Изобретение относится к новой культуральной среде для культивирования эпителиальных стволовых клеток, особенно эпителиальных стволовых клеток тонкого кишечника и толстого кишечника, и для культивирования органоидов, содержащих указанные стволовые летки. Кроме того, изобретение относится к потомкам клеток и органоидов, культивированных с использованием культуральной среды по изобретению, и к применению указанных потомков в анализах токсичности и в регенеративной медицине.
Самообновляющийся эпителий тонкого кишечника упорядочен в крипты и ворсинки (Gregorieff and Clevers, 2005. Genes Dev 19, 877-90). Клетки, вновь образующиеся в криптах, утрачиваются путем апоптоза на вершинах ворсинок, причем общее время обновления эпителия у мышей составляет 5 суток. Давно известно, что самообновляющиеся стволовые клетки располагаются вблизи основания крипты и образуют быстро пролиферирующие переходные делящиеся (TA) клетки, способные дифференцироваться во все ростки. Оцененное количество стволовых клеток составляет от 4 до 6 на крипту (Bjerknes and Cheng, 1999. Gastroenterology 116, 7-14). Из TA-клеток образуется три типа дифференцированных клеток: энтероциты, бокаловидные клетки и энтероэндокринные клетки, и они продолжают их миграцию в последовательно расположенные слои вдоль оси крипта-ворсинка. В каждую ворсинку поступают клетки из множества различных крипт. Четвертый из основных типов дифференцированных клеток, клетки Панета, находится в основании крипты.
Недавно был идентифицирован ген Lgr5, который специфично экспрессируется в пятом типе клеток, циклирующих обновляющихся столбчатых клетках основания крипты (CBC), которые представляют собой небольшие клетки, распределенные между клетками Панета (указаны черными стрелками на фиг. 8b) (Barker et al., 2007. Nature 449: 1003-1007). С использованием мыши, у которой в локус Lgr5 встроена кассета индуцируемой GFP/тамоксифеном рекомбиназы Cre, путем прослеживания родословной было показано, что Lgr5+ CBC-клетки представляют собой мультипотентные стволовые клетки, которые образуют все типы клеток эпителия, даже при оценке через 14 месяцев после индукции Cre.
Недавно было открыто, что помимо Lgr5 уникальным маркером для взрослых стволовых клеток также явлется Lgr6, но не Lgr4. В то время как Lgr5 экспрессируется в стволовых клетках головного мозга, почки, печени, сетчатки, желудка, кишечника, поджелудочной железы, молочной железы, волосяного фолликула, яичника, мозгового вещества надпочечников и кожи, Lgr6 экспрессируется в стволовых клетках головного мозга, легких, молочной железы, волосяного фолликула и кожи.
В общем, полагают, что для заякоривания и поддержания эпителиальных стволовых клеток и для обеспечения правильной ориентации, необходимой для получения правильным образом полярно ориентированной трехмерной структуры требуется тесный контакт между эпителиальными стволовыми клетками и субэпителиальными фибробластами.
Хотя описано множество культуральных систем для культивирования первичных эпителиальных стволовых клеток, включая стволовые клетки эпителия кишечника (Bjerknes and Cheng, 2006. Methods Enzymol. 419: 337-83), к настоящему времени не получено длительной культуральной системы, которая сохраняет плюрипотентность эпителиальных стволовых клеток. Более того, не известно культуральной системы, которая сохраняет основную физиологию крипта-ворсинка в криптах, выделенных из толстого или тонкого кишечника, или которая сохраняет основную физиологию выделенных фрагментов поджелудочной железы или фрагментов ткани желудка.
Таким образом, изобретение относится к способу культивирования эпителиальных стволовых клеток, выделенных фрагментов эпителиальной ткани, содержащих указанные эпителиальные стволовые клетки, или клеток аденомы, причем способ включает предоставление внеклеточного матрикса, инкубацию эпителиальной стволовой клетки, выделенного фрагмента ткани, содержащего указанные эпителиальные стволовые клетки, или клетки аденомы с внеклеточным матриксом, культивирование стволовой клетки, выделенного фрагмента ткани или клетки аденомы в присутствии клеточной культуральной среды, содержащей базальную среду для клеток животного или человека, в которую добавлен ингибитор морфогенетического белка кости (BMP), от 5 до 500 нграмм/мл или по меньшей мере 5 и не более чем 500 нграмм/мл митогенного фактора роста, и, если культивируют эпителиальные стволовые клетки и выделенные фрагменты ткани, агонист Wnt.
Авторы настоящего изобретения неожиданно открыли, что способ по изобретению позволяет культивирование эпителиальных стволовых клеток, выделенных фрагментов из тонкого кишечника, толстого кишечника, желудка и поджелудочной железы, содержащих указанные стволовые клетки, и клеток аденомы, сохраняя наличие стволовых клеток, которые сохраняют недифференцированный фенотип и способность к самоподдержанию. Например, выделенные крипты, которые культивируют способом по изобретению, развиваются в органоиды крипта-ворсинка, содержащие центральный просвет, который выстилает эпителий, подобный эпителию ворсинок. Рост выделенных крипт обеспечивался стволовыми клетками, которые присутствуют в криптах. Полученные органоиды претерпевают многократные деления крипт. Еще более удивительным было наблюдение, что способ по изобретению позволяет выращивание единичных выделенных эпителиальных стволовых клеток в органоиды крипта-ворсинка в отсутствие ниши стволовых клеток. Выделенные фрагменты желудка из пилорической области желудка ведут себя как органоиды кишечных крипт. Открытая верхняя часть элемента закрывалась и просвет заполнялся апоптотическими клетками. Вновь образованные органоиды желудка претерпевали непрерывное почкование (напоминающее деление железы) при сохранении их полярности и центрального просвета. Более того, культивирование фрагментов поджелудочной железы приводило к появлению структур, подобных островкам поджелудочной железы, которые экспрессируют инсулин и другие специфические маркеры островков поджелудочной железы, напоминающих островки Лангерганса поджелудочной железы.
Эпителий, выстилающий пилорическую область тонкого и толстого кишечника, охватывает выпячивания в просвет, ворсинки и инвагинации, крипты. Каждая клетка вдоль оси крипта-ворсинка является полярной, причем клетки на вершине кишечных ворсинок или в основании крипт толстого кишечника являются наиболее дифференцированными и постоянно утрачиваются в просвет. Непрерывная пролиферация стволовых клеток, находящихся в основании крипт, и массивная пролиферация клеток-предшественников, находящихся в середине крипт, обеспечивает надлежащую замену утрачиваемых клеток.
Стволовые клетки найдены во многих органах взрослых людей и мышей. Хотя может точные характеристики взрослых стволовых клеток могут значительно варьировать в отдельных тканях, взрослые стволовые клетки имеют следующие общие характеристики. Они сохраняют недифференцированный фенотип, их потомки могут дифференцироваться во все ростки, присутствующие в данной ткани, они сохраняют способность к самоподдержанию на протяжении жизни и они способны регенерировать данную ткань после повреждения. Стволовые клетки располагаются в специализированной области, нише стволовых клеток, которая обеспечивает надлежащие контакты между клетками и сигналы для поддержания указанной популяции стволовых клеток.
Эпителиальные стволовые клетки способны образовывать отдельные типы клеток, из которых состоит эпителий. Некоторые типы эпителия, такие как эпителий кожи и тонкого кишечника, демонстрируют быстрое обновление клеток, указывая на то, что существующие в них стволовые клетки должны непрерывно пролиферировать. Другие типы эпителия, такие как эпителий печени или поджелудочной железы, демонстрируют очень медленное обновление в нормальных условиях.
Крипты можно выделять из двенадцатиперстной кишки, тонкого и толстого кишечника, включая тощую кишку, подвздошную кишку и толстую кишку, и пилорической области желудка, с помощью протоколов, которые известны специалисту в данной области. Например, крипты можно выделять путем инкубации выделенной ткани с хелатирующими агентами, которые освобождают клетки от их кальций- и магний-зависимых взаимодействий с базальной мембраной и стромальными типами клеток. После промывания ткани слой эпителиальных клеток счищают с подслизистой оболочки с помощью предметного стекла и измельчают. После этого проводят инкубацию в трипсине или, более предпочтительно, EDTA и/или EGTA и отделение нерасщепленных фрагментов ткани и отдельных клеток от крипт с использованием, например, стадий фильтрации и/или центрифугирования. Вместо трипсина можно использовать другие протеолитические ферменты, такие как коллагеназа и/или диспаза I. Сходные способы используют для выделения фрагментов поджелудочной железы и желудка.
Способы выделения стволовых клеток из эпителиальной ткани известны в данной области. Предпочтительный способ основан на том факте, что стволовые клетки экспрессируют Lgr5 и/или Lgr6 на их поверхности, которые относятся к крупному суперсемейству сопряженных с G белком рецепторов (GPCR). Подсемейство Lgr является уникальным тем, что оно несет крупный богатый лейцином эктодомен, важный для связывания лиганда. Лиганды для Lgr5 и Lgr6 пока не описаны в литературе. Таким образом, предпочтительный способ включает получение суспензии клеток из указанной эпителиальной ткани, контактирование указанной суспензии клеток с соединением, связывающим Lgr5 и/или 6, выделение указанного соединения, связывающего Lgr5 и/или 6, и отделение стволовых клеток от указанного связывающего соединения. Предпочтительно суспензию отдельных клеток из выделенных крипт формировать механически, содержащую эпителиальные стволовые клетки, поскольку было выявлено, что эта стадия путем обработки эпителиальных стволовых клеток трипсином приводит к значительно более низким уровням выживаемости.
Предпочтительные связывающие Lgr5 и/или 6 соединения содержат антитела, такие как моноклональные антитела, которые специфично распознают и связывают внеклеточный домен либо Lgr5, либо Lgr6, такие как моноклональные антитела, включая моноклональные антитела мыши и крысы. С использованием такого антитела стволовые клетки, экспрессирующие Lgr5 и/или Lgr6, можно выделять, например, с помощью магнитных гранул и путем активированной флуоресценцией сортировки клеток, как понятно специалисту в данной области.
В предпочтительном способе по изобретению указанные эпителиальные стволовые клетки выделяют из крипт, фрагментов желудка или фрагментов поджелудочной железы. Например, указанные эпителиальные стволовые клетки выделяют из крипт, которые выделяют из кишечника. Предпочтительные эпителиальные стволовые клетки выделяют из тонкого кишечника, включая двенадцатиперстную кишку, тощую кишку и подвздошную кишку, поджелудочной железы или желудка.
Выделенные стволовые клетки предпочтительно культивируют в микроокружении, которое по меньшей мере частично имитирует клеточную нишу, в которой указанные стволовые клетки находятся в природе. Указанную клеточную нишу имитируют путем культивирования указанных стволовых клеток в присутствии биоматериалов, таких как матрицы, каркасы и культуральные субстраты, которые соответствуют ключевым регуляторным сигналам, контролирующим судьбу стволовых клеток. Указанные биоматериалы содержат природные, полусинтетические и синтетические биоматериалы и/или их смеси. Каркас обеспечивает двухмерную или трехмерную сеть. Пригодные синтетические материалы для указанного каркаса включают полимеры, выбранные из пористых твердых веществ, нановолокон и гидрогелей, например, такие как пептиды, включающие самособирающиеся пептиды, гидрогели, состоящие из фосфата полиэтиленгликоля, фумарата полиэтиленгликоля, полиакриламида, полигидроксиэтилметакрилата, ацетата полицеллюлозы и/или их сополимеров (см., например, Saha et al., 2007. Curr Opin Chem Biol. 11(4): 381-387, Saha et al., 2008. Biophysical Journal 95: 4426-4438, Little et al., 2008. Chem Rev. 108, 1787-1796). Как известно специалисту в данной области, механические свойства, например, такие как эластичность каркаса, влияют на пролиферацию, дифференцировку и миграцию стволовых клеток. Предпочтительный каркас содержит биодеградируемые (со)полимеры, которые заменяются природным компонентами после трансплантации индивидууму, например, для стимуляции регенерации ткани и/или заживления ран. Более того, предпочтительно, чтобы указанный каркас по существу не индуцировал иммуногенный ответ после трансплантации индивидууму. Указанный каркас дополняют природными, полусинтетическими или синтетическими лигандами, которые обеспечивают сигналы, требуемые для пролиферации и/или дифференцировки, и/или миграции стволовых клеток. В предпочтительном варианте осуществления указанные лиганды содержат определенные аминокислотные фрагменты. Примеры указанных синтетических полимеров включают поверхностно-активное вещество блок-сополимер Pluronic® F127 (BASF) и Ethisorb® (Johnson and Johnson).
Клеточная ниша представляет собой часть, определяемую стволовыми клетками и окружающими клетками и внеклеточным матриксом (ECM), который продуцируется клетками в указанной нише. В предпочтительном способе по изобретению, выделенные крипты или эпителиальные стволовые клетки прикреплены к ECM. ECM состоит из различных полисахаридов, воды, эластина и гликопротеинов, где гликопротеины включают коллаген, энтактин (нидоген), фибронектин и ламинин. ECM секретируется клетками соединительной ткани. Известны различные типы ECM, обладающие различным составом, включая различные типы гликопротеинов и/или различную комбинацию гликопротеинов. Указанный ECM может быть предоставлен посредством культивирования продуцирующих ECM клеток, например, таких как фибробластные клетки, в емкости, с последующим удалением этих клеток и добавлением выделенных крипт или эпителиальных стволовых клеток. Примерами клеток, продуцирующих внеклеточный матрикс, являются хондроциты, продуцирующие, главным образом, коллаген и протеогликаны, фибробластные клетки, продуцирующие, главным образом, коллаген типа IV, ламинин, интерстициальные проколлагены и фибронектин, и миофибробласты толстой кишки, продуцирующие, главным образом, коллагены (тип I, III и V), протеогликан хондроитинсульфат, гиалуроновую кислоту, фибронектин и тенасцин-C. Альтернативно указанный ECM предоставляют коммерчески. Примерами коммерчески доступных внеклеточных матриксов являются белки внеклеточного матрикса (Invitrogen) и MatrigelTM (BD Biosciences). Применение ECM для культивирования стволовых клеток повышало длительное выживание стволовых клеток и сохраняло наличие недифференцированных стволовых клеток. В отсутствие ECM культуры стволовых клеток нельзя было культивировать в течение длительных периодов времени, и не наблюдали сохраняющегося наличия недифференцированных стволовых клеток. Кроме того, присутствие ECM позволяло культивирование трехмерных тканевых органоидов, которые не было возможно культивировать в отсутствие ECM.
Предпочтительный ECM для применения в способе по изобретению содержит по меньшей мере два различных гликопротеина, таких как два различных типа коллагена или коллаген и ламинин. Указанный ECM представлять собой синтетический внеклеточный матрикс на основе гидрогеля или встречающийся в природе ECM. Наиболее предпочтительным предоставляемым ECM является MatrigelTM (BD Biosciences), который содержит ламинин, энтактин и коллаген IV.
Клеточная культуральная среда, которую используют в способе по изобретению, включает любую клеточную культуральную среду. Предпочтительная клеточная культуральная среда представляет собой синтетическую среду с определенным химическим составом, забуференную до pH 7,4 (предпочтительно от 7,2 до 7,6 или по меньшей мере 7,2 и не более чем 7,6) с помощью карбонатного буфера, при этом клетки культивируют в атмосфере, содержащей от 5% до 10% CO2, или по меньшей мере 5% и не более чем 10% CO2, предпочтительно 5% CO2. Предпочтительную клеточную культуральную среду выбирают из DMEM/F12 и RPMI1640, дополненных глутамином, инсулином, пенициллином/стрептомицином и трансферрином. В следующем предпочтительном варианте осуществления используют Advanced DMEM/F12 или Advanced RPMI, которые оптимизированы для бесклеточной культуры и уже включают инсулин. В этом случае указанные среды Advanced DMEM/F12 или Advanced RPMI предпочтительно дополняют глутамином и пенициллином/стрептомицином. Более того, предпочтительно, чтобы указанная клеточная культуральная среда была дополнена очищенным, природным, полусинтетическим и/или синтетическим фактором роста и не содержала компонента с неопределенным химическим составом, такого как эмбриональная телячья сыворотка. Добавки, например, такие как B27 (Invitrogen), N-ацетилцистеин (Sigma) и N2 (Invitrogen) стимулируют пролиферацию некоторых клеток и могут быть дополнительно добавлены в среду, если потребуется.
Компонентом, который добавляют в базальную культуральную среду, является ингибитор BMP. BMP связываются в качестве димерного лиганда с рецепторным комплексом, состоящим из двух различных рецепторных серин/треонин-киназ, рецепторов типа I и типа II. Рецептор типа II фосфорилирует рецептор типа I, что приводит к активации этой рецепторной киназы. Рецептор типа I впоследствии фосфорилирует специфические рецепторные субстраты (SMAD), что приводит к каскаду передачи сигнала, ведущему к транскрипционной активности.
Указанный ингибитор BMP определяют как вещество, которое связывается с молекулой BMP с образованием комплекса, где активность BMP нейтрализуется, например, путем предотвращения или ингибирования связывания молекулы BMP с рецептором BMP. Альтернативно указанный ингибитор представляет собой вещество, которое действует в качестве антагониста или обратного агониста. Этот тип ингибитора связывается с рецептором BMP и препятствует связыванию BMP с указанным рецептором. Примером последнего вещества является антитело, которое связывает рецептор BMP и препятствует связыванию BMP со связанным антителом рецептором.
Указанный ингибитор BMP ингибирует BMP-зависимую активность в клетке до уровня самое большее 90%, более предпочтительно, самое большее 80%, более предпочтительно, самое большее 70%, более предпочтительно, самое большее 50%, более предпочтительно, самое большее 30%, более предпочтительно, самое большее 10%, более предпочтительно 0%, относительно уровня активности BMP в отсутствие указанного ингибитора. Как известно специалисту в данной области, активность BMP можно определять путем измерения транскрипционной активности BMP, например, как показано в Zilberberg et al., 2007. BMC Cell Biol. 8:41.
Известно несколько классов природных связывающих BMP белков, включая Noggin (Peprotech), хордин и хордин-подобные белки (R&D sytems), содержащие домены хордина, фоллистатин и фоллистатин-связанные белки (R&D sytems), содержащие домен фоллистатина, DAN и DAN-подобные белки (R&D sytems), содержащие домен цистеинового узла DAN, склеростин/SOST (R&D sytems), декорин (R&D sytems) и альфа-2-макроглобулин (R&D systems).
Предпочтительный ингибитор BMP для применения в способе по изобретению выбран из Noggin, DAN и DAN-подобных белков, включая Cerberus и Gremlin (R&D sytems). Эти диффундирующие белки способны связывать лиганд BMP с различной степенью аффинности и ингибировать их доступ к рецепторам передачи сигнала. В дополнение к любому из этих ингибиторов BMP, базальная культуральная среда препятствует утрате стволовых клеток, которая в ином случае происходит приблизительно после 2-3 недель культивирования.
Наиболее предпочтительным ингибитором BMP является Noggin. Noggin предпочтительно добавляют к базальной культуральной среде в концентрации по меньшей мере 10 нг/мл, более предпочтительно по меньшей мере 20 нг/мл, более предпочтительно по меньшей мере 50 нг/мл, более предпочтительно по меньшей мере 100 нг/мл. Наиболее предпочтительная концентрация составляет приблизительно 100 нг/мл или 100 нг/мл. В процессе культивирования стволовых клеток указанный ингибитор BMP предпочтительно добавляют в культуральную среду раз в двое суток, в то время как культуральную среду обновляют предпочтительно раз в четверо суток.
Следующим компонентом, который добавляют в базальную культуральную среду, является агонист Wnt. Каскад передачи сигнала Wnt определяется серией событий, которые происходят, когда белок Wnt связывается с рецептором клеточной поверхности, представителем семейства рецепторов Frizzled. Это приводит к активации белков семейства Dishevelled, которые ингибируют комплекс белков, которые включают аксин, GSK-3 и белок APC, деградирующих внеклеточный β-катенин. В результате, увеличенное содержание β-катенина усиливает транскрипцию с помощью факторов транскрипции семейства TCF/LEF.
Агонист Wnt определяют как средство, которое активирует опосредуемую TCF/LEF транскрипцию в клетке. Таким образом, агонисты Wnt выбирают из истинных агонистов Wnt, которые связывают и активируют представителя семейства рецепторов Frizzled, включая любые и все из белков семейства Wnt, ингибитора внутриклеточной деградации β-катенина и активаторов TCF/LEF. Указанный агонист Wnt стимулирует активность Wnt в клетке по меньшей мере на 10%, более предпочтительно по меньшей мере на 20%, более предпочтительно по меньшей мере на 30%, более предпочтительно по меньшей мере на 50%, более предпочтительно по меньшей мере на 70%, более предпочтительно по меньшей мере на 90%, более предпочтительно по меньшей мере на 100%, относительно уровня указанной активности Wnt в отсутствие указанной молекулы. Как известно специалисту в данной области, активность Wnt можно определять путем измерения транскрипционной активности Wnt, например посредством люциферазных репортерных конструкций pTOPFLASH и pFOPFLASH Tcf (Koπnek et al., 1997. Science 275: 1784-1787).
Агонист Wnt включает секретируемый гликопротеин, включающий Wnt-1/Int-1; Wnt-2/Irp (Int-1-связанный белок); Wnt-2b/13; Wnt-3/Int-4; Wnt-3a (R&D sytems); Wnt-4; Wnt-5a; Wnt-5b; Wnt-6 (Kirikoshi H et al. 2001. Biochem Biophys Res Com 283: 798-805), Wnt-7a (R&D sytems); Wnt-7b, Wnt-8a/8d, Wnt-8b, Wnt-9a/14, Wnt-9b/14b/15; Wnt-10a, Wnt-10b/12, Wnt-11 и Wnt-16. Обзор белков Wnt человека представлен в "THE WNT FAMILY OF SECRETED PROTEINS", R&D Systems Catalog, 2004. Следующие агонисты Wnt включают семейство секретируемых белков R-спондинов, которое вовлечено в активацию и регуляцию каскада передачи сигнала Wnt и которое состоит из 4 представителей (R-спондин 1 (NU206, Nuvelo, San Carlos, CA), R-спондин 2 ((R&D sytems), R-спондин 3 и R-спондин-4), и Norrin (также называемым белком болезни Норри или NDP) (R&D sytems), который представляет собой секретируемый регуляторный белок, который функционирует как белок Wnt в том, что он связывается с высокой аффинностью с рецептором Frizzled-4 и индуцирует активацию каскада передачи сигнала Wnt (Kestutis Planutis et al. (2007) BMC Cell Biol 8: 12). Недавно был идентифицирован низкомолекулярный агонист каскада передачи сигнала Wnt, производное аминопиримидина, и он также прямо включен в качестве агониста Wnt (Liu et al. (2005) Angew Chem Int Ed Engl. 44, 1987-90).
Известные ингибиторы GSK включают малые интерферирующие РНК (siRNA, Cell Signaling), литий (Sigma), кенпауллон (Biomol International, Leost, M et al. (2000) Eur J Biochem 267, 5983-5994), 6-броминдирубин-30-ацетоксим (Meijr, L. et al. (2003) Chem. Biol. 10, 1255-1266), SB 216763 и SB 415286 (Sigma-Aldrich) и представители семейства FRAT и происходящие из FRAT пептиды, которые препятствуют взаимодействию GSK-3 с аксином. Обзор предоставлен в Meijer et al., (2004) Trends in Pharmacological Sciences 25, 471-480, который включен в настоящий документ в качестве ссылки. Способы и анализы для определения уровня ингибирования GSK-3 известны специалисту в данной области и включают, например, способы и анализ, описанные Liao et al 2004, Endocrinology, 145(6): 2941-9).
В предпочтительном варианте осуществления указанный агонист Wnt выбирают из одного или нескольких представителей семейства Wnt, R-спондинов 1-4, Norrin, и ингибитора GSK. Авторами настоящего изобретения было открыто, что добавление по меньшей мере одного агониста Wnt к базальной культуральной среде является необходимым для пролиферации эпителиальных стволовых клеток или выделенных крипт.
В следующем предпочтительном варианте осуществления, указанный агонист Wnt содержит R-спондин 1 или состоит из него. R-спондин 1 предпочтительно добавляют в базальную культуральную среду в концентрации по меньшей мере 50 нг/мл, более предпочтительно по меньшей мере 100 нг/мл, более предпочтительно по меньшей мере 200 нг/мл, более предпочтительно по меньшей мере 300 нг/мл, более предпочтительно по меньшей мере 500 нг/мл. Наиболее предпочтительная концентрация R-спондина 1 составляет приблизительно 500 нг/мл или 500 нг/мл. В процессе культивирования стволовых клеток указанный представитель семейства Wnt предпочтительно добавляют в культуральную среду раз в двое суток, а культуральную среду предпочтительно обновляют раз в четверо суток.
В предпочтительном варианте осуществления агонист Wnt выбран из группы, состоящей из R-спондина, Wnt-3a и Wnt-6. Более предпочтительно, в качестве агониста Wnt используют как R-спондин, так и Wnt-3a. Эта комбинация является особенно предпочтительной, поскольку эта комбинация неожиданно обладает синергическим эффектом на образование органоида. Предпочтительные концентрации составляют приблизительно 500 нг/мл или 500 нг/мл для R-спондина и приблизительно 100 нг/мл или 100 нг/мл для Wnt3a.
Другим компонентом, который добавляют в базальную культуральную среду, является митогенный фактор роста, выбранный из семейства факторов роста, включающего эпидермальный фактор роста (EGF, (Peprotech), трансформирующий фактор роста-альфа (TGF-альфа, Peprotech), основный фибробластный фактор роста (bFGF, Peprotech), происходящий из головного мозга нейротрофический фактор (BDNF, R&D Systems) и фактор роста кератиноцитов (KGF, Peprotech). EGF представляет собой сильнодействующий митогенный фактор для множества культивируемых эктодермальных и мезодермальных клеток и он обладает выраженным эффектом на дифференцировку определенных клеток in vivo и in vitro и некоторых фибробластов в клеточной культуре. Предшественник EGF существует в качестве связанной с мембраной молекулы, которая протеолитически расщепляется с образованием пептидного гормона из 53 аминокислот, который стимулирует клетки. Предпочтительным митогенным фактором роста является EGF. EGF предпочтительно добавляют в базальную культуральную среду в концентрации от 5 до 500 нг/мл или по меньшей мере 5 и не более чем 500 нг/мл. Предпочтительная концентрация составляет по меньшей мере 10, 20, 25, 30, 40, 45 или 50 нг/мл и не более чем 500, 450, 400, 350, 300, 250, 200, 150 или 100 нг/мл. Более предпочтительная концентрация составляет по меньшей мере 50 и не более чем 100 нг/мл. Еще более предпочтительная концентрация составляет приблизительно 50 нг/мл или 50 нг/мл. Те же концентрации можно использовать для FGF, предпочтительно для FGF10 или FGF7. Если используют более одного FGF, например FGF7 и FGF10, концентрация FGF является такой, как определено выше, и она относится к общей используемой концентрации FGF. В процессе культивирования стволовых клеток, указанный митогенный фактор роста предпочтительно добавляют в культуральную среду раз в двое суток, а культуральную среду обновляют предпочтительно раз в четверо суток. Можно использовать любой представитель семейства bFGF. Предпочтительно, используют FGF7 и/или FGF10. FGF7 также известен как KGF (фактор роста кератиноцитов). В следующем предпочтительном варианте осуществления в базальную культуральную среду добавляют комбинацию митогенных факторов роста, например, таких как EGF и KGF, или EGF и BDNF. В следующем предпочтительном варианте осуществления в базальную культуральную среду добавляют комбинацию митогенных факторов роста, например, таких как EGF и KGF, или EGF и FGF10.
Следующий вариант осуществления способа по изобретению включает культуральную среду, содержащую ингибитор Rock (Rho-киназы). Было выявлено, что добавление ингибитора Rock препятствует аноикису, особенно при культивировании единичных стволовых клеток. Указанный ингибитор Rock предпочтительно выбирают из моногидрата дигидрохлорида (R)-(+)-транс-4-(1-аминоэтил)-N-(4-пиридил)циклогексанкарбоксамида (Y-27632, Sigma-Aldrich), 5-(1,4-диазепан-1-илсульфонил)изохинолина (фасудил или HA1077, Cayman Chemical), и дигидрохлорида (S)-(+)-2-метил-1-[(4-метил-5-изохинолинил)сульфонил]-гексагидро-1H-1,4-диазепина (H-1152, Tocris Bioschience). Указанный ингибитор Rho-киназы, например Y-27632, предпочтительно добавляют в культуральную среду раз в двое суток в ходе первых семи суток культивирования указанных стволовых клеток. Предпочтительная концентрация для Y27632 составляет 10 мкМ.
В следующем варианте осуществления способ по изобретению включает культуральную среду, дополнительно содержащую агонист Notch. Было показано, что передача сигнала Notch играет важную роль в определении судьбы клетки, также как и в выживании и пролиферации клетки. Рецепторные белки Notch могут взаимодействовать с рядом связанных с поверхностью или секретируемых лигандов, включая, но не ограничиваясь ими, Delta 1, Jagged 1 и 2, и Delta-подобный 1, Delta-подобный 3, Delta-подобный 4. При связывании лиганда рецепторы Notch активируются серией событий расщепления, вовлекающей представителей семейства протеаз ADAM, а также внутримембранным расщеплением, регулируемым гамма-секретазой пресинилином. В результате происходит транслокация внутриклеточного домена Notch в ядро, где он транскрипционно активирует последующие гены. Предпочтительный агонист Notch выбирают из Jagged 1 и Delta 1, или их активного фрагмента или производного. Наиболее предпочтительным агонистом Notch является пептид DSL (Dontu et al., 2004. Breast Cancer Res 6: R605-R615) с последовательностью CDDYYYGFGCNKFCRPR. Указанный пептид DSL (ANA spec) предпочтительно используют в концентрации от 10 мкМ до 100 нМ или по меньшей мере 10 мкМ и не более чем 100 нМ. Добавление агониста Notch, особенно на первой неделе культивирования, увеличивает эффективность культивирования с коэффициентом 2-3. Указанный агонист Notch предпочтительно добавляют в культуральную среду раз в двое суток в течение первых семи суток культивирования указанных стволовых клеток.
Агонист Notch определяют как молекулу, которая стимулирует активность Notch в клетке по меньшей мере на 10%, более предпочтительно по меньшей мере на 20%, более предпочтительно по меньшей мере на 30%, более предпочтительно по меньшей мере на 50%, более предпочтительно по меньшей мере на 70%, более предпочтительно по меньшей мере на 90%, более предпочтительно по меньшей мере на 100%, относительно уровня активности Notch в отсутствие указанной молекулы. Как известно специалисту в данной области, активность Notch можно определять путем измерения транскрипционной активности Notch, например с помощью люциферазной репортерной конструкции 4xwtCBF1, как описано (Hsieh et al., 1996. Mol. Cell. Biol. 16, 952-959).
Кроме того, изобретение относится к клеточной культуральной среде, содержащей базальную среду для клеток животного или человека, в которую добавлен ингибитор морфогенетического белка кости (BMP), агонист Wnt; и от 5 до 500 нанограмм/мл или по меньшей мере 5 и не более чем 500 нанограмм/мл митогенного фактора роста, выбранного из группы, состоящей из EGF, TGFβ, KGF, FGF10 и FGF. Предпочтительно, митогенный фактор выбран из групп, состоящих из EGF, TGFβ и KGF, или из EGF, TGF-β и FGF7, или из EGF, TGFβ и FGF, или из EGF и KGF, или из EGF и FGF7, или из EGF и FGF, или из TGFβ и KGF, или из TGFβ и FGF7, или из TGFβ и FGF. EGF может быть заменен на TGFβ. Позднее идентифицировано несколько предпочтительных культуральных сред, в зависимости от органоида, подлежащего получению. Клеточная культуральная среда согласно изобретению позволяет выживание и/или пролиферацию и/или дифференцировку эпителиальных стволовых клеток или выделенных крипт на внеклеточном матриксе. Термин "клеточная культуральная среда" является синонимом терминам "среда", "культуральная среда" или "клеточная среда".
В предпочтительном способе по изобретению, первая культуральная среда содержит Noggin в качестве ингибитора BMP, как эпидермальный фактор роста, так и фактор роста кератиноцитов, в качестве митогенных факторов роста, и R-спондин 1 в качестве агониста Wnt, дополненные B27, N2 и N-ацетилцистеином. KGF может быть заменен на FGF, или на FGF10. Также в эту первую среду можно добавлять [Leu15]-гастрин I, экзендин и/или никотинамид.
В другом предпочтительном варианте осуществления указанная культуральная среда, называемая второй культуральной средой, идентична указанной первой среде, за исключением того, что в ней нет Noggin и предпочтительно нет [Leu15]-гастрина I, экзендина и/или никотинамида. Таким образом, указанная вторая культуральная среда содержит как эпидермальный фактор роста, так и фактор роста кератиноцитов в качестве митогенных факторов роста, и R-спондин 1 в качестве агониста Wnt, дополненные B27, N2 и N-ацетилцистеином. KGF может быть заменен на FGF или FGF10. Эти две клеточные культуральные среды поддерживают фрагменты поджелудочной железы, содержащие стволовые клетки, которые выращивают в этих средах во внеклеточном матриксе Matrigel, с образованием органоидов поджелудочной железы, содержащих подобные островкам поджелудочной железы структуры на внеклеточном матриксе. Вторая среда без Noggin представляет собой минимальную среду, в то время как первая среда с Noggin дает улучшенную среду для роста фрагментов поджелудочной железы. Среда для роста представляет собой среду, которая предпочтительно обеспечивает выживание и/или пролиферацию клеток в процессе культивирования в течение по меньшей мере двух суток.
Была разработана третья среда, которая способна стимулировать, индуцировать дифференцировку клеток в направлении органоидов поджелудочной железы в течение по меньшей мере 5 суток. Одним из предпочтительных маркеров образования органоида поджелудочной железы является нейрогенин-3, экспрессию которого можно выявлять способом RT-PCR или иммуногистохимией. Среду для дифференцировки, как например, третью или четвертую среду называют функциональной, когда нейрогенин-3 может быть выявлен способом RT-PCR или иммуногистохимией после по меньшей мере пяти суток культивирования в указанной среде. Эту стадию дифференцировки предпочтительно проводят после первой стадии выращивания в среде, такой как первая или вторая среда, как определено выше. Эта третья среда идентична второй среде, идентифицированной выше, за исключением того, что в ней нет FGF или KGF или FGF10. Эта третья среда содержит эпидермальный фактор роста и R-спондин 1 в качестве агониста Wnt, дополненные B27, N2 и N-ацетилцистеином.
Была разработана четвертая среда, которая идентична указанной первой среде, где четвертая среда также дополнена [Leu15]-гастрином I и/или экзендином. Указанная третья среда представляет собой минимальную среду для дифференцировки, в то время как указанная четвертая среда представляет собой улучшенную среду для дифференцировки. Среда для дифференцировки представляет собой среду, которая предпочтительно индуцирует или стимулирует специфическую дифференцировку клеток в течение по меньшей мере пяти суток в культуре. В случае органоида поджелудочной железы, дифференцировку можно определять путем детекции присутствия специфического маркера, ассоциированного с ростком поджелудочной железы, как определено в настоящем документе выше. Примеры других маркеров, ассоциированных с ростком поджелудочной железы, включают секрецию инсулина, которая поддается детекции посредством RT-PCR или иммуногистохимии после культивирования в течение по меньшей мере 7, 8, 9, 10 суток в среде для дифференцировки.
Таким образом, в предпочтительном способе получения и/или культивирования органоида поджелудочной железы, эпителиальные стволовые клетки, выделенные фрагменты ткани, содержащие указанные эпителиальные стволовые клетки, или клетки аденомы культивируют на первой стадии либо в первой, либо во второй среде, с последующей второй стадией либо в третьей, либо в четвертой среде. Первая стадия может длиться в течение по меньшей мере двух недель и более. Первую стадию можно проводить в течение более чем 1, 2, 3, 4, 5, 6, 7, 8, 9 или более чем 10 месяцев. Вторая стадия может длиться 8, 9, 10, 11, 12, 13, 14, 15, 16 суток или более. Каждую стадию предпочтительно проводят с использованием внеклеточного матрикса, как определено в настоящем документе. Предпочтительные концентрации каждого соединения, присутствующего в каждой среде, уже определены в настоящем документе в описании или в примерах. В предпочтительном варианте осуществления, если органоид поджелудочной железы подлежит применению в регенеративной медицине, начинают с эпителиальных клеток или с выделенного фрагмента поджелудочной железы. В другом предпочтительном варианте осуществления, если органоид поджелудочной железы подлежит применению в качестве системы для разработки лекарственных средств, начинают с аденомы. Таким образом, органоид поджелудочной железы, получаемый способом по изобретению, представляет собой следующий аспект изобретения. Таким образом, в следующем аспекте изобретение относится к первой, второй, третьей, четвертой среде, как определено в настоящем документе.
Насколько известно авторам изобретения, впервые был получен органоид поджелудочной железы, который является функциональным и живым после по меньшей мере десяти месяцев культивирования (см. экспериментальную часть). Функциональность предпочтительно охарактеризовывают по секреции инсулина. Поскольку конечное количество полученных органоидов поджелудочной железы коррелирует с длительностью культивирования, специалисту в данной области понятно, что изобретение представляет собой пионерское изобретение и потенциально открывает новые возможности, например, в регенеративной медицине.
Таким образом, в предпочтительном способе получения и/или культивирования органоида поджелудочной железы на первой стадии эпителиальные стволовые клетки, выделенные фрагменты ткани, содержащие указанные эпителиальные стволовые клетки, или клетки аденомы культивируют в контакте с внеклеточным матриксом в среде, содержащей EGF, KGF или FGF, и R-спондин 1 в качестве агониста Wnt, дополненной B27, N2 и N-ацетилцистеином, а затем на второй стадии в среде, содержащей EGF и R-спондин 1 в качестве агониста Wnt, дополненной B27, N2 и N-ацетилцистеином.
В следующем предпочтительном способе по изобретению культуральная среда содержит Noggin в качестве ингибитора BMP, эпидермальный фактор роста в качестве митогенного фактора роста, R-спондин 1 и/или Wnt3a в качестве агониста Wnt. Эта клеточная культуральная среда поддерживает культивирование выделенных крипт тонкого кишечника в 3-мерных культурах, содержащих Matrigel в качестве внеклеточного матрикса.
