Область техники, к которой относится изобретение
Изобретение относится к биотехнологии и может найти применение в области биомедицинских исследований и разработок для поиска новых терапевтических подходов к терапии нейродегенеративных заболеваний. Изобретение позволяет получить глиальные производные индуцированные плюрипотентные стволовые клетки с повышенной экспрессией фактора роста нервов (NGF).
Уровень техники
В последнее время стали появляться исследования, которые свидетельствуют о репаративных процессах в постнатальном периоде, которые также происходят в головном мозге, но с меньшей интенсивностью. Выяснилось, что в головном мозге существуют тканеспецифические нейральные стволовые клетки (НСК), которые сосредоточены в субвентрикулярной зоне боковых желудочков и зубчатой извилине гиппокампа [1,2]. Повреждение головного мозга оказывает стимулирующее воздействие на пролиферативную активность и миграцию НСК в поврежденную зону и последующей их дифференцировкой в нейрональные и глиальные клетки, заместительная способность которых составляет только 0,2% от числа погибших клеток [3,4]. Следующим механизмом репаративного процесса является реорганизация нейрональной сети за счет пластичности нейронов. Данный процесс обусловлен изменением количества и длины нейрональных отростков и установление новых синаптических связей [5]. Неотъемлемыми частью репаративных процессов является также восстановление ГЭБ и неоваскуляризация [6,7]. Данные факты о способности ЦНС к репарационным процессам позволяют сделать заключение, что регенерация является важным механизмом неврологического восстановления и является новой мишенью терапевтических воздействий. Поэтому поиск новых средств для стимуляции регенерации поврежденной нервной ткани является актуальной медицинской задачей.
К настоящему времени известно несколько аналогичных по своему назначению изобретений, представляющих собой нейропептидные лекарственные препараты, полученные из тканей головного мозга млекопитающих, которые представляют собой композицию биологически-активных полипептидов, стимулирующих регенерацию нервной ткани. Для получения активных фармкомпозиций используются ткани мозга сельскохозяйственных животных. Эти препараты отличаются по источнику сырья, срокам гестации при заборе, молекулярным массам входящих в него белково - полипептидных фракций, их качественному составу, по способу применения (парентерально, интратекально, интраназально, субконъюктивально), по биологической и фармакологической активности. К таким препаратам относят: церебролизин - средство для лечения нейродегенеративных заболеваний фирмы «Ebewe» (Австрия) - является продуктом обработки мозга интактных свиней; цереброкурин - лечебное средство при заболеваниях, сопровождающихся нарушением функций центральной нервной системы, фирмы ООО «НИР» (Украина) - является продуктом обработки мозга эмбрионов животных; кортексин - препарат полипептидной природы, получаемый путем экстракции из коры головного мозга крупного рогатого скота и свиней, фирмы ООО «Герофарм» (Россия), обладающий ноотропным, нейропротекторным, противосудорожным и антиоксидантным действием; церебролизат - препарат, повышающий устойчивость ткани головного мозга к интоксикации, гипоксии, гипогликемии, механической травме, фирмы «Микроген» НПО ФГУП (Иммунопрепарат), представляющий собой гидролизат коры головного мозга крупного рогатого скота. Однако ксеногенное происхождение препаратов ограничивает их применение из-за их высокой иммуногенности.
На сегодняшний день стволовые клетки стали ключевым элементом регенеративной медицины благодаря способности к дифференцировке в различные типы клеток. На их применении основана терапия многих нейродегенеративных заболеваний [8-10]. Недавно появилось предположение, что положительные эффекты от трансплантации стволовых клеток не могут быть ограничены лишь способностью дифференцироваться и замещать поврежденные и погибшие клетки реципиента [11-13]. В последние годы была продемонстрирована способность стволовых клеток секретировать биологически активные вещества, которые положительно влияют на течение многих неврологических заболеваний [13].
При регенеративных процессах нервной ткани важную роль играют нейротрофические факторы. Было показано, что наибольшим нейропротективным потенциалом обладают нейротрофины, такие как BDNF, CNTF, NGF и GDNF [14-17].
NGF является наиболее изученным нейротрофином, регулирующим выживание, развитие и трофику специфических популяций нейронов в периферической и центральной нервной системе (ЦНС) [18]. Во взрослом мозге самые высокие уровни NGF обнаруживаются в гиппокампе, коре и обонятельных областях, которые представляют собой мишени для холинергических нейронов базального переднего мозга [19, 20]. Первоначально NGF был описан как белок, действующий исключительно на симпатические периферические нейроны, и лишь намного позже было обнаружено, что он также активен на нейронах центральной нервной системы и на различные типы иммунных клеток [21]. В целом, присутствие NGF в тех лимбических областях ЦНС, которые участвуют в формировании настроения и познания, а также в нейроэндокринных реакций, а также в клетках иммунной и эндокринной системы, указывает на гораздо более широкую роль этого нейротрофина [22, 23]. NGF необходим для развития и поддержания сенсорных нейронов [24], а также со своими рецепторами (p75 и TrkA) играет решающую трофическую роль в холинергических нейронах переднего мозга, которые дегенерируют при старении мозга и нейродегенеративных расстройствах [25,26]. Во время развития и NGF, и BDNF регулируют естественную гибель клеток, синаптическую связь, направление волокон и морфологию дендритов [27]. Кроме того, они участвуют в пластичности мозга, участвуя в зависимых от активности функциях нейронов [28]. Известен способ, который является ближайшим аналогом настоящего изобретения - NGF-сверхэкспрессирующие мезенхимальные стволовые клетки (МСК) человека, которые секретируют NGF в концентрации 190±41.34 пг/мл [29]. Его основным недостатком является тип трансдуцированных клеток. Способность глиальных предшественников к секреции не только NGF, но и других нейротрофических факторов, характерных для ранних этапов формирования ЦНС, делают эту культуры более подходящим объектом для клеточной терапии нейродегенеративных заболеваний по сравнению с МСК. Главным отличием и преимуществом настоящего изобретения является более повышенная секреция NGF - 970±25,4 пг/мл, что в 6 раза выше существующего аналога.
