Область техники, к которой относится изобретения
Настоящее изобретение относится к способу получения белка о5-интерферона (ИФНа5) путем экспрессии в продуцирующей ИФНа5 клетке-хозяине Escherichia coli, причем инкорпорация дополнительного остатка метионина в N-концевую часть полипептидной цепи минимизирована, равно как генерация его окисленных молекул. Белок ИФНа5 можно очистить эффективным способом с получением биологически активного ИФНа5.
Уровень техники
Интерфероны (ИФН) представляют собой группу образующихся в естественных условиях плейотропных гликопротеинов, известных как цитокины, секретируемых различными видами клеток (эпителиальными клетками, фибробластами, лимфоцитами, макрофагами) путем индукции рядом стимуляций (вирус, бактерии, клетки, опухоли и макромолекулы) и обеспечивающих антивирусные, антипролиферативные и иммуномодулирующие свойства, а также обезболивающее действие. После эндогенной продукции или введения интерферон взаимодействует со специфическими рецепторами на клеточной поверхности и начинает трансдукцию сигнала через цитоплазму в ядро, индуцируя экспрессию генов, которые кодируют специфические белки, обладающие антивирусной и иммуностимулирующей активностью. Признаны лечебные возможности ИФН, что продемонстрировано одобрением нескольких различных типов ИФН в плане применения у человека, например, ИФН1а (ребиф, авонекс), ИФН1b (бетасерон), в качестве лекарственных препаратов для лечения рассеянного склероза, и рекомбинантного человеческого ИФНа2а (роферон А) и ИФНа2b (интрон А) в качестве лекарственных препаратов для лечения злокачественных (рака) и вирусных заболеваний.
Основываясь на типе рецептора, посредством которого проходит сигнал ИФН, человеческие ИФН разделены на три основных типа, а именно (i) ИФН типа I [все ИФН типа I связываются со специфическим комплексом рецептора поверхности клетки, известным как рецептор альфа-ИФН (ИФНАР) - ИФН типа I у человека представляют собой альфа-ИФН (α-ИФН), бета-ИФН (β-ИФН) и омега-ИФН (ω-ИФН)], (ii) ИФН типа II [ИФН типа II связывается с рецептором гамма-ИФН (ИФНГР) - ИФН типа II у человека представляет собой гамма-ИФН (γ-ИФН)], и ИФН типа III [ИФН типа III передают сигнал через комплекс рецептора, состоящий из IL10R2 и ИФНLР1]. Альфа- и бета-ИФН, известные как ИФН типа I, структурно коррелируют, стабильны при кислом рН и конкурируют за один и тот же клеточный рецептор (ИФНАР).
В настоящее время альфа-, бета- и гамма-ИФН могут быть получены в рекомбинантной форме с двойным преимуществом в виде получения значительно больших количеств продукта по сравнению с количествами, полученными путем выделения из природных источников (лейкоцитов, фибробластов, лимфоцитов), и снижения сложности способов очистки и проверки безопасности продукта. В действительности большую часть имеющегося в продаже рекомбинантного ИФН фармацевтического качества получают и очищают из Escherichia coli.
Системой выбора для получения ИФН. Действительно гены ИФН не имеют интронов, и белковые продукты, как правило, негликозилированы. Кроме того, Е. coli может быстро расти до высокой густоты клеток, и штаммы, используемые для рекомбинантной продукции белка, генетически модифицированы, так что их, как правило, считают безопасными для крупномасштабной ферментации.
Экспрессия кДНК ИФН достигнута непосредственно в Е. coli вскоре после того, как она была впервые клонирована [см. статьи Goedell et al. Nature., 287, 411-416, 1980; Pestka, S. Arch. Biochem. Biophys., 221 (1), 1-37, 1983; Mizoguchi et al. DNA., 4, 221-32, 1985; Pestka et al. Ann. Rev. Biochem., 56, 727-777, 1987; Baron and Narula. Critical reviews in Biotechnology, 10 (3), 179-190, 1990]. В действительности альфа-ИФН (ИФНа) представляет собой один из первых белков, получаемых с помощью Е. coli при использовании технологии рекомбинантной ДНК [см. статьи Derynck et al., Nature, 287, 193-197, 1980; Nagata et al., Nature, 284, 316-320, 1980].
Однако экспрессия ИФН в Е. coli демонстрирует ряд проблем. ИФН, экспрессируемые в большом количестве в Е. coli, часто осаждаются в виде нерастворимых агрегатов, называемых тельцами включения (IB) [см. статьи Swaminathan et al., Prot Express. Purif., 15, 236-242, 1999; Bedarrain et al., Biotechnol. Appl. Biochem., 33, 173-182, 2001; Srivasta etal. Prot. Express. Purif 41, 313-322, 2005], которые, как правило, представляют собой неправильно уложенные и, вследствие этого, биологически неактивные белки [см. статью Villaverde and Carrio, Biotechnol. Lett., 25, 1385-1395, 2003]. Для получения данных белков в нативной форме необходимо подвергнуть указанные IB фазе денатурации с последующей фазой ренатурации с окислением в случае, когда необходимо формирование дисульфидных мостиков, как в природном белке. Кроме того, инкорпорация дополнительного остатка метионина в N-концевую часть последовательности целевого белка (например, ИФН) является характерным признаком экспрессии белка в Е. coli. Как известно, остатки метионина, распределенные внутри последовательности белка, чувствительны к окислению. Данный процесс может иметь место в способе получения белка или фармацевтической композиции, содержащей данный белок (если он может быть использован в качестве лекарственного препарата, например, ИФН) и является более выраженным при длительном хранении при повышенных температурах.
Хотя разработан ряд способов получения и очистки ИФН в бактериях в виде 1 В [например, см. статью Thatcher and Panayotatos, Methods Enzymol. 119, 166-177, 1986; патентные заявки US 4511502; US 4765903; US 4845032; EP 1310559 или ЕР 1990349], существуют другие факторы, которые могут представлять собой препятствия для успешного получения и очистки ИФН, а именно ИФНа терапевтического качества, такие как инкорпорация дополнительного остатка метионина в N-конец целевого ИФН и генерация его окисленных молекул, которые следует удалять (если продукт предназначен для применения в качестве лекарственного препарата), уменьшая, таким образом, общий выход при получении ИФН, повышая сложность способа очистки и делая способ получения и очистки альфа-ИФН терапевтического качества трудоемким процессом.
Соответственно, сохраняется потребность в способе, который обеспечивает получение ИФНа, в частности, ИФНа5 терапевтического качества, из клеток-хозяев Е. coli с высоким выходом и рентабельным образом.
Раскрытие изобретения
В настоящее время неожиданно обнаружено, что концентрация микроэлементов (следовых элементов) в ферментационной среде играет важную роль в пост-трансляционных модификациях ИФНа5. Таким образом, контролируя концентрацию микроэлементов в ферментационной среде возможно минимизировать инкорпорацию дополнительного остатка метионина в N-концевую часть ИФНа5, образуемого в клетке-хозяине Е. coli, продуцирующей ИФНа5, а также минимизировать генерацию его окисленных молекул, что повышает выход продукции и упрощает способ очистки, делая, таким образом, способ получения и очистки ИФНа5, а именно ИФНа5 терапевтического качества, образуемого в клетке-хозяине Е. coli, продуцирующей ИФНа5, рентабельным менее сложным способом.
В сущности в Примере 4 показывают, что формирование окисленной метионилированной формы человеческого альфа 5-ИФН (чИФНа5) элиминируется и количество ацетилированных форм чИФНа5 снижается в два раза (т.е. наполовину), когда 1 литр (л) углеродного раствора для подпитки содержит от приблизительно 3,0 мл до приблизительно 3,7 мл исходного раствора микроэлементов, и предпочтительно, когда средняя специфическая скорость роста культуры (µ) после индукции равна или превышает 0,17.
Вследствие этого в одном аспекте изобретение относится к способу получения белка альфа 5-интерферона (ИФНа5) путем экспрессии в клетке-хозяине Е. coli, продуцирующей ИФНа5, который предусматривает:
a) получение клетки-хозяина Е. coli, продуцирующей ИФНа5;
b) культивирование клетки-хозяина Е. coli, продуцирующей ИФНа5, в условиях, эффективных в плане экспрессии указанного белка ИФНа5 указанной рекомбинантной клеткой-хозяином Е. coli, продуцирующей ИФНа5, в ферментационной среде с добавлением углеродного раствора для подпитки, причем
- указанная ферментационная среда не содержит компоненты животной природы или дрожжевой природы и
- указанный углеродный раствор для подпитки содержит источник углерода и от приблизительно 3,0 до приблизительно 3,7 мл раствора микроэлементов на литр добавленного углеродного раствора для подпитки;
и
с) выделение и необязательная очистка экспрессированного белка ИФНа5.
В конкретном варианте осуществления указанная клетка-хозяин Е. coli представляет собой штамм Е. coli с дефицитом протеазы, такой как штамм-хозяин Е. coli lon-/ompT- с дефицитом протеазы, предпочтительно штамм Е. coli BL21, наиболее предпочтительно штамм Е. coli BL21 (DE3).
В другом конкретном варианте осуществления, когда штамм Е. coli представляет собой штамм Е. coli BL21 (DE3), условия, описанные для стадии b), включают индукцию с помощью IPTG.
В другом конкретном варианте осуществления стадия с) выделения и очистки экспрессированного белка ИФНа5 включает последовательно после лизиса клеток-хозяев Е. coli выделение указанного бека ИФНа5 в форме телец включения (IB), подвергая указанные IB солюбилизации и полученную смесь окислительной ренатурации и последовательной серии хроматографической очистки, включающей:
1) проведение гидрофобной хроматографии смеси, содержащей ренатурированный ИФНа5;
2) проведение анионообменной хроматографии раствора, полученного на стадии 1);
3) проведение первой катионообменной хроматографии раствора,
полученного на стадии 2); и
4) проведение второй катионообменной хроматографии раствора, полученного на стадии 3), причем указанный раствор необязательно разводят буфером, содержащим метионин, с целью получения очищенного ИФНа5.
В конкретном варианте осуществления предпочтительно, когда указанный ИФНа5 представляет собой человеческий ИФНа5 (чИФНа5).
Краткое описание чертежей
На Фигуре 1 показывают схему конструирования штамма, продуцирующего ИФНа5.
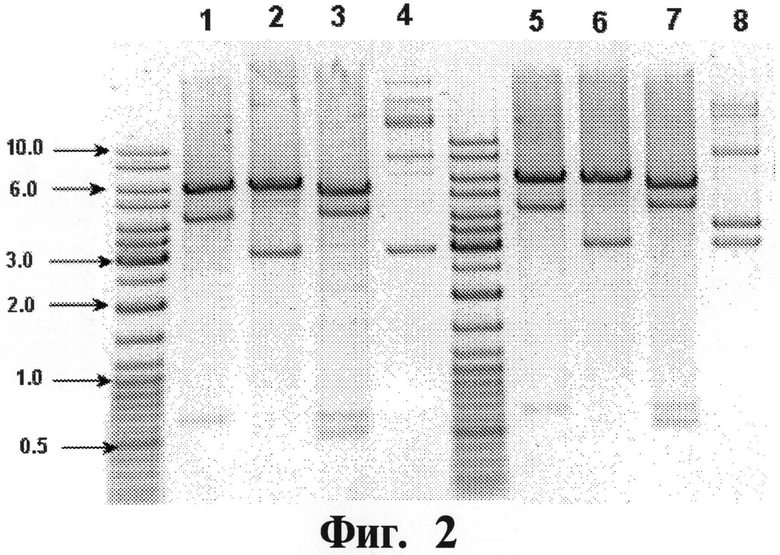
На Фигуре 2 показывают результаты рестрикционного анализа первичной плазмидной ДНК рЕТ28-альфа5-ИФН. Дорожки 1-3 и 5-7: данные рестрикционного анализа рЕТ28-альфа5-ИФН; дорожки 1 и 5: рЕТ28-альфа5-ИФН/ВаmНI; дорожки 2 и 6: рЕТ28-альфа5-ИФН/BamHI; дорожки 3 и 7: рЕТ28-альфа5-ИФН/Ndel+ВаmНI; дорожки 4 и 8: рЕТ28-альфа5-ИФН, нерасщепленный. Размер полос маркерной ДНК (Gene Ruler DNA Ladder Mix, Fermentas, Lithuania) указан в килопарах нуклеотидов (т.п.н.).
