Родственные заявки
По настоящей заявке по пункту 35 U.S.C. 119 испрашивается приоритет соответствующих Предварительных Патентных заявок США No. 61/309623 и 61/316634, поданных 2 марта 2010 г. и 23 марта 2010 г., соответственно, полное содержание которых приведено в настоящем документе в качестве ссылки.
Область изобретения
Описание относится к продуктам и способам для уменьшения концентрации акриламида в пище, так же как к пищевым продуктам, обладающим сниженным содержанием акриламида. В частности, описание относится к микроорганизмам, генетически модифицированным для их способности уменьшать количество акриламида.
Предпосылки изобретения
Акриламид представляет собой кристаллическое твердое вещество без цвета и без запаха, которое является важным промышленным мономером, общеупотребительным в качестве связующего для цемента и для синтеза полимеров и гелей. На основании различных исследований in vivo и in vitro получено убедительное доказательство канцерогенных и генотоксических эффектов акриламида и его метаболита глицидамида (Wilson et al., 2006; Rice, 2005). Акриламид оценен Международным агентством онкологических исследований (IARC) в 1994 г. и классифицирован как «возможно канцерогенный для человека» на основании положительных биологических исследований на мышах и крысах, что поддерживается доказательством того, что акриламид подвергается биологической трансформации в тканях млекопитающих до генотоксического метаболита глицидамида (IARC, 1994). Известно, что биологическая трансформация акриламида в глицидамид эффективно происходит в тканях как человека, так и грызунов (Rice, 2005). В дополнение к классификации IARC, как «The Scientific Committee on Toxicity, Ecotoxicity and the Environment» Европейского Союза, так и независимый «Committee on Carcinogenicity of Chemicals in Food, Consumer Products and the Environment» в Великобритании, советуют контролировать воздействие акриламида на людей на уровне, настолько низком, насколько возможно, из-за его по существу токсичных свойств, включающих нейротоксичность и генотоксичность как для соматических, так и для зародышевых клеток, канцерогенность и репродуктивную токсичность.
Что касается эпидемиологических исследований на человеке воздействия акриламида, поступающего с пищей, не существует доказательств какого-либо канцерогенного эффекта этого химического вещества; однако известно также, что эти эпидемиологические исследования акриламида могут не являться достаточно чувствительными для выявления потенциальных опухолей у человека после воздействия акриламида (Rice, 2005; Wilson et al., 2006).
В 2002 г. Шведская Национальная Пищевая Организация опубликовала подробное описание концентраций акриламида, обнаруженного в общепринятых пищевых продуктах, особенно подвергнутых тепловой обработке богатых углеводами пищевых продуктах, таких как картофель фри и картофельные чипсы. В настоящее время этот список расширен с включением пищевых продуктов на основе зерна, пищевых продуктов на основе овощей, пищевых продуктов на основе бобовых, напитков, таких как кофе или заменители кофе; в таблице 1 показаны данные FDA по концентрациям акриламида в различных пищевых продуктов.
В настоящее время установлено, что акриламид образуется в ходе приготовления пищевых продуктов в первую очередь посредством реакции Майларда между аминокислотой аспарагином и восстанавливающими сахарами, такими как глюкоза, где аспарагин является лимитирующим предшественником (Amrein et al., 2004; Becalski et al., 2003; Mustafa et al., 2005; Surdyk et al., 2004; Yaylayan et al., 2003).
Существовал также ряд попыток уменьшения содержания акриламида в пищевых продуктах, включая добавление коммерческих препаратов фермента аспарагиназы (Acrylaway®, Novozymes, Denmark и PreventASe, DSM, Netherlands), продолжительную ферментацию дрожжей в течение 6 часов (Fredriksson et al., 2004), применение глицина в тесте до брожения (Brathen et al., 2005; Fink et al., 2006), погружение картофеля в хлорид кальция перед обжариванием во фритюре (Gokmen and Senyuva, 2007), замену восстанавливающих сахаров сахарозой (Amrein et al., 2004), общую оптимизацию условий переработки, таких как температура, pH и влагосодержание (Claus et al., 2007; Gokmen et al., 2007) и исследования, касающиеся различного выбора сырья (Claus et al., 2006). Все из этих перечисленных способов являются до некоторой степени неадекватными или им присущи ограничения, делающие их непрактичными во время изготовления пищевых продуктов, включая стоимость, эффект на органолептические свойства пищевых продуктов и/или неэффективное снижение количества акриламида в условиях переработки пищевых продуктов.
Подобно многим микроорганизмам, Saccharomyces cerevisiae способны естественным образом потреблять/деградировать предшественники акриламида аспарагин и восстанавливающие сахара. Это может являться причиной наблюдаемого снижения количества содержания акриламида в хлебе после продолжительного времени брожения в течение 6 часов (Fredriksson et al., 2004). Однако такое продолжительное время брожения для эффективного снижения количества акриламида является непрактичным для современных способов переработки пищевых продуктов.
В S. cerevisiae генами, ответственными за аспарагин, являются ASP1 и ASP3, кодирующие аспарагиназу цитозоля и аспарагиназу клеточной стенки, соответственно. Существуют также по меньшей мере 41 ген в S. cerevisiae, аннотированные термином «транспорт аминокислот», и известно, что шесть из этих транспортеров способны транспортировать аспарагин в клетку [«Saccharomyces Genome Database» http://www.yeastgenome.org/ (10/01/09)]. Названия генов для этих шести транспортеров аспарагина в S. cerevisiae представляют собой GAP1, AGP1, GNP1, DIP5, AGP2 и AGP3. Хорошо известно также, что S. cerevisiae способны использовать широкое множество источников азота для роста и что в культурах на смешанных субстратах они последовательно выбирают от хороших до плохих источников азота (Cooper, 1982). Это последовательное использование контролируют молекулярные механизмы, состоящие из сенсорной системы и механизма регуляции транскрипции, известного как азотная катаболитная репрессия (NCR). Как правило, NCR относится к различиям экспрессии генов пермеаз и катаболических ферментов, необходимых для деградации источников азота. Экспрессию в путях катаболитов азота регулируют четыре регулятора, известные как Gln3p, Gat1p, Dal80p и Gzf3p, которые связываются с вышестоящей активирующей консенсусной последовательностью 5'-GATAA-3'. Gln3p и Gat1p положительно действуют на экспрессию генов, в то время как Dal80p и Gzf3p действуют отрицательно. В присутствии хорошего источника азота Gln3p и Gat1p фосфорилируются посредством TOR киназ Tor1p и Tor2p; затем образуют цитозольные комплексы с Ure2p и таким образом являются ингибированными против активации NCR-чувствительной транскрипции. В присутствии плохих источников азота или при азотном голодании Gln3p и Gat1p становятся дефосфорилированными, диссоциируют из Ure2p, накапливаются в ядре и активируют NCR-чувствительную транскрипцию.
Хорошо документировано также, что конкретная мутация URE2 приводит к доминантной мутации, обозначаемой [URE3]. [URE3] представляет собой прион дрожжей, образуемый посредством автокаталитического превращения Ure2p в инфекционные, устойчивые к протеазам амилоидные волокна (Wickner, 1994). Фенотипы клеток S. cerevisiae с отсутствием функционального Ure2p и инфицированных [URE3] клеток являются сходными, поскольку они больше не отвечают на NCR (Wickner, 1994; Wickner et al., 1995). Как указано выше, в ответ на хороший источник азота, Ure2p вовлекается в понижающую регуляцию активности Gln3p и Gat1p.
Краткое изложение сущности изобретения
Настоящее описание относится к микроорганизму, трансформированному по меньшей мере одной молекулой нуклеиновой кислоты для уменьшения азотной катаболитной репрессии в условиях приготовления/переработки пищевых продуктов. Настоящее описание относится также к микроорганизму, трансформированному по меньшей мере одной молекулой нуклеиновой кислоты для сверхэкспрессии гена, кодирующего внеклеточный белок, вовлеченный в деградацию аспарагина, и/или гена, кодирующего белок, вовлеченный в транспорт аспарагина в условиях приготовления/переработки пищевых продуктов. Настоящее описание относится также к микроорганизму, трансформированному по меньшей мере одной молекулой нуклеиновой кислоты для снижения азотной катаболитной репрессии и/или для сверхэкспрессии гена, кодирующего внеклеточный белок, вовлеченный в деградацию аспарагина, и/или гена, кодирующего белок, вовлеченный в транспорт аспарагина в условиях приготовления/переработки пищевых продуктов.
В одном варианте осуществления микроорганизм трансформируют молекулой нуклеиновой кислоты, кодирующей внеклеточную аспарагиназу, такую как связанная с клеточной стенкой аспарагиназа, Asp3p. В другом варианте осуществления микроорганизм трансформируют молекулой нуклеиновой кислоты, кодирующей транспортер аминокислот, такой как транспортер аминокислоты аспарагина, например, Gap1p, Agp1p, Gnp1p, Dip5p, Agp2p и/или Agp3p.
В другом варианте осуществления микроорганизм трансформируют молекулой нуклеиновой кислоты, кодирующей как Asp3p, так и Gap1p или как Asp3p, так и Gat1p. В другом варианте осуществления микроорганизм трансформируют первой и второй молекулой нуклеиновой кислоты, где первая молекула нуклеиновой кислоты кодирует Asp3p, и вторая молекула нуклеиновой кислоты кодирует Gap1p или Gat1p.
В другом варианте осуществления микроорганизм трансформируют молекулой нуклеиновой кислоты, модифицирующей активность регуляторного фактора азотной катаболитной репрессии транспорта/деградации аспарагина, такого как Ure2p, Dal80p, Gzf3p, Gln3p, Gat1p, Tor1p и/или Tor2p. В другом варианте осуществления микроорганизм трансформируют молекулой нуклеиновой кислоты, модифицирующей активность обоих регуляторных факторов азотной катаболитной репрессии Gln3p и Ure2p. В другом варианте осуществления микроорганизм трансформируют первой и второй молекулой нуклеиновой кислоты, модифицирующей азотную катаболитную репрессию, где первая молекула нуклеиновой кислоты кодирует Gln3p, и вторая молекула нуклеиновой кислоты модифицирует экспрессию Ure2p.
В одном варианте осуществления микроорганизм представляет собой гриб или бактерию. Гриб может представлять собой любой гриб, включая дрожжи, такие как Saccharomyces cerevisiae, Saccharomyces bayanus, Saccharomyces carlsbergensis, Candida albicans, Candida kefyr, Candida tropicalis, Cryptococcus laurentii, Cryptotoccous neoformans, Hansenula anomala, Hansenula polymorpha, Kluyveromyces fragilis, Kluyveromyces lactis, Kluyveromyces marxianus var lactis, Pichia pastoris, Rhodotorula rubra, Schizosaccharomyces pombe, Yarrowia lipolyitca или любые виды дрожжей, принадлежащих к царству грибов. Другие грибы, которые можно использовать, включают в себя, но без ограничения, виды из родов Aspergillus, Penicillium, Rhizopus и Mucor. Бактерии могут представлять собой любые бактерии, включая Erwinia sp., Lactobacillus sp., Lactococcus sp., Bacillus sp., Pediococcus sp., Pseudomonas sp., Brevibacterium sp., и Leuconostoc sp. В одном варианте осуществления микроорганизм является неактивным, таким как неактивные дрожжи.
В одном варианте осуществления по меньшей мере одна молекула нуклеиновой кислоты является функционально связанной с констутивно активным промотором. В другом варианте осуществления по меньшей мере одна молекула нуклеиновой кислоты является функционально связанной с промотором, не подверженным азотной катаболитной репрессии.
В настоящем документе представлен также способ снижения количества акриламида в пищевом продукте, включающий в себя добавление микроорганизма, описанного в настоящем документе, в пищевой продукт в условиях приготовления или переработки; где микроорганизм снижает азотную катаболитную репрессию или сверхэкспрессирует ген, вовлеченный в транспорт и/или деградацию аспарагина в условиях приготовления или переработки; таким образом снижая количество акриламида в пищевом продукте.
Кроме того, в настоящем документе представлен способ снижения количества акриламида в пищевом продукте, включающий в себя (a) трансформацию микроорганизма по меньшей мере одной молекулой нуклеиновой кислоты для снижения азотной катаболитной репрессии или для сверхэкспрессии гена, кодирующего внеклеточный белок, вовлеченный в деградацию аспарагина и/или гена, кодирующего белок, вовлеченный в транспорт аспарагина; (b) добавление микроорганизма в пищевой продукт в условиях приготовления или переработки; где микроорганизм снижает азотную катаболитную репрессию или сверхэкспрессирует ген, кодирующий внеклеточный белок, вовлеченный в деградацию аспарагина, и/или ген, кодирующий белок, вовлеченный в транспорт аспарагина, таким образом снижая количество акриламида в пищевом продукте.
В другом варианте осуществления представлен пищевой продукт, обладающий сниженной концентрацией акриламида, полученный с использованием трансформированного микроорганизма, описанного в настоящем документе. В другом варианте осуществления представлен пищевой продукт, обладающий сниженной концентрацией акриламида, полученный с использованием способа, описанного в настоящем документе.
В одном варианте осуществления пищевой продукт представляет собой пищевой продукт на основе зерна, включая, без ограничения, бисквиты, хлеб и крекеры, пищевой продукт на основе овощей включая, без ограничения, картофельные продукты, напиток, включая, без ограничения, кофе и заменители кофе, фруктовый, бобовый, молочный или мясной продукт.
Другие признаки и преимущества по настоящему описанию становятся очевидными из следующего подробного описания. Следует понимать, однако, что подробное описание и конкретные примеры, в то время как в них указаны предпочтительные варианты осуществления, приведены только для иллюстрации, поскольку различные изменения и модификации в пределах содержания и объема изобретения очевидны специалистам в данной области из этого подробного описания.
Краткое описание фигур
Описание поясняется фигурами, на которых:
Фиг.1 представляет собой схематическое представление сконструированной генетической кассеты ASP3 и последующие стадии утраты маркера kanMX после интеграции в локус LEU2 или URA3 штаммов S. cerevisiae. Маркер kanMX удаляют посредством рекомбинации прямых повторов промотора PGK1, что приводит к самоклонирующемуся штамму, содержащему только нативные последовательности ДНК.
Фиг.2 представляет собой схематическое представление сконструированной генетической кассеты GAP1 и последующие стадии утраты маркера kanMX после интеграции в локус URA3 штаммов S. cerevisiae. Маркер kanMX удаляют посредством рекомбинации прямых повторов промотора PGK1, что приводит к самоклонирующемуся штамму, содержащему только нативные последовательности ДНК.
Фиг.3 представляет собой схематическое представление сконструированной генетической кассеты AGP3 и последующие стадии утраты маркера kanMX после интеграции в локус LEU2 штаммов S. cerevisiae. Маркер kanMX удаляют посредством рекомбинации прямых повторов промотора PGK1, что приводит к самоклонирующемуся штамму, содержащему только нативные последовательности ДНК.
Фиг.4 представляет собой схематическое представление сконструированной генетической кассеты AGP2 и последующие стадии утраты маркера kanMX после интеграции в локус LEU2 штаммов S. cerevisiae. Маркер kanMX удаляют посредством рекомбинации прямых повторов промотора PGK1, что приводит к самоклонирующемуся штамму, содержащему только нативные последовательности ДНК.
Фиг.5 представляет собой схематическое представление сконструированной генетической кассеты GNP1 и последующие стадии утраты маркера kanMX после интеграции в локус LEU2 штаммов S. cerevisiae. Маркер kanMX удаляют посредством рекомбинации прямых повторов промотора PGK1, что приводит к самоклонирующемуся штамму, содержащему только нативные последовательности ДНК.