На сегодняшний день не существует препаратов, которые в полной мере восстанавливали поврежденные участки головного мозга. Для решения данной проблемы предложено получение глиальных производных индуцированных плюрипотентных стволовых клеток человека, модифицированных путем введения гена NGF посредством лентивирусной трансдукции. Модифицированные клетки отличаются повышенной секрецией NGF по сравнению со стандартной культурой. При этом клеточная культура модифицированных глиальных предшественников демонстрирует экспрессию глиального маркера s100β и состоит из 97±6% s100β+ - клеток. Данные клетки усиливают выживание мозжечковых нейронов в модели глутаматной эксайтотоксичности в большей степени, чем глиальные предшественники без модификаций. Также данный тип клеток обладает более выраженным антиамнестическим действием за счет повышенной секреции нейротрофина NGF.
Раскрытие изобретения
Технической задачей изобретения является разработка способа получения клеточных культур человека с повышенной экспрессией нейротрофического фактора NGF, демонстрирующих высокую эффективность в моделях глутаматной эксайтотоксичности in vitro и амнезии, вызванной скополамином, in vivo.
Техническим результатом, на достижение которого направлено заявленное изобретение, является возможность получения глиальных производных индуцированные плюрипотентные стволовые клетки с выраженными нейропротективными свойствами за счет повышенной экспрессии NGF. На всех стадиях получения глиальных производных индуцированных плюрипотентных стволовых клеток человека, модифицированных путем введения гена NGF, используются только культуральные бессывороточные среды полностью охарактеризованного состава, не содержащие компонентов ксеногенного происхождения. Видовая специфичность используемых клеточных культур позволяет избежать иммунологических конфликтов, развитие которых возможно при введении пептидов ксеногенного происхождения. Кроме того, использование клеточных культур человека позволит получать не только NGF в повышенной концентрации, но и другие полноразмерные белковые ростовые и нейротрофические факторы. Эффективность действия такой композиции потенциально выше, чем у коротких пептидов, а иммуногенность отсутствует.
Для достижения указанного технического результата предложено изобретение, описываемое следующей формулой: из первичной культуры фибробластов получают индуцированные плюрипотентные стволовые клетки человека (ИПСК) с помощью генетических конструкций на основе вируса Сендай, культивируют ИПСК человека в количестве 40000 клеток/см2, проводят лентивирусную трансдукцию ИПСК геном NGF, затем осуществляют дифференцировку трансдуцированных ИПСК в глиальном направлении и культуру глиальных предшественников стандартизуют по количественному анализу s100β+ - клеток, которых должно быть не менее 97±6% в культуре. В итоге получают глиальные производные индуцированных плюрипотентных стволовых клеток с повышенной экспрессией NGF - 970±25,4 пг/мл.
Осуществление изобретения
Изобретение осуществляется следующим образом.
В процессе проведения исследований была установлена значимость следующих принципиальных моментов на различных этапах реализации изобретения:
1. Получение первичных культур фибробластов кожи человека
Для получения культуры фибробластов проводят биопсию небольшого участка кожи за ушной раковиной (0,5-1см2). Биоптат переносят в чашку Петри, промывают раствором Хэнкса с цефазолином (ПанЭко, РФ), затем добавляют раствор Трипсин/Версен 1:1 (ПанЭко, РФ) и инкубируют 1,5 часа при 37°C. Затем суспензию пипетируют и проводят центрифугирование при 1100 об/мин 10 минут. Супернатант удаляют, а осадок ресуспедируют в среде DMEM/F12 с 10% эмбриональной телячьей сыворотки, 2 mM L-глутамина, пенициллина-стрептомицина 100 мг/л (ПанЭко, РФ) и рассевают на чашки Петри диаметром 35 мм (Corning, США) в плотности 1500-2000 клеток/см2. Смена культуральной среды происходит раз в три дня. При достижении 70-80% конфлюентности клетки снимали раствором Трипсин/Версен 1:1 и рассеивали в плотности 1500-2000 клеток/см2.
