Все документы, цитированные в настоящем описании, включены в настоящее описание в качестве ссылок в полном объеме.
ОБЛАСТЬ ТЕХНИКИ
Изобретение относится к области культуральных сред и способов для стволовых клеток, в частности, к культуральным средам и способам для увеличения в количестве популяций стволовых клеток, например, эпителиальных стволовых клеток человека.
УРОВЕНЬ ТЕХНИКИ
Существует значительный интерес в культуральных средах и способах увеличения в количестве популяций стволовых клеток. Популяции стволовых клеток имеют множество применений. Например, стволовые клетки и их дифференцированных потомков можно использовать в клеточных анализах, скрининге лекарственных средств и анализах токсичности. Стволовые клетки также могут быть перспективными для клеточных способов терапии, как например, в регенеративной медицине для лечения поврежденной ткани. Также они могут действовать в качестве источника дифференцированных клеток для целей трансплантации, например, трансплантации бета-клеток поджелудочной железы для лечения диабета и т.д. Более того, эффективные среды для культивирования клеток важны для предоставления и поддержания популяций клеток для научных целей.
Также существует интерес в культуральных средах и способах культивирования стволовых клеток для образования, поддержания и увеличения в количестве органоидов, таких как органоиды крипта-ворсинка, органоиды желудка или поджелудочной железы. Органоид содержит стволовые клетки, такие как эпителиальные стволовые клетки, которые сохраняют их недифференцированный фенотип и свойства самообновления, но также имеет дифференцирующихся потомков, которые растут в подобные тканям структуры. Аналогично популяциям родственных или идентичных клеток, органоиды крипта-ворсинка, органоиды желудка или поджелудочной железы, которые в наибольшей степени имитируют основную физиологию их ткани-источника, можно использовать в анализах токсичности или анализах лекарственных средств или пищевых добавок. Также они могут подходить для культивирования патогенов, для которых в настоящее время отсутствуют подходящие модели в культуре тканей или модели на животных. Более того, такие органоиды могут быть подходящими в регенеративной медицине, например, при репарации кишечного эпителия после облучения и/или после хирургической операции, или при репарации кишечного эпителия у пациентов, страдающих воспалительным заболеванием кишечника.
Понятно, что существует множество клинических и научных применений для стволовых клеток и их дифференцированных потомков. Для всех этих применений воспроизводимые способы культивирования стволовых клеток имеют крайнюю важность для обеспечения достаточных количеств клеток приемлемого качества. Например, для эффективного скрининга лекарственных средств, условия необходимо тщательно контролировать, что требует точных способов культивирования для контроля дифференцировки и пролиферации клеток, так чтобы можно было получить чистые популяции фенотипически и кариотипически идентичных клеток. Аналогично, для клеточных способов терапии, где культивированные клетки можно прямо предоставлять пациентам, клетки должны быть генетически и фенотипически подходящими, так чтобы избежать нежелательных иммунных ответов или нежелательной судьбы клеток при предоставлении пациенту.
Хотя описано множество культуральных систем для культивирования первичных эпителиальных стволовых клеток, включая эпителиальные стволовые клетки кишечника (Bjerknes and Cheng, 2006. Methods Enzymol. 419: 337-83), на настоящий момент не было получено долговременной системы культивирования, которая сохраняет потенциал к дифференцировке и фенотипическую и геномную целостностью эпителиальных стволовых клеток человека.
В международной патентной заявке WO2010/090513 описан способ культивирования эпителиальных стволовых клеток или выделенных фрагментов тканей. Способ оптимизирован для культивирования крипт толстого кишечника и кишечных крипт человека путем добавления Wnt-3a в среду. Это был первый раз, когда культуры стволовых клеток кишечника человека культивировали в течение длительного периода времени (вплоть до 3 месяцев), и была предоставлена первая воспроизводимая система культивирования стволовых клеток кишечника человека. Однако все еще существует потребность в улучшенных культуральных средах и способах для культивирования стволовых клеток, в частности, средах и способах для культивирования стволовых клеток человека, которые повышают скорость пролиферации, время выживания и фенотипическую и геномную целостность стволовых клеток, выращенных в культуре.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Изобретение относится к улучшенным средам и способам для культивирования стволовых клеток, в частности, эпителиальных стволовых клеток человека, и органоидов, содержащих указанные стволовые клетки, которые обеспечивают значительные преимущества над известными средами и способами для культивирования. Изобретение также относится к связанным с ними добавкам в культуральную среду, композициям и применениям.
Таким образом, изобретение относится к культуральной среде для увеличения в количестве популяции стволовых клеток, где культуральная среда содержит по меньшей мере один или несколько ингибиторов, которые связываются и снижают активность одной или нескольких мишеней в виде серин/треониновых протеинкиназ. Это имеет эффект, позволяющий непрерывный рост в течение по меньшей мере 3 месяцев при скорости увеличения в количестве, составляющей увеличение в количестве приблизительно в пять раз за неделю. Серин/треониновая протеинкиназа, предпочтительно, выбрана из группы, включающей: киназу 1 рецептора TGF-бета, ALK4, ALK5, ALK7, p38. Неожиданно, авторы изобретения обнаружили, что включение ингибиторов определенных серин/треониновых киназ в культуральную среду значительно увеличивает эффективность культуральной среды в отношении увеличения в количестве популяции стволовых клеток. Популяция стволовых клеток может представлять собой нормальные (здоровые) клетки или пораженные заболеванием клетки (например, злокачественные стволовые клетки). В частности, было показано, что ингибиторы p38 и ALK обеспечивают наибольшее улучшение из всех исследованных соединений. Это является неожиданном, поскольку не существует известного механизма, предсказывающего то, как эти конкретные ингибиторы могут действовать. Действительно, несколько низкомолекулярных ингибиторов, которые были выбраны для исследования и функционируют в сходных каскадах, не имели эффекта на способ. Таким образом, квалифицированный специалист не мог предсказать, что ингибиторы этих конкретных киназ могут обеспечить такое выраженное усовершенствование культуральной среды. Следующее усовершенствование наблюдали, когда два ингибитора, например, ингибитор p38, такой как SB202190, и ингибитор ALK, такой как A83-01, добавляли в культуральную среду вместе.
Для осознания этого авторы изобретения исследовали каскады передачи сигналов, которые известны тем, что они нарушаются при определенных злокачественных опухолях, например, при раке ободочной и прямой кишки. Они предположили, что эти каскады, которые влияют на клеточную судьбу при злокачественной опухоли, также могут играть роль в определении клеточной судьбы в условиях культивирования. Однако следует подчеркнуть, что эта гипотеза является абсолютно новой; учитывая уровень техники, не было возможным предсказать эффект любого из этих дополнительных соединений на культуральную среду и нельзя было, в частности, ожидать, что любое из этих соединений может в действительности иметь благоприятный эффект.
В первом скрининговом эксперименте исследовали серию витаминов, гормонов и факторов роста в комбинации со стандартной средой для культивирования стволовых клеток. Первоначально были идентифицированы гастрин и никотинамид в качестве обеспечивающих значительное улучшение условий культивирования. Проводили второй скрининговый эксперимент с включением этих факторов в стандартные условия культивирования, в котором исследовали низкомолекулярные ингибиторы, связанные с соответствующими каскадами передачи сигнала, такими как каскады ERK, p38, JNK, PTEN, ROCK и Hedgehog. Эти каскады были выбраны, поскольку известно, что они нарушаются при определенных злокачественных опухолях.
Предшествующие попытки культивировать стволовые клетки кишечника человека с помощью ранее описанной среды для культивирования стволовых клеток (содержащей эпидермальный фактор роста (EGF или ("E"), Noggin ("N") и R-спондин ("R"), обозначаемой в настоящем описании как среда "ENR"), оптимизированной с помощью Wnt-3A ("W") (обозначаемой в настоящем описании как среда "WENR"), привели к дезинтеграции большинства клеток в течение 7 суток, причем очень мало клеток выжило более 1 месяца. Для таких попыток также были характерны медленное время пролиферации, хромосомные нарушения и морфологические изменения от почкования до кистозных структур. Под "кистозным" подразумевают, что органоид является по большей части сферическим. Под "почкованием" подразумевают, что органоид имеет множество участков, растущих из основной структуры. Наличие почкующихся структур не всегда является преимуществом, хотя почкующиеся структуры обычно имеют увеличенную площадь поверхности и, как правило, в большей степени напоминают соответствующую ткань in vivo.
Авторы изобретения показали, что улучшенный способ позволил непрерывный рост стволовых клеток в течение по меньшей мере семи месяцев.
Новый способ также увеличил скорость пролиферации клеток в увеличенной в количестве популяции. Это, несомненно, в высокой степени применимо при выращивании клеток для коммерческих и терапевтических целей.
Новый способ также увеличил качество клеток в увеличенной в количестве популяции. Это является значительным преимуществом, поскольку для клинических и научных применений стволовых клеток и их дифференцированных потомков требуются воспроизводимые способы культивирования стволовых клеток, которые обеспечивают популяции клеток высокого качества. Главным образом, увеличение стволовых клеток в количестве in vitro нацелено на предоставление популяции клеток, которые напоминают их аналоги in vivo настолько, насколько это возможно. Это свойство называют в настоящем описании "геномной и фенотипической целостностью" клеток.
Впервые авторы изобретения открыли, что можно увеличивать в количестве эпителиальные стволовые клетки человека в культуре без утраты геномной и фенотипической целостности в течение по меньшей мере 7 месяцев (см. пример 1). В улучшенных условиях культивирования по изобретению органоиды кишечника человека проявляли почкование структур органоидов, а не кистозные структуры, наблюдаемые в предшествующих условиях культивирования. В метафазных пластинках органоидов возрастом более 3 месяцев выявляли 46 хромосом в каждой из 20 клеток, взятых от трех различных доноров. Более того, анализ на микрочипах выявил, что стволовые клетки в культуре обладали сходными молекулярными профилями с клетками кишечных крипт, включая гены стволовых клеток кишечника.
Авторы изобретения также продемонстрировали, что органоиды кишечника человека, полученные с помощью сред и способов по настоящему изобретению, имитировали решение о клеточной судьбе in vivo в ответ на внешние факторы. Например, ранее было показано, что ингибирование Notch в стволовых клетках кишечника завершает пролиферацию эпителия кишечника и индуцирует гиперплазию бокаловидных клеток in vivo. Авторы изобретения были способны показать, что органоиды кишечника по изобретению после обработки ингибитором Notch прекращали пролиферировать, и большинство клеток преобразовывалось в бокаловидные клетки в течение 3 суток.
Сходные преимущества наблюдали при включении ингибитора TGF-бета и/или ингибитора p38 в культуральную среду для увеличения в количестве стволовых клеток или органоидов из других эпителиальных тканей, таких как желудок, поджелудочная железа, печень и предстательная железа (см. примеры). Ткани могут представлять собой нормальные (здоровые) ткани или пораженные заболеванием ткани, например, злокачественные ткани или ткани, демонстрирующие фенотип кистозного фиброза.
Эти результаты демонстрируют значительное увеличение геномной и фенотипической целостности стволовых клеток и органоидов, полученных с помощью способов и сред по настоящему изобретению, по сравнению с предшествующими способами и средами.
Таким образом, изобретение относится к культуральной среде для увеличения в количестве и/или дифференцировки популяции взрослых стволовых клеток, где указанная культуральная среда содержит:
i. любой из R-спондина 1-4 и/или миметика R-спондина; и
ii. один или несколько ингибиторов, которые прямо или непрямо отрицательно регулируют передачу сигнала TGF-бета.
Изобретение также относится к композиции, содержащей культуральную среду согласно изобретению и внеклеточный матрикс или 3D-матрикс, который имитирует внеклеточный матрикс благодаря его взаимодействию с белками клеточной мембраны, такими как интегрины, например, содержащий ламинин внеклеточный матрикс, такой как MatrigelTM (BD Biosciences).
Изобретение также относится к герметично закрытой емкости, содержащей культуральную среду или композицию согласно изобретению.
Изобретение также относится к применению культуральной среды согласно изобретению для увеличения в количестве и/или дифференцировки стволовой клетки, популяции стволовых клеток, фрагмента ткани или органоида.
Изобретение также относится к способам увеличения в количестве единичной стволовой клетки, популяции стволовых клеток или фрагмента ткани, предпочтительно, для получения органоида, где способ включает культивирование единичной стволовой клетки или популяции стволовых клеток в культуральной среде согласно изобретению.
Изобретение также относится к органоидам или популяциям клеток, получаемым способами по изобретению.
Изобретение также относится к органоиду, предпочтительно, получаемому способами по изобретению, который представляет собой трехмерный органоид, содержащий эпителиальные клетки, окружающие центральный просвет, где необязательно эпителиальные клетки существуют в отдельных делящихся доменах и дифференцирующихся доменах.
Изобретение также относится к органоиду, предпочтительно, получаемому способами по изобретению, который представляет собой трехмерный органоид, содержащий эпителиальные клетки, расположенные в областях монослоев, необязательно сложенных монослоев, и в областях распределенных на слои клеток, и предпочтительно, который представляет собой трехмерный органоид, содержащий эпителиальные клетки, окружающие центральный просвет, где необязательно эпителиальные клетки существуют в отдельных делящихся доменах и дифференцирующихся доменах.
Изобретение также относится к композиции, содержащей:
i) один или несколько органоидов или одну или несколько популяций клеток по изобретению; и
ii) культуральную среду по изобретению и/или внеклеточный матрикс.
Изобретение также относится к органоиду, популяции клеток или композиции согласно изобретению для применения в скрининге лекарственных средств, подтверждении мишени, выявлении мишени, токсикологии, токсикологическом скрининге, персонализированной медицине, регенеративной медицине или в моделях клеток/органов ex vivo, например, для применения в качестве модели заболевания.
Изобретение также относится к органоиду, популяции клеток или композиции согласно изобретению для применения в трансплантации указанного органоида, популяции клеток или композиции млекопитающему, предпочтительно, человеку.
Изобретение также относится к популяции стволовых клеток или органоидам, содержащим указанные стволовые клетки, которые получены или могут быть получены с использованием культуральной среды по изобретению. Стволовые клетки или органоиды, содержащие указанные стволовые клетки, можно использовать, например, для целей трансплантации или других терапевтических применений. Например, стволовые клетки или органоиды, содержащие указанные стволовые клетки, можно использовать для скрининга лекарственных средств, подтверждения мишени, выявления мишени, токсикологии и токсикологического скрининга, персонализированной медицины, регенеративной медицины и моделей клеток/органов ex vivo, например, моделей заболевания.
Изобретение также относится к композициям, содержащим культуральную среду по изобретению.
Изобретение также относится к добавкам в культуральную среду, содержащим ингибитор согласно изобретению.
Изобретение также относится к герметично закрытой емкости, содержащей культуральную среду и/или добавку в культуральную среду согласно изобретению.
Конкретные ингредиенты культуральных сред, добавок и композиций по изобретению могут варьировать в зависимости от конкретных потребностей и применений. Аналогично, точные стадии способов по изобретению могут варьировать в зависимости от конкретных потребностей и применений.
Культуральные среды, добавки, способы, композиции и применения согласно настоящему изобретению также можно оптимизировать путем стандартного экспериментирования. Например, если культуральная среда, добавка или композиция не обеспечивает желаемый уровень увеличения стволовых клеток в количестве, в последующих экспериментах можно изменять такие переменные, как количество каждого ингредиента в культуральной среде или добавке, плотность посева, условия культивирования, периоды культивирования и т.д. Количество каждого из ингредиентов, описанных в настоящем описании, можно оптимизировать независимо от других ингредиентов путем стандартных способов оптимизации или один или несколько ингредиентов можно добавлять или устранять. Культуральную среду можно исследовать в отношении ее способности поддерживать увеличение стволовых клеток в количестве путем исследования ее вместе или вместо известной культуральной среды или способа.
Культуральные среды, добавки, способы, композиции и применения по изобретению более подробно описаны ниже. Для осуществления на практике настоящего изобретения можно использовать, если нет иных указаний, общепринятые способы культивирования клеток, молекулярной биологии и микробиологии, которые находятся в пределах квалификации специалистов в данной области.
Доступны многочисленные справочники, в которых предоставлено руководство по средам и способам для культивирования клеток млекопитающих, включая справочники, посвященные культуральным средам и способам для культивирования стволовых клеток. Такие справочники включают "Basic Cell Culture Protocols", J. Pollard and J. M. Walker (1997), "Mammalian Cell Culture: Essential Techniques", A. Doyle and J. B. Griffiths (1997), "Culture of Animal Cells: A Manual of Basic Technique", R. I. Freshney (2005), "Basic Cell Culture Protocols", C. Helgason and C. L. Miller (2005), "Stem Cells: From Bench to Bedside", A. Bongso (2005), "Human Stem Cell Manual: A Laboratory Guide", J. F. Loring, R. L. Wesselschmidt and P. H. Schwartz (2007).
Стволовые клетки и реагенты для культивирования клеток и устройство для применения в рамках изобретения являются коммерчески доступными, например, от Cellartis AB (Göteborg, Швеция), VitroLife AB (Kungsbacka, Швеция), GIBCO® (Invitrogen), Millipore Corporation (Billerica, Массачусетс), Sigma® (St. Louis, Миссури) и Biomol International L.P. (Exeter, Великобритания).
ПОДРОБНОЕ ОПИСАНИЕ
Согласно изобретению, предусмотрена культуральная среда для увеличения в количестве популяции стволовых клеток, где культуральная среда содержит по меньшей мере один или несколько ингибиторов, которые связываются с одной или несколькими мишенями в виде серин/треониновых протеинкиназ и снижают их активность, где культуральная среда обладает эффектом обеспечения непрерывного роста популяции стволовых клеток в течение по меньшей мере 3 месяцев, предпочтительно, по меньшей мере 4 месяцев, по меньшей мере 5 месяцев, по меньшей мере 6 месяцев, по меньшей мере 7 месяцев, по меньшей мере 9 месяцев или по меньшей мере 12 месяцев или более.
Ингибиторы
Культуральная среда, используемая в соответствии с первым аспектом изобретения, содержит любой ингибитор, который прямо или непрямо отрицательно регулирует передачу сигнала TGF-бета или p38. В предпочтительном варианте осуществления культуральная среда по изобретению содержит ингибитор, который прямо или непрямо отрицательно регулирует передачу сигнала TGF-бета. В некоторых вариантах осуществления культуральная среда по изобретению содержит ингибитор, который прямо или непрямо отрицательно регулирует передачу сигнала TGF-бета, и ингибитор, который прямо или непрямо отрицательно регулирует передачу сигнала p38. В следующем варианте осуществления культуральная среда по изобретению дополнительно содержит R-спондин или миметик R-спондина.
Один или несколько ингибиторов, предпочтительно, нацелены на серин/треониновую протеинкиназу, выбранную из группы, включающей: киназу 1 рецептора TGF-бета, ALK4, ALK5, ALK7, p38. Ингибитор любой из этих киназ представляет собой ингибитор, который обеспечивает снижение ферментативной активности любой одной (или нескольких) из этих молекул. Ранее было показано, что ингибирование киназ ALK и p38 связано с B-клеточной лимфомой (Bakkebø M Huse K, Hilden VI, Smeland EB, Oksvold MP, "TGF-beta-induced growth inhibition in B-cell limphoma correlates with Smad1/5 signalling and constitutively active p38 MAPK", BMC Immunol. 11:57, 2010). В этой публикации было выявлено, что чувствительные к TGF-бета клеточные линии экспрессировали более высокие уровни на клеточной поверхности ALK-5 и что конститутивное фосфорилирование p38 было ограничено чувствительными к TGF-бета клеточными линиями. Ингибирование p38 MAPK приводило к сниженной чувствительности к TGF-бета, что указывает на то, что фосфорилирование Smad1/5 является важным для антипролиферативных эффектов TGF-бета в B-клеточной лимфоме. Результаты указывают на роль p38 MAPK в регуляции индуцируемых TGF-бета антипролиферативных эффектов.
Без связи с теорией, авторы настоящего изобретения полагают, что ALK и p38 принадлежат каскаду, который отрицательно регулирует долговременное поддержание стволовых клеток, в частности, эпителиальных стволовых клеток человека. Авторы изобретения полагают, что ингибиторы, которые действуют на любом уровне этого каскада, в том числе, например, путем ингибирования передачи сигнала Smad1/5, также являются благоприятными для культуры стволовых клеток. Smad играют ключевую роль в передаче сигнала TGF-бета.
В некоторых вариантах осуществления ингибитор по изобретению связывает и снижает активность серин/треониновой протеинкиназы, выбранной из группы, включающей: киназу 1 рецептора TGF-бета, ALK4, ALK5, ALK7, p38.
В некоторых вариантах осуществления изобретения культуральная среда содержит ингибитор TGF-бета, что означает любой ингибитор, который прямо или непрямо отрицательно регулирует передачу сигнала TGF-бета. В некоторых вариантах осуществления культуральная среда по изобретению содержит один или несколько ингибиторов TGF-бета, которые связывают и снижают активность одной или нескольких серин/треониновых протеинкиназ, выбранных из группы, состоящей из ALK5, ALK4, киназы 1 рецептора TGF-бета и ALK7.
Все из ALK4, ALK5 и ALK7 являются близкородственными рецепторами суперсемейства TGF-бета. ALK4 имеет номер GI 91; ALK5 (также известная как киназа 1 рецептора TGF-бета) имеет номер GI 7046; и ALK7 имеет номер GI 658. В одном из вариантов осуществления ингибитор согласно изобретению связывает и снижает активность ALK4, ALK5 (киназа 1 рецептора TGF-бета) и/или ALK7. В другом варианте осуществления рецептор TGF-бета связывает и снижает активность белка Smad, например, R-SMAD или SMAD1-5 (т.е. SMAD 1, SMAD 2, SMAD 3, SMAD 4 или SMAD 5). В предпочтительном варианте осуществления культуральная среда по изобретению содержит ингибитор ALK5.
Известны различные способы для определения того, является ли вещество ингибитором TGF-бета. Например, можно использовать клеточный анализ, в котором клетки стабильно трансфицированы репортерной конструкцией, содержащей промотор PAI-1 человека или участки связывания Smad, регулирующие репортерный ген люциферазы. Ингибирование активности люциферазы относительно контрольных групп можно использовать в качестве меры активности соединения (De Gouville et al., Br J Pharmacol. 2005 May; 145(2): 166-177). Другим примером является анализ с помощью AlphaScreen® phosphosensor для измерения активности киназы (Drew A E et al., Comparison of 2 Cell-Based Phosphoprotein Assays to Support Screening and Development of an ALK Inhibitor, J Biomol Screen. 16(2) 164-173, 2011).
Различные ингибиторы TGF-бета известны в данной области (например, см. таблицу 1). В некоторых вариантах осуществления ингибитор, который прямо или непрямо отрицательно регулирует передачу сигнала TGF-бета, выбран из группы, состоящей из A83-01, SB-431542, SB-505124, SB-525334, SD-208, LY-36494 и SJN-2511.
В некоторых вариантах осуществления изобретения культуральная среда содержит ингибитор p38, что означает любой ингибитор, который прямо или непрямо отрицательно регулирует передачу сигнала p38. В некоторых вариантах осуществления ингибитор согласно изобретению связывает и снижает активность p38 (номер GI 1432). Протеинкиназы p38 являются частью семейства митоген-активируемых протеинкиназ (MAPK). MAPK представляют собой серин/треонин-специфические протеинкиназы, которые отвечают на внеклеточные стимулы, такие как воздействие внешних условий и воспалительные цитокины, и регулируют различные клеточные виды активности, такие как экспрессия генов, митоз, дифференцировка, пролиферация и выживание клеток/апоптоз. p38 MAPK существуют в качестве изоформ α, β, β2, γ и δ. Ингибитор p38 представляет собой средство, которое связывает и снижает активность по меньшей мере одной изоформы p38. Известны различные способы определения того, является ли вещество ингибитором p38, и их можно использовать применительно к изобретению. Примеры включают: выявление с помощью фосфо-специфического антитела фосфорилирования по Thr180/Tyr182, которое обеспечивает общепризнанный показатель активации или ингибирования клеточного p38; биохимические анализы с рекомбинантными киназами; анализы секреции фактора некроза опухоли альфа (TNFα); и высокопроизводительную платформу для скрининга DiscoverRx для ингибиторов p38 (см. http://www.discoverx.com/kinases/literature/biochemical/collaterals/DRx_poster_p38%20KBA.pdf). Также существует несколько наборов для анализа активности p38 (например, Millipore, Sigma-Aldrich).
Авторы изобретения полагают, что в некоторых вариантах осуществления высокие концентрации (например, более 100 нМ или более 1 мкМ, более 10 мкМ, или более 100 мкМ) ингибитора p38 могут иметь эффект ингибирования TGF-бета. Однако авторы изобретения не хотели бы быть ограниченными этой гипотезой, и в других вариантах осуществления ингибитор p38 не ингибирует передачу сигнала TGF-бета.
В данной области известны различные ингибиторы p38 (например, см. таблицу 1). В некоторых вариантах осуществления ингибитор, который прямо или непрямо отрицательно регулирует передачу сигнала p38, выбран из группы, состоящей из SB-202190, SB-203580, VX-702, VX-745, PD-169316, RO-4402257 и BIRB-796. В следующем варианте осуществления изобретения культуральная среда содержит оба из: a) ингибитора, который связывает и снижает активность любой одной или нескольких киназ из группы, состоящей из: ALK4, ALK5 и ALK7; и b) ингибитора, который связывает и снижает активность p38. В предпочтительном варианте осуществления культуральная среда содержит ингибитор, который связывает и снижает активность ALK5, и ингибитор, который связывает и снижает активность p38.
В одном из вариантов осуществления ингибитор согласно изобретению связывает и снижает активность его мишени (например, TGF-бета или p38) более чем на 10%; более чем на 30%; более чем на 60%; более чем на 80%; более чем на 90%; более чем на 95%; или более чем на 99% по сравнению с контролем при оценке с помощью клеточного анализа. Примеры клеточных анализов для измерения ингибирования мишени известны в данной области, как описано выше.
Ингибитор согласно изобретению может иметь величину IC50, равную или меньшую 2000 нМ; меньшую 1000 нМ; меньшую 100 нМ; меньшую 50 нМ; меньшую 30 нМ; меньшую 20 нМ или меньшую 10 нМ. Величина IC50 относится к эффективности ингибитора в отношении ингибирования его биологический или биохимической функции его мишени. IC50 указывает на то, сколько конкретного ингибитора требуется для ингибирования киназы на 50%. Величины IC50 можно вычислять в соответствии со способами анализа, описанными выше.
Ингибитор согласно изобретению может действовать конкурентно, неконкурентно, внеконкурентно или путем смешанного ингибирования. Например, в определенных вариантах осуществления, ингибитор может представлять собой конкурентный ингибитор кармана связывания ATP в киназе-мишени.
Ингибиторы согласно изобретению могут существовать в различных формах, включая природные или модифицированные субстраты, ферменты, рецепторы, низкомолекулярные органические молекулы, такие как низкомолекулярные природные или синтетические органические молекулы массой вплоть до 2000 Да, предпочтительно, 800 Да или менее, пептидомиметики, неорганические молекулы, пептиды, полипептиды, антисмысловые олигонуклеотиды, аптамеры, и их структурные или функциональные миметики, включая низкомолекулярные соединения. Ингибитор согласно изобретению также может представлять собой аптамер. Как используют в рамках изобретения, термин "аптамер" относится к цепям олигонуклеотидов (ДНК или РНК), которые могут принимать высоко специфические трехмерные конформации. Аптамеры конструируют так, чтобы они обладали высокой аффинностью связывания и специфичностью в отношении определенных молекул-мишеней, включая внеклеточные и внутриклеточные белки.
Например, ингибитор может представлять собой низкомолекулярную синтетическую молекулу с молекулярной массой от 50 до 800 Да, от 80 до 700 Да, от 100 до 600 Да или от 150 до 500 Да.
В некоторых вариантах осуществления низкомолекулярный ингибитор содержит пиридинилимидазол или 2,4-дизамещенный птеридин или хиназолин, например, содержит:
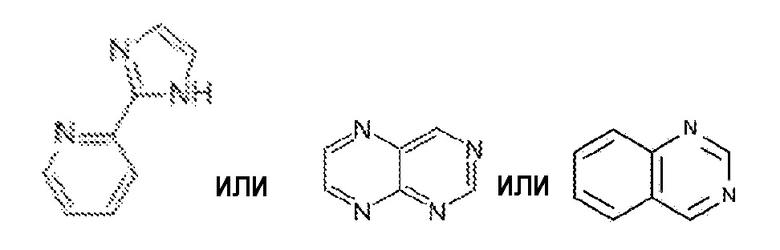
Конкретные примеры ингибиторов, которые можно использовать в соответствии с изобретением, включают, но не ограничиваются ими: SB-202190, SB-203580, SB-206718, SB-227931, VX-702, VX-745, PD-169316, RO-4402257, BIRB-796, A83-01 SB-431542, SB-505124, SB-525334, LY 364947, SD-208, SJN 2511 (см. таблицу 1). Культуральная среда по изобретению может содержать один или несколько любых из ингибиторов, приведенных в таблице 1. Культуральная среда по изобретению может содержать любую комбинацию одного приведенного ингибитора с другим приведенным ингибитором. Например, культуральная среда по изобретению может содержать SB-202190 или SB-203580 или A83-01; или культуральная среда по изобретению может содержать SB-202190 и A83-01; или культуральная среда по изобретению может содержать SB-203580 и A83-01. Квалифицированному специалисту будет понятно, что в соответствии с изобретением в культуральную среду или добавку в культуральную среду могут быть включены другие ингибиторы и комбинации ингибиторов, которые связывают и снижают активность мишеней согласно изобретению.
Ингибиторы согласно изобретению можно добавлять в культуральную среду до конечной концентрации, которая является подходящей, учитывая значение IC50 ингибитора.
Например, SB-202190 можно добавлять в культуральную среду в концентрации от 50 нМ до 100 мкМ, или от 100 нМ до 50 мкМ, или от 1 мкМ до 50 мкМ. Например, SB-202190 можно добавлять в культуральную среду в концентрации приблизительно 10 мкМ.
SB-203580 можно добавлять в культуральную среду в концентрации от 50 нМ до 100 мкМ, или от 100 нМ до 50 мкМ, или от 1 мкМ до 50 мкМ. Например, SB-203580 можно добавлять в культуральную среду в концентрации приблизительно 10 мкМ.
VX-702 можно добавлять в культуральную среду в концентрации от 50 нМ до 100 мкМ, или от 100 нМ до 50 мкМ, или от 1 мкМ до 25 мкМ. Например, VX-702 можно добавлять в культуральную среду в концентрации приблизительно 5 мкМ.
VX-745 можно добавлять в культуральную среду в концентрации от 10 нМ до 50 мкМ, или от 50 нМ до 50 мкМ, или от 250 нМ до 10 мкМ. Например, VX-745 можно добавлять в культуральную среду в концентрации приблизительно 1 мкМ.
PD-169316 можно добавлять в культуральную среду в концентрации от 100 нМ до 200 мкМ, или от 200 нМ до 100 мкМ, или от 1 мкМ до 50 мкМ. Например, PD-169316 можно добавлять в культуральную среду в концентрации приблизительно 20 мкМ.
RO-4402257 можно добавлять в культуральную среду в концентрации от 10 нМ до 50 мкМ, или от 50 нМ до 50 мкМ, или от 500 нМ до 10 мкМ. Например, RO-4402257 можно добавлять в культуральную среду в концентрации приблизительно 1 мкМ.
BIRB-796 можно добавлять в культуральную среду в концентрации от 10 нМ до 50 мкМ, или от 50 нМ до 50 мкМ, или от 500 нМ до 10 мкМ. Например, BIRB-796 можно добавлять в культуральную среду в концентрации приблизительно 1 мкМ.
A83-01 можно добавлять в культуральную среду в концентрации от 10 нМ до 10 мкМ, или от 20 нМ до 5 мкМ, или от 50 нМ до 1 мкМ. Например, A83-01 можно добавлять в культуральную среду в концентрации приблизительно 500 нМ.
SB-431542 можно добавлять в культуральную среду в концентрации от 80 нМ до 80 мкМ, или от 100 нМ до 40 мкМ, или от 500 нМ до 10 мкМ. Например, SB-431542 можно добавлять в культуральную среду в концентрации приблизительно 1 мкМ.
SB-505124 можно добавлять в культуральную среду в концентрации от 40 нМ до 40 мкМ, или от 80 нМ до 20 мкМ, или от 200 нМ до 1 мкМ. Например, SB-505124 можно добавлять в культуральную среду в концентрации приблизительно 500 нМ.
SB-525334 можно добавлять в культуральную среду в концентрации от 10 нМ до 10 мкМ, или от 20 нМ до 5 мкМ, или от 50 нМ до 1 мкМ. Например, SB-525334 можно добавлять в культуральную среду в концентрации приблизительно 100 нМ.
LY 36494 можно добавлять в культуральную среду в концентрации от 40 нМ до 40 мкМ, или от 80 нМ до 20 мкМ, или от 200 нМ до 1 мкМ. Например, LY 36494 можно добавлять в культуральную среду в концентрации приблизительно 500 нМ.
Иллюстративные ингибиторы согласно изобретению
SD-208 можно добавлять в культуральную среду в концентрации от 40 нМ до 40 мкМ, или от 80 нМ до 20 мкМ, или от 200 нМ до 1 мкМ. Например, SD-208 можно добавлять в культуральную среду в концентрации приблизительно 500 нМ.
LY364947 можно добавлять в культуральную среду в концентрации от 40 нМ до 40 мкМ, или от 80 нМ до 20 мкМ, или от 200 нМ до 1 мкМ. Например, LY364947 можно добавлять в культуральную среду в концентрации приблизительно 500 нМ.
SJN 2511 можно добавлять в культуральную среду в концентрации от 20 нМ до 20 мкМ, или от 40 нМ до 10 мкМ, или от 100 нМ до 1 мкМ. Например, SJN 2511 можно добавлять в культуральную среду в концентрации приблизительно 200 нМ.
Таким образом, в некоторых вариантах осуществления ингибитор, который прямо или непрямо отрицательно регулирует передачу сигнала TGF-бета или p38, добавляют в культуральную среду в концентрации от 1 нМ до 100 мкМ, от 10 нМ до 100 мкМ, от 100 нМ до 10 мкМ, или приблизительно 1 мкМ, например, где общая концентрация одного или нескольких ингибиторов составляет от 10 нМ до 100 мкМ, от 100 нМ до 10 мкМ или приблизительно 1 мкМ.
Дополнительно к ингибитору среда для культивирования клеток, как правило, содержит ряд компонентов, которые необходимы для продолжения поддержания и/или увеличения в количестве культивируемых клеток. Среда для культивирования клеток по изобретению, таким образом, обычно содержит множество других компонентов в дополнение к ингибитору согласно изобретению. Подходящие комбинации компонентов могут быть легко составлены квалифицированным специалистом, учитывая представленное ниже описание. Культуральная среда согласно изобретению, как правило, представляет собой питательный раствор, содержащий стандартные компоненты для культивирования клеток, такие как аминокислоты, витамины, неорганические соли, углеродный источник энергии и буфер, как более подробно описано ниже. Другие стандартные компоненты для культивирования клеток, которые могут быть включены в культуру, включают гормоны, такие как прогестерон, белки, такие как альбумин, каталаза, инсулин и трансферрин. Эти другие стандартные компоненты для культивирования клеток составляют "базальную" культуральную среду.
Культуральную среду согласно изобретению можно получать путем модификации существующей среды для культивирования клеток. Квалифицированному специалисту из общих знаний будут понятны типы культуральных сред, которые можно использовать для культуры стволовых клеток. Потенциально подходящие среды для культивирования клеток являются коммерчески доступными и включают, но не ограничиваются ими, модифицированную способом Дульбекко среду Игла (DMEM), минимальную поддерживающую среду (MEM), нокаут-DMEM (KO-DMEM), минимальную поддерживающую среду Глазго (G-MEM), базальную среду Игла (BME), DMEM/среду Хэма F12, расширенную DMEM/среду Хэма F12, модифицированную способом Искова минимальную необходимую среду (MEM), среду Хэма F-10, среду Хэма F-12, среду 199 и среду RPMI 1640. Таким образом, в некоторых вариантах осуществления одну из этих уже существующих сред для культивирования клеток используют в качестве базальной культуральной среды, в которую добавляют ингибитор, который прямо или непрямо отрицательно регулирует передачу сигнала TGF-бета или p38, и необязательно в которую добавляют один или несколько других компонентов, как описано в настоящем описании.
В некоторых вариантах осуществления, культуральная среда по изобретению содержит один или несколько дополнительных компонентов, выбранных из: ингибитора BMP, агониста Wnt, лиганда рецепторной тирозинкиназы, ингибитора Rock, никотинамида и гастрина. В некоторых вариантах осуществления культуральная среда по изобретению содержит любой из R-спондина 1-4 и/или миметика R-спондина, ингибитора TGF-бета, ингибитора BMP (например, Noggin) и агониста Wnt (например, Wnt(3a)).
В некоторых вариантах осуществления культуральная среда по изобретению содержит любой из R-спондина 1-4 и/или миметика R-спондина, ингибитор BMP (например, Noggin), ингибитор TGF-бета, лиганд рецепторной тирозинкиназы (например, EGF), никотинамид, агонист Wnt (например, Wnt(3a)) и необязательно один или несколько дополнительных компонентов, выбранных из: ингибитора p38, гастрина, FGF10, HGF и ингибитора Rock. Можно добавлять необязательные дополнительные компоненты для оптимизации культуральной среды для культивирования клеток, происходящих из конкретных тканей, как более подробно объяснено ниже.
Культуральные среды по изобретению могут содержать один или несколько ингибиторов морфогенетического белка кости (BMP). Лиганды BMP передают сигнал в качестве димеров путем сборки четырехкомпонентного комплекса трансмембранного серин/треонинового киназного рецептора, состоящего из двух рецепторов типа I и двух рецепторов типа II. Сборка комплекса инициирует каскад фосфорилирования, активирующий отвечающие на BMP Smads1/5/8 и приводящий к изменениям в активности транскрипции. Преимущественно авторы настоящего изобретения показали, что ингибиторы BMP стимулируют экспрессию Lgr5, и, таким образом, присутствие ингибитора BMP в культуральной среде по изобретению, вероятно, приведет к более способным к пролиферации органоидам, чем когда ингибитор BMP отсутствует (например, см. пример 3). Таким образом, ингибиторы BMP являются преимущественным компонентом среды для увеличения в количестве по изобретению. Таким образом, применение ингибитора BMP является преимущественным при применении среды для увеличения в количестве, когда желательно культивировать клетки в течение по меньшей мере 3 месяцев (например, по меньшей мере 4, 5, 6, 7, 8 или 9 месяцев) без дифференцировки клеток.
Известно несколько классов природных связывающих BMP белков, включая Noggin (Peprotech), хордин и хордин-подобные белки (R&D sytems), содержащие домены хордина, фоллистатин и фоллистатин-связанные белки (R&D sytems), содержащие домен фоллистатина, DAN и DAN-подобные белки (R&D sytems), содержащие домен цистеинового узла DAN, склеростин/SOST (R&D sytems), декорин (R&D sytems) и альфа-2-макроглобулин (R&D systems). Ингибитор BMP представляет собой средство, которое связывается с молекулой BMP с образованием комплекса, где активность BMP снижена, например, путем предотвращения или ингибирования связывания молекулы BMP с рецептором BMP. Альтернативно ингибитор может представлять собой средство, которое связывается с рецептором BMP и предотвращает связывание лиганда BMP с рецептором, например, антитело, которое связывает рецептор. Ингибитор BMP может представлять собой белок или низкомолекулярное соединение и он может быть встречающимся в природе, модифицированным и/или частично или полностью синтетическим. Ингибитор BMP культуральной среды по изобретению может представлять собой Noggin, DAN, или DAN-подобных белков, включая Cerberus и Gremlin (R&D sytems). Эти диффундирующие белки способны связывать лиганд BMP с различной степенью аффинности и ингибировать их доступ к рецепторам передачи сигнала. Предпочтительным ингибитором BMP для применения в культуральной среде по изобретению является Noggin. Noggin можно использовать в любой подходящей концентрации. В некоторых вариантах осуществления базальная среда для культуральной среды по изобретению может содержать от приблизительно 10 нг/мл до приблизительно 100 нг/мл Noggin. Например, культуральная среда может содержать по меньшей мере 10 нг/мл Noggin, по меньшей мере 20 нг/мл Noggin, по меньшей мере 50 нг/мл Noggin, по меньшей мере 100 нг/мл Noggin, приблизительно 100 нг/мл Noggin или 100 нг/мл Noggin. В некоторых вариантах осуществления культуральная среда может содержать менее чем 200 нг/мл Noggin, менее чем 150 нг/мл Noggin, менее чем 100 нг/мл Noggin, менее чем 75 нг/мл Noggin, менее чем 50 нг/мл Noggin или менее чем 30 нг/мл Noggin. Ингибитор BMP можно добавлять в культуральную среду раз в двое суток в процессе культивирования или каждые сутки в процессе культивирования, или раз в трое суток, раз в четверо суток, раз в пятеро суток или при необходимости. Ингибиторы BMP являются особенно преимущественными компонентами среды для увеличения в количестве, например, для увеличения в количестве стволовых клеток поджелудочной железы, тонкого кишечника, толстого кишечника, печени, предстательной железы. Однако было показано, что Noggin препятствует некоторой дифференцировке (например, см. пример 3). Таким образом, в некоторых вариантах осуществления ингибитор BMP исключен из среды для дифференцировки по изобретению.
В некоторых вариантах осуществления клетки, культивируемые с ингибитором BMP, имеют активированную экспрессию Lgr5 по сравнению с клетками, культивируемыми без ингибитора BMP. Таким образом, добавление ингибитора BMP, как правило, приводит к более способным к пролиферации органоидам. Это является неожиданным, поскольку в литературе описано, что активность BMP пригодна для дифференцировки клеток поджелудочной железы как в протоковые (см. экспрессию кератина 7 и 19), так и в эндокринные клетки. Таким образом, квалифицированный специалист может предполагать, что включение ингибитора BMP, такого как Noggin, снижает пролиферацию и увеличивает дифференцировку. Однако авторы изобретения неожиданно открыли, что применение ингибитора BMP является преимущественным, поскольку оно приводило к более способным к пролиферации органоидам и к более высокой экспрессии Lgr5. Культуральные среды по изобретению могут содержать один или несколько агонистов Wnt. Каскад передачи сигнала Wnt определяется серией событий, которые происходят, когда белок Wnt связывается с рецептором клеточной поверхности, представителем семейства рецепторов Frizzled. Это приводит к активации белков семейства Dishevelled, которые ингибируют комплекс белков, которые включают Axin, GSK-3 и белок APC, деградирующих внеклеточный бета-катенин. В результате, увеличенное содержание бета-катенина усиливает транскрипцию с помощью факторов транскрипции семейства TCF/LEF. Агонист Wnt определяют как средство, которое активирует опосредуемую TCF/LEF транскрипцию в клетке. Таким образом, агонисты Wnt выбирают из истинных агонистов Wnt, которые связывают и активируют представителя семейства рецепторов Frizzled, включая любые и все из белков семейства Wnt, ингибитора внутриклеточной деградации бета-катенина и активаторов TCF/LEF. Указанный агонист Wnt стимулирует активность Wnt в клетке по меньшей мере на 10%, более предпочтительно, по меньшей мере на 20%, более предпочтительно, по меньшей мере на 30%, более предпочтительно, по меньшей мере на 50%, более предпочтительно, по меньшей мере на 70%, более предпочтительно, по меньшей мере на 90%, более предпочтительно, по меньшей мере на 100%, относительно уровня указанной активности Wnt в отсутствие указанной молекулы. Как известно специалисту в данной области, активность Wnt можно определять путем измерения транскрипционной активности Wnt, например, посредством люциферазных репортерных конструкций pTOPFLASH и pFOPFLASH Tcf (Korinek et al., 1997. Science 275: 1784-1787).
В некоторых вариантах осуществления агонист Wnt содержит секретируемый гликопротеин, включающий Wnt-1/Int-1; Wnt-2/Irp (InM-связанный белок); Wnt-2b/13; Wnt-3/Int-4; Wnt-3a (R&D sytems); Wnt-4; Wnt-5a; Wnt-5b; Wnt-6 (Kirikoshi H et al. 2001. Biochem Biophys Res Com 283: 798-805), Wnt-7a (R&D sytems); Wnt-7b, Wnt-8a/8d, Wnt-8b, Wnt-9a/14, Wnt-9b/14b/15; Wnt-10a, Wnt-10b/12, WnM 11 и Wnt-16. Обзор белков Wnt человека представлен в "THE WNT FAMILY OF SECRETED PROTEINS", R&D Systems Catalog, 2004. Следующие агонисты Wnt включают семейство секретируемых белков R-спондинов, которое вовлечено в активацию и регуляцию каскада передачи сигнала Wnt и которое состоит из 4 представителей (R-спондин 1 (NU206, Nuvelo, San Carlos, CA), R-спондин 2 ((R&D sytems), R-спондин 3 и R-спондин-4), и Norrin (также называемым белком болезни Норри или NDP) (R&D sytems), который представляет собой секретируемый регуляторный белок, который функционирует как белок Wnt в том, что он связывается с высокой аффинностью с рецептором Frizzled-4 и индуцирует активацию каскада передачи сигнала Wnt (Kestutis Planutis et al. (2007) BMC Cell Biol 8: 12). В некоторых вариантах осуществления один или несколько агонистов Wnt для применения в рамках изобретения представляет собой миметик R-спондина, например, агонист Lgr5, такой как антитело против Lgr5. Недавно был идентифицирован низкомолекулярный агонист каскада передачи сигнала Wnt, производное аминопиримидина, и он также прямо включен в качестве агониста Wnt (Lm et al. (2005) Angew Chem Int Ed Engl. 44, 1987-90).
В некоторых вариантах осуществления агонист Wnt представляет собой ингибитор GSK. Известные ингибиторы GSK включают малые интерферирующие РНК (siRNA, Cell Signaling), литий (Sigma), кенпауллон (Biomol International, Leost, M et al. (2000) Eur J Biochem 267, 5983-5994), 6-броминдирубин-30-ацетоксим (Meyer, L et al. (2003) Chem Biol 10, 1255-1266), SB 216763 и SB 415286 (Sigma-Aldrich) и представители семейства FRAT и происходящие из FRAT пептиды, которые препятствуют взаимодействию GSK-3 с Axin. Обзор предоставлен в Meijer et al., (2004) Trends in Pharmacological Sciences 25, 471-480, который включен в настоящий документ в качестве ссылки. Способы и анализы для определения уровня ингибирования GSK-3 известны специалисту в данной области и включают, например, способы и анализ, описанные Liao et al 2004, Endocrinology, 145(6): 2941-2949.
В некоторых вариантах осуществления агонист Wnt представляет собой ингибитор RNF43 или ZNRF3. Авторы изобретения открыли, что Rnf43 и ZNRF3 располагаются в клеточной мембране и отрицательно регулируют уровни рецепторного комплекса Wnt в мембране, возможно посредством убиквитинилирования Frizzled. Таким образом, авторы изобретения полагают, что ингибирование RNF43 или ZNRF3 посредством антагонистических антител, РНК-i или низкомолекулярных ингибиторов может непрямо стимулировать каскад Wnt. Rnf43 и ZNRF3 имеют каталитический кольцевой домен (с активностью убиквитинилирования), на который можно осуществлять нацеливание с помощью низкомолекулярного соединения. Несколько антител против Rnf43 и несколько антител против ZNRF3 являются коммерчески доступными. В некоторых вариантах осуществления такие антитела являются подходящими агонистами Wnt в контексте изобретения.
В некоторых вариантах осуществления указанный агонист Wnt выбран из группы, состоящей из Wnt-3a, ингибитора GSK (такого как CHIR99021), Wnt 5, Wnt-6a, Norrin и любого другого белка семейства Wnt.
В некоторых вариантах осуществления указанный агонист Wnt содержит или состоит из любого из R-спондина 1, R-спондина 2, R-спондина 3 или R-спондина 4. В предпочтительном варианте осуществления указанный агонист Wnt выбирают из одного или нескольких представителей семейства Wnt, R-спондинов 1-4, Norrin и ингибитора GSK. В некоторых вариантах осуществления указанный агонист Wnt представляет собой ингибитор GSK-3, такой как CHIR99021 (Stemgent 04-0004). В некоторых вариантах осуществления CHIR99021 добавляют в культуральную среду до конечной концентрации от 50 нМ до 100 мкМ, например, от 100 нМ до 50 мкМ, от 1 мкМ до 10 мкМ, от 1 мкМ до 5 мкМ, или 3 мкМ. В некоторых вариантах осуществления, в которых используют ингибитор GSK-3, ингибитор GSK-3 не является BIO (6-броминдирубин-3’-оксим, Stemgent 04-0003). Авторами настоящего изобретения было открыто, что добавление по меньшей мере одного агониста Wnt к базальной культуральной среде является необходимым для пролиферации эпителиальных стволовых клеток или выделенных крипт.
В следующем предпочтительном варианте осуществления указанный агонист Wnt содержит R-спондин 1 или R-спондин 4, или состоит из него. R-спондин 1, R-спондин 2, R-спондин 3 или R-спондин 4, предпочтительно, добавляют в базальную культуральную среду в концентрации по меньшей мере 50 нг/мл, более предпочтительно, по меньшей мере 100 нг/мл, более предпочтительно, по меньшей мере 200 нг/мл, более предпочтительно, по меньшей мере 300 нг/мл, более предпочтительно, по меньшей мере 500 нг/мл. Наиболее предпочтительная концентрация R-спондина 1, R-спондина 2, R-спондина 3 или R-спондина 4 составляет приблизительно 500 нг/мл или 500 нг/мл. В некоторых вариантах осуществления R-спондин 1, R-спондин 2, R-спондин 3 или R-спондин 4 добавляют в культуральную среду в концентрации по меньшей мере 500 нг/мл, по меньшей мере 600 нг/мл, по меньшей мере 700 нг/мл, по меньшей мере 800 нг/мл, по меньшей мере 900 нг/мл, по меньшей мере 1 мкг/мл, по меньшей мере 1,5 мкг/мл или по меньшей мере 2 мкг/мл. В другом предпочтительном варианте осуществления R-спондин 1, R-спондин 2, R-спондин 3 или R-спондин 4 добавляют в культуральную среду в концентрации приблизительно 1 мкг/мл или 1 мкг/мл. В некоторых вариантах осуществления R-спондин 1, R-спондин 2, R-спондин 3 или R-спондин 4 добавляют в базальную культуральную среду в концентрации менее 1000 нг/мл, например, менее 800 нг/мл, менее 600 нг/мл, менее 550 нг/мл, менее 500 нг/мл, менее 400 нг/мл, менее 300 нг/мл или менее 200 нг/мл или менее 100 нг/мл. В некоторых вариантах осуществления в среду добавляют два или более (например, 2, 3 или 4) из R-спондина 1, R-спондина 2, R-спондина 3 и R-спондина 4 ("R-спондин 1-4"). Предпочтительно, когда добавляют два или более из R-спондинов 1-4, общая концентрация R-спондинов достигает концентраций, описанных выше. Когда указано, что культуральные среды, описанные в настоящем описании, содержат "R-спондин 1-4", подразумевают, что среда содержит любой один или несколько из R-спондина 1, R-спондина 2, R-спондина 3 и R-спондина 4. Когда указано, что культуральные среды, описанные в настоящем описании, содержат "R-спондин", подразумевают, что среда содержит любой один или несколько из R-спондина 1, R-спондина 2, R-спондина 3, R-спондина 4 и миметика R-спондина.
В процессе культивирования стволовых клеток указанный представитель семейства Wnt, предпочтительно, добавляют в культуральную среду раз в двое суток, а культуральную среду, предпочтительно, обновляют раз в четверо суток.
В предпочтительном варианте осуществления агонист Wnt выбран из группы, состоящей из R-спондина, Wnt-3a и Wnt-6. Более предпочтительно, в качестве агониста Wnt используют как R-спондин, так и Wnt-3a. Эта комбинация является особенно предпочтительной, поскольку эта комбинация неожиданно обладает синергическим эффектом на образование органоида. Предпочтительные концентрации составляют приблизительно 500 нг/мл или 500 нг/мл для R-спондина и приблизительно 100 нг/мл или 100 нг/мл для Wnt3a.
В предпочтительном варианте осуществления агонист Wnt выбран из группы, состоящей из R-спондина, Wnt-3a и Wnt-6. Более предпочтительно, оба из R-спондина и Wnt-3a используют в качестве агониста Wnt. Эта комбинация является особенно предпочтительной, поскольку эта комбинация неожиданно имеет синергический эффект на образование органоида. Предпочтительные концентрации составляют приблизительно 500 нг/мл или 500 нг/мл для R-спондина и приблизительно 100 нг/мл или 100 нг/мл для Wnt3a.
Культуральные среды по изобретению могут содержать один или несколько лигандов рецепторной тирозинкиназы. Примером лиганда рецепторной тирозинкиназы для применения в рамках изобретения является EGF, который является лигандом для рецепторной тирозинкиназы EGFR. Многие лиганды рецепторных тирозинкиназ также являются митогенными факторами роста.
Культуральные среды по изобретению могут содержать один или ненсколько митогенных факторов роста. Один или несколько митогенных факторов роста могут быть выбраны из семейства факторов роста, включающего эпидермальный фактор роста (EGF, (Peprotech), трансформирующий фактор роста-альфа (TGF-альфа, Peprotech), основный фибробластный фактор роста (bFGF, Peprotech), происходящий из головного мозга нейротрофический фактор (BDNF, R&D Systems) и фактор роста кератиноцитов (KGF, Peprotech). EGF представляет собой сильнодействующий митогенный фактор для множества культивируемых эктодермальных и мезодермальных клеток, и он обладает выраженным эффектом на дифференцировку определенных клеток in vivo и in vitro и некоторых фибробластов в клеточной культуре. Предшественник EGF существует в качестве связанной с мембраной молекулы, которая протеолитически расщепляется с образованием пептидного гормона из 53 аминокислот, который стимулирует клетки. Предпочтительным митогенным фактором роста является EGF. EGF, предпочтительно, добавляют в базальную культуральную среду в концентрации от 5 до 500 нг/мл или по меньшей мере 5 и не более чем 500 нг/мл. Предпочтительная концентрация составляет по меньшей мере 10, 20, 25, 30, 40, 45 или 50 нг/мл и не более чем 500, 450, 400, 350, 300, 250, 200, 150 или 100 нг/мл. Более предпочтительная концентрация составляет по меньшей мере 50 и не более чем 100 нг/мл. Еще более предпочтительная концентрация составляет приблизительно 50 нг/мл или 50 нг/мл. Те же концентрации можно использовать для FGF, предпочтительно, для FGF10 или FGF7. Если используют более одного FGF, например, FGF7 и FGF10, концентрация FGF является такой, как определено выше, и она относится к общей используемой концентрации FGF. В процессе культивирования стволовых клеток, указанный митогенный фактор роста, предпочтительно, добавляют в культуральную среду раз в двое суток, а культуральную среду обновляют, предпочтительно, раз в четверо суток. Можно использовать любой представитель семейства FGF. Предпочтительно, используют FGF7 и/или FGF10. FGF7 также известен как KGF (фактор роста кератиноцитов). В следующем предпочтительном варианте осуществления в базальную культуральную среду добавляют комбинацию митогенных факторов роста, например, таких как EGF и KGF, или EGF и BDNF. В следующем предпочтительном варианте осуществления в базальную культуральную среду добавляют комбинацию митогенных факторов роста, например, таких как EGF и KGF, или EGF и FGF10. Митогенный фактор роста можно добавлять в культуральную среду в концентрации от 5 до 500 нанограмм/мл или по меньшей мере 5 и но не более чем 500 нанограмм/мл, например, по меньшей мере 10, 20, 25, 30, 40, 45 или 50 нг/мл и не более чем 500, 450, 400, 350, 300, 250, 200, 150 или 100 нг/мл. Митогенный фактор роста может быть выбран из группы, состоящей из EGF, TGF-альфа, KGF, FGF7 и FGF. Предпочтительно, митогенный фактор выбран из группы, состоящей из EGF, TGF-альфа и KGF, или из EGF, TGF-альфа и FGF7 или из EGF, TGF-альфа и FGF, или из EGF и KGF, или из EGF и FGF7, или из EGF и FGF, или из TGF-альфа и KGF, или из TGF-альфа и FGF7, или из TGF-альфа и FGF. EGF может быть заменен на TGF-альфа. В некоторых вариантах осуществления митогенный фактор роста представляет собой фактор роста гепатоцитов (HGF). В некоторых вариантах осуществления в культуральную среду добавляют HGF.
В некоторых вариантах осуществления лиганд рецепторной тирозинкиназы представляет собой митогенный фактор роста, например, выбранный из семейства факторов роста, состоящего из эпидермального фактора роста (EGF), трансформирующего фактора роста-альфа (TGF-альфа), основного фибробластного фактора роста (bFGF), происходящего из головного мозга нейротрофического фактора (BDNF), фактора роста гепатоцитов (HGF) и фактора роста кератиноцитов (KGF).
В любую из описанных сред могут быть включены ингибиторы ROCK, такие как Y-27632 (10 мкМ; Sigma), в частности, в течение первых нескольких суток культивирования перед проведением экспериментов по сортировке клеток, поскольку известно, что они препятствуют аноикозу (форма запрограммированной клеточной гибели, которая индуцируется откреплением субстратзависимых клеток от окружающего внеклеточного матрикса). Таким образом, любая из сред, определенных в настоящем описании, дополнительно может содержать ингибитор ROCK в течение первых нескольких суток. В некоторых вариантах осуществления культуральные среды по изобретению дополнительно содержат ингибитор ROCK, такой как Y-27632, например, в течение первых нескольких суток культивирования перед проведением экспериментов по сортировке клеток.
Следующий вариант осуществления способа по изобретению включает культуральную среду, содержащую ингибитор Rock (Rho-киназы). Было выявлено, что добавление ингибитора Rock препятствует аноикису, особенно при культивировании единичных стволовых клеток. Указанный ингибитор Rock, предпочтительно, выбирают из моногидрата дигидрохлорида (R)-(+)-транс-4-(1-аминоэтил)-N-(4-пиридил)циклогексанкарбоксамида (Y-27632, Sigma-Aldrich), 5-(1,4-диазепан-1-илсульфонил)изохинолина (фасудил или HA1077, Cayman Chemical), и дигидрохлорида (S)-(+)-2-метил-1-[(4-метил-5-изохинолинил)сульфонил]-гексагидро-1H-1,4-диазепина (H-1 152, Tocris Bioschience). Указанный ингибитор Rho-киназы, например, Y-27632, предпочтительно, добавляют в культуральную среду раз в двое суток в ходе первых семи суток культивирования указанных стволовых клеток. Ингибитор Rock, предпочтительно, включают в среду в течение первых нескольких суток, например, в течение первых 1, 2, 3, 4, 5, 6 или 7 суток культивирования после посева отдельных клеток или после разделения. Можно использовать любую подходящую концентрацию ингибитора Rock, например, 1-200 мкМ, 1-100 мкМ, 5-50 мкМ или приблизительно 10 мкМ. Предпочтительная концентрация для Y27632 составляет 10 мкМ. Таким образом, в некоторых вариантах осуществления изобретение относится к способу культивирования стволовых клеток и/или способу получения органоида, где ингибитор Rock добавляют в культуральную среду в течение первых 1, 2, 3, 4, 5, 6 или 7 суток, необязательно раз в двое суток. В некоторых вариантах осуществления ингибитор Rock не добавляют в культуральную среду после первых 2, 3, 4, 5, 6, 7, 8, 9 или 10 суток.
Добавление ингибитора Rock является особенно важным при культивировании единичных стволовых клеток (как упоминалось выше), т.е. когда исходным материалом для органоида является единичная стволовая клетка. Таким образом, в некоторых вариантах осуществления изобретение относится к способу получения органоида, где способ включает культивирование стволовых клеток, необязательно единичных стволовых клеток, где ингибитор Rock добавляют в культуральную среду в течение первых 1, 2, 3, 4, 5, 6 или 7 суток, необязательно раз в двое суток и необязательно не добавляют ингибитор Rock в культуральную среду после первых 2, 3, 4, 5, 6, 7, 8, 9 или 10 суток.
Ингибитор Rock является менее важным, и иногда не является необходимым, при культивировании множества клеток, например, когда исходным материалом для органоида является фрагмент ткани. Таким образом, в некоторых вариантах осуществления изобретение относится к способу получения органоида, где способ включает культивирование стволовых клеток, необязательно фрагмента ткани, где ингибитор Rock не добавляют в культуральную среда либо совсем, либо после первых 2, 3, 4, 5, 6, 7, 8, 9 или 10 суток.
После разделения клеток на множество культур, ингибитор Rock можно добавлять в культуральную среду аналогичным образом, что означат в течение первых 1, 2, 3, 4, 5, 6 или 7 суток, необязательно, раз в двое суток, после разделения, в частности, когда разделение вовлекает взятие единичных стволовых клеток из первой культуры и помещение их во вторую культуру. Если разделение вовлекает взятие множества стволовых клеток из первой культуры и помещение их во вторую культуру, тогда добавление ингибитора Rock является менее важным, и иногда необязательным. Таким образом, в некоторых вариантах осуществления, когда способ получения органоидов или культивирования стволовых клеток вовлекает разделение, необязательно где единичная клетка вовлечена в разделение, ингибитор Rock добавляют в новую культуральную среду в течение первых 1, 2, 3, 4, 5, 6 или 7 суток, необязательно раз в двое суток, после разделения. В некоторых вариантах осуществления, где способ получения органоидов или культивирования стволовых клеток вовлекает разделение, необязательно где в разделение вовлечено множество клеток, его не добавляют в культуральную среду либо совсем, либо после первых 2, 3, 4, 5, 6, 7, 8, 9 или 10 суток.
В следующем варианте осуществления способ по изобретению включает культуральную среду, дополнительно содержащую агонист Notch. Было показано, что передача сигнала Notch играет важную роль в определении судьбы клетки, также как и в выживании и пролиферации клетки. Рецепторные белки Notch могут взаимодействовать с рядом связанных с поверхностью или секретируемых лигандов, включая, но не ограничиваясь ими, Delta 1, Jagged 1 и 2, и Delta-подобный 1, Delta-подобный 3, Delta-подобный 4. При связывании лиганда рецепторы Notch активируются серией событий расщепления, вовлекающей представителей семейства протеаз ADAM, а также внутримембранным расщеплением, регулируемым гамма-секретазой пресинилином. В результате происходит транслокация внутриклеточного домена Notch в ядро, где он транскрипционно активирует последующие гены. Предпочтительный агонист Notch выбирают из Jagged 1 и Delta 1, или их активного фрагмента или производного. Наиболее предпочтительным агонистом Notch является пептид DSL (Dontu et al., 2004. Breast Cancer Res 6. R605-R615) с последовательностью CDDYYYGFGCNKFCRPR. Указанный пептид DSL, предпочтительно, используют в концентрации от 10 мкм до 100 нМ или по меньшей мере 10 мкм и не более чем 100 нМ. Добавление агониста Notch, особенно на первой неделе культивирования, увеличивает эффективность культивирования с коэффициентом 2-3. Указанный агонист Notch, предпочтительно, добавляют в культуральную среду раз в двое суток в течение первых семи суток культивирования указанных стволовых клеток. Таким образом, в некоторых вариантах осуществления изобретение относится к способу культивирования стволовых клеток и/или к способу получения органоида, где агонист Notch добавляют в культуральную среду в течение первых 1, 2, 3, 4, 5, 6 или 7 суток, необязательно раз в двое суток. В некоторых вариантах осуществления агонист Notch не добавляют в культуральную среду после первых 2, 3, 4, 5, 6, 7, 8, 9 или 10 суток.
Агонист Notch определяют как молекулу, которая стимулирует активность Notch в клетке по меньшей мере на 10%, более предпочтительно, по меньшей мере на 20%, более предпочтительно, по меньшей мере на 30%, более предпочтительно, по меньшей мере на 50%, более предпочтительно, по меньшей мере на 70%, более предпочтительно, по меньшей мере на 90%, более предпочтительно, по меньшей мере на 100%, относительно уровня активности Notch в отсутствие указанной молекулы. Как известно специалисту в данной области, активность Notch можно определять путем измерения транскрипционной активности Notch, например, с помощью люциферазной репортерной конструкции 4xwtCBF1, как описано (Hsieh et al., 1996. Mol. Cell. Biol. 16, 952-959).
В следующем варианте осуществления среда для культивирования клеток дополнена ингибитором гамма-секретазы, таким как DAPT или DBZ. Ингибиторы гамма-секретазы могут влиять на решения о судьбе клеток в процессе дифференцировки. Например, в некоторых вариантах осуществления ингибиторы гамма-секретазы могут влиять на судьбу клеток в направлении секреторных клеток, таких как бокаловидные клетки. Можно использовать любую подходящую концентрацию ингибитора гамма-секретазы, например, от 1 нМ до 10 мкМ, от 1 нМ до 1 мкМ, от 1 до 100 нМ, или, предпочтительно, от 1 до 20 нМ. Например, ингибитор гамма-секретазы можно добавлять в культуральную среду до конечной концентрации приблизительно 1 нМ.
В следующем варианте осуществления среда для культивирования клеток дополнена гастрином (или подходящей альтернативой, такой как Leu15-гастрин). Гастрин (или подходящую альтернативу) можно добавлять в культуральную среду до конечной концентрации от 1 нМ до 10 мкМ, от 1 нМ до 1 мкМ, от 5 до 100 нМ или, предпочтительно, от 10 до 50 нМ. Например, Leu15-гастрин можно добавлять в культуральную среду до конечной концентрации приблизительно 10 нМ. Гастрин не является необходимым для некоторых культуральных сред по изобретению. Таким образом, в некоторых вариантах осуществления культуральная среда по изобретению не содержит гастрина. В частности, гастрин не требуется для культивирования стволовых клеток кишечника или для получения органоидов кишечника (крипта-ворсинка или крипта толстого кишечника). Однако даже когда гастрин не требуется, его, тем не менее, можно добавлять в культуральную среду без отрицательных эффектов.
В следующем варианте осуществления культуральная среда по изобретению дополнена никотинамидом. Было обнаружено, что добавление никотинамида повышает эффективность культивирования и продолжительность жизни органоидов толстого кишечника человека. Никотинамид можно добавлять в культуральную среду до конечной концентрации от 1 до 100 мМ, от 5 до 50 мМ или, предпочтительно, от 5 до 20 мМ. Например, никотинамид можно добавлять в культуральную среду до конечной концентрации приблизительно 10 мМ.
В предпочтительном варианте осуществления изобретения культуральная среда дополнена никотинамидом и гастрином (или подходящей альтернативой, такой как Leu15-гастрин), где никотинамид и гастрин добавляют в культуральную среду в любой из концентраций, описанных выше.
В некоторых вариантах осуществления культуральная среда дополнена активатором каскада передачи сигнала простагландинов (см. фиг.24, Antagonism of the prostaglandin D2 receptors DP1 and CRTH2 as an approach to treat allergic diseases. Roy Pettipher, Trevor T. Hansel & Richard Armer Nature Reviews Drug Discovery 6, 313-325 (April 2007)). Например, культуральная среда дополнена любым одним или несколькими из соединений, выбранных из перечня, содержащего: фосфолипиды, арахидоновую кислоту (AA), простагландин E2 (PGE2), простагландин G2 (PGG2), простагландин F2 (PGF2), простагландин H2 (PGH2), простагландин D2 (PGD2). Например, в некоторых вариантах осуществления культуральная среда дополнена PGE2 и/или AA. В некоторых вариантах осуществления PGE2 добавляют в среду до конечной концентрации по меньшей мере 10 нМ, например, по меньшей мере 20 нМ, по меньшей мере 30 нМ, по меньшей мере 40 нМ, по меньшей мере 45 нМ, от 10 нМ до 500 нМ, от 10 нМ до 400 нМ, от 10 нМ до 300 нМ, от 10 нМ до 200 нМ, от 10 нМ до 100 нМ, от 20 нМ до 50 нМ. В предпочтительном варианте осуществления PGE2 добавляют в среду до конечной концентрации 50 нМ. В некоторых вариантах осуществления AA добавляют в среду до конечной концентрации по меньшей мере 1 мкг/мл, по меньшей мере 5 мкг/мл, по меньшей мере 8 мкг/мл, по меньшей мере 9 мкг/мл, по меньшей мере 10 мкг/мл, например, от 1 мкг/мл до 1000 мкг/мл, от 1 мкг/мл до 500 мкг/мл, от 1 мкг/мл до 100 мкг/мл, от 1 мкг/мл до 50 мкг/мл, или от 5 мкг/мл до 20 мкг/мл. В предпочтительном варианте осуществления AA добавляют в среду до конечной концентрации 10 мкг/мл. AA и PGE2 являются взаимозаменяемыми в контексте культуральных сред по изобретению. Таким образом, когда указано, что культуральная среда, описанная в настоящем описании, включает PGE2, она альтернативно может включать AA (в соответствующей концентрации) вместо PGE2. Напротив, когда указано, что культуральная среда, описанная в настоящем описании, включает AA, она альтернативно может включать PGE2 (в соответствующей концентрации) вместо AA. Более того, квалифицированному специалисту будет понятно, что когда PGE2 и/или AA включены в культуральную среду по изобретению, культуральная среда может вместо этого содержать одно или несколько из соединений, выбранных из следующего списка, вместо или в дополнение к PGE2 и/или AA: фосфолипиды, простагландин G2 (PGG2), простагландин F2 (PGF2), простагландин H2 (PGH2) и простагландин D2 (PGD2).
В следующем варианте осуществления культуральная среда по изобретению дополнена RANK-лигандом (также обозначаемым в настоящем описании как RANKL). RANK-лиганд может быть подходящим для направления дифференцировки в сторону конкретной клеточной судьбы. Например, когда RANK-лиганд включен в культуральную среду для клеток тонкого кишечника, предпочтительно, в среду для дифференцировки клеток тонкого кишечника, он обеспечивает большую долю клеток, дифференцированных в M-клетки. Таким образом, в некоторых вариантах осуществления изобретение относится к культуральной среде, содержащей RANKL. В частности, изобретение относится к культуральной среде для культивирования, предпочтительно, для дифференцировки, клеток тонкого кишечника, где культуральная среда содержит RANKL. Можно использовать любую подходящую концентрацию RANKL, например, от 10 нг/мл до 1000 нг/мл, от 10 до 500 нг/мл, или от 50 до 100 нг/мл. Например, RANKL можно добавлять в культуральную среду до конечной концентрации приблизительно 100 нг/мл.
Культуральную среду, содержащую EGF, Noggin и R-спондин, обозначают в настоящем описании как "среда ENR". Культуральную среду, содержащую среду ENR и агонист Wnt, такой как Wnt-3a, обозначают в настоящем описании как "среда WENR". В предпочтительном варианте осуществления изобретения культуральная среда содержит среду WENR. В наиболее предпочтительном варианте осуществления изобретения культуральная среда содержит среду WENR, дополненную гастрином и/или никотинамидом (т.е. WENRg или WENR+никотинамид или WENRg+никотинамид).
pH среды может находиться в диапазоне приблизительно от 7,0 до 7,8, в диапазоне приблизительно от 7,2 до 7,6, или приблизительно составляет 7,4. pH можно поддерживать с использованием буфера. Походящий буфер может без труда выбрать специалист в данной области. Буферы, которые можно использовать, включают карбонатные буферы (например, NaHCO3), и фосфаты (например, NaH2PO4). Эти буферы обычно используют в количестве от приблизительно 50 до приблизительно 500 мг/л. Также можно использовать другие буферы, такие как N-[2-гидроксиэтил]пиперазин-N'-[2-этансульфоновая кислота] (HEPES) и 3-[N-морфолино]пропансульфоновая кислота (MOPS), обычно в количестве от приблизительно 1000 до приблизительно 10000 мг/л. Культуральная среда может содержать индикатор pH, такой как феноловый красный, для обеспечения легкого мониторинга pH-статуса среды (например, от приблизительно 5 до приблизительно 50 мг/литр).
Культуральная среда для применения в рамках изобретения может содержать одну или несколько аминокислот. Квалифицированному специалисту понятны соответствующие типы и количества аминокислот для применения в культуральных средах для стволовых клеток. Аминокислоты, которые могут присутствовать, включают L-аланин, L-аргинин, L-аспарагин, L-аспарагиновую кислоту, L-цистеин, L-цистин, L-глутаминовую кислоту, L-глутамин, L-глицин, L-гистидин, L-изолейцин, L-лейцин, L-лизин, L-метионин, L-фенилаланин, L-пролин, L-серин, L-треонин, L-триптофан, L-тирозин, L-валин и их комбинации. Некоторые культуральные среды содержат все из этих аминокислот. Как правило, каждая аминокислота, когда она присутствует, присутствует в количестве от приблизительно 0,001 до приблизительно 1 г/л среды (обычно от приблизительно 0,01 до приблизительно 0,15 г/л), за исключением L-глутамина, который присутствует в количестве от приблизительно 0,05 до приблизительно 1 г/л (обычно от приблизительно 0,1 до приблизительно 0,75 г/л). Аминокислоты могут иметь синтетическое происхождение.
Культуральная среда для применения в рамках изобретения может содержать один или несколько витаминов. Квалифицированному специалисту понятны соответствующие типы и количества витаминов для применения в средах для культивирования стволовых клеток. Витамины, которые могут присутствовать, включают тиамин (витамин B1), рибофлавин (витамин B2), ниацин (витамин B3), D-кальций пантотенат (витамин B5), пиридоксаль/пиридоксамин/пиридоксин (витамин B6), фолиевую кислоту (витамин B9), цианкобаламин (витамин B12), аскорбиновую кислоту (витамин C), кальциферол (витамин D2), DL-альфа токоферол (витамин E), биотин (витамин H) и менадион (витамин K).
Культуральная среда для применения в рамках изобретения может содержать одну или несколько неорганических солей. Квалифицированному специалисту понятны соответствующие типы и количества неорганических солей для применения в средах для культивирования стволовых клеток. Неорганические соли обычно включают в культуральные среды для способствования поддержанию осмотического равновесия клеток и для способствования регуляции мембранного потенциала. Неорганические соли, которые могут присутствовать, включают соли кальция, меди, железа, магния, калия, натрия, цинка. Соли обычно используют в форме хлоридов, фосфатов, сульфатов нитратов и бикарбонатов. Конкретные соли, которые можно использовать, включают CaCl2, CuSO4-5H2O, Fe(NO3)-9H2O, FeSO4-7H2O, MgCl, MgSO4, KCl, NaHCO3, NaCl, Na2HPO4, Na2HPO4-H2O и ZnSO4-7H2O.
Осмолярность среды может находиться в диапазоне от приблизительно 200 до приблизительно 400 мОсм/кг, в диапазоне от приблизительно 290 до приблизительно 350 мОсм/кг, или в диапазоне от приблизительно 280 до приблизительно 310 мОсм/кг. Осмолярность среды может составлять менее чем приблизительно 300 мОсм/кг (например, приблизительно 280 мОсм/кг).
Культуральная среда для применения в рамках изобретения может содержать углеродный источник энергии в форме одного или нескольких сахаров. Квалифицированному специалисту понятны соответствующие типы и количества сахаров для применения в средах для культивирования стволовых клеток. Сахара, которые могут присутствовать, включают глюкозу, галактозу, мальтозу и фруктозу. Предпочтительно, сахаром является глюкоза, в частности, D-глюкоза (декстроза). Углеродный источник энергии обычно присутствует в количестве от приблизительно 1 до приблизительно 10 г/л.
Культуральная среда по изобретению может содержать сыворотку. Можно использовать сыворотку, полученную из любого подходящего источника, включая эмбриональную телячью сыворотку (FBS), сыворотку козы или сыворотку человека. Предпочтительно, используют сыворотку человека. Сыворотку можно использовать в количестве от приблизительно 1% до приблизительно 30% по объему среды, в соответствии с общепринятыми способами.
В других вариантах осуществления культуральная среда по изобретению может содержать заместитель сыворотки. Различные составы заместителей сыворотки являются коммерчески доступными и известны квалифицированному специалисту. Когда используют заместитель сыворотки, его можно использовать в количестве от приблизительно 1% до приблизительно 30% по объему среды, в соответствии с общепринятыми способами.
В других вариантах осуществления культуральная среда по изобретению может быть бессывороточной и/или не содержащей заместителей сыворотки. Бессывороточная среда представляет собой среду, которая не содержит сыворотки животного любого типа. Бессывороточная среда может быть предпочтительной для избежания возможной ксеноконтаминации стволовых клеток. Не содержащая заместителей сыворотки среда представляет собой среду, которая не дополнена каким-либо коммерческим составом заместителя сыворотки.
В предпочтительном варианте осуществления среда для культивирования клеток дополнена очищенным, природным, полусинтетическим и/или синтетическим фактором роста и не содержит неопределенного компонента, такого как эмбриональная бычья сыворотка или эмбриональная телячья сыворотка. Например, добавки, такие как B27 (Invitrogen), N-ацетилцистеин (Sigma) и N2 (Invitrogen) стимулируют пролиферацию некоторых клеток. В некоторых вариантах осуществления среда для культивирования клеток дополнена одной или несколькими из этих добавок, например, одной, любыми двумя или всеми тремя из этих добавок.
В других вариантах осуществления среда для культивирования клеток дополнена эксендином-4. Эксендин-4, пептид из 39 аминокислот, активирует рецепторы GLP-1 (глюкагон-подобный пептид 1), увеличивая внутриклеточный cAMP в ацинарных клетках поджелудочной железы и не имеет эффекта на рецепторы VIP (вазоактивный кишечный пептид).
Культуральная среда для применения в рамках изобретения может содержать один или несколько микроэлементов, таких как ионы бария, брома, кобальта, йода, марганца, хрома, меди, никеля, селена, ванадия, титана, германия, молибдена, силикона, железа, фтора, серебра, рубидия, олова, циркония, кадмия, цинка и/или алюминия.
Среда может содержать восстановитель, такой как бета-меркаптоэтанол, в концентрации приблизительно 0,1 мМ.
Культуральная среда по изобретению может содержать одно или несколько дополнительных средств, таких как питательные вещества или факторы роста, о которых ранее сообщалось, что они улучшают культуру стволовых клеток, такие как холестерин/трансферрин/альбумин/инсулин/прогестерон, путресцин, селенит/другие факторы.
Культуральная среда по изобретению может быть диффундирована во внеклеточный матрикс (ECM). В предпочтительном способе по изобретению, выделенные фрагменты ткани или выделенные эпителиальные стволовые клетки прикреплены в ECM. ECM состоит из различных полисахаридов, воды, эластина и гликопротеинов, где гликопротеины включают коллаген, энтактин (нидоген), фибронектин и ламинин. ECM секретируется клетками соединительной ткани. Известны различные типы ECM, обладающие различным составом, включая различные типы гликопротеинов и/или различную комбинацию гликопротеинов. Указанный ECM может быть предоставлен посредством культивирования продуцирующих ECM клеток, например, таких как фибробластные клетки, в емкости, с последующим удалением этих клеток и добавлением выделенных крипт или эпителиальных стволовых клеток. Примерами клеток, продуцирующих внеклеточный матрикс, являются хондроциты, продуцирующие, главным образом, коллаген и протеогликаны, фибробластные клетки, продуцирующие, главным образом, коллаген типа IV, ламинин, интерстициальные проколлагены и фибронектин, и миофибробласты толстой кишки, продуцирующие, главным образом, коллагены (тип I, III и V), протеогликан хондроитинсульфат, гиалуроновую кислоту, фибронектин и тенасцин-C. Альтернативно указанный ECM предоставляют коммерчески. Примерами коммерчески доступных внеклеточных матриксов являются белки внеклеточного матрикса (Invitrogen) и препараты базальной мембраны из клеток саркомы мыши Engelbreth-Holm-Swarm (EHS) (например, MatrigelTM (BD Biosciences)). Можно использовать синтетический материал внеклеточного матрикса, такой как ProNectin (Sigma Z378666). Если желательно, можно использовать смеси материалов внеклеточного матрикса. Применение ECM для культивирования стволовых клеток повышало длительное выживание стволовых клеток и сохраняло наличие недифференцированных стволовых клеток. В отсутствие ECM культуры стволовых клеток нельзя было культивировать в течение длительных периодов времени, и не наблюдали сохраняющегося наличия недифференцированных стволовых клеток. Кроме того, присутствие ECM позволяло культивирование трехмерных тканевых органоидов, которые не было возможно культивировать в отсутствие ECM. Материал внеклеточного матрикса обычно представляет собой каплю на дне чашки, в которой клетки суспендированы. Как правило, когда матрикс застывает при 37ºC, добавляют среду, и она диффундирует в ECM. Клетки в среде прилипают к ECM путем взаимодействия с его поверхностной структурой, например, путем взаимодействия с интегринами. Для покрытия емкости для культивирования клеток можно использовать раствор фибронектина приблизительно 1 мг/мл (исходный раствор), используемый в количестве приблизительно 1 мкг/см2, или от приблизительно 1 мкг/см2 до приблизительно 250 мкг/см2, или от приблизительно 1 мкг/см2 до приблизительно 150 мкг/см2. В некоторых вариантах осуществления емкость для культивирования клеток покрыта фибронектином в количестве от 8 мкг/см2 до 125 мкг/см2.
Пример ECM для применения в способе по изобретению включает по меньшей мере один гликопротеин, такой как ламинин.
Предпочтительный ECM для применения в способе по изобретению содержит по меньшей мере два различных гликопротеина, таких как два различных типа коллагена или коллаген и ламинин. ECM может представлять собой синтетический внеклеточный матрикс на основе гидрогеля или природный ECM. Следующим предпочтительным предоставляемым ECM является MatrigelTM (BD Biosciences), который содержит ламинин, энтактин и коллаген IV. В некоторых вариантах осуществления внеклеточный матрикс представляет собой содержащий ламинин внеклеточный матрикс, такой как MatrigelTM (BD Biosciences).
В некоторых вариантах осуществления единичная стволовая клетка, популяция клеток или фрагмент ткани погружены в Matrigel, в котором необязательно снижено содержание факторов роста и/или отсутствует феноловый красный.
В некоторых вариантах осуществления культуральную среду помещают сверху ECM. Затем культуральную среду можно удалять и восполнять, если и когда это потребуется. В некоторых вариантах осуществления культуральную среду восполняют каждые 1, 2, 3, 4, 5, 6 или 7 суток. Если компоненты "добавляют" в среду или "удаляют" из среды, тогда в некоторых вариантах осуществления это может означать, что среду саму удаляют из ECM, а затем на ECM помещают новую среду, содержащую "добавленный" компонент или исключающую "удаленный" компонент.
В некоторых вариантах осуществления культуральная среда по изобретению контактирует с внеклеточным матриксом или 3D-матриксом, который имитирует внеклеточный матрикс посредством его взаимодействия с белками клеточной мембраны, такими как интегрины.
В некоторых вариантах осуществления базальная культуральная среда содержит или состоит из расширенной DMEM/F12, дополненной пенициллином/стрептомицином, 10 мМ HEPES, Glutamax, 1× N2, 1× B27 (все от Invitrogen) и 1 мМ N-ацетилцистеином (Sigma)).
Примеры культуральных сред по изобретению
В одном из вариантов осуществления среда для культивирования клеток содержит ингибитор TGF-бета, который связывает и снижает активность ALK5, и ингибитор p38, который связывает и снижает активность p38. Например, в одном из вариантов осуществления среда для культивирования клеток содержит A83-01 и/или SB202190, предпочтительно, A83-01+ SB202190. Неожиданно было обнаружено, что использование A83-01+SB202190 вместе в культуральной среде по изобретению синергично увеличивает количество пассажей органоидов толстого кишечника человека. В одном из вариантов осуществления среда для культивирования клеток содержит WENR+A83-01+SB202190. В одном из вариантов осуществления среда для культивирования клеток содержит WENR+A83-01+SB202190+никотинамид. В одном из вариантов осуществления среда для культивирования клеток содержит WENRg+никотинамид +A83-01+SB202190 (где "g" обозначает гастрин). В одном из вариантов осуществления среда для культивирования клеток содержит WENR+A83-01+никотинамид+FGF10. В одном из вариантов осуществления среда для культивирования клеток содержит WENRg+A83-01+никотинамид+FGF10. В одном из вариантов осуществления среда для культивирования клеток содержит WENRg+A83-01+никотинамид+FGF10+SB202190. В одном из вариантов осуществления среду для культивирования клеток используют для получения органоидов толстого кишечника. Также предусмотрен органоид толстого кишечника, который можно получать путем культивирования эпителиальных клеток с использованием среды для культивирования клеток, как описано для этого варианта осуществления.
Например, в одном из вариантов осуществления среда для культивирования клеток содержит WENRg+A83-01+FGF10, где агонист Wnt представляет собой R-спондин, но не присутствует никакого другого агониста Wnt и не присутствует никотинамид. Например, в некоторых вариантах осуществления среда для культивирования клеток содержит EGF (например, 50 нг/мл), R-спондин (например, 10% или 1 мкг/мл), Noggin (например, 100 нг/мл), FGF10 (например, 100 нг/мл), A8301 (например, 500 нМ) и гастрин (например, 10 мкМ) и необязательно SB202190. Эти компоненты можно добавлять в базальную среду, такую как среда DMEM/F12. В некоторых вариантах осуществления базальная среда далее дополнена любым одним или несколькими (например, 1, 2, 3, 4 или 5) или всеми из компонентов, выбранных из перечня компонентов, включающего: P/S, Glutamax, 10 мМ Hepes, B27, N2 и N-ацетилцистеин. Было обнаружено, что применение такой среды для культивирования клеток является пригодным для получения органоидов поджелудочной железы. Также предусмотрен органоид поджелудочной железы, полученный путем культивирования эпителиальных клеток с использованием среды для культивирования клеток, как описано для этого варианта осуществления. В некоторых вариантах осуществления гастрин или никотинамид, или гастрин и никотинамид исключены из культуральной среды.
Тканеспецифическая культуральная среда по изобретению
Особенно предпочтительные культуральные среды описаны в разделе "Примеры" настоящего описания. Культуральную среду по изобретению можно адаптировать для применения для различных тканей, например, как описано ниже.
Культуральная среда для кишечника
В некоторых вариантах осуществления культуральная среда для крипт тонкого кишечника, таких как крипты тонкого кишечника мыши, содержит или состоит из базальной среды, например, как описано выше, дополнительно содержащей: EGF, такой как EGF мыши; ингибитор BMP, такой как Noggin мыши; и R-спондин, такой как R-спондин 1 или 4 человека. В некоторых вариантах осуществления эта культуральная среда дополнительно содержит ингибитор TGF-бета (такой как A83-01) и/или ингибитор p38 (такой как SB202190). В некоторых вариантах осуществления культуральная среда для крипт толстого кишечника, таких как крипты толстого кишечника мыши, содержит или состоит из базальной среды, например, как описано выше, дополнительно содержащей: агонист Wnt, такой как рекомбинантный Wnt-3A человека или кондиционированная Wnt-3A среда; EGF, такой как EGF мыши; ингибитор BMP, такой как Noggin мыши; и R-спондин, такой как R-спондин-1 или 4 человека. В некоторых вариантах осуществления эта культуральная среда дополнительно содержит ингибитор TGF-бета (такой как A83-01) и/или ингибитор p38 (такой как SB202190).
В некоторых вариантах осуществления культуральная среда для стволовых клеток кишечника человека, крипт тонкого кишечника человека или крипт толстого кишечника человека (также известная как культуральная среда HISC), содержит или состоит из базальной среды, например, как описано выше, дополнительно содержащей: агонист Wnt, такой как рекомбинантный Wnt-3A человека или кондиционированная Wnt-3A среда; EGF; ингибитор BMP, такой как Noggin; R-спондин, такой как R-спондин-1 человека; ингибитор TGF-бета, такой как A83-01; ингибитор p38, такой как SB202190; гастрин и никотинамид. В некоторых вариантах осуществления ингибитор p38 и/или гастрин могут быть исключены из культуральной среды HISC.
В некоторых вариантах осуществления изобретение относится к культуральной среде для культивирования клеток кишечника, содержащей или состоящей из базальной среды, Wnt-3a, EGF, Noggin, любого из R-спондина 1-4, ингибитора TGF-бета, никотинамида и, предпочтительно, ингибитора p38.
В некоторых вариантах осуществления культуральная среда для увеличения в количестве клеток тонкого кишечника или толстого кишечника, например, клеток тонкого кишечника или толстого кишечника человека, содержит или состоит из базальной среды (например, содержащей расширенную DMEM/F12, B27 (50x), n-ацетилцистеин (1 мМ) и глутамин/glutamax), Wnt3A (необязательно кондиционированная среда), любой из R-спондина 1-4 (предпочтительно, 1 мкг/мл), Noggin (предпочтительно, 50-100 нг/мл), никотинамид (предпочтительно, 10 мМ), EGF (предпочтительно, 10-50 нг/мл), гастрин (предпочтительно, 10 нМ), ингибитор TGF-бета, например, A83-01 (предпочтительно, 500 нМ). В следующем варианте осуществления эта культуральная среда дополнительно содержит ингибитор p38, например, SB202190 (предпочтительно, 100 нМ). В следующем варианте осуществления эта культуральная среда дополнительно содержит ингибитор Rock, например, LY2157299.
В некоторых вариантах осуществления изобретение относится к культуральной среде для дифференцировки клеток кишечника, содержащей или состоящей из базальной среды, EGF, Noggin, ингибитора TGF-бета и ингибитора p38.
В некоторых вариантах осуществления культуральная среда для дифференцировки стволовых клеток тонкого кишечника или толстого кишечника, например, стволовых клеток тонкого кишечника или толстого кишечника человека, содержит или состоит из базальной среды (например, содержащей расширенную DMEM/F12, B27 (50x), n-ацетилцистеин (1 мМ) и глутамин/glutamax), Noggin (предпочтительно, 50-100 нг/мл), EGF (предпочтительно, 10-50 нг/мл), гастрина (предпочтительно, 10 нМ), ингибитора TGF-бета, например, A83-01 (предпочтительно, 500 нМ) и ингибитора p38, например, SB202190 (предпочтительно, 100 нМ). В некоторых вариантах осуществления гастрин может быть исключен из этой среды для дифференцировки. В некоторых вариантах осуществления в среду для дифференцировки можно добавлять ингибитор гамма-секретазы (предпочтительно, в концентрации 1 мкМ). Ингибиторы гамма-секретазы могут влиять на решения о судьбе клетки в процессе дифференцировки, например, в направлении секреторных клеток, таких как бокаловидные клетки. В некоторых вариантах осуществления в среду для дифференцировки можно добавлять RANKL (например, в концентрации 100 нг/мл). RANKL может влиять на решения о судьбе клетки в процессе дифференцировки, например, в направлении M-клеток.
Среды для культивирования клеток злокачественной опухоли
В некоторых вариантах осуществления культуральная среда для клеток рака толстого кишечника содержит или состоит из базальной среды, например, как описано выше, дополнительно содержащей: агонист Wnt, такой как рекомбинантный Wnt-3A человека или кондиционированная Wnt-3A среда; EGF; ингибитор BMP, такой как Noggin; R-спондин, такой как R-спондин 1 человека; ингибитор TGF-бета, такой как A83-01; ингибитор p38, такой как SB202190; гастрин и никотинамид.
В одном из вариантов осуществления культуральная среда для карциномы толстого кишечника, например, карциномы толстого кишечника человека, содержит базальную среду (например, содержащую расширенную DMEM/F12, B27 (50x), n-ацетилцистеин (1 мМ), примоцин и/или P/S (антибиотики) (500x) и hepes), R-спондин (необязательно кондиционированная среда) (предпочтительно, 1 мкг/мл), Noggin (предпочтительно, 100 нг/мл), никотинамид (предпочтительно, 10 мМ), EGF (предпочтительно, 50 нг/мл), гастрин (предпочтительно, 50 нМ), ингибитор TGF-бета, например, A83-01 (предпочтительно, 500 нМ), ингибитор p38, такой как SB202190 (предпочтительно, 10 мкМ), необязательно PGE2 (предпочтительно, 10 нМ) и/или ингибитор Rock (предпочтительно, 10 мкМ).
В некоторых вариантах осуществления клетки рака толстого кишечника также можно выращивать в культуральной среде HISC. В некоторых вариантах осуществления клетки рака толстого кишечника можно культивировать в культуральной среде HISC, где один или несколько или все из следующих компонентов исключены из среды: EGF, Noggin, R-спондин, ингибитор TGF-бета и ингибитор p38. Злокачественные клетки могут иметь мутации, которые конститутивно активируют или инактивируют определенные каскады роста. Например, многие типы рака толстого кишечника приводят к конститутивной активации каскада Wnt. В таких случаях для культуральной среды не требуется агонист Wnt. Другие мутации могут позволить не включать в среду другие факторы, как описано выше. В культуральных средах по изобретению также можно выращивать другие эпителиальные злокачественные опухоли (карциномы). В предпочтительном варианте осуществления органоид злокачественной опухоли, полученный из злокачественных стволовых клеток, выращивают в культуральной среде, которая пригодна для роста соответствующего органоида с нормальной тканью, полученного из нормальных стволовых клеток, необязательно с исключением определенных факторов из среды. Например, органоид рака желудка, полученный путем культивирования стволовых клеток рака желудка, можно выращивать в тех же условиях культивирования в качестве нормального органоида желудка, полученного культивированием стволовых клеток желудка, необязательно с исключением определенных факторов из среды. В другом примере органоид рака поджелудочной железы, полученный культивированием стволовых клеток рака поджелудочной железы, можно выращивать в тех же условиях культивирования, что и нормальный органоид поджелудочной железы, полученный культивированием стволовых клеток поджелудочной железы, необязательно с исключением определенных факторов из среды. В другом примере органоид рака предстательной железы, полученный культивированием стволовых клеток рака предстательной железы, можно выращивать в тех же условиях культивирования, что и нормальный органоид предстательной железы, полученный путем культивирования стволовых клеток предстательной железы, необязательно с исключением определенных факторов из среды. В другом примере органоид рака печени, полученный культивированием стволовых клеток рака печени, можно выращивать в тех же условиях культивирования, что и нормальный органоид печени, полученный культивированием стволовых клеток печени, необязательно с исключением определенных факторов из среды. Во многих случаях может быть предпочтительным (или по меньшей мере более удобным) выращивать органоиды злокачественной опухоли в среде для нормальной ткани (без исключения каких-либо факторов). Среда для нормальной ткани должна позволять злокачественным опухолям с любым генетическим фоном расти без исключения каких-либо конкретных злокачественных мутаций.
Таким образом, в некоторых вариантах осуществления изобретение относится к культуральной среде для культивирования злокачественных клеток, например, стволовых клеток злокачественной опухоли, таких как клетки аденокарциномы или карциномы, из представляющего интерес типа ткани, где культуральная среда содержит или состоит из компонентов культуральной среды, используемых для культивирования клеток из соответствующего представляющего интерес незлокачественного типа ткани, необязательно где один или несколько из следующих исключены из среды, которую используют для культивирования незлокачественных клеток представляющего интереса типа ткани: Wnt-3a, EGF, Noggin, R-спондин, ингибитор TGF-бета, ингибитор p38, никотинамид, гастрин, FGF10 и HGF.
Культуральная среда для клеток аденомы
В некоторых вариантах осуществления культуральная среда для аденом кишечника, таких как аденомы кишечника мыши, содержит базальную среду, например, как описано выше, дополнительно содержащую EGF, такой как EGF мыши.
Культуральные среды для желудка
В некоторых вариантах осуществления изобретение относится к культуральной среде для культивирования клеток желудка, содержащей или состоящий из базальной среды, Wnt-3a, EGF, Noggin, любого из R-спондина 1-4, ингибитора TGF-бета, гастрин, никотинамид, FGF-10, и предпочтительно, ингибитор p38.
В некоторых вариантах осуществления культуральная среда для стволовых клеток желудка, например, стволовых клеток желудка человека, содержит или состоит из базальной среды (например, содержащей расширенную DMEM/F12, B27 (50x), n-ацетилцистеин (1 мМ), примоцин и/или P/S (антибиотики) (500x) и глутамин/glutamax), любого из R-спондинов 1-4 (необязательно кондиционированная) (предпочтительно, 1 мкг/мл), Noggin (необязательно кондиционированная среда) (предпочтительно, 100 нг/мл), Wnt3A (необязательно кондиционированная среда), никотинамида (предпочтительно, 5 мМ), EGF (предпочтительно, 50 нг/мл), FGF10 (предпочтительно, 200 нг/мл), гастрина (предпочтительно, 1 нМ), ингибитора TGF-бета, например, A83-01 (предпочтительно, 2 мкМ). Культуральная среда для стволовых клеток желудка необязательно дополнительно содержит ингибитор p38, например, SB202190 (предпочтительно, 10 нМ). Культуральная среда для стволовых клеток желудка необязательно дополнительно содержит PGE2 (предпочтительно, 500 нМ). Культуральная среда для стволовых клеток желудка необязательно дополнительно содержит ингибитор Rock (предпочтительно, 10 мкМ).
В некоторых вариантах осуществления культуральная среда для стволовых клеток желудка, например, клеток желудка мыши, содержит или состоит из базальной среды (например, содержащей расширенную DMEM/F12, B27 (50x), n-ацетилцистеин (1 мМ), примоцин и/или P/S (антибиотики) (500x) и глутамин/glutamax), любой из R-спондина 1-4 (необязательно кондиционированная среда) (предпочтительно, 1 мкг/мл), Noggin (необязательно кондиционированная среда) (предпочтительно, 100 нг/мл), Wnt3A (необязательно кондиционированная среда), EGF (предпочтительно, 50 нг/мл), FGF10 (предпочтительно, 200 нг/мл), гастрина (предпочтительно, 1 нМ) и ингибитора Rock (предпочтительно, 10 мкМ). В некоторых вариантах осуществления эта культуральная среда дополнительно содержит ингибитор TGF-бета (такой как A83-01) и/или ингибитор p38 (такой как SB202190).
Культуральные среды для предстательной железы
В некоторых вариантах осуществления культуральная среда для увеличения в количестве стволовых клеток предстательной железы содержит тестостерон, необязательно дигидротестостерон (также обозначаемый в настоящем описании как DHT). Тестостерон представляет собой стероидный гормон из группы андрогенов. У человека большой процент тестостерона претерпевает 5α-восстановление с образованием более мощного андрогена дигидротестостерона. Тестостерон, дигидротестостерон или миметик тестостерона (например, молекула, которая имитирует активность тестостерона, связывающаяся с рецептором андрогена) можно добавлять в культуральную среду по изобретению. Таким образом, когда используют термин "тестостерон", он всегда может быть заменен терминами "дигидротестостерон" или "миметик тестостерона". Авторы изобретения показали, что добавление тестостерона в культуральную среду для стволовых клеток предстательной железы приводит к увеличению дифференцировки, но также к продолжению увеличения в количестве популяции стволовых клеток (например, см. фиг.41-45). Это является в высокой степени неожиданным, поскольку в литературе описано, что тестостерон играет важную роль в дифференцировке клеток, действуя путем подавления пролиферации и поддержания терминальной дифференцировки (Mirochnik et al. PLoS One, 7(3), e31052, 2012; Niu et al. Oncogene 29, 3593-3604, 2010). Квалифицированный специалист может ожидать, что добавление тестостерона в культуральную среду для клеток предстательной железы может привести к полностью дифференцированным органоидам без дальнейшего потенциала к увеличению в количестве. Это может быть сходным с тем, что наблюдают при дифференцировке органоидов толстого кишечника, поджелудочной железы и печени в среде для дифференцировки. Однако, напротив, авторы настоящего изобретения открыли, что, хотя тестостерон усиливает дифференцировку, он также обеспечивает продолжение увеличения в количестве стволовых клеток. Таким образом, органоиды, выращенные в культуральной среде, содержащей тестостерон, неожиданно содержат стволовые клетки и дифференцированные клетки, т.е. люминальные и базальные клетки.
В некоторых вариантах осуществления культуральная среда для получения органоида предстательной железы содержит базальную среду и тестостерон, необязательно дигидротестостерон и любой из R-спондина 1-4 или миметика R-спондина. В некоторых вариантах осуществления культуральная среда дополнительно содержит ингибитор BMP, например, Noggin. В некоторых вариантах осуществления культуральная среда дополнительно содержит лиганд тирозиновой рецепторной киназы, необязательно где лиганд тирозиновой рецепторной киназы представляет собой митогенный фактор роста, такой как EGF, FGF, KGF или HGF. В некоторых вариантах осуществления культуральная среда для получения органоида предстательной железы содержит EGF, Noggin, или любой из R-спондинов 1-4 и тестостерон.
В предпочтительном варианте осуществления культуральная среда для клеток предстательной железы содержит ингибитор TGF-бета. В некоторых вариантах осуществления культуральная среда для клеток предстательной железы содержит EGF, Noggin, любой из R-спондинов 1-4, ингибитор TGF-бета и тестостерон. В некоторых вариантах осуществления культуральная среда для клеток предстательной железы дополнительно содержит ингибитор p38. В некоторых вариантах осуществления культуральная среда для получения органоида предстательной железы не содержит ингибитора по изобретению, например, ингибитора TGF-бета и/или ингибитора p38. В некоторых вариантах осуществления культуральная среда для стволовых клеток предстательной железы не содержит тестостерона. В некоторых вариантах осуществления культуральная среда содержит базальную среду, EGF, Noggin и любой из R-спондинов 1-4 и необязательно ингибитор TGF-бета, и не содержит тестостерона.
В некоторых вариантах осуществления изобретение относится к культуральной среде для культивирования клеток предстательной железы, содержащей или состоящей из базальной среды, EGF, любого из R-спондинов 1-4, Noggin, никотинамида, ингибитора TGF-бета, и, предпочтительно, Wnt-3a и FGF-10. В некоторых вариантах осуществления культуральная среда для культивирования клеток предстательной железы дополнительно содержит тестостерон, например, (дигидро)тестостерон. В некоторых вариантах осуществления культуральная среда дополнительно содержит ингибитор p38. В некоторых вариантах осуществления культуральная среда для клеток предстательной железы, например, мыши, человека, нормальных или из карциномы, содержит базальную среду (например, содержащую расширенную DMEM/F12, B27 (50x), n-ацетилцистеин (1 мМ) и глутамин/glutamax), любой из R-спондинов 1-4 (необязательно кондиционированная среда) (предпочтительно, 1 мкг/мл), Noggin (необязательно кондиционированная среда) (предпочтительно, 100 нг/мл), никотинамид (предпочтительно, 10 мМ), EGF (предпочтительно, 50 нг/мл), FGF10 (предпочтительно, 100 нг/мл), ингибитор TGF-бета, например, A83-01 (предпочтительно, 500 нМ), (дигидро)тестостерон (предпочтительно, 1 нМ) 10 нМ и необязательно Wnt-3a. В некоторых вариантах осуществления эта культуральная среда дополнительно содержит ингибитор Rock (предпочтительно, 10 мкМ). В некоторых вариантах осуществления культуральная среда для клеток предстательной железы дополнительно содержит ингибитор p38, например, SB202190. В некоторых вариантах осуществления, где культивируют клетки предстательной железы мыши, ингибитор TGF-бета может быть исключен из культуральной среды. В других вариантах осуществления никотинамид, FGF10 и/или ингибитор Rock могут быть исключены из культуральной среды.
Культуральная среда для поджелудочной железы
В некоторых вариантах осуществления изобретение относится к культуральной среде для увеличения в количестве клеток поджелудочной железы, содержащей или состоящей из базальной среды, любого из R-спондина 1-4, Noggin, EGF, FGF10, гастрина, ингибитора TGF-бета и, предпочтительно, эксендина 4 и Wnt-3a.
В некоторых вариантах осуществления культуральная среда для увеличения в количестве стволовых клеток поджелудочной железы, например, стволовых клеток поджелудочной железы человека, содержит или состоит из базальной среды (например, содержащей расширенную DMEM/F12, B27 (50x), n-ацетилцистеин (1 мМ) и глутамин/glutamax), любого из R-спондинов 1-4 (необязательно кондиционированная среда) (предпочтительно, 1 мкг/мл), Noggin (необязательно кондиционированная среда) (предпочтительно, 100 нг/мл), никотинамида (предпочтительно, 10 мМ), EGF (предпочтительно, 50 нг/мл), FGF10 (предпочтительно, 100 нг/мл), гастрина (предпочтительно, 100 нМ) и ингибитора TGF-бета, например, A83-01 (предпочтительно, 2 мкМ). В следующем варианте осуществления эта культуральная среда дополнительно содержит Wnt-3a. В следующем варианте осуществления эта культуральная среда дополнительно содержит ингибитор p38, например, SB202190 (предпочтительно, 100 нМ). В следующем варианте осуществления эта культуральная среда дополнительно содержит ингибитор Rock, например, LY2157299 (предпочтительно, 10 мкМ). В следующем варианте осуществления эта культуральная среда дополнительно содержит эксендин 4 (предпочтительно, 50 нг/мл).
В некоторых вариантах осуществления культуральная среда для увеличения в количестве стволовых клеток поджелудочной железы, например, стволовых клеток поджелудочной железы мыши, содержит или состоит из базальной среды (например, содержащей расширенную DMEM/F12, B27 (50x), n-ацетилцистеин (1 мМ), примоцин и/или P/S (антибиотики), Hepes и глутамин/glutamax), любого из R-спондина 1-4 (необязательно кондиционированная среда) (предпочтительно, 1 мкг/мл), Noggin (необязательно кондиционированная среда) (предпочтительно, 100 нг/мл), никотинамида (предпочтительно, 10 мМ), EGF (предпочтительно, 50 нг/мл), FGF10 (предпочтительно, 100 нг/мл), гастрина (предпочтительно, 100 нМ) и ингибитора TGF-бета, например, A83-01 (предпочтительно, 2 мкМ). В следующем варианте осуществления эта культуральная среда дополнительно содержит ингибитор Rock, например, LY2157299 (предпочтительно, 10 мкМ). В некоторых вариантах осуществления культуральная среда для клеток поджелудочной железы дополнительно содержит ингибитор p38, например, SB202190.
В некоторых вариантах осуществления изобретение относится к культуральной среде для дифференцировки клеток поджелудочной железы, содержащей или состоящий из базальной среды, Noggin, EGF, FGF10, гастрина, ингибитора TGF-бета, ингибитора гамма-секретазы и предпочтительно, эксендина 4.
В некоторых вариантах осуществления культуральная среда для дифференцировки стволовых клеток поджелудочной железы, например, стволовых клеток поджелудочной железы человека, содержит или состоит из базальной среды (например, содержащей расширенную DMEM/F12, B27 (50x), n-ацетилцистеин (1 мМ) и глутамин/glutamax), Noggin (предпочтительно, 100 нг/мл), EGF (предпочтительно, 50 нг/мл), FGF10 (предпочтительно, 10 нМ), гастрина (предпочтительно, 100 нМ), ингибитора TGF-бета, например, A83-01 (предпочтительно, 50 нМ) и ингибитора гамма-секретазы (DAPT/DBZ) (предпочтительно, 10 мкМ). В следующем варианте осуществления эта культуральная среда дополнительно содержит эксендин 4 (предпочтительно, 50 нг/мл). В некоторых вариантах осуществления культуральная среда для клеток поджелудочной железы дополнительно содержит ингибитор p38, например, SB202190.
В некоторых вариантах осуществления культуральная среда для дифференцировки стволовых клеток поджелудочной железы, например, стволовых клеток поджелудочной железы мыши, содержит или состоит из базальной среды (например, содержащей расширенную DMEM/F12, B27 (50x), n-ацетилцистеин (1 мМ) и глутамин/glutamax), EGF (предпочтительно, 50 нг/мл) и ингибитора гамма-секретазы (например, DAPT/DBZ) (предпочтительно, 10 мкМ).
Культуральная среда для пищевода Барретта
В некоторых вариантах осуществления культуральная среда для пищевода Барретта содержит или состоит из базальной среды, например, как описано выше, дополнительно содержащей: агонист Wnt, такой как рекомбинантный Wnt-3A человека или кондиционированная Wnt-3A среда; EGF; ингибитора BMP, такого как Noggin; R-спондина, такого как R-спондин 1 человека; ингибитора TGF-бета, такого как A83-01; ингибитора p38, такого как SB202190; гастрина; никотинамида; и FGF, такого как FGF10 человека (т.е. HISC+FGF). В некоторых вариантах осуществления из этой культуральной среды исключен гастрин.
В некоторых вариантах осуществления изобретение относится к способу получения органоида пищевода Барретта, где способ включает культивирование выделенного эпителия из пищевода Барретта в течение 1, 2, 3, 4, 5, 6, 7 или более суток в культуральной среде HISC, необязательно дополнительно содержащей FGF10; и удаление никотинамида и SB202190 после первых 1, 2, 3, 4 или более суток. В некоторых вариантах осуществления культуральная среда дополнительно содержит ингибитор Notch, такой как DBZ. В некоторых вариантах осуществления органоид пищевода Барретта, культивируемый в присутствии ингибитора Notch, практически не содержит или не содержит пролиферирующих клеток, и содержит больше бокаловидных клеток относительно органоида, культивируемого в отсутствие ингибитора Notch (см. фиг.5).
Культуральная среда для печени
В некоторых вариантах осуществления клетки печени можно выращивать в первой культуральной среде для "увеличения в количестве" (также обозначаемой в настоящем описании как EM), с последующим предпочтительным культивированием клеток во второй культуральной среде для "дифференцировки" (также обозначаемой в настоящем описании как DM). Однако в некоторых вариантах осуществления стадию дифференцировки в среде DM не проводят, например, в некоторых способах клетки трансплантируют и позволяют им дифференцироваться in vivo. Аналогично, существуют культуральные среды для увеличения в количестве и культуральные среды для дифференцировки для других тканей, таких как поджелудочная железа, тонкий кишечник и толстый кишечник (см. выше).
В одном из вариантов осуществления среда для увеличения в количестве клеток печени содержит EGF, агонист Wnt, FGF и никотинамид. Предпочтительно, агонист Wnt представляет собой R-спондин 1-4 (например, любой один или несколько из R-спондинов 1, 2, 3 и 4) и, таким образом, среду для увеличения в количестве обозначают как "ERFNic". Особенно предпочтительная среда для увеличения в количестве дополнительно содержит HGF и ее обозначают как "ERFHNic".
В некоторых вариантах осуществления среда для увеличения в количестве клеток печени дополнена ингибитором TGF-бета. В некоторых вариантах осуществления TGF-бета присутствует в количестве по меньшей мере 5 нМ, например, по меньшей мере 50 нМ, по меньшей мере 100 нМ, по меньшей мере 300 нМ, по меньшей мере 450 нМ, по меньшей мере 475 нМ, например, 5 нМ-500 мМ, 10 нМ-100 мМ, 50 нМ-700 мкМ, 50 нМ-10 мкМ, 100 нМ-1000 нМ, 350-650 нМ или, более предпочтительно, 500 нМ. Присутствие ингибитора TGF-бета в среде для увеличения в количестве является особенно предпочтительным для вариантов осуществления с клетками человека.
В некоторых вариантах осуществления изобретение относится к культуральной среде для увеличения в количестве клеток печени, содержащей или состоящей из базальной среды, любого из R-спондинов 1-4, Noggin, никотинамида, EGF, FGF10, HGF, гастрина, ингибитора TGF-бета и PGE2, и предпочтительно Wnt-3a.
В некоторых вариантах осуществления среда для увеличения в количестве клеток печени дополнительно содержит ингибитор p38.
