ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Настоящая заявка испрашивает приоритет в соответствии со статьей 35 § 119(e) Свода законов США на основании предварительной заявки на патент США номер 61/556090, поданной 4 ноября 2011 г.; предварительной заявки на патент США номер 61/557262, поданной 8 ноября 2011 г.; и предварительной заявки на патент США номер 61/645547, поданной 10 мая 2012 г., содержание каждой из которых полностью включено в данную заявку посредством отсылок.
ОБЛАСТЬ ТЕХНИКИ
В настоящем изобретении в целом предложены гетеродимеры полипептидов, содержащие их композиции и способы получения и применения таких гетеродимеров полипептидов. В частности, настоящее изобретение относится к термоустойчивым мультиспецифическим, включая биспецифические, антителам, содержащим гетеродимерный домен Fc.
УРОВЕНЬ ТЕХНИКИ
Биспецифические лекарственные средства представляют собой молекулы на основе антител, которые могут одновременно связывать две отдельные и различные мишени или различные эпитопы одного и того же антигена. Биспецифические антитела состоят из соединений на основе домена иммуноглобулина и стремятся структурно и функционально имитировать компоненты молекулы антитела. Одним из применений биспецифических антител является перенацеливание эффекторных цитотоксических клеток иммунной системы на усиленное уничтожение опухолевых клеток, например, посредством антителозависимой клеточной цитотоксичности (АЗКЦ). В данном случае, одно плечо биспецифического антитела связывает антиген на поверхности опухолевой клетки, а другое плечо связывает детерминанту, экспрессированную на эффекторных клетках. Осуществляя перекрестное связывание опухолевых и эффекторных клеток, биспецифическое антитело не только сближает эффекторные клетки с опухолевыми клетками, но также одновременно запускает активацию первых, что приводит к ффективному уничтожению опухолевых клеток. Биспецифические антитела также применяют для обогащения опухолевых тканей химио- или радиотерапевтическими агентами, чтобы минимизировать вредное действие на нормальную ткань. В данном контексте, одно плечо биспецифического антитела связывает антиген, экспрессированный на поверхности клетки, которую нацеливают на разрушение, а другое плечо доставляет химиотерапевтическое средство, радиоактивный изотоп или токсин. Выходя за рамки биспецифичности, существует потребность в достижении эффективности белковых лекарственных средств путем нацеливания на несколько механизмов одновременно. Такое комплексное и новое биологическое действие белкового лекарственного средства можно получить путем разработки белка, который связывает множество мишеней и обладает множеством функций.
Для разработки такого мультифункционального и связывающего множество мишеней лекарственного средства необходим крепкий каркас, который обеспечит основу для слияния с другими функциональными реакционными группами или доменами, связывающими целевой белок. В идеале каркас должен не только обеспечить основу, но также обеспечить доступность для разработанного терапевтического средства множества других терапевтически важных и полезных свойств. Основным препятствием для общей разработки биспецифических и мультифункциональных лекарственных средств на основе антител служит сложность получения материалов достаточного качества и в достаточном количестве как для доклинических, так и для клинических исследований. В данной области сохраняется потребность в полипептидных конструкциях, которые содержат отдельные вариабельные домены в качестве связывающих белок доменов, которые связаны с вариантом Fc-области, где указанный вариант Fc содержит домены СН3, которые модифицированы с целью выбора гетеродимеров с повышенной стабильностью и чистотой.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Согласно одному аспекту настоящего изобретения предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный гетеродимерный домен СН3, где указанный модифицированный домен СН3 включает: первый модифицированный полипептид домена СН3, содержащий по меньшей мере три модификации аминокислот по сравнению с полипептидом домена СН3 дикого типа, и второй модифицированный полипептид домена СН3, включающий по меньшей мере три модификации аминокислот по сравнению с полипептидом домена СН3 дикого типа; причем по меньшей мере один из указанных первого и второго полипептидов домена СН3 включает модификацию аминокислоты K392J, где J выбран из L, I или аминокислоты с объемом боковой цепи, значительно не превышающим объем боковой цепи K; при этом указанные первый и второй модифицированные полипептиды домена СН3 предпочтительно образуют гетеродимерный домен СН3 с температурой плавления (Tm), составляющей по меньшей мере приблизительно 74°С, и чистотой по меньшей мере 95%; и при этом по меньшей мере одна модификация аминокислоты не затрагивает аминокислоту, которая находится на границе между указанным первым и указанным вторым полипептидами домена СН3. В некоторых вариантах реализации предложена гетеромультимерная конструкция Fc, описанная в