Данная заявка претендует на преимущества в соответствии с 35 U.S.С. §1 19(е) предварительной заявки на патент США No. 61/410746, поданной 5 ноября 2010 года; предварительной заявки на патент США No.61/425375, поданной 21 декабря 2010 года; предварительной заявки на патент США No. 61/439341, поданной 03 февраля 2011 года; предварительной заявки на патент США No. 61/475614, поданной 14 апреля 2011 года; предварительной заявки на патент США No. 61/491846, поданной 31 мая 2011 года, и предварительной заявки на патент США No. 61/497861, поданной 16 июня 2011 года, каждая из которых включена в данное изобретение посредством отсылки.
Данная заявка включает, посредством отсылки, Список последовательностей, представленный в виде текстового файла Zymeworks V84467WO.txt, созданного 4 ноября 2011 года и имеющего объем 15 килобайт.
Область техники, к которой относится изобретение
Настоящая заявка, в целом, предусматривает гетеродимерные полипептиды, их композиции и способы получения и применения таких полипептидных гетеродимеров. Более конкретно, настоящее изобретение относится к термоустойчивым мультиспецифическим, включая биспецифические, антителам, содержащим гетеродимерный Fc домен.
Сведения о предшествующем уровне техники
Биспецифические антитела представляют собой молекулы антител, которые могут одновременно связывать два отдельных различных антигена (или различные эпитопы одного и того же антигена). Одним из применений биспецифических антител является перенацеливание цитотоксических иммунных эффекторных клеток на повышенный лизис опухолевых клеток, такой как при использовании антителозависимой клеточной цитотоксичности (ADCC). В данном случае одно плечо биспецифического антитела связывает антиген на опухолевой клетке, а другое плечо связывает детерминанту, экспрессирующуюся на эффекторных клетках. При перекрестном слиянии опухолевой и эффекторной клеток биспецифическое антитело не только обеспечивает сближение эффекторных и опухолевых клеток, но также одновременно включает их активацию, что приводит к эффективному киллингу опухолевых клеток. Биспецифические антитела также применялись для обогащения химио- или радиотерапевтических агентов в опухолевых тканях с целью минимизации вредного действия на здоровые ткани. В этом случае одно плечо биспецифического антитела связывает антиген, экспрессирующийся на клетке, нацеленной на разрушение, а второе плечо доставляет химиотерапевтическое лекарство, радиоизотоп или токсин.
Основным препятствием в общей разработке биспецифических антител была трудность получения материалов в достаточном количестве и удовлетворительного качества как для преклинических, так и для клинических исследований.
Обычное получение полноразмерных биспецифических антител основано на совместной экспрессии двух пар тяжелая цепь-легкая цепь иммуноглобулина, где две цепи имеют разные специфичности (Millstein et al., 1983, Nature, 305:537-539). Свойственное участку Fc молекулы антитела стремление димеризоваться приводит к образованию сложных смесей, содержащих до 10 различных молекул IgG, состоящих из различных комбинаций тяжелых и легких цепей, из которых только одна комбинация имеет корректную биспецифическую структуру. Очистка корректной молекулы, которую обычно проводят на стадиях аффинной хроматографии, является достаточно трудоемкой, а выходы продукта низкие. Аналогичные методы раскрываются в Международной патентной заявке WO 93/08829 и в статье Traunecker et al., 1991, EMBO J., 10:3655-3659. Таким образом, получение биспецифического антитела с двумя Fab плечами, выбранного с целью связывания с двумя различными мишенями, с применением традиционного метода гибридом является трудной задачей [Segal DM et al. (2001) J Immunol Methods. 248, 1-6.]. Трифункциональное антитело, Катумаксомаб (Catumaxomab), является биспецифическим mAb, полученным при использовании квадрогибридомы крысы/мыши, а очистку этого антитела проводят хроматографией на колонке с белком А с применением рН-зависимой элюции [Lindhofer Н. et al. (1995) J Immunol 155, 219-225].
Другим традиционным методом получения биспецифического антитела является химическая конъюгация двух антител или их фрагментов, имеющих разные специфичности. Однако этот метод также является сложным, а процесс химической модификации может инактивировать антитело или вызвать агрегацию. Так как очистка от нежелательных продуктов остается сложной, низкий выход и плохое качество биспецифического антитела делает этот процесс неподходящим для крупномасштабного производства, необходимого для клинических разработок. Кроме того, эти молекулы могут не сохранить конформацию обычного антитела.
В последнее время для усовершенствования получения биспецифических антител были применены различные методы гетеродимеризации. Однако, слияние доменов простой гетеродимеризацией, подобное слиянию доменов Jun/Fos суперспираль с scFv доменами, дает смесь гомо- и гетеродимеров и требует сборки рефолдингом (de Kruif and Logtenberg, J. Biol. Chem. 271:7630-4, 1996). Слияние фрагментов scFv с целыми антителами также использовали в качестве механизма димеризации (Coloma and Morrison, Nat. Biotechnol. 15:159-63, 1997). Однако, такое слияние дает большую молекулу, обладающую плохой способностью проникать в однородные ткани. Слияние вместе двух фрагментов scFv также применяли для получения биспецифических белков (например, антител BITE® от Micromet Inc., Bethesda, MD, патент США No. 7635472). Однако, такие белки не содержат областей Fc и, следовательно, не позволяют манипулировать с их активностями, используя области Fc. Кроме того, эти белки являются низкомолекулярными (~55 кДа) и, следовательно, имеют относительно короткий период полужизни в сыворотке.
В других методах гетеродимеризации биспецифические антитела состоят из тяжелой цепи гибридного иммуноглобулина с первой специфичностью связывания в одном плече и парой тяжелая цепь-легкая цепь гибридного иммуноглобулина (предоставляющей вторую специфичность связывания) в другом плече. Было найдено, что такая асимметричная структура способствует отделению заданного биспецифического соединения от нежелательных комбинаций, так как присутствие легкой цепи иммуноглобулина только в одной половине биспецифической молекулы обеспечивает легкий способ отделения. Этот метод раскрывается в Международной заявке на патент WO 94/04690. Более подробно получение биспецифических антител описано, например, в Suresh et al., 1986, Methods in Enzymology, 121:210.
Согласно другому методу, описанному в Международной заявке на патент WO96/27011, можно сконструировать пару молекул антитела, чтобы достичь максимального процентного содержания гетеродимеров, которые регенерируются из рекомбинантной клеточной культуры. Этим методом одну или более низкомолекулярных аминокислотных боковых цепей из CH3 интерфейса (области контакта) первой молекулы антитела заменяли на более высокомолекулярные боковые цепи (например, тирозин или триптофан). Компенсирующие "полости" размера, идентичного или аналогичного размеру более высокомолекулярной(-ых) боковой(-ых) цепей(-и), создают в области контакта (на интерфейсе) второй молекулы антитела, заменяя аминокислотную(-ые) боковую(-ые) цепь(-и) большего размера на меньшие (например, аланин или треонин). Это обеспечивает механизм повышения выхода гетеродимера по сравнению с другими нежелательными конечными продуктами, такими как гомодимеры [патентная заявка США US005731168A, патент США US007183076B2, Ridgway JB, Presta LG, Carter P. Protein Eng 1996 Jul; 9(7):617-21; Atwell S, Ridgway JB, Wells JA, Carter P. J Mol Biol 1997 Jul 4; 270(1):26-35.]. Недавно Gunasekaran и сотрудники [Gunasekaran K. et al. (2010) J Biol Chem. 285, 19637-46] применили стратегию комплементарного электростатического взаимодействия для осуществления селективной гетеродимеризации. Davis и сотрудники [Davis, JH. et al. (2010) Prot Eng Des Sel; 23(4):195-202] создали CH3 домены, используя домен, полученный обменом нитей (SEED), который состоит из чередующихся сегментов последовательностей CH3 человеческих IgA и IgG, и они, преимущественно, ассоциированы в виде гетеродимеров. Однако, во всех этих методах получают антитела, содержащие гетеродимерные области Fc, которые значительно менее устойчивы, чем исходная молекула или молекула дикого типа.
Таким образом, в области техники остается потребность в альтернативных мультиспецифических вариантных Fc гетеродимерах, конкретно, в вариантах CH3 доменов, которые были бы модифицированы таким образом, чтобы выбрать гетеродимеры с повышенной устойчивостью и чистотой.
РАСКРЫТИЕ ИЗОБРЕТЕНИЯ
Согласно одному аспекту изобретения предусматривается выделенный гетеродимер, содержащий гетеродимерную Fc область, при этом гетеродимерная Fc область содержит вариантную форму CH3 домена, включающую мутации аминокислотных остатков, содействующие образованию гетеродимера, при этом гетеродимерная Fc область дополнительно содержит вариантную форму СН2 домена, включающую асимметричные аминокислотные модификации, стимулирующие селективное связывание Fc-гамма-рецептора. Согласно одному варианту СН2 домен селективно связывает Fc-гамма IIIa-рецепторы по сравнению с СН2 доменом дикого типа. Согласно одной модификации вариантная форма CH3 домена имеет температуру плавления (Tm) около 70°С или выше.
В другом аспекте изобретения предусматривается выделенный гетеромультимер, содержащий гетеродимерную Fc область, причем гетеродимерная Fc область включает вариантную форму домена CH3, содержащую аминокислотные мутации, стимулирующие образование гетеродимера с повышенной устойчивостью, и этот вариантный CH3 домен имеет температуру плавления (Tm) около 70°С или выше. Согласно одному варианту гетеродимерная Fc область не содержит дополнительной дисульфидной связи в CH3 домене по сравнению с Fc областью дикого типа, более конкретно, гетеродимерная Fc область не содержит дополнительной дисульфидной связи в CH3 домене по сравнению с Fc областью дикого типа. Согласно альтернативному варианту изобретения гетеродимерная Fc область содержит дополнительную дисульфидную связь в вариантном CH3 домене по сравнению с Fc областью дикого типа, при условии, что температура плавления (Tm) равна около 70°С или выше в отсутствие дополнительной дисульфидной связи. Согласно еще одному варианту гетеродимерная Fc область содержит дополнительную дисульфидную связь в вариантной форме CH3 домена по сравнению с Fc областью дикого типа, и при этом вариантный CH3 домен имеет температуру плавления (Tm) около 77.5°С или выше.
Согласно одному варианту предусматривается выделенный гетеромультимер, содержащий гетеродимерную область Fc, причем гетеродимерная область Fc включает вариантную форму CH3 домена, содержащую аминокислотные мутации, стимулирующие образование гетеродимера с повышенной устойчивостью, при этом вариантная форма домена CH3 имеет температуру плавления (Tm) около 70°С или выше, а чистота гетеродимерной области Fc равна более, примерно, 90%, или чистота гетеродимерной области Fc равна более, примерно, 95% или выше, или чистота гетеродимерной области Fc равна более, примерно, 98% или выше.
Согласно одному варианту изобретения предусматривается также выделенный гетеромультимер, содержащий гетеродимерную область Fc, причем гетеродимерная область Fc включает вариантную форму CH3 домена, содержащую аминокислотные мутации, стимулирующие образование гетеродимера с повышенной устойчивостью, при этом вариантная форма домена CH3 имеет температуру плавления (Tm) около 70°С или выше, или Tm около 71°С или выше, или Tm около 74°С или выше. Согласно другому варианту чистота гетеродимерной области Fc равна более, примерно, 98% или выше, а Tm равна около 73°С, или чистота гетеродимерной области Fc равна более, примерно, 90% или выше, а Tm равна около 75°С.
Согласно некоторым вариантам изобретения предусматривается выделенный гетеромультимер, содержащий гетеродимерную область Fc, причем гетеродимерная область Fc включает первый полипептид CH3 домена, содержащий аминокислотные модификации L351Y и Y407A, и второй полипептид CH3 домена, содержащий аминокислотные модификации Т366А и K409F. В одном аспекте первый полипептид CH3 домена или второй полипептид CH3 домена содержит дополнительную аминокислотную модификацию в положении Т411, D399, S400, F405, N390 или K392. Аминокислотная модификация в положении Т411 выбрана из T411N, T411R, T411Q, T411K, T411D, T411E или T411W. Аминокислотная модификация в положении D399 выбрана из D399R, D399W, D399Y или D399K. Аминокислотная модификация в положении S400 выбрана из S400E, S400D, S400R, или S400K. Аминокислотная модификация в положении F405 выбрана из F405I, F405M, F405T, F405S, F405V или F405W. Аминокислотная модификация в положении N390 выбрана из N390R, N390K или N390D. Аминокислотная модификация в положении K392 выбрана из K392V, K392M, K392R, K392L, K392F или K392E.
Согласно некоторым вариантам изобретения предусматривается выделенный гетеромультимер, содержащий гетеродимерную область Fc, причем гетеродимерная область Fc включает первый полипептид CH3 домена, содержащий аминокислотные модификации L351Y и Y407A, и второй полипептид CH3 домена, содержащий аминокислотные модификации Т366А и K409F.
Согласно другому варианту предусматривается выделенный гетеромультимер, содержащий гетеродимерную область Fc, причем гетеродимерная область Fc включает первый полипептид CH3 домена, содержащий аминокислотную модификацию Y407A, и второй полипептид CH3 домена, содержащий аминокислотные модификации Т366А и K409F. В одном аспекте первый полипептид CH3 домена или второй полипептид CH3 домена содержит дополнительные аминокислотные модификации K392, Т411, D399 и S400. В другом аспекте первый полипептид CH3 домена содержит аминокислотную модификацию D399R, S400R и Y407A, а второй полипептид CH3 домена содержит аминокислотную модификацию Т366А, K409F, K392E и Т411Е. Согласно другому варианту изобретения вариантная форма CH3 домена имеет температуру плавления (Tm) около 74°С или выше, а чистота гетеродимера составляет около 95% или выше.
Согласно другому варианту предусматривается выделенный гетеромультимер, содержащий гетеродимерную область Fc, причем гетеродимерная область Fc включает первый полипептид CH3 домена, содержащий аминокислотную модификацию в положении L351 и аминокислотную модификацию Y407A, и второй полипептид CH3 домена, содержащий аминокислотную модификацию в положении Т366 и аминокислотную модификацию K409F. В одном аспекте аминокислотная модификация в положении L351 выбрана из L351Y, L351I, L351D, L351R или L351F. В другом аспекте аминокислотная модификация в положении Y407 выбрана из Y407A, Y407V или Y407S. Еще в одном аспекте аминокислотная модификация в положении Т366 выбрана из Т366А, T366I, T366L, Т366М, T366Y, T366S, Т366С, T366V или T366W. Согласно одному варианту изобретения вариантная форма CH3 домена имеет температуру плавления (Tm) около 75°С или выше, а чистота гетеродимера составляет около 90% или выше.
Согласно другому варианту предусматривается выделенный гетеромультимер, содержащий гетеродимерную область Fc, причем гетеродимерная область Fc включает первый полипептид CH3 домена, содержащий аминокислотную модификацию в положении F405 и аминокислотные модификации L351Y и Y407V, а второй полипептид CH3 домена содержит аминокислотную модификацию в положении T394W. В одном аспекте первый полипептид CH3 домена или второй полипептид CH3 домена содержит дополнительные аминокислотную модификацию в положениях K392, Т411, Т366, L368 или S400. Аминокислотная модификация в положении F405 представляет собой F405A, F405I, F405M, F405T, F405S, F405V или F405W. Аминокислотная модификация в положении K392 представляет собой K392V, K392M, K392R, K392L, K392F или K392E. Аминокислотная модификация в положении Т411 представляет собой T411N, T411R, T411Q, T411K, T411D, Т411Е или T411W. Аминокислотная модификация в положении S400 представляет собой S400E, S400D, S400R или S400K. Аминокислотная модификация в положении Т366 представляет собой Т366А, T366I, T366L, Т366М, T366Y, T366S, Т366С, T366V или T366W. Аминокислотная модификация в положении L368 представляет собой L368D, L368R, L368T, L368M, L368V, L368F, L368S и L368A.
Согласно другому варианту предусматривается выделенный гетеромультимер, содержащий гетеродимерную область Fc, причем гетеродимерная область Fc включает первый полипептид CH3 домена, содержащий аминокислотные модификации L351Y, F405A и Y407V, а второй полипептид CH3 домена содержит аминокислотную модификацию в положении T394W. В одном аспекте второй полипептид CH3 домена содержит аминокислотную модификацию T366L или T366I.
Согласно еще одному варианту предусматривается выделенный гетеромультимер, содержащий гетеродимерную область Fc, причем гетеродимерная область Fc включает первый полипептид CH3 домена, содержащий аминокислотные модификации F405A и Y407V, а второй полипептид CH3 домена содержит аминокислотные модификации в T366I, K392M и T394W.
Согласно некоторым вариантам предусматривается выделенный гетеромультимер, содержащий гетеродимерную область Fc, причем гетеродимерная область Fc включает первый полипептид CH3 домена, содержащий аминокислотные модификации F405A и Y407V, а второй полипептид CH3 домена содержит аминокислотные модификации T366L, K392M и T394W.
Согласно другому варианту изобретения предусматривается выделенный гетеромультимер, содержащий гетеродимерную область Fc, причем гетеродимерная область Fc включает первый полипептид CH3 домена, содержащий аминокислотные модификации F405A и Y407V, а второй полипептид CH3 домена содержит аминокислотные модификации T366L и T394W.
Согласно другому варианту изобретения предусматривается выделенный гетеромультимер, содержащий гетеродимерную область Fc, причем гетеродимерная область Fc включает первый полипептид CH3 домена, содержащий аминокислотные модификации F405A и Y407V, а второй полипептид CH3 домена содержит аминокислотные модификации T366I и T394W.
Согласно некоторым вариантам гетеромультимера предусматривается биспецифическое антитело или мультиспецифическое антитело.
Согласно другому варианту предусматривается композиция, содержащая гетеромультимер по изобретению и фармацевтически приемлемый носитель.
Согласно другому варианту предусматривается клетка-хозяин, содержащая нуклеиновую кислоту, кодирующую гетеромультимер по изобретению.
Согласно некоторым вариантам изобретения предусматривается мультимер, который содержит, по меньшей мере одно терапевтическое антитело. В одном аспекте терапевтическое антитело выбрано из группы, состоящей из абаговомаба, адалимумаба, алемтузумаба, аурограба, бапинеузумаба, базиликсимаба, белимумаба, бевацизумаба, бриакинумаба, канакинумаба, катумаксомаба, цертолизумаба пегол, цетуксимаба, даклизумаба, деносумаба, эфализумаба, галиксимаба, гемтузумаба озогамицина, голимумаба, ибритумомаба тиуксетана, инфликсимаба, ипилимумаба, лумиликсимаба, меполизумаба, мотавизумаба, муромонаба, микограба, натализумаба, нимотузумаба, окрелизумаба, офатумумаба, омализумаба, паливизумаба, панитумумаба, пертузумаба, ранибизумаба, реслизумаба, ритуксимаба, теплизумаба, тоцилизумаба/атлизумаба, тозитумомаба, трастузумаба, ProxiniumTM, RencarexTM, устекинумаба и залутумумаба.
Согласно другому варианту гетеромультимера по изобретению предусматривается способ лечения рака у пациента, больного раком, характеризующимся антигеном рака, причем указанный способ включает введение указанному пациенту терапевтически эффективного количества гетеромультимера.
Согласно другому варианту гетеромультимера по изобретению предусматривается способ лечения иммунных расстройств (нарушений) у пациента, страдающего иммунным нарушением, характеризующимся иммунным антигеном, причем указанный способ включает введение указанному пациенту терапевтически эффективного количества гетеромультимера.
Согласно еще одному варианту предусматривается выделенный гетеромультимер, содержащий гетеродимерную область Fc, причем гетеродимерная область Fc включает вариантную форму CH3 домена, содержащую аминокислотные мутации, стимулирующие образование гетеродимера с повышенной устойчивостью, а вариантная форма CH3 доменов выбрана из вариантов, перечисленных в Таблице 1, Таблице 6 или Таблице 7.
КРАТКОЕ ОПИСАНИЕ ФИГУР
На Фигуре 1 графически представлена 3-D структура антитела дикого типа, показывающая области CH3 (вверху), СН2 (в середине) и рецепторную области. Пунктирный прямоугольник слева в увеличенном виде представлен справа и показывает две области, Область 1 и Область 2, целевой области CH3;
На Фигуре 2 дано графическое 3-D изображение остатка дикого типа в положении 368;
На Фигуре 3 дано графическое 3-D изображение Области 1, показывающее изменившееся (мутировавшее) положение 368;
На Фигуре 4 дано графическое 3-D изображение дополнительных мутаций в Области 2;
На Фигуре 5 представлена таблица расчетов компьютерного моделирования (in silico) оценки столкновений, разницы площади области контакта (интерфейса), разницы упаковки, разницы электростатической энергии и общей "оценки аффинности" для первых трех вариантов AZ1, AZ2 и AZ3;
На Фигуре 6 дано графическое 3-D изображение вариантов AZ2 и AZ3, которые "основаны на" варианте AZ1;
На Фигуре 7 дано графическое 3-D изображение вариантов AZ2 и AZ3.
На Фигуре 8 представлена таблица, аналогичная таблице на Фигуре 5, но для AZ1, AZ2 и AZ3 гетеродимеров и гомодимеров. Оценка аффинности для гомодимеров не является релевантной, поэтому отсутствует оценка этого аспекта для гомодимеров;
На Фигуре 9 дано графическое 3-D изображение дикого типа (слева) и мутантного AZ4 (справа);
На Фигуре 10 представлена таблица, аналогичная таблице на Фигуре 5, показывающая компьютерные (in silico) расчеты для моделирования AZ4 гетеродимера и гомодимеров;
На Фигуре 11 дано графическое изображение CH3 вариантов AZ5 (слева) и AZ6 (справа);
На Фигуре 12 представлена таблица, аналогичная таблице на Фигуре 5, показывающая in silico данные для AZ4, AZ5 и AZ6;
На Фигуре 13 дано графическое 3-D изображение антитела слева с изображением обоснования особенностей связывания в рецепторной области с применением гетеродимерного подхода;
На Фигуре 14 дано схематическое изображение молекулы IgG;
На Фигуре 15 показано множественное выравнивание последовательностей Fcγ рецепторов. ID последовательностей в Genbank/Uniprot: FcγRIIA (sp P12318), FcγRIIB (sp P31994), FcγRIIC (gi 126116592), FcγRIIIA (sp P08637), FcγRIIIB (sp 075015);
На Фигуре 16 схематически представлена структура кристалла комплекса Fc-FcγRIIIb Complex [PDB ID: 1T83, Radaev & Sun]. В полученном комплексе 1:1 Fc и рецептора Fcγ наблюдается асимметричный контакт между двумя цепями Fc и FcγR;
На Фигуре 17 схематически представлены альтернативные мультифункциональные молекулы на основе асимметричной Fc каркасной структуры:
Асимметричный Fc каркас и Асимметричный Fc- плечо мономерного IgG;
На Фигуре 18 схематически представлены альтернативные мультифункциональные молекулы на основе асимметричной Fc каркасной структуры:
Асимметричный Fc- плечи моноспецифического IgG и Асимметричный Fc- плечи биспецифического IgG (общая легкая цепь);
На Фигуре 19 схематически представлены альтернативные мультифункциональные молекулы на основе асимметричной Fc каркасной структуры:
Асимметричный Fc- плечи биспецифического IgG и функциональная молекула, такая как токсин;
На Фигуре 20 схематически представлены альтернативные мультифункциональные молекулы на основе асимметричной Fc каркасной структуры:
Асимметричный Fc- одно плечо scFv и Асимметричный Fc- биспецифические scFv плечи.
На Фигуре 21 схематически представлены альтернативные мультифункциональные молекулы на основе асимметричной Fc каркасной структуры: Асимметричный Fc-триспецифические scFv плечи и Асимметричный Fc-тетраспецифические scFv плечи.
На Фигуре 22 представлен асимметричный дизайн мутаций на одной стороне Fc, который с целью повышения FcγR селективности вводит благоприятную сторону для FcγR взаимодействий и неблагоприятную сторону для взаимодействий, подобных взаимодействиям дикого типа. Мутации на неблагоприятной стороне Fc можно вводить с целью блокады взаимодействий с FcR и полярности смещения Fc с тем, чтобы взаимодействие происходило только с благоприятной стороны.
На Фигуре 23 показана аминокислотная последовательность для дикого типа человеческого IgG1.
На Фигуре 24 показан итеративный процесс дизайна Fc гетеродимера, объединяющий стратегию позитивного и негативного дизайна, подробнее описанную ниже.
На Фигуре 25 показан in vitro анализ, применяемый для определения чистоты гетеродимера. Анализ проводили с использованием структуры полноразмерного моноспецифического антитела с двумя Fc тяжелыми цепями различной молекулярной массы. Тяжелая цепь А имеет С-концевую метку HisTag (His), а тяжелая цепь В имеет C-концевую отщепляемую метку mRFP Tag (RFP). Две тяжелых цепи A (His) и В (RFP) экспрессируются в различных относительных пропорциях вместе с фиксированным количеством легкой цепи, что приводит в результате к 3 возможным видам димеров с различной молекулярной массой: а) Гомодимер Цепь A (His)/ Цепь A (His) (~150 кДа); б) Гетеродимер Цепь A (His)/ Цепь В (RFP) (~175 кДа); в) Гомодимер Цепь В (RFP)/ Цепь В (RFP) (~200 кДа). После экспрессии, описанной в Примере 2, соотношение гетеродимер: два гомодимера определяли методом SDS-PAGE гель-электрофореза (электрофазу в полиакриламидном геле в присутствии додецилсульфата натрия) в невосстанавливающих условиях, который позволяет разделить 3 вида димера в зависимости молекулярной массы. Гели SDS-PAGE окрашивали Кумасси бриллиантовым голубым.
Фигура 25А. Проверяли следующие варианты: WT цепь A (His) только; WT цепь В (RFP) только; WT цепь A (His) плюс цепь В (RFP); Контроль 1 цепь A (His) плюс цепь В (RFP) с подтвержденной чистотой гетеродимера >95%. Состав полос димеров проверяли Вестерн-блоттингом с использованием антител, специфических к IgG-Fc (anti-Fc), mRFP Tag (anti-mRFP) и к HisTag (anti-His), показанных выше. Анализ методом SDS-PAGE гель-электрофореза показывает одну полосу для гомодимера His/His, двойную полосу для гетеродимера His/RFP и несколько полос для гомодимера RFP. Несколько полос является свойством, признаком mRFP Tag и, как было подтверждено, не влияет на физические свойства гетеродимера Fc.
Фигура 25B. Анализ методом SDS-PAGE гель-электрофореза подтверждали с использованием в качестве контроля опубликованных гетеродимерных Fc вариантов Контроль 1-4, См., Таблицу А. Варианты экспрессировали с различными соотношениями цепи A (His) и цепи В (RFP): Конкретно, соотношение 1:3 эквивалентно соотношению LC,HC_His,HC_mRFP 25%,10%,65%; соотношение 1:1 эквивалентно 25%,20%,55% и соотношение 3:1 эквивалентно 25%, 40%,35%, соответственно (как было определено, кажущееся соотношение 1:1 экспрессии цепи A (His) к цепи В (RFP) близко к 20%/55% (His/RFP) для WT Fc).
На Фигуре 25С. показан анализ методом SDS-PAGE гель-электрофореза в невосстанавливающих условиях по определению чистоты вариантов гетеродимера с каркасной структурой 1. Fc варианты экспрессировали с различными соотношениями цепи A (His) к цепи В (RFP) и анализировали методом SDS-PAGE гель-электрофореза в невосстанавливающих условиях, как описано на Фигуре 2. Конкретно, соотношение 1:3 эквивалентно соотношению LC,HC_His,HC_mRFP 25%,10%,65%; соотношение 1:1 эквивалентно 25%,20%,55% и соотношение 3:1 эквивалентно 25%, 40%,35%, соответственно как было определено, кажущееся соотношение 1:1 экспрессии цепи А (His) к цепи В (RFP) близко к 20%/55% (His/RFP) для WT Fc).
На Фигуре 26 показаны варианты гетеродимеров Fc, экспрессирующихся с конкретным соотношением цепи A (His) к цепи В (RFP) (См. Таблицу 2), очищенные аффинной хроматографией на колонке с Белком и проанализированные методом SDS-PAGE гель-электрофореза в невосстанавливающих условиях, как описано на Фигуре 25.
На Фигуре 26А показано, как различные варианты были сгруппированы в зависимости от категории чистоты на основании визуального изучения результатов SDS-PAGE гель-электрофореза. Для сравнения на гель наносили эквивалентное количество продукта, очищенного на колонке с Белком А. Это определение степени чистоты методом SDS-PAGE гель-электрофореза в невосстанавливающих условиях было подтверждено методом LC/MS для выбранных вариантов (см. Фигуру 28).
На Фигуре 26B показаны типичные результаты SDS-PAGE гель-электрофореза для выбранных гетеродимерных вариантов с каркасной структурой 1 и 2, очищенных на колонке с Белком A (AZ94, AZ86, AZ70, AZ33 и AZ34).
На Фигуре 27 проиллюстрирован анализ методом DSC (дифференциальной сканирующей калориметрии, ДСК) по определению температуры плавления домена СН3-CH3, при этом применяли два независимых метода.
Фигура 27А. Термограммы были построены по 4 независимым переходам "non-two-state" и оптимизированы, чтобы получить значения для СН2 и Fab переходов, близкие литературным данным для герцептина ~72°С (СН2) и ~82°С (Fab).
Фигура 27B. Нормализованные и скорректированные относительно базовой линии термограммы для гетеродимерных вариантов вычитали из WT, получая пик положительной и отрицательной разницы только для CH3 перехода.
На Фигуре 28 иллюстрируется ЖХ/МС анализ типичного варианта AZ70, описанного в примере 2. Указаны предполагаемые (вычисленные средние) массы гликозилированных гетеродимера и гомодимеров. Область, соответствующая массе гетеродимера, содержит основные пики, соответствующие потере глицина (-57 Да) и добавлению 1 или 2 гексоз (+162 Да и +324 Да, соответственно). Чистоту гетеродимера определяли как >90%, если отсутствовали заметные пики, соответствующие любому из гомодимеров.
На Фигуре 29 показана область контакта CH3 Фиг.29А WT Fc; Фиг.29B AZ6; Фиг.29С AZ33; Фиг.29D AZ19. Полный in silico анализ, описанный в разделе "Осуществление изобретения", и сравнение вариантов с WT показало, что одной из причин более низкой устойчивости исходного гетеродимера AZ33, чем WT, является отсутствие взаимодействия ядра/упаковки Y407 и Т366. В исходном AZ33 наблюдается неоптимальная упаковка в этом гидрофобном ядре, показанная на Фиг.29B, это означает, что оптимизация этой области, в частности, в положении Т366, должна повысить устойчивость AZ33. Это иллюстрируется на Фиг.29С и Фиг.29D введением T366I и T366L. Экспериментальные данные коррелируют с этим структурным анализом и показывают, что T366L дает наибольшее увеличение Tm. См., Пример 5.
На Фигуре 30 (А, В-увеличение части фигуры А) иллюстрируется полезность и важность анализа конформационной динамики, поясняемая вариантом AZ8 с исходным каркасом 1. Структура после in silico мутагенеза (конформация скелета близка к конформации WT) совмещается с характерной структурой 50 не моделирования методами молекулярной динамики. На Фигуре четко показана большая разница между конформациями в области петли D399-S400 варианта AZ8 по сравнению с WT, что, в свою очередь, делает доступным гидрофобное ядро для растворителя и служит причиной пониженной устойчивости AZ8 гетеродимера.
На Фигуре 31 показано, как сведения, полученные с помощью полного in silico анализа и моделирования MD, применялись для описания стратегии позитивного дизайна. Как показано на Фигуре 30, одной из причин пониженной устойчивости AZ8 по сравнению с WT является более слабое взаимодействие петли 399-400 с 409, вызванное, главным образом, утратой взаимодействия с F405 упаковки (см. сравнение Фиг.31А (WT) с Фиг.31B (AZ8)). Одним из подходов позитивного дизайна явилась оптимизация гидрофобной упаковки области с целью стабилизации конформации петли 399-400. Эту оптимизацию осуществляли с помощью мутации K392M, показанной на Фиг.31С. На Фиг.31С показан гетеродимер AZ33, который имеет Tm 74° по сравнению с Tm 68° начального варианта негативного дизайна AZ8.
На Фигуре 32 представлена динамика Fc молекулы, полученная Методом главных компонент, позволяющим рассчитать основные движения из траектории молекулярной динамики. На Фиг.32А показан чертеж скелета Fc структуры в качестве эталона. На Фиг.32B и С показано наложение динамики, наблюдаемой в соответствии с 2 самыми главными способами движения в Fc структуре. Наблюдается заметное движение раскрытие/закрытие СН2 доменов цепей А и В относительно друг друга, тогда как CH3 домены являются сравнительно неизменными. Мутации в области контакта (интерфейсе) CH3 оказывают воздействие на относительную гибкость и динамику этого движения открыть/закрыть в СН2 доменах.
На Фигуре 33 показана упаковка гидрофобного ядра двух вариантов с каркасной структурой -2 по сравнению с WT. Фиг.33А WT Fc; Фиг.33B AZ63; и Фиг.33С AZ70. Полный анализ in-silico исходного варианта с каркасной структурой -2 показал, что утрата взаимодействий ядра WT Y407-T366 является одной из причин более низкой устойчивости исходных вариантов с каркасом-2 по сравнению с WT. Утрата Y407- Т366 частично компенсируется мутациями K409F, но, как показано на Фиг.33B, в особенности мутация Т366А, оставляет в гидрофобном ядре впадину, которая дестабилизирует вариант по сравнению с WT. Оказалось, что нацеливание на это гидрофобное ядро дополнительных мутаций T366VJL351Y, как показано на примере Fc варианта AZ70 на Фиг.33С, является успешным; экспериментально определенная Tm AZ70 равна 75.5°С. См. Таблицу 4 и Пример 6.
На Фигуре 34 показаны взаимодействия петли 399-400 двух вариантов каркасной структуры-2 по сравнению с WT: Фиг.34А WT Fc; Фиг.34В AZ63; и Фиг 34С AZ94. Полный анализ in-silico исходного варианта с каркасной структурой -2 показал, что утрата WT солевого мостика K409-D399 (Фиг.34А) вследствие мутации K409F и, следовательно, ненасыщенного остатка D399 (Фиг.34B) является причиной более ′открытой′ конформации петли 399-400. Помимо всего прочего, это увеличивает доступ растворителя к гидрофобному ядру и дополнительно повышает дестабилизацию варианта по сравнению с WT. Одним из методов, применяемых для стабилизации петли 399-400 и компенсации утраты взаимодействия K409-D399, являлось создание дополнительных солевых мостиков D399R-T411E и S400R-K392E, как показано на Фиг.34С для варианта AZ94. Экспериментально доказано, что степень чистоты >95% и Tm равна 74°С. См. Таблицу 4 и Пример 6. Далее, хотя AZ94 имеет значительно более высокую степень чистоты и более устойчив по сравнению с начальным вариантом с каркасом-2 (чистота <90%, Tm 71°С), мутации в гидрофобном ядре AZ94 являются менее предпочтительными, чем ′наилучшие′ мутации в гидрофобном ядре, идентифицированные в варианте AZ70 (Фигура 33). Так как мутации в гидрофобном ядре в AZ70 (T366V_L351Y) отдалены от мутаций солевых мостиков AZ94 в петле 399-400, предполагается, что комбинация аминокислотных мутаций AZ70 и дополнительных мутаций AZ94 даст более высокую температуру плавления, чем температура плавления AZ70 или AZ94. Эту комбинацию проверяли, как описано в Примерах 1-4.
На Фигуре 35 показаны константы ассоциации (Ka(М-1)) для процессов связывания гомодимерного IgG1 Fc, гетеродимерного вариантов het1 (Контроль 1): A:Y349C_T366S_L368A_Y407V/B:S354C_T366W и het2 (Контроль 4): A:K409D_K392D/B:D399K_D356K с шестью Fc гамма рецепторами. Для гетеродимерных Fc вариантов наблюдается тенденция к слегка измененному связыванию с Fc гамма рецепторами по сравнению с дикого типа IgG1 Fc. См. Пример 7.
На Фигуре 36А показана относительная прочность связывания (связывающая способность) дикого типа IgG1 Fc и его различных гомодимерных и асимметричных мутантных форм с рецепторами IIbF, IIBY и IIaR, прочность связывания для дикого типа применяли в качестве эталона. (Homo Fc + S267D) относится к связывающей способности гомодимерного Fc с мутацией S267D в обеих цепях. (Het Fc + asym S267D) относится к прочности связывания гомодимерного Fc с мутацией S267D, введенной в одну из двух цепей в Fc. Приводится средняя величина прочности связывания способности, полученная при введении мутации в любую из двух цепей Fc. Введение этой мутации в одну цепь уменьшает прочность связывания, приблизительно, вдвое по сравнению с прочностью связывания для той же самой мутации в гомодимерном продукте. (Het Fc + asym S267D + asym E269K) относится к прочности связывания гомодимерного Fc как с мутацией S267D, так и с мутацией and E269K, введенными асимметрично в одну из двух цепей Fc. Мутация E269K блокирует взаимодействие FcgR с одной из сторон Fc и способна снижать прочность связывания, приблизительно, вдвое по сравнению с прочностью связывания, наблюдаемой для самого асимметричного S267D варианта (Het Fc + S267D). В данном случае Het Fc состоит из CH3 мутаций, указанных для варианта het2 (Контроль 4) на Фигуре 35.
На Фигуре 36B приводится константа ассоциации (Ka(M-1)) различных Fc и их вариантов с несколькими FcgRIIa, FcgRIIb и FcgRIIIa аллотипами. Ka дикого типа IgG1 Fc с различными Fcg рецепторами представлена в виде столбцов со штриховкой горизонтальными линиями. Столбцы со штриховкой вертикальными линиями (homodimer base2) изображают Ka гомодимерного Fc с мутациями S239D/D265S/I332E/S298A. Столбцы со штриховкой наклонными линиями изображают Ka гетеродимерного Fc с асимметричными мутациями A:S239D/D265S/I332E/E269K и B:S239D/D265S/S298A в СН2 домене. Введение ассиметричных мутаций позволяет достичь повышенной селективности при выборе между IIIa и IIa/IIb рецепторами. В данном случае гетеродимерный Fc состоит из CH3 мутаций, указанных для варианта het2 (Контроль 4) на Фигуре 35.
На Фигуре 36С показана константа ассоциации (Ka(М-1)) для дикого типа IgG1 и трех других вариантов, включающих гомодимерные или асимметричные мутации в СН2 домене области Fc. Значения Ka дикого типа Fc представлены столбцом, заштрихованным в виде сетки. Значения Ka варианта Fc с основной мутацией S239D/K326E/A330L/I332E/S298A, введенной "в виде гомодимера" (homodimer basel) в обе цепи Fc, показаны в виде столбца со штриховкой наклонными линиями. Асимметричное введение родственных мутаций в цепи А и В гетеродимерного Fc (hetero basel) показано в виде столбца с горизонтальной штриховкой. Столбец со штриховкой вертикальными линиями изображает асимметричный вариант, включающий мутацию E269K (hetero base 1+PD). В данном случае гетеродимерный Fc состоит из CH3 мутаций, указанных для варианта het2 (Контроль 4) на Фигуре 35.
На Фигуре 37 изображена Таблица 6, в которой представлен список вариантов CH3 доменов на основании третьей фазы дизайна, описанной в Примере 5 для каркасной структуры (Scaffold) 1.
На Фигуре 38 изображена Таблица 7, в которой представлен список вариантов CH3 доменов на основании третьей фазы дизайна, описанной в Примере 6 для каркасной структуры (Scaffold) 2.
ОСУЩЕСТВЛЕНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение предусматривает модифицированные CH3 домены, содержащие специфические аминокислотные модификации, стимулирующие образование гетеродимера. Согласно одному варианту изобретения модифицированные CH3 домены содержат специфические аминокислотные модификации, стимулирующие образование гетеродимера (См., например. Таблицу 1). Согласно другому варианту изобретения модифицированные CH3 домены содержат специфические аминокислотные модификации, стимулирующие образование гетеродимера с повышенной устойчивостью (См., например, Таблицу 4, Таблицу 6 и Таблицу 7). Устойчивость (стабильность) определяют количественно как температуру плавления (Tm) домена CH3, а понятие "повышенная устойчивость" относится к Tm около 70°С или выше. Домены CH3 образуют участок области Fc гетеромультимерного или биспецифического антитела. Таким образом, согласно одному варианту настоящего изобретения предусматриваются гетеромультимеры, содержащие гетеродимерную Fc область, причем гетеродимерная Fc область содержит модифицированный или вариантный CH3 домен, включающий аминокислотные мутации, стимулирующие образование гетеродимера, а вариантные CH3 домены выбраны из вариантов, перечисленных в Таблице 1. Согласно второму варианту изобретения предусматриваются гетеромультимеры, содержащие гетеродимерную область Fc, причем гетеродимерная Fc область содержит модифицированный или вариантный CH3 домен, включающий аминокислотные мутации, стимулирующие образование гетеродимера с повышенной устойчивостью, и вариантный CH3 домен имеет температуру плавления (Tm) около 70°С или выше.
Аминокислотные модификации, которые можно применять для получения модифицированного домена CH3, включают, но без ограничения, аминокислотные инсерции, делеции, замены и перестановки. Модификации домена CH3 и модифицированные CH3 домены в настоящем описании совместно называются "CH3 модификации", "модифицированные CH3 домены", "вариантные CH3 домены" или "CH3 варианты". Эти модифицированные CH3 домены могут включаться в предпочтительную молекулу. Соответственно, согласно одному варианту изобретения предусматриваются молекулы, в частности, полипептиды, более конкретно, иммуноглобулины (например, антитела) и другие связывающие белки, содержащие область Fc (термины "область Fc" и аналогичные термины охватывают любой домен константной области тяжелой цепи, содержащий по меньшей мере участок домена CH3), включающую модифицированный CH3 домен. Молекулы, содержащие области Fc, включающие модифицированный CH3 домен (например, домен CH3, содержащий одну или более аминокислотных инсерции, делеции, замен или перестановок), называются в данном описании "Fc вариантами", "гетеродимерами" или "гетеромультимерами". Fc варианты по настоящему изобретению содержат CH3 домен, асимметрично модифицированный с целью получения гетеродимерных Fc вариантов или областей. Fc область состоит из двух полипептидов константных доменов тяжелых цепей - Цепи А и Цепи В, которые можно применять взаимозаменяемо при условии, что каждая Fc область содержит один полипептид Цепи А и Цепи В. Аминокислотные модификации вводятся в CH3 асимметрично, в результате получают гетеродимер, если два модифицированных CH3 домена образуют Fc вариант (См., например. Таблицу 1). Асимметричные аминокислотные модификации по настоящему описанию представляют собой любую модификацию, в которой аминокислота в конкретном положении одного полипептида (например, "Цепи А") отличается от аминокислоты во втором полипептиде (например, "Цепи В") в том же самом положении гетеродимера или Fc варианта. Это может явиться результатом модификации только одной или двух аминокислот или модификации обеих аминокислот в две различные аминокислоты из Цепи А и Цепи В Fc варианта. Понятно, что вариантные CH3 домены содержат одну или более асимметричных аминокислотных модификаций.
Следует понимать, что в настоящей заявке любой интервал концентраций, любой интервал величин в процентах, любой интервал соотношений или любая область целочисленных значений включает любое целое значение внутри указанного интервала и, при необходимости, если не указано иначе, его части (такие как одна десятая и одна сотая целого числа). Термин "около" ("приблизительно", "примерно") означает ±10% от указанных интервала, значения, последовательности или структуры, если не указано иначе. Следует понимать, что термины в единственном числе по данному описанию относятся к "одному или более" перечисленных компонентов, если не указано или из контекста не следует иное. Следует понимать, что употребление альтернативы, выбора (например, "или") означает либо одну, обе или любую комбинацию альтернатив. Термины "включать" и "содержать" употребляются в качестве синонимов. Кроме того, следует понимать, что отдельные одноцепочечные полипептиды или гетеродимеры, полученные с применением различных комбинаций структур и заместителей (например, вариантные CH3 домены) по данному описанию раскрываются в настоящей заявке в той же самой степени, как если бы каждый одноцепочечный полипептид или гетеродимер был представлен индивидуально. Таким образом, отбор конкретных компонентов для образования отдельных одноцепочечных полипептидов или гетеродимеров входит в объем настоящей заявки.
"Первый полипептид" означает любой полипептид, который должен ассоциироваться со вторым полипептидом, также называемый в данном описании "Цепь А". Первый и второй полипептид встречаются в "области контакта" ("интерфейса"). "Второй полипептид" означает любой полипептид, который должен ассоциироваться с первым полипептидом в "области контакта" (через "интерфейс"), также в данном описании его называют "Цепь В". "Область контакта", "интерфейс" содержит те "контактные" аминокислотные остатки в первом полипептиде, которые взаимодействуют с одним или более "контактных" аминокислотных остатков в области контакта (в интерфейсе) второго полипептида. Область контакта, интерфейс по данному описанию включает CH3 домен Fc области, который, предпочтительно, получен из IgG антитела, и наиболее предпочтительно, из человеческого IgG1 антитела.
Употребляемый в данном описании термин "выделенный" гетеромультимер означает гетеромультимер, который был идентифицирован и выделен из и/или отделен от компонента его естественной клеточной культуральной среды. Примесные компоненты в его естественной среде представляют собой материалы, которые могли бы мешать применению гетеромультимера в диагностике или терапии, и включают ферменты, гормоны и другие белковые или небелковые растворы.
Вариантные Fc гетеродимеры, как правило, очищают практически (по существу) до гомогенности. Выражения "по существу гомогенный", "по существу гомогенная форма" и "по существу гомогенность" употребляют, чтобы показать, что продукт по существу не содержит побочных продуктов, образующихся при нежелательных комбинациях полипептидов (например, гомодимеров). Выражение "по существу гомогенность", относящееся к степени чистоты, означает, что количество побочных продуктов не превышает 10%, и, предпочтительно, составляет менее 5%, более предпочтительно, менее 1%, наиболее предпочтительно, менее 0.5%, где процентное содержание дается в весовых процентах.
Каждый термин, понятный специалистам по технологии антител, имеет значение, принятое в данной области техники, если в данном описании специально не указано иное. Известно, что антитела имеют вариабельные области, шарнирную область и константные домены. Структура и функция иммуноглобулинов обсуждаются, например, в книге Harlow et al, Eds., Antibodies: A Laboratory Manual, Chapter 14 (Cold Spring Harbor Laboratory, Cold Spring Harbor, 1988).
Создание вариантных Fc гетеродимеров из дикого типа гомодимеров иллюстрируется методом позитивного и негативного дизайна в условиях белковой инженерии путем регулирования соотношения устойчивость/специфичность, причем мутации вводят с целью стимулировать преимущественное образование гетеродимера по сравнению с образованием гомодимера, когда полипептиды экспрессируются в клеточной культуре. Методы негативного дизайна максимально повышают невыгодные для образования гомодимеров взаимодействия, либо за счет введения объемных боковых цепей в одной цепи и небольших побочных цепей в противоположную цепь, например, стратегия выступ-в-углубление (knobs-into-holes), разработанная в Genentech (Ridgway JB, Presta LG, Carter P. ′Knobs-into-holes′ engineering of antibody CH3 domains for heavy chain heterodimerization. Protein Eng. 1996 Jul; 9(7):617-21; Atwell S, Ridgway JB, Wells JA, Carter P. Stable heterodimers from remodeling the domain interface of a homodimer using a phage display library. J Mol Biol. 270(1):26-35 (1997)), либо за счет использования электростатических взаимодействий, которые приводят к исключению ("отталкиванию") образования гомодимера, например, электростатическое управление, разработанное фирмой Amgen (Gunaskekaran K, et al. Enhancing antibody Fc heterodimer formation through electrostatic steering effects: applications to bispecific molecules and monovalent IgG. JBC 285 (25):19637-19646 (2010)). В этих двух примерах асимметричные точковые мутации негативного дизайна вводили в CH3 домен дикого типа для стимуляции образования гетеродимеров. До настоящего времени для разработки Fc гетеродимеров применялись только методы негативного дизайна. Опубликованные результаты показывают, что гетеродимеры, созданные методом негативного дизайна, дают высокую специфичность в случае >95% гетеродимеров, но значительно дестабилизируют комплекс (Supra). В случае этих димеров, полученных методами негативного дизайна, температура плавления модифицированного CH3 домена равна 69°С или менее, отсутствуют дополнительные дисульфидные связи по сравнению с антителом дикого типа. См. ниже Таблицу А.
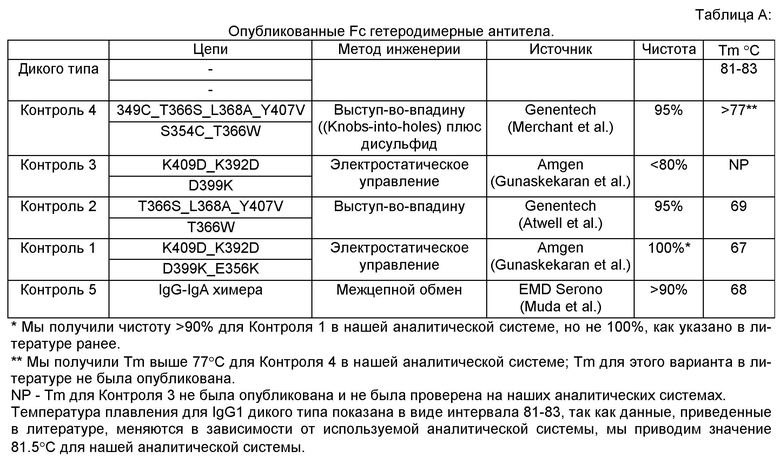
В отличие от негативного дизайна, общей методологией, применяемой для белковой инженерии, является позитивный дизайн. В данных примерах аминокислотные модификации вводили в полипептиды, чтобы получить максимальное количество выгодных взаимодействий внутри белков и между белками. Эта стратегия предполагает, что при введении нескольких мутаций, которые специфически стабилизируют заданный гетеродимер и в то же время не действуют на гомодимеры, суммарным эффектом будет более высокая специфичность взаимодействий заданного гетеродимера по сравнению с гомодимерами и, следовательно, более высокая специфичность гетеродимера. Понятно, что в контексте белковой инженерии методы позитивного дизайна позволяют оптимизировать устойчивость нужных белок-белковых взаимодействий, но редко позволяют достичь специфичности >90% (Havranek JJ & Harbury PB. Automated design of specificity in molecular recognition. Nat Struct Biol. 10(1):45-52 (2003); Bolon DN, Grant RA, Baker ТА, Sauer RT. Specificity versus stability in computational protein design. Proc Natl Acad Sci USA. 6; 102(36):12724-9 (2005); Huang PS, Love JJ, Mayo SL. A de novo designed protein protein interface Protein Sci. 16(12):2770-4 (2007)). И как следствие, до настоящего времени методы позитивного дизайна не применялись для создания Fc гетеродимеров, так как для получения и разработки терапевтических антител специфичность была более важна, чем устойчивость. Кроме того, выгодные с точки зрения позитивного дизайна мутации могут быть труднопредсказуемы. Были попытки применить другие методы повышения устойчивости, такие как дополнительные дисульфидные связи для повышения устойчивости Fc гетеродимеров, но они имели ограниченный успех. (См. Таблицу А) Это может быть вызвано тем, что все дисульфидные связи созданных Fc CH3 доменов доступны для растворителя, что определяет короткий период существования дисульфидной связи и, следовательно, оказывает значительное влияние на долговременную устойчивость гетеродимера - в особенности, если созданный CH3 домен имеет Tm ниже 70°С без дополнительной сульфидной связи (как в Контроле 4, который имеет Tm 69°С без дисульфидной связи (см. Контроль 2). Предполагалось, что другие методы повышения устойчивости, такие как дисульфидные связи, можно также применять с Fc вариантами по настоящему изобретению при условии, что собственная устойчивость (количественно определяемая температурой плавления) CH3 домена равна 70°С или выше без дисульфидной связи, в частности, когда собственная устойчивость (количественно определяемая температурой плавления) CH3 домена равна 72°С или выше без дисульфидной связи.
По этой причине мы раскрываем в данном описании новый метод конструирования Fc гетеродимеров, который позволяет получать как устойчивые, так и высокоспецифические гетеродимеры. Этот метод конструирования (дизайна) объединяет стратегию как негативного, так и позитивного дизайна наряду с методами белковой инженерии на основе структурного и компьютерного моделирования. Этот мощный метод позволил нам создать новые комбинации мутаций в IgG1 CH3 домене, причем при использовании только стандартных клеточных культур были получены гетеродимеры с чистотой 90% по сравнению с гомодимерами, и полученные гетеродимеры имели температуру плавления 70°С или выше. Согласно типичным вариантам изобретения Fc вариантные гетеродимеры имеют температуру плавления 73°С или выше и чистоту выше 98%. Согласно другим типичным вариантам изобретения Fc вариантные гетеродимеры имеют температуру плавления 75°С или выше и чистоту выше 90%.
Согласно некоторым вариантам изобретения предусматривается выделенный гетеромультимер, содержащий гетеродимерную Fc область, причем гетеродимерная Fc область содержит вариантный CH3 домен, включающий аминокислотные мутации, стимулирующие образование гетеродимера с повышенной устойчивостью, а вариантный CH3 домен имеет температуру плавления (Tm) 70°С или выше. Выражения "повышенная устойчивость", "повышенная стабильность" или "устойчивый гетеродимер" относятся к вариантному CH3 домену, в гетеродимерной конструкции, с температурой плавления около 70°С или выше. Кроме того, понятно, что выражение "стимулировать образование гетеродимера" относится в данном описании к аминокислотным мутациям в CH3 домене, которые способствуют получению более 90% гетеродимерной конструкции по сравнению с гомодимерной конструкцией.
Согласно другому варианту изобретения эта повышенная устойчивость заключается в отсутствии дополнительной дисульфидной связи. Конкретно, повышенная устойчивость заключается в отсутствии дополнительной дисульфидной связи в CH3 домене. Согласно одному варианту изобретения вариантный CH3 домен не содержит дополнительную дисульфидную связь по сравнению с дикого типа CH3 доменом. Согласно альтернативному варианту изобретения вариантный CH3 домен содержит по меньшей мере одну дисульфидную связь по сравнению с дикого типа CH3 доменом, при условии, что вариантный CH3 домен имеет температуру плавления 70°С или выше в отсутствие дисульфидной связи. Согласно одному варианту изобретения вариантный CH3 домен содержит по меньшей мере одну дисульфидную связь по сравнению с дикого типа CH3 доменом, и вариантный CH3 домен имеет температуру плавления (Tm) около 77.5°С или выше. Согласно одному варианту изобретения вариантный CH3 домен содержит по меньшей мере одну дисульфидную связь по сравнению с дикого типа CH3 доменом, и вариантный CH3 домен имеет температуру плавления (Tm) выше, чем, примерно, 78°С, или выше. Согласно другому варианту изобретения вариантный CH3 домен содержит по меньшей мере одну дисульфидную связь по сравнению с дикого типа CH3 доменом, и вариантный CH3 домен имеет температуру плавления (Tm) около 78°С или выше, примерно, 78.5°С, или выше, примерно, 79°С, или выше, примерно, 79.5°С, или выше, примерно, 80°С, или выше, примерно, 80.5°С, или выше, примерно, 81°С.
Согласно одному варианту изобретения вариантный CH3 домен имеет температуру плавления выше, примерно, 70°С, или выше, примерно, 70.5°С, или выше, примерно, 71°С, или выше, примерно, 71.5°С, или выше, примерно, 72°С, или выше, примерно, 72.5°С, или выше, примерно, 73°С, или выше, примерно, 73.5°С, или выше, примерно, 74°С, или выше, примерно, 74.5°С, или выше, примерно, 75°С, или выше, примерно, 75.5°С, или выше, примерно, 76°С, или выше, примерно, 76.5°С, или выше, примерно, 77°С, или выше, примерно, 77.5°С, или выше, примерно, 78°С, или выше, примерно, 78.5°С, или выше, примерно, 79°С, или выше, примерно, 79.5°С, или выше, примерно, 80°С, или выше, примерно, 80.5°С, или выше, примерно, 81°С. Согласно другому варианту изобретения вариантный CH3 домен имеет температуру плавления около 70°С, или около 70.5°С, или около 71°С, или около 71.5°С, или около 72°С, или около 72.5°С, или около 73°С, или около 73.5°С, или около 74°С, или около 74.5°С, или около 75°С, или около 75.5°С, или около 76°С, или около 76.5°С, или около 77°С, или около 77.5°С, или около 78°С, или около 78.5°С, или около 79°С, или около 79.5°С, или около 80°С, или около 80.5°С, или около 81°С. Согласно еще одному варианту изобретения вариантный CH3 домен имеет температуру плавления от около 70°С до около 81°С, или от около 70.5°С до около 81°С, или от около 71°С до около 81°С, или от около 71.5°С до около 81°С, или от около 72°С до около 81°С, или от около 72.5°С до около 81°С, или от около 73°С до около 81°С, или от около 73.5°С до около 81°С, или от около 74°С до около 81°С, или от около 74.5°С до около 81°С, или от около 75°С до около 81°С, или от около 75.5°С до около 81°С, или от около 76°С до около 81°С, или от около 76.5°С до около 81°С, или от около 77°С до около 81°С, или от около 77.5°С до около 81°С, или от около 78°С до около 81°С, или от около 78.5°С до около 81°С, или от около 79°С до около 81°С. Согласно еще одному варианту изобретения вариантный CH3 домен имеет температуру плавления от около 71°С до около 76°С, или от около 72°С до около 76°С, или от около 73°С до около 76°С, или от около 74°С до около 76°С.
Помимо повышенной устойчивости, гетеродимерная Fc область содержит вариантный CH3 домен, включающий аминокислотные мутации, стимулирующие образование гетеродимера. Понятно, что эти аминокислотные мутации стимулируют образование гетеродимера по сравнению с образованием гомодимера. Это образование гетеродимера по сравнению с образованием гомодимера вместе называется "чистота", или "специфичность", или "чистота гетеродимера", или "специфичность гетеродимера ". Понятно, что чистота гетеродимера относится к процентному содержанию образующегося заданного гетеродимера по сравнению с гомодимером, образующимся в условиях стандартной клеточной культуры, до селективной очистки гетеродимера. Например, чистота димера 90% показывает, что 90% димеров в растворе составляет заданный гетеродимер. Согласно одному варианту изобретения Fc вариантные гетеродимеры имеют чистоту выше, примерно, 90%, или выше, примерно, 91%, или выше, примерно, 92%, или выше, примерно, 93%, или выше, примерно, 94%, или выше, примерно, 95%, или выше, примерно, 96%, или выше, примерно, 97%, или выше, примерно, 98%, или выше, примерно, 99%. Согласно другому варианту изобретения Fc вариантные гетеродимеры имеют чистоту около 90%, или около 91%, или около 92%, или около 93%, или около 94%, или около 95%, или около 96%, или около 97%, или около 98%, или около 99%, или около 100%.
Согласно конкретному варианту изобретения выделенный гетеромультимер, содержит гетеродимерную Fc область, причем гетеродимерная Fc область содержит вариантный CH3 домен, включающий аминокислотные мутации, стимулирующие образование гетеродимера с повышенной устойчивостью, при этом вариантный CH3 домен имеет температуру плавления (Tm) 70°С или выше, а чистота полученного в результате гетеродимера превышает 90%. В одном аспекте полученный Fc вариантный гетеродимер имеет чистоту выше 90%, а вариантный CH3 домен имеет температуру плавления выше, примерно, 70°С или выше, примерно, 71°С или выше, примерно, 72°С или выше, примерно, 73°С или выше, примерно, 74°С или выше, примерно, 75°С или выше, примерно, 76°С или выше, примерно, 77°С или выше, примерно, 78°С или выше, примерно, 79°С или выше, примерно, 80°С или выше, примерно, 81°С. В другом аспекте вариантный CH3 домен имеет температуру плавления (Tm) 70°С или выше, а полученный Fc вариантный гетеродимер имеет чистоту выше, примерно, 90%, или выше, примерно, 91%, или выше, примерно, 92%, или выше, примерно, 93%, или выше, примерно, 94%, или выше, примерно,95%, или выше, примерно, 96%, или выше, примерно, 97%, или выше, примерно, 98%, или выше, примерно, 99%.
Для создания этих Fc вариантов с повышенной устойчивостью и чистотой мы применяли итеративный процесс компьютерного дизайна и экспериментального скрининга с целью выбрать наиболее успешные комбинации стратегий позитивного и негативного дизайна (См. Фигуру 24).
Конкретно, в начальной фазе дизайна получали различные Fc вариантные гетеродимеры с помощью негативного дизайна и проверяли экспрессию и устойчивость этих гетеродимеров как описано в Примерах 1-3. Начальная фаза дизайна включала Fc вариантные гетеродимеры AZ1-AZ16 (См. Таблицу 1). Из этого исходного набора Fc вариантных гетеродимеров негативного дизайна, которые, как предполагалось, имели низкую устойчивость (например, Tm ниже 71°С), для дальнейшей разработки выбирали Fc вариантные гетеродимеры с чистотой выше 90% и температурой плавления около 68°С или выше. Эти Fc вариантные гетеродимеры включали AZ6, AZ8 и AZ15. Во второй фазе дизайна эти выбранные Fc вариантные гетеродимеры дополнительно модифицировали, чтобы управлять как устойчивостью, так и чистотой с применением стратегии позитивного дизайна с последующим подробным компьютерным и структурным анализом. Каждый выбранный Fc вариантный гетеродимер (AZ6, AZ8 и AZ15) анализировали с помощью компьютерных методов и полного структурно-функционального анализа с целью идентифицировать структурные причины, почему эти Fc варианты имели пониженную устойчивость по сравнению с дикого типа Fc гомодимером, где Tm равна 81°С для IgG1. См. Список Fc вариантных гетеродимеров и значения Tm в Таблице 4.
Согласно некоторым вариантам изобретения вариантный CH3 домен выбран из AZ1, или AZ2, или AZ3, или AZ4, или AZ5, или AZ6, или AZ7, или AZ8, или AZ9, или AZ10, или AZ11, или AZ12, или AZ13, или AZ14, или AZ15, или AZ16. В выбранных вариантах изобретения вариантный CH3 домен представляет собой AZ6, или AZ8 или AZ15.
Вычислительные средства и структурно-функциональный анализ включали, но без ограничения, молекулярно-динамический расчет (MD, МД), "переупаковку" боковых цепей/каркаса, потенциал базы знаний (Knowledge Base Potential, KBP), анализ впадин и (гидрофобной) упаковки (LJ, CCSD, SASA, dSASA (углерод/все-атомы)), расчеты электростатических-GB взаимодействий и анализ связывания. (См. на Фигуре 24 общий обзор методологии расчетов).