В некоторых вариантах осуществления среда для увеличения в количестве клеток печени дополнена активатором каскада передачи сигнала простагландинов (также называемым активатором каскада простагландинов) (см. фиг.24). Например, среда для увеличения в количестве клеток печени может быть дополнена одним или несколькими соединениями, выбранными из списка, содержащего: фосфолипиды, арахидоновую кислоту (AA), простагландин E2 (PGE2), простагландин G2 (PGG2), простагландин F2 (PGF2), простагландин H2 (PGH2), простагландин D2 (PGD2). Например, в некоторых вариантах осуществления среда для увеличения в количестве клеток печени дополнена PGE2 и/или AA. В некоторых вариантах осуществления PGE2 добавляют в среду для увеличения в количестве клеток печени до конечной концентрации по меньшей мере 10 нМ, по меньшей мере 30 нМ, по меньшей мере 40 нМ, по меньшей мере 45 нМ, по меньшей мере 50 нМ, например, от 10 нМ до 500 нМ, от 10 нМ до 400 нМ, от 10 нМ до 300 нМ, от 10 нМ до 200 нМ, от 10 нМ до 100 нМ, от 20 нМ до 50 нМ. В предпочтительном варианте осуществления PGE2 добавляют в среду для увеличения в количестве клеток печени до конечной концентрации 50 нМ. В некоторых вариантах осуществления AA добавляют в среду для увеличения в количестве клеток печени до конечной концентрации по меньшей мере 1 мкг/мл, например, по меньшей мере 3 мкг/мл, по меньшей мере 5 мкг/мл, по меньшей мере 8 мкг/мл, по меньшей мере 9 мкг/мл, по меньшей мере 10 мкг/мл, от 1 мкг/мл до 1000 мкг/мл, от 1 мкг/мл до 500 мкг/мл, от 1 мкг/мл до 100 мкг/мл, от 1 мкг/мл до 50 мкг/мл, или от 5 мкг/мл до 10 мкг/мл. В предпочтительном варианте осуществления AA добавляют в среду до конечной концентрации 10 мкг/мл.
В предпочтительном варианте осуществления среда для увеличения в количестве клеток печени дополнена как ингибитором TGF-бета, так и активатором каскада передачи сигналов простагландинов (например, PGE2 и/или AA) и необязательно ингибитором p38.
В предпочтительных вариантах осуществления среда для увеличения в количестве клеток печени дополнительно содержит гастрин.
В одном из вариантов осуществления среда для дифференцировки клеток печени содержит EGF, ингибитор TGF-бета, FGF (например, FGF10, FGF2 или любой другой подходящий представитель семейства FGF) и ингибитор Notch. В одном из вариантов осуществления ингибитор TGF-бета представляет собой A83-01 и/или ингибитор Notch представляет собой DAPT. Эту среду для дифференцировки обозначают в настоящем описании как "EAFD" и она является предпочтительной средой для дифференцировки по изобретению. FGF необязательно может быть заменен на HGF или альтернативно как FGF, так и HGF могут присутствовать или отсутствовать в среде для дифференцировки. В некоторых вариантах осуществления EGF может быть заменен на HGF или другой лиганд рецепторной тирозинкиназы. Также можно добавлять дексаметазон, например, в концентрации от 10 нМ до 10 мкМ. Среда для дифференцировки клеток печени необязательно может включать активатор каскада простагландинов, такой как PGE2 или AA. Однако этот компонент может быть исключен из среды для дифференцировки. В некоторых вариантах осуществления также можно добавлять онкостатин M, например, в диапазоне концентраций от 1 нг/мл до 1 мг/мл, для способствования судьбе дифференцировки в гепатоциты.
В некоторых вариантах осуществления изобретение относится к культуральной среде для дифференцировки клеток печени, содержащей или состоящей из базальной среды, Noggin, EGF, гастрина, ингибитора TGF-бета, ингибитора гамма-секретазы, такого как DAPT или DBZ, и предпочтительно Wnt-3a.
В некоторых вариантах осуществления клетки печени можно первоначально культивировать в среде для увеличения в количестве, которая дополнительно содержит Wnt и Noggin, например, среде "ENRW", содержащей EGF, Noggin, R-спондин и Wnt (например, Wnt-3A), необязательно активатор каскада простагландинов, таких как PGE2 или AA, необязательно ингибитор TGF-бета и необязательно FGF, HGF, никотинамид. В предпочтительном варианте осуществления среда для увеличения в количестве клеток печени дополнена одним или, более предпочтительно, обоими из ингибитора TGF-бета и активатора каскада передачи сигнала простагландинов (например, PGE2 и/или AA).
В некоторых вариантах осуществления среда для увеличения в количестве клеток печени содержит EGF, Noggin, гастрин, FGF, никотинамид, ингибитор TGF-бета, такой как A83-01, HGF, R-спондин 1-4 (например, любой один или несколько из R-спондинов 1, 2, 3 и 4) и PGE2.
В предпочтительном варианте осуществления клетки печени можно сначала культивировать в среде для увеличения в количестве, которая содержит EGF, noggin, гастрин, FGF10, никотинамид, A8301, HGF и любой из R-спондинов 1-4, дополненных PGE2 и/или AA. R-спондин 1-4 может быть предоставлен в форме кондиционированной Rspo среды. Например, среда для увеличения в количестве может содержать EGF (100 нг/мл, Invitrogen); noggin (25 нг/мл, peprotech); гастрин (10 нМ, sigma); FGF10 (например, 100 нг/мл, peprotech); никотинамид (10 мМ, sigma); A8301 (500 нМ, Tocris); HGF (50 нг/мл, peprotech); кондиционированную Rspo среду (10%, например, 1 мкг/мл), дополненную PGE2 (50 нМ) и/или AA (10 мкг/мл). Среда для увеличения в количестве также может содержать ингибитор Rock.
При увеличении в количестве клеток печени один или несколько из следующих компонентов может быть исключен из культуральной среды, описанной выше: ингибитор TGF-бета (например, A83-01) и PGE2.
Авторы изобретения открыли, что эта среда является оптимальной для стимуляции первоначального увеличения в количестве клеток в течение первых нескольких суток. Таким образом, эту первую среду для увеличения в количестве иногда обозначают в настоящем описании как EM1. В некоторых вариантах осуществления Wnt и Noggin удаляют приблизительно через 1 сутки, 2 суток, 3 суток, 4 суток, 5 суток, 6 суток, 7 суток или более, например, 2 недели, 1 месяц, 5 недель, 8 недель, 2 месяца, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 месяцев или более, 14 месяцев или более. В некоторых вариантах осуществления затем клетки можно увеличивать в количестве в среде для увеличения в количестве по изобретению, как описано выше, которая не содержит Wnt или Noggin. Эту вторую среду для увеличения в количестве иногда обозначают в настоящем описании как EM2. В некоторых вариантах осуществления клетки культивируют в EM2 в течение приблизительно 1 суток, 2 суток, 3 суток, 4 суток, 5 суток, 6 суток, 7 суток или более длительного периода времени, такого как 3, 4, 5, 10, 20 или более недель. Затем культуральную среду можно заменять на оптимизированную среду для дифференцировки, как описано выше, которая содержит ингибитор TGF-бета и ингибитор Notch. Как правило, среда для дифференцировки не содержит агонист Wnt, R-спондин или никотинамид. В некоторых вариантах осуществления среда для дифференцировки не содержит активатор каскада простагландинов, такой как PGE2 или AA. Это способствует дифференцировке клеток в направлении зрелых гепатоцитов и холангиоцитов. Эти клетки подходят для трансплантации человеку или животным.
Среда для увеличения в количестве (EM2) клеток печени:
В одном аспекте настоящего изобретения предусмотрена среда для культивирования клеток, которая содержит или состоит из базальной среды для клеток животного или человека, в которую добавляют: эпидермальный фактор роста, FGF, способный связываться с FGFR2 или FGFR4, предпочтительно, FGF10, в качестве митогенного фактора роста, никотинамид, и предпочтительно, агонист Wnt, предпочтительно, R-спондин 1-4. Эту среду обозначают как EM2. Эта среда "EM2" является предпочтительной для увеличения в количестве клеток печени.
В некоторых вариантах осуществления EM2 содержит активатор каскада простагландинов, такой как PGE2 и/или AA.
В некоторых вариантах осуществления EM2 содержит ингибитор TGF-бета, такой как A83-01.
Предпочтительно, агонист Wnt представляет собой R-спондин 1-4. Среду, содержащую EGF, R-спондин 1-4, FGF и никотинамид, обозначают в настоящем описании как ERFNic.
В некоторых вариантах осуществления среда EM2 не содержит noggin и, более предпочтительно, не содержит ингибитор BMP. В некоторых вариантах осуществления среда EM2 не содержит Wnt, например, Wnt-3a.
В некоторых вариантах осуществления HGF присутствует в дополнение к FGF. Предпочтительной средой, содержащей HGF в дополнение к FGF, является ERFHNic (EGF + R-спондин (предпочтительно, R-спондин 1-4) + FGF (предпочтительно, FGF10) + HGF + никотинамид + ингибитор каскада простагландинов, такой как PGE2 и/или AA и ингибитор TGF-бета. Авторы изобретения обнаружили, что среда ERFHNic, содержащая ингибитор TGF-бета и активатор каскада простагландинов, является оптимальной средой для долговременного увеличения в количестве клеток. В отсутствие HGF клетки не оставались жизнеспособными в культуре в течение более чем трех месяцев. Кроме того, в отсутствие HGF после 10 пассажей клетки продемонстрировали недостаточный рост по сравнению с клетками, культивированными в присутствии HGF, о чем свидетельствует низкое соотношение пролиферации. В частности, после 15 пассажей клетки не являлись растущими органоидами с тем же соотношением скоростей в отсутствие HGF, что и в присутствии HGF. Таким образом, было обнаружено, что HGF необходим для поддержания хорошей скорости пролиферации в ходе долговременного культивирования. Таким образом, изобретение относится к применению среды ERFHNic по изобретению для культивирования клеток в течение по меньшей мере 2 недель, по меньшей мере 1 месяца, по меньшей мере 2 месяцев, более предпочтительно, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5, по меньшей мере 6, по меньшей мере 7, по меньшей мере 8, по меньшей мере 9, по меньшей мере 10, по меньшей мере 15, по меньшей мере 20, по меньшей мере 24, по меньшей мере 25, по меньшей мере 30 или более месяцев, например, 3 или более лет. На практике, некоторые варианты осуществления изобретения включают применение EM2 на протяжении приблизительно 20-50 пассажей клеток. Например, клетки можно разделять 4-8 раз один раз в неделю в течение 7-10 недель подряд. Предпочтительно, клетки увеличиваются в количестве со скоростью приблизительно 4-5 раз в неделю или более двух удвоений популяций в неделю. Кроме того, изобретение относится к применению среды ERFHNic по изобретению для культивирования клеток на протяжении по меньшей мере 8 пассажей, например, по меньшей мере 9, по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 15, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 40, по меньшей мере 50, по меньшей мере 60 пассажей или на протяжении 15-35 пассажей, например, приблизительно 20-30 пассажей. В некоторых вариантах осуществления ингибитор TGF-бета, такой как A83-01 дополнительно присутствует в среде EM2. Это является особенно пригодным, когда культивируют клетки или органоиды человека. В некоторых вариантах осуществления A83-01 присутствует в концентрации 400-600 нМ, например, 450-550 нМ, 470-530 нМ или приблизительно 500 нМ. В вариантах осуществления, в которых ингибитор TGF-бета присутствует в EM2, ингибитор Notch, предпочтительно, не присутствует. В некоторых вариантах осуществления EM2 дополнительно содержит ингибитор p38.
Среда для увеличения в количестве (EM1) клеток печени:
В одном аспекте изобретение относится к среде для культивирования клеток, содержащей или состоящей из базальной среды для клеток животного или человека, в которую добавлен EGF, ингибитор BMP, R-спондин Wnt. Предпочтительно, ингибитор BMP представляет собой Noggin и среду EM1 обозначают как "ENRW" (EGF, Noggin, R-спондин и Wnt (например, Wnt3A)). Эту среду обозначают как EM1. В некоторых вариантах осуществления EM1 дополнительно содержит активатор каскада простагландинов, такой как PGE2 и/или AA. В некоторых вариантах осуществления EM1 содержит ингибитор TGF-бета, такой как A83-01. Более предпочтительно, EM1 дополнительно содержит активатор каскада простагландинов и ингибитор TGF-бета. Авторы изобретения открыли, что среда, содержащая Wnt и Noggin, является идеальной для стимуляции первоначального увеличения в количестве клеток. Таким образом, в некоторых вариантах осуществления среду EM1 используют на протяжении только 1 пассажа или 1 недели, однако также предусматривается, что среду EM1 можно использовать в течение приблизительно одного года, поскольку она не является вредоносной для клеток. Таким образом, в некоторых вариантах осуществления среду EM1 используют для культивирования клеток с 0 суток по 10 сутки, например, на сутки 0-7 сутки, 0-6 сутки, 0-5 сутки, 0-4 сутки, 0-3 сутки, 0-2 сутки, 0-1 сутки, где сутки 0 представляют собой сутки, когда клетки выделяют из ткани, из которой они происходят, и 1 сутки являются последующими сутками, или ее используют в течение 1 или нескольких недель, например, 2, 3, 4 или более недель или 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 или более месяцев. В некоторых вариантах осуществления среду EM1 используют только на протяжении первых суток или первых двух суток культивирования. В некоторых вариантах осуществления среду EM1 используют на протяжении 1 или нескольких пассажей, например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30 или более пассажей, например, 20-30 пассажей, 30-35 пассажей, 32-40 пассажей или более. В некоторых вариантах осуществления среду EM1 используют после стадии замораживания или любой другой стадии транспортировки, вовлекающей изменение среды или температуры, которое не сочетается с оптимальным ростом. Эта среда "EM1" является предпочтительной для увеличения в количестве клеток печени.
Среда EM1 дополнена одним или несколькими соединениями, выбранными из группы, состоящей из FGF, HGF, никотинамида, гастрина, B27, N-ацетилцистеина и N2. В случае начала культур из замороженного исходного раствора или из единичной клетки, среду EM1, предпочтительно, дополняют ингибитором ROCK. Y27632 является предпочтительным ингибитором ROCK для применения в рамках изобретения.
Таким образом, в одном из вариантов осуществления предусмотрена среда для культивирования клеток, которая содержит или состоит из базальной среды для клеток животных или человека, в которую добавлены: эпидермальный фактор роста, FGF (например, FGF, способный связываться с FGFR2 или FGFR4), предпочтительно, FGF10 и HGF в качестве митогенных факторов роста,
активатор каскада простагландинов, такой как PGE2 и/или AA;
ингибитор TGF-бета;
гастрин, никотинамид, B27, N2 и N-ацетилцистеин, и предпочтительно;
ингибитор BMP, предпочтительно, Noggin; и
агонист Wnt, предпочтительно, R-спондин 1 и/или Wnt-3a.
Также в среду, определенную выше, добавляют B27 (Invitrogen), N-ацетилцистеин (Sigma) и N2 (Invitrogen), гастрин (Sigma) и никотинамид (Sigma) и полагают, что они контролируют пролиферацию клеток и способствуют стабильности ДНК. В контексте изобретения, никотинамид также обозначают в настоящем описании как "Nic".
"Добавка N2" доступна от Invitrogen, Carlsbad, CA; www.invitrogen.com; каталожный номер № 17502-048; и от PAA Laboratories GmbH, Pasching, Австрия; www.paa.com; каталожный номер № F005-004; Bottenstein & Sato, PNAS, 76(1):514-517, 1979. Добавка N2 поставляется PAA Laboratories GmbH в качестве 100x жидкого концентрата, содержащего 500 мкг/мл трансферрина человека, 500 мкг/мл бычьего инсулина, 0,63 мкг/мл прогестерона, 1611 мкг/мл путресцина и 0,52 мкг/мл селенита натрия. Добавку N2 можно добавлять в культуральную среду в качестве концентрата или разбавлять перед добавлением в культуральную среду. Ее можно использовать в 1x конечной концентрации или при других конечных концентрациях. Использование добавки N2 является удобным способом включения трансферрина, инсулина, прогестерона, путресцина и селенита натрия в культуральную среду по изобретению. В некоторых вариантах осуществления, в которых среда содержит B27, она также не содержит N2. Таким образом, варианты осуществления настоящего изобретения можно адаптировать для исключения N2, когда присутствует B27, если желательно.
"Добавку B27" (доступная от Invitrogen, Carlsbad, CA; www.invitrogen.com; в настоящее время каталожный номер № 17504-044; и от PAA Laboratories GmbH, Pasching, Austria; www.paa.com; каталожный номер № F01-002; Brewer et al., J Neurosci Res., 35(5):567-76, 1993) можно использовать для составления культуральной среды, которая содержит биотин, холестерин, линолеиновую кислоту, линоленовую кислоту, прогестерон, путресцин, ретинол, ретинилацетат, селенит натрия, трийодтиронин (T3), DL-альфатокоферол (витамин E), альбумин, инсулин и трансферрин. Добавка B27 поставляется PAA Laboratories GmbH в качестве жидкого 50x концентрата, содержащего среди прочих ингредиентов биотин, холестерин, линолеиновую кислоту, линоленовую кислоту, прогестерон, путресцин, ретинол, ретинилацетат, селинит натрия, трийодтиронина (T3), DL-альфа токоферол (витамин E), альбумин, инсулин и трансферрин. Среди этих ингредиентов по меньшей мере линоленовая кислота, ретинол, ретинилацетат и трийодтиронин (T3) являются агонистами ядерных рецепторов гормонов. Добавку B27 можно добавлять в культуральную среду в качестве концентрата или разбавлять перед добавлением в культуральную среду. Ее можно использовать в 1x конечной концентрации или в других конечных концентрациях. Применение добавки B27 является удобным способом включения биотина, холестерина, линолеиновой кислоты, линоленовой кислоты, прогестерона, путресцина, ретинола, ретинилацетата, селинита натрия, трийодтиронина (T3), DL-альфа токоферола (витамин E), альбумина, инсулина и трансферрина в культуральную среду по изобретению.
Например, среда для культивирования клеток может содержать или состоять из базальной среды, в которую добавлены: EGF и R-спондин 1, дополненные FGF10, HGF и никотинамидом; например, EGF (50 нг/мл) и R-спондин 1 (1 мкг/мл), дополненные FGF10 (100 нг/мл), HGF (25-50 нг/мл), никотинамид (1-10 мМ), активатор каскада простагландинов, такой как PGE2 (50 нМ) и/или AA (10 мкг/мл) и ингибитор TGF-бета, такой как A83-01 (500 нМ). В некоторых вариантах осуществления среда дополнительно содержит ингибитор p38. Авторы изобретения открыли, что эту среду можно использовать для длительного увеличения в количестве клеток. Таким образом, эта среда для культивирования клеток является предпочтительной для применения в качестве EM2 по изобретению. Базальная среда, предпочтительно, дополнена B27, N2 и 200 нг/мл N-ацетилцистеина. В некоторых вариантах осуществления базальная среда представляет собой расширенную DMEM/F12. Однако можно использовать любую другую подходящую базальную среду.
Другой пример среды для культивирования клеток и способа с использованием этой среды содержит или состоит из расширенной DMEM/F12, предпочтительно, дополненной B27, N2, 200 нг/мл N-ацетилцистеина, 50 нг/мл EGF, 1 мкг/мл R-спондина 1, 10 нМ гастрина, 100 нг/мл FGF10, 10 мМ никотинамида, 50 нг/мл HGF, 50% кондиционированной Wnt среды, активатора каскада простагландинов, такого как PGE2 (50 нМ) и/или AA (10 мкг/мл), и ингибитора TGF-бета, такого как A83-01 (500 нМ) и, предпочтительно, 10-100 нг/мл Noggin. Кондиционированная Wnt среда содержит расширенную DMEM, P/S, B27, N2 и также FCS. Клетки 293T, трансфицированные экспрессирующей Wnt3A плазмидой, продуцируют Wnt. Цельную среду отбирают через несколько суток (т.е. с секретированным Wnt) и используют в качестве источника Wnt.
Таким образом, изобретение относится к среде для культивирования клеток, содержащей или состоящий из базальной среды для клеток животного или человека, в которую добавлены:
эпидермальный фактор роста, FGF, способный связываться с FGFR2 или FGFR4, предпочтительно, FGF10 и HGF в качестве митогенных факторов роста,
активатор каскада простагландинов, такой как PGE2 и/или AA,
ингибитор TGF-бета;
гастрин, никотинамид, B27, N2 и N-ацетилцистеин и предпочтительно
ингибитор BMP, более предпочтительно, Noggin и
агонист Wnt, более предпочтительно, R-спондин 1 и/или Wnt-3a.
Таким образом, изобретение относится к первой предпочтительной культуральной среде, содержащей или состоящей из базальной среды для клеток животного или человека, в которую добавлены:
эпидермальный фактор роста, FGF10 и HGF в качестве митогенных факторов роста,
активатор каскада простагландинов, такой как PGE2 и/или AA,
ингибитор TGF-бета;
гастрин, никотинамид, B27, N2 и N-ацетилцистеин,
ингибитор BMP, более предпочтительно, Noggin, и
агонист Wnt, более предпочтительно, R-спондин 1 и Wnt-3a.
В некоторых вариантах осуществления ингибитор p38 добавляют в среду для увеличения в количестве.
Эту среду можно использовать в качестве среды для культивирования клеток EM1 по изобретению для стимуляции первоначального увеличения в количестве клеток. В некоторых вариантах осуществления среда, используемая в качестве среды для культивирования клеток EM1, содержит все компоненты культуральной среды EM2 по изобретению и дополнительно содержит Wnt-3a и Noggin.
В вариантах осуществления, в которых базальная среда дополнена N-ацетилцистеином, B27 и N2, следующие компоненты, предпочтительно, добавляют в культуральные среды: EGF, R-спондин 1, гастрин, FGF10, никотинамид и HGF и кондиционированную Wnt среду и активатор каскада простагландинов, такой как PGE2 и/или AA. Предпочтительно, базальная среда дополнена N-ацетилцистеином, EGF, R-спондином 1, гастрином, FGF10, никотинамидом и HGF и кондиционированной Wnt средой и активатором каскада простагландинов, таким как PGE2 и/или AA, в соответствии с количествами, описанными в настоящем описании выше. Предпочтительно, ингибитор TGF-бета также присутствует в количествах, описанных в настоящем описании.
Например, в некоторых вариантах осуществления базальная среда может быть дополнена от 150 нг/мл до 250 нг/мл N-ацетилцистеина; предпочтительно, базальная среда дополнена, приблизительно или точно, 200 нг/мл N-ацетилцистеина. Например, в некоторых вариантах осуществления базальная среда может быть дополнена от 40 нг/мл до 60 нг/мл EGF; предпочтительно, базальная среда дополнена, приблизительно или точно, 50 нг/мл EGF. Например, в некоторых вариантах осуществления базальная среда может быть дополнена от 0,5 мкг/мл до 1,5 мкг/мл R-спондина 1; предпочтительно, базальная среда дополнена, приблизительно или точно, 1 мкг/мл R-спондина 1. Например, в некоторых вариантах осуществления базальная среда может быть дополнена от 5 нМ до 15 нМ гастрином; предпочтительно, базальная среда дополнена, приблизительно или точно, 10 нМ гастрином. Например, в некоторых вариантах осуществления базальная среда может быть дополнена 25-200 нг/мл FGF10, например, от 70 нг/мл до 130 нг/мл FGF10; предпочтительно, базальная среда дополнена, приблизительно или точно, 100 нг/мл FGF10. Например, в некоторых вариантах осуществления базальная среда может быть дополнена от 5 мМ до 15 мМ никотинамидом; предпочтительно, базальная среда дополнена, приблизительно или точно, 10 мМ никотинамидом. Например, в некоторых вариантах осуществления базальная среда может быть дополнена от 25 нг/мл до 100 нг/мл HGF, например, от 35 нг/мл до 65 нг/мл HGF; предпочтительно, базальная среда дополнена, приблизительно или точно, 50 нг/мл HGF. Например, в некоторых вариантах осуществления базальная среда может быть дополнена 35%-65% кондиционированной Wnt средой; предпочтительно, базальная среда дополнена, приблизительно или точно, 50% кондиционированной Wnt средой.
Например, в некоторых вариантах осуществления среда для увеличения в количестве клеток печени дополнена активатором каскада передачи сигнала простагландинов (см. фиг.24). Например, среда для увеличения в количестве клеток печени может быть дополнена любым одним или несколькими из соединений, выбранных из перечня, содержащего: фосфолипиды, арахидоновую кислоту (AA), простагландин E2 (PGE2), простагландин G2 (PGG2), простагландин F2 (PGF2), простагландин H2 (PGH2), простагландин D2 (PGD2). Например, в некоторых вариантах осуществления среда для увеличения в количестве клеток печени дополнена PGE2 и/или AA. В некоторых вариантах осуществления PGE2 добавляют в среду для увеличения в количестве клеток печени до конечной концентрации по меньшей мере 10 нМ, например, от 10 нМ до 500 нМ, от 10 нМ до 400 нМ, от 10 нМ до 300 нМ, от 10 нМ до 200 нМ, от 10 нМ до 100 нМ, от 20 нМ до 50 нМ. В предпочтительном варианте осуществления PGE2 добавляют в среду для увеличения в количестве клеток печени до конечной концентрации 50 нМ. В некоторых вариантах осуществления AA добавляют в среду для увеличения в количестве клеток печени до конечной концентрации по меньшей мере 1 мкг/мл, например, от 1 мкг/мл до 1000 мкг/мл, от 1 мкг/мл и 500 мкг/мл, от 1 мкг/мл до 100 мкг/мл, от 1 мкг/мл до 50 мкг/мл, или от 5 мкг/мл до 10 мкг/мл. В предпочтительном варианте осуществления AA добавляют в среду до конечной концентрации 10 мкг/мл.
В некоторых вариантах осуществления один или оба из гастрина и N2 не присутствуют в среде для культивирования клеток.
Предпочтительно, базальная среда представляет собой расширенную DMEM/F12.
Первую культуральную среду (например, EM1, EM2 или обе из EM1 и EM2), предпочтительно, используют на протяжении первых двух недель способа культивирования по изобретению. Однако ее можно использовать в течение более короткого периода времени, такого как в течение 1, 2, 3, 5, 7 или 10 суток, или в течение более длительного периода времени, такого как 3, 4, 5, 10, 20 или более недель, 5 месяцев или более, например, 6, 7, 8, 9, 10, 11, 12 месяцев или более.
Среда для дифференцировки (DM) клеток печени:
В другом аспекте предусмотрена вторая среда для культивирования клеток, которая содержит или состоит из базальной среды для клеток животного или человека, в которую добавлены: EGF, ингибитор TGF-бета, ингибитор Notch и активатор каскада простагландинов, такой как PGE2 и/или AA. Авторы изобретения открыли, что эта среда подходит для дифференцировки клеток. Среда, используемая для дифференцировки клеток, может быть обозначена в настоящем описании как DM.
Предпочтительно, вторая среда для культивирования клеток также содержит FGF и/или HGF.
В одном из вариантов осуществления вторая культуральная среда содержит или состоит из базальной среды для клеток животного или человека, в которую добавлены:
эпидермальный фактор роста, FGF10 и HGF в качестве митогенных факторов роста;
ингибитор Notch;
ингибитор TGF-бета; и
активатор каскада простагландинов, такой как PGE2 и/или AA.
В одном из вариантов осуществления ингибитор TGF-бета представляет собой A83-01 и/или ингибитор Notch представляет собой DAPT. В другом варианте осуществления среда для культивирования клеток DM дополнительно содержит дексаметазон. В другом варианте осуществления среда для культивирования клеток DM дополнительно содержит онкостатин M. В другом варианте осуществления среда для культивирования клеток DM дополнительно содержит гастрин.
Предпочтительная вторая среда для культивирования клеток и способ использования этой среды описаны в примерах, и она содержит или состоит из базальной среды, в которую добавлены: 50 нг/мл EGF, 100 нг/мл FGF10, 50 нМ A8301 и 10 мкМ DAPT. В качестве базальной среды можно использовать расширенную DMEM/F12, а также можно использовать любую другую подходящую базальную среду.
В некоторых вариантах осуществления среда для дифференцировки клеток печени, например, клеток печени человека, содержит или состоит из базальной среды (например, содержащей расширенную DMEM/F12, B27 (50x), n-ацетилцистеин (1 мМ), глутамин/glutamax), Noggin (предпочтительно, 100 нг/мл), EGF (предпочтительно, 50 нг/мл), гастрина (предпочтительно, 10 нМ), ингибитора TGF-бета, такого как A83-01 (предпочтительно, 50 нМ) и ингибитора гамма-секретазы (например, DAPT/DBZ) (предпочтительно, 10 мкМ).
В некоторых вариантах осуществления среда для дифференцировки клеток печени, например, клеток печени мыши, содержит или состоит из базальной среды (например, содержащей расширенную DMEM/F12, B27 (50x), n-ацетилцистеин (предпочтительно, 1 мМ), глутамин/glutamax), EGF (предпочтительно, 50 нг/мл), FGF10 (предпочтительно, 100 нг/мл) гастрина (предпочтительно, 10 нМ), ингибитора TGF-бета, такого как A83-01 (предпочтительно, 50 нМ) и ингибитора гамма-секретазы (например, DAPT/DBZ) (предпочтительно, 10 мкМ).
В некоторых вариантах осуществления вторая среда для культивирования клеток не содержит R-спондина или Wnt. В некоторых вариантах осуществления вторая среда для культивирования клеток не содержит агониста Wnt. В некоторых вариантах осуществления вторая среда для культивирования клеток не содержит никотинамида. В некоторых вариантах осуществления вторая среда для культивирования клеток не содержит ингибитора BMP. В некоторых вариантах осуществления вторая среда для культивирования клеток не содержит активатора каскада простагландинов, такого как PGE2 и/или AA.
Авторы изобретения обнаружили, что оба из R-спондина 1 и никотинамида ингибируют экспрессию маркера зрелых гепатоцитов CYP3A11 и, тем не менее, стимулируют экспрессию маркера гепатобластов альбумина. Таким образом, для увеличения дифференцировки клеток в более зрелые клетки печени, авторы изобретения удалили R-спондин и никотинамид из клеточной культуры. Авторы изобретения также открыли, что экспрессия специфических билиарных факторов транскрипции в высокой степени активирована в культурах для увеличения в количестве, содержащих R-спондин 1, что указывает на то, что экспрессия генов в культуре является несбалансированой в большей степени в сторону билиарных клеток. Каскады передачи сигнала Notch и TGF-бета вовлечены в судьбу билиарных клеток in vivo. Действительно, делеция Rbpj (необходимого для достижения активной передачи сигнала Notch) приводит к аномальному тубулогенезу (Zong Y. Development 2009) и добавление TGF-бета к эксплантатам печени способствует билиарной дифференцировке in vitro (Clotman F. Genes and Development 2005). Поскольку каскады как Notch, так и TGF-бета были в высокой степени активированы в культурах печени (фиг.22) авторы изобретения обоснованно предположили, что ингибирование судьбы клеток желчных протоков может запускать дифференцировку клеток в направлении более гепатоцитарного фенотипа. Было выявлено, что добавление ингибитора TGF-бета (такого как A8301) и ингибитора Notch (такого как DAPT) в среду для дифференцировки, которая, предпочтительно, не содержит R-спондина или Wnt, повышает экспрессию маркеров зрелых гепатоцитов и увеличивает количество подобных гепатоцитам клеток (например, см. пример 5).
Общая культуральная среда
Среда для культивирования клеток согласно изобретению обеспечивает выживание и/или пролиферацию и/или дифференцировку эпителиальных стволовых клеток или выделенных крипт на внеклеточном матриксе. В некоторых вариантах осуществления среда для культивирования клеток согласно изобретению позволяет выживание и/или пролиферацию и/или дифференцировку органоида по изобретению, такого как органоид крипта-ворсинка, органоид толстого кишечника, органоид поджелудочной железы, органоид желудка, органоид пищевода Барретта, органоид аденокарциномы или органоид карциномы толстого кишечника, на внеклеточном матриксе. В некоторых вариантах осуществления среда для культивирования клеток согласно изобретению позволяет выживание и/или пролиферацию и/или дифференцировку органоида по изобретению, такого как органоид тонкого кишечника (крипта-ворсинка), органоид толстого кишечника, органоид поджелудочной железы, органоид желудка, органоид пищевода Барретта, органоид аденокарциномы, органоид карциномы, органоид карциномы толстого кишечника, органоид предстательной железы или органоид карциномы предстательной железы, на внеклеточном матриксе. Предпочтительно, в вариантах осуществления, в которых присутствует ингибитор TGF-бета и/или ингибитор p38, среда для культивирования клеток позволяет выживание и/или пролиферацию, предпочтительно, выживание и пролиферацию популяции клеток или органоида по изобретению. Предпочтительно, варианты осуществления, в которых ингибитор TGF-бета и/или ингибитор p38 первоначально присутствует в среде для культивирования клеток, но впоследствии удален из среды (например, вследствие того, что его не добавляли, когда среду обновляли), позволяют выживание и/или дифференцировку, предпочтительно выживание и дифференцировку популяции клеток или органоида по изобретению.
В некоторых вариантах осуществления ингибитор p38 добавляют в любую из сред, описанных в настоящем описании.
Термин "среда для культивирования клеток" является синонимичным для среды, культуральной среды или клеточной среды.
Использование культуральных сред по изобретению
Также изобретение относится к использованию культуральной среды по изобретению для увеличения в количестве и/или дифференцировки стволовой клетки, популяции стволовых клеток, фрагмента ткани или органоида.
В некоторых вариантах осуществления стволовая клетка, популяция стволовых клеток, фрагмент ткани или органоид выбраны из группы, состоящей из одной или нескольких стволовых клеток кишечника, крипт тонкого кишечника, крипт толстого кишечника, стволовых клеток желудка, стволовых клеток печени, стволовых клеток поджелудочной железы и стволовых клеток предстательной железы.
В некоторых вариантах осуществления стволовую клетку, популяцию стволовых клеток, фрагмент ткани или органоид можно получать из нормальной ткани.
В некоторых вариантах осуществления стволовую клетку, популяцию стволовых клеток, фрагмент ткани или органоид можно получать из пораженной заболеванием ткани, например, из аденомы, карциномы, аденокарциномы, кишечника пациента, имеющего кистозный фиброз, или кишечника пациента, имеющего воспалительное заболевание кишечника.
Стволовые клетки, культивируемые согласно изобретению
Стволовые клетки находятся во многих органах взрослых людей и мышей. Хотя может существовать значительное варьирование в точных характеристиках взрослых стволовых клеток в отдельных тканях, взрослые стволовые клетки обладают по меньшей мере следующими общими характеристиками: они сохраняют недифференцированный фенотип; их потомство может дифференцироваться в направлении всех ростков, присутствующих в соответствующей ткани; они сохраняют способность к самоподдержанию на протяжении жизни; и они способны регенерировать соответствующую ткань после повреждения. Стволовые клетки располагаются в специализированной области, нише стволовых клеток, которая обеспечивает соответствующие контакты клетка-клетка и сигналы для поддержания указанной популяции стволовых клеток. Стволовые клетки согласно изобретению, предпочтительно, экспрессируют Lgr5.
В одном из вариантов осуществления изобретение относится к популяции клеток или одному или нескольким органоидам, содержащим указанные стволовые клетки, которые получены или можно получать культивированием стволовых клеток или фрагментов тканей согласно изобретению, которые культивируют в течение по меньшей мере 3 месяцев, предпочтительно, по меньшей мере 4 месяцев, по меньшей мере 5 месяцев, по меньшей мере 6 месяцев, по меньшей мере 7 месяцев, по меньшей мере 9 месяцев, или по меньшей мере 12 месяцев или более.
"Популяция" клеток представляет собой любое количество клеток, превышающее 1, однако предпочтительно, она составляет по меньшей мере 1×103 клеток, по меньшей мере 1×104 клеток, по меньшей мере 1×105 клеток, по меньшей мере 1×106 клеток, по меньшей мере 1×107 клеток, по меньшей мере 1×108 клеток или по меньшей мере 1×109 клеток.
Стволовые клетки по изобретению, культивируемые согласно изобретению, могут представлять собой стволовые клетки человека. Стволовые клетки по изобретению, культивируемые согласно изобретению, могут представлять собой эпителиальные стволовые клетки.
В некоторых вариантах осуществления стволовые клетки по изобретению и/или культивируемые согласно изобретению, не являются эмбриональными стволовыми клетками. В некоторых вариантах осуществления стволовые клетки по изобретению и/или культивируемые согласно изобретению не являются эмбриональными стволовыми клетками человека. Предпочтительно, стволовые клетки по изобретению представляют собой взрослые стволовые клетки.
В предпочтительном варианте осуществления стволовые клетки могут представлять собой эпителиальные стволовые клетки человека. Эпителиальные стволовые клетки человека включают стволовые клетки, происходящие из эпителиальной ткани человека. Они включают, но не ограничиваются ими, ткани поджелудочной железы, тонкого кишечника, толстого кишечника, роговицы, ткани органов обоняния, респираторные ткани, ткани желудка, печени и кожи, ткани молочной железы и/или предстательной железы, например, ткань, выбранную из группы, состоящей из тканей поджелудочной железы, тонкого кишечника, толстого кишечника, роговицы, органов обоняния и респираторных тканей. Эпителиальные стволовые клетки способны образовывать различные типы клеток, из которых состоит эпителий. Некоторые типы эпителия, такие как эпителий кожи и кишечника, проявляют быстрое обновление клеток, что указывает на то, что находящиеся в них стволовые клетки должны непрерывно пролиферировать. Другие типы эпителия, такие как эпителий печени и поджелудочной железы, проявляют очень медленное обновление в нормальных условиях.
Стволовые клетки кишечника
Самообновляющийся эпителий тонкого кишечника упорядочен в крипты и ворсинки (Gregoreff and Clevers, 2005 Genes Dev 19, 877-90). Каждая клетка вдоль оси крипта-ворсинка является полярной, причем клетки на вершине кишечных ворсинок или в верхних положениях крипт толстого кишечника являются наиболее дифференцированными и постоянно утрачиваются в просвет вследствие апоптоза. Непрерывная пролиферация стволовых клеток, находящихся в основании крипт, и массивная пролиферация клеток-предшественников, находящихся в середине крипт, обеспечивает надлежащую замену утрачиваемых клеток. У мыши конечное время обновления эпителия составляет 5 суток. Давно известно, что самообновляющиеся стволовые клетки располагаются вблизи основания крипты и образуют быстро пролиферирующие переходные делящиеся (TA) клетки, способные дифференцироваться во все ростки. Оцененное количество стволовых клеток составляет от 4 до 6 на крипту (Bjerknes and Cheng, 1999. Gastroenterology 116, 7-14). Из TA-клеток образуется три типа дифференцированных клеток: энтероциты, бокаловидные клетки и энтероэндокринные клетки, и они продолжают их миграцию в последовательно расположенные слои вдоль оси крипта-ворсинка. В каждую ворсинку поступают клетки из множества различных крипт. Четвертый из основных типов дифференцированных клеток, клетки Панета, находится в основании крипты.
Толстый кишечник напоминает тонкий кишечник, однако с плоским поверхностным эпителием вместо ворсинок. Крипты толстого кишечника организованы подобно криптам тонкого кишечника. Клетки Панета не присутствуют в криптах толстого кишечника; вместо этого существуют так называемые "глубокие секреторные клетки крипт". Плоская поверхность эпителия содержит дифференцированные клетки (колоноциты и секреторные клетки). На протяжении крипты встречаются дифференцированные бокаловидные клетки, также смешанные с переходными делящимися клетками.
Выделение фрагментов тканей и стволовых клеток
Крипты можно выделять из тонкого и толстого кишечника, включая двенадцатиперстную кишку, тощую кишку, подвздошную кишку и толстую кишку, и пилорической области и области тела желудка, с помощью протоколов, которые известны специалисту в данной области. Например, крипты можно выделять путем инкубации выделенной ткани с хелатирующими агентами, которые освобождают клетки от их кальций- и магний-зависимых взаимодействий с базальной мембраной и стромальными типами клеток. После промывания ткани слой эпителиальных клеток счищают с подслизистой оболочки с помощью предметного стекла и измельчают. После этого проводят инкубацию в трипсине или, более предпочтительно, EDTA и/или EGTA и отделение нерасщепленных фрагментов ткани и отдельных клеток от крипт с использованием, например, стадий фильтрации и/или центрифугирования. Вместо трипсина можно использовать другие протеолитические ферменты, такие как коллагеназа и/или диспаза I. Сходные способы используют для выделения фрагментов поджелудочной железы и желудка. Сходные способы можно использовать для выделения фрагментов других тканей, описанных в настоящем описании. Культуральные среды по изобретению подходят для культивирования таких фрагментов тканей (см. пример 1).
Культуральная среда согласно изобретению позволяет установление условий долговременного культивирования, при которых единичные крипты претерпевают множество событий деления крипт при одновременном формировании эпителиальных доменов, подобных ворсинкам, в которых присутствуют все дифференцированные типы клеток. Культивируемые крипты претерпевают значительные морфологические изменения после извлечения их из культуры. Верхнее отверстие свежевыделенных крипт закрывается, и эта область постепенно расширяется и заполняется апоптотическими клетками, подобно тому, как апоптотические клетки отделяются на вершине ворсинки. Область крипты претерпевает непрерывное почкование, которое создает дополнительные крипты, и этот процесс напоминает деление крипт. В предпочтительном варианте осуществления изобретения органоиды содержат подобные криптам расширения, которые содержат все типы дифференцированных эпителиальных клеток, включая пролиферативные клетки, клетки Панета, энтероциты и бокаловидные клетки. В органоидах не было идентифицировано миофибробластов или других неэпителиальных клеток ни на одной из стадий.
Расширение почкующихся структур крипты обеспечивает органоиды, содержащие подобные криптам структуры, окружающие центральный просвет, выстилаемый эпителием, подобным эпителию ворсинок, и заполненный апоптотическими клеточными тельцами. Органоиды крипта-ворсинка содержат центральный просвет, выстилаемый эпителием, подобным эпителию ворсинок. Просвет открывается через последовательные временные интервалы, высвобождая содержимое в среду.
Сходная структура органоида крипта-ворсинка образуется, когда культивируют единичные эпителиальные стволовые клетки. После приблизительно одной недели образуются структуры, которые высоко сходны со структурами органоидов крипта-ворсинка, которые получают с помощью целых крипт.
Способы выделения стволовых клеток известны и специалист в данной области может выбрать подходящие способы для применения в рамках настоящего изобретения, в зависимости от используемого типа стволовых клеток. Например, выделение эпителиальных стволовых клеток можно проводить с использованием соединений, которые связываются с Lgr5 и/или Lgr6, которые являются уникальными маркерами клеточной поверхности на эпителиальных стволовых клетках. Примерами таких соединений являются антитела против Lgr5 и против Lgr6.
В некоторых вариантах осуществления изобретения единичные Lgr5+ эпителиальные стволовые клетки, например, из толстого кишечника, тонкого кишечника или поджелудочной железы, можно использовать для формирования органоидов, таких как органоиды толстого кишечника, органоиды крипта-ворсинка или органоиды поджелудочной железы, соответственно.
В следующем примере единичные Lgr5+ эпителиальные стволовые клетки из печени, предстательной железы или желудка можно использовать для получения органоидов, таких как органоиды печени, предстательной железы или желудка, соответственно.
В альтернативном варианте осуществления фрагменты ткани, такие как культивируемые крипты из желудочно-кишечного тракта, содержащие Lgr5+ стволовые клетки, можно использовать для получения органоидов с использованием способов и культуральных сред, описанных в настоящем описании.
В некоторых вариантах осуществления единичная Lgr5+ эпителиальная стволовая клетка или фрагмент ткани могут представлять собой злокачественную стволовую клетку или фрагмент злокачественной ткани, например, из карциномы или аденокарциномы. В некоторых вариантах осуществления единичная Lgr5+ эпителиальная клетка может представлять собой стволовую клетку или фрагмент ткани из ткани, пораженной неопластической патологией или заболеванием, например, пищеводом Барретта, кистозным фиброзом или аденомой. Органоиды, полученные из злокачественного, неопластического или пораженного заболеванием исходного материала имеют характеристики, напоминающие исходный материал in vivo, и, таким образом, пригодны в качестве исследовательского инструмента для скрининга лекарственных средств, подтверждения мишени, выявления мишени, токсикологии и токсикологических скринингов, персонализированной медицины, регенеративной медицины и моделей клеток/органов ex vivo, например, моделей заболевания. В одном из вариантов осуществления изобретение относится к органоидам, создаваемым или получаемым путем культивирования стволовых клеток или фрагментов тканей человека в соответствии со способом по изобретению. В одном из вариантов осуществления изобретение относится к органоидам крипта-ворсинка, или органоидам желудка, или органоидам поджелудочной железы, или органоидам толстого кишечника, или органоидам пищевода Барретта, или органоидам аденокарциномы, или органоидам карциномы толстого кишечника, создаваемым или получаемым путем культивирования стволовых клеток или фрагментов тканей человека в соответствии со способом по изобретению. В одном из вариантов осуществления изобретение относится к органоидам предстательной железы, создаваемым или получаемым путем культивирования стволовых клеток или фрагментов тканей человека в соответствии со способом по изобретению. Каждая из таких популяций органоидов, например, органоидов крипта-ворсинка, органоидов желудка или органоидов поджелудочной железы, создаваемых или получаемых путем культивирования стволовых клеток или фрагментов тканей человека в соответствии со способом по изобретению, может содержать более 10, предпочтительно, более 20, более предпочтительно, более 40 органоидов. Указанный набор органоидов, предпочтительно, содержит по меньшей мере 10% жизнеспособных клеток, более предпочтительно, по меньшей мере 20% жизнеспособных клеток, более предпочтительно, по меньшей мере 50% жизнеспособных клеток, более предпочтительно, по меньшей мере 60% жизнеспособных клеток, более предпочтительно, по меньшей мере 70% жизнеспособных клеток, более предпочтительно, по меньшей мере 80% жизнеспособных клеток, более предпочтительно, по меньшей мере 90% жизнеспособных клеток. Жизнеспособность клеток можно оценивать с использованием окрашивания красителем Хехста или окрашивания йодидом пропидия в FACS.
Авторы изобретения показали, что культуральные среды и способы по изобретению можно использовать для культивирования злокачественных клеточных линий, включая рак ободочной и прямой кишки и аденокарциному (см. пример 1). Как пояснено в примере 1, технология культивирования широко применима в качестве исследовательского инструмента для инфекционных, воспалительных и неопластических патологий. Таким образом, стволовые клетки согласно изобретению могут представлять собой стволовые клетки злокачественной опухоли. В некоторых вариантах осуществления изобретения стволовые клетки злокачественной опухоли могут образовывать органоиды аденомы или рака толстого кишечника. В некоторых вариантах осуществления эти органоиды содержат клетки, которые являются Ki67+ (Thermo Scientific* Cellomics, Millipore).
Аналогично, авторы изобретения показали, что культуральные среды и способы по изобретению можно использовать для культивирования стволовых клеток с другими связанными с заболеванием генотипами и/или фенотипами. Например, стволовые клетки кишечника, взятые от пациентов с кистозным фиброзом, можно увеличивать в количестве с использованием культуральных сред и способов по изобретению. Эти стволовые клетки сохраняют генотип и фенотип кистозного фиброза. Таким образом, в некоторых вариантах осуществления изобретения проводят взятие стволовых клеток от пациента с заболеванием, например, кистозным фиброзом, воспалительным заболеванием кишечника (таким как болезнь Крона), карциномой, аденомой, аденокарциномой, раком толстого кишечника, диабетом (таким как диабет типа I или типа II), пищеводом Барретта, болезнью Гоше, дефицитом альфа-1-антитрипсина, синдромом Леш-Найхена, анемией, синдромом Швахмана-Бодиана-Даймонда, истинной полицитемией, первичным миелофиброзом, болезнью накопления гликогена, семейной гиперхолестеринемией, синдромом Криглера-Найяра, наследственной тирозинемией, болезнью Помпе, прогрессирующим семейным холестазом, синдромом Харлера, SCID или пропускающим SCID, синдромом Оменна, хрящево-волосяной гипоплазией, обусловленным вирусом простого герпеса энцефалитом, склеродермией, несовершенным остеогенезом, мышечной дистрофией Беккера, мышечной дистрофией Дюшенна, врожденным дискератозом и т.д. В некоторых вариантах осуществления изобретения пораженные заболеванием органоиды можно получать путем культивирования стволовых клеток, взятых от человека или животного с заболеванием. Пораженные заболеванием органоиды все еще имеют характеристики ткани, из которой они получены. Таким образом, органоид тонкого кишечника с кистозным фиброзом, выращенный из крипты тонкого кишечника, соответствует определению органоида тонкого кишечника. Аналогично, органоид карциномы толстого кишечника относится к определению органоида толстого кишечника.
Существует некоторая неразбериха в литературе в отношении определения стволовой клетки злокачественной опухоли. В настоящем описании авторы следуют консенсусу, достигнутому на недавнем семинаре AACR (Clarke et al., 2006. Cancer Res. 66:9339-44), согласно которому стволовая клетка злокачественной опухоли "представляет собой клетку в опухоли, которая обладает способностью к самообновлению и обеспечению гетерогенных ростков злокачественных клеток, которые содержат опухоль. Таким образом, стволовые клетки злокачественные клетки могут быть определены только экспериментально по их способности повторять образование непрерывно растущей опухоли". Альтернативно термины в литературе включают инициирующую опухоль клетку и опухолеродную клетку. Анализы активности стволовых клеток злокачественной опухоли должны быть направлены на потенциал к самообновлению и размножению опухоли. В настоящее время золотым стандартом анализа является серийная ксенотрансплантация иммунодефицитным мышам. Кроме того, злокачественные стволовые клетки в контексте настоящего изобретения обычно экспрессируют Lgr5. Однако в некоторых вариантах осуществления инициирующие злокачественную опухоль/увеличивающие злокачественную опухоль/стволовые клетки, которые не экспрессируют Lgr5, также можно культивировать с помощью культуральных сред и способов по изобретению.
Геномная и фенотипическая целостность стволовых клеток и органоидов, содержащих указанные стволовые клетки
Клинические и научные применения для стволовых клеток и их дифференцированного потомства требуют воспроизводимых способов культивирования стволовых клеток, которые обеспечивают популяции клеток подходящего качества. Как правило, увеличение стволовых клеток в количестве in vitro нацелено на предоставление популяции клеток, которые напоминают их аналоги in vivo настолько, насколько это возможно. Это свойство называют в настоящем описании "геномной и фенотипической целостностью" клеток. Органоиды, полученные путем культивирования пораженных заболеванием клеток, таких как злокачественные клетки или клетки кистозного фиброза, также напоминают их аналоги in vivo, т.е. они сохраняют их обусловленный заболеванием генотип и/или фенотип и, таким образом, также сохраняют их "геномную и фенотипическую целостность" в том смысле, что они сохраняют генетическую или фенотипическую нестабильность, характерную для заболевания, которая сходна с ситуацией in vivo. Таким образом, в некоторых вариантах осуществления изобретение относится к "нормальным" органоидам, полученным из здоровой ткани. В других вариантах осуществления изобретение относится к "пораженным заболеванием" органоидам, таким как органоиды злокачественной опухоли (например, органоиды карциномы толстого кишечника или органоиды аденокарциномы) или органоиды тонкого кишечника с кистозным фиброзом, полученные из пораженной заболеванием ткани.
Впервые авторы изобретения открыли, что можно увеличить в количестве эпителиальные стволовые клетки человека в культуре с минимальной утратой геномной и фенотипической целостности в течение по меньшей мере 3 месяцев, предпочтительно, по меньшей мере 4 месяцев, по меньшей мере 5 месяцев, по меньшей мере 6 месяцев, по меньшей мере 7 месяцев, по меньшей мере 9 месяцев, или по меньшей мере 12 месяцев или более (см. пример 1). В усовершенствованных условиях культивирования по изобретению органоиды кишечника человека проявляли почкование структур органоидов, а не кистозные структуры, наблюдаемые в предшествующих условиях культивирования. Метафазные пластинки органоидов возрастом более 3 месяцев стабильно демонстрировали 46 хромосом в каждых из 20 клеток, взятых от трех различных доноров. Более того, анализ на микрочипах показал, что стволовые клетки в культуре обладали сходными молекулярными признаками с клетками кишечных крипт, включая гены стволовых клеток кишечника.
Таким образом, в некоторых вариантах осуществления изобретение относится к органоидам, которые выращивали в течение по меньшей мере 3 месяцев, предпочтительно, в течение по меньшей мере 4 месяцев, в течение по меньшей мере 5 месяцев, в течение по меньшей мере 6 месяцев, в течение по меньшей мере 7 месяцев, в течение по меньшей мере 9 месяцев, или в течение по меньшей мере 12 месяцев или более с минимальной утратой геномной и фенотипической целостности.
В некоторых вариантах осуществления изобретение относится к органоидам кишечника человека, содержащим почкующиеся структуры. В некоторых вариантах осуществления изобретения органоиды кишечника человека не содержат кистозных структур. В некоторых вариантах осуществления изобретения органоиды кишечника человека содержат больше почкующихся структур, чем кистозных структур. Авторы изобретения также продемонстрировали, что органоиды кишечника человека, полученные с помощью сред и способов по настоящему изобретению, имитируют решения о судьбе клеток in vivo в ответ на внешние факторы. Например, ранее было показано, что ингибирование Notch в стволовых клетках кишечника завершает пролиферацию эпителия кишечника и индуцирует гиперплазию бокаловидных клеток in vivo. Авторы изобретения были способны продемонстрировать, что органоиды кишечника по изобретению, когда их обрабатывали ингибитором Notch, прекращали пролиферировать, и большинство клеток преобразовывалось в бокаловидные клетки в пределах 3 суток.
Эти результаты демонстрируют значительное увеличение геномной и фенотипической целостности стволовых клеток и органоидов, полученных с помощью способов и сред по настоящему изобретению, по сравнению с предшествующими способами и средами.
Геномную целостность стволовых клеток по изобретению можно подтверждать с помощью анализа кариотипа. Кариотипирование стволовых клеток и их потомков можно проводить с использованием известных способов, как описано в Sato, T et al., Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature 459, 262-265, 2009.
"Нормальным кариотипом" является кариотип, когда все хромосомы присутствуют (т.е. эуплоидия) без заметных изменений. Таким образом, в предпочтительных вариантах осуществления изобретения более 50%; более 70%; более 80%; более 90%; более 95%; или более 99% стволовых клеток и дифференцированных клеток в увеличенной в количестве популяции проявляют нормальные кариотипы после 1, 2, 3, 4, 5, 6, 9, 12 или более месяцев. Термин "увеличенная в количестве популяция" охватывает органоиды.
"Нормальный фенотип" относится к клеткам, которые проявляют, в первом приближении, те же внешние характеристики, экспрессию генов и поведение, что и средняя клетка-аналог in vivo. В предпочтительных вариантах осуществления изобретения более 50%; более 70%; более 80%; более 90%; более 95%; или более 99% стволовых клеток в увеличенной в количестве популяции, культивированных согласно изобретению, проявляют нормальные фенотипы после 1, 2, 3, 4, 5, 6, 9, 12 или более месяцев.
Например, визуально нормальный фенотип можно определять по количеству погибших клеток вне органоида, величине "почкования" органоида по сравнению с кистозным ростом (почкующиеся структуры являются предпочтительными), и общей целостности отдельного слоя эпителиальных клеток (например, столбчатый-плоский фенотип). Кроме того, присутствующие типы клеток могут помочь решить, является ли органоид визуально "нормальным".
Предпочтительные свойства стволовых клеток и органоидов по изобретению представлены ниже.
Маркеры стволовых клеток
Когда гены мыши упоминаются в настоящем описании, органоид человека по изобретению может иметь сходный профиль генов, но гены-аналоги человека заменяют гены мыши. Таким образом, также изобретение относится к органоиду человека, имеющему профиль экспрессии генов, как описано в настоящем описании, но в отношении соответствующих генов человека. Аналоги из человека для генов мыши, приведенных в настоящем описании, хорошо доступны специалисту в данной области.
В одном из вариантов осуществления изобретение относится к популяции взрослых стволовых клеток, характеризующейся природной экспрессией Lgr5. В предпочтительном варианте осуществления изобретение относится к популяции взрослых стволовых клеток, характеризующейся природной экспрессией по меньшей мере Lgr5 и одного или нескольких (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22 или 23) маркеров стволовых клеток из группы, состоящей из: LGR4, epcam, Cd24a, Cdca7, Axin, CK19, нестина, соматостатина, CXCR4+, CD133+, DCAMKL-1, CD44, Sord, Sox9, CD44, Prss23, Sp5, Hnf1α, Hnf4a, Sox9, KRT7 и KRT19, Tnfrsf19. Маркеры стволовых клеток могут быть тканеспецифическими. Например, стволовые клетки или органоиды поджелудочной железы могут характеризоваться природной экспрессией одного или нескольких (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15, например, 1, 2, 3 или 4) из: CK19, нестина, соматостатина, инсулина, глюкагона, CXCR4+, Ngn3, Pdx1, NeuroD, Nkx2.2, Nkx6.1, Pax6, Mafa, Hnf1b, необязательно, Tnfrsf19 на значительном уровне; органоиды желудка могут характеризоваться природной экспрессией одного или нескольких (например, 1, 2, 3 или 4) из: CD133+, DCAMKL-1, CD44, необязательно Tnfrsf19 на значительном уровне; и органоиды крипта-ворсинка могут характеризоваться экспрессией одного или нескольких или всех органоиды (например, 1 или 2) из: Sord и/или Prss23, на значительном уровне или всех генов, приведенных в таблице/на фиг.14, например, на значительном уровне.
Термин "значительный уровень", как используют в рамках изобретения в контексте экспрессии маркера, используют синонимично с термином "поддающийся выявлению уровень", как описано ниже.
Популяции клеток органоидов тонкого кишечника и желудка также экспрессируют маркеры популяций-предшественников, общих для тонкого кишечника и желудка, такие как один или оба из Cd44 и Sox9 (Barker & Huch et al. Cell stem cell 2010). Они на высоком уровне экспрессируются в стволовых клетках согласно изобретению. В клетках в соответствии с этим аспектом изобретения также могут активироваться гены-мишени Wnt, включая, например, один, два или все из MMP7, Sp5 Tnfrs19 и axin2. Это обеспечивает убедительное доказательство потребности в активной и стабильной активности канонической передачи сигнала Wnt для поддержания способности этих культур к самообновлению.
Авторы изобретения обнаружили, что экспрессия генов "стволовых клеток" присутствует в ранних органоидах на уровне, значительно превышающем уровень в дифференцированных клетках, которые становятся потомками этих стволовых клеток. Например, гены LGR5, LGR4, Epcam, CD44, Tnfrsf19, Sox9, Cd24a, Sp5, Prom1/CD133, Cdca7, предпочтительно, экспрессируются в органоидах по изобретению, но предпочтительно, значительно подавляются при дифференцировке органоидов поджелудочной железы, печени, тонкого кишечника и толстого кишечника. Кроме того, гены RNF43 и ZNRF3, предпочтительно, экспрессируются в органоидах по изобретению.
Под "природной экспрессией" подразумевают, что клетки не были подвергнуты рекомбинантным манипуляциям никоим образом, т.е. клетки не индуцировали искусственным образом для экспрессии этих маркеров или для модулирования экспрессии этих маркеров путем введения экзогенного генетического материала, такого как введение гетерологичных (неприродных) или более сильных промоторов или других регуляторных последовательностей, функционально связанных либо с эндогенными генами, либо с экзогенно введенными формами генов. Природная экспрессия осуществляется с геномной ДНК в клетках, включая интроны между кодирующими экзоны последовательностями, где они существуют. Природная экспрессия осуществляется не с кДНК. Природная экспрессия может быть подтверждена любым из различных способов, таких как секвенирование в пределах рамки считывания гена для проверки того, не присутствует ли какая-либо внешняя гетерогенная последовательность. "Взрослый" означает постэмбриональный. Что касается стволовых клеток по настоящему изобретению, термин "взрослая стволовая клетка" означает, что стволовая клетка выделена из ткани или органа животного на стадии роста позднее эмбриогенной стадии.
Эта популяция стволовых клеток также может характеризоваться отсутствием природной экспрессии определенных маркеров на каком-либо значительном уровне, многие из которых связаны клеточной дифференцировкой. В частности, клетки популяции выделенных взрослых стволовых клеток не экспрессируют естественным образом один или несколько из Cd11b, CD13, CD14, AFP, Pdx1, любого представителя CYP (например, CYP3A11, CYP 11A1) на значительном уровне. Как определено в настоящем описании, эти маркеры называют отрицательными маркерами.
Выявление маркеров и выделение клеток
Термин "экспрессируемый" используют для описания присутствия маркера в клетке. Чтобы считаться экспрессируемым, маркер должен присутствовать на поддающемся выявлению уровне. Под "поддающимся выявлению уровнем" подразумевают, что маркер может быть выявлен с использованием одной из стандартных лабораторных методологий, таких как ПЦР, блоттинг или FACS-анализ. Ген считается экспрессируемым клеткой из популяции по изобретению, если экспрессия может быть надлежащим образом выявлена после 30 циклов ПЦР, что соответствует уровню экспрессии в клетке по меньшей мере приблизительно 100 копий на клетку. Термины "экспрессирует" и "экспрессия" имеют соответствующие значения. При уровне экспрессии ниже этого порога маркер считается не экспрессируемым. Сравнение между уровнем экспрессии маркера в клетке по изобретению и уровнем экспрессии того же маркера в другой клетке, например, такой как эмбриональная стволовая клетка, предпочтительно, можно проводить путем сравнения двух типов клеток, выделенных из одного и того же вида. Предпочтительно, этот вид представляет собой млекопитающее и, более предпочтительно, этот вид представляет собой человека. Такое сравнение можно удобным образом проводить с использованием эксперимента с полимеразной цепной реакцией с обратной транскриптазой (ОТ-ПЦР).
В некоторых вариантах осуществления популяцию клеток или органоид по изобретению считают экспрессирующими маркер, если по меньшей мере приблизительно 5% (например, по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100%) клеток в популяции клеток или органоиде согласно изобретению демонстрируют экспрессию маркера.
В некоторых вариантах осуществления клетки экспрессируют клеточный маркер на значительном уровне, если они содержат количество копий РНК, кодирующей клеточный маркер, в от 1×102 до 1×105, например, в от 5×102 до 1×104 или в от 1×103 до 1×104 раз большее, чем количество копий мРНК гена домашнего хозяйства GADPH.
В некоторых вариантах осуществления экспрессия гена в органоиде или клетке по изобретению при культивировании в среде для увеличения в количестве в несколько раз (например, по меньшей мере в 1,5 раза, в 2 раза, в 3 раза, в 4 раза, в 5 раз) превышает экспрессию гена, когда органоид или клетку культивируют в среде для дифференцировки или в полностью дифференцированной взрослой ткани. В некоторых вариантах осуществления клетка или органоид по изобретению при культивировании в условиях дифференцировки проявляет увеличение экспрессии генов, которые известны в качестве генов дифференцировки, по сравнению с клеткой или органоидом по изобретению, когда их культивируют в условиях увеличения в количестве, и также могут демонстрировать снижение уровня экспрессии по меньшей мере одного или нескольких генов стволовой клетки/предшественника по сравнению с клеткой или органоидом по изобретению при культивировании в среде для увеличения в количестве.
Любой из ряда физических способов разделения, известных в данной области, можно использовать для выбора клеток согласно этому аспекту изобретения и отличения их от других типов клеток. Такие физические способы могут вовлекать FACS и различные иммуноаффинные способы на основе маркеров, специфично экспрессируемых клетками по изобретению. Как описано выше, Lgr5, CD44 и Sox9 являются тремя клеточными маркерами, экспрессируемыми на высоких уровнях в стволовых клетках по изобретению. Таким образом, только в качестве иллюстрации, стволовые клетки по изобретению можно выделять рядом физических способов разделения, которые основаны на присутствии этих маркеров.
В одном из вариантов осуществления клетки по изобретению можно выделять с помощью FACS с использованием антитела, например, против одного из этих маркеров. Активированную флуоресценцией сортировку клеток (FACS) можно использовать для выявления маркеров, характерных для конкретного типа клеток или ростка. Как будет понятно специалисту в данной области, этого можно достигать с помощью флуоресцентно меченного антитела или с помощью флуоресцентно меченного вторичного антитела, обладающего специфичностью связывания с первичным антителом. Примеры подходящих флуоресцентных меток включают, но не ограничивается ими, FITC, Alexa Fluor® 488, GFP, CFSE, CFDA-SE, DyLight 488, PE, PerCP, PE-Alexa Fluor® 700, PE-Cy5 (TRI-COLOR®), PE-Cy5.5, PI, PE-Alexa Fluor® 750, и PE-Cy7. Этот список предоставлен только в качестве примера и не предназначен для ограничения.
Специалисту в данной области будет понятно, что FACS-анализ с использованием антитела против Lgr5 обеспечит популяцию очищенных стволовых клеток. Однако в некоторых вариантах осуществления может быть предпочтительной дальнейшая очистка популяции клеток путем проведения последующего раунда FACS-анализа с использованием одного или нескольких из других идентифицируемых маркеров.
Также можно использовать иммуногистохимию для понимания распределения и локализации биомаркеров и дифференциально экспрессируемых белков в различных частях клеточной популяции или органоида. Визуализацию взаимодействия антитело-антиген можно проводить рядом способов, которые хорошо известны в данной области, такими как способы, которые описаны в Barker et al, Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature, 2007 Oct 25;449(7165):1003-7.
В другом варианте осуществления клетки по изобретению можно выделять с помощью иммуноаффинной очистки, которая представляет собой способ разделения, хорошо известный в данной области. Только в качестве иллюстрации, клетки по изобретению можно выделять с помощью иммуноаффинной очистки, направленной на c-kit. Как будет понятно специалисту в данной области, этот способ основан на иммобилизации антител на колонке для очистки. Затем образец клеток наносят на колонку, позволяя соответствующим клеткам связываться с антителами, и, таким образом, связываться с колонкой. После стадии промывания клетки элюируют с колонки с использованием конкурента, который связывается, предпочтительно, с иммобилизованным антителом против c-kit, и обеспечивает высвобождение клеток с колонки. Специалисту в данной области будет понятно, что иммуноаффинная очистка с использованием иммобилизованного антитела обеспечит популяцию очищенных клеток. Однако в некоторых вариантах осуществления может быть предпочтительной дальнейшая очистка популяции клеток путем проведения следующего раунда иммуноаффинной очистки с использованием одного или нескольких других идентифицируемых маркеров, и использования аликвоты выделенных клонов для установления экспрессии других соответствующих внутриклеточных маркеров.
Специалисту в данной области будет очевидно, что очистке по LGR5 или очистке стволовых клеток может предшествовать любое количество стадий очистки, таких как очистка эпителия с помощью способов, известных в данной области, например, очистки с помощью EDTA или FACS-сортировки эпителия по Epcam.
Специалисту в данной области будет очевидно, что последовательные стадии очистки не обязательно требуют вовлечения одного и того же физического способа разделения. Таким образом, будет понятно, например, что клетки можно очищать с помощью стадии FACS с использованием антитела против Lgr5, с последующей стадией иммуноаффинной очистки с использованием аффинной колонки SSEA-1. В определенных вариантах осуществления клетки можно культивировать после выделения в течение по меньшей мере приблизительно 15, по меньшей мере приблизительно 20 суток, по меньшей мере приблизительно 25 суток, или по меньшей мере приблизительно 30 суток. В некоторых аспектах клетки увеличивают в количестве в культуре в течение более длительного периода времени для повышения однородности фенотипа клеток в клеточной популяции.
Анализ на микрочипах, кластерный анализ и сравнительное определение профиля экспрессии генов можно использовать для сравнения фенотипа популяции с фенотипом исходных родительских клеток или соответствующих аналогов in vivo (Sato T et al., Paneth cells constitute the niche for Lgr5 stem cells in intestinal crypts. Nature 469 415-418).
Отслеживание ростка Lgr5+ стволовых клеток демонстрирует сохранение характеристик крипт-ворсинок в органоидах.
В другом варианте осуществления одновременный многопараметрический анализ можно использовать для оценки фенотипической целостности стволовых клеток. Например, существует ряд наборов и платформ для одновременного многопараметрического скрининга, таких как 4 color ImageXpress ULTRA для точечного сканирования (Molecular Devices, Union City, США), вращающийся диск (диск Нипкова) Pathway 855 и 435 от BD Biosciences (formerly Atto Biosciences, Rockville, Maryland), Opera (PerkinElmer Inc., Waltham, MA) и IN Cell 3000 для щелевого сканирования (GE/Amersham Biosciences, Cardiff, Великобритания), Arrayscan VTI (Cellomics (Cellomics)), IN Cell Analyzer 2000 (GE Healthcare Piscataway, New Jersey, США), Acumen eX3 (TTP LabTech Ltd (Acumen eX3)), Scanalyzer (Scanalyzer LemnaTec, Aachen, Германия) и ImageXpress MICRO (Molecular Devices, Union City, США), IN Cell 1000 (GE/Amersham Biosciences Piscataway, New Jersey, США), Pathway HT (Becton Dickinson Biosciences) и ImageXpress MICRO (Molecular Devices, Union City, США), Scan^R (Olympus Soft Imaging Solutions, Германия).
Плотность посева
В некоторых вариантах осуществления изобретения обычно проводят посев суспензий отдельных клеток или небольших кластеров клеток (2-50 клеток/кластер) вместо крупных кластеров клеток, как известно в данной области. По мере их деления такие клетки будут высевать на подложку с плотностью, которая обеспечивает пролиферацию клеток. Как правило, когда выделяют отдельные клетки, используют плотность посева по меньшей мере 1-500 клеток/лунка, где поверхность лунки составляет 0,32 см2. Когда кластеры высевают, плотность посева, предпочтительно, составляет 250-2500 клеток/см2. Для повторного посева можно использовать плотность от приблизительно 2500 клеток/см2 до приблизительно 5000 клеток/см2. В ходе повторного посева обычно проводят посев суспензий единичных клеток или небольшого кластера клеток вместо крупных кластеров клеток, как известно в данной области.
Дальнейшая дифференцировка
В некоторых вариантах осуществления изобретения определенные компоненты среды для увеличения в количестве можно устранять для изменения клеточной судьбы культивируемых клеток в направлении дифференцировки. Из культуральной среды можно устранять любые компоненты культуральной среды, которые ответственны за поддержание недифференцированного состояния и/или активацию генетических программ стволовых клеток или предшественников.
В некоторых вариантах осуществления изобретения устранение ингибиторов по изобретению может обеспечить дифференцировку клеток органоидов в зрелые клетки, такие как зрелые бокаловидные или энтероэндокринные клетки в органоидах крипта-ворсинка. Таким образом, в некоторых вариантах осуществления изобретение относится к способу дальнейшей дифференцировки органоидов с использованием второй культуральной среды, которая не содержит ингибитора по изобретению. Например, см. пример 1.
Например, в некоторых вариантах осуществления ингибитор TGF-бета и/или ингибитор p38 устраняют из среды для культивирования клеток для обеспечения дифференцировки клеток. Под "устраненным" или "устранением" компонента из среды для культивирования клеток подразумевают, что, когда клетки повторно высевают и среду заменяют, компонент не добавляют в свежую среду.
В некоторых вариантах осуществления Wnt присутствует в среде для увеличения в количестве, но не в среде для дифференцировки. Например, некоторые варианты осуществления включают устранение Wnt для дифференцировки органоидов толстого кишечника в зрелые энтероциты. Wnt также можно устранять для обеспечения дифференцировки органоидов крипта-ворсинка.
В некоторых вариантах осуществления R-спондин присутствует в среде для увеличения в количестве, но не в среде для дифференцировки. Например, некоторые варианты осуществления включают устранение R-спондина для дифференцировки органоидов толстого кишечника в зрелые энтероциты. R-спондин также можно устранять для обеспечения дифференцировки органоидов крипта-ворсинка. В некоторых вариантах осуществления R-спондин и Wnt можно устранять для обеспечения дифференцировки органоидов крипта-ворсинка.
В некоторых вариантах осуществления никотинамид присутствует в среде для увеличения в количестве, но не в среде для дифференцировки. Таким образом, в некоторых вариантах осуществления никотинамид и SB202190 (или другой ингибитор p38) устраняют из среды для культивирования клеток для обеспечения дифференцировки клеток, например, в органоиды крипта-ворсинка или органоиды толстого кишечника.
Таким образом, способ получения дифференцированных клеток или органоидов может включать культивирование эпителиальных клеток в способе культивирования по изобретению, который включает ингибитор TGF-бета и/или p38 для обеспечения выживания и/или пролиферации клеток (т.е. среда для увеличения в количестве), а затем продолжение культивирования клеток и восполнение среды, где восполненная среда не содержит ингибитора TGF-бета и/или p38 (т.е. среда для дифференцировки).
В некоторых вариантах осуществления среда для дифференцировки содержит дополнительные компоненты. Например, в некоторых вариантах осуществления среда для дифференцировки содержит ингибитор гамма-секретазы, например, DAPT или DBZ. В некоторых вариантах осуществления среда для дифференцировки содержит RANK-лиганд (также обозначаемый в настоящем описании как RANKL). Как упоминалось выше, добавление ингибитора гамма-секретазы может контролировать дифференцировку клеток кишечных органоидов, таких как клетки органоидов тонкого кишечника, в направлении секреторных клеток, таких как бокаловидные клетки. Добавление RANKL в культуральную среду может направлять дифференцировку клеток кишечных органоидов, таких как клетки органоидов тонкого кишечника, в направлении M-клеток.
В некоторых вариантах осуществления изобретение относится к культуральной среде для дифференцировки стволовых клеток из представляющей интерес ткани, где культуральная среда содержит или состоит из компонентов культуральной среды, используемых для увеличения в количестве стволовых клеток из представляющего интерес типа ткани, но где один или несколько из следующих компонентов исключены из среды для дифференцировки стволовых клеток: Wnt, R-спондин, ингибитор BMP, ингибитор TGF-бета, лиганд рецепторной тирозинкиназы, ингибитор p38 и никотинамид.
Более того, изобретение относится к способу увеличения в количестве единичной стволовой клетки или популяции стволовых клеток, предпочтительно, для получения органоида, где способ включает культивирование единичной стволовой клетки или популяции стволовых клеток в культуральной среде согласно изобретению, где способ включает:
культивирование стволовой клетки, популяции стволовых клеток или фрагментов тканей в первой среде для увеличения в количестве;
продолжение культивирования стволовой клетки, популяции стволовых клеток или фрагментов ткани и восполнение среды средой для дифференцировки, где среда для дифференцировки не содержит одного или нескольких, предпочтительно, всех из факторов, выбранных из: ингибитора TGF-бета, ингибитора p38, никотинамида и Wnt.
Как правило, когда компонент описывают как "удаленный" из среды, подразумевают, что компонент не добавляют, когда среду восполняют, т.е. компонент исключают из восполненной среды. Когда среда является "восполненной", это означает, что среда физически удалена из внеклеточного матрикса, а затем заменена свежей средой.
В случае толстого кишечника, печени и поджелудочной железы очень мало дифференцированных клеток присутствует в среде для увеличения в количестве. Только после замены среды для увеличения в количестве средой для дифференцировки клетки начинают дифференцироваться. На этой стадии органоиды также начинают утрачивать их стволовые клетки. Дифференцированные органоиды могут быть пригодными для определенных применений, таких как (но не ограничиваясь ими) трансплантация, скрининг лекарственных средств от метаболических заболеваний, токсикология (например, с использованием органоидов печени, содержащих гепатоциты) и для исследования антибактериальных функций тонкого кишечника. Увеличение в количестве органоидов обычно может быть более пригодным для других применений, таких как (но не ограничиваясь ими) регенеративная медицина и скрининг лекарственных средств, например, от злокачественной опухоли или кистозного фиброза. Увеличивающиеся в количестве органоиды, как правило, имеют больший потенциал к росту (и, таким образом, больший срок жизни), чем дифференцированные органоиды. В некоторых вариантах осуществления органоиды толстого кишечника, печени и поджелудочной железы далее не дифференцируют.
Органоиды тонкого кишечника и предстательной железы отличаются от органоидов толстого кишечника, печени и поджелудочной железы тем, что они сохраняют увеличивающуюся в количестве популяцию стволовых клеток, одновременно также дифференцируясь. Нет необходимости в культивировании их в отдельной среде для дифференцировки для того, чтобы присутствовали дифференцированные типы клеток. Можно считать, что они имеют свойства как увеличивающегося в количестве, так и дифференцированного органоида. Однако для обеспечения полной дифференцировки органоидов тонкого кишечника их можно культивировать в отдельной среде для дифференцировки, которая, предпочтительно, не содержит Wnt3a и которая предпочтительно содержит ингибитор гамма-секретазы и/или RANK-лиганд (также обозначаемый в настоящем описании как RANKL). Под "полной" дифференцировкой подразумевают, что присутствуют все дифференцированные типы клеток, включая бокаловидные клетки, нейроэндокринные клетки, щеточные клетки, M-клетки, энтероциты и клетки Панета. Некоторые из этих дифференцированных типов клеток, например, клетки Панета, также присутствуют (иногда в меньших количествах) в увеличивающихся в количествах органоидах.
Органоиды
Клетки, описанные выше, вырастают в органоиды. Таким образом, органоид, который можно получить способом по изобретению, является следующим аспектом. Также предусмотрен органоид, как описано в настоящем описании. Органоид, предпочтительно, представляет собой органоид человека. Насколько известно авторам настоящего изобретения, на этот раз впервые были получены органоиды человека, которые являются функциональными и живыми после такого длительного периода времени (т.е. по меньшей мере 3 месяцев, предпочтительно, по меньшей мере 4 месяцев, по меньшей мере 5 месяцев, по меньшей мере 6 месяцев, по меньшей мере 7 месяцев, по меньшей мере 9 месяцев, или по меньшей мере 12 месяцев или более в культуре; см. примеры, включенные в настоящее описание). Функциональность, предпочтительно, характеризуется присутствием тканеспецифических маркеров и/или структурой указанного органоида, как определено в настоящем описании. Поскольку конечное количество полученных органоидов коррелирует с длительностью культивирования, квалифицированному специалисту будет понятно, что изобретение является пионерским изобретением и потенциально открывает новые возможности, например, в регенеративной медицине. Таким образом, предусматривается органоид, как описано в настоящем описании, который является функциональным и живым после по меньшей мере 3 месяцев (например, по меньшей мере 4, 5, 6, 7, 8 или более месяцев) культивирования. Например, предусматривается органоид, как описано в настоящем описании, который сохраняет по меньшей мере одно или несколько (например, 1, 2 или 3) из его структуры, экспрессии маркеров и функции после по меньшей мере 3 месяцев (например, по меньшей мере 4, 5, 6, 7, 8 или более месяцев) в культуре.
Например, органоид в соответствии с настоящим изобретением может содержать популяцию клеток, состоящую из по меньшей мере 1×103 клеток, по меньшей мере 1×104 клеток, по меньшей мере 1×105 клеток, по меньшей мере 1×106 клеток, по меньшей мере 1×107 клеток или более. Каждый органоид содержит приблизительно от 1×103 клеток до 5×103 клеток. Авторы изобретения показали, что можно выращивать органоиды из единичных Lgr5+ стволовых клеток в органоиды, содержащие популяцию клеток, как описано выше, или содержащие популяцию клеток размером приблизительно 104 клеток. Например, в рамках настоящего изобретения для мыши было показано, что можно начать рост органоида из единичных стволовых клеток. Таким образом, изобретение относится к способу получения органоида из единичной стволовой клетки. В некоторых вариантах осуществления органоид содержит приблизительно 104 клеток. В некоторых вариантах осуществления 10-20, или 20-30 или 30-40 или 40-50 органоидов можно выращивать вместе в одной ячейке планшета с 24 ячейками.
В некоторых вариантах осуществления изобретение относится к органоиду или популяции клеток, которые способны выживать в культуре в течение по меньшей мере 3 месяцев, например, в течение по меньшей мере 4 месяцев, в течение по меньшей мере 5 месяцев, в течение по меньшей мере 6 месяцев, в течение по меньшей мере 7 месяцев, в течение по меньшей мере 9 месяцев, или в течение по меньшей мере 12 месяцев или более при культивировании в культуральной среде по изобретению.
В некоторых вариантах осуществления изобретение относится к органоиду или популяции клеток, где органоид или популяция клеток увеличиваются в количестве со скоростью по меньшей мере в 3 раза, по меньшей мере в 4 раза, по меньшей мере в 5 раз, по меньшей мере в 6 раз, по меньшей мере в 7 раз, по меньшей мере в 8 раз, по меньшей мере в 9 раз или по меньшей мере в 10 раз в неделю.
Предпочтительно, популяция клеток или органоидов увеличивается в количестве со скоростью приблизительно 4-5 раз в неделю или более двух удвоений популяции в неделю. Таким образом, в некоторых вариантах осуществления популяция клеток или органоидов увеличивается в количестве со скоростью по меньшей мере в 3 раза, по меньшей мере в 4 раза, по меньшей мере в 5 раз, по меньшей мере в 6 раз, по меньшей мере в 7 раз, по меньшей мере в 8 раз, по меньшей мере в 9 раз или по меньшей мере в 10 раз в неделю.
Органоиды по изобретению можно получать с использованием клеток, выделенных из любого подходящего источника. Как правило, клетки, используемые для получения органоида, можно выделять из того же типа ткани, что и органоид, который получают. Органоиды, предпочтительно, представляют собой органоиды млекопитающего, например, мыши, крупного рогатого скота, свиньи или человека. Наиболее предпочтительно, органоиды представляют собой органоиды человека.
В некоторых вариантах осуществления изобретение относится к органоиду или популяция клеток, где органоид или популяция клеток являются нормальными (здоровыми) органоидом или популяцией клеток или пораженными заболеванием органоидом или популяцией клеток, например, полученными путем культивирования стволовых клеток, взятых от человека или животного с заболеванием.
В некоторых вариантах осуществления изобретение относится к органоиду или популяции клеток, которые замораживают и хранят при температуре ниже -5°C, ниже -10°C, ниже -20°C, ниже -40°C, ниже -60°C или ниже -80°C, ниже -100°C, ниже -150°C или приблизительно -180°C. Органоид или популяцию клеток по изобретению можно хранить в жидком азоте. Таким образом, в некоторых вариантах осуществления изобретение относится к органоиду или популяции клеток, которые хранят в жидком азоте.
В некоторых вариантах осуществления изобретение относится к органоиду по изобретению, где органоид представляет собой органоид тонкого кишечника, органоид толстого кишечника, органоид желудка, органоид поджелудочной железы, органоид печени или органоид предстательной железы.
Структура и морфология органоидов
Органоиды по изобретению, которые можно получать путем увеличения в количестве стволовых клеток, обеспечивают популяцию клеток, которые напоминают их аналоги in vivo.
Для оценки характеристик клеток в культуре, таких как морфология клеток; клеточные структуры; признаки апоптоза или лизиса клеток; и состав и структура органоида, можно использовать визуализирующий анализ. В данной области хорошо известно множество типов визуализирующих анализов, таких как электронная микроскопия, конфокальная микроскопия, стереомикроскопия, флуоресцентная микроскопия. Гистологический анализ может выявить основную архитектуру и типы клеток.
Иллюстративные примеры органоидов, полученных согласно изобретению, приведены на прилагаемых чертежах. Можно видеть, что органоиды согласно изобретению могут обладать слоем клеток с по меньшей мере одной почкой и центральным просветом. Органоиды снаружи Matrigel имеют тенденцию к тому, чтобы быть больше органоидов в центре Matrigel, возможно поскольку они имеют лучший доступ к необходимым факторам роста. Структурно органоиды согласно изобретению часто имеют удлиненную форму. Они могут включать одну или несколько почкующихся структур - эпителиальный слой толщиной в одну клетку, обладающий сходством с протоками или островками. При конфокальной микроскопии структуры могут окрашиваться положительно на кератин. Они могут включать клетки с полярными ядрами и небольшой цитоплазмой. Органоиды могут иметь срез, который образован из множества слоев; такие клетки часто имеют тенденцию к тому, чтобы их ядра располагались более центрально в клетках, т.е. не были полярными. Клетки в многослойном срезе могут самоорганизовываться так, чтобы включать щель или просвет между клетками.
В некоторых вариантах осуществления органоиды по изобретению содержат или состоят из эпителиальных клеток. В некоторых вариантах осуществления органоиды содержат или состоят из одного слоя эпителиальных клеток. В некоторых вариантах осуществления неэпителиальные клетки отсутствуют в органоидах. В некоторых вариантах осуществления органоиды по изобретению содержат все дифференцированные типы клеток, которые существуют в их соответствующем аналоге ткани in vivo.
В некоторых вариантах осуществления органоиды кишечника человека проявляют почкующиеся структуры органоида вместо кистозных структур, наблюдаемых в предшествующих условиях культивирования. Метафазные пластинки органоидов возрастом более 3 месяцев стабильно демонстрировали 46 хромосом в каждой из 20 клеток, взятых от трех различных доноров.
В некоторых вариантах осуществления органоиды по изобретению содержат один монослой клеток. В некоторых вариантах осуществления органоиды по изобретению имеют срез, который образован из множества слоев. Множество слоев клеток также называют в настоящем области как области "распределенных на слои" клеток. Под "распределенными на слои" подразумевают, что существует множество (более одного) слоев клеток. В некоторых вариантах осуществления органоиды по изобретению содержат единичные монослои, которые являются сложенными (или инвагинированными) с образованием двух или более слоев. Иногда трудно различить сложенные (или инвагинированные) монослои и области распределенных на слои клеток. В некоторых вариантах осуществления органоид содержит как области распределенных на слои клеток, так и сложенных монослоев. В некоторых вариантах осуществления органоиды по изобретению имеют срез, который образован множеством монослоев, и срез, содержащий один монослой клеток. Морфологически клетки выглядят подобно их соответствующему аналогу ткани in vivo.
Таким образом, в некоторых вариантах осуществления изобретение относится к органоиду, предпочтительно, получаемому с использованием культуральных сред и способов по изобретению, который представляет собой трехмерный органоид, содержащий эпителиальные клетки, окружающие центральный просвет, где необязательно эпителиальные клетки существуют в отдельных делящихся доменах и дифференцирующихся доменах. В некоторых вариантах осуществления органоид по изобретению представляет собой трехмерный органоид, содержащий эпителиальные клетки, расположенные в областях монослоев, необязательно сложенных монослоев и областях распределенных на слои клеток. В некоторых вариантах осуществления неэпителиалььные клетки отсутствуют в указанном органоиде. В некоторых вариантах осуществления в указанном органоиде присутствуют все дифференцированные типы клеток нормальной ткани in vivo.
Органоиды крипта-ворсинка
В органоидах крипта-ворсинка тонкого кишечника структурная организация органоидов в высокой степени сходна со структурой крипты-ворсинки in vivo: Lgr5+ стволовая клетка и клетки ниши (клетки Панета) расположены рядом друг с другом в основании крипты, за ними следуют переходные делящиеся клетки, находящиеся непосредственно выше основания крипты и ведущие в боковые области ворсинок и, наконец, дифференцированные клетки, такие как энтероциты, которые составляют остальную часть ворсинок и становятся более дифференцированными в направлении верхней части ворсинка. Можно видеть, что органоиды согласно изобретению могут иметь слой клеток с по меньшей мере одной почкой и центральным просветом. Органоиды снаружи Matrigel имеют тенденцию к тому, чтобы быть больше органоидов в центре Matrigel, возможно потому, что они обладают лучшим доступом к необходимым факторам роста. Структурно органоиды согласно изобретению часто имеют удлиненную форму. При конфокальной микроскопии структуры могут иметь положительную окраску на кератин. Они могут включать клетки с полярными ядрами и небольшой цитоплазмой. Органоиды крипта-ворсинка обычно являются однослойными.
В некоторых вариантах осуществления, например, для органоида крипта-ворсинка мыши, органоид крипта-ворсинка представляет собой трехмерный органоид, содержащий подобные криптам домены, окружающие центральный просвет, выстилаемый подобными ворсинкам эпителиальными доменами, которые представляют собой эпителиальные домены, содержащие дифференцированные типы клеток. В некоторых вариантах осуществления неэпителиальные клетки в указанном органоиде отсутствуют.
В некоторых вариантах осуществления, например, для органоида крипта-ворсинка человека, органоид крипта-ворсинка представляет собой трехмерный органоид, содержащий подобные криптам домены, окружающие центральный просвет. В некоторых вариантах осуществления делящиеся клетки ограничены почкующимися структурами. Не присутствует или присутствует мало дифференцированных клеток. В условиях дифференцировки образуются дифференцированные клетки кишечника. В некоторых вариантах осуществления неэпителиальные клетки отсутствуют в указанном органоиде. В некоторых вариантах осуществления, когда органоид является увеличивающимся в количестве, например, когда он находится в культуральной среде для увеличения в количестве согласно изобретению, органоид имеет мало или не имеет дифференцированных клеток.
В некоторых вариантах осуществления органоид тонкого кишечника по изобретению, культивируемый в культуральной среде по изобретению, содержащей RANKL, содержит M-клетки. В некоторых вариантах осуществления изобретения органоид тонкого кишечника по изобретению, культивируемый в культуральной среде по изобретению, содержащей ингибитор гамма-секретазы, содержит бокаловидные клетки. В некоторых вариантах осуществления органоид тонкого кишечника, культивируемый в среде для дифференцировки (например, где среда для дифференцировки содержит базальную среду, Noggin, EGF, ингибитор TGF-бета и ингибитор p38, ингибитор гамма-секретазы и RANKL) содержит все типы дифференцированных клеток, включая, например, бокаловидные клетки, нейроэндокринные клетки, щеточные клетки, M-клетки, энтероциты и клетки Панета. Некоторые из этих типов дифференцированных клеток, например, клетки Панета, также присутствуют (иногда в меньших количествах) в увеличивающихся в количестве органоидах.
Органоиды кишечника человека проявляют почкующиеся структуры органоидов, а не кистозные структуры, наблюдаемые в предшествующих условиях культивирования. Верхнее отверстие свежевыделенных крипт закрывается, и эта область постепенно расширяется и заполняется апоптотическими клетками, подобно тому, как апоптотические клетки отделяются на вершине ворсинки. Таким образом, в некоторых вариантах осуществления органоиды крипта-ворсинка имеют подобную крипте структуру, окружающую центральный просвет, выстилаемый подобным ворсинке эпителием и заполненный апоптотическими клеточными тельцами. В некоторых вариантах осуществления просвет открывается в последующие интервалы времени для высвобождения содержимого в среду.
В некоторых вариантах осуществления область крипты претерпевает непрерывные события почкования, которые создают дополнительные крипты, что является процессом, напоминающим деление крипты.
Авторы изобретения также продемонстрировали, что органоиды кишечника человека, полученные с помощью сред и способов по настоящему изобретению, имитируют решение о клеточной судьбе in vivo в ответ на внешние факторы. Например, ранее было показано, что ингибирование Notch в стволовых клетках кишечника завершает пролиферацию эпителия кишечника и индуцирует гиперплазию бокаловидных клеток in vivo. Таким образом, в некоторых вариантах осуществления, когда органоид крипта-ворсинка по изобретению обрабатывают ингибитором Notch, пролиферация прекращается и большинство клеток (например, более 50%, более 60%, более 70%, более 80%, более 90%, более 95%, более 98%) преобразуются в бокаловидные клетки в течение 3 суток.
В метафазных пластинках органоидов возрастом более 3 месяцев выявляли 46 хромосом в каждой из 20 клеток, взятых от трех различных доноров. Более того, анализ на микрочипах выявил, что стволовые клетки в культуре обладали сходными молекулярными профилями с клетками кишечных крипт, включая гены стволовых клеток кишечника.
Органоиды толстого кишечника
Органоиды толстого кишечника проявляют сходный клеточный состав с органоидами крипта-ворсинка. Таким образом, комментарии для органоидов крипта-ворсинка выше применимы к органоидам толстого кишечника с соответствующими изменениями. Например, см. фиг.1 и 2.
Главным образом, различие между органоидами толстого и тонкого кишечника состоит в том, что крипты являются более мелкими в толстом кишечнике, что делает их несколько более похожими на "футбольный мяч", чем на сферу с выступами. Как органоиды тонкого кишечника, так и органоиды толстого кишечника имеют домены, которые содержат стволовые клетки и переходные делящиеся (TA) клетки, и другие домены, содержащие дифференцирующиеся и/или дифференцированные клетки. Для органоидов тонкого кишечника дифференцированные домены иногда называют "подобными ворсинке". Дифференцированные домены органоидов толстого кишечника, как правило, сходны по клеточному составу с "подобными ворсинке" доменами тонкого кишечника, однако сам по себе толстый кишечник не имеет ворсинок.
Количество присутствующего Wnt может влиять на размер почкующихся структур (т.е. глубину крипт) в органоидах. Большее количество Wnt уменьшает почкование. Толстый кишечник продуцирует больше Wnt, чем тонкий кишечник и, таким образом, требует меньше дополнительного Wnt в культуральной среде и, как правило, имеет более мелкие крипты, чем тонкий кишечник. Те же отличия наблюдают в органоидах.
В некоторых аспектах изобретение относится к органоидам толстого кишечника. Авторы изобретения открыли, что органоиды толстого кишечника мыши можно получать путем культивирования крипт толстого кишечника в клеточных культуральных средах ENR + Wnt3A (WENR). Таким образом, в некоторых вариантах осуществления изобретение относится к органоиду толстого кишечника, получаемому путем культивирования крипт толстого кишечника в средах WENR.
Авторы изобретения также неожиданно открыли, что органоиды толстого кишечника человека можно поддерживать с использованием культуральной среды, содержащей WENR вместе с гастрином вместе с никотинамидом. В некоторых вариантах осуществления органоид толстого кишечника человека по изобретению можно получать с использованием сред, содержащих WENR вместе с гастрином вместе с никотинамидом и также содержащих ингибитор TGF-бета. Например, в некоторых вариантах осуществления следующие среды для культивирования клеток можно использовать для получения органоида толстого кишечника человека: WENR+гастрин+никотинамид+A8301+SB202190. В других вариантах осуществления следующую среду для культивирования клеток можно использовать для получения органоида толстого кишечника человека: WENR+никотинамид+A83-01.
В некоторых вариантах осуществления органоид толстого кишечника мыши имеет максимальный диаметр приблизительно 200-700 мкм, например, 250-600 мкм, 300-500 мкм, 320-450 мкм, 340-400 мкм, 300-380 мкм, например, приблизительно 360 мкм. В некоторых вариантах осуществления органоид толстого кишечника имеет минимальный диаметр приблизительно 100-400 мкм, например, 150-350 мкм, 170-300 мкм, 190-280 мкм, 195-250 мкм, например, приблизительно 235 мкм. В следующем варианте осуществления органоиды могут иметь диаметр вплоть до 1 мм. В некоторых вариантах осуществления органоид толстого кишечника человека имеет максимальный диаметр приблизительно 300-800 мкм, например, 350-700 мкм, 400-600 мкм, 450-550 мкм, 475-540 мкм, 500-530 мкм, например, приблизительно 500 мкм. В некоторых вариантах осуществления органоид толстого кишечника имеет минимальный диаметр приблизительно 200-500 мкм, например, 250-450 мкм, 300-415 мкм, 350-400 мкм, 325-380 мкм, например, приблизительно 375 мкм. В следующем варианте осуществления органоиды могут иметь диаметр вплоть до 1 мм. В некоторых вариантах осуществления органоид толстого кишечника по изобретению содержит почкующиеся структуры. Они могут быть видны при использовании краски EdU для визуализации пролиферирующих клеток.
Органоиды толстого кишечника человека сохраняют их характерную почкующуюся структуру в условиях культуры стволовых клеток кишечника человека ("HISC") (WENRg+никотинамид+ингибитор TGF-бета (например, A83-01)+ингибитор p38 (например, SB202190)). В некоторых вариантах осуществления органоид толстого кишечника представляет собой трехмерный органоид, содержащий почкующиеся структуры, которые являются пролиферирующими и содержат стволовые клетки. Эти домены стволовых клеток окружают центральный просвет. Делящиеся клетки, главным образом, ограничены почкующимися структурами. В некоторых вариантах осуществления отсутствуют или присутствует мало дифференцирующихся клеток. В условиях дифференцировки образуются дифференцированные клетки кишечника, например, зрелые энтероциты. В некоторых вариантах осуществления неэпителиальные клетки отсутствуют в указанном органоиде.
Органоиды поджелудочной железы
Органоиды поджелудочной железы по изобретению, предпочтительно, проявляют почкование. В некоторых вариантах осуществления органоиды поджелудочной железы имеют диаметр 100-1000 микрометров, например, 200-900 микрометров, 300-1000 микрометров, 400-700 микрометров. Органоиды поджелудочной железы, предпочтительно, являются однослойными. Структуры островков и протоков имеются только на начальной стадии. Почкующаяся структура указывает на нормальное состояние пролиферации и поддержание стволовых клеток.
В некоторых вариантах осуществления, например, когда органоиды поджелудочной железы выращивают в культуральной среде по изобретению (и в отсутствие ингибиторов TGF-бета), органоиды поджелудочной железы, главным образом, представляют собой кистозные структуры с небольшим количеством почкующихся структур или подобных протокам доменов. Кистозные структуры содержат, главным образом, монослои, но иногда могут присутствовать области распределенных на слои клеток. Клетки экспрессируют маркеры стволовых клеток и предшественников (протоков). В органоидах присутствуют дифференцированные клетки, такие как бета-клетки. Киста, главным образом, образуется монослоем, но существуют распределенные на слои части. Типы клеток напоминают стволовые клетки / предшественника (экспрессия генов клеток протоков). Отсутствуют дифференцированные клетки (β-клетки).
В других вариантах осуществления, например, когда органоиды поджелудочной железы выращивают в присутствии ингибитора TGF-бета, такого как A83-01, например, в культуральной среде по изобретению, органоиды поджелудочной железы содержат больше почкующихся структур/подобных протокам доменов (это означает, что клетки являются подобными протоковым клеткам в большей степени, чем структура является подобной протоку), как показано по окрашиванию на Krt19 (например, см. фиг.31). Можно идентифицировать монослои полярных клеток, но также можно идентифицировать области с распределенными на слои клетками.
Органоиды аденокарциномы и рака толстого кишечника
Органоиды аденокарциномы и рака толстого кишечника, как правило, образуют кистозные структуры вместо почкующихся структур. Это напоминает случай отсутствия хорошей поддерживающей ниши для клеток. Крипты аденомы, культивируемые с EFG+Noggin, демонстрируют приблизительно 16x увеличение в количестве в течение первых 10 суток. Органоиды адено(карцино)мы и рака толстого кишечника могут обеспечить полезные исследовательские инструменты и модели для скрининга лекарственных средств.
Органоиды карциномы, аденомы и аденокарциномы являются по большей части кистозными (например, см. фиг.4 и 9). Однако в некоторых вариантах осуществления они также содержат структуры, которые напоминают их нормальные аналоги органоидов тканей.
Органоиды пищевода Барретта (BE)
Органоид BE по изобретению содержит почкующиеся структуры (например, см. фиг.5).
Морфологически клетки в органоидах по изобретению выглядят подобно их соответствующему аналогу ткани in vivo.
Пищевод Барретта представляет собой заболевание, для которого характерно присутствие стоблчатого эпителия в нижнем отделе пищевода, заменяющего нормальный плоский эпителий в результате метаплазии. Гистологическим отличительным признаком пищевода Барретта является присутствие бокаловидных клеток кишечника в пищеводе. Исследуя сходства между пищеводом Барретта и эпителием кишечника, авторы изобретения показали, что культуральные среды и способы по изобретению можно использовать для поддержания эпителия пищевода Барретта в течение вплоть до 1 месяца. Авторы изобретения также впервые продемонстрировали, что добавление FGF10 в культуральную среду по изобретению обеспечивает образование органоидами пищевода Барретта почкующихся структур и значительно продлевает длительность культивирования вплоть до трех месяцев. Таким образом, органоид пищевода Барретта является примером органоида по изобретению. В некоторых вариантах осуществления органоид пищевода Барретта имеет кистозную структуру. В некоторых вариантах осуществления органоид пищевода Барретта по изобретению содержит клетки Панета. В некоторых вариантах осуществления органоид пищевода Барретта по изобретению экспрессирует лизоцим.
Таким образом, авторы изобретения также описывают культуральную среду согласно изобретению, содержащую FGF10, для культивирования эпителия пищевода Барретта.
В некоторых вариантах осуществления изобретения органоиды пищевода Барретта можно выращивать с использованием культуральной среды по изобретению, также содержащей FGF10. В некоторых вариантах осуществления эти органоиды пищевода Барретта экспрессируют Ki67 и имеют минимальное количество, предпочтительно, менее 10%, менее 5% или менее 1% PAS-положительных клеток и муцин-положительных клеток. В некоторых вариантах осуществления органоиды пищевода Барретта содержат лизоцим-положительные клетки Панета.
Органоиды желудка (гастральные органоиды) (например, см. фиг.46)
Органоиды желудка мыши, выращенные в культуральной среде по изобретению, представляют собой трехмерные органоиды, содержащие или состоящие из однослойного эпителия, который содержит домены, подобные основаниям желез желудка (образованные стволовыми клетками и клетками-предшественниками), окружающими центральный просвет, выстилаемый эпителиальными доменами, содержащими типы дифференцированных клеток, и необязательно где неэпителиальные клетки отсутствуют в указанном органоиде.
Органоиды желудка человека, выращенные в культуральной среде по изобретению, содержат кистозные структуры. Кистозная структура представляет собой монослой полярных клеток. Эти органоиды желудка человека, выращенные в присутствии ингибитора TGF-бета, напоминают органоиды желудка мыши значительно в большей степени, чем органоиды человека, выращенные в отсутствие ингибитора TGF-бета.
Органоиды предстательной железы (см. фиг.41-43)
В условиях культивирования, включающих EGF, Noggin, R-спондин, органоиды предстательной железы мыши образуют трехмерные кистозные структуры с просветом. Со временем слои складываются внутрь, образуя 3-4 слоя (распределенных на слои) эпителиальных клеток. Наружный слой состоит по большей части из CK5+ базальных эпителиальных клеток, в то время как внутренние слои состоят по большей части из CK8+ люминальных эпителиальных клеток. Не было идентифицировано компартмента стволовых клеток; все домены содержали делящиеся клетки. Таким образом, в некоторых вариантах осуществления органоид предстательной железы, выращенный в отсутствие тестостерона, содержит стратифицированные слои делящихся эпителиальных клеток. В следующем варианте осуществления органоид предстательной железы содержит наружный слой клеток, содержащий CK5+ базальные эпителиальные клетки и внутренний слой, содержащий CK8+ люминальные эпителиальные клетки. В некоторых вариантах осуществления органоид предстательной железы, выращенный в отсутствие тестостерона, не содержит никаких стволовых клеток.
Добавление тестостерона в среду для культивирования клеток предстательной железы
Авторы изобретения показали, что добавление (дигидро)тестостерона к условиям культивирования для органоидов предстательной железы приводит к тому, что большинство клеток дифференцируются в CK8+ люминальные клетки, которые образуют единственный слой эпителия, который сам по себе складывается в два слоя. Органоиды предстательной железы, выращенные в присутствии тестостерона, состоят по большей части из клеток протоков со вторым слоем базальных клеток или без него. Структура напоминает структуру in vivo. Присутствуют как дифференцированные, так и делящиеся клетки, а также стволовые клетки и предшественники. Таким образом, в некоторых вариантах осуществления, например, при культивировании в среде, содержащей тестостерон, органоид предстательной железы представляет собой трехмерный органоид, содержащий кистозные структуры и просвет. В некоторых вариантах осуществления органоид предстательной железы содержит CK8+ люминальные клетки, которые образуют монослой эпителия. В некоторых вариантах осуществления монослой складывается в два или более слоев. В других вариантах осуществления органоид может содержать области распределенных на слои клеток. В некоторых вариантах осуществления органоид предстательной железы содержит дифференцированные клетки при сохранении популяции делящихся стволовых клеток. В некоторых вариантах осуществления форма органоида определяется происхождением клеточного или тканевого исходного материала (т.е. положением в предстательной железе до выделения). Предстательная железа состоит из различных долей или областей, которые обладают различными эпителиальными структурами, описанными выше (распределенными на слои и сложенными), После культивирования in vitro органоиды, по-видимому, сохраняют отличительную макроскопическую структуру (распределенную на слои или сложенную) той части предстательной железы, из которой она происходит.
Органоиды печени
Структурно большинство органоидов печени мыши согласно изобретению часто имеют удлиненную форму. Они могут включать одну или несколько почкующихся структур - одноклеточный эпителиальный слой, который имеет структуру, не являющуюся непохожей на желчный проток. При конфокальной микроскопии структуры могут окрашиваться положительно на кератин. Они могут включать клетки с полярным ядрами и небольшой цитоплазмой. Органоиды могут иметь срез, который образован множеством слоев; такие клетки часто имеют тенденцию к тому, чтобы их ядра были расположены более центрально в клетках, т.е. не являются полярными. Клетки в многослойном срезе могут самоорганизовываться так, чтобы они включали щель или просвет между клетками. В некоторых вариантах осуществления органоиды печени человека по изобретению имеют в общем кистозную структуру.
В некоторых вариантах осуществления органоид печени представляет собой трехмерный органоид с кистозной структурой (например, см. фиг.30). В условиях увеличения в количестве органоид может состоять из стволовых клеток и клеток-предшественников, где определены два домена: (1) подобный протокам домен, образованный из однослойного кубического эпителия (положительного на маркер протоков Krt19) с клетками, выстилающими центральный просвет; и (2) псевдо-распределенный на слои эпителиальный домен, где выявляются положительные по krt19 клетки и рассеянные положительные по альбумину клетки. Эта архитектура (области с однослойным эпителием вместе с областями псевдо-распределенного на слои эпителия) напоминает эмбринальную почку печени. В условиях увеличения в количестве полностью дифференцированные клетки не присутствуют, хотя специфичные для гепатоцитов/гепатобластов маркеры в некоторых вариантах осуществления могут выявляться. Условия дифференцировки приводят к образованию кистозного органоида, где подобный протоку домен (однослойный эпителий) утрачивается и вся структура становится псевдо-распределенным на слои эпителием, содержащим >50% полярных гепатоцитов.
Органоид печени, предпочтительно, содержит гепатоцит и холангиоцит (хотя гепатоциты, главным образом, наблюдаются после дифференцировки в DM и не требуются для увеличения в количестве), более предпочтительно, где может быть выявлен по меньшей мере один из следующих маркеров: по меньшей мере один маркер гепатоцитов, такой как альбумин, транстиретин, B-1 интегрин и глутаминсинтетаза и/или по меньшей мере один из CYP3A11, FAH, tbx3, TAT и Gck, и/или по меньшей мере один маркер холангиоцитов, такой как кератин 7 и 19. Квалифицированному специалисту известно, как выявить каждый из этих маркеров (т.е. ОТ-ПЦР и/или иммунофлуоресценция). Предпочтительно, экспрессию каждого из этих маркеров оценивают так, как описано в экспериментальной части. Каждый из этих маркеров обычно экспрессирутся после культивирования по меньшей мере две недели, три недели или один месяц с использованием способа по изобретению. Анализ на микрочипах органоидов в обоих условиях культивирования показал, что органоиды печени напоминают ткань печени взрослых.
Предпочтительно, все клетки в органоиде печени экспрессируют поверхностные маркеры гепатоцитов. Например, в некоторых вариантах осуществления по меньшей мере 50% (например, 50-60%), по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 99% или 100% клеток в органоиде печени экспрессируют маркеры гепатоцитов. В некоторых вариантах осуществления приблизительно 35% клеток в органоиде печени экспрессируют поверхностный маркер гепатоцитов, например, 25-45%, 30-40%, 33-37%, 35% или менее, или 15-35% клеток. В некоторых вариантах осуществления фаза увеличения в количестве может иметь меньше гепатоцитов, например, менее 20%, менее 10%, менее 5% клеток, менее 2%, менее 1%, предпочтительно, 0% клеток. Предпочтительно, клетки и органоиды, полученные согласно изобретению, также обладают функциями гепатоцитов, такими как экспрессия или положительное окрашивание на зрелые маркеры печени альбумин, интегрин B-1, CK-8, CK-18, транстиретин (TTR), глюкозу 6P, Met, глутаминсинтазу (Glul), трансферрин, Fahd1, Fahd2a, K7, K19 и изоформы цитохрома P450 3A13 (CYP3A13), 51 (CYP51) 2D10 (CYP2D10), 2j6 (CYP2j6), 39A1 (CYP39A1), 4A10 (CYP4A10), 4F13 (CYP4F13) 4F16 (CYP4F16), CYP4B1 и 20A1(CYP20A1). Также ген печени эмбриона AFP в некоторых вариантах осуществления не выявляется ни в одних из обоих условий культивирования, как во взрослой печени. В некоторых вариантах осуществления экспрессия альфа-фетопротеина немного превышает фоновую экспрессию генов.
Также, хорошо известные факторы транскрипции печени, такие как HNF1a, HNF1b и HNF4a, на высоком уровне экспрессируются в обоих условиях.
