Настоящее изобретение относится к вирусным вакцинам. В частности, настоящее изобретение относится к генетически сконструированным мутантным вирусам для использования в качестве вакцин; к вакцинам, содержащим, мутантные вирусы; к рекомбинантным клеткам; к способам получения вакцин.
Вирусные вакцины обычно делятся на два типа. К первому типу относятся так называемые "убитые" вакцины, которые представляют собой вирусные препараты, обезвреженные путем обработки соответствующими химическими веществами, такими, как бета-пропиолактон. Ко второму типу относятся живые "ослабленные" вакцины, представляющие собой вирусы, патогенность которых по отношению к хозяину была ослаблена с помощью специфической генетической модификации вирусного генома, либо более обычным способом - путем пассажа в систему тканевой культуру определенного типа. Однако, каждый из этих двух типов вакцин имеет свои собственные недостатки. Поскольку убитые вакцины не реплицируются в хозяине, то они должны быть введены путем инъекции, а поэтому они могут продуцировать неподходящий вид иммунного ответа. Например, вакцина Солка, представляющая собой убитый препарат полиовируса, продуцирует образование иммуноглобулина (Ig)G, но не стимулирует образование IgА в кишке, природном очаге первичной инфекции. Поэтому, хотя эта вакцина может защищать от неврологических осложнений полимиелита, однако она не блокирует первичную инфекцию и не сообщает "популяцинный иммунитет". Кроме того, убитые вирусы не могут внедряться и реплицироваться внутри хозяйских клеток. Поэтому получить с их использованием какой-либо желательный иммунологический ответ против неструктурных белков, продуцируемых во время репликации, не представляется возможным. Эти вирусы также не могут способствовать продуцированию цитотоксичных Т-клеток, направленных против вирусных антигенов. "Погибшие" антигены могут быть захвачены антиген-презентирующими клетками и презентированы Т-клетками. Однако эта презентация происходит посредством молекул МНС класса II и стимулирует активацию Т-херперных клеток. В свою очередь, Т-херперные клетки помогают В-клеткам продуцировать специфические антитела против антигена.
Для стимуляции продуцирования цитотоксических Т-клеток, вирусные антигены должны быть процессированы посредством партикулярного метаболизма внутри инфецированной клетки и презентированы в виде расщепленных пептидных фрагментов на молекулах МНС класса 1. Очевидно, что этот путь разложения наиболее эффективно работает для белков, синтезированных внутри инфекцированной клетки, а следовательно, только вирус, который вводится в хозяйские клетки и экспрессирует иммуногенный вирусный белок, способен генерировать вирус-специфические цитотоксичные Т-клетки. Поэтому убитые вакцины являются плохими индукторами клеточного иммунитета против вирусной инфекции. Отсюда явствует, что использование живых аттенюированных вакцин является более предпочтительным.
До настоящего времени живые ослабленные вирусы получали путем удаления несущественного гена или путем частичного повреждения одного или нескольких существенных генов (в этом случае повреждение проводят таким образом, что гены остаются функциональными, но девствуют не так эффективно). Однако живые ослабленные вирусы часто остаются патогенными и могут неблагоприятным обрезом воздействовать на хозяина. Кроме того, в этих вакцинах, если только их ослабление не вызвано путем специфической делении, не исключена возможность реверсии к более вирулентной форме. Тем не менее, тот факт, что в хозяине имеет место продуцирование некоторого количества вирусного белка означает, что указанные вакцины являются более эффективными, чем убитые вакцины, которые не могут продуцировать этот вирусный белок.
Живые ослабленные вирусы, которые сами по себе используются в качестве вакцин, могут быть также использованы в качестве "вакцинных векторов" для других генов, т.е., другими словами, в качестве носителей генов от второго вируса (или другого патогена), против которого необходимо выработать иммунитет. Обычно в качестве вакцинных векторов используют вирусы группы оспы, например, вирус коровьей оспы. Если вирус используется в качестве вакцинного вектора, то очень важно, чтобы этот вирус не имел патогенного действия. Другими словами, может потребоваться его ослабление тем же способом, которым ослабляют простую вирусную вакцину. Следовательно, и в этом случае имеют место недостатки, описанные выше.
Было обнаружено, что можно удалить ген из вирусного генома и получить так называемую "комплементирующую" клетку, которая имеет вирус, полученный в результате делеции гена. Указанная операция была осуществлена для некоторых вирусов, например аденовирусов, вирусов герпеса и ретровирусов. В случае аденовирусов клеточная линия человека была трансформирована фрагментами ДНК аденовируса типа 5 (Graham, Smiley, Russ ell&Nairn, J.Gen. Virol., 36, 59-72, 1977). Эта клеточная линия экспрессировала некоторые вирусные гены и, как было установлено, она способна поддерживать развитие вирусных мутантов, имеющих делегированные иди инактивированные гены (Harrison, Graham&Williams, Virology 77, 319-329, 1977). Хотя этот вирус хорошо развивался на этой клеточной линии (линии комплементирующей клетки) и продуцировал стандартные вирусные частицы, однако он абсолютно не развивался на нормальных клетках человека. Клетка, экспрессирующие Т-антиген-кодирующую область генома вируса SV40 (паповавируса), обладают также способностью поддергивать репликацию вирусов, специфически делегированных в этой области (Gluzman, Cell, 23, 182-195, 1981). Для вируса простого герпеса были продуцированы клеточные линии, экспрессирующие гликопротеин gB (Саi и др., J.Virol., 62, 714-721, 1987), гликопротеин gD (Ligas и Johnson, J.Virol., 62, 1486, 1988) и предранний белок ICP4 (Deluca и др., J.Virol., 56, 558, 1985), и эти клеточные линии обнаружили способность поддерживать репликацию вирусов со специфически инактивированными копиями соответствующих генов.
Настоящее изобретение относится к мутантному вирусу для использования его в качестве вакцины, где вирусный ген, кодирующий белок, ответственный за продуцирование инфекционного вируса, является делегированным или инактивированным; и где указанный вирус может быть культивирован в клетке, которая имеет гетерологичную нуклеотидную последовательность, позволяющую указанной клетке экспрессировать необходимый белок, кодированный указанным делегированным или инактивированным вирусным геном.
Настоящее изобретение также относится к получению вакцины, которая содержит вышеуказанный вирус в сочетании с одним или несколькими наполнителями и/или адъювантами. Вирусный геном может сам по себе продуцировать иммуноген, либо он может содержать вставку гетерологичного гена, экспрессирующую иммуногенный белок.
Настоящее изобретение также относится к комплементирующей клетке, трансфецированной ослабленным вирусом, описанным выше и предназначенным для получения вакцины.
Настоящее изобретение также относится к способу, который заключается в использовании описанного выше вируса для получения вакцины, предназначенной для применения в терапевтических или профилактических целях.
Настоящее изобретение, кроме того, относится к способу получения вакцины, который заключается в том, что культивируют клетку, инфицированную вирусом, имеющим делегированный или инактивированный вирусный ген, кодирующий белок, ответственный за продуцирование инфекционного вируса, где указанная хозяйская клетка имеет гетерологичную нуклеотидную последовательность, содержащую вирусный ген, и обладает способностью экспрессировать основной белок, кодированный указанным геном; собирают продуцированный таким образом вирус и используют его для вакцины.
Этот вирус может быть получен от вируса простого герпеса (HSV), в которой, например, делегирован или инактивирован ген, кодирующий гликопротеин Н(gН). Мутантный вирус может также содержать гетерологичную последовательность, кодирующую иммуноген, происходящий от патогена. Хозяйская клетка соответственно должна быть рекомбинантной эукариотической клеточной линией, содержащей ген, кодирующий гликопротеин Н HSV. В качестве другого примера можно использовать вирус, происходящий из ортопоксвируса, например вируса коровьей оспы, который также может содержать гетерологичную последовательность, кодирующую иммуноген, происходящий от патогена.
