Изобретение относится к биотехнологии, в частности к генной инженерии и микробиологической промышленности, и касается нового рекомбинантного штамма бактерий Escherichia coli (E.coli), который может быть использован для получения высокоактивного препарата метилзависимой сайт-специфической ДНК-эндонуклеазы ElmI, расщепляющей обе цепи последовательности ДНК 5′-GC^NGC-3′/3′-CGN^CG-5′, в которой не менее трех цитозинов метилированы в положении С5 с образованием 5′-выступающих однонуклеотидных концов.
Главными биотехнологически значимыми параметрами сайт-специфических метилзависимых ДНК-эндонуклеаз (MD-эндонуклеаз) являются узнаваемая последовательность, узнаваемый узор метилирования, позиция гидролиза относительно этой последовательности, необходимость присутствия кофакторов (например, Mg2+ или АТФ) и активность препарата фермента, выделенного из соответствующего штамма-продуцента.
Наиболее близким к заявляемому штамму - прототипом, является штамм Planomicrobium koreense 78K, продуцирующий метилзависимую эндонуклеазу PkrI [1], которая узнает последовательность нуклеотидов 5′-GCN^GC-3′/3′-CG^NCG-5′ со сходным узором метилирования, требует для эффективного расщепления наличия в узнаваемой последовательности не менее трех 5-метитиозинов (5 mC), но расщепляет эту последовательность после центрального нуклеотида N.
Недостатком известного штамма является то, что продуцируемая им сайт-специфическая эндонуклеаза не расщепляет узнаваемую последовательность перед центральным нуклеотидом N с образованием 5′-выступающих концов. Это свойство оказывается существенно важным при проведении генно-инженерных манипуляций, когда необходимо с помощью сайт-специфической ДНК-эндонуклеазы получать одноцепочечные выступающие (липкие) концы ДНК, которые затем могут быть сшиты с фрагментами, образованными в результате гидролиза другими эндонуклеазами и имеющими такую же «липкость».
В настоящее время не описаны штаммы бактерий, являющиеся продуцентами метилзависимых сайт-специфических ДНК-эндонуклеаз, узнающих и расщепляющих последовательность 5′-GC^NGC-3′/3′-CGN^CG-5′ перед центральным нуклеотидом N, в которой вместо цитозина присутствуют три или четыре 5-метилцитозина.
Задачей изобретения является получение рекомбинантного штамма E. coli, продуцирующего MD-эндонуклеазу, которая узнает и расщепляет обе цепи нуклеотидной последовательности ДНК 5′-GC^NGC-3′/3′-CGN^CG-5′, при метилировании в ней не менее трех цитозинов в положении С5 и не расщепляет данную последовательность, если число метилированных цитозинов оказывается меньше трех.
Поставленная задача решается путем получения рекомбинантного штамма бактерий Escherichia coli N42 (pElmI) - продуцента MD-эндонуклеазы, узнающей и расщепляющей последовательности нуклеотидов:
5′-G(5mC)^NG(5mC)-3′/3′-(5mC)GN^G(5mC)-5′ (четыре 5-метилцитозина), G(5mC)^NG(5mC)-3′/3′-(5mC)GN^GC-5′ (три 5-метилцитозина), G(5mC)^NG(5mC)-3′/3′-CGN^G(5mC)-5′ (три 5-метилцитозина), где знаком «^» указаны позиции расщепления ДНК.
Техническим результатом изобретения является получение рекомбинантного штамма бактерий Escherichia coli N42 (pElmI), продуцирующего MD-эндонуклеазу заданной специфичности с более высоким выходом по сравнению с исходным природным штаммом Escherichia coli N17.
Предлагаемый штамм-продуцент получен путем трансформации клеток Escherichia coli штамма ER2267 плазмидной ДНК pElmI. Данная плазмида, полученная на основе вектора pMTL22 [2], включает участок хромосомной ДНК штамма Escherichia coli N17, являющегося продуцентом метилзависимой ДНК-эндонуклеазы ElmI, выделенного из природного материала (воды) в результате целенаправленного систематического поиска, но не являющегося высокопродуктивным (активность препарата фермента, выделенного из Escherichia coli N17, составляла всего 100 е.а./мл). Благодаря активации промотора гена lacZ в pMTL22 путем добавления индуктора (0.5 мМ IPTG) в процессе выращивания предлагаемого штамма Escherichia coli N42 (pElmI) удалось добиться повышения удельной активности целевого фермента в биомассе в 20 раз по сравнению с природным штаммом.