Один аспект нашего метода белковой инженерии опирается на общие структурные данные о Fc IgG белке, полученные методами рентгеноструктурного анализа с применением компьютерного моделирования дикого типа и вариантных форм CH3 домена. Эти методы позволили нам получить новые структурные и физико-химические данные о возможной роли отдельных аминокислот и их совместного действия. Эти сведения о структуре и физико-химических свойствах, полученные при изучении многих вариантных CH3 доменов, наряду с полученными эмпирическими данными, относящимися к их устойчивости и чистоте, помогли нам углубить понимание связи между чистотой и устойчивостью Fc гетеродимера по сравнению с Fc гомодимерами и воспроизведенными структурными моделями. Чтобы осуществить наше моделирование, мы начали с построения полных и правдоподобных моделей и уточнения свойств дикого типа Fc структуры IgG1 антитела. Структуры белков, полученные методом рентгеноструктурного анализа, не содержат подробностей, относящихся к некоторым свойствам белка в водной среде в физиологических условиях, и наши методы уточнения относились к этим ограничениям. Эти методы включают построение отсутствующих областей белковой структуры, зачастую гибких участков белка, таких как петли и боковые цепи некоторых остатков, оценку и определение состояний протонирования нейтральных и заряженных остатков и расположение потенциальных функционально релевантных молекул воды, ассоциированных с белком.
Алгоритмы молекулярной динамики (MD) являются одним из инструментов, который мы использовали при моделировании структуры белка, чтобы оценить характерную динамическую природу Fc гомодимера и вариантного CH3 домена в водной среде. Имитационные модели молекулярной динамики повторяют траекторию движения молекулы в результате взаимодействий и сил, действующих между всеми атомами в белке и их локальным окружением, в данном случае межу атомами, составляющими Fc, и окружающими их молекулами воды. После моделирования методами молекулярной динамики анализировались различные аспекты траекторий для лучшего понимания сущности структурных и динамических характеристик Fc гомодимера и вариантного Fc гетеродимера, которые мы применяли для идентификации специфических аминокислотных мутаций с целью повысить как чистоту, так и устойчивость молекулы.
По этой причине полученные MD траектории изучались такими методами, как анализ главных компонентов, с целью выявить внутренние низкочастотные колебания в Fc структуре. Это позволяет понять возможные конформационные "подсостояния" белка (См. Фигуру 32). Хотя важнейшие белок-белковые взаимодействия между цепью A и цепью B в Fc области происходят в области контакта CH3 доменов, наши имитационные модели показали, что эта область контакта (интерфейс) ведет себя как шарнир при движении, которое включает "открытие" и "закрытие" N-концов СН2 доменов относительно друг друга. СН2 домен взаимодействует с FcgR на этом конце, что видно на Фигуре 16. Итак, без связи с какой-либо теорией, можно сказать, что, по-видимому, введение аминокислотных мутаций в область контакта CH3 влияет на величину и характер движения открыть/закрыть на N-конце Fc и, следовательно, на то, как Fc взаимодействует с FcgR′s. См. Пример 4 и Таблицу 5.
Полученные MD траектории также изучали с целью определения мутируемости положений конкретных аминокислотных остатков в Fc структуре на основании анализа профиля их гибкости и их окружения. Этот алгоритм позволил нам идентифицировать остатки, которые могли бы повлиять на структуру и функцию белка, что дало оригинальное представление о характеристиках и мутабильности остатков, необходимое для последующих фаз дизайна вариантных CH3 доменов. Этот анализ позволил также сравнить несколько имитационных моделей и оценить изменчивость мутируемости остатков на основании показателей последующего анализа профиля.
Полученные MD траектории изучали также с целью определить взаимосвязанные, согласованные движения остатков в белке и образование сети остатков в результате осуществления связей между ними. Открытие динамических корреляций и сетей остатков в структуре Fc является важнейшей стадией для понимания белка как динамической структуры для проникновения в суть влияния мутаций в дальних сайтах. См., например, Пример 6.
Итак, мы подробно изучили влияние мутаций на локальное окружение сайта мутации. Образование плотно упакованного ядра в области контакта CH3 между цепью А и цепью В чрезвычайно важно для спонтанного спаривания двух цепей в устойчивой Fc структуре. Плотная упаковка является результатом строгого структурного соответствия между взаимодействующими молекулярными партнерами, связанными с помощью благоприятных взаимодействий между контактирующими группами. Благоприятные взаимодействия являются результатом скрытых гидрофобных контактов, весьма удаленных от воздействия растворителя, и/или результатом комплементарных электростатических взаимодействий между гидрофильными полярными группами. Эти гидрофобные и гидрофильные контакты вносят энтропийный и энтальпийный вклад в свободную энергию образования димера в области контакта CH3. Мы применяли разные алгоритмы для того, чтобы создать точную модель упаковки в области контакта CH3 между цепью А и цепью В, а потом оценить термодинамические свойства области контакта сравнением релевантных физико-химических свойств.
Мы применяли несколько методов упаковки белков, включая гибкие скелеты, с целью оптимизировать и получить модельные структуры для большого числа вариантов, компьютерный скрининг которых мы проводили. После упаковки мы оценивали ряд показателей, включая плотность контактов, вклад нежелательных контактов, водородные связи и электростатику. Использование моделей сольватации позволило нам более точно направлять действие растворителя на различные типы остатков и сопоставлять разницу свободной энергии после мутации в конкретных положениях в белке с этими различными типами остатков. Плотность контактов и вклад нежелательных контактов дают оценку комплементарности, важного аспекта эффективной упаковки белка. Эти методы скрининга основаны на применении возможностей базы знаний или схем анализа связывания, опираясь на вычисления энергии попарного взаимодействия остатков и энтропии.
Этот полный компьютерный (in-silico) анализ дал всестороннее представление об отличии каждого Fc варианта от дикого типа в отношении "горячих точек" (гипервариабельных участков), сайтов асимметрии, впадин и областей с неплотной упаковкой, структурной динамики отдельных сайтов локального развертывания в области контакта. Эти совместные результаты описанного компьютерного анализа позволили идентифицировать специфические остатки, последовательность/структурные мотивы и впадины, которые не были оптимизированы и в комбинации отвечают за пониженную устойчивость (например, Tm 68°С) и/или пониженную специфичность <90% чистоты. Во второй фазе дизайна мы применяли целевой позитивный дизайн с целью конкретно доказать эту гипотезу с помощью дополнительных точковых мутаций и тестировали их с использованием компьютерной (in-silico) инженерии вышеописанными методами и анализом (См. Фигуру 24). Правильность выбора Fc вариантных гетеродимеров, созданных с целью повысить устойчивость и чистоту, в каждом случае целевого дизайна во второй фазе (Fc вариантные гетеродимеры AZ17-AZ101) подтверждали экспериментально, проверяя экспрессию и устойчивость, как описано в Примерах 1-4.
Согласно некоторым вариантам изобретения предусматриваются выделенные гетеромультимеры, содержащие гетеродимерную Fc область, причем гетеродимерная Fc область содержит вариантный CH3 домен, включающий аминокислотные мутации, стимулирующие образование гетеродимера с повышенной устойчивостью, где вариантный CH3 домен представляет собой AZ17, или AZ18, или AZ19, или AZ20, или AZ21, или AZ22, или AZ23, или AZ24, или AZ25, или AZ26, или AZ27, или AZ28, или AZ29, или AZ30, или AZ21, или AZ32, или AZ33, или AZ34, или AZ35, или AZ36, или AZ37, или AZ38, или AZ39, или AZ40, или AZ41, или AZ42, или AZ43, или AZ44, или AZ45, или AZ46, или AZ47, или AZ48, или AZ49, или AZ50, или AZ51, или AZ52, или AZ53, или AZ54, или AZ55, или AZ56, или AZ57, или AZ58, или AZ59, или AZ60, или AZ61, или AZ62, или AZ63, или AZ64, или AZ65, или AZ66, или AZ67, или AZ68, или AZ69, или AZ70, или AZ71, или AZ72, или AZ73, или AZ74, или AZ75, или AZ76, или AZ77, или AZ78, или AZ79, или AZ80, или AZ81, или AZ82, или AZ83, или AZ84, или AZ85, или AZ86, или AZ87, или AZ88, или AZ89, или AZ90, или AZ91, или AZ92, или AZ93, или AZ94, или AZ95, или AZ96, или AZ97, или AZ98, или AZ99, или AZ100, или AZ101. Согласно типичному варианту изобретения CH3 домен представляет собой AZ17, или AZ18, или AZ19, или AZ20, или AZ21, или AZ22, или AZ23, или AZ24, или AZ25, или AZ26, или AZ27, или AZ28, или AZ29, или AZ30, или AZ21, или AZ32, или AZ33, или AZ34, или AZ38, или AZ42, или AZ43, или AZ 44, или AZ45, или AZ46, или AZ47, или AZ48, или AZ49, или AZ50, или AZ52, или AZ53, или AZ54, или AZ58, или AZ59, или AZ60, или AZ61, или AZ62, или AZ63, или AZ64, или AZ65, или AZ66, или AZ67, или AZ68, или AZ69, или AZ70, или AZ71, или AZ72, или AZ73, или AZ74, или AZ75, или AZ76, или AZ77, или AZ78, или AZ79, или AZ81, или AZ82, или AZ83, или AZ84, или AZ85, или AZ86, или AZ87, или AZ88, или AZ89, или AZ91, или AZ92, или AZ93, или AZ94, или AZ95, или AZ98, или AZ99, или AZ100, или AZ101. Согласно одному конкретному варианту изобретения вариантный CH3 домен представляет собой AZ33 или AZ34. Согласно другому варианту изобретения вариантный CH3 домен представляет собой AZ70 или AZ90.
Согласно одному типичному варианту CH3 домен содержит первый и второй полипептид (также в данном описании обозначаемые как Цепь А и Цепь В), причем первый полипептид содержит аминокислотные модификации L351Y, F405A и Y407V, а второй полипептид содержит аминокислотные модификации T366I, K392M и T394W. Согласно другому варианту изобретения первый полипептид содержит аминокислотные модификации L351Y, S400E, F405A и Y407V, а второй полипептид содержит аминокислотные модификации T366I, N390R, K392M и T394W.
Этот итеративный процесс компьютерного структурно-функционального анализа, целевой инженерии и экспериментального подтверждения был применен для конструирования остальных Fc вариантов, перечисленных в Таблице 1, в последующих фазах дизайна, и в результате получены Fc вариантные гетеродимеры с чистотой выше 90% и повышенной устойчивостью с температурой плавления CH3 домена выше 70°С. Согласно некоторым вариантам изобретения Fc варианты содержат аминокислотные мутации, выбранные из AZ1 до AZ 136. Согласно другим вариантам изобретения Fc варианты содержат аминокислотные мутации, выбранные из Fc вариантов, перечисленных в Таблице 4.
В результате первой и второй фаз дизайна были идентифицированы две ядерные каркасные структуры, Каркас 1 и Каркас 2, причем дополнительные аминокислотные модификации вводились в эти каркасы для точной регулировки чистоты и устойчивости Fc вариантных гетеродимеров. См. в Примере 5 подробное описание разработки Каркаса 1, включая AZ8, AZ17-62 и варианты, перечисленные в Таблице 6. См. в Примере 6 подробное описание разработки Каркаса 2, включая AZ15 и AZ63-101 и варианты, перечисленные в Таблице 7.
Ядерные мутации Каркаса 1 включают L351Y_F405A_Y407V/T394W. Каркас 1а содержит аминокислотные мутации T366I_K392M_T394W/F405A_Y407V, а Каркас 1b содержит аминокислотные мутации T366L_K392M_T394W/F405A_Y407V. См. Пример 5. Согласно некоторым вариантам изобретения вариантный CH3 домен содержит первый и второй полипептид (также в данном описании называемый Цепь А и Цепь В), причем первый полипептид содержит аминокислотные модификации L351Y, F405A и Y407V, а второй полипептид содержит аминокислотную модификацию T394W. В одном аспекте вариантный CH3 домен дополнительно содержит точковые мутации в положениях F405 и/или K392. Эти мутации в положении K392 включают, но без ограничения, K392V, K392M, K392R, K392L, K392F или K392E. Эти мутации в положении F405 включают, но без ограничения, F405I, F405M, F405S, F405S, F405V или F405W. В другом аспекте вариантный CH3 домен дополнительно содержит точковые мутации в положениях Т411 и/или S400. Эти мутации в положении Т411 включают, но без ограничения, T411N, T411R, T411Q, T411K, T411D, Т411Е или T411W. Эти мутации в положении S400 включают, но без ограничения, S400E, S400D, S400R или S400K. Согласно еще одному варианту изобретения вариантный CH3 домен содержит первый и второй полипептид, причем первый полипептид содержит аминокислотные модификации L351Y, F405A и Y407V, а второй полипептид содержит аминокислотную модификацию T394W, и первый и/или второй полипептид содержит дополнительные аминокислотные модификации в положениях Т366 и/или L368. Эти мутации в положении Т366 включают, но без ограничения, Т366А, T366I, T366L, Т366М, T366Y, T366S, Т366С, T366V или T366W. Согласно одному типичному варианту изобретения аминокислотная мутация в положении Т366 представляет собой мутацию T366I. Согласно другому типичному варианту изобретения аминокислотная мутация в положении Т366 представляет собой мутацию T366L. Мутации в положении L368 включают, но без ограничения, L368D, L368R, L368T, L368M, L368V, L368F, L368S и L368A.
Согласно некоторым вариантам изобретения вариантный CH3 домен содержит первый и второй полипептид (также в данном описании называемый Цепь А и Цепь В), причем первый полипептид содержит аминокислотные модификации L351Y, F405A и Y407V, а второй полипептид содержит аминокислотные модификации T366L и T394W. Согласно другому варианту изобретения вариантный CH3 домен содержит первый и второй полипептид, причем первый полипептид содержит аминокислотные модификации L351Y, F405A и Y407V, а второй полипептид содержит аминокислотные модификации T366I и T394W.
Согласно некоторым другим вариантам изобретения вариантный СИЗ домен содержит первый и второй полипептид (также в данном описании называемый Цепь А и Цепь В), причем первый полипептид содержит аминокислотные модификации L351Y, F405A и Y407V, а второй полипептид содержит аминокислотные модификации T366L, K392M и T394W. Согласно другим вариантам изобретения вариантный CH3 домен содержит первый и второй полипептид, причем первый полипептид содержит аминокислотные модификации L351Y, F405A и Y407V, а второй полипептид содержит аминокислотные модификации T366I, K392M и T394W.
Согласно еще одному варианту изобретения вариантный CH3 домен содержит первый и второй полипептид (также в данном описании называемый Цепь А и Цепь В), причем первый полипептид содержит аминокислотные модификации F405A и Y407V, а второй полипептид содержит аминокислотные модификации T366L, K392M и T394W. Согласно другому варианту изобретения вариантный CH3 домен содержит первый и второй полипептид, причем первый полипептид содержит аминокислотные модификации F405A и Y407V, а второй полипептид содержит аминокислотные модификации T366I, K392M и T394W.
Согласно некоторым вариантам изобретения вариантный CH3 домен содержит первый и второй полипептид (также в данном описании называемый Цепь А и Цепь В), причем первый полипептид содержит аминокислотные модификации F405A и Y407V, а второй полипептид содержит аминокислотные модификации T366L и T394W. Согласно другому варианту изобретения вариантный CH3 домен содержит первый и второй полипептид, причем первый полипептид содержит аминокислотные модификации F405A и Y407V, а второй полипептид содержит аминокислотные модификации T366I и T394W.
В одном типичном варианте изобретения предусматриваются выделенные гетеродимеры, содержащие гетеродимерную Fc область, причем гетеродимерная Fc область содержит вариантный CH3 домен, содержащий аминокислотные мутации, стимулирующие образование гетеродимера с повышенной устойчивостью, при этом вариантный CH3 домен имеет температуру плавления (Tm) около 74°С или выше.
Согласно другому варианту изобретения предусматриваются выделенные гетеромультимеры, содержащие гетеродимерную Fc область, при этом гетеродимерная Fc область включает вариантный CH3 домен, содержащий аминокислотные мутации, стимулирующие образование гетеродимера с повышенной устойчивостью, где вариантный CH3 домен имеет температуру плавления (Tm) выше 70°С, а вариантные CH3 домены выбраны из Таблицы 6.
Ядерные мутации в Каркасе 2 включают L351Y_Y407A/T366A_K409F. Каркас 2а включает аминокислотные мутации L351Y_Y407A/T366V_K409F, а Каркас 2b включает аминокислотные мутации Y407A/T366A_K409F. См. Пример 6.
Согласно некоторым вариантам изобретения вариантный CH3 домен содержит первый и второй полипептид (также в данном описании называемый Цепь А и Цепь В), причем первый полипептид содержит аминокислотные модификации L351Y и Y407A, а второй полипептид содержит аминокислотные модификации Т366А и K409F. В одном аспекте вариантный CH3 домен дополнительно содержит точковые мутации в положениях Т366, L351 и Y407. Эти мутации в положении Т366 включают, но без ограничения, мутации T366I, T366L, Т366М, T366Y, T366S, Т366С, T366V или T366W. В одном конкретном варианте изобретения мутация в положении Т366 представляет собой мутацию T366V. Мутации в положении L351 включают, но без ограничения, мутации L351I, L351D, L351R или L351F. Мутации в положении Y407 включают, но без ограничения, мутацию Y407V или Y407S. См. CH3 варианты AZ63-AZ70 в Таблице 1 и Таблице 4 и в Примере 6.
Согласно одному типичному варианту изобретения вариантный CH3 домен содержит первый и второй полипептид (также в данном описании называемый Цепь А и Цепь В), причем первый полипептид содержит аминокислотные модификации L351Y и Y407A, а второй полипептид содержит аминокислотную модификацию T366V и K409F.
Согласно одному типичному варианту в настоящем изобретении предусматриваются выделенные гетеромультимеры, содержащие гетеродимерную Fc область, причем гетеродимерная Fc область включает вариантный CH3 домен, содержащий аминокислотные мутации, стимулирующие образование гетеродимера с повышенной устойчивостью, при этом вариантный CH3 домен имеет температуру плавления (Tm) около 75.5°С или выше. Согласно другому варианту в настоящем изобретении предусматриваются выделенные гетеромультимеры, содержащие гетеродимерную Fc область, причем гетеродимерная Fc область включает CH3 домен, содержащий аминокислотные мутации, стимулирующие образование гетеродимера с повышенной устойчивостью, при этом вариантный CH3 домен имеет температуру плавления (Tm) около 75°С или выше, а гетеродимер имеет чистоту около 90% или выше.
Согласно другим вариантам изобретения вариантный CH3 домен содержит первый и второй полипептид (также в данном описании называемый Цепь А и Цепь В), причем первый полипептид содержит аминокислотные модификации L351Y и Y407A, а второй полипептид содержит аминокислотные модификации Т366А и K409F, при этом вариантный CH3 домен содержит одну или более аминокислотных модификаций в положениях Т411, D399, S400, F405, N390 и/или K392. Эти мутации в положении D399 включают, но без ограничения, D399R, D399W, D399Y или D399K. Мутации в положении Т411 включают, но без ограничения, мутации T411N, T411R, T411Q, T411K, T411D, Т411Е или T411W. Мутации в положении S400 включают, но без ограничения, S400E, S400D, S400R или S400K. Мутации в положении F405 включают, но без ограничения, F405I, F405M, F405S, F405S, F405V или F405W. Мутации в положении N390 включают, но без ограничения, N390R, N390K или N390D. Мутации в положении K392 включают, но без ограничения, K392V, K392M, K392R, K392L, K392F или K392E. См. CH3 варианты AZ71-101 в Таблице 1 и Таблице 4 и в Примере 6.
Согласно одному типичному варианту изобретения вариантный CH3 домен содержит первый и второй полипептид (также в данном описании называемый Цепь А и Цепь В), причем первый полипептид содержит аминокислотную модификацию Y407A, а второй полипептид содержит аминокислотную модификацию Т366А и K409F. В одном аспекте этот вариантный CH3 домен дополнительно содержит аминокислотные модификации K392E, Т411Е, D399R и S400R. Согласно другому варианту изобретения вариантный CH3 домен содержит первый и второй полипептид, причем первый полипептид содержит аминокислотную модификацию D399R, S400R и Y407A, а второй полипептид содержит аминокислотную модификацию Т366А, K409F, K392E и Т411Е.
Согласно одному типичному варианту в настоящем изобретении предусматриваются выделенные гетеромультимеры, содержащие гетеродимерную Fc область, причем гетеродимерная Fc область включает вариантный CH3 домен, содержащий аминокислотные мутации, стимулирующие образование гетеродимера с повышенной устойчивостью, при этом вариантный CH3 домен имеет температуру плавления (Tm) около 74°С или выше. Согласно другому варианту в настоящем изобретении предусматриваются выделенные гетеромультимеры, содержащие гетеродимерную Fc область, причем гетеродимерная Fc область включает CH3 домен, содержащий аминокислотные мутации, стимулирующие образование гетеродимера с повышенной устойчивостью, при этом вариантный CH3 домен имеет температуру плавления (Tm) около 74°С или выше, а гетеродимер имеет чистоту около 95% или выше.
Согласно некоторым вариантам в настоящем изобретении предусматриваются выделенные мультимеры, содержащие гетеродимерную Fc область, причем гетеродимерная Fc область включает вариантный CH3 домен, содержащий аминокислотные мутации, стимулирующие образование гетеродимера с повышенной устойчивостью, при этом вариантный CH3 домен имеет температуру плавления (Tm) выше 70°С и вариантные CH3 выбраны из Таблицы 7.
Кроме того, этот новый метод конструирования Fc вариантных гетеродимеров с повышенной устойчивостью и чистотой можно применять для других классов и изотипов Fc областей. Согласно другим вариантам изобретения Fc область представляет собой Fc область человеческого IgG. Согласно другим вариантам изобретения Fc область человеческого IgG представляет собой Fc область человеческого IgG1, IgG2, IgG3 или IgG4. Согласно некоторым вариантам изобретения Fc области получены из иммуноглобулина, выбранного из группы, состоящей из IgG, IgA, IgD, IgE и IgM. Согласно некоторым вариантам изобретения IgG относится к подклассу, выбранному из группы, состоящей из IgG1, IgG2a, IgG2b, IgG3 и IgG4.
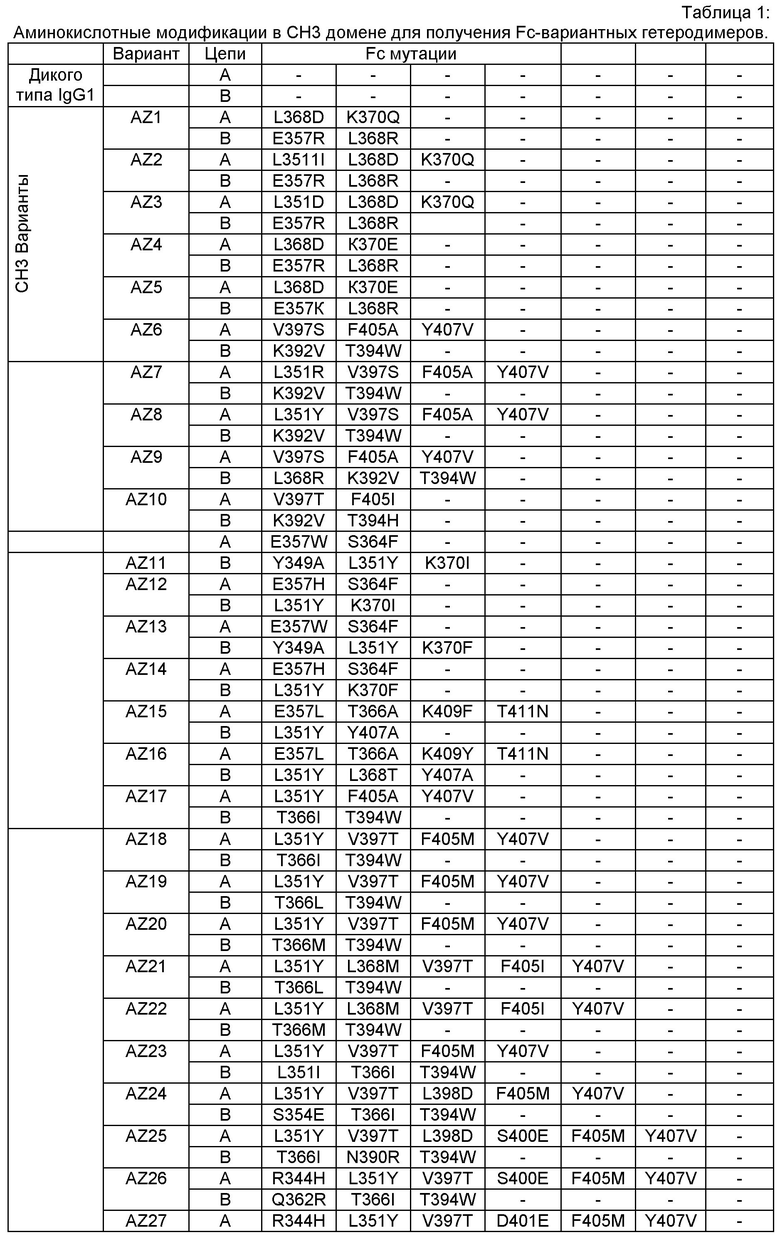
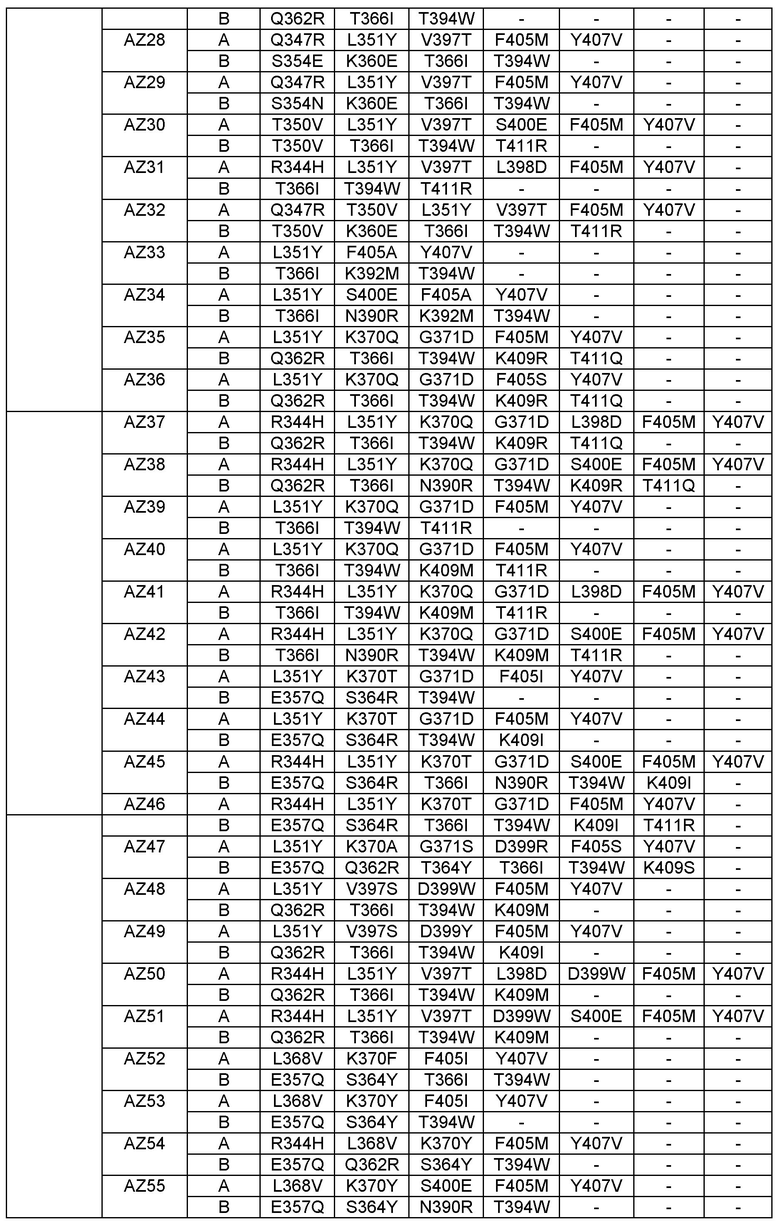
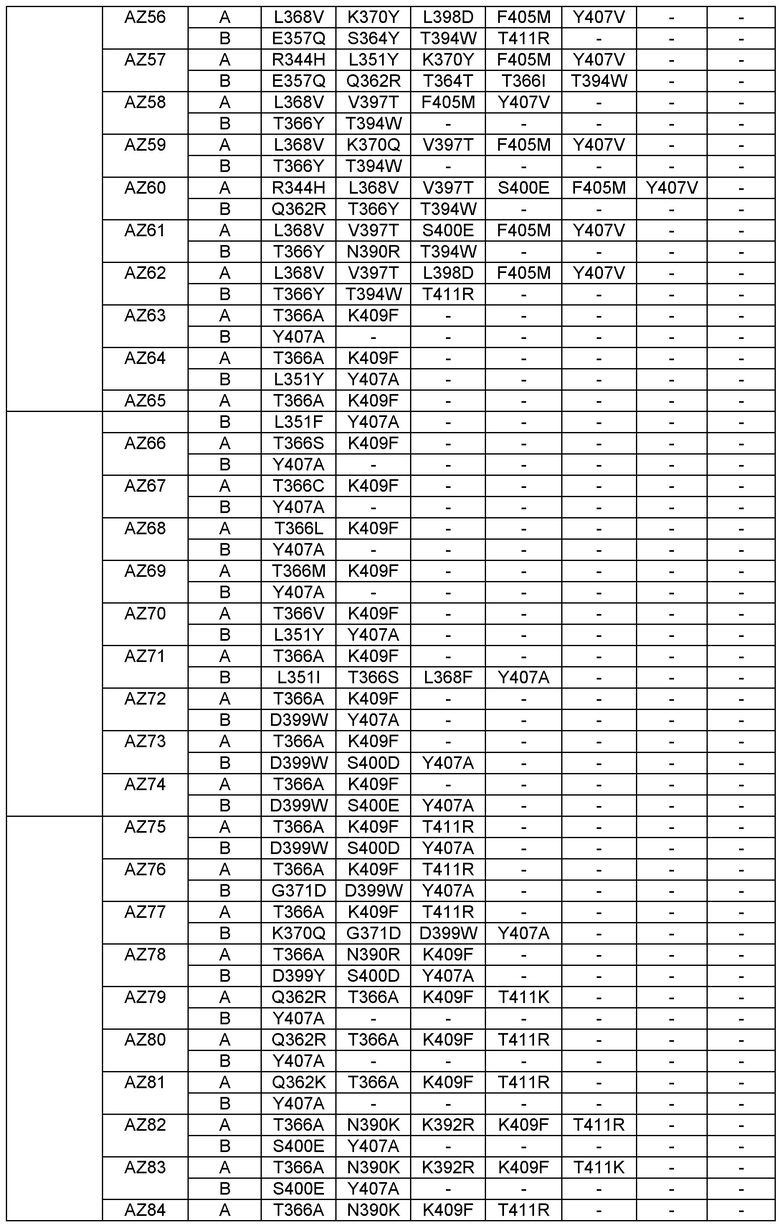
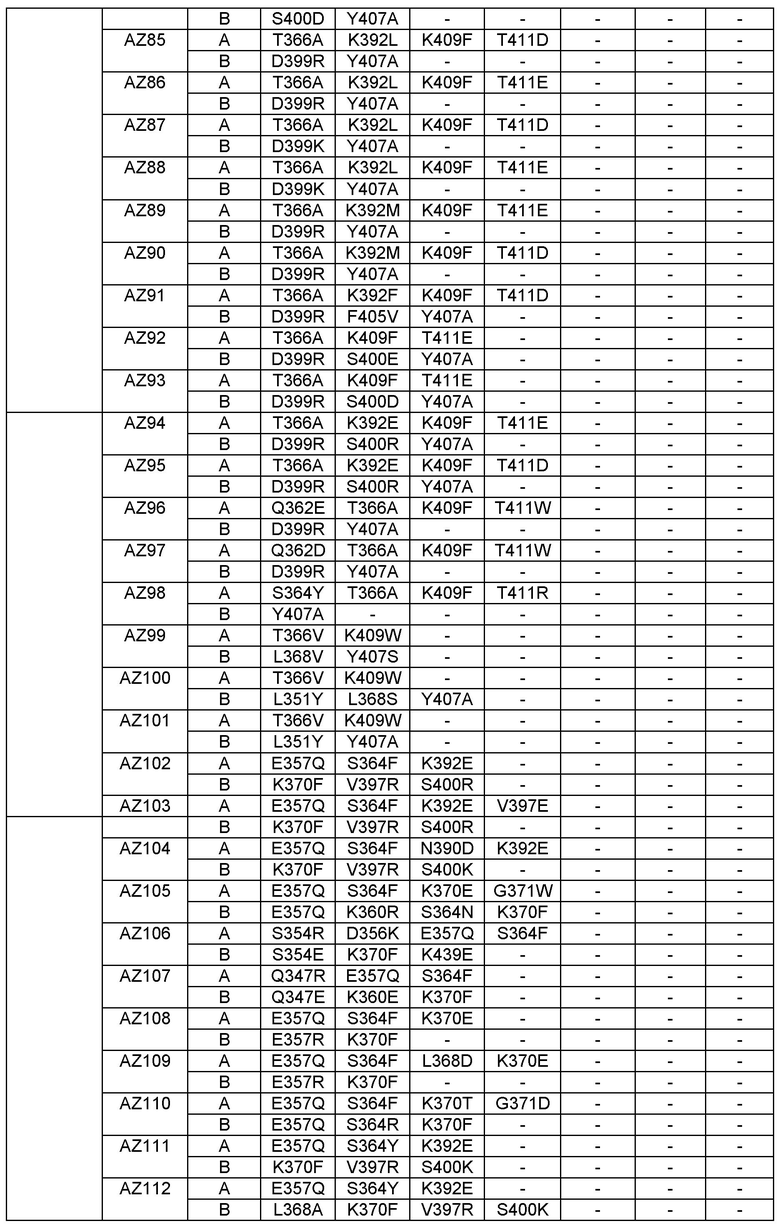
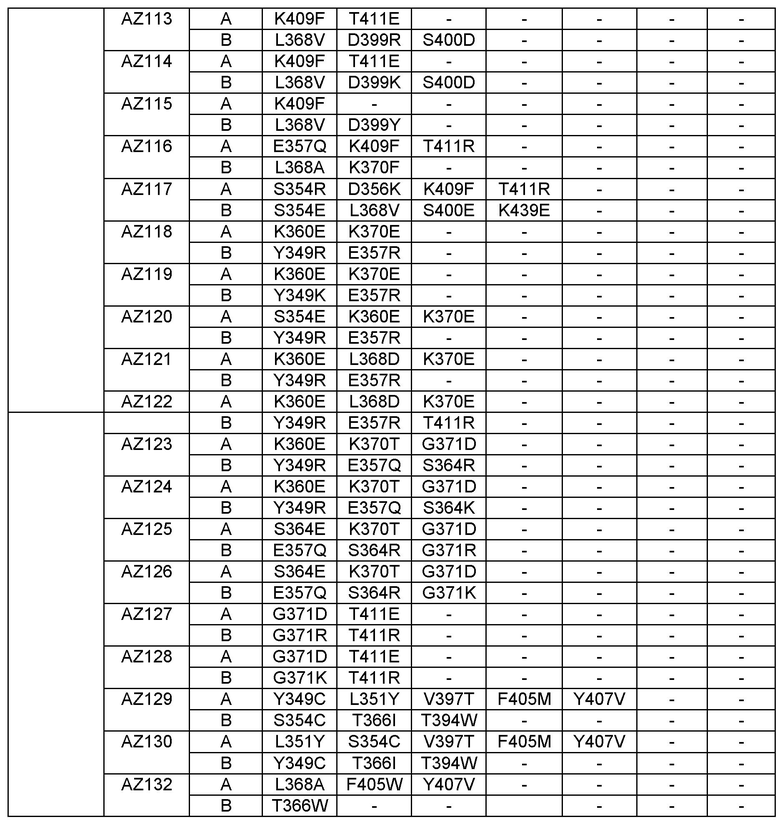
Область Fc по определению в данной заявке содержит CH3 домен или его фрагмент и, кроме того, может содержать в константной области один или более дополнительных доменов, или их фрагментов, включая шарнирную область, СН1 или СН2. Понятно, что нумерация Fc аминокислотных остатков соответствует системе нумерации EU-индекс по Kabat et al., 1991, NIH Publication 91-3242, National Technical Information Service, Springfield, Va. "EU-индекс no Kabat" относится к нумерации в системе EU-индекс для человеческого IgG1 по Kabat. Для удобства в Таблице В приводятся аминокислоты с нумерацией в соответствии с системой EU-индекс по Kabat СН2 и CH3 домена человеческого IgG1.
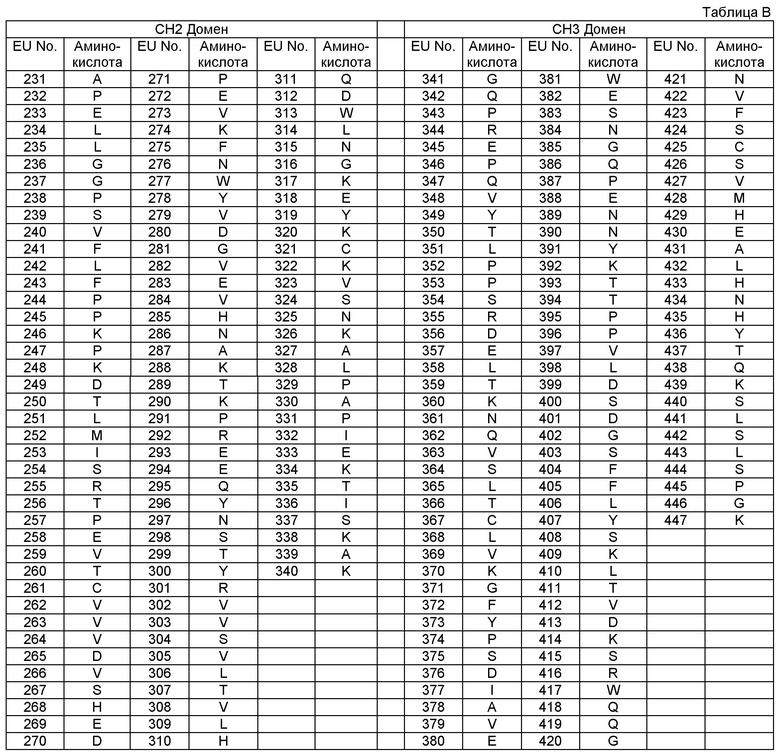
Согласно некоторым вариантам изобретения Fc вариант содержит СН2 домен. Согласно некоторым вариантам изобретения СН2 домен представляет собой вариантный СН2 домен. Согласно некоторым вариантам изобретения вариантный СН2 домен содержит асимметричные аминокислотные замены в первой и/или второй полипептидной цепи. Согласно некоторым вариантам изобретения гетеромультимер содержит асимметричные аминокислотные замены в СН2 домене таким образом, что одна цепь указанного гетеромультимера селективно связывает Fc рецептор.
Согласно некоторым вариантам изобретения, гетеромультимер селективно связывает Fc рецептор. Согласно некоторым вариантам изобретения Fc рецептор является членом семейства Fcγ рецепторов. Согласно некоторым вариантам изобретения рецептор выбран из FcγRI, FcγRIIa, FcγRIIb, FcγRIIc, FcγRIIIa и FcγRIIIb. Согласно одному варианту СН2 домен содержит асимметричные аминокислотные модификации, стимулирующие селективное связывание с Fc гамма рецепторами.
Согласно некоторым вариантам изобретения гетеромультимер селективно связывается с FcγRIIIa. Согласно некоторым вариантам изобретения гетеромультимер содержит асимметричные аминокислотные замены, выбранные из S267D, K392D и K409D. Согласно некоторым вариантам изобретения гетеромультимер селективно связывается с FcγRIIa. Согласно некоторым вариантам изобретения гетеромультимер содержит асимметричные аминокислотные замены, выбранные из S239D, K326E, A330L и I332E. Согласно некоторым вариантам изобретения гетеромультимер селективно связывается с FcγRIIb. Согласно некоторым вариантам изобретения гетеромультимер содержит асимметричные аминокислотные замены, выбранные из S239D, D265S, E269K и I332E. Согласно некоторым вариантам изобретения гетеромультимер селективно связывается с FcγRIIIa и FcγRIIa. Согласно некоторым вариантам изобретения гетеромультимер содержит асимметричные аминокислотные замены, выбранные из S239D, D265S и S298A. Согласно некоторым вариантам изобретения гетеромультимер селективно связывается с FcγRIIIa and FcγRIIb. Согласно некоторым вариантам изобретения гетеромультимер содержит асимметричные аминокислотные замены, выбранные из S239D, S298A, K326E, A330L и I332E. Согласно некоторым вариантам изобретения гетеромультимер селективно связывается с FcγRIIa и FcγRIIb. Согласно некоторым вариантам изобретения гетеромультимер содержит асимметричные аминокислотные замены, выбранные из S239D, D265S, S298A и I332E.
Согласно некоторым вариантам изобретения предусматривается метод конструирования (разработки, создания) многофункциональных лекарственных средств, содержащих гетеромультимер по данному описанию. Согласно некоторым вариантам изобретения предусматривается метод разработки бифункциональных лекарственных средств, содержащих вариантный Fc гетеродимер. Согласно некоторым вариантам изобретения предусматривается метод создания асимметричных мутаций в СН2 домене вариантного Fc гетеродимера, полученного с использованием мутаций в CH3 домене. Согласно некоторым вариантам изобретения предусматривается метод достижения селективности по отношению к различным Fc гамма рецепторам на основе мутаций в асимметричном Fc. Согласно некоторым вариантам изобретения предусматривается метод дизайна мутаций, которые смещают связывание Fc гамма рецепторов с одной стороной молекулы Fc. Согласно некоторым вариантам изобретения предусматривается метод дизайна факторов полярности, которые сдвигают взаимодействие Fcγ рецептора в одну сторону асимметричного Fc каркаса гетеромультимера по данному описанию.
Согласно некоторым вариантам изобретения предусматривается полипептид, содержащий мутации в СН2 домене асимметричного Fc, которые предопределяют предпочтительные профили селективности по отношению к Fc гамма рецепторам. Согласно некоторым вариантам изобретения мутации в CH3 домене предопределяют предпочтительное образование гетеродимерного Fc. Согласно некоторым вариантам изобретения предусматривается метод дизайна биспецифических терапевтических структур на основе асимметричных Fc по данному описанию. Согласно некоторым вариантам изобретения предусматривается метод разработки (дизайна) мультиспецифических терапевтических структур на основе асимметричных Fc по данному описанию.
Моноклональные антитела, такие как IgG, представляют собой симметричные молекулы, состоящие из двух эквивалентных тяжелых и двух легких полипептидных цепей (Фигура 14), каждая из которых содержит несколько структурных доменов иммуноглобулина (Ig). Класс IgG моноклональных антител (mAb) существует в виде одной из четырех изоформ, IgG1, IgG2, IgG3 или IgG4. Тяжелая цепь состоит из четырех (VH, CH1, СН2 и CH3), а легкая цепь из двух (VL и CL) Ig доменов, соответственно. Домены VH и CH1 каждой тяжелой цепи связываются с доменами VL и CL легкой цепи с образованием двух Fab ("антигенсвязывающий фрагмент") плеч mAb. CH3 и СН2 домены двух тяжелых цепей взаимодействуют через посредство белок-белковых связей на всем протяжении CH3 доменов и гликозилирование в СН2 доменах с образованием гомодимерной Fc ("кристаллизующийся фрагмент") области. Линкерная область между CH1 и СН2 доменами антитела образует шарнирную область молекулы антитела. Помимо связывания областей Fab и Fc mAb, шарнир также содержит дисульфидные связи поперек двух тяжелых цепей и удерживает их вместе. Число аминокислот и дисульфидных связей в шарнирной области четырех изотипов IgG заметно различается. Паттерн гликозилирования может очень различаться в молекулах IgG, в которых обнаружено около 30 различных углеводных остатков IgG [Arnold J.N.; Wormald M.R.; Sim R.B.; Rudd P.M. and Dwek R.A. (2007) Annual Reviews of Immunology 25, 21-50].
Симметричный характер структуры моноклональных антител обусловливает то, что оба Fab плеча, обладающие способностью к связыванию с антигеном, в результате созревания аффинности распознают один и тот же эпитоп. На другом конце Fc участок молекулы антитела участвует во взаимодействиях с различными рецепторными молекулами на иммунных или "эффекторных" клетках, и некоторые из этих взаимодействий отвечают за опосредование эффекторных функций, таких как антителозависимая клеточная цитотоксичность (ADCC, АЗКЦ), антителозависимый клеточный фагоцитоз (ADCP, АЗКЦ) и активация комплемента. Как правило, эффекторная функция включает иммунные ответы, приводящие к нейтрализации или исключению патогена или токсина, активации комплемента и фагоцитарного ответа под влиянием системы гуморального иммунитета. Молекулы Fcγ рецептора (FcγR) на эффекторных клетках связываются с Fc активированного IgG антитела, участвующего в целостном иммунном комплексе антитело-антиген, опосредующем и регулирующем эффекторный ответ. Оптимизация взаимодействия белковых терапевтических агентов на основе моноклонального антитела с этими Fcγ рецепторами может привести к повышению эффективности этих потенциальных лекарств.
У человека имеется три известных класса FcγR с дополнительными полиморфными типами в каждом классе. Известно, что Fc в молекуле IgG1 связывает FcγRI (CD64) с константами диссоциации в наномолярной области, тогда как связывание FcγRII (CD32) и fcγRIII (CD16) происходит в микромолярных концентрациях [Bruhns P.; Iannascoli В.; England P.; Mancardi D.A.; Fernandez N.; Jorieux S. and Daeron M. (2009) Blood 113:3716-25]. Высокоаффинные FcγRI могут связывать IgG в мономерной форме, тогда как низкоаффинные FcγRII и FcγRIII рецепторы могут связывать иммунные комплексы или IgG агрегаты антиген-антитело только в результате эффектов авидности. Различные формы IgG обладают различной аффинностью к различным FcγR′s; в частности, IgG1 и IgG3 проявляют повышенную активность. Fcγ рецепторы представляют собой внеклеточные домены трансмембранных белков и имеют цитоплазматические домены, которые участвуют в регуляции сигнальных путей внутри клетки. Будучи кластеризованы на поверхности иммунной клетки при ассоциации с антитело-опосредованными иммунными комплексами, в зависимости от характера сигнальных элементов, связанных с the FcγR′s на цитоплазматическом конце этих рецепторов клеточной поверхности, эти молекулы регулируют эффекторный ответ [Nimmerjahn F. and Ravetch J.V. (2008) Nature Immu Rev 8(1):34-47].
На хромосомном уровне у человека три гена кодируют FcγRI (FcγRIA, FcγRIB, FcγRIC) и FcγRII (FcγRIIA, FcγRIIB, FcγRIIC) и два гена кодируют FcγRIII (FcγRIIIA, FcγRIIIB). Было показано, что среди связывающих IgG человеческих рецепторов Fcγ рецепторы типов FcγRIA, FcγRIC и FcγRIIIA являются мембраносвязанными с адаптерным белком сигнального каскада с общей γ-цепью, который содержит иммунорецепторный тирозиновый активационный мотив (ITAM) в его цитоплазматическом домене, который приводит к активации эффекторной функции. FcγRIIA и FcγRIIC также содержат цитоплазматический домен ITAM, но без адаптерного белка сигнального каскада с общей γ-цепью. В то же время FcγRIIB связан с иммунорецепторным тирозиновьм ингибирующим мотивом (ITIM). Активация FcγRIIB, вызывающая фосфорилирование ITIM, приводит к ингибированию активирующего сигнального каскада. FcγRIIIB, при отсутствии любого из иммуномодулирующих мотивов на основе тирозина на цитоплазматических хвостах, содержит GPI якорь (гликозил-фосфатидил-инозитол) и, как было показано, вносит вклад в активацию некоторых гранулоцитов в присутствии FcγRIIA.
В то время как функциональная роль мотивов ITAM и ITIM и ассоциированных рецепторных молекул известна, природа и механизмы модуляции передачи сигнала в комбинации не совсем понятна, особенно в сочетании с активностью хозяина других иммуннорецепторов на поверхности клетки и адаптерных молекул (например, BCR, CD22, CD45 и т.д.), участвующих в сигнальной трансдукции. В этой ситуации дизайн Fc-подобных молекул, которые могут взаимодействовать с этими Fcγ рецепторами с профилями с исключительной селективностью, представляет собой ценный каркас при любой попытке провести деконволюцию и модулировать эффект таких рецепторных молекул с тонкой регуляторной активностью.
В процессе конструирования молекул антитела, которые могут различать FcγR′, попытки усложняются тем фактом, что во внеклеточных Fc-связывающих отрезках рецепторов типа FcγRII и FcγRIII наблюдается высокая степень подобия последовательностей (Фигура 15), что можно отнести, по меньшей мере частично, за счет сегментной дупликации одного предкового генома. Последовательности двух основных типов FcγRII рецепторов, А и В, идентичны на 69%, тогда как идентичность последовательностей FcγRIIA и FcγRIIIA составляет около 44%. Последовательности FcγRIIB и FcγRIIC отличаются только 2 остатками во внеклеточной области, хотя они значительно различаются во внутриклеточной области, а именно, присутствием мотивов ITIM и ITAM, соответственно. В результате можно ожидать, что молекулы терапевтического антитела, необходимые для связывания одного рецептора, теоретически, будут также связываться с рецепторами других классов, что, вероятно, вызовет непредусмотренный терапевтический эффект.
Кроме того, осложняющим моментом является то, что каждый класс рецепторов имеет несколько однонуклеотидных полиморфизмов (SNPs, ОНП) и вариации числа копий (CNVs). Обусловленное этим разнообразие рецепторов по-разному влияет на их аффинность к IgG′s и на механизм действия. Эти генетические вариации могли бы повлиять на аффинность конкретных подклассов IgG к Fcγ рецепторам, изменить последующие эффекторные события или повлиять на механизмы, которые изменяют уровни экспрессии рецепторов, приводя к функционально релевантным фенотипам, нефункциональным вариантам рецепторов или к вариантам рецепторов с неизвестной функцией (Bournazos S.; Woof J.M.; Hart S.P. and Dransfield I. (2009) Clinical and Experimental Immunology 157(2):244-54). Они, теоретически, могут вызвать сложные эффекты, изменяющие баланс между активацией и ингибированием передачи сигнала рецептором, что приведет в результате к созданию фенотипов, восприимчивых к заболеванию.
Некоторые из этих аллельных вариантов перечислены в Таблице С. В частности, вариант R131 в FcγRIIa с высокой эффективностью взаимодействуют с IgG1 (высокореактивный вариант), тогда как альтернативные H131 варианты более эффективно взаимодействуют с IgG2 и IgG3. В случае FcγRIIIa доноры, гомозиготные по V в положении 158, проявляют повышенную NK клеточную активность по сравнению с гомозиготными F/F158 индивидуумами вследствие повышенной аффинности первого аллотипа к человеческим IgG1, IgG3 и IgG4. Аллельные варианты NA1 и NA2 рецептора FcγRIIIb - это результат замены четырех аминокислот, что, в свою очередь, приводит к различию в гликозилировании рецептора. В случае NA1 аллеля наблюдается усиленное связывание и повышенный фагоцитоз иммунного комплекса нейтрофилами. FcγRIIB имеет два известных аллельных варианта, 232I и 232Т. Известно, что у варианта 232Т очень понижена активность негативной регуляции. Сообщалось о частоте встречаемости FcγR полиморфизмов и их связи с неодинаковой ответной реакцией на инфекции или предрасположенностью к болезненным состояниям, таким как системная красная волчанка (SLE), ревматоидный артрит (RA, РА), васкулит, иммуноопосредованная тромбоцитопеническая пурпура (ITP), тяжелая псевдопаралитическая миастения, рассеянный склероз (MS) и иммунные заболевания нервной системы (синдром Гийена-Барре (GBS)).
Была продемонстрирована вариация числа копий в локусе FcγR генов, в частности для FcγRIIIB, FcγRIIc и FcγRIIIA, и наблюдалась дополнительная корреляция этих различий с экспрессией этих рецепторов на клеточной поверхности. Напротив, в случае FcγRIIa и FcγRIIb не наблюдается вариации числа копий гена. На самом деле низкое число копий FcγRIIIb было связано с гломерулонефритом при аутоиммунном заболевании системной красной волчанкой (SLE) [Aitman TJ et al. (2006) Nature 16; 439(7078):851-5]. Это в особенности интересно, принимая во внимание тот факт, что несигнальный GPI модуль заякоривает FcγRIIIb рецептор. Можно предположить, что эти FcγRIIIb, в случае их присутствия, теоретически могли бы вести себя как конкурентные ингибиторы Fc взаимодействия с другими сигнальными FcγR. Эффект вариации числа копий в FcγRIIc также особенно интересен. С/Т SNP в положении 202 в FcγRIIc преобразует глутаминовый остаток в стоп-кодон, предупреждающий образование функционального белка. Функциональная открытая рамка считывания FcγRIIc экспрессируется у 9% здоровых людей (представителей белой расы), и в ITP популяции наблюдается заметное доминирование (19%) этого аллеля, что свидетельствует о предрасположенности этих фенотипов к ITP [Breunis WB et al. (2008) Blood 111(3):1029-38]. Было показано, что у отдельных людей, в организме которых экспрессируются функциональные FcγRIIc на NK клетках, достигнутая ADCC опосредуется этими рецепторами в большей степени, нежели FcγRIIIa. Такие сложности, обусловленные этими полиморфизмами и генетическими вариациями, указывают на необходимость персонализованной стратегии лечения, требующей высокоиндивидуализированных лекарственных средств.
Презентация этих Fcγ рецепторов различными эффекторными клетками, а также их гуморальное и тканевое распределение различны, тем самым внося вклад в вариации их активации и действия [Таблица D]. Тонкая регулировка селективности терапевтических антител для распознавания специфических FcγR типов и модуляция действия определенных классов эффекторных клеток приводит к оптимизации эффекторного механизма применительно к конкретным болезненным состояниям. Под этим понимают селективную активацию или селективное ингибирование специфических эффекторных функций в зависимости от подлежащего лечению болезненного состояния.
Кроме того, FcγR также экспрессируются фолликулярными дендритными клетками, эндотелиальными клетками, микроглиальными клетками, остеокластами и мезангиальными клетками. В настоящее время функциональная значимость экспрессии FcγR на этих других клетках неизвестна.
Высокоаффинный FcγRI состоит из трех доменов С-типа суперсемейства иммуноглобулинов (IgSF), тогда как низкоаффинные FcγRII и FcγRIII состоят из двух IgSF доменов C-типа каждый. Структура рецепторных белков FcγRIIa, FcγRIIb, FcγRIIIa и FcγRIIIb была решена кристаллографическими методами. Два IgSF домена в этих структурах расположены под углом 50-55 градусов относительно друг друга и связаны шарниром.
Общедоступная структура совместного комплекса Fc-FcγR представляет собой структуру системы Fc-FcγRIIIb, и геометрия FcγR в комплексе сохраняется очень близкой к геометрии, наблюдаемой в состоянии апобелка [Sondermann P.; Huber R.; Oosthuizen V. and Jacob U. (2000) Nature 406, 267-273; Radaev S.; Motyaka S.; Fridman W.; Sautes-Fridman C. and Sun P.D. (2001) J Biol Chem 276, 16469-16477; Sondermann P. et al. Biochem Soc Trans. 2002 Aug; 30(4):481-6; Sondermann P, Oosthuizen V. Immunol Lett. 2002 Jun 3; 82(1-2):51-6; Radaev S, Sun P. Mol Immunol. 2002 May; 38(14):1073-83.] [Фигура 16]. Большое подобие последовательностей и большое структурное подобие двух рецепторов создает основу для моделей сравнения Fc, связанного с другими рецепторами. С другой стороны, подобие последовательностей и структур этих рецепторных молекул также делает дизайн Fc с исключительной селективностью между рецепторами и их разнообразными изотипами трудным, но интересным.
Прежде, чем оценивать структуру комплекса Fc-FcγR кристаллографическими методами, нужно было решить вопрос, действительно ли ось симметрии второго порядка в молекуле Fc означает два возможных сайта связывания и эффективную 2:1 стехиометрию для комплекса Fc-FcγR. Структурные исследования взаимодействий Fc-FcγR методом ядерного магнитного резонанса (NMR, ЯМР) показывают, что связывание одной молекулы Fc с одной молекулой FcγR на одной стороне молекулы вызывает конформационное изменение, которое препятствует связыванию второй молекулы FcγR с Fc той же самой молекулы антитела [Kato K. et al (2000) J Mol Biol. 295(2):213-24]. Геометрия имеющегося сокристаллического комплекса Fc-FcγRIIIb подтверждает связь (ассоциацию) FcγR с Fc в асимметричной ориентации со стехиометрией 1:1. Как показано на Фигуре 16, FcγR связывается с углублением (расселиной) на одном конце подковообразной молекулы Fc и контактирует с СН2 доменами обеих цепей.
Аланин-сканирующий мутагенез [Shields RL et al. (2001) JBC 276(9):6591-604] дает представление об остатках области контакта Fc с различными типами рецепторов и, следовательно, участвующих во взаимодействии и узнавании Fc-FcγR. Обычно при оптимизации терапевтических антител внимание сосредоточено на мутациях, которые проявляют более прочное связывание с активирующими рецепторами FcγRIII [патент США No. 6737056] или пониженную аффинность к FcγRIIb [заявка на патент США US2009/0010920A1]. Во всех этих альтернативных вариантах мутации вводят одновременно в обе цепи.
Моноклональные антитела часто проявляют терапевтическую активность, индуцируя пространственную локализацию целевых (клеток-мишеней) и эффекторных иммунных клеток. Природное антитело опосредует это событие, взаимодействуя с мишенью с помощью своих Fab доменов и с эффекторной клеткой с использованием Fc домена. Они способны расположить иммунный комплекс непосредственно напротив эффекторной клетки (наложить его на эффекторную клетку) таким образом, чтобы можно было индуцировать клеточно-опосредованный ответ. Эффекты авидности, включающие нацеливание на одну мишень нескольких молекул антитела, является другим примером значимости пространственно-временной организации работы иммунной системы.
Есть также пространственно-временной аспект клеточной сигнализации, которая индуцируется как часть эффекторной активности молекул mAb. Клеточная сигнализация, такая как клеточная сигнализация за счет активации молекулы FcγR, включает локализацию соответствующих рецепторных молекул в области мембранного домена, называемого липидным рафтом. Липидные рафты обогащены гликосфинголипидом и холестерином и некоторыми классами upstream сигнальных трансдукторов, включая киназы семейства Src. В результате клеточной стимуляции рекрутируются различные сигнальные молекулы, адаптерные белки и сигнальные киназы, а также фосфатазы. Молекулярная сборка в липидных рафтах важна для сигнальной трансдукции.
Стратегия не-природного дизайна, объединяющего различные антигенные специфичности и повышенную авидность для улучшения связывающих свойств, лежит в основе дизайна биспецифического терапевтического антитела. Биспецифические антитела или другие формы биспецифических или многофункциональных белковых лекарственных средств конструируют с целью опосредовать взаимодействия между мишенью и различными эффекторными клетками [Muller & Kontermann (2010) BioDrugs 24(2):89-98]. Мультиспецифические терапевтические молекулы создавали для того, чтобы перенацелить хелперные Т-клетки или другие иммунные эффекторные клетки на специфические клетки-мишени.
Согласно другому варианту изобретение относится к методу компьютерной (in silico) идентификации Fc вариантных полипептидов на основании вычисленных величин аффинностей связывания с FcγRIIa, FcγRIIb и/или FcγRIIIa. Согласно другому варианту изобретения метод дополнительно включает компьютерный (in silico) расчет электростатики, сольватации, упаковки, плотности упаковки, связывания водородной связью и энтропийного эффекта указанных Fc вариантных полипептидов. Согласно еще одному варианту метод по настоящему изобретению дополнительно включает конструирование Fc вариантных полипептидов и экспрессию указанных полипептидов применительно к терапевтическому антителу и последующую экспрессию указанного антитела в клетках млекопитающих. Согласно еще одному варианту метод по настоящему изобретению включает конструирование Fc вариантных полипептидов, идентифицированных in silica с использованием сайт-направленного мутагенеза, PCR (ПЦР)-мутагенеза, кассетного мутагенеза или синтезом de novo.
Факторы, принимаемые в расчет при дизайне синтетического Fc каркаса, включают компьютерные (in silico) расчеты пространственного отталкивания, изменения скрытой области контакта, относительной плотности контакта, относительной сольватации и электростатического эффекта. Все эти матрицы применялись, чтобы прийти к оценке аффинности.
В одном аспекте данной заявки описывается молекулярный дизайн с целью получения профилей с исключительной селективностью по отношению к FcγR посредством создания асимметричного каркаса на основе гетеродимерного Fc. Этот каркас позволяет осуществлять асимметричные мутации в СН2 домене для получения целого ряда новых профилей селективности. Кроме того, этот каркас имеет характерные признаки для инженерии мультифункциональных (би-, три-, тетра- или пентафункциональных) терапевтических молекул.
Асимметричный каркас можно оптимизировать по рН-зависимым свойствам связывания с неонатальным Fc рецептором (FcRn) с целью содействовать возвращению молекулы в цикл и повысить ее период полужизни и связанные с этим фармакокинетические свойства.
Асимметричный каркас можно оптимизировать по связыванию с функционально релевантными аллотипами FcγRI рецептора. FcγRI является известным маркером на макрофагах, которые участвуют в хронических воспалительных расстройствах, таких как ревматоидных артрит, атопический дерматит, псориаз и ряд легочных заболеваний.
Асимметричный каркас можно оптимизировать по связыванию с Белком А. Связывание с Белком А часто используют для выделения и очистки молекул антитела. В асимметричный каркас можно вводить мутации, чтобы избежать агрегации лекарственного средства при хранении.
По этой причине конкретно рассматривается, чтобы Fc варианты по изобретению могли содержать, наряду с прочим, одну или более дополнительных аминокислотных замен, мутаций и/или модификаций, в результате которых получается антитело с предпочтительными характеристиками, включая, но без ограничения: повышенный период полужизни в сыворотке, повышенную аффинность связывания, пониженную иммуногенность, повышенное продуцирование, повышенную или пониженную ADCC или CDC активность, измененные гликозилирование и/или дисульфидные связи и модифицированную специфичность связывания.
