Изобретение относится к медицине, в частности к онкологии, и может быть использовано для лечения асцитных форм злокачественных новообразований.
Онкологические заболевания занимают второе место по общей смертности после сердечно-сосудистых заболеваний, а страх перед возможностью возникновения злокачественного новообразования имеет глубокий социальный резонанс. Статистические данные свидетельствуют о том, что заболеваемость онкологическими заболеваниями ежегодно растет, и при этом прослеживается явная тенденция к «омоложению рака».
Известны способы лечения онкологических заболеваний, основанные на применении химиотерапевтических препаратов, применяемых для лечения злокачественных заболеваний различных групп: алкилирующие препараты, антиметаболиты, противоопухолевые антибиотики, алкалоиды и другие биологические активные вещества растительного происхождения, а также ферментные и гормональные препараты [1].
В мировой практике разработаны и переведены в стандарты основные способы лечения больных со злокачественными новообразованиями.
Общепринятой терапией, вне зависимости от происхождения рака и его формы, является использование различных схем циторедуцирующего воздействия. Циторедуцирующая терапия с применением цитостатиков, являющаяся основой лечения раков, направлена на удаление видимой массы опухоли. Как следует из огромной клинической практики, угроза рецидивов в ближайшие 5 лет при такой терапии достигает 30-50%. Этот факт связан с присутствием в неотрансформированной ткани популяции клеток, обладающих потенциалом стволовых клеток [2, 3].
К одной из наиболее труднолечимых форм рака, связанной с терминальной стадией развития заболевания, относится асцитная форма. Злокачественный (малигнизированный) асцит - это патологическое накопление жидкости в брюшной или плевральной полости, развивающееся вследствие опухолевого поражения брюшины или легких. Асцит вызывает значительный дискомфорт и приводит к различным функциональным нарушениям у пациентов в запущенных стадиях онкологического заболевания и является сложной клинической проблемой. Малигнизированный асцит составляет примерно 10% среди всех случаев асцита и наиболее часто развивается при раке молочной железы, яичников, желудка, поджелудочной железы и толстой кишки [4].
В онкологической клинике асцитные формы рака являются грозным осложнением и, в подавляющем большинстве случаев, приводят к гибели пациентов. Смерть таких больных наступает даже при отсутствии отдаленных метастазов и прогрессии опухоли.
Для лечения асцитных форм рака используется несколько общепринятых способов лечения, позволяющих продлить и улучшить "качество" жизни пациентов [5].
Один из известных способов [6-8] основан на том, что производят пункцию с последующим удалением экссудата и вводят в полость противоопухолевые препараты и стероидные гормоны, наиболее эффективными из которых признаны цисплатин, адриамицин, тиоТЭБ и циклофосфан (ЦФ) [6-8].
Недостатком такого способа является относительно низкая эффективность, в результате чего для большинства больных он дает лишь кратковременный паллиативный эффект [9], что определяет ограниченность их применения.
Известен также способ лечения асцита, основанный на комбинированном применении двух препаратов, диоксадэта и цисплатина [10].
Сравнительное изучение противоопухолевой активности цитостатиков диоксадэта, цисплатина и их комбинации проводилось на модели асцитной опухоли яичника у 38 крыс-самок Вистар. Асцитная опухоль яичника перевивалась внутрибрюшинно в количестве 5×106 клеток на крысу. Диоксадэт и цисплатин вводили внутрибрюшинно однократно через 48 ч после перевивки опухоли яичника в максимально переносимых дозах: 1,5 и 4,0 мг/кг массы тела соответственно. В комбинации диоксадэт и цисплатин вводили по той же схеме, но в дозах, составляющих половину от максимально переносимой дозы. Противоопухолевые эффекты препаратов оценивали по увеличению продолжительности жизни животных. В контрольной группе крыс, не получавших лечения, средняя продолжительность жизни составила 16±6,3 дня. Диоксадэт и цисплатин проявили сравнимую противоопухолевую активность, увеличив среднюю продолжительность жизни на 63,1% и 48,1% соответственно. Средняя продолжительность жизни при введении комбинации препаратов значимо увеличилась на 107,5%, что равно сумме эффектов каждого препарата в отдельности.
Недостатком этого способа лечения также является относительно низкая его эффективность, поскольку, как подтвердили результаты экспериментальных исследований, при его применении не удается полностью остановить развитие асцита, а лишь тормозится его рост, увеличивая при этом продолжительность жизни животных примерно в два раза [10].
Кроме того, известен способ внутрибрюшинной химиотерапии больных асцитным раком яичников III-IV стадий [11, 12], согласно которому в предоперационном периоде амбулаторно проводят иммунокоррекцию путем внутрибрюшинного введения Т-активина по 1 ампуле в день - 100 мкг, в течение 10 дней с одномоментным внутрибрюшинным введением химиопрепаратов: в 1-й день 1-2 г ЦФ, во 2-й день 150-200 мкг препарата, производного платины, в зависимости от веса больной.
Этот способ характеризуется меньшей токсичностью и большей эффективностью, что позволяет значительно увеличить процент регрессии, операбельности и резектабельности опухоли, а также продолжительность жизни больных асцитной формой рака яичников III-IV стадии заболевания (на 16-26%) по сравнению с другими методами химиотерапии.
Однако способ направлен только на предоперационную подготовку больных, позволяющий создать условия большей операбельности и резектабельности опухоли, а также увеличить продолжительность жизни пациентов на 16-20%, и не обеспечивает надежно полную эрадикацию первичного асцита и не предотвращает его повторное появление.
Поэтому описанный выше способ лечения также недостаточно эффективен.
Помимо указанных выше, известен способ лечения распространенного рака яичника [13], сущность которого заключается в том, что в послеоперационном периоде инкубируют асцитическую жидкость больного с различными сочетаниями цитостатиков в присутствии рихлокаина в концентрации 0,005 мг на 1 мл инкубационной смеси и при обнаружении резкого увеличения числа мертвых клеток в асцитической жидкости выбирают эту комбинацию цитостатиков для дальнейшей внутрибрюшинной химиотерапии с предварительным внутрибрюшинным введением рихлокаина в суммарной дозе 10-30 мл 0,5% раствора.
Способ позволяет повысить противоопухолевое действие внутрибрюшинно введенных цитостатиков в сочетании с рихлокаином, осуществляется индивидуальный подбор схемы химиотерапии, способствующий улучшению состояния больных, снижению токсичности, удлинению срока стойкой ремиссии, нормализации ряда структурно-функциональных показателей крови, а также исключается использование неэффективных лекарственных средств.
Однако недостаточная эффективность и ограниченность области применения этого способа связана с тем, что оценка и выбор эффективности цитостатического лекарственного средства определяется при ex vivo обработке асцита прямым добавлением цитостатика, значительно сужая тем самым спектр лекарственных препаратов цитостатического ряда, часть которых проявляет свою противораковую активность в форме метаболитов, генерируемых в печени, например, ЦФ. Поэтому и этот способ недостаточно эффективен и не решает задачу полной эрадикации первичного асцита и предотвращения его повторного появления.
Известны также способы лечения раков, в том числе асцитных, основанных на проведении химиотерапией сверхмалыми дозами противоопухолевых цитостатиков [14, 15]. Для феномена сверхмалых доз установлен ряд общих закономерностей, присущих разным биологически активным веществам, в том числе лекарственным препаратам. Эти закономерности обнаружены при исследовании зависимостей «доза/эффект» в большом диапазоне доз. В наиболее простом, схематичном виде, эти зависимости являются бимодальными: эффект постоянно регистрируется при определенной «обычной» (для эндогенных веществ - «физиологической», для лекарственных препаратов - «терапевтической») дозе (обычно порядка 10-2-10-6 М, т.е. мг/кг, г/кг); при постепенном уменьшении дозы эффект снижается и в определенном интервале низких доз он становится небольшим или не регистрируется совсем («мертвая зона»). При дальнейшем уменьшении дозы и переходе в область сверхмалых доз вновь начинает регистрироваться эффект, который достигает максимума при определенной, характерной для данного вещества, сверхмалой дозе. В определенных ситуациях этот эффект может быть близким к эффекту «терапевтической дозы», меньше или даже больше его, иметь тот же или противоположный знак (например, ингибирование при «терапевтической дозе» может смениться на стимулирование при сверхмалой дозе). Во многих случаях наблюдаются более сложные полимодальные зависимости эффекта от дозы в интервале от терапевтических до сверхмалых доз. В этих случаях на кривой «доза/эффект» регистрируются несколько пиков эффекта и несколько «мертвых зон». Перечень биологически активных веществ различной природы, для которых уже доказана эффективность в сверхмалых дозах, составляет более сотни наименований, и он постоянно увеличивается. В этот список входят внутренние биорегуляторы (различные пептиды, гормоны), токсины, а также разнообразные синтетические соединения и вещества природного происхождения. Для целого ряда веществ, в том числе стандартных противоопухолевых препаратов, показано наличие противоопухолевой активности при использовании в сверхмалых дозах. Способность сверхмалых дох биологически активных веществ модифицировать противоопухолевый эффект терапевтических доз цитостатиков показана на примере совместного применения синтетического антиоксиданта фенозана и доксорубицина у мышей с асцитной лейкемией Р-388. Установлено, что комбинация фенозана в разных дозах (10-7-10-17 М) с терапевтической (1,4×10-3 М, 8 мг/кг) дозой доксорубицина приводит к увеличению эффекта по сравнению с использованием доксорубицина в виде монотерапии. Выживаемость животных, получавших фенозан в дозах 10-7 и 10-9 М, а также в значительно меньших (10-12 и 10-17 М) дозах, была существенно выше выживаемости мышей, получавших только доксорубицин. При использовании промежуточной (10-11 М) дозы фенозана не только не наблюдалось усиление эффекта доксорубицина, но даже имело место его полное отсутствие. Эти данные хорошо согласуются с отмеченными выше общими закономерностями действия сверхмалых доз биологически активных веществ.
Однако этот способ лечения также имеет ограниченную область применения, поскольку противоопухолевая активность сверхмалых доз показана не для всех изученных противоопухолевых препаратов и реализовывалась не на всех опухолевых моделях.
Известен и способ лечения опухолевых плевритов [16], включающий виутриплевральное введение противоопухолевых препаратов, причем у больного производят забор аутокрови в количестве 100-150 мл в стерильный флакон с гемоконсервантом "Глюгицир", центрифугируют при Vоб=1500 об/мин в течение 30 мин, отбирают 20 мл надосадочной плазмы, растворяют в ней разовую дозу цитостатика для внутриплеврального введения, инкубируют при 37°C в течение 30 мин, оставшиеся во флаконе клеточную часть и плазму реинфузируют, затем под местной анестезией производят пункцию плевральной полости, максимально эвакуируют плевральный выпот и вводят приготовленную аутоплазму с цитостатиком внутриплеврально, при необходимости процедуру повторяют.
При внутриплевральном введении цитостатиков на аутоплазме возможно сочетание с системной химиотерапией.
Указанный способ недостаточно эффективен, поскольку, обеспечивая стабильную на протяжении 8 месяцев ремиссию в развитии первичного асцита, не гарантирует от развития вторичного асцита в отдаленной (1-3 года) перспективе.
Наиболее близким к предложенному является способ [17], основанный на совместном воздействии цитостатика циклофосфана и препарата фрагментированной ДНК человека, при котором мышам линии СВА весом 20-35 г, несущим развитый асцит, парентерально вводится циклофосфан в дозе 200 мг/кг и далее в течение 12 часов каждый час после введения циклофосфана вводят интраперитонеально препарат ДНК, состоящий из смеси препаратов фрагментированной ДНК человека и кросс-линкированной ДНК человека, обработанной нитроген-мустардом.
Особенностью способа является то, что в качестве препарата ДНК используют смесь препаратов фрагментированной ДНК человека и кросс-линкированной ДНК человека размером 300-6000 пар оснований, а в препарате кросс-линкированной ДНК на 1000 пар оснований приходится 80-120 межцепочечных сшивок.
Наиболее близкое техническое решение основано на воздействии на стволовые инициирующие раковые клетки асцитной формы опухоли (на примере Кребс-2), при котором происходит эрадикация (уничтожение) популяции раковых клеток, обладающих инициирующими свойствами (стволовые-туморогенные раковые клетки). Для этого используется эффект совместного воздействия цитостатика циклофосфана (ЦФ) и препарата двуцепочечной ДНК.