Поскольку печень и поджелудочная железа являются в высокой степени сходными органами, авторы настоящего изобретения исследовали, экспрессируют ли их культуры печени также специфические для поджелудочной железы гены. Поджелудочную железу функционально подразделяют на эндокринную и экзокринную поджелудочную железу. Эндокринная поджелудочная железа, главным образом, характеризуется экспрессией инсулина, глюкагона и соматостатина. Экспрессия этих гормонов строго контролируется набором эндокринных специфичных для поджелудочной железы факторов транскрипции, наиболее важными из которых являются Pdx1 и NeuroD. Экзокринная поджелудочная железа образована ацинарными и протоковыми компартментами, ответственными за продукцию пищеварительных ферментов амилазы, липазы поджелудочной железы и химотрипсина, среди прочих. Экспрессия этих генов также регулируется специфическими экзокринными генами поджелудочной железы, такими как Ptf1.
Специфичные для поджелудочной железы гены Ptf1a, амилаза поджелудочной железы (Amy2a4), липаза поджелудочной железы (Pnlip), инсулин (ins1 и ins2), глюкагон (Gcg), химотрипсин (cela1), Pdx1 и NeuroD отсутствовали в культурах печени, описанных в настоящем описании.
В некоторых вариантах осуществления один или несколько или все из следующих генов экспрессируются в органоидах печени на сходном уровне с соответствующими генами в гепатоцитах взрослой печени: Aqp1, Bmp2, Apo3, Apol7a, Sord, C3, Ppara, Pparg, tbx3, lgf1, ll17rb, ll1b, Tgfbi, Apoa1, Apoa4, Apob, Cyp26b1, Cyp27a1, Cyp2b13, Cyp2b9, Cyp2c37, Cyp2f2, Cyp2g1, Cyp2j13, Cyp3a11, Cyp4a10 и Cypf14. Например, см. фиг.27A.
В некоторых вариантах осуществления один или несколько из следующих генов экспрессируется в органоидах печени на аналогичном уровне выключения по сравнению с соответствующими генами в гепатоцитах взрослой печени: Ccl2, Osmr, Icam1 и Cxcl2.
В некоторых вариантах осуществления один или оба из следующих генов дифференциально экспрессируются как в органоиде печени, так и в печени новорожденных: mKi67 и cdkn3, что означает, что экспрессия этих генов является более высокой в органоидах, чем в дифференцированных органоидах или целом органе.
В некоторых вариантах осуществления один, два или все из следующих генов экспрессируются на сходном уровне в органоиде печени и в печени новорожденных: cyp2j6, olfm4 и Lefty 1. Например, см. фиг.27B.
В некоторых вариантах осуществления органоид печени по изобретению имеет фенотип протока при культивировании в среде для увеличения в количестве по изобретению (например, EM1 или EM2).
В некоторых вариантах осуществления органоид печени по изобретению экспрессирует маркеры взрослой печени при культивировании в среде для дифференцировки по изобретению.
В одном из вариантов осуществления органоид печени по изобретению имеет профиль экспрессии генов, как показано на фиг.27C.
В особенно предпочтительном варианте осуществления популяция клеток или органоид печени мыши по изобретению имеют профиль экспрессии генов, как показано на фиг.28. Например, в одном предпочтительном варианте осуществления популяция клеток или органоид печени мыши по изобретению:
a) экспрессируют по меньшей мере один (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11), предпочтительно, все из следующих маркеров стволовых клеток: lgr5, lgr4, epcam, Cd44, Tnfrsf19, Sox9, Sp5, Cd24a, Prom1, Cdca7 и Elf3; и/или
b) не экспрессируют следующий маркер стволовых клеток: lgr6; и/или
с) экспрессируют по меньшей мере один (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19), предпочтительно, все из следующих маркеров гепатоцитов или холангиоцитов при выращивании в среде для увеличения в количестве по изобретению: Hnf1a, Hnf1b, Hnf4a, Hhex, Onecut1, Onecut2, Prox1, Cdh1, Foxa2, Gata6, Foxm1, Cebpa, Cebpb, Cebpd, Cebpg, Glul, Krt7, Krt19 и Met; и/или
d) не экспрессируют по меньшей мере один (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17) из следующих генов при выращивании в среде для увеличения в количестве по изобретению: afp, Ins1, Ins2, Gcg, Ptf1a, Cela1, Cela2a, Cela3b, Neurod1, Neurod2, Neurog1, Neurog2, Neurog3, Amy2a4, Igf1r, Igf2 и Cd34; и/или
e) экспрессируют по меньшей мере один (например, 1, 2 или 3) из следующих генов перепрограммирования: Klf4, Myc и Pou5f1, и/или
f) не экспрессируют следующий ген перепрограммирования: Sox2.
где экспрессию генов, предпочтительно, выявляют путем измерения экспрессии на уровне мРНК, например, с использованием микрочипа.
Более предпочтительно, популяция клеток или органоид печени мыши имеют следующие признаки a)-f) выше.
В некоторых вариантах осуществления профиль экспрессии генов, описанный выше для популяции клеток печени или органоида печени мыши по изобретению, представляет собой профиль экспрессии генов для популяции клеток или органоида мыши, культивируемых в среде для увеличения в количестве клеток печени по изобретению.
В некоторых вариантах осуществления предусмотрена популяция клеток или органоид печени человека по изобретению, которые имеют признак экспрессии генов, представленный на фиг.29. Например, популяция клеток или органоид печени человека, культивируемые в EM1 по изобретению, предпочтительно, экспрессируют гены, указанные на фиг.29 в качестве экспрессируемых в среде для культивирования клеток EM1. Например, популяция клеток или органоид печени человека, культивируемые в EM2 по изобретению, предпочтительно, экспрессируют гены, указанные на фиг.29, в качестве экспрессируемых в среде EM2 для культивирования клеток. Например, популяция клеток или органоид печени человека, культивируемые в DM по изобретению, предпочтительно, экспрессируют гены, указанные на фиг.29 в качестве экспрессируемых в среде DM для культивирования клеток.
Например, в одном предпочтительном варианте осуществления популяция клеток или органоид печени человека по изобретению:
a) экспрессируют по меньшей мере один (например, 1, 2, 3, 4, 5, 6, 7, 8, 9), предпочтительно, все из следующих генных профилей стволовых клеток: LGR4, TACSTD1/Epcam, CD44, SOX9, SP5, CD24, PROM1, CDCA7 и ELF3; и/или
b) экспрессируют по меньшей мере один (например, 1, 2, 3, 4), предпочтительно, все из следующих перепрограммирующих генов: KLF4, MYC, POU5F1 и SOX2; и/или
c) экспрессируют по меньшей мере один (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19), предпочтительно, все из следующих специфичных для гепатоцитов/холангиоцитов генов: HNF1A, HNF1B, HNF4A, HHEX, ONECUT1, ONECUT2, PROX1, CDH1, FOXA2, GATA6, FOXM1, CEBPA, CEBPB, CEBPD, CEBPG, GLUL, KRT7, KRT19 и MET; и/или
d) не экспрессируют по меньшей мере один (например, 1, 2, 3, 4, 5, 6), предпочтительно, все из следующих генов, специфичных для гепатоцитов/холангиоцитов: NEUROG2, IGF1R и CD34, AFP, GCG и PTF1A, например, они не экспрессируют NEUROG2, IGF1R и CD34; и/или
e) экспрессируют по меньшей мере один (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18), предпочтительно, все из следующих генов, специфичных для гепатоцитов: TTR, ALB, FAH, TAT, CYP3A7, APOA1, HMGCS1, PPARG, CYP2B6, CYP2C18, CYP2C9, CYP2J2, CYP3A4, CYP3A5, CYP3A7, CYP4F8, CYP4V2 и SCARB1;
где экспрессию генов, предпочтительно, выявляют путем измерения экспрессии на уровне мРНК, например, с использованием микрочипа.
Более предпочтительно, популяция клеток или органоид печени человека по изобретению имеют все из признаков a)-e) выше.
В некоторых вариантах осуществления гены в популяции клеток или органоиде печени человека по изобретению активированы или подавлены относительно экспрессии эталонной РНК, как показано на фиг.29. Предпочтительно, эталонная РНК представляет собой универсальную эталонную РНК человека (Stratagene, каталожный номер #740000). В некоторых вариантах осуществления ген активирован или подавлен относительно эталонной РНК, если он также показан на фиг.29 как активированный или подавленный относительно эталонной РНК, однако степень активации или подавления не должна быть одинаковой. В других вариантах осуществления степень активации или подавления составляет +/-35%, +/-30%, +/-25%, +/-20%, +/-20%, +/-15%, +/-10%, +/-5%, +/-3%, или более предпочтительно, отличается в +/-1,5 раза, +/-2 раза, +/-3 раза, +/- 5 раз или является приблизительно такой же, как показано на фиг.29. В других вариантах осуществления абсолютный уровень экспрессии генов в органоиде человека по изобретению составляет +/-35%, +/-30%, +/-25%, +/-20%, +/-15%, +/-10%, +/-5%, +/-3%, или отличается в +/-1,5 раза, +/-2 раза, +/-3 раза, +/- 5 раз или является приблизительно таким же, как показано на фиг.29.
Популяция клеток или органоиды печени человека также, предпочтительно, экспрессируют Lgr5 и/или Tnfrsf19, предпочтительно, оба из них. В некоторых вариантах осуществления популяция клеток или органоиды печени человека при культивировании в среде для увеличения в количестве по изобретению экспрессируют Lgr5 и/или Tnfrsf19, предпочтительно, оба из них. Предпочтительно, экспрессию Lgr5 и/или Tnfrsfr19 выявляют с помощью ОТ-ПЦР. В некоторых вариантах осуществления Lgr5 и/или Tnfrsf19 присутствуют на значительно более низких уровнях экспрессии в органоидах или клетках при культивировании в среде для дифференцировки по сравнению с их уровнем экспрессии в органоидах или клетках при культивировании в среде для увеличения в количестве (например, по меньшей мере в 2 раза, по меньшей мере в 3 раза, по меньшей мере в 4 раза, по меньшей мере в 5 раз, по меньшей мере в 10 раз, по меньшей мере в 15 раз более низкие уровни).
Клетки и органоиды печени в соответствии с настоящим изобретением, предпочтительно, могут быть способны секретировать альбумин, например, со скоростью от приблизительно 1 мкг в час на 106 клеток до 10 мкг в час на 106 клеток, предпочтительно, от 2 мкг до 6 мкг в час на 106 клеток.
Более того, такие клетки и органоиды печени могут секретировать мочевину. Например, в 35-мм чашках с клетками активность синтеза мочевины может составлять от 1 мкг до 50 мкг в течение 48 часов, предпочтительно, от 5 мкг до 30 мкг.
Клетки и органоиды печени согласно изобретению могут демонстрировать видимые запасы гликогена, например, при окрашивании. Способность клеток и органоидов согласно изобретению активно синтезировать гликоген можно исследовать по переключению культуральной среды со среды для дифференцировки с низким содержанием глюкозы на DMEM с высоким содержанием глюкозы, дополненную 10% FBS и 0,2 мкМ дексаметазоном, в течение двух суток.
Клетки и органоиды печени согласно изобретению могут обладать активностью индуцибельного цитохрома P450 (например, CYP1A). Такую активность можно исследовать, например, с использованием анализа с этоксирезоруфин-O-деэтилазой (EROD) (Cancer Res, 2001, 61: 8164-8170). Например, клетки или органоиды можно подвергать воздействию субстрата P450, такого как 3-метилхолантрен и уровни активности EROD сравнивать с контрольными клетками.
Морфологически клетки органоида печени выглядят похожими на гепатоциты.
Предпочтительный органоид печени содержит или состоит из кистозной структуры с почками на наружном слое клеток и с центральным просветом, как представлено на фиг.30. Этот органоид печени может иметь одну или несколько (например, 2, 3 или все 4) из следующих характеристик: (a) наличие плотности клеток >5×105 клеток/см3, предпочтительно, >10×105 клеток/см3; (b) наличие толщины, эквивалентной 2-30 слоям клеток, предпочтительно, толщины, эквивалентной 2-15 слоям клеток; (c) клетки взаимно контактируют в трех измерениях, (d) демонстрируют функцию, присущую здоровой ткани печени, (e) имеют удлиненную форму с 2 определенными доменами, т.е. однослойным эпителиальным доменом, где выявляются высоко полярные клетки и экспрессируются маркеры кератина (этот домен напоминает домен желчного протока), и другой домен составляет основную массу органоида и образован многослойным эпителием с неполярными клетками, где может выявляться экспрессия альбумина. Квалифицированному специалисту понятно, что такой органоид печени, предпочтительно, не является фрагментом печени и/или не содержит кровеносного сосуда, и/или не содержит долю печени или желчный проток.
В контексте изобретения фрагмент печени является частью взрослой печени, предпочтительно, взрослой печени человека. Предпочтительно, органоид печени, идентифицированный в настоящем описании, таким образом, не является фрагментом печени. Органоид печени, предпочтительно, получают с использованием клетки из взрослой печени, предпочтительно, эпителиальной стволовой клетки из взрослой печени, более предпочтительно, экспрессирующей Lgr5 эпителиальной стволовой клетки из взрослой печени. Органоид печени также можно получать из любой клетки, которая при повреждении или культивировании экспрессирует Lgr5 и, таким образом, экспрессирующей Lgr5 осуществляющей цикл стволовой клетки.
В некоторых вариантах осуществления органоид печени содержит клетки, которые экспрессируют Lgr5. Например, в некоторых вариантах осуществления по меньшей мере 2%, более предпочтительно, по меньшей мере 5%, по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95% клеток в органоиде печени экспрессируют Lgr5. Аналогично, изобретение относится к клетке или популяции клеток, которые экспрессируют Lgr5, где указанные клетки получены из органоида печени по изобретению. Также изобретение охватывает потомков таких клеток.
В одном из вариантов осуществления органоид печени представляет собой органоид печени, который все еще культивируют с использованием способа по изобретению и, таким образом, он контактирует с внеклеточным матриксом. Предпочтительно, органоид печени погружен в немезенхимный или мезенхимный внеклеточный матрикс. В контексте изобретения, "в контакте" означает физический или механический или химический контакт, что означает, что для отделения указанного органоида печени от указанного внеклеточного матрикса необходимо применить силу.
В предпочтительном варианте осуществления органоид печени можно культивировать в течение по меньшей мере 2, 3, 4, 5, 6, 7, 8, 9, 10 недель или 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 месяцев или дольше. В некоторых вариантах осуществления органоид печени увеличивают в количестве или поддерживают в культуре в течение по меньшей мере 3 месяцев, предпочтительно, по меньшей мере 4 месяцев, по меньшей мере 5 месяцев, по меньшей мере 6 месяцев, по меньшей мере 7 месяцев, по меньшей мере 9 месяцев, или по меньшей мере 12 месяцев или более. Предпочтительно, органоид печени, культивируемый с использованием среды для увеличения в количестве по изобретению, содержащей ингибитор TGF-бета, можно культивировать в течение по меньшей мере 4 недель, более предпочтительно, по меньшей мере 5 недель с увеличением в количестве, составляющим в 5 раз в неделю или два или более удвоений популяций в неделю (например, в течение по меньшей мере 10 удвоений, по меньшей мере 20 удвоений, более предпочтительно, по меньшей мере 25 удвоений, например, по меньшей мере 30 удвоений). Предпочтительно, органоид печени, культивируемый с использованием среды для увеличения в количестве по изобретению, содержащей активатор каскада простагландинов в дополнение к ингибитору TGF-бета, можно культивировать в течение по меньшей мере 7 недель, более предпочтительно, по меньшей мере 8 недель, при 2 или более удвоений (например, 2-3 удвоениях) в неделю (т.е. по меньшей мере 15 удвоений, по меньшей мере 25 удвоений, по меньшей мере 30 удвоений, по меньшей мере 32 удвоений, по меньшей мере 35 удвоений, например, 32-40 удвоений или по меньшей мере 40 удвоений, например, по меньшей мере 50 удвоений). Таким образом, предпочтительно, органоид печени по изобретению, например, органоид печени человека, получают с использованием среды для увеличения в количестве по изобретению.
В другом предпочтительном варианте осуществления органоид печени происходит из единичной клетки, предпочтительно, экспрессирующей Lgr5, более предпочтительно, где единичная клетка содержит конструкцию нуклеиновой кислоты, содержащую представляющую интерес молекулу нуклеиновой кислоты.
Состав органоида и экспрессия генов
Органоиды крипта-ворсинка, крипты толстого кишечника и поджелудочной железы, как правило, содержат стволовые клетки и/или клетки-предшественники и, таким образом, эти органоиды обладают определенными общими профилями экспрессии генов. В некоторых вариантах осуществления можно выявлять один или несколько (например, 1, 2, 3, 4, 5, 6 или 7) или все из следующих маркеров: LGR5, LGR4, epcam, Cd44, Sox9, Cd24a и CD133/Prom1 и необязательно Tnfrsf19. В другом варианте осуществления можно выявлять экспрессию одного или двух или всех из следующих генов предшественников: Pdx1, Nkx2.2 и Nkx6.1. Ожидается, что после дифференцировки профили экспрессии генов органоида крипта-ворсинка, крипты толстого кишечника и поджелудочной железы станут отличаться, поскольку дифференцированные органоиды экспрессируют тканеспецифические взрослые маркеры, например, такие как инсулин в поджелудочной железе.
Органоиды крипта-ворсинка
В некоторых вариантах осуществления изобретения органоиды содержат подобные криптам-ворсинкам удлинения, которые содержат все типы дифференцированных эпителиальных клеток, включая пролиферативные клетки, клетки Панета, энтероциты и бокаловидные клетки. В некоторых вариантах осуществления органоиды крипта-ворсинка по изобретению не содержат миофибробластов или других неэпителиальных клеток. Органоид крипта-ворсинка по изобретению предпочтительно, содержит энтероциты, включая осуществляющие всасывание энтероциты, бокаловидные клетки, энтероэндокринные клетки и клетки Панета, в структурах, подобных структурам крипта-ворсинка. Предпочтительно, может быть выявлен по меньшей мере один (например, 2, 3, 4, 5 или 6) из следующих маркеров: SMOC2, CDCA7, OLFM4, ASCL2, AXIN2 и/или Lgr5 Tnfrsf19, CD24a, Sox9, CD44, Prom1 (см. фиг.2e и фиг.14). В некоторых вариантах осуществления могут быть выявлены маркеры RNF43 и ZNRF3. В некоторых вариантах осуществления один или несколько (например, 1, 2, 3, 4 или 5) или все из SMOC2, CDCA7, OLFM4, ASCL2, AXIN2 и/или Lgr5 по меньшей мере в 2 раза, в 3 раза или 4 раза активированы в криптах, в то время как маркеры, которые по меньшей мере в 2 раза, в 3 раза или в 4 раза подавлены в криптах, включают по меньшей мере один или несколько (например, 1, 2, 3 или 4) или все из ABCG1, ENPP3, CSTE, MUC17 и/или APOA1. В этом контексте "активация" представляет собой активацию относительно ворсинки кишечника или верхнего отдела крипты толстого кишечника. Анализ на микрочипах, сравнивающий экспрессию генов дифференцированных клеток органоидов со стволовыми клетками показал, что органоиды крипта-ворсинка тонкого кишечника и органоиды толстого кишечника обладают сравнимыми молекулярными профилями кишечных крипт, включающими экспрессию генов стволовых клеток кишечника. Таким образом, изобретение также относится к органоиду толстого кишечника, имеющему молекулярный профиль, описанный выше для органоидов крипта-ворсинка. Органоиды, культивируемые in-vitro, отчетливо проявляют сходный профиль экспрессии со свежевыделенными криптами тонкого кишечника и экспрессируют известные маркеры стволовых клеток.
В некоторых вариантах осуществления мРНК, кодирующая один или несколько генов (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25), приведенных на фиг.14 (например, все из генов, выделенных затемнением на фиг.14) в качестве активируемых в органоидах крипта-ворсинка или органоидах толстого кишечника, соответственно, активированы в органоиде крипта-ворсинка или в органоиде толстого кишечника по изобретению по сравнению со свежевыделенными ворсинками тонкого кишечника, при определении с помощью микрочипа. В некоторых вариантах осуществления мРНК, кодирующая один или несколько (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25) генов, приведенных на фиг.14 (например, все из генов, выделенных затемнением на фиг.14) в качестве подавленных в органоидах крипта-ворсинка или органоидах толстого кишечника, соответственно, подавлены в органоиде крипта-ворсинка или органоиде толстого кишечника по изобретению по сравнению со свежевыделенными ворсинками тонкого кишечника при определении с помощью микрочипа. В некоторых вариантах осуществления кратность активации или подавления является такой, как указано на фиг.14 + /- 25%, например, +/ - 20%, +/ - 15%, + /- 10%, +/-5%, +/-3% или приблизительно как указано на фиг.14. Например, органоид крипта-ворсинка по изобретению может иметь ADORA2B, активированный в 9,54 раз +/- 25% по сравнению со свежевыделенными ворсинками тонкого кишечника. То же самое применимо, с соответствующими изменениями, к другим генам, приведенным в таблице 14.
В некоторых вариантах осуществления органоиды крипта-ворсинка демонстрируют природную экспрессию Lgr5. В некоторых вариантах осуществления органоиды крипта-ворсинка демонстрируют природную экспрессию по меньшей мере Lgr5 и одного или нескольких (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16) или всех из маркеров стволовых клеток из группы, состоящей из: CK19, нестина, соматостатина, CXCR4+, CD133+, DCAMKL-1, CD44, Sord, Sox9, CD44, Prss23, Sp5, Hnf1α, Hnf4a, Sox9, KRT7 и KRT19. Дополнительно или альтернативно, органоиды крипта-ворсинка могут быть охарактеризованы экспрессией одного или нескольких или всех (например, 1 или 2) из: Sord и/или Prss23. Дополнительно или альтернативно, органоиды крипта-ворсинка могут быть охарактеризованы экспрессией CD44 и/или Sox9. В другом варианте осуществления органоиды крипта-ворсинка демонстрируют экспрессию одного или нескольких (например, 1, 2, 3, 4, 5, 6, 7, 8, 9) или всех из маркеров из группы, состоящей из: lgr5, lgr4, epcam (tacstd1), Cd44, Tnfrsf19, Sox9, Sp5, Cd24a, Prom1 и Cdca7.
В некоторых вариантах осуществления органоид крипта-ворсинка содержит клетки Панета, экспрессирующие лизоцим.
Органоиды толстого кишечника
В некоторых вариантах осуществления органоид толстого кишечника содержит энтероэндокринные клетки (например, выявляемые с использованием окрашивания на хроматогранин A), бокаловидные клетки (выявляемые с использованием окрашивания на муцин 2). В некоторых вариантах осуществления менее 10% клеток в органоиде толстого кишечника представляют собой энтероэндокринные клетки (например, 0,01-5%, 0,1-3%). В некоторых вариантах осуществления менее 30% клеток в органоиде толстого кишечника представляют собой бокаловидные клетки (например, 1-25%, 1-15%, 5-10%). В некоторых вариантах осуществления распределение энтероэндокринных клеток и/или бокаловидных клеток является таким, как показано на фиг.1d.
В некоторых вариантах осуществления органоид толстого кишечника содержит зрелые энтероциты (например, визуализируемые путем окрашивания на щелочную фосфатазу). В некоторых вариантах осуществления менее 10% клеток органоида толстого кишечника представляют собой зрелые энтероциты (например, менее 5%, менее 3%, 0,01-5%, 0,1-3%, 0,1-5%).
В предпочтительных вариантах осуществления органоид толстого кишечника не содержит клетки Панета, поскольку в естественных условиях в толстом кишечнике in vivo клетки Панета отсутствуют.
В некоторых вариантах осуществления органоиды толстого кишечника демонстрируют природную экспрессию Lgr5.
В некоторых вариантах осуществления органоид толстого кишечника экспрессирует один или несколько (например, 1, 2, 3 или 4) из Villin1, Alpi, ChgA и Muc2. В некоторых вариантах осуществления относительное количество мРНК Villin1, экспрессируемой органоидом толстого кишечника по изобретению по сравнению со свежевыделенной криптой толстого кишечника, составляет по меньшей мере 3% (например, по меньшей мере 5%, по меньшей мере 8%, по меньшей мере 10%), например, 5-15%. В некоторых вариантах осуществления относительное количество мРНК Alpi, экспрессируемое органоидом толстого кишечника по изобретению по сравнению со свежевыделенной криптой толстого кишечника, составляет по меньшей мере 0,5% (например, по меньшей мере 1%, по меньшей мере 2%), например, 0,5-5%. В некоторых вариантах осуществления относительное количество мРНК ChgA, экспрессируемой органоидом по изобретению по сравнению со свежевыделенной криптой толстого кишечника, составляет по меньшей мере 15% (например, по меньшей мере 20%, по меньшей мере 22%), например, 15-30%. В некоторых вариантах осуществления относительное количество мРНК Muc2, экспрессируемое органоидом толстого кишечника по изобретению по сравнению со свежевыделенной криптой толстого кишечника, составляет по меньшей мере 20% (например, по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 35%), например, 25-37%.
В некоторых вариантах осуществления органоид толстого кишечника человека по изобретению экспрессирует известные маркеры стволовых клеток.
Органоиды поджелудочной железы
Поджелудочная железа содержит три класса типов клеток: клетки протоков, ацинарные клетки и эндокринные клетки. Эндокринные клетки продуцируют гормоны глюкагон, инсулин, соматостатин и панкреатический полипептид (PP), которые секретируются в кровоток и помогают организму регулировать метаболизм сахаров. Ацинарные клетки являются частью экзокринной системы, которая продуцирует пищеварительные ферменты, и клетки протоков образуют протоки поджелудочной железы, которые соединяют ацинарные клетки с органами пищеварения. Полагают, что в процессе развития островки Лангерганса происходят из эндокринных клеток-предшественников, которые возникают из протока поджелудочной железы и после дифференцировки агрегируют с образованием островков Лангерганса. Островки Лангерганса содержат α-клетки, β-клетки, δ-клетки и PP-клетки.
Клетки органоида поджелудочной железы могут иметь профиль экспрессии, который напоминает маркеры клеток протоков, такие как один или несколько (например, 1, 2 или все) из K7, K19 и Hnf1b, и/или один или несколько маркеров стволовых клеток, таких как Sox9 и/или Onecut1. Это, вероятно, является частью их профиля стволовых клеток. Как правило, наблюдают меньшее количество маркеров дифференцировки. В некоторых вариантах осуществления, в которых клетку выделяют из протока поджелудочной железы для получения органоида поджелудочной железы по изобретению, тип клетки, который дает начало органоиду поджелудочной железы по изобретению не является клеткой протока (что означает эпителиальные клетки, положительные по кератину 7 и кератину 19, которые образуют трубку протока), а представляет собой клетку, прикрепленную к протоку поджелудочной железы, что означает клетку, которая расположена в следующем слое клеток после протока в контакте с тканью поджелудочной железы (т.е. не обращена в просвет протока). Таким образом, в вариантах осуществления, в которых тип клетки, который дает начало органоиду поджелудочной железы не является клеткой протока, органоид поджелудочной железы не экспрессирует K7 или K19. Однако такой органоид поджелудочной железы все еще предпочтительно экспрессирует один или несколько основных маркеров предшественников стволовых клеток, таких как Sox9.
Органоид поджелудочной железы по изобретению, предпочтительно, содержит α-клетки, β-клетки, δ-клетки и PP-клетки. В следующем предпочтительном варианте осуществления органоид поджелудочной железы содержит бета-клетки. Например, органоид поджелудочной железы может содержать более 1%, более 5%, более 10%, более 15% или более 20% бета-клеток. Экспрессию инсулина можно использовать в качестве маркера для бета-клеток. В альтернативном варианте осуществления органоид поджелудочной железы содержит типы клеток-предшественников, необязательно происходящие из протоков, которые могут дать начало дифференцированным типам клеток при трансплантации человеку или животному. В предпочтительном варианте осуществления типы клеток-предшественников могут дать начало секретирующим инсулин бета-клеткам при трансплантации человеку или животному. Авторы изобретения показали, что органоиды поджелудочной железы человека, выращенные с помощью сред и способов по изобретению, можно трансплантировать мышам и стимулировать инсулин-секретирующие клетки в пределах одного месяца (см. пример 4). Можно без труда понять, что это может обеспечить революционные способы лечения для пациентов с диабетом и дефицитом инсулина.
В некоторых вариантах осуществления органоид поджелудочной железы по изобретению может содержать клетки протоков, ацинарные клетки и эндокринные клетки. В некоторых вариантах осуществления в качестве маркера для клеток протоков используют K19.
В некоторых вариантах осуществления в островках поджелудочной железы или островках Лангерганса существует бета-клетка. Островок, как правило, содержит in vivo приблизительно 1500 клеток, например, 1300-1700 клеток. В одном из вариантов осуществления органоид поджелудочной железы содержит по меньшей мере 0,5%, по меньшей мере 1%, по меньшей мере 1,5%, по меньшей мере 2%, по меньшей мере 3%, по меньшей мере 5%, по меньшей мере 10%, по меньшей мере 15%, по меньшей мере 20%, по меньшей мере 25%, по меньшей мере 30% или более островков Лангерганса по массе. В некоторых вариантах осуществления островки Лангерганса органоида поджелудочной железы состоят приблизительно на 65-90% из бета-клеток, приблизительно на 15-20% из альфа-клеток, приблизительно на 3-10% из дельта-клеток и приблизительно на 1% из PP-клеток. Однако это ни в коем случае не является исключительным. Например, в некоторых вариантах осуществления желательно иметь множество бета-клеток в органоиде по изобретению. Альтернативно органоид может содержать клетки-предшественники, которые могут быть трансплантированы так, чтобы они дифференцировались in vivo.
В некоторых вариантах осуществления органоид поджелудочной железы экспрессирует один, два или все три из Pdx1, Nkx2.2 и Nkx6.1. Органоид поджелудочной железы может экспрессировать один, два, три или все четыре из NeuroD, Pax6, Pax4 и Mafa. Pax4 служит в качестве маркера на присутствие продуцирующих инсулин клеток, поскольку он является необходимым фактором транскрипции для дифференцировки продуцирующих инсулин клеток из эндокринных клеток-предшественников в ходе эмбрионального развития. Органоид поджелудочной железы может экспрессировать Ngn3.
В некоторых вариантах осуществления по меньшей мере один (например, 1, 2, 3, 4, 5) из следующих маркеров может быть выявлен в органоиде поджелудочной железы по изобретению: инсулин (ins1 и/или ins2), глюкагон (Gcg), соматостатин, Pdx1 и NeuroD. В некоторых вариантах осуществления по меньшей мере один (например, 1, 2, 3, 4, 5) из следующих маркеров может быть выявлен в органоиде поджелудочной железы по изобретению: инсулин (ins1 и/или ins2), глюкагон (Gcg), соматостатин, Pdx1 и NeuroD, и следующие маркеры не выявляются: ptf1a, amy2a4, Pnlip и cela1. В некоторых вариантах осуществления по меньшей мере один (например, 1, 2, 3, 4, 5, 6, 7, 8 или 9) из следующих маркеров может выявляться в органоиде поджелудочной железы по изобретению: Ptf1a, панкреатическая амилаза (Amy2a4), панкреатическая липаза (Pnlip), инсулин (ins1 и/или ins2), глюкагон (Gcg), соматостатин, химотрипсин (cela1), Pdx1 и NeuroD.
В некоторых вариантах осуществления органоиды демонстрируют природную экспрессию Lgr5. В некоторых вариантах осуществления органоиды поджелудочной железы демонстрируют природную экспрессию по меньшей мере Lgr5 и одного или нескольких (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20) маркеров стволовых клеток, выбранных из группы, состоящей из: CK19, нестина, CXCR4+, CD133+, DCAMKL-1, CD44, Sord, Sox9, CD44, Prss23, Sp5, Hnf1α, Hnf4a, Sox9, KRT7 и KRT19, prom1, Cd24a, Lgr4, epcam. Альтернативно или дополнительно, в некоторых вариантах осуществления органоиды поджелудочной железы могут характеризоваться природной экспрессией одного или нескольких (например, 1, 2, 3 или 4) из: CK19, нестина, (инсулин, глюкагон) и CXCR4+.
В некоторых вариантах осуществления органоиды поджелудочной железы или клетки по изобретению экспрессируют сматостатин. Соматостатин представляет собой гормон, экспрессируемый в дифференцированных дельта-клетках, и также он может служить в качестве маркера для дельта-клеток.
Альтернативно или дополнительно, в некоторых вариантах осуществления органоиды поджелудочной железы демонстрируют природную экспрессию одного или нескольких ранних эндокринных маркеров, например, по меньшей мере одного или нескольких (например, 1, 2, 3, 4, 5, 6 или 7) из следующих ранних эндокринных маркеров: Sox9, Hnf1b, Hnf6, Hnf1a, Nkx2.2, Nkx6.1 и Pdx1.
Альтернативно или дополнительно, в некоторых вариантах осуществления органоиды поджелудочной железы демонстрируют природную экспрессию одного или нескольких ранних эндокринных маркеров, например, по меньшей мере одного или нескольких (например, 1, 2, 3 или 4) из следующих эндокринных маркеров: Foxa2, Hnf6, Hnf1b и Sox9. В некоторых вариантах осуществления, хотя органоиды поджелудочной железы проявляют природную экспрессию одного или нескольких (например, 1, 2, 3 или 4) из следующих маркеров: Foxa2, Hnf6, Hnf1b и Sox9, они не демонстрируют экспрессию Ngn3.
Альтернативно или дополнительно, в некоторых вариантах осуществления органоиды поджелудочной железы демонстрируют природную экспрессию одного или нескольких маркеров протоков, например, один или оба из кератина 7 и кератина 19. В некоторых вариантах осуществления органоиды поджелудочной железы демонстрируют природную экспрессию одного или нескольких маркеров протоков на значительном или поддающемся выявлению уровне. Таким образом, в некоторых вариантах осуществления органоиды поджелудочной железы имеют фенотип протоков. В некоторых вариантах осуществления органоиды поджелудочной железы демонстрируют экспрессию одного или нескольких (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15) или всех из следующих маркеров, выбранных из группы: Hnf1A, Hnf1B, Hnf4A, HHEX, ONECUT1, ONECUT2, CDH1, FOXA2, GATA6, CEBPB, CEBPD, CEBPG, Glul, Krt7, Krt19 и MET.
Однако органоиды поджелудочной железы могут иметь некоторые признаки протоков в комбинации с признаками продуцирующих инсулин клеток-предшественников. Например, они могут экспрессировать один или несколько маркеров протоков, как показано на фиг.16B. В некоторых вариантах осуществления органоид поджелудочной железы проявляет профиль экспрессии генов относительно органоидов взрослой поджелудочной железы или печени, приблизительно как показано на фиг. 16B. Например, в некоторых вариантах осуществления эти гены активированы или подавлены в органоидах поджелудочной железы по сравнению с органоидами взрослой поджелудочной железы или печени приблизительно до того же кратного соотношения, что и на фиг.16B, например, менее +/- 3%, менее +/- 5%, менее +/- 10%, менее +/- 20%.
В некоторых вариантах осуществления инсулин-положительные клетки появляются из выстилки протоков органоидов поджелудочной железы.
В некоторых вариантах осуществления один или несколько (например, 1, 2, 3, 4, 5, 6 или 7), предпочтительно, все из следующих генов активированы в органоидах поджелудочной железы по сравнению с органоидами печени: Aaas, Rps4y2, Atp2c2, Akap2, Uts2, Sox17, Agr2. Например, в некоторых вариантах осуществления эти гены активированы в органоидах поджелудочной железы по сравнению с органоидами печени до приблизительно того же кратного соотношения, что и на фиг.19, например, менее +/- 3%, менее +/- 5%, менее +/- 10%, менее +/- 20%.
В одном из вариантов осуществления органоид поджелудочной железы содержит всего по меньшей мере 103, по меньшей мере 104, по меньшей мере 105 или более клеток. В одном из вариантов осуществления органоид поджелудочной железы содержит более 50%, более 60%, более 70% или более 80% подобных протоковым эндокринных клеток-предшественников. Однако также предусматриваются более низкие проценты подобных протоковым эндокринных клеток-предшественников.
Органоиды пищевода Барретта (BE)
Органоид BE по изобретению является Ki67+.
Предпочтительно, органоид BE имеет минимальное количество (например, менее 25%, менее 20%, менее 10%, менее 5%, менее 2%, менее 1% клеток) PAS+ и муцин+ клеток через 4 суток после устранения никотинамида и SB202190 из среды для увеличения в количестве, требуемом для преобразования ее в среду для дифференцировки.
В некоторых вариантах осуществления органоид BE содержит бокаловидные клетки. Они могут быть индуцированы обработкой среды для дифференцировки ингибитором гамма-секретазы, таким как DBZ (например, в количестве 10 мкМ), например, в течение 4 суток.
В некоторых вариантах осуществления органоид пищевода Барретта по изобретению содержит клетки Панета.
В некоторых вариантах осуществления органоид пищевода Барретта по изобретению экспрессирует лизоцим.
Органоиды желудка
В некоторых вариантах осуществления органоиды желудка по изобретению демонстрируют природную экспрессию Lgr5. В некоторых вариантах осуществления органоиды желудка по изобретению демонстрируют природную экспрессию по меньшей мере Lgr5 и одного или нескольких (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16 или 17) из маркеров стволовых клеток из группы, состоящей из: CK19, нестина, соматостатина, CXCR4+, CD133+, DCAMKL-1, CD44, Sord, Sox9, CD44, Prss23, Sp5, Hnf1α, Hnf4a, Sox9, KRT7 и KRT19. Альтернативно или дополнительно, в некоторых вариантах осуществления органоиды желудка могут характеризоваться природной экспрессией одного или нескольких (например, 1, 2 или 3) из: CD133+, DCAMKL-1 и CD44. Альтернативно или дополнительно, органоиды желудка могут характеризоваться CD44 и Sox9.
Органоиды предстательной железы
В некоторых вариантах осуществления органоиды предстательной железы по изобретению, такой как предстательная железа мыши, демонстрируют природную экспрессию Lgr5. В некоторых вариантах осуществления органоиды предстательной железы демонстрируют природную экспрессию маркеров просвета предстательной железы, таких как цитокератин 18 (CK18) и цитокератин 8 (CK8). В некоторых вариантах осуществления органоиды предстательной железы по изобретению демонстрируют природную экспрессию рецептора андрогена (AR). В некоторых вариантах осуществления органоиды предстательной железы экспрессируют базальные маркеры, такие как p63 и/или цитокератин 5 (CK5). В некоторых вариантах осуществления, когда тестостерон (например, DHT) добавляют в среду, экспрессия базальных маркеров Lgr5 и Tnfrsf19 подавляется по сравнению с органоидами, выращенными в отсутствие тестостерона (например, DHT). Специфический для предстательной железы фактор транскрипции NKX3.1 экспрессируется во всех условиях. Таким образом, в некоторых вариантах осуществления органоиды предстательной железы по изобретению демонстрируют природную экспрессию специфического для предстательной железы маркера Nkx3.1.
В некоторых вариантах осуществления органоиды предстательной железы по изобретению, например, нормальные ли злокачественные органоиды предстательной железы человека, демонстрируют природную экспрессию маркеров просвета, таких как CK18, CK8 и/или B-MSP. В некоторых вариантах осуществления органоиды предстательной железы демонстрируют природную экспрессию AR. В некоторых вариантах осуществления экспрессируются маркеры базального эпителия, такие как CK14, CK5 и/или p63. В некоторых вариантах осуществления экспрессирутся TNFRSF19. Специфический для предстательной железы фактор транскрипции NKX3.1 экспрессируется во всех условиях. Таким образом, в некоторых вариантах осуществления органоиды предстательной железы по изобретению демонстрируют природную экспрессию специфического для предстательной железы маркера Nkx3.1.
Добавление тестостерона (например, DHT) в культуральную среду согласно изобретению позволяет расти органоидам предстательной железы, которые сохраняют популяцию стволовых клеток, позволяющую вплоть до в 3 раза более быстрый рост (чем без тестостерона), и также присутствует большинство (или даже все) дифференцированных типов клеток предстательной железы (как базальные клетки, так и клетки просвета). Эти условия позволяют неограниченное увеличение в количестве клеток (на настоящий момент 9 месяцев при 2,5 удвоениях популяций в неделю). Таким образом, в некоторых вариантах осуществления органоид предстательной железы содержит все дифференцированные типы клеток предстательной железы, например, как базальные клетки, так и клетки просвета. В предпочтительном варианте осуществления органоид предстательной железы содержит все дифференцированные типы клеток, например, как базальные клетки, так и клетки протоков, и стволовые клетки.
В нормальной ткани добавление тестостерона (например, DHT) увеличивает экспрессию AR во всех условиях культивирования. В некоторых вариантах осуществления органоиды предстательной железы имеют активированную экспрессию AR по сравнению с клетками предстательной железы, выращенными в отсутствие тестостерона. В опухолевой ткани на экспрессию AR добавление тестостерона (например, DHT) не влияет. Таким образом, в некоторых вариантах осуществления органоид рака предстательной железы не имеет увеличенную экспрессию AR относительно клеток рака предстательной железы in vivo. Маркер стволовых клеток LGR5 экспрессируется в условиях ENRF в органоидах предстательной железы из нормальной ткани. В органоидах предстательной железы, полученных из опухолевой ткани, экспрессия LGR5 индуцируется добавлением тестостерона (например, DHT). В некоторых вариантах осуществления органоиды предстательной железы экспрессируют LGR5.
Функции органоидов
В некоторых вариантах осуществления органоиды, получаемые с помощью сред и способов по настоящему изобретению, имитируют решения о клеточной судьбе in vivo в ответ на внешние факторы. Предпочтительно, клетки и органоиды, полученные согласно изобретению, также обладают тканеспецифическими функциями.
Органоиды поджелудочной железы
Органоид поджелудочной железы, предпочтительно, обладает эндокринными и экзокринными функциями поджелудочной железы, такими как экспрессия одного или нескольких (например, 1, 2 или всех 3) из инсулина, глюкагона и соматостатина. Экспрессия этих гормонов строго регулируется набором эндокринных специфических для поджелудочной железы факторов транскрипции, наиболее из важным из которых являются Pdx1 и NeuroD. Экзокринная поджелудочная железа образована ацинарным и протоковым компартментами, ответственными за продукцию пищеварительных ферментов амилазы, панкреатической липазы и химотрипсина, среди прочих. Экспрессия этих генов также регулируется определенными генами экзокринной поджелудочной железы, такими как Ptf1a.
Клетки и органоиды поджелудочной железы в соответствии с настоящим изобретением, предпочтительно, могут быть способны секретировать инсулин, например, со скоростью от приблизительно 1 мкг в час на 106 клеток до 10 мкг в час на 106 клеток, например, от 2 мкг до 6 мкг в час на 106 клеток. Уровень секреции инсулина можно выявлять способами, хорошо известными в данной области, например, вестерн-блоттингом по сравнению с эталоном или посредством Elisa для C-пептида. Предпочтительным способом продемонстрировать, что органоиды поджелудочной железы могут секретировать инсулин, является исследование продукции C-пептида. C-пептид проинсулина служит в качестве важного линкера между A- и B-цепями инсулина и способствует эффективной сборке, сворачиванию и процессингу инсулина в эндоплазматической сети. Затем эквимолярные количества C-пептида и инсулина запасаются в секреторных гранулах бета-клеток поджелудочной железы и оба из них в конечном итоге высвобождаются в кровообращение в системе воротной вены. Таким образом, C-пептид является предпочтительным маркером секреции инсулина.
Таким образом, в одном из вариантов осуществления предусматривается органоид поджелудочной железы, который секретирует инсулин после трансплантации in vivo. В некоторых вариантах осуществления после трансплантации in vivo органоид поджелудочной железы секретирует инсулин со скоростью по меньшей мере 1 мкг в час на 106 клеток, например, по меньшей мере 2 мкг в част на 106 клеток, по меньшей мере 4 мкг в час на 106 клеток, по меньшей мере 6 мкг в час на 106 клеток, по меньшей мере 8 мкг в час на 106 клеток или по меньшей мере 10 мкг в час на 106 клеток. В некоторых вариантах осуществления клетки в органоиде поджелудочной железы не способны секретировать инсулин и/или не экспрессируют инсулин в качестве маркера при культивировании in vitro. Однако клетки из органоида поджелудочной железы по настоящему изобретению, предпочтительно, способны секретировать инсулин in vivo при трансплантации пациенту, например, в поджелудочную железу пациента. В некоторых вариантах осуществления способность секретировать инсулин может не присутствовать сразу после трансплантации, но присутствует приблизительно через один месяц после трансплантации, например, через 6 недель, 2 месяца или 3 месяца после трансплантации.
Если из органоида поджелудочной железы по изобретению получают образец, обогащенный эндокринными клетками, в некоторых вариантах осуществления 75-85% клеток в обогащенном эндокринными клетками образце представляют собой секретирующие инсулин клетки.
В некоторых вариантах осуществления изобретение относится к органоидам поджелудочной железы для применения при лечении диабета. В некоторых вариантах осуществления органоиды поджелудочной железы представляют собой увеличивающиеся в количестве органоиды, в то время как в других вариантах осуществления они могут представлять собой дифференцированные органоиды. В некоторых вариантах осуществления один или несколько (например, 1, 2, 3, 4, 5, 6, 7 и т.д.) целых органоидов трансплантируют животному или пациенту, в то время как в других вариантах осуществления образец клеток трансплантируют пациенту.
Органоиды крипта-ворсинка
Органоид крипта-ворсинка, предпочтительно, обладает секреторной функцией и функцией самообновления. Например, органоид крипта-ворсинка, предпочтительно, секретирует муцин, ферментативные и гормональные секреты, такие как лизоцим, холицистокинин, секретин и желудочный ингибиторный пептид, и другие гликопротеины.
Органоиды желудка
Желудок человека анатомически и функционально подразделяется на две основных области. Привратниковая пещера вблизи кишечника, главным образом, продуцирует защитную слизь и секретирует гормоны, такие как гастрин. Тело желудка секретирует хлористоводородную кислоту и желудочные ферменты, такие как пепсиноген. Эпителий обеих областей желудка организован в виде инвагинаций, называемых железами. Эти железы содержат стволовые клетки желудка, клетки-предшественники и дифференцированные клетки. Точный состав дифференцированных клеток варьирует в зависимости от функции анатомической области. В привратниковой пещере железы, главным образом, состоят из клеток, продуцирующих муцин 6, и продуцирующих гормоны эндокринных клеток. В теле продуцирующие пепсиноген главные клетки и секретирующие кислоту париетальные клетки распределены между продуцирующими слизь клетками и редкими эндокринными клетками. Поверхностная область между железами желудка занята продуцирующими слизь клетками, которые, главным образом, продуцируют поверхностный муцин 5.
Органоиды желудка по структуре и функции напоминают эпителий желудка. Хотя они по большей части являются сферическими, они могут иметь домены с инвагинациями, которые в наибольшей степени напоминают железистые структуры. Окрашивание муцинов и пепсиногена показывает, что наиболее распространенными типами клеток в органоидах желудка являются слизистые клетки, продуцирующие муцин 6, и продуцирующие пепсиноген главные клетки (и/или их предшественники). Таким образом, ОТ-ПЦР указывает на экспрессию пепсиногена и муцина 6. Далее, экспрессия гастрина указывает на присутствие эндокринных клеток, и экспрессия Lgr5 указывает на присутствие стволовых клеток.
В некоторых вариантах осуществления органоиды желудка имеют природную экспрессию одного или нескольких (например, 1, 2, 3 или 4) из гастрина, пепсиногена, муцина 6 и/или Lgr5. В некоторых вариантах осуществления органоиды желудка содержат слизистые клетки, продуцирующие муцин 6, и продуцирующие пепсиноген главные клетки и необязательно Lgr5+ стволовые клетки. В некоторых вариантах осуществления органоиды желудка содержат эндокринные клетки. В некоторых вариантах осуществления органоиды желудка по большей части являются сферическими, но имеют домены с инвагинациями, которые напоминают железистые структуры.
Органоиды предстательной железы
В некоторых вариантах осуществления органоиды предстательной железы содержат или состоят из двух различных эпителиальных ростков: базальных клеток и клеток протоков. В некоторых вариантах осуществления базальные клетки и клетки протоков секретируют жидкости предстательной железы.
In vivo эпителий предстательной железы является в значительной степени сложенным, обеспечивая максимальную площадь поверхности. Два эпителиальных ростка образуют простой распределенный на слои эпителий с базальными эпителиальными клетками, образующими базальный/наружный слой, и строго полярными эпителиальными клетками просвета, расположенными сверху, образующими внутренний слой/слой просвета. Компартмент просвета необходим для секреторной функции предстательной железы. Жидкость предстательной железы является щелочной и состоит из нескольких белков, таких как простатспецифический антиген (PSA), калликреин человека 2 (KLK2) и β-микросеминопротеин (β-MSP). Основными функциями жидкости предстательной железы являются: 1) подготовка среды в матке для спермы, которая осуществляется благодаря щелочным свойствам жидкости и паракринным функциям β-MSP, и 2) увеличение текучести семенной жидкости, позволяющей сперматозоидам свободно плыть, которое осуществляется благодаря протеазам PSA и KLK2, которые разрушают семиногелины.
Экспрессия секреторных белков строго регулируется рецептором андрогенов (AR), который связывается с тестостероном, а затем перемещается в ядро и активирует транскрипцию. Нарушение функции AR демонстрирует сильное подавление секреторных белков на транскрипционном уровне и на уровне белков.
На фиг.43 показано расслоение органоидов предстательной железы, выращенных в условиях ENR+дигидротестостерон (DHT), отчетливо демонстрирующих базальные клетки цитокератин 5+, образующие внешний слой клеток, и люминальные клетки цитокератин 8+, образующие строго полярный внутренний слой. В некоторых вариантах осуществления органоид предстательной железы содержит базальные клетки цитокератин 5+ и люминальные клетки цитокератин 8+, необязательно где базальные клетки цитокератин 5+ образуют внешний слой клеток и люминальные клетки цитокератин 8+ образуют строго полярный внутренний слой. В некоторых вариантах осуществления органоид предстательной железы содержит уложенные слои клеток, необязательно имеющие устойчивую укладку. Такая укладка максимизирует площадь поверхности секреторных клеток, демонстрируя, что на морфологическом уровне органоиды предстательной железы напоминают предстательную железу in vivo. В некоторых вариантах осуществления морфология органоида предстательной железы напоминает морфологию предстательной железы in vivo.
Органоиды предстательной железы, культивируемые в условиях ENR, не секретируют никакой жидкости предстательной железы в просвет. Напротив, добавление тестостерона (например, DHT) в среду приводит к секреции жидкостей, например, жидкости предстательной железы, в просвет органоида. Это является следствием активации AR-зависимой программы транскрипции в органоидах предстательной железы тестостероном (например, DHT), что приводит к секреции жидкостей CK8+ клетками просвета. Данные демонстрируют, что органоиды предстательной железы, как морфологически, так и функционально, напоминают эпителий предстательной железы in vivo.
Таким образом, в некоторых вариантах осуществления, например, где органоиды предстательной железы культивируют в культуральной среде, содержащей тестостерон (например, DHT), органоид предстательной железы секретирует жидкость, например, жидкость предстательной железы, в просвет органоида. В некоторых вариантах осуществления, например, где органоиды предстательной железы культивируют в культуральной среде, содержащей тестостерон (например, DHT), функциональность органоида предстательной железы напоминает функциональность предстательной железы in vivo.
Фрагменты тканей
В контексте изобретения фрагмент ткани является частью взрослой ткани, предпочтительно, взрослой ткани человека, такой как часть взрослого тонкого кишечника, толстого кишечника или поджелудочной железы человека. Следующие примеры взрослой ткани человека в контексте настоящего изобретения включают желудок, печени и предстательную железу. Ткань может представлять собой нормальную (здоровую) ткань или она может представлять собой пораженную заболеванием или инфицированную ткань. Таким образом, предпочтительно, органоид, как определено в настоящем описании, не является фрагментом ткани. Органоид, предпочтительно, получают с использованием клетки из взрослой ткани, предпочтительно, эпителиальной стволовой клетки из взрослой ткани, необязательно из фрагмента взрослой ткани, более предпочтительно, эпителиальной стволовой клетки из взрослой ткани или фрагмента взрослой ткани, экспрессирующих Lgr5. Таким образом, в контексте настоящего изобретения фрагмент ткани, предпочтительно, содержит Lgr5+ стволовые клетки.
В одном из вариантов осуществления органоид представляет собой органоид, который все еще культивируют с использованием способа по изобретению (предпочтительно, с использованием культуральной среды по изобретению) и, таким образом, он находится в контакте с внеклеточным матриксом. Предпочтительно, органоид погружен в немезенхимный внеклеточный матрикс. В контексте изобретения, "в контакте" означает физический или механический или химический контакт, что означает, что для отделения указанного органоида от указанного внеклеточного матрикса необходимо применить силу. В некоторых вариантах осуществления внеклеточный матрикс представляет собой гелеобразную смесь белков, секретируемую клетками саркомы мыши Engelbreth-Holm-Swarm (EHS), такую как Matrigel (BD Biosciences). В других вариантах осуществления изобретения органоиды можно удалять из культуры и использовать для целей трансплантации и регенерации. Таким образом, изобретение относится к органоиду по изобретению для применения в трансплантации млекопитающему, предпочтительно, человеку.
Уровень выживаемости
В рамках настоящего изобретения авторы изобретения впервые показали, что добавление ингибитора ALK4, ALK5, ALK7 или киназы p38, в ранее описанную среду для культивирования стволовых клеток, улучшает эффективность посева по меньшей мере на 50% и в некоторых случаях более чем на 100% (см. таблицу 2). Авторы изобретения также показали, что включение обоих ингибиторов (ингибитор ALK и ингибитор p38, например, A83-01 и SB-202190) в культуральную среду синергично продлевает период культивирования.
Таким образом, в одном из вариантов осуществления изобретения стволовые клетки выживают в течение по меньшей мере 3 месяцев, предпочтительно, по меньшей мере 4 месяцев, по меньшей мере 5 месяцев, по меньшей мере 6 месяцев, по меньшей мере 7 месяцев, по меньшей мере 9 месяцев, или по меньшей мере 12 месяцев или более.
Скорость пролиферации
Скорость пролиферации можно оценивать в значениях уровня удвоения клеточной популяции. Уровень удвоения популяции относится к общему количеству раз удвоений клеток в популяции с момента их первичного выделения in vitro. Уровень удвоения популяции можно определять путем подсчета клеток. Альтернативно скорость пролиферации можно оценивать с помощью анализа пролиферации клеток, например, в котором специфический флуоресцентный зонд измеряет активность синтеза ДНК по включению BrdU и состояние пролиферации клеток по экспрессии Ki67 (Thermo Scientific* Cellomics, Millipore).
Следующие примеры анализов пролиферации клеток для стволовых клеток свободно доступны и могут быть найдены через Интернет или в журналах, таких как Current Protocols. Одним примером из многих является:
http://products.invitrogen.com/ivgn/en/US/adirect/invitrogen?cmd=catDisplayStyle&catKey=101&filterDispName=Cellular Proloferation Assays for Stem Cells&filterType=1&OP=фильтр&фильтр=ft_1101%2Ff_494303*&_bcs_=H4sIAAAAAAAAAH2NsQrDMAxEv0ZTsEkdKFmzZC70C4IjakFsGVuOfz%2FK0I6F4x284c48YJxfhffm%0ApQ7gnsMby0ke6x8fRDJMC7hV03u3lE6Swh9M1nNUWUlQq1UFJkXgeIvvotnSbn6Lbl1yPshvQpyq%0ADRIPfQE33RlnKQ21Lvuql7CrAAAA.
Авторы изобретения выявили, что с использованием культуральных сред по изобретению клетки могут увеличиваться в количестве вплоть до в среднем 5 раз в неделю. Например, выращивание единичной клетки в течение двух недель может обеспечить в среднем приблизительно 25 клеток. Квалифицированному специалисту будет понятно, что среднее время удвоения популяции стволовых клеток по изобретению может варьировать, в зависимости от нескольких факторов, таких как количество пассажей, условия культивирования, плотность посева и т.д.
В одном из вариантов осуществления среднее время удвоения популяции может составлять от 6 до 48 часов, от 12 до 36 часов, от 18 до 30 часов, или приблизительно 24 часов. Например, можно ожидать, что популяция стволовых клеток, культивированная с использованием культуральной среды по изобретению, будет удваиваться приблизительно 4-7 раз, или приблизительно 5 раз в неделю.
В другом варианте осуществления среднее время удвоения популяции составляет от 12 до 96 часов, от 24 до 72 часов, или приблизительно 72 часов. В другом варианте осуществления популяция клеток удваивается в среднем более одного раза, более двух раз, более трех раз, более четырех раз или более пяти раз в неделю.
Другие свойства органоидов по изобретению
В предпочтительном варианте осуществления органоид можно культивировать в течение по меньшей мере 2, 3, 4, 5, 6, 7, 8, 9, 10 недель или 1, 2, 3, 4, 5, 6 месяцев или более. В предпочтительном варианте осуществления органоид можно культивировать в течение по меньшей мере 2, 3, 4, 5, 6, 7, 8, 9, 10 недель или 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 месяцев или более. В другом предпочтительном варианте осуществления органоид происходит из единичной клетки, предпочтительно, экспрессирующей Lgr5, более предпочтительно, где единичная клетка содержит конструкцию нуклеиновой кислоты, содержащую представляющую интерес молекулу нуклеиновой кислоты.
Более того, изобретение относится к органоиду, предпочтительно, содержащему по меньшей мере 50% жизнеспособных клеток, более предпочтительно, по меньшей мере 60% жизнеспособных клеток, более предпочтительно, по меньшей мере 70% жизнеспособных клеток, более предпочтительно, по меньшей мере 80% жизнеспособных клеток, более предпочтительно, по меньшей мере 90% жизнеспособных клеток. Жизнеспособность клеток можно оценивать с использованием окрашивания красителем Хехста или окрашивания йодидом пропидия в FACS.
Жизнеспособные клетки, предпочтительно, обладают тканеспецифическими функциями, или характеристиками тканеспецифических функций, как описано выше.
Авторы изобретения также показали, что органоиды, полученные с помощью сред и способов по настоящему изобретению, можно замораживать и хранить при -80°C или ниже, как например, в жидком азоте. Замороженные органоиды можно размораживать и помещать в культуру без утраты их 3D-структуры и целостности и без значительной гибели клеток. Таким образом, в одном из вариантов осуществления изобретение относится к замороженным органоидам, хранящимся при температуре ниже -5°C, ниже -10°C, ниже -20°C, ниже -40°C, ниже -60°C или ниже -80°C.
Клетки и органоиды по настоящему изобретению отличаются от любых клеток и органоидов, которые были получены ранее (WO2009/022907 и WO2010/016766) тем, что они имеют лучшую фенотипическую (лучший профиль дифференцировки, включая преобразование в бокаловидные клетки при добавлении ингибиторов гамма-секретазы для органоидов крипта-ворсинка) и кариотипическую целостность при определении способами, описанными выше, лучшие показатели выживаемости и более высокие скорости пролиферации клеток. Таким образом, для вариантов осуществления, относящихся к тонкому кишечнику, толстому кишечнику и поджелудочной железе, органоид по настоящему изобретению отчетливо демонстрирует эпителий тонкого кишечника, толстого кишечника и поджелудочной железы человека с полным сохранением фенотипической и кариотипической целостности и поддержанием пролиферации и дифференцировки.
Применение стволовых клеток или органоидов по изобретению
Изобретение относится к применению органоида или увеличенной в количестве популяции клеток по изобретению для применения в скрининге лекарственных средств, подтверждения мишени (лекарственного средства), выявления мишени (лекарственного средства), токсикологии и токсикологическом скрининге, персонализированной медицине, регенеративной медицине и/или в качестве моделей клеток/органов ex vivo, таких как модели заболевания.
Полагают, что клетки и органоиды, культивируемые с помощью сред и способов по изобретению, правдоподобно отражают ситуацию in vivo. Это является справедливым для увеличенных в количестве популяций клеток и органоидов, выращенных из нормальной ткани, и для увеличенных в количестве популяций клеток и органоидов, выращенных из пораженной заболеванием ткани. Таким образом, помимо обеспечения нормальных моделей клеток/органов ex vivo, органоиды или увеличенную в количестве популяцию клеток по изобретению можно использовать в качестве моделей заболеваний ex vivo.
Органоиды по изобретению также можно использовать для культивирования патогена и, таким образом, можно использовать в качестве моделей инфекции ex vivo. Примеры патогенов, которые можно культивировать с использованием органоида по изобретению, включают вирусы, бактерии, прионы или грибы, которые вызывают заболевание в их животном-хозяине. Таким образом, органоид по изобретению можно использовать в качестве модели заболевания, которая отражает инфицированное состояние. В некоторых вариантах осуществления изобретения органоиды можно использовать для разработки и/или получения вакцин.
Заболевания, которые можно исследовать с помощью органоидов по изобретению, таким образом, включают генетические заболевания, метаболические заболевания, патогенные заболевания, воспалительные заболевания и т.д., например, включая, но не ограничиваясь ими: кистозный фиброз, воспалительное заболевание кишечника (такое как болезнь Крона), карциному, аденому, аденокарциному, рак толстого кишечника, диабет (такой как тип I или тип II), пищевод Барретта, болезнь Гоше, дефицит альфа-1-антитрипсина, синдром Леш-Найхена, анемию, синдром Швахмана-Бодиана-Даймонда, истинную полицитемию, первичный миелофиброз, болезнь накопления гликогена, семейную гиперхолестеринемию, синдром Криглера-Найяра, наследственную тирозинемию, болезнь Помпе, прогрессирующий семейный холестаз, синдром Харлера, SCID или пропускающий SCID, синдром Оменна, хрящево-волосяную гипоплазию, обусловленный вирусом простого герпеса энцефалит, склеродермию, несовершенный остеогенез, мышечную дистрофию Беккера, мышечную дистрофию Дюшенна, врожденный дискератоз и т.д.
Традиционно, клеточные линии и позднее клетки iPS использовали в качестве моделей клеток/органов ex vivo и/или моделей заболеваний (например, см. Robinton et al. Nature 481, 295, 2012). Однако эти способы имеют проблемы и недостатки. Например, клеточные линии нельзя получить от всех пациентов (только определенные виды биопсии приводят к успешно полученным клеточным линиям) и, таким образом, клеточные линии нельзя использовать в персонализированной диагностике и медицине. В случае клеток iPS обычно требуется некоторый уровень генетического манипулирования для перепрограммирования клеток на определенную клеточную судьбу. Альтернативно они являются объектом условий культивирования, которые влияют на кариотипическую целостность и, таким образом, время в культуре должно быть минимальным (это также справедливо для эмбриональных стволовых клеток человека). Это означает, что клетки iPS не могут точно отражать ситуацию in vivo, а вместо этого представляют собой попытку имитировать поведение клеток in vivo. Клеточные линии и клетки iPS также обладают генетической нестабильностью.
Напротив, органоиды по изобретению обеспечивают генетически стабильную платформу, которая правдоподобно отражает ситуацию in vivo. Органоиды по изобретению также можно непрерывно увеличивать в количестве, что обеспечивает хороший источник генетически стабильных клеток. В частности, увеличивающуюся в количестве популяцию можно "разделять", что означает, что органоид разделяют на части и все клетки органоида распределяют в новые культуральные чашки или флаконы. Отделенные клетки извлекают из органоида, а затем их сами можно культивировать и увеличивать в количестве с получением новых органоидов, содержащих далее увеличенные в количестве популяции, которые затем можно вновь разделять. Случаи разделения также называют в настоящем описании "пассажами". Органоид по изобретению можно культивировать в ходе 1 или нескольких пассажей, например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30 или более пассажей, например, 20-30 пассажей, 30-35 пассажей, 32-40 пассажей или более. В некоторых вариантах осуществления увеличивающуюся в количестве популяцию клеток или органоид разделяют один раз в месяц, один раз в две недели, один раз в неделю, два раза в неделю, три раза в неделю, четыре раза в неделю, пять раз в неделю, шесть раз в неделю или каждые сутки. Таким образом, органоиды по изобретению могут обеспечить непрерывный источник генетически стабильного клеточного материала. В некоторых вариантах осуществления увеличивающиеся в количестве органоиды по изобретению содержат все дифференцированные типы клеток, которые присутствуют в соответствующей ситуации in vivo. В других вариантах осуществления органоиды по изобретению могут быть дифференцированными для обеспечения всех дифференцированных типов клеток, которые присутствуют in vivo. Таким образом, органоиды по изобретению можно использовать для получения механистического представления о различных заболеваниях и терапевтических средствах, для осуществления скрининга лекарственных средств in vitro, для оценки потенциальных лекарственных средств, для идентификации возможных мишеней (например, белков) для дальнейшей разработки новой (лекарственной) терапии и/или для исследования репарации генов, сопряженной с клеточно-заместительной терапией.
Органоиды по изобретению можно замораживать и размораживать и помещать в культуру без утраты их генетической целостности и фенотипических характеристик и без утраты пролиферативной способности. Таким образом, органоиды можно легко хранить и транспортировать.
По этой причине органоиды или увеличенные в количестве популяции клеток по изобретению могут представлять собой инструмент для скрининга лекарственных средств, подтверждения мишени, выявления мишени, токсикологии и токсикологического скрининга и персонализированной медицины.
Таким образом, в следующем аспекте изобретение относится к применению увеличенной в количестве популяции стволовых клеток или органоида, таких как кишечные органоиды крипта-ворсинка или органоиды поджелудочной железы согласно изобретению, в скрининге для выявления лекарственных средств, анализа токсичности или в медицине, такой как регенеративная медицина. Например, любые из органоидов тонкого кишечника, толстого кишечника, поджелудочной железы, желудка, печени или предстательной железы можно использовать в скрининге для выявления лекарственных средств, анализе токсичности или в медицине, такой как регенеративная медицина.
Мукозальные вакцины
Дополнительным важным применением органоидов является разработка мукозальной вакцинации. Мукозальные вакцины представляют собой вакцины, которые вводят через слизистую оболочку. Она может представлять собой любую слизистую поверхность, такую как слизистая поверхность носа, рта или прямой кишки. Их можно вводить через ингалятор, спрей или другие внешние вспомогательные средства. Это имеет несколько очевидных преимуществ над инъекциями, таких как отсутствие необходимости в медицинском персонале для введения вакцины, что может быть важно, например, в развивающихся странах.
В кишечнике M-клетки (или "микроскладчатые клетки") представляют собой клетки, встречающиеся в связанном с фолликулами эпителии агрегированных лимфоидных узлов в подвздошной кишке. Они транспортируют организмы и частицы из просвета кишечника в иммунные клетки через эпителиальный барьер, и, таким образом, важны для стимуляции иммунитета слизистых оболочек. Они обладают уникальной способностью захватывать антиген из просвета тонкого кишечника через эндоцитоз или фагоцитоз, а затем доставлять его через трансцитоз в дендритные клетки (антигенпредставляющие клетки) и лимфоциты (а именно, T-клетки), расположенные в уникальной подобной карману структуре на их базально-латеральной стороне.
На фиг.48 показано, что органоиды мыши могут развиваться в M-клетки при стимуляции RANK-лигандом. На фиг.49 показано, что также можно получать M-клетки в кишечных органоидах человека. Таким образом, в некоторых вариантах осуществления изобретения увеличенная в количестве популяция клеток содержит M-клетки. В некоторых вариантах осуществления изобретения органоид, например, органоид тонкого кишечника, содержит M-клетки.
Эффективность мукозальных вакцин может существенно увеличиваться, когда они нацелены на M-клетки. Таким образом, увеличенную в количестве популяцию стволовых клеток или органоид по изобретению можно использовать для исследования способности M-клеток захватывать патогены или антигены и представлять их иммунной системе. Таким образом, в некоторых вариантах осуществления изобретение относится к применению увеличенной в количестве популяции стволовых клеток или органоида по изобретению для скрининга лекарственных средств, например, для разработки вакцин и/или получения вакцин. Например, в некоторых вариантах осуществления увеличенную в количестве популяцию стволовых клеток или органоид можно использовать для разработки или получения вакцин против вирусных, бактериальных, грибковых других паразитарных инфекций, например, (но не ограничиваясь ими) холеры, респираторно-синцитиального вируса (RSV), ротавируса и ВИЧ. В конкретном варианте осуществления изобретение относится к органоидам тонкого кишечника, дифференцированных в культуральной среде по изобретению, содержащей RANKL, для применения в разработке мукозальных вакцин.
Скрининг лекарственных средств
Предпочтительно, для высокопроизводительных целей, указанную увеличенную в количестве популяцию стволовых клеток или органоид по изобретению, такой как органоиды крипта-ворсинка или органоиды поджелудочной железы, культивируют в многолуночных планшетах, например, таких как 96-луночные планшеты или 384-луночные планшеты. Для идентификации молекул, которые влияют на указанные органоиды, используют библиотеки молекул. Предпочтительные библиотеки включают библиотеки фрагментов антител, библиотеки пептидного фагового дисплея, пептидные библиотеки (например, LOPAPTM, Sigma Aldrich), библиотеки липидов (BioMol), библиотеки синтетических соединений (например, LOP ACTM, Sigma Aldrich) или библиотеки природных соединений (Specs, TimTec). Более того, можно использовать генетические библиотеки, которые индуцируют или подавляют экспрессию одного или нескольких генов в потомстве стволовых клеток. Эти генетические библиотеки включают библиотеки кДНК, антисмысловые библиотеки и миРНК (siRNA) или библиотеки других некодирующих РНК. Клетки, предпочтительно, подвергают воздействию множества концентраций исследуемого вещества в течение определенного периода времени. В конце периода воздействия культуры оценивают. Термин "влияющий" используют для охвата любого изменения в клетке, включая, но не ограничиваясь ими, снижение или утрату пролиферации, морфологическое изменение и гибель клеток. Указанную увеличенную в количестве популяцию стволовых клеток или органоид по изобретению, такой как органоиды крипта-ворсинка или органоиды поджелудочной железы, также можно использовать для идентификации лекарственных средств, которые специфически нацелены на эпителиальные клетки карциномы, но не на указанную увеличенную в количестве популяцию стволовых клеток или органоид по изобретению, такие как органоиды крипта-ворсинка или органоиды поджелудочной железы.
Авторы изобретения показали, что можно взять биоптат из тонкого кишечника и увеличивать его в количестве в течение только 7-14 суток и получить органоид, который готов для осуществления скрининга лекарственных средств. Возможность получения пригодного органоида по изобретению за такой короткий период времени демонстрирует, что органоиды могут быть в высокой степени полезными для исследования индивидуальных ответов пациентов на лекарственные средства и коррекции лечения в соответствии со способностью к ответу. В некоторых вариантах осуществления, где органоид получают из биоптата от пациента, органоид культивируют в течение менее чем 21 суток, например, менее чем 14 суток, менее чем 13 суток, менее чем 12 суток, менее чем 11 суток, менее чем 10 суток, менее чем 9 суток, менее чем 8 суток, менее чем 7 суток (и т.д.).
Органоиды также пригодны для более широких целей выявления лекарственных средств. Например, на фиг.32-40 показано, что органоиды тонкого кишечника, взятые от здоровых пациентов и от пациентов с кистозным фиброзом, можно использовать для исследования лекарственных средств против кистозного фиброза. В частности, на фиг.32-40 показано, что индуцируемое форсколином набухание нормальных органоидов тонкого кишечника зависит от регулятора трансмембранной проводимости при кистозном фиброзе (CFTR), и, таким образом, можно исследовать коррекцию функции CFTR с использованием индуцируемого форсколином набухания в качестве положительного показателя. Таким образом, в некоторых вариантах осуществления органоиды по изобретению можно использовать для скрининга лекарственных средств от кистозного фиброза. Однако специалисту в данной области будет понятно, что органоиды по изобретению могут быть широко применимыми в качестве инструментов для скрининга лекарственных средств от инфекционных, воспалительных и неопластических патологий желудочно-кишечного тракта человека и других заболеваний желудочно-кишечного тракта, и инфекционных, воспалительных и неопластических патологий и других заболеваний других тканей, описанных в настоящем описании, включая поджелудочную железу, печень и предстательную железу. В некоторых вариантах осуществления органоиды по изобретению можно использовать для скрининга лекарственных средств от злокачественной опухоли.
В некоторых вариантах осуществления увеличенные в количестве популяции клеток, например, органоиды по изобретению или органоиды, полученные с использованием сред и способов по изобретению, можно использовать для исследования библиотек химических реагентов, антител, природного продукта (растительных экстрактов), и т.д. в отношении пригодности для применения в качестве лекарственных средств, косметических средств и/или профилактических лекарственных средств. Например, в некоторых вариантах осуществления представляющий интерес биоптат клеток пациента, такой как клетки опухоли от пациента со злокачественной опухолью, можно культивировать с использованием культуральных сред и способов по изобретению, а затем обрабатывать химическим соединением или химической библиотекой. Затем можно определить, какие соединения эффективно модифицируют, уничтожают и/или лечат клетки пациента. Это позволяет исследовать способность к ответу у конкретного пациента на конкретное лекарственное средство, таким образом, позволяя адаптацию лечения для конкретного пациента. Таким образом, это предусматривает подход персонализированной медицины.
Дополнительным преимуществом использования органоидов для идентификации лекарственных средств таким образом является то, что можно осуществлять скрининг нормальных органоидов (органоиды, происходящие из здоровой ткани) для проверки того, какие лекарственные средства и соединения имеют минимальный эффект на здоровую ткань. Это обеспечивает скрининг лекарственных средств с минимальной нецелевой активностью или нежелательными побочными эффектами.
Таким образом, можно проводить скрининг лекарственных средств от любого количества заболеваний. Например, органоиды по изобретению можно использовать для скрининга лекарственных средств от кистозного фиброза, пищевода Барретта, карцином, аденокарцином, аденом, воспалительного заболевания кишечника (такого как болезнь Крона), болезни печени и т.д. Параметры исследования зависят от представляющего интерес заболевания. Например, при скрининге лекарственных средств от злокачественной опухоли обычно конечной целью является гибель злокачественных клеток. В случае кистозного фиброза интерес представляет измерение увеличения в количестве органоидов в ответ на лекарственные средства и стимулы CFTR. В других вариантах осуществления метаболиты и экспрессию генов можно оценивать для исследования эффектов соединений и лекарственных средств при скрининге на представляющих интерес клетках или органоидах.
Таким образом, изобретение относится к способу скрининга терапевтического или профилактического лекарственного средства или косметического средства, где способ включает:
культивирование увеличенной в количестве популяции клеток (например, органоида) по изобретению, например, с помощью культуральной среды по изобретению, необязательно в течение менее 21 суток;
воздействие на указанную увеличенную в количестве популяцию клеток (например, органоид) по изобретению одной или библиотеки молекул-кандидатов;
оценку указанных увеличенных в количестве популяций клеток (например, органоидов) в отношении каких-либо эффектов, например, какого-либо изменения в клетке, такого как снижение или утрата пролиферации, морфологическое изменение и/или гибель клеток;
идентификацию молекулы-кандидата, которые вызывает указанные эффекты, в качестве потенциального лекарственного средства или косметического средства.
В некоторых вариантах осуществления используют способы культивирования и сбора данных с помощью компьютера или робота для увеличения производительности скрининга.
В некоторых вариантах осуществления увеличенную в количестве популяцию клеток (например, органоид) получают из биоптата пациента. В некоторых вариантах осуществления молекулу-кандидат, которая обеспечивает желаемый эффект на культивируемую увеличенную в количестве популяцию клеток (например, органоид), вводят указанному пациенту.
Таким образом, в одном из аспектов предусматривается способ лечения пациента, включающий:
(a) получение биоптата из представляющей интерес пораженной заболеванием ткани у пациента;
(b) скрининг подходящего лекарственного средства с использованием способа скрининга по изобретению; и
(с) лечение указанного пациента лекарственным средством, полученным на стадии (b).
В некоторых вариантах осуществления лекарственное средство или косметическое средство используют для лечения, профилактики или смягчения симптомов генетических заболеваний, метаболических заболеваний, патогенных заболеваний, воспалительных заболеваний и т.д., например, включающих, но не ограничивающихся ими: кистозный фиброз, воспалительное заболевание кишечника (такое как болезнь Крона), карциному, аденому, аденокарциному, рак толстого кишечника, диабет (такой как тип I или тип II), пищевод Барретта, болезнь Гоше, дефицит альфа-1-антитрипсина, синдром Леш-Найхена, анемию, синдром Швахмана-Бодиана-Даймонда, истинную полицитемию, первичный миелофиброз, болезнь накопления гликогена, семейную гиперхолестеринемию, синдром Криглера-Найяра, наследственную тирозинемию, болезнь Помпе, прогрессирующий семейный холестаз, синдром Харлера, SCID или пропускающий SCID, синдром Оменна, хрящево-волосяную гипоплазию, обусловленный вирусом простого герпеса энцефалит, склеродермию, несовершенный остеогенез, мышечную дистрофию Беккера, мышечную дистрофию Дюшенна, врожденный дискератоз и т.д.
Выявление мишени
В некоторых вариантах осуществления органоиды по изобретению или клетки, выращенные с использованием культуральных сред и способов по изобретению, можно использовать для выявления мишени. Клетки органоидов, происходящие из здоровой или пораженной заболеванием ткани, можно использовать для идентификации мишени. Органоиды по изобретению можно использовать для выявления мишеней лекарственных средств от кистозного фиброза, воспалительного заболевания кишечника (такого как болезнь Крона), карциномы, аденомы, аденокарциномы, рака толстого кишечника, диабета (такого как тип I или тип II), пищевода Барретта, болезни Гоше, дефицита альфа-1-антитрипсина, синдрома Леш-Найхена, анемии, синдрома Швахмана-Бодиана-Даймонда, истинной полицитемии, первичного миелофиброза, болезни накопления гликогена, семейной гиперхолестеринемии, синдрома Криглера-Найяра, наследственной тирозинемии, болезни Помпе, прогрессирующего семейного холестаза, синдрома Харлера, SCID или пропускающего SCID, синдрома Оменна, хрящево-волосяной гипоплазии, обусловленного вирусом простого герпеса энцефалита, склеродермии, несовершенного остеогенеза, мышечной дистрофии Беккера, мышечной дистрофии Дюшенна, врожденного дискератоза и т.д. Полагают, что клетки и органоиды, культивируемые с помощью сред и способов по изобретению, правдоподобно отражают ситуацию in vivo. По этой причине они могут быть инструментом для поиска новых (молекулярных) мишеней при конкретных заболеваниях.
Для поиска новых мишеней лекарственных средств можно использовать библиотеку соединений (таких как миРНК) для трансдукции клеток и инактивации определенных генов. В некоторых вариантах осуществления летки трансдуцируют посредством миРНК для ингибирования функции (большой) группы генов. Для определения того, имеет ли мишень значение для исследования, можно использовать любой функциональный показатель группы генов или конкретной клеточной функции. Специфический для заболевания показатель можно определять с использованием анализов, хорошо известных в данной области. Например, пролиферацию клеток анализируют для исследования генов, вовлеченных в злокачественную опухоль. Например, анализ Topflash, как описано в настоящем описании, можно использовать для выявления изменений в активности Wnt, вызываемых ингибированием с помощью миРНК. Когда происходит снижение роста или гибель клеток, соответствующие родственные миРНК гены можно идентифицировать способами, известными в данной области. Эти гены являются возможными мишенями для ингибирования роста этих клеток. При идентификации необходимо определять специфичность идентифицируемой мишени в отношении клеточного процесса, который исследуют, с помощью способов, хорошо известных в данной области. С использованием этих способов можно идентифицировать новые молекулы в качестве возможных мишеней лекарственных средств для терапии.
Скрининг для подтверждения мишени и лекарственного средства
Органоиды конкретного пациента, полученные из пораженной заболеванием и/или нормальной ткани, можно использовать для подтверждения мишени молекул, идентифицированных в высокопроизводительном скрининге. То же самое справедливо для подтверждения соединений, которые были идентифицированы в качестве возможных терапевтических лекарственных средств в высокопроизводительном скрининге. Использование первичного материала пациента, увеличенного в количестве в культуральной системе органоида, может быть пригодным для исследования на ложноположительные результаты и т.д. в высокопроизводительных исследованиях клеточных линий по выявлению лекарственных средств.
В некоторых вариантах осуществления увеличенную в количестве клеточную популяцию (например, органоид по изобретению), такую как органоиды крипта-ворсинка или органоиды поджелудочной железы, можно использовать для проверки соединений, которые были идентифицированы в высокопроизводительном скрининге в качестве возможных лекарственных средств или косметических средств.
Анализ токсичности
Указанная увеличенная в количестве популяция стволовых клеток (например, органоид по изобретению), такая как органоиды крипта-ворсинка или органоиды поджелудочной железы может далее заменять применение клеточных линий, таких как клетки Caco-2, в анализах токсичности потенциальных новых лекарственных средств или известных или новых пищевых добавок.
Токсикологический скрининг осуществляют аналогично скринингу лекарственных средств (как описано выше), однако он исследует только токсические эффекты лекарственных средств, но не терапевтические эффекты. Таким образом, в некоторых вариантах осуществления эффекты соединений-кандидатов являются токсическими.
Культивирование патогенов
Более того, указанную увеличенную в количестве популяцию стволовых клеток (например, органоид по изобретению), такую как органоиды крипта-ворсинка или органоиды поджелудочной железы, можно использовать для культивирования патогена, такого как норовирус, который в настоящее время лишен подходящей тканевой культуры или модели на животных.
Регенеративная медицина и трансплантация
Культуры, содержащие увеличенную в количестве популяцию стволовых клеток (например, органоид по изобретению), такую как органоиды крипта-ворсинка или органоиды поджелудочной железы, пригодны в регенеративной медицине, например, в репарации кишечного эпителия после облучения и/или после хирургической операции, в репарации кишечного эпителия у пациентов, страдающих воспалительным заболеванием кишечника, таким как болезнь Крона и язвенный колит, и в репарации кишечного эпителия у пациентов, страдающих синдромом короткой кишки. Дальнейшим применением является применение в репарации кишечного эпителия у пациентов с наследственными заболеваниями тонкого кишечника/толстого кишечника. Культуры, содержащие органоиды поджелудочной железы, также пригодны в регенеративной медицине, например, в качестве имплантатов после резекции поджелудочной железы или ее части и для лечения диабета, такого как диабет I и диабет II.
В альтернативном варианте осуществления увеличенные в количестве эпителиальные стволовые клетки перепрограммируют на судьбу родственной ткани, например, такой как клетки поджелудочной железы, включая бета-клетки поджелудочной железы. К настоящему времени, невозможно регенерировать клетки поджелудочной железы из взрослых стволовых клеток. Способы культивирования по настоящему изобретению позволят анализировать факторы, которые обеспечивают транс-дифференцировку родственной эпителиальной стволовой клетки в клетку поджелудочной железы, включая бета-клетку поджелудочной железы.
Квалифицированному специалисту будет понятно, что генную терапию, кроме того, можно использовать в способе, направленном на репарацию поврежденной или пораженной заболеванием ткани. Например, можно использовать аденовирусный или ретровирусный носитель для доставки генов, чтобы доставлять генетическую информацию, такую как ДНК и/или РНК, в стволовые клетки. Квалифицированный специалист может заменять или осуществлять репарацию конкретных генов, на которые направлена генная терапия. Например, нормальный ген можно встраивать в неспецифическую область в геноме для замены нефункционального гена. В другом примере, аномальной последовательностью гена можно заменять нормальную последовательность гена путем гомологичной рекомбинации. Альтернативно, селективная обратная мутация может возвращать ген к его нормальной функции. Следующим примером является изменение регуляции (степени, с которой ген включается или выключается) конкретного гена. Предпочтительно, стволовые клетки обрабатывают ex vivo с помощью подхода генной терапии, а затем переносят млекопитающему, предпочтительно, человеку, нуждающемуся в лечении.
Поскольку биоптаты тонкого кишечника, взятые от взрослых доноров, можно увеличивать в количестве без какого-либо явного предела или генетического вреда, эта технология может служить для получения трансплантируемого эпителия для регенеративных целей. Тот факт, что органоиды можно замораживать и размораживать и помещать в культуру без утраты их 3D-структуры и целостности и без существенной гибели клеток, далее увеличивает применимость органоидов для целей трансплантации. Более того, в некоторых вариантах осуществления органоиды, погруженные в ECM или находящиеся в контакте с ECM, можно трансплантировать млекопитающему, предпочтительно, человеку. В другом варианте осуществления органоиды и ECM можно трансплантировать одновременно млекопитающему, предпочтительно, человеку.
Квалифицированному специалисту будет понятно, что ECM можно использовать в качестве 3D-каркаса для получения подобных тканям структур, содержащих увеличенные в количестве популяции клеток или органоиды согласно изобретению. Затем такие структуры можно трансплантировать пациенту способами, хорошо известными в данной области. Каркас ECM может быть получен синтетически с использованием белков ECM, таких как коллаген и/или ламинин, или альтернативно каркас ECM может быть получен "децеллюляризацией" выделенного органа или фрагмента ткани, чтобы остался каркас, состоящий из ECM (например, см. Macchiarini et al. The Lancet, Volume 372, Issue 9655, Pages 2023 - 2030, 2008). В некоторых вариантах осуществления каркас ECM можно получать путем децеллюляризации органа или фрагмента ткани, где необязательно указанный орган или фрагмент ткани происходит из поджелудочной железы, печени, кишечника, желудка или предстательной железы.
Как упоминалось выше, изобретение относится к органоиду или популяции клеток по изобретению для применения в трансплантации млекопитающему, предпочтительно, человеку. Также предусматривается способ лечения пациента, нуждающегося в трансплантации, включающий трансплантацию органоида или популяции клеток по изобретению указанному пациенту, где указанным пациентом является млекопитающее, предпочтительно, человек.
Преимущественно, изобретение позволяет брать небольшой биоптат от взрослого донора и увеличивать его в количестве без явного предела или генетического вреда, и, таким образом, технология, описанная в настоящем описании, может служить для получения трансплантируемого эпителия для регенеративных целей.
Существенно, что авторы изобретения открыли, что когда органоиды поджелудочной железы человека по изобретению трансплантируют под периренальную капсулу у мышей, эти клетки дифференцируются с образованием зрелых бета-клеток, которые секретируют инсулин. Это является существенным, поскольку это означает, что даже если популяция клеток или органоид по изобретению не секретируют инсулин на поддающемся выявлению уровне, пока клетки или органоиды культивируют in vitro, эти клетки могут быть пригодными для трансплантации пациенту для лечения нарушения с дефицитом инсулина, такого как диабет.
Таким образом, изобретение относится к способу лечения пациента с нарушением с дефицитом инсулина, такого как диабет, или пациента, имеющего дисфункцию поджелудочной железы, включающему трансплантацию органоида поджелудочной железы по изобретению или клеток из органоида поджелудочной железы по изобретению пациенту.
В некоторых вариантах осуществления клетки или органоид не экспрессируют или не секретируют инсулин при трансплантации пациенту, но дифференцируются у пациента так, что они секретируют инсулин. Например, способность секретировать инсулин может не выявляться сразу после трансплантации, но может присутствовать приблизительно через один месяц после трансплантации, например, через 6 недель, 2 месяца или 3 месяца после трансплантации.
Пациентом, предпочтительно, является человека, но альтернативно им может являться не являющееся человеком млекопитающее, такое как кошка, собака, лошадь, корова, свинья, овца, кролик или мышь.
Таким образом, в объем изобретения включены способы лечения пациента, представляющего собой человека или не являющегося человеком животного, с помощью клеточной терапии. Такая клеточная терапия охватывает использование стволовых клеток или органоидов по изобретению у пациента с помощью любых подходящих средств. В частности, такие способы лечения вовлекают регенерацию поврежденной ткани. В соответствии с изобретением пациента можно лечить с помощью аллогенных или аутологичных стволовых клеток или органоидов. "Аутологичные клетки" представляют собой клетки, которые происходят из того же организма, в который их повторно вводят для клеточной терапии, например, для обеспечения регенерации ткани. Однако клетки не обязательно выделяют из той же ткани, что и ткань, в которую их вводят. В случае аутологичной клетки не требуется подбор пациента для преодоления проблем отторжения. "Аллогенные клетки" представляют собой клетки, которые произошли из индивидуума, который отличается от индивидуума, которому клетки вводят для клеточной терапии, например, для обеспечения регенерации ткани, хотя они представляют собой клетки того же вида. Для предупреждения проблемы отторжения все еще может потребоваться некоторая степень подбора пациента.
Как правило, клетки или органоиды по изобретению вводят в организм пациента путем инъекции или имплантации. Как правило, клетки прямо инъецируют в ткань, в которой предполагается их действие. Альтернативно, клетки можно инъецировать через воротную вену. Шприц, содержащий клетки по изобретению и фармацевтически приемлемый носитель, входит в объем изобретения. Катетер, подсоединенный к шприцу, содержащему клетки по изобретению и фармацевтически приемлемый носитель, входит в объем изобретения.
Квалифицированный специалист способен выбрать подходящий способ и путь введения, в зависимости от материала, который трансплантируют (т.е. популяция клеток, отдельные клетки в суспензии, органоиды или фрагменты органоидов), а также от органа, который лечат.
Как рассмотрено выше, клетки по изобретению можно использовать для регенерации ткани. Для обеспечения этой функции клетки можно инъецировать или имплантировать прямо в поврежденную ткань, где они могут увеличиваться в количестве и в конечном итоге дифференцироваться в требуемый тип клеток, в соответствии с их положением в организме. Альтернативно органоид можно инъецировать или имплантировать прямо в поврежденную ткань. Ткани, которые являются чувствительными к лечению, включают все поврежденные ткани, в частности, включая ткани, которые могут быть повреждены вследствие заболевания, повреждения, травмы, аутоиммунной реакции или вирусной или бактериальной инфекции. В некоторых вариантах осуществления изобретения клетки или органоиды по изобретению используют для регенерации толстого кишечника, тонкого кишечника, поджелудочной железы, пищевода или желудочной системы.
Например, в одном из вариантов осуществления клетки или органоиды по изобретению инъецируют пациенту с использованием шприца Hamilton.
Квалифицированному специалисту будет известно, какими будут соответствующие дозировки клеток или органоидов по изобретению для конкретного состояния, подвергаемого лечению.
В одном из вариантов осуществления клетки или органоиды по изобретению, либо в растворе, либо в микросферах, либо в микрочастицах различного состава, вводят в артерию, снабжающую ткань или часть поврежденного органа, нуждающегося в регенерации. Как правило, такое введение проводят с использованием катетера. Катетер может представлять собой один из большого множества баллонных катетеров, используемых для ангиопластики и/или доставки клеток, или катетер, сконструированный для конкретной цели доставки клеток в конкретную область организма. Для определенных применений клетки или органоиды можно инкапсулировать в микросферы, изготовленные из ряда различных биодеградинуемых соединений и с диаметром приблизительно 15 мкм. Этот способ может позволить введенным внутрисосудистым путем клеткам или органоидам оставаться в области повреждения и не проходить через капиллярную сеть и в системный кровоток при первом прохождении. Удержание в артериальной части капиллярной сети также может способствовать их перемещению во внесосудистое пространство.
В другом варианте осуществления клетки или органоиды можно ретроградно инъецировать в сосудистую сеть, либо через вену для доставки всему организму, либо локально в конкретную вену, которая осуществляет отток в ткань или часть организма, на которые направлены клетки или органоиды. Для этого варианта осуществления можно использовать множество препаратов, описанных выше.
В другом варианте осуществления клетки или органоиды по изобретению можно имплантировать в поврежденную ткань, прикрепленную к биосовместимому имплантату. В рамках настоящего варианта осуществления клетки могут быть прикреплены к биосовместимому имплантату in vitro перед имплантацией пациенту. Как будет понятно специалисту в данной области можно использовать любой из множества прикрепляющих материалов для прикрепления клеток к имплантату перед имплантацией. Только в качестве примера, такой прикрепляющий материал может включать фибрин, один или несколько представителей семейства интегринов, один или несколько представителей семейства кадгеринов, один или несколько представителей семейства селектинов, одну или несколько молекул клеточной адгезии (CAM), один или несколько представителей семейства иммуноглобулинов и один или несколько искусственных прикрепляющих материалов. Этот перечень предоставлен только в качестве иллюстрации и подразумевается, что он не является ограничивающим. Специалисту в данной области будет понятно, что можно использовать комбинацию одного или нескольких прикрепляющих материалов.
В другом варианте осуществления клетки или органоиды по изобретению можно погружать в матрикс перед имплантацией матрикса пациенту. Как правило, матрикс имплантируют в поврежденную ткань пациента. Примеры матриксов включают матриксы на основе коллагена, матриксы на основе фибрина, матриксы на основе ламинина, матриксы на основе фибронектина и искусственные матриксы. Этот перечень предоставлен только в качестве иллюстрации и подразумевается, что он не является ограничивающим.
В следующем варианте осуществления клетки или органоиды по изобретению можно имплантировать или инъецировать пациенту вместе с образующим матрикс компонентом. Это может позволить клеткам образовать матрикс после инъекции или имплантации, обеспечивая удержание клеток или органоидов в соответствующей области у пациента. Примеры образующих матрикс компонентов включают жидкий фибриновый клей, мономеры алкилцианоакрилатов, пластификаторы, полисахариды, такие как декстран, содержащие оксид этилена олигомеры, блок-сополимеры, такие как полоксамер и Pluronics, неионные поверхностно-активные вещества, такие как Tween и Triton"8", и компоненты, образующие искусственный матрикс. Этот перечень предоставлен только в качестве иллюстрации и подразумевается, что он не является ограничивающим. Специалисту в данной области будет понятно, что можно использовать любую комбинацию одного или нескольких образующих матрикс компонентов.
В следующем варианте осуществления клетки или органоиды по изобретению могут содержаться в микросфере. В этом варианте осуществления клетки могут быть инкапсулированы в центр микросферы. Также в этом варианте осуществления клетки могут быть погружены в материал матрикса микросферы. Материал матрикса может включать любой подходящий биодеградируемый полимер, включая, но не ограничиваясь ими альгинаты, полиэтиленгликоль (PLGA) и полиуретаны. Этот перечень предоставлен только в качестве иллюстрации и подразумевается, что он не является ограничивающим.
В следующем варианте осуществления клетки или органоиды по изобретению могут быть прикреплены к медицинскому устройству, предназначенному для имплантации. Примеры таких медицинских устройств включают стенты, спицы, швы, разделители, кардиостимуляторы, суставные протезы, искусственную кожу и стержни. Этот перечень предоставлен только в качестве иллюстрации и подразумевается, что он не является ограничивающим. Специалисту в данной области будет понятно, что клетки могут быть прикреплены к медицинскому устройству различными способами. Например, клетки или органоиды могут быть прикреплены к медицинскому устройству с использованием фибрина, одного или нескольких представителей семейства интегринов, одного или нескольких представителей семейства кадгеринов, одного или нескольких представителей семейства селектинов, одной или нескольких молекул клеточной адгезии (CAM), одного или нескольких представителей семейства иммуноглобулинов и одного или нескольких искусственных прикрепляющих материалов. Этот перечень предоставлен только в качестве иллюстрации и подразумевается, что он не является ограничивающим. Специалисту в данной области будет понятно, что можно использовать комбинацию одного или нескольких прикрепляющих материалов.
Способы по изобретению
Изобретение также относится к способу увеличения в количестве популяции стволовых клеток, где способ включает:
a) предоставление популяции стволовых клеток;
b) предоставление культуральной среды согласно изобретению;
c) контактирование стволовых клеток с культуральной средой; и
d) культивирование клеток в соответствующих условиях.
Более того, изобретение относится к способу увеличения в количестве выделенных фрагментов тканей, где способ включает:
a) предоставление выделенного фрагмента ткани;
b) предоставление культуральной среды согласно изобретению;
c) контактирование выделенного фрагмента ткани с культуральной средой; и
d) культивирование клеток в соответствующих условиях.
Способ "увеличения в количестве" популяции клеток или выделенных фрагментов ткани представляет собой способ, который вовлекает поддержание или увеличение количества стволовых клеток в первоначальной популяции для получения увеличенной в количестве популяции стволовых клеток, которые сохраняют их недифференцированный фенотип и свойства самообновления. Однако также он может включать получение дифференцирующегося потомства, которое может, например, образовывать подобные тканям структуры, участвующие в образовании органоида. Таким образом, в настоящем описании предусматриваются способы получения органоида, такого как органоид тонкого кишечника (крипта-ворсинка), органоид толстого кишечника, органоид поджелудочной железы, органоид желудка, органоид предстательной железы, органоид печени, органоид аденокарциномы, органоид карциномы или органоид пищевода Барретта, включающие культивирование стволовых клеток или фрагментов тканей, содержащих указанные стволовые клетки, в культуральной среде по изобретению. Изобретение относится к способу увеличения в количестве единичной стволовой клетки или популяции стволовых клеток, предпочтительно, для получения органоида, где способ включает культивирование единичной стволовой клетки, популяции стволовых клеток или фрагмента ткани в культуральной среде согласно изобретению. В некоторых вариантах осуществления способ получения органоида включает культивирование стволовых клеток или фрагментов тканей с первой средой для "увеличения в количестве", а затем культивирование стволовых клеток и фрагментов тканей со второй средой для "дифференцировки". В некоторых вариантах осуществления среда для дифференцировки не содержит определенных компонентов среды для увеличения в количестве, например, среда для дифференцировки не содержит Wnt, R-спондина, никотинамида, ингибитора TGF-бета и/или ингибитора p38.
В некоторых вариантах осуществления способ увеличения в количестве единичной стволовой клетки или популяции стволовых клеток, предпочтительно, для получения органоида, включает увеличение в количестве единичной стволовой клетки, популяции стволовых клеток или фрагмента ткани в первой культуральной среде согласно изобретению, и необязательно, дифференцировку увеличенных в количестве клеток или фрагментов тканей во второй культуральной среде согласно изобретению.
Таким образом, изобретение относится к способу увеличения в количестве единичной стволовой клетки или популяции стволовых клеток, предпочтительно, для получения органоида, где способ включает:
предоставление стволовой клетки, популяции стволовых клеток или выделенного фрагмента ткани;
предоставление культуральной среды согласно изобретению;
контактирование стволовых клеток с культуральной средой;
культивирование клеток в соответствующих условиях.
В некоторых вариантах осуществления способ включает приведение в контакт стволовой клетки, популяции стволовых клеток или выделенного фрагмента ткани и культуральной среды с внеклеточным матриксом или 3D-матриксом, который имитирует внеклеточный матрикс вследствие его взаимодействия с белками клеточной мембраны, такими как интегрины, например, содержащим ламинин внеклеточным матриксом, таким как MatrigelTM (BD Biosciences). В некоторых вариантах осуществления культуральная среда диффундирует во внеклеточный матрикс.
В некоторых вариантах осуществления изобретение относится к способу увеличения в количестве единичной стволовой клетки или популяции стволовых клеток или фрагмента ткани, предпочтительно, для получения органоида, где способ включает:
культивирование стволовой клетки, популяции стволовых клеток или фрагмента ткани в первой среде для увеличения в количестве;
продолжение культивирования стволовой клетки, популяции стволовых клеток или фрагмента ткани и восполнение среды средой для дифференцировки, где среда для дифференцировки не содержит одного или нескольких, предпочтительно, всех из факторов, выбранных из: ингибитора TGF-бета, ингибитора p38, никотинамида и Wnt.
В некоторых вариантах осуществления изобретение относится к способу увеличения в количестве единичной стволовой клетки или популяции стволовых клеток, предпочтительно, для получения органоида представляющей интерес ткани, включающему:
увеличение в количестве стволовых клеток или фрагментов ткани из указанной представляющей интерес ткани в культуральной среде по изобретению, которая пригодна для указанной представляющей интерес ткани; и необязательно
дифференцировку увеличенных в количестве стволовых клеток или фрагментов ткани в культуральной среде по изобретению, которая пригодна для указанной представляющей интерес ткани.
Выделенные стволовые клетки, предпочтительно, культивируют в микроокружении, которое имитирует по меньшей мере частично клеточную нишу, в которой указанные стволовые клетки находятся в природе. Указанную клеточную нишу имитируют путем культивирования указанных стволовых клеток в присутствии биоматериалов, таких как матриксы, каркасы и культуральные субстраты, которые предоставляют ключевые регуляторные сигналы, контролирующие судьбу стволовых клеток. Указанные биоматериалы включают природные, полусинтетические и синтетические биоматериалы и/или их смеси. Каркас обеспечивает двухмерную или трехмерную сеть. Подходящие синтетические материалы для указанного каркаса включают полимеры, выбранные из пористых твердых веществ, нановолокон и гидрогелей, например, таких как пептиды, включающие осуществляющие самосборку пептиды, гидрогели, состоящие из полиэтиленгликоля фосфата, полиэтиленгликоля фумарата, полиакриламида, полигидроксиэтилметакрилата, полицеллюлозы ацетата и/или их сополимеров (см., например, Saha et al., 2007 Curr Opin Chem Biol 1 1(4) 381-387, Saha et al., 2008 Biophysical Journal 95 4426-4438, Little et al., 2008 Chem Rev 108, 1787-1796). Как известно специалисту в данной области, механические свойства, например, такие как эластичность каркаса, влияют на пролиферацию, дифференцировку и миграцию стволовых клеток. Предпочтительный каркас содержит биодеградируемые (со)полимеры, которые заменяются встречающимся в природе компонентами после трансплантации индивидууму, например, для обеспечения регенерации ткани и/или заживления раны. Более того, предпочтительно, чтобы указанный каркас по существу не индуцировал иммуногенного ответа после трансплантации индивидууму. Указанный каркас дополняют природными, полусинтетическими или синтетическими лигандами, которые обеспечивают сигналы, требуемые для пролиферации и/или дифференцировки, и/или миграции стволовых клеток. В предпочтительном варианте осуществления указанные лиганды содержат определенные фрагменты аминокислот. Примеры указанных синтетических полимеров включают поверхностно-активное вещество на основе блок-сополимера Pluronic® F 127 (BASF), и Ethisorb® (Johnson and Johnson).
Клеточная ниша является частью, определяемой стволовыми клетками и окружающими клетками, и внеклеточным матриксом (ECM), который продуцируется клетками в указанной нише. В одном способе по изобретению, выделенные крипты или эпителиальные стволовые клетки прикреплены к ECM. ECM состоят из различных полисахаридов (по большей части гепаринсульфатных протеогликанов), воды, эластина, и гликопротеинов, где гликопротеины включают коллаген, энтактин (нидоген), фибронектин и ламинин. ECM секретируется клетками соединительной ткани. Известны различные типы ECM, обладающие различным составом, включая различные типы гликопротеинов и/или различную комбинацию гликопротеинов. Указанный ECM может быть предоставлен посредством культивирования продуцирующих ECM клеток, например, таких как фибробластные клетки, в емкости, с последующим удалением этих клеток и добавлением выделенных крипт или эпителиальных стволовых клеток. Примерами клеток, продуцирующих внеклеточный матрикс, являются хондроциты, продуцирующие, главным образом, коллаген и протеогликаны, фибробластные клетки, продуцирующие, главным образом, коллаген типа IV, ламинин, интерстициальные проколлагены и фибронектин, и миофибробласты толстой кишки, продуцирующие, главным образом, коллагены (тип I, III и V), протеогликан хондроитинсульфат, гиалуроновую кислоту, фибронектин и тенасцин-C. Альтернативно указанный ECM предоставляют коммерчески. Примерами коммерчески доступных внеклеточных матриксов являются белки внеклеточного матрикса (Invitrogen) и MatrigelTM (BD Biosciences). Использование ECM для культивирования стволовых клеток увеличивало долговременную выживаемость стволовых клеток и сохраняло присутствие недифференцированных стволовых клеток.
Пример ECM для применения в способе по изобретению включает по меньшей мере два различных гликопротеина, таких как два различных типа коллагена или коллаген и ламинин. Указанный ECM может представлять собой внеклеточный матрикс на основе синтетического гидрогеля или встречающийся в природе ECM. Наиболее предпочтительный ECM обеспечивается с помощью MatrigelTM (BD Biosciences), который содержит ламинин, энтактин и коллаген IV. Таким образом, в некоторых вариантах осуществления ECM для применения в способе по изобретению представляет собой 3D-матрикс, который имитирует внеклеточный матрикс путем его взаимодействия с белками клеточной мембраны, такими как интегрины.
Таким образом, в некоторых вариантах осуществления способ по изобретению включает приведение в контакт стволовой клетки, популяции стволовых клеток или выделенного фрагмента ткани и культуральной среды в контакт с внеклеточным матриксом, например, содержащим ламинин внеклеточным матриксом, таким как MatrigelTM (BD Biosciences). В некоторых вариантах осуществления культуральная среда диффундирует во внеклеточный матрикс.
Композиции и другие формы по изобретению
Изобретение относится к композиции, содержащей культуральную среду согласно изобретению и стволовые клетки. Изобретение также относится к композиции, содержащей культуральную среду согласно изобретению и органоиды. Более того, изобретение относится к композиции, содержащей культуральную среду согласно изобретению и внеклеточный матрикс.
Изобретение также относится к композиции, содержащей культуральную среду по изобретению, внеклеточный матрикс и стволовые клетки по изобретению. Изобретение также относится к композиции, содержащей культуральную среду по изобретению, внеклеточный матрикс и один или несколько органоидов по изобретению. Изобретение также относится к добавке в культуральную среду, которую можно использовать для получения культуральной среды, как описано в настоящем описании. "Добавка в культуральную среду" представляет собой смесь ингредиентов, которая сама по себе не может поддерживать стволовые клетки, но которая делает возможной или улучшает культуру стволовых клеток при комбинировании с другими ингредиентами клеточной культуры. Таким образом, добавку можно использовать для получения функциональной среды для культивирования клеток по изобретению путем комбинирования ее с другими ингредиентами клеточной культуры для обеспечения соответствующего состава среды. Применение добавок в культуральную среду хорошо известно в данной области.
Изобретение относится к добавке в культуральную среду, которая содержит ингибитор согласно изобретению. Добавка может содержать любой ингибитор (или комбинацию ингибиторов), описанный в настоящем описании. Добавка также может содержать один или несколько дополнительных ингредиентов клеточной культуры, как описано в настоящем описании, например, один или несколько ингредиентов клеточной культуры, выбранных из группы, состоящей из аминокислот, витаминов, неорганических солей, углеродных источников энергии и буферов.
Культуральная среда или добавка в культуральную среду может представлять собой концентрированную жидкую культуральную среду или добавку (например, от 2x до 250x концентрированную жидкую культуральную среду или добавку) или она может представлять собой сухую культуральную среду или добавку. Как жидкие, так и сухие культуральные среды и добавки хорошо известны в данной области. Культуральная среда или добавка может быть лиофилизированной.
