Данная заявка притязает на льготы предварительной заявки США 60/271033, поданной 23 февраля 2001 года, описание которой включено в его полном объеме путем ссылки.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ДАННОЕ ИЗОБРЕТЕНИЕ
Данное изобретение относится к области медицины и биотехнологии, в частности к области обработки и получения белков с желательной биологически активной конформацией, а также к составлению рецептур и фармацевтическим композициям.
УРОВЕНЬ ТЕХНИКИ
Высокие уровни экспрессии многих белков эукариотического происхождения были достигнуты в прокариотических экспрессионных хозяевах. Такие эукариотические белки часто неправильно укладываются и накапливаются в виде нерастворимых телец включения в прокариотическом хозяине. Для получения биологически активного белка белки, заключенные в тельцах включения, должны быть развернуты и повторно уложены при жестких условиях, включающих в себя хаотропные агенты и восстановление тиолов.
Экспрессия белков эукариотического происхождения в эукариотических хозяевах позволяет избежать этих проблем. При условии, что экспрессирующий вектор был сконструирован правильно (например, с секреторными сигнальными пептидами и т.д.), линии эукариотических клеток склонны правильно процессировать и секретировать внеклеточные эукариотические белки в виде растворимых продуктов.
Однако в то время как экспрессионные системы и векторы были улучшены для максимизации уровней экспрессии в эукариотических клетках, не весь рекомбинантный белок, экспрессируемый и секретируемый из этих хозяев, находится в желательной, наиболее активной конформации. Данное изобретение предназначено для преодоления таких проблем экспрессии и максимизации выходов биологически активного белка.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение основывается частично на открытии, что не весь препарат рекомбинантного белка, который экспрессируется эукариотическими клетками-хозяевами, уложен в нативную третичную конформацию. Кроме того, было обнаружено, что районы или домены рекомбинантных белков могут быть правильно уложены, тогда как другие районы или домены могут иметь нежелательные конформации. Таким образом, в одном аспекте данное изобретение обеспечивает способ контактирования препарата рекомбинантного белка, который содержит смесь по меньшей мере двух изомеров рекомбинантного белка с реагентом для сопряженного окисления-восстановления в течение времени, достаточного для увеличения относительной доли желательного конформационного изомера, и определения относительной доли желательного конформационного изомера в этой смеси. В другом аспекте данное изобретение описывает контактирование препарата рекомбинантного белка, который был продуцирован клетками млекопитающего с реагентом для сопряженного окисления-восстановления при рН от приблизительно 7 до приблизительно 11 и выделение фракции этого препарата рекомбинантного белка с желательной конформацией. Предпочтительными рекомбинантными белками являются гликозилированные рекомбинантные белки, такие как, например, белки, продуцируемые эукариотическими клетками. Данное изобретение относится также к способам приготовления препаратов в стерильной дозированной форме и композициям, получаемым способами данного изобретения.
КРАТКОЕ ОПИСАНИЕ ФИГУР
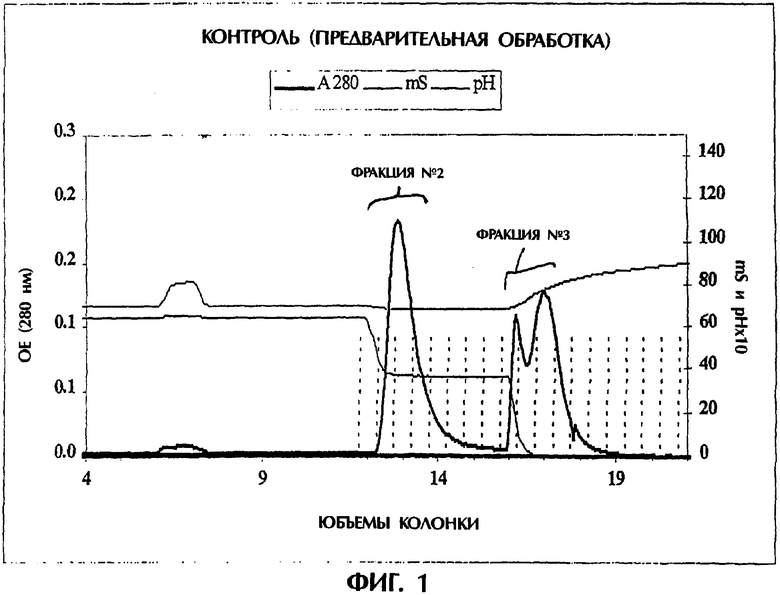
Фигура 1. Гидрофобная хроматография (HIC) TNFR:Fc. Этот препарат TNFR:Fc элюируется во время HIC в виде трех отдельных пиков, собираемых во фракцию №2 и фракцию №3, как указано.
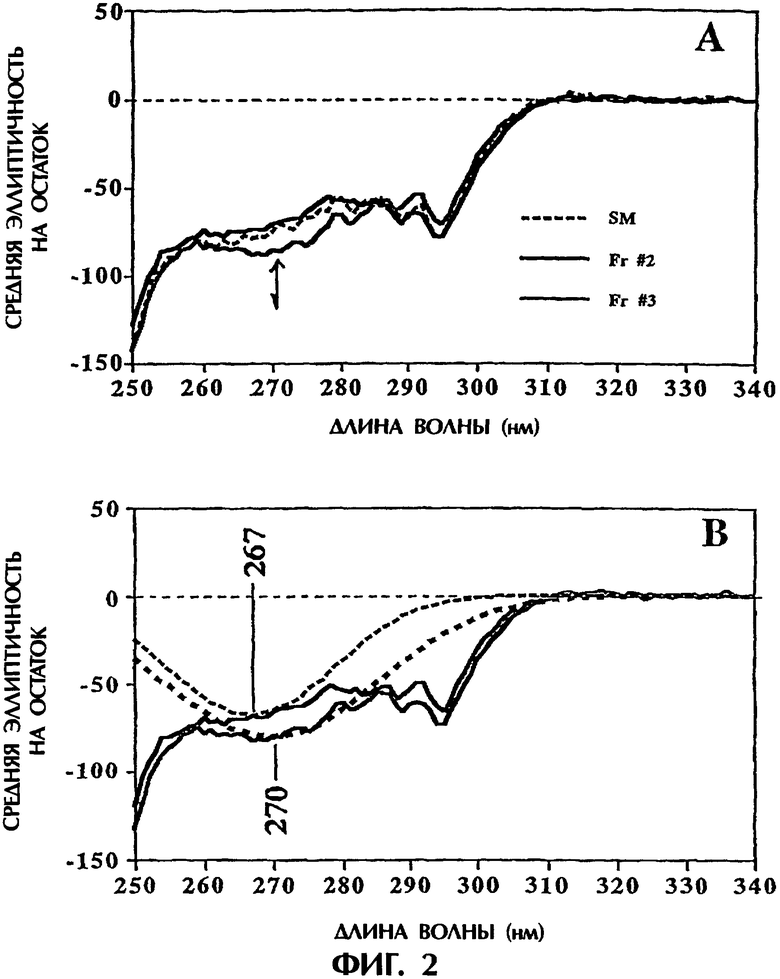
Фигура 2. Анализ кругового дихроизма фракций №2 и №3. Измерения кругового дихроизма в ближней УФ-области спектра, выраженные в виде средней эллиптичности на остаток, показаны на фигуре 2. Фигура 2А представляет спектральные данные; линия для фракции №3 является наиболее близкой к указанному стрелкой отрицательному сдвигу при приблизительно 270 нм, приписываемому вкладу дисульфидных связей, а линией для фракции №2 является более темная сплошная линия. Фигура 2В представляет данные в виде сглаженной кривой для фракции №2 (мелкий пунктир) и фракции №3 (более крупный пунктир).
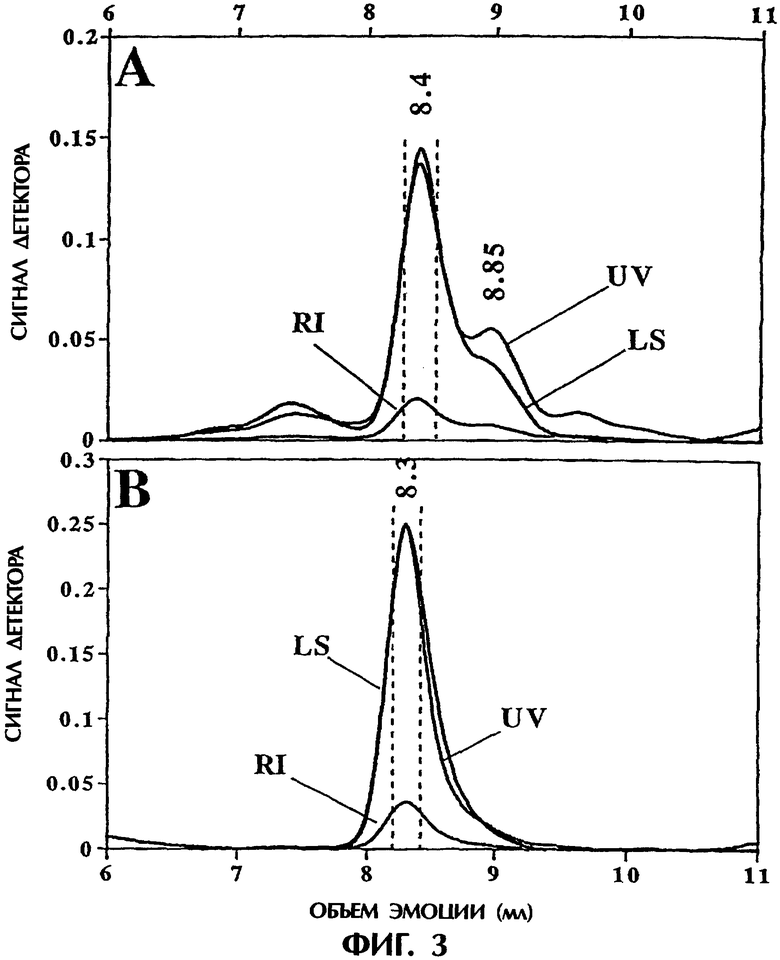
Фигура 3. Определение молекулярной массы в режиме on-line методом гель-фильтрационной хроматографии (SEC), с последовательной детекцией в ультрафиолете (UV), по рассеянию света (LS) и показателю преломления (RI) (On-line SEC/UV7LS/RI). Фигура ЗА представляет фракцию №3, а фигура 3В - фракцию №2. Вертикальные пунктирные линии указывают срезы, которые оценивались для определения молекулярной массы в районе, окружающем основной пик.
