Изобретение относится к области биотехнологии и может быть использовано для осуществления исчерпывающего сайт-специфического гидролиза фаговой, плазмидной или геномной ДНК по нуклеотидным последовательностям, который невозможно получить с применением традиционно используемых эндонуклеаз рестрикции.
При проведении эпигенетического анализа ДНК, например, при сравнении статуса метилирования ДНК малигнантных и здоровых клеток человека анализируют преимущественно сайты узнавания метилзависимых сайт-специфических ДНК-эндонуклеаз (MD-эндонуклеаз), которые расщепляются с низкой эффективностью, что существенно ограничивает возможности использования данных ферментов.
В настоящее время описано чуть более 350 прототипов эндонуклеаз рестрикции (рестриктаз), отличающихся друг от друга узнаваемой нуклеотидной последовательностью (1). Для некоторых сайтов узнавания известны рестриктазы, обладающие способностью расщеплять в разных позициях гидролиза относительно сайта узнавания - гетерошизомеров. Однако по-прежнему остается актуальной задача дальнейшего расширения спектра последовательностей, которые можно избирательно гидролизовать на двухцепочечной ДНК, что важно для молекулярной биологии, генной инженерии, биотехнологии и медицины.
Известен способ сайт-специфического гидролиза С5 метилированной ДНК, включающий обработку ДНК сайт-специфической ДНК-эндонуклеазой (обычно эндонуклеазой рестрикции II типа) в реакционном буфере, содержащем 3-10 мМ Mg2+ (2). Данный способ позволяет осуществлять сайт-специфический гидролиз ДНК примерно по 360-380 последовательностям длиной от 3 до 10 п.н.
Однако данным способом невозможно осуществить гидролиз ДНК по целому ряду протяженных последовательностей длиной 6-8 п.н. из-за отсутствия MD-эндонукулеаз (из природных или рекомбинантных бактериальных штаммов-продуцентов), узнающих и сайт-специфично расщепляющих эти последовательности.
Наиболее близким аналогом к заявляемому способу - прототипом - является способ сайт-специфического гидролиза ДНК (3), включающий подготовку реакционной смеси, содержащей буферный раствор и образец исследуемой ДНК, метилирование исследуемой ДНК путем добавления к смеси адениновой ДНК-метилтрансферазы (метилазы) и инкубирования смеси в течение часа, обработку полученной Ν6-метилированной ДНК MD-эндонуклеазой DpnI, расщепляющей ее по образовавшимся в результате метилирования последовательностям 5′-G(m6A)TC-3′, с последующей остановкой реакции инактивированием ферментов при температуре 70°С и анализом результата гидролиза ДНК с помощью гель-электрофореза. Известный способ позволяет гидролизовать только два сайта: 5′-TCGATCGA-3′ и 5′-ATCGATCGAT-3′.
Недостатками данного способа являются:
1) ограниченные функциональные возможности способа, поскольку он позволяет гидролизовать только два сайта гидролиза ДНК с применением небольшого набора адениновых метилаз;
2) неполнота гидролиза по сайтам 5′-GATC-3′ в случае, если аденин метилирован только на одной из цепей ДНК;
3) отсутствие возможности определения разных вариантов позиций гидролиза из-за использования лишь одной рестриктазы DpnI с фиксированной позицией гидролиза относительно сайта узнавания.
Задачей изобретения является расширение функциональных возможностей известного способа.
Технический результат: осуществление исчерпывающего сайт-специфического гидролиза ДНК по шести- и восьминуклеотидным последовательностям, не расщепляемым традиционно используемыми эндонуклеазами рестрикции.
Поставленная задача достигается предлагаемым способом сайт-специфического гидролиза ДНК, заключающемся в следующем.