Культуральную среду или добавку по изобретению, как правило, стерилизуют перед применением для предупреждения контаминации, например, ультрафиолетовым светом, нагреванием, облучением или фильтрацией. Культуральную среду или добавку в культуральную среду можно замораживать (например, при -20°C или -80°C) для хранения или транспортировки. В некоторых вариантах осуществления культуральную среду можно хранить в качестве жидкости (например, при приблизительно 4°C). В некоторых вариантах осуществления культуральную среду можно разделять и хранить в качестве двух компонентов: замороженного компонента (например, при температуре от приблизительно -20°C до приблизительно -80°C) и жидкого компонента (например, при приблизительно 4°C). В частности, в замороженный компонент, предпочтительно, включены чувствительный к температуре или чувствительный к времени деградируемый материал, в то время как менее чувствительный материал (например, DMEM или FCS) можно хранить в жидкой форме и, таким образом, включать в жидкий компонент для хранения и транспортировки.
Изобретение также относится к герметично закрытой емкости, содержащей культуральную среду или добавку в культуральную среду по изобретению. Герметично закрытые емкости могут быть предпочтительными для транспортировки или хранения культуральных сред или добавок в культуральные среды, описанные в настоящем описании, для предотвращения контаминации. Емкость может представлять собой любую подходящую емкость, такую как колба, планшет, бутылка, банка, флакон или мешок.
Изобретение также относится к набору, содержащему культуральную среду, добавку в культуральную среду и/или композицию по изобретению. В некоторых вариантах осуществления набор, кроме того, содержит по меньшей мере один другой дополнительный компонент, например, выбранный из перечня, включающего: ECM (например, MatrigelTM), популяцию клеток и органоид.
Общая информация
Выше в настоящем описании использована нумерация "GI". Номер GI, или "идентификатор GenInfo", представляет собой серию цифр, приписанных последовательно каждой записи последовательности, обработанной в NCBI, когда последовательности добавляют в его базы данных. Номер GI не имеет сходства с номером доступа в записи о последовательности. Когда последовательность обновляют (например, для коррекции или добавления дополнительной аннотации или информации), тогда ей присваивают новый номер GI. Таким образом, последовательность, связанная с данным номером GI, никогда не меняется.
Термин "содержащий" охватывает "включающий", а также "состоящий", например, композиция, "содержащая" X может состоять исключительно из X или может включать что-либо дополнительное, например, X + Y.
Словосочетание "по существу" не исключает "полностью", например, композиция, которая "по существу свободна" от Y, может быть полностью свободна от Y. Когда это необходимо, словосочетание "по существу" может быть пропущено в определении по изобретению.
Термин "приблизительно" применительно к числовой величине x является необязательным и означает, например, x±10%.
Если конкретно не указано, способ, включающий стадию смешения двух или более компонентов, не требует какого-либо конкретного порядка смешения. Таким образом, компоненты можно смешивать в любом порядке. Когда имеется три компонента, тогда два компонента можно комбинировать друг с другом, а затем комбинацию можно комбинировать с третьим компонентом и т.д.
Различные аспекты и варианты осуществления изобретения описаны ниже более подробно в качестве примера. Будет понятно, что модификация деталей может быть осуществлена без отклонения от объема изобретения.
ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг.1. Культура толстого кишечника мыши
a. слева: экспрессия Axin2 находится под контролем каскада передачи сигнала Wnt. Органоиды крипты толстого кишечника мышей с репортером Axin2-LacZ культивировали с EGF, Noggin и R-спондином (ENR) в течение 3 суток. Отсутствие окрашивания на LacZ указывает на то, что активный сигнал Wnt не присутствует в органоидах толстого кишечника в условиях роста ENR. На вкладке показана активная передача сигнала Wnt, визуализированная посредством экспрессии LacZ (темное окрашивание) в свежевыделенных криптах толстого кишечника из репортерных мышей Axin2-LacZ. Справа: происходящие из мышей Axin2-LacZ крипты, культивированные с ENR + Wnt3A (WENR) в течение 10 суток. Темное окрашивание указывает на экспрессию LacZ в этих органоидах.
b. слева: крипты толстого кишечника Lgr5-GFP-ires-CreER, культивированные с ENR в течение 3 суток. Отсутствие флуоресценции GFP указывает на утрату экспрессии Lgr5 в органоидах толстого кишечника в условиях роста ENR. На вкладке показана экспрессия Lgr5-GFP в свежевыделенных криптах толстого кишечника из мышей Lgr5-GFP-ires-CreER. Справа: крипта толстого кишечника Lgr5-GFP-ires-CreER, культивированная с WENR в течение 10 суток демонстрирует присутствие Lgr5 стволовых клеток.
c. Эффективность культивирования определяют в этих трех различных условиях: ENR, полные крипты WENR, и крипты WENR после мягкого ферментативного расщепления (расщепленные WENR). Крипты толстого кишечника выделяли из проксимального отдела толстого кишечника (черные столбцы) или дистального отдела толстого кишечника (белые столбцы). *:p<0,05.
d, e: 4 суток после удаления Wnt3A из культуральной среды WENR приводят к дифференцировке органоида. d. Можно видеть хроматогранин A (ChA) в энтероэндокринных клетках; муцин 2 (muc2) в бокаловидных клетках и контрастное окрашивание с помощью DAPI. e. Зрелые энтероциты визуализируют с помощью окрашивания на щелочную фосфатазу.
f. Представлена относительная экспрессия мРНК зрелых маркеров эпителиальных клеток (Vil1 (Villin1), Alpi (щелочная фосфатаза), Chga (хромогранин A), Muc2 (муцин 2)). Культивируемые в WENR органоиды крипты толстого кишечника культивируют в течение 4 суток в условиях WENR (заштрихованный паттерн) или ENR (черный). Свежевыделенные крипты толстого кишечника (белый) используют в качестве контроля. Масштабная метка в a, b, d, e: 50 мкм. Планки погрешности указывают на s.e.m. n=3.
Фиг.2. Культура толстого кишечника человека
a. Эффект никотинамида на органоиды крипт толстого кишечника человека. Большинство органоидов крипты толстого кишечника погибают в течение нескольких суток в условиях WENR+гастрин (WENRg) (левая панель). Добавление никотинамида (средняя панель: WENRg+nic) повышает эффективность культуры и продолжительность жизни органоидов толстого кишечника человека. * p<0,001. nic: никотинамид.
b. Эффект низкомолекулярного ингибитора Alk4/5/7 (A83-01) и MAP-киназы p38 (SB202190) на органоиды крипт толстого кишечника человека. Левая панель: Органоиды толстого кишечника человека, культивируемые в среде, содержащей WENRg + никотинамид, образуют кистозные структуры после культивирования в течение 3-4 недель. Средняя панель: органоиды толстого кишечника человека сохранят их характерную почкующуюся структуру в условиях культивирования стволовых клеток кишечника человека ("HISC") (WENRg+никотинамид+A83-01+SB202190). Правая панель: A83-01 и SB202190 синергично увеличивают количество пассажей органоидов толстого кишечника человека. * p<0,001. N.S.= нет статистически значимых отличий. Планки погрешностей указывают на s.e.m. n=5.
c. Пролиферирующие клетки, визуализированные путем включения EdU, ограничиваются почкующимися структурами. DAPI используют в качестве контрастного красителя.
d. Типичная картина кариотипа из органоида крипты толстого кишечника человека возрастом 3 месяца. Масштаб: 100 мкм.
e. Тепловая карта профиля экспрессии культивируемых органоидов кишечника человека. Тепловая карта представляет собой сравнение крипт тонкого кишечника человека и ворсинок тонкого кишечника человека. Гены с более высокой экспрессией в крипте выделены темно-серым цветом (верхняя половина тепловой карты), гены с более высокой экспрессией в ворсинке выделены светло-серым цветом (нижняя половина тепловой карты). Органоиды, культивируемые in vitro, отчетливо проявляют сходные профили экспрессии со свежевыделенными криптами тонкого кишечника и экспрессируют известные маркеры стволовых клеток. (Дорожка 1: органоиды тонкого кишечника человека #1, дорожка 2: органоиды тонкого кишечника человека #2, дорожка 3: органоиды толстого кишечника человека, дорожка 4: свежевыделенные крипты тонкого кишечника человека. Четыре образца сравнивают с ворсинкой тонкого кишечника человека).
Фиг.3. Состав типов клеток органоида кишечника человека
(a-c) Органоиды человека дифференцируются в различные типы клеток кишечника после устранения никотинамида и SB202190. Чтобы продемонстрировать дифференцировку, использовали маркеры различных типов клеток. (a) Верхняя панель: окрашивание на щелочную фосфатазу зрелых энтероцитов, средняя панель: окрашивание на PAS бокаловидных клеток, нижняя панель: окрашивание на синаптофизин энтероэндокринных клеток. (b) В каждом случае светлая область указывает на окрашивание. Окрашивание на муцин 2 (Muc2) в средней панели отражает бокаловидные клетки и окрашивание на хромогранин A (ChgA) в левой панели отражает энтероэндокринные клетки (см. стрелку и вкладку). DAPI используют в качестве контрастного красителя (правая панель). (c) Окрашивание на лизоцим (Lysz) представлено на левой панели, чтобы показать клетки Панета. DAPI используют в качестве контрастного красителя (правая панель).
(d-f) Дифференцировка бокаловидных клеток (Muc2) блокируется обработкой посредством SB202190 органоидов (d), в то время как ингибитор Notch DBZ увеличивает количество бокаловидных клеток в органоидах человека (f). Количество пролиферирующих клеток, отображаемых по включению EdU (средняя панель), увеличено в органоидах, обработанных SB202190 (d) или снижено в органоидах, обработанных DBZ (f).
Органоиды культивируют в следующих условиях в течение 5 суток: a) сверху: ENRg+A83-01+SB202190+никотинамид, a) в середине и снизу, b), c) WENRg+A83-01, d)WENRg+A83-01+SB202190, e) WENRg+A83-01, f) WENRg+A83-01+DBZ.
Масштабная метка: 20 мкм (a), 50 мкм (b-f). a, b, d-f: органоиды крипты толстого кишечника человека, c: органоиды тонкого кишечника человека.
Фиг.4. Культуры адено(карцино)мы
a. Крипты Lgr5-GFP-ires-CreER/APCfl/fl, культивированные с EGF (E) (сверху) или EGF+Noggin (EN) (снизу) в течение 10 суток. b. Относительная экспрессия мРНК Lgr5 и Axin2. Свежевыделенные клетки аденомы (белый) культивировали с EGF (заштрихованный) или EGF+Noggin (черный).
c. Эффективность культивирования органоидов из отсортированных клеток Lgr5-GFPhi, Lgr5-GFPlo, Lgr5-GFP-ve. *p<0,01. Односторонний ANOVA. Планки погрешности указывают на s.e.m. n=3.
d. Культивирование с течением времени клеток аденокарциномы толстого кишечника человека.
Фиг.5. Культивирование пищевода Барретта и обработка ингибитором Notch.
a. Выделенный эпителий из пищевода Барретта (BE), культивированный в условиях HISC в течение 7 суток, образует кистозные структуры. b. Добавление FGF10 значительно увеличивает количество пассажей для органоидов BE. Планки погрешностей указывают на s.e.m. n=3. c. Типичная динамика органоида BE. d. Парафиновые срезы органоидов BE. Никотинамид и SB202190 устраняют в течение 4 суток с (справа) или без (слева) ингибитора Notch DBZ, добавленного в среду. Пролиферирующие клетки (краситель Ki67) исчезают и PAS+ бокаловидные клетки увеличиваются в количестве при обработке DBZ.
Фиг.6. Экспрессия мРНК Axin2 в выделенных органоидах толстого кишечника в присутствии Wnt-3A
Выделенные крипты толстого кишечника анализируют в отношении экспрессии мРНК Axin2 после культивирования в течение 3 суток или 7 суток с ENR (заштрихованный) или WENR (черный). Свежевыделенные крипты толстого кишечника использовали в качестве контроля. Планки погрешностей указывают на s.e.m. n=3
Фиг.7. Относительная экспрессия мРНК маркеров зрелых эпителиальных клеток
Свежевыделенные крипты тонкого кишечника (белый) культивировали в условиях HISC в течение 14 суток, а затем культивировали в указанных условиях культивирования в течение 4 суток. Анализировали экспрессию мРНК ALPI (щелочная фосфатаза), VIL1 (Villin1), LYZ (лизоцим), CHGB (хромогранин B) и MUC2 (муцин 2). Условия культивирования: HISC (черный), ENRg+A83-01+SB202190+никотинамид, WENRg+A83-01, ENRg+A83-01, ENRg. Свежевыделенные крипты тонкого кишечника использовали в качестве контроля (установленного как 1,0 для ALPI, VIL1 и LYZ, как 5,0 для CHGB и MUC2. Планки погрешностей указывают на s.e.m. n=3.)
Фиг.8. Отсортированные клетки Lgr5-GFP образуют Lgr5-GFP+ органоиды
Единичные отсортированные клетки аденомы Lgr5-GFP-APCfl/fl культивируют с EGF+Noggin (EN) или EGF (E) в течение 7 суток. В органоидах аденомы, происходящих из Lgr5-GFP-клеток, восстанавливалась экспрессия Lgr5-GFP в условиях EN, но не в условиях E (a, c: справа, b, d: аутофлуоресценция GFP).
Фиг.9. Гистохимический анализ органоидов аденомы/рака толстого кишечника
Органоиды аденомы тонкого кишечника мыши (левая панель) и органоиды рака толстого кишечника человека (правая панель) анализировали с указанными гистохимическими (HE, PAS и щелочная фосфатаза) или иммуногистохимическими (хромогранин A, Ki67 и каспаза 3) красителями.
Фиг.10. Клетки Панета в органоидах BE
Лизоцим + клетки Панета наблюдали в дифференцированных органоидах BE.
Фиг.11. Перечень реагентов, использованных для культивирования органоидов
Фиг.12. Перечень реагентов, использованных для оптимизации культуры органоида кишечника человека
Фиг.13. Перечень низкомолекулярных ингибиторов, использованных для оптимизации культуры органоидов кишечника человека
Фиг.14. Перечень 25 наиболее активируемых и наиболее подавляемых генов
мРНК из органоидов тонкого кишечника или органоидов толстого кишечника сравнивают с мРНК из свежевыделенных кишечных ворсинок с помощью микрочипа. Представлены 25 наиболее активируемых и наиболее подавляемых генов. Заштрихованными линиями выделены гены, которые находились среди 70 наиболее активируемых и наиболее подавляемых генов в свежевыделенных криптах тонкого кишечника относительно ворсинок.
Фиг.15. Обобщение статуса пролиферации, дифференцировки и апоптоза для условий культивирования каждого органоида
Фиг.16: Сравнение с помощью микрочипов органоидов поджелудочной железы мыши
A - Анализ кластеризации на микрочипах, сравнивающий РНК из органоидов поджелудочной железы (культивированных в условиях, описанных в примере 2) со взрослой поджелудочной железой, взрослой печенью и печенью новорожденного. Слева направо: i) органоид поджелудочной железы; ii) взрослая поджелудочная железа; iii) взрослая печень (образец 1 [S1] и образец 2 [2]); iv) взрослая печень S2; и v) печень новорожденного.
B - Исходные данные о сигнале в анализе на микрочипах, сравнивающем уровни экспрессии выбранных маркеров протоков, факторов транскрипции, необходимых для экспрессии Ngn3, и эндокринных маркеров во взрослой печени, взрослой поджелудочной железе, органоидах поджелудочной железы и органоидах печени в среде для увеличения в количестве.
Фиг.17: Эффект Noggin на увеличение в количестве органоидов поджелудочной железы
A - Гистограммы, демонстрирующие анализ экспрессии генов органоидов поджелудочной железы, культивируемых в EGFRA так, чтобы они никогда не культивировались с Noggin (черный), и органоидов, культивируемых в EGFRAN так, чтобы они всегда культивировались с Noggin (белый). Также показан эффект культивирования органоидов поджелудочной железы в EGFRA в течение 2 суток, а затем устранения Noggin и культивирования в течение следующих 2 или 4 суток (светло-серый) и эффект культивирования органоидов поджелудочной железы в EGFRA в течение 2 суток, а затем добавления Noggin и культивирования в течение последующих 2 или 4 суток (темно-серый) на экспрессию генов. Уровни мРНК (произвольные единицы) представлены на оси Y. мРНК следующих ранних эндокринных маркеров проанализированы на основной фигуре: Sox9, Hnf1b, Hnf6, Hnf1a, Nkx2.2, Nkx6.1 и Pdx1. мРНК следующих маркеров протоков проанализирована на вкладке: кератин 7 (Krt7) и кератин 19 (Krt19).
B - Гистограмма, показывающая эффект Noggin на экспрессию Lgr5 в органоидах поджелудочной железы в культуральной среде для увеличения в количестве. Данные предоставлены для органоидов поджелудочной железы, культивируемых в EGFRA, которые никогда не культивировали с Noggin (черный), и органоидов, культивируемых в EGFRAN, которые всегда культивировали с Noggin (белый). Также показан эффект культивирования органоидов поджелудочной железы в EGFRAN, а затем устранения Noggin и культивирования в течение следующих 6 суток (светло-серый) и эффект культивирования органоидов поджелудочной железы в EGFRA, а затем добавления Noggin и культивирования следующих 6 суток (темно-серый) на экспрессию гена Lgr5. Уровни мРНК (произвольные единицы) представлены на оси Y.
Фиг.18: Продуцирующие инсулин человека клетки развиваются из увеличенных в количестве ex vivo, трансплантированных in vivo клеток-предшественников
A - Рост ткани поджелудочной железы из клеток-предшественников (стволовые клетки поджелудочной железы) на P0: (1 сутки); P0: (5 сутки); P1: (12 сутки) и P3: (24 сутки), где "P" относится к количеству пассажей.
На фиг.18B и C показана трансплантация органоидов поджелудочной железы человека в периренальную капсулу мыши.
B - через 3 часа после трансплантации клеток органоидов поджелудочной железы реципиентной мыши: окрашивание DAPI (ядерный маркер) на верхнем изображении указывает на все клетки; окрашивание K19 (маркер протоков) на нижнем изображении демонстрирует все трансплантированные клетки, и инсулин (маркер бета-клеток) на нижнем изображении указывает на продуцирующие инсулин клетки.
C - через 1 месяц после трансплантации клеток органоида поджелудочной железы реципиентным мышам: окрашивание DAPI (ядерный маркер) на верхнем изображении (синий цвет) указывает на все клетки; экспрессия CK19 (маркер протоков) на среднем изображении (зеленый цвет) указывает на все трансплантированные клетки, и инсулин (маркер бета-клеток) на нижнем изображении (красный цвет) указывает на продуцирующие инсулин клетки. Группа продуцирующих инсулин клеток обведена в круг, но полагают, что все четко окрашенные клетки являются положительными по инсулину.
Фиг.19: Экспрессия генов органоида поджелудочной железы
В этой таблице показана экспрессия генов поджелудочной железы для наиболее активированных генов по сравнению с органоидами печени.
Фиг.20: Культура органоидов печени мыши демонстрирует стабильное кариотипирование после долговременного культивирования.
A - изображения DIC органоидов печени, поддерживаемых в EGF (E) и R-спондине 1 (R), дополненных FGF10, HGF и никотинамидом (левая фиг., ER), или поддерживаемых в той же комбинации, дополненной Noggin (N) и кондиционированной Wnt3A средой (W) (правая фиг., ENRW) в течение 24 месяцев.
B - Кариотипический анализ органоидов печени мыши после культивирования в течение 8 месяцев. Были выявлены нормальное количество хромосом (n=40, фиг. на левой панели) и полиплоидия, типичный признак гепатоцитов (n=80, фиг. на правой панели).
Фиг.21: Дополнительные факторы FGF10, HGF и никотинамид; эффект на рост и дифференцировку органоида печени.
A - Диаграмма, на которой представлены гены, дифференциально экспрессируемые в течение 3 стадий развития печени, от гепатобласта до зрелого гепатоцита.
B - Схема, на которой представлен используемый протокол. Культуры высевали в среде для увеличения в количестве (ERFHNic: EGF (E) и R-спондин 1 (R), дополненная FGF10, HGF и никотинамидом; ERFHNic указана как "ER" на фиг.8B) за 2 суток до эксперимента. Через двое суток культуральные среды заменяли либо на EGF (E) отдельно, либо на EGF, дополненную R-спондином 1 (ER) с или без дополнительных добавок, выбранных из FGF10 (F), или HGF (H), или никотинамидом (Nic) или их комбинацией, в концентрациях, указанных в тексте. Через пять суток культуры разделяли и повторно высевали в соотношении 1:4 для каждых условий. В этих условиях культуры разделяли и повторно высевали каждые 7 суток в течение общего периода 10 недель
C - Первые сутки после первого разделения в каждых исследованных условиях культивирования. Результаты показывают, что EGF и R-спондин 1 в комбинации с FGF10, или HGF, или никотинамидом, или их комбинации необходимы для осуществления по меньшей мере 1 пассажа.
D - Долговременное культивирование, комбинация ER, дополненная FNic, или ER, дополненная FHNic, обе приводят к высоким количествам пассажей. После 10 пассажей скорость роста является лучшей для условий культивирования, включающих 3 дополнительных фактора, ERFHNic.
E - Анализ ОТ-ПЦР, демонстрирующий экспрессию различных маркеров гепатоцитов (CYP3A11, Alb, FAH) и маркер холангиоцитов (K19) через 5 суток после устранения определенных факторов (исходная точка представляла собой ERFHNic). Следует отметить, что только условия EF продемонстрировали экспрессию всех исследованных маркеров гепатоцитов. HPRT использовали в качестве гена домашнего хозяйства для нормализации экспрессии генов.
Фиг.22: Таблица, в которой показано количественное определение различных специфических факторов транскрипции гепатоцитов и холангиоцитов в клетках из трех различных условий культивирования печени и ткани взрослой печени. Также показана экспрессия ключевых компонентов каскадов передачи сигнала Notch и TGF-бета. E=EFHNic, ER=ERFHNic, ENRW=ENRWFHNic.
Фиг.23: Протокол дифференцировки
A - Схема, демонстрирующая использованный протокол. Культуры высевали в среде для увеличения в количестве (ERFHNic: EGF (E) и R-спондин 1 (R), дополненные FGF10, HGF и никотинамидом; ERFHNic указана как "ER" на фиг.10A) за 2 суток до эксперимента. Через двое суток культуральные среды заменяли на EGF (E), дополненный либо A8301 (A), либо DAPT (D), либо FGF10 (F), либо HGF (H), либо никотинамидом (Nic) или R-спондином 1 (R) или Wnt3A или Noggin (N) или их комбинацией в показанных концентрациях. РНК выделяли в несколько моментов времени. Ткань печени мыши использовали в качестве положительного контроля (+), в то время как в качестве отрицательного контроля (-) использовали воду.
B - Анализ ОТ-ПЦР, демонстрирующий экспрессию маркеров гепатоцитов CYP3A11, Alb, FAH, tbx3, TAT и Gck через 7 суток после условий дифференцировки. Следует отметить, что только условия EADF продемонстрировали экспрессию всех исследованных маркеров гепатоцитов. HPRT использовали в качестве гена домашнего хозяйства для нормализации экспрессии генов.
C - Анализ экспрессии с течением времени после условий дифференцировки. На 2, 5 и 8 сутки после дифференцировки экспрессию маркеров гепатоцитов CYP3A11, Alb, FAH, и маркера холангиоцитов K19, анализировали с помощью ОТ-РЦР. Следует отметить, что экспрессия маркеров печени CYP3A11 и FAH начинается через 5 суток и достигает пика еще через 8 суток. HPRT использовали в качестве гена домашнего хозяйства для нормализации экспрессии. A; A8301, D; DAPT, F; FGF10, H; HGF, De; дексаметазон.
D - Эксперимент по титрованию, демонстрирующий экспрессию маркеров гепатоцитов CYP3A11, Alb, FAH, tbx3, TAT, G6P и Gck через 7 суток после обработки различными концентрациями соединений для дифференцировки A и D. HPRT использовали в качестве гена домашнего хозяйства для нормализации экспрессии гена.
E - Иммунофлуоресцентное окрашивание маркеров печени K19, альбумина и поверхностного маркера гепатоцитов.
F - Окрашивание Xgal органоидов, происходящих из печени мышей Albcreert2LacZ. Положительные по альбумину клетки (стрелки) были выявлены после дифференцировки с помощью EADF в индуцированных тамоксифеном происходящих из Albcreert2LacZ культурах.
Фиг.24: Каскад передачи сигнала простагландинов (Antagonism of the prostaglandin D2 receptors DP1 and CRTH2 as an approach to treat allergic diseases. Roy Pettipher, Trevor T. Hansel & Richard Armer Nature Reviews Drug Discovery 6, 313-325 (April 2007)).
Фиг.25: Органоиды печени, культивируемые в (A) базальной среде, содержащей hEGF (100 нг/мл, Invitrogen); noggin человека (hnoggin) (25 нг/мл, peprotech); гастрин (10 нМ, sigma); hFGF10 (peprotech); никотинамид (10 мМ, sigma); A8301 (500 нМ, Tocris); hHGF (50 нг/мл, peprotech); кондиционированую Rspo среду (10%); (B) базальной среде + PGE2 (50 нМ); (C) базальной среде + CHIR99021 (3 мкМ); (D) базальной среде + CHIR99021 (3 мкМ) + PGE2 (50 нМ).
Фиг.26: Оганоиды печени, культивируемые в базальной среде (как описано для фиг.25) с арахидоновой кислотой и без нее.
Фиг.27: Профиль экспрессии генов органоидов печени мыши в условиях дифференцировки напоминает профиль взрослой печени и печени новорожденного
A - Кластеры генов, демонстрирующие гены, сходным образом экспрессируемые (a) или сходным образом подавляемые (b) между условиями дифференцировки EADF и взрослой печенью или печенью новорожденного.
B - Кластеры генов, демонстрирующие гены, дифференциально экспрессируемые между органоидами печени и взрослой печенью или печенью новорожденного (a) и генами, сходным образом экспрессируемыми между EADF и печенью новорожденного (b).
С - Исходные данные сигналы от анализа микрочипов, сравнивая уровни экспрессии отдельных маркеров протоков, факторов транскрипции, необходимых для экспрессии Ngn3 и эндокринных маркеров во взрослой печени, поджелудочной железы взрослого, органоидов поджелудочной железы и органоидов печени в среде для роста.
Фиг.28: Генные профили печени мыши
Таблица, демонстрирующая a) маркеры, экспрессируемые в стволовых клетках печени мыши; b) маркеры, не экспрессируемые в стволовых клетках печени мыши; c) маркеры гепатоцитов и холангиоцитов, экспрессируемые в профиле стволовых клеток печени мыши для органоидов печени мыши в среде для увеличения в количестве; d) маркеры гепатоцитов и холангиоцитов, не экспрессируемые в профиле стволовых клеток печени мыши для органоидов печени мыши в среде для увеличения в количестве; e) перепрограммирование генов, экспрессируемых в ораганоидах печени мыши; f) перепрограмирование генов, не экспрессируемых в органоидах печени мыши. Результаты получали с использованием микрочипов для печени с использованием универсальной эталонной РНК мыши (Strategene, каталожный номер #740100) в качестве эталонной РНК. Если выявленные абсолютные значения составляли менее 100, ген считали не выявляемым.
Фиг.29: Генные профили печени человека
Таблица, демонстрирующая результаты для органоидов человека, полученные на микрочипе для печени. Слева направо результаты показаны для a) среды для увеличения в количестве EM1, b) среды для увеличения в количестве EM2, c) среды для дифференцировки, d) взрослой печени.
Числа (log2) в левых четырех столбцах представляют собой результат сравнения между образцом и эталонным (коммерческим) образцом РНК, который используют для всех чипов. Представлена относительная экспрессия мРНК в каждом образце по сравнению с РНК, присутствующей в эталонном образце. Использованная эталонная РНК представляла собой универсальную эталонную РНК человека (Stratagene, каталожный номер #740000). Таким образом, отрицательные числа в этих столбцах не связаны с реальными уровнями экспрессии, они означают только, что этой РНК меньше, чем в эталонном образце. В столбце 4 справа представлены абсолютные числа. Если они составляют менее 100, ее считают не выявляемой.
Фиг.30: Морфология органоидов печени. (A) Верхние панели: парафиновый срез печени мыши, демонстрирующий различные домены (PT= воротная триада, CV= центральная вена). Нижние панели: парафиновый срез органоида печени, демонстрирующий различные домены b (однослойный эпителий) и h (расслоенный эпителий) (B) Правая панель: окрашивание на E-кадгерин в органоидах печени. Может быть идентифицировано два различных домена. Домен b образован однослойным эпителием, который напоминает структуры желчных протоков в печени. Этот домен желчных протоков образован высокополярными клетками, которые демонстрируют положительное окрашивание на панцитокератин (PCK) (нижняя панель). На левой панели показано присутствие второго домена в органоидах печени. Этот домен h образован расслоенным эпителием с неполярными клетками. Клетки организованы вокруг центрального просвета и экспрессируют маркер гепатоцитов Alb. Увеличение 10x.
Фиг.31: Окрашивание H&E органоидов поджелудочной железы
Органоиды поджелудочной железы мыши культивировали в условиях увеличения в количестве EGFNRA83-01 [(EGF (50 нг/мл), гастрин (50 нМ), Noggin (10%), R-спондин (5%), FGF10 (100 нг/мл) и A8301 (50 нМ)] на протяжении 8 пассажей (~10 недель). Органоиды удаляли из Matrigel с использованием раствора BD Cell Recovery Solution согласно инструкциям изготовителя и фиксировали 4% параформальдегидом при комнатной температуре в течение 1 ч. Затем органоиды промывали три раза холодным PBS, дегидратировали возрастающими концентрациями спирта и погружали в парафин. 3-мкм парафиновые срезы окрашивали гематоксилином-эозином для анализа гистологии органоидов поджелудочной железы.
Наблюдали существенную вариабельность формы и структуры органоидов. Некоторые из органоидов представляют собой кистозные структуры, образованные монослоем полярных эпителиальных клеток. Другие органоиды демонстрируют тот же монослой эпителиальных клеток и некоторые расслоенные области, где клетки имеют меньший размер и округлую форму. В некоторых органоидах наблюдали инвагинации, занимающие внутренне пространство кистозной структуры.
Окрашивания показали, что некоторые органоиды содержат по большей части монослои эпителиальных клеток (слева снизу), в то время как другие органоиды содержат расслоенные области и/или псевдорасслоенные области и/или сложенные монослои (справа снизу). Большинство органоидов поджелудочной железы содержат области расслоенных клеток и монослои (иногда сложенные).
Фиг.32. Количественное определение индуцированного форсколином набухания органоида тонкого кишечника мыши. (a) Анализ с помощью световой микроскопии органоидов, стимулированных форсколином или DMSO. Показаны репрезентативные примеры для указанных моментов времени после начала стимуляции. Красной линией показан внутренний просвет органоида. (b) Флуоресцентное конфокальное изображение меченного кальцеином зеленым органоида с распознаванием объекта (зеленая линия) с помощью программного обеспечения для анализа изображений. (c) Репрезентативный пример стимулированного форсколином меченного кальцеином зеленым органоида. Дифференциальный контраст интерференции (DIC) и флуоресценцию визуализировали с использованием конфокальной микроскопии живых клеток. Площадь поверхности относительно t=0 указана в верхнем левом углу. (d) Площадь поверхности относительно t=0 (нормализованная площадь) для 11 отдельных органоидов в единичной лунке. Среднее значение указано черным цветом (среднее значение ± s.e.m.). (e) Дозозависимое увеличение площади поверхности благодаря форсколину (5 мкМ (количество проанализированных органоидов n=4), 5×10-2 мкМ (n=11), 5×10-4 мкМ (n=10), DMSO n=9)). Масштабные метки (a-c) 30 мкм. Все результаты являются репрезентативными по меньшей мере для трех независимых экспериментов.
Фиг.33. Индуцируемое форсколином набухание органоидов мыши является зависимым от CFTR. (a) Нормализованные кривые набухания стимулированных форсколином меченных кальцеином зеленым органоидов, предварительно инкубированных с DMSO (n=8), CFTR-inh172 (n=7), GlyH-101 (n=9) или как CFTR-inh172, так и GlyH-101 (n=11) (среднее значение ± s.e.m.). (b) Репрезентативные изображения, полученные с помощью конфокальной микроскопии, меченных кальцеином зеленым органоидов дикого типа или органоидов с дефицитом CFTR в ответ на форсколин. Масштабная метка 50 мкм. (c) Количественное определение индуцируемого форсколином набухания в органоидах дикого типа (n=6) или органоидах с дефицитом CFTR (n=11) (среднее значение ± s.e.m.) (d,e) Аналогично b,c, но для органоидов дикого типа (n=8) и CFTR-F508del (n=12). Масштабные метки 30 мкм. (f) Абсолютный размер органоидов дикого типа или органоидов с дефицитом CFTR, количественно определенных в (c), в момент времени t=0 (среднее значение ± s.e.m.). (g) Стимулированное форсколином набухание меченных кальцеином зеленым органоидов CFTR-F508del, культивируемых при 37°C, с (n=20) или без (n=15) ингибирования CFTR или культивируемых при 27°C в течение 24 часов с (n=31) или без (n=27) ингибирования CFTR (среднее значение ± s.e.m.). (h) Индуцируемое форсколином набухание меченных кальцеином зеленым органоидов CFTR-F508del, предварительно инкубированных в течение 24 часов с DMSO с (n=15) или без (n=18) ингибирования CFTR или предварительно инкубированных с корректирующим CFTR соединением VRT-325 с (n=14) или без (n=26) ингибирования CFTR (среднее значение ± s.e.m.). (i) Нормализованное индуцированное форсколином набухание органоидов CFTR-F508del, предварительно обработанных в течение 24 часов DMSO (n=16), VRT-325 (n=18), Corr-4a (n=20) или обоими корректирующими веществами (n=20) (среднее значение ± s.e.m.). Все результаты являются репрезентативным по меньшей мере для трех независимых экспериментов.
Фиг.34. Индуцируемое форсколином набухание нормальных контрольных органоидов или органоидах кистозного фиброза человека. (a) Количественное определения индуцируемого форсколином набухания органоидов, предварительно инкубированных с DMSO, CFTRinh-172, GlyH-101 или как CFTRinh-172, так и GlyH-101 (n=5, n=7, n=8, n=10) (среднее значение ± s.e.m.). (b) Стимулируемое форсколином набухание органоидов, происходящих из 4 отдельных здоровых контролей (1-4: n=30, n=18, n=13, n=42) и 1 гомозиготного по CFTR-F508del пациента с CF (n=30) (среднее значение ± s.e.m.). (c) Нормализованное набухание индуцируемых форсколином меченных кальцеином зеленым органоидов CFTR-F508del, культивируемых в течение 24 при 37°C, или при 27°C с или без ингибирования CFTR (n=10 для всех условий) (среднее значение ± s.e.m.). (d) Репрезентативные изображения, полученные с помощью конфокальной микроскопии, меченых кальцеином зеленым происходящих из HC или происходящих из пациента с CF органоидов в ответ на форсколин при фармакологическом манипулировании с помощью CFTR. Масштабные метки 60 мкм. (e) Нормализованное индуцируемое форсколином набухание органоидов CFTR-F508del, предварительно обработанных в течение 24 часов DMSO, VRT-325, Corr-4a или обоими корректирующими веществами (n=10 для всех условий) (среднее значение ± s.e.m.). (f) Набухание происходящих из пациента с CF органоидов в ответ на форсколин с или без предварительной обработки в течение 24 часов, состоящей в стимуляции корректирующим соединением VX-809, VX-770 (одновременно с форсколином) или в комбинированной обработке VX-809 и VX-770 (n=10 для всех условий) (среднее значение ± s.e.m.). (g) Индуцируемое форсколином набухание органоидов при обработке DMSO (контроль) или комбинированной обработке соединениями из e и f, по сравнению с органоидами HC (n=10 для всех условий) (среднее значение ± s.e.m.). Каждая линия в (d) соответствует набуханию органоида, усредненному по меньшей мере для трех независимых экспериментов на индивидуума. Результаты других фигур отражают по меньшей мере три независимых эксперимента.
Фиг.35. Анализ с помощью световой микроскопии органоидов мыши дикого типа, стимулированных форсколином или DMSO. Показаны репрезентативные примеры для указанных моментов времени после начала стимуляции. Индуцируемое форсколином набухание (FIS) органоидов обращалось вспять при удалении форсколина промыванием.
Фиг.36. мРНК CFTR экспрессируется в органоидах мыши и человека. Столбиками указаны пороговые величины (CT) в цикле ПЦР в реальном времени, соответствующие уровням мРНК CFTR, β2m или GAPDH, выделенной из органоидов CFTR-F508del (левый график) или CFTR-/- (средний график) и их соответствующих органоидов дикого типа, или органоидов человека.
Фиг.37. Постепенное индуцируемое форсколином набухание препятствует спадению органоида. Нормализованная площадь поверхности индивидуальных стимулированных форсколином (a) органоидов дикого типа, (b) органоидов CFTR-F508del (спасенных при помощи температуры) и (c) органоидов человека (5% кондиционированная Wnt3a среда, WCM). Усредненное индуцируемое форсколином набухание различных типов органоидов анализировали вплоть до 10 (фиг.1d,e + 2a,c,e), 20 (фиг.2g) или 40 (фиг.2h,i + 2a-c) минут (пунктирная линия).
Фиг.38. Увеличение FIS вследствие обработки корректирующими соединениями зависит от CFTR. Индуцируемое форсколином набухание меченных кальцеином зеленым органоидов CFTR-F508del человека, предварительно инкубированных в течение 24 часов с DMSO, или с обоими из VRT-325 и Corr-4a, с или без ингибирования CFTR (n=10 для всех условий) (среднее значение ± s.e.m.). Результаты являются репрезентативными по меньшей мере для трех различных экспериментов.
Фиг.39. Индуцируемое холерным токсином набухание органоидов в случае органоидов человека зависит от CFTR. Форсколин и холерный токсин индуцируют набухание происходящих из HC органоидов, но не органоидов CFTR-F508del. Ответ на холерный токсин является замедленным по сравнению с форсколином вследствие его апикальной внеклеточной функции. (n=10 для всех условий) (среднее значение ± s.e.m.). Результаты являются репрезентативными по меньшей мере для трех различных экспериментов.
Фиг.40. Органоиды человека в нормальных условиях культивирования или в условиях культивирования со сниженным содержанием Wnt3a. (a) Изображения, полученные световой микроскопией, органоидов человека, культивированных при нормальных (50%, левая панель) или сниженных (5%, правая панель) концентрациях кондиционированной Wnt3a среды (WCM). Масштабные метки - 400 мкм. (b) Репрезентативные примеры индуцированного форсколином набухания в условиях нормальной или сниженной концентрации Wnt3a. Масштабные метки 50 мкм. Пунктирной линией показан внутренний просвет органоида (c) Количественное определение индуцируемого форсколином набухания органоида при нормальных уровнях Wnt3a, предварительно инкубированного с DMSO, CFTRinh-172, GlyH-101, или как CFTRinh-172 так и GlyH-101 (n=29, n=41, n=26, n=15) (среднее значение ± s.e.m.). (d) Количественное определение индуцируемого форсколином набухания почкующихся органоидов после низкого количества пассажей, культивированных при 50% (n=9) или 5% (n=12) концентрациях кондиционированной Wnt3a среды (WCM), усредненных для двух независимых экспериментов (среднее значение ± s.e.m.). Все результаты являются репрезентативными по меньшей мере для трех независимых экспериментов.
Фиг.41. Окрашивание посредством H&E органоидов предстательной железы
Фрагменты ткани эпителия предстательной железы мыши погружали в Matrigel. Увеличивающиеся в количестве клетки разделяли каждую неделю. Культуру можно поддерживать в течение более длительного периода времени без утраты генетической стабильности или способности к пролиферации. На фиг.41 показано сравнение эпителия предстательной железы непосредственно в предстательной железе и культурах органоидов после трех месяцев. Органоиды предстательной железы мыши растут в среде, содержащей ENR (EGF, Noggin и R-спондин) в присутствии или в отсутствие тестостерона. Культура человека требует добавления ингибитора TGF-бета. Окрашивание H&E после фиксации и погружения в парафин демонстрирует различные уровни распределения на слои и складывания эпителия in vivo. Культивируемые органоиды предстательной железы демонстрируют сходное разнообразие складывания и распределения на слои.
Фиг.42. Окрашивание на CK8 (маркер дифференцировки) органоидов предстательной железы
Фрагменты ткани эпителия предстательной железы мыши погружали в Matrigel. Увеличивающиеся в количестве клетки разделяли каждую неделю. Культуру можно поддерживать в течение длительного периода времени без утраты генетической стабильности или способности к пролиферации. На фиг.42 показано присутствие экспрессирующих CK8 клеток просвета. Культуры выращивают в среде, содержащей ENR (EGF, Noggin и R-спондин) в присутствии или в отсутствие тестостерона. Культура человека требует добавления ингибитора TGF-бета. Добавление тестостерона позволяет дифференцировку в положительные по CK8 клетки просвета при одновременной стимуляции поддержания и увеличения в количестве стволовых клеток. Тестостерон также увеличивает распределение на слои и складывание эпителия.
Фиг.43. Предстательная железа мыши после 25 недель в культуре.
Органоиды предстательной железы выращивают в EGF, Noggin, R-спондине, NAC, B27, глутамин/glutamax, pen/strep, Ad-DMEM/F12 + тестостерон. Форма органоида определяется происхождением ткани (положение в предстательной железе перед выделением). Предстательная железа состоит из различных долей и областей. Различные области проявляют определенные эпителиальные структуры (распределение на слои и складывание). После культивирования in vitro органоиды, по-видимому, сохраняют особую макроскопическую структуру (распределенную на слои или сложенную) той части предстательной железы, из которой они происходят.
Фиг.44. ПЦР 3-недельной культуры предстательной железы человека
Нормальный и злокачественный эпителий предстательной железы выделяли и выращивали в течение трех недель в условиях культивирования ENR FGF10, ENRF+DHT, WENRF, WENRF + DHT, ENR, ENR + DHT. Все условия культивирования включали A83, P38i и никотинамид.
Фиг.44(a): РНК выделяли и проводили ОТ-ПЦР для маркеров эпителия предстательной железы. Как в нормальной, так и в опухолевой ткани экспрессируются люминальные маркеры CK18, CK8 и B-MSP. Во всех условиях культивирования экспрессируется AR. В нормальной ткани добавление DHT увеличивает экспрессию AR во всех условиях культивирования. В опухолевой ткани добавление DHT не влияет на экспрессию AR. Во всех условиях культивирования экспрессируются базальные эпителиальные маркеры CK14, CK5 и p63. Предполагаемый маркер стволовых клеток LGR5 экспрессируется в условиях ENRF в нормальной ткани. В опухолевой ткани экспрессия LGR5 индуцируется добавлением DHT во всех условиях культивирования. TNFRSF19, также предполагаемый маркер стволовых клеток, экспрессируется во всех условиях в нормальной и опухолевой ткани. Простатспецифический фактор транскрипции NKX3.1 экспрессируется во всех условиях. Добавление тестостерона увеличивает рост/количество удвоений при сохранении маркеров различных типов клеток предстательной железы (базальные и люминальные).
Фиг.44(b): Два репрезентативных изображения органоидов человека, выращенных в условиях ENRF+ 1 нМ DHT
Дорожка 1: линия 1, ENRF, нормальная ткань
Дорожка 2: линия 1, ENRF + 1 нМ DHT, нормальная ткань
Дорожка 3: линия 2, WENRF, нормальная ткань
Дорожка 4: линия 2, WENRF + 1 нМ DHT, нормальная ткань
Дорожка 5: линия 3, ENR, нормальная ткань
Дорожка 6: линия 3, ENR + 1 нМ DHT, нормальная ткань
Дорожка 7: целая предстательная железа, нормальная ткань
Дорожка 8: H2O
Дорожка 9: линия 1, ENRF, опухолевая ткань
Дорожка 10: линия 1, ENRF + 1 нМ DHT, опухолевая ткань
Дорожка 11: линия 2, WENRF, опухолевая ткань
Дорожка 12: линия 2, WENRF + 1 нМ DHT, опухолевая ткань
Дорожка 13: линия 3, ENR, опухолевая ткань
Дорожка 14: линия 3, ENR + 1 нМ DHT, опухолевая ткань
Дорожка 15: целая предстательная железа, опухолевая ткань
Дорожка 16: H2O
Фиг.45. ПЦР органоида предстательной железы мыши
Три биологически независимых линии культивировали в условиях ENR или ENR + 1 нМ DHT. РНК выделяли и проводили ОТ-ПЦР для маркеров эпителия предстательной железы. В обоих условиях культивирования широко экспрессируются люминальные маркеры предстательной железы цитокератин 18 (CK18) и цитокератин 8 (CK8). Рецептор андрогенов (AR) экспрессируется в обоих условиях. Базальные маркеры p63 и цитокератин 5 (CK5) экспрессируются в обоих условиях культивирования, однако при добавлении DHT базальные маркеры подавляются. Предполагаемые маркеры стволовых клеток Lgr5 и Tnfrsf19 подавляются при добавлении DHT. Однако в этих условиях стволовость сохраняется, хотя также присутствуют дифференцированные клетки. Эти условия позволяют неограниченное увеличение в количестве клеток (на настоящий момент 9 месяцев при 2,5 удвоениях популяции в неделю). Все культуры являются положительными на простатспецифический маркер Nkx3.1. Добавление тестостерона увеличивает рост/удвоения вплоть до 3 раз при сохранении маркеров для различных типов клеток предстательной железы (базальных и люминальных).
Дорожка 1: линия 1, ENR
Дорожка 2: линия 1, ENR + 1 нМ DHT
Дорожка 3: линия 2, ENR
Дорожка 4: линия 2, ENR + 1 нМ DHT
Дорожка 5: линия 3, ENR
Дорожка 6: линия 3, ENR + 1 нМ DHT
Дорожка 7: целая предстательная железа мыши
Дорожка 8: H2O
Фиг.46: Органоиды желудка (гастральные)
Органоиды желудка человека. Ткань выделяли из тела. Клетки культивировали в среде для органоидов желудка (EGF, Noggin, R-спондин, Wnt, никотинамид, FGF10, гастрин, ингибитор TGF-бета (A8301). Клетки разделяли каждую неделю.
Фиг.46(a): Это изображение было получено после культивирования в течение 2 месяцев.
Фиг.46(b): Окрашивание H&E и различными антителами после фиксации и получения среза в парафине культуры после культивирования в течение 2 недель. На ней показано присутствие следующих клеток: PAS для продуцирующих муцин клеток; Muc5Ac для поверхностных слизистых ямочных клеток; Muc6 для слизистых шеечных клеток. Окрашивание посредством H&E демонстрирует один слой поляризованного эпителия.
Фиг.47: Выделение ткани предстательной железы
См. пример 8 (фотографии от UCSF)
Фиг.48: A) Органоиды мыши культивировали в присутствии различных доз рекомбинантного RANKL мыши в течение 72 ч. Уровни экспрессии мРНК для RANK, фактора транскрипции SpiB и специфичных маркеров M-клеток GP2 и аннексина V определяли с помощью q-ПЦР. B) Конфокальный анализ экспрессии GP2 (см. стрелку) и аннексина V (см. стрелку) в органоидах мыши, культивированных с 100 нг/мл RANKL в течение 72 ч.
Фиг.49: Органоиды человека культивировали в присутствии различных доз рекомбинантного RANKL человека в течение 7 суток. Уровни экспрессии мРНК для RANK, SipB и специфичного маркера M-клеток GP2 определяли с помощью q-ПЦР. EM: Среда для увеличения в количестве; DM: среда для дифференцировки.
ПРИМЕР 1:
Для удовлетворения потребности в усовершенствованных культуральных средах и способах для эпителиальных стволовых клеток человека, авторы изобретения исследовали каскады передачи сигналы, которые известны тем, что они нарушаются при определенных злокачественных опухолях, например, раке ободочной и прямой кишки. Было предположено, что эти каскады, которые влияют на судьбу клеток при злокачественной опухоли, также могут играть роль в определении судьбы клеток в условиях культивирования клеток in vitro.
В первом скрининговом эксперименте исследовали серию витаминов, гормонов и факторов роста в комбинации со стандартными средами для культивирования стволовых клеток. Было выявлено, что гастрин и никотинамид приводят к значительному улучшению условий культивирования. Включая эти факторы в стандартные условия культивирования, проводили второй скрининговый эксперимент, в котором исследовали определенные низкомолекулярные ингибиторы, связанные с соответствующими каскадами передачи сигнала, такие как ERK, p38, JNK, PTEN, ROCK и Hedgehog. На существующем уровне техники отсутствует приемлемый путь предсказания того, каким будет результат каждого из этих дополнительных соединений для свойств культуральной среды.
Перечень реагентов, использованных для оптимизации культуры органоидов кишечника человека
0 = нет изменений; 1+ = <50% увеличение; 2+ = 50-100% увеличение; 3+ = >100% увеличение;
1- = 0-50%; 2- = 50-100% снижение; 3- = >100% снижение.
** WENR содержит EGF+Noggin+R-спондин+Wnt-3a
*** Полужирным шрифтом выделены соединения, которые продемонстрировали наибольшее улучшение культуральной среды.
В общем, авторы изобретения установили условия долговременного культивирования, при которых единичные крипты или стволовые клетки, происходящие из тонкого кишечника (SI) мыши, увеличиваются в количестве в течение длительных периодов времени. Растущие крипты претерпевают множество событий деления крипт при одновременном образовании подобных ворсинкам доменов, в которых присутствуют все дифференцированные типы клеток. Авторы изобретения далее адаптировали условия культивирования для выращивания сходных эпителиальных органоидов из толстого кишечника мыши и SI и толстого кишечника человека. Исходя из культуральной системы тонкого кишечника мыши, авторы изобретения оптимизировали культуральную систему толстого кишечника мыши и человека. Они открыли, что добавление Wnt3A к коктейлю факторов роста позволило криптам толстого кишечника мыши увеличиваться в количестве неограничено. Дальнейшее добавление никотинамида, низкомолекулярного ингибитора Alk и ингибитора p38 было предпочтительным для длительного культивирования SI и толстого кишечника человека. Культуральная система также обеспечивала рост аденом мыши Apcmin, рака ободочной и прямой кишки человека и метапластического эпителия пищевода Барретта. Технология культивирования является широко применимой в качестве исследовательского инструмента для инфекционных, воспалительных и неопластических патологий желудочно-кишечного тракта человека. Более того, могут стать возможными регенеративные применения для увеличенного в количестве эпителия кишечника ex vivo. Самообновление эпителия тонкого кишечника и толстого кишечника запускается посредством пролиферации стволовых клеток и их потомков, расположенных в криптах. Хотя описано множество культуральных систем (Evans GS et al. J Cell Sci 1992; 101 (Pt 1):219-31; Fukamachi H. J Cell Sci 1992;103 (Pt 2):511-9; Perreault N & Jean-Francois B. Exp Cell Res 1996;224:354-64; Whitehead RH et al. Gastroenterology 1999;117:858-65), только недавно стали доступными системы долговременного культивирования, которые сохраняют основную физиологию крипт. Было опубликовано два различных протокола, которые позволяют длительное увеличение в количестве эпителия тонкого кишечника мыши. Kuo и коллеги продемонстрировали долговременный рост небольших фрагментов, содержащих эпителиальные, а также стромальные элементы, независимым от факторов роста образом (Ootani A et al. Nat Med 2009;15:701-6). Авторы изобретения создали культуральную систему для единичных стволовых клеток путем комбинирования ранее определенных идей о потребностях эпителия кишечника для роста. Передача сигнала Wnt является ключевой потребностью для пролиферации крипт (Korinek V et al. Nat Genet 1998;19:379-83; Pinto D et al. Genes Dev 2003;17:1709-13; Kuhnert F et al. Proc Natl Acad Sci USA 2004;101:266-71) и агонист Wnt R-спондин 1 индуцирует значительную гиперплазию крипт in vivo (Kim KA et al. Science 2005;309:1256-9). Во-вторых, передача сигнала EGF связана с пролиферацией кишечника (Dignass AU & Sturm A. Eur J Gastroenterol Hepatol 2001;13:763-70). В-третьих, трансгенная экспрессия Noggin индуцирует увеличение в количестве крипт (Haramis AP et al. Science 2004;303:1684-6). В-четвертых, выделенные клетки кишечника претерпевают аноикоз вне условий нормальной ткани (Hofmann C et al. Gastroenterology 2007;132:587-600). Поскольку ламинин (α1 и α2) обогащен в основании крипты (Sasaki T et al. Exp Cell Res 2002;275:185), авторы изобретения исследовали обогащенный ламинином Matrigel для поддержания роста эпителия кишечника. Культуры на основе Matrigel успешно используют для выращивания эпителия молочной железы (Stingl J et al. Breast Cancer Res Treat 2001;67:93-109). В этих условиях культивирования (R-спондин 1, EGF, и Noggin в Matrigel), авторы изобретения получили постоянно увеличивающиеся в количестве органоиды тонкого кишечника, которые проявляли все отличительные признаки эпителия тонкого кишечника с точки зрения архитектуры, состава типов клеток и динамики самообновления.
Несмотря на значительные усилия, долговременное культивирование взрослых эпителиальных клеток кишечника человека остается трудным. Существуют некоторые модели долговременного культивирования, однако эти способы и клеточные линии не получили широкого признания, возможно в результате присущих им технических трудностей, связанных с извлечением и поддержанием жизнеспособных клеток (Rogler G et al. Scandinavian journal of gastroenterology 2001;36:389-98; Buset M et al. In vitro cellular & developmental biology: journal of the Tissue Culture Association 1987;23:403-12; Whitehead RH et al. In vitro cellular & developmental biology: journal of the Tissue Culture Association 1987;23:436-42; Deveney CW et al. The Journal of surgical research 1996;64:161-9; Pang G et al. Gastroenterology 1996;111:8-18; Latella G et al. International journal of colorectal disease 1996;11:76-83; Panja A. Laboratory investigation; a journal of technical способы и pathology 2000;80:1473-5; Grossmann J et al. European journal of cell biology 2003;82:262-70). Побужденные установлением культуры тонкого кишечника мыши, авторы изобретения преследовали цель приспособления условий культивирования для эпителия толстого кишечника мыши и человека. Авторы изобретения далее описали установление протоколов долговременного культивирования для эпителия толстого кишечника мыши и человека, которые могут быть адаптированы для первичной аденомы/аденокарциномы толстого кишечника и пищевода Барретта.
Результаты
Установление культуральной системы для толстого кишечника мыши
В попытках установить культуральную систему для толстого кишечника мыши, авторы изобретения исследовали их условия культивирования тонкого кишечника (в настоящем описании называемые ENR: EGF+Noggin+R-спондин). По опыту авторов настоящего изобретения, первоначальный рост эпителия толстого кишечника часто наблюдают в условиях культивирования ENR, однако он неизменно прерывается. Образование органоидов исследовали с использованием эпителия, выделенного из дистальной части толстого кишечника мыши. В условиях ENR эффективность посева единичных дистальных крипт толстого кишечника была значительно более низкой, чем для тонкого кишечника (1-3% против >90%) и эти органоиды нельзя было пассировать. Недавно авторы изобретения показали, что клетки Панета продуцируют несколько лигандов Wnt (Gregorieff A et al. Gastroenterology 2005;129:626-38), и что продукция Wnt этими клетками Панета необходима для поддержания стволовых клеток кишечника (Sato T et al. Nature;469:415-8). Для определения статуса передачи сигнала Wnt в органоидах толстого кишечника авторы изобретения культивировали крипты толстого кишечника из мышей Axin2-lacZ (достоверный репортер Wnt) (Lustig B et al. Mol Cell Biol 2002;22:1184-93) или из нокинных мышей Lgr5-GFP (Lgr5 является Wnt-зависимым маркером стволовых клеток)(Barker N et al. Nature 2007;449:1003-7).
Свежевыделенные крипты толстого кишечника безо всяких сложностей экспрессировали Axin2-LacZ или Lgr5-GFP в их основании, однако они утрачивали экспрессию репортеров Wnt вскоре после инициации культуры (фиг.1a,b и фиг.6). Напротив, органоиды тонкого кишечника конститутивно экспрессировали репортеры Wnt на их почкующихся структурах (Sato T et al. Nature;469:415-8; Sato T et al. Nature 2009;459:262-5). Эти данные показали, что органоиды толстого кишечника продуцируют достаточные количества лигандов Wnt для поддержания стволовых клеток толстого кишечника. Для преодоления этого авторы изобретения добавили рекомбинантный Wnt3a или кондиционированную Wnt3a среду в культуральную среду ENR (среда WENR). Это увеличивало эффективность посева крипт порядка в 10 раз. Крипты толстого кишечника образовывали структуры органоидов с многочисленными Axin2-LacZ (фиг.1a) или Lgr5-GFP+ (фиг.1b) почками, подразумевая, что активация Wnt восстанавливалась. Свежевыделенные крипты толстого кишечника содержат полностью зрелые клетки в их верхних частях и авторы изобретения пришли к выводу, что эти зрелые клетки могут препятствовать росту органоида. Когда авторы изобретения в мягких условиях расщепили крипты толстого кишечника на небольшие кластеры клеток, таким образом, физически отделяя пролиферативное дно крипт от дифференцированных верхних областей крипт, большинство фрагментов, происходящих из верхней части крипты, погибали, однако кластеры клеток из основания крипты толстого кишечника эффективно образовывали органоиды (фиг.1c).
Эпителий тонкого кишечника мыши, выращенный в условиях ENR, образует все дифференцированные эпителиальные типы клеток с сосуществующим самообновлением стволовых клеток. Авторы изобретения ранее показали, что добавление Wnt3A в эти культуры препятствует дифференцировке кишечника и приводит к органоидам, которые по большей части состоят из недифференцированных предшественников (Sato T et al. Nature;469:415-8). Это является неожиданным, учитывая центральную роль передачи сигнала Wnt в поддержании недифференцированного состояния предшественников крипт (van de Wetering M et al. Cell 2002;111:241-50). В соответствии с этим наблюдением, органоиды толстого кишечника в условиях WENR не дифференцировались надлежащим образом. При устранении Wnt-3A авторы изобретения выявили дифференцировку по всем эпителиальным росткам (фиг.1d-f). Следует отметить, что единичные отсортированные Lgr5+ эпителиальные стволовые клетки толстого кишечника могут образовывать органоиды при культивировании в присутствии Y-27632 в течение первых двух суток.
Установление культуральной системы для толстого кишечника человека
Вдохновленные успехом улучшенной культуры крипт толстого кишечника мыши, авторы изобретения применили эти условия культивирования для крипт толстого кишечника человека. Хотя эти крипты первоначально выжили, большинство из них впоследствии дезинтегрировали в течение 7 суток. Для увеличения эффективности посева крипт толстого кишечника человека, авторы изобретения провели скрининг являющихся кандидатами факторов роста, гормонов и витаминов (перечень на фиг.12). Среди них, авторы изобретения выявили, что гастрин и никотинамид (предшественник NAD+, и выявлено, что он подавляет активность сиртуина (Denu JM. Trends Biochem Sci 2005;30:479-83)) повышали эффективность культивирования (фиг.12). Эффект гастрина на эффективность посева был несущественным. Однако этот гормон не препятствовал дифференцировке клеток кишечника и авторы изобретения решили включить гастрин (далее сокращенно обозначаемый как "g") во все условия культивирования кишечника человека. Важно, что никотинамид (10 мМ) был необходим для продления периода культивирования за пределы первоначальных 7 суток (фиг.2a). В этих условиях культивирования органоиды толстого кишечника человека можно было увеличивать в количестве в течение по меньшей мере 1 месяца. Начиная с 1 месяца, органоиды толстого кишечника изменяли их морфологию с почкующейся структуры органоидов на кистозные структуры (фиг.2b слева). Одновременно с морфологическим преобразованием пролиферация постепенно снижалась. Иногда кистозные органоиды вновь приобретали их пролиферативный потенциал. Однако рост всех органоидов в конечном итоге прекращался в течение 3 месяцев. Двухфазную остановку роста наблюдали в других первичных культуральных системах, таких как эпителиальные клетки молочной железы или кератиноциты, и ее называли как стадия смертности 1 (M1; старение) и стадия смертности 2 (M2; кризис) (Shay et al., 2006). В органоидах кишечника человека, культивированных в этих условиях, дифференцировку множества ростков не наблюдали даже после устранения Wnt (данные не представлены).
Авторы изобретения предположили, что остановка роста происходила вследствие ненадлежащих условий культивирования, а не присущего клеткам свойства старения/репликативного устаревания. Таким образом, авторы изобретения расширили их попытки оптимизировать условия культивирования. Авторы изобретения провели скрининг различных низкомолекулярных модуляторов MAP-киназ, молекул передачи сигнала, мутантных при раке толстого кишечника, и модификаторов гистонов (фиг.12) в условиях культивирования WENR+гастрин+никотинамид. Авторы изобретения открыли, что два низкомолекулярных ингибитора A83-01 (ингибитор Alk4/5/7; нМ) и SB202190 (ингибитор p38; 10 мкМ) значительно повышали эффективность посева. Другие ингибиторы рецептора TGF-бета 1 (ALK 5), которые также исследовали и которые показали те же результаты, что и A83-01, представляли собой LY364947, SB431542, SB505124. Можно было бы ожидать, что другие ингибиторы ALK также действуют аналогичным путем. Более того, комбинация двух соединений синергично продлевала период культивирования. Авторы изобретения продемонстрировали, что все из десяти протестированных образцов увеличивались в количестве в течение по меньшей мере 6 месяцев с разделением 1:5 каждую неделю. В этих условиях культивирования органоиды толстого кишечника человека проявляли почкующиеся структуры органоидов, а не кистозные структуры, наблюдаемые в предшествующих условиях культивирования (фиг.2b). Пролиферирующие клетки были ограничены почками (фиг.2c). В метафазных пластинках органоидов возрастом более 3 месяцев неизменно выявляли 46 хромосом в каждой клетке (20 клеток от каждого из трех различных доноров; фиг.2d). Авторы изобретения отсеквенировали весь экзом (все экзоны) органоидов толстого кишечника после культивирования в течение двух месяцев. Количество мутаций в органоидах было чрезвычайно низким. В действительности, в четырех параллельных культурах органоидов, происходящих из одного клона, была выявлена только одна мутация, которая присутствовала во всех культурах и, таким образом, вероятно, происходила из исходной ткани.
Эти результаты подразумевают, что передача сигнала рецептора Alk и p38 отрицательно регулируют долговременное поддержание эпителиальных клеток кишечника человека. Авторы изобретения называют оптимизированные условия культивирования условиями HISC (культура стволовых клеток кишечника человека).
Органоиды кишечника человека имитируют дифференцировку in vivo
В условиях HISC авторы изобретения не выявили дифференцированных клеток. Как наблюдали для органоидов толстого кишечника мыши, устранение Wnt требовалось для дифференцировки зрелых энтероцитов в органоиды человека (фиг.3a, верхняя панель, и фиг.7). Однако дифференцировка бокаловидных и энтероэндокринных клеток оставалась блокированной. Авторы настоящего изобретения открыли, что никотинамид и SB202190 значительно ингибировали эту дифференцировку, в то время как устранение этих двух реагентов позволило органоидам продуцировать зрелые бокаловидные и энтероэндокринные клетки (фиг.3a (средняя и нижняя панель), 3b и фиг.7.
Те же эффекты Wnt, никотинамида и SB202190 в отношении ингибирования дифференцировки наблюдали в органоидах тонкого кишечника человека. Клетки Панета лизоцим+ наблюдали в органоидах тонкого кишечника, но не в органоидах толстого кишечника (фиг.3d). Было описано, что обработка ингибитором p38 in vivo ингибирует дифференцировку бокаловидных клеток и усиливает пролиферацию кишечного эпителия (Otsuka M. Gastroenterology 2010;138:1255-65, 1265 e1-9). Действительно, авторы изобретения наблюдали тот же фенотип в органоидах тонкого кишечника, обработанных ингибитором p38 (фиг.3d против e).
Авторы изобретения далее исследовали ответ органоидов кишечника человека на ингибирование Notch. Авторы изобретения ранее показали, что ингибирование Notch либо ингибиторами γ-секретазы (дибензазепин; DBZ) или зависимое от условий нацеливание на фактор транскрипции каскада Notch CSL истощало стволовые клетки кишечника, завершало пролиферацию эпителия кишечника и индуцировало гиперплазию бокаловидных клеток in vivo (van Es JH et al. Nature 2005;435:959-63). Действительно, при обработке DBZ органоиды кишечника прекращали их пролиферацию и большинство клеток преобразовывалось в бокаловидные клетки в пределах 3 суток (фиг.3g против f).
Установление аденомы с дефицитом APC и аденокарциномы толстого кишечника
Недавно авторы изобретения сообщили об эффективном образовании аденомы кишечника у мыши из Lgr5 стволовых клеток у мышей Lgr5-GFP-ires-CreERT2 x APCflox/flox при индуцируемой тамоксифеном активации Cre (Barker N et al. Genes Dev 2008;22:1856-64). Авторы изобретения выделили аденомы кишечника через 10 суток после индукции и оптимизировали условия культивирования. Аденомы эффективно образовывали кистозную структуру органоида без почкования. Поскольку утрата APC конститутивно активирует каскад Wnt, авторы изобретения ожидали, что R-спондин 1 может стать несущественным для роста органоида аденомы. Это действительно наблюдали. Более того, Noggin, который необходим для долговременного культивирования нормального тонкого кишечника, был несущественным для органоидов аденомы. Интересно, что авторы изобретения выявили утрату Lgr5-GFP, но не Axin2-LacZ в аденоматозных органоидах через 7 суток после устранения Noggin (фиг.4a,b и данные не представлены). Сходные наблюдения были сделаны для нормальных органоидов кишечника при выращивании в среде ER (Sato T et al. Nature 2009;459:262-5). Это указывало на то, что Noggin, наиболее вероятно через ингибирование сигналов BMP, требуется для поддержания экспрессии Lgr5, но не требуется для увеличения в количестве органоидов аденомы. Свежевыделенные клетки Lgr5hi (но не Lgr5low), выделенные из крипт кишечника, могут инициировать рост органоидов in vitro (Sato T et al. Nature 2009;459:262-5). Для определения существования сходной иерархии Lgr5 в аденомах авторы изобретения выделили клетки Lgr5-GFPhi, GFPlow и GFP-ve из культивируемых в EN органоидов и исследовали их способность образовывать органоиды. После культивирования в течение 7 суток Lgr5-GFPhi продемонстрировали наиболее высокую эффективность образования органоидов. Тем не менее, Lgr5-GFPlow или -ve также образовывали органоиды со значительной эффективностью (фиг.4c). Следует отметить, что отсортированные клетки аденомы GFP-ve могли давать начало органоидам Lgr5-GFPhi (фиг.8).
На протяжении последних четырех десятилетий было выделено множество клеточных линий рака ободочной и прямой кишки. Как правило, такие клеточные линии появляются в качестве редких клональных выростов после вхождения первичных культур опухолей толстого кишечника в кризис культуры тканей. В настоящее время не существует устойчивой культуральной системы, которая позволяет постоянное культивирование первичных образцов рака толстого кишечника человека без кризиса культуры и последующего клонального разрастания адаптированных к культуре клеток. В качестве следующей стадии авторы изобретения использовали условия культивирования аденомы для образцов рака ободочной и прямой кишки человека. Как и ожидалось, для клеток рака толстого кишечника не требовались ни R-спондин, ни Noggin. EGF был необязательным в большинстве органоидов рака толстого кишечника, в то время как пролиферация некоторых органоидов рака толстого кишечника замедлялась после устранения EGF. В отличие от аденомы кишечника мыши, органоиды рака ободочной и прямой кишки в условиях культивирования росли в качестве неправильных компактных структур, а не простых кистозных структур (фиг.4d).
Авторы изобретения исследовали состояние пролиферации/дифференцировки органоида аденомы и рака толстого кишечника. Как и ожидалось, большинство клеток представляли собой Ki67+. В соответствии со значительным ингибиторным эффектом Wnt на дифференцировку энтероцитов (фиг.1f и фиг.7), окрашивание щелочной фосфатазы не наблюдалось в обоих типах органоидов (фиг.9). Напротив, авторы настоящего изобретения иногда наблюдали PAS+ бокаловидные клетки и эндокринные клетки хромогранин A+ в органоидах аденомы и в некоторых органоидах рака толстого кишечника (фиг.9).
Культивирование метапластического эпителия Барретта человека
Пищевод Барретта характеризуется присутствием столбчатого эпителия в нижней части пищевода, заменяющий нормальный плококлеточный эпителий в результате метаплазии (Odze RD. Nat Rev Gastroenterol Hepatol 2009;6:478-90). Историческим отличительным признаком пищевода Барретта является присутствие бокаловидных клеток кишечника в пищеводе. Исследуя сходство между эпителием Барретта и кишечным эпителием, авторы изобретения подвергли небольшиен биоптаты эпителия Барретта (BE) условиям культивирования толстого кишечника человека. В этих условиях культивирования нормальные плоские клетки пищевода временно пролиферировали в течение 1 недели, однако органоиды нельзя было пассировать. Эпителий пищевода Барретта можно было поддерживать в течение вплоть до 1 месяца в условиях HISC (фиг.5a). Органоиды BE образовывали кистозные структуры органоидов, неотличимые от структур стареющих органоидов толстого кишечника, и, как правило, претерпевали остановку роста после 1 месяца культивирования. Добавление FGF10 в условия HISC позволило органоидам BE образовывать почкующиеся структуры и значительно продлевало длительность культивирования (> 3 месяцев) (фиг.5 b, c). В противоположность органоидам кишечника человека, органоиды BE оставались Ki67+ с минимальным количеством клеток PAS+ и муцин+ через 4 суток после устранения никотинамида и SB202190. Обработка ингибитором γ-секретазы DBZ (10 мкМ) в течение 4 суток после устранения блокировала пролиферацию и индуцировала дифференцировку бокаловидных клеток (фиг.5 d-g). Это подтвердило более раннее предположение авторов настоящего изобретения о том, что локальная доставка таких ингибиторов может отражать полезную терапевтическую стратегию для устранения очагов повреждения пищевода Барретта посредством дифференцирующей терапии (Menke V et al. Disease models & mechanisms 2010;3:104-10). Следует отметить, что авторы настоящего изобретения иногда выявляли клети Панета лизоцим+ (фиг.10), что указывает на то, что в органоидах BE сохраняется дифференцировка множества ростков.
Обсуждение
Протоколы, разработанные в рамках настоящего изобретения, позволяют надежное и долговременное культивирование первичных эпителиальных клеток человека, выделенных из тонкого кишечника, толстого кишечника, адено(карцин)ом и пищевода Барретта (таблица 3).
Перечень компонентов культуральных систем органоидов
В противоположность тонкому кишечнику мыши, эпителиальные клетки толстого кишечники мыши требуют лиганда Wnt в культуральной среде. Авторы изобретения ранее описали, что клетки Панета CD24hi продуцируют Wnt-3/11, которые необходимы для поддержания стволовых клеток в тонком кишечнике (Sato T, et al. Nature 2011;469:415-8). мРНК Wnt-6 и -9b экспрессируется в основании крипт кишечника (Gregorieff A, et al. Gastroenterology 2005;129:626-38). Остается неопределенным, является ли эта локальная продукция Wnt клетками основания крипты кишечника достаточной для активации канонической передачи сигнала Wnt in vivo или существует другой источник лиганда Wnt в слизистой оболочке толстого кишечника. Отличие между условиями культивирования органоидов кишечника человека и мыши были неожиданно значительными. A83-01 ингибирует ALK4/5/7, рецепторы, которые с помощью микрочипа выявляются как в криптах мыши, так и в криптах человека. Авторы изобретения в настоящее время исследуют механизм, посредством которого сигнал ALK регулирует рост органоидов человека. Авторы изобретения не выявили трансформации клеток в долговременных культурах и в оптимизированных условиях культивирования не выявлялось хромосомных изменений. Более того, органоиды могут претерпевать значительно более высокое количество делений клеток, чем сообщено для другой культуральной системы для эпителия человека (Dey D et al. PloS one 2009;4:e5329; Garraway IP et al. The Prostate 2010;70:491-501). Главным образом, полагают, что соматические клетки по своей природе ограничены по их пролиферативной способности, и это явление называют репликативным устареванием (Walen KH. In vitro cellular & developmental biology. Animal 2004;40:150-8). Полагают, что для большинства нормальных клеток человека рассчитано количество раз их делений, и в конечном итоге они претерпевают остановку роста, называемую клеточным старением. Этот процесс может запускаться укорочением теломер и последующей активацией сигналов повреждения ДНК (M1), или изнашиванием теломер (M2). В отсутствие двух низкомолекулярных ингибиторов киназ органоиды кишечника человека претерпевают остановку роста после 10-20 удвоений популяций. Напротив, репликативная способность в оптимизированных условиях культивирования была расширена по меньшей мере вплоть до 100 удвоений популяции путем добавления ингибиторов, что превышало предел Хейфлика (Hayflick L. The Journal of investigative dermatology 1979;73:8-14). Этот результат отчетливо указывает на то, что стареющий фенотип, наблюдаемый в первой культуральной системе, отражает ненадлежащие условия роста, а не присущее репликативное старение.
Технологии культивирования можно использовать для исследования основных аспектов биологии стволовых клеток и контроля дифференцировки, иллюстрируемого истощением стволовых клеток и дифференцировкой бокаловидных клеток при обработке ингибитором Notch. Более того, платформу культивирования органоидов можно использовать для фармакологических, токсикологических или микробиологических исследований патологии кишечного тракта, поскольку органоиды в большей степени напоминают эпителий кишечника, чем часто используемые клеточные линии рака толстого кишечника, такие как CaCo2 или DLD1. Наконец, поскольку небольшие биоптаты, взятые из взрослых доноров, можно увеличивать в количестве без какого-либо заметного предела или генетического вреда, эта технология может служить для получения трансплантируемого эпителия для регенеративных целей.
ПРИМЕР 2 - Культивирование органоидов поджелудочной железы мыши
Использование ингибитора TGF-бета также исследовали в культуральной среде для органоидов поджелудочной железы мыши. Среда для увеличения в количестве, которую использовали, представляла собой среду DMEM/F12 (дополненная P/S, Glutamax, 10 мМ Hepes, B27, N2 и N-ацетилцистеином), EGF (50 нг/мл), R-спондин (10%), Noggin (100 нг/мл), FGF10 (100 нг/мл), A8301 (ингибитор TGF-бета, 500 нМ) и гастрин (10 мкМ). Это несколько отличается от описанной выше культуры HISC, использованной в примере 2, тем, что не добавляют агониста Wnt (отличного от R-спондина) или никотинамида и FGF10. Однако эти культуральные среды обладают общим количеством ключевых компонентов (ENR + гастрин + ингибитор TGF-бета), в обоих случаях является преимущественным добавление ингибитора TGF-бета. Органоиды поджелудочной железы, выращенные в этих условиях, можно было увеличивать в количестве в течение > 3 месяцев и пассировать по меньшей мере 5 раз.
Эксперименты на микрочипах осуществляли для органоидов поджелудочной железы, выращенных в описанной выше среде для увеличения в количестве и результаты сравнивали с взрослой поджелудочной железой, взрослой печенью и печенью новорожденного (см. фиг.16A). Органоид поджелудочной железы отчетливо образует кластеры с образцами взрослой поджелудочной железы, а не с образцами печени, что демонстрирует хорошее фенотипическое сходство со взрослой поджелудочной железой.
На фиг.16B показан необработанный сигнал из эксперимента на микрочипах, сравнивающего уровни экспрессии в органоидах поджелудочной железы, взрослой поджелудочной железе, взрослой печени и органоидах печени в отношении маркеров протоков, эндокринных маркеров и факторов транскрипции, необходимых для экспрессии Ngn3 (Ngn3 является фактором транскрипции, который связан с характеристиками эндокринных ростков). Высокие уровни экспрессии Krt19, Krt7 и других маркеров протоков в органоидах поджелудочной железы демонстрируют, что органоиды поджелудочной железы явно имеют фенотип протоков. Эти органоиды поджелудочной железы первоначально выращивали из препаратов протоков. Также в органоидах поджелудочной железы экспрессировались все необходимые факторы транскрипции для экспрессии Ngn3 (Foxa2, Hnf6, Hnf1b, Sox9), хотя сама по себе экспрессия Ngn3 не выявлялась в условиях увеличения в количестве.
Уровни экспрессии генов, важных для получения продуцирующих инсулин клеток, являются низкими. Однако очевидно, что в среде для увеличения в количестве, профили пролиферации и экспрессии органоидов поджелудочной железы в высокой степени напоминают профили пролиферации и экспрессии, наблюдаемые в ранних эндокринных клетках-предшественниках.
Поджелудочная железа в основном образована тремя различными типами клеток: ацинарные клетки, клетки протоков и эндокринные клетки. В образце тотальной РНК взрослой поджелудочной железы 90% РНК происходит из ацинарных клеток, так что уровни экспрессии эндокринных маркеров являются значительно разреженными в общем образце поджелудочной железы. Таким образом, дальнейшие эксперименты запланированы для каждого конкретного типа клеток. Например, авторы изобретения планируют осуществить сравнение на микрочипах органоидов поджелудочной железы, обогащенного препарата ацинарных клеток, обогащенного препарата клеток протоков и обогащенного препарата эндокринных клеток, чтобы иметь лучшую оценку уровней мРНК важных генов в их органоидах поджелудочной железы по сравнению с уровнями, присутствующими в продуцирующих инсулин клетках. Например, в обогащенном образце эндокринных клеток 75-85% присутствующих клеток будут представлять собой секретирующие инсулин клетки).
ПРИМЕР 3 - Эффект Noggin на среду для увеличения в количестве
Для исследования роли ингибитора BMP Noggin в среде для увеличения в количестве авторы изобретения сравнивали уровни мРНК ранних эндокринных маркеров и маркеров протоков в органоидах поджелудочной железы, которые всегда культивировали в среде EGFRA, так чтобы они никогда не культивировались в присутствии Noggin, с уровнем экспрессии тех же маркеров в органоидах, которые всегда культивировали в среде EGFRAN (т.е. всегда в присутствии Noggin). Авторы изобретения также сравнивали уровни мРНК этих маркеров в органоидах поджелудочной железы, в которых Noggin добавляли или удаляли из культур соответственно. В частности, один образец органоидов поджелудочной железы культивировали в среде EGFRA, а затем добавляли Noggin, и органоиды культивировали в течение дальнейших 2 или 4 суток. Другой образец органоидов поджелудочной железы культивировали в среде EGFRAN, а затем Noggin удаляли и органоиды культивировали в течение дополнительных 2 или 4 суток. Экспрессию генов сравнивали, и результаты представлены на фиг.17A. Было открыто, что Noggin уменьшает экспрессию кератина 7 и кератина 19 (маркеры протоков), что демонстрирует, что Noggin блокирует дифференцировку в направлении фенотипа протоков (уровни кератина в образцах, обозначенных белым и темно-серым цветом, ниже, чем в образцах, обозначенных черным цветом). На уровни экспрессии некоторых факторов транскрипции, необходимых для получения продуцирующих инсулин клеток (т.е. Sox9, Hnf6, Hnf1a, Pdx1, Nkx2.2, Nkx6.1 и Hnf1b) Noggin не влиял. Хотя Noggin препятствует приобретению культурами полного фенотипа протоков, что вероятно будет препятствовать дальнейшей дифференцировке в продуцирующие инсулин клетки, авторы изобретения включают Noggin в среду для увеличения в количестве, поскольку он позволяет клеткам увеличиваться в количестве при сохранении некоторых признаков протоков в комбинации с признаками продуцирующих инсулин клеток-предшественников.
Эффект присутствия или отсутствия Noggin, или его добавления или устранения из среды EGFRA на экспрессию гена Lgr5 оценивали с использованием органоидов поджелудочной железы, полученных из протоков поджелудочной железы. Результаты на фиг.17B показывают, что органоиды поджелудочной железы, культивированные с Noggin, экспрессируют в 2 раза больше Lgr5, чем органоиды поджелудочной железы, культивируемые без Noggin (сравнить второй слева белый столбик с черным столбиком слева). Также было показано, что добавление (темно-серый) или устранение (светло-серый) Noggin влияет на уровни Lgr5. Неявно, является ли увеличение экспрессии гена Lgr5 в присутствии Noggin следствием увеличенного количества Lgr5+ клеток или следствием увеличенного уровня экспрессии Lgr5 на клетку. Однако авторы настоящего изобретения показали в рамках настоящего изобретения, что ингибиторы BMP, такие как Noggin, стимулируют экспрессию Lgr5 и, таким образом, обеспечивают более пролиферативные органоиды. Таким образом, показано, что ингибиторы BMP являются преимущественным компонентом в среде для увеличения в количестве.
Это является неожиданным, поскольку в литературе описано, что активность BMP является полезной для дифференцирующей культуры клеток поджелудочной железы. Это заключение основано на наблюдениях, что передача сигнала BMP требуется для дифференцировки как в клетки протоков (см. экспрессию кератина 7 и 19), так и в эндокринные клетки. Таким образом, квалифицированный специалист может ожидать, что включение ингибитора BMP, такого как Noggin, будет неблагоприятным в среде для увеличения в количестве. Однако авторы изобретения неожиданно открыли, что применение ингибитора BMP было преимущественным, поскольку оно приводило к более пролиферативным органоидам и более высокой экспрессии Lgr5.
ПРИМЕР 4 - Трансплантация органоидов поджелудочной железы человека под почечную капсулу у мышей
Органоиды поджелудочной железы, которые увеличили в количестве с использованием протокола, описанного в примере 1 (см. фиг.18A), трансплантировали под почечную капсулу иммунодефицитных мышей.
Непосредственно перед трансплантацией органоиды обрабатывали раствором для выделения клеток (BD#354253, BD Biosciences), чтобы избавиться от остатков Matrigel. Органоиды промывали несколько раз посредством PBS и осаждали.
Трансплантацию этих органоидов под почечную капсулу иммунодефицитных реципиентов осуществляли с использованием рекомендованной NIH методики трансплантации островков под почечную капсулу ("Purified Human Pancreatic Islets, In Vivo Islets Function", Document No. 3104, A04, Effective Date 7th July 2008, DAIT, NIAID, NIH). За неделю до трансплантации у реципиентных мышей (NOD/SCID/IL2RgammaKO, также известные как NSG) химически индуцировали гипергликемию посредством инъекции высокой дозы стрептозотоцина, составлявшей 130 мг/кг. Проводили мониторинг уровней глюкозы в крови и мышей, имеющих уровень глюкозы в крови выше 18 ммоль/л, считали гипергликемическими.
Для трансплантации гипергликемического реципиента подвергали анестезии и осуществляли небольшой разрез в левом боку для обнажения левой почки. Приблизительно 2,5-3,0 мм3 органоидов собирали в силиконизированную пробирку для трансплантации PE50 и трансплантировали под почечную капсулу с использованием шприца Hamilton. После каутеризации поврежденной капсулы почку помещали обратно в брюшную полость. Затем брюшину и кожу закрывали с помощью наложения швов шелковой нитью 5-0.
Одну мышь умерщвляли через три часа после трансплантации и трансплантат анализировали на маркеры зрелых бета-клеток и предшественников. У этой мыши нельзя было выявить продуцирующих инсулин клеток в периренальной капсуле мыши (фиг.18B).
Следующей мыши позволяли выздороветь в клетке с нагреваемой подстилкой под тщательным наблюдением. Мониторинг массы тела и уровней глюкозы в крови у мыши с трансплантацией проводили в течение 1 месяца. Через один месяц мышь умерщвляли и трансплантат анализировали на маркеры зрелых бета-клеток и предшественников.
Через 1 месяц после трансплантации можно было идентифицировать количество продуцирующих инсулин клеток. Все эти продуцирующие инсулин клетки являются окрашенными клетками на фиг.18C, выборка которых обведена для повышения ясности. В частности, положительные по инсулину клетки появились в выстилке протоков, в то время как в первоначальных препаратах не наблюдали положительных по инсулину клеток.
Данные о том, что продуцирующие инсулин клетки присутствуют через 1 месяц после трансплантации, но не присутствуют через 3 часа после трансплантации, демонстрируют, что продуцирующие инсулин клетки появляются главным образом или только после трансплантации.
Эти результаты демонстрируют, что клетки, взятые из органоидов поджелудочной железы по настоящему изобретению, культивированных с помощью сред и способов по настоящему изобретению, можно трансплантировать мышам, и они могут стимулировать рост продуцирующих инсулин клеток в поджелудочной железе. Удивительно, что можно трансплантировать органоиды поджелудочной железы человека. Это открывает ряд существующих возможностей применения трансплантированных органоидов для обеспечения продукции инсулина, например, для лечения диабета.
ПРИМЕР 5 - Культура органоидов печени, содержащая ингибитор TGF-бета
В условиях ER или ENRW культуры органоидов печени самообновляются, и они могут поддерживаться и увеличиваться в количестве на еженедельной основе в течение вплоть до 1 года (фиг.20A). Кариотипический анализ после 1 года демонстрирует отсутствие данных о хромосомных аберрациях. Более 66% анализированных клеток имели нормальное количество хромосом и 13% из них также демонстрировали полиплоидию, характерный признак гепатоцитов (фиг.20B).
Комбинация EGF (50 нг/мл) и R-спондина 1 (1 мкг/мл), дополненная FGF10 (100 нг/мл), HGF (25-50 нг/мл) и никотинамидом (1-10 мМ), была предпочтительной для долговременного поддержания культур. В этих условиях авторы настоящего изобретения получали длительно живущие клеточные культуры, которые экспрессируют маркеры желчных протоков и некоторые маркеры гепатобластов или незрелых гепатоцитов (Glul, альбумин). Однако количество клеток, положительных по этим маркерам гепатоцитов, было очень низким. В этих условиях культивирования, не было выявлено маркеров зрелых гепатоцитов (например, цитохромы p450). Эти результаты указывают на то, что условия культивирования, описанные в настоящем описании, способствуют увеличению в количестве предшественников печени, способных образовывать подобные гепатоцитам клетки, хотя из в более низких количествах, но не полностью зрелые гепатоциты (фиг.21A).
Для усиления гепатоцитарного характера культур и получения зрелых гепатоцитов in vitro авторы настоящего изобретения сначала определили, оказывают ли три дополняющих фактора (FGF10, HGF и никотинамид), добавленных к EGF и R-спондину 1, положительный или отрицательный эффект на экспрессию маркеров гепатоцитов, а также на самообновление культуры. Авторы настоящего изобретения получили культуры органоидов печени и культивировали их либо с EGF, либо с EGF и R-спондином 1 вместе с FGF10, или HGF, или никотинамидом, или их комбинацией, и они разделяли культуры один раз в неделю на протяжении общего периода 10 недель. В каждый момент времени авторы настоящего изобретения также анализировали экспрессию нескольких маркеров зрелых гепатоцитов (FAH, CYP3A11) и маркеров гепатобластов (альбумин) (фиг.21B).
Было выявлено, что R-спондин 1 и никотинамид в комбинации с FGF10 необходимы для роста и самообновления культур печени (фиг.21C и D). Как R-спондин 1, так и никотинамид, ингибируют экспрессию зрелого маркера CYP3A11 и, тем не менее, стимулируют экспрессию маркера гепатобластов альбумина. Добавление либо FGF10, либо HGF, в среду, содержащую только EGF (без R-спондина 1 и без никотинамида), способствовало экспрессии зрелого маркера CYP3A11, хотя и на очень низких уровнях (фиг.21E). Для идентификации дополнительных соединений, которые могут способствовать дифференцировке гепатоцитов, авторы настоящего изобретения использовали два различных подхода, оба на основе условий: EGF + HGF и/или FGF10.
Первый подход вовлекал исследование серии соединений в дополнение к условиям EGF + FGF10 или HGF. Полный перечень проанализированных соединений представлен в таблице 4.
Dexa
A 0937
Второй подход учитывал знание из опубликованных исследований, касающихся экспрессии факторов транскрипции, необходимых для обеспечения билиарной и гепатоцитарной дифференцировки in vivo. Сравнительный анализ экспрессии факторов транскрипции в органоидах в условиях E, или ER, или ENRW, дополненных FGF10, HGF и никотинамидом, представлен на фиг.21. Присутствовали все факторы транскрипции, требуемые для спецификации в гепатоциты, помимо tbx3 и prox1. Однако авторы настоящего изобретения отметили, что экспрессия специфических билиарных факторов транскрипции была в высокой степени активирована в культурах, содержащих R-спондин 1 (R), что указывает на то, что экспрессия генов в культуре была несбалансирована в направлении в большей степени билиарной судьбы клеток.
Каскады передачи сигнала Notch и TGF-бета вовлечены в судьбу билиарных клеток in vivo. В действительности, делеция Rbpj (необходимого для достижения активной передачи сигнала Notch) приводит к нарушению тубулогенеза (Zong Y. Development 2009) и добавление TGFb к эксплантатам печени способствует билиарной дифференцировке in vitro (Clotman F. Genes and Development 2005). Поскольку каскады передачи сигнала как Notch, так и TGFb, в высокой степени активированы в культурах печени (фиг.22), авторы настоящего изобретения сделали вывод, что ингибирование судьбы клеток билиарных протоков может запускать дифференцировку клеток в направлении более гепатоцитарного фенотипа. A8301 был выбран в качестве ингибитора рецептора TGFb ALK5, 4 и 7, и DAPT был выбран в качестве ингибитора гамма-секретазы, активной протеазы, необходимой для активации каскада Notch. Сначала авторы изобретения культивировали клетки в течение 2 суток в условиях увеличения в количестве (среда ER) и на 2 сутки (фиг.23A) они запустили условия дифференцировки путем добавления комбинации различных соединений. Среду заменяли раз в двое суток и экспрессию маркеров дифференцировки анализировали через 8-9 суток. Условия ER и ENRW использовали в качестве отрицательного контроля.
Комбинация EGF + FGF10 с DAPT и A8301 приводила к неожиданно высокому усилению экспрессии анализируемых маркеров гепатоцитов (CYP3A11, TAT, альбумин) (фиг.23B). Эффект уже поддавался выявлению к 5 суткам и достигал пика на 8-9 сутки (фиг.23C). Максимальная эффективность концентрации достигалась при 10 мкМ (DAPT) и 50 нМ (A8301) (фиг.23D), соответственно. Добавление дексаметазона (известная молекула для дифференцировки гепатоцитов) не приводило к какому-либо улучшению экспрессии генов. Комбинация EGF, FGF10, A8301 и DAPT не только усиливает экспрессию, но также увеличивает количество подобных гепатоцитам клеток при оценке с помощью иммунофлуоресценции против маркеров гепатоцитов альбумина и 2F8, и окрашивания Xgal на происходящие из AlbCreLacZ органоиды (фиг.23E и F). Таким образом, авторы настоящего изобретения могут заключить, что упомянутый выше протокол дифференцировки способствует получению подобных гепатоцитам клеток in vitro из культур стволовых клеток печени.
СПОСОБЫ
Реагенты
Реагенты, использованные в экспериментах по культивированию, представлены в таблице 4.
Мыши. Мышей Lgr5-EGFP-ires-creERT2 (Barker N et al. Nature 2007;449:1003-7), APCfl/fl (Sansom OJ et al. Genes Dev 2004;18:1385-90), мышей Axin2-lacZ (Lustig B et al. Mol Cell Biol 2002;22:1184-93), мышей дикого типа C57B/6 (в возрасте 6-12 недель) генотипировали, как описано ранее, и использовали для экспериментов. Мышей Lgr5-EGFP-ires-creERT2 скрещивали с мышами APCfl/fl. Активность фермента Cre индуцировали путем внутрибрюшинных инъекций тамоксифена (2 мг/мышь). Мышей умерщвляли через 4 недели после индукции тамоксифеном. Тонкие кишечники и толстые кишечники мышей вскрывали в продольном направлении, промывали холодным PBS, а затем обрабатывали для выделения крипт. Области, содержащие аденомы кишечника, идентифицировали с использованием стереомикроскопа, разрезали скальпелем и промывали холодным PBS.
Материалы тканей человека
Хирургически вырезанные ткани кишечника получали от 30 пациентов из Diaconessen Hospital Utrecht или UMCU Hospital.
Материал пациентов собирали от 20 пациентов с раком толстого кишечника (9 слепая кишка-восходящая кишка, 7 сигмовидная кишка, 4 прямая кишка; в возрасте 33-86 лет), 5 пациентов при скрининговой колоноскопии (в возрасте 33-63 лет) и 5 пациентов с пищеводом Барретта (в возрасте 45-78 лет). Для нормальной ткани сохраняли расстояние более 3 см до опухолей. Ткани кишечника промывали и отделяли от нижележащих мышечных слоев. Ткань нарезали на фрагменты размером приблизительно 5 мм, и далее промывали холодным PBS. Эндоскопические биоптаты (кишечные или пищеводные) получали из больницы UMCU. Для каждого случая собирали по меньшей мере 5 образцов биоптатов и хранили в холодном PBS. Это исследование было одобрено этическим комитетом DHU и UMCU, и все образцы получали с информированным согласием.
Выделение крипт/аденомы и диссоциация клеток
Фрагменты кишечника (нормальный толстый кишечник мыши, нормальный тонкий и толстый кишечник человека) далее промывали холодным PBS до тех пор, пока супернатант не становился прозрачным. Далее, фрагменты ткани инкубировали в 2 мМ холодном хелатирующем буфере EDTA (дистиллированная вода с 5,6 мМ Na2HPO4, 8,0 мМ KH2PO4, 96,2 мМ NaCL, 1,6 мМ KCl, 43,4 мМ сахарозой, 54,9 мМ D-сорбитом, 0,5 мМ DL-дитиотреитолом) в течение 30 мин на льду (Gregorieff A Gastroenterology 2005(129)626-638). После удаления буфера EDTA фрагменты ткани энергично ресуспендировали в холодном хелатирующем буфере с использованием 10-мл пипетки для выделения крипт кишечника. Фрагментам ткани позволяли осесть при нормальной гравитации в течение 1 мин и супернатант удаляли для исследования с помощью инвертационной микроскопии. Процедуру ресуспендирования/осаждения обычно проводили 6-8 раз, и супернатанты, не содержавшие крипт, выбрасывали. Супернатанты, содержавшие крипты, собирали в 50-мл пробирки falcon, покрытые бычьим сывороточным альбумином. Выделенные крипты осаждали, промывали холодным хелатирующим буфером и центрифугировали при 150-200 g в течение 3 мин для отделения крипт от единичных клеток.
Крипты толстого кишечника мыши осаждали и ресуспендировали с помощью TrypLE express (Invitrogen) и инкубировали в течение 15 мин при 37°C. В этих условиях диссоциации крипты толстого кишечника расщепляли в мягких условиях, тем самым физически отделяя основания крипт от верхней части крипт толстого кишечника.
Фрагменты кишечника, содержащие аденомы из индуцированных тамоксифеном мышей Lgr5-EGFPires-creERT2/APCfl/fl, инкубировали в 2 мМ хелатирующем буфере EDTA в течение 60 мин на льду. После промывания холодным хелатирующим буфером большинство нормальных эпителиальных клеток кишечника откреплялись, в то время как клетки аденомы оставались прикрепленными к мезенхиме. Далее фрагменты аденомы инкубировали в буфере для расщепления (DMEM с 2,5% эмбриональной телячьей сывороткой, пенициллин/стрептомицин (Invitrogen), 75 Е/мл коллагеназы типа IX (Sigma), 125 мкг/мл диспазы типа II (Invitrogen)) в течение 30 мин при 37°C. Фрагментам аденомы позволяли осесть при нормальной гравитации в течение 1 мин и супернатант собирали в 50-мл пробирку falcon, осаждали и промывали PBS. Выделенные клетки аденомы центрифугировали при 150-200 g в течение 3 мин для отделения аденомы от единичных клеток.
Биоптаты из образцов эпителия Барретта и рака толстого кишечника человека, измельченные на фрагменты размером 5 мм, промывали PBS несколько раз. Фрагменты ткани инкубировали в буфере для расщепления в течение 60 мин при 37°C. После расщепления фрагменты ткани вручную отбирали под микроскопом.
Для экспериментов по сортировке выделенные крипты диссоциировали с помощью TrypLE express (Invitrogen), включавшего 2000 Е/мл ДНК-азы (Sigma) в течение 60 мин при 37°C. Диссоциированные клетки пропускали через 20-мкм клеточное сито (CellTrics) и промывали PBS. Жизнеспособные единичные эпителиальные клетки отбирали по прямому рассеянию, боковому рассеянию и ширине импульса, и отрицательному окрашиванию йодидом пропидия или 7-ADD (eBioscience).
Культура крипт кишечника, аденом, эпителия Барретта и рака толстого кишечника
Выделенные крипты кишечника, клетки эпителия Барретта и рака толстого кишечника, подсчитывали с использованием гемоцитометра. Крипты, фрагменты эпителия или единичные клетки погружали в Matrigel на льду (со сниженным содержанием факторов роста, не содержащий фенолового красного; BD bioscience) и всевали в 48-луночные планшеты (500 крипт/фрагменты или 1000 единичных клеток на 25 мкл Matrigel на лунку). Matrigel полимеризовали в течение 10 мин при 37°C, и наслаивали 250 мкл/лунка базальной культуральной среды (расширенная DMEM/F12, дополненная пенициллином/стрептомицином, 10 мМ HEPES, Glutamax, 1× N2, 1× B27 (все от Invitrogen) и 1 мМ N-ацетилцистеином (Sigma)), содержавшей следующие оптимизированные комбинации факторов роста: EGF мыши для аденом кишечника мыши, ENR (EGF мыши, noggin мыши, R-спондин-1 человека) для крипт тонкого кишечника мыши, WENR (рекомбинантный Wnt-3A человека или кондиционированная Wnt-3A среда+ENR) для крипт толстого кишечника мыши, HISC (стволовые клетки кишечника человека: WENR+гастрин+никотинамид+A83-01+SB202190) для крипт тонкого кишечника/толстого кишечника человека, HISC+FGF10 человека для эпителия Барретта. Клетки рака толстого кишечника демонстрируют гетерогенное поведение и требуют либо не добавления факторов роста, либо EGF мыши и/либо, A83-01, и/либо SB202190. Для экспериментов по сортировке клеток Y-27632 (10 мкМ; Sigma) включали в среду в течение первых 2 суток, чтобы избежать аноикоза. Реагенты и концентрации каждого фактора роста указаны на фиг.12. Обзор оптимизированных комбинаций факторов роста и низкомолекулярных ингибиторов для каждого органа представлен на фиг.12.
Анализ изображений
Изображения органоидов получали либо с помощью конфокальной микроскопии с Leica SP5, либо с помощью инвертационного микроскопа (Nikon DM-IL), либо с помощью стереомикроскопа (Leica, MZ16-FA). Для иммуногистохимии образцы фиксировали с помощью 4% параформальдегида (PFA) в течение 1 ч при комнатной температуре, и парафиновые срезы обрабатывали с помощью стандартных способов. Иммуногистохимию проводили, как описано ранее. Для иммунного окрашивания тотального препарата органоиды крипт выделяли из Matrigel с использованием раствора для выделения (BD bioscience) и фиксировали с помощью 4% PFA, а затем обеспечивали проницаемость с помощью 0,1% Triton X-100. Первичные антитела представляли собой: антитело мыши против Ki67 (1:250, Monosan), антитело кролика против Muc2 (1:100, Santa Cruz), антитело кролика против лизоцима (1:1000, Dako), антитело кролика против синаптофизина (1:100, Dako) и антитела против хромогранина A (1:100, Santa Cruz). Вторичные антитела представляли собой конъюгированные с пероксидазой антитела или конъюгированные с Alexa-568 антитела. Окрашивание EdU проводили в соответствии с протоколом изготовителя (Click-IT; Invitrogen). ДНК окрашивали с помощью DAPI (Molecular Probes). Трехмерные изображения получали с помощью конфокальной микроскопии и реконструировали с помощью программного обеспечения Volocity Software (Improvision).
Анализ на микрочипах и анализ с помощью ПЦР в реальном времени
Данные были выложены в базе данных GEO под номером доступа GSE28907.
ПРИМЕР 6 - Культура органоидов печени, содержащая простагландин-2 или арахидоновую кислоту
Выживание in vitro, рост и размножение органоидов печени существенно усиливались путем добавления простагландина E2 (PGE2) или арахидоновой кислоты (AA) в базальную среду.
На фиг.25 и 26 показано, что добавление PGE2 в концентрации 50 нМ (также наблюдали, что он действует в диапазоне 10-500 нМ) или добавление AA в концентрации 10 мкг/мл (также действует в концентрации 100 мкг/мл, но не так хорошо), приводит к более высокому количеству более крупных органоидов, чем с использованием базальной среды отдельно. Важно, что добавление PGE2 или AA позволяет более длительное время увеличения в количестве. Это означает, что органоиды можно увеличивать в количестве на протяжении большего количества удвоений популяций перед снижением или замедлением их роста. Без PGE2 снижение роста наблюдают после культивирования в течение 5 недель при увеличении в количестве в 5 раз в неделю. С PGE2 не происходит снижение роста до по меньшей мере 8 недель при увеличении в количестве в 5 раз в неделю. Было выявлено, что PGE2 имеет немного больший эффект, чем AA. Использованная базальная среда представляла собой: hEGF (100 нг/мл, Invitrogen); noggin человека (hnoggin) (25 нг/мл, peprotech); гастрин (10 нМ, sigma); hFGF10 (peprotech); никотинамид (10 мМ, sigma); A8301 (500 нМ, Tocris); hHGF (50 нг/мл, peprotech); кондиционированная Rspo среда (10%).
Как PGE2, так и AA находятся на одном и том же каскаде передачи сигнала простагландинов (см. фиг.24), вместе с фосфолипидами, простагландином G2 (PGG2), простагландином F2 (PGF2), простагландином H2 (PGH2), простагландином D2 (PGD2). Можно ожидать, что добавление любого другого активирующего компонента этого каскада может иметь тот же благоприятный эффект на культуральную среду.
Добавление PGE2 или AA является особенно полезным для культуральной среды для увеличения в количестве. Однако в некоторых обстоятельствах их также можно включать в среду для дифференцировки.
ПРИМЕР 7 - Ингибиторы GSK3 являются эффективными агонистами Wnt в культуральных средах
Было показано, что CHIR99021, ингибитор GSK3, является эффективным агонистом Wnt для культуральной среды. В частности, было показано, что он является подходящей заменой для Wnt в культуральной среде для органоидов толстого кишечника и печени.
Более того, в качестве расширения примера 6, на фиг.25 показано, что клетки печени человека, выращенные в присутствии как CHIR99021 (агонист Wnt), так и PGE2, приводят к большему количеству и более крупным органоидам, чем клетки, выращенные с агонистом либо Wnt, либо PGE2, отдельно и определенно к большему количеству/более крупным органоидам, чем только в базальной среде.
Таким образом, ингибиторы GSK3 можно использовать в культуральных средах вместо или в дополнение к другим агонистам Wnt, таким как Wnt или R-спондин 1-4.
Удивительно, что CHIR99021 был такой эффективной заменой Wnt, поскольку GSK3 вовлечен в ряд различных каскадов, а не только в каскад Wnt. Это открытие открывает возможность конструирования других агонистов Wnt, нацеленных GSK3, которые могут быть пригодными в культуральных средах.
Пример 8 - Предстательная железа
Выделение эпителия предстательной железы (протокол мыши).
Пронумерованные стадии соответствуют фиг.47.
i) Умертвить самца мыши в возрасте минимум 8 недель для выделения зрелой предстательной железы; выделить мочеполовой синус из мыши.
ii) Удалить семенные пузырьки путем разрушения/разрезания кровеносных сосудов и соединительной ткани и проведения разреза в основании уретры
iii) Удалить мочевой пузырь путем разрушения/разрезания его вблизи основания уретры
iv) Удалить оставшиеся везикулы и жировую ткань путем осторожного вытягивания и отрезания. Должны остаться доли предстательной железы (их 6) и розовая структура в середине, которая представляет собой уретру;
v) Удалить уретру, легко узнаваемую по розовому цвету (окрашена темным цветом на изображении). Осторожно оттянуть доли предстательной железы, так чтобы они более не были связаны с уретрой; выделить каждую долю по отдельности, только путем разделения их растягиванием, или продолжить с целой предстательной железой.
Далее измельчить предстательную железу (доли) на небольшие фрагменты; расщепить предстательную железу в 1 мл коллагеназы II в концентрации 10 мг/мл (растворенной в ADMEM/F12) в течение 1½ часа при 37°C; после расщепления коллагеназы должны оставаться только "похожие на палец" структуры эпителиальных клеток.
- Промыть в ADMEM/F12
- Позволить фрагментам осесть и отобрать супернатант (центрифугирование при низкой скорости избавляет от большей части мезенхимы)
- Центрифугировать 50xG 5 мин 4’C
- Ресуспендировать в 1 мл трипсина (TLE) и расщеплять в течение приблизительно 30 мин 37’C.
Пипетировать вверх и вниз каждые 10 минут для обеспечения расщепления
- Промыть в ADMEM/F12
- Либо начать культивирование в ENR или ENR+ 1 нМ дигидротестостерон (сеять приблизительно 5000 клеток на лунку) (0,1 нМ-10 мкМ), авторам не известен верхний предел
- Либо продолжить выделением конкретного типа клеток с помощью FACS
Результаты
Эпителиальные клетки предстательной железы, культивированные в ENR + дигидротестостерон, в соответствии со способами, описанными выше, до настоящего времени можно было поддерживать в течение 35 недель. В присутствии тестостерона культуры увеличиваются в количестве так же, как и без тестостерона. Однако с тестостероном присутствуют все типы клеток, включая стволовые клетки, переходные делящиеся клетки и дифференцированные клетки, т.е. происходит увеличенная дифференцировка при сохранении популяции стволовых клеток. Органоиды предстательной железы, выращенные в присутствии тестостерона, также выглядят более похожими на орган in vivo (см. фиг.41 и 42). Более того, IHC и ОТ-ПЦР демонстрируют, что органоиды предстательной железы, выращенные в присутствии тестостерона, содержат как базальные, так и люминальные клетки.
Изобретение также относится к следующим пронумерованным вариантам осуществления:
1. Культуральная среда для увеличения в количестве популяции стволовых клеток, где культуральная среда содержит по меньшей мере один или несколько ингибиторов, которые связывают и снижают активность одной или нескольких мишеней в виде серин/треониновой протеинкиназы, выбранной из группы, включающей: киназу рецептора TGFβ 1, ALK4, ALK5, ALK7, p38; и где культуральная среда позволяет непрерывный рост в течение по меньшей мере 3 месяцев.
2. Культуральная среда согласно варианту осуществления 1, где по меньшей мере один или несколько ингибиторов включают:
a) ингибитор, который связывает и снижает активность ALK5; и
b) ингибитор, который связывает и снижает активность p38.
3. Культуральная среда согласно варианту осуществления 1 или варианту осуществления 2, где ингибитор представляет собой средство, которое связывает и снижает активность его мишени более чем на 95%; при оценке с помощью клеточного анализа.
4. Культуральная среда согласно любому из предшествующих вариантов осуществления, где ингибитор имеет величину IC50 менее 100 нМ.
5. Культуральная среда согласно любому из предшествующих вариантов осуществления, где ингибитор действует конкурентно; неконкурентно; внеконкурентно или путем смешанного ингибирования.
6. Культуральная среда согласно любому из предшествующих вариантов осуществления, где ингибитор действует конкурентно и связывается со связывающим ATP карманом мишени в виде серин-треониновой протеинкиназы.
7. Культуральная среда согласно любому из предшествующих вариантов осуществления, где ингибитор представляет собой: a) низкомолекулярный ингибитор; b) белок или пептид; c) антисмысловой олигонуклеотид; или d) аптамер.
8. Культуральная среда согласно любому из предшествующих вариантов осуществления, где низкомолекулярный ингибитор имеет молекулярную массу от 50 до 800 Да.
9. Культуральная среда согласно любому из предшествующих вариантов осуществления, где ингибитор представляет собой пиридинилимидазол или 2,4-дизамещенный птеридин или происходящий из хиназолина ингибитор.
10. Культуральная среда согласно любому из предшествующих вариантов осуществления, где ингибитор добавляют в концентрации от 10 нМ до 10 мкМ.
11. Культуральная среда согласно любому из предшествующих вариантов осуществления, где ингибитор выбран из группы соединений, включающей: SB-202190, SB-203580, SB-206718, SB-227931, VX-702, VX-745, PD-169316, RO-4402257, BIRB-796, A83-01, LY364947 SB-431542, SB-505124, SB-525334, LY 364947, SD-093 и SJN 2511.
12. Культуральная среда согласно любому из предшествующих вариантов осуществления, где SB-202190 или SB-203580 добавляют до концентрации от 50 нМ до 100 мкМ.
13. Культуральная среда согласно любому из предшествующих вариантов осуществления, где стволовые клетки представляют собой стволовые клетки человека.
14. Культуральная среда согласно любому из предшествующих вариантов осуществления, где стволовые клетки представляют собой эпителиальные стволовые клетки.
15. Культуральная среда согласно варианту осуществления 14, где эпителиальные стволовые клетки человека представляют собой a) стволовые клетки поджелудочной железы; b) стволовые клетки кишечника; или c) стволовые клетки толстого кишечника.
16. Культуральная среда согласно любому из предшествующих вариантов осуществления, где стволовые клетки образуют часть органоида или выделенный фрагмент ткани.
17. Культуральная среда согласно любому из предшествующих вариантов осуществления, где стволовые клетки представляют собой злокачественные стволовые клетки.
18. Культуральная среда согласно любому из предшествующих вариантов осуществления, где процент клеток в популяции стволовых клеток, имеющих нормальный кариотип после 1, 2 или 3 или более месяцев, составляет более 90%.
19. Культуральная среда согласно любому из предшествующих вариантов осуществления, где процент клеток в популяции стволовых клеток, имеющих нормальный фенотип после 1, 2 или 3 или более месяцев, составляет более 90%.
20. Культуральная среда согласно любому из предшествующих вариантов осуществления, где стволовые клетки выживают в течение более чем трех месяцев; как например, более шести месяцев.
21. Культуральная среда согласно любому из предшествующих вариантов осуществления, где стволовые клетки имеют среднее время удвоения популяции от 12 до 36 часов, от 18 до 30 часов или приблизительно 24 часа.
22. Культуральная среда согласно любому из предшествующих вариантов осуществления, где культуральная среда содержит базальную среду для клеток животного или человека и:
a) один или несколько ингибиторов морфогенетического белка кости (BMP);
b) один или несколько митогенных факторов роста; и
c) один или несколько агонистов Wnt.
23. Культуральная среда согласно любому из предшествующих вариантов осуществления, где культуральная среда содержит гастрин и/или никотинамид.
24. Способ увеличения в количестве популяции стволовых клеток, где способ включает:
a) предоставление популяции стволовых клеток;
b) предоставление культуральной среды согласно любому из предшествующих вариантов осуществления;
c) контактирование стволовых клеток с культуральной средой; и
d) культивирование клеток в соответствующих условиях.
25. Композиция, содержащая культуральную среду согласно любому из предшествующих вариантов осуществления и стволовые клетки.
26. Композиция, содержащая культуральную среду согласно изобретению и внеклеточный матрикс.
27. Добавка в культуральную среду, содержащая один или несколько ингибиторов согласно любому из предшествующих вариантов осуществления.
28. Герметично закрытая емкость, содержащая культуральную среду согласно любому из предшествующих вариантов осуществления или добавку в культуральную среду согласно варианту осуществления 27.
29. Культуральная среда согласно любому из вариантов осуществления 1-23 для культивирования эпителия пищевода Барретта, где культуральная среда дополнительно содержит FGF10.
30. Стволовые клетки или органоиды, полученные с использованием культуральной среды согласно любому из предшествующих вариантов осуществления, для применения в целях трансплантации или других терапевтических применениях.
31. Органоид поджелудочной железы, содержащий бета-клетки.
32. Органоид поджелудочной железы согласно варианту осуществления 31, дополнительно содержащий α-клетки, δ-клетки и PP-клетки.
33. Органоид поджелудочной железы согласно любому из вариантов осуществления 31 или 32, содержащий α-клетки, β-клетки, δ-клетки и PP-клетки.
34. Органоид поджелудочной железы согласно любому из вариантов осуществления 31 или 33, который экспрессирует один, два или все три из Pdx1, Nkx2.2 и Nkx6.1.
35. Органоид поджелудочной железы согласно любому из вариантов осуществления 31-34, который экспрессирует один, два или все три из NeuroD, Pax6 и Mafa.
36. Органоид поджелудочной железы согласно варианту осуществления 35, который дополнительно экспрессирует Ngn3.
37. Органоид поджелудочной железы, например, органоид поджелудочной железы согласно любому из вариантов осуществления 31-36, который способен секретировать инсулин после трансплантации органоида пациенту.
38. Органоид поджелудочной железы, как указано согласно любому из вариантов осуществления 31-37, для применения в лечении пациента, имеющего нарушение с дефицитом инсулина, такое как диабет.
39. Способ лечения пациента, имеющего нарушение с дефицитом инсулина, такое как диабет, включающий трансплантацию органоида поджелудочной железы согласно любому из вариантов осуществления 31-37, пациенту.
40. Органоид человека, выбранный из группы, состоящей из органоида крипта-ворсинка, органоида толстого кишечника, органоида поджелудочной железы, органоида желудка, органоида пищевода Барретта, органоида аденокарциномы и органоида карциномы толстого кишечника.
41. Органоид тонкого кишечника или органоид крипта-ворсинка, полученный с использованием культуральной среды согласно любому из вариантов осуществления 1-23 для применения при лечении поврежденного эпителия, например, у пациентов с болезнью включения микроворсинок (MVID).
42. Культуральная среда для печени, содержащая или состоящая из базальной среды для клеток животного или человека, в которую добавлены: один или несколько лигандов рецепторной тирозинкиназы, такой как митогенный фактор роста (например, EGF), никотинамид, и, предпочтительно, агонист Wnt, предпочтительно, R-спондин 1-4 и/или CHIR99021, и один или оба из a) активатора каскада простагландинов, такого как PGE2 и/или AA, и b) ингибитора TGF-бета, такого как A83-01.