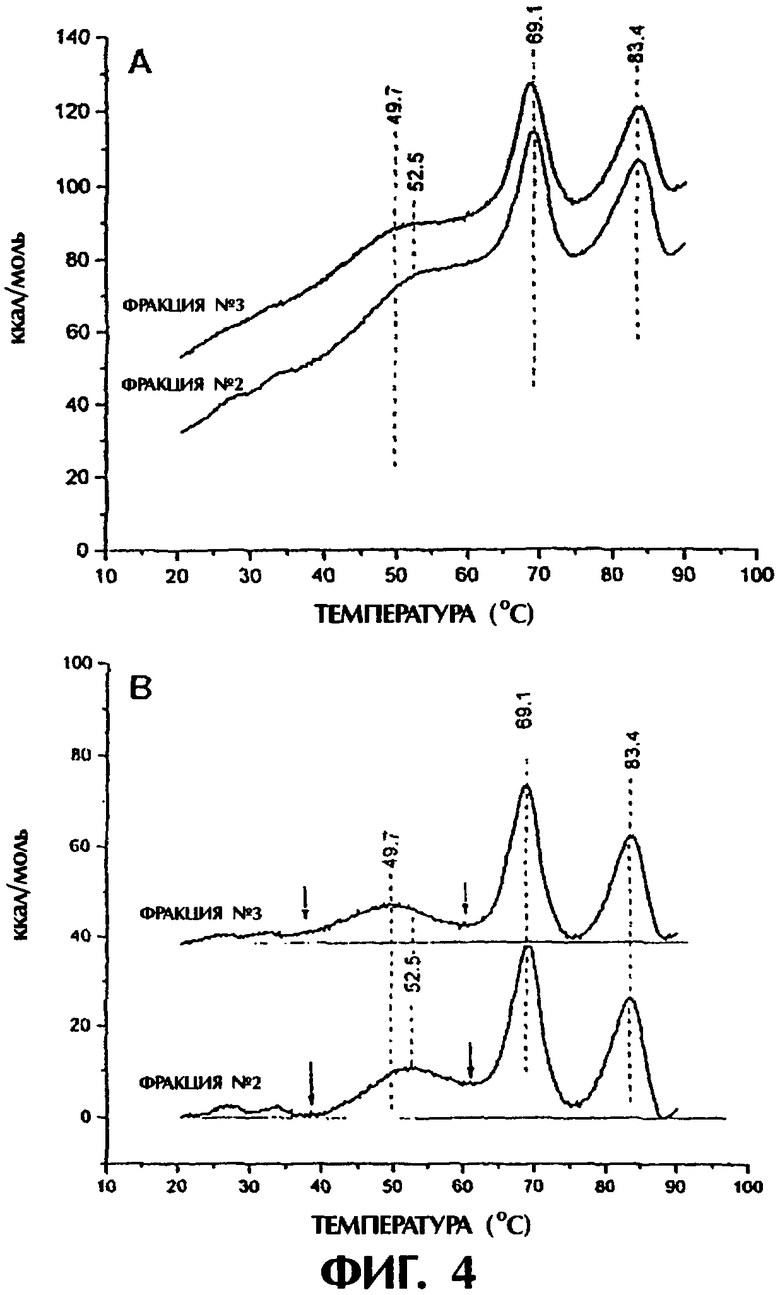
Фигура 4. Дифференциальный сканирующий калориметрический анализ фракций №2 и №3. Фигура 4А представляет нескорректированные данные, а фигура 4В представляет скорректированные в отношении фона данные. Термические переходы плавления отмечены вертикальными пунктирными линиями. Стрелки указывают сдвиг энтальпии. Горизонтальные пунктирные линии на фигуре 4В используют в качестве опорно-фонового уровня.
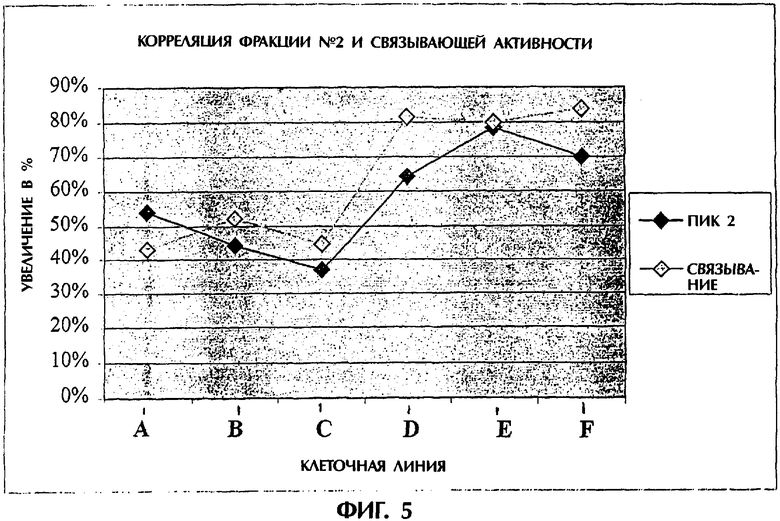
Фигура 5. Корреляция фракции №2 и связывающей активности. Шесть различных препаратов TNFR:Fc (обозначаемых A-F) из шести различных клеточных линий испытывали на корреляцию между увеличением доли фракции №2 в процентах (темные ромбы) и увеличением единиц связывания TNF-альфа в процентах (светлые ромбы).
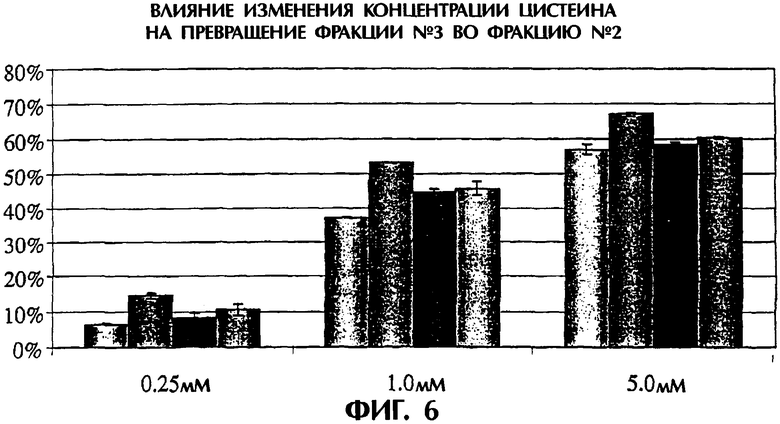
Фигура 6. Влияние изменения концентрации цистеина на превращение фракции №3 во фракцию №2. Пробы белка обрабатывали различными концентрациями цистеина (0,25-5,0 мМ) и изменения во фракции №3 оценивали с использованием HIC. Четыре разные партии TNFR:Fc обрабатывали в течение 18 часов при указанной на оси х концентрации цистеина. Процент фракции №3 в каждой партии, превратившейся во фракцию №2, показан на оси у.
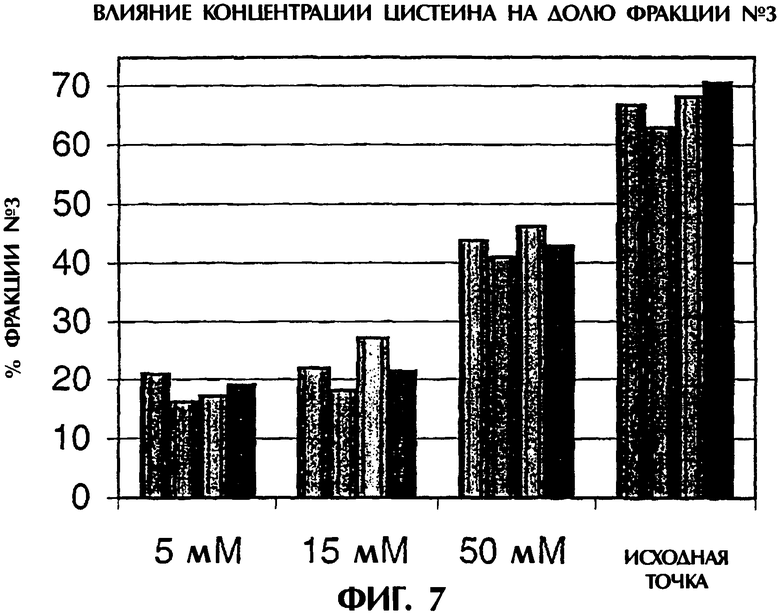
Фигура 7. Влияние концентрации цистеина на долю фракции №3. Пробы белка из четырех разных партий обрабатывали различными концентрациями цистеина (0-50 мМ) и полученный уровень фракции №3 оценивали с использованием HIC.
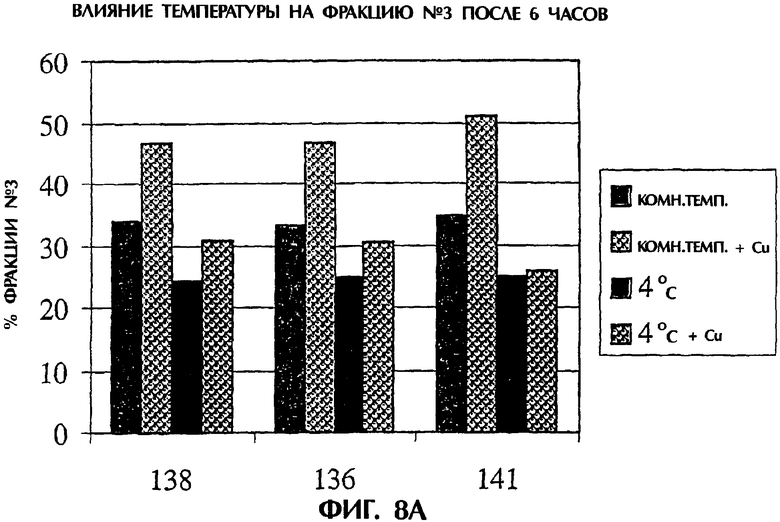
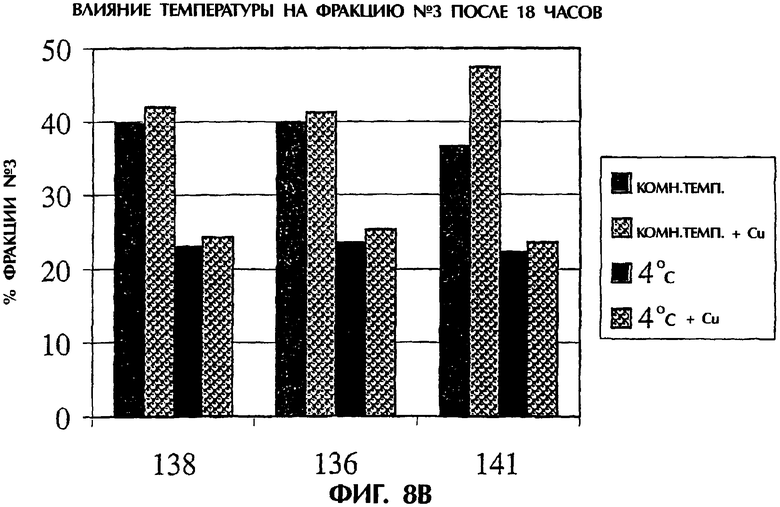
Фигура 8. Влияние температуры на дисульфидный обмен. Фракции белка обрабатывали при комнатной температуре или при 4°С в присутствии или в отсутствие меди в течение различных периодов времени. Фигура 8А представляет изменения во фракции №3 по результатам HIC после 6 часов, а фигура 8В представляет изменения во фракции №3 по результатам HIC после 18 часов.
СВЕДЕНИЯ, ПОДТВЕРЖДАЮЩИЕ ВОЗМОЖНОСТЬ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Данное изобретение обеспечивает способы увеличения выхода активных рекомбинантных белков. В частности, данное изобретение включает в себя промотирование желательной конформации белка в препаратах рекомбинантного белка. Важным моментом является то, что данное изобретение обеспечивает мягкие способы изменения структуры белка без необходимости применения жестких хаотропных обработок (таких как, например, сильные денатурирующие агенты, такие как ДСН, гуанидинийхлорид или мочевина). Использование способов данного изобретения на препаратах рекомбинантного белка приводит к более высокому проценту, или большей относительной фракции, рекомбинантного белка в препарате с желательной конформацией. Желательной конформацией для рекомбинантного белка является трехмерная структура белка, которая наиболее сходна со структурой и/или дублирует функцию природного домена этого белка. Такие мягкие способы являются особенно полезными, когда рекомбинантный белок предназначен для использования in vivo в качестве лекарственного средства или биопрепарата.