Предполагается, что Fc варианты по изобретению могут иметь другие измененные характеристики, включая повышенные периоды полужизни (например, периоды полужизни в сыворотке) в организме млекопитающего, в частности, человека, повышенную устойчивость in vivo (например, период полужизни в сыворотке) и/или in vitro (например, срок хранения) и/или повышенную температуру плавления (Tm) по сравнению с сопоставимой молекулой. Согласно одному варианту Fc вариант по изобретению имеет in vivo период полужизни более 15 дней, более 20 дней, более 25 дней, более 30 дней, более 35 дней, более 40 дней, более 45 дней, более 2 месяцев, более 3 месяцев, более 4 месяцев или более 5 месяцев. Согласно другому варианту Fc вариант по изобретению имеет in vitro период полужизни (например, жидкий или порошковый препарат) более 15 дней, более 30 дней, более 2 месяцев, более 3 месяцев, более 6 месяцев, или более 12 месяцев, или более 24 месяцев, или более 36 месяцев, или более 60 месяцев.
Специалист в данной области техники поймет также, что Fc варианты по изобретению могут обладать измененной иммуногенностью при введении субъекту. Соответственно, предполагается, что вариантный CH3 домен, который снижает иммуногенность Fc варианта до минимума, как правило, является более желательным для применения в терапии.
Fc варианты по настоящему изобретению можно комбинировать с другими Fc модификациями, включая, но без ограничения, модификации, изменяющие эффекторную функцию. Изобретение охватывает комбинирование Fc варианта по изобретению с другими Fc модификациями, чтобы получить аддитивные, синергические или новые свойства антител или Fc-слитых белков. Такие модификации могут быть в шарнирной области, в доменах СН1 или СН2 (или CH3 при условии, что они не оказывают негативного влияния на свойства устойчивости и чистоты настоящих вариантных CH3 доменов) или в их комбинации. Предполагается, что Fc варианты по изобретению повышают качество модификации, с которой они объединяются. Например, если Fc вариант по изобретению объединяется с мутантом, заведомо связывающим FcγRIIIA с более высокой аффинностью, чем сопоставимая молекула, содержащая дикого типа Fc область, комбинирование с мутантом по изобретению приводит к более многократному повышению аффинности к FcγRIIIA.
Согласно одному способу воплощения Fc варианты по настоящему изобретению можно комбинировать с другими известными Fc вариантами, такими как Fc варианты, раскрываемые в Duncan et al, 1988, Nature 332:563-564; Lund et al, 1991, J Immunol 147:2657-2662; Lund et al., 1992, Mol Immunol 29:53-59; Alegre et al, 1994, Transplantation 57:1537-1543; Hutchins et al, 1995. Proc Natl. Acad Sci USA 92:11980-11984; Jefferis et al. 1995, Immunol Lett. 44:111-117; Lund et al., 1995, Faseb J 9:115- 119; Jefferis et al, 1996, Immunol Lett 54:101-104; Lund et al, 1996, Immunol 157:4963-4969; Armour et al., 1999, Eur J Immunol 29:2613-2624; Idusogie et al, 2000, J Immunol 164:4178-4184; Reddy et al, 2000. J Immunol 164:1925-1933; Xu et al., 2000, Cell Immunol 200:16-26; Idusogie et al, 2001. J Immunol 166:2571-2575; Shields et al., 2001, J Biol Chem 276:6591-6604; Jefferis et al, 2002, Immunol Lett 82:57-65; Presta et al., 2002, Biochem Soc Trans 30:487-490); патенты США No. 5624821; 5885573; 6194551; заявки на патент США No. 60/601634 и 60/608852; опубликованные патентные заявки РСТ No. WO 00/42072 и WO 99/58572.
Специалист в данной области техники поймет, что Fc варианты по изобретению могут иметь измененные свойства связывания Fc лиганда (например, FcγR, C1q) (примеры свойств связывания включают, но без ограничения, специфичность связывания, равновесную константу диссоциации (KD), константы скорости диссоциации и ассоциации (Koff и Kon, соответственно), аффинность и/или авидность связывания) и что определенные изменения являются более или менее желательными. В данной области техники хорошо известно, что равновесная константа диссоциации (KD) определяется как koff/kon. В целом понятно, что связывающая молекула (например, антитело) с низкой KD предпочтительней связывающей молекулы (например, антитела) с высокой KD. Однако, в некоторых случаях величина kon или koff может быть более релевантной, чем величина KD. Специалист в данной области техники может определить, какой кинетический параметр наиболее важен для данного применения антитела. Например, модифицированный CH3 и/или СН2, который усиливает связывание Fc с одним или более позитивных регуляторов (например, FcγRIIIA), в то же время оставляя неизменным или даже ослабляя связывание Fc с негативным регулятором FcγRIIB, был бы более предпочтителен для повышения ADCC активности. Или же, модифицированный CH3 и/или СН2, который ослаблял связывание с одним или более позитивных регуляторов и/или усиливал связывание с FcγRIIB, был бы более предпочтителен для снижения ADCC активности. Соответственно, соотношение величин аффинности связывания (например, равновесных констант диссоциации (KD)) может показывать, является ли ADCC активность Fc варианта повышенной или пониженной. Например, уменьшение отношения FcγRIIIA/FcγRIIB равновесных констант диссоциации (KD) будет коррелировать с повышенной ADCC активностью, тогда как повышение отношения будет коррелировать со снижением ADCC активности.
В процессе определения характеристик Fc вариантов они были протестированы на аффинность связывания с FcγRIIIA (CD16a) и FcγRIIB (CD32b), описанную в виде отношения, по сравнению с дикого типа IgG1. (См. Пример 4 и Таблицу 5). В этом примере можно было оценить влияние мутаций в CH3 домене на связывание с этими активирующим и ингибирующим Fc рецепторами. Согласно одному варианту в данном изобретении предусматриваются выделенные гетеромультимеры, содержащие гетеродимерную Fc область, где гетеродимерная Fc область включает вариантный CH3 домен, содержащий аминокислотные мутации, стимулирующие образование гетеродимера с повышенной устойчивостью, при этом вариантный CH3 домен имеет температуру плавления (Tm) выше 70°С, а связывание гетеродимера с CD16a примерно такое же, как и дикого типа гомодимера. Согласно некоторым вариантам изобретения связывание гетеродимера с CD16a является более сильным по сравнению со связыванием дикого типа гомодимера. Согласно альтернативному варианту изобретения связывание гетеродимера с CD16a является более слабым по сравнению со связыванием дикого типа гомодимера.
Согласно некоторым вариантам в данном изобретении предусматриваются выделенные гетеромультимеры, содержащие гетеродимерную Fc область, где гетеродимерная Fc область включает вариантный CH3 домен, содержащий аминокислотные мутации, стимулирующие образование гетеродимера с повышенной устойчивостью, при этом вариантный CH3 домен имеет температуру плавления (Tm) выше 70°С, а связывание гетеродимера с CD32b примерно такое же, как и дикого типа гомодимера. Согласно некоторым вариантам изобретения связывание гетеродимера с CD32b является более сильным по сравнению со связыванием дикого типа гомодимера. Согласно альтернативному варианту изобретения связывание гетеродимера с CD32b является более слабым по сравнению со связыванием дикого типа гомодимера.
Специалист в данной области техники понимает, что вместо того, чтобы характеризовать Ко связывания CD16a и CD32b как отношение связывания Fc варианта и гомодимера дикого типа. Ко можно было бы описывать как отношение связывания Fc варианта с CD16a к связыванию Fc варианта с CD32b (данные не показаны). Это отношение было бы показателем влияния мутации в вариантном CH3 домене на ADCC, либо неизменное, повышенное, пониженное по сравнению с гомодимером дикого типа, более подробно описано ниже.
Аффинности и связывающие свойства Fc вариантов по изобретению по отношению к FcγR сначала определяли известными в области техники методами анализа in vitro (биохимическими и иммунологическими методами анализа) для определения Fc-FcγR взаимодействий, т.е. специфического связывания Fc области с FcγR, включая, но без ограничения, ELISA (ИФА), поверхностный плазменный резонанс, иммунопреципитацию, (раздел, озаглавленный "Определение характеристик и функциональные анализы", см. ниже) и другие методы, такие как непрямые анализы связывания, конкурентные анализы ингибирования, резонансный перенос энергии флуоресценции (FRET), гель-электрофорез и хроматография (например, гель-фильтрация). В этих и других методах может применяться метка на одном или более изучаемых компонентов, и/или могут применяться различные методы обнаружения (детекции) этих меток, включающих, но без ограничения, хромогенные, флуоресцентные, люминесцентные или изотопные метки. Подробное описание значений аффинности и кинетики связывания можно найти в книге Paul, W. E., ed., Fundamental Immunology, 4th Ed., Lippincott-Raven, Philadelphia (1999), в которой основное внимание обращается на взаимодействие антитело-иммуноген.
Предполагается, что связывающие свойства молекул по изобретению характеризуются также функциональными анализами in vitro по определению одной или более эффекторных функций, опосредуемых FcγR (раздел, озаглавленный "Определение характеристик и функциональные анализы", см. ниже). Согласно некоторым вариантам молекулы по изобретению имеют связывающие свойства в in vivo моделях (такие как описанные и раскрываемые в настоящей заявке), аналогичные связывающим свойствам анализах in vitro. Однако настоящее изобретение не исключает молекулы по изобретению, которые не проявляют нужный фенотип в анализах in vitro, но проявляют нужный фенотип in vivo.
Изобретение охватывает Fc варианты, которые связывают FcγRIIIA (CD16a) с повышенной аффинностью по сравнению с сопоставимой (адекватной) молекулой. Согласно конкретному варианту Fc варианты по изобретению связывают FcγRIIIA с повышенной аффинностью и связывают FcγRIIB (CD32b) с аффинностью связывания, либо неизмененной, либо пониженной по сравнению с сопоставимой (адекватной) молекулой. Согласно еще одному варианту изобретения Fc варианты имеют отношение FcγRIIIA/FcγRIIB равновесных констант диссоциации (KD), пониженное по сравнению с сопоставимой молекулой.
Настоящее изобретение охватывает также Fc варианты, которые связывают FcγRIIIA (CD16a) с пониженной аффинностью по сравнению с сопоставимой молекулой. Согласно одному конкретному варианту Fc варианты по изобретению связывают FcγRIIIA с пониженной аффинностью по сравнению с сопоставимой молекулой и связывают FcγRIIB (CD32b) с аффинностью связывания, либо неизмененной, либо повышенной по сравнению с сопоставимой (адекватной) молекулой.
Согласно одному варианту изобретения Fc варианты связываются с FcγRIIIA с повышенной аффинностью. Согласно одному конкретному варианту изобретения аффинность указанных Fc вариантов к FcγRIIIA по меньшей мере в 2 раза, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 7 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 30 раз, или по меньшей мере в 40 раз, или по меньшей мере в 50 раз, или по меньшей мере в 60 раз, или по меньшей мере в 70 раз, или по меньшей мере в 80 раз, или по меньшей мере в 90 раз, или по меньшей мере в 100 раз, или по меньшей мере в 200 раз превышает аффинность сопоставимой молекулы. Согласно другим вариантам изобретения аффинность Fc вариантов к FcγRIIIA повышена по меньшей мере на 10%, или по меньшей мере на 20%, или по меньшей мере на 30%, или по меньшей мере на 40%, или по меньшей мере на 50%, или по меньшей мере на 60%, или по меньшей мере на 70%, или по меньшей мере на 80%, или по меньшей мере на 90%, или по меньшей мере на 100%, или по меньшей мере на 150%, или по меньшей мере на 200% по сравнению с сопоставимой молекулой.
Согласно другому варианту изобретения равновесная константа диссоциации (KD) комплекса Fc варианта с Fc лигандом (например, FcγR, C1q) понижена, примерно, в 2-10 раз, или, примерно, в 5-50 раз, или, примерно, в 25-250 раз, или, примерно, в 100-500 раз, или, примерно, в 250-1000 по сравнению с сопоставимой молекулой.
Согласно другому варианту изобретения равновесная константа диссоциации (KD) комплекса указанных Fc вариантов с FcγRIIIA понижена по меньшей мере в 2 раза, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 7 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 30 раз, или по меньшей мере в 40 раз, или по меньшей мере в 50 раз, или по меньшей мере в 60 раз, или по меньшей мере в 70 раз, или по меньшей мере в 80 раз, или по меньшей мере в 90 раз, или по меньшей мере в 100 раз, или по меньшей мере в 200 раз, или по меньшей мере в 400 раз, или по меньшей мере в 600 раз по сравнению с сопоставимой молекулой. Согласно другому варианту изобретения равновесная константа диссоциации (KD) комплекса указанных Fc вариантов с FcγRIIIA понижена по меньшей мере на 10%, или по меньшей мере на 20%, или по меньшей мере на 30%, или по меньшей мере на 40%, или по меньшей мере на 50%, или по меньшей мере на 60%, или по меньшей мере на 70%, или по меньшей мере на 80%, или по меньшей мере на 90%, или по меньшей мере на 100%, или по меньшей мере на 150%, или по меньшей мере на 200% по сравнению с сопоставимой молекулой.
Согласно одному варианту изобретения Fc вариант связывается с FcγRIIB с аффинностью, которая является неизменной или пониженной. Согласно одному конкретному варианту изобретения указанные Fc варианты проявляют аффинность к FcγRIIB, которая является неизменной или пониженной по меньшей мере в 1 раз, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 50 раз, или по меньшей мере в 100 раз по сравнению с сопоставимой молекулой. Согласно другим вариантам изобретения Fc варианты проявляют аффинность к FcγRIIB, которая является неизменной или пониженной по меньшей мере на 10%, или по меньшей мере на 20%, или по меньшей мере на 30%, или по меньшей мере на 40%, или по меньшей мере на 50%, или по меньшей мере на 60%, или по меньшей мере на 70%, или по меньшей мере на 80%, или по меньшей мере на 90%, или по меньшей мере на 100%, или по меньшей мере на 150%, или по меньшей мере на 200% по сравнению с сопоставимой молекулой.
Согласно другому варианту изобретения равновесная константа диссоциации (KD) комплекса Fc вариантов с FcγRIIB остается неизменной или является повышенной по меньшей мере в 2 раза, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 7 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 30 раз, или по меньшей мере в 40 раз, или по меньшей мере в 50 раз, или по меньшей мере в 60 раз, или по меньшей мере в 70 раз, или по меньшей мере в 80 раз, или по меньшей мере в 90 раз, или по меньшей мере в 100 раз, или по меньшей мере в 200 раз по сравнению с сопоставимой молекулой. Согласно другому конкретному варианту изобретения равновесная константа диссоциации (KD) комплекса Fc вариантов с FcγRIIB остается неизменной или является повышенной по меньшей мере на 10%, или по меньшей мере на 20%, или по меньшей мере на 30%, или по меньшей мере на 40%, или по меньшей мере на 50%, или по меньшей мере на 60%, или по меньшей мере на 70%, или по меньшей мере на 80%, или по меньшей мере на 90%, или по меньшей мере на 100%, или по меньшей мере на 150%, или по меньшей мере на 200% по сравнению с сопоставимой молекулой.
Согласно еще одному варианту изобретения Fc варианты связывают FcγRIIIA с повышенной аффинностью по сравнению с сопоставимой молекулой и связывают FcγRIIB с аффинностью, которая остается неизменной или является пониженной по сравнению с сопоставимой молекулой. Согласно конкретным вариантам изобретения Fc варианты проявляют аффинность к FcγRIIIA, повышенную по меньшей мере в 1 раз, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 50 раз, или по меньшей мере в 100 раз по сравнению с сопоставимой молекулой. Согласно другим конкретным вариантам изобретения Fc варианты проявляют аффинность к FcγRIIB, которая является неизменной или пониженной по меньшей мере в 2 раза, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 7 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 50 раз, или по меньшей мере в 100 раз по сравнению с сопоставимой молекулой. Согласно другим конкретным вариантам изобретения Fc варианты проявляют аффинность к FcγRIIIA, повышенную по меньшей мере на 10%, или по меньшей мере на 20%, или по меньшей мере на 30%, или по меньшей мере на 40%, или по меньшей мере на 50%, или по меньшей мере на 60%, или по меньшей мере на 70%, или по меньшей мере на 80%, или по меньшей мере на 90%, или по меньшей мере на 100%, или по меньшей мере на 150%, или по меньшей мере на 200% по сравнению с сопоставимой молекулой, и Fc варианты проявляют аффинность к FcγRIIB, которая является либо неизменной, либо пониженной по меньшей мере на 10%, или по меньшей мере на 20%, или по меньшей мере на 30%, или по меньшей мере на 40%, или по меньшей мере на 50%, или по меньшей мере на 60%, или по меньшей мере на 70%, или по меньшей мере на 80%, или по меньшей мере на 90%, или по меньшей мере на 100%, или по меньшей мере на 150%, или по меньшей мере на 200% по сравнению с сопоставимой молекулой.
Согласно еще одному варианту изобретения Fc варианты имеют отношение FcγRIIIA/FcγRIIB равновесных констант диссоциации (Ко), пониженное по сравнению с сопоставимой молекулой. В конкретном варианте изобретения Fc варианты имеют отношение FcγRIIIA/FcγRIIB равновесных констант диссоциации (KD), пониженное, по меньшей мере, в 1 раз, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 50 раз, или по меньшей мере в 100 раз по сравнению с сопоставимой молекулой. Согласно другому варианту изобретения Fc варианты имеют отношение FcγRIIIA/FcγRIIB равновесных констант диссоциации (KD), пониженное по меньшей мере на 10%, или по меньшей мере на 20%, или по меньшей мере на 30%, или по меньшей мере на 40%, или по меньшей мере на 50%, или по меньшей мере на 60%, или по меньшей мере на 70%, или по меньшей мере на 80%, или по меньшей мере на 90%, или по меньшей мере на 100%, или по меньшей мере на 150%, или по меньшей мере на 200% по сравнению с сопоставимой молекулой.
Согласно другому варианту изобретения Fc варианты связывают FcγRIIIA пониженной аффинностью по сравнению с сопоставимой молекулой. Согласно конкретному варианту изобретения аффинность указанных Fc вариантов к FcγRIIIA понижена по меньшей мере в 1 раз, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 50 раз, или по меньшей мере в 100 раз по сравнению с сопоставимой молекулой. Согласно другим вариантам изобретения аффинность Fc вариантов к FcγRIIIA понижена по меньшей мере на 10%, или по меньшей мере на 20%, или по меньшей мере на 30%, или по меньшей мере на 40%, или по меньшей мере на 50%, или по меньшей мере на 60%, или по меньшей мере на 70%, или по меньшей мере на 80%, или по меньшей мере на 90%, или по меньшей мере на 100%, или по меньшей мере на 150%, или по меньшей мере на 200% по сравнению с сопоставимой молекулой.
Согласно еще одному варианту изобретения Fc варианты связывают FcγRIIIA с пониженной аффинностью по сравнению с сопоставимой молекулой и связывают FcγRIIB с аффинностью, которая остается неизменной или является повышенной по сравнению с сопоставимой молекулой. Согласно конкретному варианту изобретения Fc варианты проявляют аффинность к FcγRIIIA, пониженную по меньшей мере в 1 раз, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 50 раз, или по меньшей мере в 100 раз по сравнению с сопоставимой молекулой. Согласно другому конкретному варианту изобретения Fc варианты проявляют аффинность к FcγRIIB, которая является повышенной по меньшей мере в 2 раза, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 7 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 50 раз, или по меньшей мере в 100 раз по сравнению с сопоставимой молекулой. Согласно другим вариантам изобретения Fc варианты проявляют аффинность к FcγRIIIA, пониженную по меньшей мере на 10%, или по меньшей мере на 20%, или по меньшей мере на 30%, или по меньшей мере на 40%, или по меньшей мере на 50%, или по меньшей мере на 60%, или по меньшей мере на 70%, или по меньшей мере на 80%, или по меньшей мере на 90%, или по меньшей мере на 100%, или по меньшей мере на 150%, или по меньшей мере на 200% по сравнению с сопоставимой молекулой, и Fc варианты проявляют аффинность к FcγRIIB, которая является повышенной по меньшей мере на 10%, или по меньшей мере на 20%, или по меньшей мере на 30%, или по меньшей мере на 40%, или по меньшей мере на 50%, или по меньшей мере на 60%, или по меньшей мере на 70%, или по меньшей мере на 80%, или по меньшей мере на 90%, или по меньшей мере на 100%, или по меньшей мере на 150%, или по меньшей мере на 200% по сравнению с сопоставимой молекулой.
Согласно еще одному варианту изобретения величина равновесной константы диссоциации (KD) комплексов Fc вариантов с FcγRIIIA является повышенной по меньшей мере, в 1 раз, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 50 раз по сравнению с сопоставимой молекулой. Согласно одному конкретному варианту изобретения величина равновесной константы диссоциации (KD) комплексов Fc вариантов с FcγRIIB является повышенной по меньшей мере, в 2 раза, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 7 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 50 раз или по меньшей мере в 100 раз по сравнению с сопоставимой молекулой.
Вариации СН2 для селективности в отношении FcγR
Fc-FcγR белок-белковое взаимодействие в этом комплексе показывает, что две цепи в Fc молекуле взаимодействуют с двумя различными сайтами в молекуле FcγR. Хотя в двух тяжелых цепях в природных Fc молекулах имеется симметрия, локальное окружение остатков в одной цепи отличается от FcγR остатков, окружающих остаток в том же положении в противоположной цепи Fc. Остатки в двух симметричных родственных положениях взаимодействуют с различным набором FcγR остатков.
Если принимать во внимание асимметрию при ассоциации Fc с FcγR, конкурентные мутации в цепи А и цепи В молекулы Fc не влияют на взаимодействия с FcγR симметрично. При введении мутации с целью оптимизировать взаимодействия в одной из Fc с ее локальным FcγR окружением в гомодимерной Fc структуре, соответствующая мутация во второй цепи может быть благоприятной, неблагоприятной или не способствующей FcγR связыванию и профилю селективности.
С применением структурного и расчетного метода созданы асимметричные мутации в двух цепях Fc для того, чтобы преодолеть эти ограничения традиционных методов инженерии Fc, которые позволяют вводить одинаковые мутации в обе цепи Fc. Можно добиться более высокой селективности связывания между рецепторами, если две цепи Fc независимо оптимизировать по усиленному связыванию молекулы рецептора с их соответствующей стороной.
Например, можно сконструировать мутации в конкретном положении в одной цепи Fc, чтобы повысить селективность по отношению к конкретному остатку, позитивный дизайн, тогда как мутация остатка в том же самом положении может привести к неблагоприятному взаимодействию с локальным окружением в альтернативном типе Fcγ рецептора, негативный дизайн, таким образом достигается повышенная селективность одного из двух рецепторов. Согласно некоторым вариантам изобретения предусматривается метод дизайна в СН2 домене асимметричных аминокислотных модификаций, которые селективно связывают один Fc гамма рецептор по сравнению с другим (отличающимся) Fc гамма рецептором (например, селективно связывают FcgRIIIa вместо FcgRIIb). Согласно некоторым другим вариантам изобретения предусматривается метод дизайна (конструирования) асимметричных аминокислотных модификаций в СН2 домене вариантного Fc гетеродимера, содержащего аминокислотные модификации в CH3 домене, с целью стимулировать образование гетеродимера. Согласно другому варианту изобретения предусматривается метод дизайна селективности по отношению к различным Fc гамма рецепторам на базе вариантного Fc гетеродимера, содержащего асимметричные аминокислотные модификации в СН2 домене. Еще в одном варианте изобретения предусматривается метод создания асимметричных аминокислотных модификаций, которые смещают связывание Fc гамма рецепторов в сторону связывания с одной стороной Fc молекулы. Согласно другим вариантам изобретения предусматривается метод создания определяющего фактора полярности, который сдвигает Fc гамма рецепторы в сторону взаимодействия только с одной стороной вариантного гетеродимера, содержащего асимметричные аминокислотные модификации в СН2 домене.
Можно создать асимметричный дизайн мутаций в СН2 домене, позволяющий узнавать FcγR на одной стороне Fc молекулы. В результате этого образуется благоприятная сторона асимметричного Fc каркаса, тогда как противоположная сторона проявляет склонность к взаимодействию дикого типа и отсутствие предумышленного профиля селективности и может рассматриваться как неблагоприятная сторона. Стратегию негативного дизайна можно применять для введения мутаций на неблагоприятной стороне с целью блокировать взаимодействия FcγR с этой стороной асимметричного Fc каркаса, тем самым стимулируя склонность к целевым взаимодействиям с Fcγ рецепторами.
Настоящее изобретение относится также к слитым полипептидам, содержащим связывающий домен, слитый с Fc областью, где Fc область, включающая вариантный CH3 домен, содержит аминокислотные мутации, стимулирующие образование гетеродимера с повышенной устойчивостью, причем вариантный CH3 домен имеет температуру плавления (Tm) выше 70°С. В частности рассматривается, что молекулы, содержащие гетеродимер, включающий вариантный CH3 домен, можно получать методами, хорошо известными специалисту в данной области техники. Коротко говоря, такие методы включают, но без ограничения, объединение вариабельной области или связывающего домена с целевой специфичностью (например, вариабельной области, выделенной из библиотеки фагового дисплея или экспрессионной библиотеки или полученной из человеческого антитела или антитела организма, отличного от человека, или связывающего домена рецептора) с вариантными Fc гетеродимерами. Или же специалист в данной области техники может получать вариантный Fc гетеродимер модификацией CH3 домена в Fc области молекулы, содержащей Fc область (например, антитела).
Согласно одному варианту изобретения Fc варианты представляют собой антитела или Fc-слитые белки. Согласно конкретному варианту изобретение предусматривает антитела, содержащие Fc область, включающую вариантный CH3 домен, содержащий аминокислотные мутации, стимулирующие образование гетеродимера с повышенной устойчивостью, где вариантный CH3 домен имеет температуру плавления (Tm) выше 70°С. Такие антитела включают молекулы IgG, которые естественным образом содержат Fc область, включающую CH3 домен, который можно модифицировать с образованием Fc варианта, или производные антител, созданных таким образом, чтобы содержать Fc область, включающую вариантный CH3 домен. Fc варианты по изобретению включают любое антитело, которое связывает, предпочтительно, специфически (т.е. побеждает в конкурентном неспецифическом связывании по определению методами иммуноанализа, хорошо известными в области техники, применяемыми для анализа специфического связывания антиген-антитело), антиген, которое содержит Fc область, включающую вариантный CH3 домен. Такие антитела включают, но без ограничения, поликлональные, моноклональные, моноспецифические, биспецифические, мультиспецифические, человеческие, гуманизированные, химерные антитела, одноцепочечные антитела. Fab фрагменты, F(ab′)2 фрагменты, связанные дисульфидной связью Fvs и фрагменты, содержащие либо VL, либо VH домен или даже участок, определяющий комплементарность (гипервариабельная область, CDR), который специфически связывает антиген, в некоторых случаях созданный таким образом, чтобы содержать вариантный Fc гетеродимер или быть слитым с ним.
Термин "антителозависимая клеточная цитотоксичность" или "ADCC" относится к форме цитотоксичности, в которой секретированное антитело, связанное с Fc рецепторами (FcRs), присутствующими на некоторых цитотоксических клетках (например, естественных киллерах (NK-клетках), нейтрофилах и макрофагах), способствует специфическому связыванию этих цитотоксических эффекторных клеток с антиген-"несущей" клеткой-мишенью и последующему киллингу клетки-мишени под действием цитотоксинов. Специфические высокоаффинные IgG антитела, направленные к поверхности клеток-мишеней, "вооружают" цитотоксические клетки и абсолютно необходимы для киллинга. Лизис клеток-мишеней является внеклеточным, требует непосредственного контакта клетки с клеткой и не включает комплемента.
Способность любого специфического антитела опосредовать лизис клетки-мишени с помощью ADCC можно определить. Для определения ADCC активности целевое антитело добавляют к клеткам-мишеням в комбинации с иммунными эффекторными клетками, которые могут активироваться комплексами антиген-антитело, приводя к цитолизу клетки-мишени. Цитолиз обычно обнаруживают по высвобождению метки (например, радиоактивных субстратов, флуоресцентных красителей или природных внеклеточных белков) из лизированных клеток. Эффекторные клетки, применимые для таких анализов, включают мононуклеарные клетки периферической крови (РВМС) и естественные клетки-киллеры (NK). Конкретные примеры in vitro ADCC анализов описаны в статьях Wisecarver et al., 1985, 79:277; Bruggemann et al., 1987, J Exp Med 166:1351; Wilkinson et al., 2001, J Immunol Methods 258:183; Patel et al., 1995 J Immunol Methods 184:29 и в данной заявке (раздел "Определение характеристик и функциональные анализы", см. ниже). Или же, или помимо этого, ADCC активность целевого антитела можно определять in vivo, например, на животной модели, такой как модель, раскрываемая в статье Clynes et al., 1998, PNAS USA 95:652.
Предполагается, что Fc варианты по изобретению характеризуются in vitro функциональными анализами по определению одной или более FcγR-опосредуемых эффекторных функций. Согласно конкретным вариантам изобретения молекулы по изобретению проявляют одинаковые связывающие свойства и реализуют одинаковые эффекторные функции как на in vivo моделях (таких как модели, описанные и раскрываемые в настоящей заявке), как и в анализах in vitro. Однако настоящее изобретение не исключает молекулы по изобретению, которые не проявляют заданный фенотип в анализах in vitro, но проявляют заданный фенотип in vivo.
Кроме того, настоящее изобретение предусматривает Fc варианты с повышенной CDC функцией. Согласно одному варианту изобретения Fc варианты обладают повышенной CDC активностью. Согласно одному варианту изобретения Fc варианты проявляют CDC активность, по меньшей мере в 2 раза, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 10 раз, или по меньшей мере в 50 раз, или по меньшей мере в 100 раз выше CDC активности сопоставимой молекулы. Согласно другому варианту изобретения Fc варианты связывают C1q с аффинностью, которая по меньшей мере в 2 раза, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 7 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 50 раз, или по меньшей мере в 100 раз выше, чем аффинность сопоставимой молекулы. Согласно еще одному варианту изобретения Fc варианты обладают CDC активностью, повышенной по меньшей мере на 10%, или по меньшей мере на 20%, или по меньшей мере на 30%, или по меньшей мере на 40%, или по меньшей мере на 50%, или по меньшей мере на 60%, или по меньшей мере на 70%, или по меньшей мере на 80%, или по меньшей мере на 90%, или по меньшей мере на 100%, или по меньшей мере на 150%, или по меньшей мере на 200% по сравнению с сопоставимой молекулой. Согласно конкретному варианту изобретения Fc варианты по изобретению связывают C1q с повышенной аффинностью, имеют повышенную CDC активность и специфически связывают по меньшей мере с одним антигеном.
Настоящее изобретение предусматривает также Fc варианты с пониженной CDC функцией. Согласно одному варианту изобретения Fc варианты проявляют пониженную CDC активность. Согласно одному варианту изобретения Fc варианты проявляют CDC активность, по меньшей мере в 2 раза, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 10 раз, или по меньшей мере в 50 раз, или по меньшей мере в 100 раз ниже CDC активности сопоставимой молекулы. Согласно другому варианту изобретения Fc варианты связывают C1q с аффинностью, которая по меньшей мере в 2 раза, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 7 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 50 раз, или по меньшей мере в 100 раз ниже по сравнению с аффинностью сопоставимой молекулы. Согласно еще одному варианту изобретения Fc варианты обладают CDC активностью, пониженной по меньшей мере на 10%, или по меньшей мере на 20%, или по меньшей мере на 30%, или по меньшей мере на 40%, или по меньшей мере на 50%, или по меньшей мере на 60%, или по меньшей мере на 70%, или по меньшей мере на 80%, или по меньшей мере на 90%, или по меньшей мере на 100%, или по меньшей мере на 150%, или по меньшей мере на 200% по сравнению с сопоставимой молекулой. Согласно конкретному варианту изобретения Fc варианты по изобретению связываются с C1q с пониженной аффинностью, имеют пониженную CDC активность и специфически связывают по меньшей мере с одним антигеном.
Согласно некоторым вариантам изобретения Fc варианты содержат одну или более генно-инженерных гликоформ, т.е. углеводных структур, связанных ковалентной связью с молекулой, содержащей Fc область. Генно-инженерные гликоформы могут применяться для различных целей, включая, но без ограничения, повышение или понижение эффекторной функции. Генно-инженерные гликоформы можно получать любым методом, известным специалисту в данной области техники, например, путем экспрессии генно-инженерных или вариантных штаммов, совместной экспрессией с одним или более ферментов, например, с β(1,4)-N-ацетилглюкозаминилтрансферазой III (GnTI11), экспрессией молекулы, содержащей Fc область, в различных организмах или клеточных линиях различных организмов, или модификацией углевода(-ов) после экспрессии молекулы, содержащей Fc область. Методы получения генно-инженерных гликоформ известны в уровне техники и включают, но без ограничения, методы, описанные в статьях Umana et al, 1999, Nat. Biotechnol 17:176-180; Davies et al., 20017 Biotechnol Bioeng 74:288-294; Shields et al, 2002, J Biol Chem 277:26733-26740; Shinkawa et al., 2003, J Biol Chem 278:3466-3473); патенте США No. 6602684; заявке на патент США Регистрационный No. 10/277370 и заявке на патент США Регистрационный No. 10/113929; Международных патентных заявках РСТ WO 00/61739A1; РСТ WO 01/292246А1; РСТ WO 02/311140A1; РСТ WO 02/30954A1; Potillegent™ technology (Biowa, Inc. Princeton, N.J.); GlycoMAb™ glycosylation engineering technology (GLYCART biotechnology AG, Zurich, Switzerland). См., например. Международную патентную заявку WO 00061739; Европейскую патентную заявку ЕА01229125; заявку на патент США US 20030115614; Okazaki et al., 2004, JMB, 336:1239-49.
Предполагается, что Fc варианты включают антитела, содержащие вариабельную область и гетеродимерную Fc область, где гетеродимерная Fc область включает вариантный CH3 домен, содержащий аминокислотные мутации, стимулирующие образование гетеродимера с повышенной устойчивостью, где вариантный CH3 домен имеет температуру плавления (Tm) выше 70°С. Fc варианты, которые представляют собой антитела, можно получать "de novo" путем объединения вариабельного домена, или его фрагмента, который специфически связывает по меньшей мере один антиген с гетеродимерной Fc областью, содержащей вариантный CH3 домен. Или же гетеродимерные Fc варианты можно получать, модифицируя CH3 домен Fc область-содержащего антитела, которое связывает антиген.
Антитела по изобретению могут включать, но без ограничения, синтетические антитела, моноклональные антитела, рекомбинантные антитела, внутриклеточные антитела (интраантитела), моноспецифические антитела, мультиспецифические антитела, биспецифические антитела, человеческие антитела, гуманизированные антитела, химерные антитела, одноцепочечные FvFcs (scFvFc), одноцепочечные Fvs (scFv) и антиидиотипические (анти-Id) антитела. В частности, антитела, применяемые в методах по настоящему изобретению, включают молекулы иммуноглобулина и иммунологически активные участки молекул иммуноглобулинов. Молекулы иммуноглобулина по изобретению могут быть любого типа (например, IgG, IgE, IgM, IgD, IgA и IgY), класса (e.g., IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2) или подкласса иммуноглобулиновой молекулы.
Антитела по изобретению могут быть антителами животного любого происхождения, включая птиц и млекопитающих (например, человеческие, мышиные, ослиные, овечьи, кроличьи, козьи, антитела морской свинки, верблюда, лошади или кур). В конкретном варианте изобретения антитела представляют собой человеческие или гуманизированные моноклональные антитела, в частности, биспецифические моноклональные антитела. Термин "человеческие" антитела по данному описанию включает антитела, имеющие аминокислотную последовательность человеческого иммуноглобулина и включает антитела, выделенные из библиотек человеческих антител или взятые у мышей, экспрессирующих антитела, полученные при использовании человеческих генов.
Антитела, подобно всем полипептидам, имеют изоэлектрическую точку (pI), которая обычно определяется как значение рН, при котором полипептид не несет никакого суммарного заряда. В области техники известно, что устойчивость белка, как правило, является самой низкой, когда значение рН раствора равно изоэлектрической точке (pI) белка. Можно оптимизировать растворимость, изменяя в антителе число и локализацию ионизируемых остатков с целью корректировать pI. Например, на pI полипептида можно влиять, делая соответствующие аминокислотные замены (например, заменяя незаряженную аминокислоту, такую как аланин, на заряженную аминокислоту, такую как лизин). Не связывая себя какой-либо конкретной теорией, можно сказать, что аминокислотные замены антитела, которые приводят к изменениям pI указанного антитела, могут повысить растворимость и/или устойчивость антитела. Специалист в данной области техники поймет, какие аминокислотные замены являются наиболее подходящими для того, чтобы получить нужное значение pI для конкретного антитела, pI белка можно определять различными методами, включая, но без ограничения, изоэлектрическое фокусирование и различные компьютерные алгоритмы (см., например, Bjellqvist et al., 1993, Electrophoresis 14:1023). Согласно одному варианту изобретения значение pI Fc находится между рН 6.2 и рН 8.0. Согласно другому варианту изобретения значение pI антител по изобретению находится между рН 6.8 и рН 7.4. Согласно одному варианту изобретения замены, приводящие к изменениям pI Fc варианта по изобретению, незначительно снижают его аффинность связывания с антигеном. Предполагается, что вариантный CH3 домен с повышенной устойчивостью также может привести к изменению pI. Согласно одному варианту изобретения вариантные Fc гетеродимеры, как правило, выбраны с целью повысить как устойчивость, так и чистоту, а также вызвать какое-либо нужное изменение pI.
Антитела по изобретению могут быть моноспецифическими, биспецифическими, триспецифическими или иметь более высокую степень мультиспецифичности. Мультиспецифические антитела могут специфически связываться с различными эпитопами нужной молекулы-мишени или могут специфически связываться как с молекулой-мишенью, так и с гетерологичным эпитопом, таким как гетерологичный полипептид или твердый твердая подложка (носитель). См., например. Опубликованные международные патентные заявки No. WO 94/04690; WO 93/17715; WO 92/08802; WO 91/00360 и WO 92/05793; Tutt, et al., 1991, J. Immunol. 147:60-69; патенты США No.: 4474893; 4714681; 4925648; 5573920 и 5601819 и Kostelny et al., 1992, J. Immunol. 148:1547).
На основе этого асимметричного каркаса, показанного на Фигуре 20, можно создать различные варианты мультифункциональных нацеливающих молекул.
Мультиспецифические антитела обладают специфичностью связывания по меньшей мере с двумя различными антигенами. Хотя обычно такие молекулы связывают два антигена (т.е. биспецифические антитела, BsAbs), настоящее изобретение охватывает антитела с дополнительными специфичностями, такие как триспецифические антитела. Примеры BsAbs включают, но без ограничения, биспецифические антитела, одно плечо которых специфично к опухолевому антигену, а другое плечо специфично к цитотоксической молекуле, или оба плеча специфичны к двум различным антигенам опухолевой клетки, или оба плеча специфичны к двум различным растворимым лигандам, или одно плечо специфично к растворимому лиганду, а другое плечо специфично к рецептору клеточной поверхности, или оба плеча специфичны к двум различным рецепторам клеточной поверхности. Методы получения биспецифических антител известны в уровне техники.
Согласно другому способу вариабельные домены антител с нужными специфичностями связывания (сайты связывания антиген-антитело) "сливают" с последовательностями константных доменов иммуноглобулина. Слияние может быть с константным доменом тяжелой цепи иммуноглобулина, содержащим по меньшей мере участок шарнирной, СН2 и CH3 областей. Рассматривается, что первая константная область тяжелой цепи (СН1), содержащая сайт, необходимый для связывания легкой цепи, присутствует по меньшей мере в одном из слияний. ДНК, кодирующие слияния тяжелой цепи, и при необходимости, легкой цепи иммуноглобулина, встраивают в отдельные экспрессионные векторы и совместно трансфецируют в организм соответствующего хозяина. Это дает большую свободу действий при корректировке соотношения трех полипептидных фрагментов в вариантах изобретения, когда неравные соотношения трех полипептидных цепей, применяемых в конструкции, обеспечивают оптимальные выходы. См. Пример 1 и Таблицу 2. Возможно, однако, встраивать кодирующие последовательности для двух или всех трех полипептидных цепей в один экспрессионный вектор, когда экспрессия по меньшей мере двух полипептидных цепей в равных отношениях дает высокие выходы или когда отношения не имеют особого значения.
Биспецифические антитела включают перекрестие связанные (сшитые) или "гетероконъюгированные" ("гетероконъюгатные") антитела. Например, одно из антител в гетероконъюгате может быть связано с авидином, другое с биотином. Такие антитела были предложены, например, для нацеливания клеток иммунной системы на нежелательные клетки (патент США No. 4676980) и для лечения ВИЧ (HIV) инфекции (Международные патентные заявки WO 91/00360, WO 92/200373 и Европейский патент ЕР 03089). Гетероконъюгированные антитела можно получать любыми подходящими методами сшивания. Подходящие сшивающие агенты хорошо известны в данной области техники и раскрываются в патенте США No. 4676980 вместе с рядом методов сшивания.
Рассматриваются антитела более чем с двумя валентностями, встраивающие CH3 домены, и полученные Fc гетеродимеры по изобретению. Например, можно получать триспецифические антитела. См., например, Tutt et al. J. Immunol. 147:60 (1991).
Антитела по настоящему изобретению также охватывают антитела, имеющие период полужизни в организме (например, полужизни в сыворотке) млекопитающего (например, человека) более 15 дней, более 20 дней, более 25 дней, более 30 дней, более 35 дней, более 40 дней, более 45 дней, более 2 месяцев, более 3 месяцев, более 4 месяцев или более 5 месяцев. Повышенный период полужизни антител по настоящему изобретению в организме млекопитающего (например, человека) обусловливает повышенный сывороточный титр указанных антител или фрагментов антител у млекопитающего и, следовательно, снижает частоту введения указанных антител или фрагментов антител и/или снижает концентрацию указанных вводимых антител или фрагментов антител. Антитела, имеющие повышенные in vitro периоды полужизни, можно получать методами, известными специалистам в данной области техники. Например, антитела с повышенными in vivo периодами полужизни можно получать модификацией (например, заменой, делецией или добавлением) аминокислотных остатков, которые идентифицированы как участвующие во взаимодействии между Fc доменом и FcRn рецептором (см., например, опубликованные Международные патентные заявки No. WO 97/34631; WO 04/029207; патент США No. 6737056 и опубликованную заявку на патент США No. 2003/0190311).
Согласно конкретному варианту изобретения вариантный Fc гетеродимер, содержащий вариантный CH3 домен, представляет собой мультиспецифическое антитело (называемое в настоящей заявке антителом по изобретению), антитело по изобретению специфически связывает целевой антиген. В частности, антитело по изобретению является биспецифическим антителом. Согласно одному варианту антитело по изобретению специфически связывает полипептидный антиген. Согласно другому варианту антитело по изобретению специфически связывает неполипептидный антиген. Согласно еще одному варианту изобретения введение антитела по изобретению млекопитающему, страдающему заболеванием или нарушением, может оказать благоприятное терапевтическое действие на этого млекопитающего.
На самом деле на любую молекулу можно нацелить и/или любую молекулу можно встроить в вариантный Fc гетеродимерный белок (например, антитела, Fc-слитые белки), включая, но без ограничения, нижеприведенный перечень белков, а также субъединицы, домены, мотивы и эпитопы, относящиеся к нижеприведенному перечню белков: ренин; гормон роста, включая человеческий гормон роста и бычий гормон роста; фактор, высвобождающий гормон роста; паратиреоидный гормон; тиреоид-стимулирующий гормон; липопротеины; альфа-1-антитрипсин; А - цепь инсулина; В - цепь инсулина; проинсулин; фолликулостимулирующий гормон; кальцитонин; лютеинизирующий гормон; глюкагон; факторы свертывания крови, такие как фактор VII, фактор VIIIC, фактор IX, тканевой фактор (TF) и фактор фон Виллебранда; факторы, предупреждающие свертывание, такие как Белок С; предсердный натрийуретический пептид; легочный сурфактант; активатор плазминогена, такой как урокиназа, или человеческая моча, или тканевой активатор плазминогена (t-PA); бомбезин; тромбин; гемопоэтический фактор роста; фактор некроза опухоли-альфа и -бета; энкефалиназа; RANTES (хемокин, выделяемый Т- клетками при активации); человеческий макрофагальный воспалительный белок (MIP-1-альфа); сывороточный альбумин, такой как человеческий сывороточный альбумин; ингибирующее вещество Мюллера (антимюллеровский гормон); А - цепь релаксина; В - цепь релаксина; прорелаксин; мышиный гонад отропин-ассоциированный пептид; белок микроорганизмов, такой как бета-лактамаза; ДНК-аза; IgE; цитотоксический Т-лимфоцит-ассоциированный антиген (CTLA), такой как CTLA-4; ингибин; активин; сосудистый эндотелиальный фактор роста (VEGF); рецепторы гормонов или факторов роста, например, такие как EGFR, VEGFR; интерфероны. Такие как альфа-интерферон (α-IFN), бета-интерферон (β-IFN) и гамма-интерферон (γ-IFN); белок А или D; ревматоидные факторы; нейротрофический фактор, такой как нейротрофический фактор мозга (BDNF), нейротропин-3, -4, -5 или -6 (NT-3, NT-4, NT-5 или NT-6), или фактор роста нервной ткани; фактор роста тромбоцитов (PDGF); фактор роста фибробластов, такой как AFGF и PFGF; эпидермальный фактор роста (EGF); трансформирующий фактор роста (TGF), такой как TGF-альфа и TGF-бета, включая TGF-1, TGF-2, TGF-3, TGF-4 или TGF-5; инсулиноподобный фактор роста-I и -II (IGF-I и IGF-II); des (1-3)-IGF-I (IGF-I ткани мозга), белки, связывающие инсулиноподобный фактор роста; CD белки, такие как CD2, CD3, CD4, CD8, CD11a, CD14, CD18, CD19, CD20, CD22, CD23, CD25, CD33, CD34, CD40, CD40L, CD52, CD63, CD64, CD80 и CD147; эритропоэтин; остеоиндуктивные факторы; иммунотоксины; костный морфогенетический белок (BMP); интерферон, такой как интерферон - alpha, -beta и -гамма; колониестимулирующие факторы (CSFs), такие как М-CSF, GM-CSF и G-CSF; интерлейкины (ILs), например, от IL-1 до IL-13; TNFα, супероксиддисмутаза; Т-клеточные рецепторы; поверхностные мембранные белки; комплемент-зависимой стимулятор гемолиза; вирусные антигены, например, такие как участок оболочки AIDS (СПИД), например, gp120; транспортные белки (белки-переносчики); хоминг-рецепторы; адрессины; регуляторные белки; молекулы клеточной адгезии, такие как LFA-1, Mac 1, р150.95, VLA-4, ICAM-1, ICAM-3 и VCAM, а4/р7 интегрин и Xv/р3 интегрин, включая любые их субъединицы, альфа-субъединицы интегрина, такие как CD49a, CD49b, CD49c, CD49d, CD49e, CD49f, альфа7, альфа8, альфа9, алъфаD, CD11a, CD11b, CD51, CD11c, CD41, альфаIIb, альфаIELb; бета-субъединицы интегрина, такие как CD29, CD18, CD61, CD104, бета5, бета6, бета7 и бета8; комбинации субъединиц интегрина, включая, но без ограничения, αVβ3, αVβ5 и α4β7; участник пути апоптоза; IgE; антигены групп крови; flk2/flt3 рецептор; рецептор ожирения (OB); mp1 рецептор; CTLA-4; белок С; Eph рецептор, такой как EphA2, EphA4, EphB2 и т.д.; человеческий лейкоцитарный антиген (HLA), такой как HLA-DR; белки комплемента, такие как рецептор комплемента CR1, C1Rq и другие факторы комплемента, такие как С3 и С5; гликопротеиновый рецептор, такой как GpIbα, GPIIb/IIIa и CD200; и фрагменты любого из вышеперечисленных полипептидов.
Также рассматриваются антитела по изобретению, которые специфически связывают раковые антигены, включая, но без ограничения, ALK рецептор (рецептор плейотрофина), плейотрофин, антиген пан-карциномы (карциномы поджелудочной железы) KS 1/4; антиген рака яичника (СА125); простатическую кислую фосфатазу; простат-специфический антиген (PSA); меланома-ассоциированный антиген р97; антиген меланомы gp75; высокомолекулярный меланома-ассоциированный антиген (HMW-МАА); простатспецифический мембранный антиген; раковый эмбриональный антиген (СЕА); антиген полиморфного эпителиального муцина; антиген жировых шариков человеческого молока; антигены, ассоциированные с колоректальным раком, такие как: СЕА, TAG-72, CO17-1A, GICA 19-9, СТА-1 и LEA; антиген 38.13 лимфомы Беркитта; CD19; антиген-CD20 В-клеточной лимфомы человека; CD33; меланомо-специфические антигены, такие как ганглиозид GD2, ганглиозид GD3, ганглиозид GM2 и ганглиозид GM3; опухолеспецифический трансплантационный антиген клеточной поверхности (TSTA); индуцируемые вирусами опухолевые антигены, включая Т-антиген, опухолевые ДНК-вирусы и оболочечные антигены опухолевых РНК-вирусов; онкофетальный антиген альфа-фетопротеин, такой как СЕА толстой кишки, 5Т4 онкофетальный трофобластический гликопротеин и онкофетальный антиген опухоли мочевого пузыря; дифференцировочный антиген, такой как антигены рака легкого L6 и L20; антигены фибросаркомы; Т-клеточный антиген-Gp37 лейкоза человека; неогликопротеин; сфинголипиды; антигены рака молочной железы, такие как EGFR (рецептор эпидермального фактора роста); NY-BR-16; NY-BR-16 и HER2 антиген (p185HER2); полиморфный эпителиальный муцин (РЕМ); антиген АРО-1 злокачественных лимфоцитов человека; дифференцировочный антиген, такой как I антиген, обнаруженный на фетальных эритроцитах; антиген I зародышевой эндодермы (энтодермы) на эритроцитах у взрослых; предимплантационные зародыши; I(Ма), обнаруженные при аденокарциномах желудка; M18, М39, обнаруженные в эпителии молочной железы; SSEA-1, обнаруженный в миелоидных клетках; VEP8; VEP9; MyI; Va4-D5; D156-22, обнаруженные при колоректальном раке; TRA-1-85 (группа крови Н); SCP-1, обнаруженный при раке яичек и яичника; С14, обнаруженный при аденокарциноме толстой кишки; F3, обнаруженный при аденокарциноме легкого; АН6, обнаруженный при раке желудка; Y гаптен; Ley, обнаруженный в клетках эмбриональной карциномы; TL5 (группа крови A); EGF рецептор, обнаруженный в А431 клетках; E1 ряд (группа крови В), обнаруженный при раке поджелудочной железы; FC10.2, обнаруженный в клетках эмбриональной карциномы; антиген аденокарциномы желудка; СО-514 (группа крови Lea), обнаруженный при аденокарциноме; NS-10, обнаруженный при аденокарциномах; СО-43 (группа крови Leb); G49, обнаруженный в EGF рецепторе клеток А431; МН2 (группа крови ALeb/Ley), обнаруженный при аденокарциноме толстой кишки; 19.9, обнаруженный при раке толстой кишки; муцины рака желудка; T5A7, обнаруженный в миелоидных клетках; R24, обнаруженный при меланоме; 4.2, GD3, D1.1, OFA-1, GM2, OFA-2, GD2 и М1:22:25:8, обнаруженные в клетках эмбриональной карциномы, м SSEA-3 и SSEA-4 в эмбрионах на 4x-8-клеточных стадиях; антиген кожной Т-клеточной лимфомы; антиген MART-1; антиген Sialyl Tn (STn); антиген рака прямой кишки NY-CO-45; антиген рака легкого NY-LU-12 вариант А; антиген аденокарциномы ART1; паранеопластический антиген, ассоциированный с раком головного мозга-яичек (онконейрональный антиген МА2; паранеопластический нейрональный антиген); антиген нейроонкологических заболеваний брюшной полости 2 (NOVA2); ген 520 антигена печеночно-клеточного рака; опухолеассоциированный антиген СО-029; опухолеассоциированные антигены MAGE-C1 (раковый/тестикулярный антиген СТ7), MAGE-B1 (MAGE-XP антиген), MAGE-B2 (DAM6), MAGE-2, MAGE-4-a, MAGE-4-b аи MAGE-X2; раковый-тестикулярный антиген (NY-EOS-1) и фрагменты любого из вышеперечисленных полипептидов.
Согласно некоторым вариантам изобретения гетеромультимеры, описанные в настоящей заявке, содержат по меньшей мере одно терапевтическое антитело. Согласно некоторым вариантам изобретения терапевтическое антитело связывает целевой раковый антиген. Согласно одному варианту изобретения терапевтическое антитело может представлять собой терапевтическое антитело, выбранное из группы, состоящей из абаговомаба, адалимумаба, алемтузумаба, аурограба, бапинеузумаба, базиликсимаба, белимумаба, бевацизумаба, бриакинумаба, канакинумаба, катумаксомаба, цертолизумаба пегола, цетуксимаба, даклизумаба, деносумаба, эфализумаба, галиксимаба, гемтузумаба озогамицина, голимумаба, ибритумомаба тиуксетана, инфликсимаба, ипилимумаба, лумиликсимаба, меполизумаба, мотавизумаба, муромонаба, микограба, натализумаба, нимотузумаба, окрелизумаба, офатумумаба, омализумаба, паливизумаба, панитумумаба, пертузумаба, ранибизумаба, реслизумаба, ритуксимаба, теплизумаба, тоцилизумаба/атлизумаба, тозитумомаба, трастузумаба, ProxiniumTM, RencarexTM, устекинумаба, залутумумаба и любых других антител.
Антитела по настоящему изобретению включают производные, которые являются модифицированными (например, ковалентным связыванием молекулы любого типа с антителом). Например, но без ограничения, производные антител включают антитела, которые модифицированы, например, гликозилированием, ацетилированием, пегилированием, фосфорилированием, амидированием, дериватизацией с помощью известных защитных/блокирующих групп, протеолитическим расщеплением, связыванием с клеточным лигандом или другим белком и т.д. Любые из множества химических модификаций можно осуществить известными методами, включая, но без ограничения, специфическое химическое расщепление, ацетилирование, формилирование, метаболический синтез в присутствии туникамицина и т.д. Помимо этого, производное может содержать одну или более неклассических аминокислот.
Антитела или их фрагменты с повышенным in vivo периодом полужизни можно получать, связывая молекулы полимеров, таких как высокомолекулярный полиэтиленгликоль (ПЭГ, PEG), с антителами или фрагментами антител. ПЭГ (PEG) можно конъюгировать с антителами или фрагментами антител с помощью или без помощи многофункционального линкера, либо сайт-специфическим связыванием ПЭГ (PEG) с N- или С-концом указанных антител или фрагментов антител, либо через эпсилон-аминогруппы лизиновых остатков. Можно применять дериватизацию линейного или разветвленного полимера, которая приводит к минимальной потере биологической активности. Точный мониторинг степени конъюгации проводят методами SDS-PAGE гель-электрофореза и масс-спектрометрии, чтобы гарантировать корректную конъюгацию молекул ПЭГ (PEG) с антителами. Непрореагировавший ПЭГ (PEG) можно отделять от конъюгатов антитело-PEG, например, высокоэффективной гель-хроматографией или ионообменной хроматографией.
Далее, антитела можно конъюгировать с альбумином с целью получения антитела или фрагмента антитела, более устойчивого in vivo или с более продолжительным периодом полужизни in vivo. Эти методы хорошо известны в уровне техники, см., например, опубликованные Международные заявки No. WO 93/15199, WO 93/15200 и WO 01/77137; и Европейский патент No. EP 413622. Настоящее изобретение охватывает применение антител или их фрагментов, конъюгированных или слитых с одним или более агентов, включая, но без ограничения, пептиды, полипептиды, белки, слитые белки, молекулы нуклеиновых кислот, низкомолекулярные вещества, миметики, синтетические лекарства, молекулы неорганических веществ и молекулы органических веществ.
Настоящее изобретение охватывает применение антител или их фрагментов, связанных методами рекомбинантной ДНК или конъюгированных химическими методами (включая конъюгацию как за счет ковалентной, так и за счет нековалентной связи), с гетерологичным белком или полипептидом (или его фрагментом, например, с полипептидом из по меньшей мере 10, по меньшей мере 20, по меньшей мере 30, по меньшей мере 40, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90 или по меньшей мере 100 аминокислот), с целью получения слитых белков. Слияние необязательно должно быть непосредственным, но может происходить при помощи линкерных последовательностей. Например, антитела могут применяться для нацеливания гетерологичных полипептидов на конкретные типы клеток in vitro или in vivo, путем связывания (конденсации) или конъюгации данных антител с антителами, специфическими к конкретным рецепторам клеточной поверхности. Антитела, слитые или конъюгированные с гетерологичными полипептидами, можно также применять в in vitro иммуноанализах и способах очистки с помощью методов, известных в уровне техники. См., например, опубликованную Международную заявку No. WO 93/21232; Европейский патент No. EP 439095; Naramura et al., 1994, Immunol. Lett. 39:91-99; патент США No. 5474981; Gillies et al., 1992, PNAS 89:1428-1432; и Fell et al., 1991, J. Immunol. 146:2446-2452.
Помимо этого, настоящее изобретение включает композиции, содержащие гетерологичные белки, пептиды или полипептиды, слитые или конъюгированные с фрагментами антител. Например, гетерологичные полипептиды могут быть слиты или конъюгированы с Fab фрагментом, Fd фрагментом, Fv фрагментом, F(ab)2 фрагментом, VH доменом, VL доменом, VH CDR, VL CDR или их фрагментом. Методы слияния или конъюгации полипептидов с участками антитела хорошо известны в уровне техники. См., например, патенты США No. 5336603; 5622929; 5359046; 5349053; 5447851 и 5112946; Европейский патент No. EP 307434 и EP 367166; опубликованные Международные патентные заявки No. WO 96/04388 и WO 91/06570; Ashkenazi et al., 1991, Proc. Natl. Acad. Sci. USA 88:10535-10539; Zheng et al., 1995, J. Immunol. 154:5590-5600; и Vil et al., 1992, Proc. Natl. Acad. Sci. USA 89:11337-11341.
Другие слитые белки, например, антител, которые специфически связывают антиген (например, supra), можно получать методами шаффлинга (перетасовки) генов, шаффлинга мотивов, шаффлинга экзонов и/или шаффлинга кодонов (в целом эти методы называются "ДНК шаффлингом"). ДНК шаффлинг можно применять для изменения активности антител по изобретению или их фрагментов (например, антител или их фрагментов с повышенными значениями аффинности и пониженными скоростями диссоциации). См. в целом патенты США No. 5605793; 5811238; 5830721; 5834252 и 5837458 и Patten et al., 1997, Curr. Opinion Biotechnol. 8:724-33; Harayama, 1998, Trends Biotechnol. 16(2):76-82; Hansson, et al., 1999, J. Mol. Biol. 287:265-76; и Lorenzo and Blasco, 1998, Biotechniques 24(2):308-313. Антитела или их фрагменты, или кодированные антитела или их фрагменты, можно изменять, осуществляя перед рекомбинацией случайный мутагенез методами ПЦР пониженной точности, случайной инсерции нуклеотида или другими методами. Один или более участков полинуклеотида, кодирующего антитело или фрагмент антитела, - причем эти участки специфически связываются с антигеном, - можно рекомбинировать с использованием одного или более компонентов, мотивов, отрезков, элементов, доменов, фрагментов и т.д. одной или более гетерологичных молекул.
Помимо этого, настоящее изобретение охватывает применение вариантных Fc гетеродимеров или их фрагментов, конъюгированных с терапевтическим агентом.
Антитело или его фрагмент может быть конъюгирован с терапевтическим веществом, таким как цитотоксин, например, с цитостатическим или цитопатическим агентом, терапевтическим агентом или ионом радиоактивного металла, например, источником альфа-частиц. Примеры включают рибонуклеазу, монометилауристатин Е и F, паклитаксел, цитохалазин В, грамицидин D, этидия бромид, эметин, митомицин, этопозид, тенопозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин, дигидроксиантрациндион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропранолол, пуромицин, эпирубицин и циклофосфамид и его аналоги или гомологи. Терапевтические агенты включают, но без ограничения, антиметаболиты (например, метотрексат, 6-меркаптопурин, 6-тиогуанин, цитарабин, 5-флуорурацил декарбазин), алкилирующие агенты (например, меклорэтамин, тиотепа, хлорамбуцил, мелфалан, кармустин (BCNU) и ломустин (CCNU), циклофосфамид, бусульфан, дибромманнит, стрептозотоцин, митомицин С и цис-дихлордиаминплатина(II) (DDP), цисплатин), антрациклины (например, даунорубицин (ранее дауномицин) и доксорубицин), антибиотики (например, дактиномицин (ранее актиномицин), блеомицин, митрамицин и антрамицин (АМС)), и антимитотические агенты (например, винкристин и винбластин). Более полный перечень терапевтических агентов (лекарственных средств) можно найти в опубликованной патентной заявке РСТ WO 03/075957.
Далее, антитело или его фрагмент могут быть конъюгированы с терапевтическим агентом или лекарственным средством, которые модифицируют заданный биологический отклик. Терапевтические агенты или лекарственные вещества необязательно должны быть сконструированы только как классические химические терапевтические агенты. Например, лекарственное вещество может представлять собой белок или полипептид, обладающий заданной биологической активностью. Такие белки могут включать, например, токсин, такой как абрин, рицин А, онконазу (или другую токсическую РНК-азу), эндотоксин pseudomonas, холероген или дифтерийный токсин; белок, такой как фактор некроза опухолей, α-интерферон, β-интерферон, фактор роста нервных клеток, фактор роста тромбоцитов, тканевый активатор плазминогена, апоптотический агент, например, TNF-α, TNF-β, AIM I (см. опубликованную Международную патентную заявку No. WO 97/33899), AIM II (см. опубликованную Международную патентную заявку No. WO 97/34911), Fas лиганд (Takahashi et al., 1994, J. Immunol., 6:1567) и VEGI (см. опубликованную Международную патентную заявку No. WO 99/23105), тромбоцитарный фактор или антиангиогенный агент, например, ангиостатин или эндостатин, или модификатор биологического отклика, например, такой как лимфокин (например, интерлейкин-1 ("IL-1"), интерлейкин-2 ("IL-2"), интерлейкин-6 ("IL-6"), гранулоцитарно-макрофагальный колониестимулирующий фактор ("GM-CSF") и гранулоцитарный колониестимулирующий фактор ("G-CSF")), или фактор роста (например, гормон роста ("GH")).
Кроме того, антитело может быть конъюгировано с лекарственными агентами, такими как радиоактивные материалы или макроциклические хелатирующие агенты, применимые для конъюгации с ионами радиоактивных металлов (см. выше примеры радиоактивных материалов). Согласно некоторым вариантам изобретения макроциклическим хелатирующим агентом является 1,4,7,10-тетраазациклододекан-N,N′,N",N"-тетрауксусная кислота (DOTA), которую можно связывать с антителом через молекулу линкера. Такие линкерные молекулы общеизвестны в уровне техники и описаны в статьях Denardo et al., 1998, Clin Cancer Res. 4:2483; Peterson et al., 1999, Bioconjug. Chem. 10:553; and Zimmerman et al., 1999, Nucl. Med. Biol. 26:943.