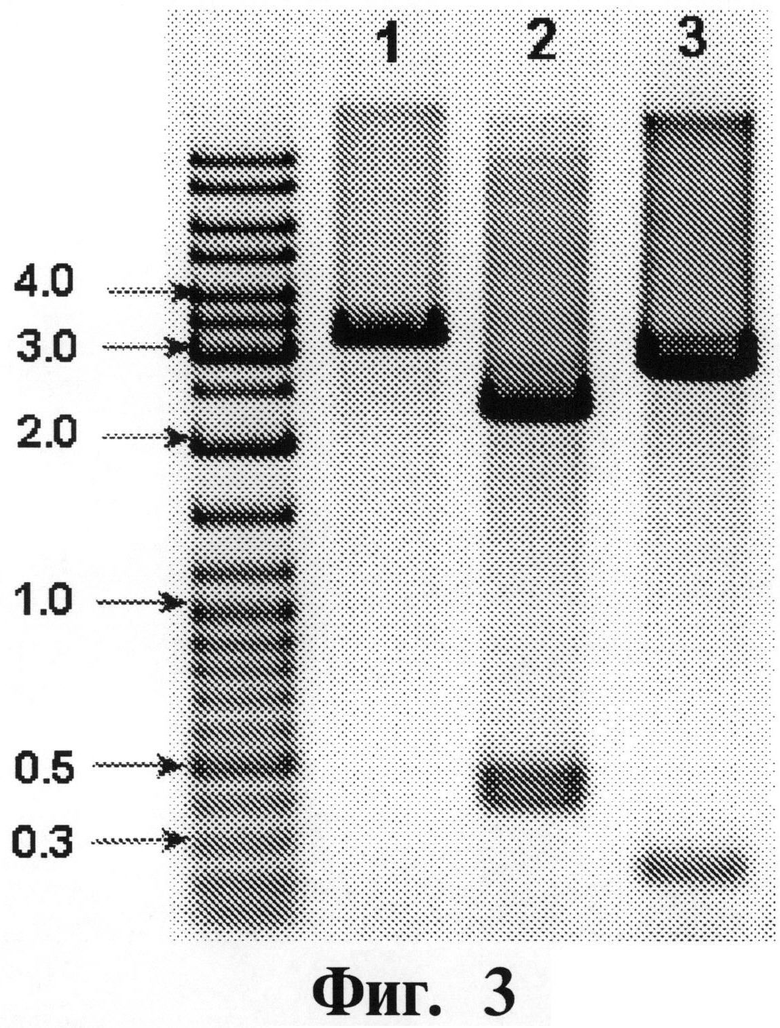
На Фигуре 3 показывают результаты рестрикционного анализа промежуточной плазмиды альфа5-ИФН. Плазмида выделена из одного клона. Размер полос маркерной ДНК (Gene Ruler DNA Ladder Mix, Fermentas, Lithuania) в т.п.н. указан. Ожидаемые размеры фрагментов (в парах нуклеотидов) приведены в скобках. Дорожка 1: рUС57-альфа5-ИФН/Рstl (28; 3197); дорожка 2: рUС57-альфа5-ИФН/Рvull (455; 406; 2364); дорожка 3: рUС57-альфа5-ИФН/Ndel (250; 2975).
На Фигуре 4 показывают фрагмент, кодирующий полную нуклеотидную последовательность человеческого альфа5-ИФН (чИФНа5), оптимизированный для экспрессии в Е. coli путем замены нескольких кодонов наименее часто встречающимися кодонами, используемыми в Е. coli. Замены нуклеотидов подчеркнуты.
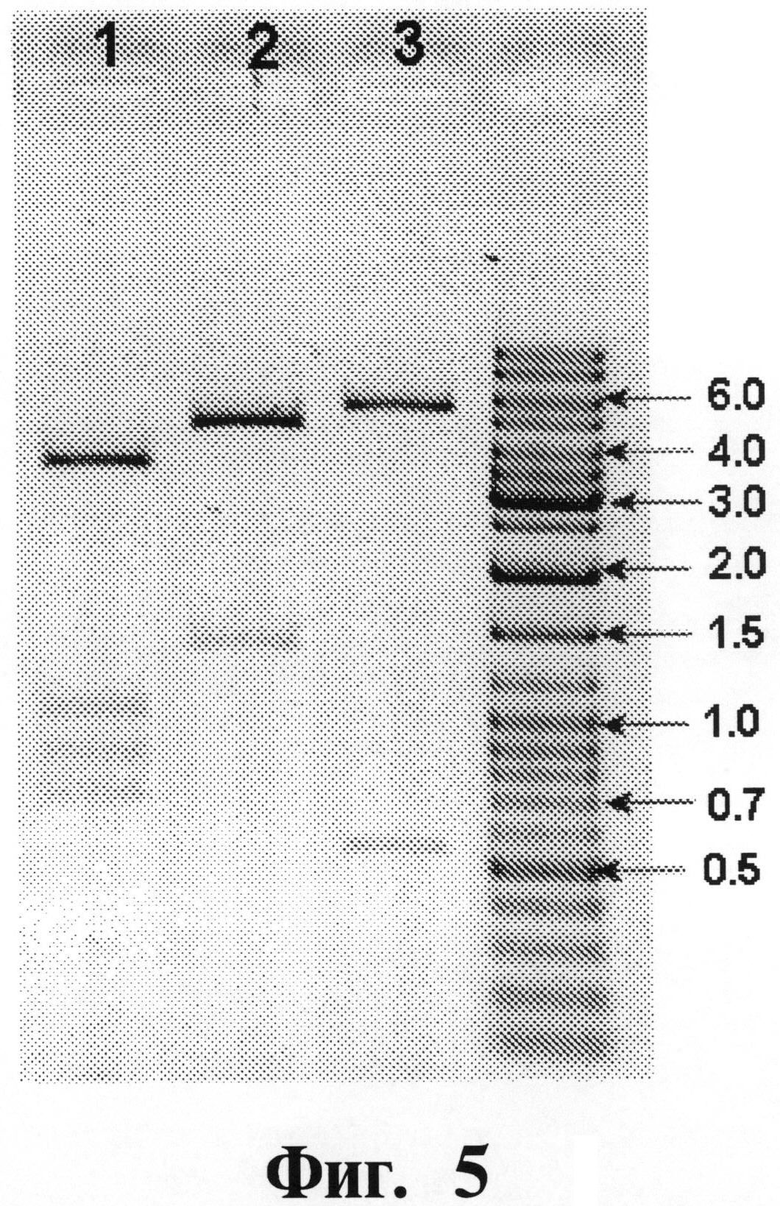
На Фигуре 5 показывают результаты рестрикционного анализа плазмиды рЕТ21-альфа5-ИФН. Плазмида выделена из одного клона. Размер полос маркерной ДНК (Gene Ruler DNA Ladder Mix, Fermentas, Lithuania) в т.п.н. указан. Ожидаемые размеры фрагментов (в парах нуклеотидов) приведены в скобках. Дорожка 1: рЕТ21-альфа5-ИФН/Раgl (673; 817; 1008; 3423); дорожка 2: рЕТ21-альфа5-ИФН/Рstl (1352; 4569); дорожка 3: рЕТ21-альфа5-ИФН/Ndel+ВаmН1 (516; 5405).
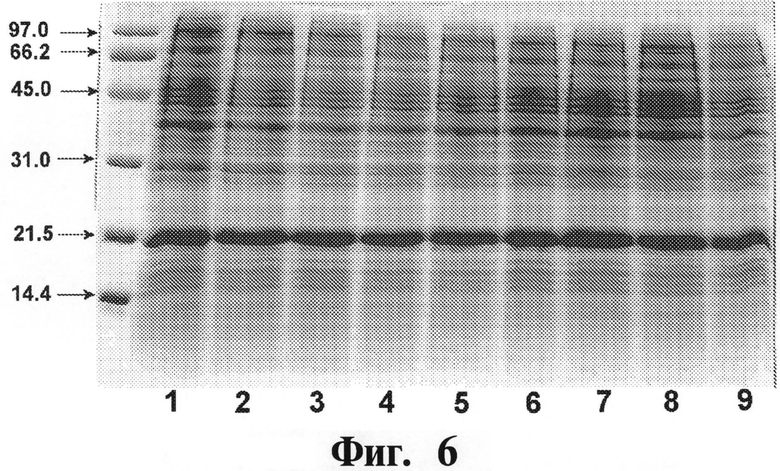
На Фигуре 6 показывают экспрессию рекомбинантного белка ИФНа5. Клеточную культуру Е. coli BL21(DE3) рЕТ21-альфа5-ИФН, пересеянную с 9 колоний (дорожки 1-9), выращивают в колбах объемом 750 мл (среда LB, объем 250 мл) при 37°С до OD600 (оптической плотности при длине волны 600) приблизительно 1,2. Экспрессию целевого белка индуцируют 1 мМ IPTG в течение 2,5 часов. Образцы общего клеточного белка проводят вместе с маркерами белка BioRad (указывают в кД слева) на 15% SDS-PAGE (электрофорез в полиакриламидном геле с использованием 15% додецилсульфата натрия) с последующим окрашиванием кумасси голубым.
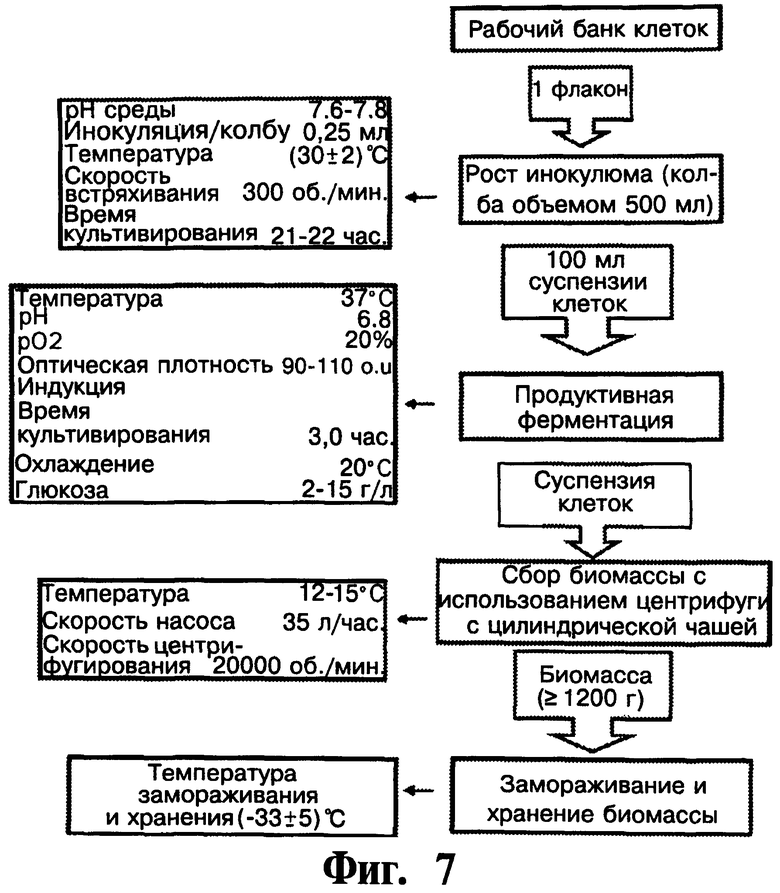
На Фигуре 7 показывают схему биосинтеза рекомбинантного чИФНа5.
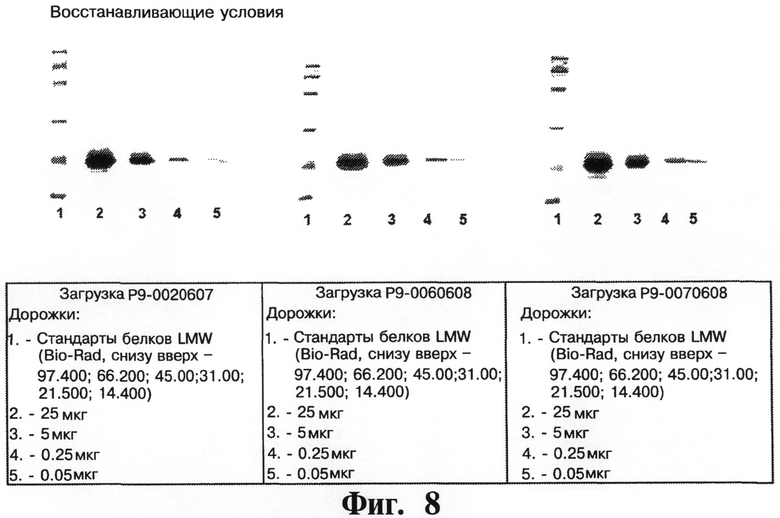
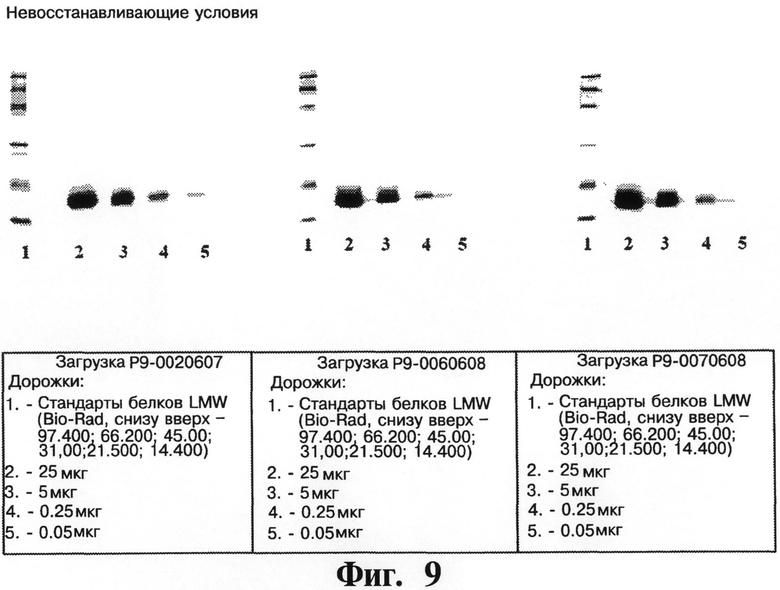
На Фигурах 8 и 9 представляют чистоту рекомбинантного чИФНа5, обработанного и изготовленного согласно данному изобретению, как определено посредством SDS-PAGE (14%) в восстанавливающих (см. Фигуру 8) и невосстанавливающих (см. Фигуру 9) условиях для белка чИФНа5 (на фигурах показывают результаты трех крупномасштабных загрузок для очистки белка чИФНа5).