2. Получение индуцированных плюрипотентных стволовых клеток человека (ИПСК) с помощью генетических конструкций на основе вируса Сендай
За день до репрограммирования фибробласты высевают в количестве 200 тысяч на лунку 6-луночного планшета. На следующий день добавляют векторы, несущие гены Oct4, Sox2, L-Myc, Klf-4 в количестве вирусных частиц на клетку (MOI) - 5, 5, 5 и 3 соответственно. Через сутки производят замену среды для фибробластов и далее среду заменяют ежедневно в течение 6 дней. На 7-ой день после репрограммирования клетки промывают дважды фосфатно-солевым буфером (ПанЭко, РФ), открепляют их реагентом TrypLE Select (Gibco, США) и центрифугируют при 1100 об/мин в течение 5 минут. Полученный супернатант отбирают, а клеточный осадок пересаживают в количестве 100 тысяч на чашку Петри диаметром 60 мм, предварительно обработав раствором витронектина в концентрации 10 мкг/мл (ThermoFisher Scientific, США). Через 24 часа после посева производят замену среды на Essential 8 Medium (Gibco, США) для культивирования плюрипотентных стволовых клеток человека. Смену среды осуществляют каждый день. Стандартизацию ИПСК проводят по экспрессии генов, специфичных для эмбриональных стволовых клеток OCT4, SOX2, NANOG, SSEA4, TRA-1-81, TRA-1-60 иммуноцихимическим методом и ПЦР с обратной транскрипцией.
3. Культивирование ИПСК человека
ИПСК культивируют в коммерческой среде Essential E8 Medium. В качестве подложки для культивирования используют рекомбинантный витронектин (Gibco, США) в концентрации 10 мкг/мл и инкубируют 1 час при комнатной температуре. Каждый клон культивируют до достижения 80% конфлюентности и затем рассевают на новые чашки в соотношении 1:3. Пассирование ИПСК осуществляют с помощью раствора Версена (ПанЭко, РФ).
4. Лентивирусная трансдукция ИПСК
В качестве матрицы для синтеза гена NGF используют коммерческие клоны от компании Евроген. Амплификацию фрагмента ДНК проводят методом полимеразной цепной реакции (ПЦР). Реакцию проводят следующим образом: первичная денатурация 95°C, 5 мин; денатурация 95° C, 20 сек; отжиг праймеров 60°C, 20 сек; элонгация 72°C, 20 секунд (25 циклов). Последовательность праймеров: for GATAGGATCCCATGCTGGACCCAAGCTCAG и rev ACTTGGGATCCAGGTTGAGGTAGGGAGGG. Далее ПЦР-продукты очищают с помощью комерческого набора для элюции ДНК из геля Wizard SV Gel and PCR Clean-Up System (Promega, Cat. № A9282) согласно протоколу фирмы-производителя. После ДНК фрагменты лигируют в p-GEM-T вектор (Promega, Cat. № A3600). Для этого берут буфер для ДНК лигазы T4, 50 нг вектора pGEM-T, 3 ед. Т4 ДНК-лигазы (SibEnzyme, Cat. № E319) и инкубируют в течение часа при комнатной температуре. Затем проводят трансформацию клеток E.coli (штаммTop10F). После их трансформации выделяют p-GEM-T векторы, содержащие фрагменты ДНК, кодирующие ген NGF человека. Полученные фрагменты секвенируют для проверки. Полученный p-GEM-T вектор со вставленным геном NGF подвергают рестрикции для выделения гена NGF.
Полученные фрагменты гена NGF подвергают кинированию с помощью фермента Т4-полинуклеотид киназы (T4 Polynucleotide Kinase, SibEnzyme, Cat. № E311). Кинированные фрагменты ДНК, содержащий ген NGF, имеющие «липкие» концы BamHI, лигируют в дефосфорилированный вектор pCMV, также рестрицированный по сайтам BamHI. Далее проводят наращивание вектора pCMV в компетентных клетках E. coli штамма Top10F. После чего клетки осаждают центрифугированием при 13000 оборотов/минуту в течение 30 секунд. Супернатант сливают, а к осажденным клеткам добавляют 200 μл лизирующего буфера (1% SDS, 0.1 H NaOH). Далее смесь центрифугируют в течение 15 минут при 13000 оборотов/минуту. К супернатанту добавляют изопропиловой спирт, после чего плазмидную ДНК высаживают центрифугированием в течение 15 минут на 12000 g при +4°С. Осадок промывают в 75% этаноле, высушивают на при комнатной температуре и растворяют в деионизованной воде (mQ). Сборку лентивирусных частиц осуществляют в специализированных клетках линии HEK293T с помощью трансфекции трех базовых плазмид, кодирующих вирусные белки, и полученной ранее плазмиды на основе вектора pCMV, содержащий ген NGF. После сборки лентивирусных частиц, их суспензию концентрируют на ультрацентрифуге для достижения рабочей концентрации.
Рабочую концентрацию лентивирусных частиц добавляют ИПСК и инкубируют в течение 24 часов. После проводят смену среды. Из полученной гетерогенной культуры получают клональные клеточные линии с оптимальным уровнем экспрессии NGF.
