ОБЛАСТЬ ТЕХНИКИ
Изобретение относится к органоиду печени, вариантам его применения и способу его получения.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Основной структурной единицей печени является печеночная долька. Каждая долька состоит из пластинок гепатоцитов, между которыми находятся синусоидальные капилляры, радиально сходящиеся в центральную выводящую вену. Печеночные дольки имеют приблизительно гексагональную форму, в которой каждый из шести углов обозначен присутствием портальной триады (портальной вены, желчного протока и печеночной артерии). Хотя гепатоциты являются главным типом паренхимных клеток печени, они функционируют совместно с холангиоцитами (клетками эпителия желчных протоков), эндотелиальными клетками, синусоидальными эндотелиальными клетками, клетками Купффера, естественными киллерами и звездчатыми клетками печени. Эта комплексная структура является критичной для печеночной функции.
Существование стволовых клеток печени остается противоречивым. С одной стороны, поддержание жизнедеятельности тканей в печени и ее регенерация при некоторых типах поражений, обеспечиваются не стволовыми клетками, а скорее делением зрелых клеток (гепатоцитов или холангиоцитов). Однако на моделях поражений печени, когда пролиферация гепатоцитов была ингибирована, также была продемонстрирована способность органа к регенерации в ответ на повреждение. Эти данные позволяют рассматривать печень как орган с факультативными стволовыми клетками.
На сегодняшний момент культуры клеток печени, полученные из гепатоцитов, либо в результате дифференцировки эмбриональных стволовых клеток (ES) или индуцированных плюрипотентных стволовых клеток, не способны размножаться и самообновляться продолжительное время.
Недавно в тонком кишечнике был идентифицирован ген Lgr5, специфично экспрессирующийся в делящихся цилиндрических клетках основания кишечных крипт (СВС-клетках), которые представляют собой мелкие клетки, находящиеся между клетками Панета (Barker et al., 2007. Nature 449: 1003-1007). С использованием мышей со встроенной в локус Lgr5 кассетой GFP-Cre-рекомбиназы, индуцируемой тамоксифеном, путем наблюдения за линиями было показано, что Lgr5+ CBC-клетки представляют собой мультипотентные стволовые клетки, из которых образуются все типы клеток эпителия, даже через 14 месяцев после индукции Cre.
Недавно было обнаружено, что кроме Lgr5, Lgr6 (но не Lgr4) также является уникальным маркером зрелых стволовых клеток. В то время как Lgr5 экспрессируется в мозге, почках, печени, сетчатке, желудке, кишечнике, поджелудочной железе, молочных железах, волосяных луковицах, яичниках, мозговом слое надпочечников и коже, Lgr6 экспрессируется в стволовых клетках мозга, легких, молочных желез, волосяных фолликул и кожи.
В настоящем изобретении авторы разработали способ культивирования предшественников зрелых клеток печени и получения органоида печени с более продолжительной жизнеспособностью, которые обладают способностью дифференцироваться в оба типа клеток - гепатоциты и холангиоциты - и сохраняют основную физиологию отдельных фрагментов печени.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Изобретение относится к способу получения органоида печени, причем способ включает в себя культивирование клеток в первой культуральной среде "для размножения" (также именуемой в настоящем документе как EM), предпочтительно, с последующим культивированием клеток во второй "дифференцирующей" культуральной среде (также именуемой в настоящем документе как DM).
Органоид печени можно получить, культивируя одну Lgr5+ стволовую клетку и/или фрагмент печени. В настоящем документе упоминание "клеток в культуральной среде" включает одну Lgr5+ стволовую клетку, популяцию клеток, содержащих по меньшей мере одну Lgr5+ стволовую клетку, и/или фрагмент печени.
В одном варианте осуществления изобретения среда для размножения содержит EGF, агонист Wnt, FGF и никотинамид. Предпочтительно, агонистом Wnt является R-спондин 1 (R-spondin 1), и поэтому среда для размножения называется "ERFNic". Особенно предпочтительная среда для размножения дополнительно содержит HGF и называется "ERFHNic".
В одном варианте осуществления изобретения дифференцирующая среда содержит EGF, ингибитор TGF-β, FGF и ингибитор Notch. В одном варианте осуществления изобретения ингибитором TGF-β является A83-01, и/или ингибитором Notch является DAPT. Эта дифференцирующая среда именуется в настоящем документе как "EAF" и является предпочтительной дифференцирующей средой по изобретению. FGF можно (необязательно) заменить HGF, или в альтернативном варианте оба, FGF и HGF, могут присутствовать или отсутствовать в дифференцирующей среде. Также может быть добавлен дексаметазон.
В предпочтительном варианте осуществления изобретения клетки можно первоначально культивировать в среде для размножения, которая дополнительно содержит Wnt и Noggin, например, в среде "ENRW", содержащей EGF, Noggin, R-спондин и Wnt, и необязательно FGF, HGF и никотинамид. Авторы изобретения обнаружили, что данная среда является оптимальной для стимуляции исходного размножения клеток в течение первых нескольких дней. Поэтому, эту первую среду для размножения иногда именуют в настоящем документе как EM1. В некоторых вариантах осуществления изобретения Wnt и Noggin удаляют приблизительно через 1 день, 2 дня, 3 дня, 4 дня, 5 дней, 6 дней, 7 дней или большее число дней, например, через 2 недели, через 1 месяц, через 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 или большее число месяцев, через 14 месяцев или после большего срока. Затем, клетки можно размножать в среде для размножения по изобретению, описанной выше, которая не содержит Wnt или Noggin. Эту вторую среду для размножения иногда именуют в настоящем документе как EM2. В некоторых вариантах осуществления изобретения клетки культивируют в EM2 приблизительно в течение 1 дня, 2 дней, 3 дней, 4 дней, 5 дней, 6 дней, 7 дней или в течение более продолжительного срока, например, в течение 3, 4, 5, 10, 20 или большего числа недель. Культуральную среду затем можно заменить на оптимизированную дифференцирующую среду, описанную выше, которая содержит ингибитор TGF-β и ингибитор Notch. Обычно, дифференцирующая среда не содержит агониста Wnt, R-спондина или никотинамида. Это усиливает дифференцировку клеток в зрелые гепатоциты и холангиоциты. Эти клетки подходят для трансплантации людям или животным.
Изобретение также относится к органоиду печени. В настоящей заявке впервые описано, что органоиды печени были выращены ex vivo.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В первом аспекте изобретение относится к способу получения и/или культивирования фрагмента печени или органоида печени, причем указанный способ включает в себя:
культивирование эпителиальных стволовых клеток и/или выделенных фрагментов ткани, включающих указанные эпителиальные стволовые клетки, в контакте с внеклеточным матриксом в присутствии среды, причем среда содержит базальную среду для клеток животных и человека, к которой добавлены: фактор роста эпидермиса, FGF, способный связывать FGFR2 или FGFR4, предпочтительно FGF10, в качестве митогенного ростового фактора, никотинамид и предпочтительно агонист Wnt, предпочтительно R-спондин 1, R-спондин 2, R-спондин 3 или R-спондин 4 и/или Wnt-3a. Предпочтительно, чтобы был добавлен HGF.
Например, в одном варианте осуществления изобретение относится к способу получения и/или культивирования фрагмента печени или органоида печени, причем указанный способ включает в себя:
культивирование эпителиальных стволовых клеток и/или выделенных фрагментов ткани, включающих указанные эпителиальные стволовые клетки, в контакте с внеклеточным матриксом в присутствии среды, причем среда содержит базальную среду для клеток животных и человека, к которой добавлены: ингибитор ВМР, агонист Wnt, фактор роста эпидермиса, FGF, способный связывать FGFR2 или FGFR4, предпочтительно FGF10, и HGF, в качестве митогенных ростовых факторов, гастрин, никотинамид, B27, N2 и N-ацетилцистеин.
Авторами настоящего изобретения неожиданно было обнаружено, что способ по изобретению позволяет культивировать эпителиальные стволовые клетки и/или выделенные фрагменты из печени, содержащие указанные стволовые клетки, одновременно поддерживая присутствие стволовых клеток, которые сохраняют недифференцированный фенотип и способность к саморазмножению. Еще более неожиданным было наблюдение, что способ по изобретению позволяет выращивать из одной выделенной эпителиальной стволовой клетки органоид печени в отсутствие "ниши стволовых клеток".
Стволовые клетки обнаружены во многих органах взрослых людей и мышей. Хотя точные характеристики зрелых стволовых клеток в индивидуальных тканях могут значительно варьировать, зрелые стволовые клетки по меньшей мере имеют следующие общие характеристики: они сохраняют недифференцированный фенотип; их потомство может дифференцироваться в различные типы клеток, присутствующих в соответствующей ткани; они сохраняют способность к саморазмножению в течение всего срока жизни; и они способны регенерировать соответствующую ткань после повреждения. Стволовые клетки имеют специализированное местоположение, нишу стволовых клеток, которая обеспечивает соответствующие межклеточные контакты и сигналы для поддержания популяции указанных стволовых клеток.
Эпителиальные стволовые клетки способны к образованию различных типов клеток, из которых состоит эпителий. Некоторый эпителий, такой как кожа или кишечный эпителий, имеет высокую скорость обновления клеток, указывая на то, что находящиеся в них стволовые клетки должны постоянно пролиферировать. Другие типы эпителия, такие как печеночный эпителий или эпителий поджелудочной железы, в нормальных условиях имеют очень низкую скорость обновления клеток.
Выделение эпителиальных стволовых клеток печени можно провести с помощью ряда различных протоколов. Хотя авторы изобретения не хотят быть связанными какой-либо определенной теорией, в настоящем документе предложена гипотеза, что за регенерацию печени после повреждения ткани отвечает популяция стволовых клеток, присутствующая в печени. Полагают, что клеточная популяция, отвечающая за данную вызванную повреждением регенерацию, экспрессирует маркер Lgr5, будучи активированной.
Путем инъекции мышам со встроенной в геном конструкцией (knock-in) Lgr5-LacZ гепатотоксичного соединения CCl4 для стимуляции повреждения печени, авторы изобретения впервые продемонстрировали, что повреждение печени вызывает экспрессию Lgr5 в печени во всех местах активной регенерации (см. пример 4). Авторы изобретения также показали, что инъекция агониста Wnt, R-спондина, вызывает повышение экспрессии, индуцируемой Wnt-сигнальным путем в печеночных протоках. Хотя авторы изобретения не хотят быть связанными какой-либо определенной теорией, в настоящем документе предложена гипотеза, что после повреждения печени регенеративные стволовые клетки/клетки-предшественники могут активироваться Wnt-сигнальным путем. Lgr5-положительные клетки затем, при необходимости, можно выделить из печени/фрагмента печени, как описано в настоящем документе. Поэтому, изобретение относится к способу получения/выделения Lgr5+ клеток из печени, включающему в себя индукцию повреждения печени или стимуляцию Wnt-сигнального пути, например, с помощью R-спондина. Это является полезным, поскольку в некоторых вариантах осуществления изобретения Lgr5+ клетки являются исходной точкой получения органоидов печени in vitro (хотя использование культуральной среды по изобретению позволяет генерировать Lgr5+ клетки путем добавления агониста Wnt, такого как R-спондин, и необязательно получать Lgr5+ клетку из печени с целью осуществления настоящего изобретения на практике). Также изобретение относится к Lgr5+ клеткам, полученным этим способом. В некоторых вариантах осуществления изобретения такие клетки предпочтительно не экспрессируют маркеры звездчатых клеток, например, SMA. Вместо этого такие клетки предпочтительно экспрессируют один или несколько маркеров печеночных клеток-предшественников или маркеров стволовых клеток, таких как Sox9. Предпочтительно, чтобы экспрессия одного или нескольких маркеров печеночных клеток-предшественников или маркеров стволовых клеток, таких как Sox9, была повышена в указанных клетках по сравнению с клетками зрелой печени.
Изобретение также относится к способу регенерации печени, включающему в себя стимулирование Wnt-сигнального пути. Такой способ может быть полезен в лечении заболеваний печени. Индукцию экспрессии Lgr5 в результате повреждения или путем стимулирования Wnt-сигнального пути можно провести in vivo, ex vivo в выделенной печени или in vitro в фрагменте печени или популяции клеток печени. В вариантах осуществления изобретения, в которых индукцию экспрессии Lgr5 проводят ex vivo или in vitro, Lgr5-положительные клетки можно ввести нуждающемуся в этом пациенту любым подходящим способом, например, с помощью инъекции или имплантации. В некоторых вариантах осуществления изобретения имплантированные клетки содержатся в биосовместимом матриксе. В некоторых вариантах осуществления изобретения клетки размножают ex vivo до использования в терапии.
В предпочтительном способе по изобретению указанные эпителиальные стволовые клетки выделяют из фрагмента печени или желчного протока печени, более предпочтительно из ткани желчного протока.
Способы выделения ткани желчных протоков известны специалистам в данной области. Например, желчный проток можно выделить из печени, как описано в примерах, изложенных в настоящем документе. Кратко, зрелую ткань печени можно промыть в холодной (4-10°C) культуральной среде, предпочтительно в среде Advanced-DMEM/F12 (Invitrogen), и затем ткань можно порубить на кусочки размером около 5 мм и дополнительно промыть холодным буфером для диссоциации (коллагеназа, диспаза, FBS в среде DMEM). Фрагменты ткани предпочтительно инкубировать около 2 ч в буфере для диссоциации при температуре около 37°C. Затем, фрагменты ткани интенсивно ресуспендируют в 10 мл холодного (4-10°C) буфера для выделения, используя пипетку на 10 мл. Первый супернатант, содержащий мертвые клетки, предпочтительно удаляют, и осадок предпочтительно ресуспендируют в буфере для диссоциации (например, в объеме 10-15 мл). После дополнительного интенсивного суспендирования фрагментов ткани супернатант обогащен желчными протоками. Таким образом, можно получить суспензию, содержащую желчные протоки, и желчные протоки собирают под микроскопом и хранят в холодной среде (DMEM + 5-10% FBS). Эту процедуру можно повторять до тех пор, пока не будут собраны по меньшей мере 10-20 желчных протоков на лунку. Затем выделенные желчные протоки можно осадить. Выделенные желчные протоки предпочтительно высевать в 50 мкл матригеля в соотношении приблизительно 20 желчных протоков на лунку.
В отличие от зрелых гепатоцитов, которые растут до конфлюэнтности в течение короткого срока, перед тем как умереть, эпителиальные стволовые клетки печени, выделенные по изобретению, способны к самообновлению и растут бесконечно. Было найдено, что самообновляющейся популяцией клеток является та, которая способна экспрессировать Lgr5 на своей поверхности. Lgr5-отрицательные клетки не самообновляются. Термин "самообновление" следует понимать, как представляющий способность клетки воспроизводить саму себя, одновременно сохраняя исходные способности клеток по изобретению к пролиферации и дифференцировке. Такие клетки пролиферируют путем деления с образованием клонов, которые дополнительно делятся на клоны, и поэтому увеличивают размер клеточной популяции без необходимости во внешнем вмешательстве, и не превращаясь в клетки с более ограниченной способностью к дифференцировке.
Предпочтительный способ основан на том факте, что стволовые клетки печени по изобретению экспрессируют Lgr5 и/или Lgr6 на своей поверхности; эти белки принадлежат к большому суперсемейству связанных с G-белками рецепторов (GPCR) (см., например, WO2009/022907, содержание которого полностью включено в настоящий документ). Подсемейство Lgr уникально тем, что оно несет крупный лейцин-богатый эктодомен, важный для связывания лиганда. Предпочтительный способ поэтому включает в себя получение суспензии клеток из указанной эпителиальной ткани, как описано выше, контактирование указанной клеточной суспензии с соединением, связывающим Lgr5 и/или 6 (таким как антитело), выделение соединения, связывающего Lgr5 и/или 6, и отделение стволовых клеток от указанного связывающего соединения.
Предпочтительно, чтобы суспензия одиночных клеток, содержащая эпителиальные стволовые клетки, была получена механически из выделенного желчного протока. Полученные таким образом в результате механического разрушения небольшие фрагменты органоида предпочтительно рассевать в соотношении приблизительно 1:6. При необходимости такие фрагменты можно кратковременно (2 или 3 минуты) инкубировать в трипсине в разведении приблизительно 1:2. Было показано, что на этой стадии эпителиальные стволовые клетки, обработанные трипсином, имеют достаточно низкую выживаемость: если клетки рассевать как индивидуальные клетки, то выживают только клетки, экспрессирующие Lgr5. Эта фракция является достаточно небольшой (приблизительно 12% от общей популяции клеток).
Предпочтительные соединения, связывающие Lgr5 и/или 6, включают в себя антитела, такие как моноклональные антитела, которые специфично связывают внеклеточный домен либо Lgr5, либо Lgr6, такие как моноклональные антитела, включая мышиные и крысиные моноклональные антитела (см., например, WO2010/016766, содержание которого полностью включено в настоящее описание). Используя такое антитело, можно выделить стволовые клетки экспрессирующие Lgr5 и/или Lgr6, например, с помощью магнитных шариков или сортинга флуоресцентно активированных клеток, как очевидно для опытного специалиста. С использованием способа по изобретению можно выделить одну единственную клетку, экспрессирующую Lgr5 и/или Lgr6, и можно применить к ней способ по изобретению. Таким образом, органоид печени можно получить из одной единственной клетки.
Ниша стволовых клеток
Выделенные стволовые клетки предпочтительно культивировать в микроокружении, которое имитирует по меньшей мере частично клеточную нишу, в которой указанные стволовые клетки находятся в природе. Эту клеточную нишу можно имитировать путем культивирования указанных стволовых клеток в присутствии биоматериалов, таких как матриксы, каркасы и культуральные субстраты, которые представляют ключевые регуляторные сигналы, контролирующие судьбу стволовых клеток. Такие биоматериалы включают в себя природные, полусинтетические и синтетические биоматериалы и/или их смеси. Каркас обеспечивает двумерную или трехмерную сеть. Подходящие синтетические материалы для такого каркаса включают полимеры, выбранные из пористых твердых веществ, нановолокон и гидрогелей, таких как, например, пептиды, включая способные к самосборке пептиды, гидрогели, состоящие из полиэтиленгликоль фосфата, полиэтиленгликоль фумарата, полиакриламида, полигидроксиэтилметакрилата, ацетата полицеллюлозы и их сополимеров (см., например, Saha et al., 2007. Curr Opin Chem Biol. 11(4): 381-387; Saha et al., 2008. Biophysical Journal 95: 4426-4438; Little et al., 2008. Chem. Rev 108, 1787-1796). Как известно опытному специалисту, механические свойства, такие как, например, эластичность каркаса, влияют на пролиферацию, дифференцировку и миграцию стволовых клеток. Предпочтительный каркас содержит биоразлагаемые (со)полимеры, которые замещаются природными компонентами после трансплантации в субъекта, например, для стимуляции регенерации ткани и/или ранозаживления. Дополнительно предпочтительно, чтобы после трансплантации субъекту такой каркас по существу не вызывал иммуногенного ответа. Такой каркас дополняется природными, полусинтетическими или синтетическими лигандами, которые обеспечивают сигналы, требуемые для пролиферации и/или дифференцировки, и/или миграции стволовых клеток. В предпочтительных вариантах осуществления изобретения указанные лиганды включают в себя определенные аминокислотные фрагменты. Примеры таких синтетических полимеров включают в себя поверхностно-активный блок-сополимер Pluronic® F127 (BASF) и Ethisorb ® (Johnson and Johnson).
Клеточная ниша частично определяется стволовыми клетками и окружающими клетками, а также внеклеточным матриксом (ЕСМ), который продуцируют клетки в указанной нише. В предпочтительном способе по изобретению выделенные фрагменты печени или выделенный желчный проток, или выделенные эпителиальные стволовые клетки прикреплены к ЕСМ. ЕСМ состоит из разнообразных полисахаридов, воды, эластина и гликопротеинов, причем гликопротеины включают в себя коллаген, энтактин (нидоген), фибронектин и ламинин. ЕСМ секретируется клетками соединительной ткани. Известны различные типы ЕСМ, содержащие различные композиции, включающие различные типы гликопротеинов и/или различные комбинации гликопротеинов. Указанный ЕСМ можно обеспечить, культивируя в носителе продуцирующие ЕСМ клетки, такие как, например, фибробластные клетки, после чего эти клетки удаляют и добавляют выделенные фрагменты печени, или выделенный желчный проток, или выделенные эпителиальные стволовые клетки. Примерами клеток, продуцирующих внеклеточный матрикс, являются хондроциты, продуцирующие в основном коллаген и протеогликаны, фибробластные клтеки, продуцирующие в основном коллаген IV типа, ламинин, интерстициальные протоколлагены и фибронектин, и миофибробласты толстого кишечника, продуцирующие в основном коллагены (I, III, и V типов), хондроитинсульфат протеогликан, гиалуроновую кислоту, фибронектин и тенасцин-С. В альтернативном варианте указанный ЕСМ доступен в продаже. Примерами доступных в продаже внеклеточных матриксов являются внеклеточные матриксные белки (Invitrogen) и препараты базальных мембран из клеток мышиной саркомы Engelbreth-Holm-Swarm (EHS) (например, Matrigel™ (BD Biosciences)). Можно использовать синтетический материал внеклеточного матрикса, такой как ProNectin (Sigma Z378666). При желании можно использовать смеси материалов внеклеточного матрикса. Применение ЕСМ для культивирования стволовых клеток увеличивало продолжительность жизни стволовых клеток и постоянное присутствие недифференцированных стволовых клеток. В отсутствие ЕСМ культуры стволовых клеток невозможно культивировать в течение продолжительного времени, и при этом не наблюдалось постоянного присутствия недифференцированных стволовых клеток. В дополнение, присутствие ЕСМ позволяло культивировать трехмерные органоиды тканей, которые невозможно культивировать в отсутствие ЕСМ. Материалом внеклеточного матрикса в норме будет покрыт сосуд для культивирования клеток, но (в дополнение или в альтернативном варианте) он может быть добавлен в раствор. Для нанесения на сосуд для культивирования клеток можно использовать раствор фибронектина с концентрацией примерно 1 мг/мл, или с концентрацией от примерно 1 мкг/см2 до примерно 250 мкг/см2, или от примерно 1 мкг/см2 до примерно 150 мкг/см2. В некоторых вариантах осуществления изобретения сосуд для культивирования клеток покрывают фибронектином с концентрацией от 8 мкг/см2 до 125 мкг/см2.
Предпочтительная ЕСМ для использования в способе по изобретению содержит по меньшей мере два разных гликопротеина, таких как два различных типа коллагена или коллаген и ламинин. Внеклеточным матриксом может быть внеклеточный матрикс из синтетического гидрогеля или природный ЕСМ. Еще один предпочтительный ЕСМ обеспечивает Matrigel™ (BD Biosciences), который содержит ламинин, энтактин и коллаген IV.
Композиции по изобретению могут содержать сыворотку или могут быть свободны от сыворотки и/или свободны от замещающих сыворотку компонентов, как описано в настоящем документе. Получения культуральной среды и клеток предпочтительно являются GMP-процессами в соответствии со стандартами, предъявляемыми FDA к биологическим продуктам для гарантии однородности продукта.
Культуральная среда
Среда для культивирования клеток, которую используют в способе по изобретению, включает в себя любую подходящую среду для культивирования клеток с ограничениями, приведенными в настоящем документе. Среда для культивирования клеток обычно содержит большое количество ингредиентов, которые необходимы для поддержки жизнедеятельности культивируемых клеток. Специалист в данной области легко сформулирует подходящие комбинации ингредиентов, принимая во внимание нижеследующее описание. Культуральной средой по изобретению, в общем, будет питательный раствор, содержащий стандартные ингредиенты для культивирования клеток, такие как аминокислоты, витамины, неорганические соли, источник углеводов и буфер, как описано более подробно в литературе и ниже.
Культуральная среда по изобретению будет в норме приготовлена на деионизированной, дистиллированной воде. Для того чтобы предупредить заражение, культуральная среда по изобретению будет, как правило, стерилизована до использования, например, ультрафиолетовым светом, нагреванием, рентгеновскими лучами или фильтрованием. Культуральная среда может быть заморожена (например, при температуре -20°C или -80°C) для хранения или для транспортировки. Среда может содержать один или несколько антибиотиков для предупреждения заражения. Содержание эндотоксинов в среде может составлять меньше 0,1 единиц эндотоксина на мл или может составлять меньше 0,05 единиц эндотоксина на мл. Способы определения содержания эндотоксинов в культуральной среде известны в данной области.
Предпочтительной клеточной культуральной средой является указанная синтетическая среда с рН 7,4 (предпочтительно с рН 7,2-7,6 или по меньшей мере 7,2, но не выше 7,6), подведенным с помощью карбонатного буфера, причем клетки культивируют в атмосфере, содержащей от 5% до 10% СО2, или по меньшей мере 5%, но не более 10% СО2, предпочтительно 5% СО2.
Исходя из общедоступной информации опытный специалист поймет, какие типы культуральной среды можно использовать в качестве базальной среды в клеточных культуральных средах по изобретению. Потенциально подходящие клеточные культуральные среды доступны в продаже и включают в себя, но не ограничены этим, модифицированную по способу Дульбекко среду Игла (DMEM), минимальную необходимую среду (MEM), Knockout™DMEM (KO-DMEM), минимальную необходимую среду Глазго (G-MEM), базальную среду Игла (BME), DMEM/среду Хэма F12, Advanced-DMEM/среду Хэма F12, модифицированную Исковым среду Дульбекко и минимальную необходимую среду, среду Хэма F-10, среду Хэма F-12, среду 199, и среду RPMI 1640.
Предпочтительная клеточная культуральная среда выбрана из DMEM/F12 и RPMI 1640 с добавлением глутамина, инсулина пенициллина/стрептомицина и трансферрина. В дополнительном предпочтительном варианте осуществления изобретения используют Advanced-DMEM/F12 или Advanced-RPMI, которые оптимизированы для культивирования бессывороточной культуры и уже включают инсулин. В этом случае в среду Advanced-DMEM/F12 или Advanced-RPMI предпочтительно добавлены глутамин и пенициллин/стрептомицин.
В некоторых вариантах осуществления изобретения базальная среда содержит Advanced-DMEM F12, Hepes, пенициллин/стрептомицин, глутамин, N-ацетилцистеин, B27, N2 и гастрин. В некоторых вариантах осуществления изобретения культивирование начинают на базальной среде, содержащей N2, и гастрин, а также пенициллин/стрептомицин, но позднее их удаляют. Например, в некоторых вариантах осуществления изобретения N2, гастрин и пенициллин/стрептомицин присутствуют в среде EM1 по изобретению, но отсутствуют в EM2 или DM. Например, в некоторых вариантах осуществления изобретения N2, гастрин и пенициллин/стрептомицин присутствуют в среде EM1 и ЕМ2 по изобретению, но не в среде DM. В особенно предпочтительных вариантах осуществления изобретения базальной средой является Advanced-DMEM/F12 или вариант DMEM с пенициллином/стрептомицином, N2, B27, глутамином и гастрином.
В предпочтительных вариантах осуществления изобретения базальная среда содержит гастрин. В некоторых вариантах осуществления изобретения базальная среда также содержит N-ацетилцистеин и/или B27.
Кроме того, предпочтительно, чтобы в указанную клеточную культуральную среду был добавлен очищенный, природный, полусинтетический и/или синтетический ростовой фактор, и она не содержала компонент с неопределенным составом, такой как фетальная бычья сыворотка или фетальная телячья сыворотка. Различные составы для замены сыворотки доступны в продаже и известны опытному специалисту в данной области. При использовании составов, заменяющих сыворотку, их можно использовать в количестве от примерно 1% до примерно 30% от объема среды в соответствии с общепринятыми методиками.
Среда для размножения (ЕМ2):
В одном аспекте настоящее изобретение относится к клеточной культуральной среде, которая содержит или состоит из базальной среды для клеток животных или человека, к которой добавлены: фактор роста эпидермиса, FGF, способный связывать FGFR2 или FGFR4, предпочтительно FGF10, в качестве митогенного ростового фактора, никотинамид, и предпочтительно агонист Wnt, предпочтительно R-спондин 1. Эта среда называется ЕМ2.
Предпочтительно, чтобы агонистом Wnt являлся R-спондин 1. В настоящем документе среда, содержащая EGF, R-спондин 1, FGF и никотинамид называется ERFNic.
