Изобретение относится к молекулярной биологии, биотехнологии, медицине и может быть использовано для повышения иммуногенности вакцин.
В настоящее время наблюдается тенденция возрастания резистентности многих патогенов, вызывающих социально значимые бактериальные и грибковые заболевания, к действию антибиотиков, в связи с чем перспективным направлением борьбы с ними представляется предупреждение развития заболевания, за счет вакцинации. Действие многих разработанных и применяющихся вакцин основано на индуцировании иммунного ответа против полисахаридных антигенов, представленных на клеточной стенке патогена. Однако полисахарид - это антиген, не связанный с реакцией Т-клеток, потому вызывает лишь краткосрочный иммунитет без иммунной памяти; вакцины, содержащие только данные вещества, неэффективны, что показано у детей до 2 лет [Greenwood В M et al., Trans R Soc Trop Med Hyg, 1980, 74:756-760; международная заявка на изобретение WO 2010120921 A1, дата приоритета 16.04.2009]. Использование полисахаридов совместно с белком-адъювантом, преимущественно в одной конструкции, позволяет задействовать все звенья иммунитета, что в результате приводит к адекватному иммунному ответу, с формированием иммунологической памяти.
Белок CRM197 является производным дифтерийного токсина, отличающимся одной мутацией, а именно заменой глицина на глутаминовую кислоту в положении 52, что элиминирует его токсичность [Uchida Т., Pappenheimer A.M., Greany R. Diphtheria toxin and related proteins. I. Isolation and properties of mutant proteins serologically related to diphtheria toxin // J. Biol. Chem. 1973. Vol. 248, №11. P. 3838-3844]. Данный белок предпочтительно применяют в противоположность химически детоксицированному токсину дифтерии, в составе вакцин против различных патогенов. Так, в настоящее время ряд вакцин производится с использованием токсоида дифтерии CRM197 в качестве иммуногенного носителя [Lepow M.L. и др., J. Infectious Diseases 150 [1984] 402-406, RU 2331435, RU 2498994, RU 2498815, RU 2325184, RU 2495049, RU 2322451, RU 2422156, RU 2471497, RU 2454244, RU 2379052, RU 2378010, RU 2378008, RU 2419628, RU 2402347, RU 2521501, RU 2518291, RU 2508122, RU 2012107657, RU 2009149359, RU 2012107480, RU 2011151781, RU 2011143398, RU 2011142774, RU 2011142747, RU 2008145859, RU 2013101418, RU 2504398, RU 2381814, RU 2012151376, RU 2012153061, RU 2360699, RU 2493870, RU 2484846, RU 2362784, RU 2435609, RU 2105568, WO 9640239, UA 96934]. Известны и конъюгаты, содержащие CRM197 [RU 2253655, RU 2385737, RU 2361609, RU 2359696, RU 2336091, RU 2333007, RU 2331435, RU 2498994, RU 2498815, RU 2495049, RU 2322451, RU 2422156, RU 2471497, RU 2454244, RU 2379052, RU 2378010, RU 2378008, RU 2419628, RU 2402347, RU 2521501, RU 2518291, RU 2412944, RU 2508122, RU 2012116119, RU 2012107657, RU 2009149359, RU 2012107480, RU 2011151781, RU 2011143398, RU 2011142774, RU 2011142747, RU 2008145859, RU 2482878, RU 2013101418, RU 2504398, RU 2381814, RU 2012151376, RU 2012153061, RU 2360699, RU 2493870, RU 2484846, RU 2362784, RU 2435609, RU 2105568, WO 9640239, UA 96934].
Вышеуказанные запатентованные в России изобретения и те, по которым планируется получить охрану в России, касаются инфекционных заболеваний, таких как менингит, вызванный Neisseria meningitidis группы A, B, C, W135 и Y, Е. coli K1-индуцированные заболевания, дифтерия, столбняк, коклюш, заболеваний, опосредованных PCSK9, Helicobacter pylori, вирусами полиомиелита, вирусом гепатита В, инфекций, вызываемых Chlamydia trachomatis, S. Pyogenes, Str. Pneumoniae, в том числе серотипов 1, 3, 4, 5, 6A, 6B, 7F, 9V, 14, 18C, 19A, 19F и 23F, H. influenzae, в том числе типа B, стафилококками, в том числе Staphylococcus aureus, преимущественно штаммов типа 8 и 5, S. epidermidis, преимущественно штаммов серотипов I, II и III, инфекций, вызываемых грамотрицательными бактериями, в том числе Neisseria gonorrhoeae, аллергических и воспалительных реакций, вызываемых взаимодействиями C3-области IgE с высокоаффинным рецептором IgE, а также тау-ассоциированных неврологических расстройств, никотиновой зависимости. Вне России защищены либо планируют получить охрану в виде патентов на изобретение технические решения для профилактики инфекций, вызываемых Streptococcus pneumoniae, Neisseria meningitidis, заболеваний, вызываемых вирусом гепатита E, вирусом гриппа и др. В основном, предложены технические решения в виде конъюгатов CRM197 с полисахаридами патогенов, причем представлены и структуры, содержащие два таких белка-носителя, конъюгированных с полисахаридом, каждый может быть конъюгирован с другим или таким же белком-носителем [RU 2004117775].
Известны и поливалентные вакцины, содержащие CRM197.
