По данной заявке испрашивается приоритет по предварительной заявке США №61/258106, поданной 4 ноября 2010, раскрытие которой включено в настоящую заявку посредством отсылки.
Уровень техники
Считается, что в ближайшие 25 лет число людей старше 65 лет в Соединенных Штатах по меньшей мере увеличится вдвое, а население людей преклонного возраста во многих других странах увеличится еще более быстрыми темпами.
С увеличением хронологического возраста существует значительно повышенный риск многочисленных истощающих заболеваний. Таким образом, существует постоянная потребность в определении стратегий для предотвращения, замедления или лечения возрастных болезней. Настоящее изобретение рассматривает данную потребность.
Раскрытие изобретения
Настоящее изобретение обеспечивает способ подавления и/или замедления клеточного старения млекопитающих. Способ включает в себя контактирование клеток млекопитающих с композицией, содержащей негенотоксичный индуктор p53 (NGIP). В некоторых вариантах осуществления NGIP представляет собой Mdm-связывающий агент или Mdm-2 антагонист. В некоторых вариантах осуществления NGIP может быть нутлином, нутлином-3A, аналогом нутлина или их комбинацией.
Способ, как ожидается, будет полезен для профилактики и/или лечения возрастных болезней и/или клеточной гипертрофии у любого человека. В одном варианте воплощения изобретения у индивидуума, подвергающегося лечению в соответствии со способом по настоящему изобретению, злокачественное заболевание не было диагностировано. В других вариантах воплощения изобретение обеспечивает способ для снижения клеточной гипертрофии в организме путем введения терапевтически эффективного количества композиции, включающей в себя антигипертрофическое соединение, в организм. Не ограничиваясь примерами, антигипертрофические соединения, которые могут быть применены при осуществлении настоящего изобретения включают нутлин, нутлин-3A, аналог нутлина, рапамицин или аналог рапамицина и их комбинации.
В различных вариантах воплощения изобретения способ по настоящему изобретению приводит к подавлению и/или замедлению клеточного старения у млекопитающих. Подавление и/или замедление клеточного старения у млекопитающих может включать соответствующее состояние покоя клеток млекопитающих.
Краткое описание фигур
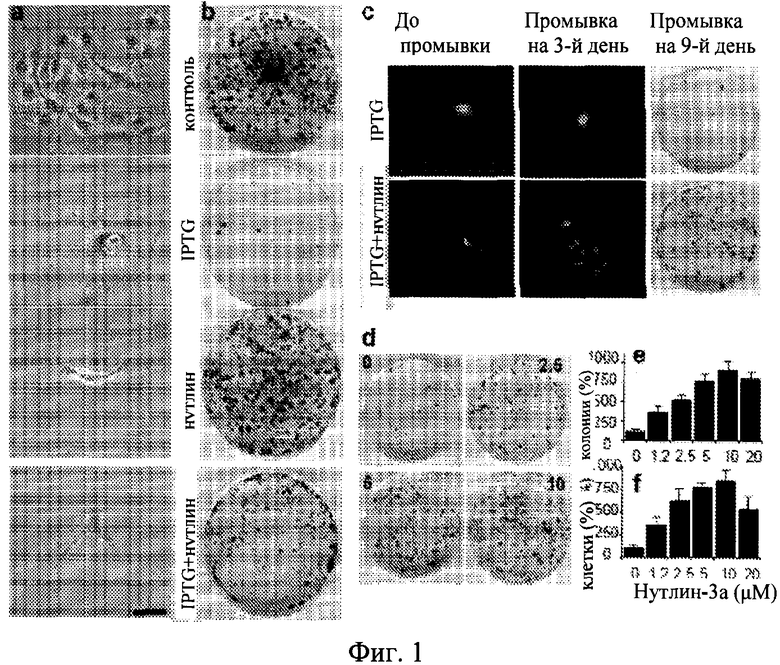
Фиг.1. Нутлин-3A преобразует старение в состояние покоя. а. Клетки HT-p21-9 обрабатывали IPTG, 10 мкМ нутлина-3a и IPTG с нутлином-3a в течение 3 дней. Клетки окрашивали на бета-Gal и фотографировали (оригинальное увеличение ×400). Масштаб столбца - 50 мкм. b. Клетки HT-p21-9 обрабатывали IPTG, 10 мкМ нутлина-3a и IPTG с нутлином-3a в течение 3 дней. После 3 дней клетки промывали для удаления IPTG и нутлина-3a. Затем клетки культивировали в свежей среде, до тех пор пока колонии становились видимыми. Планшеты окрашивали с кристаллическим фиолетовым и фотографировали на 4 день (контроль) или на 9 день (обработанные IPTG и нутлином-3a). c. Клетки HT-p21-9 обрабатывали с IPTG в присутствии или в отсутствие 10 мкм нутлина-3a в течение 3 дней до промывки. Живые клетки HT-p21-9 (экспрессирующие GFP для лучшей визуализации живых клеток) фотографировали (оригинальное увеличение ×100) под ультрафиолетовым излучением. Промывали на 3-й день. Три дня спустя препарат удаляли. Промывали на 9-й день. Девять дней спустя препарат удаляли, клетки окрашивали кристаллическим фиолетовым и фотографировали. d. Дозозависимый эффект нутлина-3a. Клетки HT-p21-9 высеивали с IPTG и 0, 2.5, 5 или 10 мкм нутлина-3a. После 3 дней планшеты промывали и клетки инкубировали в течение дополнительных 9 дней в свежей среде, окрашивали с кристаллическим фиолетовым и фотографировали. е. Колонии на планшете. Клетки HT-p21-9 высеивали с IPTG и 0, 1.2, 2.5, 5, 10 или 20 мкм нутлина-3а, как указано. После 3 дней планшеты промывали и клетки инкубировали в течение дополнительных 9 дней в свежей среде. Колонии подсчитывались, и результаты показаны как процент от контроля (только IPTG). f. Клетки на планшете. Как на панеле е. Клетки были трипсинизированы и подсчитаны. Результаты показаны как процент от контроля (только IPTG).
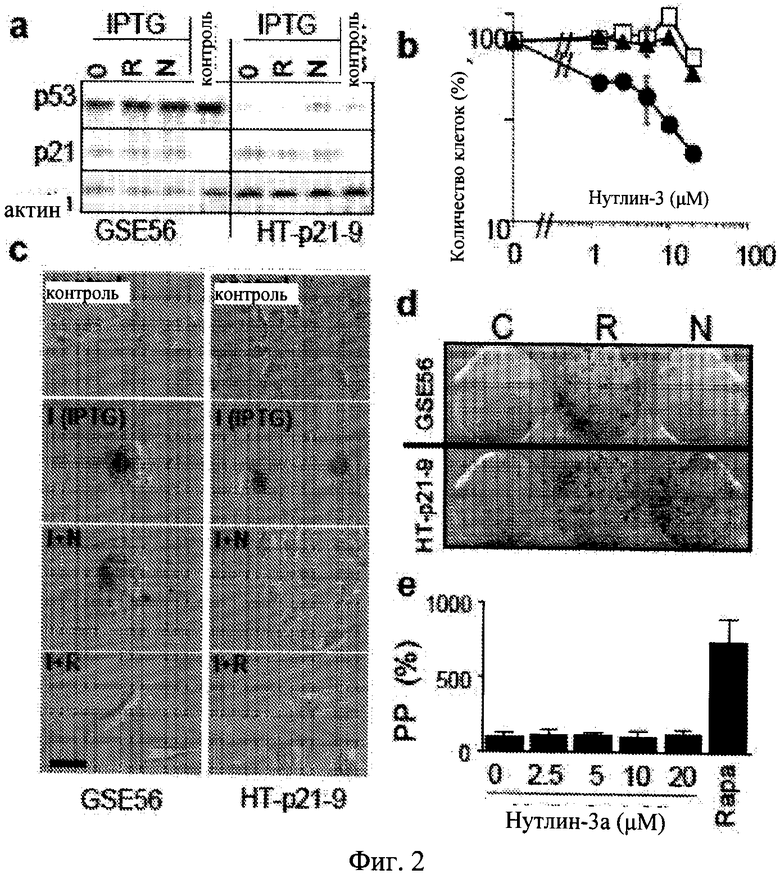
Фиг.2. p53-зависимые эффекты нутлина-3a. а. Клетки HT-p21-GSE56 и HT-p21-9 были обработаны только IPTG (0) или IPTG с рапамицином (R) и нутлином-3a (N). Контрольные клетки оставались необработанными (без IPTG). После 1 дня клетки были лизированы и производили иммуноблот. b. Клетки HT-p21-GSE56 (светлые кружки) и HT-p21-9 (темные кружки) были обработаны с нутлином-3a в течение 5 дней и затем подсчитывались. В качестве отрицательного контроля первичные клетки были обработаны с нутлином-3a (светлые квадраты). c-d. Клетки HT-p21-GSE56 и НТ-р21-9 были обработаны только IPTG или с IPTG + рапамицин (I+R) или IPTG + нутлин-3a (I+N), как указано. Контрольные клетки оставались необработанными (без IPTG). с. Морфология. После 3 дней клетки окрашивали на бета-Gal. Масштаб столбца - 50 мкм. d. Формирование колонии. После 3 дней клетки промывали и инкубировали в свежей среде без препаратов в течение дополнительных 9 дней. Планшеты окрашивали с кристаллическим фиолетовым и фотографировали. е. Пролиферативный потенциал (PP). После 3 дней, клетки HT-p21-GSE56 промывали и инкубировали в свежей среде без препаратов. Клетки были подсчитаны, и результаты показаны как процент только от IPTG.
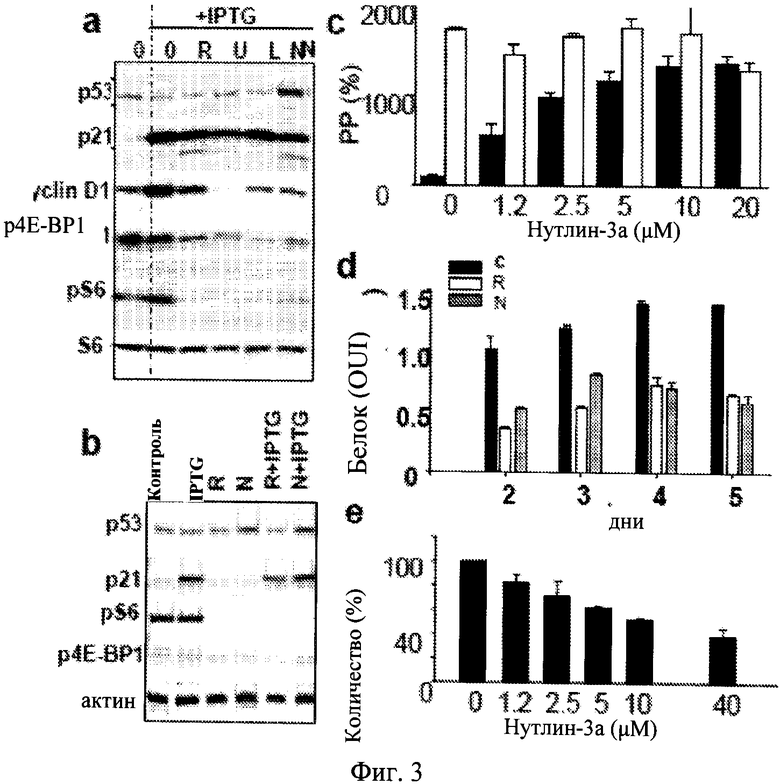
Фиг.3. Воздействия нутлина-3a на путь mTOR и синтез белка. а. Иммуноблот. Клетки HT-p21 обрабатывали только IPTG или IPTG с 500 нМ рапамицина (R), 25 мкМ LY-294002 (L), 10 мкМ U0126 (U) или 10 мкМ нутлина-3a (N) в течение 24 часов. Иммуноблот осуществляли, как описано в способах для Примера 1 ниже. b. Иммуноблот. Клетки HT-p21 были обработаны рапамицином (R) и нутлином-3a (N) в присутствии или в отсутствие IPTG в течение 18 часов. Иммуноблот осуществляли, как описано в Способах. с. Воздействия нутлина-3a на PP (пролиферативный потенциал) IPTG-обработанных клеток НТ-р21-9 в отсутствие (черные столбики) или присутствии (незакрашенные столбики) рапамицина (500 нМ). После 3 дней клетки промывали и инкубировали в свежей среде без препаратов в течение дополнительных 7 дней. Клетки подсчитывались и показаны как процент от только IPTG. d. Воздействия нутлина-3a и рапамицина на клеточную гипертрофию, вызванную IPTG. Клетки обрабатывали либо только IPTG (черные столбики), или IPTG с рапамицином (белые столбики), или с нутлином-3a (серые столбики). На 2, 3, 4 и 5 дня клетки лизировали и содержание белка в лунке измеряли. Представленные числа соответствуют содержанию белка в клетке, так как клетки не пролиферировали, и их количество оставалось неизменным в течение эксперимента. е. Воздействия нутлина-3a на синтез белка ([35S]метионин/цистеин включение). Клетки были помечены с [35S]метионин/цистеином, как описано в Способах для Примера 1.
