ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] По настоящей заявке испрашивается приоритет по предварительной заявке на патент США №62/003663, поданной 28 мая 2014 года, и предварительной заявке на патент США №62/154055, поданной 28 апреля 2015 года, которые включены в данный документ посредством ссылки в полном объеме для любых целей.
[0002] Данная заявка связана с РСТ/СА2013/050914, поданной 28 ноября 2013 года, предварительной заявке на патент США №61/730906, поданной 28 ноября 2012 года, предварительной заявке на патент США №61/761641, поданной 6 февраля 2013 года, предварительной заявке на патент США №61/818874, поданной 2 мая 2013 года, и предварительной заявке на патент США №61/869200, поданной 23 августа 2013 года, содержание которых включено в данный документ посредством ссылки в полном объеме для любых целей.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0003] Рассматриваемая заявка содержит перечень последовательностей, который представлен в электронном виде в формате ASCII и включен в данный документ посредством ссылки во всей своей полноте. Указанная копия ASCII, созданная 29 мая 2015 года, имеет название 97993-945204(000110PC)_SL.txt и размер 27012 байт.
УРОВЕНЬ ТЕХНИКИ
[0004] Биспецифические антитела способны связываться с двумя различными эпитопами. Эти эпитопы могут быть на одном и том же или на разных антигенах. Такая особенность биспецифических антител делает их привлекательным инструментом для различных терапевтических применений в случаях, если имеется терапевтическое преимущество от таргетинга или рекрутинга более чем одной молекулы при лечении болезни. Один из подходов к формированию биспецифических антител будет включать одновременную экспрессию двух уникальных тяжелых цепей антитела и двух уникальных легких цепей антитела. Правильное формирование биспецифических антител в аналогичном к дикому типу формате остается проблемой, поскольку в процессе эволюции тяжелые цепи антител приобрели свойство связывать легкие цепи антител относительно неспецифическим способом. В результате такого неспецифического спаривания одновременная экспрессия двух тяжелых цепей антитела и двух легких цепей антитела естественным образом приводит к случайной перетасовке спариваний "тяжелая цепь - легкая цепь". Такое неправильное спаривание остается серьезной проблемой для генерации биспецифических терапевтических средств, где однородное спаривание является необходимым условием хорошей технологичности и биологической эффективности.
[0005] Описано несколько подходов к получению биспецифических антител, в которых специфические легкие цепи антитела или фрагмента спариваются со специфическими тяжелыми цепями антитела или фрагментов. Обзор различных подходов к решению данной проблемы можно найти у Klein et al, (2012 год) mAbs 4:6, 1-11. Международная заявка на патент № РСТ/ЕР2011/056388 (WO 2011/131746) описывает метод получения гетеродимерного белка in vitro, в котором асимметрические мутации вводятся в области СН3 двух моноспецифических исходных белков для того, чтобы управлять направленным обменом "Fab-фрагментами" или "полумолекулами" между двумя моноспецифическими IgG4- или IgG4-подобными антителами, при инкубации в восстановительных условиях.
[0006] Schaefer et al (Roche Diagnostics GmbH) описывают способ сборки двух тяжелых и двух легких цепей, полученных из двух существующих антител, в человеческие двухвалентные биспецифические антитела IgG без использования искусственных линкеров (PNAS (2011) 108(27): 11187-11192). Способ включает обмен доменов тяжелой цепи и легкой цепи в антигенсвязывающем фрагменте (Fab) одной половины биспецифического антитела.
[0007] Strop et al (Rinat-Pfizer Inc.) описывают способ получения стабильных биспецифических антител путем экспрессии и очистки двух представляющих интерес антител по отдельности, с последующим их смешиванием при определенных окислительно-восстановительных условиях (J. Mol. Biol. (2012 год) 420:204-19).
[0008] Zhu et al (Genentech) спроектировали мутации в области контакта VL/VH конструкции диатела, состоящей из фрагментов варианта домена антитела, полностью лишенного константных областей, и синтезировали гетеродимерное диатело (Protein Science (1997 год) 6:781-788). Аналогично, Igawa et al (Chugai) также разработали мутации в области контакта VL/VH одноцепочечного диатела, чтобы способствовать избирательной экспрессии и ингибировать конформационную изомеризацию диатела (Protein Engineering, Design & Selection (2010 год) 23:667-677).
[0009] Публикация патента США №2009/0182127 (Novo Nordisk, Inc.) описывает синтез биспецифических антител путем модификации аминокислотных остатков в области контакта Fc и области контакта CH1:CL легких-тяжелых пар цепей, которые уменьшают способность легкой цепи одной пары взаимодействовать с тяжелой цепью другой пары.
[0010] Публикация патента США №2014/0370020 (Chugai) описывает регулирование связи между областями СН1 и CL антитела путем замены аминокислот, которые существуют в области контакта между этими областями, на заряженные аминокислоты.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0011] В данном документе описана выделенная полипептидная антигенсвязывающая конструкция содержащая по меньшей мере первый гетеродимер и второй гетеродимер, первый гетеродимер, содержащий первую полипептидную последовательность тяжелой цепи иммуноглобулина (H1) и первую полипептидную последовательность легкой цепи иммуноглобулина (L1); и второй гетеродимер, содержащий вторую полипептидную последовательность тяжелой цепи иммуноглобулина (Н2), и вторую полипептидную последовательность легкой цепи иммуноглобулина (L2), причем по меньшей мере одна из последовательностей H1 или L1 последовательностей первого гетеродимера отличается от соответствующей последовательности Н2 или L2 второго гетеродимера, и причем каждая из H1 и Н2 содержит по меньшей мере вариабельный домен тяжелой цепи (домен VH) и константный домен тяжелой цепи (домен CH1); каждая из L1 и L2 содержит по меньшей мере вариабельный домен легкой цепи (домен VL) и константный домен легкой цепи (домен CL); и по меньшей мере одна из H1, Н2, L1 и L2 содержит по меньшей мере одну аминокислотную модификацию по меньшей мере в одном константном домене и/или по меньшей мере одном вариабельном домене, причем H1 преимущественно спаривается с L1 по сравнению с L2, и Н2 преимущественно спаривается с L2 по сравнению с L1.
[0012] В некоторых аспектах конструкция дополнительно содержит гетеродимерный Fc, Fc, содержащий по меньшей мере две последовательности CH3, причем Fc соединен, с помощью или без помощи одного или более линкеров, с первым гетеродимером и вторым гетеродимером, причем димеризованные последовательности CH3 имеют температуру плавления (Tm) от около 68°С или выше, как определено методом дифференциальной сканирующей калориметрии (DSC), и при этом конструкция является биспецифической.
[0013] В некоторых аспектах по меньшей мере одну аминокислотную модификацию выбирают из по меньшей мере одной аминокислотной модификации, представленной в таблицах или примерах.
[0014] В некоторых аспектах H1 спаривается преимущественно с L1 по сравнению с L2, и Н2 спаривается преимущественно с L2 по сравнению с L1, в случае если H1, Н2, L1 и L2 коэкспрессируются в клетке или клетке млекопитающего, или в случае если H1, Н2, L1 и L2 коэкспрессируются в бесклеточной системе экспрессии, или в случае если H1, Н2, L1 и L2 совместно продуцируются, или в случае если H1, Н2, L1 и L2 совместно продуцируются с помощью окислительно-восстановительного способа получения.
[0015] В некоторых аспектах по меньшей мере одна из H1, Н2, L1 и L2 содержит по меньшей мере одну аминокислотную модификацию домена VH и/или VL и по меньшей мере одну аминокислотную модификацию домена CH1 и/или CL таким образом, что H1 спаривается преимущественно с L1 по сравнению с L2, и/или Н2 спаривается преимущественно с L2 по сравнению с L1.
[0016] В некоторых аспектах, если H1 содержит по меньшей мере одну аминокислотную модификацию в домене CH1, затем по меньшей мере одна из L1 и L2 содержит по меньшей мере одну аминокислотную модификацию в домене CL; и/или, если H1 содержит по меньшей мере одну аминокислотную модификацию в домене VH, то по меньшей мере одна из L1 и L2 содержит по меньшей мере одну аминокислотную модификацию в домене VL.
[0017] В некоторых аспектах H1, L1, Н2 и/или L2 содержат по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислотных мутаций. В некоторых аспектах по меньшей мере одна из H1, Н2, L1 и L2 содержит по меньшей мере 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислотных модификаций по меньшей мере одного константного домена и/или по меньшей мере одного вариабельного домена.
[0018] В некоторых аспектах, в случае, если обе L1 и L2 коэкспрессируются с по меньшей мере одной из H1 и Н2, значение относительного спаривания по меньшей мере одной из пар гетеродимеров H1-L1 и H2-L2 по отношению к соответствующей паре гетеродимеров H1-L2 или H2-L1 составляет больше, чем 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99%, и при этом значение относительного спаривания модифицированной пары гетеродимеров H1-L1 или H2-L2 больше, чем соответствующее значение относительного спаривания, наблюдаемого в соответствующей паре гетеродимеров H1-L1 или H2-L2 без по меньшей мере одной аминокислотной модификации.
[0019] В некоторых аспектах термостабильность, измеренная на основе температуры плавления (Tm), измеренной с помощью DSF по меньшей мере одного из первого и второго гетеродимеров находится в пределах около 0, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10°С от Tm соответствующего гетеродимера без по меньшей мере одной аминокислотной модификации. В некоторых аспектах термостабильность, измеренная на основе температуры плавления (Tm), измеренной с помощью DSF каждого гетеродимера, содержащего по меньшей мере одну аминокислотную модификацию, находится в пределах около 0, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10°С от Tm соответствующего гетеродимера без по меньшей мере одной аминокислотной модификации. В некоторых вариантах реализации изобретения термостабильность, измеренная с помощью температуры плавления (Tm), измеренная с помощью DSF каждого гетеродимера, содержащего по меньшей мере одну аминокислотную модификацию, находится в пределах около 0, 1, 2 или 3°С от Tm соответствующего гетеродимера без по меньшей мере одной аминокислотной модификации.
[0020] В некоторых аспектах аффинность каждого гетеродимера к антигену, с которым он связывается, находится в пределах около 1, 2, 3, 4, 5, 6, 7, 8, 9, 10-кратного значения аффинности соответствующего немодифицированного гетеродимера по отношению к тому же антигену, как измерено с помощью поверхностного плазмонного резонанса (SPR) или FACS.
[0021] В некоторых аспектах по меньшей мере одна из H1 и L1 содержит по меньшей мере один домен, содержащий по меньшей мере одну аминокислотную модификацию, что приводит к большей стерической комплементарности аминокислот, в случае если H1 спаривается с L1 по сравнению с L2. В некоторых аспектах по меньшей мере одна из Н2 и L2 содержит по меньшей мере один домен, содержащий по меньшей мере одну аминокислотную модификацию, что приводит к большей стерической комплементарности аминокислот, в случае если Н2 спаривается с L2 по сравнению с L1. В некоторых аспектах по меньшей мере одна из H1 и L1 содержит по меньшей мере один домен, содержащий по меньшей мере одну аминокислотную модификацию, что приводит к большей электростатической комплементарности между заряженными аминокислотами, в случае если H1 спаривается с L1 по сравнению с L2. В некоторых аспектах по меньшей мере одна из Н2 и L2 содержит по меньшей мере один домен, содержащий по меньшей мере одну аминокислотную модификацию, что приводит к большей электростатической комплементарности между заряженными аминокислотами, в случае если Н2 спаривается с L2 по сравнению с L1.
[0022] В некоторых аспектах по меньшей мере одна аминокислотная модификация представляет собой набор мутаций, представленный по меньшей мере в одной из таблиц или примеров.
[0023] В некоторых аспектах конструкция дополнительно содержит Fc, содержащий по меньшей мере две последовательности CH3, причем Fc соединен, с помощью или без помощи одного или более линкеров, с первым гетеродимером и вторым гетеродимером.
[0024] В некоторых аспектах Fc представляет собой человеческий Fc, Fc человеческого IgG1, Fc человеческого IgA, Fc человеческого IgG, Fc человеческого IgD, Fc человеческого IgE, Fc человеческого IgM, Fc человеческого IgG2, Fc человеческого IgG3 или Fc человеческого IgG4. В некоторых аспектах Fc представляет собой гетеродимерный Fc. В некоторых аспектах Fc содержит одну или более модификаций по меньшей мере в одной из последовательностей CH3. В некоторых аспектах температура плавления (Tm) димеризованных последовательностей CH3, измеренная с помощью DSC, составляет около 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 77,5, 78, 79, 80, 81, 82, 83, 84 или 85°С, или выше. В некоторых аспектах Fc представляет собой гетеродимер, образованный с чистотой более чем около 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% при получении; или где Fc представляет собой гетеродимер, образованный с чистотой более чем около 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% при экспрессии или при экспрессии посредством одной клетки. В некоторых аспектах Fc содержит одну или более модификаций по меньшей мере в одной из последовательностей CH3, которые способствуют образованию гетеродимерного Fc со стабильностью, сравнимой со стабильностью гомодимерного Fc дикого типа. В некоторых аспектах Fc дополнительно содержит по меньшей мере одну последовательность CH2. В некоторых аспектах последовательность(-и) CH2 Fc содержат одну или более модификаций. В некоторых аспектах Fc содержит одну или более модификаций для содействия избирательному связыванию Fc-гамма-рецепторов.
[0025] В некоторых вариантах реализации изобретения Fc содержит:
i) гетеродимерный Fc IgG1, имеющий модификации L351Y_F405A_Y407V в первом полипептиде Fc и модификации T366L_K392M_T394W во втором полипептиде Fc;
ii) гетеродимерный Fc IgG1, имеющий модификации L351Y_F405A_Y407V в первом полипептиде Fc и модификации T366LK392LT394W во втором полипептиде Fc;
iii) гетеродимерный Fc IgG1, имеющий модификации T350V_L351Y_F405A_Y407V в первом полипептиде Fc и модификации T350VT366LK392LT394W во втором полипептиде Fc;
iv) гетеродимерный Fc IgG1, имеющий модификации T350V_L351Y_F405A_Y407V в первом полипептиде Fc и модификации T350V_T366L_K392M_T394W во втором полипептиде Fc; или
v) гетеродимерный Fc IgG1, имеющий модификации T350V_L351Y_S400E_F405A_Y407V в первом полипептиде Fc и модификации T350V_T366L_N390R_K392M_T394W во втором полипептиде Fc.
[0026] В некоторых аспектах Fc спарен с гетеродимерами с помощью одного или более линкеров, или при этом Fc спарен с H1 и Н2 с помощью одного или более линкеров. В некоторых аспектах один или более линкеров представляют собой один или более полипептидных линкеров. В некоторых аспектах один или более линкеров содержит одну или более шарнирных областей антитела. В некоторых аспектах один или более линкеров содержит одну или более шарнирных областей IgG1. В некоторых аспектах один или более линкеров содержит одну или более модификаций. В некоторых аспектах одна или более модификаций в одном или более линкерах содействует избирательному связыванию Fc-гамма-рецепторов.
[0027] В некоторых аспектах по меньшей мере одна аминокислотная модификация представляет собой по меньшей мере одну аминокислотную мутацию или при этом по меньшей мере одна аминокислотная модификация представляет собой по меньшей мере одну аминокислотную замену.
[0028] В некоторых аспектах каждая из последовательностей H1, Н2, L1 и L2 является человеческой последовательностью.
[0029] В некоторых аспектах конструкция является мультиспецифической или биспецифической. В некоторых аспектах конструкция является мультивалентной или бивалентной.
[0030] В некоторых аспектах гетеродимеры, описанные в данном документе, преимущественно спариваются с образованием биспецифических антител. Например, в некоторых вариантах реализации изобретения последовательности тяжелой цепи H1 и Н2 полипептида содержат полноразмерную последовательность тяжелой цепи, содержащую константный домен тяжелой цепи (домен CH1), домен CH2 и домен CH3. В некоторых вариантах реализации изобретения процент правильно спаренных тяжелых и легких цепей в биспецифическом антителе (например, H1-L1:H2-L2) составляет более чем 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99%.
[0031] Кроме того, в данном документе описан выделенный полинуклеотид или набор выделенных полинуклеотидов, содержащий по меньшей мере одну последовательность, кодирующую конструкцию или тяжелую цепь, или легкую цепь, описанную в данном документе. В некоторых аспектах полинуклеотид или набор полинуклеотидов представляет собой кДНК. Кроме того, в данном документе описан вектор или набор векторов, содержащих один или более полинуклеотидов или наборов полинуклеотидов, описанных в данном документе. В некоторых аспектах вектор или набор векторов выбирают из группы, состоящей из плазмиды, мультицистронного вектора, вирусного вектора, неэписомального вектора млекопитающих, вектора экспрессии и рекомбинантного вектора экспрессии.
[0032] Кроме того, в данном документе описана выделенная клетка, содержащая полинуклеотид или набор полинуклеотидов, описанных в данном документе, или вектор, или набор векторов, описанных в данном документе. В некоторых аспектах клетка представляет собой гибриду, клетку яичника китайского хомячка (СНО) или клетку HEK293.
[0033] Кроме того, в данном документе описана фармацевтическая композиция, содержащая конструкцию, описанную в данном документе, и фармацевтически приемлемый носитель. В некоторых аспектах композиция дополнительно содержит одно или более веществ, выбранных из группы, состоящей из буфера, антиоксиданта, низкомолекулярного препарата, лекарственного средства, белка, аминокислоты, углевода, липида, хелатирующего агента, стабилизатора и эксципиента.
[0034] Кроме того, в данном документе описано применение конструкции или фармацевтической композиции, описанной в данном документе, для лечения заболевания или нарушения, или рака, или сосудистого заболевания у субъекта, или в получении лекарственного средства.
[0035] Кроме того, в данном документе описан способ лечения субъекта, страдающего заболеванием или нарушением, или раком, или сосудистым заболеванием, включающий введение субъекту конструкции, описанной в данном документе, или композиции, описанной в данном документе.
[0036] Кроме того, в данном документе описан способ получения конструкции, описанной в данном документе, из культуры клетки-хозяина, причем способ включает стадии: (а) получения культуры клетки-хозяина, содержащей по меньшей мере одну клетку-хозяина, содержащую одну или более последовательностей нуклеиновых кислот, кодирующих конструкцию; и (b) выделения конструкции из культуры клетки-хозяина.
[0037] Кроме того, в данном документе описан способ получения конструкции, описанной в данном документе, включающий стадии: (а) получения H1, L1, Н2 и L2; (b), позволения H1 спариваться преимущественно с L1 по сравнению с L2 и Н2 спариваться преимущественно с L2 по сравнению с L1; и (с) получения конструкции.
[0038] Кроме того, в данном документе описан способ получения конструкции, описанной в данном документе, включающий получение полинуклеотида или набора полинуклеотидов, кодирующих по меньшей мере одну конструкцию; определение оптимальных соотношений каждого из полинуклеотидов или набора полинуклеотидов для введения по меньшей мере в одну клетку-хозяина, причем оптимальные соотношения определяются путем оценки количества пар гетеродимеров H1-L1 и H2-L2, образующихся при экспрессии H1, L1, Н2 и L2 по сравнению с неправильно спаренными парами гетеродимеров H1-L2 и H2-L1, образующихся при экспрессии H1, L1, Н2 и L2; выбор предпочтительного оптимального соотношения, причем трансфекция по меньшей мере одной клетки-хозяина с предпочтительным оптимальным соотношением полинуклеотида или набора полинуклеотидов приводит к экспрессии конструкции; трансфекцию по меньшей мере одной клетки-хозяина с оптимальным соотношением полинуклеотида или набора полинуклеотидов; и культивирование по меньшей мере одной клетки-хозяина для экспрессии конструкции.
[0039] В некоторых аспектах выбор оптимального соотношения оценивается путем трансфекции в системе временной трансфекции. В некоторых аспектах трансфекция по меньшей мере одной клетки-хозяина с предпочтительным оптимальным соотношением полинуклеотида или набора полинуклеотидов приводит к оптимальной экспрессии конструкции. В некоторых аспектах конструкция содержит Fc, содержащий по меньшей мере две последовательности CH3, причем Fc соединен, с помощью или без помощи одного или более линкеров, с первым гетеродимером и вторым гетеродимером. В некоторых аспектах Fc представляет собой гетеродимер, необязательно содержащий одну или несколько аминокислотных модификаций.
[0040] Кроме того, в данном документе описан машиночитаемый носитель данных, хранящий набор данных, содержащий данные, представляющие комплементарные мутации в первом гетеродимере, содержащем первую полипептидную последовательность тяжелой цепи иммуноглобулина (H1) и первую полипептидную последовательность легкой цепи иммуноглобулина (L1); и втором гетеродимере, содержащем второю полипептидную последовательность тяжелой цепи иммуноглобулина (Н2) и вторую полипептидную последовательность легкой цепи иммуноглобулина (L2), причем каждая из H1 и Н2 содержит по меньшей мере вариабельный домен тяжелой цепи (VH домен) и константный домен тяжелой цепи (CH1 домен); причем каждая из L1 и L2 содержит по меньшей мере вариабельный домен легкой цепи (VL домен) и константный домен легкой цепи (CL домен), и при этом набор данных комплементарных мутаций содержит данные, представляющие эти мутации, перечисленные в таблицах или примерах, или подмножество этих мутаций; и исполняемый компьютером код для определения вероятности того, что H1 будет спариваться преимущественно с L1 по сравнению с L2 и/или Н2 будет спариваться преимущественно с L2 по сравнению с L1.
[0041] Кроме того, в данном документе описан компьютеризированный способ определения преимущественного спаривания, включающий: получение набора данных, содержащего данные, представляющие комплементарные мутации в первом гетеродимере, содержащем первую полипептидную последовательность тяжелой цепи иммуноглобулина (H1) и первую полипептидную последовательность легкой цепи иммуноглобулина (L1); и втором гетеродимере, содержащем второю полипептидную последовательность тяжелой цепи иммуноглобулина (Н2) и вторую полипептидную последовательность легкой цепи иммуноглобулина (L2), причем каждая из H1 и Н2 содержит по меньшей мере вариабельный домен тяжелой цепи (VH домен) и константный домен тяжелой цепи (CH1 домен); причем каждая из L1 и L2 содержит по меньшей мере вариабельный домен легкой цепи (VL домен) и константный домен легкой цепи (CL домен), и при этом набор данных комплементарных мутаций содержит данные, представляющие эти мутации, перечисленные в таблицах или примерах, или подмножество этих мутаций; и определяемая с помощью компьютерного процессора вероятность того, что H1 будет спариваться преимущественно с L1 по сравнению с L2, и/или Н2 будет спариваться преимущественно с L2 по сравнению с L1. В некоторых аспектах способ дополнительно включает получение конструкции, описанной в данном документе.
[0042] Кроме того, в данном документе описан способ получения биспецифической антигенсвязывающей полипептидной конструкции, указанная биспецифическая конструкция содержит первый гетеродимер, содержащий первую полипептидную последовательность тяжелой цепи иммуноглобулина (H1) и первую полипептидную последовательность легкой цепи иммуноглобулина (L1) из первого моноспецифического антигенсвязывающего полипептида; и второй гетеродимер, содержащий вторую полипептидную последовательность тяжелой цепи иммуноглобулина (Н2) и вторую полипептидную последовательность легкой цепи иммуноглобулина (L2) из второго моноспецифического антигенсвязывающего полипептида, причем каждая из H1 и Н2 содержит по меньшей мере вариабельный домен тяжелой цепи (VH домен) и константный домен тяжелой цепи (CH1 домен); причем каждая из L1 и L2 содержит по меньшей мере вариабельный домен легкой цепи (VL домен) и константный домен легкой цепи (CL домен), способ включает: введение одной или более комплементарных мутаций из набора данных, описанного в данном документе, в первый гетеродимер и/или второй гетеродимер; и коэкспрессию первого гетеродимера и второго гетеродимера по меньшей мере в одной клетке-хозяине для получения продукта экспрессии, содержащего биспецифическую конструкцию.
[0043] В некоторых аспектах способ дополнительно включает определение количества биспецифической конструкции в продукте экспрессии по отношению к другим полипептидным продуктам для выбора предпочтительного подмножества комплементарных мутаций. В некоторых аспектах биспецифическая конструкция продуцируется с чистотой более чем 70% (например, более чем 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) по сравнению с другими полипептидными продуктами. В некоторых аспектах набор данных представляет собой набор данных, описанный в данном документе. В некоторых аспектах способ дополнительно включает стадию добавления дополнительных аминокислотных модификаций по меньшей мере к одной из H1, Н2, L1 или L2 для увеличения чистоты биспецифической конструкции по сравнению с другими полипептидными продуктами. В некоторых аспектах конструкция содержит Fc, содержащий по меньшей мере две последовательности CH3, причем Fc соединен, с помощью или без помощи одного или более линкеров, с первым гетеродимером и вторым гетеродимером. В некоторых аспектах Fc представляет собой гетеродимер, необязательно содержащий одну или несколько аминокислотных модификаций. В некоторых аспектах антигенсвязывающий полипептид представляет собой антитело, Fab или scFv.
[0044] В некоторых вариантах реализации конструкции, H1 и/или Н2 содержит по меньшей мере одну или множество аминокислотных модификаций в L124, K145, D146, Q179 и S186, и L1 и/или L2 содержит по меньшей мере одну или множество аминокислотных модификации в Q124, S131, V133, Q160, S176, Т178 и Т180. Например, в некоторых вариантах реализации изобретения H1 и/или Н2 содержит по меньшей мере одну или множество аминокислотных модификаций, выбранных из L124R, L124E, K145M, K145T, D146N, Q179E, Q179K, S186R и S186K, и L1 и/или L2 содержит по меньшей мере одну или множество аминокислотных модификаций, выбранных из Q124E, S131R, S131K, V133G, Q160E, S176R, S176D, T178D, Т178Е и Т180Е. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации, выбранные из группы, состоящей из L124E, K145М, K145Т и Q179E, или их комбинации; L1 содержит аминокислотные модификации, выбранные из группы, состоящей из S131R, S131K, V133Gh S176R, или их комбинации; Н2 содержит аминокислотные модификации, выбранные из группы, состоящей из L124R, D146N, Q179K, S186R и S186K, или их комбинации; и L2 содержит аминокислотные модификации, выбранные из группы, состоящей из Q124E, V133G, Q160E, S176D, T178D, Т178Е и Т180Е, или их комбинации. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации L124E, K145Т и Q179E; L1 содержит аминокислотные модификации S131K, V133G и S176R; Н2 содержит аминокислотные модификации L124R и S186R; и L2 содержит аминокислотные модификации V133G, S176D и T178D.
[0045] В некоторых вариантах реализации конструкции, H1 и/или Н2 содержит по меньшей мере одну или множество аминокислотных модификаций в L124, L143, K145, D146, Q179 и S186, и L1 и/или L2 содержит по меньшей мере одну или множество аминокислотных модификации в Q124, V133, Q160, S176, Т178 и Т180. В некоторых вариантах реализации изобретения H1 и/или Н2 содержит по меньшей мере одну или множество аминокислотных модификаций, выбранных из L124E, L124R, L143E, L143D, K145T, K145M, D146N, Q179K, S186R и S186K, и L1 и/или L2 содержит по меньшей мере одну или множество аминокислотных модификаций, выбранных из Q124K, Q124E, V133G, Q160K, S176R, S176D, Т178Е, T178K, T178R, T178D и Т180Е. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации, выбранные из группы, состоящей из L124E, L143E, L143D, K145T и K145M, или их комбинации; L1 содержит аминокислотные модификации, выбранные из группы, состоящей из Q124K, V133G, Q160K, S176R, T178K и T178R, или их комбинации; Н2 содержит аминокислотные модификации, выбранные из группы, состоящей из L124R, D146N, Q179K, S186R и S186K, или их комбинации; и L2 содержит аминокислотные модификации, выбранные из группы, состоящей из Q124E, V133G, S176D, Т178Е, T178D и Т180Е, или их комбинации. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации L124E, L143E и K145T; L1 содержит аминокислотные модификации Q124K, V133G и S176R; Н2 содержит аминокислотные модификации L124R и Q179K; и L2 содержит аминокислотные модификации V133G, S176D и Т178Е. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации L124E, L143E и K145T; L1 содержит аминокислотные модификации Q124K, V133G и S176R; Н2 содержит аминокислотные модификации L124R и S186R; и L2 содержит аминокислотные модификации V133G, S176D и T178D.
[0046] В некоторых вариантах реализации конструкции, H1 и/или Н2 содержит по меньшей мере одну или множество аминокислотных модификаций в Q39, L45, L124, L143, F122 и Н172, и L1 и/или L2 содержит по меньшей мере одну или множество аминокислотных модификации в Q38, Р44, Q124, S131, V133, N137, S174, S176 и Т178. В некоторых вариантах реализации изобретения H1 и/или Н2 содержит по меньшей мере одну или множество аминокислотных модификаций, выбранных из Q39E, Q39R, L45P, F122C, L124E, L124R, L143F, Н172Т и H172R; и L1 и/или L2 содержит по меньшей мере одну или множество аминокислотных модификаций, выбранных из Q38R, Q38E, P44F, Q124C, S131T, S131E, V133G, N137K, S174R, S176R, S176K, S176D, T178Y и T178D. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации, выбранные из группы, состоящей из Q39E, L45P, F122C, L124E, L143F, Н172Т и H172R, или их комбинации; L1 содержит аминокислотные модификации, выбранные из группы, состоящей из Q38R, P44F, Q124C, S131T, V133G, N137K, S174R, S176R, S176K и T178Y, или их комбинации; Н2 содержит аминокислотные модификации, выбранные из группы, состоящей из Q39R, L124R и H172R, или их комбинации; и L2 содержит аминокислотные модификации, выбранные из группы, состоящей из Q38E, S131E, V133G, S176D и T178D, или их комбинации. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации Q39E и L124E; L1 содержит аминокислотные модификации Q38R, V133G и S176R; Н2 содержит аминокислотные модификации Q39R и L124R; и L2 содержит аминокислотные модификации Q38E, V133G и S176D. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации L45P и L124E; L1 содержит аминокислотные модификации P44F, V133G и S176R; Н2 содержит аминокислотную модификацию L124R; и L2 содержит аминокислотные модификации V133G, S176D и T178D. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации L124E и L143F; L1 содержит аминокислотные модификации V133G и S176R; Н2 содержит аминокислотную модификацию L124R; и L2 содержит аминокислотные модификации V133G, S176D и T178D. В некоторых вариантах реализации изобретения HI содержит аминокислотные модификации F122C и L124E; L1 содержит аминокислотные модификации Q124C, V133G и S176R; Н2 содержит аминокислотную модификацию L124R; и L2 содержит аминокислотные модификации V133G и S176D. В некоторых вариантах реализации изобретения HI содержит аминокислотные модификации L124E и Н172Т; L1 содержит аминокислотные модификации V133G, N137K, S174R и S176R; Н2 содержит аминокислотные модификации L124R и H172R; и L2 содержит аминокислотные модификации V133G, S176D и T178D.
[0047] В некоторых вариантах реализации конструкции, H1 и/или Н2 содержит по меньшей мере одну или множество аминокислотных модификаций в L124, А125, Н172 и K228, и L1 и/или L2 содержит по меньшей мере одну или множество аминокислотных модификации в S121, V133, N137, S174, S176 и Т178. В некоторых вариантах реализации изобретения H1 и/или Н2 содержит по меньшей мере одну или множество аминокислотных модификаций, выбранных из L124E, L124R, A125S, A125R, H172R, Н172Т и K228D; и (ii) L1 и/или L2 содержит по меньшей мере одну или множество аминокислотных модификаций, выбранных из S121K, V133G, N137K, S174R, S176K, S176R, S176D и T178D. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации, выбранные из группы, состоящей из L124E, A125S, H172R и K228D, или их комбинации; L1 содержит аминокислотные модификации, выбранные из группы, состоящей из S121K, V133G и S176R, или их комбинации; Н2 содержит аминокислотные модификации, выбранные из группы, состоящей из L124R, A125R и Н172Т, или их комбинации; и L2 содержит аминокислотные модификации, выбранные из группы, состоящей из V133G, N137K, S174R, S176D и T178D, или их комбинации. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации L124E и K228D; L1 содержит аминокислотные модификации S121K, V133G и S176R; Н2 содержит аминокислотные модификации L124R и A125R; и L2 содержит аминокислотные модификации V133G и S176D. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации L124E и H172R; L1 содержит аминокислотные модификации V133G и S176R; Н2 содержит аминокислотные модификации L124R и Н172Т; и L2 содержит аминокислотные модификации V133G, S174R и S176D.
[0048] В некоторых вариантах реализации конструкции, H1 и/или Н2 содержит по меньшей мере одну или множество аминокислотных модификаций в L124, А139 и V190, и L1 и/или L2 содержит по меньшей мере одну или множество аминокислотных модификации в F116, V133, L135 и S176. В некоторых вариантах реализации изобретения H1 и/или Н2 содержит по меньшей мере одну или множество аминокислотных модификаций, выбранных из L124E, L124R, A139W, A139G и V190A; и L1 и/или L2 содержит по меньшей мере одну или множество аминокислотных модификаций, выбранных из F116A, V133G, L135V, L135W, S176R и S176D. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации, выбранные из группы, состоящей из L124E и A139W, или их комбинации; L1 содержит аминокислотные модификации, выбранные из группы, состоящей из F116A, V133G, L135V и S176R, или их комбинации; Н2 содержит аминокислотные модификации, выбранные из группы, состоящей из L124R, A139G и V190 А, или их комбинации; и L2 содержит аминокислотные модификации, выбранные из группы, состоящей из V133G, L135W, и S176D, или их комбинации. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации L124E и A139W; L1 содержит аминокислотные модификации F116A, V133G, L135V и S176R; Н2 содержит аминокислотные модификации L124R, A139G и V190A; и L2 содержит аминокислотные модификации V133G, L135W и S176D.
[0049] В некоторых вариантах реализации конструкции, H1 и/или Н2 содержит по меньшей мере одну или множество аминокислотных модификаций в Q39, L45, K145, Н172, Q179 и S186, и L1 и/или L2 содержит по меньшей мере одну или множество аминокислотных модификации в Q38, Р44, Q124, S131, Q160, Т180 и С214. В некоторых вариантах реализации изобретения H1 и/или Н2 содержит по меньшей мере одну или множество аминокислотных модификаций, выбранных из Q39E, Q39R, L45P, K145T, H172R, Q179E и S186R; и L1 и/или L2 содержит по меньшей мере одну или множество аминокислотных модификаций, выбранных из Q38R, Q38E, P44F, Q124E, S131K, Q160E, Т180Е и C214S. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации, выбранные из группы, состоящей из Q39E, L45P, K145T, H172R и Q179E, или их комбинации; L1 содержит аминокислотные модификации, выбранные из группы, состоящей из Q38R, P44F и S131K, или их комбинации; Н2 содержит аминокислотные модификации, выбранные из группы, состоящей из Q39R, H172R и S186R, или их комбинации; и L2 содержит аминокислотные модификации, выбранные из группы, состоящей из Q38E, Q124E, Q160E, Т180Е и C214S, или их комбинации. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации Q39E, К145Т и Q179E; L1 содержит аминокислотные модификации Q38R и S131K; Н2 содержит аминокислотные модификации Q39R и S186R; и L2 содержит аминокислотные модификации Q38E, Q124E, Q160E и Т180Е. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации L45P, K145T, H172R и Q179E; L1 содержит аминокислотные модификации P44F и S131K; Н2 содержит аминокислотные модификации H172R и S186R; и L2 содержит аминокислотные модификации Q124E, Q160E и Т180Е.
[0050] В некоторых вариантах реализации конструкции, H1 и/или Н2 содержит по меньшей мере одну или множество аминокислотных модификаций в А139, L143, K145, Q179 и V190, и L1 и/или L2 содержит по меньшей мере одну или множество аминокислотных модификации в F116, Q124, L135, Q160, Т178 и Т180. В некоторых вариантах реализации изобретения H1 и/или Н2 содержит по меньшей мере одну или множество аминокислотных модификаций, выбранных из A139W, A139G, L143E, K145T, Q179E, Q179K и VI90A; и L1 и/или L2 содержит по меньшей мере одну или множество аминокислотных модификаций, выбранных из F116A, Q124R, Q124E, L135V, L135W, Q160E, T178R и Т180Е. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации, выбранные из группы, состоящей из A139W, L143E, K145T и Q179E, или их комбинации; L1 содержит аминокислотные модификации, выбранные из группы, состоящей из F116A, Q124R, L135V и T178R, или их комбинации; Н2 содержит аминокислотные модификации, выбранные из группы, состоящей из A139G, Q179K и VI90A, или их комбинации; и L2 содержит аминокислотные модификации, выбранные из группы, состоящей из Q124E, L135W, Q160E и Т180Е, или их комбинации. В некоторых вариантах реализации изобретения HI содержит аминокислотные модификации A139W, L143E, K145T и Q179E; L1 содержит аминокислотные модификации F116A, Q124R, L135V и T178R; Н2 содержит аминокислотную модификацию Q179K; и L2 содержит аминокислотные модификации Q124E, L135W, Q160E и T180E.
[0051] В некоторых вариантах реализации конструкции, H1 и/или Н2 содержит по меньшей мере одну или множество аминокислотных модификаций в Q39, L143, K145, D146, Н172 и Q179, и L1 и/или L2 содержит по меньшей мере одну или множество аминокислотных модификации в Q38, Q124, Q160, Т178 и Т180. В некоторых вариантах реализации изобретения H1 и/или Н2 содержит по меньшей мере одну или множество аминокислотных модификаций, выбранных из Q39E, Q39R, L143E, K145T, D146G, H172R, Q179E и Q179K; и L1 и/или L2 содержит по меньшей мере одну или множество аминокислотных модификаций, выбранных из Q38R, Q38E, Q124R, Q124E, Q160K, Q160E, T178R и Т180Е. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации, выбранные из группы, состоящей из Q39E, L143E, К145Т, H172R и Q179E, или их комбинации; L1 содержит аминокислотные модификации, выбранные из группы, состоящей из Q38R, Q124R, Q160K и T178R, или их комбинации; Н2 содержит аминокислотные модификации, выбранные из группы, состоящей из Q39R, H172R и Q179K, или их комбинации; и L2 содержит аминокислотные модификации, выбранные из группы, состоящей из Q38E, Q124E, D146G, Q160E и Т180Е, или их комбинации. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации Q39E, L143E, K145T и Q179E; L1 содержит аминокислотные модификации Q38R, Q124R, Q160K и T178R; Н2 содержит аминокислотные модификации Q39R, H172R и Q179K; и L2 содержит аминокислотные модификации Q38E, Q124E, Q160E и Т180Е.
[0052] В некоторых вариантах реализации конструкции, H1 и/или Н2 содержит по меньшей мере одну или множество аминокислотных модификаций в L45, L143, K145, D146, Н172 и Q179, и L1 и/или L2 содержит по меньшей мере одну или множество аминокислотных модификации в Q38, Р44, Q124, N137, Q160, S174, Т178, Т180 и С214. В некоторых вариантах реализации изобретения H1 и/или Н2 содержит по меньшей мере одну или множество аминокислотных модификаций, выбранных из L45P, L143E, K145T, D146G, H172R, Н172Т, Q179E и Q179K; и (ii) L1 и/или L2 содержит по меньшей мере одну или множество аминокислотных модификаций, выбранных из Q38E, P44F, Q124R, Q124E, N137K, Q160K, Q160E, S174R, T178R, Т180Е и C214S. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации, выбранные из группы, состоящей из L45P, L143E, K145T, H172R и Q179E, или их комбинации; L1 содержит аминокислотные модификации, выбранные из группы, состоящей из P44F, Q124R, Q160K и T178R, или их комбинации; Н2 содержит аминокислотные модификации, выбранные из группы, состоящей из D146G, H172R, Н172Т и Q179K, или их комбинации; и L2 содержит аминокислотные модификации, выбранные из группы, состоящей из Q38E, Q124E, N137K, Q160E, S174R, Т180Е и C214S, или их комбинации. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации L45P, L143E и К145Т; L1 содержит аминокислотные модификации P44F, Q124R, Q160K и T178R; Н2 содержит аминокислотные модификации D146G и Q179K; и L2 содержит аминокислотные модификации Q38E, Q124E, Q160E и Т180Е. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации L143E, K145T и H172R; L1 содержит аминокислотные модификации Q124R, Q160K и T178R; Н2 содержит аминокислотные модификации Н172Т и Q179K; и L2 содержит аминокислотные модификации Q124E, Q160E, N137K, S174R и Т180Е.
[0053] В некоторых вариантах реализации конструкции, H1 и/или Н2 содержит по меньшей мере одну или множество аминокислотных модификаций в L124, L143, K145 и Q179, и L1 и/или L2 содержит по меньшей мере одну или множество аминокислотных модификации в Q124, S131, V133, S176, Т178 и Т180. В некоторых вариантах реализации изобретения H1 и/или Н2 содержит по меньшей мере одну или множество аминокислотных модификаций, выбранных из L124W, L124A, L143E, L143F, K145T, Q179E и Q179K; и L1 и/или L2 содержит по меньшей мере одну или множество аминокислотных модификаций, выбранных из Q124R, Q124K, Q124E, S131K, V133A, V133W, S176T, T178R, T178L, Т178Е и Т180Е. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации, выбранные из группы, состоящей из L124W, L143E, К145Т и Q179E, или их комбинации; L1 содержит аминокислотные модификации, выбранные из группы, состоящей из Q124R, Q124K, S131K, V133A, S176T, T178R и T178L, или их комбинации; Н2 содержит аминокислотные модификации, выбранные из группы, состоящей из L124A, L143F и Q179K, или их комбинации; и L2 содержит аминокислотные модификации, выбранные из группы, состоящей из Q124E, V133W, S176T, T178L, Т178Е и Т180Е, или их комбинации. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации L124W, L143E, K145Т и Q179E; L1 содержит аминокислотные модификации Q124R, V133A, S176T и T178R; Н2 содержит аминокислотные модификации L124A, L143F и Q179K; и L2 содержит аминокислотные модификации Q124E, V133W, S176T, T178L и Т180Е.
[0054] В некоторых вариантах реализации конструкции, H1 и/или Н2 содержит по меньшей мере одну или множество аминокислотных модификаций в А139, L143, K145, Q179 и S186, и L1 и/или L2 содержит по меньшей мере одну или множество аминокислотных модификации в F116, Q124, V133, Q160, Т178 и Т180. В некоторых вариантах реализации изобретения H1 и/или Н2 содержит по меньшей мере одну или множество аминокислотных модификаций, выбранных из А139С, L143E, L143D, L143R, L143K, K145T, Q179E, Q179D, Q179R, Q179K, S186K, S186R; и L1 и/или L2 содержит по меньшей мере одну или множество аминокислотных модификаций, выбранных из F116C, Q124R, Q124K, Q124E, V133E, V133D, Q160K, Q160E, T178R, Т178К, Т178Е и Т180Е. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации, выбранные из группы, состоящей из А139С, L143E, L143D, K145Т, Q179E и Q179D, или их комбинации; L1 содержит аминокислотные модификации, выбранные из группы, состоящей из F116C, Q124R, Q124K, Q160K, T178R и T178K, или их комбинации; Н2 содержит аминокислотные модификации, выбранные из группы, состоящей из L143R, L143K, Q179R, Q179K, S186K и S186R, или их комбинации; и L2 содержит аминокислотные модификации, выбранные из группы, состоящей из Q124E, V133E, V133D, Q160E, Т178Е и Т180Е, или их комбинации. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации А139С, L143E, K145T и Q179E; L1 содержит аминокислотные модификации F116C, Q124R и T178R; Н2 содержит аминокислотную модификацию Q179K; и L2 содержит аминокислотные модификации Q124E, Q160E и Т180Е. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации L143E, K145T и Q179E; L1 содержит аминокислотные модификации Q124R и T178R; Н2 содержит аминокислотную модификация S186K; и L2 содержит аминокислотные модификации Q124E, Q160E и Т178Е. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации L143E, K145T и Q179E; L1 содержит аминокислотные модификации Q124R и T178R; Н2 содержит аминокислотную модификацию L143R; и L2 содержит аминокислотные модификации Q124E и V133E.
[0055] В некоторых вариантах реализации конструкции, H1 и/или Н2 содержит по меньшей мере одну или множество аминокислотных модификаций в L124, L143, K145, D146, Q179, S186 и S188, и L1 и/или L2 содержит по меньшей мере одну или множество аминокислотных модификации в Q124, S131, V133, Q160, S176, Т178 и Т180. В некоторых вариантах реализации изобретения H1 и/или Н2 содержит по меньшей мере одну или множество аминокислотных модификаций, выбранных из L124A, L143A, L143R, L143E, L143K, K145T, D146G, Q179R, Q179E, Q179K, S186R, S186K и S188L; и L1 и/или L2 содержит по меньшей мере одну или множество аминокислотных модификаций, выбранных из Q124R, Q124E, S131E, S131T, V133Y, V133W, V133E, V133D, Q160E, Q160K, Q160M, S176L, T178R, Т178Е, T178F, T178Y и Т180Е. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации, выбранные из группы, состоящей из L143E, K145T, Q179E и S188L, или их комбинации; L1 содержит аминокислотные модификации, выбранные из группы, состоящей из Q124R, Q160K и T178R, или их комбинации; Н2 содержит аминокислотные модификации, выбранные из группы, состоящей из L124A, L143A, L143R, L143K, D146G, Q179R, Q179K, S186R и S186K, или их комбинации; и L2 содержит аминокислотные модификации, выбранные из группы, состоящей из Q124E, S131E, S131T, V133Y, V133W, V133E, V133D, Q160E, Q160M, S176L, Т178Е, T178F, T178Y и Т180Е, или их комбинации. В некоторых вариантах реализации изобретения HI содержит аминокислотные модификации L143E, K145T, Q179E и S188L; L1 содержит аминокислотные модификации Q124R и T178R; Н2 содержит аминокислотную модификацию S186K; и L2 содержит аминокислотные модификации Q124E, S176L и Т180Е. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации L143E, K145T, Q179E и S188L; L1 содержит аминокислотные модификации Q124R и T178R; Н2 содержит аминокислотную модификацию S186K; и L2 содержит аминокислотные модификации Q124E, S131T, T178Y и Т180Е. В некоторых вариантах реализации изобретения HI содержит аминокислотные модификации L143E и К145Т; L1 содержит аминокислотные модификации Q124R, Q160K и T178R; Н2 содержит аминокислотную модификацию S186K; и L2 содержит аминокислотную модификацию S131E. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации L143E и K145T; L1 содержит аминокислотную модификацию Q124R; Н2 содержит аминокислотную модификацию L143R; и L2 содержит аминокислотные модификации Q124E и V133E.
[0056] В некоторых вариантах реализации конструкции H1 содержит по меньшей мере одну или множество аминокислотных модификаций в F122 и С233, и L1 содержит по меньшей мере одну или множество аминокислотных модификации в Q124 и С214. В некоторых вариантах реализации изобретения H1 содержит по меньшей мере одну или множество аминокислотных модификаций, выбранных из F122C и C233S; и L1 содержит по меньшей мере одну или множество аминокислотных модификаций, выбранных из Q124C и C214S. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации, выбранные из группы, состоящей из F122C и C233S, или их комбинации; L1 содержит аминокислотные модификации, выбранные из группы, состоящей из Q124C и C214S, или их комбинации; Н2 содержит последовательность дикого типа или немодифицированную аминокислотную последовательность; и L2 содержит последовательность дикого типа или немодифицированную аминокислотную последовательность. В некоторых вариантах реализации изобретения H1 содержит аминокислотные модификации F122C и C233S; L1 содержит аминокислотные модификации Q124C и C214S; Н2 содержит последовательность дикого типа или немодифицированную аминокислотную последовательность; и L2 содержит последовательность дикого типа или немодифицированную аминокислотную последовательность.
[0057] В некоторых вариантах реализации изобретения конструкция содержит аминокислотные модификации, выбранные из конструкций SMCA 9561-9095_1, 9561-9095_2, 9121-9373_1, 9121-9373_2, 9116-9349_1, 9116-9349_2, 9134-9521_1, 9134-9521_2, 9286-9402_1, 9286-9402_2, 9667-9830_1, 9667-9830_2, 9696-9848_1, 9696-9848_2, 9060-9756_1, 9060-9756_2, 9682-9740_1, 9682-9740_2, 9049-9759_1, 9049-9759_2, 9820-9823_1 и 9820-9823_2 из таблиц, представленных в данном документе. В некоторых вариантах реализации изобретения конструкция содержит аминокислотные модификации, выбранные из конструкций SMCA 9327-6054_1, 9815-9825_1, 9815-9825_2, 9587-9735_1, 9587-9735_2, 3522_1, 3522_2, 35191 и 3519_2 из таблиц, представленных в данном документе.
[0058] В некоторых вариантах реализации изобретения H1 и/или Н2 не содержат аминокислотную модификацию в положении Q179. В некоторых вариантах реализации изобретения H1 не содержит аминокислотную модификацию Q179E, и/или Н2 не содержит аминокислотную модификацию Q179K. В некоторых вариантах реализации изобретения L1 не содержит аминокислотную модификацию в положении S131. В другом варианте реализации L1 не содержит аминокислотную модификацию в положении S131K. В некоторых вариантах реализации изобретения L2 не содержит аминокислотную модификацию в положении Т180. В одном варианте реализации изобретения L2 не содержит аминокислотную модификацию Т180Е. В некоторых вариантах реализации изобретения конструкция не содержит комбинации аминокислотных модификаций, причем H1 содержит Q179E, L1 содержит S131K, Н2 содержит Q179K, и L2 содержит Т180Е.
[0059] В некоторых вариантах реализации изобретения H1 не содержит аминокислотную модификацию Q39 и/или Q179. В некоторых вариантах реализации изобретения H1 не содержит аминокислотную модификацию Q39E и/или Q179E. В некоторых вариантах реализации изобретения L1 не содержит аминокислотную модификацию в положении Q160. В одном варианте реализации изобретения L1 не содержит аминокислотную модификацию Q160K. В некоторых вариантах реализации изобретения Н2 не содержит аминокислотную модификацию в положении Q179. В одном варианте реализации изобретения Н2 не содержит аминокислотную модификацию Q179K. В некоторых вариантах реализации изобретения L2 не содержит аминокислотную модификацию в положении Q38, Q160 и/или Т180. В одном варианте реализации изобретения L2 не содержит аминокислотные модификации Q38E, Q160E и/или Т180Е. В некоторых вариантах реализации изобретения конструкция не содержит комбинации аминокислотных модификаций, причем H1 содержит Q39E и/или Q179E, L1 содержит Q160K, Н2 содержит Q179K, и L2 содержит Q38E, Q160E и/или Т180Е. Например, в некоторых вариантах реализации изобретения конструкция не содержит комбинации аминокислотных модификаций, причем: (i) H1 содержит Q179E, L1 содержит Q160K, Н2 содержит Q179K, и L2 содержит Q160E и Т180Е; (ii) H1 содержит Q39E и Q179E, L1 содержит Q160K, Н2 содержит Q179K, и L2 содержит Q38E, Q160E и Т180Е; или (iii) H1 содержит Q39E, L1 содержит Q160K, Н2 содержит Q179K, и L2 содержит Q38E, Q160E и Т180Е.
[0060] В некоторых вариантах реализации изобретения H1 не содержит аминокислотную модификацию в положении Q179. В некоторых вариантах реализации изобретения H1 не содержит аминокислотную модификацию Q179K или Q179E. В некоторых вариантах реализации изобретения L1 не содержит аминокислотную модификацию в положении Q160 и/или Т180. В одном варианте реализации изобретения L1 не содержит аминокислотную модификацию Q160E, Q160K и/или Т180Е. В некоторых вариантах реализации изобретения Н2 не содержит аминокислотную модификацию в положении Q179. В одном варианте реализации изобретения Н2 не содержит аминокислотную модификацию Q179K или Q179E. В некоторых вариантах реализации изобретения L2 не содержит аминокислотную модификацию в положении Q160 и/или Т180. В одном варианте реализации изобретения L2 не содержит аминокислотные модификации Q160K, Q160E и/или Т180Е. В некоторых вариантах реализации изобретения конструкция не содержит комбинации аминокислотных модификаций, причем H1 содержит Q179K или Q179E, L1 содержит Q160E, Q160K и/или Т180Е, Н2 содержит Q179K или Q179E, и L2 содержит Q160K, Q160E и/или Т180Е.
[0061] В некоторых вариантах реализации изобретения H1 и/или Н2 не содержит аминокислотную модификацию в положении Q179. В некоторых вариантах реализации изобретения H1 не содержит аминокислотную модификацию Q179K и/или Н2 не содержит аминокислотную модификацию Q179E. В некоторых вариантах реализации изобретения L1 не содержит аминокислотную модификацию в положении Т180. В одном варианте реализации изобретения L1 не содержит аминокислотную модификацию Т180Е. В некоторых вариантах реализации изобретения L2 не содержит аминокислотную модификацию в положении S131. В одном варианте реализации L1 не содержит аминокислотную модификацию в положении S131K. В некоторых вариантах реализации изобретения конструкция не содержит комбинации аминокислотных модификаций, причем H1 содержит Q179K, L1 содержит Т180Е, Н2 содержит Q179E, и L2 содержит S131K.
[0062] В некоторых вариантах реализации изобретения H1 не содержит аминокислотную модификацию в положении Q179. В некоторых вариантах реализации изобретения H1 не содержит аминокислотную модификацию Q179E. В некоторых вариантах реализации изобретения L1 не содержит аминокислотную модификацию в положении Q160. В одном варианте реализации изобретения L1 не содержит аминокислотную модификацию Q160K. В некоторых вариантах реализации изобретения Н2 не содержит аминокислотную модификацию в положении Q179. В одном варианте реализации изобретения Н2 не содержит аминокислотную модификацию Q179K. В некоторых вариантах реализации изобретения L2 не содержит аминокислотную модификацию в положении Т180. В одном варианте реализации изобретения L2 не содержит аминокислотную модификацию Т180Е. В некоторых вариантах реализации изобретения конструкция не содержит комбинации аминокислотных модификаций, причем H1 содержит Q179E, L1 содержит Q160K, Н2 содержит Q179K, и L2 содержит Т180Е.
[0063] В некоторых вариантах реализации изобретения H1 не содержит аминокислотную модификацию в положении А139. В некоторых вариантах реализации изобретения H1 не содержит аминокислотную модификацию А139С. В некоторых вариантах реализации изобретения L1 не содержит аминокислотную модификацию в положении F116. В одном варианте реализации изобретения L1 не содержит аминокислотную модификацию F116C. В некоторых вариантах реализации изобретения конструкция не содержит комбинации аминокислотных модификаций, причем H1 содержит А139С, и L1 содержит F116C.
[0064] В некоторых вариантах реализации изобретения конструкция не содержит нативные дисульфидные связи между тяжелыми и легкими цепями. Например, в некоторых вариантах реализации изобретения, цистеин в положении 214 в L1 и/или L2 модифицируется в другую аминокислоту. В некоторых вариантах реализации изобретения L1 и/или L2 содержит аминокислотную модификацию C214S. В некоторых вариантах реализации изобретения цистеин в положении 233 в H1 и/или Н2 модифицируется в другую аминокислоту. В одном варианте реализации изобретения H1 и/или Н2 содержит аминокислотную модификацию C233S.
[0065] Варианты реализации изобретения, описанные в данном документе, применимы к конструкциям в формате Fab и формате полноразмерного антитела.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0066] Фигура 1 иллюстрирует аминокислотные последовательности тяжелой цепи и легкой цепи D3H44, выровненные против канонических последовательностей зародышевой линии человека для вариабельных, константных и J-сегментов (условные обозначения на фигурах: * идентичность последовательностей). Фигура 1А иллюстрирует подгруппы VH зародышевой линии человека (одна представительная последовательность отображается для каждого семейства). Идентичность последовательности основана на выравнивании D3H44 против VH3 и IGHJ3*02. Фигура 1В иллюстрирует подгруппы каппа VL зародышевой линии человека (одна представительная последовательность отображается для каждого семейства). Идентичность последовательности основана на выравнивании D3H44 против VKI и IGKJ1*01. Фигура 1С иллюстрирует подгруппы лямбда VL зародышевой линии человека (одна представительная последовательность отображается для каждого семейства). Идентичность последовательности основана на выравнивании D3H44 против VL1 и IGLJ1*01. Фигура 1D иллюстрирует последовательности аллелей СН1 человека. Фигура 1Е иллюстрирует последовательности аллелей каппа и лямбда человека.
[0067] Фигура 2 иллюстрирует блок-схему для идентификации критических остатков области контакта и для компьютерного моделирования конструкций с преимущественным спариванием тяжелой-легкой цепи.
[0068] Фигура 3 иллюстрирует типовый набор цепей H1, L1, Н2, L2, которые были сконструированы таким образом, что H1 преимущественно спаривается с L1 по сравнению с L2, и Н2 преимущественно спаривается с L2 по сравнению с L1. Представлено динамическое изображение 3D кристаллической структуры вариабельной области тяжелой и легкой цепи области контакта. Мутации, введенные в область контакта, обеспечивают электростатическую и стерическую комплементарность для преимущественного образования облигатных пар H1-L1 и H2-L2, соответственно. С другой стороны, существует неблагоприятное стерическое и электростатическое несоответствие в неправильных парах, что приводит к пониженной предрасположенности к спариванию несоответствующей пары, а также пониженной стабильности.
[0069] Фигура 4 иллюстрирует схематический обобщенный обзор технических требований для образования биспецифичнеских Mab (моноклональное антитело), а также требований для анализа, необходимых для количественной оценки пар легкой цепи и тяжелой цепи. Цель разработки биспецифических Mab с высокой степенью чистоты (т.е., с малым количеством или без каких-либо неправильно спаренных H-L взаимодействий) может быть достигнута за счет рациональной разработки (с помощью введения специфических аминокислотных мутаций) преимущественного спаривания двух уникальных тяжелых цепей с их уникальными когнатными легкими цепями. Этот процесс показан схематично; здесь, H1 был разработан для преимущественного спаривания с L1, а не с L2. Кроме того, Н2 был разработан для преимущественного спаривания с L2, а не с L1. Экспериментальный скрининг биспецифических Mab конструкций требует анализа, способного одновременно оценить количественно H1-L1:H1-L2 и H2-L2:H2-L1. Эти требования анализа можно упростить, если предположить, что каждый биспецифический Fab-фрагмент может быть разработан независимо от другого. В этом случае анализ необходим только для количественной оценки H1-L1:H1-L2 или H2-L2:H2-L1, а не обоих одновременно.
[0070] На фигуре 5 представлена схема, изображающая процесс мечение тяжелых цепей и легких цепей, и как определяется преимущественное спаривание. На этой схеме круг представляет собой клетку, в которой трансфицируют 3 конструкции. Продукты экспрессии секретируются из клетки, и супернатант (SPNT) пропускают через устройство обнаружения, в данном случае чип SPR. На основании уровня обнаружения двух различных меток, слитых с двумя легкими цепями, конкурирующими для спаривания с тяжелой цепью, может быть определена количественная оценка преимущественного спаривания тяжелой цепи с двумя легкими цепями.
[0071] Фигура 6 иллюстрирует диаграммы размаха, которые демонстрируют средние показатели эффективности LCCA спаренных неправильно спаренных гетеродимеров Fab по меньшей мере 86:14 для каждого кластера.
[0072] Фигура 7 иллюстрирует представительные профили UPLC-SEC для А) гетеродимера Fab ДТ, а также В) представительного сконструированного гетеродимера Fab (H1L1 Fab компонент LCCA конструкций 9735, 9737 и 9740)
[0073] Фигура 8 иллюстрирует возможные продукты, связанные с тяжелой цепью, которые прогнозируются, в случае если в клетке две различные легкие цепи коэкспрессируются с двумя различными тяжелыми цепями. Преимущественное спаривания оценивается с использованием SMCA (конкурентный анализ моноклональных антител).
[0074] Фигура 9 иллюстрирует предпочтения использования смещения/цепи в пределах биспецифических систем а) D3H44/трастузумаб, b) D3H44/цетуксимаб, и с) трастузумаб/цетуксимаб. Использование цепи оценивается в различных видах, определенных с помощью LC-MS. На оси X представлено соотношение H1:H2:L1:L2, и на оси Y показан соответствующий процент каждой цепи в пределах различных экспериментов по трансфекции. В сбалансированной системе все цепи Н и L будут составлять по 25%. Смещение в сторону использования одной легкой цепи наблюдается во всех биспецифических системах.
[0075] Фигура 10 иллюстрирует представительные профили UPLC-SEC как для гетеродимерных антител дикого типа, так и для разработанных гетеродимерных антител. Фигуры 10а и 10b относятся к D3H44/трастузумаб дикого типа и 9060-9756_1, соответственно. Фигуры 10 с и 10d относятся к D3H44/цетуксимаб дикого типа и 9820-9823_1, соответственно. Фигуры 10е и 10f относятся к трастузумаб/цетуксимаб дикого типа и 9696-9848 1, соответственно.
[0076] Фигура 11 иллюстрирует диаграммы размаха изменений в % правильно спаренного компонента Fab над всеми неправильно спаренными компонентами Fab, которые используют ту же тяжелую цепь (H1: L1 над всеми видам H1 по отношению к дикому типу для D3H44/трастузумаб и D3H44/цетуксимаб; изменение H2:L2 над всеми видам Н2 по отношении к дикому типу для трастузумаб/цетуксимаб), а также изменения в процентах от желаемого биспецифического антитела по отношению к дикому типу для разработанных образцов биспецифических антител на кластер. Изменения в % правильно спаренного компонента Fab по сравнению со всеми неправильно спаренными компонентами Fab, которые используют ту же тяжелую цепь, против кластера показаны для каждой системы а) D3H44/трастузумаб, с) D3H44/цетуксимаб и е) трастузумаб/цетуксимаб. Изменения в процентах желаемого биспецифического антитела по отношению к дикому типу против кластера показаны для каждой системы b) D3H44/трастузумаб, d) D3H44/цетуксимаб и f) трастузумаб/цетуксимаб. Во всех биспецифических системах изменения в % правильно спаренных компонентов Fab над всеми неправильно спаренными компонентами Fab, которые используют ту же тяжелую цепь, против кластера показаны на фигуре 11g, и изменения в процентах от желаемого биспецифического антитела по отношению к дикому типу против кластера показаны на фигуре 11h. Следует отметить, что указанные значения также включают оценочные изменения для разработанных образцов биспецифических антител, в которых соответствующие конструкции дикого типа не были оценены с помощью SMCA.
[0077] Фигура 12 иллюстрирует способ получения биспецифического антитела с использованием библиотеки облигатных пар мутаций, представленных в данном документе.
ПОДРОБНОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
[0078] В данном документе представлены антигенсвязывающие полипептидные конструкции (также называются гетеродимерными парами), которые могут содержать первый гетеродимер и второй гетеродимер, причем каждый гетеродимер содержит тяжелую цепь иммуноглобулина или ее фрагмент и легкую цепь иммуноглобулина. Оба гетеродимера могут содержать одну или более аминокислотных модификаций в константном домене 1 тяжелой цепи иммуноглобулина (СН1) и одну или более аминокислотных модификаций в константном домене легкой цепи иммуноглобулина (CL); одну или более аминокислотных модификаций в вариабельном домене тяжелой цепи иммуноглобулина (VH) и одну или более аминокислотных модификаций в вариабельном домене легкой цепи иммуноглобулина (VL); или комбинацию предшествующих аминокислотных модификаций в обоих константных и вариабельных доменах тяжелой и легкой цепей. Модифицированные аминокислоты, как правило, являются частью области контакта между легкой цепью и тяжелой цепью и модифицированы для создания преимущественного спаривания между каждой тяжелой цепью и желаемой легкой цепью таким образом, чтобы тяжелая цепь первого гетеродимера преимущественно спаривалась с одной из легких цепей, а не с другой. Подобным образом, тяжелая цепь второго гетеродимера может преимущественно спариваться со второй легкой цепью, а не с первой.
[0079] Как было отмечено выше, специфические комбинации аминокислотных модификаций, описанные в данном документе, способствуют преимущественному спариванию тяжелых цепей со специфическими легкими цепями, что позволяет экспрессии биспецифических моноклональных антител (Mab) происходить с незначительным или ограниченным неправильным спариванием, и сводит к минимуму необходимость в очистке желательных гетеродимеров от нежелательных или неправильно спаренных продуктов. Гетеродимеры могут проявлять термическую стабильность, сопоставимую с гетеродимерами, которые не включают аминокислотные модификации, и может также демонстрировать аффинность связывания с антигеном, сравнимую с гетеродимерами, которые не включают аминокислотные модификации. Конструкции первого и второго гетеродимеров могут быть использованы для создания биспецифических антител, предназначенных для двух различных терапевтических мишеней или предназначенных для двух различных эпитопов (перекрывающихся или не неперекрывающихся) в пределах того же антигена.
[0080] Также в данном документе представлены способы получения гетеродимерных пар.
ОПРЕДЕЛЕНИЯ
[0081] Если не указано иное, все технические и научные термины, используемые в данном документе, имеют то же значение, которое обычно понимается специалистом в области техники, к которой принадлежит заявленный объект изобретения. В случае, если в данном документе имеется множество определений терминов, имеют преимущество те, которые используются в данном разделе. Если сделана ссылка на URL или другой такой идентификатор или адрес, следует понимать, что такие идентификаторы могут изменяться, и конкретная информация в интернете может появляться и исчезать, но эквивалентную информацию можно найти с помощью функции поиска в интернете. Ссылка на них свидетельствует о доступности и открытом распространении такой информации.
[0082] Следует понимать, что общее изложенное выше описание и последующее подробное описание являются только иллюстративными и пояснительными и не ограничивают какой-либо заявленный объект изобретения. В настоящей заявке использование единственного числа включает множественное число, если специально не указано иное.
[0083] В настоящем документе любой диапазон концентраций, процентный диапазон, коэффициент амплитуды или область целочисленных значений следует понимать как включающий значение любого целого числа в пределах перечисленного диапазона и, при необходимости, его частей (например, одна десятая и одна сотая целого числа), если не указано иное. Используемый в данном документе термин "около" означает ±10% от указанного диапазона, значения, последовательности или структуры, если не указано иное. Следует понимать, что используемые в данном документе формы терминов в единственном числе включают ссылки на множественное число перечисленных конструкций, если из контекста явно не следует иное. Использование альтернативы (например, "или") следует понимать как одно и другое, или любой их комбинации из альтернатив. Используемые в данном документе термины "включают" и "содержат" используются в качестве синонимов. Кроме того, следует понимать, что отдельные одноцепочечные полипептиды или конструкции иммуноглобулина, полученные из различных комбинаций структур и заменителей, описанных в данном документе, описаны в настоящей заявке в той же степени, как если бы каждый одноцепочечный полипептид или гетеродимер был изложен в индивидуальном порядке. Таким образом, выбор конкретных компонентов для образования отдельных одноцепочечных полипептидов или гетеродимеров находится в пределах объема настоящего изобретения.
[0084] Заголовки разделов, используемые в данном документе, предназначены только организационных целей и не могут быть истолкованы как ограничивающие описанный объект. Все документы или части документов, указанные в настоящей заявке, включая, без ограничений, патенты, заявки на патенты, статьи, книги, учебные пособия и трактаты, таким образом специально включены в качестве ссылки во всей их полноте для любых целей.
[0085] Надо понимать, что настоящее изобретение не ограничивается никакими определенными протоколами; клеточными линиями, конструкциями и реагентами, описанными в данном документе, и как таковое может изменяться. Надо также понимать, что терминология использована в данном документе с целью описания только конкретных вариантов реализации изобретения, и не предназначена для ограничения объема настоящего изобретения.
[0086] Все публикации и патенты, упомянутые в данном документе, включены в данный документ посредством ссылки в целях описания и раскрытия, например, конструкций и методологий, которые описаны в публикациях, которые могут быть использованы в связи со способами, композициями и соединениями, описанными в данном документе. Публикации, рассмотренные в данном документе, приведены исключительно для их раскрытия до даты подачи настоящей заявки. Ничего в данном документе не следует трактовать как допущение того, что авторы настоящего изобретения не имеют права датировать более ранним числом такое описание вследствие предшествующего изобретения или любой другой причины. В настоящей заявке названия аминокислот и названия атомов (например, N, О, С и т.д.) используются, как определено Protein DataBank (PDB) (www.pdb.org), который основан на номенклатуре IUPAC ("Номенклатура и символика для аминокислот и пептидов IUPAC" (названия остатков, названия атомов и т.д.), Eur. J. Biochem., 138, 9-37 (1984 год) вместе с их поправками в Eur. J. Biochem., 152, 1 (1985 год). Термин "аминокислотный остаток" в первую очередь предназначен для обозначения аминокислотного остатка, содержащегося в группе, состоящей из 20 аминокислот, встречающихся в природе, то есть остатков аланина (Ala или А), цистеина (Cys, или С), аспарагиновой кислоты (Asp или D), глутаминовой кислоты (Glu или Е), фенилаланина (Phe или F), глицина (Gly или G), гистидина (His или Н), изолейцина (Не или I), лизина (Lys или K), лейцина (Leu или L), метионина (Met или М), аспарагина (Asn или N), пролина (Pro или Р), глутамина (Gin или Q), аргинина (Arg или R), серина (Ser или S), треонина (Thr или Т), валина (Val или V), триптофана (Trp или W) и тирозина (Tyr или Y).
[0087] Термины "полипептид", "пептид" и "белок" используются равноправно в данном документе для обозначения полимерных аминокислотных остатков. То есть, описание, направленное на полипептид, в равной степени относится к описанию пептида и описанию белка, и наоборот. Эти термины относятся как к встречающимся в природе аминокислотным полимерам, так и к аминокислотным полимерам, в которых один или более аминокислотных остатков представляют собой не встречающуюся в природе аминокислоту. Используемые в данном документе термины охватывают аминокислотные цепи любой длины, включая непроцессированные белки, отличающиеся тем, что аминокислотные остатки соединены ковалентной пептидной связью.
[0088] Термин "нуклеотидная последовательность" или "последовательность нуклеиновой кислоты" подразумевает обозначение последовательного отрезка из двух или более нуклеотидных молекул. Нуклеотидная последовательность может представлять собой геном, кДНК, РНК, полусинтетического или синтетического происхождения или любую их комбинацию.
[0089] "Клетка", "клетка-хозяин", "линия клеток" и "культура клеток" используются в данном документе, и, как следует понимать, данные термины включают потомство, получаемое при выращивании или культивировании клеток. Термины "трансформация" и "трансфекция" используются взаимозаменяемо для обозначения процесса введения ДНК в клетку.
[0090] Термин "аминокислота" относится к встречающимся в природе и не встречающимся в природе аминокислотам, а также аминокислотным аналогам и аминокислотным миметикам, которые функционируют аналогично встречающимся в природе аминокислотам. Природными аминокислотами являются 20 общепринятых аминокислот (аланин, аргинин, аспарагин, аспарагиновая кислота, цистеин, глутамин, глютаминовая кислота, глицин, гистидин, изолейцин, лейцин, лизин, метионин, фенилаланин, пролин, серин, треонин, триптофан, тирозин и валин) и пиролизин и селеноцистеин. Аминокислотные аналоги относятся к соединениям, которые имеют основную химическую структуру, идентичную с аминокислотой, встречающейся в природе, т.е., углерод, который связан с водородом, карбоксильная группа, амино группа, и R группа, например, гомосерин, норлейцин, метионилсульфоксид, метионинметилсульфоний. Такие аналоги могут иметь модифицированные R группы (например, норлейцин) или модифицированные пептидные остовы, но сохраняют такую же основную химическую структуру, что и аминокислота, встречающаяся в природе. Термин аминокислота включает, например, встречающиеся в природе протеогенные L-аминокислоты; D-аминокислоты, химически модифицированные аминокислоты, такие как варианты аминокислот и производные; встречающиеся в природе непротеогенные аминокислоты, такие как аланин, орнитин и т.д.; и химически синтезированные соединения, имеющие свойства, известные в данной области техники, характерные для аминокислот. Примеры не встречающихся в природе аминокислот включают, без ограничений, α-метил-аминокислоты (например, метилаланин), D-аминокислоты, гистидин-подобные аминокислоты (например, 2-амино-гистидин, гидроксигистидин, гомогистидин), аминокислоты, имеющие дополнительный метилен в боковой цепи («гомо» аминокислоты), и аминокислоты, в которых функциональная группа карбоновой кислоты в боковой цепи заменяется на группу сульфоновой кислоты (например, цистеиновая кислота). Введение неприродных аминокислот, включая синтетические ненативные аминокислоты, замещенные аминокислоты или одну или более D-аминокислот в белки по настоящему изобретению может быть эффективно в ряде различных способов. Пептиды, содержащие D-аминокислоты, демонстрируют повышенную стабильность in vitro или in vivo по сравнению с аналогами, содержащими L-аминокислоты. Таким образом, конструирование пептидов, и т.д., введение D-аминокислот может быть особенно полезным в случае, если большая внутриклеточная стабильность является желаемой или требуемой. Конкретнее, D-пептиды и т.д., резистентные к эндогенным пептидазам и протеазам, таким образом, обеспечивают улучшенную биодоступность молекулы и увеличенное время жизни in vivo, в случае если такие свойства являются желаемыми. Дополнительно, D-пептиды и т.д. не могут эффективно подвергаться рестриктированному представлению по главному комплексу гистосовместимости класса II клеткам Т-хелперам и являются, таким образом, менее подходящими для индуцирования гуморального иммунного ответа в целом организме.
[0091] Аминокислоты обозначаются в данном документе либо с помощью общеизвестных трехбуквенных символов, либо с помощью однобуквенных символов, рекомендованных Комиссией по биохимической номенклатуре IUPAC-IUB. Нуклеотиды аналогичным образом могут обозначаться общепринятым однобуквенным кодом.
[0092] Термин "консервативно модифицированные варианты" применяют как к аминокислотным, так и нуклеотидным последовательностям. Принимая во внимание конкретные последовательности нуклеиновых кислот, консервативно модифицированные варианты относятся к тем нуклеиновым кислотам, которые кодируют идентичные или преимущественно идентичные аминокислотные последовательности, или, в случае если нуклеиновая кислота не кодирует аминокислотную последовательность, преимущественно идентичные последовательности. Из-за вырожденности генетического кода любой заданный белок кодируется значительным количеством функционально идентичных нуклеиновых кислот. Например, все кодоны GCA, GCC, GCG и GCU кодируют аминокислоту аланин. Таким образом, в каждом положении, в котором аланин определяется кодоном, этот кодон может быть изменен на любой из соответствующих описанных кодонов без изменения этого кодированного полипептида. Такие вариации нуклеиновых кислот являются «молчащими вариациями», которые представляют собой один вид консервативно модифицированных вариаций. В данном документе каждая последовательность нуклеиновой кислоты, которая кодирует полипептид, также описывает любую возможную молчащую вариацию нуклеиновой кислоты. Среднему специалисту в данной области техники будет понятно, что каждый кодон в нуклеиновой кислоте (за исключением AUG, который обычно представляет собой единственный кодон для метионина, и TGG, который обычно представляет собой единственный кодон для триптофана) может быть модифицирован с получением функционально идентичной молекулы. Таким образом, в каждой описанной последовательности подразумевают каждую молчащую вариацию нуклеиновой кислоты, которая кодирует полипептид.
[0093] Что касается аминокислотных последовательностей, среднему специалисту в данной области техники будет понятно, что отдельные замены, делеции или вставки в последовательность нуклеиновой кислоты, пептида, полипептида или белка, которые приводят к изменению, вставке или делеции одной аминокислоты или небольшого количества в процентах аминокислот в кодируемой последовательности, относятся к "варианту с консервативными модификациями", причем указанное изменение приводит к делеции аминокислоты, вставке аминокислоты или замене аминокислоты на химически подобную аминокислоту. Таблицы консервативных замен, в которых приведены аминокислоты с подобными функциональными свойствами, известны среднему специалисту в данной области техники. Такие консервативно модифицированные варианты являются дополнительными и не исключают полиморфные варианты, межвидовые гомологи и аллели последовательностей данного изобретения.
[0094] Таблицы консервативных замен, в которых приведены аминокислоты с подобными функциональными свойствами, известны среднему специалисту в данной области техники. Далее представлены восемь групп, каждая из которых включает аминокислоты, которые представляют собой консервативные замены друг для друга:
Аланин (А), Глицин (G);
Аспарагиновая кислота (D), Глутаминовая кислота (Е);
Аспарагин (N), Глутамин (Q);
Аргинин (R), Лизин (К);
Изолейцин (I), Лейцин (L), Метионин (М), Валин (V);
Фенилаланин (F), Тирозин (Y), Триптофан (W);
Серин (S), Треонин (Т); и
Цистеин (С), Метионин (М)
(см., например, Creighton, Proteins: Structures and Molecular Properties (W H Freeman & Co.; 2-е издание (декабрь 1995 год).
[0095] Термины "идентичный" или процентная "идентичность" в контексте двух или более последовательностей нуклеиновых кислот или полипептидных последовательностей, относятся к двум или более последовательностям, или подпоследовательностям, которые являются одинаковыми. Последовательности являются "по существу идентичными", если они содержат определенный процент аминокислотных остатков или нуклеотидов, которые являются одинаковыми (т.е. по меньшей мере около 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность в пределах конкретной области) при сравнении и выравнивании для максимального соответствия в окне сравнения или определенной области, что измеряется с использованием одного из следующих алгоритмов сравнения последовательностей (или других алгоритмов, доступных средним специалистам в данной области техники), или путем выравнивания вручную и визуального наблюдения. Это определение также относится к последовательности, комплементарной исследуемой последовательности. Идентичность может распространяться на область, длина которой составляет по меньшей мере около 50 аминокислот или нуклеотидов, или на область, длина которой составляет 75-100 аминокислот или нуклеотидов, или, если не указано иное, на всю последовательность полинуклеотида или полипептида. Полинуклеотид, кодирующий полипептид по настоящему изобретению, включая гомологи других видов, отличных от человека, может быть получен способом, включающем стадии скрининга библиотеки в жестких условиях гибридизации с использованием меченого зонда, содержащего полинуклеотидную последовательность по данному изобретению или ее фрагмент, и выделения полноразмерной кДНК и геномных клонов, содержащих указанную полинуклеотидную последовательность. Такие технологии гибридизации хорошо известны специалисту в данной области техники.
[0096] Считается, что производное или вариант полипептида характеризуется "гомологичностью" или является "гомологичным" пептиду, если аминокислотные последовательности производного или варианта по меньшей мере на 50% идентичны аминокислотной последовательности, состоящей из 100 аминокислот из оригинального пептида. В некоторых вариантах реализации изобретение производное или вариант характеризуется сходством по меньшей мере на 75% либо с пептидом, либо фрагментом пептида, имеющим такое же число аминокислотных остатков, что и производное. В некоторых вариантах реализации изобретения производное или вариант характеризуется сходством по меньшей мере на 85% либо с пептидом, либо фрагментом пептида, имеющим такое же число аминокислотных остатков, что и производное. В некоторых вариантах реализации изобретения аминокислотная последовательность производного характеризуется сходством по меньшей мере на 90% с пептидом или фрагментом пептида, имеющим такое же число аминокислотных остатков, что и производное. В некоторых вариантах реализации изобретения аминокислотная последовательность производного характеризуется сходством по меньшей мере на 95% с пептидом или фрагментом пептида, имеющим такое же число аминокислотных остатков, что и производное. В некоторых вариантах реализации изобретения производное или вариант характеризуется сходством по меньшей мере на 99% с пептидом или фрагментом пептида, имеющим такое же число аминокислотных остатков, что и производное.
[0097] В данном контексте "выделенный" полипептид или конструкция означает конструкцию или полипептид, который был идентифицирован и отделен и/или извлечен из компонента клеточной культуры его природного окружения. Контаминирующие компоненты природного окружения представляют собой вещества, которые, как правило, мешают диагностическим или терапевтическим применениям гетеромультимера и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества.
[0098] В некоторых вариантах реализации изобретения, в данном контексте, "выделенные" антигенсвязывающие полипептидные конструкции, описанные в данном документе, содержат гетеродимерные пары или "выделенные" гетеродимерные пары, которые содержат гетеродимер или гетеродимерную пару, которая была идентифицирована и отделена и/или извлечена из компонента клеточной культуры его природного окружения. Контаминирующие компоненты природного окружения представляют собой вещества, которые мешают диагностическим или терапевтическим применениям гетеродимера или антигенсвязывающих полипептидных конструкций, и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества.
[0099] Гетеродимеры и антигенсвязывающие полипептидные конструкции и пары гетеродимеров, как правило, очищают до существенной гомогенности. Фразы "по существу гомогенный", "по существу гомогенная форма" и "существенная гомогенность" используются для указания того, что продукт по существу лишен побочных продуктов, происходящих от нежелательных полипептидных комбинаций (например, гомодимеров). В данном контексте представляющий интерес вид представляет собой гетеродимер, содержащий H1 и L1 (H1-L1) или Н2 и L2 (H2-L2). Контаминанты включают гетеродимеры, содержащие H1 и L2 (H1-L2) или Н2 и L1 (H2-L1), или гомодимеры, содержащие H1 и L1 или Н2 и L2 (независимо от того, правильно или неправильно спарен Fab-фрагмент). Выраженная в терминах чистоты, существенная гомогенность означает, что количество побочных продуктов не превышает 10%, например, составляет менее 5%, менее 1% или менее 0,5% от общей интенсивности LC-MS из всех видов, присутствующих в смеси, причем процентное содержание отражает результаты масс-спектрометрического анализа.
[0100] Фраза "избирательно (или специфически) гибридизируется с" относится к связыванию, образованию дуплексов или гибридизации молекулы только с определенной нуклеотидной последовательностью в жестких условиях гибридизации, в случае если эта последовательность содержится в сложной смеси (включая, но не ограничиваясь этим, общую клеточную ДНК или РНК или библиотеку ДНК или РНК).
[0101] В данной области техники антител каждое данное значение терминов, полученное в данной области техники, понятно специалистам, если не указано иное в данном документе. Как известно, антитела имеют вариабельные области, шарнирную область и константные домены. Структура и функции иммуноглобулина рассматриваются, например, в Harlow et al, ред., Antibodies: A Laboratory Manual ch.14 (Cold Spring Harbor Laboratory, Cold Spring Harbor, NY).
[0102] В данном контексте термины "антитело", "иммуноглобулин" или "антигенсвязывающая полипептидная конструкция" используются взаимозаменяемо. "Антигенсвязывающая конструкция" относится к полипептиду, который по существу кодируется геном иммуноглобулина или генами иммуноглобулина, или к одному или более его фрагментам, которые специфически связывают анализируемое вещество (антиген). Все распознаваемые гены иммуноглобулинов включают каппа, лямбда, альфа, гамма, дельта, эпсилон и мю константные области генов, а также множество генов вариабельной области иммуноглобулина. Легкие цепи классифицируют как каппа или лямбда. Тяжелые цепи классифицируются как гамма, мю, альфа, дельта или эпсилон, которые в свою очередь определяют изотипы иммуноглобулинов, IgG, IgM, IgA, IgD и IgE, соответственно. Дополнительно антитело может принадлежать к одному из нескольких подтипов, например, IgG может принадлежать к подклассам IgG1, IgG2, IgG3 или IgG4.
[0103] Структурная единица типового иммуноглобулина (антитела) состоит из двух пар полипептидных цепей, причем каждая пара имеет одну "легкую" (около 25 кДа) и одну "тяжелую" цепь (около 50-70 кДа). Термин "легкая цепь" включает полноразмерную легкую цепь и ее фрагменты, содержащие последовательность вариабельной области, достаточную, чтобы обеспечивать специфичность связывания. Полноразмерная легкая цепь содержит домен вариабельной области VL и домен константной области CL. Домен вариабельной области легкой цепи представляет собой амино-конец полипептида. Легкие цепи включают каппа цепи и лямбда цепи. Термин "тяжелая цепь" включает полноразмерную тяжелую цепь и ее фрагменты, содержащие последовательность вариабельной области, достаточную, чтобы обеспечивать специфичность связывания. Полноразмерная тяжелая цепь содержит домен вариабельной области VH и три домена константной области CH1, СН2 и СН3. Домен VH представляет собой амино-конец полипептида, а домены СН находятся в карбоксильном конце, при этом СН3 находится наиболее близко к карбокси-концу полипептида. Тяжелые цепи могут принадлежать любому изотипу, включая IgG (включая подклассы IgG1, IgG2, IgG3 и IgG4), IgA (включая подклассы IgA1 и IgA2), IgM и IgE. Термин "вариабельная область" или "вариабельный домен" относится к части легких и/или тяжелых цепей антитела, как правило, ответственной за распознавание антигена, как правило, включающей приблизительно от 120 до 130 аминоконцевых аминокислот в тяжелой цепи (VH) и приблизительно от 100 до 110 аминоконцевых аминокислот в легкой цепи (VL). «Определяющая комплементарность область» или "CDR" представляет собой аминокислотную последовательность, которая определяет антигенсвязывающую специфичность и аффинность. "Каркасные" области (FR) могут способствовать поддержанию правильной конформации CDR для содействия связывания между антигенсвязывающей областью и антигеном. В структурном плане каркасные области могут быть расположены в антителах между CDR. Вариабельные области, как правило, обладают аналогичной общей структурой относительно консервативных каркасных областей (FR), соединенных тремя гипервариабельными областями, CDR. CDR из двух цепей каждой пары, как правило, выравниваются по каркасным областям, которые могут обеспечивать связывание со специфическим эпитопом. От N-конца к С-концу вариабельные области как легкой, так и тяжелой цепи, как правило, содержат домены FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. Как правило, аминокислоты присваивают к каждому домену в соответствии с определениями по Kabat Sequences of Proteins of Immunological Interest (National Institutes of Health, Bethesda, Md. (1987 год и 1991 год)), если не указано иное. В некоторых вариантах реализации антигенсвязывающие полипептидные конструкции содержат по меньшей мере один домен иммуноглобулина из IgG, IgM, IgA, IgD или IgE, связанный с терапевтическим полипептидом. В некоторых вариантах реализации изобретения домен иммуноглобулина, содержащийся в антигенсвязывающий полипептидной конструкции, представленной в данном документе, происходит от конструкции на основе иммуноглобулина, такой как диатело или нанотело. В некоторых вариантах реализации изобретения антигенсвязывающие полипептидные конструкции, описанные в данном документе, содержат по меньшей мере один домен иммуноглобулина из тяжелой цепи антитела, такого как верблюжье антитело. В некоторых вариантах реализации изобретения антигенсвязывающие полипептидные конструкции, представленные в данном документе, содержат по меньшей мере один домен иммуноглобулина из антитела млекопитающего, такого как бычье антитело, антитело человека, верблюжье антитело (одиночный домен и неодиночный домен), антитело грызуна, гуманизированное антитело, негуманизированное антитело, мышиное антитело, или любое химерное антитело. В некоторых вариантах реализации изобретения антигенсвязывающие полипептидные конструкции, представленные в данном документе, содержат по меньшей мере один домен иммуноглобулина из антитела, образованного из синтетической библиотеки.
[0104] "Биспецифический", "обладающий двойной специфичностью" или "бифункциональный» антигенсвязывающий белок или антитело представляет собой гибридный антигенсвязывающий белок, содержащий два разных антигенсвязывающих участка. Биспецифические антигенсвязывающие белки и антитела являются видами мультиспецифического антигенсвязывающего белка антитела. Два связывающих участка биспецифического антигенсвязывающего белка или антитела связывают два разных эпитопа, которые могут находиться на одной и той же или на разных молекулярных мишенях. "Мультиспецифический антигенсвязывающий белок" или "мультиспецифическое антитело" нацелено на более чем один антиген или эпитоп. "Бивалентный антигенсвязывающий белок" или "бивалентное антитело" содержит два антигенсвязывающих сайта. В некоторых случаях два связывающих сайта обладают специфичностью к одному и тому же антигену. Бивалентные антигенсвязывающие белки и бивалентные антитела могут быть биспецифическими, см. ниже. В определенных вариантах реализации изобретения под бивалентным антителом, отличным от "мультиспецифического" или "мультифункционального", как правило, понимается антитело, все сайты связывания которого идентичны.
[0105] Термин "преимущественное спаривание" используется в данном документе для описания схемы спаривания первого полипептида со вторым полипептидом, например, тяжелой цепи иммуноглобулина с легкой цепью иммуноглобулина в антигенсвязывающих полипептидных конструкциях и парах гетеродимеров, описанных в данном документе. Таким образом, "преимущественное спаривание" относится к преимущественному спариванию первого полипептида со вторым полипептидом, в случае если один или более дополнительных, отличных полипептидов присутствуют в одно и то же время, когда происходит спаривание между первым и вторым полипептидом. Как правило, преимущественное спаривание происходит в результате модификации (например, модификация аминокислоты) одного из или обоих первого и второго полипептида. Как правило, преимущественное спаривание приводит к тому, что спаренный первый и второй полипептид являются наиболее распространенным димером, присутствующим после спаривания. В данной области техники известно, что тяжелая цепь иммуноглобулина (H1) при коэкспрессии с двумя различными легкими цепями иммуноглобулина (L1 и L2) по статистике будет спариваться одинаково с обеими легкими цепями, что приводит к образованию смеси приблизительно 50:50 H1, спаренной с L1, и H1, спаренной с L2. В данном контексте, "преимущественное спаривание" будет происходить между, например, H1 и L1, если количество гетеродимеров тяжелой-легкой цепи H1-L1 было больше, чем количество гетеродимеров H1-L2, в случае если H1 коэкспрессируется и с L1, и с L2. Таким образом, в данном случае, H1 преимущественно спаривается с L1 по сравнению с L2.
[0106] Тем не менее, в контексте биспецифических антител дикого типа, образованных из двух стартовых систем антител, также известно в данной области техники, что в некоторых случаях существует неустранимое смещение, где легкая цепь одной системы антител преимущественно спаривается с тяжелыми цепями обоих систем антител. Таким образом, при определении прочности конструкции в контексте биспецифической антигенсвязывающей конструкции, может потребоваться оценка степени спаривания с конструкцией по сравнению со степенью спаривания в системе дикого типа. Таким образом, в одном варианте реализации изобретения конструкция считается таковой, что демонстрирует преимущественное спаривание, если количество желаемого биспецифического антитела больше, чем количество желаемого биспецифического антитела, полученного в системах дикого типа. В другом варианте реализации изобретения конструкция считается таковой, что демонстрирует преимущественное спаривание, если количество спариваний в более слабом плече антитела больше, чем наблюдаемое в системе дикого типа.
[0107] Тяжелые цепи антител спариваются с легкими цепями антител и встречаются или контактируют друг с другом в одной или более "областями контакта". "Область контакта" включает один или более "контактных" аминокислотных остатков в первом полипептиде, которые взаимодействуют с одним или более "контактными" аминокислотными остатками второго полипептида. Например, область контакта существует между СН3 полипептидными последовательностями димеризованного домена СН3, между доменом СН1 тяжелой цепи и доменом CL легкой цепи и между доменом VH тяжелой цепи и доменом VL легкой цепи. "Область контакта" может быть получена из антитела IgG и, например, из человеческого антитела IgG1.
[0108] Термин "аминокислотные модификации» в данном контексте включает, но не ограничивается ими, аминокислотные мутации, вставки, делеции, замены, химические модификации, физические модификации и перестройки.
АНТИГЕНСВЯЗЫВАЮШИЕ ПОЛИПЕПТИДНЫЕ КОНСТРУКЦИИ И ПАРЫ ГЕТЕРОДИМЕРОВ
[0109] Антигенсвязывающие полипептидные конструкции, описанные в данном документе, могут содержать первый гетеродимер и второй гетеродимер; каждый гетеродимер получен путем спаривания тяжелой цепи иммуноглобулина с легкой цепью иммуноглобулина. Структура и организация константных и вариабельных доменов тяжелых и легких цепей иммуноглобулина хорошо известна в данной области техники. Тяжелые цепи иммуноглобулина обычно содержат один вариабельный домен (VH) и три константных домена, CH1, СН2 и СН3. Легкие цепи иммуноглобулина обычно содержат один вариабельный домен (VL) и один константный домен (CL). Могут быть произведены различные модификации этих типичных форматов.
[0110] Антигенсвязывающие полипептидные конструкции и пары гетеродимеров, описанные в данном документе, могут содержать первый гетеродимер и второй гетеродимер, каждый гетеродимер, содержащий тяжелую цепь иммуноглобулина/антитела или ее фрагмент, содержащую по меньшей мере домены VH и СН1, и легкую цепь иммуноглобулина/антитела, содержащую домен VL и домен CL. В одном варианте реализации изобретения оба гетеродимера из пары гетеродимеров и антигенсвязывающие полипептидные конструкции содержат полноразмерную тяжелую цепь иммуноглобулина. В другом варианте реализации изобретения оба гетеродимера из пары гетеродимеров и антигенсвязывающие полипептидные конструкции содержат фрагмент тяжелой цепи иммуноглобулина, которая включает по меньшей мере домен VH и домен СН1. В одном варианте реализации изобретения оба гетеродимера из пары гетеродимеров содержат амино-концевой фрагмент тяжелой цепи иммуноглобулина, который содержит по меньшей мере домен VH и домен СН1. В другом варианте реализации изобретения оба гетеродимера из пары гетеродимеров содержат карбокси-концевой фрагмент тяжелой цепи иммуноглобулина, который содержит по меньшей мере домен VH и домен СН1.
[0111] Каждый гетеродимер из пары гетеродимеров может специфически связываться с антигеном или эпитопом. В одном варианте реализации изобретения тяжелая цепь иммуноглобулина и легкая цепь иммуноглобулина каждого гетеродимера получена или разработана из известного антитела, например, терапевтическое антитела. Терапевтическое антитело представляет собой антитело, которое является эффективным при лечении заболевания или расстройства у млекопитающего с заболеванием или предрасположенного к заболеванию или нарушению. Приемлемые терапевтические антитела, из которых может быть получен каждый гетеродимер, включают, но не ограничиваясь ими, абаговомаб, адалимумаб, алемтузумаб, аурограб, бапинейзумаб, базиликсимаб, белимумаб, бевацизумаб, бриакинумаб, канакинумаб, катумаксомаб, цертолизумаб пегол, цетуксимаб, даклизумаб, деносумаб, эфализумаб, галиксимаб, гемтузумаб озогамицина, голимумаб, ибритутомаб тиуксетан, инфликсимаб, ипилимумаб, лумиликсимаб, меполизумаб, мотавизумаб, муромонаб, микограб, натализумаб, нимотузумаб, окрелизумаб, офатумумаб, омализумаб, паливизумаб, панитумумаб, пертузумаб, ранибизумаб, реслизумаб, ритуксимаб, теплизумаб, тоцилизумаб/атлизумаб, тозитумомаб, трастузумаб, ПроксиниумТМ, РенкарексТМ, устекинумаб и залутумумаб.
[0112] В одном варианте реализации изобретения тяжелая цепь иммуноглобулина и/или легкая цепь иммуноглобулина каждого гетеродимера получена или разработана из антитела, которое связывает молекулу, включая, но не ограничиваясь этим, следующий список белков, а также субъединиц, доменов, мотивов и эпитопов, принадлежащих следующему списку белков: ренин; гормон роста, включая человеческий гормон роста и бычий гормон роста; фактор высвобождения гормона роста; паратиреоидный гормон; тиреотропный гормон; липопротеины; альфа-1-антитрипсин; А-цепь инсулина; В-цепь инсулина; проинсулин; фолликулостимулирующий гормон; кальцитонин; лютеинизирующий гормон; глюкагон; факторы свертывания крови, такие как фактор VII, фактор VIIIC, фактор IX, тканевой фактор (TF) и фактор фон Виллебранда; антикоагулянты, такие как протеин С; предсердный натрийуретический фактор; легочный сурфактант; активатор плазминогена, такой как урокиназа или мочевой, или тканевой активатор плазминогена (t-PA) человека; бомбезин; тромбин; гемопоэтический фактор роста; фактор некроза опухоли-альфа и -бета; энкефалиназа; RANTES (хемокин, выделяемый Т-клетками при активации); человеческий макрофагальный воспалительный белок (MIP-1-альфа); сывороточный альбумин, такой как человеческий сывороточный альбумин; мюллерова ингибирующая субстанция; А-цепь релаксина; В-цепь релаксина; прорелаксин; мышиный гонадотропин-ассоциированный пептид; микробный белок, такой как бета-лактамаза; ДНКаза; IgE; антиген цитотоксических Т-лимфоцитов (CTLA), такой как CTLA-4; ингибин; активин; фактор роста эндотелия сосудов (VEGF); рецепторы для гормонов или факторов роста, такие как, например, EGFR, VEGFR; интерфероны, такие как альфа-интерферон (альфа-IFN), бета-интерферон (бета-IFN) и гамма-интерферон (гамма-IFN); белок А или D; ревматоидные факторы; нейротрофический фактор, такой как костный нейротрофический фактор (BDNF), нейротропин-3, -4, -5 или -6 (NT-3, NT-4, NT-5 или NT-6) или фактор роста нервов; фактор роста, полученный из тромбоцитов (PDGF); фактор роста фибробластов, такой как AFGF и PFGF; эпидермальный фактор роста (EGF); трансформирующий фактор роста (TGF), такой как TGF-альфа и TGF-бета, включая TGF-1, TGF-2, TGF-3, TGF-4 или TGF-5; инсулиноподобный фактор роста-I и -II (IGF-I и IGF-II); des (l-3)-IGF-I (мозговой IGF-I), белки, связывающие инсулиноподобный фактор роста; CD протеины, такие как CD2, CD3, CD4, CD8, CD14, CD11a, CD18, CD19, CD20, CD22, CD23, CD25, CD33, CD34, CD40, CD40L, CD52, CD63, CD64, CD80 и CD147; эритропоэтин; остеоиндуктивные факторы; иммунотоксины; костный морфогенетический белок (BMP); интерферон, такой как интерферон альфа, бета и гамма; колониестимулирующие факторы (CSF), такие как M-CSF, GM-CSF и G-CSF; интерлейкины (IL), например, от IL-1 до IL-13; TNF-альфа, супероксиддисмутаза; рецепторы Т-клеток; поверхностные мембранные белки; фактор ускорения распада; вирусный антиген, такой как, например, часть оболочки СПИДа, например, gp120; транспортные белки; "хоминг" рецепторы; аддрессины; регуляторные белки; молекулы клеточной адгезии, таких как LFA-1, Mac 1, р150.95, VLA-4, ICAM-1, ICAM-3 и VCAM, интегрин а4/р7 и интегрин Xv/рЗ, включая как а, так и его субъединицы, субъединицы интегрина альфа, такие как CD49a, CD49b, CD49c, CD49d, CD49e, CDIELb; субъединицы интегрина бета, такие как CD29, CD18, CD61, CD 104, бета5, бета6, бета7 и бета8; комбинации субъединиц интегрина, включая, но не ограничиваясь этим, альфаVбета3, альфаVбета5 и альфа4бета7; элементы пути апоптоза; IgE; антигены групп крови; рецептор flk2/flt3; рецептор ожирения (OB); mp1 рецептор, CTLA-4; протеин С; Eph рецептор, такой как EphA2, EphA4, EphB2 и т.д.; антиген лейкоцитов человека (HLA), такой как HLA-DR, белки комплемента, такие как рецептор комплемента CR1, CIRq и другие факторы комплемента, таких как С3 и С5; рецептор гликопротеина, такой как GpIb.alpha., GPIIb/IIIa и CD200; и фрагменты любого из перечисленных выше полипептидов.
[0113] В варианте реализации изобретения тяжелые и/или легкие цепи иммуноглобулина каждого гетеродимера получены или разработаны из антител, которые специфически связываются с раковыми антигенами, включая, но не ограничиваясь этим, рецептор ALK (рецептор плеотрофина), плеотрофин, KS 1/4 карциноидный антиген; антиген карциномы яичников (СА125); фосфатный простатический кислый фосфат; простат-специфический антиген (PSA); связанный с меланомой антиген р97; антиген меланомы gp75; высокомолекулярный антиген меланомы (HMW-MAA); простатический специфический мембранный антиген; карциноэмбриональный антиген (СЕА); антиген полиморфного эпителиального муцина; антиген жировых глобул грудного молока человека; антигены, связанные с колоректальной опухолью, такие как: СЕА, TAG-72, С017-1А, GICA 19-9, СТА-1 и LEA; антиген-38.13 лимфомы Беркитта; CD19; антиген-CD20 В-лимфомы человека; CD33; антигены, специфические к меланоме, такие как ганглиозид GD2, ганглиозид GD3, ганглиозид GM2 и ганглиозид GM3; антиген опухоли конкретного трансплантационного типа клеточной поверхности (TSTA); антигены опухолей, вызванных вирусом, включая Т-антиген, вирусы опухолевых ДНК и вирусы капсульных антигенов опухолевых РНК; онкофетальный антиген-альфа-фетопротеин, такой как СЕА толстой кишки, 514 онкофетальный трофобластный гликопротеин и онкофетальный антиген опухоли мочевого пузыря; дифференцировочный антиген, такой как антиген карциномы легких человека L6, L20; антигены фибросаркомы; Т-клеточный антиген лейкемии человека-Gp37; неогликопротеин; сфинголипиды; опухолевый антиген рака груди, такой как EGFR (эпидермальный фактор роста рецептора), NY-BR-16; NY-BR-16 и HER2 антиген (p185HER2), полиморфный эпителиальный муцин (РЕМ); злокачественный лимфоцитарный антиген человека - АРО-1; дифференцировочный антиген, такой как I антиген, обнаруженный в фетальных эритроцитах; I антиген первичной энтодермы, обнаруженный в зрелых эритроцитах; предимплантационные эмбрионы; I(Ма), обнаруженный в аденокарциномах желудка; M18, М39, обнаруженные в эпителии груди; SSEA-1, обнаруженный в миелоидных клетках; VEP8; VEP9; Myl; Va4-D5; D156-22, обнаруженный в колоректальной злокачественной опухоли; TRA-1-85 (кровь группы Н); SCP-1, обнаруженный в карциноме яичек и яичника; С14, обнаруженный в аденокарциноме толстой кишки, F3, обнаруженный в аденокарциноме легких; АН6, обнаруженный в злокачественной опухоли желудка; Y гаптен; Ley, обнаруженный в клетках эмбриональной карциномы; TL5 (кровь группы A); EGF рецептор, обнаруженный в А431 клетках; Е1 серии (кровь группы В), обнаруженные в злокачественной опухоли поджелудочной железы; FC10.2, обнаруженный в клетках эмбриональной карциномы; антиген желудочной аденокарциномы; СО-514 (группа крови Lea), обнаруженный в аденокарциноме; NS-10, обнаруженный в аденокарциномах; СО-43 (группа крови Leb); G49, обнаруженный в EGF рецепторе клеток А431; МН2 (группа крови ALeb/Ley), обнаруженный в аденокарциноме толстого кишечника; 19.9, обнаруженный в злокачественной опухоли толстого кишечника; муцинах злокачественной опухоли желудка; Т5А7, обнаруженный в миелоидных клетках; R24, обнаруженный в меланоме; 4.2, GD3, D1.1, OFA-1, GM2, OFA-2, GD2 и М1:22:25:8, обнаруженные в клетках эмбриональной карциномы, и SSEA-3 и SSEA-4, обнаруженные в эмбрионе от 4 до 8 клеточной стадии эмбриона; антиген кожной Т-клеточной лимфомы; MART-1 антиген; Sialy Tn (STN) антиген; антиген NY-CO-45 рака толстой кишки; вариант А антигена NY-LU-12 рака легких; антиген ART1 аденокарциномы; паранеопластический антиген, связанный с раком мозга и яичек (онконейрональный антиген МА2; паранеопластический нейронный антиген); нейроонкологический вентральный антиген 2 (NOVA2); ген 520 антигена гепатоклеточной карциномы; опухолеассоциированными антиген СО-029; опухолеассоциированные антигены MAGE-C1 (раково-тестикулярный антиген СТ7), MAGE-B1 (MAGE-XP антиген), MAGE-B2 (DAM6), MAGE-2, MAGE-4-а, MAGE-4-б и MAGE-X2; раково-тестикулярный антиген (NY-EOS-1) и фрагменты любого из перечисленных выше полипептидов.
[0114] Антитела человека могут быть сгруппированы в изотипы, включая IgG, IgA, IgE, IgM и IgD. В одном варианте реализации изобретения Fc получен из изотипа IgG. В другом варианте реализации изобретения Fc получен из изотипа IgA. В другом варианте реализации изобретения Fc получен из изотипа IgE. В другом варианте реализации изобретения Fc получен из изотипа IgM. В другом варианте реализации изобретения Fc получен из изотипа IgD.
[0115] Человеческие антитела IgG также можно разделить на подклассы IgG1, IgG2, IgG3 и IgG4. Таким образом, в некоторых вариантах реализации изобретения предполагается, что Fc может быть получен из IgG1, IgG2, IgG3 или IgG4 подкласса антител.
[0116] Каждый гетеродимер из пары гетеродимеров может специфически связываться с эпитопом или антигеном. В одном варианте реализации изобретения каждый гетеродимер из пары гетеродимеров может специфически связываться с тем же эпитопом. В другом варианте реализации изобретения первый гетеродимер из пары гетеродимеров специфически связывается с эпитопом на одном антигене, и второй гетеродимер из пары гетеродимеров специфически связывается с другим эпитопом на том же антигене. В другом варианте реализации изобретения первый гетеродимер из пары гетеродимеров специфически связывается с эпитопом на первом антигене, и второй гетеродимер из пары гетеродимеров специфически связывается с эпитопом на втором антигене, который отличается от первого антигена. Например, в одном варианте реализации изобретения первый гетеродимер специфически связывается с тканевым фактором, в то время как второй гетеродимер специфически связывается с антигеном Her2(ErbB2), или наоборот. В альтернативном варианте реализации изобретения первый гетеродимер специфически связывается с тканевым фактором, в то время как второй гетеродимер специфически связывается с EGFR, или наоборот. В еще одном варианте реализации изобретения первый гетеродимер специфически связывается с EGFR, в то время как второй гетеродимер специфически связывается с антигеном Her2 или наоборот. В другом варианте реализации изобретения первый гетеродимер специфически связывается с молекулой или раковым антигеном, описанным выше. В другом варианте реализации изобретения второй гетеродимер специфически связывается с молекулой или раковым антигеном, описанным выше.
[0117] Как было указано выше, в некоторых вариантах реализации изобретения тяжелая цепь иммуноглобулина и легкая цепь иммуноглобулина каждого гетеродимера может быть получена или разработана из известного терапевтического антитела, или из антитела, которое связывается с различными молекулами-мишенями или раковыми антигенами. Аминокислотные и нуклеотидные последовательности таких многочисленных молекул легко доступны (см., например, GenBank: AJ308087.1 (домен CL и вариабельная область легкой цепи гуманизированного антитела D3H44 к человеческому тканевому фактору); GenBank: AJ308086.1 (домен СН1 и вариабельная область тяжелой цепи гуманизированного антитела D3H44 к человеческому тканевому фактору); GenBank: НС359025.1 (генетический модуль легкой цепи Fab пертузумаба); GenBank: НС359024.1 (генетический модуль тяжелой цепи Fab пертузумаба); GenBank: GM685465.1 (антитело трастузумаб (=Герцептин) - дикий тип; легкая цепь); GenBank: GM685463.1 (антитело трастузумаб (=Герцептин) - дикий тип; тяжелая цепь); GenBank: GM685466.1 (антитело трастузумаб (=Герцептин) - GC-оптимизированная легкая цепь); и GenBank: GM685464.1 (антитело трастузумаб (=Герцептин) - GC-оптимизированная тяжелая цепь. Последовательности каждого номера GenBank, описанные в данном документе, доступны на сайте NCBI по состоянию на 28 ноября 2012 года, и их содержание включено в данный документ посредством ссылки в полном объеме для любых целей. Аминокислотные и нуклеотидные последовательности для цетуксимаба также известны в данной области техники, см., например, веб-сайт Drug Bank при поддержке Канадского института исследований в области здравоохранения, Alberta Innovates - Health Solutions и Инновационного центра метаболомики (TMIC), номер доступа DB00002.
[0118] В некоторых аспектах выделенная антигенсвязывающая полипептидная конструкция содержит аминокислотную последовательность, которая по меньшей мере на 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентична аминокислотной последовательности или ее фрагменту, которая изложена в таблицах или используется под номерами доступа, описанными в данном документе. В некоторых аспектах выделенная антигенсвязывающая полипептидная конструкция содержит аминокислотную последовательность, кодированную полинуклеотидом, который по меньшей мере на 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен нуклеотидной последовательности или ее фрагменту, которая изложена в таблицах или используется под номерами доступа, описанными в данном документе.
Аминокислотные модификации тяжелых и легких цепей иммуноглобулина
[0119] По меньшей мере один из гетеродимеров из пары гетеродимеров может содержать одну или более таких аминокислотных модификаций их тяжелых и/или легких цепей иммуноглобулина, что тяжелая цепь первого гетеродимера преимущественно образует пару с одной из легких цепей, а не другой. Подобным образом, тяжелая цепь второго гетеродимера может преимущественно образовывать пару со второй легкой цепью, а не с первой. Данное преимущественное спаривание одной тяжелой цепи с одной из двух легких цепей, может быть основано на наборах конструкции, содержащих одну тяжелую цепь иммуноглобулина и две легкие цепи иммуноглобулина (обозначаемого как набор конструкции LCCA), где тяжелая цепь иммуноглобулина образовывает пару с одной из двух легких цепей иммуноглобулина, имеющей преимущество над другой, в случае если тяжелая цепь иммуноглобулина коэкспрессируется с обоими легкими цепями иммуноглобулина. Таким образом, набор конструкции LCCA может содержать одну тяжелую цепь иммуноглобулина, первую легкую цепь иммуноглобулина и вторую легкую цепь иммуноглобулина.
[0120] В одном варианте реализации изобретения одна или более модификаций аминокислот содержат одну или более аминокислотных замен.
[0121] В одном варианте реализации изобретения преимущественное спаривание, продемонстрированное в наборе конструкции LCCA, устанавливается путем модификации одной или более аминокислот, которые являются частью области контакта между легкой цепью и тяжелой цепью. В одном варианте реализации изобретения преимущественное спаривание, продемонстрированное в наборе конструкции LCCA, устанавливается путем модификации одной или более аминокислот в по меньшей мере одном из доменов СН1 тяжелой цепи иммуноглобулина, доменов CL первой легкой цепи иммуноглобулина и доменов CL второй легкой цепи иммуноглобулина.
[0122] В одном варианте реализации изобретения одна или несколько модификаций аминокислот ограничиваются консервативными остатками каркаса вариабельных (VH, VL) и константных (CH1, CL) доменов, как обозначено в нумерации остатков по Kabat. Например, Almagro [Frontiers In Bioscience (2008 год) 13: 1619-1633] предоставляет определение остатков каркасной области на основе схем нумерации Kabat, Chothia и FMGT.
[0123] В одном варианте реализации изобретения по меньшей мере один из гетеродимеров содержит одну или более мутаций, введенных в тяжелые цепи иммуноглобулина и легкие цепи иммуноглобулина, которые являются комплементарными друг к другу. Комплементарность в области контакта тяжелой и легкой цепи может быть достигнута на основе стерических и гидрофобных контактов, электростатических/зарядовых взаимодействий или комбинации различных взаимодействий. Комплементарность между белковыми поверхностями широко описана в литературе в терминах соответствия замка и ключа, выступа-во-впадину, выступа и полости, донора и акцептора и т.д., всего, что подразумевает характер структурной и химической совместимости между двумя взаимодействующими поверхностями. В одном варианте реализации изобретения по меньшей мере один из гетеродимеров содержит одну или более мутаций, где мутации, введенные в тяжелые цепи иммуноглобулина и легкие цепи иммуноглобулина, вносят новую водородную связь на поверхности контакта между легкой и тяжелой цепью. В одном варианте реализации изобретения по меньшей мере один из гетеродимеров содержит одну или более мутаций, где мутации, введенные в тяжелые цепи иммуноглобулина и легкие цепи иммуноглобулина, вносят новый соляной мостик на поверхности контакта между легкой и тяжелой цепью.
[0124] Не имеющие ограничительного характера примеры подходящих наборов конструкции LCCA описаны в Примерах, Таблицах и на Фигурах. В одном варианте реализации изобретения набор конструкции LCCA содержит тяжелую цепь иммуноглобулина с по меньшей мере одной аминокислотной модификацией в домене СН1, первую легкую цепь иммуноглобулина с по меньшей мере одной аминокислотной модификацией в домене CL и вторую легкую цепь иммуноглобулина без каких-либо модификаций аминокислот в домене CL. В другом варианте реализации изобретения набор конструкции LCCA содержит тяжелую цепь иммуноглобулина с по меньшей мере одной аминокислотной модификацией в домене СН1, первую легкую цепь иммуноглобулина с по меньшей мере одной аминокислотной модификацией в домене CL и вторую легкую цепь иммуноглобулина с по меньшей мере одной аминокислотной модификацией в домене CL. В другом варианте реализации изобретения набор конструкции LCCA содержит тяжелую цепь иммуноглобулина с по меньшей мере одной аминокислотной модификацией в домене СН1, первую легкую цепь иммуноглобулина с по меньшей мере двумя аминокислотными модификациями в домене CL и вторую легкую цепь иммуноглобулина с по меньшей мере двумя аминокислотными модификациями в домене CL. В другом варианте реализации изобретения набор конструкции LCCA содержит тяжелую цепь иммуноглобулина с по меньшей мере одной аминокислотной модификацией в домене СН1, первую легкую цепь иммуноглобулина с по меньшей мере двумя аминокислотными модификациями в домене CL и вторую легкую цепь иммуноглобулина с по меньшей мере одной аминокислотной модификацией в домене CL.
[0125] В одном варианте реализации изобретения набор конструкции LCCA содержит тяжелую цепь иммуноглобулина без каких-либо аминокислотных модификаций в домене СН1, первую легкую цепь иммуноглобулина без каких-либо аминокислотных модификаций в домене CL и вторую легкую цепь иммуноглобулина с по меньшей мере одной аминокислотной модификацией в домене CL. В одном варианте реализации изобретения набор конструкции LCCA содержит тяжелую цепь иммуноглобулина без каких-либо аминокислотных модификаций в домене СН1, первую легкую цепь иммуноглобулина без каких-либо аминокислотных модификаций в домене CL и вторую легкую цепь иммуноглобулина с по меньшей мере двумя аминокислотными модификациями в домене CL.
[0126] В одном варианте реализации изобретения набор конструкции LCCA содержит тяжелую цепь иммуноглобулина с по меньшей мере двумя аминокислотными модификациями в домене СН1, первую легкую цепь иммуноглобулина без каких-либо аминокислотных модификаций в домене CL и вторую легкую цепь иммуноглобулина с по меньшей мере одной аминокислотной модификацией в домене CL. В одном варианте реализации изобретения набор конструкции LCCA содержит тяжелую цепь иммуноглобулина с по меньшей мере двумя аминокислотными модификациями в домене СН1, первую легкую цепь иммуноглобулина с по меньшей мере одной аминокислотной модификацией в домене CL и вторую легкую цепь иммуноглобулина с по меньшей мере одной аминокислотной модификацией в домене CL. В одном варианте реализации изобретения набор конструкции LCCA содержит тяжелую цепь иммуноглобулина с по меньшей мере двумя аминокислотными модификациями в домене СН1, первую легкую цепь иммуноглобулина с по меньшей мере одной аминокислотной модификацией в домене CL и вторую легкую цепь иммуноглобулина с по меньшей мере двумя аминокислотными модификациями в домене CL. В одном варианте реализации изобретения набор конструкции LCCA содержит тяжелую цепь иммуноглобулина с по меньшей мере двумя аминокислотными модификациями в домене СН1, первую легкую цепь иммуноглобулина с по меньшей мере двумя аминокислотными модификациями в домене CL и вторую легкую цепь иммуноглобулина с по меньшей мере двумя аминокислотными модификациями в домене CL. В одном варианте реализации изобретения набор конструкции LCCA содержит тяжелую цепь иммуноглобулина с по меньшей мере двумя аминокислотными модификациями в домене СН1, первую легкую цепь иммуноглобулина с по меньшей мере тремя аминокислотными модификациями в домене CL и вторую легкую цепь иммуноглобулина с по меньшей мере двумя аминокислотными модификациями в домене CL.
[0127] В одном варианте реализации изобретения набор конструкции LCCA содержит тяжелую цепь иммуноглобулина с по меньшей мере тремя аминокислотными модификациями в домене СН1, первую легкую цепь иммуноглобулина без каких-либо аминокислотных модификаций в домене CL и вторую легкую цепь иммуноглобулина с по меньшей мере одной аминокислотной модификацией в домене CL. В одном варианте реализации изобретения набор конструкции LCCA содержит тяжелую цепь иммуноглобулина с по меньшей мере тремя аминокислотными модификациями в домене СН1, первую легкую цепь иммуноглобулина с по меньшей мере одной аминокислотной модификацией в домене CL и вторую легкую цепь иммуноглобулина с по меньшей мере одной аминокислотной модификацией в домене CL. В одном варианте реализации изобретения набор конструкции LCCA содержит тяжелую цепь иммуноглобулина с по меньшей мере тремя аминокислотными модификациями в домене СН1, первую легкую цепь иммуноглобулина с по меньшей мере тремя аминокислотными модификациями в домене CL и вторую легкую цепь иммуноглобулина с по меньшей мере двумя аминокислотными модификациями в домене CL. В одном варианте реализации изобретения набор конструкции LCCA содержит тяжелую цепь иммуноглобулина с по меньшей мере тремя аминокислотными модификациями в домене СН1, первую легкую цепь иммуноглобулина с по меньшей мере четырьмя аминокислотными модификациями в домене CL и вторую легкую цепь иммуноглобулина с по меньшей мере тремя аминокислотными модификациями в домене CL. В одном варианте реализации изобретения преимущественное спаривание, продемонстрированное в наборе конструкции LCCA, устанавливается путем модификации одной или более аминокислот в по меньшей мере одном из доменов VH тяжелой цепи иммуноглобулина, доменов VL первой легкой цепи иммуноглобулина и доменов VL второй легкой цепи иммуноглобулина. Не имеющие ограничительного характера примеры подходящих наборов конструкции LCCA приведены в Таблицах и Примерах ниже.
[0128] В одном варианте реализации изобретения набор конструкции LCCA содержит тяжелую цепь иммуноглобулина без каких-либо аминокислотных модификаций в домене VH, первую легкую цепь иммуноглобулина без каких-либо аминокислотных модификаций в домене VL и вторую легкую цепь иммуноглобулина с по меньшей мере одной аминокислотной модификацией в домене VL. В одном варианте реализации изобретения набор конструкции LCCA содержит тяжелую цепь иммуноглобулина без каких-либо аминокислотных модификаций в домене VH, первую легкую цепь иммуноглобулина без каких-либо аминокислотных модификаций в домене VL и вторую легкую цепь иммуноглобулина с по меньшей мере двумя аминокислотными модификациями в домене VL.
[0129] В одном варианте реализации изобретения набор конструкции LCCA содержит тяжелую цепь иммуноглобулина с по меньшей мере одной аминокислотной модификацией в домене VH, первую легкую цепь иммуноглобулина без каких-либо аминокислотных модификаций в домене VL и вторую легкую цепь иммуноглобулина с по меньшей мере одной аминокислотной модификацией в домене VL. В одном варианте реализации изобретения набор конструкции LCCA содержит тяжелую цепь иммуноглобулина с по меньшей мере одной аминокислотной модификацией в домене VH, первую легкую цепь иммуноглобулина с по меньшей мере одной аминокислотной модификацией в домене VL и вторую легкую цепь иммуноглобулина с по меньшей мере одной аминокислотной модификацией в домене VL. В одном варианте реализации изобретения набор конструкции LCCA содержит тяжелую цепь иммуноглобулина с по меньшей мере одной аминокислотной модификацией в домене VH, первую легкую цепь иммуноглобулина с по меньшей мере двумя аминокислотными модификациями в домене VL и вторую легкую цепь иммуноглобулина с по меньшей мере двумя аминокислотными модификациями в домене VL.
[0130] В одном варианте реализации изобретения набор конструкции LCCA содержит тяжелую цепь иммуноглобулина с по меньшей мере двумя аминокислотными модификациями в домене VH, первую легкую цепь иммуноглобулина без каких-либо аминокислотных модификаций в домене VL и вторую легкую цепь иммуноглобулина с по меньшей мере одной аминокислотной модификацией в домене VL. В одном варианте реализации изобретения набор конструкции LCCA содержит тяжелую цепь иммуноглобулина с по меньшей мере двумя аминокислотными модификациями в домене VH, первую легкую цепь иммуноглобулина с по меньшей мере двумя аминокислотными модификациями в домене VL и вторую легкую цепь иммуноглобулина с по меньшей мере одной аминокислотной модификацией в домене VL. В одном варианте реализации изобретения набор конструкции LCCA содержит тяжелую цепь иммуноглобулина с по меньшей мере двумя аминокислотными модификациями в домене VH, первую легкую цепь иммуноглобулина с по меньшей мере одной аминокислотной модификацией в домене VL и вторую легкую цепь иммуноглобулина с по меньшей мере одной аминокислотной модификацией в домене VL.
[0131] В одном варианте реализации изобретения наборы конструкции LCCA могут быть объединены для обеспечения комбинации, содержащей две разные тяжелые цепи (H1 и Н2) иммуноглобулина и две разные легкие цепи (L1 и L2) иммуноглобулина, где H1 преимущественно спаривается с L1, и Н2 преимущественно спаривается с L2, если H1, Н2, L1 и L2 коэкспрессируются. В некоторых вариантах реализации изобретения аминокислотные модификации описываются в данном документе в контексте конструкции биспецифических антител. Например, наборы конструкции, описанные в данном документе, могут быть включены в полноразмерные тяжелые цепи иммуноглобулина таким образом, что полноразмерные тяжелые цепи преимущественно спариваются с легкими цепями иммуноглобулина. В некоторых вариантах реализации изобретения полноразмерные тяжелые цепи иммуноглобулина содержат аминокислотные модификации, которые способствуют димеризации в фрагменте Fc, как описано в примерах.
Переносимость конкретных аминокислотных модификаций, определенных в настоящем описании, в отношении других антител:
[0132] Несмотря на то, что указанные выше конкретные аминокислотные модификации тяжелых и легких цепей иммуноглобулина были описаны относительно антитела D3H44 к внеклеточному домену тканевого фактора, тяжелых и легких цепей иммуноглобулина трастузумаба и цетуксимаба, предполагается и показано в данном документе (см., примеры, фигуры и таблицы), что эти аминокислотные модификации могут быть перенесены на другие тяжелые и легкие цепи иммуноглобулина, что приводит к аналогичным наборам преимущественного спаривания одной тяжелой цепи иммуноглобулина с одной из двух легких цепей иммуноглобулина, с учетом нижеприведенного.
[0133] Остатки области контакта VH:VL и CH1:CL в области контакта между тяжелыми и легкими цепями иммуноглобулина сохраняются относительно хорошо (Padlan et al, 1986, Mol. Immunol. 23(9): 951-960). Данная консервативность последовательности, результат эволюционных ограничений, увеличивает вероятность того, что функционально активные домены связывания антител будут формироваться во время комбинаторного спаривания легких и тяжелых цепей. В результате данной консервативности последовательности следует, что модификации последовательности в конкретных примерах, указанных выше для D3H44, которые запускают преимущественное спаривание, могут быть переданы другой паре тяжелой и легкой цепи гетеродимеров с приблизительно эквивалентными результатами, полученными с учетом преимущественного спаривания, поскольку этот участок показывает высокую консервативность последовательности среди антител; Кроме того, в случае если происходят различия в последовательности, обычно они находятся дистально по отношению к области контакта CH1:CL. Это особенно относится к доменам СН1 и CL. Существует, однако, некоторая вариабельность последовательности антигенсвязывающего сайта по отношению к CDR (комплементарно определяемым областям) остаткам петли (и длине), в частности, для CDR-H3. Таким образом, в одном варианте реализации изобретения, описанные в данном документе пары гетеродимеров включают гетеродимеры, где по меньшей мере один гетеродимер содержит одну или более аминокислотных модификаций в доменах VH и/или VL, которые находятся дистально по отношению к петлям CDR, в случае если аминокислотная последовательность антигенсвязывающего сайта значительно отличается от таковой в антителе D3H44. В другом варианте реализации изобретения, описанные в данном документе пары гетеродимеров включают гетеродимеры, где по меньшей мере один гетеродимер содержит одну или более аминокислотных модификаций в доменах VH и/или VL, которые находятся проксимально или дистально по отношению к петлям CDR, в случае если аминокислотная последовательность антигенсвязывающего сайта по существу является такой как в антителе D3H44.
[0134] В одном варианте реализации изобретения аминокислотные модификации, описанные в данном документе, могут быть перенесены на тяжелые и легкие цепи иммуноглобулина антител на основе IgG1 человека или гуманизированного IgG1/κ. Неограничивающие примеры таких цепей IgG1/κ содержат афатумумаб (для человека) или трастузумаб, пертузумаб или бевацизумаб (для гуманизированного).
[0135] В другом варианте реализации изобретения аминокислотные модификации, описанные в данном документе, являются переносимыми на тяжелые и легкие цепи иммуноглобулина антител с использованием широко используемых подгрупп VH и VL. Неограничивающие примеры таких антител содержат пертузумаб.
[0136] В одном варианте реализации изобретения аминокислотные модификации, описанные в данном документе, являются переносимыми на тяжелые и легкие цепи иммуноглобулина антител, имеющих каркас, близкий к эмбриональному. Примеры таких антител включают обинутузумаб.
[0137] В одном варианте реализации изобретения аминокислотные модификации, описанные в данном документе, являются переносимыми на тяжелые и легкие цепи иммуноглобулина антител, имеющих междоменный угол VH:VL, близкий к среднему значению, наблюдаемому для пар тяжелой и легкой цепи. Пример такого типа антител содержит, но без ограничений, пертузумаб. В другом варианте реализации изобретения аминокислотные модификации, описанные в данном документе, являются переносимыми на тяжелые и легкие цепи иммуноглобулина антител, имеющих канонические домены CL и СН1. Подходящие примеры таких антител содержат, но без ограничений, трастузумаб.
[0138] В некоторых вариантах реализации изобретения определенные подмножества аминокислотных модификаций, описанные в настоящем документе, используются в качестве варианта доменов в антигенсвязывающих конструкциях, приведенных выше.
[0139] Примеры, фигуры и таблицы демонстрируют, что аминокислотные модификации (например, в пределах одного или более Fab-фрагментов, содержащих вариабельную область и константную область) являются переносимыми на другие тяжелые и легкие цепи иммуноглобулина, что приводит к аналогичным наборам преимущественного спаривания одной тяжелой цепи иммуноглобулина с одной из двух легких цепей иммуноглобулина.
Преимущественное спаривание
[0140] Как было описано выше по меньшей мере один гетеродимер антигенсвязывающей конструкции/пар гетеродимеров, описанных в данном документе, может содержать одну или несколько аминокислотных модификаций к их тяжелым цепям иммуноглобулина и/или легким цепям иммуноглобулина, таким образом, что тяжелая цепь одного гетеродимера, например H1, преимущественно спаривается с одной из легких цепей, например L1, а не с другой легкой цепью, L2, и тяжелая цепь другого гетеродимера, Н2, преимущественно спаривается с легкой цепью, L2, а не легкой цепью L1. Другими словами, желаемое, преимущественное спаривание считается спаиванием между H1 и L1, а также между Н2 и L2. Преимущественное спаривание между, например, H1 и L1 считается произошедшим, если выход гетеродимера H1-L1 является большим, чем выход неправильно спаренного гетеродимера H1-L2, в случае если H1 объединяется с L1 и L2, по отношению к соответствующему спариванию соответствующей пары H1/L1 к паре H2/L2 без одной или более аминокислотных модификаций. Преимущественное спаривание между, например, Н2 и L2 считается произошедшим, если выход гетеродимера H2-L2 является большим, чем выход неправильно спаренного гетеродимера H2-L1, когда Н2 объединяется с L1 и L2, по отношению к соответствующему спариванию соответствующей пары H1-L1 к паре H2-L2 без одной или более аминокислотных модификаций. В этом контексте гетеродимер, содержащий H1 и L1 (H1-L1) или Н2 и L2 (H2-L2), упоминается в данном документе как преимущественно спаренный, корректно спаренный, обязательно спаренный или желаемый гетеродимер, в то время как гетеродимер, содержащий H1 и L2 (H1-L2) или Н2 и L1 (H2-L1), упоминается здесь как неправильно спаренный гетеродимер. Набор мутаций, соответствующий двум тяжелым цепям и двум легким цепям, предназначен для достижения избирательного спаривания H1-L1 и L2-H2 и упоминается как набор конструкции.
[0141] Таким образом, в одном варианте реализации изобретения, в случае если одна тяжелая цепь иммуноглобулина гетеродимера коэкспрессируется с двумя легкими цепями иммуноглобулина, относительный выход желаемого гетеродимера превышает 55%. В другом варианте реализации изобретения, в случае если одна тяжелая цепь иммуноглобулина гетеродимера коэкспрессируется с двумя легкими цепями иммуноглобулина, относительный выход желаемого гетеродимера превышает 60%. В другом варианте реализации изобретения, в случае если одна тяжелая цепь иммуноглобулина гетеродимера коэкспрессируется с двумя легкими цепями иммуноглобулина, относительный выход желаемого гетеродимера превышает 70%. Таким образом, в одном варианте реализации изобретения, в случае если одна тяжелая цепь иммуноглобулина гетеродимера коэкспрессируется с двумя легкими цепями иммуноглобулина, относительный выход желаемого гетеродимера превышает 80%. В другом варианте реализации изобретения, в случае если одна тяжелая цепь иммуноглобулина гетеродимера коэкспрессируется с двумя легкими цепями иммуноглобулина, относительный выход желаемого гетеродимера превышает 90%. В другом варианте реализации изобретения, в случае если одна тяжелая цепь иммуноглобулина гетеродимера коэкспрессируется с двумя легкими цепями иммуноглобулина, относительный выход желаемого гетеродимера превышает 95%.
[0142] В приведенном выше примере, преимущественное спаривание между H1-L1 считается произошедшим, если количество желаемого гетеродимера H1-L1 является большим, чем количество неправильно спаренного гетеродимера H1-L2, в случае если H1 коэкспрессируется с L1 и L2. Подобным образом, преимущественное спаривание между H2-L2 считается происшедшим, если количество желаемого гетеродимера H2-L2 является большим, чем количество неправильно спаренного гетеродимера H2-L2, в случае если Н2 коэкспрессируется с L1 и L2. Таким образом, в одном варианте реализации изобретения, в случае если одна тяжелая цепь иммуноглобулина гетеродимера коэкспрессируется с двумя легкими цепями иммуноглобулина, соотношение желаемого гетеродимера к неправильно спаренному гетеродимеру является большим, чем 1,25:1. В одном варианте реализации изобретения, в случае если одна тяжелая цепь иммуноглобулина гетеродимера коэкспрессируется с двумя легкими цепями иммуноглобулина, соотношение желаемого гетеродимера к неправильно спаренному гетеродимеру является большим, чем 1,5:1. В другом варианте реализации изобретения, в случае если одна тяжелая цепь иммуноглобулина гетеродимера коэкспрессируется с двумя легкими цепями иммуноглобулина, соотношение желаемого гетеродимера к неправильно спаренному гетеродимеру является большим, чем 2:1. В другом варианте реализации изобретения, в случае если одна тяжелая цепь иммуноглобулина гетеродимера коэкспрессируется с двумя легкими цепями иммуноглобулина, соотношение желаемого гетеродимера к неправильно спаренному гетеродимеру является большим, чем 3:1. В другом варианте реализации изобретения, в случае если одна тяжелая цепь иммуноглобулина гетеродимера коэкспрессируется с двумя легкими цепями иммуноглобулина, соотношение желаемого гетеродимера к неправильно спаренному гетеродимеру является большим, чем 5:1. В другом варианте реализации изобретения, в случае если одна тяжелая цепь иммуноглобулина гетеродимера коэкспрессируется с двумя легкими цепями иммуноглобулина, соотношение желаемого гетеродимера к неправильно спаренному гетеродимеру является большим, чем 10:1. В другом варианте реализации изобретения, в случае если одна тяжелая цепь иммуноглобулина гетеродимера коэкспрессируется с двумя легкими цепями иммуноглобулина, соотношение желаемого гетеродимера к неправильно спаренному гетеродимеру является большим, чем 25:1. В другом варианте реализации изобретения, в случае если одна тяжелая цепь иммуноглобулина гетеродимера коэкспрессируется с двумя легкими цепями иммуноглобулина, соотношение желаемого гетеродимера к неправильно спаренному гетеродимеру является большим, чем 50:1.
[0143] В некоторых вариантах реализации изобретения гетеродимеры, описанные в данном документе, преимущественно спариваются с образованием биспецифических антител. В некоторых вариантах реализации изобретения конструкции содержат гетеродимер, который преимущественно спаривается с образованием биспецифического антитела, выбранного из из D3H44/трастузумаб, D3H44/цетуксимаб и трастузумаб/цетуксимаб. В некоторых вариантах реализации изобретения биспецифическое антитело содержит аминокислотные модификации, описанные в Таблицах 28а-28с.
[0144] В некоторых вариантах реализации изобретения две конструкции полноразмерных тяжелых цепей коэкспрессируются с двумя уникальными конструкциями легких цепей, что дает десять возможных видов антител: H1-L1:H1-L1, H1-L2:H1-L2, Hl-L1:H1-L2, H2-L1:H2-L1, H2-L2:H2-L2, H2-L1:H2-L2, H1-L1:H2-L1, H1-L2:H2-L2, H1-L2:H2-L1 и H1-L1:H2-L2. Вид H1-L1:H2-L2 считается корректно спаренным биспецифическим видом антитела. В некоторых вариантах реализации изобретения подбирались соотношения ДНК с целью получить наибольшее количество корректно спаренных биспецифических видов антител. Например, в некоторых вариантах реализации изобретения соотношение H1:H2:L1:L2 составляет 15:15:53:17. В некоторых вариантах реализации изобретения соотношение H1:H2:L1:L2 составляет 15:15:17:53.
[0145] В некоторых вариантах реализации изобретения процент правильно спаренных биспецифических видов составляет по меньшей мере 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%, относительно всех видов (см., например, Таблицы 29а-29с и 30а-30с). В некоторых вариантах реализации изобретения процент корректно спаренных биспецифических видов является большим, чем процент корректно спаренных биспецифических видов, полученных с помощью коэкспрессии соответствующего H1, Н2, L1 и L2 дикого типа, без аминокислотных модификаций, описанных в Таблицах 28а-28 с. В некоторых вариантах реализации изобретения процент корректно спаренных биспецифических видов увеличивается по меньшей мере на 5%, 10%, 15%, 20%, 30%, 40%, 50%, 60%, 70% или 75% по сравнению с процентом корректно спаренных биспецифических видов, полученных с помощью коэкспрессии соответствующего H1, Н2, L1 и L2 дикого типа, без аминокислотных модификаций, описанных в Таблицах 28а-28 с. Термостабильность гетеродимеров
[0146] Помимо содействия преимущественному спариванию, аминокислотные замены были выбраны таким образом, чтобы мутации не дестабилизировали гетеродимеры Fab. Таким образом, в большинстве случаев, измерения устойчивости гетеродимеров Fab были очень близки к таковым у Fab дикого типа (например, в пределах 3°С Fab дикого типа).
[0147] Таким образом, в некоторых вариантах реализации изобретения каждый гетеродимер пары гетеродимеров, описанный в данном документе, имеет термическую стабильность, которая сравнима с таковой у гетеродимера, содержащего те же тяжелые и легкие цепи иммуноглобулина, но без аминокислотных модификаций в доменах CH1, CL, VH или VL, описанных в данном документе. В одном варианте реализации изобретения термическая стабильность определяется путем измерения температуры плавления или Тт.Таким образом, в одном варианте реализации изобретения термическая стабильность гетеродимера, описанного в настоящем документе, находится в пределах около 10°С, что и у гетеродимера, содержащего те же тяжелые и легкие цепи иммуноглобулина без аминокислотных модификаций в доменах CH1, CL, VH или VL, описанных в данном документе. Таким образом, в одном варианте реализации изобретения термическая стабильность гетеродимера, описанного в настоящем документе, находится в пределах около 5°С, что и у гетеродимера, содержащего те же тяжелые и легкие цепи иммуноглобулина без аминокислотных модификаций в доменах CH1, CL, VH или VL, описанных в данном документе. В другом варианте реализации изобретения термическая стабильность гетеродимера, описанного в настоящем документе, находится в пределах около 3°С, что и у гетеродимера, содержащего те же тяжелые и легкие цепи иммуноглобулина без аминокислотных модификаций в доменах CH1, CL, VH или VL, описанных в данном документе. В другом варианте реализации изобретения термическая стабильность гетеродимера, описанного в настоящем документе, находится в пределах около 2°С, что и у гетеродимера, содержащего те же тяжелые и легкие цепи иммуноглобулина без аминокислотных модификаций в доменах CH1, CL, VH или VL, описанных в данном документе. В другом варианте реализации изобретения термическая стабильность гетеродимера, описанного в настоящем документе, находится в пределах около 1,5°С, что и у гетеродимера, содержащего те же тяжелые и легкие цепи иммуноглобулина без аминокислотных модификаций в доменах CH1, CL, VH или VL, описанных в данном документе. В другом варианте реализации изобретения термическая стабильность гетеродимера, описанного в настоящем документе, находится в пределах около 1°С, что и у гетеродимера, содержащего те же тяжелые и легкие цепи иммуноглобулина без аминокислотных модификаций в доменах CH1, CL, VH или VL, описанных в данном документе. В другом варианте реализации изобретения термическая стабильность гетеродимера, описанного в настоящем документе, находится в пределах около 0,5°С, что и у гетеродимера, содержащего те же тяжелые и легкие цепи иммуноглобулина без аминокислотных модификаций в доменах CH1, CL, VH или VL, описанных в данном документе. В другом варианте реализации изобретения термическая стабильность гетеродимера, описанного в настоящем документе, находится в пределах около 0,25°С, что и у гетеродимера, содержащего те же тяжелые и легкие цепи иммуноглобулина без аминокислотных модификаций в доменах CH1, CL, VH или VL, описанных в данном документе.
[0148] Кроме того, в некоторых вариантах реализации изобретения термическая стабильность гетеродимера, описанного в данном документе, неожиданно улучшается (т.е., увеличивается) относительно таковой у гетеродимера, содержащего те же тяжелые и легкие цепи иммуноглобулина без аминокислотных модификаций в доменах CH1, CL, VH или VL, описанных в данном документе. Таким образом, в одном варианте реализации изобретения термическая стабильность гетеродимера, описанного в данном документе, увеличивается на около 0,1, 0,2, 0,3, 0,4, 0,5, 0,6, 0,7, 0,8. 0,9, 1,0, 1,1, 1,2, 1,3, 1,4, 1,5, 1,6, 1,7, 1,8, 1,9, 2,0, 2,1, 2,2, 2,3, 2,4, 2,5, 3,0, 3,1, 3,2, 3,3, 3,4, 3,5, 3,6, 3,7, 3,8, 3,9, 4,0, 4,5, 5,0°С или более по сравнению с гетеродимером, содержащим те же тяжелые и легкие цепи иммуноглобулина без аминокислотных модификаций в доменах CH1, CL, VH или VL, описанных в данном документе.
Аффинность гетеродимеров к антигену
[0149] В одном варианте реализации изобретения каждый гетеродимер пары гетеродимеров обладает аффинностью к соответствующему ему антигену, которая является такой же или сравнима с таковой у гетеродимера, содержащего те же тяжелые и легкие цепи иммуноглобулина, но без аминокислотных модификаций в доменах CH1, CL, VH или VL, описанных в данном документе. В одном варианте реализации изобретения гетеродимер пары гетеродимеров обладает аффинностью к соответствующему ему антигену, которая находится в около 50-кратных пределах таковой у гетеродимера, содержащего те же тяжелые и легкие цепи иммуноглобулина, но без аминокислотных модификаций в доменах CH1, CL, VH или VL, описанных в данном документе. В одном варианте реализации изобретения гетеродимер пары гетеродимеров обладает аффинностью к соответствующему ему антигену, которая находится в около 25-кратных пределах таковой у гетеродимера, содержащего те же тяжелые и легкие цепи иммуноглобулина, но без аминокислотных модификаций в доменах CH1, CL, VH или VL, описанных в данном документе. В одном варианте реализации изобретения гетеродимер пары гетеродимеров обладает аффинностью к соответствующему ему антигену, которая находится в около 10-кратных пределах таковой у гетеродимера, содержащего те же тяжелые и легкие цепи иммуноглобулина, но без аминокислотных модификаций в доменах CH1, CL, VH или VL, описанных в данном документе. В другом варианте реализации изобретения гетеродимер пары гетеродимеров обладает аффинностью к соответствующему ему антигену, которая находится в около 5-кратных пределах таковой у гетеродимера, содержащего те же тяжелые и легкие цепи иммуноглобулина, но без аминокислотных модификаций в доменах CH1, CL, VH или VL, описанных в данном документе. В другом варианте реализации изобретения гетеродимер пары гетеродимеров обладает аффинностью к соответствующему ему антигену, которая находится в около 2,5- кратных пределах таковой у гетеродимера, содержащего те же тяжелые и легкие цепи иммуноглобулина, но без аминокислотных модификаций в доменах CH1, CL, VH или VL, описанных в данном документе. В другом варианте реализации изобретения гетеродимер пары гетеродимеров обладает аффинностью к соответствующему ему антигену, которая находится в около 2-кратных пределах таковой у гетеродимера, содержащего те же тяжелые и легкие цепи иммуноглобулина, но без аминокислотных модификаций в доменах CH1, CL, VH или VL, описанных в данном документе. В другом варианте реализации изобретения гетеродимер пары гетеродимеров обладает аффинностью к соответствующему ему антигену, которая находится в около 1,5-кратных пределах таковой у гетеродимера, содержащего те же тяжелые и легкие цепи иммуноглобулина, но без аминокислотных модификаций в доменах CH1, CL, VH или VL, описанных в данном документе. В другом варианте реализации изобретения гетеродимер пары гетеродимеров обладает аффинностью к соответствующему ему антигену, которая является примерно такой же, как и у гетеродимера, содержащего те же тяжелые и легкие цепи иммуноглобулина, но без аминокислотных модификаций в доменах CH1, CL, VH или VL, описанных в данном документе.
Дополнительные необязательные модификации
[0150] В одном варианте реализации изобретения тяжелые и легкие цепи иммуноглобулина пар гетеродимеров, описанные в данном документе, могут быть дополнительно модифицированы (то есть, путем ковалентного присоединения различных типов молекул) таким образом, что ковалентное присоединение не препятствует преимущественному спариванию между тяжелой цепью и легкими цепями или влиянию на способность гетеродимера связываться с его антигеном или влиянию на его стабильность. Такая модификация включает, например, но не в качестве исчерпывающего перечня, гликозилирование, ацетилирование, пегилирование, фосфорилирование, амидирование, дериватизацию известных защитных/блокирующих групп, протеолитическое расщепление, связывание с клеточным лигандом или другим белком и т.д. Любая из многочисленных химических модификаций может быть осуществлена известными методами, в том числе, но без ограничений, специфическим химическим расщеплением, ацетилированием, формилированием, метаболическим синтезом туникамицина и т.д.
[0151] В другом варианте реализации изобретения тяжелые и легкие цепи иммуноглобулина пар гетеродимеров, описанные в данном документе, могут быть конъюгированы (прямо или опосредованно) с терапевтическим средством или молекулой лекарственного средства, модифицирующего данную биологическую реакцию. Терапевтические средства или молекулы лекарственного средства не должны быть истолкованы как ограниченные лишь классическими химическими терапевтическими средствами. Например, молекула лекарственного средства может быть белком или полипептидом, обладающим желаемой биологической активностью. Такие белки могут включать, например, токсин, такой как абрин, рицин А, Онконазу (или другую цитотоксическую РНКазу), экзотоксин синегнойной палочки, холерный токсин или дифтерийный токсин; белок, такой как фактор некроза опухоли, альфа-интерферон, бета- интерферон, нейрональный ростовой фактор, тромбоцитарный фактор роста, тканевый активатор плазминогена, апоптотический агент, например, TNF-альфа, TNF-бета, АГМ I (см., международная публикация № WO 97/33899), AIM II (см., международная публикация №WO 97/34911), лиганд Fas (Takahashi et al, 1994, J. Immunol., 6:1567) и VEGI (см., международная публикация №WO 99/23105), противотромботическое средство или ингибитор ангиогенеза, например, ангиостатин или эндостатин; или биологический модификатор, такой как, например, лимфокин (например, интерлейкин-1 ("IL-1"), интерлейкин-2 ("IL-2"), интерлейкин-6 ("IL-6"), гранулоцитарно-макрофагальный колониестимулирующий фактор ("GM-CSF"), и гранулоцитарный колониестимулирующий фактор ("G-CSF")), или фактор роста (например, гормон роста ("GH")).
[0152] Более того, в альтернативном варианте реализации изобретения антитело может быть конъюгировано с терапевтическими молекулами, такими как радиоактивные вещества или макроциклические хелатирующие агенты, используемые для конъюгации ионов радиоактивного металла (см. выше примеры радиоактивных материалов). В определенных вариантах реализации изобретения макроциклическим хелатирующим агентом является 1,4,7,10-тетраазациклододекан-N,N',N'',N'''-тетрауксусная кислота (DOTA), которая может быть присоединена к антителу с помощью линкерной молекулы. Такие линкерные молекулы, как правило, известны в данной области техники и описаны в Denardo et al, 1998, Clin Cancer Res. 4:2483; Peterson et al, 1999, Bioconjug. Chem. 10:553; и Zimmerman et al, 1999, Nucl. Med. Biol. 26:943.
[0153] В некоторых вариантах реализации изобретения тяжелые и легкие цепи иммуноглобулина гетеродимеров экспрессируются в виде слитых белков, содержащих метку для облегчения очистки и/или тестирования и т.д. Согласно изложенному в настоящем документе "метка" представляет собой любую дополнительную последовательность аминокислот, которая содержится в белке на С-конце и на N-конце или внутри последовательности, что способствует идентификации или очистке данного белка. Подходящие метки включают, но не ограничиваются метками, известными специалистам в данной области техники, которые являются полезным в очистке и/или тестировании, такие метки, как альбумин-связывающий домен (ABD), гистидиновую метку, FLAG-метку, глутатион-Б-трансферазу, гемагглютинин (НА) и связывающий мальтозу белок. Такие меченые белки также могут быть разработаны таким образом, чтобы содержать сайт расщепления, такой как сайт расщепления тромбином, энтерокиназой или фактором X, для простоты удаления метки до, во время или после очистки.
[0154] В некоторых вариантах реализации изобретения один или более остатков цистеина в нижней части домена Fab в легкой (позиция 214, нумерация по Kabat) и тяжелой (позиция 233, нумерация по Kabat) цепи, образующие межцепьевую дисульфидную связь, могут быть модифицированы на серии или аланин или на нецистеиновую или отличающуюся аминокислоту.
[0155] Следует иметь в виду, что дополнительные аминокислотные модификации могут быть проведены для тяжелых цепей иммуноглобулина с целью повышения уровня преимущественного спаривания и/или термической стабильности пар гетеродимеров. Например, дополнительные аминокислотные модификации могут быть сделаны для домена Fc тяжелой цепи иммуноглобулина с целью управления преимущественным спариванием между парами гетеродимера по отношению к парам гомодимера. Такие аминокислотные модификации известны в данной области техники и включают, например, те, которые описаны в патентной публикации США №2012/0149876. В альтернативном варианте также могут быть использованы альтернативные стратегии для управления преимущественным спариванием между парами гетеродимера по отношению к парам гомодимера, такие как, например, технологии "выступы-во-впадины", заряженные остатки с ионными взаимодействиями и разработанный домен обмена цепями (SEED). Последние стратегии были описаны в данной области техники и рассмотрены выше в Klein et al. Дальнейшее обсуждение доменов Fc следует ниже.
Домены Fc
[0156] В вариантах реализации изобретения, где антигенсвязывающая полипептидная конструкция содержит полноразмерные тяжелые цепи иммуноглобулина, конструкция будет содержать Fc. В некоторых аспектах Fc содержит по меньшей мере одну или две последовательности домена СН3. В некоторых аспектах, где антигенсвязывающая полипептидная конструкция содержит гетеродимеры, которые содержат только область Fab тяжелой цепи, Fc является соединенным, с помощью одного или более линкеров или без линкеров, с первым гетеродимером и/или вторым гетеродимером. В некоторых аспектах Fc представляет собой человеческий Fc. В некоторых аспектах Fc представляет собой IgG или IgG1 Fc человека. В некоторых аспектах Fc представляет собой гетеродимерный Fc. В некоторых аспектах Fc содержит по меньшей мере одну или две последовательности домена СН2.
[0157] В некоторых аспектах Fc содержит одну или более модификаций по меньшей мере в одной из последовательностей СH3. В некоторых аспектах Fc содержит одну или более модификаций по меньшей мере в одной из последовательностей СH2. В некоторых аспектах Fc состоит из одного полипептида. В некоторых аспектах Fc состоит из множества пептидов, например, двух полипептидов.
[0158] В некоторых аспектах Fc содержит одну или более модификаций по меньшей мере в одной из последовательностей СH3. В некоторых аспектах Fc содержит одну или более модификаций по меньшей мере в одной из последовательностей СH2. В некоторых аспектах Fc состоит из одного полипептида. В некоторых аспектах Fc состоит из множества пептидов, например, двух полипептидов.
[0159] В некоторых аспектах Fc представляет собой Fc, описанный в патентных заявках РСТ/СА2011/001238, поданной 4 ноября 2011 года, или РСТ/СА2012/050780, поданной 2 ноября 2012 года, причем полное описание каждой из них включено в данный документ в полном объеме посредством ссылки для всех целей.
[0160] В некоторых аспектах конструкция, описанная в данном документе, содержит гетеродимерный Fc, содержащий модифицированный СН3-домен, который был асимметрично модифицирован. Гетеродимерный Fc может содержать два полипептида константных доменов тяжелой цепи: первый полипептид тяжелой цепи и второй полипептид тяжелой цепи, которые могут быть использованы взаимозаменяемо при условии, что Fc содержит один первый полипептид тяжелой цепи и один второй полипептид тяжелой цепи. Как правило, первый полипептид тяжелой цепи содержит первую последовательность СН3, и второй полипептид тяжелой цепи содержит вторую последовательность СН3.
[0161] В результате введения одной или более асимметричных аминокислотных модификаций в две последовательности СН3, как правило, образуется гетеродимерный Fc, а не гомодимер, при этом происходит димеризация двух последовательностей СН3. Используемый в данном документе термин "асимметричные аминокислотные модификации" относится к любой модификации, при которой аминокислота в определенном положении в первой последовательности СН3 отличается от аминокислоты, находящейся во второй последовательности СН3 в том же самом положении, и первая и вторая последовательность СН3 преимущественно соединяются с образованием гетеродимера, а не гомодимера. Эта гетеродимеризация может происходить в результате модификации только одной из двух аминокислот в одном и том же положении соответствующей аминокислоты в каждой последовательности; или модификации обеих аминокислот в каждой последовательности в одном и том же соответствующем положении в каждой первой и второй последовательности СН3. Первая и вторая последовательность СН3 гетеродимерного Fc могут содержать одну или более чем одну асимметричную аминокислотную модификацию.
В таблице X представлена аминокислотная последовательность Fc-домена IgG1 человека, соответствующая аминокислотам от 231 до 447 тяжелой цепи полноразмерного IgG1 человека. Последовательность СН3 содержит аминокислоты 341-447 тяжелой цепи полноразмерного IgG1 человека.
[0162] Как правило, Fc может включать две непрерывные последовательности тяжелой цепи (А и В), которые способны к димеризации. В некоторых аспектах, одна или обе последовательности Fc включают одну или несколько мутаций или модификаций в следующих местах: L351, F405, Y407, Т366, К392, Т394, Т350, S400 и/или N390 согласно нумерации EU. В некоторых аспектах Fc включает мутантную последовательность, проиллюстрированную в таблице X. В некоторых аспектах Fc содержит мутации Варианта 1 А-В. В некоторых аспектах Fc содержит мутации Варианта 2 А-В. В некоторых аспектах Fc содержит мутации Варианта 3 А-В. В некоторых аспектах Fc содержит мутации Варианта 4 А-В. В некоторых аспектах Fc содержит мутации Варианта 5 А-В.
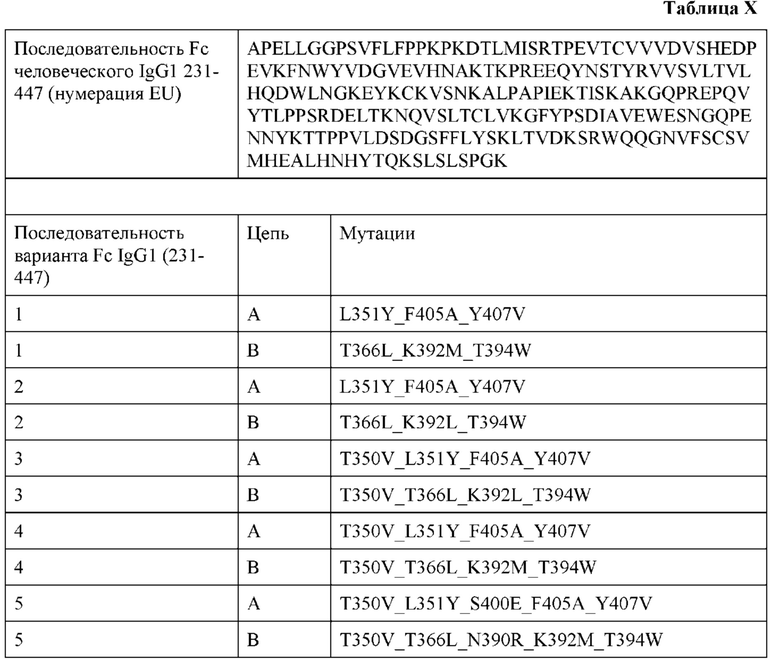
[0163] Первая и вторая последовательности СН3 могут содержать аминокислотные мутации, которые описаны в данном документе, по отношению к аминокислотам от 231 до 447 тяжелой цепи полноразмерного IgG1 человека. В одном варианте реализации изобретения гетеродимерный Fc содержит модифицированный домен СН3, причем в первой последовательности СН3 содержатся аминокислотные модификации в положениях F405 и Y407, и во второй последовательности СН3 содержатся аминокислотные модификации в положении Т394. В одном варианте реализации изобретения гетеродимерный Fc содержит модифицированный домен СН3, причем в первой последовательности СН3 содержится одна или более аминокислотных модификаций, выбранных из L351Y, F405A и Y407V, и во второй последовательности СН3 содержится одна или более аминокислотных модификаций, выбранных из T366L, T366I, K392L, K392М и T394W.
[0164] В одном варианте реализации изобретения гетеродимерный Fc содержит модифицированный домен СН3, причем в первой последовательности СН3 содержатся аминокислотные модификации в положениях L351, F405 и Y407, и во второй последовательности СН3 содержатся аминокислотные модификации в положениях Т366, K392 и Т394, и в одной из первой или второй последовательности СН3 дополнительно содержатся аминокислотные модификации в положении Q347, и в другой последовательности СН3 дополнительно содержится аминокислотная модификация в положении K360. В другом варианте реализации изобретения гетеродимерный Fc содержит модифицированный домен СН3, причем в первой последовательности СН3 содержатся аминокислотные модификации в положениях L351, F405 и Y407, и во второй последовательности СН3 содержатся аминокислотные модификации в положении Т366, K392 и Т394, причем в одной из первой или второй последовательности СН3 дополнительно содержатся аминокислотные модификации в положении Q347, и в другой последовательности СН3 дополнительно содержится аминокислотная модификация в положении K360, и в одной или обеих из указанных последовательностей СН3 дополнительно содержится аминокислотная модификация T350V.
[0165] В одном варианте реализации изобретения гетеродимерный Fc содержит модифицированный домен СН3, причем в первой последовательности СН3 содержатся аминокислотные модификации в положениях L351, F405 и Y407, и во второй последовательности СН3 содержатся аминокислотные модификации в положениях Т366, КЗ 92 и Т394, и в одной из указанных первой и второй последовательности СН3 дополнительно содержится аминокислотная модификация D399R или D399K, и причем в другой последовательности СН3 содержится одна или более модификаций из Т411Е, T411D, K409Е, K409D, K92Е и K392D. В другом варианте реализации изобретения гетеродимерный Fc содержит модифицированный домен СН3, причем в первой последовательности СН3 содержатся аминокислотные модификации в положениях L351, F405 и Y407, и во второй последовательности СН3 содержатся аминокислотные модификации в положениях Т366, К392 и Т394, причем в одной из указанных первой и второй последовательностей СН3 дополнительно содержится аминокислотная модификация D399R или D399K, и в другой последовательности СН3 содержится одна или более модификаций из Т411Е, T411D, K409Е, K409D, K392Е и K392D, и в одной или обеих из указанных последовательностей СН3 дополнительно содержится аминокислотная модификация T350V.
[0166] В одном варианте реализации изобретения гетеродимерный Fc содержит модифицированный домен СН3, причем в первой последовательности СН3 содержатся аминокислотные модификации в положениях L351, F405 и Y407, и во второй последовательности СН3 содержатся аминокислотные модификации в положениях Т366, K392 и Т394, причем в одной или обеих из указанных последовательностей СН3 дополнительно содержится аминокислотная модификация T350V.
[0167] В одном варианте реализации изобретения гетеродимерный Fc-фрагмент содержит модифицированный домен СН3, содержащий следующие ниже аминокислотные модификации, где "А" представляет собой аминокислотную модификацию в первой последовательности СН3, и "В" представляет собой аминокислотную модификацию во второй последовательности СН3: A:L351Y_F405A_Y407V, B:T366L_K392M_T394W, A:L351Y_F405A_Y407V, B:T366L_K392L_T394W, A:T350V_L351Y_F405A_Y407V, B:T350V_T366L_K392L_T394W, A:T350V_L351 Y_F405A_Y407V, B:T350V_T366L_K392M_T394W, A:T350V_L351Y_S400E_F405A_Y407V и/или В: Т350V_T3 66L_N3 90R_K3 92M_T3 94W.
[0168] Одна или более асимметричных аминокислотных модификаций могут способствовать образованию гетеродимерного Fc, причем гетеродимерный домен СН3 обладает стабильностью, которая сравнима с гомодимерным доменом СН3 дикого типа. В варианте реализации изобретения одна или более асимметричных аминокислотных модификаций способствуют образованию гетеродимерного домена Fc, причем гетеродимерный домен Fc обладает стабильностью, которая сравнима с гомодимерным доменом Fc дикого типа. В варианте реализации изобретения одна или более асимметричных аминокислотных модификаций способствуют образованию гетеродимерного домена Fc, причем гетеродимерный домен Fc обладает стабильностью, определяемой по температуре плавления (Тm) в исследовании методом дифференциальной сканирующей калориметрии, и причем температура плавления находится в пределах 4°С по отношению к температуре, наблюдаемой для соответствующего симметричного гомодимерного домена Fc дикого типа. В некоторых аспектах Fc содержит одну или более модификаций по меньшей мере в одной из последовательностей СH3, которые способствуют образованию гетеродимерного Fc со стабильностью, сравнимой со стабильностью гомодимерного Fc дикого типа.
[0169] В одном варианте реализации изобретения стабильность домена СН3 может быть оценена путем измерения температуры плавления домена СН3, например, с помощью дифференциальной сканирующей калориметрии (DSC). Таким образом, в дополнительном варианте реализации изобретения домен СН3 имеет температуру плавления около 68°С или выше. В другом варианте реализации изобретения домен СН3 имеет температуру плавления около 70°С или выше. В другом варианте реализации изобретения домен СН3 имеет температуру плавления около 72°С или выше. В другом варианте реализации изобретения домен СН3 имеет температуру плавления около 73°С или выше. В другом варианте реализации изобретения домен СН3 имеет температуру плавления около 75°С или выше. В другом варианте реализации изобретения домен СН3 имеет температуру плавления около 78°С или выше. В некоторых аспектах температура плавления (Tm) димеризованных последовательностей СH3 составляет около 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 77,5, 78, 79, 80, 81, 82, 83, 84 или 85°С, или выше.
[0170] В некоторых вариантах реализации изобретения гетеродимерный Fc, содержащий модифицированные последовательности СН3, может образоваться с показателем чистоты по меньшей мере около 75% по сравнению с гомодимерным Fc в экспрессированном продукте. В другом варианте реализации гетеродимерный Fc образуется с показателем чистоты, большим, чем около 80%. В другом варианте реализации гетеродимерный Fc образуется с показателем чистоты, большим, чем около 85%. В другом варианте реализации гетеродимерный Fc образуется с показателем чистоты, большим, чем около 90%. В другом варианте реализации гетеродимерный Fc образуется с показателем чистоты, большим, чем около 95%. В другом варианте реализации гетеродимерный Fc образуется с показателем чистоты, большим, чем около 97%. В некоторых аспектах Fc представляет собой гетеродимер, вырабатываемый с чистотой более чем около 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% при экспрессии. В некоторых аспектах Fc представляет собой гетеродимер, вырабатываемый с чистотой более чем около 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% при экспрессии на основе генетического материала одной клетки.
[0171] Дополнительные способы модификации мономерных полипептидов Fc для обеспечения образования гетеродимерных Fc, описаны в международной патентной публикации № WO 96/027011 (выступы-во-впадины), в Gunasekaran et al (Gunasekaran K. et al (2010 год) J Biol Chem. 285, 19637-46, электростатическая конструкция для достижения селективной гетеродимеризации), в Davis et al (Davis, JH. et al (2010 год) Prot Eng Des Sel; 23(4): 195-202, технология SEED (разработанный домен обмена цепями), и в Labrijn et al [Эффективное образование стабильного биспецифического IgG1 путем контролируемого обмена Fab-областей. Labrijn AF, Meesters JI, de Goeij BE, van den Bremer ET, Neijssen J, van Kampen MD, Strumane K, Verploegen S, Kundu A, Gramer MJ, van Berkel PH, van de Winkel JG, Schuurman J, Parren PW. Proc Natl Acad Sci USA. 26 марта 2013 года; 110(13):5145-50. В некоторых вариантах реализации изобретения изолированная конструкция, описанная в данном документе, содержит антигенсвязывающую конструкцию, которая связывается с антигеном; и димерную конструкцию полипептида Fc, которая имеет превосходные биофизические свойства, такие как стабильность и простота получения по отношению к антигенсвязывающей конструкции, которая не содержит такой же полипептид Fc. Ряд мутаций в последовательности тяжелой цепи Fc является известным в данной области техники для селективного изменения аффинности антитела Fc к различным Fc-гамма рецепторам. В некоторых аспектах Fc содержит одну или более модификаций для содействия избирательному связыванию Fc-гамма- рецепторов.
[0172] Домен СН2 представляет собой аминокислоты от 231 до 340 последовательности, проиллюстрированной в Таблице X. Примерные мутации перечислены ниже:
[0173] S298A/E333A/K334A, S298A/E333A/K334A/K326A (Lu Y, Vernes JM, Chiang N, et al J Immunol Methods. 28 февраля 2011 года; 365(1-2): 132-41); F243L/R292P/Y300L/V305I/P396L, F243L/R292P/Y300L/L235V/P396L (Stavenhagen JB, Gorlatov S, Tuaillon N, et al Cancer Res. 15 сентября 2007 года; 67(18):8882-90; Nordstrom JL, Gorlatov S, Zhang W, et al Breast Cancer Res. 30 ноября 2011 года; 13(6):R123); F243L (Stewart R, Thorn G, Levens M, et al Protein Eng Des Sel. сентябрь 2011 года; 24(9):671-8.), S298A/E333A/K334A (Shields RL, Namenuk AK, Hong K, et al J Biol Chem. 2 марта 2001 года; 276(9):6591-604); S239D/I332E/A330L, S239D/I332E (Lazar GA, Dang W, Karki S, et al Proc Natl Acad Sci USA. 14 марта 2006 года; 103(11):4005-10); S239D/S267E, S267E/L328F (Chu SY, Vostiar I, Karki S, et al Mol Immunol, сентябрь 2008 года; 45(15):3926-33); S239D/D265S/S298A/I332E, S239E/S298A/K326A/A327H, G237F/S298А/А330L/I332E, S239D/I332E/S298A, S239D/K3 26E/A330L/I332E/S298A, G236A/S239D/D270L/I332E, S239E/S267E/H268D, L234F/S267E/N325L, G237F/V266L/S267D и другие мутации, перечисленные в WO 2011/120134 и WO2011/120135 включены в данный документ посредством ссылки. Therapeutic Antibody Engineering (по William R. Strohl и Lila M. Strohl, Woodhead Publishing series in Biomedicine №11, ISBN 1907568379, октябрь 2012 года) мутации перечислены на странице 283.
[0174] В некоторых вариантах реализации изобретения домен СН2 содержит одну или более асимметричных аминокислотных модификаций. В некоторых вариантах реализации изобретения домен СН2 содержит одну или более асимметричных аминокислотных модификаций для содействия селективному связыванию FcγR. В некоторых вариантах реализации изобретения домен СН2 предусматривает возможность разделения и очистки изолированной конструкции, описанной в данном документе. Связывание FcRn и параметры ФК
[0175] Как известно специалистам в данной области техники, связывание с FcRn приводит к рециркуляции подвергшегося эндоцитозу антитела из эндосомы назад в кровоток (Raghavan et al, 1996, Annu Rev Cell Dev Biol 12:181-220; Ghetie et al, 2000, Annu Rev Immunol 18:739-766). Этот процесс в сочетании с уменьшением почечной фильтрации, обусловленным большим размером полноразмерной молекулы, приводит к более продолжительному периоду полувыведения антитела из сыворотки крови в пределах от одной до трех недель. Связывание Fc с FcRn также играет ключевую роль в транспорте антитела. Таким образом, в одном варианте реализации изобретения конструкции по настоящему изобретению способны связываться с FcRn.
Дополнительные модификации для улучшения эффекторной функции
[0176] В некоторых вариантах реализации изобретения конструкция, описанная в данном документе, может быть модифицирована с целью улучшения ее эффекторной функции. Такие модификации известны в данной области техники и включают афукозилирование или разработку аффинности части Fc антител к активирующему рецептору, главным образом FCGR3a для ADCC и к C1q для CDC. В приведенной ниже Таблице Y обобщены различные конструкции, описанные в литературе для разработки эффекторной функции.
[0177] Таким образом, в одном варианте реализации изобретения, описанные в данном документе конструкции могут содержать димерный Fc, который содержит одну или более аминокислотных модификаций, как указано в приведенной выше таблице, придающих улучшенную эффекторную функцию. В других вариантах реализации изобретения конструкция может быть афукозилирована с целью улучшения эффекторной функции.
Линкеры
[0178] Конструкции, описанные в данном документе, могут содержать один или более гетеродимеров, описанных в данном документе, функционально спаренных с Fc, описанным в данном документе. В некоторых аспектах Fc соединен с одним или более гетеродимерами с помощью или без одного или более линкеров. В некоторых аспектах Fc непосредственно спарен с одним или более гетеродимерами. В некоторых аспектах Fc спарен с одним или более гетеродимерами с помощью одного или более линкеров. В некоторых аспектах Fc спарен с тяжелой цепью каждого гетеродимера с помощью линкера.
[0179] В некоторых аспектах один или более линкеров представляют собой один или более полипептидных линкеров. В некоторых аспектах один или более линкеров содержат одну или более шарнирных областей антитела. В некоторых аспектах один или более линкеров содержат одну или более шарнирных областей IgG1.
Способы получения пар гетеродимеров
[0180] Как описано выше, пары гетеродимеров, описанные в данном документе, могут содержать первый гетеродимер и второй гетеродимер, каждый гетеродимер содержит тяжелую цепь иммуноглобулина или ее фрагмент, содержащую по меньшей мере домены VH и CH1, и легкую цепь иммуноглобулина, содержащую домен VL и домен CL. Тяжелые цепи иммуноглобулина и легкие цепи иммуноглобулина гетеродимера могут быть без труда получены с использованием технологии рекомбинантной ДНК, известной в данной области техники. Стандартные методы, такие как, например, те, которые описаны в Sambrook and Russell, Molecular Cloning: A Laboratory Manual (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 3-е изд., 2001 год); Sambrook et al, Molecular Cloning: A Laboratory Manual (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 2-е изд., 1989 год); Short Protocols in Molecular Biology (Ausubel et al, John Wiley and Sons, New York, 4-е изд., 1999 год); и Glick and Pasternak, Molecular Biotechnology: Principles and Applications of Recombinant DNA (ASM Press, Washington, D.C., 2-е изд., 1998 год) могут быть использованы для рекомбинантных методов нуклеиновой кислоты, синтеза нуклеиновых кислот, культивировании клеток, инкорпорации трансгена и экспрессии рекомбинантных белков. В альтернативном варианте гетеродимеры и пары гетеродимеров, описанные в данном документе, могут быть синтезированы химическим путем.
[0181] Последовательности нуклеиновых кислот и аминокислотные последовательности иммуноглобулиновых тяжелых и легких цепей антител, из которых получены гетеродимеры, либо известны в данной области техники, либо могут быть легко определены с использованием методов секвенирования нуклеиновых кислот и/или белка. Описанные в данном документе способы генетического слияния меток с тяжелыми и/или легкими цепями иммуноглобулина известны в данной области техники, и некоторые из них описаны ниже и в примерах.
[0182] Например, способы экспрессии и коэкспрессии тяжелых и легких цепей иммуноглобулина в клетке-хозяине хорошо известны в данной области техники. Кроме того, способы мечения тяжелых цепей и/или легких цепей с использованием технологии рекомбинантной ДНК также хорошо известны в данной области техники. Векторы экспрессии и клетки-хозяева, подходящие для экспрессии тяжелых и легких цепей, также хорошо известны в данной области техники, как описано ниже.
[0183] Способы получения биспецифические антител, которые не основаны на использовании только одной клональной или транзиторной клеточной линии, экспрессирующей все четыре цепи, известны в данной области техники (Gramer et al (2013 год) mAbs 5, 962; Strop et al (2012 год) J Mol Biol 420, 204.). Эти способы основаны на обмене плечами двух пар тяжелой и легкой цепи после получения, участвующих в образовании биспецифического антитела в окислительно-восстановительных условиях (окислительно-восстановительное получение). При таком подходе пары Н1:L1 и H2:L2 могут быть экспрессированы в двух различных клеточных линиях независимо друг от друга для образования двух плеч Fab. Впоследствии эти два плеча Fab смешивают в подобранных окислительно-восстановительных условиях для достижения повторного соединения двух уникальной тяжелых цепей H1 и Н2 с образованием биспецифического антитела, содержащего L1:H1:H2:L2 цепи. Можно себе представить использование библиотеки/набора данных конструкций, описанных в данном документе, в производстве биспецифических антител с использованием способа окислительно-восстановительного производства или модифицированных версий этого способа.
[0184] В некоторых вариантах реализации изобретения внеклеточные системы экспрессии белков используются для коэкспрессии полипептидов (например, полипептидов тяжелой и легкой цепи), без использования живых клеток. Вместо этого все компоненты, необходимые для транскрипции ДНК в РНК и трансляции РНК в белок (например, рибосомы, тРНК, ферменты, кофакторы, аминокислоты), предоставляются в растворе для использования in vitro. В некоторых вариантах реализации изобретения экспрессия in vitro требует (1) генетическую матрицу (мРНК или ДНК), кодирующую тяжелую и легкую цепь полипептидов и (2) реакционный раствор, содержащий необходимую для транскрипции и трансляции молекулярную машинерию. В некоторых вариантах реализации изобретения клеточные экстракты по существу поставляют компоненты реакционного раствора, например: РНК-полимеразы для транскрипции мРНК, рибосомы для трансляции полипептида, тРНК, аминокислоты, ферментные кофакторы, источник энергии и клеточные компоненты, необходимые для правильного сворачивания белков. Бесклеточные системы экспрессии белка могут быть получены с использованием лизатов, полученных из бактериальных клеток, дрожжевых клеток, клеток насекомых, клеток растений, клеток млекопитающих, человеческих клеток или их комбинации. Такие клеточные лизаты могут обеспечить правильный состав и пропорции ферментов, и строительных блоков, необходимых для трансляции. В некоторых вариантах реализации изобретения клеточные мембраны удаляются, чтобы оставить только цитозольные компоненты и органеллы клетки.
[0185] Несколько бесклеточных систем экспрессии белков известны в данной области техники, как рассмотрено у Carlson et al (2012 год) Biotechnol. Adv. 30:1185-1194. Например, бесклеточные системы экспрессии белка доступны на основе прокариотических или эукариотических клеток. Примеры бесклеточных прокариотических систем экспрессии включают те, которые получены из E.coli. Бесклеточные системы экспрессии белка доступны на основе экстрактов из, например, ретикулоцитов кролика, зародышей пшеницы и клеток насекомых. Такие бесклеточные прокариотические и эукариотические системы экспрессии белка коммерчески доступны от таких компаний, как Roche, Invitrogen, Qiagen и Novagen. Специалист в данной области техники легко сможет выбрать подходящие бесклеточные системы экспрессии белка, которые будут продуцировать полипептиды (например, полипептиды тяжелой цепи и легкой цепи), которые способны к спариванию друг с другом. Кроме того, бесклеточная система экспрессии белка также может быть дополнена шаперонами (например, BiP) и изомеразами (например, дисульфидизомераза), чтобы повысить эффективность фолдинга IgG.
[0186] В некоторых вариантах реализации изобретения внеклеточные системы экспрессии используются для коэкспрессии полипептидов тяжелой и легкой цепи из ДНК матриц (транскрипция и трансляция) или мРНК матриц (только трансляция).
Векторы и клетки-хозяева
[0187] Рекомбинантная экспрессия тяжелых и легких цепей требует создания вектора экспрессии, содержащего полинуклеотид, который кодирует тяжелую или легкую цепь (например, антитела или слитого белка). После того, как полинуклеотид, кодирующий тяжелую или легкую цепь, был получен, вектор для продуцирования тяжелой или легкой цепи может быть получен с помощью технологии рекомбинантной ДНК с использованием способов, хорошо известных в данной области техники. Таким образом, способы получения белка путем экспрессии полинуклеотида, содержащего кодирующую нуклеотидную последовательность тяжелой или легкой цепи, описаны в данном документе. Способы, которые хорошо известны специалистам в данной области техники, могут быть использованы для конструирования векторов экспрессии, содержащих кодирующую последовательность тяжелой или легкой цепи и соответствующие транскрипционные и трансляционные сигналы контроля. Эти методы включают, например, технологии рекомбинантной ДНК in vitro, технологии синтеза и генетическую рекомбинацию in vivo. Таким образом, изобретение относится к векторам репликации, содержащим нуклеотидную последовательность, кодирующую тяжелые или легкие цепи, функционально связанную с промотором.
[0188] Вектор экспрессии переносят в клетку-хозяина с помощью классических технологий, и трансфицированные клетки затем культивируют с помощью классических технологий для получения модифицированных тяжелых или легких цепей для использования в способе по изобретению. В конкретных вариантах реализации изобретения тяжелые и легкие цепи для использования в способе коэкспрессируются в клетке-хозяине для экспрессии всей молекулы иммуноглобулина, как подробно описано ниже.
[0189] Разнообразие векторных систем экспрессии в клетке-хозяине может быть использовано для экспрессии модифицированных тяжелых и легких цепей. Такие системы экспрессии в клетке-хозяине представляют собой носители, с помощью которых кодирующие последовательности, представляющие интерес, могут быть получены и затем очищены, но также представляют собой клетки, которые могут, при трансформировании или трансфицировании соответствующими кодирующими нуклеотидными последовательностями, экспрессировать модифицированные тяжелые и легкие цепи in situ. Они включают, но не ограничиваются этим, микроорганизмы, такие как бактерии (например, E.coli и В. subtilis), трансформированные рекомбинантной ДНК бактериофага, плазмидной ДНК или ДНК космидных векторов экспрессии, содержащей модифицированные кодирующие последовательности тяжелой и легкой цепи; дрожжи (например, Saccharomyces Pichia), трансформированные рекомбинантными дрожжевыми векторами экспрессии, содержащими модифицированные кодирующие последовательности тяжелой и легкой цепи; системы клеток насекомых, инфицированные рекомбинантными вирусными векторами экспрессии (например, бакуловирус), содержащими модифицированную кодирующую последовательность тяжелой и легкой цепи; системы растительных клеток, инфицированные рекомбинантными вирусными векторами экспрессии (например, вирус мозаики цветной капусты, CaMV; вирус табачной мозаики, TMV) или трансформированные рекомбинантными плазмидными векторами экспрессии (например, Ti плазмида), содержащими модифицированные кодирующие последовательности тяжелой и легкой цепи; или системы клеток млекопитающих (например, COS, СНО, ВНК, НЕК-293, NSO и 3Т3 клетки), содержащие рекомбинантные экспрессионные конструкции, содержащие промоторы, полученные из генома клеток млекопитающих (например, промотор металлотионеина) или из вирусов млекопитающих (например, поздний промотор аденовируса; 7.5К промотор вируса коровьей оспы). В некоторых вариантах реализации изобретения бактериальные клетки, такие как Escherichia coli, или эукариотические клетки используются для экспрессии модифицированных тяжелых и легких цепей, которые является рекомбинантными молекулами антитела или слитого белка. Например, клетки млекопитающих, такие как клетки яичника китайского хомячка (СНО) в сочетании с вектором, таким как основной промоторный элемент предраннего гена цитомегаловируса человека, являются эффективной системой экспрессии антител (Foecking et al, 1986, Gene 45:101; и Cockett et al, 1990, Bio/Technology 8:2). В конкретном варианте реализации изобретения экспрессия нуклеотидных последовательностей, кодирующих тяжелые и легкие цепи иммуноглобулина каждого гетеродимера, регулируется конститутивным промотором, индуцибельным промотором или тканеспецифичным промотором.
[0190] В клетках-хозяевах млекопитающих может быть использован ряд систем экспрессии на основе вирусов. В тех случаях, где аденовирус используется в качестве вектора экспрессии, представляющие интерес модифицированные кодирующие последовательности тяжелой и легкой цепи могут быть лигированы с аденовирусным комплексом контроля за транскрипцией/трансляцией, например, поздним промотором и трехкомпонентной лидерной последовательностью. Затем этот химерный ген можно вставить в геном аденовируса путем рекомбинации in vitro или in vivo. Вставка в неосновную область вирусного генома (например, область Е1 или Е3) приведет к получению рекомбинантного вируса, который является жизнеспособным и способен экспрессировать модифицированные тяжелые и легкие цепи в зараженных хозяевах (например, см. Logan & Shenk, 1984, Proc. Natl. Acad. Sci. USA 81:355-359). Специфические сигналы инициации также могут быть необходимы для эффективной трансляции вставленных кодирующих последовательностей антител. Эти сигналы включают инициирующий кодон ATG и смежные последовательности. Кроме того, инициирующий кодон должен быть синхронизирован с рамкой считывания требуемой кодирующей последовательности для обеспечения трансляции всей вставки. Эти экзогенные сигналы контроля трансляции и кодоны инициации могут иметь различное происхождение как природное, так и синтетическое. Эффективность экспрессии может быть повышена за счет включения соответствующих энхансерных элементов транскрипции, терминаторов транскрипции и т.д. (см., например, Bittner et al, 1987, Methods inEnzymol. 153:516-544).
[0191] Экспрессией тяжелых и легких цепей иммуноглобулина гетеродимеров можно управлять с помощью любого промоторного или энхансерного элемента, известного в данной области техники. Промоторы, которые могут быть использованы для контроля экспрессии гена, кодирующего модифицированные тяжелые и легкие цепи (например, антитело или слитый белок), включают, но не ограничиваются этим, SV40 раннюю промоторную область (Chambon, 1981, Nature 290: 304-310), промотор, содержащий в 3'-направлении длинный терминальный повтор вируса саркомы Рауса (Yamamoto et al, 1980, Cell 22:787-797), промотор тимидинкиназы герпеса (Wagner et al, 1981, Proc. Natl. Acad. Sci. U.S.A. 78.1441-1445), регуляторные последовательности гена металлотионеина (Brinster et al, 1982, Nature 296:39-42), промотор тетрациклина (Tet) (Gossen et al, 1995, Proc. Nat. Acad. Sci. USA 89:5547-5551); прокариотические векторы экспрессии, такие как промотор [3-лактамазы (Villa-Kamaroff et al, 1978, Proc. Natl. Acad. Sci. U.S.A. 75:3727-3731), или tac-промотор (DeBoer et al, 1983, Proc. Natl. Acad. Sci. U.S.A. 80:21-25; см. также "Useful proteins from recombinant bacteria" в Scientific American, 1980, 242:74-94); векторы экспрессии растений, содержащие промоторную область нопалин синтетазы (Herrera-Estrella et al, Nature 303:209-213) или промотор 35S-PHK вируса мозаики цветной капусты (Gardner et al, 1981, Nucl. Acids Res. 9:2871), и промотор фотосинтетического фермента рибулозобифосфаткарбоксилазы (Herrera-Estrella et al, 1984, Nature 310:115-120); промоторные элементы из дрожжей или других грибов, такие как промотор Gal 4, промотор ADC (алкогольдегидрогеназы), промотор PGK (глицерофосфат киназы), промотор щелочной фосфатазы, и следующие контрольные области транскрипции животных, которые проявляют тканевую специфичность, и использовались в трансгенных животных: контрольная область гена эластазы I, которая является активной в ацинарных клетках поджелудочной железы (Swift et al, 1984, Cell 38:639-646; Ornitz et al, 1986, Cold Spring Harbor Symp. Quant. Biol. 50:399-409; MacDonald, 1987, Hepatology 7:425-515); контрольная область гена инсулина, которая является активной в бета-клетках поджелудочной железы (Hanahan, 1985, Nature 315:115-122), контрольная область гена иммуноглобулина, которая является активной в лимфоидных клетках (Grosschedl et al, 1984, Cell 38:647-658; Adames et al, 1985, Nature 318:533-538; Alexander et al, 1987, Mol. Cell. Biol. 7:1436-1444), контрольная область вируса опухоли молочной железы мышей, которая является активной в клетках яичек, молочной железы, лимфоидных и тучных клетках (Leder et al., 1986, Cell 45:485-495), контрольная область гена альбумина, которая является активной в печени (Pinkert et al, 1987, Genes and Devel. 1:268-276), контрольная область гена альфа-фетопротеина, которая является активной в печени (Krumlauf et al, 1985, Mol. Cell. Biol. 5:1639-1648; Hammer et al, 1987, Science 235:53-58); контрольная область гена альфа-1-антитрипсина, которая является активной в печени (Kelsey et al, 1987, Genes and Devel. 1:161-171), контрольная область гена бета- глобина, которая является активной в миелоидных клетках (Mogram et al, 1985, Nature 315:338-340; Kollias et al, 1986, Cell 46:89-94); контрольная область основного белка миелина, которая является активной в олигодендроцитных клетках головного мозга (Readhead et al, 1987, Cell 48:703-712); контрольная область гена легкой цепи-2 миозина, которая является активной в скелетной мышце (Sani, 1985, Nature 314:283-286); нейрон-специфическая енолаза (НСЕ), которая является активной в нейронах (Morelli et al, 1999, Gen. Virol. 80:571-83); контрольная область гена нейротрофического фактора головного мозга (BDNF), которая является активной в нейронах (Tabuchi et al, 1998, Biochem. Biophysic. Res. Com. 253:818-823); промотор глиального фибриллярного кислого белка (GFAP), который является активным в астроцитах (Gomes et al, 1999, Braz J Med Biol Res 32(5): 619-631; Morelli et al, 1999, Gen. Virol. 80:571-83), и контрольная область гена гонадотропин-рилизинг гормона, которая является активной в гипоталамусе (Mason et al, 1986, Science 234:1372-1378).
[0192] Кроме того, штамм клетки-хозяина может быть выбран таким образом, чтобы модулировать экспрессию вставленных последовательностей или модифицировать и процессировать продукт гена желаемым определенным образом. Экспрессия от определенных промоторов может быть усилена в присутствии определенных индукторов; таким образом, экспрессию слитого белка, созданного методами генетической инженерии, можно контролировать. Кроме того, различные клетки-хозяева имеют типичные и специфические механизмы для трансляционного и пост-трансляционного процессинга и модификации (например, гликозилирование, фосфорилирование белков). Соответствующие клеточные линии или системы хозяев могут быть выбраны для обеспечения желаемой модификации и процессинга экспрессированного чужеродного белка. Например, экспрессия в бактериальной системе приведет к созданию негликозилированного продукта, и экспрессия в дрожжах приведет к созданию гликозилированного продукта. Могут быть использованы эукариотические клетки- хозяева, которые обладают клеточным механизмом для правильного процессинга первичного транскрипта (например, гликозилирование и фосфорилирование) продукта гена. Такие клетки-хозяева млекопитающих включают, но не ограничиваются этим, СНО, VERY, ВНК, Hela, COS, MDCK, НЕК-293, 3Т3, WI38, NSO, и в частности, нейрональные клеточные линии, такие как, например, клеточные линии нейробластомы человека SK-N- AS, SK-N-FI, SK-N-DZ (Sugimoto et al, 1984, J. Natl. Cancer Inst. 73 51-57), нейробластомы человека SK-N-SH (Biochim. Biophys. Acta, 1982, 704: 450-460), церебральной медуллобластомы человека Daoy (He et al, 1992, Cancer Res. 52: 1144-1148), клетки глиобластомы DBTRG-05MG (Kruse et al, 1992, In Vitro Cell. Dev. Biol. 28A: 609-614), нейробластомы человека IMR-32 (Cancer Res., 1970, 30: 2110-2118), астроцитомы человека 1321 N1 (Proc. Natl. Acad. Sci. USA, 1977, 74: 4816), астроцитомы человека MOG-G-CCM (Br. J. Cancer, 1984, 49: 269), глиобластомы-астроцитомы человека U87MG (Acta Pathol. Microbiol. Scand., 1968, 74: 465-486), глиобластомы человека A172 (Olopade et al, 1992, Cancer Res. 52: 2523-2529), клетки глиомы крысы C6 (Benda et al, 1968, Science 161: 370-371), нейробластомы мыши Neuro-2a (Proc. Natl. Acad. Sci. USA, 1970, 65: 129-136), нейробластомы мыши NB41A3 (Proc. Natl. Acad. Sci. USA, 1962, 48: 1184-1190), хориоидного сплетения овцы (SCP) (Bolin et al, 1994, J. Virol. Methods 48: 211-221), нормальных астроцитов кота G355-5, PG-4 (Haapala et al, 1985, J. Virol. 53: 827-833), мозга африканского хорька Mpf (Trowbridge et al, 1982, In Vitro 18: 952-960), и нормальные клеточные линии, такие как, например, нормальные клетки коры головного мозга крысы CTX TNA2 (Radany et al, 1992, Proc. Natl. Acad. Sci. USA 89: 6467-6471), такие как, например, CRL7030 и Hs578Bst. Кроме того, различные системы экспрессии вектор/хозяин могут в разной степени воздействовать на реакции процессинга.
[0193] Для долгосрочного, высокопроизводительного продуцирования рекомбинантных белков предпочтительной является стабильная экспрессия. Например, могут быть сконструированы клеточные линии, которые стабильно экспрессируют модифицированные тяжелые и легкие цепи по изобретению (например, антитело или слитый белок). Вместо того, чтобы использовать векторы экспрессии, которые содержат вирусные сайты инициации репликации, клетки-хозяева могут быть трансформированы с помощью ДНК, контролируемой подходящими регуляторными элементами экспрессии (например, например, промоторной, энхансерной последовательностями, терминаторами транскрипции, сайтами полиаденилирования и т.д.), и селектируемым маркером. После введения чужеродной ДНК сконструированные клетки могут расти в течение 1-2 дней в обогащенной среде, а затем их перемещают на селективную среду. Селектируемый маркер в рекомбинантной плазмиде придает устойчивость к отбору и позволяет клеткам стабильно интегрировать эту плазмиду в их хромосомы и расти с образованием очагов, которые, в свою очередь, могут быть клонированы и размножены в клеточные линии.
[0194] Могут быть использованы ряд систем селекции, в том числе, но не ограничиваясь этим, ген тимидинкиназы вируса простого герпеса 1 типа (Wigler et al, 1977, Cell 11:223), ген гипоксантингуанинфосфорибозилтрансферазы (Szybalska & Szybalski, 1962, Proc. Natl. Acad. Sci. USA 48:2026), и ген аденинфосфорибозилтрансферазы (Lowy et al, 1980, Cell 22:817) могут быть использованы в tk-, hgprt- или aprt-клетках, соответственно. Кроме того, устойчивость к антиметаболитам может быть использована в качестве основы селекции на ген dhfr, который придает устойчивость к метотрексату (Wigler et al, 1980, Natl. Acad. Sci. USA 77:3567; O'Hare et al, 1981, Proc. Natl. Acad. Sci. USA 78:1527); ген gpt, который придает устойчивость к микофеноловой кислоте (Mulligan & Berg, 1981, Proc. Natl. Acad. Sci. USA 78:2072); ген neo, который придает устойчивость к аминогликозиду G-418 (Colberre-Garapin et al, 1981, J. Mol. Biol. 150:1); и ген hygro, который придает устойчивость к гигромицину (Santerre et al, 1984, Gene 30:147).
Коэкспрессия тяжелых цепей и легких цепей
[0195] Тяжелые цепи иммуноглобулина и легкие цепи иммуноглобулина пар гетеродимеров, описанных в данном документе, могут коэкспрессироваться в клетках млекопитающих, как было отмечено выше. В одном варианте реализации изобретения одна тяжелая цепь коэкспрессируется с двумя разными легкими цепями в наборе конструкции LCCA, как описано выше, при этом тяжелая цепь преимущественно спаривается с одной из двух легких цепей. В другом варианте реализации изобретения две уникальные тяжелые цепи коэкспрессируется с двумя уникальными легкими цепями, при этом каждая тяжелая цепь преимущественно спаривается с одной из легких цепей. Испытание пар гетеродимеров
[0196] Как было описано выше по меньшей мере один гетеродимер из пар гетеродимеров, описанных в данном документе, может содержать одну или более аминокислотных модификаций в их тяжелых и/или легких цепях иммуноглобулина, таких, что, в случае если две уникальные тяжелые цепи и две уникальные легкие цепи пары гетеродимеров коэкспрессируются в клетке млекопитающего, тяжелая цепь первого гетеродимера преимущественно спаривается с одной из легких цепей, а не с другой. Аналогичным образом, тяжелая цепь второго гетеродимера преимущественно спаривается со второй легкой цепью, а не с первой. Степень преимущественного спаривания может быть оценена, например, с помощью способов, описанных ниже. Аффинность каждого гетеродимера из пары гетеродимеров к его соответствующему антигену может быть испытана, как описано ниже. Термическая стабильность каждого гетеродимера из пары гетеродимеров также может быть испытана, как описано ниже.
Способы измерения преимущественного спаривания
LCCA
[0197] В одном варианте реализации изобретения преимущественное спаривания между тяжелыми и легкими цепями иммуноглобулина определяется путем выполнения анализа конкуренции легкой цепи (LCCA). В одновременно находящейся на рассмотрении заявке на патент PCT/US2013/063306 того же заявителя, поданной 3 октября 2013 года, описаны различные варианты реализации LCCA, и она включена в данный документ посредством ссылки во всей своей полноте и во всех отношениях. Способ позволяет провести количественный анализ спаривания тяжелых цепей с определенными легкими цепями внутри смеси коэкспрессируемых белков, и может быть использован для определения того, может ли одна конкретная тяжелая цепь иммуноглобулина селективно связывается с одной из двух легких цепей иммуноглобулина, в случае если тяжелая цепь и легкие цепи коэкспрессируются. Способ можно кратко описать следующим образом: по меньшей мере одна тяжелая цепь и две различные легкие цепи коэкспрессируются в клетке в таких соотношениях, что тяжелая цепь является лимитирующим реактантом для спаривания; необязательное отделение секретируемых белков из клетки; отделение полипептидов легкой цепи иммуноглобулина, связанных с тяжелой цепью, от остальных секретируемых белков для получения изолированной спаренной фракции тяжелой цепи; определение количества каждой различной легкой цепи в изолированной фракции тяжелой цепи; и анализ относительного количества каждой различной легкой цепи в изолированной фракции тяжелой цепи, чтобы определить способность по меньшей мере одной тяжелой цепи селективно спариваться с одной из легких цепей.
[0198] Способ обеспечивает достаточную производительность и является надежным (т.е. не чувствителен к небольшим изменениям в работе, вносимыми пользователем или скоростью потока) и точным. Способ обеспечивает чувствительный анализ, который может измерять эффекты малых вариаций в белковых последовательностях. Взаимодействия промискуитетный белок-белок; домен-домен; цепь- цепь на больших площадях поверхности, как правило, требуют множественных мутаций (свопов) для внедрения селективности. Белковые продукты не нуждаются в выделении и очистке, что позволяет осуществлять более эффективный скрининг. Дополнительные подробности, касающиеся вариантов реализации изобретения по этому способу, описаны в примерах.
Альтернативные способы определения преимущественного спаривания
[0199] Альтернативные способы обнаружения преимущественного спаривания включают использование LC-MS (жидкостная хроматография - масс-спектрометрия) для количественного определения относительных совокупностей гетеродимеров, включая каждую легкую цепь, с использованием различий в их молекулярной массе для идентификации каждого отдельного вида. Анализ на антигенную активность также может быть использован для количественного определения относительных совокупностей гетеродимеров, содержащих каждую легкую цепь, в результате чего измеренная степень связывания (по отношению к контролю) будет использоваться для оценки каждой соответствующей совокупности гетеродимеров.
[0200] Дополнительные способы, например, SMCA, описаны в примерах, фигурах и таблицах.
Термостабильность
[0201] Термостабильность гетеродимеров может быть определена с помощью любого способа, известного в данной области техники. Температура плавления каждого гетеродимера свидетельствует о его термостабильности. Температура плавления гетеродимера может быть измерена с помощью таких методов, как дифференциальная сканирующая калориметрия (Chen et al (2003) Pharm Res 20:1952-60; Ghirlando et al (1999) Immunol Lett 68:47-52). В альтернативном варианте термостабильность гетеродимеров может быть измерена с помощью кругового дихроизма (Murray et al (2002) J. Chromatogr Sci 40:343-9).
Аффинность к антигену
[0202] Аффинность связывания гетеродимеров к соответствующим антигенам и скорость диссоциации взаимодействия может быть определена с помощью конкурентно- связывающих анализов в соответствии со способами, хорошо известными в данной области техники. Один из примеров конкурентно-связывающего анализа представляет собой радиоиммуноанализ, включающий инкубацию меченого антигена (например, 3Н или 125I с молекулой, представляющей интерес, (например, гетеродимеры по настоящему изобретению) в присутствии возрастающего количества немеченого антигена и обнаружения молекулы, связанной с меченым лигандом. Аффинность гетеродимера по настоящему изобретению к антигену, и связывающая скорость диссоциации может быть определена по данным насыщения с помощью анализа Скатчарда.
[0203] Кинетические параметры гетеродимеров, описанные в данном документе, также могут быть определены с помощью анализов на основе поверхностного плазмонного резонанса (SPR), известных в данной области техники (например, кинетический анализ BIAcore). Для обзора технологии на основе SPR см. Mullet et al, 2000, Methods 22: 77-91; Dong et al, 2002, Review in Mol. Biotech. 82 303-23; Fivash et al, 1998, Current Opinion in Biotechnology 9: 97-101; Rich et al, 2000, Current Opinion in Biotechnology 11: 54-61. Кроме того, любой из SPR инструментов и способов на основе SPR для измерения белок-белковых взаимодействий, описанных в патенте США №6,373,577; 6,289,286; 5,322,798; 5,341,215; 6,268,125, рассматриваются в способах по настоящему изобретению. FACS также может быть использован для измерения аффинности, как известно в данной области техники.
Образование биспецифических антител при наличии заданных Mab1 и Mab2 с использованием библиотеки конструкций мутаций биспецифических антител
[0204] В одном варианте реализации изобретения описанный в данном документе набор конструкций мутаций биспецифических антител направлен на селективно образующиеся биспецифические антитела, начиная с двух канонических антител Mab1 и Маb2, содержащих антигенсвязывающие фрагменты Fab1 и Fab2, соответственно. Набор конструкций состоит из когнатных мутаций, соответствующих Fab1, Fab2 и Fc, соответственно. В одном варианте реализации изобретения библиотеки наборов конструкций представлены наборами конструкций, включенных в таблицу 5, таблицу 12 или любую из таблиц 15-17. Мутации введены в область контакта легкой и тяжелой цепи Fab1 для достижения селективного спаривания между двумя облигатными цепями в присутствии конкурирующих легкой и тяжелой цепи Fab2. Селективное спаривание достигается путем введения благоприятных комплементарных мутаций в две облигатные легкие и тяжелые цепи на основе стерической, гидрофобной или электростатической комплементарности в область контакта между определенными горячими каркасными остатками с одновременным участием этих мутантных остатков в неблагоприятном взаимодействии областей контакта в случае необлигатных пар цепей. В каждом наборе конструкций мутации для селективного спаривания могут также быть введены в область контакта легкой и тяжелой цепи Fab2 для достижения селективного спаривания между двумя облигатными цепями в присутствии конкурирующих легкой и тяжелой цепи Fab1. Мутации направлены на снижение неправильного спаривания легкой цепи из Fab1 с тяжелой цепью Fab2, и наоборот. Мутации вводятся в область контакта Fc с целью достижения селективного спаривания тяжелых цепей для образования асимметрических молекул антител, содержащих две различные тяжелые цепи. Манипуляции в определенных позициях остатков области контакта легких и тяжелых цепей антитела часто могут привести к неблагоприятным эффектам, таким как потеря антигенсвязывающей аффинности, стабильности, растворимости, предрасположенности к агрегации и т.д. этого антитела. Может быть затронут ряд связанных свойств, например, значение константы ассоциации и константа диссоциации, температура плавления (Tm), устойчивость к таким стрессовым условиям, как кислоты, основания, окисление, замораживание/оттаивание, взбалтывание, давление и т.д. Это часто зависит от гипервариабельных участков (CDR) антитела, представляющего интерес. Учитывая, что CDR различных антител, как правило, не идентичны, влияние набора конструкций мутаций на свойства, описанные выше, не могут быть одинаковыми для всех антител. Представленное в данном документе представляет собой способ создания биспецифического антитела указанной чистоты относительно других загрязнителей, содержащих неправильно спаренные антитело-подобные структуры, при наличии заданных двух любых доступных антител Mab1 и Маb2. Легкие и тяжелые цепи Mab1 и Маb2 коэкспрессируются после введения когнатных мутаций из каждого набора конструкций мутаций, и экспрессированный продукт антитела аналитически скринируется для оценки чистоты предпочтительного биспецифического антитела по отношению к другим Mab-подобным видам, экспрессированным в белковом продукте. В некоторых вариантах реализации изобретения аналитическая процедура скрининга может быть основана на методике LC-MS. В некоторых вариантах реализации изобретения процедура аналитического скрининга может быть основана на разделении, например, методом капиллярного изоэлектрического фокусирования (cIEF) или хроматографическим методом. Пример метода скрининга, основанного на процедуре SMCA, представлен в примере 9. В некоторых вариантах реализации изобретения указанная чистота биспецифического антитела определяется как большая, чем 70% от всех полученных Mab- подобных видов в экспрессированном белковом продукте. В некоторых вариантах реализации изобретения указанная чистота биспецифического антитела определяется как большая, чем 90% от всех полученных Mab-подобных видов в экспрессированном белковом продукте. Процедура подготовки и отбора биспецифического набора конструкций Mab при наличии заданных Маb1 и Маb2 схематически показана на фигуре 12.
Фармацевтические композиции
[0205] В настоящем изобретении также предложены фармацевтические композиции, содержащие гетеродимеры или пары гетеродимеров, описанные в данном документе. Такие композиции содержат терапевтически эффективное количество гетеродимера или пары гетеродимеров, и фармацевтически приемлемый носитель. В конкретном варианте реализации изобретения термин "фармацевтически приемлемый" означает одобренный регуляторным ведомством Федерального правительства или правительства штата, или приведенный в Фармакопее США или другой общепризнанной фармакопее с целью использования на животных и, более конкретно, на людях. Термин "носитель" относится к разбавителю, адъюванту, вспомогательному веществу или носителю, с которым терапевтическое средство вводится. Такие фармацевтические носители могут быть стерильными жидкостями, такими как вода и масла, включая нефтяные масла, масла животного, растительного или синтетического происхождения, такие как ореховое масло, соевое масло, минеральное масло, кунжутное масло и тому подобное. Вода является предпочтительным носителем, в случае если фармацевтическая композиция вводится внутривенно. Солевые растворы и водные растворы декстрозы и глицерина также могут быть использованы в качестве жидких носителей, особенно для инъекционных растворов. Подходящие фармацевтические эксципиенты включают крахмал, глюкозу, лактозу, сахарозу, желатин, солод, рис, муку, мел, силикатный гель, стеарат натрия, глицеролмоностеарат, тальк, хлорид натрия, сухое обезжиренное молоко, глицерол, пропилен, гликоль, воду, этанол и тому подобное. При желании композиция может также содержать незначительные количества увлажнителей или эмульгаторов, или рН-буферных веществ. Эти композиции могут быть в виде растворов, суспензий, эмульсии, таблеток, драже, капсул, порошков, препаратов длительного высвобождения и т.п. Композиция может быть составлена в виде суппозитория с традиционным связующими веществами и носителями, такими как триглецириды. Состав для перорального введения может включать стандартные носители, такие как фармацевтические виды маннита, лактозы, крахмала, стеарата магния, натриевой соли сахарина, целлюлозы, карбоната магния и т.д. Примеры подходящих фармацевтических носителей описаны в "Remington's Pharmaceutical Sciences" by E.W. Martin. Такие композиции содержат терапевтически эффективное количество соединения, преимущественно в очищенном виде, вместе с подходящим количеством носителя для получения требуемой формы для введения пациенту. Препарат должен соответствовать способу введения.
[0206] В некоторых вариантах реализации изобретения композицию, содержащую гетеродимер или пару гетеродимеров, получают стандартными методиками в виде фармацевтической композиции, адаптированной для внутривенного введения человеку. Как правило, композиции для внутривенного введения представляют собой растворы в стерильном изотоническом буфере на водной основе. При необходимости композиция также может содержать солюбилизирующее средство и местный анестетик, такой как лидокаин, для облегчения боли в участке инъекции. Как правило, ингредиенты поставляются либо по отдельности, либо смешаны друг с другом в виде единичной лекарственной формы, например, в виде сухого лиофилизированного порошка или безводного концентрата в герметичной емкости, такой как ампула или пакет-саше, с указанием количества действующего вещества. Если композицию вводят путем инфузии, ее можно распределить с использованием инфузионного флакона, содержащего стерильную воду фармацевтической степени чистоты или солевой раствор. Если композицию вводят путем инъекции, может предоставляться ампула со стерильной водой для инъекций или физиологическим раствором, для того, чтобы можно было смешать ингредиенты до введения.
[0207] В некоторых вариантах реализации изобретения композиции, описанные в данном документе, составляют в форме нейтральных солей. Фармацевтически приемлемые соли включают соли, образующиеся с анионами, такие как, например, соли производные соляной, фосфорной, уксусной, щавелевой, винной кислот и т.д., и соли, образующиеся с катионами, такие как, например, соли производные натрия, калия, аммония, кальция, гидроксида железа, изопропиламина, триэтиламина, 2- этиламиноэтанола, гистидина, прокаина и т.д.
[0208] Количество композиции, описанное в данном документе, которое будет эффективно в лечении, ингибировании и профилактике заболевания или нарушения, связанного с неправильной экспрессией и/или активностью терапевтического белка, можно определить стандартными клиническими методами. В дополнение, анализы in vitro могут быть выборочно использованы для определения оптимального диапазона доз. Точная доза используется в препарате также в зависимости от способа введения и тяжести заболевания или нарушения, и решения в соответствии с оценкой практикующего врача и обстоятельств каждого пациента. Эффективные дозы экстраполированы из кривой зависимости доза-эффект, полученные от in vitro или тестовых систем экспериментальной модели на животном.
Использование пар гетеродимеров
[0209] Как было описано выше, пары гетеродимеров, описанные в данном документе, могут содержать первый гетеродимер и второй гетеродимер, при этом тяжелая цепь иммуноглобулина и/или легкая цепь иммуноглобулина каждого гетеродимера содержит одну или более модификаций из известного терапевтического антитела или из известного антитела, которое связывается с молекулой. Таким образом, предполагается, что гетеродимеры, содержащие модификации этих антител, могут быть использованы для лечения или профилактики того же заболевания, нарушения или инфекции, что и известное терапевтическое антитело или известное антитело.
[0210] В другом варианте реализации изобретения пары гетеродимеров, описанные в данном документе, также могут быть с успехом использованы в комбинации с другими терапевтическими средствами, известными в данной области техники, для лечения или профилактики рака, аутоиммунных заболеваний, воспалительных заболеваний или инфекционных заболеваний. В конкретном варианте реализации изобретения, пары гетеродимеров, описанные в данном документе, могут быть использованы в комбинации с моноклональными или химерными антителами, лимфокинами или гемопоэтическими факторами роста (такими как, например, IL-2, IL-3 и IL-7), которые, например, служат для увеличения количества или активности эффекторных клеток, которые взаимодействуют с молекулами, и увеличения иммунного ответа. Пары гетеродимеров, описанные в данном документе, также могут быть с успехом использованы в комбинации с одним или более лекарственными средствами, используемыми для лечения заболевания, нарушения или инфекции, такими как, например, противораковые агенты, противовоспалительные агенты или противовирусные агенты.
Наборы
[0211] В настоящем изобретении также предложены наборы, содержащие одну или более пар гетеродимеров. Отдельные компоненты набора упакованы в отдельные емкости, и вместе с такими емкостями может прилагаться уведомление в форме, установленной правительственным учреждением, регулирующим производство, использование или сбыт фармацевтической или биологической продукции, в котором отражены разрешения этого учреждения для производства, использования или сбыта препарата. Набор может необязательно содержать инструкции или указания, в которых изложен способ применения или схема применения пар гетеродимеров.
[0212] Если один или более компонентов набора предоставляются как растворы, например, водные растворы или стерильные водные растворы, сам контейнер может представлять собой ингалятор, шприц, пипетку, глазную капельницу или другие тому подобные приспособления, из которых раствор может вводиться субъекту или применяться и смешиваться с другими компонентами набора.
[0213] Компоненты набора также могут быть предоставлены в сухой или лиофилизированной форме, а набор может дополнительно содержать подходящий растворитель для восстановления лиофилизированных компонентов. Независимо от количества или типа емкостей наборы по изобретению также могут содержать устройство, помогающее пациенту с введением композиции. Такое устройство может представлять собой ингалятор, дозатор для назального спрея, шприц, пипетку, пинцет, мерную ложку, глазную капельницу или подобные средства доставки, одобренные для применения в лечебных целях.
Реализация с помощью компьютера
[0214] В одном варианте реализации изобретения компьютер содержит по меньшей мере один процессор, соединенный с чипсетом. Также соединенными с чипсетом являются запоминающее устройство, устройство для хранения данных, клавиатура, графический адаптер, указательное устройство и сетевой адаптер. Дисплей соединен с графическим адаптером. В одном варианте реализации изобретения функциональность чипсета обеспечивается с помощью контроллера концентратора памяти и контроллера концентратора ввода/вывода. В другом варианте реализации изобретения запоминающее устройство соединено непосредственно с процессором, а не с чипсетом.
[0215] Устройство для хранения данных представляет собой любое устройство, способное хранить данные, например, жесткий диск, компакт-диск "только для чтения" (CD-ROM), DVD или твердотельное запоминающее устройство. Запоминающее устройство хранит команды и данные, используемые процессором. Указывающее устройство может быть мышью, трекболом или другим типом указывающего устройства и используется в комбинации с клавиатурой для ввода данных в компьютерную систему. Графический адаптер отображает изображения и другую информацию на дисплее. Сетевой адаптер соединяет компьютерную систему с локальной или глобальной сетью.
[0216] Как известно специалистам в данной области техники, компьютер может иметь различные и/или другие компоненты, отличные от описанных ранее. Кроме того, в компьютере могут отсутствовать определенные компоненты. Кроме того, устройство для хранения данных может быть локальным и/или удаленным от компьютера (например, встроенным в сетевую систему хранения данных (SAN)).
[0217] Как известно специалистам в данной области техники, компьютер применяется для выполнения компьютерных программных модулей для обеспечения функциональных возможностей, описанных в данном документе. В данном контексте термин "модуль" относится к логике компьютерной программы, используемой для обеспечения установленной функциональности. Таким образом, модуль может быть реализован в аппаратном обеспечении, встроенном программном обеспечении и/или программном обеспечении. В одном варианте реализации изобретения программные модули хранятся на устройстве хранения данных, загружаются в запоминающее устройство и выполняются процессором.
Понятно, что примеры и варианты реализации, описанные в данном документе, приводится только в качестве иллюстрации и что различные модификации или изменения в их свете будут предложены специалистам в данной области техники и должны быть включены в сущность и сферу действия настоящей заявки и объема прилагаемой формулы изобретения.
ПРИМЕРЫ
[0218] Ниже рассматриваются конкретные варианты реализации настоящего изобретения. Примеры приводятся исключительно в иллюстративных целях и никоим образом не направлены на ограничение объема настоящего изобретения. Были предприняты усилия для того, чтобы обеспечить точность в отношении использованных числовых значений (например, количеств, температур и т.д.), однако безусловно необходимо делать поправку на некоторые экспериментальные ошибки и отклонения.
[0219] Если не указано иное, при практической реализации настоящего изобретения применяются традиционные методы химии белков, биохимии, технологии рекомбинантных ДНК и фармакологии, которые соответствуют данной области техники. Такие технологии подробно описаны в литературе. См., например, Т.Е. Creighton, Proteins: Structures and Molecular Properties (W.H. Freeman and Company, 1993); A.L. Lehninger, Biochemistry (Worth Publishers, Inc., current addition); Sambrook, et al., Molecular Cloning: A Laboratory Manual (2nd Edition, 1989); Methods In Enzymology (S. Colowick and N. Kaplan eds., Academic Press, Inc.); Remington's Pharmaceutical Sciences, 18th Edition (Easton, Pennsylvania: Mack Publishing Company, 1990); Carey and Sundberg Advanced Organic Chemistry 3rd Ed. (Plenum Press) Vols A and B(1992).
Пример 1: Молекулярное моделирование и компьютерная инженерия области контакта Fab
[0220] С использованием методов структурного и компьютерного молекулярного моделирования была создана библиотека мутантных конструкций тяжелых и легких цепей, которую можно подвергнуть скринингу в контексте других антител (AT) или их фрагментов с целью выявления мутаций, проявляющих необходимую специфичность в антителах, представляющих интерес. Стратегия инженерии преимущественного спаривания тяжелой (Н) и легкой (L) цепей включала, во-первых, выявление репрезентативного Fab (то есть, D3H44).
[0221] Как указано в таблице 1, такой Fab должен был соответствовать следующим ключевым критериям: быть человеческим/гуманизированным, иметь широко используемые подгруппы VH и VL и содержать минимальные мутации в каркасной области. Кроме того, структурные соображения предусматривали, что междоменный угол VH:VL должен был приближаться к среднему значению, наблюдаемому для антител. После отбора Fab D3H44 был выполнен in silico анализ области контакта Fab, чтобы выявить и изучить остатки, значимые для взаимодействия между тяжелыми и легкими цепями, с использованием подхода по двум направлениям.
[0222] Первое направление предусматривало общий анализ консервативности последовательностей в вариабельных и константных областях контакта Fab, который выполнялся путем выравнивания последовательностей и структур известных антител. Выравнивание последовательностей константных и вариабельных доменов различных подгрупп антител представлено на фигуре 1. Фигура 1А иллюстрирует выравнивание репрезентативных подгрупп VH зародышевой линии человека. Фигура 1А иллюстрирует выравнивание репрезентативных подгрупп каппа VL зародышевой линии человека. Фигура 1С иллюстрирует выравнивание репрезентативных подгрупп лямбда VL зародышевой линии человека. Фигура 1D иллюстрирует выравнивание последовательностей аллелей СН1 человека. Фигура 1Е иллюстрирует выравнивание последовательностей аллелей каппа и лямбда человека. Второе направление предусматривало анализ кристаллической структуры области контакта D3H44 с использованием ряда инструментов молекулярного моделирования, представленных на фигуре 2 (например, ResidueContacts™). Эти анализы привели к составлению списка "горячих" положений для инженерии преимущественного спаривания H-L. "Горячие" положения, определенные по результатам данных анализов, перечислены в таблице 2. Эти положения и аминокислоты представляют собой преимущественно каркасные остатки (за исключением нескольких остатков, расположенных в петлях CDR3), и они, по большей части, также являются консервативными остатками в цепях лямбда L. Аминокислоты в исходных последовательностях D3H44, пронумерованные по Kabat, приведены в таблицах 3а-3b.
[0223] Затем путем мутагенеза in silico и упаковки/моделирования с использованием программного обеспечения Zymepack™ были сымитированы и определены потенциальные мутации в "горячих" положениях, а также в положениях по соседству с "горячими" положениями, представляющими интерес, в трехмерной кристаллической структуре. Zymepack™ представляет собой пакет программного обеспечения, который, при наличии вводной структуры и набора мутаций, изменяет типы остатков во вводной структуре согласно заданным мутациям и создает новую структуру, представляющую собой аппроксимацию физической структуры мутантного белка. Кроме того, Zymepack позволяет оценить свойства мутантного белка, вычисляя ряд количественных показателей. Эти показатели включают измерения стерической и электростатической комплементарности, которые могут коррелировать со стабильностью, аффинностью связывания или гетеродимерной специфичностью мутантного белка.
[0224] На фигуре 3 представлен набор "горячих" положений в области контакта тяжелых и легких цепей в вариабельных доменах, и показано, каким образом в эти положения области контакта можно вводить мутации, чтобы способствовать селективному спариванию облигатных цепей, препятствуя при этом образованию неправильных пар цепей. С использованием компьютерных методов, включая программное обеспечение Zymepack™, была смоделирована стерическая комплементарность, которая также рассчитывалась на основании энергетических факторов, таких как упаковка с учетом ван-дер-ваальсовых взаимодействий, эффекты кавитации и близкие контакты гидрофобных групп. Аналогичным образом, были смоделированы энергии электростатического взаимодействия и оценены на основании кулоновских взаимодействий между зарядами, водородных связей и эффектов десольватации. Чтобы рассчитать относительные показатели стерической и электростатической комплементарности, имитировались как модели предпочтительных пар тяжелых и легких цепей, например, Н1:L1 (или H2:L2), так и модели неправильных пар, например, H1:L2 (и Н2:L1), полученные путем ввода мутаций, представляющих интерес. Это позволило определить, приводил ли конкретный набор мутаций к благоприятным энергиям, то есть, большей стерической и/или электростатической комплементарности для облигатных пар тяжелой-легкой цепей относительно некорректных (необлигатных) пар. Рассчитанные стерические и электростатические энергии являются компонентами свободной энергии, ассоциированной со спариванием легкой и тяжелой цепей. Следовательно, большая стерическая и электростатическая комплементарность свидетельствует о большем изменении свободной энергии, ассоциированном со спариванием облигатной пары, относительно спаривания необлигатной пары. Большая стерическая или электростатическая комплементарность приводить к преимущественному (селективному) спариванию облигатной пары тяжелой и легкой цепей относительно необлигатной пары.
Пример 2: Отбор и описание конструкций
[0225] Подход, описанный в Примере 1, использовали, чтобы сконструировать пары гетеродимеров тяжелая цепь-легкая цепь (т.е. H1-L1 и H2-L2), которые демонстрируют избирательное или преимущественное спаривание. Гетеродимеры были сконструированы в парах, называемых «конструкциями» или «наборами конструкций», и содержали ряд замен в цепях H1, L1, Н2 и L2, которые стимулируют преимущественное спаривание (Таблица 5). Чтобы оценить относительное спаривание, сначала исследовали наборы конструкций в виде «LCCA-конструкций» (Таблица 4), в которых одна тяжелая цепь коэкспрессировалась с двумя легкими цепями. Аминокислотные замены определены согласно Таблицам 3а, 3b с использованием системы нумерации Кабата.
[0226] Библиотеку конструкций, описанную в Таблице 30 из Международной патентной заявки номер РСТ/СА2013/050914, использовали в качестве отправной точки для определения некоторых конструкций LCCA, приведенных в Таблице 4, и наборов конструкций, приведенных в Таблице 5. Некоторые из конструкций в Таблице 4 и Таблице 5 являются новыми независимыми конструкциями. Основные конструкции приведены в Таблице 6 вместе со своими уникальными идентификационными номерами. Большинство конструкций затрагивает только константную область, и только несколько конструкций также содержат модификации в вариабельной области. Предполагалось, что эти конструкции дополнительно обеспечивают специфичность спаривания и в то же время способствуют переносу в другие системы антител.
[0227] Для полученных конструкций в качестве отправной использовали библиотеку конструкций, описанную в Таблице 30 из Международной патентной заявки номер РСТ/СА2013/050914, причем конструкции сгруппированы по структурному сходству и упорядочены на основании силы специфичности спаривания, воздействия на связывание антигена и стабильности согласно данным, полученным методом дифференциальной сканирующей калориметрии (DSC). Затем конструкции комбинировали (см. пример в Таблице 7) и/или оптимизировали (см. примеры в Таблице 8 и Таблице 9), чтобы получить соответствующие конструкции. В случае комбинаций, по меньшей мере одна из конструкций демонстрировала высокую специфичность спаривания, в то время как другая конструкция(-и) демонстрировала диапазон благоприятных значений специфичности спаривания. Все конструкции, отобранные для комбинирования и/или оптимизации, демонстрировали отсутствие воздействия/минимальное воздействие на связывание антигена и отсутствие воздействия/минимальное воздействие на температуру плавления (Tm).
[0228] Независимые конструкции исследовали по отдельности (классифицированные как независимые в колонке с типом конструкции, Таблица 5), а также в комбинации с полученными конструкциями (классифицированные как независимые/комбинированные в колонке с типом конструкции, Таблица 5; также см. пример в Таблице 10).
[0229] Конструкции аппроксимировали молекулярной моделью D3H44 и рассчитывали показатели (как описано в Примере 1). Затем отбирали лучшие конструкции на основании риска (возможное влияние на стабильность, а также иммуногенность) и воздействия (которое учитывает предполагаемую силу движущей специфичности спаривания). Такие лучшие конструкции приведены в Таблице 5.
Пример 3: Получение конструкций Fab, кодирующих тяжелые цепи D3H44 IgG и легкие цепи D3H44 IgG
[0230] Тяжелые и легкие цепи Fab дикого типа антитела D3H44 против тканевого фактора получали следующим образом. Последовательности легкой (AJ308087.1) и тяжелой (AJ308086.1) цепи Fab D3H44 получали из GenBank (Таблица 3с), проводили синтез генов и оптимизацию кодонов для экспрессии в организме млекопитающего. Вставки в вектор легкой цепи, состоящие из участка надреза 5'-EcoRI - сигнального пептида HLA-A - метки НА или FLAG - клона легкой цепи Ig - «стоп-кодона TGA» - участка надреза-3' BamH1, лигировали в вектор рТТ5 (Durocher Y et al., Nucl. Acids Res. 2002; 30, No.2 e9). Проводили секвенирование результирующей конструкции вектор + вставка, чтобы подтвердить правильность рамки считывания и последовательность кодирующей ДНК. Аналогично, вставки в вектор тяжелой цепи, состоящие из участка надреза 5'-EcoR1 - сигнального пептида HLA-A - клона легкой цепи (с терминацией в Т238; см. Таблицу 3а) - метки ABD2-His6 - стоп-кодона TGA - участка надреза-3' BamH1, лигировали в вектор рТТ5 (ABD; альбумин-связывающий домен). Также проводили секвенирование результирующей конструкции вектор + вставка, чтобы подтвердить правильность рамки считывания и последовательность кодирующей ДНК. Различные конструкции Fab D3H44, содержащие аминокислотные замены, предназначенные для наборов конструкций, получали методом синтеза генов или методом сайт-направленного мутагенеза (Braman J, Papworth С & Greener A., Methods Mol. Biol. (1996) 57:31-44).
[0231] Тяжелые и легкие цепи метили в С- и N-конце, соответственно, чтобы облегчить оценку преимущественного спаривания при помощи скрининга методом конкурентного SPR. Метка тяжелой цепи ABD2-His6 специфическим образом делала возможным захват комплексов H-L на поверхности анти-his SPR-чипа, в то время как метки легкой цепи FLAG и НА давали возможность количественно оценить относительные популяции L1 и L2.
Пример 4: Оценка преимущественного спаривания гетеродимеров Fab, содержащих модификации константного домена или комбинацию модификаций константного и вариабельного домена в легкой и/или тяжелой цепях D3H44 IgG
[0232] Конструкции, кодирующие тяжелые и легкие цепи D3H44 IgG в формате Fab, содержащие аминокислотные модификации в соответствии с наборами конструкций LCCA в Таблице 12, получали так, как описано в Примере 3. Способность конструкций к преимущественному спариванию с образованием необходимого гетеродимера H1-L1 в контексте набора конструкций LCCA (H1, L1, L2) определяли при помощи конкурентного анализа для легкой цепи (LCCA).
[0233] LCCA позволяет осуществлять количественную оценку относительного спаривания одной тяжелой цепи с по меньшей мере двумя уникальными легкими цепями и вкратце может быть описан следующим образом. Одну Fab-конструкцию тяжелой цепи D3H44 коэкспрессировали с двумя уникальными Fab-конструкциями легкой цепи D3H44 и определяли относительную специфичность спаривания легких цепей (например, Н1-L1:H1-L2) по результатам конкурентного SPR-анализа, проводимого в двух повторах. Для выявления сильных драйверов изменяли соотношение LCCA-скрининга, снижая количество L1 (сконструированной для преимущественного спаривания с Н-цепью) по сравнению с L2, (например, L1:L2=1:3, по массе), оставляя при этом ограниченное количество тяжелой цепи (т.е. H1-L1+L2, составляющее 1:3). Количество каждого из образуемых гетеродимеров (т.е. H1-L1 и H1-L2) определяли по связыванию тяжелых цепей с SPR-чипом посредством осаждения his-метки с последующим определением количества каждой из меток легкой цепи (НА или FLAG) при помощи антител, специфических в отношении этих меток. После этого проводили верификацию отобранных гетеродимеров путем верификации методом конкурентного анализа для легкой цепи, в котором соотношения ДНК L1:L2 во время трансфекции варьировали от 1:3 до 1:9, оставляя при этом ограниченное количество тяжелой цепи. Также следует отметить, что метки легкой цепи (НА или FLAG) не влияли на LCCA-спаривание в системе D3H44 (см. пример 10 из Международной патентной заявки номер РСТ/СА2013/050914). Схема, представляющая дизайн анализа, приведена на Фигуре 4. Фигура 5 иллюстрирует, каким образом метят тяжелые цепи и легкие цепи и как оценивают преимущественное спаривание. Подробности эксперимента LCCA приведены ниже.
Способ трансфекции
[0234] LCCA-конструкции, содержащие одну тяжелую цепь и две легких цепи, полученные, как описано в Примере 3, трансфицировали в клетки СНО-3Е7 следующим образом. Клетки СНО-3Е7 с плотностью 1,7-2×106 клеток/мл культивировали при 37°С в среде FreeStyle™ F17 (Invitrogen cat# А-1383501), дополненной 4 мМ глутамина и 0,1% KoliphorP188 (Sigma #K4894). Общий объем, составляющий 2 мл, трансфицировали общим количеством ДНК, составляющим 2 мкг, при помощи PEI-pro (Polyplus transfection # 115-375) с соотношением ДНК:РЕI 1:2,5. Через двадцать четыре часа после добавления смеси ДНК-РЕI клетки переносили в 32°С. На 7 день проводили исследование супернатантов в отношении экспрессии при помощи невосстанавливающего SDS-PAGE-анализа с последующим окрашиванием голубым кумасси для визуализации полос. Соотношения H:L указаны в Таблице 11.
SPR-метод конкурентного анализа
[0235] Степень преимущественного спаривания легкой цепи D3H44 с тяжелой цепью D3H44 в LCCA-конструкциях оценивали при помощи SPR-считывания уникальных эпитопных меток, расположенных в N-конце каждой легкой цепи.
[0236] Оборудование и реагенты для поверхностного плазмонного резонанса (SPR). Сенсорные чипы GLC, набор для аминного сопряжения Biorad ProteOn (1-этил-3-(3- диметиламинопропил) карбодиимид гидрохлорид (EDC), N-гидроксисульфосукцинимид (sNHS) и этаноламин) и 10 мМ буфера ацетата натрия приобретали у Bio-Rad Laboratories (Canada) Ltd. (Mississauga, ON). Буфер 4-(2-гидроксиэтил)-7-пиперазинэтансульфоновой кислоты (HEPES), этилендиаминтетрауксусную кислоту (EDTA) и NaCl приобрели у Sigma-Aldrich (Oakville, ON). 10% раствор Твин 20 приобрели у Teknova (Hollister, СА).
[0237] SPR-биосенсорный анализ. Все анализы методом поверхностного плазмонного резонанса проводили на инструменте BioRad ProteOn XPR36 (Bio-Rad Laboratories (Canada) Ltd. (Mississauga, ON)) с подвижным буфером PBST (PBS от Teknova Inc с 0,05% Твин20) при температуре 25°С. Анти-пента His захватывающую поверхность генерировали, используя сенсорный чип GLM, активируемый разведенными 1:5 стандартными растворами sNHS/EDC от BioRad, инжектируемыми в течение 140 сек при 100 мкл/мин в направлении аналита (горизонтально). Непосредственно после активации инжектировали 25 мкг/мл раствора анти-пента His антитела (Qiagen Inc.) в 10 мМ NaOAc, рН 4,5, в направлении аналита (вертикально) при скорости потока 25 мкл/мин до иммобилизации приблизительно 3000 резонансных единиц (RU). Оставшиеся активные группы тушили посредством 140 с инжекции 1 М этаноламина при 100 мкл/мин в направлении аналита, что также гарантировало создание ложно-активированных участков между областями локализации белка для контрольного сравнения.
[0238] Скрининг гетеродимеров в отношении связывания с анти-FLAG (Sigma Inc.) и анти-НА (Roche Inc.) моноклональными антителами осуществляли в две стадии: непрямой захват гетеродимеров на анти-пента His поверхности в направлении лиганда с последующей инжекцией анти-FLAG и анти-НА в направлении аналита. Сначала для стабилизации исходного уровня использовали одну инжекцию PBST в течение 30 сек при 100 мкл/мин в направлении лиганда. Для проведения каждого захвата гетеродимеров неочищенные гетеродимеры в клеточной культуральной среде разводили до 4% в PBST. В отдельные лигандные каналы одновременно инжектировали от одного до пяти гетеродимеров или контрольных образцов (т.е. контрольных образцов, содержащих 100% НА-легкой цепи или 100% FLAG-легкой цепи) в течение 240 сек при потоке 25 мкл/мин, что приводило к насыщающему захвату гетеродимеров, соответствующему приблизительно от 300 до 400 RU, на анти-пента His поверхности. Первый лигандный канал оставляли пустым, чтобы в случае необходимости использовать в качестве контроля. За стадией захвата гетеродимеров незамедлительно проводили две инжекции буфера в направлении аналита, чтобы стабилизировать базовый уровень, а затем в двух репликах инжектировали 5 нМ анти-FLAG и 5 нМ анти-НА при 50 мкл/мин в течение 120 сек с фазой диссоциации в 180 сек, получая набор сенсограмм связывания с буферным эталоном для каждого захваченного гетеродимера. Антиген тканевого фактора (TF), с которым связывается гетеродимер, также инжектировали через последний оставшийся аналитный канал в качестве контроля активности. Гетеродимеры регенерировали при помощи впрыскивания 0,85% фосфорной кислоты в течение 18 сек при 100 мкл/мин, чтобы подготовить анти-пента His поверхность к следующему циклу инжекций. Проводили выравнивание и двойной контроль сенсограмм, используя инжекцию с пустым буфером и участки между областями локализации белка, а полученные в результате сенсограммы анализировали при помощи программного обеспечения ProteOn Manager v3.0.
Результаты
[0239] Результаты LCCA приведены в Таблицах 12, 13а и 14а. Следует отметить, что в Таблицах 13 и 14 «уникальный идентификационный номер» может точно не соответствовать Таблице 5, так как уникальные идентификационные номера двух составляющих LCCA могут относиться к любой ориентации ((нa6op#H1L1L2-нa6op#H2L2L1) или (нa6op#H2L2L1-нa6op#H1L1L2)). Оценка преимущественного спаривания для каждой конструкции LCCA приведена в последних 3 колонках Таблицы 12. Те же данные включены в контексте пар конструкций в Таблицах 13а и 14а, в колонках 5, 6 и 8 или 10, 11 и 13. Каждому уникальному набору мутаций H1, L1 и L2 (конструкция LCCA) приписывали уникальный номер или «набор #» (например, 9567 или 9087). Если данные представлены в формате H1 L1 Н2 L2 (формат пары Fab или набора конструкций), такому набору конструкций последовательно приписывали «уникальный идентификационный номер», состоящий из набора цифр для двух составляющих LCCA (например, 9567-9087). Следует отметить, что большинство экспериментов LCCA проводили на конструкциях, содержащих межцепьевую дисульфидную связь(-и) Fab, находящуюся в константном домене (H/C233-L/C214, нумерация Кабата). В Таблицах 13(а и b) и 14(а и b) в целях выделения успешности конкретной конструкции в отношении преимущественного связывания два комплементарных набора LCCA (H1, L1, L2 и Н2, L2, L1) представлены в формате пары Fab. Присутствие меток (L-цепь: НА и FLAG и Н-цепь: ABD2-His6) не влияло на ожидаемое нейтральное спаривание -50%: 50% для D3H44 ДТ.
[0240] Приведенные в таблицах данные LCCA представляют средние величины в формате соотношения (H1-L1:H1-L2 и H2-L2:H2-L1), нормированные относительно соотношения ДНК L1:L2 в 1:1. Кроме того, данные LCCA нормировали относительно 100%, так как для некоторых вариантов наблюдаемое общее количество L1 и L2 существенно отличалось от 100%. Предполагается, что это расхождение в общем процентном содержании легких цепей частично связано с появлением вариабельного неспецифического связывания во время исходного захвата гетеродимеров на SPR-чипе. Так как эксперименты LCCA проводили с 2 разными соотношениями ДНК L1:L2 (L1:L2, составляющие 1:3 и 1:9, соответственно), оба нормированные соотношения LCCA приведены в таблицах. Следует отметить, что для некоторых экспериментов LCCA не приведены данные LCCA, так как полученные экспериментальные данные не соответствуют критерию включения (например, захват Fab на SPR-чипе составлял менее 100 или общее количество L1 и L2 при LCCA выпадало из диапазона от 60 до 140).
[0241] В Таблице 12 перечислены все конструкции LCCA (530), для которых были получены данные. Из 530 конструкций LCCA 490 (92,5%) конструкций LCCA характеризовались по меньшей 60% правильного спаривания (при нормированном соотношении ДНК L1:L2 в 1:1), учитывая оба соотношения ДНК L1:L2 1:3 и 1:9. Оставшиеся конструкции LCCA включали конструкции LCCA, которые были преимущественно нейтральными (32/530 или 6,0%), а также небольшую часть, для которой были получены противоречивые результаты (8/530 или 1,5%). Конструкции, приведенные в Таблице 12, были преимущественно электростатическими (на основании драйверов специфичности, которые задействуют водородное связывание или заряд-зарядовые взаимодействия), при этом некоторые конструкции также включали стерическую комплементарность и/или внутрицепьевые ковалентные дисульфидные связи. Некоторые конструкции также содержали мутации для образования новых дисульфидных связей в отсутствие природной внутрицепьевой дисульфидной связи (образуемой H/C233-L/C214).
[0242] В Таблицах 13 (а и b) и 14(а и b) перечислены 447 конструкций, для которых данные LCCA были представлены для обоих гетеродимеров набора конструкций. Таблицы 13а и 14а демонстрируют, что подход конструирования in silico, описанный в Примере 1, приводил к достижению преимущественного спаривания H1-L1 по сравнению с H1-L2 и H2-L2 по сравнению с H2-L1 среди разнообразных наборов конструкций и их вариаций.
[0243] В Таблицах 13 (а и b) перечислены те конструкции, которые имеют средние показатели LCCA (средние по медианным величинам, нормированным к соотношению L1:L2 в 1:1 для H1-L1:H1-L2 и H2-L2:H2-L1) для спаренных неправильно спаренных гетеродимеров Fab, составляющие по меньшей мере 86:14, в то время как в Таблицах 14(а и b) перечислены те конструкции, которые имеют средние показатели LCCA для спаренных неправильно спаренных гетеродимеров Fab меньше 86:14. Показатели для каждого LCCA нормировали относительно 100%, а также относительно соотношения ДНК L1:L2 в 1:1 (как описано выше в этом примере), и описывали скалярными величинами ((ln(r1/f1) или ln(r2/f2)), где r1 и r2 соответствуют медианным величинам H1L1:H1L2 и H2L2:H2L1 в экспериментальных соотношениях, соответственно, a f1 и f2 относятся к соответствующим экспериментальным значениям), а также соотношением между спаренными и неправильно спаренными гетеродимерами Fab. Каждая конструкция также характеризуется связанной медианной скалярной величиной показателя LCCA (0,5(ln(r1/f1)+ln(r2/f2))), которую также нормировали относительно 100%, а также относительно соотношения ДНК L1:L2 в 1:1 (как описано выше в этом примере). Кроме того, показан скалярный диапазон для каждой LCCA конструкции (КАЛЦ1 и КАЛЦ2, соответствующие экспериментам H1L1:H1L2 и H2L2:H2L1, соответственно). Из 447 конструкций Mab 354 (79,2%) демонстрировали по меньшей мере средние показатели LCCA, составляющие 86:14 (Таблица 13 а и b). Конструкции в Таблицах 13 (а и b) дополнительно распределили по 13 кластерам на основании сходства конструкций. Конструкции в каждом кластере выстраивали от наивысшей до низшей скалярной величины показателя LCCA.
[0244] Кроме того, данные LCCA в Таблице 13а также графически представлены на Фигуре 7. На Фигуре 7 представлены блочные графики размаха, которые иллюстрируют величины показателей LCCA для спаренных неправильно спаренных гетеродимеров Fab, составляющие по меньшей мере 86:14 для каждого кластера. Нижняя часть каждого блока отображает первый квартиль (Q1), который представляет среднюю величину показателя LCCA между наименьшей величиной и медианной величиной, таким образом, величины ниже 1-го квартиля отображают наинизшие 25% данных. Горизонтальная планка внутри блока обозначает второй квартиль, который представляет медианную среднюю величину показателя LCCA для кластера. Верхняя часть каждого блока отображает третий квартиль (Q3), который представляет среднюю величину показателя LCCA между наибольшей величиной и медианной величиной, таким образом, величины выше 3-го квартиля отображают наивысшие 25% данных. Межквартильная область представляет собой разницу между Q3 и Q1. Вертикальные усы в обоих направлениях указывают диапазон данных для величин, находящихся в пределах Q1-(1,5*IQR) или Q3+(1,5*IQR). Горизонтальные планки на концах усов указывают наибольшие и наименьшие величины в диапазоне. Данные, находящиеся за пределами блочных графиков, и усы относятся к выбросам, при этом умеренные выбросы обозначены точкой (отличаются от Q1 или Q3 на 1,5*IQR-3*IQR), а большие выбросы обозначены знаком плюс (отличаются от Q1 или Q3 более чем на 3*IQR).
Пример 5: Масштабирование для получения биофизических характеристик
[0245] Правильно спаренные гетеродимеры, указанные в ряде уникальных идентификационных номеров (Таблица 5), масштабировали (как правило, до 20 мл) и очищали описанным ниже способом, чтобы исследовать термостабильность и связывание антигена. Тяжелые и легкие цепи каждого гетеродимера экспрессировали в 20 мл культурах клеток СНО-3Е7. Клетки СНО-3Е7 с плотностью 1,7-2×106 клеток/мл культивировали при 37°С в среде FreeStyle™ F17 (Invitrogen cat# А-1383501), дополненной 4 мМ глутамина и 0,1% Koliphor Р188 (Sigma #К4894). Общий объем, составляющий 20 мл, трансфицировали общим количеством ДНК, составляющим 20 мкг, при помощи PEI-pro (Polyplus cat# 115-375) с соотношением ДНКРЕ1 1:2,5. Через двадцать четыре часа после добавления смеси ДНК-PEI клетки переносили в 32°С.
[0246] Через 7 дней после трансфекции клетки центрифугировали, а гетеродимеры очищали от супернатанта при помощи очистки методом высокопроизводительной аффинной хроматографии на никеле следующим образом. Супернатанты разводили до 20- 25% клеточного культур ал ьного супернатанта в равновесном буфере (фосфатно-солевой буфер Дульбекко (DPBS) без кальция, магния и фенолового красного (HyClone™ # SH30028.02)), а затем инкубировали с перемешиванием в течение 12 часов со смолой HisPur® Ni-NTA (Thermo Scientific # PI-88222), также предварительно уравновешенной равновесным буфером. Затем смолу собирали путем центрифугирования, переносили в 96- луночный пористый планшет, три раза промывали равновесным буфером и элюировали, используя элюирующий буфер HIS-Select® (Sigma-Aldrich # Н5413).
[0247] После очистки экспрессию гетеродимеров оценивали методом высокопроизводительного анализа экспрессии белка в невосстанавливающих условиях с использованием LabChip GXII (Perkin Elmer #760499). Процедуры проводили в соответствии с руководством для пользователя НТ Protein Express LabChip, версия 2, LabChip GXII User Manual, со следующими модификациями. Образцы гетеродимеров в количестве 2 мкл или 5 мкл (диапазон концентраций 5-2000 нг/мкл) добавляли, чтобы разделить лунки в 96-луночных планшетах (BioRad # HSP9601), вместе с 7 мкл буфера для образцов НТ Protein Express (Perkin Elmer #760328). Затем образцы гетеродимеров денатурировали при 70°С в течение 15 мин. Управление инструментом LabChip осуществляли при помощи чипа НТ Protein Express Chip (Perkin Elmer #760499) и аналитической установки Ab-200. После использования чип очищали водой MilliQ и хранили при 4°С.
Пример 6: Определение термостабильности гетеродимеров Fab методом DSF
[0248] Для оценки термостабильности применяли дифференциальную сканирующую флуориметрию (DSF) как высокопроизводительный способ для исследования всех правильно спаренных гетеродимеров по сравнению с гетеродимерами дикого типа, т.е. немодифицированными парами тяжелая цепь-легкая цепь. Гетеродимеры получали так, как описано в Примере 5.
Определение термостабильности
[0249] Термостабильность всех пар гетеродимеров определяли при помощи DSF следующим образом. Каждый гетеродимер очищали, как описано в Примере 5, и разводили до 0,5 мг/мл в DPBS (HyClone Cat # SH30028.02). Для большинства образцов рабочий раствор гелевого красителя Sypro Orange (Life Technologies Cat # S-6650) готовили путем разведения 4 мкл гелевого красителя Sypro Orange до 2 мл DPBS. Образцы DSF готовили, добавляя 14 мкл белка в концентрации 0,5 мг/мл в 60 мкл разведенного рабочего раствора гелевого красителя Sypro Orange. При этом для белков, концентрация которых составляла менее 0,5 мг/мл, каждый образец DSF готовили, добавляя 14 мкл неразведенного белка в 60 мкл рабочего раствора красителя Sypro Orange (который был разведен до 1:1500 в DPBS). Затем проводили анализ DSF в двух репликах с 20 мкл аликвотами при помощи инструмента для qPCR Rotor-Gene 6000 (QiaGen Inc). Каждый образец сканировали от 30°С до 94°C с интервалами в 1°C с 10-секундным равновесием между каждой стадией и 30-секундной задержкой на старте. Применяли фильтр возбуждения на 470 нМ и фильтр испускания на 610 нМ с коэффициентом усиления 9. Данные анализировали при помощи программного обеспечения Rotor-Gene 6000, используя максимальную величину первой производной кривой денатурации в качестве Тт.Оставшиеся образцы DSF готовили и анализировали аналогичным образом со следующими модификациями в протоколе, которые не изменяют определенные величины Tm: 1) рабочий раствор готовили путем разведения 1 мкл гелевого красителя Sypro Orange до 2 мл DPBS, 2) анализировали 30 мкл аликвоты и 3) использовали коэффициент усиления 10.
[0250] Результаты DSF приведены в Таблицах 12, 13b и 14b. Термостабильность H1:L1 Fab в контексте конструкции LCCA (величина DSF и изменение величины DSF по сравнению с диким типом) приведена в колонках 3 и 4 Таблицы 12. Те же самые величины DSF включены в контексте пар конструкций в Таблицах 13b м 14b в колонках 7 и 8. Для каждого гетеродимера Fab в случае проведения повторов указанная величина Tm представляет собой медианную величину. Сравнение величин Tm гетеродимера Fab с величиной Tm гетеродимера Fab дикого типа (конструкция Fab дикого типа, содержащая метку НА с медианным значением Tm в 81,0°С) приведено в колонке H1L1_dTm_dsf. Следует отметить, что для нескольких гетеродимеров Fab, в которых отсутствует природная внутрицепьевая дисульфидная связь (между С233 Н-цепи и С214 L-цепи), величины H1L1_dTm_dsf не определяли, так как не оценивали соответствующий Fab дикого типа, в котором отсутствует природная внутрицепьевая дисульфидная связь. Также следует отметить, что для некоторых гетеродимеров Fab не приведены величины Tm (17/230 или 7,4% гетеродимеров Fab) в связи с качеством соответствующих экспериментов (например, низкий выход, низкая интенсивность, частично перекрывающиеся пики и вариабельность между повторами для гетеродимеров Fab, превышающая 1°С). Для некоторых из этих гетеродимеров Fab вместо этого показаны оценочные величины Tm, соответствующие величинам Tm для сходных гетеродимеров Fab, которые отличаются только присутствием/отсутствием или идентичностью присоединенной метки L-цепи (НА или FLAG). В случае оценочных величин Tm соответствующая величина Tm для дикого типа (81,2°С) представляет собой медианную величину, полученную по всем гетеродимерным конструкциям Fab дикого типа (т.е. конструкциям Fab, содержащим метку НА или метку FLAG). Метка НА или FLAG существенно не влияет на величины Tm гетеродимеров Fab дикого типа. В общем случае гетеродимеры Fab демонстрировали сходные величины Tm по сравнению с ДТ. Среди гетеродимеров Fab, содержащих природную внутрицепьевую дисульфидную связь, а также тех, для которых доступны данные DSF, 93% (195/209) гетеродимеров Fab демонстрировали снижение на 3°С или менее по сравнению с ДТ. Кроме того, наиболее подверженный влиянию гетеродимер Fab демонстрировал снижение на 6,5°С по сравнению с ДТ. В Таблице 12 перечислены конструкции LCCA в порядке снижения Тт.
[0251] Кроме того, было определено тринадцать аминокислотных замен, которые в общем случае улучшали стабильность гетеродимеров Fab (см. Таблицу 34). Стабилизирующие мутации были определены после сравнения гетеродимеров Fab, которые содержат стабилизирующую мутацию, со сходными гетеродимерами Fab, которые отличаются отсутствием стабилизирующей мутации. Стабилизирующие мутации в тяжелой цепи включают A125R, F1172R, L143F, Q179D, Q179E, Q39R, S188L и V190F. Стабилизирующие мутации в легкой цепи включают Q124E, Q124R, Q160F, S176L и Т180Е. В общем случае стабилизирующие мутации повышали стабильность на 0,4°С-2,1°С.Стабилизирующие мутации в тяжелой цепи A125R, H172R, L143F, Q179D, Q179E, Q39R, S188L и V190F повышали стабильность на от 0,4°С до 0,6°С, от 0,4°С до 2,ГС, 0,4°С, от 0,5°С до 0,6°С, от 0,5°С до 0,8°С, от 1,1°С до 1,6°С, от 0,4°С до 1,2°С и 1°С, соответственно. Стабилизирующие мутации в легкой цепи Q124E, Q124R, Q160F, S176L и Т180Е повышали стабильность на от 0,4°С до 0,5°С, от 0,8°С до 0,9°С, 0,6°С, от 0,4°С до 1,0°С и 0,5°С, соответственно.
Пример 7: Определение аффинности в отношении антигена для гетеродимеров Fab
[0252] Оценивали способность гетеродимеров Fab связывать тканевой фактор, чтобы определить, оказывают ли аминокислотные замены какое-либо влияние на способность гетеродимера связываться с антигеном. Аффинность каждого гетеродимера Fab в отношении тканевого фактора определяли методом SPR следующим образом.
[0253] Оборудование и реагенты для SPR. Сенсорные чипы GLC, набор для аминного сопряжения Biorad ProteOn (EDC, sNHS и этаноламин) и 10 мМ буфера ацетата натрия приобрели у Bio-Rad Laboratories (Canada) Ltd. (Mississauga, ON). Подвижный буфер PBS с 0,05% Твин 20 (PBST) приобрели у Teknova (Hollister, CA).
[0254] Партия гетеродимеров Fab. Очищенные гетеродимеры Fab исследовали в 3 партиях - А, В и С. Партии А и В хранили при 4°С приблизительно в течение 1 месяца перед проведением анализа SPR, в то время как очищенные гетеродимеры Fab из партии С хранили при 4°С приблизительно в течение 2 месяцев перед проведением анализа SPR. Гетеродимеры Fab из партии С обозначены «+» рядом с соответствующими величинами KD в Таблице 12.
[0255] Все эксперименты методом поверхностного плазмонного резонанса проводили при помощи инструмента BioRad ProteOn XPR36 (Bio-Rad Laboratories (Canada) Ltd. (Mississauga, ON)) с подвижным буфером PBST при температуре 25°С. Анти-пента His захватывающую поверхность генерировали, используя сенсорный чип GLM, активируемый разведенными 1:5 стандартными растворами sNHS/EDC от BioRad, инжектируемыми в течение 140 сек при 100 мкл/мин в направлении аналита (горизонтально). Непосредственно после активации инжектировали 25 мкг/мл раствора анти-пента His антитела (Qiagen Inc.) в 10 мМ NaOAc, рН 4,5, в направлении аналита (вертикально) при скорости потока 25 мкл/мин до иммобилизации приблизительно 3000 резонансных единиц (RU). Оставшиеся активные группы гасили посредством 140 сек инжекции 1 М этаноламина при 100 мкл/мин в направлении аналита, что также гарантировало создание ложно-активированных участков между областями локализации белка для контрольного сравнения.
[0256] Скрининг гетеродимеров в отношении связывания с антигеном TF осуществляли в две стадии: непрямой захват гетеродимеров на поверхности анти-пента His антитела в направлении лиганда с последующей одновременной инжекцией 5 концентраций очищенного антигена и одного пустого буфера для двойного контроля в направлении аналита. Сначала стабилизировали исходный уровень одной инжекцией буфера в течение 30 сек при 100 мкл/мин в направлении лиганда. От одного до пяти гетеродимеров или контрольных образцов в концентрации 3,4 мкг/мл в PBST одновременно инжектировали в отдельные лигандные каналы в течение 240 сек при потоке 25 мкл/мин. Это приводило к среднему захвату, соответствующему приблизительно 1000 RU, на анти-пента His поверхности для серий А и В, и среднему захвату, соответствующему приблизительно 600 RU, на анти-пента His поверхности для серии С.Первый лигандный канал оставляли пустым, чтобы в случае необходимости использовать в качестве контроля. За стадией захвата гетеродимеров незамедлительно проводили две инжекции буфера при 100 мкл/мин в течение 30 сек каждая в направлении аналита, чтобы стабилизировать исходный уровень, а затем одновременно инжектировали 60 нМ, 20 нМ, 6,7 нМ, 2,2 нМ и 0,74 нМ антигена (TF) вместе с пустым буфером при 50 мкл/мин в течение 120 сек с фазой диссоциации в 600 сек. Поверхности с захваченными антителами регенерировали при помощи впрыскивания 0,85% фосфорной кислоты в течение 18 сек при 100 мкл/мин, чтобы подготовить к следующему циклу инжекции. Проводили выравнивание и двойной контроль сенсограмм, используя инжекцию с пустым буфером и участки между областями локализации белка, а полученные в результате сенсограммы анализировали при помощи программного обеспечения ProteOn Manager v3.1. После проведения двойного контроля сенсограммы аппроксимировали 1:1 моделью связывания. Величины Rmax для каждого антигена нормировали относительно уровней захвата антител для каждого варианта и сравнивали с 100% контролями.
[0257] Величины аффинности в отношении антигена (KD) для образцов гетеродимеров Fab приведены в Таблицах 12, 13b и 14b. Величины KD для H1:L1 Fab в контексте конструкции LCCA (KD, диапазон величин KD и изменение медианных величин KD по сравнению с диким типом) приведены в колонках 5, 6 и 7, соответственно, Таблицы 12. Те же самые величины KD также включены в контексте пар конструкций в Таблицы 13b и 14b в колонки 3 (KD гетеродимера Fab H1-L1), 4 (изменение KD гетеродимера Fab H1-L1 по сравнению с диким типом), 5 (KD гетеродимера Fab H2-L2) и 6 (изменение KD гетеродимера Fab H2-L2 по сравнению с диким типом). Величины KD определяли только для образцов гетеродимеров Fab, которые демонстрировали захват гетеродимеров Fab, соответствующий по меньшей мере 100 RU. Контрольная KD для дикого типа (0,157 нМ) отображает медианное значение гетеродимера Fab, в котором легкая цепь содержит метку FLAG. Гетеродимеры Fab дикого типа (содержащие метку FLAG или НА), демонстрировали сходные величины KD так, что между максимальной и минимальной величинами наблюдали 2,6-кратную разницу. В Таблицах 12, 13b и 14b показана разница в KD по сравнению с аффинностью связывания антигена дикого типа с использованием расчета -(log(KD)конструкция - log(KD)дт), таким образом, положительные значения указывают на более низкие величины KD, в то время как отрицательные значения указывают на повышенные величины KD гетеродимера Fab по сравнению с аффинностью связывания дикого типа в отношении антигена. Следует отметить, что для некоторых гетеродимеров Fab отсутствуют измеренные величины KD. В некоторых из этих случаев оценку гетеродимеров Fab проводили, но эксперименты SPR продемонстрировали низкий захват гетеродимеров Fab (т.е. менее 100 RU), и, следовательно, точное определение величин KD было невозможным. Для тех гетеродимеров Fab, которые демонстрируют сходство с другими гетеродимерами Fab (т.е. отличаются только присутствием/отсутствием или идентичностью присоединенной метки L-цепи (НА или FLAG)), вместо этого приведены оценочные величины KD (как отмечено в Таблице 12, 13b и 14b), соответствующие величинам KD для сходных гетеродимеров Fab. Соответствующая оценочная величина KD дикого типа (0,15 нМ) представляла собой медианную величину, полученную по всем конструкциям гетеродимеров Fab дикого типа (т.е. конструкциям Fab, содержащим метку НА или метку FLAG). В общем случае результаты свидетельствуют о том, что правильно спаренные гетеродимеры (с точки зрения конструкции) демонстрируют сходную с диким типом аффинность связывания в отношении антигена (в пределах приблизительно 2,3 раз относительно контрольной аффинности дикого типа).
Пример 8. Профили сверхэффективной жидкостной хроматографии и эксклюзионной хроматографии (UPLC-SEC) меченых гетеродимеров D3H44 дикого типа и преимущественно спаренных гетеродимеров
[0258] Гетеродимеры D3H44 дикого типа (одна тяжелая цепь и одна легкая цепь) с С-концевой меткой ABD2-His6 на тяжелой цепи и N-концевой меткой (FLAG в одной конструкции и НА в другой конструкции) на легкой цепи экспрессировали и очищали в соответствии с известными в данной области техники способами, аналогичными описанным в Примере 5. Преимущественно или правильно спаренные гетеродимеры масштабировали в индивидуальном порядке и очищали при помощи аффинной очистки с применением метки His, как описано в Примере 5.
[0259] UPLC-SEC проводили, используя SEC-колонку Waters ВЕН200 (2,5 мл, 4,6×150 мм, нержавеющая сталь, 1,7 мкм частицы), установленную на 30°С и присоединенную к UPLC-системе Waters Acquity с PDA-детектором. Время прогона составляло 7 мин с общим объемом на инжекцию, составляющим 2,8 мл, с подвижным буфером Hyclone DPBS/модифицированный -кальций -магний (часть № SH30028.02) при 0,4 мл/мин. Элюирование отслеживали по УФ-поглощению в диапазоне 200-400 нм, а хроматограммы получали на 280 нм. Интегрирование пиков проводили при помощи программного обеспечения Empower 3.
[0260] На Фигуре 6 приведены профили UPLC-SEC для типовых пар гетеродимеров Fab ДТ (содержащих метку FLAG на L-цепи), а также типовых (компонент H1L1 Fab конструкций LCCA 9735, 9737 и 9740) сконструированных пар гетеродимеров Fab. В общем случае сконструированные пары гетеродимеров Fab демонстрировали сходные с ДТ профили UPLC-SEC.
Пример 9: Оценка преимущественного спаривания гетеродимеров в коэкспрессионных группах, содержащих модификации константного домена или константного и вариабельного домена в формате биспецифического антитела
[0261] Оценивали конструкции гетеродимеров, чтобы определить, обеспечивают ли они возможность преимущественного спаривания в формате биспецифического антитела. В этом примере для стимуляции гетеродимеризации уникальных тяжелых цепей ассиметрично модифицировали Fc-область полноразмерной тяжелой цепи каждого гетеродимера так, чтобы одна тяжелая цепь содержала мутации T350V, L351Y, F405A и Y407V, а другая тяжелая цепь содержала мутации T350V, T366L, K392L и T394W (нумерация EU).
Получение конструкций:
[0262] Гетеродимерные конструкции исследовали в контексте следующих биспецифических антител: а) D3Н44/трастузумаб, b) D3Н44/цетуксимаб и с) трастузумаб/цетуксимаб. Следует отметить, что D3H44 является человеческим антителом, трастузумаб является гуманизированным антителом, а цетуксимаб является химерным антителом, состоящим из областей человеческого IgG1 и мышиного Fv. Конструкции, кодирующие тяжелые и легкие цепи IgG D3H44, трастузумаба и цетуксимаба, содержащие аминокислотные модификации в соответствии с конструкцией, получали следующим образом. Базовые последовательности ДНК для тяжелых и легких цепей D3H44, трастузумаба и цетуксимаба приведены в Таблице 3С. Последовательности легких цепей D3H44, трастузумаба и цетуксимаба получали, как описано в Примере 3, за исключением того, что в некоторых последовательностях отсутствовала метка, в то время как другие последовательности содержали метку FLAG или НА. Последовательности тяжелых цепей D3H44, трастузумаба и цетуксимаба получали, как описано в Примере 3, за исключением того, что полноразмерные тяжелые цепи создавали путем введения последовательности ДНК IgG1*01, кодирующей домены шарнирной области-СН2-СН3 и модифицированной, чтобы стимулировать гетеродимеризацию, на С-конце домена СН1 тяжелых цепей Fab. Следует отметить, что канонический С-концевой остаток лизина в тяжелой цепи был удален для предотвращения гетерогенности сигнала LC-MS вследствие отсечения С- концевого лизина (Lawrence W. Dick Jr. et al., Biotechnol. Bioeng. (2008) 100:1132-43).
Формат анализа (SMCA)
[0263] Способность наборов конструкций с коэкспрессией гетеродимеров к преимущественному спариванию с образованием биспецифического антитела оценивали, как описано ниже. Данный анализ основан на коэкспрессии четырех цепей (цепи H1 и L1 от одного антитела, а цепи Н2 и L2 от другого антитела) и выявлении наличия правильно образованного биспецифического антитела методом масс-спектрометрии (LC-MS). На Фигуре 8 приведена схема, иллюстрирующая четыре стартовые полипептидные цепи и потенциальные продукты, получаемые в результате коэкспрессии этих стартовых полипептидных цепей в отсутствие преимущественного спаривания между тяжелыми и легкими цепями пар гетеродимеров. Две полноразмерные конструкции тяжелых цепей коэкспрессировали с двумя уникальными конструкциями легких цепей, что приводило к получению десяти возможных видов антител: H1-L1:H1-L1, H1-L2:H1-L2, H1-L1:H1-L2, H2-L1:H2-L1, H2-L2:H2-L2, H2-L1:H2-L2, H1-L1:H2-L1, H1-L2:H2-L2, H1-L2:H2-L1 и Н1-L1:H2-L2. Вид H1-L1:H2-L2 представляет собой правильно спаренное биспецифическое антитело (см. Фигуру 8). После очистки на рА и дегликозилирования при помощи LC-MS определяли относительную специфичность спаривания в терминах количества предпочтительного вида H1-L1:H2-L2 по сравнению с другими. По возможности цепи оставляли немечеными, при условии, что все виды Mab и полу-Ab отличались друг от друга по меньшей мере на 50 Да. Когда разница в массе исключала такую возможность, к легким цепям добавляли N-концевые метки (НА или FLAG), чтобы обеспечить достаточную массовую дифференциацию между видами.
[0264] Этот метод анализа, включающий стадии экспрессии и скрининга биспецифического антитела, называется SMCA.
Метод масс-спектрометрии
[0265] Степень преимущественного спаривания легкой цепи D3H44 с тяжелой цепью D3H44 в группах коэкспрессии оценивали при помощи масс-спектрометрии после очистки на протеине А и неденатурирующего дегликизилирования. Так как гетеродимеры D3Н44/трастузумаб содержали только N-связанные гликаны Fc, эту систему обрабатывали только одним ферментом, N-гликозидазой F (PNGазой-F). Очищенные образцы дегликозилировали при помощи PNGазы-F следующим образом: 0,2 Е PNGазы-F/мкг антитела в 50 мМ Трис-HCl, рН 7,0, инкубация в течение ночи при 37°С, конечная концентрация белка 0,5 мг/мл. В случае систем D3Н44/цетуксимаб и трастузумаб/цетуксимаб вследствие наличия дополнительного N-связанного гликана в области Fab цетуксимаба, эти системы обрабатывали N-гликозидазой F плюс некоторым количеством экзогликозидаз. Как правило, в этих целях использовали смесь из четырех ферментов: N-гликозидазы F, β-галактозидазы (Prozyme), β-N-ацетилглюкозаминидазы (New England Biolabs) и нейраминидазы. N-гликозидаза F удаляет N-связанные гликаны Fc, в то время как экзогликозидазы упорядочивают N-связанные гликаны Fab в равномерную коровую структуру M3F (GlcNAc2Man3Fuc1). Очищенные образцы дегликозилировали при помощи смеси из четырех ферментов следующим образом: 0,2 Е PNGазы-F/MKr антитела, 0,002 Е α-нейраминидазы/мкг антитела, 0,0001 Е β- галактозидазы/мкг антитела и 0,2 Е β-N-ацетилглюкозаминидазы/мкг антитела в 50 мМ Трис-HCl, рН 7,0, инкубация в течение ночи при 37°С, конечная концентрация белка 0,5 мг/мл. При этом в некоторых случаях преимущественной являлась обработка тремя ферментами (N-гликозидазой F, β-галактозидазой и нейраминидазой), чтобы избежать перекрытия масс компонентов образца при проведении анализа LC-MS. В этих случаях гликаны Fab окисляли до немного более крупной структуры G0F (Man3GlcNAc2Fuc1GlcNAc2). Очищенные образцы дегликозилировали при помощи смеси из трех ферментов, используя такие же концентрации и условия, что описаны для смеси из четырех ферментов. После дегликозилирования образцы хранили при 4°С перед проведением анализа LC-MS.
[0266] Образцы дегликозилированного белка анализировали методом интактной LC-MS с использованием ВЭЖХ-системы Agilent 1100, соединенной с масс-спектрометром LTQ-Orbitrap XL (ThermoFisher Scientific) посредством источника ионов для электрораспыления Ion Max (ThermoFisher). Образцы (5 мкг) инжектировали в обращенно-фазовую колонку Poros R2, 2,1×30 мм (Applied Biosystems), и разделение проводили, используя следующие градиентные условия: 0-3 мин: 20% растворитель В; 3-6 мин: 20-90% растворитель В; 6-7 мин: 90-20% растворитель В; 7-9 мин: 20% растворитель В. Растворитель А представлял собой дегазированный 0,1% раствор муравьиной кислоты, а растворитель В представлял собой дегазированный ацетонитрил. Скорость потока составляла 3 мл/мин. После прохождения колонки поток разделяли, чтобы направить 100 мкл на поверхность раздела для электрораспыления. Колонку нагревали до 82,5°С, а растворители нагревали перед подачей в колонку до 80°С для улучшения формы белковых пиков. LTQ-Orbitrap XL калибровали при помощи калибровочного раствора ThermoFisher Scientific's LTQ Positive Ion ESI (кофеин, MRFA и Ultramark 1621) и настраивали, используя 10 мг/мл растворы С si. Напряжение на конусе (настройки для фрагментации источника) составляло 40 В, разрешение FT составляло 7500, а диапазон сканирования соответствовал соотношению масса/заряд 400-4000. LTQ-Orbitrap XL настраивали для оптимальной фиксации крупных белков (>50 кДа).
[0267] Деконволюцию диапазонов, содержащих множество заряженных ионов из полноразмерных антител (масса/заряд 2000-3800) и полуантител (масса/заряд 1400-2000), в профили молекулярной массы проводили, используя модуль MaxEnt 1 MassLynx, программное обеспечение для управления инструментом и анализа данных (Waters). Вкратце, исходные данные LC-MS по белку сначала открывали в QualBrower, модуле для просмотра спектров в Xcalibur (Thermo Scientific) и конвертировали для совместимости с MassLynx, используя Databridge, программу для конвертации файлов, предоставляемую Waters. Переконвертированные спектры белка просматривали в спектральном модуле MassLynx и проводили деконволюцию, используя MaxEnt 1. Количество разных видов антител в каждом образце определяли непосредственно из полученных профилей молекулярной массы.
Типовые конструкции для анализа SMCA
[0268] Всего из кластеров 1-12 было отобрано 25 типовых конструкций со средними величинами показателей LCCA для исследования в формате SMCA. Типовые конструкции выбирали на основании соответствующих наборов конструкций, занимающих аналогичное пространство, использующих аналогичные драйверы, а также содержащих аналогичные мутации. Из каждого кластера была выбрана по меньшей мере одна типовая конструкция. Некоторые кластеры были представлены одной типовой конструкцией (т.е. кластеры 1, 5, 7, 8, 10). Оставшиеся кластеры имели более одной типовой конструкции, так как кластеры либо были крупными (т.е. кластер 2), либо состояли из меньших кластеров (т.е. кластеры 3, 4, 6, 9, 11 и 12). Хотя конструкции в пределах каждого кластера характеризовались сходством последовательностей, меньшие кластеры в пределах кластера отличались по меньшей мере одним набором драйверных мутаций. В случае кластеров, которые состояли из меньших кластеров, из каждого меньшего кластера отбирали дополнительные типовые конструкции.
[0269] В Таблицах 15-27 перечислены аминокислотные замены для каждого из кластеров и указаны соответствующие типовые представители для каждого кластера/меньшего кластера. В случае кластера 1, кластер представляет только одна конструкция (9134-9521), так как в этих конструкциях используются аналогичные электростатические драйверы, занимающие аналогичное пространство (см. Таблицу 15). Для всех представителей этого кластера HI конструировали так, чтобы обеспечить возможность образования отрицательно заряженными заменами (L124E и Q179E) солевых мостиков с положительно заряженными заменами LI (S176R и любой из S131K или S131R). Н2 конструировали так, чтобы обеспечить возможность образования положительно заряженными заменами (L124R и любой из Q179K или S186K) солевых мостиков с отрицательно заряженными заменами L2 (S176D и любой из T178D или Т178Е и/или Т180Е). Неправильное спаривание H1L2 и H2L1 будет невыгодным главным образом вследствие электростатического отталкивания.
[0270] В случае кластера 2 были выбраны две типовые конструкции (9279-9518 и 9286-9402), чтобы представлять крупный кластер (см. Таблицу 16). В конструкциях в пределах этого кластера используются аналогичные электростатические драйверы, занимающие аналогичное пространство. Для всех представителей этого кластера H1 конструировали так, чтобы обеспечить возможность образования отрицательно заряженными заменами (L124E и L143E или L143D) солевых мостиков с положительно заряженными заменами LI (S176R и комбинации любых из (Q124K и/или Т178K) или (Q124K и Q160K)). Н2 конструировали так, чтобы обеспечить возможность образования положительно заряженными заменами (L124R и Q179K или S186K или S186R) солевых мостиков с отрицательно заряженными заменами L2 (S176D и T178D или Т178Е и/или Т180Е). Неправильное спаривание H1L2 и H2L1 будет невыгодным главным образом вследствие электростатического отталкивания.
[0271] В случае кластера 3 были выбраны пять типовых конструкций (9338-9748, 9815-9825, 6054-9327, 9066-9335 и 9121-9373), чтобы представлять каждый из пяти меньших кластеров (см. Таблицу 17). Во всех представителях этого кластера используются аналогичные электростатические драйверы в H1 (L124E), L1 (S176R), Н2 (L124R) и L2 (S176D), которые обеспечивают возможность образования солевых мостиков в преимущественно спаренных гетеродимерах, в то время как неправильные пары будут невыгодны главным образом вследствие электростатического отталкивания. Чтобы представить те конструкции, в которых используются главным образом данные драйверы константной области, для представления данного меньшего кластера была выбрана конструкция 6054-9327. Кроме этих электростатических взаимодействий один меньший кластер также содержал стерический драйвер вариабельной области (H1 L45P и L1 P44F), следовательно, для представления этого меньшего кластера был выбран типовой представитель, содержащий этот драйвер вариабельной области, (9338-9748). Другой меньший кластер также содержал электростатические драйверы вариабельной области в обоих гетеродимерах Fab (H1 Q39E, L1 Q38R, Н2 Q39R, L2 Q38E), следовательно, для представления этого меньшего кластера был выбран типовой представитель, содержащий этот драйвер вариабельной области, (9815-9825). Кроме того, один меньший кластер состоял из одного представителя, и, следовательно, одна типовая конструкция (9066-9335) содержит сконструированную дисульфидную связь между H1 F122C и L1 Q124C. В оставшемся меньшем кластере, представляемом 9121-9373, использовались главным образом драйверы константной области с дополнительными заменами Н172Т в H1 и S174R в L1, чтобы немного модифицировать взаимодействие драйверов константной области H1L1, а также исследовать влияние H172R в НС2.
[0272] В случае кластера 4 были выбраны две типовые конструкции (9168-9342 и 9118-6098), чтобы представлять каждый их двух меньших кластеров (см. Таблицу 18). Во всех представителях этого кластера используются аналогичные электростатические драйверы в H1 (L124E), L1 (S176R или S176K), Н2 (L124R) и L2 (S176D), которые обеспечивают возможность образования солевых мостиков в преимущественно спаренных гетеродимерах, в то время как неправильные пары будут невыгодны главным образом вследствие электростатического отталкивания. В одном меньшем кластере, представляемом 9118-6098, главным образом используются общие электростатические драйверы для преимущественного спаривания, в то время как в другом меньшем кластере, представляемом 9168-9342, дополнительно используются замены в H1 (K228D) и L1 (S121K), которые обеспечивают возможность образования дополнительного солевого мостика.
[0273] Кластер 5, представляемый уникальным идентификационным номером 9116-9349, состоит только из 1 представителя (см. Таблицу 19). В этой конструкции используются как электростатические драйверы в H1 (L124E), L1 (S176R), Н2 (L124R) и L2 (S176D), так и стерические драйверы в H1 (A139W), L1 (F116A_V133G_L135V), Н2 (A139G_V190A) и L2 (V133G_L135W). В результате, в случае преимущественно спаренных гетеродимеров заряженные замены также будут обеспечивать возможность образования солевых мостиков. Образование неправильно спаренных гетеродимеров Fab будет невыгодным вследствие электростатического отталкивания, а также дополнительных стерических эффектов.
[0274] В случае кластера 6 были выбраны две типовые конструкции, чтобы представлять каждый из двух меньших кластеров (см. Таблицу 20). Во всех представителях этого кластера используются аналогичные электростатические драйверы в константной области (Q179E в H1, S131K в L1, S186R в Н2 и Q124E, Q160E и Т180Е в L2), которые обеспечивают возможность образования солевых мостиков в преимущественно спаренных гетеродимерах, в то время как неправильные пары будут невыгодны главным образом вследствие электростатического отталкивания. Кроме того, меньшие кластеры также состояли из разных драйверов вариабельной области. В одном меньшем кластере, представляемом уникальным идентификационным номером 9814-9828, используются электростатические драйверы в вариабельных областях (Q39E в H1, Q38R в L1, Q39R в Н2 и Q38E в L2). В другом меньшем кластере используется стерический драйвер вариабельной области, состоящий из L45P в H1 и P44F в L1. В результате, в случае этого меньшего кластера неправильные пары будут дополнительно невыгодны вследствие привнесенных стерических эффектов. Следует отметить, что этот меньший кластер представлен конструкцией, полученной из уникального идентификационного номера 9745-9075, которая отличается только отсутствием Q38E в L2.
[0275] В случае кластера 7 была выбрана только одна конструкция (9060-9756), чтобы представлять кластер, так как в этих конструкциях используются аналогичные электростатические и стерические драйверы (см. Таблицу 21). Общие электростатические драйверы включают L143E и Q179E в H1, Q124R в L1, Q179K в Н2 и Q124E, Q160E и Т180Е в L2. Общие стерические драйверы включают A139W в H1, F116A_L135V в L1 и L135W в L2. В результате, в случае преимущественно спаренных гетеродимеров заряженные замены в областях Fab будут обеспечивать возможность образования солевых мостиков. Образование неправильно спаренных гетеродимеров будет невыгодным вследствие электростатического отталкивания, а также дополнительных стерических эффектов.
[0276] В случае кластера 8 была выбрана только одна конструкция (9820-9823), чтобы представлять кластер, так как в этих конструкциях используются аналогичные электростатические драйверы (см. Таблицу 22). В вариабельной обрасти используются Q39E в H1, Q38R в L1, Q39R в Н2 и Q38E в L2. В константной области используются L143E в H1, Q124R, Q160K и T178R в L1, Q179K в Н2 и Q124E, Q160E и Т180Е в L2. В случае преимущественно спаренных гетеродимеров заряженные замены в областях Fab будут обеспечивать возможность образования солевых мостиков, в то время как образование неправильно спаренных гетеродимеров будет невыгодным главным образом вследствие электростатического отталкивания.
[0277] В случае кластера 9 были выбраны две типовые конструкции, чтобы представлять каждый из двух меньших кластеров (см. Таблицу 23). Во всех представителях этого кластера используются аналогичные электростатические драйверы в константной области (L143E в H1, Q124R в L1, Q179K в Н2 и Q124E, Q160E и Т180Е в L2), которые обеспечивают возможность образования солевых мостиков в преимущественно спаренных гетеродимерах, в то время как неправильные пары будут невыгодны главным образом вследствие электростатического отталкивания. Кроме того, меньшие кластеры отличаются присутствием/отсутствием драйвера вариабельной области (L45P в H1 и P44F в L1). В результате, для меньшего кластера, содержащего драйвер вариабельной области, неправильные пары будут дополнительно невыгодны вследствие привнесенных стерических эффектов. Типовая конструкция для этого меньшего кластера, содержащая драйвер вариабельной области, была получена из уникального идентификационного номера 9751-9065, который отличается только отсутствием Q38E в L2. Типовой конструкцией для меньшего кластера, в которой отсутствуют замены в вариабельной области, является 9611-9077.
[0278] В случае кластера 10 была выбрана только одна конструкция (9561-9095), чтобы представлять кластер, так как в этих конструкциях используются аналогичные электростатические и стерические драйверы, занимающее аналогичное пространство (см. Таблицу 24). Общие электростатические драйверы включают L143E и Q179E в H1, аналогично расположенные Q124R, Q124K или S131K в L1, Q179K в Н2 и Q124E и Т180Е в L2. Общие стерические драйверы включают L124W BH1.V133ABL1 и V133WBL2. B результате, в случае преимущественно спаренных гетеродимеров заряженные замены в областях Fab будут обеспечивать возможность образования солевых мостиков. Образование неправильно спаренных гетеродимеров будет невыгодным вследствие электростатического отталкивания, а также дополнительных стерических эффектов.
[0279] В случае кластера 11 были выбраны три конструкции (9049-9759, 9682-9740 и 9667-9830), чтобы представлять каждый из трех меньших кластеров (см. Таблицу 25). Во всех представителях этого кластера использовались электростатические замены для стимуляции преимущественного спаривания гетеродимеров. В результате, в случае преимущественно спаренных гетеродимеров заряженные замены в областях Fab будут обеспечивать возможность образования солевых мостиков. Образование неправильно спаренных гетеродимеров будет невыгодным главным образом вследствие электростатического отталкивания. В случае меньшего кластера, представляемого уникальным идентификационным номером 9667-9830, общие замены включают отрицательно заряженные замены (L143E или L143D и Q179E или Q179D) в H1, положительно заряженные замены (T178R или T178K) в L1, положительно заряженные замены (S186K или S186R или Q179K или Q179R) в Н2 и отрицательно заряженные замены (Q124E) в L2. Другой меньший кластер, представляемый единственным представителем этого меньшего кластера (уникальный идентификационный номер 9049-9759), дополнительно содержал замены для образования сконструированной дисульфидной связи. В оставшемся кластере, представляемом уникальным идентификационным номером 9682-9740, использовались такие же драйверы для H1 и L1, что и в других двух меньших кластерах; при этом использовались разные драйверы константной области Н2 и L2. В Н2 использовалась L143R или L143K, а в L2, кроме замены Q124E (общей с другими двумя меньшими кластерами), использовалась V133E или V133D.
[0280] В случае кластера 12 были выбраны четыре конструкции (9696-9848, 9986-9978, 9692-9846 и 9587-9735), чтобы представлять каждый из четырех меньших кластеров (см. Таблицу 26). Во всех представителях этого кластера использовались электростатические замены для стимуляции преимущественного спаривания гетеродимеров. В некоторых представителях дополнительно использовались стерические драйверы. В меньшем кластере, представляемом уникальным идентификационным номером 9696-9848, использовались как электростатические, так и стерические драйверы. Общие электростатические замены в пределах этого меньшего кластера состояли из L143E в H1, Q124R и T178R в L1, аналогично расположенных S186K или S186R, или Q179K, или Q179R в Н2 и Q124E и Т180Е в L2. Общие стерические замены в пределах этого меньшего кластера состояли из S188L в H1 и любой из S176L или V133Y, или V133W в L2; в случае конструкций, в которых использовались V133Y или V133W в L2, в Н2 также присутствовали L143A или L124A для размещения объемных мутаций. В случае меньшего кластера, представляемого уникальным идентификационным номером 9692-9846, использовались такие же электростатические драйверы, что в и меньшем кластере, представляемом уникальным идентификационным номером 9696-9848; в случае некоторых представителей вместо Т180Е использовалась аналогично расположенная замена Т178Е в L2. Кроме того, в подгруппе из этого меньшего кластера также использовались аналогичные стерические драйверы с аналогично расположенной заменой T178Y или T178F вместо S176L в L2. В меньшем кластере, представляемом уникальным идентификационным номером 9986-9978, использовались только электростатические драйверы для стимуляции преимущественного спаривания. Аналогичные общие замены использовались для H1, L1 и Н2; при этом использовалась отличная замена в L2 (S131E). В оставшемся меньшем кластере, представляемом уникальным идентификационным номером 9587-9735, использовались аналогичные электростатические драйверы в H1 и L1 (за исключением того, что во всех представителях в пределах этого меньшего кластера не использовалась T178R в L1); при этом разные электростатические драйверы использовались для Н2 (L143R или L143K) и L2 (Q124E и V133E или Q124E и V133D). В паре представителей этого меньшего кластера также использовались аналогичные стерические драйверы, состоящие из S188L в H1 и S176L в L2. В целом, в случае преимущественно спаренных гетеродимеров заряженные замены в областях Fab будут обеспечивать возможность образования солевых мостиков. Образование неправильно спаренных гетеродимеров будет невыгодным главным образом вследствие электростатического отталкивания. Кроме того, в случае конструкций, в которых также использовались стерические драйверы, их образование будет дополнительно невыгодным вследствие стерических эффектов. Кластер 13 состоит из одного представителя, 9122-9371 (см. Таблицу 27). В этой конструкции используется сконструированный дисульфидный мостик между H1 F122C и L1 Q124C в качестве ковалентного драйвера для преимущественного спаривания гетеродимеров. Кроме того, так как в конструкции также отсутствует природная внутрицепьевая дисульфидная связь, образование дисульфидной связи подтверждали при помощи невосстанавливающего и восстанавливающего SDS-PAGE. Эту конструкцию не исследовали в формате SMCA; при этом сконструированную дисульфидную связь исследовали в присутствии природной внутрицепьевой дисульфидной связи и в комбинации с дополнительными драйверами константной области (кластер 3, типовая конструкция 9066-9335).
Метод трансфекции
[0281] Коэкспрессионные наборы, содержащие конструкции с двумя тяжелыми цепями и двумя легкими цепями, трансфицировали в клетки СНО-3Е7 следующим образом. Клетки СНО-3Е7 с плотностью 1,7-2×106 клеток/мл культивировали при 37°С в среде FreeStyle(TM) F17 (Invitrogen cat# A-1383501), дополненной 4 мМ глутамина и 0,1% плюроника F-68 (Invitrogen cat# 24040-032). Общий объем, составляющий 50 мл, трансфицировали общим количеством ДНК, составляющим 50 мкг, при помощи PEI-pro (Polyplus cat# 115-010) с соотношением ДНК:PEI 1:2,5. Через двадцать четыре часа после добавления смеси ДНК-PET клетки переносили в 32°С и инкубировали в течение 7 дней перед сбором. Культуральную среду собирали путем центрифугирования и подвергали вакуумной фильтрации при помощи 0,2 мкМ фильтра Steriflip. Затем отфильтрованную культуральную среду очищали при помощи смолы с протеином A Mab Select SuRe (GE Healthcare #17-5438-02) следующим образом. Отфильтрованную культуральную среду применяли к колонке (Ну clone DPBS/модифицированный, без кальция, без магния, #SH-300028.02), которая была предварительно уравновешена PBS. Затем молекулы гетеродимерных антител промывали PBS и элюировали 100 мМ цитрата, рН 3,6, в центрифужном фильтре Amicon ultra 15 Ultracel 10K (Millipore #SCGP00525). Затем буфер меняли на ФБС, а образцы оценивали при помощи калипера перед проведением дегликозилирования и LC-MS.
[0282] Чтобы оценить ошибки биспецифической системы, свойственные системам биспецифических Ab дикого типа, в которых легкая цепь одной системы преимущественно связывается с тяжелыми цепями обоих Ab-систем, исследовали ряд соотношений ДНК H1:H2:L1:L2 в экспрессионных системах СНО. Эти соотношения предназначены для компенсации естественной разницы в уровнях экспрессии и/или природных ошибок спаривания между тяжелыми и легкими цепями двух разных антител. Для всех биспецифических Ab-систем наблюдали ошибки среди всех исследуемых соотношений (Фигура 9). В случае системы D3H44/трастузумаб наблюдали ошибку в отношении трастузумаба, т.е. тяжелая цепь D3H44 преимущественно спаривалась с легкой цепью трастузумаба (см. Фигуру 9а). В случае D3H44/цетуксимаб наблюдали ошибку в отношении цетуксимаба, т.е. тяжелая цепь D3H44 преимущественно спаривалась с легкой цепью цетуксимаба (см. Фигуру 9b). В случае системы трастузумаб/цетуксимаб наблюдали ошибку в отношении трастузумаба, т.е. тяжелая цепь цетуксимаба преимущественно спаривалась с легкой цепью трастузумаба (см. Фигуру 9с).
[0283] При исследовании каждой из 25 типовых конструкций в пределах каждой биспецифической Ab-системы соотношения ДНК H1:H2:L1:L2 представляли собой соотношения для соответствующих биспецифических систем дикого типа, которые приводили к получению наибольшего количества молекул биспецифических Ab, характеризуясь при этом небольшим количеством полу-Ab (см. Таблицы 32а, b и с). В случае системы D3H44/трастузумаб применяемое соотношение соответствовало 15 (H1), 15 (Н2), 53 (L1), 17 (L2), при этом H1 и L1 относятся к D3H44, а Н2 и L2 относятся к трастузумабу. В случае системы трастузумаб/цетуксимаб применяемое соотношение соответствовало 15 (H1), 15 (H2), 17 (L1), 53 (L2), при этом H1 и L1 относятся к трастузумабу, а H2 и L2 относятся к цетуксимабу. В случае системы D3H44/цетуксимаб применяемое соотношение соответствовало 15 (H1), 15 (H2), 53 (L1), 17 (L2), при этом H1 и L1 относятся к D3H44, а H2 и L2 относятся к цетуксимабу.
[0284] Кроме этого, конструкции исследовали в обеих ориентациях в отношении биспецифических систем D3H44/цетуксимаб и трастузумаб/цетуксимаб так, что в одной ориентации замены, присутствующие в H1L1 и H2L2, исследовали на антителе 1 (Ab1) и антителе 2 (Ab2) биспецифической Ab-системы, соответственно, а в «зеркальной» ориентации замены, присутствующие в H1L1 и H2L2, исследовали на Ab2 и Ab1, соответственно (см. Таблицу 28а и b). «_1», приписанная к уникальному идентификационному номеру указывает на те конструкции, в которых замены в тяжелой цепи и ассоциированной с ней легкой цепи, которые приводят к более высокому результату LCCA для преимущественного спаривания (см. Таблицу 13а), вносили в антитело, в котором тяжелая цепь слабо конкурирует за ассоциированную с ней легкую цепь по сравнению с легкой цепью от другого антитела. «_2», приписанная к уникальному идентификационному номеру указывает на «зеркальную» ориентацию, в которой замены в тяжелой цепи и ассоциированной с ней легкой цепи, которые приводят к более высокому результату LCCA для преимущественного спаривания (см. Таблицу 13а), вносили в антитело, в котором тяжелая цепь сильнее конкурирует за ассоциированную с ней легкую цепь по сравнению с легкой цепью от другого антитела. В случае системы D3H44/трастузумаб конструкции исследовали только в ориентации «_1» (см. Таблицу 28с).
Результаты SMCA
[0285] Систему D3H44/трастузумаб обрабатывали только одним ферментом (PNGаза-F) и полностью дегликозилировали. В случае обработки несколькими ферментами присоединенные сахара в области Fab в общем случае были усечены до корового M3F (путем обработки четырьмя ферментами) или G0F (путем обработки 3 ферментами). В целом, в большинстве случаев дегликозилирующая обработка обеспечивала возможность выявления всех возможных разных молекул, определяемых LC-MS. Во многих случаях каждая молекула была представлена одним пиком LC-MS. Исключения составляли боковые пики, которые, вероятно, также соответствуют желаемым биспецифическим молекулам (возможные аддукты или гетерогенность, связанные с отщеплением лидерного пептида); однако, вследствие неопределенности боковых пиков вклад этих боковых пиков в биспецифические молекулы не учитывали. Кроме того, для некоторых конструкций в системах D3H44/цетуксимаб (3519_1, 3522_1) и трастузумаб/цетуксимаб (9748-9338_1) для молекул требовалось учитывать несколько пиков вследствие вариабельности присоединенной крупной маннозы. Все эти конструкции вносили участок гликозилирования в легкую цепь цетуксимаба. Следует отметить, что в некоторых случаях было невозможно различить некоторые малочисленные виды молекул (составляющие менее 5% от всех видов) вследствие малого различия в массе между видами (т.е. различия менее 50 Да). Кроме того, желаемые биспецифические молекулы, H1-L1_H2-L2, в общем случае невозможно экспериментально отличить на базе ЖХ/МС от неправильно спаренного типа: H1-L2_H2-L1. Следовательно, когда в таблицах приводится содержание биспецифических молекул, нельзя полностью исключить, что они не содержат этот тип неправильно спаренных молекул. При этом очень низкое содержание, наблюдаемое для таких видов, как Н1-L2_H1-L2 и H2-L1_H2-L1, а также полуантител H1-L2 и H2-L1, свидетельствует о незначительных, в случае наличия, примесях в биспецифических молекулах.
[0286] Данные LC-MS представлены в Таблицах 29а, 29b и 29с. Для сравнения, данные для дикого типа представлены в Таблицах 33а, 33b и 33с и обозначены как «NA» в колонке «уникальный идентификационный номер SMCA», а также в колонке «Кластер». Все три биспецифические системы дикого типа демонстрировали сдвиговые ошибки, когда наблюдалось доминирование одной легкой цепи в отношении обоих тяжелых цепей (см. Таблицы 33 и Фигуру 9). Кроме того, по меньшей мере в системе трастузумаб/цетуксимаб расположение метки также оказывало существенное влияние на спаривание H1L1 и H2L2. Следовательно, чтобы оценить влияние конструкций на переносимость и процентное содержание желаемых биспецифических молекул по сравнению с диким типом, проводили сравнение с соответствующей биспецифической конструкцией дикого типа при том же соотношении ДНК H1:H2:L1:L2, а результаты представляли в колонках «Изменение в % спаривания H1L1 (среди всех молекул H1) по отношению к дикому типу», «Изменение в % спаривания H2L2 (среди всех молекул Н2) по отношению к дикому типу» и «Изменение в % H1:H2:L1:L2 по отношению к дикому типу» (с учетом только полноразмерных молекул антител) (см. Таблицу 29). Следует отметить, что для оценки % спаривания H1L1 (среди всех молекул H1) или % спаривания H2L2 (среди всех молекул Н2), оценивали все молекулы в отношении спаривания в области Fab. Когда соответствующую биспецифическую конструкцию дикого типа не оценивали при помощи SMCA, сравнение проводили со сходной конструкцией дикого типа. Оценочные значения обозначены «***» рядом с приведенным значением. Сходную конструкцию дикого типа для проведения сравнения выбирали следующим образом. Чтобы оценить возможность переноса, каждая конструкция дикого типа была представлена экспериментом SMCA (проводимым при разных соотношениях), который демонстрировал самый высокий «% спаривания H1L1 и % спаривания H2L2 (среди всех молекул)». Чтобы оценить влияние конструкций на процентное содержание желаемых биспецифических молекул по сравнению с диким типом, каждая конструкция дикого типа была представлена экспериментом SMCA (проводимым при разных соотношениях), который демонстрировал самый высокий % H1:H2:L1:L2 (с учетом только полноразмерных молекул антител). Затем в обоих случаях из всех конструкций дикого типа в биспецифической системе выбирали медианные величины в качестве величин дикого типа для сравнения.
[0287] Переносимость для каждой конструкции оценивали, фиксируя повышение в общем спаривании H:L среди всех молекул по отношению к ДТ, в частности, в %H1L1/все молекулы H1 и/или %H2L2/Bce молекулы Н2. Кроме того, также оценивали влияние на процентное содержание желаемых биспецифических молекул, акцентируя внимание на сравнении только полноразмерных молекул антител, так как полуантитела, в случае наличия, могут быть удалены/минимизированы при помощи препаративной SEC или путем дополнительного титрования ДНК H1:H2:L1:L2. Таблицы 30a, b и с показывают, что препаративная SEC может быть эффективной в отношении удаления/минимизации молекул полу-Ab. Таблицы 32а, b и с показывают, что процентное содержание молекул полу-Ab также можно изменять во время трансфекции, используя разные титровальные соотношения ДНК.
[0288] В случае системы D3H44/цетуксимаб (Таблица 29а) происходил перенос всех конструкций за исключением одной (9327-6054_2) согласно оценке спаривания H1L1 среди всех молекул относительно дикого типа. Большинство конструкций (за исключением 9327-6054_2 и 9134-9521_2) также демонстрировали повышенное процентное содержание желаемых биспецифических антител с учетом только полных молекул Ab. Кроме того, за исключением одной конструкции, для которой не наблюдали перенос (9327-6054_2), конструкции снижали количество изначально неправильно спаренных молекул антител (H1H2L2L2), наблюдаемое для дикого типа. Кроме того, за исключением 9327-6054_2 и соответствующей конструкции 9327-6054_1 в другой ориентации, наблюдали перенос конструкций в обеих ориентациях, при этом большинство конструкций демонстрировали одинаково эффективное спаривание H:L в обеих ориентациях.
[0289] В случае системы D3H44/трастузумаб (Таблица 29b) происходил перенос всех конструкций согласно оценке спаривания H1L1 среди всех молекул относительно дикого типа. Кроме того, все конструкции демонстрировали повышенное процентное содержание желаемых биспецифических антител (с учетом только полных молекул Ab). Кроме того, большинство конструкций существенно снижали количество изначально неправильно спаренных молекул антител (H1H2L2L2), наблюдаемое для дикого типа. При этом следует отметить, что вследствие отсутствия экспрессии отсутствуют данные по 9611-9077_1 (Таблица 28с).
[0290] В случае системы трастузумаб/цетуксимаб (Таблица 29с) по меньшей мере 35 из 49 конструкций демонстрировали переносимость согласно оценке спаривания H2L2 среди всех молекул Н2 (положительные значения в колонке «Изменение % спаривания H2L2 (среди всех молекул Н2) относительно дикого типа»). Конструкции, которые не продемонстрировали переносимость, включают 9279-9518_2, 3522_2, 9815-9825_2, 9327-6054_2, 9118-6098_2, 9748-9338_2, 9692-9846_2, 9587-9735_2, 9814-9828_2, 3519_2, 9986-9978_2, 9168-9342_2 и 9066-9335_1 (отрицательные значения в колонке «Изменение % спаривания H2L2 (среди всех молекул Н2) относительно дикого типа»); при этом конструкции в другой ориентации не демонстрировали переносимость (следует отметить, что 9279-9518_1 не исследовали вследствие отсутствия образца). Все конструкции, которые демонстрировали переносимость, демонстрировали сниженное процентное содержание изначально неправильно спаренных молекул антител (H1H2L2L2), наблюдаемое в экспериментах для дикого типа. Кроме того, из конструкций, для которых наблюдали переносимость, только 2 конструкции (9134-9521_1 и 9279-9518_2) демонстрировали сниженное процентное содержание желаемых биспецифических Ab с учетом только полных молекул антител по сравнению с диким типом.
[0291] В общем случае большинство конструкций, которые повышали спаривание H:L более слабого конкурирующего антитела, приводили к повышению процентного содержания желаемых биспецифических антител (с учетом только полноразмерных антител). Что касается ориентации, большинство конструкций в ориентации «_1» демонстрировали сходную или лучшую переносимость, судя по спариванию H:L, по сравнению с ориентацией «_2» (за исключением изначально наблюдаемых в системе трастузумаб/цетуксимаб). Кроме того, в Таблице 35а перечислены те конструкции, для которых наблюдали переносимость в обеих ориентациях для всех 3 исследуемых биспецифических систем (D3H44/цетуксимаб, D3H44/трастузумаб и трастузумаб/цетуксимаб). В Таблице 35b перечислены те конструкции, для которых наблюдали переносимость в одной ориентации для всех 3 исследуемых биспецифических систем (D3H44/цетуксимаб, D3H44/трастузумаб и трастузумаб/цетуксимаб) и переносимость в другой ориентации только для одной биспецифической системы. Кроме того, в конкретной ориентации в тяжелой цепи и более слабой конкурирующей когнатной цепи присутствуют одинаковые мутации во всех 3 биспецифических системах, а использование легкой цепи превосходит по меньшей мере 10%.
[0292] Что до переносимости и показателей кластеров, в случае биспецифической системы D3H44/трастузумаб, все представители во всех кластерах демонстрировали переносимость (см. Фигуру 11 а) и повышенное процентное содержание желаемого биспецифического антитела (с учетом только полноразмерных антител) (см. Фигуру 11b). В случае биспецифической системы D3H44/цетуксимаб, все кластеры демонстрировали переносимость, и только один представитель из кластера 3 демонстрировал сниженное спаривание H1L1 среди всех молекул H1, по сравнению с диким типом (см. Фигуру 11 с). Также все кластеры содержали представителей, которые демонстрировали повышение процентного содержания желаемого биспецифического антитела по сравнению с диким типом (с учетом только полноразмерных антител); при этом 3 кластера (кластеры 1, 3 и 4) также содержали представителей, которые демонстрировали снижение процентного содержания желаемого биспецифического антитела по сравнению с диким типом (с учетом только полноразмерных антител) (см. Фигуру 11d). В случае биспецифической системы трастузумаб/цетуксимаб, все кластеры содержали варианты, которые демонстрировали переносимость конструкций; при этом только несколько кластеров (1, 5, 7, 8, 10, 11) содержали варианты, в которых все соответствующие представители демонстрировали переносимость (см. Фигуру 11е). Кроме того, все кластеры содержали представителей, которые демонстрировали повышенное процентное содержание желаемого биспецифического антитела по сравнению с диким типом (с учетом только полноразмерных антител) (см. Фигуру 11f). Для тех кластеров, в которых все представители демонстрировали переносимость, все представители в кластерах 5, 7, 8, 10 и 11 также демонстрировали повышение процентного содержания желаемого биспецифического антитела по сравнению с диким типом (с учетом только полноразмерных антител).
[0293] В целом, с учетом всех 3 биспецифических систем, все представители в кластерах 1, 5, 7, 8, 10 и 11 демонстрировали переносимость (см. Фигуру 11g); кластеры 5, 7, 8, 10 и 11 содержали представителей, среди которых все демонстрировали повышение процентного содержания желаемого биспецифического антитела по сравнению с диким типом (с учетом только полноразмерных антител) (см. Фигуру 11h).
[0294] В целом, большинство конструкций, которые повышали спаривание H:L более слабого конкурирующего антитела, приводили к повышению процентного содержания желаемых биспецифических антител (с учетом только полноразмерных антител). Что касается ориентации, большинство конструкций в ориентации «_1» демонстрировали сходную или лучшую переносимость, судя по спариванию H:L, по сравнению с ориентацией «_2» (за исключением изначально наблюдаемых в системе трастузумаб/цетуксимаб).
Пример 10: Препаративная эксклюзионная хроматография (SEC) выбранных биспецифических гетеродимерных антител SMCA и родительских Mab для получения биофизических характеристик
[0295] Подгруппа образцов SMCA была выбрана для получения дополнительных биофизических характеристик. Большинство из этих образцов SMCA обычно демонстрировали высокую степень спаривания (более ~80% спаривания в колонке H1L1+H2L2/Bce молекулы) и низкое количество молекул полуантител (менее ~30% с учетом всех молекул полуантител). Препаративную SEC проводили следующим образом. Проводили разделение образцов гетеродимерных антител, используя колонку Superdex 200 10/300 GL (GE Healthcare), установленную на системе Pharmacia (GE Healthcare)  Purifier. Образцы гетеродимерных антител (0,3-0,5 мл) в PBS (Hyclone DPBS/модифицированный, без кальция, без магния, номер в каталоге SH-300028.02) вручную загружали в 0,5 мл петлю, заполненную PBS. Затем образцы автоматически инжектировали в колонку и проводили разделение при 0,5 мл/мин с элюирующим объемом 1 CV. Элюирование белка отслеживали при ОП280 и собирали в 0,5 мл фракции. Для каждого образца SMCA объединяли те фракции, которые содержали основной пик, и получали дополнительные биофизические характеристики.
Purifier. Образцы гетеродимерных антител (0,3-0,5 мл) в PBS (Hyclone DPBS/модифицированный, без кальция, без магния, номер в каталоге SH-300028.02) вручную загружали в 0,5 мл петлю, заполненную PBS. Затем образцы автоматически инжектировали в колонку и проводили разделение при 0,5 мл/мин с элюирующим объемом 1 CV. Элюирование белка отслеживали при ОП280 и собирали в 0,5 мл фракции. Для каждого образца SMCA объединяли те фракции, которые содержали основной пик, и получали дополнительные биофизические характеристики.
Пример 11: Оценка преимущественного спаривания биспецифических гетеродимеров в формате антитела после препаративной эксклюзионной хроматографии
[0296] После препаративной SEC отобранные образцы анализировали в отношении преимущественного спаривания биспецифических гетеродимерных антител, используя метод LC-MS, как описано в Примере 9. Все эти образцы демонстрируют увеличение процентного содержания желаемых молекул биспецифических антител, а также снижение процентного содержания молекул полуантител (Таблицы 29 и 30).
Пример 12: Термостабильность биспецифических гетеродимерных антител SMCA
[0297] После препаративной SEC измеряли термостабильность отобранных биспецифических гетеродимерных антител SMCA и сравнивали с показателями родительских моноклональных антител D3H44 и трастузумаба, а также антитела с одним плечом - цетуксимаба. В общем случае антитела с одним плечом относятся к конструкциям, состоящим из одной полноразмерной тяжелой цепи, одной усеченной тяжелой цепи, в которой отсутствует область Fab (и внесена замена C233S), и одной легкой цепи, при этом гетеродимеризация тяжелой цепи достигается так, как описано в Примере 9.
Измерение термостабильности
[0298] Определяли термостабильность отобранных биспецифических гетеродимерных антител и контрольных антител дикого типа методом дифференциальной сканирующей калориметрии (DSC). После проведения препаративной SEC 400 мкл образцов с исходной концентрацией 0,2 мг/мл или 0,4 мг/мл в PBS использовали для анализа DSC с VP-Capillary DSC (GE Healthcare). В начале каждого эксперимента DSC проводили 5 инжекций пустого буфера, чтобы стабилизировать базовый уровень, а также для контроля инжекцию буфера проводили перед инжекцией каждого образца. Каждый образец исследовали от 20 до 100°С при скорости 60°С/ч, с низкой обратной связью, 8 сек фильтр, 5 мин preTstat и давление азота 70 фунтов/кв. дюйм. Полученные в результате термограммы обрабатывали и анализировали при помощи программного обеспечения Origin 7.
[0299] Результаты приведены в Таблицах 31a, b и с. Представленные в таблицах величины Tm для Fab дикого типа были получены для гомодимерных антител для D3H44 (79°С) и трастузумаба (81°С) и для антитела с одним плечом цетуксимаба (72°С). Для гетеродимерных антител ДТ D3H44/цетуксимаб и трастузумаб/цетуксимаб наблюдали только 2 пика, соответствующие Tm Fab. Отчетливые пики не наблюдали для СН2 (вследствие перекрытия с Fab цетуксимаба) или СН3 (вследствие перекрытия с величинами Tm для Fab D3H44 и трастузумаба). Для гетеродимерного антитела ДТ D3H44/трастузумаб величины Tm для двух Fab из D3H44 и трастузумаба одинаковы, пик на 81°С, вероятно, соответствует обоим Fab, в то время как пик приблизительно на 72°С, вероятно, соответствует СН2.
[0300] В Таблице 31а, b и с приведены только величины Tm для пиков, соответствующих обоим Fab, если не указано иное. Также следует отметить, что для некоторых гетеродимерных образцов концентрация белка была низкой (ниже 0,4 мг/мл), что приводило к повышенным шумам в базовом уровне. В результате в системе D3H44/трастузумаб некоторые образцы давали кривые DSC с низкими пиковыми интенсивностями, поэтому было затруднительно сделать различие между пиком СН2 и возможно дестабилизированным Fab. В этих случаях также приведены величины Tm для 70-72°С (Таблица 31а). В целом, большинство гетеродимерных антител демонстрировали термостабильность, сходную с соответствующими молекулами дикого типа (3°С или менее). Кроме того, большинство гетеродимерных антител не демонстрировали дополнительные пики, позволяющие предположить значительную дестабилизацию пиков СН2 или СН3. Единственное исключение составляет сконструированное гетеродимерное антитело 9611-9077_2 из системы трастузумаб/цетуксимаб, которое демонстрирует дополнительный пик на 60°С, который может появляться вследствие дестабилизации СН2.
Пример 13: Измерения антигенной аффинности биспецифических гетеродимерных антител
[0301] Способность биспецифических антител связываться с ассоциированными антигенами оценивали для определения, оказывали ли аминокислотные замены какое-либо влияния на связывание с антигеном. Аффинность связывания антигена определяли с помощью SPR следующим образом.
Биосенсорные анализы SPR
[0302] EDC: 1-этил-3-(3-диметиламинопропил) карбодиимида гидрохлорид; NHS: N-гидроксисукцинимид; SPR: поверхностный плазмонный резонанс; EDTA: этилендиаминтетрауксусная кислота; TF: тканевой фактор; EGFR ECD: внеклеточный домен рецептора эпидермального фактора роста; Her2 ECD: внеклеточный домен рецептора 2 человеческого эпителиального фактора роста.
[0303] Материалы для SPR. Сенсорный чип СМ5 S серии, набор Biacore для связывания амина (NHS, EDC и 1 М этаноламин) и буферы 10 мМ натрий ацетатный буферы были приобретены у компании GE Healthcare Life Science (Миссиссауга, штат Онтарио). Рекомбинантный белок Her2 внеклеточного домена (ECD) был приобретен у компании eBioscience (Сан Диего, штат Калифорния). Электродный буфер PBS с 1% Tween 20 (PBST) был приобретен у компании Teknova Inc. (Холлистер, штат Калифорния). Козьи поликлональные антитела к человеческому Fc были приобретены у компании Jackson Immuno Research Laboratories Inc. (Вест Гроув, штат Пенсильвания). EDTA была приобретена у компании Bishop (Берлингтон, штат Онтарио).
[0304] Все анализы поверхностного плазмонного резонанса проводились с использованием инструмента Biacore T200 Surface Plasmon Resonance (GE Healthcare Life Science (Миссиссауга, штат Онтарио)) с электродным буфером PBST (с добавлением маточного раствора 0,5 М EDTA до конечной концентрации 3,4 мМ) при температуре 25°С. Поверхность захвата к человеческому Fc была создана с использованием сенсорного чипа СМ5 S серии с использованием параметров по умолчанию в Мастере Иммобилизации в программном обеспечении системы управления Biacore T200, которое было установлено на целевые 2000 резонансных единиц (RU). Скрининг вариантов антитела для связывания с целевыми антигенами Her2 ECD, TF или EGFR ECD происходил в две стадии: непрямой захват вариантов антител на поверхности проточной ячейки с антителом к человеческому Fc с последующей инъекцией 5 концентраций очищенного антигена для кинетического анализа с использованием методологии одного цикла кинетики. Варианты или контроль для захвата вводили по 1 мкг/мл для отдельной проточной ячейки в течение отдельных 60 сек при скорости потока 10 мкл/мин. В целом, это привело к захвату приблизительно от 50 до 100 RU на поверхности с антителом к Fc. Первая проточная ячейка остается пустой для использования в качестве холостой пробы. За этой стадией захвата сразу следует введение пяти концентраций антигена (либо 5 нМ, 2,5 нМ, 1,25 нМ, 0,63 нМ и 0,31 нМ для TF или EGFR ECD антигенов, или 40 нм, 20 нм, 10 нм, 5 нм и 2,5 нм для антигена Her2 ECD), которые последовательно вводили во все четыре проточные ячейки по 100 мкл/мин в течение 180 сек с фазой диссоциации 300 сек для EGFR ECD, 1800 сек для Her2 ECD и 3600 сек для TF. Поверхности с захваченными антителами регенерируют с помощью 10 мМ глицина, рН 1,5 в течение 120 сек при 30 мкл/мин с целью подготовки к следующему циклу инъекции. Выполняются по меньшей мере две пробные буферные инъекции для каждой инъекции анализируемого вещества, которое будет использоваться для сравнения. Полученные в результате одного цикла кинетики сенсограммы были проанализированы с помощью программного обеспечения Biacore T200 BiaEvaluation и пригодны к модели связывания 1:1.
[0305] Антигенную аффинность гетеродимерных антител оценивали с учетом соответствующего контроля дикого типа: МАт для D3H44, трастузумаб ОАА и цетуксимаб ОАА. Антигенная аффинность также была получена для биспецифических антител дикого типа; вместе с тем, SPR захват биспецифических антител дикого типа может быть гетерогенным (например, включать захват неправильно спаренных гетеродимеров), тем самым препятствуя определению KD (см. таблицы 31а и с). Для гетеродимерных антител, которые имели антигенную связываемость, измеренную в системе D3H44/цетуксимаб, антигенная аффинность была аналогична соответствующим образцам контроля дикого типа (таблица 31b). Для большинства гетеродимерных антител, которые имели антигенную связываемость, измеренную в системах D3H44/трастузумаб и трастузумаб/цетуксимаб, антигенная аффинность была аналогична соответствующим образцам контроля дикого типа (см. таблицы 31а и с). Исключение включают одиннадцать разработанных антител, которые не проявляют Her2 связывания. В обоих системах D3H44/трастузумаб и трастузумаб/цетуксимаб her2 связывание не наблюдалось для шести разработанных гетеродимерных антител 9049-9759_1 и 9682-9740_1 и 3522_1. Кроме того, для системы трастузумаб/цетуксимаб пять дополнительно разработанных антител 9696-9848_1, 9561-9095_2, 9611-9077_2, 9286-9402_2 и 9060-9756_2 также не проявляли связывания с Her2. Десять из этих одиннадцати разработанных антител имеют общие мутации в константной области на Н-цепи (L143E_K145T) и L-цепи (Q124R_T178R). Другие разработанные антитела 9286-9402_2 имеют общие мутации в константной области на Н-цепи (L143E_K145T) и подобные мутации на L-цепи (Q124K и S176R).
Пример 14: Профили сверхэффективной жидкостной хроматографии - эксклюзионной хроматографии (UPLC-SEC) для разработанных гетеродимерных антител, а также для гетеродимерных антител и гомодимерных антител дикого типа
[0306] После препаративной SEC разработанных гетеродимерных антител, а также контроля биспецифических и гомодимерных антител дикого типа, UPLC-SEC анализ осуществляли с использованием колонки Waters BEH200 SEC (2,5 мл, 4,6×150 мм, нержавеющая сталь, 1,7 мкм частицы) с заданной температурой 30°С и монтировали на системе Waters Acquity UPLC с детектором PDA. Время разгона составляло 7 мин, и общий вводимый объем составлял 2,8 мл с электродным буфером из либо PBS и 0,02% полисорбата 20, либо 20 мМ NaPO4, 50 мМ KCl, 0,02% полисорбата 20, 10% ацетонитрила, рН 7 при 0,4 мл/мин. Элюирование контролировали с помощью УФ-поглощения в диапазоне 210-400 нм, и хроматограммы экстрагируют при длине волны 280 нм. Интегрирование пика осуществляли с помощью программного обеспечения Empower 3.
[0307] Фигура 10 иллюстрирует репрезентативные профили UPLC-SEC представителей разработанных гетеродимерных антител, а также представительных гетеродимерных антител дикого типа. В большинстве случаев разработанные гетеродимерные антитела демонстрируют профили UPLC-SEC, аналогичные соответствующим профилям гетеродимерных антител дикого типа, со средним процентом мономеров 99,18%, 98,70% и 98,77% для D3H44/трастузумаб, D3H44/цетуксимаб и трастузумаб/цетуксимаб, соответственно (см. таблицы 31а, 31b и 31с).
[0308] Несмотря на то, что изобретение было конкретно проиллюстрировано и описано со ссылкой на предпочтительный вариант реализации изобретения и различные альтернативные варианты реализации изобретения, специалистам в данной области техники понятно, что могут быть внесены различные изменения в отношении формы и деталей без отступления от сущности и объема данного изобретения.
[00253] Все ссылки, выданные патенты, патентные публикации и номера доступа последовательностей, описанные в данном документе, включены в него посредством ссылки в полном объеме для любых целей.
Таблицы