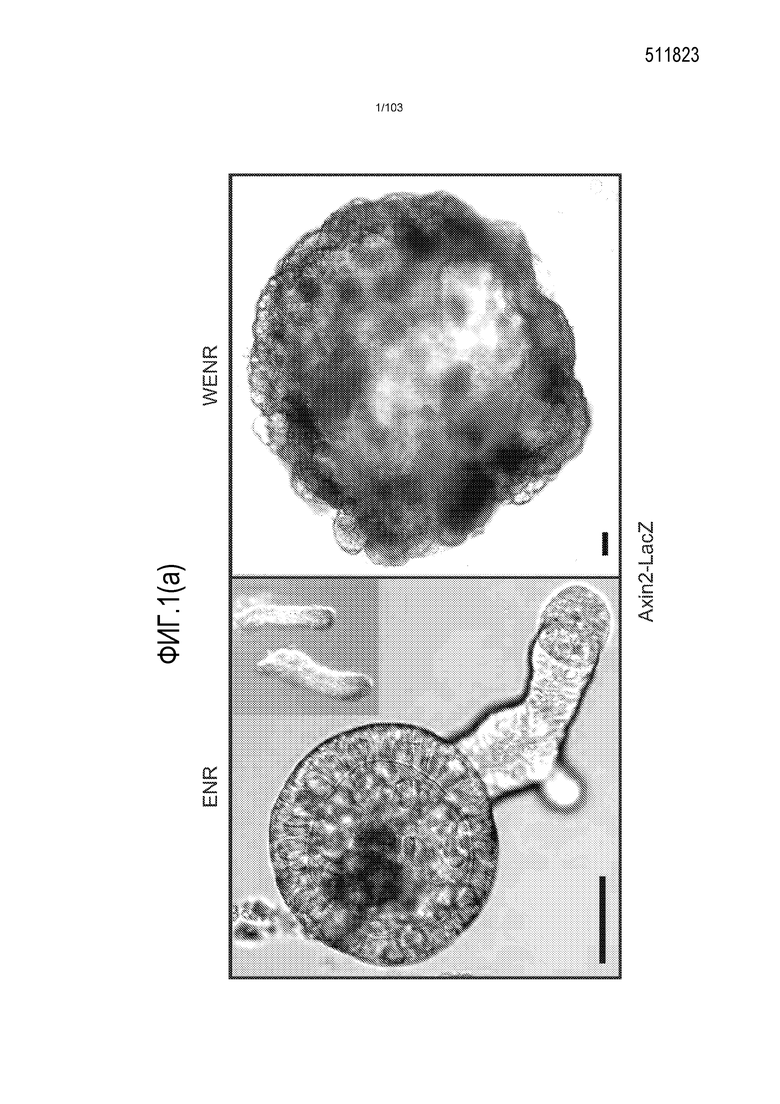
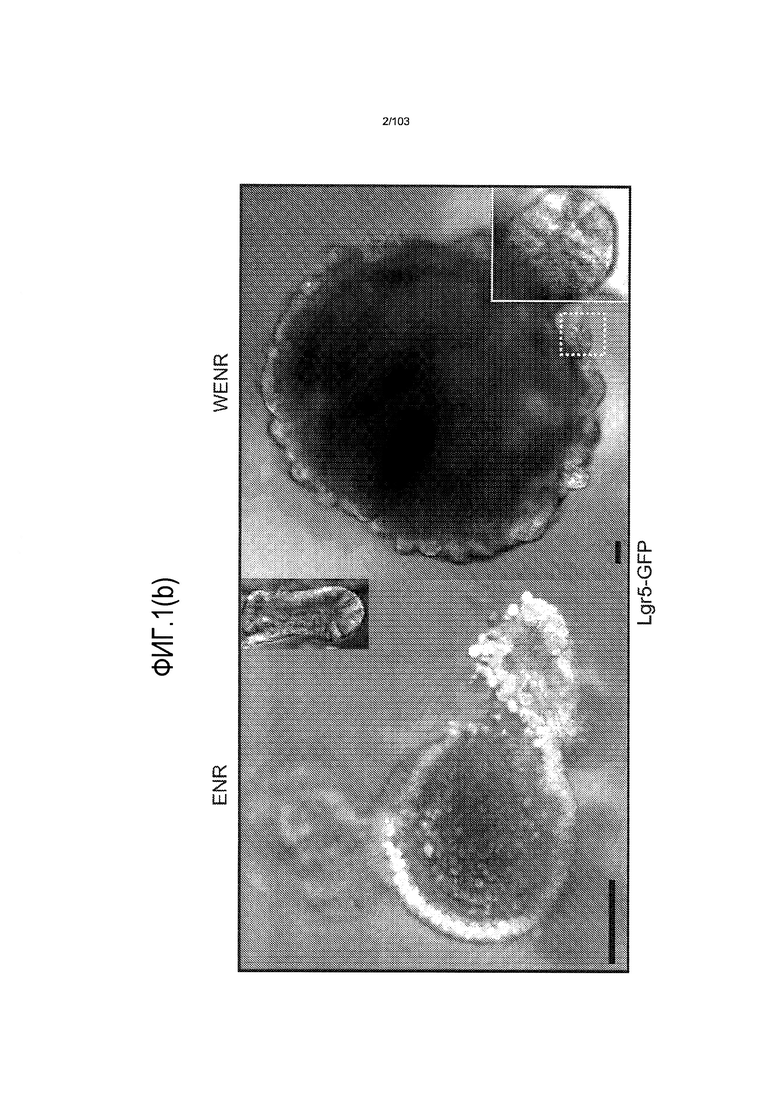
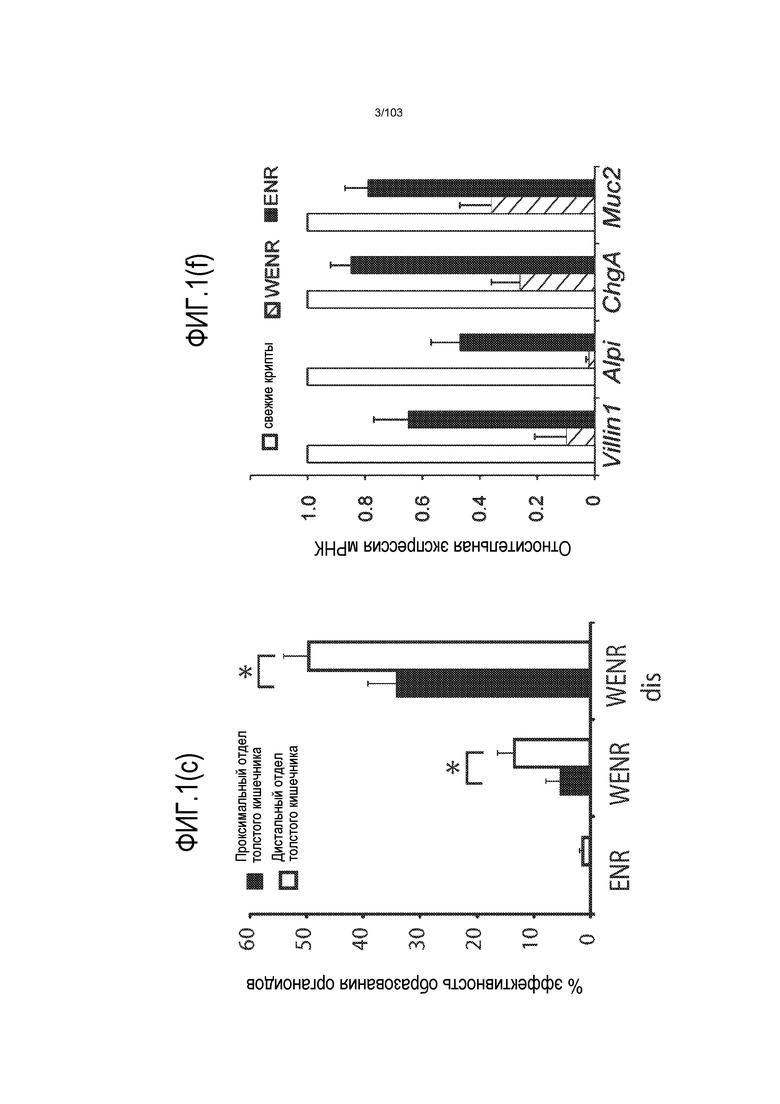
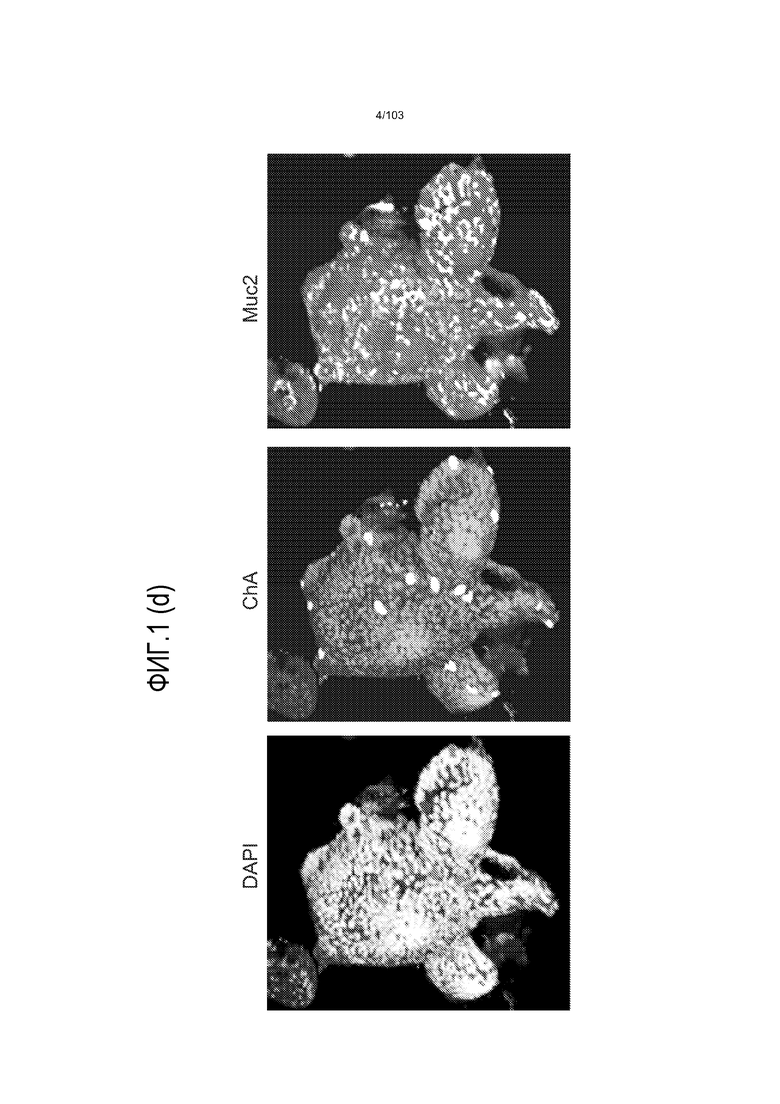
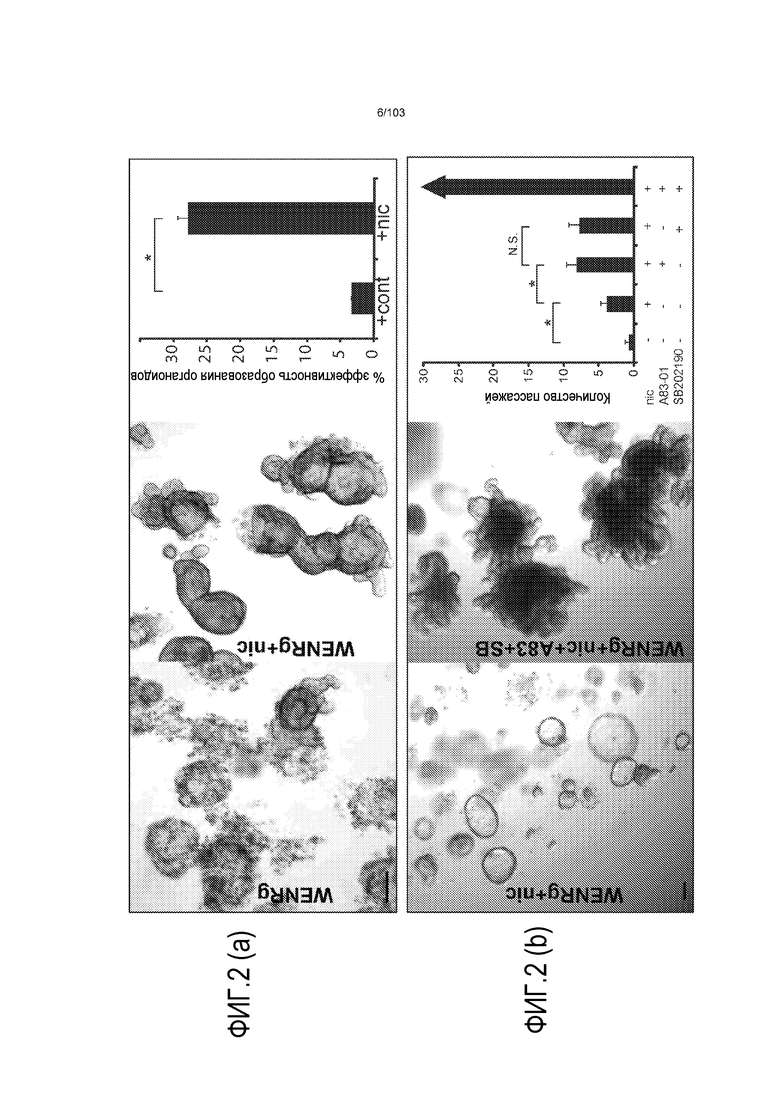