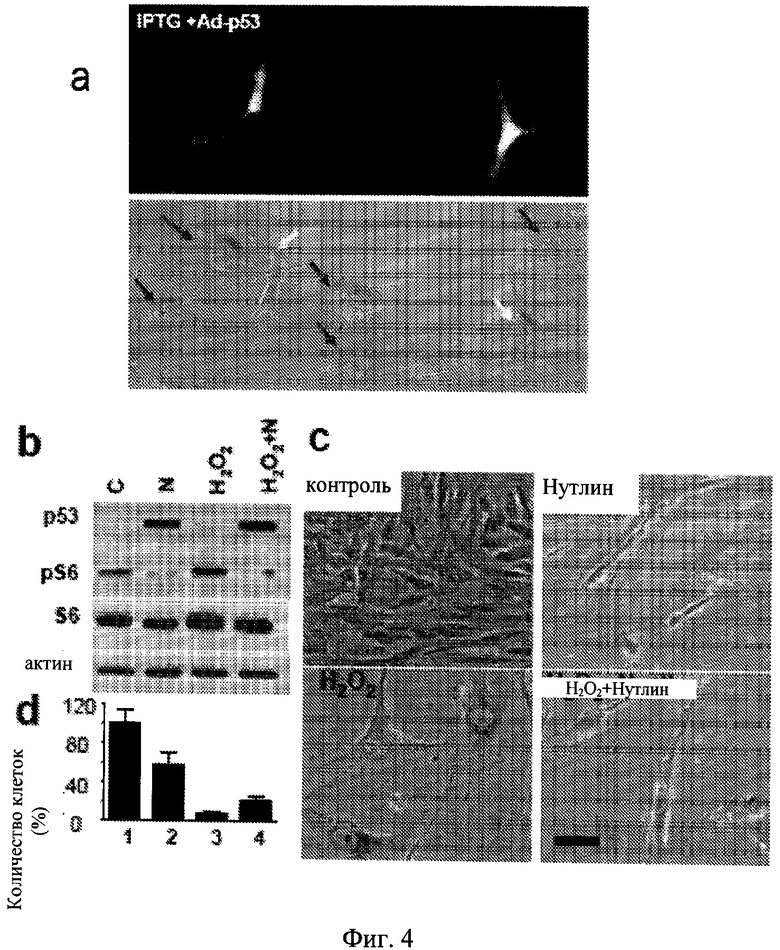
Фиг.4. Воздействия эктопического и эндогенного p53 на старение в HT-p21-9 и WI-38-tert. а. Аденовирус, экспрессирующий p53 (Ad-p53), замедляет стареющую морфологию, вызванную IPTG в клетках HT-p21-a. Клетки HT-p21-a обрабатывали IPTG и инфицировали Ad-p53. После 3 дней клетки фотографировали (оригинальное увеличение ×200): Верхняя панель. Под ультрафиолетовым облучением для визуализации клеток, экспрессирующих p53 (зеленые клетки). Нижняя панель. Под видимым светом для визуализации всех клеток. Красные стрелки указывают клетки, лишенные p53 экспрессии. Все эти клетки показывали большую, плоскую клеточную морфологию. Зеленые стрелки указывают клетки, экспрессирующие p53. b-d. Воздействия нутлина-3a на клеточное старение в фибробластах WI-38-tert, клетки WI-38-tert были обработаны с 200 мкМ Н2О2 в течение 30 минут в среде без сыворотки. Затем среда была заменена на полную среду (10% сыворотки) с или без 10 мкМ нутлина-3a. b. После 1 дня, клетки лизировали и осуществляли иммуноблот, как описано в Способах для Примера 1. с. После 3 дней клетки промывали (нутлин-3а был удален) и выращивали в течение 3 дополнительных дней в свежей полной среде. Клетки были затем окрашивали на активность бета-Gal и микрофотографировали. Масштаб столбца - 50 мкм. d. После 3 дней клетки промывали (нутлин-3a был удален) и выращивали в течение 6 дополнительных дней в свежей полной среде. Клетки затем трипсинизировали и подсчитывали. В контроле клетки достигали конфлюэнтности на 5 день и не пролиферировали дальше. Результаты показаны как процент от контроля.
Фиг.5. Морфология старения в сравнении с состоянием покоя. Клетки HT-p21 обрабатывали IPTG, нутлином-3a (мкМ) и IPTG с нутлином-3a в течение 3 дней или оставались необработанными (контроль). Живые клетки визуализировали с GFP (×100). В контроле клетки подвергали 3 разделениям, формированию микроколонии. Клетки, обработанные IPTG (большие и плоские), не подвергались никаким делениям. Клетки, обработанные нутлином-3a, подавляли после одного деления с клетками нормальной морфологии.
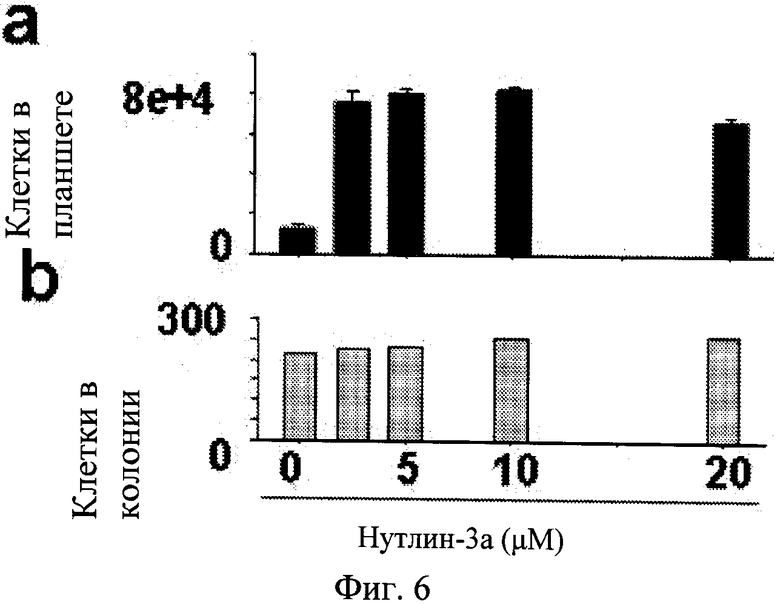
Фиг.6. Клетки HT-p21-9 высеивали в 10 мм планшеты и обрабатывали с IPTG в присутствии или в отсутствие нутлина-3a в течение 3 дней. Девять дней спустя препарат удаляли.
а. Количество клеток на планшет. Клетки на планшете были подсчитаны. b. Количество клеток на колонию. Количество клеток на колонию было подсчитано. Количество клеток на колонию было 200-250 (приблизительно равно 8 делениям) на 9 день. Таким образом, клетки в состоянии покоя характеризуются нормальным пролиферативным потенциалом после освобождения от IPTG + нутлин-3a.
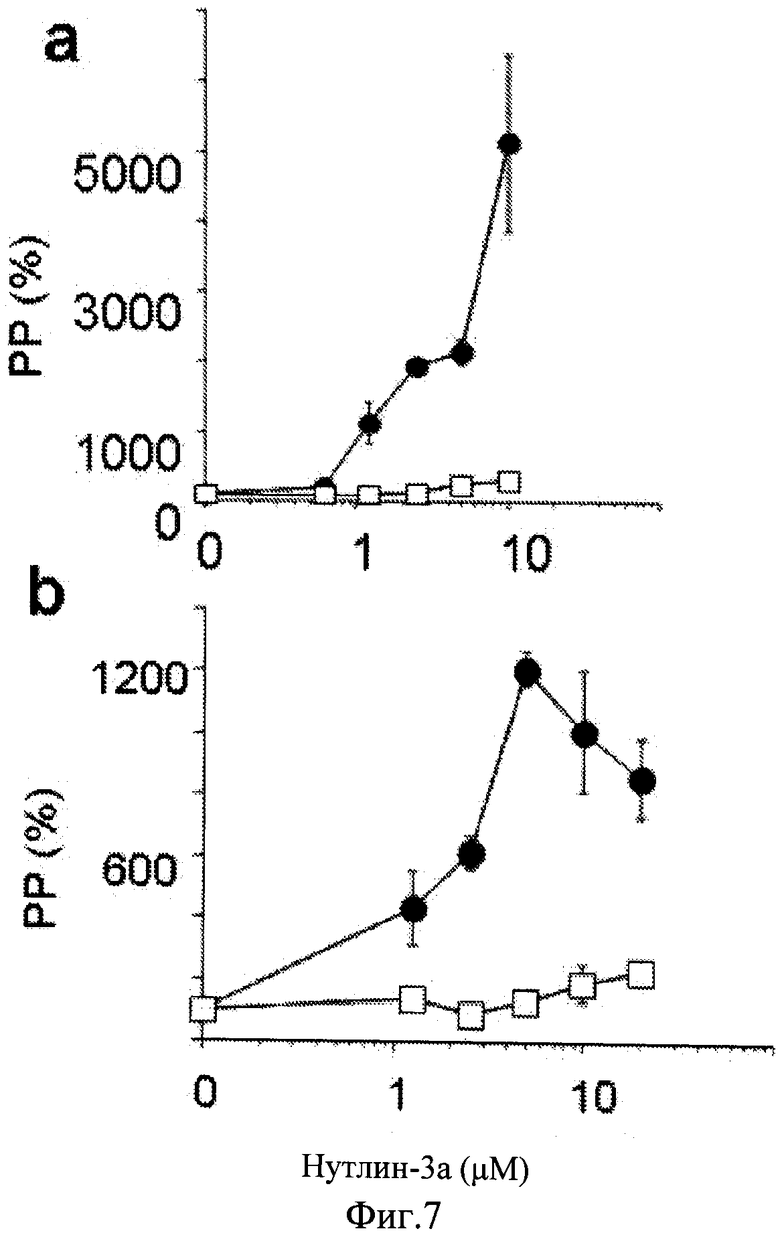
Фиг.7. Сохранение пролиферативного потенциала нутлином-3a. а. Сравнение нутлина-3a и нутлина-3b в клетках HT-p21-a. Клетки HT-p21-a обрабатывали IPTG в присутствии указанных концентраций нутлина-3a (темные кружки) и нутлина-3b (светлые квадраты) в течение 6 дней. Затем среду заменяли и клетки подсчитывали после 8 дней. b. Сравнение нутлина-3a и нутлина-3b в клетках HT-p16. Клетки HT-p16 обрабатывали в IPTG в присутствии указанных концентраций нутлина-3a (темные кружки) и нутлина-3b (светлые квадраты) в течение 3 дней. Затем среду сменяли и клетки подсчитывались после 5 дней.
Фиг.8. Воздействия IPTG и 500 нМ белка ([35S]метионин/цистеин включение). Клетки обрабатывали, как указано, в течение 24 часов и затем помечали с [35S]метионин/цистеином, как описано в Способах для Примера 1.
Фиг.9. а. Воздействия Ad-p21 и Ad-p53 на клеточную морфологию. Клетки p53, производные клеток HT-1080, были инфицированы с либо аденовирусом, экспрессирующим p21 (верхняя панель: Ad-p21), или аденовирусом, экспрессирующим p53 (нижняя панель: Ad-p53). Ad-p21 (верхняя панель) вызывал большую, плоскую морфологию. Ad-p53 не вызывал большой, плоской морфологии клеток. Клетки фотографировали при ×200. b. Ad-p53 замедлял стареющую морфологию, вызванную Ad-p21. Клетки p16-5, производные клеток HT-1080, были инфицированы с Ad-p21 и Ad-p53. Верхняя панель. Под ультрафиолетовым облучением для визуализации клеток, экспрессирующих p53 (зеленые клетки) (×200). Нижняя панель. Под видимым светом для визуализации всех клеток (×200). Красная стрелочка указывает на клетки со слабой экспрессией p53. Все другие клетки не приобретали большую, плоскую клеточную морфологию.
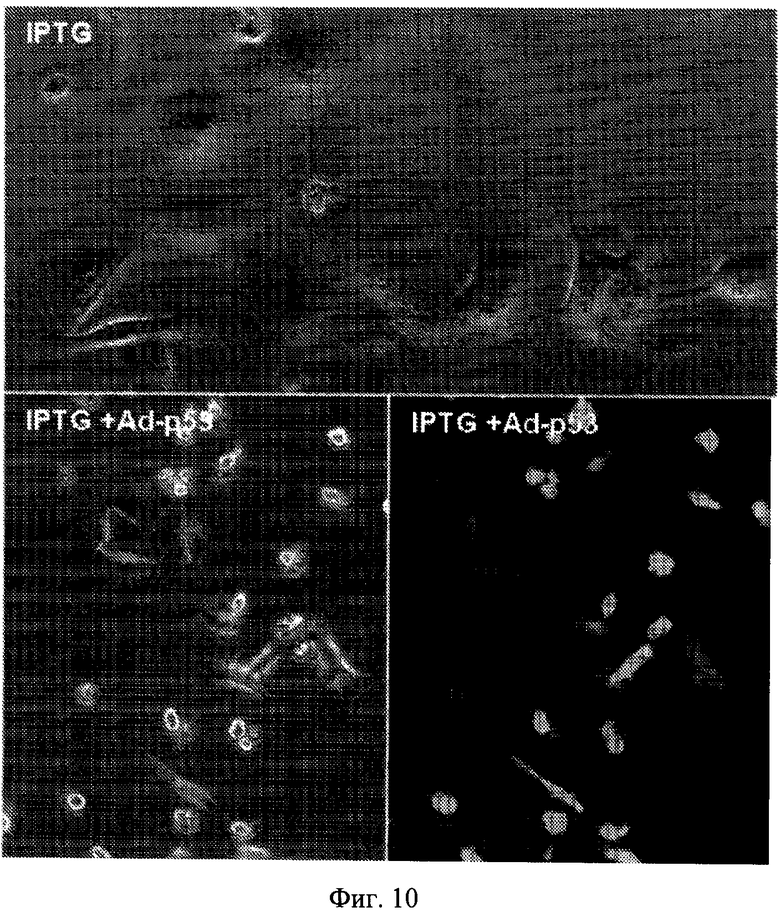
Фиг.10. Воздействия Ad-p53 на стареющую морфологию, вызванную p16. Клетки p16-5, производные клеток HT-1080, обрабатывали IPTG (верхняя панель) и IPTG с Ad-p53 (нижняя панель). IPTG (верхняя панель) вызывал большую, плоскую морфологию. Ad-p53 предотвращал данную морфологию. Клетки фотографировали при видимом свете и ультрафиолетовом облучении (×200) для визуализации клеток, экспрессирующих p53.
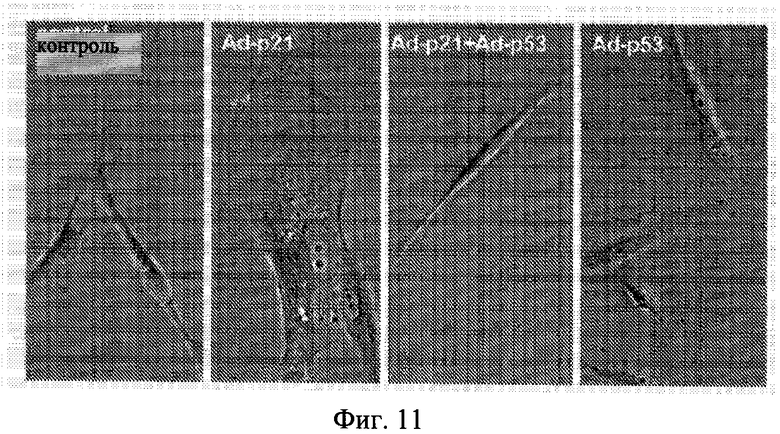
Фиг.11. Воздействия Ad-p21 и Ad-p53 на стареющую морфологию в фибробластах WI-38-tert. Клетки WI-38-tert были инфицированы либо аденовирусом, экспрессирующим p-21 (Ad-p21), или аденовирусом, экспрессирующим p53 (Ad-p53), или обоими. После 3 дней клетки окрашивали на бета-Gal.
Фиг.12. Воздействия нутлина-3a на уровень p53 и фосфорилирование S6/S6K в фибробластах WI-38-tert. Клетки WI-38-tert обрабатывали с указанными концентрациями нутлина-3а и 500 нМ рапамицина (Rapa), как указано, в течение 24 часов. Иммуноблот для р53, p-S6K, S6 и актина осуществляли, как описано в Способах для Примера 1.