данной заявке, которая включает по меньшей мере одну модификацию Т350Х, где X представляет собой природную или неприродную аминокислоту, выбранную из валина, изолейцина, лейцина, метионина и производных или вариантов указанных аминокислот. В некоторых вариантах реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, которая включает по меньшей мере одну модификацию T350V. В одном варианте реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой модифицированный домен СН3 имеет температуру плавления (Tm), составляющую по меньшей мере приблизительно 75°С или более. В одном варианте реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой модифицированный домен СН3 имеет Tm, составляющую приблизительно 77°С или более. В некоторых вариантах реализации, модифицированный домен СН3 имеет Tm, составляющую приблизительно 80°С или более. В некоторых вариантах реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой по меньшей мере один полипептид домена СН3 представляет собой модифицированный полипептид домена СН3, включающий модификацию по меньшей мере одной из аминокислот L351, F405 и Y407. В некоторых вариантах реализации предложена изолированная гетеромультимерная конструкция Fc, в которой по меньшей мере один полипептид домена СН3 представляет собой модифицированный полипептид домена СН3, дополнительно включающий модификацию аминокислоты Т366. В некоторых вариантах реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой первый полипептид домена СН3 представляет собой модифицированный полипептид домена СН3, включающий модификации аминокислот в положениях L351, F405 и Y407, и второй полипептид домена СН3 представляет собой модифицированный полипептид домена СН3, включающий модификации аминокислот в положениях Т366, K392 и Т394. В одном варианте реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой указанный первый полипептид домена СН3 включает модификации аминокислот L351Y, F405A и Y407V, и указанный второй полипептид домена СН3 включает модификации аминокислот T366L, K392M и T394W. В некоторых вариантах реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой указанный первый полипептид домена СН3 включает модификации аминокислот L351Y, F405A и Y407V, и указанный второй полипептид домена СН3 включает модификации аминокислот T366L, K392L и T394W. В дополнительном варианте реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой указанный первый полипептид домена СН3 включает модификации аминокислот L351Y, F405A и Y407V, и указанный второй полипептид домена СН3 включает модификации аминокислот T366I, K392M и T394W. В некоторых вариантах реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой указанный первый полипептид домена СН3 включает модификации аминокислот L351Y, F405A и Y407V, и указанный второй полипептид домена СН3 включает модификации аминокислот T366I, K392L и T394W. В некоторых вариантах реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой по меньшей мере один из указанных первого и второго полипептидов домена СН3 представляет собой модифицированный полипептид домена СН3, включающий модификацию аминокислоты в положении S400. В дополнительном варианте реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, включающая модификацию S400Z, в которой Ζ выбран из положительно заряженной аминокислоты и отрицательно заряженной аминокислоты. В некоторых вариантах реализации, положительно заряженная аминокислота представляет собой лизин или аргинин и отрицательно заряженная аминокислота представляет собой аспарагиновую кислоту или глутаминовую кислоту. В некоторых вариантах реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой указанный первый полипептид домена СН3 включает модификацию аминокислоты, выбранную из S400E и S400R. В некоторых вариантах реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой по меньшей мере один из указанных первого и второго полипептидов домена СН3 представляет собой модифицированный полипептид домена СН3, включающий модификацию аминокислоты в положении N390. В некоторых вариантах реализации, модификация N390 представляет собой N390Z, где Ζ выбран из положительно заряженной аминокислоты и отрицательно заряженной аминокислоты. В одном варианте реализации, N390Z представляет собой N390R. В некоторых вариантах реализации изолированной гетеромультимерной конструкции Fc, описанной в данной заявке, указанный первый полипептид домена СН3 представляет собой модифицированный полипептид домена СН3, включающий модификацию аминокислоты S400E, и указанный второй полипептид домена СН3 представляет собой модифицированный полипептид домена СН3, включающий модификацию аминокислоты N390R. В некоторых вариантах реализации изолированной гетеромультимерной конструкции Fc, описанной в данной заявке, каждый из первого и второго полипептидов домена СН3 представляет собой модифицированный полипептид домена СН3, один указанный модифицированный полипептид домена СН3 включает модификацию аминокислоты Q347R, а другой модифицированный полипептид домена СН3 включает модификацию аминокислоты K360E.