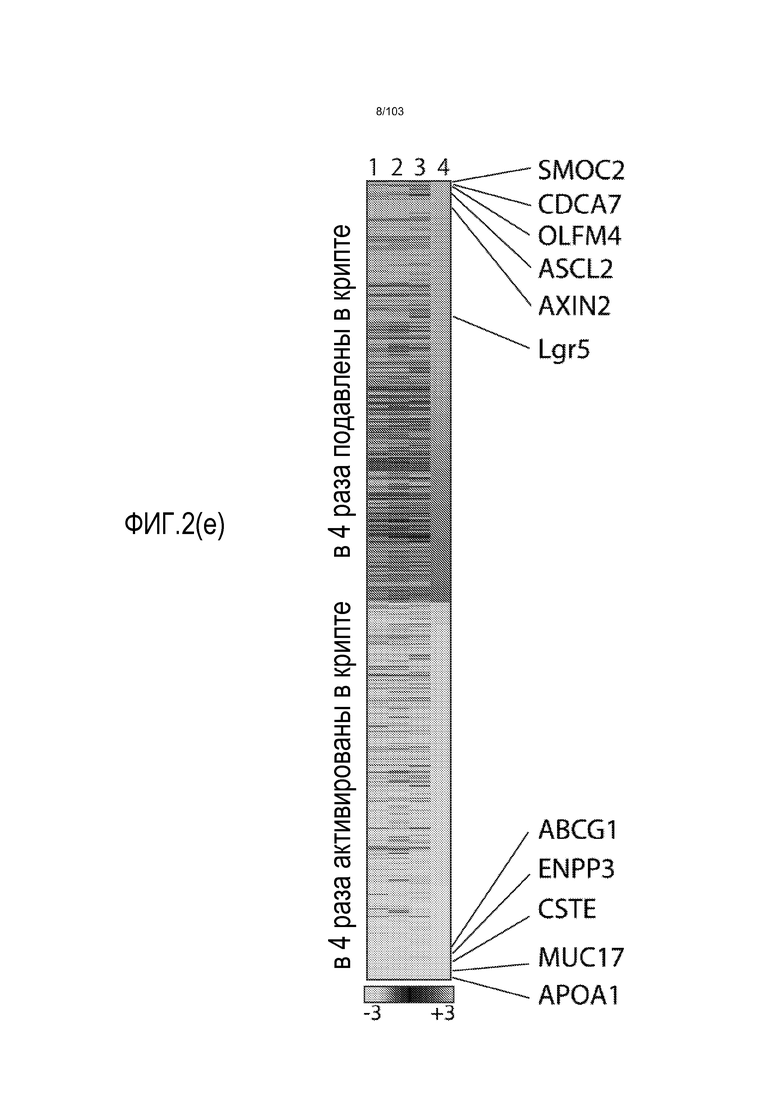
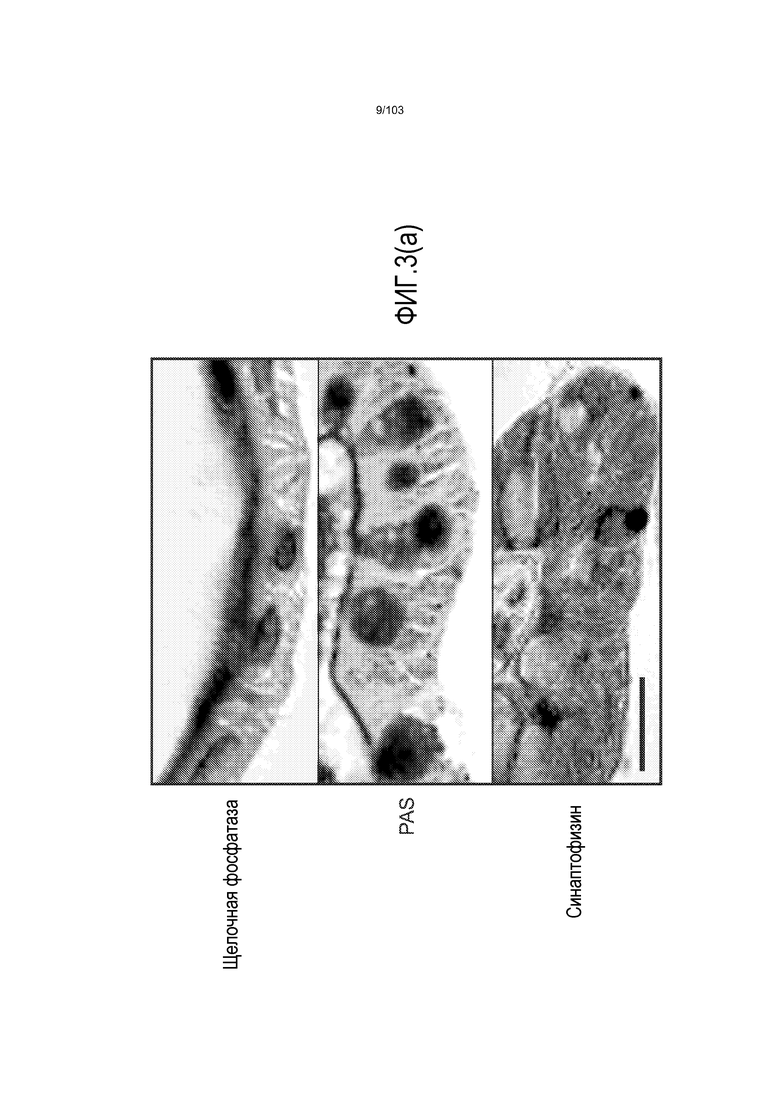
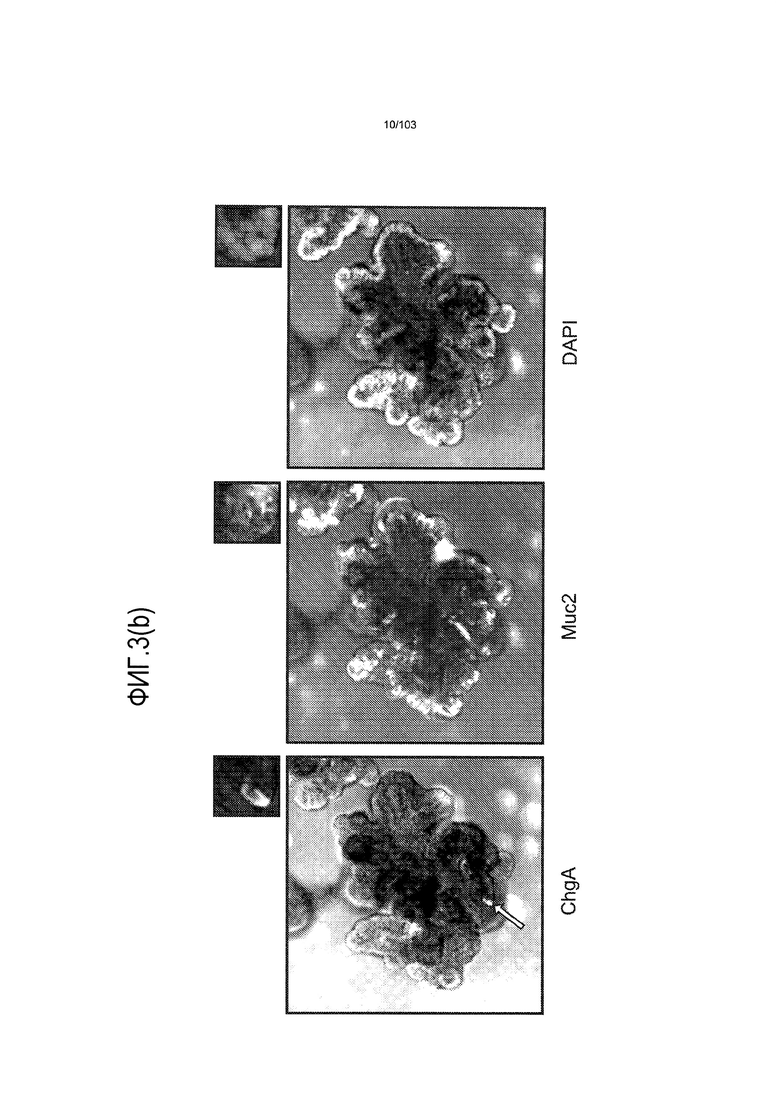
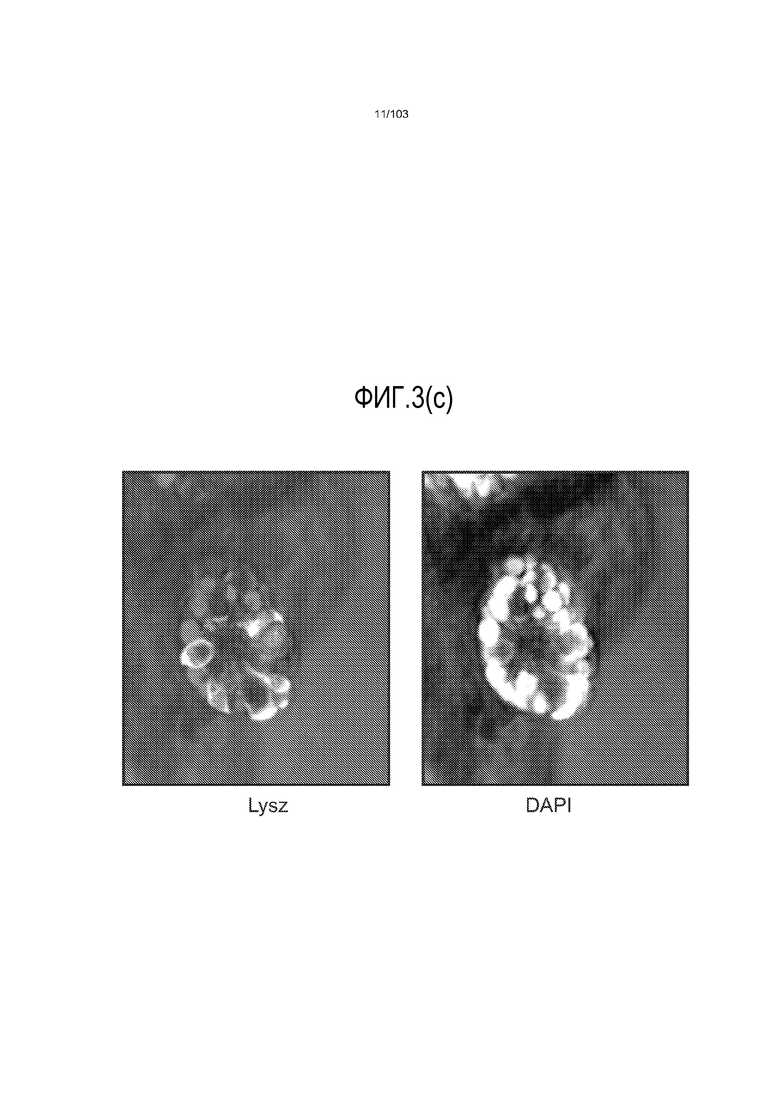
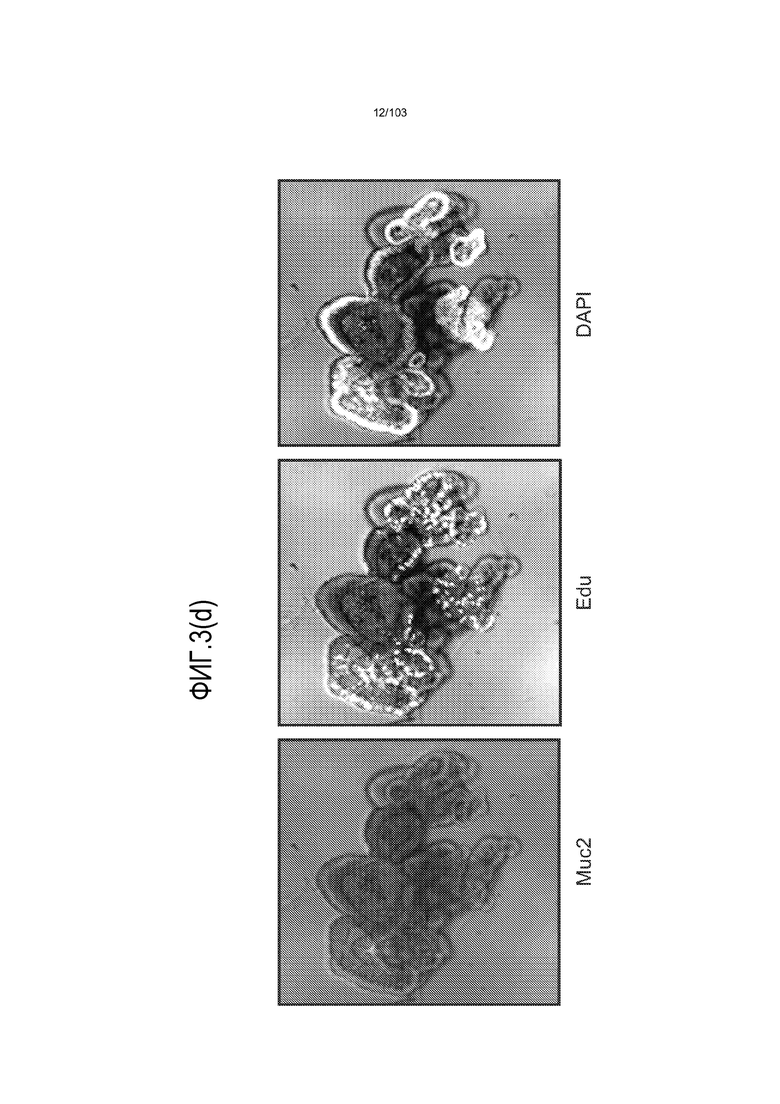
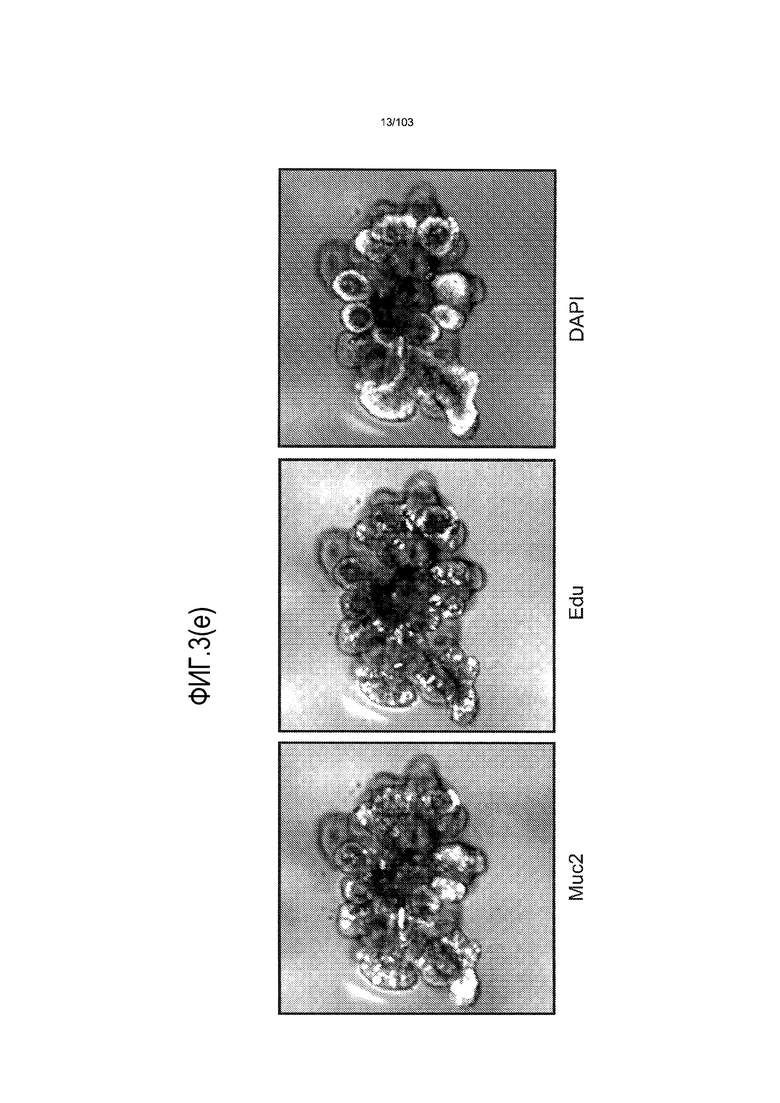
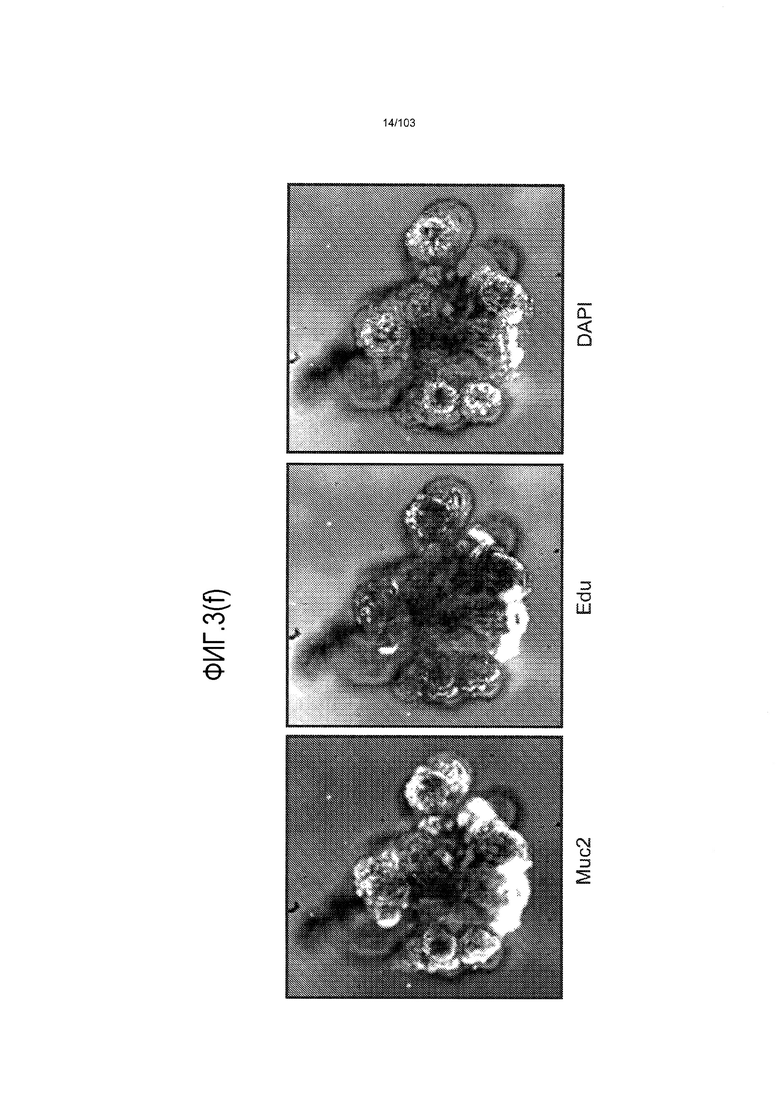
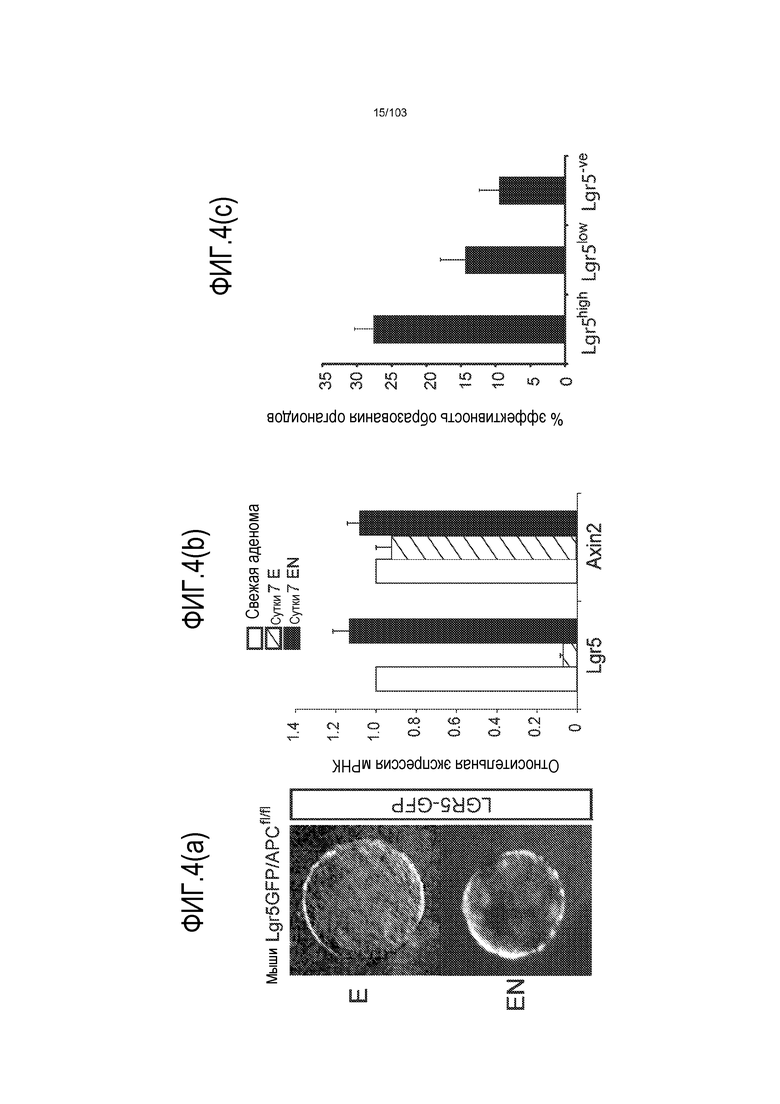
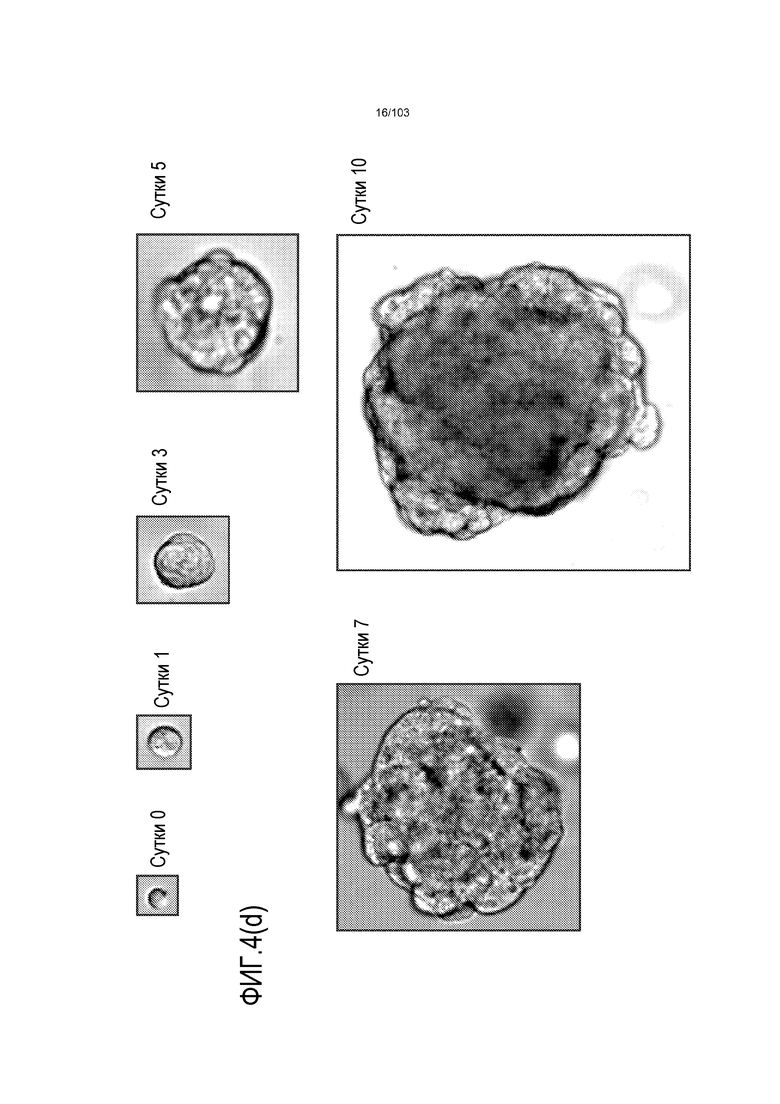
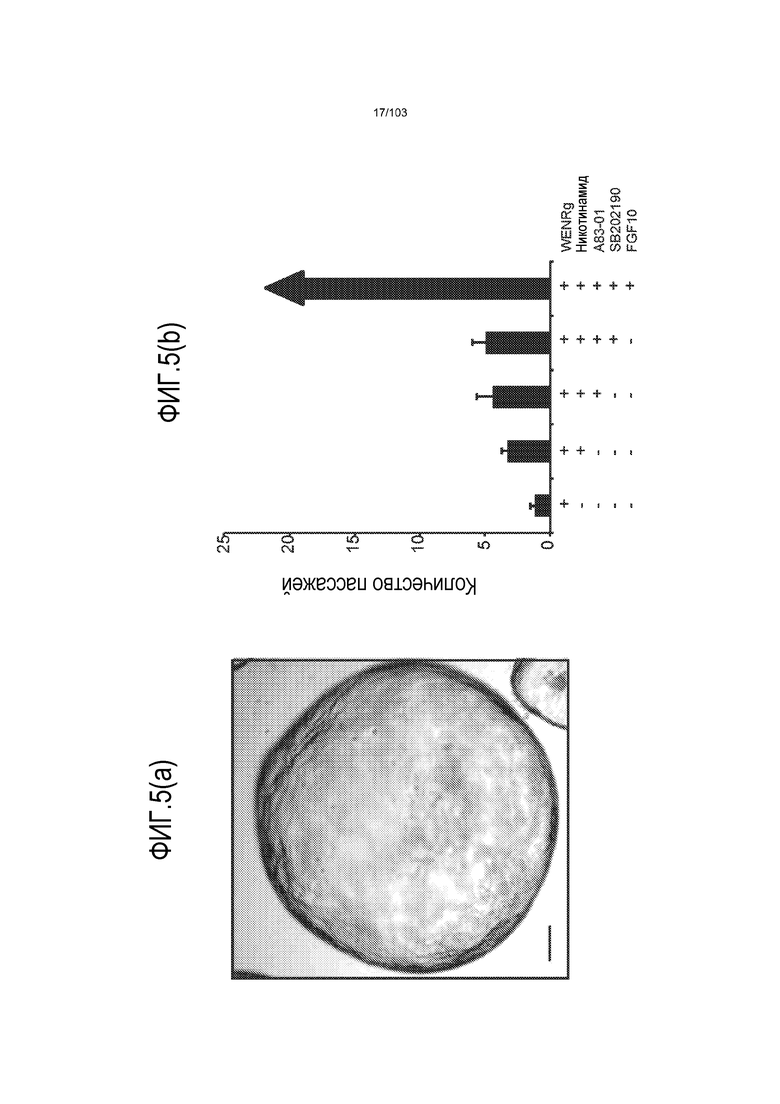
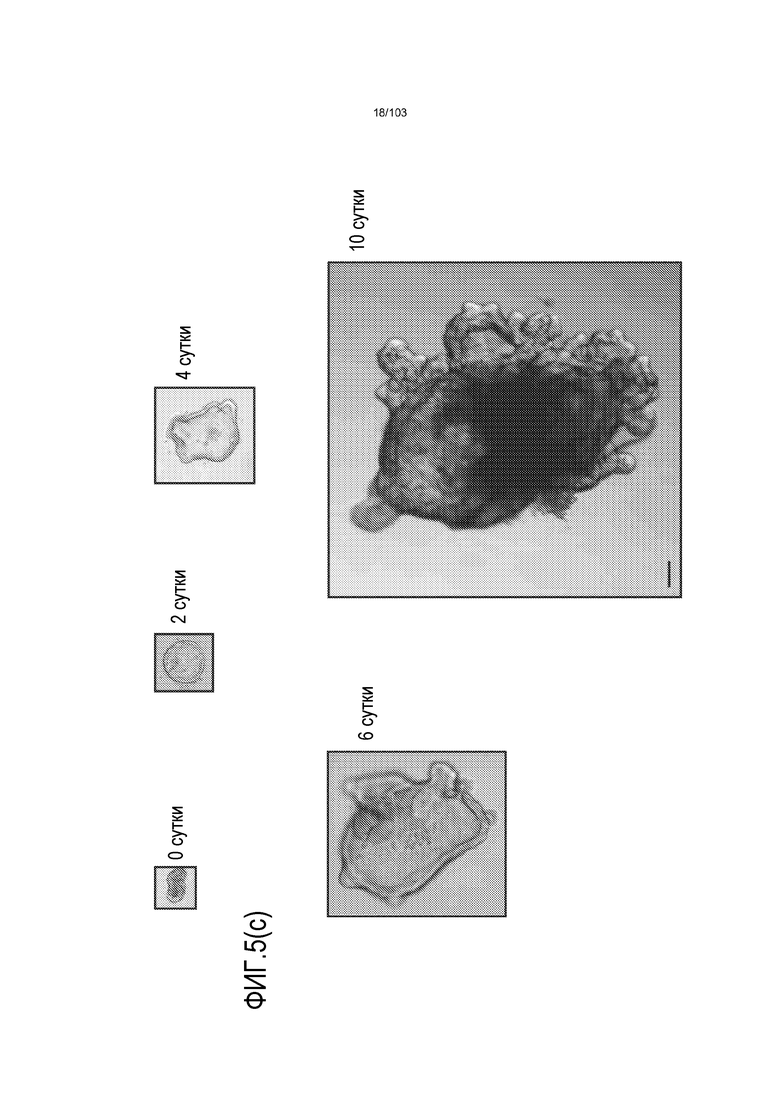
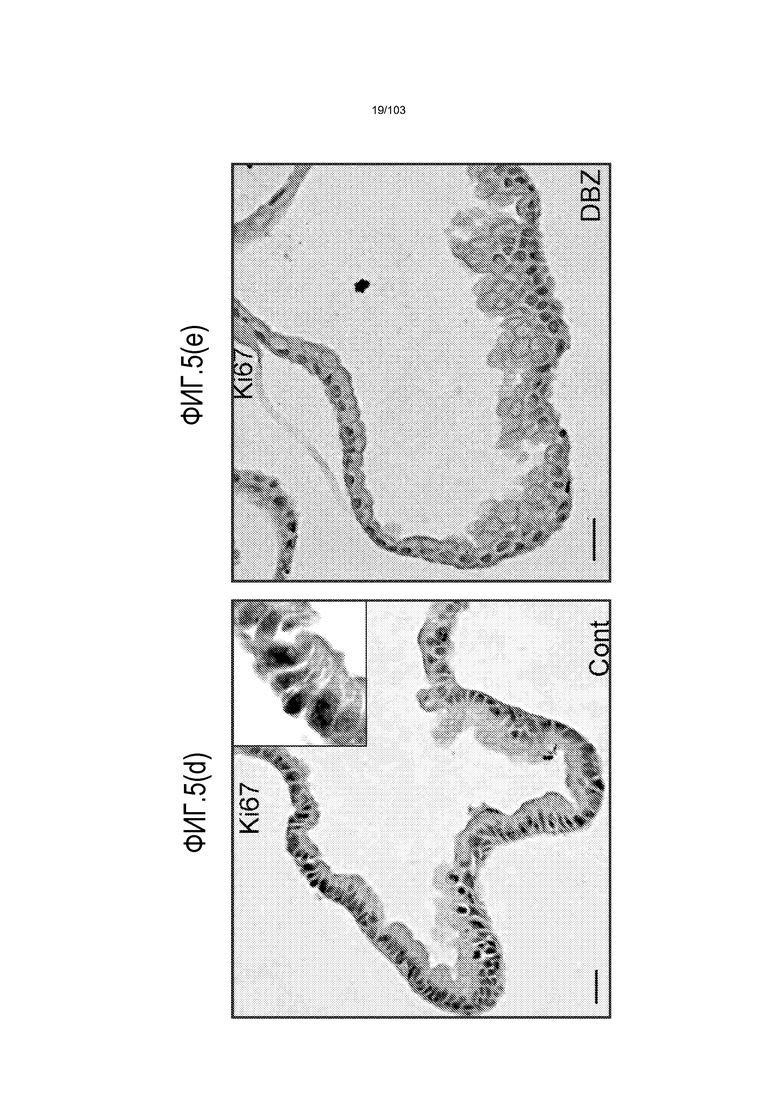
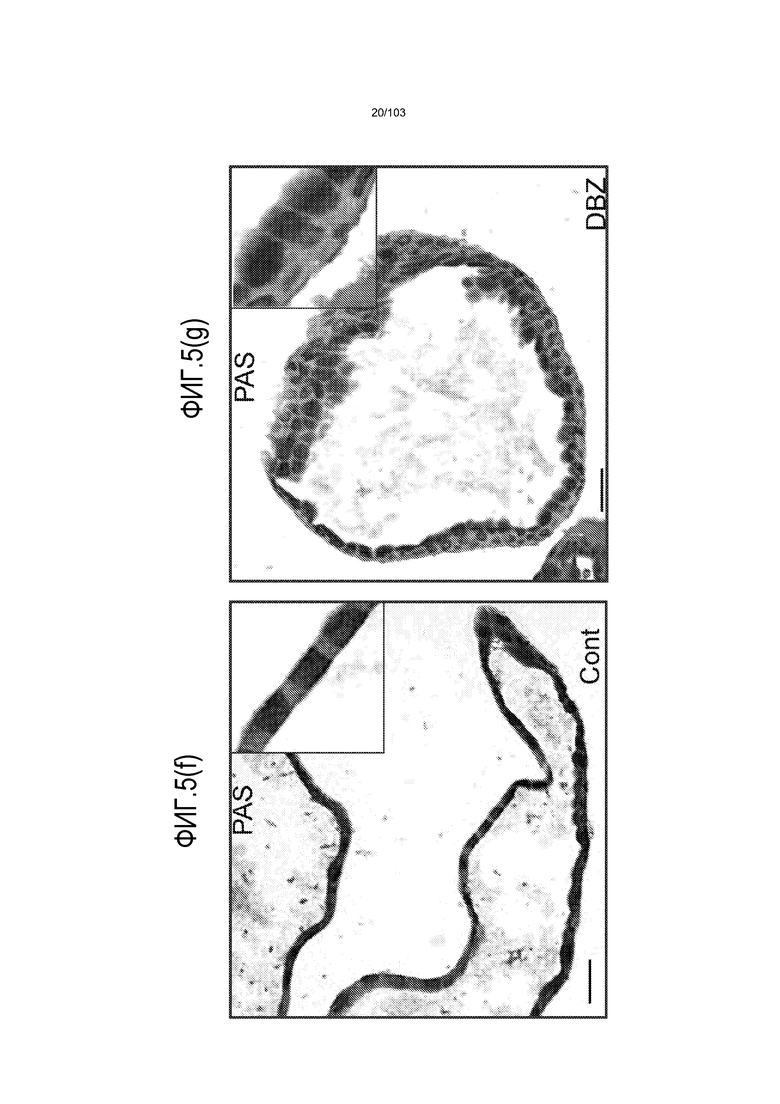
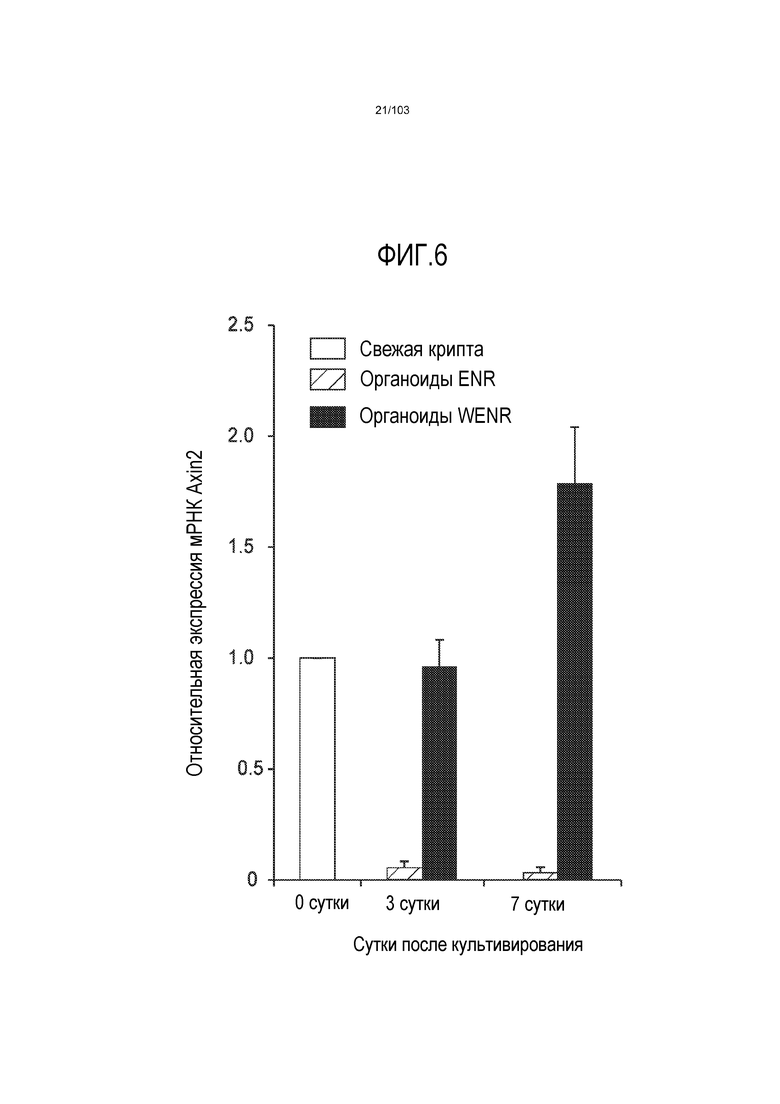
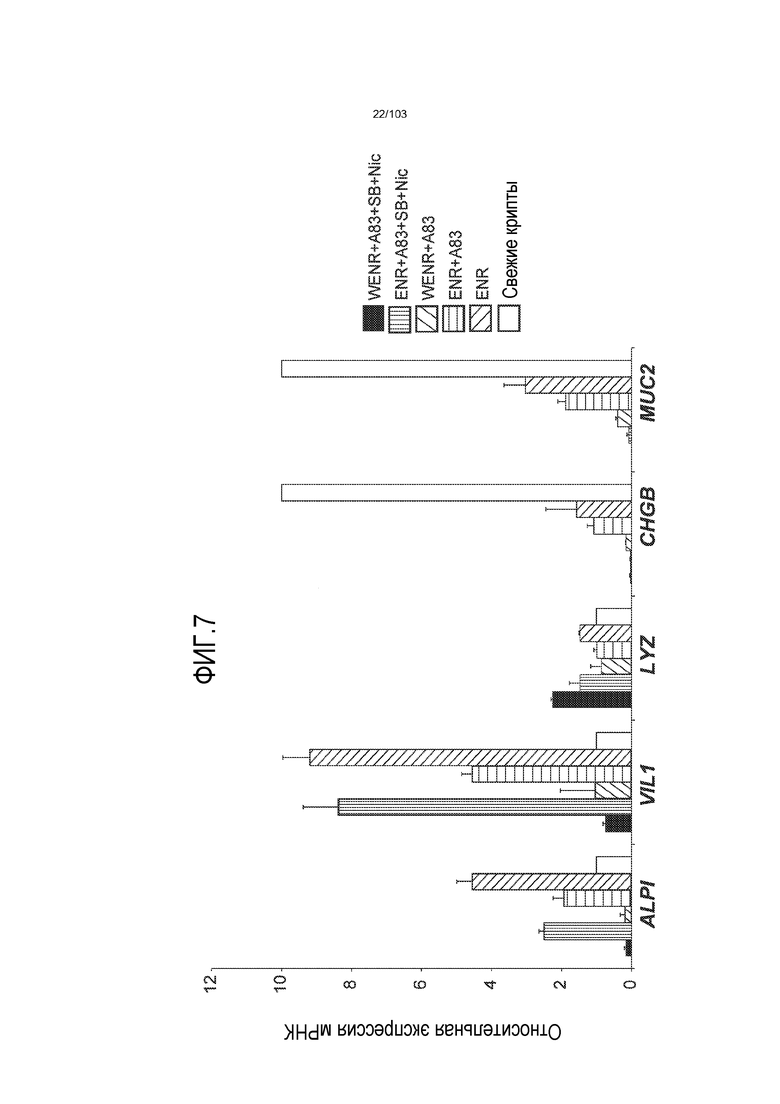