Однако наиболее близкое техническое решение также обладает относительно низкой эффективностью лечения, поскольку при его использовании применяются многоразовые инъекции препарата ДНК, что вызывает токсический эффект, при этом временной интервал действия препарата не ориентирован на интерференцию репаративного процесса гомологичной рекомбинации в стволовых инициирующих раковых клетках, что в значительной степени ограничивает терапевтический эффект от такой терапии и не предполагает полного вылечивания от асцитной формы рака.
Задача, решаемая в настоящем изобретении, направлена на повышение эффективности лечения путем увеличения эрадикации стволовых инициирующих раковых клеток с целью полной элиминации первичного ракового асцита и уменьшения возможности появления вторичного асцита, или полного вылечивания от асцитной формы рака.
Требуемый технический результат заключается в повышении эффективности лечения асцитной формы рака путем повышения эрадикации стволовых инициирующих раковых клеток, элиминации первичного ракового асцита и предотвращения появления вторичного асцита на основе использования теоретически обоснованных и экспериментально подтвержденных авторами четких временных интервалов применения препаратов ЦФ и ДНК в соответствии с вновь выявленными и описанными биологическими свойствами стволовых инициирующих раковых клеток.
Поставленная задача решается, а требуемый технический результат достигается в способе лечения асцитной формы рака, основанном на совместном воздействии цитостатика циклофосфана и препарата двуцепочечной ДНК, при котором, согласно изобретению, мышам-опухоленосам линии СБА весом 20-35 г, несущим развитый асцит, парентерально вводят цитостатик циклофосфан в 0, 36, 72 часа каждый раз в дозе 100 мг/кг, а через 18, 54, 90 часов после каждой инъекции циклофосфана, соответственно, мышам-опухоленосам вводят интраперитонеально препарат ДНК, состоящий из композиции препаратов нативной двуцепочечной ДНК человека в дозе 3.0 мг и смеси нативной и кросс-линкированной двуцепочечной ДНК спермы лосося в соотношении 5:3 в дозе 3 мг суммарно в количестве 6.0 мг на инъекцию, независимо от веса мышей-опухоленосов.
Кроме того, требуемый технический результат достигается тем, что для достижения максимального терапевтического действия на восьмые сутки после первой инъекции циклофосфана вводят дополнительно циклофосфан в дозе 100 мг/кг и через 18 часов препарат ДНК суммарно в количестве 6 мг.
Кроме того, требуемый технический результат достигается тем, что интраперитонеально препарат ДНК в количестве 6 мг на инъекцию вводят однократно в объеме 500 мкл из расчета половины объема четырехсуточного асцита, соответствующего 1 мл и 1/20 веса животного, содержащего ~200 млн раковых клеток.
Сущность предложенного способа заключается в следующем.
Вначале проводят разрушение комметированных раковых клеток возбуждением трех последовательных без перерыва полных циклов репарации межцепочечных сшивок, индуцируемых инъекциями циклофосфана в 0, 36, 72 часа в дозе по 100 мг/кг. При этом эрадикация стволовых инициирующих раковых клеток осуществляется одновременно на фоне инъекций циклофосфана путем инъекции препарата ДНК в 18, 54, 90 часов (через 18 часов после каждой инъекции циклофосфана), соответственно, при которых вторая и третья инъекции циклофосфана будут индуцировать вторую и третью волну двуцепочечных разрывов в раковых клетках, находящихся в этот момент в поздней S/G2/M фазе клеточного цикла, а так же в раковых клетках, находящихся в момент первой инъекции циклофосфана в G2/M фазах клеточного цикла и не чувствительных, по этой причине, к обработке ДНК, т.е. вторая и третья обработки ЦФ будут индуцировать вторую и третью волну двуцепочечных разрывов в уже арестованных стволовых инициирующих раковых клетках, повторно запирая их выход из состояния ареста, и одновременно будут индуцироваться межцепочечные сшивки и арест клеточного цикла у другой части стволовых инициирующих раковых клеток, находившихся при первой обработке цитостатиком в нечувствительной G2/M фазе клеточного цикла и перешедших в G1/S чувствительную фазу.
Таким образом, к концу цикла лечения индуцируется апоптоз всех коммутированных раковых клеток и происходит синхронизация в поздней S/G2/M всех стволовых раковых клеток, успешно преодолевших терапевтические процедуры. При этом для каждой последующей инъекции препарата ДНК используется точка 18 часов после каждой инъекции циклофосфана, представляющая собой точку разделения двух фаз процесса репарации межцепочечных сшивок, индуцированных циклофосфаном, в которой закладывается процесс разрушения стволовых инициирующих раковых клеток по двум независимым модам в форме интерференции конечных интермедиатов репаративной фазы NER одной составляющей препарата ДНК, и интерференции начальных интермедиатов репаративной фазы гомологичной рекомбинации второй составляющей препарата ДНК. После проведения первого цикла лечения проводят аналогичный первому второй цикл через 7-9 суток после начала первого цикла, т.е. в момент начала масштабного разрушения комметированных раковых клеток и синхронизации в G2/M, и переходу в следующую по очередности S фазу TAMRA+ стволовых инициирующих раковых клеток.
В целом, предложенный способ основан на естественной способности свободно расположенных в асцитном бульоне не объединенных в компартменты стволовых (инициирующих) раковых клеток Кребс-2 интернализовать экзогенные экстраклеточные фрагменты двуцепочечной ДНК, на участии интернализованных фрагментов в репаративном процессе при репарации межцепочечных сшивок в фазе NER и гомологичной рекомбинации после действия цитостатических препаратов кросс-линкирующего ряда (ЦФ), а также на определенной временной интервальной схеме, определяемой вновь открытыми биологическими свойствами стволовых инициирующих раковых клеток Кребс-2.
В современной литературе отсутствуют указания на предлагаемый способ убийства стволовых раковых клеток и эрадикации развитого асцита и возможности предотвращения развития вторичного асцита на протяжении 190 суток (на модели асцитной формы рака Кребс-2) после начала заболевания. Следовательно, предложение отвечает критериям новизны и изобретательского уровня.
Ниже приводятся теоретические и экспериментальные данные, подтверждающие, что изобретение отвечает критерию практической (промышленной) применимости.
На чертеже представлены следующие иллюстрирующие материалы.
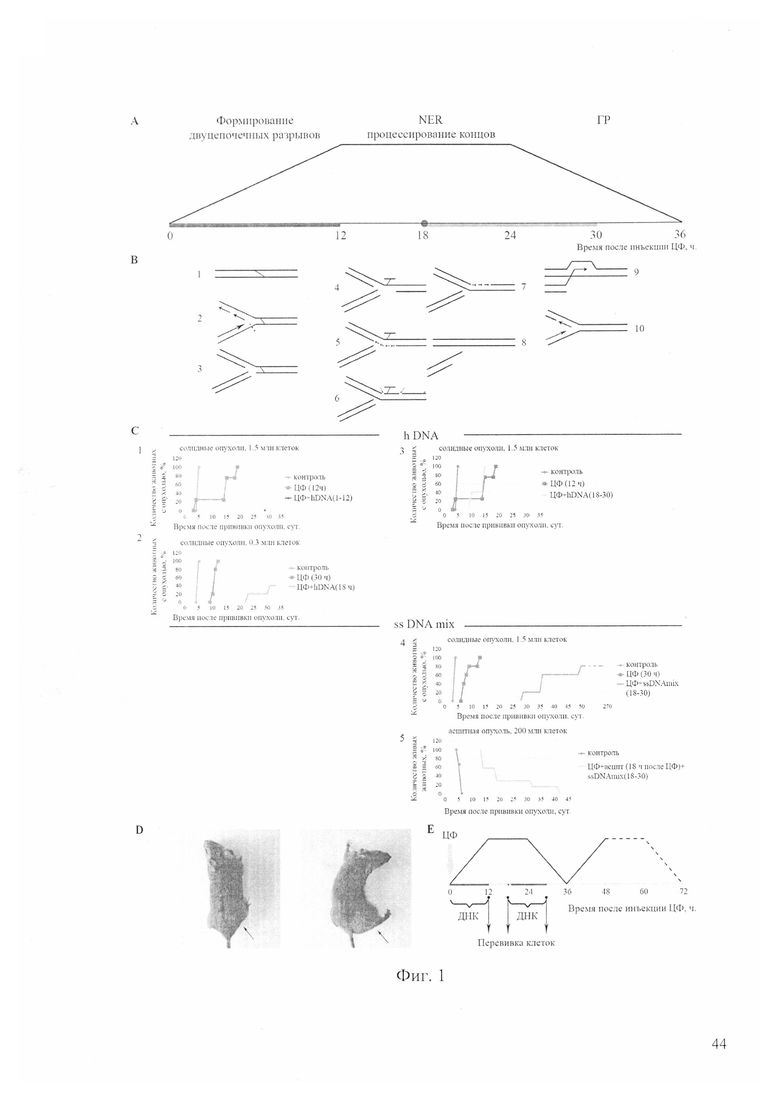
На фиг. 1 - корреляция противоракового эффекта совместных обработок ЦФ и препаратами ДНК со стадией репарации межцепочечных сшивок, индуцированных действием ЦФ, полученная на клетках асцитной формы опухоли Кребс-2 (данные на графиках приведены в сравнении с контролем и действием только ЦФ, где А - схематическое изображение стадий репаративного процесса в клетках асцита Кребс-2 и наложение на него режимов введения препаратов двуцепочечной ДНК (линия от 0 до 12 часов и от 18 до 30 часов, соответственно); линия от 0 до 12 часов - введение нативного препарата ДНК человека, линия от 18 до 30 часов - введение смеси кросс-линкированной и нативной ДНК лосося (ssDNAmix) в соотношении 3/5; В - схематическое изображение интермедиатов репарации межцепочечных сшивок, расположенных в соответствии с фазами формирования и репарации двуцепочечных разрывов: 1 - появление межцепочечной сшивки, 2 - остановка репликативной вилки в месте повреждения, 3, 4 - обнаружение межцепочечной сшивки NER системой клетки и формирование двуцепочечного разрыва, 5 - транслокация аддукта и репаративный синтез прямой цепи ДНК, осуществляемый либо путем TLS, либо по гомологичной матрице, 6 - второй раунд NER, вырезание аддукта, 7 - репаративный синтез обратной цепи ДНК, 8 - процессинг 3′-конца двуцепочечного разрыва, 9, 10 - инвазия процессированного 3′-конца двуцепочечного разрыва в гомологичный участок цепи ДНК и восстановление репликативной вилки; С - 1, 2, 3, 4 - динамика появления опухолей при перевивке в виде солидных внутримышечных трансплантатов клеток, обработанных in vivo ЦФ и препаратами ДНК, где количество перевиваемых клеток указано над графиком, 5 - динамика гибели экспериментальных животных с асцитной формой опухоли Кребс-2 после проведенных обработок, где количество исходно привитых клеток асцита указано над графиком, 1-3 - экспериментальные группы животных, получавшие в сочетании с ЦФ препарат ДНК человека (hDNA), 4, 5 - экспериментальные группы животных, получавшие в сочетании с ЦФ препарат ДНК спермы лосося (ssDNAmix); D - слева мышь из группы «CP + ssDNAmix (18-30)» (фиг. 2, С4, кривая «Hp + ssDNAmix (18-30)»), у которой через 270 дней после прививки графта трансплантат так и не развился, справа - мышь из контрольной группы на 20-й день после прививки опухоли, стрелками указано место прививки трансплантата; Е - обобщенная схема обработки экспериментальных мышей ЦФ и препаратами ДНК и точки забора клеток для последующей трансплантации здоровым животным в виде солидных опухолей.
