клиренса PCSK9 у пациента, скорости клиренса вводимого вещества у пациента и рассудительности лечащего врача. Обычно врач будет назначать антагонистическое антитело, пептид или аптамер против PCSK9 до тех пор, пока не достигнет дозы, при которой получают желаемый результат. Доза и/или частота могут изменяться во время курса лечения. Эмпирические расчеты, такие как время полужизни, как правило, способствуют определению дозы. Например, антитела, которые совместимы с иммунной системой человека, такие как гуманизированные антитела или полностью человеческие антитела, можно использовать для удлинения времени полужизни антитела и предотвращения атаки на антитело со стороны иммунной системы организма-хозяина. Частота введения может быть определена и адаптирована на протяжении курса лечения, и, как правило, но необязательно, исходя из лечения и/или супрессии и/или облегчения и/или отсрочки симптомов, например, гиперхолестеринемии. Альтернативно может быть адекватно замедленное непрерывное высвобождение композиций антител-антагонистов против PCSK9. Различные композиции и устройства для достижения замедленного высвобождения известны в уровне техники.
В одном варианте осуществления дозы для антагонистического антитела, пептида или аптамера могут быть определены эмпирически у индивидуумов, у которых было одно или несколько введений антагонистического антитела, пептида или аптамера. Индивидуумам вводят постепенно увеличивающиеся дозы антагонистического антитела, пептида или аптамера против PCSK9. Для оценки эффективности можно отслеживать индикатор данного заболевания.
Введение антагонистического антитела пептида или аптамера против PCSK9 в соответствии со способом по настоящему изобретению может быть непрерывным или периодическим, в зависимости, например, от физиологического состояния реципиента, является ли данное введение профилактическим или терапевтическим, и других факторов, известных квалифицированным практикующим врачам. Введение антагонистического антитела, пептида или аптамера против PCSK9 может быть, по существу, непрерывным в течение заранее установленного периода времени или может быть сериями разделенных по времени доз.
В некоторых вариантах осуществления могут присутствовать более чем одно антагонистическое антитело, пептид или аптамер. Могут присутствовать по меньшей мере одно, по меньшей мере два, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять различных, или более, антагонистических антител и/или пептидов. Как правило, эти антагонистические антитела или пептиды против PCSK9 могут иметь взаимодополняющие действия, которые не оказывают негативного влияния друг на друга. Антагонистическое антитело, пептид или аптамер против PCSK9 также можно использовать в сочетании с другими антагонистами против PCSK9 или антагонистами против рецептора PCSK9. Например, можно использовать один или несколько из следующих антагонистов против PCSK9: антисмысловая молекула, направленная на PCSK9 (в том числе антисмысловая молекула, направленная на нуклеиновую кислоту, кодирующую PCSK9), соединение, ингибирующее PCSK9, и структурный аналог PCSK9. Антагонистическое антитело, пептид или аптамер против PCSK9 также можно применять в сочетании с другими веществами, которые предназначены для усиления или дополнения эффективности данных агентов.
Приемлемые носители, наполнители или стабилизаторы являются нетоксичными в используемых дозах и концентрациях и могут включать буферы, такие как фосфатный, цитратный и других органических кислот; соли, как например, хлорид натрия; антиоксиданты, в том числе аскорбиновая кислота и метионин; консерванты (как например, октадецилдиметилбензил хлорид аммония; хлорид гексаметония; бензалконий хлорид, бензетония хлорид; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехин; резорцин; циклогексанол; 3-пентанол; и м-крезол); низкомолекулярные (менее чем приблизительно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин, или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, в том числе глюкоза, манноза или декстрины; хелатообразующие вещества, как, например, ЭДТА; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, как например, натрий; комплексные соединения металлов (например, комплексы Zn-белок); и/или неионогенные поверхностно-активные вещества, такие как TWEENTM, PLURONICSTM или полиэтиленгликоль (PEG).
Липосомы, содержащие антагонистическое антитело, пептид или аптамер против PCSK9, получены способами, известными в уровне техники, такими как описаны у Epstein, et al., 1985, Proc. Natl. Acad. Sci. USA 82:3688; Hwang, et al., 1980, Proc. Natl Acad. Sci. USA 77:4030; и патентах США №№ 4485045 и 4544545. Липосомы с увеличенным временем циркулирования раскрыты в патенте США № 5013556. Чрезвычайно эффективные липосомы можно получить методом обращенно-фазового выпаривания с липидной композицией, содержащей фосфатидилхолин, холестерин и PEG-дериватизированный фосфатидилэтаноламин (PEG-PE). Липосомы экструдированы через фильтры с определенным размером пор для получения липосом с необходимым диаметром.
Действующие вещества также могут быть заключены в микрокапсулы, полученные, например методом коацервации или посредством полимеризации на границе раздела фаз, например гидроксиметилцеллюлозные или желатиновые микрокапсулы и поли(метилметакрилатные) микрокапсулы, соответственно, в коллоидных системах доставки лекарственного средства (например, липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы), или в микроэмульсиях. Такие способы описаны у Remington, The Science and Practice of Pharmacy, 20th Ed., Mack Publishing (2000).
Могут быть получены препараты с замедленным высвобождением. Подходящие примеры препаратов с замедленным высвобождением включают полупроницаемые матрицы твердых гидрофобных полимеров, содержащих антитело, каковые матрицы представляют собой форму профилированных изделий, например пленок или микрокапсул. Примеры матриц замедленного высвобождения включают полиэфиры, гидрогели (например, поли(2-гидроксиэтил-метакрилат), или поли(виниловый спирт)), полилактиды (патент США № 3773919), сополимеры L-глютаминовой кислоты и 7 этил-L-глутамата, неразлагающийся сополимер этилена и винилацетата, разлагающиеся сополимеры молочная кислота-гликолевая кислота, такие как LUPRON DEPOTTM (инъецируемые микросферы, состоящие из сополимера молочная кислота-гликолевая кислота и лейпролид ацетата), изобутират ацетата сахарозы и поли-D-(-)-3-гидроксимасляную кислоту.
Композиции, используемые для применения in vivo, должны быть стерильными. Это требование без труда выполняется, например, с помощью фильтрования через стерильные фильтрационные мембраны. Терапевтическую композицию антагонистического антитела, пептида или аптамера против PCSK9 обычно помещают в контейнер, имеющий стерильное входное отверстие, например мешок для внутривенного раствора или флакон с пробкой, протыкающейся с помощью иглы для подкожных инъекций.
Подходящие эмульсии могут быть приготовлены с использованием коммерчески доступных жировых эмульсий, таких как IntralipidTM, LiposynTM, InfonutrolTM, LipofundinTM и LipiphysanTM. Действующее вещество может быть либо разведено в предварительно смешанной эмульсионной композиции, либо альтернативно может быть разведено в масле (например, масло соевых бобов, сафлоровое масло, масло из семян хлопка, кунжутное масло, кукурузное масло и миндальное масло) и эмульсия готовится при смешивании фосфолипида (например, фосфолипиды яиц, фосфолипиды соевых бобов или лецитин соевых бобов) и воды. Следует принимать во внимание, что могут быть добавлены другие ингредиенты, например глицерин или глюкоза, для коррекции тоничности данной эмульсии. Пригодные эмульсии обычно будут содержать до 20% масла, например между 5 и 20%. Жировые эмульсии могут содержать капли жира от 0,1 до 1,0 мкм, в частности от 0,1 до 0,5 мкм, и имеют значение рН в диапазоне от 5,5 до 8,0.
Данными эмульсионными композициями могут быть композиции, полученные в результате смешивания антагонистического антитела, пептида или аптамера против PCSK9 с IntralipidTM или его компонентами (масло соевых бобов, фосфолипиды яйца, глицерин и вода).
Композиции для ингаляции или инсуффляции включают растворы и суспензи в фармацевтически пригодных водных или органических растворителях, или их смеси, и порошки. Жидкие или твердые композиции могут содержать соответствующие фармацевтически приемлемые наполнители, как представлено выше. В некоторых вариантах осуществления данные композиции вводятся перорально или через носовой дыхательный путь для локального или системного эффекта. Композиции в предпочтительно стерильных фармацевтически приемлемых растворителях могут быть распылены путем использования газов. Распыленные растворы можно вдыхать непосредственно из распыляющего устройства, или распыляющее устройство может быть присоединено к маске для лица, тенту или аппарату для дыхания с перемежающимся положительным давлением. Композиции растворов, суспензий или порошков могут быть введены, преимущественно, перорально или назально, с помощью устройств, которые доставляют данную композицию надлежащим образом.
B. Антагонисты PCSK9
В способах по данному изобретению используют антагонистическое антитело, пептид или аптамер против PCSK9, которые относятся к любому пептиду или молекуле нуклеиновой кислоты, которая блокирует, подавляет или уменьшает (в том числе значительно уменьшает) биологическую активность PCSK9, в том числе нисходящий метаболический путь, опосредованный сигнальным путем PCSK9, например выявление клеточного ответа на PCSK9.
Антагонистическое антитело, пептид или аптамер против PCSK9 должно проявлять одно или несколько из следующих свойств: (a) связываться с PCSK9; (b) блокировать взаимодействие PCSK9 с LDLR; (c) блокировать или уменьшать PCSK9-опосредованную угнетающую регуляцию LDLR; (d) ингибировать PCSK9-опосредованное снижение клиренса LDL, (e) повышать клиренс LDL в среде посредством культивированных гепатоцитов, (f) повышать клиренс LDL крови с помощью печени in vivo, (g) повышать чувствительность к статинам, и (h) блокировать взаимодействие PCSK9 с другими, даже идентичными факторами.
Для целей этого изобретения антитело, пептид или аптамер, предпочтительно реагирует с PCSK9 таким образом, чтобы ингибировать сигнальную функцию PCSK9 и взаимодействие с LDLR. В некоторых вариантах осуществления антитело-антагонист против PCSK9 специфически распознает PCSK9 приматов. В некоторых вариантах осуществления антитело-антагонист против PCSK9 связывается с PCSK9 приматов и грызунов.
Антитела, эффективные в настоящем изобретении, могут охватывать моноклональные антитела, поликлональные антитела, фрагменты антител (например, Fab, Fab', F(ab')2, Fv, Fc и тому подобное), гибридные антитела, биспецифические антитела, гетероконъюгатные антитела, одиночную цепь (ScFv), их мутантов, слитые белки, включающие часть антитела (например, домен антитела), антитела человека, гуманизированные антитела и любую другую модифицированную конфигурацию молекулы иммуноглобулина, которая содержит участок распознавания антигена требуемой специфичности, в том числе гликозилированные варианты антител, варианты аминокислотных последовательностей антител и ковалентно модифицированные антитела. Антитела могут быть мышиными, крысиными, человеческими или любого другого происхождения (в том числе гибридные или гуманизированные антитела).
В некоторых вариантах осуществления антитело-антагонист против PCSK9 представляет собой моноклональное антитело. Антитело-антагонист против PCSK9 также может быть гуманизированным. В других вариантах осуществления данным антителом является антитело человека.
В некоторых вариантах осуществления данное антитело содержит модифицированную константную область, такую как константная область, которая иммунологически инертна, то есть имеющая сниженный потенциал для провокации иммунной реакции. В некоторых вариантах осуществления данная константная область модифицирована, как описано в Eur. J. Immunol., 1999, 29:2613-2624; PCT публикации № WO99/58572; и/или патентной заявке Великобритании № 9809951.8. Fc может представлять собой IgG2 человека или IgG4 человека. Fc может представлять собой IgG2 человека, содержащий мутацию A330P331 в S330S331 (IgG2∆a), в которой аминокислотные остатки пронумерованы в соответствии с последовательностью IgG2 дикого типа. Eur. J. Immunol., 1999, 29:2613-2624. В некоторых вариантах осуществления данное антитело содержит константную область из IgG4, содержащую следующие мутации (Armour et al., 2003, Molecular Immunology 40 585-593): E233F234L235 в P233V234A235 (IgG4∆c), с нумерацией в соответствии с IgG4 дикого типа. В еще одном варианте осуществления Fc представляет собой IgG4 человека E233F234L235 в P233V234A235 с делецией G236 (IgG4∆b). В другом варианте осуществления Fc представляет собой Fc любого IgG4 человека(IgG4, IgG4∆b или IgG4∆c), содержащий стабилизирующую мутацию шарнирного участка из S228 в P228 (Aalberse et al., 2002, Immunology 105, 9-19). В другом варианте осуществления данная Fc может представлять собой дегликозилированную Fc.
В некоторых вариантах осуществления константная область является дегликозилированной в результате видоизменения остатка присоединения олигосахарида (такого как Asn297) и/или фланкирующих остатков, которые являются частью последовательности узнавания гликозилирования в данной констатной области. В некоторых вариантах осуществления данная константная область является ферментативно дегликозилированной для N-сопряженного гликозилирования. Константная область может быть дегликозилирована для N-сопряженного гликозилирования ферментативно или в результате экспрессии в клетке-хозяине с недостаточным гликозилированием.
Аффинность к связыванию (KD) антитела-антагониста против PCSK9 с PCSK9 (например, PCSK9 человека) может составлять приблизительно от 0,002 до приблизительно 200 нМ. В некоторых вариантах осуществления аффинность к связыванию может быть любой из приблизительно 200 нМ, приблизительно 100 нМ, приблизительно 50 нМ, приблизительно 10 нМ, приблизительно 1 нМ, приблизительно 500 пМ, приблизительно 100 пМ, приблизительно 60 пМ, приблизительно 50 пМ, приблизительно 20 пМ, приблизительно 15 пМ, приблизительно 10 пМ, приблизительно 5 пМ или приблизительно 2 пМ. В некоторых вариантах осуществления аффинность к связыванию меньше чем приблизительно 250 нМ, приблизительно 200 нМ, приблизительно 100 нМ, приблизительно 50 нМ, приблизительно 10 нМ, приблизительно 1 нМ, приблизительно 500 пМ, приблизительно 100 пМ, приблизительно 50 пМ, приблизительно 20 пМ, приблизительно 10 пМ, приблизительно 5 пМ или приблизительно 2 пМ.
Одним способом определения аффинности к связыванию антител с PCSK9 является определение аффинности к связыванию монофункциональных фрагментов Fab данного антитела. Для того чтобы получить монофункциональные фрагменты Fab, антитело (например, IgG) должно быть расщеплено с помощью папаина или экспрессировано рекомбинантно. Аффинность к PCSK9 фрагмента Fab антитела может быть определена с помощью поверхностно-плазмонного резонанса (система поверхностно-плазмонного резонанса (SPR) Biacore3000TM, Biacore, INC, Piscataway NJ), снабженного предиммобилизированными стрептавидиновыми сенсорными чипами (SA) с использованием подвижного буфера HBS-EP (0,01M HEPES, pH 7,4, 0,15 NaCl, 3 мМ ЭДТА, 0,005% (объем/объем) сурфактант P20). Биотинилированный PCSK9 человека (или любой другой PCSK9) может быть разведен в буфере HBS-EP до концентрации менее чем 0,5 мкг/мл и впрыснут через отдельные каналы чипа в различное время контакта, для достижения двух диапазонов плотности антигена, либо 50-200 реакционных единиц (RU) для подробных кинетических исследований, либо 800-1000 RU для скринингового анализа. Исследования регенерации показали, что 25 мМ NaOH в 25% (объем/объем) этаноле эффективно удаляет связь Fab, тогда как сохраняет активность PCSK9 на чипе для более 200 впрыскиваний. Обычно серийные разведения (переркрывающиеся концентрации 0,1-10× расчетной KD) очищенных образцов Fab впрыскивают в течение 1 мин при 100 мкл/минуту и разрешенные сроки диссоциации составляют до 2 часов. Концентрации белков Fab определяют с помощью ELISA и/или электрофореза в SDS-PAGE с использованием Fab известной концентрации (как определено по аминокислотному анализу) в качестве стандарта. Кинетические скорости ассоциации (kon) и скорости диссоциации (koff) получены одновременно, путем аппроксимации данных в целом к модели связывания Langmuir 1:1 (Karlsson, R. Roos, H. Fagerstam, L. Petersson, B., 1994. Methods Enzymology 6. 99-110), используя программу BIAevaluation. Значения равновесной константы диссоциации (KD) определены как koff/kon. Этот протокол является пригодным для использования при определении аффинности к связыванию антитела с любым PCSK9, в том числе PCSK9 человека, PCSK9 другого млекопитающего (как например, PCSK9 мыши, PCSK9 крысы, PCSK9 приматов), а также различными формами PCSK9 (например, α и β формы). Аффинность антитела к связыванию обычно измеряют при 25°C, но также можно измерять при 37°C.
Антитела-антагонисты против PCSK9 могут быть получены любым способом, известным в уровне техники, в том числе способом, представленным в Примере 1. Метод и план иммунизации животного-хозяина обычно соответствуют разработанным и общепринятым технологическим приемам для стимулирования и получения антител, как описано здесь далее. Общие методики для производства антител человека и мыши известны в уровне техники и/или описаны здесь. Предпочтительный в настоящее время способ получения антител включает иммунизацию нокаут-животных PCSK9- (PCSK9 -/-), как раскрыто здесь.
Предусматривается, что любое млекопитающее, в том числе человек, или его антителопродуцирующие клетки могут быть обработаны таким образом, чтобы служить в качестве основы для получения гибридомных клеточных линий млекопитающего, в том числе человека. В основном вакцинацию животного-хозяина проводят внутрибрюшинно, внутримышечно, перорально, подкожно, внутриподошвенно и/или внутрикожно с количеством иммуногена, в том числе, как описано здесь.
Гибридомы можно получить из лимфоцитов и иммортализованных клеток миеломы, используя обычную технологию гибридизации соматических клеток по Kohler, B. и Milstein, C, 1975, Nature 256:495-497, или модификацию по Buck, D. W., et al., 1982, In Vitro, 18:377-381. В данной гибридизации можно использовать доступные линии миеломы, в том числе, но не ограничиваясь ими, X63-Ag8.653 и миеломные линии, доступные в Salk Institute, Cell Distribution Center, San Diego, Calif., USA. Обычно данный способ вовлекает слияние миеломных клеток и лимфоидных клеток с использованием вещества, способствующего слиянию клеток, такого как полиэтиленгликоль, или с помощью электрических способов, хорошо известных специалисту в данной области. После слияния клетки отделяют от среды, в которой произошло слияние, и выращивают в селективной питательной среде, такой как среда гипоксантин-аминоптерин-тимидин (HAT), для устранения негибридизованных родительских клеток. Любая среда, описанная здесь, дополненная или не дополненная сывороткой, может быть использована для культивирования гибридом, которые секретируют моноклональные антитела. В качестве другой альтернативы технологии слияния клеток можно использовать EBV-иммортализованные В клетки для продукции моноклональных антител против PCSK9 по данному изобретению. Гибридомы увеличены и сублокнированы, при желании, и супернатанты исследуют на активность против иммуногена посредством общепринятых процедур иммунологического анализа (например, радиоиммуноанализ, иммуноферментный анализ или флуоресцентный иммуноанализ).
Гибридомы, которые можно использовать в качестве источника антител, охватывают все производные, клетки-потомство родительских гибридом, которые продуцируют специфические моноклональные антитела против PCSK9, или их части.
Гибридомы, которые продуцируют такие антитела, можно выращивать in vitro или in vivo, используя известные процедуры. Данные моноклональные антитела можно изолировать из культуральной среды или жидкостей организма, используя общепринятые процедуры очищения иммуноглобулинов, такие как фракционирование сульфатом аммония, гель-электрофорез, диализ, хроматография и ультрафильтрация, если необходимо. Нежелательная активность, если присутствует, может быть устранена, например, прогоняя данный препарат через адсорбирующие вещества, изготовленные из данного иммуногена, прикрепленного к твердой фазе, и элюируя или высвобождая требуемые антитела от иммуногена. Иммунизация животного-хозяина с помощью PCSK9 человека, или фрагмента, содержащего целевую аминокислотную последовательность, конъюгированную с белком, который является иммуногенным для биологических видов, подвергающихся иммунизации, например гемоцианин лимфы улитки, сывороточный альбумин, тиреоглобулин быка или соевый ингибитор трипсина, с использованием бифункционального или дериватизирующего агента, например малеимидобензол сульфосукцинимид эфир (конъюгация посредством остатков цистеина), N-гидроксисукцинимид (посредством остатков лизина), глутаральдегид, янтарный ангидрид, SOCl2 или R1N=C=NR, где R и R1 представляют собой разные алкильные группы, может привести к продукции популяции антител (например, моноклональных антител).
При необходимости может быть определена последовательность представляющего интерес антитела-антагониста против PCSK9 (моноклонального или поликлонального), и данная полинуклеотидная последовательность может затем быть клонирована в вектор для экспрессии или воспроизводства. Последовательность, кодирующая представляющее интерес антитело, может поддерживаться в векторе в клетке-хозяине, и данную клетку-хозяина затем можно размножить и заморозить для использования в будущем. Производство рекомбинантных моноклональных антител в клеточной культуре можно осуществлять посредством клонирования генов антитела из В-клеток, используя известные в уровне техники способы. Смотри, например, Tiller et al., 2008, J. Immunol. Methods 329, 112; патент США № 7314622.
Альтернативно полинуклеотидную последовательность можно использовать для генетических манипуляций для «гуманизации» антитела или для улучшения аффинности или других характеристик данного антитела. Например, константная область может быть сконструирована таким образом, чтобы больше походить на константные области человека, во избежание иммунной реакции, если антитело используют в клинических испытаниях и лечении человека. Может быть необходимо проведение генетических манипуляций с последовательностью антитела для получения большей аффинности к PCSK9 и большей эффективности при ингибировании PCSK9. Квалифицированному специалисту в данной области будет очевидно, что можно изменить один или несколько полинуклеотидов в данном антителе-антагонисте против PCSK9 и все же поддерживать его аффинность к связыванию с PCSK9.
Существуют четыре основных этапа гуманизации моноклонального антитела. Это: (1) определение нуклеотидной и предвычисленной аминокислотной последовательности вариабельных доменов легкой и тяжелой цепей исходного антитела; (2) проектирование гуманизированного антитела, то есть принятие решения, какая каркасная область антитела будет использоваться в процессе гуманизации; (3) фактические методы/технологии гуманизации; и (4) трансфекция и экспрессия гуманизированного антитела. Смотри, например, патенты США №№ 4816567; 5807715; 5866692; 6331415; 5530101; 5693761; 5693762; 5585089; и 6180370.
Был описан целый ряд молекул «гуманизированных» антител, содержащих участок связывания антигена, полученный из нечеловеческого иммуноглобулина, в том числе химерные антитела, имеющие V области грызунов, или модифицированные V области грызунов, и их соответствующие CDR, слитые с константными доменами человека. Смотри, например, Winter et al., 1991, Nature 349:293-299; Lobuglio et al., 1989, Proc. Nat. Acad. Sci. USA 86:4220-4224; Shaw et al., 1987, J Immunol. 138:4534-4538; и Brown et al., 1987, Cancer Res. 47:3577-3583. В других ссылочных материалах описаны CDR грызунов, встроенные в несущую каркасную область человека (FR) перед слиянием с константным доменом подходящего антитела человека. Смотри, например, Riechmann et al.,1988, Nature 332:323-327; Verhoeyen et al., 1988, Science 239:1534-1536; и Jones et al., 1986, Nature 321:522-525. В другом ссылочном материале описаны CDR грызунов, усиленные рекомбинантно сконструированными каркасными областями грызунов. Смотри, например, европейскую патентную публикацию № 0519596. Эти «гуманизированные» молекулы сконструированы для сведения к минимуму нежелательной иммунологической реакции по отношению к молекулам античеловеческого антитела грызунов, которая ограничивает продолжительность и эффективность терапевтического применения этих веществ у реципиента-человека. Например, константная область антитела может быть сконструирована таким образом, чтобы быть инертной (например, не запускать комплементарный лизис). Смотри, например, PCT публикацию № WO99/58572; патентную публикацию Великобритании № 9809951.8. Другие способы гуманизации антител, которые также можно использовать, раскрыты у Daugherty et al., 1991, Nucl. Acids Res. 19:2471-2476 и патентах США №№ 6180377; 6054297; 5997867; 5866692; 6210671; и 6350861; и в PCT публикации № WO 01/27160.
В еще одном варианте полностью человеческие антитела могут быть получены, используя коммерчески доступные линии мышей, которые были выведены для того, чтобы экспрессировать специфические иммуноглобулины человека. Трансгенные животные, выведенные для выработки более целесообразной или более активной иммунной реакции, также могут быть использованы для выработки гуманизированных антител или антител человека. Примерами таких систем являются XenomouseTM, предоставленные фирмой Abgenix, Inc. (Fremont, CA), HuMAb-Mouse® и TC Mouse TM, предоставленные фирмой Medarex, Inc. (Princeton, NJ), и мыши Veloclmmune® фирмы Regeneron Pharmaceuticals, Inc. (Tarrytown, NY).
Альтернативно антитела могут быть получены рекомбинантно и экспрессируются с использованием любого способа, известного в уровне техники. В другом варианте антитела могут быть получены рекомбинантно с помощью технологии фагового дисплея. Смотри, например, патенты США №№ 5565332; 5580717; 5733743; и 6265150; и Winter et al., 1994, Annu. Rev. Immunol. 12:433-455. Альтернативно, технологию фагового дисплея (McCafferty et al., 1990, Nature 348:552-553) можно использовать для продукции антител человека и фрагментов антител in vitro, из различных библиотек генов вариабельного (V) домена иммуноглобулина иммунизированных доноров. По этой технологии гены домена V антитела клонированы внутри рамки считывания в ген либо главного, либо минорного белка оболочки нитевидного бактериофага, такого как М13 или fd, и воспроизводятся в виде функциональных фрагментов антитела на поверхности фаговой частицы. Из-за того что нитевидные частицы содержат копию одноцепочечной ДНК фагового генома, отбор на основании функциональных свойств данного антитела также приводит в результате к отбору гена, кодирующего антитело, проявляющее эти свойства. Таким образом, фаг имитирует некоторые из свойств В-клетки. Фаговый дисплей может быть осуществлен в различных видах; смотри, например, Johnson, Kevin S. and Chiswell, David J., 1993, Current Opinion in Structural Biology 3:564-571. Для фагового дисплея можно использовать несколько источников V-сегментов гена. Clackson et al., 1991, Nature 352:624-628 изолировали разнотипную группу антител против оксазолона из небольшой рандомизированной комбинаторной библиотеки V генов, полученных из селезенок иммунизированных мышей. Антигенный репертуар генов V от неиммунизированных людей-доноров может быть сконструирован, и антитела к разнотипной группе антигенов (в том числе, аутоантигенов) могут быть выделены, в общем и целом, следуя методам, описанным у Mark et al., 1991, J. Mol. Biol. 222:581-597, или Griffith et al., 1993, EMBO J. 12:725-734. При естественной иммунной реакции в генах антител с высокой скоростью накапливаются мутации (соматическая сверхмутация). Некоторые из введенных изменений будут обеспечивать более высокую аффинность, и В-клетки, воспроизводящие высокоаффинный поверхностный иммуноглобулин, предпочтительно реплицируются и дифференцируются во время последующей сенсибилизации антигеном. Этот естественный процесс можно смоделировать, используя технологию, известную как «перетасовка цепей» (Marks et al., 1992, Bio/Technol. 10:779-783). В этом методе аффинность «первичных» антител человека, полученных с помощью фагового дисплея, может быть усилена посредством последовательной замены генов V областей тяжелой и легкой цепей на репертуар встречающихся в природе вариантов (иммунный репертуар) генов V доменов, полученных от неиммунизированных доноров. Этот метод делает возможной выработку антител и фрагментов антител с аффиностями в диапазоне пМ-нМ. Стратегия получения очень большого репертуара фаговых антител (также известно, как «основательница всех библиотек») была описана у Waterhouse et al., 1993, Nucl. Acids Res. 21:2265-2266. Перетасовку генов также можно использовать для извлечения антител человека из антител грызунов, где антитело человека имеет схожие аффинности и специфичности с исходным антителом грызуна. В соответствии с этим способом, который также называют «импринтиг антигенной детерминанты», гены V домена тяжелой или легкой цепи антигенов грызунов, полученные посредством технологии фагового дисплея, замещены иммунным репертуаром генов V доменов человека, получая в результате гибриды грызун-человек. Отбор на антиген приводит в результате к выделению человеческих вариабельных областей, способных к восстановлению функционального участка связывания с антигеном, то есть эпитоп обусловливает (фиксирует) выбор партнера. Когда процесс повторяют для того, чтобы заменить оставшийся V-домен грызуна, получают антитело человека (смотри PCT публикацию № WO 93/06213). В отличие от традиционной гуманизации антител грызунов посредством введения CDR, этот способ обеспечивает полностью человеческие антитела, которые не имеют ни остатков каркаса, ни CDR, происходящих от грызуна.
Совершенно очевидно, что хотя вышеприведенное обсуждение относится к гуманизированным антителам, обсуждаемые общие принципы применимы при получении антител для использования, например, у собак, кошек, приматов, лошадей и жвачных животных. Дополнительно очевидно, что один или несколько аспектов описанной здесь гуманизации антитела могут быть скомбинированы, например введение CDR, мутация каркаса и мутация CDR.