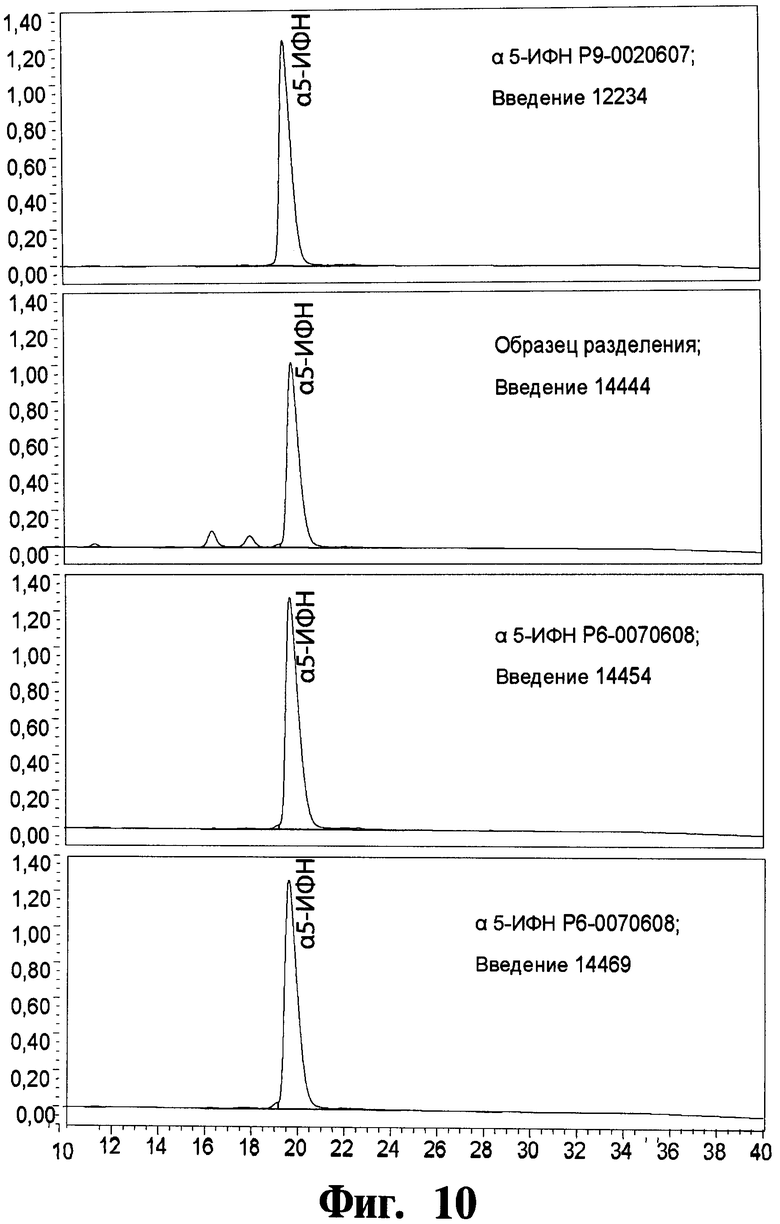
На Фигуре 10 показывают чистоту рекомбинантного чИФНа5, обработанного и изготовленного согласно данному изобретению, как определено посредством анализа с использованием "высокоэффективной жидкостной хроматографии с обращенной фазой" (ОФ-ВЭЖХ) (на фигуре показывают результаты трех крупномасштабных загрузок рекомбинантного чИФНа5).
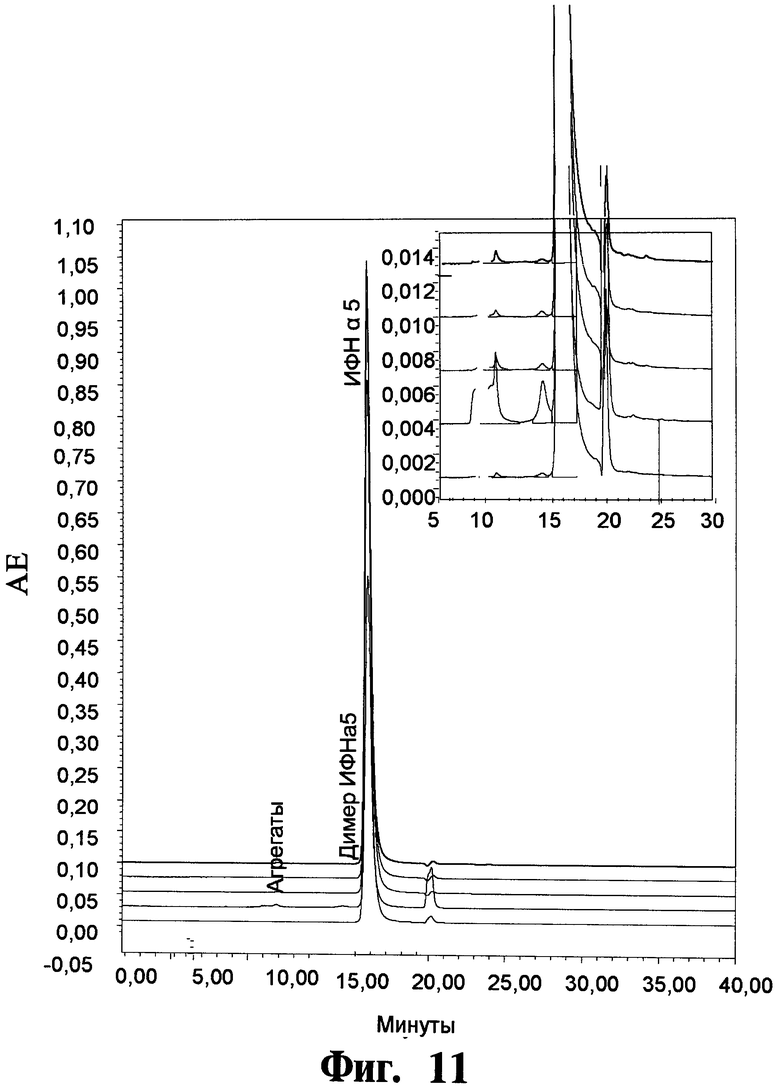
На Фигуре 11 показывают чистоту рекомбинантного чИФНа5, обработанного и изготовленного согласно данному изобретению, как определено посредством анализов с использованием "эксклюзионной по размеру ВЭЖХ" (ЭР-ВЭЖХ) (на фигуре показывают результаты трех крупномасштабных загрузок рекомбинантного чИФНа5).
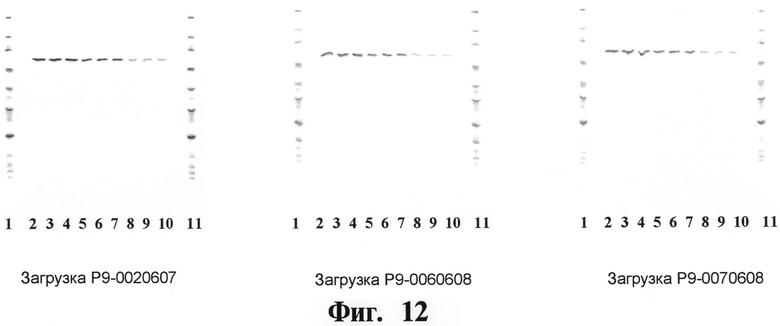
На Фигуре 12 показывают чистоту рекомбинантного чИФНа5, обработанного и изготовленного согласно данному изобретению, как определено посредством анализов с использованием изоэлектрического фокусирования (на фигуре показывают результаты трех крупномасштабных загрузок рекомбинантного чИФНа5). Дорожки 1,11: стандарты pi (Amersham Pharmacia); дорожки 2, 3, 4: рекомбинантный чИФНа5, 15 мкг; дорожки 5, 6, 7: рекомбинантный чИФНа5, 5 мкг; дорожки 8, 9, 10: рекомбинантный чИФНа5, 1 мкг.
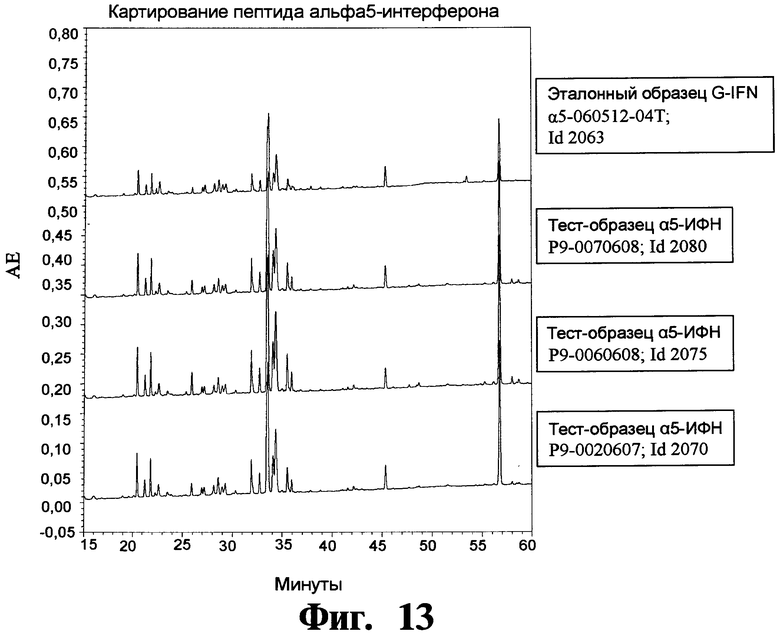
На Фигуре 13 показывают хроматограммы картирования пептидов трех крупномасштабных загрузок препаратов рекомбинантного чИФНа5, обработанного и изготовленного согласно данному изобретению.
Осуществление изобретения
Определения
Для облегчения понимания настоящего изобретения настоящим представлены значения некоторых терминов и выражений, как используют в контексте изобретения.
Как используют в данном контексте, термин "альфаб-интерферон" (или "альфа5-ИФН" или "ИФНа") относится к белку, продуцируемому лейкоцитами, который по всей видимости главным образом участвует во врожденном иммунном ответе в отношении вирусной инфекции, способен связываться со специфическим комплексом рецептора поверхности клетки, известным как рецептор альфа-ИФН (ИФНАР).
Белки ИФНа5 описаны, например, в WO 83/02459 (чИФНа5).
Термин "ИФНа5" включает белки, имеющие (i) последовательность аминокислот, которая по меньшей мере в существенной степени идентична последовательности аминокислот нативного белка ИФНа5 и (ii) биологическую активность, которая свойственна нативному ИФНа5. Выражение в существенной степени идентичная последовательность аминокислот означает, что последовательности идентичны или отличаются одним или более изменениями аминокислот (т.е. делениями, добавлениями, заменами), которые не приводят к вредному функциональному различию между синтетическим белком и нативным ИФНа5, например, белки ИФНа5, имеющие идентичность по меньшей мере 70% с одним из приведенных белков ИФНа5. Х% идентичности между белком ИФНа5 (Р) и эталонным белком ИФНа5 (R) означает, что при элайнменте данных двух последовательностей Х% аминокислот Р идентичны соответствующей аминокислоте в последовательности R или заменены аминокислотой той же группы, такой как:
- аминокислоты с неполярными группами R: аланин, валин, лейцин, изолейцин, пролин, фенилаланин, триптофан, метионин;
- аминокислоты с незаряженными полярными группами R: глицин, серин, треонин, цистеин, тирозин, аспарагин, глутамин;
- аминокислоты с заряженными полярными группами R (отрицательно заряженными при рН 6,0); аспарагиновая кислота, глутаминовая кислота;
- основные аминокислоты (положительно заряженные при рН 6,0): лизин, аргинин, гистидин (при рН 6,0);
- аминокислоты, имеющие фенильные группы: фенилаланин, триптофан, тирозин.
