Изобретение относится к области молекулярной биологии, биотехнологии и микробиологической промышленности и касается нового рекомбинантного штамма бактерий Escherichia coli, который может быть использован для получения высокоактивного препарата ДНК-метилтрансферазы M.AgsI, переносящей метальную группу с S-аденозил-L-метионина (SAM) на 6-й атом азота внешнего аденинового основания сайта узнавания, с образованием метилированной последовательности 5′-TTSA(m6A)-3′/3′-(m6A)ASTT-5′ на обеих цепях ДНК (где символ «S» означает гуанин или цитозин).
Главными биотехнологически значимыми параметрами ДНК-метилтрансфераз являются узнаваемая последовательность ДНК, узнаваемые нуклеотиды этой последовательности, метилируемые атомы оснований этих нуклеотидов, а также активность препарата фермента, выделенного из соответствующего штамма-продуцента.
ДНК-метилтрансферазы переносят метальную группу SAM на основания ДНК и классифицируются на цитозин (С5)-ДНК-метилтрансферазы, цитозин (N4)-ДНК-метилтрансферазы и аденин (N6)-ДНК-метилтрансферазы. Метилированные основания обозначают соответственно m5C, m4C и m6A (1). Наиболее разнообразна специфичность ДНК-метилтрансфераз у бактерий, в которых открыто около 300 прототипов этих ферментов (2).
Известен штамм бактерии Sporosarcina species 9D, продуцирующий ДНК-метилтрансферазу M.Sse9I, образующую метилированную последовательность 5′-А(m6A)ТТ-3′ (3). Недостатком этого штамма является то, что M.Sse9I может образовывать 5′-TTSA(m6A)-3′ только в составе последовательности 5′-AATTSA(m6A)TT-3′.
В настоящее время не описаны штаммы бактерий, являющиеся продуцентами ДНК-метилтрансфераз, узнающих последовательность 5′-TTSAA-3′ и метилирующих внешний остаток аденина этой последовательности с образованием 5′-TTSA(m6A)-3′.
Наиболее близким к заявляемому штамму - прототипом, является рекомбинантный штамм бактерии Escherichia coli, который продуцирует ДНК-метилтрансферазу M.MseI, образующую последовательность 5′-ТТА(m6A)-3′ (4). Для получения данного рекомбинантного штамма, несущего клонированные гены системы рестрикции-модификации MseI, узнающей последовательность 5′-ТТАА-3′, используется метод клонирования системы рестрикции-модификации MseI, заключающийся в том, что благодаря наработке целевой модифицирующей метилазы в клетке поддерживается полная защита ДНК во время всех фаз роста, в которых присутствует родственный фермент рестрикции.
Недостатком известного штамма является то, что M.MseI не образует метилированную последовательность 5′-TTSA(m6A)-3′.
Задачей изобретения является расширение ассортимента высокопродуктивных штаммов - продуцентов ДНК-метилтрансфераз.
Поставленная задача решается путем получения штамма бактерий Escherichia coli N106 (pM.AgsI) - продуцента ДНК-метилтрансферазы, узнающей последовательность нуклеотидов 5′-TTSAA-3′ и метилирующей в присутствии SAM внешний остаток аденина этой последовательности с образованием 5′-TTSA(m6A)-3′.
Техническим результатом изобретения является получение рекомбинантного штамма бактерий Escherichia coli N106 (pM.AgsI), продуцирующего высокоактивную ДНК-метилтрансферазу заданной специфичности.
Предлагаемый штамм-продуцент получен трансформацией плазмидной ДНК pM.AgsI-106 клеток штамма Escherichia coli RR1. Плазмида pM.AgsI-106 получена в результате клонирования в E.coli участка хромосомной ДНК штамма Agrococcus species 25, являющегося продуцентом ферментов системы рестрикции-модификации AgsI, и выделенного из природного материала (почвы) в результате целенаправленного систематического поиска. Однако этот штамм не является высокопродуктивным для сопряженной ДНК-метилтрансферазы (активность M.AgsI в клетках природного штамма составляет 3000 е.а./г).
Плазмида pM.AgsI-106 является рекомбинантной, содержит слитые гены, кодирующие две субъединицы ДНК-метилтрасферазы M.AgsI (А+В), метилирующей внешний аденин в положении N6 в последовательности 5′-TTSAA-3′.