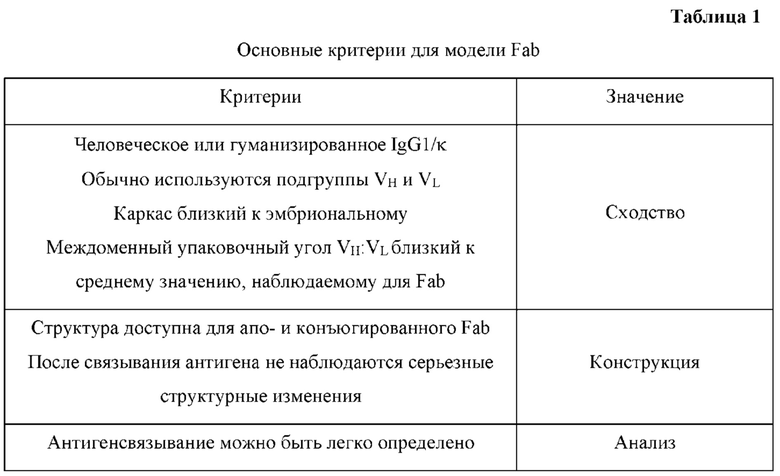
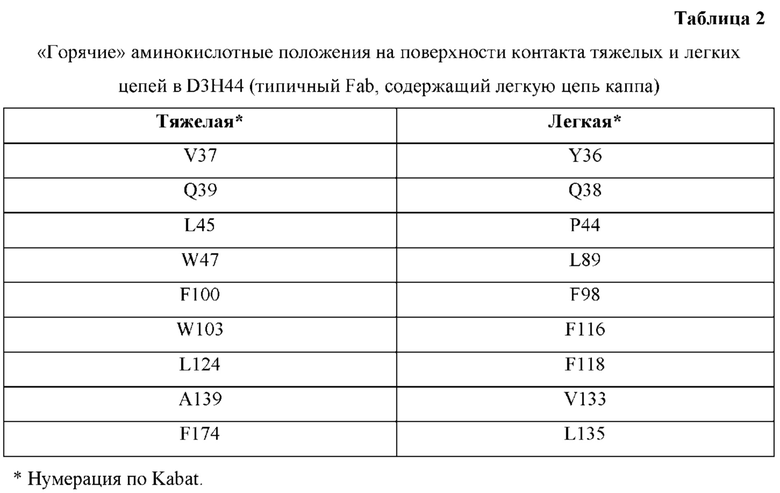
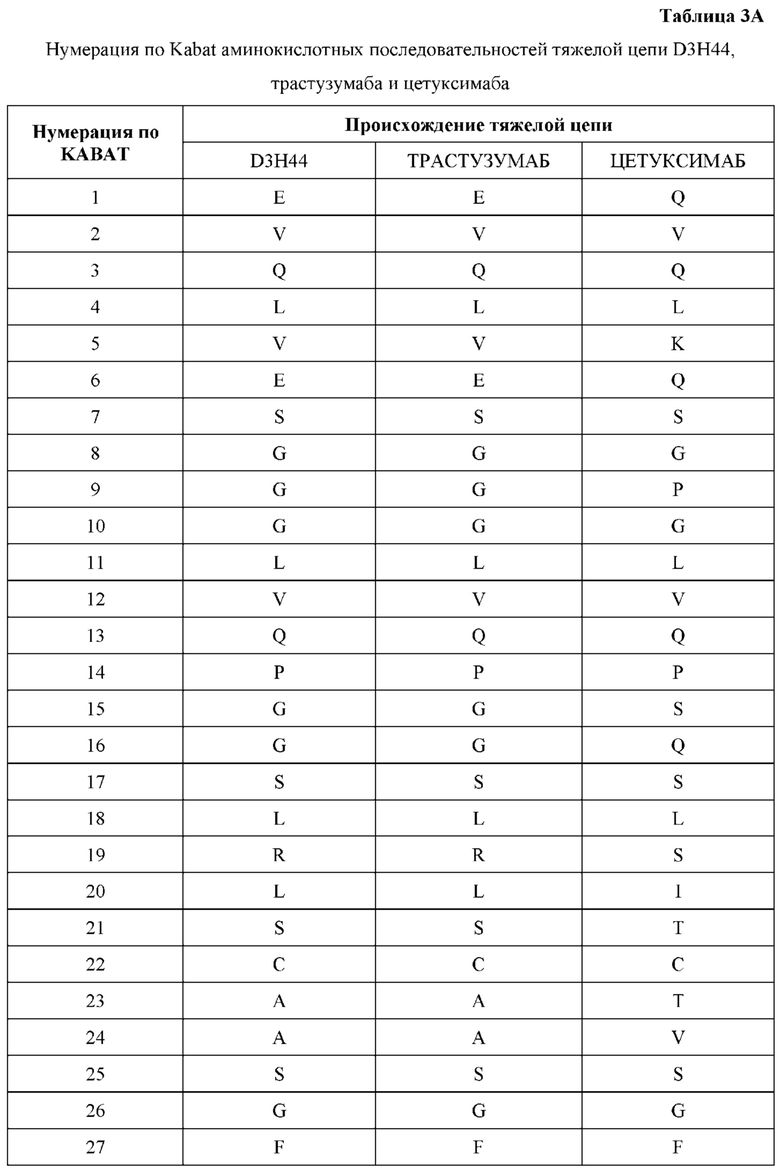
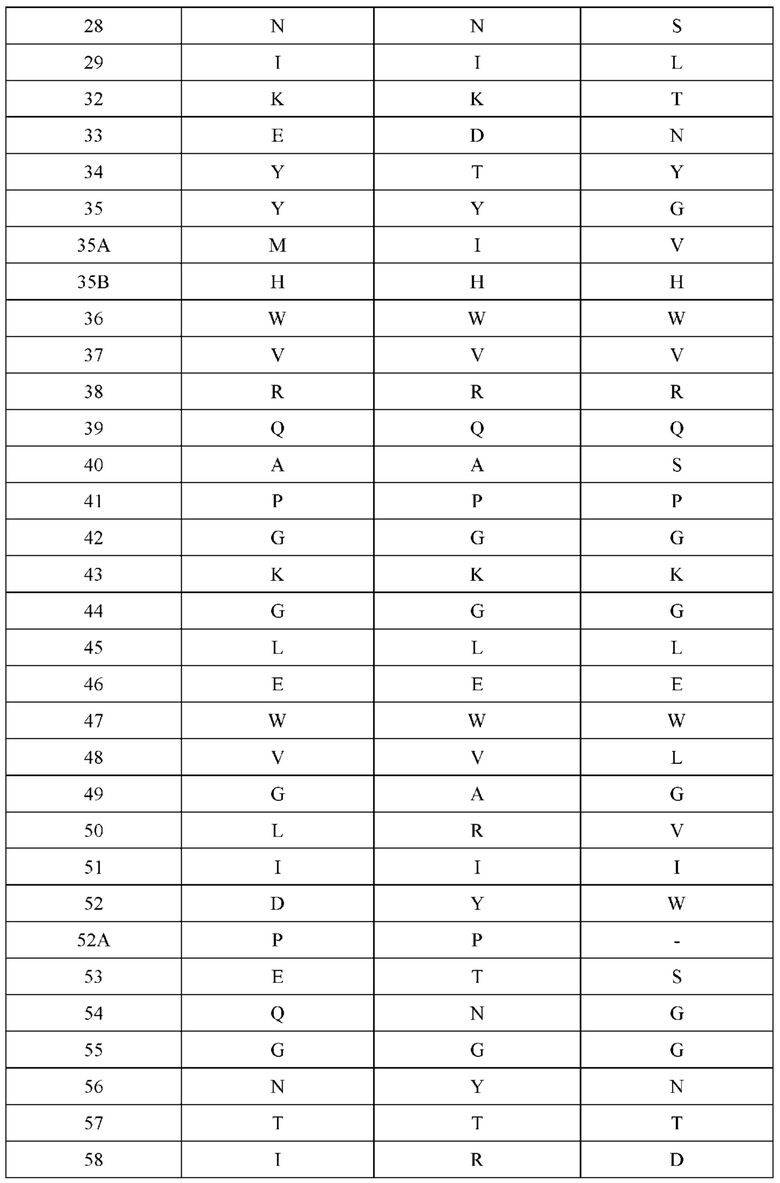
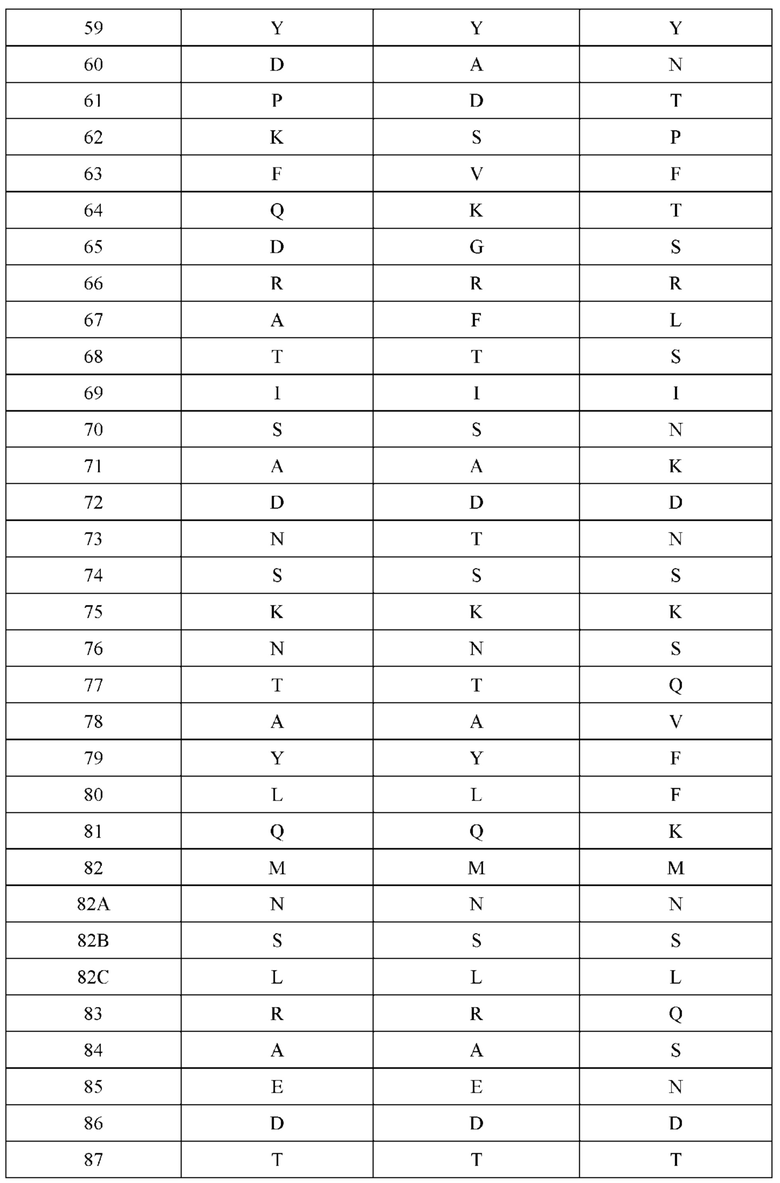
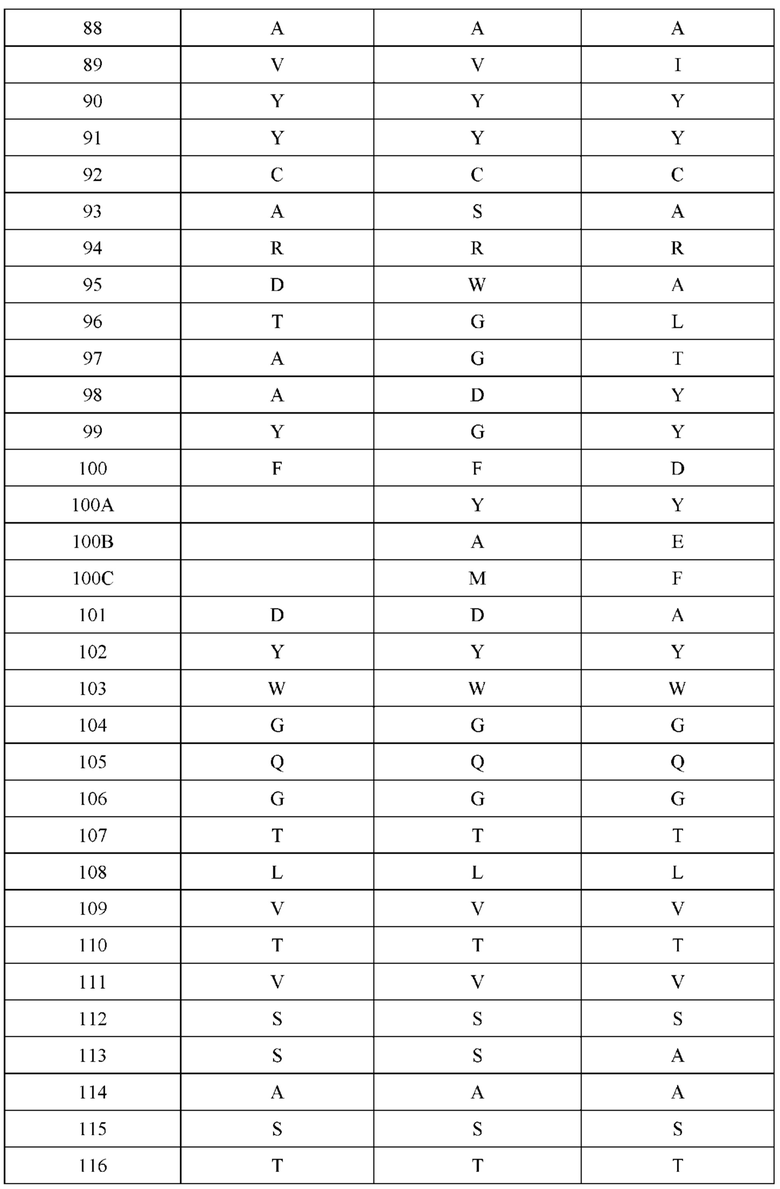
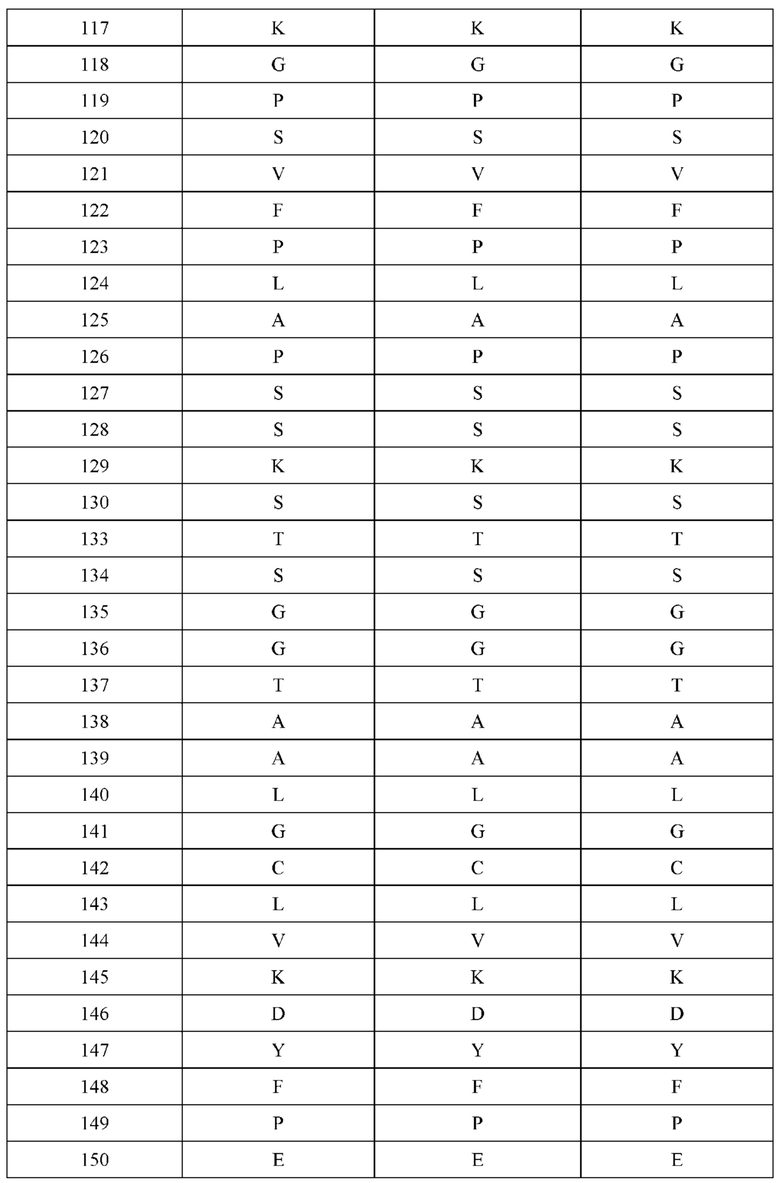
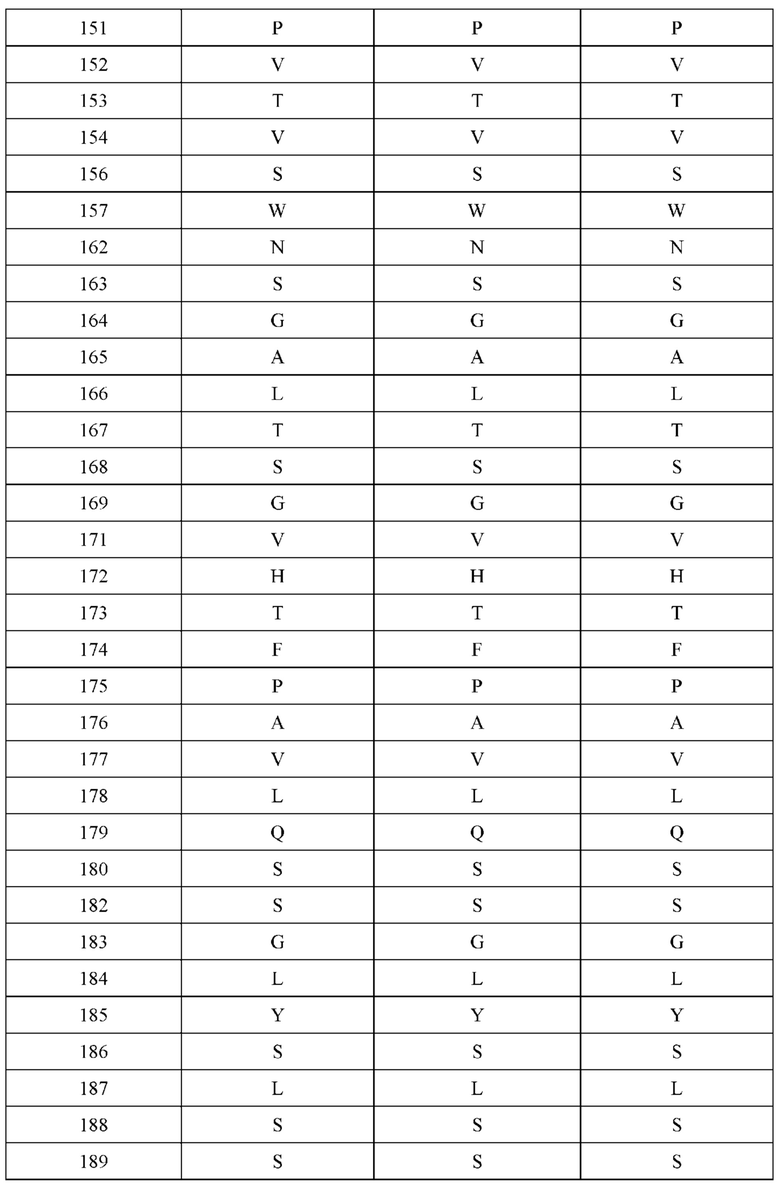
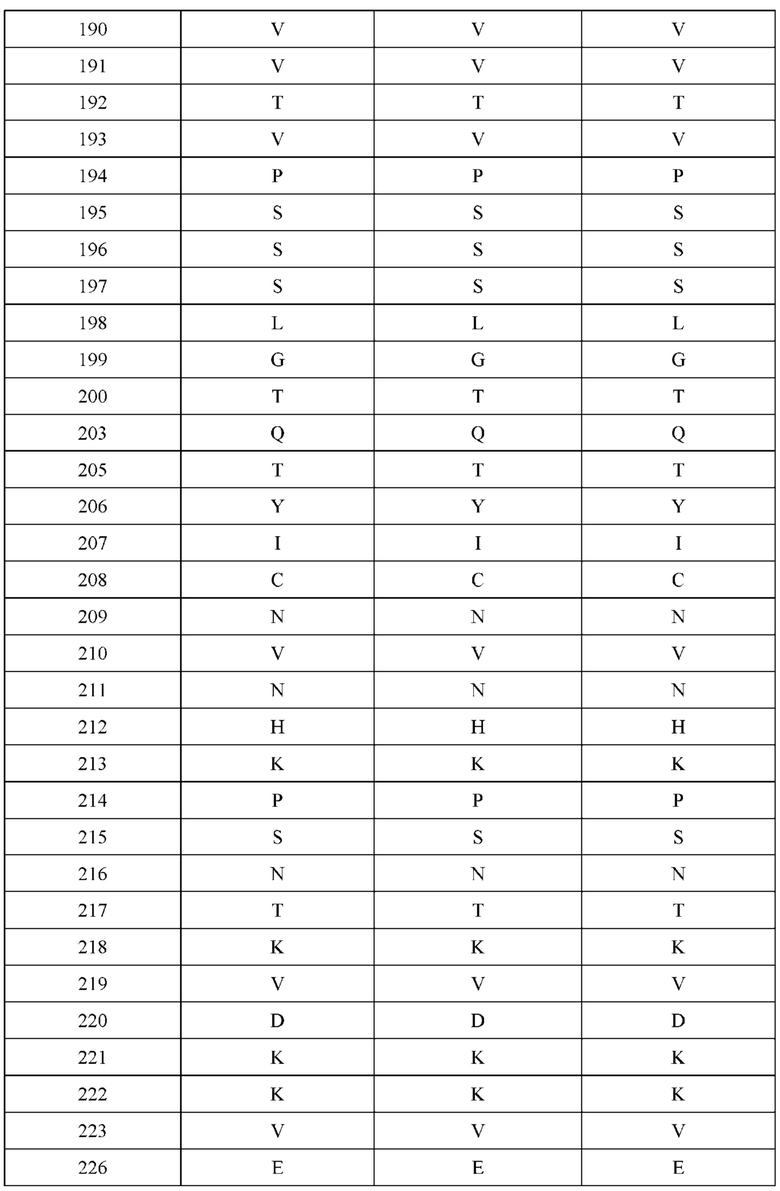
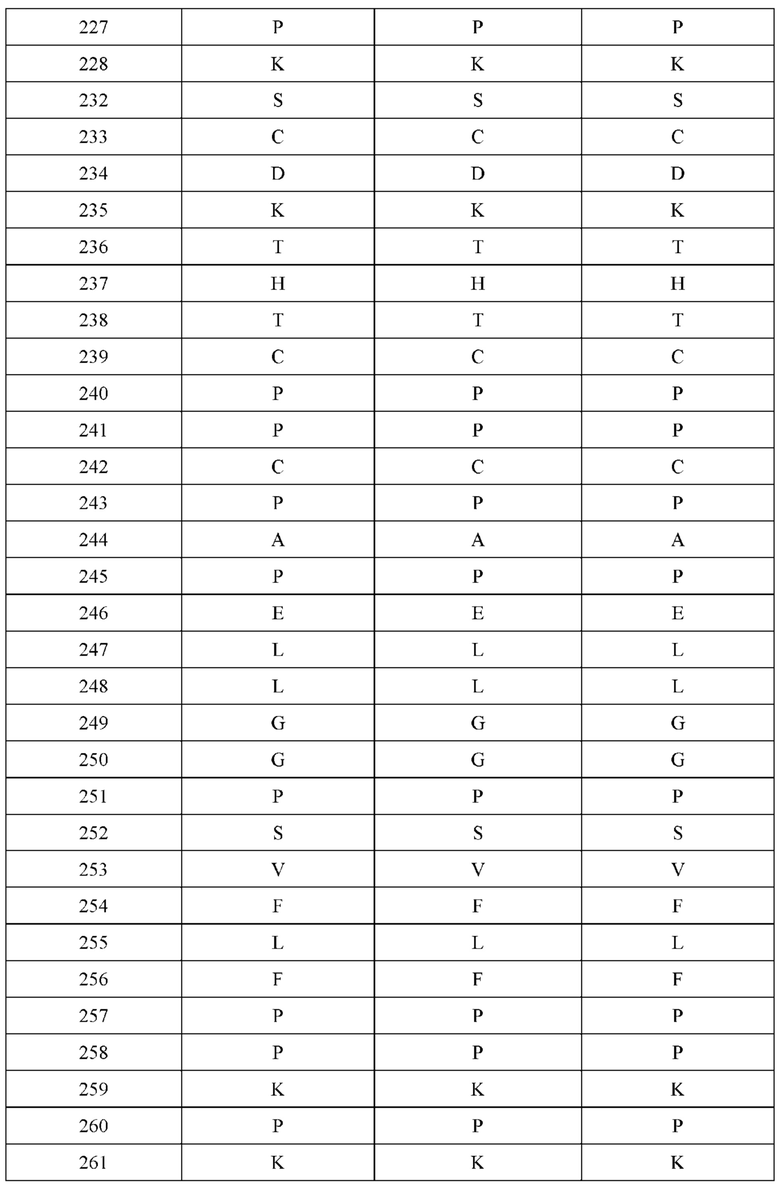
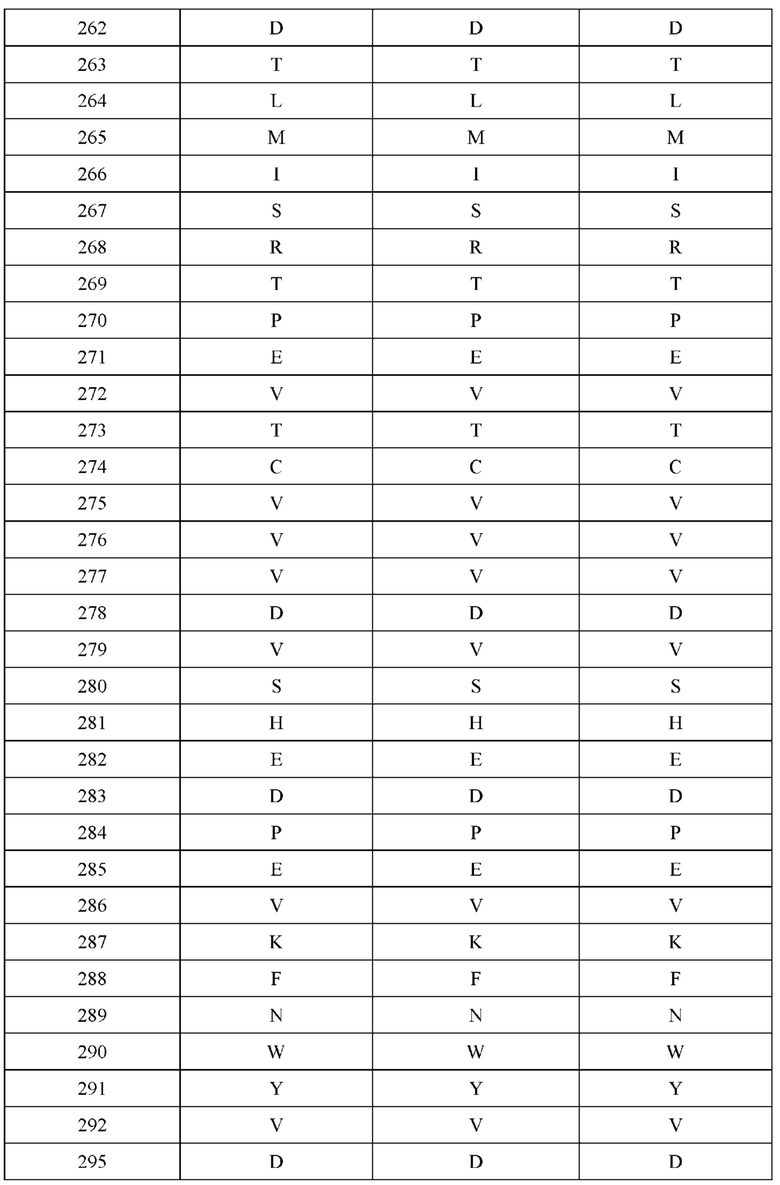
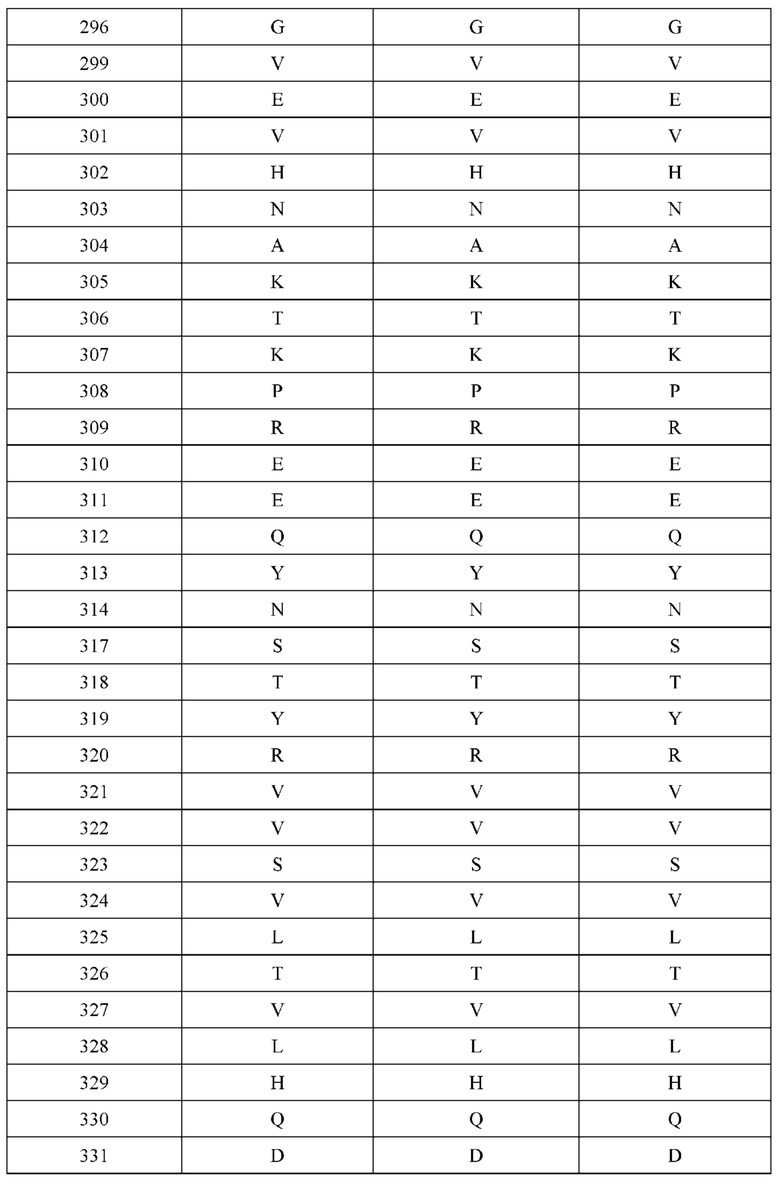
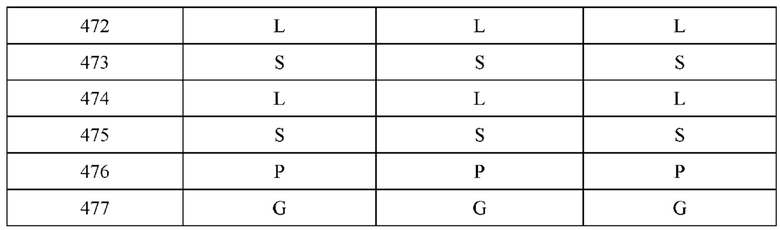
Вариабельные области: HFR1; 1-30, CDR-H1; 31-35, HFR2; 36-49, CDR-H2; 50-65, HFR3; 66-94, CDR-H3; 95-102, HFR4; 103-113 (Ссылка:, Molecular Immunology. Том 45, Номер 14 Август 2008 года, Страницы 3832-3839).
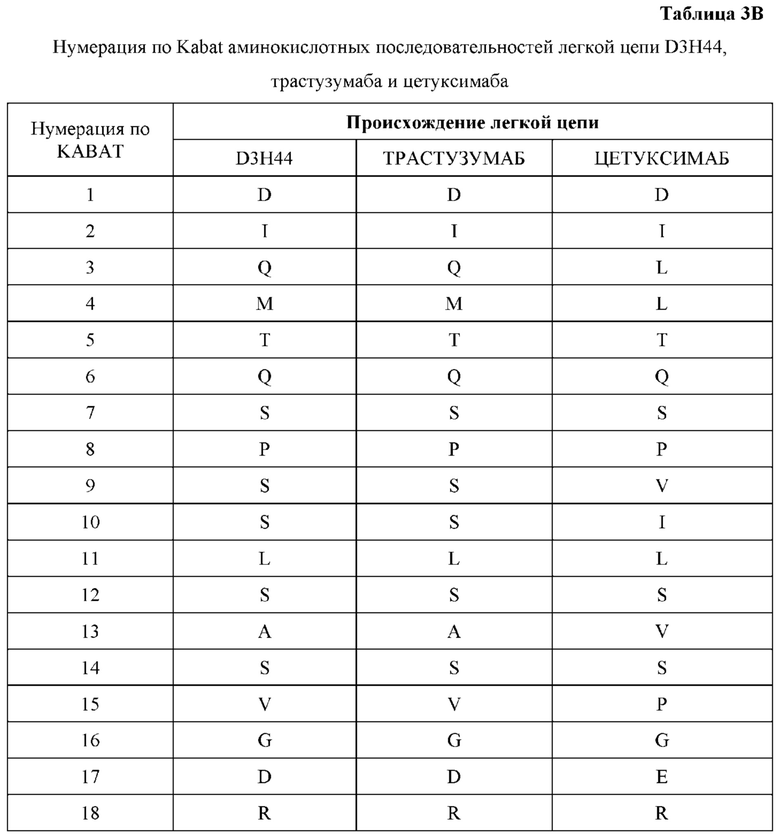
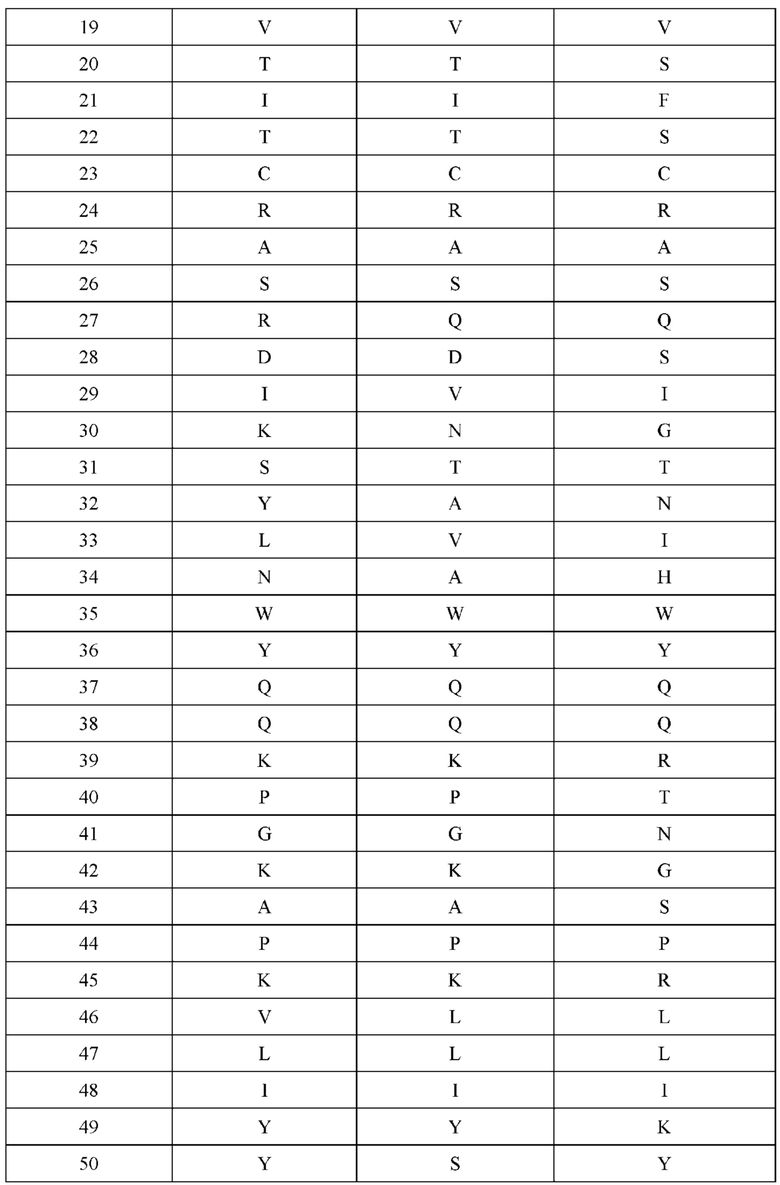
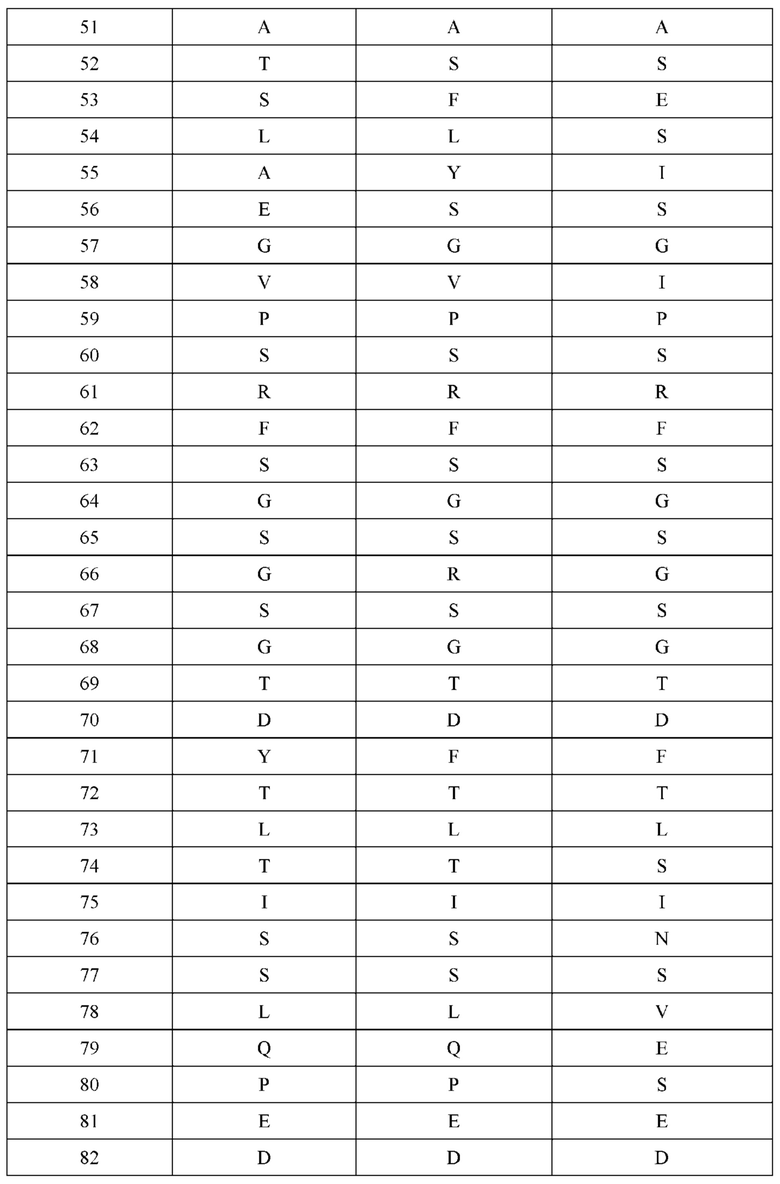
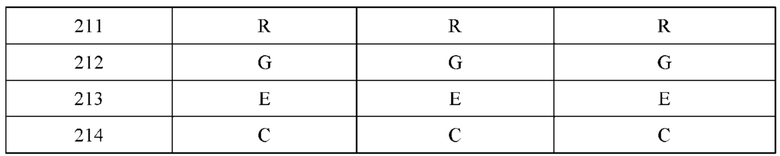
Вариабельные области: LFR1; 1-23, CDR-L1; 24-34, LFR2; 35-49, CDR-L2; 50-56, LFR3; 57-88, CDR-L3; 89-97, LFR4; 98-110 (Ссылка: Molecular Immunology. Том 45, Номер 14, Август 2008 года, Страницы 3832-3839).
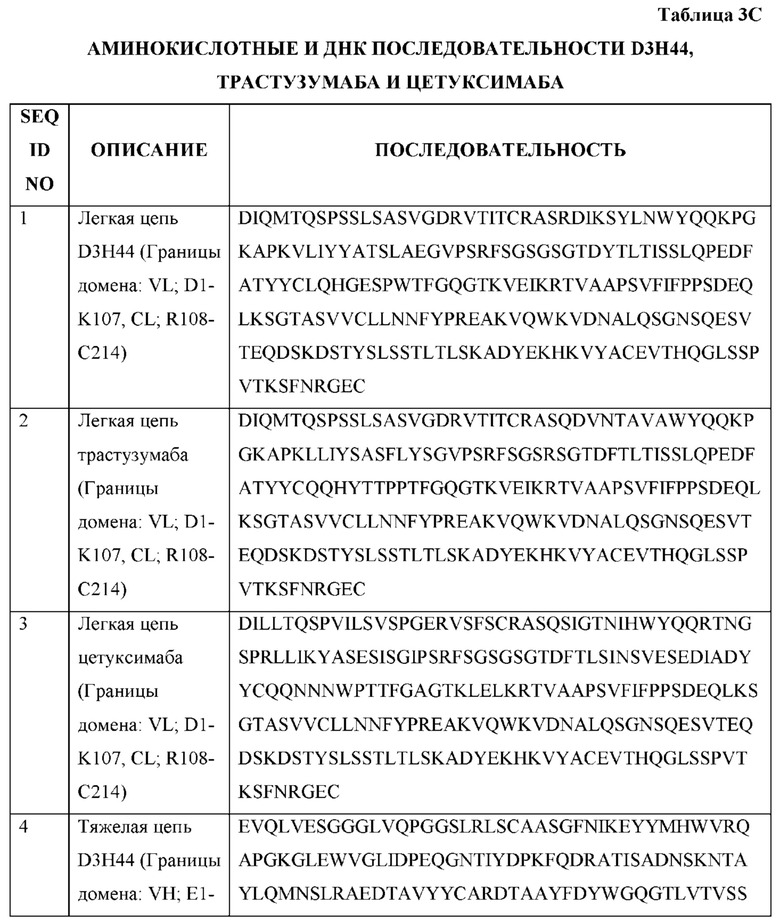