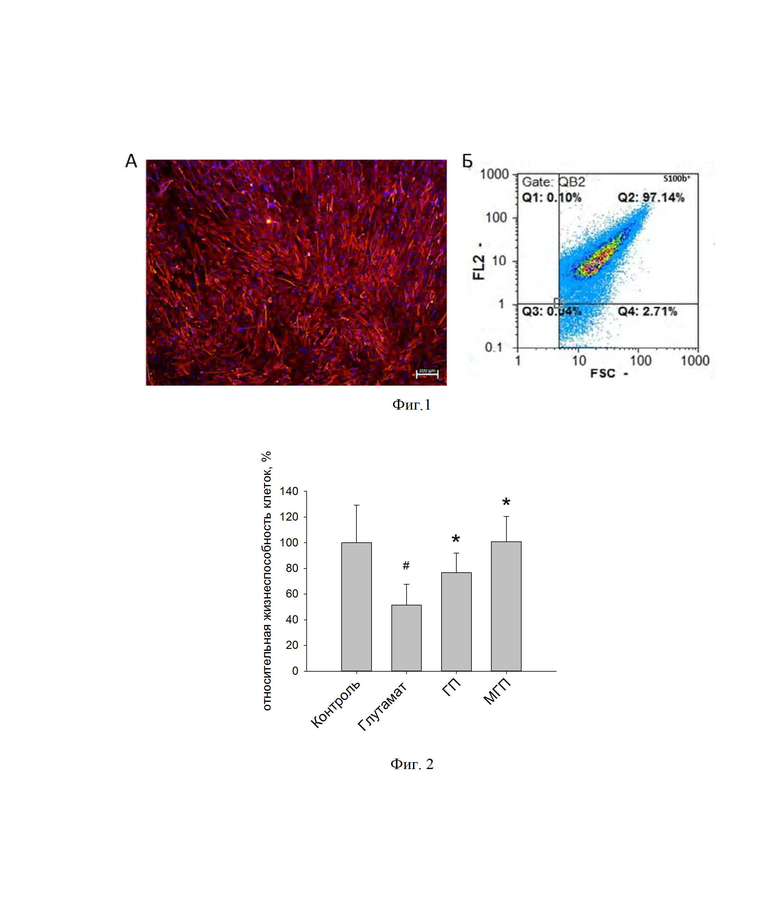
Далее ИПСК со сверэкспрессией NGF дифференцируют в нейральном направлении. Для этого клетки культивируют в среде DMEM/F12 (Gibco, США) с 1% N2 (Gibco, США), 2мМ глутамина (Панэко, РФ), 100 мг/л пенициллин-стрептомицина (Панэко, РФ), 10 мкМ SB431542 (Stemcell Technologies, США), 2 мкМ дорсоморфина (Stemcell Technologies, США), 200 нМ LDN193189 (Sigma-Aldrich, США). Образовавшиеся нейральные стволовые клетки дифференцируют в глиальные предшественники путем добавления в среду DMEM/F12 следующих компонентов: 1% заменителя сыворотки, 1% N2, 2мМ глутамина, 100 мг/л пенициллин-стрептомицина (Панэко, РФ), 10 нг/мл FGF-2, 20 нг/мл EGF (ProSpec, Великобритания), 20нг/мл CNTF (PeproTech, США). Следующие три дня культивировали клетки в среде DMEM/F12, 1% заменителя сыворотки, 1% N2 добавка, 2мМ глутамин, 100 мг/л пенициллин-стрептомицина (Панэко, РФ), 20 нг/мл EGF (ProSpec, Великобритания), 20 нг/мл CNTF (PeproTech, США). Стандартизацию глиальных предшественников проводят по количественному анализу s100β+ - клеток, которых должно быть не менее 97±6% в культуре (фиг. 1 А, Б).
Пример 1. Эффективность лентивирусной трансдукции
Эффективность лентивирусной продукции оценивается опосредованно содержанием фактора NGF в кондиционированных средах, полученных при культивировании глиальных предшественников. Для оценки секреции проводят иммуноферментный анализ согласно инструкции производителя, концентрация NGF выражается в пг/мл. Было исследовано 10 образцов кондиционированных сред для каждой культуры глиальных предшественников. Средняя концентрация NGF после лентивирусной трансдукции составляет 970±25,4 пг/мл, что в 16 раз больше чем в культуре глиальных предшественников без модификаций (табл. 1).
Пример 2.Оценка эффективности кондиционированных сред на модели глутаматной эксайтотоксичности in vitro
Оценку нейропротективного действия кондиционированных сред, полученных при культивировании глиальных предшественников со сверхэкспрессией NGF и без модификаций, проводят на модели глутаматной эксайтотоксичности, выполненной на мозжечковых нейронах. Для этого у крысят линии Wistar (Ро) производят декапитацию в соответствии с этическими требованиями по работе с животными. Суспензию мозжечковых нейронов получают путем обработки ткани мозга трипсином (0,05%, Gibco, США), и дальнейшим осаждением в центрифуге (3000 rpm). Клеточный осадок ресуспендируют в соответсвующем объеме культуральной среды Neurobasal Medium (Gibco, США), содержащей 2% добавки B27 (Gibco, США), и переносят в лунки 48-луночных планшетов (Corning, США), предварительно покрытые поли-DL-лизином (ПанЭко, РФ) в концентрации 0,1 мг/мл. В каждую лунку вносят по 250 тысяч клеток в 500 мкл среды. На второй-третий день добавляют 10 мкМ арабинозинмоноцитозида (Sigma-Aldrich, США), который удаляют спустя сутки. В течение культивирования каждые три дня производят смену среды. Культуру мозжечковых нейронов инкубируют в течение 7 сут. (37°C, 5% СО2, относительная влажность 98%) для достижения необходимой экспрессии глутаматных рецепторов. Добавление кондиционированной среды (20 мкл), предварительно сконцентрированной в 25 раз на фильтрах 3 кДа (Amicon Ultra), проводят за 16-18 часов до моделирования глутаматной эксайтотоксичности. Для моделирования глутаматной эксайтотоксичности культуральную среду собирают в отдельные пробирки, а культуру мозжечковых нейронов дважды промывают раствором DPBS без Ca2+ и Mg2+ (ПанЭко, РФ), затем клетки инкубируют 1 ч. в присутствии глутамата (100 мкМ) в буферном растворе следующего состава (мМ): NaCl - 140, KCl - 5, CaCl2 - 2, глицин - 10, HEPES - 20, глюкоза - 5, (рН 7,4, доводили 1М NaOH) (Sigma-Aldrich, США). После добавления глутамата клетки на 1 час оставляют при комнатной температуре. Затем дважды промывают буфером, содержащим (мМ): NaCl - 140, KCl - 5, MgCl2 - 2, HEPES - 20, глюкоза - 5, (рН 7,4) (Sigma-Aldrich, США) и заливают исходной культуральной средой. МТТ анализ проводят на следующий день после моделирования. МТТ вносят в культуральную среду до конечной концентрации 0,5 мг/мл и инкубируют при 37°С в течение 1,5 ч с последующим растворением кристаллов формазана в ДМСО (ПанЭко, РФ). Оптическую плотность измеряют при длине волны 570/620 нм на планшетном ридере (PerkinElmer, США). Результаты оценивают по отношению к контролю, который принимают за 100%.
