Предложена рекомбинантная плазмида pPICZαA-PLA2-K4, содержащая синтетический ген фосфолипазы А2 (SEQ ID NO: 1), состоящего из измененной последовательности фосфолипазы А2, оптимизированной для экспрессии гена в Pichia pastoris. Плазмида имеет размер 3859 п. о. Использован штамм дрожжей Pichia pastoris X-33/pPICZαA-PLA2-K4, являющийся продуцентом рекомбинантной секреторной фосфолипазы А2 PLA2 Streptomyces violaceoruber и полученный путем трансформации дрожжей Pichia pastoris Х-33 линеаризованной плазмидой pPICZαA-PLA2-K4. Изобретение обеспечивает получение активной рекомбинантной фосфолипазы А2 Streptomyces violaceoruber, секретируемой в культуральную среду с высоким выходом.
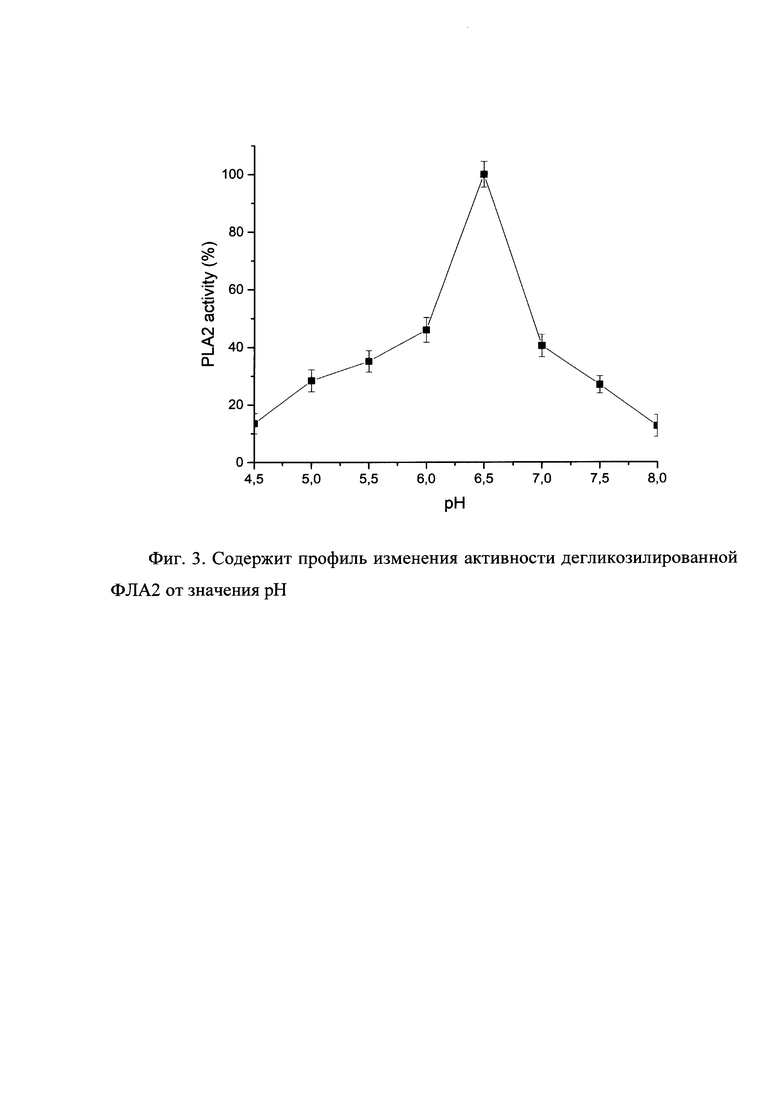
Изобретение включает в себя изменения в последовательности гена, замещающие аминокислоты в сайте гликозилирования, позволяющие после очистки получать гомогенный белковый продукт, обладающий видоизмененной ферментативной активностью. Так, нам удалось изменить оптимум рН фермента фосфолипазы А2 с 6.0 до 6.5 за счет удаления сайтов гликозилирования.
Изобретение относится к генной инженерии и биотехнологии и может быть использовано для получения фосфолипазы А2. Предложена рекомбинантная плазмида pPICZαA-PLA2-K4, содержащая синтетический ген фосфолипазы А2 (SEQ ID NO: 1), состоящего из измененной последовательности фосфолипазы А2, оптимизированной для экспрессии гена в Pichia pastoris.
