РОДСТВЕННЫЕ ЗАЯВКИ
Данная заявка испрашивает приоритет по заявке США 61/419,635, поданной 3 декабря 2010 и 61/510,620, поданной 22 июля 2011. Содержание данных заявок включено в настоящую заявку во всей полноте посредством отсылки.
ОБЛАСТЬ ТЕХНИКИ
Данная заявка относится к иммунологии, в частности к способам иммунизации против Streptococcus pneumoniae.
УРОВЕНЬ ТЕХНИКИ
Отит среднего уха является распространенным заболеванием у детей. Термин «отит среднего уха» объединяет ряд клинических расстройств, включающий мирингит, экссудативный отит среднего уха (ОМЕ), гнойный отит среднего уха и острый отит среднего уха (АОМ) (24). Острый отит среднего уха (АОМ) является симптоматическим заболеванием ассоциированным с симптомами в верхних респираторных путях, болью, лихорадкой и отореей. Острый отит среднего уха является наиболее распространенным инфекционным заболеванием в мире. Данное заболевание приводит к избыточному потреблению антибиотиков детьми в развитых странах и к значительной распространенности нарушений слуха и других осложнений в развивающихся странах (1-3).
АОМ распространен достаточно широко и около 60-70% детей переносят, по крайней мере, один эпизод АОМ в течение первых трех лет жизни (4, 5). Часть детей страдает от повторяющегося отита. Те, у кого количество эпизодов АОМ за 6 месяцев 3 и более или 4 эпизода за год, считаются восприимчивыми к отиту. Количество восприимчивых детей составляет 10-30% от общего количества детей в популяции (4, 5).
Для развития АОМ необходима назофарингеальная инфекция одним или несколькими организмами-отопатогенами. Наиболее широко распространенными отопатогенами являются Streptococcus pneumoniae (Spn), нетипируемый штамм Haemophilus influenzae (NTHi) и Moraxella Catarrhalis, среди которых доминирует Spn (6). Отмечена прямая взаимосвязь между частотой колонизации NTHi и частотой возникновения АОМ (J. Infect Dis 170:862-866).
При лечении рецидивирующего отита в настоящее время применяются различные антибиотики по мере увеличения эффективности их действия. Такой подход основан на предположении, что рецидивирующие инфекции вызываются размножением антибиотико-резистентных штаммов бактерий. Если частота рецидивов составляет 3 и более раз за 6 месяцев или 4 и более раз за 12 месяцев, часто применяется тимпанотомия, с или без сопутствующей аденэктомии и/или тонзилэктомии.
В качестве профилактических мер в настоящее время доступны два типа вакцин на основе пневмококка. Первая включает капсульные полисахариды 23 типов S. pneumoniae, которые вместе представляют собой капсульные полисахариды около 90% штаммов, вызывающих пневмококковые инфекции. Однако данная вакцина не является высокоиммуногенной у маленьких детей (Fedson, and Musher 2004, "Pneumococcal Polysaccharide Vaccine", pp. 529-588; In Vaccines. S.A. Plotikin and W.A. Orenstein (eds.), W.B. Saunders and Co., Philadelphia, PA; Shapiro et. al., N. Engl. J. Med. 325:1453-1460 (1991)), поскольку у них не развивается выраженного иммунного ответа на антигены полисахаридной природы до достижения 2-летнего возраста. Данная вакцина не рекомендована для предотвращения развития отита среднего уха.
Вторым доступным типом вакцин на основе пневмококка являются конъюгированные вакцины. Данные вакцины вызывают серотип-специфическую защиту и включают специфические для серотипа капсульные полисахаридные антигены, конъюгированные с белком-носителем. В настоящее время доступны 7 и 13-валентные конъюгированные вакцины: 7-валетная включает 7 полисахаридных антигенов (из капсулы серотипов 4, 6В, 9V, 14, 18С, 19F и 23F), 13-валентная - 13 полисахаридных антигенов (из капсулы серотипов 1, 3, 5, 6А, 7F, и 19А, а также серотипов, включенных в 7-валетную вакцину). Также разработаны 9 и 11-валентные конъюгированные вакцины, каждая из которых включает дополнительные по сравнению с 7-валетной вакциной серотип-специфические полисахариды (т.е. серотипы 1 и 5 в 9-влетной и типы 3 и 7F в 11-валентной вакцинах).
В то же время существуют ограничения для применения конъюгированных вакцин. Например, поскольку такие вакцины повышают серотип-специфическую защиту, для защиты от других серотипов, включая доминирующие серотипы в развивающихся странах, в состав вакцин должны быть включены дополнительные серотип-специфические полисахариды, что усложняет производство таких вакцин (Di Fabio et al., Pediatr. Infect. Dis. J. 20:959-967 (2001); Mulholland, Trop.Med. Int. Health 10:497-500 (2005)). Применение 7-ми валентной конъюгированной вакцины также привело к более частой колонизации и развитию заболевания штаммами, полисахариды капсулы которых не включены в состав данной вакцины (Bogaert et al., Lancet Infect. Dis. 4:144-154 (2004); Eskola et al., N. Engl. J. Med. 344-403-409 (2001); Mbelle et al., J. Infect. Dis. 180:1171-1176 (1999)). Что касается отита среднего уха, вызываемого пневмококковыми инфекциями, доступные в настоящее время конъюгированные вакцины не обладают эффективным защитным действием, как это обычно бывает по отношению к инфекционным заболеваниям. Кроме того, развитие рецидивов АОМ возможно после вакцинации; например, у части детей, особенно восприимчивых к рецидивам АОМ, несмотря на иммунизацию конъюгированной вакциной, развивается ряд рецидивов и такие дети становятся восприимчивы к отиту.
Таким образом, существует потребность в создании композиций для применения в качестве вакцин, разработке способов профилактики и лечения рецидивирующего пневмококкового АОМ.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Описаны способы профилактики и лечения рецидивирующего АОМ, возникающего при инфекции S. pneumoniae, у пациентов группы риска. Группа риска включает, например, младенцев и детей с рецидивами АОМ (например, восприимчивых к отиту), а также пациентов с АОМ, у которых терапия оказалась не успешной. Например, описаны способы профилактики и лечения рецидивов острого отита среднего уха вызванного инфекцией Streptococcus pneumoniae у пациентов группы риска развития рецидива пневмококкового АОМ, способ включающий, по крайней мере, однократное введение таким пациентам терапевтически эффективного количества композиции, содержащей, по меньшей мере, один выделенный и очищенный иммуногенный пептид, выбранный из группы, состоящей из полипептидов Streptococcus pneumoniae PhtD, PhtE, PcpA, LytB и детоксифицированного пневмолизина, или иммуногенного фрагмента данных полипептидов. В определенных вариантах исполнения, пациент предварительно мог перенести, по крайней мере, один эпизод острого отита среднего уха. В определенных вариантах исполнения пациент мог перенести 3 и более эпизодов острого отита среднего уха в течение 6 месяцев или перенести 4 и более эпизодов острого отита среднего уха в течение 12 месяцев. В некоторых вариантах исполнения пациент может страдать от острого отита среднего уха.
Также описаны композиции для применения при профилактике и лечении рецидива АОМ. Данные композиции содержат, по крайней мере, один иммуногенный полипептид S. pneumoniae выбранный из группы, состоящей из PhtD, PhtE, РсрА, LytB и детоксифицированного пневмолизина, или их иммуногенных фрагментов. Основное содержание данной заявки характеризует несколько преимуществ. Например, способы, описанные в данной заявке, могут быть использованы для инициации или интенсификации образования антиген-специфических CD4+ Т-клеток.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Описание изобретения может быть более ясным из следующего описания со ссылкой на чертежи.
Фигура 1. На данной фигуре представлена доля субпопуляций CD45RALow CD4+ Т-клеток памяти, продуцирующих различные цитокины (ИФН-γ, (b) ИЛ-4, (с) ИЛ-2, (d) ИЛ-17а) в ответ на стимуляцию 6 пневмококковыми антигенами, в периферической крови восприимчивых и невосприимчивых к отиту детей. Столбики на диаграмме представляют собой средние значения процента CD69+CD4+ Т-клеток после стимуляции антигеном. Планки погрешностей построены для стандартной ошибки среднего (SEM), значения Р определены с использованием критерия Манн-Уитни. *Р<0.05; **Р<0.005.
Фигура 2. На данной фигуре приведены результаты сравнения уровня IgG против пневмококковых антигенов (PhtD, LytB, РсрА, PhtE и Ply) в сыворотке пациентов двух групп: восприимчивых и невосприимчивых к отиту детей. *Р<0.05; **Р<0.005; ***Р<0.0005. По оси Y отложены значения геометрического среднего титра, планки погрешностей обозначают 95% доверительный интервал.
Фигура 3. На данной фигуре приведены результаты исследования ответа CD4+ Т-лимфоцитов на стимуляцию суперантигеном (SEB). Клетки мононуклеарной фракции периферической крови восприимчивых и невосприимчивых к отиту детей были стимулированы SEB, после чего оценивали продукцию цитокинов субпопуляцией CD45RALow CD4+ Т-лимфоцитов.
Фигура 4. На данной фигуре приведены данные по измерению уровня сывороточных IgG у детей во время АОМ (образцы собирали во время визита к врачу по поводу развившегося АОМ): 35 пациентов из группы восприимчивых к отиту, 25 из группы AOMTF и 34 пациента из группы невосприимчивых к отиту. Примечание: все значения концентраций антител, специфичных к пяти белкам, представляют собой титр в конечной точке. Линиями отмечены значительные отличия между двумя группами пациентов. ***р value<0.0001, **р value<0.001, и *р value<0.05.
Фигура 5. На данной фигуре приведены результаты измерения уровня IgG против пяти белков S. pneumoniae у пациентов из групп восприимчивых и невосприимчивых к отиту, в зависимости от возраста (6-24 мес). Образцы собирали в возрасте 6, 9, 12, 15, 18 и 24 месяца. 107, 88, 65, 61, 55, и 44, соответственно, образца сыворотки было проанализировано от невосприимчивых к отиту детей, и 10, 10, 9, 10, 10 и 4 образца сыворотки было проанализировано от восприимчивых к отиту детей. Обнаружены значимые различия концентрации IgG против всех пяти белков, за исключением LytB (р<0,07), сопровождаемые относительным увеличением концентрации сывороточных IgG со временем, у пациентов из группы невосприимчивых к отиту. Показанные различия не были статистически достоверны у пациентов группы восприимчивых к отиту (р=0.40 для белка PhtD, р=0.39 для LytB, р=0.11 для РсрА, р=0.09 для PhtE и р=0.42 для Ply).
Фигура 6. Фигура состоит из трех панелей: А, В, С.На Фигуре 6А приведены результаты измерения представленности В-клеток памяти; на Фигуре 6В приведены результаты измерения уровня IgG-ответа на пять пневмококковых антигенов й сыворотке восприимчивых и невосприимчивых к отиту детей (по оси Y отложено геометрическое среднее титра, планки погрешностей обозначают 95% доверительный интервал); на Фигуре 6С показана корреляция между количеством PhtD-специфических В-клеток памяти в периферической крови (по оси X) с концентрацией PhtD-специфических IgG в сыворотке (по оси Y).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Описаны способы профилактики и/или лечения рецидивирующего острого отита среднего уха, возникающего в результате инфекции S. pneumoniae у пациентов группы риска (например, детей). Также описаны композиции, используемые при данных способах профилактики и/или лечения рецидивирующего острого отита среднего уха. Данные композиции содержат, по крайней мере, один иммуногенный полипептид S. pneumoniae выбранный из группы, состоящей из полипептидов PhtD, PhtE, РсрА, LytB и детоксифицированного пневмолизина, или их иммуногенные фрагменты. Данные способы и композиции описаны далее.
Описанные профилактические и терапевтические способы включают, по меньшей мере, однократное введение терапевтически эффективного количества композиции (например, фармацевтической композиции), содержащей, по меньшей мере, один выделенный и очищенный иммуногенный пептид, выбранный из группы, состоящей из полипептидов Streptococcus pneumoniae PhtD, PhtE, РсрА, LytB и деактивированного пневмолизина, или иммуногенный фрагмент данных пептидов, пациентам с высоким риском развития рецидива пневмококкового АОМ (т.е. симптоматической инфекции S. pneumoniae приводящей к рецидиву АОМ).
Группа риска включает, например, младенцев и детей, перенесших один, два, три, четыре или больше эпизодов АОМ на протяжении жизни; восприимчивых к отиту детей и младенцев (т.е. перенесших 3 или более эпизодов АОМ в течение месяцев или 4 или более эпизодов АОМ в течение года); младенцев и детей, у которых терапия АОМ была неудачна (т.е. в случае не достижения ликвидации бактериальной инфекции и/или разрешения симптомов после, по меньшей мере, 48 часов соответствующей антибиотикотерапии; или младенцев и детей у которых симптомы АОМ возникли снова в течение 14 дней после завершения курса антибиотикотерапии). Группа риска также включает, например, младенцев и детей: с генетической предрасположенностью к рецидивирующему АОМ (Casselbrant ML et al JAMA 1999; 282:2125-2130); посещающих детские дошкольные учреждения; посещающих детские сады семейного типа или уход на дому у няни; в семье которых курит один или более родителей/опекунов; использующих соску-пустышку; получающих преимущественно искусственное вскармливание; перенесших АОМ в первые 6 месяцев жизни (Bentdal et al Int. J. Ped. Otorhinolaryngol. 2007; 71:1251-1259). По мере взросления дети становятся менее восприимчивы к АОМ в следствие анатомических изменений в евстахиевой трубе. Обычно дети, восприимчивые к отиту «перерастают» восприимчивость в возрасте 3-5 лет (40, 48-51). В определенных вариантах исполнения пациенты имеют пневмококковый АОМ или находятся в группе риска. Как описано в Примерах в данной заявке, дети, восприимчивые к развитию отита (т.е. пациенты группы риска) имеют сниженную иммунореактивность по отношению к антигенам Spn (например, PhtD, PhtE, РсрА, LytB, Ply) в отличие от невосприимчивых в отиту детей. Например, по сравнению с невосприимчивыми детьми, у восприимчивых детей наблюдается отсутствие или сниженное количество специфических к пневмококковому антигену функциональных CD4+ Т-клеток памяти (например, функциональных CD4+ Т-клеток памяти специфичных к PhtD, PhtE, РсрА, LytB или Ply) и сниженный уровень IgG, специфичных к пневмококковым антигенам (например, к PhtD, PhtE, РсрА, LytB или Ply), в сыворотке. У таких детей, однако, нет дефицита по функциональным Т-клеткам памяти вообще или детектируемому иммунному ответу за счет В-клеток и антител против антигенов после вакцинации. Иммунологические реакции детей после неудачной терапии АОМ (AOMTF) схожи с таковыми у восприимчивых к отиту детей. Пациентами группы риска являются те пациенты, у которых наблюдается описанная сниженная иммунореактивность против антигенов Spn, таких как; например, PhtD, PhtE, РсрА, LytB и/или Ply.
При использовании в данной заявке, профилактика рецидива АОМ у пациента подразумевает введение пациенту терапевтически эффективного количества композиции, описанной в данной заявке, для профилактики развития рецидива острого пневмококкового отита среднего уха у пациента.
При использовании в данной заявке, лечение рецидива АОМ (или восприимчивого к отиту пациента или пациента с рецидивирующим АОМ) подразумевает введение терапевтически эффективного количества композиции, описанной в данной заявке, пациентом с АОМ вызванным S. pneumoniae или инфицированным S. pneumoniae и имевшем прежде АОМ, где целью является излечение, способствование излечению, облегчение состояния, смягчение симптомов, изменение состояния, освобождение от заболевания, улучшение или изменение состояния (например, АОМ) или симптомов заболевания (т.е. АОМ).
Терапевтически эффективным количеством называется количество, которое обеспечивает терапевтический эффект при данных условиях и режиме приема. Терапевтически эффективное количество может быть определено медицинским работником обычной квалификации на основе характеристик пациента (возраст, вес, пол, состояние, наличие осложнений и т.д.). Также значение терапевтически эффективного количества зависит от способа введения композиции.
В отдельных примерах введение композиции вызывает или усиливает образование антиген-специфических CD4+ Т-клеток. Образовавшиеся антиген-специфические CD4+ Т-клетки могут продуцировать, например, ИФН-γ, ИЛ-4, ИЛ-2 и/или ИЛ-17а. Например, в одном варианте исполнения введение композиции вызывает или усиливает образование антиген-специфических CD4+ Т-клеток, продуцирующих IFN-γ. При использовании в данной заявке выражение «вызывает или усиливает образование антиген-специфических CD4+ Т-клеток» подразумевает, что происходит увеличение количества или доли (%) антиген-специфических CD4+ Т-клеток. Количество клеток может увеличиваться, например, на 25%, 30%, 35%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 100%, 150%, 200%, 300%, 400% или более по сравнению с таковым непосредственно перед введением композиции.
В одном варианте исполнения введение композиции вызывает или усиливает продукцию антиген-специфических антител (например, IgG). При инициации или усилении выработки антител возрастает суммарная концентрация (титр) антиген-специфических IgG относительно концентрации (титра) непосредственно перед введением. Титр конечного разведения может возрастать, например, на 25%, 30%; 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 100%, 150%; 200% и более по сравнению с титром непосредственно перед введением композиции. В одном варианте исполнения, титр антиген-специфических IgG возрастает, например, в 2, 3 или 4 раза по сравнению с титром непосредственно перед введением композиции.
Также описан способ снижения риска развития рецидива острого отита среднего уха у пациента (например, ребенка), включающий введение пациенту композиции, содержащей один или более описанных в данной заявке иммуногенных полипептидов. Риск такого рецидива может быть снижен с помощью способов описанных в данной заявке.
В определенных вариантах исполнения реализован способ профилактики или лечения состояния восприимчивости к отиту у пациента группы риска (т.е. пациента, перенесшего по меньшей мере один или более рецидивирующих эпизодов АОМ).
Настоящая заявка также описывает способ выработки иммунного ответа у пациента группы риска с помощью введения композиций описанных в данной заявке. Выработка иммунного ответа может быть достигнута введением в виде фармацевтически приемлемой лекарственной формы композиции пациенту для презентации, по меньшей мере, одного иммуногенного полипептида иммунной системе пациента.
Данная заявка также описывает использование одного или более иммуногенных полипептидов S. pneumoniae в композициях, таких как, например, вакцинные композиции. Такие композиции при введении в организм (например, млекопитающего) вызывают или усиливают иммунный ответ направленный против иммуногенного полипептида (т.е. антигена) включенного в состав композиции. Данный ответ может включать выработку антител (например, за счет стимуляции В клеток) или Т-клеточный иммунный ответ (например, цитотоксический). Такой иммунный ответ может быть защитным или нейтрализующим, или нет. Нейтрализующим или защитным иммунным ответом является негативный иммунный ответ по отношению к инфицирующему организму соответствующему антигену (например, организму из которого был получен антиген) и благоприятный по отношению к организму, в который была введена композиция (например, за счет снижения или предотвращения развития инфекции). При использовании в данной заявке защитные и нейтрализующие антитела могу взаимодействовать с соответствующим полипептидом S. pneumoniae дикого типа и могут снижать или ингибировать летальность соответствующего организма или соответствующего полипептида S. pneumoniae дикого типа для организма (например, млекопитающего). Иммунологическая композиция, приводящая к защитному или нейтрализующему иммунному ответу при введении в организм может рассматриваться как вакцина. Композиции описанные в данной заявке используются в способах профилактики или лечения рецидивов АОМ у пациентов группы риска, которые, как определено выше, имеют риск быть инфицированы S. pneumoniae с развитием рецидива АОМ. Данная композиция также используется в способах профилактики и лечения рецидивирующего АОМ.
