Область техники настоящего изобретения
Изобретение относится к области микробиологии, молекулярной биологии и биохимии, в частности к получению штамма генномодифицированного микроорганизма, используемого для получения рекомбинантного фактора роста эндотелия сосудов (VEGF).
Предшествующий уровень техники настоящего изобретения
Наличие факторов, стимулирующих рост кровеносных сосудов и тем самым привлекающих вновь образуемые сосуды для питания растущей опухоли, было постулировано несколько десятилетий назад, хотя идентификация и выделение этих факторов долгое время оставались недостижимыми. В настоящее время фактор роста эндотелия сосудов (VEGF), который был идентифицирован в 1980-е годы, считается ключевым регулятором нормального и аномального роста кровеносных сосудов. В 1993 году было показано, что моноклональные антитела против VEGF вызывают резкое подавление опухолевого роста in vivo. Это наблюдение было положено в основу разработки противоракового средства бевацизумаба (авастина (Avastin; Genentech)), гуманизированного варианта этого антитела против VEGF. Недавнее одобрение бевацизумаба US FDA в качестве терапевтического средства первой очереди при метастатическом колоректальном раке подтверждает представления о том, что VEGF является ключевым медиатором опухолевого ангиогенеза и что блокирование ангиогенеза является эффективной стратегией лечения злокачественных опухолей у человека.
Наблюдение того, что опухолевый рост может сопровождаться усиленной васкуляризацией, было опубликовано более века назад (с обзором можно ознакомиться в ссылке (1). Однако лишь в 1939 году Ide et al. впервые постулировали наличие опухолевого фактора, стимулирующего рост кровеносных сосудов, способствующего привлечению кровоснабжения к растущей опухоли (2). Несколько лет спустя на основании наблюдения, что быстрому росту опухоли предшествует локальное увеличение плотности сосудистой сети, Algire et al. предположили, что быстрый рост опухолевых трансплантатов зависит от формирования разветвленной сосудистой сети (3). Затем новых сообщений в этой области техники не появлялось до 1960-х годов, когда в ходе экспериментов Greenblatt и Shubik (4) и Ehrmann и Knoth (5) были получены первые доказательства того, что опухолевый ангиогенез опосредован способными к диффузии факторами, продуцируемыми опухолевыми клетками.
В 1971 году Folkman предположил, что подавление ангиогенеза могло бы являться эффективной противораковой стратегией (6). На основании этой революционной гипотезы в начале 1970-х годов Folkman et al. предприняли усилия, направленные на выделение "фактора опухолевого ангиогенеза" из опухолей человека и животных (7). В 1978 году Gullino также предположил, что блокирование ангиогенеза способно предотвратить развитие злокачественной опухоли (8). Впоследствии были описаны ангиогенные эффекты множества факторов (например, эпидермального фактора роста (EGF), трансформирующего фактора роста (TGF)-α, TGF-β, фактора-α некроза опухоли (TNF-α) и ангиогенина) (9). Однако, хотя эти факторы усиливали ангиогенез в ходе некоторых биологических анализов, ни один из них не действовал физиологически (10).
Львиная доля внимания была уделена двум широко распространенным и высокоэффективным митогенам эндотелиоцитов и ангиогенным факторам - кислому и основному факторам роста фибробластов (aFGF и bFGF). В середине 1980-х годов были проведены очистка до состояния гомогенности, секвенирование и клонирование кДНК этих FGF (11). Неожиданная находка состояла в том, что гены как aFGF, так и bFGF не кодируют стандартный секреторный сигнальный пептид. Однако, как указано выше, более ранние исследования позволяли предположить, что опухолевый ангиогенез опосредован способными к диффузии молекулами (4, 5). Кроме того, некоторые исследования свидетельствовали о том, что иммунонейтрализация FGF не оказывала эффекта или оказывала слабый эффект на опухолевый ангиогенез (12, 13), что позволяло предположить, что ключевые регуляторы ангиогенеза все еще не были идентифицированы.