В одном аспекте предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный гетеродимерный домен СН3, где указанный модифицированный домен СН3 содержит: первый модифицированный полипептид домена СН3, включающий по меньшей мере три модификации аминокислот по сравнению с полипептидом домена СН3 дикого типа, и второй модифицированный полипептид домена СН3, включающий по меньшей мере три модификации аминокислот по сравнению с полипептидом домена СН3 дикого типа; при этом по меньшей мере один из указанных первого и второго полипептидов домена СН3 включает модификацию аминокислоты K392J, где J выбран из L, I или аминокислоты с объемом боковой цепи, значительно не превышающим объем боковой цепи K; при этом указанные первый и второй модифицированные полипептиды домена СН3 предпочтительно образуют гетеродимерный домен СН3 с температурой плавления (Tm), составляющей по меньшей мере приблизительно 74°С, и чистотой по меньшей мере 95%; и при этом по меньшей мере одна модификация аминокислоты не затрагивает аминокислоту, которая находится на границе между указанным первым и указанным вторым полипептидами домена СН3. В некоторых вариантах реализации предложена гетеромультимерная конструкция Fc, описанная в данной заявке, которая включает по меньшей мере одну модификацию Т350Х, где X представляет собой природную или неприродную аминокислоту, выбранную из валина, изолейцина, лейцина, метионина и производных или вариантов указанных аминокислот. В некоторых вариантах реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, которая включает по меньшей мере одну модификацию T350V. В одном варианте реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой модифицированный домен СН3 имеет температуру плавления (Tm), составляющую по меньшей мере приблизительно 75°С или более. В одном варианте реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой модифицированный домен СН3 имеет Tm, составляющую приблизительно 77°С или более. В некоторых вариантах реализации, модифицированный домен СН3 имеет Tm, составляющую приблизительно 80°С или более. В одном варианте реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой по меньшей мере один полипептид домена СН3 представляет собой модифицированный полипептид домена СН3, включающий модификацию по меньшей мере одной из аминокислот K409 и Т411. В некоторых вариантах реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, включающая по меньшей мере одну из модификаций K409F, Т411Е и T411D. В некоторых вариантах реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой по меньшей мере один полипептид домена СН3 представляет собой модифицированный полипептид домена СН3, включающий модификацию аминокислоты D399. В некоторых вариантах реализации, модификация аминокислоты D399 представляет собой по меньшей мере одну из модификаций D399R и D399K.
В одном аспекте предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный гетеродимерный домен СН3, где указанный модифицированный домен СН3 включает: первый модифицированный полипептид домена СН3, включающий по меньшей мере три модификации аминокислот по сравнению с полипептидом домена СН3 дикого типа, и второй модифицированный полипептид домена СН3, включающий по меньшей мере три модификации аминокислот по сравнению с полипептидом домена СН3 дикого типа; причем по меньшей мере один из указанных первого и второго полипептидов домена СН3 включает модификацию аминокислоты K392J, где J выбран из L, I или аминокислоты с объемом боковой цепи, не превышающим значительно объем боковой цепи K; при этом указанные первый и второй модифицированные полипептиды домена СН3 предпочтительно образуют гетеродимерный домен СН3 с температурой плавления (Tm), составляющей по меньшей мере приблизительно 74°С, и чистотой по меньшей мере 95%; и при этом по меньшей мере одна модификация аминокислоты не затрагивает аминокислоту, которая находится на границе между указанным первым и указанным вторым полипептидами домена СН3. В некоторых вариантах реализации предложена гетеромультимерная конструкция Fc, описанная в данной заявке, которая включает по меньшей мере одну модификацию Т350Х, где X представляет собой природную или неприродную аминокислоту, выбранную из валина, изолейцина, лейцина, метионина и производных или вариантов указанных аминокислот. В некоторых вариантах реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, которая включает по меньшей мере одну модификацию T350V. В одном варианте реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой модифицированный домен СН3 имеет температуру плавления (Tm), составляющую по меньшей мере приблизительно 75°С или более. В одном варианте реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой модифицированный домен СН3 имеет Tm, составляющую приблизительно 77°С или более. В некоторых вариантах реализации, модифицированный домен СН3 имеет Tm, составляющую приблизительно 80°С или более. В некоторых вариантах реализации изолированной гетеромультимерной конструкции Fc, описанной в данной заявке, в которой первый полипептид домена СН3 представляет собой модифицированный полипептид домена СН3, включающий по меньшей мере одну модификацию аминокислоты, выбранную из K409F, Т411Е и T411D, и второй полипептид домена СН3 представляет собой модифицированный полипептид домена СН3, включающий по меньшей мере одну модификацию аминокислоты, выбранную из Y407A, Y407I, Y407V, D399R и D399K. В некоторых вариантах реализации предложена любая из изолированных гетеромультимерных конструкций Fc, описанных в данной заявке, дополнительно содержащая первый модифицированный домен СН3, включающий одну из модификаций аминокислот T366V, T366I, Т366А, Т366М и T366L; и второй модифицированный домен СН3, включающий модификацию аминокислоты L351Y. В некоторых вариантах реализации предложена любая из изолированных гетеромультимерных конструкций Fc, описанных в данной заявке, содержащая первый модифицированный домен СН3, включающий одну из модификаций аминокислот K392L или K392E; и второй модифицированный домен СН3, включающий одну из модификаций аминокислот S400R или S400V.