В некоторых вариантах осуществления изобретения среда ЕМ2 не содержит Noggin и более предпочтительно не содержит ингибитора BMP. В некоторых вариантах осуществления изобретения среда ЕМ2 не содержит Wnt, например, Wnt-3a.
В некоторых вариантах осуществления изобретения присутствует HGF дополнительно к FGF. Предпочтительной средой, содержащей HGF дополнительно к FGF, является ERFHNic (EGF + R-спондин (предпочтительно R-спондин 1) + FGF (предпочтительно FGF10) + HGF + никотинамид). Авторы изобретения обнаружили, что среда ERFHNic является оптимальной для размножения клеток в течение длительного времени. В отсутствие HGF клетки не выживали в культуре более чем три месяца. Кроме того, в отсутствие HGF через 10 пассажей у клеток наблюдалось отставание роста по сравнению с клетками, культивируемыми в присутствии HGF, о чем свидетельствовала меньшая скорость пролиферации. В частности, через 15 пассажей из клеток не вырастали органоиды с такой же скоростью роста в отсутствие HGF как в присутствии HGF. Следовательно, было найдено, что HGF необходим для поддержания хорошей скорости пролиферации при длительном культивировании. Следовательно, изобретение относится к применению среды ERFHNic по изобретению для культивирования клеток в течение по меньшей мере 2 недель, по меньшей мере 1 месяца, по меньшей мере 2 месяцев, более предпочтительно, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5, по меньшей мере 6, по меньшей мере 7, по меньшей мере 8, по меньшей мере 9, по меньшей мере 10, по меньшей мере 15, по меньшей мере 20, по меньшей мере 24, по меньшей мере 25, по меньшей мере 30 или большего числа месяцев, например, в течение 3 или более лет. На практике, некоторые варианты осуществления изобретения включают в себя применение Е2 примерно на протяжении 20-30 пассажей клеток. Например, клетки можно рассевать 20-30 раз, обычно один раз в неделю. Предпочтительно, количество клеток увеличивается примерно в 4 раза за неделю, или происходит два умножения популяции за неделю. Изобретение дополнительно относится к применению среды ERFHNic по изобретению для культивирования клеток в течение по меньшей мере 10 пассажей, например, по меньшей мере 11, по меньшей мере 12, по меньшей мере 15, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 40, по меньшей мере 50, по меньшей мере 60 пассажей или в течение 15-35 пассажей, например, приблизительно 20-30 пассажей. В некоторых вариантах осуществления изобретения в среде ЕМ2 дополнительно присутствует ингибитор TGF-β, такой как A83-01. Это особенно полезно при культивировании клеток или органоидов человека. В некоторых вариантах осуществления изобретения A83-01 присутствует в концентрации 400-600 нМ, например, 450-550 нМ, 470-530 нМ или приблизительно 500 нМ. В вариантах осуществления изобретения, в которых ингибитор TGF-β присутствует в среде ЕМ2, предпочтительно, чтобы в среде отсутствовал ингибитор Notch.
Среда для размножения (ЕМ1):
В одном аспекте настоящее изобретение относится к клеточной культуральной среде, которая содержит или состоит из базальной среды для клеток животных или человека, к которой добавлены: EGF, ингибитор BMP, R-спондин и Wnt. Предпочтительно, чтобы ингибитором ВМР являлся Noggin и среда называлась "ENRW" (EGF, Noggin, R-спондин и Wnt). Эта среда называется ЕМ1. Авторы изобретения обнаружили, что среда, содержащая Wnt и Noggin, является идеальной для первоначального размножения клеток. Следовательно, в некоторых вариантах осуществления изобретения среду ЕМ1 используют всего для 1 пассажа или в течение 1 недели, но также предусмотрено, что среду ЕМ1 можно использовать около года, поскольку она не вредна для клеток. Следовательно, в некоторых вариантах осуществления изобретения среду ЕМ1 используют для культивирования клеток от 0-го дня до 10-го дня, например, в течение 0-7 дней, 0-6 дней, 0-5 дней, 0-4 дней, 0-3 дней, 0-2 дней, 0-1 дней, причем 0-й день является днем выделения клеток из соответствующей ткани, а 1-й день является следующим днем, или среду ЕМ1 используют в течение 1 или большего количества недель, например, в течение 2, 3, 4 или большего количества недель, или в течение 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 или большего количества месяцев. В некоторых вариантах осуществления изобретения среду ЕМ1 используют только в течение первого дня или первых двух дней культивирования. В некоторых вариантах осуществления изобретения среду ЕМ1 используют в течение 1 или большего числа пассажей, например, в течение 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30 или большего числа пассажей, например, в течение 20-30 пассажей. В некоторых вариантах осуществления изобретения среду ЕМ1 используют после стадии замораживания или любой другой стадии транспортировки, включающей в себя смену температуры или среды, которые не соответствуют оптимальному росту.
Среду ЕМ1 дополняют одним или несколькими соединениями, выбранными из группы, состоящей из FGF, HGF, никотинамида, гастрина, N-ацетилцистеина и N2. В случае начала культур из замороженного источника или из единственной клетки, среду ЕМ1 предпочтительно дополнить ингибитором ROCK. Y27632 является предпочтительным ингибитором ROCK для использования в изобретении.
Следовательно, в одном варианте осуществления изобретение относится к клеточной культуральной среде, которая содержит или состоит из базальной среды для клеток животных или человека, в которую добавлены: фактор роста эпидермиса, FGF, способный связывать FGFR2 или FGFR4, предпочтительно FGF10, и HGF в качестве митогенных ростовых факторов,
Гастрин, никотинамид, В27, N2 и N-ацетилцистеин и предпочтительно;
ингибитор ВМР, предпочтительно Noggin; и
агонист Wnt, предпочтительно R-спондин 1 и/или Wnt-3a.
B27 (Invitrogen), N-ацетилцистеин (Sigma) и N2 (Invitrogen), Гастрин (Sigma) и никотинамид (Sigma) также добавляют в описанную выше среду, и, полагают, что они стимулируют пролиферацию клеток. В контексте изобретения никотинамид также называют "Nic".
"Добавка N2" доступна от Invitrogen, Carlsbad, CA; www.invitrogen.com; каталожный № 17502-048; и от PAA Laboratories GmbH, Pasching, Austria; www.paa.com; каталожный № F005-004; Bottenstein & Sato, PNAS, 76(1):514-17, 1979. Добавка N2 поставляется PAA Laboratories GmbH в виде 100× жидкого концентрата, содержащего 500 мкг/мл трансферрина человека, 500 мкг/мл бычьего инсулина, 0,63 мкг/мл прогестерона, 1611 мкг/мл путресцина и 0,52 мкг/мл селенита натрия. Добавку N2 можно добавлять в культуральную среду в виде концентрата или разведенную в культуральной среде перед добавлением. Ее можно использовать в 1× конечной концентрации или других конечных концентрациях. Использование добавки N2 является удобным путем включения трансферрина, инсулина, прогестерона, путресцина и селенита натрия в культуральную среду по изобретению.
"Добавку В27" (доступна от Invitrogen, Carlsbad, CA; www.invitrogen.com; текущий каталожный № 17504-044; и от PAA Laboratories GmbH, Pasching, Austria; www.paa.com; каталожный № F01-002; Brewer et al., J Neurosci Res., 35(5):567-76, 1993) можно использовать для составления культуральной среды, которая содержит биотин, холестерин, линолевую кислоту, линоленовую кислоту, прогестерон, путресцин, ретинол, ретинилацетат, селенит натрия, трийодтиронин (Т3), DL-α-токоферол (витамин Е), альбумин, инсулин и трансферрин. Добавка В27 поставляется PAA Laboratories GmbH в виде 50× концентрата, содержащего помимо прочих ингредиентов биотин, холестерин, линолевую кислоту, линоленовую кислоту, прогестерон, путресцин, ретинол, ретинилацетат, селенит натрия, трийодтиронин (Т3), DL-α-токоферол (витамин Е), альбумин, инсулин и трансферрин. Из этих ингредиентов по меньшей мере линоленовая кислота, ретинол, ретинилацетат и трийодтиронин (Т3) являются агонистами ядерных рецепторов гормонов. Добавку В27 можно добавлять к культуральной среде в виде концентрата или разведенную перед добавлением в культуральную среду. Ее можно использовать в 1× конечной концентрации или в других конечных концентрациях. Использование добавки N2 является удобным путем включения биотина, холестерина, линолевой кислоты, линоленовой кислоты, прогестерона, путресцина, ретинола, ретинилацетата, селенита натрия, трийодтиронина (Т3), DL-α-токоферола (витамина Е), альбумина, инсулина и трансферрина в культуральную среду по изобретению.
Ингибиторы ВМР
Компонентом, который предпочтительно добавлять к базальной культуральной среде, является ингибитор ВМР. ВМР связываются в виде димерного лиганда с рецепторным комплексом, состоящим из двух различных рецепторных сериновых/треониновых киназ, рецепторов I и II типов. Рецептор II типа фосфорилирует рецептор I типа, приводя к активации этой рецепторной киназы. Рецептор I типа далее фосфорилирует специфичные субстраты рецептора (SMAD), обеспечивая передачу сигнала, приводящую к транскрипционной активности.
Ингибитор ВМР определяют как агент, который связывает молекулу ВМР, образуя комплекс, причем активность ВМР нейтрализуется, например, в результате предупреждения или ингибирования связывания молекулы ВМР с рецептором ВМР. В альтернативном варианте указанный ингибитор является агентом, который действует в качестве антагониста или обратного агониста. Этот тип ингибитора связывается с рецептором ВМР и предупреждает связывание ВМР с указанным рецептором. Примером последнего агента является антитело, которое связывает рецептор и предупреждает связывание ВМР со связанным с антителом рецептором.
Ингибитор ВМР можно добавлять к среде в количестве, эффективном для ингибирования ВМР-зависимой активности в клетке самое большее до 90%, более предпочтительно, самое большее до 80%, более предпочтительно, самое большее до 70%, более предпочтительно, самое большее до 50%, более предпочтительно, самое большее до 30%, более предпочтительно, самое большее до 10%, более предпочтительно до 0%, относительно уровня активности ВМР в отсутствие указанного ингибитора, оцениваемой на том же типе клеток. Опытному специалисту в данной области известно, что активность ВМР можно определить, измеряя его транскрипционную активности, например, как описано в Zilberberg et al., 2007. BMC Cell Biol. 8:41.
Известны несколько классов ВМР-связывающих белков, включая Noggin (Peprotech), Chordin и Chordin-подобные белки (R&D systems), содержащие Chordin-домены, фоллистатин и родственные фоллистатину белки (R&D systems), содержащие фоллистатиновый домен, DAN и DAN-подобные белки (R&D systems), домен "цистеиновый узел" DAN, склеростин/SOST (R&D systems), декорин (R&D systems) и α-2-макроглобулин (R&D systems).
Предпочтительный ингибитор ВМР для использования в способе по изобретению выбирают из Noggin, DAN и DAN-подобных белков, включая Cerberus и Gremlin (R&D systems). Эти диффундирующие белки способны связывать ВМР-лиганды с различной степенью аффинности и ингибировать их доступ к сигнальным рецепторам. Добавление любого из этих ингибиторов ВМР к базальной культуральной среде предупреждает потерю стволовых клеток.
Предпочтительным ингибитором ВМР является Noggin. В контексте культуральной среды по изобретению Noggin также именуется в настоящем документе как "N". Noggin предпочтительно добавлять к базальной культуральной среде в концентрации по меньшей мере 10 нг/мл, более предпочтительно, по меньшей мере 20 нг/мл, более предпочтительно, по меньшей мере 50 нг/мл, более предпочтительно, по меньшей мере 100 нг/мл. Еще более предпочтительно, чтобы концентрация составляла приблизительно 100 нг/мл или точно 100 нг/мл. В ходе культивирования стволовых клеток указанный ингибитор ВМР можно добавлять к культуральной среде по необходимости, например, ежедневно или через день. Ингибитор ВМР предпочтительно добавлять к культуральной среде через день. Культуральную среду можно обновлять при необходимости, например, ежедневно или через день, хотя предпочтительно обновлять среду на каждый четвертый день.
Агонисты Wnt
Дополнительным компонентом, который можно добавить к базальной культуральной среде, является агонист Wnt. В контексте культуральной среды по изобретению агонист Wnt также именуется в настоящем документе как "W". Wnt-сигнальный путь определяется серией событий, которые происходят после связывания белком Wnt рецептора на клеточной поверхности, принадлежащего к семейству рецепторов Frizzled. Это приводит в результате к активации белков из семейства Dishevelled, которые ингибируют деградацию β-катенина комплексом белков, который включает в себя Axin, GSK-3 и белок АРС. В результате накопление ядерного β-катенина усиливает транскрипцию, опосредованную транскрипционными факторами семейства TCF/LEF.
Агонист Wnt определяют как агент, который активирует TCF/LEF-опосредованную транскрипцию в клетке. Поэтому, агонистов Wnt выбирают из действительных агонистов Wnt, которые связывают и активируют члена семейства рецепторов Frizzled, включая все до одного белки семейства Wnt, ингибитор внутриклеточной деградации β-катенина и активаторы TCF/LEF. Указанный агонист Wnt добавляют к среде в количестве, эффективном для стимуляции активности Wnt в клетке по меньшей мере на 10%, более предпочтительно, по меньшей мере на 20%, более предпочтительно, по меньшей мере на 30%, более предпочтительно, по меньшей мере на 50%, более предпочтительно, по меньшей мере на 70%, более предпочтительно, по меньшей мере на 90%, более предпочтительно, по меньшей мере на 100% относительно уровня активности указанного Wnt в отсутствие указанной молекулы, оцениваемой в том же типе клеток. Как известно опытному специалисту в данной области, активность Wnt можно определять путем измерения транскрипционной активности Wnt, например, с помощью Tcf-люциферазных репортерных конструкций, pTOPFLASH и pFOPFLASH (Korinek et al., 1997. Science 275:1784-1787).
Агонист Wnt может включать в себя секретируемый гликопротеин, включая Wnt-1/Int-1; Wnt-2/Irp (Int-1-родственный белок); Wnt-2b/13; Wnt-3/Int-4; Wnt-3a (R&D systems); Wnt-4; Wnt-5a; Wnt-5b; Wnt-6 (Kirikoshi H et al. 2001. Biochem Biophys Res Com 283: 798-805); Wnt-7a (R&D systems); Wnt-7b; Wnt-8a/8d; Wnt-8b; Wnt-9a/14; Wnt-9b/14b/15; Wnt-10a; Wnt-10b/12; Wnt-11 и Wnt-16. Обзор белков Wnt человека приведен в "THE WNT FAMILY OF SECRETED PROTEINS", R&D Systems Catalog, 2004.
Дополнительные агонисты Wnt включают: семейство секретируемых белков R-спондинов, которое участвует в активации и регуляции Wnt-сигнального пути и которое состоит из 4 членов (R-спондин 1 (NU206, Nuvelo, San Carlos, CA), R-спондин 2 (R&D systems), R-спондин 3 и R-спондин-4); и Norrin (также называемый белком болезни Норри или NDP) (R&D systems), который представляет собой секретируемый регуляторный белок, функционирующий аналогично белку Wnt в том, что он с высокой аффинностью связывает рецептор Frizzled-4 и вызывает активацию Wnt-сигнального пути (Kestutis Planutis et al. (2007) BMC Cell Biol. 8: 12).
Соединения, которые имитируют активность R-спондина, можно использовать в качестве агонистов Wnt по изобретению. Недавно было найдено, что R-спондин взаимодействует с Lgr5. Поэтому, агонисты Lgr5, такие как антитела к Lgr5, являются примерами агонистов Wnt, которые можно использовать в изобретении.
В контексте культуральной среды по изобретению R-спондин также именуется в настоящем документе как "R".
Низкомолекулярный агонист Wnt-сигнального пути, аминопиримидиновое соединение, было недавно идентифицировано и также однозначно включено в качестве агониста Wnt (Liu et al. (2005) Angew Chem Int Ed Engl. 44, 1987-90).
Известные ингибиторы GSK включают в себя малые интерферирующие РНК (миРНК; Cell Signaling), литий (Sigma), кенпауллон (Biomol International; Leost, M. et al. (2000) Eur. J. Biochem. 267, 5983-5994), 6-бромоиндирубин-30-ацетоксим (Meijer, L. et al. (2003) Chem. Biol. 10, 1255-1266), SB 216763 и SB 415286 (Sigma-Aldrich), и члены FRAT-семейства и полученные из FRAT пептиды, которые предупреждают взаимодействие GSK-3 с Axin. Обзор приведен в статье Meijer et al., (2004) Trends in Pharmacological Sciences 25, 471-480, которая, таким образом, включена в настоящий документ путем ссылки. Способы и анализы для определения уровня ингибирования GSK-3 известны опытному специалисту и включают в себя, например, способы и анализы, описанные в Liao et al 2004, Endocrinology, 145(6): 2941-9.
В предпочтительном варианте осуществления изобретения указанный агонист Wnt выбирают из одного или нескольких из членов семейства Wnt, R-спондина 1-4 (например, R-спондин 1 или R-спондин 4), Norrin и ингибитора GSK. Авторы изобретения обнаружили, что добавление по меньшей мере одного агониста Wnt в базальную культуральную среду важно для пролиферации эпителиальных стволовых клеток печени или выделенного желчного протока или выделенных фрагментов печени.
В дополнительном предпочтительном варианте осуществления изобретения указанный агонист Wnt включает в себя или состоит из R-спондина 1. R-спондин 1 предпочтительно добавлять к базальной культуральной среде в концентрации по меньшей мере 200 нг/мл, более предпочтительно, по меньшей мере 300 нг/мл, более предпочтительно, по меньшей мере 500 нг/мл. Еще более предпочтительная концентрация R-спондина 1 составляет приблизительно 500 нг/мл или точно 500 нг/мл. В ходе культивирования стволовых клеток указанный член семейства Wnt можно добавлять к культуральной среде по необходимости, например, ежедневно или через день. Белок из семейства Wnt предпочтительно добавлять к культуральной среде через день. Культуральную среду можно обновлять при необходимости, например, ежедневно или через день, хотя предпочтительно обновлять среду на каждый четвертый день.
В предпочтительном варианте осуществления изобретения агонист Wnt выбран из группы, состоящей из: R-спондина, Wnt-3a и Wnt-6. Более предпочтительно использовать оба, R-спондин и Wnt-3a, в качестве агониста Wnt. Эта комбинация является особенно предпочтительной, поскольку она неожиданно оказывает синергетический эффект на образование органоида. Предпочтительные концентрации составляют 1-500 нг/мл, например, 1-10 нг/мл, 10-100 нг/мл, 1-50 нг/мл, 10-200 нг/мл, от 200 до 500 нг/мл, 30 нг/мл для R-спондина и от 100 нг/мл до 1000 нг/мл, например, 100 нг/мл или 1000 нг/мл для Wnt3a.
Агонистом Wnt предпочтительно является Wnt-лиганд, такой как, например, Wnt3a, и его можно добавлять свежим в культуральную среду. В альтернативном варианте Wnt-лиганд экспрессируется в клеточной линии в результате трансфекции или инфекции клеточной линии соответствующей экспрессионной конструкцией, экспрессирующей указанный Wnt-лиганд. Указанную клеточную линию культивируют, и культуральную среду, содержащую секретируемый Wnt-лиганд, собирают через соответствующие временные интервалы. Например, клетки будут продуцировать Wnt3a, как только они достигнут конфлюентности и перестанут расти. В качестве контроля используют культуральную среду от клеток, которые не были трансфицированы или инфицированы указанной экспрессионной конструкцией. Кондиционную среду собирают и тестируют, например, методом анализа, в котором экспрессия люциферазы находится под контролем TCF-отвечающих элементов для тестирования на присутствие агониста, такого как Wnt3a (Korinek et al., 1997. Science 275: 1784-1787). Среду разводят при использовании в культурах для регенерации ткани. Опытному специалисту известно, что добавление избытка Wnt-лиганда иногда вредно для культуры, также как и добавление слишком малого количества Wnt-лиганда. Поэтому, фактическое разведение кондиционной среды будет зависеть от количества Wnt-лиганда, которое определяют в тесте.
Митогенные ростовые факторы
Еще одним дополнительным компонентом, который добавляют в базальную культуральную среду, является комбинация митогенных ростовых факторов, выбранных из группы фактора роста эпидермиса (EGF; (предпочтительно от Peprotech)), фактора роста фибробластов (FGF), способного связываться с FGFR2 или FGFR4, и фактора роста гепатоцитов (HGF) (также предпочтительно от Peprotech). FGF, способный связываться с FGFR2 (рецептором FGF) или FGFR4, предпочтительно являются FGF4, FGF7 или FGF10 (предпочтительно от Peprotech), наиболее предпочтительно FGF10. Предпочтительно использовать все три фактора - EGF, FGF и HGF. В контексте культуральной среды по изобретению EGF также именуется в настоящем документе как "Е", FGF также именуется в настоящем документе как "F", а HGF также именуется в настоящем документе как "Н". EGF является сильным митогенным фактором для ряда культивируемых эктодермальных и мезодермальных клеток и обладает выраженным эффектом на дифференцировку определенных клеток in vivo и in vitro и некоторых фибробластов в клеточной культуре. Предшественник EGF существует в виде мембраносвязанной молекулы, которая протеолитически отщепляется с образованием 53-аминокислотного пептидного гормона, стимулирующего клетки. EGF проявляет свои эффекты на клетки-мишени путем связывания с расположенной на плазматической мембране рецептором EGF. Рецептор EGF является трансмембранной протеинтирозинкиназой. Связывание EGF с рецептором вызывает активацию киназы и последующее аутофосфорилирование рецептора. Аутофосфорилирование необходимо для взаимодействия рецептора с его субстратами. Сигнальные пути, активируемые EGF, включают фосфатидилинозитольный путь, приводящий к активации протеинкиназы С и увеличению концентрации внутриклеточного Са2+, и ras-сигнальный путь, приводящий к активации МАР-киназы. Эти пути участвуют в регуляции клеточной пролиферации, дифференцировки и выживания (Herbst, 2004, Int Journal of radiation oncology, 59, 2, S21-S26).
EGF предпочтительно добавляют к базальной культуральной среде в концентрации от 5 до 500 нг/мл, или составляющей по меньшей мере 5 и не выше 500 нг/мл. Предпочтительная концентрация составляет по меньшей мере 10, 20, 25, 30, 40, 45, или 50 нг/мл и не превышает 500, 450, 400, 350, 300, 250, 200, 150 или 100 нг/мл. Более предпочтительная концентрация составляет по меньшей мере 50 и не превышает 100 нг/мл. Еще более предпочтительная концентрация составляет примерно 50 нг/мл или 50 нг/мл.
FGF10 является белком, который принадлежит к семейству белков факторов роста фибробластов (FGF). Члены FGF-семейства обладают широким спектром митогенных функций и клеточной выживаемости, и участвуют в ряде биологических процессов, включая эмбриональное развитие, клеточный рост, морфогенез, восстановление тканей, рост и инвазию опухолей. FGF стимулируют клетки путем взаимодействия с тирозинкиназными рецепторами (FGFR) на клеточной поверхности. Идентифицированы четыре близкородственных рецептора (FGFR1-FGFR4). Было показано, что гены FGFR1-FGFR3 кодируют множественные изоформы, и эти изоформы могут быть критичны для определения лиганд-специфичности. Большинство FGF связывают более одного рецептора (Ornitz J Biol Chem. 1998 Feb 27;273 (9):5349-57). Однако FGF10 и FGF7 являются уникальными среди других FGF в том, что они взаимодействуют только с определенной изоформой FGFR2, обозначаемой FGFR2b, которая экспрессируются исключительно в эпителиальных клетках (Igarashi, J Biol Chem. 1998 273(21):13230-5). FGF10 является предпочтительным FGF, способным связывать FGFR2 или FGFR4.
Фактор роста гепатоцитов/рассеивающий фактор (HGF/SF) является морфогенным фактором, который регулирует клеточный рост, клеточное движение и морфогенез путем активации тирозинкиназного сигнального каскада после связывания протоонкогенного c-Met-рецептора.
Для FGF10 и HGF можно использовать одинаковые концентрации. Предпочтительные концентрации для FGF10 составляют 20, 50, 100, 500 нг/мл, не превышая 500 нг/мл. Предпочтительные концентрации для HGF составляют 1, 10, 20, 50 нг/мл, не превышая 50 нг/мл. В ходе культивирования стволовых клеток указанную комбинацию митогенных ростовых факторов (а именно, EGF, FGF10 и HGF) предпочтительно добавлять к культуральной среде по необходимости, например, ежедневно или через день. Предпочтительно добавление через день. Культуральную среду можно обновлять при необходимости, например, ежедневно или через день, хотя предпочтительно обновлять среду на каждый четвертый день.
В некоторых вариантах осуществления изобретения в среде EM1 дополнительно присутствует ингибитор TGF-β, такой как A83-01. Это особенно полезно при культивировании клеток или органоидов человека. В некоторых вариантах осуществления изобретения A83-01 присутствует в концентрации 400-600 нМ, например, 450-550 нМ, 470-530 нМ или приблизительно 500 нМ. В вариантах осуществления изобретения, в которых в среде EM1 присутствует ингибитор TGF-β, предпочтительно, чтобы отсутствовал ингибитор Notch.
Предпочтительная среда для размножения по изобретению
Предпочтительная культуральная среда и способы ее использования описаны в разделе "Примеры".
Например клеточная культуральная среда может содержать или состоять из базальной среды, к которой добавлены: EGF и R-спондин 1, дополненные FGF10, HGF и никотинамидом; например, EGF (50 нг/мл) и R-спондин 1 (1 мкг/мл), дополненные FGF10 (100 нг/мл), HGF (25-50 нг/мл) и никотинамидом (1-10 мМ). Авторами изобретения было найдено, что данная среда предпочтительна для продолжительного размножения клеток. Поэтому, эта клеточная культуральная среда предпочтительна для использования в качестве ЕМ2 по изобретению. Базальную среду предпочтительно дополнять B27, N2 и 200 нг/мл N-ацетилцистеина. В некоторых вариантах осуществления изобретения базальной средой является Advanced-DMEM/F12. Однако можно использовать любую другую подходящую базальную среду.
Другая предпочтительная клеточная культуральная среда и способ ее использования описаны в примерах, и она содержит или состоит из Advanced-DMEM/F12, предпочтительно дополненной B27, N2, 200 нг/мл N-ацетилцистеина, 50 нг/мл EGF, 1 мкг/мл R-спондина 1, 10 нМ гастрином, 100 нг/мл FGF10, 10 мМ никотинамидом и 50 нг/мл HGF, и 50% Wnt-кондиционной средой, а также предпочтительно 10-100 нг/мл Noggin. Wnt-кондиционная среда содержит Advanced-DMEM, P/S, B27, N2, а также FCS. Клетки 293T, трансфицированные Wnt3A-экспрессионной плазмидой, продуцируют Wnt. Полную среду собирают через несколько дней (то есть, с секретированным Wnt) и используют в качестве источника Wnt.
Изобретение, таким образом, относится к клеточной культуральной среде, содержащей или состоящей из базальной среды для клеток животных или человека, к которой добавлены:
Фактор роста эпидермиса, FGF, способный связывать FGFR2 или FGFR4, предпочтительно FGF10, и HGF, в качестве митогенных ростовых факторов,
гастрин, никотинамид, B27, N2 и N-ацетилцистеин и предпочтительно
ингибитор ВМР, более предпочтительно Noggin, и
агонист Wnt, более предпочтительно R-спондин 1 и/или Wnt-3a.
Следовательно, изобретение охватывает первую предпочтительную культуральную среду, содержащую или состоящую из базальной среды для клеток животных или человека, к которой добавлены:
Фактор роста эпидермиса, FGF10, и HGF, в качестве митогенных ростовых факторов,
гастрин, никотинамид, B27, N2 и N-ацетилцистеин,
ингибитор ВМР, более предпочтительно Noggin, и
агонист Wnt, более предпочтительно R-спондин 1 и/или Wnt-3a.
Эту среду можно использовать в качестве клеточной культуральной среды ЕМ1 по изобретению для стимуляции первичного размножения клеток.
В некоторых вариантах осуществления изобретения среда, используемая в качестве клеточной культуральной среды ЕМ1, содержит все компоненты культуральной среды ЕМ2 по изобретению и дополнительно содержит Wnt-3a и Noggin.