Фиг.6 представляет собой схематическое представление сконструированной генетической кассеты AGP1 и последующие стадии утраты маркера kanMX после интеграции в локус URA3 штаммов S. cerevisiae. Маркер kanMX удаляют посредством рекомбинации прямых повторов промотора PGK1, что приводит к самоклонирующемуся штамму, содержащему только нативные последовательности ДНК.
Фиг.7 представляет собой схематическое представление сконструированной генетической кассеты GAT1 и последующие стадии утраты маркера kanMX после интеграции в локус LEU2 штаммов S. cerevisiae. Маркер kanMX удаляют посредством рекомбинации прямых повторов промотора PGK1, что приводит к самоклонирующемуся штамму, содержащему только нативные последовательности ДНК.
Фиг.8 представляет собой схематическое представление интеграции самоклонирующейся кассеты ure2∆ в локус URE2 штаммов S. cerevisiae с использованием маркера kanMX и последующей утраты маркера посредством рекомбинации части фланкирующих последовательностей 5'URE2, действующих как прямые повторы. Полученная трансформация делетирует ген URE2 из генома.
На фиг.9 показана карта сконструированной плазмиды pAC1, использованной в клонируемых генетических кассетах для интеграции в локус LEU2.
На фиг.10 показаны карты сконструированной плазмиды pAC2, использованной в клонируемых генетических кассетах для интеграции в локус URA3.
На фиг.11 показано потребление аспарагина в хлебном тесте с использованием коммерческих хлебных дрожжей (BY) со сверхэкспрессией гена ASP1 или ASP3.
На фиг.12 показаны концентрации акриламида в образце хлебного теста, взятого во временной точке 5 час из эксперимента, изображенного на фиг.11.
На фиг.13 показано потребление аспарагина в хлебном тесте с использованием коммерческих хлебопекарных дрожжей (BY) со сверхэкспрессией ASP3 или GAP1 и комбинации ASP3/GAP1.
На фиг.14 показано потребление аспарагина в хлебном тесте с использованием лабораторных (LY) с нокаутом гена DAL80 или URE2.
На фиг.15 показаны концентрации акриламида в образце хлебного теста, взятого во временной точке 5 час из эксперимента, изображенного на фиг.14.
На фиг.16 показано потребление аспарагина в комплексной среде с использованием коммерческих хлебопекарных дрожжей (BY) со сверхэкспрессией AGP2 или AGP3 после 5 часов роста.
На фиг.17 показано потребление аспарагина в синтетической среде, содержащей аспарагин и аммиак, с использованием коммерческих хлебопекарных дрожжей (BY) со сверхэкспрессией GAT1 или ASP3 и комбинации GAT1/ASP3.
На фиг.18 показано потребление аспарагина в синтетической среде, содержащей аспарагин и аммиак, с использованием коммерческих хлебопекарных дрожжей (BY) со сверхэкспрессией GNP1.
На фиг.19 показано потребление аспарагина в синтетической среде, содержащей аспарагин и аммиак, с использованием лабораторных дрожжей (LY) со сверхэкспрессией ASP3 или делецией TOR1 и с комбинацией tor1∆/ASP3.
На фиг.20 показано потребление аспарагина в синтетической среде, содержащей аспарагин, с использованием коммерческих хлебопекарных дрожжей (BY) со сверхэкспрессией AGP1 и лабораторных дрожжей (LY) с нокаутом GZF3 после 5 часов роста.
Подробное описание
Авторы настоящего изобретения получили штаммы дрожжей, обладающие увеличенной способностью к потреблению и/или деградации аспарагина, который является лимитирующим предшественником, получаемым в ходе переработки или приготовления пищевого продукта, который приводит к продукции акриламида.
Микроорганизмы
В одном варианте осуществления представлен микроорганизм, трансформированный по меньшей мере одной молекулой нуклеиновой кислоты для снижения азотной катаболитной репрессии и/или для сверхэкспрессии гена, кодирующего внеклеточный белок, вовлеченный в деградацию аспарагина, и/или гена, кодирующего белок, вовлеченный в транспорт аспарагина, в условиях приготовления/переработки пищевых продуктов.
В другом варианте осуществления микроорганизм трансформируют по меньшей мере двумя, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5 или более молекулами нуклеиновой кислоты.
Фраза «сверхэкспрессия гена, кодирующего внеклеточный белок, вовлеченный в деградацию аспарагина, и/или гена, кодирующего белок, вовлеченный в транспорт аспарагина», как применяют в настоящем документе, относится к увеличенной экспрессии мРНК или белков, которые транспортируются к мембране клетки или секретируются к клеточной стенке и которые вовлечены в транспорт и/или деградацию аминокислоты аспарагина по сравнению с контролем, не трансформированным молекулой нуклеиновой кислоты.
Молекула нуклеиновой кислоты может представлять собой любую молекулу нуклеиновой кислоты, кодирующую белок, вовлеченный, напрямую или опосредованно, в транспорт аспарагина, и/или внеклеточный белок, вовлеченный напрямую или опосредованно в деградацию аспарагина. В одном варианте осуществления молекула нуклеиновой кислоты кодирует аспарагиназу клеточной стенки или ее фрагмент, обладающий активностью деградации аспарагина. Внеклеточные аспарагиназы представляют собой ферменты, известные в данной области, и включают в себя, без ограничения, внеклеточные аспарагиназы, такие как аспарагиназы клеточной стенки из любого источника, способные превращать аспарагин в аспартат, такого как дрожжи Asp3p, или их гомологи, и могут быть кодированы любыми генами аспарагиназы, кодирующие аспарагиназы клеточной стенки, включая, без ограничения, ASP3 или их гомологи. В одном варианте осуществления аспарагиназа клеточной стенки является кодированной молекулой нуклеиновой кислоты ASP3, как показано на SEQ ID NO:2 или ее гомолог или фрагмент, или содержит аминокислотную последовательность Asp3p, как показано на SEQ ID NO:1, или ее гомолог или фрагмент. Микроорганизмы, содержащие молекулы нуклеиновой кислоты, кодирующие внеклеточные аспарагиназы, способные деградировать аспарагин в условиях приготовления и переработки пищевых продуктов.
В другом варианте осуществления молекула нуклеиновой кислоты кодирует транспортер аминокислот или его фрагмент, обладающий способностью транспортировать аспарагин в клетку. Транспортеры аминокислот известны в данной области и включают в себя, без ограничения, транспортеры аминокислот из любого источника, способные к активному транспорту аспарагина в микроорганизм, такой как дрожжи Gap1p, Agp1p, Gnp1p, Dip5p, Agp2p и Agp3p (NP_012965, NP_009905, NP_010796, NP_015058, NP_009690 и NP_116600), или их гомолог, и могут быть кодированы любым геном транспортера аминокислот, включая, без ограничения, GAP1, AGP1, GNP1, DIP5, AGP2 и AGP3 (SGD:S000001747, SGD:S000000530, SGD:S000002916, SGD:S000006186, SGD:S000000336 и SGD:S000001839) или их гомолог. Соответственно, в одном варианте осуществления транспортер аминокислот кодирован молекулой нуклеиновой кислоты GAP1, AGP3, AGP2, GNP1, AGP1 или DIP5, как показано на SEQ ID NO:4, 6, 8, 10, 12 или 30 соответственно, или ее гомологом или фрагментом, или содержит аминокислотную последовательность Gap1p, Agp3p, Agp2p, Gnp1p, Agp1p или Dip5p, как показано на SEQ ID NO:3, 5, 7, 9, 11 или 29 соответственно, или ее гомолог или фрагмент. Микроорганизмы, содержащие молекулы нуклеиновой кислоты, кодирующие транспортеры аминокислот, способны потреблять или поглощать аспарагин в условиях приготовления и переработки пищевых продуктов.
В другом варианте осуществления микроорганизм трансформируют нуклеиновой кислотой, кодирующей аспарагиназу клеточной стенки, и нуклеиновой кислотой, кодирующей транспортер аминокислот. В таком варианте осуществления микроорганизм является способным потреблять и деградировать аспарагин.
Фраза «снижать азотную катаболитную репрессию (NCR)» транспорта/деградации аспарагина, как применяют в настоящем документе, относится к действительному снижению репрессии генов для NCR-чувствительных генов или относится к увеличенной эндогенной экспрессии или гетерологичной экспрессии NCR-чувствительных генов. Например, молекула нуклеиновой кислоты для снижения NCR может представлять собой регуляторный фактор, который модифицирует выражение азотной катаболитной репрессии или может представлять собой сверхэкспрессию NCR-чувствительного гена.
В другом варианте осуществления молекула нуклеиновой кислоты модифицирует активность регуляторного фактора азотной катаболитной репрессии. Регуляторные факторы для азотной катаболитной репрессии известны в данной области и включают в себя, без ограничения, регуляторные факторы из любого источника, такого как дрожжи Gat1p, Ure2p, Tor1p, Dal80p, Gzf3p, Tor2p, или Gln3p, как показано на SEQ ID NO:13, 15, 17, 19, 21, 33 или 31, или их гомолог или фрагмент, и могут быть кодированы любым геном, кодирующим регуляторный фактор, такой как GAT1, URE2, TOR1, DAL80, GZF3, TOR2, или GLN3, как показано на SEQ ID NO:14, 16, 18, 20, 22, 34 или 32. Например, можно получить микроорганизм, больше не обладающий функциональным отрицательным регулятором, таким как Ure2p, Tor1p, Tor2p Dal80p или Gzf3p. Этого можно достигать, например, посредством молекулы нуклеиновой кислоты, приводящей к делеции гена URE2, выделения и экспрессии мутантного фенотипа ure2, так что он больше не осуществляет понижающей регуляции активности Gln3p и Gat1p, посредством скрещивания штамма дикого типа со штаммом [URE3], или индукции фенотипа [URE3] любыми способами молекулярной биологии, включая цитодукию и сверхэкспрессию URE2. Последствия отсутствия у клеток функционального Ure2p приводят к тому, что чувствительные к NCR гены, такие как гены, вовлеченные в транспорт и утилизацию аспарагина (т.е. ASP3, AGP1, GAP1, GAT1, DAL80 и GZF3), больше не репрессированы в присутствии хорошего источника азота, такого как аммиак или глутамин. Соответственно, в одном варианте осуществления молекула нуклеиновой кислоты содержит делеционную кассету URE2, TOR1, TOR2, DAL80 и/или GZF3. Микроорганизмы с отсутствием функционального Ure2p, Tor1p, Dal80p и/или Gzf3p способны потреблять и деградировать аспарагин в условиях приготовления и переработки пищевых продуктов. Альтернативно, этого можно достигать посредством молекулы нуклеиновой кислоты, приводящей к сверхэкспрессии функционального положительного регулятора, такого как Gat1p и/или Gln3p.
Термин «ген», как применяют в настоящем документе, находится в соответствии с его обычным определением, для обозначения функционально связанной группы последовательностей нуклеиновой кислоты. Модификация гена в контексте настоящего описания может включать в себя модификацию любой из различных последовательностей, функционально связанной с геном. Под «функционально связанными» понимают, что конкретные последовательности взаимодействуют напрямую или опосредованно для выполнения предназначенной им функции, такой как опосредование или модуляция экспрессии гена. Взаимодействие функционально связанных последовательностей может быть, например, опосредовано белками, которые, в свою очередь, взаимодействуют с последовательностями нуклеиновой кислоты.
Последовательности различных генов и нуклеиновых кислот по описанию могут представлять собой рекомбинантные последовательности. Термин «рекомбинантный», как применяют в настоящем документе, относится к чему-либо, что подвергали рекомбинации, так что по отношению к конструкции нуклеиновой кислоты термин относится к молекуле, содержащей последовательности нуклеиновой кислоты, соединенные вместе в некоторой точке или полученные способами молекулярной биологии. Термин «рекомбинантный» по отношению к белку или полипептиду обозначает молекулу белка или полипептида, которую экспрессируют с использованием рекомбинантной конструкции нуклеиновой кислоты, полученной способами молекулярной биологии. Термин «рекомбинантный» по отношению к генетической композиции относится к гамете или потомству, или клетке, или геному с новыми комбинациями аллелей, не встречающимися в природных родительских геномах. Рекомбинантные конструкции нуклеиновых кислот могут включать в себя нуклеотидную последовательность, лигированную, или подвергнутую манипуляциям для лигирования с последовательностью нуклеиновой кислоты, с которой она не лигирована в природе, или с которой она лигирована в природе в другой локализации. Обозначение конструкции нуклеиновой кислоты как «рекомбинантной» таким образом, обозначает, что с молекулой нуклеиновой кислоты манипулировали посредством вмешательства человека с использованием генной инженерии.
Молекулы нуклеиновой кислоты можно химически синтезировать с использованием таких способов, как описанные, например, в Itakura et al., Патент США No. 4598049; Caruthers et al., Патент США No. 4458066; и Itakura, Патенты США No. 4401796 и 4373071. Такие синтетический нуклеиновые кислоты по их природе являются «рекомбинантными», как этот термин применяют в настоящем документе (являясь продуктом последовательных стадий объединения составляющих частей молекулы).
Степень гомологии между последовательностями (такими как природные аминокислотные последовательности Asp3p, Gap1p, Dip5p, Gnp1p, Agp1p, Agp2p, Agp3p, Tor1p, Tor2p, Gat1p, Gln3p, Dal80p, Gzf3p или Ure2p, или природные последовательности нуклеиновой кислоты ASP3, GAP1, DIP5, GNP1, AGP1, AGP2, AGP3, TOR1, TOR2, GAT1, GLN3, DAL80, GZF3 или URE2, и последовательность гомолога) можно выражать как процент идентичности, когда последовательности оптимально выровнены, подразумевая существование точных совпадений между последовательностями. Оптимальное выравнивание последовательностей для сравнений идентичности можно проводить с использованием множества алгоритмов, таких как алгоритм поиска локальных гомологий Смита и Уотермана, 1981, Adv. Appl. Math 2: 482, алгоритм выравнивания по гомологии Нидлемана и Вунша, 1970, J. Mol. Biol. 48:443, способ поиска сходства по Пирсону и Липману, 1988, Proc. Natl. Acad. Sci. USA 85: 2444, и компьютеризированные реализации этих алгоритмов (такие как GAP, BESTFIT, FASTA и TFASTA в пакете программного обеспечения Wisconsin Genetics Software Package, Genetics Computer Group, Madison, WI, U.S.A.). Выравнивание последовательностей можно проводить также с использованием алгоритма BLAST, описанного в Altschul et al., 1990, J. Mol. Biol. 215:403-10 (с использованием опубликованных параметров по умолчанию). Программное обеспечение для проведения анализа BLAST может быть доступно через Национальный центр биотехнологической информации (через Интернет на http://www.ncbi.nlm.nih.gov/). Алгоритм BLAST включает в себя первичную идентификацию пар последовательностей с высокой степенью сходства (HSP) посредством идентификации коротких слов длиной W в запрашиваемой последовательности, которые либо совпадают, либо удовлетворяют некоторому положительно оцениваемому пороговому показателю T при выравнивании со словом той же длины в последовательности из базы данных. T обозначает пороговую оценку сходства соседних слов. Первоначальное попадание в соседнее слово является источником для инициации поисков с целью найти более длинные HSP. Попадания в слова расширяются в обоих направлениях вдоль каждой последовательности, пока кумулятивная оценка выравнивания может увеличиваться. Расширение попадания в слова в каждом направлении прекращается, когда встречаются следующие параметры: кумулятивная оценка выравнивания уменьшается на количество Х от ее максимально достигнутого значения; кумулятивная оценка стремится к нулю или ниже из-за накопления одного или нескольких отрицательно оцениваемых выравниваний остатков; или достигнут конец какой-либо из последовательностей. Параметры алгоритма BLAST W, T и X определяют чувствительность и скорость выравнивания. Программы BLAST могут использовать по умолчанию длину слова (W), оценочную матрицу BLOSUM62 (Henikoff and Henikoff, 1992, Proc. Natl. Acad. Sci. USA 89: 10915-10919) выравнивания (B) 50, ожидание (E) 10 (которое может меняться в альтернативных вариантах осуществления до 1 или 0,1 или 0,01 или 0,001 или 0,0001; хотя значения E, намного превышающие 0,1, могут не идентифицировать функционально сходные последовательности, полезно проверять попадания с более низкой значимостью, значениями E между 0,1 и 10 по коротким областям сходства), M=5, N=4, для сравнения обеих цепей нуклеиновых кислот. Для сравнения белков BLASTP можно использовать со следующими параметрами по умолчанию: G=11 (штраф за открытие пропуска); E=1 (штраф за расширение пропуска); E=10 (величина ожидания, в этом параметре предполагают ожидание того, что 10 попаданий с показателями, равными или лучшими, чем определенный показатель выравнивания, S, появляются случайно в базе данных того же размера, что и та, в которой ведется поиск; значение E можно увеличивать или уменьшать для изменения строгости поиска.); и W=3 (длина слова, по умолчанию 11 для BLASTN, 3 для других программ blast). Матрица BLOSUM определяет оценку вероятности для каждого положения при выравнивании, которая основана на частоте, с которой, как известно, эта замена встречается среди консенсусных блоков в пределах родственных белков. Матрицу замен BLOSUM62 (штраф за существование пропуска = 11; штраф за пропуск остатка = 1; соотношение лямбда = 0,85) используют по умолчанию в BLAST 2.0. Множество других матриц может быть использовано в качестве альтернативных для BLOSUM62, включая: PAM30 (9,1,0.87); PAM70 (10,1,0.87) BLOSU 80 (10,1,0.87); BLOSUM62 (1,1,0.82) и BLOSUM45 (14,2,0.87). Одним из измерений статистического сходства между двумя последовательностями с использованием алгоритма BLAST является наименьшая сумма вероятностей (P(N)), которая указывает на вероятность, с которой совпадение двух нуклеотидных или аминокислотных последовательностей может происходить случайно. В альтернативных вариантах осуществления нуклеотидные или аминокислотные последовательности считают по существу идентичными, если наименьшая сумма вероятностей при сравнении тестируемых последовательностей составляет менее приблизительно 1, менее приблизительно 0,1, менее приблизительно 0,01 или менее приблизительно 0,001. Сходство между последовательностями можно также выражать как процентную идентичность.