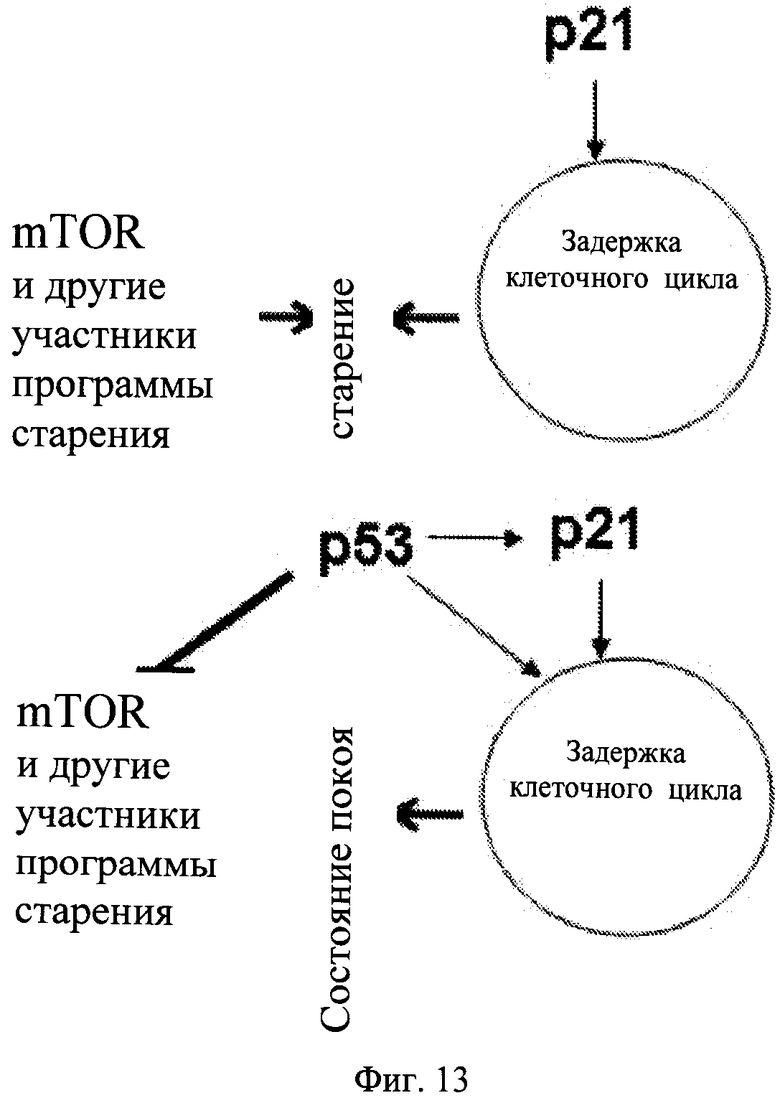
Фиг.13. Схема: Замедление старения p53. а. p21 вызывал подавление клеточного цикла, что приводило к старению. b. p53 вызывал подавление клеточного цикла и одновременно ингибировал программу старения, что приводило к состоянию покоя.
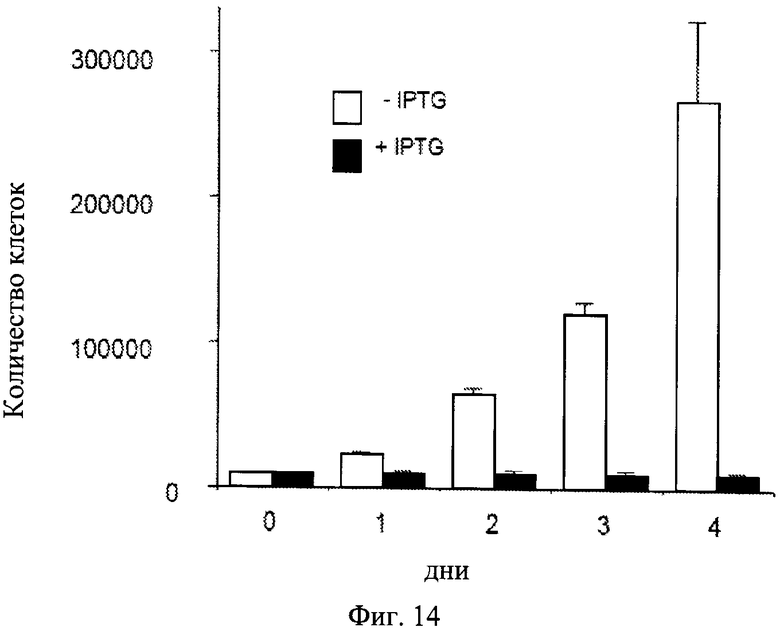
Фиг.14. Ингибирование пролифирации клеток IPTG.
Темные столбики: клетки HT-p21 обрабатывали IPTG (+IPTG). Клетки не пролиферировали. Светлые столбики: Необработанные клетки HT-p21. Экспоненциально пролиферирующие клетки. Клетки подсчитывали ежедневно.
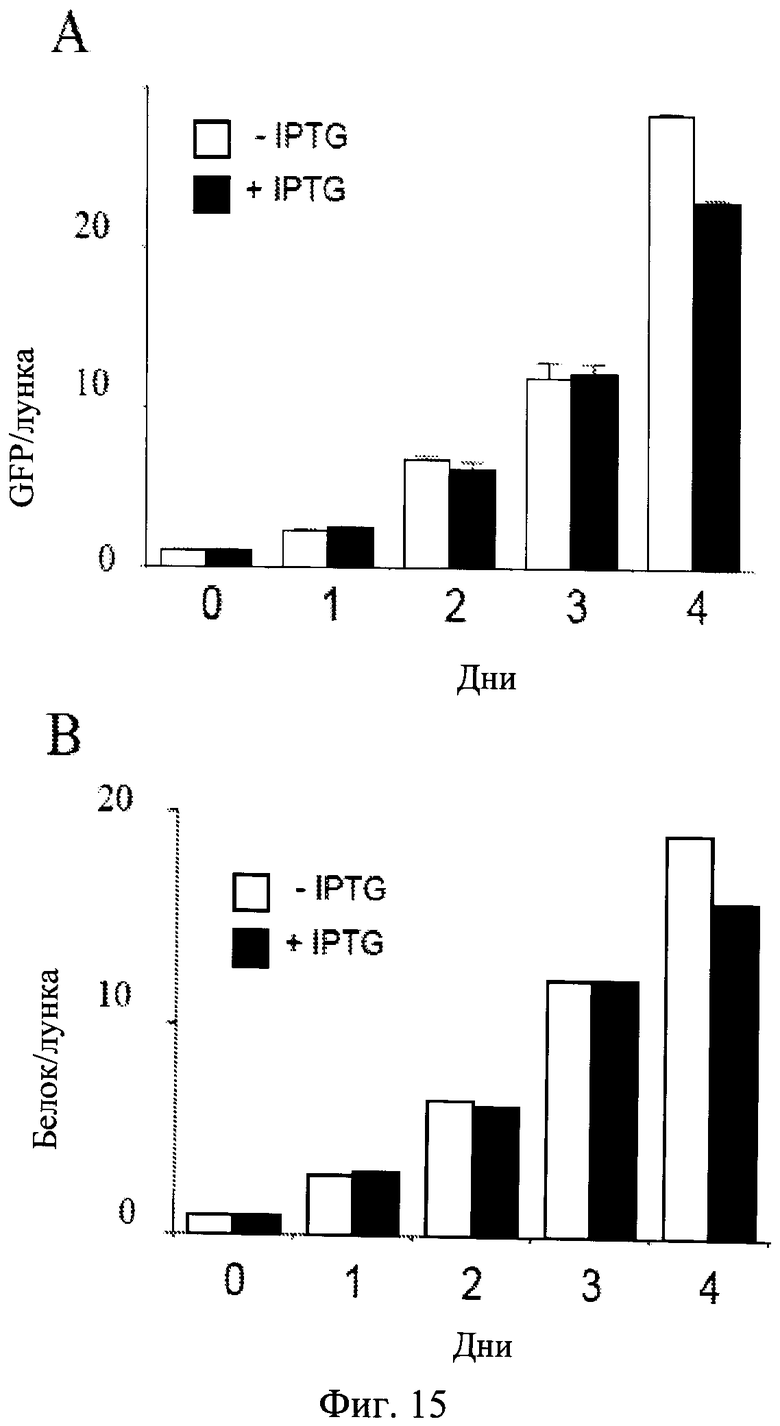
Фиг.15. Общее возрастание клеточной массы в процессе индукции старения.
Клетки HT-p21 росли в 60 мм лунках, и растворенный белок и GFP измеряли ежедневно.
Темные столбики: клетки HT-p21 обрабатывали с IPTG (+IPTG). Светлые столбики: Необработанные клетки HT-p21 (-IPTG). В обоих: пролиферирующих (-IPTG) и непролиферирующих (+IPTG) условиях, белок в лунке и GFP в лунке увеличивались. В панели В белок измеряли в двух повторностях, и показаны без стандартных отклонений, поэтому статистическое расхождение между -IPTG и +IPTG не должно рассматриваться. Панель только показывает экспоненциальную фазу роста в обоих условиях.
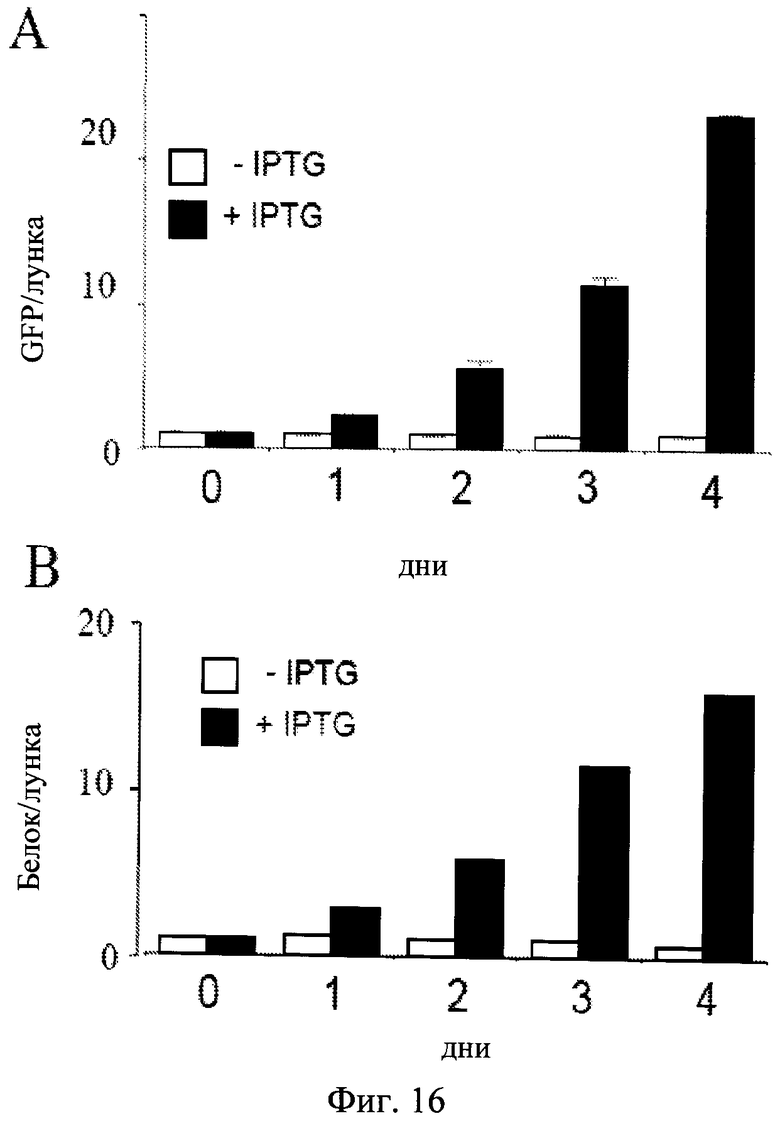
Фиг.16. Клеточная гипертрофия в процессе индукции старения.
Клетки HT-p21 росли в 60 мм лунках и количество клеток, растворимого белка и GFP измеряли ежедневно. Темные столбики: клетки HT-p21 обрабатывали IPTG (+IPTG). Светлые столбики: Необработанные клетки HT-p21 (-IPTG). Белок в клетке и GFP в клетке оставались без изменений в пролиферирующих (-IPTG) клетках. Белок в клетке и GFP в клетке увеличиваются экспоненциально в непролиферирующих (+IPTG) клетках.
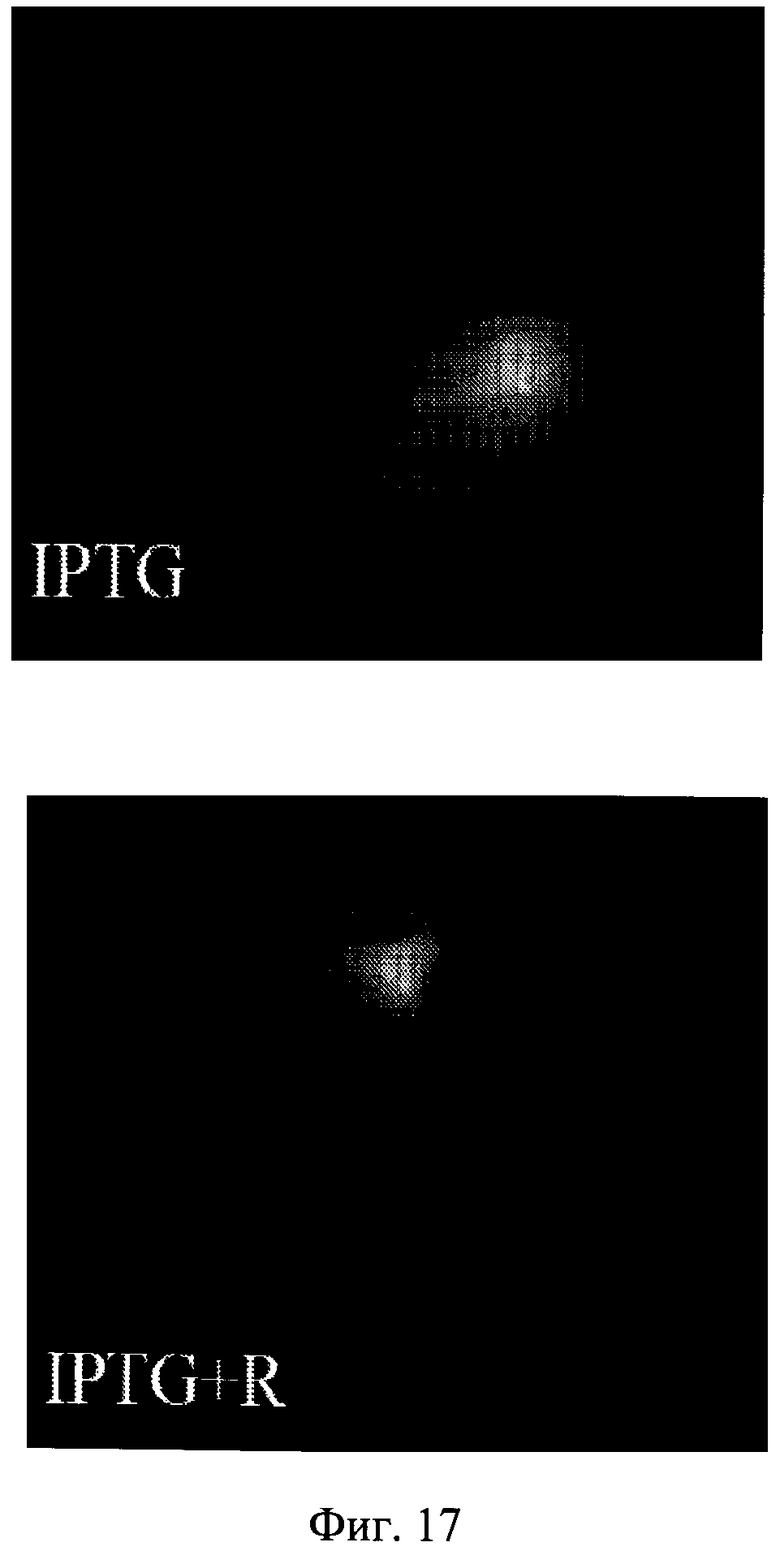
Фиг.17. Визуализация клеточной гипертрофии.