Готовят реакционную смесь, содержащую буферный раствор, содержащий 10 мМ Трис-HCl (рН 7,5 при 25°С), 0,1 мМ S-аденозил-L-метионин, 1 мМ дитиотреитол (ДТТ) и 1 мкг образца ДНК. Для метилирования образца исследуемой ДНК к смеси добавляют 1 мкл препарата ДНК-метилтрансферазы с активностью 2 е.а./мкл и смесь инкубируют в течение 1 часа при комнатной температуре. В зависимости от того, какой сайт необходимо расщепить, ДНК-метилтраснферазу выбирают из группы: Fsp4HI, НаеIII, HspAI, AluI. Для сайт-специфического гидролиза полученной С5-метилированной ДНК в смесь добавляют воду и 10-кратный реакционный SE-буфер «Y» (330 мМ Трис-ацетат (рН 7,9 при 25°С), 100 мМ магния ацетат, 660 мМ калия ацетат, 10 мМ ДТТ). Полученную реакционную смесь тщательно перемешивают, после чего добавляют к ней препарат MD-эндонуклеазы PkrI (4) с активностью 2 е.а./мкл или препарат MD-эндонуклеазы ElmI с активностью 2 е.а./мкл в зависимости от того, в какой позиции необходимо гидролизовать ДНК, и повторно инкубируют при температуре 37°С в течение 1 ч с последующей остановкой реакции инактивированием ферментов при температуре 70°С в течение 5 минут и анализом результата гидролиза С5-метилированной последовательности ДНК с помощью гель-электрофореза (5).
Конкретное количество реагентов и объем реакционной смеси зависят от количества исследуемого образца ДНК, которую необходимо подвергнуть исчерпывающему сайт-специфическому гидролизу.
Предлагаемый способ позволяет провести исчерпывающий сайт-специфический гидролиз по трем последовательностям нуклеотидов: 5′-GCNGCNGC-3′, 5′-GGCCGCGC-3′ и 5′-AGCTGCGC-3′, причем каждая из последовательностей может быть отдельно гидролизована в двух различных позициях в зависимости от того, какая из MD-эндонуклеаз используется - PkrI или ElmI:
1) 5′-GCN↓GCN↓GC-3′
3′-CG↑NCG↑NCG -5′
2) 5′-GC↓NGC↓NGC-3′
3′-CGN↑CGN↑CG -5′
3) 5′-GGC↓CGCGC-3′
3′-CCGG↑CGCG -5′
4) 5′-GGCC↓GCGC-3′
3′-CCG↑GCGCG -5′
5) 5′-AGC↓TGCGC-3′
3′-TCGA↑CGCG -5′
6) 5′-AGCT↓GCGC-3′
3′-TCG↑ACGCG -5′
Определяющими отличиями заявляемого способа по сравнению с прототипом являются:
1. Для метилирования образца ДНК используют ДНК-метилтрансферазу, модифицирующую не аденин в положении N6, а цитозин в положении С5, что обеспечивает 100% метилирование ДНК по сайтам узнавания ДНК-метилтрансфераз и образование сайтов узнавания MD-эндонуклеаз, с помощью которых осуществляется гидролиз ДНК.
2. По окончании метилирования ДНК в реакционную смесь добавляют препарат MD-эндонуклеазы, предназначенной для сайт-специфического гидролиза, при этом в зависимости от того, какая позиция гидролиза относительно сайта узнавания является предпочтительной, выбирают одну из MD-эндонуклеаз - PkrI или ElmI, позволяющих расщеплять последовательность ДНК: 5′-GCNGC-3′ перед (для ElmI) или после (для PkrI) центрального нуклеотида.
Таким образом, предлагаемый способ обеспечивает сайт-специфический гидролиз ДНК по позициям, не расщепляемым традиционно используемыми эндонуклеазами рестрикции.
При использовании способа-прототипа ни одна из традиционно используемых для этой цели эндонуклеаз рестрикции не способна расщеплять ДНК по вышеуказанным последовательностям, что ограничивает применение этих ферментов в молекулярной биологии, генной инженерии и медицине.
Изобретение иллюстрируется следующими примерами конкретного выполнения.
Пример 1. Сайт-специфический гидролиз ДНК плазмиды pUC19 по последовательности 5′-GCNGCNGC-3′ с помощью MD-эндонуклеазы PkrI
Создание сайта узнавания PkrI и ElmI возможно при метилировании метилазой Fsp4HI. В этом случае образуется разнесенная палиндромная шестинуклеотидная метилированная последовательность 5′-G(5mC)NG(5mC)NGC-3′/3′-CGN(5mC)GN(5mC)G(5mC)G-5′, которая представляет собой два метилированных пересекающихся сайта узнавания PkrI и ElmI (две пересекающихся последовательности 5′-GCNGC-3′, содержащих на обеих цепях по три 5-метилцитозиновых остатка).
В качестве объекта для проверки возможности сайт-специфического расщепления по сайту 5′-GCNGCNGC-3′ выбиралась ДНК плазмиды pUC19, содержащая две таких последовательности.