В следующем предпочтительном способе по изобретению культуральная среда содержит Noggin в качестве ингибитора BMP, эпидермальный фактор роста в качестве митогенного фактора роста, R-спондин 1 в качестве агониста Wnt, пептид Jagged-DSL в качестве агониста Notch и ингибитор Rho-киназы Y-27632. Эта клеточная культуральная среда поддерживает культивирование выделенных единичных эпителиальных стволовых клеток в 3-мерных культурах, содержащих Matrigel в качестве внеклеточного матрикса.
В следующем предпочтительном способе по изобретению культуральная среда содержит Noggin в качестве ингибитора BMP, эпидермальный фактор роста и/или BDNF в качестве митогенных факторов роста, R-спондин 1 и/или Wnt-3a в качестве агонистов Wnt, дополненные по меньшей мере одним из B27, N2 и N-ацетилцистеина. Wnt-3a представляет собой предпочтительный агонист Wnt в этом предпочтительном способе. Эта среда для культивирования клеток поддерживает культивирование выделенных крипт толстого кишечника в 3-мерных культурах, содержащих Matrigel в качестве внеклеточного матрикса. Эта среда способна стимулировать выживание и/или пролиферацию и/или дифференцировку клеток в течение по меньшей мере двух суток культивирования. Предпочтительный маркер дифференцировки в сторону образования крипты толстого кишечника может быть выбран из следующей группы: щелочная фосфатаза, указывающая на присутствие энтероцитов, Muc2, указывающий на присутствие бокаловидных клеток, и нейрогенин-3 или хромогранин, указывающие не присутствие эндокринных клеток. Экспрессию каждого из этих маркеров можно выявлять с помощью RT-PCR или иммуногистохимии. Среда, функциональная для стимуляции выживания и/или пролиферации и/или дифференцировки клеток для получения крипты толстого кишечника является такой, что по меньшей мере один из идентифицированных маркеров может быть выявлен после культивирования в течение меньшей мере 2, 3, 4, 5, 6, 7, 8, 9, 10 суток или более. Предпочтительная среда содержит Noggin в качестве ингибитора BMP, эпидермальный фактор роста в качестве митогенного фактора роста, и R-спондин 1 и/или Wnt-3a в качестве агонистов Wnt, дополненные по меньшей мере одним из B27, N2 и N-ацетилцистеина. Эту среду называют пятой средой по изобретению, которая соответствует следующему аспекту изобретения.
Таким образом, в предпочтительном способе получения и/или культивирования крипты толстого кишечника, эпителиальные стволовые клетки, выделенные фрагменты ткани, содержащие указанные эпителиальные стволовые клетки, или клетки аденомы культивируют в среде, как указано выше, предпочтительно в пятой среде. Этот способ предпочтительно проводят с использованием внеклеточного матрикса, как определено в настоящем документе. Предпочтительные концентрации каждого соединения, присутствующего в указанной среде, уже были определены в настоящем документе в описании или в примерах. Таким образом, крипта толстого кишечника, получаемая способом по изобретению, является следующим аспектом изобретения. Насколько известно авторам изобретения, впервые была получена крипта толстого кишечника, которая является функциональной и живой после культивирования в течение по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 месяцев (см. экспериментальную часть). Функциональность предпочтительно характеризуется присутствием по меньшей мере одного из маркеров, указанных выше. Изобретение является пионерским изобретением и потенциально открывает новые возможности, например, в регенеративной медицине.
Таким образом, в предпочтительном способе получения и/или культивирования крипты толстого кишечника, эпителиальные стволовые клетки, выделенные фрагменты ткани, содержащие указанные эпителиальные стволовые клетки, или клетки аденомы культивируют в контакте с внеклеточным матриксом в среде, содержащей Noggin, EGF, и R-спондин 1 и/или Wnt-3 в качестве агониста Wnt, дополненной B27, N2 и N-ацетилцистеином.
В следующем предпочтительном способе по изобретению культуральная среда содержит Noggin в качестве ингибитора BMP, эпидермальный фактор роста в качестве митогенного фактора роста, R-спондин 1 в качестве агониста Wnt, дополненные либо Wnt-3a, либо KGF, и, кроме того, содержит B27, N2, N-ацетилцистеин. Эту среду называют шестой средой и, соответственно, она соответствует следующему аспекту изобретения. KGF можно заменять на FGF или FGF10. Эта среда предпочтительно содержит Noggin в качестве ингибитора BMP, эпидермальный фактор роста и FGF10 в качестве митогенного фактора роста, R-спондин 1 и Wnt-3a в качестве агониста Wnt, и, кроме того, содержит B27, N2, N-ацетилцистеин. FGF10 является предпочтительным в качестве FGF, поскольку он дает лучшие результаты, чем, например, FGF7 (фиг. 32). Эта клеточная культуральная среда поддерживает культивирование выделенных фрагментов желудка или органоида желудка в 3-мерных культурах, содержащих Matrigel в качестве внеклеточного матрикса. Эта шестая среда представляет собой среду для роста фрагмента желудка. Среда для роста представляет собой среду, которая предпочтительно стимулирует выживание и/или пролиферацию клеток в процессе культивирования в течение по меньшей мере двух суток. Была разработана дополнительная среда, т.е. седьмая среда, которая способна стимулировать, индуцировать дифференцировку клеток в направлении органоида желудка или фрагмента желудка в течение по меньшей мере 2 суток. Эта седьмая среда идентична шестой среде, за исключением того, что концентрация Wnt-3a снижена по сравнению с его концентрацией, присутствующей в шестой среде. Концентрация является сниженной по меньшей мере на 25%, 50%, 100%, 200%, 300%, 400%, 500%, 600% или более по сравнению с концентрацией Wnt-3a, присутствующей в шестой среде. Эта седьмая среда содержит эпидермальный фактор роста и R-спондин 1 и Wnt-3a в качестве агониста Wnt, Noggin и FGF10, дополненные B27, N2, N-ацетилцистеином и гастрином. Гастрин предпочтительно используют в концентрации 1 нМ.
Указанная седьмая среда представляет собой среду для дифференцировки. Среда для дифференцировки представляет собой среду, которая предпочтительно индуцирует или стимулирует специфическую дифференцировку клеток в процессе культивирования в течение по меньшей мере двух, 3, 4, 5, 6, 7, 8, 9, 10 суток или более. В случае органоида желудка или фрагмента желудка, дифференцировку можно измерять путем детекции присутствия специфического маркера, ассоциированного с ростком желудка. Примеры маркеров, ассоциированные с ростком желудка, включают MUC5AC (маркер пит-клеток), гастрин и/или соматостатин (оба маркеры эндокринных клеток). Присутствие по меньшей мере одного из указанных маркеров предпочтительно определяют с использованием RT-PCR и/или иммуногистохимии или иммунофлуоресценции. Присутствие по меньшей мере одного из этих маркеров предпочтительно поддается детекции после по меньшей мере 6 суток в условиях дифференцировки, более предпочтительно по меньшей мере 10 суток. Среду для дифференцировки, как например, седьмую среду, называют функциональной, когда по меньшей мере один из указанных выше маркеров может быть выявлен с помощью RT-PCR или иммуногистохимии после культивирования в течение по меньшей мере шести суток в указанной среде. Эту стадию дифференцировки предпочтительно проводят после первой стадии выращивания в среде, такой как шестая среда, как определено выше.
Таким образом, в предпочтительном способе получения и/или культивирования фрагмента желудка, эпителиальные стволовые клетки, выделенные фрагменты ткани, содержащие указанные эпителиальные стволовые клетки, или клетки аденомы культивируют на первой стадии в такой шестой среде, а затем на второй стадии в такой седьмой среде. Каждую стадию предпочтительно проводят с использованием внеклеточного матрикса, как определено в настоящем документе. Первая стадия может длиться по меньшей мере 3 суток и более. Первую стадию можно проводить в течение более чем 3, 4, 5, 6, 7, 8, 9, суток. Вторая стадия может длиться 6, 7, 8, 9, 10, 11, 12, 13, 14, 15. 16 суток или более. Предпочтительные концентрации каждого соединения, присутствующего в каждой среде, уже были определены в настоящем документе в описании или в примерах. Таким образом, фрагмент желудка, получаемый способом по изобретению, представляет собой следующий аспект изобретения.
Таким образом, в предпочтительном способе получения и/или культивирования фрагмента желудка эпителиальные стволовые клетки, выделенные фрагменты ткани, содержащие указанные эпителиальные стволовые клетки, или клетки аденомы культивируют в контакте с внеклеточным матриксом на первой стадии в среде, содержащей Noggin в качестве ингибитора BMP, эпидермальный фактор роста и FGF10 в качестве митогенного фактора роста, R-спондин 1 и Wnt-3a в качестве агониста Wnt, и, кроме того, содержащей B27, N2, N-ацетилцистеин, а затем на второй стадии в среде, содержащей эпидермальный фактор роста и R-спондин 1 и Wnt-3a в качестве агониста Wnt, Noggin и FGF10, дополненные B27, N2 и N-ацетилцистеином, где концентрация Wnt-3 на второй стадии снижена по сравнению с концентрацией Wnt-3a на первой стадии.
В следующем предпочтительном способе по изобретению культуральная среда содержит Noggin в качестве ингибитора BMP и эпидермальный фактор роста в качестве митогенного фактора роста. Эта клеточная культуральная среда поддерживает культивирование выделенных фрагментов аденомы или выделенных отдельных клеток аденомы в 3-мерных культурах, содержащих Matrigel в качестве внеклеточного матрикса.
Лиганд, например, такой как Wnt3a, может быть добавлен в культуральную среду извне. Альтернативно, лиганд экспрессируют в клеточной линии путем трансфекции или инфицирования клеточной линии пригодной конструкцией, экспрессирующей указанный лиганд. Указанную клеточную линию культивируют и культуральную среду, содержащую секретированный лиганд, собирают через подходящие промежутки времени. Например, клетки начинают продуцировать Wnt3a, как только они достигают смыкания монослоя и перестают расти. В качестве контроля используют культуральную среду от клеток, которые не были трансфицированы или инфицированы указанной экспрессирующей конструкцией. Кондиционированную среду собирают и тестируют, например в анализе, где экспрессия люциферазы контролируется отвечающими элементами TCF для тестирования в отношения присутствия агониста Wnt, такого как Wnt3a (Korinek et al., 1997. Science 275: 1784-1787). При использовании для регенерации ткани среду разбавляют. Как известно специалистам в данной области, добавление избытка лиганда иногда является настолько же вредоносным для культуры, как и добавление недостаточного количества лиганда. Таким образом, истинное разведение кондиционированной среды будет зависеть от количества лиганда, которое определено в тесте.
Кроме того, изобретение относится к применению культуральной среды по изобретению для культивирования эпителиальных стволовых клеток или выделенных структур органоидов, которые содержат стволовые клетки на внеклеточном матриксе, причем указанные стволовые клетки предпочтительно не содержат эмбриональных стволовых клеток человека. Предпочтительными являются взрослые стволовые клетки человека. Более того, отдельные отсортированные эпителиальные стволовые клетки из тонкого кишечника, толстого кишечника и желудка также способны инициировать эти 3-мерные органоиды в культуральной среде по изобретению. Кроме того, изобретение относится к применению культуральной среды по изобретению для культивирования фрагментов поджелудочной железы, содержащих стволовые клетки, которые образуют органоиды поджелудочной железы, содержащие структуры, подобные островкам поджелудочной железы.
Предпочтительно, чтобы указанные стволовые клетки представляли собой стволовые клетки поджелудочной железы, желудка, кишечника или толстого кишечника, причем наиболее предпочтительными стволовыми клетками являются стволовые клетки тонкого кишечника.
Культуральная среда по изобретению позволила установить условия длительного культивирования, в которых единичные крипты претерпевают множество делений крипт, одновременно образуя подобные ворсинкам эпителиальные домены, в которых присутствуют все типы дифференцированных клеток. Использование способа культивирования по изобретению сделало возможными периоды культивирования по меньшей мере семь месяцев, по меньшей мере восемь месяцев, по меньшей мере девять месяцев, по меньшей мере десять месяцев.
Культивированные крипты претерпевают значительные морфологические изменения после извлечения их из культуры. Верхнее отверстие свежевыделенных крипт закрывается, и эта область постепенно расширяется и заполняется апоптотическими клетками, подобно тому, как апоптотические клетки отделяются на ворсинке. Было выявлено, что область крипты претерпевает непрерывное почкование, которое создает дополнительные крипты, и этот процесс напоминает деление крипт. Подобные криптам расширения содержат все типы дифференцированных эпителиальных клеток, включая пролиферативные клетки, клетки Панета, энтероциты и бокаловидные клетки. В органоидах не было идентифицировано миофибробластов или других неэпителиальных клеток ни на одной из стадий.
Расширение почкующихся структур крипты привело к органоидам, содержащим >40 подобных криптам структур, окружающих центральный просвет, выстилаемый эпителием, подобным эпителию ворсинок, и заполненный апоптотическими клеточными тельцами. Органоиды крипта-ворсинка содержат центральный просвет, выстилаемый эпителием, подобным эпителию ворсинок. Просвет открывается через последовательные временные интервалы, высвобождая содержимое в среду. Органоиды можно пассировать и поддерживать в культуре в течение по меньшей мере 6 месяцев без утраты важных характеристик. Пассирование предпочтительно вовлекает фрагментацию органоидов вручную.
Сходная структура органоида крипта-ворсинка образуется, когда культивируют отдельные эпителиальные стволовые клетки. После приблизительно одной недели образуются структуры, которые высоко сходны со структурами органоидов крипта-ворсинка, которые получают с помощью целых крипт. Гистологический анализ этих органоидов также показал сохранение основной структуры крипта-ворсиснки, присутствие всех типов дифференцированных клеток и отсутствие неэпителиальных элементов.
Таким образом, в одном аспекте изобретение относится к органоидам крипта-ворсинка, содержащим центральный просвет, выстилаемый эпителием, подобным эпителию ворсинок, которые получают культивированием эпителиальных стволовых клеток или выделенных крипт в культуральной среде по изобретению. Предпочтительно, указанный органоид крипта-ворсинка получают с использованием способа по изобретению.
В следующем аспекте изобретение относится к органоидам поджелудочной железы, полученным или получаемым культивированием фрагментов поджелудочной железы способом по изобретению.
Приблизительно 20% органоидов поджелудочной железы образуют почкующиеся структуры через 7 суток после начала культивирования. Протоки поджелудочной железы быстро пролиферируют, в противоположность ацинозной ткани, которая растет очень медленно. После пассирования органоидов поджелудочной железы наблюдают структуры, подобные островкам поджелудочной железы, которые секретируют инсулин и которые напоминают островки Лангерганса поджелудочной железы, которые присутствуют в здоровой ткани поджелудочной железы. Кроме того, изобретение относится к органоиду желудка, содержащему центральный просвет. Предпочтительно указанный органоид желудка можно получать способом по изобретению.
Другие факторы роста, которые можно добавлять в культуральную среду, например для увеличения присутствия островков поджелудочной железы в органоидах или для дальнейшего поддержания культивирования выделенных фрагментов, таких как фрагменты желудка, включают циклопамин (ингибитор Sonic-hedgehog, Tocris Bioscience), активин, GLP (глюкагон-подобный пептид) и его производное (экзендин 4, California Peptide Research), гастрин (Genscript), агонист Notch (пептид Jagged, Ana Spec), никотинамид и агонист Wnt, такой как Wnt-3a. Wnt-3a является привлекательным для применения, когда культуру начинают с единичной клетки.
Кроме того, изобретение относится к коллекциям органоидов крипта-ворсинка, желудка или поджелудочной железы, каждая из которых содержит более 10, предпочтительно более 20, более предпочтительно более 40 органоидов. Органоиды крипта-ворсинка окружают центральный просвет, выстилаемый эпителием, подобным эпителию ворсинок. Просвет заполнен апоптотическими клеточными тельцами. Клетки в органоидах крипта-ворсинка являются полярными, причем стволовые клетки располагаются в основании структур. Верхняя часть подобных криптам структур содержит апоптотические клетки, которые попадают в просвет. Указанная коллекция органоидов крипта-ворсинка предпочтительно содержит по меньшей мере 10% жизнеспособных клеток, более предпочтительно по меньшей мере 20% жизнеспособных клеток, более предпочтительно по меньшей мере 50% жизнеспособных клеток, более предпочтительно по меньшей мере 60% жизнеспособных клеток, более предпочтительно по меньшей мере 70% жизнеспособных клеток, более предпочтительно по меньшей мере 80% жизнеспособных клеток, более предпочтительно по меньшей мере 90% жизнеспособных клеток. Жизнеспособность клеток можно оценивать с использованием окрашивания Hoechst или окрашивания йодидом пропидия в FACS.
В следующем аспекте изобретение относится к применению органоидов крипта-ворсинка, органоидов желудка или органоидов поджелудочной железы по изобретению в скрининге при разработке лекарственных средств, в анализе токсичности или в регенеративной медицине.
Для целей высокой производительности указанные органоиды крипта-ворсинка, желудка или поджелудочной железы культивируют в многолуночных планшетах, например, таких как 96-луночные планшеты или 384-луночные планшеты. Для идентификации молекулы, которая влияет на указанные органоиды, используют библиотеки молекул. Предпочтительные библиотеки включают библиотеки фрагментов антител, библиотеки пептидного фагового дисплея, пептидные библиотеки (например LOPAPTM, Sigma Aldrich), библиотеки липидов (BioMol), библиотеки синтетических соединений (например LOP ACTM, Sigma Aldrich) или библиотеки природных соединений (Specs, TimTec). Более того, можно использовать генетические библиотеки, которые индуцируют или подавляют экспрессию одного или нескольких генов в потомках клеток аденомы. Эти библиотеки включают библиотеки кДНК, библиотеки антисмысловых молекул, и библиотеки siRNA или других некодирующих РНК. Клетки предпочтительно подвергают воздействию множества концентраций тестируемого вещества в течение определенного периода времени. В конце периода воздействия культуры оценивают. Термин "влияние" используют для обозначения любого изменения в клетке, включая, но не ограничиваясь ими, снижение или утрату пролиферации, морфологическое изменение и гибель клетки. Указанные органоиды крипта-ворсинка, желудка или поджелудочной железы также можно использовать для идентификации лекарственных средств, которые специфично нацелены на клетки эпителиальной карциномы, но не на указанные органоиды крипта-ворсинка, желудка или поджелудочной железы.
Указанные органоиды крипта-ворсинка, желудка или поджелудочной железы могут, кроме того, заменить применение клеточных линий, таких как клетки Caco-2, в анализах токсичности потенциальных новых лекарственных средств или известных или новых пищевых добавок.
Более того, указанные органоиды крипта-ворсинка, желудка или поджелудочной железы можно использовать для культивирования патогена, такого как норовирус, для которого в настоящее время не существует подходящей тканевой культуры или модели на животных.
Культуры, содержащие органоиды крипта-ворсинка, пригодны в регенеративной медицине, например при восстановлении эпителия кишечника после облучения и/или хирургической операции, при восстановлении эпителия кишечника у пациентов, страдающих воспалительным заболеванием кишечника, таким как болезнь Крона и язвенный колит, и при восстановлении эпителия кишечника у пациентов, страдающих синдромом укороченной тонкой кишки. Следующим применением является применение в восстановлении эпителия кишечника у пациентов с наследственными заболеваниями тонкого кишечника/толстого кишечника. Культуры, содержащие органоиды поджелудочной железы, также пригодны в регенеративной медицине, например в качестве имплантатов после резекции поджелудочной железы или ее части и для лечения диабета, такого как диабет I и диабет II.
В альтернативном варианте осуществления размножившиеся эпителиальные стволовые клетки перепрограммируют на путь родственных тканей, например, клеток поджелудочной железы, включая β-клетки поджелудочной железы, и клеток печени. К настоящему времени, невозможно регенерировать клетки поджелудочной железы или клетки печени из взрослых стволовых клеток. Способы культивирования по настоящему изобретению обеспечивают анализ факторов, которые осуществляют транс-дифференцировку близко родственных эпителиальных стволовых клеток в клетки поджелудочной железы, включая β-клетки поджелудочной железы, и клетки печени.
Специалисту будет понятно, что в способе, направленном на репарацию поврежденной или пораженной заболеванием ткани, кроме того, можно использовать генную терапию. В целях доставки генетической информации, такой как ДНК и/или РНК, в стволовые клетки можно использовать, например, аденовирусный или ретровирусный носитель для доставки генов. Специалист в данной области может заменить или восстановить конкретные гены, на которые нацелена генная терапия. Например, нормальный ген можно встраивать в неспецифическую область в геноме для замены нефункционального гена. В другом примере неправильную последовательность гена можно заменять нормальной последовательностью гена посредством гомологичной рекомбинации. Альтернативно селективная возвратная мутация может возвращать ген к его нормальному функционированию. Следующим примером является изменение регуляции (степени, с которой ген включается или выключается) конкретного гена. Предпочтительно, стволовые клетки обрабатывают ex vivo с помощью подхода генной терапии и впоследствии переносят млекопитающему, предпочтительно человеку, нуждающемуся в лечении.
В другом аспекте изобретение относится к способу культивирования эпителиальных клеток аденомы, включающий предоставление внеклеточного матрикса, прикрепление эпителиальных клеток аденомы к внеклеточному матриксу, культивирование клеток в присутствии клеточной культуральной среды, содержащей базальную среду для клеток животного или человека, в которую добавлен ингибитор морфогенетического белка кости (BMP), и от 5 до 500 нграмм/мл или по меньшей мере 5 и не более чем 500 нграмм/мл митогенного фактора роста, выбранного из EGF, TGF-альфа и KGF. KGF можно заменять на FGF или FGF10.
Эпителиальная клетка аденомы толстого кишечника содержит изменение в гене, кодирующем белок APC, которое приводит к менее эффективной деградации внутриклеточного β-катенина комплексом белков, содержащим APC. Другие мутации, распространенные в аденомах толстого кишечника, включают мутации в β-катенине или Axin2. Общим результатом является усиление передачи сигнала TCF/LEF вследствие повышенного количества β-катенина в ядре. Было выявлено, что культуральная среда без агониста Wnt является достаточной для пролиферации клеток аденомы.
Указанные клетки аденомы можно выделять из эпителиальной аденомы способами, известными в данной области, включающими применение диссоциирующих средств, таких как EDTA. Альтернативно единичные положительные по Lgr5 или Lgr-6 клетки можно выделять из аденомы с использованием Lgr5-связывающего соединения, а затем магнитных гранул или FACS-анализа.
Кроме того, изобретение относится к потомкам эпителиальных клеток аденомы, которые культивировали в присутствии клеточной культуральной среды, содержащей базальную среду для клеток животного или человека, в которую добавлен ингибитор морфогенетического белка кости (BMP) и от 5 до 500 нграмм/мл или по меньшей мере 5 и не более чем 500 нграмм/мл эпидермального фактора роста (EGF). Культивированные клетки аденомы неспособны развивать полярную трехмерную структуру, такую как структура, подобная структуре крипта-ворсинка. Вместо этого, клетки аденомы формируют подобные сфере структуры, в которых клетки случайным образом ориентированы в направлении либо периферии, либо центрального просвета. Отсутствуют признаки дифференцировки в другие типы эпителиальных клеток. Этот результат указывает на роль APC в трехмерной организации структуры, подобной структуре крипта-ворсинка.
Кроме того, изобретение относится к применению потомков клеток аденомы для направленного скрининга при разработке лекарственных средств в целях идентификации лекарственного средства, которое специфично влияет на клетки аденомы по сравнению с размножившимися нормальными эпителиальными клетками, которые культивируют в той же культуральной среде. Для целей высокой производительности, потомков клеток аденомы культивируют в многолуночных планшетах, например, таких как 96-луночные планшеты или 384-луночные планшеты. Для идентификации молекулы, которая влияет на указанных потомков, используют библиотеки молекул. Предпочтительные библиотеки включают библиотеки фрагментов антител, библиотеки пептидного фагового дисплея, пептидные библиотеки (например LOPAPTM, Sigma Aldrich), библиотеки липидов (BioMol), библиотеки синтетических соединений (например LOP ACTM, Sigma Aldrich) или библиотеки природных соединений (Specs, TimTec). Более того, можно использовать генетические библиотеки, которые индуцируют или подавляют экспрессию одного или нескольких генов в потомках клеток аденомы. Эти библиотеки включают библиотеки кДНК, библиотеки антисмысловых молекул, и библиотеки siRNA или других некодирующих РНК. Соединение, которое влияет на клетки аденомы, впоследствии или параллельно тестируют в отношении влияния на размножившиеся нормальные эпителиальные клетки. Термин "влияние" используют для обозначения любого изменения в клетке, включая, но не ограничиваясь ими, снижение или утрату пролиферации, морфологическое изменение и гибель клетки. Указанных потомков также можно использовать для идентификации лекарственных средств, которые специфично нацелены на эпителиальные клетки карциномы, по сравнению с эпителиальными клетками аденомы, включая обратное развитие клеток карциномы.
Понятно, что указанных потомков также можно использовать в высокопроизводительном подходе для определения in vitro метаболической стабильности и метаболических профилей лекарственных средств-кандидатов.
Более того, изобретение относится к применению потомков клеток аденомы по изобретению, органоидов поджелудочной железы, органоидов желудка и органоидов крипта-ворсинка по изобретению, в анализах токсичности. Указанных потоков и органоиды крипта-ворсинка легко культивировать и они более сходны с первичными эпителиальными клетками, чем, например, эпителиальные клеточные линии, такие как Caco-2 (ATCC HTB-37), I-407 (ATCC CCL6) и XBF (ATCC CRL 8808), которые в настоящее время используют в анализах токсичности. Ожидается, что результаты анализов токсичности, получаемые с помощью первичных культур аденомы или с помощью органоидов крипта-ворсинка, будут более сходны с результатами, получаемыми у пациентов. Для определения органоспецифической цитотоксичности используют клеточный тест токсичности. Соединения, которые тестируют в указанном тесте, включают химиопрофилактические средства от злокачественной опухоли, химические соединения, действующие на окружающую среду, пищевые добавки и потенциальные токсические вещества. Клетки подвергают воздействию множества концентраций тестируемого вещества в течение определенного периода времени. Диапазоны концентраций для тестируемых средств в анализе определяют в предварительном анализе с использованием воздействия в течение пяти суток и логарифмических разведений от наиболее высокой концентрации растворимого вещества. В конце периода воздействия культуры оценивают в отношении ингибирования роста. Данные анализируют для определения концентрации, которая ингибирует конечный результат на 50 процентов (TC50).
В этом документе и в формуле изобретения глагол "содержать" и его спряжения используют в его неограничивающем значении для обозначения, что объекты, следующие за этим словом, включены, но объекты, которые конкретно не указаны, не исключены. Кроме того, глагол "состоять" может быть заменен на "по существу состоять", означающий, что продукт, как определено в настоящем документе, может содержать дополнительный компонент(ы) относительно продуктов, конкретно указанных, причем указанный дополнительный компонент(ы) не изменяет уникальных характеристик изобретения. Кроме того, способ, как определено в настоящем документе, может включать дополнительную стадию(и) по сравнению со способами, конкретно указанными, причем указанная дополнительная стадия(и) не изменяет уникальных характеристик изобретения. Кроме того, указание на элемент с помощью единственного числа не исключает возможности, что присутствует более одного элемента, если контекст явно не требует, чтобы был один и только один из элементов. Таким образом, форма единственного числа означает "по меньшей мере один". Слово "приблизительно" или "примерно", когда его используют применительно к числовой величине (приблизительно 10) предпочтительно означает, что величина может представлять собой данную величину 10 плюс или минус 1% величины.
Все патентные и литературные ссылки, цитированные в настоящем описании, включены в настоящий документ в качестве ссылок в полном объеме.
Следующие примеры предназначены только для иллюстративных целей и не предназначены для ограничения объема настоящего изобретения никоим образом.
Описание фигур
Фигура 1. Потребность культуры крипты в факторах роста.
a: 500 крипт высевали с EGF (E, 0-50 нг/мл) и R-спондином 1 (R, 0-500 нг/мл) в трех экземплярах; органоиды крипт подсчитывали через 7 суток после посева. b: 500 Крипты/органоиды крипт культивировали с EGF (50 нг/мл) и R-спондином 1 (500 нг/мл) с указанными количествами Noggin и проводили 3 пассирования. Органоиды крипт подсчитывали при каждом пассировании. Эксперимент повторили три раза со сравнимыми результатами.
Фигура 2. Установление культуральной системы крипт кишечника.
a: Выращивание с течением времени выделенной единичной крипты в органоид. Контрастное изображение с дифференциальной интерференцией выявило клетки Панеты, содержащие гранулы, в основании крипт (стрелки). b, c: Единичные выделенные крипты эффективно образуют органоиды крипт. На протяжении повторяющегося деления крипт структуры образуют многочисленные подобные осьминогу органоиды на 14 сутки. d: 3D-реконструированное конфокальное изображение отдельного органоида после культивирования в течение 3 недель. Lgr5- GFP+ стволовые клетки (светло-серые) локализуются на верхушки подобных криптам доменов. Контрастное окрашивание на ДНК: ToPro-3 (темно-серый). e: Схематическое представление органоида крипты. Органоид состоит из центрального просвета, выстилаемого эпителием, подобным эпителию ворсинок, и ряда окружающих подобных криптам доменов. Темно-серые клетки на вершине домена крипты указывают на положение Lgr5+ стволовых клеток, которые присутствуют в каждом домене крипты. Измерительная линейка показывает 50 мкм.
Фигура 3. Кластерный анализ профиля экспрессии генов
Кластерный анализ уровней экспрессии с использованием свежевыделенных крипт толстого и тонкого кишечника, а также органоидов тонкого кишечника показал высокую степень сходства между органоидами тонкого кишечника, и тканью, из которой они происходили, криптами тонкого кишечника. Крипты толстого кишечника кластеризовались на отдельной ветви, указывая на отличающийся паттерн экспрессии генов этой высоко сходной ткани. Следует отметить, что была увеличена экспрессия только 1,2% всех экспрессированных генов относительно крипт тонкого кишечника, в то время как, наоборот, в криптах тонкого кишечника была увеличена экспрессия 2% генов. Анализ каскада этих отличающихся генов авторами изобретения выявил специфическое присутствие признака лимфоцитов в свежевыделенных криптах, в то время как нельзя было идентифицировать выраженного каскада в небольшом количестве генов, экспрессия которых была увеличена в органоидах (не показано). Авторы настоящего изобретения сделали вывод, что последняя группа представляет собой биологический шум, в то время как признак лимфоцитов происходит из контаминирующих внутриэпителиальных иммунных клеток, утраченных при культивировании.
Фигура 4. Органоиды крипт сохраняют основные характеристики крипты-ворсинок.
a-e: Код активации Wnt сохраняется в доменах крипт. a: Ядерный β-катенин (темно-серый, стрелки) наблюдали только в доменах крипт. Изображение более высокого разрешения представлено на фиг. 5. Звездочка, matrigel; Lu, просвет. b: EphB2 (светло-серый) экспрессируется градиентно на CBC-клетках и TA-клетках. Следует отметить, что Lgr5-GFP+ стволовые клетки указаны белой стрелкой. c: Каспаза-3+ апоптотические клетки (темно-серый, стрелки), выходящие в центральный просвет, выстилаемый энтероцитами. d: 40 хромосом в материале клеток из культуры крипты возрастом >3 месяцев. e-g: Прослеживание происхождения Lgr5+ стволовых клеток in vitro. e: Крипты из репортерных мышей Lgr5-EGFP-ires-CreERT2/Rosa26-LacZ стимулировали тамоксифеном in vitro в течение 12 ч, и культивировали в течение указанного количества суток. Окрашивание по LacZ (темно-серый) показывает, что рассеянные единичные LacZ+ клетки (сутки 1) создавали целые LacZ+ крипты in vitro (сутки 2-14). На вставках показано более высокое увеличение окрашенных органоидов крипт. f: Гистологиеский анализ демонстрирует, что целый домен LacZ+ крипты (темно-серый/черный) снабжает домен ворсинки. g: Процент органоидов крипт с LacZ+ клетками оставался постоянным с течением времени, указывая на то, что клетки Lgr5+ обладают длительной активностью стволовых клеток. 500 крипт высевали в трех экземплярах и подсчитывали ораноиды LacZ+ крипт. Планки погрешностей представляют собой стандартное отклонение для трех экземпляров. Эксперимент повторили три раза со сходными результатами.
Фигура 5. Более высокое разрешение изображения фиг. 4a, фиг. 11m и 11p.
Фигура 6. Нет данных о субэпителиальных фибробластах в органоидах крипт. a: Иммунное окрашивание на актин гладких мышц (SMA, темно-серый, примеры указаны черными стрелками) демонстрирует присутствие субэпителиальных фибробластов под эпителиальным слоем. b: Отсутствие SMA+ клеток в matrigel (звездочка) указывает на отсутствие субэпителиальных фибробластов в культуральной системе. Измерительная линейка, 50 мкм.
Фигура 7. a-c: Крипту из репортерной мыши Lgr5-EGFP-ires-CreERT2/Rosa26-YFP стимулировали тамоксифеном in vitro в течение 12 ч, и визуализировали на указанные сутки. Lgr5+ клетки являются светло-серыми и указаны белыми стрелками. d: Органоиды возрастом семь суток, происходящие из крипт Lgr5-EGFP-ires-CreERT2/Rosa26-YFP, стимулировали тамоксифеном in vitro в течение 12 ч, и культивировали и визуализировали на указанные сутки. Флуоресценция YFP (светло-серый) показывает, что рассеянные единичные YFP+ клетки (сутки 1) давали множество потомков in vitro в течение последующих пяти суток. Домен ворсинки разрывался в течение суток 1-1,5, с последующим образованием новой ворсинки (белая окружность). Следует отметить, что клетки YFP+ мигрируют в направлении домена ворсинки.
Фигура 8. Единичные отсортированные Lgr5+ стволовые клетки образуют целые структуры крипта-ворсинка. a: Lgr5-GFP+ клетки, полученные из кишечника Lgr5-EGFP-ires-CreERT2 (нижняя часть), по сравнению с однопометными животными дикого типа (верхняя часть). GFP+ клетки подразделяли на две популяции; GFPhi и GFPlow. b: Анализ конфокальной микроскопией свежевыделенной крипты демонстрирует GFPhi в CBC-клетках (черные вершины стрелок) и GFPlow выше CBC (белые вершины стрелок). c: Отсортированные клетки GFPhi. d: 1000 отсортированных клеток GFPhi (слева) и клеток GFPlow (справа) после культивирования в течение 14 суток. e-f: Четырнадцать суток после сортировки, единичные клетки GFPhi образуют органоиды крипт, причем клетки Lgr5-GFP+ (светло-серые клетки) и клетки Панета (белые стрелки) расположены в основаниях крипт. Измерительная линейка, 50 мкм. f: Более высокое увеличение основания крипты из e. g: Для визуализации пролиферирующих клеток органоиды культивировали с аналогом тимидина EdU (светло-серый, примеры указаны белыми стрелками) в течение 1 ч, после чего их фиксировали. Следует отметить, что только домены крипт включали EdU. Контрастное окрашивание: DAPI (темно-серый).
Фигура 9. a: Эффективность отдельных клеток, отсортированных в отдельные лунки, в отношении образования колоний. Среднее значение приведено для 4 отдельных экспериментов, среди которых в каждом эксперименте 100 клеток визуально проверяли, а затем наблюдали за их ростом. b: Пример успешно выращенной единичной клетки GFPhi. c: Количества клеток на единичный органоид, усредненные для 5 выращенных органоидов. d: Суспензию отдельных клеток, полученную из органоида, происходящего из единичной клетки, пересевали и выращивали в течение 2 недель.
Фигура 10. Эффективность отдельных клеток, отсортированных в отдельные лунки, в отношении образования колоний. Пример успешно выращенных единичных клеток GFPhi. Стрелки указывают на частицы пыли в качестве ориентира. Измерительная линейка: 50 мкм.
Фигура 11. Состав органоидов, происходящих из единичных стволовых клеток. a-d: Трехмерное реконструированное конфокальное изображение для a: ворсинки указаны светло-серым (вершина энтероцитов, выстилающих центральный просвет), b: окрашивание на Muc2 указано белыми стрелками (бокаловидные клетки), c: лизоцим указан светло-серым цветом (клетки Панета), d: хромогранин A указан светло-серым цветом (энтероэндокринные клетки). Ядро подвергали контрастному окрашиванию с помощью DAPI. e-g: Окрашивание парафинового среза. e: Щелочная фосфатаза указана черным цветом (вершины энтероцитов, выстилающих центральный просвет), f: PAS указан темно-серым цветом (бокаловидные клетки), g: лизоцим указан темно-серым цветом (клетки Панета), h: синаптофизин указан темно-серым цветом (энтероэндокринные клетки). i-p: Срезы для электронной микроскопии органоидов крипт демонстрируют присутствие энтероцитов (i), бокаловидных клеток (j), клеток Панета (k) и энтероэндокринных клеток, (l) m/o: Изображение крипты при низком увеличении иллюстрирует отсутствие стромальных клеток. n-o: Более высокое увеличение m. n: созревание щеточной каемки в направлении просветного компартмента органоида, как указано по различию в длине микроворсинок (черные стрелки). p: Изображение при низком увеличение ворсинки. Lu, просвет органоида крипты, заполненного апоптотическими тельцами и выстланного полярными энтероцитами. G, бокаловидные клетки; EC, энтероэндокринные клетки; P, клетки Панета; звездочка, matrigel. Измерительная линейка: 5 мкм (m, p), 1 мкм (n, o).
Фигура 12. Сравнение электронно-микроскопических изображений крипты in vivo с культивированной криптой in vitro. a, b: Нормальный кишечник в основании крипты с соединительной тканью под ней (стрелки). Для сравнения см. c-g для органоидов, также снятых в основании крипты. d: Изображение апикальной мембраны при высоком увеличении. Между мембранами двух соседних клеток существуют внутриклеточные щели (стрелки). Следует отметить десмосому (вершина стрелки), за которой следует внутриклеточная щель. e: Высокое увеличение базальной области, где за мембраной двух соседних клеток могут следовать внутриклеточные щели. Эти изображения сравнимы с a и b из кишечника нормальной мыши. Причиной этих внутриклеточных щелей может быть осмотических шок в процессе фиксации альдегидом. f, g: Все клетки, которые образуют органоид, находятся в здоровом состоянии и лишены крупных вакуолей или других признаков стресса. Можно наблюдать картины митоза (c) и в каждой клетке множество ядерных пор (f, стрелки) и целые митохондрии. ER и аппарат Гольджи (g) можно видеть без признаков набухания. Отсутствуют признаки кариорексиса, кариолизиса или кариопикноза. Таким образом, не наблюдают признаков лизиса клеток или апоптоза. Клетки в просвете органоида демонстрируют ожидаемые апоптотические признаки, как можно наблюдать в кишечнике нормальной мыши. f: показан другой пример энтероэндокринной клетки. Mi: митотические клетки, Lu: просвет, EC: энтероэндокринные клетки, G: аппарат Гольджи.