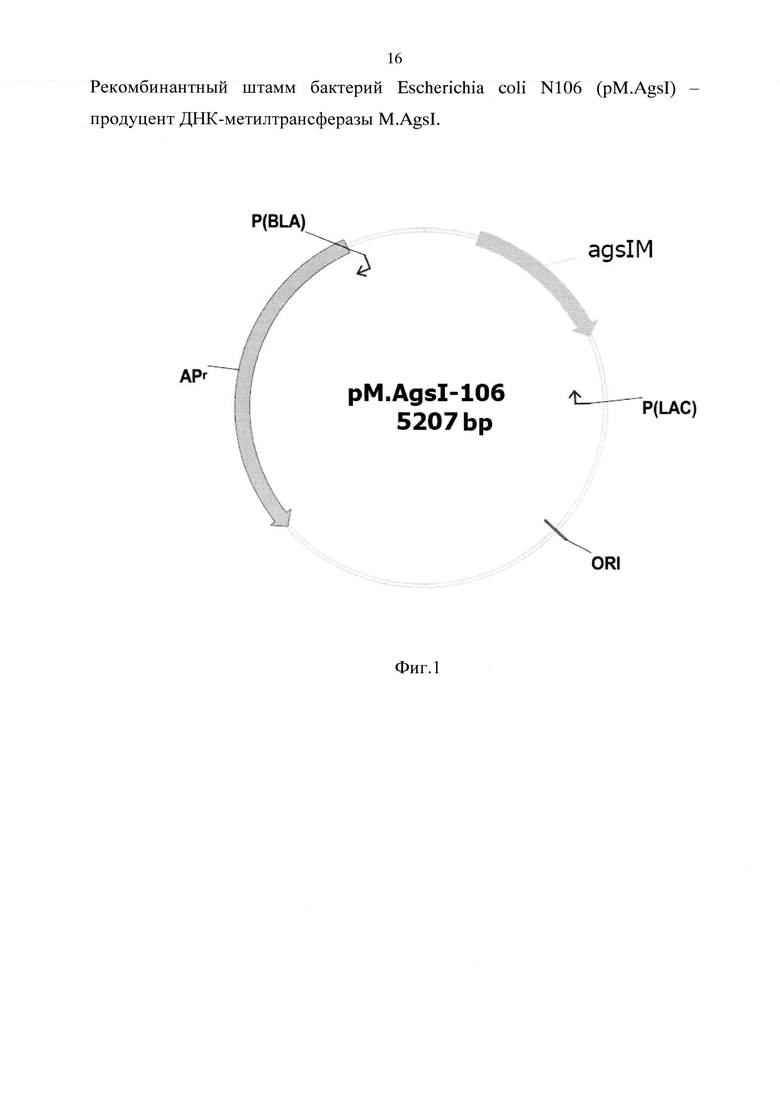
На фигуре 1 представлена физическая карта плазмиды pM.AgsI-106,
где:
ORI - точка начала репликации;
Apr - ген bla, обеспечивающий устойчивость к ампициллину;
P(BLA) - промотор гена bla;
P(LAC) - промотор гена lacZ;
agsIM - слитые гены, кодирующие ДНК-метилтрансферазу AgsI (субъединицы А+В).
Сконструированной плазмидой pM.AgsI-106 трансформируют клетки E.coli штамма RR1. Трансформантов отбирают на селективной агаризованной питательной среде, содержащей 100 мкг/мл ампициллина, по резистентности к антибиотику, т.е. способности к росту в его присутствии. Содержание рекомбинантной ДНК-метилтрасферазы M.AgsI составляет 130000 е.а./г влажной биомассы.
Преимуществом заявляемого штамма по сравнению с наиболее близким аналогом является то, что продуцируемый им фермент - M.AgsI, по своей специфичности является абсолютно новым. Этот фермент, модифицирующий ДНК по сайту 5′-TTSAA-3′, предоставляет новые возможности для молекулярно-генетических работ.
Штамм Escherichia coli N106 (pM.AgsI) имеет следующие характеристики.
Культурально-морфологические признаки
Клетки мелкие, палочковидной формы, грамотрицательные, 1×3,5 мкм, подвижные. Штамм хорошо растет на обычных питательных средах (МПА, МПБ, LB-бульон, LB-агар, минимальная среда с глюкозой). При росте на агаризованной среде LB колонии круглые, гладкие, полупрозрачные, блестящие, серые. Край ровный, диаметр колоний 1 -3 мм, консистенция пастообразная. Рост в жидких средах (LB, минимальная среда с глюкозой) характеризуется ровным помутнением, осадок легко седиментирует.