Последовательности нуклеиновой кислоты и белка, описанные в настоящем документе, могут в некоторых вариантах осуществления являться по существу идентичными, например, по существу идентичными аминокислотным последовательностям Asp3p, Gap1p, Gnp1p, Agp1p, Agp2p, Agp3p, Gat1p, Tor1p, Tor2p, Dip5p, Gln3p, Dal80p, Gzf3p или Ure2p, или последовательностям нуклеиновой кислоты ASP3, GAP1, GNP1, AGP1, AGP2, AGP3, TOR1, TOR2, DIP5, GLN3, GAT1, DAL80, GZF3 или URE2. Значительная идентичность таких последовательностей может быть отражена в проценте идентичности при оптимальном выравнивании, который может составлять, например, более 50%, 80%-100%, по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 95%, что в случае нацеливаемых на гены субстратов может относиться к идентичности части нацеливаемого на ген субстрата части последовательности-мишени, где степень идентичности может облегчать гомологичное спаривание и рекомбинацию и/или репарацию. Альтернативным указанием на то, что две последовательности нуклеиновой кислоты являются, по существу, идентичными, является то, что эти две последовательности гибридизуются друг с другом в умеренно строгих или высоко строгих условиях. Гибридизацию со связанными с фильтром последовательностями в умеренно строгих условиях можно проводить, например, в 0,5M NaHPO4, 7% додецилсульфате натрия (SDS), 1 мМ ЭДТА при 65°C, а отмывку в 0,2×SSC/0,1% SDS при 42°C (см. Ausubel, et al. (eds), 1989, Current Protocols in Molecular Biology, Vol. 1, Green Publishing Associates, Inc., и John Wiley & Sons, Inc., New York, p. 2.10.3). Альтернативно, гибридизацию со связанными с фильтром последовательностями в высоко строгих условиях можно проводить, например, в 0,5M NaHPO4, 7% SDS, 1 мМ ЭДТА при 65°C, а отмывку в 0,1×SSC/0,1% SDS при 68°C (см. Ausubel, et al. (eds), 1989, выше). Условия гибридизации можно модифицировать в соответствии с известными способами в зависимости от интересующей последовательности (см. Tijssen, 1993, Laboratory Techniques in Biochemistry and Molecular Biology - Hybridization with Nucleic Acid Probes, Часть I, Глава 2 «Обзор принципов гибридизации и стратегии анализов с использованием нуклеотидных зондов», Elsevier, New York). Как правило, выбирают строгие условия, которые приблизительно на 5°C ниже, чем точка плавления специфической последовательности при определенной ионной силе и pH. Отмывки для строгой гибридизации могут, например, составлять по меньшей мере 15 минут, 30 минут, 45 минут, 60 минут, 75 минут, 90 минут, 105 минут или 120 минут.
В данной области хорошо известно, что некоторые модификации и изменения можно вносить в структуру полипептида, такого как Asp3p, Gap1p, Gnp1p, Agp1p, Agp2p, Agp3p, Gatp, Tor1p, Tor2p, Dip5p, Gln3p, Dal80p, Gzf3p или Ure2p, без существенного изменения биологической функции этого пептида, для получения биологически эквивалентного полипептида. В одном аспекте белки, обладающие активностью транспорта аспарагина, могут включать в себя белки, отличающиеся от аминокислотных последовательностей природных Gap1p, Gnp1p, Dip5p, Agp1p, Agp2p, Agp3p или других транспортеров консервативными заменами аминокислот. Подобным образом, белки, обладающие активностью аспарагиназы, могут включать в себя белки, отличающиеся от последовательностей природного Asp3p, или других аспарагиназ клеточной стенки консервативными аминокислотными заменами. Как применяют в настоящем документе, термин «законсервированные или консервативные аминокислотные замены» обозначает замену одной аминокислоты на другую в заданном положении в белке, где замену можно осуществлять без существенной потери соответствующей функции. При осуществлении таких изменений, замены сходных аминокислотных остатков можно осуществлять на основании относительного сходства заместителей боковых цепей, например, их размера, заряда, гидрофобности, гидрофильности и тому подобного, и можно анализировать эффект таких замен на функцию белка посредством общепринятого тестирования.
В некоторых вариантах осуществления можно выполнять консервативные замены аминокислот, где аминокислотный остаток замещен на другой аминокислотный остаток, имеющий сходную величину гидрофильности (например, в пределах значения плюс или минус 2,0), где нижеследующее может представлять собой аминокислоту, обладающую индексом гидропатичности приблизительно -1,6, например, Tyr (-1,3) или Pro (-1,6), приписанным аминокислотным остаткам (как подробно описано в Патенте США No. 4554101, содержание которого приведено в настоящем документе в качестве ссылки): Arg (+3,0); Lys (+3,0); Asp (+3,0); Glu (+3,0); Ser (+0,3); Asn (+0,2); Gln (+0,2); Gly (0); Pro (-0,5); Thr (-0,4); Ala (-0,5); His (-0,5); Cys (-1,0); Met (-1,3); Val (-1,5); Leu (-1,8); Ile (-1,8); Tyr (-2,3); Phe (-2,5); и Trp (-3,4).
В альтернативных вариантах осуществления можно выполнять консервативные замены аминокислот, где аминокислотный остаток замещен на другой аминокислотный остаток, имеющий сходную величину гидропатичности (например, в пределах значения плюс или минус 2,0). В таких вариантах осуществления каждому аминокислотному остатку можно приписать индекс гидропатичности на основании характеристик его гидрофобности и заряда следующим образом: Ile (+4,5); Val (+4,2); Leu (+3,8); Phe (+2,8); Cys (+2,5); Met (+1,9); Ala (+1,8); Gly (-0,4); Thr (-0,7); Ser (-0,8); Trp (-0,9); Tyr (-1,3); Pro (-1,6); His (-3,2); Glu (-3,5); Gln (-3,5); Asp (-3,5); Asn (-3,5); Lys (-3,9); и Arg (-4,5).
В альтернативных вариантах осуществления можно выполнять консервативные замены аминокислот, где аминокислотный остаток замещен на другой аминокислотный остаток того же класса, где аминокислоты подразделяются на классы неполярных, кислых, основных и нейтральных аминокислот, следующим образом: неполярные: Ala, Val, Leu, Ile, Phe, Trp, Pro, Met; кислые: Asp, Glu; основные: Lys, Arg, His; нейтральные: Gly, Ser, Thr, Cys, Asn, Gln, Tyr.
В альтернативных вариантах осуществления консервативные замены аминокислот включают в себя замены, основанные на представлениях о гидрофильности или гидрофобности, размере, или объеме, или заряде. Как правило, аминокислоты могут быть охарактеризованы как гидрофобные или гидрофильные главным образом в зависимости от свойств боковой цепи аминокислот. Гидрофобная аминокислота имеет гидрофобность больше нуля, а гидрофильная аминокислота имеет гидрофильность менее нуля, на основании нормализованной консенсусной шкалы гидрофобности Eisenberg et al. (J. Mol. Bio. 179:125-142, 184). Генетически кодируемые гидрофобные аминокислоты включают в себя Gly, Ala, Phe, Val, Leu, Ile, Pro, Met и Trp, и генетически кодируемые гидрофильные аминокислоты включают в себя Thr, His, Glu, Gln, Asp, Arg, Ser и Lys. Негенетически кодируемые гидрофобные аминокислоты включают в себя т-бутилаланин, в то время как негенетически кодируемые гидрофильные аминокислоты включают в себя цитруллин и гомоцистеин.
Гидрофобные или гидрофильные аминокислоты можно дополнительно подразделять на основании характеристик их боковых цепей. Например, ароматическая аминокислота представляет собой гидрофобную аминокислоту с боковой цепью, содержащей по меньшей мере одно ароматическое или гетероароматическое кольцо, которое может содержать один или несколько заместителей, таких как -OH, -SH, -CN, -F, -Cl, -Br, -I, -NO2, -NO, -NH2, -NHR, -NRR, -C(O)R, -C(O)OH, -C(O)OR, -C(O)NH2, -C(O)NHR, -C(O)NRR и т.д., где R независимо представляет собой (C1-C6) алкил, замещенный (C1-C6) алкил, (C1-C6) алкенил, замещенный (C1-C6) алкенил, (C1-C6) алкинил, замещенный (C1-C6) алкинил, (C5-C20) арил, замещенный (C5-C20) арил, (C6-C26) алкарил, замещенный (C6-C26) алкарил, 5-20-членный гетероарил, замещенный 5-20-членный гетероарил, 6-26-членный алкгетероарил или замещенный 6-26-членный алкгетероарил. Генетически кодируемые ароматические аминокислоты включают в себя Phe, Tyr и Tryp.
Неполярная аминокислота представляет собой гидрофобную аминокислоту с боковой цепью, которая является незаряженной при физиологическом pH, и которая имеет связи, в которых пара электронов, обобществленная двумя атомами, как правило, в равной степени удерживается каждым из двух атомов (т.е. боковая цепь является неполярной). Генетически кодируемые неполярные аминокислоты включают в себя Gly, Leu, Val, Ile, Ala и Met. Неполярные аминокислоты можно дополнительно подразделять с включением алифатических аминокислот, которые представляют собой гидрофобные аминокислоты, имеющие алифатическую углеводородную боковую цепь. Генетически кодируемые алифатические аминокислоты включают в себя Ala, Leu, Val и Ile.
Полярная аминокислота представляет собой гидрофильную аминокислоту с боковой цепью, которая является незаряженной при физиологическом pH, но имеет одну связь, в которой пара электронов, обобществленная двумя атомами, находится ближе к одному из атомов. Генетически кодируемые полярные аминокислоты включают в себя Ser, Thr, Asn и Gln.
Кислая аминокислота представляет собой гидрофильную аминокислоту со значением pKa боковой цепи менее 7. Кислые аминокислоты, как правило, имеют отрицательно заряженные боковые цепи при физиологическом pH из-за потери иона водорода. Генетически кодируемые кислые аминокислоты включают в себя Asp и Glu. Основная аминокислота представляет собой гидрофильную аминокислоту со значением pKa боковой цепи более 7. Основные аминокислоты, как правило, имеют положительно заряженные боковые цепи при физиологическом pH из-за ассоциации с ионом гидроксония. Генетически кодируемые основные аминокислоты включают в себя Arg, Lys и His.
Специалисту в данной области понятно, что вышеупомянутые классификации не являются абсолютными, и что аминокислота может быть классифицирована в более чем одной категории. Кроме того, аминокислоты можно классифицировать на основании известного поведения и/или характерных химических, физических или биологических свойств на основании специфических анализов или по сравнению с предварительно идентифицированными аминокислотами.
Микроорганизм может представлять собой любой микроорганизм, пригодный для добавления в пищевые продукты, включая, без ограничения, грибы и/или бактерии. Грибы, пригодные по настоящему описанию, включают в себя, без ограничения, Aspergillus niger, Aspergillus oryzae, Neurospora crassa, Neurospora intermedia var. oncomensis, Penicillium camemberti, Penicillium candidum, Penicillium roqueforti, Rhizopus oligosporus, Rhizopus oryzae. В другом варианте осуществления грибы представляют собой дрожжи, такие как Saccharomyces cerevisiae, Saccharomyces bayanus, Saccharomyces carlsbergensis, Candida albicans, Candida kefyr, Candida tropicalis, Cryptococcus laurentii, Cryptotoccous neoformans, Hansenula anomala, Hansenula polymorpha, Kluyveromyces fragilis, Kluyveromyces lactis, Kluyveromyces marxianus var lactis, Pichia pastoris, Rhodotorula rubra, Schizosaccharomyces pombe, Yarrowia lipolyitca или любой штамм, принадлежащий к царству грибов. Существует множество коммерческих источников штаммов дрожжей, таких как Lallemand Inc. (Canada), AB Mauri (Australia) и Lesaffre (France). В другом варианте осуществления бактерии могут представлять собой любые бактерии, включая Erwinia sp., Lactobacillus sp., Lactococcus sp., Bacillus sp., Pediococcus sp., Pseudomonas sp., Brevibacterium sp. и Leuconostoc sp.
В одном варианте осуществления микроорганизм является неактивным, таким как неактивные дрожжи. Термин «неактивный», как применяют в настоящем документе, относится к композиции неактивных, нежизнеспособных и/или мертвых микроорганизмов, которые все еще сохраняют их содержание питательных веществ и другие свойства. Например, дрожжи можно выращивать в условиях, позволяющих сверхэкспрессию желательного белка или белков. Затем дрожжи можно использовать для получения неактивных дрожжей, например, посредством множества способов пастеризации, включая, без ограничения, высокотемпературную пастеризацию и кратковременную пастеризацию, множества способов стерилизации, включая, без ограничения, стерилизацию влажным паром и облучение, множества способов инактивации, включая, без ограничения, инактивацию под высоким давлением, фотокаталитическую инактивацию и инактивацию импульсным излучением, фотосенсибилизацию, инактивацию электрическими полями, включая RF и пульсирующие поля, разрушение клеток, обработку ультразвуком, гомогенизацию, автолиз и инактивацию на основе химических веществ, включая, без ограничения, формальдегид, тимеросол, хлорамины, диоксид хлора, иод, серебро, медь, антибиотики и озон.
Рекомбинантные конструкции нуклеиновых кислот можно, например, вводить в клетку-хозяина микроорганизма посредством трансформации. Такие рекомбинантные конструкции нуклеиновых кислот могут включать последовательности, полученные из того же самого вида клетки-хозяина или из других видов клеток-хозяев, выделенные и повторно введенные в клетки вида-хозяина.