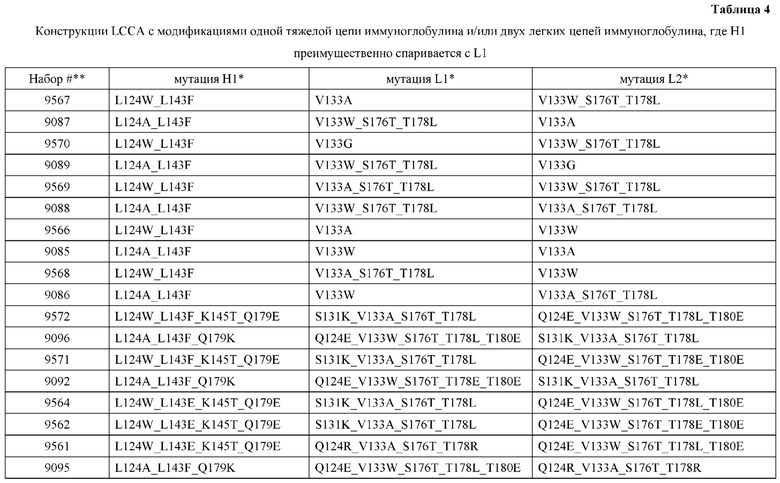
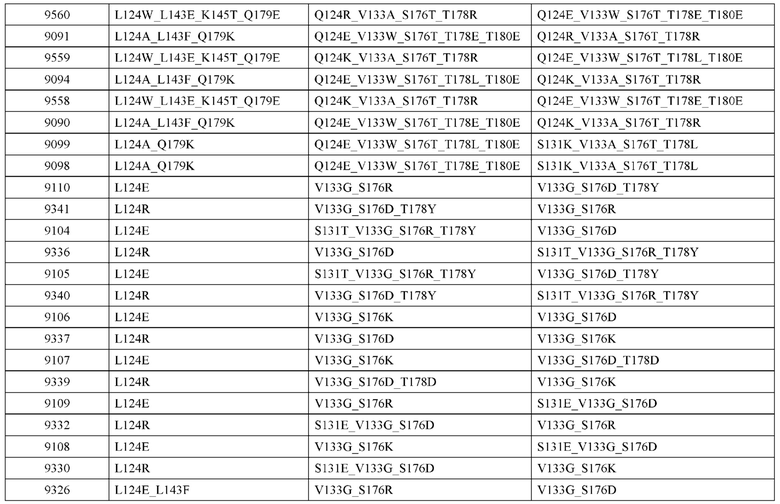
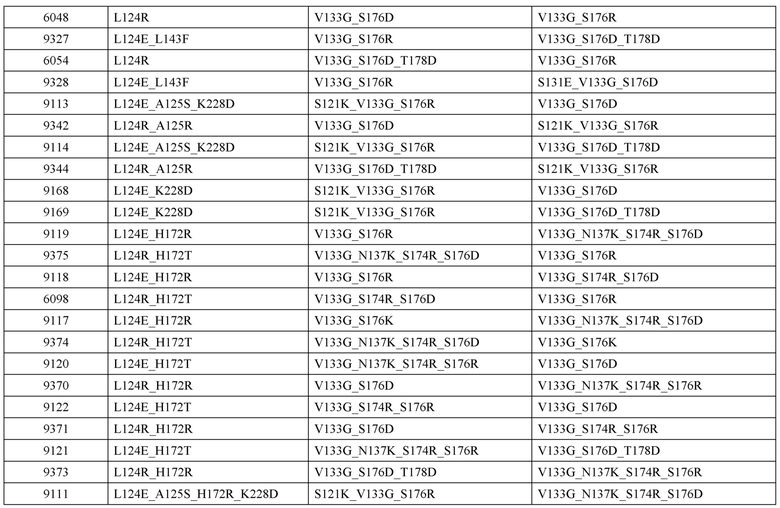
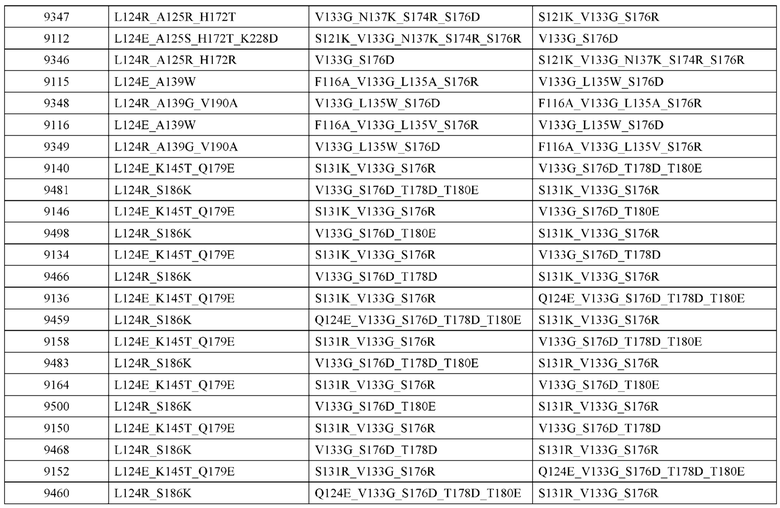
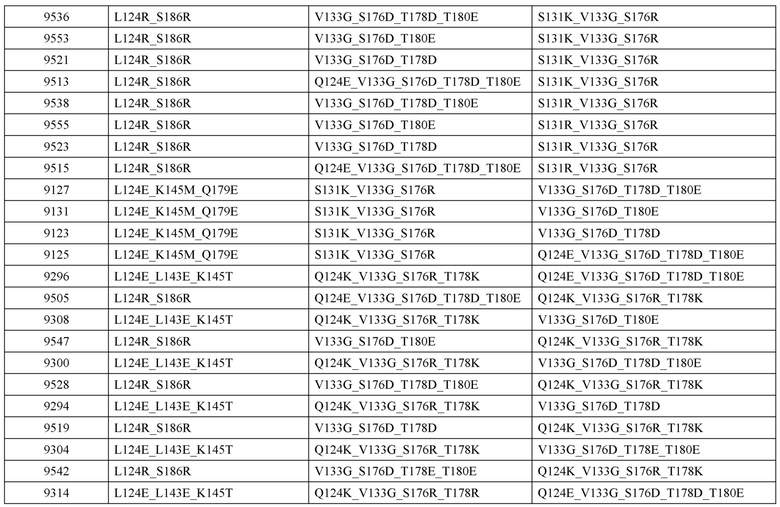
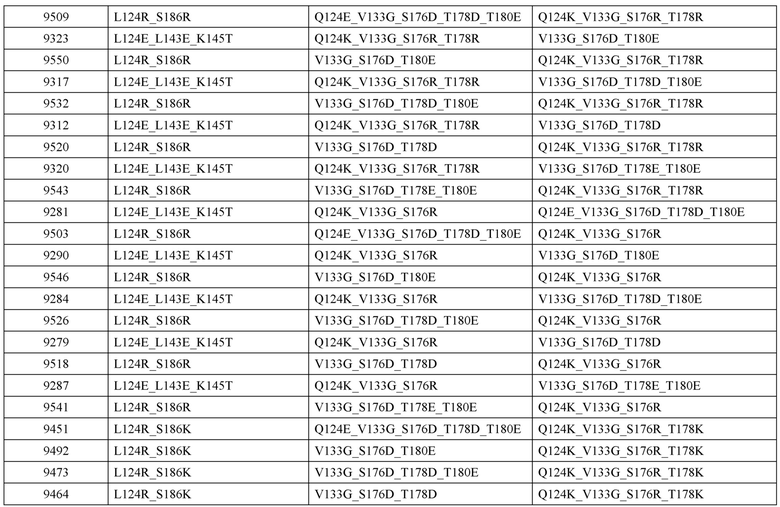
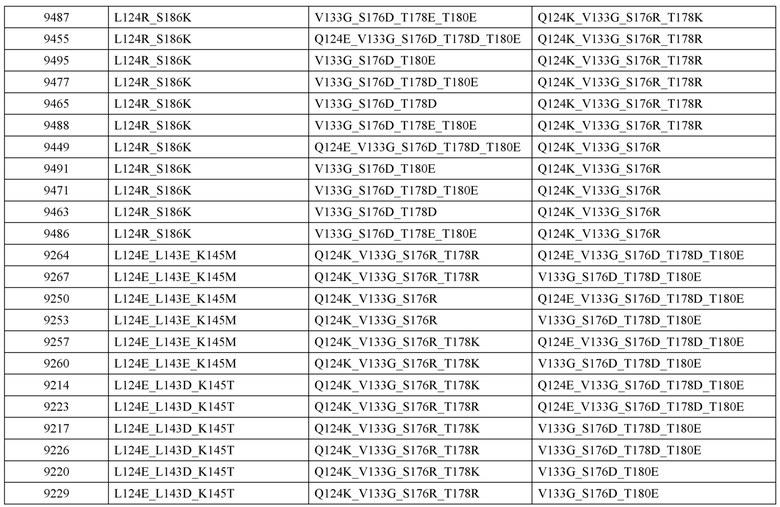
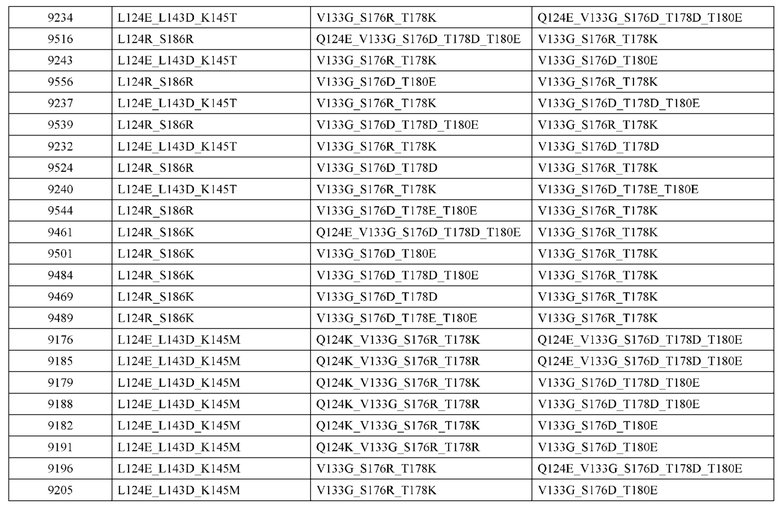
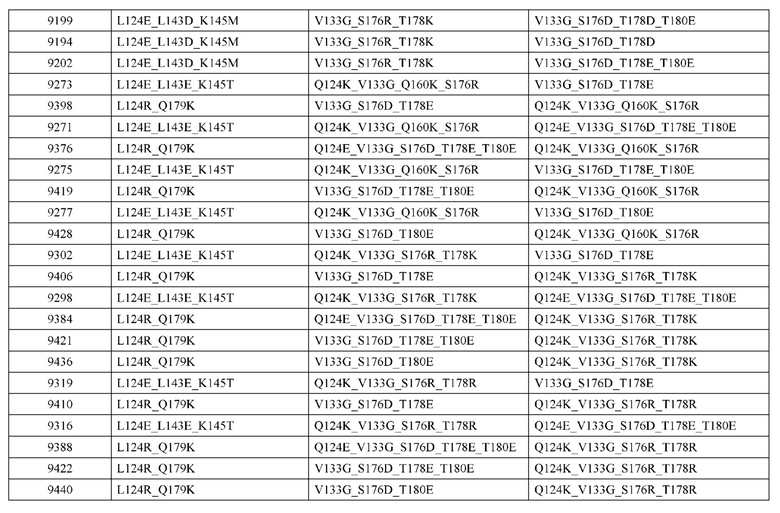
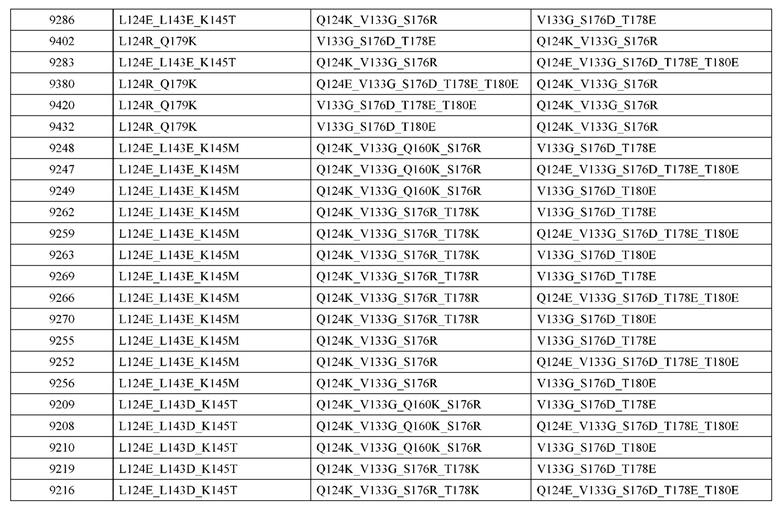
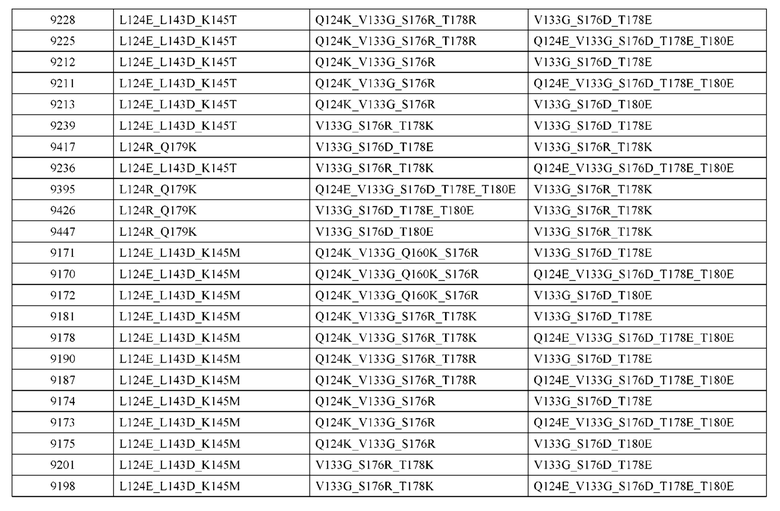
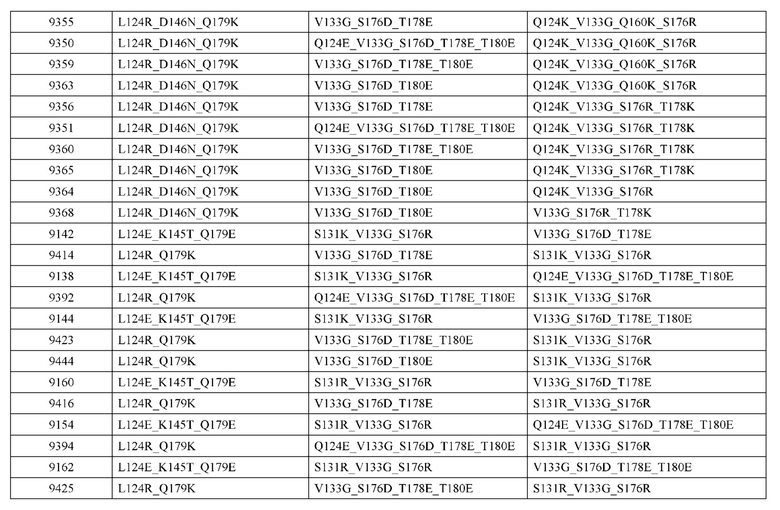
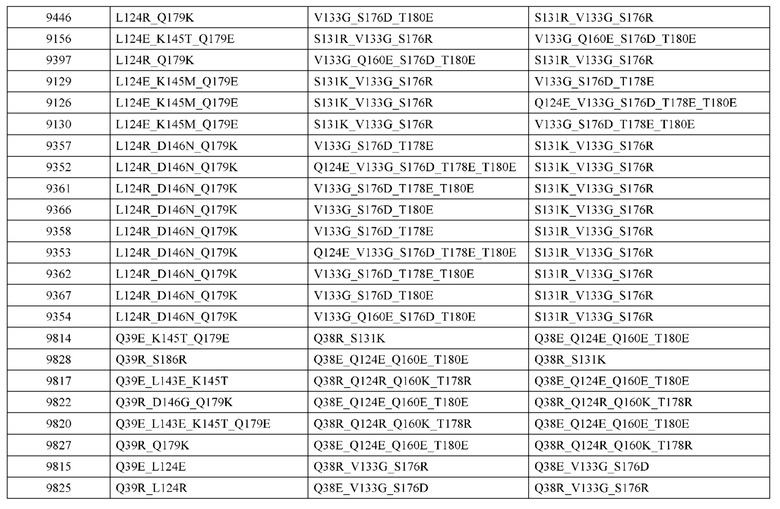
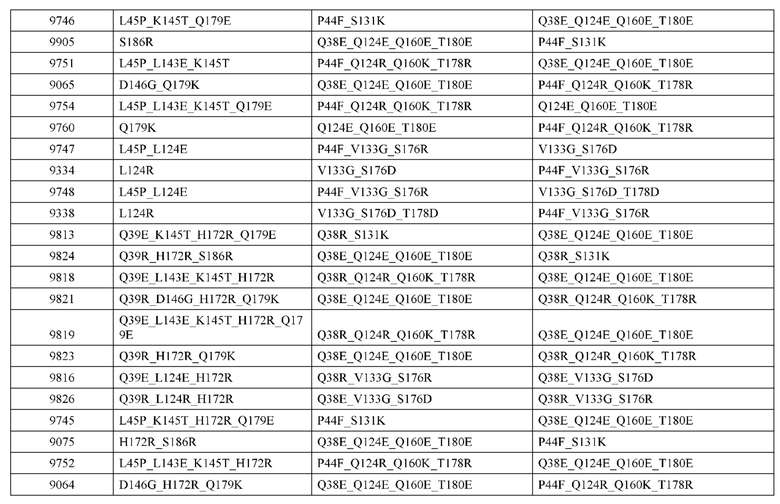
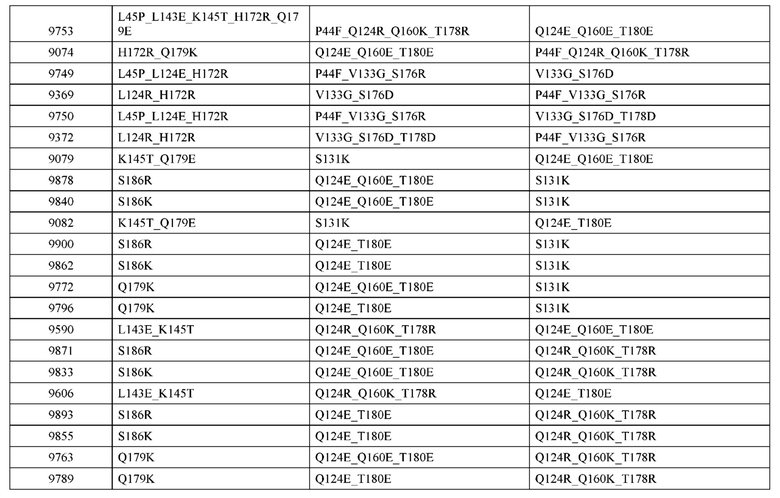
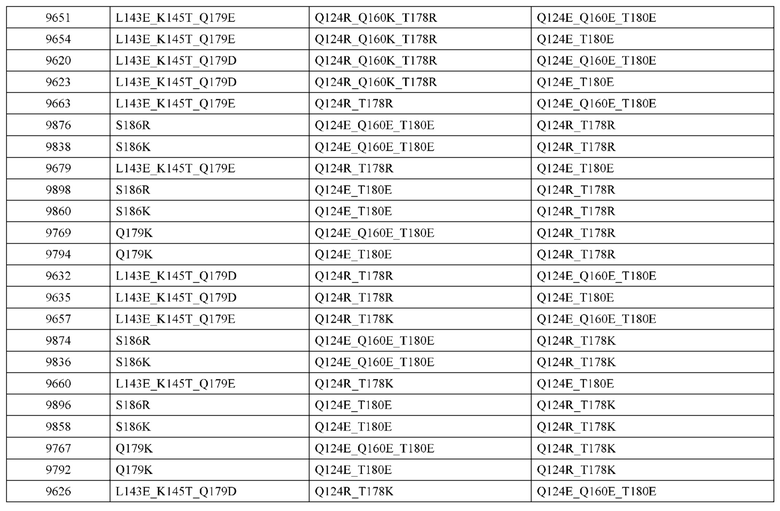
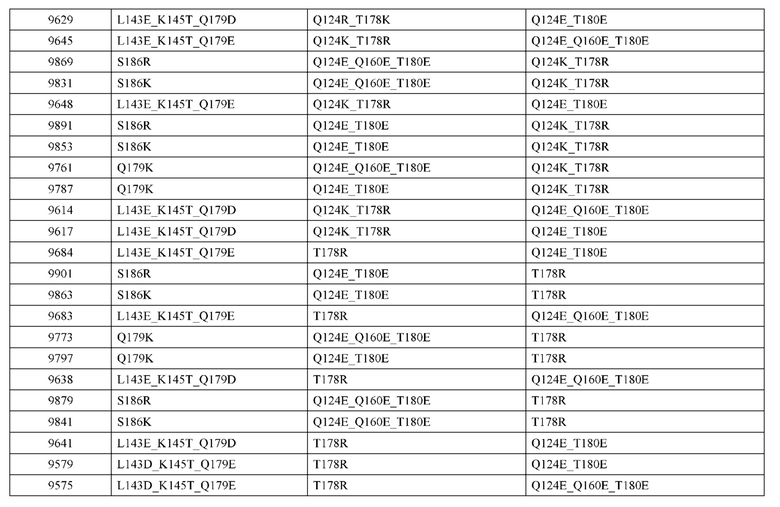
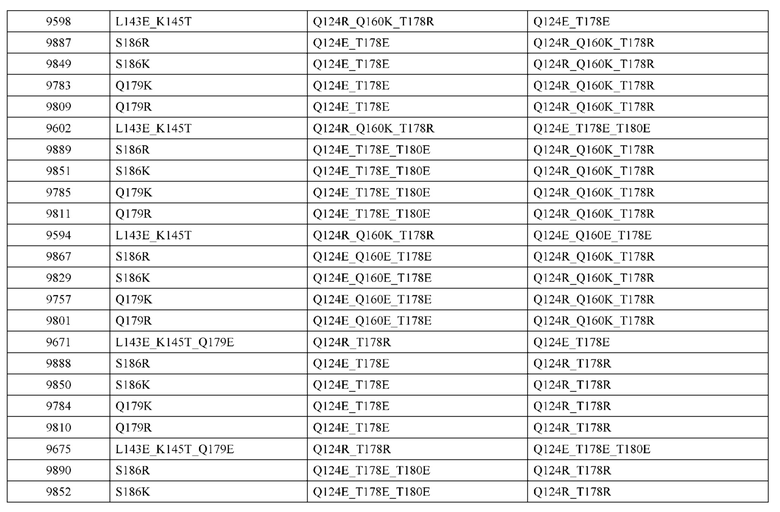
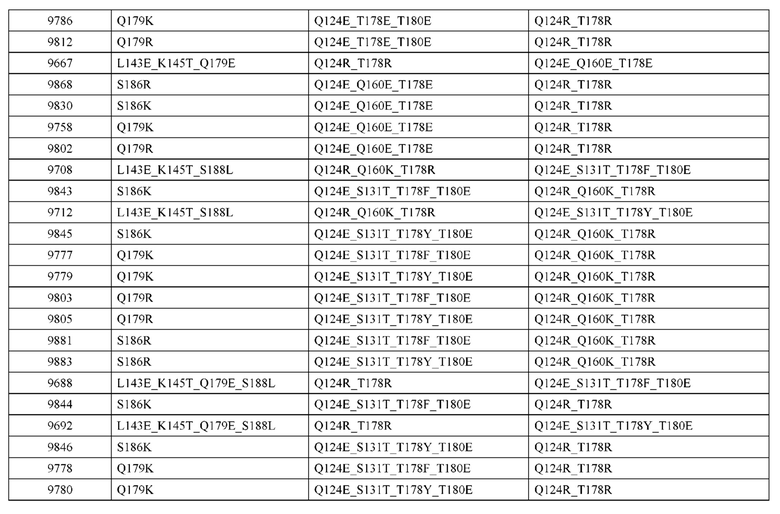
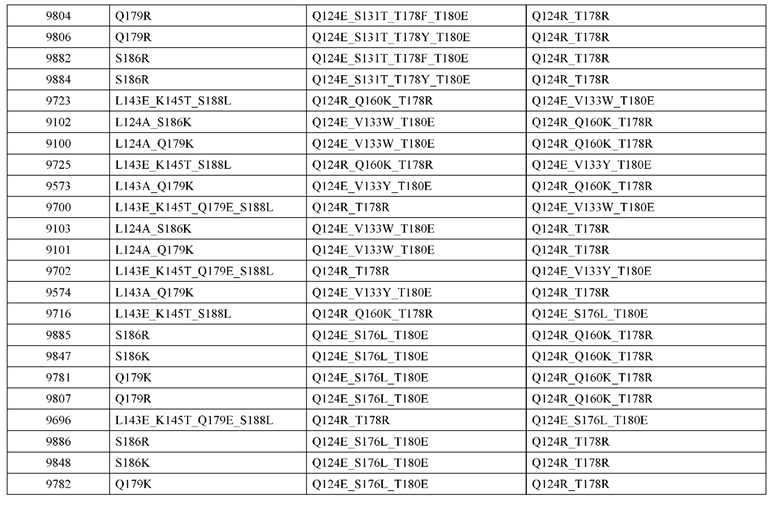
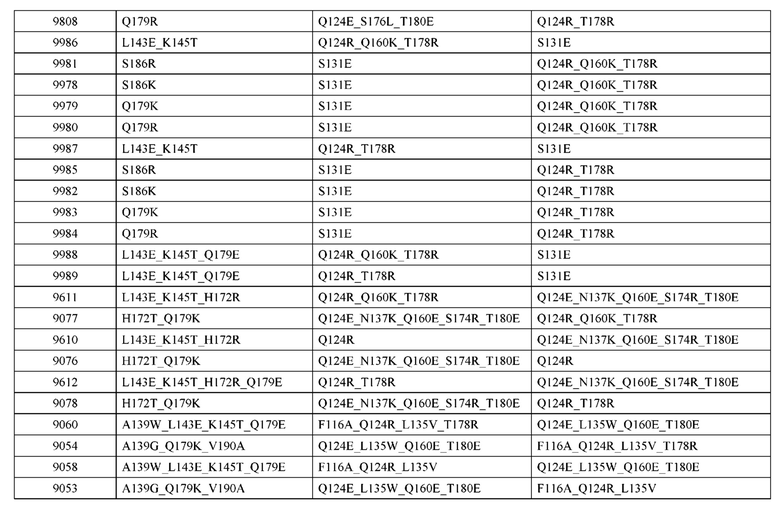
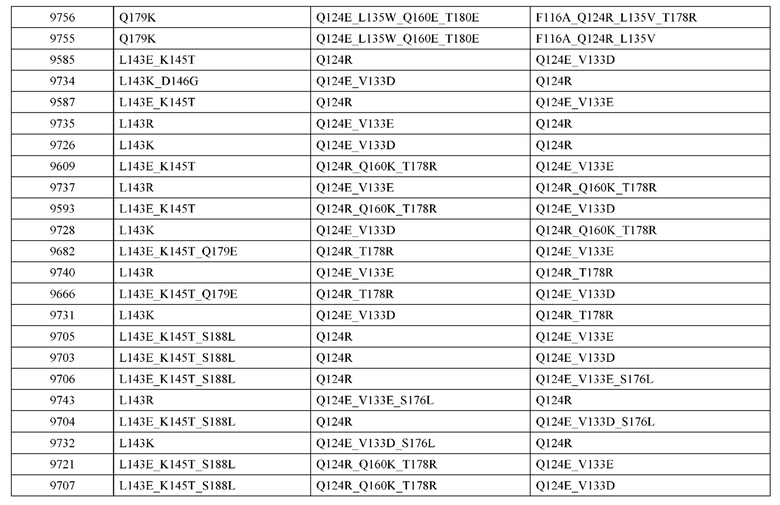
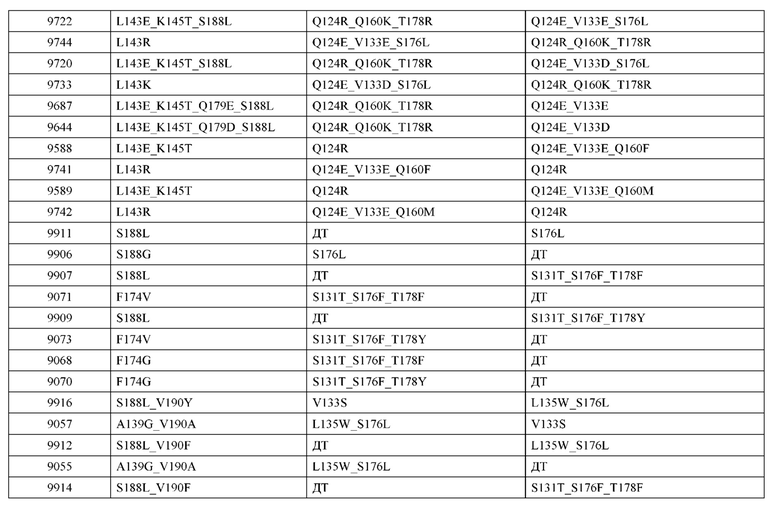
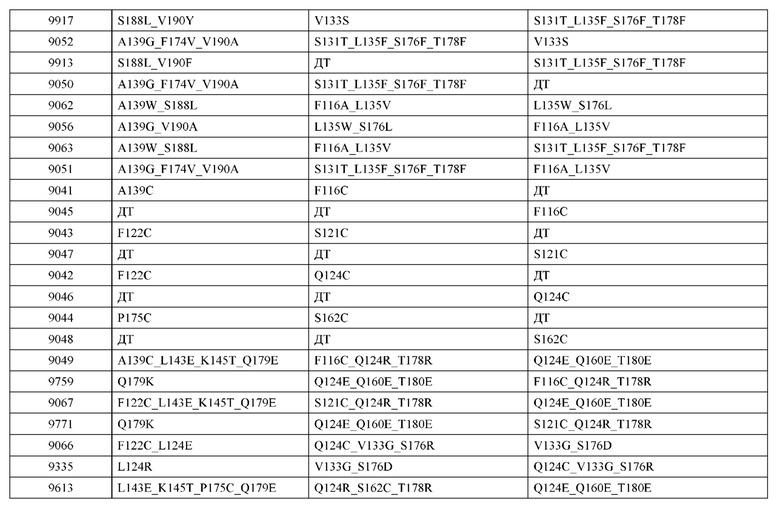