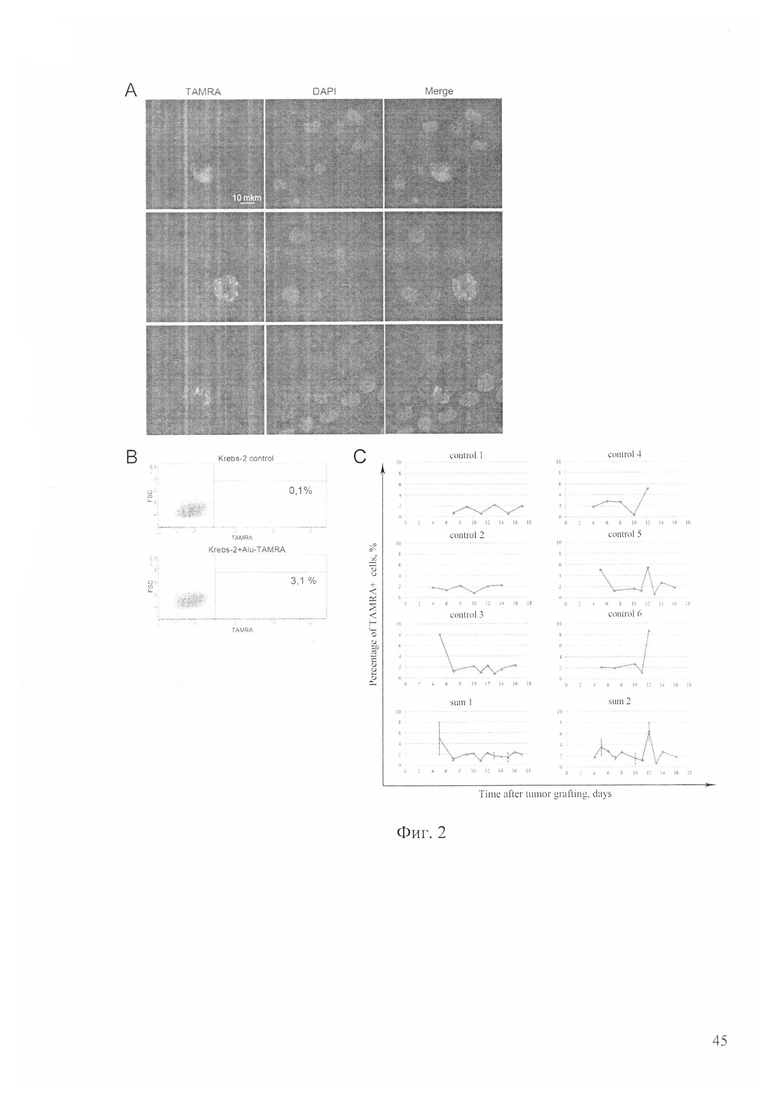
На фиг. 2 - цитофлуоресцентный (А) и проточноцитометрический (В) анализ интервализации Alu-TAMRA ДНК в стволовые инициирующие раковые клетки асцитной опухоли Кребс-2, красный сигнал (TAMRA) - интернализация меченного ДНК зонда, синим окрашены ядра клеток флуоресцентным красителем DAPI; Merge - объединенные (TAMRA/DAPI) имиджи. С - анализ содержания TAMRA+ ДНК в асците у индивидуальных мышей (control 1-6) на протяжении его развития до 15 суток, где нижние графики (sum 1, sum 2) - объединенные результаты по трем животным.
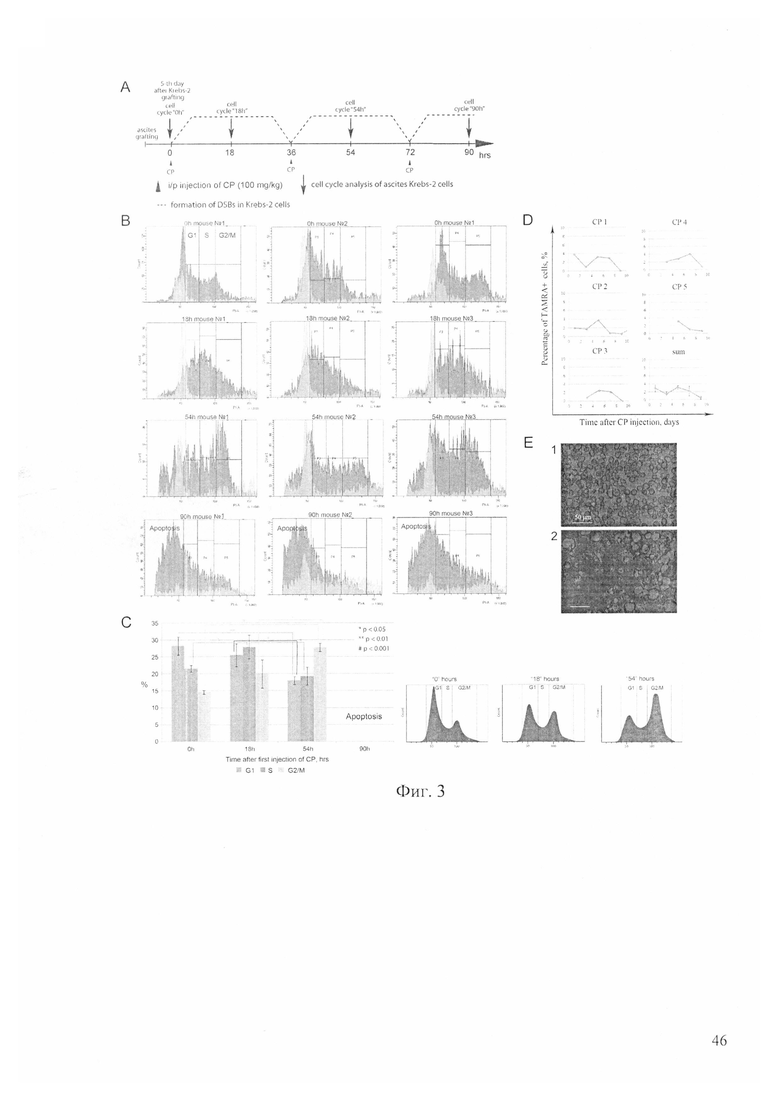
На фиг. 3 - анализ клеточного цикла асцитных клеток у мышей после обработки исследуемым терапевтическим режимом; А - схема эксперимента, где кривые соответствуют фазам репаративного процесса NER и гомологичной рекомбинации при репарации межцепочечных сшивок, индуцированных ЦФ, 0, 36 и 72 часа - точки введения цитостатика, 18, 54 и 90 часов - точки введения препарата двуцепочечной ДНК. Пунктиром обозначена принципиальная схема репарации МЦС, подробно рассмотренная на фигуре 1. В - плоты, полученные на основе анализа асцитных раковых клеток 3-х индивидуальных мышей, демонстрирующие изменения в клеточном цикле; С - суммарный график по трем животным (слева), графическое изображение изменений в клеточном цикле (справа); D - графики изменения процента TAMRA+ клеток в асците после проведенных обработок циклофосфаном, где показаны изменения у индивидуальных животных (СР1-5; n=5) и суммарный обобщающий график (sum); Е - цитологический анализ состояния клеток на 7-е сутки от начала терапии (2) по сравнению с контролем (1).
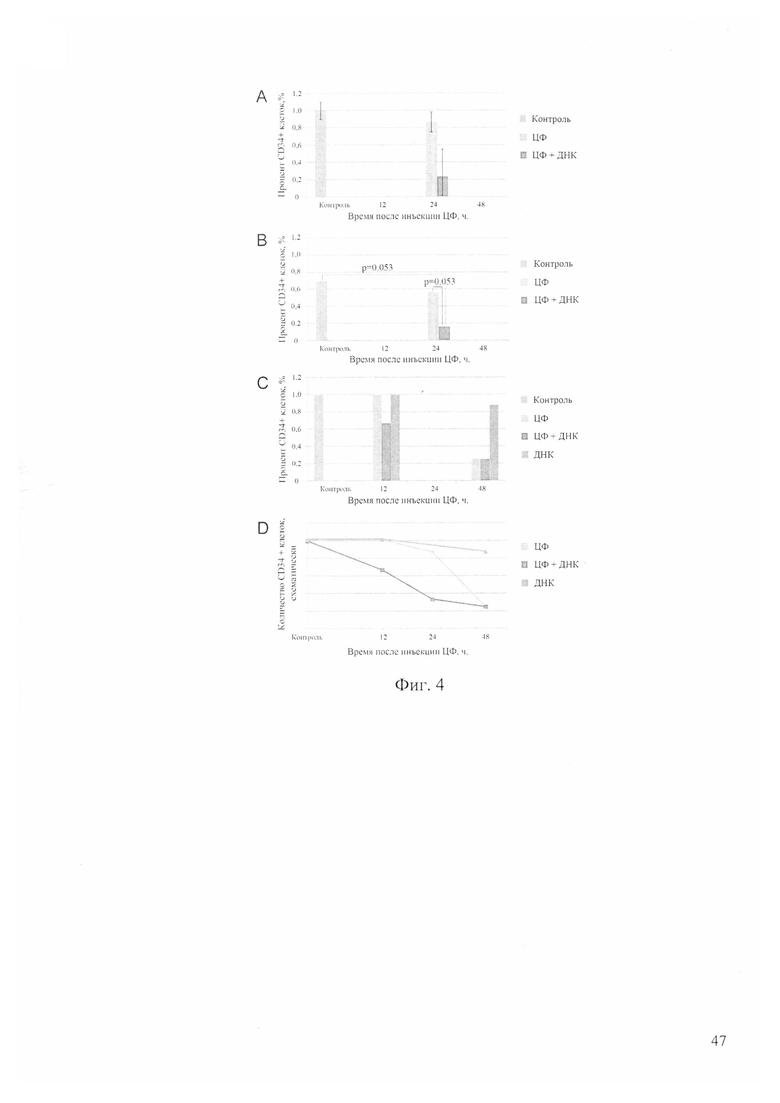
На фиг. 4 - анализ динамики количества CD34+ клеток в асцитном варианте мышиной опухоли Кребс-2 после обработки ЦФ и препаратом двуцепочечной ДНК, где «ЦФ» - однократная инъекция ЦФ (200 мг/кг), «ЦФ+ДНК» - инъекция ЦФ (200 мг/кг) и 12 инъекций препарата ДНК через каждый час после инъекции ЦФ по 0,5 мг за инъекцию, «ДНК» - 12 ежечасных инъекций препарата ДНК по 0,5 мг на инъекцию, «Контроль» - интактный асцит до обработок, 0 - время инъекции ЦФ или первой инъекции ДНК; А - FACS анализ количества CD34+ клеток в асците Кребс-2, развивающемся в мыши, в контроле (до инъекций) и через 24 часа после инъекций ЦФ или ЦФ+ДНК; В - анализ количества CD34+ клеток в асците Кребс-2, развивающемся в мыши, в контроле и через 24 часа после инъекций ЦФ, ЦФ+ДНК или ДНК при помощи флуоресцентной микроскопии; С - FACS анализ количества CD34+ клеток в асците Кребс-2, развивающемся в мыши, в контроле (до инъекций) и через 12 и 48 часов после инъекций ЦФ или ЦФ+ДНК; D - схематичное изображение изменения количества CD34+ клеток в течение двух суток в асците Кребс-2 на фоне инъекций ЦФ, ЦФ+ДНК и ДНК, где график построен с учетом результатов трех независимых экспериментов (Л-С).