Методы слияния или конъюгации антител с полипептидными компонентами известны в уровне техники. См., например, патенты США No. 5336603; 5622929; 5359046; 5349053; 5447851 и 5112946; европейский патент ЕР 307434; ЕР 367166; опубликованные Международные патентные заявки РСТ WO 96/04388 и WO 91/06570; Ashkenazi et al., 1991, PNAS USA 88:10535; Zheng et al., 1995, J Immunol 154:5590; и Vil et al., 1992, PNAS USA 89:11337. Слияние антитела с агентом необязательно должно быть непосредственным, но может происходить через линкерные последовательности. Такие линкерные последовательности общеизвестны в уровне техники и описаны в Denardo et al., 1998, Clin Cancer Res 4:2483; Peterson et al., 1999, Bioconjug Chem 10:553; Zimmerman et al., 1999, Nucl Med Biol 26:943; Gamett, 2002, Adv Drug Deliv Rev 53:171.
Рекомбинантная экспрессия Fc варианта, его производного, аналога или фрагмента (например, антитела или слитого белка по изобретению), требует конструирование экспрессионного вектора, содержащего полинуклеотид, который кодирует Fc вариант (например, антитело или слитый белок). После получения полинуклеотида, кодирующего Fc вариант (например, антитело или слитый белок), вектор для продуцирования Fc варианта (например, антитела или слитого белка) можно получать методом рекомбинантной ДНК по методикам, хорошо известным в уровне техники. Так, в настоящей заявке описаны методы получения белка экспрессией полинуклеотида, содержащего нуклеотидную последовательность, кодирующую Fc вариант, (например, антитело или слитый белок). Методы, которые общеизвестны специалистам в данной области техники, можно применять для конструирования экспрессионных векторов, содержащих последовательности, кодирующие Fc вариант (например, антитело или слитый белок), и соответствующие сигналы регуляции транскрипции и трансляции. Эти методы включают, например, in vitro методы рекомбинантной ДНК и in vivo генетическую рекомбинацию. Соответственно, изобретение предусматривает реплицируемые векторы, содержащие нуклеотидную последовательность, кодирующую Fc вариант по изобретению, функционально связанную с промотором. Такие векторы могут включать нуклеотидную последовательность, кодирующую константную область молекулы антитела (см., например, опубликованную Международную патентную заявку No. WO 86/05807; опубликованную Международную патентную заявку No. WO 89/01036; и патент США No. 5122464) и вариабельный домен антитела, или полипептид для получения Fc варианта можно клонировать в такой вектор для экспрессии цепи полноразмерного антитела (например, тяжелой или легкой цепи), или полного Fc варианта, содержащего слияние полипептида, происходящего не из антитела, и Fc области, включающей по меньшей мере вариантный CH3 домен.
Экспрессионный вектор переносят в клетку-хозяина обычными методами, а затем трансфецированные клетки культивируют обычными же методами, получая Fc вариант по изобретению. Соответственно, изобретение включает клетки-хозяева, содержащие полинуклеотид, кодирующий Fc вариант по изобретению, функционально связанный с гетерологичным промотором. Согласно конкретным вариантам изобретения для экспрессии Fc вариантов, содержащих двухцепочечные антитела, векторы, кодирующие как тяжелую, так и легкую цепи, можно совместно экспрессировать в клетке-хозяине с целью экспрессии полной молекулы иммуноглобулина, как подробно описано ниже.
Для экспрессии Fc вариантов по изобретению (например, молекул антитела или слитого белка) можно использовать различные экспрессионные системы хозяин-экспрессионный вектор (см., например, патент США No. 5807715). Такие системы хозяин-экспрессионный вектор представляют собой носители, с помощью которых можно продуцировать, а затем очищать нужные кодирующие последовательности, но также представляют собой клетки, которые, будучи трансформированы или трансфецированы с помощью соответствующих нуклеотидных кодирующих последовательностей, могут экспрессировать Fc вариант по изобретению in situ. Эти системы включают, но без ограничения, микроорганизмы, такие как бактерии (например, Е.coli и В.subtilis), трансформированные с использованием векторов экспрессии на основе рекомбинантной бактериофагальной ДНК, плазмидной ДНК или космидной ДНК, содержащих последовательности, кодирующие Fc вариант; дрожжи (например, Saccharomyces Pichia), трансформированные с использованием рекомбинантных дрожжевых экспрессионных векторов, содержащих последовательности, кодирующие Fc вариант; системы экспрессии в клетках насекомых, инфицированных с помощью рекомбинантных вирусных экспрессионных векторов (например, бакуловирусных векторов), содержащих последовательности, кодирующие Fc вариант; системы экспрессии в клетках растений, инфицированных при помощи рекомбинантных вирусных экспрессионных векторов (например, вируса мозаики цветной капусты, CaMV; вируса табачной мозаики, TMV) или трансформированных с помощью рекомбинантных плазмидных экспрессионных векторов (например, Ti плазмиды), содержащих последовательности, кодирующие Fc вариант; или системы экспрессии в клетках млекопитающих (например, клетки COS, CHO, BHK, 293, NS0 и 3Т3), содержащих рекомбинантные экспрессионные конструкции, включающие промоторы из генома клеток млекопитающих (например, промотор гена металлотионеина) или промоторы вирусов, инфицирующих млекопитающих (например, поздний промотор аденовируса; 7.5K промотор вируса коровьей оспы). Согласно некоторым вариантам изобретения бактериальные клетки, такие как Escherichia coli, или клетки эукариот, применяются для экспрессии Fc варианта, который представляет собой рекомбинантное антитело или слитый белок. Например, клетки млекопитающего, такие как клетки китайского хомячка (CHO), в сочетании с вектором, таким как основной элемент промежуточного промотора раннего гена человеческого цитомегаловируса, представляют собой эффективную систему экспрессии антител (Foecking et al., 1986, Gene 45:101; и Cockett et al., 1990, Bio/Technology 8:2). Согласно конкретному варианту изобретения экспрессия нуклеотидных последовательностей, кодирующих Fc вариант по изобретению (например, антитело или слитый белок), регулируется конститутивным промотором, индуцибельным промотором или тканеспецифическим промотором.
В бактериальных системах можно, предпочтительно, выбрать целый ряд экспрессионных векторов в зависимости от предполагаемого применения экспрессируемого Fc варианта (например, антитела или слитого белка). Например, если требуется продуцировать большое количество такого белка для получения фармацевтических композиций Fc варианта, могут быть желательны векторы, которые управляют высокоуровневой экспрессией продуктов, содержащих слитый белок, которые легко подвергаются очистке. Такие векторы включают, но без ограничения, экспрессионный вектор pUR278 E.coli (Ruther et al., 1983, EMBO 12:1791), в котором последовательность, кодирующая Fc вариант, может индивидуально лигироваться с вектором в рамке считывания с областью, кодирующей lac Z, таким образом, что продуцируется lac Z-слитый белок; pIN векторы (Inouye & Inouye, 1985, Nucleic Acids Res. 13:3101-3109; Van Heeke & Schuster, 1989, J. Biol. Chem. 24:5503-5509); и т.п. Векторы pGEX также можно применять для экспрессии чужеродных полипептидов в виде белков, слитых с глутатион 5-трансферазой (GST). Как правило, такие слитые белки являются растворимыми и их легко очистить от лизированных клеток, адсорбируя на глутатион-агарозных гранулах в качестве матрицы и связывая с этими гранулами, с последующей элюцией в присутствии свободного глутатиона. Дизайн векторов pGEX таков, что он включает сайты расщепления тромбином или протеазой фактора Ха, с тем, чтобы клонированный целевой генный продукт можно было освободить от GST фрагмента.
В системе насекомых вирус ядерного полиэдроза Autographa californica (AcNPV) применяют в качестве вектора для экспрессии чужеродных генов. Вирус выращивают в культуре клеток бабочки Spodoptera frugiperda. Последовательность, кодирующую Fc вариант (например, антитело или слитый белок), можно клонировать индивидуально в несущественные области (например, ген полиэдрина) вируса и помещать под контроль AcNPV промотора (например, промотора полиэдрина).
В клетках хозяина-млекопитающего можно использовать различные вирусные экспрессионные системы. В тех случаях, когда в качестве экспрессионного вектора применяют аденовирус, нужную последовательность, кодирующую Fc вариант (например, антитело или слитый белок), можно лигировать в комплекс регуляции транскрипции/ трансляции гена аденовируса, например, поздний промотор и трехкомпонентную лидерную последовательность. Этот химерный ген можно затем встраивать в геном аденовируса рекомбинацией in vitro или in vivo. Инсерция в несущественную область вирусного генома (например, область Е1 или Е3) дает рекомбинантный вирус, жизнеспособный и способный экспрессировать Fc вариант (например, антитело или слитый белок) в инфицированных клетках-хозяевах (например, см. Logan & Shenk, 1984, Proc. Natl. Acad. Sci. USA 81:355-359). Специфические сигналы инициации также могут потребоваться для эффективной трансляции встроенных последовательностей, кодирующих антитело. Эти сигналы включают инициирующий ко дон ATG и прилегающие последовательности. Кроме того, инициирующий кодон должен быть в фазе с рамкой считывания заданной кодирующей последовательности, чтобы гарантировать трансляцию целого инсерционного сегмента. Эти экзогенные сигналы регуляции трансляции и инициирующие кодоны могут быть различного происхождения, как естественного, так и синтетического. Эффективность экспрессии можно повысить, включая соответствующий элемент, повышающий транскрипцию, элементы терминации транскрипции и т.д. (см., например, Bittner et al., 1987, Methods in Enzymol. 153:516-544).
Экспрессию Fc варианта (например, антитела или слитого белка) можно регулировать с помощью любого промоторного или энхансерного элемента, известного в уровне техники. Промоторы, которые можно применять для регуляции экспрессии гена, кодирующего Fc вариант (например, антитело или слитый белок), включают, но без ограничения, область раннего промотора SV40 (Bernoist and Chambon, 1981, Nature 290:304-310), промотор, входящий в состав 3′ длинного концевого повтора вируса саркомы Рауса (Yamamoto, et al., 1980, Cell 22:787-797), промотор гена тимидинкиназы вируса герпеса (Wagner et al., 1981, Proc. Natl. Acad. Sci. U.S.A. 78:1441-1445), регуляторные последовательности гена металлотионеина (Brinster et al., 1982, Nature 296:39-42), тетрациклин ((Tet))-регулируемый промотор (Gossen et al., 1995, Proc. Nat. Acad. Sci. USA 89:5547-5551); векторы для прокариотической экспрессии, такие как промотор β-лактамазы (Villa-Kamaroff et al, 1978, Proc. Natl. Acad. Sci. U.S.A. 75:3727-3731), или tac промотор (DeBoer et al., 1983, Proc. Natl. Acad. Sci. U.S.A. 80:21-25; см. также "Useful proteins from recombinant bacteria" in Scientific American, 1980, 242:74-94); векторы для экспрессии в растительных клетках, содержащие промоторную область гена нопалин-синтетазы (Herrera-Estrella et al., Nature 303:209-213) или промотор 35S RNA вируса мозаики цветной капусты (Gardner et al., 1981, Nucl. Acids Res. 9:2871), и промотор фотосинтетического фермента рибулоза- бифосфат карбоксилазы (Herrera-Estrella et al., 1984, Nature 310:115-120); промоторные элементы дрожжей и других грибов, такие как промотор Gal 4, промотор ADC (алкогольдегидрогеназы), промотор PGK (фосфоглицерин-киназы), промотор щелочной фосфатазы, и нижеприведенные области регуляции транскрипции в клетках животных, которые проявляют тканеспецифичность и использовались для трансгенных животных: область контроля гена эластазы I, являющаяся активной в панкреатических ацинарных клетках (Swift et al., 1984, Cell 38:639-646; Ornitz et al., 1986, Cold Spring Harbor Symp. Quant. Biol. 50:399-409; MacDonald, 1987, Hepatology 7:425-515); область контроля гена инсулина, активную в панкреатических бета-клетках (Hanahan, 1985, Nature 315:115-122), область контроля гена иммуноглобулина, активную в лимфоидных клетках (Grosschedl et al., 1984, Cell 38:647-658; Adames et al., 1985, Nature 318:533-538; Alexander et al., 1987, Mol. Cell. Biol. 7:1436-1444), область контроля вируса опухолей молочных желез у мышей, активную в тестикулярных клетках, клетках молочной железы, в лимфоидных и тучных клетках (Leder et al., 1986, Cell 45:485-495), область контроля гена альбумина, активную в печени (Pinkert et al., 1987, Genes and Devel. 1:268-276), область контроля гена альфа-фетопротеина, активную в печени (Krumlauf et al., 1985, Mol. Cell. Biol. 5:1639-1648; Hammer et al., 1987, Science 235:53-58; область контроля гена альфа 1-антитрипсина, активную в печени (Kelsey et al., 1987, Genes and Devel. 1:161-171), область контроля гена бета-глобина, активную в миелоидных клетках (Mogram et al., 1985, Nature 315:338-340; Kollias et al., 1986, Cell 46:89-94); область контроля гена основного белка миелина, активную в клетках головного мозга (Readhead et al., 1987, Cell 48:703-712); область контроля гена легкой цепи-2 миозина, активную в скелетных мышцах (Sani, 1985, Nature 314:283-286); нейрон-специфическую енолазу (NSE), активную в нервных клетках (нейронах) (Morelli et al., 1999, Gen. Virol. 80:571-83); область контроля гена нейротропного фактора головного мозга (BDNF), активную в нейронах (Tabuchi et al., 1998, Biochem. Biophysic. Res. Corn. 253:818-823); промотор глиального фибриллярного кислого белка (GFAP), активный в астроцитах (Gomes et al., 1999, Braz J Med Biol Res 32(5):619-631; Morelli et al., 1999, Gen. Virol. 80:571-83) и область контроля гена гонадропин-рилизинг гормона, активную в гипоталамусе (Mason et al., 1986, Science 234:1372-1378).
Экспрессионные векторы, содержащие инсерции гена, кодирующего Fc вариант по изобретению (например, антитело или слитый белок), можно идентифицировать тремя общими методами: (а) гибридизацией нуклеиновых кислот, (б) по наличию или отсутствию функций "маркерного" гена, и (в) экспрессией инсерционных последовательностей. В соответствии с первым методом присутствие в экспрессионном векторе гена, кодирующего пептид, полипептид, белок или слитый белок, можно детектировать гибридизацией нуклеиновых кислот с применением зондов, содержащих последовательности, гомологичные встроенному гену, кодирующему пептид, полипептид, белок или слитый белок, соответственно. Согласно второму методу систему рекомбинантный вектор/(клетка-)хозяин можно идентифицировать и выбрать в зависимости от наличия или отсутствия определенных функций "маркерного" гена (например, тимидинкиназной активности, резистентности к антибиотикам, трансформации фенотипа, образования телец включения бакуловируса и т.д.), вызванных инсерцией в вектор нуклеотидной последовательности, кодирующей антитело или слитый белок. Например, если нуклеотидная последовательность, кодирующая Fc вариант (например, антитело или слитый белок), встроена в последовательность маркерного гена вектора, рекомбинанты, содержащие инсерцию гена, кодирующего антитело или слитого белка, можно идентифицировать по отсутствию функции маркерного гена. Согласно третьему методу векторы экспрессии рекомбинантной ДНК можно идентифицировать, анализируя генный продукт (например, антитело или слитый белок), экспрессированный рекомбинантами. Такие анализы могут базироваться на физических или функциональных свойствах слитого белка в in vitro аналитических системах, например, связывании с антителом к биоактивной молекуле.
Кроме того, можно выбрать штамм клетки-хозяина, который модулирует экспрессию инсерционных последовательностей, или модифицирует и процессирует генный продукт нужным специфическим образом. Экспрессию с использованием определенных промоторов можно повысить в присутствии некоторых индуцирующих факторов; таким образом, экспрессию генно-инженерного слитого белка можно регулировать. Далее, в различных клетках-хозяевах наблюдаются характерные и специфические механизмы трансляционных и посттрансляционных процессирования и модификации (например, гликозилирование, фосфорилирование белков). Можно выбрать соответствующие линии клеток или системы клеток-хозяев, чтобы гарантировать заданные модификацию и процессирование чужеродных экспрессированных белков. Например, экспрессия в бактериальной системе будет давать негликозилированный продукт, а экспрессией в дрожжах получают гликозилированный продукт. Можно использовать эукариотические клетки-хозяева, которые имеют клеточные механизмы соответствующего процессирования первичного транскрипта (например, гликозилирования и фосфорилирования) генного продукта. Такие клетки млекопитающих-хозяев включают, но без ограничения, СНО, VERY, BHK, Hela, COS, MDCK, 293, 3T3, WI38, NS0, и в частности, линии нервных клеток, например, таких как линий нервных клеток, как например, таких как SK-N-AS, SK-N-FI, SK-N-DZ человеческой нейробластомы (Sugimoto et al., 1984, J. Natl. Cancer Inst. 73:51-57), SK-N-SH человеческой нейробластомы (Biochim. Biophys. Acta, 1982, 704:450-460), Daoy человеческой медуллобластомы мозжечка (Не et al., 1992, Cancer Res. 52:1144-1148) клетки глиобластомы DBTRG-05MG (Kruse et al., 1992, In Vitro Cell. Dev. Biol. 28A:609-614), IMR-32 человеческой нейробластомы (Cancer Res., 1970, 30:2110-2118), 1321N1 человеческой астроцитомы (Proc. Natl. Acad. Sci. USA, 1977, 74:4816), MOG-G-CCM человеческой астроцитомы (Br. J. Cancer, 1984, 49:269), U87MG человеческой глиобластомы-астроцитомы (Acta Pathol. Microbiol. Scand., 1968, 74:465-486), A172 человеческой глиобластомы (Olopade et al., 1992, Cancer Res. 52:2523-2529), C6 клетки крысиной глиомы (Benda et al., 1968, Science 161:370-371), Neuro-2a мышиной нейробластомы (Proc. Natl. Acad. Sci. USA, 1970, 65:129-136), NB41A3 мышиной нейробластомы (Proc. Natl. Acad. Sci. USA, 1962, 48:1184- 1190), SCP клетки хороидного сплетения овец (Bolin et al., 1994, J. Virol. Methods 48:211-221), G355-5, PG-4 нормальные кошачьи астроцитомы (Haapala et al., 1985, J. Virol. 53:827-833), Mpf клетки мозга хорька (Trowbridge et al., 1982, In Vitro 18:952-960), и линии нормальных клеток, например, таких как СТХ TNA2 нормальные клетки коры головного мозга крыс (Radany et al., 1992, Proc. Natl. Acad. Sci. USA 89:6467-6471), например, такие как CRL7030 и Hs578Bst. Кроме того, различные экспрессионные системы вектор/клетка-хозяин могут в различной степени влиять на реакции процессирования.
Для долговременного высокоэффективного продуцирования рекомбинантных белков стабильная экспрессия часто является предпочтительной. Например, можно получить линии (генно-инженерных) клеток, которые стабильно экспрессируют Fc вариант по изобретению (например, антитело или слитый белок). Вместо использования экспрессионных векторов, которые содержат ориджины репликации вируса, можно трансформировать клетки-хозяева с помощью ДНК, контролируемой соответствующими элементами регуляции экспрессии (например, промоторной, энхансерной последовательностями, терминаторами транскрипции, сайтами полиаденилирования и т.п.) и селективным маркером. После введения чужеродной ДНК можно оставить генно-инженерные клетки расти в течение 1-2 дней в обогащенной среде, а затем "переключить" на селективную среду. Селективный маркер в рекомбинантной плазмиде сообщает резистентность к отбору и способствует тому, что клетки стабильно интегрируют плазмиду в свои хромосомы и растут, образуя очаги, которые, в свою очередь, можно клонировать и распространять в линии клеток. Этот метод можно, предпочтительно, применять для создания линий клеток, экспрессирующих Fc вариант, который специфически связывается с антигеном. Такие генно-инженерные линии клеток могут быть особенно применимы при скрининге и оценке соединений, влияющих на активность Fc варианта (например, полипептида или слитого белка), который специфически связывается с антигеном.
Можно применять целый ряд систем отбора, включая, но без ограничения, гены тимидинкиназы вируса простого герпеса (Wigler et al., 1977, Cell 11:223), гипоксантин-гуанин фосфорилбозилтрансферазы (Szybalska & Szybalski, 1962, Proc. Natl. Acad. Sci. USA 48:2026) и аденин фосфорилбозилтрансферазы (Lowy et al., 1980, Cell 22:817), которые можно применять в tk-, hgprt- или aprt-клетках, соответственно. Также можно использовать антиметаболитную устойчивость как основу для отбора на dhfr, который сообщает устойчивость к гену метотрексата (Wigler et al., 1980, Natl. Acad. Sci. USA 77:3567; O′Hare et al., 1981, Proc. Natl. Acad. Sci. USA 78:1527); gpt, который сообщает устойчивость к гену микофенольной кислоты (Mulligan & Berg, 1981, Proc. Natl. Acad. Sci. USA 78:2072); neo, который сообщает устойчивость к гену аминогликозида G-418 (Colberre-Garapin et al., 1981, J. Mol. Biol. 150:1); и hygro, который сообщает устойчивость к гену гигромицина (Santerre et al., 1984, Gene 30:147).
После продуцирования Fc варианта (например, антитела или слитого белка) по изобретению при использовании экспрессии рекомбинантных ДНК его можно очистить любым методом очистки белка, известным в уровне техники, например, хроматографией (например, ионообменной, аффинной, в частности, аффинной по специфическому антигену к Белку А, и колоночной гель-хроматографией), центрифугированием, за счет различной растворимости или любым другим стандартным методом очистки белков.
Обычно Fc вариант извлекают из культуральной среды в виде секретированного полипептида, хотя его можно также извлекать из лизата клеток-хозяев, когда они продуцируются непосредственно без секреторного сигнала. Если Fc вариант является мембраносвязанным, его может высвобождать из мембраны с помощью раствора подходящего детергента (например, Triton-X 100).
Если Fc вариант продуцируется в рекомбинантной клетке, отличной от клетки человеческого происхождения, она совершенно не содержит белков или полипептидов человеческого происхождения. Однако необходимо очищать Fc вариант от рекомбинантных клеточных белков или полипептидов, чтобы получать препараты, по существу гомогенные в отношении Fc варианта. На первой стадии культуральную среду или лизат обычно центрифугируют для удаления крупнодисперсного клеточного детрита.
Fc гетеродимеры, имеющие константные домены антитела, удобно очищать хроматографией на колонке с гидроксилапатитом, гель-электрофорезом, диализом или аффинной хроматографией, причем предпочтительным методом очистки является аффинная хроматография. В зависимости от выделяемого полипептида можно также применять другие методы очистки белков, такие как фракционирование на ионообменной колонке, осаждение этанолом, обращенно-фазовая ВЭЖХ хроматография, хроматография на силикагеле, хроматография на гепарин-сефарозе, хроматография на анионо- или катионообменной смоле (например, на колонке с полиаспарагиновой кислотой), хроматофокусирование, SDS-PAGE гель-электрофорез и осаждение сульфатом аммония. Возможность применения Белка А в качестве лиганда для аффинной хроматографии зависит от вида и изотипа используемого Fc домена иммуноглобулина. Белок А можно применять для очистки Fc областей иммуноглобулинов на основе человеческих γ1, γ2 или γ4 тяжелых цепей (Lindmark et al., J. Immunol. Meth. 62:1-13 (1983)). Белок G рекомендуется для всех мышиных изотипов и для γ3 человека (Guss et al., EMBO J. 5:15671575 (1986)). Матрицей, с которой связывается аффинный лиганд, чаще всего является агароза, но применимы также другие матрицы. Механически устойчивые матрицы, такие как стекло с контролируемым размером пор или сополимер стирола с дивинилбензолом, обеспечивают более быстрые скорости потока и более короткое время обработки, чем агароза. Условия связывания иммуноадгезина с Белком А или G на колонке для аффинной хроматографии диктуются исключительно характеристиками Fc домена; а именно, его видом и изотипом. Как правило, если выбран подходящий лиганд, эффективное связывание происходит непосредственно из некондиционированной культуральной среды. Связанные вариантные Fc гетеродимеры можно эффективно элюировать либо при кислом рН (при рН 3.0 или выше), или при нейтральном рН буфером, содержащим слабохаотропную соль. Эта стадия аффинной хроматографии позволяет получать препарат вариантного Fc гетеродимера с чистотой >95%.
Уровни экспрессии Fc варианта (например, антитела или слитого белка) можно повысить с помощью амплификации векторов (обзор см. Bebbington and Hentschel, The use of vectors based on gene amplification for the expression of cloned genes in mammalian cells in DNA cloning, Vol.3. (Academic Press, New York, 1987)). Например, если маркер в векторной системе, экспрессирующей антитело или слитый белок, способен амплифицировать, повышение уровня ингибитора в культуре клетки-хозяина повысит число копий маркерного гена. Так как амплифицируемая область ассоциирована с геном антитела, продуцирование антитела или слитого белка также повышается (Crouse et al., 1983, Mol. Cell. Biol. 3:257).
Клетку-хозяина можно трансфецировать совместно с использованием двух экспрессионных векторов по изобретению. Например, с использованием первого вектора, кодирующего полипептид тяжелой цепи, и второго вектора, кодирующего полипептид легкой цепи. Два вектора могут содержать идентичные селективные маркеры, которые способствуют равной экспрессии полипептидов тяжелой и легкой цепи. Или же можно использовать один вектор, который кодирует и способен экспрессировать слитый белок или полипептиды как тяжелой, так и легкой цепи. Кодирующие последовательности для слитого белка или тяжелой и легкой цепей могут содержать кДНК или геномную ДНК.
Определение характеристик и функциональные анализы
Fc варианты (например, антитела или слитые белки) по настоящему изобретению можно характеризовать различными способами. Согласно одному варианту изобретения чистоту вариантных Fc гетеродимеров определяют хорошо известными в уровне техники методами, включая, но без ограничения, SDS-PAGE гель-электрофорез, вестерн-блоттинг, денситометрию или масс-спектроскопию. Устойчивость белка можно охарактеризовать многими методами, включая, но без ограничения, высокоэффективную гель-хроматографию, УФ спектроскопию и спектроскопию в видимой области, CD-спектроскопию кругового дихроизма, масс-спектрометрию, дифференциальное светорассеяние, анализ стабильности пробоподготовки, метод замораживания-оттаивания в сочетании с другими методами определения характеристик, дифференциальную сканирующую калориметрию, дифференциальную сканирующую флуориметрию, хроматографию гидрофобного взаимодействия, изоэлектрическое фокусирование, анализы рецепторного связывания или относительные уровни экспрессии белков. Согласно одному типичному варианту изобретения устойчивость вариантных Fc гетеродимеров определяют по температуре плавления вариантного CH3 домена по сравнению с дикого типа CH3 доменом, используя методы, хорошо известные в уровне техники, такие как дифференциальная сканирующая калориметрия или дифференциальная сканирующая флуориметрия.
Fc варианты по настоящему изобретению можно также анализировать, определяя их способность специфически связываться с лигандом (например, FcγRIIIA, FcγRIIB, C1q). Такой анализ можно проводить в растворе (например, Houghten, Bio/Techniques, 13:412-421, 1992), на гранулах (Lam, Nature, 354:82-84, 1991), на чипах (Fodor, Nature, 364:555-556, 1993), на бактериях (патент США No. 5223409), на плазмидах (Cull et al., Proc. Natl. Acad. Sci. USA, 89:1865-1869, 1992), на фаге (Scott and Smith, Science, 249:386-390, 1990; Devlin, Science, 249:404-406, 1990; Cwirla et al., Proc. Natl. Acad. Sci. USA, 87:6378-6382, 1990; и Felici, J. Mol. Biol., 222:301-310, 1991). Молекулы, идентифицированные как специфически связывающиеся с лигандом (например, FcγRIIIA, FcγRIIB, C1q или с антигеном), можно затем проверять на их аффинность к лиганду.
Fc варианты по изобретению можно проверить на специфическое связывание с такими молекулами, как молекула антигена (например, ракового антигена, и перекрестную реактивность к другим антигенам) или лиганда (например, FcγR) любым известным в уровне техники методом. Методы иммуноанализа, которые можно применять для определения специфического связывания и перекрестной реактивности, включают, но без ограничения, конкурентные и неконкурентные методы анализа, такие как вестерн-блоттинг, радиоиммуноанализы, ELISA (твердофазный иммуноферментный анализ), "сэндвич"-иммуноанализы, иммунопреципитационные методы анализа, реакции с преципитинами, реакции диффузионной преципитации в геле, иммунодиффузионные методы, агглютационные тесты, фиксацию комплемента, иммунорадиометрические методы анализа, флуоресцентный иммуноанализ, иммуноанализы с Белком А. Такие анализы являются стандартными и хорошо известны в уровне техники (см., например, Ausubel et al., eds, 1994, Current Protocols in Molecular Biology, Vol.1, John Wiley & Sons, Inc., New York).
Аффинность связывания Fc вариантов по настоящему изобретению с антигеном или лигандом (например, FcγR) и скорость диссоциации при взаимодействии можно определять методами конкурентного связывания. Одним из примеров анализа конкурентного связывания является радиоиммуноанализ, включающий инкубацию меченого лиганда, такого как FcγR (например, целевой молекулы (например, Fc варианта по настоящему изобретению), меченной 3Н или 125I), в присутствии увеличивающихся количеств немеченого лиганда, такого как FcγR, и детекцию молекулы, связанной с меченым лигандом. Аффинность молекулы по настоящему изобретению к лиганду и скорости диссоциации при связывании можно определять по результатам насыщения по методу Скэтчарда.
Кинетические параметры Fc варианта можно определять также известными в уровне техники методами анализа, основанными на явлении поверхностного плазменного резонанса (SPR) (например, анализ в системе BIAcore). Обзор методов на основе SPR см. В Mullet et al., 2000, Methods 22:77-91; Dong et al., 2002, Review in Mol. Biotech., 82:303-23; Fivash et al., 1998, Current Opinion in Biotechnology 9:97-101; Rich et al., 2000, Current Opinion in Biotechnology 11:54-61. Кроме того, некоторые приборы и методы на основе SPR для количественного определения белок-белковых взаимодействий, описанные в патентах США No. 6373577; 6289286; 5322798; 5341215; 6268125, рассматриваются в способах по изобретению.
Флуоресцентно-активированную сортировку клеток (FACS) любым методом, известным специалистам в данной области техники, можно применять для характеристики связывания Fc вариантов с молекулой, экспрессированной на поверхности клетки (например, FcγRIIIA, FcγRIIB). Проточные сортеры позволяют быстро проверить большое число отдельных клеток, которые содержат вставки из библиотек (например, 10-100 миллионов клеток в час) (Shapiro et al., Practical Flow Cytometry, 1995). Проточные цитометры для сортировки и изучения биологических клеток хорошо известны в уровне техники. Известные проточные цитометры описаны, например, в патентах США No. 4347935; 5464581; 5483469; 5602039; 5643796; и 6211477. Другими известными проточными цитометрами являются система FACS Vantage™, выпускаемая Becton Dickinson and Company, и система COPAS™, выпускаемая компанией Union Biometrica.
Fc варианты по изобретению можно характеризовать по их способности опосредовать FcγR-опосредуемую функцию эффекторных клеток. Примеры функций эффекторных клеток, которые можно проанализировать, включают, но без ограничения, антителозависимую клеточную цитотоксичность (ADCC), фагоцитоз, опсонизацию, опсонофагоцитоз, связывание C1q и комплемент-зависимую цитотоксичность (CDC). Для определения активности функции эффекторных клеток можно использовать любой известный специалистам в данной области техники анализ на основе клеточных или бесклеточных систем (об анализе эффекторных клеток см. Perussia et al., 2000, Methods Mol. Biol. 121:179-92; Baggiolini et al., 1998 Experientia, 44(10):841-8; Lehmann et al., 2000 J. Immunol. Methods, 243(1-2):229-42; Brown E J. 1994, Methods Cell Biol., 45:147-64; Munn et al., 1990 J. Exp. Med., 172:231-237, Abdul-Majid et al., 2002 Scand. J. Immunol. 55:70-81; Ding et al., 1998, Immunity 8:403-411).
В частности, Fc варианты по изобретению можно анализировать на FcγR-опосредованную ADCC активность в эффекторных клетках (например, естественных киллерах) любым из стандартных методов, известных специалистам в данной области техники (см., например, Perussia et al., 2000, Methods Mol. Biol. 121:179-92). Типичный метод определения ADCC активности молекул по изобретению основан на анализе высвобождения 51Cr, включающем: мечение клеток-мишеней с помощью [51Cr]Na2CrO4 (эту проникающую через клеточную мембрану молекулу обычно используют для мечения, так как она связывает цитоплазматические белки, и хотя она самопроизвольно медленно высвобождается из клетки, она полностью высвобождается после некроза клетки-мишени); опсонизацию клеток-мишеней с применением Fc вариантов по изобретению; объединение опсонизированных меченных радиоактивной меткой клеток-мишеней с эффекторными клетками в микротитрационном планшете при соответствующем соотношении клеток-мишеней и эффекторных клеток; инкубацию смеси клеток в течение 16-18 часов при 37°С.; сбор супернатантов; и анализ радиоактивности. Цитотоксичность молекул по изобретению можно определять, например, по следующей формуле: лизис%=(экспериментальное число имп./мин-прогнозное значение имп./мин утечки)/(имп./мин при лизисе детергентом-прогнозное значение имп./мин утечки)×100%. Или же, лизис%=(ADCC-AICC)/(максимальное высвобождение - спонтанное высвобождение). Специфический лизис можно рассчитать по формуле: специфический лизис=лизис % под действием молекул по изобретению - лизис% в отсутствие молекул по изобретению. Можно построить кривую, меняя либо соотношение клетки-мишени: эффекторные клетки, либо концентрацию антитела.
Метод определения способности Fc вариантов связывать C1q и опосредовать комплемент-зависимую цитотоксичность (CDC) хорошо известны в уровне техники. Например, для определения связывания C1q можно провести анализ ELISA связывания C1q. Типичный анализ может включать следующее: планшеты для анализа можно покрывать в течение ночи при 4°С вариантом полипептида или исходным полипептидом (контроль) в буфере для сенсибилизации поверхностей. Затем планшеты можно отмывать и блокировать. После отмывки в каждую лунку можно добавлять аликвоту человеческого C1q и инкубировать в течение 2 часа при комнатной температуре. После дополнительной отмывки в каждую лунку можно добавлять 100 мкл антитела овцы против компонента комплемента C1q, конъюгированного с пероксидазой хрена пероксидазы, и инкубировать в течение 1 часа при комнатной температуре. Планшет можно снова отмывать буфером и в каждую лунку можно добавить 100 мкл субстратного буфера, содержащего OPD (O-фенилендиамин дихлорид (Sigma)). Реакцию окисления, наблюдаемую по появлению желтой окраски, можно проводить в течение 30 минут, ее прекращают, добавляя 100 мкл 4.5 N H2SO4. Затем считывают оптическую плотность при (492-405) нм.
Для определения активации комплемента можно провести анализ комплемент-зависимой цитотоксичности (CDC) (например, как описано в Gazzano-Santoro et al., 1996, J. Immunol. Methods 202:163). Коротко говоря, различные концентрации Fc варианта и человеческого комплемента можно развести в буфере. Клетки, экспрессирующие антиген, с которым связывается Fc вариант, можно разводить до плотности около 1×106 клеток/мл. Смеси Fc варианта, разведенного человеческого комплемента и клеток, экспрессирующих антиген, можно добавлять в 96-луночный планшет для тканевых культур с плоскодонными лунками и оставить инкубироваться в течение 2 часов при 37°С и 5% CO2, чтобы стимулировать опосредованный комплементом лизис клеток. Затем в каждую лунку можно добавить 50 мкл alamar blue (Accumed International) и инкубировать в течение ночи при 37°С. Оптическую плотность определяют с использованием флуориметра в 96-лун очном формате с возбуждением при 530 нм и испусканием при 590 нм. Результаты можно выражать в относительных единицах флуоресценции (RFU). Концентрации образца можно рассчитать по стандартной кривой, и активность целевого Fc варианта приводят в процентах по сравнению с сопоставимой молекулой (т.е. молекулой, содержащей Fc область с немодифицированным или дикого типа CH3 доменом).
Анализ комплемента можно проводить с использованием сыворотки морской свинки, кроличьей или человеческой сыворотки. Комплемент-опосредованный лизис клеток-мишеней можно детектировать, проводя мониторинг высвобождения внутриклеточных ферментов, таких как лактатдегидрогеназа (LDH), как описано в Korzeniewski et al., 1983, Immunol. Methods 64(3):313-20; и Decker et al., 1988, J. Immunol Methods 115(1):61-9; или высвобождения внутриклеточной метки, такой как европий, хром 51 или индий 111, которыми мечены клетки.
Методы
Настоящее изобретение охватывает введение одного или более Fc вариантов по изобретению (например, антител) животному, в частности, млекопитающему, конкретно, человеку, с целью предупреждения, лечения или уменьшения интенсивности одного или более симптомов, ассоциированных с заболеванием, нарушением или инфекцией. Fc варианты по изобретению применимы, в частности, для лечения или предупреждения заболевания или нарушения, при котором желательна измененная эффективность функции эффекторных клеток (например, ADCC, CDC). Fc варианты и их композиции применимы, в частности, для лечения или предупреждения первичного или метастатического неопластического заболевания (т.е. рака) и инфекционных заболеваний. Молекулы по изобретению можно предоставлять в фармацевтически приемлемых композициях, известных в уровне техники или описанных в настоящей заявке. Как подробно описано ниже, молекулы по изобретению можно применять в методах лечения или предупреждения рака (в частности, при пассивной иммунотерапии), аутоиммунного заболевания, воспалительных нарушений или инфекционных заболеваний.
Fc варианты по изобретению можно также применять, предпочтительно, в комбинации с другими терапевтическими агентами, известными в уровне техники, применяемых для лечения или предупреждения рака, аутоиммунного заболевания, воспалительных нарушений или инфекционных заболеваний. В одном конкретном варианте Fc варианты изобретения можно применять в комбинации с моноклональными или химерными антителами, лимфокинами или гемопоэтическими факторами роста (например, такими как IL-2, IL-3 и IL-7), которые служат, например, для увеличения числа или повышения активности эффекторных клеток, взаимодействующих с молекулами, и повышают иммунный ответ. Fc варианты по изобретению можно также, преимущественно, применять в комбинации с одним или более лекарств, применяемых для лечения заболевания, нарушения или инфекции, например, таких как противораковые агенты, противовоспалительные агенты и антивирусные агенты.
Соответственно, настоящее изобретение предусматривает методы предупреждения, лечения или уменьшения интенсивности одного или более симптомов, ассоциированных с раком или родственными состояниями, путем введения одного или более Fc вариантов по изобретению. Без связи с каким-либо механизмом действия можно сказать, что Fc вариант по изобретению, который связывает FcγRIIIA и/или FcγRIIA с более высокой аффинностью, чем сопоставимая молекула, и, кроме того, связывает FcγRIIB с более низкой аффинностью, чем сопоставимая молекула, и/или указанный Fc вариант, который проявляет повышенную эффекторную функцию, например, ADCC, CDC, фагоцитоз, опсонизация и т.д., в результате обеспечивает селективное нацеливание и эффективную деструкцию раковых клеток.
Помимо этого, изобретение охватывает введение одного или более Fc вариантов по изобретению в комбинации с другими средствами лечения, которые, как известно специалистам в данной области техники, применяются для лечения или предупреждения рака, включая, но без ограничения, современные стандартные и экспериментальные химиотерапевтические средства, гормональные препараты, биологические лекарственные средства, иммунотерапевтические средства, лучевую терапию или хирургию. Согласно некоторым вариантам молекулы по изобретению можно вводить в комбинации с терапевтически или профилактически эффективным количеством одного или более противораковых агентов, терапевтических антител или других агентов, применяемых, как известно специалистам в данной области техники, для лечения и/или предупреждения рака. Примеры режимов дозирования и методов терапии (и терапевтических средств), которые можно применять в комбинации с Fc вариантами по изобретению, хорошо известны в уровне техники и подробнее описаны в другом документе (см., например, опубликованные Международные заявки РСТ WO 02/070007 и WO 03/075957).
Раковые заболевание и родственные нарушения, которые можно лечить или предупредить методами и композициями по настоящему изобретению, включают, но без ограничения, следующие: лейкозы, лимфомы, множественные миеломы, саркомы костей и соединительной ткани, опухоли мозга, рак молочной железы, рак надпочечника, рак щитовидной железы, рак поджелудочной железы, опухоли гипофиза, рак глаза, рак влагалища, рак вульвы, рак шейки матки, рак матки, рак яичника, рак пищевода, рак желудка, рак толстой кишки, рак прямой кишки, рак печени, рак желчного пузыря, холангиокарциному, рак легкого, рак яичек, рак предстательной железы, рак пениса; рак полости рта, рак слюнных желез, рак ротоглотки, рак кожи, рак почки, рак мочевого пузыря (обзор, относящийся к этим заболеваниям, см. Fishman et al., 1985, Medicine, 2d Ed., J.B. Lippincott Co., Philadelphia and Murphy et al., 1997, Informed Decisions: The Complete Book of Cancer Diagnosis, Treatment, and Recovery, Viking Penguin, Penguin Books U.S.A., Inc., United States of America).
Помимо этого, изобретение рассматривает получение методами генной инженерии любого из антител, известных в уровне техники, для лечения и/или предупреждения рака и родственных заболеваний, таким образом, чтобы антитела содержали Fc область, включающую вариантный CH3 домен по изобретению.
Согласно конкретному варианту молекула по изобретению (например, антитело, содержащее вариантный Fc гетеродимер, ингибирует или снижает рост первичной опухоли или метастазирование раковых клеток по меньшей мере на 99%, по меньшей мере на 95%, по меньшей мере на 90%, по меньшей мере на 85%, по меньшей мере на 80%, по меньшей мере на 75%, по меньшей мере на 70%, по меньшей мере на 60%, по меньшей мере на 50%, по меньшей мере на 45%, по меньшей мере на 40%, по меньшей мере на 45%, по меньшей мере на 35%, по меньшей мере на 30%, по меньшей мере на 25%, по меньшей мере на 20% или по меньшей мере на 10% по сравнению с ростом первичной опухоли или метастазированием в отсутствие указанной молекулы по изобретению.
Настоящее изобретение охватывает применение одного или более Fc вариантов по изобретению для предупреждения, лечения или контроля одного или более симптомов, обусловленных воспалительным нарушением у субъекта. Не будучи связанными с каким-либо механизмом действия, можно сказать, что Fc варианты с повышенной аффинностью к FcγRIIB приводят к угнетению активирующих рецепторов и, следовательно, к угнетению иммунного ответа и обладают терапевтической эффективностью при лечении и/или предупреждении аутоиммунного расстройства. Помимо этого, антитела, связывающие более одной мишени, такие как биспецифические антитела, содержащие вариантный Fc гетеродимер, ассоциированные с воспалительным нарушением, могут оказывать синергическое действие, более сильное, чем действие одновалентного антитела.
Помимо этого, настоящее изобретение охватывает введение Fc вариантов по изобретению в комбинации с терапевтически или профилактически эффективным количеством одного или более противовоспалительных агентов. Изобретение также предусматривает методы предупреждения, лечения или контроля одного или более симптомов, обусловленных аутоиммунным заболеванием, дополнительно включающие введение указанному субъекту Fc варианта по изобретению в комбинации с терапевтически или профилактически эффективным количеством одного или более иммуномодулирующих агентов. Примеры аутоиммунных нарушений, которые можно лечить с помощью Fc вариантов по изобретению, включают, но без ограничения, гнездную алопецию, анкилозирующий спондилоартрит, антифосфолипидный синдром, аутоиммунная болезнь Аддисона, аутоиммунные заболевания надпочечника, аутоиммунную гемолитическую анемию, аутоиммунный гепатит, аутоиммунный оофорит и орхит, аутоиммунную тромбоцитопению, болезнь Бехчета, буллезный пемфигоид, кардиомиопатию, целиакию, синдром хронической усталости и иммунной дисфункции (CFIDS), хроническую воспалительную демиелинизирующую полинейропатию, синдром Черджа-Стросса, рубцовый пемфигоид, CREST синдром, болезнь холодовых агглютининов, болезнь Крона, дискоидную красную волчанку, эссенциальную смешанную криоглобулинемию, фибромиалгию-фибромиоцитоз, гломерулонефрит, болезнь Грейвса, болезнь Гийенна-Барре, тиреоидит Хашимото, идиопатический легочный фиброз, идиопатическую тромбоцитопеническую пурпуру (ITP), IgA-нефропатию, ювенильный артрит, красный плоский лишай, красную волчанку, смешанное заболевание соединительной ткани, рассеянный склероз, типа 1 или иммуноопосредованный сахарный диабет, тяжелую псевдопаралитическую миастению, обыкновенную пузырчатку, пернициозную анемию, узелковый полиартериит, полихондрит, полигландулярный синдром, ревматическую полимиалгию, полимиозит и дерматомиозит, первичную (а-)гаммаглобулинемию, первичный билиарный цирроз печени, псориаз, псориатический артрит, феномен Рейно, синдром Рейтера, ревматоидный артрит, саркоидоз, склеродермию, синдром Шегрена, синдром мышечной скованности, системную красную волчанку, артериит Такаясу, системную красную волчанку, красную волчанку, артериит Такаясу, височный артериит/гигантоклеточный артериит, язвенный колит, увеит, васкулиты, такие как герпетиформный дерматит, витилиго и гранулематоз Вегенера. Примеры воспалительных нарушений включают, но без ограничения, астму, энцефалит, воспалительное заболевание кишечника, хроническую обструктивную болезнь легких (ХОБЛ), аллергические нарушения, септический шок, фиброз легких, недифференцированную спондилоартропатию, недифференцированную артропатию, артрит, воспалительный остеолиз и хроническое воспаление в результате хронических вирусных или бактериальных инфекций. Некоторые аутоиммунные заболевания ассоциированы с воспалительным состоянием, т.е. происходит взаимное наложение заболевания, рассматриваемого как аутоиммунное нарушение, и воспалительного нарушения. Примеры воспалительных нарушений, которые можно предупредить, лечить или контролировать в соответствии с методами по изобретению, включают, но без ограничения, астму, энцефалит, воспалительное заболевание кишечника, хроническую обструктивную болезнь легких (ХОБЛ), аллергические нарушения, септический шок, фиброз легких, недифференцированную спондилоартропатию, недифференцированную артропатию, артрит, воспалительный остеолиз и хроническое воспаление в результате хронических вирусных или бактериальных инфекций.
Fc варианты по изобретению можно применять также для уменьшения воспаления, испытываемого животными, в частности, млекопитающими, страдающими воспалительными нарушениями. Согласно конкретному варианту Fc по изобретению уменьшает воспаление у животного по меньшей мере на 99%, по меньшей мере на 95%, по меньшей мере на 90%, по меньшей мере на 85%, по меньшей мере на 80%, по меньшей мере на 75%, по меньшей мере на 70%, по меньшей мере на 60%, по меньшей мере на 50%, по меньшей мере на 45%, по меньшей мере на 40%, по меньшей мере на 45%, по меньшей мере на 35%, по меньшей мере на 30%, по меньшей мере на 25%, по меньшей мере на 20% или по меньшей мере на 10% по сравнению с воспалением у животного, которому не вводили указанную молекулу.
Помимо этого, изобретение рассматривает получение методами генной инженерии любого из антител, известных в уровне техники, для лечения и/или предупреждения аутоиммунного заболевания или воспалительного заболевания, таким образом, чтобы антитела содержали вариантный Fc гетеродимер по изобретению.
Изобретение охватывает также методы лечения или предупреждения инфекционного заболевания у субъекта, включающие введение терапевтически или профилактически эффективного количества одного или более Fc вариантов по изобретению. Инфекционные заболевания, которые можно лечить или предупреждать с помощью Fc вариантов по изобретению, вызываются инфекционными агентами, включая, но без ограничения, вирусы, бактерии, грибы и простейшие.
Вирусные заболевания, которые можно лечить или предупреждать с помощью Fc вариантов в сочетании с методами по настоящему изобретению, включают, но без ограничения, заболевания, которые вызывают вирусы гепатита А, гепатита В, гепатита С, гриппа, ветряной оспы, аденовирус, вирусы простого герпеса типа I (HSV-I), простого герпеса типа II (HSV-II), чумы рогатого скота, риновирус, эховирус, ротавирус, респираторно-синцитиальный вирус, вирус папилломы, вирус Попова (папова вирус), цитомегаловирус, эхиновирус, арбовирус, хунтавирус, коксаки вирус, вирус свинки, вирус кори, вирус краснухи, полиовирус, вирус натуральной оспы, вирус Эпштейна-Барр, вирус иммунодефицита человека типа I (HIV-I, ВИЧ-I), вирус иммунодефицита человека типа II (HIV-II, ВИЧ II) и агенты, вызывающие вирусные заболевания, такие как вирусный менингит, энцефалит, лихорадку денге или натуральную оспу.
Бактериальные заболевания, которые можно лечить или предупредить с помощью Fc вариантов по изобретению в сочетании с методами по настоящему изобретению, вызываются бактериями, включая, но без ограничения, микобактерии, риккетсии, микоплазмы, нейссерию, S.pneumonia, Borrelia burgdorferi (возбудитель болезни Лайма), Bacillus antracis (возбудитель сибирской язвы), возбудители столбняка, стрептококк, стафилококк, возбудители коклюша, холеры, чумы, дифтерии, хламидии, золотистый стафилококк (S.aureus) и легионеллы. Протозойные заболевания, которые можно лечить или предупредить с помощью молекул по изобретению в сочетании с методами по настоящему изобретению, вызываемые простейшими, включают лейшманиоз, коксидиоз, трипасономоз или малярию. Паразитарные заболевания, которые можно лечить или предупредить с помощью молекул по изобретению в сочетании с методами по настоящему изобретению, вызываются простейшими, включающими, но без ограничения, хламидии и риккетсии.
Согласно некоторым вариантам Fc варианты по изобретению можно вводить в комбинации с терапевтически или профилактически эффективным количеством одного или дополнительных лекарственных веществ, известных специалистам в данной области техники, для лечения и/или предупреждения инфекционного заболевания. Изобретение рассматривает применение молекул по изобретению в комбинации с другими молекулами, известными специалистам в данной области техники, для лечения и/или предупреждения инфекционного заболевания, включая, но без ограничения, антибиотики, противогрибковые агенты и противовирусные агенты.
Изобретение предусматривает методы и фармацевтические композиции, содержащие Fc варианты по изобретению (например, антитела, полипептиды). Также изобретение предусматривает методы лечения, профилактики и уменьшения интенсивности одного или более симптомов, ассоциированных с заболеванием, нарушением или инфекцией, путем введения субъекту эффективного количества по меньшей мере одного Fc варианта по изобретению или фармацевтической композиции, содержащей по меньшей мере один Fc вариант по изобретению. В одном аспекте изобретения Fc вариант является по существу чистым (т.е. по существу не содержащим веществ, которые ограничивают его действие или дают нежелательные побочные эффекты, они включают гомодимеры и другой клеточный материал). Согласно одному конкретному варианту изобретения субъектом является животное, такое как млекопитающее, включая животных, отличных от приматов (например, коров, свиней, лошадей, кошек, собак, крыс и т.д.), и приматов (например, обезьян, таких как обезьяна циномолгус, и человека). Согласно одному конкретному варианту изобретения субъектом является человек. Согласно еще одному варианту изобретения антитело по изобретению получено из того же вида, к которому относится субъект.
Способ введения композиции зависит от состояния, подлежащего лечению. Например, внутривенная инъекция может быть предпочтительна для лечения системного нарушения, такого как рак лимфатической системы или метастазированная опухоль. Дозу вводимых композиций специалист в данной области техники может определить без неоправданных экспериментов параллельно со стандартными исследованиями доза-эффект. Важные обстоятельства, которые следует рассматривать при проведении этих определений, включают заболевание или заболевания, которые следует лечить, выбор композиции, которую следует вводить, возраст, вес и реакцию отдельного пациента, и тяжесть симптомов у пациента. В зависимости от заболевания композицию можно вводить пациенту перорально, парентерально, интраназально, вагинально, ректально, транслингвально, подъязычно (сублингвально), трансбуккально, интрабуккально и/или трансдермально.
Соответственно, композиции, предназначенные для перорального, транслингвального, подъязычного, трансбуккального и интрабуккального введения, можно приготовить без неоправданных экспериментов способами, хорошо известными в уровне техники, например, с инертным разбавителем или с пищевым (съедобным) носителем. Композиция может быть заключена в желатиновые капсулы или может быть в виде прессованных таблеток. Для перорального применения фармацевтические композиции по настоящему изобретению могут включать эксципиенты и выпускаться в виде таблеток, пастилок, капсул, эликсиров, суспензий, сиропов, облаток, жевательных резинок и т.п.
Таблетки, пилюли, капсулы, пастилки и т.п. могут также содержать связующие, recipients, дезинтегранты, смазки, подсластители и/или корригенты. Некоторые примеры связующих включают микрокристаллическую целлюлозу, камедь трагаканта и желатин. Примеры эксципиентов включают крахмал и лактозу. Некоторые примеры дезинтегрантов включают альгиновую кислоту, кукурузный крахмал и т.п. Примеры смазок включают стеарат магния и стеарат калия. Примером вещества, способствующего проглатыванию, является коллоидный диоксид кремния. Некоторые примеры подсластителей включают сахарозу, сахарин и т.п. Примеры корригентов включают перечную мяту, метилсалицилат, корригент со вкусом апельсина и т.п. Материалы, применяемые для приготовления этих различных композиций, должны быть фармацевтической степени чистоты и нетоксическими в применяемых количествах.
Фармацевтические композиции по настоящему изобретению можно вводить парентерально, например, в виде внутривенной, внутримышечной, интратекальной и/или подкожной инъекции. Парентеральное введение можно осуществлять, применяя композиции по настоящему изобретению в виде раствора или суспензии. Такие растворы или суспензии могут включать также стерильные разбавители, такие как вода для инъекций, физиологический раствор, нелетучие масла, полиэтиленгликоль, глицерин, пропиленгликоль и/или другие синтетические растворители. Препараты для парентерального введения могут включать также антибактериальные агенты, например, такие как бензиловый спирт и/или бензилпарабены, антиоксиданты, например, такие как аскорбиновая кислота и/или бисульфит натрия, и хелатирующие агенты, такие как, ЭДТА. Можно также добавлять буферы, такие как ацетатный, цитратный и фосфатный, и агенты для корректировки тоничности, такие как хлорид натрия и декстроза. Препараты для парентерального введения могут быть помещены в ампулы, одноразовые шприцы и/или многодозовые флаконы из стекла или пластика. Ректальное введение включает введение композиции в прямую кишку и/или толстый кишечник. Это можно осуществлять, используя суппозитории и/или клизмы. Препараты в виде суппозиториев можно готовить методами, известными в уровне техники. Трансдермальное введение включает впитывание композиции по изобретению через кожу. Трансдермальные препараты включают пластыри, мази, кремы, гели, бальзамы и т.п. Композиции по настоящему изобретению можно вводить пациенту через нос. Назальное введение по настоящему изобретению включает введение композиций через слизистую оболочку носовых проходов и/или полости носа пациента.
Фармацевтические композиции по изобретению можно применять в соответствии с методами по изобретению для предупреждения, лечения или уменьшения интенсивности одного или более симптомов, ассоциированных с заболеванием, нарушением или инфекцией. Предполагается, что фармацевтические композиции являются стерильными и находятся в виде, пригодном для введения субъекту.
Согласно одному варианту композиции по изобретению являются апирогенными препаратами, практически не содержащими эндотоксинов и/или родственных пирогенных веществ. Эндотоксины включают токсины, которые находятся внутри микроорганизма и высвобождаются, когда микроорганизмы разрушаются или гибнут. Пирогенные вещества включают также вызывающие лихорадочное состояние термоустойчивые вещества (гликопротеины) из внешней мембраны бактерий и других микроорганизмов. И те, и другие вещества, при введении их человеку, могут вызвать лихорадку, гипотензию и шок. Ввиду возможного неблагоприятного действия предпочтительно удалять даже малые количества эндотоксинов из лекарственных растворов для внутривенного введения. Управление по контролю за пищевыми продуктами и лекарственными средствами ("FDA") установило верхний предел при внутривенном введении лекарств 5 эндотоксиновых единиц (EU) на дозу на килограмм массы тела при одноразовом введении за время в течение одного часа (The United States Pharmacopeial Convention, Pharmacopeial Forum 26 (1):223 (2000)). Если фармацевтические белки вводят в количествах от нескольких сотен до тысячи миллиграмм на килограмм массы тела, как в случае моноклональных антител, предпочтительно удалять даже следовые количества эндотоксина. Согласно одному конкретному варианту изобретения уровни эндотоксинов и пирогенных веществ в композиции составляют менее 10 EU/мг, или менее 5 EU/мг, или менее 1 EU/мг, или менее 0.1 EU/мг, или менее 0.01 EU/ мг, или менее 0.001 EU/мг.
Изобретение предусматривает метод предупреждения, лечения или уменьшения интенсивности одного или более симптомов, ассоциированных с заболеванием, нарушением или инфекцией, который включает: (а) введение нуждающемуся в этом субъекту дозы, включающей профилактически или терапевтически эффективное количество композиции, содержащей один или более Fc вариантов, и (б) введение одной или более последующих доз указанных Fc вариантов для поддержания концентрации в плазме Fc варианта, который непрерывно связывается с антигеном, на заданном уровне (например, от около 0.1 мкг/мл до около 100 мкг/мл). Согласно одному специфическому варианту изобретения концентрация Fc варианта в плазме поддерживается на уровне 10 мкг/мл, 15 мкг/мл, 20 мкг/мл, 25 мкг/мл, 30 мкг/мл, 35 мкг/мл, 40 мкг/мл, 45 мкг/мл или 50 мкг/мл. Согласно конкретному варианту изобретения указанное вводимое эффективное количество Fc варианта составляет по меньшей мере от 1 мг/кг до 8 мг/кг на дозу. Согласно другому конкретному варианту изобретения указанное вводимое эффективное количество Fc варианта составляет по меньшей мере от 4 мг/кг до 5 мг/кг на дозу. Согласно другому конкретному варианту изобретения указанное вводимое эффективное количество Fc варианта составляет по меньшей мере от 50 мг до 250 мг на дозу. Согласно еще одному конкретному варианту изобретения указанное вводимое эффективное количество Fc варианта составляет по меньшей мере от 100 мг до 200 мг на дозу.
Настоящее изобретение включает также протоколы предупреждения, лечения или уменьшения интенсивности одного или более симптомов, ассоциированных с заболеванием, нарушением или инфекцией, в которых Fc вариант применяется в комбинации с препаратом (например, профилактическим или терапевтическим агентом), отличным от Fc варианта и/или варианта слитого белка. Изобретение основано, частично, на признании того факта, что Fc варианты по изобретению усиливают действие других противораковых препаратов и совместно с ними вызывают синергический эффект, повышают эффективность других противораковых препаратов, повышают толерантность к другим противораковым препаратам и/или снижают побочные эффекты, вызываемые другими противораковыми препаратами, включая современные стандартные и экспериментальные химиотерапевтические средства. Комбинированные препараты по изобретению проявляют аддитивную активность, вызывают аддитивный терапевтический эффект или синергический эффект. Комбинированные препараты по изобретению позволяют вводить субъекту, у которого наблюдается заболевание, нарушение или инфекция, пониженные дозы препарата (например, профилактических или терапевтических агентов) в сочетании с Fc вариантами для предупреждения, лечения или уменьшения интенсивности одного или более симптомов, ассоциированных с заболеванием, нарушением или инфекцией, и/или реже вводить указанному субъекту такие профилактические или терапевтические агенты для повышения качества жизни указанного субъекта и/или для достижения профилактического или терапевтического эффекта. Кроме того, комбинированные препараты по изобретению ослабляют нежелательные или вредные побочные эффекты или не допускают этих эффектов, обусловленных введением современных средств монотерапии и/или современных комбинированных препаратов, что, в свою очередь, повышает приверженность пациента соблюдению требований протокола лечения. Многочисленные соединения (молекулы), которые можно применять в комбинации с Fc вариантами по изобретению, хорошо известны в уровне техники. См., например, опубликованные Международные патентные заявки РСТ WO 02/070007; WO 03/075957 и опубликованная заявка на патент США 2005/064514.
Настоящее изобретение предусматривает наборы, содержащие один или более Fc вариантов с измененной аффинностью к FcγRs и/или C1q и измененной ADCC и/или CDC активностью, которые специфически связываются с антигеном, конъюгированным или слитым с детектируемым агентом, терапевтическим агентом или лекарством, в одном или более контейнеров для применения при мониторинге, в диагностике, для предупреждения, лечения или уменьшения интенсивности одного или более симптомов, обусловленных заболеванием, нарушением или инфекцией.
5. ПРИМЕРЫ
Приведенные ниже примеры даются для иллюстрации практического применения настоящего изобретения. Не предполагается, что они ограничивают или определяют полный объем настоящего изобретения.
Пример 1: Получение двухвалентных моноспецифических антител с гетеродимерными Fc доменами.
Гены, кодирующие тяжелую и легкую цепи антитела, получали синтезом генов с применением последовательностей, кодон-оптимизированных для экспрессии в клетках человека/млекопитающего. Последовательности Fab получали с применением известного АЬ, связывающего Her2/neu (Carter P. et al. (1992) Humanization of an anti P185 Her2 antibody for human cancer therapy. Proc Natl Acad Sci 89, 4285.), a Fc был IgG1 изотипа (SEQ ID NO:1). Конечные генные продукты субклонировали в вектор рТТ5 (NRC-BRI, Canada) для экспрессии в клетках млекопитающих (Durocher, Y., Perret, S. & Kamen, A. High-level and high-throughput recombinant protein production by transient transfection of suspension-growing human HEK293-EBNA1 cells. Nucleic acids research 30, E9 (2002)). Мутации в СИЗ домен вводили с помощью сайт-направленного мутагенеза с использованием векторов на матрице рТТ5. См. в Таблице 1, Таблице 6 и Таблице 7 список мутаций, осуществленных в вариантном CH3 домене.
Для того чтобы оценить образование гетеродимеров и определить отношение гомодимеров к гетеродимерам, были созданы две тяжелых цепи гетеродимера с C-концевыми удлинениями различной протяженности (конкретно, цепь А с C-концевым HisTag и цепь В с C-концевым mRFP плюс StrepTagII). Эти различия молекулярных масс позволяют дифференцировать гомодимеры и гетеродимер методом SDS-PAGE гель-электрофореза в невосстанавливающих условиях, как показано на Фигуре 25А.