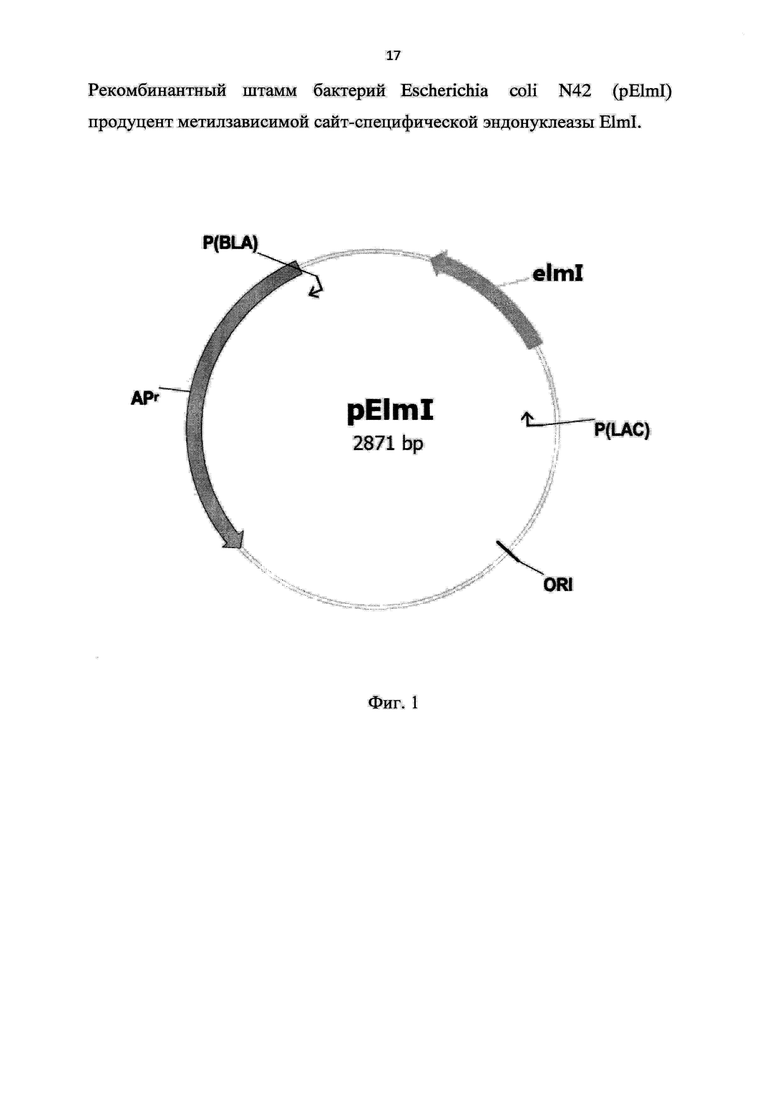
На фигуре 1 представлена физическая карта плазмиды pElmI, где
ORI - точка начала репликации плазмиды pElmI;
Apr - ген bla, обеспечивающий устойчивость к ампициллину;
P(BLA) - промотор гена bla;
P(LAC) - промотор гена lacZ;
elmI - ген, кодирующий метилзависимую эндонуклеазу ElmI;
Сконструированной плазмидой pElmI трансформируют клетки E.coli штамма ER2267 и подращивают при 37°C в течение ночи. Трансформантов отбирают на селективной агаризованной питательной среде, содержащей 100 мкг/мл ампициллина, по устойчивости к антибиотику. Содержание рекомбинантной MD-эндонуклеазы ElmI составляет не менее 4000 е.а./г влажной биомассы.
Преимуществом заявляемого штамма, по сравнению с наиболее близким аналогом, является то, что при расщеплении метилированной ДНК продуцируемой им метилзависимой ДНК-эндонуклеазой образуются однонуклеотидные 5′-выступающие «липкие» концы, сходные с концами, получающимися при расщеплении ДНК такими эндонуклеазами рестрикции, как, например, Fsp4HI (сайт узнавания 5′-GC^NGC-3′), Bst2UI (5′-CC^WGG-3′), AsuC2I (5′-CC^SGG-3′), MspR9I (5′-CC^NGG-3′), AclWI (5′-GGATC-3′ (4/5)) и других [3], а также рядом метилзависимых ДНК-эндонуклеаз, например, MteI (сайт узнавания 5′-G(5mC)G(5mC)^NG(5mC)G(5mC)-3′), что позволяет эффективно применять фермент ElmI в разнообразных генно-инженерных манипуляциях.