Композиции, описанные в данной заявке могут быть введены соответствующим образом, например, через кожу (например, внутримышечно, внутривенно, внутрибрюшинно или подкожно), трансдермально, через слизистую оболочку (например, интраназально) или местно, в количествах и в режиме определенном специалистами в данной области. Например, 100-500 мкг, 1-240 мкг, 10-100 мкг, 5-50 мкг или 10-25 мкг иммуногенного полипептида может быть введено в качестве одной дозы. С целями профилактики или терапии, данные вакцины могут вводиться однократно или многократно. Например, данные вакцины могут вводиться 1, 2, 3 или 4 раза. В одном примере, однократное или многократные введения могут применяться как часть так называемого «prime-boost» протокола. При многократном введении, дозы могут быть разделены между собой, например, неделей, месяцем или несколькими месяцами.
Иммуногенные полипептиды, описанные в данной заявке, обладают иммуногенный активностью. Термин «иммуногенная активность» означает способность полипептида вызывать иммунный ответ у организма (например, у млекопитающего). Иммунологический ответ на полипептид подразумевает развитие у животного клеточного и/или антитело-опосредованного иммунного ответа на полипептид. Как правило, иммунологический ответ включает, но не ограничивается одним или более из следующих эффектов: продукцию антител, В-клеток, хелперных Т-клеток, супрессорных Т-клеток и/или цитотоксических Т-клеток, направленных против эпитопа или эпитопов полипептида. Термин «эпитоп» означает участок антигена по отношению к которому В- и Т-клетки реагируют так, что происходит выработка антител. Иммуногенная активность может быть защитной. Термин «защитная иммуногенная активность» означает способность полипептида вызывать иммунологический ответ у пациента, предотвращающий или подавляющий инфекцию S. pneumoniae (например, подавление инфекции S. pneumoniae вызывающей рецидив АОМ).
В определенных вариантах исполнения, может быть приготовлена многокомпонентная композиция, включающая два, три, четыре или более иммуногенных пептида для защиты против рецидива АОМ, возникающего после инфекции S. pneumoniae. Предпочтительный вариант исполнения такой композиции включает иммуногенные полипептиды PhtD и РсрА. Дополнительный предпочтительный вариант исполнения включает полипептиды PhtD и РсрА и детоксифицированный пневмолизин. Отдельные, предпочтительные для применения в соответствии с описанным здесь, многокомпонентные композиции описаны в заявке WO 2011/075823 (поданной 20.12.2010 и озаглавленной «Иммуногенные композиции» / «Immunogenic Compositions))).
Компоненты многокомпонентной композиции предпочтительно совместимы и смешаны в соответствующих соотношениях с целью избежать интерференции антигенов и оптимизировать любые возможные положительные взаимодействия. Например, количества каждого компонента могут быть в диапазоне от, примерно, 5 до, примерно, 500 мкг на дозу, от 5 мкг до, примерно, 10 мкг на дозу, от 25 мкг до, примерно, 50 мкг на дозу, или от 50 до, примерно, 100 мкг на дозу. Наиболее предпочтительным диапазоном может быть диапазон, примерно, от 10 мкг до 50 мкг каждого антигенного компонента на дозу.
Иммуногенные полипептиды
Нуклеиновые кислоты, кодирующие иммуногенные полипептиды, могут быть выделены, например, но не ограничиваясь этим примером, из клеток дикого или мутантного штамма S. pneumoniae, а также могут быть получены прямо из ДНК штамма S. pneumoniae, несущего соответствующие гены (например, рсрА или phtD), с помощью полимеразной цепной реакции (ПЦР) или с использованием альтернативных стандартному способов, определяемых специалистами в данной области. Возможными штаммами S. pneumoniae могут являться TIGR4 и 14453. В предпочтительных вариантах исполнения полипептиды получают с помощью технологии рекомбинантной ДНК из клеток S. pneumoniae штамма 14453.
Полипептиды, описанные в данной заявке, могут быть получены с помощью стандартных молекулярно-биологических способов и систем экспрессии (см. например, Molecular Cloning: A Laboratory Manual, Third Edition by Sambrook et. al., Cold Spring Harbor Press, 2001). Например, фрагмент гена, кодирующего иммуногенный полипептид, может быть выделен, после чего данный полинуклеотид, кодирующий иммуногенный полипептид, может быть клонирован в любой коммерчески доступный вектор для экспрессии (такой как, например, pBR322 и pUC векторы (New England Biolabs, Inc., Ipswich, MA)) или вектор для экспрессии/очистки (такие как, например, векторы с GST-тэгом (Pfizer, Inc., Piscataway, N.J.)) и, затем, экспрессирован в подходящей про- или эукариотической или вирусной системах. Очистка продукта может производиться традиционными способами или, в случае коммерческих систем экспрессии/очистки, в соответствии с инструкциями производителя.
Также, описанные в данной заявке иммуногенные полипептиды, включая варианты, могут быть получены путем химического синтеза с использованием коммерчески доступных автоматизированных способов, таких как, например, эксклюзионный твердофазный синтез, способов частично твердофазного синтеза, конденсации фрагментов или синтеза в растворе.
Иммуногенные полипептиды РсрА содержат полноразмерную аминокислотную последовательность РсрА (включая или нет сигнальную последовательность), его фрагменты или варианты. Полипептиды РсрА, подходящие для использования в составе композиций, описанных в данной заявке, включают, например, такие, как указанные в базе данных GenBank под номерами САВ 04758, YP 817353, ААК 76194, NP 359536, ZP 01835022 и ZP 01833419, а также описанные здесь и в разделе «Примеры» ниже, и другие. Аминокислотная последовательность РсрА в одном из вариантов исполнения приведена здесь в виде последовательностей SEQ ID NO: 1 или 2.
Аминокислотная последовательность полноразмерного РсрА, кодируемого геномом S. pneumoniae 14453, приведена в виде последовательности SEQ ID NO. 1. Предпочтительные полипептиды РсрА могут обладать аминокислотной последовательностью, имеющей 50% и более идентичности (например, 60, 65, 70, 75, 80, 85, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 99.5% или более) с последовательностями SEQ ID NO.: 1, 2 или 3. Предпочтительные полипептиды могут содержать фрагмент, как минимум, из 8, 9, 10, 12, 14, 16, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250 или более, например, последовательных аминокислотных остатков последовательностей SEQ ID NO.: 1, 2 или 3. Последовательности предпочтительных фрагментов содержат эпитоп из последовательностей SEQ ID NO.: 1, 2 или 3. Другие последовательности предпочтительных фрагментов не содержат один или более аминокислотных остатков N-конца последовательностей SEQ ID NO.: 1 или 2 (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или более) и/или один или более аминокислотных остатков С-конца последовательностей SEQ ID NO.: 1 или 2 и, при этом, содержат, по крайней мере, один эпитоп последовательностей SEQ ID NO.: 1 или 2. Последовательности других предпочтительных фрагментов не содержат сигнальную последовательность N-конца последовательностей SEQ ID NO.: 1 или 2. Предпочтительная последовательность полипептида РсрА приведена в последовательности SEQ ID NO.: 3.



Последовательность иммуногенного полипептида РсрА может не содержать последовательность холин-связывающего якорного домена, в норме присутствующую в природном зрелом белке РсрА. Природная последовательность якорного холин-связывающего домена зрелого белка РсрА приведена в заявке WO 2008/022302 как последовательность SEQ ID NO: 52. Конкретнее, последовательность иммуногенного полипептида содержит N-концевую область природного РсрА с одной или более аминокислотными заменами и обладает 60-99% (или любым значением между, например 80, 85, 90, 95%) идентичности с природной последовательностью РсрА. N-концевая область может содержать последовательность SEQ ID NO.: 1 или 2 (или SEQ ID NO.: 1, 2, 3, 4, 41 или 45 заявки WO 2008/022302) в присутствии или отсутствии одной или более замен консервативных аминокислотных остатков, а также в присутствии или отсутствии сигнальной последовательности. N-концевой участок может содержать аминокислотную последовательность, обладающую 60-99% идентичности (или любой другой в диапазоне 80-99%) с последовательностями SEQ ID NO.: 1, 2 или 3 (приведены в Списке последовательностей в данной заявке) или последовательностями SEQ ID NO: 1, 2, 3, 4 или 41 заявки WO 2008/022302.
Иммуногенные фрагменты последовательностей SEQ ID NO.: 1, 2 или 3 могут содержать, например, 5, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 160, 170, 180, 190 и 191 аминокислотный остаток последовательностей SEQ ID NO.: 1, 2 или 3 или любое другое число аминокислотных остатков между 5 и 191. Примеры иммуногенных фрагментов описаны в заявке WO 2008/022302.
Варианты иммуногенных полипептидов, описанные в данной заявке, могут содержать одну или более замен консервативных аминокислотных остатков. Варианты иммуногенных полипептидов РсрА включают аминокислотные последовательности, обладающие идентичностью последовательности в 50-99% (или любой другой в диапазоне 50-99%) с последовательностями SEQ ID NO.: 1, 2 или 3 или их фрагментами. Варианты отбираются в соответствии с их иммуногенами свойствами с помощью способов, хорошо известных в данной области.
Последовательности иммуногенных полипептидов PhtX, применимых для описанных в данной заявке композиций, включают, например, полноразмерные аминокислотные последовательности PhtD или PhtE (с или без сигнальной последовательности), их иммуногенные фрагменты, их варианты и слитые белки на их основе. Полипептиды PhtD применимые для использования в композициях, описанных в данной заявке, включают, например, такие, последовательности которых приведены в базе данных GenBank под номерами ААК 06760, YP 816370 и NP 35851 и другие. Аминокислотная последовательность полноразмерного PhtD, кодируемая геномом S. pneumoniae 14453 приведена в виде последовательности SEQ ID NO.: 4, а кодируемая геномом штамма TIGR4 в виде последовательности SEQ ID NO.: 5. Предпочтительная последовательность полипептида PhtD (кодируемая геномом S. pneumoniae 14463) приведена в виде последовательности SEQ ID NO: 6. Полипептиды PhtE, пригодные для использования в композициях, описанных в данной заявке, включают те, например, чьи последовательности приведены в базе данных GenBank под номерами ААК 06761, YP 816371 и NP 358502 и другими. Полноразмерная последовательность PhtE, кодируемая геномом S. pneumoniae 14453, приведена в виде последовательности SEQ ID NO.: 7. Предпочтительная последовательность полипептида PhtE (на основе генома S. pneumoniae 14453) приведена в виде последовательности SEQ ID NO.: 8.





Иммуногенные полипептиды PhtX (например, PhtD или PhtE) могут включать полноразмерную последовательность белка с присоединенной сигнальной последовательностью, последовательность зрелого полноразмерного белка с удаленной последовательностью сигнального пептида (например, около 20 аминокислотных остатков на N-конце), варианты PhtX (природные или искусственные, например, синтетического происхождения) и иммуногенные фрагменты PhtX (например, фрагменты содержащие, по меньшей мере, 15 или 20 последовательных аминокислотных остатков естественной последовательности зрелого белка PhtX). Данные иммуногенные фрагменты и варианты полипептидов PhtX способны вызывать иммунный ответ по отношению к соответствующей зрелой полноразмерной аминокислотной последовательности. Примеры иммуногенных фрагментов PhtD описаны в заявке РСТ WO 2009/012588.
Предпочтительные для применения полипептиды PhtD могут содержать аминокислотную последовательность, обладающую 50% или более идентичности (например, 60, 65, 70, 75, 80, 85, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 99.5% или более) с последовательностями SEQ ID NO: 4, 5 или 6. Предпочтительные для применения полипептиды могут содержать фрагмент из, по меньшей мере, 8, 9, 10, 12, 14, 16, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250 или более последовательных аминокислотных остатков последовательностей SEQ ID NO: 4, 5 или 6. Предпочтительные для применения фрагменты содержат эпитоп последовательностей SEQ ID NO: 4, 5 или 6. Другие предпочтительные последовательности не содержат одного или более аминокислотных остатков N-конца последовательностей SEQ ID NO: 4, 5 или 6 (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или более) и/или один или более аминокислотных остатков С-конца последовательностей SEQ ID NO: 4, 5 или 6, и содержат, по меньшей мере, один эпитоп последовательностей SEQ ID NO: 4, 5 или 6. Другие предпочтительные фрагменты не содержат сигнальную последовательность N-конца последовательностей SEQ ID NO: 4 или 5. Последовательность предпочтительного полипептида PhtD приведена в виде последовательности SEQ ID NO: 6.
Предпочтительные для применения полипептиды PhtE могут содержать аминокислотную последовательность обладающую 50% или более идентичности (например, 60, 65, 70, 75, 80, 85, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 99.5% или более) с последовательностями SEQ ID NO: 7 или SEQ ID NO: 8. Предпочтительные для применения полипептиды могут содержать фрагмент из, по меньшей мере, 8, 9, 10, 12, 14, 16, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250 или более последовательных аминокислотных остатков последовательностей SEQ ID NO: 7 или SEQ ID NO: 8. Предпочтительные для применения фрагменты содержат эпитоп последовательностей SEQ ID NO: 7 или SEQ ID NO: 8. Другие предпочтительные последовательности не содержат одного или более аминокислотных остатков N-конца последовательностей SEQ ID NO: 7 или SEQ ID NO: 8 (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или более) и/или один или более аминокислотных остатков С-конца последовательностей SEQ ID NO: 7 или SEQ ID NO: 8, и содержат, по меньшей мере, один эпитоп последовательностей SEQ ID NO: 7 или SEQ ID NO: 8. Другие предпочтительные фрагменты не содержат сигнальную последовательность N-конца последовательностей SEQ ID NO: 7. Последовательность предпочтительного полипептида PhtE приведена в виде последовательности SEQ ID NO: 8
Иммуногенные полипептиды LytB включают полноразмерную последовательность белка с присоединенной сигнальной последовательностью, полноразмерную последовательность зрелого белка без сигнального пептида, варианты последовательности LytB (природные или искусственные, например, синтетического происхождения) и иммуногенные фрагменты LytB (например, фрагменты содержащие, по меньшей мере, 15 или 20 последовательных аминокислотных остатков естественной последовательности зрелого белка LytB). Иммуногенные варианты и фрагменты иммуногенных полипептидов LytB, описанные в данной заявке, способны вызывать иммунный ответ специфичный к соответствующей зрелой полноразмерной аминокислотной последовательности. Последовательности полипептидов LytB, пригодных для использования в композициях описанных в данной заявке включают, например, последовательности базы данных GenBank с номерами САА 09078, YP 816335, ABJ 55408, ААК 19156, NP 358461 и ААК 75086, а также другие.
Предпочтительные для применения полипептиды LytB могут содержать аминокислотную последовательность, обладающую 50% или более идентичности (например, 60, 65, 70, 75, 80, 85, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 99.5% или более) с последовательностями SEQ ID NO: 9, 10 или 11. Предпочтительные для применения полипептиды могут содержать фрагмент, по меньшей мере, из 8, 9, 10, 12, 14, 16, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250 или более последовательных аминокислотных остатков последовательностей SEQ ID NO: 9, 10 или 11. Предпочтительные для применения фрагменты содержат эпитоп последовательностей SEQ ID NO: 9, 10 или 11. Другие предпочтительные последовательности не содержат одного или более аминокислотных остатков N-конца последовательностей SEQ ID NO: 9, 10 или 11 (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или более) и/или один или более аминокислотных остатков С-конца последовательностей SEQ ID NO: 9 или 10, и содержат, по меньшей мере, один эпитоп последовательностей SEQ ID NO: 9 или SEQ ID NO: 10. Другие предпочтительные фрагменты не содержат сигнальную последовательность N-конца последовательности SEQ ID NO: 10. Предпочтительная последовательность полипептида LytB приведена в виде последовательности SEQ ID NO: 11.


SEQ ID NO: 11 (LytB конструкция, полученная на основе Spn штамм 14453; отсутствуют сигнальная последовательность и области связывающие холин; последовательность вектора подчеркнута)

Пневмолизин (Ply) является активирующим цитолиз токсином, участие которого предполагается на многих этапах развития пневмококковой инфекции, включая угнетение деятельности ресничного эпителия и нарушение плотных контактов между эпителиальными клетками (Hirst et al. Clinical and Experimental Immunology (2004)). Известно несколько пневмолизинов, которые (после деактивации) могут быть использованы для композиций описанных в данной заявке, включая, например, указанные в базе данных GenBank последовательности Q04IN8, P0C2J9, Q7ZAK5 и АВО 21381 и другие. В одном варианте исполнения Ply имеет аминокислотную последовательность, приведенную здесь как последовательность in SEQ ID NO.12.
Иммуногенные полипептиды пневмолизинов могут включать полноразмерную последовательность белка с присоединенной сигнальной последовательностью, полноразмерную последовательность зрелого белка с удаленной последовательностью сигнального пептида, варианты пневмолизина (природные или искусственные, например, синтетические) и иммуногенные фрагменты пневмолизина (например, фрагменты содержащие, по меньшей мере, 15 или 20 последовательных аминокислотных остатков из последовательности зрелого природного пневмолизина). Иммуногенные варианты и фрагменты иммуногенных полипептидов пневмолизинов могут обладать способностью вызывать специфичный иммунный ответ по отношению к соответствующей полноразмерной аминокислотной последовательности зрелого белка. Данные иммуногенные полипептиды пневмолизинов, как правило, детоксифицированы, т.е. не обладают или обладают сниженной токсичностью по сравнению со зрелым пневмолизином дикого типа, полученным из клеток S. pneumoniae. Детоксификация таких иммуногенных полипептидов пневмолизинов может быть, например, химической (например, с использованием обработки формальдегидом) или генетической (например, полученные с помощью технологии рекомбинантных ДНК мутантные формы). Предпочтительные примеры иммуногенного детоксифицированного пневмолизина описаны в заявке РСТ No. WO 2010/071986. В одном из вариантов исполнения иммуногенный детоксифицированный пневмолизин имеет аминокислотную последовательность приведенную как SEQ ID NO. 13.
Предпочтительные для применения полипептиды пневмолизина могут содержать аминокислотную последовательность обладающую 50% или более идентичности (например, 60, 65, 70, 75, 80, 85, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 99.5% или более) с последовательностями SEQ ID NO: 12 или 13. Предпочтительные полипептиды могут содержать фрагмент, по меньшей мере, из 8, 9, 10, 12, 14, 16, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250 или более последовательных аминокислотных остатков последовательностей SEQ ID NO: 12 или 13. Предпочтительные фрагменты могут содержать эпитоп последовательностей SEQ ID NO: 12 или 13. Другие предпочтительные фрагменты не содержат одного или более аминокислотных остатков N-конца последовательностей SEQ ID NO: 12 или 13 (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или более) и/или одного или более аминокислотных остатков С-конца последовательностей SEQ ID NO: 12 или 13 и содержат, по меньшей мере, один эпитоп последовательностей SEQ ID NO: 12 или 13. Другие предпочтительные фрагменты не содержат сигнальную последовательность N-конца последовательности SEQ ID NO: 12. Предпочтительная последовательность иммуногенного и детоксифицированного полипептида пневмолизина приведена в виде последовательности SEQ ID NO: 13.