Антитела можно получить рекомбинантно, сначала выделяя антитела и антителопродуцирующие клетки из животных-хозяев, получая последовательность гена и используя данную последовательность гена для экспрессии данного антигена рекомбинантно в клетках-хозяевах (например, клетки CHO). Другим способом, который можно применять, является экспрессия последовательности антитела в растениях (например, табак) или трансгенном молоке. Методы экспрессии антител рекомбинантно в растениях или молоке были описаны. Смотри, например, Peeters, 2001, et al. Vaccine 19:2756; Lonberg, N. and D. Huszar, 1995, Int. Rev. Immunol 13:65; и Pollock, et al., 1999, J Immunol Methods 231:147. Способы получения производных антител, например гуманизированных, одноцепочечных и тому подобного, известны в уровне техники.
Также для выделения антител, которые являются специфическими для PCSK9, можно использовать методы иммунологического анализа и методы сортинга с помощью проточной цитометрии, такие как клеточный сортинг с возбуждением флуоресценции (FACS).
Антитела могут быть связаны с различными носителями. Носители могут быть активными и/или инертными. Примеры хорошо известных носителей включают полипропилен, полистирол, полиэтилен, декстран, нейлон, амилазы, стекло, природную или модифицированную целлюлозу, полиакриламиды, агарозы и магнетит. Природа носителя может быть либо растворимой, либо нерастворимой, в зависимости от целей данного изобретения. Квалифицированному в данной области специалисту будут известны другие носители для связывания антител, или он будет в состоянии определить таковые с помощью рутинной экспериментальной работы. В некоторых вариантах осуществления данные носители содержат вещества, нацеленные на миокард.
ДНК, кодирующую данные моноклональные антитела, легко выделить и секвенировать, используя общепринятые методы (например, используя олигонуклеотидные зонды, которые способны специфически связываться с генами, кодирующими тяжелую и легкую цепи данных моноклональных антител). Гибридомные клетки служат в качестве предпочтительного источника такой ДНК. После выделения ДНК можно поместить в экспрессирующие векторы (такие как экспрессирующие векторы, раскрытые в PCT публикации № WO 87/04462), которые затем трансфицируют в клетки-хозяев, такие как клетки E. coli, клетки COS обезьян, клетки яичника китайского хомячка (CHO), или клетки миеломы, которые иначе не продуцируют иммуноглобулиновый белок, для достижения синтеза моноклональных антител в рекомбинантных клетках-хозяевах. Смотри, например, PCT публикацию № WO 87/04462. Данная ДНК также может быть модифицирована, например, подставляя кодирующую последовательность константных доменов легкой и тяжелой цепей человека вместо гомологичных последовательностей мыши, Morrison et al., 1984, Proc. Nat. Acad. Sci. 81:6851, или посредством ковалентного присоединения к кодирующей последовательности иммуноглобулина всей или части кодирующей последовательности неиммуноглобулинового полипептида. В этом случае получены «химерные» или «гибридные» антитела, которые имеют специфичность связывания моноклонального антитела против PCSK9.
Антитела-антагонисты против PCSK9 и полипептиды, полученные из антител, могут быть идентифицированы или охарактеризованы с использованием методов, известных в уровне техники, посредством чего уменьшение, улучшение или нейтрализация биологического действия PCSK9 определяется и/или измеряется. В некоторых вариантах осуществления антагонистическое антитело или полипептид против PCSK9 идентифицируют, инкубируя возможный агент с PCSK9 и отслеживая связывание и/или присутствующее уменьшение или нейтрализацию биологической активности PCSK9. Анализ связывания может быть осуществлен с очищенным PCSK9 полипептидом (полипептидами) или с клетками, естественным образом секретирующими или трансфицированными, чтобы экспрессировать PCSK9 полипептид(ы). В одном варианте осуществления анализ связывания представляет собой анализ конкурентного связывания, в котором оценивают способность возможного антитела конкурировать с известным антагонистом PCSK9 за связывание с PCSK9. Этот анализ можно провести в различных видах, в том числе ELISA. В других вариантах осуществления антитело-антагонист против PCSK9 было определено в результате инкубирования возможного агента с PCSK9 и отслеживания связывания и сопровождающего ингибирования экспрессии LDLR и/или клиренса холестерина крови.
После первичной идентификации активность кандидатного антитела-антагониста против PCSK9 может быть дополнительно подтверждена и улучшена в результате биологических анализов, которые известны для тестирования целенаправленных биологических активностей. Альтернативно, биоанализ можно использовать для непосредственного скрининга кандидатных веществ. Некоторые из способов идентификации и описания антагонистических антител, пептидов и аптамеров против PCSK9 описаны подробно в Примерах.
Антитела-антагонисты против PCSK9 могут быть охарактеризованы с использованием методов, хорошо известных в уровне техники. Например, один способ предназначен для выявления эпитопа, с которым связывается данное антитело, или «картирование эпитопов». В уровне техники известно множество методов картирования и характеризации местоположения эпитопов на белках, в том числе определение кристаллической структуры комплекса антитело-антиген, конкурентные анализы, анализ экспрессии фрагментов гена, синтетический анализ на основе пептидов, как описано, например, в Главе 11 у Harlow and Lane, Using Antibodies, a Laboratory Manual, (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1999). В дополнительном примере, картирование эпитопов можно использовать для определения последовательности с которой связывается антитело-антагонист против PCSK9. Картирование эпитопов доступно на коммерческой основе из различных источников, например система Pepscan Systems (Edelhertweg 15, 8219 PH Lelystad, The Netherlands). Эпитоп может представлять собой линейный эпитоп, то есть содержащий единственный аминокислотный участок, или конформационный эпитоп, образованный в результате трехмерного взаимодействия аминокислот, которые необязательно должны содержаться в единственном участке. Пептиды различной длины (например, по меньшей мере длиной 4-6 аминокислот) могут быть изолированы и синтезированы (например, рекомбинантно) и использованы в анализе связывания с антителом-антагонистом против PCSK9. В другом примере эпитоп, с которым связывается антитело-антагонист против PCSK9, может быть определено при систематическом скрининге, используя перекрывающиеся пептиды, происходящие из последовательности PCSK9, и определяя связывание с помощью антитела-антагониста против PCSK9. В соответствии с анализом экспрессии фрагментов гена открытая рамка считывания, кодирующая PCSK9, фрагментирована либо случайным образом, либо с помощью специфических генетических конструкций и определяется реактивность экспрессированных фрагментов PCSK9 с тестируемым антителом. Фрагменты гена могут быть получены, например, в результате ПЦР и затем транскрибированы и транслированы в белок in vitro, в присутствии радиоактивных аминокислот. Связывание данного антитела с радиоактивно маркированными фрагментами PCSK9 определяют затем с помощью иммунопреципитации и гель-электрофореза. Некоторые эпитопы также могут быть идентифицированы с использованием больших библиотек случайных пептидных последовательностей, выраженных на поверхности фаговых частиц (фаговые библиотеки). Альтернативно, определенная библиотека перекрывающихся пептидных фрагментов может быть протестирована на предмет связывания с тестируемым антителом с помощью простых анализов связывания. В дополнительном примере мутагенез антигенсвязывающего домена, эксперименты с обменом доменами и сканирующий аланином мутагенез могут быть осуществлены для идентификации остатков, необходимых, достаточных и/или обязательных для связывания эпитопа. Например, эсперименты по обмену доменами могут быть осуществлены с использованием мутанта PCSK9, в котором различные фрагменты данного полипептида PCSK9 были замещены (обменены) последовательностями из PCSK9 других видов, или близкородственного, но антигенно отличающегося белка (как например, другой член семейства пропротеин конвертазы). Оценивая связывание антитела с данным мутантным PCSK9, может быть определена значимость конкретного фрагмента PCSK9 для связывания с антителом.
Еще одним методом, который можно применять для описания антитела-антагониста против PCSK9, является использование конкурентного анализа с другими антителами, которые, как известно, связывают этот же антиген, то есть различные фрагменты на PCSK9, для определения того, связывает ли данное антитело-антагонист против PCSK9 тот же самый эпитоп, как и другие антитела. Конкурентный анализ хорошо известен квалифицированному специалисту в данной области.
Для описания антитела также можно использовать кристаллическую структуру комплекса антитело:антиген. Остатки определяют, рассчитывая разницу между областью доступной поверхности между кристаллической структурой L1L3:PCSK9 и структурой одного PCSK9. Остатки PCSK9, которые демонстрируют скрытую поверхность в результате формирования комплекса с антителом L1L3, включены как часть эпитопа. Данная доступная для растворения поверхность белка определена как локус в центре зондовой сферы (представляет молекулу растворителя радиусом 1,4 Å), поскольку она переворачивает ван-дер-ваальсову поверхность белка. Доступная для растворения поверхность расчитывается посредством создания поверхностных точек на расширенной сфере около каждого атома (на расстоянии от центра атома, эквивалентном сумме радиусов данного атома и зонда) и исключая те, которые находятся внутри эквивалентных сфер, ассоциированных с соседними атомами, как реализовано в программе AREAIMOL (Briggs, P.J., 2000, CCP4 Newsletter No. 38, CCLRC, Daresbury).
Эспрессирующий вектор может быть использован для прямой экспрессии антитела-антагониста против PCSK9. Специалист в данной области знаком с применением экспрессирующих векторов для получения экспрессии экзогенных белков in vivo. Смотри, например, патенты США №№ 6436908; 6413942; и 6376471. Применение экспрессирующих векторов включает локальное или системное применение, в том числе инъекции, пероральное применение, генную пушку или введение посредством катетера, и местное применение. В другом варианте осуществления данный экспрессирующий вектор вводят непосредственно в симпатический ствол или ганглий, или в коронарную артерию, предсердие, желудочек сердца или перикард.
Также можно использовать направленную доставку терапевтических композиций, включающих экспрессирующий вектор, или субгеномные полинуклеотиды. Рецептор-опосредованные технологии доставки ДНК описаны, например, у Findeis et al., 1993, Trends Biotechnol. 11:202; Chiou et al., 1994, Gene Therapeutics: Methods And Applications Of Direct Gene Transfer (J.A. Wolff, ed.); Wu et al., 1988, J. Biol. Chem. 263:621; Wu et al., 1994, J. Biol. Chem. 269:542; Zenke et al., 1990, Proc. Natl. Acad. Sci. USA 87:3655; Wu et al., 1991, J. Biol. Chem. 266:338. Терапевтические композиции, содержащие полинуклеотид, применяются в диапазоне приблизительно от 100 нг до приблизительно 200 мг ДНК для местного применения в протоколе генотерапии. Также в протоколе генотерапии можно использовать диапазон концентраций ДНК приблизительно от 500 нг до приблизительно 50 мг, приблизительно от 1 мкг до приблизительно 2 мг, приблизительно от 5 мкг приблизительно до 500 мкг и приблизительно от 20 мкг приблизительно до 100 мкг. Терапевтические полинуклеотиды и полипептиды могут быть доставлены с помощью систем доставки генов. Данная система доставки генов может быть вирусного или невирусного происхождения (смотри в общих чертах, Jolly, 1994, Cancer Gene Therapy 1:51; Kimura, 1994, Human Gene Therapy 5:845; Connelly, 1995, Human Gene Therapy 1:185; и Kaplitt, 1994, Nature Genetics 6:148). Экспрессия таких кодирующих последовательностей может быть индуцирована с помощью эндогенных промоторов млекопитающих или гетерологичных промоторов. Экспрессия кодирующей последовательности может быть конститутивной или регулируемой.
Векторы на вирусной основе для доставки необходимого полинуклеотида и экспрессии в желаемой клетке хорошо известны в уровне техники. Примеры носителей на вирусной основе включают, но не ограничиваются ими, векторы на основе рекомбинантных ретровирусов (смотри, например, PCT публикации №№ WO 90/07936; WO 94/03622; WO 93/25698; WO 93/25234; WO 93/11230; WO 93/10218; WO 91/02805; патенты США №№ 5219740 и 4777127; патент Великобритании № 2200651; и EP патент № 0 345 242), векторы на основе альфавирусов (например, векторы вируса Синдбис, вирус леса Симлики (ATCC VR-67; ATCC VR-1247), вирус реки Росс (ATCC VR-373; ATCC VR-1246) и вирус венесуэльского энцефалита лошадей (ATCC VR-923; ATCC VR-1250; ATCC VR-1249; ATCC VR-532)) и векторы на основе аденоассоциированного вируса (AAV) (смотри, например, PCT публикации №№ WO 94/12649, WO 93/03769; WO 93/19191; WO 94/28938; WO 95/11984 и WO 95/00655). Также можно использовать введение ДНК, связанной с убитым аденовирусом, как описано у Curiel, 1992, Hum. Gene Ther. 3:147.
Также можно использовать невирусные системы доставки и методы, включающие, но не ограничивающиеся ими, поликатионно конденсированную ДНК, связанную или не связанную с убитым аденовирусом, отдельно (смотри, например, Curiel, 1992, Hum. Gene Ther. 3:147); лигандсвязанную ДНК (смотри, например, Wu, J., 1989, Biol. Chem. 264:16985); систему доставки эукариотических клеток (смотри, например, патент США № 5814482; PCT публикации №№ WO 95/07994; WO 96/17072; WO 95/30763; и WO 97/42338) и нуклеиновую нейтрализацию заряда или слияние с клеточными мембранами. Также можно использовать оголенную ДНК. Примеры способов введения оголенной ДНК описаны в PCT публикации № WO 90/11092 и патенте США № 5580859. Липосомы, которые выступают в качестве системы доставки генов, описаны в патенте США № 5422120; PCT публикации №№ WO 95/13796; WO 94/23697; WO 91/14445; и EP 0524968. Дополнительные подходы описаны у Philip, 1994, Mol. Cell Biol., 14:2411 и у Woffendin, 1994 Proc. Natl. Acad. Sci. 91:1581.
Это изобретение охватывает композиции, в том числе фармацевтические композиции, содержащие антитела, описанные здесь, или полученные описанными здесь способами, и имеющие характеристики, описанные здесь. Используемые здесь композиции включают одно или несколько антител, пептидов или аптамеров, которые противодействуют взаимодействию PCSK9 с LDLR, и/или один или несколько полинуклеотидов, содержащих последовательности, кодирующие одно или несколько этих антител или пептидов. Эти композиции могут дополнительно содержать подходящие наполнители, такие как фармацевтически приемлемые наполнители, в том числе буферы, которые хорошо известны в уровне техники.
Антагонистическим антителам и пептидам против PCSK9 по данному изобретению свойственны любая (одна или несколько) из следующих характеристик: (a) связь с PCSK9; (b) блокировка взаимодействия PCSK9 с LDLR; (c) снижение PCSK9-опосредованной угнетающей регуляции данного LDLR; и (d) подавление PCSK9-опосредованного ингибирования клиренса LDL крови. Предпочтительно антитела против PCSK9 имеют два или более из этих свойств. Более предпочтительно данные антитела имеют три или более из этих свойств. Наиболее предпочтительно данные антитела имеют все четыре свойства.
Соответственно, данное изобретение обеспечивает любое из следующего, или композиции (в том числе фармацевтические композиции), содержащие любое антитело, имеющее неполную последовательность легкой цепи и неполную последовательность тяжелой цепи, как представлено в Таблице 1. Подчеркнутые последовательности представляют собой последовательности CDR по Кабату (Kabat), а выделенные жирным шрифтом - в соответствии с Хотиа (Chothia).


Данное изобретение также обеспечивает участки CDR антител против PCSK9 (в том числе CDR по Кабату (Kabat) и Хотиа (Chothia)). Установление областей CDR входит в число навыков в данной области. Подразумевается, что в некоторых вариантах осуществления CDR могут представлять собой комбинацию CDR по Кабату (Kabat) и по Хотиа (Chothia) (также называемые «комбинированные CDR» или «удлиненные CDR»). В некоторых вариантах осуществления данные CDR представляют собой CDR по Хотиа (Chothia). Другими словами, в вариантах осуществления с более чем одной CDR данные CDR могут быть любыми, по Кабату (Kabat), по Хотиа (Chothia), комбинированными CDR или их комбинациями.
Данное изобретение также обеспечивает способ получения любого из этих антител или полипептидов. Антитела по этому изобретению можно создать, используя методы, известные в уровне техники. Полипептиды можно получить в результате протеолитической или другой деградации антител, рекомбинантными методами (а именно отдельные или слитые полипептиды), как описано выше, или в результате химического синтеза. Полипептиды данных антител, особенно более короткие полипептиды приблизительно до 50 аминокислот, удобно получать в результате химического синтеза. Способы химического синтеза хорошо известны в уровне техники и доступны на коммерческой основе. Например, антитело может быть получено с помощью автоматизированного синтезатора полипептидов с применением твердофазного метода. Смотри также патенты США №№ 5807715; 4816567 и 6331415.
В другом варианте данные антитела и полипептиды можно получить рекомбинантно, используя методы, хорошо известные в уровне техники. В одном варианте осуществления полинуклеотид содержит последовательность, кодирующую вариабельные области тяжелой цепи и/или легкой цепи антитела 4A5, 5A10, 6F6, 7D4 или L1L3. Последовательность, кодирующую представляющее интерес антитело, можно поддерживать в векторе в клетке-хозяине, и данную клетку-хозяина можно размножить и заморозить для дальнейшего применения. Векторы (в том числе экспрессирующие векторы) и клетки-хозяева описаны здесь далее.
Данное изобретение также охватывает scFv антител по данному изобретению. Одноцепочечные фрагменты вариабельной области получают посредством соединения вариабельных областей тяжелой и/или легкой цепи, используя короткий сшивающий пептид. Bird et al., 1988, Science 242:423-426. Примером сшивающего пептида является (GGGGS)3 (SEQ ID NO: 24), который образует сшивку приблизительно 3,5 нм между карбоксильным концом одной вариабельной области и аминоконцом другой вариабельной области. Могут быть сконструированы и использованы линкеры с другими последовательностями. Bird et al., 1988, supra. Линкеры должны быть короткими, гибкими полипептидами и предпочтительно содержащими менее чем приблизительно 20 аминокислотных остатков. Линкеры, в свою очередь, могут быть модифицированы для выполнения дополнительных функций, как например фиксация лекарственных средств или фиксация с твердыми подложками. Одноцепочечные варианты можно получить либо рекобминантно, либо синтетически. Для синтетического получения scFv можно использовать автоматизированный синтезатор. Для рекомбинантного получения scFv подходящая плазмида, содержащая полинуклеотид, который кодирует данный scFv, может быть встроена в подходящую клетку-хозяина либо эукариотическую, например клетки дрожжей, растений, насекомых или млекопитающих, или прокариотическую, например E.coli. Полинуклеотиды, кодирующие представляющий интерес scFv, можно получить с помощью рутинных манипуляций, таких как лигирование полинуклеотидов. Полученный scFv можно изолировать, используя стандартные способы очистки белков, известные в уровне техники.
Также охвачены другие формы одноцепочечных антител, такие как диатела. Диатела представляют собой двухвалентные, биспецифические антитела, в которых домены VH и VL экспрессируются на единственной полипептидной цепи, но используя линкер, который является слишком коротким для обеспечения образования пар между данными двумя доменами на той же цепи, тем самым принуждая данные домены образовывать пары с комплементарными доменами другой цепи и образуя два участка связывания антегена (смотри, например, Holliger, P., et al., 1993, Proc. Natl. Acad Sci. USA 90:6444-6448; Poljak, R.J., et al., 1994, Structure 2:1121-1123).
Например, биспецифические антитела, моноклональные антитела, которые имеют специфичность связывания, по меньшей мере, с двумя различными антигенами, могут быть получены с использованием раскрытых здесь антител. Способы получения биспецифических антител хорошо известны в уровне техники (смотри, например, Suresh et al., 1986, Methods in Enzymology 121:210). Традиционно рекомбинантное получение биспецифических антител было основано на совместной экспрессии двух пар легкая цепь-тяжелая цепь иммуноглобулина, где данные две тяжелые цепи имеют различные специфичности (Millstein and Cuello, 1983, Nature 305, 537-539).
В соответствии с одним способом получения биспецифических антител вариабельные домены антитела с желаемой специфичностью связывания (антигенсвязывающие активные центры антитела) слиты с последовательностями константных доменов иммуноглобулина. Данное слияние предпочтительно с константным доменом тяжелой цепи иммуноглобулина, содержащим, по меньшей мере, часть шарнира, области CH2 и CH3. Предпочтительно иметь первую константную область тяжелой цепи (СН1), включающую участок, необходимый для связывания легкой цепи, присутствующей, по меньшей мере, в одном из слияний. ДНК, кодирующие слияния тяжелых цепей иммуноглобулина и, при необходимости, легкой цепи иммуноглобулина, встроены в отдельные экспрессирующие векторы и совместно трансфицированы в подходящий организм-хозяин. Это обеспечивает значительную гибкость при коррекции общего количественного соотношения данных трех полипептидных фрагментов в вариантах осуществления, в которых неравные соотношения данных трех полипептидных цепей, использованных в конструкции, обеспечивают оптимальную продуктивность. Тем не менее, возможно встраивание кодирующих последовательностей двух или все трех полипептидных цепей в один экспрессирующий вектор, в случае когда экспрессия по меньшей мере двух полипептидных цепей в одинаковых соотношениях в результате приводит к высокой продуктивности или когда соотношения не имеют определенной значимости.
В одном способе биспецифические антитела состоят из тяжелой цепи гибридного иммуноглобулина с первой специфичностью связывания в одном плече и пары тяжелая цепь-легкая цепь гибридного иммуноглобулина (обеспечивает вторую специфичность связывания) в другом плече. Эта асимметрическая структура, с легкой цепью иммуноглобулина только в одной половине данной биспецифической молекулы, облегчает отделение желаемого биспецифического соединения от нежелательных комбинаций иммуноглобулиновых цепей. Этот способ описан в PCT публикации № WO 94/04690.
Гетероконъюгатные антитела, состоящие из двух ковалентно связанных антител, также входят в диапазон данного изобретения. Такие антитела использовали для нацеливания клеток иммунной системы против нежелательных клеток (патент США № 4676980) и для лечения ВИЧ-инфекции (PCT публикации №№ WO 91/00360 и WO 92/200373; EP 03089). Гетероконъюгатные антитела можно создать, используя любые удобные способы перекрестного сшивания. Подходящие перекрестносшивающие агенты и способы перекрестного сшивания хорошо известны в уровне техники, и они описаны в патенте США № 4676980.
Химерные или гибридные антитела также можно создать in vitro, используя известные способы синтетической химии белка, в том числе те, в которых используют перекрестносшивающие агенты. Например, иммунотоксины могут быть сконструированы с использованием реакции обмена дисульфидов или посредством образования тиоэфирной связи. Примеры подходящих реагентов для этой цели включают иминотиолат и метил-4-меркаптобутиримидат.
Гуманизированные антитела, содержащие одну или несколько CDR антител 5A10 или 7D4, или одну или несколько CDR, полученных из антител 5A10 или 7D4, могут быть созданы, например, с использованием любого способа, известного в уровне техники. Например, четыре общих этапа можно использовать для гуманизации моноклонального антитела. Это: (1) определение нуклеотидной и предвычисленной аминокислотной последовательности вариабельных доменов легкой и тяжелой цепей исходного антитела; (2) проектирование гуманизированного антитела, то есть принятие решения, какая каркасная область антитела будет использоваться в процессе гуманизации; (3) фактические методы/технологии гуманизации; и (4) трансфекция и экспрессия гуманизированного антитела. Смотри, например, патенты США №№ 4816567; 5807715; 5866692; 6331415; 5530101; 5693761; 5693762; 5585089 и 6180370.
В рекомбинантных гуманизированных антителах участок Fc может быть модифицирован во избежание взаимодействия с рецептором Fсγ и комплементом и имунными системами. Способы получения таких антител описаны в WO 99/58572. Например, константную область можно сконструировать более походящей на константные области человака во избежание иммунной реакции, если данное антитело используют в клинических испытаниях и лечении человека. Смотри, например, патенты США №№ 5997867 и 5866692.
Гуманизированные антитела, содержащие вариабельные области легкой или тяжелой цепи, или одну или несколько CDR антитела или его вариантов, представленных в Таблице 1, или одну или несколько CDR, полученных из данного антитела или его вариантов, представленных в Таблице 2, можно создать, используя любые способы, известные в уровне техники.
Гуманизированные антитела можно создать любым способом, известным в уровне техники.
Данное изобретение включает модификации антител и полипептидов по вариантам изобретения, представленным в Таблице 1, в том числе, функционально эквивалентные антитела, которые не затрагивают в значительной степени их свойства и варианты, которые имеют увеличенную или сниженную активность и/или аффинность. Например, аминокислотная последовательность может быть изменена для получения антитела с требуемой аффинностью к связыванию с PCSK9. Модификация полипептидов представляет собой установившуюся методику в уровне техники и не требует подробного описания здесь. Модификация полипептидов проиллюстрирована в Примерах. Примеры модифицированных полипептидов включают полипептиды с консервативными заменами аминокислотных остатков, одну или несколько делеций или вставок аминокислот, которые не вызывают значительного вредного изменения функциональной активности или которые доводят до зрелости (усиливают) аффинность данного полипептида к его лиганду, или применение химических аналогов.
Вставки аминокислотных последовательностей включают амино- и/или карбоконцевые слияния с диапазоном длины от одного остатка до полипептидов, содержащих сто или более остатков, а также инсерции одного или множественных аминокислотных остатков внутри последовательности. Примеры концевых вставок включают антитело с N-концевым остатком метионила или антитело, слитое с эпитопной меткой. Другие инсерционные варианты молекулы антитела включают слияние с N- или C-концом антитела, фермента или полипептида, что повышает время полужизни анитела в кровотоке.
Замещенные варианты имеют по меньшей мере один удаленный аминокислотный остаток в молекуле антитела и другой остаток, вставленный на его место. Участки, представляющие собой большой интерес для замещающего мутагенеза, включают гипервариабельные участки, но изменения в FR также предусмотрены. Консервативные замены представлены в Таблице 2 под заголовком «консервативные замены». Если такие замены в результате приводят к изменению биологической активности, то более существенные замены, обозначенные в Таблице 2 как «иллюстративные замены» или как описанные далее в применении к классам аминокислот, могут быть введены и проведен скрининг полученных продуктов.
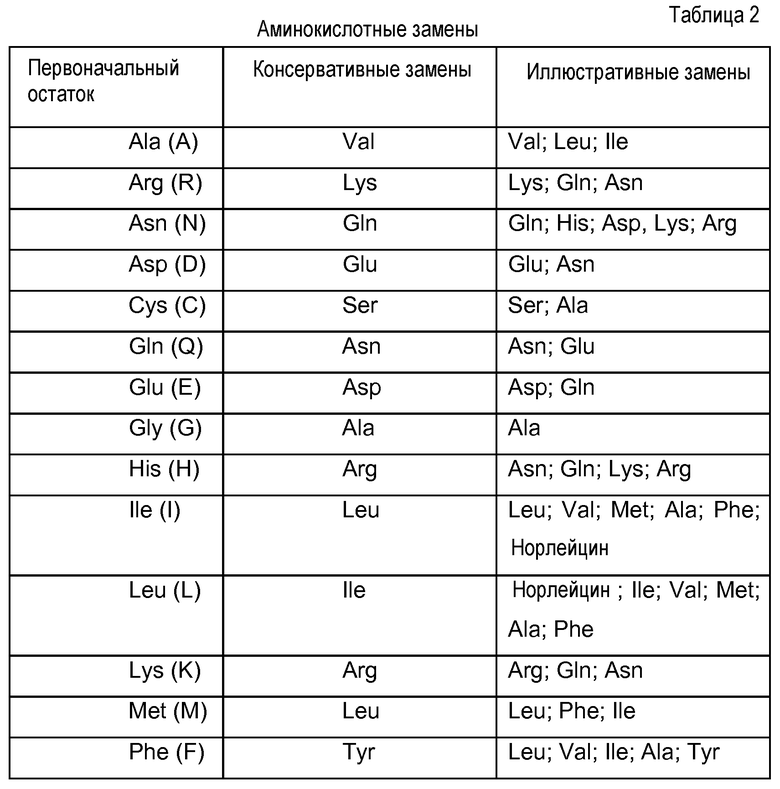
Значительные изменения биологических свойств антитела сопровождаются отбором замен, которые отличаются значительно по своему эффекту на поддержание (а) структуры полипептидного остова в области замены, например, складчатой или спиральной конформации, (b) заряда или гидрофобности молекулы в данном сайте-мишени, или (с) внутреннюю структуру боковой цепи. Встречающиеся в природе остатки разделены на группы, исходя из общих свойств боковой цепи:
(1) неполярные: Норлейцин, Met, Ala, Val, Leu, Ile;
(2) полярные незаряженные: Cys, Ser, Thr, Asn, Gln;
(3) кислые (отрицательно заряженные): Asp, Glu;
(4) основные (положительно заряженные): Lys, Arg;
(5) остатки, оказывающие влияние на ориентацию цепей: Gly, Pro; и
(6) ароматические: Trp, Tyr, Phe, His.
Неконсервативные замены происходят в результате замены члена одного из этих классов на другой класс.
Любой остаток цистеина, не вовлеченный в поддержание правильной конформации данного антитела, также может быть заменен, обычно на серин, для улучшения окислительной устойчивости данной молекулы, и для предотвращения измененного перекрестного сшивания. И наоборот, цистеиновая связь (связи) может быть введена в данное антитело для улучшения его стабильности, в частности, когда это антитело представляет собой фрагмент антитела, например фрагмент Fv.