В результате сокультивирования модифицированных глиальных предшественников (МГП) с мозжечковыми нейронами увеличивает выживаемость последних на 50,4±3,2%. При этом инкубация глиальных предшественников без модификации (ГП) приводит к увеличению количества жизнеспособных клеток на 19,4±5,2% (фиг.2).
Пример 3. Изучение антиамнестических свойств веществ на модели амнезии УРПИ, вызванной скополамином
Выработку условного рефлекса пассивного избегания (УРПИ) осуществляют согласно методическим указаниям [30]. Исследования проводят на животных беспородных крысах-самцах в возрасте 2-3 месяца, весом 220-240 г. Выработка УРПИ у крыс проводят в установке Passive avoidance фирмы "Lafayette Instrument Co" (США) по Адеру [31]. Конструктивно установка состоит из маленькой платформы, расположенной на расстоянии 1м от пола, освещенной специальной лампой и темной камеры с электродным полом. Крыс помещают на освещенную платформу перед входом в темную камеру установки хвостом к входному отверстию и регистрируют латентное время рефлекса. В эксперименте не используют животных, которые при обучении показывают латентное время рефлекса более минуты. Когда крыса оказывается в темном отсеке, отверстие закрывают и наносят животному неизбегаемое электроболевое раздражение через электродный пол (сила обучающего тока 0,35 мА, длительность каждого импульса составляет 1 с, интервал между последовательными импульсами - 2 с, 8 ударов). Тест на воспроизведение УРПИ осуществляют через 24 часа после обучения и проводят при повторном помещении животных в установку с регистрацией в течение 180 с. латентного периода первого захода крысы в темную экспериментальную камеру, и количества животных, не зашедших в темный отсек камеры в %. Кроме того, с целью анализа влияния веществ на угашение рефлекса процедуру воспроизведения навыка проводят через 7 и 14 суток после обучения.
Для получения экспериментальной амнезии крысам за 15 минут до обучения УРПИ вводят внутрибрюшинно блокатор М-холинергических рецепторов - скополамин в дозе 1,5 мг/кг, затем проводят процедуру обучения УРПИ и через 24 часа, 7 и 14 суток воспроизводят рефлекс. Последнее (5-е) введение исследуемых веществ осуществляют за 60 минут до обучения.
В экспериментах in vivo в контрольной группе крыс, получавших физраствор в течение 5 дней в объеме 1 мл на крысу, при однократном внутрибрюшинном (в/б) введении скополамина в дозе 1,5 мг/кг показано, что через 24 часа после обучения 14% (серия 1 и 2) животных помнили об ударе током и не заходили в темный отсек камеры (табл. 2 и 3). При воспроизведении УРПИ через 7 и 14 дней после обучения наблюдался низкий уровень реконсолидации памятного следа, что выражалось в коротком латентном периоде захода в темный отсек камеры и небольшом количестве животных 14%, помнящих о токе.
Изучение антиамнестической активности клеточной терапии на модели УРПИ, вызванной скополамином показало, что МГП при внутриартериальном (в/а) введении в течение 5 дней в объеме 500 тыс. кл./мл на крысу обладают выраженным антиамнестическим эффектом. При этом воспроизведение УРПИ через 24 часа после обучения выявляется до 60% животных, помнящих о токе. Через 7 и 14 суток наблюдается 30% и 60%, соответственно, животных, воспроизводивших рефлекс после скополаминовой амнезии (табл. 2).
ГП при аналогичном режиме введения обладают антиамнестическим эффектом при сравнении с группой животных контроля. При этом воспроизведение УРПИ через 24 часа после обучения выявляет до 50% животных, помнящих о токе. Через 7 и 14 суток наблюдается до 30% животных, воспроизводивших рефлекс после скополаминовой амнезии (табл. 3).
Таким образом, модификация глиальных предшественников приводит к более выраженному нейропротективному и антиамнестическому эффекту за счет повышенной секреции нейротрофина NGF клетками.