Физиолого-биохимические признаки
Клетки растут при 4-42°С, оптимум pH 6,8-7,6. В качестве источника азота используют как минеральные соли аммония, так и органические соединения: аминокислоты, пептон, триптон, дрожжевой экстракт. В качестве источника углерода при росте на минимальной среде используют глицерин, углеводы, аминокислоты.
Генетические признаки, устойчивость к антибиотикам.
Штамм-хозяин Esherichia coli RR1 (F- Δ(gpt-proA)62 leuB6 supE44 ara-14 galK2 lacY1 Δ(mcrC-mrr) rpsL20(Strr) xyl-5 mtl-1 recA+). Проявляет устойчивость к стрептомицину (25 мкг/мл).
Штамм Escherichia coli N106 (pM.AgsI) дополнительно проявляет устойчивость к ампициллину (до 300 мкг/мл), обусловленную наличием гена устойчивости в ДНК рекомбинантной плазмиды pM.AgsI-106.
Условия хранения штамма
Штамм бактерий Escherichia coli N106 (pM.AgsI) хранят на агаризованной среде LB со 100 мкг/мл ампициллина на чашках Петри или в стеклянных пробирках со скошенным агаром при 4°С. Пересевы на свежие среды делают один раз в месяц. Может храниться не менее года в среде LB, содержащей 30% глицерин, при -50-70°С.
Длительное хранение штамма осуществляется в лиофильно высушенном состоянии или в растворе 30% глицерина при температуре -70°С.
Для культивирования штамма применяют среду следующего состава (г/л): пептон - 10, дрожжевой экстракт - 5, NaCl - 5, ампициллин 0,1. Культивирование проводят при 30°С с аэрацией до достижения стационарной стадии роста.
Выход препарат фермента составляет ~2 мл/г сырой биомассы с концентрацией 50000 е.а./мл.
Рекомбинантная ДНК-метилтрансфераза M.AgsI характеризуется следующими свойствами:
1. Узнает и, в присутствии SAM, метилирует последовательность 5′-TTSAA-3′ на обеих цепях ДНК, с образованием 5′-TTSA(m6A)-3′/3′-(m6A)ASTT-5′.
2. Оптимальная температура реакции 30°С.
3. Фермент проявляет высокую активность в буфере следующего состава: 100 мкМ SAM, 33 мМ Трис-ацетат (рН 7,9 при 25°С), 66 мМ калия ацетат, 1 мМ ЭДТА, 1 мМ дитиотреитол, 100 мкг/мл бычий сывороточный альбумин.
Определяющим отличием предлагаемого штамма от всех известных в настоящее время штаммов-продуцентов ДНК-метилтрансфераз является то, что он позволяет получать коммерчески доступный препарат фермента M.AgsI, не имеющего аналогов, узнающего и, в присутствии SAM, метилирующего последовательность 5′-TTSAA-3′ на обеих цепях ДНК, с образованием 5′ -TTSA(m6A)-3′/3′-(m6A)ASTT-5′.
Поскольку предлагаемый штамм получен впервые и для выделения ДНК-метилтрансферазы, способной узнавать и метилировать вышеназванную последовательность нуклеотидов в указанной позиции, никогда не использовался, можно сделать вывод о соответствии предлагаемого штамма критериям изобретения «новизна» и «изобретательский уровень».
Изобретение иллюстрируется примерами конкретного выполнения.
Пример 1. Конструирование рекомбинантной плазм иды pM.AgsI-106
Получение библиотеки клонов E.coli
Штамм Agrococcus species 25 выращивали в LM-бульоне (г/л): триптон "Organotechnie" (Франция) - 10, дрожжевой экстракт (той же фирмы) - 5, NaCl - 5, MgCl2 - 0,5, тиамин - 0,01, рН 7,0-7,4 при 30°С со встряхиванием в колбах 2 дня. Из 3 г клеток выделяли фенольным методом (5) 4 мг хромосомной ДНК. Далее хромосомную ДНК расщепляли отдельно эндонуклеазами рестрикции PstI и Zsp2I и сшивали с помощью Т4 ДНК-лигазы по стандартной методике (6) с ДНК вектора pUC19 (7), линеаризованного PstI в соотношении вектор/хромосомная ДНК - 1/9. Лигазной смесью трансформировали клетки E.coli RR1 методом электропорации на «Easyject Prima» (Великобритания) и выращивали в 100 мл бульона LM с 0,1 г/л ампициллина. С использованием набора «QIAGEN Plasmid Maxi Kit» (Германия) из клеток выделяли 0,6 мл ДНК суммарной плазмиды ΣpAgs/PZ с концентрацией 0,3 мг/мл, представляющей библиотеку из 5000 рекомбинантов.