Рекомбинантная плазмида для экспрессии в дрожжах Pichia pastoris гена фосфолипазы А2 и штамм дрожжей Pichia pastoris - продуцент рекомбинантного белка фосфолипаза А2.
Экспрессия чужеродных генов в метилотрофных дрожжах Pichia pastoris позволяет получать не загрязненные эндотоксинами рекомбинантные белки с более высоким выходом, чем в клетка Е. coli. Поскольку дрожжи Pichia pastoris являются эукариотами, они производят растворимые белки, обеспечивая фолдинг и посттрансляционные модификации, необходимые для функциональной активности белка. Однако способность к специфическому гликозилированию рекомбинантных белков также может менять их свойства. По причине этого, в последовательность ДНК, кодирующую фосфолипазу А2, были введены замены, не позволяющие гликозилировать белок.
Из уровня техники известы генетические конструкции, позволяющие экспрессировать ген секреторной фосфолипазы А2 в системе экспрессии Pichia pastoris:
1) Плазмида pPIC9K-PLA2. Ген фосфолипазы А2 GenBank No. AY359866.1 без сигнальной последовательности был амлифицирован и клонирован в плазмиде pP1C9K по сайтам рестирикции в EcoRI/NotI для создания экспрессионной плазмиды pPIC9K-PLA2. При использовании данной плазмиды удалось получить штамм-продуцент гликозилированной секреторной фосфолипазы А2 с рН оптимумом ферментативной активности в 6.0 [Патент №CN104328095A Phospholipase А2 with most appropriate pH being in acid range and application thereof. Epub 2014].
2) Наиболее близким аналогом является плазмида pPICZαA-PhoA2-StV. Ген фосфолипазы А2 GenBank No. AY359866.1 без сигнальной последовательности был амлифицирован и клонирован в плазмиде pPICZαA по сайтам рестирикции в EcoRI/SalI для создания экспрессионной плазмиды pPICZαA-PhoA2-StV. При использовании данной плазмиды удалось получить штамм-продуцент гликозилированной секреторной фосфолипазы А2 с рН оптимумом ферментативной активности в 6.0 [Патент №RU2676321C1. Способ получения ферментного препарата фосфолипазы А2 с применением рекомбинантного штамма-продуцента Pichia pastoris Х-33/ pPICZαA-PhoA2-StV].
Таким образом, созданные на данный момент плазмиды для экспрессии гена фосфолипазы А2 позволяют получать штаммы-продуценты гликозилированной фосфолипазы А2, что затрудняет ее очистку до гомогенного состояния, а также сдвигает рН оптимум ферментативной активности с 6.5 до 6.0.
Фосфолипазы А2 активно применяются в различных отраслях биотехнологической и пищевой промышленности, в частности для ферментативного дегуммирования растительных масел и обработки яичного желтка. Создание новых штамм-продуцентов ФЛА2 с измененным рН оптимумом на основе разработанных генетических конструктов позволяет расширить спектр ферментов для биотехнологической индустрии.
Технический результат заявленного изобретения заключается в получении плазмиды, которая позволяет экспрессировать ген секреторной фосфолипазы А2 с измененным рН-оптимумом 6,5 в системе экспрессии Pichia pastoris, получение максимальной гомогенности фермента на стадии очистки.