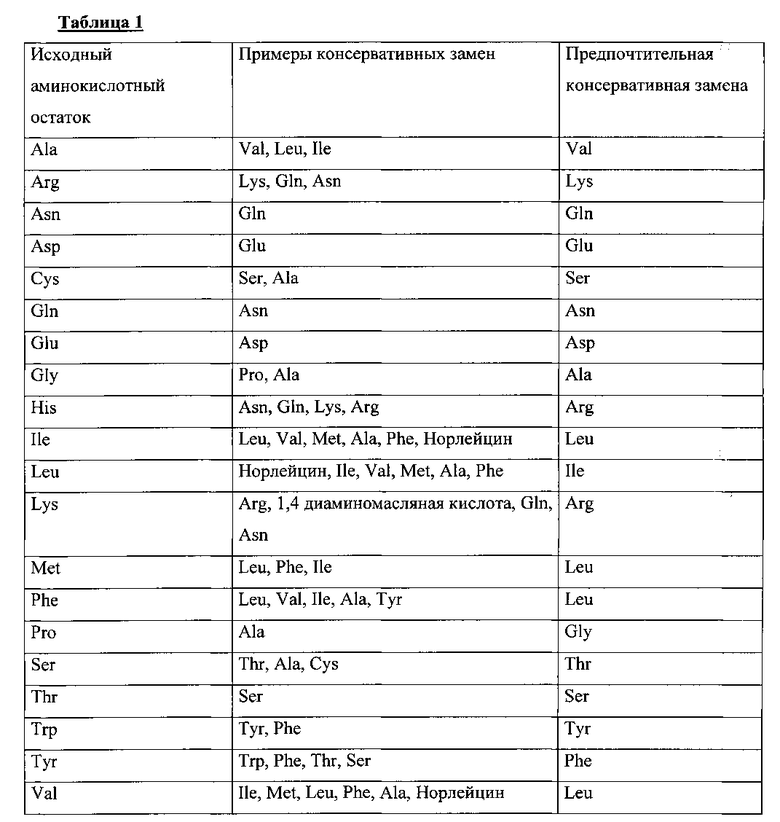
Описанные в данной заявке варианты иммуногенных полипептидов выбраны на основе их иммуногенной активности с применением способов, хорошо известных в соответствующей области. Такие варианты могут содержать аминокислотные модификации. Например, модификации аминокислотной последовательности включают замены, делеции и вставки. Замены, делеции и вставки или любые комбинации данных модификаций могут быть комбинированы в одном варианте полипептида такой же длины как длина данного варианта иммуногенного полипептида. Вставки включают присоединение на N и/или С-конец, а также встраивание внутрь последовательности одного или нескольких аминокислотных остатков. Обычно вставки внутрь последовательности меньше, чем С- или N-концевые, например в пределах от одного до четырех аминокислотных остатков. Делеции представляют собой удалением одного или более аминокислотных остатков из последовательности белка. Как правило, удаляется не более 2-6 аминокислотных остатков в любом месте белковой последовательности. Такие варианты, как правило, получают с помощью сайт-направленного мутагенеза нуклеотидов последовательности ДНК кодирующей данный белок, получая таким образом ДНК, кодирующую конкретный вариант белка, с последующей экспрессией данной последовательности ДНК в культуре рекомбинантных клеток. Способы введения замен в определенные сайты последовательности ДНК с известной последовательностью хорошо известны и включают, но не ограничиваются, мутагенез с помощью праймеров для фага М13 и ПЦР-мутагенез. Аминокислотные замены обычно состоят из одного аминокислотного остатка, но могут быть одновременно в нескольких разных позициях. Варианты полипептида с заменами, это варианты в которых по меньшей мере один аминокислотный остаток удален и другой аминокислотный остаток встроен на это место. Такие замены в основном делаются в соответствии со приведенной ниже Таблицей и называются консервативными заменами. Другие являются хорошо известными специалистам в данной области техники.
Как указано в данной заявке, аминокислотные замены могут быть консервативными и неконсервативными. Консервативные аминокислотные замены могут включать замены природного аминокислотного остатка на неприродный, такие, которые оказывают очень слабое или не оказывают вовсе никакого влияния на размер, полярность, заряд, гидрофобность или гидрофильность аминокислотного остатка в данной позиции и, в частности, не снижают иммуногенности. Применимые консервативные аминокислотные замены приведены в Таблице 1 ниже.
Специалист в данной области способен определить применимые варианты полипептидов и/или фрагментов, описанных в данной заявке, используя известные способы.
Аналоги могут отличаться от природных полипептидов S. pneumoniae последовательностью аминокислотных остатков или модификациями, не затрагивающими последовательность. Такие модификации включают изменения в ацетилировании, метилировании, фосфорилировании, карбоксилировании или гликозилировании. «Модификации» полипептидов могут включать полипептиды (или их аналоги, такие как, например, их фрагменты) с химически или энзиматически измененными одним или более последовательными аминокислотными остатками. Такие модификации могут включать, например, модификации боковых цепей, модификации основной цепи и N- и С-концевые модификации, такие как, например, ацетилирование, гидроксилирование, метилирование, амидирование и присоединение углеводных или липидных молекул, кофакторов, и подобных молекул, а также их комбинации. Модифицированные полипептиды, описанные в данной заявке могут сохранять биологическую активность немодифицированных полипептидов или иметь сниженную или повышенную биологическую активность.
Структурное сходство двух полипептидов может быть определено с помощью выравнивания аминокислотных последовательностей данных полипептидов (например, кандидатного полипептида и, например, последовательности SEQ ID NO. 2) с целью определения протяженности идентичных участков последовательностей; для оптимизации определения протяженности участков идентичных аминокислотных остатков, в одну или в обе последовательности могут вводиться бреши, при этом исходный порядок аминокислотных остатков в обеих последовательностях не должен меняться. Кандидатным является полипептид, последовательность которого должна быть сопоставлена с последовательностью референсного полипептида. Кандидатный полипептид может быть выделен, например, из бактерии или получен с помощью технологии рекомбинантных ДНК, или синтезирован химически или энзиматически.
Попарное сравнение последовательностей аминокислотных остатков может быть осуществлено с использованием глобального алгоритма, например алгоритма Needleman-Wunsch. Одновременно последовательности полипептидов могут быть сравнены с использованием локальных алгоритмов выравнивания, такого как алгоритм Blastp программы BLAST2 Search, как описано Tatiana et al., (FEMS Microbiol. Lett, 174 247-250 (1999), доступного на веб-сайте National Centre for Biotechnology Information (NCBI). Для всех параметров BLAST2 Search могут быть использованы значения по умолчанию, включая: matrix = BLOSUM62; open gap penalty = 11, extension gap penalty = 1, gap x dropoff = 50, expect 10, wordsize = 3 и filter on. Другим локальным инструментом для построения и анализа выравнивания является алгоритм Smith and Waterman (1988).
При сравнении двух аминокислотных последовательностей структурное сходство может быть выражено как «процент идентичности» или «процент сходства». «Идентичность» означает присутствие идентичных аминокислотных остатков. «Сходство» означает присутствие не только идентичных аминокислотных остатков, но также и присутствие консервативных замен. Варианты аминокислотных остатков для консервативных замен в полипептиде, описанном в данной заявке, могут быть выбраны среди аминокислот того же класса, к которому принадлежит заменяемый аминокислотный остаток, и показаны в Таблице 1.
Композиции
Композиции (например, вакцинные композиции)могут вводиться в присутствие или в отсутствие адъюванта. Адъювантами обычно являются вещества, способные повышать иммуногенность антигенов. Адъюванты могут оказывать влияние и на систему приобретенного иммунитета, и на систему врожденного (например, Toll-подобные рецепторы) различными путями, часть которых не изучена.
В качестве адъювантов могут выступать многие вещества, как природные, так и синтетические. Например, адъюванты могут включать, но не ограничиваются ими, минеральные соли, скваленовые смеси, мурамил-пептид, производные сапонина, препараты клеточных стенок микобактерий, определенные эмульсии; монофосфорил липид А, производные миколовой кислоты, неионные ПАВ на основе блоковых сополимеров, Quil А, субъединица холерного токсина В, полифосфазен и его производные, иммуностимулирующие комплексы (ISCOM), цитокиновые адъюванты, адъювант MF59, липидные адъюванты, мукозные адъюванты, определенные бактериальные эндотоксины и другие компоненты, определенные олигонуклеотиды, PLG и другие. Такие адъюванты могут быть использованы в составе композиций и в способах, описанных в данной заявке.
В определенных вариантах исполнения композиция вводится в присутствии адъюванта, который содержит эмульсию масло-в-воде включающую, по меньшей мере, сквален, водный растворитель, полиоксиэтилен-алкил эфир, гидрофильное неионное ПАВ, гидрофобное неионное ПАВ, где указанная эмульсия масло-в-воде может быть получена путем процесса обращения фаз при нагревании, и в которой 90% капель масла по объему имеют размер менее 200 нм, и, дополнительно, менее 150 нм. Такой адъювант описан в заявке WO 2007006939 (Vaccine Composition Comprising a Thermoinversable Emulsion), которая включена в данную заявку во всей полноте. Данная композиция также может включать продукт Е6020 (номер CAS 287180-63-6) в дополнение к, или вместо описанной эмульсии сквалена типа масло-в-воде. Продукт Е6020 описан в заявке US 2007/0082875 (которая включена в данную заявку во всей полноте посредством отсылки).
В определенных вариантах исполнения композиция включает агонисты TLR (например, агонист TLR4), отдельно или вместе, в комбинации с адъювантом. Например, адъювант может содержать агонист TLR4 (например, TLA4), сквален, водный растворитель, неионное гидрофильное ПАВ на основе полиоксиэтилен-алкильной эфирной группы, неионное гидрофобное ПАВ, и являться термообратимым. Примеры таких адъювантов описаны в заявке WO 2007080308 (Thermoreversible Oil-in-Water Emulsion), которая включена в данную заявку во всей полноте. В одном варианте исполнения данная композиция используется с адъювантом, состоящим из комбинации CpG и соли алюминия (например, Alum). В способах, описанных в данной заявке, одними из используемых адъювантов являются адъюванты на основе соли алюминия (или вещества). Примеры используемых адъювантов на основе солей алюминия включают гидроксид алюминия (например, кристаллический гидроксид алюминия AlO(ОН) и гидроксид алюминия Al(ОН)3. Гидроксид алюминия представляет собой алюминий содержащее вещество, содержащее ионы Al3+ и гидроксильные группы (-ОН). Смеси гидроксида алюминия с другими алюминий-содержащими веществами (например, гидроксифосфатом или гидроксисульфатом) могут также использоваться в случае, если в результате смешивания образующееся алюминий-содержащее вещество содержит гидроксильные группы. В отдельных вариантах исполнения, адъювант на основе алюминия представляет собой оксигидроксид алюминия (например, Alhydrogel®). В соответствующей области хорошо известно что композиции содержащие адъюванты на основе солей алюминия не должны подвергаться воздействию экстремальных температур, т.е. замораживаться (ниже 0°C) или нагреваться до высокой температуры (например, ≥70°C), поскольку воздействие экстремальных температур может негативно влиять на стабильность и иммуногенность как адъюванта, так и антигена.
В отдельном варианте исполнения алюминий-содержащее вещество (например, адъювант на основе гидроксида алюминия) взаимодействует с фосфатом.
В предпочтительно варианте исполнения фосфат добавляется к адъюванту на основе гидроксида алюминия в форме соли. Предпочтительно, ионы фосфата вводятся посредством буферного раствора содержащего ди и мононатрий фосфат.
Предпочтительно, как в приведенных в данной заявке примерах, что данное алюминий-содержащее вещество сочетается с фосфатом (например, с помощью процесса, описанного в заявке WO 2011/075822 (поданной 20.12.2010 и озаглавленной immunogenic Compositions and Related Methods»). В этом процессе водная суспензия гидроксида алюминия (приблизительно 20 мг/мл) смешивается с фосфатным буферным раствором (например, приблизительно 400 моль/л). Предпочтительная конечная концентрация фосфата составляет от 2 до 20 мМ. Затем смесь разбавляется буферным раствором (например, Tris-HCl, Tris-HCl с солями, HEPES) для получения суспензии гидроксида алюминия и фосфата (РО4) Предпочтительным буферным раствором является раствор: 10 mM Tris-HCl и 150 mM NaCl, при pH около 7.4. Данная суспензия затем смешивается в течение приблизительно 24 ч при комнатной температуре. Предпочтительная концентрация алюминия в конечной суспензии составляет от 0,28 мг/мл до 1,68 мг/мл. Более предпочтительной концентрацией является 0,56 мг/мл.
Иммуногенные полипептиды (например, РсрА, PhtD) по отдельности или в комбинации могут быть затем адсорбированы с подготовленным гидроксидом алюминия.
Данные композиции предпочтительно могут быть в жидкой форме, однако могут быть лиофилизированы (посредством стандартных способов) или высушены во вспененном состоянии (как описано в заявке WO 2009012601, Antigen-Adjuvant Compositions and Methods). Композиция в соответствии с одним из вариантов исполнения находится в жидкой форме. Доза для иммунизации может быть в диапазоне от 0,5 до 1 мл. Жидкие лекарственные формулы могут быть представлены в любой форме, пригодной для введения, включая, например, раствор или суспензию. Таким образом, данные композиции могут включать жидкую среду (например, солевой раствор или воду), которая может содержать буферный раствор. pH данных лекарственных формул (и композиций) предпочтительно может быть в диапазоне от 6,4 до 8,4. Более предпочтительным является pH 7,4. Ориентировочный диапазон pH данных композиций составляет 5-10, например, 5-9, 5-8, 5.5-9, 6-7.5 или 6.5-7. Указанный pH может поддерживаться с помощью буферного раствора.
Фармацевтические составы иммуногенных композиций также могут дополнительно включать одно или более вспомогательных веществ (например, разбавители, загустители, буферные растворы, консерванты, поверхностно-активные агенты, адъюванты, детергенты и/или иммуностимуляторы), хорошо известные в соответствующей области. Применимые вспомогательные вещества должны быть совместимы с антигеном (антигенами) и с адъювантом настолько, насколько это известно в соответствующей области. Примеры разбавителей включают, связывающие агенты, разрыхлители или диспергирующие агенты, такие как крахмал, производные целлюлозы, фенол, полиэтиленгликоль, пропиленгликоль или глицерин. Фармацевтические составы могут также включать один или более активных компонентов, таких как антимикробные агенты, противовоспалительные агенты и обезболивающие. Примеры детергентов включают Tween (полисорбат), такой как Tween 80. Применимые для включения в состав композиций вспомогательные вещества известны в соответствующей области. В одном варианте исполнения иммунизации с адъювантом, например, иммуногенные полипептиды и/или их фрагменты могут быть ковалентно связаны с бактериальными полисахаридами с образованием полисахаридных конъюгатов. Такие конъюгаты могут быть использованы как иммуногенные вещества вызывающие Т-зависимый иммунный ответ направленный против бактериального полисахарида, конъюгированного с полипептидами и/или их фрагментами. Иммуногенные композиции могут выпускаться в форме набора, содержащего иммуногенную композицию и адъювант или раствор для восстановления, содержащий один или более фармацевтический приемлемых разбавителей для ускорения восстановления композиции перед введением млекопитающему с использованием традиционных или других приспособлений. Такой набор может дополнительно включать устройство для введения жидкой формы композиции (например, шприц для подкожных инъекций, набор микроигл) и/или инструкции по применению.
ПРИМЕРЫ
Вышеприведенное описание в основном относится к вариантам выполнения изобретения. Более полное понимание может быть получено после изучения приведенных ниже конкретных Примеров. Данные Примеры описаны только с целью иллюстрации и не ограничивают область данной заявки. Изменения формы или замена эквивалентов рассматриваются как условия, которые могут предлагать или предоставлять средство достижения цели. Специфические термины, использованы в данной заявке для описания и не с целью ограничений.
Использованные, но не описанные подробно, способы молекулярной генетики, белковой биохимии и иммунологии достаточно подробно описаны в научной литературе и могут применяться специалистами в соответствующей области.
Иммунный ответ
CD4+ Т-клетки считаются крайне важными для иммунного ответа против внеклеточных патогенов, таких как, например, S. pneumonaie. При взаимодействии с антигеном в контексте молекул МНС класса II с помощью антиген-презентирующих клеток (АРС), наивные CD4+ Т-клетки могут дифференцироваться в функционально различные Т-хелперные (Th) субпопуляции. Такая дифференцировка на различные Th-популяции зависит от комплексного взаимодействия с АРС в определенных, способствующих дифференцировке, условиях, включая тип и количество антигена, взаимодействие костимуляторных молекул и цитокиновые сигналы (7-9). Например, Th1-клетки, характеризующиеся продукцией интерлейкина-2 (ИЛ2), интерферона-γ (ИФН-γ) и фактора некроза опухоли-β (ФНО-β), являются наиболее важными для элиминации внутриклеточных патогенов. Th2-клетки необходимы для элиминации внеклеточных патогенов и продуцируют ИЛ-4, ИЛ-5, ИЛ-6, ИЛ-10, ИЛ-13 и ИЛ-25. Недавно описанные клетки Th17 секретируют ИЛ-17, ИЛ-21 и ИЛ-22 (10).
Иммунный ответ за счет клеток памяти развивается либо в ходе иммунного ответа эффекторных клеток (линейная модель или ассиметричное деление), либо являются остатками большего пула эффекторных клонотипов, который сокращается и присутствует в организме после удаления антигена (11). Иммунологическая память, с ее быстрым развитием ответа и высоким уровнем продукции цитокинов, представляет высокоэффективный механизм обеспечения быстрой защиты против распространенной инфекции и служит первичным защитным механизмом против повторной инфекции патогеном в местах, доступных для инфицирования, таких как слизистая оболочка респираторного тракта (12, 13). Быстрый ответ за счет Т- и В-клеток памяти развивается при естественной инфекции, а также при вакцинации, за счет лимфоцитов памяти населяющих лимфоидные и нелимфоидные органы (14-16). Однажды образовавшиеся Т-клетки памяти могут детектироваться в кровотоке на протяжении долгого периода времени (15; 17-18). Современные гипотезы развития иммунного ответа против Spn основываются на исследованиях мышиной модели, определяющих основную роль для CD4+ Th-субпопуляций клеток памяти (Th-1, Th-2 и Th-17) (19-21). На животных моделях иммунный ответ за счет CD4+ Т-клеток играет значительную роль при защите от отопатогенов и также может обеспечивать антитело-зависимый иммунный ответ (20, 22, 23). Однако, в настоящее время нет данных, относительно защитной роли Th-субпопуляций клеток памяти у людей страдающих АОМ.