Вообще, когда рекомбинантный белок содержит домен рецепторного белка, желательная конформация будет иметь более высокую аффинность связывания (и, следовательно, более низкую константу диссоциации) в отношении родственного лиганда данного рецептора. Например, желательная конформация TNF-связывающей молекулы будет иметь более высокую аффинность связывания и меньшую константу диссоциации в отношении TNF (например, TNF-альфа).
Кроме того, желательная конформация рекомбинантного белка предпочтительно является более термостабильной, чем нежелательная конформация (при измерении в условиях того же самого раствора). Термостабильность может быть измерена любым из способов, таких как, например, переход в точке плавления (Tm). Желательная конформация рекомбинантного белка может иметь или может не иметь отличное расположение дисульфидных связей, хотя предпочтительно эта конформация содержит нативные дисульфидные связи. Желательная конформация рекомбинантного белка может иметь другие характеристики третичной структуры. Например, желательная конформация может быть мономером, димером, тримером, тетрамером или некой другой формой белка более высокого порядка. Для целей данного изобретения «конформацией» белка является его трехмерная структура. Две различные структуры полипептида с одной и той же первичной аминокислотной последовательностью являются «конформерами» друг друга, когда они имеют различные конформации, соответствующие минимумам энергии, и они отличаются друг от друга только тем, как их атомы ориентированы в пространстве. Конформеры могут быть взаимопревращающимися (что касается свободы вращения вокруг связей за исключением разрыва связей). Две различные структуры полипептида с одной и той же первичной аминокислотной последовательностью являются «конфигурационными изомерами», когда они имеют различные конформации, соответствующие минимумам энергии, они отличаются друг от друга тем, как их атомы ориентированы в пространстве, и они являются невзаимопревращающимися без разрыва ковалентной связи. В практике данного изобретения конфигурационные изомеры могут быть взаимопревращаемыми, например, посредством разрыва и при необходимости повторного образования дисульфидных связей.
Таким образом, данное изобретение предусматривает контактирование гликозилированного рекомбинантного белка, который содержит смесь по меньшей мере двух конфигурационных изомеров этого рекомбинантного белка, с реагентом для сопряженного окисления-восстановления в течение периода времени, достаточного для увеличения относительной доли желательного конфигурационного изомера, и определение относительной доли желательного конфигурационного изомера в данной смеси. В другом аспекте данное изобретение предусматривает контактирование препарата рекомбинантного белка, который был продуцирован клетками млекопитающего, с реагентом для сопряженного окисления-восстановления при рН от около 7 до около 11 и выделение фракции препарата этого рекомбинантного белка с желательной конформацией. Предпочтительными рекомбинантными белками являются гликозилированные рекомбинантные белки, такие как, например, белки, продуцируемые эукариотическими клетками.
Данное изобретение может быть использовано для обработки почти любого белка для промотирования желательной конформации. Под термином «белок» подразумевают обычно полипептид из по меньшей мере 10 аминокислот, более предпочтительно по меньшей мере приблизительно 25 аминокислот, еще более предпочтительно по меньшей мере приблизительно 75 аминокислот и наиболее предпочтительно по меньшей мере из приблизительно 100 аминокислот. Способы данного изобретения находят особенное применение в обработке белков, которые имеют по меньшей мере приблизительно 3 остатка цистеина, более предпочтительно по меньшей мере приблизительно 8 остатков цистеина, еще более предпочтительно по меньшей мере приблизительно 15 остатков цистеина, даже еще более предпочтительно по меньшей мере приблизительно 30 и еще более предпочтительно по меньшей мере приблизительно 50-150 остатков цистеина.
Обычно способы данного изобретения применимы для улучшения способов получения рекомбинантных белков. Рекомбинантными белками являются белки, получаемые способом генетической инженерии. Термин «генетическая инженерия» относится к любому способу рекомбинантых ДНК или РНК, используемому для создания клетки-хозяина, которая экспрессирует ген при повышенных уровнях, при пониженных уровнях и/или продуцирует мутантную форму данного гена. Другими словами, эта клетка была трансфицирована, трансформирована или трансдуцирована рекомбинантной полинуклеотидной молекулой и посредством этого изменена таким образом, чтобы заставить эту клетку изменить экспрессию желательного белка. Способ и векторы для генетически сконструированных клеток и/или клеточных линий для экспрессии представляющего интерес белка хорошо известны специалистам в данной области; например, многочисленные способы иллюстрируются в Current Protocols in Molecular Biology, Ausubel et al., eds. (Wiley & Sons, New York, 1988 и ежеквартальные обновления) и Sambrook et al., Molecular Cloning: A Laboratory Manual, (Cold Spring Laboratory Press, 1989). Способы генетической инженерии включают в себя, но не ограничиваются ими, экспрессионные векторы, нацеленную гомологичную рекомбинацию и активацию генов (см., например, Патент США № 5272071, выданный Chapel) и транс-активацию генов сконструированными факторами транскрипции (см., например, Segal et al., 1999, Proc. Natl. Acad. Sci. USA 96(6): 2758-63).
Данное изобретение находит особенное применение в улучшении продуцирования белков, которые являются гликозилированными. Конкретно белки, которые секретируются системами на основе клеток грибов (например, дрожжей, нитчатых грибов) и системами на основе клеток млекопитающих, будут гликозилированными. Предпочтительно эти белки секретируются продукционными клетками млекопитающих, адаптированными к росту в клеточной культуре. Примерами таких клеток, обычно используемых в промышленности, являются клетки СНО, VERO, BHK, HeLa, СV1 (в том числе Cos), MDCK, 293, ЗТЗ, линии клеток миеломы (в частности, мышиной), PC 12 и WI38. Особенно предпочтительными клетками-хозяевами являются клетки яичника китайского хомячка (СНО), которые широко используются для получения нескольких сложных рекомбинантных белков, например, цитокинов, факторов свертывания и антител (Brasel et al., 1996, Blood 88:2004-2012; Kaufinan et al., 1988, J Biol Chem 263:6352-6362; McKinnon et al., 1991, J Mol Endocrinol 6:231-239; Wood et al., 1990, J Immunol 145:3011-3016). Недостаточная по дигидрофолатредуктазе (DHFR) мутантная клеточная линия (Uriaub et al., 1980, Proc Natl Acad Sci USA 77:4216-4220), DXB11 и DG-44 являются предпочтительными линиями клеток-хозяев СНО, так как эффективная DHFR-селектируемая и амплифицируемая система экспрессии делает возможным экспрессию высокого уровня рекомбинантного белка в этих клетках (Kaufinan R.J., 1990 Meth En2ymol 185:527-566). Кроме того, с этими клетками легко обращаться в виде прикрепленных или суспензионных культур, и они проявляют относительно хорошую генетическую стабильность. Клетки СНО и экспрессируемые в них рекомбинантные белки были основательно охарактеризованы и одобрены для применения в клиническом производстве регулирующими ведомствами.
Было обнаружено, что данное изобретение является мягким и эффективным способом для улучшения способа получения белков, которые могут принимать множественные конформации и/или содержат более одного домена. «Домен» является непрерывным районом полипептидной цепи, который принимает определенную третичную структуру и/или имеет определенную активность, которая может быть локализована в этом районе полипептидной цепи. Например, один домен белка может иметь аффинность связывания в отношении одного лиганда и один домен белка может иметь аффинность связывания в отношении другого лиганда. В смысле термостабильности доменом может называться единица кооперативного разворачивания белка. Такие белки, которые содержат более чем один домен, могут встречаться в природе в виде одного белка или могут быть генетически сконструированы в виде слитого белка. Кроме того, домены полипептида могут иметь суб домены.
В одном аспекте способы данного изобретения могут быть использованы на препаратах рекомбинантных белков, в которых по меньшей мере один домен белка имеет стабильную конформацию и по меньшей мере один домен белка имеет нестабильную конформацию. Термины «стабильная» и «нестабильная» используются в виде сравнительных терминов. Домен белка со стабильной конформацией будет иметь, например, более высокую температуру плавления (Tm), чем нестабильный домен этого белка при измерении в одном и том же растворе. Домен является стабильным в сравнении с другим доменом, когда различие в Tm составляет по меньшей мере приблизительно 2°С, более предпочтительно приблизительно 4°С, еще более предпочтительно приблизительно 7°С, даже еще более предпочтительно приблизительно 10°С, еще более предпочтительно приблизительно 15°С, еще более предпочтительно приблизительно 20°С, даже еще более предпочтительно приблизительно 25°С и наиболее предпочтительно приблизительно 30°С при измерении в одном и том же растворе.
Данное изобретение также обычно применимо к белкам, которые имеют Fc-домен и другой домен (например, антителам и Fc-слитым белкам). Например, в одном из неограничительных воплощений, иллюстрированных ниже, относящихся к TNFR:Fc, Tm для Fc-части этой молекулы равны 69,1 и 83,4°С, тогда как Tm для TNFR-части этой молекулы находится в диапазоне от 52,5°С (в более желательной конформации) до Tm 49,7°С (в менее желательной конформации).
Особенно предпочтительными белками являются лекарственные средства на основе белков, также известные как биопрепараты. Предпочтительно эти белки экспрессируются в виде внеклеточных продуктов. Белки, которые могут быть получены с использованием способов данного изобретения, включают в себя, но не ограничиваются ими, flt3-лиганд (описанный в WO 94/28391, который включен здесь в его полном виде путем отсылки), СD40-лиганд (описанный в US6087329, который включен здесь в его полном виде путем отсылки), эритропоэтин, тромбопоэтин, кальцитонин, Fas-лиганд, лиганд для рецептора активатора NF-каппа В (RANKL), родственный фактору некроза опухолей (TNF) индуцирующий апоптоз лиганд (TRAIL, описанный в WO 97/01633, включенном здесь во всей полноте путем отсылки), полученный из стромы тимуса лимфопоэтин, гранулоцитарный колониестимулирующий фактор, гранулоцитарно-макрофагальный колониестимулирующий фактор (GM-CSF, описанный в Австралийском патенте № 588819, который включен здесь в полном виде путем отсылки), фактор роста тучных клеток, фактор роста стволовых клеток, эпидермальный фактор роста, RANTES (цитокин подсемейства С-С, продуцируемый Т-клетками), гормон роста, инсулин, инсулинотропин, инсулин-подобные факторы роста, паращитовидный гормон, интерфероны, факторы роста нервов, глюкагон, интерлейкины 1-18, колониестимулирующие факторы, лимфотоксин-р, фактор некроза опухолей (TNF), ингибиторный фактор лейкоза, онкостатин-М и различные лиганды для молекул поверхности клеток ELK и Hek (такие как лиганды для eph-родственных киназ (протеинтирозинкиназ) или LERK.S). Описания белков, которые могут быть очищены в соответствии со способами данного изобретения, могут быть найдены, например, в Human Cytokines: Handbook for Basic and Clinical Research, Vol. II (Aggarwal and Gutterman, eds. Blackwell Sciences, Cambridge, MA, 1998); Growth Factors: A Practical Approach (McKay and Leigh, eds., Oxford University Press Inc., New York, 1993); и The Cytokines Handbook (A.W. Thompson, ed.. Academic Press, San Diego, CA, 1991).