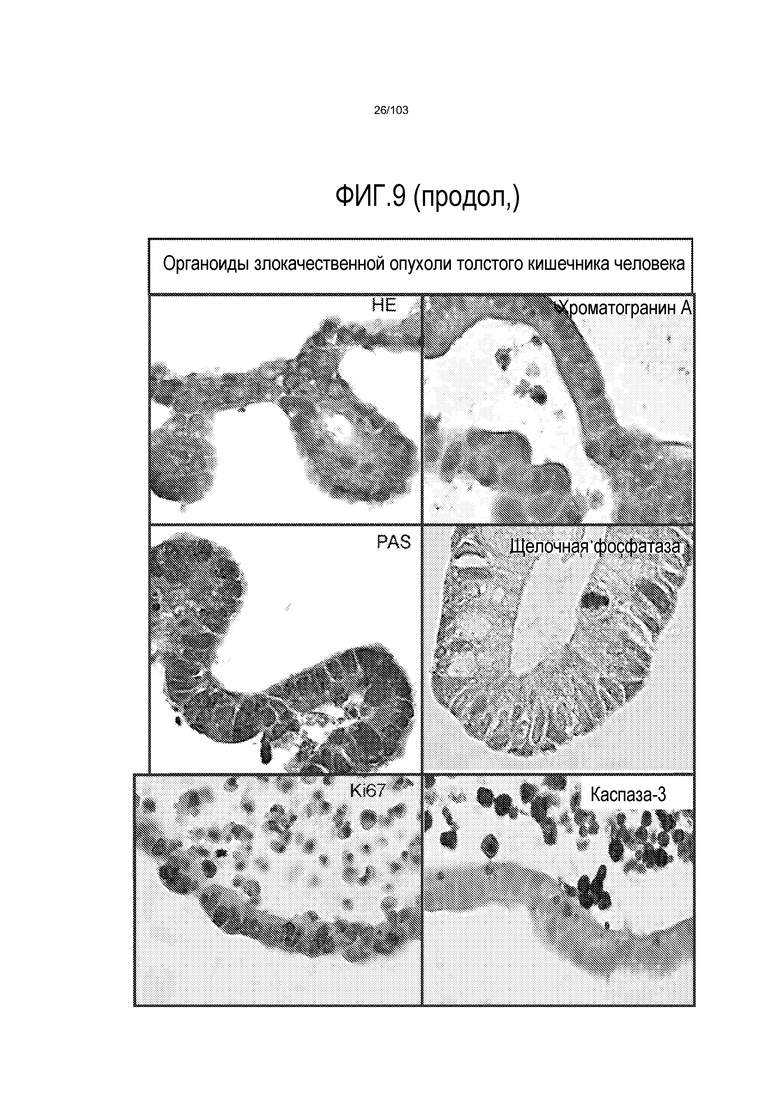

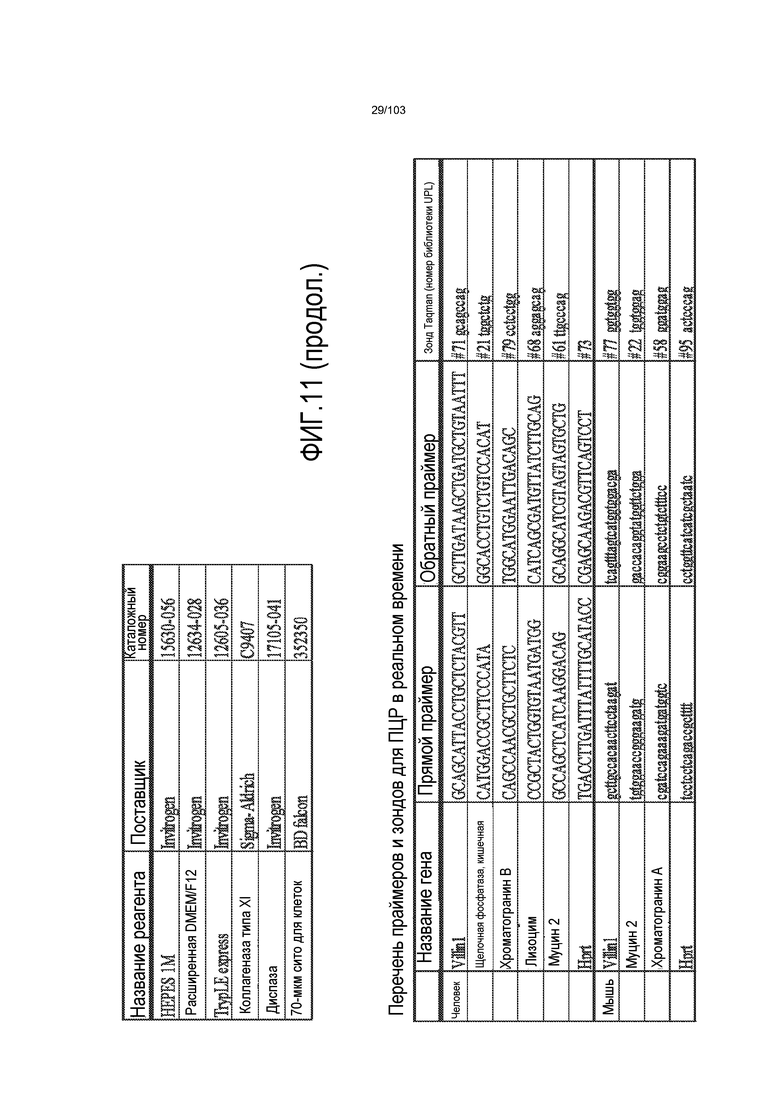

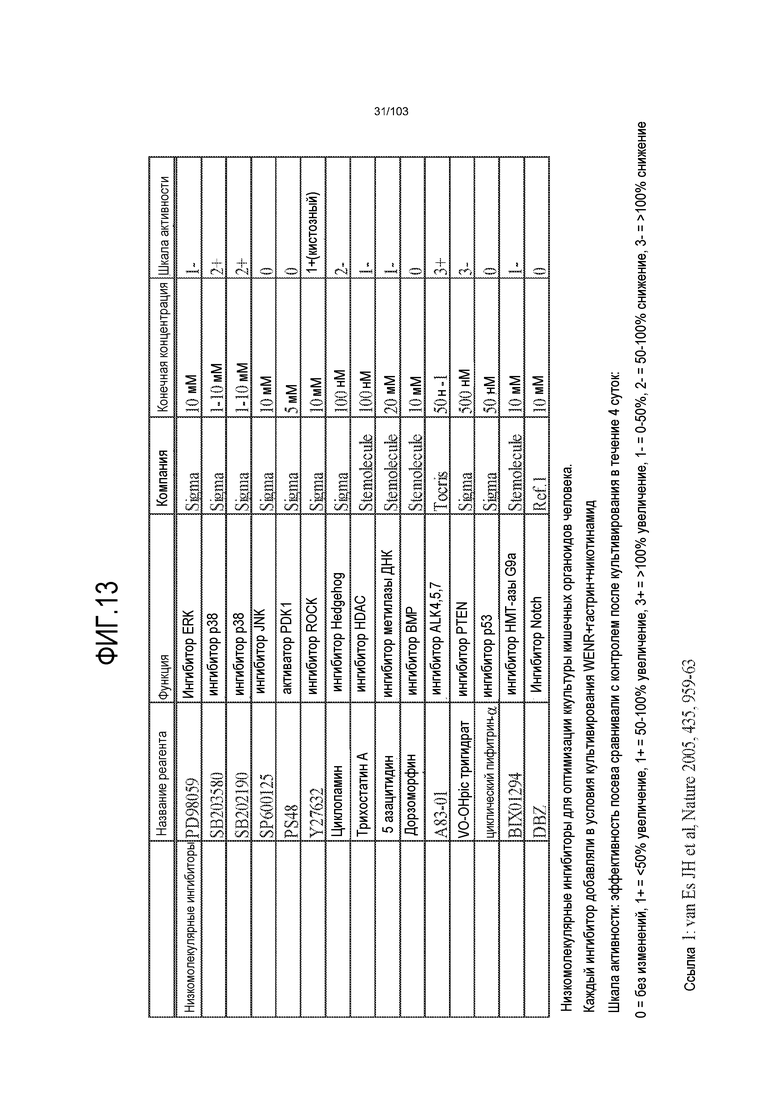
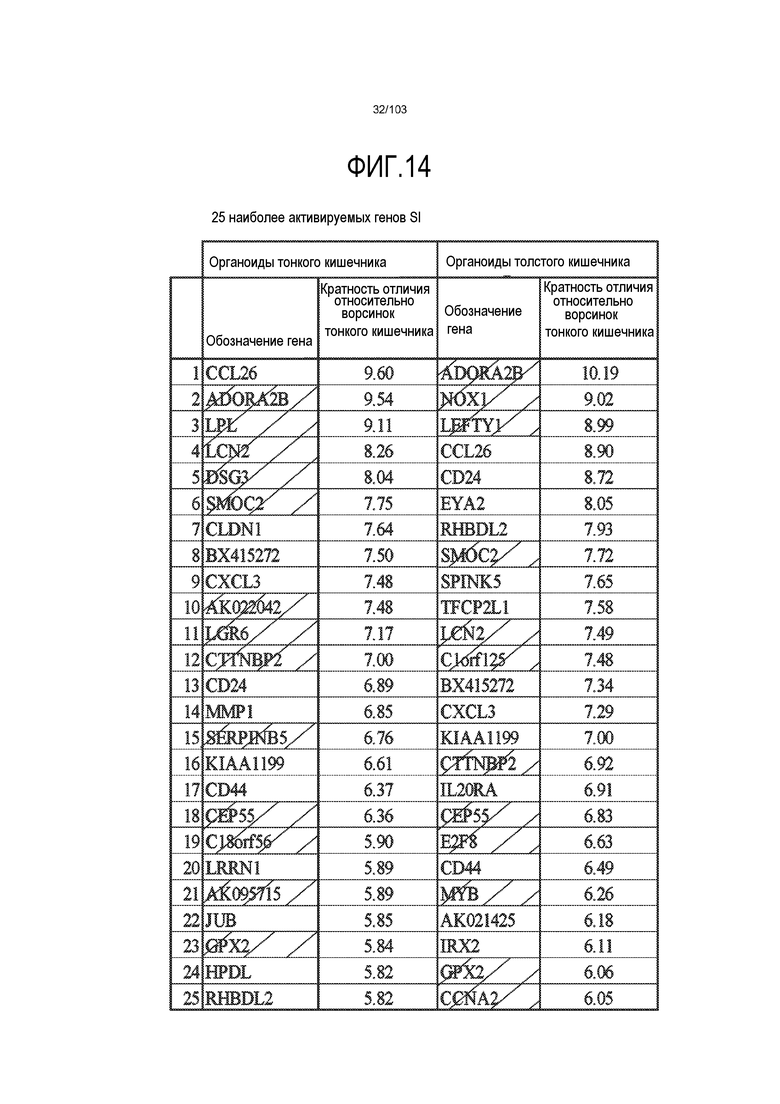

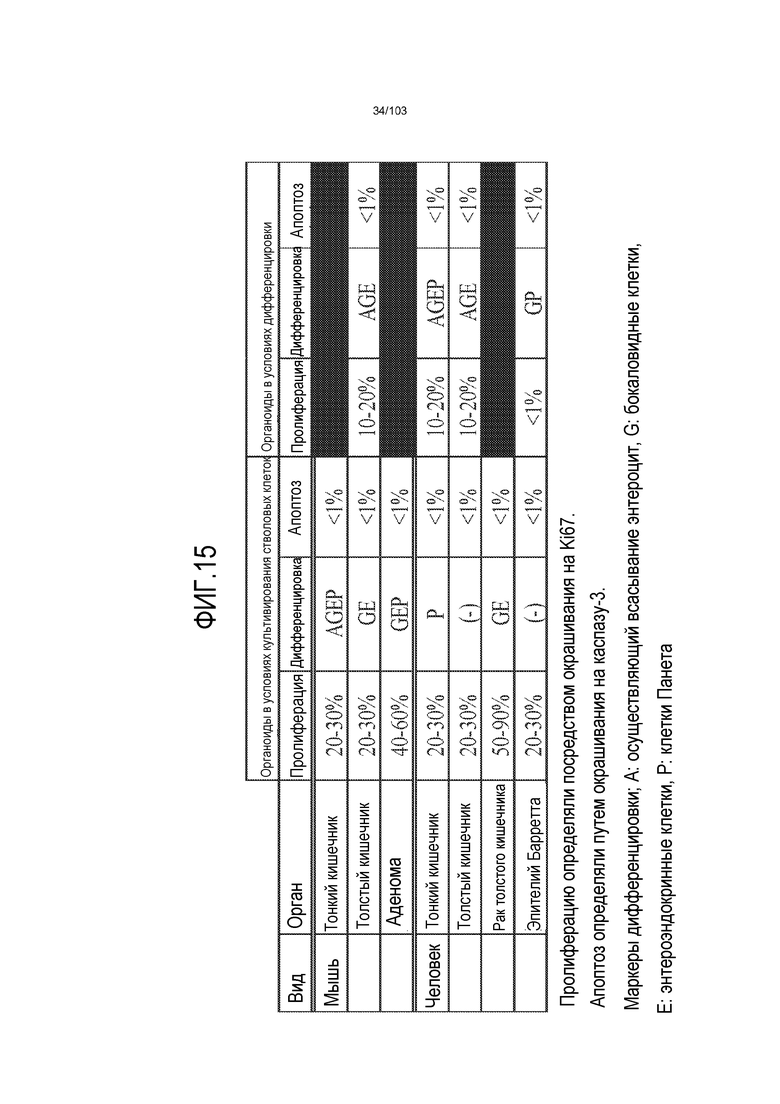



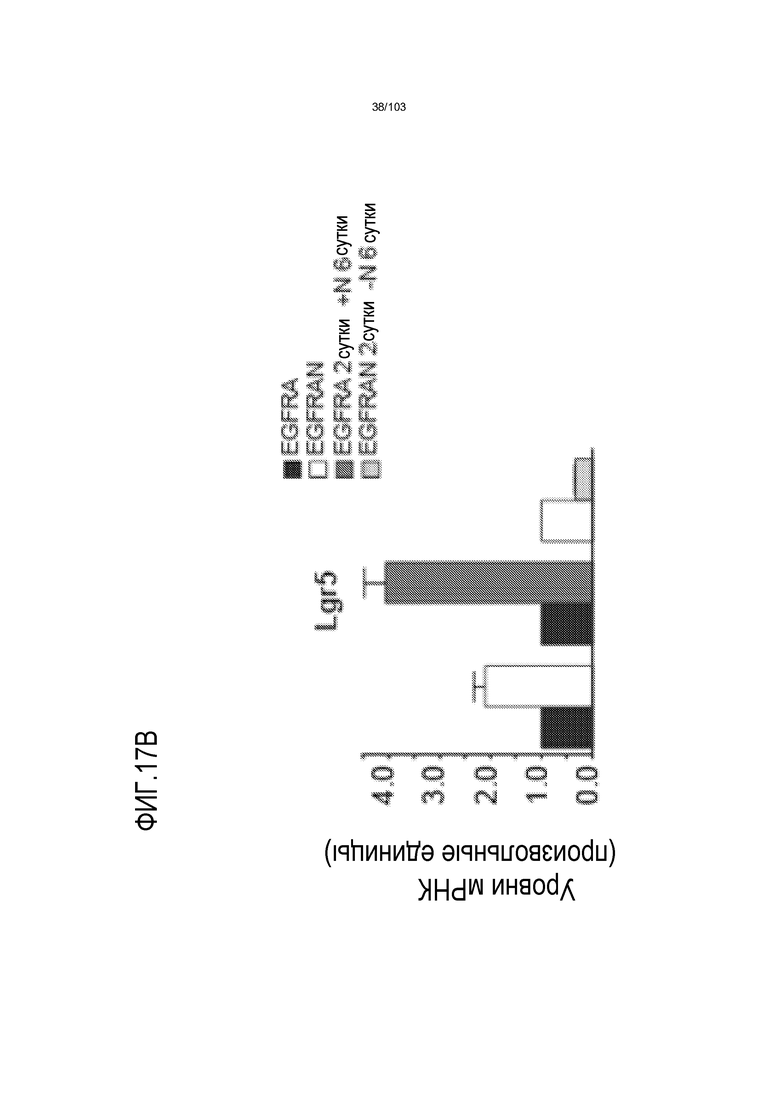
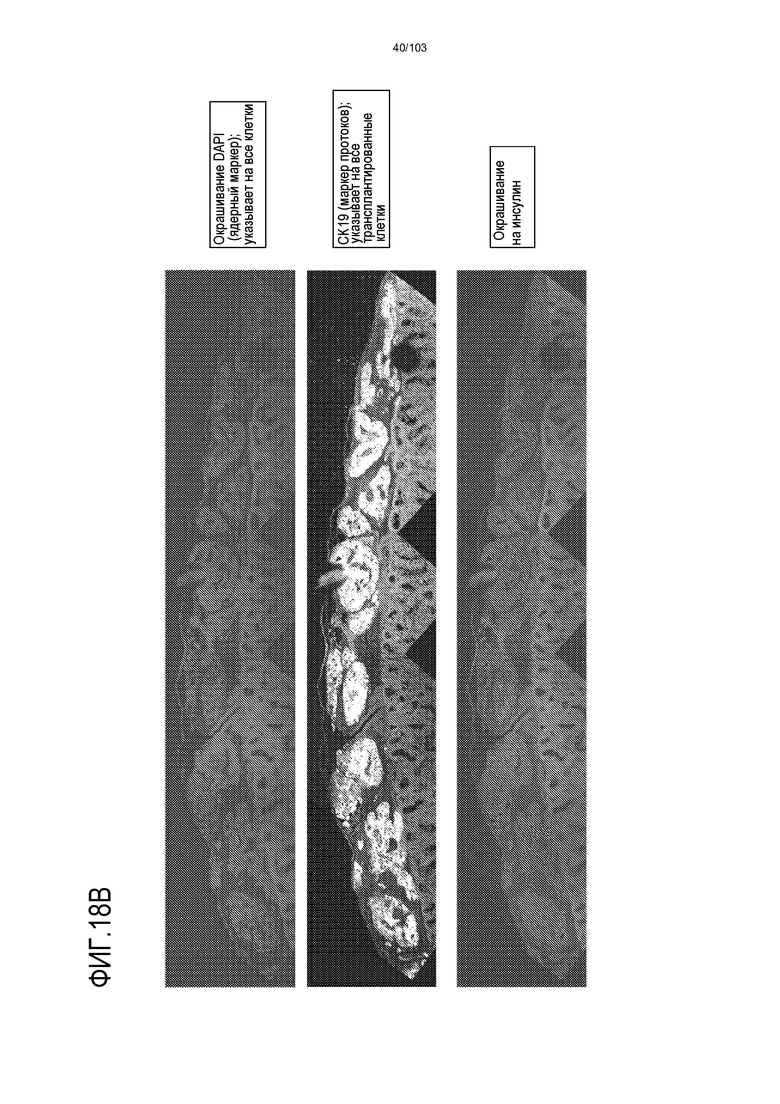
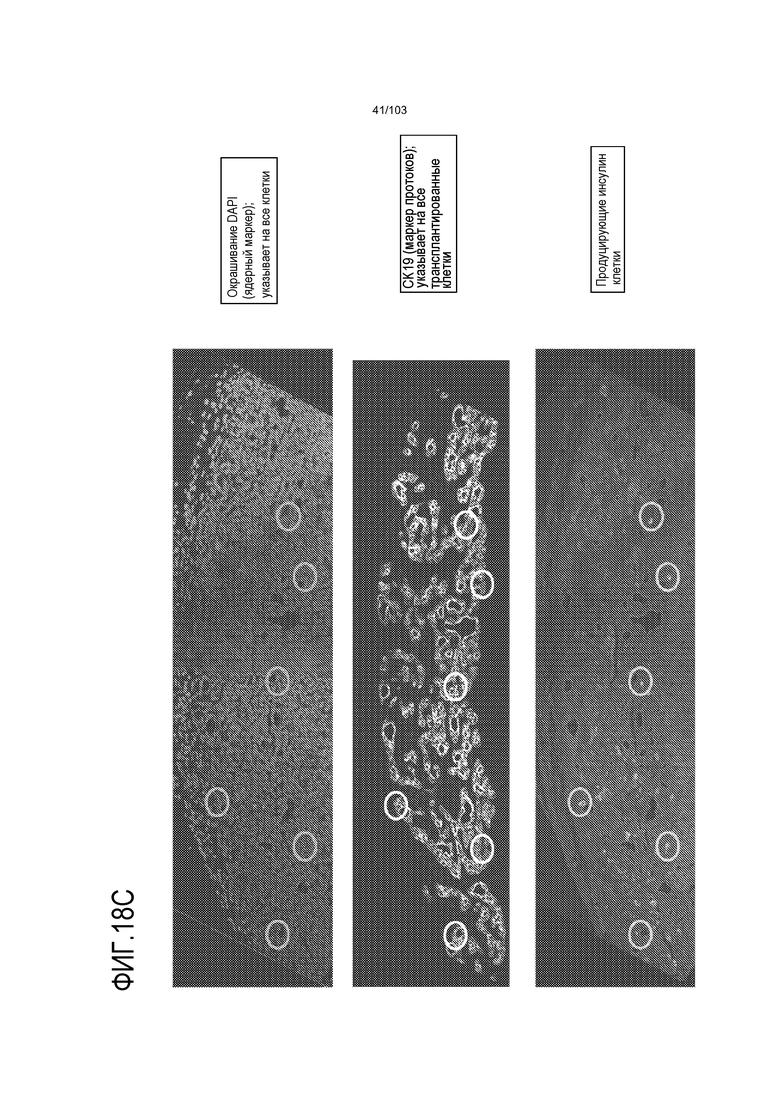

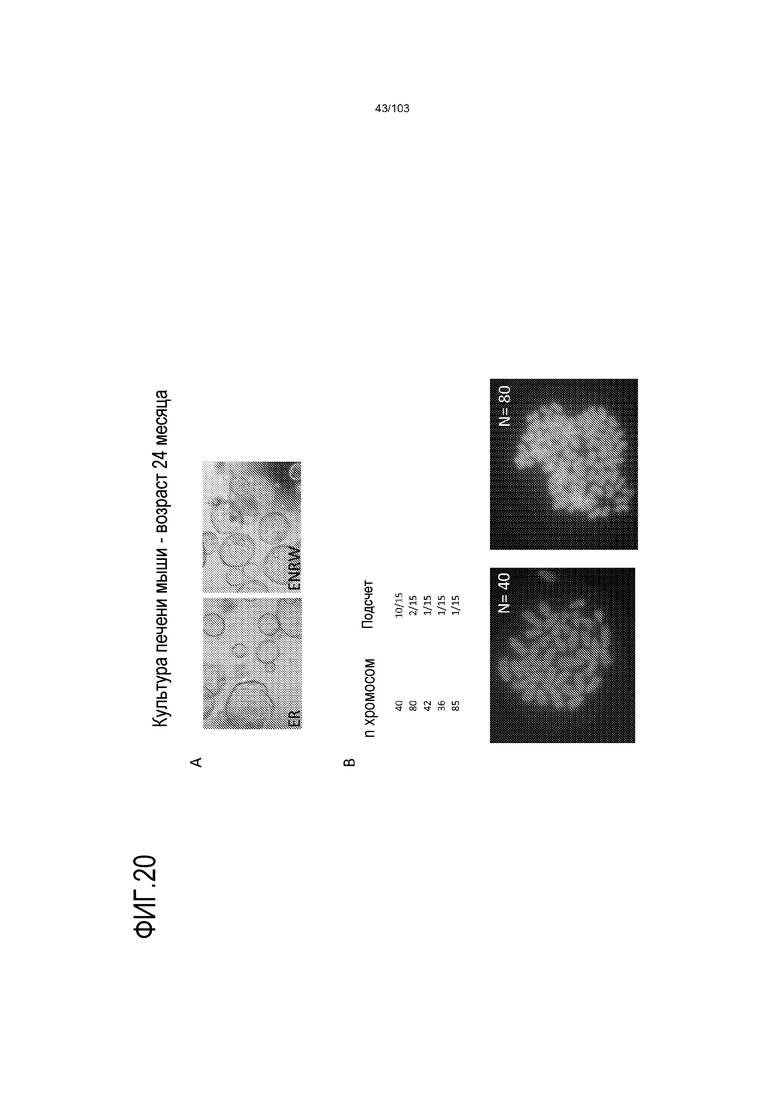


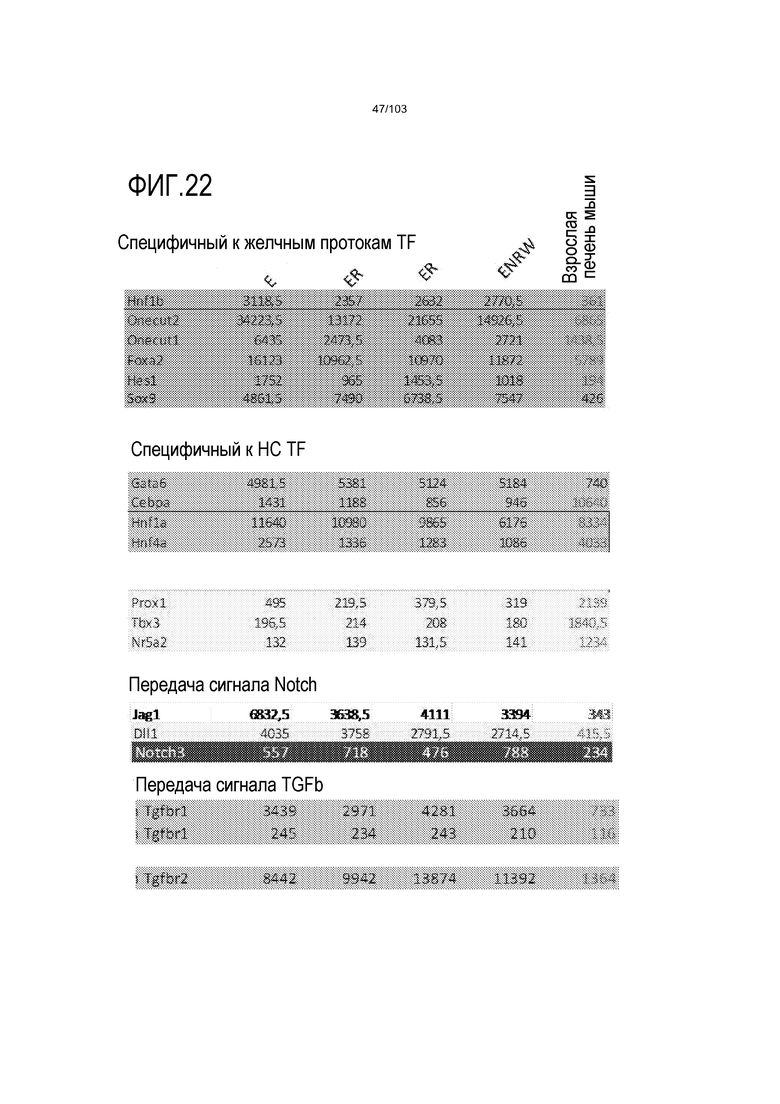

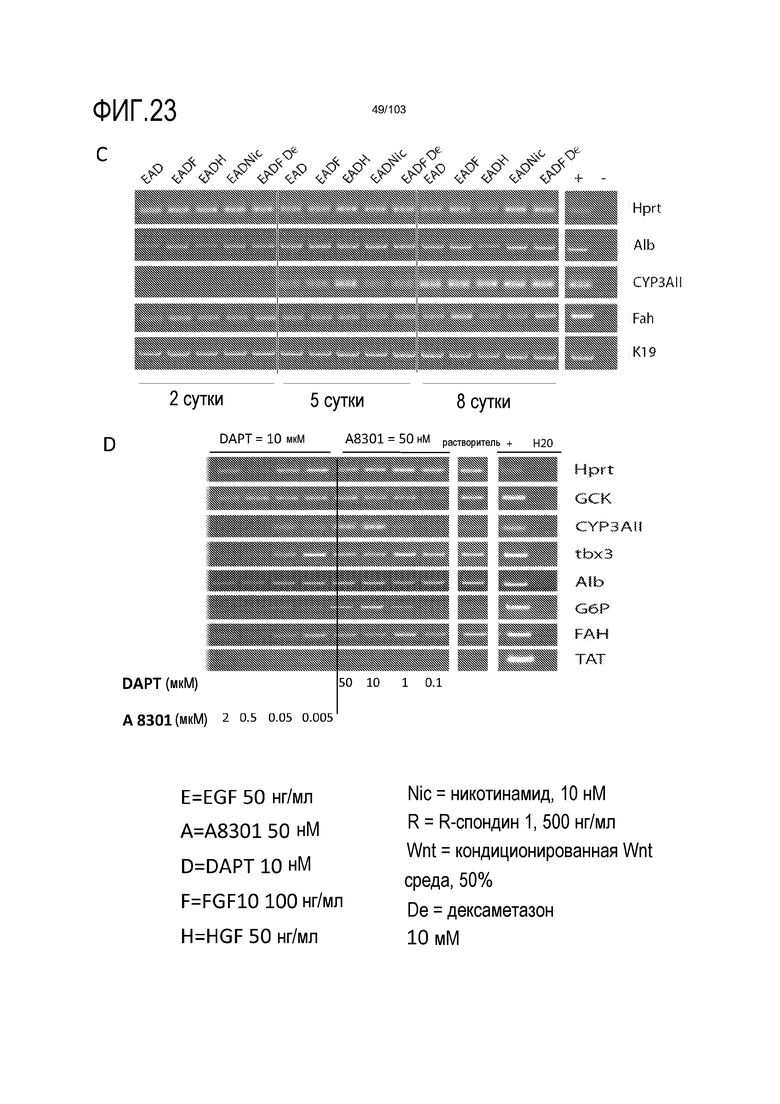
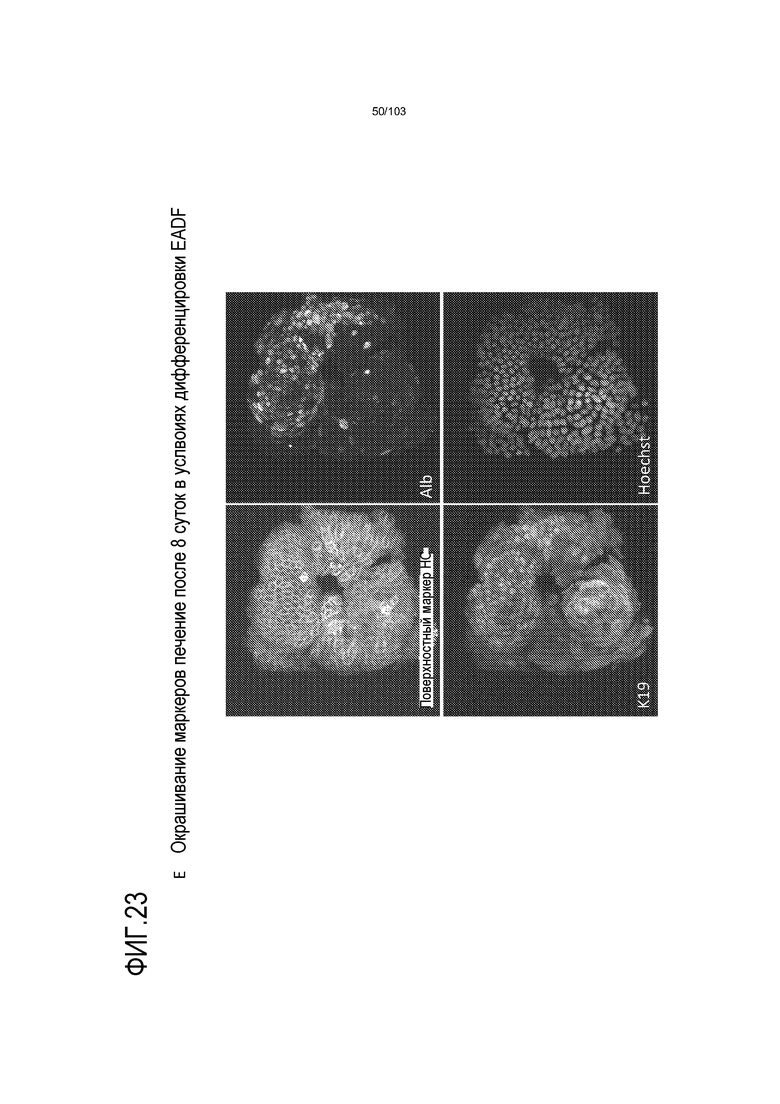
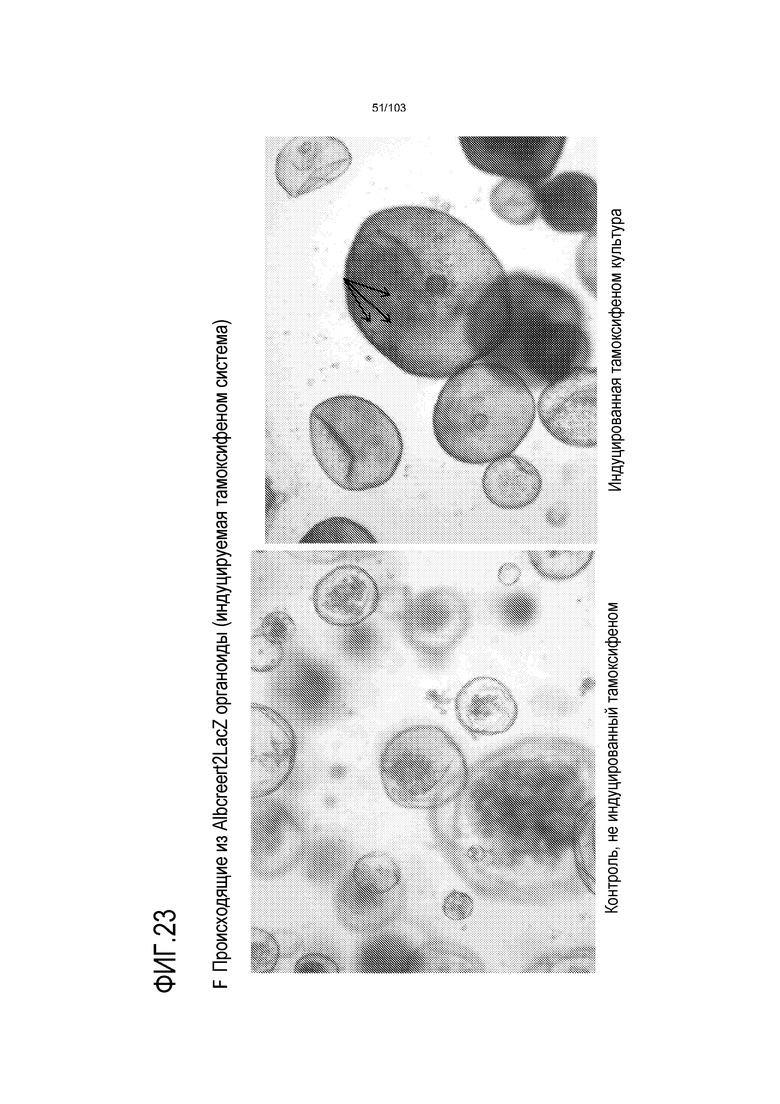
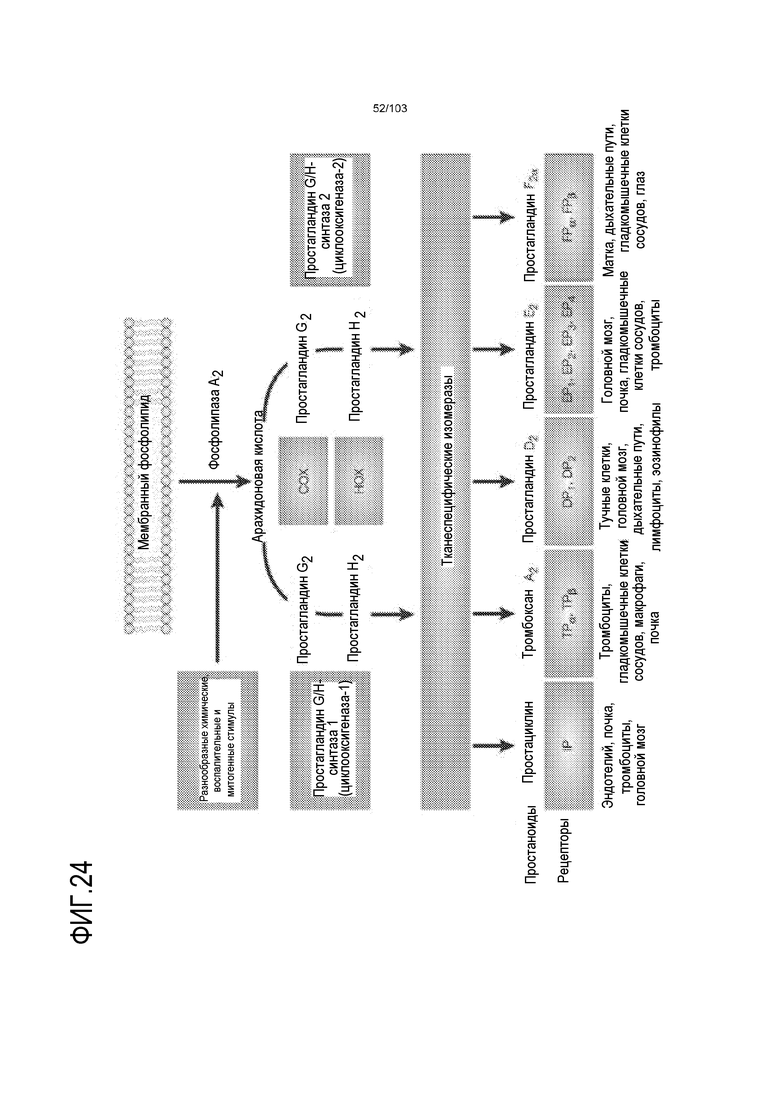
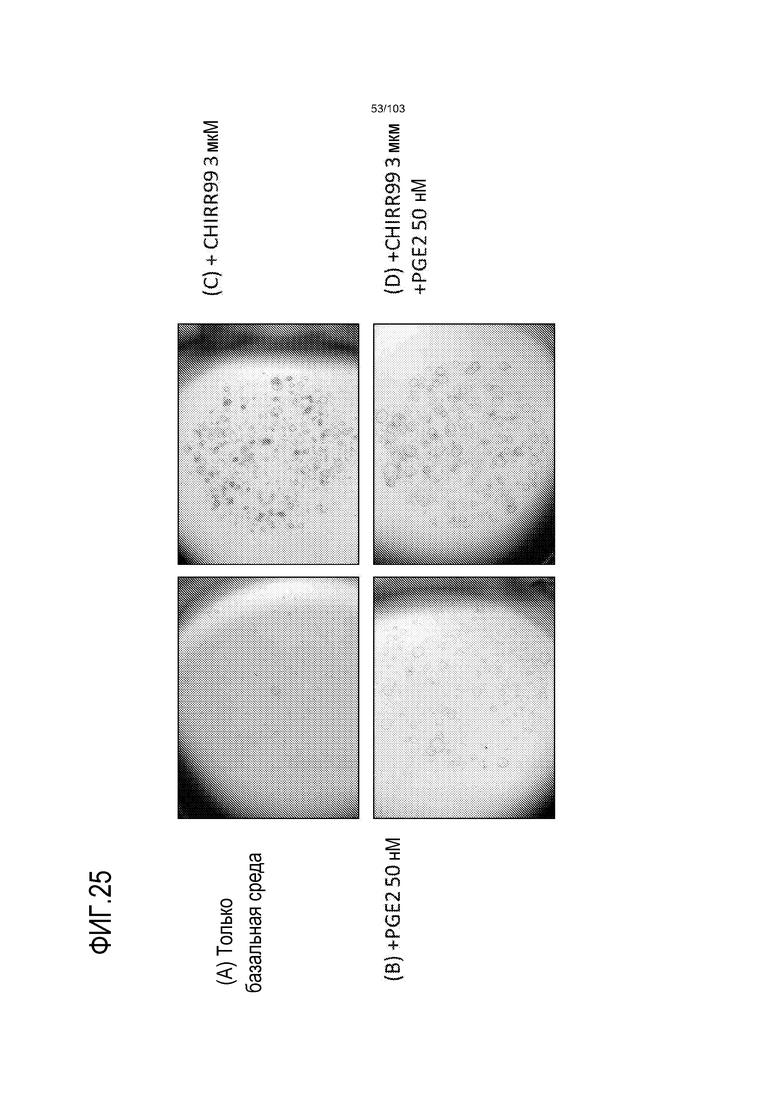
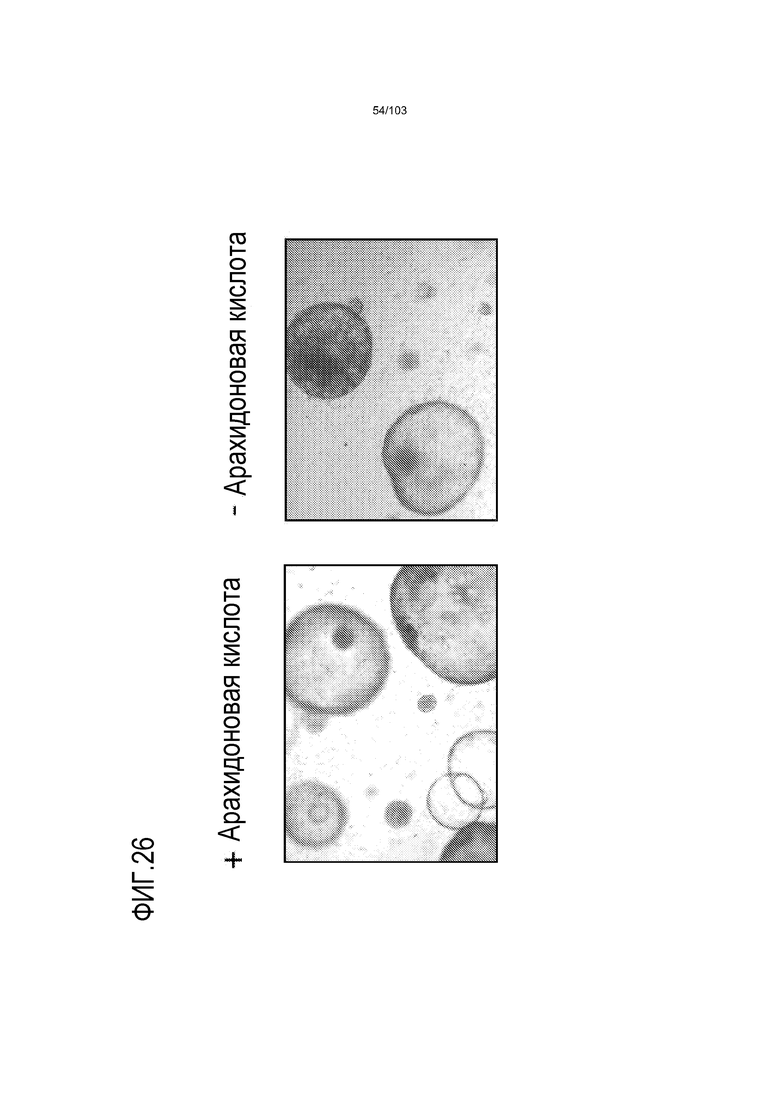







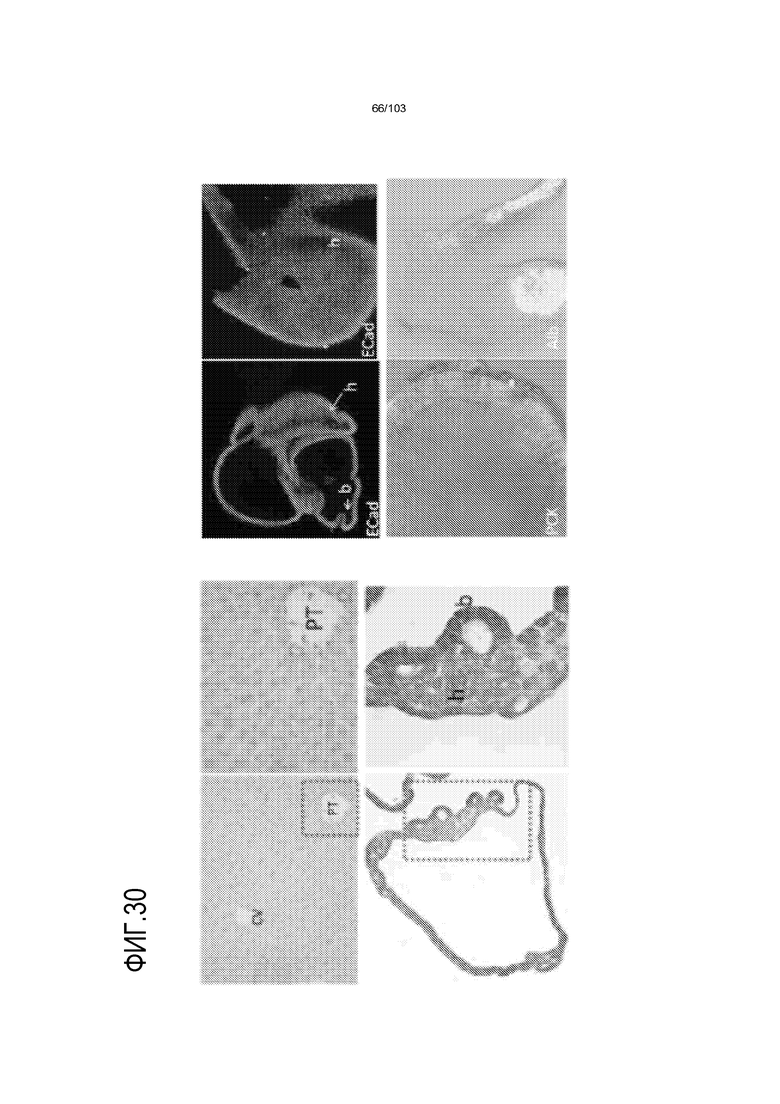
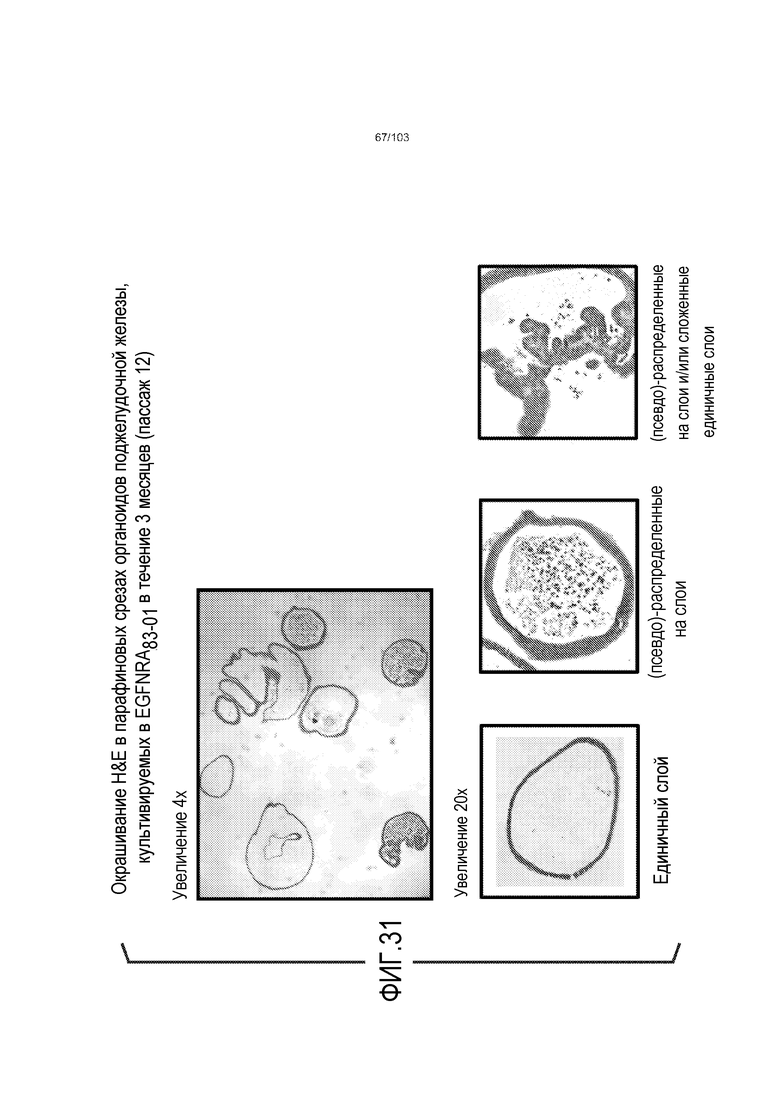