Известна композиция вакцины, включающая анатоксин дифтерии, ′D′; анатоксин столбняка, T; клеточный антиген коклюша, ′wP′; поверхностный антиген вируса гепатита B, ′HbsAg′; капсулярный сахарид Haemophilus influenzae типа b (Hib), конъюгированный с белком-носителем, и по меньшей мере один капсулярный сахарид Neisseria meningitidis, конъюгированный с белком-носителем, где каждый из капсулярного сахарида Haemophilus influenzae типа b и капсулярного сахарида (сахаридов) Neisseria meningitidis является конъюгированным с белком-носителем в виде анатоксина столбняка [RU 2420315]. Известна мультивалентная иммуногенная композиция, включающая HBV компонент, дифтерийный анатоксин и один или несколько адъювантов, вариантом также являются конъюгаты данных компонентов [RU 2442825].
Известен способ получения комбинированной вакцины, которая содержит неионогенное поверхностно-активное вещество, поверхностный антиген (HBsAg) вируса гепатита В (HBV) и антиген C. diphtheriae - дифтерийный анатоксин, по которому осуществляют очистку и объединение HBsAg-компонента [RU 2444374]. Предложены и способы получения конъюгатов, с CRM197, за счет активированных функциональных групп на аминокислотных остатках носителя или, необязательно, через линкерную молекулу [WO 2005058940].
Классическим способом получения дифтерийного токсина и его нетоксичных производных является продукция в клетках Corynebacterium diphtheria линии PW8, инфицированных частицами β фага, геном которого несет мутантный ген tox, кодирующий дифтерийный токсин [RU 2394914, RU 94007083]. При этом необходимо выращивать культуру клеток в течение 36-48 часов для достижения максимальной концентрации дифтерийного токсина или его производных. Эти белки секретируются в культуральную среду, которая, наряду с целевым белком, содержит неспецифические примеси аминокислот, пептидов и белков. Целевой белок затем выделяют центрифугированием и последующей ультрафильтрацией, либо преципитацией, и далее очищают с использованием хроматографических методов. Однако для получения большего выхода целевого белка процесс ферментации требует большого количества лизогенных мутантов С. diphtheria и четко контролируемых условий роста (температура, перемешивание, аэрация, концентрация железа). Более того, используется патогенный организм, что требует особых условий производства.
Известен способ получения мутантного дифтерийного токсина CRM197 с использованием клеток Escherichia coli и гена, функционально связанного с промотором и хотя бы одной идеальной палиндромной операторной последовательностью, в векторе, преимущественно в периплазме [WO 2013178974].
Известен способ получения CRM197 с использованием клеток Escherichia coli и гена, его кодирующего, при котором синтезируется гибридный белок, в котором CRM197-фрагмент связан с гетерологичной сигнальной последовательностью для доставки в периплазму и содержит сайт разрезания между этими двумя фрагментами, при этом собственный сигнал для транспорта в периплазму может быть удален, причем гетерологичная последовательность функционально связана с промотором из araBAD promoter (PBAD), lac promoter, 1-rhamnose - индуцибельный rhaP BAD promoter, T7 RNA polymerase promoter, trc и tac promoter, lambda phage promoter p L, anhydro tetracycline-inducible tetA promoter/operator, клонирована в высококопийный экспрессионный вектор, из pEC415, pBR322, pBAD, серии pET, серии pUC, pACT3, pEXT22, pEXT20, серии pBLUESCRIPT, серии pGEM, хотя бы 50% продуцируемого белка растворима, индукция экспрессии происходит от 20°C до 37°C, N-конец экспрессируемого белка представлен на 50-100% последовательностями ADDV, GADDV, либо MGADDV, у того же количества белка имеется дисульфидная связь между Cys186 и Cys201 [WO 2014102265].
Известна экспрессионная система, в том числе для получения белка CRM197, в которой ген, кодирующий белок CRM197, на 5′ конце содержит сигнальную последовательность для направления в периплазматическое пространство бактерии, возможно, гетерологичную, на 3′ конце содержит последовательность, кодирующую, по меньшей мере, один B- или T-клеточный эпитоп, клетка-хозяин выбрана из группы, состоящей из E. coli, Acinetobacter, Actinobacillus, Bordetella, бруцелл, бактерии Campylobacter, Цианобактерии, Enterobacter, Erwinia, Franciscella, Helicobacter, Hemophilus, Klebsiella, Legionella, Moraxella, Neisseria, Pasteurella, Proteus, Salmonella, Serratia, Shigella, Treponema, Vibrio, и Yersinia [WO 2011042516]. Также предложен способ получения данного белка в ферментере, по которому на разных стадиях различается температура(первая стадия - 20-40°C, вторая стадия - 20-28°C) и pH (первый этап - 6,0-7,0 или 7,0, второй этап - 6.5, 6.5-10.0, 7,0-8,0, 6.5-9,0 или 7,0-8,5), pH поддерживают с использованием буфера из группы, состоящей из фосфатного буфера, трис-буфера и гистидинового буфера, в концентрации 10-200 мМ, 50-100 мМ, 10-50 мМ, 100-200 мМ или 50-150 мМ, фосфатного буфера - 80-100 мМ, увеличение pH на второй стадии достигается путем добавления гидроксида натрия или аммиака, причем затем pH поддерживают между 6.5-10.0, 7,0-8,0, 9,0 6.5- или 7,0-8,5 путем добавления гидроксида натрия или аммиака, также различается скорость подачи субстрата, экспрессия индуцируется на второй стадии добавлением ИПТГ до конечной концентрации от 0,1 до 10 мМ.