В настоящее время известно, что одним таким ключевым регулятором ангиогенеза является фактор роста эндотелия сосудов (VEGF, также называемый VEGF-A), и роль семейства генов VEGF в регуляции ангиогенеза интенсивно исследуется уже на протяжении более десятилетия (1). Семейство VEGF включает в себя прототипный член VEGF-A, плацентарный фактор роста (P1GF) (14), VEGF-B (15), VEGF-C (16) и VEGF-D (17). Убедительные доказательства свидетельствуют, что тогда как построение и созревание сосудистой стенки представляют собой весьма сложные процессы, требующие согласованных воздействий ангиопоэтинов, тромбоцитарного фактора роста В (PDGF-B) и других факторов (18), действие VEGF-A представляет собой лимитирующую скорость стадию нормального и патологического роста кровеносных сосудов (19). Важным представляется то, что VEGF-C и VEGF-D регулируют ангиогенез лимфатических сосудов (20), что подчеркивает уникальную роль этого семейства генов в контролировании роста и дифференцировки некоторых анатомических компонентов сосудистой системы.
В 1983 году Senger et al. сообщили о частичном выделении из среды, кондиционированной опухолевой клеточной линией морской свинки, "фактора проницаемости сосудов" (VPF), белка, который индуцировал увеличение проницаемости сосудов кожи (21). Однако, поскольку VPF не был выделен и секвенирован, на тот момент молекулярная структура фактора осталась неизвестной. В 1989 году появилось сообщение о выделении из среды, кондиционированной фолликулярными клетками гипофиза коровы, "фактора роста эндотелия сосудов" (VEGF), специфичного в отношении эндотелиоцитов митогена (22). Аминоконцевая аминокислотная последовательность VEGF не совпадала ни с одним известным белком в доступных базах данных (22). Затем работа Connolly et al., выполненная на основе работы Senger et al., независимо привела к выделению и секвенированию VPF (23). Клонирование кДНК VEGF (24) и VPF (25) показало, что VEGF и VPF представляют собой одну и ту же молекулу. Этот результат был неожидан, учитывая, что другие митогены эндотелиоцитов, такие как FGF, не усиливают проницаемость сосудов.
VEGF характеризуется значительной гомологией с цепями А и В PDGF (24). Ген, кодирующий VEGF-A человека, состоит из восьми экзонов, разделенных семью интронами (26, 27). Альтернативный сплайсинг экзонов приводит к образованию четырех основных изоформ - VEGF121, VEGF165, VEGF189 и VEGF206, - состоящих после отщепления сигнальной последовательности соответственно из 121, 165, 189 и 206 аминокислот (24). Альтернативный сплайсинг регулирует биологическую доступность VEGF (28, 29). К настоящему времени скопилась масса доказательств того, что VEGF165 является наиболее физиологически компетентной изоформой (30, 31). Также в регуляции биологической доступности VEGF важную роль играет внеклеточный протеолиз. Плазмин способен расщеплять VEGF165 и VEGF 189 и высвобождать биологически активный продукт, состоящий из первых 110 аминоконцевых аминокислот (32). Учитывая важность активации плазминогена в ходе физиологического и патологического ангиогенеза (33), этот механизм может иметь особое значение для регуляции активности и биологической доступности VEGF при развитии ремоделирования и в ответ на сигналы микроокружения. Кроме того, в некоторых опухолях протеолиз VEGF, опосредованный мактриксной металлопротеиназой-9 (ММР9), может быть ответствен за запуск ангиогенеза (34).
Достоверно установленным действием VEGF является ускорение роста эндотелиоцитов артерий, вен и лимфатических сосудов (обзор приведен в ссылке 35). VEGF индуцирует мощный ангиогенный ответ во множестве моделей in vivo (24, 36). Кроме того, как указано выше, VEGF увеличивает проницаемость сосудов, и это свойство лежит в основе важной роли этой молекулы в воспалении и других патологических процессах (37). В этом контексте VEGF также индуцирует экспрессию эндотелием некоторых молекул адгезии, регулирующих адгезию лейкоцитов при воспалении (38).
In vitro VEGF предотвращает апоптоз эндотелиоцитов, индуцируемый истощением сыворотки, что опосредуется каскадом фосфатидилинозитол-3'-киназы (PI3K)/Akt (39). Также, VEGF индуцирует экспрессию в эндотелиоцитах антиапоптотических белков BCL2 и А1 (40). Зависимость от VEGF была показана на эндотелиоцитах вновь образованных, но не до конца сформировавшихся сосудов опухолей (41, 42). Образование выстилки перицитами, предположительно, является одним из ключевых событий, приводящих к утрате эндотелиоцитами зависимости от VEGF (42).