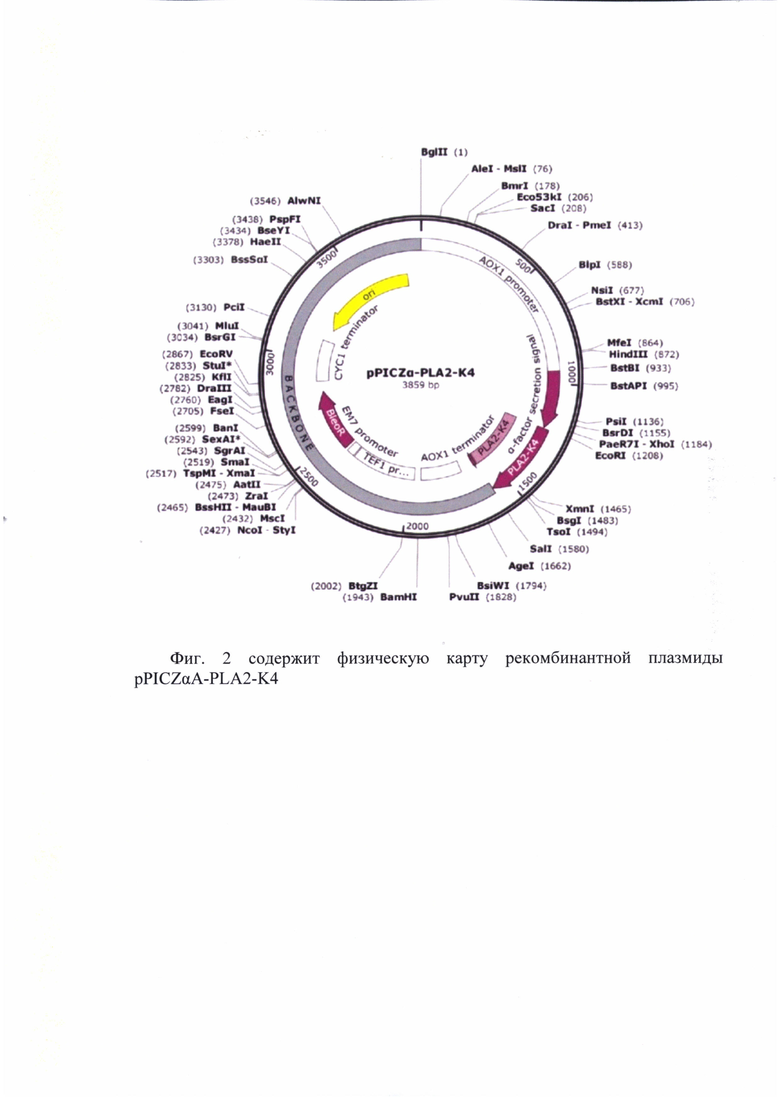
Технический результат достигается за счет плазмиды pPICZαA-PLA2-К4, обеспечивающей биосинтез и секрецию модифицированной фосфолипазы А2 Streptomyces violaceoruber (SEQ ID NO: 1), имеющей конкретное конструктивное исполнение, размер 3859 п. о. и состоящей из следующих элементов (Фиг. 1):
а) АОХ1 promoter - 5’концевая область промотора алкоголь оксидазы: 1-940;
б) Alpha factor signal peptide - участок, кодирующий N-концевой сигнальный пептид из Saccharomyces cerevisiae: 941-1212;
в) Модифицированная последовательность гена фосфолипазы А2, оптимизированная для экспрессии в Pichia pastoris, с удаленными сайтами гликозилирования, 1213-1585 п. о.;
г) АОХ1 transcription terminator - терминатор транскрипции: 1683-1929 п. о.;
д) TEF1 promoter - эукариотический промотор: 1969-2359 п. о.;
е) ЕМ7 promoter - бактериальный промотор: 2363-2410 п. о.;
ж) Zeo(R) - ген BleoR устойчивости к селективному антибиотику зеоцину: 2429-2803 п.о.;
з) CYC1 transcription terminator - терминатор транскрипции гена цитохрома С1: 2869-3116 п. о.;
и) pUC origin - бактериальная точка начала репликации pUC: 3191-3779
п. о. Получение максимальной гомогенности фермента на стадии очистки происходит за счет удаления сайтов гликозилирования.
Технический результат также достигается получением штамма дрожжей Pichia pastoris X-33/pPICZαA-PLA2-K4 - продуцента рекомбинантной дегликозилированной фосфолипазы А2, путем трансформации дрожжей Pichia pastoris Х-33 линеаризованной плазмидой pPICZαA-PLA2-K4.
Для получения рекомбинантной плазмиды pPICZαA-PLA2-K4 сначала был спроектирован модифицированный синтетический ген фосфолипазы А2 без сайтов гликозилирования.
Далее проводится оптимизация нуклеотидной последовательности синтетического гена по составу кодонов, для экспрессии в дрожжах Pichia pastoris. Оптимизацию кодонового состава осуществляют с использованием компьютерной программы «ExpOptimizer» (NovoPro Inc, China) удаляя при этом ненужные сайты рестрикции путем синонимических замен. Кроме этого, используют синонимические замены наиболее часто встречаемых пар кодонов в геноме Pichia pastoris, так как в соответствующих участках затормаживается трансляция мРНК.
Из последовательности гена PLA2-K4 были удалены сайты гликозилирования. Нуклеотидная последовательность полученного гена PLA2-K4 указана в SEQ ID NO 1, его синтез осуществляют в составе плазмиды pPICZαA-PLA2-K4. Рекомбинантную плазмиду pPICZαA-PLA2-K4 получают, проводя лигирование фрагмента плазмиды pPICZαA, гидролизованной рестриктазами EcoRI и Sall, и фрагмента ампликона гена PLA2-K4, гидролизованного рестриктазами EcoRI и Xhol, при помощи ДНК-лигазы фага Т4, как в примерах 1 и 2.