Настоящее изобретение иллюстрирует уникальный способ, в котором сочетаются эффективность и безопасность убитой вакцины с высоким иммунологическим ответом, индуцируемым посредством in vivo - продуцирования вирусного белка аттенюированной вакциной. Предпочтительный вариант осуществления настоящего изобретения имеет два отличительных признака. Во-первых, выбранный ген инактивируют в вирусном геноме обычно путем осуществления специфической делеции. Этот ген будет участвовать в продуцировании инфекционного вируса, предпочтительно не препятствуя репликации вирусного генома. Таким образом, инфицированная клетка может продуцировать большее количество вирусного белка из реплицированного генетического материала, а в некоторых случаях могут быть продуцированы новые вирусные частицы, которые не должны быть инфекционными. Это означает, что вирусная инфекция не сможет распространяться от места инокуляции.
Вторым отличительным признаком настоящего изобретения является клетка, которая дает вирус с продуктом делетированного гена, что делает возможным культивировать этот вирус в тканевой культуре. Поэтому, хотя в этом вирусе будет отсутствовать ген, кодирующий важный белок, при культивировании в подходящей хозяйской клетке, однако он будет размножаться и продуцировать вирусные частицы, которые по своему внешнему виду будут неотличимы от исходного вируса. Этот мутантный вирусный препарат является неактивным в том смысле, что поскольку он имеет дефектный геном и не может продуцировать инфекционный вирус в нормальном хозяине, то его введение в количествах, необходимых для непосредственного генерирования гуморального ответа в хозяине, является абсолютно безопасным. Таким образом, мутантный вирус необязательно должен быть инфекционным для протектируемой хозяйской клетки, и в основном он может просто действовать таким же образом, что и стандартная убитая или ослабленная вирусная вакцина. Однако предпочтительно, чтобы иммунизирующий вирус еще сам по себе был инфекционным в том смысле, чтобы он мог связываться с клеткой, внедряться в нее и инициировать цикл вирусной репликации, т.е. чтобы он был способен инициировать инфекцию внутри хозяйской клетки, принадлежащей к протектируемым видам, и продуцировать в ней некоторое количество вирусного антигена. Таким образом, представляется еще одна благоприятная возможность стимулировать клеточный арсенал иммунной системы хозяина.
Предпочтительно, чтобы делегированный или инактивированный ген участвовал как можно в более поздней фазе вирусного цикле, с тем, чтобы продуцировать как можно большее количество вирусного белке in vivo для индуцирования иммуногенного ответа. Например, этот ген может быть геном, участвующим в упаковке или в другой пострепликативной стадии, например, таким, как ген гликопротеина gН вируса НSV. Однако выбранный ген может быть гоном, участвующим в репликации вирусного генома, и тогда количество экспрессируемого in vivo белка будет зависеть от стадии, при которой обычно экспрессируется этот ген. В случае использования человеческого цитомегаловируса (НСМV), выбранным геном может быть ген (кроме предраннего гена), который будет сильно препятствовать репликации вирусного генома, поскольку предранний ген, который продуцируется до репликации вирусного генома (и фактически является главным для нее), является высокоиммуногенным.
Настоящее изобретение может бить применено к любому вирусу, в котором один или несколько главных генов могут быть идентифицированы и делегированы из вирусного генома или инактивированы в вирусном геноме. Для ДНК-вирусов, таких, как аденовирусы, вирусы герпеса, паповавирусы, вирусы папилломы, и парвовирусы, эта процедура может быть осуществлена непосредственно путем I) in vivo - модификации клонированных ДНК-копий выбранного гена в целях получения специфических ДНК-мутаций, и II) повторного введения модифицированного варианта в вирусной геном с помощью стандартной техники рекомбинации и "спасения" генетического маркера.
Однако в настоящем изобретении могут быть также использованы и РНК-вирусы. Затем может быть применена любая техника, которая позволяет манипулировать о комплементарными ДНК-копиями генома РНК-вируса in vitro с помощью стандартной генетической технологии, с последующим превращением в РНК путем in vitro-транскрипции. Полученные РНК могут быть затем снова введены в вирусный геном. Эта техника была использована для получения специфических мутаций в геноме как (+) РНК-вируса, так и (-) РНК-вируса, например полиовируса (Racaniello и Baltimore, Science, 214, 916-919, 1981), и вируса гриппа (Lutyes и др., Cell, 59, 1107-113, 1989).
Теоретически для получения аттениюрованного вируса может быть выбран любой ген, кодирующим главный белок. Однако, на практике, выбор такого гена должен проводиться исходя из следующих соображений:
1. Этот ген должен быть предпочтительно геном, который является необходимым в более поздней фазе инфекции. Таким образом, репликация аттениюрованного вируса не прерывается в ранней фазе. Это означает, что большинство, а возможно и вое другие вирусные антигены будут продуцированы в инфицированной клетке и презентованы иммунной системе хозяина совместно о молекулами МНС классе I хозяйской клетки. Такая презентация способствует развитию клеточного иммунитета против вирусной инфекции посредством продуцирования цитотоксичных Т-клеток. Цитотоксичные Т-клетки могут узнавать указанные антигены и, следовательно, нейтрализовать инфицированные вирусом клетки. Возможно, что делегированный ген должен быть таким геном, который совсем необязателен для сборки вируса, но который является необходимым для того, чтобы сборный вирус был способен инфицировать новые клетки. Примером такого белка является белок gН вируса НSV. В отсутствие этого белка вирионы НSV еще будут продуцироваться, но при этом они будут неинфекционными.
2. В идеальном случае продукт выбранного гена не должен быть сам по себе токсичным по отношению к эукаристической клетке, так, чтобы комплементирующая клетка могла относительно легко продуцироваться. Однако это требование не является категоричным, поскольку этот ген может быть помещен под контроль индуцируемого промотора в комплементирующей клетке, так, чтобы его экспрессия могла бы включаться, когда это потребуется.
Природа мутаций, создаваемые в целевой гене, также является предметом выбора. Любая мутация, которая продуцирует нефункциональный генный продукт, является вполне приемлемой, если только при этом не минимизирован риск реверсии к структуре дикого типа. Такими мутациями могут быть встраивание в целевой ген чужеродных последовательностей и создание специфических делений. Однако наиболее предпочтительной мутацией в изготовлении вакцины, предназначенной для использования в терапевтических и/или профилактических целях, является делеция, которая охватывает всю доследовательность, вводимую в комплементирующую клетку. Эта методика позволяет минимизировать риск регенерации вируса дикого типа посредством рекомбинации между вирусом и клеточной ДНК в комплементирующей клетке.
Хотя в настоящее время имеется несколько примеров комбинаций специфически инактивированных вирусов и комплементирующих клеток (см. вышеприведенную дискуссию), однако они были использованы либо для фундаментальных исследовании вирусов, либо, в случае ретровирусов, для получения безопасного вектора в целях продуцирования траногенных животных. Указанные комбинации не использовались для производства вакцин и, насколько известно заявителям, не было сделано даже никаких предположений относительно возможности такого их использования.