Клетки HT-p21 экспрессируют повышенный зеленый флуоресцентный белок (GFP) в соответствии с вирусным CMV промотором. Экспрессия GFP в клетке является маркером клеточной гипертрофии.
Низкая клеточная плотность - 2 тысячи клеток высеивали в 100 мм планшет и обрабатывали либо IPTG, либо IPTG + рапамицин.
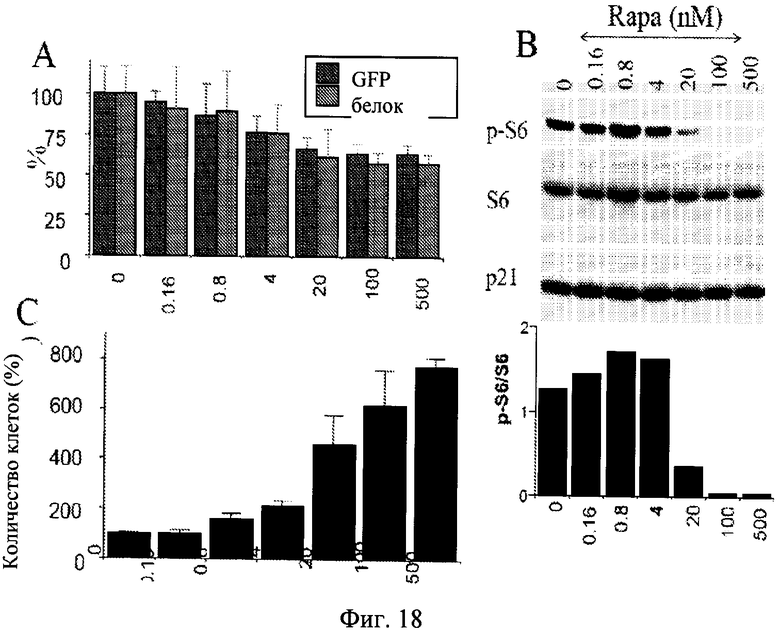
Фиг.18. Соответствие между фосфорилированием 86, гипертрофией и потерей пролиферативного потенциала в стареющих клетках. Клетки HT-p21 высеивали в 6 луночные планшеты и обрабатывали IPTG с увеличивающимися концентрациями рапамицина (от 0.16 до 500 нМ). При концентрации 0 клетки обрабатывали только IPTG. А. Клеточная гипертрофия: белок и GFP. После 3 дней растворимый белок и GFP изменяли в лунке. [Примечание: в непролиферирующих клетках белок/лунка является измерением белок/клетка]. Результаты показаны как процент от только IPTG (0) без рапамицина. В. После 3 дней клетки лизировали и иммублот для p-S6, S6 и p21. С. PC: сохранение пролиферативной способности. После 3 дней клетки промывали для удаления IPTG и RAPA. Клетки инкубировали в течение дополнительных 5 дней в свежей среде и затем подсчитывали. Результаты показаны как процент от только IPTG (0) без рапамицина.
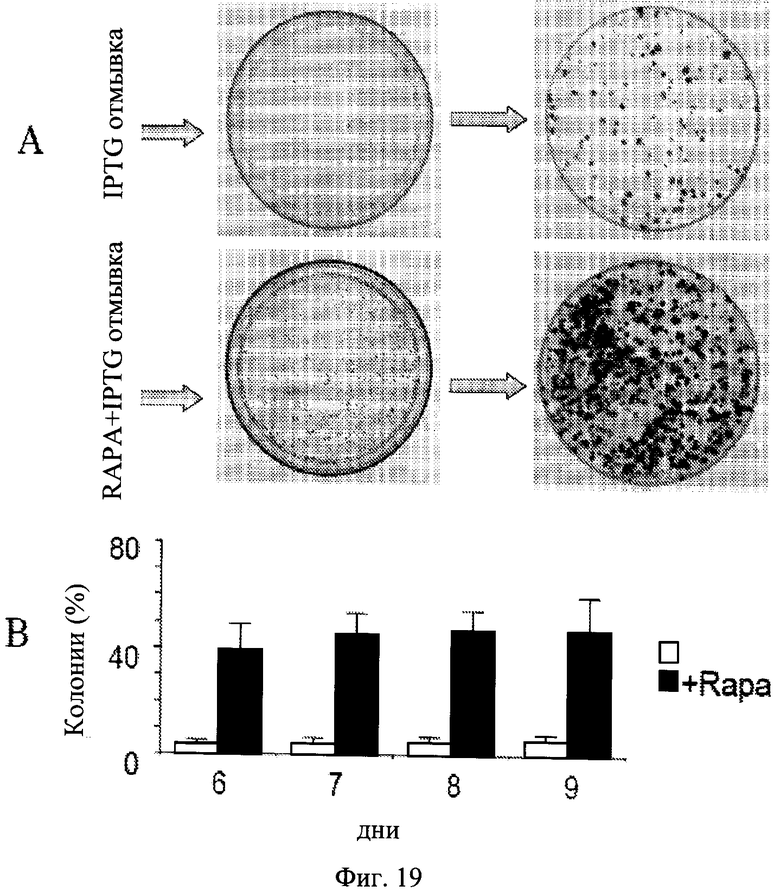
Фиг.19. Клональная пролиферация компетентных клеток. Клетки HT-p16 высеивали в 100-мм планшеты. На следующий день 50 мкМ IPTG с или без рапамицина, если указанный (RAPA) был добавлен. После 3 дней планшеты промывали для удаления IPTG и RAPA. А. Фотографии. Верхняя панель: На 5 и 8 дни (после удаления IPTG) планшеты фиксировали, окрашивали и фотографировали. Нижняя панель: На 5 и 8 дни (после удаления IPTG) планшеты фиксировали, окрашивали и фотографировали. В. Количество колоний. На 6, 7, 8 и 9 дни (после удаления IPTG), планшеты фиксировали, окрашивали и фотографировали. Количество колоний подсчитывали, и результаты показаны как процент от посеянных клеток в логарифмической шкале.
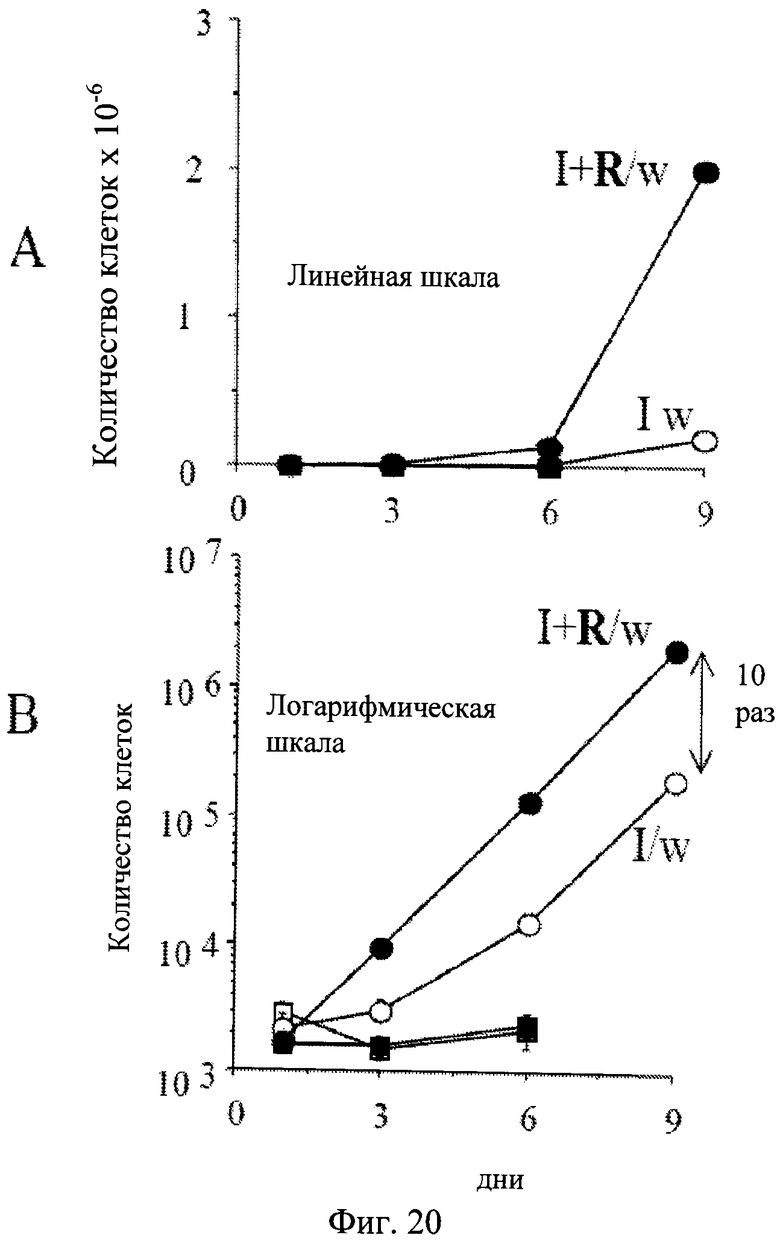
Фиг.20. Динамика количества клеток. 500 клеток HT-p21 высеивали в 12-луночные планшеты. На следующий день либо только IPTG или IPTG с рапамицином (I+R) были добавлены. После 3 дней планшеты промывали (I/w и I+R/w) или оставляли непромытыми. Клетки подсчитывались на 1, 3, 6 и 9 дни. Верхняя панель: линейная шкала. Нижняя панель: логарифмическая шкала. Светлые и темные квадраты: IPTG и IPTG с Rapa, соответственно. Светлые и темные кружки: IPTG промытые (I/w) и IPTG с Rapa промытые (I+R/w) соответственно. В присутствии IPTG (светлые квадраты) и IPTG с рапамицином (темные квадраты) клетки не пролиферируют.
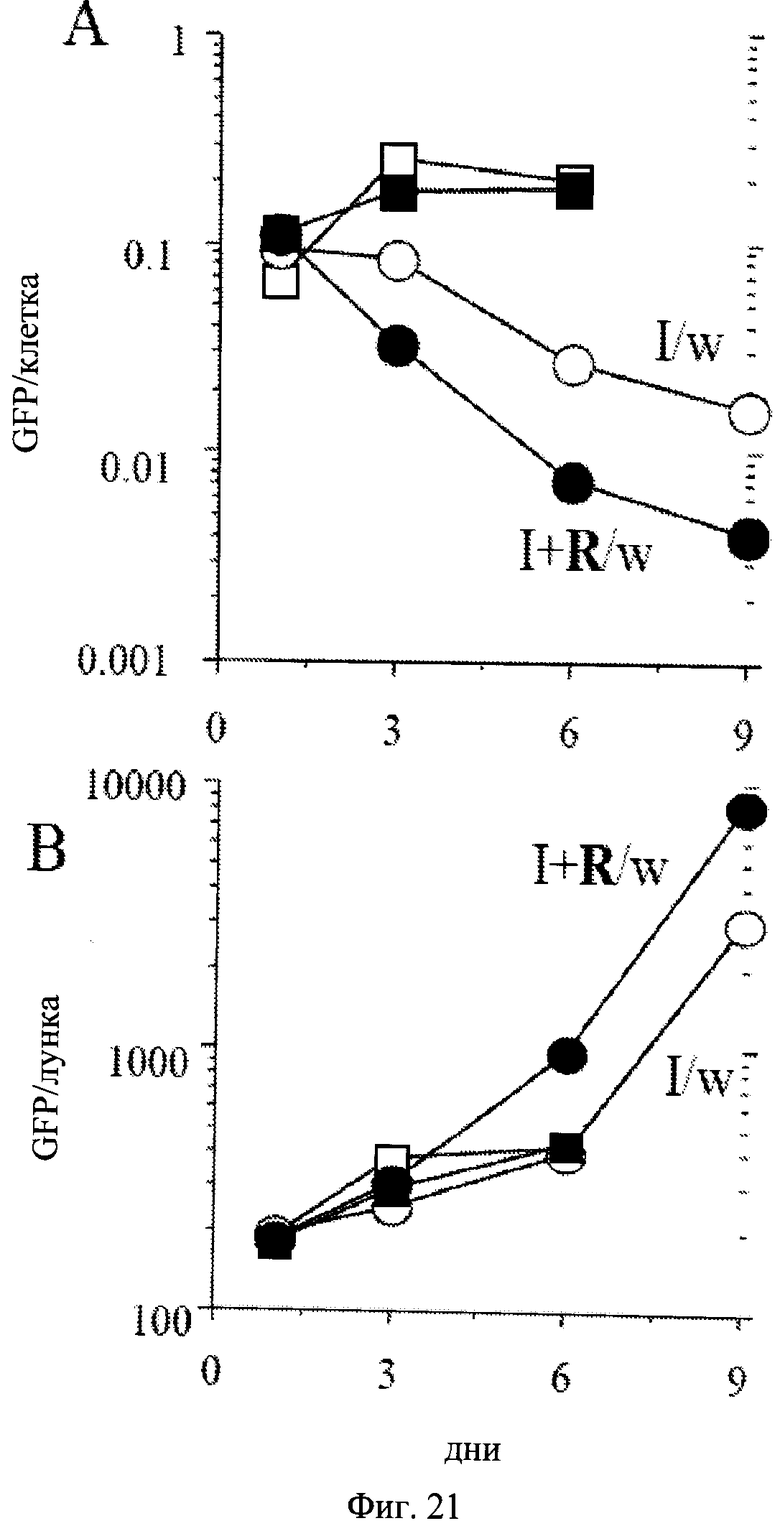
Фиг.21. Потеря гипертрофии в процессе пролиферации компетентных клеток. 500 клеток HT-p21 высеивали в 21-луночные планшеты. На следующий день добавляли либо только IPTG, или IPTG с рапамицином. После 3 дней планшеты промывали (I/w и I+RAv) или оставляли непромытыми. GFP в лунке измеряли и клетки подсчитывали на 1, 3, 6 и 9 дни. GFP в клетке подсчитывали (верхняя панель). Результаты показаны в произвольных единицах измерения (M±m). Светлые и темные квадраты: IPTG и IPTG с Rapa соответственно. Светлые и темные кружки: IPTG промытые (I/w) и IPTG с Rapa промытые (I+R/w) соответственно. Когда клетки возобновили экспоненциальную пролиферацию, GFP в клетке понизился до нормального уровня. В связи с сильной пролиферацией, имеется возрастание GFP в лунке.