Штамм дрожжей Pichia pastoris Х-33/ pPICZαA-PLA2-K4 - продуцент рекомбинантной модифицированной фосфолипазы А2, получают путем трансформации компетентных клеток нативного штамма дрожжей Pichia pastoris Х-33 линеаризованной по сайту Sad в области 5’-АОХ1 промотора плазмидой pPICZαA-PLA2-K4 методом электропорации, как в примере 3.
Биосинтез рекомбинантной фосфолипазы А2 проводят в клетках заявляемого штамма дрожжей Pichia pastoris X-33/pPICZαA-PLA2-K4 при культивировании и последующей индукции метанолом, как в примерах 4.
Функциональную активность рекомбинантной фосфолипазы А2 при различных рН определяют с помощью флуоресцентного субстрата как в примере 5.
Предложенная группа технических решений позволяет значительно увеличить выход гомогенной фосфолипазы А2 в сравнении с прототипами и другими аналогами. При культивировании клеток рекомбинантного штамма Pichia pastoris Х-33/ pPICZαA-PLA2-K4 и последующей индукции метанолом в культуральную среду секретируется растворимая фосфолипаза А2 с выходом около 125 мг на 1 л среды.
Достижение заявленного технического результата обусловлено следующим:
1. Получением рекомбинантного штамма дрожжей Pichia pastoris путем трансформации дрожжей Pichia pastoris Х-33 линеаризованной плазмидой pPICZαA-PLA2-K4, что обеспечивает экспрессию гена модифицированной фосфолипазы А2, а также секрецию и процессинг негликозилированного белка, что позволяет получить растворимую гомогенную фосфолипазу А2 с измененным рН-оптимумом фосфолипазной активности.
2. Использованием в составе линеаризованной плазмиды pPICZαA-PLA2-K4 синтетического гена фосфолипазы А2, полученного с оптимизацией состава кодонов и с учетом синонимичных замен пар кодонов, наиболее часто встречаемых пар кодонов в геноме Pichia pastoris, приводящих к торможению трансляции мРНК в соответствующих участках, что в совокупности способствовало увеличению экспрессии гена фосфолипазы А2.
3. Изобретение позволяет измененить рН-оптимума секретируемого фермента фосфолипаза А2 с 6,0 до 6,5 в системе экспрессии Pichia pastoris за счет удаления сайтов гликозилирования. Удаление сайтов гликозилирования также позволяет достичь максимальной гомогенности фермента на стадии очистки.
Сущность заявляемого изобретения далее поясняется на чертежах.
Фиг. 1 содержит физическую карту рекомбинантной плазмиды pPICZαA-PLA2-K4.
АОХ1 promoter - 5’-концевая область промотора алкоголь оксидазы; Alpha factor signal peptide - участок, кодирующий N-концевой сигнальный пептид из Saccharomyces cerevisiae; PLA2-K4 фрагмент, кодирующий измененный ген фосфолипазы А2; АОХ1 transcription terminator - терминатор транскрипции; TEF1 promoter - эукариотический промотор; ЕМ7 promoter -бактериальный промотор; BleoR - ген устойчивости к зеоцину; CYC1 transcription terminator - терминатор транскрипции гена цитохрома C1; pUC origin - бактериальная точка начала репликации.
Фиг. 2. Содержит профиль изменения активности дегликозилированной ФЛА2 от значения рН.
Осуществление изобретения пояснено на примерах.
Пример 1. Оптимизация нуклеотидной последовательности проектируемого синтетического гена фосфолипазы А2 для его эффективной экспрессии в клетках P. pastoris. Оптимизированную нуклеотидную последовательность гена фосфолипазы А2 получают путем синонимичных замен кодонов на наиболее часто встречаемые в геноме P. pastoris кодоны и удалением возникающих сайтов рестрикции при помощи программы «ExpOptimizer» (NovoPro Inc, China). Кроме того, производят синонимичную замену наиболее часто встречающихся пар кодонов, а именно TTCAAG/TTTAAG, AAGAAG/AAGAAA и TTTGAA/TTCGAA, при помощи собственной программы.
Также проверяют наличие в мРНК, кодируемой синтетическим геном, нежелательных участков вторичной структуры.