Однако выделение белка из телец включения является более простым, менее трудоемким и позволяет получить больший выход белка из 1 л культуры клеток.
Известен способ получения белка CRM197, слитого с тагом, с использованием клеток Escherichia coli [WO 2010150230]. В тексте описания указано, что использовали плазмидную ДНК pET9a и штамм E. coli BL21AI TM и BL21 (DE3). Также авторы указывают как возможные для использования векторы pET3a, pET3b, pET3c, pET5a, pET5b, pET5c, pET9b, pET9c, pET12a, pET12b, pET12c, pET17b (Novagen), pRSETA, B and C (Invitrogen) и pTYB1, pTYB2, рТУВ3 и pTYB4 (New England Biolabs), все векторы, которые имеют сильный промотор T7; штаммы E. coli, содержащие копию гена, кодирующего Т7 РНК-полимеразу, в том числе BL21StarTM(DE3), BL21-Gold(DE3), BL21(DE3)pLys, производные штамма ER2566, ER2566, ER2833, ER3011, ER3012, BL21 Al TM BL21(DE3), C41(DE3), C43(DE3) и все модифицированные штаммы Е. coli В - BLR(DE3), B834(DE3 TunerTM(DE3) или все модифицированные штаммы Е. coli K-12, HMS174(DE3), AD494(DE3), OrigamiTM(DE3), NovaBlue(DE3), RosettaTM(DE3). Предложена полинуклеотидная последовательность, кодирующая CRM197, оптимизированная для экспрессии в клетках Е. coli, которая может быть соединена с последовательностью, кодирующей любой таг.
Однако целесообразно использование продуцента, который обеспечивает высокий уровень синтеза CRM197, не слитого с какой-либо последовательностью, для уменьшения количества работы с белком и увеличения вероятности прохождения препаратом доклинических и клинических исследований.
Известен способ получения мутантного дифтерийного токсина CRM197 с использованием клеток Escherichia coli, включающий стадии получения телец включения, рефолдинг, разделение и концентрирование ультрафильтрацией раствора для рефолдинга, замену среды на Tris-HCl буфер, концентрирование раствора для рефолдинга, очистки с использованием анионообменной хроматографии, причем для хроматографии используют уравновешивающий буфер (Tris-HCl или фосфатный буфер 10 ммоль/л-1 моль/л, pH 6.0-11.0), добавление NaCl (10 ммоль/л-1 моль/л), элюирование белка в режиме вымывания, либо прерывистым элюционным градиентом [CN 101265288]. Для хроматографии используют агарозу или полистирол, с четвертичной аммониевой группой, DEAE-Sepharose FF, Q-Sepharose FF, SOURCE30-Q or Q-Big-Beds, причем может быть использована диэлектрическая среда с DEAE - Sepharose FF. В данном патенте не описан используемый штамм E. coli, в этом отношении приведена ссылка на документ CN 200610042194.7, однако такой номер не существует.
Известен способ получения мутантного дифтерийного токсина CRM197 с использованием клеток Escherichia coli, включающий стадии получения телец включения, денатурацию, центрифугирование, диализ супернатанта, помещение в буферную проточную систему и рефолдинг при перемешивании при 4°C, осаждение белка сульфатом аммония (от 20% до 40%), центрифугрование, концентрирование ультрафильтрацией с использованием ионообменной хроматографии, разделение с иерархическим использованием молекулярных сит [CN 103266125]. Используют ген, кодирующий CRM197, кодонно оптимизированный. В описании изобретения указано, что использовали клетки E. coli DH5α, а также плазмидную ДНК pBAD-DEST49, затем клетки E. coli BL21. Использование штамма E. coli, в котором возможна контролируемая экспрессия целевого белка, является более предпочтительным.
Известен способ получения CRM197 с использованием клеток Escherichia coli, в тельцах включения, который характеризуется следующими шагами: (1) клонирование гена, кодирующего CRM197 (дизайн последовательности CRM197 и ДНК-последовательностей праймеров; с учетом синтетической рекомбинантной плазмиды); (2) создание рекомбинантной экспрессионной плазмиды (разрезание последовательностей ДНК, кодирующих CRM197 и векторную ДНК (вектора серии PET, PACYC PMBP), очистка и лигирование, трансформация, скрининг и идентификация); (3) трансформация клеток E. coli (E. coli BL21(DE3), JM109(DE3), OrigmaB(DE3) или Rosseta(DE3)) полученным экспрессионным вектором, скрининг (индукция экспрессии целевого белка, анализ белка-мишени электрофорезом в полиакриламидном геле ДСН-ПААГ; секвенирование ДНК); (4) очистка CRM197 (извлечение телец включения, отмывка, рефолдинг, очистка на анионообменнике на колонке (Q или DEAE) и гель-фильтрация белка, анализ очищенного белка с использованием ДСН-ПААГ [CN 100999548]. Описанное в данном патенте на изобретение техническое решение является наиболее близким аналогом предлагаемого изобретения - прототипом.