Клетки HEK293 трансфецировали в фазе экспоненциального роста (1.5-2 миллиона клеток/мл) с использованием водного раствора полиэтиленимина 25 кДа (PEI, Poly sciences) с концентрацией 1 мг/мл при соотношении PEI: ДНК 2.5: 1. (Raymond С. et al. A simplified polyethylenimine-mediated transfection process for large-scale and high-throughput applications. Methods. 55(1):44-51 (2011)). Для определения интервала концентраций, оптимального для образования гетеродимеров, ДНК трансфецировали при трех различных соотношениях плазмид для двух тяжелых цепей. Например, этот процесс проводили в культуре объемом 2 мл и ДНК для трансфекции, содержавшей 5% GFP, 45% ДНК из молок лососевых рыб, 25% плазмиды для легкой цепи и 25% в совокупности плазмид для тяжелых цепей, где образцы плазмиды тяжелой цепи А (с C-концевым HisTag) и плазмиды тяжелой цепи В (с C-концевым StrepTagII плюс RFP) в соотношении 65%/55%/35% или 10%/20%/40%) брали в 3 различных соотношениях (цепь_A(His)/цепь_B(mRFP)) 10%/65%; 20%/55%; 40%/35% (было найдено, что кажущееся соотношение 1:1 экспрессии WT_His/WT_mRFP гетеродимера близко к соотношению ДНК 20%/55%). Через 4-48 часов после трансфекции в F17 бессывороточной среде (Gibco) добавляли TN1 пептон до конечной концентрации 0.5%. Экспрессированное антитело анализировали методом SDS-PAGE гель-электрофореза для определения наилучшего соотношения тяжелой и легкой цепей для оптимального образования гетеродимера (См. фигуру 25В и С).
Выбранное соотношение ДНК, например, 50% ДНК-плазмиды для легкой цепи, 25% ДНК-плазмиды для тяжелой цепи А, 25% ДНК-плазмиды для тяжелой цепи В AZ33 и AZ34, с 5% GFP и 45% ДНК молок лососевых рыб применяли для трансфекции 150 мл клеточной культуры как описано выше. Трансфецированные клетки собирали через 5-6 дней, культуральную среду объединяли после центрифугирования при 4000 об/мин и осветляли фильтрованием через фильтр 0.45 мкм. См. ниже в Таблице 2 перечень процентного содержания плазмид для легкой и тяжелых цепей А и В, применяемых в анализах трансфекции в увеличенном масштабе для каждого из антител с CH3 мутациями, полученных для дальнейших анализов, включая определение чистоты и температуры плавления.
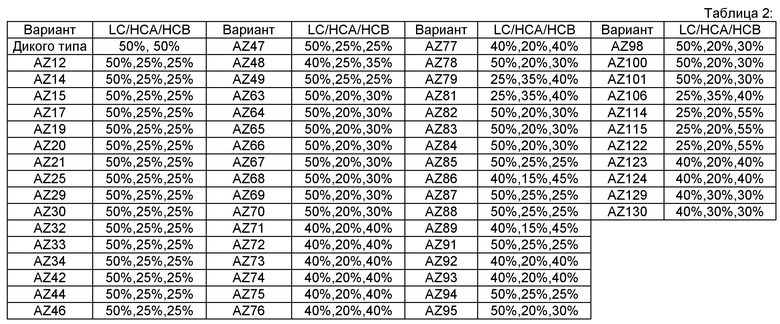
Пример 2: Очистка двухвалентных моноспецифических антител с гетеродимерными Fc доменами.
Осветленную культуральную среду загружали в колонку с хроматографической средой MabSelect SuRe (GE Healthcare) с Белком А и промывали буфером PBS при рН 7.2 в количестве, равном 10 объемам колонки. Антитело элюировали цитратным буфером при рН 3.6 (в количестве, равном 10 объемам колонки), фракции, содержащие антитело, объединяли и нейтрализовали, добавляя TRIS с рН 11. Наконец, проводили обессоливание белка на колонке Econo-Pac 10DG (Bio-Rad). C-концевой хвост mRFP в тяжелой цепи В удаляли, инкубируя антитело с энтерокиназой (NEB) в соотношении 1:10000 в течение ночи в PBS при 25°С. Антитело в смеси очищали гель-фильтрацией. Для гель-фильтрации 3.5 мг смеси, содержащей антитело, концентрировали до 1.5 мл и загружали в колонку с Sephadex 200 HiLoad 16/600 200 пг (GE Healthcare) с использованием AKTA Express FPLC при скорости потока 1 мл/мин. PBS буфер с рН 7.4 применяли при скорости потока 1 мл/мин. Фракции, соответствующие очищенному антителу, собирали, концентрировали до ~1 мг/мл и хранили при -80°С.
Образование гетеродимеров, по сравнению с гомодимерами, анализировали методами SDS-PAGE гель-электрофореза в невосстанавливающих условиях и масс-спектрометрии. Антитело, очищенное с использованием белка А, подвергали электрофорезу в градиенте 4-12% SDS-PAGE, невосстанавливающего геля, с целью определить процентное содержание образованных гетеродимеров, а потом обрабатывали энтерокиназой (EK) (см. Фигуру 26). Для масс-спектрометрии все эксперименты по жидкостной хроматографии масс-спектрометрии Trap LC/MS (ESI-TOF) проводили на Agilent 1100 ВЭЖХ системе, соединенной с масс-спектрометром Waters Q-TOF2. Пять мкг антитела, очищенного гель-фильтрацией, вводили в микроловушку для белка Protein MicroTrap (1.0 на 8.0 мм), промывали 1% раствором ацетонитрила в течение 8 минут, в градиенте от 1 до 20% ацетонитрила/0.1% муравьиной кислоты в течение 2 минут, затем элюировали в градиенте от 20 до 60% ацетонитрила/0.1% муравьиной кислоты в течение 20 минут. Элюат (30-50 мкл/мин) направляли в спектрометр, спектр снимали каждую секунду (m/z от 800 до 4000) (см. Фигуру 28). Варианты, содержавшие более 90% гетеродимеров, отбирали для последующего анализа, за исключением AZ12 и AZ14, каждый из которых содержал более 85% гетеродимеров.
Пример 3: Определение устойчивости двухвалентных моноспецифических антител с гетеродимерными Fc доменами методом дифференциальной сканирующей калориметрии (ДСК).
Все ДСК эксперименты проводили на приборе GE VP-Capillary. Заменяли буфер для белков на PBS (рН 7.4) и разводили до концентрации 0.4-0.5 мг/мл, 0.137 мл помещали в ячейку для образца и проводили измерения при скорости сканирования 1°С/мин от 20°С до 100°С. Результаты анализировали с применением программы Origin (GE Healthcare), вычитая фоновые результаты для PBS буфера. (См. Фигуру 27). В Таблице 3 представлен список тестированных вариантов и полученные температуры плавления. В Таблице 4 представлен список тестированных вариантов с температурой плавления 70°С и выше и конкретное значение Tm для каждого варианта.
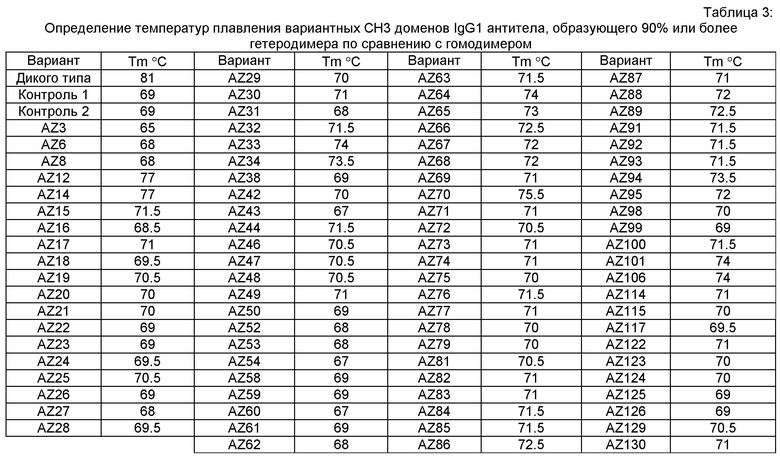
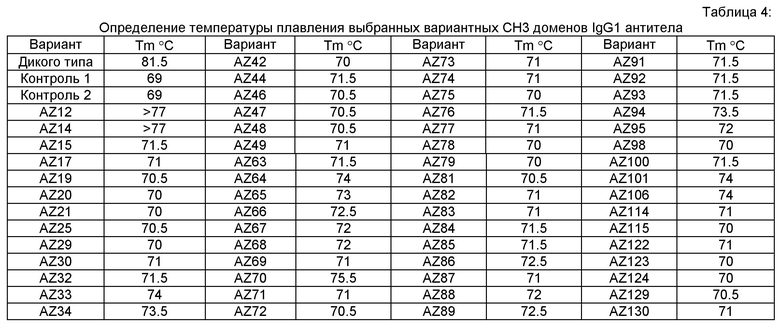
Пример 4: Определение связывания Fc гаммаR с применением поверхностного плазменного резонанса
Все эксперименты по связыванию проводили на приборе BioRad ProteOn XPR36 при 25°C с растворами, содержащими 10 мМ HEPES, 150 мМ NaCl, 3.4 мМ ЭДТА и 0.05% Tween 20 при рН 7.4. Рекомбинантный HER-2/neu (p185, ErbB-2 (Biosciences, Inc.)) улавливали на активированном GLM сенсорном чипе инжекцией 4.0 мкг/мл в 10 мМ NaOAc (рН 4.5) при скорости 25 мкл/мин, до иммобилизации, примерно, 3000 резонансных единиц (RU) при том, что остальные активные группы блокировались. 40 мкг/мл очищенных антител против HER-2/neu, содержащих вариантные CH3 домены, опосредованно улавливали на сенсорном чипе путем связывания белка Her-2/neu при инжекции со скоростью 25 мкл/мин в течение 240 сек (что дает в результате 500 RU) с последующей инжекцией буфера для установления стабильной базовой линии. Fc гамма R (CD16a (f аллотип) и CD32b), с концентрацией 6000, 2000, 667, 222 и 74.0 нМ, инъецировали со скоростью 60 мкл/мин в течение 120 сек, фаза диссоциации составляла 180 сек, получали ряд сенсограмм связывания. Значения KD определяли из изотерм связывания, используя равновесную модель, приведены средние значения из трех независимых опытов. Дано сравнение с дикого типа IgG1 Fc доменом, а связывание выражается как отношение WT KD к KD варианта (См. Таблицу 5).
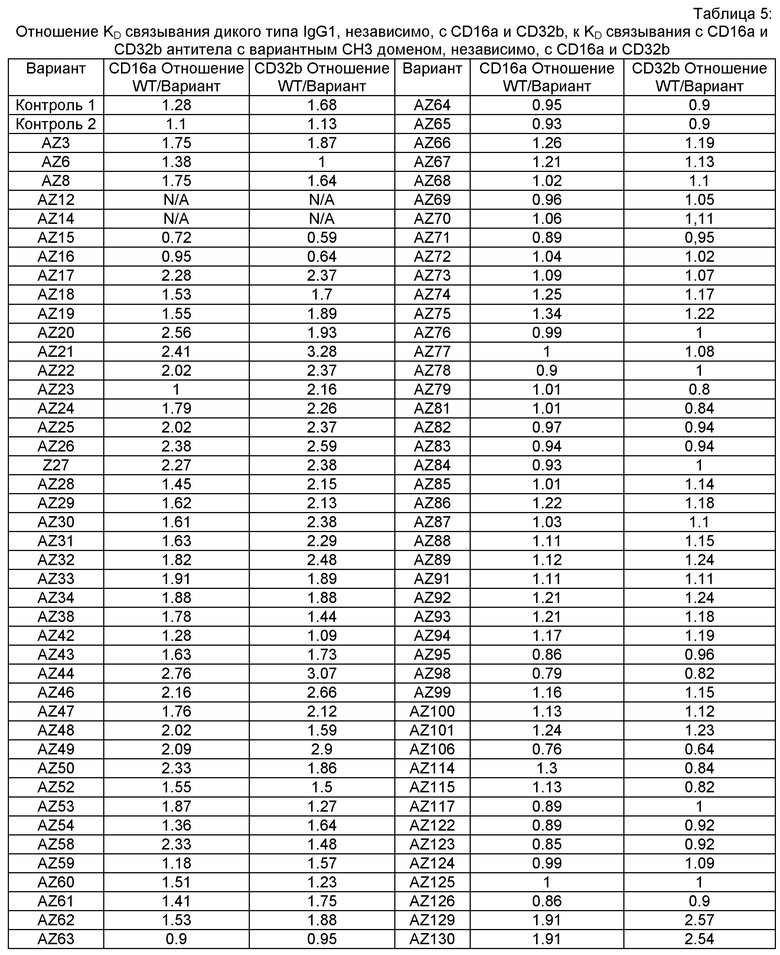
Пример 5: Рациональный дизайн Fc вариантов методами Fc_CH3 генной инженерии - Каркас 1 (1a и 1b) разработка AZ17-62 и AZ133-AZ2438
Для повышения устойчивости и чистоты первоначально полученного с помощью негативного дизайна Fc варианта AZ8 применяли описанные выше структурные и вычислительные методы. (См. Фигуру 24) Например, тщательный структурно-функциональный анализ AZ8 дал глубокое понимание роли каждой из вводимых мутаций AZ8,L351Y_V397S_F405A_Y407V/K392V_T394W по сравнению с дикого типа человеческим IgG1 и показал, что важными мутациями ядерного гетеродимера являются мутации L351Y_F405A_Y407V/T394W, тогда как мутации V397S, K392V не являются значимыми для образования гетеродимера. Ядерные мутации (L351Y_F405A_Y407V/T394W) в данной заявки называются мутациями "Каркаса 1". Кроме того, анализ выявил, что важными гипервариабельными участками в области контакта, которые утрачиваются при образовании гомодимера дикого типа (WT), являются участки взаимодействия WT-F405-K409, Y407-Т366 и упаковки Y407-Y407 и -F405 (См. Фигуру 29). Это отражается в анализе упаковки, полости и MD, которые показали большое различие между конформациями в области петли D399-S400-D401 (См. Фигуру 30) и связанных с ней β-складок в K370. Это привело в результате к утрате взаимодействия между цепями K409-D399 (См. Фигуру 30) и ослаблению сильной водородной связи K370 с Е357 (K370 больше не находится в непосредственном контакте с S364 и Е357, но полностью доступен для растворителя). В WT IgG1 CH3 домене эти области ограничивали область контакта по периферии, защищая ядро от конкурентного взаимодействия с большой массой растворителя и повышая частоту предпочтительных гидрофобных ван-дер-Ваальсовых взаимодействий. Следствием этого явилась более низкая площадь неэкспансированной поверхности AZ8 по сравнению с WT и большая доступность гидрофобного ядра для растворителя. Это показывает, что наиболее важными факторами пониженной устойчивости AZ8 по сравнению с устойчивостью WT являются а) утрата взаимодействия WT-F405-K409 и упаковки F405, и б) утрата сильного взаимодействия в упаковке Y407-Y407 и Y407-Т366. См. Фигуру 29.
Следовательно, мы идентифицировали важнейшие остатки/мотивы в последовательности, отвечающие за низкую устойчивость AZ8 по сравнению с WT. Для повышения устойчивости и специфичности гетеродимера AZ8 последующие усилия по инжинирингу с помощью позитивного дизайна мы сосредоточили конкретно на стабилизации конформации петли в положениях 399-401 в виде более ′закрытой′, WT-подобной конформации (См. Фигуру 30) и компенсации общей немного менее плотной упаковки гидрофобного ядра в положениях Т366 и L368 (См. Фигуру 29).
Для осуществления этой стабилизации конформации петли в положениях 399-401 мы применяли описанные выше компьютерные методы, чтобы оценить наши различные предположения о целевом дизайне. Конкретно, три различные независимые возможности для Fc варианта AZ8 были проанализированы с целью оптимизации идентифицированных ключевых областей для повышения устойчивости. Во-первых, полость близ положения K409 и F405A оценивали с точки зрения более плотной упаковки в гидрофобном ядре как для защиты гидрофобного ядра, так и для стабилизации конформации петли 399-400 (См. Фигуру 30). Здесь рассматривались, но без ограничения, дополнительные точковые мутации в положениях F405 и K392. Во-вторых, оценивались возможности повышения электростатических взаимодействий в положениях 399-409 для стабилизации конформации петли 399-400 и защиты гидрофобного ядра. Эти возможности включали, но без ограничения, дополнительные точковые мутации в положениях Т411 и S400. В-третьих, оценивали полость и упаковку гидрофобного ядра в положениях Т366, T394W и L368 с точки зрения улучшения упаковки гидрофобного ядра (См. Фигуру 29). Эти возможности включали, но без ограничения, дополнительные точковые мутации в положениях Т366 и L368. Различные независимые идеи позитивного дизайна были проверены in silico (на компьютерной модели) и лучшие из рассмотренных на компьютерной модели вариантов (AZ17-AZ62) оценивали экспериментально с точки зрения экспрессии и устойчивости, как описано в Примерах 1-4. См. В Таблице 4 список Fc вариантов, полученных в этой фазе дизайна, с температурой плавления 70°С или выше.
Fc вариант AZ33 представляет собой пример создания Fc варианта, в котором Каркас 1 был модифицирован, результатом чего явились мутации в Каркасе 1а для повышения устойчивости и чистоты. Этот Fc вариант был создан на базе AZ8 с целью усовершенствования упаковки гидрофобного ядра в положениях 392-394-409 и 366 как для защиты гидрофобного ядра, так и для стабилизации конформации петли 399-400. Этот Fc вариантный AZ33 гетеродимер содержит две дополнительные точковые мутации, отличные от мутаций ядра AZ8, K392M и T366I. Мутации T366I_K392M_T394W/F405A_Y407V в данной заявке называются мутациями "Каркаса 1а". Мутация K392M создана с целью улучшить упаковку в полости близ положения K409 и F405A для защиты гидрофобного ядра и стабилизации конформации петли 399-400 (См. Фигуру 31). Мутация T366I была осуществлена для улучшения упаковки гидрофобного ядра и для исключения возможности образования гомодимеров T394W цепи (См. Фигуру 29). Экспериментальные данные для AZ33 показали значительно более высокую устойчивость по сравнению с исходным, полученным с помощью негативного дизайна, Fc вариантом AZ8 (Tm 68°С), так AZ33 имел Tm 74°С и содержание гетеродимера >98%. (См. Фигуру 25С)
Усовершенствование Fc вариантов с помощью мутаций в Каркасе 1 в фазе три дизайна Fc вариантных гетеродимеров
Хотя AZ33 показывает значительное повышение устойчивости и специфичности (или чистоты) по сравнению с исходным вариантом AZ8, наш анализ показал, что можно сделать дальнейшие улучшения Fc вариантного гетеродимера с дополнительными аминокислотными модификациями, используя экспериментальные данные для AZ33 и вышеописанные методы дизайна. Различные идеи дизайна были независимо проверены с точки зрения экспрессии и устойчивости, но независимые положения дизайна могут "переноситься", и наиболее успешный гетеродимер может содержать комбинацию конструкций различного дизайна. Конкретно, для оптимизации упаковки AZ8 мутации в полости близ K409-F405A-K392 оценивали независимо от мутаций, которые оптимизируют упаковку ядра в остатках L366T-L368. Эти две области 366-368 и 409-405-392 расположены далеко друг от друга и полагаются независимыми. Например, Fc вариант AZ33 оптимизировали по упаковке в остатках 409-405-392, но не в 366-368, так как эти "оптимизирующие мутации" оценивали отдельно. Сравнение мутаций в положениях 366-368 показывает, что вариант T366L имеет повышенную устойчивость по сравнению с Т366, а также T366I, точковой мутацией, использовавшейся при разработке Fc варианта AZ33. Соответственно, экспериментальные данные в настоящей заявке прямо подсказывали, что, например, нужна дополнительная оптимизация AZ33 путем введения T366L вместо T366I. Поэтому аминокислотные мутации в CH3 домене T366L_K392M_T394W/F405A_Y407V в данной заявке называются мутациями "Каркаса 1b".
Аналогичным образом были проанализированы все экспериментальные данные, чтобы определить точковые мутации, которые можно использовать для дальнейшего усовершенствования имеющегося Fc вариантного гетеродимера AZ33. Эти идентифицированные мутации анализировали вышеописанным компьютерным методом, располагали в определенном порядке и получили список дополнительных Fc вариантных гетеродимеров на основе AZ33, приведенный в Таблице 6.
Пример 6: Рациональный дизайн Fc вариантов методами Fc_CH3 инженерии - Каркас 2 (a и b) и создание AZ63-101 и AZ2199-AZ2524
Для повышения устойчивости и чистоты исходного Fc варианта, полученного в фазе негативного дизайна, AZ15, были применены описанные выше структурные и вычислительные (компьютерные) методы (См. Фигуру 24). Например, тщательный структурно-функциональный анализ Fc варианта AZ15 дал возможность подробно изучить и осмыслить каждую из вводимых мутаций AZ15, L351Y_Y407A/E357L_T366A_K409F_T411N по сравнению с дикого типа (WT) человеческим IgG1 и показал, что важными мутациями ядерного гетеродимера были мутации L351Y_Y407A/T366A_K409F, тогда как мутации E357L, T411N не были достаточно значимы для образования и устойчивости гетеродимера. Мутации ядра (L351Y_Y407A/T366A_K409F) в данной заявке называются мутациями "Каркаса 2". Кроме того, анализ выявил, что важными гипервариабельными участками в области контакта, которые утрачиваются по сравнению с образованием дикого типа (WT) гомодимера, являются: солевой мостик D399-K409, водородная связь Y407-T366 и упаковка Y407-Y407. В приведенном ниже нашем подробном анализе описано, как мы усовершенствовали устойчивость нашего первоначального Fc варианта AZ15, и положения и аминокислотные модификации, осуществленные с целью получить эти Fc варианты с повышенной устойчивостью.
Усовершенствование Fc вариантов с помощью мутаций в Каркасе 2 и дальнейшая разработка мутаций Каркаса 2а.
Наш in silico анализ показал неоптимальную упаковку мутаций Fc варианта AZ15 K409F_T366A_Y407A и общую менее плотную упаковку гидрофобного ядра вследствие утраты взаимодействий WT-Y407-Y407. Усилия позитивного дизайна в следующей фазе инженерии были сосредоточены на точковых мутациях с целью компенсировать эти недостатки упаковки в исходном Fc варианте AZ15. Целевые остатки включали остатки в положениях Т366, L351 и Y407. Различные их комбинации были проверены на компьютерной модели (in silico) и наилучшие полученные компьютерными методами Fc варианты (AZ63-AZ70) оценивали экспериментально с точки зрения экспрессии и устойчивости, как описано в Примерах 1-4.
Fc вариант AZ70 является примером усовершенствования Fc варианта, в котором Каркас 2 был модифицирован, в результате получены мутации Каркаса 2а с повышенной устойчивостью и чистотой. Этот Fc вариант был создан на основе AZ15 с целью получить более плотную упаковку в гидрофобном ядре как описано выше. Fc вариант AZ70 содержит те же самые мутации в ядре Каркаса 2 (L351Y_Y407A/T366A_K409F), описанные выше, за исключением того, что остаток Т366 мутировал в T366V вместо Т366А (Фигура 33). Мутация L351Y повысила температуру плавления варианта 366A_409F/407A с 71.5°С до 74°С, а дополнительная замена 366А на 366V повысила Tm до 75.5°С.(См. AZ63, AZ64 и AZ70 в Таблице 4, с Tm 71.5°С, 74°С и 75.5°С, соответственно). Мутации в ядре (L351Y_Y407A/T366V_K409F) в данной заявке называются мутациями "Каркаса 2а". Экспериментальные данные для Fc варианта AZ70 показали значительно повышенную устойчивость по сравнению с исходным Fc вариантом AZ15, полученным в результате негативного дизайна (Tm 71°С), AZ70 имеет Tm of 75.5°C, а содержание гетеродимера составляет >90% (Фигуры 33 и 27).