Штамм Escherichia coli N42 (pElmI) имеет следующие характеристики.
Культурально-морфологические признаки.
Клетки мелкие палочковидной формы, грамотрицательные, 1×3,5 мкм, подвижные. Штамм хорошо растет на обычных питательных средах (ΜΠΑ, МПБ, LB-бульон, LB-arap, минимальная среда с глюкозой). При росте на агаризованной среде LB колонии круглые, гладкие, полупрозрачные, блестящие, серые. Край ровный, диаметр колоний 1-3 мм, консистенция пастообразная. Рост в жидких средах (LB, минимальная среда с глюкозой) характеризуется ровным помутнением, осадок легко седиментирует.
Физиолого-биохимические признаки.
Клетки растут при 4-42°C, оптимум рН 6,8-7,6. В качестве источника азота используют как минеральные соли аммония, так и органические соединения: аминокислоты, пептон, триптон, дрожжевой экстракт. В качестве источника углерода при росте на минимальной среде используют глицерин, углеводы, аминокислоты.
Генетические признаки, устойчивость к антибиотикам.
Штамм-хозяин Escherichia coli ER2267 (F′ proA+B+lacIqΔ(lacZ)M15 zzf::mini-Tn10 (KanR)/Δ(argF-lacZ)U169 glnV44 e14-(McrA-)Δ(mcrC-mrr)). Проявляет устойчивость к канамицину (до 25 мкг/мл).
Штамм Escherichia coli N42 (pElmI) дополнительно проявляет устойчивость к ампициллину (до 300 мкг/мл), обусловленную наличием гена устойчивости в ДНК рекомбинантной плазмиды pElmI.
Условия хранения штамма.
Штамм Escherichia coli N42 (pElmI) хранят на агаризованной среде LB со 100 мкг/мл ампициллина на чашках Петри или в стеклянных пробирках со скошенным агаром при 4°C. Пересевы на свежие среды делают один раз в месяц. Может храниться не менее года в среде LB, содержащей 30% глицерин, при -50-70°C. Длительное хранение штамма осуществляют в лиофильно высушенном состоянии или в растворе 30% глицерина при температуре -70°C.
Для культивирования штамма применяют среду следующего состава (г/л): триптон - 10, дрожжевой экстракт - 5, NaCl - 5, с добавлением 100 мкг/мл ампициллина. Культивирование проводят при 37°C с аэрацией до достижения стационарной стадии роста. Выход препарата фермента ElmI составляет ~0,5 мл/г сырой биомассы с концентрацией 2000 е.а./мл.
Продуцируемая новым рекомбинантным штаммом метилзависимая ДНК-энднуклеаза ElmI характеризуется следующими свойствами: 1. Узнает и расщепляет последовательность 5′-GC^ΝGC-3′/3′-CGN^CG-5′ на обеих цепях ДНК перед центральным нуклеотидом N при условии метилирования не менее трех из четырех возможных цитозинов в ней в положении С5.
3. Не расщепляет вышеприведенную последовательность, если количество метилированных цитозинов в ней меньше трех, а также неметилированную последовательность 5′-GCNGC-3′/3′-CGNCG-5′.
4. Оптимальная температура реакции 37°C.
5. Фермент проявляет высокую активность в буфере следующего состава: 10 мМ Трис-HCl (рН 8,5 при 25°C), 10 мМ магния ацетат, 10 мМ NaCl, 1 мМ дитиотреитол.
Определяющим отличием предлагаемого штамма от всех известных в настоящее время штаммов-продуцентов сайт-специфических эндонуклеаз является то, что он позволяет получать коммерчески доступный препарат, не имеющий аналогов, метилзависимой сайт-специфической ДНК-эндонуклеазы ElmI, которая узнает и расщепляет обе цепи последовательности 5′-GC^NGC-3′/3′-CGN^CG-5′ перед центральным нуклеотидом N при условии метилирования в этой последовательности не менее трех цитозинов в положении С5.
Новая сайт-специфическая ДНК-эндонуклеаза ElmI является неошизомером метилзависимой ДНК-эндонуклеазы PkrI, то есть узнает ту же последовательность ДНК, но расщепляет ее в другой позиции. В отличие от ElmI MD-эндонуклеаза PkrI расщепляет узнаваемый сайт 5′-GCN^GC-3′ после центрального нуклеотида с образованием 3′-выступающих однонуклеотидных концов.