Однако показано, что рекомбинантные белки, получаемые с использованием экспрессионных систем, в том числе бактериальных, которые сохраняют N-концевой метионин, в некоторых случаях отличаются от таковых, синтезируемых в клетке-хозяине, что может приводить к негативным последствиям при применении таких белков, включая индукцию формирования нежелательных антител. Использование метионинаминопептидазы для модификации рекомбинантного белка позволяет осуществить способ недорогой генерации терапевтических белков, имеющих структуру, имитирующую таковую местных белков клетки-хозяина, которые используются для борьбы с причинами различных заболеваний [Sandman et al., Biotechnology (NY) 13:504-6 (1995), US 6638750 (B1)]. Так, известен способ получения безметионинового интерферона альфа с использованием вектора, кодирующего метиониновую аминопептидазу и ген интерферона альфа человека, в клетках E. coli BL21 [CN 102367441 (B)]. Использование такой конструкции позволяет получить белок с уменьшенной иммуногенностью и уменьшить количество побочных реакций при использовании у человека.
Авторами настоящего изобретения впервые для CRM197, для белка-адъюванта, предложено использование подхода, при котором в процессе его синтеза в клетках E. coli отщепляется N-концевой метионин.
Технический результат выражается в увеличении эффективности вакцинации. Данный технический результат достигается тем, что формируются более специфичные антитела к вводимой конструкции, к полисахариду, за счет того, что уменьшается формирование антител на данный специфичный для экспрессии в клетках E. coli элемент, что позволит акцентировать внимание на конструкции целиком.
Технический результат также выражается в увеличении эффективности вакцинации разными вакцинами, содержащими одинаковый адъювант. Данный технический результат достигается тем, что конструкция не подвергается блокированию и быстрой элиминации, индуцируется ответ на всю конструкцию, благодаря тому, что уменьшается вероятность специфического ответа на N-концевой метионин конструкции, полученной в клетках E. coli, в результате появляется возможность использования данного адъюванта в различных вакцинах, вводимых одному пациенту.
Технический результат также выражается в увеличении безопасности вводимой вакцины за счет уменьшения побочных эффектов, связанных с N-концевым метионином CRM197.
Сущность изобретения
Предложен штамм-продуцент CRM197 на основе клеток Escherichia coli BL21(DE3), содержащих плазмидную ДНК pETDuet-1, с которой одновременно синтезируются белок CRM197 и метиониновая аминопептидаза E. coli, гены, кодирующие данные белки, кодонно оптимизированы для экспрессии в клетках Е. coli, в 7 результате, синтезируемый CRM197 - безметиониновый. Использование данного штамма позволяет получить более безопасный компонент вакцины - безметиониновый CRM197, при этом не требуются дополнительные стадии работы с белком, для отделения метионина, дезактивации фермента и дополнительной очистки белка, поскольку отделение происходит внутри клетки, фермент также работает внутри клетки. При этом метиониновая аминопептидаза синтезируется в растворимом виде, за счет того, что использовали видоспецифичный фермент, а также за счет кодонной оптимизации, в результате является активным ферментом, способным к котрансляционной модификации синтезируемого CRM197, при этом CRM197 переходит в нерастворимую форму (в тельца включения), в результате стоит задача выделения белка исключительно из нерастворимой фракции - из телец включения, что облегчает очистку белка.
Результаты исследований проиллюстрированы примерами 1-3 и фиг. 1.
Краткое описание чертежей.
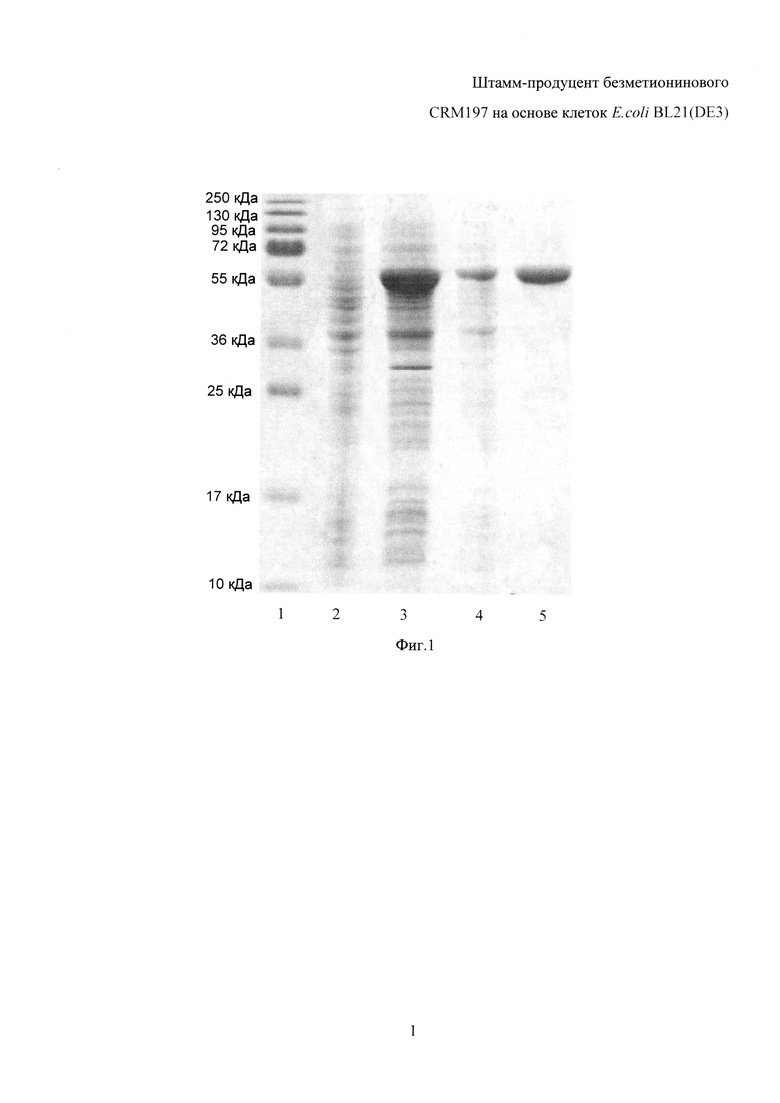
Фиг. 1. Электрофореграмма результатов индукции экпрессии гена crm197 с использованием 0,5 мМ ИПТГ, а также очистки белка CRM197, 12,5% ПААГ, 0,1% ДДС-Na. 1 - маркер молекулярного веса, 2 - лизат клеток штамма-продуцента до индукции, 3 - лизат клеток штамма-продуцента после индукции экспрессии гена 0,5 мМ ИПТГ, 4 - препарат рекомбинантного белка CRM197, полученный после проведения гидрофобной хроматографии, 5 - высокоочищенный препарат рекомбинантного белка CRM197, полученный после проведения гель-фильтрации.