Усовершенствование Fc вариантов с помощью мутаций Каркаса 2 и дальнейшее усовершенствование мутаций Каркаса 2b.
Моделирование методами молекулярной динамики (MD) и анализ упаковки продемонстрировали предпочтительно более ′открытую′ конформацию петли 399-400, возникшую, по-видимому, в результате потери WT солевого мостика K409-D399. Результатом этого является также ненасыщенный D399, который, в свою очередь, предпочел компенсирующее взаимодействие с K392, что и обусловило более ′открытую′ конформацию петли. Эта более ′открытая′ конформация петли имеет следствием общую более рыхлую упаковку и повышенную доступность для растворителя остатков ядра домена CH3 в области контакта, что, в свою очередь, в значительной степени дестабилизировало комплекс гетеродимера. По этой причине одной из целенаправленных мер позитивного дизайна было ограничение подвижности этой петли в более ′закрытой′, WT-подобной конформации с помощью дополнительных точковых мутаций, которые компенсируют утрату солевого мостика D399-K409 и взаимодействий упаковки остатка K409. Целевые остатки включали остатки в положениях Т411, D399, S400, F405, N390, K392 и их комбинации. Стратегии позитивного дизайна по инженерии различной упаковки, гидрофобных и электростатических взаимодействий проверяли in silico в отношении вышеуказанных положений и наилучшие Fc варианты, определенные компьютерным моделированием (AZ71-AZ101), определяли экспериментально с точки зрения экспрессии и устойчивости, как описано в Примерах 1-4.
Fc вариант AZ94 является примером усовершенствования Fc варианта, в котором Каркас 2 модифицировали, что привело к повышению устойчивости и чистоты в результате мутаций Каркаса 2b наряду с дополнительными точковыми мутациями. Этот Fc вариант был создан на базе AZ15 с целью ограничить эту петлю в более ′закрытой′ WT-подобной конформации и компенсировать утрату солевого мостика D399-K409 как описано выше. Fc вариант AZ94 содержит четыре дополнительных точковых мутации в Каркасе 2 (L351Y_Y407A/T366A_K409F) и возвращает L351Y в дикого типа L351, оставляя (Y407A/T366A_K409F) как мутации в ядре для этого Fc варианта. Мутации ядра Y407A/T366A_K409F в данной заявке называются мутациями "Каркаса 2b". Четырьмя дополнительными точковыми мутациями AZ94 являются K392E_Т411Е/D399R_S400R. Мутации Т411Е/D399R осуществляли с целью создать дополнительный солевой мостик и компенсировать утрату взаимодействия K409/D399 (Фигура 34). Кроме того, этот солевой мостик был образован с целью предупредить образование гомодимера, сделав невыгодными взаимодействий заряд-заряд в обоих потенциальных гомодимерах. Предполагалось, что дополнительные мутации K392E/S400R образуют другой солевой мостик и, следовательно, дополнительно ограничат петлю 399_400 в более ′закрытой′, WT-подобной конформации (Фигура 34). Экспериментальные данные показали повышенную устойчивость и чистоту AZ94 по сравнению с Fc вариантом AZ15 (Tm 71°С, чистота >90%), Fc variant AZ94 имеет Tm 74°С, а содержание гетеродимера или чистота составляет >95%.
Усовершенствование Fc вариантов с использованием мутаций в Каркасе 2 в фазе три дизайна Fc вариантных гетеродимеров
В случае обеих Fc вариантов, AZ70 и AZ94, наблюдается значительное повышение устойчивости и чистоты по сравнению с исходным Fc вариантом AZ15, полученным в результате негативного дизайна, но наш анализ и сравнение AZ70 и AZ94 прямо указывает, что дальнейшее повышение устойчивости Fc вариантного гетеродимера можно осуществить с помощью дополнительных модификаций аминокислот. Например, Fc варианты AZ70 и AZ94 были созданы в результате нацеливания на две различных неоптимизированные области в исходном варианте AZ15, усовершенствование осуществляли путем уплотнения упаковки в гидрофобном ядре и за счет мутаций вне области контакта остатков ядра, что привело в результате к образованию дополнительного солевого мостика и водородной связи и стабилизации конформации петли в положениях 399-401. Дополнительные точковые мутации Fc вариантов AZ70 и AZ94 расположены далеко друг от друга и, следовательно, являются независимыми и взаимозаменяемыми с другими Fc вариантами, усовершенствованными с помощью тех же самых мутаций в ядре Каркаса 2, включая мутации 2а и 2b. Конкретно, AZ70 несет только оптимизированные мутации в ядре L351Y_Y407A/T366A_K409F, но не несет никаких солевых мостиков, тогда как AZ94 несет четыре дополнительных мутации, участвующие в электростатических взаимодействиях, (K392E_Т411Е/D399R_S400R), но содержит на одну мутацию меньше в области контакта гидрофобного ядра (Y407A/T366A_K409F). Эти мутации Каркаса 2b сообщают более низкую устойчивость, чем устойчивость AZ70 (см. например AZ63, который содержит мутации в ядре, эквивалентные мутациям в AZ94, и Tm 72°С), но компенсируются добавлением мутаций K392E_Т411Е/D399R_S400R. Представленные экспериментальные характеристики устойчивости и чистоты показывают, что сочетание мутаций AZ70, которые оптимизируют гидрофобное ядро, и мутаций, участвующих в электростатических взаимодействиях (вариант AZ94), должны еще более повысить устойчивость и чистоту Fc вариантных гетеродимеров. Аналогичным образом были проанализированы все экспериментальные данные для Fc вариантов Каркаса 2 (AZ63-101) с целью проанализировать точковые мутации, которые можно использовать для дальнейшего усовершенствования Fc вариантных гетеродимеров AZ70 и AZ94. Эти идентифицированные мутации были дополнительно проанализированы вышеописанным компьютерным методом и классифицированы, в результате в Таблице 7 представлен список дополнительных Fc вариантных гетеродимеров на основе AZ70 и AZ94.
Пример 7: Влияние гетеродимерного CH3 на связывание Fc гамма R (FcgR)
В качестве типового примера активности гетеродимерного Fc по отношению к FcgR мы проверили два варианта антител с гетеродимерной Fc областью A:K409D_K392D/B:D399K_D356K (Контроль 1 (het 1 на Фигуре 35)) и A:Y349C_T366S_L368A_Y407V/B:S354C_T366W (Контроль 4 (het 2 на Фигуре 35)) с помощью связывания Her2 с Fab фрагментами (плечами) в анализе SPR, описанном в Примере 4 для связывания FcgR. Как показано на Фигуре 35, мы нашли, что обе гетеродимерных Fc области связывают различные Fc гамма рецепторы с той же самой относительной прочностью, что и дикого типа IgG1 Fc область, но суммарно гетеродимерная Fc область связывала каждый из FcgR несколько прочнее, чем антитело дикого типа. Это показывает, что мутации в CH3 области контакта Fc могут влиять на прочность связывания Fc области с Fc гамма рецепторами по всем СН2 доменам, как мы наблюдали при моделировании методом молекулярной динамики и с помощью анализов.
Пример 8: Влияние асимметричных мутаций в СН2 домене гетеродимерной области Fc на связывание FcgR.
Известно, что мутация серина в положении 267 домена СН2 области Fc на аспарагиновую кислоту (S267D) усиливает связывание с Fc гамма IIbF, IIbY и IIaR рецепторами при введении в две цепи СН2 домена гомодимерной Fc области. Как показывают данные, представленные на Фигуре 36А, эту мутацию можно вводить только в один из СН2 доменов в гетеродимерной Fc молекулы, чтобы выиграть, примерно, половину от повышения прочности связывания, полученного при введении этой мутации в СН2 домен гомодимерной Fc области. С другой стороны, мутация E269K в гомодимерном СН2 домене Fc области препятствует связыванию Fc области с FcgR. Мы представляем схему более тонкого воздействия на прочность связывания Fc области с Fcg рецепторами с помощью асимметричного введения этих выгодных и невыгодных мутаций в одну из двух цепей СН2 домена Fc области. Асимметричное введение мутации E269K в одну цепь СН2 домена в гетеродимерной Fc области действует как фактор, определяющий полярность, блокируя связывание FcgR с той стороны, где эта мутация находится, и в то же время давая возможность другой стороне Fc взаимодействовать с FcgR обычным образом. Результаты этого эксперимента представлены на Фигуре 36А. Возможность селективно изменять прочность связывания посредством обеих цепей Fc, независимо, предоставляет повышенные возможности управлять прочностью и селективностью связывания Fc и Fcg рецепторов. Таким образом, такой асимметричный дизайн мутаций в СН2 домене позволил нам осуществлять стратегии позитивного и негативного дизайна для того, чтобы содействовать или противодействовать определенным моделям связывания, что повышало возможность селективности.
В следующем эксперименте мы изменяли профиль селективности базового Fc мутантного варианта S239D_D265S_I332E_S298A, который продемонстрировал повышенную прочность связывания с Fc гамма IIIaF и IIIaV рецепторами, в то же время сохраняя более слабую способность связывания с Fc гамма IIaR, IIbF и IIbY рецепторами. Это видно на профиле связывания, показанном на Фигуре 36В. Вводя асимметричную мутацию E269K в цепь А и не допуская мутации I332E в цепи В, мы смогли получить новый профиль связывания FcgR, который дополнительно ослабляет связывание IIa и IIb рецепторов и делает Fc более специфическим по отношению к связыванию IIIa рецептора.
В другом примере, показанном на Фигуре 36С, особо отмечены асимметричные мутации по сравнению с гомодимерным Fc вариантом, включающие мутацию S239D/K326E/A330L/I332E/S298A в СН2 домене. По сравнению с дикого типа IgG1 Fc этот вариант продемонстрировал повышенное связывание с IIIa рецептором, но он также связывался с IIa и IIb рецепторами немного сильнее (прочнее), чем дикого типа Fc. Асимметричное введение этих мутаций A:S239D/K326E/A330L/I332E и B:S298A хотя снижало (ослабляло) связывание с IIIa рецептором, также повышало IIa/IIb рецепторное связывание, т.е. процесс утрачивал селективность. Вводя в этот гетеродимерный вариант асимметричную мутацию E269K, а именно. А:S239D/K326E/A330L/I332E/E269K и B:S298A, мы смогли понизить уровни связывания IIa/IIb обратно до уровней дикого типа. Это демонстрирует, что применение асимметричных мутаций в СН2 домене Fc может предоставить большие возможности для достижения повышенной селективности по отношению к Fc гамма R.
Реагенты, применяемые в примерах, имеются в продаже или их можно получить на доступном оборудовании или доступными методами, или реагенты известны в уровне техники. Приведенные выше примеры иллюстрируют различные аспекты изобретения и практическое применение методов по изобретению. Не предполагается, что эти примеры описывают многие отличные варианты изобретения. Так, хотя вышеописанное изобретение описано подробно для наглядности и на примерах для лучшего понимания, специалисты в данной области техники легко поймут, что в нем можно сделать многие изменения и модификации, не отходя от сущности и объема прилагаемой Формулы изобретения.
Все публикации, патенты и патентные заявки, упоминаемые в настоящем описании, вводятся в данную заявку в той степени, как если бы конкретно и индивидуально указывалось, что каждая отдельная публикация, каждый отдельный патент или каждая патентная заявка вводится в данное описание в качестве ссылки.
| название | год | авторы | номер документа |
|---|---|---|---|
| РАЗРАБОТКА СТАБИЛЬНОГО ГЕТЕРОДИМЕРНОГО АНТИТЕЛА С МУТАЦИЯМИ В ДОМЕНЕ Fc | 2012 |
|
RU2810540C2 |
| РАЗРАБОТКА СТАБИЛЬНОГО ГЕТЕРОДИМЕРНОГО АНТИТЕЛА С МУТАЦИЯМИ В ДОМЕНЕ Fc | 2012 |
|
RU2675319C2 |
| БИСПЕЦИФИЧЕСКИЕ АСИММЕТРИЧНЫЕ ГЕТЕРОДИМЕРЫ, СОДЕРЖАЩИЕ АНТИ-CD3 КОНСТРУКЦИИ | 2013 |
|
RU2650868C2 |
| ГЕТЕРОМУЛЬТИМЕРЫ С УМЕНЬШЕННОЙ ИЛИ ПОДАВЛЕННОЙ ЭФФЕКТОРНОЙ ФУНКЦИЕЙ | 2014 |
|
RU2655439C2 |
| СКОНСТРУИРОВАННЫЕ МУЛЬТИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА И ДРУГИЕ МУЛЬТИМЕРНЫЕ БЕЛКИ С АСИММЕТРИЧНЫМИ МУТАЦИЯМИ В ОБЛАСТИ CH2-CH3 | 2018 |
|
RU2804031C2 |
| МОДИФИЦИРОВАННЫЕ АСИММЕТРИЧНЫЕ АНТИТЕЛА, СВЯЗЫВАЮЩИЕ Fc-РЕЦЕПТОР, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2687043C2 |
| ВАРИАНТЫ Fc-ОБЛАСТИ С МОДИФИЦИРОВАННЫМИ СВОЙСТВАМИ СВЯЗЫВАНИЯ FcRn И БЕЛКА А | 2015 |
|
RU2713131C1 |
| ВАРИАНТЫ Fc-ОБЛАСТИ С МОДИФИЦИРОВАННЫМ СВЯЗЫВАНИЕМ FcRn И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2714116C2 |
| ВАРИАНТЫ FC-ОБЛАСТИ С УЛУЧШЕННОЙ СПОСОБНОСТЬЮ СВЯЗЫВАТЬСЯ С БЕЛКОМ А | 2015 |
|
RU2698969C2 |
| МОДИФИЦИРОВАННЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ ПОЛИПЕПТИДНЫЕ КОНСТРУКЦИИ И ИХ ПРИМЕНЕНИЕ | 2015 |
|
RU2832921C1 |







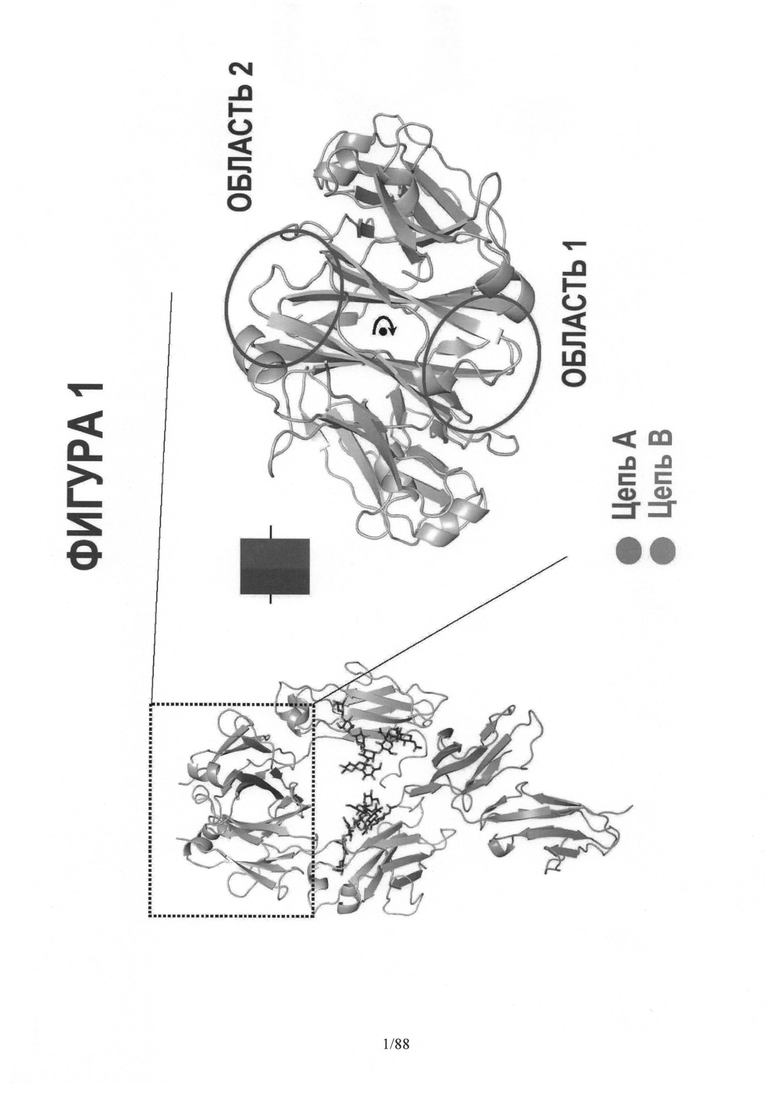

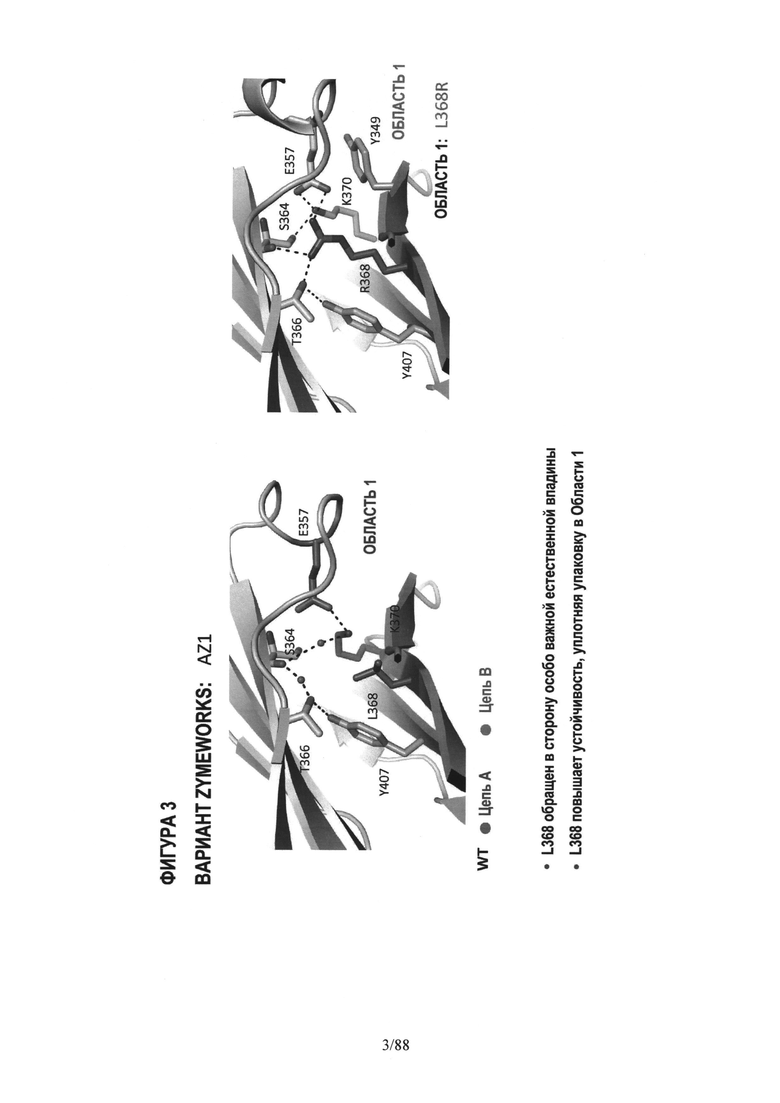
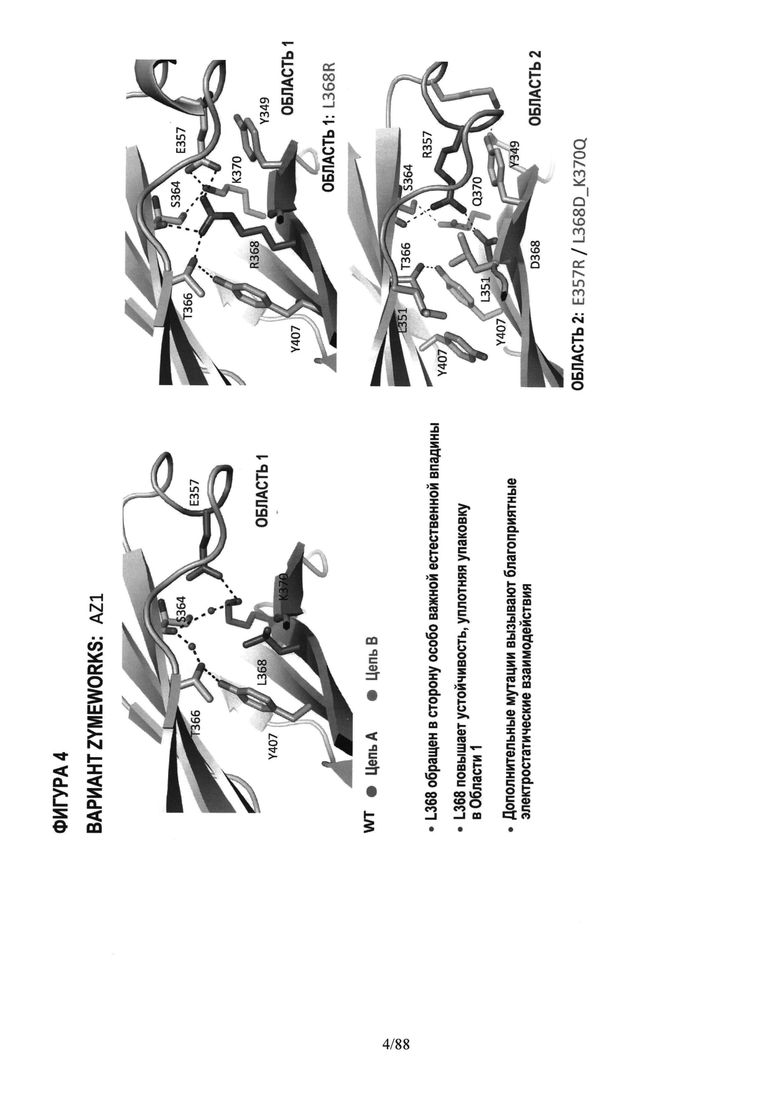
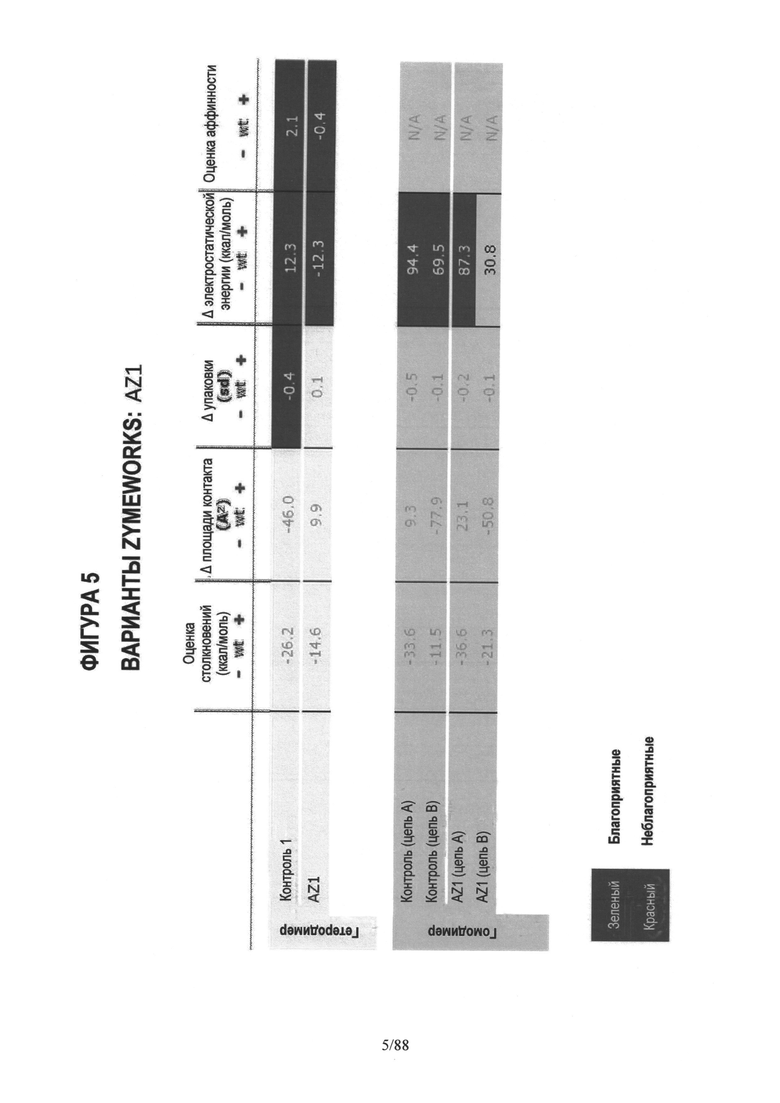




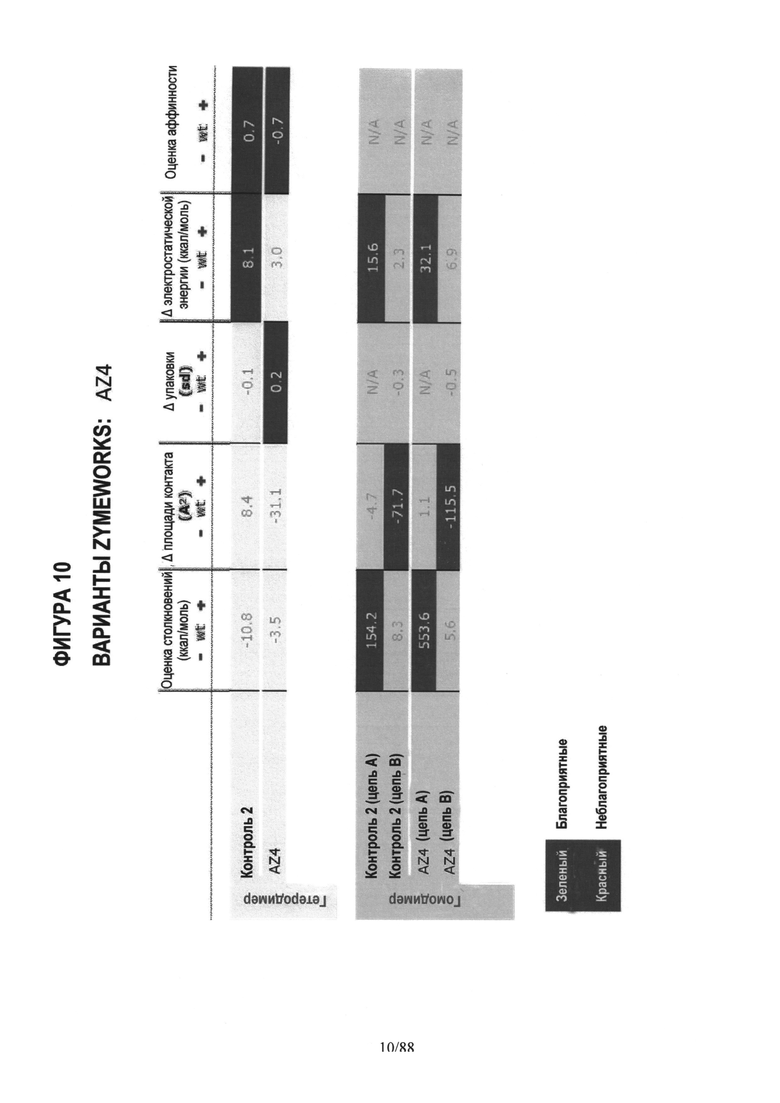
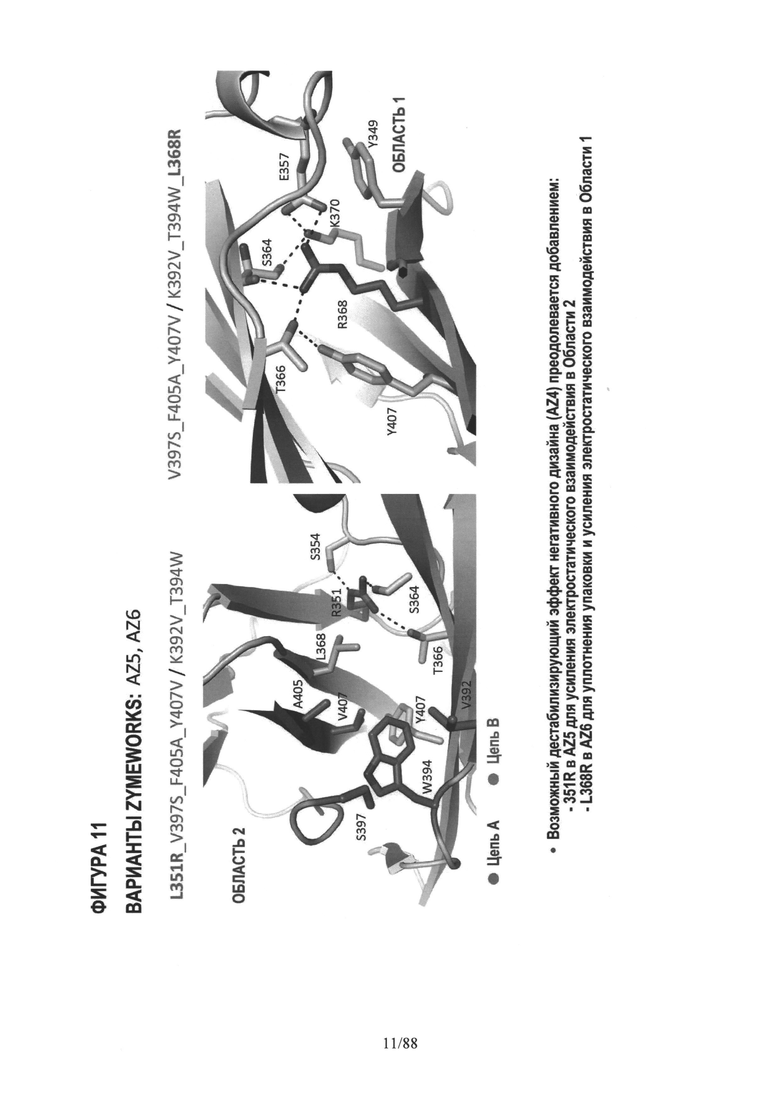
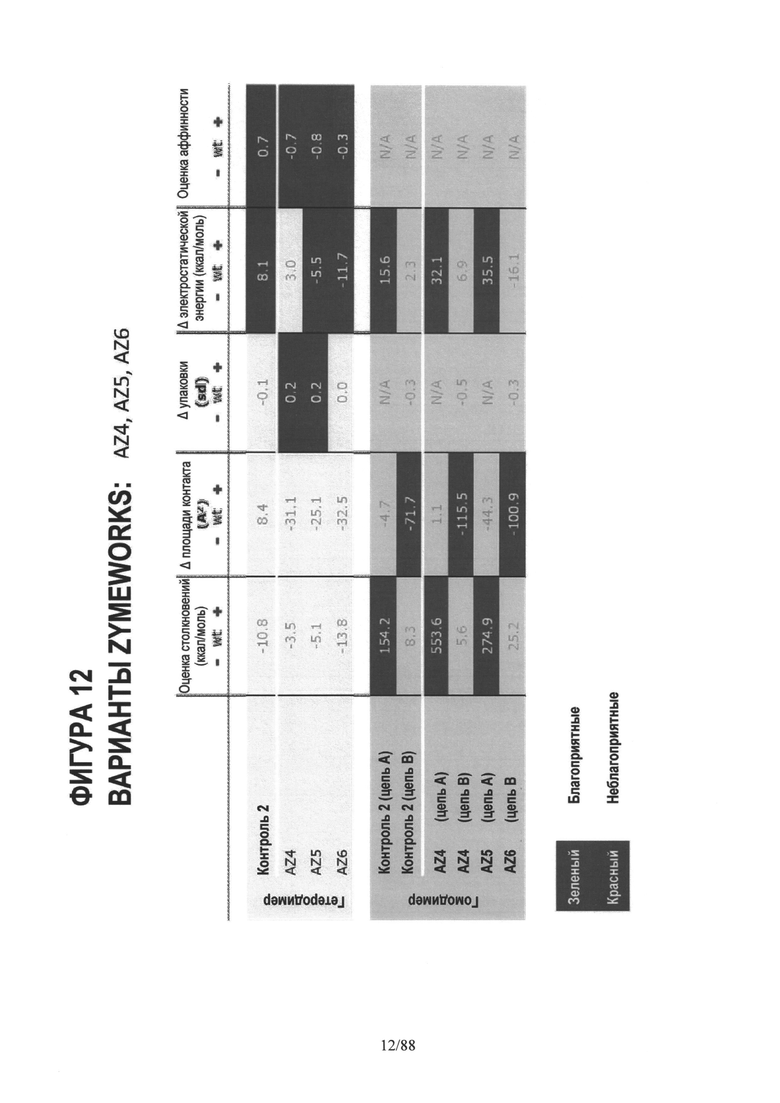
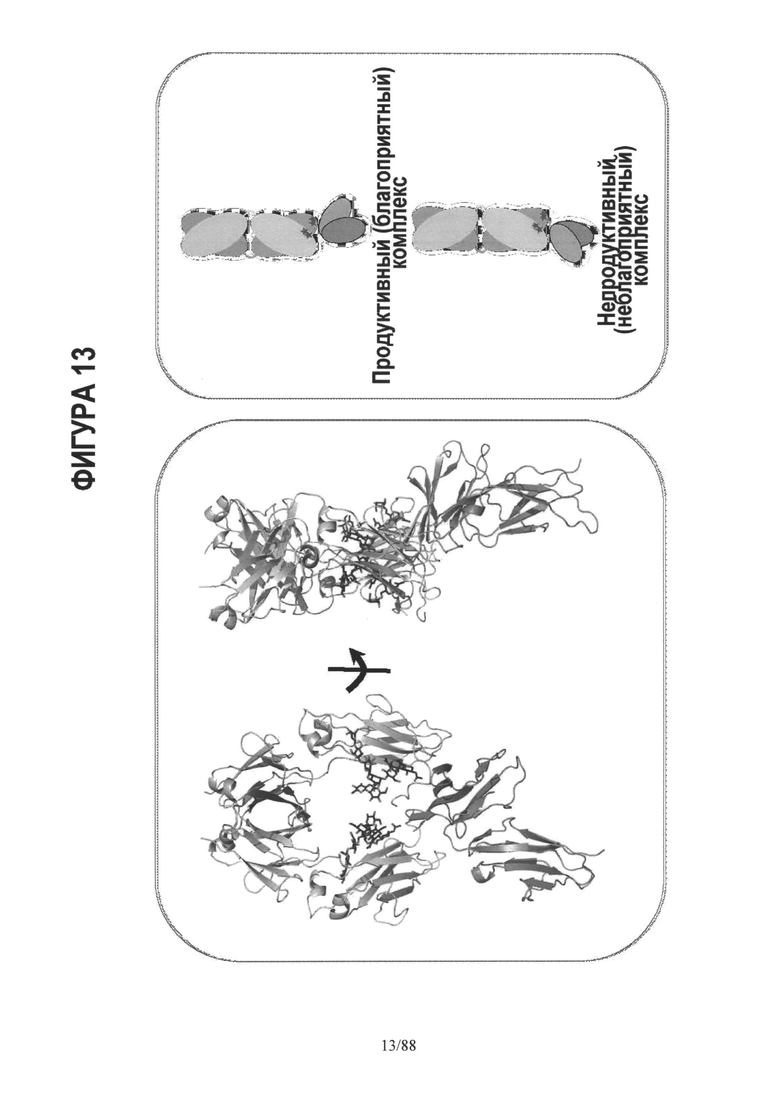
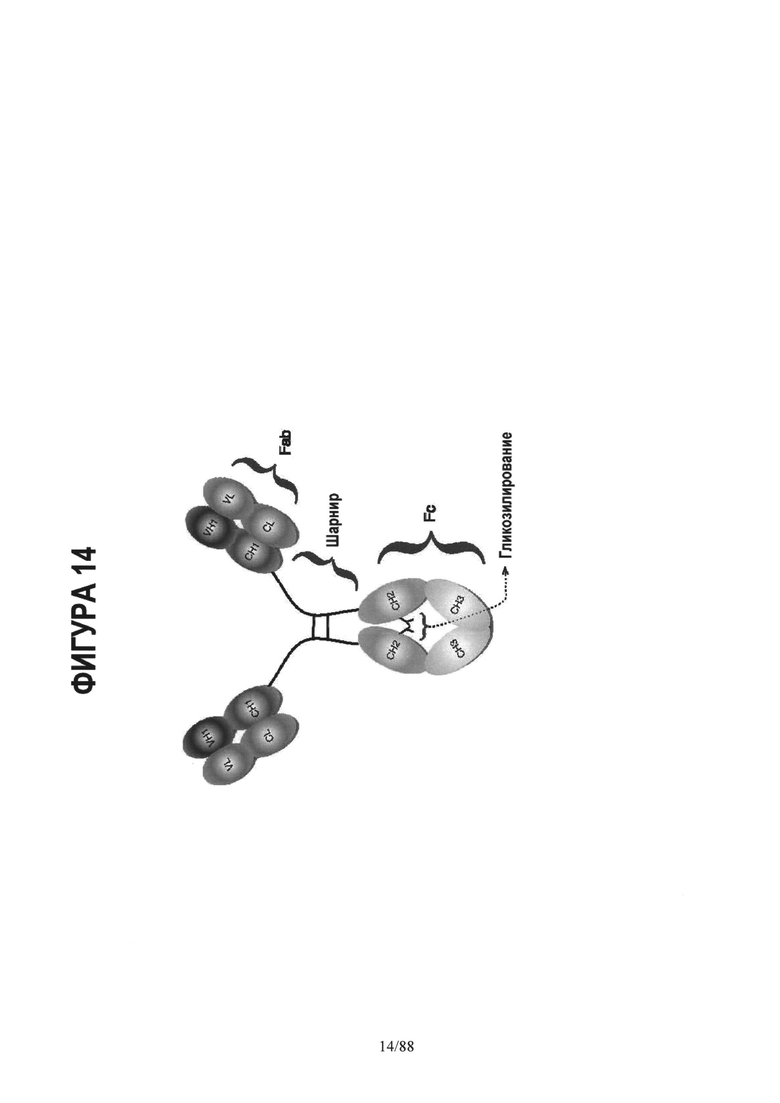


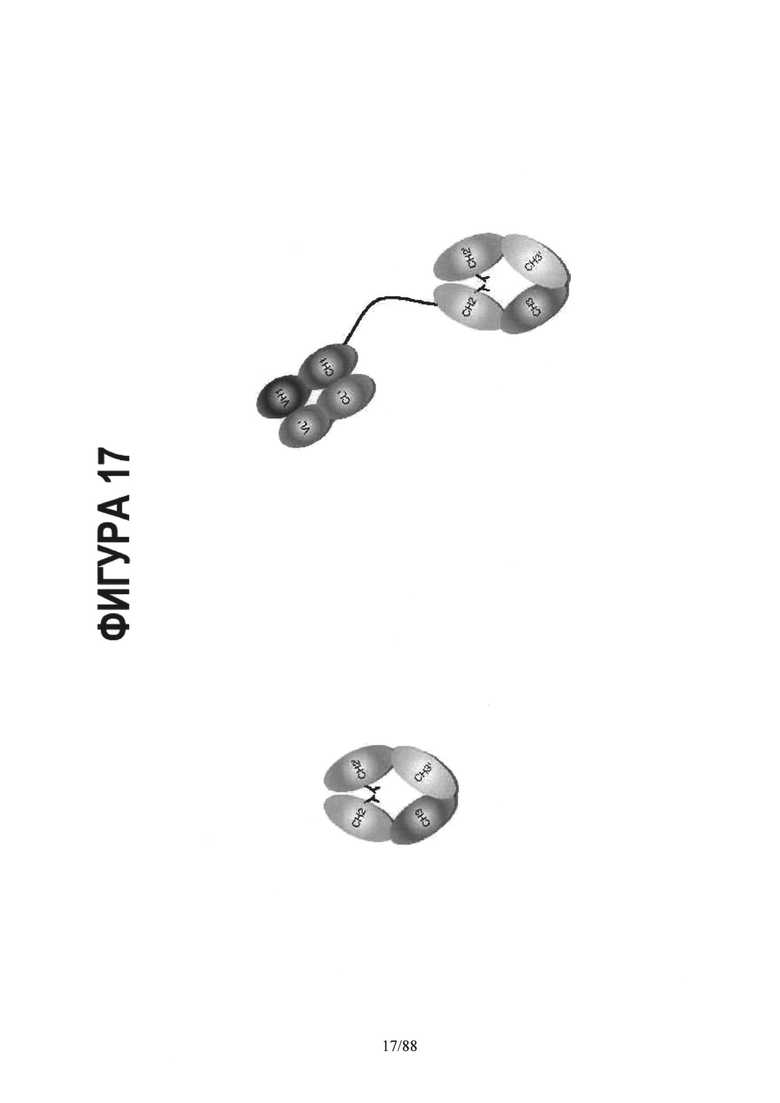
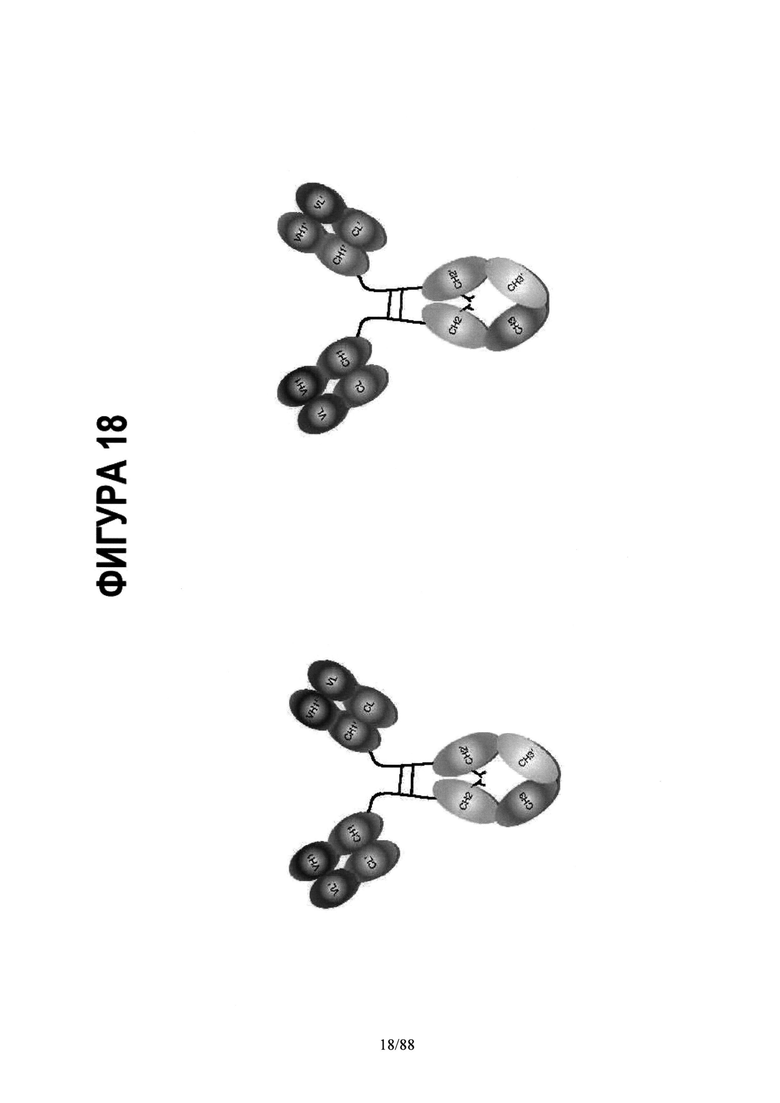
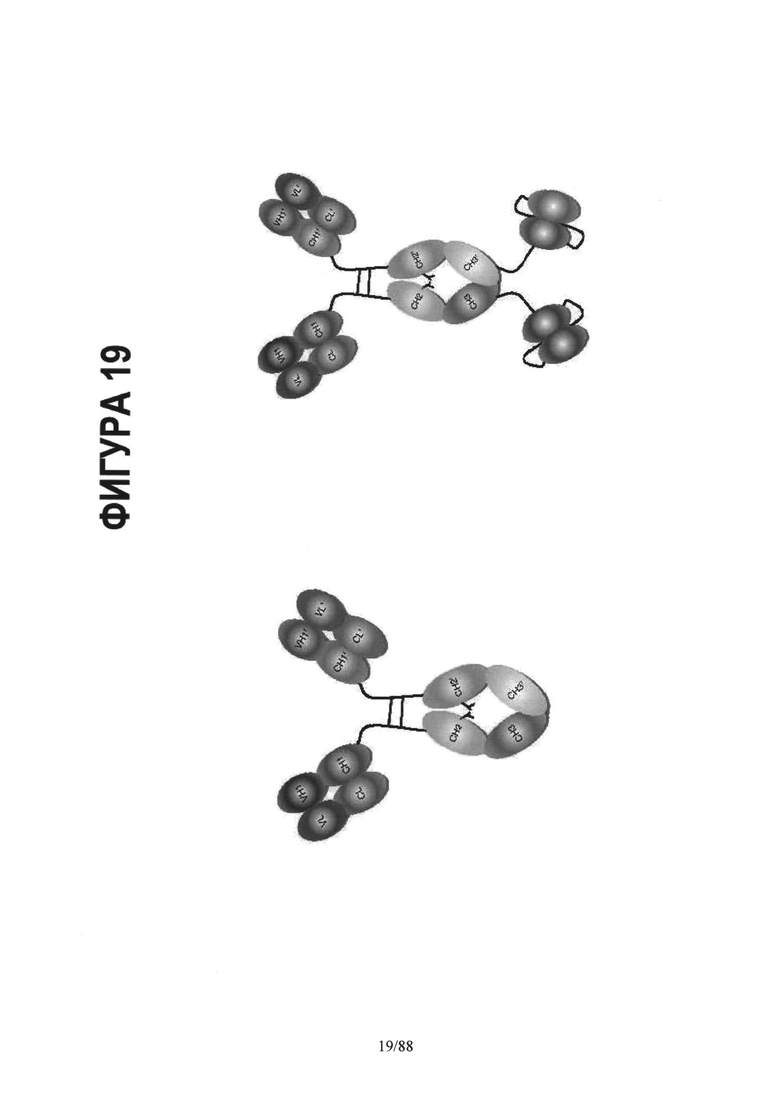
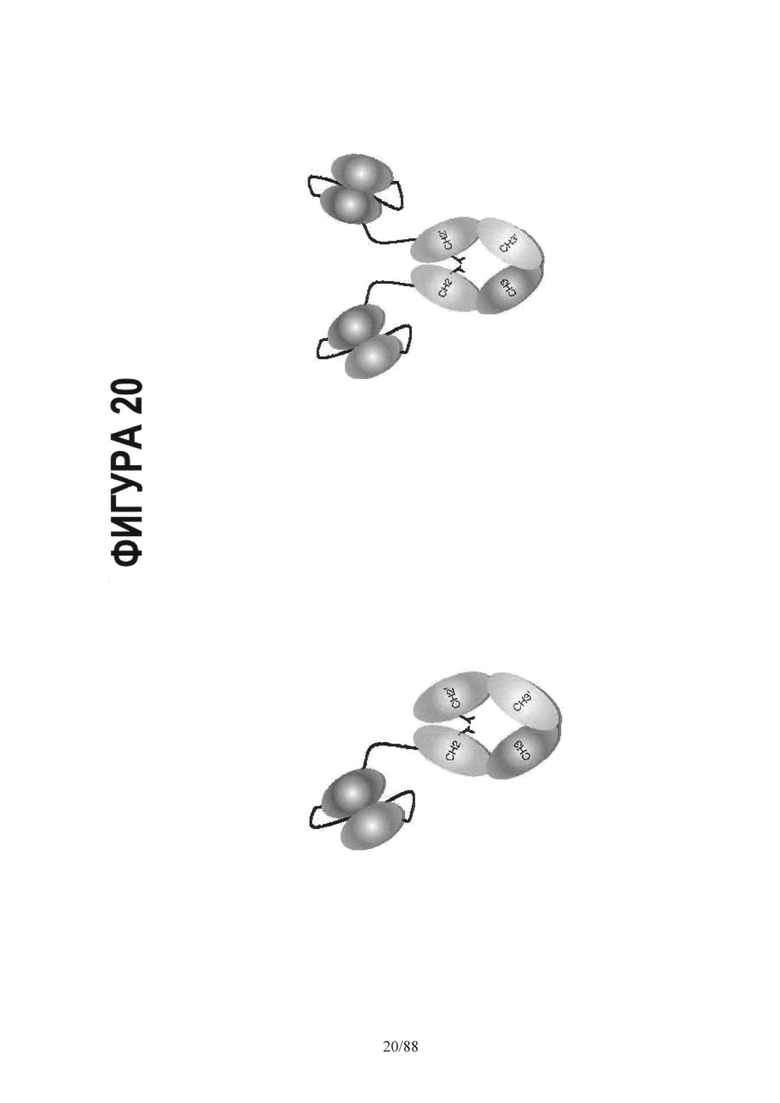

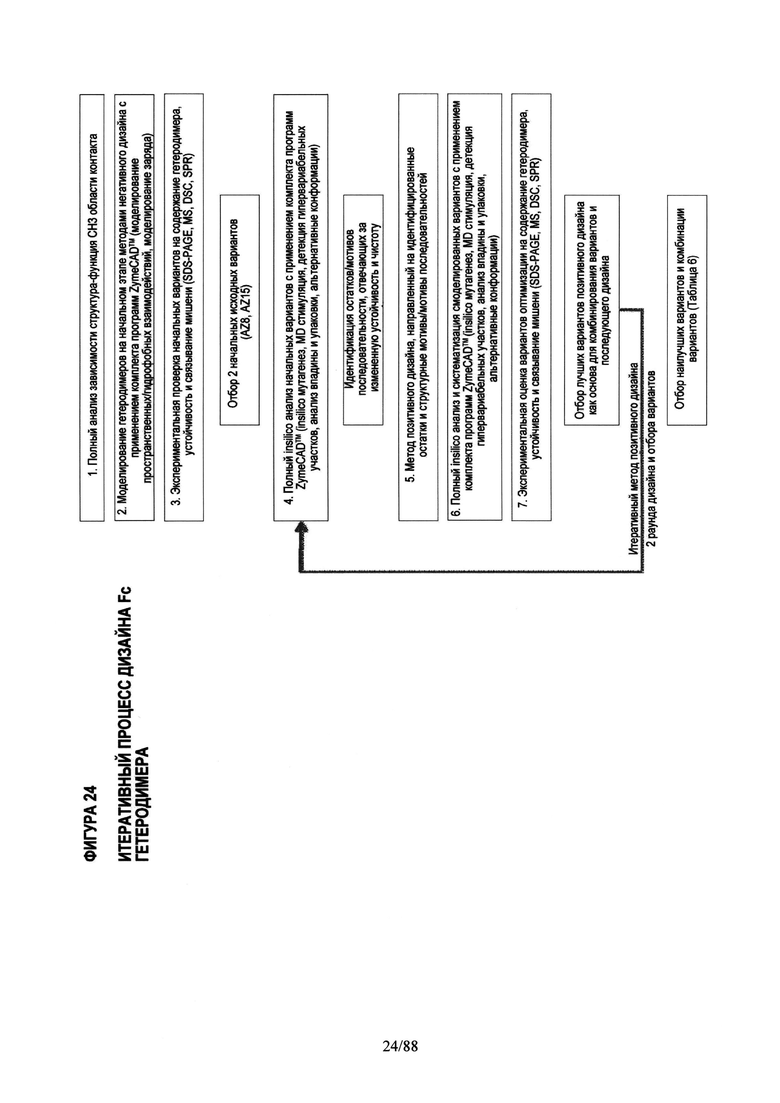
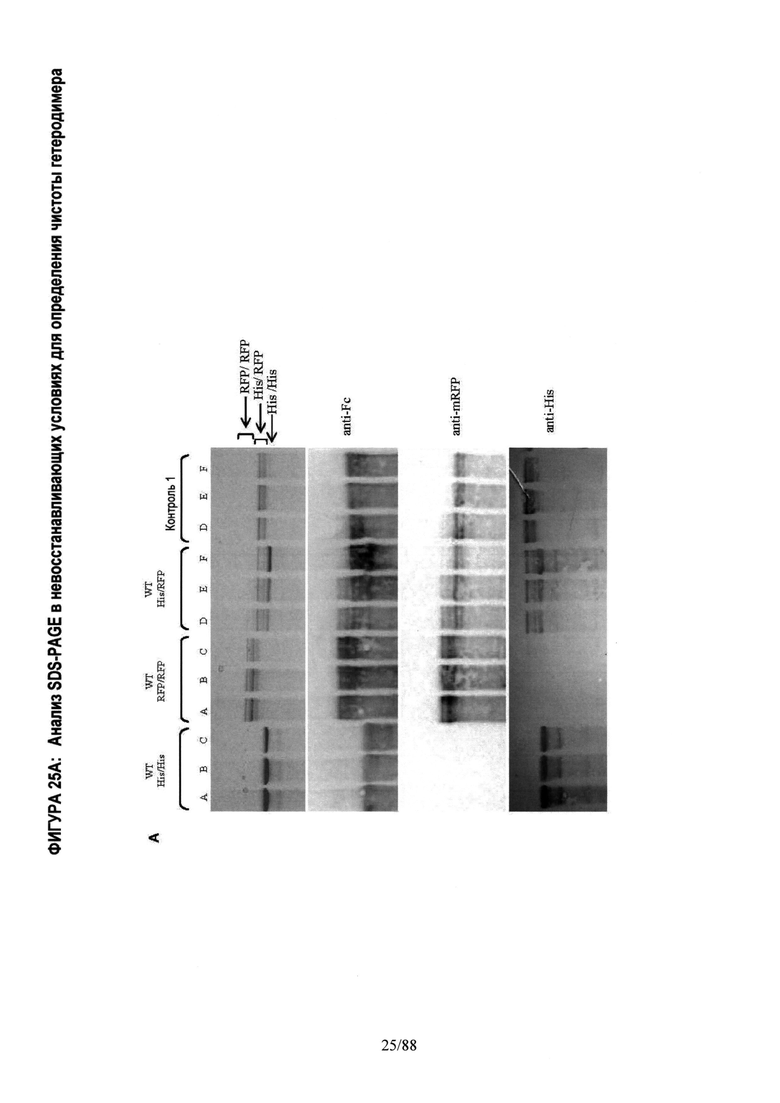
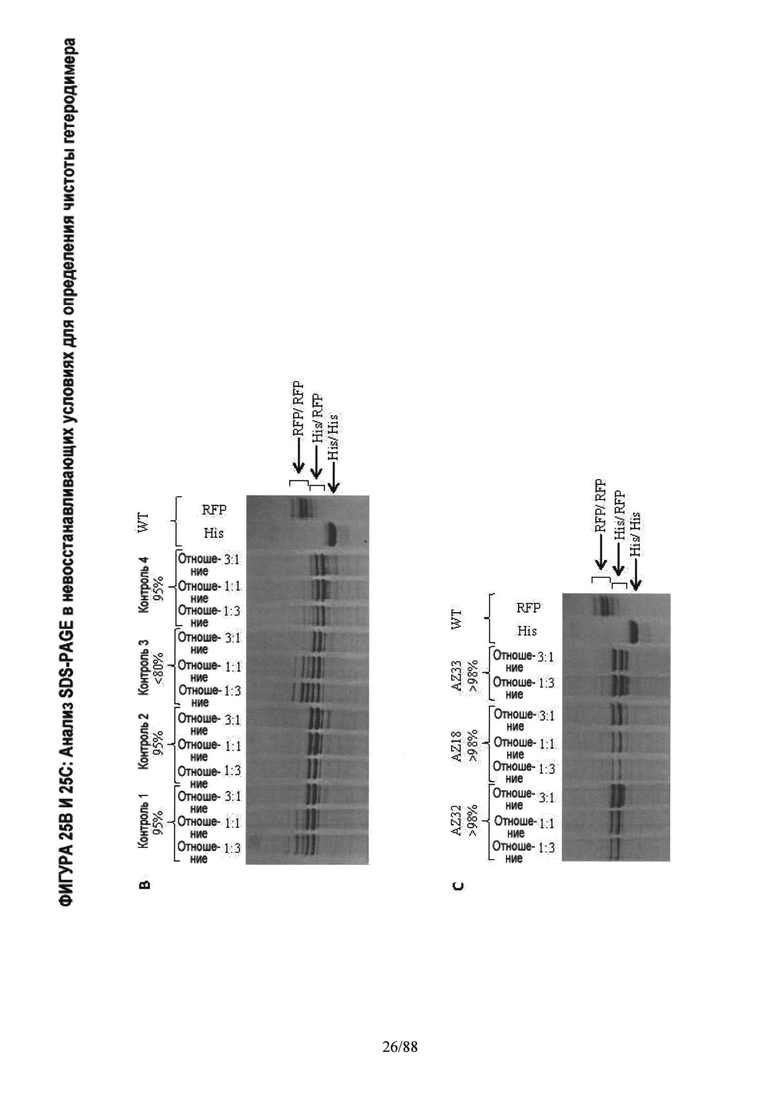
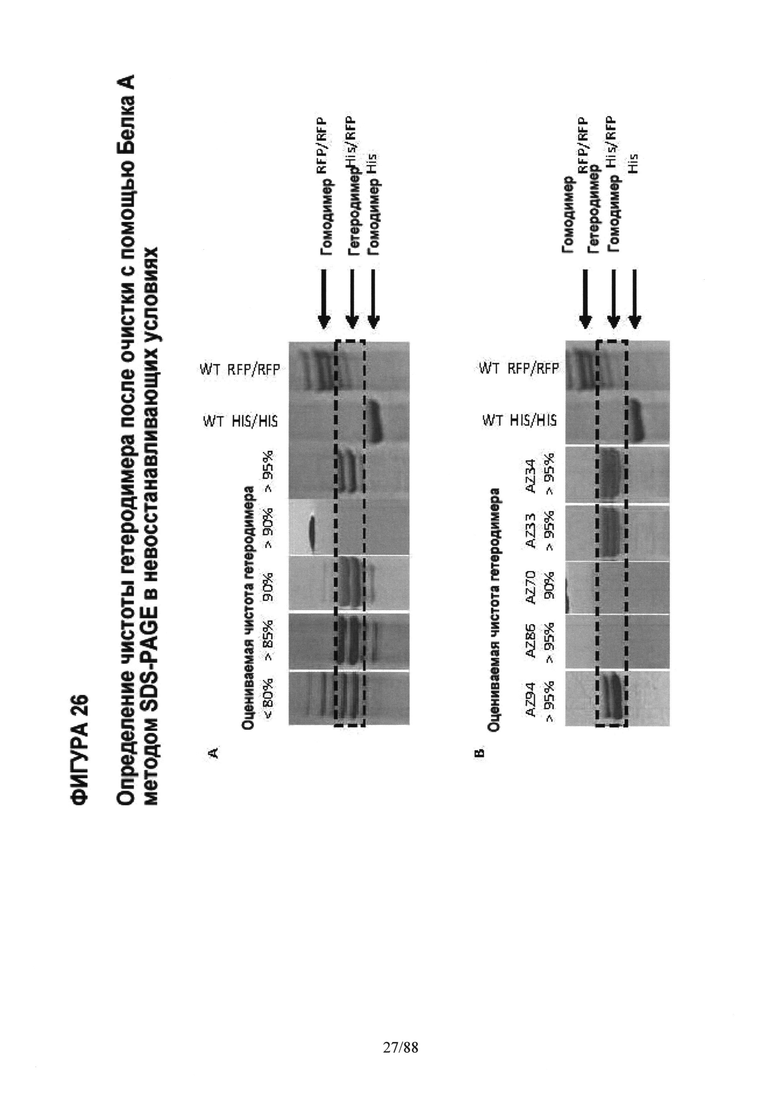
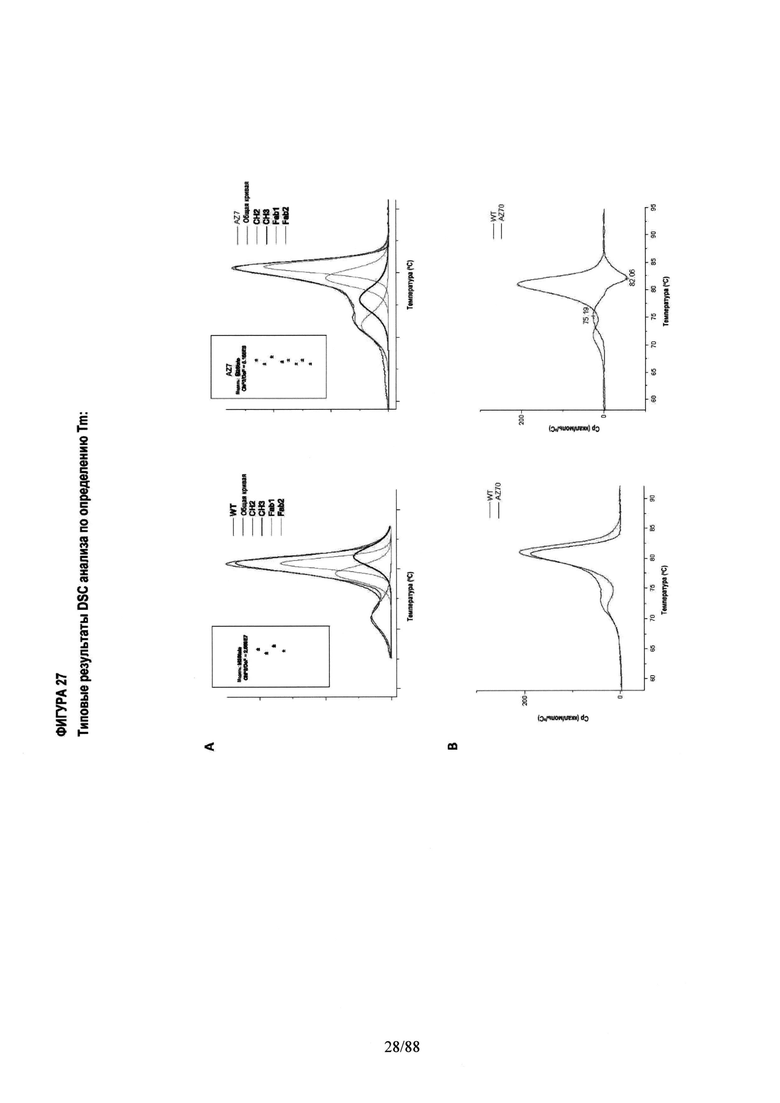




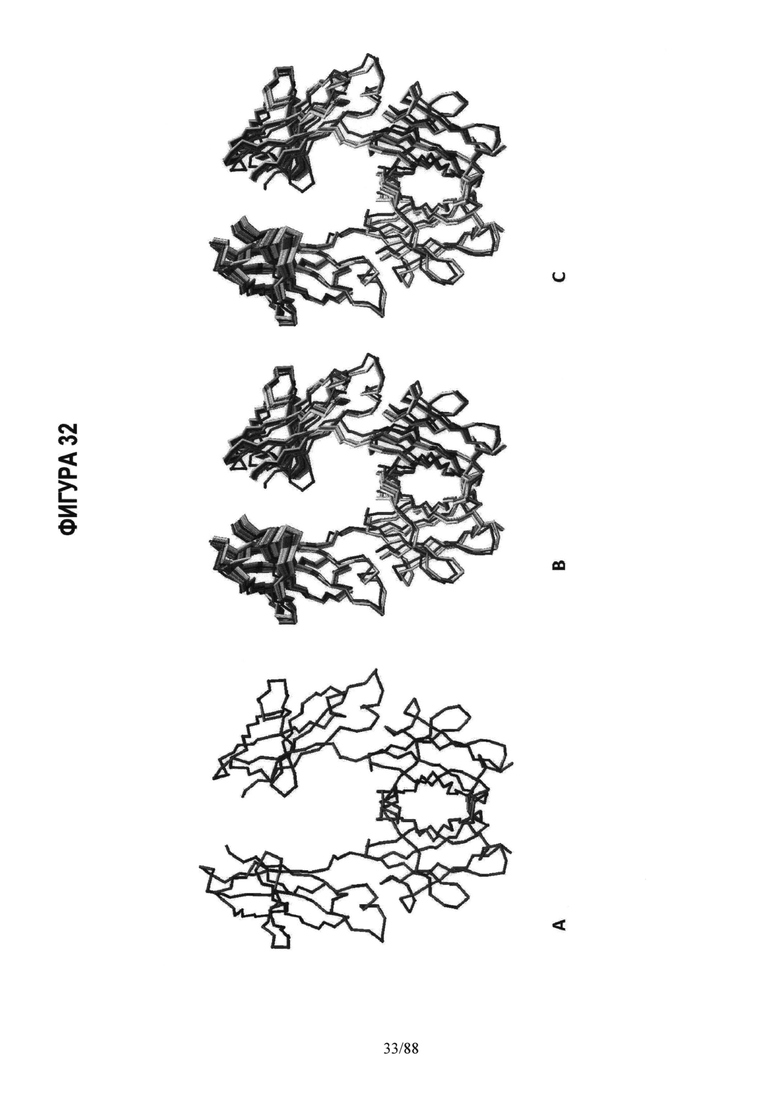
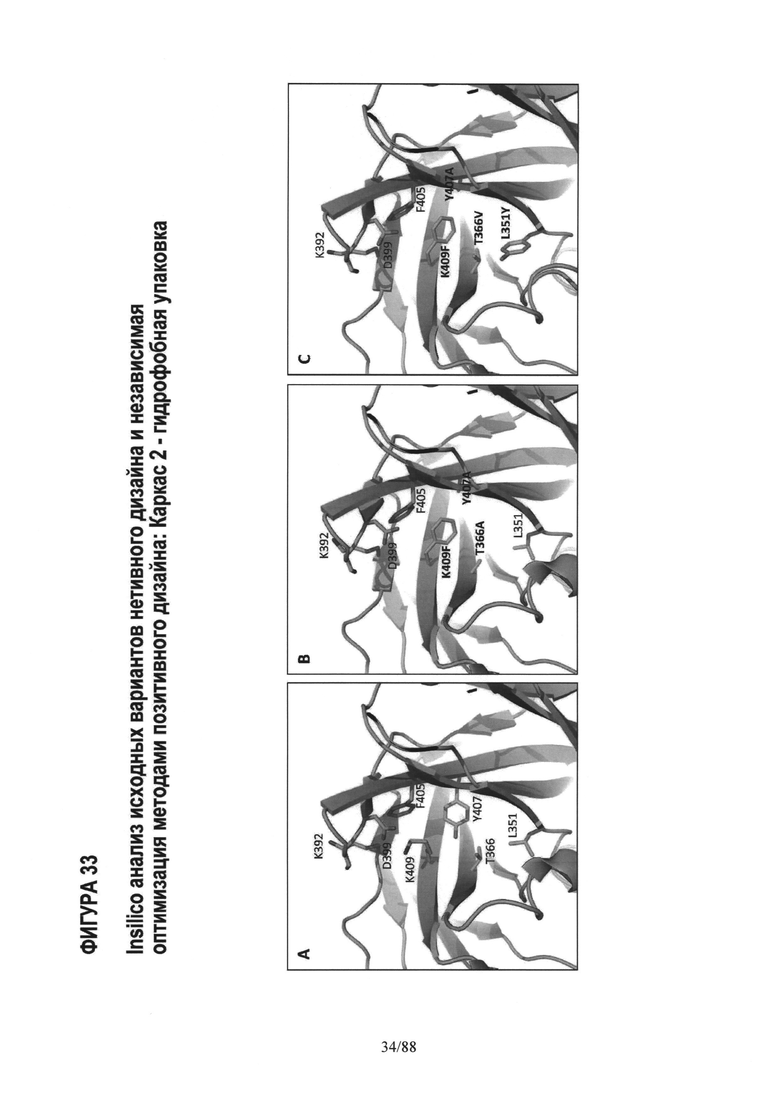
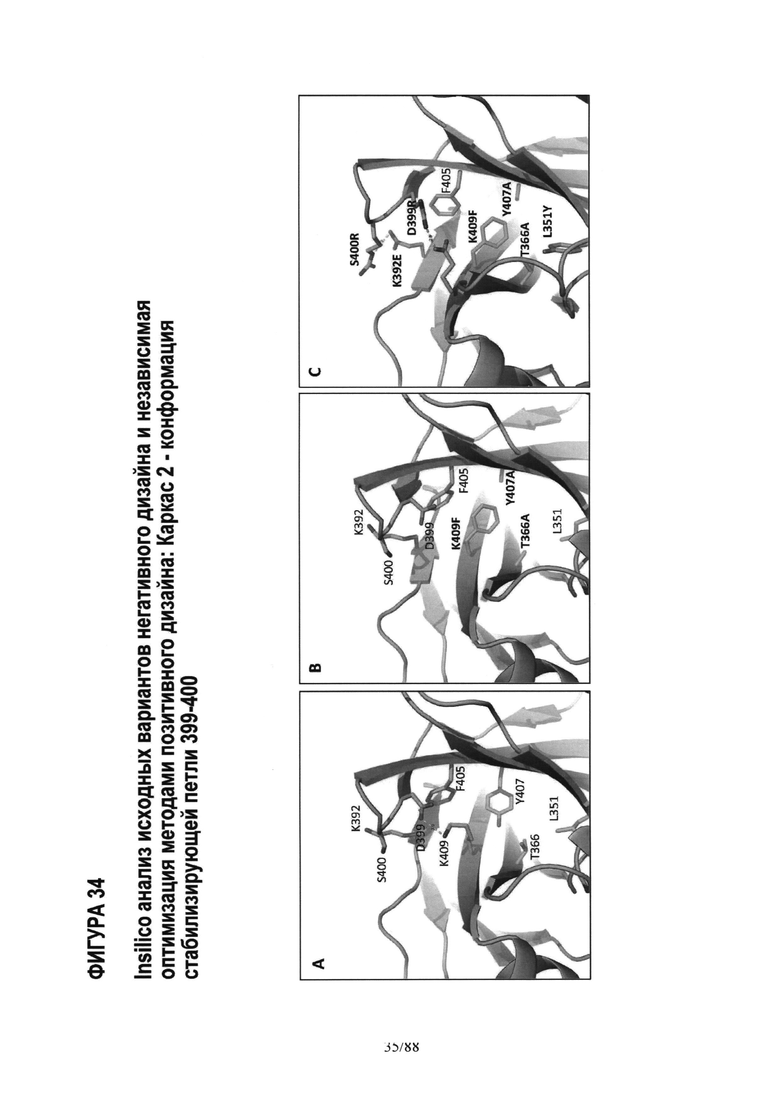
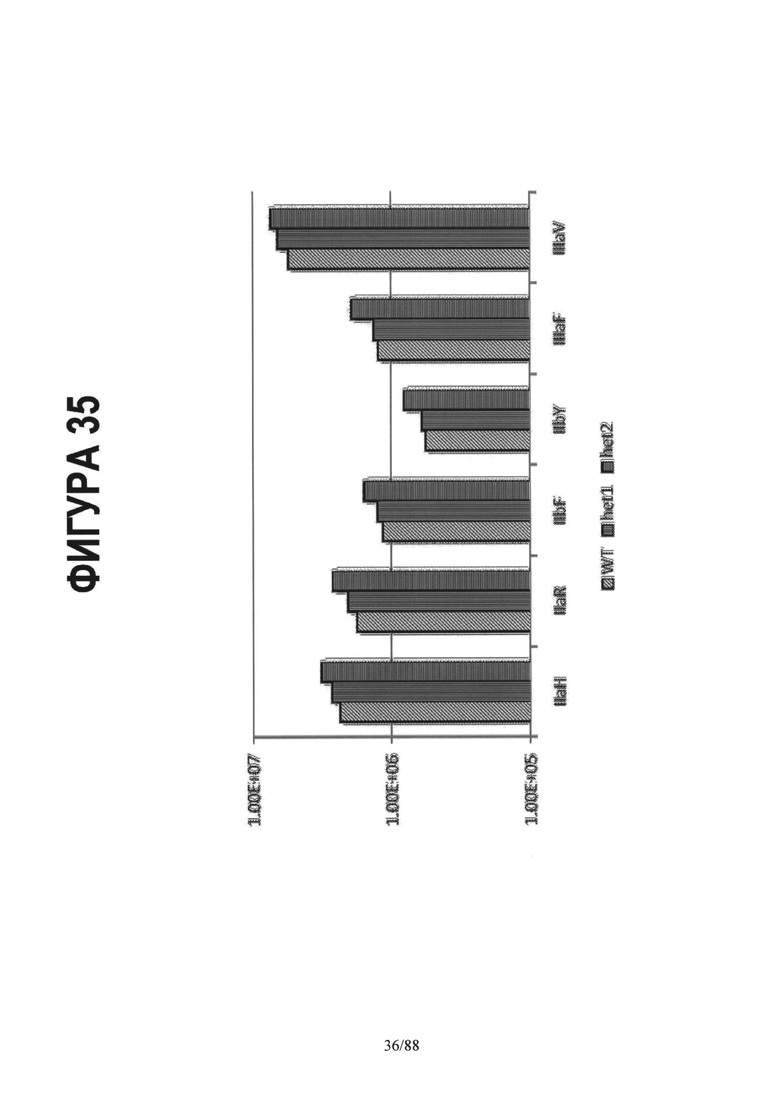
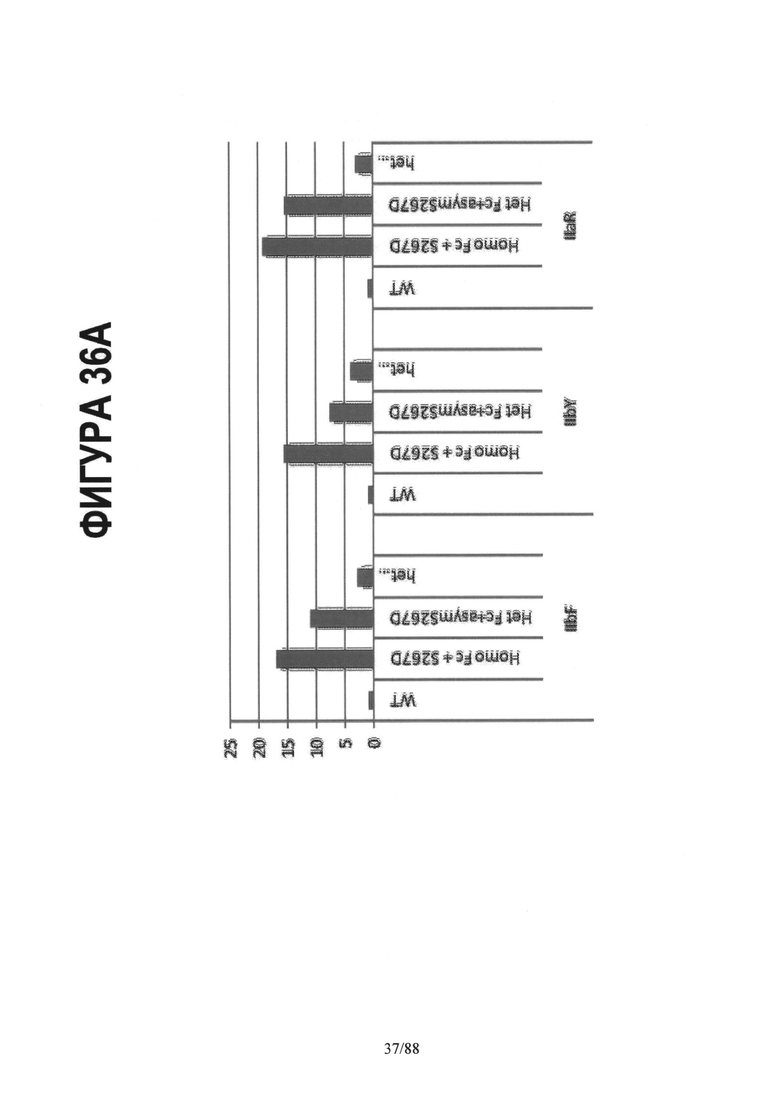
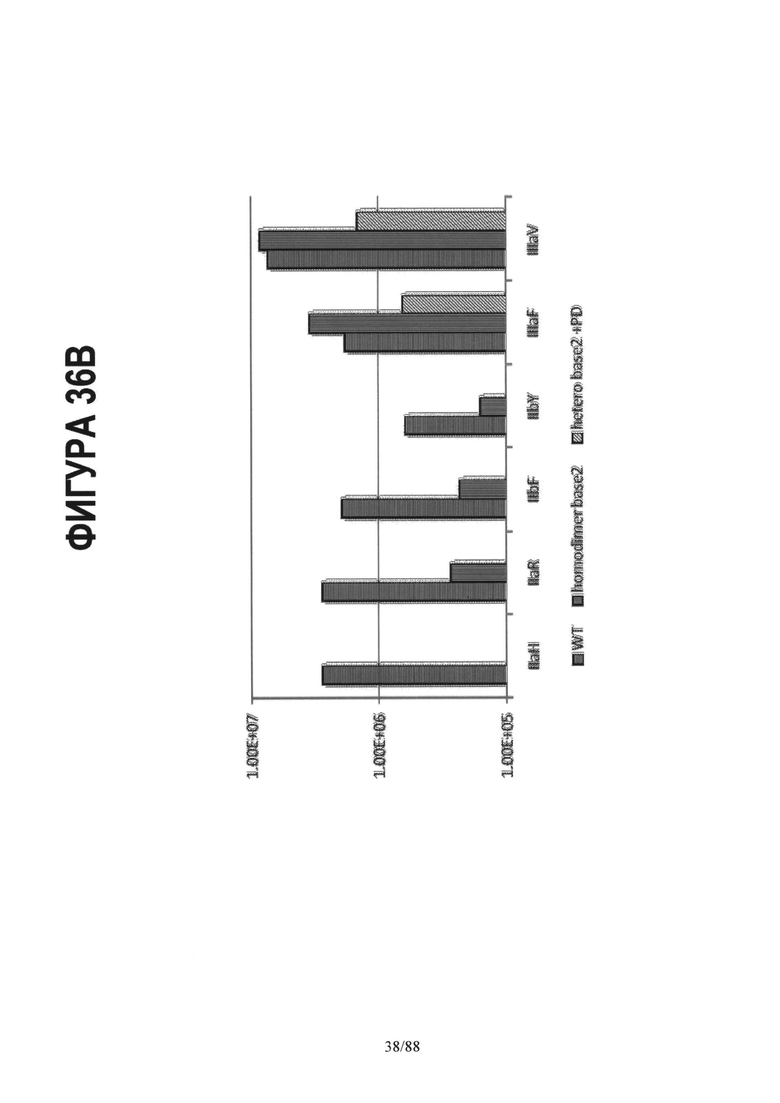
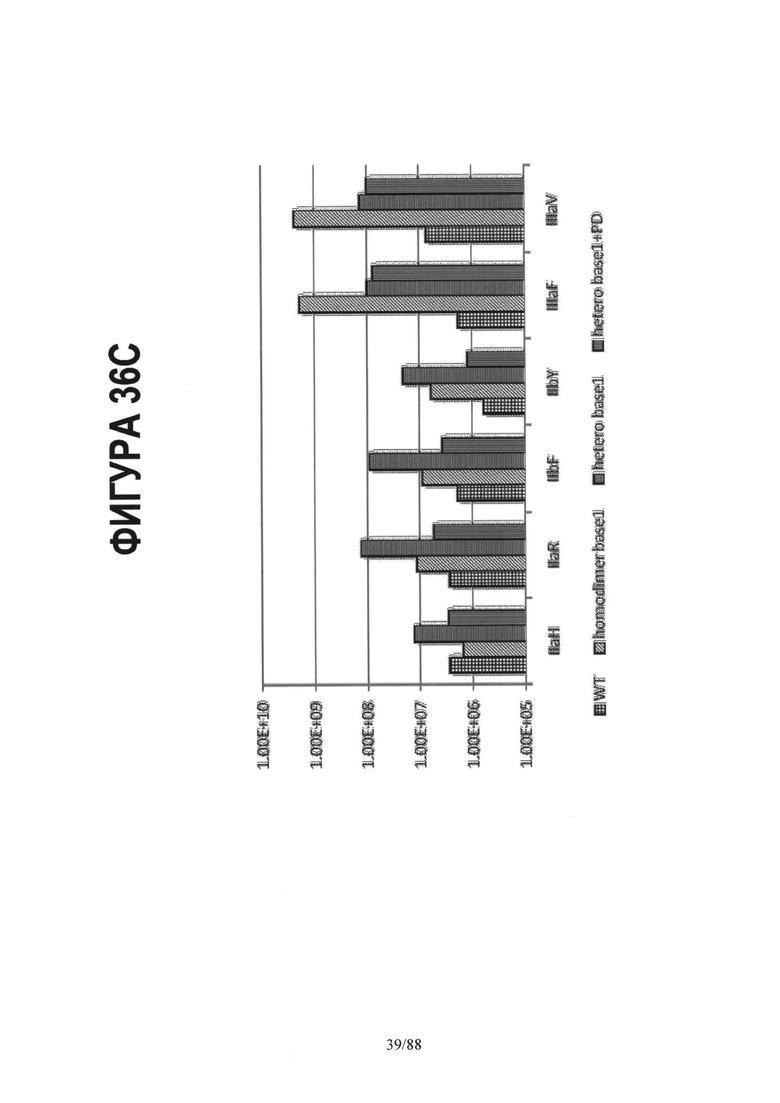
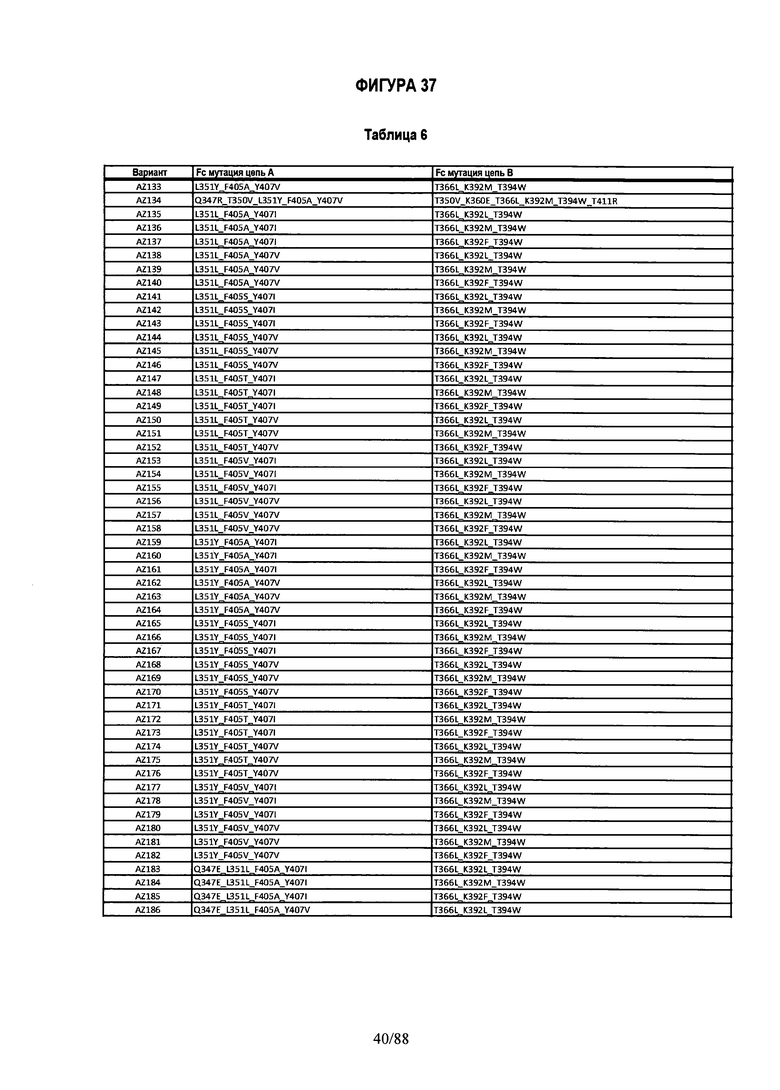
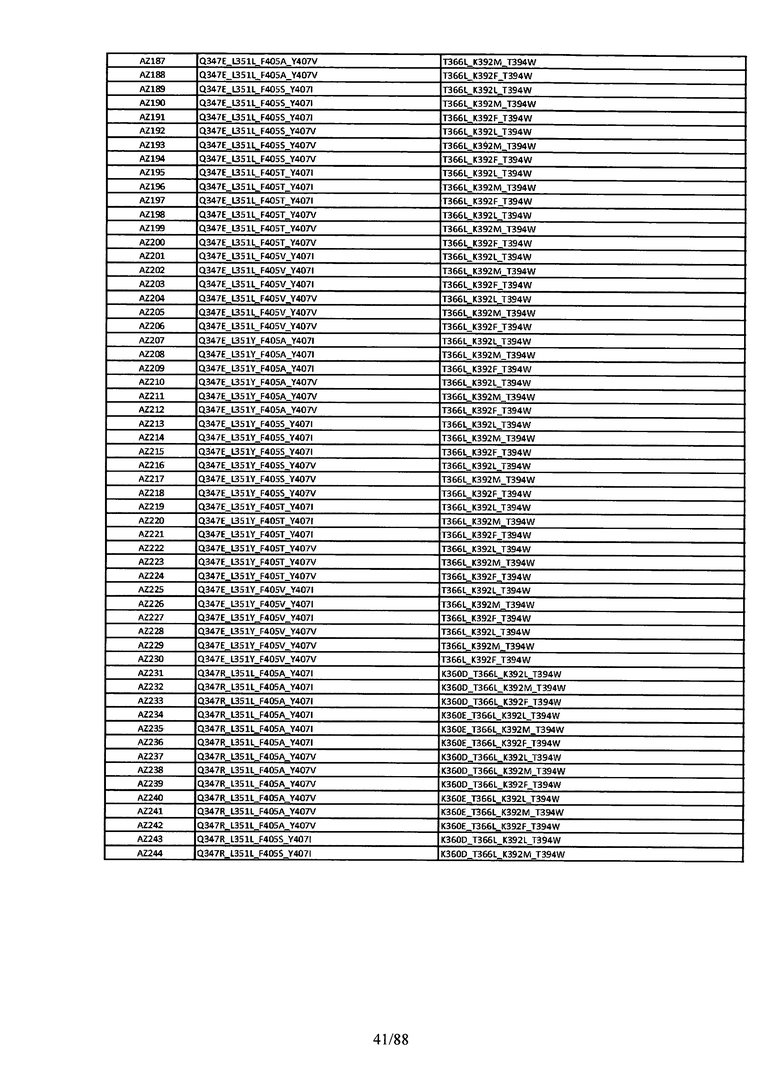
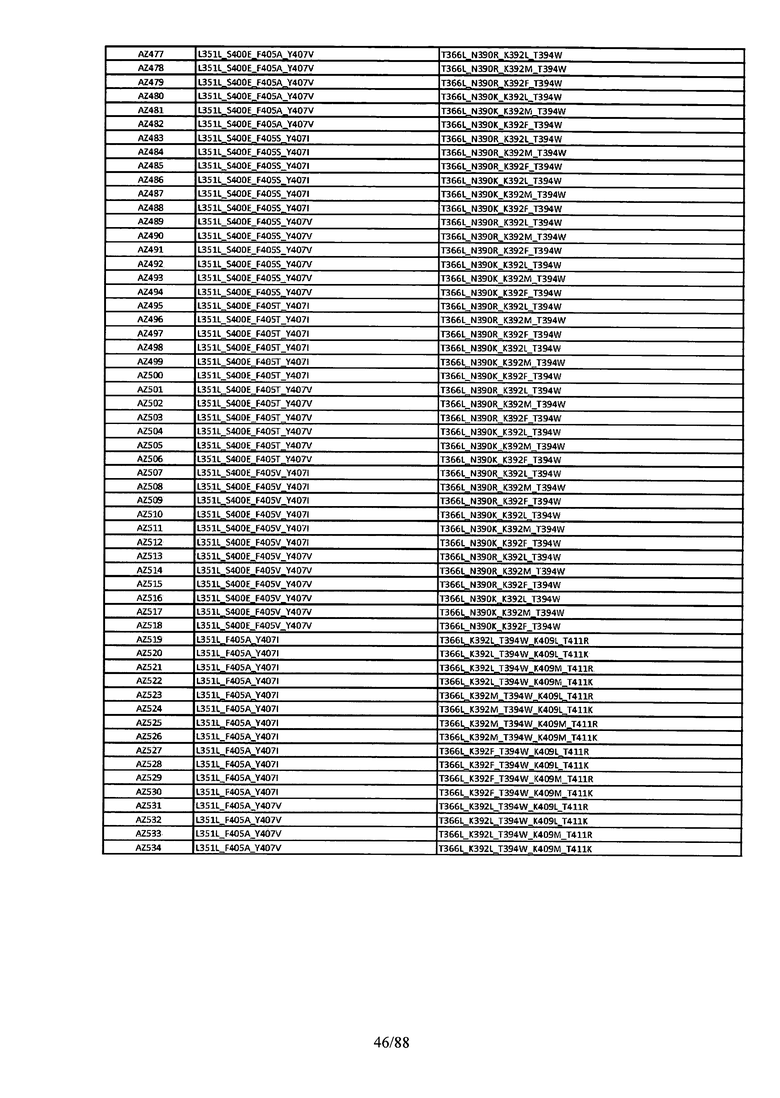
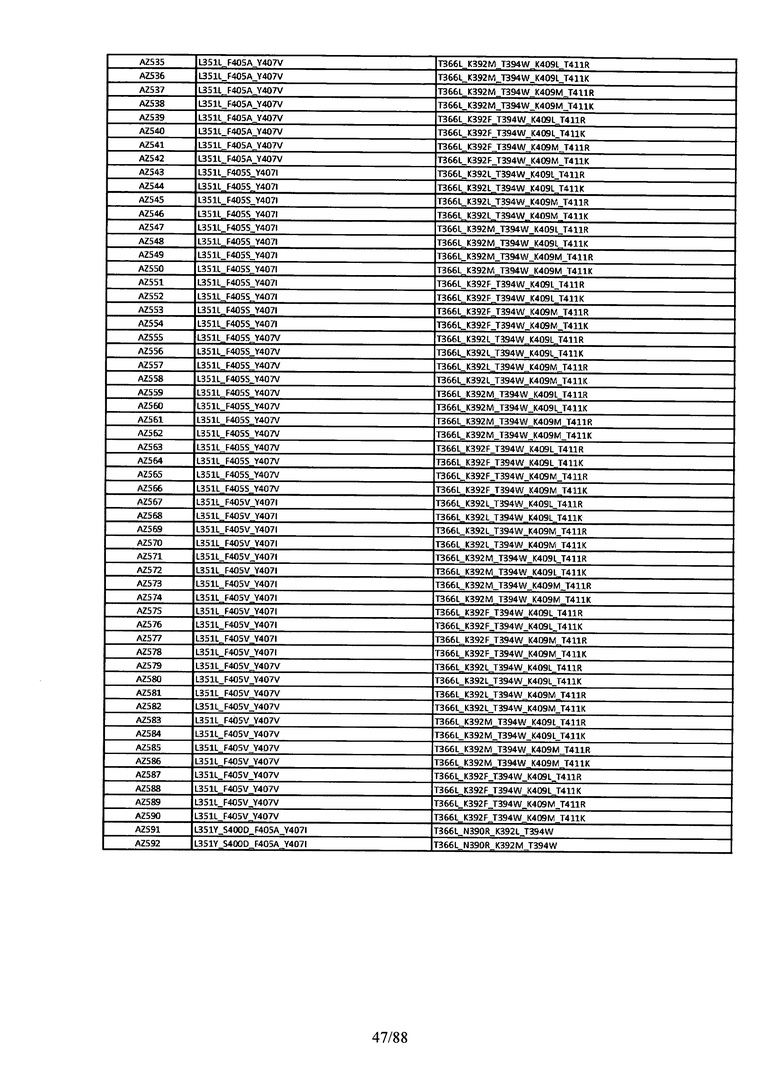
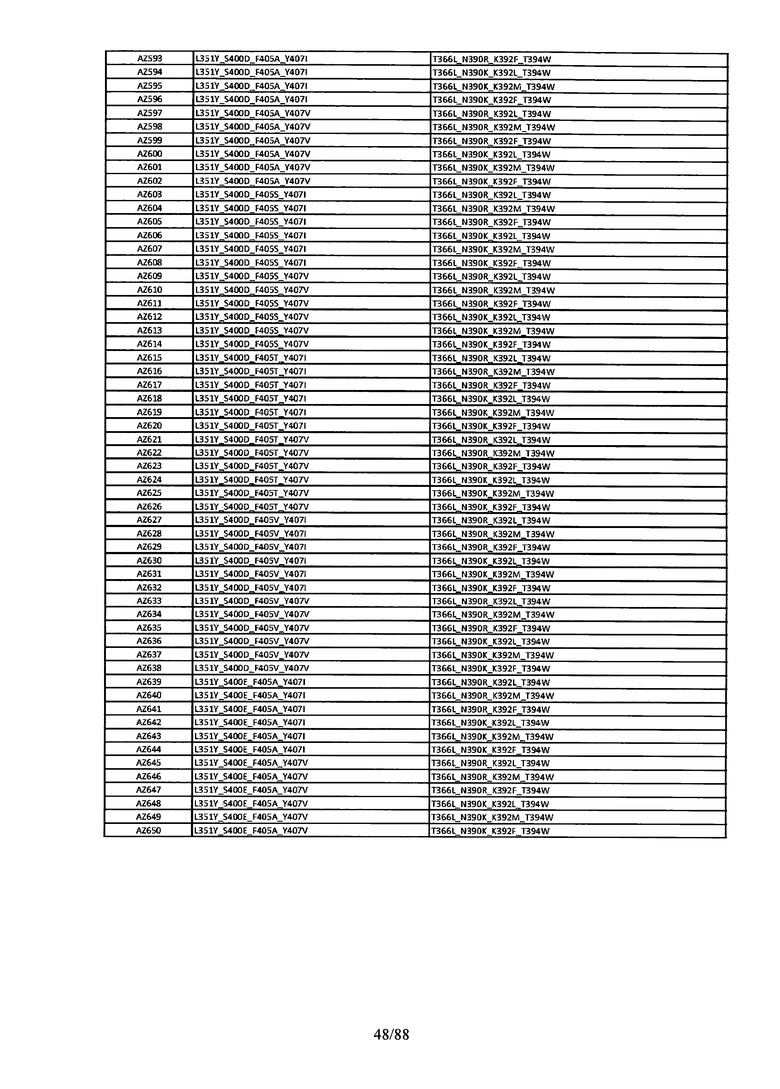
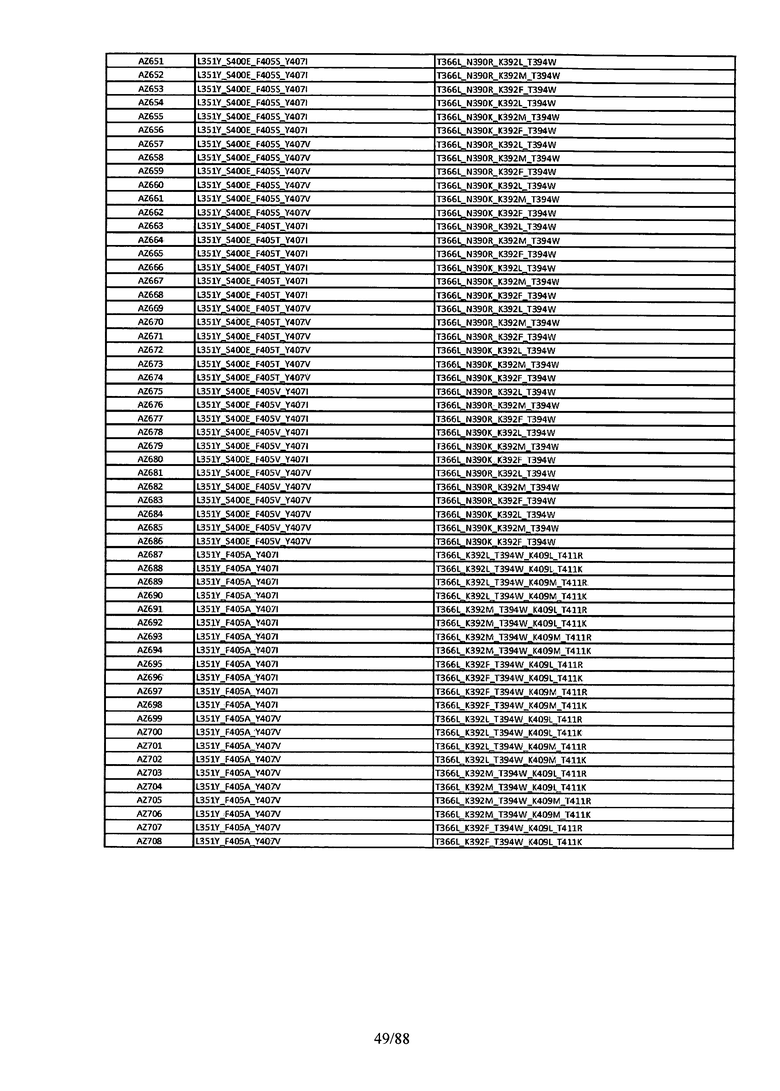
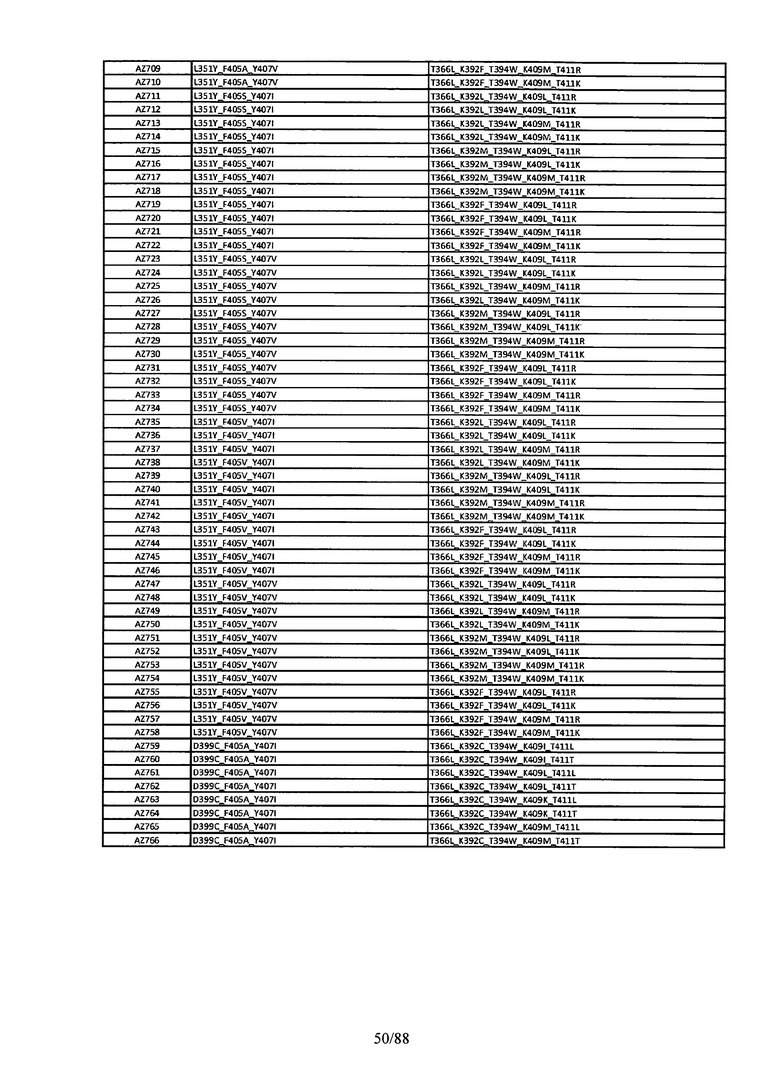
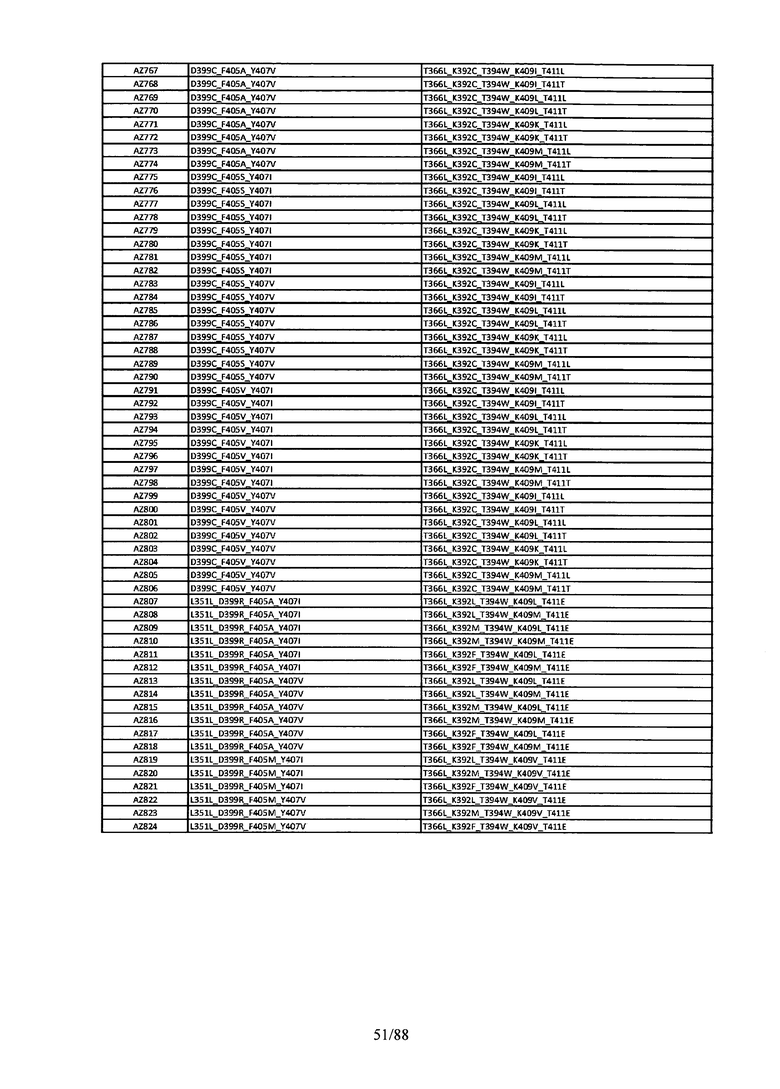
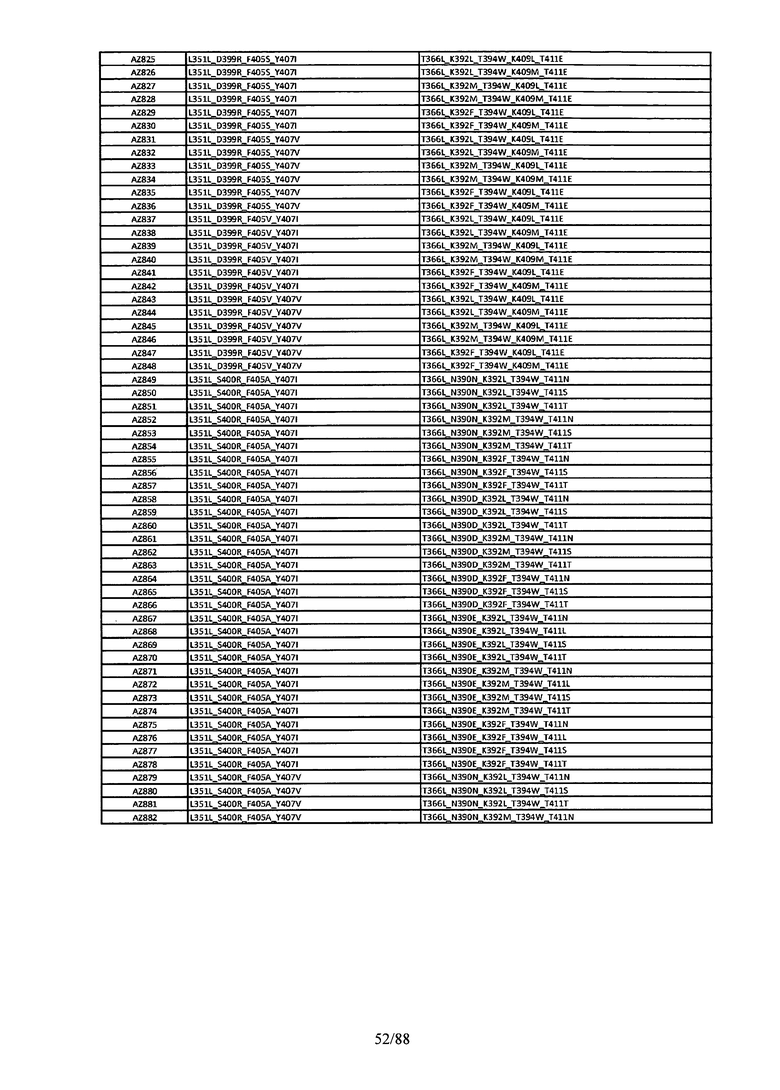
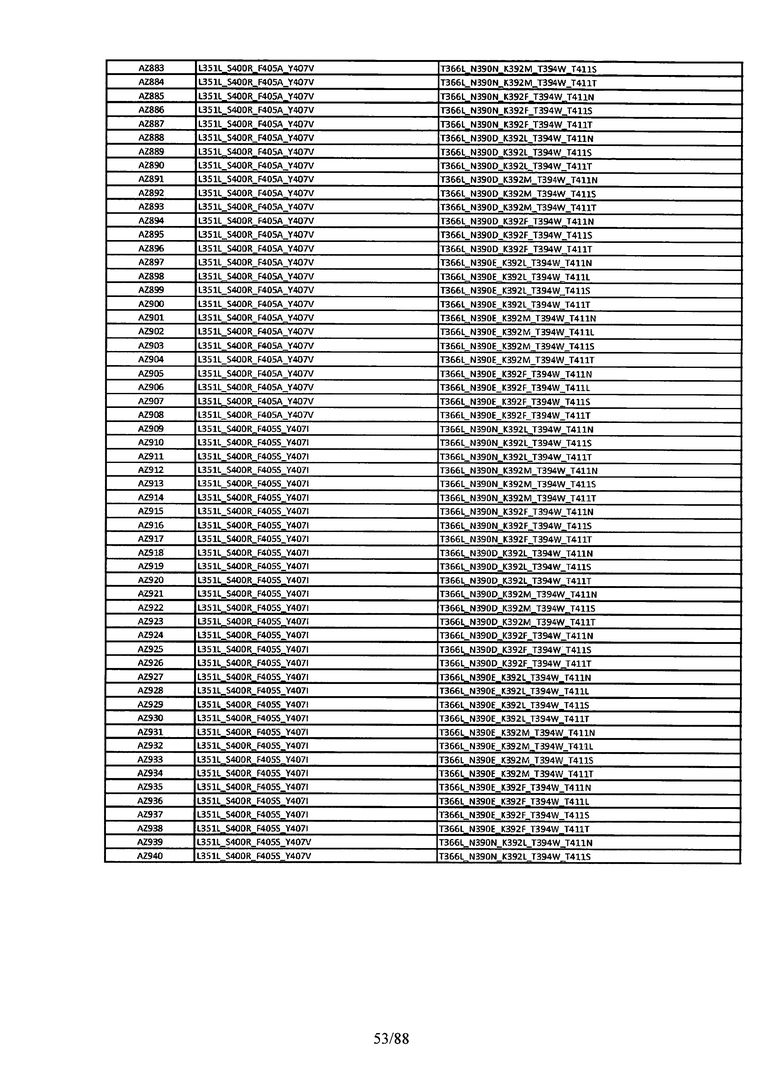
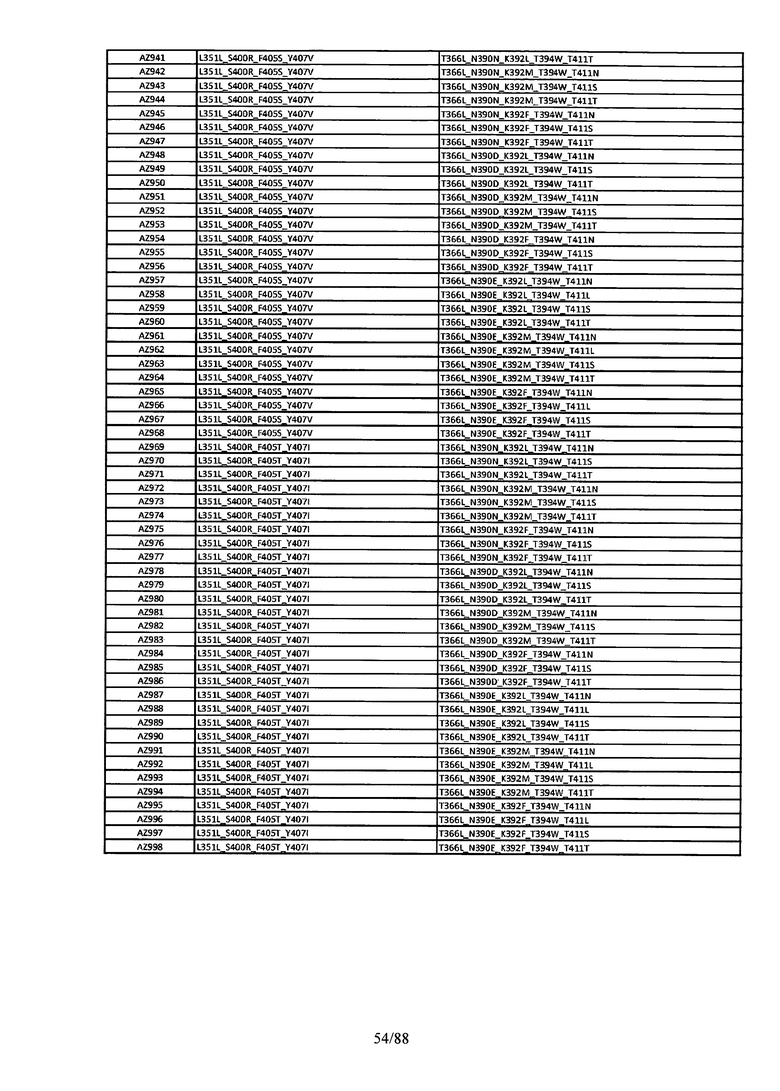
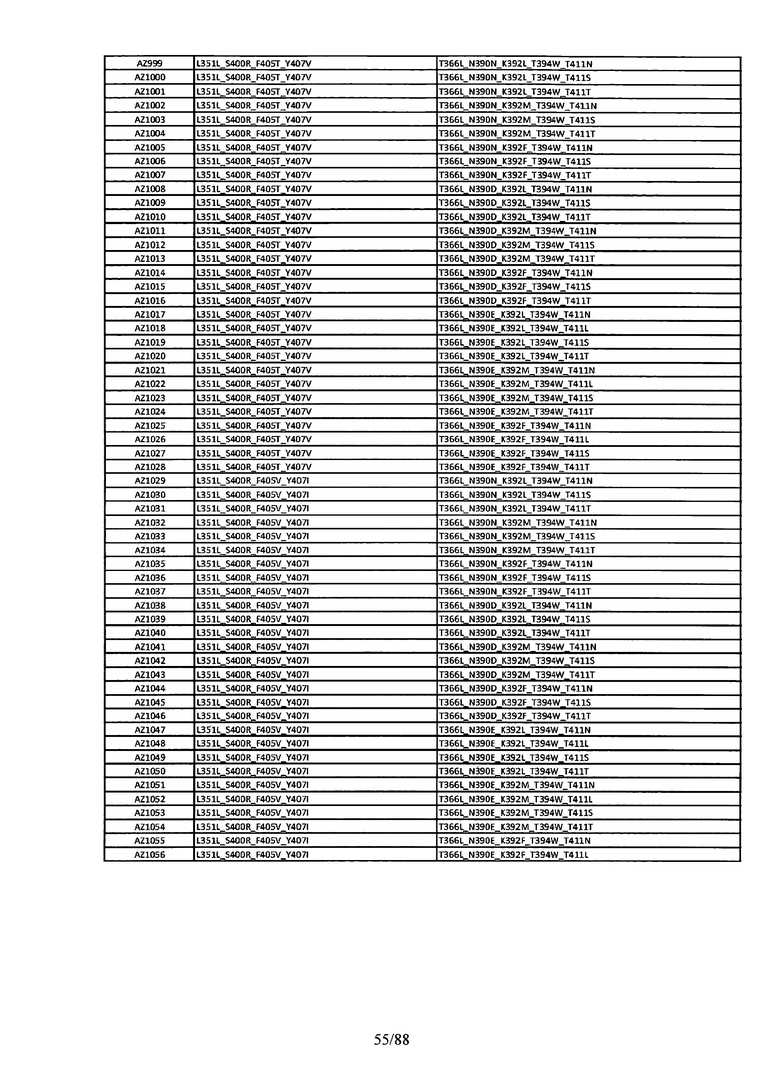
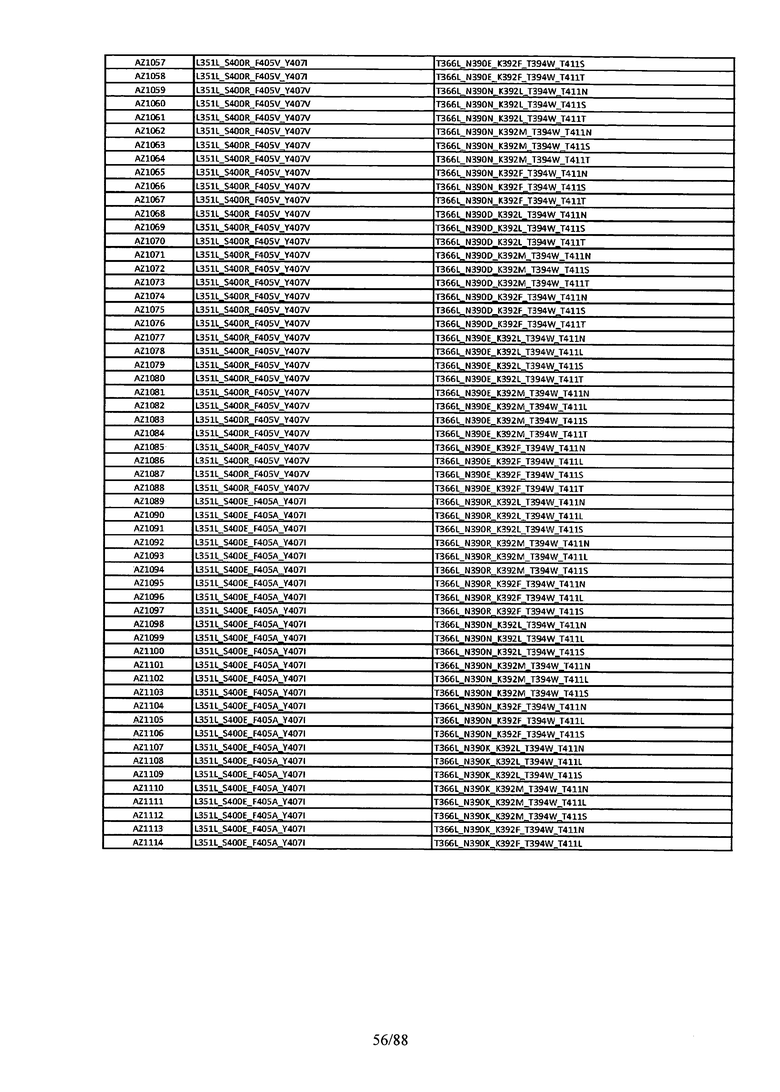
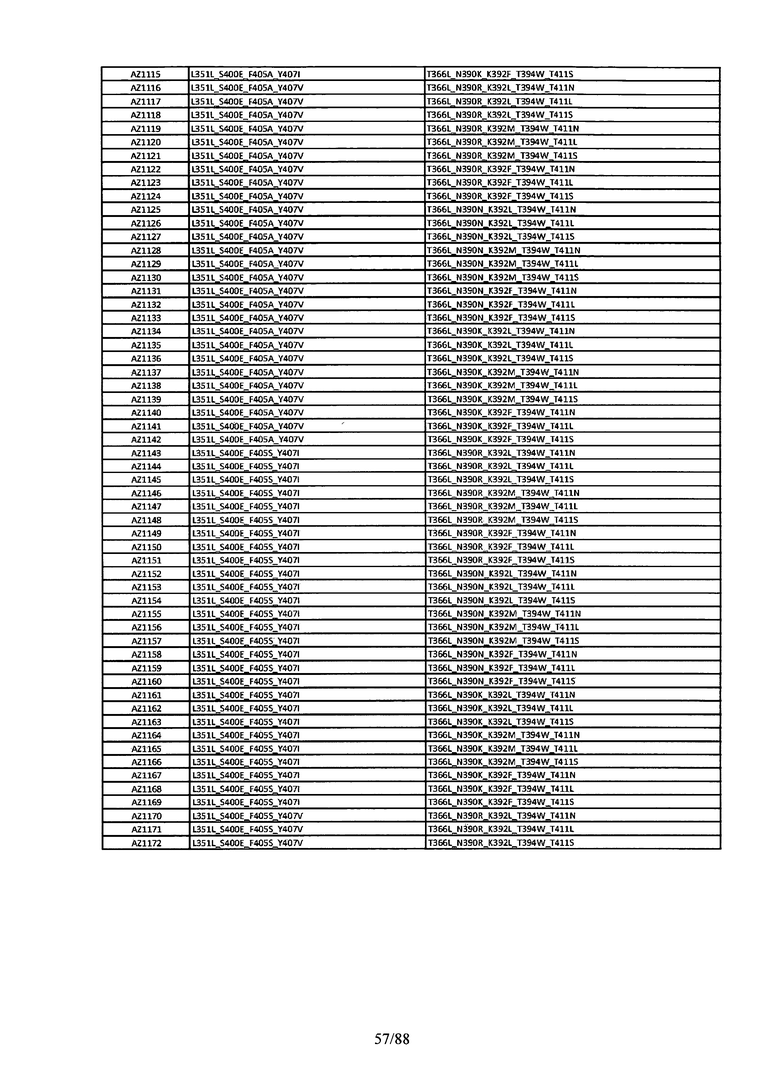
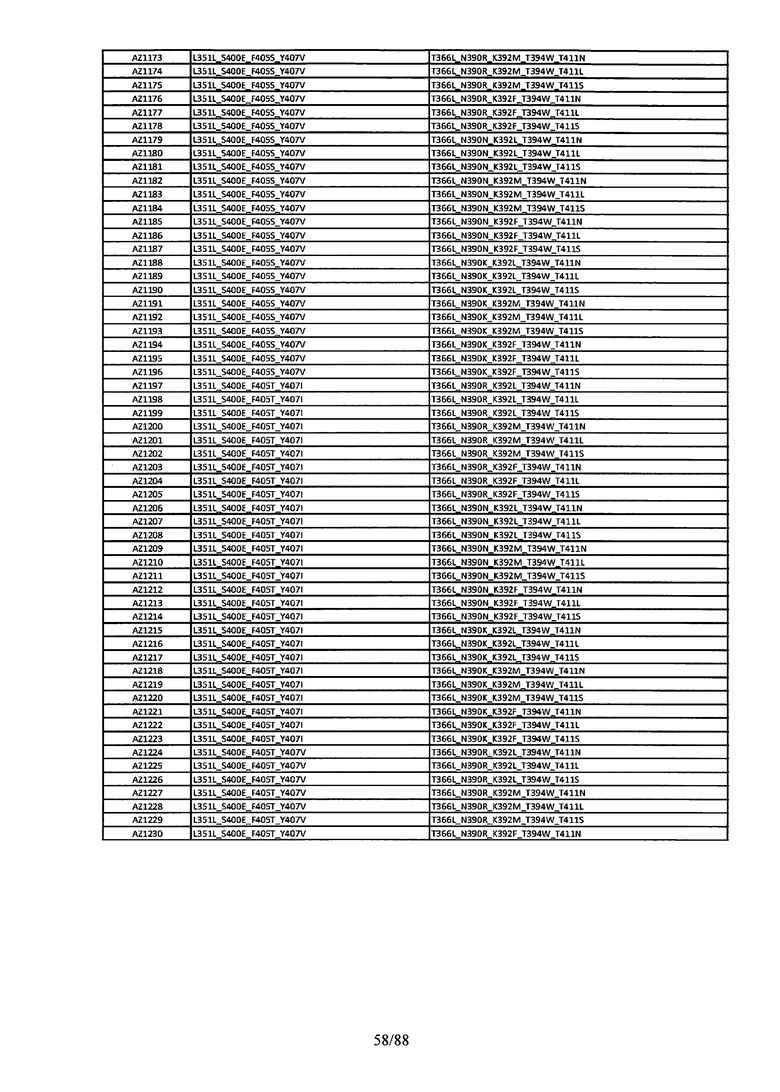
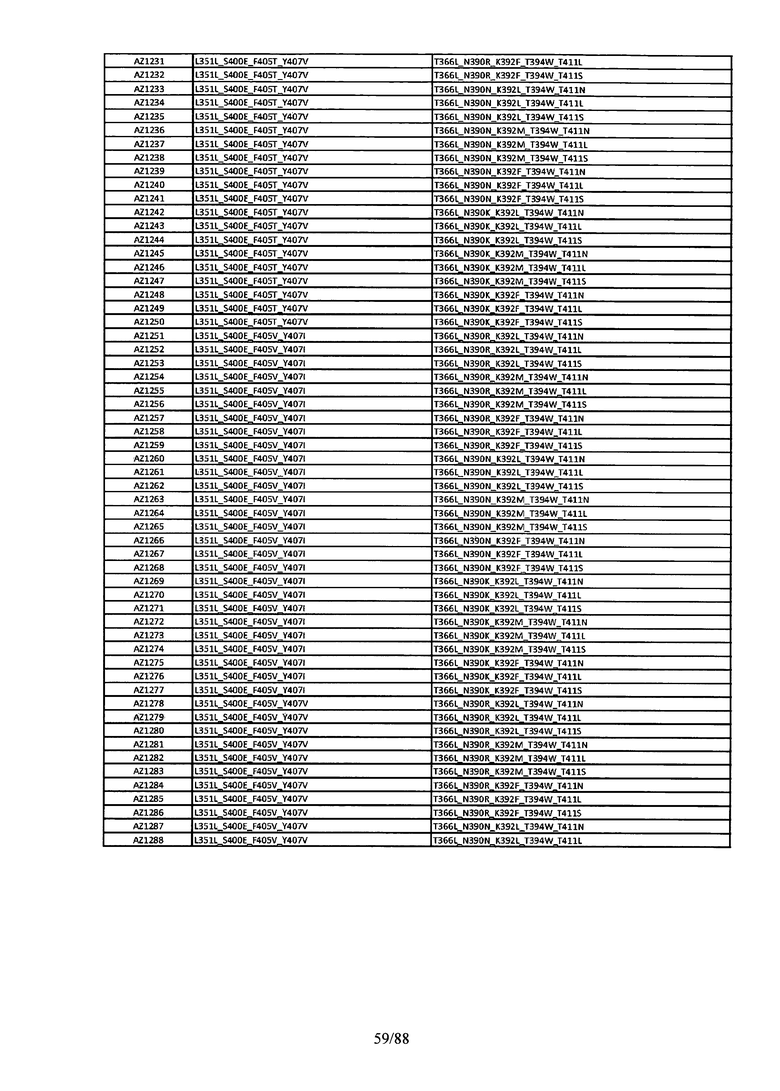
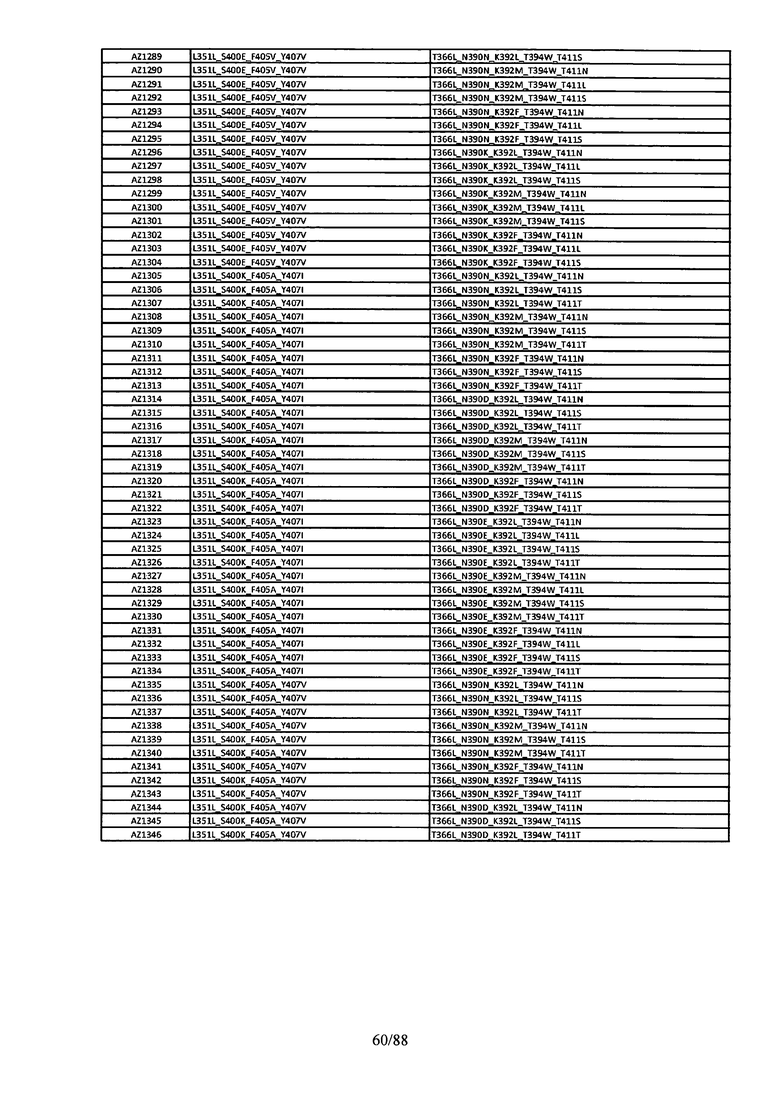
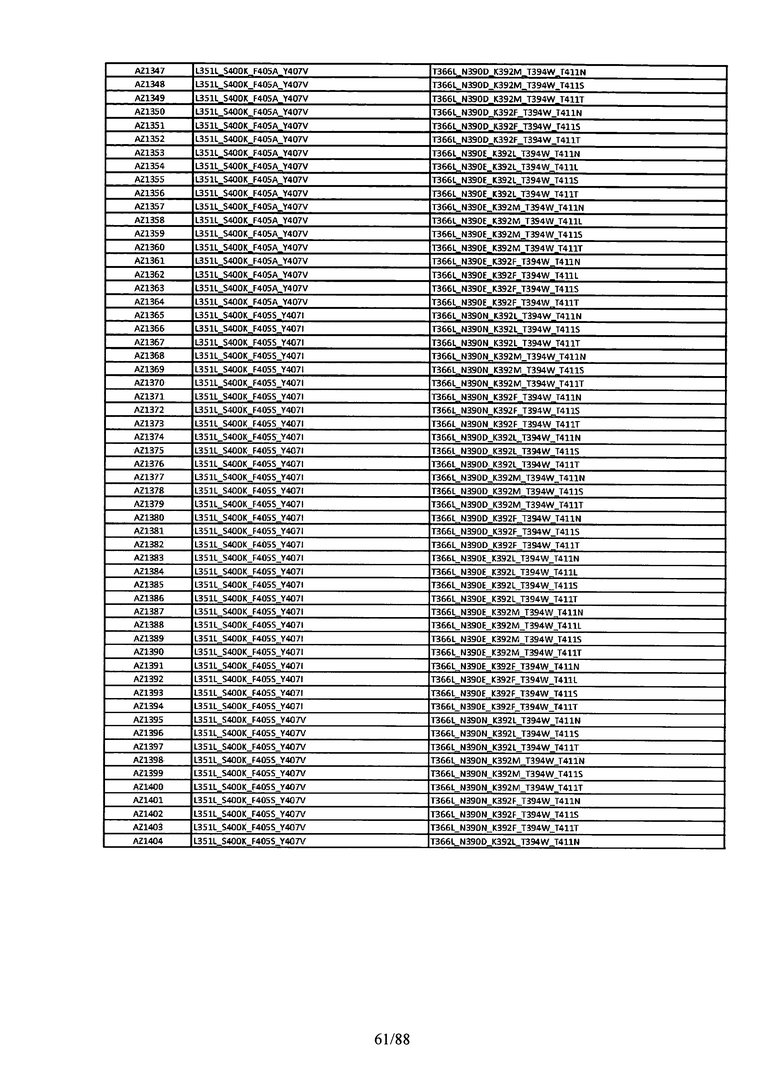
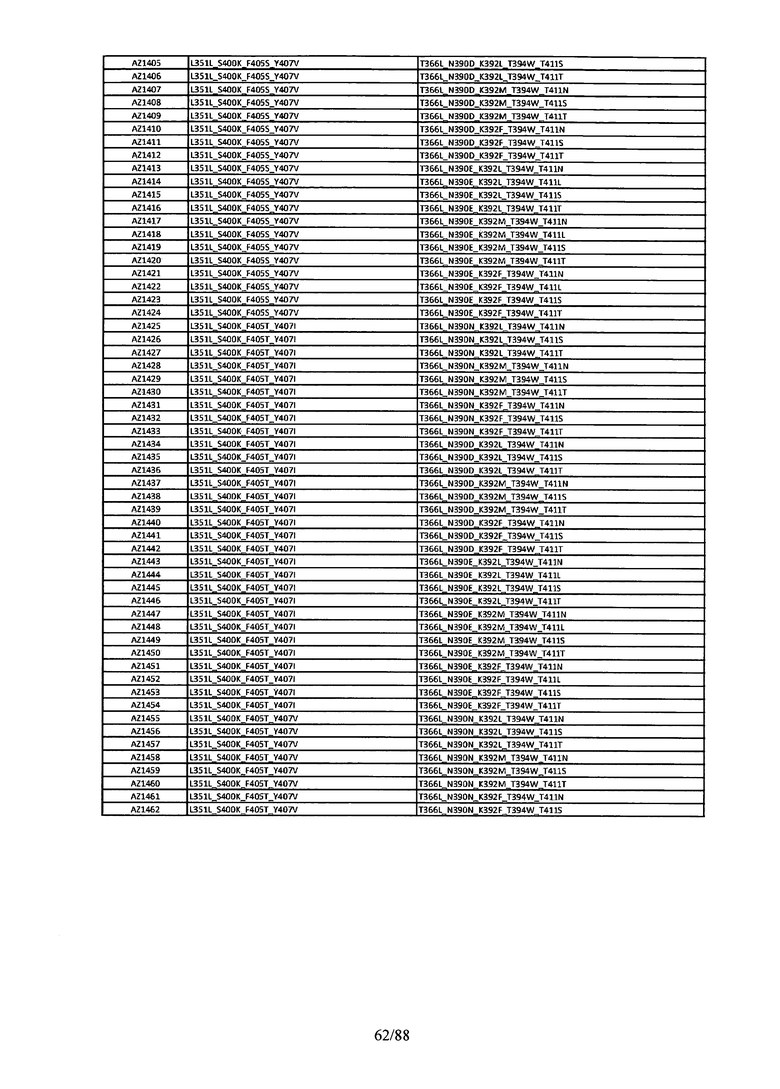
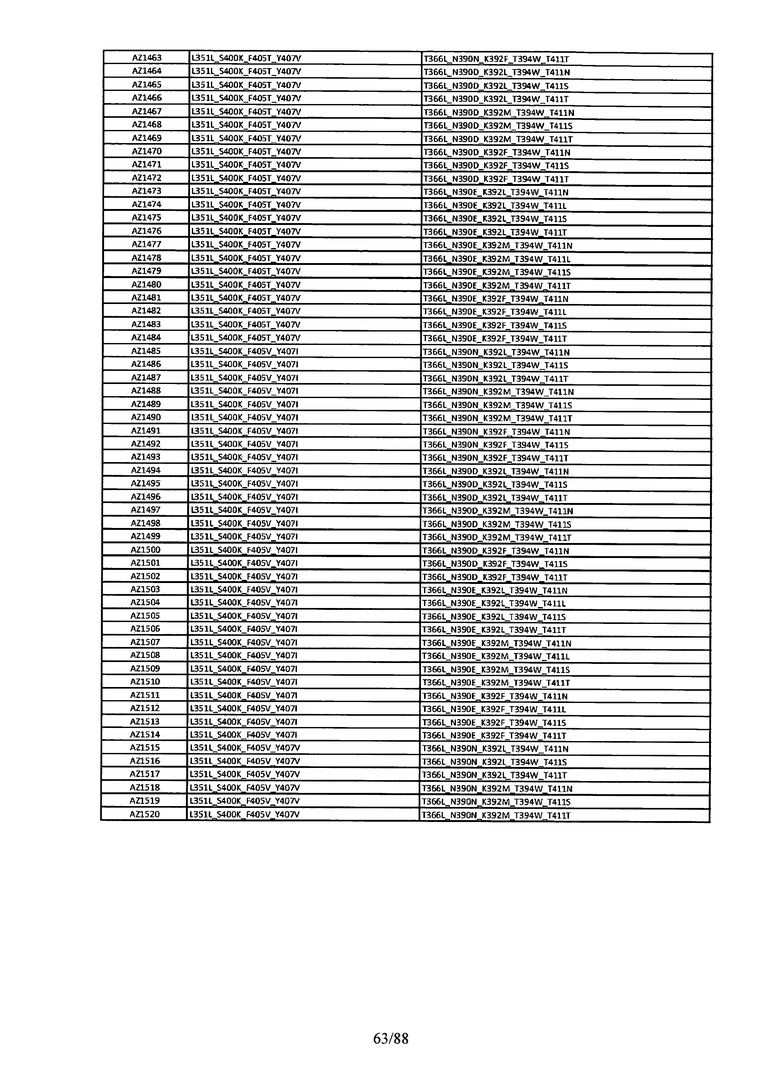
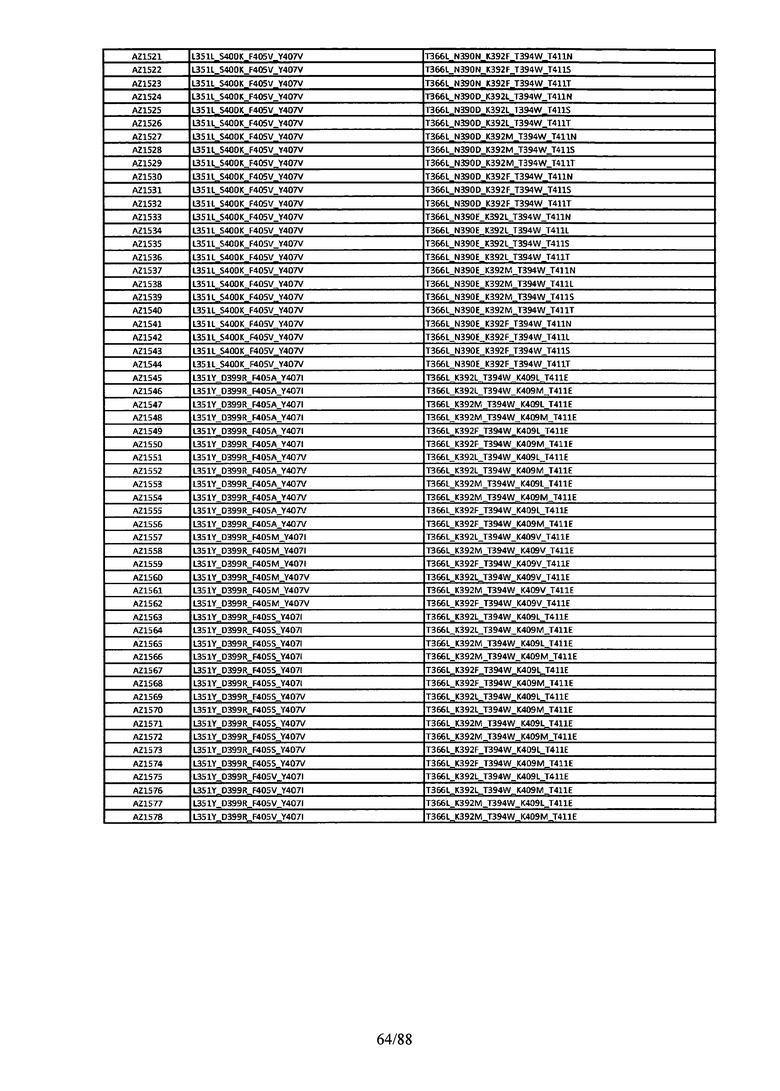
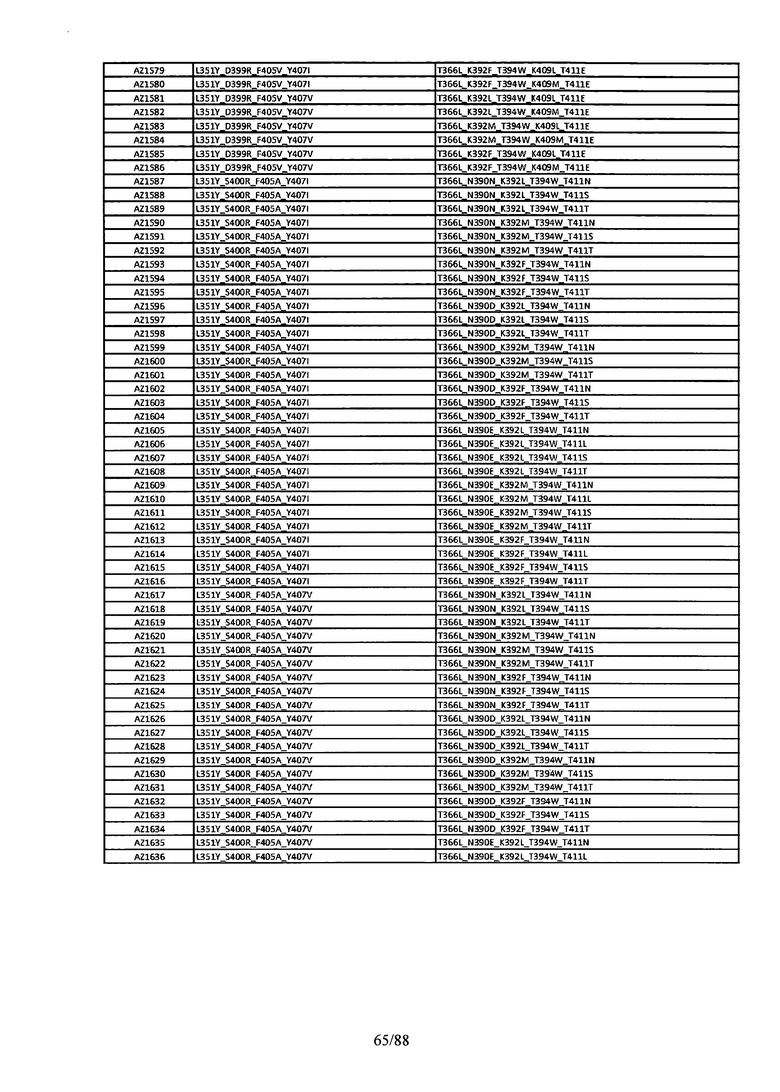
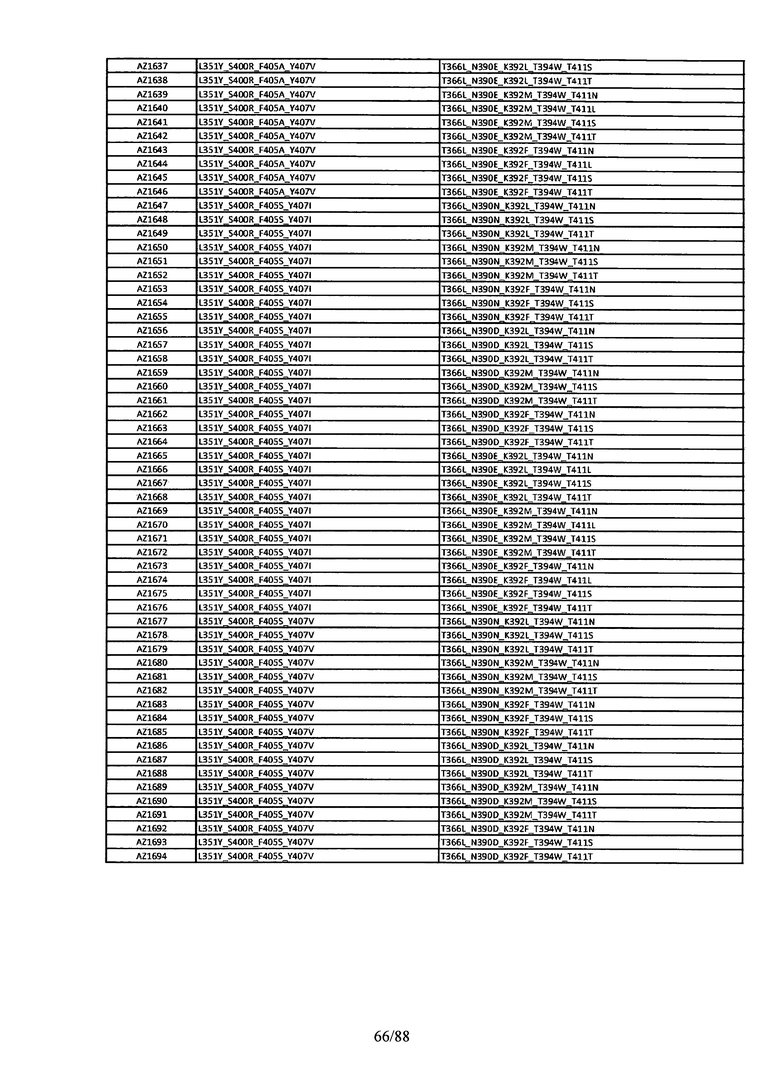
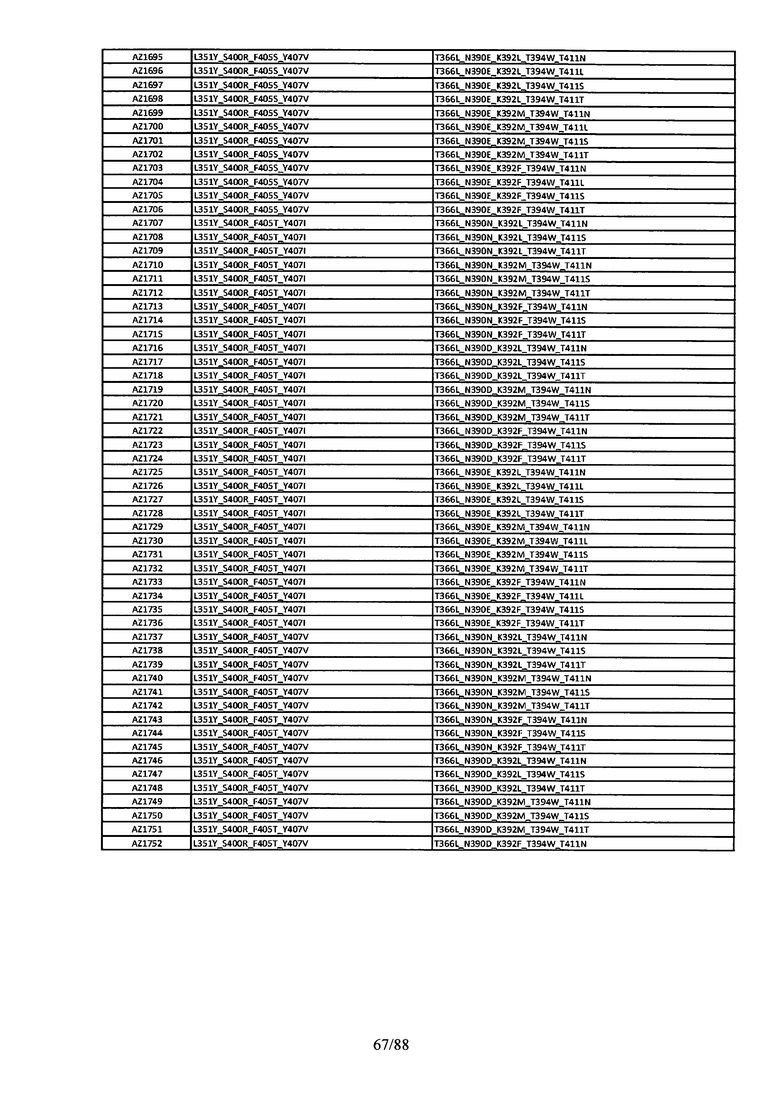
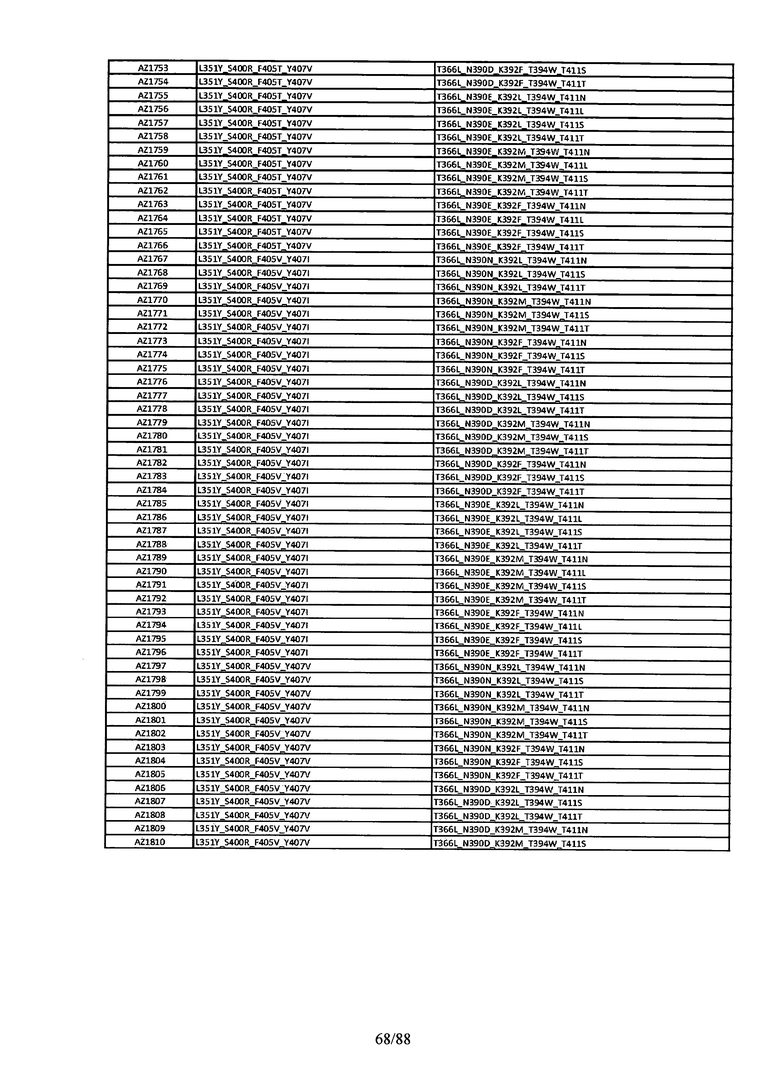
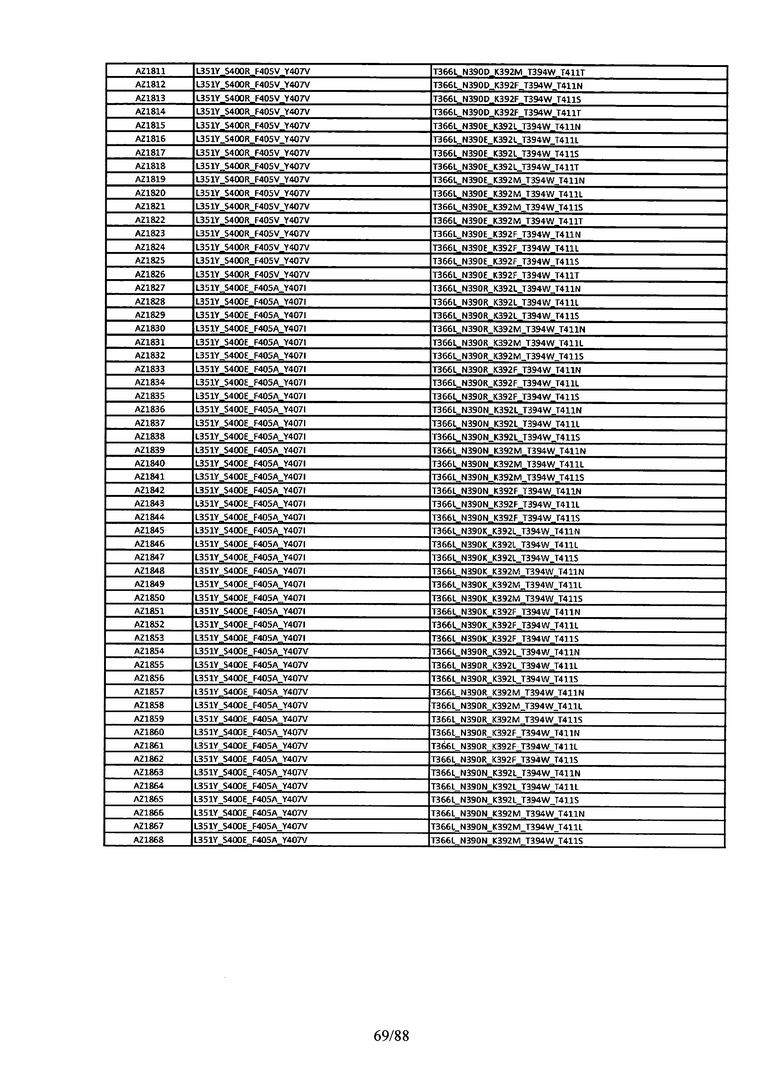
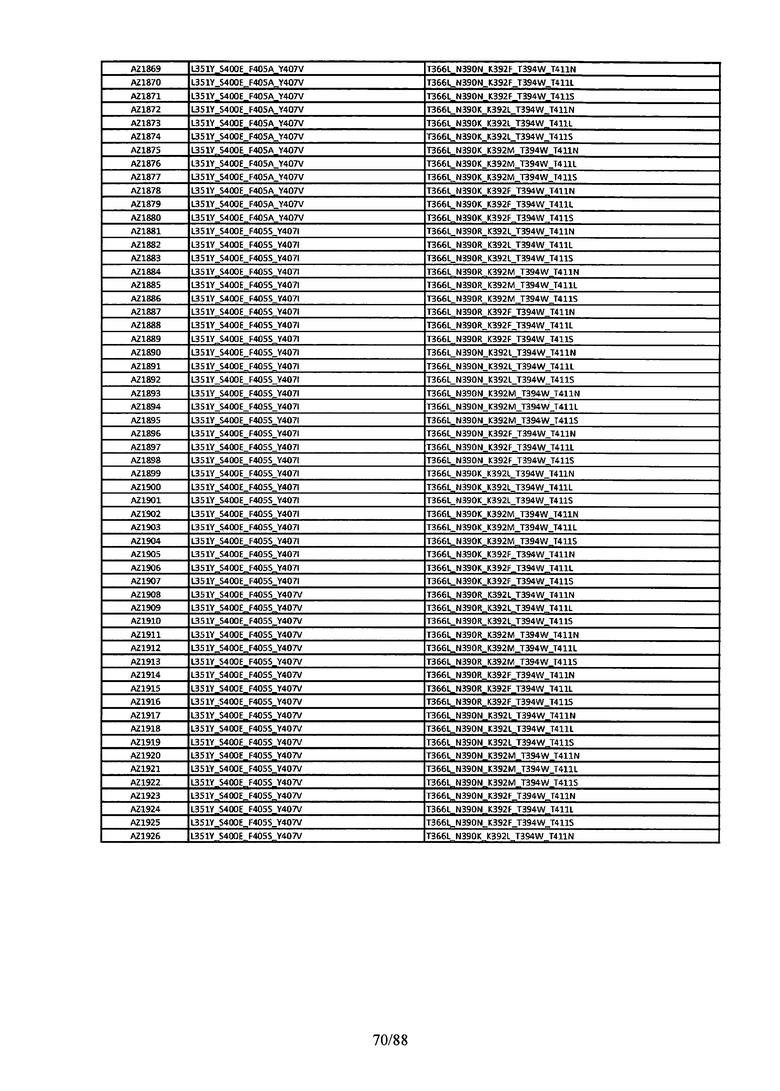
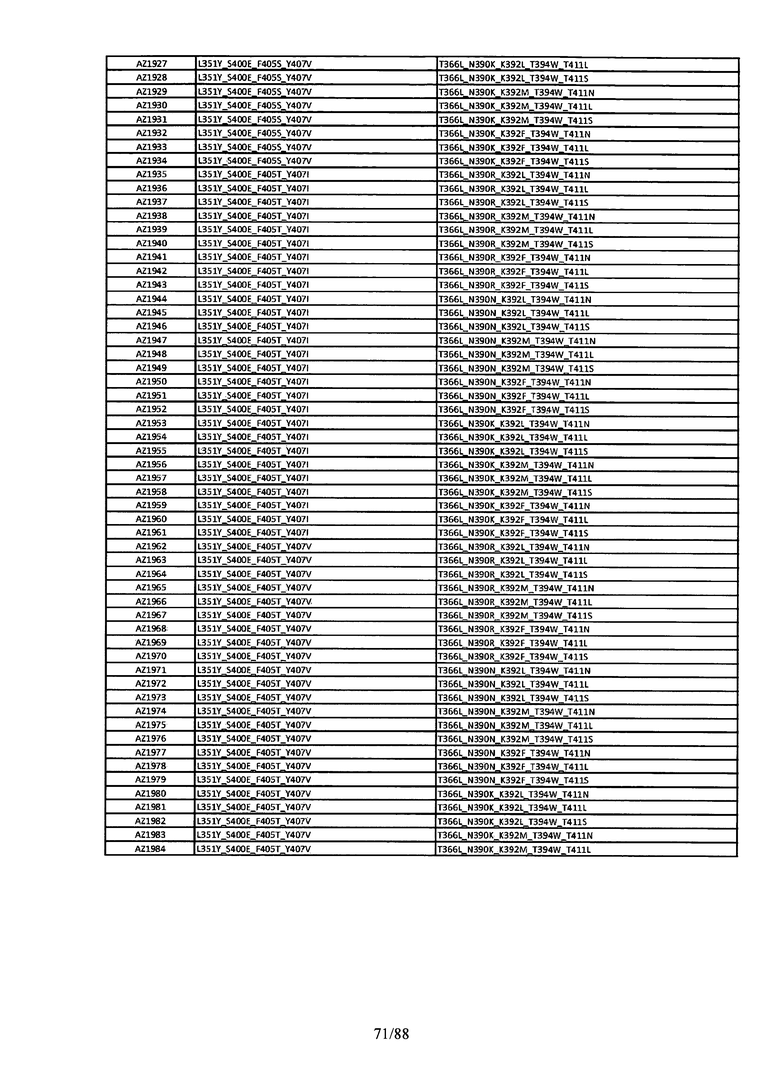
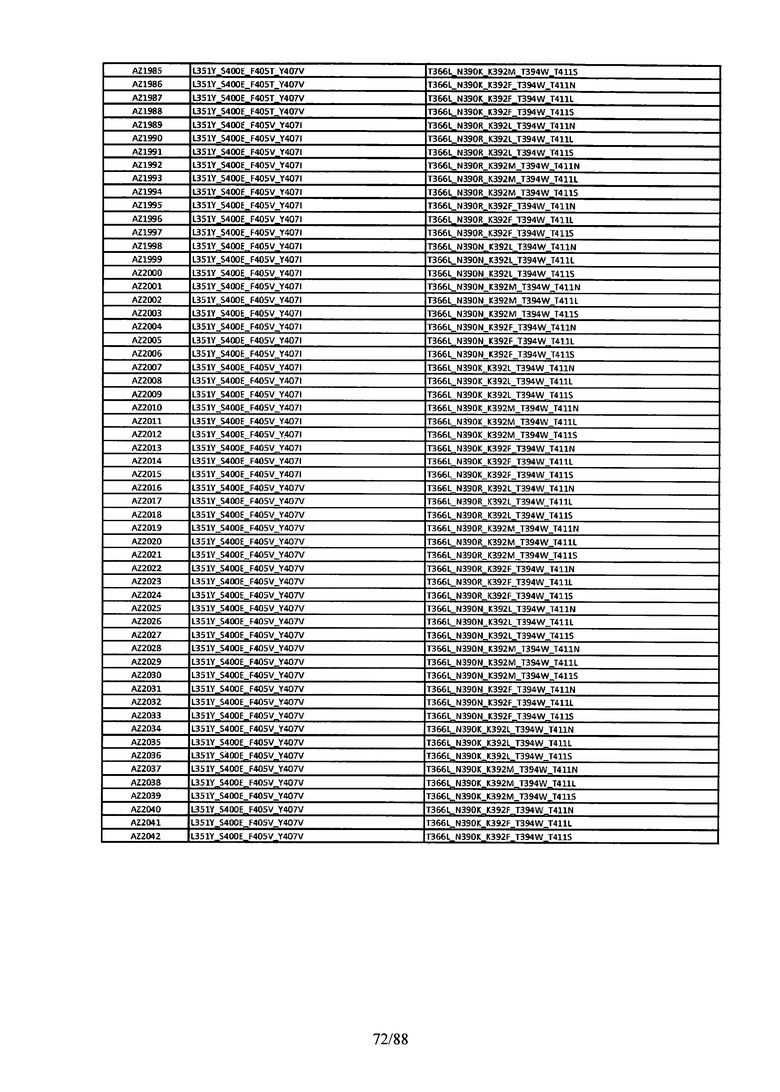
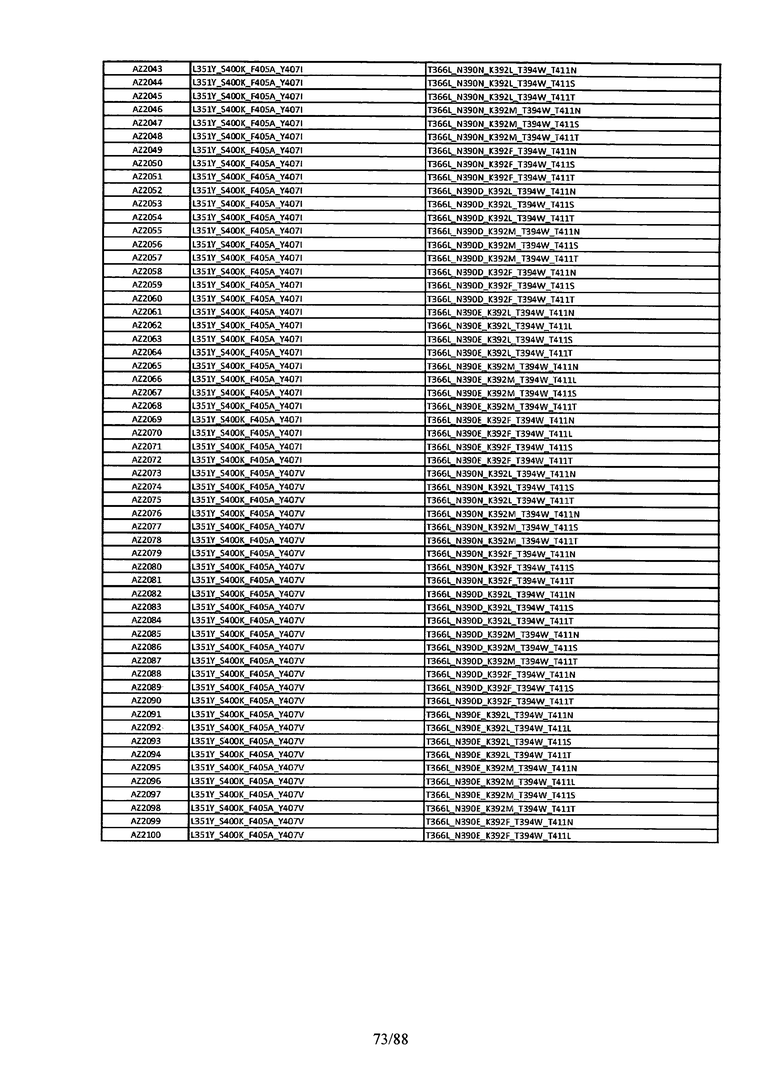
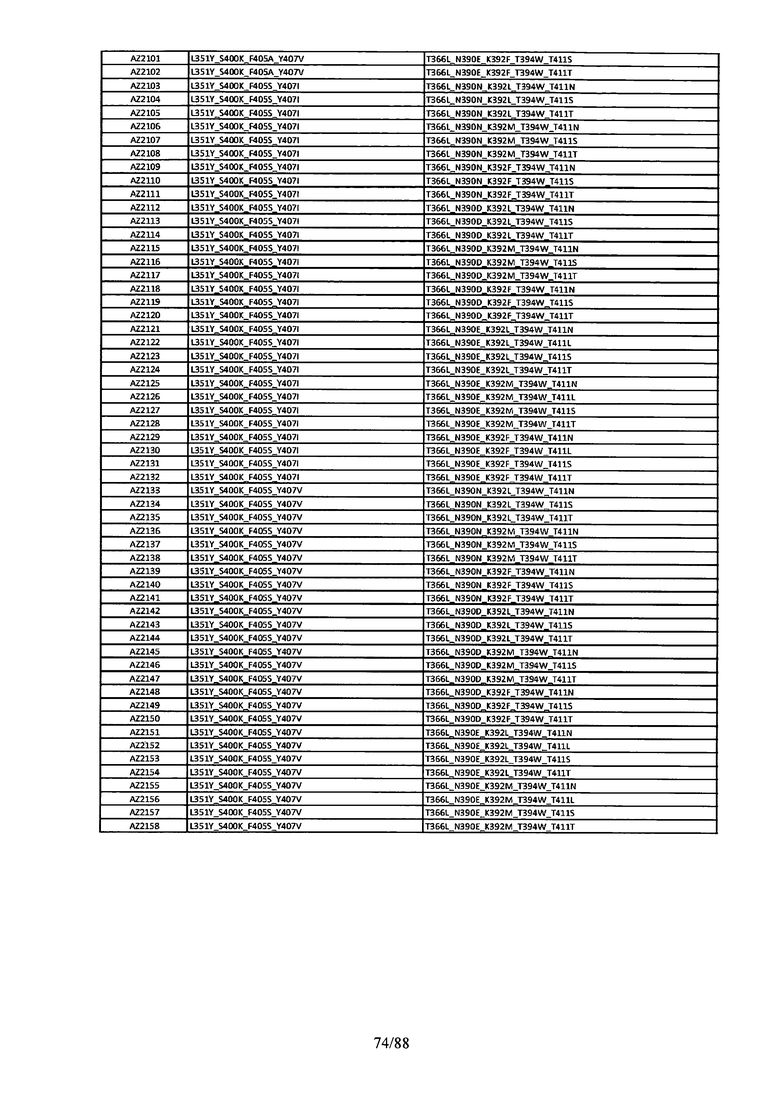
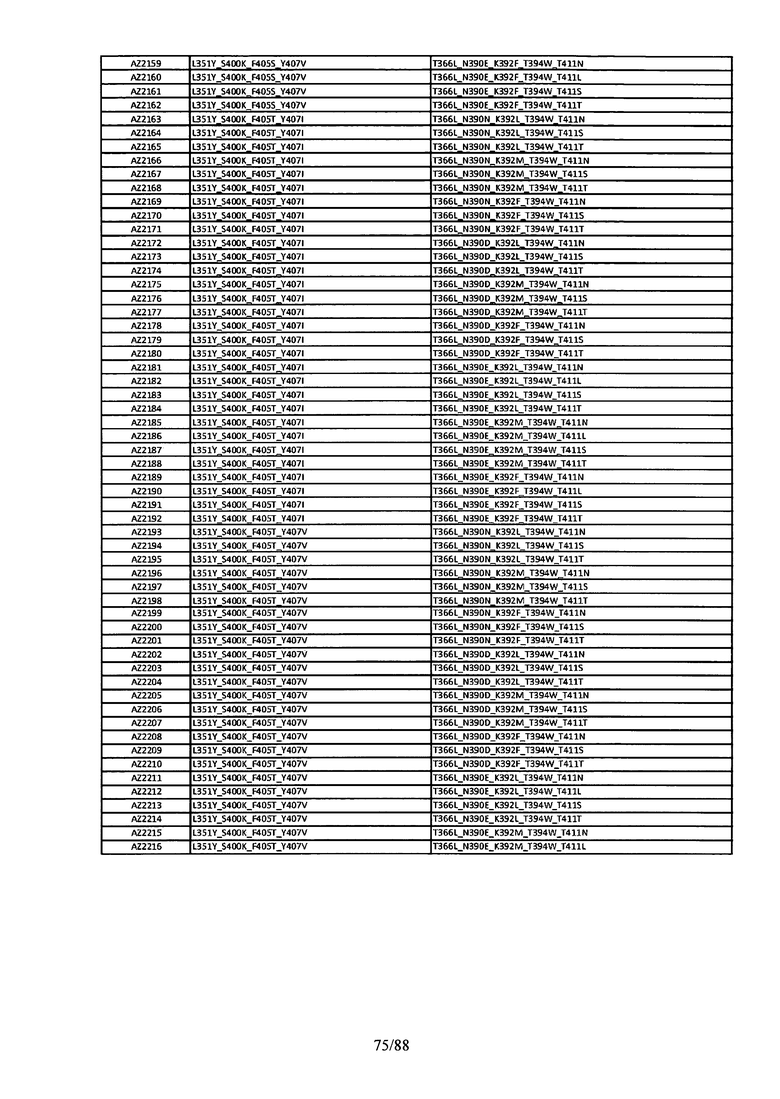
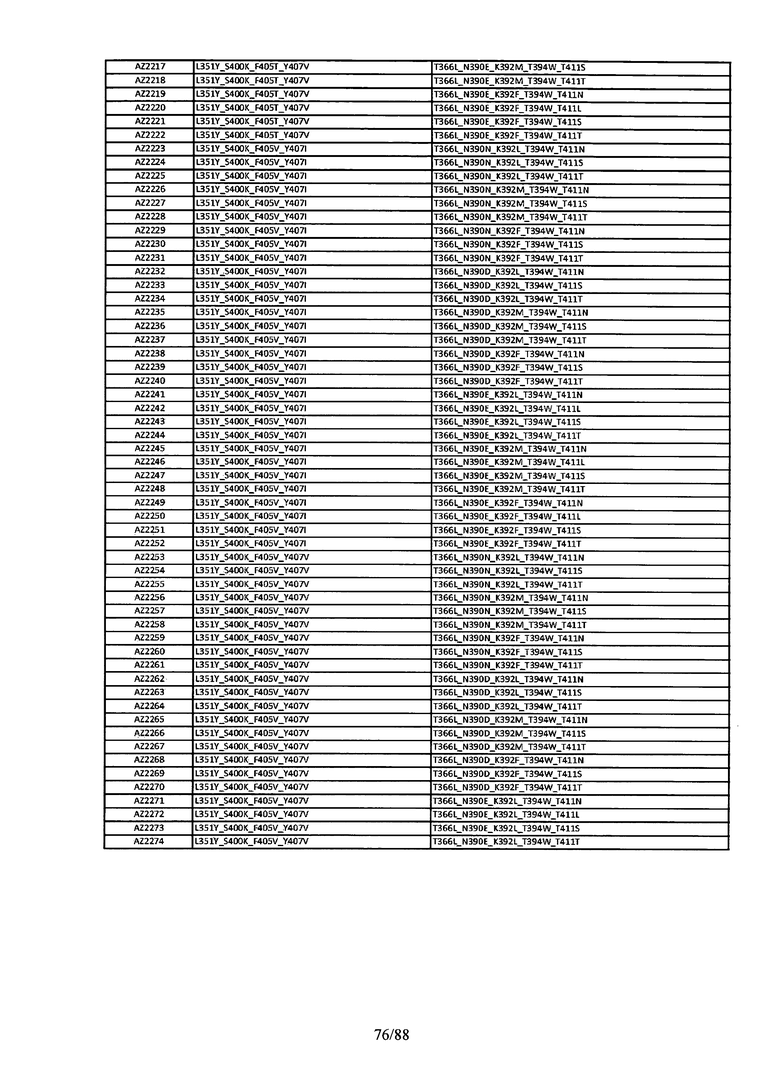
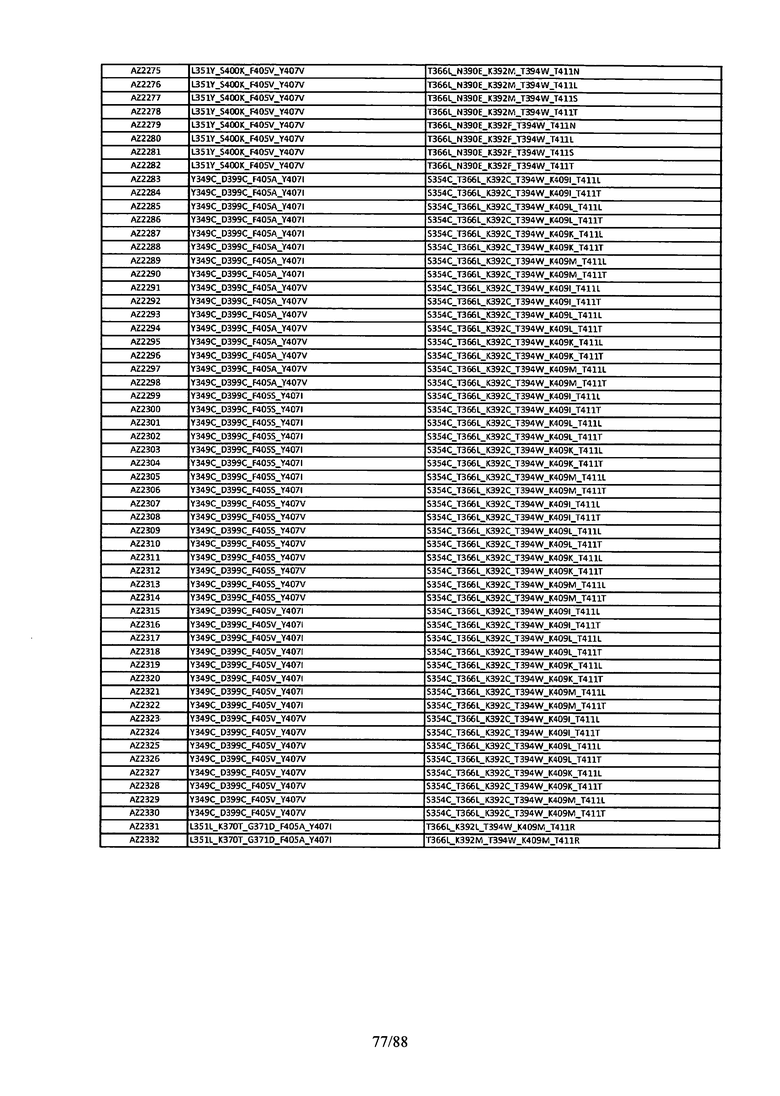
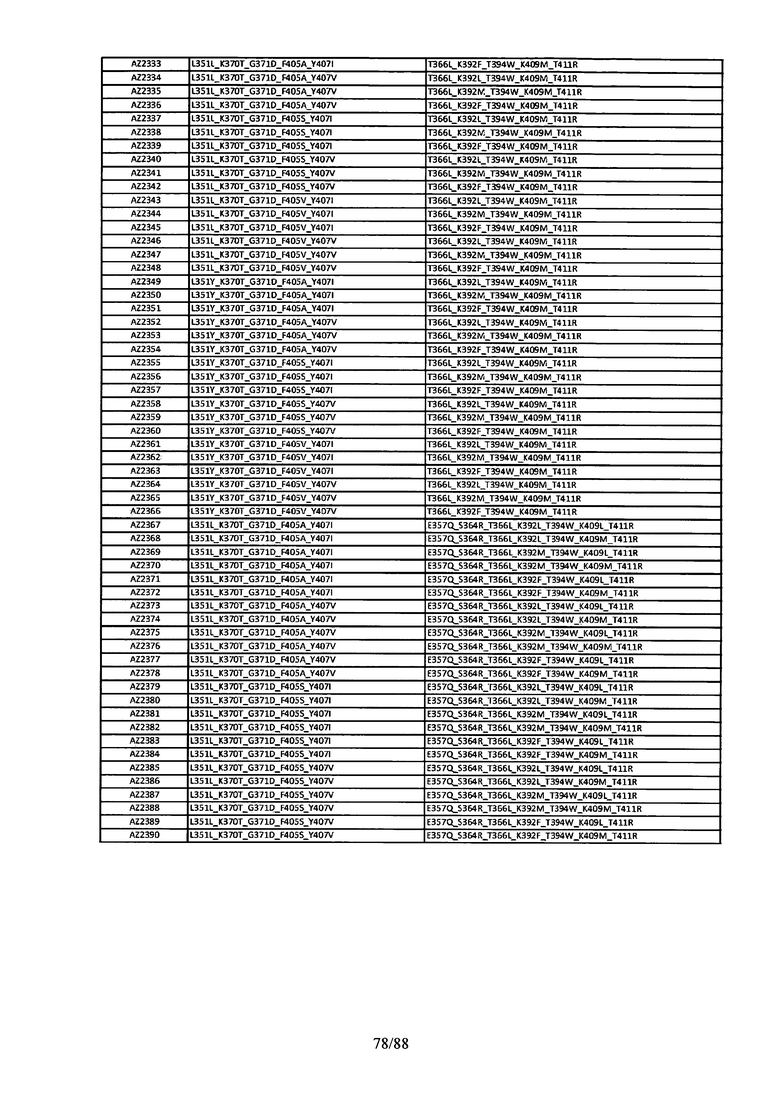
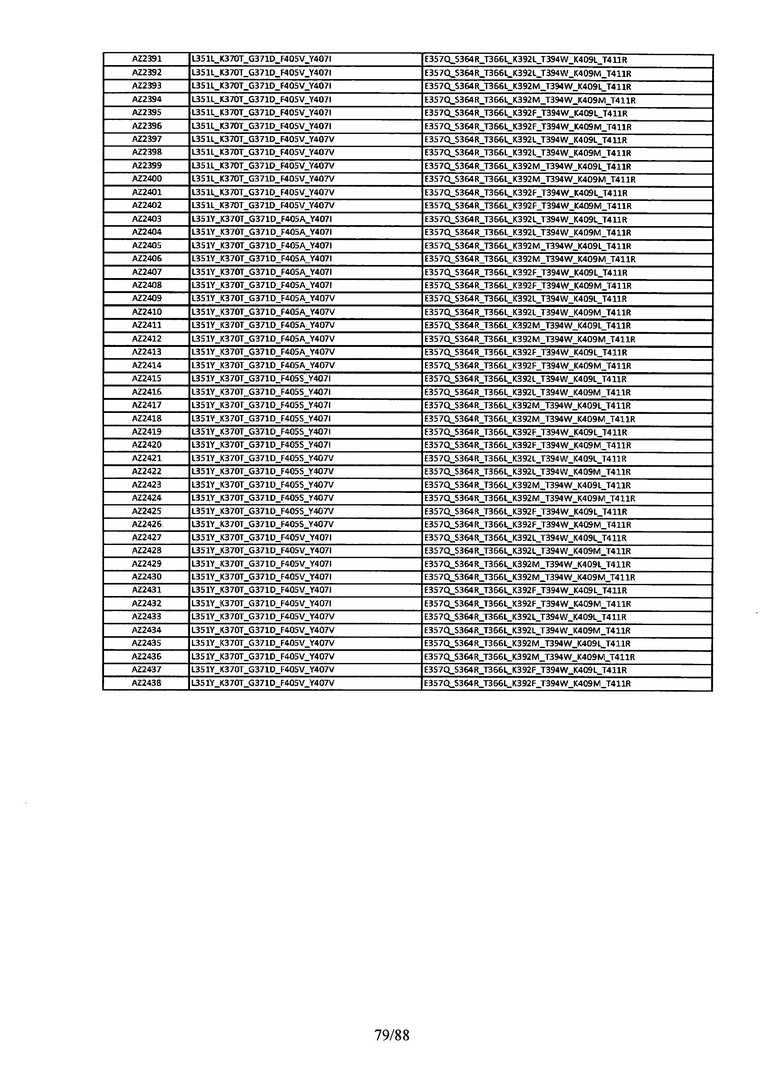
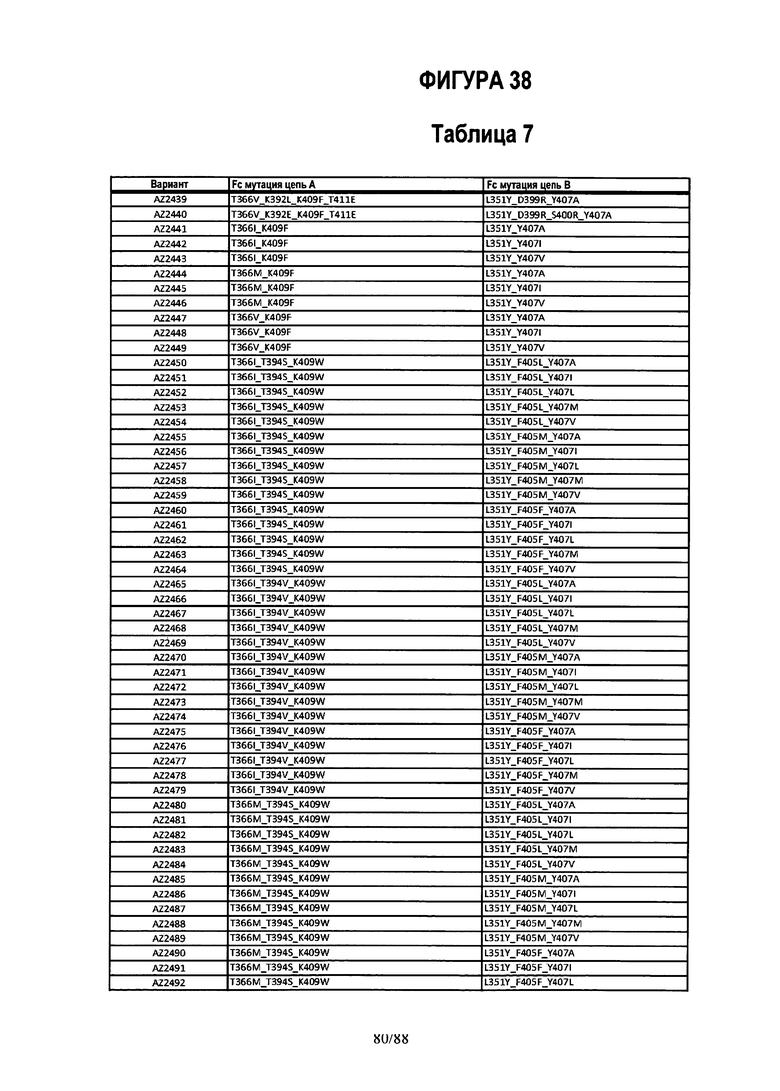
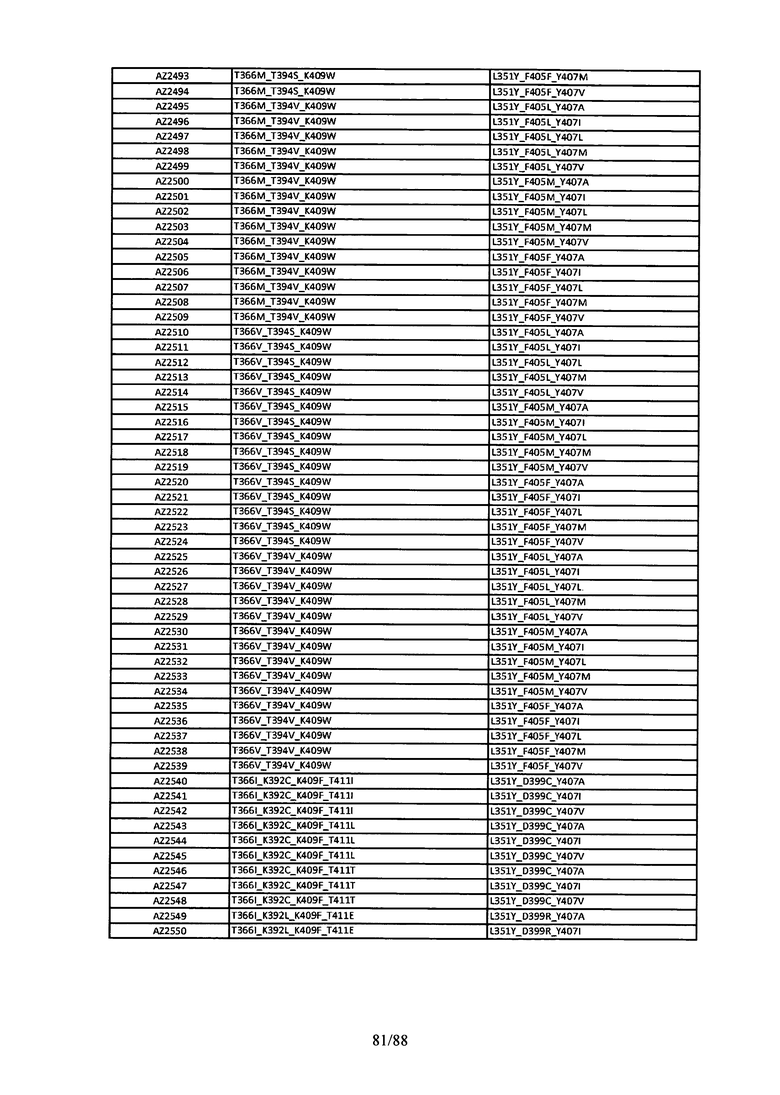
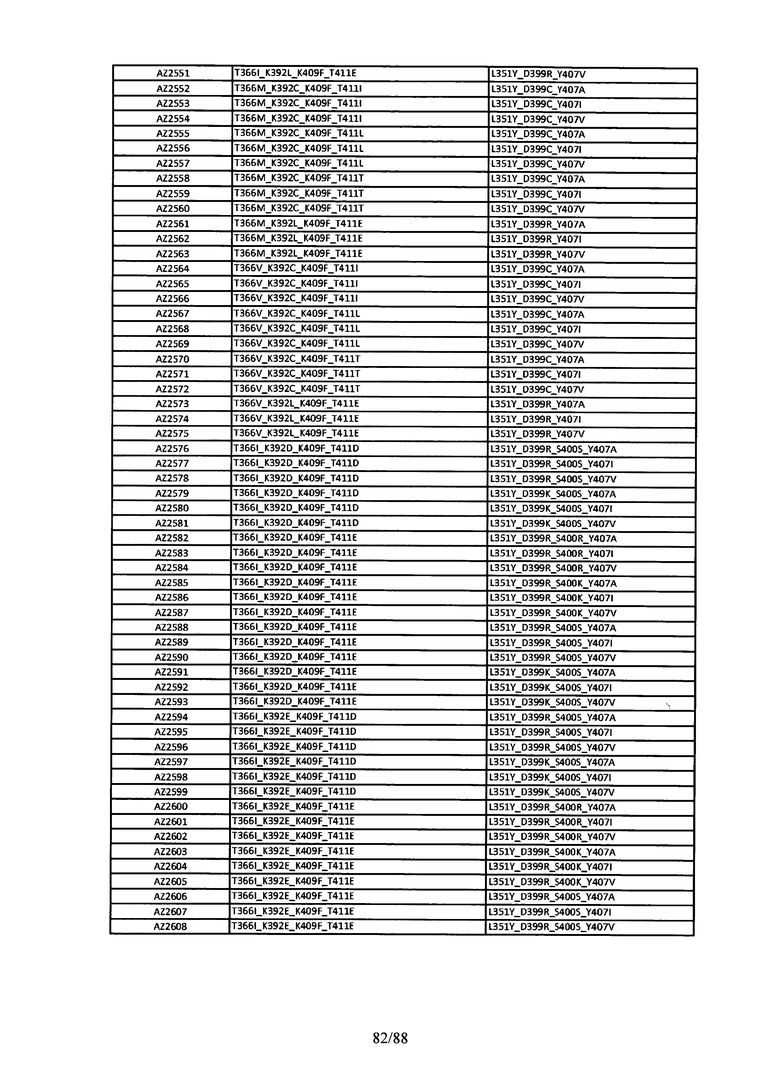
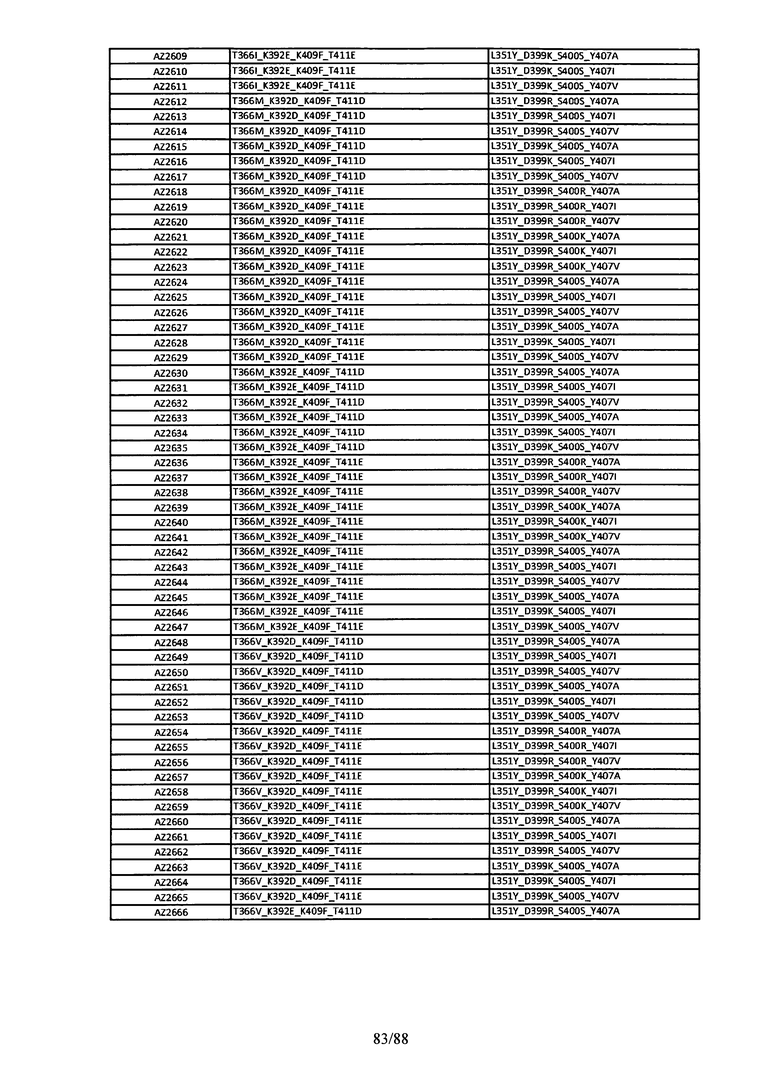
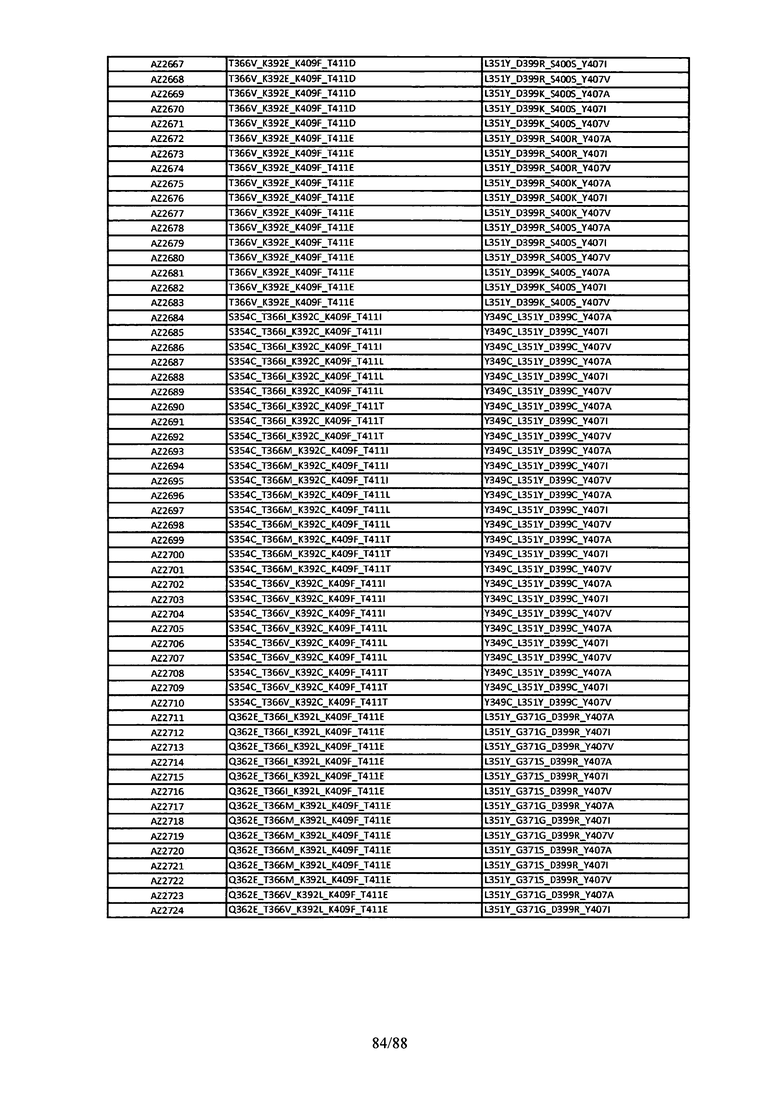
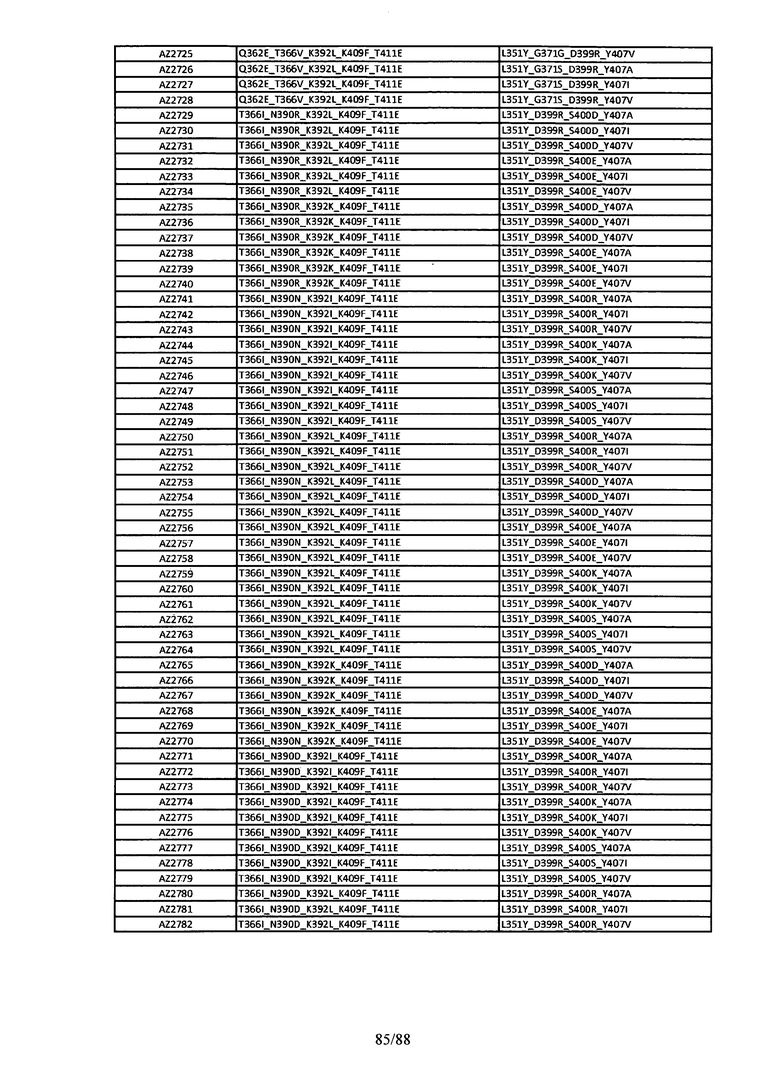
Изобретение относится к биохимии. Описаны каркасы, которые содержат тяжелые цепи, являющиеся асимметричными в различных доменах (например, СН2 и СН3), для того, чтобы достичь селективности между различными Fc рецепторами, участвующими в модуляции эффекторной функции, селективности более высокой, чем селективность, достигаемая при использовании натуральной гомодимерной (симметричной) Fc молекулы, и повышенной устойчивости и чистоты полученных в результате вариантных Fc гетеродимеров. Эти новые молекулы содержат комплексы гетерогенных компонентов, созданные с целью изменить естественное поведение антител и использовать их в лекарственных средствах. 14 н. и 66 з.п. ф-лы, 38 ил., 12 табл., 8 пр.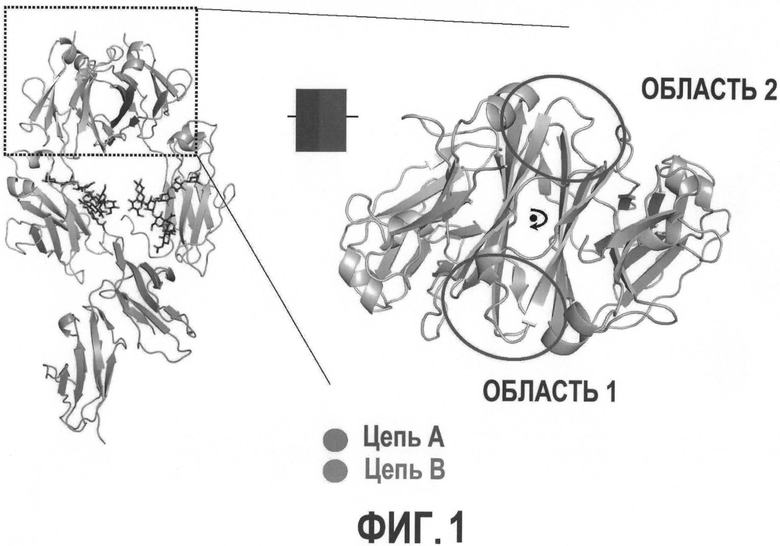
1. Гетеродимерная Fc область для получения гетеромультимера, содержащего данную гетеродимерную Fc область, причем гетеродимерная Fc область содержит вариантный СН3 домен, включающий первый полипептид СН3 домена и второй полипептид СН3 домена, где первый и второй полипептиды СН3 домена каждый независимо содержат аминокислотные модификации,
где гетеродимерная Fc область имеет чистоту больше, чем около 90%,
где вариантный СН3 домен имеет температуру плавления (Tm) около 70°C или выше,
где аминокислотные модификации содержат:
(a) модификации в положениях Т366, Т394, F405 и Y407 или
(b) модификации в положениях Т366, Y407 и K409,
и где
модификация в положении Т366 представляет собой Т366А, T366I, T366L, Т366М или T366V;
модификация в положении Т394 представляет собой T394W;
модификация в положении F405 представляет собой F405A, F405I, F405M, F405T, F405S, F405V или F405W;
модификация в положении Y407 представляет собой Y407A, Y407V, Y407L, Y407I или Y407S;
модификация в положении K409 представляет собой K409F, K409I, K409M, K409S или K409W,