Рекомбинантные последовательности нуклеиновой кислоты могут быть интегрированы в геном клетки-хозяина, либо в результате первичной трансформации клеток-хозяев, либо в результате последующих событий рекомбинации и/или репарации. Альтернативно, рекомбинантные последовательности могут сохраняться в виде внехромосомных элементов. Такие последовательности можно репродуцировать, например, с использованием организма, такого как трансформированный штамм дрожжей, в качестве исходного штамма для способов улучшения штамма, осуществляемых посредством мутации, массового скрещивания или слияния протопластов. Полученные штаммы, сохраняющие рекомбинантную последовательность по изобретению, сами считаются «рекомбинантными», как этот термин применяют в настоящем документе.
Трансформация представляет собой способ, посредством которого генетический материал, переносимый клеткой, изменяют посредством включения одной или нескольких экзогенных нуклеиновых кислот в клетку. Например, дрожжи можно трансформировать с использованием множества способов (Gietz et al., 1995). Такая трансформация может происходить посредством включения экзогенной нуклеиновой кислоты в генетический материал клетки или посредством изменения эндогенного генетического материала клетки, происходящего в результате воздействия на клетку экзогенной нуклеиновой кислоты. Трансформанты или трансформированные клетки являются клетками или потомками клеток, которые были функционально улучшены посредством поглощения экзогенной нуклеиновой кислоты. Как эти термины применяют в настоящем документе, их применяют к потомкам трансформированных клеток, где желательное генетическое изменение сохранено на протяжении последующих поколений клеток, независимо от других мутаций или изменений, которые также могут присутствовать в клетках последующих поколений.
В одном варианте осуществления может быть представлен вектор, содержащий рекомбинантную молекулу нуклеиновой кислоты, обладающую кодирующей последовательностью аспарагиназы или транспортера аминокислот или положительного регуляторного фактора NCR, или мутантного отрицательного регуляторного фактора NCR, или их гомологов, под контролем гетерологичной промоторной последовательности, опосредующей регулируемую экспрессию полипептида. Для предоставления таких векторов открытую рамку считывания (ORF), например, открытую рамку считывания, полученную из микроорганизма-хозяина, можно вставлять в плазмиду, содержащую экспрессирующую кассету, которая может регулировать экспрессию рекомбинантного гена. Альтернативно, молекула нуклеиновой кислоты может представлять собой делеционную кассету для делеции отрицательного регуляторного фактора NCR. Рекомбинантную молекулу можно вводить в избранный микроорганизм для предоставления трансформированного штамма, обладающего измененным транспортом аспарагина и деградирующей активностью. В альтернативных вариантах осуществления на экспрессию природной кодирующей последовательности аспарагиназы или транспортера аминокислот или регуляторного фактора NCR или гомолога у хозяина можно влиять также посредством замены природного промотора на другой промотор. Дополнительные регуляторные элементы также можно использовать для конструирования рекомбинантных экспрессирующих кассет с использованием эндогенной кодирующей последовательности. Рекомбинантные гены или экспрессирующие кассеты можно интегрировать в хромосомную ДНК хозяина.
В одном варианте осуществления микроорганизмы трансформируют для постоянной деградации и/или поглощения молекул аспарагина в условиях приготовления/переработки пищевых продуктов. Например, молекулу нуклеиновой кислоты можно функционально связывать с конститутивно активным промотором. Конститутивно активные промоторы известны в данной области и включают в себя, без ограничения, промотор PGK1, промотор TEF, усеченный промотор HXT7. Альтернативно, молекулу нуклеиновой кислоты можно функционально связывать с промотором, который не подвергается азотной катаболитной репрессии, таким как ADH1, GAL1, CUP1, PYK1 или CaMV 35S.
Термин «промотор», как применяют в настоящем документе, относится к нуклеотидной последовательности, способной опосредовать или модулировать транскрипцию интересующей нуклеотидной последовательности с желательной пространственной или временной картиной и до желательной степени, когда регулирующая транскрипцию область функционально связана с интересующей последовательностью. Регулирующая транскрипцию область и интересующая последовательность являются «функционально или оперативно связанными», когда последовательности функционально соединены, так чтобы позволять опосредование или модуляцию транскрипции интересующей последовательности регулирующей транскрипцию областью. В некоторых вариантах осуществления, чтобы являться функционально связанной, регулирующая транскрипцию область может быть локализована на той же цепи, что и интересующая последовательность. Регулирующая транскрипцию область может в некоторых вариантах осуществления быть локализована на 5' от интересующей последовательности. В таких вариантах осуществления регулирующая транскрипцию область может находится непосредственно на 5' от интересующей последовательности, или могут существовать промежуточные последовательности между этими областями. Регулирующие транскрипцию последовательности могут в некоторых вариантах осуществления быть локализованы на 3' от интересующей последовательности. Для функциональной связи регулирующей транскрипцию области и интересующей последовательности может являться необходимым связывание соответствующих молекул (таких как белки - активаторы транскрипции) с регулирующей транскрипцию областью, таким образом, описание включает варианты осуществления, в которых представлены такие молекулы, in vitro или in vivo.
Промоторы для использования включают в себя, без ограничения, промоторы, выбранные из подходящих природных промоторов S. cerevisiae, таких как промотор PGK1. Такие промоторы можно использовать с дополнительными регуляторными элементами, такими как терминатор PGK1. Можно использовать множество природных или рекомбинантных промоторов, где промоторы выбирают или конструируют для опосредования проявления активности деградации аспарагина, такой как активность Asp3p, в выбранных условиях, таких как условия приготовления и переработки пищевых продуктов. Множество конститутивных промоторов могут быть, например, функционально связаны с кодирующей последовательностью.
В одном варианте осуществления молекула нуклеиновой кислоты содержит генетическую кассету ASP3 или GNP1, или AGP2, или AGP3, или GAT1 (фиг.1, 3, 4, 5 или 7), которую вставляют в локус LEU2. В другом варианте осуществления молекула нуклеиновой кислоты содержит кассету GAP1 или AGP1 или ASP3, которую вставляют в локус URA3 (фиг.1, 2 и 6). В другом варианте осуществления молекула нуклеиновой кислоты содержит кассету ure2Δ, которую вставляют в локус URE2 (фиг.8).
Методы
В другом аспекте представлен способ снижения количества аспарагина во время приготовления или переработки пищевых продуктов, включающий в себя добавление микроорганизма, описанного в настоящем документе, в пищевой продукт в условиях приготовления или переработки; где микроорганизм снижает азотную катаболитную репрессию и/или сверхэкспрессирует ген, кодирующий внеклеточный белок, вовлеченный в деградацию аспарагина, и/или ген, кодирующий белок, вовлеченный в транспорт аспарагина, таким образом снижая количество аспарагина в пищевом продукте. Также в настоящем документе представлено использование микроорганизмов, описанных в настоящем документе, для снижения количества аспарагина в условиях приготовления или переработки пищевых продуктов.
В другом варианте осуществления представлен способ снижения количества аспарагина во время приготовления или переработки пищевых продуктов, включающий в себя
a) трансформацию микроорганизма по меньшей мере одной молекулой нуклеиновой кислоты для снижения азотной катаболитной репрессии и/или для сверхэкспрессии гена, кодирующего внеклеточный белок, вовлеченный в деградацию аспарагина, и/или гена, кодирующего белок, вовлеченный в транспорт аспарагина;
b) добавление микроорганизма в пищевой продукт в условиях приготовления или переработки пищевого продукта;
где микроорганизм снижает азотную катаболитную репрессию и/или сверхэкспрессирует ген, кодирующий внеклеточный белок, вовлеченный в деградацию аспарагина, и/или ген, кодирующий белок, вовлеченный в транспорт аспарагина, таким образом, снижая количество аспарагина.
Аспарагин является лимитирующим предшественником в реакции, образующей акриламид во время приготовления или переработки пищевого продукта. Соответственно, в другом варианте осуществления представлен способ снижения количества акриламида в пищевом продукте, включающий в себя добавление микроорганизма, описанного в настоящем документе, в пищевой продукт в условиях приготовления или переработки; где микроорганизм снижает азотную катаболитную репрессию и/или сверхэкспрессирует ген, кодирующий внеклеточный белок, вовлеченный в деградацию аспарагина и/или ген, кодирующий белок, вовлеченный в транспорт аспарагина, таким образом снижая количество акриламида в пищевом продукте. В настоящем документе представлено также использование микроорганизмов, описанных в настоящем документе, для снижения концентрации акриламида в условиях приготовления или переработки пищевых продуктов.
В другом варианте осуществления представлен способ снижения количества акриламида в пищевом продукте, включающий в себя
a) трансформацию микроорганизма по меньшей мере одной молекулой нуклеиновой кислоты для снижения азотной катаболитной репрессии и/или для сверхэкспрессии гена, кодирующего внеклеточный белок, вовлеченный в деградацию аспарагина, и/или гена, кодирующего белок, вовлеченный в транспорт аспарагина;
b) добавление микроорганизма в пищевой продукт в условиях приготовления или переработки пищевого продукта;
где микроорганизм снижает азотную катаболитную репрессию и/или сверхэкспрессирует ген, кодирующий внеклеточный белок, вовлеченный в деградацию аспарагина и/или ген, кодирующий белок, вовлеченный в транспорт аспарагина, таким образом, снижая количество акриламида в пищевом продукте.
В одном варианте осуществления молекула нуклеиновой кислоты кодирует аспарагиназу клеточной стенки, как описано в настоящем документе, и в условиях приготовления или переработки пищевого продукта микроорганизм экспрессирует аспарагиназу, например, посредством конститутивной экспрессии. В другом варианте осуществления молекула нуклеиновой кислоты кодирует транспортер аминокислот, как описано в настоящем документе, и в условиях приготовления или переработки пищевого продукта экспрессирует транспортер аминокислот, например, посредством конститутивной экспрессии. В другом варианте осуществления, молекула нуклеиновой кислоты кодирует как аспарагиназу клеточной стенки, так и транспортер аминокислот. В другом варианте осуществления нуклеиновая кислота модифицирует регуляторный фактор азотной катаболитной репрессии, как описано в настоящем документе, и в условиях приготовления или переработки пищевого продукта регуляторный фактор не экспрессируется, так что NCR-чувствительные гены экспрессируются в присутствии хороших источников азота. В другом варианте осуществления, после трансформации микроорганизм выращивают в условиях, позволяющих сверхэкспрессию желаемых белков, и затем микроорганизм инактивируют и перерабатывают для добавления в пищевой продукт в условиях приготовления или переработки пищевого продукта. В таком варианте осуществления белки в неактивном микроорганизме обладают активностью деградации аспарагина, таким образом, снижая количество акриламида в пищевом продукте.
В одном варианте осуществления условия приготовления или переработки пищевого продукта включают в себя ферментацию. Например, способы и применения в настоящем документе являются пригодными для ферментации пищевого продукта, включая, без ограничения, углеводы в процессе выпечки хлеба, переработки картофеля, изготовления печенья, изготовления кофе или изготовления закусочных пищевых продуктов.
В другом варианте осуществления в описании представлен способ отбора природных мутантов ферментирующего организма, обладающих желательным уровнем активности деградации аспарагина в условиях приготовления и переработки пищевых продуктов. Например, можно отбирать штаммы с отсутствием NCR транспортера аминокислот или аспарагиназы клеточной стенки, например, ASP3, GAP1, GNP1, AGP1, AGP2, AGP3, TOR1, TOR2, DIP5, GLN3, GAT1, DAL80, GZF3 или URE2. Пример мутации и протоколов отбора для дрожжей, см. в Патенте Соединенных Штатов No. 6140108, выданном Mortimer et al. 31 октября 2000 г., содержание которого приведено в настоящем документе посредством ссылки. В таких способах штамм дрожжей можно обрабатывать мутагеном, таким как этилметансульфонат, азотистая кислота, или гидроксиламин, образующим мутанты с помощью замен пар оснований. Можно проводить скрининг мутантов с измененной активностью деградации аспарагина, например, посредством рассева на подходящую среду.
В другом варианте осуществления можно использовать сайт-специфический мутагенез для изменения уровня транспорта аспарагина или активности деградации аспарагина у хозяина. Например, сайт-специфический мутагенез можно использовать для удаления опосредующих NCR элементов из промотора, такого как промотор дрожжей AGP1, ASP3, GAP1, DIP5, GAT1, TOR2, DAL80 или GZF3. Например, GATAA(G) боксы в природных последовательностях промоторов AGP1, ASP3, GAP1, DIP5, GAT1, TOR2, DAL80 или GZF3, как показано на SEQ ID NO:23-28, 35 и 36, соответственно, можно делетировать или модифицировать посредством замены. В одном варианте осуществления, например, один или все из GATAA боксов можно модифицировать посредством замены T вместо G, так что последовательность становится TATAA. Способы сайт-специфического мутагенеза описаны, например, в: Rothstein, 1991; Simon and Moore, 1987; Winzeler et al., 1999; и Negritto et al., 1997. Отобранные или модифицированные промоторы с отсутствием NCR можно затем функционально связывать с кодирующей последовательностью аспарагиназы или транспортера аминокислот, для опосредования экспрессии белка в условиях приготовления и переработки пищевых продуктов. В альтернативных вариантах осуществления гены, кодирующие Gln3p, Gat1p, Ure2p, Tor1/2p, Dal80p или Gzf3p, которые опосредуют NCR в S. cerevisiae, также можно подвергать мутации для модуляции NCR.
Относительный транспорт аспарагина или деградирующую ферментативную активность штамма микроорганизма можно измерять относительно нетрансформированного исходного штамма. Например, можно отбирать трансформированные штаммы по наличию большего транспорта аспарагина или деградирующей активности, чем исходный штамм, в условиях приготовления и переработки пищевых продуктов, или активности, которая составляет несколько больший процент, чем активность исходного штамма в тех же условиях ферментации, например, по меньшей мере 150%, 200%, 250%, 300%, 400% или 500% активности исходного штамма. Подобным образом, активность ферментов, экспрессируемых или кодируемых рекомбинантными нуклеиновыми кислотами по описанию, можно определять относительно нерекомбинантных последовательностей, из которых они получены, с использованием сходных коэффициентов активности.
В одном варианте осуществления способов и применений, описанных в настоящем документе, микроорганизм представляет собой любой активный или неактивный микроорганизм, пригодный для добавления в пищевые продукты, включая, без ограничения, грибы и/или бактерии. Как описано в настоящем документе, грибы, пригодные для настоящих способов и применений, включают в себя, без ограничения, Aspergillus niger, Aspergillus oryzae, Neurospora crassa, Neurospora intermedia var. oncomensis, Penicillium camemberti, Penicillium candidum, Penicillium roqueforti, Rhizopus oligosporus, Rhizopus oryzae. В другом варианте осуществления грибы представляют собой дрожжи, такие как Saccharomyces cerevisiae, Saccharomyces bayanus, Saccharomyces carlsbergensis, Candida albicans, Candida kefyr, Candida tropicalis, Cryptococcus laurentii, Cryptotoccous neoformans, Hansenula anomala, Hansenula polymorpha, Kluyveromyces fragilis, Kluyveromyces lactis, Kluyveromyces marxianus var lactis, Pichia pastoris, Rhodotorula rubra, Schizosaccharomyces pombe, Yarrowia lipolyitca или любой штамм, принадлежащий к царству грибов. Бактерии могут представлять собой любые бактерии, включая Erwinia sp., Lactobacillus sp., Lactococcus sp., Bacillus sp., Pediococcus sp., Pseudomonas sp., Brevibacterium sp. и Leuconostoc sp.
Пищевые продукты
В другом аспекте настоящее описание относится к пищевому продукту, обладающему сниженной концентрацией акриламида, полученному с использованием трансформированного микроорганизма, описанного в настоящем документе.
В другом варианте осуществления настоящее описание относится к пищевому продукту, обладающему сниженной концентрацией акриламида, полученному с использованием способов, описанных в настоящем документе.