Краткое описание чертежей
Изобретение поясняется таблицами, где
В табл. 1 представлены данные о эффективности лентивирусной трансдукции, оцениваемой опосредованно содержанием фактора NGF в кондиционированных средах, полученных при культивировании глиальных предшественников,
Табл. 2 - результаты интиамнестического эффекта кондиционированных сред: влияние МГП на латентное время рефлекса и количество животных, не зашедших в темный отсек камеры (%) после амнезии УРПИ, вызванной скополамином.
Влияние ГП на латентное время рефлекса и количество животных, не зашедших в темный отсек камеры (%) после амнезии УРПИ, вызванной скополамином (серия 2) и фигурами, где
Фиг. 1. Характеристика модифицированных глиальных предшественников, где
А - морфология и иммуноцитохимический анализ на маркер глиальных предшественников S100b (красный). Ядра окрашены DAPI (синий).
Б - данные проточной цитометрии, отражающие количество S100b+ -клеток в культуре модифицированных глиальных предшественников.
Фиг. 2. Относительная жизнеспособность клеток культуры мозжечковых нейронов. МТТ-тест. *p≤0,05 достоверность отличия от глутамата; #p≤0,05 от контроля. Данные проанализированы с помощью однофакторного дисперсионного анализа с поправкой теста Холма-Сидака и представлены в виде средних значений и стандартных отклонений.
Список литературы
[1] Dorsky R.I., Moon R.T., Raible D.W. Control of neural crest cell fate by the Wnt signalling pathway // Nature. - 1998. - Т. 396. - №. 6709. - С. 370.
[2] Eminli S. et al. Differentiation stage determines potential of hematopoietic cells for reprogramming into induced pluripotent stem cells // Nature genetics. - 2009. - Т. 41. - №. 9. - С. 968.
[3] Fusaki N. et al. Efficient induction of transgene-free human pluripotent stem cells using a vector based on Sendai virus, an RNA virus that does not integrate into the host genome // Proceedings of the Japan Academy, Series B. - 2009. - Т. 85. - №. 8. - С. 348-362.
[4] Garcia J.H. Experimental ischemic stroke: a review // Stroke. - 1984. - Т. 15. - №. 1. - С. 5-14.
[5] Gerrard L., Rodgers L., Cui W. Differentiation of human embryonic stem cells to neural lineages in adherent culture by blocking bone morphogenetic protein signaling // Stem cells. - 2005. - Т. 23. - №. 9. - С. 1234-1241.
[6] Ginsberg M.D., Busto R. Combating hyperthermia in acute stroke: a significant clinical concern // Stroke. - 1998. - Т. 29. - №. 2. - С. 529-534.
[7] Feuerstein G.Z., Rozanski D.G. proteins and heart failure: Is Gαq a novel target for heart failure. - 2000.
[8] Kim S.U., De Vellis J. Stem cell-based cell therapy in neurological diseases: a review // Journal of neuroscience research. - 2009. - Т. 87. - №. 10. - С. 2183-2200.
[9] Krencik R., Zhang S. C. Directed differentiation of functional astroglial subtypes from human pluripotent stem cells //Nature protocols. - 2011. - Т. 6. - №. 11. - С. 1710.
[10] Lindvall O., Kokaia Z. Martinez Serrano A //Stem cell therapy for. - 2004.
[11] Baraniak P.R., McDevitt T. C. Stem cell paracrine actions and tissue regeneration // Regenerative medicine. - 2010. - Т. 5. - №. 1. - С. 121-143.
[12] Gnecchi M. et al. Paracrine mechanisms in adult stem cell signaling and therapy // Circulation research. - 2008. - Т. 103. - №. 11. - С. 1204-1219.
[13] Shimada I.S., Spees J.L. Stem and progenitor cells for neurological repair: Minor issues, major hurdles, and exciting opportunities for paracrine-based therapeutics // Journal of cellular biochemistry. - 2011. - Т. 112. - №. 2. - С. 374-380.
[14] Almeida R.D. et al. Neuroprotection by BDNF against glutamate-induced apoptotic cell death is mediated by ERK and PI3-kinase pathways // Cell Death & Differentiation. - 2005. - Т. 12. - №. 10. - С. 1329-1343.
[15] Sofroniew M.V., Howe C.L., Mobley W.C. Nerve growth factor signaling, neuroprotection, and neural repair // Annual review of neuroscience. - 2001. - Т. 24. - №. 1. - С. 1217-1281.
[16] Sauvage M.A.C. et al. The neuroprotective agent CNTF decreases neuronal metabolites in the rat striatum: an in vivo multimodal magnetic resonance imaging study // Journal of Cerebral Blood Flow & Metabolism. - 2015. - Т. 35. - №. 6. - С. 917-921.
[17] Duarte E.P. et al. Neuroprotection by GDNF in the ischemic brain // Growth factors. - 2012. - Т. 30. - №. 4. - С. 242-257.
[18] Levi-Montalcini R., Angeletti P.U. Nerve growth factor // Physiological Reviews. - 1968. - Т. 48. - №. 3. - С. 534-569.
[19] Korsching S. et al. Levels of nerve growth factor and its mRNA in the central nervous system of the rat correlate with cholinergic innervation // The EMBO journal. - 1985. - Т. 4. - №. 6. - С. 1389-1393.
[20] Large T.H. et al. Nerve growth factor gene expression in the developing rat brain //Science. - 1986. - Т. 234. - №. 4774. - С. 352-355.