Выделение целевой плазмиды
Плазмиду, кодирующую целевую ДНК-метилтрансферазу, отбирали по методике, описанной ранее (8). При этом ДНК ΣpAgs/PZ расщепляли рестриктазой AgsI и повторно трансформировали клетки E.coli RR1. На LM-агаре с ампициллином получали 120 клонов (pAgs/PZ №1-120). Из них выделяли плазмиды с использованием набора «QIAGEN Plasmid Miniprep Kit» (Германия). В плазмидах определяли наличие метилирования и размеры вставок путем их гидролиза рестриктазами AgsI и VspI, соответственно, с последующим электрофорезом в 1% агарозном геле.
На фиг. 2 представлены результаты электрофореза после обработки одной из отобранных плазмид, названной pM.AgsI-106, рестриктазами VspI и AgsI, где дорожки:
1 - исходная ДНК pM.AgsI-106;
2 - pM.AgsI-106+VspI;
3 - pM.AgsI-106+AgsI;
M - маркер молекулярного веса ДНК 1 kb (производства ООО «СибЭнзайм»).
Данная плазмида содержит встроенный фрагмент длиной около 2500 п. н., что видно по гидролизу VspI (дорожка 2), и защищена от гидролиза AgsI (дорожка 3), то есть содержит функционально активный ген ДНК-метилтрансферазы M.AgsI. Этой плазмидой повторно трансформировали клетки Escherichia coli RR1 и получили продуцент Escherichia coli N106 (pM.AgsI).
Измерение активности фермента M.AgsI в клетках Escherichia coli N106 (pM.AgsI)
Штамм выращивали в 300 мл LM-бульона с добавлением 100 мкг/мл ампициллина в 0,5 л колбах со встряхиванием при 30°С в течение двух суток. Культура при плотности 2,2 о.е. при 540 нм/см давала 3 г/л биомассы. 1 мл центрифугировали в пробирке «Eppendorf» при 12000 об/мин в течение 3 мин. Осадок из 1 мл суспендировали в 0,2 мл лизирующего буфера: 50 мкг/мл лизоцим, 0,4% Тритон Х-100, 10 мМ Трис-HCl (рН 8,1) и 1 мМ ЭДТА, и лизировали клетки при 22°С в течение 30 мин. 25 мкл лизата вносили в 100 мкл SM-смеси, содержащей: 50 мкг/мл ДНК фага λ (dam- dcm-), 100 мкМ SAM, М-буфер «Y′» (33 мМ Трис-ацетат (рН 7,9), 1 мМ ЭДТА, 66 мМ ацетат калия и 1 мМ дитиотреитол), и 100 мкг/мл бычий сывороточный альбумин. Делали 10 дополнительных 2-кратных разбавлений в аликвотах по 50 мкл SM-смеси. Метилировали ДНК X при 30°С в течение 2 ч. Прогревали смесь при 65°С в течение 10 мин для инактивации ДНК-метилтрансферазы и охлаждали до комнатной температуры. Добавляли по 5 мкл 1000 е.а./мл эндонуклеазы рестрикции AgsI в SE-буфере В100 (10 мМ Трис-HCl (рН 7,6), 50 мМ KCl, 0,1 мМ ЭДТА, 200 мкг/мл бычий сывороточный альбумин, 1 мМ дитиотреитол), дополненном 111 мМ ацетатом магния. Инкубировали реакционную смесь при 37°С в течение 1 ч и анализировали электрофорезом в 1%-ном агарозном геле. За единицу активности ДНК-метилтрансферазы принимали количество фермента, достаточное для защиты 1 мкг ДНК фага X за 1 ч от гидролиза сопряженной эндонуклеазы рестрикции AgsI. Для простого выявления активности 5 мкл лизата либо 5 мкл пробы хроматографии (разбавленной в 100 раз в SE-буфере В100) вносили в 50 мкл SM-смеси, метилировали ДНК 30 мин при 30°С, расщепляли сопряженной рестриктазой и проводили электрофорез в 1% агарозном геле.