Пример 1. Создание штамма-продуцента безметионинового белка CRM197 на основе клеток Escherichia coli.
1.1. Создание нуклеотидной последовательности, кодирующей белок CRM197, и нуклеотидной последовательности, кодирующей метионинаминопептидазу E. coli. Использовали транслированный по нуклеотидной последовательности NCBI Reference Sequence: NC_012971.2 белок метионинаминопептидазу E. coli BL21(DE3), аминокислотную последовательность которого переводили в нуклеотидную с одновременной кодонной оптимизацией для экспрессии в клетках E. coli с использованием программы на сайте molbiol.ru и добавлением сайтов рестрикции Nco I и Hind III, а также стоп-кодона.
Использовали транслированный по нуклеотидной последовательности NCBI PRF: 224021 белок CRM197, аминокислотную последовательность которого переводили в нуклеотидную с одновременной кодонной оптимизацией для экспрессии в клетках E. coli с использованием программы на сайте molbiol.ru и добавлением сайтов рестрикции Nde I и Avr II, а также старт- и стоп-кодона.
Рассчитанные нуклеотидные последовательности синтезировали химическим методом с помощью синтезатора ДНК ASM-800 (БИОССЕТ, Россия).
1.2. Создание плазмидной ДНК, кодирующей белки CRM197 и метионинаминопептидазу E. coli.
Полученные гены клонировали в плазмиде pETDuet-1 для последующей экспрессии. Для этого проводили реакцию рестрикции, а затем лигирования генов и вектора pETDuet-1, последовательно для каждого гена, с использованием соответствующего буфера и лигазы, при +20°C в течение 2 часов.
Смесь прогревали при +95°C в течение 10 мин и очищали от солей диализом на нитроцелюлозных фильтрах с диаметром пор 0,025 мкм (Millipore, США). Диализ проводили против раствора, содержащего 0,5 мМ ЭДТА в 10% глицерине, в течение 10 мин.
1.3. Создание штамма Е. coli для амплификации полученной плазмидной ДНК.
Полученной плазмидой трансформировали клетки Е. coli штамма DH10B/R (Gibko BRL, США) с генотипом F-mcrA Δ(mrr-hsdRMS-mcrBC) φ80dlacZΔM 15 ΔlacX74 deoR recA1 endA1 araD139 Δ(ara, leu)769 galU galKλ-rpsL nupG методом электропорации.
После трансформации клетки инкубировали в SOC-среде (2% бакто-триптон, 0.5% дрожжевой экстракт, 10 мМ NaCl, 2.5 мМ KCl, 10 мМ MgCl2, 10 мМ MgSO4, 20 мМ глюкоза) в течение 40 мин при +37°C.
Проводили скрининг клеток E. coli на наличие плазмид на селективной среде, содержащей LB-arap, 50 мкг/мл ампициллина. Выросшие на селективной среде колонии клеток E. coli проверяли на наличие вставки таргетных генов.
Из выросших клонов выделяли плазмидную ДНК с использованием набора Wizard Minipreps DNA Purification System (Promega, США). Очищенную плазмидную ДНК проверяли с помощью секвенирования с использованием стандартных праймеров для секвенирования для используемой плазмидной ДНК. В ходе работы были отобраны клоны, содержащие требуемые фрагменты ДНК в составе плазмиды, - штамм E. coli для амплификации плазмидной ДНК, содержащей таргетные гены, - из которого такие плазмиды были выделены для дальнейшего создания штамма-продуцента.
1.4. Создание штамма-продуцента безметионинового белка CRM197 на основе клеток Escherichia coli BL21 (DE3).
Для экспрессии генов, кодирующих белки метионинаминопептидазу и CRM197, с полученной плазмиды использовали клетки E. coli штамма BL21 (DE3) (Invitrogen, USA), с генотипом F-ompT hsdSB (rB-mB-) gal dcm rne 131 (DE3), содержащие в геноме λDe3 лизоген и мутацию rne131. Мутированный ген rne (rne131) кодирует усеченную форму РНКазы Е, что уменьшает внутриклеточное разрушение мРНК, приводя к увеличению ее ферментативной стабильности. lon- и ompT-мутации по генам протеаз позволяют получать непротеолизированные рекомбинантные белки в больших количествах.