Препараты рецепторов, в частности растворимых форм этих рецепторов, для любого из вышеупомянутых белков могут быть также улучшены с использованием способов данного изобретения, в том числе для обеих форм TNFR (обозначаемых р55 и р75), рецепторов типов I и II интерлейкина-1 (как описано в ЕР 0460846, US 4968607 и US 5767064, которые включены здесь в их полном виде путем отсылки), рецептора интерлейкина-2, рецептора интерлейкина-4 (как описано в ЕР 0367566 и US 5856296, которые включены здесь в их полном виде путем отсылки), рецептора интерлейкина-15, рецептора интерлейкина-17, рецептора интерлейкина-18, рецептора гранулоцитарно-макрофагального колониестимулирующего фактора, рецептора гранулоцитарного колониестимулирующего фактора, рецепторов для онкостатина-М и ингибиторного фактора лейкоза, рецептора активатора NF-каппа В (RANK, описанного в US 6271349, включенном здесь в его полном виде путем отсылки), рецепторов для TRAIL (в том числе рецепторов 1, 2, 3 и 4 TRAIL) и рецепторов, которые содержат домены смерти, таких как Fas-рецептор или апопотоз-индуцирующий рецептор (AIR).
Другие белки, способ получения которых может быть улучшен с использованием способов данного изобретения, включают в себя кластер антигенов дифференцировки (обозначаемых как CD-белки), например, описанных в Leukocyte Typing VI (Proceedings of VIth International Workshop and Conference; Kixhimoto, Kikutani et al., eds.; Kobe, Japan, 1996), или CD-молекулы, раскрытые на последующих симпозиумах. Примеры таких молекул включают в себя CD27, CD30, CD39, CD40 и лиганды к ним (CD27-лиганд, CDЗ0-лиганд и CD40-лиганд). Некоторые из них являются членами семейства рецепторов TNF, которое включает в себя также 41ВВ и 0Х40; эти лиганды являются часто членами семейства TNF (например, лиганд 41ВВ и лиганд 0Х40); соответственно, члены семейств TNF и TNFR могут быть также получены с использованием данного изобретения.
Белки, которые являются ферментативно активными, могут быть также получены в соответствии с данным изобретением. Примеры включают в себя члены семейства металлопротеиназ-дезинтегринов (разрушающих интегрины), различные киназы, глюкоцереброзидазу, супероксиддисмутазу, тканевый активатор плазминогена, фактор VIII, фактор IX, аполипопротеин Е, аполипопротеин A-I, антагонист IL-2, альфа-1-антитрипсин, TNF-α-превращающий фермент и многочисленные другие ферменты. Лиганды для ферментативно активных белков могут также экспрессироваться с использованием данного изобретения.
Композиции и способы данного изобретения применимы также для получения других типов рекомбинантных белков, в том числе молекул иммуноглобулинов или их частей и химерных антител (например, антител, имеющих константную область человека, сопряженную с антигенсвязывающим участком мыши) или их фрагментов. Известны многочисленные способы, при помощи которых можно манипулировать с ДНК, кодирующей молекулы иммуноглобулинов, для получения ДНК, способных кодировать рекомбинантные белки, такие как одноцепочечные антитела, антитела с повышенной аффинностью или другие полипептиды на основе антител (см., например, Larrick et al., 1989, Biotechnology 7:934-938; Reichmann et al., 1988, Nature 332:323-327; Roberts et al., 1987, Nature 328:731-734; Verhoeyen et al., 1988, Science 239:1534-1536; Chaudhary et al., 1989, Nature 339:394-397). Препараты полностью человеческих антител (такие как препараты, получаемые с использованием трансгенных животных, и при необходимости затем модифицированные in vitro), а также гуманизированные антитела могут быть также использованы в данном изобретении. Термин гуманизированное антитело включает в себя также одноцепочечные антитела См., например, Cabilly et al., U.S. Pat. No. 4816567; Cabilly et al., European Patent No. 0125023 Bl; Boss et al., U.S. Pat. No. 4816397; Boss et al., European Patent No. 0120694 Bl; Neuberger, M.S. et al., WO 86/1533; Neuberger, M.S. et al., European Patent No. 0194276 B1; Winter, U.S. Patent No. 5225539; Winter, European Patent No. 0239400 Bl; Queen et al., European Patent No. 0451216 Bl и Padlan, E.A. et al., EP 0519596 Al. Способ данного изобретения может быть также использован во время получения конъюгатов, содержащих антитело и цитотоксическое или люминесцентное вещество. Такие вещества включают в себя: производные майтанзина (такие как DM1); энтеротоксины (такие как стафилококковый энтеротоксин); изотопы иода (такие как иод-125); изотопы технеция (такие как Тс-99т); цианиновые флуорохромы (такие как Су5.5.18); и инактивирующие рибосомы белки (такие как буганин, гелонин или сапорин-86).
Примеры антител или конъюгатов антитело/цитотоксин или антитело/люминофор, предусматриваемые данным изобретением, включают в себя такие антитела и конъюгаты, которые узнают любой вышеупомянутый белок или комбинацию вышеупомянутых белков и/или следующие антигены: CD2, CD3, CD4, CD8, CDlla, CD14, CD18, CD20, CD22, CD23, CD25, CD33, CD40, CD44, CD52, CD80 (В7.1), CD86 (В7.2), CD147, IL-ld. IL-lβ, IL-4, IL-5, IL-8, IL-10, рецептор IL-2, рецептор IL-4, рецептор IL-6, рецептор IL-13, субъединицы рецептора IL-18, PDGF-p, VEGF, TGF, TGF-p2, TGF-pl, рецептор EGF, рецептор VEGF, С 5-комплемент, IgE, опухолевый антиген СА125, опухолевый антиген MUC1, антиген РЕМ, LCG (который является продуктом гена, который экспрессируется при раке легкого), HER-2, ассоциированный с опухолью гликопротеин TAG-72, антиген SK-1, ассоциированные с опухолью эпитопы, которые присутствуют в повышенных количествах в сыворотках пациентов с раком толстой кишки и/или поджелудочной железы, ассоциированные с раком эпитопы или белки, экспрессируемые на раковых клетках молочной железы, толстой кишки, на плоских клетках, раковых клетках предстательной железы, поджелудочной железы, легкого и/или почки и/или на клетках меланомы, глиомы или нейробластомы, некротическом ядре опухоли, интегрин альфа 4 бета 7, интегрин VLA-4, интегрины В2, рецепторы TRAIL 1, 2, 3 и 4, RANK, лиганд RANK, TNF-α, молекулу адгезии VAP-1, молекулу адгезии эпителиальных клеток (ЕрСАМ), молекулу (фактор) межклеточной адгезии 3 (ICAM-3), адгезин лейкоинтегрин, гликопротеин тромбоцитов gp IIb/IIIa, тяжелую цепь миозина сердца, паращитовидный гормон, rNAPc2 (который является ингибитором тканевого фактора Vila), MHC I, раково-эмбриональный антиген (СЕА), альфа-фетопротеин (AFP), фактор некроза опухолей (TNF), CTLA-4 (который является ассоциированным с цитотоксическими Т-лимфоцитами антигеном), рецептор Fc-γ-1, HLA-DR 10 бета, антиген HLA-DR, L-селектин, IFN-y, респираторно-синцитиальный вирус, вирус иммунодефицита человека (ВИЧ), вирус гепатита В (HBV), Streptococcus mutans и Staphylococcus aureus.
С использованием способов данного изобретения могут быть получены препараты различных слитых белков. Примеры таких слитых белков включают в себя белки, экспрессируемые в виде слитых с частью молекулы иммуноглобулина белков, белки, экспрессируемые в виде слитых компонентом типа «застежки-молнии» (zipper) белков, и новые полифункциональные белки, такие как слитые белки цитокина и фактора роста (т.е. GM-CSF и IL-3, MGF и IL-3). В WO 93/08207 и WO 96/40918 описано получение различных растворимых олигомерных форм молекулы, называемой CD40L, включающей в себя слитый с иммуноглобулином белок и слитый с «застежкой-молнией» белок соответственно; обсуждаемые здесь способы применимы и к другим белкам. Любая из вышеупомянутых молекул может быть экспрессирована в виде слитого белка, включающего в себя, но не ограничивающегося ими, внеклеточный домен клеточной рецепторной молекулы, фермент, гормон, цитокин, часть молекулы иммуноглобулина, домен «застежки-молнии» и эпитоп.
Препаратом рекомбинантного белка может быть супернатант клеточной культуры, экстракт клеток, но предпочтительно частично очищенная фракция из них. Под «частично очищенной» имеют в виду, что были проведены некоторая процедура или некоторые процедуры фракционирования, но присутствуют дополнительные виды полипептидов (по меньшей мере 10%), иные, чем желательный белок или желательная конформация белка. Одним из преимуществ способов данного изобретения является то, что препарат рекомбинантного белка может быть с довольно высокой концентрацией. Предпочтительные диапазоны концентраций составляют 0,1-20 мг/мл, более предпочтительно от 0,5 до 15 мг/мл и еще более предпочтительно от 1 до 10 мг/мл.
Препарат рекомбинантного белка может быть первоначально получен культивированием рекомбинантных клеток-хозяев в условиях культивирования, подходящих для экспрессии данного полипептида. Полипептид может также экспрессироваться в виде продукта трансгенных животных, например в виде компонента молока трансгенных коров, коз, свиней или овец, которые характеризуются соматическими или зародышевыми клетками, содержащими нуклеотидную последовательность, кодирующую данный полипептид. Затем полученный экспрессированный полипептид может быть очищен или частично очищен из такой культуры или ее компонента (например, из культуральной среды или экстрактов клеток или жидкой среды организма) с использованием известных способов. Процедуры фракционирования могут включать в себя, но не ограничиваются ими, одну или несколько стадий фильтрования, центрифугирования, осаждения, разделения фаз, аффинной очистки, гель-фильтрационной хроматографии, ионообменной хроматографии, гидрофобной хроматографии (HIC; с использованием таких смол, как простой фениловый эфир, бутиловый эфир или пропиловый эфир), ВЭЖХ или какой-либо комбинации вышеперечисленного.
Например, очистка полипептида может включать в себя аффинную колонку, содержащую агенты, которые будут связываться с этим полипептидом; одну или несколько колоночных стадий с использованием таких аффинных смол, как конканавалин А-агароза, гепарин-toyopearl® или Cibacron blue ЗОА-Sepharose®; одну или несколько стадий, включающих в себя элюцию; и/или иммуноаффинную хроматографию.