Следует подчеркнуть, что хотя эндотелиоциты являются главными мишенями VEGF, в ходе нескольких исследований были показаны также его эффекты на митотическую активность/выживаемость некоторых неэндотелиальных клеточных типов, включая нейроны (43).
VEGF принципиально важен для нормального эмбрионального васкулогенеза и ангиогенеза. Инактивация отдельного аллеля VEGF у мышей приводит к эмбриональной смертности (44, 45). Ингибирование VEGF в раннем неонатальном периоде приводит к остановке роста, апоптозу эндотелиоцитов и летальному исходу главным образом в результате почечной недостаточности (46, 47). VEGF также необходим для внутрихрящевого окостенения, фундаментального механизма роста трубчатых костей. Ингибиторы VEGF подавляют эти процессы у грызунов и приматов (48, 49). Важно заметить, что этот эффект полностью обратим при прекращении анти-VEGF обработки (48, 49). Ангиогенез является ключевым аспектом нормального овариально-менструального цикла и функционирования эндометрия (50). Экспрессия мРНК VEGF связана во времени и пространстве с пролиферацией кровеносных сосудов в яичниках множества видов (51, 52). Введение ингибиторов VEGF замедляет развитие фолликулов (53) и подавляет ангиогенез в желтом теле грызунов (54) и приматов (49, 55, 56).
Исследования гибридизации in situ свидетельствуют об экспрессии мРНК VEGF во многих опухолях человека, в том числе карциномах легкого (57), молочной железы (58), желудочно-кишечного тракта (59), почки (60) и яичника (61). Однако экспрессия VEGF варьирует не только между различными типами опухолей, но также и в пределах одной опухоли. В случае полиморфной глиобластомы и других опухолей с существенным некротическим компонентом экспрессия мРНК VEGF наиболее интенсивна в гипоксических опухолевых клетках, соседствующих с зонами некроза (62, 63). Опухолью с особенно сильной экспрессией VEGF является почечно-клеточная карцинома. Положительная регуляция экспрессии VEGF может быть связана с целым рядом факторов, среди которых можно отметить гипоксию и некоторые мутации.
Клиническая применимость ингибирования VEGF в онкологии не ограничена солидными опухолями. Появляется все больше доказательств, что VEGF и его рецепторы экспрессируются множеством злокачественных опухолей системы крови, что позволяет предположить перспективность ингибирования VEGF или передачи сигнала VEGFR для лечения таких состояний (64).
Раскрытие настоящего изобретения
Сущностью настоящего изобретения является создание нового штамма клеток Escherichia coli BL21 (pVEGF-A165), продуцирующих рекомбинантный фактор роста эндотелия сосудов - белок GST-VEGF-A165. Способность клеток Escherichia coli BL21 (pVEGF-A165) секретировать рекомбинантный VEGF достигается при помощи генной модификации клеток Escherichia coli плазмидой pGEX-VEGF-A165 (SEQ ID NO: 1).
Технический результат заключается в получении штамма клеток Escherichia coli BL21 (pVEGF-A165), стабильно трансфицированных плазмидой, кодирующей VEGF, и секретирующих этот фактор во внеклеточное пространство при культивировании in vitro.
Целью настоящего изобретения является расширение коллекции уникальных клеточных штаммов, которые можно использовать для получения VEGF. Эта задача является особенно актуальной для современной фармакологической биотехнологии. Наработанный указанным штаммом VEGF может применяться во многих сферах биологических и медицинских исследований, в частности при создании и изучении лекарственных средств и других соединений, являющихся агонистами или антагонистами VEGF.