На фиг. 3 представлены результаты определения активности M.AgsI в лизате клеток Escherichia coli N106 (pM.AgsI), где дорожки:
1 - метилирование 50 мкг/мл X ДНК лизатом, разбавленным в реакционной смеси до исходной плотности клеток культуры + AgsI;
2 - метилирование лизатом, разбавленным в 2 раза + AgsI;
3 - метилирование лизатом, разбавленным в 4 раза + AgsI;
4 - метилирование лизатом, разбавленным в 8 раз + AgsI;
5 - метилирование лизатом, разбавленным в 16 раз + AgsI;
6 - метилирование лизатом, разбавленным в 32 раза + AgsI;
7 - метилирование лизатом, разбавленным в 64 раза + AgsI;
8 - метилирование лизатом, разбавленным в 128 раз + AgsI;
9 - метилирование лизатом, разбавленным в 250 раз + AgsI;
10 - метилирование лизатом, разбавленным в 500 раза + AgsI;
11 - метилирование лизатом, разбавленным в 1000 раз + AgsI;
12 - контроль без лизата + AgsI;
М - маркер молекулярного веса ДНК λ/BssT1I.
На фиг. 3 видно, что метилирование защищает ДНК от гидролиза рестриктазой AgsI до 5-го разбавления лизата, что соответствует активности M.AgsI 400 е.а./мл культуры или 130000 е.а./г клеток. Учитывая низкую активность ДНК-метилтрансфераз природных штаммов, заявляемый штамм Escherichia coli N106 (pM.AgsI) является суперпродуцентом M.AgsI.
Анализ ДНК плазмиды pM.AgsI-106
Секвенирование встроенного фрагмента в ДНК pM.AgsI-106 было проведено методом Сэнгера (9) на автоматическом секвенаторе ABI 3130xI Genetic Analyzer ("Applied Biosystems", США). Анализ первичной структуры показал, что Pstl-фрагмент ДНК A.species 25, клонированый в pUC19, состоит из 2527 п.н. Фрагмент содержит две однонаправленные пересекающиеся открытые рамки трансляции, которые являются генами двух субъединиц ДНК-метилтрансферазы M.AgsI: рамка субъединицы А, длиной 918 п. н., и рамка субъединицы В, длиной 561 п. н. Область пересечения генов составляет 41 п. н.
На фиг. 4 приведена нуклеотидная последовательность гена, кодирующего субъединицу А ДНК-метилтрансферазы M.AgsI, а также соответствующая ей аминокислотная последовательность.
На фиг. 5 приведена нуклеотидная последовательность гена, кодирующего субъединицу В ДНК-метилтрансферазы M.AgsI, а также соответствующая ей аминокислотная последовательность.
Выравнивание аминокислотных последовательностей субъединиц M.AgsI с помощью программы BLAST (http://blast.ncbi.nlm.nih.gov/Blast.cgi) выявило консервативные мотивы, характерные для аденин (N6)-ДНК-метилтрансфераз (КФ 2.1.1.72) (10).
Пример 2. Выращивание штамма и выделение рекомбинантной ДНК-метилтрансферазы M.AgsI
Получение биомассы
Клетки Escherichia coli N106 (pM.AgsI) выращивали в LM-бульоне с ампициллином в 20-литровом ферментере "New Brunswick Scientific" (США) при 30°С, аэрации 15 л/мин, перемешивании 200 об/мин 8 ч до оптической плотности 2,2-2,5 ед/см при 540 нм. Центрифугировали на центрифуге «Весктап J2-M1» (США) в роторе JA-10 при 8000 об/мин в течение 20 минут. За два выращивания получали 124 г клеток.
Условия выделения фермента
Фермент M.AgsI выделяли при 4°С с использованием буферных растворов:
буфер А - 10 мМ Трис-HCl рН 7,5, 0,1 мМ ЭДТА и 7 мМ β-меркаптоэтанол;
буфер Б - 10 мМ К-фосфат рН 7,2, 0,1 мМ ЭДТА и 7 мМ β-меркаптоэтанол.