Основная роль антиген-специфических CD4+ Т-клеток в адаптивном иммунном ответе заключается в обеспечении продукции антител В-клетками с одной стороны и в выполнении их собственных эффекторных функций с другой (7, 9, 23, 27). Далее, в условиях цитокинового окружения, обеспечиваемого Th-клетками и в ответ на антигенную стимуляцию, специфические В-клетки подвергаются клональной экспансии, переключению класса антител и соматической гипермутации, приводящей к отбору антител с большей аффинностью (28, 29). Размножившиеся В-клетки дифференцируют в плазматические клетки, с высокой скоростью секретирующие антитела и персистирующие в костном мозге и подобных местах, в то время как часть В-клеток дифференцирует в В-клетки памяти (29, 30). Данные В-клетки памяти могут быстро отвечать на повторную стимуляцию тем же антигеном и могут вносить вклад в поддержание пула плазматических клеток и, таким образом, сывороточного уровня антител в течение продолжительного периода времени при наличии постоянных сигналов от CD4+ Т-клеток (31).
ПРИМЕР 1
Для определения состояния восприимчивости детей к отиту, используя белковые пневмококковые антигены, оценивали количество Spn-специфичных субпопуляций CD4+ Т-лимфоцитов в периферической крови групп невосприимчивых и восприимчивых к отиту детей. Также у детей этих групп оценивали уровень антител IgG против тех же антигенов.
Пациенты в исследовании были участниками 5-летнего проспективного долгосрочного исследования АОМ, поддержанного NIH США (26). Те дети, которые переносили три эпизода АОМ в течение 6 месяцев или четыре эпизода на протяжении года, были отнесены к группе восприимчивых к отиту детей. Те дети, которые страдали от АОМ реже, были отнесены к группе невосприимчивых к отиту детей. Пациенты, включенные в данное исследование являются представителями среднего класса, проживающими в пригороде Рочестера, штат Нью-Йорк (Rochester, NY). Также в исследование были включены здоровые дети в возрасте 6 месяцев, не страдавшие от АОМ прежде. Для таких детей были получены семь посевов из сыворотки, назо-(NP) и отофарингеальных(ОР) мазков, в возрасте 6, 9, 12, 15, 18, 24 и 30. Обе группы включали детей различного возраста не старше двух лет. Образцы жидкости среднего уха были получены с помощью тимпаноцентеза при АОМ у пациентов. Оценку колонизации NP/OP Streptococcus pneumoniae и Haemophilus influenzae обычно проводили с помощью микробиологических способов при культивировании мазков NP и ОР и жидкости среднего уха. Мононуклеарные клетки периферической крови (РВМС) из полученных образцов замораживали перед дальнейшей обработкой. Образцы, исследованные в настоящем исследовании, были получены при визитах в связи с АОМ от восприимчивых детей и при колонизации или визите в связи с АОМ от детей из группы невосприимчивых. Дети были иммунизированы против S. pneumoniae в соответствии с календарем прививок по возрасту, соответствующими дозировками доступных конъюгированных вакцин. Антигены
Использовали следующие белковые антигены: PhtD (SEQ ID NO: 6), PhtE (SEQ ID NO: 8), LytB (SEQ ID NO: 11), PcpA (SEQ ID NO: 3) и PlyD1 (SEQ ID NO: 13) - детоксифицированное производное пневмолизина. В качестве контроля также использовали PspA. Последовательность каждого белка была получена из клеток штамма S. pneumoniae серотип 6 В и экспрессирована в клетках Е. Coli, с получением белка в растворимой форме, после чего препараты белков очищали с помощью различных сочетаний ионообменной хроматографии. По результатам анализа с помощью ДСН-ПААГ электрофореза и ВЭЖХ с обращенной фазой препараты белков имели степень очистки ≥90%.
Оптимальное дозирование для стимуляции определяли по отсутствию детектируемой токсичности для клеток, применяя окрашивание триптановым синим и/или проточную цитофлуориметрию после окрашивания йодида пропидия (данные не приведены).
Стимуляция Т-клеток
Мононуклеарные клетки периферической крови, полученные от восприимчивых к отиту детей, а также невосприимчивых детей, положительных по анализу NP-мазка или имеющих АОМ, вызванный инфекцией Spn, стимулировали тремя антигенами NTHi. Перед стимуляцией пробирки с замороженными клетками быстро нагревали до 37C на водяной бане, после чего к клеткам медленно добавляли полную культуральную среду (RPMI 1640, содержащую 10% ФБС (фетальной бычьей сыворотки), 2 мМ L-глутамина, 0,1 мМ пирувата натрия, заменимые аминокислоты, 100 ед/л пенициллина, 100 мкг/мл стрептомицина). Затем клетки отмывали и инкубировали с полной средой в течение ночи в 24-луночных планшетах. Затем клетки стимулировали с помощью стандартного протокола, описанного в предыдущих работах (35, 36), который был адаптирован. Коротко: подсчитывали количество клеток и переносили клетки в 96-луночные культуральные планшеты с плоским дном, после чего стимулировали клетки добавлением различных белковых антигенов в концентрации 1 мкг/мл или стафилококкового энтеротоксина В (SEB) в концентрации 1 мкг/мл. Для обеспечения костимуляторных сигналов и облегчения детекции антиген-специфического ответа в клеточную культуру добавляли анти-CD28 и анти-CD94d антитела (клоны L293 и L25 соответственно; BD Biosciences). анти-CD28 и анти-CD94d антитела широко используются для костимуляции не влияя на уровень фонового сигнала (18, 37). После этого клетки инкубировали в течение 2 часов при 37С в присутствии 5% СО2 для процессинга антигена. Через 2 часа к клеткам добавляли ингибитор транспорта через аппарат Гольджи для сохранения цитокинов внутри клетки, после чего инкубировали клетки еще 4 часа.
Определение уровня экспрессии цитокинов
Для детекции функциональных CD4+ Т-клеток, реагирующих на взаимодействие с белками Spn, циркулирующих в крови пациентов исследуемых групп после АОМ или назофарингеальной колонизации применяли многопараметрическую проточную цитофлуориметрию. Для определения принадлежности антиген-специфических CD4+ Т-лимфоцитов к определенным субпопуляциям (Th-1, Th-2 и Th17) применяли внутриклеточное окрашивание (intracellular cytokine staining assay (ICCS)). После стимуляции клетки переносили в 96-луночные планшеты с V-образным дном и однократно промывали буфером для цитофлуориметрии (PBS с 5% ФБС) и инкубировали с антителами против различных поверхностных маркеров клеток. Применяли следующие антитела: анти-CD4 АРС Alexafluor 750 (клон RPA Т4, eBiosciences), PE-Texas Red анти- CD45RA (клон МЕМ56, Invitrogen), анти-CCR7 PerCP/Су5.5 конъюгат (клоны TG8/CCR7, Biolegend). После чего мембраны клеток пермеабилизировали с помощью раствора для пермеабилизации и фиксации клеток (BD Biosciences) в течение 20 мин и три раза промывали 1х буферным раствором для пермеабилизации (BD Biosciences). Для окрашивания накопленных внутри клеток в результате стимуляции цитокинов использовали смесь различных цитокин-специфических антител. Использовали следующие антитела: конъюгированные с РЕ-Су7 анти-ИФН-γ (клон В27, BD biosciences), конъюгированные с Pacific blue анти- ИЛ17А (клон BL168, Biolegend), конъюгированные с Alexa fluor 700 анти-ИЛ-2 (клон MQ1-17H12, Biolegend), конъюгированные с РЕ анти-ИЛ-4 (клон 8D4-8, BD Biosciences), конъюгированные с AF 488 анти-ФНО-α, анти-СО3 Qdot 605 (клон UCHT1, Invitrogen) и РЕ-Су5 анти-CD69 (клон FN50, BD biosciences). После внутриклеточного окрашивания клетки промывали 3 раза 1х буфером для пермеабилизации, после чего промывали клетки один раз буфером для цитофлуориметрии, перед перенесением клеток в пробирки для цитофлуориметрии. Для анализа использовали оригинальный проточный флуориметр BD LSR II, способный анализировать 12 флуоресцентных меток, и программное обеспечение FLOW JO (Tree Star) для анализа. Для каждого образца определяли 2-5×105 событий. Для исключения клеточного дебриза и агрегатов использовали показания прямого и бокового светорассеяния и собирали фракцию CD4+CD45RALowCD69+клеток. Для подтверждения результатов собрали также фракцию ФНО-α+ клеток, сравнивая с результатами для других цитокинов. При низком уровне экспрессии цитокинов клетками, результаты подтверждали с помощью так называемого «обратного гейтирования». Для устранения фонового сигнала при выделении антиген-специфичных клеток использовали анти-CD28/CD49d антитела, как описано ранее (37). Вся способность анализа была стандартизирована и проведено сравнение с коммерчески доступным набором для анализа уровня экспрессии цитокинов на основе цепочек гранул (СВА, BD Biosciences)
Гуморальный иммунный ответ
Для определения уровня антител в исследуемых образцах применяли ИФА, как описано ранее (26, 38). Коротко, в лунки 96-луночных планшетов (Nunc-Immulon) сорбировали антигены из концентрации 0,25 мкг/мл (100 мкл/лунку) в буфере для сорбции (бикарбонатный буферный раствор (pH 9,4)) в течение ночи при +4°C. После отмывки, блокировали сайты неспецифического связывания 3% раствором обезжиренного молока (200 мкл/лунку), инкубируя планшеты 1 час при 37°C. Отмывали планшеты пять раз, после чего в лунки добавляли 100 мкл сыворотки в стартовом разведении 1:100 (в буфере PBS с 3% обезжиренного молока), с последующей серией последовательных разведений с шагом 2. Смесь инкубировали при комнатной температуре в течение 1 часа с последующим добавлением аффинно очищенных козьих антител против IgG человека, конъюгированных с пероксидазой корней хрена (Bethyl Laboratories, Inc, Montgomery, TX), в качестве вторичного антитела. Продукт реакции детектировали с помощью набора ТМВ Microwell Peroxidase Substrate System (KPL, Gaithersburg, MD), останавливая реакцию добавлением 1,0 M раствора фосфорной кислоты. Измерение оптической плотности проводили с помощью стандартного автоматического спектрофотометра для ИФА при длине волны 450 нм. Для количественного определения концентрации специфических антител, сигнал сравнивали с сигналом от контрольной сыворотки (смешанный образец сыворотки человека с высоким уровнем специфических антител). Количественно уровень IgG в контрольной сыворотке определяли с помощью ИФА-набора для количественного определения IgG человека (Bethyl laboratories). Для построения кривых для исследуемого и контрольного образцов применяли четырехпараметрическую log-логистическую функцию. Данный ИФА был валидирован с полном объеме в соответствии с Руководством ICH.
Для статистического анализа полученных данных использовали пакет программ Graph Pad Prism. Для проверки достоверности различий применяли критерий Манн-Уитни с двусторонним значением Р.
Результаты
В исследование в обе группы (восприимчивых и невосприимчивых к отиту) были включены дети сходного возраста. Также обе группы были сходны по половому составу, дню начала лечения, уровню пассивного курения в месте проживания, числу сибсов младше 8 лет, применению грудного вскармливания.
После стимуляции мононуклеарной фракции периферических клеток крови специфическими антигенами, сравнивали количество различных субпопуляций Th-клеток памяти специфичных против Spn у восприимчивых и невосприимчивых к отиту детей. Для этого рассчитывали количество CD45RALow CD4+ Т-клеток памяти, продуцирующих ИФН-γ, ИЛ-4, ИЛ-2 или ИЛ-17, собирая активированные CD69+ Т-клетки. Полученные данные нормализовали на количество, полученное для нестимулированных мононуклеарных клеток или стимулированных неспецифическим антигеном (гемоцианин брюхоногого моллюска).
На фигуре 1 приведены сводные результаты, демонстрирующие уровень различных субпопуляций CD45RALow CD4+ Т-клеток памяти, специфических по отношению ко всем антигенам Spn, использованным для стимуляции, у невосприимчивых к отиту детей (n=15) с АОМ (n=6) или с назофарингеальной колонизацией Spn (n=9). В отличие от этой группы, у детей группы восприимчивых к отиту (n=13) показано заметно сниженное количество Spn-специфичных CD4+ Т-клеток памяти в периферической крови после АОМ (n=10) и назофарингеальной колонизации (n=3). В частности, обнаружено полное отсутствие CD4+ Т-клеток памяти продуцирующих ИФН-γ против LytB, PhtE и PspA. Кроме того, клетки таких пациентов продуцировали значительно меньшее количество ИФН-γ при стимуляции PhtD, РсрА и PspA (Р<0,02) (Фиг. 1а). В группе восприимчивых к отиту детей показано значительное снижение уровня продукции CD4+ Т-клетками памяти ИЛ-4 при стимуляции PhtD и LytB (Р<0.02) антигенами (Фиг. 1b). Уровень продукции ИЛ-2 CD4+ Т-клетками памяти детей этой группы был значимо ниже при стимуляции PhtD (Р<0.05), РсрА (Р<0.005), PhtE (Р<0.05), Ply (Р<0.005) и PspA (0.02) (Фиг. 1с). Кроме того, у восприимчивых к отиту детей было показано сниженное количество клеток продуцирующих ИЛ-17а при стимуляции антигенами PhtD, РсрА и PhtE (Р<0.05) (Фиг. 1d).
Поскольку отсутствие антиген-специфических Th-клеток памяти может приводить к нарушению антиген-специфического В-клеточного ответа (9), оценивали титр антиген-специфических антител у детей групп восприимчивых и невосприимчивых к отиту. Сывороточный уровень IgG против антигенов пневмококка у детей соответствующих групп приведен на Фигуре 2. Как предполагалось, при высоком уровне Т-клеток памяти титр IgG против PhtD, LytB, PhtE, Ply значимо выше у детей группы невосприимчивых к отиту по сравнению с детьми 3 группы восприимчивых к отиту (Р<0.05; 0.0005; 0.0005; 0.005 соответственно) (Фиг. 2). Также отмечена разница титра IgG против РсрА, однако не являющаяся статистически достоверной (Фиг. 3).
Поскольку иммунная система маленьких детей не полностью сформирована в отношении В- и Т-клеточного ответа (39, 40), анализировали В- и Т-клеточный ответ для оценки связи сниженного иммунного ответа за счет CD4+ Т-клеток памяти у восприимчивых к отиту детей, с нарушениями Т- или В-клеточного звена в целом. Клетки мононуклеарной фракции периферической крови стимулировали с помощью SEB, антигена, стимулирующего Т-клетки независимо от презентации на АПК (антиген-презентирующих клетках) (41).На Фигуре 3 показано, что доля CD4+ Т-клеток памяти продуцирующих ИФН-γ, ИЛ-4, ИЛ-2 ли ИЛ17а одинакова у детей обеих групп - восприимчивых и невосприимчивых к отиту. Для проверки отсутствия общего иммунодефицитного состояния у детей восприимчивых к отиту оценивали титр IgG против антигенов дифтерии, столбняка и коклюша, входивших в состав вакцины DTaP (АКДС), полученной детьми. Значимых различий в концентрации IgG против дифтерийного токсина, токсина столбняка, токсина коклюша, филаментозного гемагглютинина или пертактина не найдено между двумя исследованными группами (данные не приведены).
В сумме полученные данные демонстрируют отсутствие или значительно сниженное количество CD4+ Т-клеток памяти, специфичных против антигенов пневмококка, у детей, восприимчивых к отиту, по сравнению с детьми из группы невосприимчивых. Данный эффект ассоциирован со сниженным количеством IgG специфических к антигенам взятым в исследование. Как показано в нашем исследовании, у восприимчивых к отиту детей не происходит иммунного ответа за счет специфических к Spn функциональных CD45RALow Th-клеток памяти. Выработка антител против белковых антигенов Spn у таких детей снижена. Однако, такие пациенты не имеют общей недостаточности функциональной активности Т-клеток памяти или В-клеточного гуморального иммунного ответа (например, выработки IgG) против антигенов, в отношении которых была проведена вакцинация.
Несмотря на тот факт, что CD4+ хелперные Т-клетки грают роль в элиминации инфекций вызванных Spn или NTHi, в настоящее время не опубликовано исследований, демонстрирующих прямую связь между CD4+ Т-клетками памяти и развитием Spn- или NTHi-опосредованного АОМ у детей. Очевидно, что сниженная активность образования CD4+ Т-клеток памяти у детей приведет к сниженному В-клеточному гуморальному иммунному ответу. Отсутствие иммунологической памяти, т.о. может привести к повторяющейся восприимчивости к инфекциям уха. В данном исследовании впервые показано, что у восприимчивых к отиту детей отсутствуют/снижено количество Th-клеток памяти специфических к патогенам уха (например, S. pneumoniae) в периферической крови после АОМ и/или NP-колонизации. У детей невосприимчивых к отиту напротив, происходит образование специфических CD4+ Т-клеток после АОМ и/или назофарингеальной колонизации патогенами уха.
По видимому, у восприимчивых к отиту детей происходит развитие краткосрочного В-клеточного ответа, поскольку некоторые антитела могут быть детектированы у таких детей после АОМ или назофарингеальной колонизации S. pneumoniae. Однако, при отсутствии Т-клеток памяти, после того, как уровень антител снижается, такие дети быстро становятся восприимчивы к новым инфекциям АОМ. Таким образом, основа иммунодефицита у восприимчивых к отиту детей, по-видимому, заключается в неспособности к образованию Т-клеток памяти. Поскольку у восприимчивых к отиту детей иммунный ответ на антигены, не требующие презентации АПК (SEB), а также на антигены, вводимые парентерально в форме вакцины DTaP, схож с таковым у невосприимчивых к отиту детей, возможно, что нарушение иммунного ответа у восприимчивых детей против Spn и NTHi связано с нарушениями процессинга и презентации их антигенов АПК слизистой носа.
В предыдущей работе показана роль антигенов Spn и NTHi при пролиферативном ответе CD4+ Т-клеток (в течение 5-7 дней) у детей и взрослых (42, 43). В более раннем исследовании, при анализе пролиферации субпопуляции CD4+ Т-клеток из клеток аденоидов или миндалин восприимчивых к отиту детей, показано отсутствие пролиферации при стимуляции белком Р6 NTHi (44).Такие исследования позволяют получить информацию об антиген-специфической пролиферации Т-клеток, однако в них никак не исследуются антиген-специфические CD4+ Т-клетки памяти.
Тогда как CD4+Th-2 клетки в основном обеспечивают гуморальный иммунный ответ, способствующий элиминации бактериальных патогенов из организма, в недавних исследованиях на мышиной модели показан антитело-независимый иммунный ответ на назофарингеальную колонизацию Spn, осуществляемый CD4+ Т-клетками, продуцирующими ИЛ-17а (Th17-клетки) (20). В нашем исследовании впервые показано повышенное количество Spn-специфичных Th-клеток памяти продуцирующих ИЛ-17а в периферической крови невосприимчивых к отиту детей, по сравнению с восприимчивыми детьми. Таким образом, Spn-специфические Th-клетки памяти, продуцирующие ИЛ 17а, могут защищать от развития состояний восприимчивости к отиту.