Помимо использования указанной комбинации инактивированного вируса и комплементирующей клетки в целях продуцирования безопасных вакцин против вируса дикого типа, настоящее изобретение также относится к использованию указанной системы для получения безопасных вирусных векторов, предназначенных для использования в качестве вакцин против чужеродных патогенов. В качестве примера может служить вектор, полученный на основе HSV. Геном вируса является достаточно большим, чтобы вместить значительное дополнительное количество генетической информации, и несколько примеров рекомбинантных вирусов HSV, несущих и экспрессируюших чужеродный генетический материал, было описано в литературе (например, Ligas и Johnson, J.Virol. 1988, см. выше). Так, например, вирус, имеющий делецию в главной вирусном гене, описанном выше, а также несущем и экспрессирующем определенный чужеродный ген, может быть использован в качестве безопасного вектора для вакцинации в целях генерирования иммунного ответа против чужеродного белка.
Отличительной особенностью вируса HSV является тот факт, что он может быть латентным в нейронах инфицированного организма, но иногда он снова реактивируется, вызывая тем самым очаговое поражение. Такими образом, вирус НSV, имеющий делецию в главном вирусе гена и экспрессирующий чужеродный ген, может быть использован для продуцирования желательной латентной инфекции нейронов у подвергающегося лечению пациента. Реактивация такой латентной инфекции не должна вызывать поражения, поскольку вирусный вектор не способен полностью реплицироваться, но может инициировать начальную цикла репликации вируса. В течение этого периода времени может иметь место экспрессия чужеродного антигена, генерируя тем самым иммунный ответ. В том случае, когда делетированный ген вируса HSV определяет белок, который является необязательным для сборки вируса, а необходим лишь для инфекционности сборного вируса, чужеродный антиген может быть введен в сборные вирусные частицы в целях усиления иммуногенного действия. Эта экспрессия чужеродного гена и введение его белке в вирусную частицу может, конечно, также происходить на той стадии, где мутантный вирус сначала продуцируется в комплементирующей клетке, и в этом случае мутантный вирус при использовании его в качестве вакцины будет сразу представлять чужеродный белок обрабатывамым видам.
В другом примере вирус коровьей оспы, поксвирус, может иметь и экспрессировать гены от различных патогенов, и это свидетельствует о том, что такие вакцины могут быть эффективными при использовании их в живых экспериментальных системах. Возможности использовать этот вирус для человека довольно широки, но из-за известных побочных эффектов, связанных с широко распрастраненным использованием вируса коровьей оспы в качестве вакцины против натуральной оспы, использовать в широком масштабе немодифицированный вирус коровьей оспы на человеке нежелательно. Были сделаны попытки ослабить вирус коровьей оспы путем делеции несущественных генов, напримэр, гена фактора роста коровьей оспы (Buller, Chakrabarti, Cooper, Twardzik & Moss, J.Virolooy, 62, 866-874, 1988). Однако такие ослабленные вирусы еще способны раплицироваться in vivo, хотя и при более низком уровне. Но пока еще не было получено вируса коровьей оспы с делецией в главном гене, хотя именно такой вирус, т.е. вирус с делецией в главном гене, описанный выше, позволил бы получить безопасный вариант этого вакцинного вектора.
Еще одно преимущество описываемой стратегии иммунизации против гетерологичных белков заключается в том, что с одним и тем же вирусным вектором можно осуществлять многократные эффективные вакцинации, что невозможно сделать, используя стандартные живые вирусные векторы. Поскольку эффективность стандартной живой вирусной вакцины основана, вероятно, на ее способности к репликации в животном-хозяине посредством многих циклов инфицирования, то эта эффективность будет сильно сокращаться в организме, где вырабатывается иммунитет против этого вируса. Так, например, весьма вероятно, что второе контрольное заражение тем же вирусом, независимо от того, проводится ли эта бустериммунизация против этого же белка, либо с целью вызвать новый ответ против другого белка, будет неэффективным. Однако использование вирусного вектора с делецией в главном гене в том случае, где многоциклическая репликация является нежелательной или необязательной, дает эффективную реакцию почти сразу после иммунизации. Доза мутантного вируса может быть относительно большой, поскольку этот вирус абсолютно безопасен, и поэтому маловероятно, что такое раннее воздействие мутантного вируса будет блокироваться иммунным ответом хозяина, для мобилизации которого требуется некоторое время.
Хотя все сказанное выше относилось к мутантному вирусу, имеющему делению в главном гене и необязательно содержащему ген для иммуногенного белка-патогена, однако этот мутант может быть дефектным в более чем одном главном гене и/иди содержать более чем один ген для иммуногенного белка-патогена. Так, например, мутантный вирус может включать в себя ген для gp 120 ВИЧ, в результате чего вакцина будет действовать предполагаемым выше способом, а также ген для gag-белка ВИЧ для того, чтобы этот белок экспрессировался в вакцинированном хозяине и был презентирован на поверхности хозяйской клетки вместе о MHC-I в целях стимуляции Т-клеточного ответа в хозяине.
Для более наглядной иллюстрации настоящего изобретения нижe представлены примеры, которые, однако, не должны рассматриваться как некое ограничение изобретения; а также следующие чертежи:
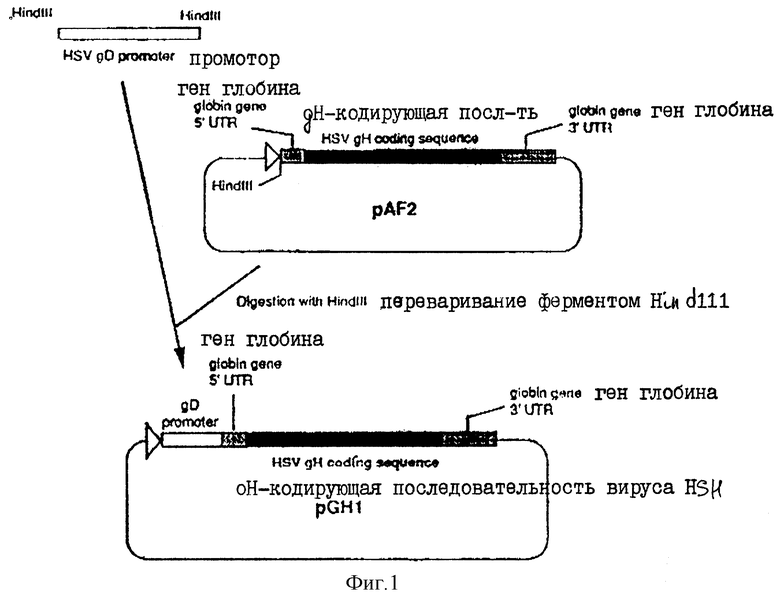
На фиг.1 показано продуцирование плазмиды pCHl.
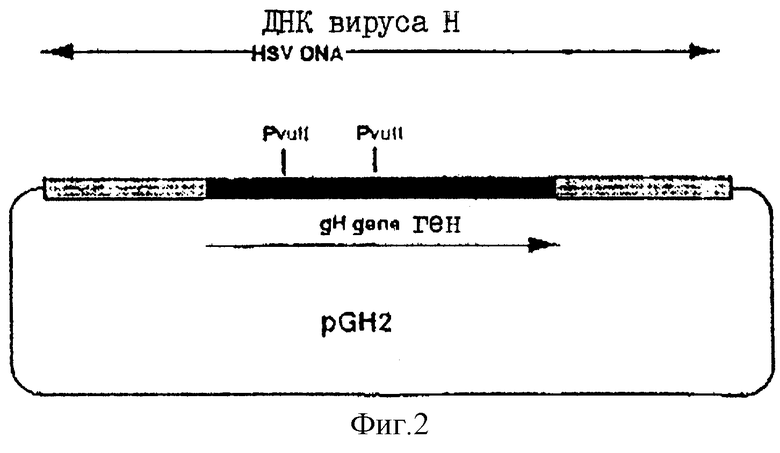
На фиг.2 показано продуцирование плазмиды рGH2.