Поставленная задача решается тем, что получен новый штамм клеток Escherichia coli BL21 (pVEGF-A165), трансформированных плазмидой pGEX-VEGF-A165 (SEQ ID NO: 1) и продуцирующих рекомбинантный белок GST-VEGF-A165 в количестве не менее 2 мкг очищенного рекомбинантного белка на 1 мл ростовой среды за 24 часа (по данным ИФА (ELISA)). Полученный штамм обладает стабильными культуральными и морфологическими свойствами и в результате генной модификации обладает способностью секретировать рекомбинантный фактор роста эндотелия сосудов. Штамм клеток Escherichia coli BL21 (pVEGF-A165) депонирован во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) ФГУП ГосНИИГенетика под номером ВКПМ В-10042.
Краткое описание чертежей
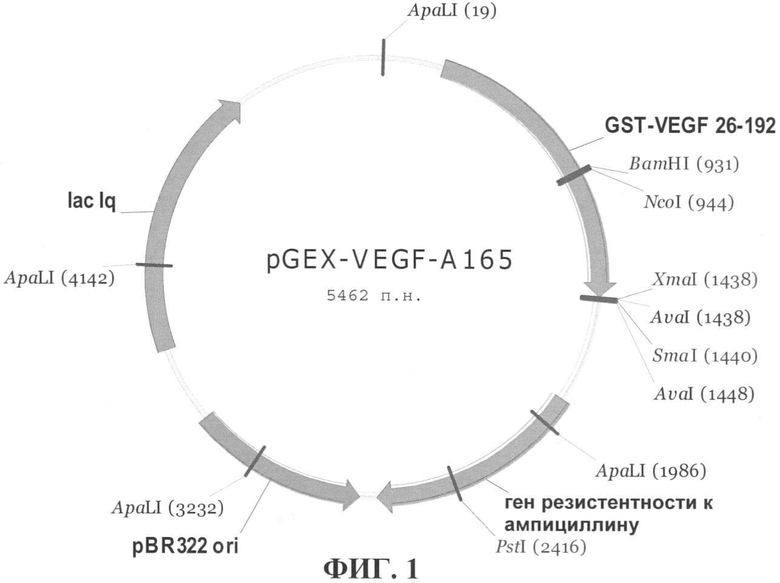
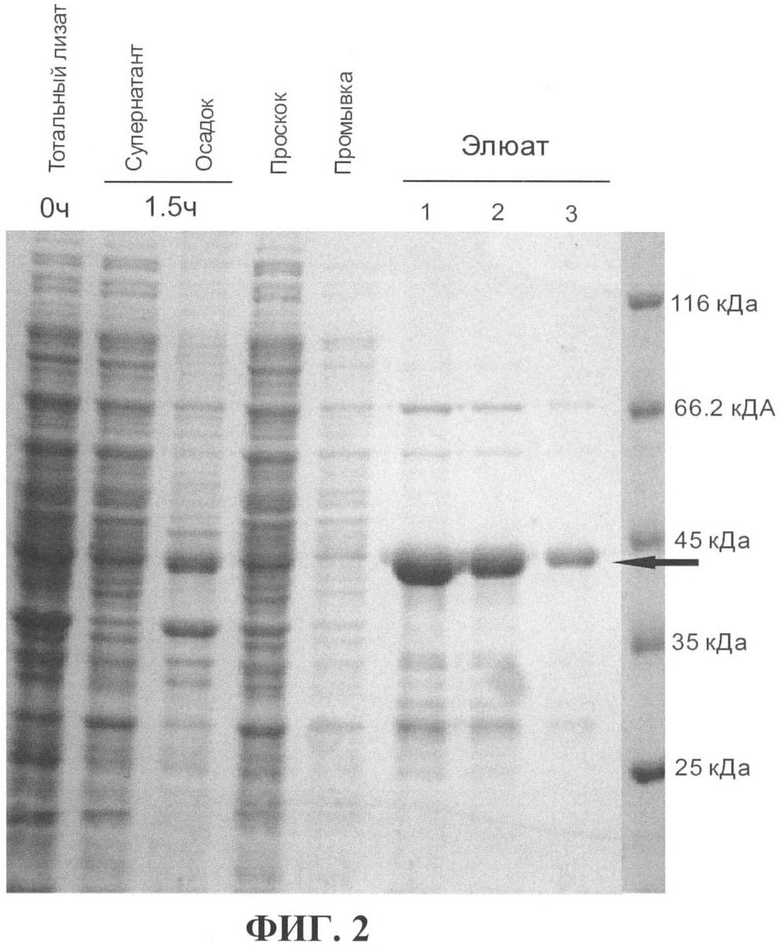
На фиг.1 приведена генетическая карта плазмиды pGEX-VEGF-A165. На фиг.2 показан результат анализа фракций, собранных в процессе очистки рекомбинантного белка GST-VEGF-A165. Пропорциональные количества белка из каждой фракции анализировали с помощью ДСН-ПАГЭ в 10%-ном акриламидном геле с последующим окрашиванием геля красителем Coomassie Brilliant Blue (Serva). Для оценки молекулярной массы белков использовали маркер молекулярной массы Unstained Protein Molecular Weight Marker производства (Fermentas), крайняя дорожка справа. Стрелка указывает на положение в геле рекомбинантного белка GST-VEGF-A165. Кажущаяся молекулярная масса белка GST-VEGF-A165 составляет около 45 кДа и соответствует теоретически рассчитанной (45526 Да).