100 г биомассы суспендировали в 500 мл буфера А, содержащего 5% глицерин, 0,5% Тритон Х-100, 1 мг/мл лизоцим, 0,1 мМ PMSF (фенилметилсульфонилфторид - ингибитор протеаз), и перемешивали 1 ч. По 100 мл суспензии разрушали ультразвуком на дезинтеграторе Soniprep 150 («MSE» Великобритания) с насадкой диаметром 2 см при амплитуде 22-24 мкм пятью импульсами по 1 мин с охлаждением во льду по 1 мин. Добавляли NaCl до 0,2 М и центрифугировали на «Beckman J2-M1» в роторе JA-20 при 18000 об/мин 30 минут.
560 мл полученного экстракта наносили на колонку (15 см × 50 мм) с фосфоцеллюлозой Р-11 («Whatman» Великобритания) объемом 300 мл, уравновешенной буфером А с 0,2 М NaCl. Промывали 600 мл буфера А с 0,2 М NaCl. Элюировали линейным градиентом 0,2 М-0,6 М NaCl в буфере А объемом 3 л. Собирали 200 фракций по 15 мл. Фракции 68-89, содержащие M.AgsI с максимальной активностью, объединяли.
Объединенные фракции диализовали 4 ч против 3 л буфера А с 0,05 М NaCl и наносили на колонку (8 см × 25 мм) с гепарин-сефарозой («Bio-Rad» США) объемом 40 мл. Промывали 80 мл буфера А с 0,05 М NaCl и элюировали градиентом 0,05 М - 0,5 М NaCl в буфере А объемом 800 мл.
Собирали 50 фракций по 16 мл. Фракции 18-20, содержащие пик активности M.AgsI, объединяли.
Объединенные фракции наносили на колонку (205 см × 25 мм) с 1000 мл Сефакрила S-200 («Bio-Rad» США), уравновешанного в буфере А с 0,8 М NaCl. Элюировали 1000 мл буфера А с 0,8 М NaCl. Собирали 100 фракций по 10 мл, из которых фракции 65-72 содержали пик целевой активности.
Объединенные фракции наносили на колонку (10 см × 25 мм) с 50 мл гидроксиапатита («Bio-Rad» США) в буфере Б. Промывали 100 мл буфера Б и элюировали линейным градиентом 0,01-0,2 М K-PO4 (рН 7,2) в буфере Б объемом 1000 мл. Собирали 60 фракций по 16,7 мл. Фракции 27-32, содержащие пик активности M.AgsI, объединяли и диализовали против 1 л буфера, содержащего: 50% глицерин, 10 мМ Трис-HCl (рН 7,5), 0,05 М NaCl, 0,1 мМ ЭДТА и 7 мМ β-меркаптоэтанол. 22 мл полученного препарата ДНК-метилтрансферазы M.AgsI хранили при -20°С.
Пример 3. Определение специфичности ДНК-метилтрансферазы M.AgsI
Основание, метилируемое M.AgsI, определяли методом IIS-кассеты (11). Олигодезоксирибонуклеотидный дуплекс, синтезированный на автоматическом синтезаторе ДНК/РНК ASM-800 («БИОССЕТ» Россия), содержал сайт TTSAA, метилируемый M.AgsI (выделен жирным шрифтом), и вне метилирования сайт GGATG, узнаваемый FokI, эндонуклеазой рестрикции IIS-типа (выделен рамкой). При этом точки гидролиза FokI (показаны стрелками) приходились на метилируемый сайт.
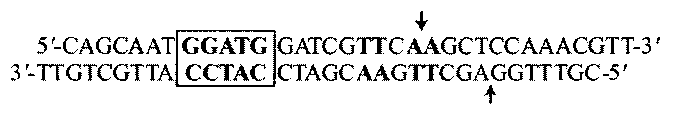
Дуплекс метилировали M.AgsI с использованием 3H-SAM, расщепляли FokI, фрагменты разделяли электрофорезом в 20% полиакриламидном геле, для каждого из них измеряли радиоактивность. Распределение метки во фрагментах олигонуклеотидного дуплекса, меченного M.AgsI в присутствии 3H-SAM, после его гидролиза FokI представлено в таблице. Из табл. видно, что метка включалась преимущественно во фрагменты длиной 13 и 27 нуклеотидов. Если больший фрагмент содержал весь сайт, узнаваемый M.AgsI, то меньший только его внешний аденин. Следовательно, M.AgsI метилировала внешний аденин узнаваемой последовательности, образуя 5′-TTSA(3Hm6A)-3′.