Аминокислотные модификации могут колебаться от изменений или модификации одной или нескольких аминокислот до полного изменения области, например вариабельной области. Изменения в вариабельной области могут изменять аффинность и/или специфичность связывания. В некоторых вариантах осуществления не более чем от одной до пяти замен проведено в пределах домена CDR. В других вариантах осуществления не более чем от одной до трех консервативных аминокислотных замен проведено в пределах домена CDR. В других вариантах осуществления домен CDR представляет собой CDR Н3 и/или CDR L3.
Модификации также включают гликозилированные и негликозилированные полипептиды, а также полипептиды с другими посттрансляционными модификациями, такими как, например, гликозилирование с различными сахарами, ацетилирование и фосфорилирование. Антитела являются гликозилированными в консервативных положениях в их константных областях (Jefferis and Lund, 1997, Chem. Immunol. 65:111-128; Wright and Morrison, 1997, TibTECH 15:26-32). Олигосахаридные боковые цепи иммуноглобулинов оказывают действие на функцию белка (Boyd et al., 1996, Mol. Immunol. 32:1311-1318; Wittwe and Howard, 1990, Biochem. 29:4175-4180) и внутримолекулярные взаимодействия между участками гликопротеина, что может оказывать воздействие на конформацию и представляет трехмерную поверхность данного гликопротеина (Jefferis and Lund, supra; Wyss and Wagner, 1996, Current Opin. Biotech. 7:409-416). Олигосахариды могут также способствовать нацеливанию данного гликопротеина на некоторые молекулы на основании специфических распознающих структур. Также сообщалось, что гликозилирование антител воздействует на антителозависимую клеточноопосредованную цитотоксичность (ADCC). В частности, клетки СНО с регулируемой тетрациклином экспрессией β(1,4)-N-ацетилглюкозаминилтрансферазы III (GnTIII), гликозилтрансферазы, катализирующей образование делящейся пополам GlcNAc, как сообщалось, имеют улучшенную активность ADCC (Umana et al., 1999, Nature Biotech. 17:176-180).
Гликозилирование антител является обычно либо N-сопряженным, либо О-сопряженным. N-сопряженное относится к присоединению молекулы углевода к боковой цепи остатка аспарагина. Трипептидные последовательности аспарагин-Х-серин, аспарагин-Х-треонин и аспарагин-Х-цистеин, где Х представляет собой любую аминокислоту, кроме пролина, являются распознаваемыми последовательностями для ферментативного присоединения молекулы углевода к аспарагину боковой цепи. Таким образом, наличие любой из этих трипептидных последовательностей в полипептиде создает потенциальный участок гликозилирования. O-сопряженное гликозилирование относится к присоединению одного из сахаров N-ацетилгалактозамин, галактоза или ксилоза к гидроксиаминокислоте, чаще всего к серину или треонину, хотя 5-гидроксипролин или 5-гидроксилизин также могут быть использованы.
Добавление участков гликозилирования в антитело удобно выполняется посредством изменения аминокислотной последовательности, так чтобы она содержала одну или несколько из вышеупомянутых трипептидных последовательностей (для N-сопряженных участков гликозилирования). Изменение также может быть осуществлено в результате добавления, или замены на, одного или нескольких остатков серина или треонина в последовательность исходного антитела (для O-сопряженных участков гликозилирования).
Характер гликозилирования антитела также может быть изменен без изменения лежащей в основе нуклеотидной последовательности. Характер гликозилирования в значительной степени зависит от клетки-хозяина, используемой для экспрессии антитела. Поскольку клеточный тип, используемый для экспрессии рекомбинантных гликопротеинов, например антител, в качестве потенциальных терапевтических средств редко является нативной клеткой, вариации в характере гликозилирования данных антител могут быть ожидаемы (смотри, например, Hse et al., 1997, J. Biol. Chem. 272:9062-9070).
Кроме выбора клетки-хозяина, факторы, которые влияют на гликозилирование во время рекомбинантного получения антител, включают способ выращивания, состав культуральной среды, насыщенность культуральной среды, насыщение кислородом, pH, системы очистки и тому подобное. Различные способы были предложены для изменения характера гликозилирования, выполненного в определенном организме-хозяине, в том числе введение или сверхпродуцирование некоторых ферментов, вовлеченных в выработку олигосахаридов (патенты США №№ 5047335; 5510261 и 5278299). Гликозилирование, или некоторые типы гликозилирования, может быть ферментативно удалено из гликопротеина, например, используя эндогликозидазу Н (Endo H), N-гликозидазу F, эндогликозидазу F1, эндогликозидазу F2 и эндогликозидазу F3. Кроме того, рекомбинантная клетка-хозяин может быть генетически сконструирована таким образом, чтобы быть непригодной для обработки некоторых типов полисахаридов. Эти и схожие способы хорошо известны в уровне техники.
Другие методы модификации включают использование методов присоединения, известных в уровне техники, в том числе, но не ограничиваясь ими, ферментативные способы, окислительное замещение и хелатообразование. Можно использовать модификации, например, для присоединения меток для иммунологического анализа. Модифицированные полипептиды получают, используя общепринятые в данной области методы, и их скрининг может быть проведен с помощью стандартных испытаний, известных в уровне техники, некоторые из которых описаны ниже в Примерах.
В некоторых вариантах осуществления данного изобретения данное антитело содержит модифицированную константную область, такую как константная область, которая иммунологически инертна или частично инертна, например не запускает комплемент-опосредованный лизис, не стимулирует ADCC или не активирует микроглию; или имеют сниженные активности (по сравнению с немодифицированным антителом) в одном или нескольких из следующих процессов: запуск комплемент-опосредованного лизиса, стимулирование ADCC или активирование микроглии. Различные модификации константной области можно использовать для достижения оптимального уровня и/или комбинации эффекторных функций. Смотри, например, Morgan et al., 1995, Immunology 86:319-324; Lund et al., 1996, J. Immunology 157:4963-9 157:4963-4969; Idusogie et al., 2000, J. Immunology 164:4178-4184; Tao et al., 1989, J. Immunology 143:2595-2601; и Jefferis et al., 1998, Immunological Reviews 163:59-76. В некоторых вариантах осуществления константная область модифицирована, как описано в Eur. J. Immunol., 1999, 29:2613-2624; PCT публикации № WO99/58572; и/или патентной заявке Великобритании № 9809951.8. В других вариантах осуществления данное антитело содержит константную область тяжелой цепи IgG2 человека, включающую следующие мутации: A330P331 в S330S331 (нумерация аминокислот в соответствии с последовательностью IgG2 дикого типа). Eur. J. Immunol., 1999, 29:2613-2624. В других вариантах осуществления данная константная область является дегликозилированной для N-сопряженного гликозилирования. В некоторых вариантах осуществления данная константная область является дегликозилированной для N-сопряженного гликозилирования посредством изменения гликозилируемых аминокислотных остатков или фланкирующих остатков, которые являются частью распознаваемой последовательности N-гликозилования в данной константной области. Например, участок N-гликозилирования N297 может быть изменен в A, Q, K или H. Смотри, Tao et al., 1989, J. Immunology 143:2595-2601; и Jefferis et al., 1998, Immunological Reviews 163:59-76. В некоторых вариантах осуществления данная константная область является дегликозилированной для N-сопряженного гликозилирования. Данная константная область может быть дегликозилирована для N-сопряженного гликозилирования ферментативно (например, удаляя углевод с помощью фермента PNGase) или в результате экспрессии в клетке-хозяине с дефицитом гликозилирования.
Другие модификации антител включают антитела, которые были модифицированы, как описано в PCT публикации № WO 99/58572. Эти антитела содержат, в дополнение к домену связывания, направленного на молекулу-мишень, эффекторный домен, имеющий аминокислотную последовательность, по существу гомологичную всему или части константного домена тяжелой цепи иммуноглобулина человека. Эти антитела способны связывать молекулу-мишень без запуска значительного комплементопосредованного лизиса, или клеточноопосредованного разрушения данной мишени. В некоторых вариантах осуществления данный эффекторный домен способен специфически связывать FcRn и/или FcγRIIb. Они обычно основаны на химерных доменах, происходящих от двух или нескольких доменов СН2 тяжелой цепи иммуноглобулина человека. Антитела, модифицированные таким образом, являются чрезвычайно подходящими для использования в длительном лечении антителами, во избежание воспаления и других нежелательных реакций на традиционное лечение антителами.
Данное изобретение включает варианты доведения аффинности до созревания. Например, антитела с созревшей аффинностью могут быть получены с помощью процедур, известных в уровне техники (Marks et al., 1992, Bio/Technology, 10:779-783; Barbas et al., 1994, Proc Nat. Acad. Sci, USA 91:3809-3813; Schier et al., 1995, Gene, 169:147-155; Yelton et al., 1995, J. Immunol., 155:1994-2004; Jackson et al., 1995, J. Immunol., 154(7):3310-9; Hawkins et al., 1992, J. Mol. Biol., 226:889-896; и PCT публикация № WO2004/058184).
Следующие способы можно использовать для регулирования аффинности антитела и для описания характеристик CDR. Один способ описания характеристик CDR антитела и/или изменения (например, улучшение) связывающей аффинности полипептида, такого как антитело, называется «сканирующий мутагенез библиотеки». Как правило, сканирующий мутагенез библиотеки работает следующим образом. Аминокислоты в одном или нескольких положениях в данной CDR замещают на две или несколько (например, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20) аминокислот, используя методы, признанные в уровне техники. В результате этого генерируются небольшие библиотеки клонов (в некоторых вариантах осуществления, одна для каждого анализируемого положения аминокислоты), каждая со сложностью два или несколько членов (если две или несколько аминокислот замещены в каждом положении). Как правило, данная библиотека также включает клон, содержащий нативную (незамещенную) аминокислоту. На небольшом количестве клонов, например приблизительно 20-80 клонов (в зависимости от сложности данной библиотеки), из каждой библиотеки проводят скрининг на аффинность к связыванию с полипептидом-мишенью (или другой мишенью связывания), и выявляются кандидаты с повышенной, такой же, пониженной аффинностью или без связывания. Способы определения аффинности к связыванию хорошо известны в уровне техники. Аффинность к связыванию может быть определена в результате анализа поверхностного плазмонного резонанса Biacore, который выявляет 2-кратное или более различие в аффинности к связыванию. Biacore является чрезвычайно эффективным, когда исходное антитело уже связывает с относительно высокой аффинностью, например с KD приблизительно 10 нМ или ниже. Скрининг с использованием поверхностного плазмонного резонанса Biacore описан здесь в Примерах.
Аффинность к связывнию может быть определена с использованием Kinexa Biocensor, сцинтилляционного анализа сближения, ELISA, иммунологического анализа ORIGEN (IGEN), гашения флуоресценции, переноса флуоресценции, и/или дрожжевого дисплея. Также может быть проведен скрининг аффинности к связыванию с использованием подходящего биологического анализа.
В некоторых вариантах осуществления каждое аминокислотное положение в CDR замещено (в некоторых вариантах осуществления, по одному) всеми 20 встречающимися в природе аминокислотами с использованием способов мутагенеза, признанных в уровне техники (некоторые из которых описаны здесь). В результате этого генерируются небольшие библиотеки клонов (в некоторых вариантах осуществления, одна для каждого анализируемого положения аминокислоты), каждая со сложностью два или несколько членов (если все 20 аминокислот замещены в каждом положении).
В некоторых вариантах осуществления данная библиотека для скрининга содержит замены в двух или нескольких положениях, которые могут быть в той же самой CDR, или в двух или нескольких CDR. Таким образом, данная библиотека может содержать замещения в двух или нескольких положениях в одной CDR. Данная библиотека может содержать замещение в двух или нескольких положениях в двух или нескольких CDR. Данная библиотека может содержать замещение в 3, 4, 5 или более положениях, указанные положения обнаружены в двух, трех, четырех, пяти или шести CDR. Данное замещение может быть получено с использованием кодонов с низкой избыточностью. Смотри, например, Таблицу 2 у Balint et al., 1993, Gene 137(1):109-18.
Данная CDR может представлять собой CDRH3 и/или CDRL3. Данная CDR может быть одной или несколькими из CDRL1, CDRL2, CDRL3, CDRH1, CDRH2 и/или CDRH3. Данная CDR может быть CDR по Кабату (Kabat), CDR по Хотиа (Chothia) или расширенной CDR.
Можно провести секвенирование кандидатных соединений с улучшенным связыванием и тем самым выявить мутационную замену в CDR, приводящую к усиленной аффинности (также называемая «повышенная» замена). Кандитатные соединения, которые связаны, также могут быть секвенированы, тем самым выявляют замены CDR, сохраняющие связывание.
Могут быть проведены многократные циклы скрининга. Например, кандидатные соединения (каждое содержит аминокислотную замену в одном или нескольких положениях в одной или нескольких CDR) с улучшенным связыванием также могут быть полезны для конструирования второй библиотеки, содержащей, по меньшей мере, исходную и замещенную аминокислоту в каждом улучшенном положении CDR (то есть аминокислотное положение в данной CDR, в котором замещающая мутация проявляет улучшенное связывание). Получение, скрининг и отбор этой библиотеки обсуждается далее.
Сканирующий мутагенез библиотеки также обеспечивает способы описания характеристик CDR, поскольку частота клонов с улучшенным связыванием, таким же связыванием, сниженным связыванием и без связывания также обеспечивают информацию, относящуюся к значимости каждого аминокислотного положения для стабильности комплекса антитело-антиген. Например, если положение CDR сохраняет связывание при изменении всех 20 аминокислот, данное положение определено как положение, которое маловероятно необходимо для связывания антигена. С другой стороны, если положение CDR сохраняет связывание только в малом проценте замен, это положение распознается как положение, важное для функционирования CDR. Таким образом, способы сканирующего мутагенеза библиотеки вырабатывают информацию, относящуюся к положениям в CDR, которые могут быть изменены на многие разные аминокислоты (в том числе все 20 аминокислот), и положениям в CDR, которые не могут быть изменены или могут быть заменены только на некоторые из аминокислот.
Кандидатные соединения, которые улучшают аффинность, могут быть скомбинированы во вторую библиотеку, которая включает улучшенные аминокислоты и может дополнительно включать дополнительные замены в этом положении, в зависимости от сложности желаемой библиотеки, или допустимого использования методов желаемого скрининга или отбора. Кроме того, при необходимости, и смежное аминокислотное положение может быть рандомизировано для по меньшей мере двух или нескольких аминокислот. Рандомизация смежных аминокислот может обеспечить дополнительную конформационную гибкость в мутантной CDR, что, в свою очередь, может обеспечить или облегчить введение большего числа улучшающих мутаций. Данная библиотека также может содержать замены в положениях, которые не проявляют улучшенной аффинности в первом цикле скрининга.
Проводится скрининг или отбор в данной второй библиотеке для выявления членов библиотеки с улучшенной и/или измененной аффинностью к связыванию, с использованием любого способа, известного в уровне техники, в том числе скрининг с использованием анализа поверхностного плазмонного резонанса Biacore, и отбор с использованием любого метода отбора, известного в уровне техники, в том числе фаговый дисплей, дрожжевой дисплей и рибосомный дисплей.
Данное изобретение также охватывает слитые белки, содержащие один или несколько фрагментов или областей антител или полипептидов по данному изобретению. В одном варианте осуществления, обеспечен слитый полипептид, который состоит по меньшей мере из 10 заменимых аминокислот вариабельной области легкой цепи, представленной в SEQ ID NO: 53, 16, 17, 18 или 19, и/или по меньшей мере 10 аминокислот вариабельной области тяжелой цепи, представленной в SEQ ID NO: 54, 20, 21, 22 или 23. В других вариантах осуществления обеспечен слитый полипептид, который состоит из по меньшей мере приблизительно 10, по меньшей мере приблизительно 15, по меньшей мере приблизительно 20, по меньшей мере приблизительно 25 или по меньшей мере приблизительно 30 заменимых аминокислот вариабельной области легкой цепи и/или по меньшей мере приблизительно 10, по меньшей мере приблизительно 15, по меньшей мере приблизительно 20, по меньшей мере, приблизительно 25 или по меньшей мере приблизительно 30 заменимых аминокислот вариабельной области тяжелой цепи. В другом варианте осуществления данный слитый полипептид включает вариабельную область легкой цепи и/или вариабельную область тяжелой цепи, как представлено в любой из пар последовательностей, выбранных из SEQ ID NO: 53 и 54, 16 и 20, 17 и 21, 18 и 22 и 19 и 23. В другом варианте осуществления данный слитый полипептид содержит одну или несколько CDR. В других вариантах осуществления данный слитый полипептид содержит CDR H3 (VH CDR3) и/или CDR L3 (VL CDR3). Для целей данного изобретения слитый белок содержит одно или несколько антител и другую аминокислотную последовательность, с которой нет соединения в нативной молекуле, например гетерологичную последовательность или гомологичную последовательность из другой области. Примеры гетерологичных последовательностей включают, но не ограничиваются этим, «метку», такую как метка FLAG или метка 6His. Метки хорошо известны в уровне техники.
Слитый полипептид можно получить способами, известными в уровне техники, например, синтетически или рекомбинантно. В основном слитые белки по этому изобретению создают в результате получения экспрессии кодирующего их полинуклеотида, используя рекомбинантные способы, описанные здесь, хотя они могут быть получены любыми другими способами, известными в уровне техники, в том числе, например, с помощью химического синтеза.
Данное изобретение также обеспечивает композиции, содержащие антитела или полипептиды, конъюгированные (например, сопряженные) с агентом, который облегчает контакт с твердой подложкой (такой как биотин или авидин). Для упрощения ссылки в основном будут сделаны на антитела с пониманием, что эти методы применимы к любому связывающемуся с PCSK9 и/или антагонистическому против PCSK9 варианту осуществления, описанному здесь. Конъюгация, как правило, относится к сопряжению этих компонентов, как описано здесь. Сопряжение (которое обычно фиксирует эти компоненты в непосредственной близости, по меньшей мере, для введения) может быть достигнуто любым количеством способов. Например, непосредственная реакция между агентом и антителом возможна, когда каждый обладает заместителем, способным реагировать с другим. Например, нуклеофильная группа, такая как амино- или сульфгидрильная группа, на одном соединении может быть способна к реагированию с карбонилсодержащей группой, такой как ангидрид или галогенангидрид, или с алкильной группой, содержащей легко уходящую группу (например, галоген) на другом соединении.
Антитело или полипептид по данному изобретению может быть сопряжено с агентом-меткой, таким как флуоресцентная молекула, радиоактивная молекула или любые другие метки, известные в уровне техники. В уровне техники известны метки, которые в большинстве случаев обеспечивают (либо непосредственно, либо косвенно) сигнал.
Данное изобретение также обеспечивает композиции (в том числе, фармацевтические композиции) и наборы, содержащие, как разъясняет данное описание изобретения, любое или все антитела и/или полипептиды, описанные здесь.
Данное изобретение также обеспечивает выделенные полинуклеотиды, кодирующие антитела и пептиды по данному изобретению, и векторы и клетки-хозяева, содержащие данный полинуклеотид.
Соответственно, данное изобретение обеспечивает полинуклеотиды (или композиции, в том числе фармацевтические композиции), включающие полинуклеотиды, кодирующие любое из следующего: антитела 4A5, 5A10, 6F6, 7D4, L1L3, или любой их фрагмент или часть, имеющие способность вызывать антагонизм к PCSK9.
В другом аспекте данное изобретение обеспечивает полинуклеотиды, кодирующие любые антитела (в том числе фрагменты антител) и полипептиды, описанные здесь, такие как антитела и полипептиды, имеющие ослабленную эффекторную функцию. Полинуклеотиды могут быть получены и экспрессированы с помощью процедур, известных в уровне техники.
В другом аспекте данное изобретение обеспечивает композиции (такие как фармацевтические композиции), содержащие любой из полинуклеотидов по данному изобретению. В некоторых вариантах осуществления данная композиция содержит экспрессирующий вектор, содержащий полинуклеотид, кодирующий антитело, как описано здесь. В другом варианте осуществления данная композиция содержит экспрессирующий вектор, содержащий полинуклеотид, кодирующий любое из антител или полипептидов, описанных здесь. В других вариантах осуществления данная композиция содержит любой или оба полинуклеотида, представленные в SEQ ID NO: 25 и SEQ ID NO: 26. Далее здесь описаны экспрессирующие векторы и применение полинуклеотидных композиций.
В другом аспекте данное изобретение обеспечивает способ получения любого из описанных здесь полинуклеотидов.
Также настоящим изобретением охвачены полинуклеотиды, комплементарные любым таким последовательностям. Полинуклеотиды могут быть одноцепочечными (кодирующими или антисмысловыми) или двухцепочными и могут представлять собой молекулы ДНК (геномная, кДНК или синтетическая) или РНК. Молекулы РНК включают молекулы гяРНК, которые содержат интроны и соответствуют молекуле ДНК взаимно однозначно, и молекулы мРНК, которые не содержат интронов. Дополнительные кодирующие или некодирующие последовательности могут, но необязательно, присутствовать внутри полинуклеотида по настоящему изобретению, и полинуклеотид может, но не обязан быть сопряжен с другими молекулами и/или подложками.
Полинуклеотиды могут содержать нативную последовательность (а именно эндогенная последовательность, которая кодирует антитело или его часть) или может содержать вариант такой последовательности. Полинуклеотидные варианты содержат одну или несколько замен, дополнений, делеций и/или инсерций, так что иммунореактивность кодируемого полипептида не уменьшена по отношению к нативной иммунореактивной молекуле. Эффект на иммунореактивность кодируемого полипептида можно в большинстве случаев оценить, как описано здесь. Варианты предпочтительно проявляют по меньшей мере 70% идентичность, более предпочтительно по меньшей мере приблизительно 80% идентичность, еще более предпочтительно приблизительно 90% идентичность и наиболее предпочтительно приблизительно 95% идентичность с полинуклеотидной последовательностью, которая кодирует нативное антитело или его часть.
Две полинуклеотидные или полипептидные последовательности называются «идентичными», если данная последовательность нуклеотидов или аминокислот в данных двух последовательностях является одной и той же, при выравнивании для максимального соответствия, как описано далее. Сравнения между двумя последовательностями обычно осуществляют посредством сравнения данных последовательностей через окно сравнения для идентификации и сравнения локальных областей схожести последовательностей. «Окно сравнения», как используется здесь, относится к участку из по меньшей мере приблизительно 20 соприкасающихся положений, обычно от 30 до приблизительно 75 или 40 до приблизительно 50, в котором последовательность может быть сравнена с референсной последовательностью с таким же числом прилежащих положений после оптимального выравнивания данных двух последовательностей.
Оптимальное выравнивание последовательностей для сравнения может быть проведено с помощью программы Megalign в блоке Lasergene программного обеспечения для биоинформатики (DNASTAR, Inc., Madison, WI), используя параметры по умолчанию. Эта программа объединяет несколько схем выравнивания, описанных в следующих справочных документах: Dayhoff, M.O., 1978, A model of evolutionary change in proteins - Matrices for detecting distant relationships. In Dayhoff, M.O. (ed.) Atlas of Protein Sequence and Structure (National Biomedical Research Foundation, Washington DC), Vol. 5, Suppl. 3, pp. 345-358; Hein J., 1990, Unified Approach to Alignment and Phylogenes pp. 626-645 Methods in Enzymology vol. 183, (Academic Press, Inc., San Diego, CA); Higgins, D.G. and Sharp, P.M., 1989, CABIOS 5:151-153; Myers, E.W. and Muller W., 1988, CABIOS 4:11-17; Robinson, E.D., 1971, Comb. Theor. 11:105; Santou, N., Nes, M., 1987, Mol. Biol. Evol. 4:406-425; Sneath, P.H.A. and Sokal, R. R., 1973, Numerical Taxonomy the Principles and Practice of Numerical Taxonomy (Freeman Press, San Francisco, CA); Wilbur, W.J. and Lipman, D.J., 1983, Proc. Natl. Acad. Sci. USA 80:726-730.
Предпочтительно «процент идентичности последовательности» определен посредством сравнения двух оптимально выровненных последовательностей через окно сравнения из по меньшей мере 20 положений, где данный участок полинуклеотидной или полипептидной последовательности в окне сравнения может содержать дополнения или делеции (то есть разрывы) на 20 процентов или менее, обычно от 5 до 15 процентов или от 10 до 12 процентов по сравнению с референсными последовательностями (которые не содержат дополнений или делеций) для оптимального выравнивания данных двух последовательностей. Процент рассчитывается, определяя количество положений, в которых идентичные основания нуклеиновых кислот или аминокислотные остатки встречаются в обеих последовательностях, чтобы получить число совпадающих положений, деля данное число совпадающих положений на общее количество положений в референсной последовательности (то есть размер окна) и умножая полученные результаты на 100, для получения процента идентичности последовательности.
Варианты могут также, или альтернативно, быть по существу гомологичными нативному гену, или его части, или комплемента. Такие полинуклеотидные варианты являются способными к гибридизации в умеренно строгих условиях с природной последовательностью ДНК, кодирующей нативное антитело (или комплементарной последовательностью).
Соответствующие «умеренно строгие условия» включают предварительную отмывку в растворе 5× SSC, 0,5% SDS, 1,0 мM ЭДТА (pH 8,0); гибридизацию при 50°C-65°C, 5× SSC, в течение ночи; с последующей промывкой два раза при 65°C, в течение 20 минут каждый раз, с использованием 2×, 0,5× и 0,2× SSC, содержащей 0,1% SDS.
«Чрезвычайно строгие условия» или «условия с высокой строгостью» представляют собой такие, при которых: (1) используется низкая ионная сила и высокая температура для промывки, например 0,015 M хлорид натрия/0,0015 M цитрат натрия/0,1% додецилсульфат натрия при 50°C; (2) во время гибридизации используется денатурирующее вещество, такое как формамид, например 50% (объем/объем) формамид с 0,1% альбумином бычьей сыворотки/0,1% фиколл/0,1% поливинилпирролидон/50 мM натриево-фосфатный буфер при pH 6,5 с 750 мM хлорида натрия, 75 мM цитрат натрия при 42°C; или (3) используется 50% формамид, 5× SSC (0,75 M NaCl, 0,075 M цитрат натрия), 50 мM фосфат натрия (pH 6,8), 0,1% пирофосфат натрия, 5× раствор Денхардта, озвученная ультразвуком ДНК из спермы лососевых (50 мкг/мл), 0,1% SDS, и 10% сульфат декстрана при 42°C, с промывками при 42°C в 0,2× SSC (хлорид натрия/цитрат натрия) и 50% формамиде при 55°C, с последующей промывкой при 55°С в условиях высокой строгости с использованием 0,1× SSC, содержащим ЭДТА. Квалифицированный специалист в данной области поймет, как подобрать температуру, ионную силу и тому подобное, по мере необходимости, чтобы учесть факторы, такие как длина пробы и тому подобное.
Специалисту с обычными навыками в данной области будет понятно, что в результате вырожденности генетического кода существует множество нуклеотидных последовательностей, которые кодируют полипептид, как описанный здесь. Некоторые из этих полинуклеотидов несут минимальную гомологичность нуклеотидной последовательности любого нативного гена. Тем не менее, настоящим изобретением, в частности, предусмотрены полинуклеотиды, которые отличаются в результате различий использования кодонов. Дополнительно, аллели данных генов, содержащие полинуклеотидные последовательности, предоставленные здесь, входят в диапазон настоящего изобретения. Аллели представляют собой эндогенные гены, которые изменены в результате одной или нескольких мутаций, таких как делеции, дополнения и/или замены нуклеотидов. Полученные мРНК и белок могут, но необязательно, иметь измененную структуру или функцию. Аллели можно идентифицировать с помощью стандартных процедур (таких как гибридизация, амплификация и/или сравнение баз данных последовательностей).
Полинуклеотиды по данному изобретению можно получить, используя химический синтез, рекомбинантные методы или ПЦР. Способы химического синтеза хорошо известны в уровне техники и не нуждаются в подробном описании здесь. Квалифицированный специалист в данной области может использовать последовательности, обеспеченные здесь, и коммерческие ДНК-синтезаторы для получения необходимой последовательности ДНК.
Для получения полинуклеотидов с помощью рекомбинантных методов, полинуклеотид, содержащий требуемую последовательность, может быть встроен в подходящий вектор, и данный вектор, в свою очередь, может быть введен в подходящую клетку-хозяина для репликации и размножения, как обсуждается далее здесь. Полинуклеотиды могут быть встроены в клетки-хозяев любыми способами, известными в уровне техники. Клетки могут быть трансформированы в результате введения экзогенного полинуклеотида посредством прямого поглощения, эндоцитоза, трансфекции, F-скрещивания или электропорации. После встраивания экзогенный полинуклеотид может поддерживаться в данной клетке как неинтегрированный вектор (такой как плазмида) или интегрированный в геном клетки-хозяина. Амплифицированный таким образом полинуклеотид может быть выделен из клетки-хозяина способами, хорошо известными в уровне техники. Смотри, например, Sambrook et al., 1989, supra.
Альтернативно ПЦР делает возможным воспроизведение последовательностей ДНК. Метод ПЦР хорошо известен в уровне техники и описан в патентах США №№ 4683195, 4800159, 4754065 и 4683202, а также в PCR: The Polymerase Chain Reaction, MuIMs et al., 1994, eds. (Birkauswer Press, Boston, MA).