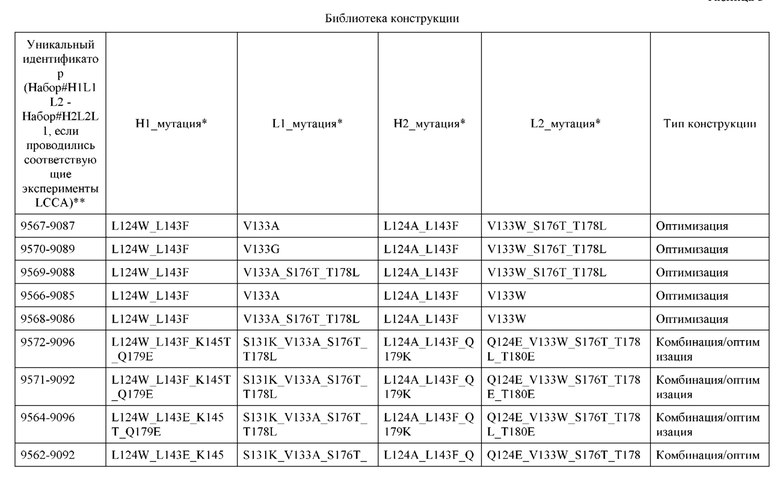
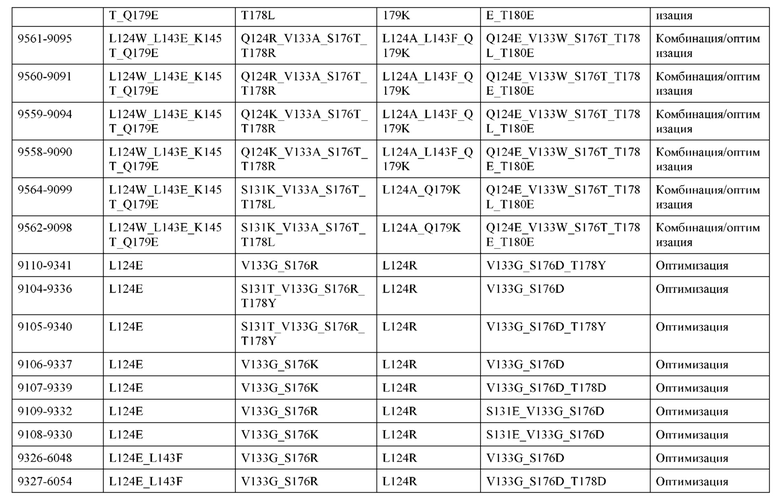
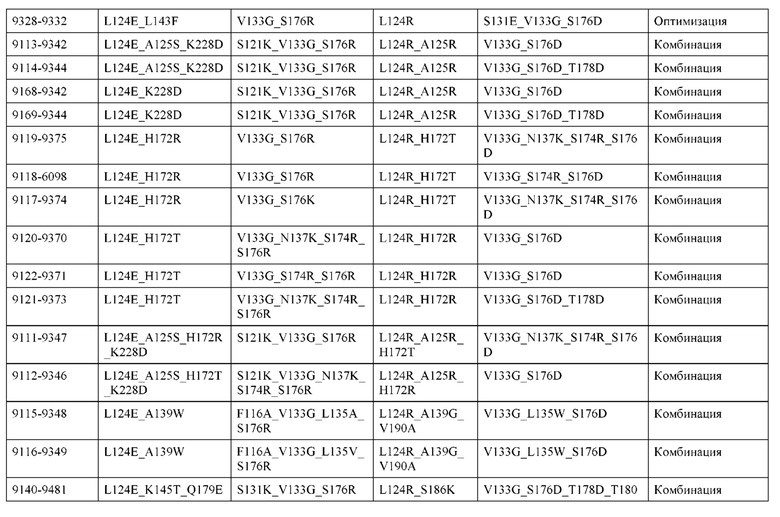
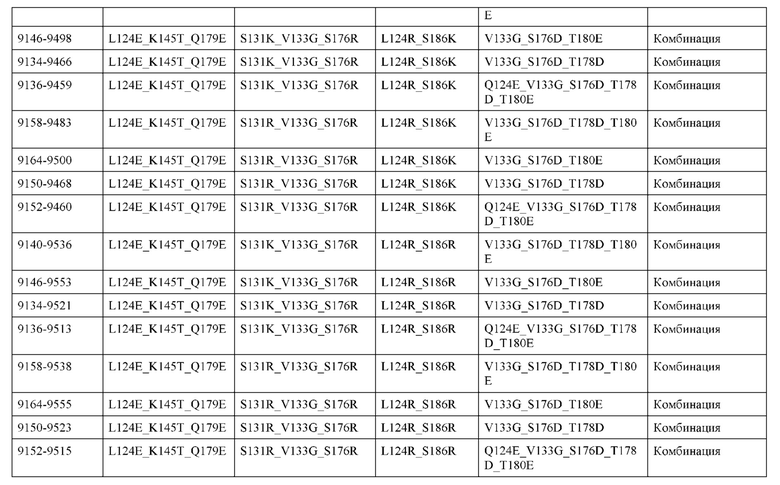
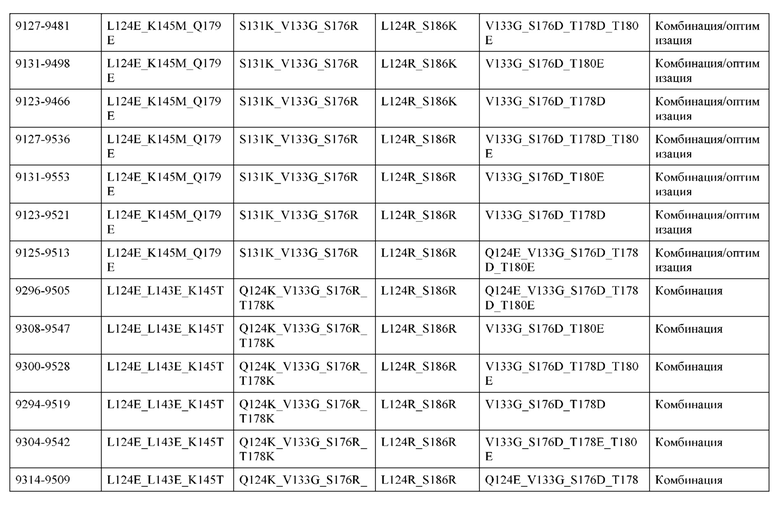
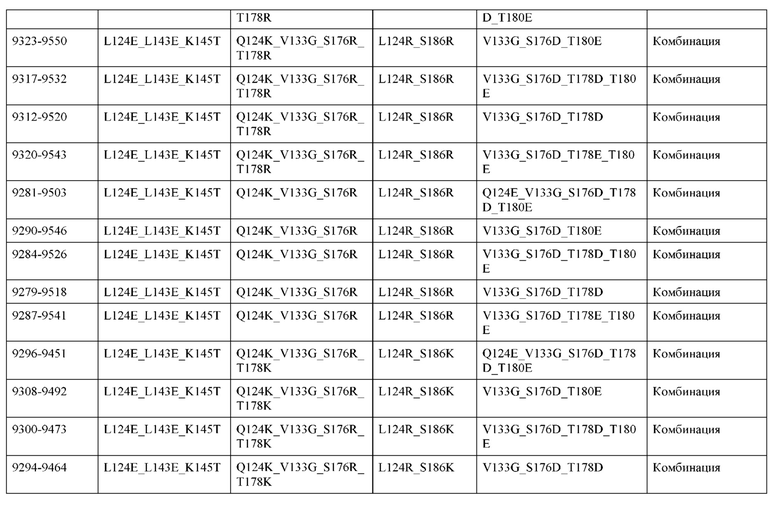
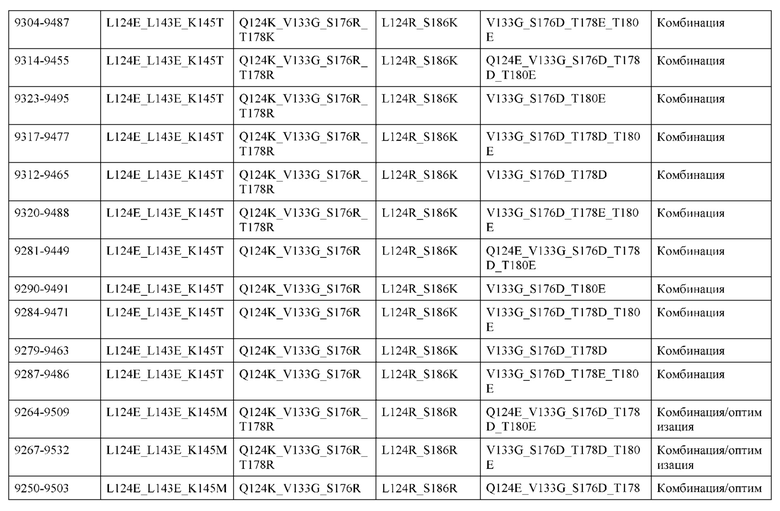
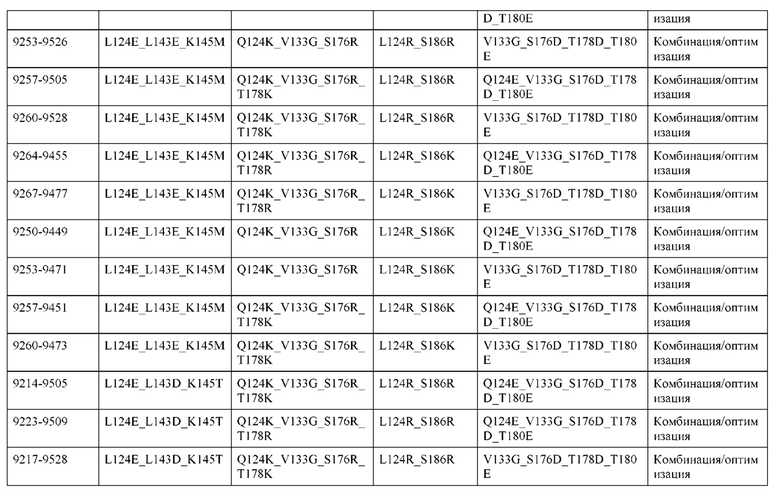
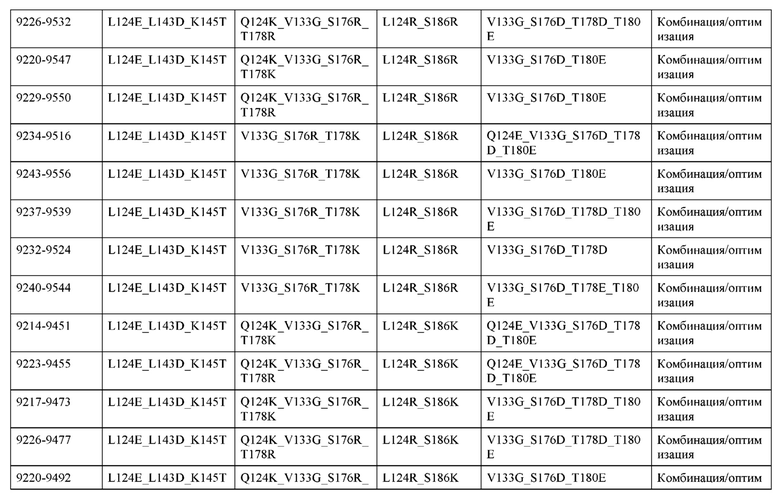
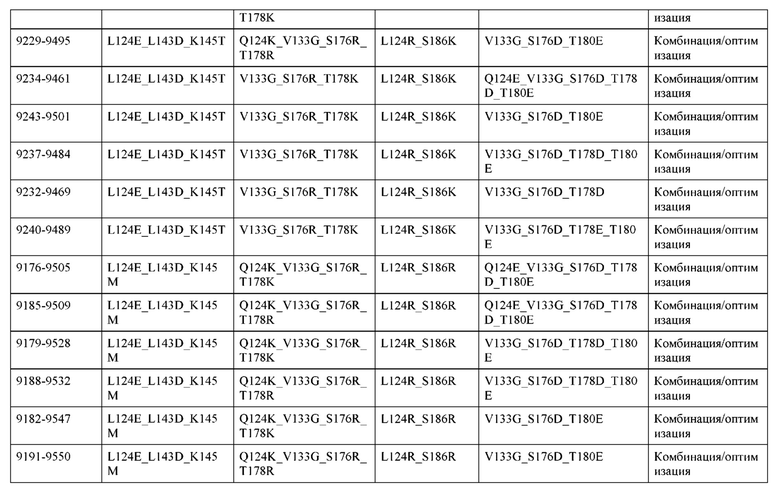