На фиг. 1 представлено сравнение теоретически рассчитанной с помощью программы Vector NTI картины гидролиза ДНК pUC19 по разнесенной палиндромной шестинуклеотидной последовательности 5′-GCNGCNGC-3′ с экспериментальными данными и приведена картина расщепления ДНК pUC19, метилированной метилазой Fsp4HI, с помощью PkrI после проведения электрофореза в 1% агарозном геле, где дорожки:
1 - исходная ДНК pUC19 (кольцевая форма);
2 - ДНК pUC19, метилированная ДНК-метилтрансферазой Fsp4HI и обработанная MD-эндонуклеазой PkrI;
М - маркер молекулярного веса ДНК 1 kb.
Справа - теоретически смоделированная (в программе Vector NTI) картина расщепления ДНК pUC19 по сайту 5′-GCNGCNGC-3′.
Как видно из фиг. 1, теоретически рассчитанная картина расщепления ДНК и результаты эксперимента совпадают.
Таким образом, полученные результаты свидетельствуют о возможности получения полного (100%) сайт-специфического гидролиза ДНК по шестинуклеотидной последовательности 5′-GCNGCNGC-3′ с помощью предварительного метилирования ДНК метилазой Fsp4HI и последующим расщеплением ее MD-эндонуклеазой PkrI.
Пример 2. Сайт-специфический гидролиз ДНК аденовируса типа 2 по последовательности 5′-GGCCGCGC-3′ с помощью эндонуклеазы ElmI
Создание сайта узнавания PkrI и ElmI возможно при совместном метилировании метилазами НаеIII и HspAI. В этом случае образуется непалиндромная восьминуклеотидная метилированная последовательность
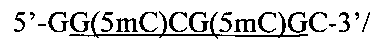
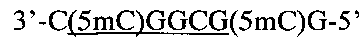
внутри которой содержится сайт узнавания PkrI и ElmI, содержащий три 5-метилцитозиновых остатка (подчеркнут).
Удобным объектом для проверки возможности сайт-специфического расщепления по сайту 5′-GGCCGCGC-3′ является ДНК аденовируса типа 2 - Ad2 (производство «New England Biolabs», США), так как она содержит 7 таких сайтов.
На фиг. 2 представлено сравнение теоретически рассчитанной с помощью программы Vector NTI картины гидролиза аденовирусной ДНК по непалиндромной восьминуклеотидной последовательности 5′-GGCCGCGC-3′ с экспериментальными данными и приведена картина расщепления метилированной метилазами НаеIII и HspAI аденовирусной ДНК с помощью ElmI после проведения электрофореза в 1% агарозном геле, где дорожки:
1 - ДНК Аd2;
2 - ДНК Ad2, метилированная ДНК-метилтрансферазами НаеIII и HspAI и обработанная MD-эндонуклеазой ElmI;
М - маркер молекулярного веса ДНК 1 kb.
Справа - теоретически смоделированная (в программе Vector NTI) картина расщепления ДНК фага лямбда по сайту 5′-GGCCGCGC-3′.
Как видно из фиг. 2, теоретически рассчитанная картина расщепления ДНК и результаты эксперимента совпадают.
Таким образом, полученные результаты свидетельствуют о возможности получения полного (100%) сайт-специфического гидролиза ДНК по восьминуклеотидной последовательности 5′-GGCCGCGC-3′ с помощью предварительного метилирования ДНК метилазами НаеIII и HspAI и последующим расщеплением ее MD-эндонуклеазой ElmI.
Пример 3. Сайт-специфический гидролиз ДНК фага лямбда по последовательности 5′-AGCTGCGC-3′ с помощью эндонуклеазы ElmI
Создание сайта узнавания PkrI и ElmI возможно при совместном метилировании метилазами AluI и HspAI. В этом случае образуется непалиндромная восьминуклеотидная метилированная последовательность:
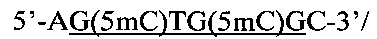
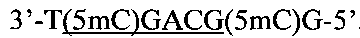
внутри которой содержится сайт узнавания ElmI и PkrI, содержащий три 5-метилцитозиновых остатка (подчеркнут).
Удобным объектом для проверки возможности сайт-специфического расщепления по сайту 5′-AGCTGCGC-3′ является ДНК фага лямбда, так как она содержит 7 таких сайтов.