Фенотипирование клеточных популяций на месте инфекции при АОМ (слизистая и жидкость среднего уха) позволяет предполагать значительную миграцию CD45ROHigh/CD45RALow CD4+ Т-клеток не несущих рецепторов хоминга - L-селектинов (45). В других исследованиях показано накопление в основном CD4+ Т-клеток памяти в жидкости среднего уха при АОМ (45-47). Местные вторичные лимфоидные органы, такие как аденоиды, являются основными местам праймирования Т-клеток при инфекциях верхних дыхательных путей, таких как бактериальная колонизация. После миграции АПК с антигеном в локальные лимфоидные органы (аденоиды) происходит дифференцировка лимфоцитов (CD4+ Т-клеток). После выхода в кровоток такие CD4+ Т-клетки немедленно мигрируют к слизистой среднего уха (в случае АОМ) и/или верхних дыхательных путей (в течение назофарингеальной колонизации).
Хотя теоретические основания отсутствуют, считается, что задержка иммунологического созревания вызывает отсутствие функциональных Т-клеток у восприимчивых к отиту детей (48). По мере взросления такие дети становятся менее восприимчивы к АОМ вследствие анатомических изменений евстахиевой трубы, а также по мере созревания иммунной системы. Полноценный ответ на основе Т-клеток памяти развивается у детей в возрасте 3-5 лет. Обычно восприимчивые к отиту дети «перерастают» эту склонность в течение данного возрастного периода. CD4+ Т-клетки памяти могут играть ключевую роль при иммунном ответе при АОМ у человека. Таким образом, специфические против Spn CD4+ Т-клетки памяти, в случае их образования, будут эффективны при профилактике повторяющихся АОМ инфекций.
ПРИМЕР 2
В данном исследовании изучалось образование антител IgG против PhtD, PhtE, LytB, РсрА и Ply в сыворотке у детей с АОМ трех групп в возрасте от 3 до 36 месяцев: 1) восприимчивые к отиту дети, группа которых включала детей, кто перенес 3 и более эпизодов АОМ за шесть месяцев или четыре или более эпизодов за 12 месяцев; 2) группа детей, у которых лечение АОМ оказалось неудачным(АОМТР), включающая пациентов, у которых не произошло ликвидации бактерий и/или разрешения симптомов после, по меньшей мере, 48 часов соответствующей антибиотикотерапии (70, 71), а также пациентов, у которых признаки и симптомы АОМ появились вновь в течение 14 дней после завершения курса антибиотикотерапии; 3) группа невосприимчивых к отиту детей, перенесших только один или два эпизода АОМ.
Собранные и проанализированные образцы были получены в ходе проспективного исследования, описанного в Примере 1. Здоровые дети в возрасте 6 месяцев, не переносившие АОМ, были включены в исследование и наблюдались до 30-месячного возраста. Были получены по 7 образцов сыворотки, назофарингиальные (NP) и отофарингиальные культуры (OP) от каждого участника исследования в возрасте 6, 9, 12, 15, 18, 24, и 30 месяцев. Образцы, полученные для возраста 30 месяцев, были исключены из анализа, так как очень небольшое число участников эксперимента продолжало участвовать в исследовании на этот период. На протяжении всего периода исследования, во время АОМ от каждого пациента получали образцы сыворотки, NP и ОР-культур, а также жидкость среднего уха с помощью тимпаноцентеза. Образцы от выздоравливающих пациентов собирали 3 недели спустя. У большинства детей не было АОМ (около 70%), поэтому они были включены в группу №3 (невосприимчивые к отиту дети). Некоторые дети были включены группу восприимчивых к отиту (около 5%, группа №1), а некоторые в группу AOMTF (около 5%, группа №2). Для увеличения численности групп восприимчивых к отиту детей и группы AOMTF, в исследование были включены дополнительные пациенты, безотносительно возраста, в котором они стали удовлетворять критериям группы 1 или 2, на протяжении периода от 6 месячного возраста, до 36 месячного. Во время АОМ немедленно собирали образцы сыворотки, NP и ОР-культур, MEF-образцы, а три недели спустя собирали такие же образцы от выздоравливающих участников.
Для подтверждения диагноза АОМ дети обследовались детскими врачами-отоларингологами опираясь на Диагностическое руководство по АОМ от Американской Академии Педиатров. Тимпаноцентез проводили с целью подтверждения присутствия отопатогена в MEF. Образцы MEF и NP и ОР-мазки инокулировали в триптиказо-соевый бульон, высевали на чашки с триптиказо-соевым агаром с 5% овечьей крови, а также на чашки с шоколадным агаром. Бактерии выделяли в соответствии со стандартными культуральными способами CLSI.
ИФА: белки PhtD, LytB, РсрА, PhtE и PlyD1 S. pneumoniae, описанные в Примере 1, также использовали для этого исследования. Титр специфических к данным белкам антител определяли с помощью ИФА, используя очищенные рекомбинантные белки. В лунки 96-луночных планшетов Nunc-Immulon 4 сорбировали антигены из концентрации 0,5 мкг/мл (100 мкл/лунку) в бикарбонатном буферном растворе (pH 9,4) в течение ночи при 4°C. После отмывки, блокировали сайты неспецифического связывания 3% раствором обезжиренного молока (200 мкл/лунку), инкубируя планшеты 1 час при 37°C. Отмывали планшеты пять раз, после чего в лунки добавляли 100 мкл сыворотки в стартовом разведении 1:100 (в буфере PBS с 3% обезжиренного молока), с последующим), с последующей серией последовательных разведений с шагом 2. Смесь инкубировали при комнатной температуре в течение 1 часа с последующим добавлением аффинно очищенных козьих антител против IgG человека, конъюгированных с пероксидазой корней хрена (Bethyl Laboratories, Inc, Montgomery, TX), в качестве вторичного антитела. Продукт реакции детектировали с помощью набора ТМВ Microwell Peroxidase Substrate System (KPL, Gaithersburg, MD), останавливая реакцию добавлением 1,0 M раствора фосфорной кислоты. Измерение оптической плотности проводили с помощью спектрофотометра для планшетов Spectra max (Molecular Devices, Sunnyvale, Calif.) при длине волны 450 нм, используя протокол разбавления в конечной точке Softmax.
Статистическую обработку результатов проводили с использованием программного обеспечения GraphPad Prism 5. Непарный t-критерий использовали для сравнения различий в уровне IgG между тремя группами. Парный t-критерий применяли для оценки достоверности различий между образцами, полученными в остром состоянии и образцами в процессе выздоровления. Односторонний тест ANOVA применяли для увеличения концентрации антитела с течением времени. Значения р<0,05 рассматривали как статистически значимое.
Специфический титр антител против белков PhtD, LytB, РсрА, PhtE и Ply у пациентов трех групп во время АОМ
Титр антител IgG против белков PhtD, LytB, РсрА, PhtE и Ply Spn измеряли во время острого АОМ у 35 детей из группы восприимчивых к отиту, 25 детей с AOMTF и 34 детей из группы невосприимчивых, у которых АОМ был второй или третий раз в жизни (Фигура 4).
Титр IgG против белка PhtD у детей 3 группы восприимчивых к отиту был значимо ниже по сравнению с детьми из группы невосприимчивых к отиту (р<0,05). Уровень IgG против PhtD у детей из группы AOMTF также был ниже такового у невосприимчивых к отиту детей, однако разница не достигла статистически значимого значения. Титр IgG против белка LytB у восприимчивых к отиту детей и у детей с AOMTF был значимо ниже такового у невосприимчивых к отиту детей (р<0,001 для обоих сравнений). GMT иммуноглобулинов IgG против белка РсрА у детей из группы восприимчивых к отиту и у детей из группы AOMTF был почти в три раза ниже такового по сравнению с детьми из группы невосприимчивых к отиту.
Однако, различие не было статистически значимым при сравнении трех групп, вследствие большой вариабельности уровня антител. Титр IgG против белка PhtE у детей из группы восприимчивых к отиту и группы AOMTF был значимо ниже такового у невосприимчивых к отиту детей (р<0,001). Титр IgG против белка PlyD1 у детей из группы восприимчивых к отиту и группы AOMTF был значимо ниже такового у невосприимчивых к отиту детей (р=0,001; р=0,02, соответственно).
Уровень антител против белков PhtD, LytB, РсрА, PhtE и Ply S. pneumoniae в образцах пациентов при остром АОМ и при выздоровлении у детей трех групп Были получены парные образцы сыворотки крови при остром АОМ (во время АОМ) и на стадии выздоровления (3 недели спустя) у 20 детей из группы восприимчивых к отиту, 13 детей из группы AOMTF и 20 детей из группы невосприимчивых к отиту. У пациентов всех трех групп уровень антител IgG против 4 из 5 белков при остром АОМ в сравнении с периодом выздоровления не отличался в сторону значимого повышения антител в острой фазе заболевания (за исключением антител против белка PhtE у детей из группы AOMTF, р=0,04) (Таблица 1).Однако, были обнаружены значительные индивидуальные различия в уровне антител в острой стадии и на стадии выздоровления, поскольку у некоторых детей из всех трех групп уровень антител к одному или нескольким антигенам был в два раза выше (Таблица 2).
Возрастное изменение уровня антител у восприимчивых и невосприимчивых к отиту детей
На Фигуре 5 показаны данные по уровню IgG-антител, специфических против белков PhtD, LytB, РсрА, PhtE и Ply, полученные при плановых, не связанных с АОМ, визитах участвующих в проспективном исследовании восприимчивых и невосприимчивых к отиту детей в возрасте от 6 до 24 месяцев. Приведенные данные получены от 150 невосприимчивых и 10 восприимчивых к отиту детей. У невосприимчивых к отиту детей уровень антител IgGnpoTHB всех белков со временем повышался (р<0,001), кроме Lyt В (р=0,075). У детей восприимчивых к отиту такой тенденции не наблюдалось для антител против ни одного из пяти белков (р=0,40 для белка PhtD, р=0,39 для белка LytB, р=0,11 для белка РсрА, р=0,09 для белка PhtE и р=0,42 для белка Ply).
Эти данные демонстрируют, что восприимчивые к отиту и дети с AOMTF имеют значимо более низкий уровень антител к белкам Spn при начале АОМ, чем невосприимчивые к отиту дети, что позволяет предполагать, что предыдущая инфекция Spn не вызвала, или вызвала менее мощный адаптивный иммунный ответ, что отражается на сывороточном уровне антител. Эти данные позволяют предполагать, что иммунологически восприимчивые к отиту дети и дети с AOMTF схожи, однако их иммунный ответ отличается от такового у невосприимчивых к отиту детей. Кроме того, уровень сывороточных антител против 5 антигенов Spn нарастает значимо медленнее у восприимчивых к отиту детей, по сравнению с невосприимчивыми. Более медленное нарастание уровня антител после естественного контакта с патогеном при назофарингеальной колонизации у восприимчивых к отиту детей согласуется с данными о сниженном иммунном ответе у таких детей при контакте с отопатогеном. Также полученные данные свидетельствуют о том, что восприимчивые к отиту дети и дети с AOMTF не отличаются от невосприимчивых к отиту детей по уровню сывороточных антител при АОМ. По-видимому, АОМ не является иммунизирующим событием для большинства детей из любой из трех исследованных групп, по крайней мере, до 3-летнего возраста (в соответствии с данными полученными в настоящем исследовании).
Полученные данные относительно антигенспецифического иммунного ответа против Spn подтверждают наблюдения других авторов для восприимчивых к отиту детей и не согласуются с некоторыми более ранними работами, представляя собой совершенно новые данные. Так Freijd с соавт. (72) сравнивал уровень антител против полисахаридов Spn серотипов 3, 6А и 23 у 15 у восприимчивых к отиту детей в возрасте 30 месяцев с таковым у контрольной группы детей такого же возраста и у группы взрослых. Был показан значимо более низкий уровень антител против антигенов серотипов 6А и 23 у детей, восприимчивых к отиту. Prellner с соавт. (73) проводил измерение уровня сывороточных антител против полисахаридов Spn серотипов 6А, 19 и 23 у 15 восприимчивых к отиту детей. Было показано, ч то 60% детей не имели детектируемых антител. Даже в возрасте 6 лет уровень антител против полисахаридов серотипа 6А у восприимчивых к отиту детей был ниже такового у невосприимчивых детей. Hotomi с соавт. (74) оценивал уровень сывороточных антител против полисахаридов NTHi ОМР Р6 и Spn (используя 23-валентую вакцину в которой Spn выступает в роли антигена) у 36 восприимчивых (средний возраст - 18 месяцев)и 20 невосприимчивых к АОМ детей. У 55% восприимчивых к отиту детей уровень антительного ответа против Р6 и у 48% против полисахаридов Spn был ниже. В исследованиях Yamanaka и Faden (75;76) и Bernstein с соавт. (77) показан схожим образом сниженный уровень сывороточных/или антител слизистой против другого отопатогена, NTHi, у восприимчивых к отиту детей. Насколько нам известно, наше исследование -первые данные о сывороточном уровне антител к антигенам Spn у восприимчивых к отиту детей и детей с AOMTF.
Полученные данные относительно ассоциированной с АОМ выработки антител против белков PhtD, LytB, РсрА, PhtE и Ply у восприимчивых к отиту детей подтверждают общепринятую гипотезу, объясняющую восприимчивое к отиту состояние: дети, восприимчивые к отиту обладают специфической недостаточностью антительного ответа против Spn и других отопатогенов в случае, если контакт с патогеном осуществляется естественным образом через назофарингеальный путь.
Как описано в Примере 1, восприимчивые к отиту дети демонстрируют недостаточную активность функциональных хелперных Т-клеток и Т-клеток памяти в ответ на инфекцию Spn и NTHi (неопубликованные данные) (82). У таких детей выработка антител при парентеральной вакцинации вакциной против дифтерии, столбняка и коклюша не снижена, что согласуется с данными Prellner с соавт. (83) и Wiertsma с соавт. (84), которые также показали, что у восприимчивых к отиту детей вырабатывается нормальное количество антител при вакцинации против кори и других детских заболеваний. Таким образом, показанная дисфункция иммунной системы у восприимчивых к отиту детей связана с естественным контактом с отопатогеном и не связана с парентеральной вакцинацией. Данные о развитии нормального иммунного ответа против конъюгированных Spn-вакцин у восприимчивых к отиту детей, подтверждают данное заключение (85, 86).
При сравнении уровня антител против исследуемых антигенов Spn при остром состоянии и при выздоровлении, общий уровень GMT не увеличивается значимо ни у восприимчивых к отиту детей, ни у детей с AOMTF или невосприимчивых к отиту. В основном это связано с высокой вариабельностью индивидуального иммунного ответа ребенка. Действительно, у некоторых детей при выздоровлении наблюдается более высокий титр антител, тогда как у других титр антител ниже или остается на том же уровне. Наиболее вероятно, что такие различия связаны с длительностью назофарингеального носительства Spn перед развитием АОМ. У пациентов с длительным периодом носительства пик продукции антител может быть до развития АОМ, таким образом, у таких пациентов график уровня антител в сыворотке в остром состоянии и при выздоровлении может быть ровным или падающим. У других пациентов, с коротким периодом назофарингеального носительства перед развитием АОМ, график уровня антител в сыворотке в остром состоянии и при выздоровлении может быть нарастающим. Полученные результаты демонстрируют тот факт, что различные антигены вызывают антительный ответ разного профиля, что возможно является отражением их различной антигенности для маленьких детей, в случае, когда белок попадает в организм естественным путем при бессимптомной колонизации или с развитием АОМ. Схожие наблюдения были сделаны при анализе антительного ответа против белков NTHi другими группами, показавшими такую же вариабельность при остром состоянии и выздоровлении при АОМ (87-89). Soininen с соавт. исследовали процесс естественной выработки антител против полисахаридов Spn типов 1, 6В, 11А, 14, 19F и 23F, ассоциированную с назофарингеальной колонизацией и развитием АОМ группы из 329 детей в течение первых 2-х лет жизни (90). Уровень антител нарастал со временем медленно, однако значимо; серотипы 11A и 14 являются более иммуногенными в более молодом возрасте. В работе показано, что уровень антител был одинаков при назофарингеальной колонизации и при АОМ. Однако в более позднем исследовании тех же выборок детей Soininen с соавт. описали данные явления нарастания уровня антител более чем в 2 раза, как относительно нечастые события после АОМ, связанные с возрастом ребенка и серотипом Spn (89).
В соответствующем исследовании было отмечено постепенное накопление антител против аналогичных исследованным в нашей работе пяти белков Spn, а также трем белкам NTHi (Белок D, Р6 и ОМР26) у здоровых детей (69, 87). В данном исследовании у восприимчивых к отиту детей не происходило или было в значительно меньшей степени выражено нарастание с возрастом уровня антител против пяти белков Spn.
Заключая, полученные результаты вносят вклад в понимание иммунологического ответа у восприимчивых к отиту детей. Показан сниженный иммунный ответ у восприимчивых к отиту людей против антигенов Spn. Иммунный ответ у детей с AOMTF был схож с таковым у детей, восприимчивых к отиту. Введение вакцинной композиции содержащей, по меньшей мере, один или более антигенов из PhtD, PhtE, РсрА, LytB и детоксифицированного пневмолизина (например, PlyD1) парентеральным путем (дополнительно, с адъювантом) может быть использовано для минимизации сниженной иммунореактивности при естественном контакте с S. pneumoniae.
ПРИМЕР 3
Сравнивали количество Spn антиген-специфичных В-клеток в периферической крови восприимчивых и невосприимчивых к отиту детей, описанных в Примере 1. От общего количества 387 детей, участвующих в исследовании, в данное исследование были вовлечены 10 восприимчивых к отиту детей (на основе доступности образцов клеток мононуклеарной фракции периферической крови) и 12 случайно выбранных невосприимчивых к отиту детей, перенесших 1-2 АОМ, схожего с группой восприимчивых детей возраста в качестве контрольной группы. Клинические данные для детей, участвующих в данном исследовании, приведены в Таблице 3.
Количество клеток продуцирующих специфические IgG (к белкам PhtD, PhtE, LytB, РсрА, Ply) и общее количество IgG-продуцирующих клеток определяли с помощью анализа ELISPOT (внутрилабораторно стандартизированного), где В-клетки памяти были стимулированы in vitro для дифференцировки в антитело-секретирующие клетки (АСК). Вкратце, 1 миллион размороженных клеток мононуклеарной фракции периферической крови помещали в каждую лунку 24-луночного планшета, содержащие по 1 мл полной среды или по 1 мл полной среды с митогеном лаконоса (1 мкг/мл). Клетки инкубировали при 37°C в течение 3 дней для дифференцировки, затем отмывали полной средой, определяли количество и помещали в лунки 96-луночного планшета для ELISPOT (Milipore) с сорбированным антигеном (10 мкг/мл), после чего инкубировали в течение ночи. Определение плазматических клеток проводили с помощью анализа дифференцированных клеток на проточном цитофлуориметре (результаты не приведены). Для определения общего количества IgG-секретирующих клеток в лунки планшета было сорбировано моноклональное антитело против IgG человека (МТ91/145; Mabtech) из концентрации 10 мкг/мл в PBS. В качестве негативного контроля использовали лунки, в которые ничего не сорбировали или сорбировали бычий сывороточный альбумин (БСА) из такой же концентрации. Сайты неспецифического связывания блокировали инкубацией с 10% ФБС в RPMI1640 в течение 30 мин при 37°C. Определяли количество стимулированных клеток мононуклеарной фракции периферической крови и 5×105 клеток ресуспендировали в 200 мкл свежей полной среды RPMI перед внесением их в лунки с сорбированным антигеном или контрольные лунки. Планшеты инкубировали при 37°C и 5% СО2 в течение ночи и, затем, отмывали лунки PBS 5 раз. Затем добавляли в лунки 100 мкл раствора 1 мкг/мкл биотинилированных антител против IgG человека (МТ78/145; Mabtech) и инкубировали планшеты в течение часа. Затем отмывали лунки и вносили конъюгат стрептавидин-щелочная фосфатаза (в разведении 1:1000) и инкубировали планшеты в течение часа при 37°C. Затем лунки отмывали 5 раз PBS перед внесением субстрата (BCIP/NBT; Mabtech). Вследствие низкого количества антиген-специфических АСК, пятна анализировали вручную, с использованием микроскопа для микродиссекций. Долю антиген-специфических В-клеток памяти определяли на миллион клеток мононуклеарной фракции периферической крови по следующей формуле: % антиген-специфических В-клеток памяти = (число антиген-специфических пятен / общее число пятен антител)×100.