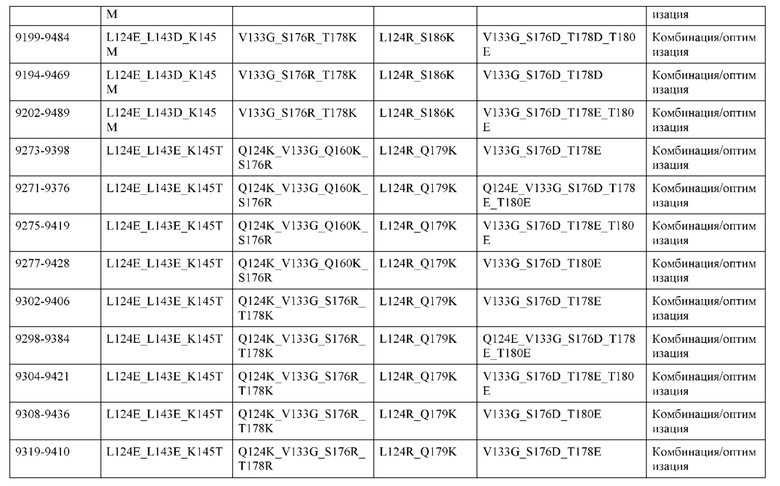
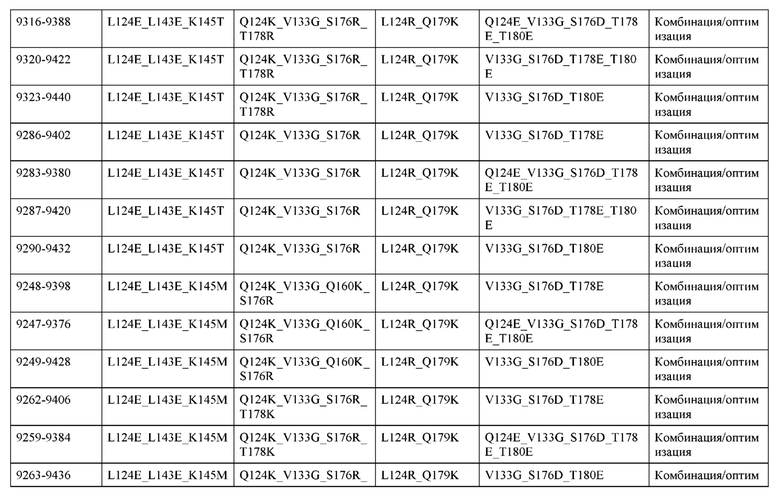
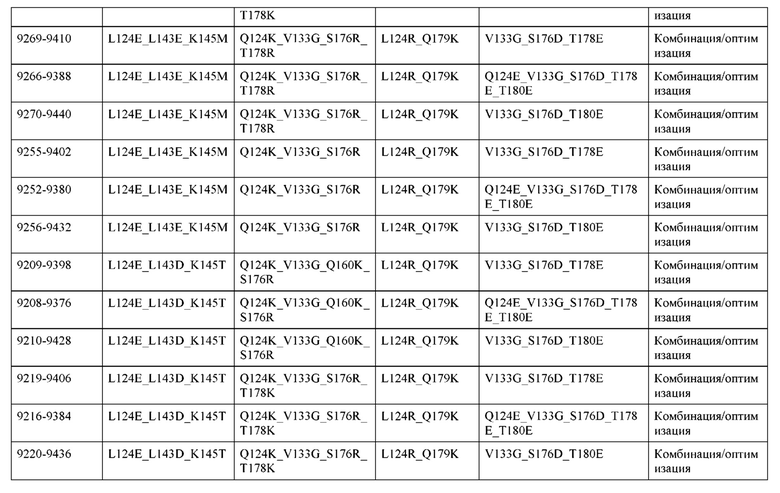
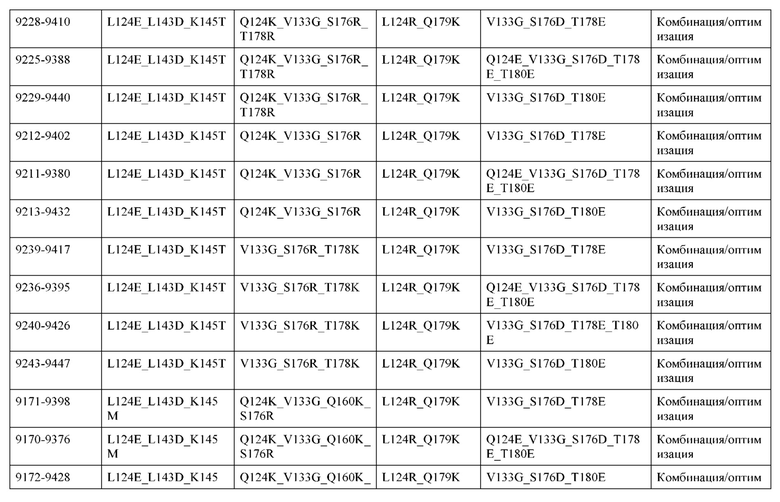
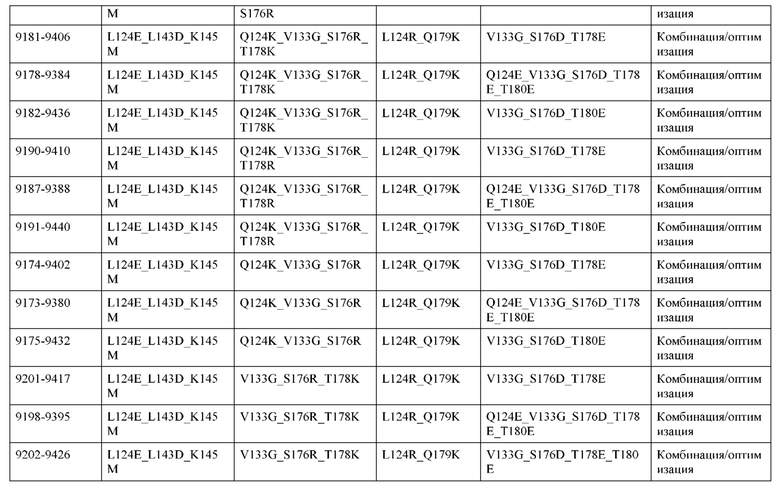
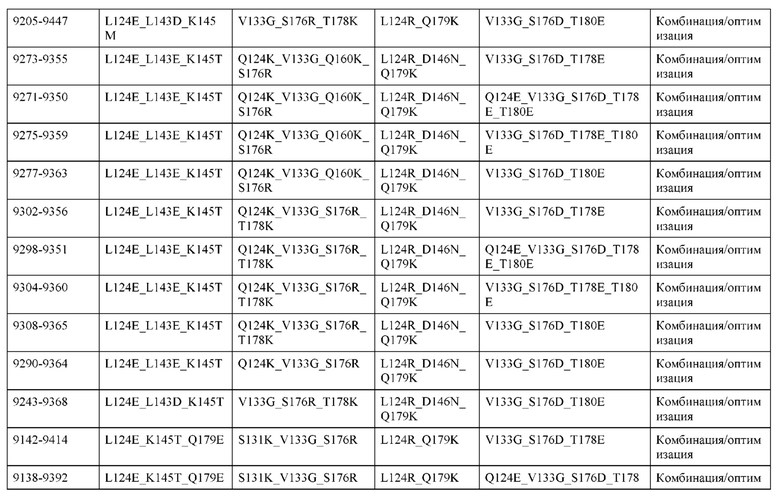
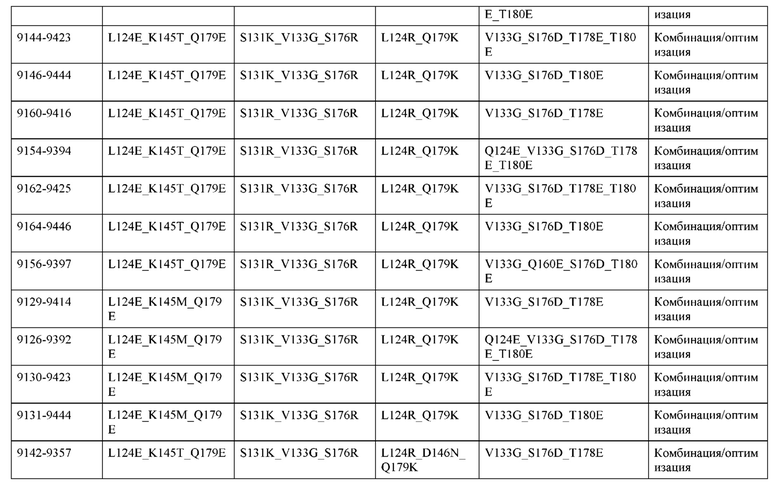
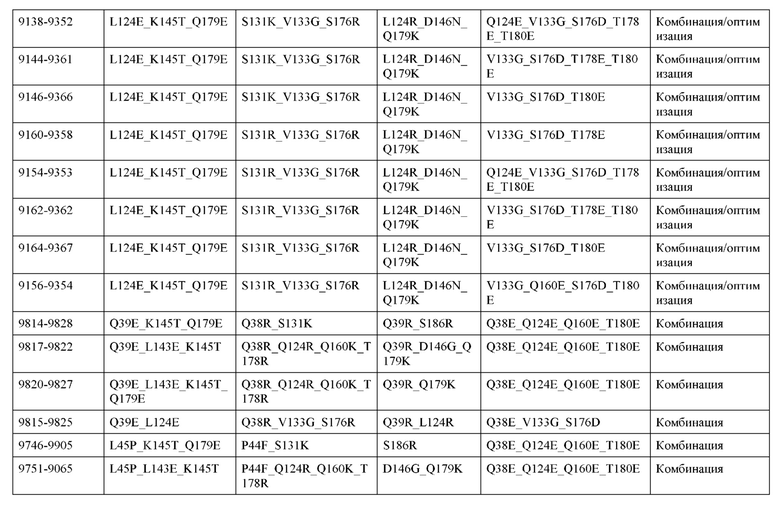
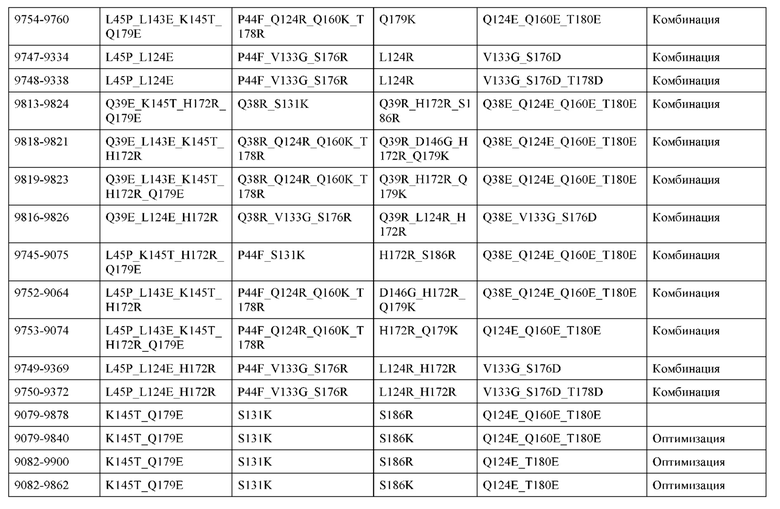
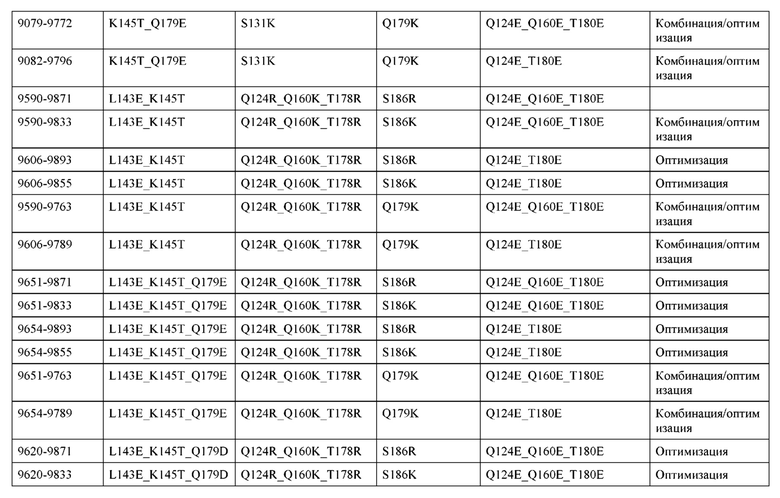
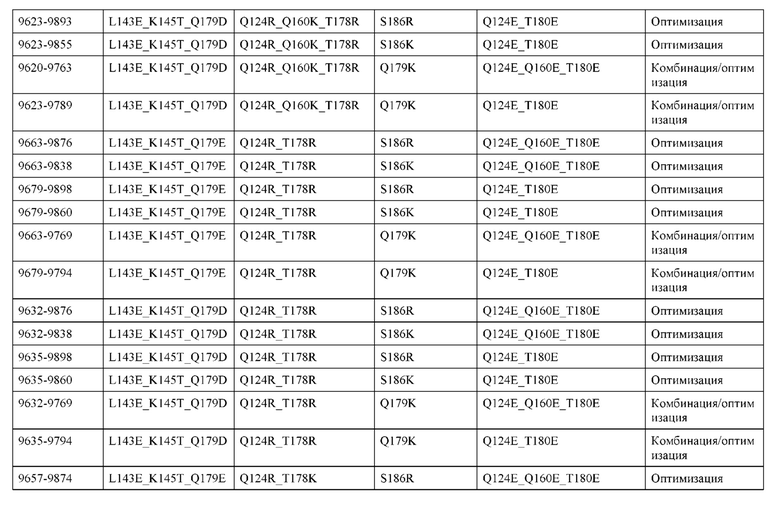
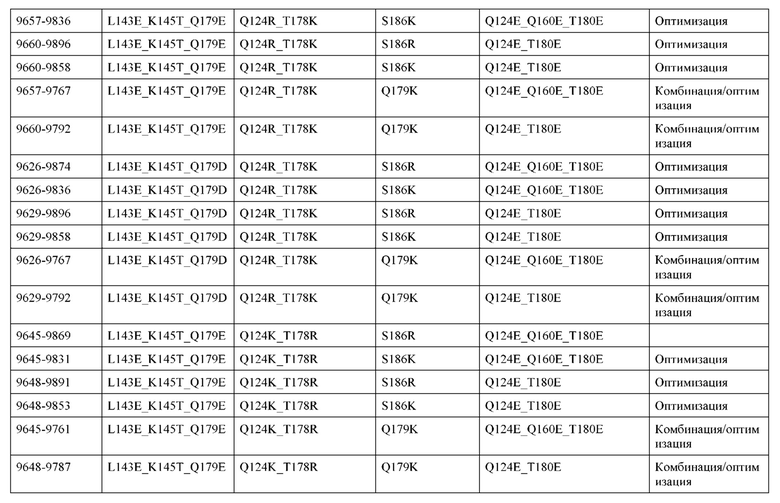
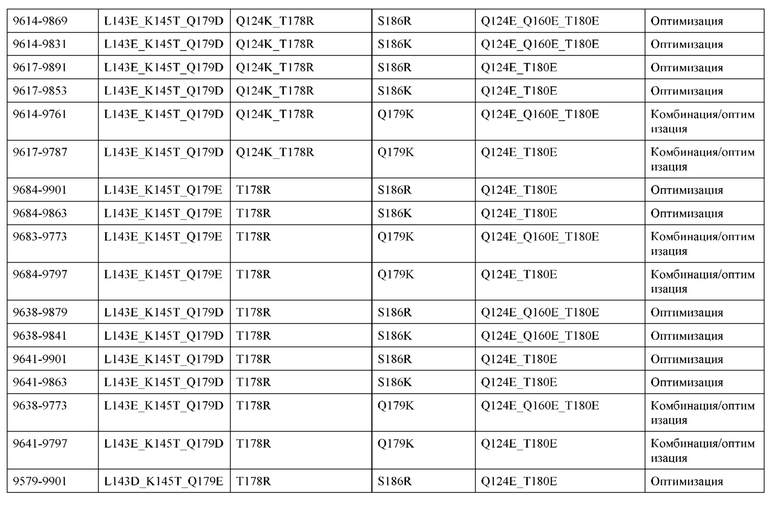
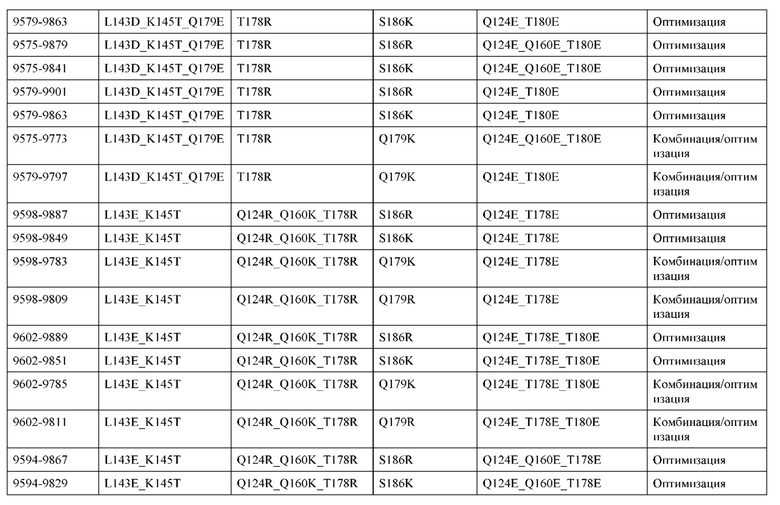
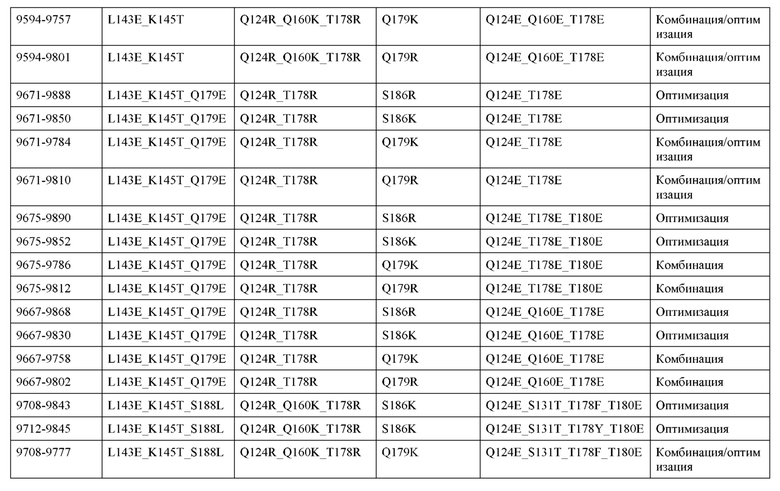
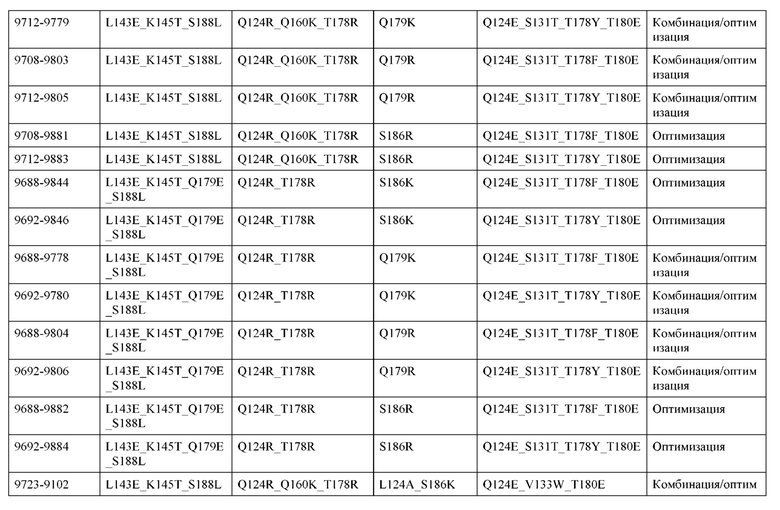
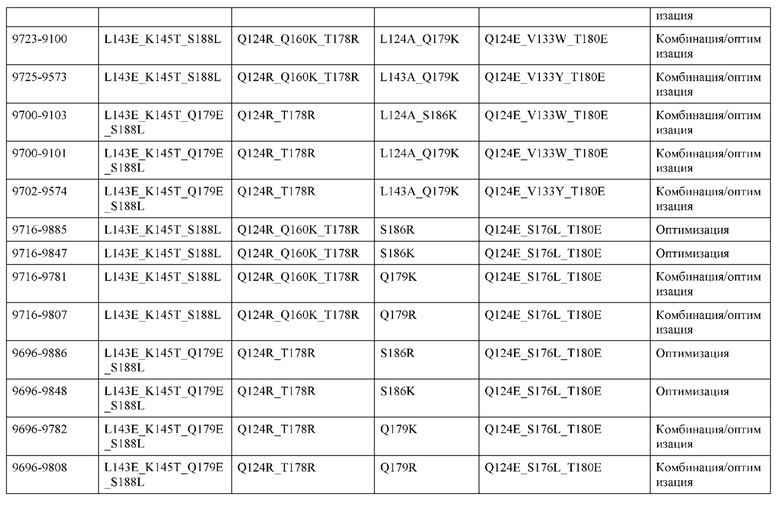
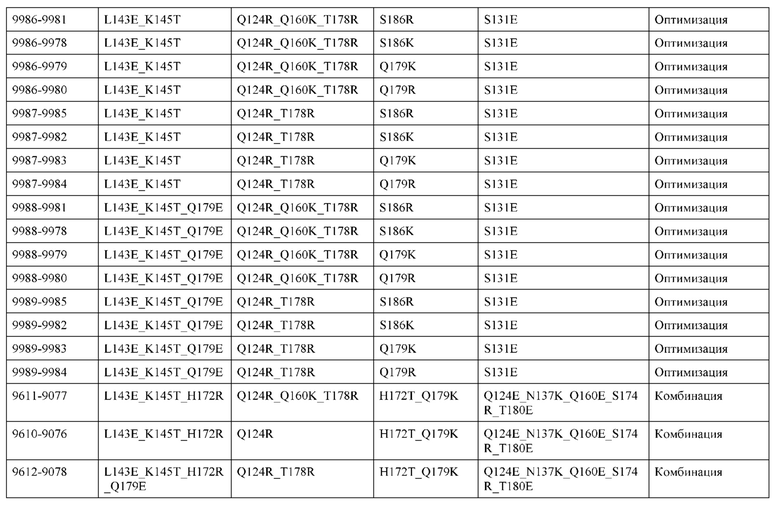
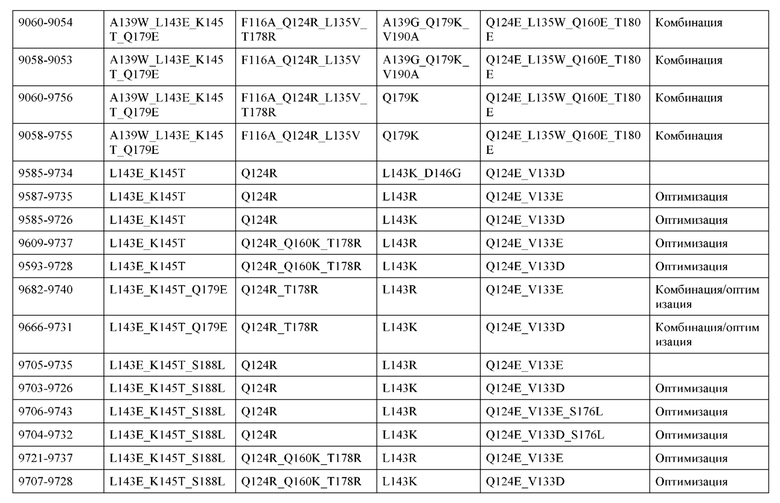
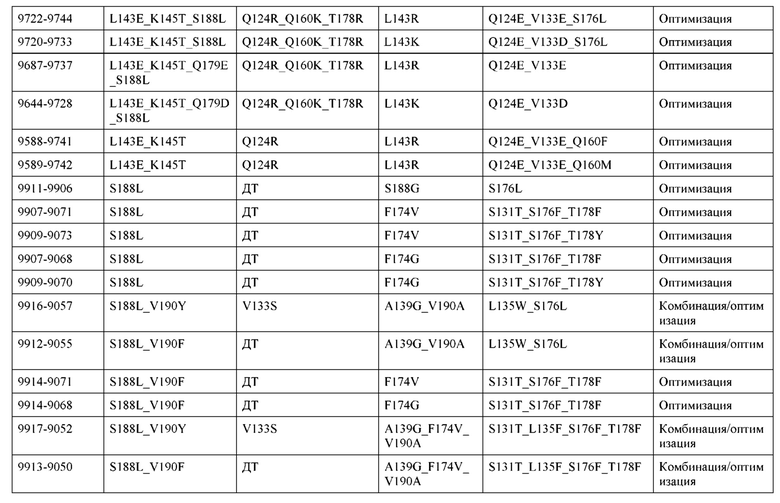
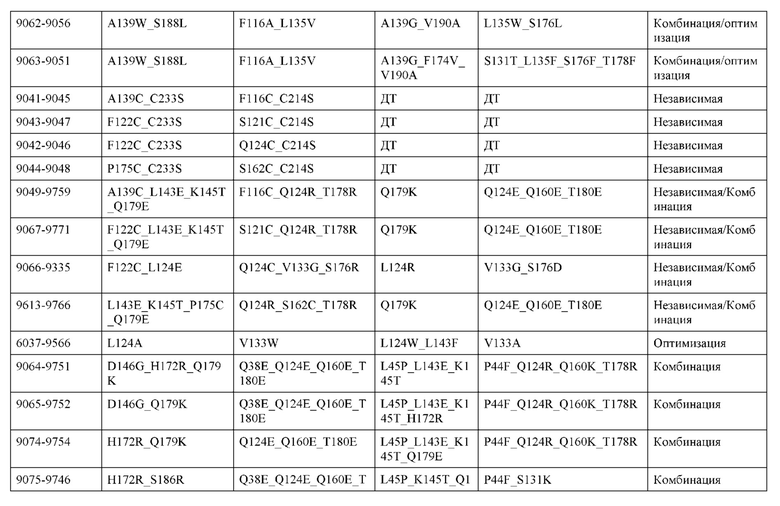
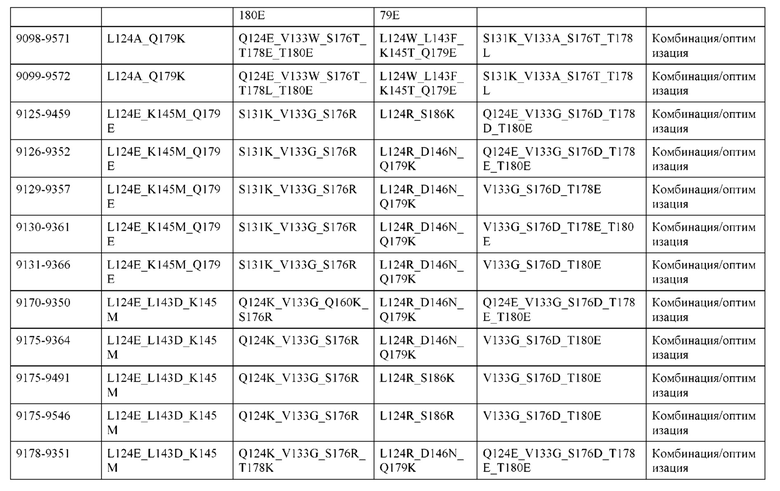
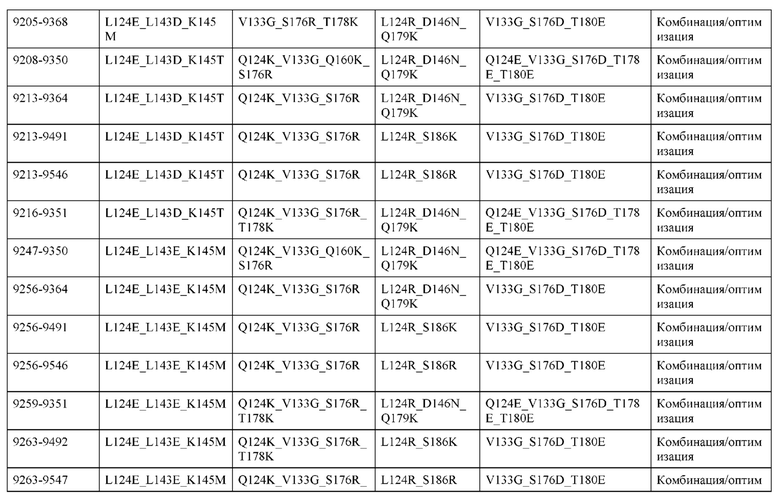
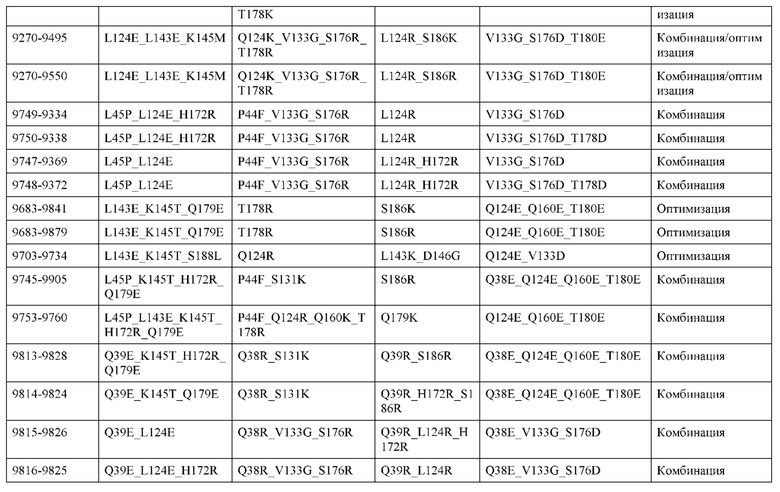
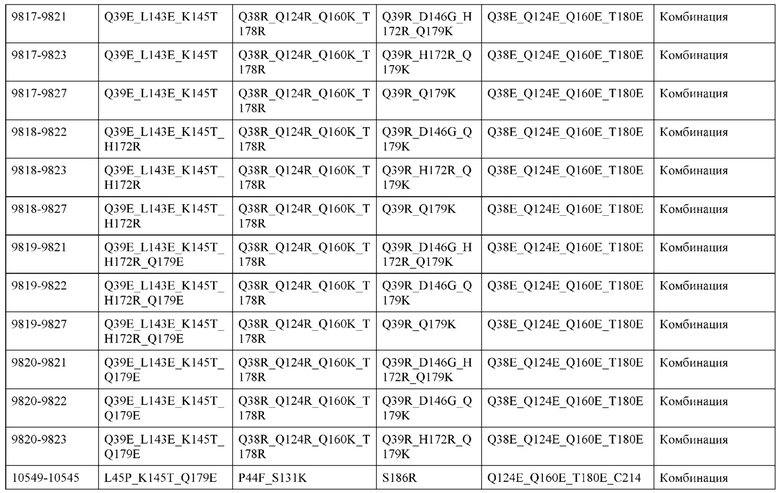
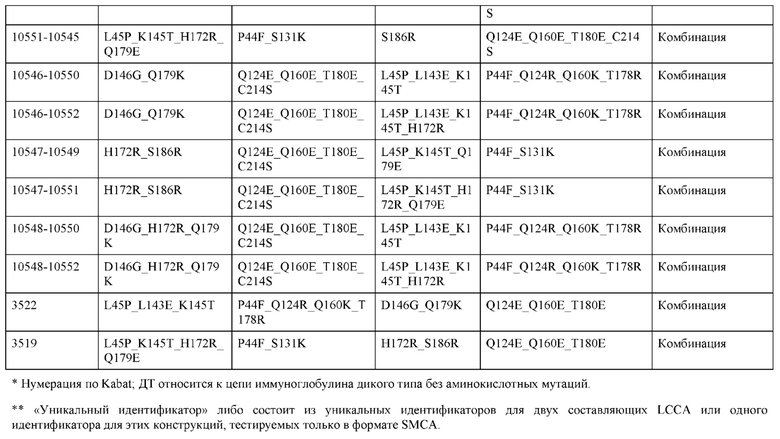
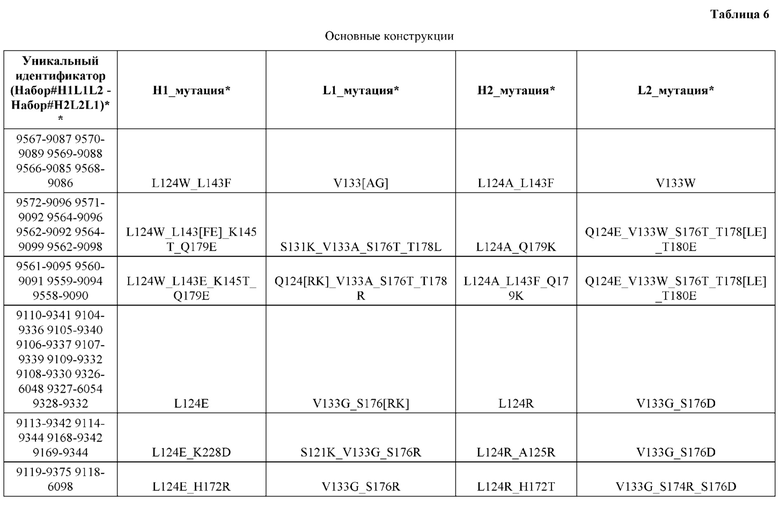
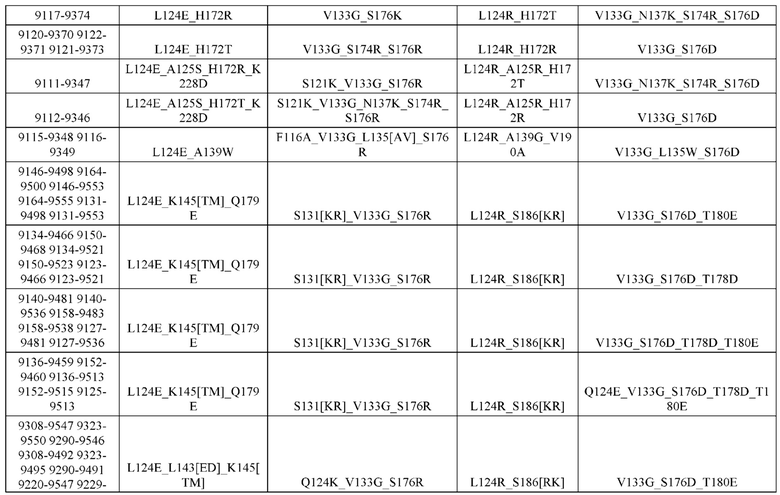
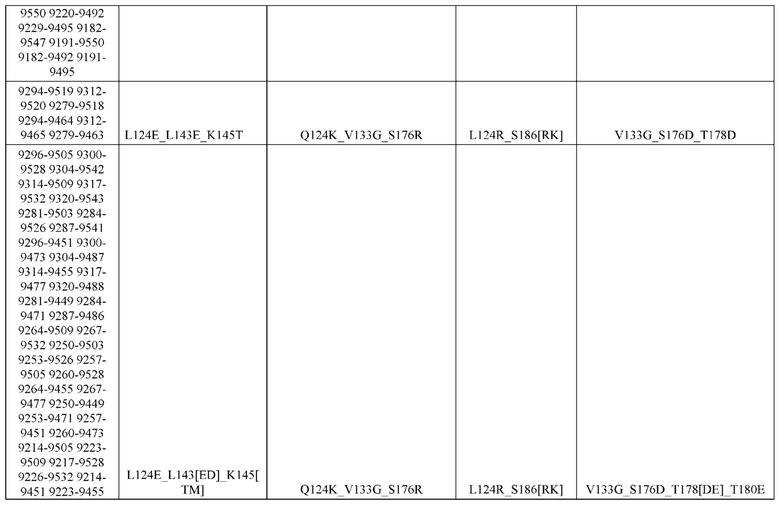
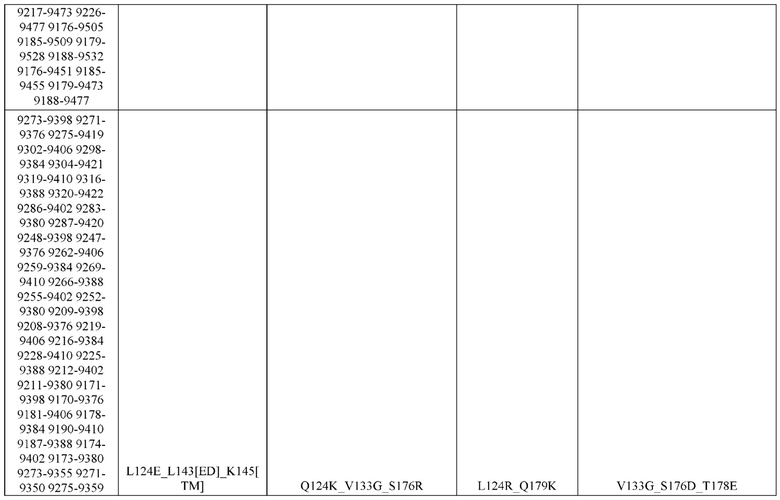
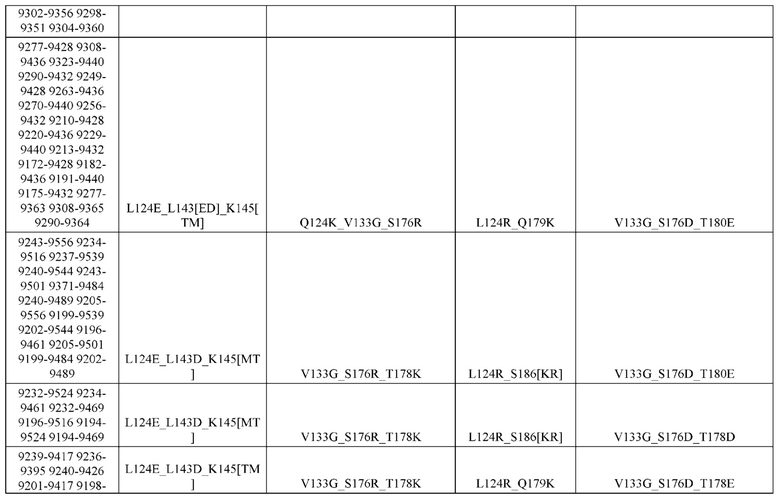
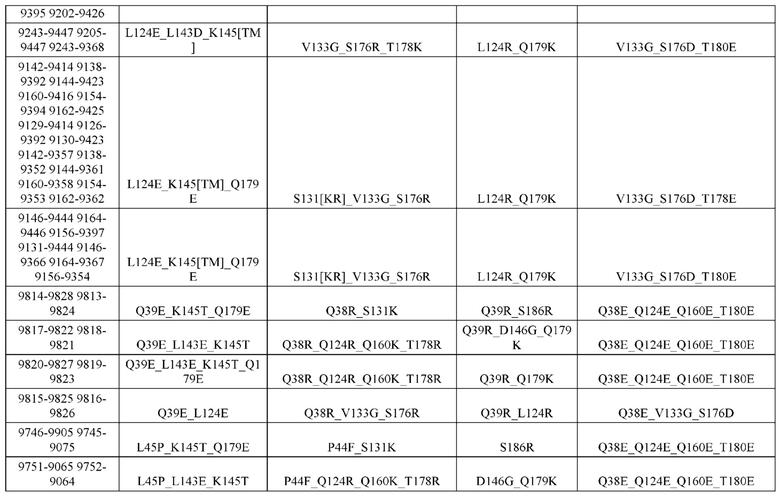
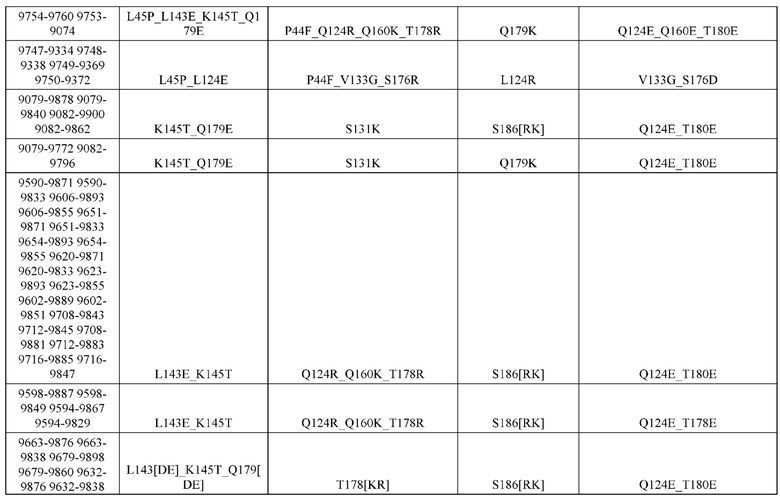
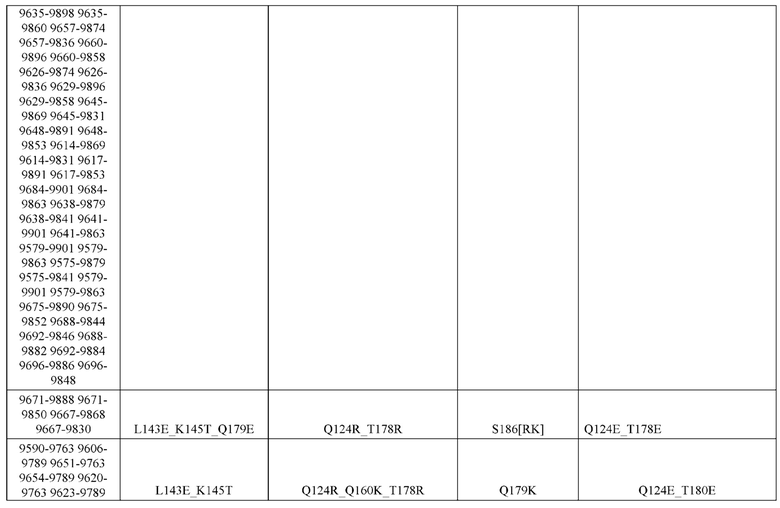
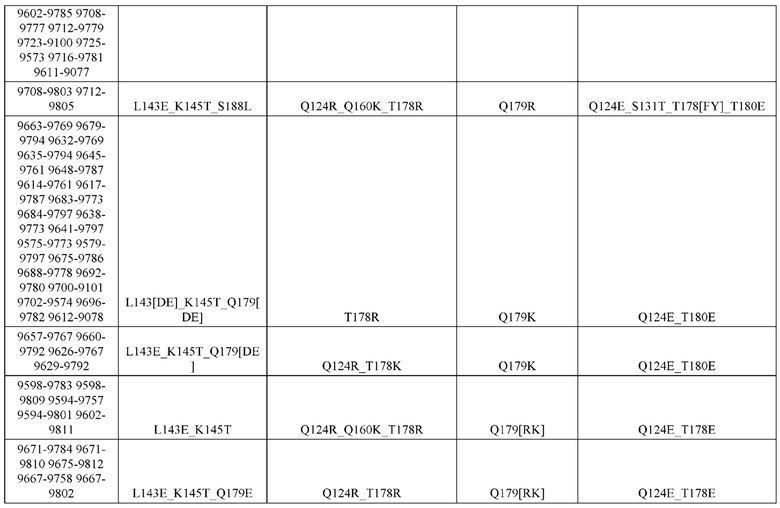
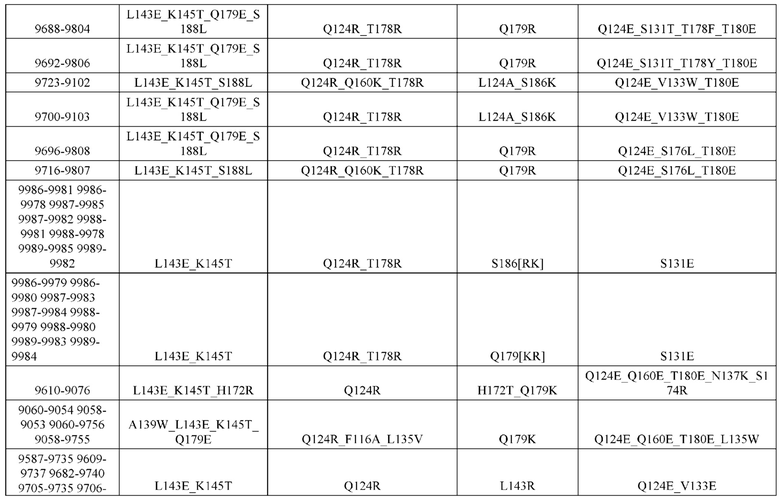
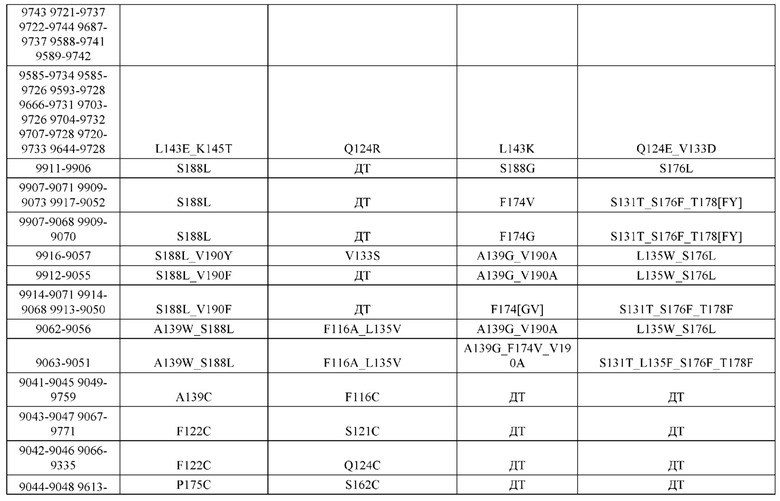