[21] Black I.B. Trophic molecules and evolution of the nervous system //Proceedings of the National Academy of Sciences. - 1986. - Т. 83. - №. 21. - С. 8249-8252.
[22] Aloe L. et al. The expanding role of nerve growth factor: from neurotrophic activity to immunologic diseases //Allergy. - 1997. - Т. 52. - №. 9. - С. 883-994.
[23] Levi-Montalcini R. A role for nerve growth factor in nervous, endocrine and immune systems //Prog Neuroendocrinimmunol. - 1990. - Т. 3. - С. 1-10.
[24] Taiwo Y. O. et al. Hyperalgesia induced in the rat by the amino-terminal octapeptide of nerve growth factor // Proceedings of the National Academy of Sciences. - 1991. - Т. 88. - №. 12. - С. 5144-5148.
[25] Fischer W. et al. Amelioration of cholinergic neuron atrophy and spatial memory impairment in aged rats by nerve growth factor //Nature. - 1987. - Т. 329. - №. 6134. - С. 65-68.
[26] Sofroniew M.V., Howe C.L., Mobley W.C. Nerve growth factor signaling, neuroprotection, and neural repair //Annual review of neuroscience. - 2001. - Т. 24. - №. 1. - С. 1217-1281.
[27] Thoenen H. Neurotrophins and neuronal plasticity //Science. - 1995. - Т. 270. - №. 5236. - С. 593-598.
[28] Castrén E. Neurotrophins as mediators of drug effects on mood, addiction, and neuroprotection //Molecular neurobiology. - 2004. - Т. 29. - С. 289-301.
[29] Zhu S.X. et al. Growth and expression of rat bone marrow mesenchymal stem cells modified by nerve growth factor in diabetic rat bladders //Molecular medicine reports. - 2013. - Т. 7. - №. 6. - С. 1791-1799.
[30] Воронина Т.А. и соавт., «Руководство по проведению доклинических исследований лекарственных средств», 2012 г.
[31] Ader R., Weijnen J., Moleman P. Retention of a passive avoidance response as a function of the intensity and duration of electric shock // Psychonomic Science. - 1972. - Т. 26. - №. 3. - С. 125-128.
Способ получения глиальных производных индуцированных плюрипотентных стволовых клеток с повышенной экспрессией NGF для терапии нейродегенеративных заболеваний
* - от группы контроля, при Р≤0,05 (критерий Манна-Уитни);
Способ получения глиальных производных индуцированных плюрипотентных стволовых клеток с повышенной экспрессией NGF для терапии нейродегенеративных заболеваний
* - от группы контроля, при Р≤0,05 (критерий Манна-Уитни)
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3" fileName="GPCs-NGF.xml"
softwareName="WIPO Sequence" softwareVersion="2.3.0"
productionDate="2024-06-07">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>2023129052/10(064568)</ApplicationNumberText>
<FilingDate>2023-11-09</FilingDate>
</ApplicationIdentification>
<ApplicantFileReference>100407</ApplicantFileReference>
<EarliestPriorityApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>2023129052/10(064568)</ApplicationNumberText>
<FilingDate>2023-11-09</FilingDate>
</EarliestPriorityApplicationIdentification>
<ApplicantName languageCode="ru">Dmitry Goldshtein</ApplicantName>
<InventionTitle languageCode="ru">Способ получения глиальных
производных индуцированных плюрипотентных стволовых клеток с
повышенной экспрессией NGF для терапии нейродегенеративных
заболеваний</InventionTitle>
<SequenceTotalQuantity>2</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>30</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..30</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>gataggatcccatgctggacccaagctcag</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>29</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..29</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q4">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>acttgggatccaggttgaggtagggaggg</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ получения глиальных производных индуцированных плюрипотентных стволовых клеток с повышенной экспрессией BDNF для терапии ишемического инсульта | 2023 |
|
RU2829361C1 |
| Ноотропная композиция на основе полипептидных комплексов, выделенных из глиальных прогениторных клеток в условиях теплового шока, и способ её получения | 2019 |
|
RU2732599C1 |
| УЛУЧШЕННЫЙ СПОСОБ ДИФФЕРЕНЦИРОВКИ ИНДУЦИРОВАННЫХ ПЛЮРИПОТЕНТНЫХ СТВОЛОВЫХ КЛЕТОК ЧЕЛОВЕКА В ДОФАМИНЭРГИЧЕСКИЕ НЕЙРОНЫ | 2023 |
|
RU2823290C1 |
| Ноотропная композиция на основе полипептидных комплексов, выделенных из нейрональных прогениторных клеток в условиях теплового шока, и способ ее получения | 2019 |
|
RU2752906C2 |
| Ноотропная композиция на основе полипептидных комплексов, выделенных из нейрональных прогениторных клеток в условиях гипоксии, и способ ее получения | 2019 |
|
RU2732600C1 |
| СПОСОБ ПОЛУЧЕНИЯ ЭМБРИОНАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК МЛЕКОПИТАЮЩИХ, МОДИФИЦИРОВАННЫХ ГЕНОМ ФАКТОРА РОСТА НЕРВОВ ЧЕЛОВЕКА | 2011 |