В вариантах осуществления изобретения в которых базальная среда дополнена N-ацетилцистеином, B27 и N2, в культуральную среду предпочтительно добавлять следующие компоненты: EGF, R-спондин 1, гастрин, FGF10, никотинамид и HGF, а также Wnt-кондиционную среду. Предпочтительно дополнять базальную среду N-ацетилцистеином, EGF, R-спондином 1, гастрином, FGF10, никотинамидом и HGF, а также Wnt-кондиционной средой в соответствии с количествами, описанными в настоящем документе выше.
Например, в некоторых вариантах осуществления изобретения базальная среда может быть дополнена от 150 нг/мл до 250 нг/мл N-ацетилцистеина; предпочтительно, чтобы базальная среда была дополнена примерно или точно 200 нг/мл N-ацетилцистеина. Например, в некоторых вариантах осуществления изобретения базальная среда может быть дополнена EGF в концентрации от 40 нг/мл до 60 нг/мл; предпочтительно, чтобы базальная среда была дополнена примерно или точно 50 нг/мл EGF. Например, в некоторых вариантах осуществления изобретения базальная среда может быть дополнена от 0,5 мкг/мл до 1,5 мкг/мл R-спондина 1; предпочтительно, чтобы базальная среда была дополнена примерно или точно 1 мкг/мл R-спондина 1. Например, в некоторых вариантах осуществления изобретения базальная среда может быть дополнена от 5 нМ до 15 нМ Гастрином; предпочтительно, чтобы базальная среда была дополнена примерно или точно 10 нМ Гастрином. Например, в некоторых вариантах осуществления изобретения базальная среда может быть дополнена 25-200 нг/мл FGF10, например, от 70 нг/мл до 130 нг/мл FGF10; предпочтительно, чтобы базальная среда была дополнена примерно или точно 100 нг/мл FGF10. Например, в некоторых вариантах осуществления изобретения базальная среда может быть дополнена от 5 мМ до 15 мМ никотинамидом; предпочтительно, чтобы базальная среда была дополнена примерно или точно 10 мМ никотинамидом. Например, в некоторых вариантах осуществления изобретения базальная среда может быть дополнена от 25 нг/мл до 100 нг/мл HGF, например, от 35 нг/мл до 65 нг/мл HGF; предпочтительно, чтобы базальная среда была дополнена примерно или точно 50 нг/мл HGF. Например, в некоторых вариантах осуществления изобретения базальная среда может быть дополнена от 35% до 65% Wnt-кондиционной среды; предпочтительно, чтобы базальная среда была дополнена примерно или точно 50% Wnt-кондиционной среды.
В некоторых вариантах осуществления изобретения любой или оба из гастрина и N2 не присутствуют в клеточной культуральной среде.
Предпочтительно, чтобы базальной средой была Advanced-DMEM/F12.
Данную первую культуральную среду (например, ЕМ1, ЕМ2 или обе ЕМ1 и ЕМ2) предпочтительно использовать в течение первых двух недель способа культивирования по изобретению. Однако ее можно использовать в течение более короткого времени, например, в течение 1, 2, 3, 5, 7 или 10 дней, или более длительного времени, например, в течение 3, 4, 5, 10, 20 или большего количества недель, 5 месяцев или большего времени, например, 6, 7, 8, 9, 10, 11, 12 месяцев или большего времени.
Дифференцирующая среда (DM):
В другом аспекте изобретение относится к второй клеточной культуральной среде, которая содержит или состоит из базальной среды для клеток животных или человека, к которой добавлены: EGF, ингибитор TGF-β и ингибитор Notch. Авторами изобретения было найдено, что эта среда полезна для дифференцирующихся клеток. Среда, используемая для дифференцировки клеток, в настоящем документе может именоваться как DM.
Предпочтительно, чтобы вторая клеточная культуральная среда также содержала FGF и/или HGF. В одном варианте осуществления изобретения вторая клеточная культуральная среда содержит или состоит из базальной среды для клеток животных или человека, к которой добавлены:
Фактор роста эпидермиса, FGF 10 и HGF в качестве митогенных ростовых факторов;
ингибитор Notch; и
ингибитор TGF-β.
В одном варианте осуществления изобретения ингибитором TGF-β является A83-01, и/или ингибитором Notch является DAPT. В другом варианте осуществления изобретения клеточная культуральная среда DM дополнительно содержит дексаметазон.
Предпочтительная вторая клеточная культуральная среда и способ ее использования описаны в примерах, и она содержит или состоит из базальной среды, к которой добавлены: 50 нг/мл EGF, 100 нг/мл FGF10, 50 нМ A8301 и 10 мкМ DAPT. В качестве базальной среды можно использовать Advanced-DMEM/F12, а также любую другую подходящую базальную среду.
В некоторых вариантах осуществления изобретения вторая клеточная культуральная среда не содержит R-спондина или Wnt. В некоторых вариантах осуществления изобретения вторая клеточная культуральная среда не содержит агониста Wnt. В некоторых вариантах осуществления изобретения вторая клеточная культуральная среда не содержит никотинамида. В некоторых вариантах осуществления изобретения вторая клеточная культуральная среда не содержит ингибитора BMP.
Авторы изобретения открыли, что R-спондин 1 и никотинамид, оба ингибируют экспрессию маркера зрелых гепатоцитов, CYP3A11, и при этом усиливают экспрессию маркера гепатобластов, альбумина. Поэтому, для усиления дифференцировки клеток до более зрелых клеток печени, авторы изобретения удалили R-спондин и никотинамид из клеточной культуры. Авторы изобретения также открыли, что экспрессия определенных билиарных транскрипционных факторов сильно повышается в размножающихся культурах, содержащих R-спондин 1, что указывает на то, что баланс экспрессии генов в культуре смещен в сторону дифференцировки в билиарные клетки. Сигнальные пути от Notch и TGF-β вовлечены в дифференцировку в билиарные клетки in vivo. В действительности, удаление Rbpj (необходим для достижения действующей передачи сигнала от Notch) приводит к аномальному тубулогенезу (Zong Y. Development 2009), а добавление TGF-β к эксплантатам печени способствует дифференцировке в билиарные клетки in vitro (Clotman F. Genes and Development 2005). Поскольку оба, Notch- и TGF-β-сигнальные пути, были сильно активированы в культурах клеток печени (фиг.9), то авторы изобретения предположили, что ингибирование дифференцировки в билиарные клетки может запустить дифференцировку клеток в клетки с более гепатоцитарным фенотипом. Было найдено, что добавление ингибитора TGF-β (такого как A8301) и ингибитора Notch (такого как DAPT) к дифференцирующей среде, которая предпочтительно не содержит R-спондина или Wnt, усиливает экспрессию маркеров зрелых гепатоцитов и увеличивает число гепатоцит-подобных клеток (например, см. пример 2).
Notch является трансмембранным поверхностным рецептором, который может активироваться в результате множества вариантов протеолитического расщепления, одним из которых является расщепление комплексом белков с протеазной активностью, называемых гамма-секретазой. Гамма-секретаза представляет собой протеазу, которая осуществляет свою протеолитическую активность внутри мембраны. Гамма-секретаза является многокомпонентным ферментом и состоит по меньшей мере из четырех различных белков, а именно, презенелинов (презенилина 1 или 2), никастрина, PEN-2 и APH-I. Презенилин является каталитическим центром гамма-секретазы. При связывании с лигандом, Notch-рецептор претерпевает конформационное изменение, которое позволяет "осыпание" эктодомена в результате действия ADAM-протеазы, которая является металлопротеазой. За этим немедленно следует действие гамма-секретазного комплекса, которое приводит к высвобождению внутриклеточного домена Notch (NICD). NICD перемещается в ядро, где взаимодействует с CSL (C-промотор-связывающим фактором/белком, связывающим последовательность сигнала рекомбинации, Jκ/подавителем гена отсутствия волосяного покрова/Lag1). Связывание NICD превращает CSL из транскрипционного репрессора в активатор, который в результате обеспечивает экспрессию генов-мишеней Notch. Примерами предпочтительных ингибиторов Notch, которые можно использовать в контексте настоящего изобретения, являются: ингибиторы гамма-секретазы, такие как DAPT или дибензазепин (DBZ) или бензодиазепин (BZ), или LY-41 1575, ингибитор, способный снижать лиганд-опосредованную активацию Notch (например, посредством доминантно-негативного лиганда Notch, или посредством доминантно-негативного Notch, или посредством антитела, способного по меньшей мере частично блокировать взаимодействие между Notch-лигандом и Notch), или ингибитор ADAM-протеаз.
TGF-β-сигнальный путь вовлечен во многие клеточные функции, включая клеточный рост, клеточную дифференцировку и апоптоз. Сигнальный путь, как правило, начинается со связывания лиганда из суперсемейства TGF-β с рецептором II типа, который рекрутирует и фосфорилирует рецептор I типа. Рецептор I типа затем фосфорилирует белки SMAD, которые действуют как транскрипционные факторы в ядре и регулируют экспрессию генов-мишеней. В альтернативном варианте, TGF-β-сигнальный путь может активировать МАР-киназные сигнальные пути, например, через МАР-киназу, р38. Лиганды суперсемейства TGF-β включают в себя костные морфогенные белки (BMP), факторы роста и дифференцировки (GDF), антимюллеров гормон (AMH), активин, нодал и белки TGF-β.
Ингибитором TGF-β является агент, который снижает активность TGF-β-сигнального пути. Существует множество возможностей нарушения TGF-β-сигнального пути, которые известны в данной области, и которые можно использовать вместе с изобретением. Например, TGF-β-сигнальный путь можно нарушить: ингибируя экспрессию TGF-β с использованием стратегии малых интерферирующих РНК; ингибируя фурин (протеазу, активирующую TGF-β); ингибируя путь с помощью физиологических ингибиторов, например, ингибируя ВМР с помощью Noggin, DAN или DAN-подобных белков; нейтрализуя TGF-β моноклональным антителом; ингибируя низкомолекулярными соединениями TGF-β-рецепторную киназу (также известную как активинрецептор-подобная киназа, ALK5), ALK4, ALK6, ALK7 или другие родственные TGF-β-рецепторным киназам; ингибируя Smad2- и Smad3-сигнальный путь посредством сверхэкспрессии их физиологического ингибитора, Smad7, или используя тиоредоксин, в качестве "якоря" для Smad, блокирующего активацию Smad (Fuchs, O. Inhibition of TGF- Signaling for the Treatment of Tumor Metastasis and Fibrotic Diseases. Current Signal Transduction Therapy, Volume 6, Number 1, January 2011, pp. 29-43(15)).
Различные способы определения того, является ли вещество ингибитором TGF-β, известны и могли бы использоваться вместе с изобретением. Например, можно использовать клеточный метод анализа, в котором клетки стабильно трансфицированы репортерной конструкцией, содержащей промотор PAI-1 человека или сайты связывания Smad, управляющие люциферазным репортерным геном. Ингибирование люциферазной активности относительно контрольных групп можно использовать для измерения активности соединения (De Gouville et al., Br J Pharmacol. 2005 May; 145(2): 166-177).
Ингибитором TGF-β по настоящему изобретению может быть белок, пептид, низкомолекулярные соединения, малые интерферирующие РНК, антисмысловой олигонуклеотид, аптамер или антитело. Ингибитор может быть природным или синтетическим. Примеры предпочтительных низкомолекулярных ингибиторов TGF-β, которые можно использовать в контексте этого изобретения, включают низкомолекулярные ингибиторы, перечисленные в таблице 1:
Низкомолекулярные ингибиторы TGF-β, мишенью которых являются рецепторные киназы
(нМ)
Способ по изобретению может включать в себя использование одного или нескольких ингибиторов, перечисленных в таблице 1. Способ по изобретению может включать в себя использование любой комбинации одного ингибитора с другим перечисленным ингибитором. Например, способ по изобретению может включать в себя использование SB-525334 или SD-208, или A83-01; либо способ по изобретению может включать в себя использование SD-208 и A83-01; либо способ по изобретению может включать в себя использование SD-208 и A83-01. Опытному специалисту в данной области будет очевидно, что существует ряд других низкомолекулярных ингибиторов, которые исходно были разработаны для других киназ, но при высоких концентрациях могут также ингибировать TGF-β-рецепторные киназы. Например, SB-203580 является ингибитором МАР-киназы, р38, который, как полагают, при высоких концентрациях (например, приблизительно 10 мкМ или выше) ингибирует ALK5. Любой такой ингибитор, который ингибирует TGF-β-сигнальный путь, также можно использовать в контексте настоящего изобретения.
A83-01 можно добавлять в культуральную среду в концентрации от 10 нМ до 10 мкМ, или от 20 нМ до 5 мкМ, или от 50 нМ до 1 мкМ. Например, A83-01 можно добавлять в культуральную среду в концентрации приблизительно 500 нМ. При использовании в ЕМ, A83-01 можно добавлять в культуральную среду в концентрации 350-650 нМ, например, 450-550 нМ, более предпочтительно приблизительно 500 нМ. При использовании в DM, A83-01 можно добавлять в культуральную среду в концентрации 25-75 нМ, например, 40-60 нМ или приблизительно 50 нМ.
SB-431542 можно добавлять в культуральную среду в концентрации от 80 нМ до 80 мкМ, или от 100 нМ до 40 мкМ, или от 500 нМ до 10 мкМ. Например, SB-431542 можно добавлять в культуральную среду в концентрации, приблизительно составляющей 1 мкМ.
SB-505124 можно добавлять в культуральную среду в концентрации от 40 нМ до 40 мкМ, или от 80 нМ до 20 мкМ, или от 200 нМ до 1 мкМ. Например, SB-505124 можно добавлять в культуральную среду в концентрации, составляющей приблизительно 500 нМ.
SB-525334 можно добавлять в культуральную среду в концентрации от 10 нМ до 10 мкМ, или от 20 нМ до 5 мкМ, или от 50 нМ до 1 мкМ. Например, SB-525334 можно добавлять в культуральную среду в концентрации, составляющей приблизительно 100 нМ.
LY 364947 можно добавлять в культуральную среду в концентрации от 40 нМ до 40 мкМ, или от 80 нМ до 20 мкМ, или от 200 нМ до 1 мкМ. Например, LY 364947 можно добавлять в культуральную среду в концентрации, составляющей приблизительно 500 нМ.
SD-208 можно добавлять в культуральную среду в концентрации от 40 нМ до 40 мкМ, или от 80 нМ до 20 мкМ, или от 200 нМ до 1 мкМ. Например, SD-208 можно добавлять в культуральную среду в концентрации, составляющей приблизительно 500 нМ.
SJN 2511 можно добавлять в культуральную среду в концентрации от 20 нМ до 20 мкМ, или от 40 нМ до 10 мкМ, или от 100 нМ до 1 мкМ. Например, A83-01 можно добавлять в культуральную среду в концентрации, составляющей приблизительно 200 нМ.
В дополнительном аспекте изобретение относится к альтернативной второй культуральной среде, содержащей или состоящей из базальной среды для клеток животных или человека, к которой добавлены:
Фактор роста эпидермиса, FGF10 и HGF в качестве митогенных ростовых факторов;
Гастрин, никотинамид, B27, N2 и N-ацетилцистеин.
Эта альтернативная вторая культуральная среда полезна в качестве дифференцирующей среды (DM). Предпочтительно использовать эту альтернативную вторую культуральную среду через две недели культивирования. На этой стадии, по-видимому, больше нет необходимости в ингибиторе ВМР и лиганде Wnt. Следовательно, в некоторых вариантах осуществления изобретения альтернативная вторая культуральная среда не содержит ингибитора ВМР или лиганда Wnt. В некоторых вариантах осуществления изобретения альтернативная вторая культуральная среда не содержит ингибитора ВМР или агониста Wnt.
Как и в случае первой культуральной средs (ЕМ, а именно, ЕМ1 и ЕМ2, либо ЕМ1 или ЕМ2) базальная среда в некоторых вариантах осуществления изобретения может быть дополнена N-ацетилцистеином, EGF, гастрином, FGF10, никотинамидом и HGF согласно количествам, описанным в настоящем документе выше. Например, предпочтительно, чтобы базальная среда была дополнена примерно или точно 200 нг/мл N-ацетилцистеина. Предпочтительно, чтобы базальная среда была дополнена примерно или точно 50 нг/мл EGF. Предпочтительно, чтобы базальная среда была дополнена примерно или точно 10 нМ гастрином. Предпочтительно, чтобы базальная среда была дополнена примерно или точно 100 нг/мл FGF10. Предпочтительно, чтобы базальная среда была дополнена примерно или точно 10 мМ никотинамидом. Предпочтительно, чтобы базальная среда была дополнена примерно или точно 50 нг/мл HGF.
Первая и вторая клеточная культуральная среда, используемая по изобретению, позволяет выживание и/или пролиферацию, и/или дифференцировку эпителиальных стволовых клеток или эпителиальных стволовых клеток печени, или выделенного желчного протока или выделенных фрагментов печени на внеклеточном матриксе.
Средой, позволяющей выживание и/или пролиферацию, является среда, которая предпочтительно индуцирует или способствует выживанию и/или пролиферации клеток в течение по меньшей мере 5, 10, 15, 20, 30, 40, 50, 60, 70, 80, 90, 100, 120, 140, 160, 180, 200 дней культивирования. Пролиферацию можно оценить, используя методики, известные в данной области, такие как окрашивание BrdU, окрашивание Edu, окрашивание Ki67, и можно использовать методы анализа кривых роста. Например, авторами изобретения было показано, что 10 желчных протоков через 6 дней можно разделить на 6-лунок с 10 органоидами на лунку (60 новых органоидов). На каждом 6-дневном пассаже авторы могли получить около 360 органоидов. Путем проведения 1 пассажа в неделю в течение 32 недель авторы были способны генерировать больше 11000 (11520) новых органоидных структур в течение всего 7 месяцев. Это является важным для промышленности, поскольку доступность клеток и органоидов для трансплантации представляет значительную проблему. Например, для мышиного трансплантата как минимум требуется 105 клеток. Вероятно, что для успешной пересадки человеческого трансплантата может быть необходимо 106 или 10×106 клеток.
Иными словами, среда, используемая по изобретению, обеспечивает возможность размножения популяции стволовых клеток с образованием органоидов печени в течение по меньшей мере 3, по меньшей мере 4, по меньшей мере 5, по меньшей мере 6, по меньшей мере 7, по меньшей мере 8, по меньшей мере 9, по меньшей мере 10, по меньшей мере 20, по меньшей мере 30, по меньшей мере 40, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90 или по меньшей мере 100 пассажей в соответствующих условиях.
Вторая среда, как было показано выше, предпочтительно индуцирует или способствует специфичной дифференцировке клеток в течение по меньшей мере пяти дней культивирования. Дифференцировку можно измерить, детектируя присутствие определенного маркера, ассоциированного с клетками печени, указанного в настоящем документе. Дифференцировку можно измерить, детектируя присутствие определенного маркера, ассоциированного с клетками печени, как указано в настоящем документе. В зависимости от маркера его экспрессию можно оценить с помощью ОТ-ПЦР или иммуногистохимии по меньшей мере через 5, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16 дней культивирования в первой среде или после культивирования в первой и впоследствии во второй среде, как указано в настоящем документе.
Дифференцирующей средой является среда, которая предпочтительно вызывает или способствует специфичной дифференцировке клеток в течение по меньшей мере пяти дней культивирования. Дифференцировку можно измерить, детектируя присутствие определенного маркера, ассоциированного с клетками печени, как указано в настоящем документе.
В контексте изобретения термин клеточная культуральная среда является синонимом среды, культуральной среды или клеточной среды, или базальной среды, или базальной клеточной среды, или базальной клеточной культуральной среды, или среды для размножения, или дифференцирующей среды.
Соответственно, в еще одном дополнительном аспекте изобретение относится к герметично закрытому сосуду, содержащему культуральную среду по изобретению. В некоторых вариантах осуществления изобретения культуральной средой является среда для размножения. В некоторых вариантах осуществления изобретения культуральной средой является дифференцирующая среда. Герметично закрытые сосуды могут быть предпочтительны для транспортировки или хранения культуральной среды с целью предупреждения загрязнения. Сосудом может быть любой подходящий сосуд, такой как колба, планшет, флакон, банка, пузырек или пакет.
Способы получения и/или культивирования стволовых клеток
Способ получения и/или культивирования фрагмента печени или органоида печени включает в себя культивирование эпителиальных стволовых клеток и/или выделенных фрагментов ткани, содержащих указанные эпителиальные стволовые клетки, в контакте с внеклеточным матриксом в присутствии одной или нескольких клеточных культуральных сред по изобретению.
В некоторых вариантах осуществления изобретения способ получения и/или культивирования фрагмента печени или органоида печени включает в себя культивирование эпителиальных стволовых клеток и/или выделенных фрагментов ткани, содержащих указанные эпителиальные стволовые клетки, в контакте с внеклеточным матриксом в присутствии среды ЕМ1 и затем среды ЕМ2, а затем среды DM.
В некоторых вариантах осуществления изобретения способ получения и/или культивирования фрагмента печени или органоида печени включает в себя культивирование эпителиальных стволовых клеток и/или выделенных фрагментов ткани, содержащих указанные эпителиальные стволовые клетки, в контакте с внеклеточным матриксом в присутствии среды ЕМ2 и затем среды DM, без использования среды ЕМ1.
В некоторых вариантах осуществления изобретения способ получения и/или культивирования фрагмента печени или органоида печени включает в себя культивирование эпителиальных стволовых клеток и/или выделенных фрагментов ткани, содержащих указанные эпителиальные стволовые клетки, в контакте с внеклеточным матриксом в присутствии среды ЕМ1 и затем среды DM, без использования среды ЕМ2.
Способ получения и/или культивирования фрагмента печени или органоида печени может включать в себя:
i) культивирование эпителиальных стволовых клеток и/или выделенных фрагментов ткани, содержащих указанные эпителиальные стволовые клетки, в контакте с внеклеточным матриксом в присутствии среды, причем среда содержит базальную среду для клеток животных или человека, к которой добавлены: фактор роста эпидермиса, FGF, способный связывать FGFR2 или FGFR4, предпочтительно FGF10, и HGF в качестве митогенных ростовых факторов, никотинамид и предпочтительно агонист Wnt, предпочтительно любой один из R-спондина 1-4; и впоследствии
ii) культивирование стволовых клеток и/или выделенных фрагментов ткани, содержащих указанные эпителиальные стволовые клетки, в контакте с внеклеточным матриксом в присутствии второй клеточной культуральной среды, указанной в настоящем документе (среды DM).
В некоторых вариантах осуществления изобретения перед стадией (i) способ включает в себя культивирование эпителиальных стволовых клеток и/или выделенных фрагментов ткани, содержащих указанные эпителиальные стволовые клетки, в контакте с внеклеточным матриксом в присутствии среды, содержащей или состоящей из базальной среды для клеток животных или человека, к которой добавлены: EGF, ингибитор BMP, R-спондин и Wnt. Предпочтительно, чтобы ингибитором ВМР являлся Noggin, и среда ЕМ1 называлась "ENRW" (EGF, Noggin, R-спондин и Wnt).
В одном варианте осуществления изобретение относится к способу получения и/или культивирования фрагмента печени или органоида печени, причем указанный способ включает в себя:
культивирование эпителиальных стволовых клеток и/или выделенных фрагментов ткани, содержащих указанные эпителиальные стволовые клетки, в контакте с внеклеточным матриксом в культуральной среде ЕМ2, описанной в настоящем документе, например, среде, содержащей агонист Wnt, такой как R-спондин, фактор роста эпидермиса, FGF, способный связывать FGFR2 или FGFR4, предпочтительно FGF 10, и никотинамид. Предпочтительно, чтобы среда также содержала HGF.
В дополнительном варианте осуществления изобретение относится к способу получения и/или культивирования фрагмента печени или органоида печени, причем указанный способ включает в себя:
культивирование эпителиальных стволовых клеток и/или выделенных фрагментов ткани, содержащих указанные эпителиальные стволовые клетки, в контакте с внеклеточным матриксом в среде, содержащей ингибитор ВМР, агонист Wnt, фактор роста эпидермиса, FGF, способный связывать FGFR2 или FGFR4, предпочтительно FGF10, и HGF в качестве митогенных ростовых факторов, гастрин, никотинамид, B27, N2 и N-ацетилцистеин.
В другом предпочтительном способе после по меньшей мере двух недель культивирования в среде ЕМ1, указанной в настоящем документе, культивирование продолжают в среде, которая не содержит ингибитора ВМР и агониста Wnt. В некоторых вариантах осуществления изобретения культивирование продолжают в среде ЕМ2 по изобретению. В альтернативных вариантах осуществления изобретения культивирование продолжают во второй среде по изобретению (DM), которая дополнительно содержит ингибитор TGF-β, такой как A83-01, и ингибитор Notch, такой как DAPT. В некоторых вариантах осуществления изобретения это культивирование продолжают в ЕМ2, а затем в среде DM по изобретению.
Поэтому, в предпочтительном способе получения и/или культивирования органоида печени, эпителиальные стволовые клетки, и/или выделенные фрагменты ткани печени, содержащие указанные эпителиальные стволовые клетки, культивируют на первой стадии в первой среде (ЕМ), затем последовательно на отдельной второй стадии - во второй среде (DM). Длительность первой стадии культивирования может составлять по меньшей мере две недели и может быть длиннее, например, может составлять 3 недели или больше, 4 недели или больше, или может составлять 1, 2, 3, 4, 5, 6, 7, 8 месяцев или больше. Длительность второй стадии предпочтительно составляет 6 недель или меньше, более предпочтительно 1 месяц или меньше, например, 3 недели или меньше, 2 недели или меньше, 1 неделю или меньше. Было найдено, что многие клетки начинают умирать после двух недель в DM, и, таким образом, предпочтительно, чтобы вторая стадия составляла 2 недели или меньше, или самое большее 1 месяц. Поэтому, только в менее предпочтительных вариантах осуществления изобретения продолжительность второй стадии может составлять 8, 9, 10, 11, 12, 13, 14, 15, 16 дней или 3 недели или 1, 2, 3, 4, 5, 6 месяцев или больше. Каждую стадию предпочтительно проводить, используя внеклеточный матрикс, указанный в настоящем документе, и используя культуральную среду, подробно описанную в настоящем документе.
В качестве альтернативы, в другом варианте осуществления изобретения указанный способ проводят в первой среде, не переключаясь на вторую среду. Продолжительность этого способа может составлять 8, 9, 10, 11, 12, 13, 14, 15, 16 дней или 3 недели, либо 1, 2, 3, 4, 5, 6, 7, 8 месяцев или больше.
В некоторых вариантах осуществления изобретения стадия культивирования в первой среде по изобретению включает в себя как культивирование в среде ЕМ1, так и затем культивирование в среде ЕМ2. Следовательно, описанные выше периоды времени могут относиться к общему периоду, за который клетки или фрагменты культивируют в ЕМ1 и впоследствии в среде ЕМ2. В некоторых вариантах осуществления изобретения продолжительность стадии культивирования в среде ЕМ1 может составлять меньше 10 дней, например, меньше 9, меньше 8, меньше 7, меньше 6, меньше 5, меньше 4, меньше 3, меньше 2 дней или меньше 1 дня. В некоторых вариантах осуществления изобретения стадия культивирования в среде ЕМ1 может иметь продолжительность от 1 до 10 дней, например, 1-7 дней, 2-6 дней, 3-5 дней, например, может составлять 4 или 5 дней. В некоторых вариантах осуществления изобретения продолжительность стадии культивирования в среде ЕМ2 может составлять 10 дней или больше, например, 11, 12, 13, 14, 15, 16 дней или больше, либо 1-3 недели или 2-4 недели, или 3-6 недель, или 1, 2, 3, 4, 5, 6 месяцев или больше.
Предпочтительные концентрации каждого соединения, присутствующего в каждой среде, уже были указаны в настоящем документе в описании или в примерах.
Описанные в настоящем документе способы получения и/или культивирования фрагмента печени или органоида печени также можно использовать для получения и/или культивирования популяции клеток, экспрессирующих Lgr5, и такие способы входят в объем изобретения. Следовательно, изобретение также относится к популяции клеток, экспрессирующих Lgr5, полученных способом по изобретению.