Фигура 13. Крипты, происходящие из толстого кишечника, также можно поддерживать в культуре. Единичные выделенные крипты, происходящие из толстого кишечника, эффективно образуют органоиды крипт с использованием тех же условий культивирования, которые использовали для крипт тонкого кишечника. Путем многократного слияния крипт, на 14 сутки структуры образуют многочисленные подобные осьминогу органоиды крипт.
Фигура 14. Добавление BDNF увеличивает эффективность культивирования. Единичные выделенные крипты толстого кишечника культивировали в присутствии EGF, Noggin, R-спондина и BDNF. Изображения органоидов крипт толстого кишечника, полученные на 0, 4 и 14 сутки после начала культивирования.
Фигура 15. Добавление Wnt3a далее увеличивает эффективность культивирования органоидов крипт толстого кишечника. Отдельные выделенные крипты толстого кишечника культивировали в присутствии EGF, Noggin, R-спондина. Применение кондиционированной среды Wnt3a (+Wnt3a) увеличивало эффективность культивирования вплоть до 30% по сравнению с культивированием органоидов толстой кишки в контрольной среде (-Wnt3a).
Фигура 16. Аденома, выделенная из мышей APC-/-, может расти in vitro. Единичные выделенные аденомы из мышей APC-/- диссоциировали и культивировали с использованием условий, как описано выше, за исключением того, что R-спондин не был включен в культуральные среды. a: Органоиды аденомы, как показано здесь на 4 сутки, главным образом, растут в качестве простого пузыря, содержащего один центральный просвет, содержащий апоптотические клетки. b: Более высокое увеличение одного органоида аденомы. c: Один органоид аденомы окрашивали на β-катенин (темно-серый) и гематоксилином (светло-серый в просвете). Наружный слой органоида состоит из эпителиальных клеток с окрашиванием ядерного β-катенина. Внутренний просвет содержит погибшие клетки, которые захватили гематоксилин, окрашивающий темно-серым цветом. d: Более высокое увеличение наружного слоя эпителиальных клеток, демонстрирующее светлый ядерный β-катенин.
Фигура 17. Добавление Wnt3a увеличивает эффективность образования органоида. a: Клетки Lgr5-GFPhi отсортировывали и культивировали с Wnt3a (100 нг/мл) или без него, в дополнение к общепринятым условиям для культивирования отдельных клеток (EGF, noggin, R-спондин, Notch-лиганд и Y-27632, как описано выше для отдельных клеток). Эти изображения чашек с культивированными органоидами в присутствии и в отсутствие Wnt3a являются типичными. b: 100 клеток/лунка высевали и количество органоидов определяли через 14 суток после посева. На этом графике представлено количество органоидов/чашка.
Фигура 18. Модель функции R-спондина 1
Передача сигнала Wnt/β-катенин инициируется связыванием канонического лиганда Wnt с Frizzled и ассоциацией с рецепторами LRP5/6. В отсутствие R-спондина 1, передача сигнала Wnt ограничена количеством LRP6 на поверхности клеток, которая поддерживается на низком уровне опосредуемой DKK1/Kremen1 интернализацией. R-спондин 1 усиливает передачу сигнала Wnt путем антагонизма опосредуемому DKK1/Kremen1 кругообороту LRP6, что приводит к повышенным уровням LRP6 на поверхности клеток. Эта фигура взята из PNAS 104: 14700, 2007.
Фигура 19. Клетки Панета расположены рядом с Lgr5+ стволовыми клетками в тонком кишечнике. Крипты выделяли из тонкого кишечника мышей со вставкой Lgr5-EGFP-ires-CreERT2. Здесь представлены примеры типичных крипт. GFP+ клетки являются Lgr5+ (светло-серый, указаны черными стрелками) и они, как правило, расположены рядом с клетками Панета (указанными *).
Фигура 20. В отсутствие жизнеспособных клеток Панета, эффективность образования органоидов снижается.
Выделенные крипты инкубировали с 1 мкМ Newport Green-DCF (Molecular probe) в PBS + 0,1% Pluronic 127 (Sigma) в течение 3 мин при комнатной температуре, а затем промывали PBS. После этого крипты погружали в Matrigel и культивировали с использованием стандартных условий, как описано выше.
Фигура 21. Эффективность культуры органоида желудка. (a) GFP (стрелки, указывающие GFP-положительные клетки) и изображение DIC выделенных желез желудка из пилорической области желудка мышей Lgr5-GFP. Ядра окрашены DAPI. Увеличение 63x (b) 100 желез желудка/лунка высевали в двух экземплярах с EGF(E), R-спондином 1 (R), Noggin (N), EGF + R-спондин 1 (ER), EGF + Noggin (EN), EGF + R-спондин 1 + Noggin (ERN), EGF + R-спондин 1 + Noggin + Wnt3A (ERNW) или EGF + R-спондин 1 + Noggin + Wnt3A + KGF (ERNWK). Количество органоидов желудка подсчитывали через 2, 5 и 7 суток. Результаты представлены в качестве среднего значения ±SEM для 2 независимых экспериментов. (b) 100 желез желудка/лунка высевали в двух экземплярах с рекомбинантным белком Wnt3A (ENRWK) или кондиционированной Wnt3A средой (ENRWCMK), дополненной другими факторами роста, описанными в a. Количество почкующихся органоидов подсчитывали на 7 сутки после посева и на 2 сутки после первого пассирования.
Фигура 22. Образование органоидов желудка in vitro. (a) Выделенные железы желудка, выращенные в органоиды. Контрастные изображения с дифференциальной интерференцией на 1, 2, 5 и 7 сутки после посева. Увеличение 10x (сутки 1, 2, 5). Сутки 7, увеличение 4x, вставка - 10x. (b) Культуры пассировали каждые 4-7 суток путем механической диссоциации. Культуры выращивали в течение по меньшей мере одного месяца. Типичные изображения, демонстрирующие почкующиеся структуры, выходящие из органоидов при различных пассажах. Пассаж 1 (P1), пассаж 2 (P2) и пассаж 4 (P4) соответствуют суткам 8, 11, 20, соответственно.
Фигура 23. Маркеры желез желудка (a) культуры желудка от мышей Lgr5-LacZ. Детекцию экспрессии LacZ проводили в почкующемся желудке на 5 сутки после высевания (см. стрелку, указывающую на LacZ-положительные (темно-серые) клетки), показавшую наличие положительных по Lgr5 клеток. Увеличение 20x. (b) Окрашивание Ki67 (черный) демонстрируют положительные пролиферирующие клетки в основании подобной железе структуры. (c) Апоптотические клетки с каспазой-3 (темно-серый) присутствуют внутри просвета органоида. (d) В органоидах желудка присутствуют клетки, положительные по муцину желудка 5AC (темно-серый). Lu, просвет органоида. Увеличение 20x.
Фигура 24. Протоки поджелудочной железы могут образовывать органоиды, подобные поджелудочной железе, in vitro. Свежевыделенные протоки поджелудочной железы культивировали в присутствии EGF, Noggin, R-спондина 1 и KGF. Контрастные изображения с дифференциальной интерференцией на 0, 4 и 14 сутки после посева.
Фигура 25. Структуры, подобные островкам поджелудочной железы, образуются приблизительно после 3 недель культивирования in vitro. Контрастные изображения с дифференциальной интерференцией на 21 сутки после посева.
Фигура 26
Мышам Axin-LacZ инъецировали носитель отдельно (A) или R-спондин (B). Через 2 суток поджелудочную железу извлекали и определяли наличие экспрессии LacZ путем окрашивания с помощью X-gal. На средней панели B представлено более высокое увеличение протока, который демонстрирует положительное окрашивание на LacZ, указывая на экспрессию Axin-LacZ вдоль выстилки протока поджелудочной железы. На левой панели показано, что небольшие клетки протоков в центроацинозных клетках или клетках вставочного протока, экспрессировали Axin2-LacZ (примеры которых указаны черными стрелками). В углу каждого изображения показаны увеличения. Лигирование протока поджелудочной железы проводили у мышей дикого типа. В различные моменты времени после PDL поджелудочные железы выделяли и срезы тканей, полученные из областей PDL и не PDL, окрашивали H&E. Показаны увеличения для каждого момента времени. (C) Лигирование протока поджелудочной железы проводили у мышей wt и Axin2-LacZ через 7 суток PDL, поджелудочную железу выделяли и определяли экспрессию Axin2-LacZ окрашиванием с помощью X-gal фиксированных срезов тканей (D) или целых препаратов фрагментов органов. (E) Белые окружности указывают на лигированную часть поджелудочной железы. Экспрессия Ki67 (примеры, указанные стрелками) в срезах ткани поджелудочной железы через 5 суток после PDL. Увеличения показаны. (F) Включение BrdU (примеры, указанные стрелками) в ткань поджелудочной железы через 2 суток после обработки in vivo R-спондином. Увеличения показаны. (G) Экспрессию мРНК Lgr5 определяли посредством Q-PCR в ткани поджелудочной железы, полученной от мышей, подвергнутых PDL или имитирующей операции. В поджелудочной железе поле PDL, область PDL и область не PDL подвергали Q-PCR. Показана кратность увеличения экспрессии Lgr5 по сравнению с геном домашнего хозяйства белка, связывающего TATA-бокс (tbp). (H) Через 13 суток после PDL, поджелудочную железу выделяли и экспрессию Lgr5-LacZ определяли посредством окрашивания X-gal фиксированных срезов тканей. Примеры окрашенных клеток указаны черными стрелками (I).
Фигура 27. Изображения фрагментов протоков поджелудочной железы, выращенных in vitro в EM, полученные в различные моменты времени после выделения из мыши дикого типа (A, верхняя панель). Центроациозные клетки не росли в течение периодов, превышающих 7 суток, после чего они распадались (A, нижняя панель). Фрагменты поджелудочной железы выращивали в присутствии или в отсутствие EGF (50 нг/мл), R-спондина (1 мкг/мл), FGF10 (100 нг/мл) или Noggin (100 нг/мл). Изображения культур получали через 7 и 14 суток после начала культивирования со свежевыделенных фрагментов поджелудочной железы. Культуры без EGF не выживали в течение более чем 10 суток. (B) Фрагменты поджелудочной железы, выделенные из мышей Axin2-LacZ, культивировали в отсутствие или в присутствии R-спондина (1 мкг/мл) в течение 3 суток. Окрашивание с помощью X-gal продемонстрировало экспрессию отвечающего на Wnt Axin-LacZ в клетках протоках после 3 и 14 суток только в присутствии R-спондина (примеры, указанные белыми стрелками). Не было выявлено окрашивания посредством X-gal в ацинозных или островковых клетках. (C) Фрагменты протоков выделяли из мышей Lgr5-LacZ и культивировали в течение 3 суток в отсутствие или в присутствии R-спондина. Экспрессия Lgr5-LacZ, как показало окрашивание X-gal, продемонстрировала Lgr5+ клетки на вершинах почек, аналогично его экспрессии после PDL. (D) Окрашивание посредством FACS клеток, полученных из фрагментов поджелудочной железы, культивированных в присутствии агониста Wnt, R-спондина. Клетки окрашивали в отношении EpCAM, общего эпителиального клеточного маркера, и LacZ (флуоресцеиндигалактопиранозид, FDG). Процент клеток Lgr5+ значимо возрастал, когда фрагменты поджелудочной железы культивировали в присутствии сигнала Wnt (E).
Фигура 28
Поджелудочную железу выделяли из мышей через 7 суток после обработки PDL и клетки поджелудочной железы окрашивали посредством EpCAM-APC и флуоресцентного субстрата для LacZ (набор FluoroReporter kit), сортировали и культивировали в EM, включающем 50% кондиционированную Wnt3A среду и 10 мМ Y-27632, в течение 4 суток. Через 4 суток культуральную среду заменяли на среду EM без Wnt и Y-27632. Изображения получали на указанные сутки, и представлено увеличение 40x.
Фигура 29
Органоиды поджелудочной железы переносили из EM в DM. Эффектом удаления FGF10 из среды для размножения, приводящего к DM, была индукция дифференцировки в островки. Органоиды поджелудочной железы культивировали в течение 10 суток в DM после чего структуры, подобные островкам, можно было выявить in vitro. Представлены изображения культур в присутствии и в отсутствие FGF10 (A) и показана повышенная экспрессия определенных маркеров дифференцировки, Ngn3 и соматостатина, при измерении посредством PCR. Hprt представляет собой ген домашнего хозяйства (B). Через несколько моментов времени после переноса в DM, экспрессию ряда маркеров оценивали посредством PCR (C). Изменение морфологии с панкреатических пузырей на структуры, подобные β-клеткам (D) сопровождалось появлением определенных β-клеточных маркеров, таких как инсулин и C-пептид при детекции иммунофлуоресценцией (E). Присутствие R-спондина в DM является необходимым для регенерации предшественников β-клеток, как показано посредством положительного иммунофлуоресцентного окрашивания на Ngn3 (примеры указаны белыми стрелками) (F).
Фигура 30.
Фрагменты поджелудочной железы человека выделяли свежими и культивировали в EM. Изображения культур получали в указанные моменты времени после начала культивирования.
Фигура 31. Культуры крипт in vitro продуцируют лиганд(ы) Wnt
(A) Схематическое представление каскада Wnt. Когда лиганды Wnt секретируются, они могут аутокринно или паракринно активировать каскад передачи сигнала. Поркупин важен для надлежащей секреции лигандов Wnt. Ингибиторы IWP приводят к ингибированию секреции лиганда Wnt.
(B) Органоиды мыши, культивированные в нормальных условиях, как указано в примере 1.
(C) Инкубация культур органоидов мыши с 1 мкм IWP приводит к гибели клеток культур органоидов.
(D) Добавление среды, кондиционированной Wnt3a, улучшает культуры органоидов мыши.
(E) Индуцируемой IWP гибели органоидов препятствует добавление среды, кондиционированной Wnt3a.
Показано увеличение 10x (B-E).
Фигура 32. Установление культуры крипты кишечника человека. Органоиды человека, культивированные из тонкого кишечника (A-D) и толстого кишечника (E-H) после 3 (A, C, E, G) и 5 (B, D, F, H) суток в среде, дополненной EGF, Noggin и R-спондином со средой, кондиционированной Wnt3a (A, B, E, F), и без нее (C, D, G, H).
Фигура 33. Установление культуры органоидов желудка.
(A) Всего 100 желез желудка/лунка высевали в двух экземплярах с EGF (E), R-спондин 1 (R), Noggin (N), EGF + R-спондин 1 (ER), EGF + Noggin (EN), EGF + R-спондин 1 + Noggin (ERN), EGF + R-спондин 1 + Noggin + Wnt3A (ERNW), EGF + R-спондин 1 + Noggin + Wnt3A + FGF10 (ERNWF), EGF + R-спондин 1 + Noggin+ контрольная кондиционированная среда + FGF10 (ERNCCMF) или EGF + R-спондин 1 + Noggin + среда, кондиционированная Wnt3A, + FGF10 (ERNWCMF). Количество органоидов желудка подсчитывали через 2, 5 и 7 суток. Результаты представлены в качестве среднего значения ±SEM для 2 независимых экспериментов.
(B) Всего 100 желез желудка/лунка высевали в двух экземплярах со средой, кондиционированной Wnt3A (ENRWCM), или со средой, кондиционированной Wnt3A, дополненной FGF10 (ENRWCMF). Количество почкующихся органоидов подсчитывали через 7, 15 (пассаж 2) и 60 суток (пассаж 10) в культуре.
(C) Всего 100 желез желудка/лунка высевали в среду, кондиционированную Wnt3A, (WCM) + EGF + Noggin и R-спондин, дополненную либо FGF7/KGF (K), либо FGF10 (F), тестировали как 100, так и 1000 нг/мл. Количество почкующихся органоидов подсчитывали после 4 суток (пассаж 7) в культуре. Показан репрезентативный эксперимент.
(D) Выделенные железы желудка, развивающиеся в органоиды. Контрастные изображения с дифференциальной интерференцией на 1, 2, 3, 4, 7 сутки после посева. Через одну неделю культуры требовали разделения 1:5 или 1:6. Субкультивирование и поддержание проводили, как описано в разделе "Дополнительные материалы и способы". Типичные изображения культур после культивирования в течение 15 суток, 3 месяцев, 4,5 и 6 месяцев; (увеличение 10x).
(E) Пример культуры, выращиваемой в течение 5 суток в контрольной кондиционированной среде. Следует отметить, что культура не является растущей и не образует домены желез. В этих условиях культура выживала не более 7 суток.
(F) Окрашивание на E-кадгерин тотального препарата органоида желудка возрастом 3 месяца.
Фигура 34. Единичные Lgr5+ клетки образуют длительно живущие органоиды желудка in vitro.
(A) Конфокальный анализ свежевыделенного пилорического элемента желудка мыши Lgr5-EGFP-ires-CreERT2. Стрелками показаны различные популяции GFPhi (серый), GFPlo (черный) GFP- (белый).
(B) Lgr5-EGFP+ клетки отличаются от популяций GFPlo и GFP- их уровнем экспрессии GFP. FSC, рассеяние вперед.
(C) Типичный пример растущего органоида, происходящего из единичной Lgr5+ клетки. Стрелками показано образование почек подобных железам доменов на 7. Исходные увеличения: 1-4 сутки, увеличение 40x; 5-6 сутки, увеличение 20x; 7-8 сутки, увеличение 10x; и 9 сутки, увеличение 5x.
(D) Органоиды, происходящие из единичных Lgr5+ клеток, диссоциировали и разделяли каждые 5-7 суток. Типичные изображения культуры возрастом 3 месяца. Исходные увеличения: левая панель, увеличение 4x; правая панель, увеличение 10x.
(E) Конфокальный анализ экспрессирующих Lgr5 EGFP клеток в культуре желудка возрастом 14 суток, выращенной из единичной клетки GFPhi. Следует отметить, что Lgr5-GFP+ клетки расположены в основании доменов желез (белая стрелка; увеличение 10x).
(F) Органоиды, культивированные с аналогом тимидина EdU (красный) в течение 1,5 ч. Только домены желез включают EdU (белые стрелки, увеличение 20x). Контрастное окрашивание 4,6-диамидино-2-фенилиндол (DAPI; ядерный).
(G) Культуру возрастом 2 недели из культуры единичных клеток репортерной мыши Lgr5-EGFP-ires-CreERT2/Rosa26-YFP стимулировали тамоксифеном in vitro в течение 20 ч, и визуализировали на указанные сутки. Флуоресценция YFP (желтый) показывает, что рассеянные единичные желтые клетки (сутки 1,5) образуют множество потомков in vitro. Следует отметить, что YFP+ клетки мигрируют в направлении центрального просвета (белая пунктирная окружность).
(H) Анализ экспрессии специфических генов желудка из культур возрастом 2 месяца, происходящим из Lgr5+ единичных клеток. Культуры поддерживали в среде с высоким (левая панель) или низким (средняя панель) уровнем Wnt3A. Следует отметить, что происходящие из желудка культуры являются отрицательными в отношении специфических генов кишечника (правая панель).
(I) Культуры поддерживали в среде с низким уровнем Wnt3A в течение по меньшей мере 10 суток. Верхняя панель: конфокальное изображение окрашивания по ECad (красный, происходящие из эпителия органоиды). Контрастное окрашивание: Hoescht 33345 (синий). Нижние панели: парафиновые срезы, окрашенные на Tff2 (коричневый, слизистые клетки шейки), перйодная кислота Шиффа (красный, пит-клетки), MUC5AC (коричневый, пит-клетки) и хроматогранин A (коричневый, энтероэндокринные клетки).
Примеры
Пример 1. Культивирование крипт и ворсинок тонкого кишечника in vitro
Материалы и способы
Мыши: использовали аутбредных мышей в возрасте 6-12 недель. Получение и генотипирование аллеля1 Lgr5-EGFP-Ires-CreERT2 описано ранее1. Репортерные мыши Rosa26-lacZ или YFP Cre были получены в Jackson Labs.
Выделение крипт, диссоциация клеток и культивирование. Крипты высвобождали из тонкого кишечника мыши посредством инкубации в 2 мМ EDTA/PBS в течение 30 мин при 4°C. Выделенные крипты подсчитывали и осаждали. 500 крипт смешивали с 50 мкл Matrigel (BD Bioscience) и высевали в 24-луночные планшеты. После полимеризации Matrigel добавляли 500 мкл культуральной среды для крипт (Advanced DMEM/F12 с факторами роста (10-50 нг/мл EGF (Peprotech), 500 нг/мл R-спондина 111 и 100 нг/мл Noggin (Peprotech)). Для экспериментов по сортировке, выделенные крипты инкубировали в культуральной среде в течение 45 мин при 37°С, а затем ресуспендировали стеклянной пипеткой. Диссоциированные клетки пропускали через 20-мкм сито для клеток. Клетки GFPhi, GFPlow или GFP сортировали проточной цитометрией (MoFlo, Dako). Отдельные жизнеспособные эпителиальные клетки отбирали с помощью параметра рассеяния вперед, бокового рассеяния и длительности импульса, и отрицательного окрашивания йодидом пропидия. Отсортированные клетки собирали в культуральную среду для крипт и погружали в Matrigel, включающий пептид Jagged-1 (Ana Spec, 1 мкМ) в количестве 1 клетка/лунка (в 96 луночном планшете, 5 мкл Matrigel). Наслаивали культуральную среду для крипт (250 мкл для 48-луночного планшета, 100 мкл для 96-луночного планшета), включая Y-27632 (10 мкМ). Факторы роста добавляли раз в двое суток и всю среду заменяли раз в 4 суток. Для пассирования органоиды удаляли из Matrigel и механически диссоциировали на отдельные домены крипт и переносили в новый Matrigel. Пассирование проводили каждые 1-2 недели с соотношением для разделения 1:5.
Реагенты: рекомбинантный EGF и Noggin мыши приобретали от Peprotech. Для экспериментов по культивированию использовали рекомбинантный R-спондин 1 человека11, Y-27632 (Sigma), 4-гидрокситамоксифен (Sigma) и Edu (Invitrogen). Для иммунного окрашивания использовали следующие антитела: антитело против лизоцима (Dako), антитело против синаптофизина (Dako), антитело против BrdU (Roche), антитело против β-катенина (BD Bioscience), антитело против E-кадгерина (BD Bioscience), антитело против актина гладки мышц (Sigma), антитело против EphB2 и B3 (R&D), антитело против виллина, антитело против Muc2, антитело против хромогранина A (Santa Cruz), антитело против каспазы-3 (Cell Signaling).
Выделение крипт: Выделенные тонкие кишечники открывали продольно и промывали холодным PBS. Ткань измельчали на фрагменты приблизительно 5 мм, и дополнительно промывали холодным PBS. Фрагменты ткани инкубировали в 2 мМ EDTA с PBS в течение 30 мин на льду. После удаления среды с EDTA, фрагменты ткани энергично суспендировали с помощью 10-мл пипетки с холодным PBS. Супернатант представлял собой фракцию ворсинок и его удаляли, осадок ресуспендировали с помощью PBS. После дальнейшего энергичного суспендирования и центрифугирования супернатант был обогащен криптами. Эту фракцию пропускали через 70-км сито (BD bioscience) для удаления остаточного материала ворсинок. Выделенные крипты центрифугировали при 300 об./мин. в течение 3 мин для отделения крипт от отдельных клеток. Конечная фракция состояла по существу из чистых крипт и ее использовали для культивирования и диссоциации на единичные клетки.
Индукция тамоксифеном и окрашивание с помощью X-gal: Для активации CreERT2 крипты инкубировали с низкой дозой 4-гидрокситамоксифена (100 нМ) в течение 12 ч и культивировали в культуральной среде для крипт. Окрашивание с помощью X-gal проводили, как описано ранее1. Без обработки 4-гидрокситамоксифеном окрашивания не наблюдали.
Анализ электронной микроскопией: Как описано ранее1, Matrigel, включающий органоиды крипт, фиксировали в фиксаже Karnovsky (2% параформальдегид, 2,5% глутаральдегид, 0,1 M Na-какодилат, 2,5 мМ CaCl2 и 5 мМ MgCl2, pH 7,4) в течение 5 ч при комнатной температуре. Образцы погружали в смолу Epon и исследовали с помощью микроскопа Phillips CM10 (Eindhoven, Нидерланды).
Анализ на микрочипах: Анализ экспрессии генов крипт толстого кишечника, крипт тонкого кишечника и органоидов. Свежевыделенные крипты тонкого кишечника от двух мышей разделяли на две части. Из одной части сразу выделяли РНК (набор RNeasy Mini Kit, Qiagen), другую часть культивировали в течение одной недели, с последующим выделением РНК. Авторы изобретения получили меченную кРНК в соответствии с инструкциями изготовителя (Agilent Technologies). Дифференциально меченную кРНКиз крипт и органоидов тонкого кишечника гибридизовали по отдельности для двух мышей на микрочипах 4X44k Agilent Whole Mouse Genome dual colour Microarrays (G4122F) в экспериментах с обменом двумя красителями, что приводило к четырем индивидуальным чипам. Кроме того, выделенные крипты толстого кишечника гибридизовали против дифференциально меченных крипт тонкого кишечника в экспериментах с обменом двумя красителями, что приводило к четырем индивидуальным чипам. Сигнал микрочипа и фоновую информацию получали с использованием Feature Extraction (V 9.5.3, Agilent Technologies). Все анализы данных проводили с использованием ArrayAssist (5.5.1, Stratagene Inc.) и Microsoft Excel (Microsoft Corporation). Необработанные данные об интенсивности сигнала корректировали, вычитая локальный фон. Отрицательные величины заменяли на положительную величину, близкую к нулю (стандартное отклонение локального фона) для обеспечения вычисления соотношений интенсивностей для признаков, присутствующих только в одном канале (крипты тонкого кишечника или органоиды) или (крипты тонкого кишечника или крипты толстого кишечника). Нормализацию проводили с использованием алгоритма Lowess и отдельные признаки фильтровали, если изменялись обе (крипты тонкого кишечника или органоиды) или (крипты тонкого кишечника или крипты толстого кишечника) интенсивности или если обе интенсивности менее чем в два раза превышали фоновый сигнал. Более того, отфильтровывали неоднородные признаки. Данные доступны в GEO (Gene Expression Omnibus, номер GSE 14594) после публикации. На нормализованных интенсивностях (обработанный сигнал с Feature Extraction) крипт тонкого кишечника/толстого кишечника и органоидов проводили неуправляемую иерархическую кластеризацию с использованием Cluster 3 (расстояние: city block, корреляция: среднее сцепление) и визуализировали с помощью TreeView. Гены считали значимо измененными, если их экспрессия согласованно в трех чипах более чем в 3 раза более высокой в органоидах или криптах.
Анализ изображений: изображения органоидов крипт получали либо с помощью конфокальной микроскопии (Leica, SP5), либо с помощью инвертационного микроскопа (Nikon DM-IL) или стереомикроскопа (Leica, MZ16-FA). Для иммуногистохимии образцы фиксировали с помощью 4% параформальдегида (PFA) в течение 1 ч при комнатной температуре, и парафиновые срезы обрабатывали стандартным способом1. Иммуногистохимию проводили, как описано ранее1. Для иммунного окрашивания цельного препарата органоиды крипт выделяли из matrigel с использованием диспазы (Invitrogen), и фиксировали с помощью 4% PFA, с последующим обеспечением проницаемости с помощью 0,1% Triton-X. Окрашивание с помощью EdU проводили в соответствии с протоколом изготовителя (Click-IT, Invitrogen). ДНК окрашивали с помощью DAPI или ToPro-3 (Molecular Probe). Изображения 3D получали с помощью конфокальной микроскопии (Leica, SP5) и реконструировали с помощью программного обеспечения Volocity Software (Improvision).
Результаты
Эпителий кишечника является наиболее быстро самообновляющейся тканью у взрослых млекопитающих. Авторы изобретения недавно продемонстрировали присутствие приблизительно шести циклирующих Lgr5+ стволовых клеток в основании крипт тонкого кишечника1. Авторы настоящего изобретения установили условия длительного культивирования, в которых единичные крипты претерпевают множество делений крипт при одновременном образовании подобных ворсинкам эпителиальных доменов, в которых присутствуют все дифференцированные типы клеток. Единичные отсортированные Lgr5+ стволовые клетки также могут инициировать эти органоиды крипта-ворсинка. Эксперименты по наблюдению указывают на то, что в органоидах поддерживается иерархия Lgr5+ стволовых клеток. Авторы настоящего изобретения заключили, что элементы кишечника крипта-ворсинка являются самоорганизующимися структурами, которые могут образовываться из единичной стволовой клетки в отсутствие неэпителиальной клеточной ниши.
Самообновляющийся эпителий тонкого кишечника упорядочен в крипты и ворсинки2. В криптах клетки вновь образуются и утрачиваются посредством апоптоза на вершинах ворсинок, причем время кругооборота у мышей составляет 5 суток. Давно известно, что самообновляющиеся стволовые клетки находятся вблизи основания крипт и образуют быстро пролифирирующие переходные делящиеся (TA) клетки. Оцененное количество стволовых клеток составляет от 4 до 6 на крипту. Из TA-клеток образуется три дифференцированных типа клеток: энтероциты, бокаловидные клетки и энтероэндокринные клетки, и они продолжают их миграцию в последовательные слои вдоль оси крипта-ворсинка. В каждую ворсинку поступают клетки из множества различных крипт. Четвертый из основных типов дифференцированных клеток, клетки Панета, находится в основании крипты. Авторы изобретения недавно идентифицировали ген Lgr5, который специфично экспрессируется в пятом типе клеток, циклирующих обновляющихся столбчатых клетках основания крипты, которые распределены между клетками Панета. С использованием мыши, у которой в локус Lgr5 встроена кассета индуцируемой GFP/тамоксифеном рекомбиназы Cre, путем прослеживания происхождения авторы настоящего изобретения показали, что Lgr5+ CBC-клетки представляют собой мультипотентные стволовые клетки, которые образуют все типы клеток эпителия, даже при оценке через 14 месяцев после индукции Cre3.
Хотя описано множество культуральных систем4-7, не было получено длительной культуральной системы, которая поддерживает основную физиологию крипта-ворсинка2.
Препараты крипт мыши суспендировали Matrigel. Рост крипт требовал EGF и R-спондина 1 (фиг. 1a). Пассирование выявило потребность в Noggin (фиг. 1b). Культивируемые крипты имели стереотипическое поведение (фиг. 2a). Верхнее отверстие быстро закрывалось и просвет заполнялся апоптотическими клетками. Область крипты претерпевала непрерывное почкование, напоминающее деление крипт17. В области почки всегда присутствовали клетки Панета. Можно было культиваровать большую часть крипт (фиг. 2b). Последующее размножение приводило к органоидам, содержащим >40 доменов крипт, окружающих центральный просвет, выстилаемый эпителием, подобным эпителию ворсинки ("домен ворсинки") (фиг. 2c-e). Окрашивание на E-кадгерин выявило одноклеточный слой (данные не представлены). Раз в неделю органоиды механически диссоциировали и реплицировали в количестве 1/5 от плотности до высевания. Органоиды культивировали в течение >6 месяцев без утраты характеристик, описанных ниже. Анализ экспрессии с помощью микрочипов выявил, что органоиды оставались высоко сходными со свежевыделенными криптами тонкого кишечника по сравнению, например, со свежими криптами толстого кишечника (фиг. 3).
Культивирование крипт Lgr5-EGFP-ires-CreERT2 выявило Lgr5-GFP+ стволовые клетки, смешанные с клетками Панета, в основании крипты. Активация Wnt, о которой свидетельствует ядерный β-катенин (фиг. 4a, фиг. 5) и экспрессия генов-мишеней Wnt, Lgr5 (фиг. 2d) и EphB2 18 (фиг. 4b) была ограничена криптами. Апоптотические клетки утрачивались в центральный просвет в процессе, напоминающем утрачивание апоптотических клеток на вершинах ворсинок in vivo (фиг. 4c). Метафазные пластинки органоидов возрастом >3 месяцев выявили 40 хромосом/клетка (n=20) (фиг. 4d). Неожиданно, авторы настоящего изобретения не выявили данных о наличии миофибробластов и других неэпителиальных клеток (фиг. 6).
Авторы настоящего изобретения культивировали крипты от мышей Lgr5-EGFP-ires-CreERT2, скрещенных с мышами с Cre-активируемым репортером Rosa26-LacZ для обеспечения прослеживания происхождения. Непосредственно после индукции тамоксифеном в низкой дозе авторы настоящего изобретения заметили единичные меченые клетки (фиг. 4e, g). Более 90% этих полученных полностью синих крипт (фиг. 4e-g) предполагают, что Lgr5-GFP+ клетки действительно сохраняли свойства стволовых клеток. Крипты из мышей с Cre-активируемым репортером Rosa26-YFP 19 позволяли прослеживание происхождения с помощью конфокального анализа. Непосредственно после обработки тамоксифеном авторы настоящего изобретения отметили единичные меченые клетки, которые обеспечивали прослеживание происхождения в течение последующих суток, как в свежевыделенных криптах (фиг. 7a-c), так и в установившихся органоидах (фиг. 7d).
Недавно были получены эпителиальные структуры молочной железы из единичных стволовых клеток in vitro 21. Когда единичные Lgr5-GFPhi отсортировывали, они сразу погибали. Ингибитор Rho-киназы Y-27632 значительно уменьшал эту гибель клеток. Было выявлено, что пептид-агонист24 способствует поддержанию пролифиративных крипт23. В этих условиях выживали значительные количества Lgr5-GFPhi клеток и они образовывали крупные органоиды крипт. Органоиды образовывались редко, когда высевали дочерние клетки GFPlow (фиг. 8d). Множество клеток LgrS-GFPhi было смешано с клетками Панета в основаниях крипт (фиг. 8e-f). Включение EdU (аналог тимидина) выявило клетки S-фазы в криптах (фиг. 8g).
Авторы настоящего изобретения отсортировали клетки в количестве 1 клетка/лунка, визуально проверили присутствие единичных клеток и наблюдали за последующим ростом. В каждом из четырех отдельных экспериментов авторы изобретения идентифицировали и наблюдали 100 единичных клеток. В среднем приблизительно 6% из клеток Lgr5-GFPhi вырастали в органоиды, в то время как остальные клетки, как правило, погибали в пределах первых 12 часов, возможно вследствие физического и/или биологического стресса, свойственного процедуре выделения. Клетки GFPlow редко вырастали (фиг. 9a). На фиг. 9b и фиг. 10 проиллюстрирован рост органоида из единичной клетки Lgr5-GFPhi. К четвертым суткам культивирования структуры состояли приблизительно из 100 клеток, что согласовывалось с 12-часовым клеточным циклом пролиферативных клеток крипт25 (фиг. 9c). Через две недели органоиды диссоциировали на единичные клетки и пересевали для образования новых органоидов (фиг. 9d). Эту процедуру можно было повторять в течение по меньшей мере четырех раз в течение 2 недель без заметного снижения эффективности пересевания.
Органоиды, происходящие из единичных стволовых клеток, выглядели неотличимыми от органоидов, происходящих из целых крипт. Клетки Панета и стволовые клетки были расположены в основаниях крипт (фиг. 8e, f, фиг. 11c, g). Полностью полярные энтероциты, на которые указывали виллин+-зрелые щеточные каемки и апикальная щелочная фосфатаза, выстилали центральный просвет (фиг. 11 a, e, i). Бокаловидные клетки (Muc2+, фиг. 11b, PAS+, фиг. 11f) и энтероэндокринные клетки (хроматогранин A+, фиг. 11d, синаптофизин+, фиг. 11h) были рассеяны по структуре органоида. С помощью электронной микроскопии было распознано четыре типа клеток (фиг. 11i-l). Неэпителиальные (стромальные/мезенхимные) клетки отсутствовали, наблюдение подтверждали визуализацией EM (фиг. 11i-p, фиг. 12c-g). Как крипты (фиг. 11m, o), так и центральный эпителий просвета (фиг. 11p) состояли из одного слоя поляризованных эпителиальных клеток, покоящихся непосредственно на подложке из matrigel. Изображения высокого разрешения для этих изображений EM приведены на фиг. 5. Органоид, окрашенный на E-кадгерин красным цветом и контрастно окрашенный на ядра синим цветом, демонстрируют однослойный характер эпителия органоида (данные не представлены).
Хорошо известно, что эпителиальные крипты находятся в тесном контакте с субэпителиальными миофибробластами26-28 и, главным образом, полагают, что последние клетки создают специализированную клеточную нишу в основании крипты27,29,30. Такая ниша может создавать уникальное окружение для заякоривания и поддержания стволовых клеток кишечника. Авторы изобретения показали, что самообновляющийся эпителий можно получить с помощью ограниченного набора ростовых сигналов, которые присутствуют постоянно. Несмотря на это, выделенные стволовые клетки автономно создают асимметрию высоко стереотипическим образом. Это быстро приводит к образованию структур, подобных структурам крипт, с образовавшимися de novo стволовыми клетками и клетками Панета, расположенными в их основании, и заполненных TA-клетками. Эти подобные криптам структуры снабжают подобные ворсинкам домены просвета, состоящие из постмитотических энтероцитов, где апоптотические клетки отщепляются в просвет, напоминая утрату клеток на верхушках ворсинок. Парадоксальное наблюдение, что единичные клетки, на которые воздействует однородное стимулирующее рост окружение, могут создавать асимметричные структуры, особенно очевидно при изучении каскада Wnt. Хотя все клетки подвергаются воздействию R-спондина 1, только клетки в криптах проявляют признаки активной передачи сигнала Wnt, т.е. ядерный β-катенин и экспрессию генов-мишеней Wnt. Очевидно, в основе образования оси крипта-ворсинка лежит отличающаяся способность к ответу на передачу сигнала Wnt, а не отличающееся воздействие разных внеклеточных сигналов Wnt.