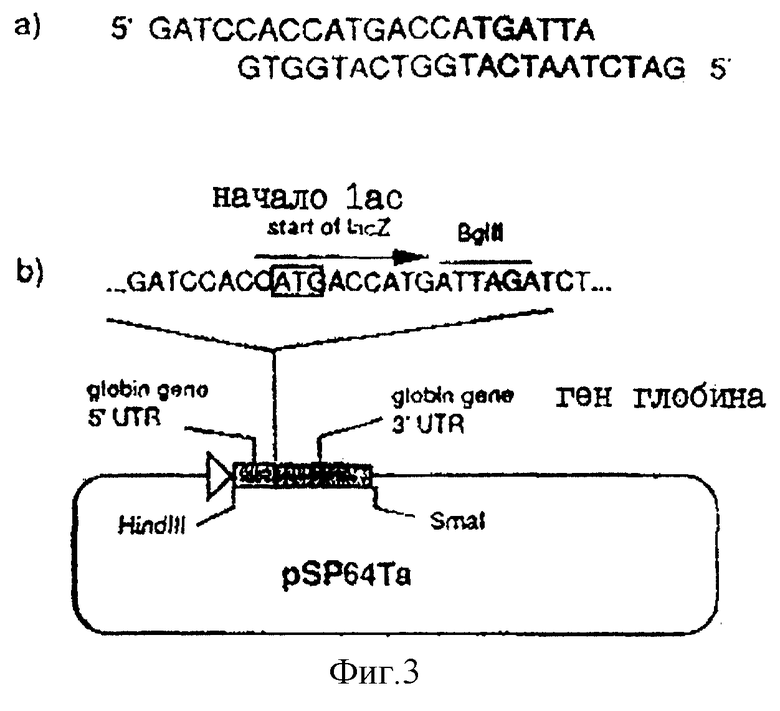
На фиг.3а показаны два комплементарных олигонуклеотида, используемых для получения плазмиды рSР64Та; На фиг.3в показано продуцирование плазмиды рSР64ТА.
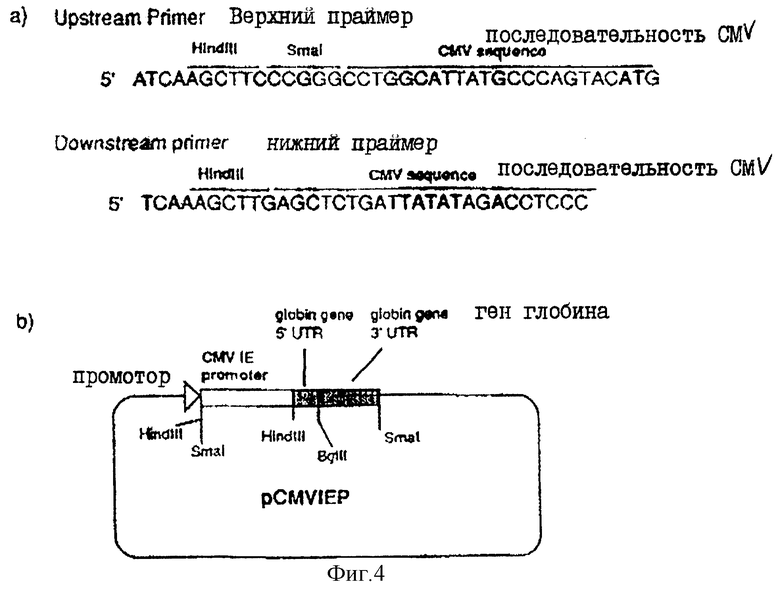
На фиг.4a показаны два олигонуклеотида, используемые для получения плазмиды рСMVIEP.
На фиг.4в показана плазмида pCMVIEP.
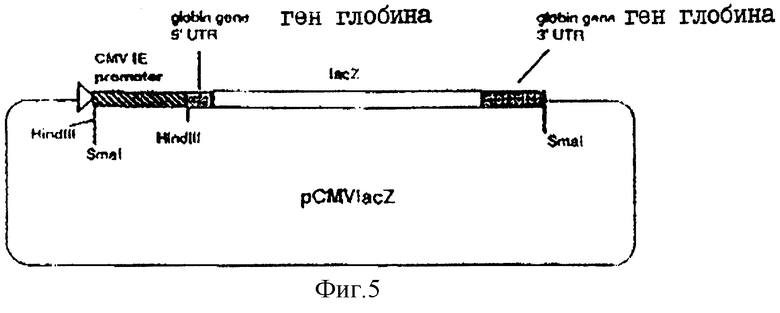
На фиг.5 показана плазмида рСMVIacz.
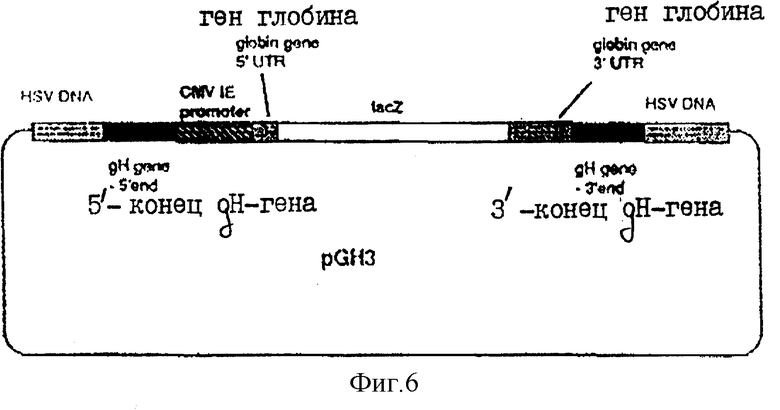
На фиг.6 показана плазмида pGH3.
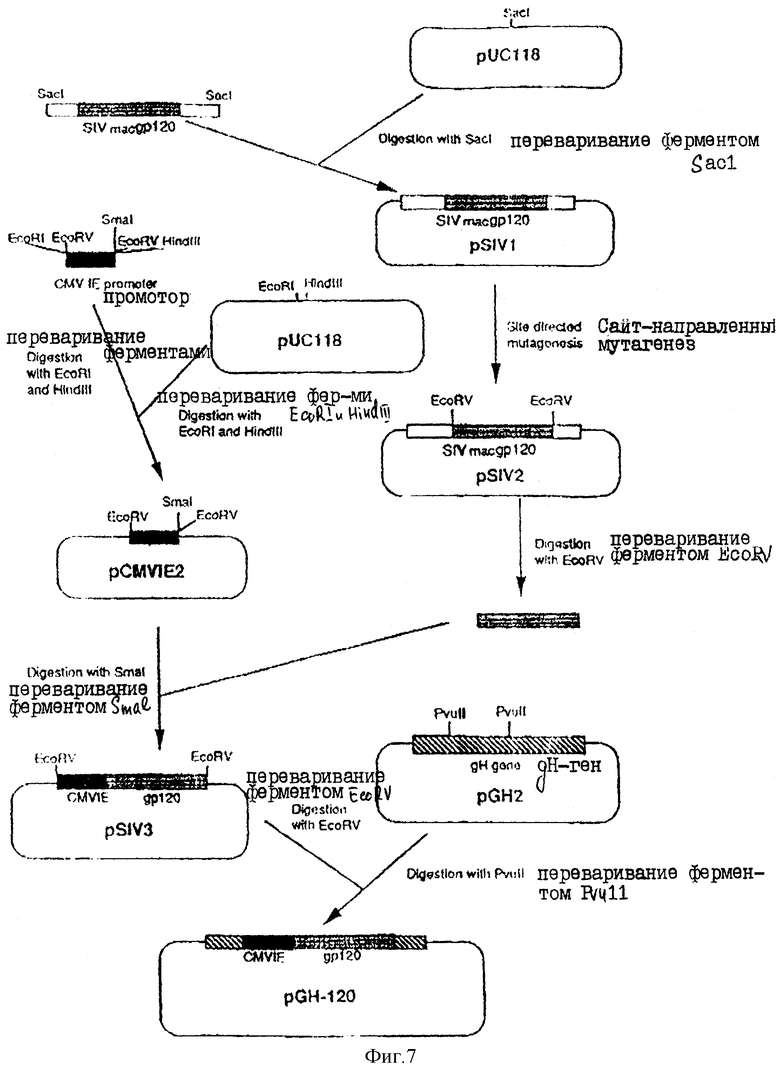
На фиг.7 показана стратегия конструирования плазмиды рGН-120.