Поскольку предлагаемый штамм получен впервые и для выделения сайт-специфической эндонуклеазы, узнающей и расщепляющей вышеназванную последовательность нуклеотидов, в указанной позиции никогда не использовался, можно сделать вывод о соответствии предлагаемого штамма критериям изобретения «новизна» и «изобретательский уровень».
Изобретение иллюстрируется примерами конкретного выполнения.
Пример 1. Конструирование рекомбинантной плазмиды pElmI.
Ген, кодирующий MD-эндонуклеазу ElmI, был амплифицирован с помощью ПЦР с использованием хромосомной ДНК дикого штамма-продуцента Escherichia coli N17, выделенного из природного источника (воды).
Для ПЦР использовались следующие праймеры:
Entero direct: 5′-CCCCCATATGAGTGCACGTGAAGCA-3′
Entero reverse 1: 5′-CGCGGATCCTTAGGGATTACACTGACTGAA-3′
На следующем этапе осуществляли встраивание амплифицированного гена, обработанного эндонуклеазами рестрикции FauNDI+BamHI в плазмиду pMTL22 по сайтам рестрикции FauNDI+BglII. Процедуру лигазной сшивки, а также дальнейшую трансформацию клеток E.coli штамма ER2267 осуществляли известным способом [4].
Из одного из полученных трансформантов выделяли рекомбинантную плазмиду, названную pElmI, и содержащую клонированный FauNDI-BamHI ПЦР-фрагмент ДНК Escherichia coli N17 в векторе pMTL22/FauNDI-BglII. Длина встроенного фрагмента составляла 439 пар нуклеотидов. Фрагмент являлся геном метилзависимой сайт-специфической ДНК-эндонуклеазы ElmI (позиция открытой рамки считывания - 183-625 по комплементарной цепи). Направление рамки считывания гена elmI в рекомбинантной плазмиде совпадало с направлением считывания гена lacZ из pMTL22 (Фиг. 1).
Первичная структура встроенного гена, кодирующего MD-эндонуклеазу ElmI, была определена методом Сэнгера [5] на автоматическом секвенаторе ABI 3130xI Genetic Analyzer ("Applied Biosystems", США).
На фиг. 2. приведена нуклеотидная последовательность гена, кодирующего MD-эндонуклеазу ElmI, и соответствующая аминокислотная последовательность белка.
Пример 2. Выращивание штамма и выделение рекомбинантной MD-эндонуклеазы ElmI.
Получение биомассы клеток штамма-продуцента.
Для получения биомассы клетки штамма-продуцента Escherichia coli N42 (pElmI) переносили в чашку Петри на агаризованную среду LB с ампициллином (100 мкг/мл) и инкубировали в течение ночи при 37°C. Свежевыращенные колонии переносили в две колбы объемом 500 мл, содержащих по 300 мл бульона (1% триптон («Organotechnie», Франция), 0,5% дрожжевой экстракт («Organotechnie», Франция), 0,5% NaCl, 0,05% MgCl2 и 0,001% тиамин, (рН 7,0), и 100 мкг/мл ампициллина) и подращивали при 37°C в течение 12 часов без встряхивания. Затем культура рассевалась по 10 мл по 20 колбам, содержащим по 300 мл бульона того же состава. Колбы встряхивали при 120-140 об/мин в инкубаторе («New Brunswick», США) при +37°С в течение 3 часов, затем добавляли IPTG до 0.5 мМ и растили еще 16 часов.
Для измерения в клетках активности ElmI из культуры отбирались по две пробы по 1 мл в пробирки «Eppendorf», клетки осаждали в настольной центрифуге «5416 Eppendorf» ("Eppendorf GmbH", Германия) при 12000 об/мин 3 минуты, супернатанты убирали, а клетки замораживали при -18°C. Биомассы культур осаждали на центрифуге J2-21 («Beckman», США) в течение 30 минут в роторе JA-10 при 8000 об/мин, замораживали и хранили при -20°C. Получали 12 г замороженных клеток, что составляло 2 г/л бульона. Содержание рекомбинантной рестриктазы ElmI составляло 4000 е.а./г влажной биомассы. Разрушение биомассы клеток, выделение и очистку фермента проводили по модификации известной методики [6]. В результате из 12 г биомассы Escherichia coli N42 (pElmI) получили -5,5 мл препарата рекомбинантной MD-эндонуклеазы ElmI с концентрацией 2000 е.а./мл.
Пример 3. Сайт-специфический гидролиз субстратных ДНК рекомбинантной MD-эндонуклеазой ElmI.