В общем, авторы изобретения заключили, что единичная Lgr5+ стволовая клетка кишечника может действовать независимо от позиционных ориентиров из ее окружения и что она может образовывать постоянно размножающуюся самоорганизующуюся эпителиальную структуру, напоминающую нормальный кишечник. Описанная культуральная система упрощает исследование запускаемой стволовыми клетками биологии крипта-ворсинка. Более того, она может открыть новые пути для регенеративной медицины и генной терапии.
Пример 2. Культивирование крипт толстого кишечника и ворсинок in vitro
Материалы и способы
Среда, кондиционированная Wnt3a
Клеточную линию, экспрессирующую лиганд Wnt3a и ту же клеточную линию без лиганда Wnt3a (контрольная среда) культивируют в течение 3-4 недель. Клетки начинают продуцировать Wnt3a, как только они прекращают расти при смыкании монослоя. Среду собирают и тестируют в анализе TOPflash, люциферазном анализе с использованием конструкции, отвечающей на TCF элементы-luc (TOP) и той же конструкции, но без мутации в отвечающих на TCF элементах (FOP). Соотношение между TOP/FOP для среды, подлежащей применению в культурах, должно составлять более 20. Среду разбавляют на 25-50% при использовании в культурах для регенерации ткани.
Свежевыделенный толстый кишечник открывали и промывали посредством PBS или DMEM, и нарезали на небольшие фрагменты. Фрагменты инкубировали с 2 мМ EDTA/PBS в течение 1 часа при 40°С при осторожном качании. После удаления раствора EDTA фрагменты ткани энергично суспендировали в 10 мл холодного PBS с помощью 10-мл пипетки. Первый супернатант, содержащий дебрис, удаляли и осадок суспендировали с помощью 10-15 мл PBS. После последующего энергичного суспендирования фрагментов тканей, супернатант был обогащен криптами толстого кишечника. Фрагменты осаждали и смешивали с matrigel и культивировали в качестве культуральной системы органоида тонкого кишечника. Matrigel инкубировали в течение 5-10 мин при 37°C. После полимеризации matrigel, добавляли 500 мкл среды для культивирования тканей (50% Advanced-DMEM/F12/50% среда, кондиционированная Wnt-3a, дополненная 200 нг/мл N-ацетилцистеина, 50 нг/мл EGF, 1 мкг/мл R-спондина 1, 100 нг/мл Noggin, 100 нг/мл BDNF (Peprotech). Всю среду заменяли каждые 2-3 суток. Для пассирования органоиды удаляли из Matrigel с использованием 1000-мкл пипетки и диссоциировали механически на небольшие фрагменты и переносили в свежий Matrigel. Пассирование проводили в соотношении для разделения 1:4 по меньшей мере один раз каждые две недели. В этих условиях культуры поддерживали в течение по меньшей мере 3 месяцев.
Результаты
Органоиды толстого кишечника росли медленнее и менее эффективно по сравнению с органоидами тонкого кишечника. При тех же условиях факторов роста, что и для тонкого кишечника, менее 5% крипт толстого кишечника, выделенных из дистального толстого кишечника, вырастало и образовывало структуру органоида (фиг. 13). Было трудно выращивать крипты толстого кишечника из проксимальной части толстого кишечника. Поскольку авторы настоящего изобретения выявили активацию trkB, рецептора BDNF (происходящего из головного мозга нейротрофического фактора), в анализе на микрочипах (клетки Lgr5-GFPhi толстого кишечника против клеток Lgr5-GFPlow толстого кишечника), авторы настоящего изобретения определили эффект BDNF в отношении органоидов толстого кишечника. Авторы настоящего изобретения постоянно наблюдали приблизительно в 2 раза более высокую эффективность культивирования в культуре BDNF+ по сравнению с культурой BDNF-. Как правило, один органоид толстого кишечника может содержать приблизительно 10 доменов крипт (фиг. 14). В соответствии с их происхождением, нельзя было выявить клеток Панета. По сравнению с органоидами тонкого кишечника, крипта толстого кишечника не обладает продуцирующими Wnt-3a клетками Панета в основании крипты, таким образом, дополнение с помощью Wnt-3 увеличивает эффективность культивирования крипт толстого кишечника, но не крипт тонкого кишечника. Главным образом, авторы изобретения достигли вплоть до 30% эффективности культивирования, когда они добавили среду, кондиционированную Wnt-3a (фиг. 15).
В заключение крипты, происходящие как из тонкого кишечника, так и из толстого кишечника, можно поддерживать и размножать in vitro с использованием описанных выше условий, что делает этот способ первым описанным способом культивирования для получения эпителия кишечника в искусственной системе.
Пример 3. Культивирование аденом in vitro
Материалы и способы (см. пример 1)
Результаты
Аденомы исторически было трудно культивировать in vitro. Поскольку описанные выше условия успешно использовали для культивирования здоровых крипт, происходящих из тонкого кишечника, а также из толстого кишечника, то определяли, могут ли сходные условия поддерживать аденомы in vitro. После выделения аденомы из мышей APC-/- с использованием 2,5 мМ EDTA, единичные аденомы культивировали в сходных условиях, как описано выше. Важно, что эти условия были подходящими для поддержания роста аденом in vitro, однако R-спондин стал ненужным. Это можно легко объяснить тем фактом, что он более не был необходим для индукции каскада передачи сигнала Wnt, поскольку отсутствие APC в этих клетках автоматически приводит к ядерному β-катенину. Это делает R-спондин, агонист Wnt, ненужным при культивировании аденом in vitro. На фиг. 16a, и на более крупном увеличении на фиг. 16b, показано, что в противоположность нормальным органоидам крипт, в которых можно видеть почкования крипт с центральным просветом, органоиды аденомы просто растут в качестве пузырей. Погибшие клетки утрачиваются в просвет, как можно заключить из присутствия большого количества погибших клеток внутри просвета. В нормальных органоидах крипт ядерный β-катенин наблюдают только в основании домена крипты (см. фиг. 4a). В органоидах аденомы (фиг. 16c и более крупное увеличение на 16d), ядерный β-катенин наблюдали в каждой эпителиальной клетке, что согласуется с генетической мутацией APC. Эти органоиды можно пассировать неопределенно долго.
Кроме того, было протестировано, способны ли единичные отсортированные Lgr5+ клетки, происходящие из аденом у мышей Lgr5-EGFP-Ires-CreERT2/APCflox/flox, образовывать сходные органоиды аденом in vitro с использованием указанных выше условий культивирования (без R-спондина). Действительно, это было так, и полученные органоиды были высоко сравнимы по структуре с органоидами, которые были получены с использованием полных аденом в качестве исходного материала для культивирования in vitro (данные не представлены).
Пример 4. Тестирование эффекта других агонистов Wnt
Для определения того, обладают ли другие агонисты Wnt тем же эффектом, что и R-спондин, а именно, образованием органоидов крипта-ворсинка in vitro, к отсортированным единичным Lgr5+ клеткам добавляли растворимый Wnt3a и оценивали эффект на образования крипты-ворсинок in vitro.
Материалы и способы
LgrS-GFPhi клетки отсортировывали и культивировали с Wnt3a (100 нг/мл) или без него в дополнение к общепринятым условиям культивирования единичных клеток (EGF, noggin, R-спондин, Notch-лиганд и Y-27632, как описано выше для единичных клеток). Авторы посеяли 100 клеток/лунка и подсчитали количество органоидов через 14 суток после посева.
Выделенные крипты инкубировали с 1 мкМ Newport Green-DCF (Molecular Probes) в PBS + 0,1% Pluronic 127 (Sigma) в течение 3 мин при комнатной температуре, а затем промывали посредством PBS. После этого крипты погружали в Matrigel и культивировали с использованием стандартных условий, как описано выше.
Результаты
Добавление Wnt3a в отсутствие R-спондина не имело какого-либо эффекта на образование колоний, в отсутствие R-спондина образовывалось от малого до нулевого количества колоний. Однако в присутствии R-спондина, увеличенную эффективность образования органоидов наблюдали только в присутствии Wnt3a (фиг. 17). Это указывает на то, что оба фактора поддерживают друг друга в их способности стимулировать и поддерживать дифференцировку стволовых клеток во все клетки, необходимые для образования полного эпителиального клеточного слоя. Современная гипотеза состоит в том, что R-спондин отвечает за ингибирование интернализации ко-рецептора Frizzled, LRP6, перед передачей сигнала через Frizzled. При связывании фактора Wnt с Frizzled и с корецептором LRP6, каскад передачи сигнала Wnt активируется31. Когда LRP6 присутствует на поверхности клеток, происходит активация Wnt (фиг. 18). Таким образом, если R-спондин не присутствует в культуральной среде, Wnt3a не способен активировать каскад передачи сигнала Wnt, поскольку LRP6 интернализуется и недоступен для передачи сигнала в комбинации с фактором Wnt, тем самым препятствуя активации каскада передачи сигнала Wnt.
Wnt3a представляет собой растворимый фактор, который в физиологических условиях продуцируется клетками Панета. Эти клетки, как правило, расположены рядом со стволовыми клетками (фиг. 19) и предполагают, что эти клетки способствуют поддержанию постоянной дифференцировки слоя эпителиальных клеток кишечника. Другими факторами Wnt, которые также секретируются клетками Панета, являются Wnt6, 9b и 11. Ожидается, что Wnt6 будет иметь тот же эффект на дифференцировку стволовых клеток, какой имеет Wnt3a. Эти данные подтверждают представление, что клетки Панета важны для образования ниши стволовых клеток. Эти данные являются неожиданными, поскольку ниша стволовых клеток широко обсуждалась, но до настоящего времени не существовало данных, подтверждающих существование такой ниши. Дополнительное подтверждение присутствия ниши стволовых клеток вытекает из эксперимента, в котором клетки Панета селективно уничтожали. Крипты выделяли из тонкого кишечника мыши и культивировали in vitro в присутствии хелатора цинка32, который специфично устраняет клетки Панета. Его использовали в таких низких концентрациях и в течение такого короткого периода времени, чтобы он влиял только на клетки Панета, но не на другие клетки в крипте. После обработки хелатором цинка оценивали образование органоида. Когда клетки Панета более не присутствовали в исходных криптах, наблюдали значительное уменьшение образования органоида (фиг. 20). В присутствии Wnt3a происходило частичное восстановление после этого уменьшения (данные не представлены). Это подтверждает роль клеток Панета в поддержании ниши стволовых клеток, которая поддерживает дифференцировку Lgr5+ стволовых клеток в крипте.
Пример 5. Условия культивирования также поддерживают рост органоидов желудка
Желудок состоит из 3 топографических областей (дно, тело и пещеры) и 2 функциональных железистых областей (секретирующей кислоту и пилорической). Секретирующая кислоту область составляет 80% органа, в то время как пилорическая область составляет 20% органа. Эпителий желудка млекопитающих организован в желудочные элементы, состоящие из плоского поверхностного эпителия, короткого углубления и длинной железы. Углубление выстилается клетками, секретирующими слизь, в то время как железа состоит из секретирующих клеток, разделенных на три области: перехват, шейка и основание. Эпителий желудка постоянно обновляется. Исследования с прослеживанием, проведенные в лаборатории авторов изобретения, показали, что LGR5-положительные клетки, расположенные в основании железы, удовлетворяют определению стволовости (Barker et al., подготавливается). До настоящего времени, однослойные культуры желудка не были способны объединять признаки элемента желудка, который образован несколькими дифференцированными желудочными клетками. Более того, описанные системы способа 3-D-культивирования реконструируют только высокодифференцированные слизистые клетки поверхности желудка, не демонстрируя каких-либо эндокринных клеток. Более того, эти культуры проводили только в течение 7 суток, таким образом, указывая на недостаток способности к самообновлению (Ootani A, Toda S, Fujimoto K, Sugihara H, Am J Pathol. 2003 Jun, 162(6): 1905-12). Здесь авторы изобретения разработали способ выделения желудочных элементов из пилорической области желудка мыши и были способны разработать систему 3D-культуры, которая демонстрирует более длительное поддержание.
Материалы и способы
Выделение желудочного элемента
Выделенные желудки открывали продольно и промывали холодной Advanced-DMEM/F12 (Invitrogen). Под стереоскопом пилорическую область вырезали и извлекали из организма и слизистую оболочку каридальной и пилорической области тщательно отделяли от мышечного слоя с помощью пинцета. Затем ткань измельчали на фрагменты размером приблизительно 5 мм и далее промывали холодным буфером для выделения (28 мМ Na2HPO4 + 40 мМ KH2PO4 + 480 мМ NaCl + 8 мМ KCl + 220 мМ сахароза + 274 мМ D-Сорбит + 2,6 мМ DL-дитиотреитол). Фрагменты ткани инкубировали в 5 мМ EDTA с буфером для выделения в течение 2 ч при 4°C при осторожном качании. После удаления раствора EDTA, фрагменты ткани тщательно суспсендировали в 10 мл холодного буфера для выделения с помощью 10-мл пипетки. Первый супернатант, содержавший погибшие клетки, удаляли и осадок суспендировали с помощью 10-15 мл холодного буфера для выделения. После дополнительного энергичного суспендирования фрагментов ткани супернатант был обогащен желудочными элементами. Каждые 10-20 суспендирований супернатант заменяли свежим холодным буфером для выделения и держали на льду и проводили проверку на наличие желудочных элементов. Эту процедуру повторяли до полного высвобождения желудочных элементов, обычно 4-5 раз. Обогащенные желудочными элементами суспензии центрифугировали при 600 об./мин. в течение 2-3 мин для отделения выделенных желудочных элементов от отдельных клеток и осадок использовали для культивирования.
Культура желудка
Целые желудочные элементы, содержавшие железу, область перехвата и пит-область, выделяли из пилорической области желудка мыши путем инкубации с 5 мМ EDTA при 4°C в течение 2 ч, как указано в предыдущем разделе. Выделенные желудочные элементы подсчитывали и осаждали. 100 желудочных элементов смешивали с 25 мкл Matrigel (BD Bioscience), высевали на 48-луночные планшеты для культивирования тканей и инкубировали в течение 5-10 мин при 37°C до полной полимеризации Matrigel. После полимеризации добавляли 250 мкл среды для культивирования тканей (Advanced-DMEM/F12, дополненная B27, N2, 200 нг/мл N-ацетилцистеина, 50 нг/мл EGF, 1 мкг/мл R-спондина 1, 100 нг/мл Noggin, 100 нг/мл Wnt3A, 50 или 100 нг/мл KGF). Всю среду заменяли каждые 2 суток. Для пассирования органоиды удаляли из Matrigel с использованием 1000-мкл пипетки и диссоциировали механически на небольшие фрагменты и переносили в свежий Matrigel. Пассирование проводили в соотношении для разделения 1:4 один раз или два раза в неделю. В этих условиях культуры поддерживали в течение по меньшей мере 1 месяца.
Реагенты
Advanced DMEM/F12 и добавки N2 и B-27. Бессывороточную добавку приобретали от Invitrogen и N-ацетилцистеин приобретали от Sigma. Рекомбинантный EGF, Noggin мыши и KGF человека приобретали от Peprotech, и рекомбинантный белок Wnt3A приобретали от Stem Cell Research. Из указанных факторов роста отличающиеся концентрации тестировали только для R-спондина 1 и KGF. При 50 нг/мл R-спондин 1 ингибирует рост культуры. KGF можно использовать в концентрации либо 50, либо 100 нг/мл, однако эффективность почкования является более высокой в условиях 100 нг/мл.
Среду, кондиционированную Wnt3A, получали, как описано ранее (Willert K, Brown JD, Danenberg E, Duncan AW, Weissman IL, Reya T, Yates JR 3rd, Nusse R. Nature 2003 May 22; 423(6938): 448-52).
Иммуногистохимия и визуализирующий анализ
Для окрашивания посредством X-gal органоиды прямо фиксировали в matrigel с 0,25% глутаральдегидом (Sigma) в 100 мМ MgCl2 в PBS, в течение 1-2 ч при комнатной температуре. После этого культуры промывали 3 раза раствором для промывания (0,01% дезоксихолат натрия + 0,02% NP40 + 5 мМ MgCl2 в PBS) и инкубировали в течение 16 ч при 37°C с 1 мг/мл X-Gal (Invitrogen) в присутствии 0,21% K4Fe(CN)6 и 0,16% K3Fe(CN)6. После промывания в PBS культуры фиксировали с помощью 2% PFA в PBS в течение 15 мин при комнатной температуре. Все реагенты были приобретены у Sigma.
Для иммуногистохимии органоиды выделяли из matrigel с использованием трипсина (Tryple Select, Invitrogen), фиксировали с помощью 4% PFA в течение 1 ч при комнатной температуре и погружали в парафин. Парафиновые срезы обрабатывали стандартными способами и иммуногистохимию проводили, как описано ранее. Использовали следующие антитела: антитело против Ki67 мыши (клон MM1, Monosan) (1:200), антитело против расщепленной каспазы-3 кролика (Cell Signaling Technology) (1:400) и антитело против желудочного муцина человека 5AC (Novocastra clone, 45 мл) (1:200). Во всех случаях проводили извлечение антигена в цитратном буфере. Срезы контрастно окрашивали гематоксилином Майера. Изображения органоидов желудка и выделенных желез желудка получали с помощью инвертационного микроскопа (Nikon DM-IL) или конфокальной микроскопии (Leica SP5).
Результаты
До настоящего времени культуры желудка выращивали монослоями. Однако монослойные культуры лишены способности повторять признаки всего желудочного элемента, который образован несколькими дифференцированными желудочными клетками (слизистые пит-клетки, энтероэндокринные клетки и пролиферирующие не содержащие слизи клетки). Недавно в лаборатории авторов изобретения было показано путем прослеживания происхождения in vivo, что положительные по Lgr5 клетки, находящиеся в основании крипт кишечника, являются истинными стволовыми клетками кишечника (Barker N, van Es JH, Kuipers J, Kujala P, van den Born M, Cozijnsen M, Haegebarth A, Korving J, Begthel H, Peters PJ, Clevers H. Nature 2007; 449: 1003-7). Как и эпителий кишечника, эпителий желудка постоянно обновляется. Положительные по Lgr5 клетки были выявлены в основании элементов пилорических желез желудка и, исследования по прослеживанию показали, что эти положительные по LGR5 клетки удовлетворяют определению стволовости, демонстрируя способность к самообновлению и мультипотентности (Barker et al. подготавливается). Поскольку авторы изобретения были способны культивировать крипты кишечника из единичных Lgr5+ клеток в 3-D-структурах, то определяли, могут ли сходные условия поддерживать рост пилорических желудочных элементов in vitro.
После выделение элементов желудочных желез с использованием 5 мМ EDTA, железы желудка (фиг. 21a) суспендировали в Matrigel. Рост культуры желудка требовал EGF (50 нг/мл), Noggin (100 нг/мл), R-спондина 1 (1 мкг/мл) и Wnt3A (100 нг/мл) (фиг. 21b). Для образования почкующихся элементов, и, таким образом, размножения культур, был необходим KGF (50 или 100 нг/мл). Таким образом, культивированные пилорические элементы имели такое же поведение, как и органоиды крипт кишечника. Открытая верхняя часть закрывается и просвет заполняется апоптотическими клетками. Вновь образованные органоиды желудка претерпевают непрерывное почкование (напоминающее деление желез) при сохранении их полярности, с почкованием желез желудка в центральный просвет. Когда использовали среду, кондиционированную Wnt3A, которая демонстрирует в 10-100 раз более высокую активность Wnt по сравнению с рекомбинантным белком Wnt3A, было выявлено значительное увеличение эффективности образования почек (фиг. 21c), указывая на дозозависимость образования и морфогенеза почек от Wnt.
Органоиды культивировали в течение по меньшей мере 1 месяца без утраты описанных свойств. Раз в неделю органоиды пассировали 1:4 путем механической диссоциации (фиг. 22). Культура Lgr5-LacZ пилорических желудочных элементов показала присутствие Lgr5-положительных стволовых клеток в желудочных органоидах (фиг. 23a). Как выявлено окрашиванием на Ki67, пролиферирующие клетки расположены в основании подобных железам структур (фиг. 23b), в то время как обнаружено, что апоптотические положительные по каспазе 3 клетки вытесняются в просвет (фиг. 23c). Муцин желудка 5AC (MUC5AC) является специфическим маркером желудочных пит-клеток, также называемых фовеолярными клетками. Положительные по MUC5AC клетки найдены в органоидах, что указывает на присутствие по меньшей мере одного дифференцированного ростка желудочных клеток (фиг. 23d). Однако не было выявлено клеток, происходящих из эндокринных желез. Таким образом, требуются дополнительные факторы. Они могут включать рилизинг-пептид гастрина, активаторы или ингибиторы семейств Hedgehog и Notch, другие активаторы каскада Wnt и другие ингибиторы семейства BMP, активаторы семейства TGF.
Пример 6a. Органоиды поджелудочной железы можно выращивать in vitro
Материалы и способы
Свежевыделенную поджелудочную железу нарезали на небольшие фрагменты и инкубировали в DMEM (Invitrogen) со смесью расщепляющих ферментов (300 Ед/мл коллагеназы типа XI (Sigma), 0,01 мг диспазы I (Roche) и 0,1 мг ДНК-азы) в течение 10 минут в орбитальном устройстве для встряхивания (80 об./мин., 37°C). После инкубации фрагменты ткани мягко диссоциировали механическим пипетированием. Нерасщепленные фрагменты осаждали в течение 1 минуты при нормальной гравитации, и супернатант переносили в новую пробирку. Супернатант пропускали через сито с размером ячеек 70 мкм, и осадок промывали DMEM. Фрагменты, оставшиеся на сите с ячейками, собирали ополаскиванием посредством DMEM перевернутого сита с ячейками, и осаждали. Фрагменты, главным образом, состояли из ацинозной ткани поджелудочной железы и включали протоки поджелудочной железы. Осадок смешивали с matrigel и культивировали в качестве культуральной системы органоида тонкого кишечника (см. материалы и способы примера 1). Matrigel инкубировали в течение 5-10 мин при 37°C. После полимеризации matrigel добавляли 500 мкл среды для культивирования тканей (Advanced-DMEM/F12, дополненная B27, N2, 200 нг/мл N-ацетилцистеина, 50 нг/мл EGF, 1 мкг/мл R-спондина 1, 100 нг/мл Noggin, 50 или 100 нг/мл KGF (Peprotech). Факторы роста добавляли каждые 2 суток. Всю среду заменяли каждые 4-6 суток. Для пассирования органоиды извлекали из Matrigel с использованием 1000-мкл пипетки и диссоциировали механически на небольшие фрагменты и переносили в свежий Matrigel. Пассирование проводили в соотношении для разделения 1:4 один раз или два раза в неделю. В этих условиях культуры поддерживали в течение по меньшей мере 2 месяцев.
Результаты
Ткань поджелудочной железы образовывала простую структуру пузыря через 3-4 суток после культивирования в присутствии EGF. Noggin и R-спондин синергично увеличивали размер структуры пузыря, но не влияли на морфогенез органоидов. KGF в значительной степени индуцировал образование почек, а также эффективность культивирования. С использованием оптимальной комбинации факторов роста (EGF, Noggin, R-спондин-1 и KGF), в наилучшей комбинации факторов роста росли более 80% поджелудочных протоков.
После помещения протоков поджелудочной железы в культуру, протоки быстро закрываются с обоих концов структуры и образуют простую структуру. Приблизительно 20% органоидов начинали образовывать почкующуюся структуру через 7 суток после начала культивирования (фиг. 24). Протоки поджелудочной железы быстро пролиферируют, в противоположность ацинозной ткани, которая растет очень медленно.
Интересно, что после пассирования органоидов, приблизительно через 2-3 недели после начала культивирования, наблюдали структуру, подобную островку поджелудочной железы (фиг. 25). Эти структуры, подобные островкам поджелудочной железы, как правило, не наблюдались перед пассированием. Островки выживают в течение по меньшей мере 7 суток, но пролиферируют очень медленно или не пролиферируют совсем. Эти структуры, подобные островкам поджелудочной железы, напоминают островки Лангенганса поджелудочной железы, которые присутствуют в здоровой ткани поджелудочной железы. Такие островки содержат, среди прочих, альфа-клетки и бета-клетки, которые продуцируют глюкагон и инсулин, соответственно. Наблюдаемые подобные островам структуры содержат клетки, которые экспрессирует инсулин, нейрогенин 3 и Pdx-1. Для определения того, увеличивают ли они наличие β-клеток поджелудочной железы в органоидах, которые происходят из ткани поджелудочной железы будет протестировано несколько факторов роста. Факторы роста-кандидаты включают циклопамин (ингибитор Sonic-hedgehog), активин, GLP (глюкагон-подобный пептид) и его производное (экзендин 4), гастрин и никотинамид.
Пример 6b. Органоиды поджелудочной железы можно выращивать in vitro
Материалы и способы
Свежевыделенную поджелудочную железу нарезали на небольшие фрагменты и инкубировали в DMEM (Invitrogen) со смесью расщепляющих ферментов (300 Е/мл коллагеназы типа XI (Sigma), 0,01 мг/мл диспазы I (Roche) и 0,1 мг/мл ДНК-азы) в течение 10 минут в орбитальном устройстве для встряхивания (80 об./мин., 37°C). После инкубации фрагменты тканей мягко диссоциировали механическим пипетированием. Нерасщепленные фрагменты осаждали в течение 1 минуты при нормальной гравитации. Нерасщепленные фрагменты далее расщепляли с помощью смеси расщепляющих ферментов в течение 10 минут. Эту процедуру расщепления повторяли до тех пор, пока нерасщепленные фрагменты, главным образом, не состояли из протоков поджелудочной железы. Структуры протоков поджелудочной железы вручную отбирали из неращепленных фрагментов под микроскопом. Протоки поджелудочных желез смешивали с matrigel и культивировали в качестве культуральной системы органоида тонкого кишечника (см. материалы и способы примера 1). Matrigel инкубировали в течение 5-10 мин при 37°C. После полимеризации matrigel добавляли 500 мкл среды для культивирования тканей (Advanced-DMEM/F12, дополненная 1x Glutamax, пенициллин/стрептомицином, 10 мМ Hepes, B27, N2, 10 мМ N-ацетилцистеином, 10 нМ [Leu15]-гастрином I, 100 нМ экзендином 4, 10 мМ никотинамидом, 50 нг/мл EGF, 1 мкг/мл R-спондина 1, 100 нг/мл Noggin, 50 или 100 нг/мл FGF7 (KGF) или FGF10 (Peprotech). Культуральную среду заменяли каждые 2 суток. Для пассирования органоиды извлекали из Matrigel с использованием 1000-мкл пипетки и диссоциировали механически на небольшие фрагменты и переносили в свежий Matrigel. Пассирование проводили в соотношении для разделения 1:4 один раз или два раза в неделю. В этих условиях культуры поддерживали в течение по меньшей мере 10 месяцев.
Результаты
Ткань поджелудочной железы образовывала простую структуру пузыря через 3-4 суток после культивирования в присутствии EGF. Noggin и R-спондин синергично увеличивали размер структур пузыря, но не влияли на морфогенез органоидов. FGF7 (KGF)/FGF10 в значительной степени индуцировали образование почек, а также эффективность культивирования. С использованием оптимальной комбинации факторов роста (EGF, Noggin, R-спондин-1 и FGF7 (KGF)/FGF10), в наилучшей комбинации факторов роста росло более чем 80% протока поджелудочной железы. После отбора протоков поджелудочной железы в культуру, протоки быстро закрывались с обоих концов структуры и образовывали простую структуру. Приблизительно 80% органоидов начинало образовываться из почкующейся структуры через 7 суток после начала культивирования (фиг. 24). Протоки поджелудочной железы быстро пролиферировали, в противоположность ацинозной ткани, которая росла только очень медленно. Интересно, что после пассирования органоидов приблизительно через 2-3 недели после начала культивирования наблюдали структуру, подобную островку поджелудочной железы (фиг. 25). Эти структуры, подобные островкам, как правило, не наблюдали перед пассированием. Островки выживают в течение по меньшей мере 14 суток, но пролиферируют очень медленно или не пролиферируют совсем. Эти подобные островкам структуры напоминают островки Лангерганса поджелудочной железы, которые присутствуют в здоровой ткани поджелудочной железы. Такие островки содержат, среди прочих, альфа-клетки и бета-клетки, которые продуцируют глюкагон и инсулин, соответственно. Наблюдаемые подобные островкам структуры содержат клетки, которые экспрессируют инсулин, нейрогенин 3 и Pdx-1. Несколько факторов роста будет протестировано для определения, увеличивают ли они присутствие β-клеток в органоидах, которые происходят из ткани поджелудочной железы. Факторы роста-кандидаты включают циклопамин (ингибитор Sonic-hedgehog), активин, GLP (глюкагон-подобный пептид) и его производное (экзендин 4), гастрин и никотинамид.
Пример 7. Беспрепятственное размножение предшественников поджелудочной железы in vitro путем запуска регенеративного ответа Wnt/Lgr5
Материалы и способы
Мыши, реагенты и ткани
Ткань поджелудочной железы получали из следующих мышей: со вставкой Axin-LacZ (Lustig et al. Mol. Cell. Biol. 2002), со вставкой Lgr5-LacZ (Barker et al, 2007), Lgr5-GFP (Barker et al, 2007). Мышам Axin-LacZ внутрибрюшинно инъецировали 100 мкг очищенного R-спондина 1 человека (любезно предоставленного A. Abo, Nuvelo Inc, CA, США) и их умерщвляли через 48 часов после этого для анализа экспрессии LacZ в поджелудочной железе.
Лигирование протоков поджелудочной железы проводили, как описано для крыс (Wang et al, 1995) с некоторыми небольшими модификациями. Процедура эксперимента по PDL была следующей: животных подвергали анестезии с помощью смеси флуанизон: фентанил: мидазолам, инъецированной внутрибрюшинно в дозировке 3,3 мг/кг, 0,105 мг/кг и 1,25 мг/кг соответственно. Животных помещали в положение лежа на спине и поверхность живота брили и очищали антисептическим раствором (раствор йода). После этого проводили срединный надрез на верхней передней стенке живота от мечевидного отростка грудины и обнажали поджелудочную железу. Под препаровальной лупой селезеночную долю локализуют и проток поджелудочной железы лигировали швом из полипропиленовой мононити 7-0 приблизительно на 1 мм дистальнее соединения с протоком желудочной доли. После хирургической операции подкожно вводили анальгетик бупренорфин в дозе 0,01-0,05 мг/кг. После этого стенку желудка и кожу закрывали швами из шелковой нити 5-0.
Свежевыделенную поджелудочную железу обрабатывали, как описано в примере 6, получая фрагменты поджелудочной железы, которые культивировали в условиях, описанных ниже. Главный проток поджелудочной железы и первую ветвь протоков механически выделяли. Эти фрагменты нарезали на небольшие фрагменты и инкубировали в DMEM (Invitrogen) со смесью расщепляющих ферментов (300 Е/мл коллагеназы типа XI (Sigma), 0,01 мг/мл диспазы I (Roche) и 0,1 мг/мл ДНК-азы) в течение 30 минут в орбитальном устройстве для встряхивания (80 об./мин., 37°C). После расщепления большинство ацинозных клеток высвобождались из фрагментов. Нерасщепленные фрагменты, главным образом, состоящие из клеток протоков поджелудочной железы, осаждали в течение 1 минуты при нормальной гравитации и супернатант удаляли. После промывания в течение трех раз с помощью PBS, нерасщепленные фрагменты инкубировали с помощью 2 мМ EDTA/PBS в течение 30 мин при комнатной температуре. Фрагменты энергично пипетировали и осаждали в течение 1 минуты при нормальной гравитации. Супернатант, обогащенный клетками протоков, переносили в новые пробирки и промывали посредством PBS три раза. Клетки протоков осаждали и смешивали с Matrigel. Matrigel инкубировали в течение 5-10 мин при 37°C. После полимеризации matrigel добавляли 500 мкл среды для размножения (Advanced-DMEM/F12, дополненная 1x Glutamax, пенициллином/стрептомицином, 10 мМ Hepes, B27, N2, 1 мМ N-ацетилцистеином, 10 нМ [Leu15]-гастрином 1, 100 нМ экзендином 4, 10 мМ никотинамидом, 50 нг/мл EGF, 1 мкг/мл R-спондина 1, 100 нг/мл Noggin, 50 или 100 нг/мл FGF7 (KGF) или FGF10 (Peprotech). Полную среду заменяли каждые 2 суток. Для пассирования органоиды удаляли из Matrigel с использованием 1000-мкл пипетки и механически диссоциировали на небольшие фрагменты и переносили в свежий Matrigel. Пассирование проводили в соотношении для разделения 1:4 один раз в неделю. В этих условиях культуры поддерживались в течение по меньшей мере 2 месяцев. Для дифференцировки среду для размножения заменяли на среду для дифференцировки (Advanced-DMEM/F12, дополненная Glutamax, пенициллином/стрептомицином, 10 мМ Hepes, B27, N2, 200 нг/мл N-ацетилцистеином, 10 нМ [Leu15]-гастрином 1, 100 нМ экзендином 4, 50 нг/мл EGF, 1 мкг/мл R-спондина 1, 100 нг/мл Noggin).
FGF10 был получен от Peprotech. BrdU был получен от Sigma.
Q-PCR
РНК выделяли с помощью набора RNA easy mini kit (Quiagen), и подвергали обратной транскрипции с использованием обратной транскриптазы вируса лейкоза мышей Молони (Promega). кДНК амплицировали в термоблоке.
Использованные праймеры представлены ниже.
mmTBP (прямой) TATTGTATCTACCGTGAATCTTGG
mmTBP (обратный) CAGTTGTCCGTGGCTCTC
Lgr5 (прямой) TCCAACCTCAGCGTCTTC
Lgr5 (обратный) TGGGAATGTGTGTCAAAG (Tm=57°C)
PCR
Все праймеры были разработаны так, чтобы они фланкировали или охватывали последовательности интронов, чтобы отличить геномную ДНК.
Hprt, (F) AAGTTTGTTGTTGGATATGC (R) CATCTTAGGCTTTGTATTTGG, (Tm) 57°C, 106 т.п.н.
Ngn3, (F) TCCTCGGAGCTTTTCTACGA (R) TGTGTCTCTGGGGACACTTG (Tm) 60°С, 239 т.п.н./373 т.п.н. (геномная полоса).
Pax6, (F) AACAACCTGCCTATGCAACC (R) ACTTGGACGGGAACTGACAC, TM 60°С, 206 т.п.н.
Глюкокиназа, (F) AAGATCATTGGCGGAAAG (R) GAGTGCTCAGGATGTTAAG, (Tm) 57°C, 193 т.п.н.
Хромогранин A, (F) GCTGACAGCAGAGAAGCGGCT (R) GACAGGCTCTCTAGCTCCTGG, (Tm) 60°С 231 т.п.н.
Glut2, (slc2a2) (F) AAGTTGGAAGAGGAAGTCAG (R) AGACCTTCTGCTCAGTCG (Tm), 57°C 124 т.п.н.
Инсулин, (F) TTTGTCAAGCAGCACCTTTG (R) TCTACAATGCCACGCTTCTG, (Tm) 57°С, 214 т.п.н.
Соматостатин, (F) GAGGCAAGGAAGATGCTGTC (R) GGGCATCATTCTCTGTCTGG, (Tm) 57°C, 214 т.п.н.
Глюкагон (F) TTACTTTGTGGCTGGATTGCTT (R) AGTGGCGTTTGTCTTCATTCA, (Tm) 57°C, 149 т.п.н.
Анализ изображений
Изображения органоидов крипт получали либо с помощью конфокальной микроскопии с использованием инвертационного микроскопа Leica SP5 (Nikon DM-IL), либо с помощью стереомикроскопа (Leica, MZ16-FA). Для иммуногистохимии образцы фиксировали 4% параформальдегидом (PFA) в течение 1 ч при комнатной температуре, и парафиновые срезы обрабатывали стандартными способами (Barker et al., Nature. 2007). Иммуногистохимию проводили, как описано ранее (Barker et al., Nature. 2007). Для иммунного окрашивания тотального препарата органоиды поджелудочной железы выделяли из Matrigel с использованием диспазы (Invitrogen) и фиксировали с помощью 4% PFA с последующим увеличением проницаемости с помощью 0,1% Triton X-100. Для иммуногистохимии использовали следующие антитела: антитело против BrdU (Amersham), антитело против Ki67 (Dako), антитело против инсулина (Sigma), антитело против C-пептида (Cell signaling), антитело против Ngn3 (Developmental hybridoma studies bank).
ДНК окрашивали с помощью DAPI или ToPro-3 (Molecular Probes). Трехмерные изображения получали с помощью конфокальной микроскопии. Окрашивание с помощью X-gal проводили, как описано в примере 5 в разделе "Иммуногистохимия" и "Анализ изображений".
FACS
Органоиды поджелудочной железы культивировали в присутствии или в отсутствие R-спондина (1 мкг/мл) и удаляли из matrigel механически или ферментативно (TrypLE). Выделенные органоиды далее расщепляли TrypLE в течение 10 минут при 37°С. Диссоциированные клетки пропускали через сито с ячейками размером 40 мкм (BD bioscience) и окрашивали конъюгированным с APC антителом против EpCAM (eBioscience). LacZ окрашивали с помощью набора FluoReporter kit (Invitrogen) в соответствии с протоколом изготовителя. Единичные жизнеспособные клетки отсортировали с помощью длительности импульса, параметра бокового рассеяния и окрашивания йодидом пропидия.
Размножение in vitro единичных положительных по Axin2-LacZ клеток поджелудочной железы. Поджелудочную железу извлекали из мышей через 7 суток после проведения PDL, и выделяли протоки поджелудочной железы, как описано выше. Выделенные протоки поджелудочной железы инкубировали с TrypLE Express (Invitrogen) в течение 20 минут при 37°С, а затем пропускали через сито с ячейками размером 40 мкм (BD bioscience). Клетки окрашивали с помощью EpCAM-APC и флуоресцентного субстрата для LacZ (набор FluoroReporter kit), как описано в примере 7. Клетки анализировали и единичные жизнеспособные эпителиальные клетки сортировали с помощью проточного цитометра (MoFlo, Dako Cytomation), и собирали в среду EM. Отсортированные клетки осаждали, смешивали с Matrigel и культивировали со средой EM, включающей 50% среду, кондиционированную Wnt, и 10 мМ Y-27632 в течение 4 суток. Через 4 суток культуральную среду заменяли на среду EM без Wnt и Y-27632.