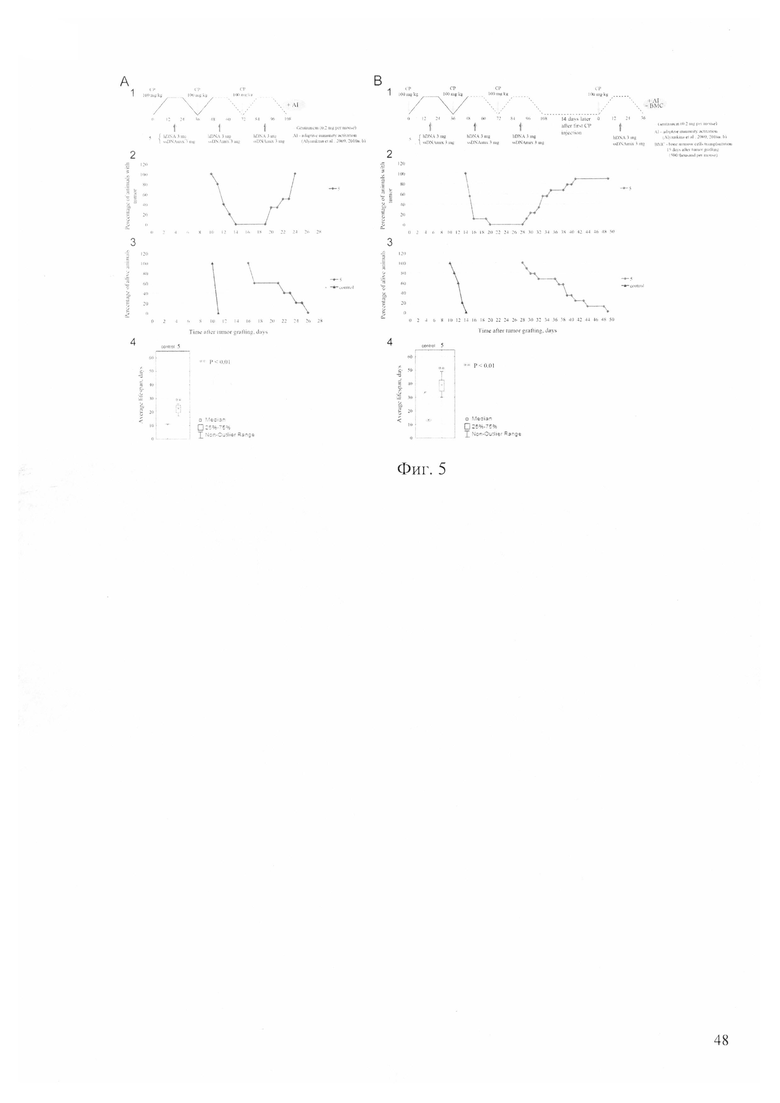
На фиг. 5 - А - синергичное действие ЦФ и препарата двуцепочечной ДНК в режиме «три ЦФ+ДНК», не предусматривающее финальной терапевтической инъекции; В - синергичное действие ЦФ и препарата двуцепочечной ДНК в режиме «три ЦФ+ДНК + четвертая инъекция ЦФ+ДНК» в момент времени полного восстановления асцитных раковых клеток от воздействия первоначальной процедуры (на 14 сутки от первой инъекции ЦФ). AI - активация адаптивного иммунного ответа препаратом «Панаген». ВМС - трансплантация костного мозга. 1 - терапевтическая схема; 2 - график резорбции асцита, ремиссии и рецидива; 3 - график гибели животных; 4 - плоты, демонстрирующие среднюю продолжительность жизни.
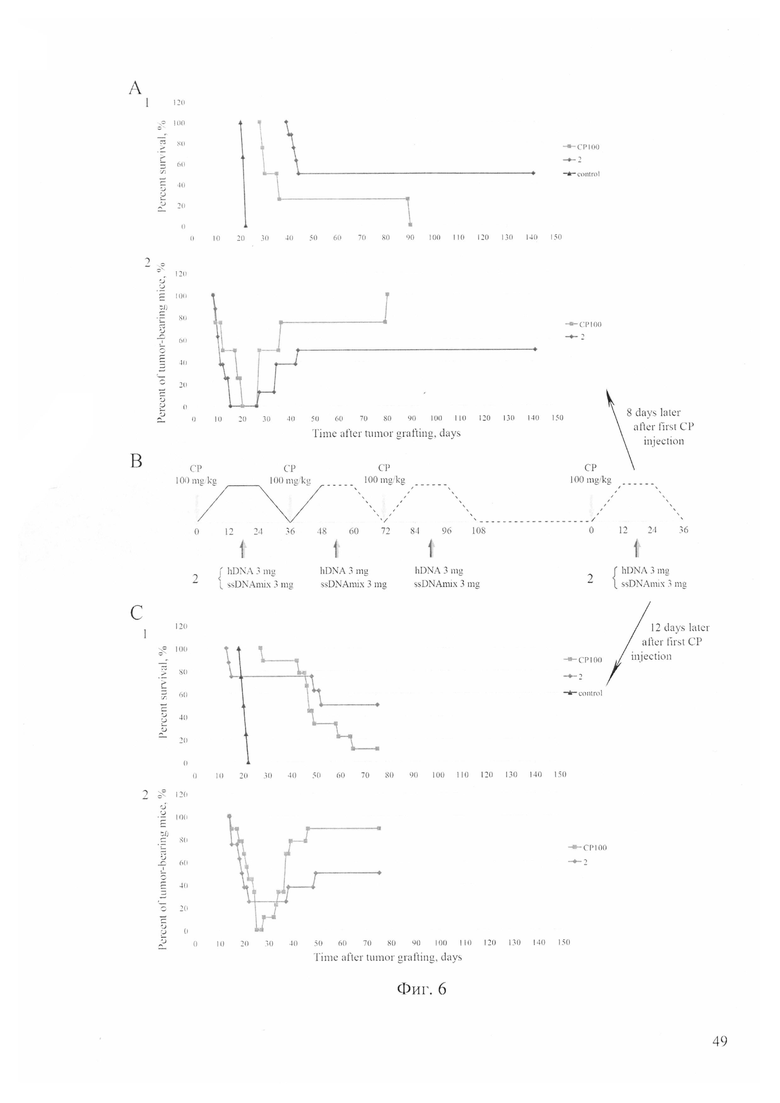
На фиг. 6 - А - синергичное действие ЦФ и препарата двуцепочечной ДНК в режиме «три ЦФ+ДНК + четвертая инъекция ЦФ+ДНК» в терминальную точку терапии в момент разрушения комметированных раковых клеток и выхода из ареста в G2/M стволовых инициирующих раковых клеток Кребс-2 на 8 сутки от первой инъекции ЦФ. (1) динамика гибели животных и (2) регрессии асцита в экспериментальных группах (ЦФ и группа после проведенной терапии); В - схема эксперимента; С - синергичное действие ЦФ и препарата двуцепочечной ДНК в режиме «три ЦФ+ДНК + четвертая инъекция ЦФ+ДНК» в терминальную точку терапии в момент разрушения комметированных раковых клеток и выхода из ареста в G2/M стволовых инициирующих раковых клеток Кребс-2 (на 12 сутки от первой инъекции ЦФ). (1) динамика гибели животных и (2) регрессии асцита в экспериментальных группах (ЦФ и группа после проведенной терапии).
На фиг. 7 - А - общая схема событий, произошедших с экспериментальными животными после проведенной терапевтической процедуры и до 150 суток эксперимента; В - демонстрация мышей до терапии (левая группа панелей), в момент регрессии асцита (на 17 сутки от первой инъекции ЦФ) и вылеченных мышей на 150 сутки эксперимента (правая группа панелей).
Актуальность данного изобретения подтверждается тем, что, несмотря на достигнутые успехи в химиотерапии злокачественных опухолей и, в частности, асцитных форм рака существует проблема, не позволяющая сформировать четкое законченное представление о правильности выбранной стратегии лечения и средств воздействия на малегнизированные клетки. Такой проблемой является отсутствие понимания всей совокупности биологических свойств опухолевых клеток и в первую очередь стволовых инициирующих раковых клеток.
В этой связи наиболее важным является вопрос чувствительности туморогенных стволовых клеток к выбранной терапии. Все цитостатики, так или иначе, разрушают комметированные раковые клетки, и при этом в настоящее время не существует адекватного, простого подхода в определении эффективности воздействия выбранного лечения на стволовые инициирующие раковые клетки.
В этой связи подходы, позволяющие контролировать процесс эрадикации стволовых инициирующих раковых клеток и строящие свою стратегическую платформу на четком знании определенных биологических закономерностей стволовых инициирующих раковых клеток, являются тем важным инструментом, позволяющим осознанно применять терапевтические процедуры, основной целью которых является эрадикация стволовых инициирующих раковых клеток и тем самым лишение опухоли туморогенного начала. Кроме этого, оценка состояния популяции стволовых инициирующих раковых клеток позволяет следить за эффективностью терапии и корректировать лечебную тактику.
В этой связи разработка новых терапевтических стратегий эрадикации туморогенного начала опухоли, основанных на знании принципиально новых биологических свойств стволовых инициирующих раковых клеток асцитных опухолей, является актуальной задачей, а нахождение адекватных методов терапии позволит сделать скачок в лечении злокачественных асцитных раков.
Ниже приводятся экспериментальные доказательства обнаружения возможности полного вылечивания от асцитной формы рака на модели асцитной формы рака мышей Кребс-2.
Используемые методики
Экспериментальные животные
В экспериментах использовались мыши линии CBA/Lac и С57В1, которые разводились в условиях вивария при Институте цитологии и генетики СО РАН. Животные содержались в клетках, по 5-10 особей в каждой, и имели свободный доступ к воде и пище.
Опухолевая модель
В экспериментах использовали мышиную опухолевую модель Кребс-2. Для получения асцитной формы опухоли клетки перевивали мышам внутрибрюшинно (в/б) по 2 млн клеток в 200 мкл физиологического раствора. Для формирования солидных опухолей Кребс-2 клетки перевивали внутримышечно в заднюю правую лапу в необходимом количестве в 100 мкл среды RPMI-1640 или PBS. В ходе экспериментов, как только опухоли начинали пальпироваться (примерно через 5-7 дней после прививки в контроле), каждые 1-2 дня проводили измерение их объема с помощью штангенциркуля. Объем опухоли рассчитывали, используя стандартную формулу: длина × высота × ширина.
Введение препаратов циклофосфана и (экзогенной) ДНК
Цитостатик ЦФ вводили мышам внутрибрюшинно в дозе 100-200 мг/кг массы тела в зависимости от эксперимента. Препараты фрагментированной ДНК (ДНК человека и ДНК спермы лосося) и кросс-линкированной ДНК, обработанной нитроген-мустардом, вводили внутрибрюшинно в дозе 0,5-1 мг на инъекцию ежечасно или каждые 2 часа (суммарно по 6 мг ДНК на мышь). Временные интервалы введения препаратов либо описаны непосредственно в тексте, либо изображены схематически на соответствующих фигурах.
Получение препарата ДНК человека и лосося
Геномную ДНК человека выделяли безфенольным методом из плацентарного материала и гидролизовали ультразвуком до фрагментов размером 200-6000 п.о. в соответствии с регламентом производства фармакопейного препарата «Панаген» (ЛСР №004429/08 от 09.06.2008). Препарат охарактеризован и не содержит стероидных гормонов, РНК, имеет отрицательные результаты ПЦР анализов на присутствие ДНК вируса гепатита В, РНК вируса гепатита С, ДНК ВИЧ, РНК ВИЧ.
Препарат ДНК спермы лосося получали аналогичным образом из молок лосося.
Подготовка препарата ДНК, кросс-линкированной нитроген мустардом.
Способ получения кросс-линкированной ДНК приведен в работе [19].
Определение количества CD34+ клеток в асцитной (форме) опухоли Кребс-2 при помощи проточной цитометрии и микроскопического анализа.
Клетки асцита Кребс-2 отбирали у животных после соответствующих обработок, промывали PBS и осаждали при 4°C, 400 g в течение 5 мин. После осаждения центрифугированием клетки ресуспендировали в PBS с 0,1% NaN3, 1% FBS (Fetal Bovine Serum) и подсчитывали в камере Горяева. К 1 млн клеток добавляли 4 мкг антител (FITC Rat anti-Mouse CD34, BD Pharmingen) и, соответственно, 4 мкг изотип-контроля (FITC Rat IgG2a, к Isotype Control, BD Pharmingen) и инкубировали в течение 40 мин при 4°C в 200 мкл среды. Процент клеток CD34+ определяли либо при помощи проточного цитофлуориметра BD FACSAria относительно результатов изотип-контроля в каждом образце, либо обработанные указанным выше способом клетки асцита Кребс-2 промывали PBS и наносили цитоспином на предметное стекло (1000 об/мин в течение 1 мин). Затем на полученный препарат наносили ~10 мкл Antifade DABCO, накрывали покровным стеклом и анализировали при помощи флуоресцентного микроскопа Axioskop 2 Plus (Zeiss) с использованием программы AxioVision.
Мечение ДНК флюорохромом TAMRA
Метод мечения ДНК флуорохромом TAMRA в реакции ПЦР подробно описан в статье [18].
Анализ перевивочного потенциала клеток асцита Кребс-2, обработанных in vivo.
Мыши с развитым асцитом Кребс-2 (4-7 суток после прививки) получали внутрибрюшинные инъекции цитостатика ЦФ (100-200 мг/кг) и препаратов ДНК (суммарно по 6 мг по схемам, приведенным в каждом конкретном эксперименте). В определенный интервал времени после проведенных обработок у животных забирали асцит, клетки осаждали при 400 g в течение 5 мин (4°C), ресуспендировали, подсчитывали в камере Горяева и перевивали интактным животным в мышцы задней правой лапы в 100 мкл среды RPMI-1640 или PBS. Количество перевиваемых клеток в экспериментах варьировалось. Животным контрольных групп перевивали аналогичное количество клеток асцита Кребс-2, не подвергнутого обработкам.