Полипептид может экспрессироваться в форме, которая облегчает очистку. Например, он может экспрессироваться в виде слитого полипептида, такого как слитый полипептид с мальтозусвязывающим полипептидом, глутатион-8-трансферазой (GST) или тиоредоксином (TRX). Наборы для экспрессии и очистки таких слитых полипептидов являются коммерчески доступными от New England BioLab (Beverly, Mass.), Pharmacia (Piscataway, N.J.) и InVitrogen соответственно. Полипептид может быть помечен эпитопом и затем очищен с использованием специфического антитела, направленного на такой эпитоп.Один такой эпитоп (FLAG®) является коммерчески доступным от Kodak (New Haven, Conn.). Можно также использовать аффинную колонку, содержащую полипептидсвязывающий полипептид, например моноклональное антитело к данному рекомбинантному белку, для аффинной очистки экспрессированных полипептидов. Другими типами аффинной очистки может быть белок А- или белок G-колонка, аффинные агенты которой связываются с белками, содержащими Fc-домены. Полипептиды могут быть удалены с аффинной колонки с использованием общепринятых способов, например, в высокосолевом буфере для элюции, и затем диализованы против низкосолевого буфера для применения или изменением рН или других компонентов в зависимости от используемого аффинного матрикса, или могут быть конкурентно удалены с использованием природного субстрата аффинного компонента. В одном воплощении данного изобретения, иллюстрированном ниже, препарат рекомбинантного белка частично очищали посредством белок А-аффинной колонки.
Некоторые или все из предыдущих стадий очистки в различных комбинациях могут быть использованы для получения подходящего препарата рекомбинантного белка для использования в способах данного изобретения и/или для дополнительной очистки рекомбинантного полипептида после контактирования препарата рекомбинантного белка с реагентом для сопряженного окисления-восстановления. Полипептид, который является по существу свободным от других полипептидов млекопитающих, определяется как «изолированный полипептид».
Полипептид может быть также получен известным общепринятым химическим синтезом. Способы конструирования полипептидов посредством синтетических способов известны специалистам в данной области. Синтетически сконструированные полипептидные последовательности могут быть гликозилированы in vitro.
Желательная степень конечной очистки зависит от предполагаемого применения данного полипептида. Например, относительно высокая степень чистоты желательна, когда полипептид должен вводиться in vivo. В подобном случае полипептид очищают таким образом, чтобы никакие полипептидные полосы, соответствующие другим полипептидам, не детектировались при анализе электрофорезом в ДСН-полиакриламидном геле (ДСН-ПААГ). Специалисту в подходящей области будет понятно, что множественные полосы, соответствующие данному полипептиду, могут быть визуализированы при электрофорезе в ДСН-ПААГ вследствие дифференциального гликозилирования, дифференциального посттрансляционного процессинга и т.п. Более предпочтительно полипептид данного изобретения очищают до существенной гомогенности, как следует из единственной полипептидной полосы при анализе электрофорезом в ДСН-ПААГ. Эта полипептидная полоса может быть визуализирована окрашиванием серебром, Кумасси голубым и/или (если полипептид является радиоактивно меченым) авторадиграфией.
Под "контактированием" имеют в виду подвергание действию, и/или экспонирование действию в растворе. Белок или полипептид также может быть подвергнут контактированию, когда он связан с твердым носителем (например, аффинной колонкой или хроматографическим матриксом). Предпочтительно этот раствор является забуференным. Для максимизации выхода белка с желательной конформацией рН раствора выбирают защищающим стабильность данного белка и оптимальным для дисульфидного обмена. В практике данного изобретения рН раствора предпочтительно не является сильно кислым. Так, предпочтительные диапазоны рН являются большими, чем рН 5, предпочтительно от около рН 6 до около рН 11, более предпочтительно от приблизительно рН 7 до приблизительно рН 10 и еще более предпочтительно от приблизительно рН 7,6 до приблизительно рН 9,6. В одном неограничительном воплощении данного изобретения с использованием TNFR:Fc, который иллюстрируется ниже, было обнаружено, что оптимальный рН должен быть около рН 8,6. Однако оптимальный рН для конкретного воплощения данного изобретения может быть легко определен экспериментально специалистами в данной области.
Реагент для сопряженного окисления-восстановления является источником восстанавливающих агентов. Предпочтительными восстанавливающими агентами являются свободные тиолы (SH). Реагент для сопряженного окисления-восстановления предпочтительно включает соединение из группы, состоящей из восстановленного и окисленного глутатиона, дитиотреитола (ДТТ), 2-меркаптоэтанола, дитионитробензоата, цистеина и цистина. Для облегчения применения и экономии может быть использован восстановленный глутатион и/или восстановленный цистеин.
Реагент для сопряженного окисления-восстановления присутствует в концентрации, достаточной для увеличения относительной доли желательной конформации. Оптимальная концентрация реагента для сопряженного окисления-восстановления зависит от концентрации белка и числа дисульфидных связей в данном белке. Например, было обнаружено с использованием белка (TNFR:Fc) с 29 дисульфидными связями в концентрации 2 мг/мл (приблизительно 14 мкМ белок или 400 мкМ дисульфид), что реагент для сопряженного окисления-восстановления с 2 мМ восстановленным тиолом работал хорошо, увеличивая относительную долю желательной конформации. Это соответствует отношению приблизительно 35 свободных тиолов на 1 дисульфидную связь. Однако было также обнаружено, что отношения от 20 до 400 свободных тиолов на дисульфид также хорошо работали. Конечно, количество тиола, используемого для конкретной концентрации, может несколько варьировать в зависимости от восстанавливающей способности данного тиола и может быть легко определено специалистом в данной области.
Таким образом, обычно концентрация свободных тиолов из реагента для сопряженного окисления-восстановления может быть от приблизительно 0,05 до приблизительно 50 мМ, более предпочтительно приблизительно 0,1 - приблизительно 25 мМ и еще более предпочтительно приблизительно 0,2 - приблизительно 20 мМ.
Кроме того, реагент для сопряженного окисления-восстановления может содержать окисленные тиолы при более высоких, равных или более низких концентрациях в сравнении с восстановленным тиоловым компонентом. Например, реагент для сопряженного окисления-восстановления может быть комбинацией восстановленного глутатиона и окисленного глутатиона. Посредством реальных рабочих примеров было обнаружено, что отношение восстановленного глутатиона к окисленному глутатиону от приблизительно 1:1 до приблизительно 100:1 (восстановленные тиолы:окисленные тиолы) может работать одинаково хорошо. Альтернативно в другом воплощении реагент для сопряженного окисления-восстановления может быть цистеином или комбинацией цистеина и цистина. Таким образом, при включении окисленных тиолов в исходный реагент для сопряженного окисления-восстановления отношение восстановленных тиолов к окисленным тиолам может быть в предпочтительном варианте от приблизительно 1:10 до приблизительно 1000:1, более предпочтительно приблизительно 1:1 - приблизительно 500:1, еще более предпочтительно приблизительно 5:1-100:1, еще более предпочтительно приблизительно 10:1.
Контактирование препарата рекомбинантного белка с реагентом для сопряженного окисления-восстановления выполняют в течение времени, достаточного для увеличения относительной доли желательной конформации. Любое относительное увеличение в этой доле является желательным, но предпочтительно по меньшей мере 10% белка с нежелательной конформацией превращается в белок с желательной конформацией. Более предпочтительно по меньшей мере 20, 30, 40, 50, 60, 70 и даже 80% этого белка превращают из нежелательной в желательную конформацию. Типичные выходы, которые достигались способами данного изобретения, были в диапазоне от 40 до 80%. Если стадию контактирования выполняют на частично очищенном или высокоочищенном препарате рекомбинантного белка, стадия контактирования может быть выполнена в течение такого короткого периода времени, как приблизительно 1 час - приблизительно 4 часа, и в течение такого длительного периода времени, как приблизительно 6 часов - приблизительно 4 дня. Было обнаружено, что стадия контактирования приблизительно 4 часа - приблизительно 16 часов или приблизительно 18 часов работает хорошо. Стадия контактирования может также иметь место во время другой стадии, например, на твердой фазе или во время фильтрования или во время любой другой стадии очистки.
Способы данного изобретения могут выполняться в широком температурном диапазоне. Например, способы данного изобретения успешно осуществлялись при температурах от приблизительно 4 до приблизительно 37°С, однако наилучшие результаты были получены при более низких температурах. Типичной температурой для контактирования частично или полностью очищенного препарата рекомбинантного белка является температура приблизительно 4 - приблизительно 25°С (температура окружающей среды), но контактирование может также выполняться при более низких температурах и при более высоких температурах.
Препарат рекомбинантного белка может контактировать с реагентом для сопряженного окисления-восстановления в различных объемах в зависимости от целесообразности. Например, способы данного изобретения проводили успешно в аналитическом лабораторном масштабе (1-50 мл), препаративном масштабе (50 мл-10 л) и промышленном масштабе (10 или более литров). Таким образом, способы данного изобретения могут проводиться как в малом масштабе, так и в крупном масштабе с воспроизводимыми результатами.
В предпочтительных аспектах стадию контактирования выполняют в отсутствие значительных количеств хаотропных агентов, таких как, например, ДСН, мочевина и гуанидин-HCl. Значительные количества хаотропных агентов требуются для наблюдения ощутимого разворачивания. Обычно, присутствует менее чем 1 М хаотропный агент, более предпочтительно менее чем 0,5 М, еще более предпочтительно менее чем 0,1 М хаотропный агент. Раствор является по существу свободным от хаотропа (например, ДСН, мочевины и гуанидин-HCl), когда хаотроп не добавляют специально к этому раствору, и могут присутствовать лишь следовые количества (например, менее чем 10 мМ) (например, из сосуда или в качестве клеточного побочного продукта).
Дисульфидный обмен может быть остановлен любым из способов, известных специалистам в данной области. Например, реагент для сопряженного окисления-восстановления может быть удален, или его концентрация может быть уменьшена посредством стадии очистки, и/или он может быть химически инактивирован, например, подкислением раствора. Обычно, когда реакцию останавливают подкислением, рН раствора, содержащего реагент для сопряженного окисления-восстановления, будет доводиться до показателя ниже рН 7. Предпочтительно рН доводят до показателя менее рН 6. Обычно, рН снижают до показателя между приблизительно рН 2 и приблизительно рН7.
Определение конформации белка и относительных долей конформации белка в смеси может быть выполнено с использованием любого из различных аналитических и/или качественных способов. Если имеется различие в активности между этими конформациями белка, определение относительной доли конформации в смеси может быть выполнено при помощи анализа активности (например, связывания с лигандом, ферментативной активности, биологической активности и т.д.). Например, в одном из неограничительных воплощений, описанных ниже, по меньшей мере две различные конформации TNFR:Fc могут быть разделены с использованием твердофазного анализа связывания TNF. Этот анализ, в основном такой, как описанный для IL-1R (Slack, et al., 1993, J. Biol. Chem. 268:2513-2524), может различать между относительными долями различных конформации белка посредством изменений получаемых констант ассоциации, диссоциации или ингибирования связывания лиганд-рецептор. Альтернативно результаты связывания могут быть выражены в виде единиц активности на мг белка.
Если эти две конформации разделяются различным образом в ходе хроматографии, электрофореза, фильтрования или других способов очистки, то относительная доля конформации в данной смеси может быть определена с использованием таких способов очистки. Например, в неограничительных воплощениях, описанных ниже, по меньшей мере две различные конформации TNFR:Fc могли быть разделены с использованием гидрофобной хроматографии. Далее, поскольку для расчета состава вторичной структуры белков использовали круговой дихроизм в дальней УФ-области спектра (Perczel et al., 1991, Protein Engrg. 4:669-679), такой способ может определить, присутствуют ли альтернативные конформации белка. Другим способом, используемым для определения конформации, является флуоресцентная спектроскопия, которая может быть использована для определения дополнительных различий в третичной структуре, могущих быть связанными с флуоресценцией триптофана и тирозина. Другими способами, которые могут быть использованы для определения различий в конформации и, следовательно, относительных долей конформации, являются SEC в режиме on-line для измерения агрегационного состояния, дифференциальная сканирующая калориметрия для измерения переходов плавления (Тm) и энтальпий компонентов и хаотропное разворачивание.