Пищевой продукт может представлять собой любой пищевой продукт, полученный в условиях приготовления или переработки, приводящих к продукции аспарагина и в конечном счете к продукции акриламида. Типичные условия приготовления и переработки, приводящие к продукции акриламида, включают в себя приготовление, включающее в себя высокие температуры кулинарной обработки (более 120°C) и включают в себя, без ограничения, обжаривание во фритюре и выпекание, поджаривание, обжаривание, приготовление на гриле, тушение и жарение на открытом огне. Акриламид, как правило, обнаруживают в высокой концентрации в полученных из картофеля продуктах, хлебобулочных изделиях и в любых продуктах из злаков или зерна (см. также таблицу 1). Соответственно, в одном варианте осуществления пищевой продукт представляет собой полученный из овощей продукт, такой как полученный из картофеля, таро или оливок продукт, хлебобулочное изделие или продукт из злаков или зерна. Продукты из картофеля включают в себя, без ограничения, картофель фри, картофельные чипсы, обжаренные во фритюре/выпеченные закуски из картофеля и формованные продукты из картофеля. Хлебобулочные изделия включают в себя, без ограничения, бисквиты, печенье, крекеры, хлеб, безопарные хлебные продукты, обжаренные в кляре продукты, тортильи из кукурузы и муки, мучные кондитерские изделия, коржи для тортов, смеси для тортов и кексов и тесто для мучных кондитерских изделий. Например, хлеб может включать в себя, без ограничения, свежие и замороженные хлеб и тесто, закваску, тесто для пиццы, сдобные булки и рулеты и различные виды хлеба, так же как родственные хлебопродукты, такие как обжаренные во фритюре или выпеченные закуски или панировочные сухари; и мучные кондитерские изделия могут включать в себя, без ограничения, сладкие сдобные булки, пончики и торты. Продукты из злаков или зерна включают в себя, без ограничения, типичные сухие завтраки, пивной солод и продукты из молочной сыворотки, кукурузные чипсы и соленые крендели. Другие пищевые продукты, которые перерабатывают при высоких температурах включают в себя, без ограничения, кофе, обжаренные орехи, обжаренную спаржу, пиво, солод и напитки из молочной сыворотки, порошок шоколада, продукты из рыбы, продукты из мяса и домашней птицы, луковый суп и смесь для макания, ореховая паста, глазированный арахис, обжаренные соевые бобы, обжаренные семена подсолнечника, обжаренные во фритюре или выпеченные пищевые продукты, такие как фалафель и кеббе и шоколадные батончики.
Настоящее описание в общем описано в приведенном выше описании. Более полное понимание можно получить по отношению к следующим ниже конкретным примерам. Эти примеры описаны единственно с целью иллюстрации и не предназначены для ограничения объема описания. Предусмотрены изменения формы и замена эквивалентов, как могут диктовать условия или как представляется целесообразным. Хотя в настоящем документе используют конкретные термины, такие термины подразумевают в описательном смысле, а не с целью ограничения.
Следующие неограничивающие примеры иллюстрируют настоящее описание:
ПРИМЕРЫ
Пример 1: Клонирование и конститутивная экспрессия генов ASP3, ASP1, GAP1, GNP1, AGP1, AGP2, AGP3 и GAT1 в штамме Saccharomyces cerevisiae и делеция URE2, TOR1, DAL80 и GZF3
Для отбора клонов использовали маркер устойчивости к антибиотику kanMX. Штамм промышленных/коммерческих хлебных дрожжей или лабораторный штамм трансформировали для конститутивной экспрессии ASP3, ASP1, GAP1, GNP1, AGP1, AGP2, AGP3 или GAT1, или комбинации ASP3 и GAP1, или комбинации ASP3 и GAT1, или для делеции гена URE2, TOR1, DAL80 или GZF3, или для комбинации tor1Δ и сверхэкспрессии ASP3. Единственными генетическими и метаболическими модификациями являлись намеченная конститутивная экспрессия ASP3, ASP1, GAP1, GNP1, AGP1, AGP2, AGP3 или GAT1, или комбинации ASP3 и GAP1, или комбинации ASP3 и GAT1, или наличие делеции гена URE2 TOR1, DAL80 и GZF3, или комбинация tor1Δ и сверхэкспрессии ASP3.
Пример 2: Трансформация дрожжей генной кассетой ASP3, ASP1, GAP1, GNP1, AGP1, AGP2, AGP3 или GAT1 или делеционной кассетой гена URE2
Дрожжи трансформировали рекомбинантной нуклеиновой кислотой, содержащей ген ASP3, ASP1, GAP1, GNP1, AGP1, AGP2, AGP3 или GAT1 под контролем промотора и сигнала терминации PGK1. Промотор PGK1 не подвержен NCR. Делеционная кассета URE2 содержала 5'- и 3'-фланкирующие последовательности URE2 для направленной делеции гена.
Пример 3: Самоклонирующаяся кассета, позволяющая удаление селективного маркера
Фиг.1-8 иллюстрируют, как разработанные генетические кассеты позволяют отбор трансформированных дрожжей и последующее удаление маркера устойчивости к антибиотику посредством рекомбинации прямых повторов, используемых в этом примере, как описано ниже. Самоклонирующуюся кассету ASP1 конструировали сходным образом, проводили трансформацию и удаление маркера устойчивости к антибиотику, как проиллюстрировано для других примеров.
Пример 4: Исследования снижения количества аспарагина и акриламида с помощью самоклонирующихся дрожжей для определения случаев снижения количества акриламида или лимитирующего предшественника аспарагина
На фиг.11-20 показано значительное снижение количества аспарагина и/или акриламида для дрожжей, трансформированных с помощью ASP3, GAP1, GNP1, AGP1, AGP2, AGP3 или GAT1, или комбинации ASP3 и GAP1, или комбинации ASP3 и GAT1, или обладающих делецией гена URE2, TOR1, DAL80 или GZF3 или комбинацией tor1Δ и сверхэкспрессии ASP3. На фигуре 11 ясно показано также, что сверхэкспрессия цитозольного ASP1 не действует по сравнению со сверхэкспрессией ASP3, кодирующего связанную с клеточной стенкой аспарагиназу.
Некоторые из трансформированных штаммов тестировали в хлебе, например, ASP3, GAP1/ASP3 и ure2∆ (фиг.11, 13 и 14). Как трансформированные штаммы, так и контрольные штаммы коммерческих хлебных дрожжей выращивали одновременно в двух отдельных ферментерах, и клетки собирали на следующие сутки для исследований теста. Аспарагин добавляли в тесто для мониторирования потребления аспарагина с использованием ферментного анализа. После смешивания трансформированных дрожжей с тестом отмечали, что уровни аспарагина немедленно начинали снижаться; в отличие от этого, заметного уменьшения количества аспарагина не измеряли при использовании контрольного штамма. После формирования теста с момента добавления дрожжей периодически отбирали образцы для тестирования концентрации аспарагин. Тесто из некоторых из этих экспериментов (с более высокими уровнями аспарагина) использовали также для получения выпеченного образца для определения концентрации акриламида в конечном хлебном продукте. Результаты для акриламида из этого эксперимента показаны на фиг.12 и 14, и выявлено, что штаммы трансформированных дрожжей снижали количество акриламида значительно больше, чем контрольные образцы дрожжей. Этот результат согласуется со снижением количества аспарагина, обнаруженным при анализе теста.
Трансформированные дрожжи тестировали также в жидкой среде для имитации промышленных условий переработки, где условия внешней среды для дрожжей могут иметь более высокое содержание влажности (т.е. производство картофеля, злаков и кофе). Равные количества клеток каждого штамма инокулировали в отдельные пробирки, содержащие комплексную среду или синтетическую лабораторную среду с точным добавлением различных уровней аспарагина. Образцы отбирали периодически, и концентрацию аспарагина определяли с использованием ферментного набора или посредством LC-MS/MS. На фиг.16-20 показаны штаммы трансформированных дрожжей с улучшенной деградацией аспарагина.
Для снижения количества акриламида в пищевых продуктах, производители сталкиваются с задачей изменения параметров процессов и/или продуктов без ухудшения вкуса, текстуры и внешнего вида их продуктов. В качестве примера изготовлены различные виды хлеба с использованием трансформированных дрожжей и контрольных коммерческих хлебных дрожжей. Для конечных продуктов не показано различий в цвете, размере или текстуре. Важно, что не потребовалось никаких изменений в способе выпечки для достижения этого значительного снижения образования акриламида в хлебе.
ЭКСПЕРИМЕНТАЛЬНЫЕ СПОСОБЫ, ИСПОЛЬЗУЕМЫЕ В ВЫШЕУКАЗАННЫХ ПРИМЕРАХ
1. Конструирование pAC1-ASP3, pAC1-AGP1, pAC1-AGP3, pAC1-GNP1 и pAC1-GAT1
Для помещения ASP3, AGP1, GNP1 и GAT1 под контроль конститутивного промотора и сигналов терминации PGK1, каждую из ORF клонировали в pAC1 (фиг.9). Каждую ORF от старта до стоп-кодона амплифицировали с геномной ДНК S. cerevisiae с использованием праймеров, содержащих участки для ферментов рестрикции Mlu1 и Bmt1 на 5'-концах.
После ПЦР, визуализации в 0,8% агарозном геле и очистки продуктов ПЦР (Qiagen, USA - PCR Purification Kit), как продукт ПЦР (вставку), так и pAC1 (вектор) расщепляли Mlu1 и Bmt1 (Fermentas, Canada). После обработки расщепленного вектора щелочной фосфатазой rAPiD (Roche, USA) для предотвращения рециркуляризации, вставку и дефосфорилированный вектор лигировали при комнатной температуре (ДНК-лигаза T4 - Roche, USA); смесь после лигирования (2 мкл) использовали для трансформации компетентных клеток DH5α™ (Invitrogen, USA), которые затем выращивали на чашках с LB (Difco, USA), дополненных 100 мкг/мл ампициллина (Sigma-Aldrich, USA). Плазмиды из случайной выборки трансформированных колоний собирали (Qiagen, USA - QIAprep Spin Miniprep kit) и расщепляли Mlu1 и Bmt1 (Fermentas, Canada) для идентификации плазмид с корректным размером вставки; секвенирование подтвердило, что вставка соответствовала AGP1, AGP3, GNP1 или GAT1.
2. Конструирование pAC2-GAP1, pAC2-AGP1 и pAC2-ASP3
Для помещения GAP1, AGP1 и ASP3 под контроль конститутивных промоторного и терминаторного сигналов PGK1, каждую ORF клонировали в pAC2 (фиг.10). Каждую ORF от старта до стоп-кодона амплифицировали из геномной ДНК S. cerevisiae с использованием праймеров, содержащих участки для ферментов рестрикции Mlu1 и Bmt1, встроенные в 5'-концы.
После ПЦР, визуализации в 0,8% агарозном геле и очистки продуктов ПЦР (Qiagen, USA - PCR Purification Kit), как продукт ПЦР (вставку), так и pAC2 (вектор) расщепляли Mlu1 и Bmt1 (Fermentas, Canada). После обработки расщепленного вектора щелочной фосфатазой rAPiD (Roche, USA) для предотвращения рециркуляризации вставку и дефосфорилированный вектор лигировали при комнатной температуре (ДНК-лигаза T4 - Roche, USA); смесь после лигирования (2 мкл) использовали для трансформации компетентных клеток DH5α™ (Invitrogen, USA), которые затем выращивали на чашках с LB (Difco, USA), дополненных 100 мкг/мл ампициллина. Плазмиды из случайной выборки трансформированных колоний собирали (Qiagen, USA - QIAprep Spin Miniprep kit) и расщепляли Mlu1 и Bmt1 (Fermentas, Canada) для идентификации плазмид с корректным размером вставки; секвенирование подтвердило, что вставка соответствовала GAP1, AGP1 или ASP3.
3. Конструирование кассеты ure2∆
Кассету ure2∆ полностью получали синтезом ДНК (MrGene, Germany).
4. Трансформация линейными кассетами S. cerevisiae и отбор трансформантов
Каждую кассету вырезали из соответствующей плазмиды с использованием Swa1 (Fermentas, Canada) и визуализовали в 0,8% агарозном геле. Полосу ожидаемого размера из геля вырезали и экстрагировали (Qiagen, USA - Gel extraction kit). После экстракции, очистки и количественного определения 500 нг линейной кассеты использовали для трансформации штаммов S. cerevisiae. Штаммы дрожжей трансформировали с использованием способа с ацетатом лития/полиэтиленгликолем/оцДНК. После трансформации клетки оставляли для восстановления в YEG при 30°C на 3 часа перед рассевом на чашки с YPD, дополненной 500 мкг/мл G418 (Sigma, USA). Чашки инкубировали при 30°C до появления колоний.
5. Трансформация линейной кассетой ure2∆ S. cerevisiae и отбор трансформантов
Кассету ure2∆ размером 3149 п.о. вырезали из pMrG-ure2∆ с использованием Pme1 (Fermentas, Canada) и визуализовали в 0,8% агарозном геле. Полосу ожидаемого размера 3149 п.о. вырезали и экстрагировали (Qiagen, USA - Gel extraction kit). После экстракции, очистки и количественного определения, 500 нг линейной кассеты использовали для трансформации штаммов PDM S. cerevisiae. Штаммы дрожжей трансформировали с использованием способа с ацетатом лития/полиэтиленгликолем/оцДНК. После трансформации клетки оставляли для восстановления в YEG при 30°C на 3 часа перед рассевом на чашки с YPD, дополненной 500 мкг/мл G418 (Sigma, USA). Чашки инкубировали при 30°C до появления колоний.
Штаммы лабораторных дрожжей, являющихся делеционными мутантами для tor1∆, dal80∆, gzf3∆ и ure2∆, также получали из коммерческого источника для комплектации некоторых тестов.
6. Исследования снижения концентрации аспарагина и акриламида
Тесто для хлеба из цельнозерновой пшеничной муки получали со следующими ингредиентами: мука из цельной пшеницы, нативная пшеничная клейковина, соль, растительное масло, меласса, вода и дрожжи (либо тестируемый штамм, либо контроль). Способ близко следовал процессу из способа «безопарного теста». Во временной точке 5 час образцы также нагревали для получения данных по акриламиду (подробности приведены ниже).
1. Охладить сосуд Дьюара для жидкого азота в морозильнике при -30°C и наполнить жидким N2.
2. В 250-мл бутыли для среды растворить L-аспарагин в 50-мл фильтрованной воды.
3. Определить содержание влаги/твердых веществ в дрожжах (влажных или сухих), подлежащих добавлению по рецептуре теста.
4. Отмерить вычисленное количество дрожжей в 200-мл коническую пробирку Falcon.
5. Определить необходимое количество очищенной с помощью обратного осмоса воды с учетом содержания влаги, поступающей с дрожжами, подлежащими добавлению. Отмерить необходимое количество очищенной с помощью обратного осмоса воды по массе на чашечных весах.
6. Ресуспендировать соответствующее количество дрожжей с помощью 2/3 оставшейся очищенной с помощью обратного осмоса воды (30°C). Использовать оставшуюся 1/3 для промывки.
7. Определить массу чаши для смешивания.
8. Отвесить сухие ингредиенты (муку, глютен и соль) в чашу для смешивания Kitchen Aid. Перемешать сухие ингредиенты с помощью лопатки в течение 20-30 с. Заменить лопатку на крюк.
9. Добавить отмеренное растительное масло и мелассу, и раствор L-аспарагина в чашу для смешивания. Перемешивать на скорости 2 до достижения равномерной консистенции теста.
10. Установить таймер на 10 минут.
11. Добавить суспензию дрожжей к перемешиваемому тесту. Немедленно запустить таймер и перемешивать на скорости 2.