Таким образом, результаты M.AgsI-3Н-метилирования сайта TTSAA олигонуклеотидного дуплекса, M.AgsI-метилирования λ ДНК, блокирующего расщепление сайта TTSAA эндонуклеазой рестрикции M.AgsI; а также выравнивание аминокислотных последовательностей M.AgsIA и M.AgsIB с доменами, характерными для аденин (N6)-ДНК-метилтрансфераз, доказывают, что ДНК-метилтрансфераза M.AgsI образует TTSA(m6A) на ДНК и относится к аденин (N6)-ДНК-метилтрансферазам (КФ 2.1.1.72).
Заявленная ДНК-метилтрансфераза M.AgsI, являясь ферментом с новой специфичностью, может представлять интерес для молекулярно-биологических и генно-инженерных исследований.
Источники информации
1. Cheng X., Blumenthal R.M. (Eds.). S-Adenosylmethionine-Dependent Methyltransferases. Structures and Functions. // Singapore, New Jersey, London, Hong Kong: World Scientific Publishing Co. Pte. Ltd. - 1999.
2. REBASE. - 2015. Available at: http://rebase.neb.com.
3. Gonchar D.A., Wolf Y.I., Degtyarev S.K. Cloning and characterization of Sse9I DNA-methyltransferase recognizing 5′-AATT-3′. // Nucleic Acids Res. - 1996. - V. 24. - P. 2790-2792.
4. Vaisvila R., Morgan R.D., Kucera R.B., Claus Т.Е., Raleigh, E.A. A Method for cloning and producing the Msel restriction endonuclease. // Patent US 6846658 B1, оп. 25.01.2005.
5. Smith C.L., Klco S.R. and Cantor C.R. In: Davies K. (ed.). Genome Analysis. A Practical Approach. // - 1987. - Oxford. UK: IRL Press.
6. Sambrook J., Fritsch E.F. and Maniatis T. Molecular Cloning. A laboratory manual, 2nd ed. // - 1989. - Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press.
7. Yanisch-Perron C, Vieira J., Messing J. Improved M13 phage cloning vectors and host strains: nucleotide sequences of the M13mp18 and pUC19 vectors. // Gene. - 1985. - 33. - P. 103-119.
8. Mann M.В., Rao R.N., Smith H.O. Cloning of restriction and modification genes in E.coli: the HhaII system from Haemophilus haemolyticus. // Gene. - 1978. - V. 3. - P. 97-112.
9. Sanger F., Nicklen S., Coulson A. R. DNA sequencing with chain-terminating inhibitors. // Proc Natl Acad Sci USA. - 1977. - V. 74. - P. 5463-5467.
10. Timinskas A., Butkus V., Janulaitis A. Sequence motifs characteristic for DNA [cytosine-N4] and DNA [adenine-N6] methyltransf erases. Classification of all DNA methyltransferases. // Gene. - 1995. - V. 157. - P. 3-11.
11. Чернухин В.А., Каширина Ю.Г., Суханова К.С., Абдурашитов М.А., Гончар Д.А., Дегтярев С.Х. ДНК-метилтрансфераза FauIA, модифицирующая второй остаток цитозина в непалиндромной последовательности 5′-CCCGC-3′ (Выделение и свойства). // Биохимия. - 2005. - Т. 70. - С. 829-837.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНЫЙ ШТАММ БАКТЕРИЙ ESCHERICHIA COLI N16 (PM.ALUBI) - ПРОДУЦЕНТ ДНК-МЕТИЛТРАНСФЕРАЗЫ M.ALUBI | 2015 |
|
RU2603086C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ БАКТЕРИЙ Escherichia coli N42 (pElmI) - ПРОДУЦЕНТ МЕТИЛЗАВИСИМОЙ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ ElmI | 2015 |
|