Особенно предпочтительными консервативными заменами являются:
Lys на Аrg и наоборот, так чтобы можно было сохранить положительный заряд;
Glu на Asp и наоборот, так чтобы можно было сохранить отрицательный заряд;
Ser на Thr, так чтобы можно было сохранить свободный -ОН; и
Gin на Asn, так чтобы можно было сохранить свободный NH2.
Данные проценты идентичности последовательности можно получить при использовании программы BLAST (blast2seq, параметры по умолчанию) [см. статью Tatutsova and Madden, FEMS Microbiol. Lett., 174, 247-250 (1990)].
Как используют в данном контексте, термин "продуцирующая ИФНа5 клетка-хозяин Е. coli" относится к клетке-хозяину Е. coli, которая генетически сконструирована таким образом, что продуцирует белок, который обладает биологической активностью, ассоциированной с ИФНа5. В конкретном варианте осуществления, клетка-хозяин Е. coli представляет собой штамм Е. coli с дефицитом протеазы, такой как штамм-хозяин Е. coli lon-/ompT- с дефицитом протеазы, предпочтительно штамм Е. coli BL21, наиболее предпочтительно штамм Е. coli BL21 (DE3).
Как используют в данном контексте, термин "биологическая активность ИФНа5" относится к любой биологической активности указанного ИФНа5, включая терапевтическую активность указанного ИФНа5. WO 83/02459 раскрывает, что ИФНа5 проявляет антивирусную активность в отношении ДНК- и РНК-вирусов, активность в отношении роста клеток и способность регулировать образование внутриклеточных ферментов и других образуемых клеткой субстанций; соответственно, предполагают, что ИФНа5 можно использовать для лечения вирусных инфекций (например, инфекции хронического гепатита В), опухолей и рака.
Как используют в данном контексте, выражение ферментационная среда "не содержит компоненты животного происхождения и дрожжевого происхождения" означает, что отсутствует риск переноса агентов, вызывающих губчатую энцефалопатию, через продукты медицинского назначения, хотя отсутствуют какие-либо доказательства заражения BSE (коровьей губчатой энцефалопатии) или случаев vCJD (нового варианта болезни Крейтцфельдта-Якоба), связанных с фармацевтическими продуктами. Продукт свободен от следовых количеств примесных белков из дрожжевых клеток.
Способ получения ИФНа5
В одном аспекте изобретение относится к способу получения белка альфа5-интерферона (ИФНа5) путем экспрессии в клетке-хозяине Escherichia coli, продуцирующей ИФНа5, далее "способ, соответствующий изобретению", который предусматривает:
a) получение продуцирующей ИФНа5 клетки-хозяина Е. coli;
b) культивирование продуцирующей ИФНа5 клетки-хозяина Е. coli в условиях, эффективных в плане экспрессии указанного белка ИФНа5 указанной рекомбинантной продуцирующей ИФНа5 клеткой-хозяином Е. coli в ферментационной среде с добавлением углеродного раствора для подпитки, причем
- указанная ферментационная среда не содержит компоненты животного происхождения или дрожжевого происхождения и
- указанный углеродный раствор для подпитки содержит источник углерода и от приблизительно 3,0 до приблизительно 3,7 мл раствора микроэлементов на литр добавленного углеродного раствора для подпитки;
и
c) выделение и необязательную очистку экспрессированного белка ИФНа5.
Рекомбинантный ИФНа5, который можно получить согласно способу, соответствующему изобретению, определен ранее. В конкретном варианте осуществления указанный рекомбинантный ИФНа5, который можно получить согласно способу, соответствующему изобретению, представляет собой белок ИФНа5, который в существенной степени идентичен нативному ИФНа5, т.е. белок, который образуется продуцирующей ИФНа5 клеткой-хозяином Е. coli, которая трансформирована геном, кодирующим ИФНа5, или его модификацией, которая кодирует белок, имеющий (1) последовательность аминокислот, которая по меньшей мере в существенной степени идентична последовательности аминокислот нативного ИФНа5, и (2) биологическую активность, которая присуща нативному ИФНа5. А предпочтительном варианте осуществления указанный ИФНа5 представляет собой чИФНа5.
В более конкретном варианте осуществления рекомбинантный ИФНа5, получаемый согласно способу, соответствующему изобретению, представляет собой ИФНа5, имеющий последовательность аминокислот, показанную в SEQ ID NO: 1 (см. Фигуру 4), которая соответствует зрелому чИФНа5, имеющему дополнительный остаток метионина в N-концевой части полипептидной цепи.
Согласно способу, соответствующему изобретению, предусматривают продуцирующую ИФНа5 клетку-хозяин Е. coli [стадия а)]. Хотя в принципе в способе, соответствующем изобретению, можно использовать любой штамм Е. coli, в предпочтительном варианте осуществления клетка-хозяин Е. coli представляет собой штамм Е. coli с дефицитом протеазы, такой как штамм-хозяин Е. coli lon-/ompT- с дефицитом протеазы, предпочтительно штамм Е. coli BL21, наиболее предпочтительно штамм Е. coli BL21 (DE3).
Продуцирующую ИФНа5 клетку-хозяин Е. coli можно получить при использовании принятых способов и протоколов клонирования и экспрессии ИФНа5 [см., например, монографии Sambrook et al. Molecular cloning: A Laboratory Manual (Лабораторное руководство по молекулярному клонированию), 2 изд., CSH Laboratory, Cold Spring Harbor, 1989; Current Protocols in Molecular Biology (Современные протоколы в молекулярной биологии), т.1-3 (под ред. Ausubel F.M. et al.) John Wiley & Sons, Inc., Brooklyn, New York, 1994-1998]. В конкретном варианте осуществления клонирование и экспрессию гена, кодирующего ИФНа5, и конструкцию бактериального штамма, продуцирующего рекомбинантный белок ИФНа5 (т.е. продуцирующей ИФНа5 клетки-хозяин Е. coli) можно осуществить согласно способу, который предусматривает клонирование кДНК гена, кодирующего ИФНа5, модификацию последовательности ДНК указанного гена с целью оптимизации его экспрессии в Е. coli, конструирование экспрессирующей плазмиды, трансформацию выбранной плазмидой подходящего штамма Е. coli и выбор условий экспрессии/индукции. В Примере 1 раскрывают конструкцию продуцирующей чИФНа5 клетки-хозяина Е. coli.
В конкретном варианте осуществления, когда клетка-хозяин Е. coli представляет собой штамм Е. coli с дефицитом протеазы, такой как штамм Е. coli BL21 (DE3), указанную клетку-хозяин трансформируют вектором, содержащим последовательность, кодирующую белок ИФНа5 под контролем индуцируемого промотора; в данном случае для экспрессии белка необходимо добавление индуктора, такого как, например, изопропил-β-D-тиогалактопиранозид (IPTG). Таким образом, согласно конкретному варианту осуществления, когда клетка-хозяин Е. coli представляет собой штамм Е. coli BL21, такой как штамм Е. coli BL21 (DE3), условия на стадии b) включают индукцию с помощью IPTG.
На стадии b) способа, соответствующего изобретению, продуцирующую ИФНа5 клетку-хозяин Е. coli культивируют в условиях, эффективных в плане экспрессии указанного белка ИФНа5 указанной рекомбинантной продуцирующей ИФНа5 клеткой-хозяином Е. coli в ферментационной среде с добавлением углеродного раствора для подпитки, причем
- указанная ферментационная среда не содержит компоненты животного происхождения и дрожжевого происхождения и
- указанный углеродный раствор для подпитки содержит источник углерода и от приблизительно 3,0 до приблизительно 3,7 мл раствора микроэлементов на литр добавляемого углеродного раствора для подпитки;
и
Эффективные условия, в которых следует культивировать продуцирующую ИФНа5 клетку-хозяин Е. coli с целью экспрессии указанного белка ИФНа5, как правило, известны компетентному специалисту в области техники. Данные условия включают ферментационную среду, содержащую источник азота, источник углерода и источник металлов, подходящие для того, чтобы культивировать клетку-хозяин Е. coli при условии, соответствующем настоящему изобретению, что ферментационная среда не содержит компоненты животного происхождения или дрожжевого происхождения, такую как синтетическая химическая ферментационная среда.
В конкретном варианте осуществления диаммонийфосфат и аммиак в виде монокомпонента или в комбинации можно использовать в качестве источника азота. В другом конкретном варианте осуществления источником углерода может являться лимонная кислота, глюкоза или их комбинации.
В Примере 2 раскрывают ферментационную среду для культивирования продуцирующего ИФНа5 штамма Е. coli BL21 (DE3), причем данная среда содержит диаммонийфосфат, сульфат магния, дигидрофосфат калия, лимонную кислоту, D(+)-глюкозы и исходный раствор микроэлементов, где данный исходный раствор микроэлементов содержит микроэлементы, выбранные из группы микроэлементов, состоящей из железа, кальция, цинка, марганца, меди, кобальта, молибдена, бора и их комбинаций. В конкретном варианте осуществления указанная ферментационная среда содержит источник микроэлементов, выбранный из группы источников микроэлементов, состоящих из хлорида железа (III), хлорида кальция, сульфат цинка (II), сульфат марганца (II), сульфат меди (II), хлорид кобальта (II), молибдат натрия, борная кислота и их комбинации. В конкретном варианте осуществления исходный раствор микроэлементов (следовых элементов) содержит (в г/л): гексагидрат хлорида железа (III) (30,0), дигидрат хлорида кальция (4,05), гептагидрат сульфата цинка (II) (6,75), моногидрат сульфата марганца (II) (1,5), пентагидрат сульфата меди (II) (3,0), гексагидрат хлорида кобальта (II) (1,14), дигидрат молибдата натрия (0,3) и борную кислоту (0,69) (см. Пример 4).
Другой признак способа, соответствующего изобретению? относится к тому факту, что углеродный раствор для подпитки содержит источник углерода и от приблизительно 3,0 до приблизительно 3,7 мл раствора микроэлементов на литр добавленного углеродного раствора для подпитки. Отдельные компоненты указанного раствора микроэлементов определены ранее. В конкретном варианте осуществления углеродный раствор для подпитки содержит источник углерода (например, лимонную кислоту и/или глюкозу), сульфат магния и, согласно изобретению, концентрированный раствор микроэлементов в концентрации от приблизительно 3,0 мл до приблизительно 3,7 мл раствора микроэлементов/литр добавляемого углеродного раствора для подпитки. Как отмечено выше, в Примерах 2 и 4 раскрывают исходный раствор микроэлементов, содержащий микроэлементы, выбранные из группы микроэлементов, состоящей из железа, кальция, цинка, марганца, меди, кобальта, молибдена, бора и их комбинаций; в конкретном варианте осуществления указанный исходный раствор микроэлементов содержит хлорид железа (III), хлорида кальция, сульфат цинка (II), сульфат марганца (II), сульфат меди (II), хлорид кобальта (II), молибдат натрия, борную кислоту и их комбинации. В конкретном варианте осуществления исходный раствор микроэлементов, включенный в концентрации от приблизительно 3,0 мл до приблизительно 3,7 мл раствора микроэлементов на литр углеродного раствора для подпитки, представляет собой исходный раствор микроэлементов, раскрытый в Примере Example 2 и 4. В Примере 2 раскрывают способ биосинтеза чИФНа5 с использованием штамм-продуцента.
Различные исследования, проведенные авторами, показывают эффект микроэлементов на пост-трансляционные модификации рекомбинантного ИФНа5 (например, чИФНа5), а именно то, что относительное количество окисленного-Met, обнаруженного в очищенном API рекомбинантного ИФНа5 (например, рекомбинантного чИФНа5), образующегося в Е. coli, находится в тесной связи с количеством "непроцессированного метионина" в N-концевой части полипептидной цепи. Для получения меньше чем 10% указанного непроцессированного метионина в N-концевой части количество окисленного-Met ИФНа5 (определенное с помощью ОФ-ВЭЖХ) после развертывания должно составлять не более 1%.
При оптимизации способа биосинтеза рекомбинантного чИФНа5 авторы наблюдают, что концентрация микроэлементов в ферментационной среде имеет важный эффект на пост-трансляционные модификации чИФНа5. В действительности определено, что образование окисленной метионилированной формы чИФНа5 (окисленный-Met чИФНа5) элиминируется и количество ацетилированных форм чИФНа5 снижается в два раза, когда 1 л углеродного раствора для подпитки содержит 3,0 мл - 3,7 мл указанного исходного раствора микроэлементов или когда он находится в пределах 0,0048 мл/Л/o.u. - 0,0070 мл/Л/o.u. [o.u. - оптические единицы].