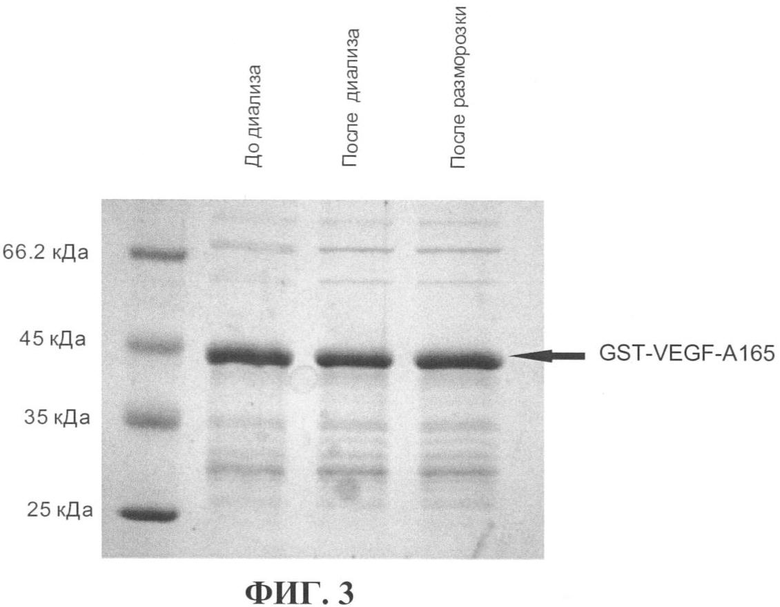
На фиг.3 показан результат анализа стабильности рекомбинантного белка GST-VEGF-A165 в процессе диализа и замораживания-оттаивания. Пропорциональные количества белка из каждой фракции анализировали с помощью ДСН-ПАГЭ в 10%-ном акриламидном геле с последующим окрашиванием геля красителем Coomassie Brilliant Blue (Serva). Для оценки молекулярной массы белков использовали маркер молекулярной массы Unstained Protein Molecular Weight Marker производства (Fermentas), крайняя дорожка слева. Стрелка указывает на положение в геле рекомбинантного белка GST-VEGF-A165. Белок GST-VEGF-A165 стабилен в процессе проведения диализа и выдерживает замораживание в жидком азоте и последующее оттаивание.
Примеры осуществления настоящего изобретения
Пример 1. Процедура получения рекомбинантной плазмидной ДНК, кодирующей кДНК белка VEGF-A165, слитого с белком глутатион-8-трансферазой (GST-VEGF-A165)
кДНК VEGF человека амплифицировали с помощью ПЦР с использованием в качестве матрицы первых цепей кДНК из плаценты человека. Праймеры подбирали таким образом, чтобы из кДНК VEGF был удален сигнальный пептид (аминокислоты 1-26), а 5'-праймер [5'-CAT GGA TCC GCT GCA CCC ATG GCA GAA GGA-3'] и 3'-праймер [5'-GTC ACC CGG GTC ACC GCC TCG GCT-3'] содержали сайты узнавания эндонуклеаз рестрикции BamHI и SmaI, соответственно. Далее BamHI-SmaI фрагмент клонировали по соответствующим сайтам в плазмидный вектор pGEX-4T2 (Amersham) таким образом, чтобы кДНК белка VEGF-A165 находилась в одной рамке считывания с кДНК белка GST.