причем нумерация аминокислотных остатков соответствует системе нумерации EU-индекс по Kabat.
2. Гетеродимерная Fc область по п. 1, дополнительно включающая вариантный СН2 домен, содержащий асимметричные аминокислотные модификации, стимулирующие селективное связывание Fc гамма рецептора.
3. Гетеродимерная Fc область по п. 2, в которой вариантный СН2 домен селективно связывает Fc гамма IIIa рецептор по сравнению с СН2 доменом дикого типа.
4. Гетеродимерная Fc область по любому из пп. 1-3, в которой гетеродимерная область Fc имеет чистоту около 95% или выше.
5. Гетеродимерная Fc область по любому из пп. 1-3, где Tm равна около 74°C или выше.
6. Гетеродимерная Fc область по п. 1, в которой аминокислотные модификации содержат модификации по подпункту 1(a), и в которой аминокислотная модификация в положении T366 представляет собой T366I, T366L, T366M или T366V; аминокислотная модификация в положении T394 представляет собой T394W; аминокислотная модификация в положении F405 представляет собой F405A, F405T, F405S или F405V, и аминокислотная модификация в положении Y407 представляет собой Y407V, Y407A, Y407L или Y407I.
7. Гетеродимерная Fc область по п. 6, в которой один из первого и второго полипептидов СН3 домена содержит аминокислотные модификации в положениях F405 и Y407, и другой полипептид СН3 домена содержит аминокислотные модификации в положениях T366 и T394.
8. Гетеродимерная Fc область по п. 7, в которой
(a) по меньшей мере один из первого и второго полипептидов СН3 домена дополнительно содержит аминокислотную модификацию в положении L351, выбранную из L351Y, L351I и L351F, и/или
(b) по меньшей мере один из первого и второго полипептидов СН3 домена дополнительно содержит аминокислотную модификацию в положении K392, выбранную из K392L, K392M, K392V и K392F.
9. Гетеродимерная Fc область по п. 1, в которой аминокислотные модификации содержат модификации по подпункту 1(b), и в которой один из первого и второго полипептидов СН3 домена содержит аминокислотную модификацию в положении Y407, и другой полипептид СН3 домена содержит аминокислотные модификации в положениях T366 и K409.
10. Гетеродимерная Fc область по п. 9, в которой по меньшей мере один из первого и второго полипептидов СН3 домена дополнительно содержит аминокислотную модификацию в положении L351, выбранную из L351Y, L351I и L351F.
11. Гетеродимерная Fc область по п. 10, в которой первый полипептид СН3 домена содержит аминокислотные модификации L351Y и Y407A, и второй полипептид СН3 домена содержит аминокислотную модификацию K409F и одну из аминокислотных модификаций T366V, T366I или T366L.
12. Гетеродимерная Fc область по п. 10, в которой первый полипептид СН3 домена или второй полипептид СН3 домена содержит дополнительную аминокислотную модификацию в одном или более из положений T411, D399, S400, F405, N390 или K392, причем
аминокислотная модификация в положении T411 представляет собой T411N, T411R, T411Q, T411K, T411D, T411E или T411W;
аминокислотная модификация в положении D399 представляет собой D399R, D399W, D399Y или D399K;
аминокислотная модификация в положении S400 представляет собой S400E, S400D, S400R или S400K;
аминокислотная модификация в положении F405 представляет собой F405I, F405M, F405T, F405S, F405V или F405W;
аминокислотная модификация в положении N390 представляет собой N390R, N390K или N390D, и
аминокислотная модификация в положении K392 представляет собой K392V, K392M, K392R, K392L, K392F или K392E.
13. Гетеродимерная Fc область по п. 12, в которой:
(a) первый полипептид СН3 домена дополнительно содержит одну из аминокислотных модификаций D399R или D399W, и второй полипептид СН3 домена дополнительно содержит одну из аминокислотных модификаций K392E или K392L и одну из аминокислотных модификаций T411E или T411D, или
(b) первый полипептид СН3 домена дополнительно содержит одну из аминокислотных модификаций D399R или D399W и одну из аминокислотных модификаций S400R или S400K, и второй полипептид СН3 домена дополнительно содержит одну из аминокислотных модификаций K392E или K392L и одну из аминокислотных модификаций T411E или T411D.
14. Гетеродимерная Fc область по п. 10, в которой первый полипептид СН3 домена содержит аминокислотные модификации L351Y и Y407A, и второй полипептид СН3 домена содержит аминокислотные модификации T366V и K409F.
15. Гетеродимерная Fc область по п. 9, в которой первый полипептид СН3 домена содержит аминокислотную модификацию Y407A, и второй полипептид СН3 домена содержит аминокислотные модификации T366A и K409F.
16. Гетеродимерная Fc область по п. 15, в которой первый полипептид СН3 домена или второй полипептид СН3 домена содержит дополнительные аминокислотные модификации K392E, T411E, D399R и S400R.
17. Гетеродимерная Fc область по п. 15, в которой первый полипептид СН3 домена дополнительно содержит аминокислотные модификации D399R и S400R, а второй полипептид СН3 домена дополнительно содержит аминокислотные модификации K392E и T411E.
18. Гетеродимерная Fc область по п. 6, в которой первый полипептид СН3 домена содержит аминокислотные модификации F405A и Y407V, и второй полипептид СН3 домена содержит одну из аминокислотных модификаций T366L или T366I и аминокислотную модификацию T394W.
19. Гетеродимерная Fc область по п. 18, в которой второй полипептид СН3 домена дополнительно содержит
одну из аминокислотных модификаций K392V, K392M, K392L или K392F.
20. Гетеродимерная Fc область по п. 18, в которой первый полипептид СН3 домена дополнительно содержит аминокислотную модификацию L351Y.
21. Гетеродимерная Fc область по п. 7, в которой первый полипептид СН3 домена или второй полипептид СН3 домена дополнительно содержит аминокислотную модификацию в одном или более из положений S400, Q347, K360 или N390, где:
аминокислотная модификация в положении S400 представляет собой S400E, S400D, S400R или S400K;
аминокислотная модификация в положении Q347 представляет собой Q347R, Q347E или Q347K;
аминокислотная модификация в положении K360 представляет собой K360D, K360E или K360R, и
аминокислотная модификация в положении N390 представляет собой N390R, N390K или N390D.
22. Гетеродимерная Fc область по п. 21, в которой первый полипептид СН3 домена дополнительно содержит по меньшей мере одну из аминокислотных модификаций S400E или Q347R, и второй полипептид СН3 домена дополнительно содержит по меньшей мере одну из аминокислотных модификаций N390R или K360E.
23. Гетеродимерная Fc область по п. 6, в которой первый полипептид СН3 домена содержит аминокислотные модификации L351Y, F405A и Y407V, и второй полипептид СН3 домена содержит одну из аминокислотных модификаций T366I или T366L, одну из аминокислотных модификаций K392L или K392M и аминокислотную модификацию T394W.
24. Гетеродимерная Fc область по п. 19, в которой
(a) первый полипептид СН3 домена содержит аминокислотные модификации F405A и Y407V, и второй полипептид СН3 домена содержит аминокислотные модификации T366I, K392M и T394W, или
(b) первый полипептид СН3 домена содержит аминокислотные модификации F405A и Y407V, и второй полипептид СН3 домена содержит аминокислотные модификации T366L, K392M и T394W.
25. Гетеродимерная Fc область по п. 18, в которой
(a) первый полипептид СН3 домена содержит аминокислотные модификации F405A и Y407V, и второй полипептид СН3 домена содержит аминокислотные модификации T366L и T394W, или
(b) первый полипептид СН3 домена содержит аминокислотные модификации F405A и Y407V, и второй полипептид СН3 домена содержит аминокислотные модификации T366I и T394W.
26. Гетеродимерная Fc область для получения гетеромультимера, содержащего данную гетеродимерную Fc область, причем гетеродимерная Fc область содержит первый полипептид СН3 домена, содержащий аминокислотную модификацию в положении F405 и аминокислотные модификации L351Y и Y407V, и второй полипептид СН3 домена, содержащий аминокислотную модификацию T394W, причем аминокислотная модификация в положении F405 представляет собой F405A, F405I, F405M, F405T, F405S, F405V или F405W, причем нумерация аминокислотных остатков соответствует системе нумерации EU-индекс по Kabat.
27. Гетеродимерная Fc область по п. 26, в которой первый полипептид СН3 домена или второй полипептид СН3 домена дополнительно содержит аминокислотную модификацию в одном или большем количестве из положений K392, T411, T366, L368 или S400, причем
аминокислотная модификация в положении К392 представляет собой K392V, K392M, K392R, K392L, K392F или K392E;
аминокислотная модификация в положении Т411 представляет собой T411N, T411R, T411Q, T411K, T411D, T411E или T411W;
аминокислотная модификация в положении S400 представляет собой S400E, S400D, S400R или S400K;
аминокислотная модификация в положении T366 представляет собой T366A, T366I, T366L, T366M, T366Y, T366S, T366C, T366V или T366W, и
аминокислотная модификация в положении L368 представляет собой L368D, L368R, L368T, L368M, L368V, L368F, L368S и L368A.
28. Гетеродимерная Fc область для получения гетеромультимера, содержащего данную гетеродимерную Fc область, причем гетеродимерная Fc область содержит первый полипептид СН3 домена, содержащий аминокислотные модификации L351Y, F405A и Y407V, и второй полипептид СН3 домена, содержащий аминокислотную модификацию T394W, причем нумерация аминокислотных остатков соответствует системе нумерации EU-индекс по Kabat.
29. Гетеродимерная Fc область по п. 28, в которой второй полипептид СН3 домена дополнительно содержит аминокислотную модификацию T366L или T366I.
30. Гетеродимерная Fc область для получения гетеромультимера, содержащего данную гетеродимерную Fc область, причем гетеродимерная Fc область содержит
(a) первый полипептид СН3 домена, содержащий аминокислотные модификации L351Y, F405A и Y407V, и второй полипептид СН3 домена, содержащий аминокислотные модификации T366I, K392M и T394W, или
(b) первый полипептид СН3 домена, содержащий аминокислотные модификации L351Y, F405A и Y407V, и второй полипептид СН3 домена, содержащий аминокислотные модификации T366L, K392M и T394W, или
(c) первый полипептид СН3 домена, содержащий аминокислотные модификации L351Y, F405A и Y407V, и второй полипептид СН3 домена, содержащий аминокислотные модификации T366L, K392L и T394W, или
(d) первый полипептид СН3 домена, содержащий аминокислотные модификации L351Y, F405A и Y407V, и второй полипептид СН3 домена, содержащий аминокислотные модификации T366I, K392L и T394W,
причем нумерация аминокислотных остатков соответствует системе нумерации EU-индекс по Kabat.
31. Гетеродимерная Fc область по п. 30, в которой
(a) первый полипептид СН3 домена дополнительно содержит аминокислотную модификацию S400E, или
(b) второй полипептид СН3 домена дополнительно содержит аминокислотную модификацию N390R, или
(c) первый полипептид СН3 домена дополнительно содержит аминокислотную модификацию S400E и второй полипептид СН3 домена дополнительно содержит аминокислотную модификацию N390R.
32. Гетеродимерная Fc область по п. 1, в которой аминокислотные модификации в подпункте 1(a) дополнительно содержат модификации в положении L351 и аминокислотные модификации в подпункте 1(b) дополнительно содержат модификации в одном или обоих положениях L351 и F405, причем
модификация в положении L351 представляет собой L351Y, L351I, L351D, L351R или L351F;
модификация в положении T366 представляет собой T366I, T366L, T366M или T366V,
модификация в положении Y407 представляет собой Y407A, Y407V или Y407S; и
модификация в положении K409 представляет собой K409F.
33. Гетеродимерная Fc область по п. 32, в которой первый полипептид СН3 домена содержит аминокислотные модификации в положениях F405 и Y407, и второй полипептид СН3 домена содержит аминокислотные модификации в положениях T366, T394 и K409.
34. Гетеродимерная Fc область по п. 32, в которой первый полипептид СН3 домена содержит аминокислотные модификации в положениях L351, F405 и Y407, а второй полипептид СН3 домена содержит аминокислотные модификации в положениях T366 и T394.
35. Гетеродимерная Fc область по п. 34, в которой первый полипептид СН3 домена содержит аминокислотные модификации L351Y, F405A и Y407V, и второй полипептид СН3 домена содержит одну из аминокислотных модификаций T366I, T366L, T366M или T366V и аминокислотную модификацию T394W.
36. Гетеродимерная Fc область по п. 32, в которой первый полипептид СН3 домена содержит аминокислотные модификации в положениях L351 и Y407, а второй полипептид СН3 домена содержит аминокислотные модификации в положениях T366 и K409.
37. Гетеродимерная Fc область по п. 36, в которой первый полипептид СН3 домена содержит аминокислотные модификации L351Y и Y407A, а второй полипептид СН3 домена содержит одну из аминокислотных модификаций T366I, T366L, Т366М или T366V и аминокислотную модификацию K409F.
38. Гетеродимерная Fc область для получения гетеромультимера, содержащего данную гетеродимерную Fc область, причем гетеродимерная Fc область содержит вариантный СН3 домен, выбранный из:
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации E357H и S364F, и второй полипептид СН3 домена, содержащий аминокислотные модификации L351Y и K370I;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации E357H и S364F, и второй полипептид СН3 домена, содержащий аминокислотные модификации L351Y и K370F;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации E357L, T366A, K409F и T411N, и второй полипептид СН3 домена, содержащий аминокислотные модификации L351Y и Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации L351Y, F405A и Y407V, и второй полипептид СН3 домена, содержащий аминокислотные модификации T366I и T394W;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации L351Y, V397T, F405M и Y407V, и второй полипептид СН3 домена, содержащий аминокислотные модификации T366L и T394W;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации L351Y, V397T, F405M и Y407V, и второй полипептид СН3 домена, содержащий аминокислотные модификации T366M и T394W;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации L351Y, L368M, V397T, F405I и Y407V, и второй полипептид СН3 домена, содержащий аминокислотные модификации T366L и T394W;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации L351Y, V397T, L398D, S400E, F405M и Y407V, и второй полипептид СН3 домена, содержащий аминокислотные модификации T366I, N390R и T394W;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации Q347R, L351Y, V397T, F405M и Y407V, и второй полипептид СН3 домена, содержащий аминокислотные модификации S354N, K360E, T366I и T394W;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T350V, L351Y, V397T, S400E, F405M и Y407V, и второй полипептид СН3 домена, содержащий аминокислотные модификации T350V, T366I, T394W и T411R;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации Q347R, T350V, L351Y, V397T, F405M и Y407V, и второй полипептид СН3 домена, содержащий аминокислотные модификации T350V, K360E, T366I, T394W и T411R;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации L351Y, F405A и Y407V, и второй полипептид СН3 домена, содержащий аминокислотные модификации T366I, K392M и T394W;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации L351Y, S400E, F405A и Y407V, и второй полипептид СН3 домена, содержащий аминокислотные модификации T366I, N390R, K392M и T394W;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации R344H, L351Y, K370Q, G371D, S400E, F405M и Y407V, и второй полипептид СН3 домена, содержащий аминокислотные модификации T366I, N390R, T394W, K409M и T411R;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации L351Y, K370T, G371D, F405M и Y407V, и второй полипептид СН3 домена, содержащий аминокислотные модификации E357Q, S364R, T394W и K409I;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации R344H, L351Y, K370T, G371D, F405M и Y407V, и второй полипептид СН3 домена, содержащий аминокислотные модификации E357Q, S364R, T366I, T394W, K409I и T411R;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации L351Y, K370A, G371S, D399R, F405S и Y407V, и второй полипептид СН3 домена, содержащий аминокислотные модификации E357Q, Q362R, T364Y, T366I, T394W и K409S;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации L351Y, V397S, D399W, F405M и Y407V, и второй полипептид СН3 домена, содержащий аминокислотные модификации Q362R, T366I, T394W и K409M;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации L351Y, V397S, D399Y, F405M и Y407V, и второй полипептид СН3 домена, содержащий аминокислотные модификации Q362R, T366I, T394W и K409I;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366A и K409F, и второй полипептид СН3 домена, содержащий аминокислотную модификацию Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366A и K409F, и второй полипептид СН3 домена, содержащий аминокислотные модификации L351Y и Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366A и K409F, и второй полипептид СН3 домена, содержащий аминокислотные модификации L351F и Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366S и K409F, и второй полипептид СН3 домена, содержащий аминокислотную модификацию Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366C и K409F, и второй полипептид СН3 домена, содержащий аминокислотную модификацию Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366L и K409F, и второй полипептид СН3 домена, содержащий аминокислотную модификацию Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366M и K409F, и второй полипептид СН3 домена, содержащий аминокислотную модификацию Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366V и K409F, и второй полипептид СН3 домена, содержащий аминокислотные модификации L351Y и Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366A и K409F, и второй полипептид СН3 домена, содержащий аминокислотные модификации L351I, T366S, L368F и Y407A,