Под термином "выделение" имеют в виду физическое отделение по меньшей мере одного компонента смеси от других компонентов смеси. Выделение компонентов или конкретных конформации белка может быть достигнуто с использованием любого способа очистки, который имеет тенденцию разделять подобные компоненты. Таким образом, можно проводить одну или несколько стадий хроматографии, в том числе, но не только, HIC, гидроксиапатитную хроматографию, ионообменную хроматографию, аффинную хроматографию и SEK. Другими способами очистки являются фильтрование (например, фильтрование с тангенциальным потоком), электрофоретические способы (например, электрофорез, электроэлюция, изоэлектрическое фокусирование) и разделение фаз (например, разделение фаз с использованием ПЭГ-декстрана), если называть некоторые из них. Кроме того, фракция препарата рекомбинантного белка, которая содержит белок в нежелательной конформации, может быть обработана снова способами данного изобретения для дальнейшей оптимизации выходов белка с желательной конформацией.
Например, после обработки могут быть приготовлены образцы белка для гидрофобной хроматографии (HIC) следующим образом. Равный объем 850 мМ цитрата натрия, 50 мМ фосфата натрия, рН 6,5, добавляют к обработанной пробе и дают уравновешиваться при комнатной температуре. После фильтрования (например, с использованием фильтра 0,22 мкМ) проводят хроматографию HIC на смоле Toyopearl® Butyl 650-М (Tosoh Biosep LLC, Montgomeryville, PA), при скорости тока 150 см/ч и нагрузке по весу 2,1 мг на мл смолы. Колонку предварительно уравновешивают 3 колоночными объемами 425 мМ цитрата натрия, 50 мМ фосфата, рН 6,5, наносят пробу и затем колонку промывают 3 колоночными объемами 425 мМ цитрата натрия, 50 мМ фосфата, рН 6,5. Элюция может выполняться с использованием градиента 425 мМ цитрат натрия, 50 мМ фосфат, рН 6,5 - 0 мМ цитрат натрия, 50 мМ фосфат, рН 6,5 в общем объеме, равном 5 объемам колонки. Во время элюции могут собираться фракции. Колонка может быть отмыта 3 колоночными объемами воды с последующим промыванием 3 колоночными объемами 0,1 М NaOH. С использованием способов данного изобретения могут быть получены таким образом препараты TNFR:Fc, которые содержат более 85, более 90 и даже более 95% TNFR:Fc, присутствующего в данном препарате, в наиболее активной конформации (фракция №2). Таким образом, композиции, в том числе фармацевтические композиции, TNFR:Fc, содержащие такие доли фракции №2, также обеспечиваются данным изобретением.
Данное изобретение также при необходимости включает в себя составление рецептуры с этими белками. Под термином «составление рецептуры» имеют в виду, что эти белки могут быть подвергнуты смене буфера, стерилизации, упакованы в валовой форме и/или упакованы для конечного потребителя. Для целей данного изобретения термин «стерильная валовая форма» означает, что эта форма является не содержащей или по существу не содержащей микробного загрязнения (в такой степени, которая приемлема для пищевых и/или лекарственных целей) и имеет определенный состав и концентрацию. Термин «стерильная дозированная форма» означает форму, которая является пригодной для введения потребителю и/или пациенту или потребления потребителем и/или пациентом. Такие композиции могут содержать эффективное количество белка в комбинации с другими компонентами, такими как физиологически приемлемые разбавитель, носитель и/или наполнитель. Термин «физиологически приемлемый» означает нетоксичный материал, который не препятствует эффективности биологической активности активного ингредиента (активных ингредиентов). Рецептуры, подходящие для введения, включают в себя водные и неводные стерильные инъекционные растворы, которые могут содержать антиоксиданты, буферы, бактериостатические агенты и растворенные вещества, которые делают эту рецептуру изотонической с кровью реципиента; и водные и неводные стерильные суспензии, которые могут содержать суспендирующие агенты или загущающие агенты. Кроме того, стерильные валовые формы и стерильные дозированные формы могут содержать небольшую концентрацию (приблизительно 1 до приблизительно 10 мМ) реагента для сопряженного окисления-восстановления (например, глутатиона, цистеина и т.д.). Эти полипептиды могут быть составлены в рецептуру в соответствии с известными способами приготовления фармацевтически применимых композиций. Они могут быть объединены в смеси, либо в виде единственного активного вещества, либо с другими известными активными веществами, пригодными для определенного медицинского показания, с фармацевтически приемлемыми разбавителями (например, солевым раствором, забуференными Трис-HCl, ацетатом или фосфатом растворами), консервантами (например, тимерозалом, бензиновым спиртом, парабенами), эмульгаторами, солюбилизаторами, адъювантами и/или носителями. Подходящие рецептуры для фармацевтических композиций включают в себя рецептуры, описанные в Remington's Pharmaceutical Sciences, 16th ed. 1980, Mack Publishing Company, Easton, PA. Кроме того, такие композиции могут быть в комплексе с полиэтиленгликолем (ПЭГ), ионами металлов, и/или могут быть включены в полимерные соединения, такие как полиуксусная кислота, полигликолевая кислота, гидрогели, декстран и т.д., или могут быть включены в липосомы, микроэмульсии, мицеллы, однослойные или многослойные везикулы, тени эритроцитов или сферобласты. Подходящие липиды для липосомной рецептуры включают в себя, без ограничения, моноглицериды, диглицериды, сульфатиды, лизолецитин, фосфолипиды, сапонин, желчные кислоты и т.п.
Приготовление таких липосомных рецептур находится в пределах уровня квалификации в данной области, как описано, например, в патентах США с номерами 4235871, 4501728, 4837028 и 4737323. Такие композиции будут влиять на физическое состояние, растворимость, стабильность, скорость высвобождения in vivo и скорость клиренса in vivo и, следовательно, могут выбираться в соответствии с предполагаемым применением, так что характеристики носителя будут зависеть от выбранного пути введения. Формы с замедленным высвобождением, пригодные для применения, включают в себя, но не ограничиваются ими, полипептиды, которые инкапсулированы в медленно растворяющемся биосовместимом полимере (таком как микрочастицы альгината, описанные в патенте США с номером 6036978), смешаны с таким полимером (в том числе наносимыми местно гидрогелями) или заключены в биосовместимом полупроницаемом имплантате.
После описания данного изобретения следующие примеры приводятся в качестве иллюстрации, но не для ограничения изобретения.
ПРИМЕР1
Биофизическая оценка фракций №2 и №3 TNFR:Fc
TNFR:Fc элюируется с колонки для гидрофобной хроматографии (HIC) в виде трех отдельных пиков, обозначенных фракция №1, фракция №2 и фракция №3 (см. фигуру 1). Фракция №2 является желательной фракцией. Фракция №3 представляла особый интерес, так как она может содержать от 20 до 60% образца, и было показано, что она проявляет низкую TNF-связывающую активность и биологическую активность A3 75 в сравнении с фракцией №2. Таким образом, в интересах понимания различий между этими двумя фракциями и для определения, какие факторы способствуют потере активности во фракции №3 в связи со структурой и конформацией, проводили биофизические исследования. В этом примере авторы изобретения анализировали фракцию №2 и фракцию №3 с использованием кругового дихроизма, флуоресценции, SEC/UV/LS/RI в режиме on-line и дифференциальной сканирующей калориметрии (DSC).
Материалы и способы:
Материалы: Исходным материалом был TNFR:Fc в TMS-буфере (10 мМ Трис, 4% маннит, 1% сахароза). Элюированные при HIC фракции этого материала получали в виде фракций №2 и №3 для описанных ниже экспериментальных исследований.
Круговой дихроизм: Исследования проводили в ближней (250-340 нм) и дальней (190-250 нм) УФ-областях спектра. Исследования в ближней УФ-области проводили для выяснения различий в третичной структуре, тогда как исследования в дальней УФ-области использовали для характеристики различий во вторичной структуре.
Измерения кругового дихроизма в ближней УФ-области проводили в TMS растворах со следующими концентрациями. Исходный материал разбавляли до 6,25 мг/мл, тогда как фракции №2 и №3 оценивали при их имеющихся концентрациях 9,4 и 5,4 мг/мл соответственно. Использовали ячейку для кругового дихроизма с длиной оптического пути 0,1 см и сканирования проводили от 340 до 250 нм.
Измерения кругового дихроизма в дальней УФ-области проводили в буфере для белка, замененном на 10 мМ фосфат натрия (рН 7,0), и затем оценивали с использованием ячейки с длиной оптического пути 0,1 см и сканирования проводили от 250 до 190 нм. Вторичную структуру оценивали с использованием анализа ограничений на выпуклость (convex constraint analysis, CCA) (Perczel et al., 1991, Protein Engrg. 4:669-679).
Флуоресцентная спектроскопия: Образцы исследовали после разбавления до приблизительно 50 микрограммов на мл с использованием двух различных длин волн возбуждения. Флуоресценцию тирозина и триптофана исследовали с использованием возбуждения при 270 нм, тогда как флуоресценцию исключительно триптофана оценивали, используя возбуждение при 295 нм (Lakowicz, J.R. in "Principles of fluorescence spectroscopy". Plenum Press, 1983. New York, N.Y., 342-343). Сканирование флуоресценции по длинам волн проводили от 300 до 440 нм для возбуждения при 270 нм и от 310 до 440 нм для возбуждения при 295 нм. Четыре последовательных сканирования усредняли по сигналу для каждого спектра. Приводятся нормализованные данные для оценки различий в флуоресценции, происходящей из разных образцов.
SEC/UV/LS/RI в режиме on-line: Молекулярные массы элюирующихся компонентов с использованием гель-фильтрационной хроматографии определяли с использованием последовательной детекции в ультрафиолете (280 нм УФ), по светорассеянию (90°С) и показателю преломления (RI). Этот способ был хорошо документирован (см. Arakawa et al., 1992, Anal. Biochem. 203:53-57 и Wen et al., 1996, Anal. Biochem. 240:155-166) и имеет преимущество, заключающееся в измерении негликозилированных молекулярных масс белков и пептидов, которые являются гликозилированными. Данные SEC и UV собирали с использованием системы ВЭЖХ Integral (PerSeptive Biosystems, Inc.) с колонкой BioSil-400-5 (от BioRad) при скорости тока 1 мл/мин. Буфер для элюции состоял из 100 мМ фосфата (рН 6,8) и 100 мМ Nad. Многоугольный детектор светорассеяния DAWN DSP и рефрактометр Optilab DSP были оба куплены у Wyatt Technology, Inc. Калибровочные стандарты для определения приборных констант включали в себя димер БСА, мономер БСА и овальбумин (фигура 2).