Полученная плазмидная конструкция имела размер 5462 п.н. (фиг.1). Рекомбинантный белок GST-VEGF-A165, кодируемый данной плазмидой, имел размер 392 аминокислоты и расчетную молекулярную массу 45526 Да.
Пример 2. Процедура получения штамма бактерий Е.coli BL21, продуцирующих рекомбинантный белок GST-VEGF-A165
Плазмидный вектор pGEX-VEGF-A165 трансформировали в штамм Е.coli BL21 с помощью стандартной методики трансформации бактерий (65) и высевали на LB агар (0,171 М NaCl, 1% триптон, 0,5% дрожжевой экстракт, 1,5% агар), содержащий селективный антибиотик ампициллин в концентрации 50 мкг/мл, и растили в течение 16-18 часов при температуре 37°С. Единичные колонии бактерий, выросших на селективной среде, пересевали на свежую селективную чашку и растили в течение 16-18 часов при температуре 37°С. С чашки Петри бактериальной петлей брали единичную колонию бактерий и инокулировали 10 мл жидкой ростовой среды LB (0,171 М NaCl, 1% триптон, 0,5% дрожжевой экстракт), содержащей селективный антибиотик ампициллин в концентрации 50 мкг/мл, и растили в течение 16-18 часов при температуре 37°С и постоянном перемешивании. Брали 2 мл бактериальной суспензии, клетки осаждали центрифугированием, супернатант удаляли, а осадок ресуспендировали в 1,5 мл среды для заморозки (жидкая среда LB, содержащая 15% глицерина). Полученный штамм-продуцент хранили при температуре -80°С.
Пример 3. Процедура получения рекомбинантного белка GST-VEGF-A165 в бактериальной системе экспрессии
Штамм-продуцент рекомбинантного белка GST-VEGF-A165 высевали штрихом на чашку Петри с LB агаром, содержащую селективный антибиотик ампициллин в концентрации 50 мкг/мл, и растили в течение 16-18 часов при температуре 37°С. С чашки Петри брали единичную колонию бактерий, инокулировали 10 мл жидкой среды LB, содержащей 50 мкг/мл ампициллина, и растили при постоянном перемешивании и температуре 37°С в течение 16-18 часов.
По 5 мл ночной культуры штамма-продуцента переносили стерильной пипеткой в две 2-литровые колбы, содержащие 200 мл жидкой среды LB, и растили при постоянном перемешивании и температуре 37°С в течение 2,5 часов, после чего индуцировали экспрессию рекомбинантного белка путем добавления изопропил-β-D-тиогалактозида (Fermentas) до конечной концентрации 0,02 мМ и бактерии растили при температуре 28°С в течение 1,5 часов.
Клетки осаждали центрифугированием при 4000 об/мин и температуре 4°С в течение 10 минут, супернатант удаляли, а осадок промывали с использованием 40 мл фосфатно-солевого буфера (0,01 М фосфат натрия, 0,138 М NaCl, 0,0027 М KCl, рН 7,4), предварительно охлажденного до 4°С, и затем клетки осаждали центрифугированием при 4000 об/мин и температуре 4°С в течение 10 минут. Супернатант удаляли, а осадок клеток ресуспендировали в 10 мл буфера (0,01 М фосфат натрия, 0,138 М NaCl, 0,0027 М KCl, 2,5 мМ MgCl2, 1 мМ ЭДТА, 0,05% NP-40, рН 7,4) и добавляли кристаллический лизоцим (Sigma) до конечной концентрации 10 мкг/мл. Далее суспензию бактериальных клеток замораживали в жидком азоте и хранили при температуре -80°С до стадии очистки белка.
Суспензию бактериальных клеток медленно оттаивали на льду и подвергали воздействию ультразвука при температуре +0°С с помощью прибора Vibracell 72434 (Bioblock Scientific) до тех пор, пока суспензия не просветлялась и вязкость не исчезала (продолжительность ультразвукового импульса составляла 60 секунд, между импульсами делали перерывы не менее 60 секунд; проводили не менее 3 импульсов).