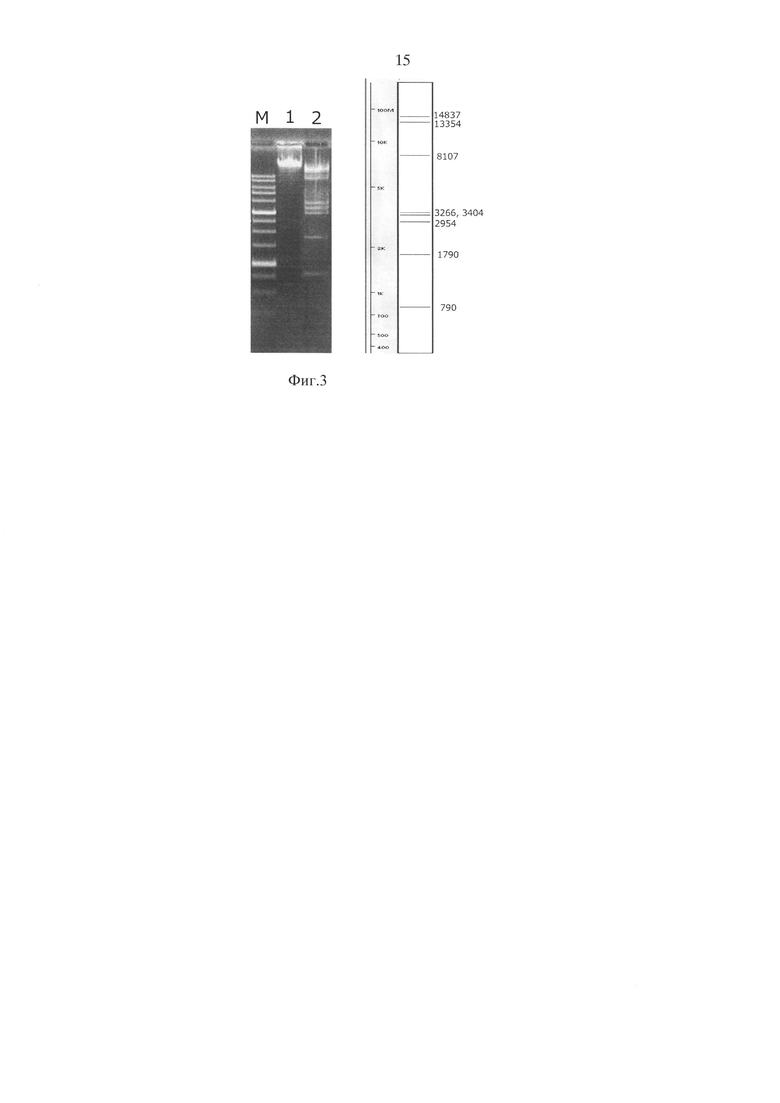
На фиг. 3 представлено сравнение теоретически рассчитанной с помощью программы Vector NTI картины гидролиза ДНК фага лямбда по непалиндромной восьминуклеотидной последовательности 5′-AGCTGCGC-3′ с экспериментальными данными и приведена картина расщепления метилированной метилазами AluI и HspAI ДНК фага лямбда с помощью ElmI после проведения электрофореза в 1% агарозном геле, где дорожки:
1 - исходная ДНК фага лямбда;
2 - ДНК фага лямбда, метилированная ДНК-метилтрансферазами AluI и HspAI и обработанная MD-эндонуклеазой ElmI;
M - маркер молекулярного веса ДНК 1 kb.
Справа - теоретически смоделированная (в программе Vector NTI) картина расщепления ДНК фага лямбда по сайту 5′-AGCTGCGC-3′.
Как видно из фиг. 3, теоретически рассчитанная картина расщепления ДНК и результаты эксперимента совпадают.
Таким образом, полученные результаты свидетельствуют о возможности получения полного (100%) сайт-специфического гидролиза ДНК по восьминуклеотидной последовательности 5′-AGCTGCGC-3′ с помощью предварительного метилирования ДНК метилазами AluI и HspAI и последующим расщеплением ее MD-эндонуклеазой ElmI.
Пример 4. Осуществление сайт-специфического гидролиза по последовательности 5′-GCNGCNGC-3′
Все расчеты количества используемых реагентов приведены для объема реакционной смеси 50 мкл, но могут быть масштабированы.