Синтетический ген фосфолипазы А2 PLA2-K4 (SEQ ID NO 1) получают химическим синтезом, клонированным в составе бактериальной плазмиды pTBA-PLA2-K4.
Пример 2. Получение рекомбинантной химерной плазмиды pPICZαA-PLA2-K4.
Ампликон гена PLA2-K4 получают методом ПЦР на матрице плазмиды pTBA-PLA2-K4 с помощью высокоточной термостабильной полимеразы Phusion (Thermo scientific, USA), с прямым праймером pla2-K4-F GCTGAATTCGCTCCAGCAGATAAACC с достройкой 5'-концевого сайта рестрикции EcoRI и обратного праймера pla2-K4-R GATGGTCGACTTAAAGAATCTTTACGGCTTGATAG с достройкой 3’-концевого сайта рестрикции SalI. Ампликон выделяют экстракцией фенолом/хлороформом с последующим осаждением изопропанолом и гидролизуют рестриктазами EcoRI и SalI. Полученный фрагмент очищают при помощи фенол/хлороформной экстракции и осаждают изопропанолом.
Плазмиду pPICZoA гидролизуют по сайтам рестрикции EcoRI и SalI, фрагмент плазмиды размером 3487 п. о. получают, выделяя из 0,8% агарозного гела при помощи набора Qiagen USA.
Полученные фрагменты лигируют с использованием Т4 ДНК лигазы (Сибэнзим, Россия) в амплификаторе «Терцик» в течение 12-ти часов по заданной программе при температуре, меняющейся циклически от 10°С до 30°С.Полученной лигазной смесью трансформировали клетки Е. coli BL21 (DE3). Отбор клонов, содержащих рекомбинантную плазмиду pPICZαA-PLA2-K4., проводили на агаризованной среде LB, содержащей 50 мкг/мл зеоцина. Наличие целевой ДНК-вставки определяли методом ПЦР колоний, с праймерами AOX1-F: GACTGGTTCCAATTGACAAGC и AOX1-R GCAAATGGCATTCTGACATCC, а также проводили рестрикционный анализ плазмид, выделенных из полученных клонов. Клон, содержащий плазмиду pPICZαA pTBA-PLA2-K4 использовали для препаративной наработки.
Пример 3. Получение рекомбинантного штамма дрожжей Pichia pastoris X-33/pPICZαA-PLA2-K4. - продуцента рекомбинантной фосфолипазы А2.
Рекомбинантную плазмиду, полученную в примере 2, обрабатывают рестриктазой SacI, находящейся в области АОХ1 промотора (Фиг. 1). При этом происходит встройка экспрессирующей кассеты, состоящей из гена устойчивости к зеоцину и синтетического гена рекомбинантной фосфолипазы А2, который слит с находящимся в плазмиде pPICZαA-PLA2-К4 альфа-сигналом секреции, путем гомологичной рекомбинации. Таким образом, трансформанты, выросшие на агаризованной среде с селективным антибиотиком зеоцином, генетически стабильны как в присутствии, так и в отсутствие в среде культивирования селективного антибиотика.
Кроме того, отбор трансформантов на высоких концентрациях зеоцина (500-1000 мкг/мл) позволяет отобрать клоны, в которых содержится несколько экспрессирующих кассет с геном устойчивости к зеоцину, таким образом, эти клоны должны продуцировать большее количество рекомбинантного белка.
Компетентные клетки Pichia pastoris Х-33 для электропорации получали по следующей методике: Выращивали 5 мл штамма Pichia pastoris Х-33 в среде YPD в 5 мл пробирке при 30°С в течение ночи; инокулировали 200 мл среды YPD в 1 литровой колбе 0,1-0,5 мл ночной культуры; выращивали в течение ночи до СЮбоо=1.3-1.5; центрифугировали клетки при 1500 хд в течение 5 мин при+4°С (ЦФ) в центрифуге Hitachi CR-22N; промывали осадок в 200 мл ледяной (0°С) стерильной деионизованной воды; ЦФ; промывали осадок в 100 мл ледяной (0°С) стерильной деионизованной воды; ЦФ; промывали осадок в 8 мл ледяного (0°С) стерильного 1М сорбита; ЦФ; ресуспендировали осадок в 400 мкл ледяного (0°С) 1М сорбитола, получив конечный объем примерно 600 мкл. Клетки использовали в аликвотах по 4 0 мкл.