Бактериальный лизат переносили в охлажденные микроцентрифужные пробирки (Costar) емкостью 1,5 мл и центрифугировали в настольной микроцентрифуге при 13000 об/минут и температуре 4°С в течение 15 минут. Осветленный лизат собирали и смешивали с 500 мкл 4Fast Flow глутатионовой сефарозы (Amersham Biosciences), предварительно уравновешенной охлажденным фосфатно-солевым буфером.
Далее в течение 1,5 ч при 4°С и постоянном перемешивании проводили связывание рекомбинантного белка GST-VEGF-A165 с глутатионовой сефарозой. После связывания лизат при 4°С наносили на колонку, механически задерживающую глутатион-сефарозу, и позволяли лизату протечь через колонку под действием силы тяжести. Затем при температуре 4°С глутатион-сефарозу один раз промывали 10 мл буфера (0,01 М фосфат натрия, 0,138 М NaCl, 0,0027 М KCl, 2,5 мМ MgCl2, 1 мМ ЭДТА, 0,05% NP-40, рН 7,4) и два раза 5 мл фосфатно-солевого буфера.
Связавшийся с глутатион-сефарозой рекомбинантный белок GST-VEGF-A165 трижды элюировали 500 мкл буфера для элюции (50 мМ Трис, рН 8,0, 10 мМ восстановленный глутатион), инкубируя глутатион-сефарозу в буфере для элюции в течение 10 минут при комнатной температуре и постоянном перемешивании с помощью ротомикса (10 об/мин).
На всех стадия очистки белка фракции собирали и анализировали с помощью ДСН-ПАГЭ в 10%-ном акриламидном геле (66). Результат анализа фракций, собранных в процессе очистки рекомбинантного белка GST-VEGF-A165, приведен на фиг.2.
Элюированный рекомбинантный белок диализовали при температуре 4°С против фосфатно-солевого буфера, замораживали в жидком азоте и хранили при -80°С в аликвотах. Для проверки стабильности белка GST-VEGF-A165 аликвоту размораживали и концентрацию рекомбинантного белка анализировали методом ДСН-ПАГЭ. Результат анализа стабильности рекомбинантного белка GST-VEGF-A165 в процессе диализа и замораживания-оттаивания приведен на фиг.3.
Пример 4. Процедура определения концентрации рекомбинантного белка GST-VEGF-A165.
Концентрацию рекомбинантного белка GST-VEGF-A165 определяли методом иммуноферментного анализа (ELISA) с использованием набора для ИФА-определения белка VEGF Human VEGF ELISA Development kit производства PeproTech в соответствии с протоколом производителя.
Антитела для связывания разводили в фосфатно-солевом буфере до конечной концентрации 0,5 мкг/мл, 100 мкл данного раствора добавляли в лунки 96-луночного планшета для ИФА и инкубировали при комнатной температуре в течение 16-18 часов. Раствор антител удаляли и лунки четырежды промывали 300 мкл буфера для промывания (фосфатно-солевой буфер, содержащий 0,05% Tween-20). Остатки буфера для промывания тщательно удаляли, в лунки добавляли 300 мкл буфера для блокирования (1% BSA в фосфатно-солевом буфере) и планшет инкубировали при комнатной температуре в течение 1 часа. Раствор для блокирования удаляли и лунки четырежды промывали 300 мкл буфера для промывания.
Белковые стандарты (в концентрации от 0 до 4 нг/мл) и образцы в объеме 100 мкл добавляли в лунки планшета по 3 образца в параллель и инкубировали при комнатной температуре в течение 2 часов.
Далее образцы удаляли, а лунки четырежды промывали 300 мкл буфера для промывания. Антитела для детекции разводили в буфере для разведения (фосфатно-солевой буфер, содержащий 0,05% Tween-20 и 0,1% BSA) до конечной концентрации 0,25 мкг/мл, 100 мкл добавляли в лунку и инкубировали при комнатной температуре в течение 2 часов.
Раствор антител удаляли, а лунки четырежды промывали 300 мкл буфера для промывания. 6 мкл авидинпероксидазы разводили в буфере для разведения из расчета 1:2000, в каждую лунку добавляли 100 мкл данного раствора и инкубировали при комнатной температуре в течение 30 минут.