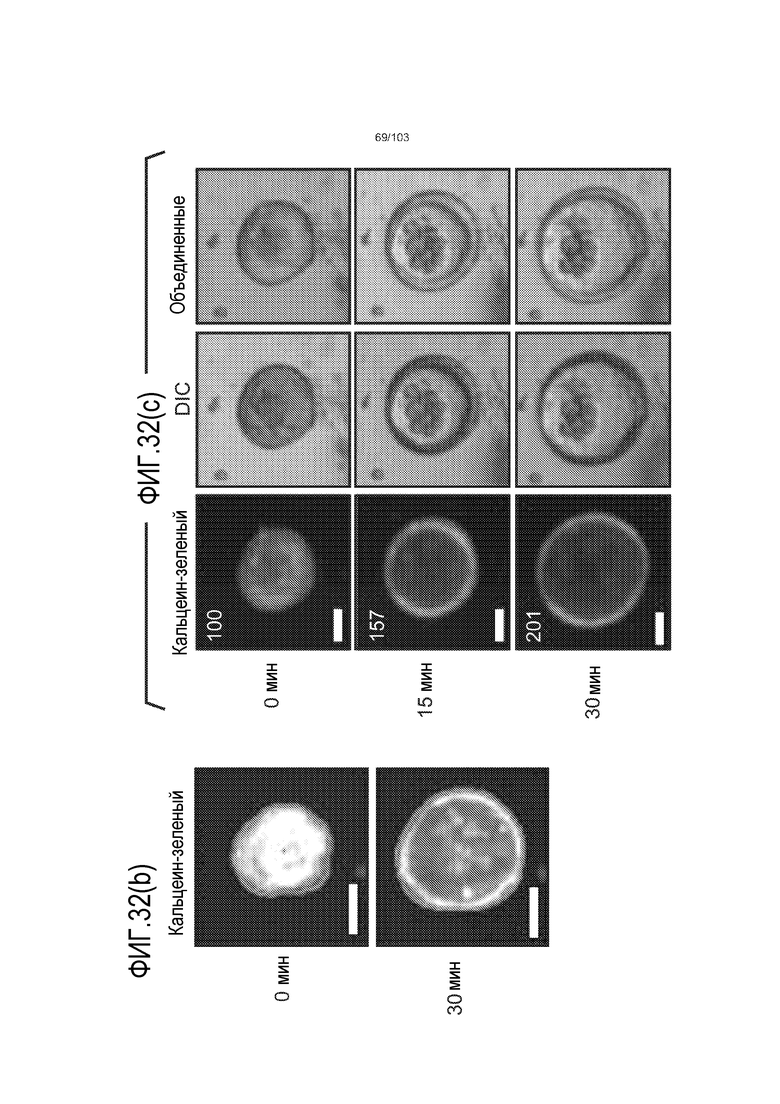
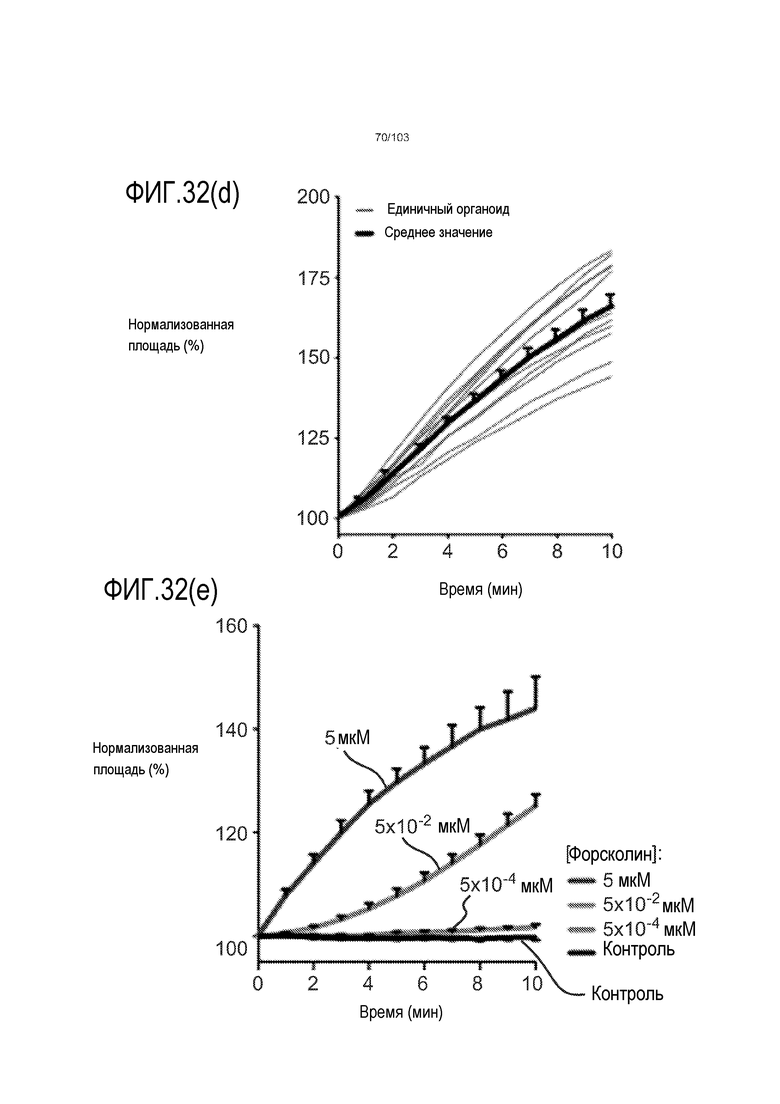
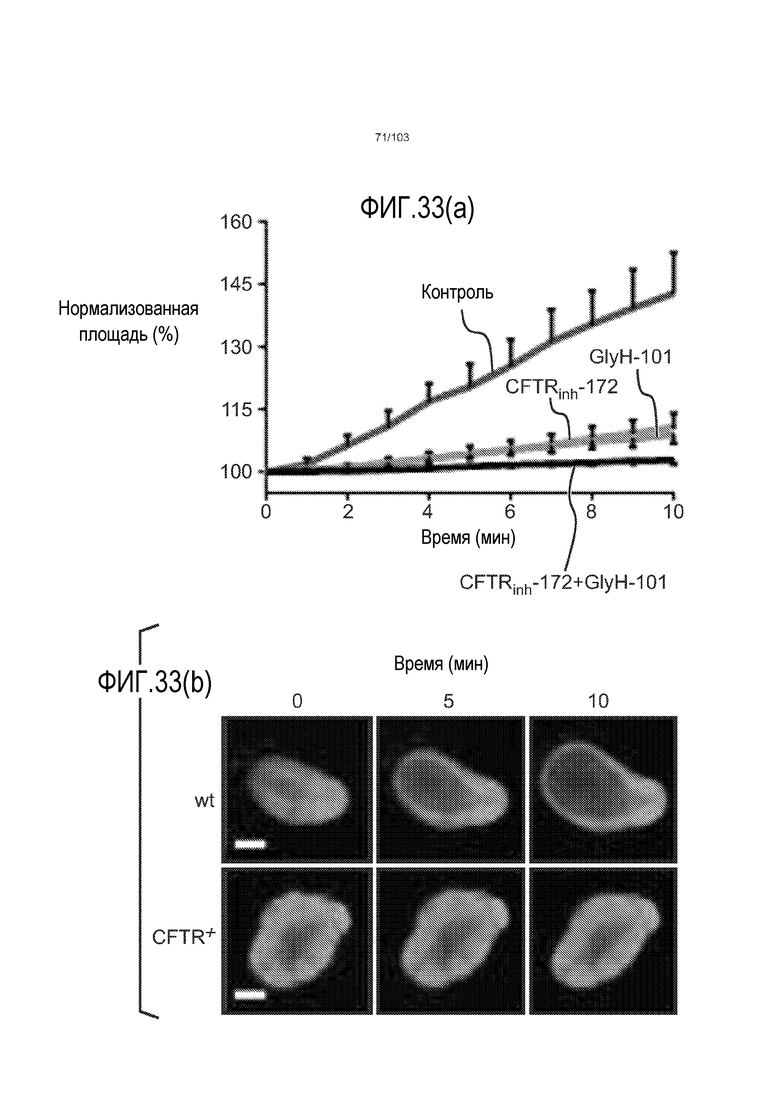
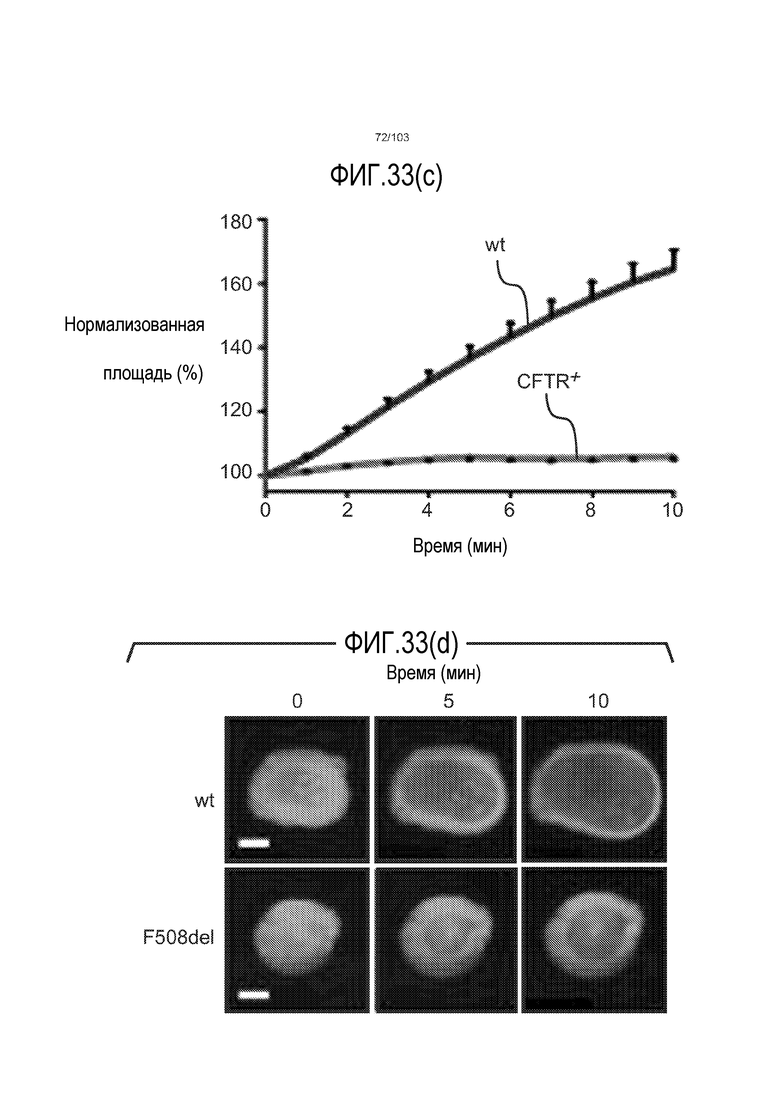
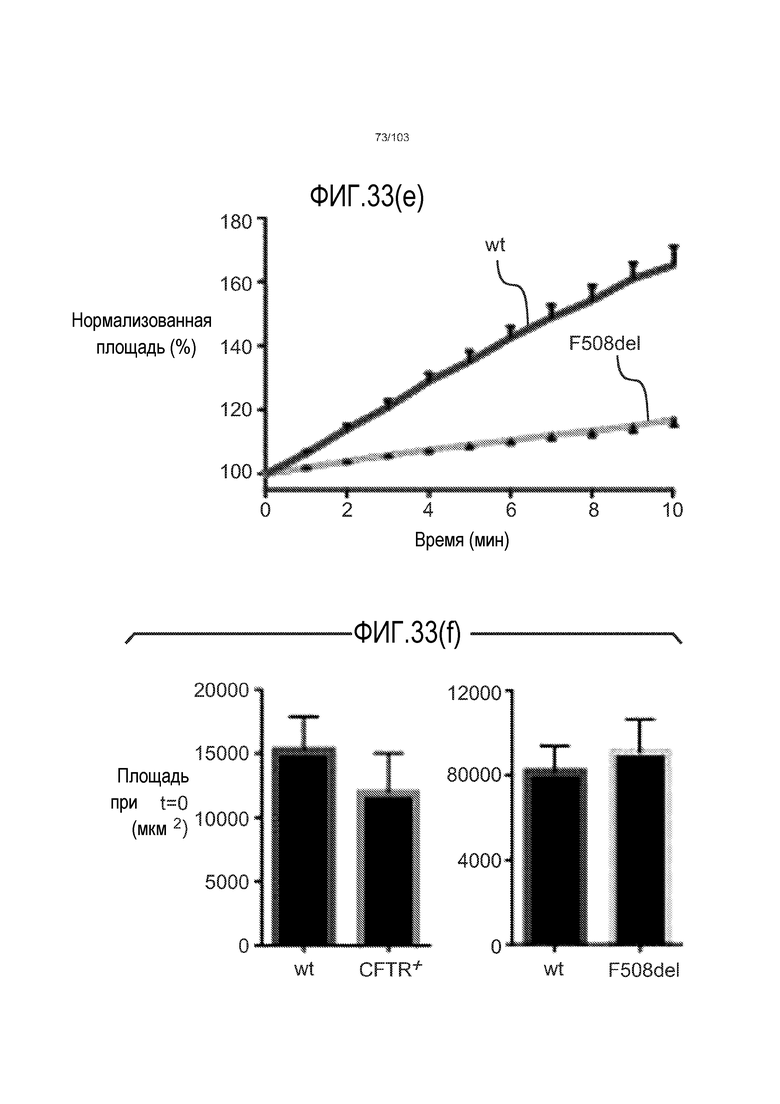
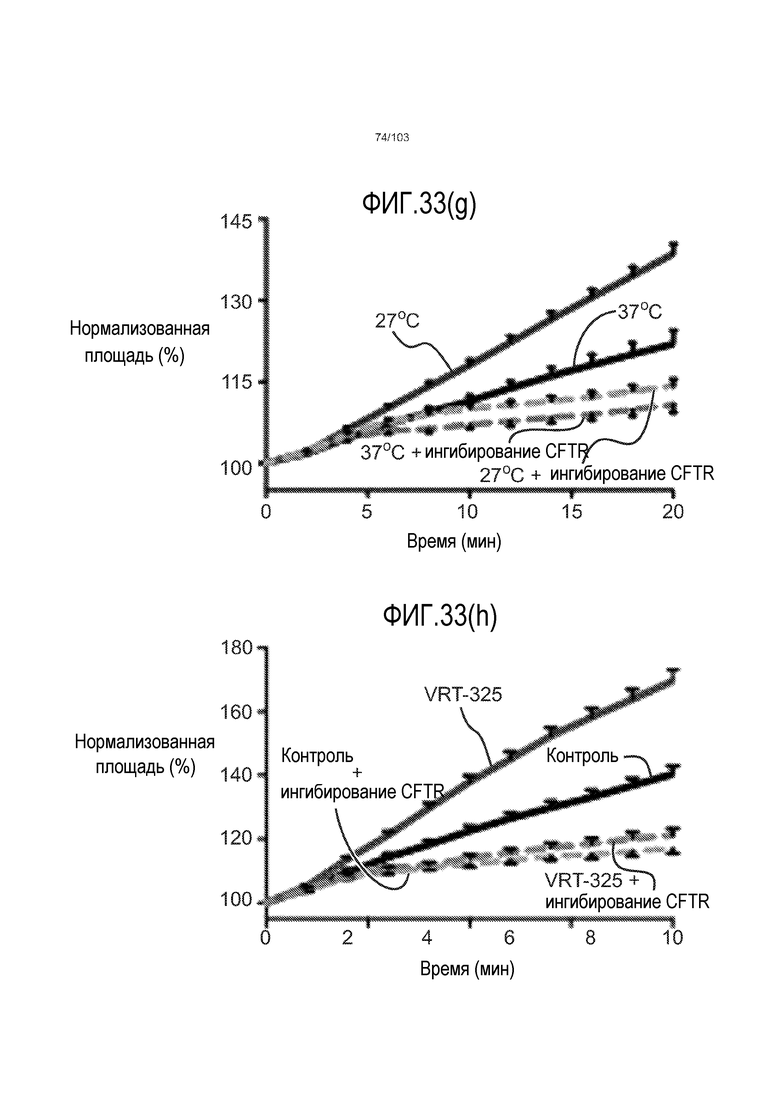
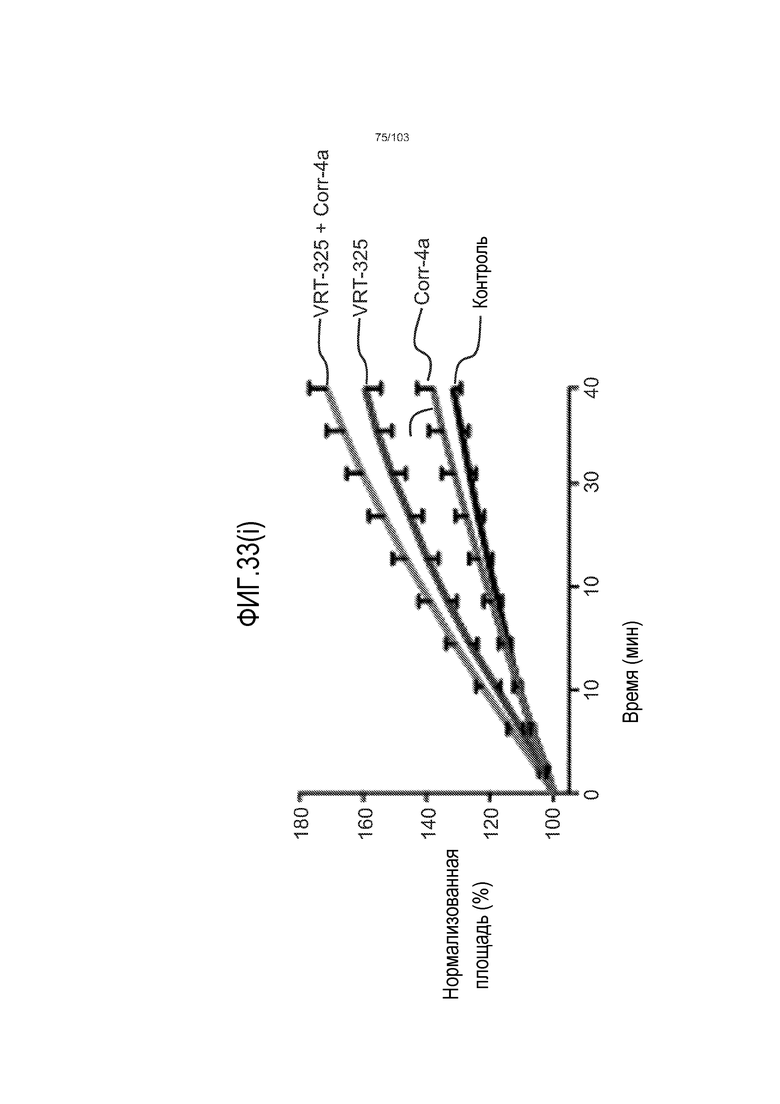
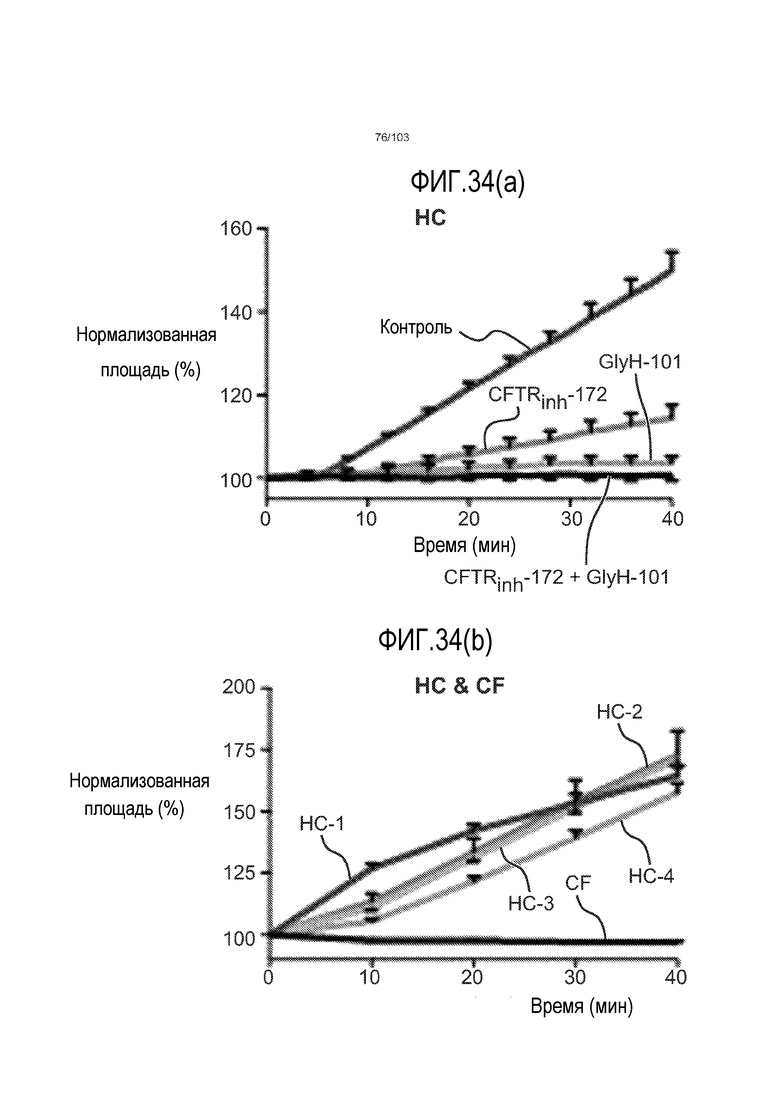
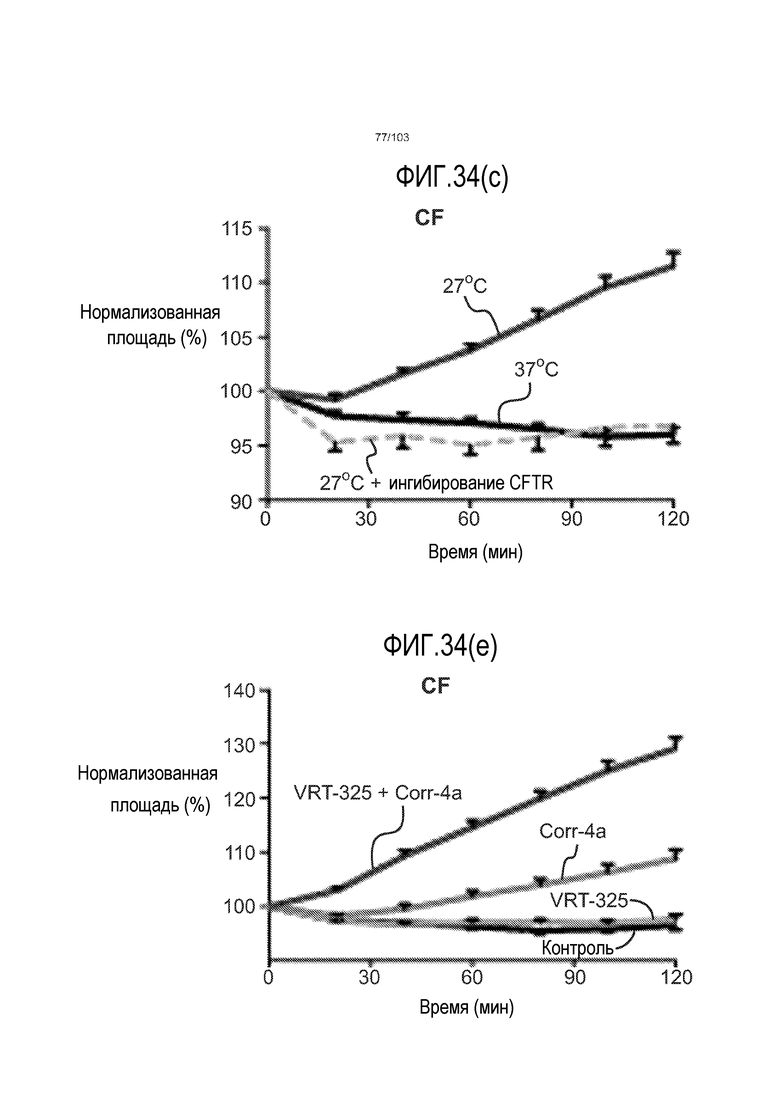

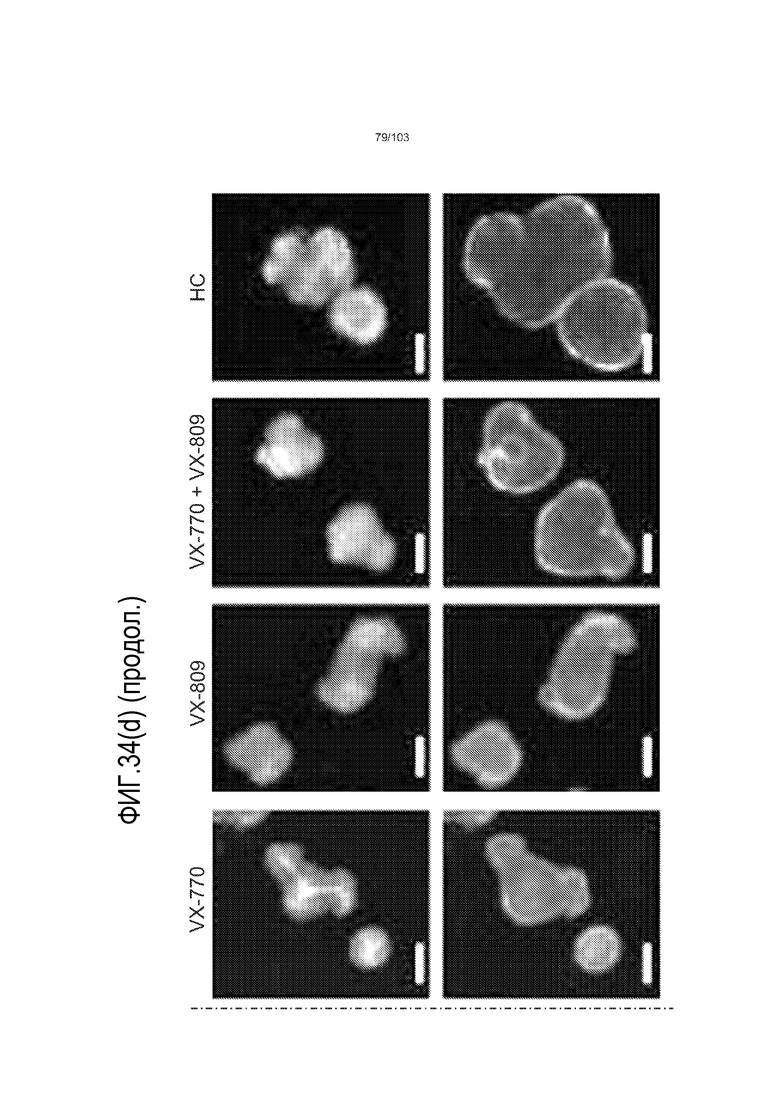

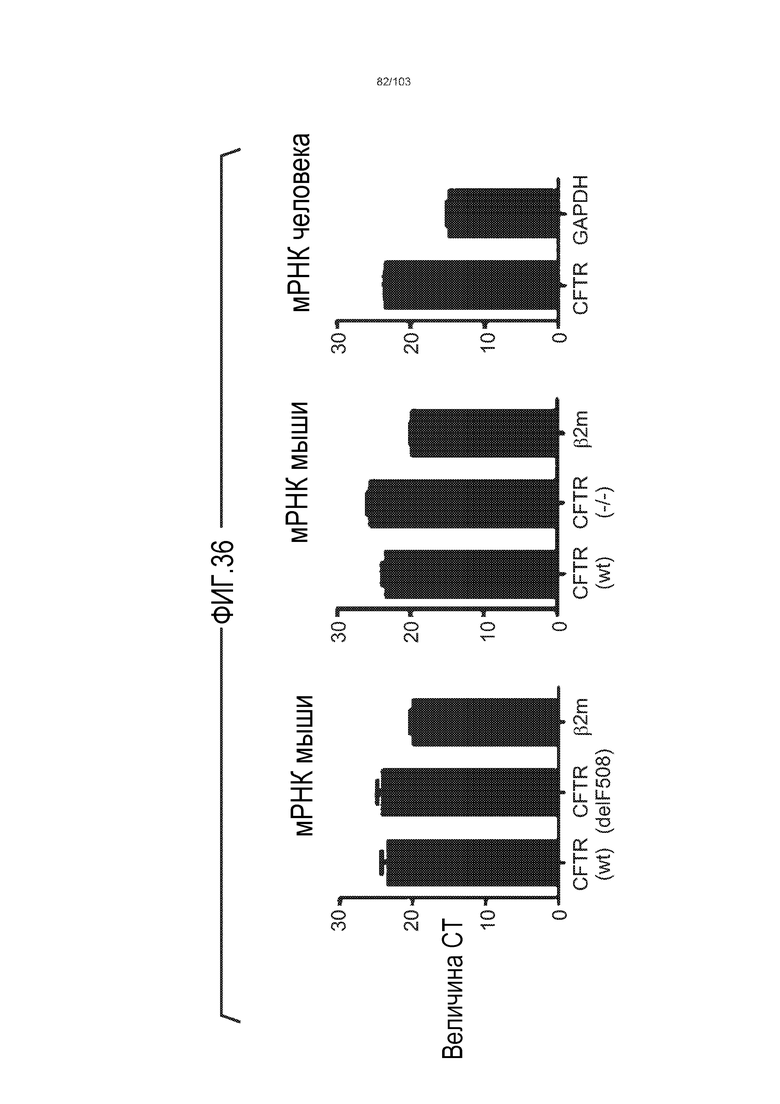
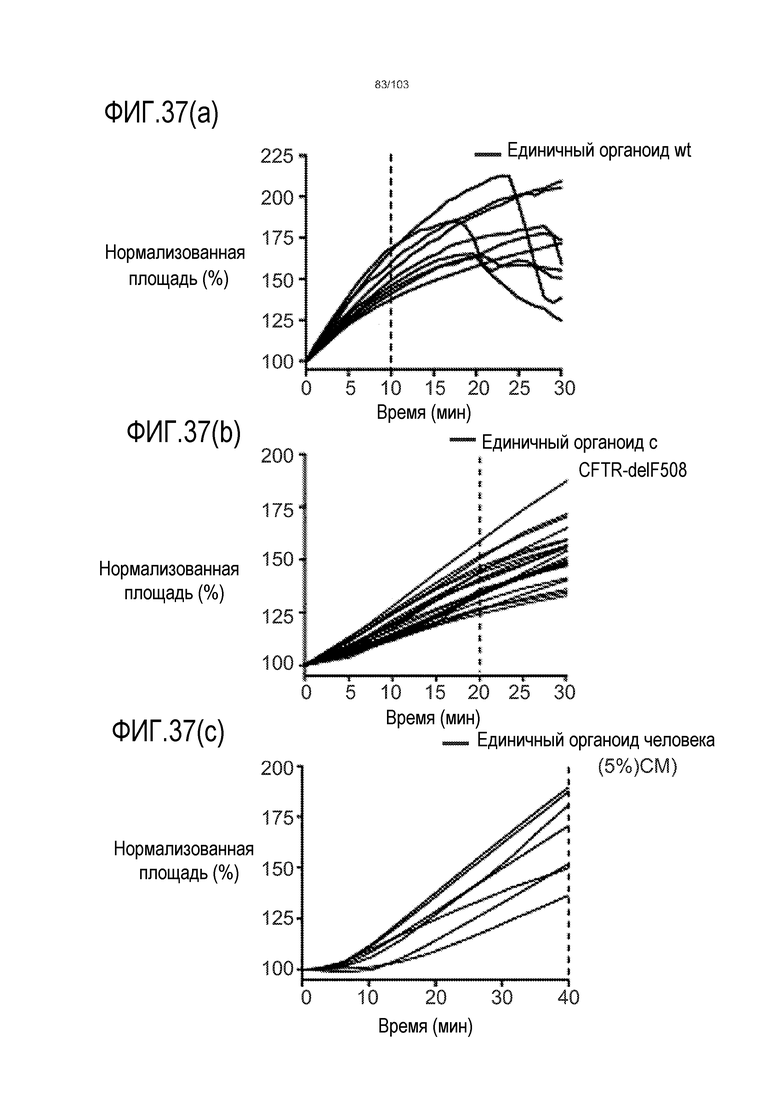
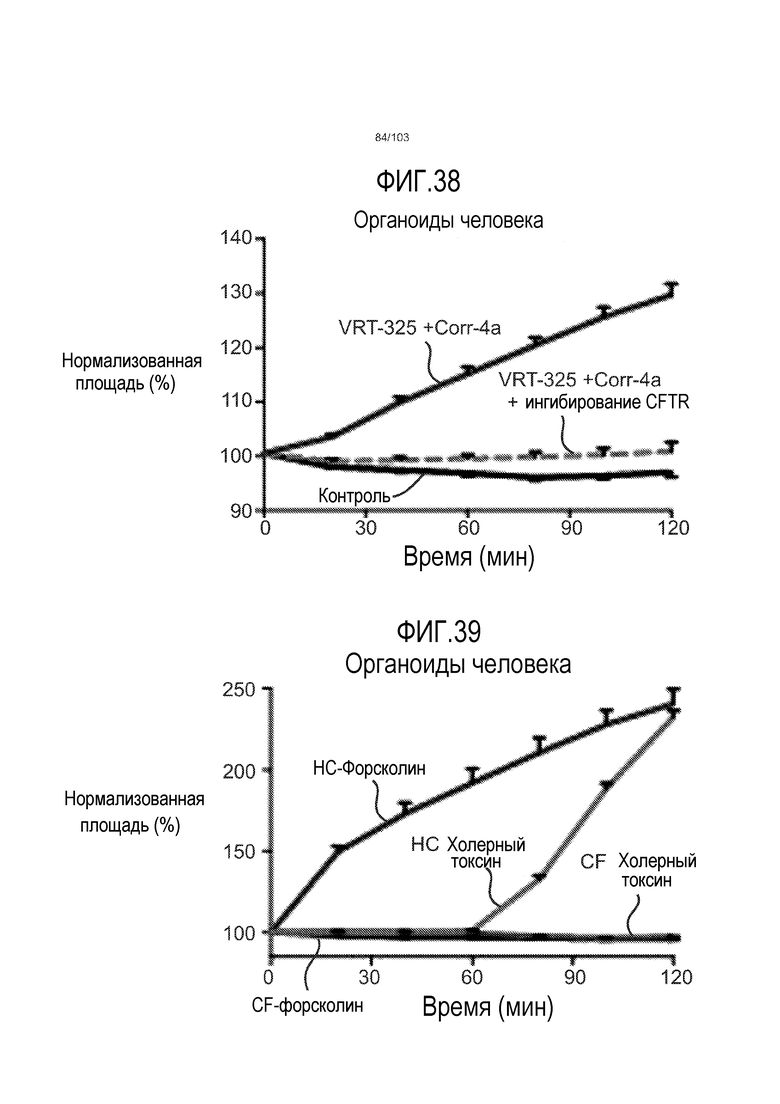

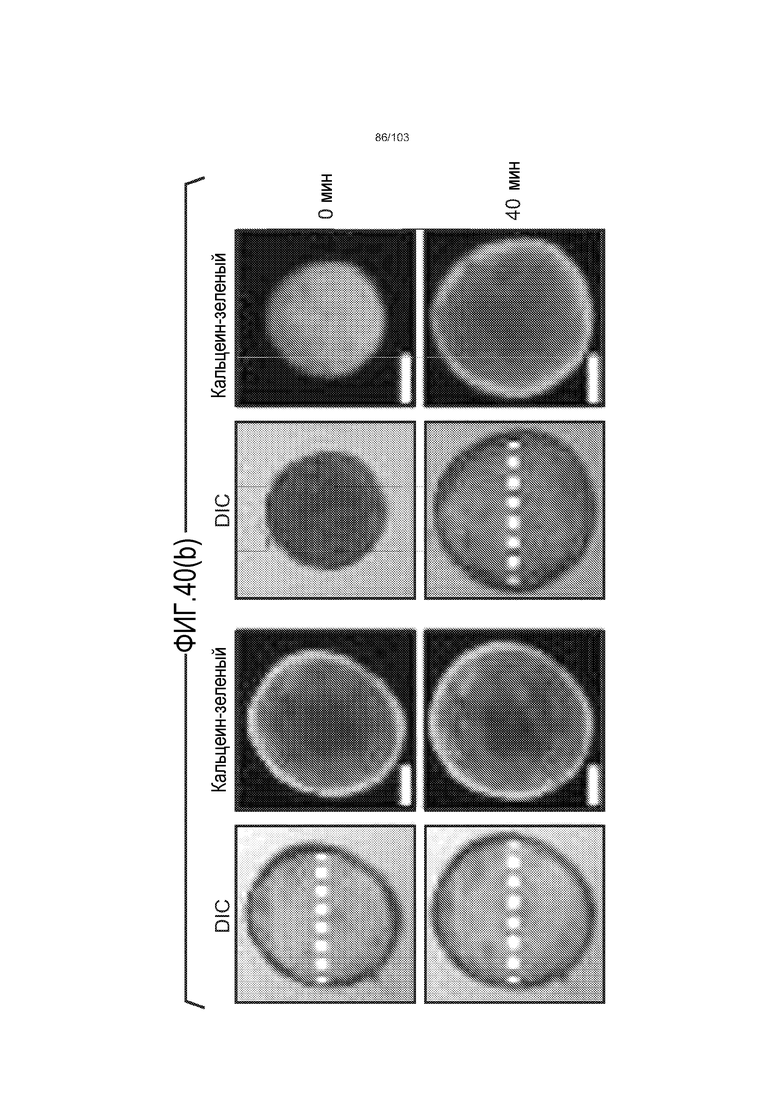
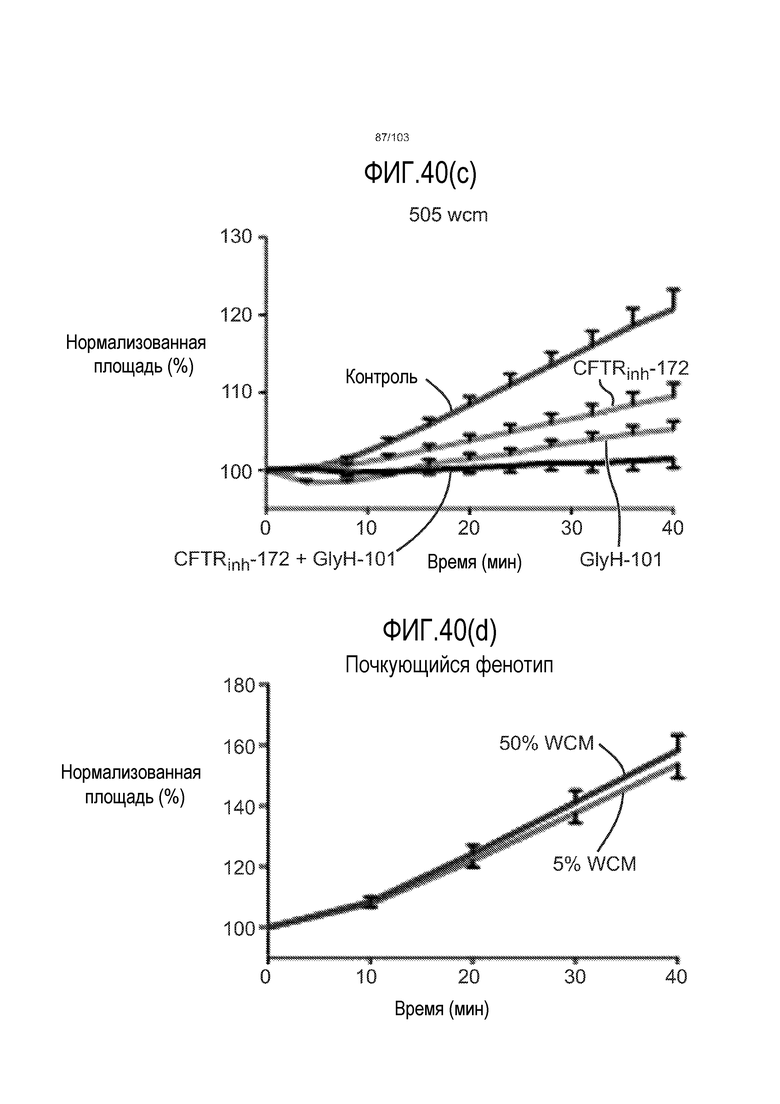

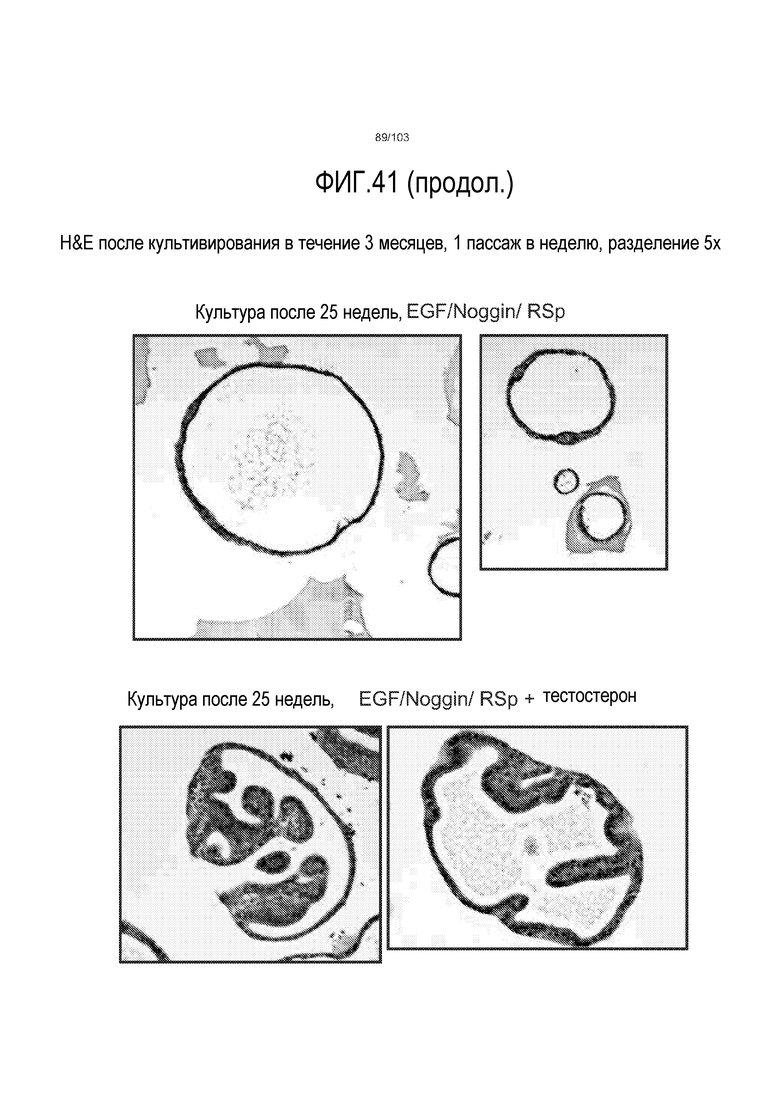
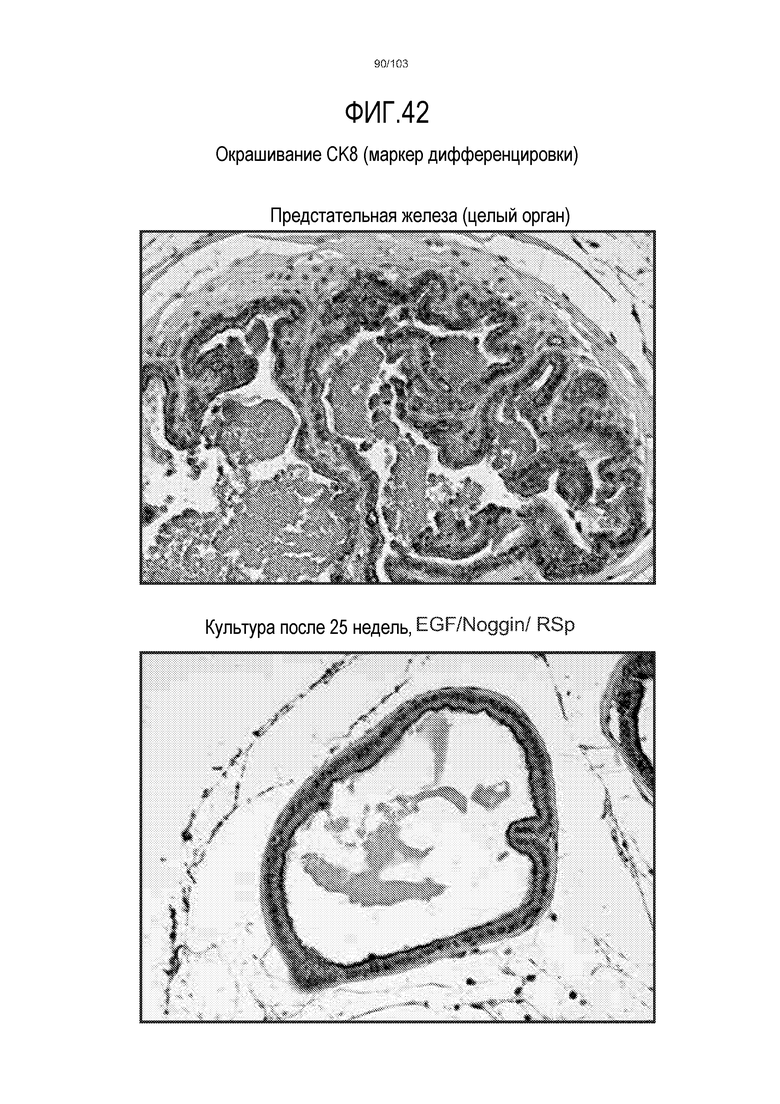
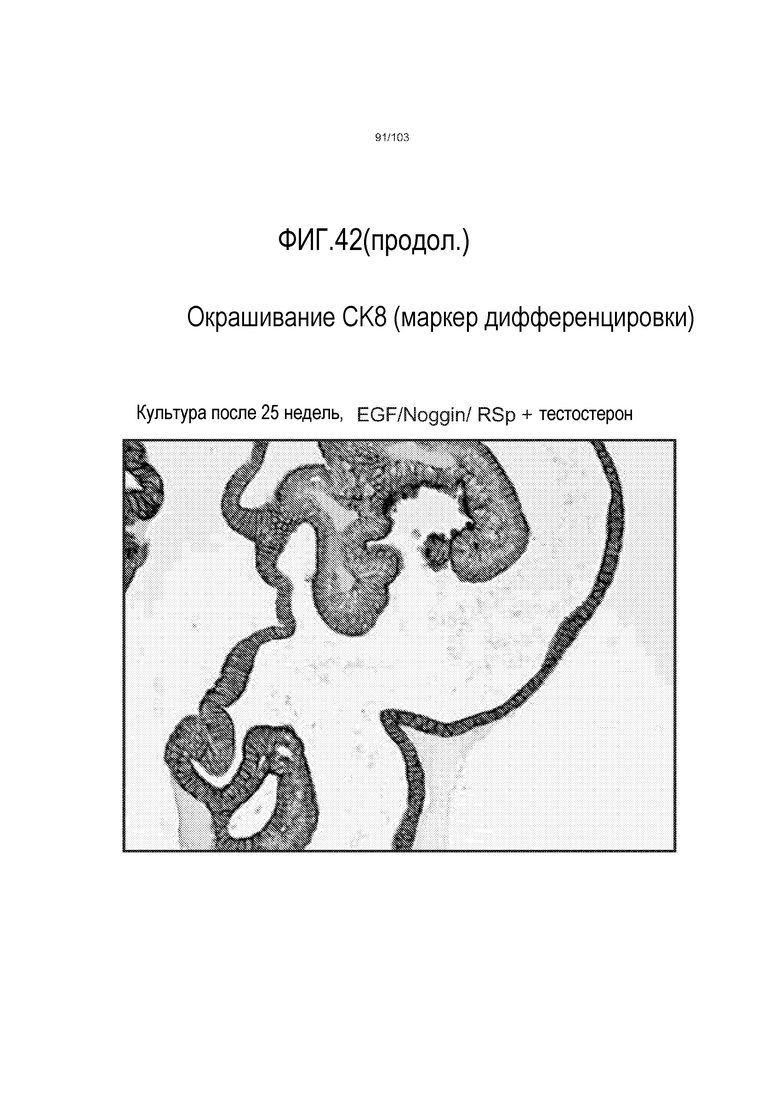
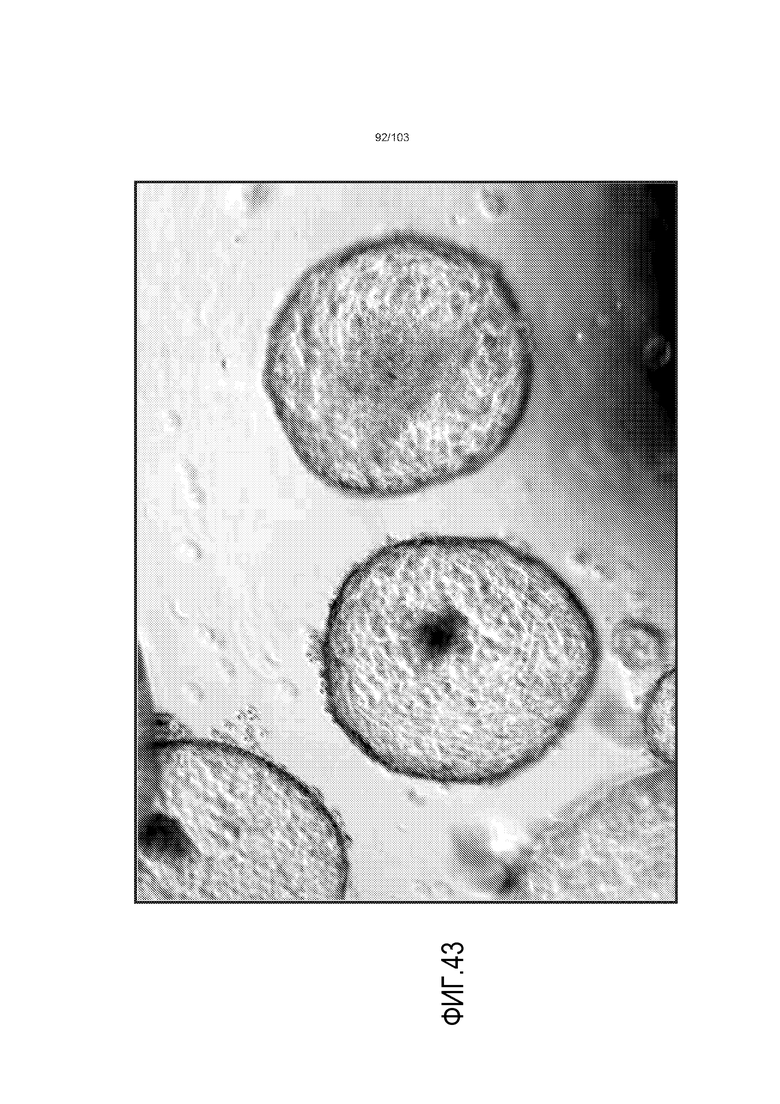
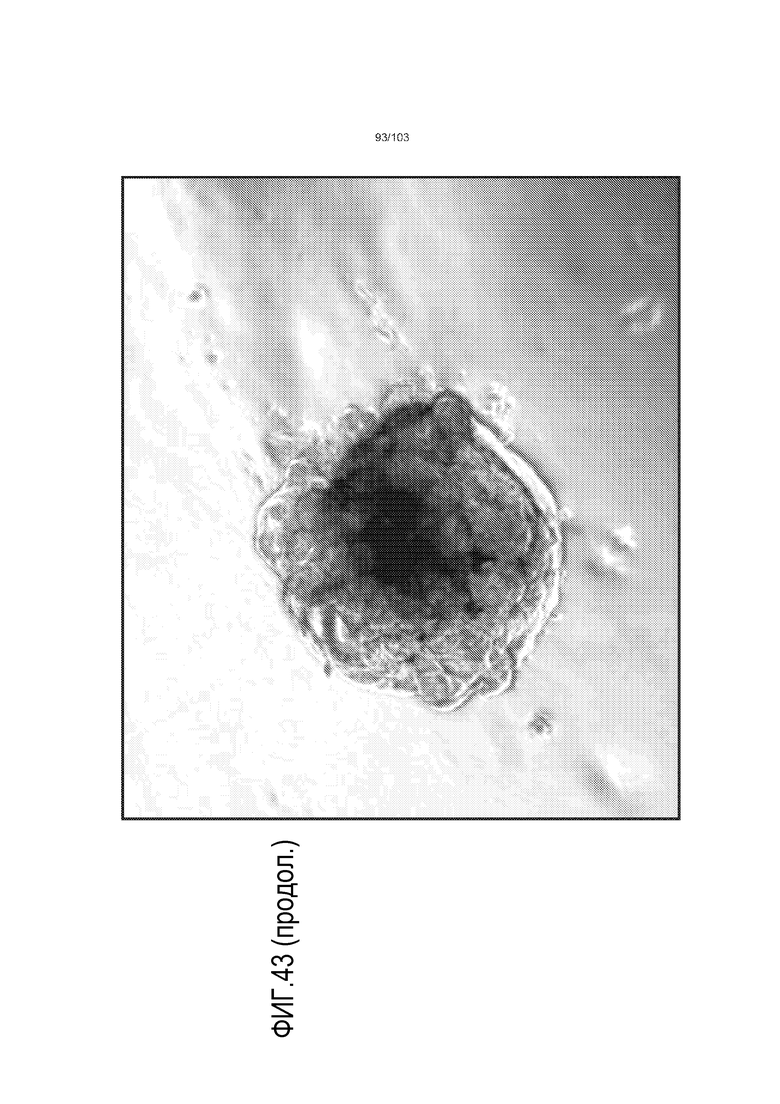
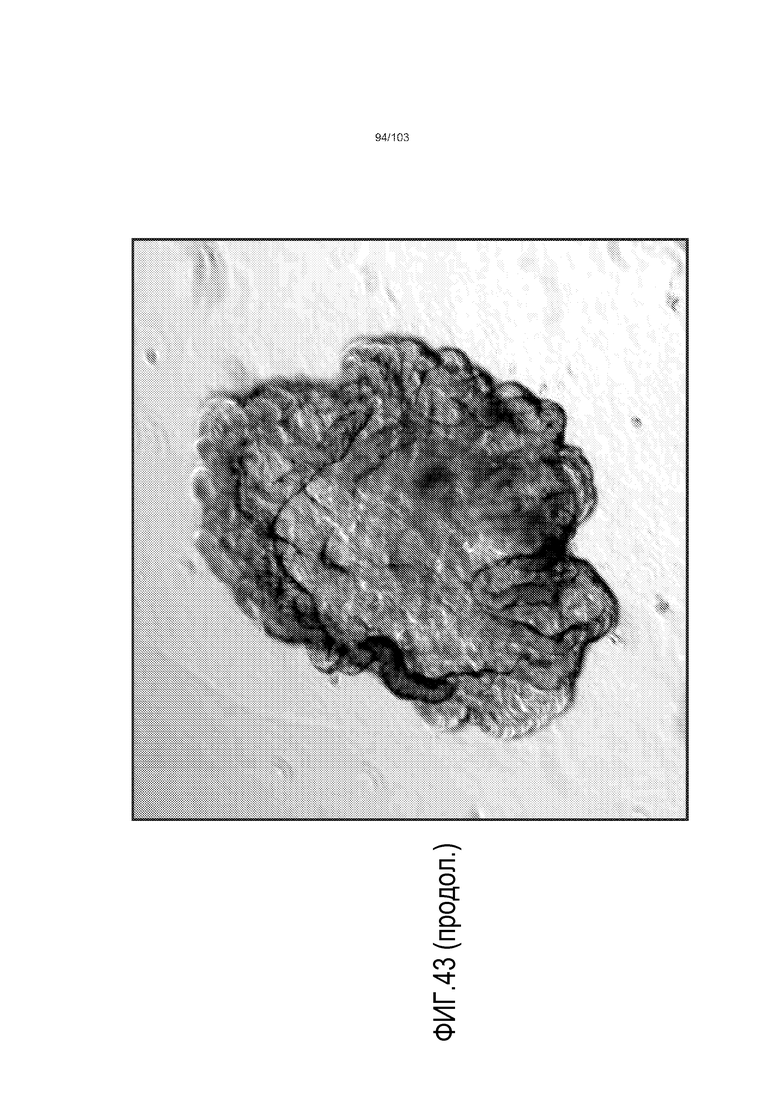


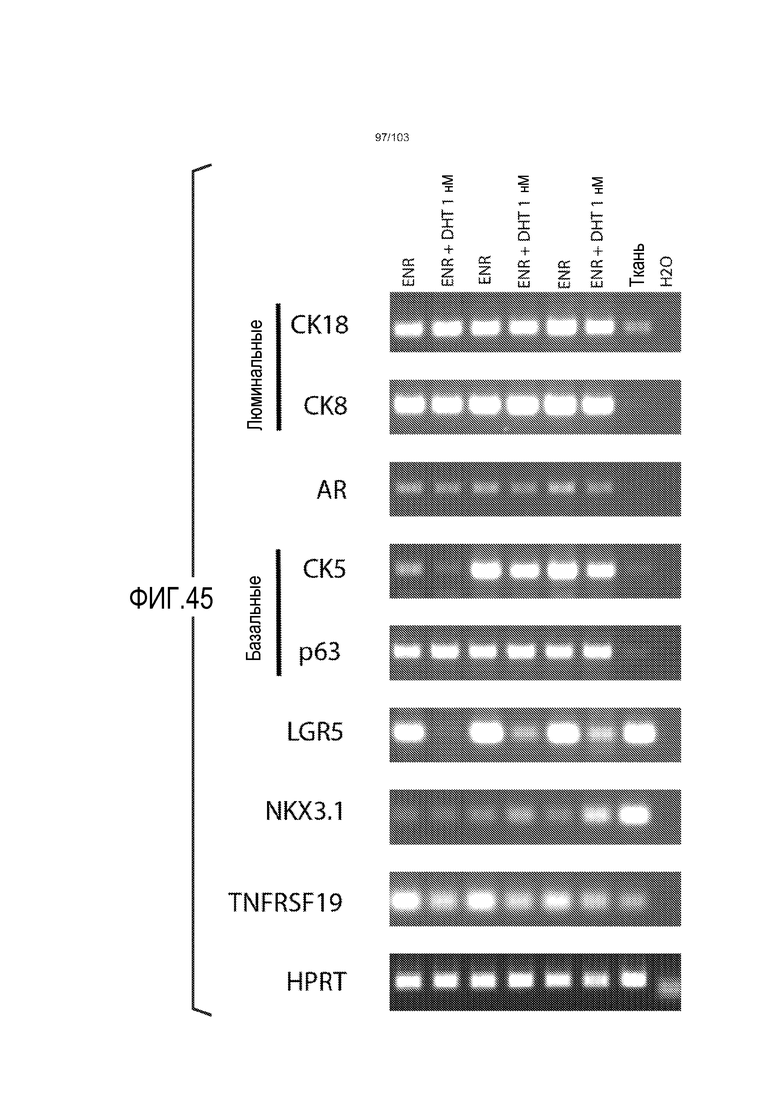
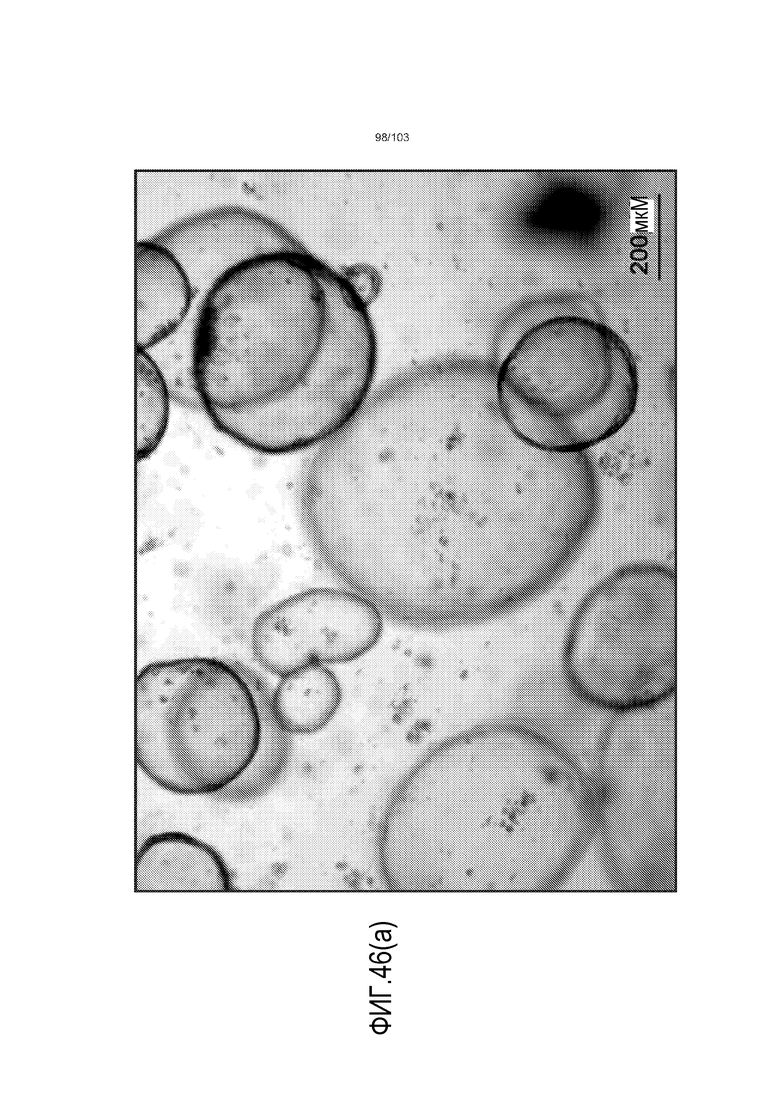
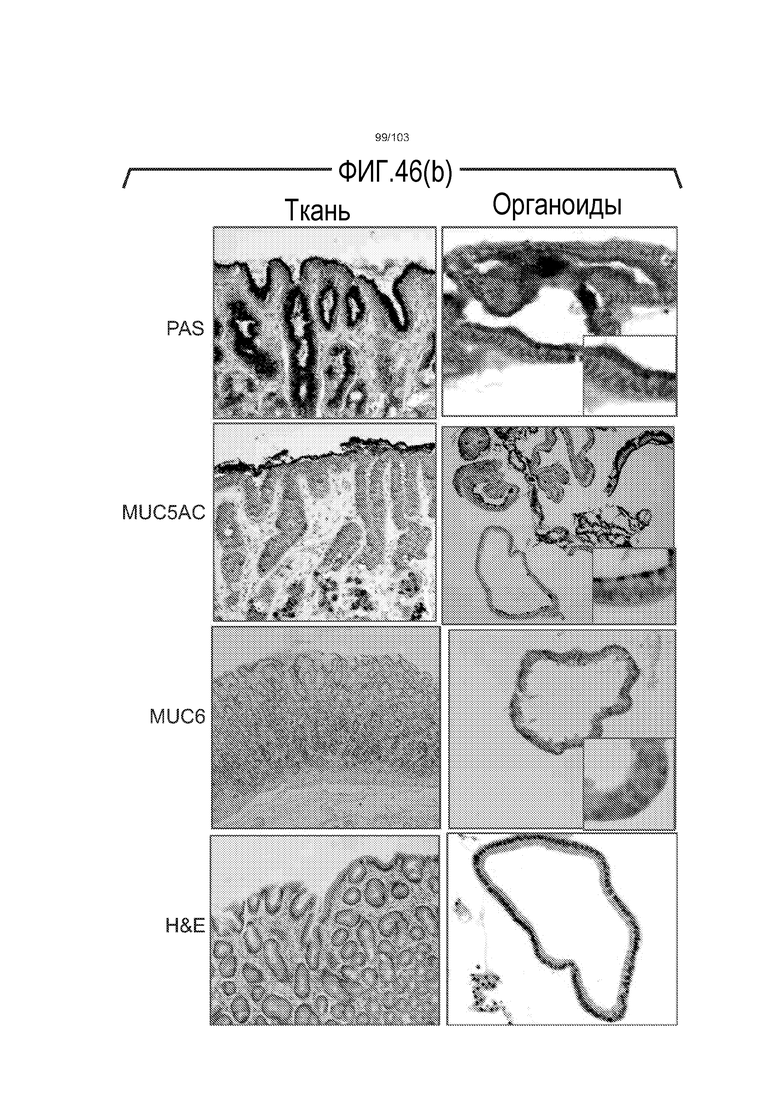

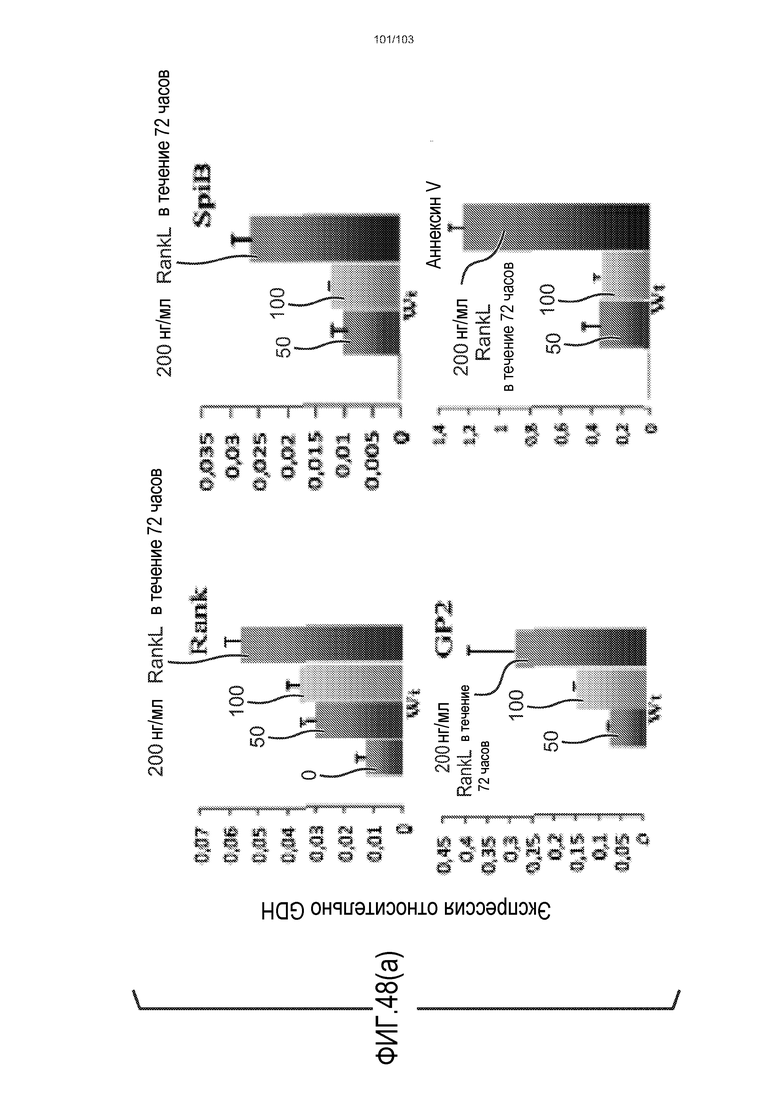
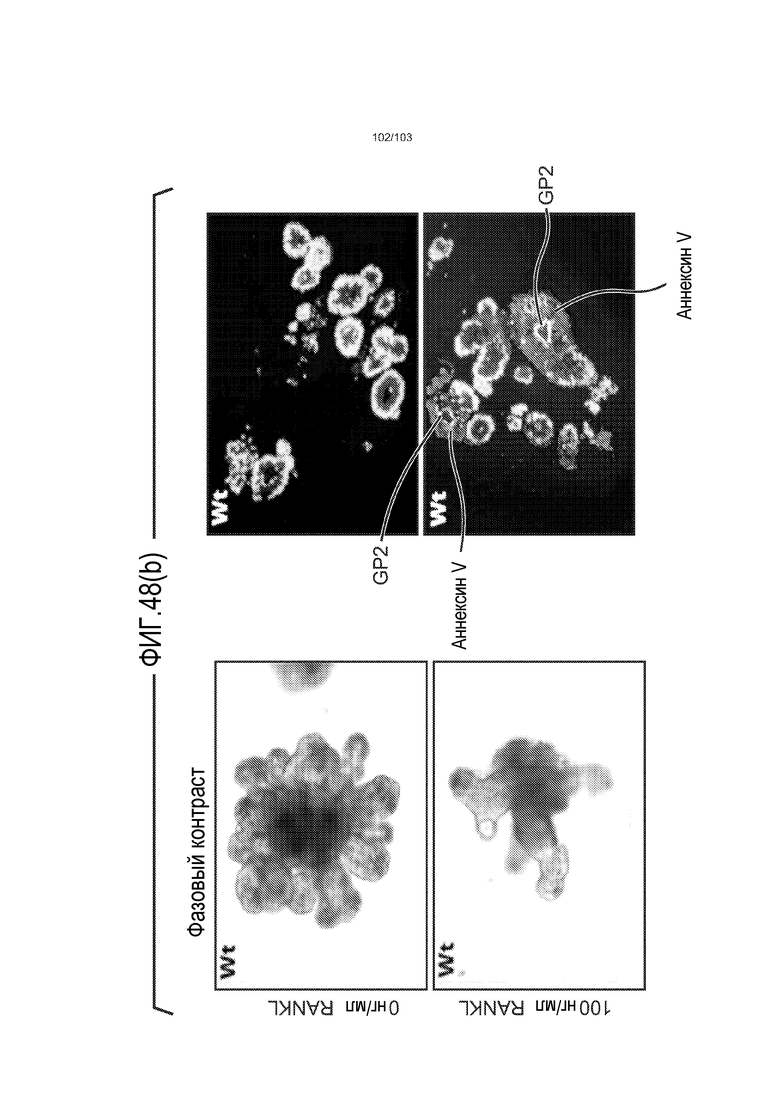
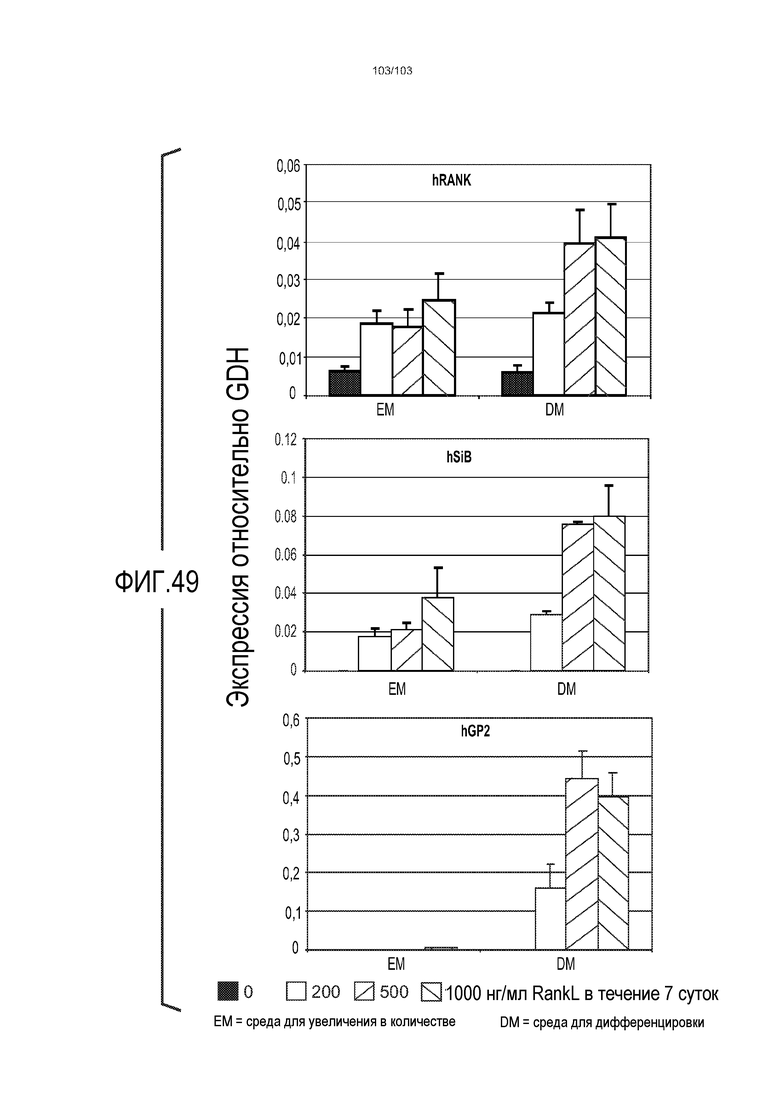
Изобретение относится к области биохимии, в частности к культуральной среде для увеличения в количестве популяции взрослых стволовых клеток, где указанная культуральная среда содержит базальную среду, к которой добавлены агонист Wnt, ингибитор BMP и один или несколько ингибиторов TGF-бета, которые представляют собой ингибитор ALK5, ALK4 и/или ALK7, а также к ее применению для увеличения в количестве стволовой клетки, популяции стволовых клеток или фрагмента ткани или органоида, содержащих стволовую клетку или популяцию стволовых клеток. Также раскрыт способ увеличения в количестве единичной стволовой клетки, популяции стволовых клеток или фрагмента ткани, содержащей стволовую клетку или популяцию стволовых клеток, для получения органоида с использованием вышеуказанной культуральной среды. Изобретение позволяет эффективно увеличивать в количестве популяцию взрослых стволовых клеток. 4 н. и 18 з.п. ф-лы, 49 ил., 4 табл., 8 пр.
1. Культуральная среда для увеличения в количестве популяции взрослых стволовых клеток, где указанная культуральная среда содержит базальную среду, к которой добавлены:
i. агонист Wnt;
ii. ингибитор BMP; и
iii. один или несколько ингибиторов TGF-бета, которые представляют собой ингибитор ALK5, ALK4 и/или ALK7.
2. Культуральная среда по п.1, где один или несколько ингибиторов TGF-бета связываются и снижают активность одной или нескольких серин/треониновых протеинкиназ, выбранных из группы, состоящей из ALK5, ALK4 и ALK7.
3. Культуральная среда по п.1 или 2, где один или несколько TGF-бета ингибиторов, которые прямо или непрямо отрицательно регулируют передачу сигнала TGF-бета, выбраны из группы, состоящей из A83-01, SB-431542, SB-505124, SB-525334, SD-208, LY-36494 и SJN-2511.
4. Культуральная среда по п.1, где ингибитор TGF-бета добавлен в концентрации от 1 нМ до 100 мкМ, от 10 нМ до 100 мкМ, от 100 нМ до 10 мкМ, или приблизительно 1 мкМ, например, где общая концентрация одного или нескольких ингибиторов составляет от 10 нМ до 100 мкМ, от 100 нМ до 10 мкМ, или приблизительно 1 мкМ.
5. Культуральная среда по п.1, где агонист Wnt является агонистом Lgr5 или любым из R-спондина 1-4.
6. Культуральная среда по п.1, где культуральная среда содержит один или несколько дополнительных компонентов, выбранных из лиганда рецепторной тирозинкиназы, никотинамида, ингибитора p38, ингибитора Rock, гастрина, активатора каскада передачи сигнала простагландинов и тестостерона.
7. Культуральная среда по п.6, где культуральная среда предназначена для культивирования стволовых клеток кишечника человека, крипт тонкого кишечника человека или крипт толстого кишечника человека и где культуральная среда содержит базальную среду и дополнительно содержит ингибитор BMP, R-спондин, ингибитор TGF-бета, агонист Wnt, EGF, ингибитор p38, гастрин и никотинамид.
8. Культуральная среда по п.6, где культуральная среда предназначена для культивирования стволовых клеток поджелудочной железы и где культуральная среда содержит базальную среду, любой из R-спондин 1-4, Noggin, ингибитор TGF-бета, EGF, FGF10, гастрин и предпочтительно экстендин 4 и Wnt-3a.
9. Культуральная среда по п.6, где культуральная среда предназначена для культивирования клеток предстательной железы и где культуральная среда содержит базальную среду, любой из R-спондин 1-4, Noggin, ингибитор TGF-бета, никотинамид, EGF, тестостерон и предпочтительно Wnt-3a и FGF10.
10. Культуральная среда по п.6, где культуральная среда предназначена для культивирования желудочных клеток и где культуральная среда содержит базальную среду, Noggin, R-спондин 1-4, ингибитор TGF-бета, Wnt-3a, EGF, гастрин, никотинамид, FGF10 и предпочтительно ингибитор p38.
11. Культуральная среда по п.6, где культуральная среда предназначена для культивирования злокачественных клеток и где из среды исключен один или несколько следующих компонентов: Wnt-3a, EGF, Noggin, R-спондин, ингибитор p38, никотинамид, гастрин, FGF10 и HGF.
12. Культуральная среда по любому из пп.1-11, где взрослые стволовые клетки представляют собой эпителиальные клетки.
13. Композиция для увеличения популяции взрослых стволовых клеток, где указанная композиция содержит культуральную среду по любому из пп.1-11 и внеклеточный матрикс или 3D-матрикс, который имитирует внеклеточный матрикс путем его взаимодействия с белками клеточной мембраны, такими как интегрины, например, содержащий ламинин внеклеточный матрикс, такой как MatrigelTM (BD Biosciences).
14. Культуральная среда по любому из пп.1-11, где культуральная среда содержится в герметично закрытой емкости.
15. Композиция по п.13, где композиция содержится в герметично закрытой емкости.
16. Применение культуральной среды по любому из пп.1-11 для увеличения в количестве стволовой клетки, популяции стволовых клеток или фрагмента ткани или органоида, содержащих стволовую клетку или популяцию стволовых клеток.
17. Способ увеличения в количестве единичной стволовой клетки, популяции стволовых клеток или фрагмента ткани, содержащей стволовую клетку или популяцию стволовых клеток, для получения органоида, где способ включает культивирование единичной стволовой клетки, популяции стволовых клеток или фрагмента ткани, содержащей стволовую клетку или популяцию стволовых клеток в культуральной среде по любому из пп.1-11.
18. Способ по п.17, где способ включает:
обеспечение наличия стволовой клетки, популяции стволовых клеток или выделенного фрагмента ткани;
обеспечение наличия культуральной среды по любому из пп.1-11;
приведение стволовых клеток в контакт с культуральной средой; и
культивирование клеток в подходящих условиях.
19. Способ по п.17, где способ включает приведение стволовой клетки, популяции стволовых клеток или выделенного фрагмента ткани и культуральной среды в контакт с внеклеточным матриксом или 3D-матриксом, который имитирует внеклеточный матрикс путем его взаимодействия с белками клеточной мембраны, такими как интегрины, например, содержащим ламинин внеклеточным матриксом, таким как MatrigelTM (BD Biosciences).
20. Способ по любому из пп.17-19, где способ включает:
культивирование стволовой клетки, популяции стволовых клеток или фрагментов ткани в первой среде для увеличения в количестве в соответствии с любым из пп.1-11;
продолжение культивирования стволовой клетки, популяции стволовых клеток или фрагментов ткани и восполнение среды средой для дифференцировки, где среда для дифференцировки не содержит одного или нескольких, предпочтительно всех, из факторов, выбранных из: ингибитора TGF-бета, ингибитора p38, никотинамида и Wnt.
21. Способ по любому из пп.17-19, где клетки представляют собой эпителиальные клетки.
22. Способ по любому из пп.17-19, где органоид представляет собой человеческий органоид.
| WO2010090513 A2, 12.08.2010 | |||
| TOSHIRO SATO ET AL, Long-term expansion of epithelial organoids from human colon, adenoma, adenocarcinoma and and Barrett`s epithelium, Gastroenterology, Elsevier, Philadelphia, vol.141, no.5, 27.07.2011 | |||
| WO2012014976 A2, 02.02.2012 | |||
| SAIYONG ZHU ET AL, Chemical strategies for stem cell biology and regenerative |