Опытному специалисту по прочтении изобретения будет очевидно, что предпочтительные способы культивирования по изобретению имеют преимущество, благодаря отсутствию необходимости в "питающих" клетках. Слои питающих клеток часто используют для поддержания культуры стволовых клеток и для ингибирования их дифференцировки. Слой питающих клеток, как правило, представляет собой монослой клеток, которые совместно культивируют с представляющими интерес клетками и которые обеспечивают поверхность, подходящую для их роста. Слой питающих клеток обеспечивает окружение, в котором могут расти клетки, представляющие интерес. Питающие клетки часто митотически инактивированы (например, облучением или обработкой митомицином С) для предупреждения их пролиферации. Использование питающих клеток является нежелательным, поскольку это осложняет пассирование клеток (клетки должны быть отделены от питающих клеток при каждом пассаже, и для каждого пассажа требуются новые питающие клетки). Использование питающих клеток также может привести к загрязнению целевых клеток питающими клетками. Очевидно, что это является проблемой для любых медицинских приложений, и даже в контексте исследований осложняет анализ результатов любых экспериментов, проведенных на клетках. Как отмечено в настоящем документе, культуральные среды по изобретению особенно предпочтительны, поскольку их можно использовать для культивирования клеток без контакта с питающими клетками, то есть способы по изобретению не требуют слоя питающих клеток для поддержки клеток, чей рост они обеспечивают.
Соответственно, композиции по изобретению могут быть композициями, свободными от питающих клеток. Композиция обычно считается свободной от питающих клеток, если клетки печени в композиции культивируются по меньшей мере в течение одного пассажа в отсутствие слоя питающих клеток. Свободная от питающих клеток композиция по изобретению будет обычно содержать меньше примерно 5%, меньше примерно 4%, меньше примерно 3%, меньше примерно 2%, меньше примерно 1% питающих клеток (выраженных в % от общего числа клеток в композиции) или предпочтительно совсем не будет содержать питающих клеток.
В одном предпочтительном варианте осуществления изобретения, например, если органоид печени предназначен для использования в регенеративной медицине, то способ может начинаться с эпителиальных клеток или с выделенного фрагмента печени. Клетки или фрагмент печени могут быть аутологичными или аллогенными. "Аутологичные" клетки является клетками, которые происходят из того же организма, в который они повторно вводятся для клеточной терапии, например, с целью обеспечения возможности регенерации ткани. Аутологичная клетка в принципе не требует проведения процедуры соответствия пациенту с целью преодоления проблем иммунного отторжения, и/или она снижает необходимость иммуносупрессивного вмешательства после трансплантации. "Аллогенные" клетки являются клетками, которые происходят из индивидуума, отличающегося от индивидуума, которому вводят эти клетки для клеточной терапии, например, с целью обеспечения возможности регенерации ткани, хотя и относящегося к тому же биологическому виду. Для предупреждения проблем отторжения еще может потребоваться проведение процедуры соответствия пациенту. Опытным специалистам в данной области будут известны методики минимизации отторжения тканей.
Предпочтительный способ по изобретению охватывает способ культивирования выделенного фрагмента печени, содержащего указанные эпителиальные стволовые клетки.
Другой предпочтительный способ охватывает способ получения органоида печени из эпителиальных стволовых клеток и/или выделенных фрагментов печени, содержащих указанные эпителиальные стволовые клетки.
Другой предпочтительный способ охватывает способ получения и культивирования органоида печени из эпителиальных стволовых клеток и/или выделенных фрагментов печени, содержащих указанные эпителиальные стволовые клетки.
В предпочтительном способе, как было показано ранее, ингибитор ВМР выбран из Noggin, DAN и DAN-подобных белков, включая Cerberus и Gremlin, предпочтительно ингибитор ВМР содержит Noggin или является им; и/или указанный агонист Wnt выбран из одного или нескольких из Wnt, предпочтительно Wnt-3a, R-спондина 1-4, Norrin и ингибитора GSK, более предпочтительно агонист Wnt содержит Wnt-3a и/или R-спондин или является Wnt-3a и/или R-спондином.
В другом предпочтительном способе органоид печени происходит от одной единственной клетки, предпочтительно клетки, экспрессирующей Lgr5, более предпочтительно, чтобы единственная клетка содержала нуклеотидную конструкцию, содержащую представляющую интерес молекулу нуклеиновой кислоты. Клетки также могут экспрессировать печеночные специфичные маркеры, такие как Hnf1α и Hnf4.
Выделение некоторых типов клеток, экспрессирующих Lgr5, было описано ранее (см., например, WO2009/022907 и WO2010/016766). Однако специфичные для печени стволовые клетки, экспрессирующие Lgr5, раскрытого в настоящем документе типа, ранее не были описаны. Соответственно, изобретение относится к популяции стволовых клеток взрослого человека, отличающихся природной экспрессией по меньшей мере Lgr5 и одного из следующих маркеров - Hnf1α, Hnf4a, Sox9, KRT7 и KRT19 - на значительном уровне. Эта клеточная популяция также экспрессирует маркеры, общие для популяций клеток-предшественников тонкого кишечника и желудка, такие как Cd44 и Sox9 (Barker & Huch et al Cell stem cell 2010). Эти маркеры экспрессируются на высоком уровне в стволовых клетках по изобретению, но не экспрессируются в зрелой печени, усиливая способность к самообновлению культур клеток печени, описанных в настоящем документе. В клетках по этому аспекту изобретения также может быть повышена экспрессия генов-мишеней Wnt, включая, например, MMP7, Sp5 и Tnfrs19. Это уверенно свидетельствует в пользу необходимости активного и сильного канонического Wnt-сигнального пути для сохранения способности этих культур к самообновлению.
Под "природной экспрессией" понимают, что с клетками не проводились никакие рекомбинантные манипуляции, то есть в клетках не была искусственно индуцирована или не модулировалась экспрессия этих маркеров путем введения экзогенного генетического материала, например, введением гетерологичных (неприродных) или более сильных промоторов, или других регуляторных последовательностей, функционально соединенных либо с эндогенными генами, либо с экзогенно введенными формами генов. Природная экспрессия осуществляется с геномной ДНК в клетках, включающей интроны между экзонными кодирующими последовательностями, если они присутствуют. Природная экспрессия не осуществляется с кДНК. Природную экспрессию при необходимости можно подтвердить любым одним из различных способов, таких как секвенирование рамки считывания гена для проверки отсутствия чужеродной гетерогенной последовательности. "Зрелый" означает пост-эмбриональный. В отношении стволовых клеток по настоящему изобретению термин "зрелая стволовая клетка" означает, что стволовая клетка выделена из ткани или органа животного на стадии роста после эмбриональной стадии.
Эта популяция стволовых клеток также отличается отсутствием природной экспрессии некоторых маркеров на любом значительном уровне, многие из которых ассоциированы с клеточной дифференцировкой. Более конкретно, клетки выделенной популяции зрелых стволовых клеток не экспрессируют природно один или несколько из Cd11b, CD13, CD14, AFP, Pdx1, любого белка из семейства CYP (например, CYP3A11, CYP11A1) на значительном уровне. Как указано в настоящем документе, полагают, что эти маркеры являются негативными маркерами.
Термин "экспрессированный" используют для описания присутствия маркера в клетке. Для того чтобы считаться экспрессированным, маркер должен присутствовать на детектируемом уровне. Под "детектируемым уровнем" понимается, что маркер можно детектировать с использованием одной из стандартных лабораторных методик, таких как ПЦР, блоттинг или FACS-анализ. Ген считается экспрессированным клеткой популяции по изобретению, если экспрессия обоснованно детектируется после 30 циклов ПЦР, что соответствует уровню экспрессии в клетке по меньшей мере примерно 100 копий на клетку. Термины "экспрессировать" и "экспрессия" имеют аналогичные значения. При уровне экспрессии ниже этого порога, считают, что маркер не экспрессируется. Сравнение между уровнем экспрессии маркера в клетке по изобретению и уровнем экспрессии того же маркера в другой клетке, такой как, например, эмбриональная стволовая клетка, можно предпочтительно проводить путем сравнения двух типов клеток, которые были выделены из одного биологического вида. Предпочтительно, чтобы этот вид относился к млекопитающим, и, более предпочтительно, чтобы этим видом был человек. Такое сравнение можно удобно проводить, используя полимеразную цепную реакцию, совмещенную с обратной транскрипцией (ОТ-ПЦР).
Любой один из ряда физических способов разделения, известных в данной области, можно использовать для отбора клеток по этому аспекту изобретения и отличить их от других типов клеток. Такие физические способы могут включать FACS и различные иммуноаффинные способы, основанные на том, что маркеры специфично экспрессируются клетками по изобретению. Как описано выше, Lgr5, Hnf1α и Hnf4 являются 3 клеточными маркерами, экспрессирующимися на высоком уровне в клетках по изобретению. Поэтому, только с целью иллюстрации, клетки по изобретению можно выделить рядом физических способов разделения, которые основаны на присутствии этих маркеров.
В одном варианте осуществления изобретения клетки по изобретению можно выделить с помощью FACS, используя антитело, например, к одному из этих маркеров. Опытному специалисту в данной области будет очевидно, что этого можно достигнуть посредством флуоресцентно меченного антитела или посредством флуоресцентно меченного вторичного антитела, специфично связывающего первичное антитело. Примеры подходящих флуоресцентных меток включают, но не ограничены этим, FITC, Alexa Fluor® 488, GFP, CFSE, CFDA-SE, DyLight 488, PE, PerCP, PE-Alexa Fluor® 700, PE-Cy5 (TRI-COLOR®), PE-Cy5.5, PI, PE-Alexa Fluor® 750 и PE-Cy7. Этот список приведен только для примера и не подразумевает ограничение объема изобретения.
Опытному специалисту в данной области будет очевидно, что FACS-анализ с использованием анти-Lgr5 антитела будет обеспечивать очищенную популяцию клеток. Однако в некоторых вариантах осуществления изобретения может быть предпочтительно очищать клеточную популяцию дополнительно, проводя дополнительный раунд FACS-анализа с использованием одного или нескольких других идентифицируемых маркеров, предпочтительно, Hnf1α и Hnf4, но можно использовать и другие маркеры.
В другом варианте осуществления изобретения клетки по изобретению можно выделить иммуноаффинной очисткой, которая представляет собой способ разделения, хорошо известный в данной области. Только с целью иллюстрации клетки по изобретению можно выделить иммуноаффинной очисткой по белку c-kit. Опытному специалисту в данной области будет очевидно, что этот способ основан на иммобилизации антител на колонке для очистки. Образец клеток затем наносят на колонку, позволяя соответствующим клеткам связаться с антителами, и, таким образом, связаться с колонкой. После стадии промывки клетки элюируют с колонки, используя конкурент, который предпочтительно связывается с иммобилизованным анти-c-kit антителом и позволяет клеткам высвободиться с колонки.
Опытному специалисту в данной области будет очевидно, что иммуноаффинная очистка с использованием иммобилизованного антитела будет обеспечивать очищенную популяцию клеток. Однако в некоторых вариантах осуществления изобретения может быть предпочтительно дополнительно очистить популяцию клеток, проводя дополнительный раунд иммуноаффинной очистки с использованием одного или нескольких других идентифицируемых маркеров, например, Hnf4, и использовать аликвоту выделенных клонов для выявления экспрессии других значимых внутриклеточных маркеров.
Опытному специалисту в данной области будет очевидно, что последовательные стадии очистки не требуют использования одного физического способа разделения. Поэтому, будет ясно, что, например, клетки можно очистить посредством FACS с использованием анти-Lgr5 антитела с последующей иммуноаффинной очисткой с использованием SSEA-1-аффинной колонки. В некоторых вариантах осуществления изобретения клетки можно культивировать после выделения в течение по меньшей мере примерно 15, по меньшей мере примерно 20 дней, по меньшей мере примерно 25 дней, или по меньшей мере примерно 30 дней. В некоторых аспектах клетки размножают в культуре в течение большего времени для улучшения гомогенности клеточного фенотипа в популяции клеток.
Другие признаки этого способа определены в части описания, посвященной определениям. Как известно в данной области, обычно высевают одноклеточные суспензии или маленькие кластеры клеток (2-50 клеток/кластер), а не большие кластеры клеток. После их деления такие клетки будут высевать на подложку с плотностью, которая способствует пролиферации клеток. Как правило, при выделении одиночных клеток используют плотность посева по меньшей мере 1-500 клеток/лунку при площади поверхности лунки, составляющей 0,32 см2. При высевании кластеров плотность посева предпочтительно составляет 250-2500 клеток/см2. При пересеве можно использовать плотность посева клеток, составляющую от примерно 2500 клеток/см2 до примерно 5000 клеток/см2. Как известно в данной области, при пересеве обычно высевают одноклеточные суспензии или маленькие кластеры клеток, а не крупные кластеры клеток.
Органоиды по изобретению
Описанные выше клетки вырастают в тельца, которые в настоящем документе называют "органоидами". Соответственно, органоид печени, который можно получить способом по изобретению, является дополнительным аспектом изобретения. Насколько известно авторам изобретения, это является первым случаем получения органоида печени, который является функциональным и живым после длительного периода времени (а именно, по меньшей мере 7 месяцев культивирования; см. примеры, включенные в настоящий документ). Функциональность предпочтительно отличается присутствием печеночного маркера, определенного в настоящем документе, и/или структурой указанного органоида, определенной в настоящем документе. Поскольку конечное количество полученных органоидов печени коррелирует с длительность культивирования, то опытному специалисту будете понятно, что изобретение является передовым изобретением и потенциально открывает новые возможности, например, в области регенеративной медицины.
Например, органоид по настоящему изобретению может содержать популяцию клеток, состоящую по меньшей мере из 1×103 клеток, по меньшей мере из 1×104 клеток, по меньшей мере из 1×105 клеток, по меньшей мере из 1×106 клеток, по меньшей мере из 1×107 клеток или большего количества клеток. В некоторых вариантах осуществления изобретения каждый органоид содержит приблизительно от 1×103 до 5×103 клеток; в общем, 10-20 органоидов могут расти совместно в одной лунке 24-луночного планшета.
Клетки и органоиды по настоящему изобретению могут принадлежать животному или человеку. Авторы изобретения впервые показали возможность выращивания и сохранения органоидов печени как животных, так и человека, in vitro, с использованием культуральной среды и способов по изобретению.
Иллюстративные примеры органоидов, полученных по изобретению, приведены в сопровождающих фигурах. Можно увидеть, что органоиды по изобретению могут иметь кистозную структуру с внешней стороны со слоем клеток и по меньшей мере с одним зачатком и центральным просветом. Органоиды по краям матригеля обычно крупнее органоидов в центре матригеля, возможно, вследствие лучшего доступа к необходимым ростовым факторам. Структурно, органоиды по изобретению часто имеют вытянутую форму. Они могут включают одну или несколько зачаточных структур - монослойный эпителий, который имеет структуру, не похожую на желчный проток. На конфокальной микроскопии структуры могут окрашиваться положительно на кератин. Они могут включать клетки с поляризованными ядрами и маленьким объемом цитоплазмы. Органоиды могут иметь отдел, который образуется множеством слоев; ядра таких клеток часто имеют тенденцию к более центральному расположению в клетках, то есть, не являются поляризованными. Клетки в многослойном отделе могут самоорганизовываться с включением промежутка или просвета между клетками.
Органоид печени предпочтительно содержит гепатоциты и холангиоциты, более предпочтительно, в которых можно детектировать по меньшей мере один из следующих маркеров: по меньшей мере один гепатоцитарный маркер, такой как альбумин, транстиретин, В-1 интегрин и глутаминсинтетазу и/или по меньшей мере один из CYP3A11, FAH, tbx3, TAT и Gck, и/или по меньшей мере один холангиоцитарный маркер, такой как кератин 7 и 19. Опытному специалисту известно, как детектировать каждый из этих маркеров (а именно, с помощью ОТ-ПЦР и/или иммуногистохимии). Предпочтительно, чтобы оценку экспрессии каждого из этих маркеров проводили, как описано в экспериментальной части. Каждый из этих маркеров обычно экспрессируется по меньшей мере через две недели, три недели или один месяц культивирования с использованием способа по изобретению. Анализ органоидов методом микроматриц в двух условиях культивирования показал, что органоиды печени похожи на ткань зрелой печени.
В некоторых вариантах осуществления изобретения приблизительно 35% клеток в органоиде печени экспрессируют поверхностный маркер гепатоцитов, например, 25-45%, 30-40%, 33-37%, 35% или меньше, или 15-35% клеток.
Предпочтительно, чтобы клетки и органоиды, полученные по изобретению, также обладали функциями гепатоцитов, такими как экспрессия или положительное окрашивание на маркеры зрелых гепатоцитов - альбумин, интегрин В-1, CK-8, CK-18, транстиретин (TTR), глюкозо-6-фосфатазу, Met, глутаминсинтетазу (Glul), трансферрин, Fahd1, Fahd2a, K7, K19 и изоформы цитохрома P450 - 3A13 (CYP3A13), 51 (CYP51) 2D10 (CYP2D10), 2j6 (CYP2j6), 39A1 (CYP39A1), 4A10 (CYP4A10), 4F13 (CYP4F13), 4F16 (CYP4F16), CYP4B1 и 20A1 (CYP20A1). Также в некоторых вариантах осуществления изобретения ген AFP, экспрессирующийся в эмбриональной печени, не детектируется ни в одном из двух условий культивирования, подобно зрелой печени. В некоторых вариантах осуществления изобретения экспрессия альфа-фетопротеина немного выше фонового уровня экспрессии.
Кроме того, хорошо известные печеночные транскрипционные факторы, такие как HNF1a, HNF1b и HNF4a, экспрессируются на высоком уровне при двух условиях культивирования.
Поскольку печень и поджелудочная железа являются близкородственными органами, авторы изобретения провели исследование возможности экспрессии полученными ими культурами клеток печени генов, специфичных для клеток поджелудочной железы. Поджелудочная железа функционально разделена на эндокринную и экзокринную части поджелудочной железы. Эндокринная часть в основном отличается экспрессией инсулина, глюкагона и соматостатина. Экспрессия этих гормонов плотно регулируется набором специфичных для эндокринной части поджелудочной железы транскрипционных факторов, наиболее важными из которых являются Pdx1 и NeuroD. Экзокринную часть поджелудочной железы образуют ацинарный и протоковый отделы, отвечающие за продукцию пищеварительных ферментов в том числе - амилазы, панкреатической липазы и химотрипсина. Экспрессия этих генов также регулируется специфичными для экзокринной части генами, например, Ptf1.
Специфичные для клеток поджелудочной железы гены Ptf1a, панкреатической амилазы (Amy2a4), панкреатической липазы (Pnlip), инсулина (ins1 и ins2), глюкагона (Gcg), химотрипсина (cela1), Pdx1 и NeuroD отсутствовали в культурах клеток печени, описанных в настоящем документе.
В некоторых вариантах осуществления изобретения один или несколько из следующих генов экспрессируются в органоидах печени на уровне, аналогичному уровню экспрессии соответствующего гена в зрелых гепатоцитах печени: Aqp1, Bmp2, Apo3, Apol7a, Sord, C3, Ppara, Pparg, tbx3, lgf1, Il17rb, Il1b, Tgfbi, Apoa1, Apoa4, Apob, Cyp26b1, Cyp27a1, Cyp2b13, Cyp2b9, Cyp2c37, Cyp2f2, Cyp2g1, Cyp2j13, Cyp3a11, Cyp4a10 и Cypf14. Например, см. фигуру 16А.
В некоторых вариантах осуществления изобретения один или несколько из следующих генов экспрессируется в органоидах печени на аналогичном минимальном уровне по сравнению с экспрессией соответствующего гена в зрелых гепатоцитах печени: Ccl2, Osmr, Icam1 и Cxcl2.
В некоторых вариантах осуществления изобретения один или оба из следующих генов дифференциально экспрессируются как в органоиде печени, так и в печени новорожденного: mKi67 и cdkn3.
В некоторых вариантах осуществления изобретения один, два или все из следующих генов экспрессируются на аналогичном уровне как в органоиде печени, так и в печени новорожденного: cyp2j6, olfm4 и Lefty 1. Например, см. фиг.16B.
В некоторых вариантах осуществления изобретения органоид печени по изобретению имеет протоковый фенотип при культивировании в среде для размножения по изобретению (например, ЕМ1 или ЕМ2).
В некоторых вариантах осуществления изобретения органоид печени по изобретению экспрессирует маркеры зрелой печени при культивировании в дифференцирующей среде по изобретению.
В одном варианте осуществления изобретения органоид печени по изобретению имеет профиль экспрессии генов, показанный на фиг.16С.
В особенно предпочтительном варианте осуществления изобретения популяция клеток или органоид печени мыши по изобретению имеет профиль экспрессии генов, показанный на фиг.18. Например, в одном предпочтительном варианте осуществления изобретения популяция клеток или органоид печени мыши по изобретению:
а) экспрессируют по меньшей мере один (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11), предпочтительно все, из следующих маркеров стволовых клеток: lgr5, lgr4, epcam, Cd44, Tnfrsf19, Sox9, Sp5, Cd24a, Prom1, Cdca7 и Elf3; и/или
b) не экспрессируют следующий маркер стволовых клеток: lgr6; и/или
с) экспрессируют по меньшей мере один (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19), предпочтительно все, из следующих маркеров гепатоцитов или холангиоцитов при выращивании в среде для размножения по изобретению: Hnf1a, Hnf1b, Hnf4a, Hhex, Onecut1, Onecut2, Pro1, Cdh1, Foxa2, Gata6, Foxm1, Cebpa, Cebpb, Cebpd, Cebpg, Glul, Krt7, Krt19 и Met; и/или
d) не экспрессируют по меньшей мере один (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17) из следующих генов при выращивании в среде для размножения по изобретению: afp, Ins1, Ins2, Gcg, Ptfla, Cela1, Cela2a, Cela3b, Neurod1, Neurod2, Neurog1, Neurog2, Neurog3, Amy2a4, Igf1r, Igf2 и Cd34; и/или
e) экспрессируют по меньшей мере один (например, 1, 2 или 3) из следующих репрограммирующих генов: Klf4, Myc и Pou5f1 и/или
f) не экспрессируют следующий репрограммирующий ген: Sox2.
Причем экспрессию генов предпочтительно детектировать путем измерения экспрессии на уровне мРНК, например, с использованием метода микроматриц.
Более предпочтительно, чтобы популяция клеток или органоид печени мыши по изобретению имели все признаки (а)-(f), перечисленные выше.
В некоторых вариантах осуществления изобретения профиль экспрессии генов, описанный выше для популяции клеток или органоида печени мыши по изобретению, относится к популяции клеток или органоиду мыши, культивируемым в среде для размножения по изобретению.
В некоторых вариантах изобретение относится к популяции клеток или органоиду печени человека по изобретению, которые имеют характеристическую экспрессию генов, показанную на фиг.19. Например, популяция клеток или органоид печени человека, культивированные в ЕМ1 по изобретению, предпочтительно экспрессируют гены, указанные на фиг.19, как экспрессирующиеся в клеточной культуральной среде ЕМ1. Например, популяция клеток или органоид печени человека, культивированные в ЕМ2 по изобретению, предпочтительно экспрессируют гены, указанные на фиг.19, как экспрессирующиеся в клеточной культуральной среде ЕМ2. Например, популяция клеток или органоид печени человека, культивированные в DM по изобретению, предпочтительно экспрессируют гены, указанные на фиг.19, как экспрессирующиеся в клеточной культуральной среде DM.
Например, в одном предпочтительном варианте осуществления изобретения популяция клеток или органоид печени человека по изобретению:
a) экспрессируют по меньшей мере один (например, 1, 2, 3, 4, 5, 6, 7, 8, 9), предпочтительно все, из следующих характеристических генов стволовых клеток: LGR4, TACSTD1/Epcam, CD44, SOX9, SP5, CD24, PROM1, CDCA7 и ELF3; и/или
b) экспрессируют по меньшей мере один (например, 1, 2, 3, 4), предпочтительно все, из следующих репрограммирующих генов: KLF4, MYC, POU5F1 и SOX2; и/или
c) экспрессируют по меньшей мере один (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19), предпочтительно все, из следующих специфичных для гепатоцитов/холангиоцитов генов: HNF1A, HNF1B, HNF4A, HHEX, ONECUT1, ONECUT2, PROX1, CDH1, FOXA2, GATA6, FOXM1, CEBPA, CEBPB, CEBPD, CEBPG, GLUL, KRT7, KRT19 и MET; и/или
d) не экспрессируют по меньшей мере один (например, 1, 2, 3, 4, 5, 6), предпочтительно все, из следующих специфичных для гепатоцитов/холангиоцитов генов: NEUROG2, IGF1R и CD34, AFP, GCG и PTF1A, например, они не экспрессируют NEUROG2, IGF1R и CD34; и/или
e) экспрессируют по меньшей мере один (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18), предпочтительно все, из следующих специфичных для гепатоцитов генов: TTR, ALB, FAH, TAT, CYP3A7, APOA1, HMGCS1, PPARG, CYP2B6, CYP2C18, CYP2C9, CYP2J2, CYP3A4, CYP3A5, CYP3A7, CYP4F8, CYP4V2 и SCARB1;
причем экспрессию генов предпочтительно детектировать путем измерения экспрессии на уровне мРНК, например, с использованием метода микроматриц.
Более предпочтительно, чтобы популяция клеток или органоид печени человека по изобретению имели все признаки (а)-(е), перечисленные выше.
В некоторых вариантах осуществления изобретения экспрессия генов в популяции клеток или органоиде печени человека по изобретению повышена или снижена относительно экспрессии референсной РНК, как показано на фиг.19. Предпочтительно, чтобы референсной РНК являлась Универсальная референсная РНК человека (Stratagene, Catalog #740000). В некоторых вариантах осуществления изобретения экспрессия гена повышена или снижена относительно референсной РНК, если она также показана на фиг.19, как повышенная или пониженная относительно референсной РНК, но степень повышения или снижения экспрессии необязательно должна быть одинаковой. В других вариантах осуществления изобретения степень повышения или снижения экспрессии составляет +/-35%, +/-30%, +/-25%, +/-20%, +/-20%, +/-15%, +/-10%, +/-5%, +/-3% или является приблизительно одинаковой с показанной на фиг.19. В других вариантах осуществления изобретения абсолютный уровень экспрессии генов в органоиде человека по изобретению составляет +/-35%, +/-30%, +/-25%, +/-20%, +/-15%, +/-10%, +/-5%, +/-3% или является приблизительно таким же, как показанный на фиг.19.
Популяция клеток печени человека или органоиды по изобретению также предпочтительно экспрессируют Lgr5 и/или Tnfrsf19, предпочтительно оба гена. В некоторых вариантах осуществления изобретения популяция клеток или органоиды печени человека при культивировании в среде для размножения по изобретению экспрессируют Lgr5 и/или Tnfrsf19, предпочтительно оба гена. Предпочтительно детектировать экспрессию Lgr5 и/или Tnfrsfr19 с помощью ОТ-ПЦР. В некоторых вариантах осуществления изобретения Lgr5 и/или Tnfrsf19 имеют значительно меньший уровень экспрессии в органоидах или клетках, культивируемых в дифференцирующей среде, по сравнению с их уровнем экспрессии в органоидах или клетках, культивируемых в среде для размножения (например, по меньшей мере в 2 раза, по меньшей мере в 3 раза, по меньшей мере в 4 раза, по меньшей мере в 5 раз, по меньшей мере в 10 раз, по меньшей мере в 15 раз меньше).
Клетки и органоиды по настоящему изобретению могут предпочтительно обладать способностью секретировать альбумин, например, со скоростью от приблизительно 1 мкг в час на 106 клеток до 10 мкг в час на 106 клеток, предпочтительно от 2 мкг до 6 мкг в час на 106 клеток.
Кроме того, такие клетки и органоиды могут секретировать мочевину. Например, для клеток в чашках Петри диаметром 35 мм интенсивность синтеза мочевины может составлять от 1 мкг до 50 мкг за 48 часов, предпочтительно от 5 мкг до 30 мкг.
В клетки и органоидах по изобретению могут наблюдаться видимые запасы гликогена, например, при окрашивании. Способность клеток и органоидов по изобретению активно синтезировать гликоген можно протестировать путем переключения культуральной среды с дифференцирующей среды с низким содержанием глюкозы на DMEM с высоким содержанием глюкозы, дополненную 10% FBS и 0,2 мкМ дексаметазоном, на два дня.