В данной заявке предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный домен СН3, содержащий первый модифицированный полипептид домена СН3 и второй модифицированный полипептид домена СН3, при этом каждый модифицированный полипептид домена СН3 включает по меньшей мере четыре мутации аминокислот, при этом по меньшей мере один из указанного первого и указанного второго модифицированных полипептидов домена СН3 включает мутацию, выбранную из N390Z и S400Z, где Ζ выбран из положительно заряженной аминокислоты и отрицательно заряженной аминокислоты, и при этом указанные первый и второй модифицированные полипептиды домена СН3 предпочтительно образуют гетеродимерный домен СН3 с температурой плавления (Tm), составляющей по меньшей мере приблизительно 70°С, и чистотой по меньшей мере 90%. В одном варианте реализации предложена изолированная гетеромультимерная конструкция Fc, в которой указанный первый модифицированный полипептид домена СН3 включает модификации аминокислот в положениях F405 и Y407 и указанный второй модифицированный полипептид домена СН3 включает модификацию аминокислоты в положении Т394. В одном варианте реализации предложена изолированная гетеромультимерная конструкция Fc, в которой первый модифицированный полипептид домена СН3 включает модификацию аминокислоты в положении L351. В некоторых вариантах реализации предложен изолированный гетеромультимер, описанный в данной заявке, в котором указанный второй модифицированный полипептид домена СН3 включает модификацию в по меньшей мере одном из положений Т366 и K392. В некоторых вариантах реализации предложен изолированный гетеромультимер, описанный в данной заявке, в котором модифицированный домен СН3 имеет температуру плавления (Tm), составляющую по меньшей мере приблизительно 75°С, и получен с чистотой по меньшей мере приблизительно 95%. В некоторых вариантах реализации предложен изолированный гетеромультимер, описанный в данной заявке, в котором по меньшей мере один модифицированный полипептид домена СН3 включает по меньшей мере одну из модификаций аминокислот N390R, S400E и S400R. В некоторых вариантах реализации предложен изолированный гетеромультимер, описанный в данной заявке, в котором один из указанных первого и второго модифицированных полипептидов домена СН3 включает модификацию аминокислоты в положении 347 и другой модифицированный полипептид домена СН3 включает модификацию аминокислоты в положении 360. В некоторых вариантах реализации предложен изолированный гетеромультимер, описанный в данной заявке, в котором по меньшей мере один из указанных первого и второго модифицированных полипептидов домена СН3 включает модификацию аминокислоты T350V. В конкретных вариантах реализации предложен изолированный гетеромультимер, описанный в данной заявке, в котором указанный первый модифицированный полипептид домена СН3 включает по меньшей мере одну модификацию аминокислоты, выбранную из L351Y, F405A и Y407V; и указанный второй модифицированный полипептид домена СН3 включает по меньшей мере одну модификацию аминокислоты, выбранную из T366L, T366I, K392L, K392M и T394W. В некоторых вариантах реализации, описанных в данной заявке, предложен изолированный гетеромультимер, в котором первый модифицированный полипептид домена СН3 включает модификации аминокислот в положениях D399 и Y407 и второй модифицированный полипептид домена СН3 включает модификации аминокислот в положениях K409 и Т411. В некоторых вариантах реализации предложен изолированный гетеромультимер, описанный в данной заявке, в котором первый полипептид домена СН3 включает модификацию аминокислоты в положении L351 и второй модифицированный полипептид домена СН3 включает модификации аминокислот в положениях Т366 и K392. В конкретных вариантах реализации предложены изолированные гетеромультимеры, описанные в данной заявке, в которых по меньшей мере один из указанных первого и второго полипептидов домена СН3 включает модификацию аминокислоты T350V. В некоторых вариантах реализации предложены изолированные гетеромультимеры, описанные в данной заявке, в которых модифицированный домен СН3 имеет температуру плавления (Tm), составляющую по меньшей мере приблизительно 75°С или более, и получен с чистотой по меньшей мере приблизительно 95%. В некоторых вариантах реализации предложены изолированные гетеромультимерные конструкции Fc, описанные в данной заявке, в которых указанный первый модифицированный полипептид домена СН3 включает модификации аминокислот, выбранные из L351Y, D399R, D399K, S400D, S400E, S400R, S400K, Y407A и Y407V; и указанный второй модифицированный полипептид домена СН3 включает модификации аминокислот, выбранные из T366V, T366I, T366L, Т366М, N390D, N390E, K392L, K392I, K392D, K392E, K409F, K409W, T411D и Т411Е.