Дифференциальная сканирующая калориметрия (DSC): Физические характеристики разворачивания измеряли с использованием прибора MicroCal MC-2 DSC в режиме восходящего сканирования. Образцы готовили заменой буфера на тот же самый TMS-буфер с рН 7,4. Пробы содержали приблизительно 4 мг/мл белка, и их оценивали против буфера (в отсутствие белка) в качестве контроля. Скорость сканирования была 67°С/час при охвате температурного диапазона от 20 до 90°С. Затем полученные данные сканирования преобразовывали в нормализованные по концентрации данные для лучшего сравнения поведения энтальпий переходов разворачивания с учетом различий в концентрации (данные приводятся в ккал/моль). Результаты:
Круговой дихроизм. Измерения кругового дихроизма в ближней УФ-области, выраженные в виде средней эллиптичности на остаток, показаны на фигуре 2. Изменения в широкой области вблизи 270 нм были очевидными между фракциями №2 и №3, как показано большей долей отрицательной эллиптичности в спектре фракции №3 (указанной стрелкой на фигуре 2А). Отмечалось, что спектральное поведение исходного материала близко совпадает со спектральным поведением фракции №2, но действительно проявляет слабый отрицательный сдвиг в той же самой области вблизи 270 нм. Этот результат представляется согласующимся, так как фракция №3 составляет малую часть исходного материала, и, следовательно, ее вклад в общую эллиптичность в этой области был сильно уменьшенным, но в том же самом направлении сдвига. Воспроизводимость спектра фракции №3 подтверждала, что наблюдаемый сдвиг для этого образца был истинным. Имея это в виду и зная, что дисульфиды приводят к появлению признака широкой отрицательной эллиптичности в этой области спектра кругового дихроизма (см. Kahn, Р.С., 1978, Methods Enzymol. 61:339-378 и Kosen et al., 1981, Biochemistry 20:5744-5754), спектр кругового дихроизма в ближней УФ-области был представлен сглаженной кривой для оценки, что означают наблюдаемые изменения в этой области с позиций третичной структуры. Результаты сглаживания данных представлены на фигуре В2, и они показали небольшое смещение в красную область (3 нм) и увеличенный отрицательный сдвиг, согласующиеся со вкладом, возникающим из изменения третичной структуры с участием дисульфидов, при сравнении фракции №3 с фракцией №2.
Круговой дихроизм в дальней УФ-области спектра использовали для оценки состава вторичной структуры белков (Perczel et al., 1991, Protein Engrg. 4:669-679). Соотношение со вторичной структурой выполняли с использованием ССА. Рассчитанные спектры, состоящие из суммы элементов вторичной структуры, сравнивали с экспериментально наблюдаемыми спектрами, и они обнаруживали хорошее совпадение. Вторичные структуры обеих фракций были сравнимыми в пределах экспериментальной точности (в пределах 10%). Таким образом, этот эксперимент не обнаружил каких-либо различий в отношении вторичной структуры для каждой из этих двух фракций.
Флуоресцентная спектроскопия. Зная, что имелись значительные различия, наблюдаемые в спектре кругового дихроизма в ближней УФ-области, использовали флуоресцентную спектроскопию для обнаружения дополнительных различий в третичной структуре, связанных с флуоресценцией триптофана и тирозина. С использованием двух длин волн возбуждения удалось определить, что спектры для всех рассматриваемых трех случаев (SM, фракция №2 и фракция №3) были накладываемыми друг на друга с максимумом флуоресценции вблизи 338 нм. Поскольку трехмерная структура определенного белка является ответственной за максимумы испускания нативных белков, эти результаты предполагают, что усредненная структура, включающая в себя естественные флуорофоры, триптофан и тирозин, была ненарушенной.
SEC/UV/LS/RI в режиме on-line. Исследования светорассеяния, выполненные в режиме on-line с SEC, дали молекулярные массы основного пика элюции, которые согласовались с молекулярной массой негликозилированного полипептида TNFR:Fc (например, 102 кД). Хотя были явные различия в составах элюируемых молекулярных форм, оцениваемых этим способом, при сравнении профиля элюции фракции №3 с профилем элюции фракции №2 (фигура ЗА и В) измерение показало, что основной пик, содержащий основной компонент, имел молекулярную массу 102,5±1,6 кДа (удерживаемый объем=8,4 мл) и 101,9±2,1 кДа (удерживаемый объем = 8,3 мл) соответственно. Точность выражали в виде стандартного отклонения для 23 срезов через пик элюции, ограниченный вертикальными пунктирными линиями на фигуре 3. Отмечалось также, что значимый сигнал нисходящего плеча для фракции №3 позволял определить, что молекулярная масса этого полипептида была 78,1±3,7 кД (эта оценка учитывала 8 срезов, окружающих пик, отмеченный на 8,85 мл). Как показала точность, связанная с определением молекулярной массы этого компонента, этот пик проявлял более высокую гетерогенность, и вследствие этого в нем предполагалась большая полидиперсность, чем в основном пике. Фракция №3 содержала также значительное количество высокомолекулярных форм молекул, что согласуется с объемом элюции преимущественно димерной формы TNFR:Fc (вблизи 7,5). Таким образом, было определено, что фракция №3 состоит из нескольких форм, включающих в себя агрегаты и фрагментированные части этой молекулы.
Дифференциальная сканирующая калориметрия. Измерения DSC, проведенные на двух фракциях, дали значимые различия в разворачивании TNFR-части молекулы TNFR:Fc (фигура 4). Как более ясно следует из скорректированных на фон данных (фигура 4В), имеется смещение 2,8°С в сторону более низкой температуре в переходе плавления (Tm) при сравнении Tm 52,5°С (фракция №2) с Tm 49,7°С (фракция №3). Этот переход является слегка более широким для фракции №3 с полушириной 8°С на половине максимума перехода в сравнении с фракцией №2, имеющей полуширину 6,5°С. Из экспериментов по термическому разворачиванию мономера TNFR:Fc было установлено, что этот низкотемпературный переход обусловлен TNFR-доменом этой молекулы. Термические переходы при 69,1 и 83,4°С были отнесены в Fc-части молекулы. Эти последние два перехода разворачивания хорошо сопоставимы и являются сравнимыми в отношении Tm и энтальпий компонентов.
Обсуждение:
Среди испытанных способов, различия наблюдались при измерениях кругового дихроизма в ближней УФ-области спектра и DSC. Данные дифференциальной сканирующей калориметрии подтверждают разрыхление структуры, которое может быть отнесено к рецепторной части этой молекулы, с малым изменением в области Fc. Результаты определения кругового дихроизма в ближней УФ-области спектра предполагают, что дисульфиды участвуют в изменениях третичной структуры, связанных с фракцией №3. Эти изменения могут возникать как следствие погруженных дисульфидов, приобретающих большую открытость для растворителя и ответственных за увеличение гидрофобности, как следует из небольшого увеличения удерживаемого объема, наблюдаемого при элюции фракции №3 при HIC. Интересно, что не было ощутимых различий, обнаруживаемых по флуоресцентным данным, которые указывали бы на такое изменение в конформационной структуре. Если проанализировать первичную структуру TNFR:Fc в связи с распределением тирозинов (Y) и триптофанов (W), становится очевидным, что район, простирающийся от С-концевой части остатка 104 домена TNFR до остатка 296 N-концевой части Fc (содержащий 40% линейной последовательности TNFR:Fc), лишен этих естественных флуорофоров. Таким образом, одним возможным объяснением, согласующимся с этими данными, может быть то, что третичная структура, удаленная от шарнирной области Fc, является относительно неизмененной, тогда как структура области приблизительно от остатка С115 до С281 может быть несколько изменена конформационно. Эта область данной молекулы содержит 10 возможных цистеинов, которые могут подвергаться воздействию с предположительно малыми последствиями структурного изменения, влияющего на локальную структуру тирозинов и триптофанов. Отмечается, что в настоящее время неизвестно, как укладывается эта молекула, и представляется вероятным, что цистеины, которые образуют дисульфидные связи, которые являются более удаленными от любого конкретного остатка триптофана или тирозина, будут логическими подозреваемыми в изменениях третичной структуры, которые приводят к наблюдаемым результатам определения кругового дихроизма в ближней УФ-области спектра, но оказывают малое влияние на виципальную структуру, включающую в себя тирозины и триптофаны. Эта идея не исключает возможности, что имеется некоторое необычное изменение в структуре в одном или обоих плечах TNFR, которое не вызывает заметного изменения в общем эффекте флуоресценции, обусловленном тирозином и триптофаном. Тот факт, что флуоресцентные данные (которые являются нечувствительными к дисульфидам) не обнаруживают изменения и ближняя УФ-область спектра (которая является чувствительной к дисульфидам, тирозинам и триптофанам) показывает небольшой отрицательный сдвиг, согласующейся с дисульфидной структурной модификацией, действительно предполагает, что дисульфиды ответственны за различия между фракциями №2 и №3.
При суммировании остальных результатов, касающихся фракции №3, было обнаружено, что все показатели, относящиеся к молекулярной массе и вторичной структуре, являются неотличимыми от показателей фракции №2.
ПРИМЕР 2
Эксперименты по дисульфидному обмену на фракции №3 TNFR:Fc с глутатионом
Этот эксперимент был предназначен для оценки способности различных обработок переводить фракцию №3 TNFR:Fc в конформацию фракции №2 в процессе, пригодном для крупномасштабной промышленной эксплуатации.
Материалы и способы:
Материалы. Исходным материалом был TNFR:Fc в виде элюата с колонки с белком А, чистого элюата фракции №3 после HIC и смеси 50:50 элюатов фракции №2 и фракции №3 после HIC. Буферами были 0,1 М цитрат или 0,1 М Трис/глицин при рН 7,6, рН 8,6 или рН 9,6. Концентрация белка TNFR:Fc была от 0,2 до 4,5 мг/мл. Сопряженную редокс-систему восстановленного глутатиона и глутатиона (GSH/GSSG в соотношении 10:1) добавляли при 0,1-5 мМ GSH. Температура инкубирования варьировалась при 4, 22 или31°С.
Способы. Дисульфидный обмен останавливали подкислением образца до рН 6 1 М уксусной кислотой. Обработанные препараты рекомбинантного белка характеризовали аналитической HIC, SEC (время удерживания, концентрация агрегатов) и твердофазным анализом связывания TNF для определения процента и выхода фракции №2.
Результаты и обсуждение:
Эффективность обработки как функция рН и концентрации GSH. Значимый% белка во фракции №3 (по меньшей мере 10%) превращался во фракцию №2, когда обработку выполняли как 0,1 мМ GSH/pH 7,6, так и 0,1 мМ GSH/pH 8,6. Однако эффективность значительно улучшалась (от 45 до почти 70%), когда обработку выполняли 0,1 мМ GSH/pH 9,6; 1 мМ GSH/pH 7,6; 1 мМ GSH/pH 8,6 и 1 мМ GSH/pH 9,6. Таким образом, хотя эффективность обработки является чувствительной к рН и концентрации свободных тиолов (SH), она может эффективно выполняться в широком диапазоне этих переменных.
Температурные эффекты. Фракцию №3 обрабатывали при трех различных температурах: 4, 22 и 31°С. Концентрацию GSH поддерживали при 1 мМ и рН 8,6. Спустя 16 часов при всех вариантах обработки обнаруживалось значительное превращение фракции №3 во фракцию №2, но это превращение казалось слегка более эффективным при двух более низких температурах.
Клональные эффекты. Шесть различных клонов клеточной линии, все из которых продуцируют TNFR:Fc, испытывали в стандартизованном протоколе, основанном на приведенных выше результатах. Конкретно элюат с колонки с белком А, содержащий 0,4-0,7 мг/мл TNFR:Fc (при приблизительно рН 4), доводили до рН 8,6 при помощи 1 М Трис/глицина (конечная концентрация 0,1 М Трис/глицин). Эти растворы доводили до 1 мМ ЭДТА и 2,5 мМ GSH/0,25 мМ GSSG и инкубировали при комнатной температуре в течение приблизительно 16 часов. Дисульфидный обмен останавливали подкислением, как описано выше.