К 10 мкл реакционной смеси, содержащей буферный раствор, содержащий 10 мМ Трис-HCl (рН 7,5 при 25°С), 0,1 мМ 5-аденозил-L-метионин, 1 мМ ДТТ и 1 мкг образца гидролизуемой ДНК, добавляли 1 мкл препарата ДНК-метилтрансферазы Fsp4HI с активностью 2 е.а./мкл. Реакционную смесь инкубировали в течение 1 часа при комнатной температуре (24-30°С), после чего в смесь добавляли 36 мкл воды и 4 мкл 10-кратного реакционного SE-буфера «Y» (330 мМ Трис-ацетат (рН 7,9 при 25°С), 100 мМ магния ацетат, 660 мМ калия ацетат, 10 мМ ДТТ). Полученную реакционную смесь объемом 50 мкл тщательно перемешивали, после чего добавляли 2 мкл препарата MD-эндонуклеазы PkrI с активностью 2 е.а./мкл - для получения гидролиза в позиции после центральных нуклеотидов N -
5′-GCN↓GCN↓GC-3′
3′-CG↑NCG↑NCG -5′
или добавляли 2 мкл препарата MD-эндонуклеазы ElmI с активностью 2 е.а./мкл - для получения гидролиза перед центральными нуклеотидами N -
5′-GC↓NGC↓NGC-3′
3′-CGN↑CGN↑CG -5′
Полученную реакционную смесь повторно инкубировали при температуре 37°С в течение 1 часа с последующей остановкой реакции инактивированием ферментов при температуре 70°С в течение 5 минут. Результаты гидролиза анализировали с помощью электрофореза в 1% агарозном геле.
Пример 5. Осуществление сайт-специфического гидролиза по последовательности 5′-GGCCGCGC-3′
Все расчеты количества используемых реагентов приведены для объема реакционной смеси 50 мкл.
К 10 мкл буферного раствора, содержащего 10 мМ Трис-HCl (рН 7,5 при 25°С), 0,1 мМ S-аденозил-L-метионин, 1 мМ ДТТ и 1 мкг образца гидролизуемой ДНК, добавляли 0,5 мкл препарата ДНК-метилтрансферазы НаеIII с активностью 2 е.а./мкл и 0,5 мкл ДНК-метилтрансферазы HspAI с активностью 2 е.а./мкл.
Реакционную смесь инкубировали в течение 1 часа при 37°С, после чего в смесь добавляли 36 мкл воды и 4 мкл 10-кратного реакционного SE-буфера «Y» (330 мМ Трис-ацетат (рН 7,9 при 25°С), 100 мМ магния ацетат, 660 мМ калия ацетат, 10 мМ ДТТ). Полученную реакционную смесь объемом 50 мкл тщательно перемешивали пипеткой, после чего для гидролиза в позиции после третьего нуклеотида на верней цепи и после четвертого нуклеотида на нижней цепи -
5′-GGC↓CGCGC-3′
3′-CCGG↑CGCG -5′
добавляли 2 мкл препарата MD-эндонуклеазы ElmI с активностью 2 е.а./мкл, а для получения гидролиза в последовательности
5′-GGCC↓GCGC-3′
3′-CCG↑GCGCG -5′
в позиции, указанной стрелочками, добавляли 2 мкл препарата MD-эндонуклеазы PkrI с активностью 2 е.а./мкл.
Полученную реакционную смесь повторно инкубировали при температуре 37°С в течение 1 часа с последующей остановкой реакции инактивированием ферментов при температуре 70°С в течение 5 минут. Результаты гидролиза анализировали с помощью электрофореза в 1% агарозном геле.
Пример 6. Осуществление сайт-специфического гидролиза по последовательности 5′-AGCTGCGC-3′
Все расчеты количества используемых реагентов приведены для объема реакционной смеси 50 мкл.