Анализ перевивочного потенциала клеток асцита Кребс-2, обработанных in vivo ЦФ и ex vivo препаратами ДНК.
Отличие настоящего подхода от предыдущего заключается в том, что экспериментальным животным единовременно прививали клетки асцита Кребс-2 в количестве, соответствующем количеству клеток, содержащихся в 4-суточном асците, развившемся в мыши после прививки 2 млн клеток (~200 млн клеток).
Мышь с развившейся асцитной опухолью получала внутрибрюшинную инъекцию ЦФ в дозе 100 мг/кг. Через 18 часов у нее отбирали асцит, клетки осаждали при 400 g в течение 5 мин, ресуспендировали и подсчитывали в камере Горяева. Далее клетки смешивали со смесью препаратов кросс-линкированной и нативной ДНК лосося в соотношении 3/5 (ssDNAmix) из расчета 0,5 мг/мышь и прививали по 200 млн клеток экспериментальным животным, получившим также предварительную инъекцию ЦФ (100 мг/кг) за 18 часов до прививки асцита. После этого мыши получили инъекции аналогичной дозы смеси препаратов ДНК еще 11 раз (режим 18-30).
Анализ доставки TAMRA-меченой ДНК в клетки асцита Кребс-2.
Клетки асцита Кребс-2 отбирали у мыши и осаждали центрифугированием в течение 5 мин, 400 g при 4°C и промывали средой RPMI-1640 один раз. Затем клетки подсчитывали в камере Горяева. 1 млн клеток инкубировали в 200 мкл этой же среды с 10 или 1000 нг TAMRA-меченой ДНК в течение часа при комнатной температуре. Затем клетки осаждали центрифугированием, промывали небольшим объемом среды и осадок ресуспендировали в конечном объеме среды. Далее количество клеток, захвативших TAMRA-меченую ДНК, анализировали при помощи проточного цитофлуориметра BD FACSAria (Becton Dickinson, США). При этом процент TAMRA+ клеток оценивали относительно контроля, т.е. клеток, инкубировавшихся в среде без добавления TAMRA-меченой ДНК.
Введение препаратов циклофосфана, экзогенной ДНК и антибиотика гентамицина.
Цитостатик ЦФ вводили мышам внутрибрюшинно в дозе 50-100 мг/кг массы тела одно-, дву-, трех- или четырехкратно в зависимости от эксперимента. Препараты фрагментированной ДНК (ДНК человека и ДНК спермы лосося) и кросс-линкированной ДНК, обработанной нитроген-мустардом, или их смесь (в соотношении кросс-линкированная/нативная 1/3 или 3/5) вводили внутрибрюшинно в дозе 0,5-1 мг на инъекцию каждые 1, 2 или 3 часа (суммарно по 6 мг ДНК на мышь за 12 часов). Временные интервалы введения препаратов либо описаны непосредственно в тексте, либо изображены схематически на соответствующих фигурах. Антибиотик гентамицин вводили животным внутрибрюшинно по 0,2 мг/мышь каждые 1-2 суток в течение месяца относительно начала эксперимента.
Активация адаптивного иммунитета
Для активации у экспериментальных животных адаптивного иммунитета вводили препарат нативной ДНК человека через 1, 2 и 3 суток после последней инъекции ЦФ по 500 мкг/мышь.
Пересадка клеток костного мозга
Для получения клеток костного мозга использовали здоровых мышей-доноров той же линии. Клетки костного мозга вымывали PBS из больших берцовых костей, промывали средой RPMI-1640, подсчитывали в камере Горяева и перевивали в хвостовую вену экспериментальным животным в количестве 200 тыс клеток на мышь.
Гистологоанатомический анализ тканей и органов мышей
Кусочки органов фиксировали в 4%-ном параформальдегиде, обезвоживали в спиртах возрастающей концентрации, просветляли ксилолом, заливали в парафин. Парафиновые срезы толщиной до 5 мкм окрашивали гематоксилином и эозином. Просмотр препаратов и микрофотосъемку проводили на световом микроскопе Axiolmager ZI (Zeiss, Германия).
Оценка ремиссии асцитной опухоли Кребс-2
О ремиссии заболевания судили визуально: «уходом» асцита считали состояние мыши, когда внешний вид живота соответствовал виду интактного животного и появлялись характерные рубцы, свидетельствующие о резком уменьшении объема жидкости, находящейся в брюшной полости.
Статистическая обработка данных
Статистическую обработку данных проводили при помощи программы Microsoft Excel, на фигурах отображено стандартное отклонение. Сравнение продолжительности жизни между группами животных проводили методом непараметрической статистики с использованием Т-критерия Вилкоксона-Манна-Уитни в программе Статистика.
Рассмотрим более подробно базовые положения предлагаемого подхода.
Работа посвящена разработке стратегии излечения мышей от злокачественной опухоли Кребс-2. Кребс-2 представляет собой недифференцированную карциному с высокой степенью пролиферативной активности. Внутримышечные трансплантаты этой опухоли приводят к гибели животных на 30-35 сутки. Асцитная форма Кребс-2 приводит к летальному исходу на 14-18 сутки. Опухоль слабо чувствительна или не чувствительна к действию цитостатиков и, в частности, к ЦФ [20].
В работе [18] охарактеризованы два общебиологических феномена. Интернализация фрагментов экстраклеточной двуцепочечной ДНК в стволовые инициирующие раковые клетки Кребс-2 и способность этих интериализованных фрагментов интерферировать процесс репарации межцепочечных сшивок таким образом, что перевиваемый трансплантат теряет свое туморогенное начало.
Экспериментальный дизайн исследования имел две принципиальных платформы. В первой части исследования использовался подход, состоящий из обработки терапевтической процедурой, включающей инъекции ЦФ и препаратов двуцепочечной ДНК мышей-асцитоносов и последующей перевивки обработанных асцитных клеток мышам той же лини в форме солидного трансплантата. Вторая часть исследования посвящена поиску и характеристике режима полного вылечивания мышей-асцитоносов от асцита Кребс-2 терапевтической процедурой, примененной in vivo.
Основные положения терапии асцита in vivo базируются на общебиологических свойствах стволовых инициирующих раковых клеток Кребс-2 и феноменах, возникающих при репарации межцепочечных сшивок, индуцированных инъекциями ЦФ, с последующими инъекциями препарата двуцепочечной ДНК, описанных в наших недавних работах [18, 19] и в настоящем исследовании.
Рассмотрим влияние синергичного действия ЦФ и препаратов двуцепочечной ДНК человека и лосося на перевивочный потенциал асцитных клеток Кребс-2.
Для оценки биологического эффекта синергичного воздействия ЦФ и ЦФ+ДНК, которое подразумевало элиминацию TAMRA+ клеток (стволовых инициирующих раковых клеток, характеризующихся захватом TAMRA меченной ДИК) или изменение их функционального состояния, были проведены эксперименты по перевивке в форме солидного трансплантата мышам той же линии обработанных непосредственно в асците опухолевых клеток. Такая тактика была связана с тем, что в начале исследования [19] в экспериментах использовался асцит в возрасте старше 7 суток от перевивки, и после обкола ЦФ и препаратом двуцепочечной ДНК наблюдалась тотальная гибель мышей. Причины токсического воздействия обработок были неизвестны. Как показали дальнейшие эксперименты, одновременное апоптотическое разрушение массы асцитных клеток, составляющей для асцита такого возраста около миллиарда, и связанные с этим патологические последствия (вторичный некроз, разрушение кишечного эпителия, системная воспалительная реакция, сепсис, коллапс иммунной системы), вызванные проведенными обработками, явились причиной смертельного токсического эффекта [19, 21]. Скоротечная гибель мышей и возможность длительного нахождения (до 14 суток) молекул ДНК в реципиентной клетке [22] не позволяли оценить состояние TAMRA+ клеток у мышей с асцитом Кребс-2, которым вводился препарат двуцепочечной ДНК.
На фиг. 1 приводятся результаты по нескольким примененным режимам, в которых был использован зрелый (>7 суток) асцит. Асцит in vivo обрабатывался препаратами нативной человеческой двуцепочечной ДНК в режиме ЦФ+ДНК (1-12), ЦФ+ДНК (18-30) или ЦФ+ДНК (18) и ДНК лосося в смеси кросс-линкированная ДНК/нативная ДНК (2-3/5) в режиме ЦФ+ДНК (18-30), после чего выделенные асцитные клетки перевивались внутримышечно мышам той же линии.
В проведенных исследованиях удалось добиться длительного, по сравнению с контролем, промежутка времени появления опухоли или полностью лишить клетки асцита туморогенного начала. Полученные результаты свидетельствуют о том, что:
- в синергизме с ЦФ в фазе репарации NER (1-12 часов после введения ЦФ) при интерференции процесса репарации межцепочечных сшивок фрагментами двуцепочечной ДНК человека, попавшими в стволовые инициирующие раковые клетки, последние теряют жизнеспособность или изменяют свой функциональный статус, при этом перевивка 40-50-кратного избытка клеток не приводит к развитию солидного трансплантата, а единственная инъекция двуцепочечной ДНК человека в 18 часов после введения ЦФ снижает туморогенный потенциал перевиваемого графта (при перевивке 300 тыс клеток у 3 из 5 опытных животных опухоль не появилась после 60 дней наблюдения (фиг. 1, С1, 2);
- в рассматриваемой серии экспериментов был установлен контроверсивный факт, что в синергизме с ЦФ в фазе репарации гомологичной рекомбинации (18-30 часов после введения ЦФ) при интерференции процесса репарации межцепочечных сшивок фрагментами двуцепочечной ДНК человека, попавшими в стволовые инициирующие раковые клетки, корректность репаративного процесса сохраняется, и стволовые инициирующие раковые клетки выживают более эффективно, чем при действии одного ЦФ, происходит стимуляция развития солидного трансплантата, который приживается в 100% перевивочных экспериментов (фиг. 1, C3);
- в синергизме с ЦФ использование кросс-линкированной ДНК в чистом виде смертельно токсично для мышей при всех вариантах обработок;
- в синергизме с ЦФ в фазе репарации гомологичной рекомбинации (18-30 часов после введения ЦФ) при использовании смеси кросс-линкированная ДНК/нативная ДНК лосося (2/5) происходят события, интерферирующие процесс репарации таким образом, что снижается туморогенный потенциал прививаемого солидного трансплантата, причем в проведенном эксперименте у одной из 5 мышей солидный трансплантат не сформировался к 270 дню наблюдения, у остальных мышей трансплантат формировался последовательно, в зависимости от степени сохранившегося туморогенного потенциала на протяжении длительного промежутка времени, но и в этом случае наблюдается ярко выраженное количественное изменение признака (разрушение способности индуцировать развитие новой опухоли), (фиг. 1, С4);
- в синергизме с ЦФ в фазе репарации гомологичной рекомбинации (18-30 часов после введения ЦФ) при использовании смеси кросс-линкированная ДНК/нативная ДНК лосося (3/5) при первичной ex vivo обработке асцитных клеток происходят события, интерферирующие процесс репарации таким образом, что в значительной степени редуцируется туморогенный потенциал прививаемого (асцитного) трансплантата (фиг. 1, С5).
В анализируемом эксперименте количество привитых клеток составляло 200 млн, что соответствовало 4-суточному асциту. Средняя продолжительность жизни в 4 раза превысила этот показатель в контроле, где мыши погибли на 4-6 сутки от прививки асцита. У 70% животных экспериментальной группы асцит развивался последовательно. Максимальная продолжительность жизни составила 42 дня.
Таким образом, экспериментально было показано, что интраперитонеальная обработка in vivo мышей с асцитом Кребс-2 препаратами двуцепочечной ДНК человека и смеси ДНК человека и модифицированной ДНК спермы лосося в различные промежутки времени и после однократной инъекции ЦФ в определенном проценте случаев приводит к полной потере туморогенного начала клетками асцита.
Сопоставляя два факта, что незначительная популяция асцитных клеток Кребс-2, обладающая свойствами стволовых инициирующих раковых клеток, захватывает экстраклеточные фрагменты двуцепочечной ДНК и что обработка раковых клеток асцита Кребс-2 ЦФ и препаратами двуцепочечной ДНК приводит к потере раковыми асцитными клетками способности давать жизнеспособный трансплантат, было сделано предположение, что основное воздействие такая обработка оказывает на стволовые инициирующие раковые клетки, от которых зависит туморогенность графта. Полученные данные предполагали, что проведенные обработки разными препаратами двуцепочечной ДНК и при разных режимах введения могут приводить к одному и тому же результату - полной эрадикации всех стволовых инициирующих раковых клеток в обработанном асците или полному изменению их функционального ракового статуса, что сопровождается потерей или значительным снижением туморогенного потенциала перевиваемого графта.