В данной заявке предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный домен СН3, содержащий первый модифицированный полипептид домена СН3 и второй модифицированный полипептид домена СН3, причем каждый модифицированный полипептид домена СН3 включает мутации по меньшей мере трех аминокислот, при этом один из указанных первого и указанного второго модифицированных полипептидов домена СН3 включает мутацию, выбранную из Т411Е и T411D, и при этом указанные первый и второй модифицированные полипептиды домена СН3 предпочтительно образуют гетеродимерный домен СН3 с температурой плавления (Tm), составляющей по меньшей мере приблизительно 70°С, и чистотой по меньшей мере 90%. В одном варианте реализации предложена изолированная гетеромультимерная конструкция Fc, в которой указанный первый модифицированный полипептид домена СН3 включает модификации аминокислот в положениях F405 и Y407 и указанный второй модифицированный полипептид домена СН3 включает модификацию аминокислоты в положении Т394. В одном варианте реализации предложена изолированная гетеромультимерная конструкция Fc, в которой первый модифицированный полипептид домена СН3 включает модификацию аминокислоты в положении L351. В некоторых вариантах реализации предложен изолированный гетеромультимер, описанный в данной заявке, в котором указанный второй модифицированный полипептид домена СН3 включает модификацию в по меньшей мере одном из положений Т366 и K392. В некоторых вариантах реализации предложен изолированный гетеромультимер, описанный в данной заявке, в котором модифицированный домен СН3 имеет температуру плавления (Tm), составляющую по меньшей мере приблизительно 75°С, и получен с чистотой по меньшей мере приблизительно 95%. В некоторых вариантах реализации предложен изолированный гетеромультимер, описанный в данной заявке, в котором по меньшей мере один модифицированный полипептид домена СН3 включает по меньшей мере одну из модификаций аминокислот N390R, S400E и S400R. В некоторых вариантах реализации предложен изолированный гетеромультимер, описанный в данной заявке, в котором один из указанных первого и второго модифицированных полипептидов домена СН3 включает модификацию аминокислоты в положении 347 и другой модифицированный полипептид домена СН3 включает модификацию аминокислоты в положении 360. В некоторых вариантах реализации предложен изолированный гетеромультимер, описанный в данной заявке, в котором по меньшей мере один из указанных первого и второго модифицированных полипептидов домена СН3 включает модификацию аминокислоты T350V. В конкретных вариантах реализации предложен изолированный гетеромультимер, описанный в данной заявке, в котором указанный первый модифицированный полипептид домена СН3 включает по меньшей мере одну модификацию аминокислоты, выбранную из L351Y, F405A и Y407V; и указанный второй модифицированный полипептид домена СН3 включает по меньшей мере одну модификацию аминокислоты, выбранную из T366L, T366I, K392L, K392M и T394W. В некоторых вариантах реализации, описанных в данной заявке, предложен изолированный гетеромультимер, в котором первый модифицированный полипептид домена СН3 включает модификации аминокислот в положениях D399 и Y407 и второй модифицированный полипептид домена СН3 включает модификации аминокислот в положениях K409 и Т411. В некоторых вариантах реализации предложен изолированный гетеромультимер, описанный в данной заявке, в котором первый полипептид домена СН3 включает модификацию аминокислоты в положении L351 и второй модифицированный полипептид домена СН3 включает модификации аминокислот в положениях Т366 и K392. В конкретных вариантах реализации предложены изолированные гетеромультимеры, описанные в данной заявке, в которых по меньшей мере один из указанных первого и второго полипептидов домена СН3 включает модификацию аминокислоты T350V. В некоторых вариантах реализации предложены изолированные гетеромультимеры, описанные в данной заявке, в которых модифицированный домен СН3 имеет температуру плавления (Tm), составляющую по меньшей мере приблизительно 75°С или более, и получен с чистотой по меньшей мере приблизительно 95%. В некоторых вариантах реализации предложены изолированные гетеромультимерные конструкции Fc, описанные в данной заявке, в которых указанный первый модифицированный полипептид домена СН3 включает модификации аминокислот, выбранные из L351Y, D399R, D399K, S400D, S400E, S400R, S400K, Y407A и Y407V; и указанный второй модифицированный полипептид домена СН3 включает модификации аминокислот, выбранные из T366V, T366I, T366L, Т366М, N390D, N390E, K392L, K392I, K392D, K392E, K409F, K409W, T411D и Т411Е.
В данной заявке предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный домен СН3, содержащий первый модифицированный полипептид домена СН3, включающий модификации аминокислот L351Y, F405A и Y407V; и второй модифицированный полипептид домена СН3, включающий модификации аминокислот T366I, K392M и T394W.
В некотором аспекте предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный домен СН3, содержащий первый модифицированный полипептид домена СН3, включающий модификации аминокислот L351Y, F405A и Y407V; и второй модифицированный полипептид домена СН3, включающий модификации аминокислот T366I, K392L и T394W.
В некотором аспекте предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный домен СН3, содержащий первый модифицированный полипептид домена СН3, включающий модификации аминокислот L351Y, F405A и Y407V; и второй модифицированный полипептид домена СН3, включающий модификации аминокислот T366L, K392M и T394W.
В некоторых аспектах предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный домен СН3, содержащий первый модифицированный полипептид домена СН3, включающий модификации аминокислот L351Y, F405A и Y407V; и второй модифицированный полипептид домена СНЗ, включающий модификации аминокислот T366L, K392L и T394W.
В некотором аспекте предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный домен СН3, содержащий первый модифицированный полипептид домена СН3, включающий модификации аминокислот T350V, L351Y, F405A и Y407V; и второй модифицированный полипептид домена СНЗ, включающий модификации аминокислот T350V, T366L, K392L и T394W.
В некотором аспекте предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный домен СН3, содержащий первый модифицированный полипептид домена СН3, включающий модификации аминокислот T350V, L351Y, S400R, F405A, Y407V; и второй модифицированный полипептид домена СНЗ, включающий модификации аминокислот T350V, T366L, K392M и T394W.
В некотором аспекте предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный домен СН3, содержащий первый модифицированный полипептид домена СН3, включающий модификации аминокислот T350V, L351Y, S400E, F405A, Y407V; и второй модифицированный полипептид домена СН3, включающий модификации аминокислот T350V, T366L, N390R, K392M и T394W.