Результаты
Единичные Wnt-зависимые Lgr5+ стволовые клетки, происходящие из тонкого кишечника, можно культивировать для получения непрерывно размножающихся подобных пищеварительному каналу органоидов (Sato et al., 2009). В здоровой взрослой поджелудочной железе каскад Wnt является неактивным и, следовательно, Lgr5 не экспрессируется. При повреждении путем частичного лигирования протока (PDL) авторы настоящего изобретения открыли, что каскад Wnt устойчиво активируется, в то время как экспрессия Lgr5 появляется на почках регенерирующих протоков. В условиях, модифицированных из культуральной системы кишечника, свежевыделенные фрагменты взрослого протока инициировали экспрессию Lgr5 и образовали почкующиеся пузыри, которые размножались в 10 раз за неделю в течение >30 недель. Удаление стимулов роста преобразует эти пузыри в структуры с незрелой морфологией островков, экспрессирующие эндокринные маркеры и маркеры β-клеток. Единичные стимулированные Wnt клетки из поврежденной поджелудочной железы также могли инициировать эти длительные культуры. Авторы настоящего изобретения заключили, что предел Хейфлика не был применим ко взрослым клеткам-предшественником, когда их культивировали в оптимизированных условиях. Таким образом, способы культивирования, способствующие размножению органоспецифических стволовых клеток, могут являться альтернативной для получения ткани на основе ES или iPS.
Хотя развитие экзокринных и эндокринных компартментов эмбриональной поджелудочной железы понятно достаточно детально (Jensen, 2004), значительно менее известно об образовании клеток островков в постнатальной поджелудочной железе (Bonner-Weir and Weir, 2005, Bouwens and Rooman, 2005). Прослеживание генетического происхождения обеспечило доказательство того, что предсуществующие β-клетки, а не стволовые клетки/клетки-предшественники, образуют новые β-клетки у взрослых мышей, как в нормальных физиологических условиях, так и после частичной панкреатоэктомии (Dor et al., 2004, Teta et al., 2007). Недавно было описано существование мультипотентных клеток-предшественников в выстилке протоков поджелудочной железы взрослых мышей, которые могут активироваться в поврежденной поджелудочной железе для увеличения массы функциональных β-клеток (Xu et al., 2008). Контролируемое повреждение обеспечивали путем проведения PDL на поджелудочной железе взрослых мышей, несущих промоторный репортер Ngn3, который кодирует главный выключатель для клеток-предшественников эмбриональных островков (Apelqvist et al., 1999, Gradwohl et al., 2000, Gu et al., 2002, Schwitzgebel et al., 2000) и который является молчащим в нормальной постнатальной поджелудочной железе (Gu et al., 2002). Дифференцировка этих предшественников β-клеток является Ngn3-зависимой и дает начало всем типам клеток островков, включая отвечающие на глюкозу β-клетки (Xu et al., 2008). В настоящее время неизвестно, какие сигналы запускают появление этих предшественников при повреждении. Такое представление кажется важным, поскольку оно может направить разработку подходов для размножения клеток in vitro.
Для определения того, играет ли передача сигнала Wnt роль в индукции предшественников β-клеток, наблюдали за экспрессией аллеля Axin2-LacZ во взрослой поджелудочной железе. Показано, что аллель Axin2-LacZ является проверенным общим репортером для передачи сигнала Wnt (Lustig et al., Mol. Cell. Biol. 2002). Как ожидалось, репортер был неактивным во взрослой поджелудочной железе (фиг. 26A). Однако когда авторы настоящего изобретения инъецировали агонист Wnt Rspol (Kim et al., 2005) мышам Axin2-LacZ для активации каскада передачи сигнала Wnt, они отметили наличие отвечающих на Wnt клеток вдоль протоков, но не в ацинусах или островках поджелудочной железы (фиг. 26B). Поскольку предшественники β-клеток ранее были выявлены только при повреждении поджелудочной железы, авторы настоящего изобретения тестировали, активировался ли ответ на Wnt физиологически в этих клетках при проведении PDL. На фигуре 26C показано окрашивание H&E срезов тканей поджелудочной железы, выделенных из области PDL и области не PDL. Как описано ранее (Abe et al. 1995), ацинозные клетки становились апоптотическими через 5 суток и заменялись вновь образованными структурами протоков с помощью механизма, понятного не полностью. Через 7 суток также наблюдали увеличение количества островков (неогенез островков), а также размера островков (как указано звездочкой). Это указывает на то, что PDL было успешным. Действительно, репортер Axin2-LacZ специфично активировался вдоль протоков лигированной части поджелудочной железы, в то время как нелигированная часть не демонстрировала этого ответа (фиг. 26D и E). Более того, пролиферативный ответ, при определении по окрашиванию на Ki67, был наиболее ограничен протоками лигированной части, в то время как в протоках нелигированной части нельзя было выявить ядерного Ki67 (фиг. 26F). Это напоминало детекцию пролиферативных положительных по BrdU клеток в поджелудочной железы после обработки R-спондином (фиг. 26 G).
Авторы изобретения ранее показали, что в кишечниках определенная популяция отвечающих на Wnt клеток представляют собой стволовые клетки (Barker et al., 2007). Маркером для этой популяции клеток был Lgr5. Ген Lgr5 представляет собой, подобно Axin2, отвечающий на Wnt ген. Кроме кишечника и кожи, он экспрессируется только в стимулированных Wnt стволовых клетках, но не в переходных делящихся клетках (Barker et al., 2007, Jaks et al., 2008)). Таким образом, он считается истинным маркером стволовых клеток. Авторы настоящего изобретения предположили, что, аналогично Lgr5+ клеткам в кишечниках, клетки Lgr5+ в поджелудочной железе также могут происходить из предшественников β-клеток, как выявлено после повреждения. Для тестирования этой гипотезы авторы настоящего изобретения провели PDL в поджелудочной железе мышей Axin-LacZ и Lgr5-LacZ и определили экспрессию мРНК Lgr5 и провели окрашивание LacZ. Интересно, что Lgr5 быстро стал поддаваться детекции посредством qPCR после PDL (фиг. 26H). Более того, PDL у мышей со вставкой Lgr5-LacZ привело к специфической активности репортера в почках регенерирующих протоков (указано звездочками), как продемонстрировано посредством окрашивания посредством X-gal (фиг. 26I). Появление экспрессии Lgr5 в областях активной регенерации показало, что Lgr5 может не только маркировать стволовые клетки при физиологическом самообновлении (например, в кишечнике желудке или волосяных фолликулах), но что его экспрессия также может предвещать активацию посредством Wnt регенеративных стволовых клеток/предшественников при повреждении.
С учетом появления Wnt-зависимого маркера стволовых клеток Lgr5, авторы изобретения заключили, что взрослые предшественники поджелудочной железы можно размножать в ранее установленных условиях культивирования органоида кишечника (Sato et al., 2009). Ранее были получены культуры гетерогенных популяций клеток поджелудочной железы и, как правило, они включают факторы роста, такие как EGF (Githens et al. In Vitro Cell Dev Biol. 1989), FGF10 (Miralles et al. Proc Natl Acad Sci USA. 1999) и HGF (Lefebvre et al. Diabetes. 1998, Suzuki et al., Diabetes 53, 2004) и добавки в сыворотку, такие как гастрин (Rooman et al. Gastroenterology. 2001), никотинамид (Rooman et al. Diabetologia. 2000) и другие. Ряд таких культур приводил к образованию in vitro клеток с фенотипами, подобными β-клеткам (Bonner-Weir et al., 2000, Seaberg et al., 2004, Suzuki et al., 2004), которые в определенных условиях были способны вызывать обратное развитие гипергликемии при трансплантации мышам с диабетом (Hao et al., 2006, Ramiya et al., 2000). Большинство из этих подходов начинаются со смешанных популяций клеток, которые подвергаются старению с течением времени. По-видимому, является справедливым, что в настоящее время не существует устойчивой длительной культуральной системы, которая поддерживает устойчивое размножение определенных нетрансформированных взрослых предшественников поджелудочной железы в течение длительных периодов времени, которые сохраняют способность дифференцироваться в эндокринный росток.
Авторы изобретения впервые предприняли попытку вырастить очищенные фрагменты протоков в среде для размножения (EM). Как показано на фиг. 27A, небольшие фрагменты протоков сразу претерпевают размножение в структуры, подобные пузырям, претерпевающие непрерывное почкование, в то время как островки (данные не представлены) и ацинусы (нижняя панель) постепенно дезинтегрируют. Культуры размножаются в 10 раз/неделя (и их часто пассируют раз в неделю) в течение более чем 30 недель. Для определения требуемых сигналов для оптимального размножения клеток поджелудочной железы in vitro было протестировано множество факторов роста (фиг. 27B). Очевидно, в отсутствие EGF, культуры дезинтегрировали через 7 суток. Также отсутствие R-спондина или FGF10 уменьшало жизнеспособность культур через 14 суток. Напротив, Noggin, ингибитор BMP, не оказывал какого-либо эффекта на непрерывный рост фрагментов поджелудочной железы. Добавление никотинамида, экзендина 4, гастрина, в среду для размножения, не было необходимым, а приводило к увеличению эффективности культивирования (данные не представлены).
Поскольку авторы изобретения продемонстрировали, что передача сигнала Wnt активировалась при PDL, определяли эффект добавления агониста Wnt на свежевыделенные фрагменты поджелудочной железы in vitro на непрерывный рост. Когда протоки выделяли из мышей Axin2-LacZ, целые почкующиеся пузыри окрашивались синим цветом только в присутствии агониста Wnt R-спондина 1 (фиг. 27C), напоминая ситуацию in vivo после PDL (фиг. 26D и E). В свежевыделенных островках или ацинусах из поджелудочной железы Axin2-LacZ не наблюдали окрашивания синим цветом. Наряду с наблюдениями in vivo, при PDL, только пучки пузырей Lgr5-LacZ окрашивались синим цветом (фиг. 27D). Более того, культивирование органоидов поджелудочной железы Lgr5-LacZ в течение 14 суток в присутствии R-спондина значимо увеличивало процент Lgr5+ клеток (фиг. 27E). Важно, что когда фрагменты поджелудочной железы культивировали в отсутствие R-спондина в EM, органоиды прекращают пролиферировать в течение 1 месяца, в то время как в присутствии R-спондина они могут размножаться в течение неограниченного периода времени. Эти наблюдения подразумевают, что отвечающие на Wnt предшественники, расположенные вблизи протоков, поддерживали рост почкующихся пузырей, которые впоследствии поддерживались экспрессирующими Lgr5 клетками со свойствами, подобными свойствам стволовых клеток.
Для непосредственного тестирования этого представления авторы изобретения отсортировали положительные по Axin2-LacZ клетки из мышей через 7 суток после PDL и выявили, что эти клетки эффективно инициировали почкующиеся пузыри, которые были неотличимы от инициируемых протоками пузырей (фиг. 28). Единичные клетки требуют присутствия Wnt3a в среде. Сравнение эффективности культивирования в присутствии или в отсутствие Wnt3A после диссоциации на единичные клетки показало, что единичные клетки, культивированные в отсутствие Wnt3A, первоначально растут в качестве небольших структур пузырей, но прекращают пролиферировать через 2-4 суток. Это не так для культур поджелудочной железы, начинаемых с выделенных фрагментов поджелудочной железы. Интересно, что Wnt3A можно удалять через 4 суток, что указывает на то, что этот сигнал более не является необходимым для стимуляции роста или что продукция Wnt3A инициировалась клетками, происходящими из единичных отсортированных клеток, с которых начинали культивирование.
Затем авторы изобретения предприняли попытку оценить потенциал почкующихся пузырей в отношении образования клеток эндокринного ростка. Для этого авторы настоящего изобретения протестировали ряд изменений EM для определения среды для дифференцировки (DM). Ряд факторов были протестированы в отношении их эффекта на дифференцировку в эндокринные ростки. Удаление FGF10 оказалось важным для индукции дифференцировки. Структуры, подобные островкам, появлялись только в отсутствие FGF10 (фиг. 29A), что соответствовало появлению экспрессии нескольких маркеров дифференцировки для предшественников β-клеток (Ngn3), β-клеток (инсулин), глюкагона (α-клетки) и соматостатина (δ-клетки) (фиг. 29B и C). Более того, начиная на 10 сутки после начала воздействия среды DM повышалась регуляция маркеров дифференцировки, таких как глюкокиназа, Pax6 и хроматогранин A. Таким образом, DM оптимально состояла по меньшей мере из EGF и R-спондина и не содержала ни FGF7, ни или FGF 10. Непрерывную экспрессию Lgr5, маркера стволовых клеток в условиях дифференцировки можно объяснить присутствием R-спондина, агониста Wnt, в DM, поскольку Lgr5 представляет собой отвечающий на Wnt ген. Когда клетки культивировали в присутствии никотинамида в EM, также было важно удалить его из среды для обеспечения полной дифференцировки. Когда почкующиеся пузыри после какого-либо периода культивирования переносили из EM в DM пузыри претерпевали процесс стереотипической "инволюции": постепенное направленное внутрь сворачивание стенки приводило к сжатию пузыря в компактное тельце меньших размеров, напоминающее островок (фиг. 29D). Подобную островкам морфологию подтверждали посредством маркеров островком β-клеток, таких как инсулин и C-пептид (фиг. 29E). Для подтверждения зависимости этой стадии процесса регенерации от передачи сигнала Wnt, фрагменты поджелудочной железы культивировали в DM в отсутствие или в присутствии R-спондина. Важно, что предшественники β-клеток, как продемонстрировано по экспрессии Ngn3, поддавались детекции только в присутствии R-спондина (фиг. 29F).
Пример 8. Размножение фрагментов поджелудочной железы in vitro
В процессе эмбрионального развития поджелудочной железы в сети протоков поджелудочной железы наблюдали клетки, экспрессирующие нейрогенин-3+ или инсулин, и было предположено, что клетки протоков поджелудочной железы дают начало эндокринным предшественникам и, следовательно, взрослым эндокринным клеткам. Было показано, что клетки протоков поджелудочной железы человека дифференцируются в отвечающие на глюкозу продуцирующие инсулин клетки in vitro (Bonner-Weir, S et al. 2000. PNAS), и это открытие сделало клетки протоков поджелудочной железы привлекательным источником для заместительной терапии бета-клетками. Однако трудно размножить клетки протоков без утраты способности к эндокринной дифференцировке. В ранее описанной культуральной системе клетки протоков поджелудочной железы утрачивали эпителиальные свойства и подвергались старению после от 2 до 5 недель (Trautmann B et al. Pancreas vol. 8: 248-254). Таким образом, не существует устойчивой культуральной системы для размножения клеток протоков поджелудочной железы, которая сохраняет способность к эндокринной дифференцировке. Пользуясь преимуществом установления культуральной системы органоидов поджелудочной железы мыши, здесь авторы настоящего изобретения предприняли попытку установить культуральную систему органоида человека.
Выращивание предшественников поджелудочной железы человека in vitro
Поджелудочную железу человека получали от Leiden University Medical Center, Нидерланды. Важно, что в тех же условиях, как описано для фрагментов поджелудочной железы выше (пример 7), также можно выращивать свежевыделенные фрагменты поджелудочной железы человека in vitro (фиг. 30).
В этих условиях размножения эффективность культивирования фрагментов поджелудочной железы составляла приблизительно 80%, что означало что 80% свежевыделенных фрагментов поджелудочной железы эффективно размножалось in vitro в течение более длительного периода времени. По сравнению с поджелудочной железой мыши, ацинозная ткань более легко образует структуры пузырей, однако эта структура прекращала пролиферировать в течение 4 недель. Клетки протоков поджелудочной железы из более крупной сети более эффективно продуцируют структуры пузырей и в конечном итоге образуют органоиды из почек. Органоиды поджелудочной железы разделяли в соотношении 1:5 один раз в неделю и поддерживали in vitro в течение по меньшей мере 5 недель без утраты способности к пролиферации.
В общем, авторы настоящего изобретения установили культуральную систему органоидов поджелудочной железы человека и успешно размножили клетки протоков поджелудочной железы по меньшей мере 3000 раз из исходного объема. Авторы изобретения оптимизируют условия культивирования для эндокринной дифференцировки клеток протоков поджелудочной железы человека, и после оптимизации этот подход in vitro может иметь важное значение для создания заместительной терапии бета-клетками, доступной для большого количества людей с сахарным диабетом 1 и 2 типа.
Ссылки



Пример 9. Культивирование крипт тонкого кишечника или толстого кишечника in vitro
Как описано в примерах 1 и 2, впервые стало возможным создание условий длительного культивирования эпителия тонкого кишечника и толстого кишечника мыши. Органоиды крипта-ворсинка растут с помощью дополнения набором определенных факторов роста и внеклеточного матрикса. Органоиды содержат стволовые клетки кишечника, которые активно делятся и дают начало всем основным росткам дифференцированных клеток, присутствующим в кишечнике. В этом примере авторы изобретения показали, что эти условия культивирования не являются уникальными для эпителия кишечника мыши, а также их можно использовать для выращивания эпителия кишечника человека.
Материалы и способы
Культуры органоидов толстого кишечника мыши
Культуры органоидов мыши культивировали, как описано в примере 1. Для ингибирования секреции Wnt использовали продукцию ингибитора Wnt (IWP-2) (Chen et al., Nat Chem Biol. 2009 Feb, 5(2): 100-7).
Культуры органоидов толстого кишечника человека
Крипты толстого кишечника человека выделяли из образца нормального подвергнутого резекции толстого кишечника и культивировали в качестве структур органоидов в течение 7 суток с использованием установленной системы культивирования органоидов (Sato et al., 2009. Nature. May 14, 459(7244): 262-5). Поскольку этот протокол был оптимизирован для происходящих из мыши культур органоидов, авторы настоящего изобретения внесли небольшое изменение путем добавления среды, кондиционированной Wnt3a, для обеспечения оптимального роста органоидов толстого кишечника человека. Для получения этой кондиционированной среды Wnt3a экспрессировали в клеточной линии путем трансфекции пригодной экспрессирующей конструкции, кодирующей указанный лиганд. Указанную клеточную линию культивируют и культуральную среду, содержащую секретированный лиганд, собирают через подходящие интервалы времени. Например, клетки начинают продуцировать Wnt3a в тот момент, когда они достигают смыкания монослоя и перестают расти. В качестве отрицательного контроля использовали культуральную среду клеток, которые не были трансфицированы или инфицированы указанной пустой экспрессирующей конструкцией. Кондиционированную среду собирали и тестировали, например в анализе, где экспрессия люциферазы контролируется отвечающими на TCF элементами, для количественного определения присутствия агониста Wnt, такого как Wnt3a (Korinek et al., 1997 Science 275: 1784-1787).
Результаты
Пролиферация эпителия кишечника зависит от каскада передачи сигнала Wnt. Однако точное расположение источника Wnt является неясным (Gregorieff and Clevers, 2005, Genes Dev Apr. 15, 19(8): 877-90). Поскольку органоиды кишечника мыши росли зависимым от ниши образом (Sato et al., 2009. Nature. May 14, 459(7244): 262-5) авторы изобретения предположили, что эти органоиды могут продуцировать их собственные лиганды Wnt. Для тестирования этого предположения авторы настоящего изобретения ингибировали секрецию Wnt посредством инкубации с ингибитором поркупина. Поркупин важен для секреции Wnt (схематическая фиг. 31A). Инкубация с 1 мкМ IWP (Chen et al, Nat Chem Biol. 2009 Feb; 5(2): 100-7) приводила к гибели органоидов (фиг. 31B и C). Органоиды можно было спасти добавлением среды, кондиционированной Wnt3a, что указывало на то, что органоиды действительно продуцируют лиганды Wnt (фиг. 31D и E).
Далее авторы изобретения предприняли попытку культивирования органоидов кишечника человека. Оказалось, что добавление Wnt3a в среду было необходимым, поскольку без него органоиды крипт никогда не образовывали почкующиеся структуры и погибали в течение 5-10 суток для тонкого кишечника и в течение 3-4 суток для толстого кишечника (фиг. 32). В целом, органоиды крипт кишечника человека росли сравнимым образом с культурами органоидов мыши. Главным образом, авторы изобретения достигали эффективности культивирования вплоть до 80%, в зависимости от активности среды, кондиционированной Wnt-3a. Культуры кишечника человека находились в культуре в течение вплоть до 3 месяцев. Эффект Wnt-3a в толстом кишечнике человека был ожидаемым, поскольку в культуре органоидов толстого кишечника мыши также было выявлено, что он усиливает эффекты. Потребность в Wnt-3a в тонком кишечнике и толстом кишечнике человека может вытекать из более низкой продукции эндогенных лигандов Wnt органоидами человека вследствие меньшего количества клеток Панета, присутствующих в кишечнике человека по сравнению с кишечником мыши. До настоящего времени не существовало воспроизводимой длительной культуральной системы для кишечника человека, и культуральная система авторов изобретения пригодна не только для понимания биологии стволовых клеток кишечника человека, но также для применения в клинически ориентированных тестах, таких как скрининг лекарственных средств.
Пример 10. Оптимизированные условия культивирования для выращивания органоидов желудка
Как описано в примере 5, была идентифицирована культуральная среда, которую можно использовать для культивирования эпителия желудка в течение длительных периодов времени. Здесь авторы изобретения описывают оптимизированные условия для этих культур органоидов желудка.
Материалы и способы
Выделение желудочного элемента, диссоциация единичных клеток и сортировка EGFP+ клеток. Железы желудка выделяли из пилорических областей мышей, как описано ранее, с некоторыми модификациями (Bjerknes and Cheng, 2002, Am J Physiol Gastrointest Liver Physiol. Sep; 283(3): G767-77). В кратком изложении под микроскопом желудок открывали вдоль большего изгиба, промывали солевым раствором и извлекали привратник желудка. Мышечный слой желудка удаляли и оставшийся эпителий разделяли на 5-мм фрагменты и инкубировали в течение 3-5 ч в забуференном солевом растворе (28 мМ Na2HPO4, 40 мМ KH2PO4, 480 мМ NaCl, 8 мМ KCl, 220 мМ сахароза, 274 мМ D-сорбит, 2,6 мМ DL-дитиотреитол), содержащем 10 мМ EDTA (для культивирования или окрашивания) или 5 мМ EGTA (для выделения РНК) при 4°C. После удаления хелатирующего агента, фрагменты ткани энергично суспендировали в забуференном растворе с использованием 10-мл пипетки. После суспендирования и центрифугирования осадок был обогащен железами желудка. После выделения желез клетки собирали и ресуспендировали в не содержащей кальций среде SMEM (Invitrogen), дополненной 10 мг/мл трипсина и 0,8 единиц/мкл ДНК-азы I (для анализа на микрочипах) или ресуспендировали в TrypleExpress (GIBCO), дополненной 0,8 единиц/мкл ДНК-азы (для целей культивирования). В обоих случаях после инкубации при 37°С в течение 20-25 минут клетки осаждали и фильтровали через сито с ячейками 40 мкМ. Клетки EGFPhi и EGFPlo сортировали проточной цитометрией (MoFlo, Beckman Coulter). Единичные жизнеспособные эпителиальные клетки отсортировывали с помощью рассеяния вперед и параметра длительности импульса. Когда было задано, клетки улавливали либо в отношении отрицательного окрашивания йодидом пропидия, собирали в Trizol LS (Invitrogen) и подвергали выделению РНК в соответствии с протоколом изготовителя, либо собирали в культуральную среду для желудка, погружали в Matrigel (BD Bioscience) и культивировали согласно протоколу, подробно описанному ниже.
Культура для желудка
Для культивирования выделенные железы желудка подсчитывали и всего 100 желез смешивали с 50 мкл Matrigel (BD Bioscience) и высевали в 24-луночные планшеты. После полимеризации Matrigel наслаивали культуральную среду желудка (Advanced DMEM/F12, дополненную B27, N2 и n-ацетилцистеином (Invitrogen), содержащую факторы роста (50 нг/мл EGF (Peprotech), 1 мкг/мл R-спондина 1, 100 нг/мл Noggin (Peprotech), 100 нг/мл FGF10 (Preprotech) и среда, кондиционированная Wnt3A). Для культивирования единичных клеток всего 100 отсортированных EGFPhi клеток/лунка собирали в культуральную среду для желудка и погружали в Matrigel (BD Bioscience). После полимеризации Matrigel наслаивали культуральную среду для желудка. В течение первых 2 суток после посева среду также дополняли 10 мкМ ингибитором ROCK Y-27632 (Sigma Aldrich), для предотвращения аноикиса. Факторы роста добавляли раз в двое суток и всю среду заменяли раз в 4 суток. Для пассирования органоиды желудка извлекали из Matrigel, механически диссоциировали и переносили в свежий Matrigel. Пассирование проводили каждые 1-2 недели с соотношением для разделения 1:5-1:8. Для подтверждения потребности в Wnt3A добавляли рекомбинантный белок Wnt3A мыши (Stem cell technologies) вместо среды, кондиционированной Wnt3A. Для экспериментов по прослеживанию in vitro, органоиды желудка возрастом 2 недели инкубировали с 100 нМ 4-гидрокситамоксифена в культуральной среде для желудка в течение 20 ч для активации Lgr5-CreERT2. Затем визуализировали YFP и регистрировали в живых органоидах с использованием конфокальной микроскопии (Leica, SP5).
Среда, кондиционированная Wnt3a
Среду с Wnt3a подготавливали в соответствии с протоколом, описанным в литературе (Willert et al., 2003, Nature May 22, 423(6938): 448-52). Для тестирования Wnt в среде, кондиционированной Wnt3a, и контрольной кондиционированной среды использовали анализ TOP/FOP, как описано Wetering и коллегами (van de Wetering et al., 2001. Cancer Res Jan 1, 61(1): 278-84). Соотношение TOP/FOP ≥50 считали средой с высоким содержанием Wnt и разбавляли ее 1:1 с помощью культуральной среды для органоидов желудка. Разведение 1:10 этой среды с высоким содержанием Wnt3a (соотношение TOP/FOP ~5) считали средой с низким содержанием Wnt и использовали для целей дифференцировки.
Иммуногистохимия органоидов желудка
Для иммуногистохимии органоиды желудка промывали один раз посредством PBS и сразу фиксировали с помощью параформальдегида 4% в течение 15-20 мин при RT. Затем указанные органоиды желудка погружали в парафин и обрабатывали с использованием стандартных способов. Для окрашивания тотального препарата увеличивали проницаемость в образцах с помощью PBS, 0,5% Triton-X100-1% BSA и инкубировали в течение ночи с первичными антителами. После нескольких промываний PBS с 0,3% Triton X100, образцы инкубировали со вторичным антителом. Окрашивание на EdU проводили в соответствии с инструкциями изготовителя (Click-IT, Invitrogen). Ядра окрашивали с помощью йода TOPRO3 или Hoescht33342. Изображения желез желудка и органоидов желудка получали с использованием конфокальной микроскопии (Leica, SP5). Трехмерную реконструкцию проводили с использованием программного обеспечения Volocity Software (Improvision).
RT-PCR
РНК экстрагировали из культур клеток желудка или свежевыделенной ткани с использованием набора для экстракции РНК RNeasy Mini RNA Extraction Kit (Qiagen) и подвергали обратной транскрипции с использованием обратной транскриптазы вируса лейкоза мышей Молони (Promega). кДНК амплифицировали в термоблоке (GeneAmp PCR System 9700; Applied Biosystems, Лондон, Великобритания), как описано ранее (Huch et al., 2009). Использованные праймеры представлены ниже (за символом гена следуют прямой (5'-3') и обратный (5'-3') праймеры).

Результаты
Для определения оптимального роста желудочных элементов in vitro авторы выделили элементы желез желудка, которые суспендировали в Matrigel и культивировали в различных условиях. Условия роста культуры желудка были сходны с условиями роста культур тонкого кишечника (включая EGF, Noggin и R-спондин 1), за исключением прямой зависимости от Wnt3A в форме кондиционированной среды. Эта потребность была подтверждена с использованием очищенного белка Wnt3a (фиг. 33A). Более того, было показано, что FGF10 является необходимым компонентом для запуска почкования и для размножения культур в органоиды из множества элементов (фиг. 33B). Для замены FGF7 (KGF), который использовали в примере 5, можно использовать FGF10 и это даже приводит к увеличению в 2 раза % почкующихся органоидов через 4 суток после начала культивирования (фиг. 33C). Вновь образующиеся органоиды желудка претерпевали постоянное почкование, при сохранении их полярности, причем почки домена желез желудка были распределены вокруг центрального просвета (фиг. 33D). В отсутствии среды, кондиционированной Wnt3A, органоиды желудка быстро портились (фиг. 33E). Каждую неделю органоиды механически диссоциировали и разделяли до одной пятой их плотности до посева. Культивированные пилорические элементы представляли собой однослойные эпителиальные структуры, на что указывало окрашивание на E-Cad (фиг. 33F). Авторы изобретения успешно культивировали органоиды желудка в течение по меньшей мере 8 месяцев без какой-либо поддающейся детекции утраты свойств, описанных выше.
Для определения того, способны ли Lgr5+ клетки желудка (фиг. 34A) образовывать и поддерживать элементы пилорических желез желудка in vitro авторы изобретения отсортировали клетки с высоким уровнем Lgr5-EGFP (фиг. 34B). Когда единичные клетки с высоким содержанием Lgr5-EGFP были отсортированы, в среднем 8% клеток вырастали в органоиды, в то время как остальные клетки погибали в течение первых 24 ч. Отсортированные клетки Lgr5-EGFPhi быстро начинали делиться и небольшие подобные пузырям структуры становились видимыми уже через 5 суток. На следующие сутки вновь образованные (подобные пузырям) структуры начинали образовывать подобные железам домены (фиг. 34C). После 9-11 суток в культуре органоиды желудка диссоциировали вручную и разделяли для получения новых органоидов. Органоиды желудка, происходящие из единичных клеток, успешно пересевали раз в неделю в течение по меньшей мере 3 месяцев без утраты описанных свойств (фигура 34D). Начиная с 7 суток экспрессия Lgr5-EGFP ограничивалась основанием подобных железам доменов (фиг. 34E). Как показано с помощью окрашивания на EdU, пролиферирующие клетки были расположены в основании этих подобных железам доменов (фиг. 34F), в то время как было выявлено, что апоптотические положительные по каспазе 3 клетки вытеснялись в просвет (данные не представлены). Прослеживание происхождения исследовали в установившихся органоидах, происходящих из единичных Lgr5+ клеток, выделенных из репортерной мыши Lgr5-EGFP-ires-CreERT2/Rosa26-YFP. После индукции тамоксифеном, в единичных Lgr5+ клетках в подобных железам доменах быстро активировался YFP+ репортерный ген. В течение последующих нескольких суток домен, экспрессирующий YFP, значительно расширялся в растущих органоидах, подтверждая вклад Lgr5+ стволовых клеток в рост органоидов in vitro (фиг. 34G). Органоиды, происходящие из одноклеточных культур, представляли собой однослойные эпителиальные структуры, на что указывало окрашивание по E-кадгерину (фиг. 34I). В дополнение к Lgr5, культуры экспрессировали маркеры эпителия желудка: внутренний фактор желудка муцин 6 и пепсиноген C. В этих условиях культивирования не наблюдали дифференцировки пит-ростков или энтероэндокринных ростков (это отличается от примера 5, где наблюдали росток пит-клеток. Однако в этом примере использовали белок Wnt3a вместо среды, кондиционированной Wnt, который является менее активным. Снижение концентрации среды, кондиционированной Wnt, приводит к дифференцировке в росток пит-клеток, см. ниже). Снижение концентрации Wnt3A в культуральной среде приводило к образованию сравнимых желудочных структур, содержащих полярные пит-клетки, на что указывала экспрессия желудочного муцина 5AC (MUC5AC) и перйодной кислоты Шиффа (PAS), слизистые шеечные клетки, на что указывала экспрессия Tff2, и некоторые рассеянные незрелые энтероэндокринные клетки (хромогранин A) (фиг. 34H, I). Добавление дополнительных факторов роста, таких как RA, IGF и экзендин 4, может приводить к более зрелой дифференцировке культур желудка в направлении различных ростком клеток. Взятые вмести, эти наблюдения in vivo и in vitro демонстрируют, что Lgr5 является маркером ранее недооцениваемой популяции самообновляющихся мультипотентных взрослых стволовых клеток в пилорическом отделе желудка.