Вирус простого герпеса, делетированный в гене гликопротеина Н (gH-HSV)
Вирус простого герпеса (НSV) является большим ДНК-вирусом, который вызывает широкий ряд патогенных симптомов у человека, включая рецидивирующие поражения лица и гениталий, и нередко энцефалиты с летальным исходом. Инфицирование этим вирусов можно до некоторой степени устранить с помощью химиотерапии с использованием лекарственного средства Ацилоквира, однако еще не найдено соответствующей вакцины, которая позволила бы предупредить первичную инфекцию. Трудность вакцинации против вируса НSV заключается в том, что этот вирус в основном распространяется в организме путем непосредственного переноса от клетки к клетке. Поэтому в этом случае гуморальный иммунитет будет малоэффективным, поскольку циркулирующие антитела могут нейтрализовать лишь внеклеточный вирус. Для борьбы о вирусной инфекцией такого рода был бы эффективен клеточный иммунитет, и при этом предпочтительной была бы такая вакцина, которая была бы способна генерировать как гуморальный, так и клеточный иммунитет, но которая также была бы безопасной.
Целевым геном, подходящим для инактивации в геноме Н является ген гликопротеина Н (gН). gН-белок представляет собой гликопротеин, присутствующий на поверхности вирусной оболочки. Этот белок, по-видимому, участвует в процессе сращивания мембраны во время внедрения вируса в инфицированную клетку. Поэтому чувствительные к температуре вирусные мутанты с дефектом в этом гене не выбрасываются из вирус-инфицированных клеток при непермиссивных температурах (desai и др., J.Gen. Vir ol., 69, 1147-1156, 1988). Если этот белок экспрессируется в поздней стадии инфицирования, а также если он отсутствует, то при этом может все же иметь место значительный синтез вирусного белка.
Все генетические манипуляции осуществляли в соответствии со стандартными методами, описанными в "Molecular Cloning", А Laboratory Manual, изд. Sambrook, Fritsch and Maniatis, Cold Spring Harbor Laboratory, 1989.
A. Получение клеточной лининии, экспрессирующей ген gН Н
Ген gН находится в уникально длинной области (UL) генома вируса НSV типа I, между нуклеотидами 46382 и 43868 (МcGееch и др., J.Gеn. Virol. 69, 1531-1574, 1988). Клонированная копия этого гена имеется в плазмиде рAF2. Эта плазмида была получена путем вырезания BglI-XhoI-фрагмента, охватывающего полную gН-кодирующую последовательность, из плазмиды pTZgН, и его клонирования в ВgII-сайт плазмиды рSР64Т, описанной Gompels и Мinson (J.Virol., 63, 4744-4755, 1989). Затем из плазмиды рSVD4 (Everett NucI. Acids Res., II, 6647-6667, 1983) вырезали HindIII-фрагмент, содержащий промоторную последовательность для гена гликопротеина D(gD) (простирающуюся от нуклеотида -392 до +11 по отношению к началу гена gD), и клонировали в уникальный HindIII-сайт рАF2 для продуцирования pGHI (фиг.1), так, чтобы промоторная последовательность была в правильной ориентации, необходимой для стимулирования экспрессии гена gН. Таким образом, эта плазмида содержит полную gН-кодирующую последовательность под контролем промотора гена gD HSV типа I. Затем эту плазмиду очищали и трансфецировали в клетки Vero вместе с плазмидой pNEO (Pharmacia LKB Biotechnology Inc.) с использованием стандартной техники совместного осаждения фосфатом кальция (Graha и Ver der Eb, Virology 52, 456-467, 1973). Клетки vero с приобретенной устойчивостью к неомищину затем отбирали путем пассирования клеток в лекарственную среду G418, а колонии этих клеток клонировали путей конечного разведения. Эти неомицин-резистентные клетки были затем амплифицированы в тканевой культуре, после чего образцы инфицировали вирусом HSV типа 2. Инфицирование вирусом HSV типа 2 способствует индуцированию транскрипции от промотора gD типа I, присутствующего в геноме комплементирующей клетки, а также стимулирует продуцирование белка gН типа I в комплементирующей клетке. Лизаты инфицированных клеток затем скринировали на экспрессию gН-белка с помощью вестерн-блоттинга, используя при этом поликлональную антисыворотку, которая, как известно, способна к специфическому узнаванию gН-белка типа I (Desei и др., 1988, см. выше). Клетки, которые экспрессируют нужный белок, затем сохраняли, и приготавливали замороженный шток. Этот материал представлял собой gН + линию комплементирующих клеток.
В. Получение вируса НSV типа I с прерванным геном gН
ВоIII-фрагмент в 6432 пар оснований, содержащий gH-кодирующую последовательность вместе с фланкирующими последовательностями Н, вырезали из плазмиды pUG102 (Gompel и Мinson, Virology 153, 230-247, 1986) и клонировали в плазмиду рАTI53 (twigg и Sherrat, Nature, 283, 216, 1980), в результате чего получали плазмиду рGН2 (фиг.2). Эту плазмиду переваривали PvuII, причем ее разрезали только в gН-кодирующей последовательности в двух положениях (в нуклеотидах 44955 и 46065) согласно схеме нумерации McGeoch и др., 1988, см. выше), и самый большой из двух (фрагментов очищали. ДНК-фрагмент, содержащий полный ген В-галактозидазы от E.coli, находящийся ниже (в прямом направлении) от промотора продранного гена, происходящего от цитомегаловируса (CMV), затем получали с помощью следующей процедуры. Прежде всего отжигали два комплементарных олигонуклеотида (фиг.3а) и лигировали с BgIII-переваренной и обработанной фосфатвной плазмидой рSР64Т Krieg и Melton, NucI. Acids Res., 12, 7057-7071, 1984), в результате чего получали плазмиду рSP64Та показанную на фиг.3в. Присоединенный линкер также содержал инициирующий кодон и первые три кодона гена В-галактозидазы (Iacz) E.coli. Затем "область серцевины" промотора предраннего гена от CMV амплифицировали из плазмиды рUG-HI (Gompels и Мinson, 1988, см. выше) с помощью полимеразно-цепной реакции (PCR-Molecular Сloning, изд. Sambrook и др., см. выше), где использовались два олигонуклеотида, показанные на фиг.4а, и соответствующие последовательностям от -302 до -288, и от -13 до -36, соответственно (нумерация проводилась по отношению к началу предраннего гена CMV, описанного Аnrigg и др., Virus Rеsеаreh, 2, 107-121, 1985). Эти олигонуклеотиды также содержат у своих 5 концов сайты рестрикции для фермента HindIII, а в случае отжига олигонуклеотида выше от промотора еще и Smal-сайт. Затем PCR-амплифицированный продукт ДНК переваривали HindlII, и клонировали в HindIII-переваренную рР64Та, в результате чего получали плазмиду pCMVIEP (фиг.4в). И, наконец, ДНК-фрагмент, содержащий полную копию В-галактозидазы, в котором отсутствует лишь крайняя 5'-концевая область кодирующей последовательности, выделяли путем переваривания плазмиды рSС8 (Chakzabarti и др., Mol. Cell. Biol., 5, 3403-3409, 1985) ферментом BamHI и клонировали в уникальный BoIII-сайт pCMVIEP, в результате чего получали pCMVlacz (фиг.5), ДНК-фрагмент, содержащий ген В-галактозидазы под контролем CMV IE-промотора, затем выделяли путем переваривания плазмиды pCMV-Iacz ферментом Smal, и лигировали с очищенным PvuIII-фрагментом плазмиды рGН2, описанной выше, в результате чего получали плазмиду рGH3, которая содержала копию gН-гена, прерванного функуиональным геном В-галактозидазы (фиг.6). Следующая стадия заключалась в замене gН-гена дикого типа в геноме HSV полученным прерванным мутантом, и эту стадию осуществаляли с помощью рекомбинации между HSV-ДНК и плазмидой рGН3, с последующим отбором тех вирусов, которые приобрели функциональный ген В-гадактозидазы. Для этого ДНК плазмиды рGН3 трансфецировали в клетки, экспрессирующие gН-ген (gН+комплементирующая клеточная линия, описанная в части А), совместно с очищенной ДНК HSV, выделенной из очищенных HSV-вирионов (Killington и PoweII "Technigues in Virology: A practical Appzoach" (изд. В.W.C.Маhу, стр.207-236, IPL Press, Оксфорд (1985)), с использованием стандартной техники осаждения фосфатом кальция (Grаhаm и Van de Eb, 1973, см. выше). Потомство вируса HSV, полученное в результате описанного эксперимента по трансфекции, затем высевали на монослои gН+ комплементирующих клеток, проводя стандартный анализ бляшкообразования и используя верхний агаровый слой для покрытия, в присутствии 5-бромохлоро-3-индолил-β-D-галактозида (Х-gаI), хромогенного субстрата, который при наличии β-галактозидазы приобретал синий цвет.