В некотором аспекте предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный домен СН3, содержащий первый модифицированный полипептид домена СН3, включающий модификации аминокислот T350V, L351Y, F405A, Y407V; и второй модифицированный полипептид домена СН3, включающий модификации аминокислот T350V, T366L, K392L и T394W.
В одном аспекте предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный домен СН3, содержащий первый модифицированный полипептид домена СН3, включающий модификации аминокислот T366V, K392L, K409F и Т411Е; и второй модифицированный полипептид домена СН3, включающий модификации аминокислот L351Y, D399R и Y407A.
В одном аспекте предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный домен СН3, содержащий первый модифицированный полипептид домена СН3, включающий модификации аминокислот T366V, K392L, K409F и Т411Е; и второй модифицированный полипептид домена СН3, включающий модификации аминокислот L351Y, D399R, S400R и Y407A.
Согласно одному аспекту настоящего изобретения предложен изолированный гетеромультимер, включающий гетеродимерную Fc-область, при этом гетеродимерная Fc-область содержит модифицированный домен СН3, включающий мутации аминокислот, способствующие образованию гетеродимера, при этом гетеродимерная Fc-область дополнительно содержит вариант домена СН2, включающий по меньшей мере одну асимметричную модификацию аминокислоты, способствующую селективному связыванию рецептора Fc-гамма. В одном варианте реализации вариант домена СН2 избирательно связывает рецептор Fc-гамма IIIa, по сравнению с доменом СН2 дикого типа. В одном варианте реализации, модифицированный домен СН3 имеет температуру плавления (Tm), составляющую приблизительно 70°С или более. В некоторых вариантах реализации, модифицированный домен СН3 имеет температуру плавления (Tm), составляющую по меньшей мере приблизительно 75°С. В некоторых вариантах реализации, модифицированный домен СН3 имеет температуру плавления (Tm), составляющую по меньшей мере приблизительно 80°С.
В другом аспекте предложен изолированный гетеромультимер, включающий гетеродимерную Fc-область, причем гетеродимерная Fc-область содержит модифицированный домен СН3, включающий мутации аминокислот, при этом модифицированный домен СН3 имеет температуру плавления (Tm), составляющую приблизительно 70°С или более, и при этом указанный модифицированный домен СН3 способствует образованию гетеродимерной Fc-области с повышенной стабильностью по сравнению с доменом СН3, который не содержит мутаций аминокислот. В одном варианте реализации, гетеродимерная Fc-область не содержит дополнительную дисульфидную связь в домене СН3 по сравнению с Fc-областью дикого типа. В альтернативном варианте реализации, гетеродимерная Fc-область содержит по меньшей мере одну дополнительную дисульфидную связь в модифицированном домене СН3 по сравнению с Fc-областью дикого типа при условии, что температура плавления (Tm) составляет приблизительно 70°С или более в отсутствии дополнительной дисульфидной связи. В другом варианте реализации, гетеродимерная Fc-область содержит по меньшей мере одну дополнительную дисульфидную связь в модифицированном домене СН3 по сравнению с Fc-областью дикого типа, и при этом модифицированный домен СН3 имеет температуру плавления (Tm), составляющую приблизительно 77,5°С или более.
В одном варианте реализации предложен изолированный гетеромультимер, включающий гетеродимерную Fc-область, при этом гетеродимерная Fc-область содержит модифицированный домен СН3, включающий мутации аминокислот, при этом модифицированный домен СН3 имеет температуру плавления (Tm), составляющую приблизительно 70°С или более, и гетеродимерная Fc-область получена с чистотой, более приблизительно 90%, или гетеродимерная Fc-область получена с чистотой, составляющей приблизительно 95% или более, или гетеродимерная Fc-область получена с чистотой, составляющей приблизительно 98% или более.
Также в некоторых вариантах реализации предложен изолированный гетеромультимер, включающий гетеродимерную Fc-область, при этом гетеродимерная Fc-область содержит модифицированный домен СН3, включающий одну или более мутаций аминокислот, которые приводят к образованию гетеродимерной Fc-области с повышенной стабильностью, по сравнению с доменом СН3, который не содержит указанную одну или более мутаций аминокислот, при этом модифицированный домен СН3 имеет температуру плавления (Tm), составляющую приблизительно 70°С или более, или Tm составляет приблизительно 71°С или более, или Tm составляет приблизительно 74°С или более. В другом варианте реализации, гетеродимерную Fc-область получают в растворе с чистотой, составляющей приблизительно 98% или более, и Tm приблизительно 73°С, или при этом гетеродимерную Fc-область получают с чистотой, составляющей приблизительно 90% или более, и Tm приблизительно 75°С.