Клетки и органоиды по изобретению могут обладать индуцируемой активностью цитохрома Р450 (например, CYP1A). Такую активность можно протестировать, например. Используя этоксирезоруфин-О-деэтилазный (EROD) метод анализа (Cancer Res, 2001, 61: 8164-8170). Например, к клеткам или органоидам можно добавить субстрат Р450, такой как 3-метилхолантрен, и сравнить уровень активности EROD с контрольными клетками.
Морфологически клетки выглядят похожими на гепатоциты.
Предпочтительный органоид печени содержит или состоит из кистозной структуры с внешней стороны со слоем клеток с зачатками и центральным просветом, показанными на фиг.2. Этот органоид печени может иметь одну (например, 2, 3 или все 4) из следующих характеристик: (а) иметь плотность клеток >5×105 клеток/см3, предпочтительно >10×105 клеток/см3; (b) иметь толщину, равную 2-30 слоям клеток, предпочтительно толщину, равную 2-15 слоям клеток; (с) клетки контактируют друг с другом во всех трех пространственных направлениях; (d) демонстрирует функцию, характерную для ткани нормальной печени; (е) имеет вытянутую форму с 2 выраженными доменами, а именно, состоящий из одного слоя клеток эпителиальный домен, в котором детектируются высоко поляризованные клетки и экспрессируются кератиновые маркеры (этот домен напоминает домен желчных протоков), а другой домен составляет основное тело органоида и образован многослойным эпителием с неполяризованными клетками, в которых можно детектировать экспрессию альбумина. Опытному специалисту в данной области ясно, что такой органоид печени предпочтительно не является фрагментом печени и/или не содержит кровеносных сосудов, и/или не содержит печеночную дольку или желчного протока.
В контексте изобретения фрагментом печени является часть зрелой печени, предпочтительно зрелой печени человека. Поэтому предпочтительно, что органоид печени, определенный в настоящем документе, не является фрагментом печени. Органоид печени, предпочтительно получать, используя клетку из зрелой печени, предпочтительно эпителиальную стволовую клетку из зрелой печени, более предпочтительно эпителиальную стволовую клетку из зрелой печени, экспрессирующую Lgr5.
В некоторых вариантах осуществления изобретения органоид печени содержит клетки, которые экспрессируют Lgr5. Например, в некоторых вариантах осуществления изобретения по меньшей мере 2%, более предпочтительно, по меньшей мере 5%, по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95% клеток в органоиде печени экспрессируют Lgr5. Аналогично, изобретение относится к клетке или популяции клеток, которые экспрессируют Lgr5, причем указанные клетки получены из органоида печени по изобретению. Изобретение также охватывает потомство таких клеток.
В варианте осуществления изобретения органоидом печени является органоид печени, который все еще культивируется с использованием способа по изобретению и поэтому находится в контакте c внеклеточным матриксом. Предпочтительно, чтобы органоид печени был заключен в немезенхимальный внеклеточный матрикс. В контексте изобретения "в контакте" означает физический, механический или химический контакт, означающий, что для отделения указанного органоида печени от указанного внеклеточного матрикса необходимо приложить усилие.
В предпочтительном варианте осуществления изобретения органоид печени можно было бы культивировать в течение по меньшей мере 2, 3, 4, 5, 6, 7, 8, 9, 10 недель или 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 месяцев или большего периода времени.
В другом предпочтительном варианте осуществления изобретения органоид печени получен из одной клетки, предпочтительно экспрессирующей Lgr5, более предпочтительно, что одна клетка содержит конструкцию нуклеиновой кислоты, содержащую представляющую интерес молекулу нуклеиновой кислоты.
Изобретение дополнительно относится к использованию культуральной среды по изобретению для культивирования эпителиальных стволовых клеток или выделенных органоидных структур, которые содержат эти стволовые клетки, на внеклеточном матриксе, в результате чего указанные стволовые клетки предпочтительно не содержат эмбриональных стволовых клеток человека. Предпочтительными являются зрелые стволовые клетки человека. Кроме того, одиночные отсортированные эпителиальные стволовые клетки из печени также способны инициировать рост этих 3-мерных органоидов в культуральной среде по изобретению. Изобретение дополнительно относится к использованию культуральной среды по изобретению для культивирования фрагментов печени, содержащих стволовые клетки.
Предпочтительно, чтобы указанные стволовые клетки являлись стволовыми клетками печени или эпителиальными стволовыми клетками. Культуральная среда по изобретению позволяла установить условия для длительного культивирования, при которых образуется органоид печени, в котором присутствуют все типы дифференцированных клеток. Использование способа культивирования по изобретению позволяло культивирование в течение по меньшей мере семи месяцев, по меньшей мере восьми месяцев, по меньшей мере девяти месяцев, по меньшей мере десяти месяцев.
Изобретение дополнительно относится к органоиду печени, предпочтительно содержащему по меньшей мере 50% жизнеспособных клеток, более предпочтительно, по меньшей мере 60% жизнеспособных клеток, более предпочтительно, по меньшей мере 70% жизнеспособных клеток, более предпочтительно, по меньшей мере 80% жизнеспособных клеток, более предпочтительно, по меньшей мере 90% жизнеспособных клеток. Жизнеспособность клеток можно оценить, используя окрашивание Хехстом или окрашивание йодистым пропидием в FACS-анализе.
Жизнеспособные клетки предпочтительно обладают гепатоцитарными функциями или характеристиками гепатоцитов, описанными выше.
Применение клеток и органоидов по изобретению
В дополнительном аспекте изобретение относится к применению клетки или органоида печени по изобретению, описанных выше, в скрининге для поиска новых лекарственных соединений, анализе токсичности или в регенеративной медицине. Изобретение дополнительно относится к применению потомства органоидов печени по изобретению в методах анализа токсичности. Такими методами анализа токсичности могут быть методы анализа in vitro с использованием клеток, полученных из органоида печени, либо органоида печени или его части. Такое потомство и органоиды печени легко культивировать, и они в большей степени похожи на первичные эпителиальные клетки, чем, например, эпителиальные клеточные линии, такие как Caco-2 (ATCC HTB-37), I-407 (ATCC CCL6) и XBF (ATCC CRL 8808), которые на сегодняшний момент используются в методах анализа токсичности. Ожидается, что результаты анализа токсичности, полученные с органоидами печени, более точно отражают результаты, полученные на пациентах. Клеточный тест на токсичность используют для определения орган-специфичной цитотоксичности. Соединения, которые проходят тестирование в указанном тесте, включают противораковые химиопрофилактические агенты, бытовую химию, пищевые добавки и потенциальные токсические соединения. На клетки воздействуют различными концентрациями тестируемого агента в течение определенного периода времени. Концентрационные диапазоны для тестируемых соединений в анализе определяют в предварительном анализе, используя время воздействия 5 дней и 10-кратные разведения от максимальной растворимой концентрации. В конце периода воздействия культуры оценивают на ингибирование роста. Данные анализируют для определения концентрации, которая ингибирует рост в конечной точке на 50 процентов (ТС50).
Например, индукция ферментов цитохромов 450 в гепатоцитах печени является ключевым фактором, который определяет эффективность и токсичность лекарственных соединений. В частности, индукция ферментов Р450 является важным механизмом в проблемном взаимодействии лекарственных соединений между собой и также важным фактором, который ограничивает эффективность лекарственного соединения и определяет его токсичность. Методы анализа индукции цитохрома Р450 тяжело разрабатывать, поскольку они требуют интактных нормальных гепатоцитов человека. Было показано, что эти клетки трудны для получения в количествах, достаточных для поддержания массового получения высокопропускных методов анализа.
Например, по этому аспекту изобретения соединение-кандидат может контактировать с клеткой или органоидом, описанными в настоящем документе, и можно контролировать любое изменение в клетках или активности клеток. Примеры других вариантов нетерапевтического применения клеток или органоидов по настоящему изобретению включают исследование эмбрионального развития печени, типов клеток печени и пути дифференцировки; исследования экспрессии генов, включая экспрессию рекомбинантных генов; исследования механизмов, вовлеченных в повреждение и восстановление печени; исследования воспалительных и инфекционных заболеваний печени; исследования патогенетических механизмов; и исследования механизмов трансформации клеток печени и этиологии рака печени.
Для высокопропускных целей указанные органоиды печени культивируют в мультилуночных планшетах, таких как, например, 96-луночные планшеты или 384-луночные планшеты. Для идентификации молекулы, которая влияет на указанные органоиды, используют библиотеки молекул. Предпочтительные библиотеки включают в себя библиотеки фрагментов антител, библиотеки пептидного фагового дисплея, пептидные библиотеки (например, LOPAP™, Sigma Aldrich), липидные библиотеки (BioMol), библиотеки синтетических соединений (например, LOP AC™, Sigma Aldrich) или библиотеки природных соединений (Specs, TimTec). Кроме того, можно использовать генетические библиотеки, которые индуцируют или репрессируют экспрессию одного или нескольких генов в потомстве клеток аденомы. Эти генетические библиотеки включают в себя кДНК-библиотеки, библиотеки антисмысловых последовательностей и библиотеки миРНК или других некодирующих РНК. На клетки предпочтительно воздействовать различными концентрациями тестируемого агента в течение определенного периода времени. В конце этого периода проводят оценку культур. Термин "влияние" используют для описания любого изменения в клетке, включая, без ограничений, снижение или потерю пролиферации, морфологические изменения и клеточную смерть. Указанные органоиды печени также можно использовать для идентификации лекарственных соединений, специфичной мишенью которых являются клетки карциномы эпителия, но не указанные органоиды печени.
Органоиды печени по изобретению могут дополнительно заменить использование клеточных линий, таких как клетки Caco-2, в методах анализа токсичности потенциальных новых лекарственных соединений, или известных или новых пищевых добавок.
Кроме того, такие органоиды печени можно использовать для культивирования патогена.
Культуры, содержащие органоиды печени, пригодны в регенеративной медицине, например, при восстановлении эпителия печени после облучения и/или после хирургического вмешательства, при восстановлении эпителия у пациентов, страдающих от хронической печеночной недостаточности или заболевания печени. Заболевания печени включают, но не ограничены этим, гепатоцеллюлярную карциному, синдром Алажиля, недостаточность альфа-1-антитрипсина, аутоиммунный гепатит, билиарную атрезию, хронический гепатит, рак печени, кистозное заболевание печени, жировую дистрофию печени, галактоземию, синдром Жильбера, первичный билиарный цирроз, гепатит А, гепатит В, гепатит С, первичный склерозирующий холангит, синдром Рейе, саркоидоз, тирозинемию, гликогеновую болезнь I типа, болезнь Вильсона, неонатальный гепатит, неалкогольный стеатогепатит, порфирию и гемохроматоз.
Генетические дефекты, которые приводят к печеночной недостаточности, могут компенсироваться клеточной терапией в форме частичного или полного замещения клеток с использованием клеток, культивированных в среде и/или по способам изобретения. Неограничивающий список генетических дефектов, которые приводят к печеночной недостаточности, включает: прогрессирующий семейный внутрипеченочный холестаз, гликогеновую болезнь III типа, тирозинемию, недостаточность дезоксигуанозинкиназы, недостаточность пируваткарбоксилазы, врожденную дизэритропоетическую анемию, поликистоз печени, поликистоз почек, недостаточность альфа-1-антитрипсина, нарушения цикла мочевины, органические ацидемии, лизосомные болезни накопления и нарушения окисления жирных кислот. Другие патологические состояния, которые также могут компенсироваться клеточной терапией, включают болезнь Вилсона и наследственный амилоидоз (FAP).
Другие не связанные с гепатоцитами причины печеночной недостаточности, которые требовали бы трансплантации всей печени для достижения полного терапевтического эффекта, могут тем не менее компенсироваться в результате некоторого временного восстановления функций с помощью клеточной терапии с использованием клеток, культивированных в среде и/или согласно способам по изобретению. Неограничивающий список примеров таких патологических состояний включает: первичный билиарный цирроз, первичный склерозирующий холангит, синдром Алажиля, гомозиготную семейную гиперхолестеринемию, гепатит В с циррозом, гепатит С с циррозом, синдром Бадда-Киари, первичную гипероксалурию, аутоиммунный гепатит и алкогольную болезнь печени.
Органоиды печени по изобретению можно использовать в способе лечения наследственного заболевания, которое связано с нарушением функций гепатоцитов. Такие заболевания могут иметь раннее или позднее начало. Заболевания с ранним началом включают связанную с метаболизмом органную недостаточность (например, недостаточность альфа-1-антитрипсина), гликогеновые болезни (например, GSD II, болезнь Помпе), тирозинемию, легкую недостаточность DGUOK, CDA I типа, нарушения цикла мочевины (например, недостаточность ОТС), органические ацидемии и нарушения окисления жирных кислот. Заболевания позднего начала включают первичную гипероксалурию, семейную гиперхолестеринемию, болезнь Вилсона, наследственный амилоидоз и поликистозную болезнь печени. Частичное или полное замещение здоровыми гепатоцитами из органоидов печени по изобретению можно использовать для восстановления функций печени или для отсрочки печеночной недостаточности.
Органоиды печени по изобретению можно использовать в способе лечения хронической печеночной недостаточности, возникающей вследствие наследственного метаболического заболевания или в результате инфекции гепатоцитов. Лечение наследственного метаболического заболевания может включать в себя введение генетически модифицированных аутологичных органоидов печени по изобретению. Лечение инфекции гепатоцитов может включать в себя введение аллогенных органоидов печени по изобретению. В некоторых вариантах осуществления изобретения органоиды печени вводят в течение 2-3 месяцев.
Органоиды печени по изобретению можно использовать для лечения острой печеночной недостаточности, например, в результате интоксикации печени, которая может быть результатом применения парацетамола, лекарственных соединений или спирта. В некоторых вариантах осуществления изобретения терапия для восстановления функции печени будет включать в себя инъекции суспензии гепатоцитов из замороженных, готовых для использования аллогенных гепатоцитов, полученных из органоидов по изобретению. Возможность замораживать подходящие органоиды означает, что органоиды могут быть доступны для немедленной доставки, то есть нет необходимости ожидать переливания крови.
В случае замещения или исправления недостаточности функции печени можно сконструировать клеточно-матриксную структуру из одного или нескольких органоидов печени, полученных по настоящему изобретению. Полагают, что только примерно 10% массы клеток печени необходимы для ее достаточного функционирования. Благодаря этому имплантация детям композиций из органоидных единиц особенно предпочтительна относительно трансплантации целого органа, вследствие относительно ограниченной доступности доноров и меньшего размера органов детей. Например, здоровая печень 8-месячного ребенка весит приблизительно 250 г. Этому ребенку, поэтому, необходимо примерно 25 г ткани. Печень взрослого человека весит приблизительно 1500 г; поэтому требуемый имплант будет составлять только примерно 1,5% от печени взрослого человека. При имплантации органоидных единиц по изобретению, необязательно прикрепленных к полимерному каркасу, они будут пролиферировать в новом хозяине, и полученная масса печеночных клеток заменит недостаточность функции печени хозяина. Авторы изобретения впервые показали возможность получения зрелых гепатоцитов из стволовых клеток зрелой печени или фрагментов ткани печени, содержащих стволовые клетки, которые подходят для трансплантации животным или человеку. Используя первый способ культивирования по изобретению авторы продемонстрировали возможность сохранения и размножения популяции стволовых клеток печени. Используя второй способ культивирования по изобретению авторы показали, что гепатобласты могут дифференцироваться in vivo в зрелые гепатоциты, подходящие для трансплантационных целей. Следовательно, авторы изобретения обеспечили новый источник гепатоцитов для регенерации печени, замещения или исправления недостаточности функции печени.
Авторы изобретения также продемонстрировали успешную трансплантацию органоидов, выращенных способами по настоящему изобретению, иммунодефицитным мышам (см. пример 7), причем из трансплантированных органоидных клеток генерируются как холангиоциты, так и гепатоциты, in vivo. Поэтому, в одном варианте осуществления изобретение относится к органоидам или органоидным клеткам по изобретению для трансплантации человеку или животным.
Использование органоидов печени человека для трансплантационных целей предпочтительнее использования зародышевых или взрослых гепатоцитов по ряду причин. Во-первых, способы культивирования по изобретению обеспечивают неограниченное размножение клеток и, следовательно, их неограниченный источник. В частности, авторы изобретения показали, что при правильных условиях культивирования (например, использовании культуральной среды для размножения по изобретению) клетки Lgr5+ могут проходить более чем через 1000 делений in vitro. Таким образом, клетки Lgr5+ можно выделить из органоидов печени и перепассировать, обеспечивая постоянный самообновляющийся источник трансплантируемых генерирующих гепатоциты и холангиоциты клеток. В отличие от этого, зародышевые и зрелые гепатоциты, полученные из донорской печени, обеспечивают только один раунд трансплантации. Кроме того, донорские клетки могут поддерживаться живыми только в течение нескольких дней, но теряют свои гепатоцитарные свойства. Это означает, что трансплантация должна проводиться как можно скорее после того, как донор стал доступен. Органоидные клетки, с другой стороны, сохраняют свой фенотип после многочисленных делений и после больших временных интервалов, что означает их готовность и доступность для трансплантации на любой стадии. Это также могло бы позволить использование органоидных клеток в качестве временного лечения печени для увеличения продолжительности жизни пациентов в случае их нахождения в списке ожидания на трансплантацию печени. Дополнительным преимуществом органоидов печени по изобретению является возможность их заморозки и последующей разморозки без потери функциональности. Это дает возможность создания банков клеток, легкого хранения и быстрой доступности при срочной необходимости использования. Это может быть полезно, например, при получении стандартного готового продукта, который можно использовать для лечения острой печеночной токсичности. Органоиды также можно выращивать из клеток или фрагментов ткани, забранных в виде небольших образцов биопсии от живых доноров, минимизируя любые этические запреты лечения. Донором может являться сам пациент, подвергающийся лечению, что может снизить любые побочные эффекты, ассоциированные с трансплантацией чужеродных клеток и органов, и уменьшить необходимость в иммуносупрессирующих лекарственных препаратах.
Соответственно, в объем изобретения включены способы лечения пациента-животного или человека посредством клеточной терапии. Термин "животное" в настоящем документе обозначает всех млекопитающих животных, предпочтительно людей. Он также включает отдельное животное на всех стадиях развития, включая эмбриональные и зародышевые стадии. Например, пациент может быть взрослым человеком, или терапия может применяться в педиатрии (например, у новорожденного, ребенка или подростка). Такая клеточная терапия охватывает введение клеток или органоидов, полученных по изобретению, пациенту любыми соответствующими средствами. Более конкретно, такие способы лечения включают регенерацию поврежденной ткани. Термин "введение", используемый в настоящем документе, относится к хорошо известным формам введения, таким как внутривенное введение или с помощью инъекции, а также к введению путем трансплантации, например, трансплантации хирургическим путем, пересадке или трансплантации печени, полученной с помощью тканевой инженерии из клеток или органоидов по настоящему изобретению. В случае клеток, возможно их системное введение индивидууму, например, инфузией в верхнюю брыжеечную артерию, брюшную артерию, подключичную вену через грудной лимфатический проток, инфузией в сердце через верхнюю полую вену, или инфузией в брюшную полость с последующей миграцией клеток через поддиафрагмальные лимфатические протоки или напрямую в участки печени посредством инфузии в источник артериальной крови печени или в воротную вену.
На инфузию можно вводить от 104 до 1013 клеток на 100 кг веса человека. Предпочтительно, на 100 кг веса человека можно вводить внутривенной инфузией от примерно 1-5×104 до 1-5×107 клеток. Более предпочтительно, на 100 кг веса человека можно вводить внутривенной инфузией от примерно 1×104 до 10×106 клеток. Некоторые варианты осуществления изобретения относятся к однократному введению клеток или органоидов. В других вариантах осуществления изобретения используется многократное введение. Многократное ведение можно осуществлять по исходной схеме лечения, например, в течение 3-7 последовательных дней, и затем повторить в другое время.
В некоторых вариантах осуществления изобретения желательно заменить популяцию/заместить 10-20% печени пациента здоровыми гепатоцитами, выращенными из органоида печени по изобретению.
Также возможно восстановить органоид печени из одной единственной клетки, экспрессирующей Lgr5, описанной в настоящем документе. Эта единственная клетка может быть модифицирована путем введения конструкции нуклеиновой кислоты, описанной в настоящем документе, например, для исправления генетически обусловленной недостаточности или мутации. Также было бы возможно при желании специфично подавить экспрессию, например, используя миРНК. Полипептидами, которые потенциально возможно экспрессировать, могли бы быть любые из полипептидов, недостаточность которых наблюдается при метаболических заболеваниях печени, включая, например, ААТ (альфа-антитрипсин). Для изучения физиологии лечения авторы также могли бы экспрессировать или инактивировать гены, вовлеченные в сигнальные пути Wnt, EGF, FGF, BMP или Notch. Также, для скрининга на токсичность лекарственных соединений большой интерес представляла бы экспрессия или инактивация генов, отвечающих за метаболизм лекарственных соединений в печени (например, генов из семейства CYP).
В одном варианте осуществления изобретения размноженные эпителиальные стволовые клетки можно репрограммировать по желательному пути дифференцировки, такому как, например, в клетки печени, включая гепатоцит и холангиоцит. До настоящего времени не было возможным регенерировать клетки печени из зрелых стволовых клеток. Способы культивирования по настоящему изобретению обеспечивают возможность анализа факторов, которые трансдифференцируют близкородственные эпителиальные стволовые клетки в клетки печени, включая гепатоцит и холангиоцит.
Опытному специалисту в данной области будет ясно, что генная терапия может дополнительно использоваться в способе, направленном на восстановление поврежденной или больной ткани. Для доставки генетической информации, например, ДНК и/или РНК, в стволовые клетки, можно использовать, например, аденовирусные или ретровирусные средства доставки генов. Опытный специалист в данной области может заместить или восстановить конкретные гены, являющиеся мишенью генной терапии. Например, можно встроить нормальный ген в неспецифичное место в геноме для замещения нефункционального гена. В другом примере, аномальную последовательность гена можно заменить на нормальную последовательность гена посредством гомологичной рекомбинации. В альтернативном варианте селективная обратная мутация может вернуть гену его нормальную функцию. Дополнительным примером является изменение регуляции (степень, до которой экспрессия гена включена или выключена) конкретного гена. Предпочтительно проводить генную терапию на стволовых клетках ex vivo, и после переносить в млекопитающее, предпочтительно человека, нуждающегося в лечении. Например, органоидные клетки можно генетически модифицировать в культуре перед трансплантацией пациенту.
Авторы изобретения обнаружили, что Lgr5 не детектируется в здоровой печени, хотя можно детектировать его остаточную экспрессию. Поэтому, изобретение дополнительно относится к способу диагностики повреждения печени, включающему в себя детекцию экспрессии Lgr5, причем экспрессия белка Lgr5 указывает на повреждение печени. Изобретение также относится к способу мониторинга восстановления или регенерации печени путем мониторинга экспрессии Lgr5 в печени. Экспрессию Lgr5 можно детектировать любым подходящим способом, например, проточной цитометрией, иммуногистохимией или используя способы ПЦР.
Изобретение также относится к композиции или клеточному культуральному сосуду, содержащему клетки и/или органоиды по любому одному из аспектов изобретения, описанных выше, и культуральную среду по любому одному из аспектов изобретения, описанных выше. Например, такая композиция или клеточный культуральный сосуд могут содержать любое число клеток или органоидов, культивированных способом по изобретению, в культуральной среде, описанной выше. Например, предпочтительная культуральная среда содержит или состоит из Advanced-DMEM/F12, дополненной B27, N2, 200 нг/мл N-ацетилцистеина, 50 нг/мл EGF, 1 мкг/мл R-спондина 1, 10 нМ гастрина, 100 нг/мл FGF10, 10 мМ никотинамида и 50 нг/мл HGF и 50% Wnt-кондиционной среды.
Определения
Нуклеотидная конструкция содержит представляющую интерес молекулу нуклеиновой кислоты и будет гарантировать экспрессию данной молекулы нуклеиновой кислоты в клетках, в которые она введена. Особенно предпочтительной нуклеотидной конструкцией является экспрессионный вектор, в котором кодирующая полипептид молекула нуклеиновой кислоты функционально связана с промотором, способным управлять экспрессией указанной молекулы нуклеиновой кислоты (например, кодирующей последовательности) в указанных клетках. Фраза "экспрессионный вектор" в общем относится к молекуле нуклеиновой кислоты, способной осуществлять экспрессию гена/молекулы нуклеиновой кислоты, которые она содержит, в клетке, совместимой с такими последовательностями. Эти экспрессионные векторы, как правило, включают в себя по меньшей мере подходящие промоторные последовательности и, необязательно, сигналы терминации транскрипции. Нуклеиновая кислота или ДНК, или нуклеотидная последовательность, кодирующая полипептид, включена в ДНК/нуклеотидную конструкцию, способную входить и экспрессироваться в клеточной культуре in vitro, указанной в способе по изобретению. ДНК-конструкция, созданная для введения в конкретную клетку, как правило, включает в себя систему репликации, распознаваемую указанной клеткой, целевой сегмент ДНК, кодирующий заданный полипептид, и регуляторные последовательности инициации и терминации транскрипции и трансляции, функционально связанные с кодирующим полипептид сегментом. Сегмент ДНК "функционально связан", когда он находится в функциональной взаимосвязи с другим сегментом ДНК. Например, промотор или энхансер функционально связаны с кодирующей последовательностью, если они стимулируют транскрипцию последовательности. ДНК сигнальной последовательности функционально связана с кодирующей полипептид ДНК, если она экспрессируется в виде пребелка, который участвует в секреции полипептида. В общем, функционально связанная ДНК-последовательность является непрерывной, и в случае сигнальной последовательности, непрерывной и с сохранением рамки считывания. Однако энхансеры не должны быть непрерывными с кодирующей последовательностью, чью транскрипцию они регулируют. Соединение выполняется с помощью лигирования по удобным сайтам рестрикции или через адаптеры или линкеры, встроенные вместо них.
Выбор соответствующей промоторной последовательности в общем зависит от клетки-хозяина, выбранной для экспрессии сегмента ДНК. Примеры подходящих промоторных последовательностей включают в себя эукариотические промоторы, хорошо известные в данной области (см., например, Sambrook and Russell, 2001, выше). Транскрипционная регуляторная последовательность, как правило, включает гетерологичный энхансер или промотор, которые распознаются клеткой. Подходящие промоторы включают в себя CMV-промотор. Экспрессионный вектор включает в себя систему репликации, и могут использоваться транскрипционные и трансляционные регуляторные последовательности вместе с сайтом для встраивания кодирующего полипептид сегмента. Примеры работающих комбинаций клеточных линий и экспрессионных векторов описаны в Sambrook and Russell (2001, выше) и в Metzger et al. (1988) Nature 334: 31-36.
Изобретение не ограничено определенным полипептидом или представляющей интерес молекулой нуклеиновой кислоты, экспрессируемой в экспрессирующей Lgr5 клетке, указанной в настоящем документе. В зависимости от цели способа может быть предусмотрена экспрессия полипептида в клетке печени и/или инактивация экспрессии полипептида в клетке печени. Потенциальными пептидами для экспрессии могли бы быть все из тех полипептидов, недостаточность которых наблюдается при метаболических заболеваниях печени, как, например, ААТ (альфа-антитрипсин). Для изучения физиологии печения авторы также могли бы экспрессировать или инактивировать гены, вовлеченные в сигнальные пути Wnt, EGF, FGF, BMP или Notch. Также, для скрининга на токсичность лекарственных соединений большой интерес представляла бы экспрессия или инактивация генов, отвечающих за метаболизм лекарственных соединений в печени (например, генов из семейства CYP).
Некоторые аспекте изобретения касаются использования конструкции нуклеиновой кислоты или экспрессионного вектора, содержащего нуклеотидную последовательность, указанную выше, причем вектором является вектор, подходящий для генной терапии. Векторы, которые подходят для генной терапии, описаны в Anderson 1998, Nature 392: 25-30; Walther and Stein, 2000, Drugs 60: 249-71; Kay et al., 2001, Nat. Med. 7: 33-40; Russell, 2000, J. Gen. Virol. 81: 2573-604; Amado and Chen, 1999, Science 285: 674-6; Federico, 1999, Curr. Opin. Biotechnol. 10: 448-53; Vigna and Naldini, 2000, J. Gene Med. 2: 308-16; Marin et al., 1997, Mol. Med. Today 3: 396-403; Peng and Russell, 1999, Curr. Opin. Biotechnol. 10: 454-7; Sommerfelt, 1999, J. Gen. Virol. 80: 3049-64; Reiser, 2000, Gene Ther. 7: 910-3; и указанных в этих статьях ссылках. Примеры включают встраиваемые или невстраиваемые векторы, такие как векторы на основе ретровирусов, аденовирусов (AdV), адено-ассоциированных вирусов (AAV), лентивирусов, вирусов оспы, альфавирусов и вирусов герпеса.