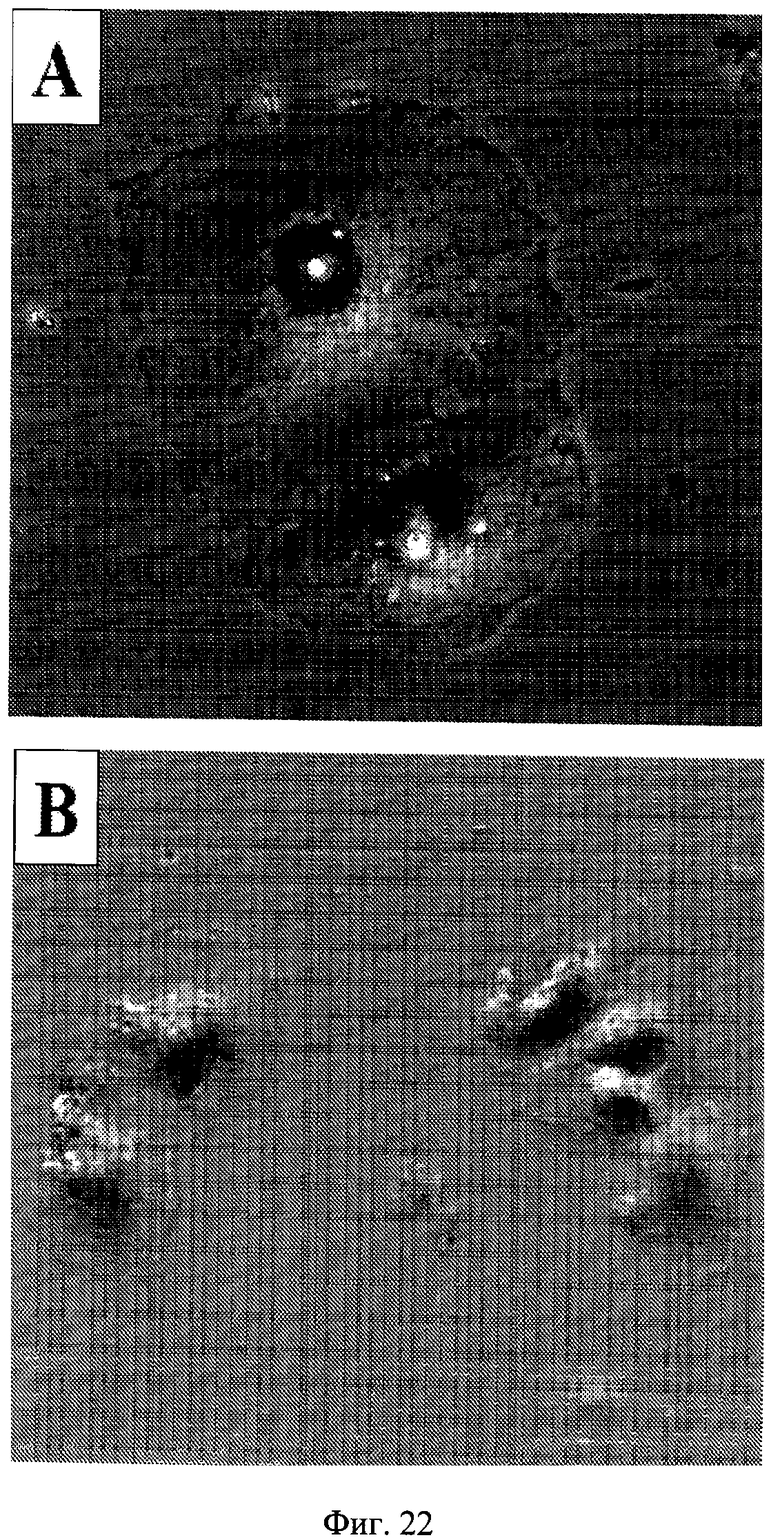
Фигура 22. Морфология клеток в процессе восстановления. 500 клеток HT-p21 высеивали в 12-луночные планшеты. На следующий день добавляли IPTG (А) или IPTG с рапамицином (В). После 3 дней планшеты промывали и микрофотографирование выполняли после дополнительных 3 дней. Клетки окрашивали на бета-Gal. А: I/w; В: I+R/w.
Фигура 23. Визуализация потери гипертрофии в процессе пролиферации компетентных клеток. 500 клеток HT-p21 (А) обрабатывали с IPTG (В) или IPTG с рапамицином (С), как указано, или оставляли необработанными. После 3 дней планшеты промывали и инкубировали без препаратов для обеспечения пролиферации. А. Нормальный размер пролиферирующих клеток. В. Клеточная гипертрофия стареющих клеток. С. Пример 1. Клональная пролиферация компетентных клеток приводит к потере гипертрофии. С. Пример 2. Клетки, которые остались угнетенными, остаются гипертрофированными.
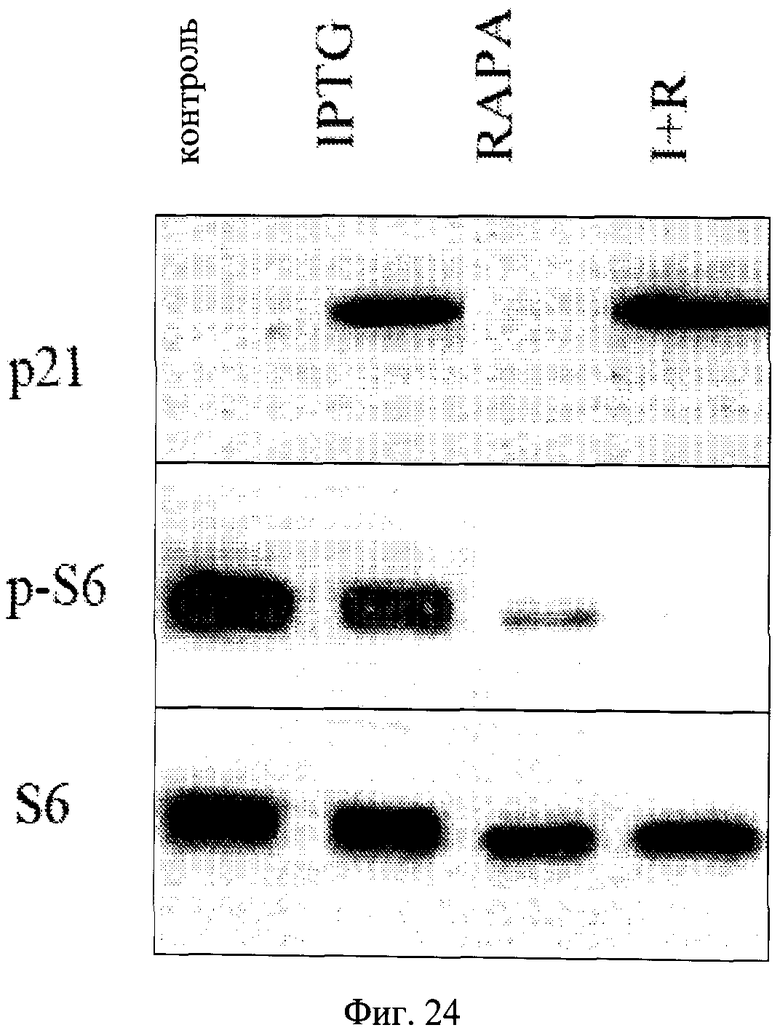
Фигура 24. Индукция p21 с помощью IPTG. Клетки HT-p21 высеивали на 6-луночные планшеты и обрабатывали с IPTG с или без рапамицина, как указано. На следующий день клетки лизировали и иммуноблот для p-S6, S и p21 осуществляли, как описано в Способах. IPTG существенно индуцировал p21, не влияя на фосфорилирование S6, тогда как рапамицин ингибировал фосфорилирование S6, не влияя на индукцию p21.
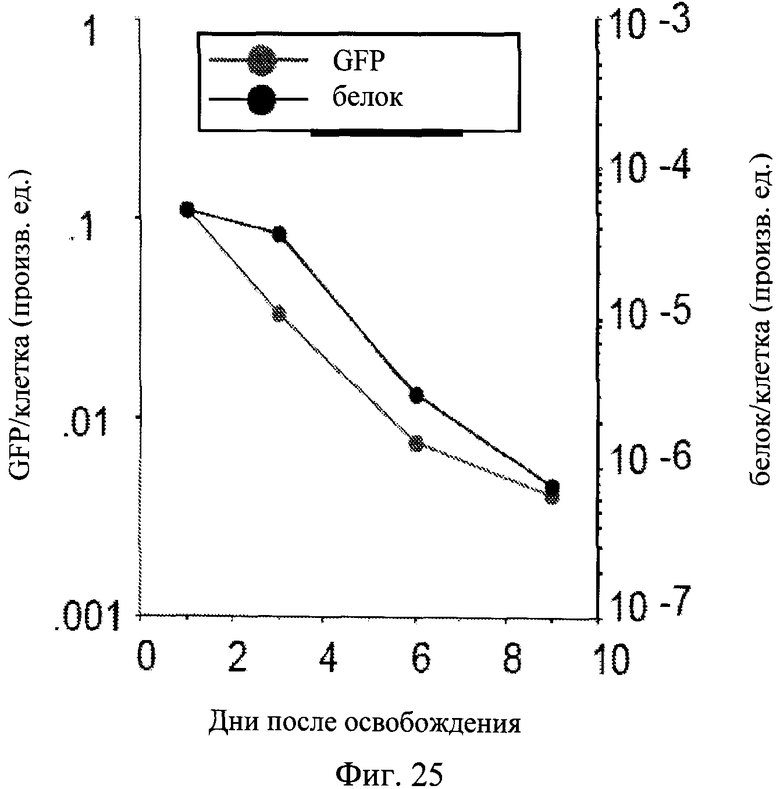
Фигура 25. Потеря гипертрофии после освобождения. Клетки HT-p21 обрабатывали IPTG с 500 нМ рапамицина в течение 3 дней. Затем клетки промывали и инкубировали в свежей среде без препаратов. В указанные дни растворимый белок, GFP и количество клеток измеряли в лунке. Белок (pr) в клетке и GFP в клетке подсчитывали и наносили на график в произвольных единицах измерения.
ОСУЩЕСТВЛЕНИЕ ИЗОБРЕНИЯ
Настоящее изобретение обеспечивает способ для профилактики и/или лечения возрастных заболеваний и/или симптомов такого рода заболеваний. Без намерения привязываться к какой-либо конкретной теории, считается, что настоящее изобретение обеспечивает данные воздействия путем подавления процесса старения.
В настоящем изобретении используется преимущество открытия, изложенного в настоящей заявке впервые, что p53, поначалу считающийся символическим индуктором клеточного старения, вместо этого принимает участие в подавлении клеточного старения. В связи с этим в предыдущих исследованиях подавление старения под действием p53 было, по всей видимости, скрыто задержкой клеточного цикла, индуцированного действием p53, которая (в случае дальнейшего продолжения) может приводить к старению. Поскольку предыдущие исследования основывались на том, что сам p53 вызывает задержку клеточного цикла, было невозможно отличить, активно ли p53 подавлял старение, или просто не удавалось индуцировать его в некоторых экспериментальных ситуациях Однако в настоящем изобретении авторы смогли различить эти два варианта развития событий с помощью тестирования воздействия p53 на старение, индуцированное p21 или p16 вместо самого p53. Авторы изобретения обнаружили, что в любых p21 или p16-подавленных клетках p53 преобразует старение (необратимую задержку со стареющей морфологией) в состояние покоя (обратимую задержку с сохранением пролиферативной способности и нестареющей морфологией). Таким образом, настоящее изобретение основывается отчасти на обнаружении авторами настоящей заявки парадоксального подавления клеточного старения p53.
Применительно к настоящему изобретению считается, что «старение» означает организменное старение и/или клеточное старение (старение). Организменное старение происходит вследствие клеточного старения и рассматривается как увеличение вероятности смерти с возрастом (временем). Подавление старения снижает вероятность смерти и, следовательно, увеличивает продолжительность жизни. Организменное старение проявляется возрастными болезнями, распространенность которых увеличивается с возрастом. Смерть от старения означает смерть от возрастных болезней. Подавление старения задерживает одно, некоторые или большинство заболеваний. Медленное старение проявляется задержкой возрастных болезней. Медленное старение рассматривается как вид здорового старения. Возрастные болезни считаются биомаркерами организменного старения. Соединение, которое задерживает возрастные болезни, увеличивает продолжительность жизни и может быть рассмотрено как лекарственное средство, замедляющее старение. Также соединение, которое подавляет старение, задерживает возрастные болезни.
Без намерения привязываться к какой-либо конкретной теории, клеточное старение (старение), считается, вызвано сверхстимуляцией и сверхактивацией путей сигнальной трансдукции, таких как mTOR путь, особенно когда клеточный цикл является блокированным, что приводит к клеточной гиперактивации и гиперфункции. В свою очередь, это вызывает сигнал вторичной резистентности и компенсирующую функциональную недостаточность. Как клеточная гиперфункция так и сигнал-резистентность являются причиной повреждения органов (включая в себя отдаленные органы), проявляются в виде старения (субклиническое повреждение) и возрастными болезнями (клиническое повреждение), в результате приводящие к организменной смерти. Не ограничиваясь примерами, маркерами клеточного старения считаются клеточная гипертрофия, постоянная потеря пролиферативного потенциала, большая, плоская клеточная морфология и бета-Gal окрашивание.
При осуществлении настоящего изобретения авторы продемонстрировали, что p53 подавляет клеточное старение и что негенотоксичные индукторы p53 (NGIP) предотвращают, замедляют и подавляют клеточное старение. Кроме того, клеточное старение характеризуется не только необратимой потерей пролиферативного потенциала, определенной морфологией, гиперсекреторным и провосполительным фенотипом, но также большим размером старой клетки (гипертрофией). Гипертрофия стареющих клеток способствует возрастным болезням, таким как увеличение простаты, гипертрофия сердца, почечная гипертрофия, утолщение стенки артерии и ожирение, при этом ожирение происходит от увеличения размера жировых клеток, а необязательно от увеличения количества клеток. Авторы настоящего изобретения показали, что оба NGIP (такой как Нутлин-3a) и ингибиторы mTOR (такие как рапамицин) снижают гипертрофию стареющих клеток. Таким образом, предполагается, что антигипертрофические агенты, такие как нутлин-3a и рапамицин, могут применяться для уменьшения размера клеток при возрастных болезнях, тем самым дополнительно способствуя антивозрастному воздействию данных соединений.