В некоторых вариантах реализации предложен изолированный гетеромультимер, включающий гетеродимерную Fc-область, при этом гетеродимерная Fc-область включает первый и второй полипептиды домена СН3, при этом по меньшей мере один из указанных первого и второго полипептидов домена СН3 включает модификацию аминокислоты T350V. В некоторых вариантах реализации предложен изолированный гетеромультимер, включающий гетеродимерную Fc-область, при этом гетеродимерная Fc-область включает первый полипептид домена СН3, включающий модификацию аминокислоты T350V, и второй полипептид домена СН3, также включающий модификацию аминокислоты T350V. В некоторых вариантах реализации предложен изолированный гетеромультимер, включающий гетеродимерную Fc-область, при этом гетеродимерная Fc-область включает первый полипептид домена СН3, включающий модификации аминокислот в положениях F405 и Y407, и второй полипептид домена СН3, включающий модификацию аминокислоты в положении Т394. Первый полипептид домена СН3 включает модификации аминокислот в положениях D399 и Y407 и второй полипептид домена СН3 включает модификации аминокислот в положениях K409 и Т411. В некоторых вариантах реализации предложен изолированный гетеромультимер, включающий гетеродимерную Fc-область, при этом гетеродимерная Fc-область включает первый полипептид домена СН3, включающий модификации аминокислот L351Y и Y407A, и второй полипептид домена СН3, включающий модификации аминокислот Т366А и K409F. В одном аспекте, первый полипептид домена СН3 или второй полипептид домена СН3 включает дополнительную модификацию аминокислоты в положении Т411, D399, S400, F405, N390 или K392. Модификацию аминокислоты в положении Т411 выбирают из T411N, T411R, T411Q, T411K, T411D, Т411Е или T411W. Модификацию аминокислоты в положении D399 выбирают из D399R, D399W, D399Y или D399K. Модификацию аминокислоты в положении S400 выбирают из S400E, S400D, S400R или S400K. Модификацию аминокислоты в положении F405 выбирают из F405I, F405M, F405T, F405S, F405V или F405W. Модификацию аминокислоты в положении N390 выбирают из N390R, N390K или N390D. Модификацию аминокислоты в положении K392 выбирают из K392V, K392M, K392R, K392L, K392F или K392E.
В некоторых вариантах реализации предусмотрен изолированный гетеромультимер, включающий гетеродимерную Fc-область, при этом гетеродимерная Fc-область включает первый полипептид домена СН3, включающий модификации аминокислот T350V и L351Y, и второй полипептид домена СН3, также включающий модификации аминокислот T350V и L351Y.
В другом варианте реализации предусмотрен изолированный гетеромультимер, включающий гетеродимерную Fc-область, при этом гетеродимерная Fc-область включает первый полипептид домена СН3, включающий модификацию аминокислоты Y407A, и второй полипептид домена СН3, включающий модификации аминокислот Т366А и K409F. В одном аспекте первый полипептид домена СН3 или второй полипептид домена СН3 включает дополнительные модификации аминокислот K392E, Т411Е, D399R и S400R. В другом аспекте, первый полипептид домена СН3 включает модификации аминокислот D399R, S400R и Y407A и второй полипептид домена СН3 включает модификации аминокислот Т366А, K409F, K392E и Т411Е. В дополнительном варианте реализации, модифицированный домен СН3 имеет температуру плавления (Tm), составляющую приблизительно 74°С или более, и гетеродимер имеет чистоту, составляющую приблизительно 95% или более.
В другом варианте реализации предложен изолированный гетеромультимер, включающий гетеродимерную Fc-область, при этом гетеродимерная Fc-область включает первый полипептид домена СН3, включающий модификацию аминокислоты в положении L351 и модификацию аминокислоты Y407A, и второй полипептид домена СН3, включающий модификацию аминокислоты в положении Т366 и модификацию аминокислоты K409F. В одном аспекте, модификацию аминокислоты в положении L351 выбирают из L351Y, L351I, L351D, L351R или L351F. В другом аспекте, модификацию аминокислоты в положении Y407 выбирают из Y407A, Y407V или Y407S. В другом дополнительном аспекте, модификацию аминокислоты в положении Т366 выбирают из Т366А, T366I, T366L, Т366М, T366Y, T366S, Т366С, T366V или T366W. В одном варианте реализации, модифицированный домен СН3 имеет температуру плавления (Tm), составляющую приблизительно 75°С или более, и гетеродимер имеет чистоту, составляющую приблизительно 90% или более.
В другом варианте реализации предложен изолированный гетеромультимер, включающий гетеродимерную Fc-область, при этом гетеродимерная Fc-область включает первый полипептид домена СН3, включающий модификацию аминокислоты в положении F405 и модификации аминокислот L351Y и Y407V, и второй полипептид домена СН3, включающий модификацию аминокислоты T394W. В одном аспекте первый полипептид домена СН3 или второй полипептид домена СН3 включает модификации аминокислот в положениях K392, Т411, Т366, L368 или S400. Модификация аминокислоты в положении F405 представляет собой F405A, F405I, F405M, F405T, F405S, F405V или F405W. Модификация аминокислоты в положении K392 представляет собой K392V, K392M, K392R, K392L, K392F или K392E. Модификация аминокислоты в положении Т411 представляет собой T411N, T411R, T411Q, T411K, T411D, Т411Е или T411W. Модификация аминокислоты в положении S400 представляет собой S400E, S400D, S400R или S400K. Модификация аминокислоты в положении Т366 представляет собой Т366А, T366I, T366L, Т366М, T366Y, T366S, Т366С, T366V или T366W. Модификация аминокислоты в положении L368 представляет собой L368D, L368R, L368T, L368M, L368V, L368F, L368S и L368A.