РНК может быть получена, используя выделенную ДНК в соответствующем векторе и встраивая ее в подходящую клетку-хозяина. После репликации клетки и транскрипции ДНК в РНК данная РНК затем может быть выделена с помощью методов, хорошо известных специалисту в данной области, как описано, например, у Sambrook et al., 1989, supra.
Подходящие векторы для клонирования могут быть сконструированы в соответствии со стандартными способами или могут быть выбраны из большого числа векторов для клонирования, доступных в уровне техники. Несмотря на то что векторы для клонирования могут различаться в зависимости от используемой клетки-хозяина, пригодные векторы для клонирования в основном имеют способность к самовоспроизводству, могут обладать единственной мишенью для определенной эндонуклеазы рестрикции и/или могут нести гены маркера, который может быть использован при отборе клонов, содержащих данный вектор. Подходящие примеры включают плазмиды и бактериальные вирусы, например, pUC18, pUC19, Bluescript (например, pBS SK+) и его производные, mp18, mp19, pBR322, pMB9, ColE1, pCR1, RP4, фаговые ДНК и челночные векторы, такие как pSA3 и pAT28. Эти и многие другие векторы для клонирования доступны у коммерческих поставщиков, таких как BioRad, Strategene и Invitrogen.
Экспрессирующие векторы, как правило, представляют собой воспроизводимые полинуклеотидные конструкции, которые содержат полинуклеотид по данному изобретению. Это означает, что экспрессирующий вектор обязан быть воспроизводимым в клетках-хозяевах либо как эписомы, либо как составляющий элемент хромосомной ДНК. Подходящие экспрессирующие векторы включают, но не ограничены ими, плазмиды, вирусные векторы, в том числе аденовирусы, аденоассоциированные вирусы, ретровирусы, космиды и экспрессирующий вектор(ы), раскрытый в PCT публикации № WO 87/04462. Векторные компоненты могут, как правило, включать, но не ограничиваясь этим, один или несколько из следующего: сигнальная последовательность; начало репликации; один или несколько маркерных генов; подходящие элементы, контролирующие транскрипцию (такие как промоторы, энхансеры и терминаторы). Для экспрессии (а именно трансляции) также обычно требуется один или несколько элементов, контролирующих транскрипцию, такие как участки связывания рибосом, участки инициации трансляции и стоп-кодоны.
Векторы, содержащие представляющие интерес полинуклеотиды, могут быть встроены в клетку-хозяина любым из многих соответствующих способов, в том числе посредством электропорации, трансфекции с использованием хлорида кальция, хлорида рубидия, фосфата кальция, DEAE-декстрана или другого вещества; бомбардировки микрочастицами; липофекции; и инфицирования (например, когда вектор представляет собой инфицирующий агент, например вирус коровьей оспы). Выбор векторов для трансформации или полинуклеотидов зачастую будет зависеть от свойств клетки-хозяина.
Данное изобретение также обеспечивает клетки-хозяева, содержащие любой из полинуклеотидов, описанных здесь. Любые клетки-хозяева, способные сверхпродуцировать гетерологичные ДНК, могут быть использованы для цели выделения генов, кодирующих данное представляющее интерес антитело, полипептид или белок. Неограничивающие примеры клеток-хозяев млекопитающих включают, но не ограничиваясь ими, клетки COS, HeLa, NSO и CHO. Смотри также PCT публикацию № WO 87/04462. Подходящие клетки-хозяева, полученные не от млекопитающих, включают прокариотов (например, E. coli или B.subtillis) и дрожжи (как например, S.cerevisae, S.pombe; или K.lactis). Предпочтительно данные клетки-хозяева экспрессируют кДНК на уровне приблизительно 5-кратно превышающем, более предпочтительно 10-кратно превышающем, даже более предпочтительно 20-кратно превышающем уровень соответствующего эндогенного представляющего интерес антитела или белка, если присутствует, в данных клетках-хозяевах. Скрининг данных клеток-хозяев на предмет специфического связывания с PCSK9 или доменом PCSK9, осуществляется посредством иммунологического анализа или FACS. Может быть определена клетка, сверхпродуцирующая представляющее интерес антитело или белок.
C. Композиции
Композиции, используемые в методах по данному изобретению, включают эффективное количество антагонистического антитела против PCSK9, полипептида, полученного из антагонистического антитела против PCSK9, или других антагонистов PCSK9, описанных здесь. Примеры таких композиций, а также способ их составления, также описаны в более раннем разделе, и далее. В одном варианте осуществления данная композиция дополнительно содержит антагонист против PCSK9. В другом варианте осуществления данная композиция содержит одно или несколько антител-антагонистов против PCSK9. В других вариантах осуществления данное антитело-антагонист против PCSK9 распознает PCSK9 человека. В других вариантах осуществления данное антитело-антагонист против PCSK9 является гуманизированным. В других вариантах осуществления данное антитело-антагонист против PCSK9 содержит константную область, которая не запускает нежелательной или непредусмотренной иммунной реакции, такой как антителоопосредованный лизис или ADCC. В других вариантах осуществления данное антитело-антагонист против PCSK9 содержит одну или несколько CDR данного антитела (как например, одну, две, три, четыре, пять или в некоторых вариантах осуществления все шесть CDR). В некоторых вариантах осуществления данное антитело-антагонист против PCSK9 является человеческим.
Подразумевается, что данные композиции могут содержать более одного антитела-антагониста против PCSK9 (например, смеси антител-антагонистов против PCSK9, которые распознают различные эпитопы на PCSK9). Другие иллюстративные композиции содержат более одного антитела-антагониста против PCSK9, которые распознают один и тот же эпитоп(ы), или различные виды антител-антагонистов против PCSK9, которые связываются с различными эпитопами на PCSK9.
Композиция, используемая в настоящем изобретении, может дополнительно содержать фармацевтически приемлемые носители, наполнители или стабилизирующие вещества (Remington: The Science and Practice of Pharmacy 20th Ed., 2000, Lippincott Williams and Wilkins, Ed. K. E. Hoover), в форме лиофилизированных составов или водных растворов. Приемлемые носители, наполнители или стабилизирующие вещества являются нетоксичными для реципиентов в данных дозировках и концентрациях и могут включать буферы, такие как фосфатные, цитратный или других органических кислот; антиоксиданты, в том числе аскорбиновую кислоту и метионин; консерванты (такие как октадецилдиметилбензил аммоний хлорид, хлорид гексаметония; хлорид бензалкония, хлорид бензетония; фенольный, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехин; резорцин; циклогексанол; 3-пентанол; и м-крезол); низкомолекулярные (менее чем приблизительно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин, или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу, или декстраны; хелатобразующие агенты, такие как ЭДТА; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, комплексы Zn-белок); и/или неионные сурфактанты, такие как TWEENTM, PLURONICSTM или полиэтиленгликоль (PEG). Фармацевтически приемлемые наполнители дополнительно описаны здесь.
В одном варианте осуществления антитело вводится в организм в композиции в виде стерильного водного раствора, имеющего значение рН в диапазоне от приблизительно 5,0 до приблизительно 6,5 и содержащего антитело приблизительно от 1 мг/мл до приблизительно 200 мг/мл, гистидиновый буфер приблизительно от 1 миллимолярного до приблизительно 100 миллимолярного, полисорбат 80 приблизительно от 0,01 мг/мл до приблизительно 10 мг/мл, трегалозу приблизительно от 100 миллимолярной до приблизительно 400 миллимолярной и двунатриевый ЭДТА дигидрат приблизительно от 0,01 миилимолярного до приблизительно 1,0 миллимолярного.
Антитело-антагонит против PCSK9 и его композиции также можно использовать в сочетании с другими агентами, которые предназначены для усиления и/или дополнения эффективности данных веществ.
D. Наборы
Данное изобретение также обеспечивает наборы для использования в экспресс-системах. Наборы по данному изобретению включают один или несколько контейнеров, содержащих антитело-антагонист против PCSK9 (например, гуманизированное антитело) или пептид, описанный здесь, и инструкции по применению в соответствии с любыми способами по данному изобретению, описанными здесь. Как правило, эти инструкции содержат описание применения антагонистического антитела, пептида или аптамера против PCSK9 для вышеописанных терапевтических воздействий.
В некоторых вариантах осуществления данное антитело представляет собой гуманизированное антитело. В некоторых вариантах осуществления данное антитело представляет собой антитело человека. В других вариантах осуществления данное антитело является моноклональным антителом. Инструкции по применению антитела-антагониста против PCSK9, как правило, включают информацию о дозировке, режиме дозирования, пути введения для планируемого лечения. Эти контейнеры могут содержать однократные дозы, многодозовые упаковки (например, упаковки с многократными дозами) или субоднократные дозы. Инструкции, которыми снабжены наборы по данному изобретению, в большинстве случаев представляют собой письменные инструкции на этикетке или листовке-вкладыше в упаковке (например, бумажный бланк, включенный в набор), но также пригодны распознаваемые машиной инструкции (например, инструкции на магнитном диске или диске оптической памяти).
Наборы по данному изобретению находятся в подходящей упаковке. Подходящая упаковка включает, но не ограничивается ими, ампулы, флаконы, баночки, гибкую тару (например, запаянные пакеты из пленки майлар или полиэтиленовые пакеты) и тому подобное. Также предусмотрены упаковки для применения в комбинации со специальным устройством, таким как ингалятор, устройство для назального введения (например, пульверизатор) или устройство для инфузии, такое как минипомпа. Набор может иметь стерильное отверстие для доступа (например, данный контейнер может представлять собой мешок для раствора для внутривенного введения или флакон, имеющий пробку, протыкаемую иглой для подкожной инъекции). Контейнер также может иметь стерильное отверстие для доступа (например, контейнер может представлять собой пакет с раствором для внутривенного введения или флакон с пробкой, прокалываемой стерильной иглой для подкожных инъекций). По меньшей мере одно действующее вещество в данной композиции представляет собой антитело-антагонист против PCSK9. Данный контейнер (например, предварительно заполненный шприц или автоинжектор) может дополнительно содержать второе фармацевтически действующее вещество.
В наборах могут быть, при желании, обеспечены дополнительные компоненты, такие как буферы, и информация для интерпретации. Обычно набор содержит контейнер и ярлык или листок(листки)-вкладыш на контейнере, или присоединенный к контейнеру.
Мутации и модификации
Для экспрессии антител против PCSK9 по настоящему изобретению фрагменты ДНК, кодирующие области VH и VL, сначала могут быть получены с использованием любого метода из описанных выше. Различные модификации, например мутации, делеции и/или дополнения, также могут быть введены в последовательности ДНК с помощью стандартных методов, известных специалисту в данной области. Например, мутагенез может быть осуществлен с использованием стандартных методов, таких как ПЦР-опосредованный мутагенез, в которых мутированные нуклеотиды введены в праймеры для ПЦР, так что ПЦР содержит требуемые мутации, или сайт-направленный мутагенез.
Одним типом замены, например, который можно провести, является замена одного или нескольких цистеинов в антителе, который может быть химически реактивным, на другой остаток, такой как, без ограничения, аланин или серин. Например, может быть произведена замена неканонического цистеина. Замена может быть осуществлена в CDR, или каркасной области вариабельного домена, или в константном домене антитела. В некоторых вариантах осуществления цистеин является каноническим.
Антитела также могут быть модифицированы, например, в вариабельных доменах тяжелой и/или легкой цепей, например, для изменения связывающих свойств данного антитела. Например, мутация может быть осуществлена в одной или нескольких областях CDR для повышения или понижения KD данного антитела для PCSK9, для повышения или понижения koff или для изменения специфичности связывания данного антитела. Способы сайт-направленного мутагенеза хорошо известны в уровне техники. Смотри, например, Sambrook et al. и Ausubel et al., выше.
Модификации или мутации также могут быть осуществлены в каркасной области или константном домене для повышения времени полужизни антитела против PCSK9. Смотри, например, PCT публикацию № WO 00/09560. Мутация в каркасной области или константном домене также может быть осуществлена для изменения иммуногенности данного антитела, для обеспечения участка для ковалентного или нековалентного связывания с другой молекулой или для изменения таких свойств, как фиксация комплемента, связывание FcR и антителозависимая клеточноопосредованная цитотоксичность. В соответствии с данным изобретением единственное антитело может содержать мутации в любой одной или нескольких CDR, или каркасных областях вариабельного домена, или в константном домене.
В процессе, называемом «процесс возвращения к зародышевому генотипу» («germlining»), некоторые аминокислоты в последовательностях VH и VL могут быть изменены для того, чтобы совпадать с таковыми, обнаруженными в природе в последовательностях VH и VL зародышевой линии. В частности, аминокислотные последовательности каркасных областей последовательностей VH и VL могут быть видоизменены для совпадения с последовательностями зародышевой линии, чтобы снизить риск иммуногенности при введении данного антитела. Последовательности ДНК зародышевой линии для генов VH и VL человека известны в уровне техники (смотри, например, базу данных последовательностей зародышевых линий человека «Vbase»; также смотри Kabat, E. A., et al. (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH публикацию № 91-3242; Tomlinson et al., 1992, J. Mol. Biol. 227:776-798; и Cox et al., 1994, Eur. J. Immunol. 24:827-836.
Другой тип аминокислотной замены, который может быть осуществлен, представляет собой удаление потенциальных протеолитических сайтов в данном антителе. Такие сайты могут встречаться в CDR или каркасной области вариабельного домена или в константном домене антитела. Замена цистеиновых остатков и удаление протеолитических сайтов могут снизить риск гетерогенности в продукте антитела и тем самым повысить его гомогенность. Другой тип аминокислотной замены устраняет пары аспарагин-глицин, которые образуют потенциальные участки дезамидирования, в результате изменения одного или обоих этих остатков. В другом примере может быть расщеплен лизин С-конца тяжелой цепи антитела против PCSK9 по данному изобретению. В различных вариантах осуществления данного изобретения тяжелые и легкие цепи антител против PCSK9 могут при желании включать сигнальную последовательность.
После получения фрагментов ДНК, кодирующих сегменты VH и VL по настоящему изобретению, эти фрагметы ДНК могут быть далее обработаны с использованием стандартных методов рекомбинантных ДНК, например, для преобразования генов вариабельной области в гены полноразмерной цепи антитела, в гены Fab-фрагмента или ген scFv. При этих манипуляциях фрагмент ДНК, кодирующий VL или VH, функционально связан с другим фрагментом ДНК, кодирующим другой белок, такой как белок константной области антитела или гибкий линкер. Используемый здесь термин «функционально связанный» означает, что два фрагмента ДНК соединены таким образом, что аминокислотные последовательности, кодируемые этими двумя фрагментами ДНК, сохраняются в рамке.
Выделенная ДНК, кодирующая область VH, может быть преобразована в ген полноразмерной тяжелой цепи посредством функционального связывания данной ДНК, кодирующей VH, с другой молекулой ДНК, кодирующей константные области тяжелой цепи (CH1, CH2 и CH3). Последовательности генов константной области тяжелой цепи человека известны в уровне техники (смотри, например, Kabat, E. A., et al., 1991, Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publ. No. 91-3242), и фрагменты ДНК, вмещающие эти области, могут быть получены с помощью стандартной ПЦР-амплификации. Константная область тяжелой цепи может представлять собой константную область IgG1, IgG2, IgG3, IgG4, IgA, IgE, IgM или IgD, но наиболее предпочтительной является константная область IgG1 или IgG2. Последовательность константной области IgG может быть любой из множества аллелей или аллотипов, встречающихся среди различных индивидуумов, например Gm(1), Gm(2), Gm(3) и Gm(17). Эти аллотипы являются представителями встречающихся в природе аминокислотных замен в константных областях IgG1. Для гена тяжелой цепи фрагмента Fab ДНК, кодирующая VH, может быть функционально связана с другой молекулой ДНК, кодирующей только константную область СН1 тяжелой цепи. Константная область тяжелой цепи СН1 может быть получена из любых генов тяжелых цепей.
Выделенная ДНК, кодирующая область VL, может быть преобразована в ген полноразмерной легкой цепи (а также в ген Fab легкой цепи) посредством функционального связывания данной ДНК, кодирующей VL, с другой молекулой ДНК, кодирующей константную область легкой цепи, CL. Последовательности генов константной области легкой цепи человека известны в уровне техники (смотри, например, Kabat, E. A., et al., 1991, Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publ. No. 91-3242), и фрагменты ДНК, вмещающие эти области, могут быть получены с помощью стандартной ПЦР-амплификации. Константная область легкой цепи может представлять собой константную область каппа или лямбда. Константная область каппа может быть любой из разных аллелей, встречающихся у различных индивидуумов, например Inv(1), Inv(2) и Inv(3). Константная область лямбда может быть получена из любого из трех генов лямбда.
Для создания гена scFv фрагменты ДНК, кодирующие VH и VL, функционально связывают с другим фрагментом, кодирующим гибкий линкер, например кодирующим аминокислотную последовательность (Gly4 -Ser)3, так что данные последовательности VH и VL могут быть экспрессированы в виде сопряженного одноцепочечного белка с областями VL и VH, соединенными данным гибким линкером (смотри, например, Bird et al., 1988, Science 242:423-426; Huston et al., 1988, Proc. Natl. Acad. Sci. USA 85:5879-5883; McCafferty et al., 1990, Nature 348:552-554). Данное одноцепочечное антитело может быть моновалентным, если используются только одиночные VH и VL, бивалентным, если использовали два VH и VL, или поливалентным, если использовали больше двух VH и VL. Могут быть созданы биспецифические или поливалентные антитела, которые специфически связываются с PCSK9 и с другой молекулой.
В другом варианте осуществления может быть получено слитое антитело или иммуноадгезин, который содержит все или часть антитела против PCSK9 по настоящему изобретению, связанное с другим полипептидом. В другом варианте осуществления только вариабельные домены антитела против PCSK9 связаны с данным полипептидом. В другом варианте осуществления домен VH антитела против PCSK9 связан с первым полипептидом, в то время как домен VL антитела против PCSK9 связан со вторым полипептидом, который соединен с первым полипептидом таким образом, что домены VH и VL могут взаимодействовать друг с другом для формирования участка связывания антигена. В другом предпочтительном варианте осуществления домен VH отделен от домена VL линкером, так что данные домены VH и VL могут взаимодействовать друг с другом. Атитело VH-линкер-VL затем соединяют с представляющим интерес полипептидом. Кроме того, могут быть созданы слитые антитела, в которых два (или несколько) одноцепочечных антитела соединены друг с другом. Это используется, если есть желание создать двухвалентное или поливалентное антитело на единственной полипептидной цепи или если есть желание создать биспецифическое антитело.
В других вариантах осуществления другие модифицированные антитела могут быть получены с использованием молекул нуклеиновых кислот, кодирующих антитело против PCSK9. Например, «каппа тела» (Ill et al., 1997, Protein Eng. 10:949-57), «минитела» (Martin et al., 1994, EMBO J. 13:5303-9), «диатела» (Holliger et al., 1993, Proc. Natl. Acad. Sci. USA 90:6444-6448) или «Janusins» (Traunecker et al., 1991, EMBO J. 10:3655-3659 and Traunecker et al., 1992, Int. J. Cancer (Suppl.) 7:51-52) могут быть получены с помощью стандартных методов молекулярной биологии, следуя идеям данного описания изобретения.
Биспецифические антитела или антигенсвязывающие фрагменты, могут быть получены различными способами, в том числе слиянием гибридом или связыванием фрагментов Fab'. Смотри, например, Songsivilai & Lachmann, 1990, Clin. Exp. Immunol. 79:315-321, Kostelny et al., 1992, J. Immunol. 148:1547-1553. Кроме того, биспецифические антитела могут быть сформированы, как «диатела» или «Janusins». В некоторых вариантах осуществления биспецифические антитела связываются с двумя различными эпитопами на PCSK9. В некоторых вариантах осуществления данные модифицированные антитела, описанные выше, получены с использованием одного или нескольких вариабельных доменов или областей CDR человеческого антитела против PCSK9, обеспеченного здесь.
Получение антигенспецифических антител
Более 500 поликлональных и моноклональных антител создано против рекомбинантного полноразмерного PCSK9 человека, рекомбинантного полноразмерного PCSK9 мыши, и была проведена оценка различных синтетических пептидов на предмет их способности к угнетающей регуляции общего белка LDLR в культивированных клетках печени человека Huh7. Среди этих антител была группа антител, выращенных для и реактивных с группой полипептидов из 12-20 аминокислотных остатков, которые, как было предсказано исходя из структуры PCSK9, покрывают большую часть поверхности белка. При наибольшей концентрации самые лучшие антитела демонстрировали только приблизительно 60% блокирующей активности.
Таким образом, был применен альтернативный и, до настоящего времени, не изученный подход, а именно выработка моноклональных антител с использованием PCSK9-«нуль» мышей, иммунизированных рекомбинантным полноразмерным белком PCSK9. Этот способ получения антитела обеспечил выход антител-антагонистов, которые демонстрируют полную блокировку связывания PCSK9 с LDLR, полную блокировку PCSK9-опосредованного снижения уровней LDLR в клетках Huh7 и снижение LDLc in vivo, в том числе у мышей, до уровней, сравнимых с таковыми, наблюдаемыми у мышей PCSK9 -/-, как показано в Примере 7.
Репрезентативные антитела (гибридомы) по настоящему изобретению были помещены на хранение в Американскую коллекцию типовых культур (АТСС) 28 февраля 2008 года, и присвоенные их номера доступа представлены в Таблице 3. Были помещены на хранение гибридомы для антител 4А5, 5А10, 6F6 и 7D4.
Примеры
Пример 1: Выработка и скрининг антител-антагонистов против PCSK9
Общие процедуры по иммунизации животных для выработки моноклональных антител:
Мышам Balb/c или 129/bl6 pcsk9 -/- делали инъекцию с 100 мкг антигена 5 раз в течение 13-дневного плана иммунизации. PCSK9 -/- (которые являются «нуль»- или нокаут-мышами) могут быть получены от, или как описано у, Rashid et al., 2005, Proc Natl Acad Sci USA 102:5374. Смотри также патент США № 7300754. Для первых 4 инъекций антиген был приготовлен посредством смешивания рекомбинантных белков с адъювантом, иммуноген был введен посредством инъекции в заднюю часть шеи, в подошву стопы или внутрибрюшинно, приблизительно каждые 3 дня в течение 11-дневного курса, с последним бустерным введением внутривенно, без адъюванта. На 13 день мышей безболезненно умерщвляли, и извлекали их селезенки. Лимфоциты были иммортализованы посредством слияния с разработанной клеточной линией для создания клонов гибридом с использованием стандартной гибридомной технологии, распределены в 96-луночных планшетах. Клоны оставляли для роста, затем проводили отбор посредством скрининга с помощью ELISA, используя иммунизирующий антиген, как представлено ниже.
ELISA скрининг антител:
Проводили скрининг супернатанта от растущих клонов гибридомы по отдельности на их способность связывать рекомбинантный PCSK9 человека или рекомбинантный PCSK9 мыши. Эти исследования проводили в 96-луночных планшетах, покрытых в течение ночи 100 мкл 1 мкг/мл раствора одного из антигенов. После каждого этапа планшеты отмывали от избыточных реагентов с помощью PBS, содержащего 0,05% Tween-20. Затем планшеты были блокированы с помощью PBS, содержащего 0,05% BSA. Супернатант добавляли в эти планшеты и инкубировали при комнатной температуре в течение 2 часов. Добавляли анти-мышиный Fc козла, конъюгированный с пероксидазой хрена (HRP), для связывания с мышиными антителами, соединенными с данным антигеном. Затем добавляли тетраметил бензидин в качестве субстрата для HRP, для того чтобы определить количество мышиного антитела, находящегося в данном супернатанте. Реакцию останавливали и относительное количество антитела вычисляли посредством считывания поглощения при 450 нм. Клоны гибридомы, которые секретировали антитела, способные к связыванию либо с мышиным, либо с человеческим PCSK9, были отобраны для дальнейшего анализа.
PCSK9-опосредованная угнетающая регуляция LDLR в клетках Huh7:
Клоны гибридомы, секретирующие антитела, связывающие человеческий или мышиный PCSK9, были размножены и супернатанты были собраны. Общие IgG были очищены из приблизительно 10 мл супернатанта с использованием бусин с протеином А, диализированы в буфере PBS, и конечный объем был уменьшен для получения растворов с 0,7-1 мг/мл антител. Очищенные антитела затем использовали для тестирования их возможности ингибировать способность PCSK9 опосредовать угнетающую регуляцию LDLR в клетках Huh7. Клетки Huh7 высевали в 96-луночные планшеты и оставляли расти до достижения 80% конфлюентности в среде RPMI, содержащей 10% FBS, 4 мМ глутамин, пенициллин и стрептавидин. Данную среду заменяли на другую, содержащую 10% делипидизированный FBS, на 8-16 часов для индукции экспрессии LDLR. Затем клетки инкубировали в течение 8-16 часов с 40 мкл/лунку среды для экспрессии 293, дополненной 6 мкг/мл человеческого (предпочтительно) или мышиного PCSK9, с или без 70-100 мкг/мл тестируемых антител. В конце инкубирования удаляли среду, содержащую PCSK9 и антитело, и клетки лизировали с помощью 17 мкл лизирующего буфера посредством встряхивания в течение часа при 4°С. Данный лизирующий буфер состоит из 50 мМ глицерин-фосфата, 10 мМ HEPES рН 7,4, 1% Triton X-100, 20 мM NaCl и смеси ингибиторов протеаз (Roche). Клеточные лизаты собирали и проводили анализ на уровни белка LDLR через окрашивание вестерн-блотов после электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия. Клоны гибридом, вырабатывающие антитела, которые могут частично или полностью сохранять уровень LDLR, были отобраны для дальнейшего анализа. Под «анализом угнетающей регуляции LDLR» подразумевают вышеописанный анализ с использованием клеток Huh7.
На Фигуре 1 продемонстрирован эффект антагонистических моноклональных антител 7D4.4, 4A5.G3, 6F6.G10.3 и 5A10.B8 против PCSK9 на способность человеческого и мышиного PCSK9 подавлять LDLR в культивированных клетках Huh7. Использовали 100 нМ мышиный или человеческий рекомбинантный PCSK9, и серийные разведения 25-800 нМ антител. A) PCSK9 мыши. B) PCSK9 человека. На фигурах представлены вестерн-блоттинг, демонстрирующие, что данные антитела, в большинстве случаев, являются более эффективными при блокировании функции PCSK9 человека, чем PCSK9 мыши. Несколько антител имеют, как правило, схожие аффинности к PCSK9 человека, но различаются своими аффинностями к PCSK9 мыши.
Пример 2: Определение связывающей аффинности антитела
Аффиности антител к PCSK9 определяли с помощью поверхностного плазмонного резонанса на биосенсоре Biacore 3000, оборудованном сенсорным чипом исследовательского качества, с использованием подвижного буфера HBS-EP (Biacore AB, Uppsala, Sweden - now GE Healthcare). Кроличьи поликлональные анти-Ms IgG были аминосоединенными насыщенно с чипом, используя стандартные химикаты N-гидроксисукцинимид/этилдиметиламинопропил карбодиимид (NHS/EDC). Буфер заменяли на HBS-EP+1 мг/мл BSA+1 мг/мл CM-декстран. Полноразмерные IgG против PCSK9 разбавляли приблизительно до 15 мкг/мл и захватывали на 1 мин при 5 мкл/мин для получения уровней приблизительно 500 RU на проточную кювету, оставляя одну пустой в качестве референсного канала. 3,73-302 нМ hPCSK9 или 2,54-206 нМ mPCSK9 впрыскивали в виде 5-членных 3-кратных серий в течение 1 мин при 100 мкл/мин. Диссоциацию отслеживали в течение 5 мин. Чип восстанавливали после последнего впрыска каждого титра с помощью двух 30-секундных стимулов 100 мМ фосфорной кислоты. Буферные циклы обеспечили бланки для двойного сравнения с эталоном данных, которые затем в целом были аппроксимированы в простую модель связывания с использованием программного обеспечения Biaevaluation v.4.1. Аффинности были выведены из соотношения кинетических констант скоростей (KD=koff/kon). Результаты Примера 2 представлены в Таблице 4. Эти результаты демонстрируют, что данные антитела имеют превосходную активность к PCSK9 мыши или PCSK9 человека, как показано.
Пример 3: Анализ эффекта антител против PCSK9 на взаимодействие PCSK9-LDLR
Было показано, что PCSK9 связывается с LDLR с аффинностью 180 нМ при нейтральном значении рН (Cunningham et al., 2007, Nat Struct Mol Biol, 14(5):413-9). Рекомбинантный белок PCSK9 человека или мыши был биотинилирован с использованием реагентов Pierce, следуя инструкциям фирмы-производителя. Планшеты для ELISA (Corning Mixisorb) покрывали раствором 1 мкг/мл внеклеточного домена рекомбинантного LDLR (R&D Systems) в каждой лунке, при 4°C в течение ночи, блокировали с помощью 2% BSA+PBS в течение 2 часов при комнатной температуре, а затем 5 раз промывали с помощью буфера для промывки (1× PBS+0,05% Tween-20). Лунки инкубировали с 50 мкл биотинилированного белка PCSK9 заданной концентрации в течение 1 ч при комнатной температуре. Связь LDLR-PCSK9 может быть стабилизирована посредством добавления 50 мкл раствора 4% FDH+4% сахароза+PBS и инкубирования в течение 5 мин. Лунки промывали 5 раз с помощью буфера для промывки, инкубировали с 1:2000 разведением HRP, конъюгированным со стрептавидином (Invitrogen) в течение 1 ч при комнатной температуре, промывали 5 раз с помощью буфера для промывки. Субстрат TMB добавляли в лунки, данный раствор инкубировали от 20 до 30 мин при комнатной температуре, и данную реакцию останавливали добавлением 1 М фосфорной кислоты. Сигналы считывали при 450 нм.