Результаты, приведенные в настоящей заявке, являются значительными потому, что p53 вызывает апоптоз, обратимую задержку клеточного цикла (состояние покоя) и необратимую задержку клеточного цикла (старение). Предполагается, что p53 активно вызывает старение.
Авторы настоящего изобретения показали, что нутлин-3a вызывает состояние покоя (обратимую задержку без морфологического старения) в клетках HT-p21 и WI-38-tert. В этих же клеточных линиях индуцируемые эктопические p21 и p16 вызывают старение. Согласно общепринятым основным положениям нутлин-3a в предыдущих исследованиях просто не активировал программу старения, потому что, например, недостаточная индукция p21. Без намерения привязываться к какой-либо конкретной теории, авторы настоящего изобретения считают, что нутлин-3a ингибирует программу старения. В настоящей заявке авторы изобретения показали, что p53 в действительности превращает старение в состояние покоя. Авторы изобретения пришли к выводу, что помимо его способности вызывать задержку клеточного цикла p53 представляет собой сильный супрессор старения. Таким образом, впервые авторы настоящего изобретения показали, что p53 подавляет клеточное старение, которое не было признано ранее, и применили полученные сведения посредством способа по настоящему изобретению. Также авторы настоящего изобретения показали, что эктопический p53 сам подавляет старение. Таким образом, предполагается, что любые p53-индуцирующие агенты также подавляют старение.
В одном из вариантов осуществления настоящего изобретения способ включает в себя контактирование клеток или введение человеку композиции, включающей в себя негенотоксический инкуктор p53 (NGIP), отличающийся тем, что контактирование и/или введение приводит к предотвращению, ингибированию или лечению возрастной болезни или симптома такого рода болезни. NGIP может применяться в количестве, эффективном для предотвращения, ингибирования или лечения возрастной болезни или ее симптома.
В одном из вариантов осуществления изобретение обеспечивает способ подавления и/или замедления клеточного старения у млекопитающих с помощью контактирования клеток с NGIP. В одном из вариантов осуществления настоящего изобретения клетки млекопитающих присутствуют в организме человека. В одном из вариантов осуществления настоящего изобретения человеку ранее не вводили NGIP.
В одном из вариантов осуществления настоящего изобретения человеку, для которого осуществляется заявленный способ, ранее не вводили NGIP. В одном из вариантов осуществления настоящего изобретения человек не имеет злокачественного заболевания.
В одном из вариантов осуществления настоящего изобретения подавление и/или замедление клеточного старения у млекопитающих характеризуется тем, что клетки млекопитающих, которые контактируют с NGIP, переходят в состояние покоя. В одном из вариантов осуществления настоящего изобретения до того, как добиться состояния покоя с помощью осуществления заявленного способа, клетки млекопитающего старели. Таким образом, в некоторых вариантах осуществления изобретение обеспечивает способы для того, что бы добиться превращения в состояние покоя клеток млекопитающих.
Другой вариант осуществления изобретения относится к профилактике и/или лечению гипертрофии стареющих клеток. Гипертрофия стареющих клеток способствует возрастным болезням, таким как увеличение простаты, гипертрофия сердца, утолщение стенки артерии и гипертрофические жировые клетки или ожирение. В связи с этим авторы настоящего изобретения показали, что NGIPs и ингибиторы mTOR снижают гипертрофию стареющих клеток. Таким образом, в одном из вариантов осуществления изобретение включает в себя способ ингибирования или снижения гипертрофии клеток путем введения человеку, нуждающемуся в этом, композиции, включающей в себя эффективное количество NGIP, ингибитора mTOR или их комбинацию. В различных вариантах осуществления настоящего изобретения человек, которому вводили ингибитор mTOR, ранее не получал ингибитор mTOR и/или человек ранее не получал пересадку органа, и/или не являлся кандидатом на пересадку органа. В одном из вариантов осуществления настоящего изобретения человек не нуждается в иммуносупрессорной терапии.
Предполагается, что заявленный способ может применяться для профилактики или лечения любых возрастных болезней и/или клеточной гипертрофии у любого человека. Не ограничиваясь примерами, возрастные болезни включают в себя доброкачественные опухоли, сердечно-сосудистые заболевания (такие как инсульт, атеросклероз, гипертензию), ангиому, остеопороз, инсулинорезистентность и диабеты II типа (диабетическую ретинопатию, нейропатию), болезнь Альцгеймера, болезнь Паркинсона, возрастную дегенерацию макулы, артрит, себорейный кератоз, актинический кератоз, фотостареющую кожу и кожную сыпь, рак кожи, системную красную волчанку, псориаз, пролиферацию гладкомышечных клеток и утолщение интимы после сосудистого повреждения, воспаление, артрит, побочное воздействие химиотерапии, доброкачественную гиперплазию предстательной железы (ДГПЖ или увеличение простаты), а также малораспространенные заболевания, которые распространены больше у лиц пожилого возраста, чем у молодых людей.
Предполагается, что любой NGIP может применяться в заявленном способе. В различных вариантах осуществления настоящего изобретения NGIP представляет собой агент, который индуцирует p53 путем блокирования взаимодействия p53 с другими белками, такими как Mdm-2, FAK, COP1 и p73/р63. Таким образом, в одном из вариантов осуществления настоящего изобретения NGIP представляет собой Mdm (Hdm)-связывающийся агент или антагонист Mdm-2. В различных вариантах осуществления настоящего изобретения Mdm-связывающийся агент представляет собой нутлин, включающий нутлин-3a и его аналоги. В одном из вариантов осуществления настоящего изобретения NGIP представляет собой нутлин-3a. Такого рода агенты могут также применяться в качестве антигипертрофических агентов.
Также предполагается, что любой ингибитор mTOR может применяться в настоящем изобретении. Ингибитором mTOR может быть любое соединение, которое является прямым или косвенным ингибитором mTOR. Подходящие косвенные ингибиторы mTOR включают в себя, но не ограничиваются ими, Mek ингибиторы, PI-3K ингибиторы или AMPK активаторы. В одном из вариантов осуществления настоящего изобретения mTOR ингибитор применяется с NGIP.
В одном из вариантов осуществления настоящего изобретения mTOR ингибитор представляет собой рапамицин или аналог рапамицина. Подходящие аналоги рапамицина включают в себя, но не ограничиваются ими, эверолимус, такролимус, CCI-779, ABT-578, АР-23675, АР-23573, АР-23841, 7-эпи-рапамицин, 7-тиометил-рапамицин, 7-эпи-триметоксифенил-рапамицин, 7-эпи-тиометил-рапамицин, 7-деметокси-рапамицин, 32-деметокси-рапамицин, 2-дезметил-рапамицин, 42-O-(2-гидроски)этил рапамицин и их комбинации. Изобретение можно также осуществлять применяя сочетания NGIPs и антигипертрофических агентов.
Для применения в профилактике и/или лечении возрастных болезней композиции, описанные в данной заявке, могут быть введены в общепринятой лекарственной форме, приготовленной путем смешивания со стандартными фармацевтически приемлемыми носителями в соответствии с известными методиками. Некоторые примеры фармацевтически приемлемых носителей приведены в: Remington: The Science and Practice of Pharmacy (2005) 21st Edition, Philadelphia, PA. Lippincott Williams & Wilkins. В различных вариантах осуществления настоящего изобретения композиции могут быть обеспечены в качестве фармацевтических препаратов, примеры которых включают в себя, но не ограничиваются ими, пилюли, таблетки, смеси, растворы, линименты, глазные капли и композиции наночастиц.
Различные способы, известные специалистам в данной области техники, могут применяться для введения заявленной композиции человеку и/или в условиях in vitro. Подходящие способы для введения композиций человеку включают в себя, но не ограничиваются этим, внутричерепной, интратекальный, внутрикожный, внутримышечный, внутрибрюшинный, внутривенный, подкожный, пероральный, интраназальный и ретроградные пути.
Специалистам в данной области будет понятно, что форма и характер конкретного режима дозирования, используемого в способе по настоящему изобретению, будет определяться путем введения и другими хорошо известными изменяемыми параметрами, такими как скорость клиренса, рост и массой тела индивидуума и стадией конкретного заболевания, подлежащего лечению. Исходя из такого рода критериев специалист в данной области техники может определить количество любых конкретных композиций, описанных в настоящей заявке, которое будет эффективно для профилактики и/или лечения возрастных болезней и/или клеточной гипертрофии у любого человека.
Способ по настоящему изобретению может быть выполнен в сочетании с общепринятыми омолаживающими и/или антивозрастными терапиями. Композиции по настоящему изобретению могут быть введены до, одновременно или после выполнения общепринятых омолаживающих и/или антивозрастных терапий. Такого рода лечения могут включать в себя, но не ограничиваются ими, химиотерапии, лучевую терапию и операции, в частности раковых опухолей. Кроме того, дополнительные соединения могут быть введены в сочетании с вводимыми композициями согласно изобретению. Например, композиция, содержащая NGIP, может быть введена со вторым соединением, предназначенным для увеличения, дополнения или обеспечения синергитического эффекта в сочетании с NGIP. Такого рода соединения включают в себя, но не ограничиваются ими, витамин D, витамин Е, витамин А, метформин, антиоксиданты, ресвератрол, нестероидное противовоспалительное средство, такое как СОХ ингибитор, mTOR ингибиторы, L-карнитин, липоевая кислота, лептин, Pgp ингибитор, ингибиторы каспазы и их комбинации. Кроме того, если антигипертрофическое соединение вводят, оно может быть введено со вторым соединением, предназначенным для увеличения, дополнения или обеспечения синергитического эффекта в сочетании с антигипетрофическим соединением. Такого рода соединения включают в себя, но не ограничиваются ими, витамин D, метформин, антиоксиданты, витамины, ресвератрол, нестероидное противовоспалительное средство, такое как СОХ ингибиторы, ингибитор Pgp/MRP (при нейродегенерации, для снижения выделения и для изменения биологической доступности) и ингибиторы метаболизирующих ферментов и комбинации вышеупомянутых.
Дополнительные соединения, которые могут применяться в сочетании в композициями, содержащими NGIP и/или антигипертрофическое соединение, могут быть введены одновременно, до или после введения композиции, содержащей NGIP и/или антигипертрофическое соединение.
Следующие Примеры предназначены для иллюстрации, но не ограничивают настоящее изобретение.
Пример 1.
Следующие Материалы и Способы были использованы для получения данных и результатов, представленных в данном Примере.
Способы.
Клеточные линии и реагенты. Клетки HT-p21-9 и HT-p21-a представляют собой производные клеток фибросаркомы человека HT1080, где экспрессия p21 может быть включена или выключена с использованием изопропил-тио-галактозидаз (IPTG) (7, 16, 28, 29, 36), Клетки HT-p21-9 экспрессируют GFP, тогда как клетки HT-p21-a нет. Клетки HT-p16 представляют собой производные клеток HT1080, в которых экспрессия p16 может быть включена или выключена с использованием IPTG (16, 36). WI-38-Tert, WI-38 представляют собой фибробласты, иммортализированные теломеразой. Клетки HT-p21-GSE56: p53 ингибирующий белок GSE56 (18) был введен в клетки HT1080 p21-9 с помощью ретровирусного вектора LXSE (37). Клетки выращивали с высоким уровнем глюкозы в DMEM с 10% FC2 сывороткой. Клетки WI-38-tert выращивали с низким уровнем глюкозы в DMEM с 10% FCS. Рапамицин получали от LC Laboratories (Wobum, МА). IPTG (конечная концентрация 50 мкг/мл) и FC2 были получены от Sigma-Aldrich (St. Louis, МО). Нутлин-3a и -b были получены от Sigma-Aldrich и La Roche, Nutley, NJ (38). p53, p21 и p53-GFP, экспрессирующихся аденовирусами (Ad-p53, Ad-p21 и Ad-p53-GFP), были описаны ранее (20, 39) и получены от Dr. Wafik El-Deiry (Univ. Penn. Philadelphia, PA).
Анализ колониеобразования. Планшеты фиксировали и окрашивали 1.0% метиленовым синим или кристаллическим фиолетовым (13).
Иммуноблот-анализ. Белки были разделены на 4-15% градиента Tris-HCl геля (Bio Rad). Использовали следующие антитела: мышиные антиактин от Santa Cruz Biotechnology, кроличьи анти-фосфо-S6 (Ser240/244) и (Ser235/236), мышиные aHra-S6, мышиные анти-фосфо-p70 S6 киназа (Thr389), мышиные анти-p21 и анти-p53, кроличьи анти-фосфо-4E-BP1 (Thr37/46) от Cell Signaling; мышиные анти-4E-BP1 от Invitrogen, мышиные анти-p53 (Ab-6) от Calbiochem.
Окрашивание на бета-галактозидазу. Бета-gal окрашивание осуществляли с использованием набора Senescence - galactosidase staining kit (Cell Signaling Technology).
Метаболическое мечение. Клетки HT-p21-9 высевали при плотности 25,000 клеток/лунка в 12-луночные планшеты. На следующий день клетки обрабатывали препаратами. После 24 часов клетки были помечены с 30 мкКи [35S]метионин/цистеином (Amersham) на мл Met/Cys-свободной Дульбекко модифицированная среда Игла (Invitrogen) в течение 1 часа при 37°С. Клетки промывали PBS и лизировали в 1% ДДCNa 0.5% БСА. Для определения 35S включения общий белок осаждали с 0.5 мл 10% ТХУ и собирали на нитроцеллюлозных фильтрах. Фильтры были высушены на воздухе и радиоактивность определяли с использованием жидкостного сцинтилляционного счетчика.
При использовании Материалов и Способов, описанных выше, были получены следующие результаты.
Результаты
Активатор p53 Нутлин-3a подавляет старение, индуцированное p21.