Каждый из шести различных клонов показал улучшение в продуцировании и выходе фракции №2. Уменьшение фракции №3 после HIC в результате обработки в различных клонах составляло 64, 72, 77, 78, 78 и 83%. Увеличение фракции №2 после HIC в тех же самых клонах было 37, 64, 78, 70, 44 и 54% соответственно. Увеличение фракции №2 после HIC, выраженное в процентах, хорошо коррелировало с увеличением единиц связывания в процентах, как показано на фигуре 5. Таким образом, эти способы оказались общеприменимыми ко всем испытанным клонам.
Анализы связывания. Три различных препарата TNFR:Fc анализировали в твердофазном анализе связывания TNF. Образцы 11-6 и 12 были элюатами с колонки с белком А. Образец 8085-47 также элюировали с колонки с белком А и затем подвергали дополнительной стадии очистки с использованием HIC; этот образец содержал исключительно фракцию №3. Образцы испытывали в анализе связывания до и после дисульфидного обмена, как описано выше. Результаты, представленные ниже в таблице 1, показывают увеличение в активности связывания лиганда после обработки всех образцов глутатионом.
TNF-связывающая активность TNFR:Fc до и после дисульфидного обмена
ПРИМЕР 3
Эксперименты по дисульфидному обмену на TNFR:Fc, обработанном L-цистеином
Этот эксперимент был предназначен для оценки цистеина/цистина в качестве реагентов для сопряженного окисления-восстановления TNFR:Fc. Эта процедура позволяет оценить изменение фракции №3 после HIC в конформацию фракции №2 в процессе, пригодном для крупномасштабной промышленной эксплуатации. Эта процедура может быть проведена на очищенной фракции №3, смеси фракций №2 и №3 и/или после других способов разделения, таких как хроматография на белке А, со сходными результатами.
Материалы и способы:
Исходным материалом был TNFR:Fc в виде чистого элюата фракции №3 после HIC или в виде элюированного с колонки с белком А TNFR:Fc, содержащего как фракцию №2, так и фракцию №3. Буферами были 0,1 М цитрат или 0,2 М Трис при рН 8,5. Концентрация белка TNFR:Fc была от 2,5-3 мг/мл.
Использовали сопряженную редокс-систему L-цистеина (в диапазоне от 0 до 50 мМ). Эта процедура оценивала также +/- L-цистин (0,025-0,5 мМ) и +/- 1 мМ ЭДТА. Температуру инкубации оценивали при 4, 15 и 22°С для 6, 18 и 48 часов. Дисульфидный обмен останавливали подкислением пробы до рН 7 NaH2PO4 или 0,85 М цитратом. Обработанные препараты рекомбинантного белка характеризовали аналитической HIC и SEC (время удерживания, концентрация агрегатов) для определения процента и выхода фракции №2 и фракции №3, анализа цистеинилирования и анализа свободных сульфгидрильных групп. Результаты и обсуждение:
Эффективность обработки как функция концентрации L-цистеина (0-5 мМ). Значительный процент белка TNFR:Fc во фракции №3 после HIC (в среднем 10%) превращался во фракцию №2, когда обработку выполняли 0,25 мМ L-цистеином в отсутствие L-цистина или ЭДТА в четырех повторностях образцов (фигура 6). Однако эффективность сильно улучшалась (с 45 до почти 70%), когда обработку выполняли 1 мМ L-цистеином или 5 мМ цистеином (фигура 6). Действие цистина в этих условиях реакции менялось в зависимости от присутствия ЭДТА (см. ниже). Для конкретной партии клеточной культуры (обрабатывали пробы из четырех различных партий клеточных культур) способ обработки был воспроизводимым.
Эффективность обработки как функция более высокой концентрации L-цистеина (5-50 мМ). Более высокие концентрации L-цистеина (5, 15 и 50 мМ L-цистеина), используемые для обработки TNFR:Fc, приводили к уменьшению фракции №3 после HIC из исходного материала в каждом случае, но 5 мМ L-цистеин был наиболее эффективным в стимуляции накопления фракции №2 (фигура 7). Считается, что более высокие концентрации L-цистеина либо значительно восстанавливали сульфгидрильные группы в молекуле, либо требовали слишком большого времени для повторного окисления.
Эффективность обработки как функция дополнительного внесения L-цистеина. В попытке увеличения эффективности дисульфидного обмена TNFR:Fc обрабатывали 5 мМ L-цистеином и инкубировали при 4°С в течение 18 часов. Затем добавляли дополнительное количество L-цистеина (0-5 мМ) и пробы инкубировали при 4°С в течение двух дополнительных дней. При этих условиях не отмечали значимого влияния на отношение фракции №3 после HIC к фракции №2 после HIC при дополнительном внесении L-цистеина.
Действие ЭДТА, цистина и L-цистеина. Действие цистина (0-0,4 мМ) в комбинации с L-цистеином (5 мМ) на TNFR:Fc оценивали в присутствии или в отсутствие 1 мМ ЭДТА. Оптимальные результаты в присутствии 1 мМ ЭДТА имели место при концентрациях цистина в диапазоне 0,1-0,2 мМ.
Внесение меди, температуры и времени. TNFR:Fc обрабатывали при 5 мМ L-цистеине при 4°С в течение 6 или 18 часов. Завершение обработки TNFR:Fc оценивали по добавлении меди с последующей HIC. После 6 часов инкубирования дисульфидный обмен является более полным при 4°С, чем при 22°С, и обработка является явно более полной после 18 часов при 4°С (фигура 8А и 8В).
Сравнение эффективности обработки L-цистеином в аналитическом масштабе с эффективностью в препаративном масштабе. На основе условий обработки, оптимизированных в малом масштабе, TNFR:Fc (2,5 мг/мл в 0,2 М Трисе, рН 8,5) в количествах либо 3 мл, либо 20 л обрабатывали 5 мМ L-цистеином (в отсутствие цистина или ЭДТА), инкубировали при 4°С в течение 18 часов, разбавляли равным объемом 850 мМ цитрата натрия, 50 мМ фосфата натрия, рН 6,5 для остановки обработки и хроматографировали на HIC. Контрольные образцы TNFR:Fc препаративного и аналитического масштабов имели 63% и 68% фракции №3, соответственно. После обработки с использованием вышеуказанных условий фракция №3 уменьшалась до 28% как в препаративном, так и в аналитическом масштабе. Таким образом, эффективность обработки была 56 и 59% для препаративного и аналитического образцов, соответственно (таблица 2). Этот эксперимент демонстрирует, что этот способ применим для крупномасштабной обработки.

Таким образом, хотя на окислительно-восстановительную эффективность обработки влияют концентрация свободных тиолов, температура и время, она может быть эффективно оптимизирована и выполнена в широком диапазоне переменных. Протоколы обработки могут также выполняться в малом и крупном масштабе с хорошей воспроизводимостью.
Данное изобретение не ограничивается в объеме описанными здесь конкретными воплощениями, которые предназначены только в качестве иллюстрации отдельных аспектов данного изобретения, и функциональные эквивалентные способы и компоненты находятся также в рамках данного изобретения. Действительно, разнообразные модификации данного изобретения, помимо показанных и описанных здесь, будут очевидными специалистам в данной области из предыдущего описания и сопутствующих чертежей. Предполагается, что такие модификации находятся в объеме прилагаемой формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ВЫДЕЛЕНИЯ ПОВТОРНО СВЕРНУТОГО РЕКОМБИНАНТНОГО БЕЛКА ИЗ КУЛЬТУРЫ ПРОКАРИОТИЧЕСКИХ КЛЕТОК (ВАРИАНТЫ) | 2007 |
|
RU2439076C2 |
| РЕКОМБИНАНТНОЕ ПОЛУЧЕНИЕ СВЯЗЫВАЮЩИХ ГЕПАРИН БЕЛКОВ | 2006 |
|
RU2385877C1 |
| СПОСОБ ОЧИСТКИ БЕЛКА СЛИЯНИЯ | 2015 |
|
RU2698654C2 |
| СПОСОБ ПОЛУЧЕНИЯ СЛИТОГО БЕЛКА TNFR-FC С ЗАДАННЫМ СОДЕРЖАНИЕМ ПРИМЕСЕЙ | 2015 |
|
RU2670166C1 |
| ОДНОЦЕПОЧЕЧНЫЕ БЕЛКИ-АГОНИСТЫ РЕЦЕПТОРА CD40 | 2016 |
|
RU2745801C2 |
| СТАБИЛЬНАЯ ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ НА ОСНОВЕ СЛИТОГО БЕЛКА TNFR:FC | 2013 |
|
RU2664691C2 |
| ЖИДКИЙ СОСТАВ ПОЛИПЕПТИДОВ, СОДЕРЖАЩИХ FC-ДОМЕН ИММУНОГЛОБУЛИНА | 2011 |
|
RU2600847C2 |
| ПОЛУЧЕНИЕ БИОЛОГИЧЕСКИ АКТИВНЫХ БЕЛКОВ | 2007 |
|
RU2441911C2 |
| КОЛИЧЕСТВЕННАЯ ОЦЕНКА НЕПРАВИЛЬНО СВЕРНУТОГО TNFR2:Fc | 2015 |
|
RU2698671C2 |
| ПОЛУЧЕНИЕ ГЕТЕРОМУЛЬТИМЕРНЫХ БЕЛКОВ | 2011 |
|
RU2624027C2 |
Изобретение относится к области медицины и биотехнологии и может быть использовано в фармацевтической промышленности. Долю наиболее активной конформации гликозилированного рекомбинантного белка, секретированного клеткой млекопитающего, увеличивают путем контактирования его с реагентом для сопряженного окисления-восстановления. Данный способ промотирования наиболее активной конформации белка используют в способе получения гликозилированного рекомбинантного белка в наиболее активной конформации. Конфигурационный изомер белка, полученный указанным способом получения гликозилированного рекомбинантного белка в наиболее активной конформации, используют в способе получения композиции белка для введения потребителю и/или пациенту или потребления потребителем и/или пациентом. Применение заявленного изобретения позволяет повысить активность гликозилированного белка, полученного методом рекомбинантных ДНК с помощью клетки млекопитающего. 3 н. и 24 з.п. ф-лы, 8 ил., 2 табл.
контактирование указанного препарата с реагентом для сопряженного окисления-восстановления в течение времени, достаточного для увеличения относительной доли желательного конфигурационного изомера, и затем определение относительной доли желательного конфигурационного изомера в смеси.
| Способ изготовления мембранных предохранительных устройств | 1975 |
|
SU553494A1 |
| СПОСОБ ВЫРАЩИВАНИЯ КЛУБЕНЬКОВБ1Х | 0 |
|
SU293785A1 |
| WO 9603141 А, 02.08.1996 | |||
| ПРОДУКТ, СОДЕРЖАЩИЙ ГРАНУЛОЦИТАРНЫЙ КОЛОНИЕСТИМУЛИРУЮЩИЙ ФАКТОР (G - CSF) И TNF-СВЯЗУЮЩИЙ ПРОТЕИН | 1995 |
|
RU2139084C1 |