Расщепление субстратных ДНК рекомбинантной MD-эндонуклеазой ElmI проводили в оптимальных условиях при температуре 37°C в реакционном буфере - SE-буфер «W» (10 мМ Трис-HCl (рН 8,5 при 25°C), 10 мМ магния ацетат, 10 мМ NaCl, 1 мМ дитиотреитол) в течение 60 мин.
Продукты расщепления ДНК разделяли путем электрофореза в 1% агарозном геле в буфере ТАЕ.
В качестве субстратов для определения специфичности фермента ElmI использовали ДНК плазмид, содержащих гены различных ДНК-метилтрансфераз. Благодаря активности этих генов в штаммах E.coli, из которых были выделены эти плазмиды, данные ДНК-субстраты оказывались модифицированными соответствующими ДНК-метилтрансферазами и несли определенный узор метилирования.
В качестве таких метилированных ДНК-субстратов были использованы следующие плазмиды:
1) pMHpaII, содержащая ген, кодирующий ДНК-метилтрансферазу HpaII, вследствие чего в данной плазмиде метилирован первый цитозин во всех последовательностях 5′-CCGG-3′ на обеих цепях ДНК [7].
2) pMHaeIII, содержащая ген, кодирующий ДНК-метилтрансферазу HaeIII, вследствие чего в данной плазмиде метилирован первый цитозин во всех последовательностях 5′-GGCC-3′ на обеих цепях ДНК [8].
3) pHspAI2, содержащая ген, кодирующий ДНК-метилтрансферазу HspAI, вследствие чего в данной плазмиде метилирован первый цитозин во всех последовательностях 5′-GCGC-3′ на обеих цепях ДНК [9]. Кроме того, данная плазмида содержит гиперметилированный участок:
5′-G(5mC)G(5mC)G(5mC)GC-3′
3′-CG(5mC)G(5mC)G(5mC)G-5′.
4) pHspAI10, также содержащая ген, кодирующий ДНК-метилтрансферазу HspAI, вследствие чего в данной плазмиде метилирован первый цитозин во всех последовательностях 5′-GCGC-3′ на обеих цепях ДНК [10]. Кроме того, данная плазмида содержит гиперметилированный участок:
5′-G(5mC)G(5mC)G CAG(5mC)G(5mC)G С-3′
3′-С G(5mC)G(5mC)GTC G(5mC)G(5mC)G-5′.
5) Та же pHspAI10, но дополнительно метилированная ДНК-метилтрансферазой Fsp4HI, в результате чего в данной плазмиде дополнительно метилирован первый цитозин во всех последовательностях 5′-GCNGC-3′ [10]. Данная плазмида содержит гиперметилированный участок:
5′-G(5mC)G(5mC)G(5mC)A G(5mC)G(5mC)G С-3′
3′-С G(5mC)G(5mC) G T(5mC)G(5mC)G(5mC)G-5′.
6) pFsp4HI3, содержащая ген, кодирующий ДНК-метилтранс-феразу Fsp4HI, в результате чего в данной плазмиде во всех последовательностях 5′-GCNGC-3′ метилирован первый цитозин [11]. Данная плазмида содержит гиперметилированный участок
5′-G(5mC)C G(5mC)G G(5mC)A G C-3′
3′-С G G(5mC) G C(5mC)G T(5mC)G-5′.
Все плазмиды были предварительно переведены из кольцевой формы в линейную путем гидролиза эндонуклеазой рестрикции DriI по уникальному (для каждой плазмиды) сайту 5′-GACNNNNNGTC-3′.
На фиг. 3 представлена электрофореграмма продуктов расщепления указанных выше ДНК-субстратов рекомбинантной метилзависимой ДНК-эндонуклеазой ElmI, где дорожки:
1 - pMHpaII/DriI;
2 - pMHpaII/DriI+ElmI;
3 - pMHaeIII/DriI;
4 - pMHaeIII/DriI+ElmI;
5 - pHspAI2/DriI;
6 - pHspAI2/DriI+ElmI;
7 - pHspAI10/DriI;
8 - pHspAI10/Dril+ElmI;
9 - pHspAI10/(M.Fsp4HI+DriI);
10 - pHspAI10/(M.Fsp4HI+DriI)+ElmI;
11 - pFsp4HI3/DriI;
12 - pFsp4HI3/DriI+ElmI
M - маркер молекулярного веса ДНК 1 kb (производства ООО «СибЭнзайм»).
Как видно из фиг. 3, эндонуклеаза ElmI не расщепляет последовательности, метилированные ДНК-метилтрансферазами HpaII (дорожка 2), HaeIII (дорожка 4), HspAI (дорожка 6).