Особенно предпочтительный вектор для генной терапии включает аденовирусный (Ad) вектор и вектор на базе адено-ассоциированного вируса (AAV). Эти векторы инфицируют широкий ряд делящихся и неделящихся типов клеток, включая клетки печени. Кроме того, аденовирусные векторы способны к высокому уровню экспрессии трансгена. Однако вследствие эписомальной природы аденовирусных и AAV векторов после вхождения в клетку, эти вирусные векторы наиболее подходят для вариантов терапевтического применения, требующих только транзиентной экспрессии трансгена (Russell, 2000, J. Gen. Virol. 81: 2573-2604; Goncalves, 2005, Virol. J. 2(1):43), указанного выше. Предпочтительные аденовирусные векторы модифицированы для снижения ответа клетки-хозяина, см. обзор Russell (2000, выше). Безопасность и эффективность переноса генов с помощью AAV интенсивно исследовались у людей с многообещающими результатами в отношении печени, мышечной ткани, ЦНС и сетчатки (Manno et al Nat medicine 2006, Stroes et al ATVB 2008, Kaplitt, Feigin, Lancet 2009; Maguire, Simonelli et al NEJM 2008; Bainbridge et al NEJM 2008). AAV2 является наиболее хорошо описанным серотипом для исследований переноса генов, как у человека, так и в экспериментальных моделях. AAV2 имеет природный тропизм в отношении скелетных мышц, нейронов, клеток сосудистой гладкой мускулатуры и гепатоцитов. Поэтому, AAV2 является хорошим выбором вектора, направленного на ткани печени. Другие примеры невстраиваемых векторов на основе адено-ассоциированных вирусов включают AAV1, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11 и псевдотипированный AAV. Использование серотипов, не характерных для человека, таких как AAV8 и AAV9, может быть полезно для преодоления иммунологических ответов у субъектов, и клинические испытания только начались (ClinicalTrials.gov Identifier: NCT00979238). Было показано, что для переноса гена в клетку печени эффективными векторами являются аденовирус 5-го серотипа или AAV 2-го, 7-го или 8-го серотипов, и поэтому они являются предпочтительными Ad или AAV серотипами (Gao, Molecular Therapy (2006) 13, 77-87).
Предпочтительным вектором для применения в настоящем изобретении является экспрессионная конструкция на основе лентивирусов. Лентивирусные векторы имеют уникальную способность инфицировать неделящиеся клетки (Amado and Chen, 1999 Science 285: 674-6). Способы конструирования и использования экспрессионных конструкций на основе лентивирусов описаны в патентах США №№ 6,165,782, 6,207,455, 6,218,181, 6,277,633 и 6,323,031 и в Federico (1999, Curr Opin Biotechnol 10: 448-53) и Vigna et al. (2000, J Gene Med 2000; 2: 308-16).
В общем, векторы для генной терапии будут как экспрессионные векторы, описанные выше, в том смысле, что они содержат нуклеотидную последовательность, кодирующую экспрессируемый полипептид по изобретению, в результате чего нуклеотидная последовательность функционально связана с соответствующими регуляторными последовательностями, как указано выше. Такая регуляторная последовательность будет по меньшей мере содержать промоторную последовательность. Подходящие промоторы для экспрессии нуклеотидной последовательности, кодирующей полипептид для векторов генной терапии, включают промежуточный ранний промотор цитомегаловируса (CMV), промоторы вирусных длинных концевых повторов (LTR), такие как промоторы LTR из вируса лейкемии Молони мышей (мМLV), вируса саркомы Рауса или HTLV-1, ранний промотор вируса обезьян 40 (SV 40) и промотор тимидин-киназы вируса герпеса. Подходящие промоторы описаны ниже.
Описаны несколько индуцибельных промоторных систем, которые можно индуцировать путем введения низкомолекулярных органических или неорганических соединений. Такие индуцибельные промоторы включают промоторы, контролируемые тяжелыми металлами, такие как металлотиониновый промотор (Brinster et al. 1982 Nature 296: 39-42; Mayo et al. 1982 Cell 29: 99-108); RU-486 (агонистом прогестерона) (Wang et al. 1994 Proc. Natl. Acad. Sci. USA 91: 8180-8184); стероидами (Mader and White, 1993 Proc. Natl. Acad. Sci. USA 90: 5603-5607), тетрациклином (Gossen and Bujard 1992 Proc. Natl. Acad. Sci. USA 89: 5547-5551; патент США № 5,464,758; Furth et al. 1994 Proc. Natl. Acad. Sci. USA 91: 9302-9306; Howe et al. 1995 J. Biol. Chem. 270: 14168-14174; Resnitzky et al. 1994 Mol. Cell. Biol. 14: 1669-1679; Shockett et al. 1995 Proc. Natl. Acad. Sci. USA 92: 6522-6526); и систему tTAER, основанную на мультихимерном трансактиваторе, состоящем из полипептида tetR, активаторного домена из VP 16 и лиганд-связывающего домена эстрогенового рецептора (Yee et al., 2002, патент США № 6,432,705).
Подходящие промоторы для нуклеотидных последовательностей, кодирующих малые РНК для выключения определенных генов с помощью РНК-интерференции (см. ниже) включают, в дополнение к указанным выше промоторам для полимеразы II, промоторы для полимеразы III. РНК-полимераза III (pol III) отвечает за синтез большого числа ядерных и цитоплазматических некодирующих РНК, включая 5S, U6, аденовирусную область VA1, Vault, теломеразную РНК и тРНК. Структуры промоторов большого числа генов, кодирующих эти РНК, были определены, и было найдено, что промоторы РНК-полимеразы III разделяются на три типа структур (обзор см. в Geiduschek and Tocchini-Valentini, 1988 Annu. Rev. Biochem. 57: 873-914; Willis, 1993 Eur. J. Biochem. 212: 1-11; Hernandez, 2001, J. Biol. Chem. 276: 26733-36). Для экспрессии миРНК особенно подходят 3-й тип промоторов для РНК-полимеразы III, в которых транскрипция управляется цис-действующими элементами, найденными только в 5'-фланкирующей области, то есть выше сайта начала транскрипции. Расположенные выше элементы последовательности включают традиционный ТАТА-бокс (Mattaj et al., 1988 Cell 55, 435-442), близкорасположенный элемент последовательности и дальний элемент последовательности (DSE; Gupta and Reddy, 1991 Nucleic Acids Res. 19, 2073-2075). Примерами генов под контролем 3-го типа промотора РНК-полимеразы III являются гены малой ядерной РНК U6 (U6-мяРНК), 7SK, Y, MRP, H1 и теломеразной РНК (см., например, Myslinski et al., 2001, Nucl. Acids Res. 21: 2502-09).
Вектор для генной терапии может необязательно включать вторую, либо одну или несколько дополнительных нуклеотидных последовательностей, кодирующих второй или дополнительный полипептид. Вторым или дополнительным полипептидом может быть (селектируемый) маркерный полипептид, который позволяет идентификацию, отбор и/или скрининг клеток, содержащих экспрессионную конструкцию. Подходящими маркерными белками для этой цели являются, например, флуоресцентный белок GFP, селектируемые маркерные гены тимидинкиназы HSV (для селекции на НАТ-среде), бактериальная гигромицин В-фосфотрансфераза (для селекции на гигромицине В), аминогликозид-фосфотрансфераза Tn5 (для селекции на G418) и дигидрофолатредуктаза (DHFR) (для селекции на метотрексате), CD20, ген низкоаффинного фактора роста нервных клеток. Источники получения указанных маркерных генов и способы их использования описаны в Sambrook и Russel (2001) "Molecular Cloning: A Laboratory Manual (3rd edition), Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, New York.
В альтернативном варианте, вторая или дополнительная нуклеотидная последовательность может кодировать полипептид, который обеспечивает безопасный механизм, позволяющий освобождать субъект от трансгенных клеток после излечения, если это необходимо. Такая нуклеотидная последовательность, часто называемая суицидным геном, кодирует белок, который способен превращать пролекарственное соединение в токсичное соединение, способное уничтожать трансгенные клетки, в которых экспрессируется полипептид. Соответствующие примеры таких суицидных генов включают, например, ген цитозиндезаминазы из E.coli или один из генов тимидинкиназы из вируса Herpes simplex, цитомегаловируса и вируса ветряной оспы, в случае которых можно использовать ганцикловир в качестве пролекарственного соединения для уничтожения IL-10 трансгенных клеток у субъекта (см., например, Clair et al., 1987, Antimicrob. Agents Chemother. 31: 844-849).
Для выключения экспрессии определенного полипептида, вектор для генной терапии или другую экспрессионную конструкцию используют для экспрессии целевой нуклеотидной последовательности, которая предпочтительно кодирует РНКи-агент, то есть молекулу РНК, способную к РНК-интерференции, или которая является частью РНК-молекулы, способной к РНК-интерференции. Такие молекулы РНК называются киРНК (короткими интерферирующими РНК, включая короткие шпилечные РНК). В альтернативном варианте молекула киРНК может вводиться напрямую, например, в фармацевтической композиции, вводимой в клетку, экспрессирующую Lrg5, или в ее окружение, или в клетку печени (а именно, гепатоцит или холангиоцит) или в органоид печени.
Желаемая нуклеотидная последовательность содержит антисмысловую кодирующую ДНК, кодирующую антисмысловую РНК, специфичную к области мРНК гена-мишени, и/или смысловую кодирующую ДНК, кодирующую смысловую РНК, специфичную к той же области мРНК гена-мишени. В ДНК-конструкции по изобретению антисмысловая и смысловая кодирующие ДНК функционально связаны с одним или несколькими промоторами, описанными в настоящем документе, которые способны экспрессировать антисмысловую и смысловую РНК, соответственно. "миРНК" предпочтительно означает малые интерферирующие РНК, которые представляют собой короткие двухцепочечные РНК, нетоксичные для клеток млекопитающих (Elbashir et al., 2001, Nature 411: 494-98; Caplen et al., 2001, Proc. Natl. Acad. Sci. USA 98: 9742-47). Длина необязательно ограничена 21-23 нуклеотидами. Для длины миРНК нет конкретных ограничений, при условии ее нетоксичности. Длина "миРНК" может составлять, например, по меньшей мере 15, 18 или 21 нуклеотид и до 25, 30, 35 или 49 нуклеотидов. В альтернативном варианте участок двухцепочечной РНК конечного продукта транскрипции экспрессируемой киРНК, может составлять, например, по меньшей мере 15, 18 или 21 нуклеотид и до 25, 30, 35 или 49 нуклеотидов в длину.
"Антисмысловая РНК" является предпочтительно цепью РНК, имеющей последовательность, комплементарную мРНК гена-мишени, и, как полагают, индуцирует РНКи путем связывания с мРНК гена-мишени. "Смысловая РНК" комплементарна по последовательности антисмысловой РНК и отжигается со своей комплементарной антисмысловой РНК, образуя киРНК. Термин "ген-мишень" в этом контексте предпочтительно относится к гену, чья экспрессия подавляется в результате киРНК, экспрессируемой настоящей системой, и может выбираться произвольно. В качестве гена-мишени предпочтительно выбирать, например, гены, чьи последовательности известны, но чьи функции остаются невыясненными, и гены, чья экспрессия, как полагают, является причиной заболеваний. Геном-мишенью может быть ген, чья геномная последовательность еще не полностью определена, при условии что определена частичная последовательность мРНК гена, имеющая в длину по меньшей мере 15 нуклеотидов или больше, которая способна связывать одну из цепей (антисмысловую РНК-цепь) миРНК. Поэтому, в качестве "гена-мишени" можно выбирать гены, маркеры экспрессированной последовательности (EST) и участки РНК, для которых были определены некоторые последовательности (предпочтительно, по меньшей мере 15 нуклеотидов), даже если их полноразмерные последовательности не определены.
Двухцепочечные участки РНК в миРНК, в которых две цепи РНК спарены, не ограничены полностью спаренными цепями и могут содержать неспаренные участки вследствие наличия несовпадения (соответствующие нуклеотиды не комплементарны), выпячивания (отсутствия соответствующего комплементарного нуклеотида в одной цепи) и т.п. Неспаренные участки могут содержаться при условии, что они не препятствуют образованию киРНК. В контексте настоящего изобретения "выпячивание" предпочтительно включает 1-2 неспаренных нуклеотида, а область двухцепочечной РНК в миРНК, на протяжении которой спарены две цепи РНК, содержит предпочтительно от 1 до 7, более предпочтительно от 1 до 5 выпячиваний. В дополнение, "несовпадение" в контексте настоящего изобретения предпочтительно содержится в области двухцепочечной РНК в киРНК, на протяжении которой спарены две цепи РНК, в количестве предпочтительно от 1 до 7, более предпочтительно от 1 до 5. В предпочтительном несовпадении одним из нуклеотидов является гуанин, а другим является урацил. Такое несовпадение является следствием мутации из С в Т, G в A или их смесей в ДНК, кодирующей смысловую РНК, но конкретно не ограничено ими. Кроме того, в настоящем изобретении область двухцепочечной РНК в миРНК, на протяжении которой спарены две цепи РНК, может содержать как выпячивание, так и несовпадения, число которых в сумме может составлять предпочтительно от 1 до 7, более предпочтительно от 1 до 5. Такие неспаренные участки (несовпадения или выпячивания и т.д.) могут подавлять вышеописанную рекомбинацию между антисмысловой и смысловой кодирующей ДНК и могут обеспечивать стабильность системе экспрессии миРНК, описанной ниже. Кроме того, поскольку трудно секвенировать структуру ДНК "стебель-петля", не содержащую неспаренных участков в области двухцепочечной РНК в миРНК, на протяжении которой спарены две цепи РНК, возможность секвенирования обеспечивают введением несовпадений или выпячиваний, описанных выше. Более того, киРНК, содержащие несовпадения или выпячивания в области спаренной двухцепочечной РНК, обладают преимуществом стабильности в E. coli или животных клетках.
Структура концов миРНК может быть либо "тупой", либо "липкой" (с выступающими концами), при условии что миРНК обеспечивает возможность подавления экспрессии целевого гена вследствие своего эффекта РНКи. Структура липких (выступающих) концов не ограничена только 3'-выступающими концами, также может быть включена 5'-выступающая структура, при условии что структура способна вызывать эффект РНКи. В дополнение, число выступающих нуклеотидов не ограничено опубликованными 2 или 3, а может составлять любое число, при условии что структура с выступающими концами способна индуцировать эффект РНКи. Например, выступающие концы могут состоять из 1-8, предпочтительно 2-4 нуклеотидов. В настоящем изобретении общая длина киРНК, имеющей структуру с липкими концами, выражена как сумма длины спаренного двухцепочечного участка и участка пары, содержащего выступающие одиночные цепи на обеих сторонах. Например, в случае 19-нуклеотидной части двухцепочечной РНК с 4-нуклеотидными выступающими концами на обеих сторонах, общая длина выражена как 23 п.о. Кроме того, поскольку эта выступающая последовательность имеет низкую специфичность к гену-мишени, она необязательно комплементарна (антисмысловая) или идентична (смысловая) последовательности гена-мишени. Кроме того, при условии что миРНК способна сохранять свой эффект подавления экспрессии в отношении гена-мишени, она может содержать низкомолекулярную РНК (которая может быть природной молекулой РНК, такой как тРНК, рРНК или вирусная РНК, или молекула синтетической РНК), например, в выступающем участке на ее одном конце.
В дополнение, структура концов "миРНК" обязательно отщепляется на обоих концах, как описано выше, и может иметь структуру "стебель-петля", в которой концы каждой стороны двухцепочечной РНК соединены линкерной РНК ("кшРНК"). Длина области двухцепочечной РНК (участка стебель-петля) может составлять по меньшей мере 15, 18 или 21 нуклеотид и до 25, 30, 35 или 49 нуклеотидов в длину. В альтернативном варианте длина области двухцепочечной РНК, которая является конечным продуктом транскрипции экспрессируемой киРНК, составляет, например, по меньшей мере 15, 18 или 21 нуклеотид и до 25, 30, 35 или 49 нуклеотидов в длину. Кроме того, нет конкретных ограничений на длину линкера, при условии что его длина не мешает образованию пар в участке стебля. Например, для образования стабильных пар в участке стебля и подавления рекомбинации между ДНК, кодирующей участок, линкерный участок может иметь тРНК-структуру "клеверного листа". Даже в том случае, когда длина линкера мешает образованию пар в участке стебля, возможно, например, сконструировать линкерный участок, чтобы он включал интроны таким образом, чтобы интроны вырезались в ходе процессинга предшественника РНК в зрелую РНК, таким образом, позволяя спаривание в участке стебля. В случае миРНК "стебель-петля", любой конец (голова или хвост) РНК без петельной структуры может иметь низкомолекулярную РНК. Как описано выше, этой низкомолекулярной РНК может быть природная молекула РНК, такая как тРНК, рРНК или вирусная РНК, или молекула синтетической РНК.
Для экспрессии антисмысловых и смысловых РНК, соответственно, с антисмысловой и смысловой кодирующей ДНК, ДНК-конструкция по настоящему изобретению содержит промотор, описанный выше. Число и расположение промоторов в конструкции могут в принципе быть произвольно выбраны, при условии что они способны экспрессировать антисмысловую и смысловую кодирующие ДНК. В качестве простого примера ДНК-конструкции по изобретению можно создать тандемную систему экспрессии, в которой промотор расположен выше как антисмысловой, так и смысловой кодирующих ДНК. Эта тандемная система экспрессии способна продуцировать миРНК, имеющие вышеуказанные отщепляемые структуры на обеих концах. В системе экспрессии миРНК "стебель-петля" (стеблевой экспрессионной системе) антисмысловая и смысловая кодирующие ДНК расположены в противоположном направлении, и эти ДНК соединены посредством линкерной ДНК для создания транскрипционной единицы. Промотор связан с одной стороной этой единицы для конструирования системы экспрессии РНК "стебель-петля". В настоящем изобретении нет конкретного ограничения на длину и последовательность линкерной ДНК, которая может иметь любую длину и последовательность, при условии что ее последовательность не является последовательностью терминации, и ее длина и последовательность не мешают образованию пар в участке стебля в ходе получения зрелой РНК, как описано выше. Например, в качестве линкерной ДНК можно использовать ДНК, кодирующую вышеуказанную тРНК и т.п.
В обоих случаях систем экспрессии - тандемной и стебель-петля - 5'-конец может иметь последовательность, способную стимулировать транскрипцию с промотора. Более конкретно, в случае тандемной киРНК, эффективность продукции миРНК можно повысить путем добавления последовательности, способной стимулировать транскрипцию с промоторов, на 5'-концы антисмысловой и смысловой кодирующих ДНК. В случае миРНК стебель-петля, такую последовательность можно добавить на 5'-конец вышеописанной единицы. Транскрипт с такой последовательности можно использовать прикрепленным к киРНК, при условии что это не препятствует подавлению экспрессии гена-мишени с помощью миРНК. Если это состояние препятствует подавлению экспрессии гена, предпочтительно проводить отщепление транскрипта, используя средства отщепления (например, рибозим, известный в данной области). Опытному специалисту в данной области будет ясно, что антисмысловая и смысловая РНК могут экспрессироваться в одном векторе или в разных векторах. Для того чтобы избежать добавления избыточных последовательностей ниже смысловой и антисмысловой РНК, предпочтительно поместить терминатор транскрипции на 3'-концы соответствующих цепей (цепей, кодирующих антисмысловую и смысловую РНК). Терминатором может быть последовательность из четырех или большего числа последовательных адениновых (А) нуклеотидов.
В этом документе и его формуле изобретения глагол "содержать/включать в себя" и их спряжения используют в неограничивающем смысле для обозначения того, что элементы, следующие за ними, включены, но элементы, специально не упомянутые, не исключены. В дополнение, глагол "состоять из" можно заменить на "состоять из по существу", означающий, что продукт, указанный в настоящем документе, может содержать дополнительный(-ые) компонент(-ы) относительно конкретно указанных, причем указанный(-ые) дополнительный(-ые) компонент(-ы) не изменяют уникальные черты изобретения. В дополнение, способ, указанный в настоящем документе, может включать в себя дополнительную(-ые) стадию(-и) относительно конкретно указанных, причем указанная(-ые) дополнительная(-ые) стадия(-и) не изменяют уникальные черты изобретения. В дополнение, упоминание элемента в единственном числе не исключает возможности присутствия более одного элемента, кроме тех случаев, когда контекст ясно требует одного и только одного элемента. Единственное число поэтому обычно означает "по меньшей мере" один. Наречие "примерно" или "приблизительно" при использовании в связи с численной величиной (примерно 10) предпочтительно означает, что величина может составлять данную величину плюс/минус 1% от величины.
Все патенты и литературные ссылки, цитируемые в настоящем описании, таким образом полностью включены в него посредством ссылки.
Нижеследующие примеры приведены только для иллюстративных целей, и не предназначены для ограничения объема настоящего изобретения никаким образом.
ОПИСАНИЕ ФИГУР
Фиг.1. Требования органоидов печени к ростовым факторам. (А) DIC-изображения органоидов печени, поддерживаемых на EGF (E), R-спондине 1 (R), Noggin (N), Wnt3A-кондиционной среде (W) или их комбинации, которые дополнены FGF10, HGF и никотинамидом. (B) Число органоидов подсчитывали еженедельно, и органоиды пассировали по необходимости. Результаты показаны как среднее±SEM для трех независимых экспериментов. (С) Анализ экспрессии генов Lgr5, кератина 7 (K7) и альбумина (Alb) с помощью ОТ-ПЦР. (D) Выделенные желчные протоки, вырастающие в органоиды. Дифференциальные интерференционно-контрастные изображения для соответствующих дней после посева. 10-кратное увеличение (дни 0, 1, 3 и 5). 4-кратное увеличение в 15-й день. Культуры пассировали каждые 4-7 дней с помощью механической диссоциации. Культуры растили по меньшей мере 8 месяцев.
Фиг.2. Морфология органоидов печени. (А) Верхние панели: парафиновый срез печени мыши, не котором показаны разные домены (РТ = портальная триада, CV = центральная вена). Нижние панели: парафиновый сред органоида печени, на котором показаны разные домены - b (однослойный эпителий) и h (многослойный эпителий) (B) Правая панель: окрашивание Е-кадхерина (Ecad) в органоидах печени. Идентифицировано два разных домена. Домен b, образованный однослойным эпителием, который напоминает структуры желчных протоков в печени. Этот домен желчных протоков образован высоко поляризованными клетками, который показывает положительное окрашивание на панцитокератин (PCK) (нижняя панель). На левых панелях показано присутствие второго домена в органоидах печени. Этот домен h образован многослойным эпителием с неполяризованными клетками. Клетки упорядочены вокруг центрального просвета и экспрессируют маркер гепатоцитов - Alb. Увеличение - 10×.
Фиг.3. Wnt-сигнальный путь в культурах печени. Экспрессию Lac Z детектировали в культурах, полученных из мышей Lgr5-LacZ или Axin2-LacZ. Никакого положительного окрашивания не детектировали в культурах печени, полученных из мышей В16. Увеличение - 4×, врезка - 20×.
Фиг.4. Экспрессия маркеров дифференцировки печени. (А) Иммуногистохимический и иммунофлуоресцентный анализ экспрессии маркера холангиоцитов, кератина 7 (K7) и маркеров гепатоцитов, кератина 8 (К8) и альбумина (Alb). (B) Анализ экспрессии генов маркеров гепатоцитов: альбумина (Alb), транстиретина (Ttr), глутаминсинтетазы (Glul), глюкозо-6-фосфатазы (G6P) и изоформы 3А11 цитохрома p450 (CYP3A11); и маркеров холангиоцитов: кератина 7 (K7) и кератина 19 (К19).
Фиг.5. Одноклеточные культуры клеток печени. (А) Диаграмма проточной цитометрии, на которой указана область отсортированных клеток. (В) Одиночная клетка, вырастающая в органоиды за указанные временные интервалы. Увеличение - 40× (день 1-3), 10× (день 16), 4× (день 21 и далее). (С и D) Репрезентативное изображение эффективности образования колоний из сортированных по Lgr5GFP одиночных клеток. 100 клеток высевали в трех повторах, и подсчитывали колонии через 10 дней.
Фиг.6. Анализ культур печени методом микроматриц. Анализ профиля экспрессии генов зрелой ткани печени и культур органоидов печени, выращиваемых в течение 1 месяца в среде ER или среде ER, дополненной Noggin (ENR) или Noggin и Wnt (ENRW). Этот генетический профиль сравнивали между различными образцами и генетическим профилем бурой жировой ткани (ВАТ), белой жировой ткани (WAT), мышечной ткани и печени новорожденного. (А) Теплокарта, показывающая, что культуры имеют профиль экспрессии, аналогичный зрелой печени, но отличающийся от профиля непеченочных тканей, таких как мышцы, BAT и WAT. (B) Список маркеров гепатоцитов и маркеров холангиоцитов при различных условиях.
Фиг.7: Культура органоидов печени мыши показывает стабильное кариотипирование после продолжительного культивирования.
A - DIC-изображения органоидов печени, культивируемых в EGF (E) и R-спондине 1 (R), дополненных FGF10, HGF и никотинамидом (левая фигура, ER), или культивируемых в той же комбинации, дополненной Noggin (N) и Wnt3A-кондиционной средой (W) (правая фигура, ENRW) в течение 24 месяцев.
B - Анализ кариотипа органоидов печени мыши через 8 месяцев в культуре. Были найдены нормальное число хромосом (n=40, левая панель) и полиплоидия, типичный признак гепатоцитов (n=80, правая панель).
Фиг.8: дополнительные факторы FGF10, HGF и никотинамид; эффект на рост и дифференцировку.
А - Диаграмма, указывающая гены, дифференциально экспрессирующиеся в течение 3 стадий развития печени, от гепатобласта до зрелого гепатоцита.
В - Схема, показывающая используемый протокол. Культуры высевали в среде для размножения EM2 (ERFHNic: EGF (E) и R-спондин 1 (R), дополненные FGF10, HGF и никотинамидом; ERFHNic указана как "ER" на фиг.8В) за 2 дня до эксперимента. Через два дня культуральную среду заменяли либо только на EGF (E), либо на EGF, дополненный R-спондином 1 (ER) с или без дополнительных добавок, выбранных из FGF10 (F) или HGF (H), или никотинамида (Nic), или их комбинации в концентрациях, указанных в тексте. Через пять дней культуры пересевали в соотношении 1:4 для каждого условия культивирования. В этих условиях культуры пересевали каждые 7 дней в течение всего 10 недель.
С - Первый день после первого пересева в каждом из тестируемых культуральных условий. Результаты показывают, что EGF и R-спондин 1, объединенные с FGF10, или HGF, или никотинамидом, или их комбинацией, необходимы для достижения по меньшей мере 1 пассажа.
D - После продолжительной культуры, комбинация ER, дополненная FNic, или ER, дополненная FHNic, обе в результате давали большое число пассажей. После 10-го пассажа скорость роста была лучше для условий культивирования, включающих 3 дополнительных фактора - ERFHNic (фиг.15A, B).
Е - ОТ-ПЦР-анализ, показывающий экспрессию различных маркеров гепатоцитов (CYP3A11, Alb, FAH), маркера холангиоцитов (K19) и маркера стволовых клеток (IGR5) через 5 дней после удаления некоторых факторов (исходной точкой была среда ERFHNic). Следует отметить, что только культуральные условия EF показали экспрессию всех тестируемых маркеров гепатоцитов. В качестве гена "домашнего хозяйства" для нормализации экспрессии генов использовали HPRT.
Фиг.9: Таблица, показывающая количественную оценку различных гепатоцит- и холангиоцит-специфичных транскрипционных факторов в клетках для трех различных условий культивирования клеток печени и в ткани зрелой печени. Также показана экспрессия ключевых компонентов Notch- и TGF-β-сигнальных путей. E=EFHNic, ER=ERFHNic, ENRW=ENRWFHNic.