Индукция p21 в HT1080-производных HT-p21-9 клеток, несущих IPTG-индуцируемую p21 экспрессию конструкции, вызывает старение. В тех же клетках индукция p53 нутлином-3a вызывает обратимую задержку клеточного цикла (состояние покоя) и клетки возобновляют пролиферацию после удаления нутлина-3a (Huang В, Deo D, Xia M, Vassilev LT (2009) Pharmacologic p53 Activation Blocks Cell Cycle Progression but Fails to Induce Senescence in Epithelial Cancer Cells. Mol Cancer Res. 7: 1497-509). Авторы настоящего изобретения применяли нутлин-3a, ингибитор связывания p53-Mdm2, в данных экспериментах, поскольку он индуцирует p53 при физиологических уровнях без повреждения ДНК и является высокоспецифическим (17). Таким образом, физиологические уровни p53 индуцировали состояние покоя, тогда как эктопическая экспрессия p21 индуцировала старение (Huang et al. 1999). Имеются две альтернативных модели, которые могли бы объяснить данные результаты. Во-первых, общепринятая модель показывает, что физиологические уровни p53, индуцированные нутлином-3a, не достаточны для индукции p21 в размере, необходимом для активации программы старения в данной клеточной линии. Таким образом, добавление нутлина-3a к IPTG может только усилить старение. Второй альтернативной моделью является то, что p53 на самом деле подавляет старение. При этом активацию p53 нутлином-3a вместе с IPTG-опосредованной индукцией p21 можно ожидать для преобразования старения в состояние покоя.
Как показано на Фигуре 1 и сообщалось ранее (Huang et al., 1999), клетки, обработанные IPTG и нутлином-3a, являются положительным контролем старения и состояния покоя соответственно. Обработка IPTG индуцировала характерную стареющую морфологию (крупные, плоские, SA-бета-Gal-позитивные клетки), а клетки, обработанные нутлином-3a, оставались маленькими, безжировыми и SA-бета-Gal-негативными (Фиг.1A). Кроме того, анализ колониеобразования показал, что обработка IPTG приводила к необратимой потере пролиферативного потенциала (только некоторые клетки формировали колонии после удаления IPTG), а обработка нутлином-3a вызывала обратимую задержку (заметное колониеобразование после удаления нутлина-3a) (Фиг.1B).
При анализе данных наблюдений авторы настоящего изобретения исследовали, будет ли влиять добавление нутлина-3a к IPTG на преобразование старения в состояние покоя. В результате, этот ключевой эксперимент показал, что обработка с нутлином-3a предотвращает стареющую морфологию, вызванную IPTG: клетки оставались маленькими, безжировыми и негативными на SA-бета-Gal окрашивание (Фиг.1a). Кроме того, такие клетки сохранили пролиферативный потенциал и клоногенность (Фиг.1б). Таким образом, авторы настоящего изобретения определили, что воздействие нутлина-3a на IPTG-индуцированное старение было доминирующим. Важно отметить, что нутлин-3a ни изменяет, ни уменьшает уровней p21 (см. иммуноблоты на Фигурах, представленных в этом Примере). Нутлин-3a не отменяет цитотоксического эффекта IPTG и IPTG вызывает моментальную задерку клеточного цикла, проявляющуюся в виде одиноких клеток со стареющей морфологией при низкой плотности клеток (Фиг.5). В присутствии только нутлина-3a клетки подвергаются одному делению и не пролиферируют дальше, как показано на колониях 2 приспособленных клетки с нестареющей морфологией (Фиг.5). В присутствии как нутлина-3a, так и IPTG клетки подавляются сразу же без единого деления, но не приобретают стареющей морфологии (Фиг.5). Таким образом, не отменяя задержку клеточного цикла, вызванного IPTG, нутлин-3a преобразует старение в обратимое состояние (состояние покоя). Когда IPTG и нутлин-3a вымыли из культур, клетки возобновили пролиферацию, формирование микроколоний (Фиг.1c) и затем макроколоний (Фиг.1c). Данные результаты показали, что нутлин-3a предохраняет клетки от подвержения IPTG-индуцированным старением. Подавление старения нутлином-3a наблюдалось в диапазоне активных концентраций нутлина-3a в зависимости от дозы (Фиг.1d-e). Лучшим количественным способом для измерения сохранения пролиферативного потенциала (PP) является общее количество клеток на планшет. Нутлин-3a сохраняет пролиферативный потенциала (PP) в зависимости от дозы (Фиг.If). Авторы настоящего изобретения измерили количество клеток в колонии в зависимости от количества колоний в планшете (Фиг.6). Таким образом, нутлин-3a увеличивает количество клеток с нормальным PP. Сохранение пролиферативного потенциала нутлином-3a в клетках с индуцированной IPTG задержкой было подтверждено в обоих, IPTG-регулируемых p16- и p21-экспрессирующих клетках (Фиг.7).
Подавление старения требует транс-активации функции p53.
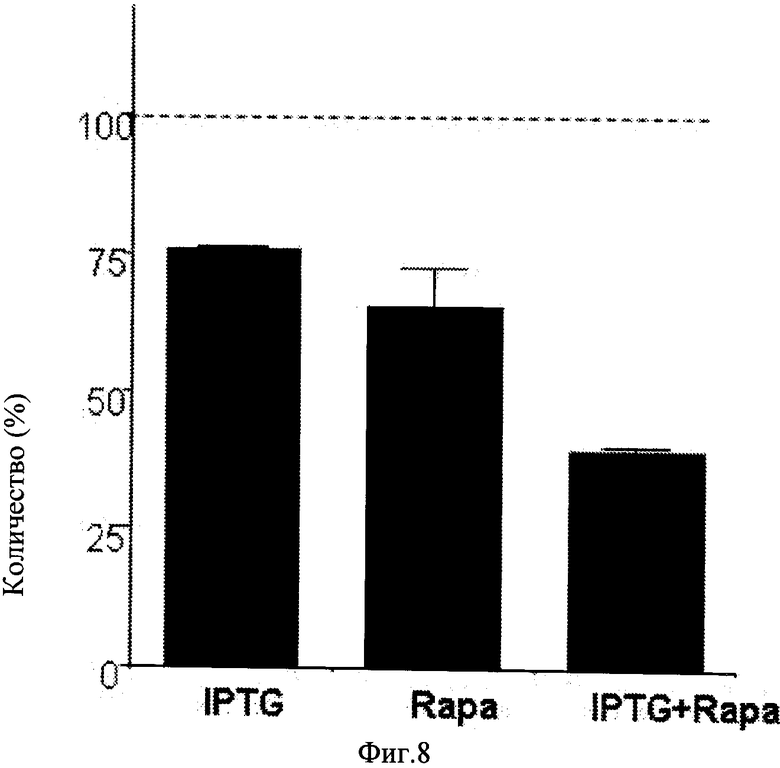
Нутлин-3a представляет собой высоко специфический активатор p53 и считается, не сообщалось о нецелевых эффектах данного соединения. Известно, что нутлин-3b, оптимер нутлина-3a, который не блокирует взаимодействие Mdm-2/p53, был не способен к преобразованию старения в состояние покоя (Фиг.7b-c). Для непосредственной проверки ингибирования нутлином-3a старения p53-зависимым механизмом авторы настоящего изобретения применяли клетки HT-p21-GSE56, производные клеточной линии HT-p21, в которых функция p53 блокирована транс-доминантным ингибитором, GSE56 (Ossovskaya VS, et al. (1996) Use of genetic suppressor elements to dissect distinct biological effects of separate p53 domains. Proc Nati Acad Sci USA 93: 10309-14.). Результаты показали, что р53 экспрессировался на очень высоком уровне в данных клетках, поскольку ингибирование его функции транс-активации приводило к стабилизации белка (по аналогии с мутантным p53). Хотя нутлин-3a индуцировал p53 в клетках HT-p21, он не влиял на уровень p53 в клетках HT-p21-GSE56 (Фиг.2a). IPTG сильно индуцировал p21 в клетках HT-p21-GSE56 и нутлин-3a, не влияя на данную индукцию (Фиг.2a). Нутлин-3a не ингибировал пролиферацию клеток HT-p21-GSE56 (Фиг.20), тем самым подтверждая, что модель подходит для исследования подавления старения нутлином-3a в зависимости от p53. Кроме того, важно использовать положительный контроль для p53-независимого подавления старения. Авторы настоящего изобретения показали, что активация mTOR (мишень рапамицина у млекопитающих) была необходимой для клеточного старения, а деактивация mTOR рапамицином предотвращала старение, вызывая взамен этому состояние покоя. Рапамицин не индуцировал p53 (Фиг.2a) согласно его p53-независимому ингибированию mTOR. Рапамицин подавлял IPTG-индуцированное старение в клетках HT-p21-GSE56 (Фиг.2c). В отличие от этого нутлин-3a подавлял старение в клетках HT-p21-9, обработанных только IPTG, а в аналогично обработанных клетках HT-p21-GSE56 - нет (Фиг.2c). Согласно с данными выводами нутлин-3a (в отличие от рапамицина) не сохранял колониеобразование и пролиферативный потенциал (PP) в клетках HT-p21-GSE56, обработанных IPTG, лишенных функционального p53 (Фиг.2d-e). Эти данные показывают, что транскрипциональная активность p53 необходима для старения нутлином-3a. В отличие от этого рапамицин ингибировал старение независимо от p53, о чем свидетельствует его способность к предотвращению морфологического старения (Фиг.2c), и сохранял пролиферативный потенциал (Фиг.3d-e) в клетках HT-p21-GSE56, обработанных IPTG.
Ингибирование mTOR пути нутлином-3a.
Авторы настоящего изобретения ранее сообщали, что ингибиторы mTOR (рапамицин), PI-3K (LY294002) и МЕК (U0126), все деактивируют mTOR путь в клетках HT-p21-9, что измеряется отсутствием фосфорилирования рибосомального белка S6, и подавляют клеточное старение (Demidenko ZN, Shtutman M, Blagosklonny MV (2009) Pharmacologic inhibition of МЕК and PI-3K converges on the mTOR/S6 pathway to decelerate cellular senescence. Cell Cycle 8: 1896-900). Как и все данные агенты, нутлин-3a ингибировал фосфорилирование S6 и частично ингибировал фосфорилирование 4E-BPI, другой нижележащей мишени mTOR пути (Фиг.3a). Нутлин-3a также нормализировал повышенный уровень циклина D1, связанного с клеточным старением. Как и рапамицин, нутлин-3a ингибировал mTOR пути как в присутствии так и в отсутствие IPTG и не предотвращал индукции p21 с помощью IPTG (Фиг.3b). Важно отметить, что IPTG-индуцируемый p21 не влиял на фосфорилирование S6 и 4E-BPI (Фиг.3a-b).
Рапамицин и нутлин-3a являются одинаково эффективными в подавлении старения (сохранение пролиферативного потенциала) в клетках HT-p21-9, обработанных IPTG (Фиг.3c). Однако в присутствии рапамицина в дозах, которые полностью ингибировали mTOR, нутлин-3a не мог бы еще больше подавлять старение, что измеряли по сохранению пролиферативного потенциала (PP) IPTG-угнетенных клеток (Фиг.3c). Это подтверждает представление того, что нутлин-3a и рапамицин влияли и тот и другой одинаково или перекрывающимися путями. mTOR путь стимулировал синтез белка. Важно отметить, что синтез белка оставался на высоком уровне в IPTG-угнетенных клетках и ингибировался рапамицином (Фиг.8), что объясняет клеточную гипертрофию, связанную со старением. Оба, нутлин-3а и рапамицин, снижали содержание белка в клетке в IPTG-обработанных клетках HT-p21-9 (Фиг.3d). Для оценки, насколько данное снижение участвовало в ингибировании синтеза белка, авторы настоящего изобретения измеряли включение 35S-метионин/цистеина в образующихся белках в присутствии нутлина-3a (Фиг.3e). Нутлин-3a ингибировал включение 35S-метионин/цистеина в клетки HT-p21-9, обработанные IPTG, в зависимости от дозы (Фиг.3e).
Подавление старения эктопической экспрессией p53.
Для того чтобы подтвердить результаты, полученные авторами настоящего изобретения без зависимости от нутлина-3a для активации p53, авторы проверяли будет ли экспрессия эндогенного p53 также приводить к подавлению p21-индуцированного старения. Авторы настоящего изобретения применяли аденовирус, который направляет конструктивную экспрессию p53 вместе с GFP (Ad-p53-GFP) (Wang W, Takimoto R, Rastinejad F, E1-Deiry WS (2003) Stabilization of p53 by CP-31398 inhibits ubiquitination without altering phosphorylation at serine 15 or 20 or MDM2 binding. Mol Cell Biol. 23: 2171-2181.) так, что инфицированные клетки могли быть легко определены путем флуоресцентной микроскопии. В данных экспериментах авторы изобретения применяли клетки HT-p21-a, которые в отличие от HT-p21-9 не экспрессировал внутренний GFP и вследствие этого не зеленые. При низких титрах Ad-p53-GFP инфицировал -20% клеток HT-p21-a; поэтому авторы изобретения имели возможность сравнить клетки, сверхэкспрессирующие p53, и неинфицированные в одном и том же микроскопическом препарате. Как и предполагалось, в неинфицированных клетках обработка IPTG вызывала стареющую морфологию (Фиг.4a, красные стрелки на нижней панели). В отличие от этого Ad-p53-GFP-инфицированные клетки не приобретали стареющей морфологии (Фиг.4a). Для исследования различных средств, вызывающих старение, авторы изобретения применяли заражение с p-21-экспрессирующим аденовирусом (Ad-p21) вместо IPTG для индукции p21. Ad-p21 инфицированные клетки быстро приобретали стареющую морфологию, тогда как Ad-p53-GFP инфицированные клетки не приобретали (Фиг.9a). Кроме того, Ad-p53-GFP подавляли старение, вызванное Ad-p21 (Фиг.9b) и IPTG-индуцированным р16 (Фиг.10).
Подавление стрессиндуцированного старения в фибробластах.
Для продолжения наблюдения авторами p53-опосредованного подавления старения клеток, не связанных с HT1080, авторы настоящего изобретения применяли теломеразо-иммортализованные WI-38 фибробласты человека (клетки WI-38-tert). Как показано на Дополнительной Фигуре 11, инфицирование данных клеток с Ad-p53 также приводило морфологию в состояние покоя (уменьшенные в весе, бета-Gal-отрицательные клетки) однако заражение с Ad-p21 индуцировало стареющую морфологию. Самое главное, коинфицирование клеток с Ad-p53 и Ad-p21 показало, что p53 подавляет p21-индуцированное старение (Фиг.11). Поскольку заражение Ad-p53 приводило к избыточному уровню p53, обнаруженный эффект ограничивался сопутствующей индукцией апоптоза. Поэтому авторы настоящего изобретения применяли нутлин-3a для индукции p53 при физиологическом уровне в данной системе. Авторы изобретения показали, что обработка клеток WI-38-tert с нутлином-3a вызывало состояние покоя. Важно отметить, что нутлин-3a (при концентрациях, которые индуцировали p53) ингибировал фосфорилирование S6K и 86 (Фиг.12). В отличие от этого доксорубицин не ингибировал mTOR. Это может объяснить, почему нутлин-3a индуцировал состояние покоя в клетках WI-38-tert, тогда как доксорубицин вызывал старение в клетках WI-38-tert. Далее авторы изобретения исследовали, может ли нутлин-3a подавлять старение, вызванное перекисью водорода (H2O2), каноническим индуктором клеточного старения в фибробластах. В клетках WI-38-tert H2O2 ингибировал клеточную пролиферацию без индукции p53 и без влияния на фосфорилирование S6 (Фиг.4b). Это приводило к стареющей морфологии (Фиг.4c). Нутлин-3a индуцировал p53, ингибировал фосфорилирование S6 (Фиг.4b) и подавлял старение, индуцированное H2O2 (Фиг.5c). Кроме того, нутлин-3a частично сохранял пролиферативный потенциал в клетках, обработанных H2O2 (Фиг.4d). Таким образом, авторы изобретения применяли различные клеточные линии, а также различные средства, индуцирующие клеточное старение и активирующие p53, для того чтобы показать, что p53 подавляет старение.