Подготавливали вышеуказанные клетки Е. coli следующим образом. Инкубировали клетки при +37°С в течение ночи в 5 мл L-бульона, содержащего 1% триптон, 1% дрожжевой экстракт и 1% натрий хлористый. Разводили культуру свежим L-бульоном в 50-100 раз и выращивали на качалке при +37°С до оптической плотности 0,2-0,3 при длине волны 590 нм. При достижении оптической плотности более 0,3 культуру разводили свежим L-бульоном до оптической плотности 0,1 и растили 30 мин. Переносили 100 мл культуры в стерильную центрифужную пробирку и осаждали клетки при +4°С на 5000 g в течение 10 мин. Супернатант сливали, клетки ресуспендировали в деионизованной воде в исходном объеме с последующим центрифугированием. Процедуру отмывки повторяли трижды. После отмывки осадок клеток ресуспендировали в малом объеме деионизованной воды и центрифугировали 30 сек. при 5000 об/мин. на микроцентрифуге.
Трансформацию компетентных клеток осуществляли методом электропорации. Для этого 1 мкл плазмидной ДНК добавляли к 12 мкл компетентных клеток, перемешивали и проводили электропорацию с использованием электропоратора MicroPulser (BioRad) в стерильных ячейках при электрическом импульсе напряженностью 10 кВ/см длительностью 4 мсек.
После трансформации клетки инкубировали в SOC-среде (2% бакто-триптон, 0,5% дрожжевой экстракт, 10 мМ NaCl, 2,5 мМ KCl, 10 мМ MgCl2, 10 мМ MgSO4, 20 мМ глюкоза) в течение 40 мин. при +37°С. 10-100 мкл клеточной суспензии высевали на селективную LB-среду (Gibko BRL, США), содержащую ампициллин (50 мкг/мл), для отбора клонов, содержащих плазмиды (штаммов-продуцентов).
Полученная после трансформации компетентных клеток штаммов Е. coli плазмида обеспечивала высокий уровень биосинтеза рекомбинантных белков, закодированных в ней.
Пример 2. Получение безметионинового CRM197 в клетках штамма-продуцента E. coli 2.1. при индукции синтеза белков 0.2% лактозой по методу Штудиера
Для культивирования полученного штамма-продуцента использовали стандартную агаризованную LB-среду, содержащую ампициллин в концентрации 50 мкг/мл и глюкозу в концентрации 1% для блокирования неспецифической экспрессии.
Индукцию экспрессии проводили при достижении культурой клеток оптической плотности 0.6-0.8 оптических единиц при длине волны 600 нм.
Для автоиндукции экспрессии по методу Штудиера использовали среду PYP-5052, состоящую из 1% пептона (Gibco, США), 0.5% дрожжевого экстракта (Gibco, США), 50 мМ Na2HPO4, 50 мМ K2HPO4, 25 мМ (NH4)2SO4, 2 мМ MgSO4, 0.5% глицерола, 0.05% глюкозы и 0.2% лактозы, в качестве индуктора использовали 0.2% лактозу [Studier FW. Protein production by auto-induction in high density shaking cultures. Protein Expr Purif. 2005 May; 41(1):207-34].
В среду PYP-5052, содержащую ампициллин в концентрации 50 мкг/мл, инокулировали единичную колонию штамма-продуцента. Ферментацию проводили при +37°C в термостатированном шейкере роторного типа при 250 об. мин. в течение 20 часов до отсутствия существенного изменения ОП600 за 1 час. Отбирали аликвоту на анализ экспрессии генов, кодирующих белки CRM197 и метиониновую аминопептидазу, методом электрофореза в ПААГ, а оставшуюся биомассу осаждали центрифугированием при 9000 g.
Клетки ресуспендировали в лизирующем буфере, содержащем 20 мМ трис-HCl pH 7,5, 5 мМ ЭДТА и 1 мМ феноксиметилсульфонилфторид, из расчета на 1 г клеток 5-7 мл буфера. Суспензию клеток обрабатывали ультразвуком 7 раз по 30 сек с интервалом в 30 сек (частота ультразвука составляет 22 кГц), отбирали пробу на анализ SDS-PAGE (PolyAcrylamide Gel Electrophoresis with Sodium dodecyl sulfate). Полученный препарат содержал около 5% метиониновой аминопептидазы и около 25% белка CRM197 от общего количества белков E. coli.
Лизат центрифугировали 10 мин. при +4°C, 5000 g. Отбирали пробу надосадочной жидкости (супернатанта) и осадка для анализа локализации белков и оценки их растворимости, с использованием SDS-PAGE. Анализ показал наличие белка массой около 29-30 кДа в супернатанте, что соответствует рассчитанной массе метионинаминопептидазы (29,3 кДа), и, таким образом, продемонстрировал растворимость полученного белка. Осадок содержал белок массой около 58-59 кДа, что соответствует рассчитанной массе CRM197 (58,5 кДа), таким образом, анализ продемонстрировал нерастворимость полученного белка.