Время добавления дрожжей:
12. Промыть пробирку Falcon с помощью оставшейся воды и добавить смыв в чашу для смешивания.
13. Продолжать перемешивание до сигнала таймера через 10 минут.
14. Определить конечную массу чаши для смешивания + тесто:
15. Немедленно раскатать тесто до толщины ~1,0 см и использовать круглый штамп для печенья для вырезания соответствующего количества образцов теста для эксперимента.
Быстро отобрать 1 образец теста и разломить, и затем налить жидкий азот в ступку для замораживания кусков теста. Это представляет собой образец «T=15 мин».
Сохранять замороженные куски теста в маркированной 50-мл пробирке Falcon при -80°C для дальнейшего анализа.
16. Поместить оставшиеся образцы теста на бумагу для выпечки и инкубировать при 30°C.
17. Отбирать образец теста в желательной для эксперимента временной точке и разламывать на мелкие куски, и замораживать с помощью жидкого азота.
Сохранять замороженные куски в маркированной 50-мл пробирке Falcon при -80°C.
18. Для некоторых экспериментов при T=5 часов вырезать дополнительное печенье и выпекать при 400°F (204°C) в течение 20 мин, и сохранять при -80°C.
Препараты жидкой среды получали общепринятым способом и точно добавляли различные количества аспарагина. Равные количества клеток каждого штамма инокулировали в отдельные пробирки, содержащие стерильную подготовленную среду и периодически отбирали образы, концентрацию аспарагина определяли с использованием ферментного набора (Megazyme, K-ASNAM) или посредством LC-MS/MS (описана ниже).
7. Количественное определение аспарагина и акриламида
Ранее полученные образцы теста обрабатывали жидким азотом во время получения, чтобы прекратить активность аспарагиназы. Затем образцы размалывали и хранили при -80 градусов Цельсия до анализа. Анализ количества аспарагина в образцах теста проводили посредством ферментного анализа (K-ASNAM - Megazyme), следуя способу экстракции производителя для хлебобулочных изделий со следующими изменениями: Гомогенизированные образцы теста (2 г) быстро взвешивали и переносили в 100 мл мерные колбы. Приблизительно 90 мл MilliQ H2O при 80 градусов Цельсия добавляли для предотвращения любого повторного появления ферментативной активности, и образцы инкубировали в водяной бане при 80 градусах Цельсия в течение 20 минут. Затем образцы оставляли охлаждаться до комнатной температуры, доводили до метки и аликвоту осаждали центрифугированием (RT, 4000×g, 15 мин.) для анализа.
Акриламид в полученных в лаборатории выпеченных образцах анализировали способом ELISA. Образцы хлеба измельчали в мельнице, что тоже обеспечивало гомогенность. Образцы хранили при -80 градусов Цельсия до анализа. 2 г гомогенатов образцов взвешивали и экстрагировали водой в течение 30 минут. Затем образцы фильтровали и центрифугировали перед очисткой твердофазной экстракцией и элюцией акриламида. Затем экстрагированный аналит анализировали посредством анализа ELISA (Abraxis).
Для анализа аспарагина посредством LC-MS/MS образцы культуры клеток, полученные в жидкой среде, анализировали с использованием следующих параметров. Колонка 2×250 мм Aquasil (Thermo) и бинарная подвижная фаза, состоящая из 12% MeOH и 1 мМ формиата аммония, мониторирование ионных переходов аспарагина 133,0→74,0 и 133,0→87,0 (MRM). Внутренний стандарт из меченного изотопом 13C - акриламида (Cambridge Isotope Laboratories) использовали в концентрации 0,01 г/л, добавленным непосредственно в осветленные супернатанты культуры клеток.
В то время как настоящее описание описано в отношении того, что в настоящее время рассматривают как предпочтительные примеры, следует понимать, что описание не является ограниченным описанными примерами. Напротив, описание предназначено, чтобы охватывать различные модификации и эквивалентные схемы, включенные в содержание и объем прилагаемой формулы изобретения.
Полное содержание всех публикаций, патентов и патентных заявок приведено в настоящем документе в качестве ссылки в такой степени, как если бы конкретно и отдельно было указано, что полное содержание каждой отдельной публикации, патента или патентной заявки приведено в качестве ссылки.
Средневзвешенная величина



















Список литературы
Amrein, T.M., Schonbachler, B., Escher, F., Amado, R., 2004. Acrylamide in gingerbread: critical factors for formation and possible ways for reduction. Journal of Agricultural and Food Chemistry 52, 4282-4288.
Becalski, A., Lau, B.P.Y., Lewis, D., Seaman, S.W., 2003. Acrylamide in foods: occurrence, sources and modeling. Journal of Agricultural and Food Chemistry 51, 802-808.
Brathen, E., Kita, A., Knutsen, S.H., Wicklund, T., 2005. Addition of glycine reduces the content of acrylamide in cereal and potato products. Journal of Agricultural and Food Chemistry 53, 3259-3264.
Claus, A., Schreiter, P., Weber, A., Graeff, S., Hermann, W., Claupein, W., Schieber, A., Carle, R., 2006. Influence of agronomic factors and extraction rate on the acrylamide contents in yeast-leavened breads. Journal of Agricultural and Food Chemistry 54, 8968-8976.
Claus, A., Mongili, M., Weisz, G., Schieber, A., Carle, R., 2007. Impact of formulation and technological factors on the acrylamide content of wheat bread and bread rolls. Journal of Cereal Science 47, 546-554.
Cooper, T.G., 1982. In The Molecular Biology of the Yeast Saccharomyces: Metabolism and Gene Expression, eds Strathern J.N., Jones E.W., Borach J. (Cold Spring Harbour Laboratory, Cold Spring Harbor, NY), pp 39-99.
Fink, M., Andersson, R., Rosen, J., Aman, P., 2006. Effect of added asparagine and glycine on acrylamide content in yeast-leavened bread. Cereal Chemistry 83, 218-222.
Fredriksson, H., Tallving, J., Rosen, J., Aman, P., 2004. Fermentation reduces free asparagine in dough and acrylamide content in bread. Cereal Chemistry 81, 650-653.
Gietz, R.D., Schiestl, R.H., 1995. Transforming Yeast with DNA. Methods in Molecular and Cellular Biology. Vol 5, #5, 255-269.
Gokmen, V., Senyuva, H.Z., 2007. Acrylamide formation is prevented by divalent cations during the Maillard reaction. Food Chemistry 103, 196-203.