На Фигуре 2 продемонстрирован эффект дозы антагонистических против PCSK9 моноклональных антител 6F6.G10.3, 7D4.4, 4A5.G3, 5A10.B8, антитела отрицательного контроля 42H7 и PBS, на блокирование связывания рекомбинантного биотинилированного PCSK9 человека и PCSK9 мыши с иммобилизированным внеклеточным доменом рекомбинантного LDLR in vitro. В части A) продемонстрировано связывание PCSK9 человека с внеклеточным доменом LDLR человека и что 7D4, 4A5, 5A10 и 6F6 являются эффективными при блокировании связывания, тогда как 42H7 и PBS не являются. В части B) продемонстрировано связывание PCSK9 мыши с внеклеточным доменом LDLR человека.
Данное взаимодействие также может быть продемонстрировано в свободном растворе при нейтральном значении рН. На Фигуре 3 представлен эффект дозы антагонистических моноклональных антител 6F6.G10.3, 7D4.4, 4A5.G3 и 5A10.B8 против PCSK9 на блокирование связывания рекомбинантного биотинилированного PCSK9 человека (30 нМ) с меченным европием внеклеточным доменом рекомбинантного LDLR (10 нМ) в растворе с нейтральным значением рН in vitro. В этом анализе измеряется связывание в свободном растворе при нейтральном значении рН.
Пример 4: Картирование эпитопов/связывание антител с использованием кристаллической структуры комплекса L1L3:PCSK9, Biacore и мутагенеза
a. Кристаллическая структура комплекса L1L3:PCSK9. Остатки были идентифицированы в результате вычисления разницы площади доступной поверхности между кристаллической структурой L1L3:PCSK9 и структурой одного PCSK9. Остатки PCSK9, которые показывают скрытую площадь поверхности при формировании комплекса с антителом L1L3, были учтены как часть эпитопа. Данная доступная для растворения поверхность белка определена как локус в центре зондовой сферы (представляет молекулу растворителя радиусом 1,4 Å), поскольку она переворачивает ван-дер-ваальсову поверхность белка. Доступная для растворения поверхность расчитывается посредством создания поверхностных точек на расширенной сфере около каждого атома (на расстоянии от центра атома, эквивалентном сумме радиусов данного атома и зонда) и исключая те, которые находятся внутри эквивалентных сфер ассоциированных с соседними атомами, как реализовано в программе AREAIMOL (Briggs, P.J., 2000, CCP4 Newsletter No. 38, CCLRC, Daresbury).
Результаты анализа кристаллической структуры представлены на Фигуре 23. На Фигуре 23A представлена кристаллическая структура PCSK9 (светло-серое изображение поверхности), связанного с антителом L1L3 (черное изображение). Эпитоп для связывания L1L3 с PCSK9 включает остатки 153-155, 194, 197, 237-239, 367, 369, 374-379 и 381 аминокислотной последовательности PCSK9 (SEQ ID NO: 53). Для сравнения, эпитоп для связывания домена EGF LDLR с PCSK9 включает остатки 153-155, 194, 238, 367, 369, 372, 374-375 и 377-381 (Kwon et al., 2008, PNAS 105:1820-1825).
b. Группы антител и эпитопов, исходя из конкуренции за связывание с PCSK9. Полноразмерные IgG были амино-соединены с сенсорным чипом СМ5 (три на чип, конечное приблизительно 7000 RU), используя EDC/NHS-опосредованную стандартную химию аминосоединения. Одну проточную кювету оставляли немодифицированной для обеспечения референсного канала. PCSK9 человека (100 нМ) предварительно смешивали со смесью IgG (конечное 500 нМ) и эти комплексы впрыскивали в чип посредством 1-минутной инъекции при 10 мкл/мин. Антитела, которые связываются с конкурирующими эпитопами, будут блокировать связывание PCSK9 с антителом, иммобилизированным на чипе. Альтернативно использовали классический сэндвич-метод, сначала впрыскивая PCSK9 человека при 50 нМ в течение 1 мин со скоростью 10 мкл/мин (для привязывания его посредством IgG на данный чип) и затем связывая смесь IgG (конечное 500 нМ каждое) в течение 2 мин каждое. Зафиксированные IgG восстанавливали с помощью слабой кислоты (мягкий буфер для элюирования Pierce+1M NaCl). Антитела, направленные против различных известных эпитопов, использовали в качестве контролей для положительного образования сэндвич в этом анализе.
c. Структурно-обусловленный мутагенез для картирования антителосвязывающих эпитопов. Основываясь на кристаллической структуре PCSK9 и возможном вовлечении D374 в связывание LDLR (Cunningham et al., 2007, Nat Struct MoI Biol, 14(5):413-419), девятнадцать мутантных поверхностных остатков PCSK9 (F379A, I369A, R194A, D374Y, D238R, T377R, K222A, R199A, F216A, R218A, R237A, D192R, D367R, R165A, R167A, A443T, A53V, I474V, H449A), поблизости или вдалеке от положения D374, были выбраны для изменения для картирования антителосвязывающих эпитопов.
d. Мутант и образование антител. Данные 19 точковых мутаций были сгенерированы из ранее описанной конструкции ДНК дикого типа (Cunningham et al., 2007, выше), используя стандартные ДНК технологии. Мутантные белки были экспрессированы с использованием неустойчивой трансфекции в клетках HEK293T, секретированы в клеточной среде. Данные мутантные белки были очищены с помощью высокопропускной системы AKTA Xpress (GE Healthcare) посредством Ni2+ гель-хроматографии, с использованием условий, схожих с описанными ранее. Концентрации белка определяли с помощью прибора LabChip (Bio-Rad). Блокирующие PCSK9 мышиные антитела 4A5, 7D4, 5A10 и 6F6 были экспрессированы посредством неустойчивой трансфекции в клетках HEK293F, очищены с помощью колонки с протеином G, элюированы буфером 0,1 М глицина при значении рН 2,8 и нейтрализованы в 1,0 М Tris при pH 9,0.
e. Области PCSK9, которые контактируют с моноклональными антителами 5A10 и 7D4 (получение описано далее) были определены посредством томографии белка (Sidec AB, Stockholm, Sweden). Петли в положениях 186-200, 371-379, 176-181, 278-283, 449-453, 402-406 и 236-245 данного PCSK9 были близки к аминокислотным остаткам данного антитела. Последовательности, соответствующие петлям, представлены в Таблице 5, и в предпочтительном варианте осуществления, антагонисты по данному изобретению связываются с одной или несколькими из этих последовательностей в PSCK9.
f. Biacore-связывание мутантов с иммобилизированным LDLR. Белок внеклеточного домена рекомбинантного LDLR был зафиксирован на чипе Biacore SA. Каждый мутантный белок был инъецирован в Biacore-3000 M) в двух копиях от 25 мМ до 0,012 мМ и при пяти концентрациях от 1°С, с подвижным буфером 50 мМ Tris pH 7,5, 2 мM CaCl2, 200 мM NaCl, 0,02% P20 и 1 мг/мл BSA. Все результаты точно соответствуют кинетической модели связывания 1:1. В соответствии с ожидаемым мутация остатков с непосредственным контактом с доменом EGF-A (F379A, R194A, I369A, T377R, D238R) значительно ослабляет (в 10-100 крат) связывание LDLR. Более того, три мутации, не контактирующие с EGF-A (R199A, R218A, K222A) продемонстрировали более слабое связывание (5-15-кратное). Эти новые данные указывают на то, что они вовлечены в связывание других доменов LDLR. В целом эти эксперименты подтверждают целостность и активность данных мутантов для последующих экспериментов по картированию эпитопов.
g. Связывание мутантов с иммобилизированными антителами 4A5, 7D4, 5A10 и 6F6. Биотинилированные антитела против PCSK9 были иммобилизированы на чипах SA с применением стандартных способов. Эксперименты по связыванию мутантов были осуществлены с помощью Biacore 3000 при 25°C с подвижным буфером 50 мM Tris-HCl pH 7,5, 150 мM NaCl и 0,02% P20. Мутанты были протестированы при концентрациях 333 нМ или 111 нМ в дупликатах с теми, которые дают ослабленное связывание по сравнению с диким типом, из-за остатков, вовлеченных в связывание mAb (перечислены ниже).
Пример 5: Клонирование и определение последовательности антител
Один миллион клеток гибридомы гомогенизировали с помощью спин-колонок QIAshredder, и общую РНК выделяли с использованием набора RNAeasy Micro фирмы QIAGEN. Используя набор Superscript III RT производства фирмы Invitrogen, синтезировали кДНК. Вариабельные области антител против PCSK9 клонировали с использованием наборов мышиный IgG-Primer Sets фирмы Novagen, которые содержат вырожденные праймеры для клонирования генов тяжелой цепи IgG мыши и каппа и лямбда легких цепей IgG мыши. Условия ПЦР были следующие: 1 цикл при 92°C в течение 2 мин; два цикла при 94°C в течение 30 с, 44°C в течение 30 с и 72°C в течение 2 мин; два цикла при 94°C в течение 30 с, 46°C в течение 30 с и 72°C в течение 2 мин; два цикла при 94°C в течение 30 с, 48°C в течение 30 ск и 72°C в течение 2 мин; два цикла при 94°C в течение 30 с, 50°C в течение 30 с и 72°C в течение 2 мин; два цикла при 94°C в течение 30 с, 52°C в течение 30 с и 72°C в течение 2 мин; с последующими 35 циклами при 94°C в течение 30 с, 54°C в течение 30 с и 72°C в течение 45 с. Полученные ПЦР-продукты клонировали в вектор для клонирования Topo-TA, фирмы Invitrogen, и секвенировали. Последовательности клонированных антител были подтверждены с помощью N-концевого секвенирования первых 10 аминокислот исходных антител, полученных из асцитов.
Пример 6: Выработка антигенов для иммунизации
Рекомбинантный белок PCSK9 человека получали, как описано у Cunningham et al., 2007, Nat Struct Mol Biol, 14(5):413-9. Для получения рекомбинантного белка PCSK9 мыши кДНК мышиного PCSK9 была клонирована в экспрессирующий вектор PRK5 млекопитающего с добавлением метки 6-His на С-конце, используя методы, известные в уровне техники, неустойчиво трансфицирована и экспрессирована в клетках HEK293. Рекомбинантный белок очищали от кондиционированной среды с помощью Ni-колонки.
Поверхностные пептиды человеческого и мышиного PCSK9 отбирали, исходя из структуры белка PCSK9, и синтезировали с помощью Elim Biopharmaceuticals.
Пример 7: PCSK9-специфические антитела в качестве антагонистов против PCSK9
1. Идентификация PCSK9-специфических антител-антагонистов
a. Идентификация антител, блокирующих PCSK9
Мышиные антитела к человеческому и/или мышиному PCSK9 были выработаны в результате иммунизации мышей с помощью синтетических пептидов PCSK9 человека и PCSK9 мыши, полученных как описано в Примере 6, или рекомбинантных белков, и проводили скрининг антител посредством ELISA-анализа, с использованием рекомбинантного белка PCSK9 человека и/или мыши в качестве антигенов, как описано в Примере 1 и других стандартных гибридомных способах. Более 500 положительных клонов было получено и выращено до конфлуентности в 6-луночных планшетах в 10 мл среды. Собирали супернатанты, и общие иммуноглобулины в кондиционированной среде были очищены с помощью mAb Select (Pierce). Способность очищенных и концентрированных мышиных IgG ингибировать функцию мышиного и человеческого PCSK9 была протестирована в клетках Huh7, используя способы, описанные в Примере 1. Клоны гибридомы, экспрессирующие IgG, которые проявляют некоторую степень блокады, были размножены и повторно протестированы. 60 перспективных клонов были субклонированы, размножены и инъецированы либо в Balb/c, либо в безтимусных мышей, для формирования асцита. Антитела, очищенные из асцитной жидкости, были повторно протестированы на их способность ингибировать угнетающую регуляцию LDLR человеческим или мышиным PCSK9 в клетках Huh7. Четыре гибридомных клона, 4A5, 5A10, 6F6 и 7D4, были идентифицированы как обладающие способностью полностью ингибировать функцию PCSK9 человека, и, по меньшей мере, частично ингибировать функцию PCSK9 мыши. Для определения IC50 для каждого из этих блокирующих антител, серийные разведения IgG использовали в данном исследовании, начиная со 100 мкг/мл до 3,125 мкг/мл, с концентрацией человеческого и мышиного PCSK9 постоянно равной 6 мкг/мл.
b. Действие антагонистов PCSK9 на связь PCSK9-LDLR
Было показано, что PCSK9 локализуется совместно с LDLR в клеточных компартментах (Lagace et al., 2006, J Clin Inv, 116(11):2995-3005). Рекомбинантный белок PCSK9 также связывается с внеклеточным доменом LDLR in vitro (Fisher et al., 2007, JBC, 282(28):20502-12). Для определения связи между ингибированием PCSK9-опосредованной угнетающей регуляции LDLR и ингибированием связи PCSK9-LDLR антителами, авторы данного изобретения протестировали антитела против PCSK9, которые частично или полностью блокируют функцию PCSK9 на LDLR, и образцы антител, которые не блокируют. Все частично антагонистические антитела также частично ингибировали связывание внеклеточного домена LDLR с PCSK9, за исключением одного. Антитела-антагонисты, которые полностью блокируют функцию PCSK9, а именно 4A5, 5A10, 6F6 и 7D4, также полностью блокировали связывание внеклеточного домена LDLR с PCSK9 (Таблица 5). Значения IC50 для этих четырех антител коррелируют с их аффинностью к связыванию с PCSK9.
c. Определение эпитопов данных блокирующих антител
На Фигуре 4 представлено связывание анти-PSCK9 антител с эпитопами. В части (A) представлена информация об эпитопе анти-PCSK9 mAb, определенная по связыванию с синтетическими 13-18-мерными пептидами или связыванию эпитопа посредством Biacore. В части (B) отображена, посредством анализа Biacore, способность иммобилизированных антител 6F6, 5A10 и 4A5 к связыванию с человеческим PCSK9, предварительно смешанным с mAb, указанными по оси y.
Другое моноклональное антитело против PCSK9, называемое 6G7, связывается с рекомбинантным PCSK9 мыши, но не с PCSK9 человека. Смотри Таблицу 6. 6G7, 4A5, 5A10, 6F6 и 7D4 взаимно исключают способность друг друга к связыванию с мышиным PCSK9. Гибридный анализ между мышиным и человеческим PCSK9 показал, что для связывания 6G7 с PCSK9 необходим каталитический домен. Смотри Таблицу 6. Таким образом, участки связывания на 4A5, 5A10, 6F6 и 7D4 перекрывают каталитический участок и/или эпитоп, связанный с 6G7.
d. Определение видоспецифических последовательностей антител против PCSK9
Для определения видоспецифичности данных антител против PCSK9 антитела инкубировали с плазмой, полученной от различных видов, и полученные комплексы были очищены и протестированы с независимым антителом против PCSK9 с помощью вестерн-блоттинга. Антитела 4A5, 5A10, 6F6 и 7D4 распознавали PCSK9 человека, человекообразных обезьян, мыши и крысы. Смотри Фигуру 5. Анитело 6G7 распознавало только мышиный PCSK9, и независимое контрольное антитело 42H7 не распознавало ни один из протестированных PCSK9. Id.
e. Определение последовательностей антител-антагонистов против PCSK9
Аминокислотные последовательности вариабельных доменов PCSK9-антител 4A5, 5A10, 6F6 и 7D4 были определены используя метод, описанный в Примере 5. Данные последовательности указывают на то, что данные антитела являются родственными, но отличными друг от друга. В Таблице 1 представлены аминокислотные последовательности вариабельных областей каждого антитела. В Таблице 7 представлены последовательности CDR легких цепей и тяжелых цепей из Таблицы 1, как определено по методам Кабата (Kabat) и Хотиа (Chotia).
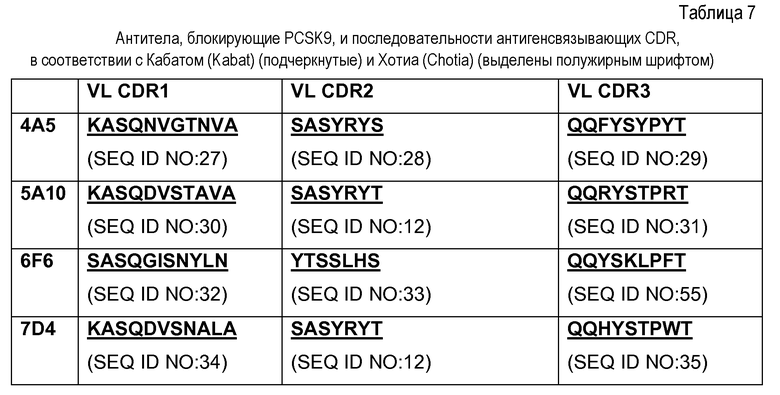

Анти-PCSK9 IgG 4A5, 5A10 и 6F6 с помощью амина были соединены с чипом Biacore. hPCSK9 (100 нМ) смешивали с 500 нМ 4A5, 5A10, 6F6 или 7D4 в различных соотношениях и впрыскивали в течение 1 мин со скоростью 10 мкл/мин. Данные четыре антитела взаимно блокируют друг друга независимо от исследованной ориентации, что указывает на то, что они все связываются с конкурирующими эпитопами. В отличие от этого они способны образовывать «сэндвич»-комплексы с другими частично блокирующими антителами, которые были картированы в специфических областях, с помощью синтетических пептидов.
2. Действие PCSK9-специфических антител в качестве антагониста против PCSK9 in vivo
a. Антитела-антагонисты против PCSK9 снижают уровень холестерина в сыворотке крови у мышей
Для определения того, могут ли моноклональные антитела-антагонисты против PCSK9 воздействовать на уровни холестерина in vivo, ингибируя функцию внеклеточного PCSK9, действие 7D4 было протестировано против мышиного PCSK9 in vitro, на холестерин сыворотки крови, после инъекции мышам. Самцов мышей C57/bl6 в возрасте 6-7 недель содержали при 12 ч светотемновом цикле, на день -7 пускали кровь для сбора приблизительно 70 мкл сыворотки крови. Антитело-антагонист 7D4 против PCSK9 и контрольное, совпадающее по изотипу, моноклональное антитело вводили 7-недельным самцам мышей C57/bl6 посредством внутрибрюшинных (i.p.) инъекций в день 0, 1, 2 и 3. Мышей безболезненно умерщвляли на день 4 без предварительного ограничения в пище и собирали образцы сыворотки крови. Все замороженные образцы сыворотки крови были отправлены в лаборатории IDEXX для измерений общего холестерина, триглицеридов, HDL-холестерина и LDL-холестерина. На Фигуре 6 показано, что 7D4 снижал уровень холестерина в сыворотке крови на 48%, в то время как контрольное антитело не имело никакого значительного эффекта. И количество, и процент сокращения были похожи с таковыми, которые сообщались для мышей PCSK9-/- (нокаут PCSK9 мыши), что указывает на то, что можно добиться полного или почти полного ингибирования функции PCSK9, блокируя только внеклеточный PCSK9, и что внутриклеточный PCSK9 играет небольшую роль, или не играет никакой, в угнетающей регуляции LDLR в нормальных физиологических условиях. Как и ожидалось, печеночные уровни LDLR были стимулированы у животных, которым было введено 7D4, по сравнению с животными, которым было введено контрольное антитело (Фигура 6).
b. Частично блокирующие антитела не влияют на уровни холестерина в крови
На Фигуре 7 показано, что частично антагонистическое поликлональное анти-PCSK9 mAb CRN6 не влияет на уровни холестерина у мышей. Двум группам 8-недельных мышей C57/bl6 (n=10 мышей/группа) пускали кровь и проводили тест уровней холестерина на день -7; мышам вводили дозы 15 мг/кг/день антитела CRN6 или контрольного антитела, посредством в/в введения в день 0, 1, 2 и 3; и затем пускали кровь и проверяли уровни холестерина через 24 часа после последней дозы. На Фигуре 7A показано, что антитело CRN6 частично блокирует опосредованную PCSK9 угнетающую регуляцию LDLR в клетках Huh7 in vitro. На Фигуре 7В показано, что введение антитела CRN6 не влияет на уровни холестерина в сыворотке крови у мышей.
c. Пролонгированный эффект mAb-антагониста против PCSK9 на уровни холестерина сыворотки крови у мышей.
Исследование периода действия было осуществлено для определения времени начала и продолжительности снижающего уровень холестерина действия антител-антагонистов против PCSK9 у мышей. mAb 7D4 или контрольный физиологический раствор вводили посредством в/в инъекции в дозе 10 мг/кг или 3 мл/кг 48 6-недельным мышам C57/bl6. Восемь мышей из каждой обрабатываемой группы были безболезненно умерщвлены на день 1, 2, 4, 7, 14 и 21 после инъекции. Однократная инъекция 7D4 вызвала быстрый и продолжительный эффект снижения уровней холестерина сыворотки крови. Снижение на 25% холестерина в сыворотке крови наблюдали через 24 ч после инъекции. Смотри Фигуру 8. Максимальное снижение холестерина сыворотки крови наблюдали на 7 день. В день 21 снижение холестерина больше не является статистически значимым. В части B) представлены уровни HDL-холестерина. Уровни LDL-холестерина были очень низкие.
На Фигуре 9 показано, что антитело-антагонист против PCSK9 mAb 7D4 дозозависимо снижает уровни общего холестерина, HDL и LDL в сыворотке крови у мышей. 8-недельным мышам C57/bl6 из шести групп (n=8/группа) пускали кровь и проводили тест базальных уровней холестерина в день -7 и вводили обозначенные дозы антител или физиологического раствора в день 0, 1, 2 и 3 посредством внутрибрюшинной (i.p.) болюсной инъекции. Образцы сыворотки крови собирали и проводили определение уровней холестерина через 24 часа после последней дозы. На Фигуре 9A представлены уровни общего холестерина, которые снижены до менее 60% от контроля после введения от 3 до 30 мг/кг/день. Максимальный эффект на общий холестерин наблюдали при дозе 10 мг/кг, и статистически значимое снижение при 1 мг/кг. На Фигуре 9B представлены уровни HDL, которые снижены до менее 70% после введения от 3 до 30 мг/кг/день. На Фигуре 9C представлены уровни LDL, которые снижены почти до нуля при всех протестированных дозах от 0,3 мг/кг/день и выше.
d. Эффект дозы антител-антагонистов специфических к PCSK9 у мышей
На Фигуре 10 показано, что анти-PCSK9 антагонистическое антитело 5А10 дозозависимо снижает уровни холестерина у мышей. На Фигуре 10A представлены шесть групп 8-недельных мышей C57/bl6 (n=8/группа), которым были введены указанные дозы антител или физиологический раствор ежедневно в день 0, 1, 2 и 3, посредством в/в болюсной инъекции. Образцы сыворотки крови собирали и проводили определение уровней холестерина через 24 часа после последней дозы, и было показано ступенчатое снижение с повышением дозы антитела. На Фигуре 10B представлены пять групп 8-недельных мышей C57/bl6 (n=8/группа), которым были введены указанные дозы антител или физиологический раствор в день 0 посредством внутрибрюшинной (i.p.) болюсной инъекции. Собирали образцы сыворотки крови и определяли уровни холестерина на 7 день, и также показали ступенчатое снижение с увеличением доз антитела.
На Фигуре 11 показано, что анти-PCSK9 антагонистические антитела 4A5 и 6F6 снижают уровни холестерина у мышей в дозозависимой манере. 8-недельным мышам C57/bl6 (n=8/группа) вводили обозначенные дозы антител или физиологического раствора в день 0 посредством внутрибрюшинной (i.p.) болюсной инъекции. Собирали образцы сыворотки крови и определяли уровни холестерина на 7 день. На Фигуре 11A антитело 4А5 демонстрирует ступенчатое снижение общего холестерина сыворотки крови с повышением дозы антитела. На Фигуре 11B антитело 6F6 показывает снижение общего холестерина сыворотки крови при дозе 10 мг/кг/день.
Анти-PCSK9 антитела-антагонисты 4A5, 5A10, 6F6 и 7D4 повышают печеночные уровни LDLR у мышей, как показано в результате вестерн-блоттинга. Смотри Фигуру 12. Для 4A5, 5A10 и 6F6, 8-недельным мышам C57/bl6 вводили 10 мг/кг антитела или физиологический раствор в день 0, посредством в/в болюсной инъекции, на 7 день животных безболезненно умерщвляли, и лизат цельной печени от 3 отдельных животных анализировали на уровни белка LDLR и GAPDH посредством вестерн-блоттинга. Для антитела 7D4, 8-недельным мышам Bl6/c57 вводили 10 мг/кг антител в день 0, 1, 2 и 3 посредством внутрибрюшинной (i.p.) болюсной инъекции, животных безболезненно умерщвляли на день 4, и лизат цельной печени от 3 отдельных животных анализировали на уровни белка LDLR и GAPDH посредством вестерн-блоттинга. У всех мышей, получивших инъекции антител, выявили высокие уровни LDLR по сравнению с контрольными мышами, получавших PBS.
На Фигуре 13 показано, что анти-PCSK9 антитело-антагонист не оказывает действие на мышей LDLR-/-. 8-недельным мышам LDLR-/- (мыши LDLR KO) вводили 10 мг/кг 4A5 или физиологический раствор в день 0, посредством внутрибрюшинной (i.p.) болюсной инъекции. Образцы сыворотки крови (от n=9-10 мышей) собирали и проводили определение уровней холестерина на день 7. Введение этого антитела не изменяло заметно уровни общего холестерина, HDL или LDL в сыворотке крови.
На Фигуре 14 показано, что комплексное лечение мышей анти-PCSK9 антителами-антагонистами может значительно снизить общий холестерин сыворотки крови. 8-недельным мышам C57/bl6 вводили обозначенные дозы антител или PBS в день 0, 7, 14 и 21 посредством в/в болюсной инъекции. В день 28 образцы сыворотки (n=5-11 мышей) собирали и проводили определение уровней холестерина.
Пример 8: Антитела-антагонисты против PCSK9 снижают уровень LDL в сыворотке крови у приматов, не принадлежащих к человеческому роду
Для определения действия in vivo антител против PCSK9 антитело 7D4 было протестировано на человекообразных обезьянах. Четырем 3-4-летним человекообразным обезьянам в день 0 инъецировали носитель (PBS+0,01% Tween 20) и антитело 7D4 10 мг/кг в день 7. Липидные профили плазмы крови анализировали в день 0, 2, 7, 9, 11, 14, 21 и 28, натощак. Однократная инъекция 10 мг/кг антитела 7D4 вызвала резкое снижение LDL в плазме крови (60%) (Фигура 15A) и числа частиц LDL (Фигура 15D) у всех 4 животных, в то время как имело минимальное действие на их уровни HDL (Фигура 15B) и числа частиц HDL (Фигура 15E). Общий холестерин (Фигура 15C) также был снижен после введения 7D4, в то время как уровень триглицеридов (Фигура 15F) не был значительно изменен. Также измеряли уровень общего 7D4 (G) и уровни общего PCSK9 (H).
На Фигуре 16 проиллюстрирован эффект дозы анти-PCSK9 антитела 7D4 на уровни холестерина сыворотки крови у человекообразных обезьян. Двум самцам и двум самкам человекообразных обезьян, возрастом 3-5 лет, в каждой группе посредством в/в болюсной инъекции вводили определенную дозу антитела 7D4 в день 7 и эквивалентный объем физиологического раствора в день 0. Образцы плазмы крови получали в обозначенные моменты времени и измеряли уровни LDL плазмы крови.
На Фигуре 17 показано сравнение влияния анти-PCSK9 антител 4A5, 5A10, 6F6 в 7D4 на уровни холестерина крови у человекообразных обезьян. Двум самцам и двум самкам человекообразных обезьян, возрастом 3-6 лет, в каждой группе посредством в/в болюсной инъекции вводили 1 мг/кг указанного антитела в день 0. Образцы плазмы крови получали в обозначенные моменты времени, измеряли уровни LDL плазмы крови и нормализовали с таковыми в день -2.
На Фигуре 18 представлен эффект анти-PCSK9 антитела-антагнониста 7D4 на уровни холестерина плазмы крови у человекообразных обезьян, чей рацион содержал 33,4% кКал жира с добавлением 0,1% холестерина. В течение 16 недель рацион шести 3-5-летних человекообразных обезьян был с высоким содержанием жира. В определенный срок трем обезьянам вводили антитело 7D4 в дозе 10 мг/кг, и трем обезьянам вводили физиологический раствор. Уровни LDL у отдельных обезьян были измерены и нормализованы с таковыми на день обработки.