Таким образом, из вышеизложенного будет понятно, что в настоящей заявке впервые описано, что состояние покоя, индуцированное p53, на самом деле является результатом подавления старения под действием p53.
Пример 2.
Следующие Материалы и Способы были использованы для получения результатов, представленных в данном Примере.
Материалы и Способы.
Клеточные линии и реагенты. В клетках HT-p21 экспрессия p21 может быть включена или выключена с применением изопропил-тио-галактозидаз (IPTG) [14, 15]. Клетки LC LaboratoriesSigma-Aldrich (St. Louis, МО). IPTG растворяли в воде до 50 мг/мл исходного раствора и применяли в культуре клеток в конечной концентрации 50 мкг/мл. Иммуноблот-анализ. Клетки лизировали и растворимые белки были собраны, как описывалось ранее [9]. Иммуноблот-анализ осуществляли с применением мышиных моноклональных анти-p21, мышиных моноклональных анти-фосфо-S6 Ser240/244 (Cell Signaling, MA, USA), кроличьих поликлональных анти-S6 (Cell Signaling, MA, USA) и мышиных моноклональных анти-тубулин Ab, как описывалось ранее [9]. Подсчет клеток. Клетки были подсчитаны на устройстве для подсчета клеток Coulter Z1 (Haileah, FL).
Анализ колониеобразования. Две тысячи клеток HT-p21 были посеяны на 100 мм планшеты. На следующий день клетки обрабатывали с 50 мкг/мл IPTG и/или 500 нМ рапамицина, как указано. После 3 дней среду удаляли; клетки промывали и культивировали в свежей среде. Когда колонии становились видимыми, планшеты фиксировали и окрашивали с 0.1% кристаллическим фиолетовым (Sigma). Планшеты фотографировали и количество колоний определяли, как описывалось ранее [9].
SA-Gal окрашивание. Клетки фиксировали в течение 5 минут в фиксаторе бета-галактозидазы (2% формальдегида; 0.2% глутаральдегида в PBS) и промывали в PBS и окрашивали в рестворе-галактозидазы (1 мг/мл 5-бромо-4-хлоро-3-индолил-бета-gal (X-gal) в 5 мМ феррицианида калия, 5 мМ ферроцианида калия, 2 мМ MgCl2 в PBS) при 37°C, до бета-Gal окрашивания становятся видимыми в любой экспериментальной или контрольной планшете. Затем клетки промывали в PBS и количество позитивных по активности - β-галактозидазы клеток (синее окрашивание) были подсчитаны при освещении в светлом поле.
Применяя Материалы и Способы, описанные выше в данном Примере, были получены следующие результаты.
Экспоненциальное возрастание массы предшествует старению.
Количество пролиферирующих клеток возрастало экспоненциально (с удвоением времени 20-24 часов). Как в Примере 1, индукция p21 с помощью IPTG вызывала угнетение G1 и G2, полностью блокируя клеточную пролиферацию (Фиг.14). p21-угнетенные клетки продолжали увеличиваться в размерах, становясь гипертрофическими. Поскольку эти клетки содержали CMV-контролируемый EGFP, авторы изобретения измерили как белок, так и GFP. На лунку количества GFP и белка увеличивалось почти в геометрической прогрессии с или без IPTG (Фиг.15). На лунку количества GFP и белка увеличивались только в IPTG-обработанных (неделящихся) клетках (Фиг.16). Для пролиферирующих клеток (без IPTG), GFP в клетке и белок в клетке оставались постоянными (Фиг.16), потому что возрастание массы уравновешивалось клеточным делением. В отличие от этого в клетках, обработанных IPTG, белок/клетка и GFP/клетка увеличивались почти в геометрической прогрессии в течение 3 дней (Фиг.16). В процессе индукции старения с помощью IPTG клеточная масса продолжала увеличиваться, но не была уравновешена клеточным делением. Во всех случаях белок и GFP взаимосвязаны (Фиг.16), образование GFP в клетке подходящий маркер клеточной гипертрофии.
Эти данные могут объяснить, как индукция p21 может индуцировать GFP без транс-активации CMV промотера: ингибированием клеточного цикла без ингибирования клеточного роста. Кроме того, представления о том, что GFP в клетке является маркером гипертрофии, дает 2 прогноза. Во-первых, мутантный p21, который не может связывать CDKs и, следовательно, не может угнетать клеточный цикл, не может индуцировать GFP. Во-вторых, антигипертрофические агенты, такие как рапамицин, уменьшают GFP в клетке без отмены угнетения клеточного цикла.
Дозозависимое подавление клеточной гипертрофии.
Затем авторы настоящего изобретения исследовали воздействия рапамицина на гипертрофию стареющих клеток. Клетки индуцировали к старению с помощью IPTG в присутствии (+R) или в отсутствие рапамицина. На 3 и 5 дни оценивали воздействия рапамицина на клеточную гипертрофию. С помощью микроскопии антигипертрофическое воздействие рапамицина было наиболее очевидным при низкой плотности клеток (такой как 1000 клеток на 10-мм планшет), потому что было достаточно пространства для клеток, обработанных IPTG, для увеличения в размере при отсутствии рапамицина (Фиг.17). Однако авторы изобретения не могли надежно измерить уровни белка при такой низкой плотности клеток. При регулярной плотности клеток рапамицин (500 нМ) снижал клеточную гипертрофию до 30%-40% (Фиг.18А, данные не представлены). Два маркера гипертрофии (белок/клетка и GFP/клетка) взаимосвязаны (Фиг.18A). Антигипертрофическое воздействие рапамицина не было статистически значимым при концентрациях рапамицина ниже 20 нМ. Сначала это приводило в замешательство, учитывая, что рапамицин ингибировал mTOR путь в низких концентрациях во многих типах клеток. Таким образом, авторы изобретения исследовали дозозависимый эффект mTOR ингибирования путем измерения фосфорилирования S6, маркера активности mTOR. В соответствии с антигипертрофическим эффектом, рапамицин ингибировал фосфорилирование S6 в концентрации 20 нМ или выше, достигал максимального эффекта при 100 нМ-500 нМ (Фиг.18B). Таким образом, ингибирование фосфорилирования S6 и ингибирование гипертрофии взаимосвязаны, объясняя необходимость высокой концентрации (100-500 нМ) рапамицина для антигипертрофического эффекта в данной клеточной линии.
Дозозависимое сохранение клеточной компетентности.
Рапамицин сохранял пролиферативный потенциал в угнетенных клетках, то есть клетки могли успешно делиться, когда подавление отменено. Но рапамицин не индуцировал пролиферацию, а наоборот, мог вызвать состояние покоя (у некоторых типов клеток). Для того чтобы четко отличать возможную пролиферацию (компетентность) и фактическую пролиферацию, авторы настоящего изобретения использовали понятие компетентности (способности к пролиферации) и некомпетентности (безвозвратной потери пролиферативного потенциала, связанной с клеточным старением). В клетках HT-1080 рапамицин сохранял компетентность в продолжении задержки клеточного цикла, вызванной p21. В отличие от стареющих клеток клетки, находящиеся в состоянии покоя, являлись компетентными.
Авторы настоящего изобретения определяли, будет ли сохранение компетентности (PC) взаимосвязано с ингибированием фосфорилирования S6 и антигипертрофическим эффектом рапамицина. Клетки обрабатывали с IPTG и увеличивающимися концентрациями рапамицина в диапазоне от 0 до 500 нМ (Фиг.18C). Через 3 дня IPTG вымывали, тем самым позволяя клеткам пролиферировать и затем после еще 5 дней клетки подсчитывали. Клетки, обработанные IPTG, становились некомпетентными, тогда как рапамицин подавлял некомпетентность (Фиг.18C). Существенно, что сохранение компетентности обнаруживалось при более низких концентрациях рапамицина, чем те, которые ингибировали либо фосфорилирование S6, либо клеточную гипертрофию. В частности, такая высокая чувствительность PC-теста по сравнению с ингибированием гипертрофии могла быть связана с относительной величиной эффектов (30% ингибирования гипертрофии против 800% PC). Возможно, что даже переходное ингибирование mTOR (не определяемое иммуноблоттингом) определяется увеличивающейся компетентностью. Согласно данному объяснению, даже когда рапамицин добавляли с задержкой, сохранение компетентности определялось.
Экспоненциальная пролиферация компетентных клеток.
В присутствии IPTG (с содержанием или без рапамицина) клетки не пролиферировали и не формировали колонии. Когда IPTG вымывали, 3-5% клеток сохраняли компетентность даже без рапамицина (Фиг.19). Колонии увеличивались в размерах, тогда как количество колоний почти не изменялось (Фиг.19). Рапамицин увеличивал количество колоний (количество компетентных клеток) почти в 10 раз. Авторы настоящего изобретения в дальнейшем сравнили качество пролиферации компетентных клеток, сохранившейся после обработки IPTG с содержанием или без рапамицина (I/w и I+R/w соответственно). В условиях I/w и I+R/w количество клеток начинало экспоненциально увеличиваться после 1 дня и 3 дней соответственно (Фиг.20). Через 6 дней обе кривые (I/w и I+R/w) стали параллельными. Кривая «I+R/w» была только сдвинута правее на приблизительно 3 дня (Фиг.20). Это соответствует 10-кратному различию в исходном количестве компетентных клеток, если их время удвоения составляло 1 день. Примечательно, что это также соответствует исходному различию в количестве компетентных клеток, что установлено колониеобразованием (Фиг.19). Также в обоих условиях, I/w и I+R/w, время удвоения компетентных клеток составляло около 20-24 часов, сходным со скоростью пролиферации необработанных клеток.
Восстановление гипертрофии во время пролиферации компетентных клеток.
Рапамицин снижал клеточную гипертрофию приблизительно на 30% в клетках, обработанных IPTG (Фиг.18A). Когда IPTG и рапамицин вымывали, наблюдался лаг-период около 24-30 часов для компетентных до вступления в первое деление. В продолжении периода задержки клетки увеличивались в размере, потому что рапамицин был вымыт. Следовательно, по измеренному GFP в клетке (Фиг.21A) клетки, обработанные рапамицином, достигали размеров клеток, обработанных только IPTG (Фиг.21A: I/w и I+R/w в первый день). Также по измеренному белку в клетке клетки, обработанные IPTG с рапамицином, становились полностью гипертрофическими на первый день после вымывания (данные не показаны). Независимо от восстановления гипертрофии клетки, обработанные IPTG + рапамицин, оставались компетентными (Фиг.19-20). Это указывает на то, что гипертрофия не была вызвана пролиферативными некомпетентными клетками, обработанными IPTG. Когда компетентные клетки разделились, GFP в клетке уменьшался (Фиг.21B). В соответствии, было отмечено существенное различие в клеточной морфологии типичных клеток в обоих условиях (Фиг.22). В условиях I/w большая часть клеток были все еще большими и плоскими, с окрашиванием на бета-Gal. В условиях I+R/w преобладающие клетки были с мелкоклеточной морфологией и бета-Gal-негативными. Эти клетки формировали колонии, указывающие на то, что они приобрели нестареющую морфологию в связи с пролиферацией (Фиг.23C). В отличие от этого стареющие клетки, которые не возобновили пролиферацию, оставались большими (Фиг.23C). Компетентные клетки, несмотря на то что пролиферировали и формировали колонии, уменьшались в размере (Фиг.23C). В результате средний размер клеток снижался до нормального уровня в условиях I+R/w, соответствующего уменьшению как количества белка/клетка, так и GFP/клетка (Фиг.24), что указывает на то, что оба являются маркерами клеточной гипертрофии. Несмотря на изменение гипертрофии и капли GFP/клетка количество общего GFP и белка в лунке увеличивалось за счет клеточной пролиферации (Фиг.21B, данные не показаны).




Изобретение относится к медицине и может быть использовано для подавления и/или замедления клеточного старения млекопитающих. Для этого применяют композицию, включающую негенотоксичный индуктор р53 (NGIP), путем контактирования клеток млекопитающего с указанной композицией. NGIP представляет собой Mdm-связывающий агент, или антагонист Mdm-2, или нутлин, или нутлин-3A, или аналог нутлина, или их комбинацию. Изобретение позволяет подавить старение клеток за счет перехода клеток в состояние покоя. 8 з.п. ф-лы, 25 ил., 2 пр.
1. Применение композиции, включающей не-генотоксичный индуктор р53 (NGIP) для подавления и/или замедления клеточного старения млекопитающего путем контактирования клеток млекопитающего с указанной композицией.
2. Применение по п. 1, отличающееся тем, что NGIP представляет собой нутлин, нутлин-3А, аналог нутлина или их комбинацию.
3. Применение по п. 2, отличающееся тем, что клетки млекопитающего присутствуют у человека.
4. Применение по п. 3, отличающееся тем, что у человека не был диагностирован рак.
5. Применение по п. 4, отличающееся тем, что NGIP представляет собой нутлин-3А.
6. Применение по п. 1, отличающееся тем, что NGIP представляет собой Mdm-связывающий агент или антагонист Mdm-2.
7. Применение по п. 4, отличающееся тем, что подавление и/замедление клеточного старения у млекопитающих включает переход клеток млекопитающего в состояние покоя.
8. Применение по п. 1, отличающееся тем, что NGIP индуцирует р53 путем блокирования взаимодействия р53 с другими белками.
9. Применение по п. 3, отличающееся тем, что человек нуждается в профилактике или лечении возрастных болезней, выбранных из группы, состоящей из доброкачественной гиперплазии предстательной железы, ангиомы, сердечнососудистых заболеваний, атеросклероза, гипертензии, остеопороза, инсулинорезистентности и диабета II типа, болезни Альцгеймера, болезни Паркинсона, возрастной макулярной дегенерации, ретинопатии, системной красной волчанки, псориаза, пролиферации гладкомышечных клеток и утолщении интимы после повреждения сосудов, воспаления, артрита, побочного действия химиотерапии, а также их комбинации.
| HUANG B | |||
| et al.,Pharmacologic p53 activation blocks cell cycle progression but fails to induce senescence in epithelial cancer cells, Mol Cancer Res | |||
| Колосоуборка | 1923 |
|
SU2009A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| MULLER CR | |||
| et al., Potential for | |||