Фиг.10: Протокол дифференцировки
А - Схема, показывающая использованный протокол. За 2 дня перед экспериментом культуры высевали в среде для размножения (ERFHNic: EGF (E) и R-спондин 1 (R), дополненные FGF10, HGF и никотинамидом; ERFHNic указана как "ER" на фиг.10А). Через два дня культуральную среду заменяли на EGF (E), дополненный либо A8301 (A), либо DAPT (D), либо FGF 10 (F), либо HGF (H), либо никотинамидом (Nic), либо R-спондином 1 (R), либо Wnt3A, либо Noggin (N), либо их комбинацией в показанных концентрациях. РНК выделяли в нескольких временных точках. В качестве положительного контроля (+) брали ткань печени мыши, в то время как в качестве отрицательного контроля использовали воду (-).
В - ОТ-ПЦР-анализ, показывающий экспрессию маркеров гепатоцитов, CYP3A11, Alb, FAH, tbx3, TAT и Gck, через 7 дней после введения условий для дифференцировки. Следует отметить, что только культивирование в EADF давало экспрессию всех тестируемых маркеров гепатоцитов. В качестве гена "домашнего хозяйства" для нормализации экспрессии генов использовали HPRT.
С - Временной анализ экспрессии после введения условий дифференцировки. Через 2, 5 и 8 дней после начала дифференцировки, экспрессию маркеров гепатоцитов, CYP3A11, Alb, FAH, и маркера холангиоцитов, К19, анализировали с помощью ОТ-ПЦР. Следует отметить, что экспрессия печеночных маркеров CYP3A11 и FAH начинается на 5-й и достигает пикового значения на 8-й день после начала дифференцировки. В качестве гена "домашнего хозяйства" для нормализации экспрессии генов использовали HPRT. A; A8301, D; DAPT, F; FGF10, H; HGF, De; дексаметазон.
D - Титрационный эксперимент, показывающий экспрессию гепатоцитарных маркеров, CYP3A11, Alb, FAH, tbx3, TAT, G6P и Gck, через 7 дней после различных концентраций дифференцирующих соединений А и D. В качестве гена "домашнего хозяйства" для нормализации экспрессии генов использовали HPRT.
Е - Иммунофлуоресцентное окрашивание печеночных маркеров, К19, альбумина и маркера поверхности гепатоцитов. Для окраски ядер использовали краситель Хехст.
F - Xgal-окрашивание органоидов печени, полученных из мышей Albcreert2LacZ. Положительные по альбумину клетки (указаны стрелками) детектировали после стимулированной EADF дифференцировки в индуцированных тамоксифеном культурах, полученных из Albcreert2LacZ.
Фиг.11: Культуры клеток печени человека при культивировании в ERFHNic
Фиг.12: Ответ печени на стимуляцию Wnt-сигнальным путем в физиологических условиях или в течение регенерации после повреждения
А: Инъекция лиганда Lgr5, R-спондин 1, мышам Axin2 LacZ показывает, что клетки печени отвечают на стимуляцию Wnt (стрелки указывают на X-gal-положительные клетки). При этом не наблюдалось экспрессии Lgr5, так что авторы изобретения предположили, что для инициации ответа использовался Lgr4.
B: Инъекция CCL4 мышам Axin2 LAcZ показала, что в течение регенерационного ответа активируется Wnt-сигнальный путь.
Фиг.13: повышение экспрессии Lgr5 после повреждения печени - модели регенерации
Взрослым мышам Lgr5-LacZ KI вводили с помощью инъекций 0,8 мг/кг гепатотоксичного соединения CCL4. На рисунках показано, что в печени мышей, не получавших инъекции (нет повреждений), Wnt-сигнальный путь активен только в клетках, выстилающих протоки. После повреждения в результате действия CCl4 в других клетках, а не только в выстилающих протоки, активировался Wnt-сигнальный путь.
А - Кинетический эксперимент, показывающий повышение экспрессии Lgr5 в поврежденной в результате действия CCL4 печени (стрелки, показывающие Х-gal-положительные клетки). В случае контрольных мышей дикого типа, которым инъецировали CCL4, и мышей Lgr5LacZ KI, которым инъецировали плацебо, никакого окрашивания не наблюдалось (правая панель).
В - Окрашивание клеток на Lgr5 совместно с маркерами клеток печени.
Фиг.14: Окрашивание выделенных протоков на К19
Выделение протоков из печени мышей Lgr5LacZ. Окрашивание на К19 подтверждает, что выделенные и высеянные структуры, несомненно, являются внутрипеченочными протоками.
Фиг.15: Требования в отношении ростовых факторов
3 дополнительных фактора (FGF10, HGF и никотинамид) необходимы для продолжительного поддержания культур клеток печени. После длительного культивирования комбинация ER, включающая FNic ($) или ERFHNic ($$), обе дают в результате большое число пассажей. После 10-го пассажа скорость роста выше в случае культуральных условий, включающих 3 дополнительных фактора; ERFHNic (см. фиг.16B).
Фиг.16: Профиль экспрессии генов органоидов печени мыши в дифференцирующих условиях похож на профиль экспрессии генов зрелой печени и печени новорожденной мыши
А - кластеры генов, показывающие гены, аналогично экспрессирующиеся (а) или аналогично "выключенные" (b), для дифференцирующих условий EADF и зрелой печени или печени новорожденного.
В - кластеры генов, показывающие дифференциально экспрессирующиеся гены для органоидов печени и зрелой печени или печени новорожденного (а) и аналогично экспрессирующихся для EADF и печени новорожденного (b).
С - Необработанные данные для анализа методом микроматриц, в которых сравнивается уровень экспрессии отдельных маркеров желчных протоков, транскрипционных факторов, необходимых для экспрессии Ngn3, и эндокринных маркеров в зрелой печени, зрелой поджелудочной железе, органоидах поджелудочной железы и органоидах печени в среде для размножения.
Фиг.17: Трансплантация клеток мышиной модели болезни печени
Органоиды трансплантировали мышиной модели: взрослым FGR-мышам (FAH-/-RAG-/-IL2R-/-). В качестве контроля мышам трансплантируют гепатоциты.
А - К19-положительные клетки (левая верхняя панель) и Fah-положительные клетки (средняя панель), полученные из органоидов печени, трансплантированных FAH-нокаутным мышам. Контролем являлись трансплантированные гепатоциты (верхняя правая панель). На нижних диаграммах проточной цитометрии показано, что % клеток, положительно окрашенных на маркер гепатоцитов, был выше в группе FAH-положительных пересаженных гепатоцитов.
C и D - Анализ методом проточной цитометрии трансплантированных клеток. (С) Клон 1, полученный из мыши Lgr5-GFP, и (D) клон 2, полученный из мыши Lgr5-lacZ. Окрашивание на маркер поверхности гепатоцитов показывает положительную субпопуляцию, которая включает в себя крупные клетки и сильно гранулированные клетки, то есть клетки, которые представляют фенотип зрелых гепатоцитов.
Е - Схема трансплантации.
Фиг.18: Гены, характеристические для печени мыши
В таблице показаны (а) маркеры, экспрессирующиеся в стволовых клетках печени мыши; (b) маркеры, не экспрессирующиеся в стволовых клетках печени мыши; (с) маркеры гепатоцитов и холангиоцитов, экспрессирующиеся в стволовых клетках мыши, характеристические для органоидов печени мыши в среде для размножения; (d) маркеры гепатоцитов и холангиоцитов, не экспрессирующиеся в стволовых клетках мыши, характеристические для органоидов печени мыши в среде для размножения; (е) репрограммирующие гены, экспрессирующиеся в органоидах печени мыши; (f) репрограммирующие гены, не экспрессирующиеся в органоидах печени мыши. Результаты были получены анализом экспрессии генов методом микроматриц с использованием Универсальной мышиной референсной РНК (Strategene, каталожный № 740100) в качестве референсной РНК. Если абсолютные значения при детекции составляли меньше 100, то ген считался недетектируемым.
Фиг.19: Гены, характеристические для печени мыши
В таблице показаны результаты анализа методом микроматриц органоидов человека. Слева направо, результаты показаны для (а) среды для размножения ЕМ1, (b) среды для размножения ЕМ2, (с) дифференцирующей среды, (d) зрелой печени.
Числа (log2) в левых четырех колонках представляют результат сравнения между образцом экспериментальной и референсной (покупной) РНК, которую использовали во всех анализах методом микроматриц. Показана относительная экспрессия мРНК в каждом образце по сравнению с РНК, присутствующей в референсном образце. Используемой референсной РНК была Универсальная референсная РНК человека (Stratagene, каталожный № 740000). Таким образом, отрицательные числа в этих колонках не связаны с реальным уровнем экспрессии, и они означают только то, что уровень РНК ниже уровня референсной РНК. 4 колонки справа представляют собой абсолютные значения. Если значения были ниже 100, то они считались недетектируемыми.
ПРИМЕРЫ
Пример 1 - Среда для размножения для роста и размножения органоидов печени
После выделения желчные протоки (см. фиг.1) ресуспендировали в матригеле и культивировали в различных культуральных условиях. Комбинация EGF (50 нг/мл) и R-спондин 1 (1 мкг/мл), дополненная FGF10 (100 нг/мл), HGF (25-50 нг/мл) и никотинамидом (1-10 мМ) (ERFHNic), была необходима для длительного поддержания культур, указывая на то, что Wnt- и EGF-сигнальные пути точно необходимы для поддержания пролиферации зрелых предшественников клеток печени in vitro. Добавление Noggin (100 нг/мл) и Wnt-кондиционной среды (50%) также обеспечивало продолжительное поддержание культур (см. фиг.1А и 1В). В этих условиях, обеспечивающих длительное поддержание культур, экспрессия Lgr5, а также маркеров гепатоцитов (альбумина) и маркеров холангиоцитов (К7), детектировались с помощью ОТ-ПЦР (см. фиг.1С). В этих условиях органоиды печени перепассировали каждую неделю с помощью механической или ферментативной диссоциации в разведении 1:8 и выращивали в течение многих месяцев (фиг.1D).
Авторы изобретения анализировали экспрессию генов-мишеней Wnt, Axin2 и Lgr5 в культуре. В культурах печени мышей, как Axin2LacZ, так и Lgr5-LacZ, было выявлено присутствие Axin2- и Lgr5-положительных клеток в органоидах печени через 1 месяц после посева, таким образом, подтверждая то, что Wnt-сигнальный путь активен и требуется для роста культуры (фиг.3). Культуры клеток печени также экспрессируют маркеры гепатоцитов (например, альбумин, транстиретин, глутаминсинтетазу) и маркеры холангиоцитов (кератин 7 и 19) (см. фиг.4).
При сортировке одиночных клеток Lgr5 из мышей Lgr5LacZ или Lgr5GFP, одиночные колонии вырастали в органоиды. Эти культуры также экспрессировали маркеры холангиоцитов и гепатоцитов и поддерживались и регулярно пересевались в соотношении 1:6-1:8 на протяжении более 4 месяцев (см. фиг.5А и В). Следует отметить, что только полученные из Lgr5-положительных клеток культуры вырастали в органоиды (фиг.5С и D). Эти данные указывают на то, что Lgr5-положительные клетки являются предшественниками этих культур и способны давать в качестве потомства 2 различных типа клеток печени.
Установив, что органоиды печени получены из Lgr5-положительных клеток авторы изобретения поставили целью определить их индивидуальные характеристические гены по сравнению с характеристическими генами зрелой печени. РНК выделяли из зрелой печени и из органоидов печени, выращиваемых в среде ER или ENRW, дополненной FGF10, никотинамидом и фактором роста гепатоцитов. Генетическую характеристику зрелой печени и клеток печени в двух условиях культивирования впоследствии получали, сравнивая профили экспрессии генов относительно экспрессии Универсальной референсной РНК. Использование одной референсной РНК для гибридизации во всех образцах позволило авторам сравнить друг с другом 3 независимых образца (зрелую печень, и культуры в ER и ENRW). Анализ методом теплокарт выявил, что профиль экспрессии генов в двух условиях культивирования очень похож на профиль экспрессии генов в зрелой ткани печени, в то же время отличаясь от профиля экспрессии генов в мышечной и жировой ткани (см. фиг.6). В аналогичных профилях экспрессии генов для зрелой печени и культуры печени были детектированы специфичные для печени гены, такие как HNF1a, HNF1b, HNF4, Alb, Glul, Met, G6P, Fahd1, Fahd2a, CYP4B1, K7 и K19. Анализ методом теплокарт выявил, что оба условия культивирования обеспечивали аналогичный профиль экспрессии генов между собой и в сравнении с образом, полученным из зрелой печени. Однако при подробном анализе данных авторы отметили, что условия культивирования без Wnt и Noggin давали более "дифференцированный" профиль экспрессии относительно условий, включающих оба ростовых фактора. Это согласуется с данными, показанными на фиг.1С, где дифференцировка в гепатоциты (выраженная экспрессией альбумина) практически отсутствовала в присутствии Wnt. Этот результат указывает на то, что Wnt способствует самообновлению культуры в ущерб дифференцировке.
Также при двух условиях культивирования, а также в зрелой печени, можно детектировать гены, неспецифичные для зрелой печени, такие как AFP, и неспецифичные для печени транскрипционные факторы, такие как Pdx1 или NeuroD.
Примечательно то, что в обоих условиях культивирования, но не в зрелой печени, маркер стволовых клеток, Lgr5, был одним из наиболее высокоэкспрессирующихся генов в характеристическом профиле экспрессии генов в культуре клеток печени. Кроме того, клеточные маркеры популяций-предшественников в тонком кишечнике и желудке, такие как Cd44 и Sox9 (Barker & Huch et al Cell stem cell 2010), экспрессировались на высоком уровне в двух условиях культивирования, но не в зрелой печени, снова указывая на способность культур клеток печени к обновлению, а также на состояние покоя клеток здоровой зрелой печени.
Дополнительно, помимо Lgr5, экспрессия многих генов-мишеней Wnt была сильно повышена в культурах клеток печени относительно зрелой печени, включая, среди прочих, MMP7, Sp5 и Tnfrs19, что обеспечивает веское доказательство необходимости активного и сильного канонического Wnt-сигнального пути для сохранения способности культур к самообновлению.
Пример 2 - Улучшенная дифференцирующая среда
При культивировании в условиях ER или ENRW культуры клеток печени обновляются и могут поддерживаться и размножаться еженедельно до 1 года (фиг.7А). Кариотипический анализ через 1 год не выявил свидетельств хромосомных аберраций. Более 66% проанализированных клеток имели нормальное число хромосом, а 13% из них были полиплоидными, что характерно для гепатоцитов (фиг.7В).
Для длительного культивирования была предпочтительна комбинация EGF (50 нг/мл) и R-спондина 1 (1 мкг/мл), дополненная FGF10 (100 нг/мл), HGF (25-50 нг/мл) и никотинамидом (1-10 мМ). В этих условиях авторы изобретения получили долгоживущие культуры клеток, которые экспрессируют маркеры клеток желчных протоков и некоторые маркеры гепатобластов или незрелых гепатоцитов (Glul, альбумин). Однако число клеток, положительных по этим маркерам гепатоцитов, было очень низким. В этих условиях культивирования не детектировались маркеры зрелых гепатоцитов (например, цитохромы р450). Эти результаты позволяют предположить, что описанные здесь условия культивирования, способствуют размножению предшественников клеток печени, способных генерировать гепатоцит-подобные клетки (хотя и в небольших количествах), но не полностью зрелые гепатоциты (фиг.8А).
Для усиления гепатоцитарной природы культур и получения зрелых гепатоцитов in vitro, авторы изобретения сначала определили, оказывают ли три дополнительных фактора (FGF10, HGF и никотинамид), добавленные к EGF и R-спондину 1, либо положительный, либо отрицательный эффект на экспрессию гепатоцит-специфичных генов, а также на самообновление культуры. Авторы получили культуры органоидов печени и культивировали их либо с EGF, либо с EGF и R-спондином 1, плюс FGF10 или HGF, или никотинамид, или их комбинация, и авторы пересевали культуры один раз в неделю на протяжении всего 10 недель. В каждой временной точке авторы также анализировали экспрессию нескольких маркеров зрелых гепатоцитов (FAH, CYP3A11) и маркеров гепатобластов (альбумин) (фиг.8В).
В согласии с данными на фиг.1 (см. пример 1) авторы наблюдали, что R-спондин 1 и никотинамид, объединенные с FGF10, необходимы для роста и самообновления культур клеток печени (фиг.8C и D). R-спондин 1 и никотинамид, оба ингибируют экспрессию маркера зрелых гепатоцитов CYP3A11 и еще стимулируют экспрессию маркера гепатобластов - альбумина. Добавление либо FGF10, либо HGF, к среде, содержащей только EGF (без R-спондина 1 и без никотинамида), способствовало экспрессии маркера зрелых гепатоцитов CYP3A11, хотя и на очень низком уровне (фиг.8Е). Для того чтобы идентифицировать дополнительные соединения, которые могли бы способствовать дифференцировке в гепатоциты, авторы использовали два различных подхода, оба основанные на базовых условиях культивирования: EGF + HGF и/или FGF10.
Первый подход включал тестирование ряда соединений в дополнение к условиям культивирования, содержащим EGF + FGF10 или HGF. Полный список проанализированных соединений приведен в таблице 2.
Второй подход основан на информации, полученной из опубликованных исследований в области биологии развития относительно экспрессии транскрипционных факторов, необходимых для дифференцировки билиарных клеток и гепатоцитов in vivo. Сравнительный анализ экспрессии транскрипционных факторов в органоидах в культуральных условиях E, ER или ENRW, дополненных FGF10, HGF и никотинамидом, показан на фиг.8. Присутствовали все транскрипционные факторы, требуемые для гепатоцитов, помимо tbx3 и prox1. Однако авторы также отметили, что экспрессия специфичных билиарных транскрипционных факторов была значительно повышена в культурах, содержащих R-спондин 1 (R), указывая на то, что экспрессия генов в культивируемых клетках была сдвинута в сторону дифференцировки в билиарные клетки.
Notch- и TGF-β-сигнальные пути вовлечены в дифференцировку по пути билиарных клеток in vivo. В действительности, делеция Rbpj (необходим для достижения действующей передачи сигнала от Notch) приводит в результате к аномальному тубулогенезу (Zong Y. Development 2009), а добавление TGF-β к эксплантатам печени способствует дифференцировке в билиарные клетки in vitro (Clotman F. Genes and Development 2005). Поскольку Notch- и TGF-β-сигнальные пути сильно активированы в культурах клеток печени (фиг.9), то авторы решили, что ингибирование дифференцировки в клетки желчных протоков может запустить дифференцировку клеток в клетки с более гепатоцитарным фенотипом. A8301 был выбран в качестве ингибитора рецепторов TGF-β - ALK5, 4 и 7, а DAPT - в качестве ингибитора гаммы-секретазы, активной протеазы, необходимой для активации Notch-сигнального пути. Авторы сначала культивировали клетки 2 дня в условиях для размножения (среде ER), а на 2-й день (фиг.10А) авторы начинали дифференцировку путем добавления комбинации различных соединений. Среду меняли через день, и экспрессию дифференцированных маркеров анализировали через 8-9 дней. В качестве отрицательного контроля использовали культивирование в ER и ENRW.
Комбинация EGF+FGF10 с DAPT и A8301 дала в результате необыкновенно большое увеличение экспрессии анализируемых гепатоцитарных маркеров (CYP3A11, TAT, альбумин) (фиг.10B). Эффект детектировался уже на 5-й день, а пик его приходился на 8-9-й дни (фиг.10С). Максимальная концентрационная эффективность достигалась, соответственно, при 10 мкМ (DAPT) и 50 нМ (A8301) (фиг.10D). Добавление дексаметазона (известной молекулы дифференцировки гепатоцитов) не приводило к какому-либо увеличению экспрессии генов. Комбинация EGF, FGF10, A8301 и DAPT не только усиливала экспрессию генов, но также увеличивала число гепатоцит-подобных клеток, оцениваемое иммунофлуоресцентным окрашиванием гепатоцитарных маркеров, альбумина и 2F8, и Xgal-окрашиванием органоидов, полученных из мышей AlbCreLacZ (фиг.10E и F). Поэтому, авторы заключили, что вышеуказанный протокол дифференцировки способствует получению гепатоцит-подобных клеток in vitro из культур стволовых клеток печени.
Пример 3 - Органоиды печени человека
Используя эти условия для размножения (среды ERFHNic и ENRWFHNic), авторы изобретения также были способны размножать культуры клеток человека, полученных из желчных протоков (фиг.11), с добавлением 500 мкМ ингибитора TGF-β (A83-01) в среду для размножения.
МАТЕРИАЛЫ И МЕТОДЫ (Для примеров 1-3)
Культивирование клеток печени - выделение желчных протоков
Выделенную ткань зрелой печени промывали в холодной среде Advanced-DMEM/F12 (Invitrogen), и затем ткань измельчали на кусочки размером около 5 мм и дополнительно промывали холодным буфером для диссоциации (коллагеназа, диспаза, FBS в среде DMEM). Фрагменты ткани инкубировали 2 ч с буфером для диссоциации при температуре 37°C. Затем, фрагменты ткани интенсивно ресуспендировали в 10 мл холодного (4-10°C) буфера для выделения, используя пипетку на 10 мл. Первый супернатант, содержащий мертвые клетки, удаляли, и осадок ресуспендировали в 10-15 мл буфера для диссоциации. После дополнительного интенсивного ресуспендирования фрагментов ткани супернатант обогащен желчными протоками. Эту процедуру повторяют до тех пор, пока не будет получено достаточное количество желчных протоков.
Выделенные желчные протоки осаждали и смешивали с 50 мкл матригеля (BD Bioscience), высевали на 24-луночные культуральные планшеты и инкубировали в течение 5-10 мин при 37°C до полной полимеризации матригеля. После полимеризации сверху наносили 500 мкл культуральной среды.
Состав среды: Advanced-DMEM/F12, дополненная B27, N2, 200 нг/мл N-ацетилцистеина, 50 нг/мл EGF, 1 мкг/мл R-спондина 1, 10 нМ гастрином, 100 нг/мл FGF10, 10 мМ никотинамидом и 50 нг/мл HGF, а также 50% Wnt-кондиционной среды.
Среду полностью меняли каждые 2 дня. Через 1 неделю исключали Wnt-кондиционную среду, и образовавшиеся органоиды удаляли из матригеля, используя пипетку на 1000 мкл, и механически диссоциировали на маленькие фрагменты и переносили в свежий матригель. Пассирование выполняли в соотношении 1:4 один или два раза в неделю. В этих условиях культуры содержали по меньшей мере 6 месяцев.
Реагенты
Фактор роста гепатоцитов человека (HGF) был приобретен в Peprotech, EGF - в Invitrogen, R-спондин - в Nuvelo, Noggin - в Peprotech, FGF10 - в Peprotech, гастрин - в Sigma Aldrich, никотинамид - в Sigma.
Анализ методом микроматриц
Для анализа экспрессии культур клеток печени, отобранных по положительной экспрессии Lgr5, выделяли РНК, используя набор Qiagen RNeasy из зрелой печени или из культур клеток печени, культивируемых в среде без Wnt-кондиционной среды и Noggin (ER) или с Wnt-кондиционной средой и Noggin (ENRW). 150 нг тотальной РНК метили с использованием набора Low RNA Input Linear Amp (Agilent Technologies, Palo Alto, CA). Универсальную мышиную референсную РНК (Agilent) дифференциально метили и гибридизовали либо с тканью зрелой печени или с культурами в среде ER или ENRW. Использовали 4X44K Agilent Whole Mouse Genome dual colour Microarrays (G4122F). Процедуры мечения, гибридизации и отмывок проводили согласно инструкциям Agilent.
Пример 4 - Экспрессия Lgr5 повышается после повреждения печени
В печени Wnt-сигнальный путь активен в области центральной вены. Авторы изобретения недавно наблюдали, что Wnt-сигнальный путь играет ключевую роль в метаболизме печени (Boj et al., персональное сообщение). В клетках печеночных протоков Wnt-сигнальный путь активируется после повреждения печени (Hu et al 2007, Gastroenterology, 133(5): 1579-91). Аналогичным образом, используя аллель Axin2-LacZ, который представляет достоверный общий репортер для Wnt-сигнального пути, авторы также наблюдали повышение активности Wnt-сигнального пути во всей паренхиме печени после инъекции агониста Wnt, Rspo1 (см. фиг.12A) или после повреждения печени гепатотоксическим соединением, четыреххлористым углеродом (CCl4) (см. фиг.12B).
Ген-мишень Wnt, Lgr5, экспрессируется в стволовых клетках в нескольких активно самообновляющихся тканях, но ранее не было опубликовано, что он экспрессируется при повреждении. На ранее описанных авторами мышах со встроенной в геном (knockin) конструкцией Lgr5-LacZ (Barker et al, 2007, Nature 449 (7165): 1003-7) показано, что Lgr5 по существу не детектируется в здоровой печени, хотя остаточная экспрессия мРНК детектируется количественной ПЦР. После инъекции CCl4 мышам со встроенной в геном конструкцией Lgr5-LacZ (см. Barker et al., 2007, выше, относительно мышей LacZ, и Furuyama K et al., Nat Genetics, 43, 34-41, 2001 относительно способа повреждения печени с использованием CCl4), авторы наблюдали явно выраженную экспрессию репортера в новообразованных зачаточных структурах в печени (см. фиг.13А) с пиком в день 6.5 после повреждения и поддерживающуюся до 9-го дня с явным падением после полной регенерации печени на 13-й день после повреждения (см. фиг.13А, верхняя правая панель). Никакой экспрессии репортерного гена не детектировали у однопометных животных дикого типа, к которым применяли такой же протокол повреждения печени (см. фиг.13А, нижняя правая панель).
Появление экспрессии Lgr5 в местах активной регенерации позволяло предположить, что Lgr5 может возвещать об активации de novo в результате действия Wnt регенеративных стволовых клеток/предшественников при повреждении. Более того, авторы изобретения обнаружили, что появляющиеся de novo Lgr5-положительные клетки не экспрессируют маркеры зрелых клеток печени (К19 или FAH) или звездчатых клеток (SMA), а вместо этого они положительны по недавно описанному маркеру предшественников клеток печени, Sox9 (фиг.13B). Это означает, что Lgr5+ клетки, которые являются исходной точкой получения органоидов in vitro, можно получать из фрагментов печени, индуцируя повреждение печени или стимулируя Wnt-сигнальный путь с помощью R-спондина. Индукцию экспрессии Lgr5 в клетках печени в результате повреждения или действия R-спондина можно проводить in vivo до получения клеток, ex vivo в выделенной печени, или in vitro в фрагменте печени или популяции клеток печени.
Пример 5 - Долговременное размножение культуры органоидов печени
В примере 1 было найдено, что комбинация EGF (50 нг/мл) и R-спондина 1 (1 мкг/мл), дополненная FGF10 (100 нг/мл), HGF (25-50 нг/мл) и никотинамидом (1-10 мМ), была предпочтительна для длительного поддержания культур. Авторы на сегодняшний момент имеют доказательство того, что три дополнительных фактора (FGF10, HGF и никотинамид), добавленные к EGF и R-спондин 1, все необходимы для размножения культур в течение более чем 3 месяцев. Для оценки этого авторы выделили желчные протоки из паренхимы печени, как показано на фиг.14 (для подтверждения идентичности выделенных структур авторы использовали окрашивание на К19), и получали культуры органоидов печени путем культивирования их с: (i) EGF; или (ii) EGF и R-спондин 1 плюс FGF10 или HGF, или никотинамид; или (iii) EGF и R-спондин 1 плюс FGF10, HGF и никотинамид (ERFHNic). Авторы пересевали культуры один раз в неделю в течение всего 14 недель. Результаты подтвердили, показано в примерах 1 и 2, что EGF, R-спондин 1 и никотинамид, объединенные с FGF10, необходимы для роста и самообновления культур печени. Через 10 пассажей, культуры, в которых отсутствовал HGF, показывали отставание в росте по сравнению с культурами, дополненными HGF. Хотя клетки все еще были жизнеспособными, скорость пролиферации снижалась до разведения 1:2-1:4 при пересеве по сравнению с разведением 1:6-1:8 при пересеве культур, дополненных полной комбинацией (FGF10, HGF и никотинамид). Через 15 пассажей культуры на ERFNic, не дополненные HGF, были уже нежизнеспособными. Таким образом, эти результаты позволяют предположить, что HGF является необходимым для поддержания хорошей скорости пролиферации после длительного размножения (фиг.15).