Ссылки



| название | год | авторы | номер документа |
|---|---|---|---|
| КУЛЬТУРАЛЬНАЯ СРЕДА СТВОЛОВЫХ КЛЕТОК | 2012 |
|
RU2714256C2 |
| УЛУЧШЕННЫЙ СПОСОБ ДИФФЕРЕНЦИРОВКИ | 2017 |
|
RU2772435C2 |
| ОРГАНОИД ПЕЧЕНИ, ВАРИАНТЫ ЕГО ПРИМЕНЕНИЯ И СПОСОБ КУЛЬТИВИРОВАНИЯ ДЛЯ ЕГО ПОЛУЧЕНИЯ | 2011 |
|
RU2579995C2 |
| ДВОЙНЫЕ ТРУБЧАТЫЕ СТРУКТУРЫ | 2017 |
|
RU2756404C1 |
| Способ получения органоидов из опухолевой ткани органов головы и шеи | 2021 |
|
RU2787378C1 |
| ДИФФЕРЕНЦИРОВКА ЭМБРИОНАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК ЧЕЛОВЕКА | 2008 |
|
RU2465323C2 |
| КУЛЬТУРАЛЬНАЯ СРЕДА ДЛЯ ПЕРВИЧНЫХ ЭПИТЕЛИАЛЬНЫХ КЛЕТОК МОЛОЧНОЙ ЖЕЛЕЗЫ, СПОСОБ ИХ КУЛЬТИВИРОВАНИЯ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2819362C1 |
| ДИФФЕРЕНЦИРОВКА ЧЕЛОВЕЧЕСКИХ ЭМБРИОНАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК | 2008 |
|
RU2473684C2 |
| ДИФФЕРЕНЦИРОВАНИЕ ПЛЮРИПОТЕНТНЫХ СТВОЛОВЫХ КЛЕТОК | 2014 |
|
RU2673750C2 |
| КУЛЬТУРАЛЬНАЯ СРЕДА ДЛЯ ЭПИТЕЛИАЛЬНЫХ КЛЕТОК ПЛОСКОКЛЕТОЧНОЙ КАРЦИНОМЫ ПИЩЕВОДА, СПОСОБ КУЛЬТИВИРОВАНИЯ И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2816529C1 |
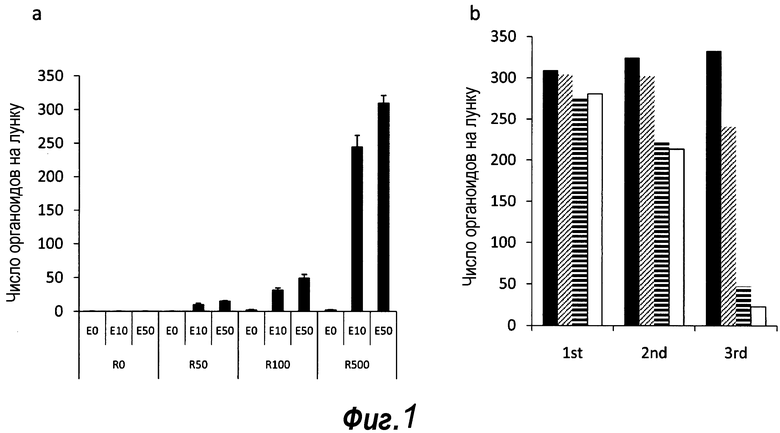
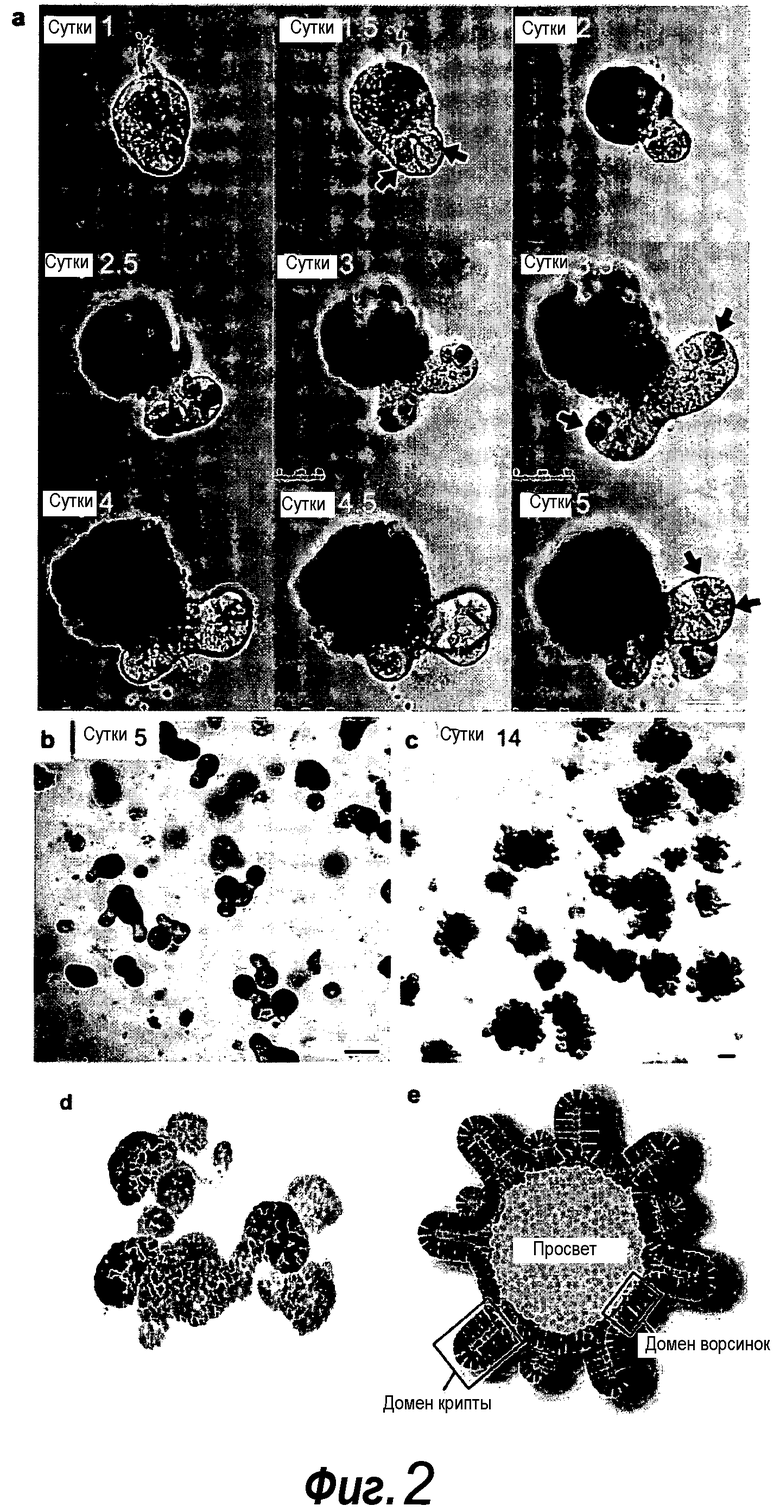
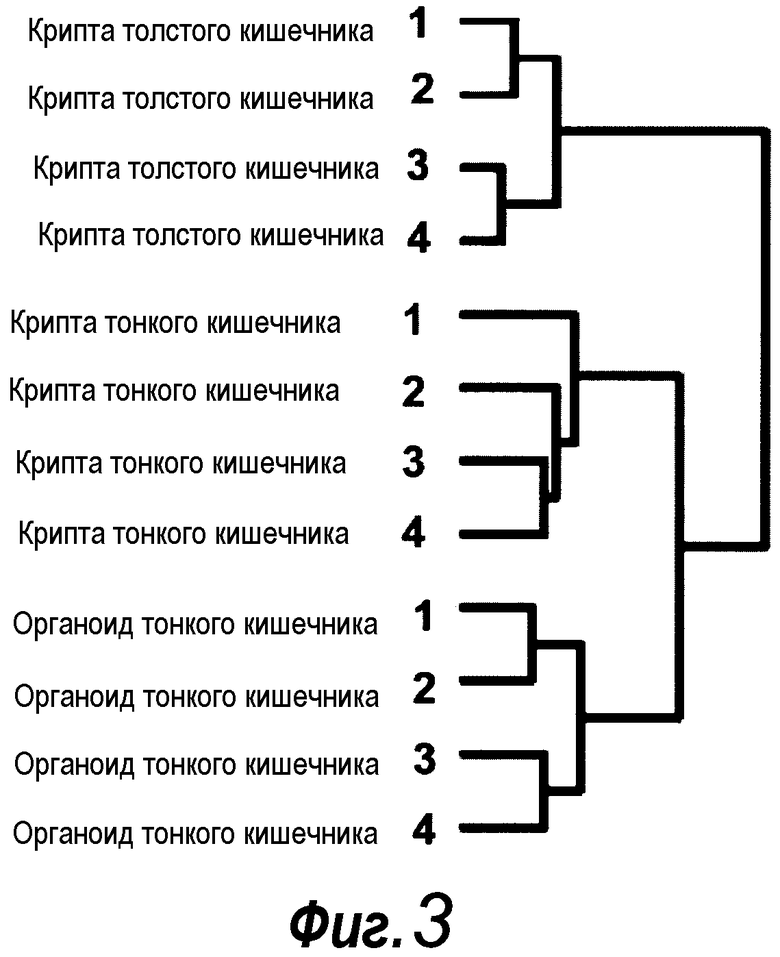
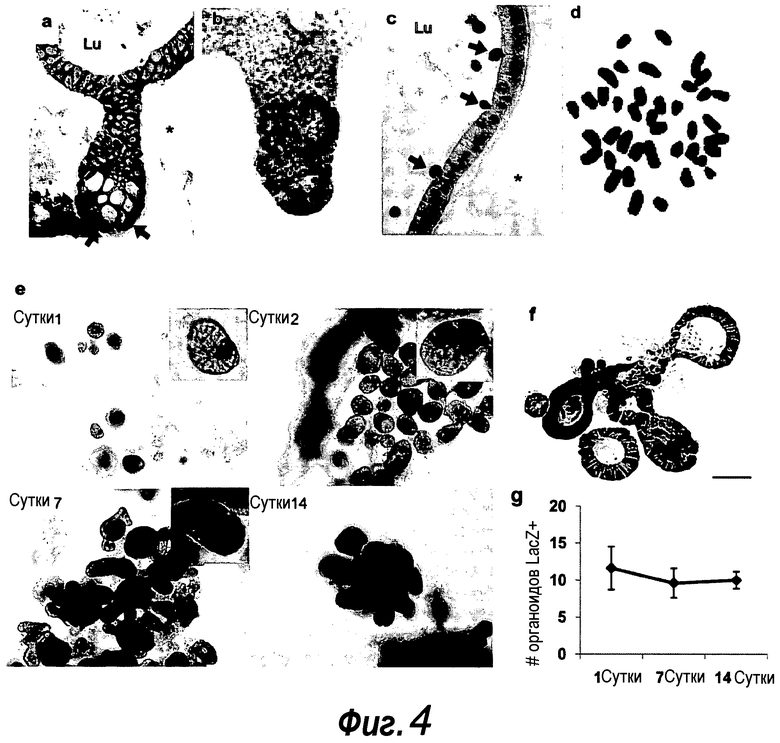
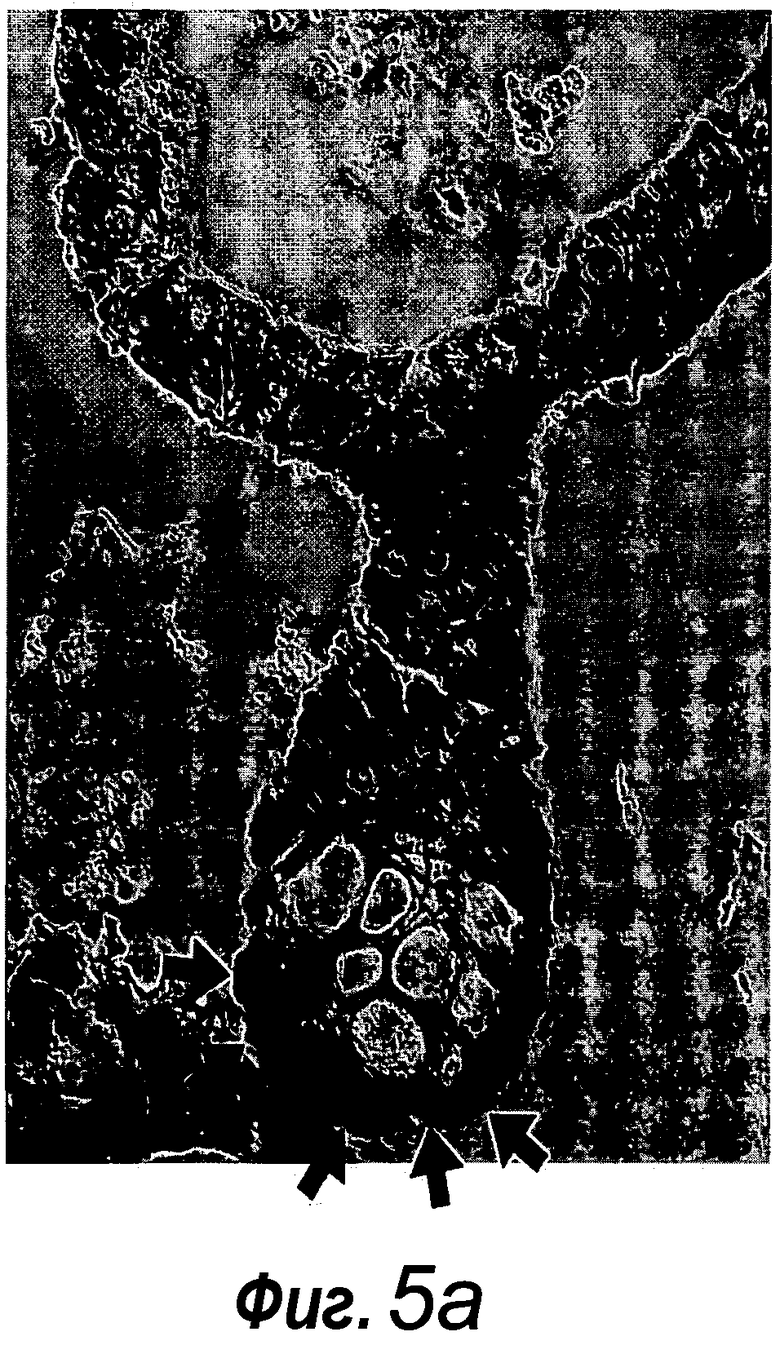

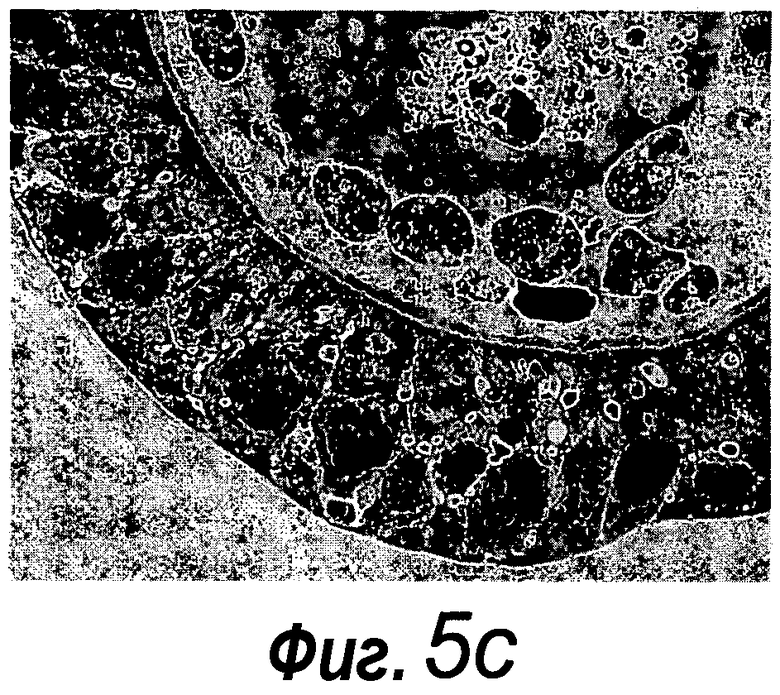
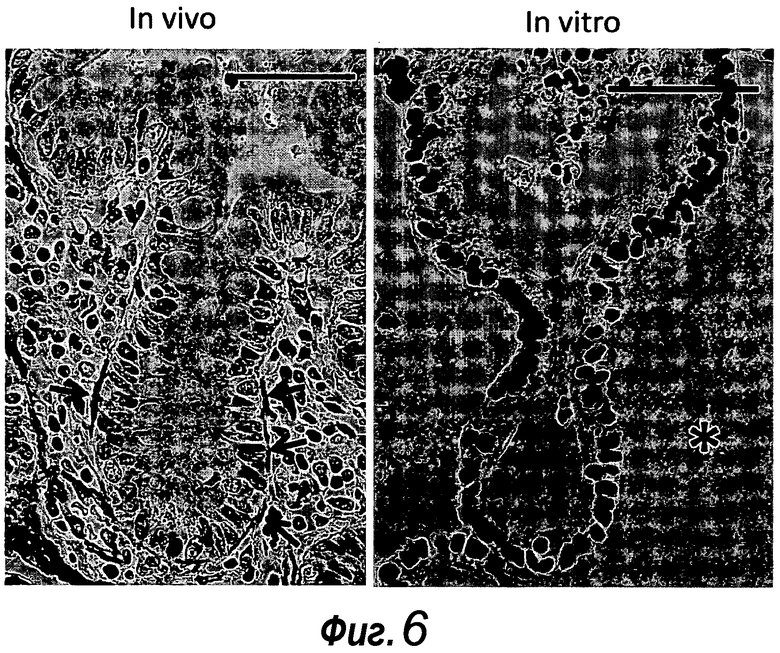
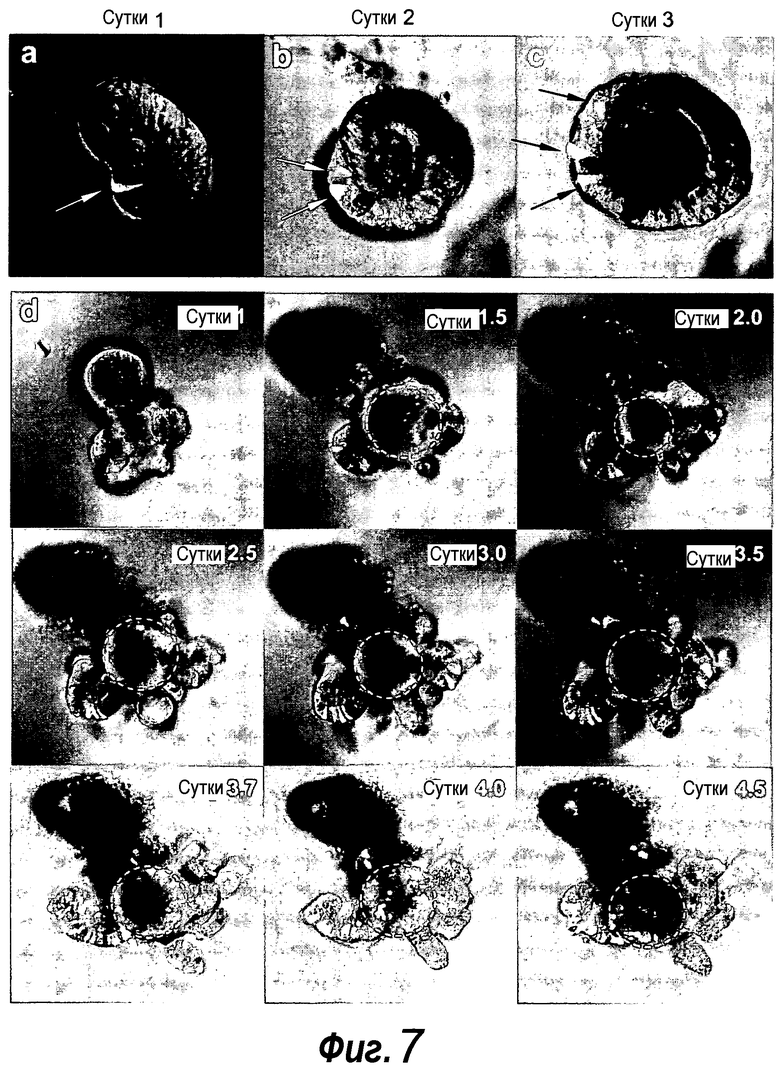
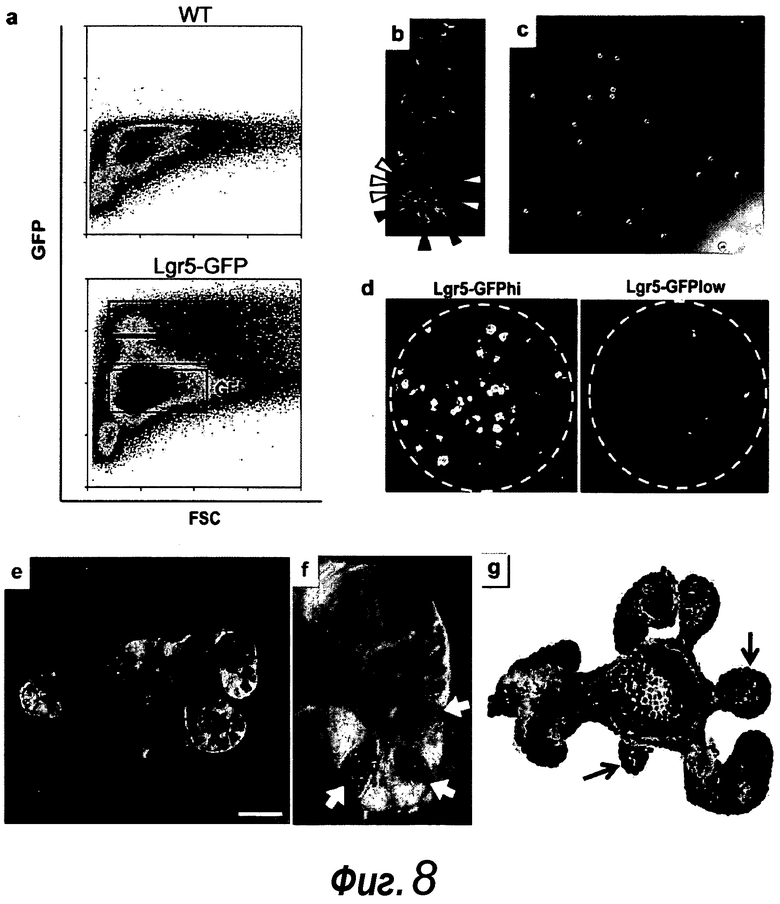
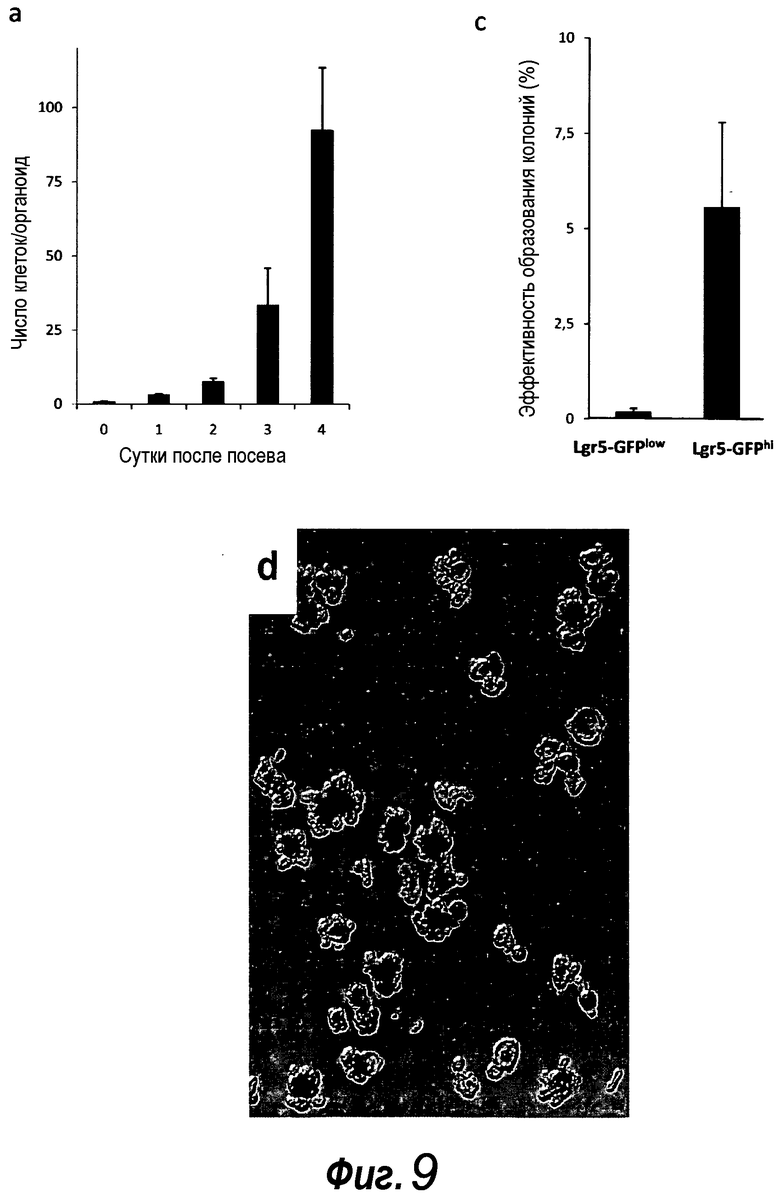
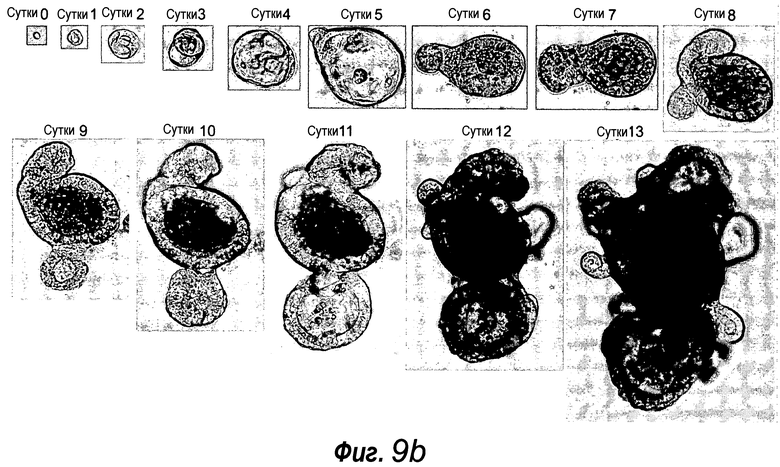
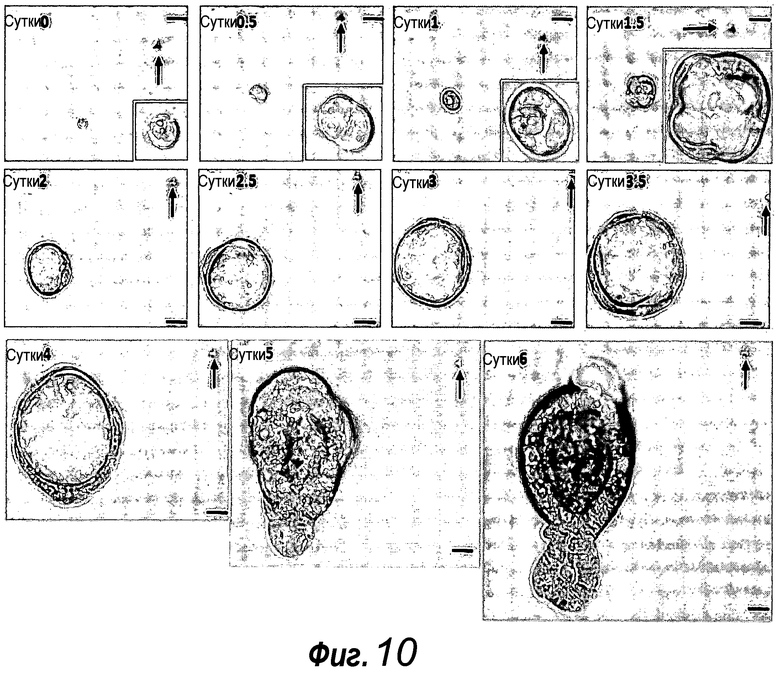
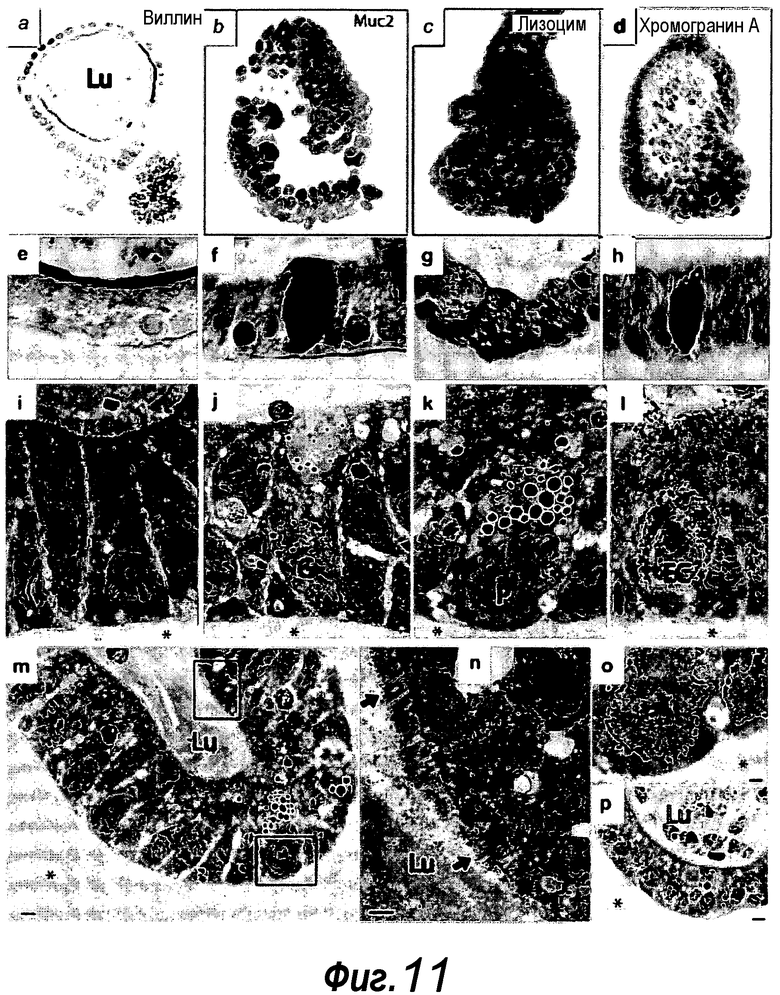
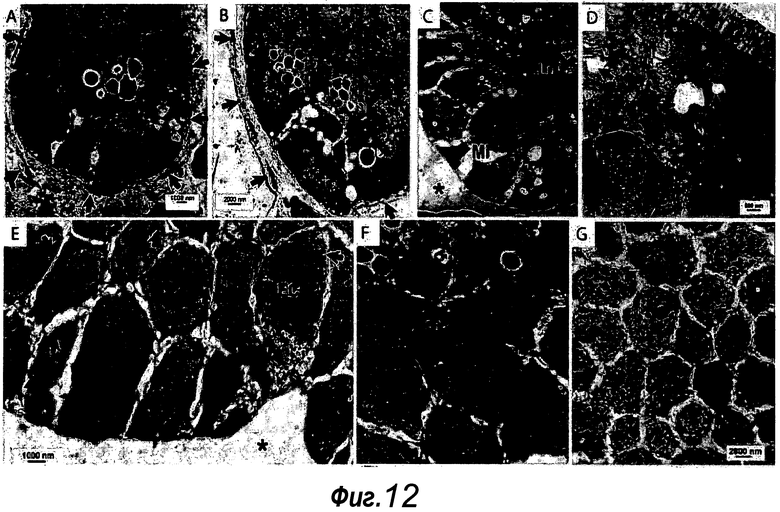
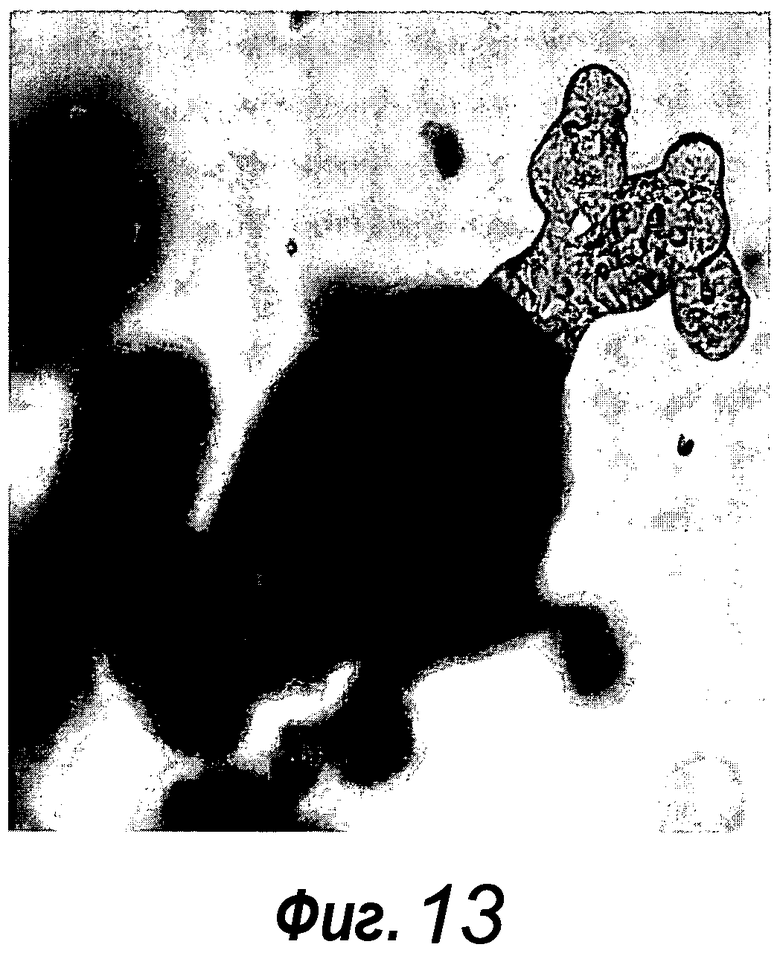


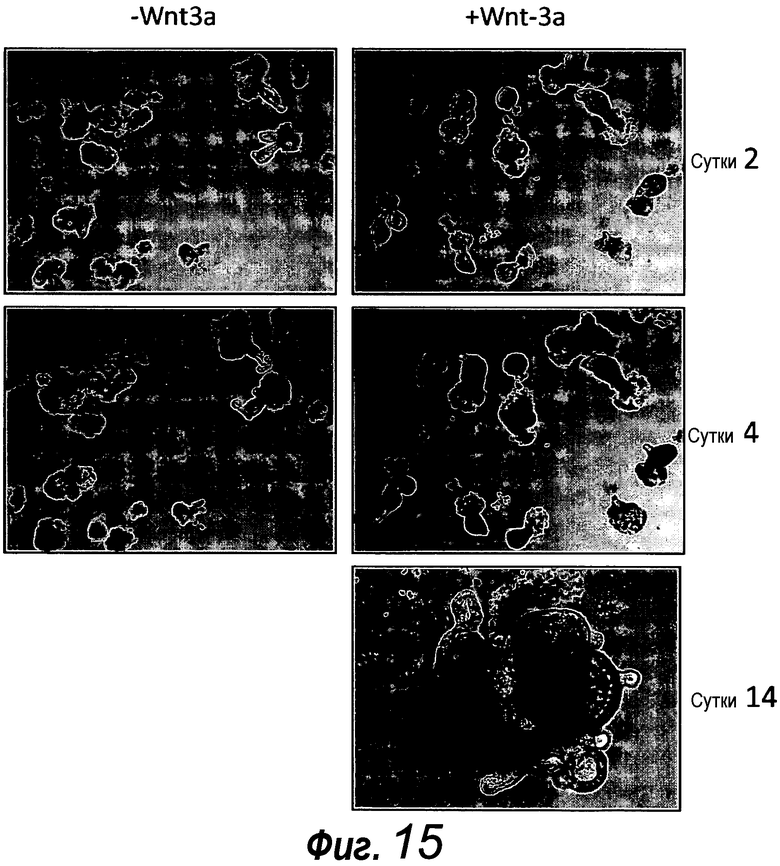
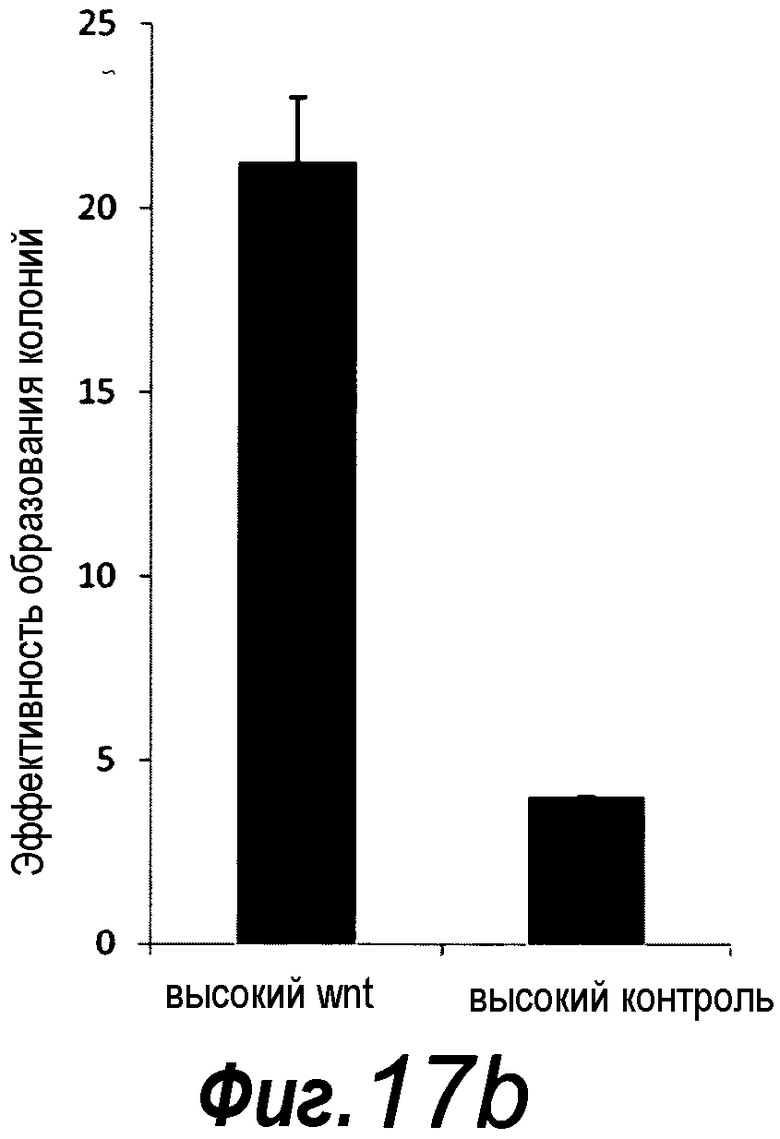
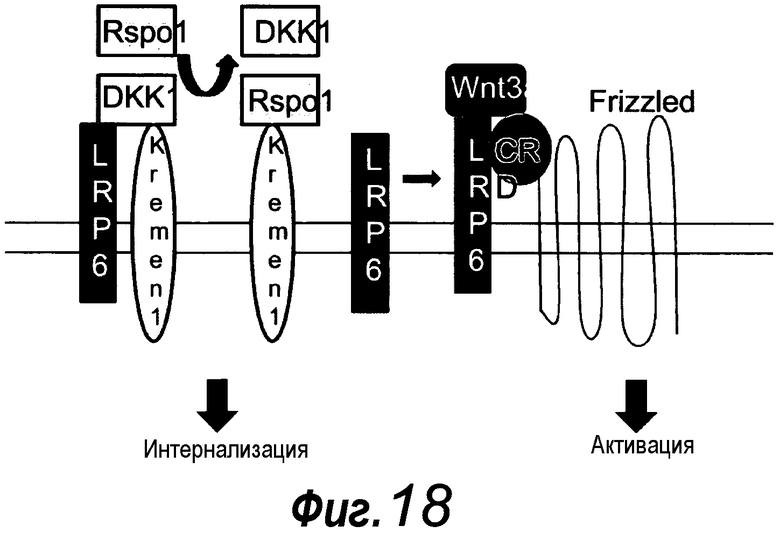
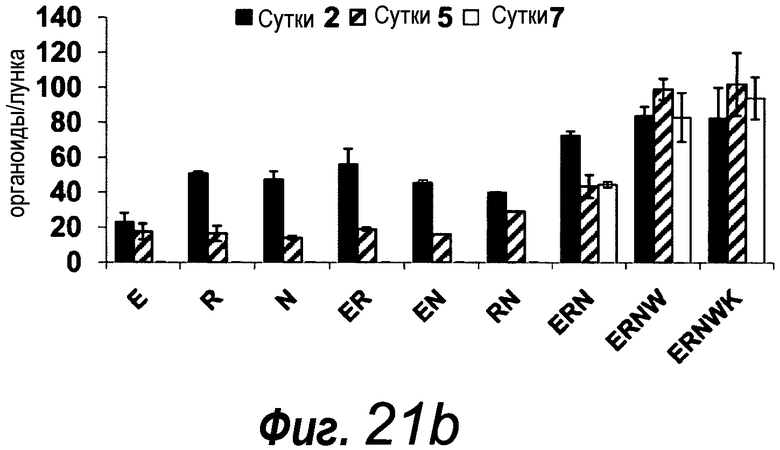
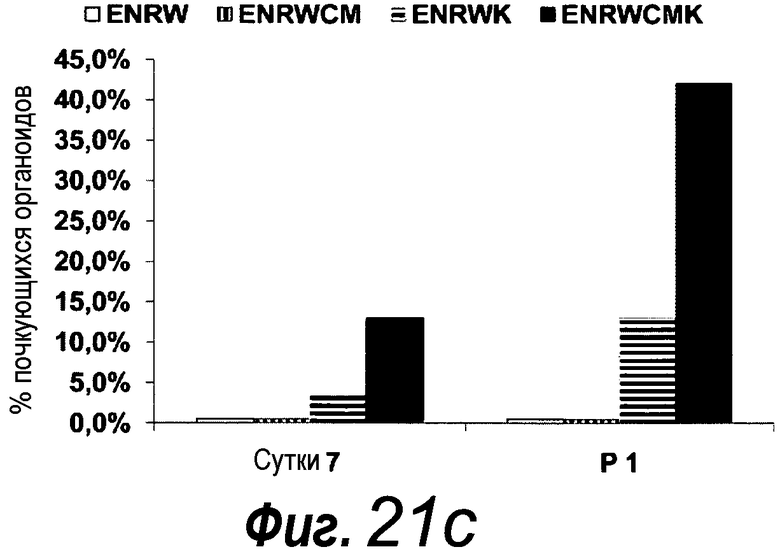
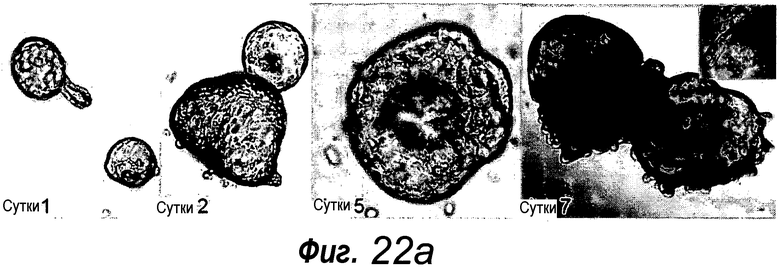

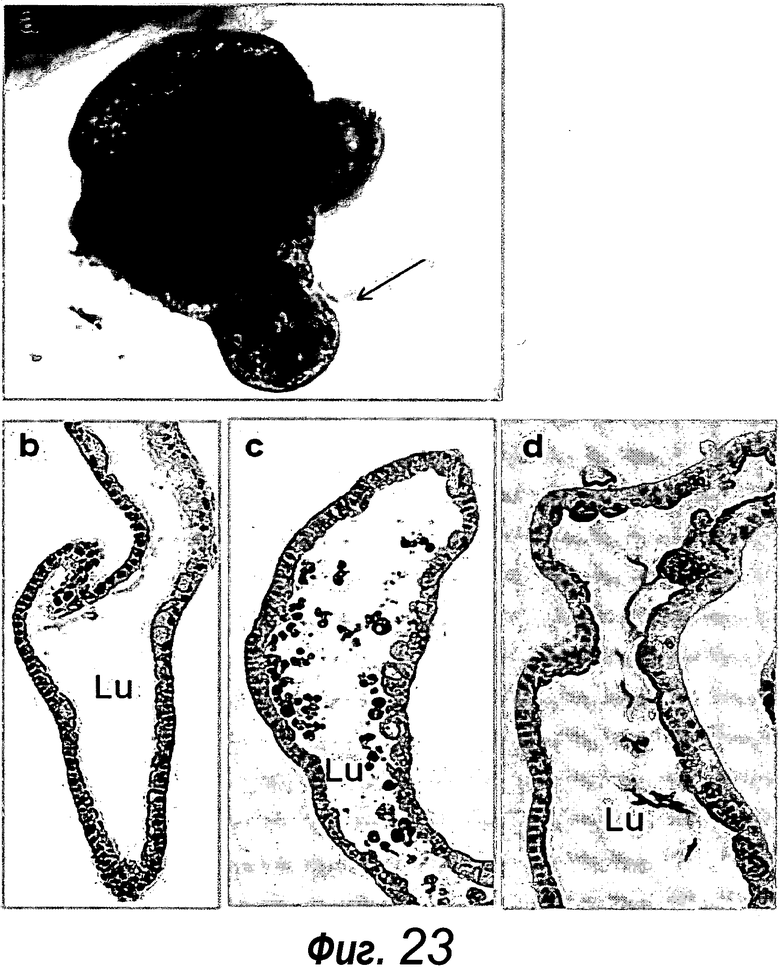
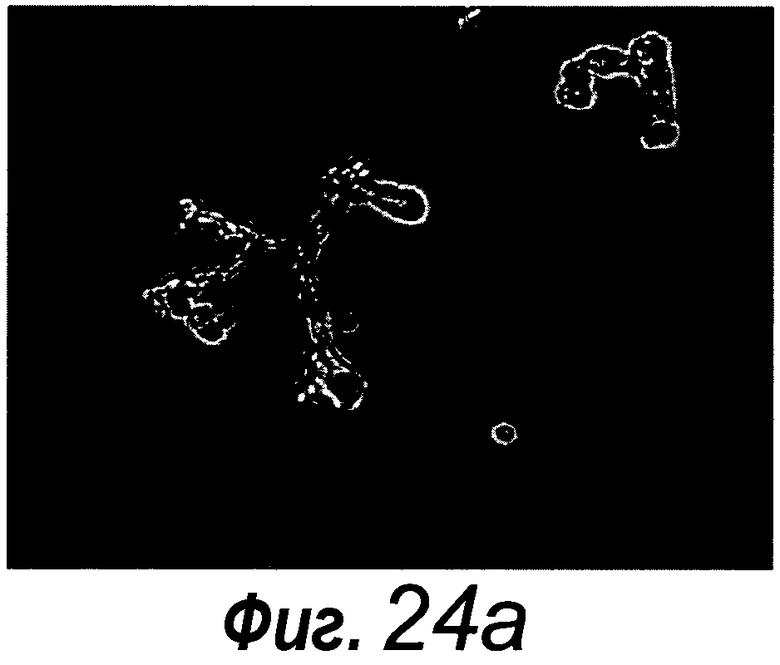
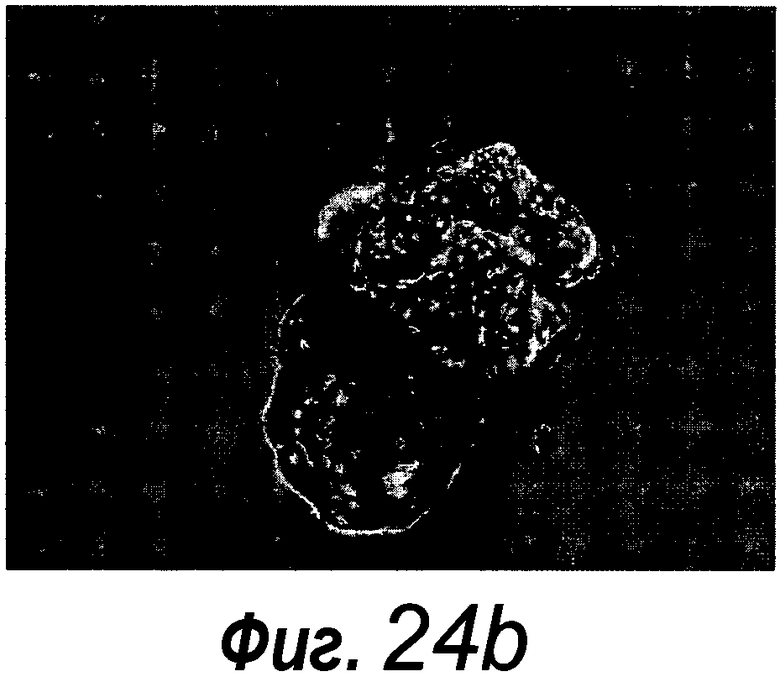

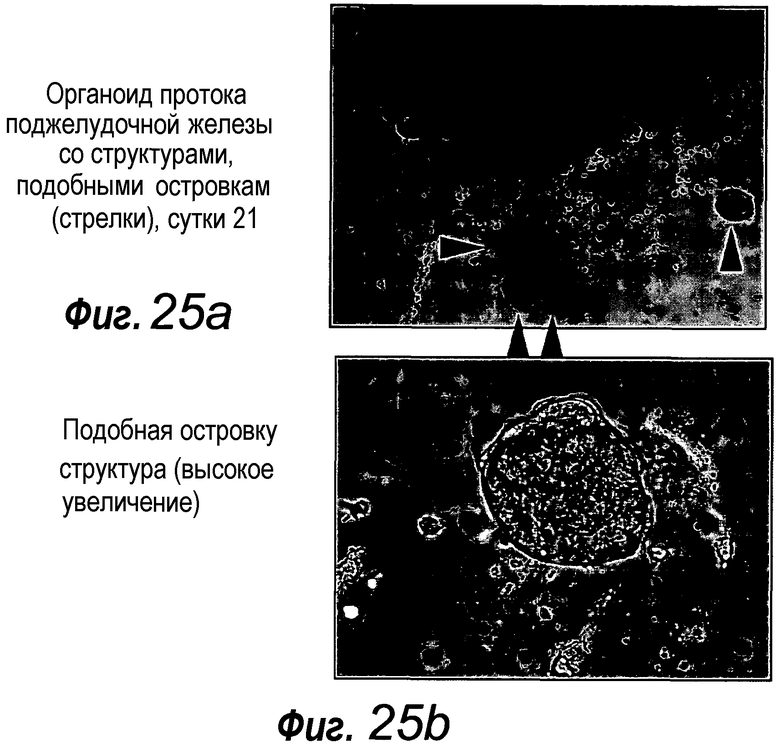
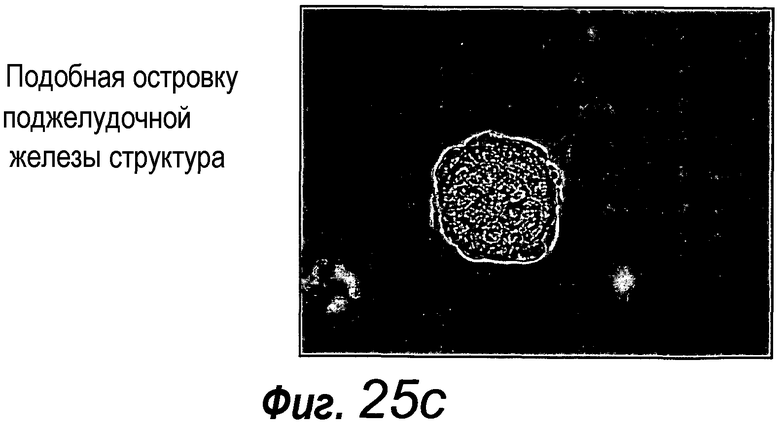
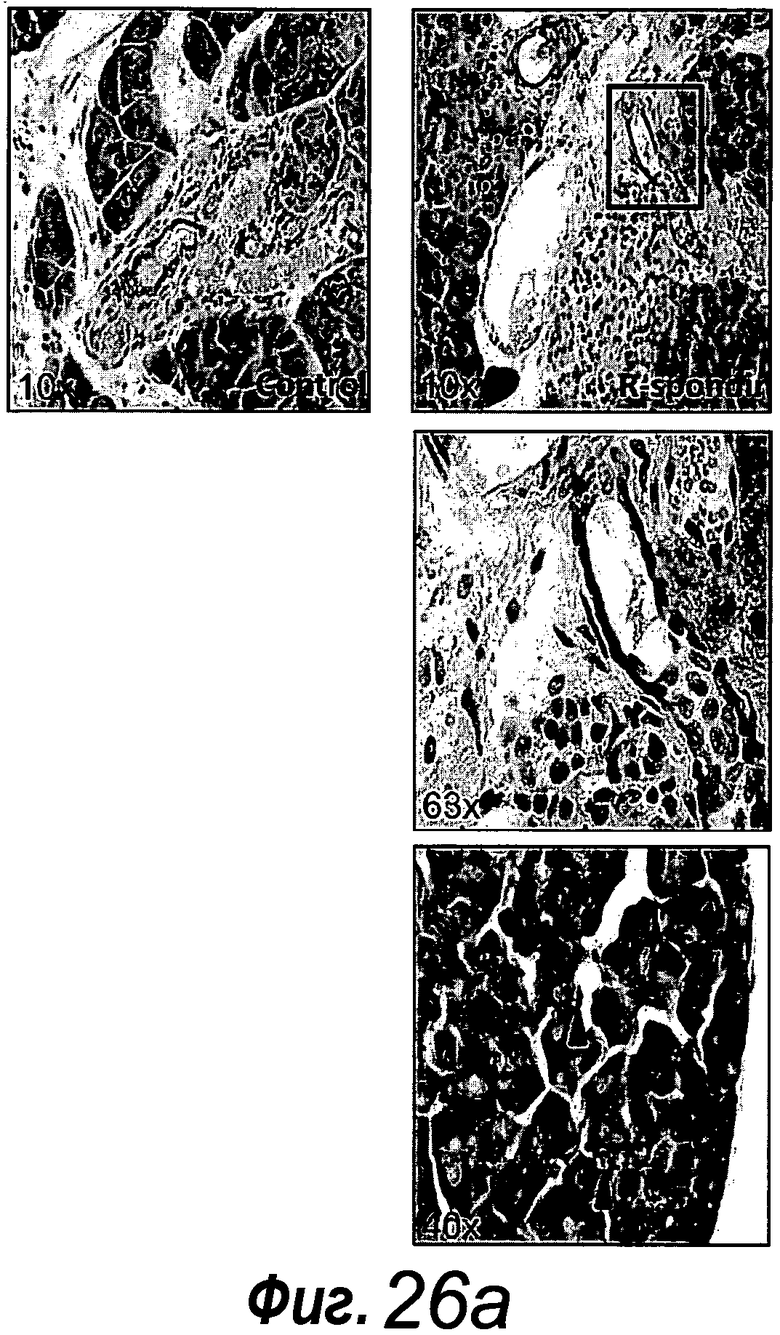
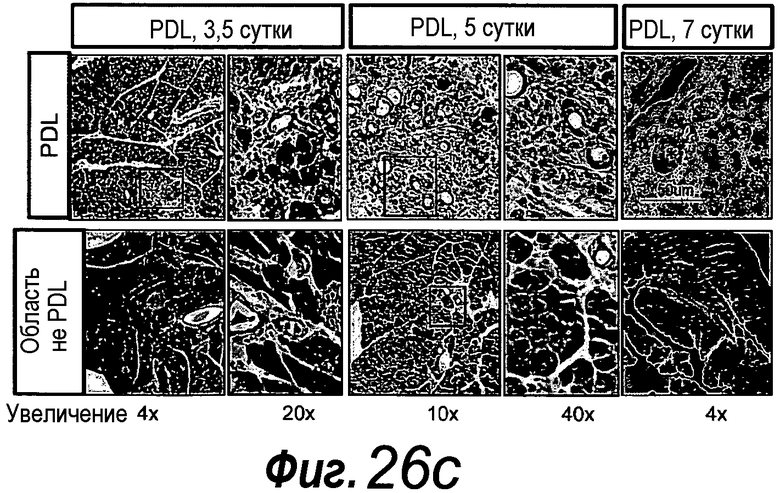
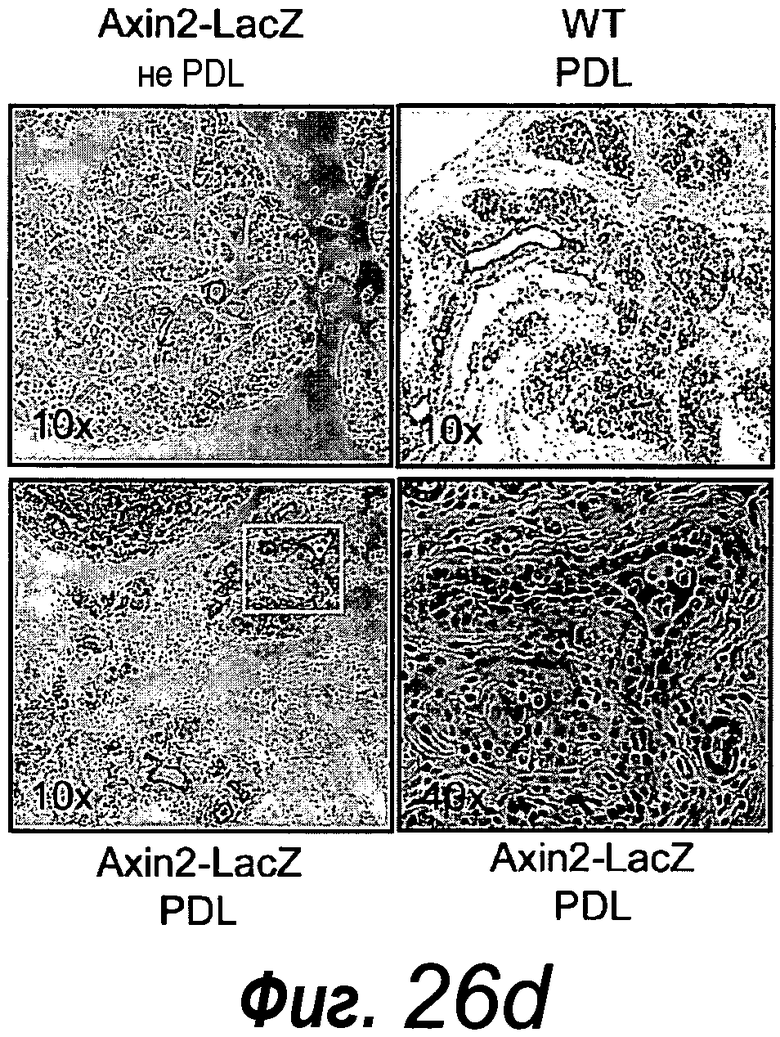
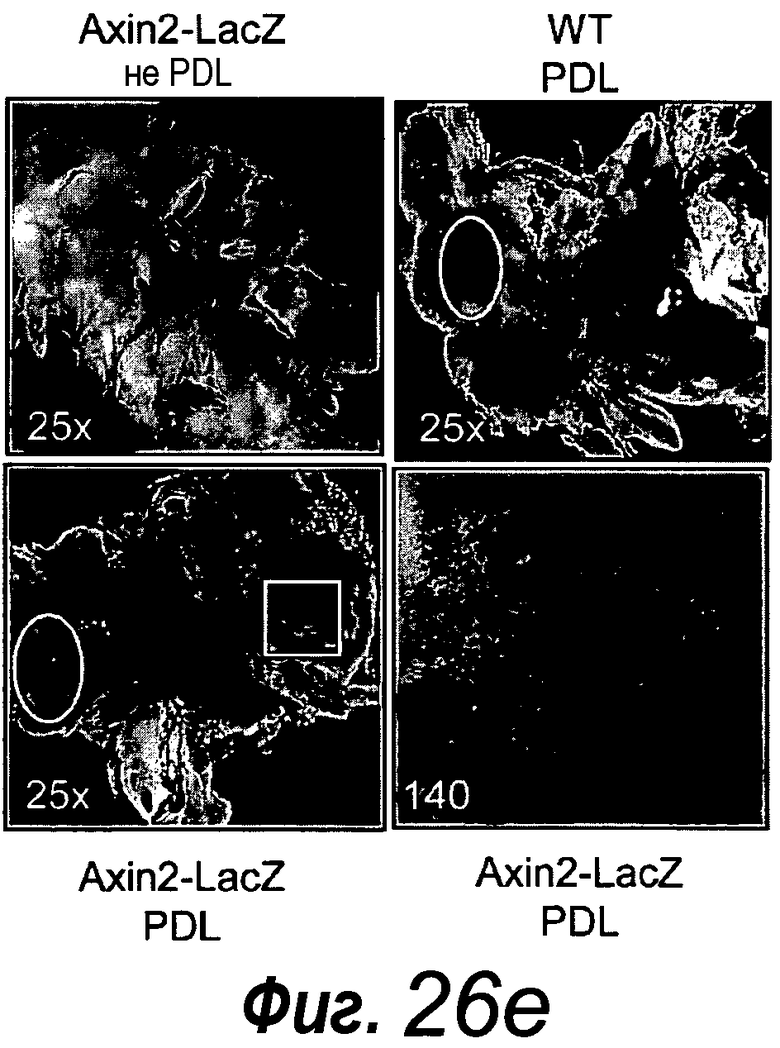
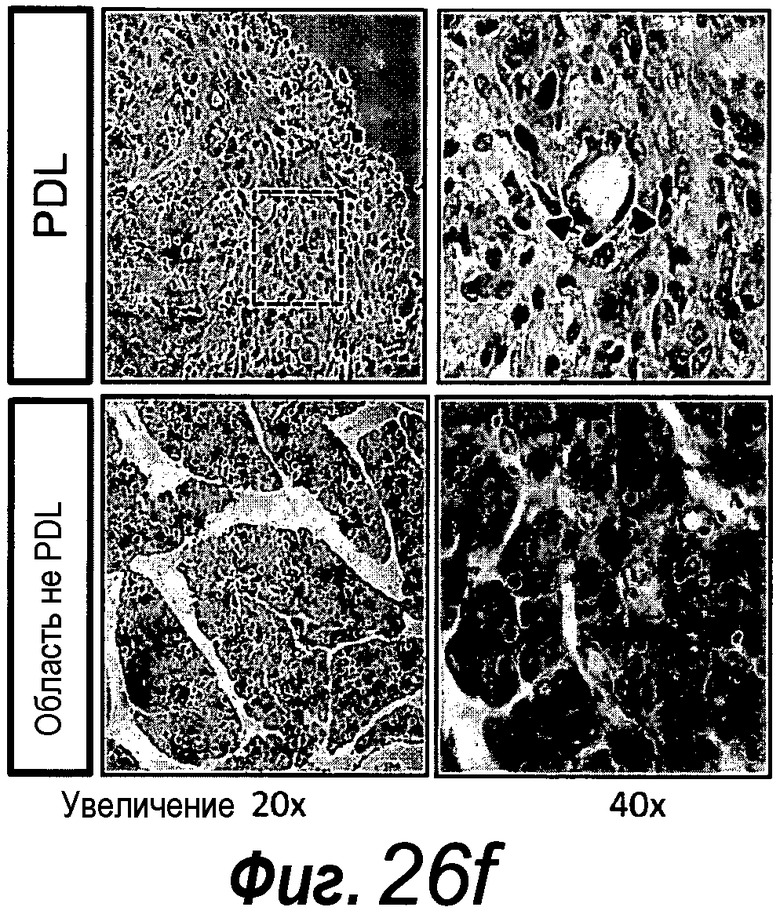

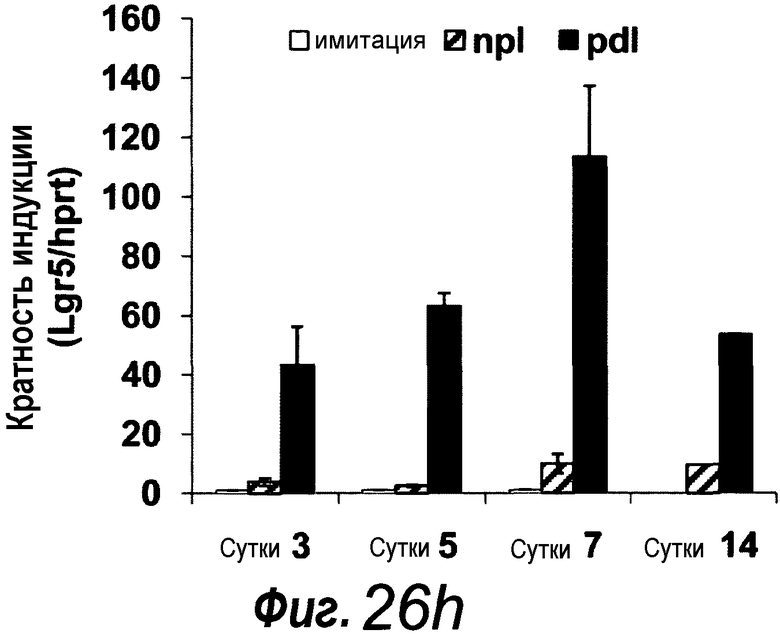
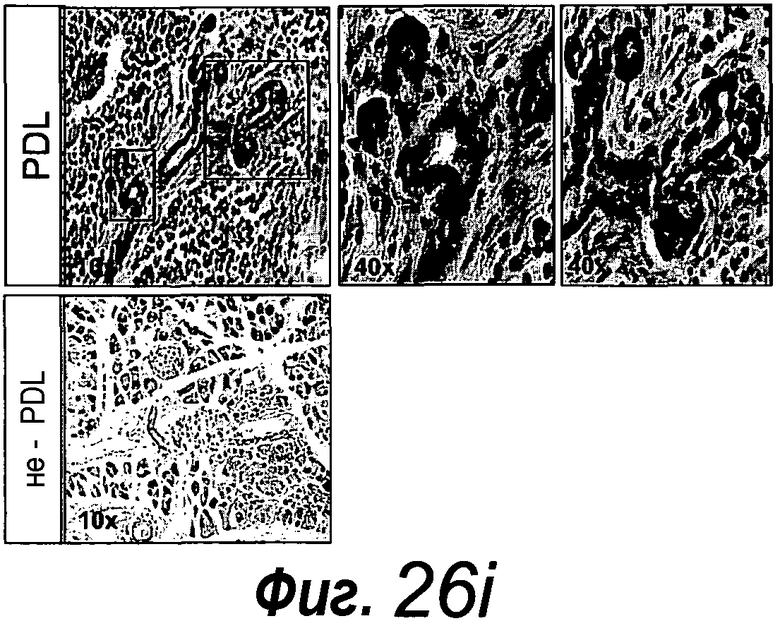
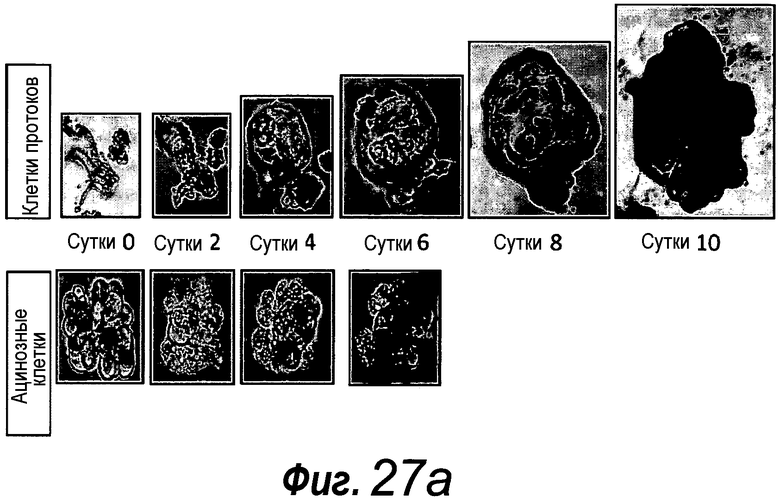
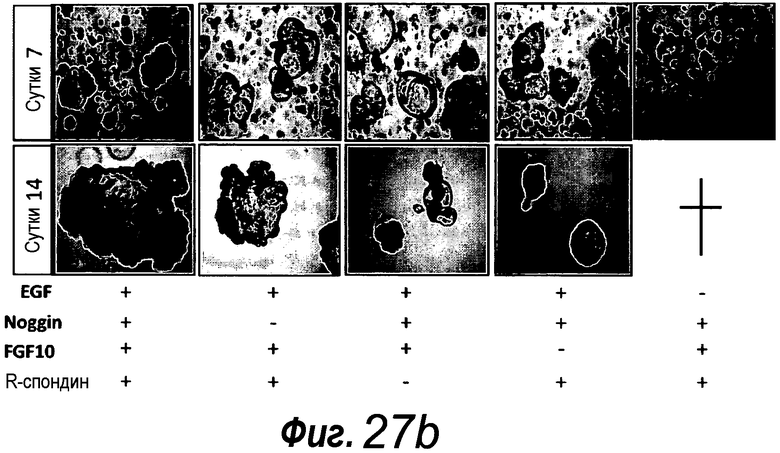
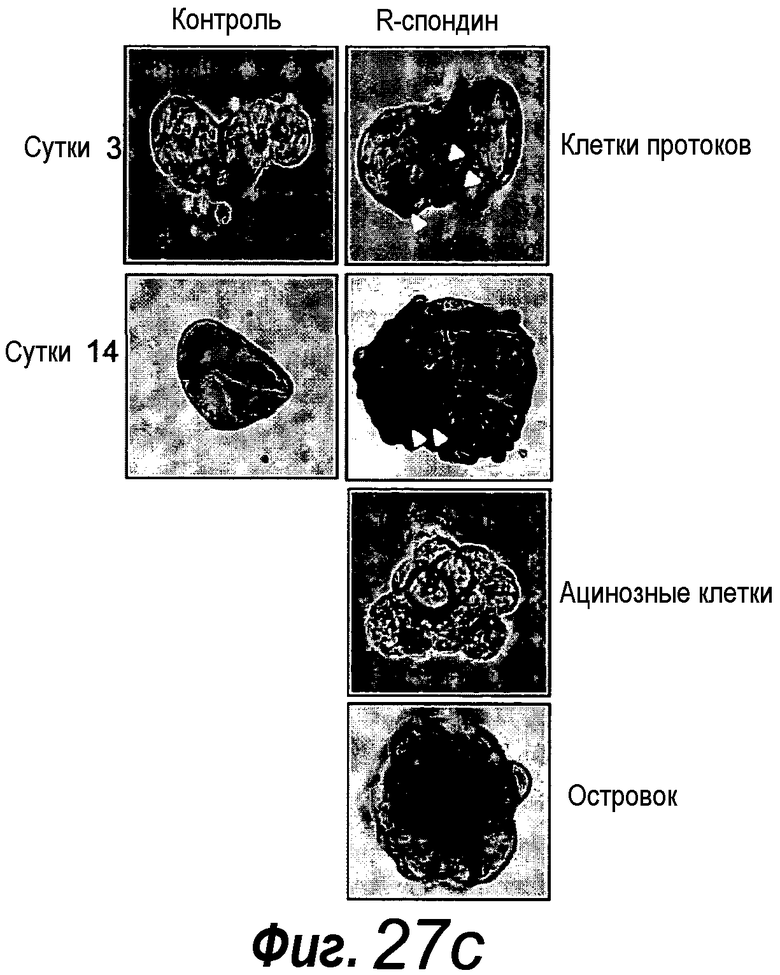

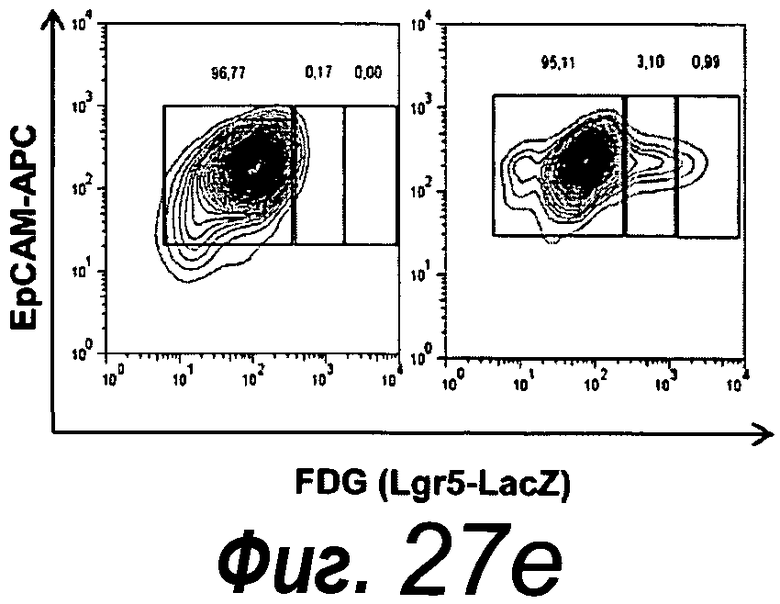
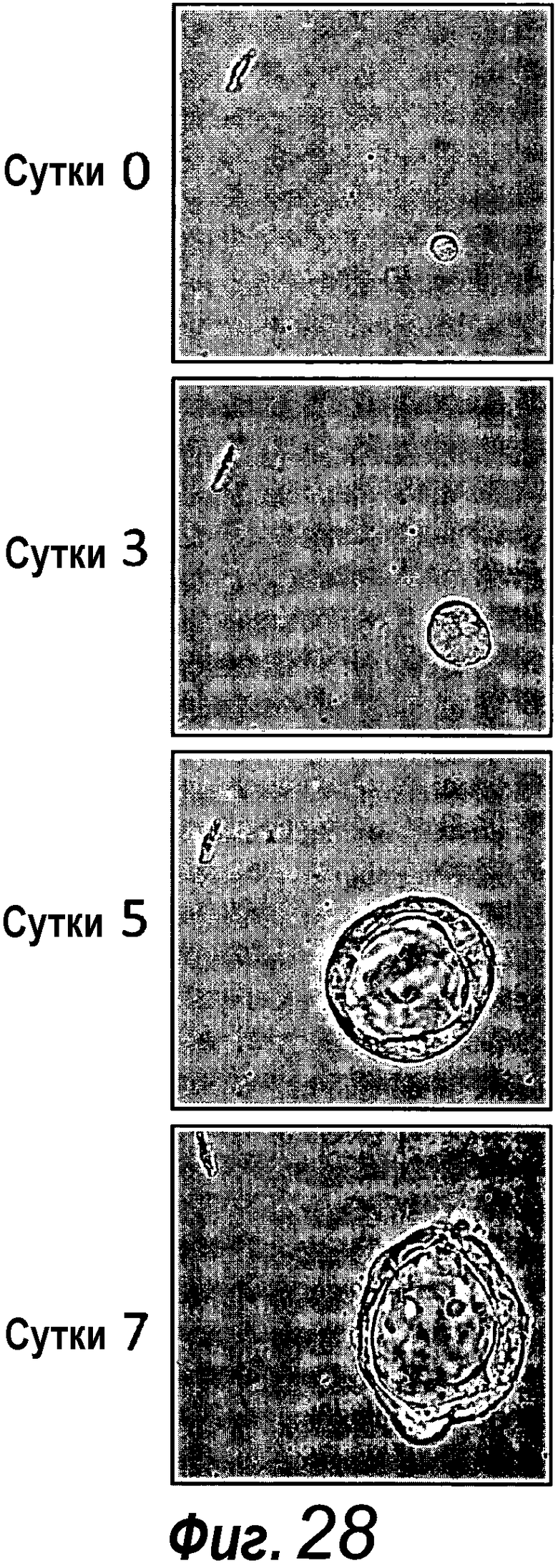
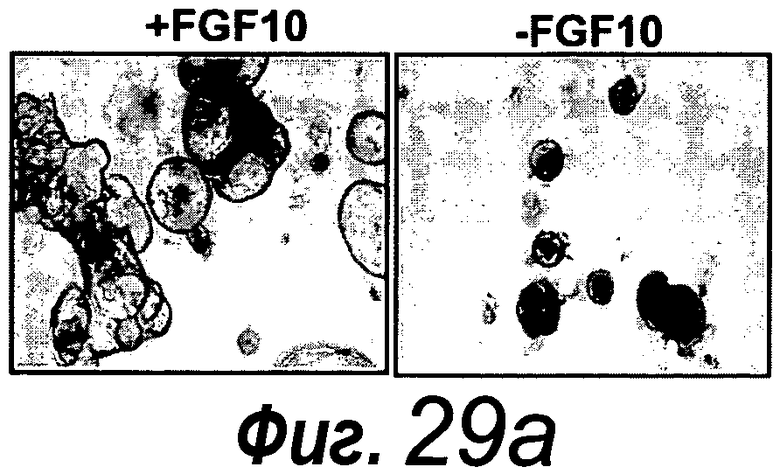
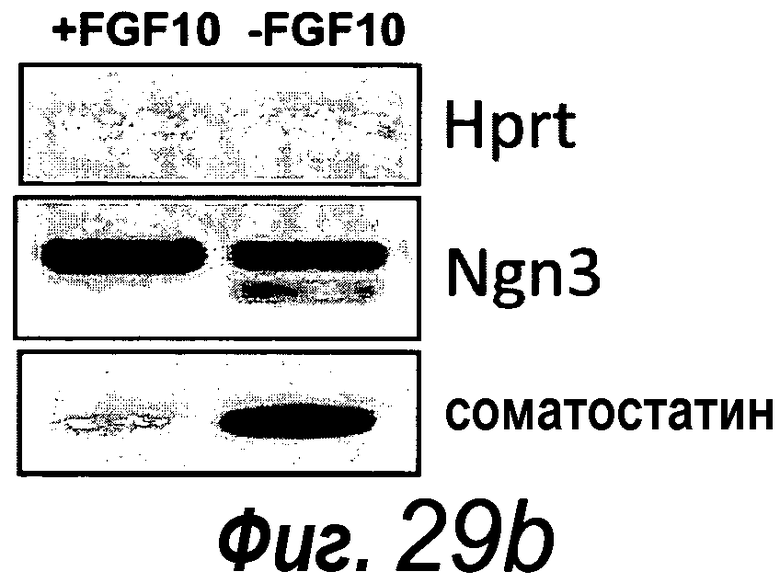
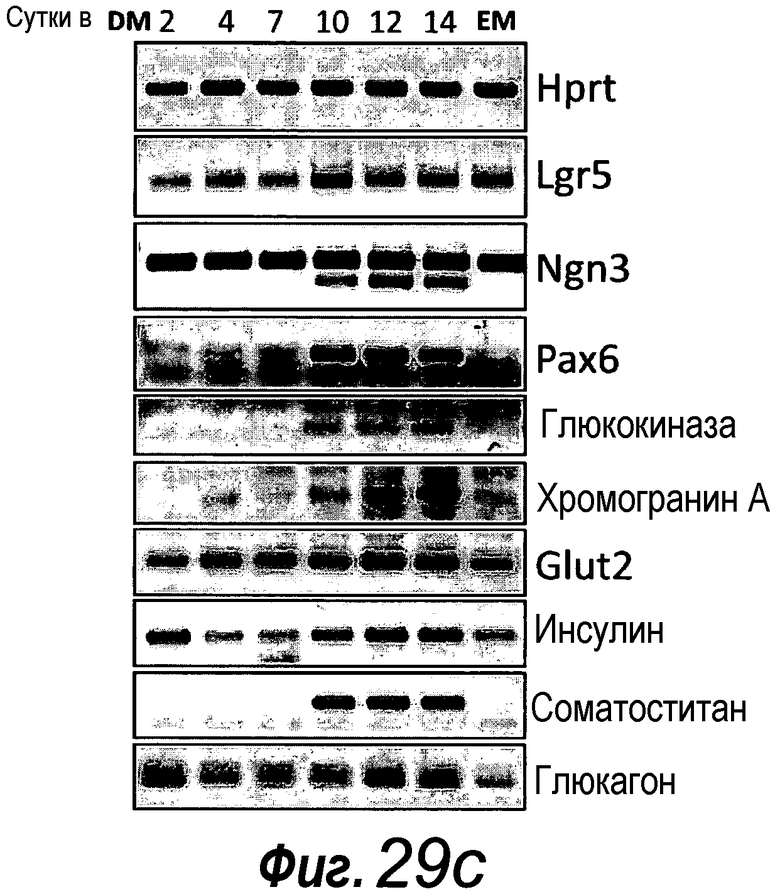
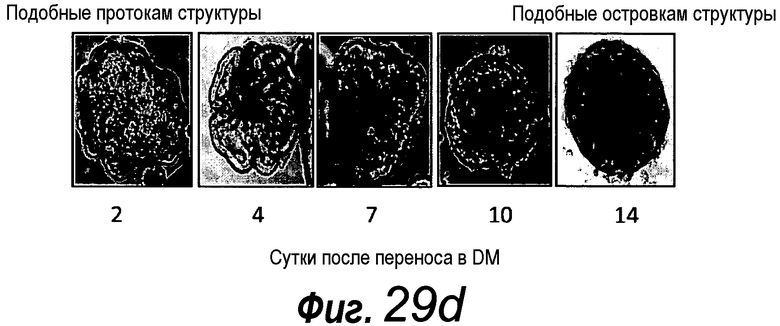

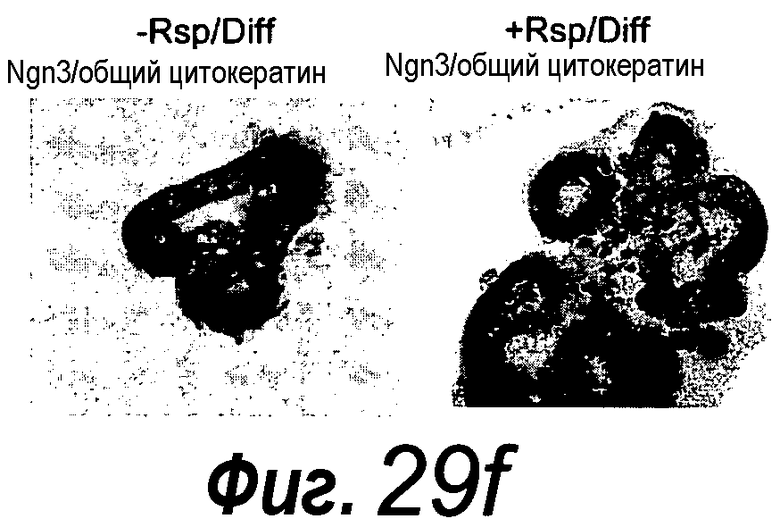
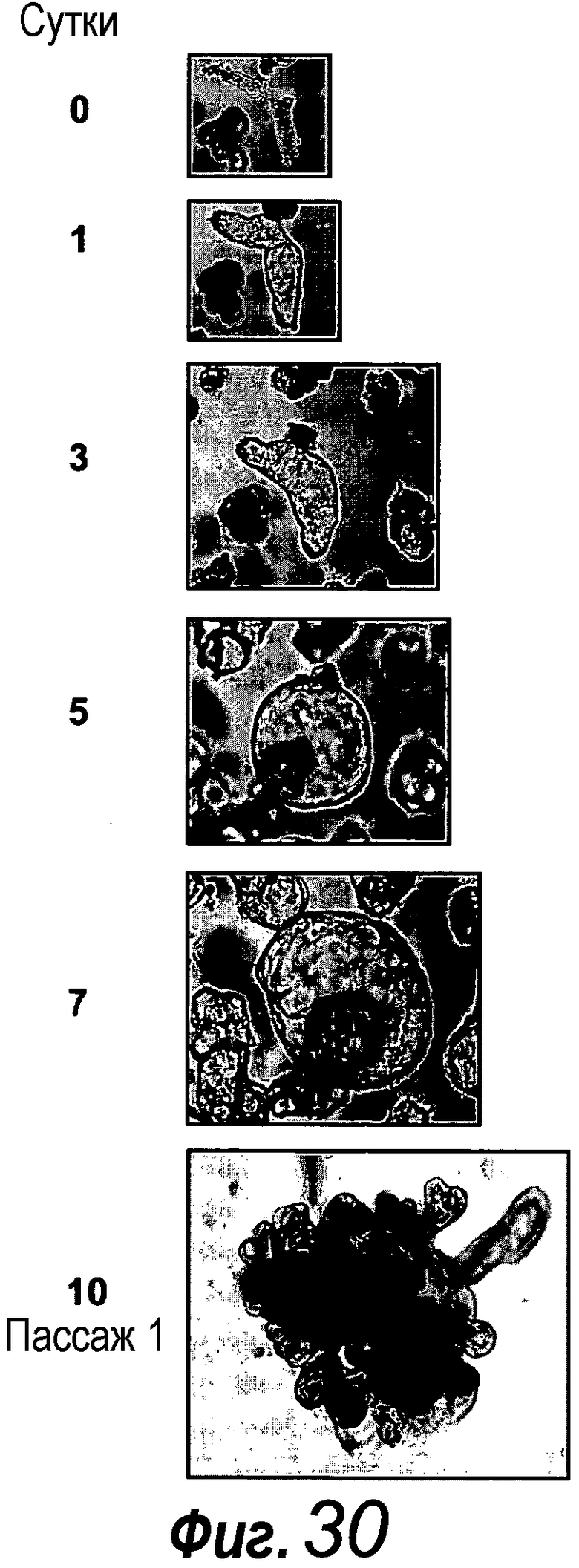
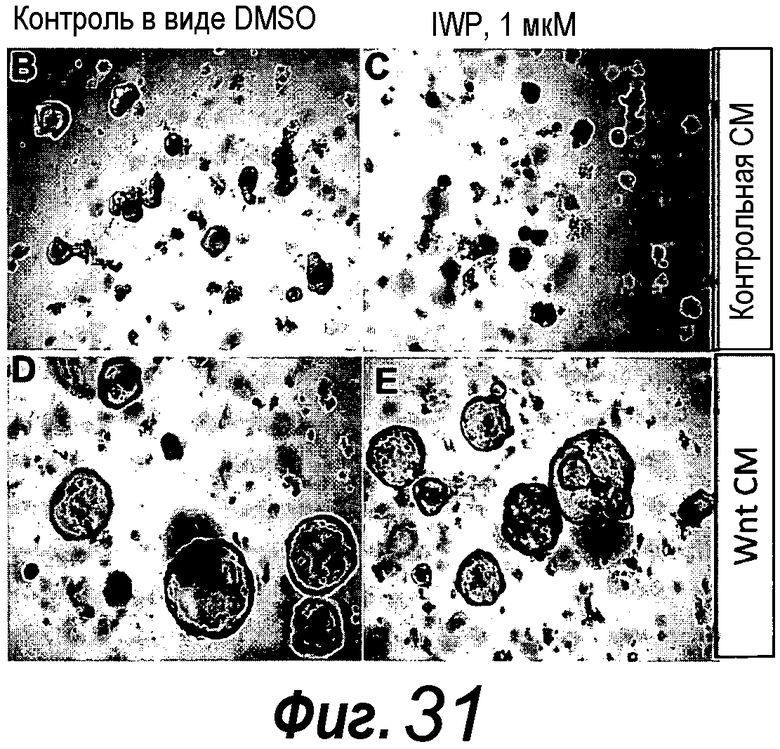
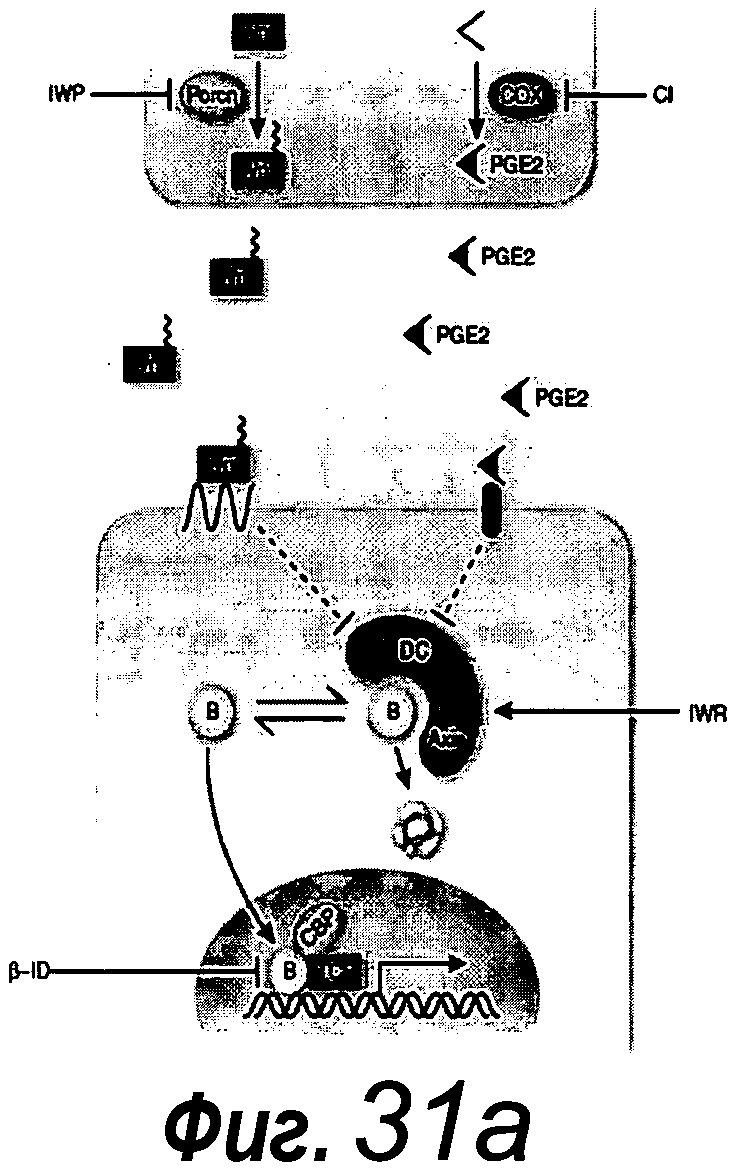
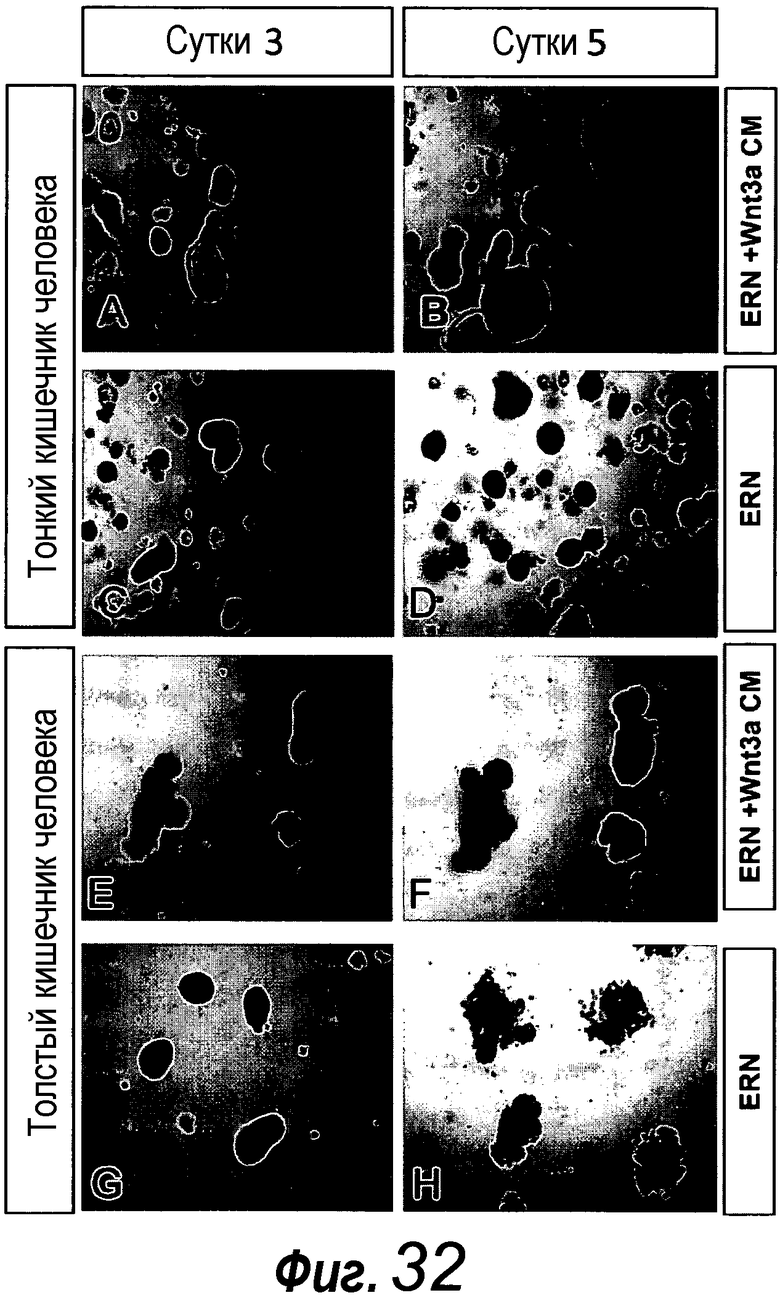
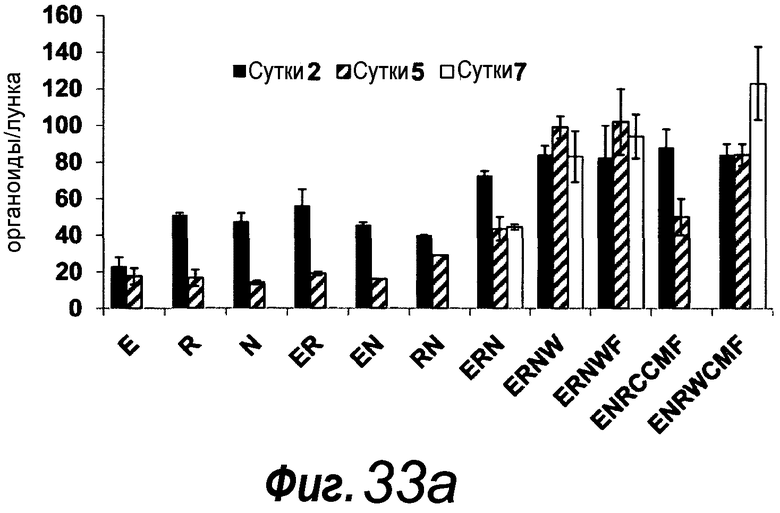
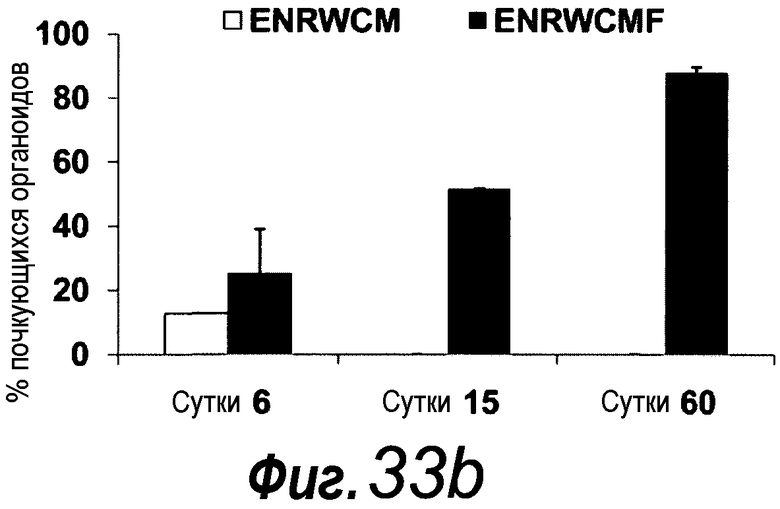
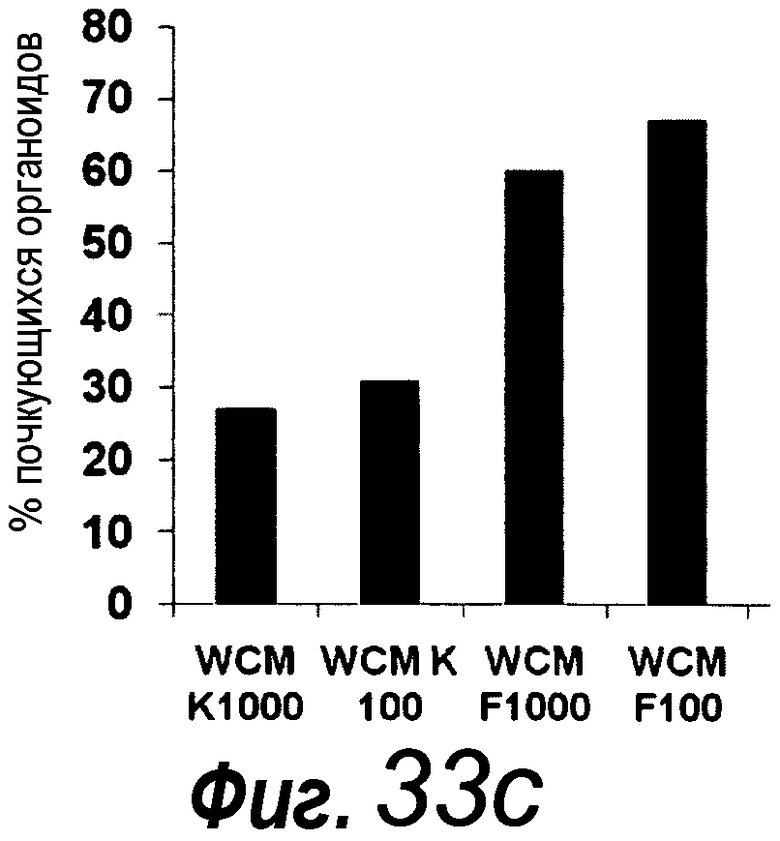
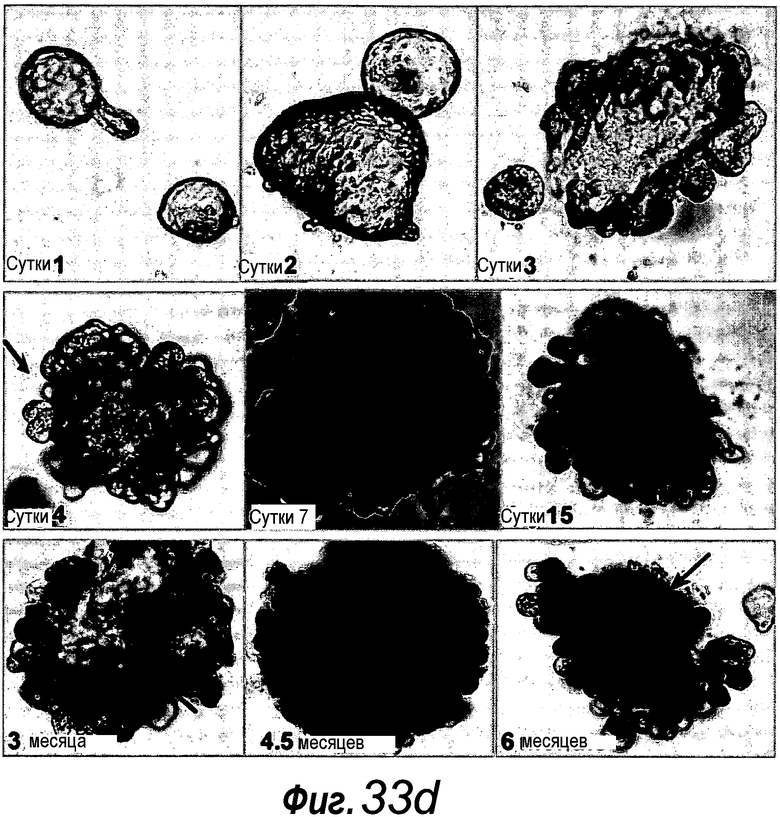
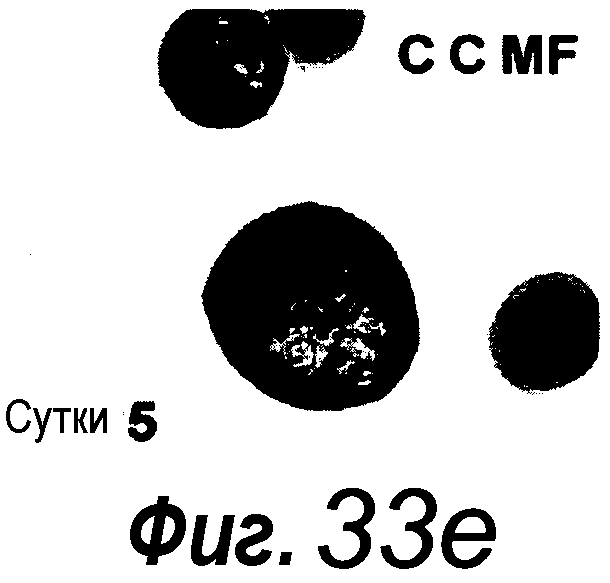

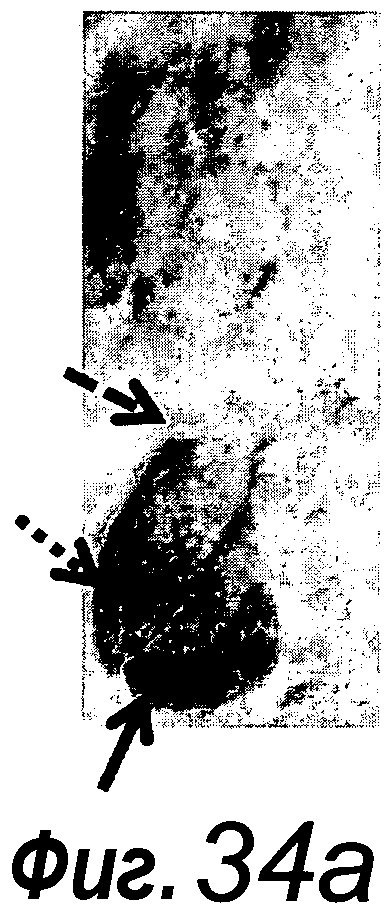
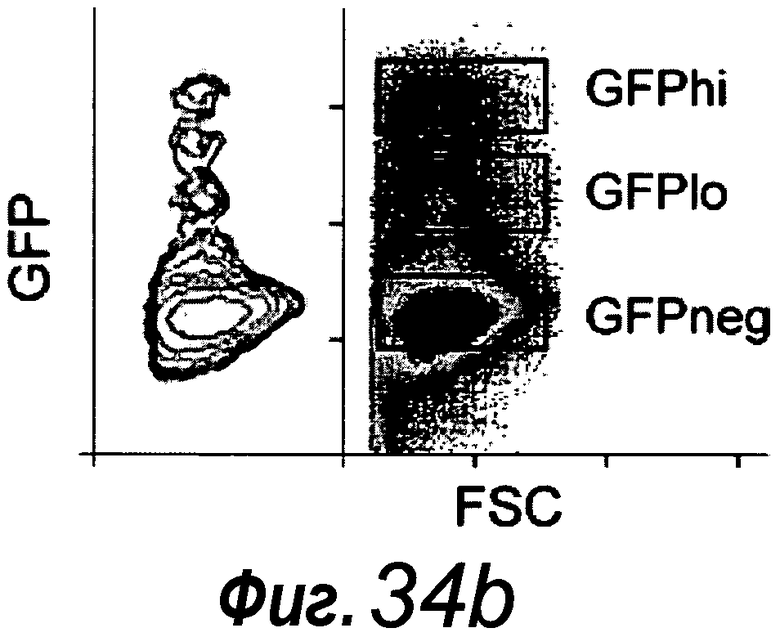
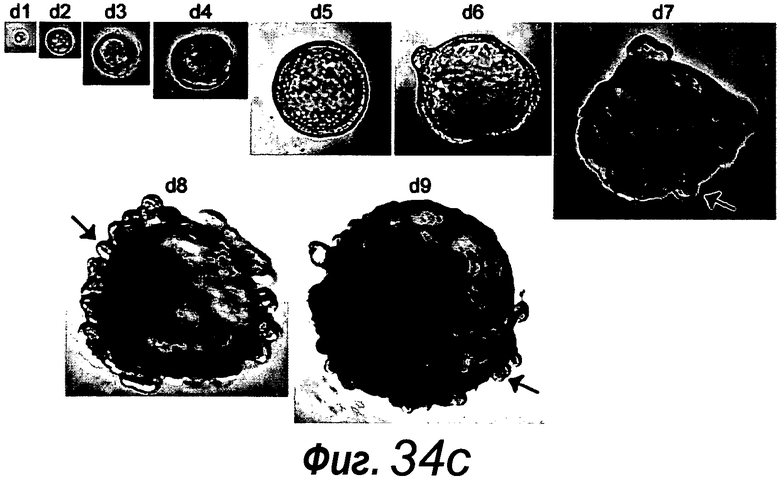

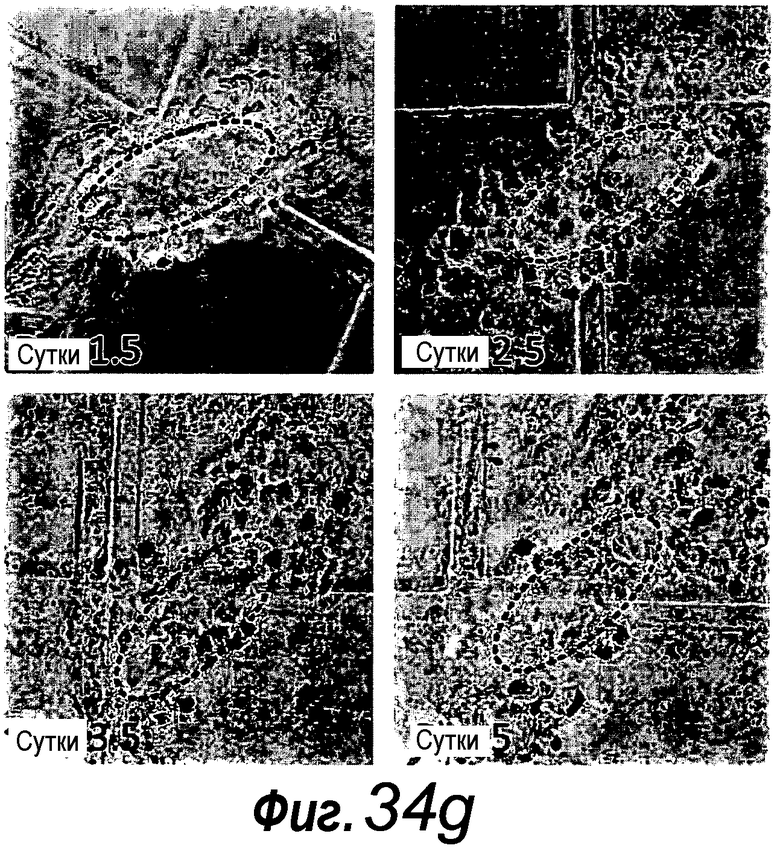
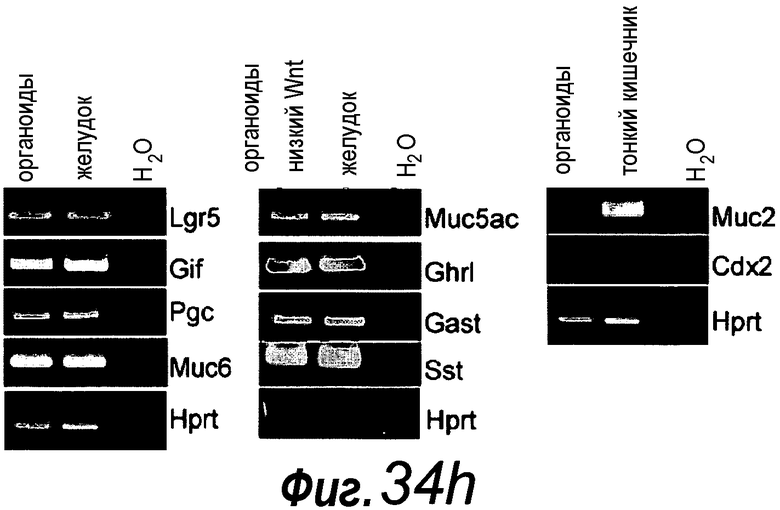
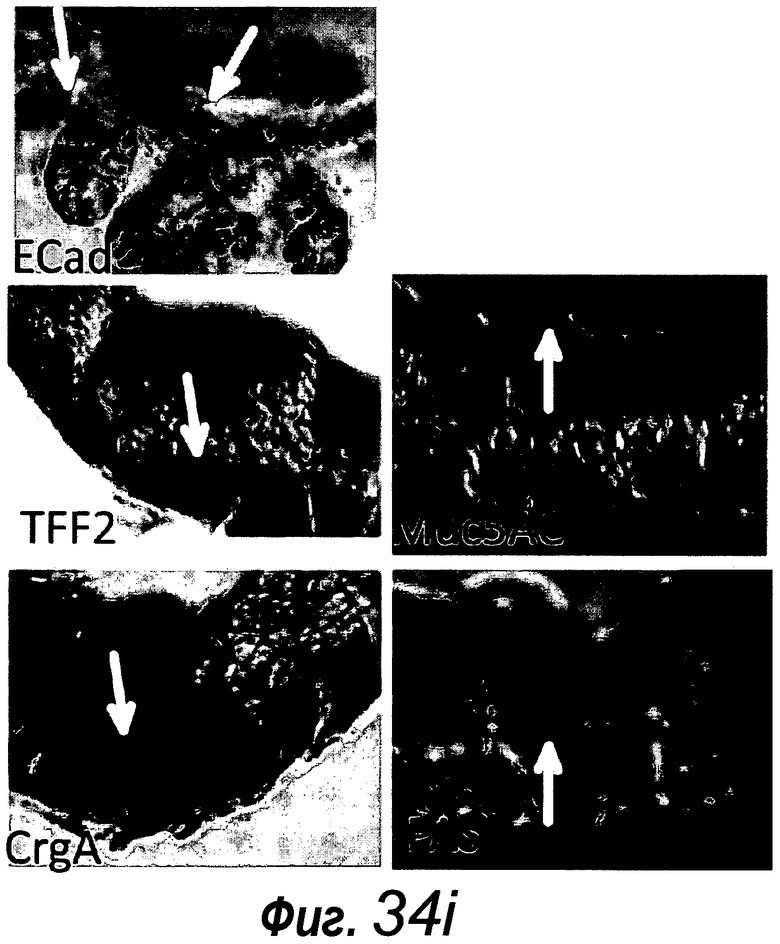
Изобретение относится к области биотехнологии. Предложен способ культивирования эпителиальных стволовых клеток или выделенных фрагментов ткани, содержащих указанные эпителиальные стволовые клетки, включающий обеспечение внеклеточного матрикса, инкубирование эпителиальной стволовой клетки или выделенного фрагмента ткани, содержащего указанные эпителиальные стволовые клетки, с внеклеточным матриксом, культивирование эпителиальной стволовой клетки или выделенного фрагмента ткани в присутствии клеточной культуральной среды, содержащей базальную среду для клеток животного или человека, в которую добавлены ингибитор морфогенетического белка кости (BMP) и 5- 500 нг/мл митогенного фактора роста, а также клеточная культуральная среда, ее применение, способ культивирования трехмерного органоида, способ его получения, трехмерный органоид и его применение. 9 н. и 23 з.п. ф-лы, 10 пр., 34 ил.