15 мкг плазмиды pPICZαA-PLA2-K4 гидролизовали 10 мкл рестриктазы SacI в течение ночи, линеаризованную плазмиду очищали фенолом/хлороформом с последующим осаждением изопропанолом, растворяли в 10 мкл ТЕ буфера. К 40 мкл компетентных клеток, находящихся на льду, добавляли 10 мкл линейной плазмиды, инкубировали 2 мин при 0°С, клетки переносили в охлажденную 0,2 см кювету, проводили электропорацию при 2000V, время импульса 1 мс, заливали в кювету 1 мл ледяной среды YPDS, клетки переносили в стерильный фалькон емкостью 15 мл инкубировали на качалке, 200 об/мин при 30°С в течение примерно 3-х часов. Высевали по 50, 100 и 200 мкл на чашки со средой YPD с концентрациями зеоцина 200 и 1000 мкг/мл. Чашки инкубировали при 30°С. На третьи сутки вырастали клоны, которые далее анализировали при помощи индукции синтеза рекомбинантного белка метанолом в 96-луночном глубоком планшете (объем лунок по 2 мл, круглодонная форма, Cellstar) следующим образом: в лунки добавляли по 300 мкл среды BMGY, стерильной зубочисткой переносили часть колонии в лунки, при этом перекалывая этой же зубочисткой колонии на 2 чашки параллельно. Выращивали при 30°С на качалке, 250 об/мин в течение 60 часов, планшет при этом накрывали 2-мя слоями стерильной марли. Добавляли по 250 мкл среды BMMY, инкубировали далее в течение 12 часов. Добавляли по 50 мкл среды BMMY через каждые последующие 24 часа. На третьи сутки индукции клетки осаждали центрифугированием при 2500×g, белки из супернатантов концентрировали в 10 раз осаждением ТХУ (добавляли 1/100 объема 1,5 раствора дезоксихолата натрия, инкубировали 10 мин при 25°С, добавляли 1/10 исходного объема насыщенного раствора ТХУ, -20°С 10 мин, 16*103 об/мин 10 мин) с промывкой ацетоном. Электрофорез по Лэммли проводили в 12.5% полиакриламидном геле с последующим окрашиванием кумасси G250.
На Фиг. 3 представлена электофореграмма белков, секретируемых в культуральную жидкость рекомбинантными клонами Pichia pastoris X-33/pPICZαA-PLA2-K4, которые выросли при концентрации зеоцина 1000 мкг/мл.
Полученный рекомбинантный штамм дрожжей Pichia pastoris X-33/pPICZαA-PLA2-K4 характеризуется следующими свойствами:
Морфологические и культуральные характеристики. При росте на агаризованной среде YPD колонии круглые, гладкие, блестящие, белые, диаметр колоний 1-5 мм; консистенция пастообразная. Рост в жидкой среде YPD характеризуется ровным помутнением.
Физиолого-биохимические характеристики. Клетки растут при температуре 10-30°С при рН 5,0-7,5 без признаков лизиса. В качестве источника азота используют как минеральные соли в виде смеси, так и органические соединения в виде пептона, дрожжевого экстракта, аминокислот. В качестве источника углерода используют глицерин, глюкозу, метанол.
Устойчивость к антибиотикам. Клетки проявляют устойчивость к зеоцину (до 1000 мкг/мл), обусловленную внесением в геном при трансформации линеаризованной плазмидой pPICZαA-PLA2-K4 гена BleoR.
Продуктивность. Заявляемый штамм обеспечивает синтез рекомбинантной модифицированной PLA2-K4 в количестве 125 мг на литр культуральной среды при наработке белка по результатам электрофоретического анализа и замера концентрации белка.
Пример №4. Очистка методом анионообменной хроматографии.
Полученный после тангенциальной фильтрации раствор ФЛА2 разбавляли 10-кратным объемом буфера 20 мМ Tris-HCl рН=8.0 и наносили на хроматографическую колонку Sepharose Q Fast Flow. Элюцию проводили 20 мМ Tris-HCl рН=8.0 в градиенте 0-1М NaCl. При этом примеси адсорбировались на хроматографическую колонку. Фосфолипаза А2 не связывалась с колонкой, при этом примеси адсорбировались на хроматографической колонке.
С помощью анионообменной хроматографии препарат ФЛА2 удалось избавить от более крупных белковых примесей. После проведения анионообменной хроматографии использовали хроматографию гидрофобных взаимодействий. Был использован носитель Butyl-Sepharose Fast Flow. Не адсорбировавшиеся фракции после анионообменной хроматографии объединяли, добавляли 8,7 г NaCl на 50 мл фракций (до 3 М NaCl) и наносили при скорости потока 1 мл/мин на хроматографическую колонку. Элюцию проводили 20 мМ Tris-HCl рН=8.0, понижая содержания хлорида натрия. Фракции собирали в диапазоне 65-100% раствора Б. Эти фракции содержали высокоочищенный препарат фосфолипазы А2.