Таким образом, бляшки, полученные в результате инфицирования вирусными геномами, содержащими и экспрессирующими ген В-галактозидазы, становились синими. Это означало, что эти вирусные геномы должны содержать вариант прерванного gН-гена. Затем вирус очищали из этих бляшек путем снятия агаровых пробок из соответствующей части планшеты, а вирусные заготовки получают путем культивирования этого вируса в gН+комплементирующей клеточной линии. Эти вирусы, поскольку они несут нефункциональные варианты gН-гена, должны быть неспособны образовывать бляшки на клетках, которые не содержат и не экспрессируют экдогенную функциональную копию gН-гена, и для подтверждения этого факта образцы анализировали на их способность образовывать бляшки на монослоях клеток дикого типа линии Vero по сравнению с gН-комплементирующими клетками. И, наконец, из этих заготовок получали вирусную ДНК и проводили тестирование на ожидаемую структуру ДНК возле gН-гена о помощью Саузерн-блоттинга. После подтверждения правильности генетической структуры приготавливали большие запасы вируса с дефицитным gН-геном путем инокуляции образца вируса в крупномасштабную культуру gН+ комлементирующей клеточной линии (множественность инфекции=0,01), а через три дня клетки собирали. Инфицированные клетки разрушали о помощью ультразвука для высвобождения ассоциированного с клетками вируса, и всю смесь, обработанную ультразвуком, хранили при -70° в виде вирусного маточного препарата. Титр этого вирусного препарата затем устанавливали с помощью анализа бляшкообразования на gН+комплементирующей клеточной линии. После этого образцы этого вирусного препарата использовали для получения рабочих препаратов, описанных выше, после чего эти рабочие препараты использовали для инфидирования лабораторных животных, как описано ниже.
С. Исследование протективного действия gН-НSV по сравнению с вирусом, обезвреженным тепловым воздействием
Для анализа иммунологического ответа хозяина на этот вирус проводили эксперименты по контрольному заражению мышей в соответствии со схемой, описанной ниже.
Затем сравнивали протективное действие живого gH--вирусного препарата с действием инактивированного препарата вируса дикого типа (WT) (штамм SCI6). Получение инактивированного вируса дикого типа для вакцинации
Вирус HSV типа I (штамм SC16) культивировали при низкой множественности заражения (0,01 б.о.е/клетка) клеток линии Vero. Через три дня вирус собирали, и цитоплазматический вирус выделяли с помощью гомогенизатора Даунса. Ядра удаляли путем центрифугирования 15 минут при 500×г, а вирус выделяли из супернатанта путем центрифугирования в градиенте сахарозы (до 40%) при 12 К в течение 60 мин, с использованием ротора Beckman SW27. Полосы вируса разводили, осаждали и очищали путем центрифугирования в градиенте сахарозы (Killington и Powell, см. выше). Полосу вируса собирали из градиента, в вирус выделяли путем центрифугирования. Затем вирус ресуспендировали в фосфатно-солевом буфере (PBS), анализировали на инфекционность путем титрования бляшек на клетках почки дитеныша хомячка (ВНК), и проводили подсчет частиц на электронном микроокопе. Отношение "частица: ифeктивнoсть" препарата составляло 110 частиц/б.о.е. Вирус разводили до 2,5×1010 б.о.е./мл в PBS, и инактивировали путем обработки β-пропиолактоном при 20°С в течение 60 минут. Полученные аликвоты затем хранили при -70°С.
Получение живого gН--вируса для вакцинации
Этот материал получали так же, как и вирус дикого типа, за исключением того, что этот вирус культивировали в gH+ комплементирующей клеточной линии, содержащей и экспрессирующей gН-ген HSV дикого типа, и не подвергали инактивации путем обработки β-пропионолактоном. Отношение "частицы: инфекционность" этого препарата составляло 150:1. Концентрацию этого препарата доводили до 2,5×1010 б.о.е./мл, и аликвоты хранили в PBS при -70°C.
Схема вакцинации
4-недельных самок мышей баlb/С (поставляемых Tucksv. k. Ltd.) подвергали вакцинации различными дозами инактивированного вируса дикого типа или живого вируса gH- в 2 мкл-объемах фосфатно-солевого буфера путем капельной аппликации и скарификации иглой правого уха по следующей схеме:
Группа А Контроль - не обрабатывали вирусом
Группа В 5×104 б.о.е. вирусной вакцины
Группа С 5×105 б.о.е. вирусной вакцины
Группа Д 5×106 б.о.е. вирусной вакцины
Группа Е 5×107 б.о.е. вирусной вакцины
Через 14 дней всех мышей подвергали контрольному заражению путем аналогичной инокуляции левого уха 2×106 б.о.е. штамма SCI6 HSV-I (вируса дикого типа). Через 5 дней мышей забивали и анализировали на вирусную инфекционность в левом ухе и в левых шеечных ганглиях СII, CIII и СIV (объединенные данные). Для исследования латентного состояния, других вакцинированных и подвергнутых контрольному заражению животных забивали через 1 месяц и испытывали из латентную инфекцию путем иссечения гинглий cII, cIII и cIV. Эта ткани инкубировали в среднем 5 дней, а затем гомогенизировали и оценивали на присутствие инфекционного вируса путем стандартного анализа бляшкообразования. Все полученные результаты выражали в б.о.е./орган (бляшкообразующих единиц не орган).