Титра антиген-специфических антител в сыворотке данных двух групп детей определяли с помощью ИФА, с использованием в общем схожего с описанным в Примере 1 протокола, с изменениями: антиген сорбировали из раствора с концентрацией 0,5 мкг/мл, в качестве вторичных антител использовали аффинно очищенные антитела козы против IgG, IgM или IgA человека, конъюгированные с пероксидазой корней хрена (Bethyl Laboratories, Inc., Montgomery, TX).
Статистическую обработку полученных данных проводили с помощью программного обеспечения Graph Pad Prism. Для оценки достоверности отличий использовали критерий Манн-Уитни с двусторонним значением Р.
Полученные результаты суммированы на Фигуре 6 (А, В, С). Доля В-клеток памяти специфичных к 5 антигенам Spn (PhtD, PhtE, LytB, РсрА, Ply) у восприимчивых и невосприимчивых к отиту детей показана на Фигуре 6А. В отличие от невосприимчивых детей, восприимчивые к отиту дети имеют заметно сниженное количество Spn-специфических В-клеток памяти в периферической крови после АОМ или назофарингеальной колонизации (Фигура 6А). В частности, значимо более низкая доля В-клеток памяти, продуцирующих антиген-специфические IgG, наблюдалась по отношению к антигенам PhtD, PhtE и PlyD1 (Р<0,02). Кроме того, у восприимчивых к отиту детей наблюдалось также сниженное общее количество антиген-специфических В-клеток памяти по отношению к антигену LytB, однако показанное различие не было статистически достоверным (р=0,01). Не показано статистически значимых различий в количестве РсрА-специфических В-клеток памяти между группами восприимчивых и невосприимчивых к отиту детей (Фигура 6А). Также между образцами, полученными для изученных групп, не различалось общее число IgG-секретирующих клеток (данные не приведены).
Сывороточный уровень IgG против Spn-антигенов у представителей изученных групп приведен на Фигуре 6 В. У детей из группы невосприимчивых к отиту титр IgG специфических к антигенам PhtD, РсрА и PhtE был значимо выше по сравнению с таковым у восприимчивых к отиту детей (Р<0,05). Титра антител против Ply был ниже и статистически не различался у изученных групп (Фигура 6В). Титр антител против LytB оказался наиболее низким по сравнению с антителами против других антигенов в данном анализе у обеих групп детей (Фигура 6В).
В этом исследовании показано сниженное количество В-клеток памяти в периферической крови восприимчивых к отиту детей с последующим АОМ и/или назофарингеальной колонизацией (Фиг. 6А). После встречи наивной В-клетки с антигеном во вторичных лимфоидных органах образуется антиген-специфическая В-клетка памяти и антитело-секретирующая клетка, которые мигрируют из периферической крови в костный мозг, селезенку и целевой орган, например дыхательный тракт (16). Поскольку уровень антител в сыворотке поддерживается В-клетками памяти (31), после анализа количества образующихся антиген-специфических В-клеток памяти, нами получено более точное иммунологическое объяснение показанному сниженному уровню антител у восприимчивых к отиту детей. Для подтверждения связи низкого количества В-клеток памяти с уровнем антител в сыворотке, измеряли титр Spn-специфических антител. Показано, что титр значимо снижен у восприимчивых к отиту детей (Фиг. 1В), что сходно с результатами, полученными в Примере 1 с использованием образцов сыворотки от различных групп невосприимчивых (n=15) и восприимчивых к отиту детей (n=13) с последующим АОМ или назофарингеальной колонизацией. Общая тенденция более высокого титра Spn-специфических антител у невосприимчивых к отиту детей согласуется с таковой, наблюдавшейся в Примере 1, хотя точные результаты относительно статистической значимости различий между группами по антиген-специфическому ответу различаются в разных случаях. Например, у небольшой группы детей, взятых в данное исследование, не было найдено различий в титре Ply-специфических антител. Хотя антительный ответ и образование В-клеток, специфичных против конкретного белкового антигена, после бактериальной колонизации и/или АОМ могут различаться у разных индивидуумов, меньшая степень различий ожидается при вакцинации.
Как показано в Примере 1, восприимчивые к отиту дети демонстрируют субоптимальный ответ CD4+ Т-клеток памяти, специфичных к пневмококковым антигенам (96). Эти данные подтверждают результаты, описанные в более ранних Примерах (т.е. что у детей восприимчивых к отиту может развиваться некоторый антительный ответ), поскольку количество антител и В-клеток памяти было на определимом уровне у данных детей после АОМ и назофарингеальной колонизации отопатогенами (Фиг. 6А-В). Однако, при отсутствии антиген-специфических В-клеток памяти или CD4+ Т-клеток памяти, уровень антител снижается, и восприимчивые к отиту дети не способны поддерживать подходящий уровень антител в сыворотке и становятся восприимчивыми к повторению инфекции АОМ. Вакцинация конъюгированной вакциной с пневмококковыми полисахаридами применима для создания защитного уровня противополисахаридных антител (86), однако вариабельность на уровне серотипов ограничивает защитную эффективность штамм-специфических противополисахаридных антител (95). Более того, несмотря на тот факт, что у восприимчивых к отиту детей могут образовываться серотип-специфические антитела к конъюгированной вакцине, повторные инфекции распространены среди представителей такой восприимчивой группы (86). Эти данные говорят о том, что развивающийся серотип-нейтрализующий иммунный ответ является неполным и непродолжительным.
Примечательно, что доля специфичных к PhtD В-клеток памяти в периферической крови коррелирует с уровнем PhtD в сыворотке (Фиг. 6С). Также наблюдались различия между долей антиген-специфических В-клеток памяти и сывороточным уровнем антител против РсрА и PlyD1 (Фиг. 6А-В).
Заключая, в отношении антигенов в данном исследовании у восприимчивых к отиту детей происходит значительно более слабое образование В-клеток памяти, которые могут дифференцироваться в антитело-секретирующие клетки (АСК). Клинические последствия этих особенностей очевидны. Антиген-специфические В-клетки памяти поддерживают на определенном уровне антитела в сыворотке и способны пролиферировать в АСК при повторной инфекции данным антигеном, что приводит к росту сывороточного уровня антител. В нашем исследовании показано, что у восприимчивых к отиту детей нет нарушений в общем количестве IgG-секретирующих клеток. Более того, в результате анализа с помощью проточного цитофлуориметра, обнаружено, что у восприимчивых к отиту детей нет нарушений при трансформации В-клеток памяти (CD19+IgD-) в плазматические антитело-секретирующие клетки (CD27+CD38+CD138+), в ответ на поликлональную стимуляцию (данные не приведены).
Эти данные демонстрируют наличие Spn антиген-специфического иммунного ответа после АОМ или назофарингеальной колонизации у детей обеих групп: восприимчивых и невосприимчивых к отиту. Хотя у восприимчивых к отиту детей наблюдается сниженный иммунный ответ, тем не менее, у таких детей наблюдается иммунный ответ после естественной инфекции или колонизации поддержанной введением вакцинной композиции содержащей, по крайней мере, один или более из PhtD, PhtE, РсрА, LytB и инактивированного пневмолизина (например, PlyD1), как это описано ранее (например, Пример 2), для снижения иммунологической нечувствительности, присутствующей после естественного контакта с S. pneumoniae.
Хотя в данной заявке описаны примеры способов, белков, композиций и другие характеристики, это не подразумевает сокращения или ограничения области данного изобретения, описания или применения. Модификации, изменения и вариации будут очевидны для специалиста в данной области техники. Таким образом, данное описание не ограничено специфическими деталями, представленными средствами и примерами, показанным и описанными в данной заявке. Список последовательностей является частью данной заявки.
Содержание всех цитированных источников включено в данную заявку посредством ссылки. Применение единственного числа в данной заявке не исключает возможности применения множественного, там, где это не противоречит контексту. Таким образом, если в формуле изобретения указано единственное число для X или Y, это может быть интерпретировано как более одного X или более одного Y, если не указано обратное. Союз «или» используемый в описании или в формуле изобретения подразумевает значение «А или В или оба» (например, в случае «А или В»). В ситуациях, когда по смыслу должно быть использовано «только А или В, но не оба» используется фраза «только А или В, но не оба». Таким образом, союз «или» в данной заявке применяется в объединительном, а не в исключительном смысле. Другие варианты исполнения даны в соответствии со следующей формулой изобретения.


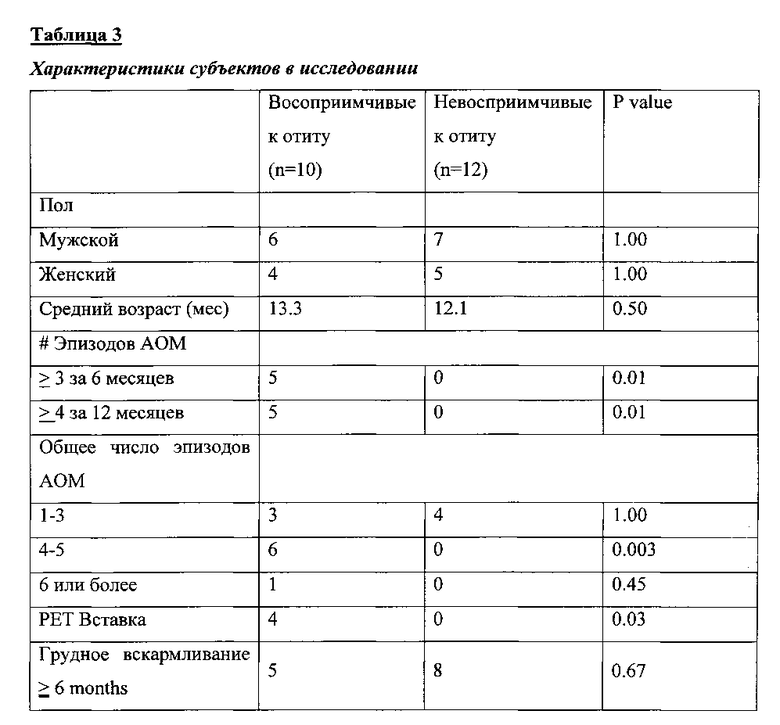
ССЫЛКИ
1. Pichichero ME. Recurrent and persistent otitis media. Pediatr. Infect. Dis.J. 2000; 19 (9):911-6.
2. Pichichero ME, Casey JR. Otitis media. Expert. Opin. Pharmacother. 2002; 3 (8):1073-90.
3. Vergison A, Dagan R, Arguedas A, Bonhoeffer J, Cohen R, Dhooge I et al. Otitis media and its consequences: beyond the earache. Lancet Infect.Dis. 2010;10(3):195-203.
4. Teele DW, Klein JO, Rosner B. Epidemiology of otitis media during the first seven years of life in children in greater Boston: a prospective, cohort study. J.Infect.Dis. 1989; 160 (1):83-94.
5. Poehling KA, Szilagyi PG, Grijalva CG, Martin SW, LaFleur B, Mitchel E et al. Reduction of frequent otitis media and pressure-equalizing tube insertions in children after introduction of pneumococcal conjugate vaccine. Pediatrics 2007; 119 (4):707-15.
6. Del Beccaro MA, Mendelman PM, Inglis AF, Richardson MA, Duncan NO, Clausen CR et al. Bacteriology of acute otitis media: a new perspective. J. Pediatr. 1992; 120 (1):81-4.
7. Fietta P, Delsante G. The effector T helper cell triade. Riv. Biol. 2009; 102 (1):61-74.
8. Mosmann TR, Sad S. The expanding universe of T-cell subsets: Th1, Th2 and more. Immunol.Today 1996; 17 (3):138-46.
9. Mosmann TR, Coffman RL. TH1 and TH2 cells: different patterns of lymphokine secretion lead to different functional properties. Annu.Rev.Immunol. 1989; 7:145-73.
10. Korn T, Bettelli E, Oukka M, Kuchroo VK. IL-17 and Th17 Cells. Annu.Rev.Immunol. 2009; 27:485-517.
11. van Leeuwen EM, Sprent J, Surh CD. Generation and maintenance of memory CD4(+) T Cells. Curr. Opin. Immunol. 2009; 21 (2):167-72.
12. McKinstry KK, Strutt TM, Swain SL. The potential of CD4 T-cell memory. Immunology 2010; 130(1):1-9.
13. Combadiere B, Siberil S, Duffy D. Keeping the memory of influenza viruses. Pathol.Biol.(Paris) 2010; 58 (2):e79-e86.
14. Lanzavecchia A, Sallusto F. Human В cell memory. Curr.Opin.Immunol. 2009; 21 (3):298-304.
15. de Bree GJ, Daniels H, Schilfgaarde M, Jansen HM, Out ТА, van Lier RA et al. Characterization of CD4+ memory T cell responses directed against common respiratory pathogens in peripheral blood and lung. J.Infect.Dis. 2007; 195 (11):1718-25.
16. Kelly DF, Pollard AJ, Moxon ER. Immunological memory: the role of В cells in long-term protection against invasive bacterial pathogens. JAMA 2005; 294 (23):3019-23.
17. Picker LJ, Singh MK, Zdraveski Z, Treer JR, Waldrop SL, Bergstresser PR et al. Direct demonstration of cytokine synthesis heterogeneity among human memory/effector T cells by flow cytometry. Blood 1995; 86 (4):1408-19.
18. Pitcher CJ, Quittner C, Peterson DM, Connors M, Koup RA, Maino VC et al. HIV-1-specific CD4+ T cells are detectable in most individuals with active HIV-1 infection, but decline with prolonged viral suppression. Nat.Med. 1999; 5 (5):518-25.
19. Kodama S, Suenaga S, Hirano T, Suzuki M, Mogi G. Induction of specific immunoglobulin A and Th2 immune responses to P6 outer membrane protein of nontypeable Haemophilus influenzae in middle ear mucosa by intranasal immunization. Infectlmmun. 2000; 68 (4):2294-300.
20. Malley R, Srivastava A, Lipsitch M, Thompson CM, Watkins C, Tzianabos A et al. Antibody-independent, interleukin-17A-mediated, cross-serotype immunity to pneumococci in mice immunized intranasally with the cell wall polysaccharide. Infect.Immun. 2006; 74 (4):2187-95.
21. van den Biggelaar AH, Richmond PC, Pomat WS, Phuanukoonnon S, Nadal-Sims MA, Devitt CJ et al. Neonatal pneumococcal conjugate vaccine immunization primes T cells for preferential Th2 cytokine expression: a randomized controlled trial in Papua New Guinea. Vaccine 2009; 27 (9): 1340-7.
22. Chen SJ, Chu ML, Wang CJ, Liao CL, Hsieh SL, Sytwu HK et al. Kinetic Th1/Th2 responses of transgenic mice with bacterial meningitis induced by Haemophilus influenzae. Clin.Sci.(Lond) 2006; 111 (4):253-63.
23. Snapper CM, Shen Y, Khan AQ, Colino J, Zelazowski P, Mond JJ et al. Distinct types of T-cell help for the induction of a humoral immune response to Streptococcus pneumoniae. Trends Immunol. 2001; 22 (6):308-11.
24. Faden H. The microbiologic and immunologic basis for recurrent otitis media in children. Eur.J.Pediatr. 2001; 160 (7):407-13.
25. Murphy TF, Yi K. Mechanisms of recurrent otitis media: importance of the immune response to bacterial surface antigens. Ann.N.Y.Acad.Sci. 1997; 830:353-60.
26. Kaur R, Casey JR, Pichichero ME. Serum Antibody Response to Three Non-typeable Haemophilus influenzae Outer Membrane Proteins During Acute Otitis Media and Nasopharyngeal Colonization in Otitis Prone and Non-Otitis Prone Children. Vaccine 2011; 29 (5):1023-8.
27. Mosmann TR, Schumacher JH, Street NF, Budd R, O′Garra A, Fong ТА et al. Diversity of cytokine synthesis and function of mouse CD4+ T cells. Immunol.Rev. 1991; 123:209-29.
28. Rajewsky K. Clonal selection and learning in the antibody system. Nature 1996; 381 (6585):751-8.
29. Manz RA, Hauser AE, Hiepe F, Radbruch A. Maintenance of serum antibody levels. Annu.Rev.Immunol. 2005; 23:367-86.
30. Slifka MK, Antia R, Whitmire JK, Ahmed R. Humoral immunity due to long-lived plasma cells. Immunity. 1998; 8 (3):363-72.
31. Bernasconi NL, Traggiai E, Lanzavecchia A. Maintenance of serological memory by polyclonal activation of human memory В cells. Science 2002; 298 (5601):2199-202.
32. De Las RB, Garcia JL, Lopez R, Garcia P. Purification and polar localization of pneumococcal LytB, a putative endo-beta-N-acetylglucosaminidase: the chain-dispersing murein hydrolase. J.Bacteriol. 2002; 184 (18):4988-5000.
33. Kirkham LA, Jefferies JM, Kerr AR, Jing Y, Clarke SC, Smith A et al. Identification of invasive serotype 1 pneumococcal isolates that express nonhemolytic pneumolysin. J. Clin. Microbiol. 2006; 44 (1): 151-9.
34. Kirkham LA, Kerr AR, Douce GR, Paterson GK, Dilts DA, Liu DF et al. Construction and immunological characterization of a novel nontoxic protective pneumolysin mutant for use in future pneumococcal vaccines. Infect. Immun. 2006; 74 (1):586-93.
35. Perfetto SP, Chattopadhyay PK, Lamoreaux L, Nguyen R, Ambrozak D, Koup RA et al. Amine reactive dyes: an effective tool to discriminate live and dead cells in polychromatic flow cytometry. J. Immunol. Methods 2006; 313 (1-2): 199-208.
36. Waldrop SL, Pitcher CJ, Peterson DM, Maino VC, Picker LJ. Determination of antigen-specific memory/effector CD4+ T cell frequencies by flow cytometry: evidence for a novel, antigen-specific homeostatic mechanism in HIV-associated immunodeficiency. J. Clin. Invest 1997; 99 (7): 1739-50.