Супернатант сливали, к осадку добавляли раствор 1 M мочевины из расчета 10 мл на 1 г клеток, интенсивно перемешивали. Повторяли центрифугирование. Супернатант сливали, осадок ресуспендировали в растворе 2 М мочевины того же объема. Повторяли центрифугирование, сливали супернатант, осадок (тельца включения) использовали для выделения белка CRM197.
2.2. При индукции синтеза белков ИПТГ.
Индукцию экспрессии генов с использованием ИПТГ осуществляли следующим образом. Инкубировали клетки единичной колонии штамма-продуцента при +37°C в термостатированном шейкере роторного типа при 250 об./мин. в течение ночи в LB среде (1% триптон, 1% дрожжевой экстракт и 1% натрий хлористый), содержащей ампициллин в концентрации 50 мкг/мл. Разводили культуру свежей LB средой в 50 раз и выращивали в термостатированном шейкере роторного типа при 250 об./мин. +37°C до достижения культурой клеток оптической плотности 0.6-0.8 оптических единиц при длине волны 600 нм. После этого осуществляли индукцию экспрессии рекомбинантных генов добавлением ИПТГ к культуре до конечной концентрации 0,1 мМ. 0,5 мМ или 1 мМ. Индукцию проводили в течение 18 часов для определения оптимальной концентрации индуктора для получения высокого уровня экспрессии таргетных генов, после чего отбирали пробу для анализа с использованием SDS-PAGE, клетки концентрировали с помощью центрифугирования.
Клетки ресуспендировали в лизирующем буфере, содержащем 20 мМ трис-HCl pH 7,5, 5 мМ ЭДТА и 1 мМ феноксиметилсульфонилфторид, из расчета на 1 г клеток 5-7 мл буфера. Суспензию клеток обрабатывали ультразвуком 7 раз по 30 сек с интервалом в 30 сек (частота ультразвука составляет 22 кГц), отбирали пробу на анализ SDS-PAGE. Полученный препарат содержал в случае индукции 0,1 мМ ИПТГ около 2% метиониновой аминопептидазы и около 18% белка CRM197 от общего количества белков E. coli, при индукции 0,5 мМ ИПТГ - около 5% метиониновой аминопептидазы и около 27% белка CRM197 от общего количества белков E. coli, при индукции 1 мМ ИПТГ около 3% метиониновой аминопептидазы и около 20% белка CRM197 от общего количества белков E. coli.
Лизат центрифугировали 10 мин. При +4°C, 5000 g. Отбирали пробу надосадочной жидкости (супернатанта) и осадка для анализа локализации белков и оценки их растворимости, с использованием SDS-PAGE. Анализ проб после индукции ИПТГ во всех исследуемых концентрациях показал наличие белка массой около 29-30 кДа в супернатанте, что соответствует рассчитанной массе метионинаминопептидазы (29,3 кДа), и, таким образом, продемонстрировал растворимость полученного белка. Осадок содержал белок массой около 58-59 кДа, что соответствует рассчитанной массе CRM197 (58,5 кДа), таким образом, анализ продемонстрировал нерастворимость полученного белка.
Таким образом, наблюдали экспрессию генов, кодирующих белок CRM197 и метионинаминопептидазу, при всех вариантах индукции, а именно - с использованием 0,1 мМ, 0,5 мМ и 1 мМ ИПТГ, и 0,2% лактозы. При всех исследованных вариантах индукции белок CRM197 получали в нерастворимой фракции (тельцах включения), метионинаминопептидазу - в растворимой фракции, что облегчает выделение и очистку белка CRM197. Подобран оптимальный протокол индукции экспрессии генов, кодирующих рекомбинантные белки CRM197 и метиониновую аминопептидазу, - индукция 0,5 мМ ИПТГ при 37°C в течение ночи. Данный способ является оптимальным по выходу белков и затратам на его осуществление (Фиг. 1).
Пример 3. Очистка рекомбинантного белка CRM197.
Осуществляли очистку белка CRM197, полученного с использованием индукции 0,5 мМ ИПТГ. Биомассу, полученную из 1 л жидкой культуры клеток штамма-продуцента после индукции экпрессии гена crm197 (4 г), ресуспендировали на ледяной бане, добавляя 3 мл на 1 г биомассы охлажденного ресуспендирующего буфера (40 мМ Tris-HCl, pH 8,0; 1 мМ ЭДТА; 1 мМ дитиотреитол - ДТТ) и разрушали 5 циклами соникации. Фрагменты клеток осаждали центрифугированием при 15000 g в течение 40 мин. Для осветления лизата к супернатанту добавляли по каплям 10%-ный Тритон Х-100 до 0,1%, перемешивали, оставляли на 30 мин при 4° и затем центрифугировали при 25000 g 30 мин [DeWalt B.W. et al. Compaction agent clarification of microbial lysates // Protein Expr. Purif. 2003. Vol. 28, №2. P. 220-223].