В конкретном варианте осуществления способ, соответствующий изобретению, осуществляют в условиях, при которых средняя "специфическая скорость роста культуры" (µ) после индукции равна или превышает 0,17 [µ-((In OD2 - In OD1)/T2-T1), где OD означает "оптическую плотность" (оптические единицы, o.u.) и Т означает "время"]. Исследования, проведенные авторами, показывают, что рост культуры и концентрация микроэлементов тесно взаимозависимы, особенно когда концентрация микроэлементов очень низкая/почти ограничивающая. Когда концентрация микроэлементов в среде для культивирования ниже чем 0,95 мл/л конечного объема суспензии (см. Пример 4, Таблица 3) или меньше чем 3,0 мл/л углеродного раствора для подпитки, среднее значение - после индукции достигает точно 0,121-0,158, т.е. меньше чем 0,17 (М-83, М-84, М-85, М-86). Однако, когда среднее значение - после индукции меньше чем 0,17, присутствие окисленной метионилированной формы ИФНа5 практически гарантировано. Концентрация микроэлементов в среде для культивирования, превышающая 1,23 мл/л конечного объема суспензии, приводит в результате к ускоренному росту (М-89, М-90), повышенному WCW и повышенному количеству ацетилированной формы ИФНа5+неизвестного белка. Однако, когда среднее значение - после индукции равно или выше 0,17 способ работает лучше, пока концентрация микроэлементов выше чем 0,123 мл/л конечного объема суспензии или больше чем 3,7 мл/л углеродного раствора для подпитки.
Стадия с) способа, соответствующего изобретению, предусматривает выделение и необязательную очистку экспрессированного белка ИФНа5. В конкретном варианте осуществления после лизирования продуцирующих ИФНа5 клеток-хозяев Е. coli белок ИФНа5 выделяют в форме телец включения (IB) посредством солюбилизации указанных IB, чтобы получить смесь, содержащую денатурированный ИФНа5, который, в свою очередь, подвергают окислительной ренатурационной обработке, чтобы получить смесь, содержащую ренатурированный ИФНа5, который затем подвергают процессу очистки с целью получения соответствующего очищенного ИФНа5. В конкретном варианте осуществления предпочтительно, когда указанный ИФНа5 представляет собой чИФНа5.
Для выделения и очистки ИФНа5, экспрессированного согласно способу, соответствующему изобретению, продуцирующие ИФНа5 клетки-хозяева Е. coli сначала лизируют с целью выделения указанного рекомбинантного ИФНа5 в форме телец включения (IB). Вкратце, в конкретном варианте осуществления клеточные мембраны продуцирующих ИФНа5 клеток-хозяев Е. coli лизируют при использовании принятых способов, таких как гомогенизация, обработка ультразвуком или циклическое изменение давления. Предпочтительные способы включают обработку ультразвуком или гомогенизацию с помощью гомогенизатора Потера (тефлон/стекло). После лизиса клеток IB, содержащие ИФНа5, отделяют от жидкой фазы лизата, например, центрифугированием, и ресуспендируют в соответствующем буферном растворе. IB необязательно можно промыть для удаления из них любых водорастворимых белков Е. coli.
Затем указанные IB солюбилизируют в присутствии солюбилизирующего вещества, такого как вещество, вызывающее диссоциацию, например, агент, денатурирующий белок, который вызывает диссоциацию водородных связей и действует на третичную и вторичную структуру белка, вызывая его развертывание, как правило, в водном буферном растворе, для того, чтобы получить смесь, содержащую денатурированый ИФНа5. Иллюстративные неограничивающие примеры веществ, вызывающих диссоциацию, включают мочевину и гидрохлорид гуанидиния (GdmHCI), предпочтительно гидрохлорид гуанидиния, сильное вызывающее диссоциацию вещество, которое препятствует карбамоилированию полипептидной цепи (которое может происходить при использовании концентрированного раствора мочевины). Концентрация вещества, вызывающего диссоциацию, будет зависеть от конкретного используемого вещества, вызывающего диссоциацию, и количества присутствующего клеточного материала. Предпочтительно, когда используют раствор гидрохлорида гуанидиния, имеющего концентрацию 6-7 М, наиболее предпочтительно 6 М. рН можно подвести добавлением подходящих буферов, и предпочтительно, когда рН будет выше 7, как правило, равным или выше приблизительно 8, предпочтительно равным или выше 8,6, более предпочтительно от 9,55 до 9,65, содержащих вещество, вызывающее диссоциацию. Как правило, в предпочтительном варианте осуществления солюбилизацию IB проводят при том же рН, что используют для стадии развертывания, избегая, таким образом, дополнительного подведения рН солюбилизата для стадии развертывания.
После солюбилизации IB, содержащих ИФНа5, нерастворимые твердые частицы отделяют и отбрасывают. Денатурированный ИФНа5, присутствующий в смеси, содержащей денатурированный ИФНа5, ренатурируют разведением указанной смеси в ренатурирующем растворе, таком как буфер для ренатурации. В конкретном варианте осуществления указанный буфер для ренатурации содержит активирующее вещество (например, L-аргинин и т.п.), окислительно-восстановительную пару (например, GSH/GSSG и т.п.) и, необязательно, хелатирующее соединение в буферной системе, имеющей значение рН выше 7,0, как правило, равное или выше приблизительно 8, предпочтительно равное или выше 8,6, более предпочтительно от 9,55 до 9,65. После ренатурации полученный в результате раствор белка, содержащий правильно уложенный ИФНа5, осветляют принятыми способами, например, центрифугированием или фильтрацией, с целью удаления любых оставшихся твердых частиц. Затем при необходимости рН осветленного раствора белка подводят до 8,0-8,20 подходящей кислотой (например, НС1) и затем смесь, содержащую ренатурированный ИФНа5 (раствор белка), подвергают любому подходящему способу очистки ИФНа5.
Хотя можно использовать практически любой способ очистки ИФНа5, изобретение далее предусматривает эффективный способ очистки ИФНа5, который включает обработку ренатурированного ИФНа5 в четырехстадийном хроматографическом способе, предусматривающем:
1) проведение гидрофобной хроматографии указанной смеси, содержащей ренатурированный ИФНа5;
2) проведение анионообменной хроматографии раствора, полученного на стадии 1);
3) проведение первой катионообменной хроматографии раствора,
полученного на стадии 2); и
4) проведение второй катионообменной хроматографии раствора, полученного на стадии 3), причем указанный раствор необязательно разводят буфером, содержащим метионин.
Вкратце, осветленный раствор белка с подведенным рН, содержащий совокупность (пул) белков, полученный после обработки с целью окислительной ренатурации, наносят на стадии 1) на колонку Фенил-Сефароза с целью отделения ренатурированного ИФНа5 от других компонентов, например, остаточных веществ, вызывающих диссоциацию, и т.п. Кроме того, контакт ренатурированного ИФНа5 с гидрофобной поверхностью абсорбента способствует созреванию ИФНа5.
Затем на стадии 2) пул белков, полученный на стадии 1), подводят до проводимости (например, 13,00-14,00 мСм/см) и значение рН подводят до 8,75-8,85 и наносят на колонку Q-Сефароза(анионообменная хроматография) с целью отделения мономера ИФНа5 от его агрегированных форм. Фракции, имеющие определенную чистоту (например, равную или превышающую 55%), могут быть объединены для дальнейшей очистки.
Затем на стадии 3) пул белков, полученный на стадии 2), подводят до проводимости (например, 6,00-7,00 мСм/см) и значение рН подводят до 5,15-5,20 и наносят на колонку SP-Сефароза (первая катионообменная хроматография) с целью отделения основной формы ИФНа5 от заряженных изоформ, таких как N-метионил-ИФНа5 и ацетилированный ИФНа5 (формы, которые представляют собой продукты пост-трансляционных модификаций). Фракции, имеющие определенную чистоту (например, равную или превышающую 70%), могут быть объединены для дальнейшей очистки.
Наконец, на стадии 4) пул белков, полученный на стадии 3), подводят до проводимости (например, 6,00-7,00 мСм/см) и значение рН подводят до 5,00-5,20 и наносят на колонку SP-Сефароза (вторая катионообменная хроматография) с целью отделения основной формы ИФНа5 от заряженных изоформ. В конкретном варианте осуществления, L-метионин добавляют к нагружаемому раствору, чтобы предотвратить окисление ИФНа5 во время хроматографии, проводимой при комнатной температуре. Фракции могут быть объединены таким образом, чтобы могла быть достигнута чистота ИФНа5, равная или выше чем (>) 95% (определено с помощью ОФ-ВЭЖХ).
При необходимости полученный таким образом ИФНа5 может быть получен с фармацевтически приемлемыми носителями и наполнителями, например, фосфатом натрия, рН 6,80-7,20, содержащим хлорид натрия. Раствор белка, если требуется, можно сконцентрировать до необходимой концентрации, например, в конкретном варианте осуществления раствор белка концентрируют до 10 мг/мл, например, до концентрации белка 1,0-1,5 мг/мл, и буфер заменяют ультрафильтрацией и стерилизуют с использованием стерильной фильтрации через устройство стерильного фильтра с максимальным размером пор 0,22 мкм.
В Примере 3 раскрывают способ выделения и очистки чИФНа5 из штамма, продуцирующего чИФНа5.
Следующие примеры служат для дальнейшей иллюстрации вариантов осуществления настоящего изобретения.
Пример 1. Конструирование штамма E.coli, экспрессирующего ИФНа5.
Данный пример раскрывает разработку и конструирование штамма Е. coli, продуцирующего рекомбинантный человеческий альфа-5 интерферон (чИФНа5). Вкратце, клонирование и экспрессию гена чИФНа5 и конструирование бактериального штамма, продуцирующего рекомбинантный белок ИФНа5, достигают, как описано ниже с использованием следующих стадий: клонирование кДНК гена, кодирующего чИФНа5, модификация последовательности ДНК данного гена с целью оптимизации его экспрессии в Е. coli, конструирование экспрессирующей плазмиды, трансформация отобранной плазмидой подходящего штамма Е. coli и выбор условий экспрессии/индукции.
Способы
При клонировании и экспрессии чИФНа5 используют принятые способы и протоколы [см. монографии Sambrook et al. Molecular cloning: A Laboratory Manual. (Лабораторное руководство по молекулярному клонированию), 2 изд., CSH Laboratory, Cold Spring Harbor, 1989; Current Protocols in Molecular Biology (Современные протоколы в молекулярной биологии), т.1-3 (под ред. Ausubel F.M. et al.) John Wiley & Sons, Inc., Brooklyn, New York, 1994-1998].
Все операции с ферментами, маркерами ДНК и белками проводят в соответствии с инструкциями изготовителей [в основном Fermentas (Lithuania)].
Генетическое конструирование
Конструирование продуцирующей чИФНа5 Е. coli осуществляют, следуя стадиям, показанным на схеме разработки генетических конструкций, представленной на Фигуре 1.
Первичная плазмида
Кодирующую чИФНа5 последовательность (без сигнального пептида) клонируют из нормальной ткани печени, полученной от анонимного пациента-донора (после информированного согласия), подвергнутого хирургическому вмешательству в брюшной полости не вследствие патологии печени, следующим образом: нормальную ткань печени гомогенизируют в 1 мл раствора Ultraspec (Biotex) и общую РНК обрабатывают ДНКазой (Gibco-BRL, Paisley, U.K.) перед обратной транскрипцией с помощью обратной транскриптазы M-MLV (Gibco- BRL) в присутствии РНКазы OUT (Gibco- BRL). Кодирующую последовательность чИФНа5 (без сигнального пептида) амплифицируют с помощью ПЦР (полимеразной цепной реакции) с предварительно полученной комплементарной ДНК(кДНК) при использовании следующих находящихся выше и ниже по ходу транскрипции праймеров (5'-3'):
GGAATTCCATATGTGTGATCTGCCTCAGACCCA (SEQ ID NO: 2) и
CGGGATCCTTGAACCAGTTTTCATTCCTTC (SEQ ID NO: 3).
Оба праймера содержат последовательность чИФНа5 (показана жирным шрифтом) и специфические последовательности рестрикционных ферментов: Ndel и BamHI (показаны подчеркиванием). Продукт ПЦР анализируют электрофорезом на агарозном геле и вырезают полосу из геля и очищают с использованием набора Gene Clean Kit (MP Biomedicals). Очищенный продукт ПЦР клонируют в плазмиду pCR 2.1 ТОРО с использованием набора для клонирования ТОРО ТА Cloning Kit (Invitogen). Клоны из инсерта секвенируют в анализаторе ABIPRISM 310 Genetic Analyzer (Perkin Elmer) с использованием набора для секвенирования с терминатором цикла красителем родамином (Perkin Elmer) для проверки того, что инсерт точно соответствует последовательности чИФНа5. После этого pCR 2.1 ТОРО- альфа5ИФН разрезают рестрикционными ферментами Ndel и BamHI и полосу 534 п.н. (соответствующую последовательности, кодирующей ИФНа5) клонируют в вектор pET28b(Novagen), предварительно разрезанный теми же ферментами. Последовательность снова проверяют при использовании того же способа.
ДНК первичной плазмиды рЕТ28-альфа5-ИФН, выделенную из разных колоний, анализируют с помощью рестрикционного анализа (см. Фигуру 2).
Плазмиду рЕТ28-альфа5-ИФН анализируют секвенированием обеих цепей ДНК с использованием анализатора последовательностей ABI Prism 377. Данный анализ подтверждает кодирующую последовательность чИФНа5. Плазмиду рЕТ28-альфа5-ИФН используют для конструирования зрелой структуры с оптимизацией кодона в качестве матрицы для ПЦР-амплификации.