ElmI также не расщепляет плазмиду pHspAI10, предварительно линеаризованную Dril (дорожка 8). В данной плазмиде содержится гиперметилированный участок, включающий последовательность 5′-GCAG(5mC)-3′/3′-(5mC)GTCG-5′, в которой метилированы вторые (внешние) цитозины на обеих цепях, и которая не расщепляется ElmI. Однако ElmI расщепляет эту же плазмиду (pHspAI10) после дополнительного ее метилирования ДНК-метилтрансферазой Fsp4HI с образованием пяти фрагментов (дорожка 10).
Поскольку ElmI узнает и расщепляет последовательность 5′-GCNGC-3′ при наличии в ней не менее трех 5-метилцитозинов, можно предположить, что сайты узнавания ElmI на данной плазмиде могут образовываться в двух случаях:
1) При перекрывании сайтов узнавания ДНК-метилтрансфераз HspAI и Fsp4HI (5′-GCGCNGC-3′) с образованием гиперметилированного сайта
5′-G(5mC)G(5mC)NGC-3′/3′-CG(5mC)GN(5mC)G-5′.
2) При перекрывании двух сайтов узнавания ДНК-метилтрансферазы Fsp4HI (5′-GCNGCNGC-3′) с образованием гиперметилированного сайта
5′-G(5mC)NG(5mC)NGC-3′/3′-CGN(5mC)GN(5mC)G-5′
В обоих случаях эти гиперметилированные сайты содержат
последовательности 5′-GCNGC-3′ с тремя 5-метилцитозинами.
Анализ первичной структуры плазмиды pHspAI10, проведенный с помощью программы Vector NTI Suite 7, показал, что при расщеплении по двум указанным выше сайтам эндонуклеазой ElmI (с учетом предварительной линеаризации плазмидной ДНК рестриктазой Dril) образуются ДНК-фрагменты следующей длины: ~1800, ~1200, ~490, ~420, ~60 п. н.
Как видно из фиг. 3, электрофоретическая подвижность ДНК-фрагментов, образованных ElmI после гидролиза pHspAI10/(M.Fsp4HI+DriI) соответствует теоретически предсказанной длине (дорожка 10).
Анализ первичной структуры плазмиды pFsp4HI3, проведенный с помощью программы Vector NTI Suite 7, показал, что при ее расщеплении по двум указанным выше сайтам эндонуклеазой ElmI (с учетом предварительной линеаризации плазмидной ДНК рестриктазой Dril) образуются фрагменты следующей длины: ~3500, ~490 (два фрагмента), -340 п. н. Как видно из фиг 3, электрофоретическая подвижность ДНК-фрагментов, образованных после гидролиза плазмиды pFsp4HI3/DriI эндонуклеазой Elm соответствует теоретически предсказанной длине (дорожка 12). Остальные же последовательности 5′-GCNGC-3′ в данной плазмиде содержат только два 5-метилцитозина и не расщепляются ElmI.
Полученные результаты свидетельствует о том, что MD-эндонуклеаза ElmI узнает и расщепляет только те варианты последовательности 5′-GCNGC-3′, которые содержат только три или четыре 5-метилцитозина.
Пример 4. Определение позиции гидролиза ДНК эндонуклеазой ElmI.
Определение места гидролиза ДНК ElmI осуществляли путем сравнения длин фрагментов, образуемых при расщеплении эндонуклеазами рестрикции ElmI и BisI олигонуклеотидного дуплекса D1/D2, образованного из олигонуклеотидов D1 и D2 (узнаваемая ElmI последовательность выделена рамочкой):

На фиг. 4 приведен радиоавтограф электрофореграммы продуктов расщепления радиоактивно меченного дуплекса D1*/D2 в 20% полиакриламидном геле с 7М мочевиной, где дорожки:
1 - дуплекс D1*/D2;
2 - дуплекс D1*/D2, обработанный эндонуклеазой ElmI;
3 - дуплекс D1*/D2, обработанный эндонуклеазой BisI;
Знаком «*» обозначена меченная цепь.
Как видно из фиг. 4, фрагменты ДНК, образованные в результате гидролиза ферментами ElmI и BisI дуплекса D1*/D2, имеют одинаковую длину, что говорит об идентичности позиций гидролиза узнаваемой последовательности этих метилзависимых эндонуклеаз. Так как BisI расщепляет последовательность 5′-GC^NGC-3′ перед центральным нуклеотидом N, то и ElmI также расщепляет ее перед центральным нуклеотидом.