Дадим характеристику некоторых свойств TAMRA+ асцитных клеток необработанного асцита и асцита после инъекций ЦФ. Определим эффект воздействия обработки ЦФ и двуцепочечной ДНК на TAMRA+/CD34+ асцитные клетки.
В настоящей части раздела были оценены некоторые биологические особенности TAMRA+ асцитных клеток в нативном асците и асците, обработанном ЦФ.
Анализируя состояние клеток по ходу развития асцита, было обнаружено, что TAMRA+ клетки сохраняли свой процент ориентировочно на одном уровне (около 3%) на протяжении всего развития асцита (до 15 суток). Для одной группы животных этот процент достоверно осциллировал в пределах 0.3-2%. Период осцилляции составлял 3 суток. Для другой группы мышей четкой осцилляции не обнаруживалось, а процент TAMRA+ клеток имел пик на 12 сутки. Для третьей группы мышей наблюдался провал в количестве и процентном отношении TAMRA+ клеток на 7 сутки развития асцита (фиг. 2, таблица 1).
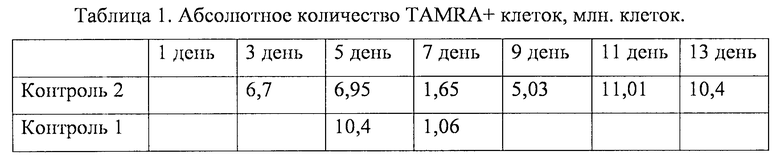
Показано, что асцит по мере старения набирает объем не прямолинейно. Общее число клеток асцита также изменяется без видимой закономерности и колебалось от 190 млн до ~500 млн клеток на протяжении развития асцита (до 15 суток). Для взятого в эксперимент животного на 3 сутки объем асцитной жидкости составлял 2 мл, а к 15 суткам увеличивался до 4.5 мл. Абсолютное количество TAMRA+ клеток менялось от 1 млн до 11 млн клеток и также не было связано видимой закономерностью (таблица 1).
В подготовительный период была проведена серия масштабных экспериментов, характеризующих влияние ЦФ как монопрепарата на асцит Кребс-2. Оказалось, что несколько последовательных инъекций ЦФ в определенные точки репаративного цикла репарации межцепочечных сшивок обладают ярко выраженным терапевтическим эффектом.
В литературе подробно, шаг за шагом, описано формирование и разрешение интермедиатов репарации межцепочечных сшивок [24-27] после инъекции ЦФ. Первоначально идет формирование двуцепочечных разрывов, затем фаза NER. Далее происходит процессинг оставшегося двуцепочечного конца и на последнем этапе осуществляется акт гомологичной рекомбинации (фиг. 1А, В). Для асцитных раковых клеток Кребс-2 эти фазы репарации разбиты на временные отрезки следующим образом. До 12 часов идет накопление двуцепочечных разрывов. Репарация двуцепочечных разрывов (фаза гомологичной рекомбинации) начинается в 24 часа и заканчивается в 36 часов после инъекции ЦФ. Между 12 и 24 часами происходят два события. А именно - осуществляется NER и формируется одноцепочечный участок двуцепочечного конца. Было сделано предположение, что повторение аналогичной терапевтической процедуры в фазу клеточного цикла (через 36 часов послед первой инъекции ЦФ), когда остаточные раковые клети, находящиеся при первой обработке в нечувствительной для них фазе клеточного цикла (G2/M), будут доступными (в следующей по порядку после первой обработки фазе G1/S) для действия ЦФ и одновременно, произойдет повторное запирание в поздней S фазе готовящихся выйти из нее клеток первой очереди воздействия, нанесет максимально эффективное деструктивное действие на раковые клетки, свободно расположенные в асцитном бульоне. Трехкратное повторение такой процедуры с большой долей вероятность охватит всю популяцию стволовых инициирующих раковых клеток.
Анализируя состояние клеток после обработки ЦФ, было установлено, что последовательные инъекции ЦФ в четко определенные временные точки приводят к аресту раковых клеток в поздней S/G2/M на 54 час эксперимента. На 90 час эксперимента четкое деление клеток по количеству хроматина исчезает, а клеточный материал уходит в зону апототического «мусора» (фиг. 3). К 7-9 суткам происходит полный коллапс всей массы асцитных раковых клеток (фиг. 3). В момент клеточного коллапса TAMRA+ клетки не обнаруживаются. Состояние коллапса продолжается несколько суток и сопровождается апототической деградацией клеток и визуально детектируемым и экспериментально подтвержденным уходом асцита. К 14 суткам состояние клеток восстанавливается до нормы, и их число возрастает до контрольного. Процент TAMRA+ клеток возвращается к норме, составляющей ~3%.
Одним из первостепенных вопросов, касающихся воздействия ЦФ и двуцепочечной ДНК на стволовые инициирующие раковые клетки, был вопрос эффективности элиминации стволовых инициирующих раковых клеток, захватывающих двуцепочечную ДНК и находящихся под воздействием ЦФ. Использование терапевтического количества двуцепочечной ДНК исключает возможность применения маркерной TAMRA ДНК, поскольку все свободное внутреннее пространство клеток будет занято терапевтическими фрагментами двуцепочечной ДНК, а применение одной TAMRA+ маркерной ДНК не будет обладать требуемым терапевтическим действием. Смесь таких ДНК не дает должного уровня чувствительности.
Были проведены эксперименты по оценке воздействия двух препаратов ЦФ и двуцепочечной ДНК на клетки асцита Кребс-2, способные интернализовать фрагменты двуцепочечной ДНК, с использованием дополнительного маркера этих клеток CD34. Было выявлено, что популяция клеток, способных интернализовать TAMRA-меченую ДНК, на 40% перекрывается с популяцией клеток, несущих маркер CD34 (причем от 40 до 90% CD34+ клеток являются TAMRA-позитивными). Если при проведенных обработках будет обнаружено снижение числа CD34+ клеток, то этот факт может с определенной долей вероятности означать, что произошла элиминация или TAMRA+ стволовых инициирующих раковых клеток или поверхностного CD34 маркера. Однако нельзя исключить возможность воздействия указанных обработок на CD34+ клетки, не способные захватывать TAMRA-меченую ДНК, что подразумевает относительность указанной оценки. Было проведено несколько экспериментов по оценке изменения числа CD34+ клеток в обработанном асците. Результаты с использованием человеческой двуцепочечной ДНК в режиме 1-12 часов после введения ЦФ представлены на фиг. 4.
Анализируя полученные результаты, можно сделать следующие выводы. К 48 часам обработка как ЦФ, так и его сочетанием с препаратом двуцепочечной ДНК человека приводит к снижению количества CD34+ клеток до значения 0.2% (фиг. 4С, ЦФ, ЦФ+ДНК). Полного исчезновения этого типа клеток не происходит. На отрезке времени 0-48 часов обнаружена разница в динамике изменения количества CD34+ клеток при обработке одним ЦФ и ЦФ совместно с двуцепочечной ДНК человека (фиг. 4А, В, С, ЦФ, ЦФ+ДНК). Можно отметить более выраженное снижение количества CD34+ клеток в случае обработки ЦФ и двуцепочечной ДНК по сравнению с действием одного ЦФ (фиг. 4D, ЦФ, ЦФ+ДНК). Такая картина могла означать, что более сильное воздействие на этот тип клеток оказывает совместная обработка ЦФ и двуцепочечной ДНК, и что клетки, интернализующие двуцепочечную ДНК и одновременно позитивные по CD34, более чувствительны к такому воздействию. Общий вывод следующий. Обработка асцита Кребс-2 препаратом ДНК человека в режиме 1-12 на фоне воздействия ЦФ элиминирует TAMRA+ клетки или меняет их функциональное состояние, что в обоих случаях сопровождается исчезновением клеток, несущих CD34 поверхностный маркер.
Таким образом, следуя полученным результатам по перевивке и анализу воздействия ЦФ на TAMRA+ стволовые инициирующие раковые клетки Кребс-2, можно сделать следующий вывод. ЦФ создает условия для терминальной эрадикации стволовых инициирующих раковых клеток из асцитной опухоли Кребс-2 дополнительными инъекциями препарата двуцепочечной ДНК.
Рассмотрим режим полного вылечивания экспериментальных мышей от асцита Кребс-2.
Согласно предположению, высказанному в первом разделе работы, точка 18 часов после введения ЦФ является пограничной точкой, разделяющей две фазы процесса репарации межцепочечных сшивок. До 18 часов идет и завершается стадия NER, с 18 и до 30 часов идет и завершается стадия гомологичной рекомбинации. Нативная двуцепочечная ДНК человека интерферирует NER, тогда как исковерканная ДНК лосося интерферирует стадию гомологичной рекомбинации. Было сделано предположение, что если в 18 часов после введения ЦФ однократно ввести препарат двуцепочечной ДНК, состоящий из смеси нативной ДНК человека и смеси кросс-линкированной и нативной ДНК лосося в определенной пропорции, то фрагменты нативной ДНК будут губительно для стволовых инициирующих раковых клеток интерферировать конечные этапы NER, а исковерканные фрагменты ДНК спермы лосося будут губительно для стволовых инициирующих раковых клеток интерферировать начальные этапы гомологичной рекомбинации. И, таким образом, можно полностью перекрыть все возможности для стволовых инициирующих раковых клеток корректно завершить репарацию межцепочечных сшивок или изменить их раковый статус, и тем самым элиминировать из асцита туморогенное начало опухоли. Повторение аналогичной процедуры после каждого введения ЦФ по схеме трех ЦФ охватит всю популяцию стволовых инициирующих раковых клеток. Проведенные эксперименты свидетельствовали, что такая терапия приводит к одновременному, акцидентальному, в течение 2-3 суток, уходу всей асцитной жидкости. Фаза ремиссии продолжается 7-9 суток. При такой терапии всегда наблюдалось возникновение вторичного асцита, что подробно обсуждается в цитируемых работах и связано с сохранением стволовых инициирующих раковых клеток в тканях, недоступных для терапевтической обработки (фиг. 5).
В клеточном сообществе асцитных раковых клеток существует две дискриминируемые по признаку интернализации TAMRA ДНК популяции клеток. Было показано, что TAMRA+ клетки обладают свойствами стволовых инициирующих раковых клеток. Было сделано предположение, что эти две популяции клеток завершают арест в поздней S/G2/M и связанную с арестом синхронизацию в этой фазе клеточного цикла по-разному. Коммутированные раковые клетки разрушаются апоптозом с последующим вторичным некрозом [19, 21]. Часть генетически экстремально пластичных стволовых инициирующих раковых клеток TAMRA+ выдерживают терапию и после завершения ареста в G2/M выходят из репаративного процесса и стремятся завершить деление. Происходит полная синхронизация всех выживших после проведенной терапии стволовых инициирующих раковых клеток, и они одновременно входят в первую S фазу после ареста в поздней S/G2/M предыдущего цикла. И именно эта точка может позволить нанести завершающий удар по всем оставшимся причинным стволовым инициирующим раковым клеткам Кребс-2. Через несколько суток произойдет естественное расхождение раковых клеток, в том числе и стволовых инициирующих раковых клеток, по фазам клеточного цикла и стволовые инициирующие раковые клетки станут не досягаемыми для однократной обработки по схеме разрабатываемого терапевтического режима (фиг. 5).
Было проведено два полномасштабных эксперимента для проверки предложенной гипотезы. Результаты проведенных экспериментов представлены на фиг. 6.