|
RU2458984C1 |
| Средство для сдерживания гибели нейронов при ишемическом инсульте головного мозга и способ клеточно-опосредованной генной терапии ишемического инсульта головного мозга средством сдерживания гибели нейронов при ишемическом инсульте головного мозга | 2017 |
|
RU2676701C1 |
| Ноотропная композиция на основе полипептидных комплексов, выделенных из нейронов и глиальных клеток, полученных методом направленной дифференцировки индуцированных плюрипотентных стволовых клеток человека | 2018 |
|
RU2690846C1 |
| СПОСОБ ОЦЕНКИ МОРФОФУНКЦИОНАЛЬНОГО СОСТОЯНИЯ ДИФЕРЕНЦИРОВАННЫХ В ДОФАМИНЕРГИЧЕСКИЕ НЕЙРОНЫ ИНДУЦИРОВАННЫХ ПЛЮРИПОТЕНТНЫХ СТВОЛОВЫХ КЛЕТОК БОЛЬНЫХ ПАРКИНСОНИЗМОМ | 2012 |
|
RU2501853C1 |
| Способ получения ноотропной композиции на основе полипептидных комплексов, выделенных из нейронов и глиальных клеток, полученных методом направленной дифференцировки индуцированных плюрипотентных стволовых клеток человека | 2018 |
|
RU2690498C1 |
Изобретение относится к области биотехнологии, в частности к cпособу получения глиальных предшественников, полученных из индуцированных плюрипотентных стволовых клеток, с повышенной экспрессией NGF для терапии нейродегенеративных заболеваний. Изобретение обеспечивает получение клеточной культуры модифицированных глиальных предшественников, которая демонстрирует экспрессию глиального маркера s100β, стимулирует повышенную выживаемость мозжечковых нейронов и обладает выраженным антиамнестическим действием за счет повышенной секреции нейротрофина NGF. 2 ил., 3 табл., 3 пр.
Способ получения глиальных предшественников, полученных из индуцированных плюрипотентных стволовых клеток, с повышенной экспрессией NGF для терапии нейродегенеративных заболеваний, характеризующийся тем, что для получения культуры фибробластов проводят биопсию небольшого участка кожи за ушной раковиной 0,5-1 см2, при достижении 70-80% конфлюентности фибробласты снимают раствором Трипсин/Версен 1:1 и рассеивают в плотности 1500-2000 клеток/см2, затем получают индуцированные плюрипотентные стволовые клетки человека с помощью генетических конструкций на основе вируса Сендай, при этом за день до репрограммирования фибробласты высевают в количестве 200 тысяч на лунку 6-луночного планшета, на следующий день добавляют векторы, несущие гены Oct4, Sox2, L-Myc, Klf-4 в количестве вирусных частиц на клетку - 5, 5, 5 и 3 соответственно, проводят культивирование ИПСК человека, при этом каждый клон культивируют до достижения 80% конфлюентности и затем рассевают на новые чашки в соотношении 1:3, осуществляют лентивирусную трансдукцию ИПСК, амплификацию фрагмента ДНК проводят методом полимеразной цепной реакции, используют последовательность праймеров: for GATAGGATCCCATGCTGGACCCAAGCTCAG и rev ACTTGGGATCCAGGTTGAGGTAGGGAGGG, полученный p-GEM-T вектор со вставленным геном NGF подвергают рестрикции для выделения гена NGF, который затем кинируют с помощью фермента Т4-полинуклеотид киназы и лигируют в дефосфорилированный вектор pCMV, сборку лентивирусных частиц осуществляют в специализированных клетках линии HEK293T с помощью трансфекции трех плазмид, кодирующих вирусные белки, и плазмиды на основе вектора pCMV, содержащего ген NGF, далее ИПСК со сверэкспрессией NGF дифференцируют в нейральном направлении, образовавшиеся нейральные стволовые клетки дифференцируют в глиальные предшественники путем добавления в среду DMEM/F12 следующих компонентов: 1% заменителя сыворотки, 1% N2, 2 мМ глутамина, 100 мг/л пенициллин-стрептомицина, 10 нг/мл FGF-2, 20 нг/мл EGF, 20 нг/мл CNTF, стандартизацию глиальных предшественников проводят по количественному анализу s100β+-клеток, которых должно быть не менее 97±6% в культуре, при этом клеточная культура модифицированных глиальных предшественников демонстрирует экспрессию глиального маркера s100β, стимулирует повышенную выживаемость мозжечковых нейронов и обладает выраженным антиамнестическим действием за счет повышенной секреции нейротрофина NGF.
| НОВОСАДОВА Е.В | |||
| и др | |||
| Получение и характеристика глиальных клеток из индуцированных плюрипотентных стволовых клеток человека, Нейрохимия, 2020, т | |||
| Пишущая машина | 1922 |
|
SU37A1 |
| САЛИХОВА Д.И | |||
| и др | |||
| Ключевые этапы дифференцировки ИПСК в нейрональные и глиальные клетки, Гены и клетки, vol | |||
| Насос | 1917 |
|
SU13A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Устройство для устранения мешающего действия зажигательной электрической системы двигателей внутреннего сгорания на радиоприем | 1922 |
|
SU52A1 |
| УСТРОЙСТВО ДЛЯ ИЗМЕРЕНИЯ МГНОВЕННЫХ ЗНАЧЕНИЙ ПЕРИОДИЧЕСКИ ПОВТОРЯЮЩИХСЯ ЭЛЕКТРИЧЕСКИХ ПРОЦЕССОВ | 1934 |
|
SU41083A1 |
| TELIAS M | |||
| Neural | |||