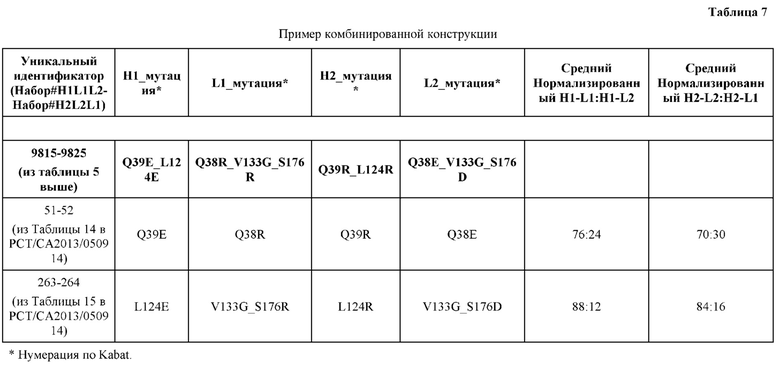
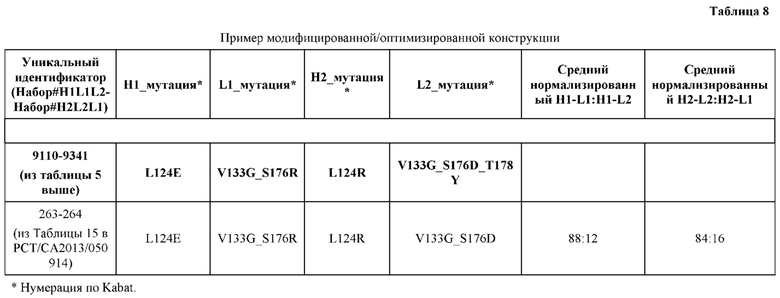
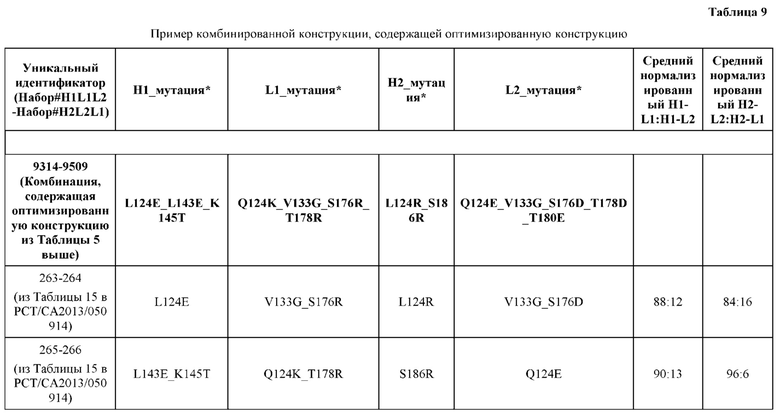
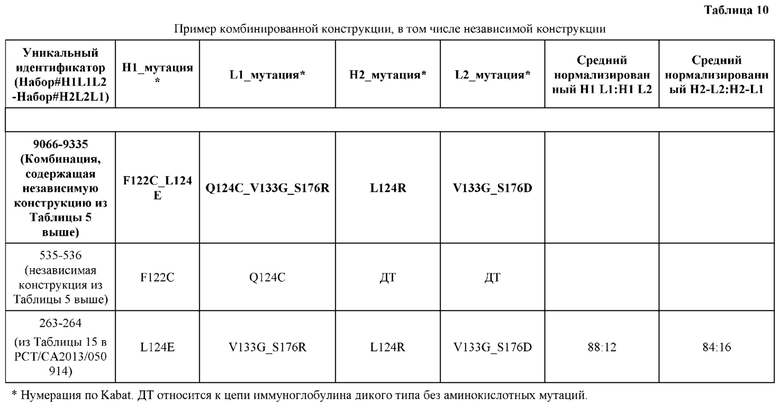
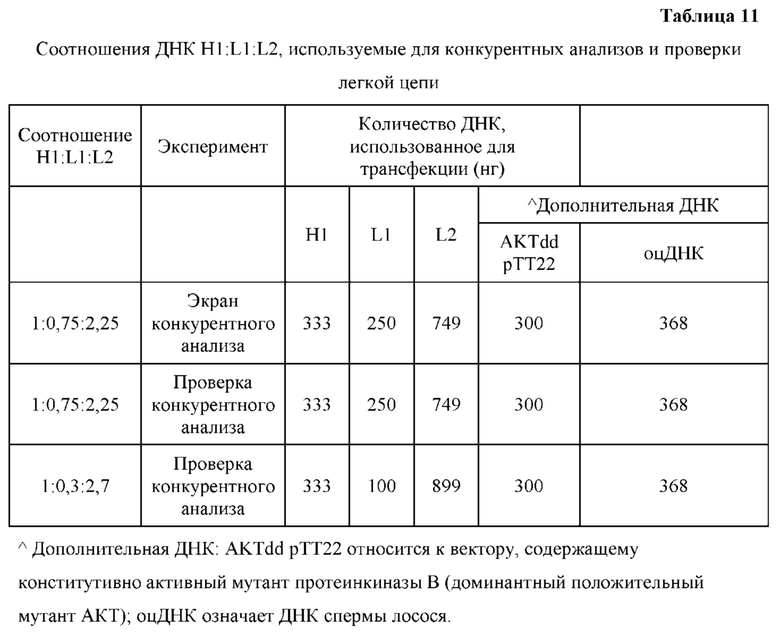
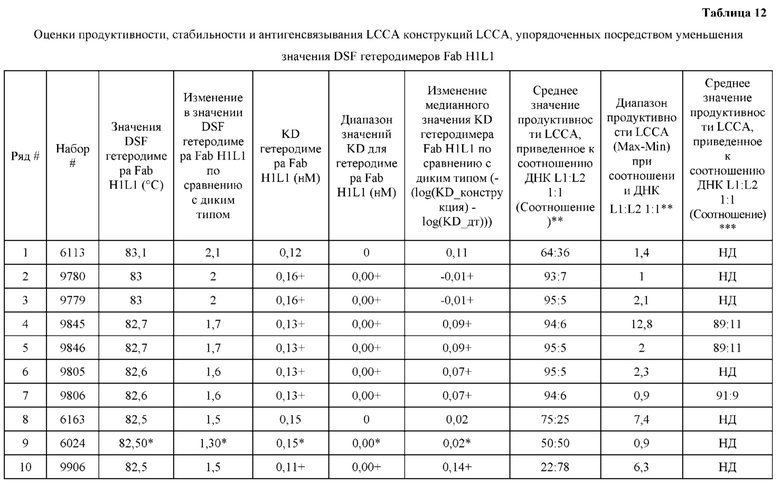
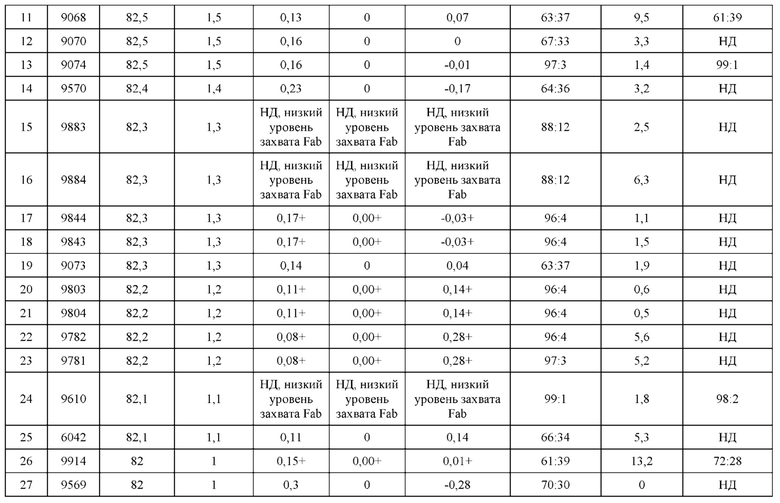
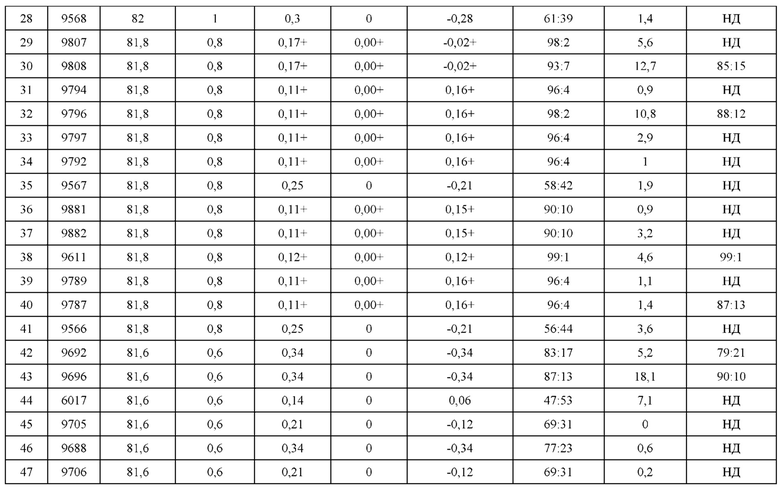
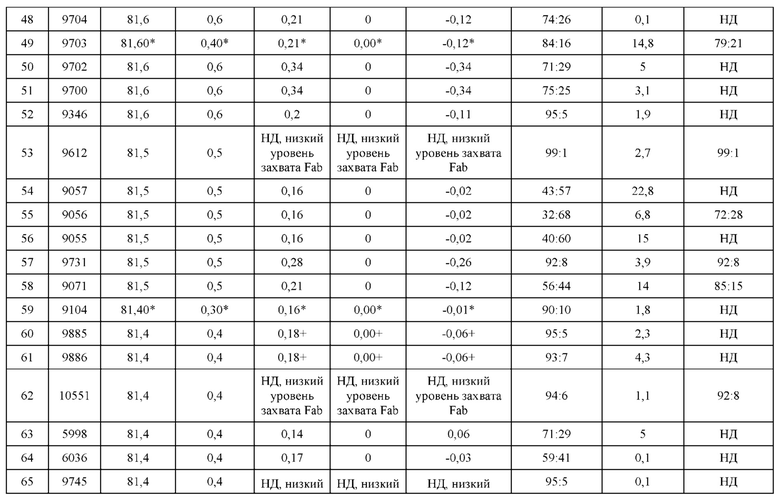
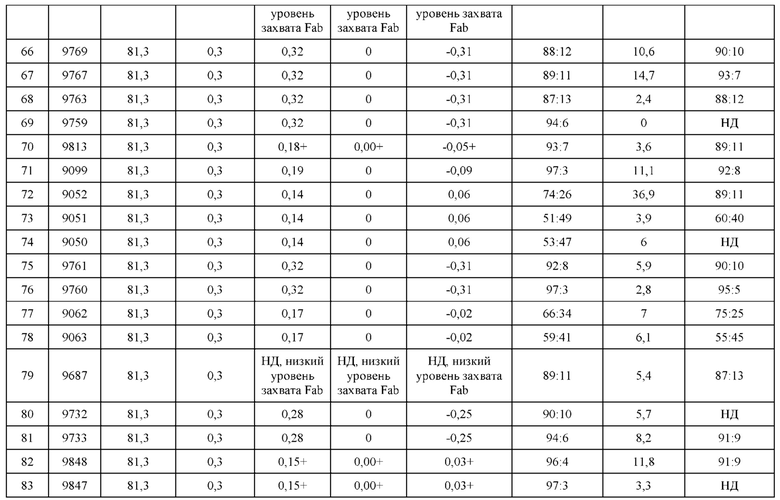
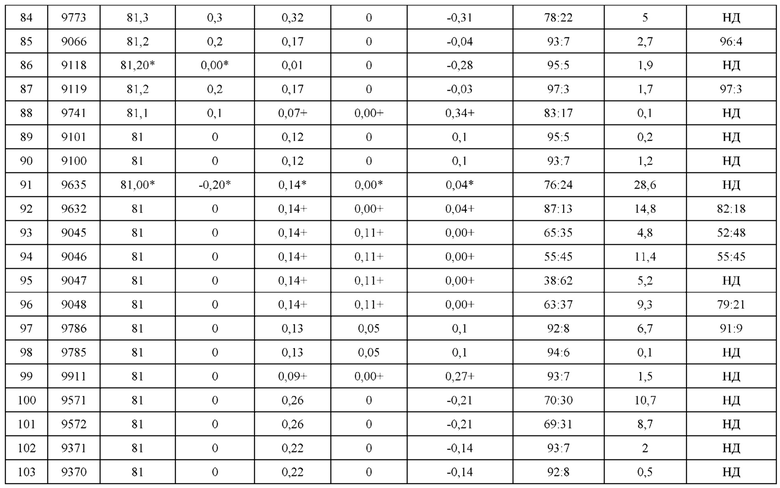
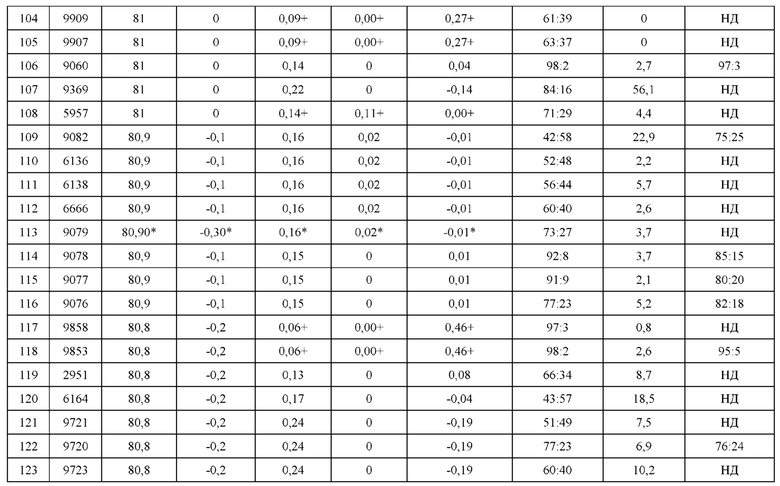
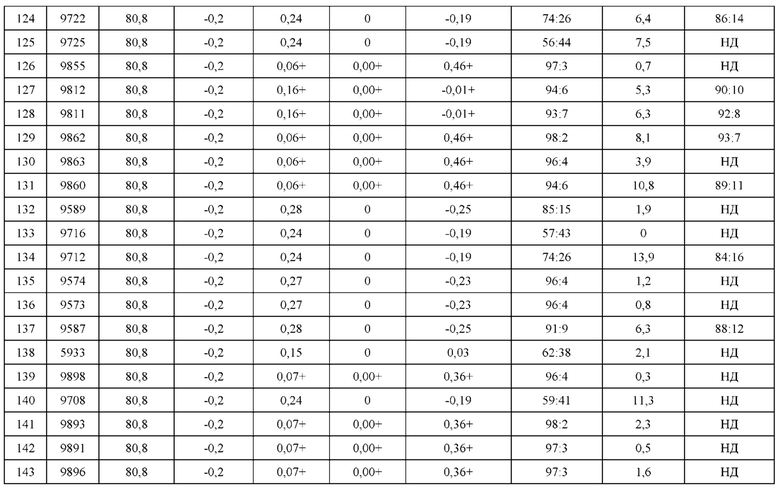
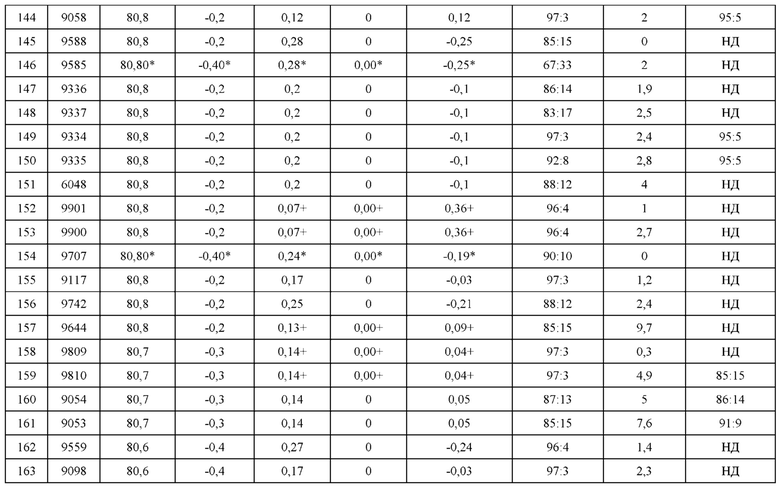
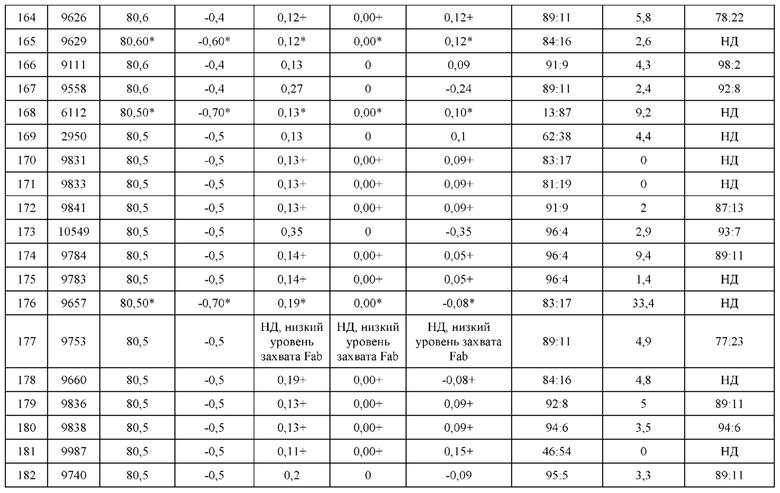
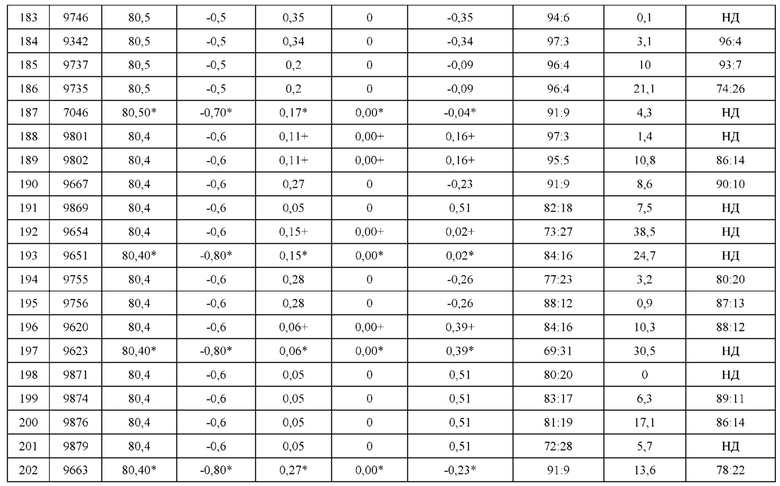
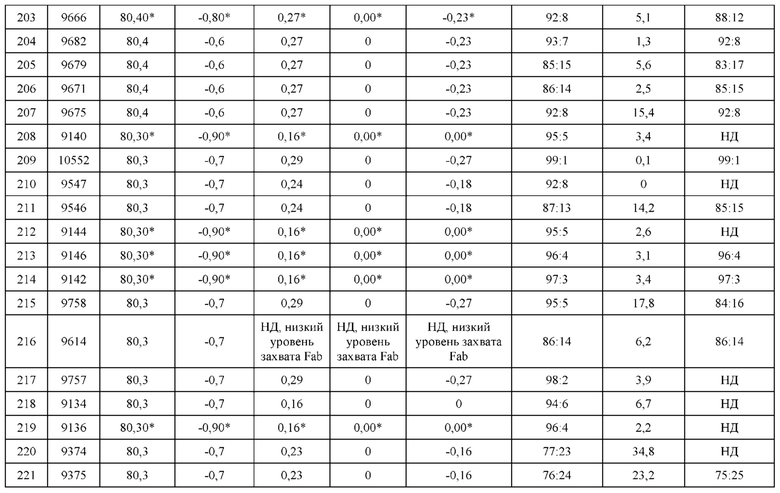
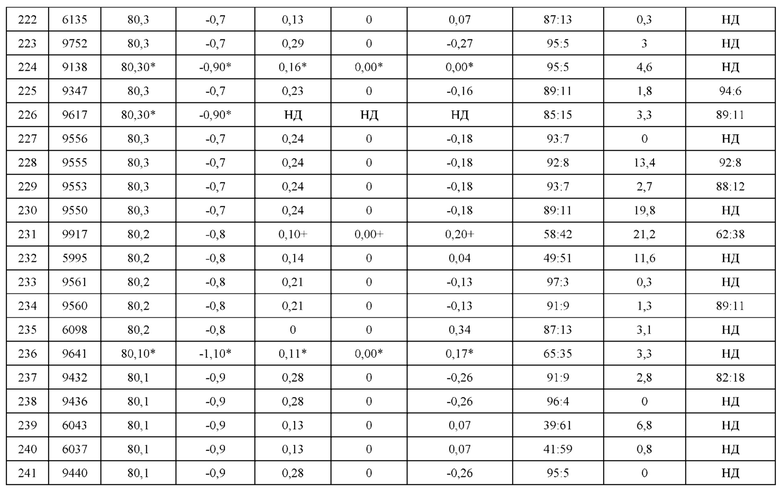
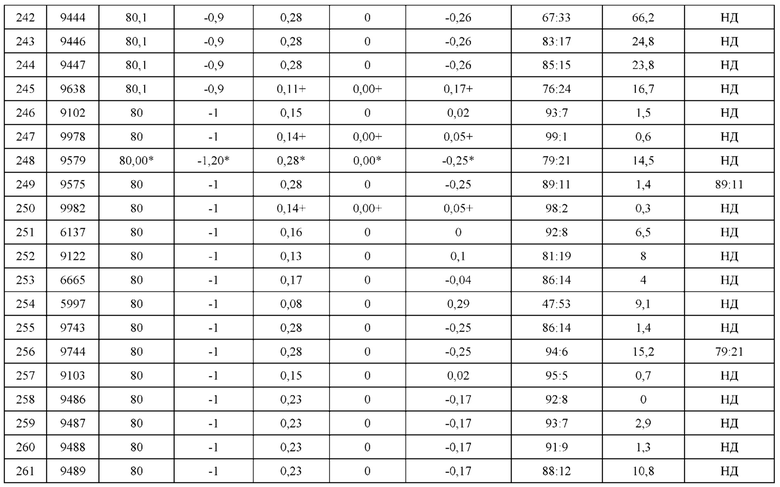
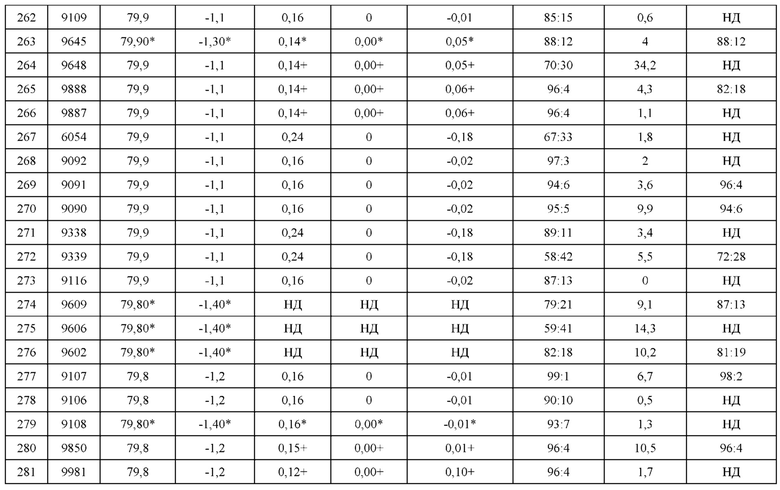
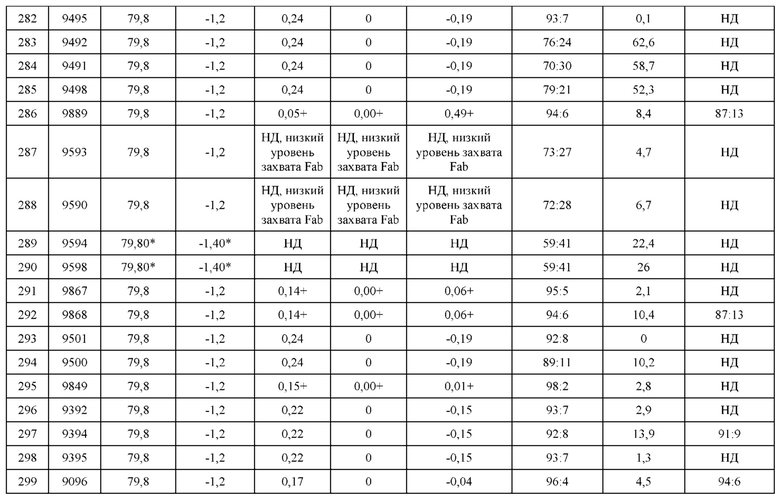
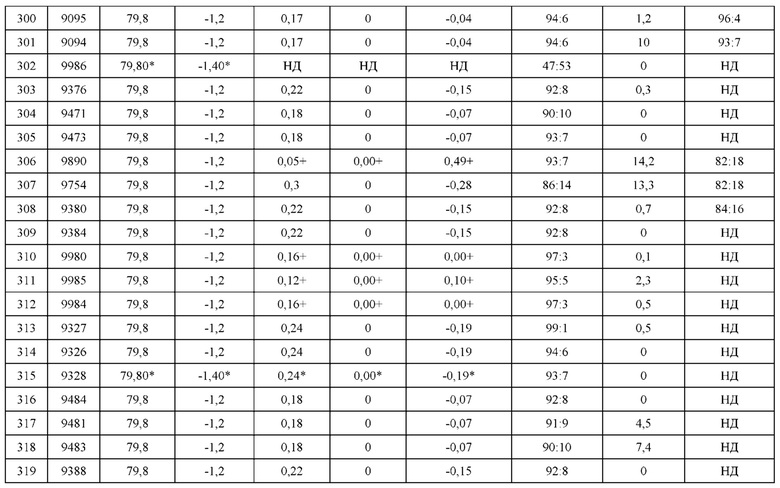
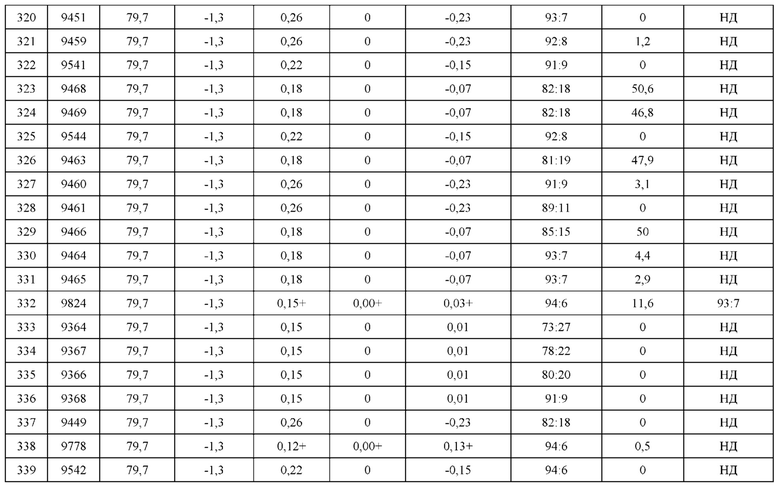
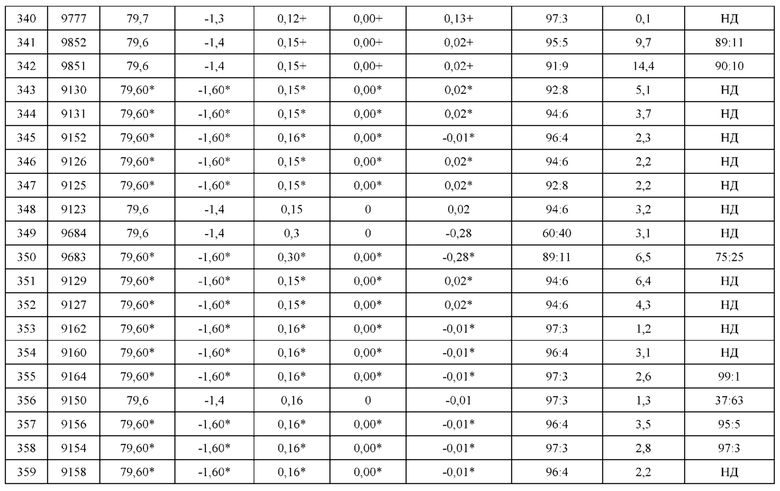
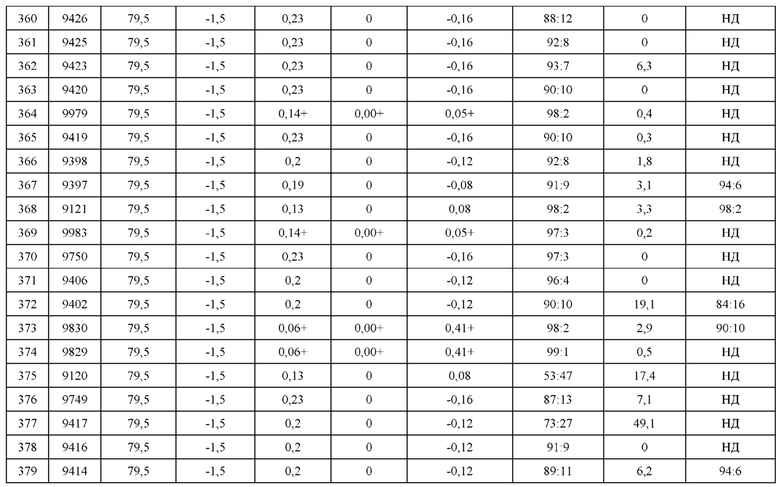
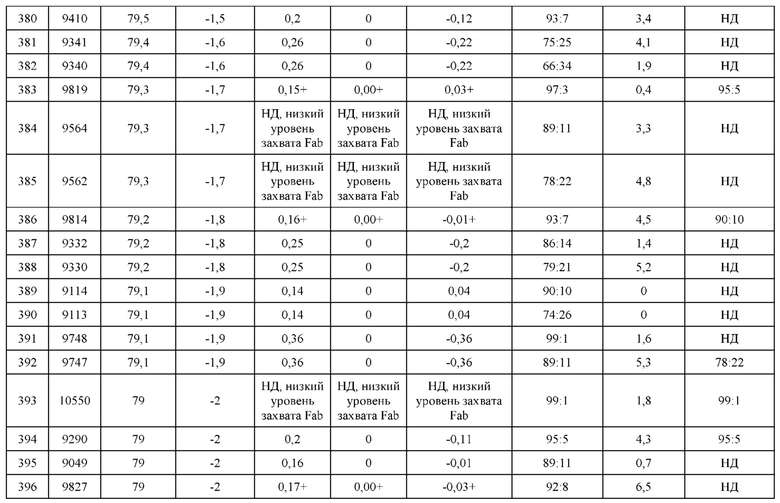
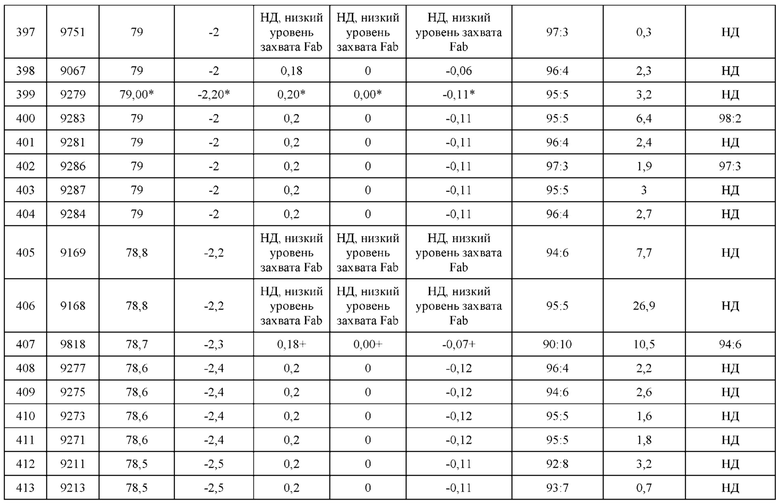
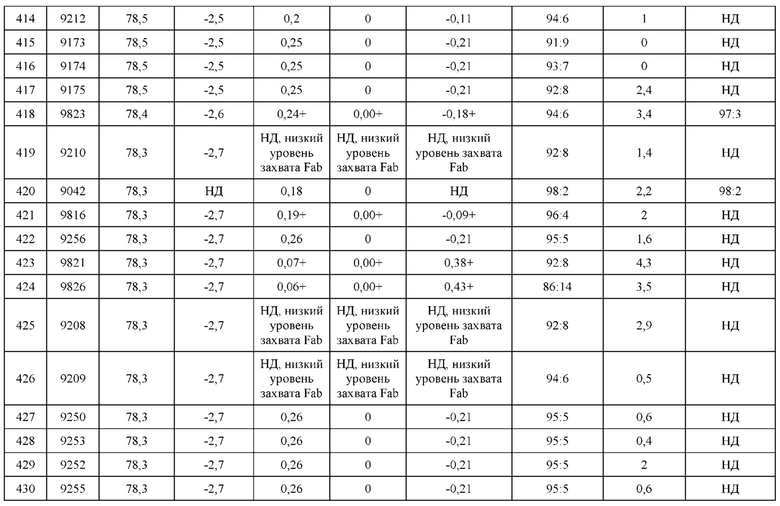
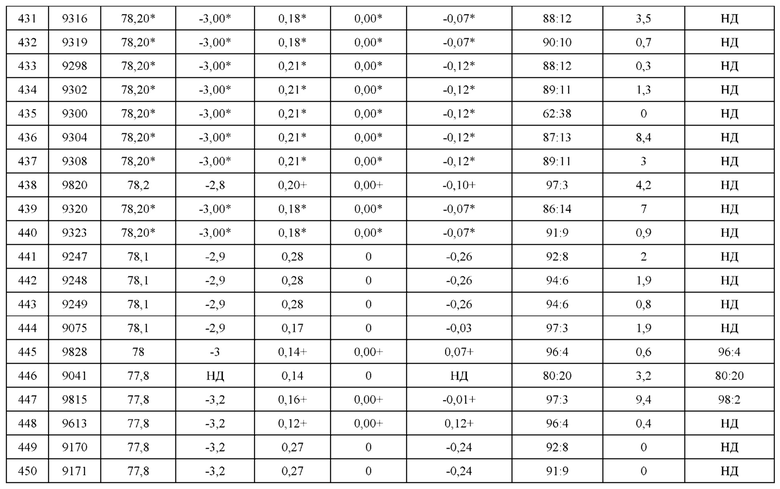
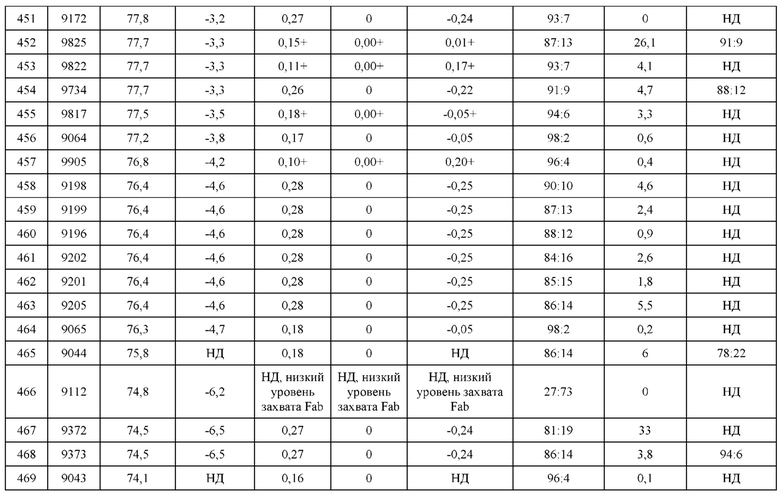
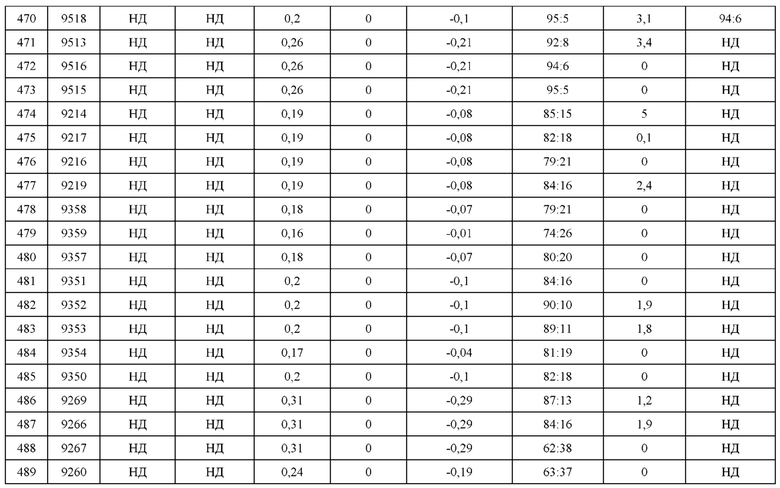
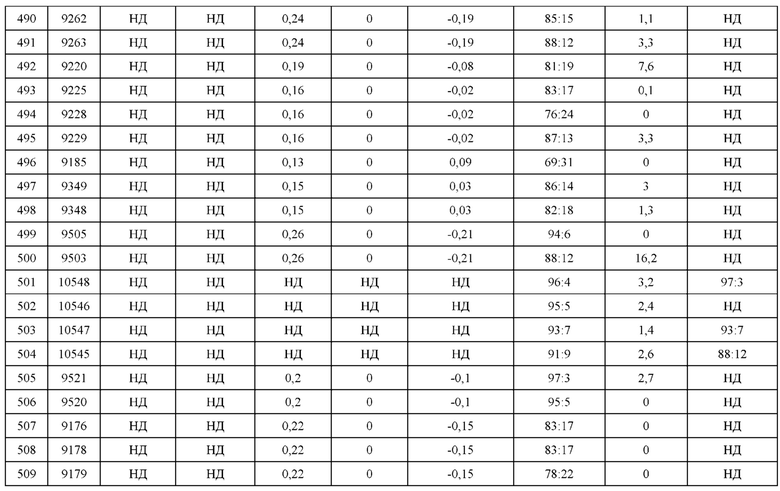
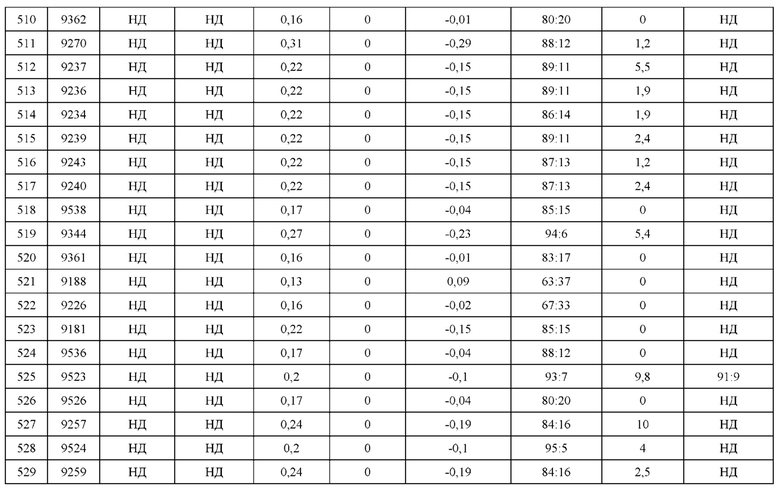

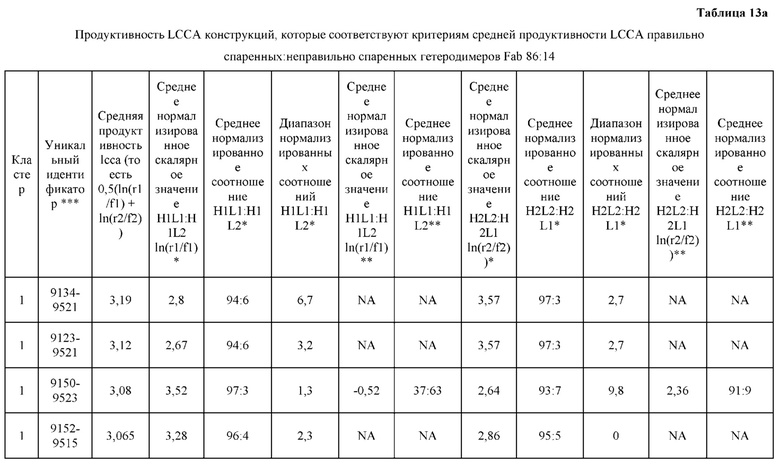
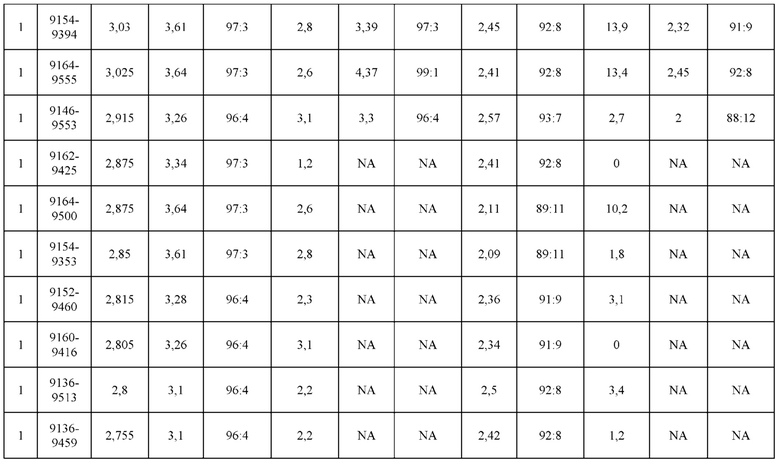
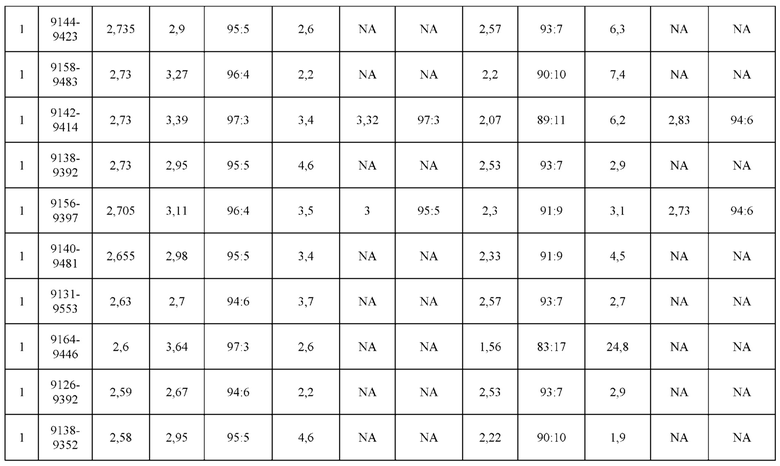
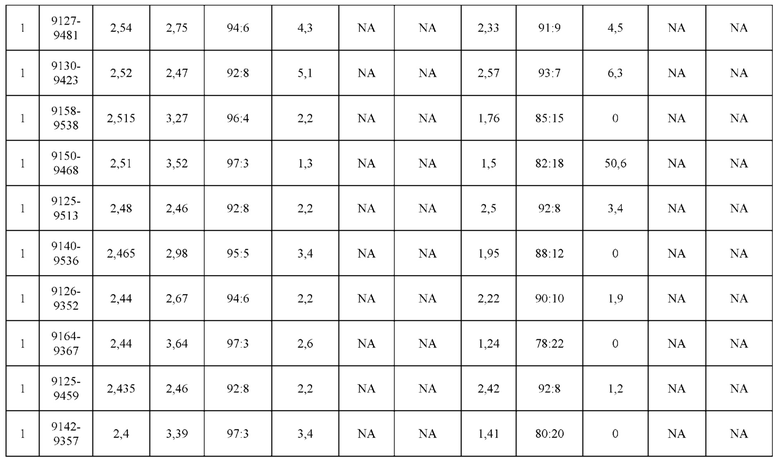
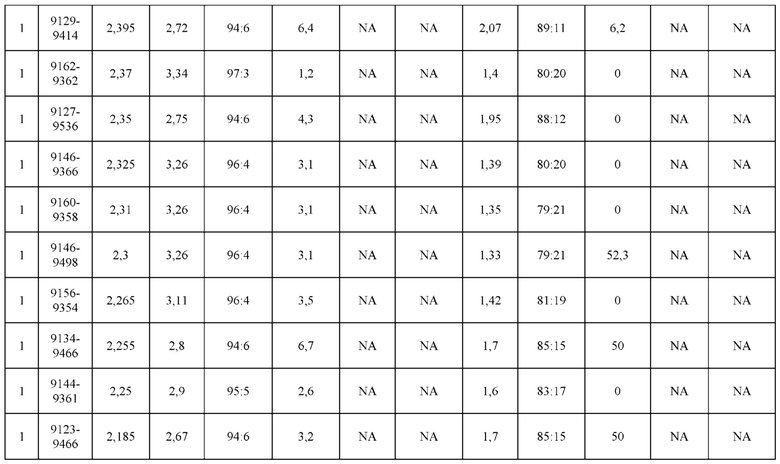
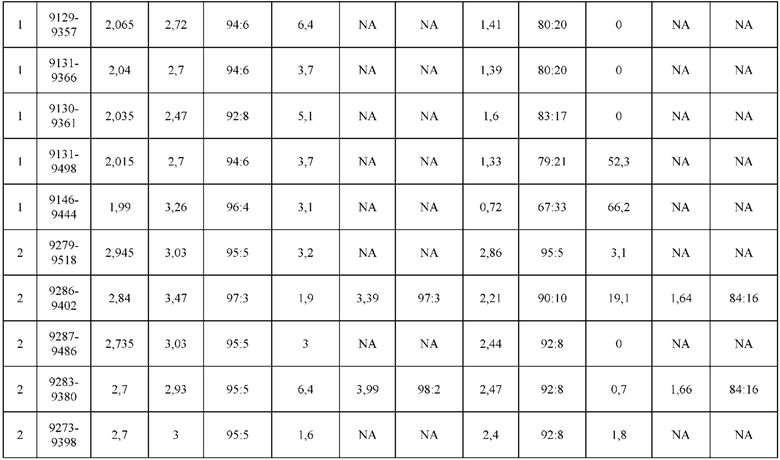
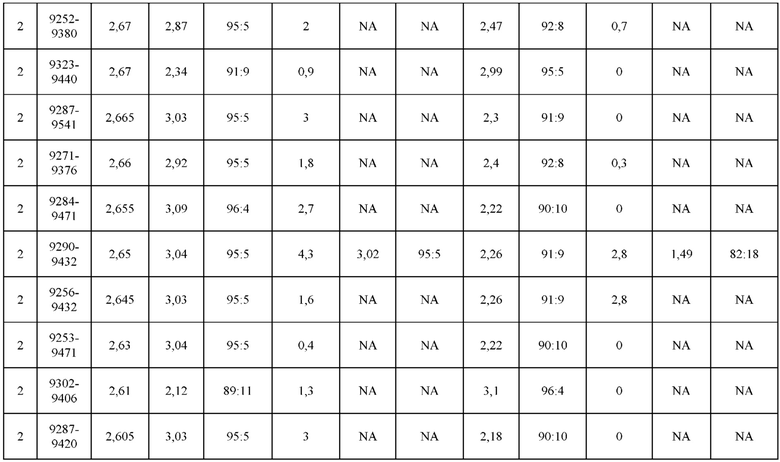
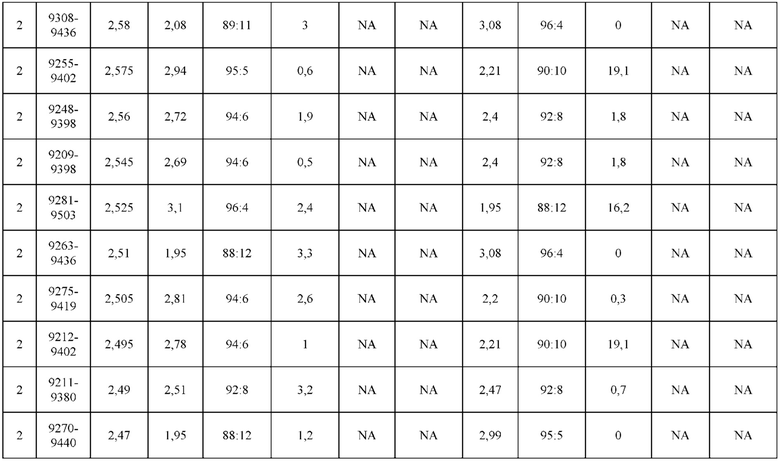
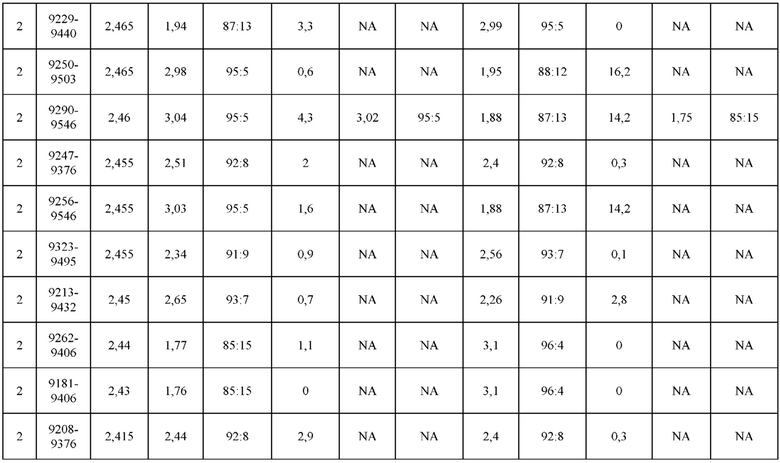
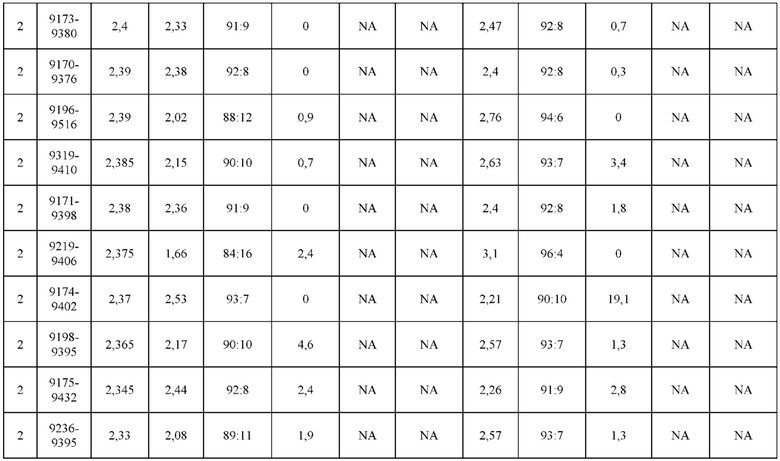
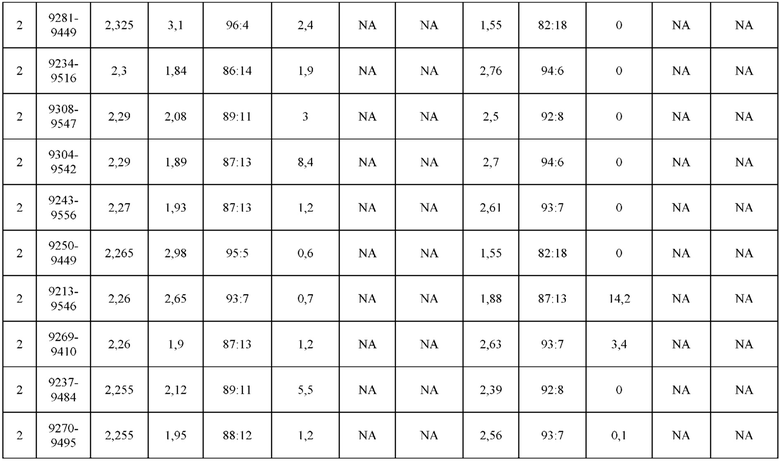
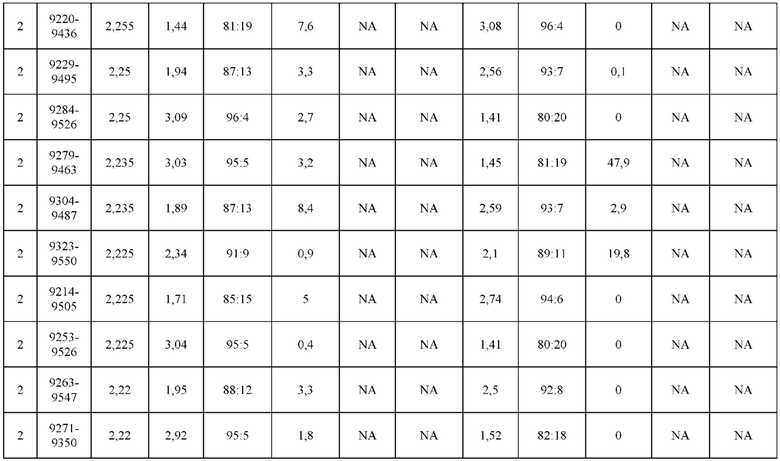
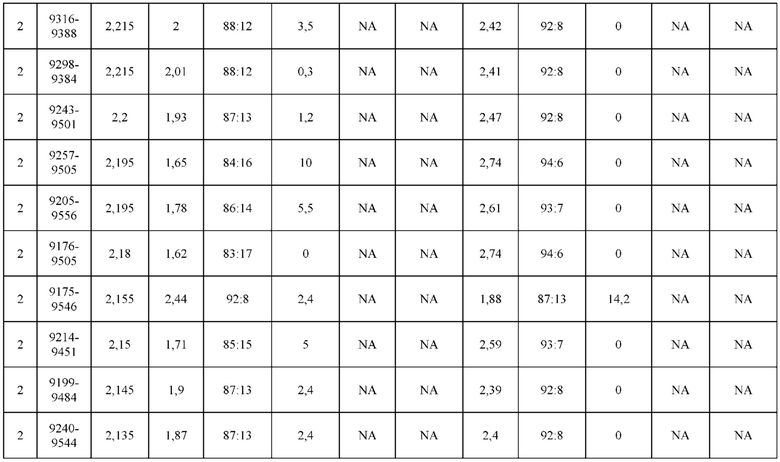
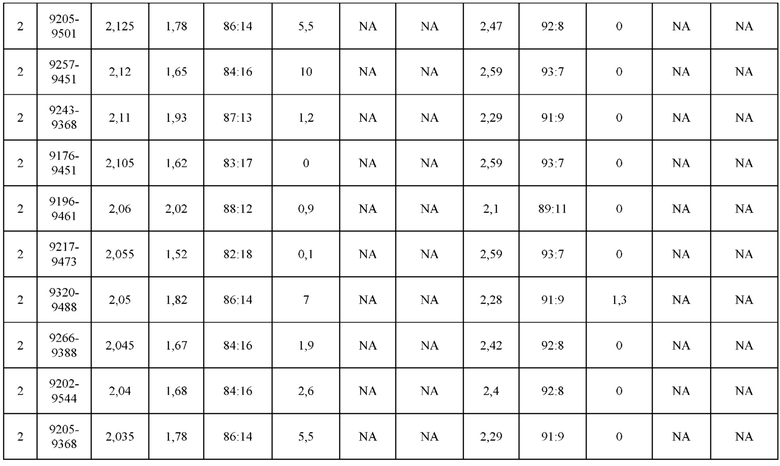
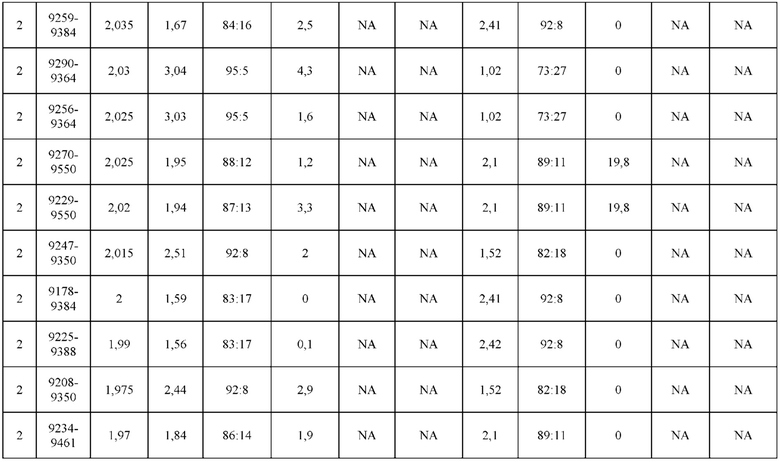
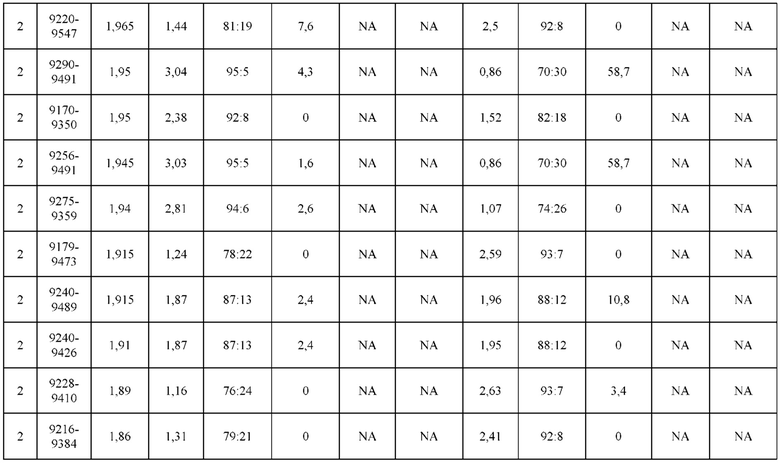
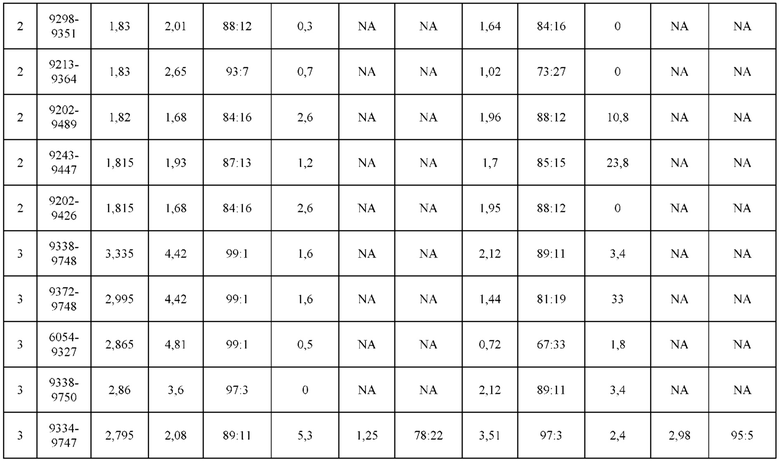
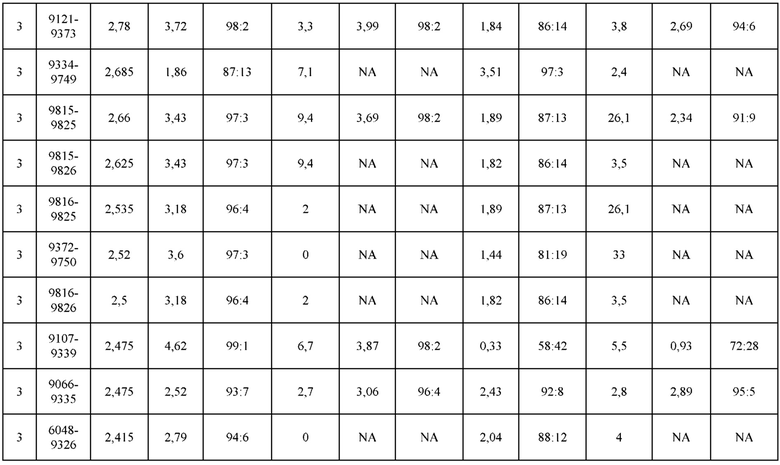
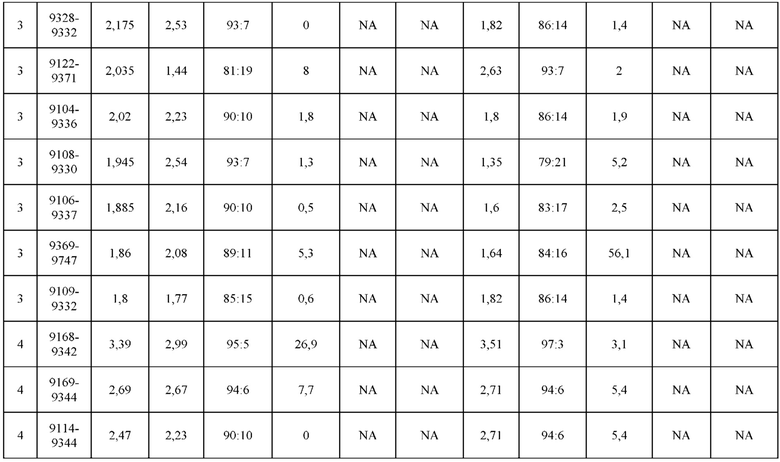
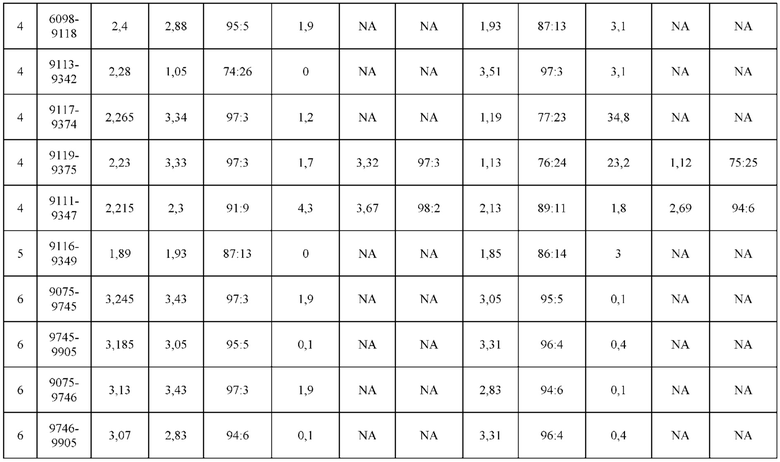
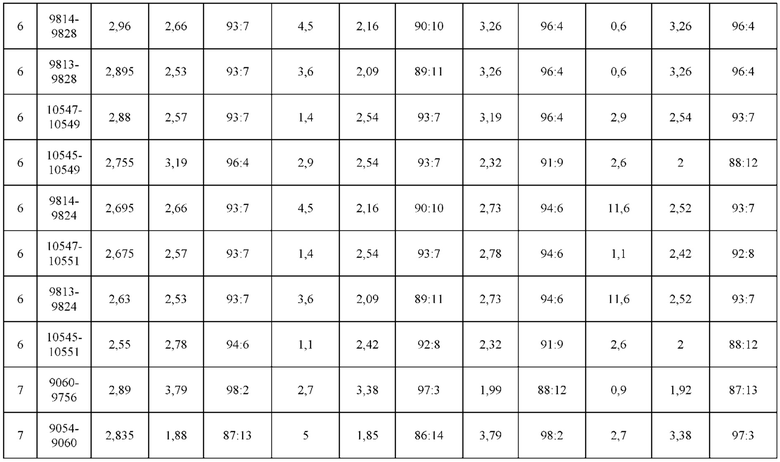
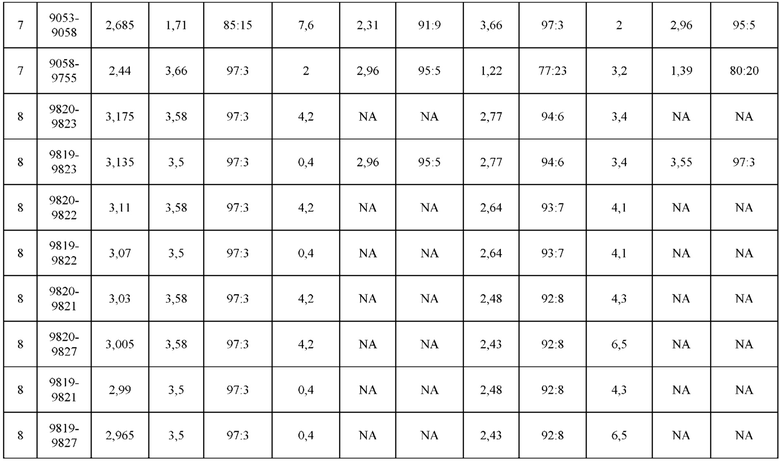
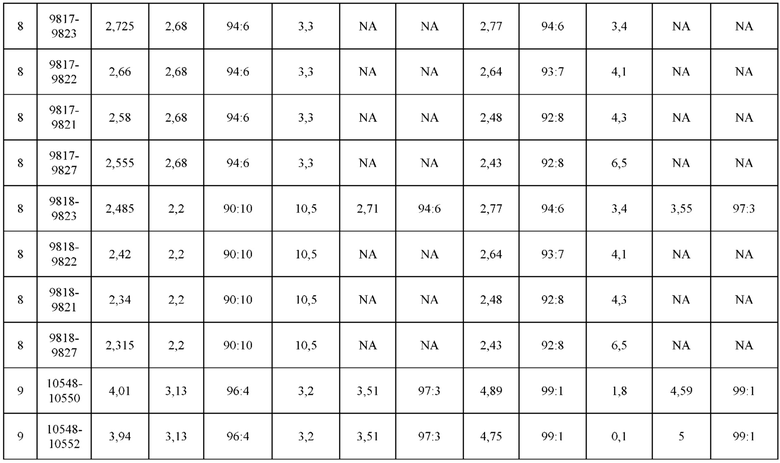
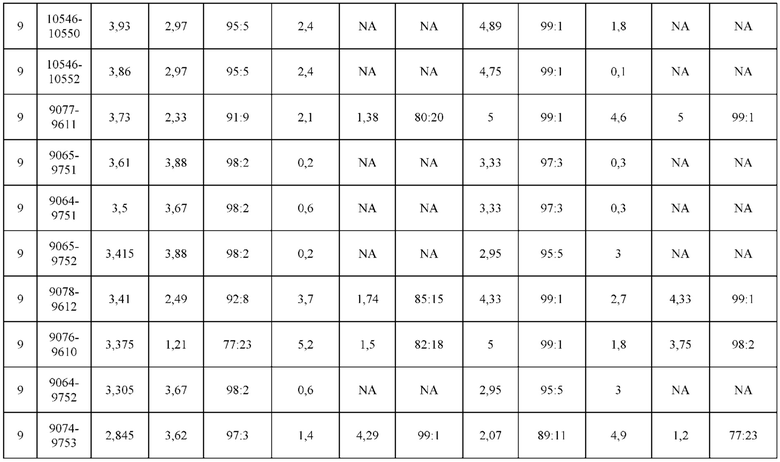
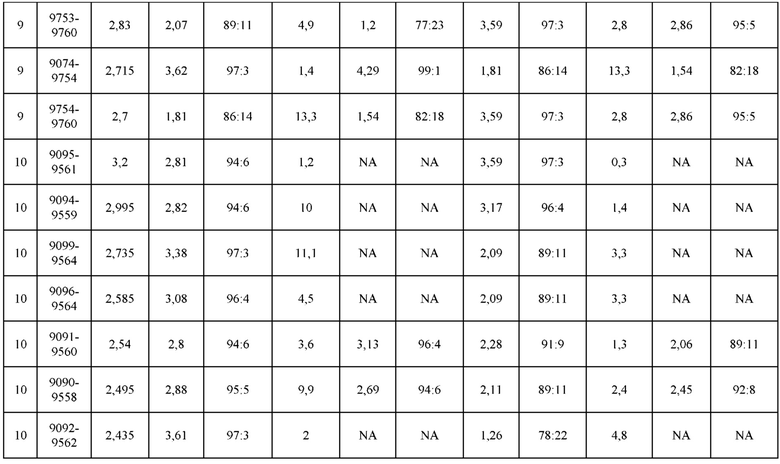
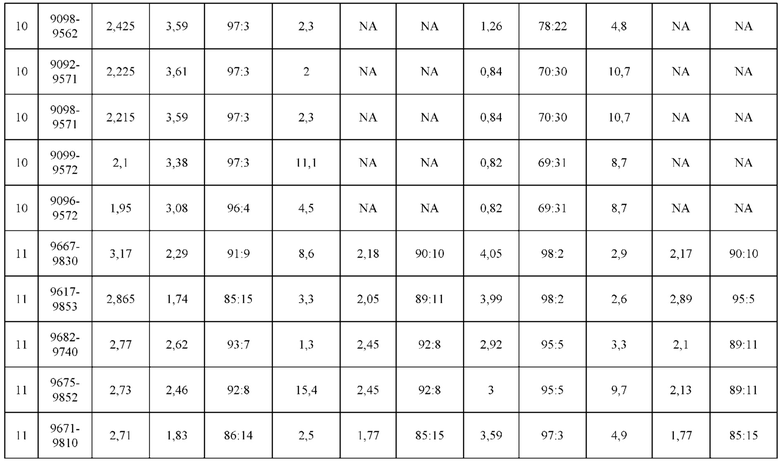
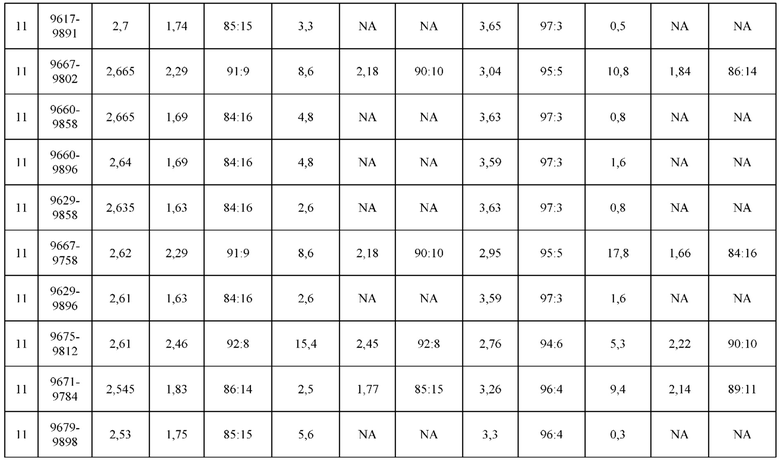
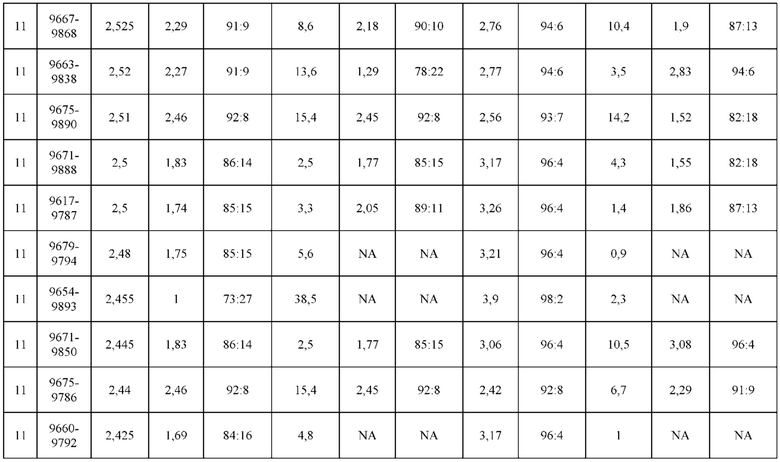
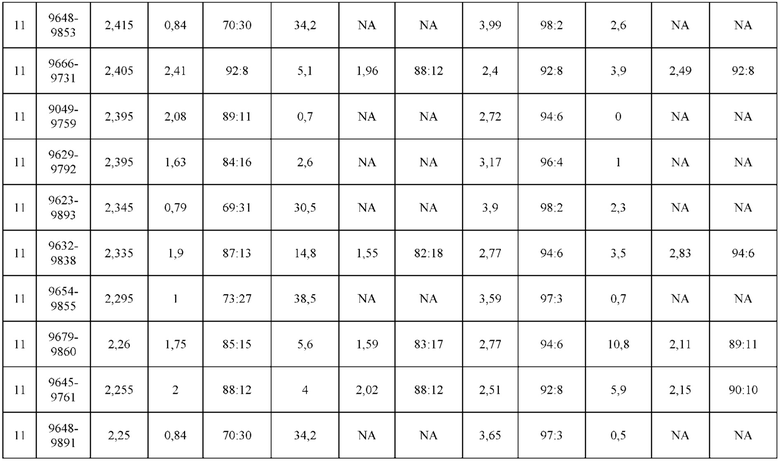
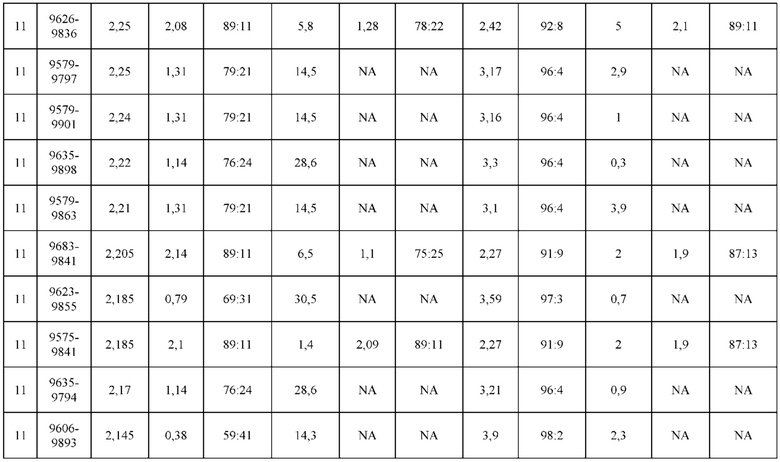
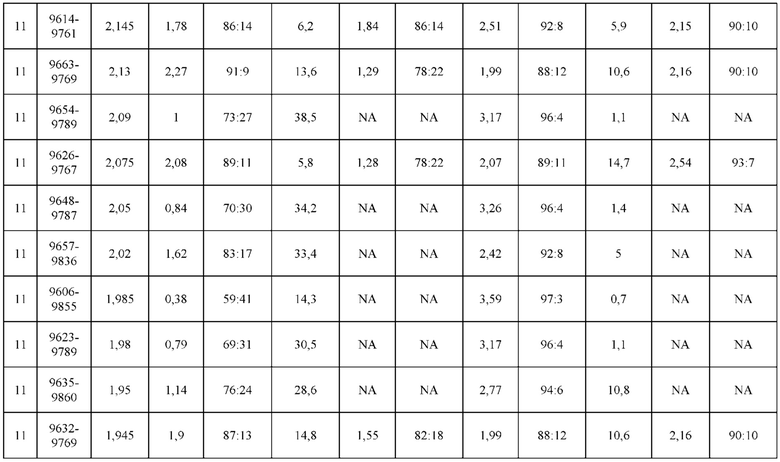
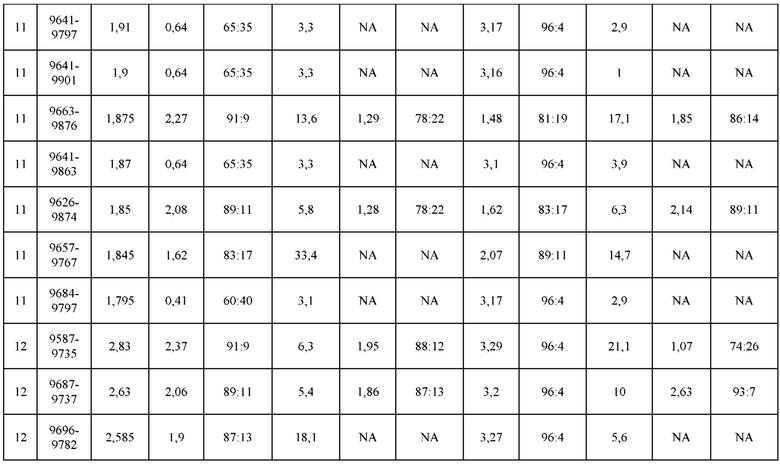
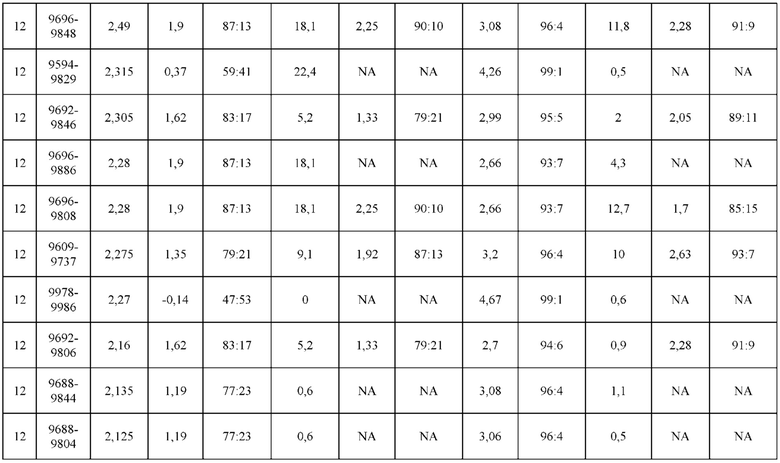
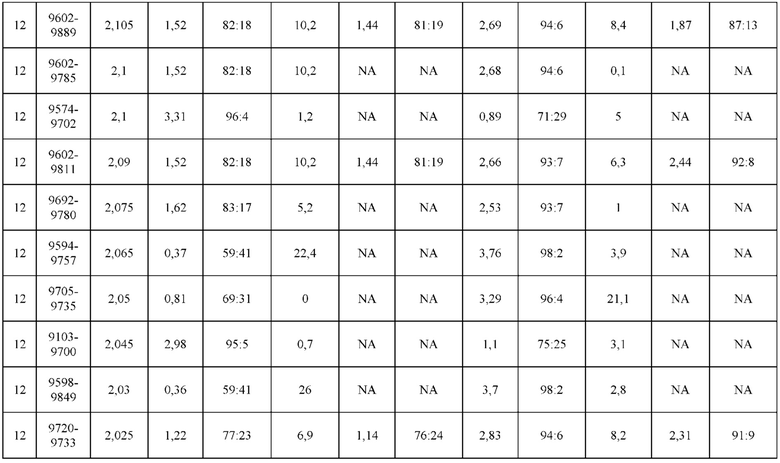
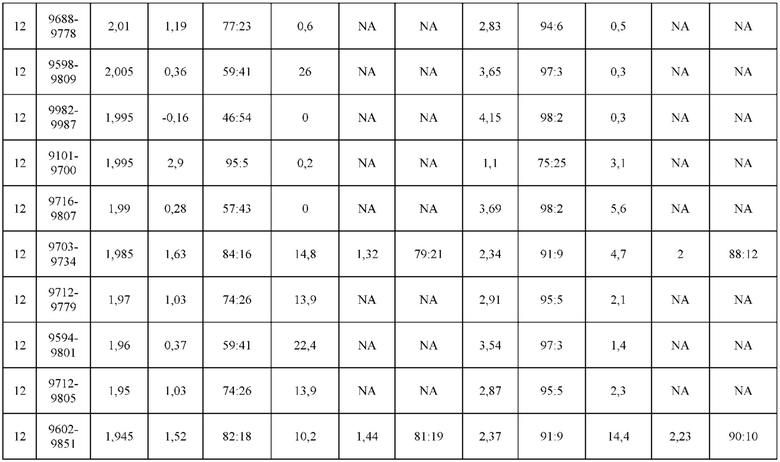
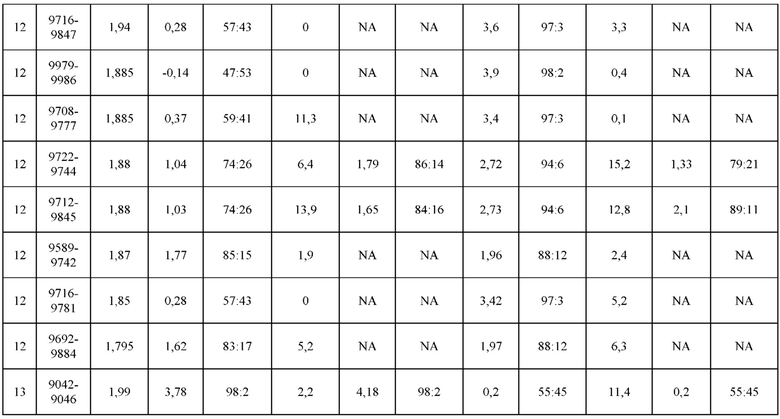

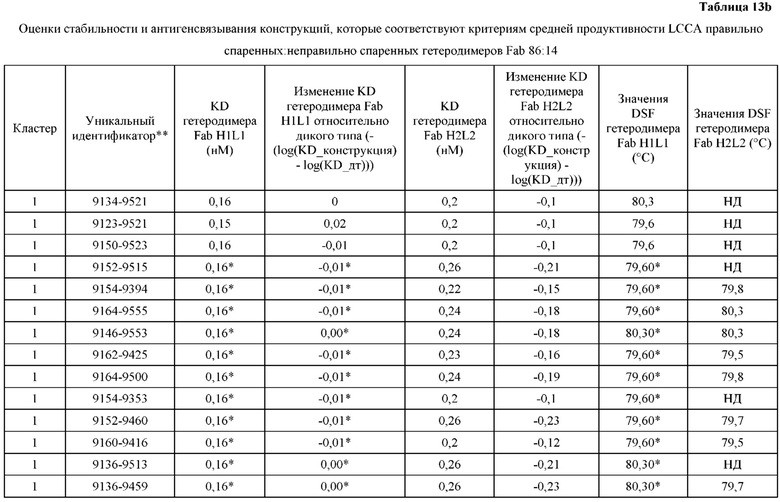
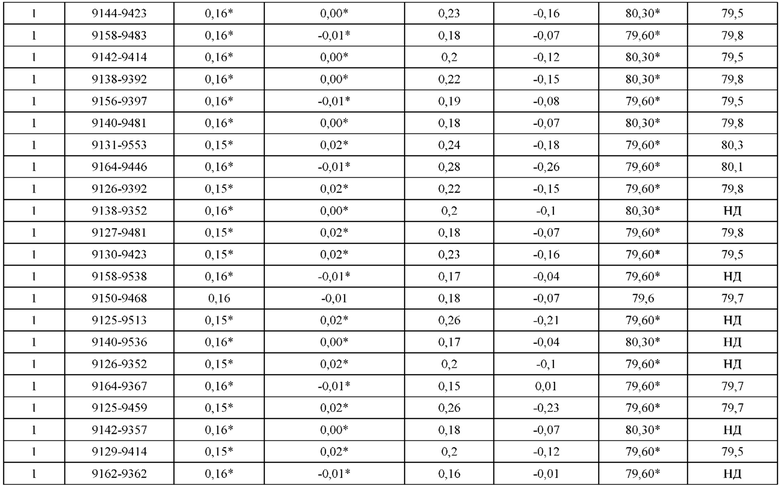
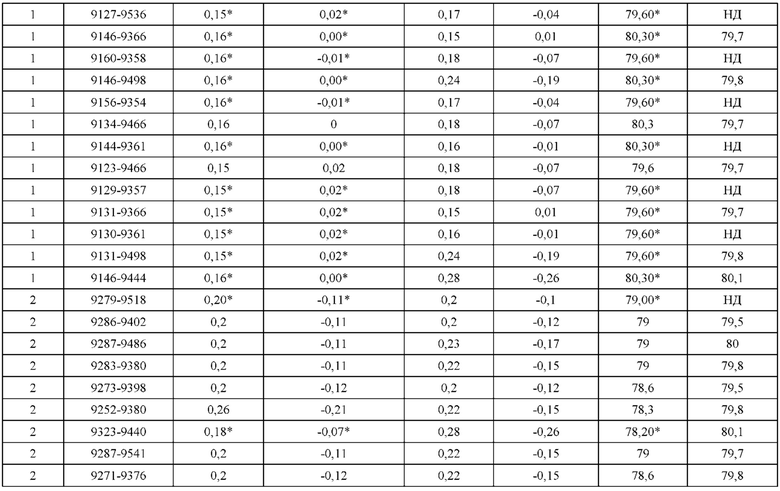
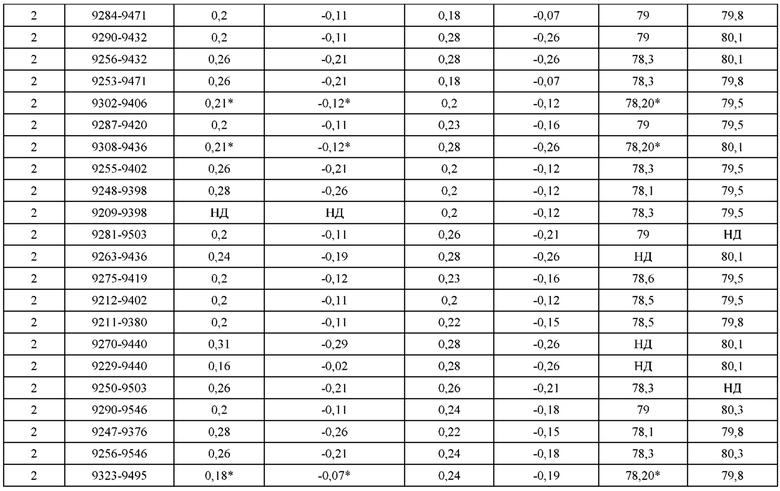
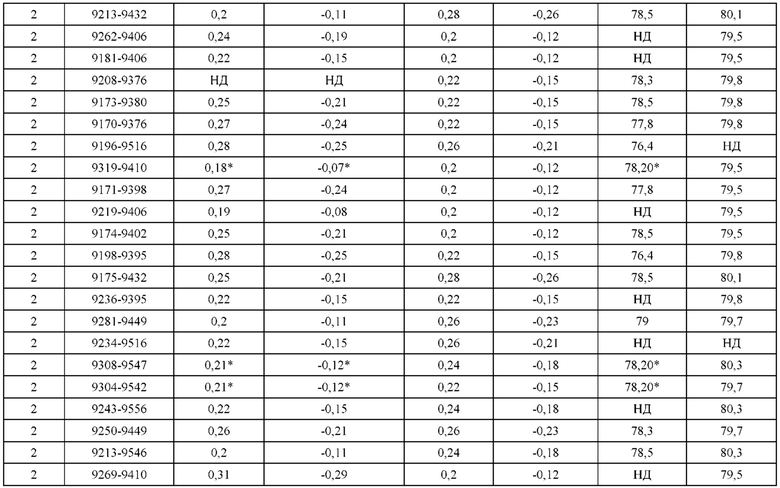
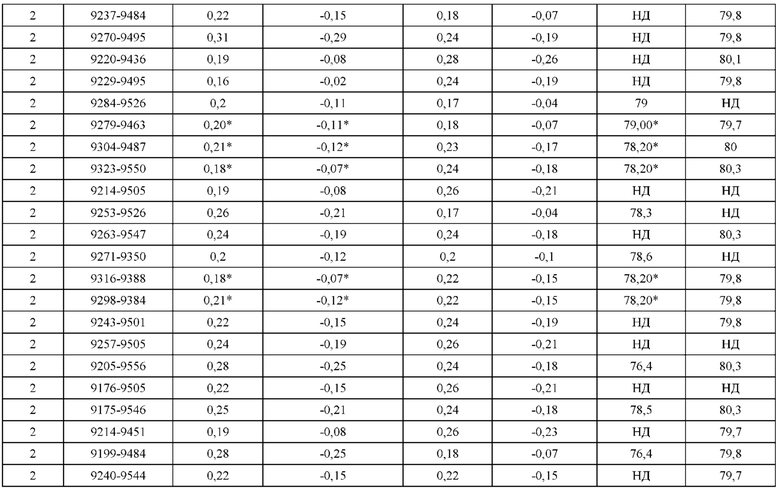
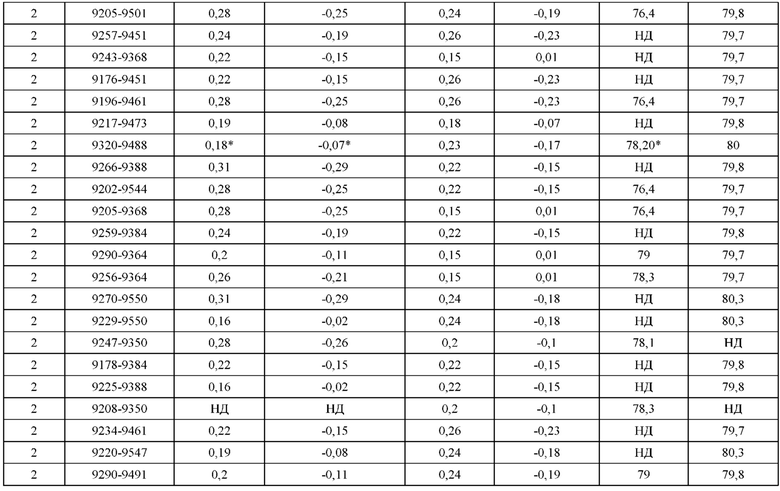
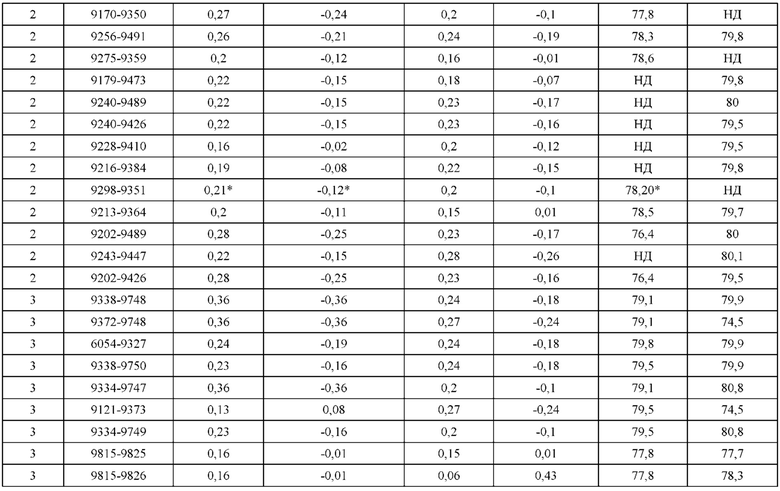
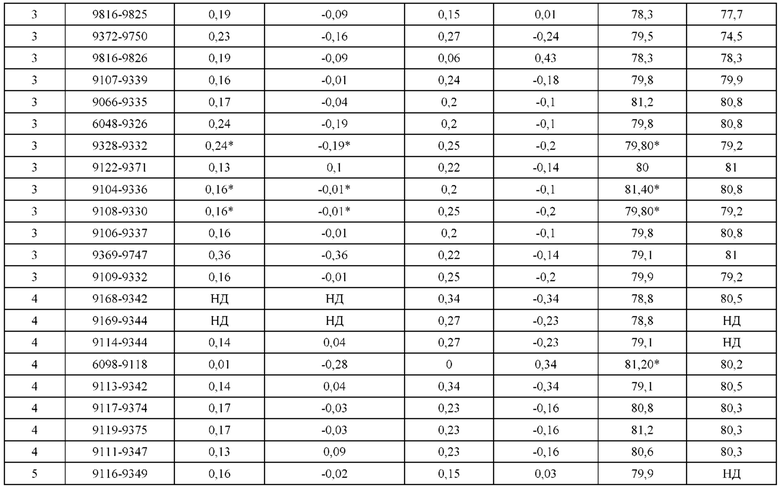
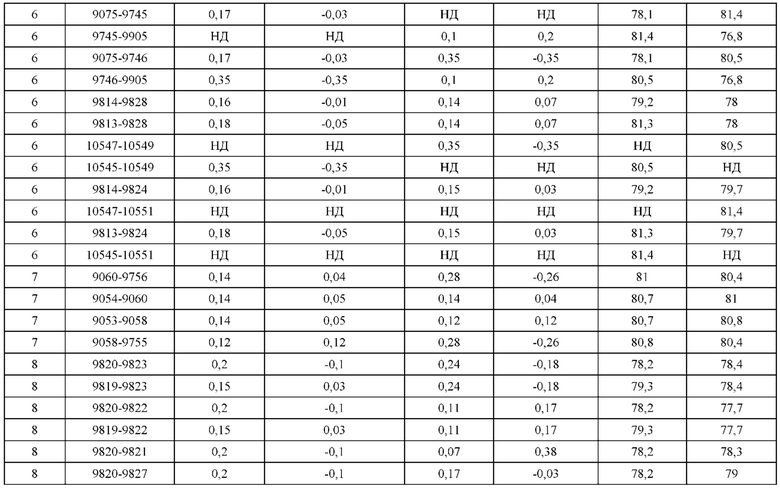
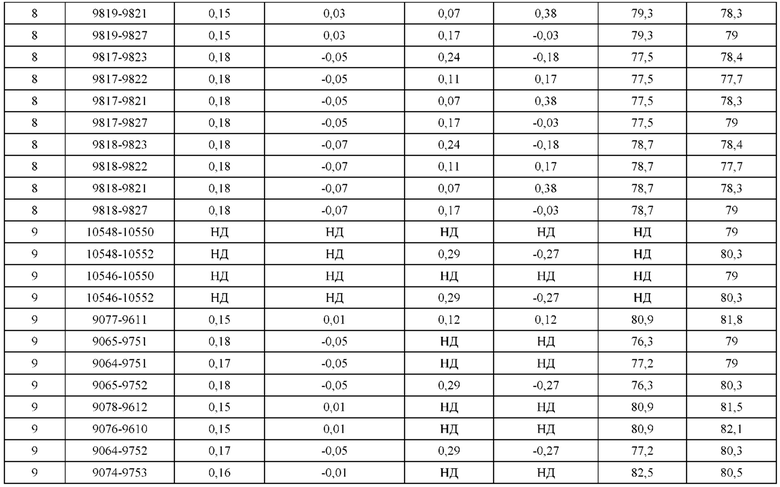
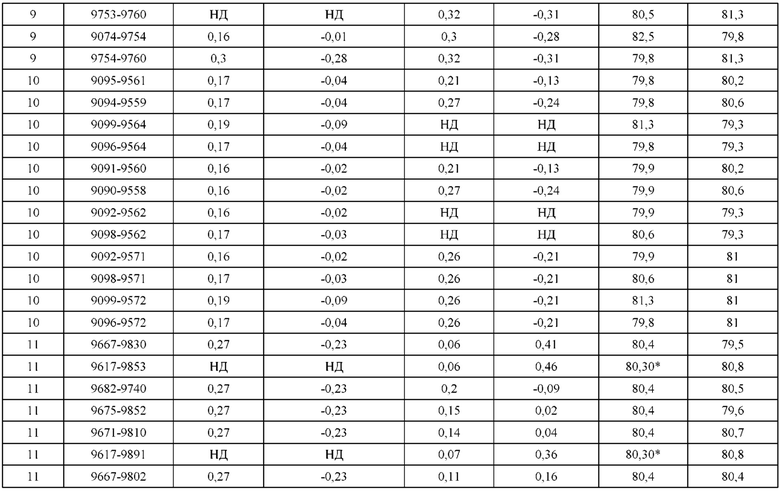
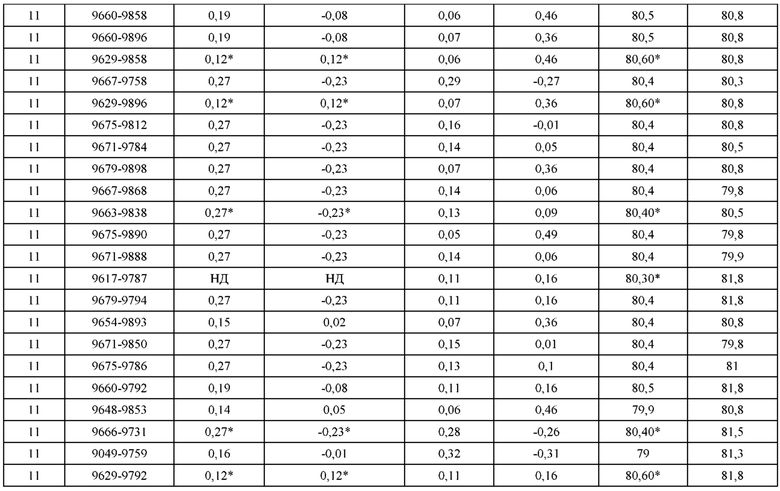
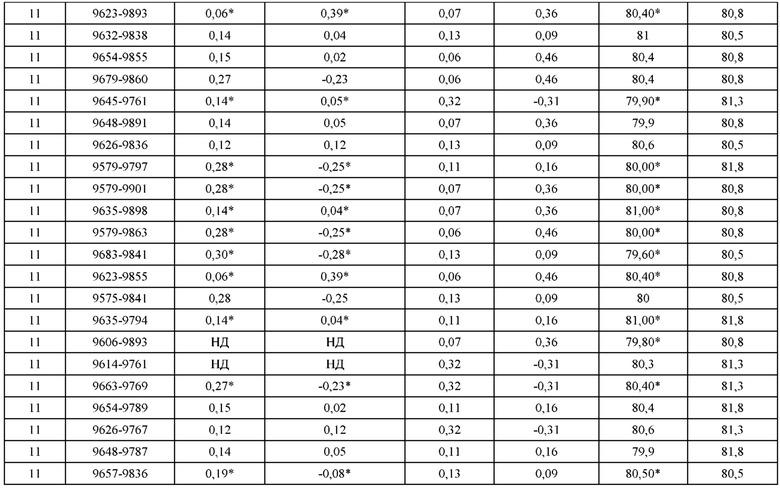
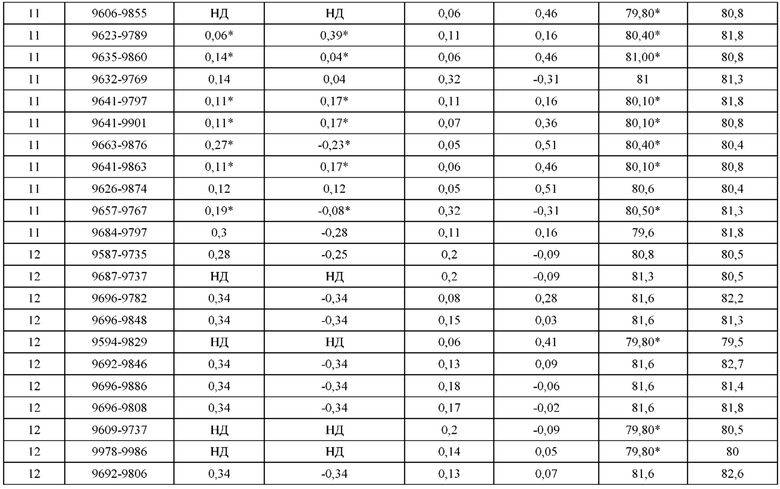
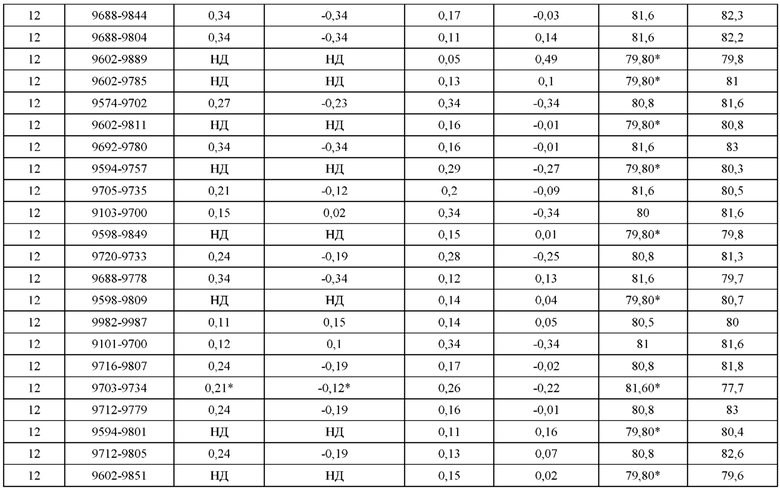
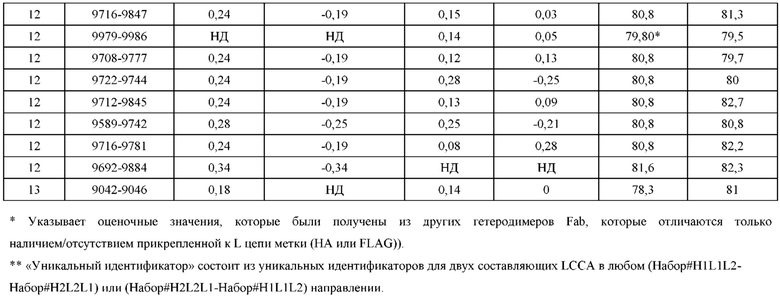
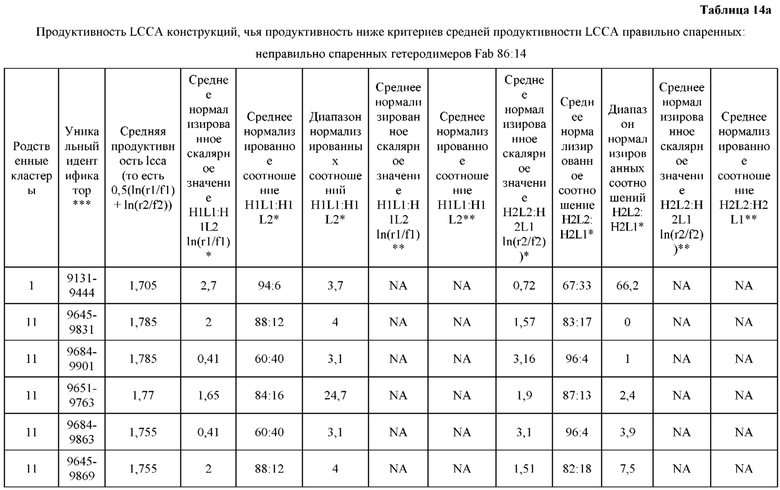
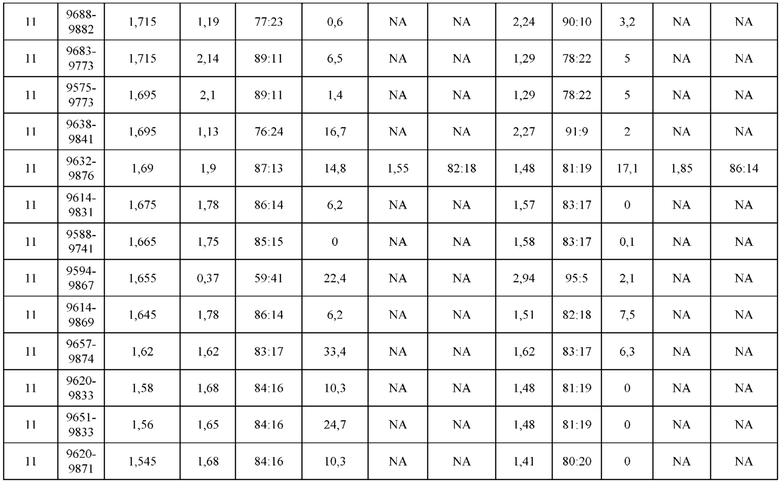
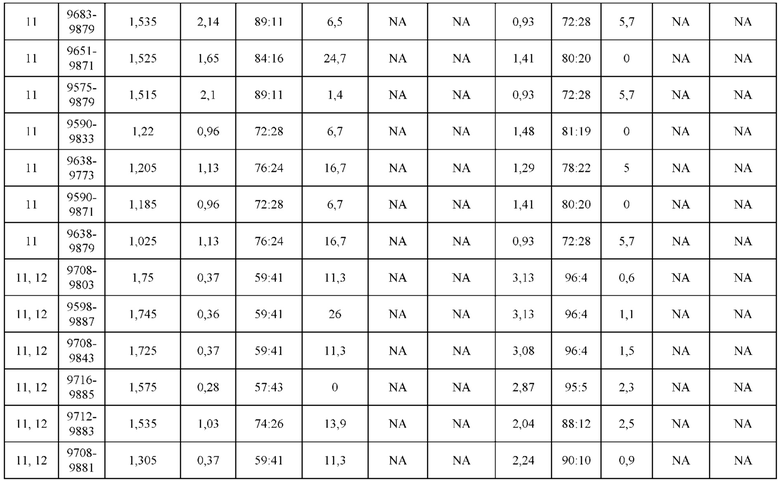
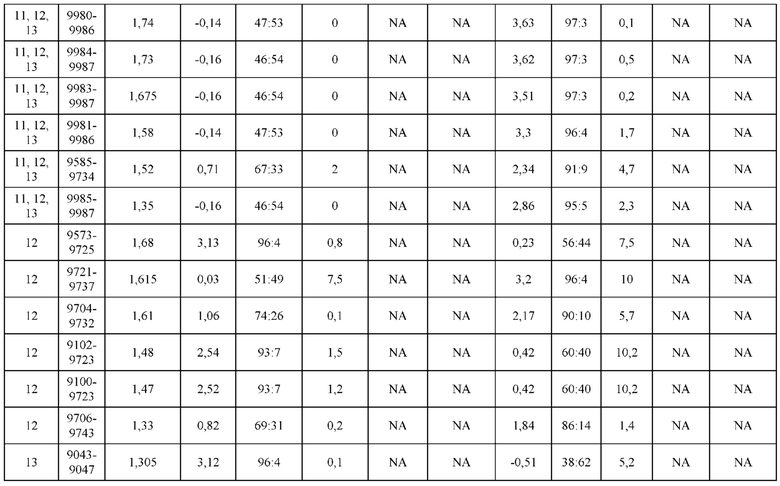
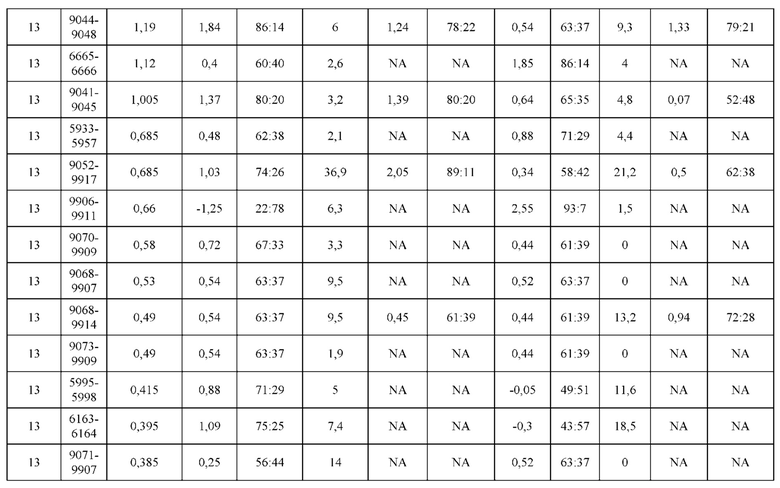
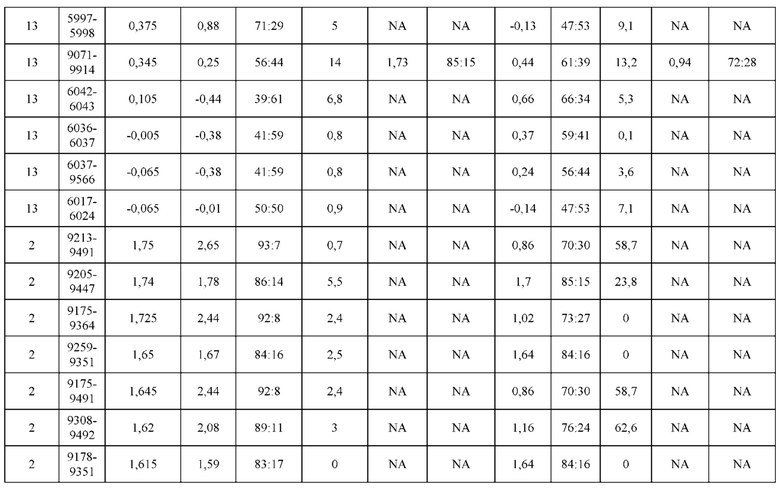
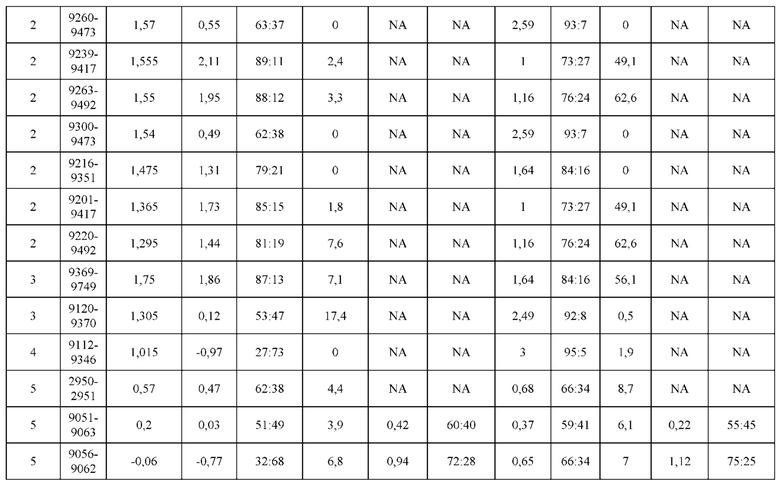
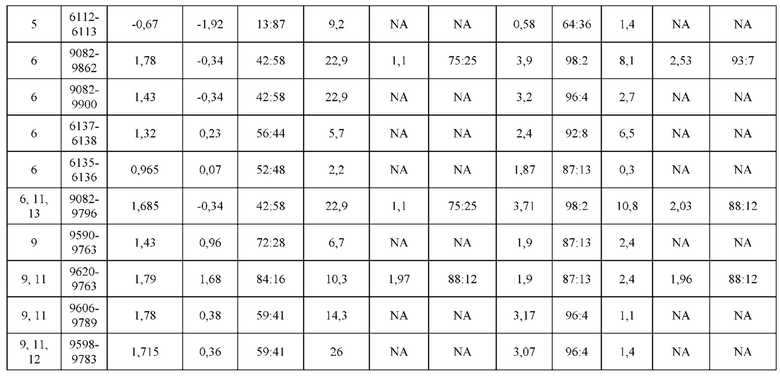