1. Способ культивирования эпителиальных стволовых клеток или выделенных фрагментов ткани, содержащих указанные эпителиальные стволовые клетки, где способ включает:
обеспечение внеклеточного матрикса;
инкубирование эпителиальной стволовой клетки или выделенного фрагмента ткани, содержащего указанные эпителиальные стволовые клетки, с внеклеточным матриксом;
культивирование эпителиальной стволовой клетки или выделенного фрагмента ткани в присутствии клеточной культуральной среды, содержащей базальную среду для клеток животного или человека, в которую добавлены:
ингибитор морфогенетического белка кости (BMP); и
от 5 до 500 нг/мл митогенного фактора роста.
2. Способ по п. 1, в котором эпителиальные стволовые клетки представляют собой нормальные эпителиальные клетки или эпителиальные клетки аденомы и в случае культивирования нормальных эпителиальных клеток добавляют агонист Wnt.
3. Способ по п. 1, где указанный ингибитор BMP выбран из Noggin, DAN и DAN-подобных белков, включая Cerberus и Gremlin.
4. Способ по п. 2, где указанный агонист Wnt выбран из одного или нескольких из Wnt, R-спондина 1-4, Norrin и ингибитора GSK.
5. Способ по п. 2, где указанный агонист Wnt содержит R-спондин 1-4 и/или Wnt-3а.
6. Способ по п. 1, где указанный митогенный фактор роста выбран из группы, состоящей из EGF, TGF-альфа, FGF, BDNF и KGF.
7. Способ по п. 6, где указанный митогенный фактор роста представляет собой EGF.
8. Способ по п. 1, где ингибитор BMP представляет собой Noggin, митогенный фактор роста представляет собой эпидермальный фактор роста (EGF), а агонист Wnt представляет собой один из R-спондинов 1-4.
9. Способ по п. 1, где указанная культуральная среда дополнительно содержит ингибитор Rock (Rho-киназа), необязательно выбранный из Y-27632, фасудила и Н-1152.
10. Способ по п. 1, где указанная культуральная среда дополнительно содержит агонист notch.
11. Клеточная культуральная среда, подходящая для культивирования эпителиальных стволовых клеток, содержащая базальную среду для культивации клеток животного или человека, в которую добавлен:
ингибитор морфогенетического белка кости (BMP); и
митогенный фактор роста в количестве, в частности, от 5 до 500 нг/мл.
12. Клеточная культуральная среда по п. 11, где митогенетический фактор роста выбран из одного или нескольких митогенных факторов роста из группы, состоящей из эпидермального фактора роста (EGF), трансформирующего фактора роста-альфа (TGF-альфа), фибробластного фактора роста (FGF), происходящего из головного мозга нейротрофического фактора (BDNF) и фактора роста кератиноцитов (KGF), в частности, где митогенетический фактор роста представляет собой EGF.
13. Клеточная культуральная среда по п. 11, которая дополнительно содержит агонист Wnt, в частности любой из R-спондинов 1-4, и/или где ингибитор BMP представляет собой Noggin, и где среда необязательно дополнена по меньшей мере В27, N2 и N-ацетилцистеином.
14. Клеточная культуральная среда по п. 13, где ингибитор BMP представляет собой Noggin, митогенетический фактор роста представляет собой EGF, а агонист Wnt представляет собой один из R-спондинов 1-4.
15. Клеточная культуральная среда по п. 11, которая дополнительно содержит внеклеточный матрикс, в частности, с внеклеточным матриксом, который содержит коллаген и ламинин, в частности с Matrigel™.
16. Клеточная культуральная среда по п. 13, которая дополнительно содержит внеклеточный матрикс, в частности, с внеклеточным матриксом, который содержит коллаген и ламинин, в частности с Matrigel™.
17. Клеточная культуральная среда по п. 15, где культуральная среда добавлена к или включена во внеклеточный матрикс.
18. Клеточная культуральная среда по п. 16, где культуральная среда добавлена к или включена во внеклеточный матрикс.
19. Применение культуральной среды по любому из пп. 11-18 для культивирования эпителиальных стволовых клеток, выделенных фрагментов тканей или выделенных органоидных структур на или во внеклеточном матриксе.
20. Применение по п. 19, где эпителиальные стволовые клетки представляют собой нормальные эпителиальные клетки или эпителиальные клетки аденомы.
21. Способ культивирования трехмерного тканевого органоида в присутствии внеклеточного матрикса, где способ включает культивирование органоида в культуральной среде по любому из пп. 11-18.
22. Способ по п. 21, где указанный трехмерный тканевой органоид культивируют в среде, содержащей Noggin, EGF и, необязательно, R-спондин 1 и/или Wnt-3а в качестве агониста Wnt.