Зрелая структура с оптимизацией кодона
ПЦР-амплификация гена, кодирующего чИФНа5, включающая оптимизацию кодона
ПЦР-амплификацию проводят при использовании плазмиды рЕТ28-альфа5-ИФН в качестве матрицы. Синтезируют следующие олигонуклеотиды:
Смысловой праймер (SP): 5'- CAT ATG TGT GAT CTG CCG CAG ACC CAC TCC CTG TCT AAC CGT CGT ACT CTG ATG АТС ATG GCA CAG ATG GGT CGT АТС TCT CCT TTC [SEQ ID NO: 4]
Антисмысловой праймер (ASP): 5'- CTG CAG TTA TTC CTT ACG ACG TAA ACG TTC TTG CAA G [SEQ ID NO: 5]
SP [SEQ ID NO: 4] и ASP [SEQ ID NO: 5] праймеры используют для замещения кодонов, которые наименее часто используют в Е. coli. Оптимизация кодонов в основном относится к кодонам аргинина AGA и AGG.
Клонирование полученного при ПНР фрагмента в промежуточную плазмиду
Очищенные продукты амплификации размером приблизительно 500 п.н. клонируют в плазмиду pUC57/T (#SD0171 Fermentas, Lithuania), используя набор для быстрого лигирования ДНК (Rapid DNA ligation Kit) (#К1421, Fermentas, Lithuania) и трансформируют E.coli JM109 (АТСС 53323, АТСС Bacteria and Bacteriophages (Американская коллекция типовых культур бактерий и бактериофагов), 19 изд., 1996). Рекомбинантные клоны отбирают на основании рестрикционного анализа (см. Фигуру 3). Отбирают два клона и секвенируют выделенные плазмиды.
Анализ последовательности промежуточной плазмиды альфа5-ИФН и реклонирование последовательности, кодирующей человеческий альфа5-ИФН, в плазмиду рЕТ21b (+)
Анализ нуклеотидной последовательности подтверждает последовательность части, кодирующей чИФНа5, и показан на Фигуре 4.
Фрагмент, кодирующий чИФНа5, разрезают Ndel+BamHI и очищенный фрагмент ДНК лигируют в разрезанный Ndel+BamHI вектор рЕТ21b(+) (Novagen), чтобы получить плазмиду рЕТ21-альФа5-ИФН. После трансформации штамма Е. coli JM109 бактерии селектируют путем добавления 100 мкг/мл ампициллина. Анализ на рекомбинантный инсерт колоний, полученных из трансформированных клеток, проводят при использовании способа тестирования колоний с помощью ПЦР. Детальный рестрикционный анализ плазмиды рЕТ21-альфа5-ИФН, очищенной из ПЦР-положительных колоний, дает в результате ожидаемый тип рестрикции (см. Фигуру 5).
Экспрессия рекомбинантного чИФНа5
После адаптации (стабилизации) плазмиды рЕТ21-альфа5-ИФН в неэкспрессирующем хозяине ею трансформируют Е. coil-хозяина BL21(DE3), несущую соответствующие генетические элементы для экспрессии целевых белков, чтобы получить штамм Е. coli BL21 (DE3) рЕТ21-ИФНа5. Экспрессию чИФНа5 индуцируют 1 мМ IPTG (изопропил-β-D-тиогалактопиранозидом), и результаты представляют на Фигуре 6.
Согласно данным SDS-PAGE молекулярная масса чИФНа5 составляет приблизительно 20 кД, что коррелирует с рассчитанным значением 19,7 кД.
Рекомбинантный чИФНа5 выявляют в нерастворимой фракции общего клеточного лизата; выход целевого белка составляет приблизительно 20% от общего клеточного белка. чИФНа5 составляет почти 40% нерастворимой фракции клеточного лизата. Получают одну колонию экспрессирующего штамма для создания банка мастер-клеток для исследований (RMCB).
Пример 2. Способ биосинтеза чИФНа5 продуцирующим штаммом.
Штамм Е. coli BL21 (DE3) рЕТ21-ИФНа5 (см. Пример 1) культивируют в средах, имеющих следующий состав (г/л):
a) для получения инокулюма (культивирования) в колбах (г/л):
гидрофосфат динатрия (17,0), дигидрофосфат калия (1,82), сульфат аммония (3,0), гептагидрат сульфата магния (0,5), моногидрат D(+)-глюкозы (15,0) и исходный раствор микроэлементов е) (0,16 мл);
b) для ферментации (г/л): диаммонийфосфат (4,0), гептагидрат сульфата магния (0,5), дигидрофосфат калия (13,3), лимонная кислота моногидрат (1,6), D(+)-глюкоза моногидрат (30,0) и исходный раствор микроэлементов е) (0,25 мл);
c) раствор для подпитки А (г/л): D(+)-глюкоза моногидрат (700,0), гептагидрат сульфата магния (20,7) и исходный раствор микроэлементов е) (3,4 мл/Л);
d) раствор для подпитки В (г/л): диаммонийфосфат (360,0) и дигидрофосфат калия (306,7); и
e) исходный раствор микроэлементов (следовых элементов) (г/л):
гексагидрат хлорида железа (III) (30,0), дигидрат хлорида кальция (4,05), гептагидрат сульфата цинка (II) (6,75), моногидрат сульфата марганца (II) (1,5), пентагидрат сульфата меди (II) (3,0), гексагидрат хлорида кобальта (II) (1,14), дигидрат молибдата натрия (0,3) и борная кислота (0,69).
На Фигуре 7 показывают полную схему процесса биосинтеза человеческого альфа5-ИФН.
Получение инокулюма: Колбу Эрленмейера, содержащую 500 мл стерильной среды для культивирования в колбах [а)], инокулируют 0,25 мл маточной культуры WCB (рабочего банка культур) Е. coli BL21 (DE3) рЕТ21-ИФНа5. Затем колбу инкубируют на ротационном шейкере при скорости встряхивания 300 об/мин, при температуре 30°С в течение 21-22 часов. Оптическая плотность после инкубирования должна быть равна или превышать 4,50 о.и. (оптические единицы) [А=595 нм].
Ферментация: Ферментер (общий объем 13,7 л), содержащий 7,0 л ферментационной среды [о)], инокулируют 1,5-1,6% культуры, полученной в колбах для инокулюма. Ферментацию проводят при автоматическом контроле температуры (37°С), рН (6,8) и рO2(20%). 25% раствор аммиака используют для подведения рН. Через 8-9 часов культивирования начинают дополнительную подпитку. Раствор для подпитки А [с)] закачивают в определенных дозах с целью поддержания концентрации глюкозы в среде для культивирования от приблизительно 3 г/л до 22 г/л. Индукцию осуществляют с помощью IPTG при 90-110 o.u. (λ=595 нм), чтобы сделать конечную концентрацию IPTG 0,5 мМ. Специфическая скорость роста культуры в точке индукции не должна быть ниже чем 0,45, чтобы иметь достаточно специфическую скорость роста после индукции. Средняя специфическая скорость роста культуры после индукции должна быть выше чем 0,17. Раствор для подпитки В [d)] закачивают в раздельных дозах: 150 мл при 60-70 o.u., 150 мл при 120-140 o.u., 75 мл через 1,5 ч(90 минут) и 75 мл через 2 часа после индукции. Ферментацию продолжают в течение 3 часов после индукции в тех же условиях. Затем клеточную суспензию охлаждают в ферментере до 12-15°С и переносят в центрифугу с помощью перистальтического насоса (35 л/час.). Клеточную суспензию центрифугируют при скорости 5000 об/мин при 4°С.
Полученную биомассу собирают в полиэтиленовую емкость и помещают в холодильник (-33±5)°С для замораживания и последующего хранения. Часть замороженной биомассы берут для оценки общего белка и экспрессии чИФНа5.
Пример 3. Способ выделения и очистки чИФНа5 из продуцирующего штамма.
1. Гомогенизация биомассы, разрушение и выделение телец включения (IB)
680,0-700,0 г биомассы, полученной в Примере 2, гомогенизируют в буфере для ресуспендирования (0,1 М Трис-HCl, рН 7,80-8,00, содержащем 2 мМ ЭДТА)этилендиаминтетрауксусной кислоты), 0,1% Тритон Х-100 и 1 мМ PMSF) в соотношении 1/10 (масс./об.), т.е. 1 г сырой биомассы/10 мл буфера для ресуспендирования.
Ресуспендирование осуществляют в гомогенизаторе Потера (Тефлон/стекло) и затем клетки разрушают в гомогенизаторе высокого давления при 600-800 бар при температуре 4-10°С.После разложения клеток тельца включения (IB) отделяют центрифугированием при 8000 об/мин в течение 30-35 мин.
2. Промывание телец включения (IB)
Предварительное промывание выделенных IB осуществляют способом из четырех последовательных стадий с использованием буферов для промывания I-III:
Буфер для промывания I: 10 мМ буфер Трис-HCl, рН 7,45-7,55, содержащий 1 М NaCl, 0,1% Полисорбата-80;
Буфер для промывания II: 10 мМ буфер Трис-HCl, рН 8,00-8,20, содержащий 6 М мочевину;
Буфер для промывания III: 10 мМ буфер Трис-HCl, рН 8,00-8,20.
Вкратце, Промывание IB осуществляют следующим образом:
a) первые два промывания (стадии 1 и 2) - IB промывают буфером для промывания I;
b) третье промывание (стадия 3) - IB промывают буфером для промывания II; и
c) четвертое промывание (стадия 4) - IB промывают буфером для промывания III.
Соотношение буфер для промывания/сырая биомасса, составляющее 10 мл буфера/1 г сырой биомассы, поддерживают на протяжении всей процедуры промывания IB.
3. Солюбилизация IB
Для солюбилизации IB используют буфер для солюбилизации (50 мМ глицин/буфер NaOH, pH 9,55-9,65, содержащий 6 М GdmHCl).
Соотношение солюбилизации: IB, выделенные из 1 г биомассы, в 6 мл буфера для солюбилизации в течение 2 час. при 2-8°С с последующим центрифугированием при 8000 об/мин в течение 25-30 мин.
4. Ренатурация
Буфер для ренатурации: 50 мМ буфер глицин/NaOH, pH 9,55-9,65, содержащий 1,2 М NaCl, 0,22 М L-аргинин, 2,85 мМ GSH, 0,285 мМ GSSG, проводимость 110-110 мСм/см при 4-10°С.
Денатурированный GdmHCl человеческий альфа5-ИФН ренатурируют добавлением по каплям солюбилизата IB к буферу для ренатурации (объемное соотношение 1:7) для достижения конечной концентрации в ренатурационной смеси 0,2М L-аргинина, 2,5/0,25 мМ GSH/GSSG. Ренатурационную смесь непрерывно перемешивают в течение 44-66 часов при 2-8°С.После ренатурации раствор белка осветляют центрифугированием при 8000 об/мин в течение 30-35 мин.
5. Хроматография
чИФНа5 очищают четырехстадийным хроматографическим способом, как указано ниже.
5.1 Хроматография на колонке Фенил-Сефароза (гидрофобная хроматография)
Колонка Фенил-Сефароза предназначена для отделения ренатурированного чИФНа5 от остаточного вещества, вызывающего диссоциацию, GdmHCl. Кроме того, контакт ренатурированного чИФНа5 с гидрофобной поверхностью абсорбента способствует созреванию чИФНа5.
Вкратце, рН осветленного раствора белка подводят до 8,00-8,20 6 М НСl и затем наносят раствор белка на хроматографическую колонку Фенил-Сефароза при следующих параметрах процесса:
Хроматографическая среда: Фенил-Сефароза быстрая (Amersham Pharmacia Biotech AB);
Используемая колонка: BPG 100×500 мм, диаметр 10 см;
Линейная скорость потока: 60 см/час;
Объем слоя хроматографической среды: 2,7±0,3 л;
Уравновешивающий буфер: 20 мМ буфер Трис-HCl, рН 8,00-8,20, содержащий 1,5 М хлорид натрия, проводимость 115-125 мСм/см при 15-25°С (110-120 мСм/см при 4-10°С);
Буфер для элюции: 10 мМ буфер Трис-HCl, рН 9,20-9,25, проводимость 0,1-0,2 мСм/см при 15-25°С;
Элюцию проводят более чем 6 объемами колонки (CV) [т.е. 6CV] буфером для элюции. Собранный раствор белка составляет 2-6 CV.
5.2 Хроматография на колонке Q-Сефароза (анионообменная хроматография)
Колонку Q-Сефароза используют для отделения мономера чИФНа5 от его агрегированных форм.