Таким образом, ElmI узнает последовательность ДНК 5′-GC^NGC-3′ и расщепляет ее перед центральным нуклеотидом N на обеих цепях ДНК, образуя 5′-выступающие однонуклеотидные «липкие» концы.
Использование предлагаемого изобретения позволит расширить ассортимент высокопродуктивных штаммов-продуцентов, позволяющих получать высокоактивный препарат MD-эндонуклеазу ElmI с активностью 2000 е.а./мл.
Данная сайт-специфическая эндонуклеаза может быть использована в молекулярной биологии, эпигенетике и генной инженерии для сайт-специфического расщепления метилированной ДНК и для получения библиотек в совокупности с ферментами, дающими в результате гидролиза ДНК такие же «липкие» концы.
Источники информации
1. Чернухин В.А., Наякшина Т.Н., Болтенгаген А.А., Тарасова Г.В., Дедков B.C., Михненкова Н.А., Дегтярев С.Х. Штамм бактерий Planomicrobium koreense 78Л - продуцент сайт-специфической эндонуклеазы PkrI // Патент RU 2475534, C1, оп. - 2011.
2. Chambers S.P., Prior S.E., Barstow D.A., Minton N.P. The pMTLnic-cloning vectors. I. Improved pUC polylinker regions to facilitate the use of sonicated DNA for nucleotide sequencing // Gene - 1988. - V.68. - P. 139-149.
3. Roberts R.J., Vineze T., Posfai J., Macelis D. REBASE - restriction enzymes and methylases // Nucl. Acids Res. - 2003. - V.31. - P.418-420.
4. Sambrook J., Fritsch E.F. and Maniatis T. / Molecular Cloning: A laboratory manual. - 2-nd ed. - Cold Spring Harbor Laboratory Press. - Cold Spring Harbor, New York. - 1989.
5. Sanger F., Nicklen S., Coulson A. R. DNA sequencing with chain-terminating inhibitors. // Proc Natl Acad Sci USA.- 1977. - V. 74. - P.5463-5467.
6. Bickle, T.A., Pirrotta, V. and Imber, R. A simple, general procedure for purifying restriction endonucleases // Nucleic Acids Res. - 1977. - V.4 - P. 561-2572.
7. Чернухин B.A., Килева E.B., Томилова Ю.Э., Болтенгаген А.А., Дедков B.C., Михненкова Н.А., Гончар Д.А., Голикова Л.Н., Дегтярев С.Х. Новая метил-зависимая сайт-специфическая эндонуклеаза KroI узнает и расщепляет последовательность ДНК 5′-G^C(5mC)GGC-3′ // Вестник биотехнологии и физико-химической биологии имени Ю. А. Овчинникова. - 2011. - Т. 7. - №1. - С. 14-20.
8. Чернухин В.А., Беличенко О.А., Тарасова Г.В., Гончар Д.А., Акишев А.Г., Дедков B.C., Михненкова Н.А., Дегтярев С.Х. Штамм бактерии Arthrobacter oxydans - продуцент сайт-специфической эндонуклеазы AoxΙ // Патент RU 2399663, C1, оп. - 2009.
9. Чернухин В.А., Наякшина Т.Н., Абдурашитов М.А., Томилова Ю.Э., Мезенцева Н.В., Дедков B.C., Михненкова Н.А., Гончар Д.А., Дегтярев С.Х. Новая эндонуклеаза рестрикции GlaI узнает метилированную последовательность 5′-G(5mC)^GC-3′ // Биотехнология. - 2006. - №4. - С. 31-35.
10. Чернухин В.А., Гончар Д.А., Килева Е.В., Соколова В.А., Голикова Л.Н., Дедков B.C., Михценкова Н.А., Дегтярев С.Х. Новая метилзависимая сайт-специфическая ДНК-эндонуклеаза MteI расщепляет девятинуклеотидную последовательность 5′-G(5mC)G(5mC)NG(5mC)GC-3′/3′-CG(5mC)GN(5mC)G(5mC)G-5′ // Вестник биотехнологии и физико-химической биологии имени Ю.А. Овчинникова. - 2012.- Т.8. - №1. -С. 16-26.