RU2597987C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ БАКТЕРИЙ Escherichia coli N41 (pBpuN4/MR)-ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ РЕСТРИКЦИИ BpuN4I | 2013 |
|
RU2529362C1 |
| НОВЫЕ ВАРИАНТЫ ПОЛИПЕПТИДА PapM БАКТЕРИЙ РОДА Streptomyces | 2003 |
|
RU2351608C2 |
| Штамм бактерий Micrococcus luteus 805 - продуцент сайт-специфической метилзависимой эндонуклеазы MluVI | 2016 |
|
RU2614262C1 |
| СПОСОБЫ АМПЛИФИКАЦИИ ДНК ДЛЯ СОХРАНЕНИЯ СТАТУСА МЕТИЛИРОВАНИЯ | 2018 |
|
RU2754038C2 |
| ШТАММ БАКТЕРИЙ Plantibacter flavus 3Kz - ПРОДУЦЕНТ МЕТИЛЗАВИСИМОЙ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ PfsI | 2015 |
|
RU2593723C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pEstPc, КОДИРУЮЩАЯ ПОЛИПЕПТИД СО СВОЙСТВАМИ ЭСТЕРАЗЫ Psychrobacter cryohalolentis K5, И ШТАММ БАКТЕРИЙ Escherichia coli - ПРОДУЦЕНТ ПОЛИПЕПТИДА СО СВОЙСТВАМИ ЭСТЕРАЗЫ Psychrobacter cryohalolentis K5 | 2011 |
|
RU2478708C1 |
| ЛИТИЧЕСКАЯ ПРОТЕАЗА AlpA БАКТЕРИИ LYSOBACTER SP. XL1, ФРАГМЕНТ ДНК, КОДИРУЮЩИЙ ЛИТИЧЕСКУЮ ПРОТЕАЗУ AlpA БАКТЕРИИ LYSOBACTER SP. XL1, И СПОСОБ ПОЛУЧЕНИЯ ЛИТИЧЕСКОЙ ПРОТЕАЗЫ AlpA БАКТЕРИИ LYSOBACTER SP. XL1 | 2009 |
|
RU2407782C2 |
| ЛИТИЧЕСКАЯ ПРОТЕАЗА AlpB БАКТЕРИИ Lysobacter sp. XLI, ФРАГМЕНТ ДНК, КОДИРУЮЩИЙ ЛИТИЧЕСКУЮ ПРОТЕАЗУ AlpB БАКТЕРИИ Lysobacter sp. XLI, И СПОСОБ ПОЛУЧЕНИЯ ЛИТИЧЕСКОЙ ПРОТЕАЗЫ AlpB БАКТЕРИИ Lysobacter sp. XLI | 2009 |
|
RU2408725C2 |





Изобретение относится к фармацевтической промышленности, а именно к способу получения рекомбинантного штамма бактерий Escherichia coli N106 (pM.AgsI) . Рекомбинантный штамм бактерий Escherichia coli N106 (pM.AgsI) получают трансформацией клеток штамма Escherichia coli RR1 плазмидой pM.AgsI-106, продуцирующий ДНК-метилтрансферазу M.AgsI, узнающую нуклеотидную последовательность ДНК 5′-TTSAA-3′ и метилирующую внешний остаток аденина этой последовательности с образованием 5′-TTSA(m6A)-3′. Вышеописанный способ позволяет получить рекомбинантный штамм бактерий Escherichia coli N106 (pM.AgsI), продуцирующий высокоактивную ДНК-метилтрансферазу заданной специфичности. 5 ил., 1 табл., 3 пр.
Рекомбинантный штамм бактерий Escherichia coli N106 (pM.AgsI), полученный трансформацией клеток штамма Escherichia coli RR1 плазмидой pM.AgsI-106, продуцирующий ДНК-метилтрансферазу M.AgsI, узнающую нуклеотидную последовательность ДНК 5′-TTSAA-3′ и метилирующую внешний остаток аденина этой последовательности с образованием 5′-TTSA(m6A)-3′.
| US 0006846658 B1 (New England Biolabs, Inc.) 25.01.2005 | |||
| Gonchar D.A., Wolf Y.I., Degtyarev S.K | |||
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| // Nucleic Acids Res | |||
| Предохранительное устройство для паровых котлов, работающих на нефти | 1922 |
|
SU1996A1 |
| - V | |||
| Пишущая машина для тюркско-арабского шрифта | 1922 |
|
SU24A1 |
| - P | |||
| Кольцевая шпонка для соединения конструктивных деревянных частей | 1925 |
|
SU2790A1 |
| РЕКОМБИНАНТНЫЙ ШТАММ БАКТЕРИЙ Escherichia coli N41 (pBpuN4/MR)-ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ РЕСТРИКЦИИ BpuN4I | 2013 |
|
RU2529362C1 |