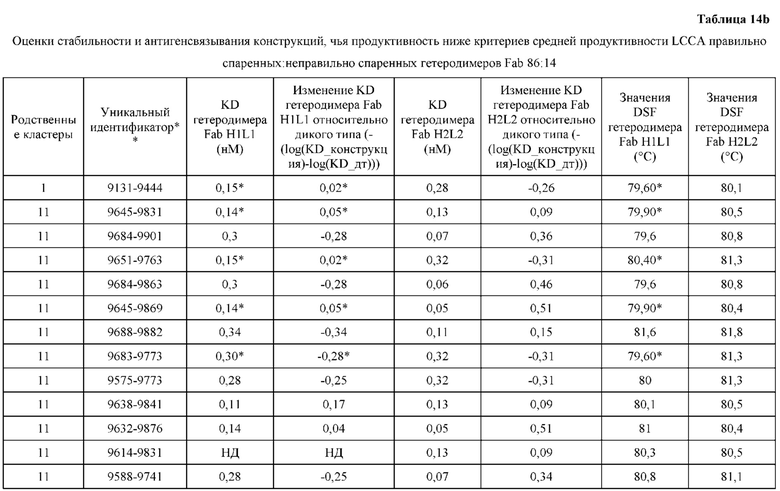
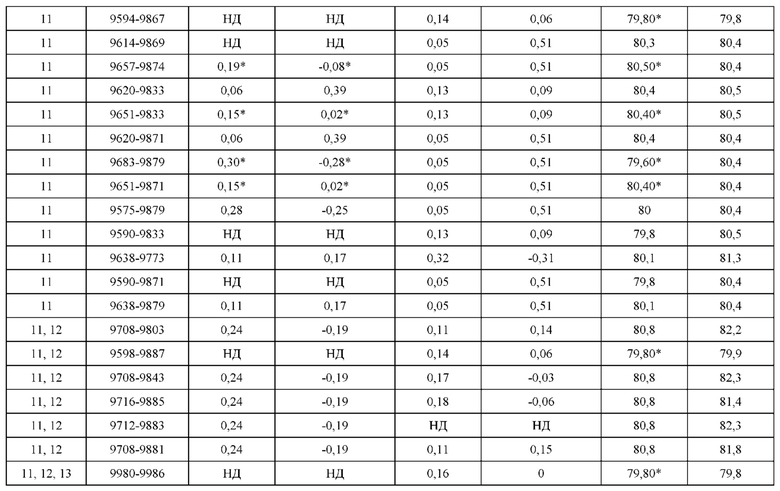
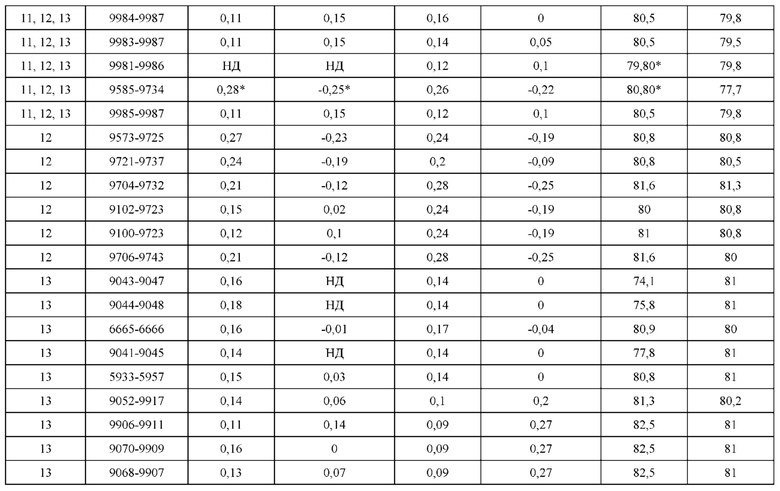
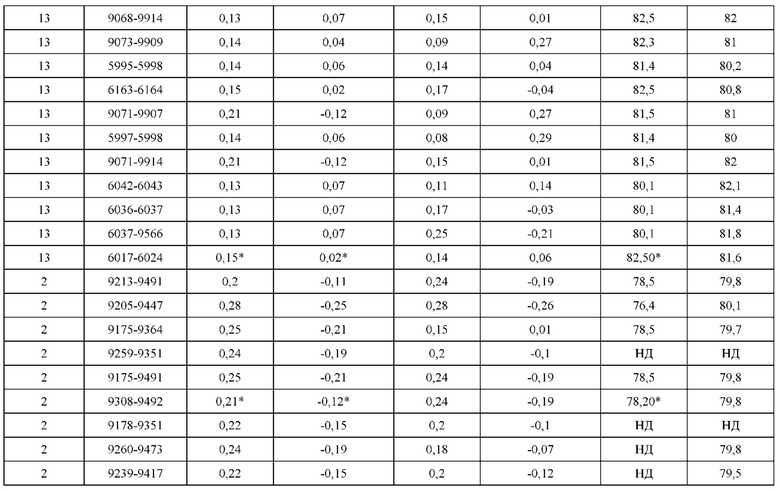
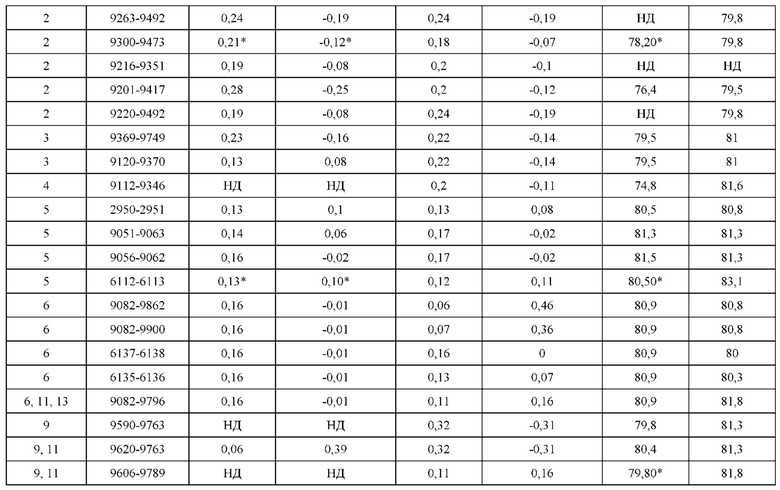

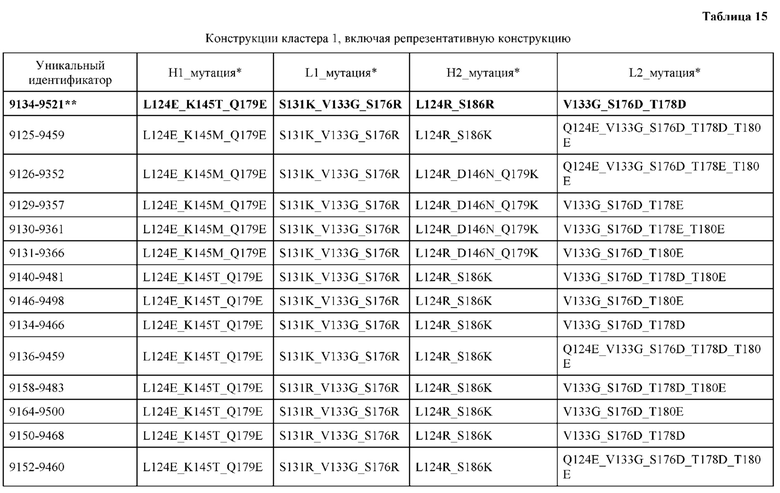
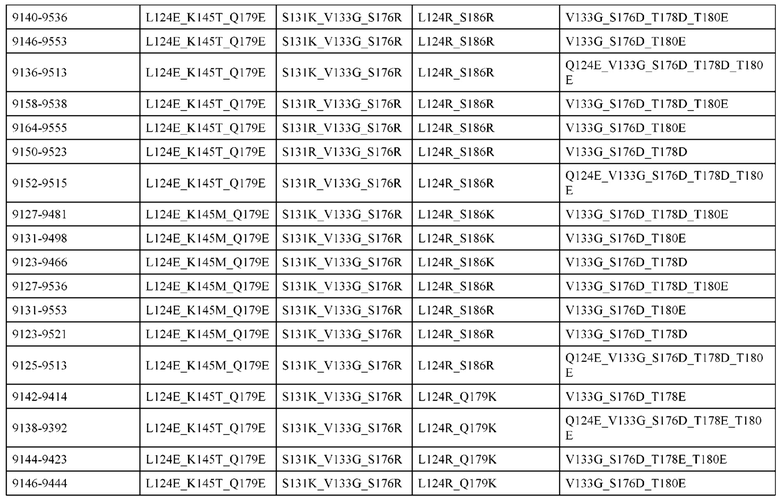
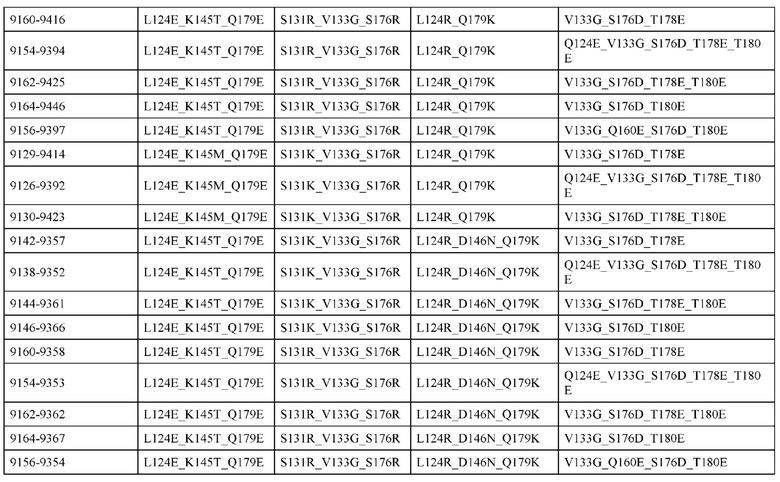

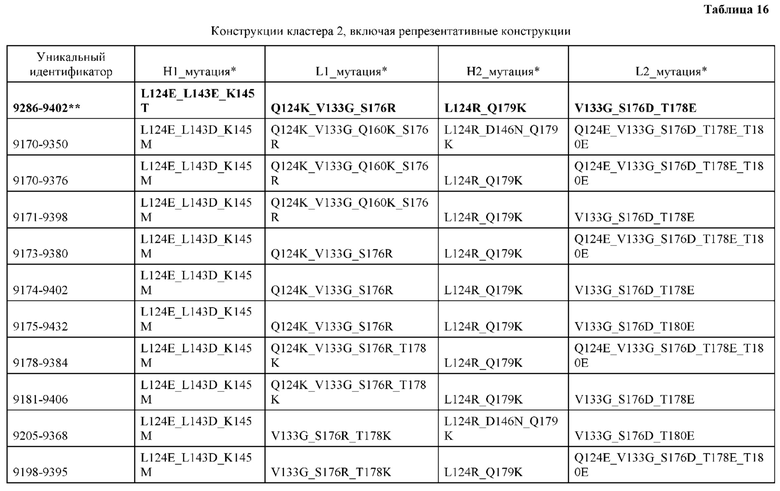
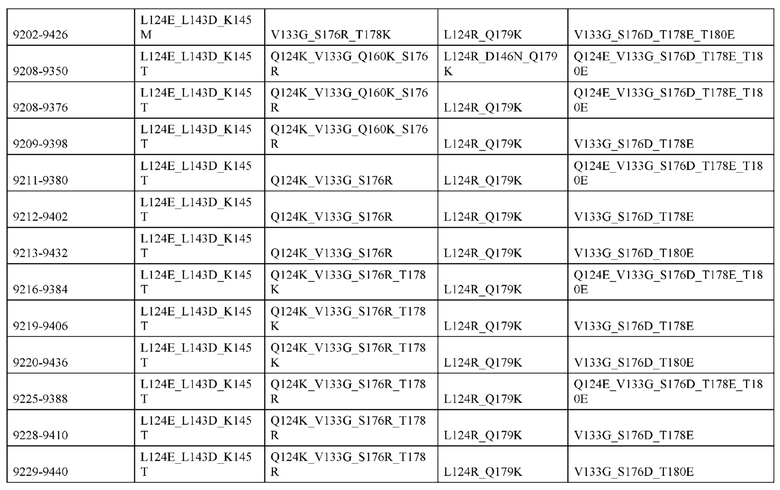
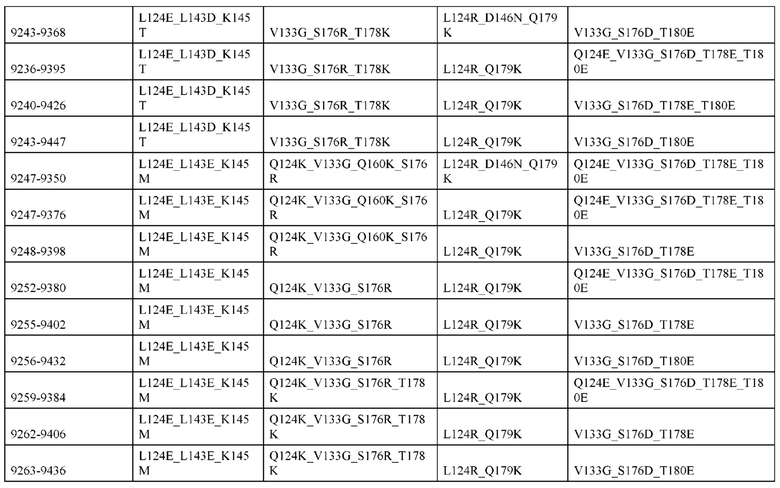
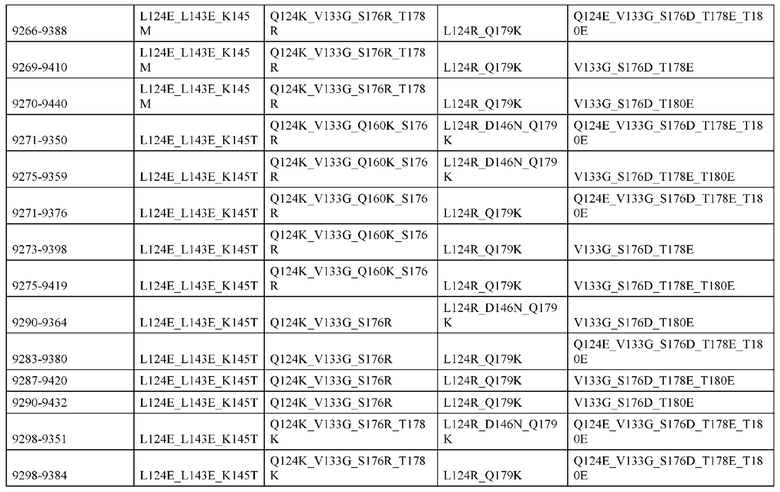
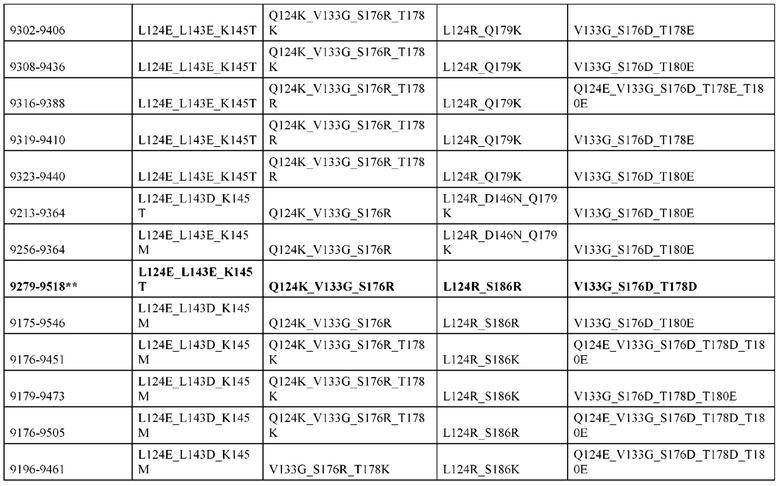
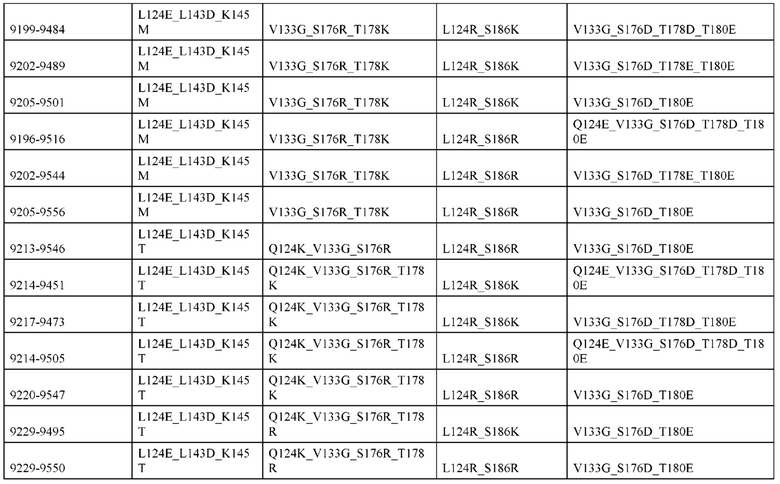
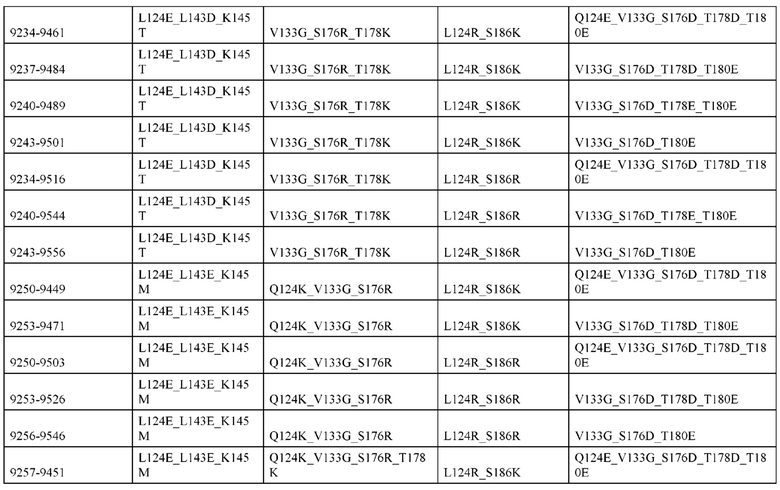
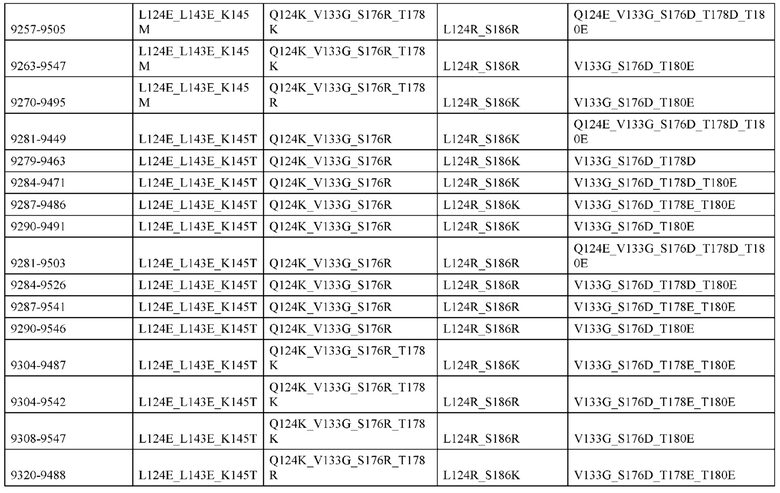

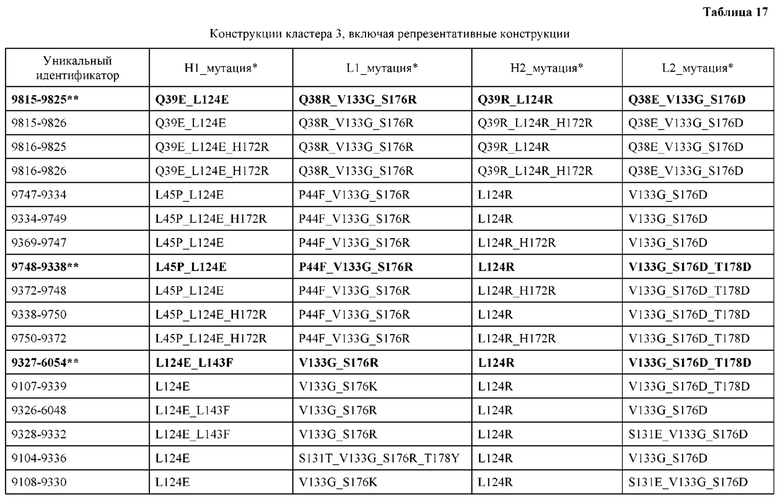

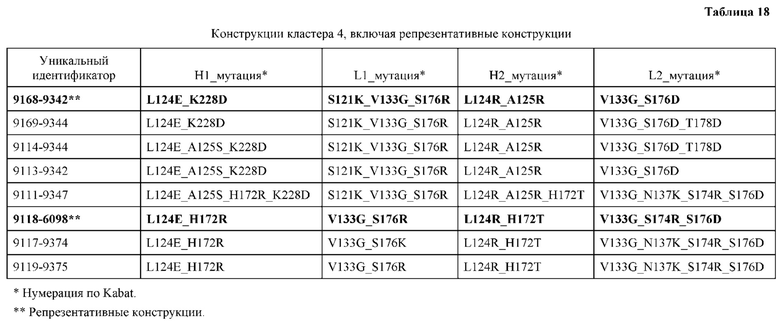

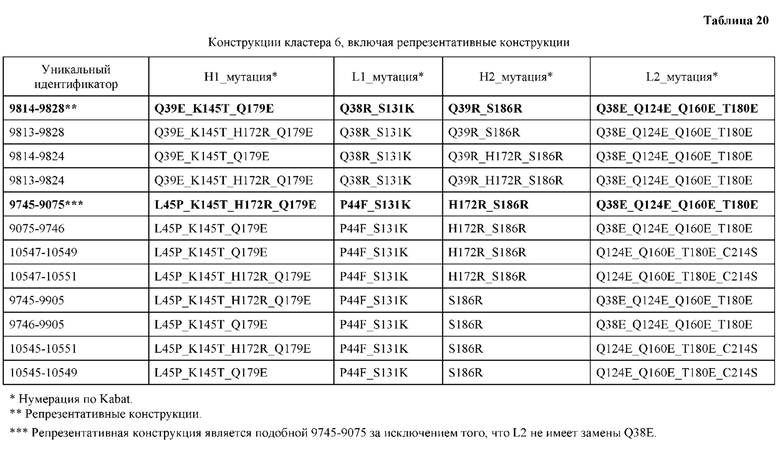
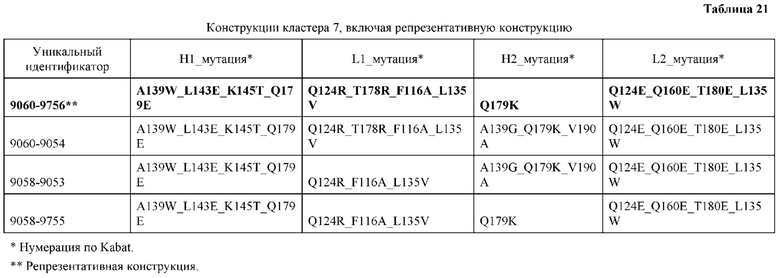
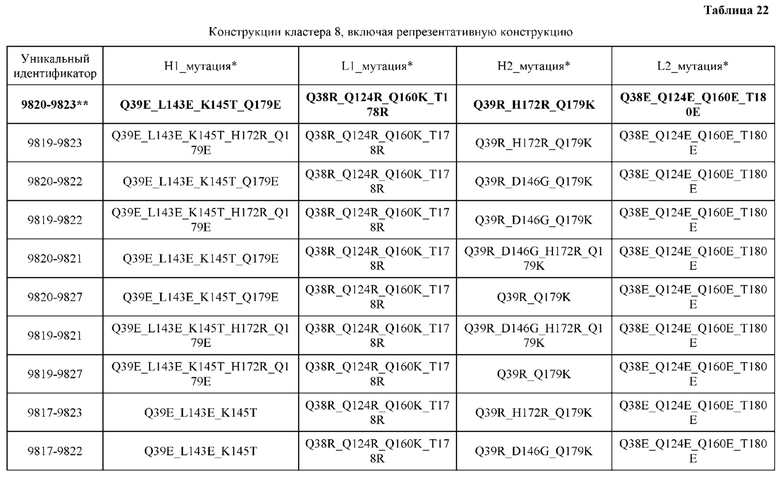
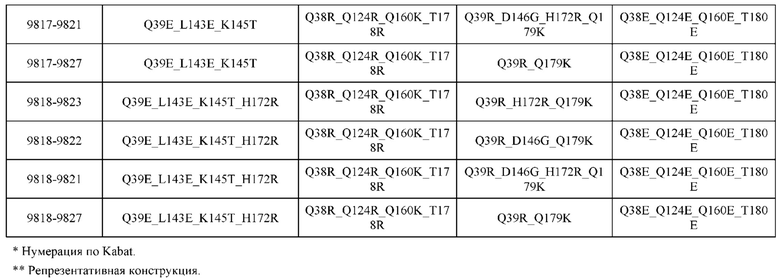
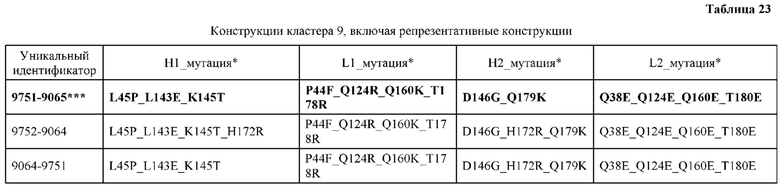
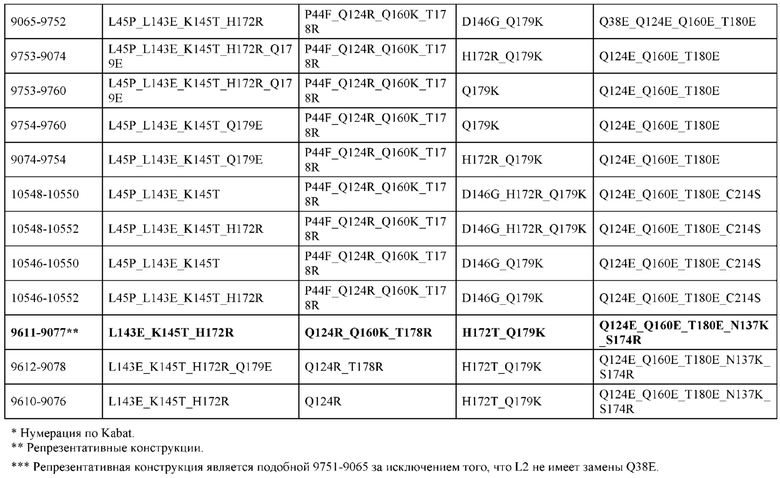
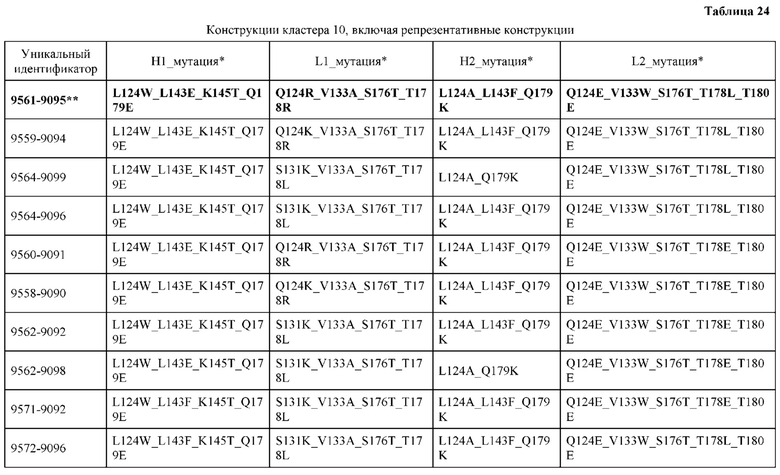

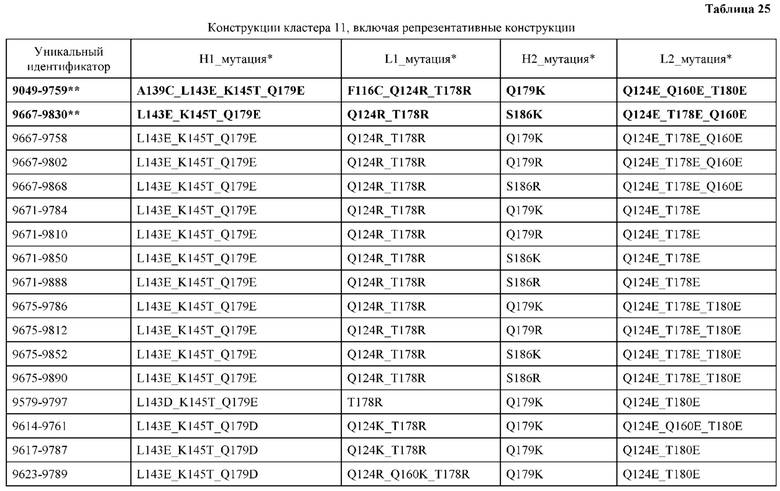
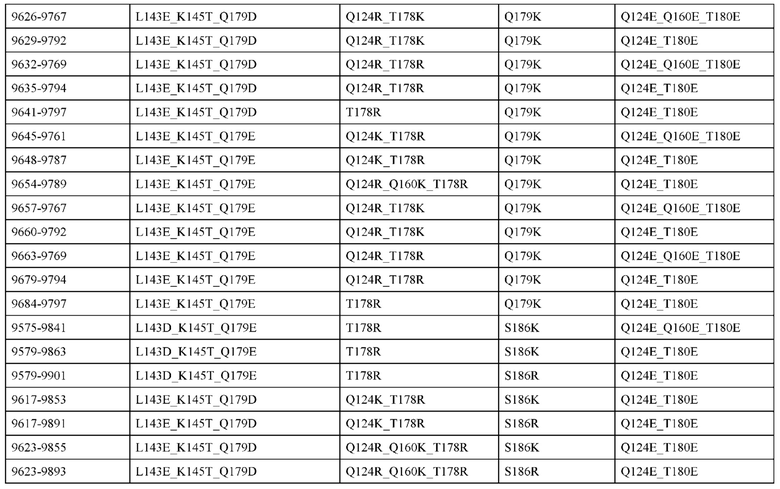
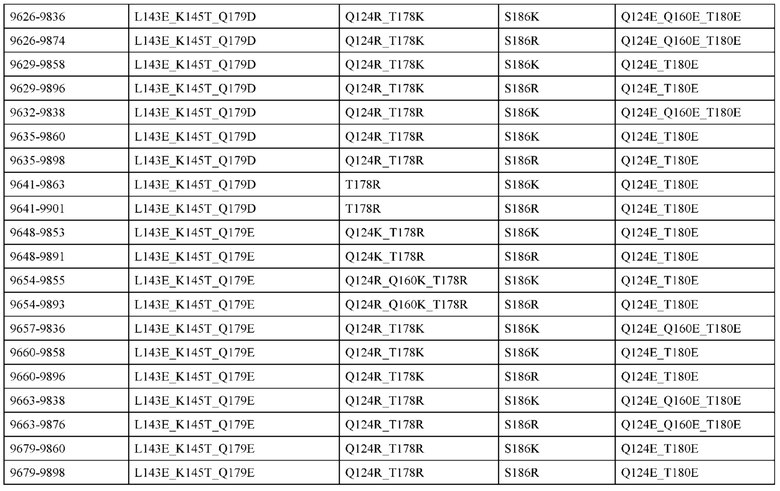
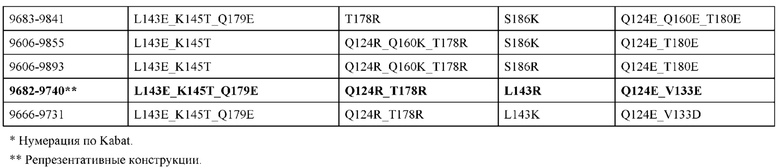
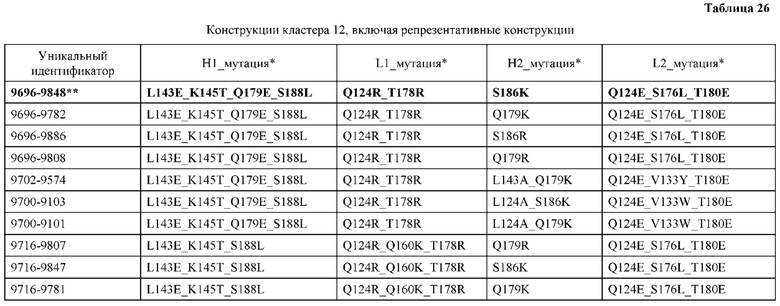
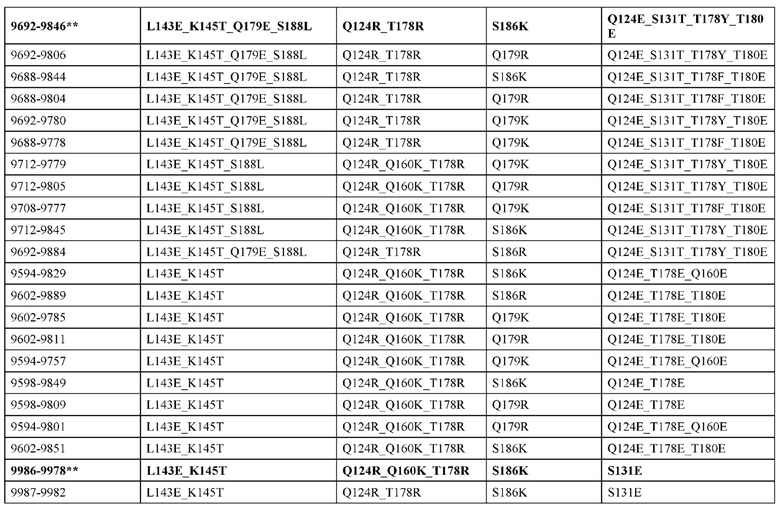
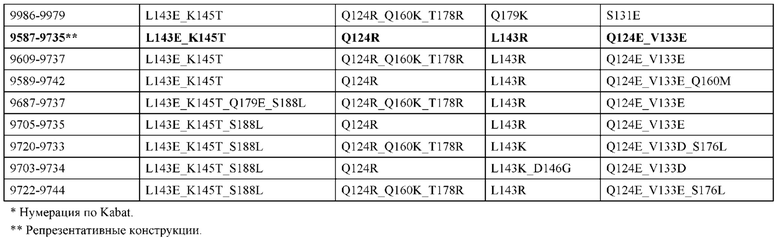

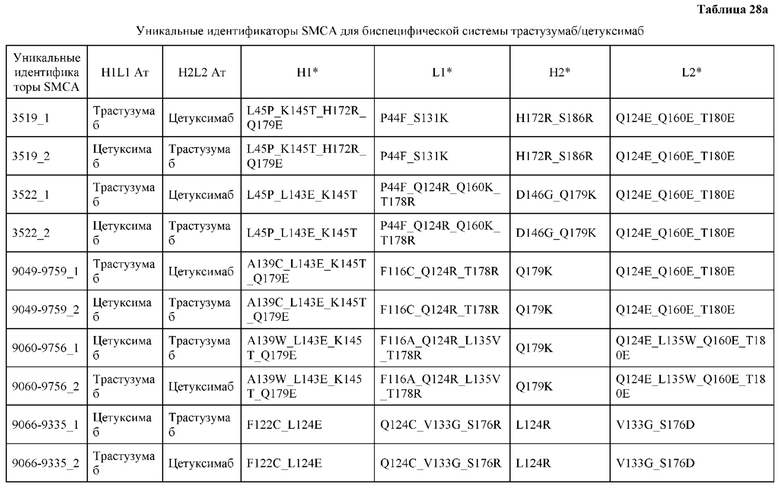
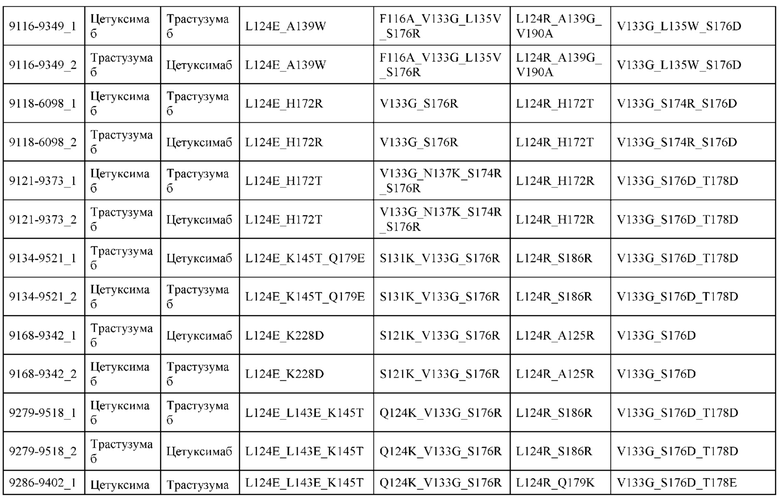
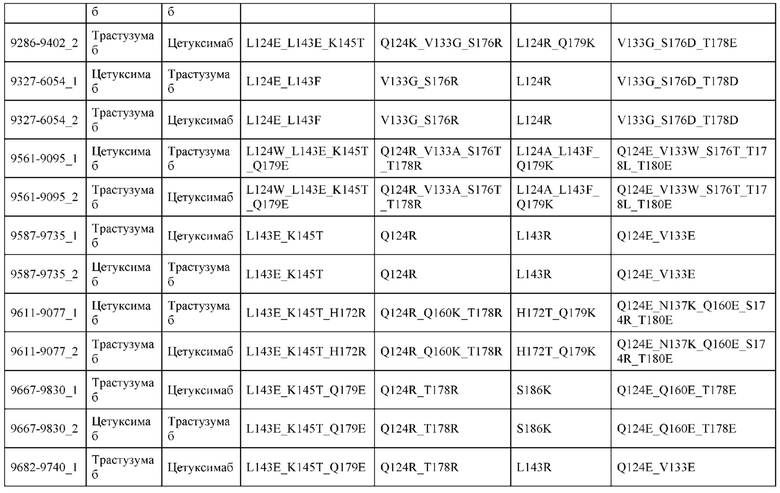
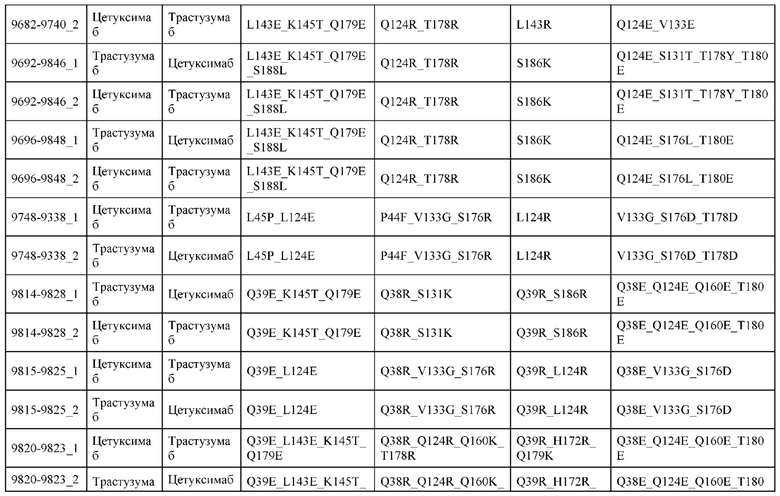
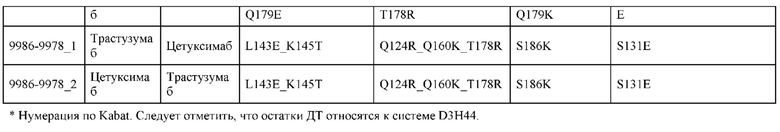
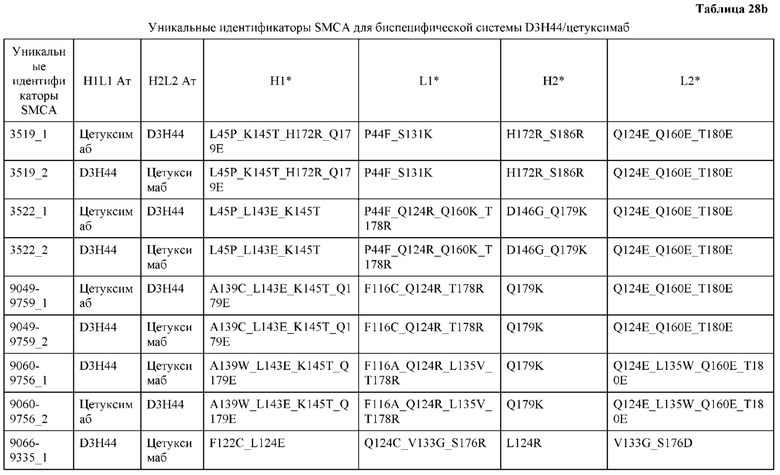
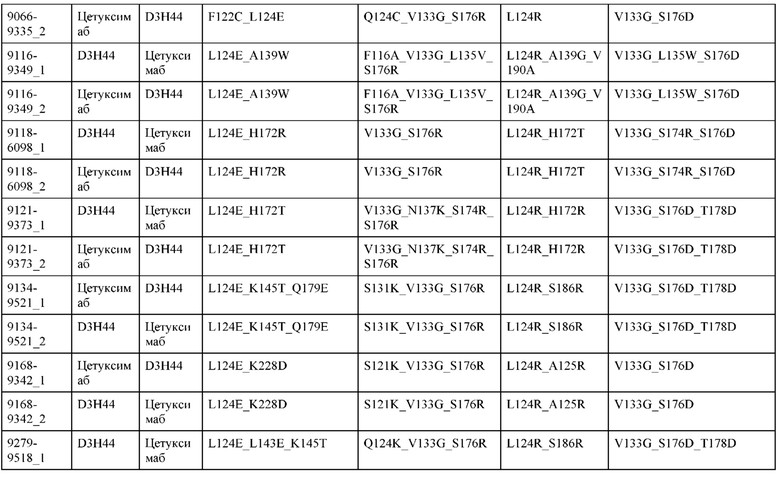
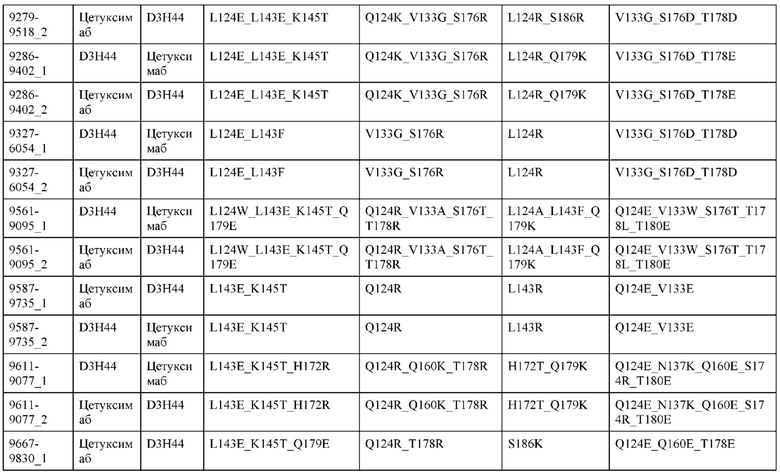
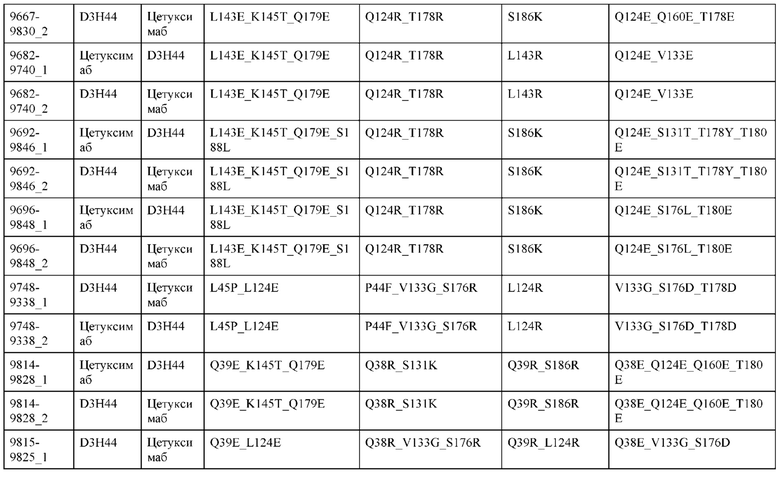
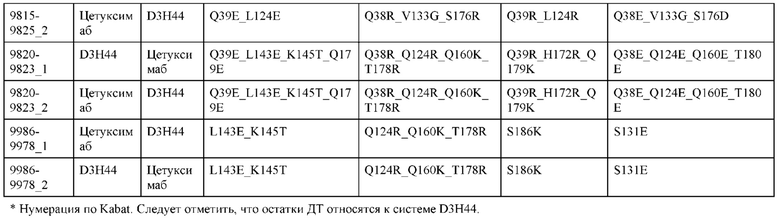
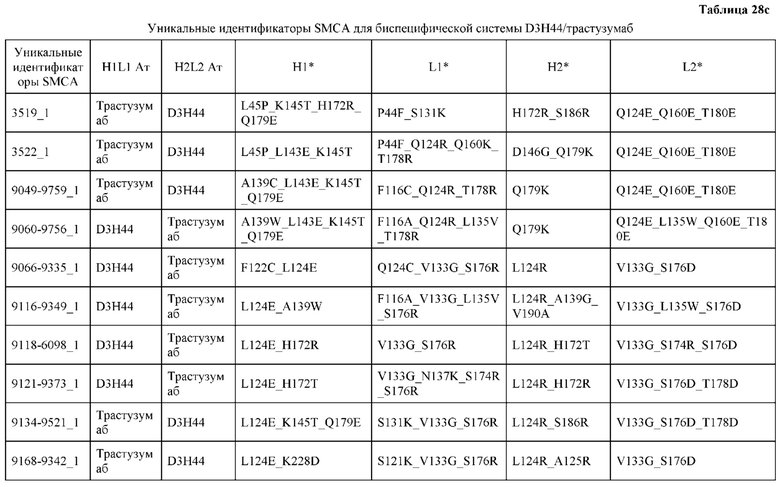
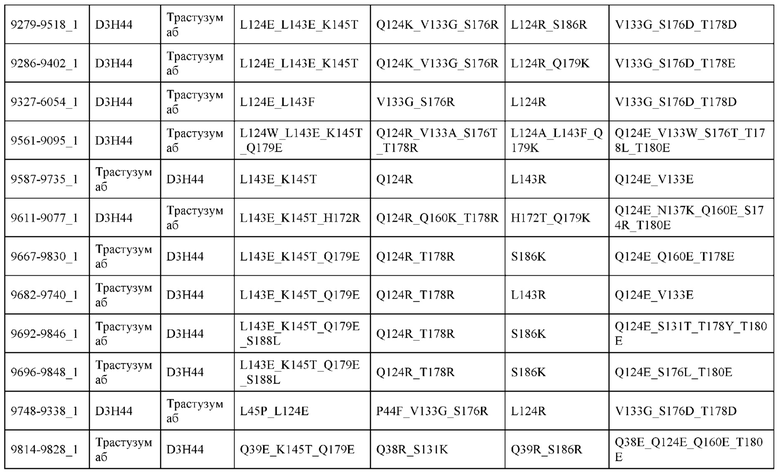
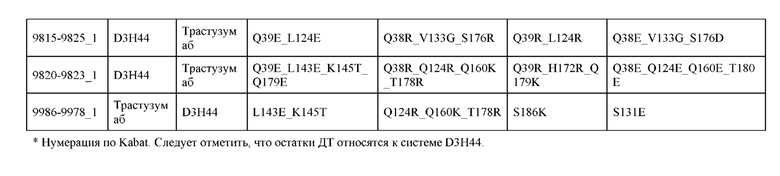
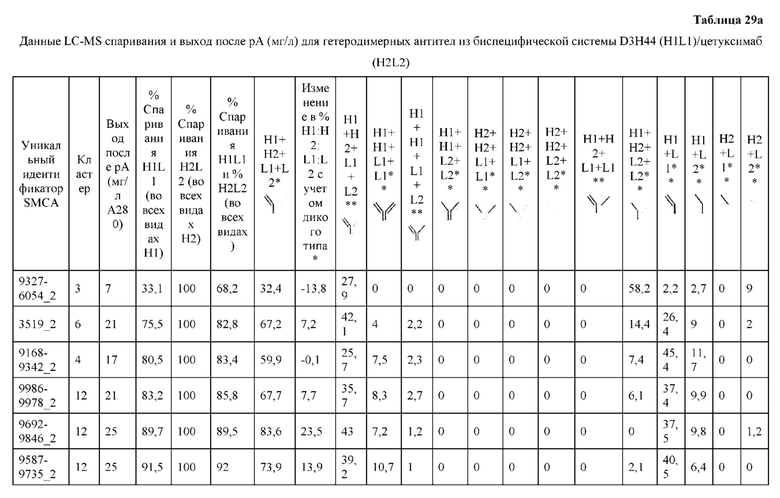
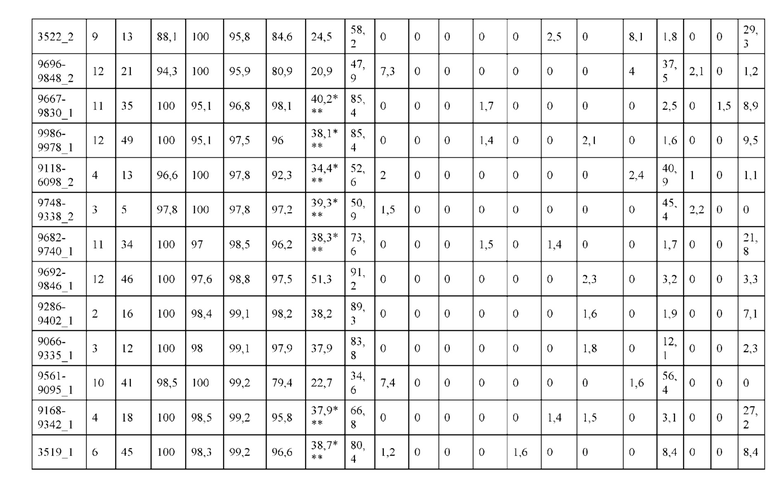
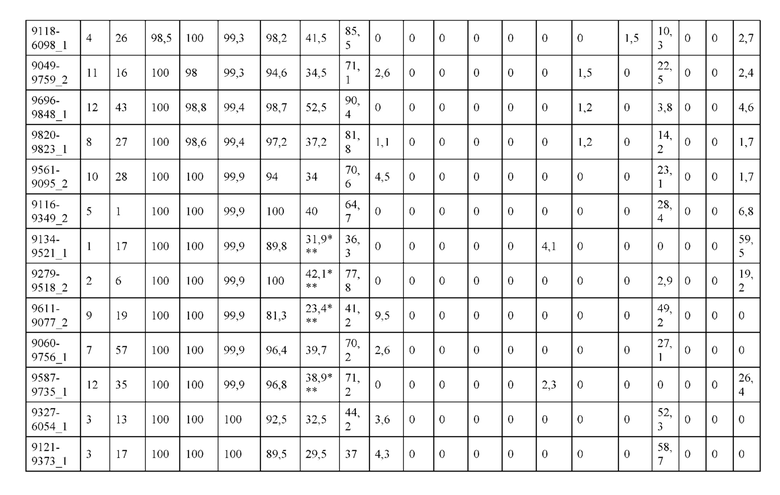
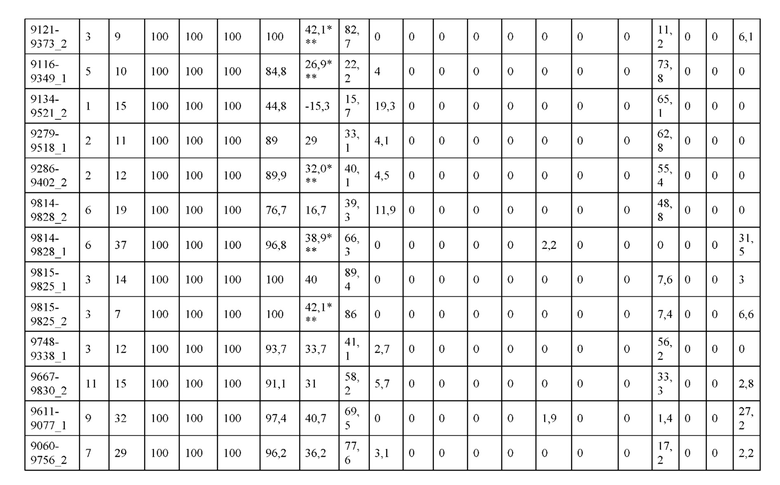
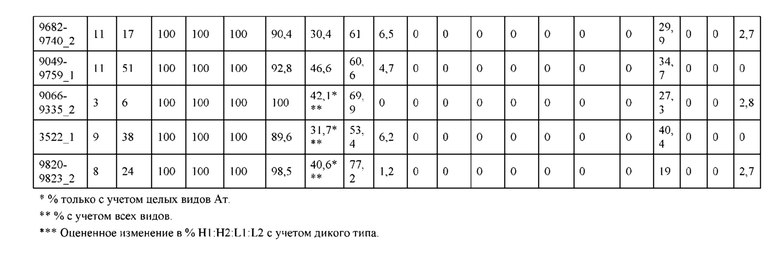
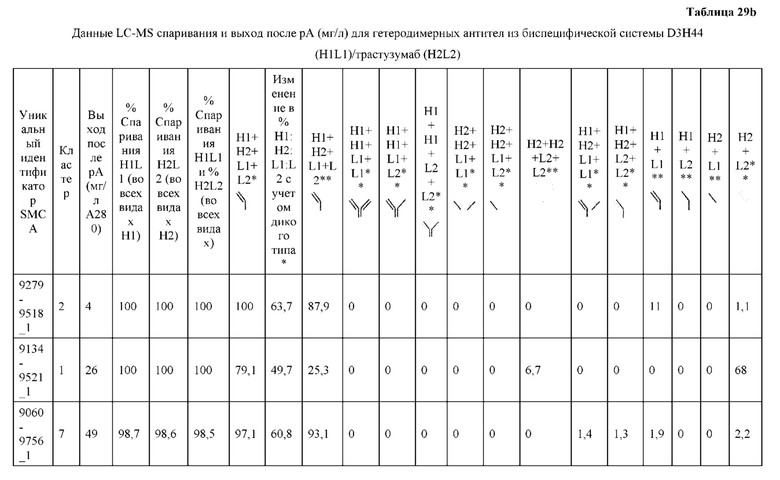
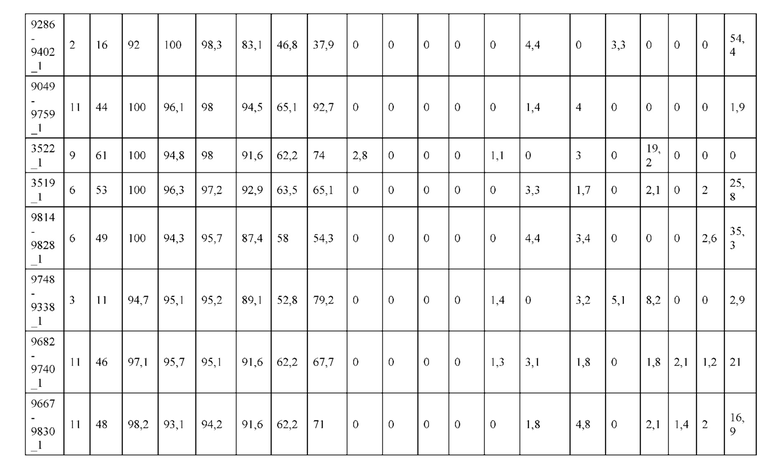
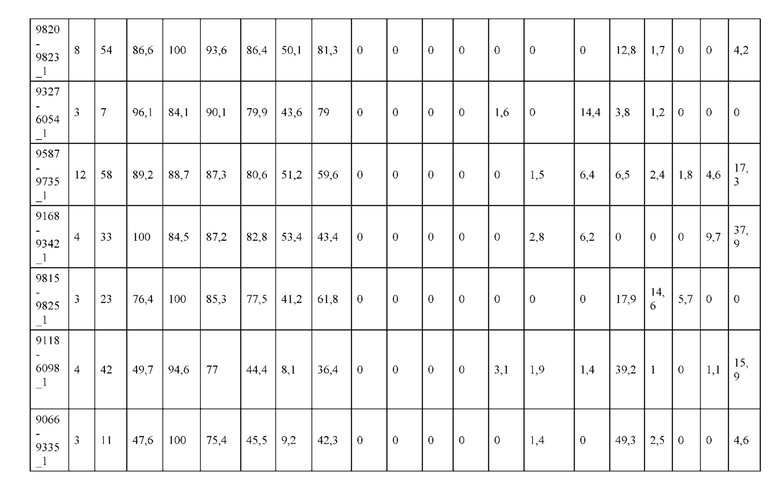
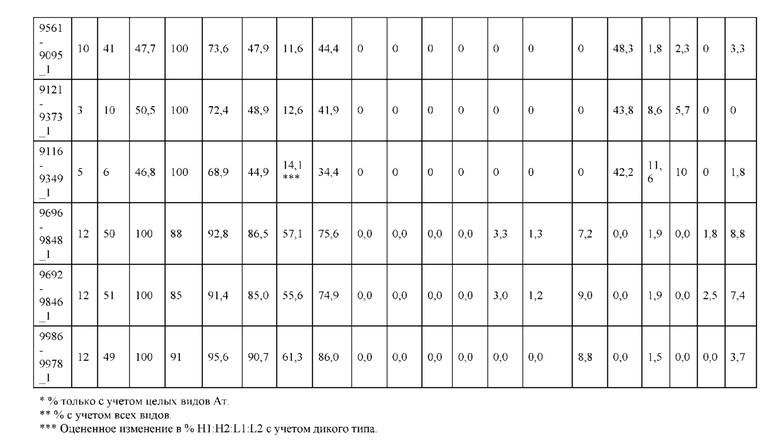
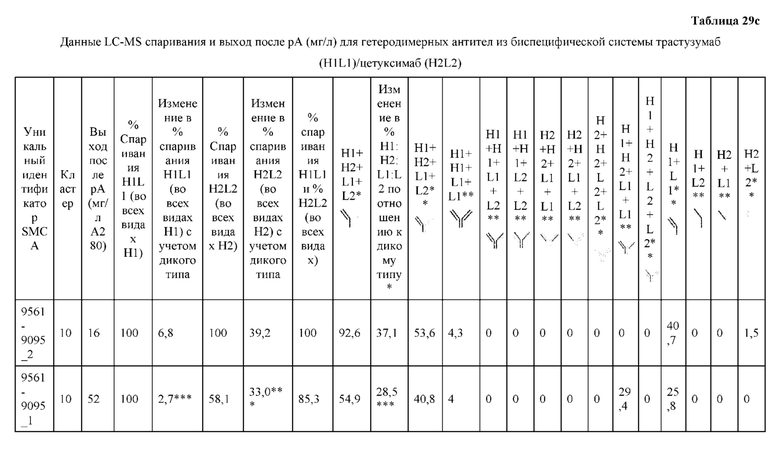
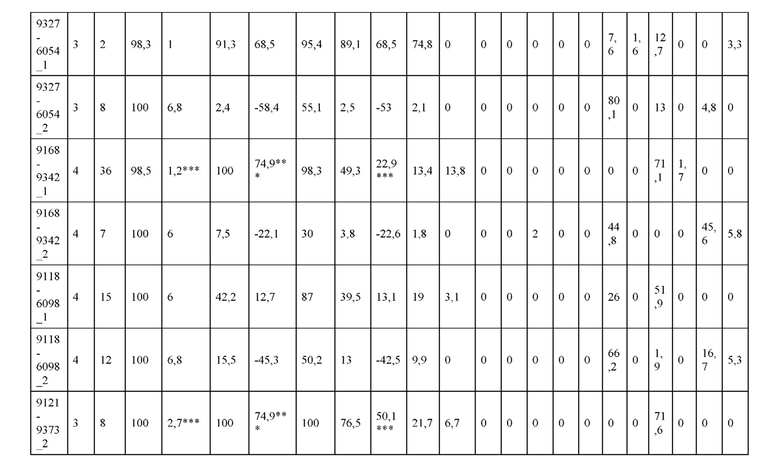
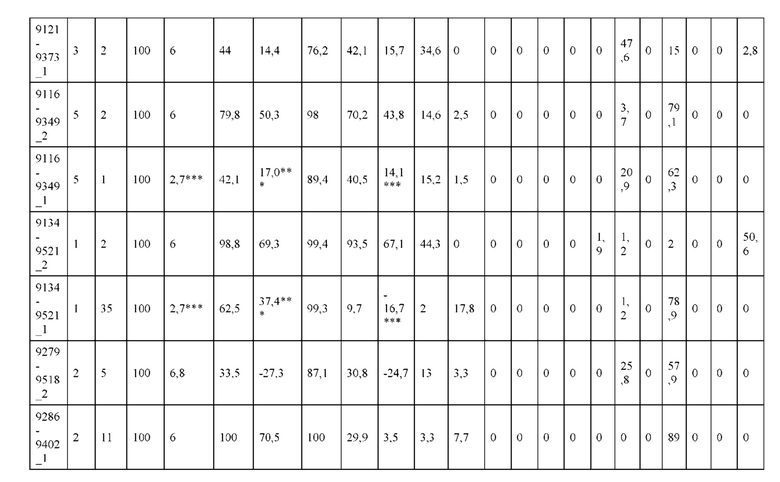
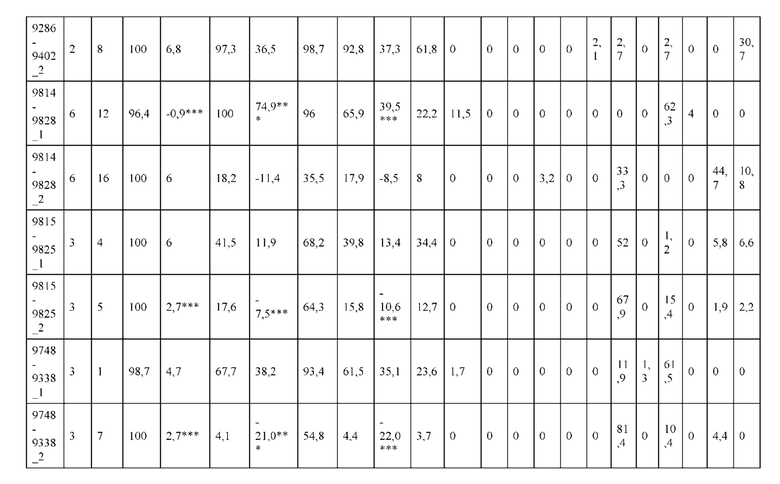
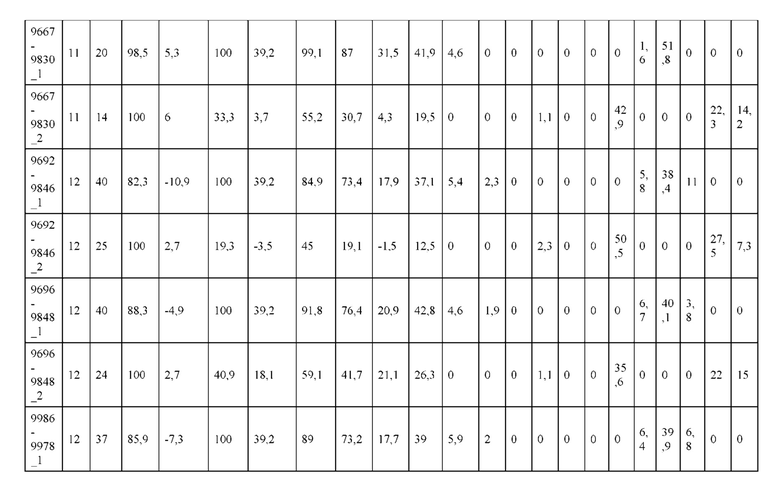
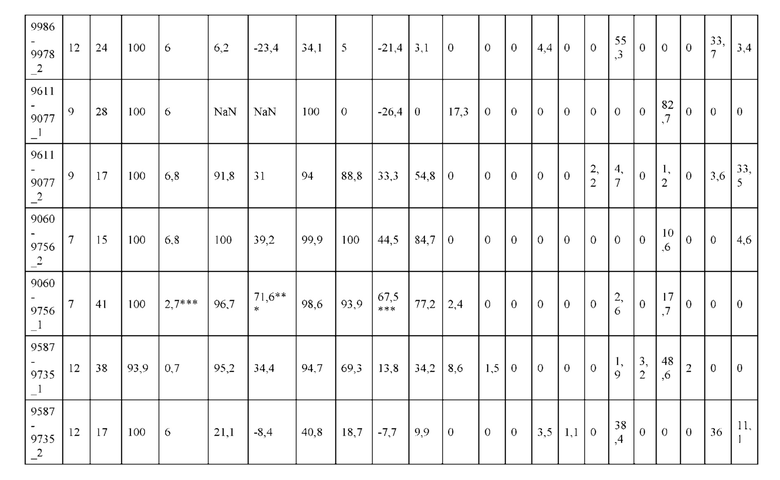
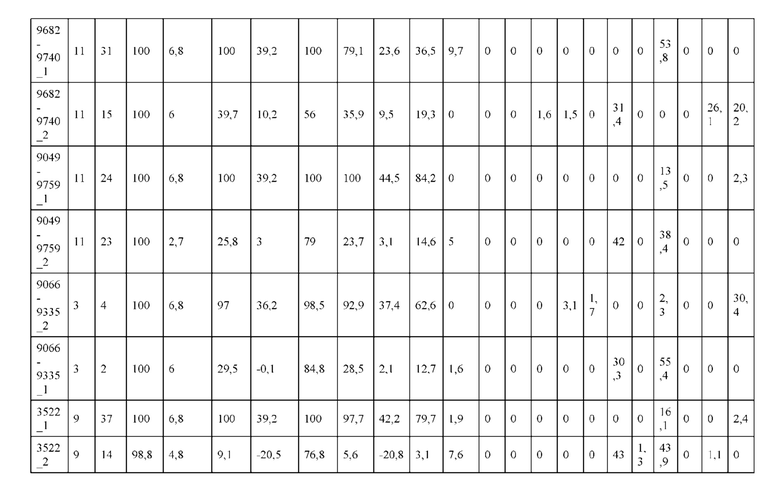
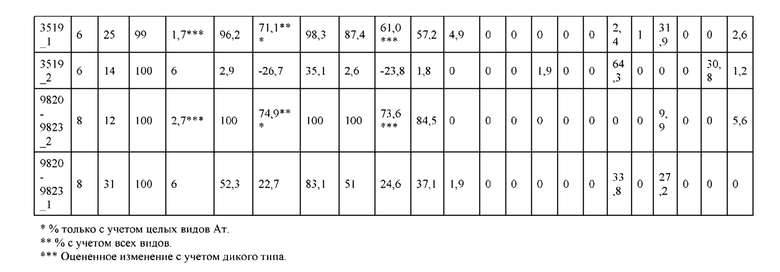

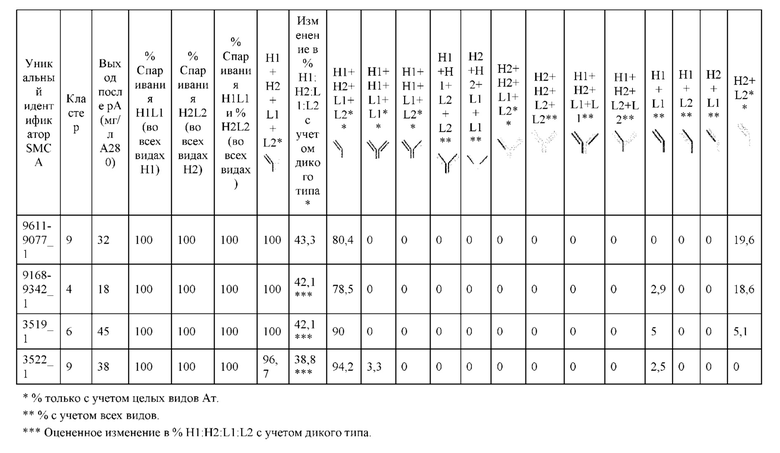
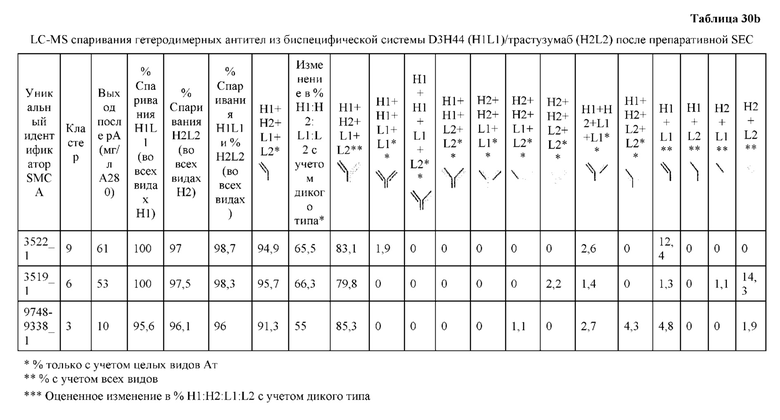
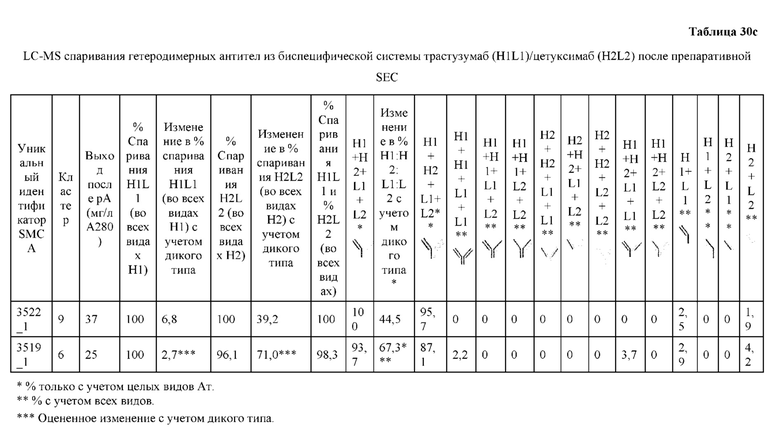
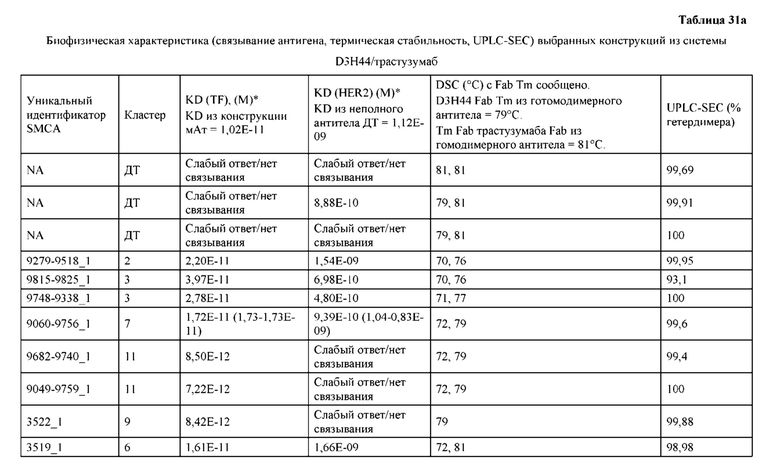

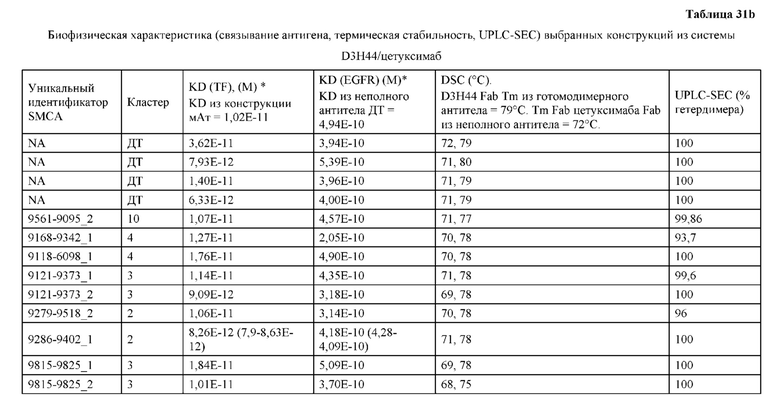
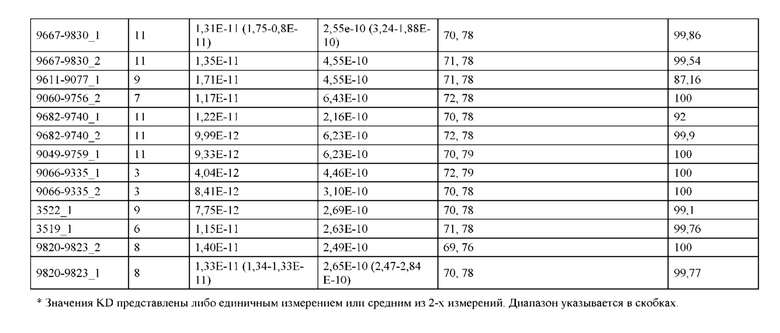
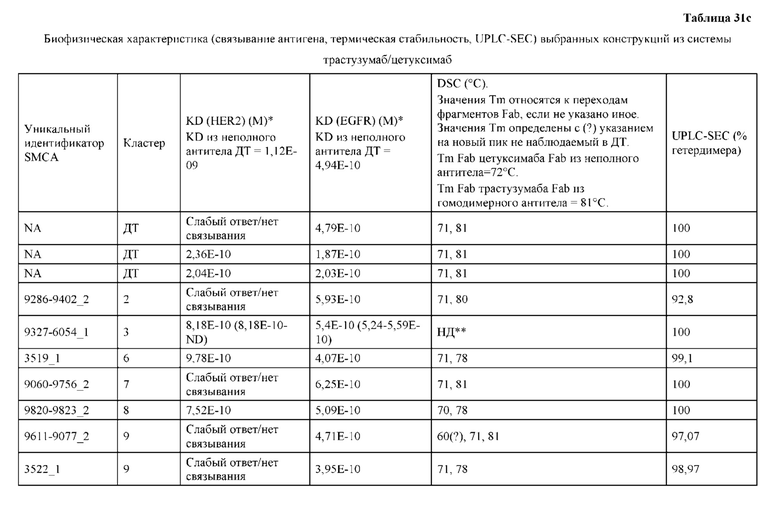
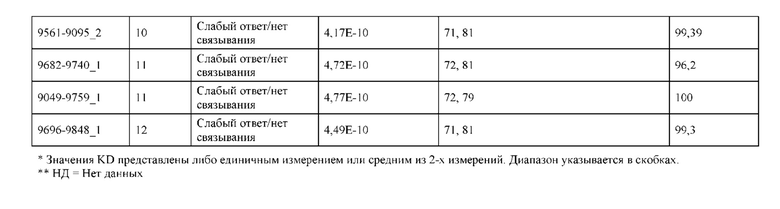
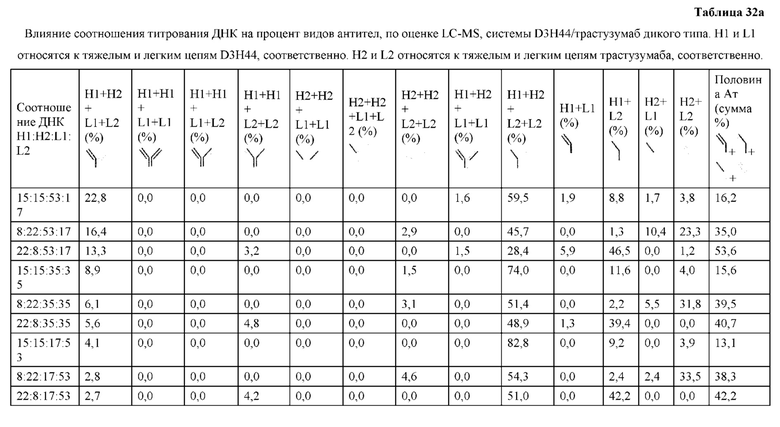
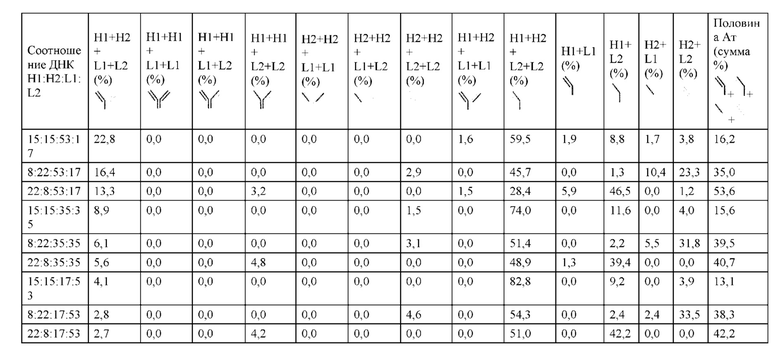
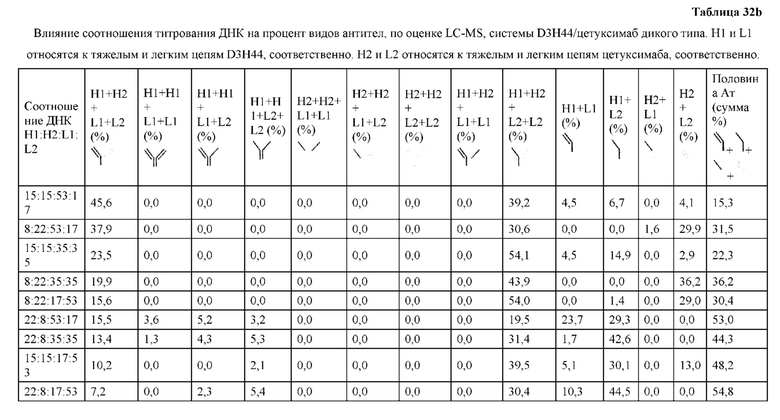
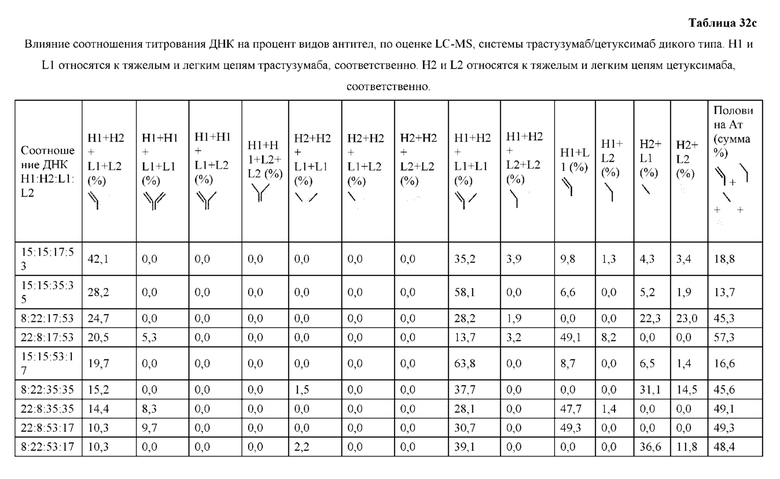
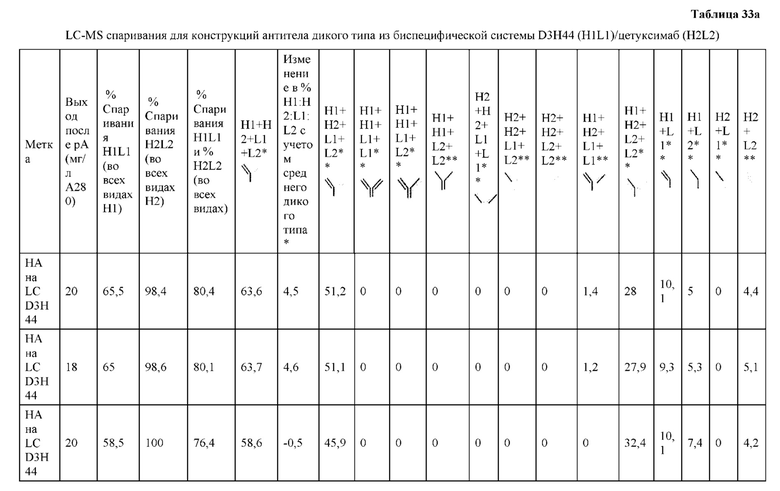
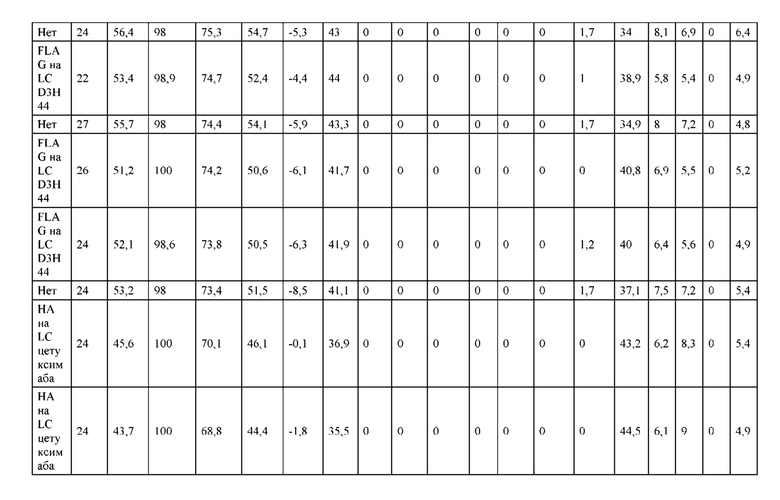
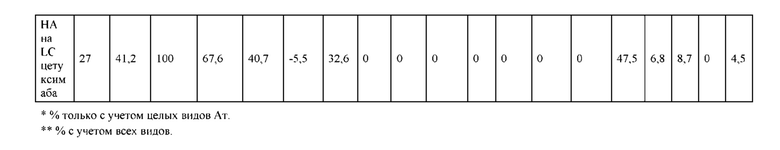
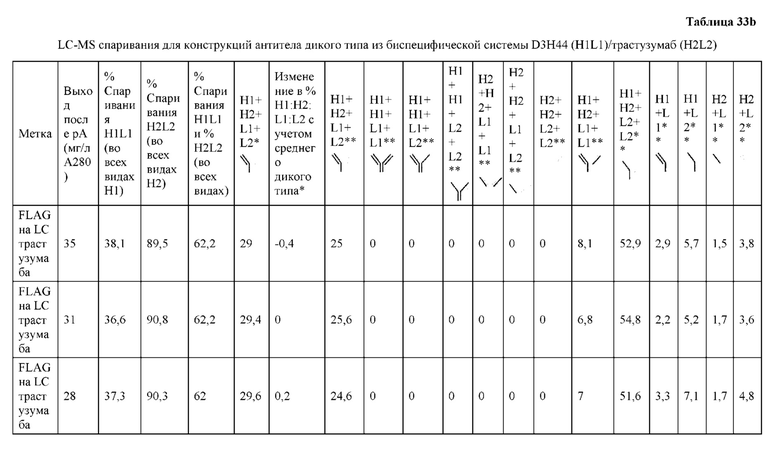
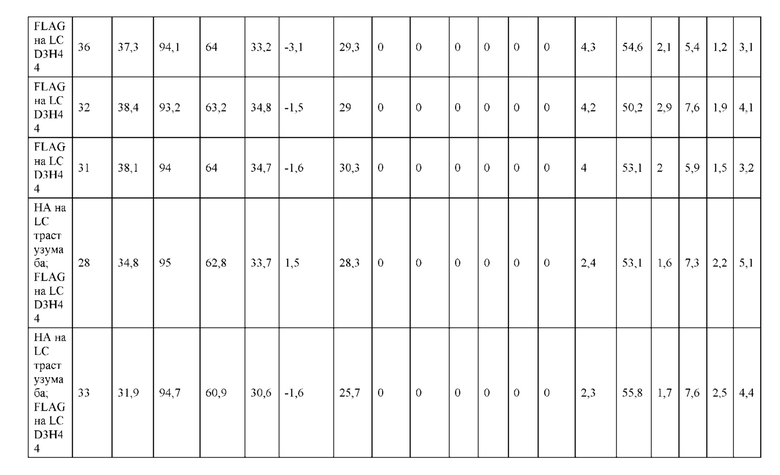
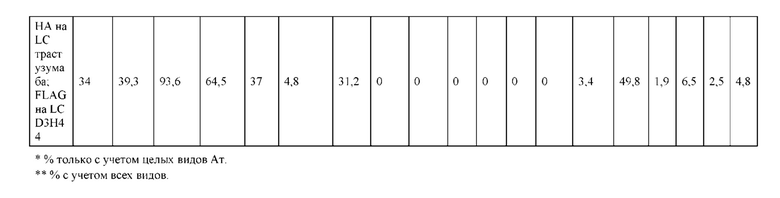
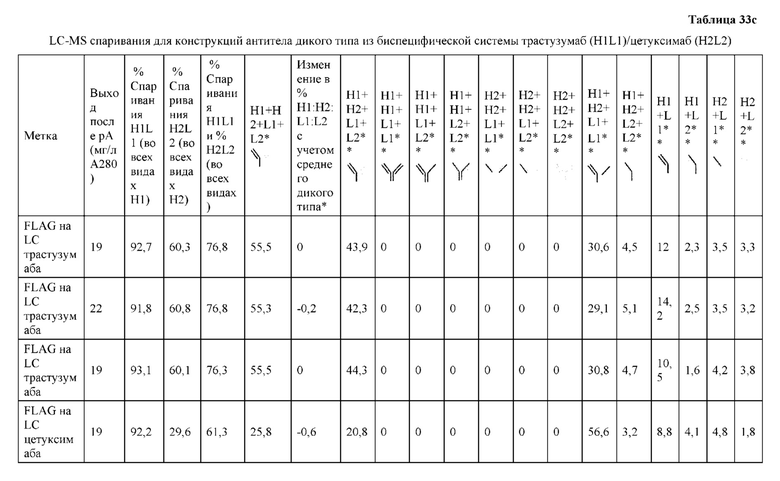
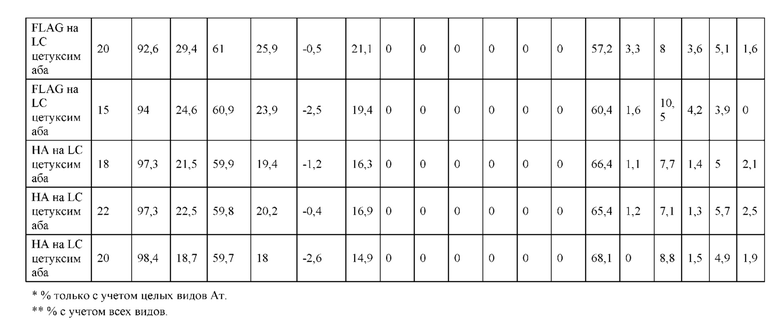