К 40 мл осветленного клеточного лизата добавляли буфер для нанесения (50 мМ Tris-HCl, 1 мМ ЭДТА, 2 мМ ДТТ, 0.2 M бензамидин-HCl, 0.2 мМ PMSF, pH 7.5), после чего наносили на колонку HiPrep 16/10 Q XL (GE Healthcare, Швеция), уравновешенную буфером для нанесения, и промывали 50 мл того же буфера при скорости 10 мл/мин. Элюцию белков, связавшихся с носителем, проводили градиентом NaCl (0% В в 5 CV, 30% В в 5 CV, 100% В в 5 CV) до 10 M NaCl. Оптический профиль элюции определяли по поглощению при 280 нм на спектрофотометре. Фракции (5 мл каждая), содержащие белок CRM197, объединяли и наносили на колонку SOURCE 15ISO (GE Healthcare, Швеция), уравновешенную буфером для нанесения (1.6 M (NH4)2SO4, 10% глицерин, 50 мМ Tris-HCl, 1 мМ ЭДТА, 2 мМ ДТТ, 0.2 мМ бензамидин-HCl, 0.2 мМ PMSF, pH 7.5). После промывания колонки 50 мл того же буфера белок элюировали буфером для элюции (50 мМ Tris-HCl, 10% глицерин, 1 мМ ЭДТА, 2 мМ ДТТ, 0.2 мМ бензамидин-HCl, 0.2 мМ PMSF, pH 7.5) при скорости 5 мл/мин. Для элюции использовали градиент - 0-16% В в 4 CV, 16-24% В в 8 CV, 24-35% В в 4 CV, 100% В в 4 CV. Объем каждой фракции составил 5 мл.
Для доочистки белка CRM197, а также перевода в нативные условия использовали гель-фильтрацию. Данный метод не только позволяет разделять молекулы по их размеру, но и обеспечивает формирование правильной пространственной структуры рекомбинантных белков [Li M., Su Z.-G., Janson J.-C. In vitro protein refolding by chromatographic procedures // Protein Expr. Purif. 2004. Vol. 33, №1. p. 1-10].
Хроматографию проводили на гель-фильтрационной колонке XK26/60 с сорбентом Superdex S-100, при скорости элюции 1,3 мл/мин, элюент - 0,01 M натрий фосфатный буфер, pH 7,2-7,4. Результаты анализировали с помощью электрофореза в 12,5%-ном полиакриламидном геле в денатурирующих условиях по Леммли [Laemmli U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4 // Nature. 1970. Vol. 227, №5259. P. 680-685], гели окрашивали Кумасси G-250. Концентрацию белка определяли по методу Бредфорд [Bradford М.М. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding // Anal. Biochem. 1976. Vol. 72. P. 248-254].
В результате был получен препарат белка CRM197 с чистотой 97%, выход белка составил 0,3 г из 1 литра жидкой культуры штамма-продуцента (Фиг. 1).
Пример 4. Свойства полученного штамма-продуцента белка CRM197.
Культуральные свойства
Грамотрицательные прямые палочки, размером 1,1-1,5-2,0-3,0 мкм, одиночные, спор и капсул не образуют. Каталазоположительные. Оксидазоотрицательные. Факультативные анаэробы. Интервал pH 5-7. Катализируют D-глюкозу и некоторые другие углеводы с образованием кислоты и газа, не сбраживают лактозу, арабинозу и галактозу. Реакция Фогес-Проскауэра отрицательная, не образуют H2S, но гидролизуют мочевину.
Ростовые характеристики Клетки растут в интервале температур от 8°C до 43°C, интервал для культивирования - 28-38°C, оптимум роста при 37°C.
Описание визуальных и цитологических наблюдений при стандартных условиях культивирования
Клетки хорошо растут на простых питательных средах, содержащих и не содержащих ампициллин. На агаризованной среде - колонии гладкие, круглые, слабо выпуклые, с ровным краем. В жидких средах образуют равномерную светорассеивающую суспензию, при хранении без перемешивания оседают на дно.
Изобретение относится к области молекулярной биологии и касается штамма-продуцента безметионинового белка CRM197. Предложенный штамм получен на основе клеток Escherichia coli BL21 (DE3) и содержит плазмидную ДНК pETDuet-1, включающую ген, с которого синтезируется белок CRM197, и дополнительно ген, с которого синтезируется метиониновая аминопептидаза E. coli, при этом гены кодонно оптимизированы для экспрессии в клетках E. coli и синтез двух белков осуществляется одновременно. Изобретение может быть использовано для повышения иммуногенности вакцин. 1 ил., 4 пр.
Штамм-продуцент безметионинового белка CRM197 на основе клеток Escherichia coli BL21 (DE3), содержащий плазмидную ДНК pETDuet-1, включающую ген, с которого синтезируется белок CRM197, и дополнительно ген, с которого синтезируется метиониновая аминопептидаза E. coli, при этом гены кодонно оптимизированы для экспрессии в клетках E. coli.
| КОНЪЮГАТ ПОЛИСАХАРИД-БЕЛОК ДЛЯ ИНДУЦИРОВАНИЯ ИММУННОГО ОТВЕТА И ЗАЩИТЫ ПРОТИВ ИНФЕКЦИИ Staphylococcus aureus, СПОСОБЫ ПОЛУЧЕНИЯ КОНЪЮГАТА (ВАРИАНТЫ), КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ КОНЪЮГАТ И СПОСОБЫ ИНДУЦИРОВАНИЯ ИММУННОГО ОТВЕТА И ПРЕДОТВРАЩЕНИЯ ИНФЕКЦИИ Staphylococcus aureus | 2010 |
|
RU2531234C2 |
| CN 100999548 A, 18.07.2007 | |||
| WO 2010150230 A1, 29.12.2010 | |||
| WO 2001087982 A2, 22.11.2001. | |||