International Agency on Research on Cancer, 1994. IARC Monographs on the Evaluation of Carcinogenic Risks to Humans. Some Industrial Chemicals, vol. 60. Acrylamid, Lyon, France, IARC 1994, pp. 389-433.
Mustafa, A., Andersson, R., Rosen, J., Kamal-Eldin, A., Aman, P., 2005. Factors influencing acylamide content and color in rye crisp bread. Journal of Agricultural and Food Chemistry 53, 5985-5989.
Negritto, M.T., Wu, X., Kuo, T., Chu, S., Bailis, A.M., 1997. Influence of DNA sequence identity on efficiency of targeted gene replacement. Mol Cell Biol 17, 278-286.
Rice, J.M., 2005. The carcinogenicity of acrylamide. Mutation Research 580, 3-20.
Rothstein, R., 1991. Targeting, disruption, replacement, and allele rescue: integrative DNA transformation in yeast. Methods Enzymol. 194, 281-301.
Simon, J.R., Moore, P.D., 1987. Homologous recombination between single-stranded DNA and chromosolmal genes in Saccharomyces cerevisiae. Mol Cell Biochem 7, 2329-2334.
Surdyk, N., Rosen, J., Andersson, R., Aman, P., 2004. Effects of asparagine, fructose and baking conditions on acrylamide content in yeast-leavened wheat bread. Journal of Agricultural and Food Chemistry 52, 2047-2051.
Wilson, K.M., Rimm, E.B., Thompson, K.M., Mucci, L.A., 2006. Dietary acrylamide and cancer risk in humans: a review. Journal fur Verbraucherschutz und Lebensmittelsicherheit 1, 19-27. Cited in Claus, A., Carle, R., Schieber, A., 2008. Acrylamide in cereal products: a review. Journal of Cereal Science 47, 118-133.
Winzeler EA, Shoemaker DD, Astromoff A, Liang H, Anderson K, Andre B, Bangham R, Benito R, Boeke JD, Bussey H, Chu AM, Connelly C, Davis K, Dietrich F, Dow SW, El Bakkoury M.Foury F, Friend SH, Gentalen E, Giaever G, Hegemann JH, Jones T, Laub M, Liao H, Liebundguth N, Lockhart DJ, Lucau-Danila A, Lussier M, M'Rabet N, Menard P, Mittmann M,Pai C, Rebischung C, Revuelta JL, Riles L, Roberts CJ, Ross-MacDonald P, Scherens B, Snyder M, Sookhai-Mahadeo S, Storms RK, Veronneau S, Voet M, Volckaert G, Ward TR.Wysocki R, Yen GS, Yu K, Zimmermann K, Philippsen P, Johnston M, Davis RW., 1999. Functional characterization of the S. cerevisiae genome by gene deletion and parallel analysis. Science 285, 901-906.
Yaylayan, V.A., Wnorowski, A., Locas Perez, C, 2003. Why asparagine needs carbohydrates to generate acrylamide. Journal of Agricultural and Food Chemistry 51, 1753-1757.
Wickner, R.B., 1994. [URE3] as an altered URE2 protein: evidence for a prion analog in Saccharomyces cerevisiae. Science 264(5158), 566-9.
Wickner, R.B., Masison, D.C., Edskes, H.K., 1995. [PSI] and [URE3] as yeast prions. Yeast 11(16), 1671-85.










































































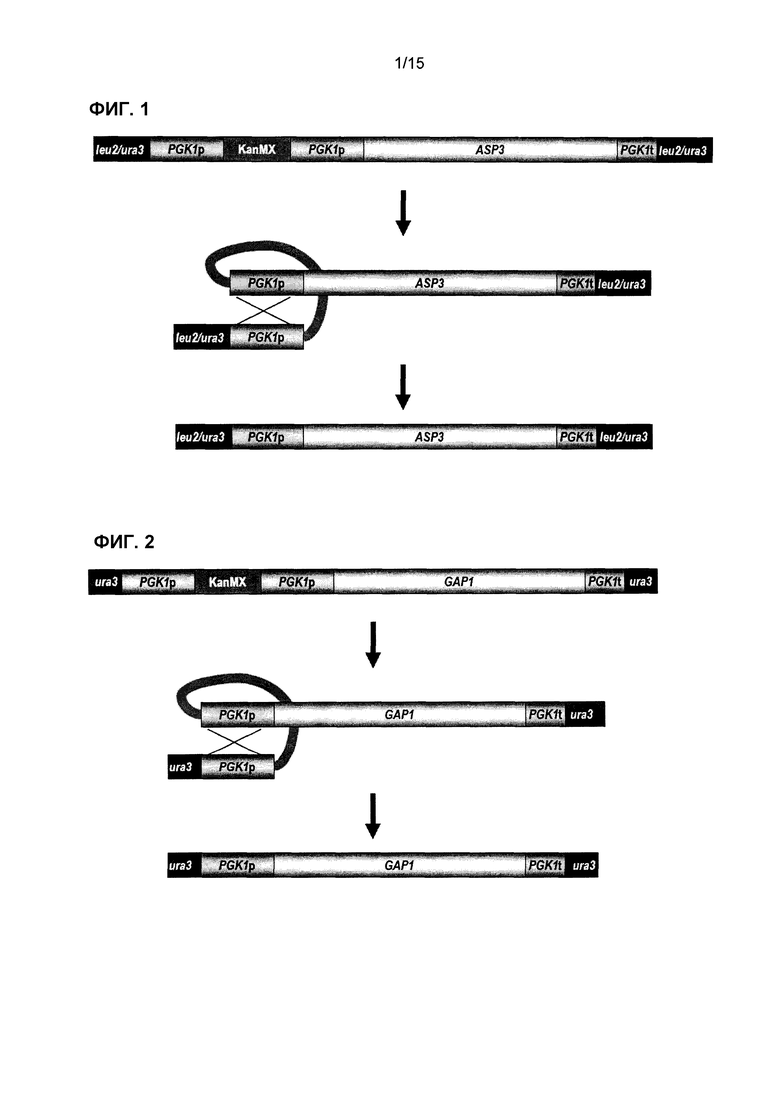
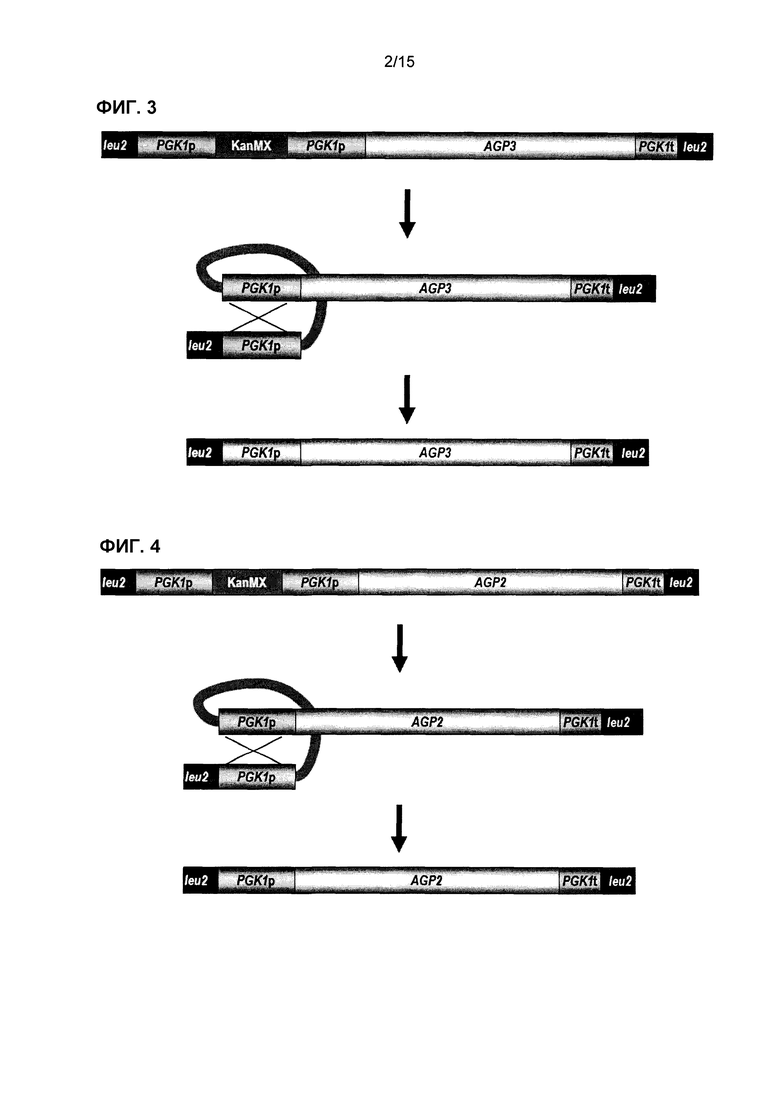

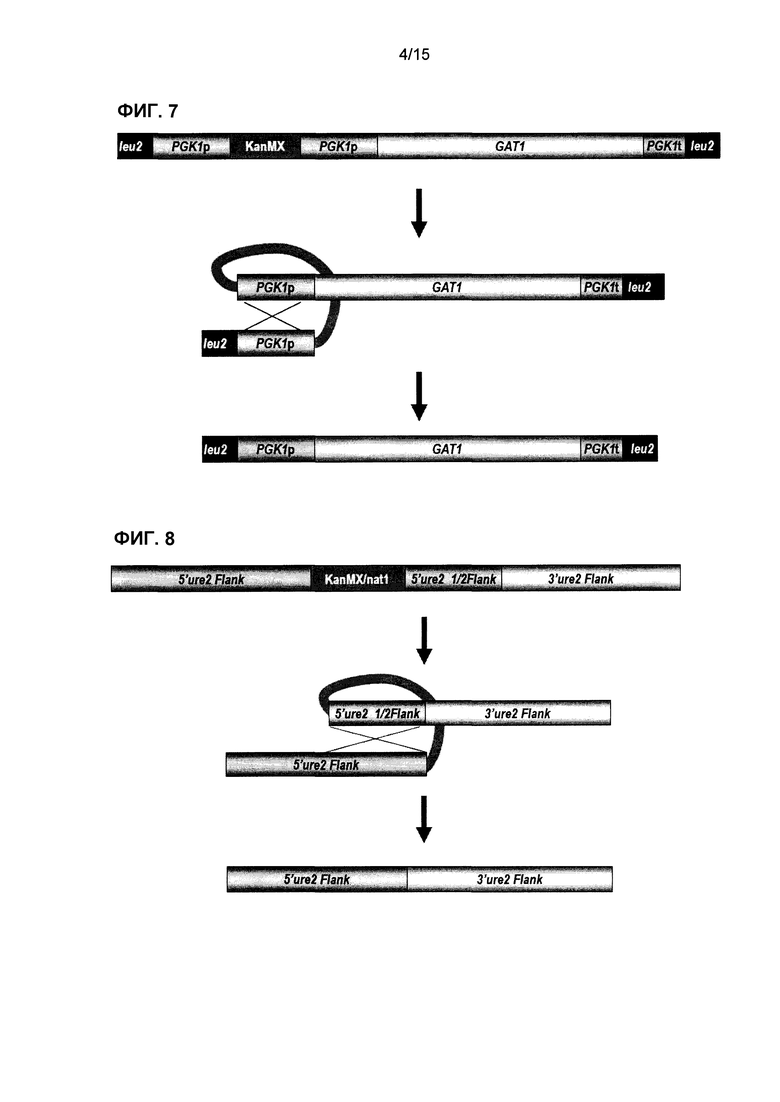


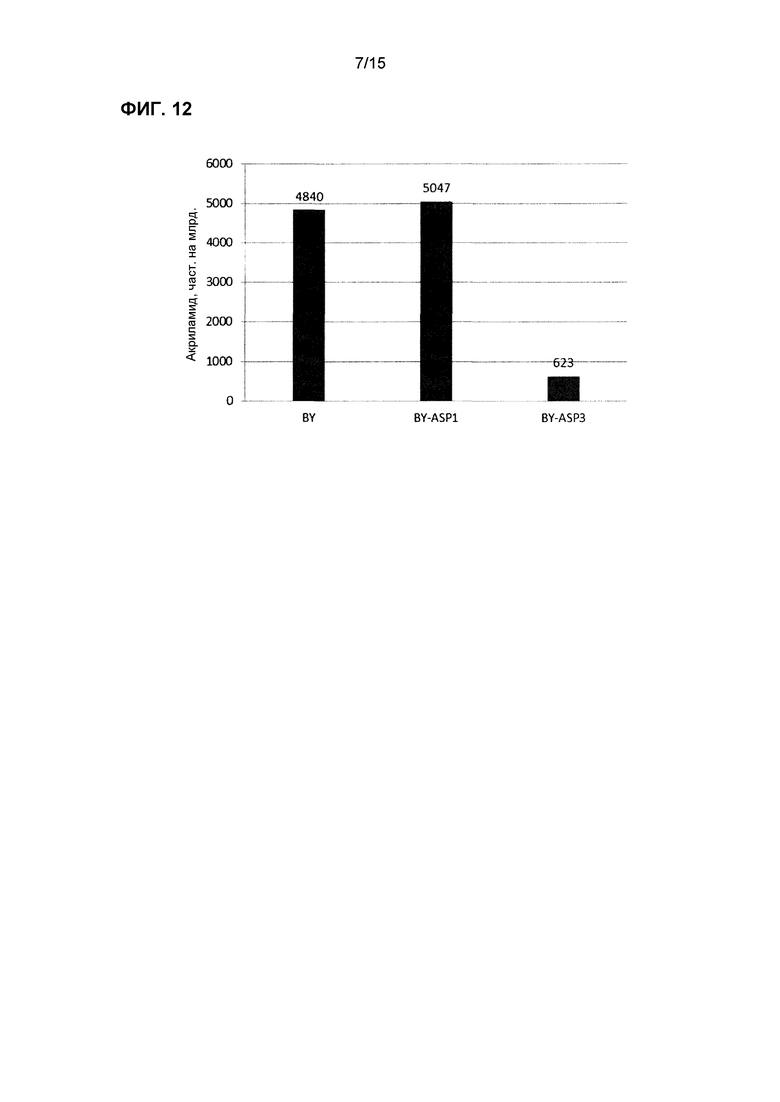
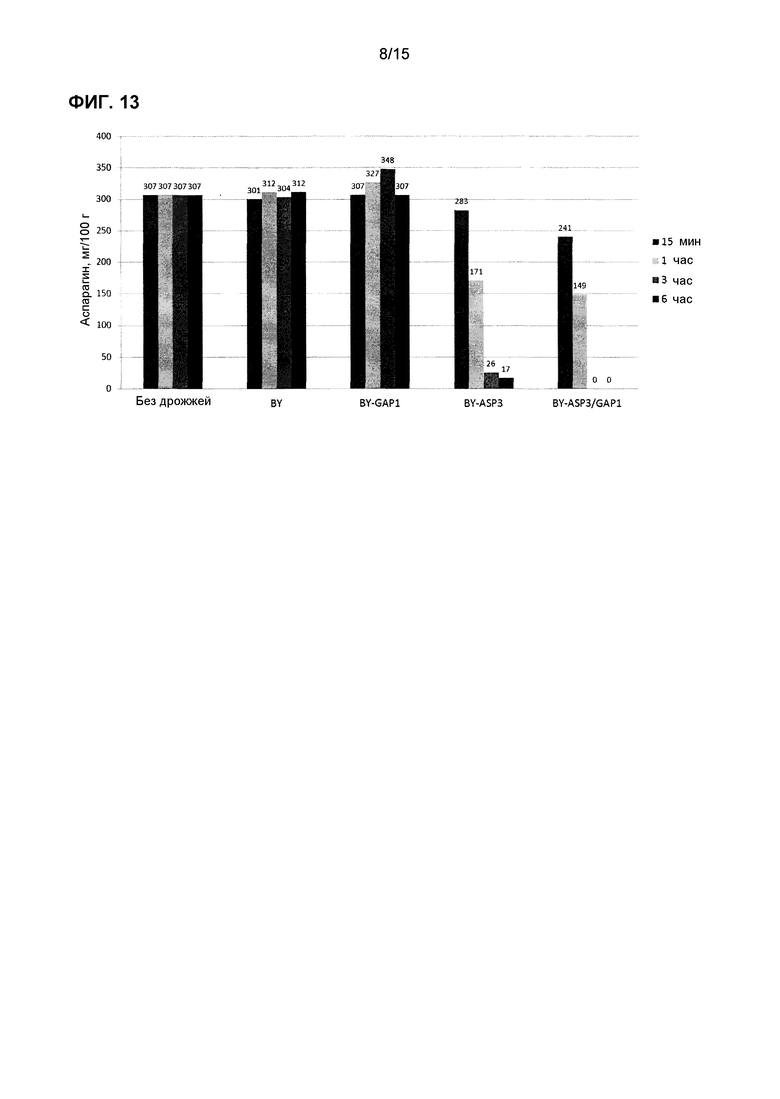

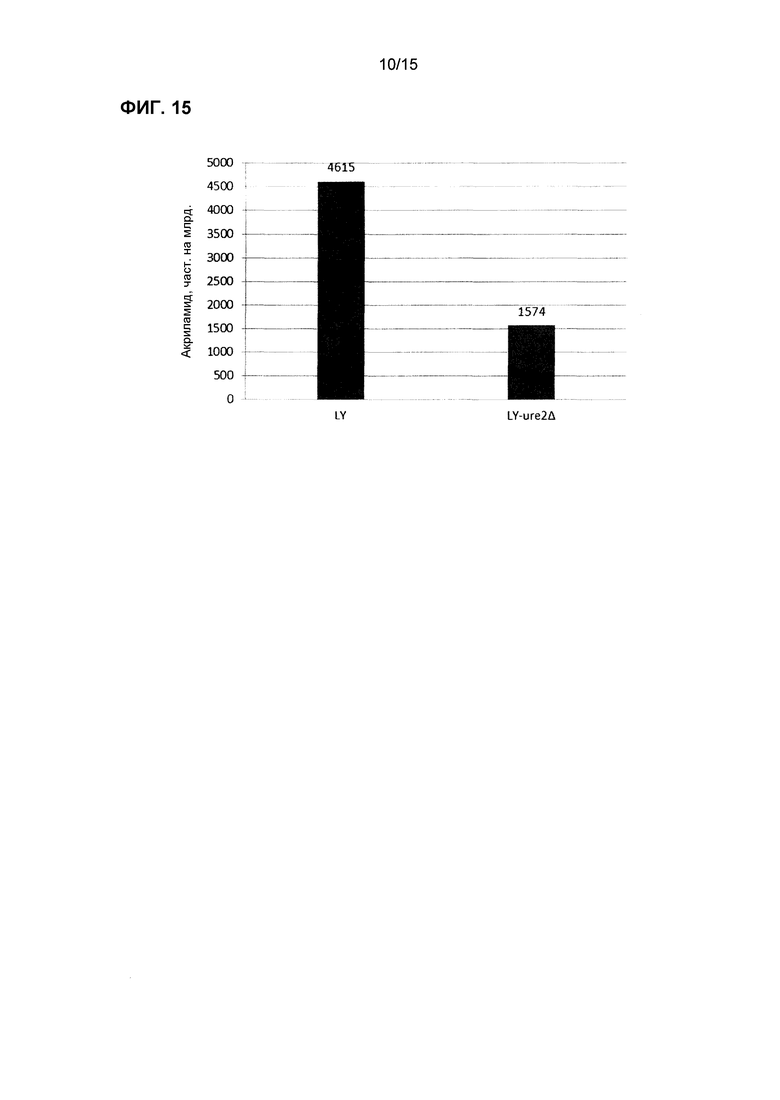

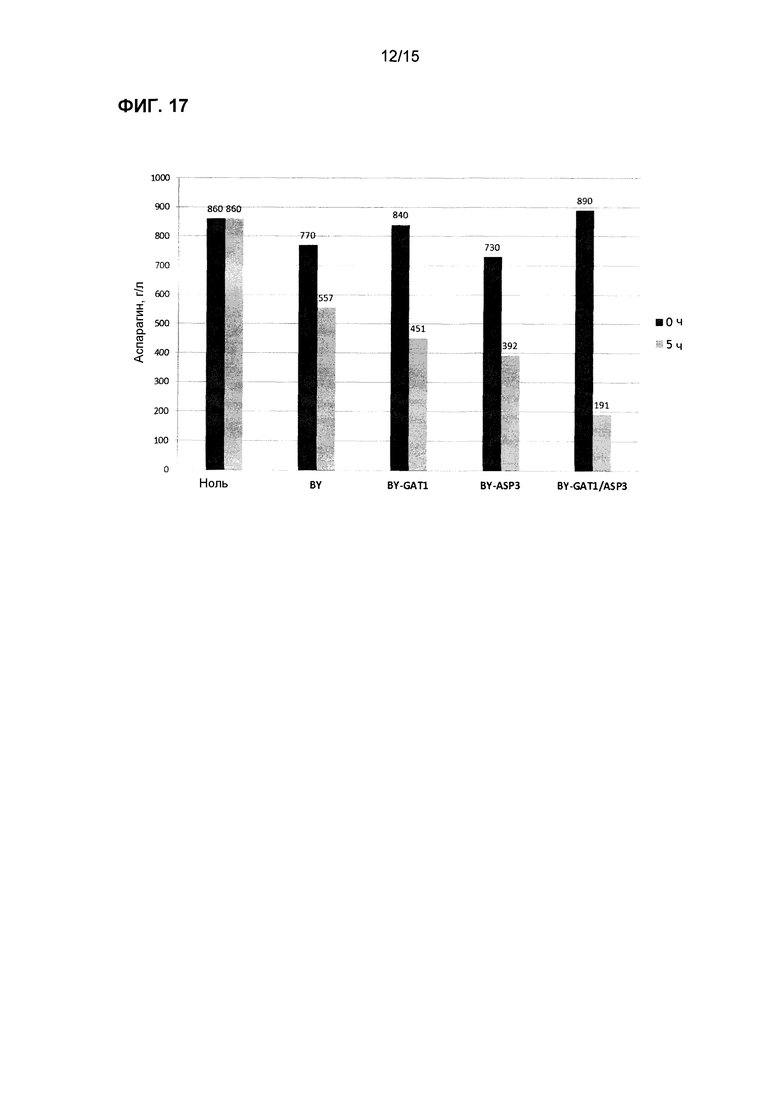
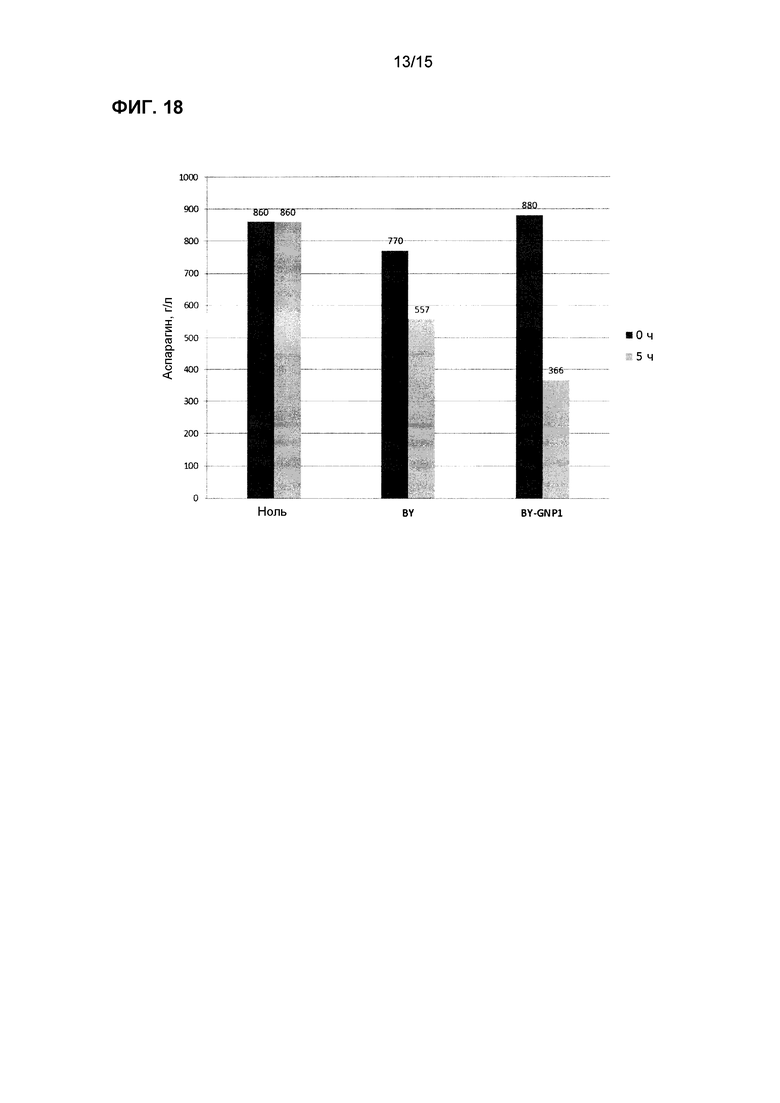
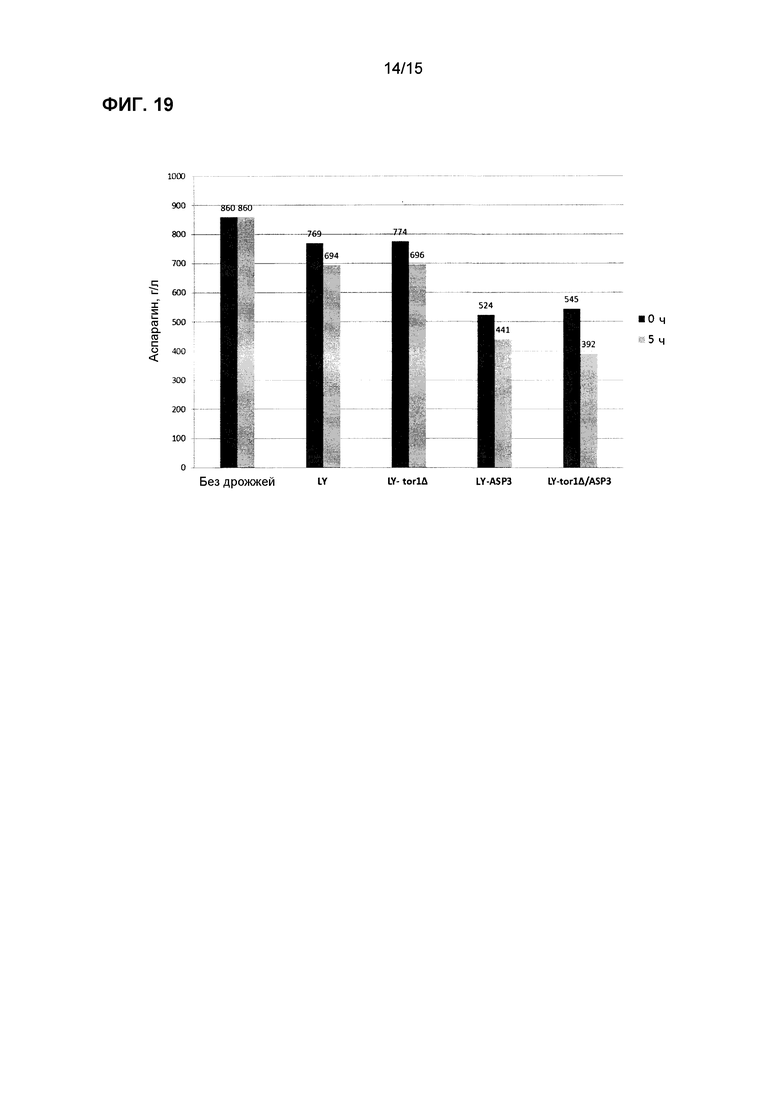
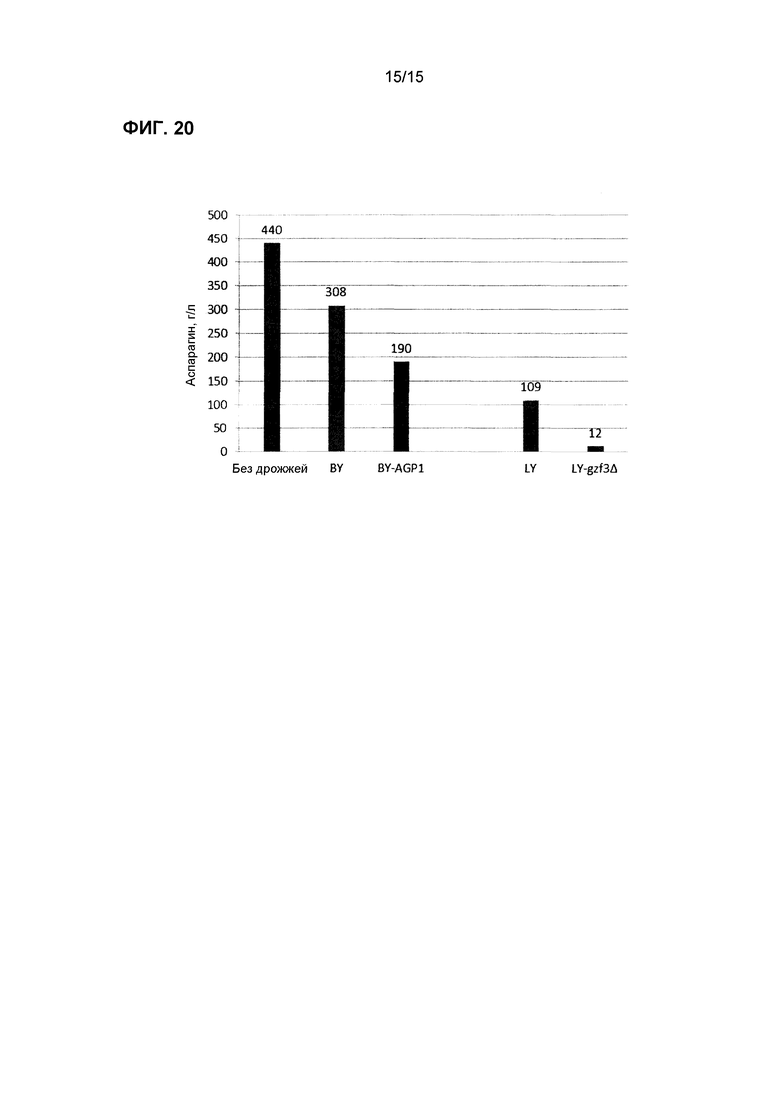
Изобретение относится к дрожжам для снижения акриламида или аспарагина в пищевом продукте. Дрожжи трансформированы по меньшей мере двумя из следующих модификаций: а) делеция или инактивация гена, кодирующего отрицательный регуляторный фактор, регулируемого азотной катаболитной репрессией, или введение молекулы нуклеиновой кислоты, которая сверхэкспрессирует положительный регуляторный фактор генов, регулируемых азотной катаболитной репрессией, для снижения азотной катаболитной репрессии; b) введение молекулы нуклеиновой кислоты для сверхэкспрессии гена, кодирующего аспарагиназу клеточной стенки, которая расщепляет аспарагин; и c) введение молекулы нуклеиновой кислоты, кодирующей транспортер аминокислот, который транспортирует аспарагин в условиях приготовления/переработки пищевых продуктов. Предложены также варианты способа снижения количества аспарагина или акриламида в пищевых продуктах и пищевые продукты, обладающие сниженным содержанием акриламида, полученные с использованием указанных дрожжей. Изобретение обеспечивает улучшенную способность снижать концентрацию акриламида в пищевых продуктах, приготовленных посредством нагревания. 7 н. и 26 з.п. ф-лы, 20 ил., 2 табл., 4 пр.
1. Дрожжи для снижения акриламида или аспарагина в пищевом продукте, где упомянутые дрожжи имеют по меньшей мере две из следующих модификаций:
a) делецию или инактивацию гена, кодирующего отрицательный регуляторный фактор, регулируемого азотной катаболитной репрессией, или молекулу нуклеиновой кислоты, которая сверхэкспрессирует положительный регуляторный фактор генов, регулируемых азотной катаболитной репрессией, для снижения азотной катаболитной репрессии;
b) молекулу нуклеиновой кислоты для сверхэкспрессии гена, кодирующего аспарагиназу клеточной стенки, которая расщепляет аспарагин; и
c) молекулу нуклеиновой кислоты, кодирующей транспортер аминокислот, который транспортирует аспарагин в условиях приготовления/переработки пищевых продуктов.
2. Дрожжи по п.1, где аспарагиназу кодирует ASP3.
3. Дрожжи по п.1, где аспарагиназа представляет собой Asp3p.
4. Дрожжи по п.1, где транспортер аминокислот кодируют GAP1, AGP1, GNP1, DIP5, AGP2 или AGP3.
5. Дрожжи по п.1, где транспортер аминокислот представляет собой Gap1p, Agp1p, Gnp1p, Dip5p, Agp2p или Agp3p.
6. Дрожжи по п.1, где положительный регуляторный фактор кодируют GAT1 или GLN3 или где отрицательный регуляторный фактор кодируют URE2, TOR1, TOR2, DAL80, или GZF3.
7. Дрожжи по п.1, где положительный регуляторный фактор представляет собой Gat1p или Gln3p, или где отрицательный регуляторный фактор представляет собой Ure2p, Tor1p, Tor2p, Dal80p, или Gzf3p.
8. Дрожжи по п.1, где по меньшей мере одна молекула нуклеиновой кислоты подпункта (а) содержит делеционную кассету URE2.
9. Дрожжи по п.1, где по меньшей мере одна молекула нуклеиновой кислоты подпункта (а) кодирует [URE3].
10. Дрожжи по любому из пп.1-9, где дрожжи являются неактивными.
11. Дрожжи по пп.1-10, где дрожжи представляют собой S. cerevisiae.
12. Дрожжи по любому из пп.1-11, трансформированные для непрерывной деградации и/или поглощения молекул аспарагина в условиях приготовления/переработки пищевых продуктов.