Пример 9: Гуманизированное анти-PCSK9 антитело
Мышиное моноклональное антитело 5А10 было гуманизировано и его аффинность доведена до зрелости, для обеспечения антитела L1L3. Аффинность антитела L1L3 к мышиному PCSK9 составляет 200 пM, и аффинность к PCSK9 человека составляет 100 пM, при измерении посредством Biacore. L1L3 полностью ингибирует PCSK9-опосредованную угнетающую регуляцию LDLR в культивированных клетках Huh7, при инкубировании со 100 нМ человеческого или мышиного антитела против PCSK9. Смотри Фигуру 19.
На Фигуре 20 представлен эффект дозы L1L3, мышиного предшественника 5A10 и антитела отрицательного контроля 42H7 на блокаду связывания рекомбинантного биотинилированного PCSK9 человека и PCSK9 мыши с иммобилизированным внеклеточным доменом рекомбинантного LDLR in vitro. На Фигуре 20A показан PCSK9 человека, связанный с внеклеточным доменом LDLR человека при pH 7,5. На Фигуре 20B представлен PCSK9 человека, связанный с внеклеточным доменом LDLR человека при pH 5,3. На фигуре 20C показан PCSK9 мыши, связанный с внеклеточным доменом LDLR человека при pH 7,5. На фигуре 20D показан PCSK9 мыши, связанный с внеклеточным доменом LDLR человека при pH 5,3.
На Фигуре 21 представлен эффект применения 10 мг/кг L1L3 на уровень холестерина сыворотки крови у мышей. 8-недельным мышам C57/bl6 в двух группах (n=8/группа) в день 0 посредством внутрибрюшинной (i.p.) инъекции вводили 10 мг/кг антитела L1L3 или эквивалентное количество физиологического раствора. Образцы сыворотки крови собирали в день 2, 4 и 7 и определяли уровни холестерина. L1L3 снижало общий холестерин сыворотки крови приблизительно на 40% в день 2 и 4. В другом исследовании, когда 10 мг/кг L1L3 вводили в виде однократной внутрибрюшинной дозы (IP) мышам C57BL/6, получавшим нормальный рацион (n=10), уровни холестерина сыворотки крови были снижены на 47% по сравнению с группой, обработанной физиологическим раствором, на 4 день после обработки. Когда в испытании эффекта дозы L1L3 вводили как однократную IP дозу 0, 0,1, 1, 10 и 80 мг/кг (n=6/группа) самцам крыс Sprague-Dawley, получавших нормальный рацион, уровни холестерина были дозозависимо снижены, с максимальным эффектом 50% при дозах 10 и 80 мг/кг, через 48 часов после введения. Продолжительность подавления холестерина также была дозозависима в диапазоне от 1 до 21 дней.
Аминокислотная последовательность полностью гуманизированной тяжелой цепи L1L3 (SEQ ID NO: 15) представлена в Таблице 8. Последовательность вариабельной области подчеркнута (SEQ ID NO: 54).

Аминокислотная последовательность полностью гуманизированной легкой цепи L1L3 (SEQ ID NO: 14) представлена в Таблице 9. Последовательность вариабельной области подчеркнута (SEQ ID NO: 53).
rfsgsgsgtd
ftftisslqp
ediatyycqq
ryslwrtfgq
gtkleikrtv aapsvfifpp 120
sdeqlksgta svvcllnnfy preakvqwkv dnalqsgnSq esvteqdskd styslsstlt 180
lskadyekhk vyacevthqg lsspvtksfn rgec 214
На Фигуре 22 представлено действие внутривенного введения эффективной дозы (3 мг/кг) антитела 5A10 (черные кружочки на графике) или антитела L1L3 (черные квадратики) на каждую из четырех человекообразных обезьян в день ноль. Изменение HDL сыворотки крови (Фигура 22A) и LDL сыворотки крови (Фигура 22B) измеряли в промежутке от -2 до +28 дня. Введение обоих антител привело в результате к более чем 70% снижению уровней LDL в сыворотке крови приблизительно на семь дней, эффект, который в значительной степени сохранялся приблизительно еще шесть дней у животных, получивших L1L3. У всех животных наблюдали нормальную функцию печени и почек и почти нормальный гематокрит.
L1L3 дозозависимо снижало LDL-C, с максимальным эффектом, наблюдаемым в группе, получившей 10 мг/кг, что поддерживало 70% снижение уровня LDL-C до 21 дней после введения дозы, и полностью восстановился на 31 день. Введение L1L3 не повлияло на уровни HDL-C во всех дозируемых группах. Животным в группе с дозой 3 мг/кг (n=4) также вводили две дополнительные в/в дозы 3 мг/кг антитела L1L3 в день 42 и 56 (с интервалом 2 недели). Эти две дополнительные дозы снова снизили LDL-C и поддерживали уровни LDL-C ниже 50% в течение 4 недель. Уровни LDL-C вернулись к нормальным через две недели. Уровни HDL-C сыворотки крови оставались неизменными на всем протяжении исследования.
Были исследованы эффективность L1L3 у страдающих гиперхолестеринемией приматов, не являющихся человеком, и фармакодинамические взаимодействия между L1L3 и статинами, ингибирующими HMG-CoA редуктазу. Перед началом данного исследования, в течение более 18 месяцев группе человекообразных обезьян (n=12) давали рацион, содержащий 35% жира (вес/вес) и 600 частей на миллион (ppm) холестерина, чем подняли уровни LDL-C в среднем до 120 мг/дл по сравнению с нормальными средними уровнями 50 мг/дл. Как ни удивительно, не наблюдался эффект на уровень общего холестерина или уровни LDL-C сыворотки крови после ежедневного введения средней дозы (10 мг/животное) Crestor® (розувастатин кальция) в течение 6 недель, и после последующего ежедневного введения высоких доз (20 мг/кг) в течение 2 недель. Однократное введение 3 мг/кг L1L3 с применением