Гидрофобная хроматография позволила избавиться от подавляющего большинства примесных продуктов, добиться высокой (~90% по данным денситометрического анализа) степени очистки, а также сконцентрировать образец. После очистки активность полученной фосфолипазы А2 соответствовала ферментативной активности в 210 ед./мл.
Пример №5. Определение фосфолипазной активности.
Фосфолипазную активность определяли с использованием флуоресцентного субстрата 1 -пальминтоил-2-(10-пиренилдеканоил)-8п-глицеро-3-фосфорилхолина (10-pyrene PC, Molecular probes, Голландия). Для измерения флуоресценции к 1 мл буфера, содержащего 50 мМ Tris-HCl, рН=7.5, 100 мМ NaCl, 1 мМ EDTA добавляли 10 мкл субстрата (конечная концентрация 2 мкМ), 10 мкл 10% раствора БСА (конечная концентрация 0,1%), и 6 мкл 1 М CaCl2 (конечная концентрация 6 мМ). Реакцию инициировали добавлением 5 мкл раствора 0.5 мг/мл ФЛА2. В качестве положительного контроля использовали кислую ФЛА2 СМ2 из яда кобры N. kaouthia. Флуоресценцию детектировали на флуоресцентном спектрофотометре Fluoromax-4 (HORIBA Scientfic, UK) (λexmax exтam exтam =345 нм, λемmах=395 нм).
Активность фосфолипаз А2 рассчитывали по формуле A=2x10-4x(S-S0)xV/Fmax и приводили в мг белка, где S -увеличение интенсивности флуоресценции в минуту в присутствии фосфолипазы; SO -увеличение интенсивности флуоресценции в минуту в контроле; V - объем добавляемого 0,2 мМ 10-pyrene PC, мкл; Fmax - максимальная интенсивность флуоресценции при добавлении 5 мкг кислую ФЛА2 СМ2 из яда кобры N. kaouthia.
--->
Перечень последовательностей
<110> Федеральное государственное учреждение «Федеральный
исследовательский центр «Фундаментальные основы биотехнологии»
Российской академии наук» (ФИЦ Биотехнологии РАН)
Российская Федерация, г. Москва, Ленинский проспект д. 33,
корп. 2, 119071
<120> Получение гена фосфолипазы А2 с измененным оптимумом рН
путем удаления сайтов гликозилирования
<160> 1
<210> 1
<211> 363
<212> DNA
<213> Artificial sequence for the synthesis of recombinant
phospholipase A2 of Streptomyces violaceoruber
<220>
<221> CDS
<222> (l)-(363)
<223> Sequence encodes modified recombinant phospholipase A2 of
Streptomyces violaceoruber
<400> 1
gcccctgctgataaacctcaggtactagcatcattcacccaaacatctgcttctt
cacagaatgcctggttggcagcaaacagggcccagtctgcatgggccgcctacga
atttgactggtctactgatctgtgtacccaagctccagataaccctttcggtttc
ccatttaatacggcttgtgctagacacgacttcggttaccgtaactataaagccg
ctggaagtttcgacgcagccaaatcaagaattgattctgctttttatgaagatat
gaaacgtgtatgtacaggatacactggtgagaaaaacaccgcttgtgcaagtact
gcttggacatactatcaggccgttaagattcta
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ получения штамма-продуцента фосфолипазы А2 Komagataella phaffii (Pichia pastoris) YIB Δleu2_PLA2S | 2020 |
|
RU2746817C1 |
| Способ получения ферментного препарата фосфолипазы А2 с применением рекомбинантного штамма-продуцента Pichia pastoris X-33/ pPICZαA-PhoA2-StV | 2018 |
|
RU2676321C1 |
| Фосфолипаза А2 для экспрессии в дрожжах (варианты) | 2018 |
|
RU2716087C1 |
| Способ очистки рекомбинантного ферментного препарата фосфолипазы А2 из штамма продуцента Pichia pastoris | 2019 |
|
RU2746563C1 |
| Рекомбинантная плазмида для экспрессии в дрожжах Pichia pastoris гена химерного белка ангиогенина человека и штамм дрожжей Pichia pastoris - продуцент рекомбинантного химерного белка ангиогенина человека | 2017 |