К 10 мкл буферного раствора, содержащего 10 мМ Трис-HCl (рН 7,5 при 25°С), 0,1 мМ S-аденозил-L-метионин, 1 мМ ДТТ и 1 мкг образца гидролизуемой ДНК, добавляли 0,5 мкл препарата ДНК-метилтрансферазы AluI с активностью 2 е.а./мкл и 0,5 мкл ДНК-метилтрансферазы HspAI с активностью 2 е.а./мкл. Реакционную смесь инкубировали в течение 1 часа при 37°С, после чего в смесь добавляли 36 мкл воды и 4 мкл 10-кратного реакционного SE-буфера «Y» (330 мМ Трис-ацетат (рН 7,9 при 25°С), 100 мМ магния ацетат, 660 мМ калия ацетат, 10 мМ ДТТ). Полученную реакционную смесь объемом 50 мкл тщательно перемешивали пипеткой, после чего для получения гидролиза в позиции после третьего нуклеотида на верхней цепи и после четвертого нуклеотида на нижней цепи -
5′-AGC↓TGCGC-3′
3′-TCGA↑CGCG -5′
добавляли 2 мкл препарата MD-эндонуклеазы ElmI с активностью 2 е.а./мкл, а для получения гидролиза в последовательности
5′-AGCT↓GCGC-3′
3′-TCG↑ACGCG -5′
в позиции, указанной стрелочками, добавляли 2 мкл препарата MD-эндонуклеазы PkrI с активностью 2 е.а./мкл.
Полученную реакционную смесь повторно инкубировали при температуре 37°С в течение 1 часа. По окончании инкубации все ферменты в реакционной смеси инактивировали инкубацией пробирки с реакционной смесью при температуре 70°С в течение 5 минут. Результаты гидролиза анализировали с помощью электрофореза в 1% агарозном геле.
Предлагаемый способ обеспечивает возможность получить исчерпывающий гидролиз ДНК, предварительно метилированной подобранными для этих целей ДНК-метилтрансферазами, а затем обработанной MD-эндонуклеазой PkrI или ElmI, каждая из которых обеспечивает гидролиз ДНК по шести- и восьминуклеотидным последовательностям, не расщепляемым традиционно используемыми эндонуклеазами рестрикции.
ИСТОЧНИКИ ИНФОРМАЦИИ
1. Pingoud Α., Wilson G.G., Wende W.Type II restriction endonucleases - a historical perspective and more // Nucleic Acids Res. - 2014. - V. 42. - P. 7489-527.
2. Pingoud Α., Jeltsch A. Structure and function of type II restriction endonucleases // Nucleic Acids Research. - 2001. - V. 29. - P. 705-3727
3. McClelland M, Kessler LG, Bittner M. Site-specific cleavage of DNA at 8- and 10-base-pair sequences // Proc. Natl. Acad. Sci. USA. - 1984. - V.81. - P. 983-987.
4. Чернухин Β.Α., Наякшина Т.Н, Гончар Д.Α., Томилова Ю.Э., Тарасова М.В., Дедков B.C., Михненкова Н.А., Дегтярёв С.Х. Новая сайт-специфическая метилзависимая ДНК эндонуклеаза PkrI узнает и расщепляет метилированную последовательность 5`-GCN∧AGC-3`/3`-CG∧NCG-5`, содержащую не менее 3-х 5-метилцитозинов // Вестник биотехнологии и физико-химической биологии имени Ю.А. Овчинникова. - 2011. - Т. 7. - №3, С. 35-42.
5. Maniatis Т., Fritsch E.F. and Sambrook J. Molecular cloning: A Laboratory Manual. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press,1982. 545 pages.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНЫЙ ШТАММ БАКТЕРИЙ Escherichia coli N42 (pElmI) - ПРОДУЦЕНТ МЕТИЛЗАВИСИМОЙ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ ElmI | 2015 |
|
RU2597987C1 |
| ШТАММ БАКТЕРИЙ Plantibacter flavus 3Kz - ПРОДУЦЕНТ МЕТИЛЗАВИСИМОЙ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ PfsI | 2015 |
|
RU2593723C1 |
| Штамм бактерий Micrococcus luteus 805 - продуцент сайт-специфической метилзависимой эндонуклеазы MluVI | 2016 |
|
RU2614262C1 |
| ШТАММ БАКТЕРИЙ Planomicrobium koreense 78K - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ PkrI | 2011 |
|
RU2475534C1 |
| СПОСОБ САЙТ-СПЕЦИФИЧЕСКОГО ГИДРОЛИЗА С5-МЕТИЛИРОВАННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ ДНК | 2015 |
|