Вкратце, пул белков, полученный на предшествующей стадии (5.1), подводят до проводимости 13,00-14,00 мСм/см добавлением 20 мМ буфера Трис-HCl, рН 8,75-8,85, содержащего 5 М NaCI и подводят рН до 8,75-8,85 с помощью 6 М HCI. Затем раствор белка наносят на хроматографическую колонку Q-Сефароза при следующих параметрах процесса:
Хроматографическая среда: Q-Сефароза быстрая (Amersham Pharmacia Biotech AB);
Используемая колонка: BPG 140 X 500 мм, диаметр 14 см;
Линейная скорость потока: 60 см/час;
Объем слоя хроматографической среды: 3,3±0,3 л
Уравновешивающий буфер: 20 мМ буфер Трис-HCl, рН 8,75-8,85, содержащий 0,12 М хлорид натрия, проводимость 13,00-14,00 мСм/см при 15-25°С;
Буфер для элюции: 20 мМ буфер Трис-HCl, рН 8,75-8,85, содержащий 0,23 М хлорид натрия, проводимость 23,00-25,00 мСм/см при 15-25°С;
Элюция: линейный градиент до 100% буфер для элюции более чем 5 объемами колонки (5CV) и 5CV 100% буфера для элюции.
Объем фракции составляет 400-1000 мл.
Для дальнейшей очистки объединяют только фракции, равные или превышающие по чистоте (≥) чистоту 55% чИФНа5 (как определено посредством ОФ-ВЭЖХ).
5.3 Первая хроматография на колонке SP-Сефароза (катионообменная хроматография I)
Колонку SP-Сефароза используют на третьей и четвертой стадиях хроматографии с целью отделения основной формы чИФНа5 от заряженных изоформ, таких как N-метионил-чИФНа5 и ацетилированные чИФНа5 (формы, которые представляют собой продукты пост-трансляционных модификаций).
Вкратце, пул белков, полученный на предшествующей стадии (5.2), подводят до проводимости 6,00-7,00 мСм/см добавлением 10 мМ буфера на основе ацетата натрия, рН 4,95-5,05 и рН подводят до 5,15-5,20 4 М уксусной кислотой. Затем раствор белка наносят на хроматографическую колонку SP-Сефароза при следующих параметрах процесса:
Хроматографическая среда: SP-Сефароза быстрая (Amersham Pharmacia Biotech AB);
Используемая колонка: BPG 100×500 мм, диаметр 10 см;
Линейная скорость потока: 60 см/час;
Объем слоя хроматографической среды: 3,3±0,3 л;
Уравновешивающий буфер: 20 мМ буфер на основе ацетата натрия, рН 5,15-5,20, содержащий 50 мМ хлорид натрия, проводимость 6,00-7,00 мСм/см при 15-25°С;
Буфер для элюции: 20 мМ буфер на основе ацетата натрия, рН 5,15-5,20, содержащий 2 мМ L-метионин и 0,1 М NaCI, проводимость 11,00-13,00 мСм/см при 15-25°С;
Элюцию осуществляют более чем 10 объемами колонки (10CV) буфера для элюции.
Объем фракции составляет 400-2000 мл.
Для дальнейшей очистки объединяют только фракции, равные или превышающие по чистоте (≥) чистоту 70% чИФНа5 (как определено посредством ОФ-ВЭЖХ).
5.4 Вторая хроматография на колонке SP-Сефароза (катионообменная хроматография II)
Вкратце, нагружаемый раствор [пул фракции белка, выделенный из первой колонки SP-Сефароза (стадия 5.3)] разводят 5 мМ буфером на основе ацетата натрия, рН 5,00-5,20, содержащим 2 мМ L-метионин, проводимость 0,200-0,800 мСм/см при 15-25°С до проводимости 6,00-7,00 мСм/см при 15-25°С. Введение 2 мМ L-метионина в нагружаемый раствор проводят с целью предупреждения окисления чИФНа5 во время хроматографии, осуществляемой при комнатной температуре. Нагружаемый раствор наносят на хроматографическую колонку SP-Сефароза при следующих параметрах процесса:
Хроматографическая среда: SP-Сефароза быстрая (Amersham Pharmacia Biotech AB);
Используемая колонка: BPG 100×500 мм, диаметр 10 см;
Линейная скорость потока: 60 см/час;
Объем слоя хроматографической среды: 3,0+0,3 л;
Уравновешивающий буфер: 20 мМ буфер на основе ацетата натрия, рН 5,15-5,20, содержащий 50 мМ хлорид натрия, проводимость 6,00-7,00 мСм/см при 15-25°С;
Буфер для элюции: 20 мМ буфер на основе ацетата натрия, рН 5,15-5,20, содержащий 0,1 М NaCl, проводимость 11,00-13,00 мСм/см при 15-25°С;
Линейная скорость потока при элюции: 45 см/час;
Элюция: линейный градиент до 100% буфера для элюции в 20 объемах колонки (20CV).
Объем фракции составляет 400-2000 мл.
Фракции объединяют таким образом, что чистота чИФНа5 по ОФ-ВЭЖХ равна или выше чем (≥) 95%.
Препарат, концентрированно, стерильная фильтрация
Буфер для препарата: 25 мМ фосфат натрия, рН 6,80-7,20, содержащий 0,1 М хлорид натрия, проводимость 10,00-14,00 мСм/см при 15-25°С.
Замену буфера/концентрирование раствора белка осуществляют диафильтрацией/концентрированием через мембрану Biomax 10 кД с получением равной или выше чем (≥) 1,00 мг/мл, стерильно фильтруют через стерильный фильтр 0,22 мкм (Millipak 20) и наполняют стеклянные флаконы.
На Фигурах 8 и 9 показывают чистоту рекомбинантного чИФНа5, обработанного и изготовленного согласно данному изобретению, как определено посредством SDS-PAGE (14%), как в восстанавливающих, так и в невосстанавливающих условиях для рекомбинантного белка чИФНа5 по трем крупномасштабным загрузкам очистки.
На Фигуре 10 показывают чистоту рекомбинантного чИФНа5, обработанного и изготовленного согласно данному изобретению, как определено посредством "анализа с использованием высокоэффективной жидкостной хроматографии с обращенной фазой" (ОФ-ВЭЖХ) (на фигурах показывают результаты по трем крупномасштабным загрузкам рекомбинантного чИФНа5).
На Фигуре 11 показывают чистоту рекомбинантного чИФНа5, обработанного и изготовленного согласно данному изобретению, как определено посредством анализов с использованием "эксклюзионной хроматографии по размеру - ВЭЖХ" (ЭР-ВЭЖХ) по трем крупномасштабным загрузкам рекомбинантного чИФНа5.
На Фигуре 12 показывают чистоту рекомбинантного чИФНа5, обработанного и изготовленного согласно данному изобретению, как определено посредством анализов с использованием изоэлектрического фокусирования по трем крупномасштабным загрузкам рекомбинантного чИФНа5. Дорожки 1,11: стандарты pl (Amersham Pharmacia); дорожки 2, 3, 4: рекомбинантный чИФНа5, 15 мкг; дорожки 5, 6, 7: рекомбинантный чИФНа5, 5 мкг; дорожки 8, 9, 10: рекомбинантный чИФНа5, 1 мкг.
На Фигуре 13 представлены хроматограммы картирования пептидов трех крупномасштабных загрузок препаратов рекомбинантного чИФНа5, обработанного и изготовленного согласно данному изобретению.
Пример 4. Оценка эффекта микроэлементов и концентрации глюкозы на биосинтез рекомбинантного человеческого альфа5-ИФН.
Целью данного примера является подтверждение эффекта микроэлементов на пост-трансляционные модификации рекомбинантного человеческого альфа5-ИФН (чИФНа5) с целью установления пределов критической концентрации микроэлементов в углеродном растворе для подпитки и оценки эффекта концентрации глюкозы на биосинтез рекомбинантного чИФНа5. Таким образом, в данном примере описывают проведенный анализ и результаты, полученные при исследовании, и он служит обоснованием концентрации микроэлементов в углеродном растворе для подпитки и обоснованием концентрации глюкозы в процессе биосинтеза рекомбинантного чИФНа5.
При оптимизации процесса биосинтеза чИФНа5 устанавливают, что концентрация металлов или микроэлементов (следовых элементов) в ферментационной среде обладает эффектом на пост-трансляционные модификации белка чИФНа5. Таким образом, цель данного исследования состоит в подтверждении эффекта указанных следовых элементов на пост-трансляционные модификации белка чИФНа5 и установлении пределов критической концентрации микроэлементов с углеродном растворе для подпитки. Кроме того, решают оценить концентрацию глюкозы (17 г/л и 22 г/л) в ферментационной среде после введения каждой дозы углеродной подпитки с целью изучения, оказывает ли она какой-либо эффект на рост культуры (т.е. включает ли она ограничение роста культуры, воздействуя тем самым на выход биомассы, или она не имеет эффекта на рост культуры), а также на количество целевого белка (рекомбинантного чИФНа5).
Материалы и способы
План экспериментов разрабатывают и мониторируют с помощью руководства "Разработка экспериментов" (DOE), и он представлен с Таблице 1
Включенные эксперименты (*), которые предварительно проведены ранее в тех же условиях.
Ферментации с подпиткой с высокой густотой клеток проводят в химически определенной среде на основе минеральных солей/глюкозы.
- Композиция среды на основе минеральных солей/глюкозы для культивирования в колбе (г/л): гидрофосфат динатрия -17,0, дигидрофосфат калия - 1,82, сульфат аммония - 3,0, гептагидрат сульфата магния - 0,5, D(+)-глюкоза моногидрат -15,0, исходный раствор микроэлементов - 0,16 мл.
- Исходный раствор микроэлементов (следовых элементов или металлов) (г/л): гексагидрат хлорида железа (III) - 30,0, дигидрат хлорида кальция - 4,05, гептагидрат сульфата цинка (II) - 6,75, моногидрат сульфата марганца (II) - 1,5, пентагидрат сульфата меди (II) - 3,0, гексагидрат хлорида кобальта (II) - 1,14, дигидрат молибдата натрия - 0,3, борная кислота - 0,69.
- Композиция среды на основе минеральных солей/глюкозы для ферментации (г/л): дигидрофосфат аммония - 4,0, гептагидрат сульфат магния - 0,5, дигидрофосфат калия - 13,3, лимонная кислота моногидрат -1,6, D(+)-глюкоза моногидрат - 30,0, исходный раствор микроэлементов -0,25 мл.
- Углеродный раствор для подпитки: 70,0% глюкоза, 2,1% MgSO4×7H2O и исходный раствор микроэлементов, соответствующий плану эксперимента.
Концентрацию глюкозы, начиная с 8 часа культивирования, измеряют каждые (15-30) мин и после этого добавляют рассчитанную (для достижения верхнего предела концентрации глюкозы 17 г/л или 22 г/л согласно плану эксперимента) дозу углеродной подпитки.
Кроме того, добавляют 70 мл раствора обоих фосфатов (дигидрофосфат аммония - 25,0 г, дигидрофосфат калия - 21,0 г, разделенные на 3 дозы в соотношении 2:2:1 или 28:28:14 мл) в раздельных дозах при 60-75, 120-135 и 170-180 o.u.
При необходимости поступающий воздух автоматически обогащают чистым кислородом (до 60% на литр общего объема ферментера), что поддерживать концентрацию растворенного кислорода на уровне 20%.
0,25 мл маточной культуры Е. coli BL21 (DE3) рЕТ21-ИФНа5 из WCB (хранение при -70°С) [см. Пример 2] размножают в 500 мл среды на основе минеральных солей/глюкозы (рН приблизительно 7,7) при инкубировании на планетарном шейкере (300 об/мин) в течение 22 часов при 30°С.
Инокулюм составляет приблизительно 1,0% (20 мл) рабочего объема ферментера, что означает 1,54% реального объема.
Ферментации проводят в ферментере общего объема 3 л/рабочего объема 2 л "Biostat В" при рН 6,8, pO2 - 20%, температура - 37°С. Автоматически контролируемые переменные процесса ферментации в режиме реального времени (температура, перемешивание, рН, pO2, потребление кислот/оснований) и автономные переменные - оптическая плотность, регистрируют на графиках MFCS/win.
Для коррекции рН используют 35% ортофосфорную кислоту и 25% раствор аммиака.
Пену гасят 20% Pluronic® 31 R1.
Индукцию осуществляют при OD 91,6-103,6 o.u. (λ=600 нм) с помощью IPTG, чтобы создать конечную концентрацию IPTG 0,5 мМ для конечного рабочего объема. Ферментацию продолжают в течение 3 дополнительных часов.
Специфическую скорость роста культуры (µ), входящие (дополнительная подпитка) и выходящие (отбор проб для измерения OD и глюкозы) объемы точно рассчитывают на протяжении всех ферментации.
Результаты и анализ данных
Проведенный анализ и результаты, полученные во время данного исследования, убеждают в том, что пост-трансляционные модификации рекомбинантного белка чИФНа5, полученного в Е. coli, зависят от концентрации микроэлементов в среде для культивирования.
Окисленный метионилированный относящийся к чИФНа5 белок элиминируется, когда концентрация микроэлементов (исходного раствора) равны или больше чем 3,0 мл/л углеродного раствора для подпитки или равна или выше чем 0,95 мл/л реального конечного объема суспензии и средняя специфическая скорость роста культуры (µ) после индукции равна или выше чем (≥) 0,17 (см. Таблицы 2-3).