|
RU2658758C1 |
| Рекомбинантная плазмида, способ её конструирования и штамм дрожжей Komagataella pastoris - продуцент иммунного интерферона-гамма собаки | 2020 |
|
RU2756852C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА ДЛЯ ЭКСПРЕССИИ В ДРОЖЖАХ PICHIA PASTORIS ГЕНА ФОСФОЛИПАЗЫ, ШТАММ ДРОЖЖЕЙ PICHIA PASTORIS - ПРОДУЦЕНТ ФОСФОЛИПАЗЫ | 2009 |
|
RU2409671C1 |
| ИНТЕГРАТИВНЫЙ ПЛАЗМИДНЫЙ ВЕКТОР ДЛЯ ЭКСПРЕССИИ ГЕНОВ В ДРОЖЖАХ | 2008 |
|
RU2388823C1 |
| РЕКОМБИНАНТНЫЙ ХИМЕРНЫЙ БЕЛОК SUMO3-apoA-I ДЛЯ ПОЛУЧЕНИЯ ЗРЕЛОГО АПОЛИПОПРОТЕИНА A-I ЧЕЛОВЕКА, ШТАММ ДРОЖЖЕЙ Pichia pastoris - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО ХИМЕРНОГО БЕЛКА SUMO3-apoA-I И СПОСОБ ПОЛУЧЕНИЯ ЗРЕЛОГО АПОЛИПОПРОТЕИНА A-I ЧЕЛОВЕКА | 2015 |
|
RU2605326C1 |
| Штамм Komagataella phaffii T07/pPZL-4x-OA-xyl-AsOr, обладающий способностью продуцировать ксиланазу из грибов вида Aspergillus oryzae | 2022 |
|
RU2796447C1 |

Предложена плазмида pPICZaA-PLA2-K4, обеспечивающая биосинтез и секрецию гомогенной дегликозилированной рекомбинантной фосфолипазы А2 Streptomyces violaceoruber с рН оптимумом 6,5. Плазмида имеет размер 3859 п.н. Плазмида содержит модифицированную последовательность гена фосфолипазы А2 с SEQ ID NO: 1, оптимизированную для экспрессии в Pichia pastoris, с удаленными сайтами гликозилирования. Изобретение обеспечивает получение активной рекомбинантной фосфолипазы А2 Streptomyces violaceoruber, секретируемой в культуральную среду с высоким выходом. 3 ил., 5 пр.
Плазмида pPICZaA-PLA2-K4, обеспечивающая биосинтез и секрецию гомогенной дегликозилированной рекомбинантной фосфолипазы А2 Streptomyces violaceoruber, с рН оптимумом 6,5, имеющая размер 3859 п.н. и состоящая из следующих элементов:
а) АОХ1 promoter - 5'-концевая область промотора алкоголь оксидазы: 1-940;
б) Alpha factor signal peptide - участок, кодирующий N-концевой сигнальный пептид из Saccharomyces cerevisiae: 941-1212;
в) модифицированная последовательность гена фосфолипазы А2 (SEQ ID NO: 1), оптимизированная для экспрессии в Pichia pastoris, с удаленными сайтами гликозилирования: 1213-1585 п.о.;
г) АОХ1 transcription terminator - терминатор транскрипции: 1683-1929 п.о.;
д) TEF1 promoter - эукариотический промотор: 1969-2359 п.о.;
е) ЕМ7 promoter - бактериальный промотор: 2363-2410 п.о.;
ж) Zeo(R) - ген BleoR устойчивости к селективному антибиотику зеоцину: 2429-2803 п.о.;
з) CYC1 transcription terminator - терминатор транскрипции гена цитохрома С1: 2869-3116 п.о.;
и) pUC origin - бактериальная точка начала репликации pUC: 3191–3779 п.о.
| Способ получения ферментного препарата фосфолипазы А2 с применением рекомбинантного штамма-продуцента Pichia pastoris X-33/ pPICZαA-PhoA2-StV | 2018 |
|
RU2676321C1 |
| CN 104328095 B, 29.12.2017 | |||
| LIU A | |||
| ET AL | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Appl Biochem Biotechnol | |||
| Устройство для закрепления лыж на раме мотоциклов и велосипедов взамен переднего колеса | 1924 |
|
SU2015A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| YU X.W | |||
| ET AL | |||
| Identification of novel factors enhancing recombinant | |||