RU2597985C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ БАКТЕРИЙ ESCHERICHIA COLI N16 (PM.ALUBI) - ПРОДУЦЕНТ ДНК-МЕТИЛТРАНСФЕРАЗЫ M.ALUBI | 2015 |
|
RU2603086C1 |
| ШТАММ БАКТЕРИЙ GLACIAL ICE BACTERIUM - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ GLU I | 2006 |
|
RU2322492C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ НУКЛЕОТИДНОЙ ПОСЛЕДОВАТЕЛЬНОСТИ Pu(5mC)GPy В ЗАДАННОМ ПОЛОЖЕНИИ ПРОТЯЖЕННОЙ ДНК | 2013 |
|
RU2525710C1 |
| ШТАММ БАКТЕРИЙ Arthrobacter luteus B-ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ Alu BI | 2007 |
|
RU2340670C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ГИПЕРМЕТИЛИРОВАННЫХ CpG ОСТРОВКОВ В ОБЛАСТИ ГЕНОВ-СУПРЕССОРОВ ОПУХОЛЕВОГО РОСТА В ДНК ЧЕЛОВЕКА | 2009 |
|
RU2413773C1 |

Изобретение относится к области биотехнологии. Способ предусматривает приготовление реакционной смеси, содержащей буферный раствор и исследуемый образец ДНК. Добавляют к смеси ДНК-метилтрансферазу с активностью 2 е.а./мкл, модифицирующую цитозин в положении С5, выбранную из группы: Fsp4HI, НаеIII, HspAI, AluI, в зависимости от того, какой сайт необходимо расщепить, смесь инкубируют в течение часа, полученную С5-метилированную ДНК обрабатывают MD-эндонуклеазой ElmI с активностью 2 е.а./мкл или PkrI с активностью 2 е.а./мкл в зависимости от того, в какой позиции необходимо гидролизовать ДНК. С помощью данного способа можно осуществить исчерпывающий сайт-специфический гидролиз ДНК по шести- и восьминуклеотидным последовательностям, не расщепляемым традиционно используемыми эндонуклеазами рестрикции. 2 з.п. ф-лы, 3 ил., 6 пр.
1. Способ сайт-специфического гидролиза ДНК, включающий подготовку реакционной смеси, содержащей буферный раствор и образец исследуемой ДНК, метилирование ДНК путем добавления к смеси препарата ДНК-метилтрансферазы и инкубирования смеси в течение часа при комнатной температуре, добавление в смесь реакционного буфера и препарата MD-эндонуклеазы и повторное инкубирование полученной реакционной смеси при температуре 37°С в течение 1 ч с последующей остановкой реакции инактивированием ферментов при температуре 70°С и анализом результата гидролиза ДНК с помощью гель-электрофореза, отличающийся тем, что для метилирования образца ДНК используют ДНК-метилтрансферазу с активностью 2 е.а./мкл, модифицирующую цитозин в положении С5, выбранную из группы: Fsp4HI, HaeIII, HspAI, AluI, в зависимости от того, какой сайт необходимо расщепить, а для сайт-специфического гидролиза полученной С5-метилированной ДНК в реакционную смесь добавляют препарат MD-эндонуклеазы PkrI с активностью 2 е.а./мкл или препарат ElmI с активностью 2 е.а./мкл в зависимости от того, в какой позиции необходимо гидролизовать ДНК.
2. Способ по п. 1, отличающийся тем, что для метилирования ДНК в качестве реакционного буфера используют раствор, содержащий 10 мМ Трис-HCl (рН 7,5), 0,1 мМ S-аденозил-L-метионин, 1 мМ дитиотреитол.
3. Способ по п. 1, отличающийся тем, что для сайт-специфического гидролиза С5-метилированной ДНК используют 10-кратный SE-буфер «Y», содержащий 330 мМ Трис-ацетат (рН 7,9 при 25°С), 100 мМ магния ацетат, 660 мМ калия ацетат, 10 мМ дитиотреитол.
| MCCLELLAND M et al., Site-specific cleavage of DNA at 8- and 10-base-pair sequences | |||
| Proc | |||
| Natl | |||
| Acad | |||
| Sci | |||
| USA | |||
| Колосниковая решетка с чередующимися неподвижными и движущимися возвратно-поступательно колосниками | 1917 |
|
SU1984A1 |
| Горный компас | 0 |
|
SU81A1 |
| - p | |||
| Регистрационный ящик | 1918 |
|
SU983A1 |
| ЧЕРНУХИН В.А | |||
| и др | |||
| Кипятильник для воды | 1921 |
|
SU5A1 |