Раствор авидинпероксидазы удаляли, а лунки четырежды промывали 300 мкл буфера для промывания. В лунки добавляли 100 мкл жидкого субстрата ABTS и инкубировали при комнатной температуре до развития окраски.
Измерение оптической плотности образцов в лунках проводили при длине волны 405 нм и коррекции длины волны, равной 650 нм, с использованием микропланшетного ридера Multiscan EX (Labsystems).
Основные характеристики клеток (по данным паспорта штамма)
1. Родовое и видовое название штамма-хозяина (реципиента): Escherichia coli
2. Номер или наименование штамма: BL21 (pVEGF-A165)
3. Способ получения штамма: получен трансформацией.
4. Продукт, синтезируемый штаммом: GST-VEGF-A165.
5. Активность (продуктивность) штамма: 2 мкг очищенного рекомбинантного белка на 1 мл ростовой среды по данным ИФА (ELISA).
6. Способ, условия и состав сред для длительного хранения штамма: среда LB+15% глицерина в ампулах при -80°С.
7. Способ, условия и состав сред для размножения штамма: среда LB, содержащая 50 мкг/мл ампициллина).
8. Группа уровня риска штамма: I
9. Генетические особенности штамма:
а. Генотип штамма хозяина (мутации, делеции, инверсии, наличие плазмид или профагов, устойчивость (чувствительность) к антибиотикам, фагам и т.д., прочие генетические особенности):
[F-, ompT, hsdSB (r-B, m-B), dcm, gal].
б. Описание рекомбинантной плазмиды:
Название плазмиды: pGEX-VEGF-A165.
Размер плазмиды: 5462 пар нуклеотидов.
в. Сведения о векторе, на основе которого сконструирована плазмида:
Название вектора: pGEX-4T2.
Размер вектора: 4970 п.н.
Полная нуклеотидная последовательность вектора pGEX-4T2: SEQ ID NO: 2.
Происхождение вектора и его основных генетических элементов:
вектор содержит
- ген устойчивости к ампициллину (bla);
- ген, кодирующий фермент глутатион S-трансферазу из Schistosoma japonicum,
- ген lad репрессора из Е.coli;
- точку начала репликации из плазмиды pBR322 из Е.coli.
г. Сведения о клонированной ДНК:
Видовая принадлежность донорного организма: Homo sapiens
Размер клонированного фрагмента и включенные в его состав гены: 501 п.н.;
фрагмент кДНК VEGF (GenBank № NM_003376).
Литература








Изобретение относится к биотехнологии. В частности, к штамму Escherichia coli BL21 (pVEGF-A165) и может быть использовано для продукции фактора роста эндотелия сосудов - белка GST-VEGF-A165. Получают новый штамм клеток Escherichia coli BL21 (pVEGF-A165), трансформированный плазмидой pGEX-VEGF-A165. Данный штамм продуцирует рекомбинантный белок GST-VEGF-A165. Изобретение позволяет получить штамм Escherichia coli BL21 (pVEGF-A165), стабильно трансфицированный плазмидой, кодирующей VEGF, и секретирующих этот фактор во внеклеточное пространство при культивировании in vitro. 3 ил.
Штамм клеток Escherichia coli BL21 (pVEGF-A165), трансформированных плазмидой pGEX-VEGF-A165 (SEQ ID NO: 1) и продуцирующих рекомбинантный белок GST-VEGF-A165, депонированный во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) под номером ВКПМ В-10042.
| РЕКОМБИНАНТНЫЙ ПЛАЦЕНТАРНЫЙ ФАКТОР РОСТА И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2002 |
|
RU2295535C2 |
| KECK P.J | |||
| et al | |||
| Vascular permeability factor, an endothelial cell mitogen related to PDGF, Science, 1989, v.246, n.4935, p.1309-12 | |||
| HOUCK K.A | |||
| et al | |||
| The vascular endothelial growth factor family: identification of a fourth molecular species and characterization of alternative splicing of RNA, Mol Endocrinol., 1991, v.5, n.12, p.1806-14. | |||