Пример 6 - Маркеры, экспрессирующиеся в органоидах печени в дифференцирующих условиях
Используя протокол дифференцировки, описанный в примере 2, авторы были способны детектировать маркер гепатобластов (альбумин) и маркер поверхности гепатоцитов в органоидах печени. Для количественной оценки числа этих гепатоцит-подобных клеток авторы провели анализ методом культур проточной цитометрии с использованием маркера поверхности гепатоцитов. Авторы наблюдали, что хотя в среде для размножения клетки, положительные по маркеру поверхности гепатоцитов, практически не детектировались, после дифференцировки до 35% клеток были положительными по маркеру поверхности гепатоцитов (см. фиг.17B и C).
Авторы затем проанализировали профиль экспрессии генов органоидов печени мыши в данных условиях дифференцировки (фиг.16 и фиг.1; авторы наблюдали сильное повышение экспрессии, например, генов Alb, FAH, TAT и Cyp3). Авторы обнаружили, что экспрессия генов в органоидах печени мыши похожа на профиль экспрессии генов в зрелых гепатоцитах и/или печени мыши.
Пример 7 - Трансплантация органоидов печени мышам
Клетки были взяты из органоидов, которые выращивались с использованием условий для размножения ERFHNic и дифференцирующих условий EAFD, и трансплантировали иммунодефицитному штамму мышей с недостаточностью фермента катаболизма тирозина, фумарилацетоацетат-гидролазы (FAH), мышиной модели тирозинемии I типа человека (Azuma et al. 2007, Nature Biotech, 25(8), 903-910). Схема трансплантации показана на фиг.17D. Предварительные результаты показывают, что рассеянные FAH-положительные клетки можно обнаружить в паренхиме печени мышей с недостаточностью FAH, указывая на то, что клетки печени, полученные из культур органоидов, прижились в реципиентной печени (см. фиг.17А, правую сторону). Кроме того, в печени мышей-реципиентов также детектировали значительно увеличенное число К19-положительных клеток. Это позволяет предположить, что полученные из органоидов трансплантированные клетки способны генерировать оба типа клеток in vivo: гепатоциты (показаны маркером FAH) и холангиоциты (показаны маркером К19) (см. фиг.17А, левая верхняя панель). Эти данные дополнительно были подтверждены анализом посредством метода проточной цитометрии трансплантированных клеток, которые произошли от двух различных клонов из двух отдельных культур (фиг.17В и 17С, соответственно). Lgr5+ клетки были трансдуцированы вирусом, содержащим GFP, и после дифференцировки проводили анализ методом проточной цитометрии. Клетки, положительные по маркеру поверхности гепатоцитов, имели большее рассеяние, указывающее на более крупные клетки, которые имеют гранулярность и зрелость, то есть, клетки зрелых гепатоцитов. Клетки, отрицательные по маркеру поверхности гепатоцитов, имели меньшее рассеяние, указывающее на меньшие клетки, то есть, менее зрелые предшественники. Таким образом, в дифференцированной культуре присутствуют все типы клеток (зрелые и клетки-предшественники). Остаток клеток, то есть клетки, не используемые для FACS-анализа, использовали для экспериментов по трансплантации.
Пример 8
Органоиды из печени мышей, культивированные согласно способу по изобретению, анализировали, используя анализ методом микроматриц, для того чтобы определить, какие гены экспрессируются, а какие не экспрессируются.
Пример 9
Органоиды из печени человека, культивированные с использованием среды ЕМ1, ЕМ2 и DM по изобретению, анализировали, используя анализ методом олигонуклеотидных микроматриц, для того чтобы определить, какие гены экспрессируются, а какие не экспрессируются. Профили экспрессии значительно отличались для генов, экспрессируемых в среде для размножения, в дифференцирующей среде и зрелой печени. Общая закономерность для экспрессии генов гепатоцитов была примерно одинаковой, как и для мышиных клеток, но дифференцировка органоидов была менее выраженной по сравнению с органоидами печени мышей. Это могло быть следствием использования клеток человека.
Как часто случает в анализе с использованием метода олигонуклеотидных микроматриц, Lgr5 и Tnfrsf19 не детектировались. Однако было найдено, что они присутствуют в органоидах, культивированных в среде для размножения.
МАТЕРИАЛА И МЕТОДЫ (для примеров 4-7)
Воздействие на животных
Двухмесячные Lgr5LacZ или Axin2-LacZ или однопометные дикого типа мыши BL6/Balbc F1 получали внутрибрюшинные инъекции 0,8 мл/кг CCL4, растворенного в кукурузном масле (n= ), или просто кукурузного масла (n= ). Мышей умерщвляли через 2, 5, 9 или 13 дней, и выделяли печень и далее обрабатывали ее для выделения РНК или окрашивания на β-галактозидазу.
Окрашивание на β-галактозидазу (lacZ)
Ткани печени выделяли и немедленно инкубировали 2 часа в 20-кратном объеме ледяного фиксирующего буфера (1% формальдегида; 0,2% глутарового альдегида; 0,02% NP40 в PBS0) при 4°C на вращающейся платформе. Фиксирующий буфер удаляли и ткани дважды промывали в промывающем буфере (PBS0; 2 мМ MgCl2; 0,02% NP40; 0,1% деоксихолата натрия) в течение 20 минут при комнатной температуре на вращающейся платформе. Затем добавляли субстрат β-галактозидазы (5 мМ K3Fe(CN)6; 5 мМ K4Fe(CN)6·3H2O; 2 мМ MgCl2; 0,02% NP40; 0,1% деоксихолата Na; 1 мг/мл X-gal в PBS0), и ткани инкубировали 2 часа в темноте при 37°C и в течение ночи при комнатной температуре. Субстрат удаляли, и ткани промывали дважды PBS0 по 20 минут при комнатной температуре на вращающейся платформе. Затем ткани фиксировали в течение ночи в 20-кратном объеме 4%-го параформальдегида (PFA) в PBS0 при 4°C в темноте на вращающейся платформе. Удаляли PFA, и ткани промывали дважды в PBS0 по 20 минут при комнатной температуре на вращающейся платформе.
Окрашенные ткани переносили в кассеты, и приготавливали парафиновые блоки, используя стандартные способы. Приготавливали среды тканей (4 мкМ) и окрашивали нейтральным красным.
Обработка R-спондином 1
6-8-недельным мышам Axin2-lacZ вводили внутрибрюшинно с помощью инъекций 100 мкг очищенного R-спондина 1 и умерщвляли через 48 часов для анализа экспрессии LacZ в печени.
ОТ-ПЦР
РНК экстрагировали из культур клеток кишечника или свежевыделенной ткани, используя набор RNeasy Mini RNA Extraction Kit (Qiagen), и использовали в реакции обратной транскрипции с использованием обратной транскриптазы из вируса лейкоза мышей Молони (Promega). кДНК амплифицировали в термоциклере (GeneAmp PCR System 9700; Applied Biosystems, London, UK), как было описано ранее (Huch et al., 2009). Праймеры показаны в таблице 3 ниже.
Праймеры для ОТ-ПЦР
Иммуногистохимия
Используемая в настоящем изобретении процедура иммуноокрашивания была описана ранее в Huch et al. 2009. Кратко, пяти-микрометровые срезы депарафинировали, регидрировали, и срезы ткани пермеабилизовали, используя PBS-T (PBS; Tween20 0,1%). При необходимости среды обрабатывали 10 мМ цитратным буфером (рН 6,0) для извлечения антигена, блокировали, используя универсальный блокирующий буфер (BioGenex), и инкубировали с первичным антителом. Затем срезы дважды промывали PBS и инкубировали с вторичными антителами, конъюгированными с пероксидазой. В качестве хромогенного субстрата использовали DAB+ (DAKO). Срезы окрашивали гематоксилином Майера и визуализировали на микроскопе Leica DMR. Использовали следующие первичные антитела: кроличьи анти-Sox9 (в разведении 1:600; 1 ч при комнатной температуре, Millipore), мышиные анти-SMA (в разведении 1:1000, в течении ночи при 4°C, Sigma), кроличьи анти-FAH (в разведении 1:5000; в течении ночи при 37°C, дар от M. Grompe), кроличьи анти-K19 (в разведении 1:500; в течении ночи при 4°C, дар от M. Grompe). Используемыми конъюгированными с пероксидазой вторичными антителами были анти-мышиные или анти-кроличьи антитела Brightvision (ImMunologic).
Иммунофлуоресценция
Для окрашивания тотальных препаратов, органоиды или выделенные желчные протоки фиксировали ацетоном (органоиды) или 4%-м PFA (желчные протоки) в течение 30 мин, промывали один раз PBS, пермеабилизовали PBS + 0,3% Triton-X100 в течение 5 мин, блокировали с использованием универсального блокирующего раствора (Power block HK085-5KE BioGenex) и инкубировали в течение ночи с первичными антителам, разведенными в PBS + 1% FBS. После нескольких промывок в PBS образцы инкубировали с вторичным антителом. Ядра окрашивали Hoescht 33342. Изображения получали с помощью конфокальной микроскопии (Leica, SP5). Трехмерную реконструкцию проводили, используя Volocity Software (Improvision). Используемыми первичными антителами были: кроличье анти-K19 (в разведении 1:500; дар от M. Grompe), крысиное антитело к маркеру поверхности гепатоцитов (в разведении 1:50, дар от M. Grompe), козье антитело к альбумину (в разведении 1:50, Santa Cruz). Используемыми вторичными антителами были во всех случаях ослиные антитела, конъюгированные с различными флуорофорами Alexa (ослиные анти-козьи Alexa-568, ослиные анти-крысиные Alexa-488, ослиные анти-кроличьи Alexa-647, Molecular probes).
Проточная цитометрия
Диссоциированные клетки ресуспендировали в концентрации 1×104 клеток на миллилитр в 1 мл DMEM + 2% FBS перед добавлением супернатанта гибридомы MIC1-1C3 в разведении 1:20 или супернатанта гибридомы OC2-2F8 в разведении 1:50, и инкубировали 30 мин при 4°C. После промывки холодным фосфатно-солевым буфером Дульбекко (DPBS) клетки ресуспендировали в DMEM + 2% FBS, содержащей в разведении 1:200 APC-конъюгированное козье анти-крысиное вторичное антитело, сорбированное против мышиных сывороточных белков (Jackson Immunoresearch). Окрашивание йодистым пропидием использовали для мечения мертвых клеток с целью их исключения из анализа. Клетки анализировали и сортировали на Cytopeia inFluxV-GS (Becton-Dickenson).
Трансплантационный метод анализа
Инъекцию отсортированных клеточных популяций в селезенку и удаление NTBC для индукции селекции гепатоцитов проводили, как описано ранее (Overturf et al. 1996). Удаление лекарственного соединения осуществляли на протяжении 3 недель с последующим восстановлением введения до восстановления нормального веса у животных-реципиентов.
| название | год | авторы | номер документа |
|---|---|---|---|
| КУЛЬТУРАЛЬНАЯ СРЕДА СТВОЛОВЫХ КЛЕТОК | 2012 |
|
RU2714256C2 |
| КУЛЬТУРАЛЬНАЯ СРЕДА ДЛЯ ЭПИТЕЛИАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК И ОРГАНОИДОВ, СОДЕРЖАЩИХ УКАЗАННЫЕ СТВОЛОВЫЕ КЛЕТКИ | 2010 |
|
RU2555545C2 |
| УЛУЧШЕННЫЙ СПОСОБ ДИФФЕРЕНЦИРОВКИ | 2017 |
|
RU2772435C2 |
| ДИФФЕРЕНЦИРОВКА ЧЕЛОВЕЧЕСКИХ ЭМБРИОНАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК | 2008 |
|
RU2473685C2 |
| ДИФФЕРЕНЦИРОВКА ЭМБРИОНАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК ЧЕЛОВЕКА | 2008 |
|
RU2465323C2 |
| МУЛЬТИПОТЕНТНЫЕ СТВОЛОВЫЕ КЛЕТКИ ВНЕПЕЧЕНОЧНЫХ ЖЕЛЧНЫХ ПУТЕЙ И СПОСОБЫ ИХ ВЫДЕЛЕНИЯ | 2010 |
|
RU2580246C2 |
| КОМПОЗИЦИИ ТРАНСПЛАНТАТА ДЛЯ ПЕРЕСАДКИ КЛЕТОК | 2018 |
|
RU2828021C2 |
| СПОСОБ ТРАНСПЛАНТАЦИИ КЛЕТОК ИЗ ПЛОТНЫХ ТКАНЕЙ | 2011 |
|
RU2574364C2 |
| Способ получения, культивирования и характеристики бронхиальных и легочных органоидов из индуцированных плюрипотентных стволовых клеток человека | 2022 |
|
RU2821564C2 |
| СПОСОБ ПОЛУЧЕНИЯ КЛЕТОК ДЛЯ ЗАМЕСТИТЕЛЬНОЙ КЛЕТОЧНОЙ ТЕРАПИИ ПАТОЛОГИЙ ПЕЧЕНИ | 2012 |
|
RU2510276C1 |




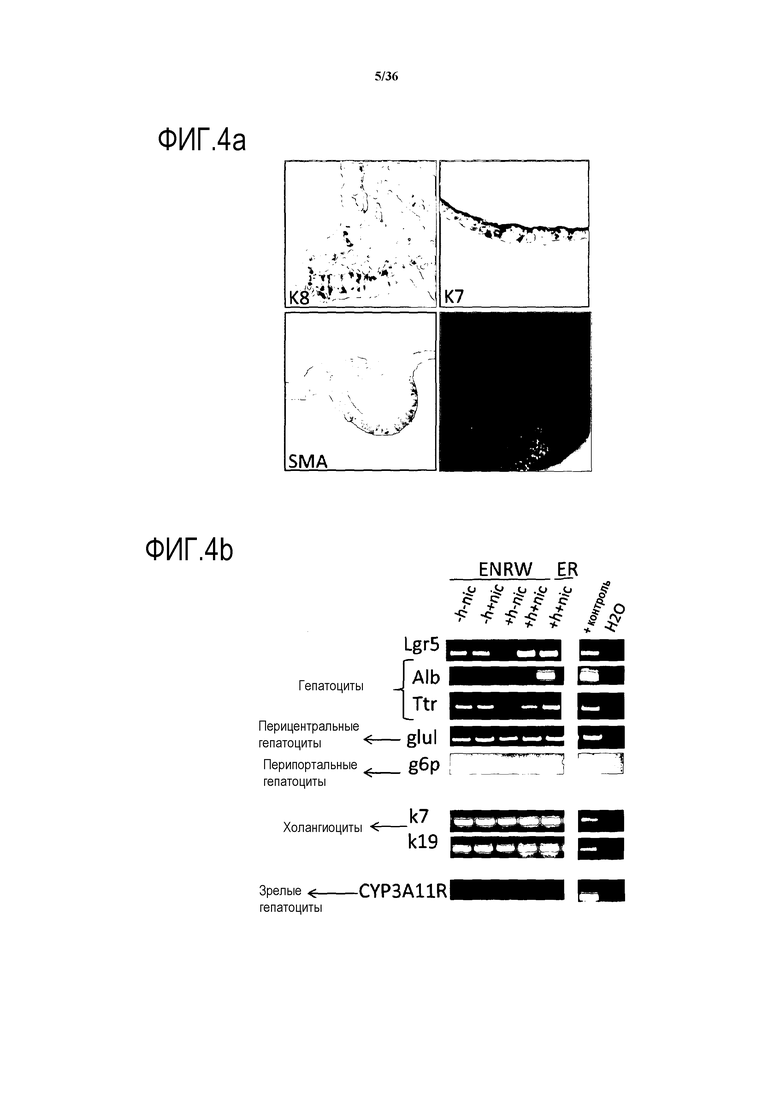

















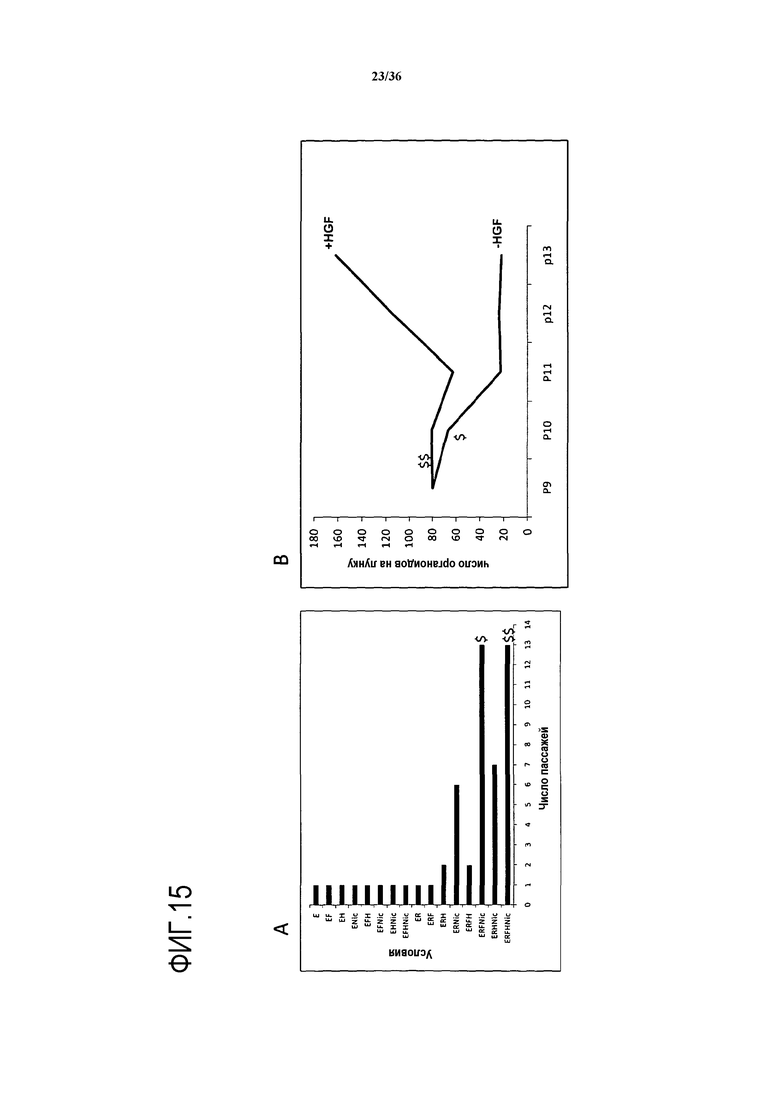










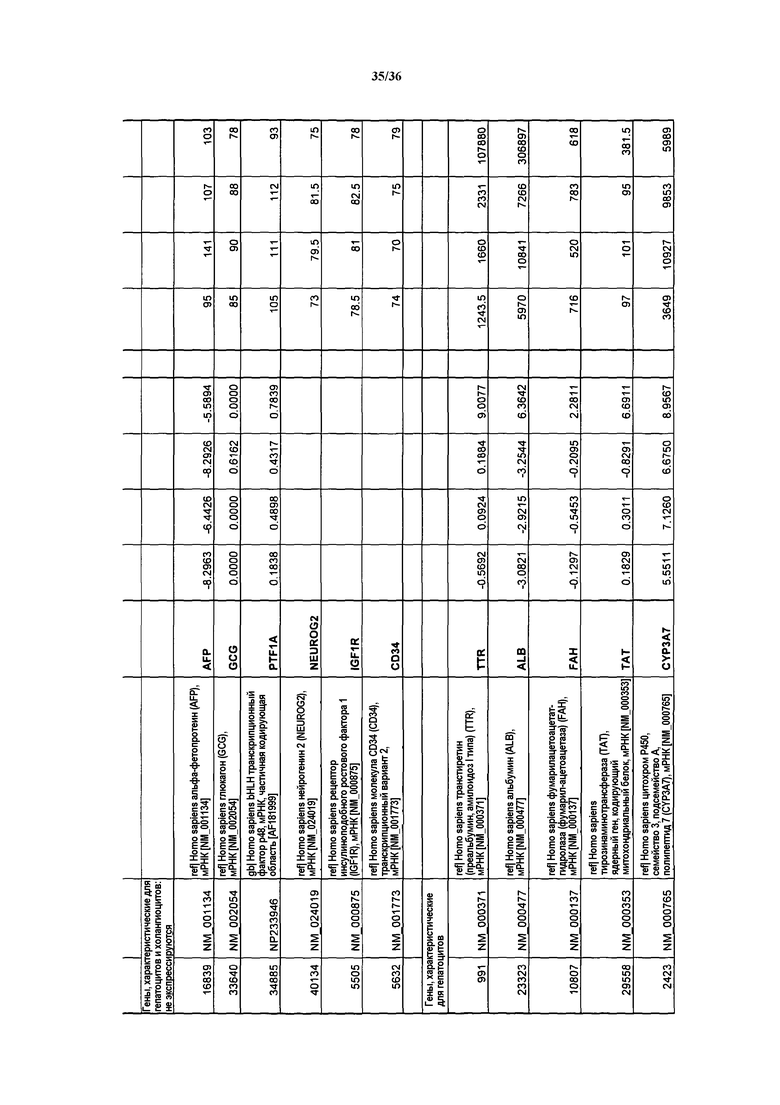






Изобретение относится к гистологии. Раскрыт способ получения органоида печени. Культивируют по меньшей мере одну зрелую эпителиальную стволовую клетку из фрагмента печени или желчного протока печени в контакте с внеклеточным матриксом в присутствии среды. Причем среда содержит базальную среду для клеток животных или человека, к которой добавлены: фактор роста эпидермиса (EGF), агонист Wnt, фактор роста фибробластов (FGF) и никотинамид. Описаны соответствующие культуральные среды. Также описан органоид печени и его применение для скрининга новых лекарственных средств или в анализе токсичности, а также в разнообразных исследованиях, для получения лекарственных средств для лечения печени и в регенеративной медицине. 9 н. и 17 з.п. ф-лы, 19 ил., 3 табл., 9 пр.
1. Способ получения органоида печени, где указанный способ содержит:
культивирование по меньшей мере одной зрелой эпителиальной стволовой клетки из фрагмента печени или желчного протока печени в контакте с внеклеточным матриксом в присутствии среды, причем среда содержит базальную среду для клеток животных или человека, к которой добавлены: фактор роста эпидермиса (EGF), агонист Wnt, фактор роста фибробластов (FGF) и никотинамид.
2. Способ по п. 1, где одно, два или три соединения, выбранные из группы, состоящей из ингибитора BMP, ингибитора TGF-β и HGF, добавляют в среду.
3. Способ по п. 1 или 2, в котором по меньшей мере через день среду меняют на вторую среду, где второй средой является среда для дифференциации, содержащая базальную среду для клеток животных или человека, к которой добавлены EGF, FGF и/или HGF, ингибитор TGF-β и ингибитор Notch.
4. Способ по п. 3, где способ содержит культивирование указанных эпителиальных стволовых клеток в клеточной культуральной среде, содержащей или состоящей из базальной среды для клеток животных или человека, к которой добавлены EGF, ингибитор BMP, R-спондин и Wnt; и впоследствии во второй среде для размножения, которая содержит или состоит из базальной среды для клеток животных или человека, к которой добавлены: EGF, FGF и HGF, никотинамид и R-спондин; и впоследствии в дифференцирующей среде, которая содержит или состоит из базальной среды для клеток животных или человека, к которой добавлены: EGF, FGF и/или HGF, ингибитор TGF-β и ингибитор Notch.
5. Способ по любому из пп. 1-4, где агонист Wnt выбран из члена семейства Wnt, R-спондина 1-4, Norrin и ингибитора GSK.
6. Способ по любому из пп. 2-4, где TGF-бета ингибитор представляет собой низкомолекулярный ингибитор, выбранный из группы, состоящей из: А83-01, SB-431542, SB-505124, SB-525334, LY 364947, SD-208, SJN 2511.
7. Способ по любому из пп. 1-4, где FGF представляет собой FGF, способный связываться с FGFR2 или FGFR4, и предпочтительно является FGF10.
8. Способ по любому из пп. 1-4, где одно, два, три, четыре или более соединений, выбранных из группы, содержащей гастрин, В27, N2 и N-ацетилцистеин, добавляют в базальную среду.
9. Культуральная среда для культивирования зрелых эпителиальных стволовых клеток, содержащая базальную среду для клеток животных и человека, к которой добавлены: EGF, агонист Wnt, FGF и никотинамид.
10. Культуральная среда по п. 9, в которую добавлены одно, два или три соединения, выбранных из группы, содержащей: ингибитор BMP, ингибитор TGF-β и HGF.
11. Культуральная среда для культивирования зрелых эпителиальных стволовых клеток, содержащая базальную среду для клеток животных и человека, к которой добавлены EGF, FGF и/или HGF, ингибитор TGF-β и ингибитор Notch.
12. Культуральная среда по п. 11, в которой ингибитором Notch является ингибитор гамма-секретазы, такой как DAPT или дибензазепин (DBZ), или бензодиазепин (BZ), или LY-411575.
13. Культуральная среда по п. 9 или 11, в которой среда дополнительно содержит дексаметазон.
14. Культуральная среда по п. 9 или 11, где агонист Wnt выбран из члена семейства Wnt, R-спондина 1-4, Norrin и ингибитора GSK.
15. Культуральная среда по п. 9 или 11, где ингибитором TGF-β является низкомолекулярный ингибитор, выбранный из группы, состоящей из А83-01, SB-431542, SB-505124, SB-525334, LY 364947, SD-208, SJN 2511.
16. Культуральная среда по п. 9 или 11, где FGF является FGF, способный связываться с FGFR2 или FGFR4, и предпочтительно является FGF10.
17. Культуральная среда по п. 9 или 11, где одно, два, три, четыре или более соединений, выбранных из группы, содержащей гастрин, В27, N2 и N-ацетилцистеин, добавляют в базальную среду.
18. Органоид печени, который получают способом по любому одному из пп. 1-8, который содержит кистозную структуру и с внешней стороны слой клеток по меньшей мере с одним зачатком и центральным просветом.
19. Органоид печени по п. 18, который имеет одну или несколько (например, 2, 3, 4 или все 5) из следующих характеристик: (а) плотность клеток >5×105 клеток/см3, предпочтительно >10×105 клеток/см3; (b) толщину, равную 2-30 слоям клеток, предпочтительно толщину, равную 2-15 слоям клеток; (с) клетки контактируют друг с другом в трех пространственных направлениях; (d) демонстрирует функцию, присущую здоровой ткани печени, (е) имеет вытянутую форму с двумя выраженными доменами.
20. Органоид печени по п. 18, содержащий монослойный эпителиальный домен, где детектируются поляризованные клетки и экспрессируются кератиновые маркеры, и второй домен, составляющий основное тело органоида и образованный многослойным эпителием с неполяризованными клетками, необязательно экспрессирующими альбумин.
21. Органоид печени по п. 18, который происходит от клетки, экспрессирующей Lgr5, и который содержит популяцию по меньшей мере от 1×103 до 5×104 клеток, например по меньшей мере от 1×103 до 5×103 клеток.
22. Применение органоида печени по любому одному из пп. 18-21 в скрининге для поиска новых лекарственных соединений.
23. Применение органоида печени по любому одному из пп. 18-21 в анализе токсичности.
24. Применение органоида печени по любому одному из пп. 18-21 в исследованиях в области эмбриогенеза печени, типов клеток печени и путей дифференцировки; исследованиях экспрессии генов, включая рекомбинантную экспрессию генов; исследованиях механизмов, вовлеченных в повреждение и восстановление печени; исследованиях воспалительных и инфекционных заболеваний печени; исследованиях патогенетических механизмов; или исследованиях механизмов трансформации клеток печени и этиологии рака печени.
25. Применение органоида печени по любому из пп. 18-21 для получения лекарственного средства для лечения нарушения, состояния или заболевания печени.
26. Применение органоида печени по любому из пп. 18-21 в регенеративной медицине.
| WO2004087896 A2, 14.10.2004 | |||
| КОМПОЗИЦИИ ПОДВЕРГНУТЫХ РЕСТРИКЦИИ КЛЕТОК, СПОСОБНЫХ К БЫСТРОМУ РОСТУ, КОТОРЫЕ ПРОДУЦИРУЮТ ВЕЩЕСТВА, ПОДАВЛЯЮЩИЕ ПРОЛИФЕРАЦИЮ КЛЕТОК, И ИХ ПРИМЕНЕНИЕ | 1999 |
|
RU2236855C2 |