11. Чернухин В.А., Чмуж Е.В., Томилова Ю.Э., Наякшина Т.Н., Гончар Д.А., Дедков B.C., Дегтярев С.Х. Новая сайт-специфическая эндонуклеаза GluI узнает метилированную последовательность ДНК 5′-G(5mC)^NG(5mC)-3′/3′-(5mC)GN^(5mC)G-5′. // Вестник биотехнологии и физико-химической биологии имени Ю.А. Овчинникова. - 2007. - Т3. - №2. - С. 13-17.
| название | год | авторы | номер документа |
|---|---|---|---|
| Штамм бактерий Micrococcus luteus 805 - продуцент сайт-специфической метилзависимой эндонуклеазы MluVI | 2016 |
|
RU2614262C1 |
| ШТАММ БАКТЕРИЙ Plantibacter flavus 3Kz - ПРОДУЦЕНТ МЕТИЛЗАВИСИМОЙ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ PfsI | 2015 |
|
RU2593723C1 |
| СПОСОБ САЙТ-СПЕЦИФИЧЕСКОГО ГИДРОЛИЗА ДНК | 2015 |
|
RU2597984C1 |
| ШТАММ БАКТЕРИИ Paracoccus carotinifaciens 3K - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ PcsI. | 2008 |
|
RU2377294C1 |
| ШТАММ БАКТЕРИЙ Microbacterium testaceum 17B - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ MteI | 2011 |
|
RU2475533C1 |
| СПОСОБ САЙТ-СПЕЦИФИЧЕСКОГО ГИДРОЛИЗА С5-МЕТИЛИРОВАННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ ДНК | 2015 |
|
RU2597985C1 |
| ШТАММ БАКТЕРИИ Arthrobacter oxydans - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ AoxI | 2009 |
|
RU2399663C1 |
| ШТАММ БАКТЕРИИ Kocuria rosea - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ KroI | 2009 |
|
RU2394099C1 |
| ШТАММ БАКТЕРИЙ Planomicrobium koreense 78K - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ PkrI | 2011 |
|
RU2475534C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ БАКТЕРИЙ ESCHERICHIA COLI N16 (PM.ALUBI) - ПРОДУЦЕНТ ДНК-МЕТИЛТРАНСФЕРАЗЫ M.ALUBI | 2015 |
|
RU2603086C1 |



Изобретение относится к биотехнологии и представляет собой рекомбинантный штамм бактерий Escherichia coli N42 (pElmI), являющийся продуцентом метилзависимой сайт-специфической ДНК-эндонуклеазы ElmI, расщепляющей обе цепи последовательности ДНК 5′-GC^NGC-3′/3′-CGN^CG-5′, в которой не менее трех цитозинов метилированы в положении С5 с образованием 5′-выступающих однонуклеотидных концов. Штамм получен путем трансформации клеток Escherichia coli ER2267 плазмидой pElmI. Данная плазмида сконструирована на основе вектора pMTL22 и содержит ген, кодирующий метилзависимую сайт-специфическую ДНК-эндонуклеазу ElmI из природного штамма Escherichia coli N17. Изобретение позволяет получить ДНК-эндонуклеазы ElmI заданной специфичности с более высоким выходом. 4 ил., 4 пр.
Рекомбинантный штамм бактерий Escherichia coli N42 (pElmI) - продуцент метилзависимой сайт-специфической эндонуклеазы ElmI, узнающей последовательность ДНК 5′-GC^ANGC-3′, в которой не менее трех цитозинов метилированы в положении С5, и расщепляющей обе ее цепи перед центральным нуклеотидом N с образованием 5′-выступающих однонуклеотидных концов, полученный путем трансформации штамма Escherichia coli ER2267 плазмидой pElmI, сконструированной на основе вектора pMTL22 и содержащей ген, имеющий нуклеотидную последовательность SEQ ID NO: 1, кодирующий метилзависимую сайт-специфическую эндонуклеазу ElmI.
| ШТАММ БАКТЕРИЙ Planomicrobium koreense 78K - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ PkrI | 2011 |
|
RU2475534C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ БАКТЕРИЙ Escherichia coli N41 (pBpuN4/MR)-ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ РЕСТРИКЦИИ BpuN4I | 2013 |
|
RU2529362C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pBinPLUS-ARS-EPSPS, ОБЕСПЕЧИВАЮЩАЯ ЭКСПРЕССИЮ ГЕНА АГРОБАКТЕРИАЛЬНОЙ 5-ЕНОЛПИРУВИЛ-ШИКИМАТ-3-ФОСФАТ-СИНТЕТАЗЫ В ТРАНСГЕННЫХ МИКРОВОДОРОСЛЯХ РОДА ХЛОРЕЛЛА | 2009 |
|
RU2407794C1 |