Оказалось, что предложенное решение позволяет полностью вылечить мышей от асцитной формы рака Кребс-2. И ЦФ, и комбинированное действие ЦФ и препарата двуцепочечной ДНК демонстрировали ярко выраженный терапевтический эффект на развитие асцита. Последняя обработка для первого эксперимента была осуществлена на 8 сутки от введения ЦФ. В первом эксперименте в группе ЦФ у 50% мышей асцит не развился к 32 суткам эксперимента. К 42 суткам эксперимента осталось 25% живых и без асцита животных. К 80 дню эксперимента последнее животное было с раковым асцитом и погибло в течение недели. В экспериментальной группе ЦФ+ДНК к 42 суткам эксперимента осталось живых 70% мышей, без асцита 60% животных. К 80 суткам эксперимента у 50% (4) животных экспериментальной группы вторичный асцит не проявился. Животные этого эксперимента продолжают оставаться живыми до настоящего времени (190 суток). Эксперимент был повторен. Дополнительная терапия была проведена на 12 сутки после первой инъекции ЦФ, то есть на четверо суток позже, чем в первом эксперименте. Такой подход позволил частично оценить продолжительность терминального исчерпывающего терапевтического отрезка времени. Результаты повторного эксперимента представлены на фиг. 6. Так же, как и в первом эксперименте, обе экспериментальные группы демонстрировали высокое терапевтическое действие. Во втором эксперименте в группе ЦФ к 50 суткам эксперимента 87.5% мышей были с асцитом или погибли. У большинства животных рецидив асцита произошел в промежуток времени 30-46 сутки эксперимента. К 80 суткам эксперимента осталась одна мышь (12.5%) без асцита. В экспериментальной группе ЦФ+ДНК к 50 суткам эксперимента 50% мышей погибли, остальные мыши (50%) продолжали жить без признаков заболевания до 90 суток наблюдения.
В обоих экспериментах была достигнута 50% выживаемость мышей для группы ЦФ+ДНК. Гибель животных в экспериментальных группах обуславливалась двумя причинами. Первое, это развитие вторичной опухоли. Второе - развитие неуправляемой полиорганной недостаточности [21, 23]. Из 50% погибших животных для первого эксперимента одна мышь погибла от полиорганной недостаточности без признаков асцита, три мыши погибли с развившимся вторичным асцитом. Для второго эксперимента две мыши погибли от полиорганной недостаточности без признаков асцита, два животных погибло с асцитом (фиг. 7).
Дадим общебиологическую оценку некоторым свойствам TAMRA+ стволовых инициирующих раковых клеток опухоли Кребс-2.
TAMRA+ клетки, как было определено, являются туморогенными раковыми клетками Кребс-2. В результате проведенных исследований установлено, что в среднем процент TAMRA+ клеток не превышает 3% с определенными колебаниями на протяжении всего времени развития асцита. Для одних животных процентное содержание этого типа клеток по ходу развития асцита осциллирует с периодом ~3 суток, при этом соотношение TAMRA+ клеток изменяется (для наиболее показательного животного) от 0.5% до 2.0% и обратно. Для другой группы, на 12 сутки развития асцита, % этих клеток резко возрастает до 7%, а затем также резко падает. Для третьей группы животных наблюдается ярко выраженный спад TAMRA+ клеток на 7 сутки развития асцита до значений 0.3%-0.8% и такой же выраженный подъем количества TAMRA+ клеток (Таблица 1). Сохранение на определенном уровне процента активно делящихся TAMRA+ клеток предполагает, что после определенного числа делений эти клетки теряют свою способность интернализовать TAMRA ДНК зонд. Это означает что, для поддержания статус-кво сообщества раковых клеток Кребс-2 требуется около 3% TAMRA+ стволовых инициирующих раковых клеток, и что на определенную массу раковых клеток должно присутствовать определенное количество «клеток-операторов». По-видимому, TAMRA+ клетки и являются такими операторами, поддерживающими гомеостаз асцитного сообщества клеток.
Известно, что стволовая инициирующая раковая клетка может делиться как симметрично, давая такую же плюрипотентную клетку, так и не симметрично, давая коммутированного потомка. Последняя, обладающая конечным, но значительным пролиферативным потенциалом, продуцирует множество дифференцированных раковых клеток [29-32].
Как следует из оценки динамики процентного соотношения TAMRA+ клеток, не все TAMRA позитивные клетки обладают одними и теми же пролиферативными характеристиками. Исходными, истинными стволовыми инициирующими раковыми клетками можно считать те TAMRA+ клетки и то их минимальное количество, составляющее менее 1% и 1 млн в численном выражении для различных животных, до которого падает их число в определенных точках наблюдения. Оставшиеся TAMRA+ клетки представляют собой комметированные потомки первого эшелона с высокой пролиферативной активностью. Именно они после определенного числа делений в очередной раз делятся, теряя свойство захватывать ДНК зонд, и переходят в статут дифференцированных клеток. Именно этот переход детектируется как резкое падение процентного содержания и абсолютного числа TAMRA+ клеток до минимального, а также наблюдаемые падения при осцилляции. Далее идет фаза несимметричного деления истинных стволовых инициирующих раковых клеток (0.3-0,8%), дающих прослойку TAMRA+ потомков первого эшелона комметирования, увеличивающая процент TAMRA+ клеток в асците до ~3%. Такое соотношение поддерживается на протяжении всего периода развития асцита. Можно полагать, что TAMRA+ стволовые инициирующие раковые клетки в процессе становления и первых дней развития асцита претерпевают серию симметричных делений, образуют необходимое базовое количество «клеток-операторов».
Подведем общий итог по охарактеризованной стратегии вылечивания асцита Кребс-2.
Что происходит при использовании режима 3+1 инъекции ЦФ+ДНК? Первые три инъекции ЦФ одна за другой уменьшают количество клеток, не подверженных воздействию ЦФ. Каждый раз при введении ЦФ в 36 часов после предыдущей инъекции ЦФ новая обработка попадает на время, когда часть популяции TAMRA интернализующих клеток, находившаяся в G2/M фазе клеточного цикла и не чувствительная к ЦФ, переходит в G1/S фазу и становится чувствительной к обработке цитостатитком. Одновременно каждая следующая инъекция ЦФ повторно запирает в поздней S/G2/M фазе готовящиеся выйти из нее клетки первой очереди воздействия, то есть очередная обработка ЦФ индуцирует новые кросс-линки в клетках, уже залечивших сшивки от предыдущей обработки, и не дает им перейти в митоз. Происходит полномасштабная синхронизация как комметированных раковых клеток, так и стволовых инициирующих раковых клеток Кребс-2 в поздней S/G2/M фазе. Одновременно при добавлении в реакцию ДНК в точку разделения двух репаративных фаз происходит интерференция репаративного процесса по двум векторам, NER и гомологичной рекомбинации. На 7-9 сутки после проведенных обработок детектируется тотальное масштабное разрушение асцитных раковых клеток. Комметированные клетки уходят в апоптоз. Часть генетически пластичных стволовых инициирующих раковых клеток, выживших при проведенной терапии, синхронно завершают деление и синхронно входят в первую после ареста S фазу клеточного цикла. Экспериментально было подтверждено предположение, что если в это время еще раз провести обработку ЦФ и ДНК в указанном режиме, то происходит полная эрадикация стволовых инициирующих раковых клеток и разрушение туморогенного начала опухоли. Без дополнительного вмешательства (иммунокоррекция, антисепсисная терапия) выживает 50% животных. При этом патоморфологический анализ свидетельствует, что у обработанных в указанном режиме мышей отсутствуют значимые патологии тканей и органов, что подразумевает возможность переноса данной терапии на человека.
Таким образом, охарактеризован режим полного вылечивания 50% экспериментальных животных от, как считается не излечимой, в высшей степени злокачественной асцитной формы рака Кребс-2. Охарактеризованы контрольные точки приложения терапевтических обработок, базирующиеся на общебиологических свойствах стволовых инициирующих раковых клеток Кребс-2, к которым относятся: 1) Способность стволовых инициирующих раковых клеток Кребс-2 интернализовать фрагменты двуцепочечной ДНК естественным механизмом интернализации. 2) Фрагменты двуцепочечной ДНК, доставленные во внутриклеточное пространство стволовых инициирующих раковых клеток, при репарации межцепочечных сшивок, интерферируют процесс репарации таким образом, что стволовая инициирующая раковая клетка или погибает, или лишается своего туморогенного статуса.
Факторы применимости терапии и контрольные точки приложения терапевтических обработок, определяющие возможность полного вылечивания мышей от асцитной формы рака Кребс-2.
1. Повторяемость инъекций ЦФ. Первые три инъекции ЦФ одна за другой уменьшают количество клеток, остающихся не подверженным воздействию ЦФ. Каждый раз при введении ЦФ в 36 часов после предыдущей инъекции ЦФ новая обработка попадает на время, когда часть популяции интернализующих ДНК TAMRA+ клеток, находившаяся в G2/M фазе клеточного цикла и не чувствительная к ЦФ, переходит в G1/S фазе и становится чувствительной к обработке цитостатиком. Одновременно повторная обработка повторно запирает уже обработанные асцитные клетки в той же поздней S/G2/M фазе.
2. Для полной эрадикации асцита необходимо проводить дополнительную обработку в рассматриваемом режиме в момент апоптотического разрушения комметированных потомков раковых клеток Кребс-2 и выхода стволовых инициирующих раковых клеток Кребс-2 синхронизованных в поздней S/G2/M фазе в S фазу следующего цикла деления.
3. При добавлении в реакцию сложнокомпозитной ДНК в точку демаркации NER и гомологичной рекомбинации (18 часов после любой инъекции ЦФ) происходит одновременная интерференция репаративного процесса по двум векторам, NER и гомологичной рекомбинации. В ходе трех обработок подавляющая часть популяции стволовых инициирующих раковых клеток погибает, опухоль в значительной степени теряет туморогенное начало. Композиция препарата двуцепочечной ДНК составлена таким образом, что одна составляющая препарата интерферирует NER, а вторая гомологичную рекомбинацию. Оба эффекта приводят к разрушению стволовых инициирующих раковых клеток.
Дополнительные факторы
1. В схемах применения эффективного режима в обязательном порядке должна присутствовать антибиотиковая терапия на протяжении всего времени лечения, а также особая диета, используемая при лечении сепсиса [33].
2. Для купирования последствий масштабного одновременного разрушения асцитных раковых клеток необходимо проводить терапевтические процедуры при развитом асците, характеризующимся объемом 1/20 от веса тела мыши, в котором содержится 200-300 млн асцитных раковых клеток.
3. Доза вводимого препарата должна составлять 6 мг препарата в 500 мкл физраствора на 1 мл асцитной жидкости.
Предложим генеральную схему регламента стратегии вылечивания асцитного рака на примере асцитной опухоли Кребс-2.
Для любой асцитной опухоли первоначально определяется наличие и процент TAMRA+ стволовых инициирующих раковых клеток. Далее анализируется репаративный цикл (если для человека, то асцит переносится в NOD/SKIDE мышей и там культивируется до определения параметров клеточного цикла). Находится точка демаркации NER и гомологичной рекомбинации.
Проводятся три повторных инъекции ЦФ в контрольные точки репаративного цикла репарации межцепочечных сшивок, арестующие все раковые клетки в поздней S/G2/M фазе клеточного цикла. По аресту в поздней S/G2/M определяется время синхронизации раковых клеток. Цитологически или биохимически (ЛДГ) анализируется завершение процесса разрушения раковых клеток. Выбирается день терминальной эрадикации стволовых инициирующих раковых клеток.
Поводится полный курс терапии, состоящий из следующих процедур. Проводится дополнительная обработка в определенную точку демаркации NER и гомологичной рекомбинации сложнокомпозиционным препаратом на основе двуцепочечной ДНК, одновременно интерферирующим процесс репарации NER и гомологичной рекомбинации, таким образом, что асцит теряет туморогенное начало. Проводится терминальная обработка ЦФ и препаратом двуцепочечной ДНК в указанном режиме в фазу апоптотической деструкции асцитных раковых клеток, когда стволовые инициирующие раковые клетки, избежавшие апоптотической смерти, синхронно переходят в следующего после одновременного ареста S фазу клеточного цикла.
Список использованных источников
1. Машковский М.Д. Лекарственные средства. М.: Медицина, 1984, часть 2, стр. 425-475.
2. Lapidot Т, Sirard С, Vormoor J, Murdoch В, Hoang Т, Caceres-Cortes J, Minden M, Paterson B, Caligiuri MA, Dick JE. A cell initiating human acute myeloid leukaemia after transplantation into SOD mice. Nature. 1994 Feb 17; 367(6464):645-8.
3. Bonnet D, Dick JE. Human acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cell. Nat Med. 1997 Jul; 3(7):730-7.
4. RU 2464974, C1, A61K 31/00, A61P 35/00, 27.10.2012.
5. Лайт P.У. Болезни плевры. M.: Медицина, 1986.