Crestor® или носителя в течение 2 недель эффективно снижало уровни LDL-C сыворотки крови на 56% на 5 день после обработки, и уровни LDL-C постепенно восстанавливались в течение 2,5-3 недель, в то время как не было воздействия на уровни HDL-C. После перевода животных на ежедневное применение 50 мг/кг Zocor® (симвастатин), их уровни LDL-C достигли максимального снижения на 43% на день 5 и стабилизировались впоследствии. После 3 недель введения Zocor® 50 мг/кг/день этим животным вводили однократную дозу 3 мг/кг L1L3, в то время как им все еще продолжали вводить Zocor® 50 мг/кг/день. Введение L1L3 привело в результате к дополнительному 65% снижению LDL-C, вдобавок к 43% снижению посредством Zocor®, на день 5, и уровни вернулись к изначальным в течение 2 недель.
Другие аминокислотные замены в CDR были осуществлены в 5А10 в процессе гуманизации и доведения аффинности до зрелости и для получения специфических свойств. Последовательности модифицированных CDR и способность связываться с PCSK9 для данных антител, содержащих эти модифицированные CDR, представлены на Фигурах 24A-G. Номера, следующие за каждой последовательностью на Фигурах 24A-G, представляют собой SEQ ID NO для такой последовательности.
Описания всех процитированных справочных материалов включены здесь посредством ссылки.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТАГОНИСТЫ PCSK9 | 2014 |
|
RU2618869C2 |
| ЛЕЧЕНИЕ АНТИТЕЛАМИ ПРОТИВ PCSK9 | 2012 |
|
RU2576034C2 |
| АНТИТЕЛА С РН-ЗАВИСИМЫМ СВЯЗЫВАНИЕМ АНТИГЕНА | 2011 |
|
RU2570729C2 |
| ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ, СОДЕРЖАЩИЕ АНТИТЕЛА К PCSK9 ЧЕЛОВЕКА | 2012 |
|
RU2604139C2 |
| АНТИТЕЛА ЧЕЛОВЕКА К PCSK9 ДЛЯ ПРИМЕНЕНИЯ В СПОСОБАХ ЛЕЧЕНИЯ КОНКРЕТНЫХ ГРУПП ИНДИВИДУУМОВ | 2012 |
|
RU2603481C2 |
| Антитело против PCSK9 и его применение | 2016 |
|
RU2756012C2 |
| СПОСОБ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ, СВЯЗАННЫХ С ХОЛЕСТЕРИНОМ | 2021 |
|
RU2832333C1 |
| PCSK9 ВАКЦИНЫ | 2015 |
|
RU2698971C2 |
| АНТАГОНИСТЫ | 2019 |
|
RU2839429C2 |
| ВАКЦИНА | 2012 |
|
RU2640258C2 |

















































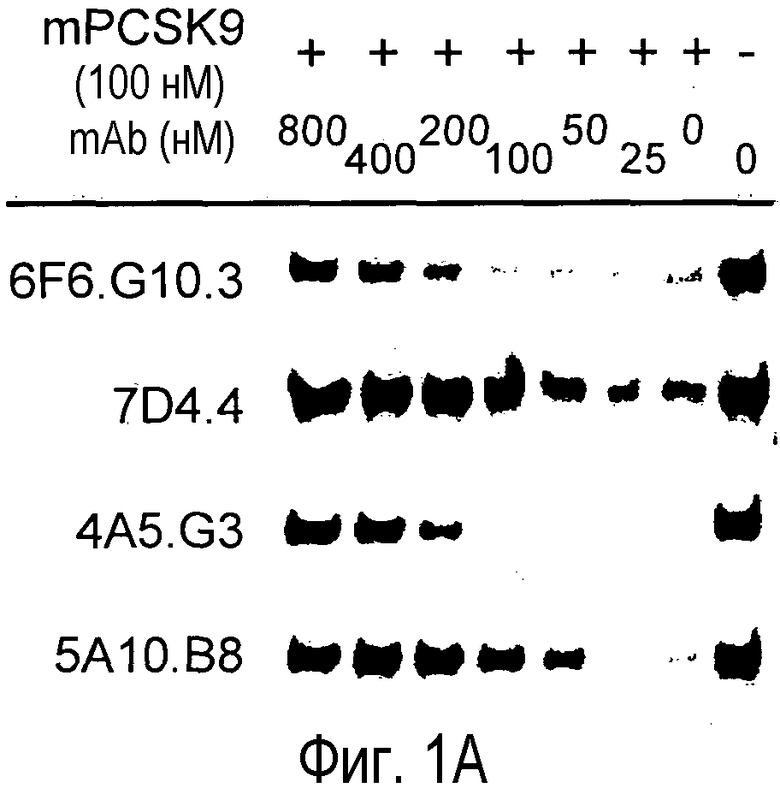
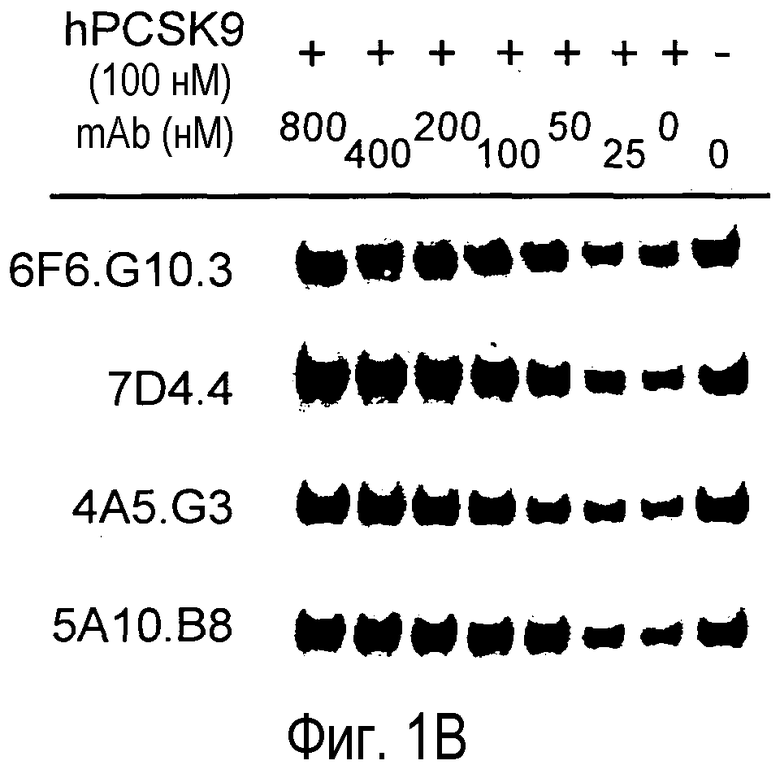
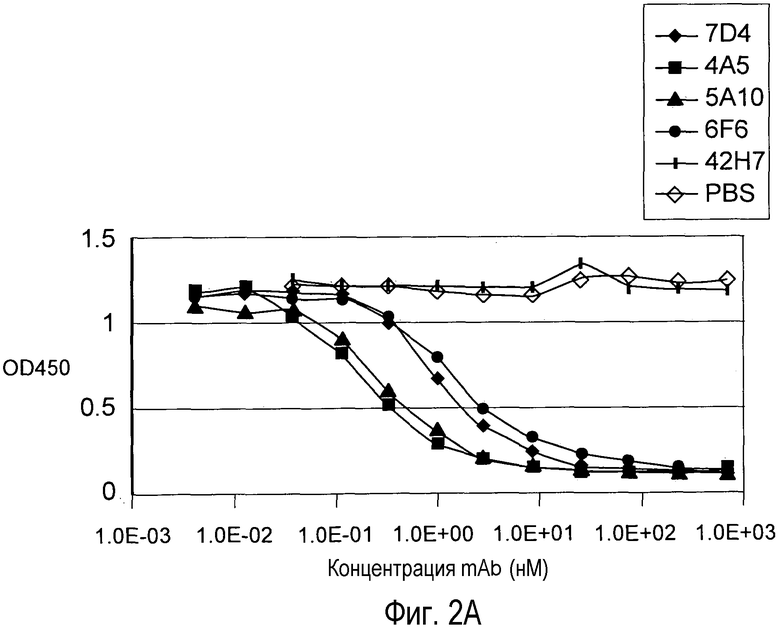
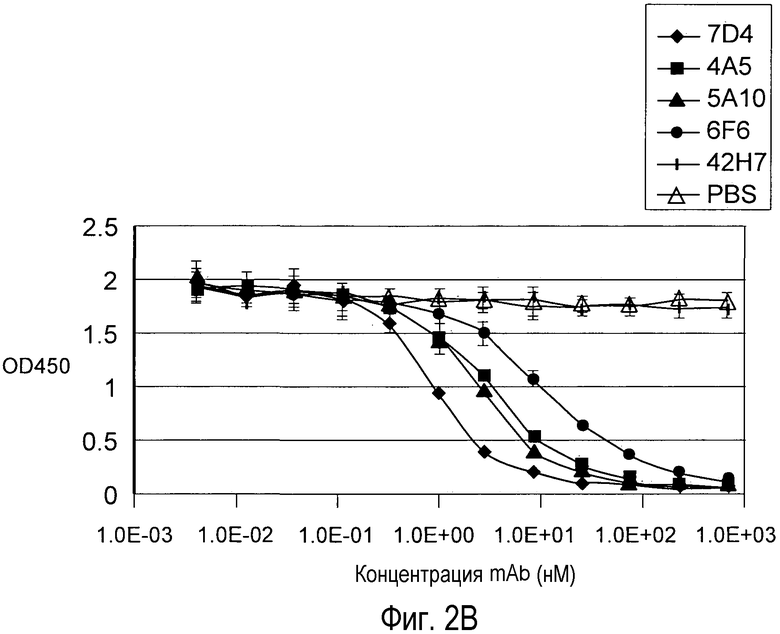
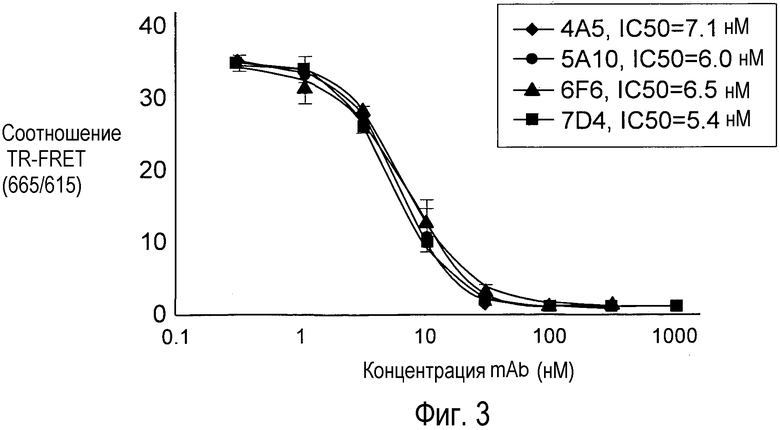
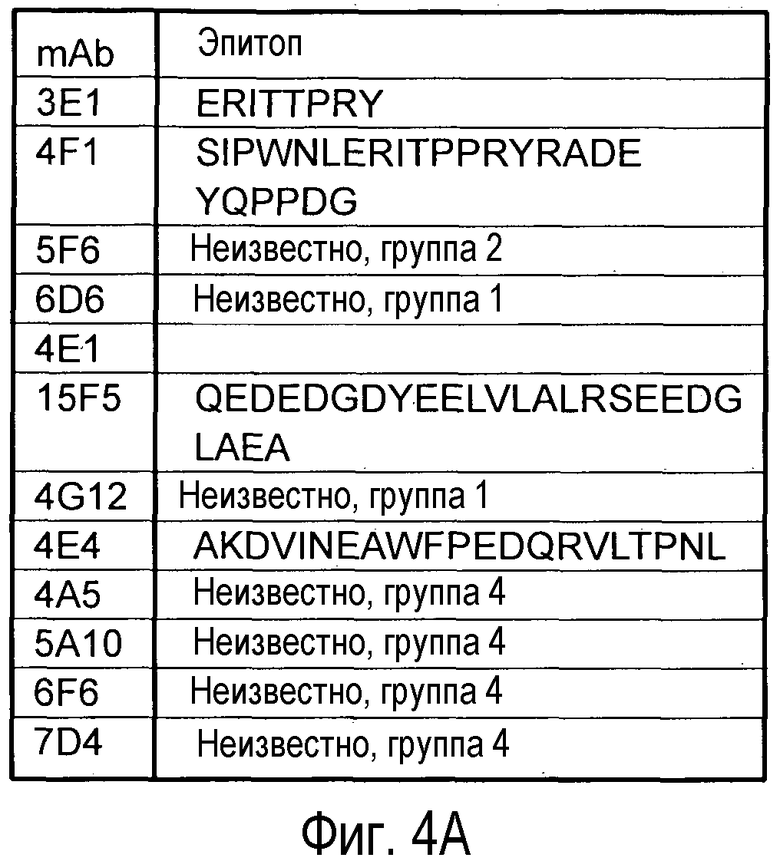
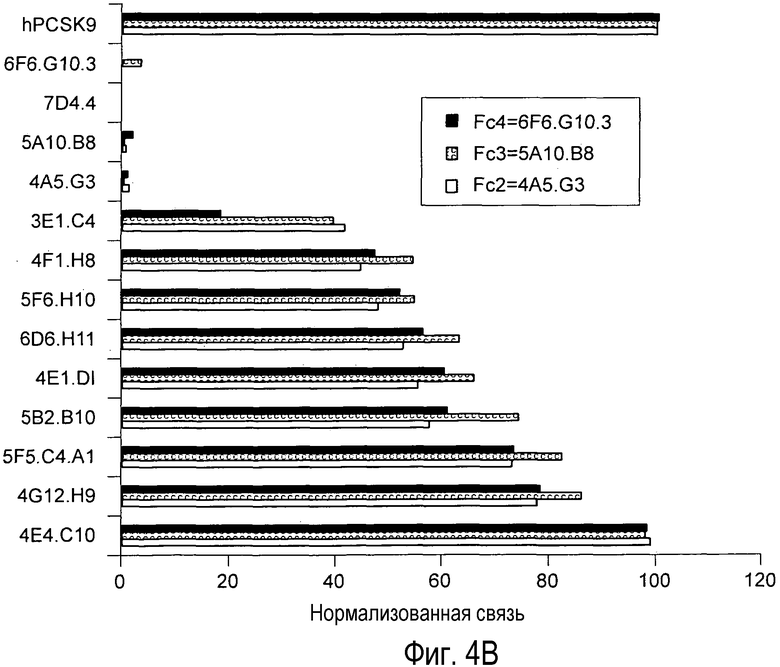
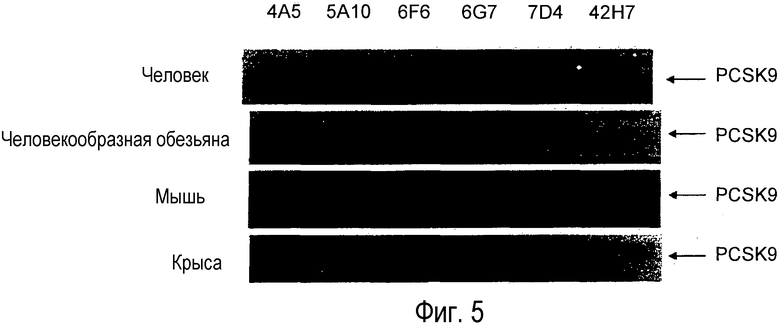
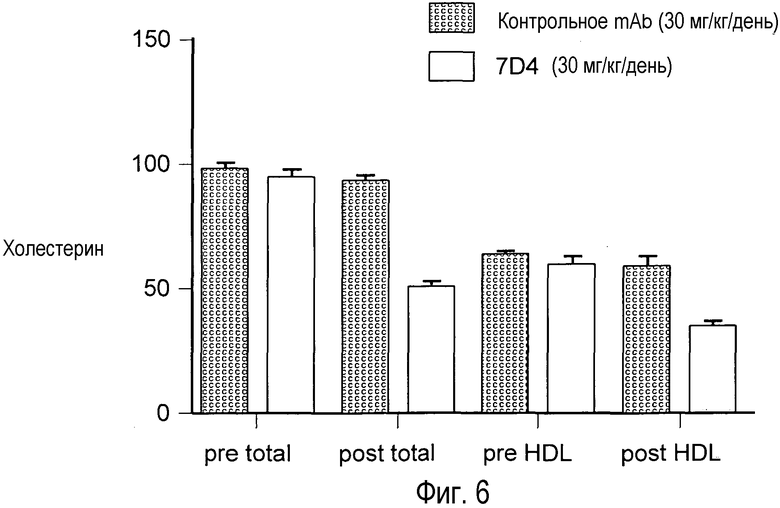
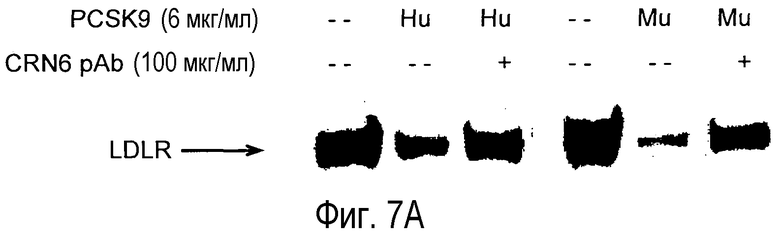
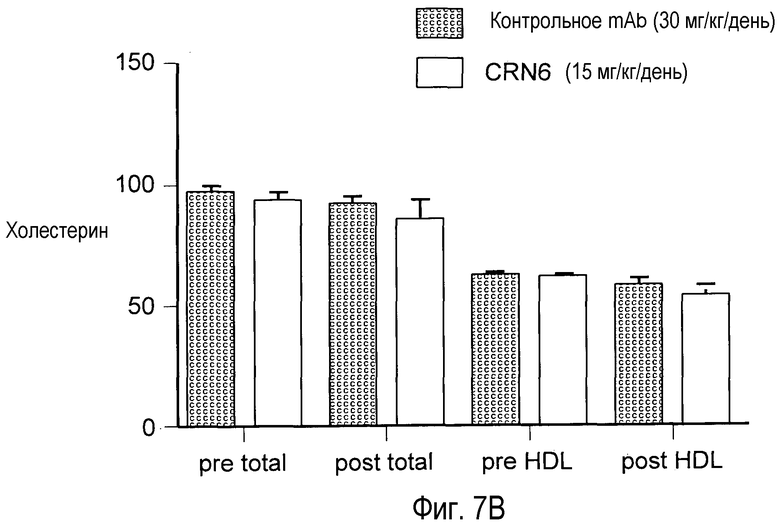
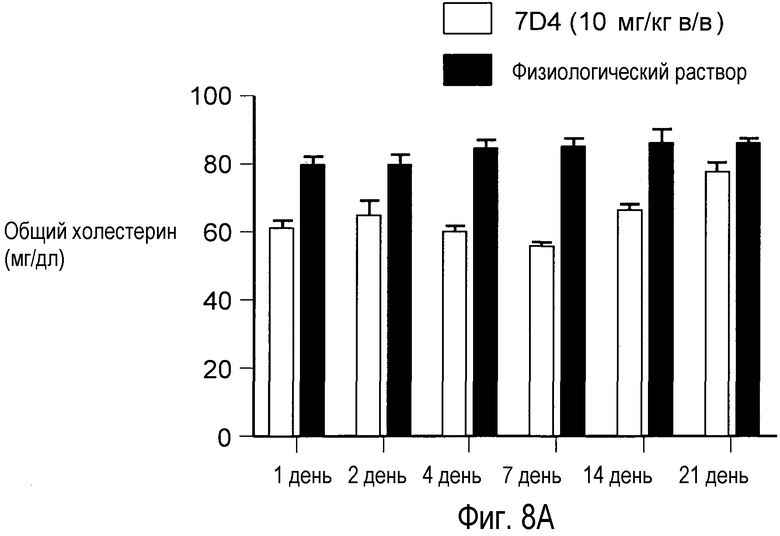
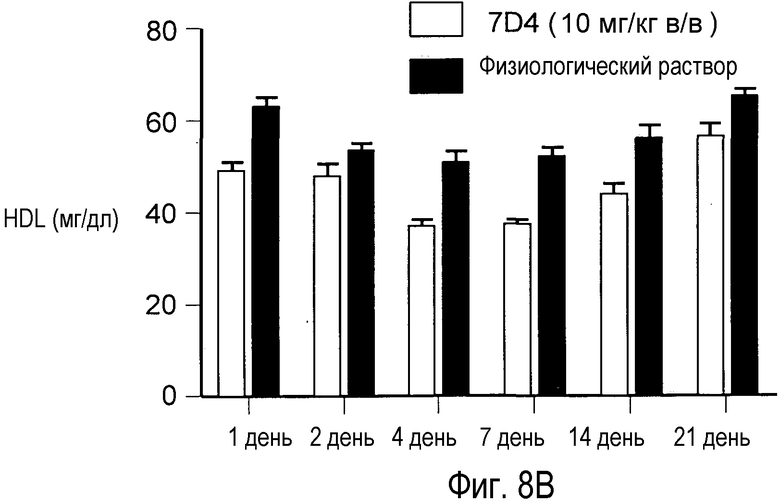
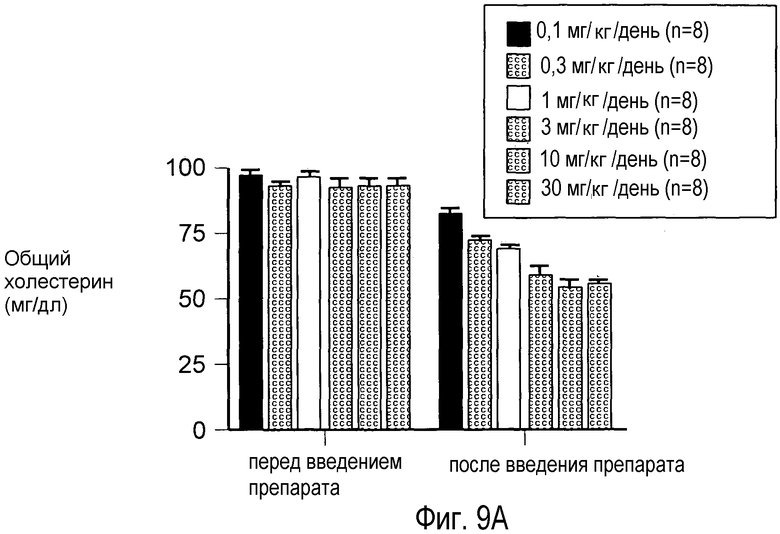
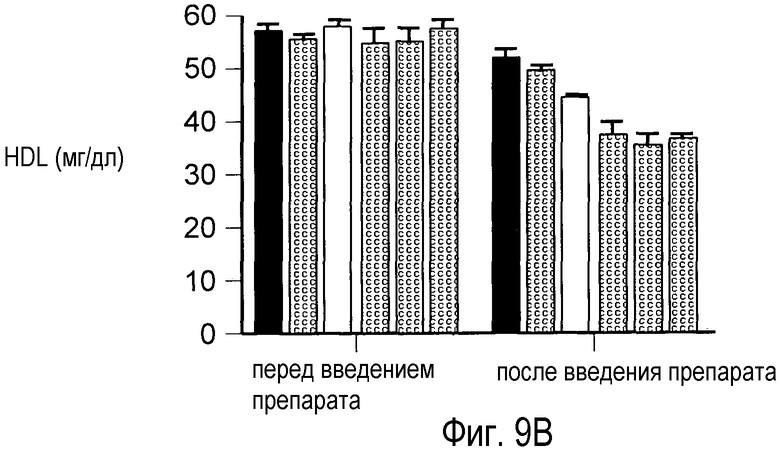
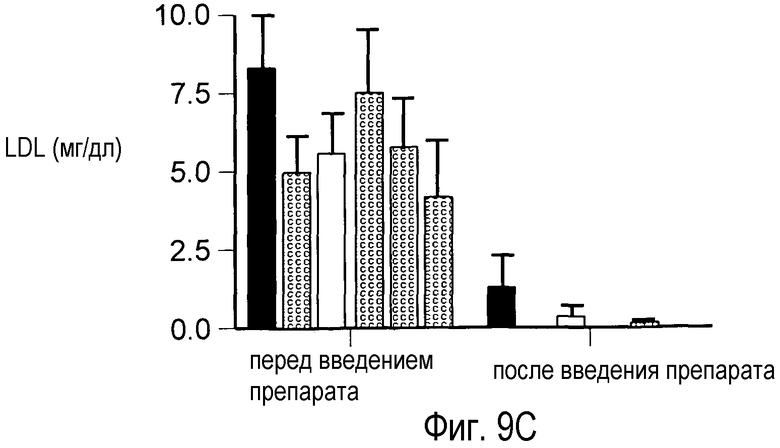
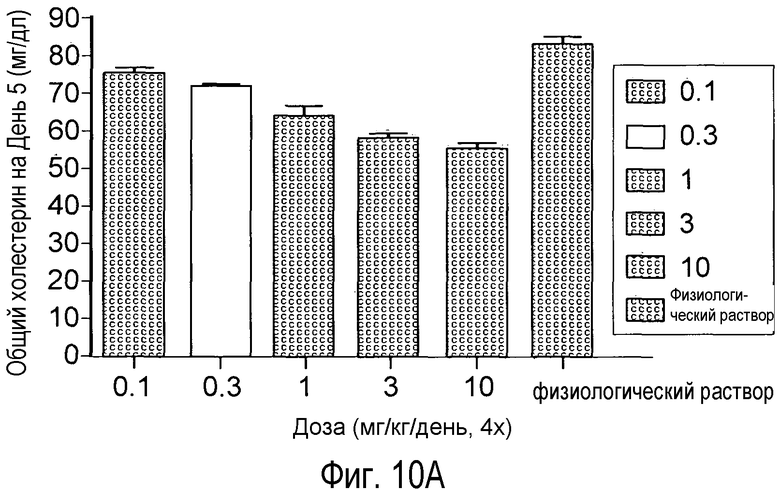
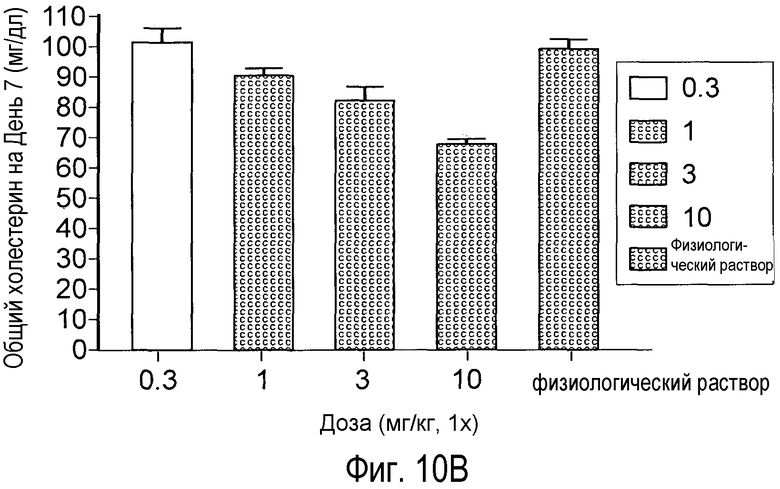
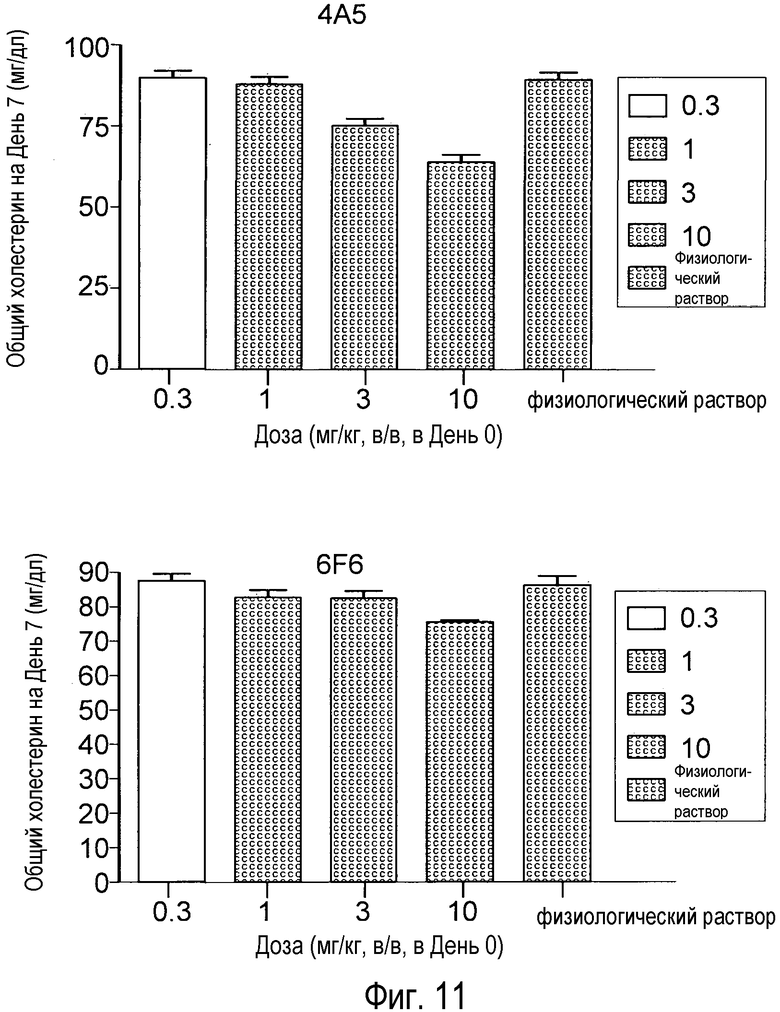
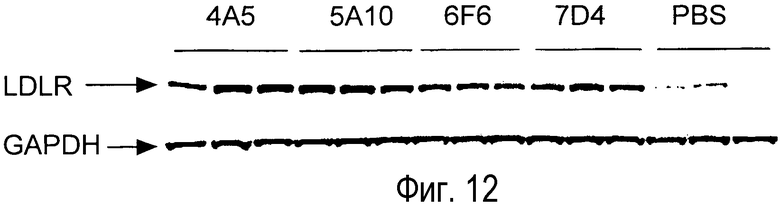
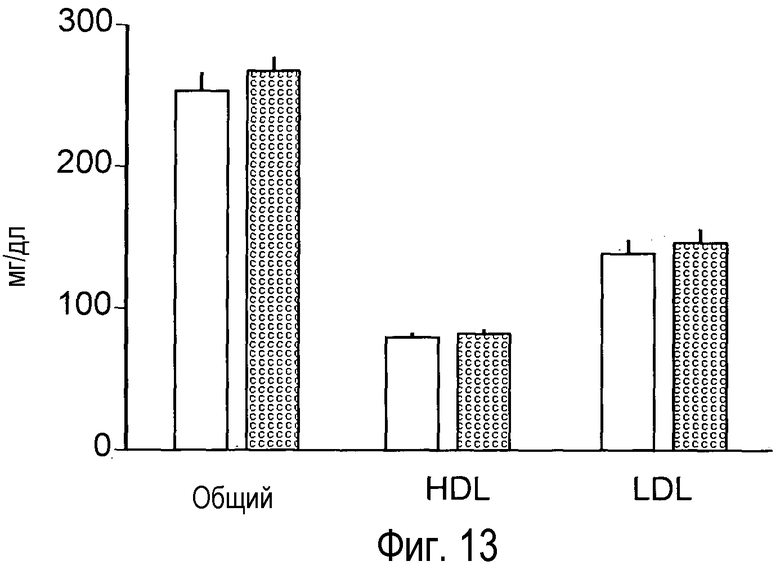
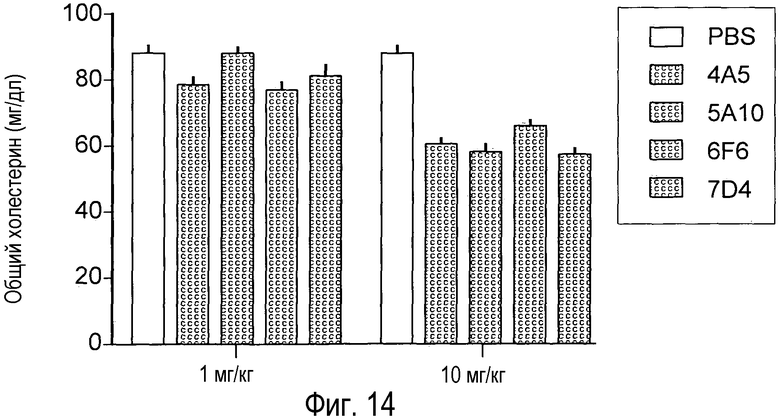
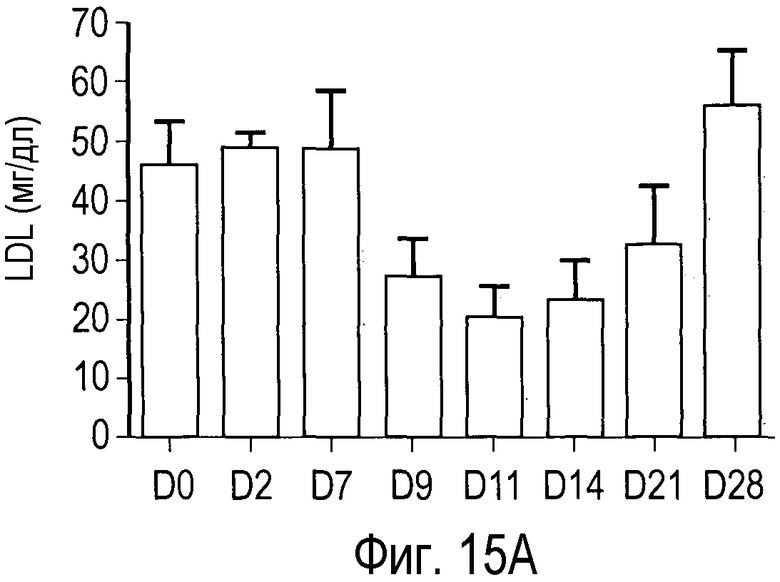
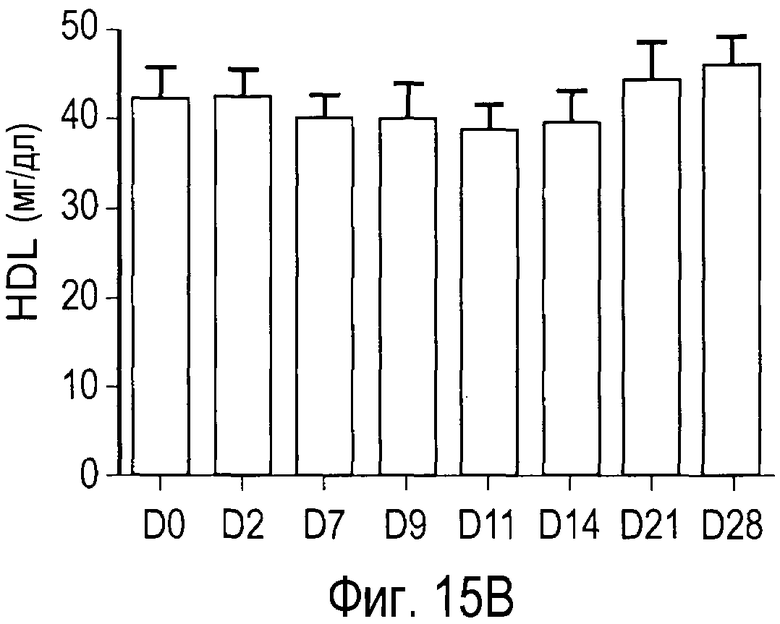
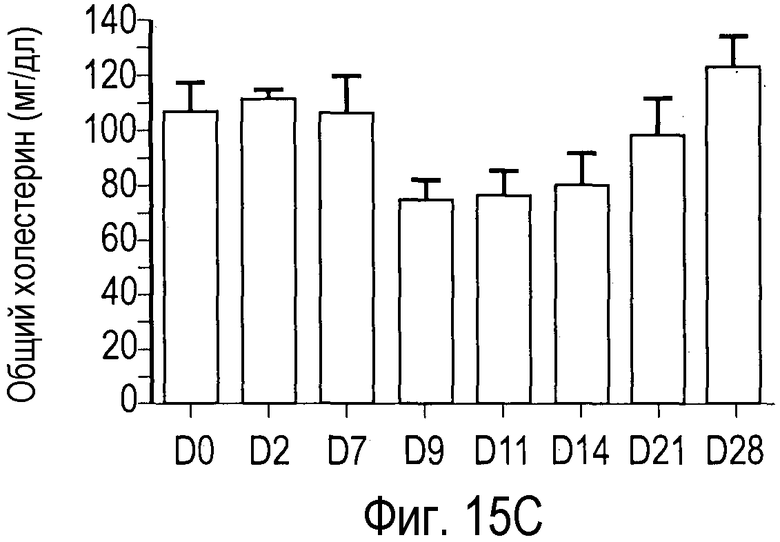
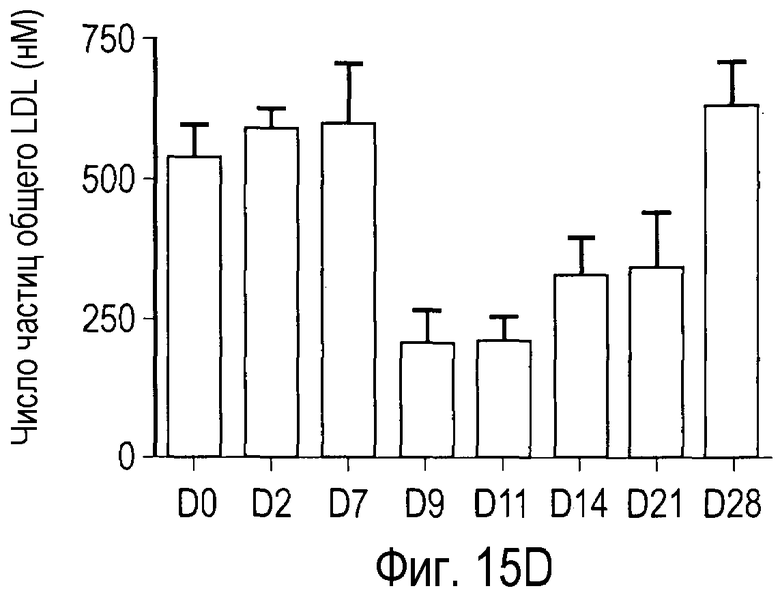
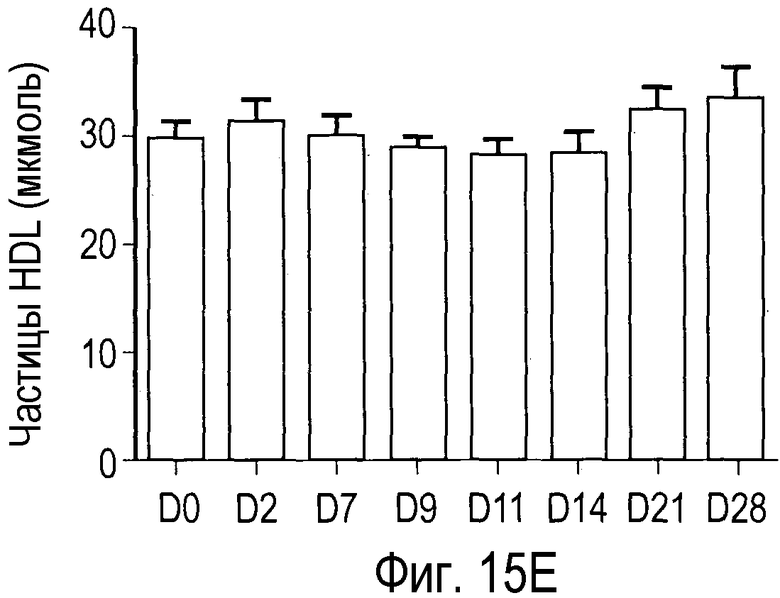
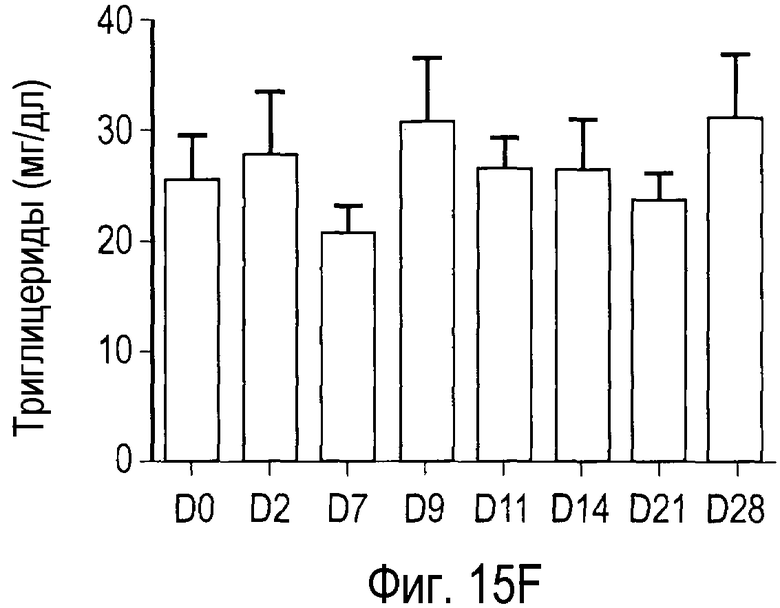
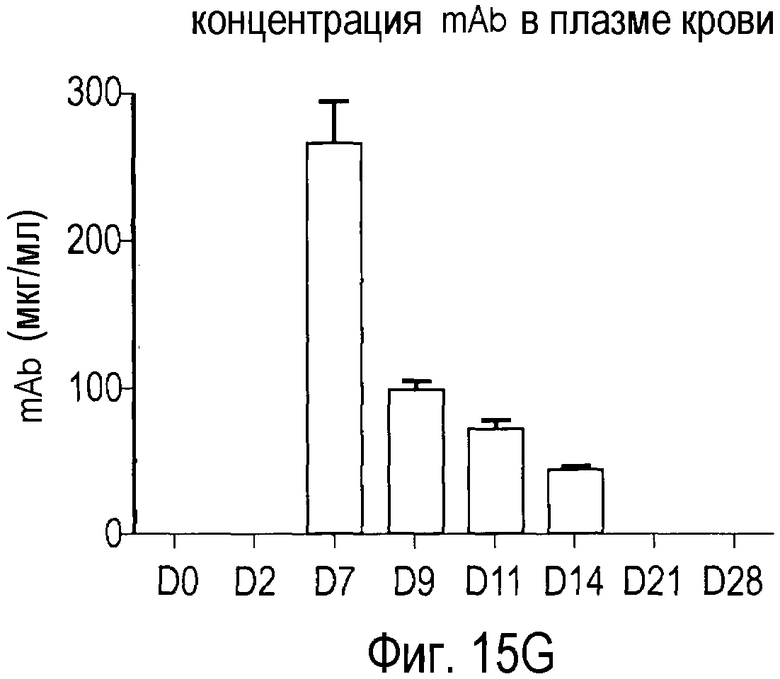
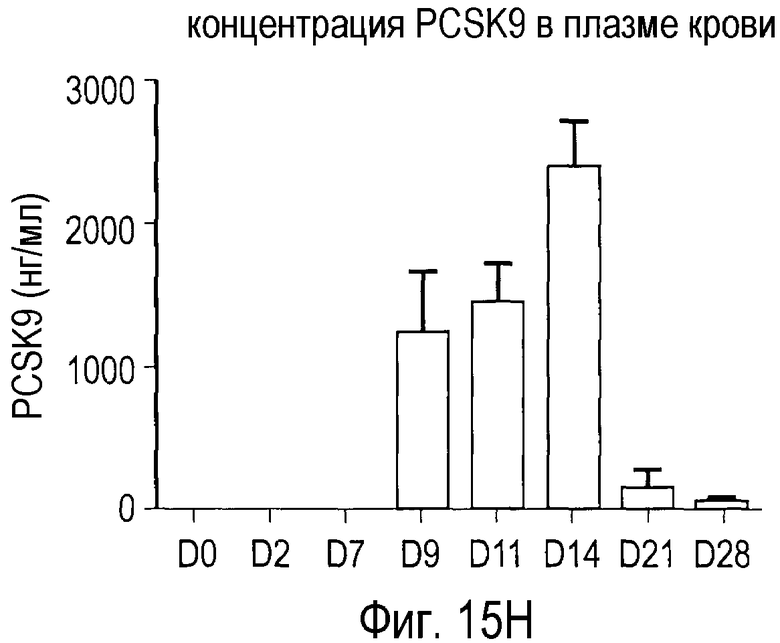
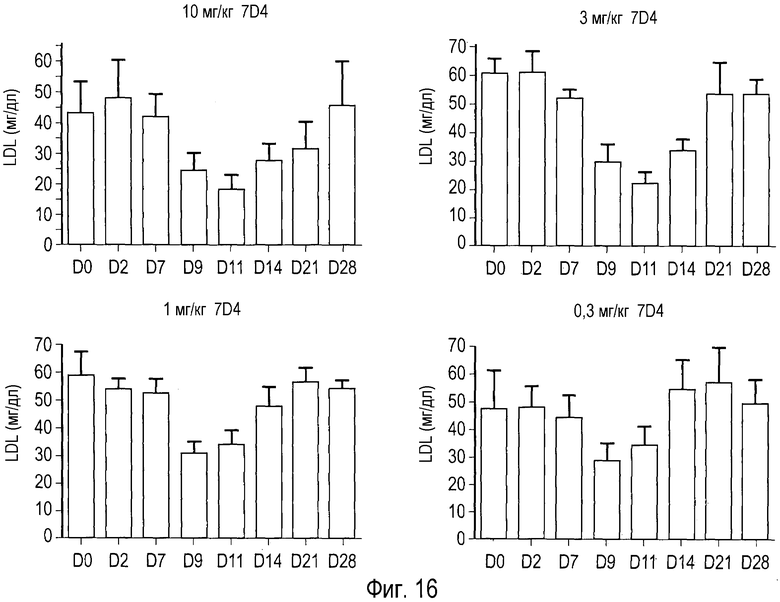
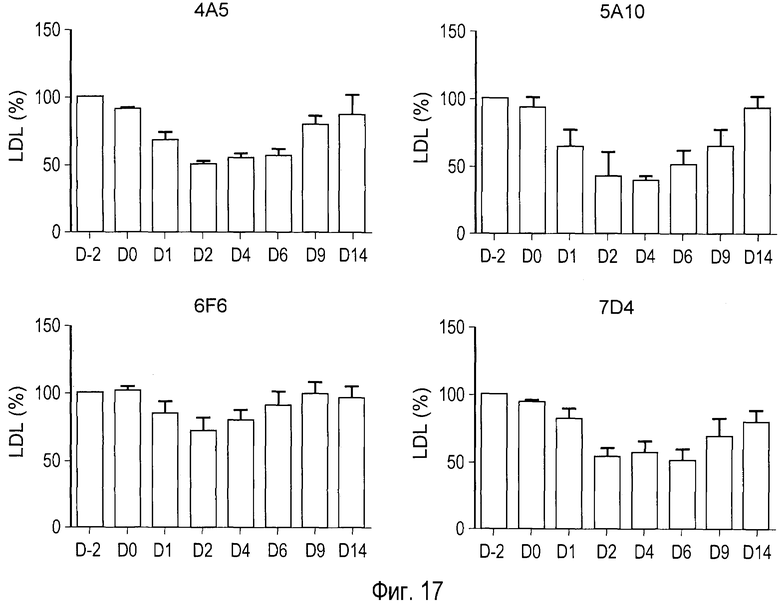
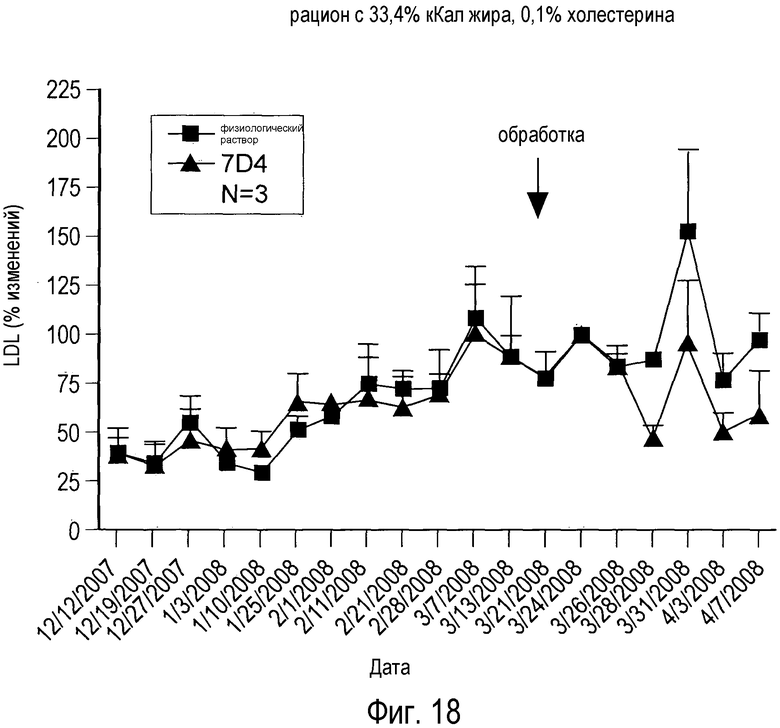
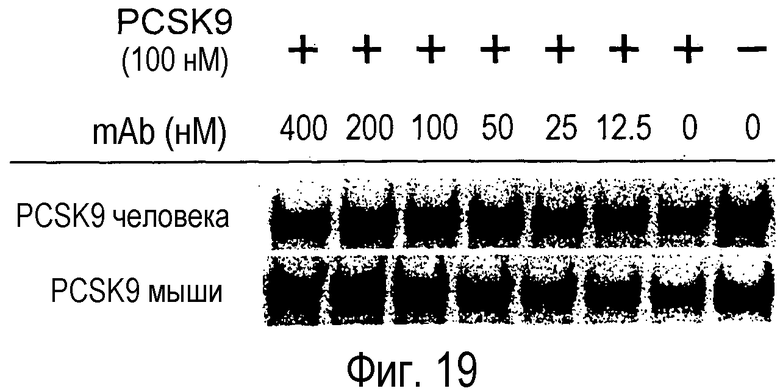
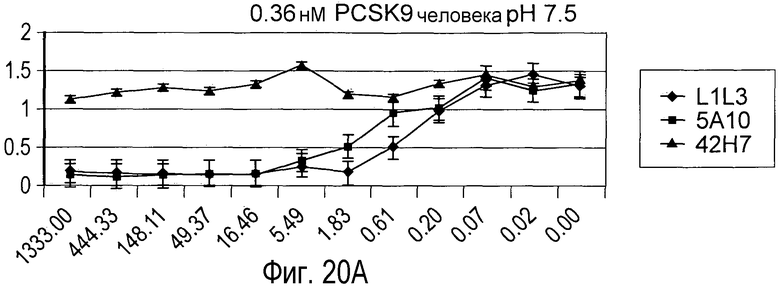
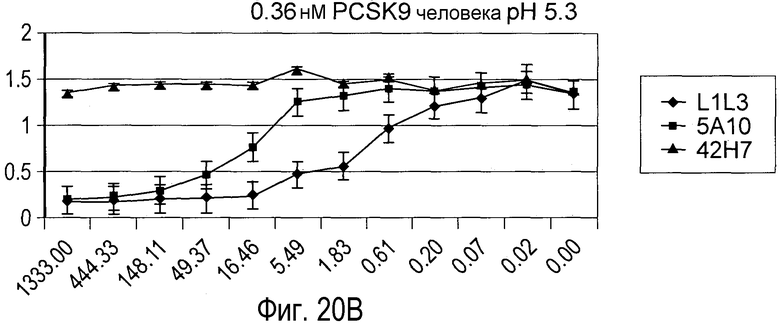
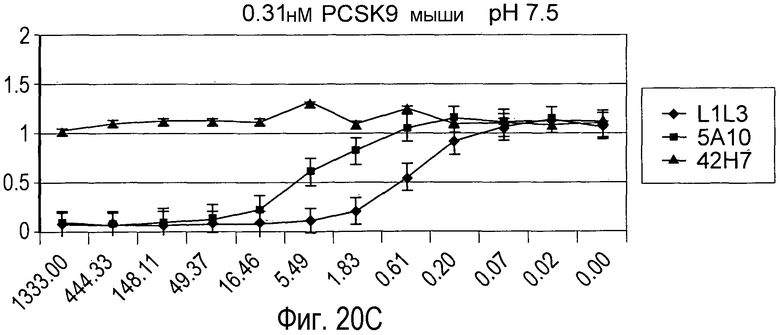
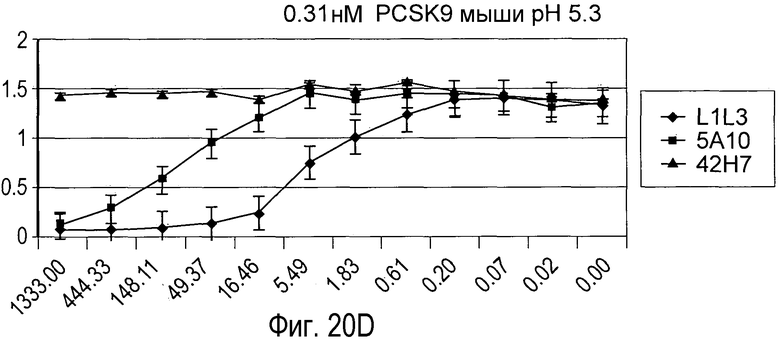
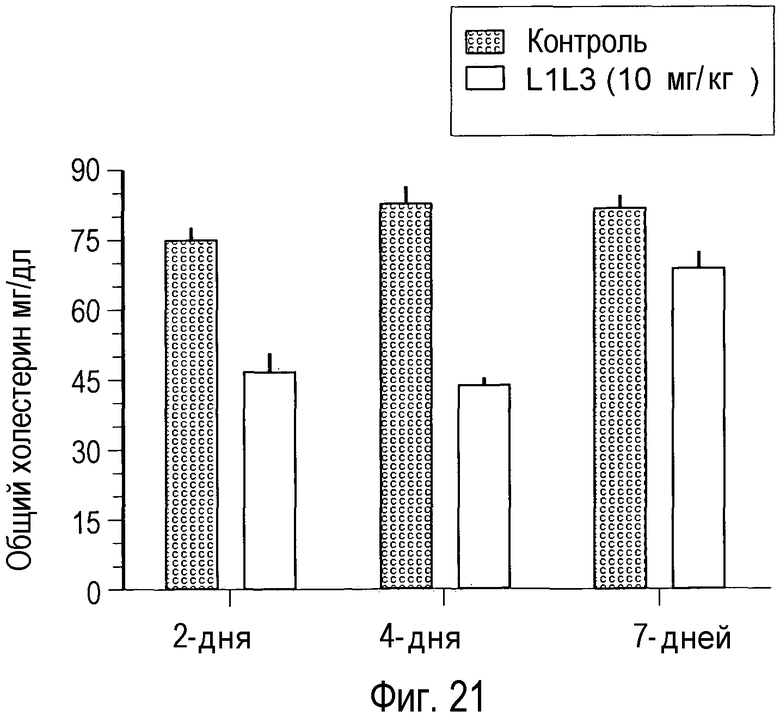
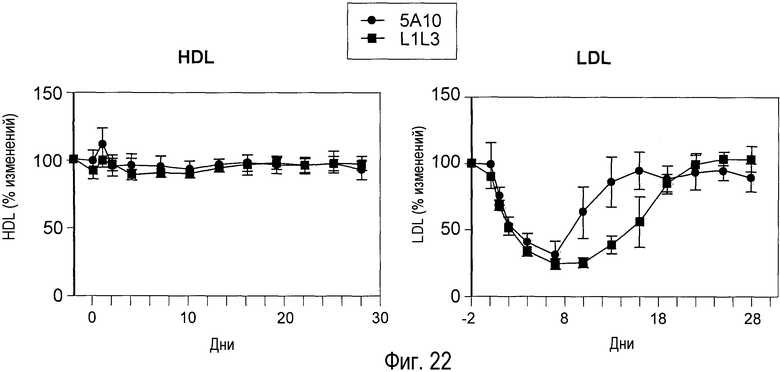
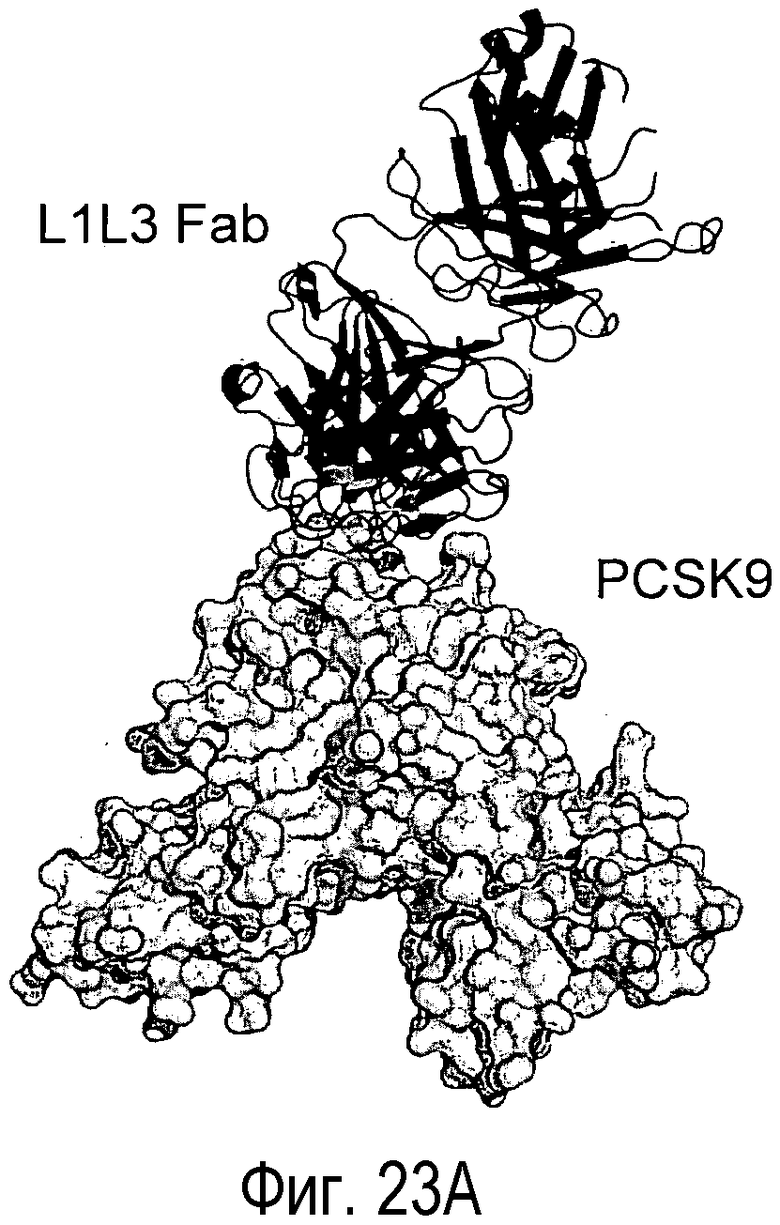
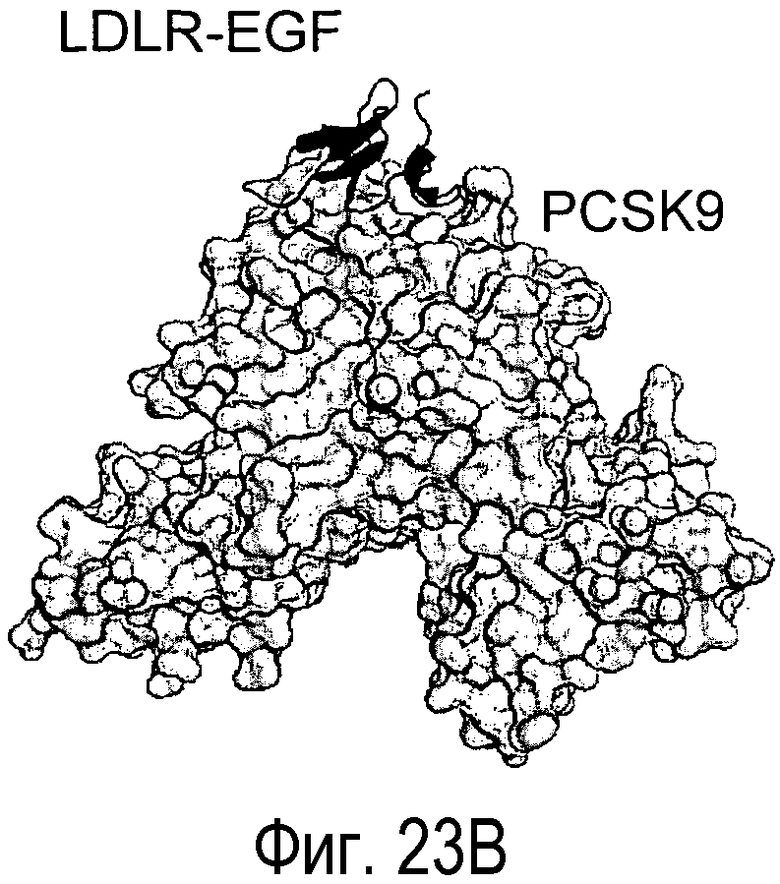
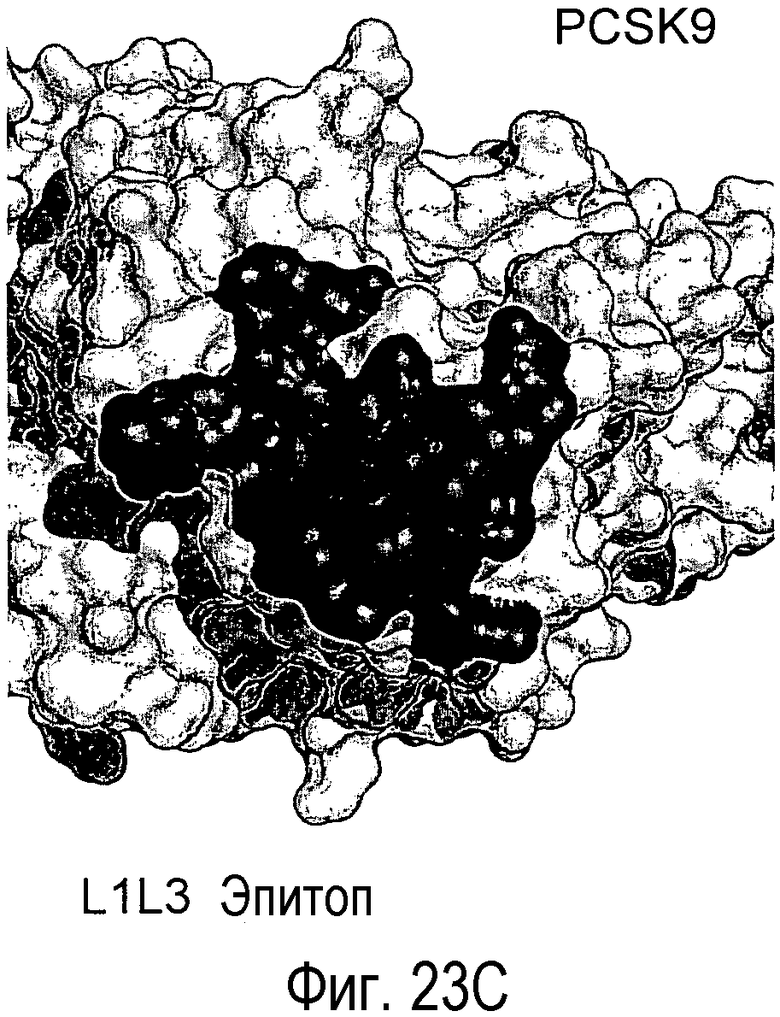
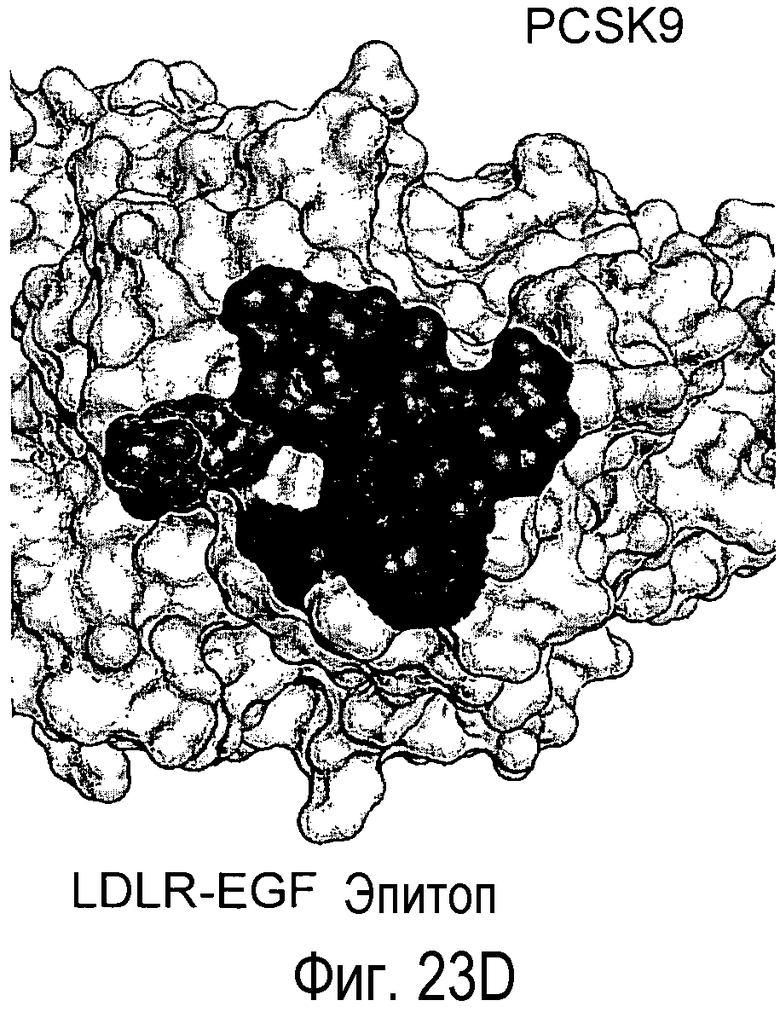
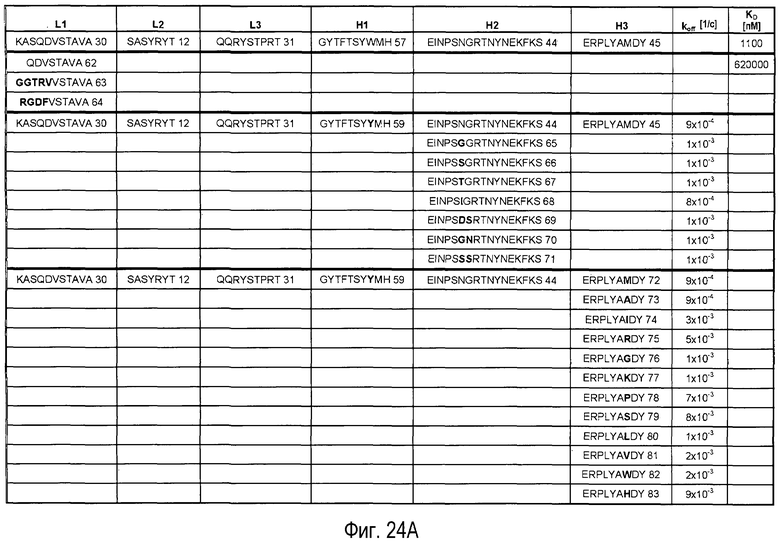
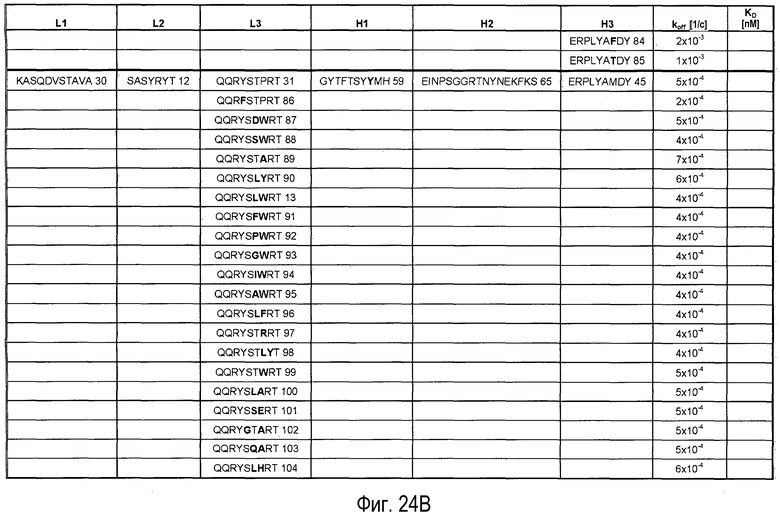
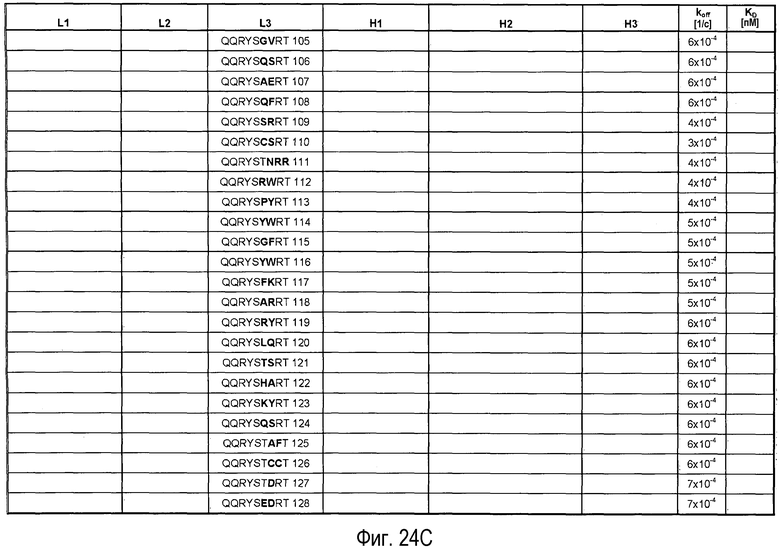
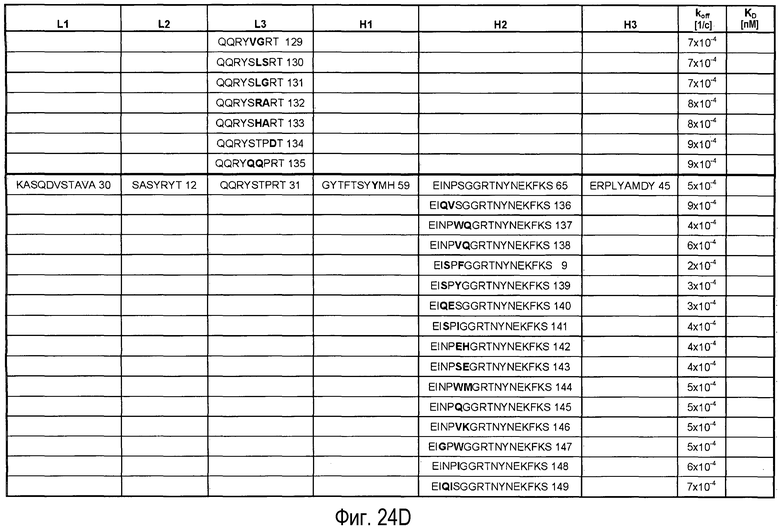
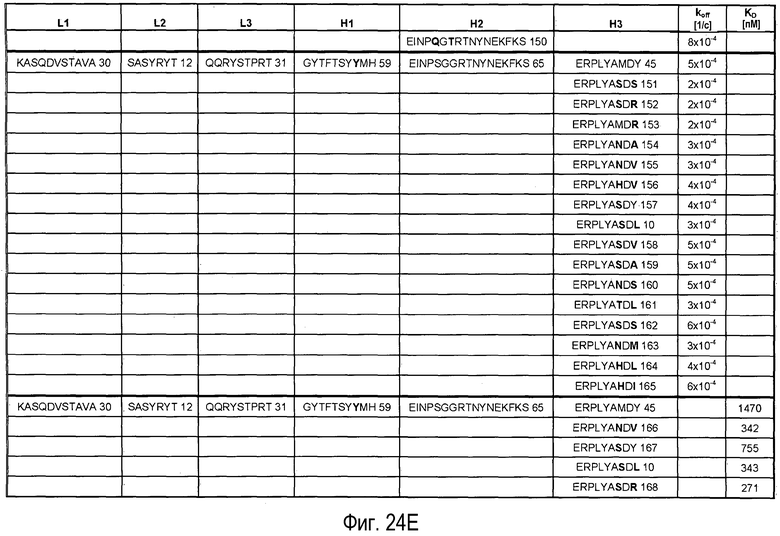
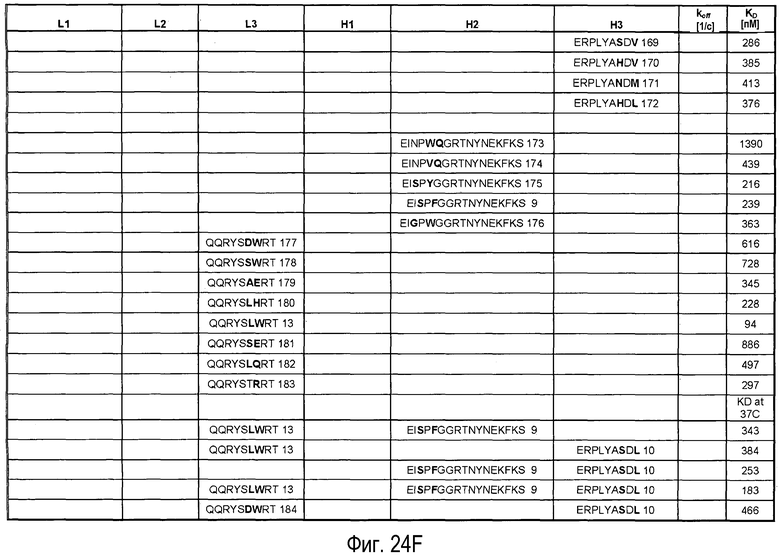
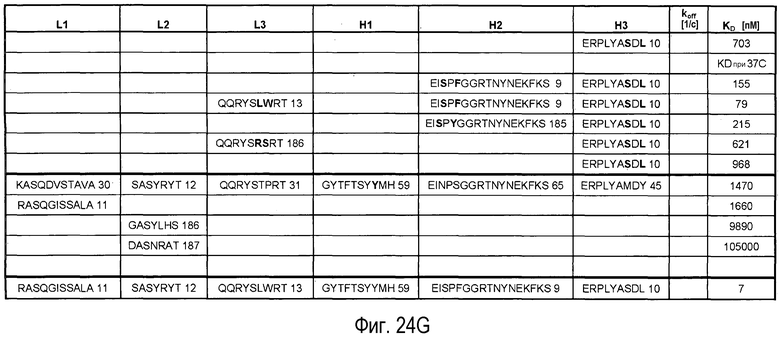
Настоящее изобретение относится к иммунологии и биотехнологии. Предложены: изолированное антитело или его вариант, специфически распознающие PCSK9, и фармацевтическая композиция для снижения уровня холестерина-LDL на их основе. Описаны: применение для снижения уровня холестерина и/или LDL крови; снижения заболеваемости и/или исправления нарушенных уровней холестерина и/или LDL; применение при получении лекарственного средства для снижения уровня холестерина и/или LDL крови; снижения заболеваемости и/или исправления нарушенных уровней холестерина и/или LDL на их основе. Раскрыты варианты клеточных линий, продуцирующие антитело против PCSK9 или его антигенсвязывающую часть и депонированные в коллекции АТСС под номерами РТА-8986, АТСС РТА-8984, АТСС РТА-8983, соответственно. Описана кодирующая нуклеиновая кислота и клетка-хозяин для продуцирования антитела на её основе. Изобретение обеспечивает антагонистические антитела против PCSK9, что может найти применение в медицине для снижения уровня холестерина. 12 н. и 6 з.п. ф-лы, 24 ил., 9 табл., 9 пр.