37. Waldrop SL, Davis KA, Maino VC, Picker LJ. Normal human CD4+ memory T cells display broad heterogeneity in their activation threshold for cytokine synthesis. J. Immunol. 1998; 161 (10):5284-95.
38. Pichichero ME, Deloria MA, Rennels MB, Anderson EL, Edwards KM, Decker MD et al. A safety and immunogenicity comparison of 12 acellular pertussis vaccines and one whole-cell pertussis vaccine given as a fourth dose in 15- to 20-month-old children. Pediatrics 1997; 100 (5):772-88.
39. Adkins B, Bu Y, Guevara P. The generation of Th memory in neonates versus adults: prolonged primary Th2 effector function and impaired development of Th1 memory effector function in murine neonates. J. Immunol. 2001; 166 (2):918-25.
40. Holt PG. Functionally mature virus-specific CD8(+) T memory cells in congenitally infected newborns: proof of principle for neonatal vaccination? J. Clin. Invest 2003; 111 (11):1645-7.
41. Fleischer B. Superantigens. APMIS 1994; 102 (1):3-12.
42. Mureithi MW, Finn A, Ota MO, Zhang Q, Davenport V, Mitchell TJ et al. T cell memory response to pneumococcal protein antigens in an area of high pneumococcal carriage and disease. J.Infect.Dis. 2009; 200 (5):783-93.
43. Zhang Q, Bagrade L, Bematoniene J, Clarke E, Paton JC, Mitchell TJ et al. Low CD4 T cell immunity to pneumolysin is associated with nasopharyngeal carriage of pneumococci in children. J.Infect.Dis. 2007; 195 (8): 1194-202.
44. Kodama H, Faden H, Harabuchi Y, Kataura A, Bernstein JM, Brodsky L. Cellular immune response of adenoidal and tonsillar lymphocytes to the P6 outer membrane protein of non-typeable Haemophilus influenzae and its relation to otitis media. Acta Otolaryngol. 1999; 119 (3):377-83.
45. Mattila PS, Nykanen A, Eloranta M, Tarkkanen J. Adenoids provide a microenvironment for the generation of CD4(+), CD45RO(+), L-selectin(-), CXCR4(+), CCR5(+) T lymphocytes, a lymphocyte phenotype found in the middle ear effusion. Intlmmunol. 2000; 12 (9): 1235-43.
46. Forseni M, Hansson GK, Bagger-Sjoback D, Hultcrantz M. Infiltration of immunocompetent cells in the middle ear during acute otitis media: a temporal study. Am. J. Otol. 1999; 20 (2): 152-7.
47. Skotnicka B, Stasiak-Barmuta A, Hassmann-Poznanska E, Kasprzycka E. Lymphocyte subpopulations in middle ear effusions: flow cytometry analysis. Otol.Neurotol. 2005; 26 (4):567-71.
48. Zaghouani H, Hoeman CM, Adkins B. Neonatal immunity: faulty T-helpers and the shortcomings of dendritic cells. Trends Immunol. 2009; 30 (12):585-91.
49. Adkins B, Leclerc C, Marshall-Clarke S. Neonatal adaptive immunity comes of age. NatRev.Immunol. 2004; 4 (7):553-64.
50. Siegrist CA. Neonatal and early life vaccinology. Vaccine 2001; 19 (25-26):3331-46.
51. Tu W, Chen S, Sharp M, Dekker C, Manganello AM, Tongson EC et al. Persistent and selective deficiency of CD4+ T cell immunity to cytomegalovirus in immunocompetent young children. J.Immunol. 2004; 172 (5):3260-7.
52. Klein NP, Gans HA, Sung P, Yasukawa LL, Johnson J, Sarafanov A et al. Preterm infants′ T cell responses to inactivated poliovirus vaccine. J.Infect.Dis. 2010; 201 (2):214-22.
53. Geiger R, Duhen T, Lanzavecchia A, Sallusto F. Human naive and memory CD4+ T cell repertoires specific for naturally processed antigens analyzed using libraries of amplified T cells. J.Exp.Med. 2009; 206 (7): 1525-34.
54. Upham JW, Rate A, Rowe J, Kusel M, Sly PD, Holt PG. Dendritic cell immaturity during infancy restricts the capacity to express vaccine-specific T-cell memory. Infectlmmun. 2006; 74 (2):1106-12.
55. Bluestone CD, Stephenson JS, Martin LM. Ten-year review of otitis media pathogens. Pediatr Infect Dis J 1992; 11:S7-11.
56. Luotonen J, Herva E, Karma P, Timonen M, Leinonen M, Makela PH. The bacteriology of acute otitis media in children with special reference to Streptococcus pneumoniae as studied by bacteriological and antigen detection methods. Scand.J Infect Dis 1981; 13:177-83.
57. Berkley JA, Lowe BS, Mwangi I et al. Bacteremia among children admitted to a rural hospital in Kenya. N.Engl.J Med. 2005; 352:39-47.
58. Denny FW, Loda FA. Acute respiratory infections are the leading cause of death in children in developing countries. Am.J Trop.Med.Hyg. 1986; 35:1-2.
59. Huang SS, Piatt R, Rifas-Shiman SL, Pelton SI, Goldmann D, Finkelstein JA. Post-PCV7 changes in colonizing pneumococcal serotypes in 16 Massachusetts communities, 2001 and 2004. Pediatrics 2005; 116:e408-e413.
60. Kadioglu A, Weiser JN, Paton JC, Andrew PW. The role of Streptococcus pneumoniae virulence factors in host respiratory colonization and disease. Nat.Rev.Microbiol. 2008; 6:288-301.
61. Paton JC, Andrew PW, Boulnois GJ, Mitchell TJ. Molecular analysis of the pathogenicity of Streptococcus pneumoniae: the role of pneumococcal proteins. Annu. Rev. Microbiol. 1993; 47:89-115.
62. Ogunniyi AD, Grabowicz M, Mahdi LK et al. Pneumococcal histidine triad proteins are regulated by the Zn2+-dependent repressor AdcR and inhibit complement deposition through the recruitment of complement factor H. FASEB J 2009; 23:731-8.
63. Adamou JE, Heinrichs JH, Erwin AL et al. Identification and characterization of a novel family of pneumococcal proteins that are protective against sepsis. Infect Immun. 2001; 69:949-58.
64. Garcia P, Gonzalez MP, Garcia E, Lopez R, Garcia JL. LytB, a novel pneumococcal murein hydrolase essential for cell separation. Mol. Microbiol. 1999; 31:1275-81.
65. Rosenow C, Ryan P, Weiser JN et al. Contribution of novel choline-binding proteins to adherence, colonization and immunogenicity of Streptococcus pneumoniae. Mol. Microbiol. 1997; 25:819-29.
66. Glover DT, Hollingshead SK, Briles DE. Streptococcus pneumoniae surface protein PcpA elicits protection against lung infection and fatal sepsis. Infect Immun. 2008; 76:2767-76.
67. Walker JA, Allen RL, Falmagne P, Johnson MK, Boulnois GJ. Molecular cloning, characterization, and complete nucleotide sequence of the gene for pneumolysin, the sulfhydryl-activated toxin of Streptococcus pneumoniae. Infect Immun. 1987; 55:1184-9.
68. Musher DM, Phan HM, Baughn RE. Protection against bacteremic pneumococcal infection by antibody to pneumolysin. J Infect Dis 2001; 183:827-30.
69. Pichichero ME, Kaur R, Casey JR, Xu Q, Almudevar A, Ochs M. Antibody Response to Streptococcus pneumoniae Vaccine Targets PhtD, LytB, PcpA, PhtE and PlyDl After Nasopharyngeal Colonization and Acute Otitis Media in Children. 10th International Symposium on Recent Advances in Otitis Media. New Orleans LA, USA, Abstract J03, page 105, June 5-9, 2011
70. Pelton SI, Leibovitz E. Recent advances in otitis media. Pediatr Infect Dis J 2009; 28:S133-S137.
71. Pichichero ME, Casey JR, Hoberman A, Schwartz R. Pathogens causing recurrent and difficult-to-treat acute otitis media, 2003-2006. Clin.Pediatr (Phila) 2008; 47:901-6.
72. Freijd A, Hammarstrom L, Persson MA, Smith CI. Plasma anti-pneumococcal antibody activity of the IgG class and subclasses in otitis prone children. Clin.Exp.Immunol. 1984; 56:233-8.
73. Prellner K, Kalm O, Pedersen FK. Pneumococcal antibodies and complement during and after periods of recurrent otitis. Int. J Pediatr Otorhinolaryngol. 1984; 7:39-49.
74. Hotomi M, Yamanaka N, Saito T et al. Antibody responses to the outer membrane protein P6 of non-typeable Haemophilus influenzae and pneumococcal capsular polysaccharides in otitis-prone children. Acta Otolaryngol. 1999; 119:703-7.
75. Yamanaka N, Faden H. Antibody response to outer membrane protein of nontypeable Haemophilus influenzae in otitis-prone children. J Pediatr 1993; 122:212-8.
76. Yamanaka N, Faden H. Local antibody response to P6 of nontypable Haemophilus influenzae in otitis-prone and normal children. Acta Otolaryngol. 1993; 113:524-9.
77. Bernstein JM, Bronson PM, Wilson ME. Immunoglobulin G subclass response to major outer membrane proteins of nontypable Haemophilus influenzae in children with acute otitis media. Otolaryngol.Head Neck Surg. 1997; 116:363-71.
78. Giebink GS, Mills EL, Huff JS, Cates KL, Juhn SK, Quie PG. Polymorphonuclear leukocyte dysfunction in children with recurrent otitis media. J Pediatr 1979; 94:13-8.
79. Freijd A, Oxelius VA, Rynnel-Dagoo B. A prospective study demonstrating an association between plasma IgG2 concentrations and susceptibility to otitis media in children. Scand. J Infect Dis 1985; 17:115-20.
80. Veenhoven R, Rijkers G, Schilder A et al. Immunoglobulins in otitis-prone children. Pediatr Res. 2004; 55:159-62.
81. Berman S, Lee B, Nuss R, Roark R, Giclas PC. Immunoglobulin G, total and subclass, in children with or without recurrent otitis media. J Pediatr 1992; 121:249-51.
82. Sharma S, Kaur R, Casey JR, Pichichero ME. Lack of Generation of T Cell Memory to Pneumococcal and Haemophilus influenzae Proteins in Early Childhood Explains Immunologic Susceptibility to the Otitis Prone Condition, - (unpublished results).
83. Prellner K, Harsten G, Lofgren B, Christenson B, Heldrap J. Responses to rubella, tetanus, and diphtheria vaccines in otitis-prone and non-otitis-prone children. Ann. Otol. Rhinol. Laryngol. 1990; 99:628-32.
84. Wiertsema SP, Sanders EA, Veenhoven RH et al. Antibody levels after regular childhood vaccinations in the immunological screening of children with recurrent otitis media. J Clin. Immunol. 2004; 24:354-60.
85. Breukels MA, Rijkers GT, Voorhorst-Ogink MM, Zegers В J, Sanders LA. Pneumococcal conjugate vaccine primes for polysaccharide-inducible IgG2 antibody response in children with recurrent otitis media acuta. J Infect Dis 1999; 179:1152-6.
86. Barnett ED, Pelton SI, Cabral HJ et al. Immune response to pneumococcal conjugate and polysaccharide vaccines in otitis-prone and otitis-free children. Clin. Infect Dis 1999; 29:191-2.
87. Pichichero ME, Kaur R, Casey JR, Sabirov A, Khan MN, Almudevar A. Antibody Response to Haemophilus influenzae Outer Membrane Protein D, P6, and OMP26 After Nasopharyngeal Colonization and Acute Otitis Media in Children. Vaccine 2010; 28:7184-92.
88. Rapola S, Kilpi T, Lahdenkari M, Makela PH, Kayhty H. Antibody response to the pneumococcal proteins pneumococcal surface adhesin A and pneumolysin in children with acute otitis media. Pediatr Infect Dis J 2001; 20:482-7.
89. Soininen A, Lahdenkari M, Kilpi T, Makela PH, Kayhty H. Antibody response to pneumococcal capsular polysaccharides in children with acute otitis media. Pediatr Infect Dis J 2002; 21:186-92.
90. Soininen A, Pursiainen H, Kilpi T, Kayhty H. Natural development of antibodies to pneumococcal capsular polysaccharides depends on the serotype: association with pneumococcal carriage and acute otitis media in young children. J Infect Dis 2001; 184:569-76.
91. Zhang Q, Bernatoniene J, Bagrade L et al. Serum and mucosal antibody responses to pneumococcal protein antigens in children: relationships with carriage status. Eur. J Immunol. 2006; 36:46-57.
92. Obaro SK, Adegbola RA, Tharpe JA et al. Pneumococcal surface adhesin A antibody concentration in serum and nasopharyngeal carriage of Streptococcus pneumoniae in young African infants. Vaccine 2000; 19:411-2.
93. Holmlund E, Simell B, Jaakkola T et al. Serum antibodies to the pneumococcal surface proteins PhtB and PhtE in Finnish infants and adults. Pediatr Infect Dis J 2007; 26:447-9.
94. Holmlund E, Quiambao B, Ollgren J et al. Antibodies to pneumococcal proteins PhtD, CbpA, and LytC in Filipino pregnant women and their infants in relation to pneumococcal carriage. Clin. Vaccine Immunol. 2009; 16:916-23.
95. Casey JR, Adlowitz DG, Pichichero ME. New patterns in the otopathogens causing acute otitis media six to eight years after introduction of pneumococcal conjugate vaccine. Pediatr. Infect. DisJ. 2010; 29 (4):304-9.