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366A и K409F, и второй полипептид СН3 домена, содержащий аминокислотные модификации D399W и Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366A и K409F, и второй полипептид СН3 домена, содержащий аминокислотные модификации D399W, S400D и Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366A и K409F, и второй полипептид СН3 домена, содержащий аминокислотные модификации D399W, S400E и Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366A, K409F и T411R, и второй полипептид СН3 домена, содержащий аминокислотные модификации D399W, S400D и Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366A, K409F и T411R, и второй полипептид СН3 домена, содержащий аминокислотные модификации G371D, D399W и Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366A, K409F и T411R, и второй полипептид СН3 домена, содержащий аминокислотные модификации K370Q, G371D, D399W и Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366A, N390R и K409F, и второй полипептид СН3 домена, содержащий аминокислотные модификации D399Y, S400D и Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации Q362R, T366A, K409F и T411K, и второй полипептид СН3 домена, содержащий аминокислотную модификацию Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации Q362K, T366A, K409F и T411R, и второй полипептид СН3 домена, содержащий аминокислотную модификацию Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366A, N390K, K392R, K409F и T411R, и второй полипептид СН3 домена, содержащий аминокислотные модификации S400E и Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366A, N390K, K392R, K409F и T411K, и второй полипептид СН3 домена, содержащий аминокислотные модификации S400E и Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366A, N390K, K409F и T411R, и второй полипептид СН3 домена, содержащий аминокислотные модификации S400D и Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366A, K392L, K409F и T411D, и второй полипептид СН3 домена, содержащий аминокислотные модификации D399R и Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366A, K392L, K409F и T411E, и второй полипептид СН3 домена, содержащий аминокислотные модификации D399R и Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366A, K392L, K409F и T411D, и второй полипептид СН3 домена, содержащий аминокислотные модификации D399K и Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366A, K392L, K409F и T411E, и второй полипептид СН3 домена, содержащий аминокислотные модификации D399K и Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366A, K392M, K409F и T411E, и второй полипептид СН3 домена, содержащий аминокислотные модификации D399R и Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366A, K392F, K409F и T411D, и второй полипептид СН3 домена, содержащий аминокислотные модификации D399R, F405V и Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366A, K409F и T411E, и второй полипептид СН3 домена, содержащий аминокислотные модификации D399R, S400E и Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366A, K409F и T411E, и второй полипептид СН3 домена, содержащий аминокислотные модификации D399R, S400D и Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366A, K392E, K409F и T411E, и второй полипептид СН3 домена, содержащий аминокислотные модификации D399R, S400R и Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366A, K392E, K409F и T411D, и второй полипептид СН3 домена, содержащий аминокислотные модификации D399R, S400R и Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации S364Y, T366A, K409F и T411R, и второй полипептид СН3 домена, содержащий аминокислотную модификацию Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366V и K409W, и второй полипептид СН3 домена, содержащий аминокислотные модификации L351Y, L368S и Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации T366V и K409W, и второй полипептид СН3 домена, содержащий аминокислотные модификации L351Y и Y407A;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации S354R, D356K, E357Q и S364F, и второй полипептид СН3 домена, содержащий аминокислотные модификации S354E, K370F и K439E;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации K409F и T411E, и второй полипептид СН3 домена, содержащий аминокислотные модификации L368V, D399K и S400D;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотную модификацию K409F, и второй полипептид СН3 домена, содержащий аминокислотные модификации L368V и D399Y;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации K360E, L368D и K370E, и второй полипептид СН3 домена, содержащий аминокислотные модификации Y349R, E357R и T411R;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации K360E, K370T и G371D, и второй полипептид СН3 домена, содержащий аминокислотные модификации Y349R, E357Q и S364R;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации K360E, K370T и G371D, и второй полипептид СН3 домена, содержащий аминокислотные модификации Y349R, E357Q и S364K;
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации Y349C, L351Y, V397T, F405M и Y407V, и второй полипептид СН3 домена, содержащий аминокислотные модификации S354C, T366I и T394W; или
вариантного СН3 домена, который содержит первый полипептид СН3 домена, содержащий аминокислотные модификации L351Y, S354C, V397T, F405M и Y407V, и второй полипептид СН3 домена, содержащий аминокислотные модификации Y349C, T366I и T394W.
39. Гетеродимерная Fc область по любому из пп. 1-3 и 6-38, причем гетеродимерная Fc область является на основе Fc области IgG.
40. Гетеродимерная Fc область по п. 39, в которой Fc область IgG представляет собой Fc область IgG1.
41. Гетеродимерная Fc область по любому из пп. 1-3 и 6-38, причем указанная гетеродимерная Fc область содержится в гетеромультимере, который представляет собой биспецифическое антитело.
42. Гетеродимерная Fc область по любому из пп. 1-3 и 6-38, причем указанная гетеродимерная Fc область содержится в гетеромультимере, который представляет собой мультиспецифическое антитело.
43. Гетеродимерная Fc область по любому из пп. 1-3 и 6-38, причем гетеродимерная Fc область содержится в гетеромультимере, который связывается с по меньшей мере одним антигеном рака.
44. Гетеродимерная Fc область по любому из пп. 1-3 и 6-38, причем гетеродимерная Fc область содержится в гетеромультимере, который конъюгирован с терапевтическим агентом.
45. Применение гетеродимерной Fc области по любому из пп. 1-44 для получения фармацевтической композиции для лечения заболевания, расстройства или инфекции пациента.
46. Клетка-хозяин для получения гетеродимерной Fc области по любому из пп. 1-43, причем клетка-хозяин содержит нуклеиновую кислоту, кодирующую гетеродимерную Fc область по любому из пп. 1-43.
47. Нуклеиновокислотная композиция для получения гетеродимерной Fc области по любому из пп. 1-43, содержащая один или большее количество полинуклеотидов, кодирующих гетеродимерную Fc область по любому из пп. 1-43.
48. Нуклеиновокислотная композиция по п. 47, в которой один или большее количество полинуклеотидов содержатся в одном или большем количестве экспрессионных векторов.
49. Гетеродимерная Fc область по любому из пп. 1-3 и 6-38, входящая в состав гетеромультимера, который содержит вариабельный домен или его фрагмент из терапевтического антитела.
50. Гетеродимерная Fc область по п. 49, причем терапевтическое антитело выбрано из группы, состоящей из абаговомаба, адалимумаба, алемтузумаба, аурограба, бапинеузумаба, базиликсимаба, белимумаба, бевацизумаба, бриакинумаба, канакинумаба, катумаксомаба, цертолизумаба пегол, цетуксимаба, даклизумаба, деносумаба, эфализумаба, галиксимаба, гемтузумаба озогамицина, голимумаба, ибритумомаба тиуксетана, инфликсимаба, ипилимумаба, лумиликсимаба, меполизумаба, мотавизумаба, муромонаба, микограба, натализумаба, нимотузумаба, окрелизумаба, офатумумаба, омализумаба, паливизумаба, панитумумаба, пертузумаба, ранибизумаба, реслизумаба, ритуксимаба, теплизумаба, тоцилизумаба/атлизумаба, тозитумомаба, трастузумаба, ProxiniumTM, RencarexTM, устекинумаба и залутумумаба.
51. Применение терапевтически эффективного количества гетеродимерной Fc области по любому из пп. 1-44, 49 и 50 для лечения рака у пациента, имеющего рак, характеризующийся раковым антигеном.
52. Применение терапевтически эффективного количества гетеродимерной Fc области по любому из пп. 1-44 для лечения иммунного расстройства у пациента, имеющего иммунное расстройство, характеризующееся иммунным антигеном.
53. Гетеродимерная Fc область для получения гетеромультимера, содержащего данную гетеродимерную Fc область, причем гетеродимерная Fc область содержит вариантный СН3 домен, включающий первый полипептид СН3 домена и второй полипептид СН3 домена,
причем гетеродимерная Fc область имеет чистоту больше, чем около 90%,
причем вариантный СН3 домен имеет температуру плавления (Tm) около 70°C или выше,
причем первый полипептид СН3 домена содержит аминокислотные модификации в положениях D399 и Y407, и второй полипептид СН3 домена содержит аминокислотные модификации в положениях K409 и T411, причем модификация в положении D399 представляет собой D399R, D399W, D399Y или D399K;
модификация в положении Y407 представляет собой Y407A, Y407I, Y407V или Y407S;
модификация в положении K409 представляет собой K409F, K409W, K409D или K409E, и
модификация в положении T411 представляет собой T411N, T411R, T411Q, T411K, T411D, T411E или T411W,
причем нумерация аминокислотных остатков соответствует системе нумерации EU-индекс по Kabat.
54. Гетеродимерная Fc область по п. 53, в которой первый полипептид СН3 домена дополнительно содержит аминокислотную модификацию в положении L351, и второй полипептид СН3 домена дополнительно содержит аминокислотные модификации в положениях T366 и K392, причем
модификация в положении L351 представляет собой L351Y, L351I, L351D, L351R или L351F;
модификация в положении T366 представляет собой T366I, T366L, T366M или T366V, и
модификация в положении K392 представляет собой K392M, K392L, K392I, K392E или K392D.
55. Гетеродимерная Fc область по п. 53, в которой первый полипептид СН3 домена дополнительно содержит аминокислотную модификацию в положении S400, и второй полипептид СН3 домена дополнительно содержит аминокислотную модификацию в положении N390, причем
модификация в положении S400 представляет собой S400E, S400D, S400R, или S400K, и
модификация в положении N390 представляет собой N390R, N390K или N390D.
56. Гетеродимерная Fc область по п. 53, в которой вариантный СН3 домен имеет температуру плавления Tm около 74°C или выше и гетеромультимер имеет чистоту около 95% или выше.
57. Гетеродимерная Fc область по п. 53, в которой первый полипептид СН3 домена содержит одну из аминокислотных модификаций D399R или D399K и одну из аминокислотных модификаций Y407A, Y407I или Y407V и дополнительно содержит одну или большее количество из аминокислотных модификаций, выбранных из L351Y, S400K и S400R, и второй полипептид СН3 домена содержит одну из аминокислотных модификаций K409F или K409W и одну из аминокислотных модификаций T411D или T411E и дополнительно содержит одну или большее количество аминокислотных модификаций, выбранных из T366V, T366I, T366L, T366M, N390D, N390E, K392L, K392I, K392D и K392E.
58. Гетеродимерная Fc область по п. 54, в которой первый полипептид СН3 домена содержит аминокислотные модификации:
L351Y,
D399R или D399K, и
Y407A, Y407I или Y407V;
и второй полипептид СН3 домена содержит аминокислотные модификации:
T366V, T366I, T366L или T366M,
K392L, K392I, K392D или K392E,
K409F или K409W и
T411D или T411E.
59. Гетеродимерный СН3 домен для получения гетеромультимера, имеющего гетеродимерную Fc область, содержащую гетеродимерный СН3 домен, причем гетеродимерный СН3 домен содержит первый полипептид СН3 домена и второй полипептид СН3 домена, где первый и второй полипептиды СН3 домена содержат аминокислотные модификации в положениях Т366, Т394, F405 и Y407,
где аминокислотная модификация в положении T366 представляет собой T366I, T366L, T366M или T366V, аминокислотная модификация в положении T394 представляет собой T394W, аминокислотная модификация в положении F405 представляет собой F405A, и аминокислотная модификация в положении Y407 представляет собой Y407V, Y407A, Y407L или Y407I,
где гетеродимерный СН3 домен имеет температуру плавления (Tm) около 70°C или выше и чистоту выше, чем около 90%, и
причем нумерация аминокислотных остатков соответствует системе нумерации EU-индекс по Kabat.
60. Гетеродимерный СН3 домен по п. 59, в котором по меньшей мере один из первого и второго полипептидов СН3 доменов дополнительно содержит аминокислотную модификацию L351Y.
61. Гетеродимерный СН3 домен по п. 59, в котором по меньшей мере один из первого и второго полипептидов СН3 домена дополнительно содержит аминокислотную модификацию в положении K392, выбранную из K392L, K392M, K392V и K392F.
62. Гетеродимерный СН3 домен по п. 59, имеющий Tm около 74°C или выше.
63. Гетеродимерный СН3 домен по п. 59, имеющий чистоту около 95% или выше.
64. Гетеродимерный СН3 домен п. 59, в котором аминокислотная модификация в положении Т366 представляет собой T366I или T366L.
65. Гетеродимерный СН3 домен по п. 64, в котором аминокислотная модификация в положении Y407 представляет собой Y407V.
66. Гетеродимерный СН3 домен по п. 65, в котором первый полипептид СН3 домена содержит аминокислотные модификации L351Y, F405A и Y407V, а второй полипептид СН3 домена содержит одну из аминокислотных модификаций T366I или T366L; одну из аминокислотных модификаций K392L или K392M и аминокислотную модификацию T394W.
67. Гетеродимерный СН3 домен по п. 66, в котором первый полипептид СН3 домена содержит аминокислотные модификации L351Y, F405A и Y407V, а второй полипептид СН3 домена содержит аминокислотные модификации T366L, K392M и T394W.
68. Гетеродимерный СН3 домен по п. 66, в котором первый полипептид СН3 домена содержит аминокислотные модификации L351Y, F405A и Y407V, а второй полипептид СН3 домена содержит аминокислотные модификации T366L, K392L и T394W.
69. Гетеродимерный СН3 домен по п. 66, в котором первый полипептид СН3 домена содержит аминокислотные модификации L351Y, F405A и Y407V, а второй полипептид СН3 домена содержит аминокислотные модификации T366I, K392M и T394W.
70. Гетеродимерный СН3 домен по п. 66, в котором первый полипептид СН3 домена содержит аминокислотные модификации L351Y, F405A и Y407V, а второй полипептид СН3 домена содержит аминокислотные модификации T366I, K392L и T394W.
71. Гетеродимерный СН3 домен по п. 66, в котором первый полипептид СН3 домена дополнительно содержит аминокислотную модификацию S400E.
72. Гетеродимерный СН3 домен по п. 71, в котором второй полипептид СН3 домена дополнительно содержит аминокислотную модификацию N390R.
73. Гетеродимерный СН3 домен по п. 72, в котором первый полипептид СН3 домена содержит аминокислотные модификации L351Y, S400E, F405A и Y407V, а второй полипептид СН3 домена содержит аминокислотные модификации T366L, N390R, K392M и T394W.
74. Гетеродимерный СН3 домен по любому из пп. 59-73, который содержится в Fc области IgG.
75. Гетеродимерный СН3 домен по п. 74, в котором Fc область IgG представляет собой Fc область IgG1.
76. Гетеродимерный СН3 домен по любому из пп. 59-73, который содержится в гетеромультимере, представляющем собой биспецифическое или мультиспецифическое антитело.
77. Гетеродимерный СН3 домен по п. 76, где биспецифическое или мультиспецифическое антитело связано с по меньшей мере одним антигеном рака.
78. Гетеродимерный СН3 домен по любому из пп. 59-73, который содержится в гетеромультимере, конъюгированном с терапевтическим агентом.
79. Применение гетеродимерного СН3 домена по любому из пп. 59-78 для получения фармацевтической композиции для лечения заболевания, расстройства или инфекции пациента.
80. Способ экспрессии гетеромультимера, содержащего гетеродимерную Fc область по любому из пп. 1-43, 49, 50 и 53-77, в клетках млекопитающего, причем способ включает:
(a) трансфекцию по меньшей мере одной клетки млекопитающего одним или большим количеством полинуклеотидов, кодирующих гетеромультимер, содержащий гетеродимерную Fc область по любому из пп. 1-43, 49, 50 и 53-77, для получения по меньшей мере одной временно или устойчиво трансфицированной клетки млекопитающего; и
(b) культивирование по меньшей мере одной временно или устойчиво трансфицированной клетки млекопитающего в условиях, подходящих для экспрессии гетеромультимера.
| WO2009089004 A1, 16.07.2009 | |||
| Сборная бумажная гильза для дробового ружья | 1928 |
|
SU11879A1 |