* - В ранее проведенных загрузках OD измеряют с помощью BioPhotometer (Eppendorf, A=595 нм), в других - с помощью спектрометра EZ 150 (PerkinElmer, А=б00 нм); WCW: сырая масса клеток (г/л клеточной суспензии); DCW: сухая масса клеток (%).
* ферментация 10 л (рабочий объем); ** - замедленная скорость роста после индукции (разведение, поскольку датчик пеногашения вышел из строя, добавляют 84 мл раствора пеногасителя). *** - наблюдают образование дополнительной (неизвестной) формы между главным пиком и двумя пиками ацетилированных форм. НТ: Не тестируют.
Как описано в данном контексте, в Таблице 2 показывают параметры биосинтеза при различных концентрациях микроэлементов в углеродном растворе для подпитки, тогда как в Таблице 3 показывают параметры биосинтеза и развертывания при различных концентрациях микроэлементов.
Проведен ряд исследований, например i) зависимости специфической скорости роста культуры (µ) от концентрации микроэлементов в углеродном растворе для подпитки или в суспензии клеток; (ii) зависимости окисленного метионилированного чИФНа5 (OxidMet чИФНа5) после развертывания и общего ацетилированного чИФНа5 после развертывания от средней специфической скорости роста культуры (µ) после индукции; (iii) зависимости пост-трансляционных модификаций белка чИФНа5 (OxidMet чИФНа5 после развертывания, ацетилированного чИФНа5 после развертывания и правильно уложенного чИФНа5 после развертывания) от концентрации микроэлементов в углеродном растворе для подпитки и (iv) зависимости относящихся к чИФНа5 форм (ацетилированного чИФНа5 после развертывания и OxidMet чИФНа5 после развертывания) от концентрации микроэлементов в углеродном растворе для подпитки или в суспензии клеток. Кроме того, все загрузки подвергают ОФ-ВЭЖХ после развертывания. Изданных результатов очевидно, что специфическая скорость роста культуры (µ) и выход биомассы зависят от концентрации микроэлементов в среде для культивирования (см. Таблицы 2-3).
Таким образом, полученные результаты показывают, что:
- самое низкое содержание ацетилированного чИФНа5 наблюдают при самых высоких количествах окисленного метионилированного чИФНа5 и наоборот (короткие номера загрузок No. М-83, М-84, М-85 и М-86);
- Количество ацетилированного чИФНа5 составляет приблизительно 8-11%, когда концентрация микроэлементов (исходный раствор) лежит в интервале 3,0-3,7 мл/л углеродного раствора для подпитки (короткие No. загрузок М-80, М-81, М-92, М-82 и М-87) или в интервале 0,95-1,23 мл/л реального конечного объема суспензии (см. Таблицу 3); и
- Когда концентрация микроэлементов в углеродном растворе для подпитки выше чем 4 мл/л или выше чем 1,23 мл/л реального конечного объема суспензии клеток, определяют дополнительную неизвестную (неидентифицированную) форму [ее отмечают после основного пика и двух пиков ацетилированных форм (данные не показаны)].
Полученные результаты показывают также, что верхний предел (17 г/л или 22 г/л) концентрации глюкозы практически не влияет на качество целевого белка (чИФНа5) и выход биомассы, реальные полученные значения находятся в пределах ошибки (см. таблицу 2).
Заключения
- Результаты, полученные на протяжении данного исследования, служат обоснованием концентрации микроэлементов в углеродном растворе для подпитки и концентрации глюкозы в процессе биосинтеза рекомбинантного чИФНа5.
- Окисленный метионилированныйбелокчИФНа5 элиминируется, когда концентрация микроэлементов равна или выше чем 3,0 мл/л углеродного раствора для подпитки или выше чем 0,95 мл/л реального конечного объема суспензии и средняя специфическая скорость роста культуры (µ) после индукции равна или выше чем 0,17.
- Ацетилированный белок чИФНа5 составляет приблизительно 8-11%, когда концентрация микроэлементов лежит в пределах от приблизительно 3,0 до приблизительно 3,7 мл/л углеродного раствора для подпитки или от приблизительно 0,95 до приблизительно 1,23 мл/л реального конечного объема суспензии.
- Когда концентрация микроэлементов в углеродном растворе для подпитки выше чем 3,7 мл/л или больше чем 1,23 мл/л L реального конечного объема суспензии, синтезируется неизвестная (неидентифицированная) форма белка.
- Оптимальная концентрация микроэлементов, определяющая максимальный выход и наилучшее качество целевого белка чИФНа5, лежит в пределах от приблизительно 3,0 до приблизительно 3,7 мл/л углеродного раствора для подпитки.
- Концентрация глюкозы в интервале от 17 г/л до 22 г/л практически не влияет на качество целевого белка (рекомбинантного чИФНа5) и выход биомассы.
| название | год | авторы | номер документа |
|---|---|---|---|
| ФЕРМЕНТАЦИОННАЯ СРЕДА И СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНЫХ БЕЛКОВ | 2009 |
|
RU2556120C2 |
| ФЕРМЕНТАЦИОННАЯ СРЕДА ДЛЯ ПОЛУЧЕНИЯ ПРАВАСТАТИНА И СПОСОБ ПОЛУЧЕНИЯ ПРАВАСТАТИНА | 2009 |
|
RU2595380C2 |
| Рекомбинантная плазмидная ДНК pET21-IFN-γ, содержащая ген, кодирующий гамма-интерферон человека, штамм E.coli BL21/pET21-IFN-γ - продуцент рекомбинантного гамма-интерферона человека и способ получения гамма-интерферона человека | 2023 |
|
RU2827613C1 |
| ШТАММ Escherichia coli BL21(DE3)Gold/pETCYPopti - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО ЦИКЛОФИЛИНА А ЧЕЛОВЕКА | 2015 |
|
RU2603283C1 |
| ШТАММ КЛЕТОК Escherichia coli BL21 (pVEGF-A165), СЕКРЕТИРУЮЩИХ РЕКОМБИНАНТНЫЙ ФАКТОР РОСТА ЭНДОТЕЛИЯ СОСУДОВ (VEGF) | 2008 |
|
RU2395568C2 |
| ФЕРМЕНТАЦИОННАЯ СРЕДА И СПОСОБ ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНЫХ БЕЛКОВ | 2009 |
|
RU2491345C2 |
| ПОЛИНУКЛЕОТИД, КОДИРУЮЩИЙ МУТАНТНУЮ РЕКОМБИНАНТНУЮ IgA1 ПРОТЕАЗУ Neisseria meningitidis СЕРОГРУППЫ В, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, СОДЕРЖАЩАЯ УКАЗАННЫЙ ПОЛИНУКЛЕОТИД, КЛЕТКА-ХОЗЯИН, СОДЕРЖАЩАЯ УКАЗАННУЮ ПЛАЗМИДНУЮ ДНК, РЕКОМБИНАНТНАЯ IgA1 ПРОТЕАЗА Neisseria memingitidis СЕРОГРУППЫ В, СПОСОБ ПОЛУЧЕНИЯ ЗРЕЛОЙ ФОРМЫ IgA1 ПРОТЕАЗЫ | 2011 |
|
RU2486243C1 |
| СПОСОБ ПОЛУЧЕНИЯ МЕТИЛМЕТАКРИЛАТА | 2017 |
|
RU2758142C2 |
| Рекомбинантная плазмида pFM-IFN-17, обеспечивающая экспрессию интерферона альфа-2b человека, рекомбинантная плазмида pFM-АР, обеспечивающая экспрессию фермента метионинаминопептидазы E. coli, биплазмидный штамм Escherichia coli FM-IFN-АР (pFM-IFN-17, pFM-АР) - продуцент (Met-) рекомбинантного интерферона альфа-2b человека | 2016 |
|
RU2610173C1 |
| Способ выделения и очистки амброксида | 2016 |
|
RU2720094C2 |




Изобретение относится к области биотехнологии и может быть использовано для рекомбинантного получения α5-интерферона. Клетку E. coli - продуцента ИФНа5 культивируют с использованием углеродного раствора для подпитки с глюкозой в качестве источника углерода, содержащего на литр следующие компоненты, мг: 90-111 гексагидрата хлорида железа (III), 12,15-14,985 дигидрата хлорида кальция, 20,25-24,975 гептагидрата сульфата цинка (II), 4,5-5,55 моногидрата сульфата марганца (II), 9-11,1 пентагидрата сульфата меди (II), 3,42-4,218 гексагидрата хлорида кобальта (II), 0,9-1,11 дигидрата молибдата натрия и 2,07-2,553 борной кислоты. Ферментационная среда является, по существу, свободной от компонентов животного происхождения или дрожжевого происхождения и характеризуется содержанием на литр следующих компонентов, мг: 28,5-36,9 гексагидрата хлорида железа (III), 3,8475-4,9815 дигидрата хлорида кальция, 6,4125-8,3025 гептагидрата сульфата цинка (II), 1,425-1,845 моногидрата сульфата марганца (II), 2,85-3,69 пентагидрата сульфата меди (II), 1,083-1,4022 гексагидрата хлорида кобальта (II), 0,285-0,369 дигидрата молибдата натрия и 0,6555-0,8487 борной кислоты. Изобретение позволяет минимизировать инкорпорацию дополнительного остатка метионина в N-концевую часть полипептидной цепи и генерацию его окисленных молекул. 9 з.п. ф-лы, 13 ил., 3 табл., 4 пр.
1. Способ получения белка альфа5-интерферона (ИФНа5) посредством экспрессии в клетке-хозяине Escherichia coli (Е. coli), включающий использование клетки-хозяина Е. coli, продуцирующей ИФНа5, культивирование клетки-хозяина Е. coli, продуцирующей ИФНа5, в условиях, эффективных для экспрессии ИФНа5 клеткой-хозяином Е. coli, экспрессию ИФНа5 клеткой-хозяином Е. coli в ферментационной среде, которую подпитывают углеродным раствором для подпитки, выделение и необязательную очистку экспрессированного белка ИФНа5, причем углеродный раствор для подпитки содержит источник углерода, которым является глюкоза, и на литр следующие компоненты, мг:
а указанная ферментационная среда является, по существу, свободной от компонентов животного происхождения или дрожжевого происхождения и характеризуется содержанием на литр следующих компонентов, мг:
2. Способ по п. 1, в котором в качестве клетки-хозяина Е. coli используют клетку-хозяина Е. coli, трансформированную вектором, содержащим последовательность, кодирующую белок ИФНа5, под контролем индуцируемого промотора.
3. Способ по п. 1 или 2, в котором в качестве клетки-хозяина Е. coli используют клетку-хозяина Е. coli, представляющую собой штамм Е. coli с дефицитом протеазы, предпочтительно, штамм-хозяин Е. coli lon-/ompT- с дефицитом протеазы, более предпочтительно штамм Е. coli BL21, наиболее предпочтительно штамм Е. coli BL21 (DE3).
4. Способ по п. 3, в котором в качестве клетки-хозяина Е. coli используют клетку-хозяина Е. coli, представляющую собой штамм Е. coli BL21 (DE3) и экспрессию ИФНа5, индуцируют посредством изопропил-β-D-тиогалактопиранозида.
5. Способ по п. 4, который характеризуется средней специфической скоростью роста культуры (μ) после индукции, составляющей по меньшей мере 0,17.
6. Способ по п. 5, в котором выделенный ИФНа5 характеризуется содержанием окисленного-Met ИФНа5 по большей мере 1%.
7. Способ по п. 1, в котором указанный белок ИФНа5 представляет собой чИФНа5.
8. Способ по п. 1, в котором белок ИФНа5 характеризуется последовательностью, содержащей аминокислотную последовательность SEQ ID NO: 1.
9. Способ по п. 1, в котором выделение и очистку белка ИФНа5 осуществляют из смеси, содержащей указанный белок ИФНа5 в форме телец включения (IB), и включают солюбилизацию указанных телец включения с получением смеси, содержащей денатурированный ИФНа5, окислительную ренатурационную обработку указанной смеси, содержащей денатурированный ИФНа5, с формированием смеси, содержащей ренатурированный ИФНа5, и очистку указанной смеси, содержащей ренатурированный ИФНа5, с получением очищенного ИФНа5.
10. Способ по п. 9, в котором очистку смеси, содержащей ренатурированный ИФНа5, осуществляют посредством четырехстадийной хроматографической обработки, включающей последовательное проведение гидрофобной хроматографии, анионообменной хроматографии, первой катионообменной хроматографии и второй катионообменной хроматографии с необязательным разведением буфером, содержащим метионин.
| СЛАВЧЕНКО И.Ю | |||
| и др., Фенотипические проявления особенностей метаболизма клеток Escherichia coli BL21 (DE3) при выращивании на средах, содержащих различные источники углерода, Бiополiмери i клiтина, 2002, т.18, No:3, с.232-236 |