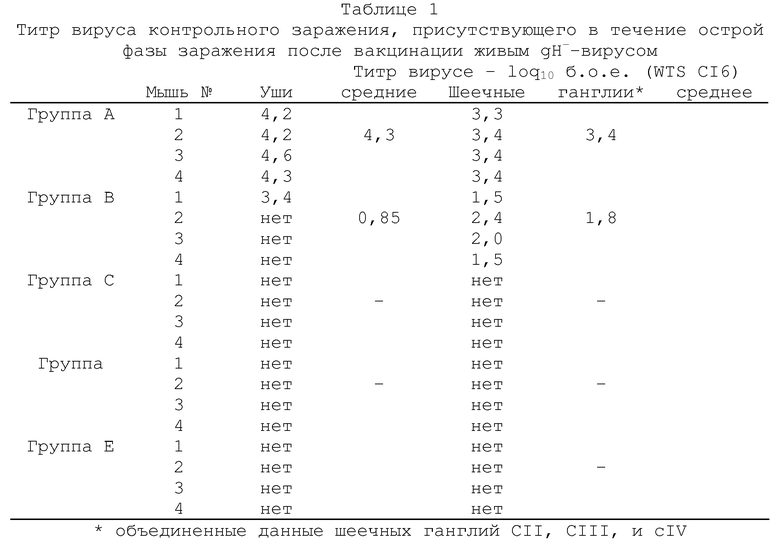
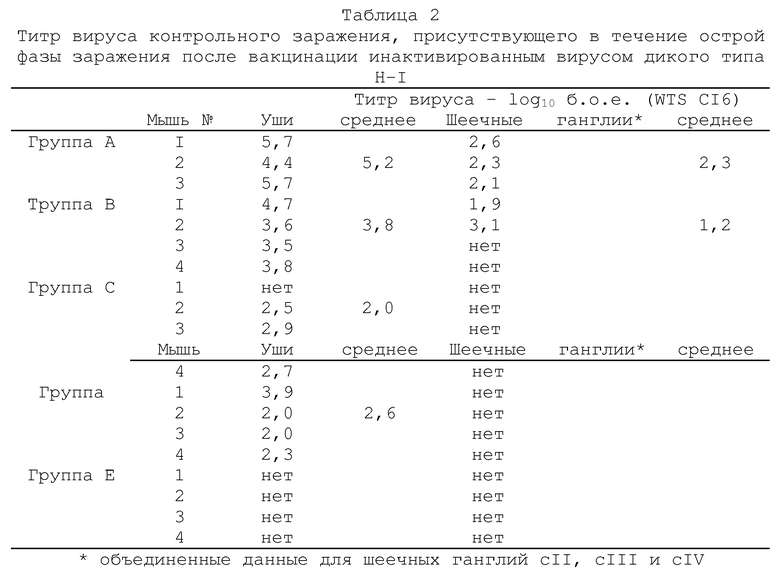
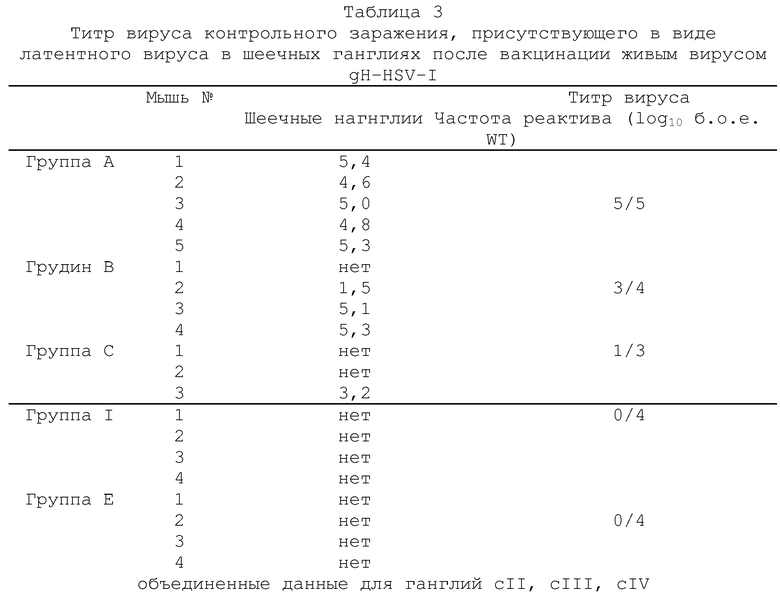
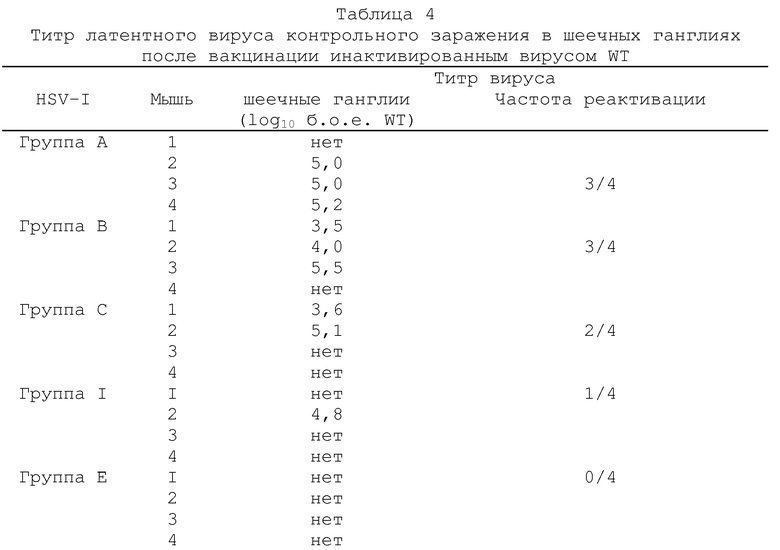
объединенные данные для шеечных ганглий cII, cIII и cIV (б.о.е. - бляшкообазующие единицы; gН--вирус с дефектным gH-геном).
Полученные результаты показывают, что титр вируса контрольного заражения дикого типа SCI6 присутствовал в ушах и шеечных ганглиях во время острой фазы заражения. Так, например, низкий титр указывает на хорошую эффективность схемы вакцинации вирусом gН-, тогда как более высокий титр указывает на более низкую эффективность. Из подученных результатов видно, что вакцинация живым gH--вирусом KSV значительно более эффективна, чем эквивалентное количество инактивированного вируса дикого типа. В случае инактивированного препарата, для предупреждения репликации в ухе вируса контрольного заражения требуется доза 5×107 б.о.е., а в случае живого qH--вируса эта доза необходимого вируса в 100-1000 раз меньше. Вакцинация живым gH--вирусом в дозе 5×105 б.о.е. и выше оказалась также способной блокировать peпликацию вируса контрольного заражения в шеечных ганглиях во время острой фазы заражения и, кроме того, явно показала протективное действие против активации латентной инфекции в шеечных ганглиях.
Вирус HSV, в котором отсутствует gН--ген и который используется в качестве вектора для иммунизации против чужеродного антигена: введение gр120-гена штамма 142SIVmac в геном gH-вируса HSV
Как было описано выше, вирусы с делениями в главных генах могут быть использованы в качестве безопасных векторов для презентации чужеродных антигенов иммунной системе, и в качестве подходящего примера такого вектора был использован gH-вирус (HIV), описанный выше. Этот вирус может быть использован для экспрессии любого нужного чужеродного антигена, но особенно привлекательной является возможность экспрессировать главные антигенные белки вируса иммунодефицита человека (ВИЧ), вызывающего СПИД. Так, например, эти последовательности могут быть введены в qH--HSV-геном так, чтобы обеспечить их экспрессию в течение заражения нормальных клеток (т.е. некомплементирующих клеток) рекомбинантным вирусом. Инфицирование организма таким вирусом приводит к латентной инфекции, которая по прошествии некоторого времени реактивируется, что способствует резкому увеличению продуцирования чужеродного антигена и тем самым стимуляции иммунного ответа к этому белку.