1. Изолированное антитело или его вариант, где антитело или его вариант специфически связывают PCSK9 и являются полным антагонистом PCSK9-опосредованного эффекта на уровни LDLR, как измерено in vitro на клетках Huh7, где антитело содержит первую определяющую комплементарность область (CDR1) вариабельной области тяжёлой цепи (VH), имеющую аминокислотную последовательность, представленную в SEQ ID NO:8, 59 или 60, VH CDR2, имеющую аминокислотную последовательность, представленную в SEQ ID NO:9 или 61, и VH CDR3, имеющую аминокислотную последовательность, представленную в SEQ ID NO:10, и CDR1 вариабельной области лёгкой цепи (VL), имеющую аминокислотную последовательность, представленную в SEQ ID NO:11, VL CDR2, имеющую аминокислотную последовательность, представленную в SEQ ID NO:12, и VL CDR3, имеющую аминокислотную последовательность, представленную в SEQ ID NO:13, а его вариант содержит CDR1 VH, CDR2 VH, CDR3 VH, CDR1 VL, CDR2 VL и CDR3 VL, где вариант отличается от антитела тем, что имеет одну, две или три консервативных аминокислотных замен в CDR1 VH, CDR2 VH, CDR3 VH, CDR1 VL, CDR2 VL и/или CDR3 VL.
2. Антитело по п.1, где вариант VH CDR1 содержит консервативную замену в аминокислотном положении 8 в SEQ ID NO:59, вариант VH CDR2 содержит консервативную замену в аминокислотных положениях 3, 4, 5, 6 или 7 в SEQ ID NO:9, и/или вариант VH CDR3 содержит консервативную замену в одном из или в обоих аминокислотных положениях 7 и 9 в SEQ ID NO:10.
3. Антитело по п.1, где область VH содержит SEQ ID NO:54 и область VL содержит SEQ ID NO:53.
4. Гуманизированное антитело, содержащее легкую цепь, имеющую SEQ ID NO:14, и тяжёлую цепь, имеющую SEQ ID NO:15, c или без C-концевого лизина в SEQ ID NO:15, где антитело специфически связывает PCSK9.
5. Антитело по п.1, где антитело дополнительно содержит иммунологически инертную константную область.
6. Антитело по п.5, где антитело имеет изотип, который выбран из группы, состоящей из IgG2, IgG4, IgG2∆a, IgG4∆c, IgG4 S228P, IgG4∆b S228P и IgG4∆c S228P.
7. Антитело по п.6, где константная область представляет собой дегликозилированный Fc.
8. Фармацевтическая композиция для снижения уровня холестерина-LDL в крови, причем фармацевтическая композиция содержит терапевтически эффективное количество антитела по любому из предшествующих пунктов.
9. Фармацевтическая композиция по п.8, дополнительно содержащая терапевтически эффективное количество статина.
10. Изолированная нуклеиновая кислота, кодирующая антитело по п.1.
11. Клетка-хозяин, продуцирующая антитело по любому из пп.1-7, содержащая нуклеиновую кислоту по п.10.
12. Способ снижения уровня холестерина-LDL в крови нуждающегося в этом пациента, включающий введение пациенту терапевтически эффективного количества антитела по любому из пп.1-7 или фармацевтической композиции по п.8 или 9.
13. Клеточная линия 4A5, имеющая № доступа ATCC PTA-8985, которая рекомбинантно продуцирует антитело против PCSK9 или его антигенсвязывающую часть.
14. Клеточная линия 5A10, имеющая № доступа ATCC PTA-8986, которая рекомбинантно продуцирует антитело против PCSK9 или его антигенсвязывающую часть.
15. Клеточная линия 6F6, имеющая № доступа ATCC PTA-8984, которая рекомбинантно продуцирует антитело против PCSK9 или его антигенсвязывающую часть.
16. Клеточная линия 7D4, имеющая № доступа ATCC PTA-8983, которая рекомбинантно продуцирует антитело против PCSK9 или его антигенсвязывающую часть.
17. Применение антитела по любому из пп.1-7, или фармацевтической композиции по п.8 или 9, для снижения уровней холестерина крови и/или уровней липопротеинов низкой плотности (LDL) крови, и/или снижения заболеваемости или исправления нарушенных уровней холестерина и/или уровней LDL, обусловленных нарушениями метаболизма холестерина и/или липопротеинов, в том числе фамильной гиперхолестеринемией, атерогенной дислипидемией, атеросклерозом и сердечно-сосудистым заболеванием.
18. Применение антитела по любому из пп.1-7, или фармацевтической композиции по п.8 или 9, при получении лекарственного средства для снижения уровней холестерина крови и/или уровней липопротеинов низкой плотности (LDL) крови, и/или снижения заболеваемости или исправления нарушенных уровней холестерина и/или уровней липопротеинов, обусловленных нарушениями метаболизма холестерина и/или липопротеинов, в том числе фамильной гиперхолестеринемией, атерогенной дислипидемией, атеросклерозом и сердечно-сосудистым заболеванием.
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| KWON et al., "Molecular basis for LDL receptor recognition by PSCK9", PNAS, 12.02.2008, vol.105, no.6, pp.1820-1825 | |||
| WO2007128121 A1, 15.11.2007 | |||
| WEDEMAYER et al., "Structural insights into the evolution of an antibody combining site", Science, 1997,Vol | |||
| СПОСОБ СОСТАВЛЕНИЯ ЗВУКОВОЙ ЗАПИСИ | 1921 |
|
SU276A1 |
| Прибор для записи работы холостого хода и простоя станков | 1925 |
|
SU5319A1 |