6. Блохин Н.Н., Переводчикова Н.И. Химиотерапия опухолевых заболеваний. М.: Медицина, 1984. 302 с.
7. Brenner DE. Intraperitoneal chemotherapy: a review. J Clin Oncol. 1986; 4 (7): 1135-1147.
8. Мещерякова Н.Г. Платиновые цитостатики в лечении опухолевых плевритов. Дисс. канд. мед. наук. М., 1993.
9. Бычков М.Б. Противоопухолевая химиотерапия. М.: Медицина, 1996.
10. Беспалов В.Г., Жабин А.А., Стуков А.Н., Беляева О.А., Муразов Я.Г., Семенов А.Л., Коньков С.А., Крылова И.М. Синергизм противоопухолевого действия диоксадэта и цисплатина на модели асцитной опухоли яичника. Сибирский онкологический журнал. 2013. №1 (55), стр. 42-46.
11. RU 2138259, C1, А61K 31/28, А61K 31/66, А61K 35/26, 27.09.1999.
12. Степура Л.А. Пролонгированная внутрибрюшинная неоадъювантная и адъювантная химиоиммунотерапия в комплексном лечении асцитной формы рака яичников III-IV стадии заболевания. Автореф. дис. канд. мед. наук. Ростов н/Д., 1998.
13. RU 2163152, С1, А61М 31/00, А61K 31/00, 20.02.2001.
14. http://is-med.com/publ/14-1-0-250
15. http://xreferat.com/66/643-1-sverhmalye-dozy-bol-shaya-zagadka-prirody.html
16. RU 2225228, С2, А61М 1/36, 10.02.2004.
17. RU 2542410, C1, А61K 31/675, А61K 31/711, А61Р 35/00, 20.02.2015.
18. Dolgova EV, Alyamkina ЕА, Efremov YR, Nikolin VP, Popova NA, Tyrinova TV, Kozel AV, Minkevich AM, Andrushkevich OM, Zavyalov EL, Romaschenko AV, Bayborodin SI, Taranov OS, Omigov VV, Shevela EY, Stupak VV, Mishinov SV, Rogachev VA, Proskurina AS, Mayorov VI, Shurdov MA, Ostanin AA, Chernykh ER, Bogachev SS. Identification of cancer stem cells and a strategy for their elimination. Cancer Biol Ther. 2014; 15(10): 1378-94.
19. Alyamkina EA, Nikolin VP, Popova NA, Minkevich AM, Kozel AV, Dolgova EV, Efremov YR, Bayborodin SI, Andrushkevich OM, Taranov OS, Omigov VV, Rogachev VA, Proskurina AS, Vereschagin EI, Kiseleva EV, Zhukova MV, Ostanin AA, Chernykh ER, Bogachev SS, Shurdov MA. Combination of cyclophosphamide and double-stranded DNA demonstrates synergistic toxicity against established xenografts. Cancer Cell Int. 2015; 15: 32. In press.
20. Parsons DF, Marko M, Braun SJ, Wansor KJ. Ascites tumor invasion of mouse peritoneum studied by high-voltage electron microscope stereoscopy. Cancer Res. 1982; 42(11): 4574-4583.
21. Поттер E.A., Долгова E.B., Минкевич A.M., Ефремов Я.Р., Таранов О.С., Омигов В.В., Николин В.П., Попова Н.А., Проскурина А.С., Верещагин Е.И., Козел А.В., Рогачев В.А., Петров Д.Б., Останин А.А., Черных Е.Р., Колчанов Н.А., Богачев С.С. Терапевтические эффекты воздействия циклофосфана, препаратов двуцепочечной ДНК и их сочетания на раковые клетки асцита Кребс-2 и различные формы трансплантатов. Часть 1. Вавиловский журнал генетики и селекции. 2015. В печати.
22. Dolgova EV, Proskurina AS, Nikolin VP, Popova NA, Alyamkina EA, Orishchenko KE, Rogachev VA, Efremov YR, Dubatolova TD, Prokopenko AV, Chernykh ER, Ostanin AA, Taranov OS, Omigov VV, Zagrebelniy SN, Bogachev SS, Shurdov MA. "Delayed death" phenomenon: A synergistic action of cyclophosphamide and exogenous DNA. Gene. 2012; 495: 134-145.
23. Поттер E.A., Долгова E.B., Минкевич A.M., Ефремов Я.Р., Таранов О.С., Омигов В.В., Николин В.П., Попова Н.А., Проскурина А.С., Верещагин Е.И., Козел А.В., Рогачев В.А., Петров Д.Б., Останин А.А., Черных Е.Р., Колчанов Н.А., Богачев С.С. Терапевтические эффекты воздействия циклофосфана, препаратов двуцепочечной ДНК и их сочетания на раковые клетки асцита Кребс-2 и различные формы трансплантатов. Часть 2. Вавиловский журнал генетики и селекции. 2015. В печати.
24. Долгова Е.В., Лихачева А.С., Орищенко К.Е., Алямкина Е.А., Богачев С.С., Шурдов М.А. Репарация межцепочечных сшивок молекулы ДНК. Информационный вестник ВОГиС. 2010. Т. 14, №2, стр. 332-356.
25.  М, Knipscheer Р, Enoiu М, Angelov Т, Sun J, Griffith JD, Ellenberger ТЕ,
М, Knipscheer Р, Enoiu М, Angelov Т, Sun J, Griffith JD, Ellenberger ТЕ,  OD, Walter JC. Mechanism of replication-coupled DNA interstrand crosslink repair. Cell. 2008; 134 (6): 969-980.
OD, Walter JC. Mechanism of replication-coupled DNA interstrand crosslink repair. Cell. 2008; 134 (6): 969-980.
26. Muniandy PA, Liu J, Majumdar A, Liu ST, Seidman MM. DNA interstrand crosslink repair in mammalian cells: step by step. Crit. Rev. Biochem. Mol. Biol. 2010; 45 (1): 23-49.
27. Deans AJ, West SC. DNA interstrand crosslink repair and cancer. Nat. Rev. Cancer. 2011; 11 (7): 467-480.
28. Долгова E.B., Поттер E.A., Ефремов Я.Р., Николин В.П., Попова Н.А., Проскурина А.С., Минкевич A.M., Козел А.В., Рогачев В.А., Петров Д.Б., Останин А.А., Черных Е.Р., Колчанов Н.А., Богачев С.С. Характеристика свойств факторов интернализации различных типов экстраклеточной ДНК в стволовые инициирующие раковые клетки опухоли мыши Кребс-2. 2015. В печати.
29. Lathia JD, Hitomi М, Gallagher J, Gadani SP, Adkins J, Vasanji A, Liu L, Eyler CE, Heddleston JM, Wu Q, Minhas S, Soeda A, Hoeppner DJ, Ravin R, McKay RDG, McLendon RE, Corbeil D, Chenn A, Hjelmeland AB, Park DM, Rich JN. Distribution of CD 133 reveals glioma stem cells self-renew through symmetric and asymmetric cell divisions. Cell Death Dis. 2011; 2(9): e200.
30.  S, Lerner RG, Petritsch C. Asymmetric cell division of stem and progenitor cells during homeostasis and cancer. Cell Mol Life Sci. 2014; 71(4): 575-597.
S, Lerner RG, Petritsch C. Asymmetric cell division of stem and progenitor cells during homeostasis and cancer. Cell Mol Life Sci. 2014; 71(4): 575-597.
31. Pine SR, Liu W. Asymmetric cell division and template DNA co-segregation in cancer stem cells. Front Oncol. 2014; 4: 226.
32. Deleyrolle LP, Ericksson G, Morrison В J, Lopez JA, Burrage K, Burrage P, Vescovi A, Rietze RL, Reynolds BA. Determination of somatic and cancer stem cell self-renewing symmetric division rate using sphere assays. PLoS One. 2011; 6(1): el5844.
33. Беркасова И.В., Верещагин Е.И., Чикинев Ю.В. Влияние глутамина на обмен нуклеиновых кислот в периоперационном периоде в реконструктивной хирургии пищевода. 2015. Подготовка к печати.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ЭРАДИКАЦИИ СТВОЛОВЫХ ИНИЦИИРУЮЩИХ РАКОВЫХ КЛЕТОК | 2013 |
|
RU2542410C1 |
| СПОСОБ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2017 |
|
RU2662354C1 |
| СПОСОБ ДЕТЕКЦИИ СТВОЛОВЫХ РАКОВЫХ КЛЕТОК | 2013 |
|
RU2530170C1 |
| СПОСОБ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2009 |
|
RU2429019C2 |
| СПОСОБ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2018 |
|
RU2675233C1 |
| Способ лечения распространенного рака молочной железы в терминальной стадии прогрессии | 2024 |
|
RU2837589C1 |
| СПОСОБ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2008 |
|
RU2387456C1 |
| СПОСОБ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2010 |
|
RU2489169C2 |
| СПОСОБ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2007 |
|
RU2345792C2 |
| СПОСОБ ТОРМОЖЕНИЯ РОСТА ПОДКОЖНОГО ТРАНСПЛАНТАТА ЭКСПЕРИМЕНТАЛЬНОЙ ГЛИОБЛАСТОМЫ ЧЕЛОВЕКА U-87, ПЕРЕВИТОГО ИММУНОДЕФИЦИТНЫМ МЫШАМ NU/J | 2019 |
|
RU2717218C1 |

Изобретение относится к медицине, а именно к онкологии, и может быть использовано для лечения асцитной формы рака. Для этого мышам-опухоленосам линии СВА весом 20-35 г, несущим развитый асцит, парентерально вводят цитостатик циклофосфан в 0, 36, 72 часа каждый раз в дозе 100 мг/кг, а через 18, 54, 90 часов после каждой инъекции циклофосфана, соответственно. Также мышам-опухоленосам вводят интраперитонеально препарат ДНК, состоящий из композиции препаратов нативной двуцепочечной ДНК человека в дозе 3.0 мг и смеси нативной и кросс-линкированной двуцепочечной ДНК спермы лосося в соотношении 5:3 в дозе 3 мг суммарно в количестве 6.0 мг на инъекцию, независимо от веса мышей-опухоленосов. Изобретение обеспечивает повышение эффективности лечения асцитной формы рака, заключающееся в предотвращении появления вторичного асцита за счет разработанной схемы введения препаратов ЦФ и ДНК. 2 з.п. ф-лы, 1 табл., 7 ил.
1. Способ лечения асцитной формы рака, основанный на совместном воздействии цитостатика циклофосфана и препарата двуцепочечной ДНК, отличающийся тем, что мышам-опухоленосам линии СВА весом 20-35 г, несущим развитый асцит, парентерально вводят цитостатик циклофосфан в 0, 36, 72 часа каждый раз в дозе 100 мг/кг, а через 18, 54, 90 часов после каждой инъекции циклофосфана, соответственно, мышам-опухоленосам вводят интраперитонеально препарат ДНК, состоящий из композиции препаратов нативной двуцепочечной ДНК человека в дозе 3.0 мг и смеси нативной и кросс-линкированной двуцепочечной ДНК спермы лосося в соотношении 5:3 в дозе 3 мг суммарно в количестве 6.0 мг на инъекцию, независимо от веса мышей-опухоленосов.
2. Способ по п. 1, отличающийся тем, что для достижения максимального терапевтического действия на восьмые сутки после первой инъекции циклофосфана вводят дополнительно циклофосфан в дозе 100 мг/кг и через 18 часов препарат ДНК суммарно в количестве 6 мг.
3. Способ по п. 1, отличающийся тем, что интраперитонеально препарат ДНК в количестве 6 мг на инъекцию вводят однократно в объеме 500 мкл из расчета половины объема четырехсуточного асцита, соответствующего 1 мл и 1/20 веса животного, содержащего ~200 млн раковых клеток.
| СПОСОБ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2010 |
|
RU2489169C2 |
| СПОСОБ ЛЕЧЕНИЯ БОЛЬНЫХ РАСПРОСТРАНЕННЫМ РАКОМ ЯИЧНИКОВ | 2007 |
|
RU2352341C1 |
| ALYAMKINA E.A | |||
| et al | |||
| Combined therapy with cyclophosphamide and DNA preparation inhibits the tumor growth in mice | |||
| Genet | |||
| Колосоуборка | 1923 |
|
SU2009A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| BESPALOV V.G., et al | |||
| Synergistic antitumor effect of dioxodate and cisplatin on the model of ovarian ascites | |||