Поскольку проводить исследования с целью проверки этого метода непосредственно на человеке не представляется возможным, то в качестве начального этапа исследования этот метод может быть проверен на обезьянах с использованием обезьяньего вируса СПИДа (SIVmac, обезьяний вирус иммунодефицита, полученный от макак). Подходящим для этой цели геном SIV является ген, кодирующий белок gp120, один из главных антигенных мишеней для этого вируса. Поэтому этот ген был введен в gH--геном HSV, после чего был проведен анализ на эффективность этого вируса в качестве вакцины в выработке иммунитета у обезьян против контрольного заражения вирусом S IV.
Прежде всего, gp120-ген SIV клонировали в середину соседнего промотора цитомегаловируса IE (Gompels и Minson, 1989, см. выше), а затем ДНК-кассету, содержащую gp120-ген и расположенный выше CMV-промотор клонировали в плазмиду рGН2 (фиг.2). Полученную плазмиду затем совместно трансфецировали в gН+ комплементирующую клеточную линию вместе с ДНК, очищенной из gH-НSV, и рекомбинантный вирус, который вместо гена β-галактозидазы, присутствующего в gН-HSV-вирусе, приобрел гэн ор120, выделяли путем скрининга на наличие гена β-галактозидазы.
А. Конструирование плазмиды для рекомбинации SIV-gp120-кодирующей последовательности в геноме НSV.
Полная схема этой процедуры показана на фиг.7. Фрагмент рестриктирующего фермента Sacl (соответствующий основаниям 5240-8721) вырезали из клонированной ДНК-копии генома SIV (Chakrabarti и др., Nature 328, 543 (1987) и клонировали в Sacl-сайт плазмиды рVС118 (Viera и Messing, Methods in Enzymology, 153, 3, 1987) в целях генерирования плазмиды pSIVI, которая может быть затем трансформирована в одноцепочечную ДНК для манипуляций с помощью сайт-направленного мутагенеза. Затем эту ДНК-область, которая включает в себя env-генSIV (расположенный между 6090-8298), модифицировали с помощью сайт-направленного мутагенеза (Brierley и др., Cell, 57, 537, 1989) в долях введения сайта для рестриктирующего фермента EcoRV в положения 6053-6058, используя при этой синтетический олигонуклеотид
5'GААGАAGGCТАТАGСТAAТАСАT
Затем вводили второй EcoRV-сайт в положения 7671-7676 внутри env-гена SIV, которые соответствуют сайту расщепления между gp120- и gp40-доменами env-генной последовательности, используя при этом синтетический олигонуклеотид
5'СAAGAАAТАААСТАТАGGTСТТТGTGС,
в результате чего получали плазмиду pSlV2. Затем путей переваривания SIV2 ферментом EcoRV получали ДНК-фрагмент (1617 пар оснований), соответствующий gp120-части env-гена SIV.
Область сердцевины промотора продранного гена СMV получали из плазмиды рVG-HI (Gompels и Minson, 1989, см. выше) при помощи полимера эко-цепной реакции с использованием следующих двух синтетических олигонуклеотидов:
праймер, расположенный выше (в направлении 5→3)
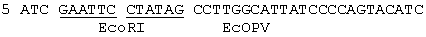
праймер, расположенный ниже
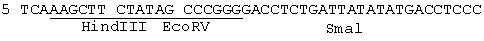
Затем продукт этой реакции гидролизовали ферментами EcoRI и HindIII с получением ДНК-фрагмента, который затем клонировали в ЕсОRI- и HindIII-переваренную плазмиду рМС118, в результате чего получали плазмиду pCMVIE2, которая имела уникальный smal-сайт, расположенный непосредственно ниже промоторной последовательности CMV. EcoRV-фрагмент, содержащий SIVmac. gp120-кодирующую последовательность, полученную в соответствии с приведенным выше описанием, затем клонировали в указанный Smal-сайт после чего отбирали плазмиду pSIV8 с SIV-кодирующей областью, правильно ориентированной, т.е. так, чтобы имела место экспрессии кодирующей последовательности от промотора. Затем эту плазмиду переваривали ферментом ЕсоRV, в результате чего получали ДНК-фрагмент с тупыми концами, который содержит SIV-последовательность вместе с CMV-промотором, и который после этого клонировали в PVuII-переваренную плаамиду pGH2 (фиг.2) с получением плазмиды рGH-120.
В. Конструирование рекомбинантного gН-HSV, несущего ген gp120S1
Из gН-вируса НSV, сконотруированного в ооответотвии с подробным описанием, приведенным выше, выделяли ДНК, которую трансфецировали в gН+комплементирующие клетки совместно с очищенной ДНК рGН-120. Вирус потомства, полученный в результате этой трансфекции, пассировали на монослои gН+комплементирующей клеточное линии с помощью стандартного анализа бляшкообразования, как описано выше, и с использованием верхнего агара в присутствии X-gal. Родительский gH-вирус содержал функциональный ген-галактозидазы, расположенный внутри остаточных gН-кодирующих последовательностях, и образовывали в присутствии Х-gal синие бляшки. Однако рекомбинантные вирусы, которые вместо гена β-галактозидазы приобрели SIV-gp120-кодирующую последовательность, продуцируют белые бляшки. Затем вирус выделяли из этих белых бляшек путем снятия агаровых пробок, и готовили вирусные препараты посредством культивирования этого вируса в gН+ комплементирующей клеточной линии. Из этих препаратов получали ДНК, которую тестировали на наличие правильной ДНК-структуры возле gН-гена посредсвом Саузерн-блоттинга с использованием соответствующих зондов, происходящих от кодирующей последовательности SIV. И, наконец, получали вирусные препараты в соответствии с приведенным выше описанием, которые могут быть затем использованы для исследований по вакцинации животных.
Вакцина, содержащая аттеннюированный вирус, может быть получена и использована в соответствии со стандартной техникой, хорошо известной специалистам. Например, эта вакцина может также содержать один или более наполнителей и/или адьвантов. Эффективная доза аттенюированного вируса, содержащегося в акцине, может быть определена традиционными способами, хорошо известными специалистам.
Изобретение относится к медицине и касается вакцины, содержащей мутантный вирус герпеса. Сущность изобретения включает мутантный вирус герпеса, содержащей геном, в котором вирусный ген, кодирующий существенный для продуцирования инфекционного вируса белок, является делегированным или инактивированным так, что вирус способен инфицировать нормальные клетки, реплицироваться и экспрессировать вирусные гены в этих клетках, но не может продуцировать нормальные инфекционные частицы, кроме тех случаев, когда вирус инфицирует комплементирующую клетку, которая имеет гетерологичную нуклеотидную последовательность, которая позволяет комплементирующей клетке экспрессировать существенный белок, кодируемый делегированным или инактивированным вирусным геном. Преимущество изобретения заключается в разработке вакцины для профилактического или терапевтического использования в выработке иммунного ответа с использованием живого, но ослабленного вируса. 17 з.п. ф-лы., 4 табл., 7 ил.
| СПОСОБ ПОЛУЧЕНИЯ ДИЭТИЛОВОГО ЭФИРА р-ХЛОРТЕТРАГИДРОФУРИЛПРОПИЛМАЛОНОВОЙ КИСЛОТЫ | 0 |
|
SU213894A1 |
| Пожарный двухцилиндровый насос | 0 |
|
SU90A1 |
| Способ размножения копий рисунков, текста и т.п. | 1921 |
|
SU89A1 |