23. Способ по п. 1, в котором получают органоид.
24. Способ получения органоида путем культивирования эпителиальных стволовых клеток толстого или тонкого кишечника или фрагментов ткани, содержащих указанные эпителиальные стволовые клетки, в частности выделенные крипты толстого или тонкого кишечника, используя способ по любому из пп. 1-10, где, в частности, стадия культивирования включает культивирование указанных эпителиальных стволовых клеток толстого или тонкого кишечника или фрагментов ткани, содержащих указанные эпителиальные стволовые клетки, в частности выделенные крипты толстого или тонкого кишечника, в контакте с внеклеточным матриксом в среде, содержащей Noggin в качестве ингибитора BMP, EGF в качестве митогенного фактора роста и дополнительно содержащей R-спондин 1 и/или Wnt-3 в качестве агониста Wnt, дополненной В27, N2 и N-ацетилцистеином.
25. Способ получения органоида поджелудочной железы путем культивирования эпителиальных стволовых клеток поджелудочной железы или фрагментов ткани, содержащих указанные эпителиальные стволовые клетки, используя способ по любому из пп. 1-10, где, в частности, стадия культивирования включает:
- культивирование указанных эпителиальных стволовых клеток поджелудочной железы или фрагментов ткани, содержащих указанные эпителиальные стволовые клетки, в контакте с внеклеточным матриксом на первой стадии в среде, содержащей Noggin в качестве ингибитора BMP, EGF, KGF или FGF в качестве митогенных факторов роста и дополнительно содержащей R-спондин 1 в качестве агониста Wnt, дополненной В27, N2 и N-ацетилцистеином, и
- затем на второй стадии в среде, содержащей EGF и R-спондин 1 в качестве агониста Wnt, дополненной В27, N2 и N-ацетилцистеином.
26. Способ получения желудочного органоида путем культивирования эпителиальных стволовых клеток желудка или фрагментов ткани, содержащих указанные эпителиальные стволовые клетки, используя способ по любому из пп. 1-10, где, в частности, стадия культивирования включает:
- культивирование указанных эпителиальных стволовых клеток желудка или фрагментов ткани, содержащих указанные эпителиальные стволовые клетки, в контакте с внеклеточным матриксом на первой стадии в среде, содержащей Noggin в качестве ингибитора BMP, EGF и FGF10 в качестве митогенных факторов роста и дополнительно R-спондин 1 и Wnt-3а в качестве агониста Wnt и дополнительно содержащей В27, N2, N-ацетилцистеин, и
- затем на второй стадии в среде, содержащей EGF и R-спондин 1 и Wnt-3а в качестве агониста Wnt, Noggin и FGF10, дополненной В27, N2 и N-ацетилцистеином, где концентрация Wnt-3 на второй стадии снижена по сравнению с концентрацией Wnt-3а, присутствующей на первой стадии.
27. Трехмерный органоид, полученный способом по любому из пп. 21-26, где органоид содержит центральный просвет и где клетки, которые не являются эпителиальными, отсутствуют в указанном органоиде.
28. Трехмерный органоид по п. 27, где органоид происходит из эпителиальных стволовых клеток тонкого кишечника, эпителиальных стволовых клеток поджелудочной железы или эпителиальных стволовых клеток желудка.
29. Трехмерный органоид по п. 27, содержащий домены, подобные криптам, окружающие центральный просвет, выстилаемый эпителиальными доменами, подобными ворсинкам, которые являются эпителиальными доменами, включающими дифференцированные типы клеток, и где в органоиде отсутствуют клетки, которые не являются эпителиальными.
30. Трехмерный органоид по п. 27, включающий структуры, подобные островкам поджелудочной железы, которые являются эпителиальными доменами, включающими дифференцированные типы клеток, и где в органоиде отсутствуют клетки, которые не являются эпителиальными.
31. Трехмерный органоид по п. 27, где органоид происходит из одной или более клеток аденомы и имеет подобную пузырю структуру, в которой клетки беспорядочно ориентированы по направлению к периферии или к центральному просвету.
32. Применение органоидов по любому из пп. 27-31 в скрининге при разработке лекарственных средств, в анализе токсичности или в регенеративной медицине.
| US 20070254359 A1, 01.11.2007 | |||
| US 20080233088 A1, 25.09.2008 | |||
| WO 2008101215 A1, 21.08.2008 | |||
| Клапан горячего дутья доменных воздухонагревателей | 1990 |
|
SU1792979A1 |
| Способ и прибор для определения коэффициента теплопередачи в котлах | 1926 |
|
SU7611A1 |
| ДИФФЕРЕНЦИРОВКА ЭМБРИОНАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК ЧЕЛОВЕКА | 2008 |
|
RU2465323C2 |