13. Дрожжи по любому из пп.1-11, где по меньшей мере одна молекула нуклеиновой кислоты функционально связана с конститутивно активным промотором.
14. Дрожжи по любому из пп.1-11, где по меньшей мере одна молекула нуклеиновой кислоты функционально связана с промотором, который не подвержен азотной катаболитной репрессии.
15. Дрожжи по п.1, трансформированные первой и второй молекулой нуклеиновой кислоты, где первая молекула нуклеиновой кислоты кодирует Asp3p и вторая молекула нуклеиновой кислоты кодирует Gap1p или Gat1p.
16. Способ снижения количества аспарагина во время приготовления или переработки пищевого продукта, включающий в себя добавление дрожжей по любому из пп. 1-15 в пищевой продукт в условиях приготовления или переработки пищевого продукта; где дрожжи снижают азотную катаболитную репрессию и/или сверхэкспрессируют ген, кодирующий аспарагиназу клеточной стенки, которая расщепляет аспарагин, и/или ген, кодирующий транспортер аминокислот, который транспортирует аспарагин во время приготовления или переработки пищевого продукта, таким образом снижая количество аспарагина во время приготовления или переработки пищевого продукта.
17. Способ снижения количества акриламида в пищевом продукте, включающий в себя добавление дрожжей по любому из пп. 1-15 в пищевой продукт в условиях приготовления или переработки пищевого продукта; где дрожжи снижают азотную катаболитную репрессию и/или сверхэкспрессируют ген, кодирующий аспарагиназу клеточной стенки, которая расщепляет аспарагин, и/или ген, кодирующий транспортер аминокислот, который транспортирует аспарагин во время приготовления или переработки пищевого продукта, таким образом, снижая количество акриламида в пищевом продукте.
18. Способ снижения количества аспарагина во время приготовления или переработки пищевого продукта, включающий в себя
а) введение в дрожжи по меньшей мере двух из следующих модификаций:
i) делеции или инактивации гена, кодирующего отрицательный регуляторный фактор, регулируемого азотной катаболитной репрессией, или молекулы нуклеиновой кислоты, которая сверхэкспрессирует положительный регуляторный фактор генов, регулируемых азотной катаболитной репрессией, для снижения азотной катаболитной репрессии;
ii) молекулы нуклеиновой кислоты для сверхэкспрессии гена, кодирующего аспарагиназу клеточной стенки, которая расщепляет аспарагин; и
iii) молекулы нуклеиновой кислоты кодирующей транспортер аминокислот, который транспортирует аспарагин;
b) добавление дрожжей в пищевой продукт в условиях приготовления или переработки пищевого продукта;
где дрожжи снижают азотную катаболитную репрессию и/или сверхэкспрессируют ген, кодирующий аспарагиназу клеточной стенки, которая расщепляет аспарагин, и/или ген, кодирующий транспортер аминокислот, который транспортирует аспарагин, таким образом, снижая количество аспарагина во время приготовления или переработки пищевого продукта.
19. Способ снижения количества акриламида в пищевом продукте, включающий в себя
а) введение в дрожжи по меньшей мере двух из следующих модификаций:
i) делеции или инактивации гена, кодирующего отрицательный регуляторный фактор, регулируемого азотной катаболитной репрессией, или молекулы нуклеиновой кислоты, которая сверхэкспрессирует положительный регуляторный фактор генов, регулируемых азотной катаболитной репрессией, для снижения азотной катаболитной репрессии;
ii) молекулы нуклеиновой кислоты для сверхэкспрессии гена, кодирующего аспарагиназу клеточной стенки, которая расщепляет аспарагин; и
iii) молекулы нуклеиновой кислоты, кодирующей транспортер аминокислот, который транспортирует аспарагин;
b) добавление дрожжей в пищевой продукт в условиях приготовления или переработки;
где дрожжи снижают азотную катаболитную репрессию и/или сверхэкспрессируют ген, кодирующий аспарагиназу клеточной стенки, которая расщепляет аспарагин, и/или ген, кодирующий транспортер аминокислот, который транспортирует аспарагин, таким образом снижая количество акриламида в пищевом продукте.
20. Способ по п.18 или 19, где аспарагиназу кодирует ASP3.
21. Способ по п.18 или 19, где аспарагиназа представляет собой Asp3p.
22. Способ по п.18 или 19, где транспортер аминокислот кодируют GAP1, AGP1, GNP1, DIP5, AGP2 или AGP3.
23. Способ по п.18 или 19, где транспортер аминокислот представляет собой Gap1p, Agp1p, Gnp1p, Dip5p, Agp2p или Agp3p.
24. Способ по п.18 или 19, где положительный регуляторный фактор кодируют GAT1 или GLN3 или где отрицательный регуляторный фактор кодируют URE2, TOR1, TOR2, DAL80, или GZF3.
25. Способ по п.18 или 19, где положительный регуляторный фактор представляет собой Gat1p или Gln3p, или где отрицательный регуляторный фактор представляет собой Ure2p, Tor1p, Tor2p, Dal80p, или Gzf3p.
26. Способ по п.18 или 19, где нуклеиновая кислота, которая модифицирует активность регуляторного фактора азотной катаболитной репрессии, содержит делеционную кассету URE2.
27. Способ по п.18 или 19, где дрожжи являются неактивными.
28. Способ по любому из пп.18-27, где дрожжи представляют собой S. cerevisiae.
29. Способ по любому из пп.16-19, где молекула нуклеиновой кислоты функционально связана с конститутивно активным промотором.
30. Способ по любому из пп.16-19, где молекула нуклеиновой кислоты функционально связана с промотором, который не подвержен азотной катаболитной репрессии.
31. Способ по любому из пп.16-19, где пищевой продукт представляет собой пищевой продукт на основе овощей, напиток, хлебобулочное изделие, продукт из зерна, фрукты, бобовые, молочный или мясной продукт.
32. Пищевой продукт, обладающий сниженной концентрацией акриламида, полученный добавлением трансформированных дрожжей по любому из пп. 1-15 в пищевой продукт в условиях приготовления или переработки.
33. Пищевой продукт, обладающий сниженной концентрацией акриламида, полученный с использованием способа по любому из пп. 16-31.
| СПОСОБ ПЕРЕРАБОТКИ НЕФТЕСОДЕРЖАЩИХ ОТХОДОВ (ШЛАМОВ) | 1998 |
|
RU2156750C2 |
| US 7666652 B2, 23.02.2010 | |||
| WO 2004032648 A1, 22.04.2004 | |||
| СПОСОБ УМЕНЬШЕНИЯ ОБРАЗОВАНИЯ АКРИЛАМИДА В ТЕРМИЧЕСКИ ОБРАБОТАННЫХ ПИЩЕВЫХ ПРОДУКТАХ | 2004 |
|
RU2335998C2 |