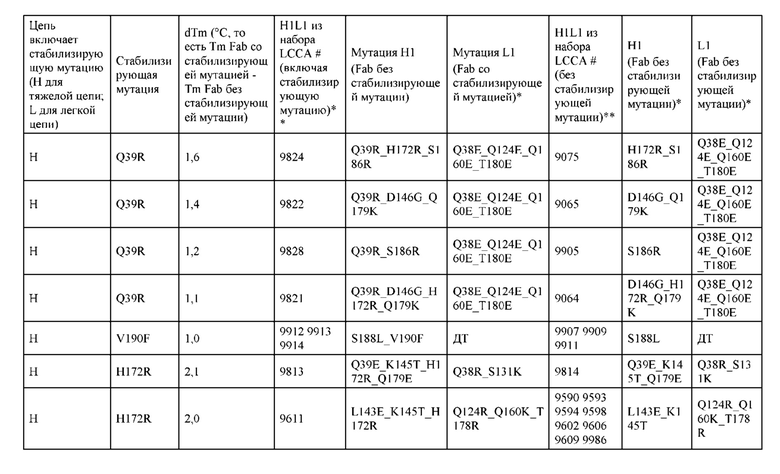
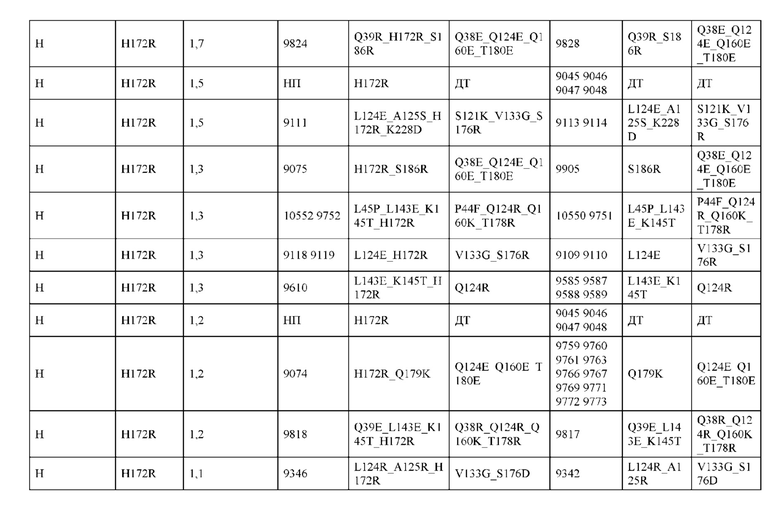
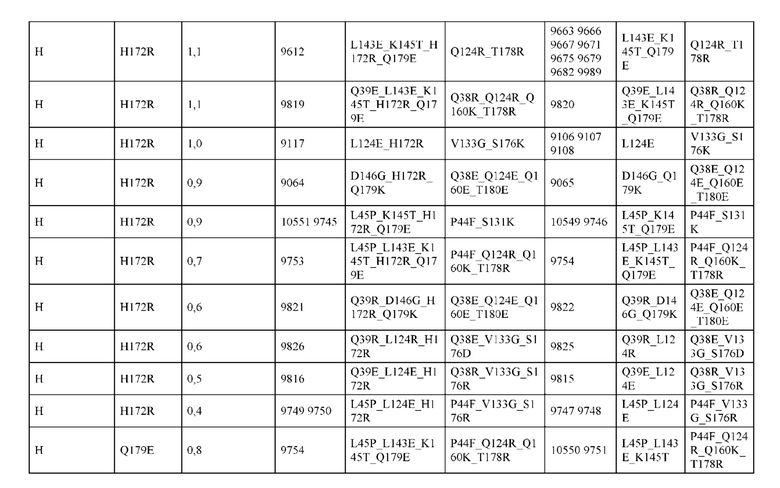
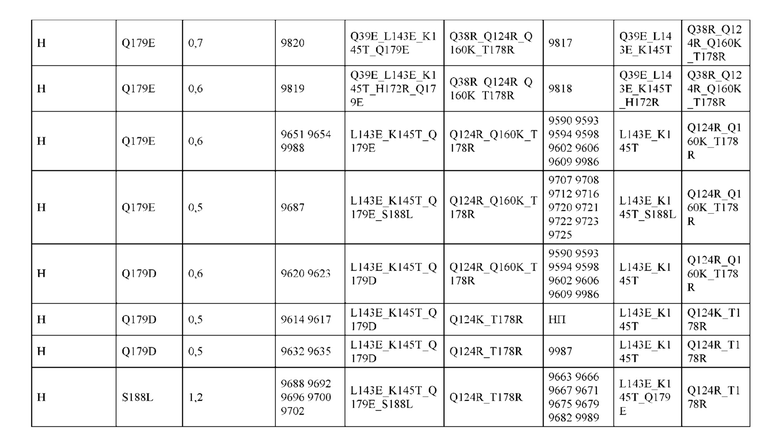
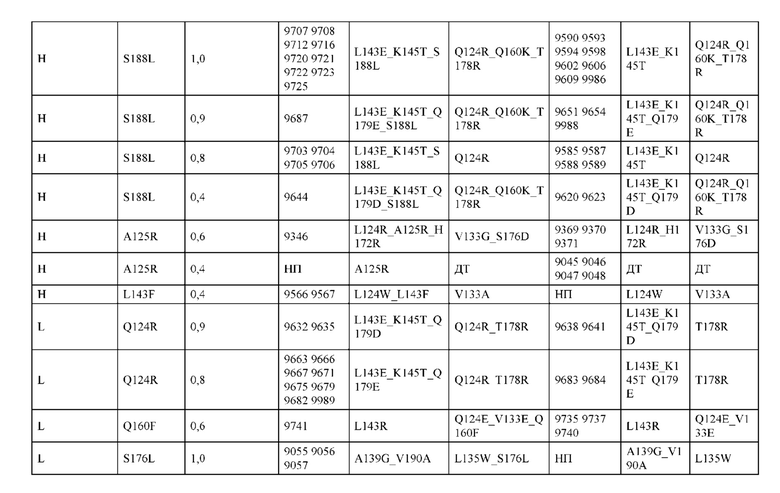
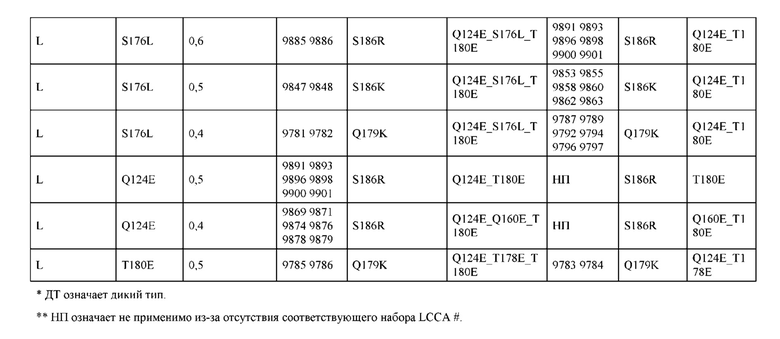
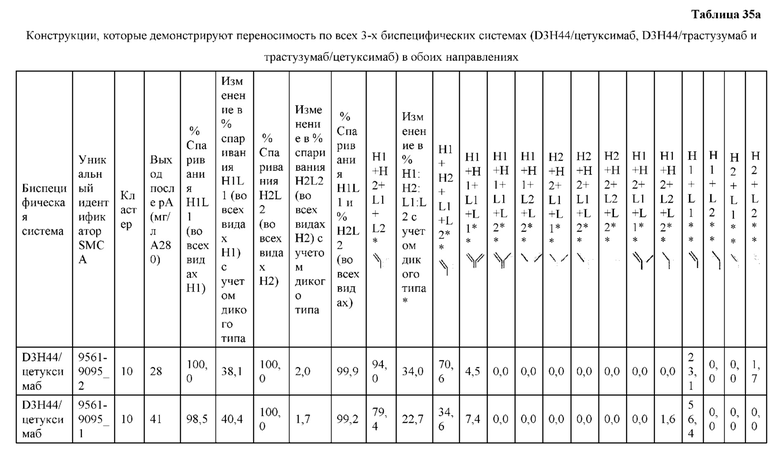
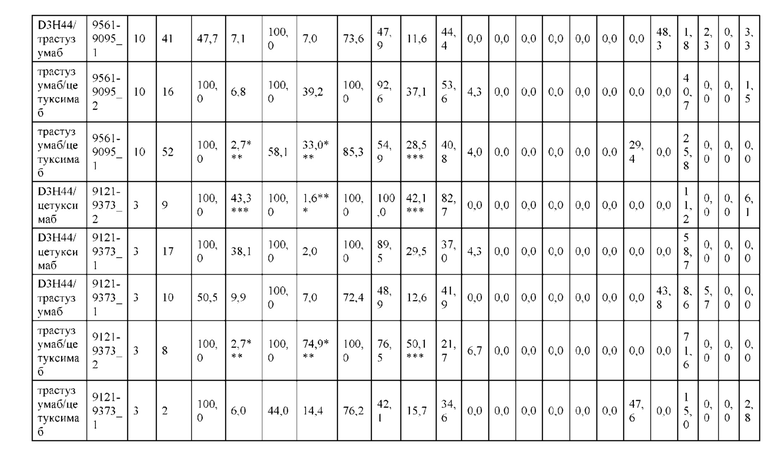
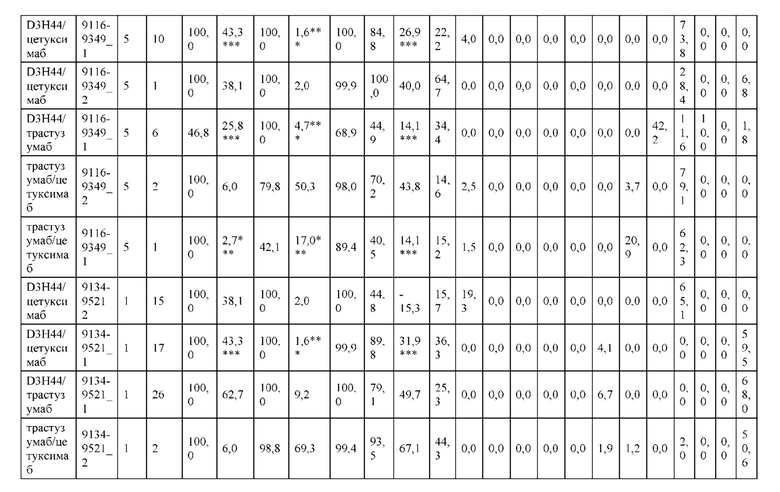
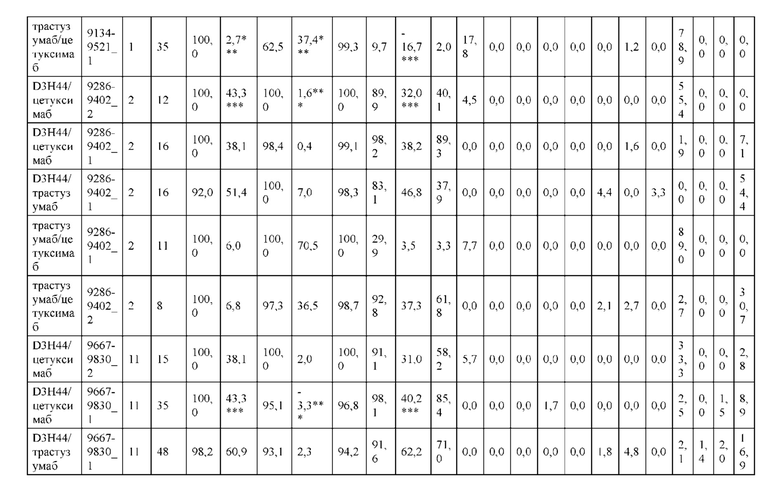
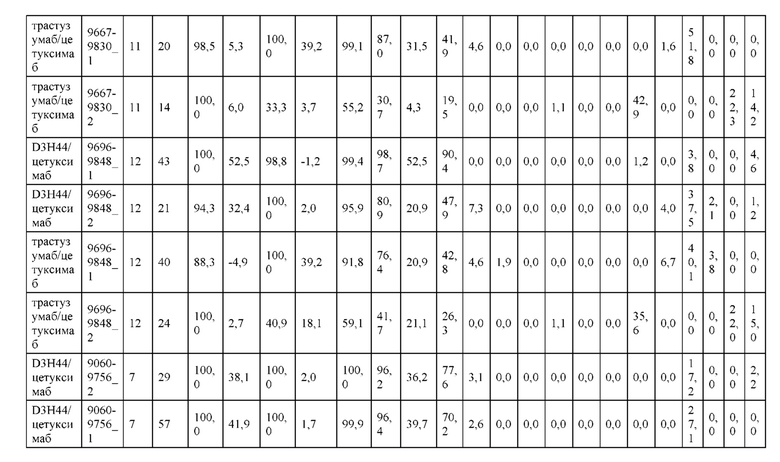
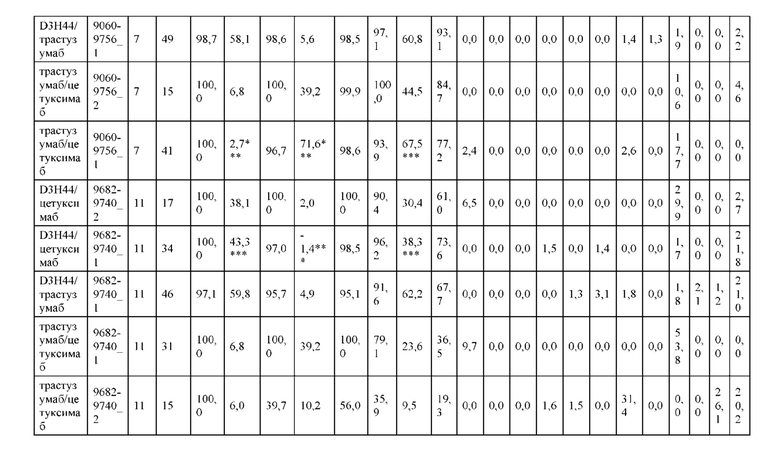
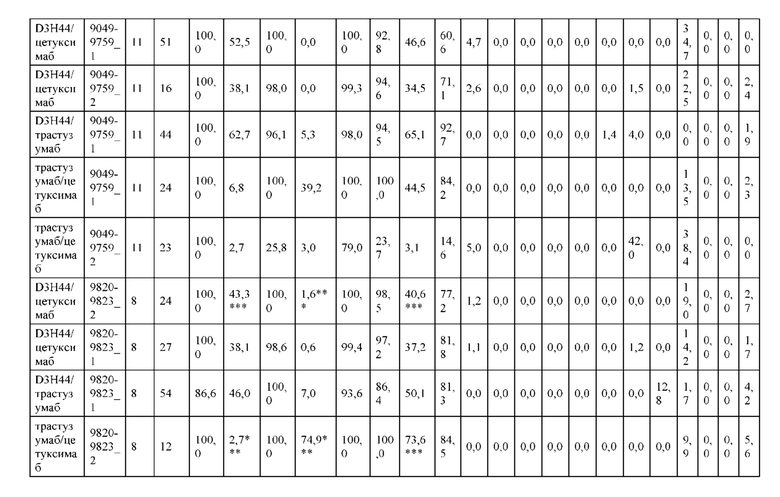
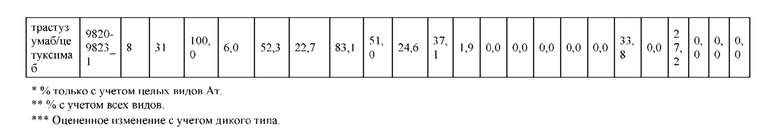
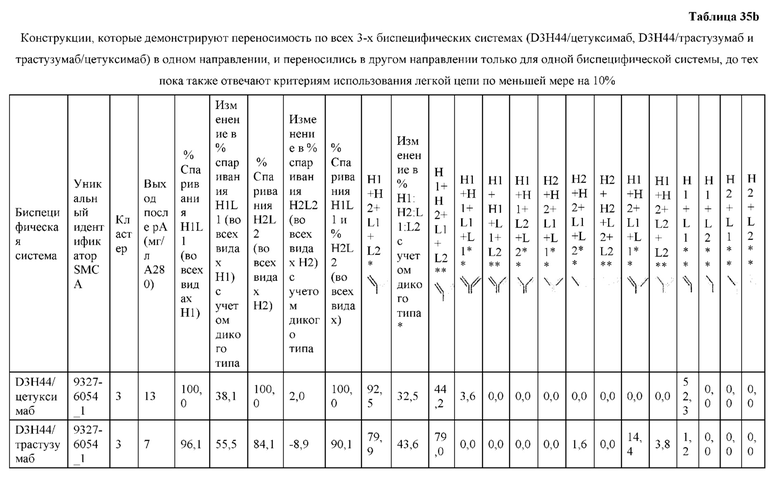
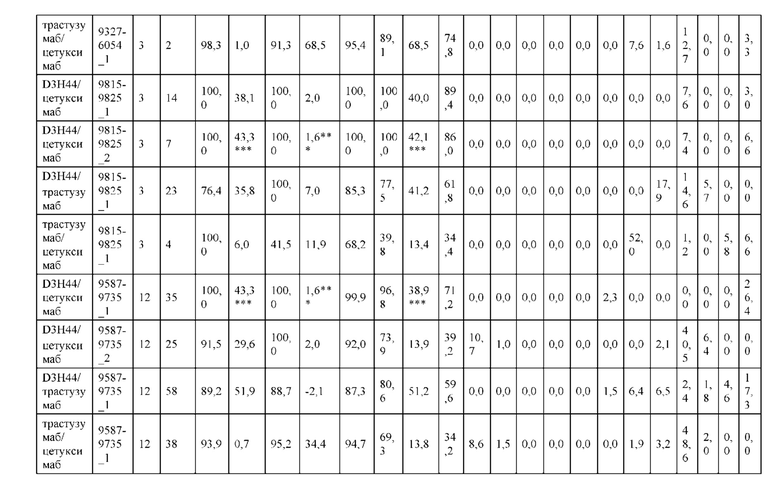
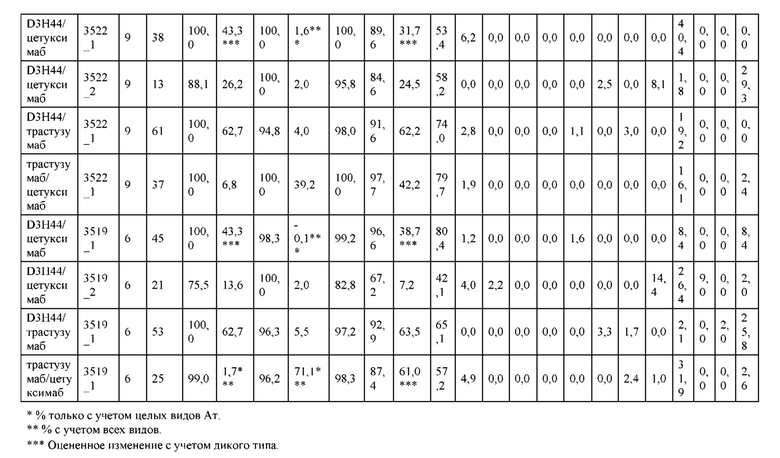
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ZYMEWORKS INC.
<120> МОДИФИЦИРОВАННЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ ПОЛИПЕПТИДНЫЕ
КОНСТРУКЦИИ И ИХ
ПРИМЕНЕНИЕ
<130> 97993-945204 (000110PC)
<140>
<141>
<150> 62/154055
<151> 2015-04-28
<150> 62/003663
<151> 2014-05-28
<160> 12
<170> PatentIn версии 3.5
<210> 1
<211> 214
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 1
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Arg Asp Ile Lys Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Val Leu Ile
35 40 45
Tyr Tyr Ala Thr Ser Leu Ala Glu Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln His Gly Glu Ser Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 2
<211> 214
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 2
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 3
<211> 214
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 3
Asp Ile Leu Leu Thr Gln Ser Pro Val Ile Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Val Ser Phe Ser Cys Arg Ala Ser Gln Ser Ile Gly Thr Asn
20 25 30
Ile His Trp Tyr Gln Gln Arg Thr Asn Gly Ser Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Ser
65 70 75 80
Glu Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Asn Asn Asn Trp Pro Thr
85 90 95
Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 4
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 4
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Glu Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Leu Ile Asp Pro Glu Gln Gly Asn Thr Ile Tyr Asp Pro Lys Phe
50 55 60
Gln Asp Arg Ala Thr Ile Ser Ala Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Thr Ala Ala Tyr Phe Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 5
<211> 449
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 5
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly
<210> 6
<211> 448
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 6
Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Asn Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Ser Gly Gly Asn Thr Asp Tyr Asn Thr Pro Phe Thr
50 55 60
Ser Arg Leu Ser Ile Asn Lys Asp Asn Ser Lys Ser Gln Val Phe Phe
65 70 75 80
Lys Met Asn Ser Leu Gln Ser Asn Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95
Arg Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 7
<211> 1347
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 7
gaggtgcagc tggtggaaag cggaggagga ctggtgcagc caggaggatc tctgcgactg 60
agttgcgccg cttcaggatt caacatcaag gacacctaca ttcactgggt gcgacaggct 120
ccaggaaaag gactggagtg ggtggctcga atctatccca ctaatggata cacccggtat 180
gccgactccg tgaaggggag gtttactatt agcgccgata catccaaaaa cactgcttac 240
ctgcagatga acagcctgcg agccgaagat accgctgtgt actattgcag tcgatgggga 300
ggagacggat tctacgctat ggattattgg ggacagggga ccctggtgac agtgagctcc 360
gcctctacca agggccccag tgtgtttccc ctggctcctt ctagtaaatc cacctctgga 420
gggacagccg ctctgggatg tctggtgaag gactatttcc ccgagcctgt gaccgtgagt 480
tggaactcag gcgccctgac aagcggagtg cacacttttc ctgctgtgct gcagtcaagc 540
gggctgtact ccctgtcctc tgtggtgaca gtgccaagtt caagcctggg cacacagact 600
tatatctgca acgtgaatca taagccctca aatacaaaag tggacaagaa agtggagccc 660
aagagctgtg ataagaccca cacctgccct ccctgtccag ctccagaact gctgggagga 720
cctagcgtgt tcctgtttcc ccctaagcca aaagacactc tgatgatttc caggactccc 780
gaggtgacct gcgtggtggt ggacgtgtct cacgaggacc ccgaagtgaa gttcaactgg 840
tacgtggatg gcgtggaagt gcataatgct aagacaaaac caagagagga acagtacaac 900
tccacttatc gcgtcgtgag cgtgctgacc gtgctgcacc aggactggct gaacgggaag 960
gagtataagt gcaaagtcag taataaggcc ctgcctgctc caatcgaaaa aaccatctct 1020
aaggccaaag gccagccaag ggagccccag gtgtacacac tgccacccag cagagacgaa 1080
ctgaccaaga accaggtgtc cctgacatgt ctggtgaaag gcttctatcc tagtgatatt 1140
gctgtggagt gggaatcaaa tggacagcca gagaacaatt acaagaccac acctccagtg 1200
ctggacagcg atggcagctt cttcctgtat tccaagctga cagtggataa atctcgatgg 1260
cagcagggga acgtgtttag ttgttcagtg atgcatgaag ccctgcacaa tcattacact 1320
cagaagagcc tgtccctgtc tcccggc 1347
<210> 8
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 8
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagagtcacc 60
atcacttgcc gggcaagtca ggacgttaac accgctgtag cttggtatca gcagaaacca 120
gggaaagccc ctaagctcct gatctattct gcatcctttt tgtacagtgg ggtcccatca 180
aggttcagtg gcagtcgatc tgggacagat ttcactctca ccatcagcag tctgcaacct 240
gaagattttg caacttacta ctgtcaacag cattacacta ccccacccac tttcggccaa 300
gggaccaaag tggagatcaa acgaactgtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccaa 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gt 642
<210> 9
<211> 1344
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 9
caggtgcagc tgaaacagag cggcccgggc ctggtgcagc cgagccagag cctgagcatt 60
acctgcaccg tgagcggctt tagcctgacc aactatggcg tgcattgggt gcgccagagc 120
ccgggcaaag gcctggaatg gctgggcgtg atttggagcg gcggcaacac cgattataac 180
accccgttta ccagccgcct gagcattaac aaagataaca gcaaaagcca ggtgtttttt 240
aaaatgaaca gcctgcagag caacgatacc gcgatttatt attgcgcgcg cgcgctgacc 300
tattatgatt atgaatttgc gtattggggc cagggcaccc tggtgaccgt gagcgcggcg 360
agcaccaaag gcccgagcgt gtttccgctg gcgccgagca gcaaaagcac cagcggcggc 420
accgcggcgc tgggctgcct ggtgaaagat tattttccgg aaccggtgac cgtgagctgg 480
aacagcggcg cgctgaccag cggcgtgcat acctttccgg cggtgctgca gagcagcggc 540
ctgtatagcc tgagcagcgt ggtgaccgtg ccgagcagca gcctgggcac ccagacctat 600
atttgcaacg tgaaccataa accgagcaac accaaagtgg ataaaaaagt ggaaccgaaa 660
agctgcgata aaacccatac ctgcccgccg tgcccggcgc cggaactgct gggcggcccg 720
agcgtgtttc tgtttccgcc gaaaccgaaa gataccctga tgattagccg caccccggaa 780
gtgacctgcg tggtggtgga tgtgagccat gaagatccgg aagtgaaatt taactggtat 840
gtggatggcg tggaagtgca taacgcgaaa accaaaccgc gcgaagaaca gtataacagc 900
acctatcgcg tggtgagcgt gctgaccgtg ctgcatcagg attggctgaa cggcaaagaa 960
tataaatgca aagtgagcaa caaagcgctg ccggcgccga ttgaaaaaac cattagcaaa 1020
gcgaaaggcc agccgcgcga accgcaggtg tataccctgc cgccgagccg cgatgaactg 1080
accaaaaacc aggtgagcct gacctgcctg gtgaaaggct tttatccgag cgatattgcg 1140
gtggaatggg aaagcaacgg ccagccggaa aacaactata aaaccacccc gccggtgctg 1200
gatagcgatg gcagcttttt tctgtatagc aaactgaccg tggataaaag ccgctggcag 1260
cagggcaacg tgtttagctg cagcgtgatg catgaagcgc tgcataacca ttatacccag 1320
aaaagcctga gcctgagccc gggc 1344
<210> 10
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 10
gacatcctgc tgactcagag cccagtgatc ctgtcagtca gcccaggaga gcgggtgtcc 60
ttctcttgca gagcaagtca gtcaatcgga acaaatattc actggtacca gcagaggact 120
aacggctccc ctcgcctgct gattaagtat gctagcgaat ccatctctgg cattccatct 180
cggttcagtg gctcagggag cggaacagac tttactctgt ccatcaattc tgtggagagt 240
gaagacattg ccgattacta ttgccagcag aacaataact ggcccaccac attcggcgct 300
gggaccaagc tggagctgaa acgaacagtg gccgctcctt ctgtcttcat ctttccccct 360
agtgacgaac agctgaaaag cggcacagcc tccgtggtct gtctgctgaa taacttttac 420
ccaagagagg caaaggtgca gtggaaagtc gataatgccc tgcagtcagg gaacagccag 480
gagtccgtga ctgaacagga ctctaaggat agtacctatt cactgagctc cactctgacc 540
ctgtccaaag ctgattacga gaagcacaaa gtgtatgcat gcgaagtcac ccatcagggg 600
ctgtctagtc ccgtgacaaa gagctttaac cggggagagt gt 642
<210> 11
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 11
gaggtgcagc tggtcgaatc tggaggagga ctggtgcagc caggagggtc actgagactg 60
agctgcgccg cttccggctt caacatcaag gagtactata tgcactgggt gaggcaggca 120
cctggcaaag gactggagtg ggtgggactg atcgacccag aacaggggaa caccatctac 180
gaccctaagt ttcaggatag ggcaaccatt tctgccgaca acagtaaaaa tacagcttat 240
ctgcagatga acagcctgag ggctgaagat actgcagtgt actattgcgc acgcgacacc 300
gcagcctact tcgattattg gggacagggc accctggtca cagtgagctc cgcatcaact 360
aagggaccca gcgtgtttcc actggccccc tctagtaaat ccacttctgg aggcaccgct 420
gcactgggct gtctggtgaa ggattacttc ccagagcccg tcacagtgag ctggaactcc 480
ggggccctga ccagcggagt ccatacattt cctgctgtgc tgcagtcaag cgggctgtac 540
tccctgtcct ctgtggtcac cgtgccaagt tcaagcctgg gaactcagac ctatatctgc 600
aacgtgaatc acaagccttc aaatacaaaa gtcgacaaga aagtggaacc aaagagctgt 660
gataaaacac atacttgccc accttgtcct gcaccagagc tgctgggagg accaagcgtg 720
ttcctgtttc cacccaagcc caaagacacc ctgatgattt cccgcacacc agaagtcact 780
tgcgtggtcg tggacgtgtc tcacgaggac cccgaagtca agttcaactg gtacgtggat 840
ggcgtcgagg tgcataatgc caagacaaaa ccccgggagg aacagtacaa ctccacatat 900
agagtcgtgt ctgtcctgac tgtgctgcac caggactggc tgaacgggaa ggagtataag 960
tgcaaagtga gtaataaggc cctgcccgct cctatcgaga aaacaattag caaggccaaa 1020
ggccagcctc gagaaccaca ggtgtacact ctgcctccat ctcgggacga gctgactaag 1080
aaccaggtca gtctgacctg tctggtgaaa ggattctatc ccagcgatat cgctgtggag 1140
tgggaatcca atggccagcc tgagaacaat tacaagacca caccccctgt gctggactct 1200
gatggcagtt tctttctgta tagtaagctg accgtcgata aatcacgatg gcagcagggg 1260
aacgtgttca gctgttcagt gatgcacgaa gccctgcaca accattacac ccagaagagc 1320
ctgagcctgt ctcccggc 1338
<210> 12
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 12
gacatccaga tgacacagtc ccctagctcc ctgagtgcct cagtggggga cagagtcact 60
atcacctgcc gggcttccag agatattaag tcttacctga actggtatca gcagaagcca 120
ggcaaagcac ccaaggtgct gatctactat gccaccagtc tggctgaagg agtgccttca 180
cggttcagcg gctccgggtc tggaactgac tacacactga ctatttctag tctgcagcct 240
gaggatttcg ctacctacta ttgcctgcag cacggcgaat ccccatggac ttttggccag 300
gggaccaaag tggagatcaa gaggacagtg gccgctccat ccgtcttcat ttttccccct 360
tctgacgaac agctgaaatc aggaactgcc agcgtggtct gtctgctgaa caatttctac 420
ccccgcgagg caaaagtgca gtggaaggtc gataacgccc tgcagagtgg caattcacag 480
gagagcgtga cagaacagga ctccaaagat tctacttata gtctgtcaag caccctgaca 540
ctgtctaagg ctgattacga gaagcacaaa gtgtatgcat gcgaagtcac ccatcagggg 600
ctgtcctctc ccgtgacaaa gagctttaat cggggagagt gt 642
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СКОНСТРУИРОВАННЫЕ ПАРЫ ТЯЖЕЛАЯ-ЛЕГКАЯ ЦЕПИ ИММУНОГЛОБУЛИНА И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2826203C1 |
| СКОНСТРУИРОВАННЫЕ ПАРЫ ТЯЖЕЛАЯ-ЛЕГКАЯ ЦЕПИ ИММУНОГЛОБУЛИНА И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2720472C2 |
| АНТИГЕНСВЯЗЫВАЮЩИЕ ПОЛИПЕПТИДНЫЕ КОНСТРУКЦИИ, СОДЕРЖАЩИЕ ЛЕГКИЕ ЦЕПИ КАППА И ЛЯМБДА, И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2824193C2 |
| АНТИТЕЛА К LY6G6D И СПОСОБЫ ПРИМЕНЕНИЯ | 2020 |
|
RU2818569C1 |
| МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕ BCMA, И ПУТИ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2812322C2 |
| АНТИТЕЛО, КОТОРОЕ СВЯЗЫВАЕТСЯ С VEGF И IL-1БЕТА, И СПОСОБЫ ПРИМЕНЕНИЯ | 2019 |
|
RU2816476C2 |
| АНТИТЕЛА К LAG3 | 2018 |
|
RU2778053C2 |
| АНТИТЕЛА К CD38 И КОМБИНАЦИИ С АНТИТЕЛАМИ К CD3 И CD28 | 2018 |
|
RU2812910C2 |
| СПОСОБЫ ЛЕЧЕНИЯ РАКА С ИСПОЛЬЗОВАНИЕМ АНТАГОНИСТОВ, СВЯЗЫВАЮЩИХСЯ С КОМПОНЕНТАМИ СИГНАЛЬНОГО ПУТИ PD-1, И ТАКСАНОВ | 2014 |
|
RU2719487C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА К CCL2 | 2020 |
|
RU2819613C1 |





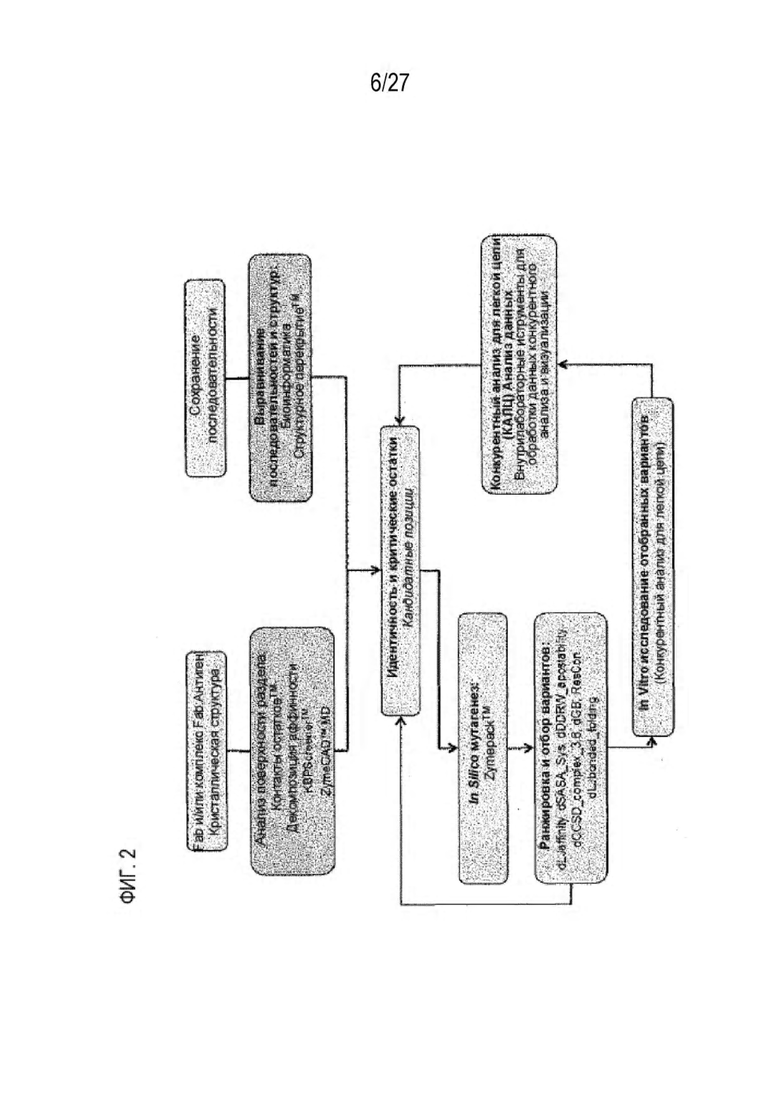
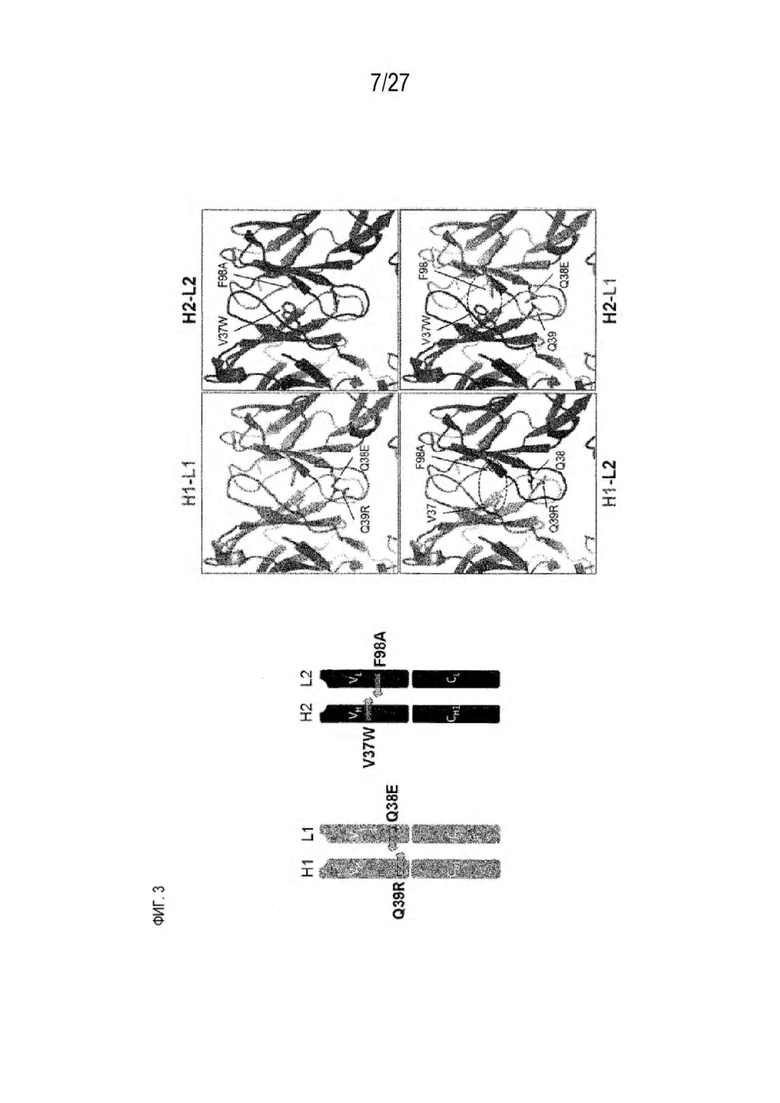

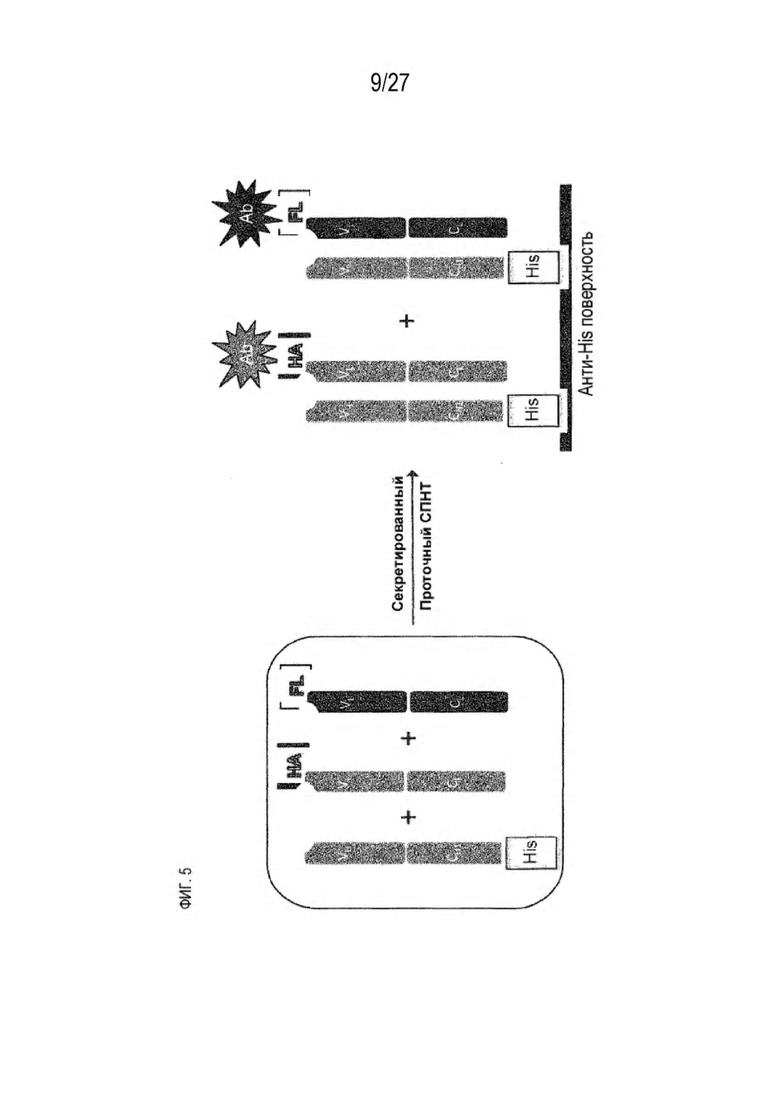

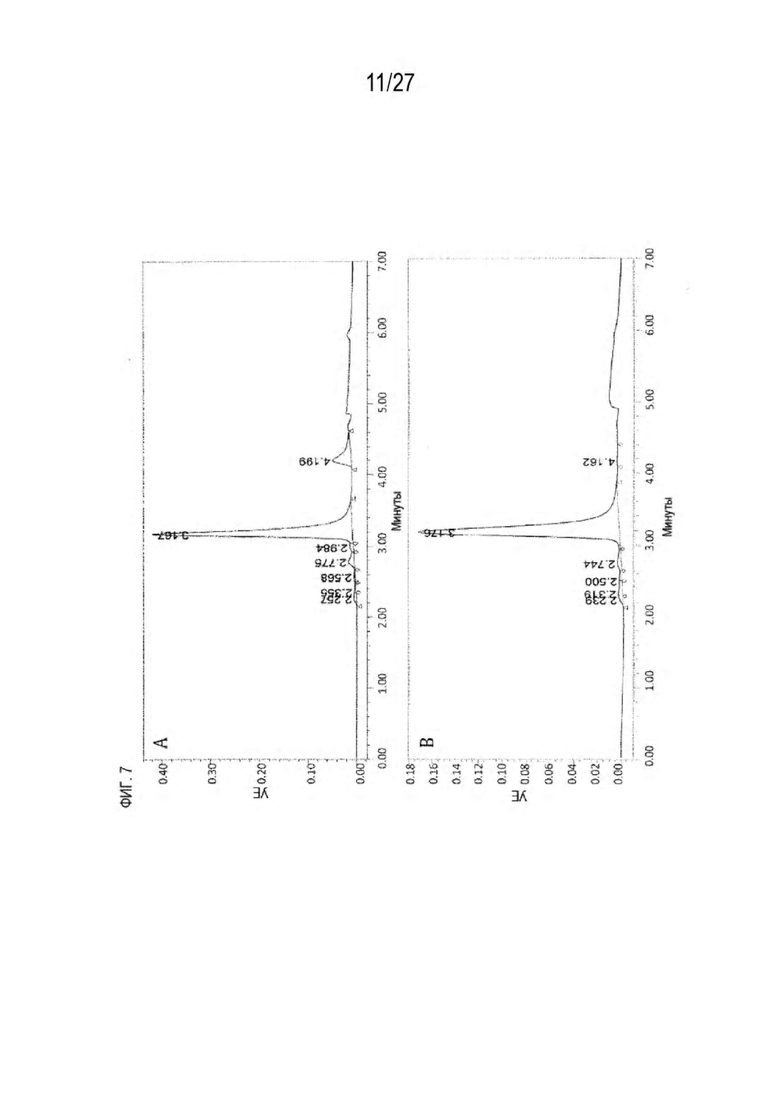
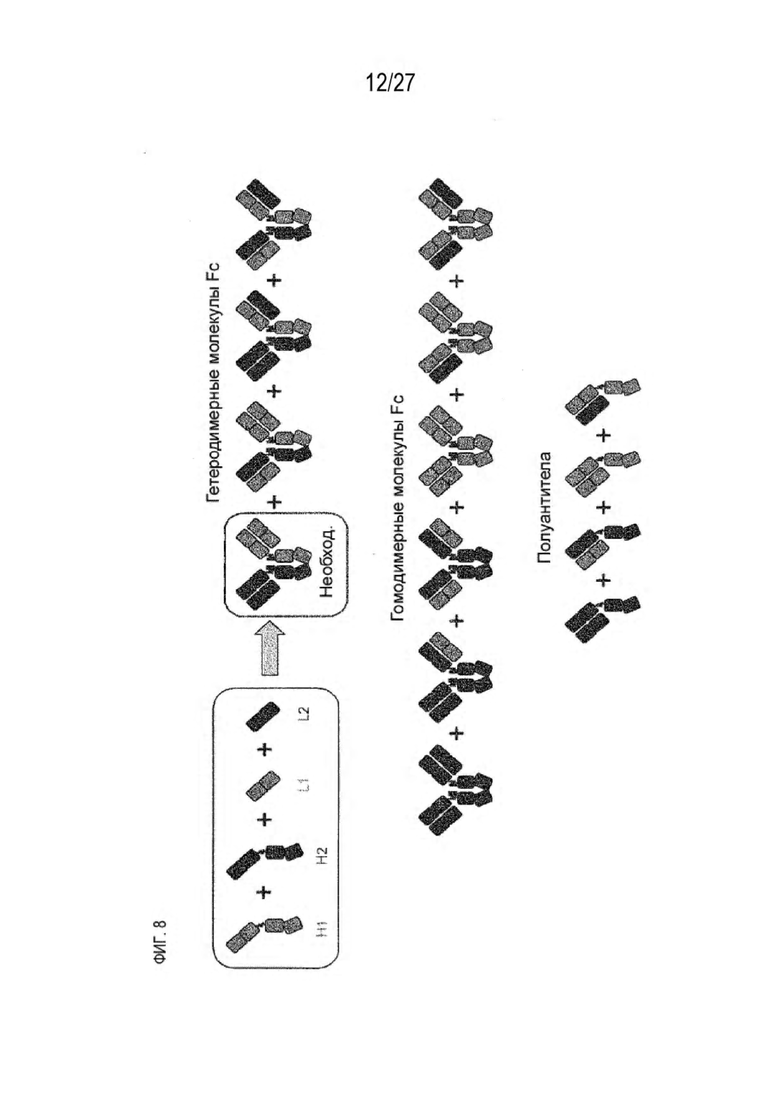
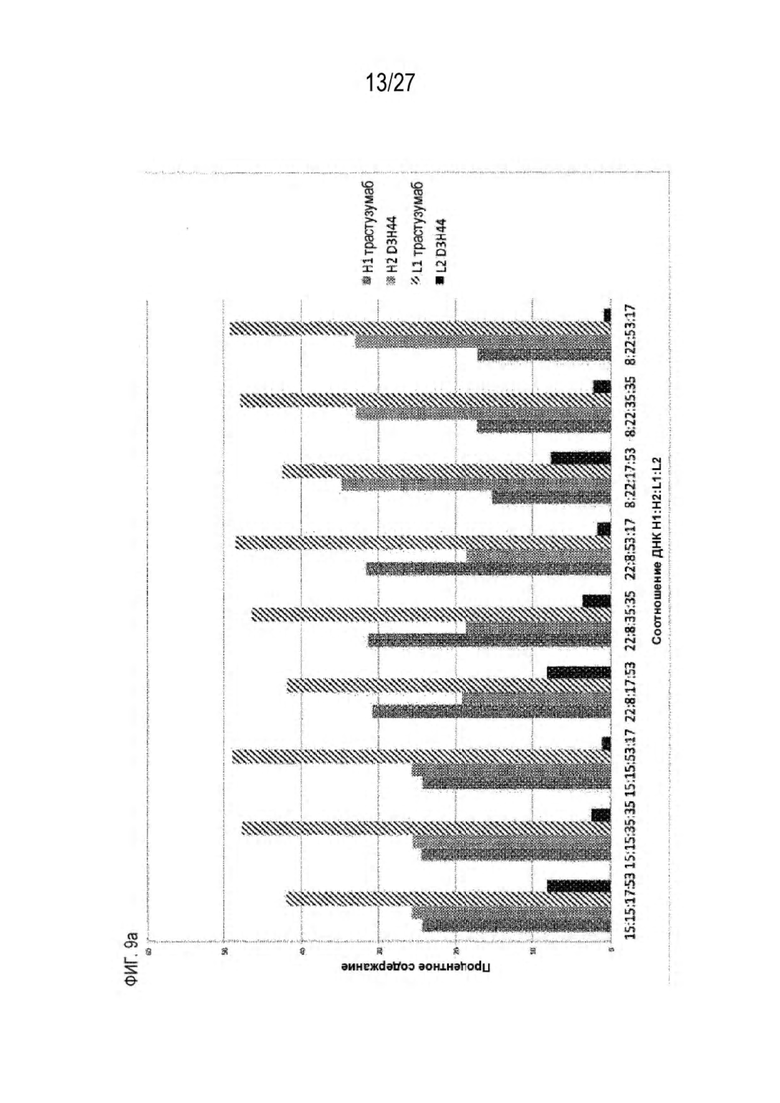
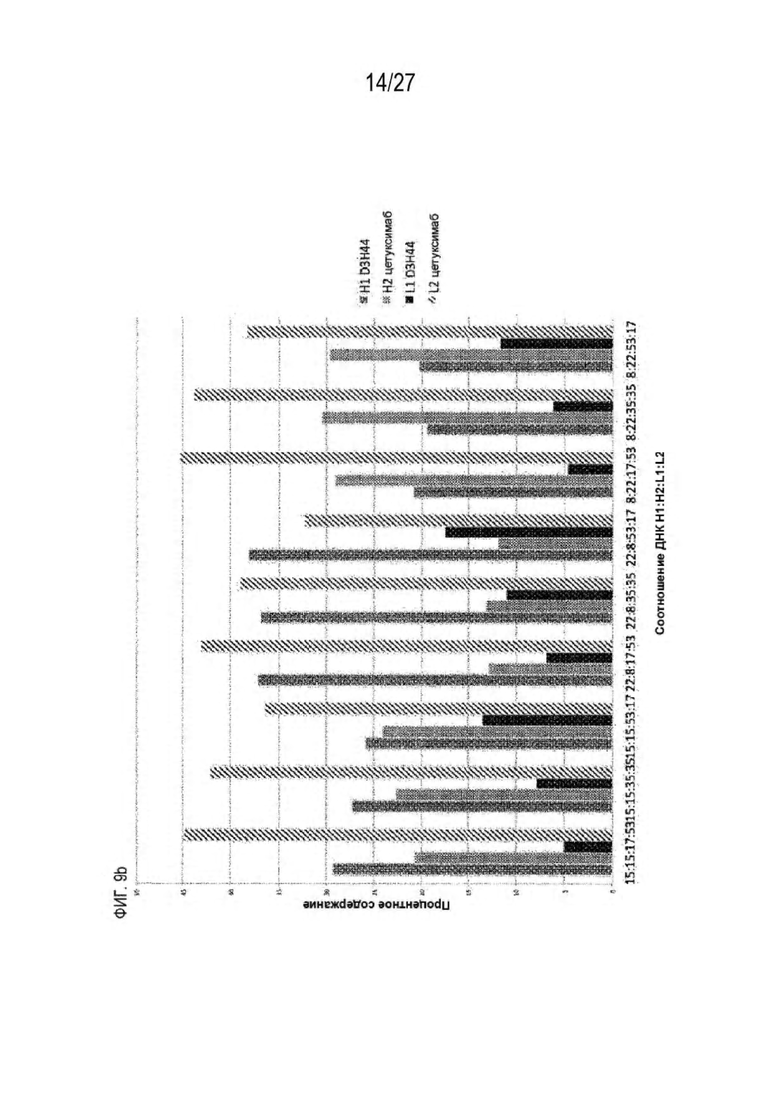

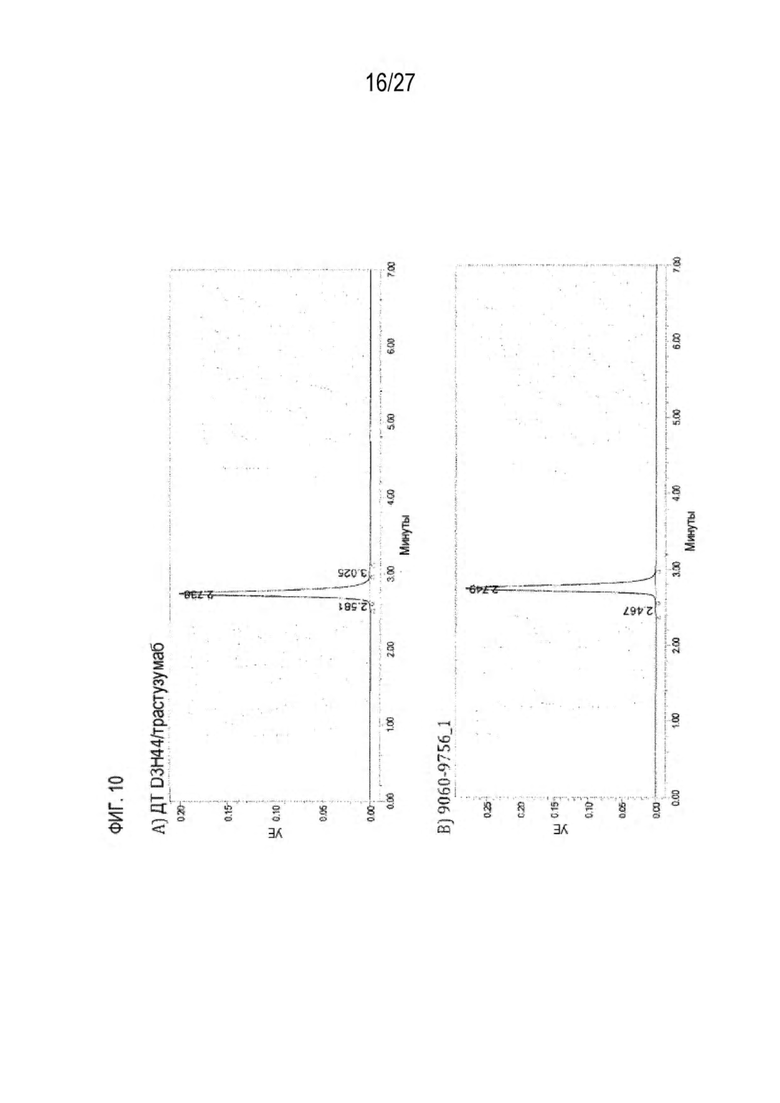
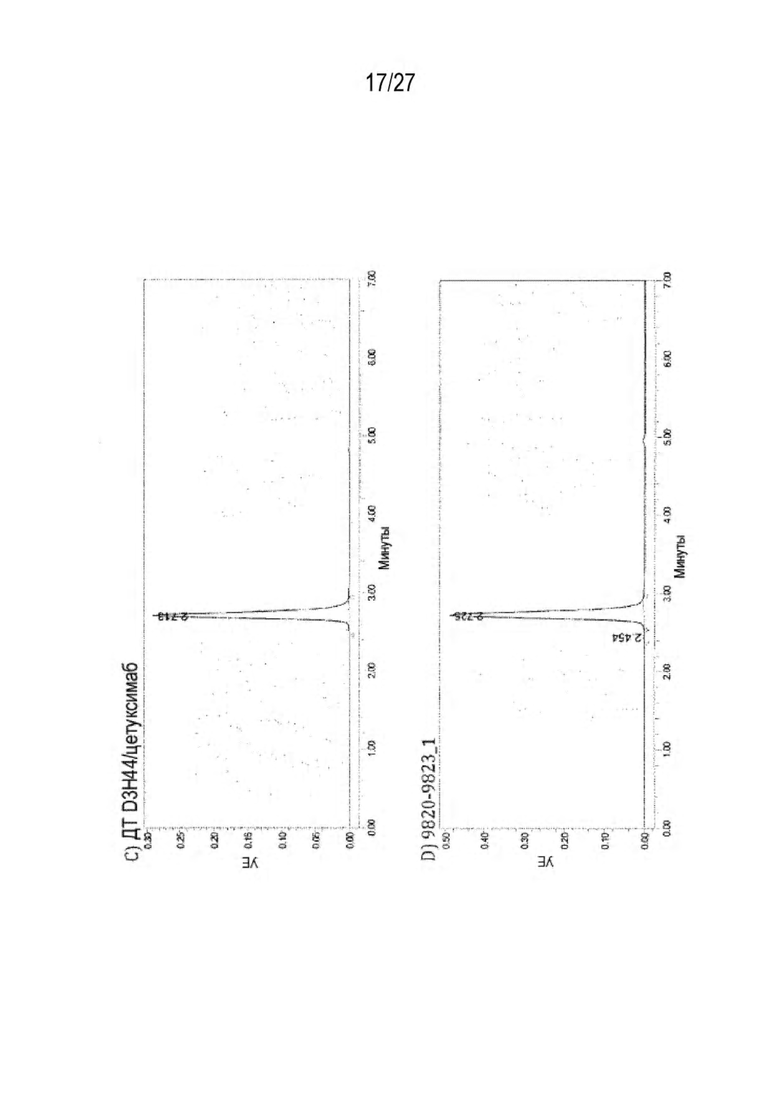
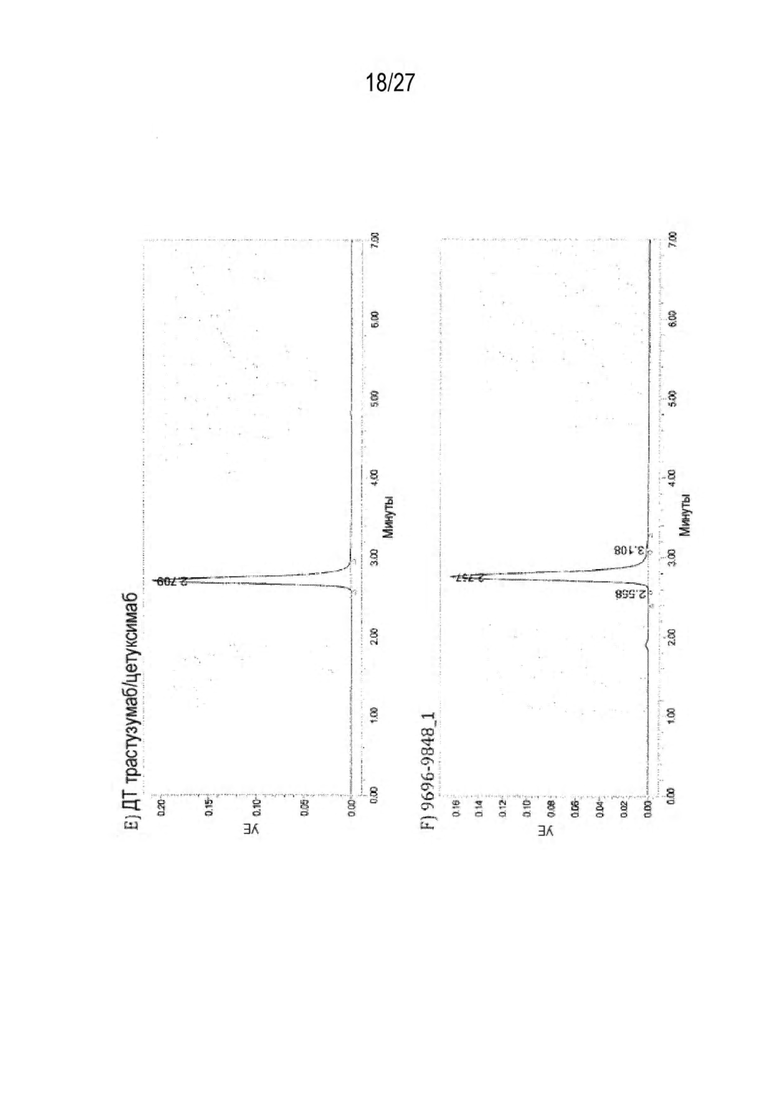
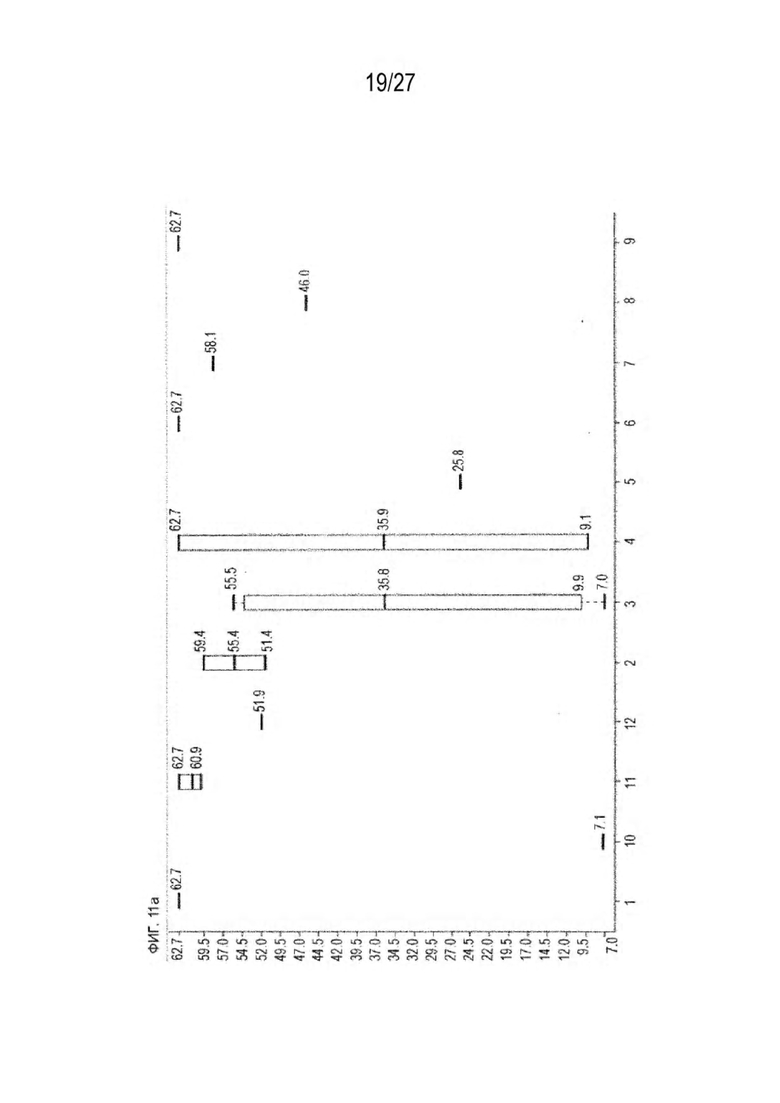
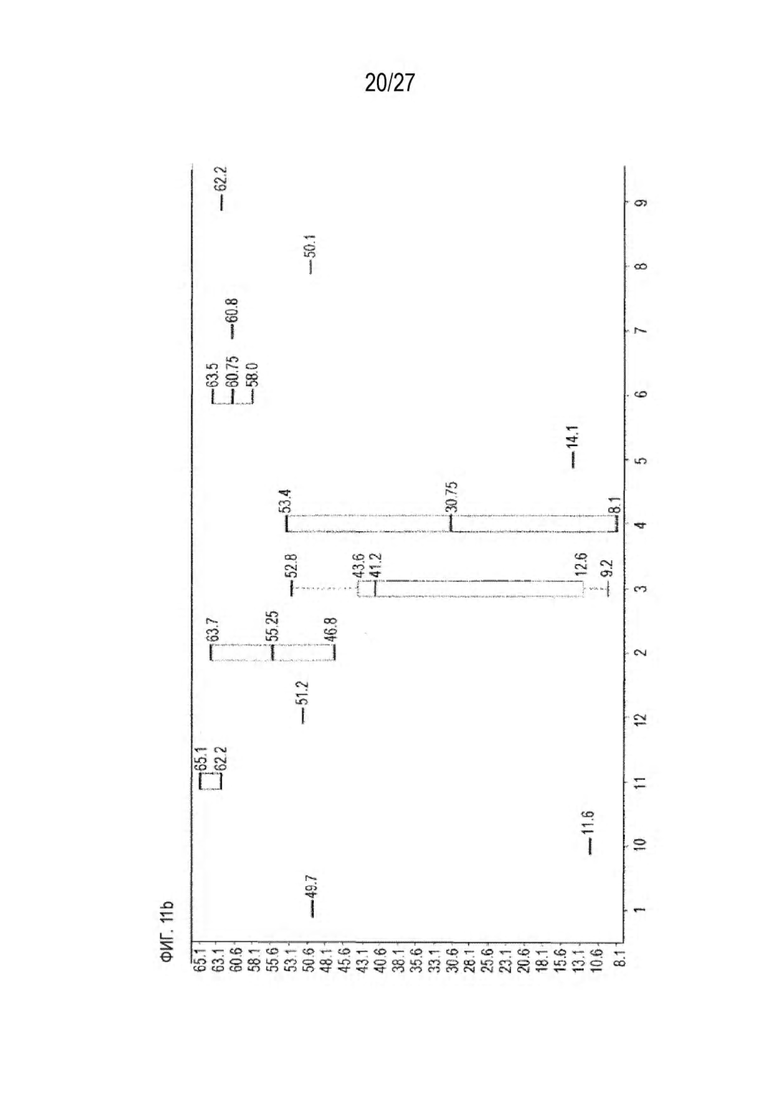


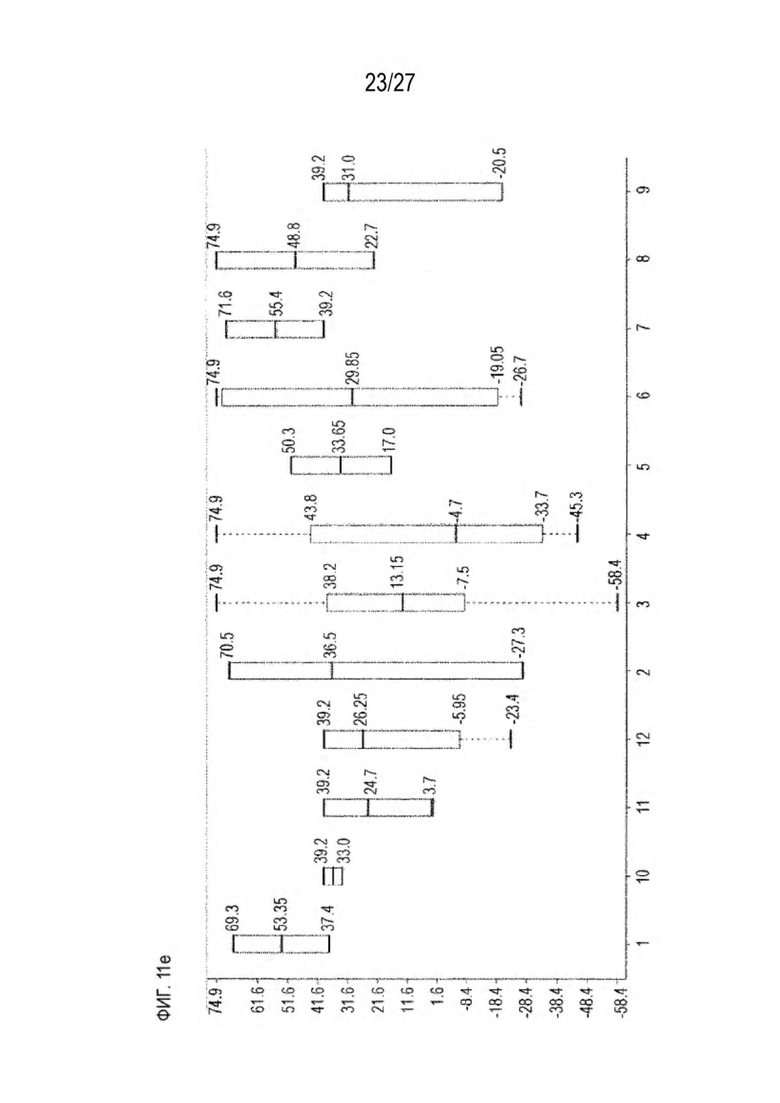
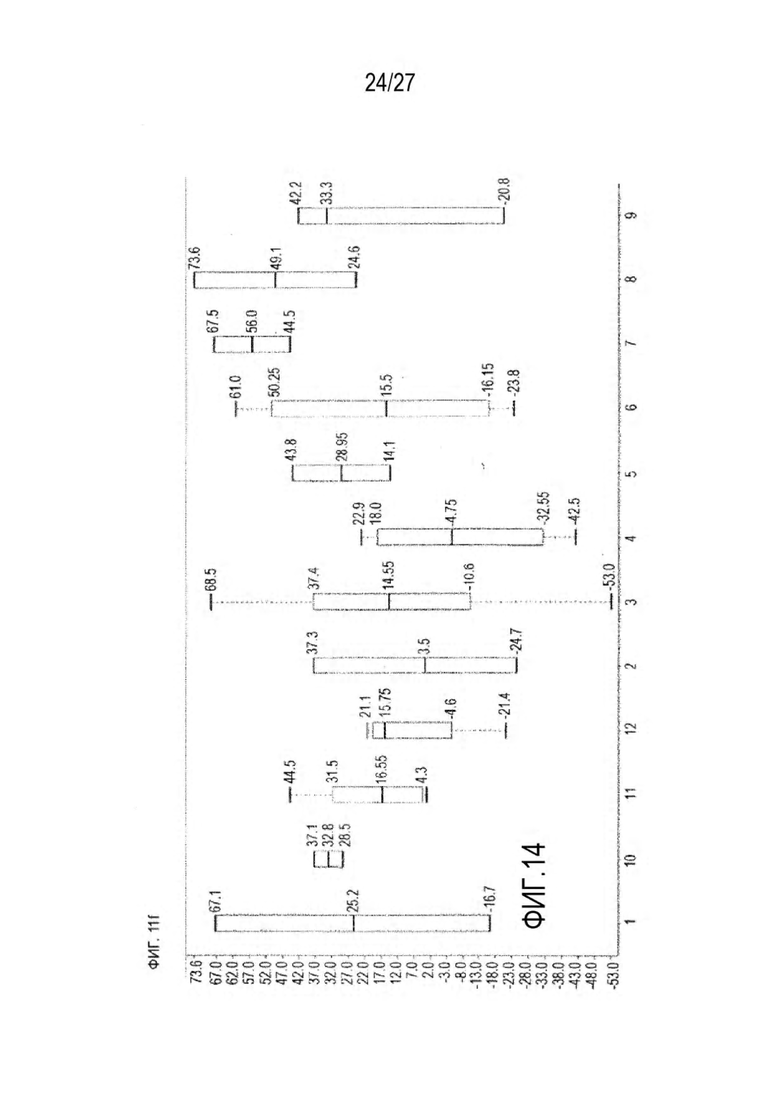

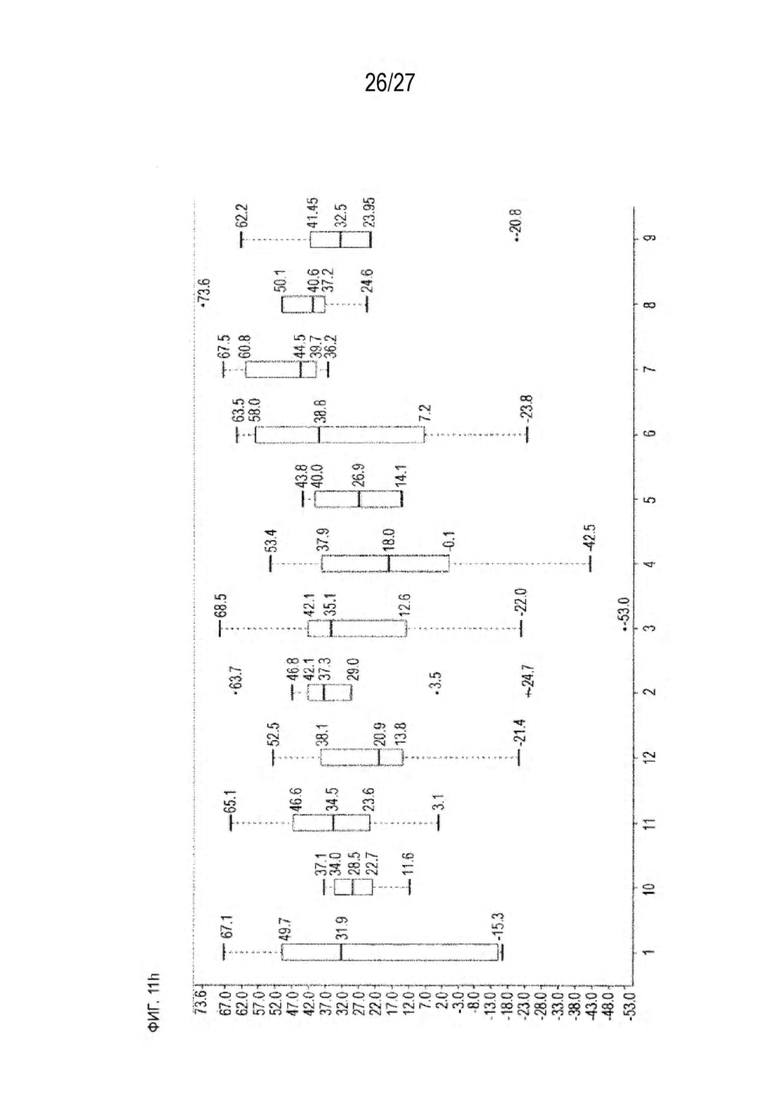
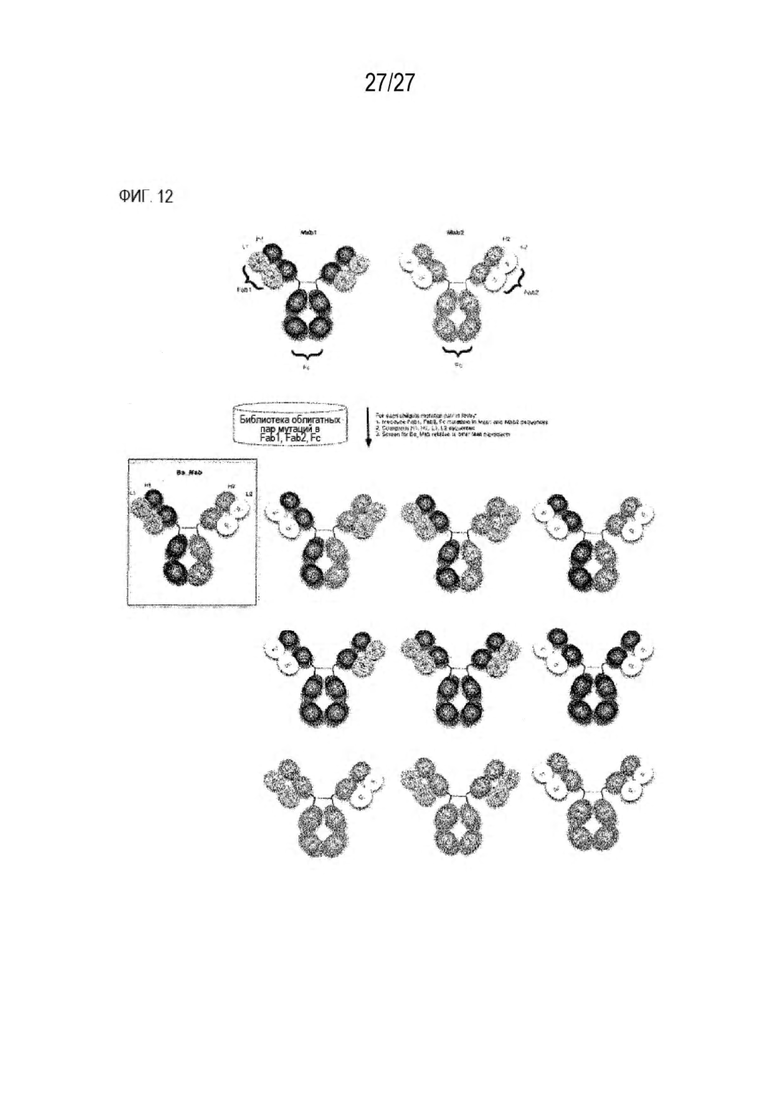
Группа изобретений относится к биотехнологии. Предложен набор полинуклеотидов для получения выделенной антигенсвязывающей полипептидной конструкции, а также набор векторов экспрессии, выделенная клетка-хозяин и способ получения антигенсвязывающей полипептидной конструкции. Группа изобретений позволяет получать антигенсвязывающие полипептидные конструкции на основе антител, обладающие повышенной стабильностью. 4 н. и 25 з.п. ф-лы, 12 ил., 35 табл., 14 пр.
1. Набор полинуклеотидов для получения антигенсвязывающей полипептидной конструкции, причем набор полинуклеотидов кодирует антигенсвязывающую полипептидную конструкцию, содержащую по меньшей мере первый гетеродимер и второй гетеродимер, где первый гетеродимер содержит первую человеческую или гуманизированную полипептидную последовательность (H1) тяжелой цепи иммуноглобулина и первую человеческую или гуманизированную полипептидную последовательность (L1) легкой цепи иммуноглобулина и имеет область Fab, связывающуюся с первым эпитопом; и второй гетеродимер содержит вторую человеческую или гуманизированную полипептидную последовательность (H2) тяжелой цепи иммуноглобулина и вторую человеческую или гуманизированную полипептидную последовательность (L2) легкой цепи иммуноглобулина и имеет область Fab, связывающуюся со вторым эпитопом, где по меньшей мере одна из последовательностей H1 или L1 первого гетеродимера отличается от соответствующей последовательности H2 или L2 второго гетеродимера и где H1 и H2 каждая содержит по меньшей мере вариабельный домен тяжелой цепи (домен VH) и константный домен тяжелой цепи (домен CH1);
L1 и L2 каждая содержит по меньшей мере вариабельный домен легкой цепи (домен VL) и константный домен легкой цепи (домен CL); и
H1, H2, L1 и L2 содержат по меньшей мере одну аминокислотную замену или совокупность аминокислотных замен, где H1 предпочтительно спаривается с L1 по сравнению с L2 и H2 предпочтительно спаривается с L2 по сравнению с L1;
где термостабильность области Fab, как измерено на основе температуры плавления (Tm) по меньшей мере первого и второго гетеродимеров, находится в пределах около 0, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10°C от Tm соответствующего гетеродимера без по меньшей мере одной аминокислотной замены или совокупности аминокислотных замен и где H1, L1, H2 и L2 содержат следующие аминокислотные замены в положениях, идентифицированных в соответствии с нумерацией Kabat:
a) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену 186K и L2 содержит аминокислотные замены 124E, 160E и 178E;
b) H1 содержит аминокислотные замены 143E, 145T и 179D, L1 содержит аминокислотные замены 124K и 178R, H2 содержит аминокислотную замену 186K и L2 содержит аминокислотные замены 124E и 180E;
c) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену 186K и L2 содержит аминокислотные замены 124E, 178E и 180E;
d) H1 содержит аминокислотные замены 143E, 145T и 179D, L1 содержит аминокислотные замены 124K и 178R, H2 содержит аминокислотную замену 186R и L2 содержит аминокислотные замены 124E и 180E;
e) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178K, H2 содержит аминокислотную замену 186K и L2 содержит аминокислотные замены 124E и 180E;
f) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178K, H2 содержит аминокислотную замену 186R и L2 содержит аминокислотные замены 124E и 180E;
g) H1 содержит аминокислотные замены 143E, 145T и 179D, L1 содержит аминокислотные замены 124R и 178K, H2 содержит аминокислотную замену 186K и L2 содержит аминокислотные замены 124E и 180E;
h) H1 содержит аминокислотные замены 143E, 145T и 179D, L1 содержит аминокислотные замены 124R и 178K, H2 содержит аминокислотную замену 186R и L2 содержит аминокислотные замены 124E и 180E;
i) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену 186R и L2 содержит аминокислотные замены 124E и 180E;
j) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену 186R и L2 содержит аминокислотные замены 124E, 160E и 178E;
k) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену 186K и L2 содержит аминокислотные замены 124E, 160E и 180E;
l) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену 186R и L2 содержит аминокислотные замены 124E, 178E и 180E;
m) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену 186R и L2 содержит аминокислотные замены 124E и 178E;
n) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R, 160K и 178R, H2 содержит аминокислотную замену 186R и L2 содержит аминокислотные замены 124E и 180E;
o) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену 186K и L2 содержит аминокислотные замены 124E и 178E;
p) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124K и 178R, H2 содержит аминокислотную замену 186K и L2 содержит аминокислотные замены 124E и 180E;
q) H1 содержит аминокислотные замены 143E, 145T и 179D, L1 содержит аминокислотные замены 124R, 160K и 178R, H2 содержит аминокислотную замену 186R и L2 содержит аминокислотные замены 124E и 180E;
r) H1 содержит аминокислотные замены 143E, 145T и 179D, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену 186K и L2 содержит аминокислотные замены 124E, 160E и 180E;
s) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R, 160K и 178R, H2 содержит аминокислотную замену 186K и L2 содержит аминокислотные замены 124E и 180E;
t) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену 186K и L2 содержит аминокислотные замены 124E и 180E;
u) H1 содержит аминокислотные замены 143E, 145T и 179D, L1 содержит аминокислотные замены 124R и 178K, H2 содержит аминокислотную замену 186K и L2 содержит аминокислотные замены 124E, 160E и 180E;
v) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124K и 178R, H2 содержит аминокислотную замену 186R и L2 содержит аминокислотные замены 124E и 180E;
w) H1 содержит аминокислотные замены 143D, 145T и 179E, L1 содержит аминокислотную замену 178R, H2 содержит аминокислотную замену 186R и L2 содержит аминокислотные замены 124E и 180E;
x) H1 содержит аминокислотные замены 143E, 145T и 179D, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену 186R и L2 содержит аминокислотные замены 124E и 180E;
y) H1 содержит аминокислотные замены 143D, 145T и 179E, L1 содержит аминокислотную замену 178R, H2 содержит аминокислотную замену 186K и L2 содержит аминокислотные замены 124E и 180E;
z) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотную замену 178R, H2 содержит аминокислотную замену 186K и L2 содержит аминокислотные замены 124E, 160E и 180E;
aa) H1 содержит аминокислотные замены 143D, 145T и 179E, L1 содержит аминокислотную замену 178R, H2 содержит аминокислотную замену 186K и L2 содержит аминокислотные замены 124E, 160E и 180E;
bb) H1 содержит аминокислотные замены 143E, 145T и 179D, L1 содержит аминокислотные замены 124R, 160K и 178R, H2 содержит аминокислотную замену 186K и L2 содержит аминокислотные замены 124E и 180E;
cc) H1 содержит аминокислотные замены 143E и 145T, L1 содержит аминокислотные замены 124R, 160K и 178R, H2 содержит аминокислотную замену 186R и L2 содержит аминокислотные замены 124E и 180E;
dd) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178K, H2 содержит аминокислотную замену 186K и L2 содержит аминокислотные замены 124E, 160E, 180E;
ee) H1 содержит аминокислотные замены 143E и 145T, L1 содержит аминокислотные замены 124R, 160K и 178R, H2 содержит аминокислотную замену 186K и L2 содержит аминокислотные замены 124E и 180E;
ff) H1 содержит аминокислотные замены 143E, 145T и 179D, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену 186K и L2 содержит аминокислотные замены 124E и 180E;
gg) H1 содержит аминокислотные замены 143E, 145T и 179D, L1 содержит аминокислотную замену 178R, H2 содержит аминокислотную замену 186R и L2 содержит аминокислотные замены 124E и 180E;
hh) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену 186R и L2 содержит аминокислотные замены 124E, 160E и 180E;
ii) H1 содержит аминокислотные замены 143E, 145T и 179D, L1 содержит аминокислотную замену 178R, H2 содержит аминокислотную замену 186K и L2 содержит аминокислотные замены 124E и 180E;
jj) H1 содержит аминокислотные замены 143E, 145T и 179D, L1 содержит аминокислотные замены 124R и 178K, H2 содержит аминокислотную замену 186R и L2 содержит аминокислотные замены 124E, 160E и 180E;
kk) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену Q179R и L2 содержит аминокислотные замены 124E и 178E;
ll) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену Q179R и L2 содержит аминокислотные замены 124E, 160E и 178E;
mm) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену 179K и L2 содержит аминокислотные замены 124E, 160E и 178E;
nn) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену 179R и L2 содержит аминокислотные замены 124E, 178E и 180E;
oo) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену 179K и L2 содержит аминокислотные замены 124E, 178E;
pp) H1 содержит аминокислотные замены 143E, 145T и 179D, L1 содержит аминокислотные замены 124K и 178R, H2 содержит аминокислотную замену 179K и L2 содержит аминокислотные замены 124E, 180E;
qq) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену 179K и L2 содержит аминокислотные замены 124E и 180E;
rr) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену 179K и L2 содержит аминокислотные замены 124E, 178E и 180E;
ss) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178K, H2 содержит аминокислотную замену 179K и L2 содержит аминокислотные замены 124E и 180E;
tt) H1 содержит аминокислотные замены 139C, 143E, 145T и 179E, L1 содержит аминокислотные замены 116C, 124R и 178R, H2 содержит аминокислотную замену 179K и L2 содержит аминокислотные замены 124E, 160E и 180E;
uu) H1 содержит аминокислотные замены 143E, 145T и 179D, L1 содержит аминокислотные замены 124R и 178K, H2 содержит аминокислотную замену 179K и L2 содержит аминокислотные замены 124E и 180E;
vv) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124K и 178R, H2 содержит аминокислотную замену 179K и L2 содержит аминокислотные замены 124E, 160E и 180E;
ww) H1 содержит аминокислотные замены 143D, 145T и 179E, L1 содержит аминокислотную замену 178R, H2 содержит аминокислотную замену 179K и L2 содержит аминокислотные замены 124E и 180E;
xx) H1 содержит аминокислотные замены 143E, 145T и 179D, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену 179K и L2 содержит аминокислотные замены 124E и 180E;
yy) H1 содержит аминокислотные замены 143E, 145T и 179D, L1 содержит аминокислотные замены 124K и 178R, H2 содержит аминокислотную замену 179K и L2 содержит аминокислотные замены 124E, 160E и 180E;
zz) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену 179K и L2 содержит аминокислотные замены 124E, 160E и 180E;
aaa) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R, 160K и 178R, H2 содержит аминокислотную замену 179K и L2 содержит аминокислотные замены 124E и 180E;
bbb) H1 содержит аминокислотные замены 143E, 145T и 179D, L1 содержит аминокислотные замены 124R и 178K, H2 содержит аминокислотную замену 179K и L2 содержит аминокислотные замены 124E, 160E и 180E;
ccc) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124K и 178R, H2 содержит аминокислотную замену 179K и L2 содержит аминокислотные замены 124E и 180E;
ddd) H1 содержит аминокислотные замены 143E, 145T и 179D, L1 содержит аминокислотные замены 124R, 160K и 178R, H2 содержит аминокислотную замену 179K и L2 содержит аминокислотные замены 124E и 180E;
eee) H1 содержит аминокислотные замены 143E, 145T и 179D, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену 179K и L2 содержит аминокислотные замены 124E, 160E и 180E;
fff) H1 содержит аминокислотные замены 143E, 145T и 179D, L1 содержит аминокислотную замену 178R, H2 содержит аминокислотную замену 179K и L2 содержит аминокислотные замены 124E и 180E;
ggg) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178K, H2 содержит аминокислотную замену 179K и L2 содержит аминокислотные замены 124E, 160E и 180E;
hhh) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотную замену 178R, H2 содержит аминокислотную замену 179K и L2 содержит аминокислотные замены 124E и 180E;
iii) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену 143K и L2 содержит аминокислотные замены 124E и 133D; или
jjj) H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену 143R и L2 содержит аминокислотные замены 124E и 133E.
2. Набор полинуклеотидов по п. 1, причем H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену 186K и L2 содержит аминокислотные замены 124E, 160E и 178E.
3. Набор полинуклеотидов по п. 1, причем H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену 179R и L2 содержит аминокислотные замены 124E, 178E и 180E.
4. Набор полинуклеотидов по п. 1, причем H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену 179K и L2 содержит аминокислотные замены 124E и 180E.
5. Набор полинуклеотидов по п. 1, причем H1 содержит аминокислотные замены 139C, 143E, 145T и 179E, L1 содержит аминокислотные замены 116C, 124R и 178R, H2 содержит аминокислотную замену 179K и L2 содержит аминокислотные замены 124E, 160E и 180E.
6. Набор полинуклеотидов по п. 1, причем H1 содержит аминокислотные замены 143E, 145T и 179E, L1 содержит аминокислотные замены 124R и 178R, H2 содержит аминокислотную замену 143R и L2 содержит аминокислотные замены 124E и 133E.
7. Набор полинуклеотидов по любому из пп. 1-6, где L1 и L2 представляют собой полипептидные последовательности легкой цепи каппа.
8. Набор полинуклеотидов по п. 7, где H1 предпочтительно спаривается с L1 по сравнению с L2 и H2 предпочтительно спаривается с L2 по сравнению с L1, в случае если H1, H2, L1 и L2 являются коэкспрессированными в клетке или клетке млекопитающего, или в случае если H1, H2, L1 и L2 являются коэкспрессированными в бесклеточной экспрессионной системе, или в случае если H1, H2, L1 и L2 являются совместно полученными, или в случае если H1, H2, L1 и L2 являются совместно полученными редокс-способом.
9. Набор полинуклеотидов по п. 8, где, в случае если обе L1 и L2 являются коэкспрессированными с по меньшей мере одной из H1 и H2, спаривание H1-L1 с H1-L2 и спаривание H2-L2 с H2-L1 является большим, чем спаривание H1-L1 с H1-L2 и спаривание H2-L2 с H2-L1 в отсутствие аминокислотных замен.
10. Набор полинуклеотидов по любому из пп. 1-9, где аффинность каждого гетеродимера к антигену, с которым он связывается, находится в пределах около 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 25, 30, 35, 40, 45 или 50 кратного значения аффинности соответствующего немодифицированного гетеродимера по отношению к тому же антигену, как измерено с помощью поверхностного плазмонного резонанса (SPR) или FACS.
11. Набор полинуклеотидов по любому из пп. 1-10, где конструкция дополнительно содержит Fc, содержащий первую последовательность CH3 и вторую последовательность CH3, где первая последовательность CH3 соединена с помощью или без помощи одного или более линкеров с первым гетеродимером, а вторая последовательность CH3 соединена с помощью или без помощи одного или более линкеров со вторым гетеродимером.
12. Набор полинуклеотидов по п. 11, где Fc представляет собой человеческий Fc, Fc человеческого IgG1, Fc человеческого IgA, Fc человеческого IgG, Fc человеческого IgD, Fc человеческого IgE, Fc человеческого IgM, Fc человеческого IgG2, Fc человеческого IgG3 или Fc человеческого IgG4.
13. Набор полинуклеотидов по п. 12, где Fc представляет собой гетеродимерный Fc.
14. Набор полинуклеотидов по п. 13, где Fc содержит одну или более модификаций в по меньшей мере одной из последовательностей CH3.
15. Набор полинуклеотидов по п. 14, где Fc содержит одну или более модификаций в по меньшей мере одной из последовательностей CH3, которые способствуют образованию гетеродимерного Fc со стабильностью, сопоставимой с гомодимерным Fc дикого типа.
16. Набор полинуклеотидов по п. 15, где Fc содержит:
i) гетеродимерный Fc IgG1, имеющий модификации L351Y_F405A_Y407V в первом полипептиде Fc и модификации T366L_K392M_T394W во втором полипептиде Fc;
ii) гетеродимерный Fc IgG1, имеющий модификации L351Y_F405A_Y407V в первом полипептиде Fc и модификации T366L_K392L_T394W во втором полипептиде Fc;
iii) гетеродимерный Fc IgG1, имеющий модификации T350V_L351Y_F405A_Y407V в первом полипептиде Fc и модификации T350V_T366L_K392L_T394W во втором полипептиде Fc;
iv) гетеродимерный Fc IgG1, имеющий модификации T350V_L351Y_F405A_Y407V в первом полипептиде Fc и модификации T350V_T366L_K392M_T394W во втором полипептиде Fc; или
v) гетеродимерный Fc IgG1, имеющий модификации T350V_L351Y_S400E_F405A_Y407V в первом полипептиде Fc и модификации T350V_T366L_N390R_K392M_T394W во втором полипептиде Fc, где нумерация остатков в полипептиде Fc дана в соответствии с нумерационной системой EU.
17. Набор полинуклеотидов по п. 16, где Fc дополнительно содержит по меньшей мере одну последовательность CH2.
18. Набор полинуклеотидов по п. 17, где последовательность(и) CH2 Fc содержит(ат) одну или более модификаций.
19. Набор полинуклеотидов по п. 18, где Fc содержит одну или более модификаций для содействия селективного связывания Fc-гамма рецепторов.
20. Набор полинуклеотидов по п. 19, где Fc соединен с гетеродимерами с помощью одного или более линкеров или где Fc спарен с H1 и H2 с помощью одного или более линкеров.
21. Набор полинуклеотидов по п. 20, где один или более линкеров содержат одну или более шарнирных областей антитела.
22. Набор полинуклеотидов по п. 21, где один или более линкеров содержат одну или более шарнирных областей IgG1.
23. Набор полинуклеотидов по п. 22, где один или более линкеров содержат одну или более модификаций.
24. Набор полинуклеотидов по п. 23, где одна или более модификаций содействуют селективному связыванию Fc-гамма рецепторов.
25. Набор полинуклеотидов по п. 1, где конструкция является мультиспецифической или биспецифической.
26. Набор векторов экспрессии для получения антигенсвязывающей полипептидной конструкции, причем набор векторов экспрессии содержит набор полинуклеотидов по любому из пп. 1-25.
27. Выделенная клетка-хозяин для получения антигенсвязывающей полипептидной конструкции, причем выделенная клетка-хозяин содержит набор полинуклеотидов по любому из пп. 1-25.
28. Способ получения или создания антигенсвязывающей полипептидной конструкции из культуры клеток-хозяев, причем способ включает стадии приготовления:
(a) получение культуры клеток-хозяев, содержащей по меньшей мере одну клетку-хозяина, содержащую набор полинуклеотидов по любому из пп. 1-25; и (b) выделение конструкции из культуры клеток-хозяев.
29. Способ по п. 28, дополнительно включающий стадию конъюгирования антигенсвязывающей полипептидной конструкции с терапевтическим агентом или лекарственным средством.
| LEWIS S.M | |||
| et al | |||
| "Generation of bispecific IgG antibodies by structure-based design of an orthogonal Fab interface." Nature biotechnology, 2014 (published online 26 January 2014), 32(2): 191- 198 | |||
| WO 2012131555 A2, 04.10.2012 | |||
| WO 2007147901 A1, 27.12.2007 | |||
| US 2006136184 A1, 22.06.2006 | |||
| ЯРИЛИН А.А., "Основы иммунологии", М.: Медицина, 1999, |