96. Sharma SK, Casey JR, Pichichero ME. Reduced memory CD4+ T Cell generation in the circulation of young children may contribute to the Otitis Prone Condition. J.I nfect. Dis. 2011.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ОЧИСТКИ БАКТЕРИАЛЬНОГО ЦИТОЛИЗИНА | 2004 |
|
RU2340627C2 |
| ИММУНОБИОЛОГИЧЕСКОЕ СРЕДСТВО НА ОСНОВЕ РЕКОМБИНАНТНОГО БЕЛКА ДЛЯ ПРОФИЛАКТИКИ ПНЕВМОКОККОВОЙ ИНФЕКЦИИ | 2024 |
|
RU2823752C1 |
| ВАКЦИНЫ И КОМПОЗИЦИИ ПРОТИВ STREPTOCOCCUS PNEUMONIAE | 2012 |
|
RU2623174C2 |
| ВАКЦИНЫ И КОМПОЗИЦИИ, НАПРАВЛЕННЫЕ ПРОТИВ STREPTOCOCCUS PNEUMONIAE | 2010 |
|
RU2580299C2 |
| КОМБИНАЦИИ, ВКЛЮЧАЮЩИЕ САХАРИД ПНЕВМОКОККА СЕРОТИПА 14 | 2010 |
|
RU2549438C2 |
| КОМБИНАЦИИ МЕНИНГОКОККОВОГО ФАКТОР Н-СВЯЗЫВАЮЩЕГО БЕЛКА И ПНЕВМОКОККОВЫХ САХАРИДНЫХ КОНЪЮГАТОВ | 2010 |
|
RU2555757C2 |
| ПОЛИВАЛЕНТНЫЕ ПНЕВМОКОККОВЫЕ ВАКЦИНЫ | 2019 |
|
RU2815390C2 |
| Капсульные полисахариды Streptococcus pneumoniae и их конъюгаты | 2015 |
|
RU2688831C2 |
| СОЕДИНЕНИЯ И СПОСОБЫ ДЛЯ ЛЕЧЕНИЯ И ДИАГНОСТИКИ ХЛАМИДИЙНОЙ ИНФЕКЦИИ | 2001 |
|
RU2410394C2 |
| Капсульные полисахариды Streptococcus pneumoniae и их конъюгаты | 2015 |
|
RU2743793C1 |
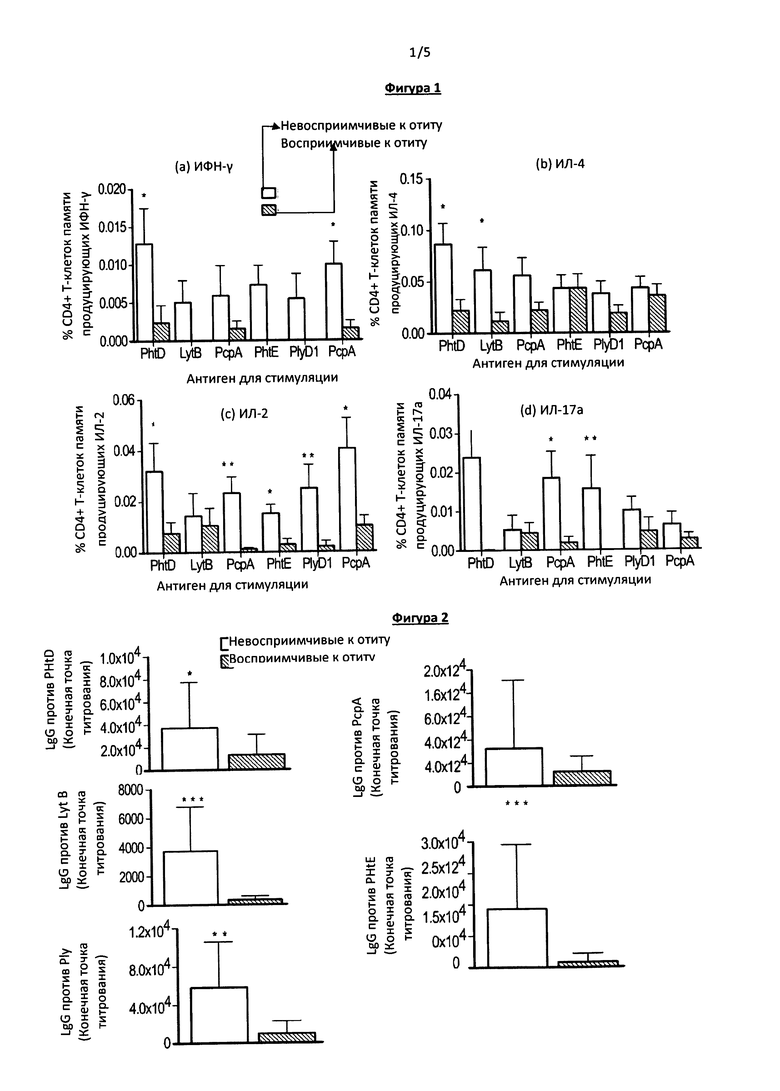
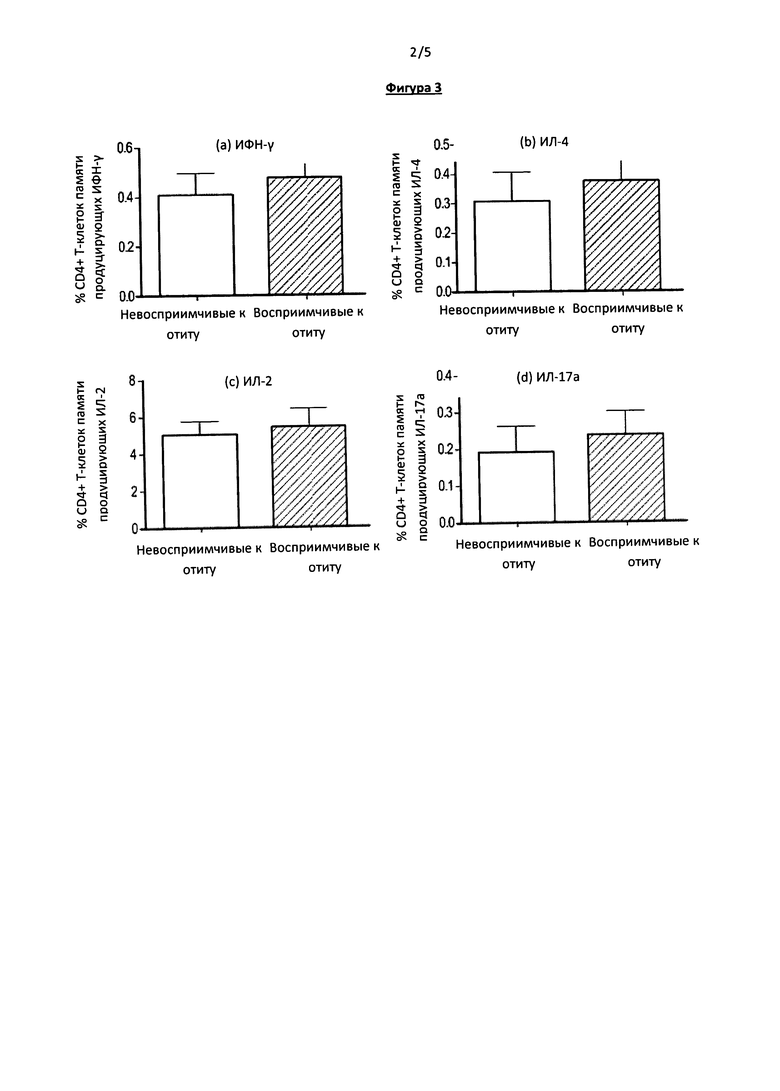
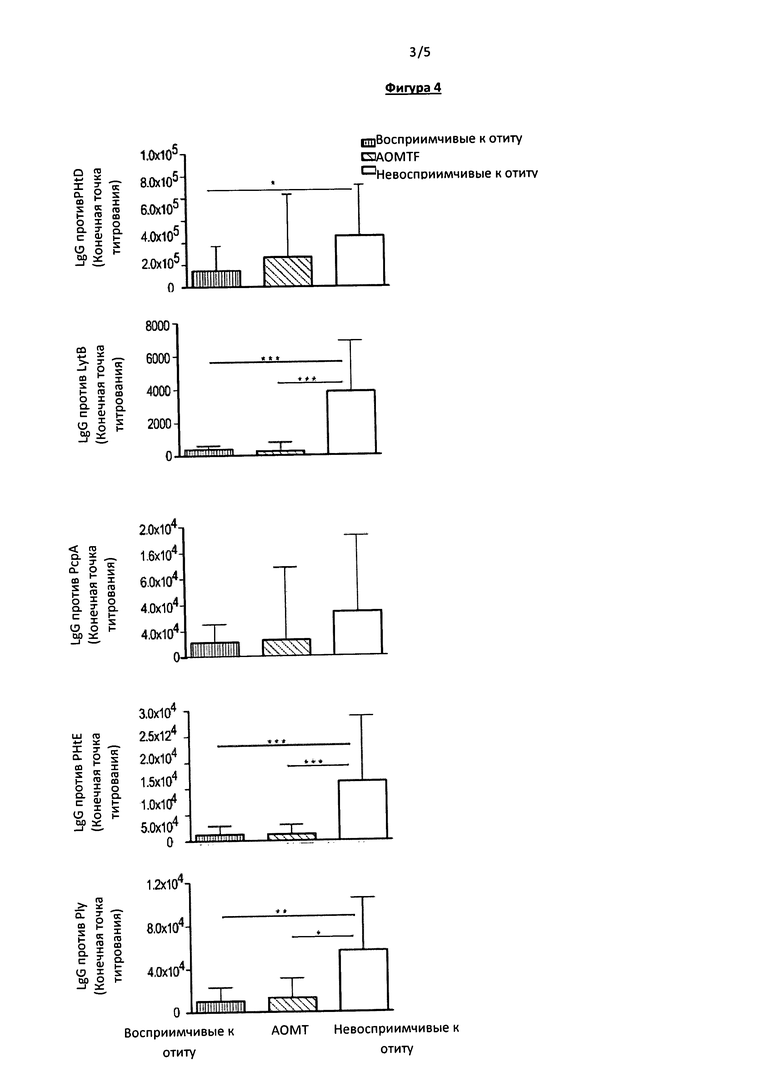
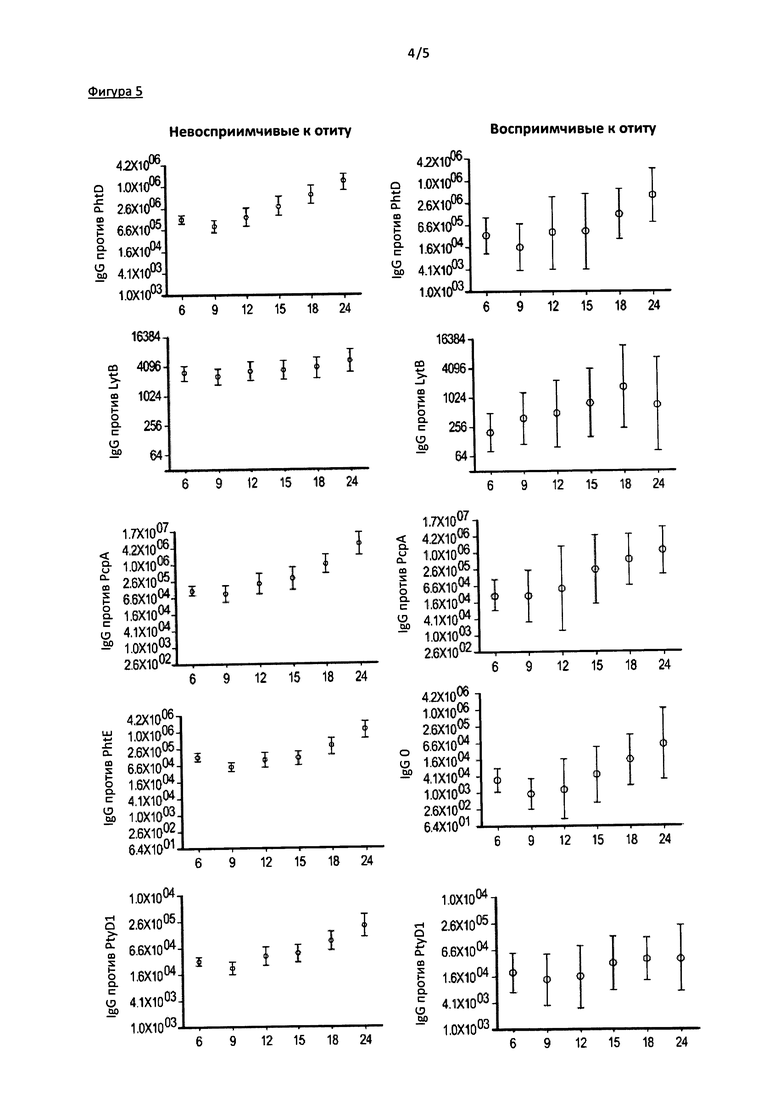
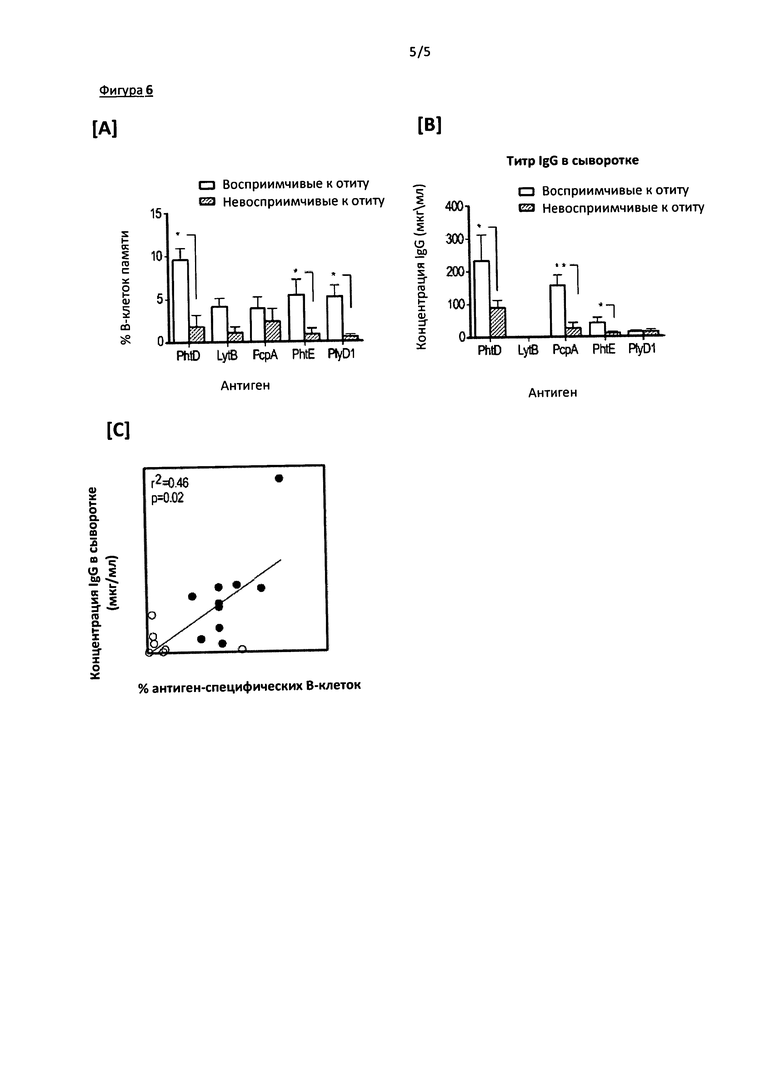




























Группа изобретений относится к способу профилактики или лечения рецидива острого отита среднего уха, возникающего в результате инфекции Streptococcus pneumoniae, у пациентов группы риска, включающему введение терапевтически эффективного количества композиции, по меньшей мере, однократно, пациенту. Вводимая композиция содержит, по меньшей мере, один иммуногенный полипептид, выбранный из группы, состоящей из полипептидов S.pneumoniae PhtD, PhtE, PcpA, LytB и деактивированного пневмолизина. Заявленная группа изобретений обладает эффективным защитным действием. 3 н. и 34 з.п. ф-лы, 6 ил., 3 табл., 3 пр.
1. Способ профилактики или лечения рецидива острого отита среднего уха, возникающего в результате инфекции Streptococcus pneumoniae, у пациента группы риска развития рецидива пневмококкового острого отита среднего уха, включающий введение данному пациенту, по меньшей мере, однократно, терапевтически эффективного количества композиции, включающей, по меньшей мере, один выделенный и очищенный иммуногенный полипептид, выбранный из группы, состоящей из полипептидов Streptococcus pneumoniae PhtX, РсрА, LytB и детоксифицированного пневмолизина или их иммуногенных фрагментов.
2. Способ по п. 1, в котором пациент предварительно перенес, по меньшей мере, один эпизод острого отита среднего уха.
3. Способ по п. 2, в котором пациент перенес 3 или более эпизодов острого отита среднего уха за период шесть месяцев или перенес 4 или более эпизодов острого отита среднего уха за период 12 месяцев.
4. Способ по п. 1, в котором у пациента развился острый отит среднего уха или он находится в группе риска развития острого отита среднего уха.
5. Способ по п. 4, в котором у пациента развился острый отит среднего уха.
6. Способ по п. 1, в котором введение композиции вызывает или усиливает образование антиген-специфических CD4+ Т-клеток.
7. Способ по п. 6, в котором введение композиции вызывает или усиливает образование антиген-специфических CD4+ Т-клеток, продуцирующих ИФН-γ, ИЛ-4, ИЛ-2 и/или ИЛ-17а.
8. Способ по п. 7, в котором доля антиген-специфических CD4+ Т-клеток, продуцирующих ИФН-γ, ИЛ-2, ИЛ-4 и/или ИЛ-17а, увеличивается относительно их доли непосредственно перед введением композиции.
9. Способ по п. 1, в котором введение стимулирует продукцию цитокинов ИФН-γ, ИЛ-2, ИЛ-4 и/или ИЛ-17а.
10. Способ по п. 1, в котором композиция также содержит адъювант.
11. Способ по п. 1, в котором композиция содержит, по меньшей мере, два выделенных и очищенных иммуногенных полипептида, выбранных из группы, состоящей из полипептидов Streptococcus pneumoniae PhtD, PhtE, РсрА, LytB и детоксифицированного пневмолизина или их иммуногенных фрагментов.
12. Способ по п. 1, в котором композиция содержит, по меньшей мере, три выделенных и очищенных иммуногенных полипептида, выбранных из группы, состоящей из полипептидов Streptococcus pneumoniae PhtD, PhtE, РсрА, LytB и детоксифицированного пневмолизина или их иммуногенных фрагментов.
13. Способ по п. 1, в котором композиция содержит, по меньшей мере, четыре выделенных и очищенных иммуногенных полипептида, выбранных из группы, состоящей из полипептидов Streptococcus pneumoniae PhtD, PhtE, РсрА, LytB и детоксифицированного пневмолизина или их иммуногенных фрагментов.
14. Способ по п. 1, в котором композиция содержит, по меньшей мере, пять выделенных и очищенных иммуногенных полипептидов, выбранных из группы, состоящей из полипептидов Streptococcus pneumoniae PhtD, PhtE, РсрА, LytB и детоксифицированного пневмолизина или их иммуногенных фрагментов.
15. Способ по одному из пп. 1-14, в котором детоксифицированный пневмолизин представляет собой мутантный белок пневмолизин, содержащий аминокислотные замены в позициях 65, 293 и 428 относительно последовательности дикого типа.
16. Способ по п. 15, в котором данные три аминокислотные замены включают Т65→С, G293→C и С428→А.
17. Композиция, содержащая, по меньшей мере, один выделенный и по существу очищенный иммуногенный полипептид, выбранный из группы, состоящей из полипептидов Streptococcus pneumoniae PhtX, РсрА, LytB и детоксифицированного пневмолизина для применения для профилактики или лечения рецидива острого отита среднего уха, возникающего в результате инфекции Streptococcus pneumoniae у пациента.
18. Композиция по п. 17, где пациент предварительно перенес, по меньшей мере, один эпизод острого отита среднего уха.
19. Композиция по п. 17, где пациент перенес 3 или более эпизодов острого отита среднего уха за период шесть месяцев или перенес 4 или более эпизодов острого отита среднего уха за период 12 месяцев.
20. Композиция по п. 17, где у пациента развился острый отит среднего уха или он находится в группе риска развития острого отита среднего уха.
21. Композиция по п. 20, где у пациента развился острый отит среднего уха.
22. Композиция по п. 17, где введение композиции вызывает образование антиген-специфических CD4+ Т-клеток.
23. Композиция по п. 22, где введение композиции вызывает образование антиген-специфических CD4+ Т-клеток, продуцирующих ИФН-γ, ИЛ-4, ИЛ-2 и/или ИЛ-17а.
24. Композиция по п. 22, где введение композиции вызывает увеличение доли антиген-специфических CD4+ Т-клеток, продуцирующих ИФН-γ, ИЛ-2, ИЛ-4 и/или ИЛ-17а, относительно введения композиции, не содержащей, по меньшей мере, один, выделенный и очищенный иммуногенный полипептид.
25. Композиция по п. 17, где введение стимулирует продукцию цитокинов ИФН-γ, ИЛ-2, ИЛ-4 и/или ИЛ-17а.
26. Композиция по п. 17, где композиция также содержит адъювант.
27. Композиция по п. 17, где композиция содержит, по меньшей мере, два выделенных и очищенных иммуногенных полипептида, выбранных из группы, состоящей из полипептидов Streptococcus pneumoniae PhtD, PhtE, РсрА, LytB и детоксифицированного пневмолизина или их иммуногенных фрагментов.
28. Композиция по п. 17, где композиция содержит, по меньшей мере, три выделенных и очищенных иммуногенных полипептида, выбранных из группы, состоящей из полипептидов Streptococcus pneumoniae PhtD, PhtE, РсрА, LytB и детоксифицированного пневмолизина или их иммуногенных фрагментов.
29. Композиция по п. 17, где композиция содержит, по меньшей мере, четыре выделенных и очищенных иммуногенных полипептида, выбранных из группы, состоящей из полипептидов Streptococcus pneumoniae PhtD, PhtE, РсрА, LytB и детоксифицированного пневмолизина или их иммуногенных фрагментов.
30. Композиция по п. 17, где композиция содержит, по меньшей мере, пять выделенных и очищенных иммуногенных полипептидов, выбранных из группы, состоящей из полипептидов Streptococcus pneumoniae PhtD, PhtE, РсрА, LytB и детоксифицированного пневмолизина или их иммуногенных фрагментов.
31. Композиция по одному из пп. 17-30, где детоксифицированный пневмолизин представляет собой мутантный белок пневмолизин, содержащий аминокислотные замены в позициях 65, 293 и 428 относительно последовательности дикого типа.
32. Композиция по п. 31, где данные три аминокислотные замены включают Т65→С, G293→C и С428→А.
33. Композиция по одному из пп. 17-30, 32, где композиция является вакциной.
34. Композиция по п. 17, где пациент перенес эпизод острого отита среднего уха в результате инфекции S. pneumoniae и у которого не произошло ликвидации бактерий и/или разрешения симптомов после, по меньшей мере, 48 часов соответствующей антибиотикотерапии.
35. Композиция по п. 17, где пациент перенес эпизод острого отита среднего уха (АОМ) в результате инфекции S. pneumoniae и у которого в течение 14 дней после завершения курса лечения АОМ антибиотиком симптомы АОМ возникли снова.
36. Способ снижения риска рецидива острого отита среднего уха (АОМ), возникающего в результате инфекции S. pneumoniae у пациента группы риска развития рецидива пневмококкового АОМ, включающий введение пациенту терапевтически эффективного количества иммуногенной композиции, содержащей, по меньшей мере, один выделенный и по существу очищенный иммуногенный полипептид, выбранный из группы, состоящей из полипептидов S. Pneumoniae PhtX, РсрА, LytB и детоксифицированного пневмолизина или их иммуногенных фрагментов.
37. Способ по п. 36, где иммуногенный полипептид PhtX представляет собой PhtD или PhtE.
| ДВИГАТЕЛЬНЫЙ ЭНЕРГОАГРЕГАТ | 0 |
|
SU222168A1 |
| WO2007001200 A1, 04.01.2007 | |||
| WO2010071986 A1, 01.07.2010 | |||
| I | |||
| J | |||
| DHOOGE et al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| International Journal of Pediatric Otorhinolaryngology | |||
| Vol | |||
| Нефтяной конвертер | 1922 |
|
SU64A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Топочная решетка для многозольного топлива | 1923 |
|
SU133A1 |