В другом варианте реализации предусмотрен изолированный гетеромультимер, включающий гетеродимерную Fc-область, при этом гетеродимерная Fc-область включает первый полипептид домена СН3, включающий модификации аминокислот L351Y, F405A и Y407V, и второй полипептид домена СН3, включающий модификацию аминокислоты T394W. В одном аспекте, второй полипептид домена СН3 включает модификацию аминокислоты T366L или T366I.
В еще одном варианте реализации предусмотрен изолированный гетеромультимер, включающий гетеродимерную Fc-область, при этом гетеродимерная Fc-область включает первый полипептид домена СН3, включающий по меньшей мере одну из модификаций аминокислот Y349C, F405A и Y407V, и второй полипептид домена СН3, включающий модификации аминокислот T366I, K392M и T394W.
В некоторых вариантах реализации предусмотрен изолированный гетеромультимер, включающий гетеродимерную Fc-область, при этом гетеродимерная Fc-область включает первый полипептид домена СН3, включающий модификации аминокислот L351Y, F405A и Y407V, и второй полипептид домена СН3, включающий модификации аминокислот K392M и T394W, и одну из T366L и T366I.
В другом варианте реализации предусмотрен изолированный гетеромультимер, включающий гетеродимерную Fc-область, при этом гетеродимерная Fc-область включает первый полипептид домена СН3, включающий модификации аминокислот F405A и Y407V, и второй полипептид домена СН3, включающий модификации аминокислот T366L и T394W.
В другом варианте реализации предусмотрен изолированный гетеромультимер, включающий гетеродимерную Fc-область, при этом гетеродимерная Fc-область включает первый полипептид домена СН3, включающий модификации аминокислот F405A и Y407V, и второй полипептид домена СН3, включающий модификации аминокислот T366I и T394W.
В некоторых вариантах реализации гетеромультимера предложено биспецифическое антитело или мультиспецифическое антитело.
В другом варианте реализации предложена композиция, содержащая гетеромультимер согласно настоящему изобретению и фармацевтически приемлемый носитель.
В другом варианте реализации предложена клетка-хозяин, содержащая нуклеиновую кислоту, кодирующую гетеромультимер согласно настоящему изобретению.
В некоторых вариантах реализации предусмотрен гетеромультимер, при этом указанный гетеромультимер включает по меньшей мере одно терапевтическое антитело. В одном аспекте, терапевтическое антитело выбирают из группы, состоящей из абаговомаба, адалимумаба, алемтузумаба, аурограба, бапинейзумаба, базиликсимаба, белимумаба, бевацизумаба, бриакинумаба, канакинумаба, катумаксомаба, цертолизумаба пегола, цетуксимаба, даклизумаба, деносумаба, эфализумаба, галиксимаба, гемтузумаба озогамицина, голимумаба, ибритумомаба тиуксетана, инфликсимаба, ипилимумаба, лумиликсимаба, меполизумаба, мотавизумаба, муромонаба, микограба, натализумаба, нимотузумаба, окрелизумаба, офатумумаба, омализумаба, паливизумаба, панитумумаба, пертузумаба, ранибизумаба, реслизумаба, ритуксимаба, теплизумаба, тоцилизумаба/атлизумаба, тозитумомаба, трастузумаба, Проксиниума™, Ренкарекса™, устекинумаба и залутумумаба.
В другом варианте реализации гетеромультимера согласно настоящему изобретению предложен способ лечения рака у пациента, страдающего раком, характеризующимся наличием ракового антигена, включающий введение указанному пациенту терапевтически эффективного количества гетеромультимера.
В другом варианте реализации гетеромультимера согласно настоящему изобретению предложен способ лечения иммунологических расстройств у пациента, страдающего иммунологическим расстройством, характеризующимся наличием иммунного антигена, включающий введение указанному пациенту терапевтически эффективного количества гетеромультимера.
В некоторых вариантах реализации, модифицированная Fc-область, используемая в гетеромультимерных конструкциях, описанных в данной заявке, включает иммуноглобулины типа G, например, иммуноглобулины, которые называют иммуноглобулинами класса 2 (IgG2) или иммуноглобулинами класса 3 (IgG3). В некоторых вариантах реализации, модифицированная Fc-область, используемая в гетеромультимерных конструкциях, описанных в данной заявке, включает иммуноглобулин М, или IgM. В некоторых вариантах реализации, модифицированная Fc-область, используемая в гетеромультимерных конструкциях, описанных в данной заявке, включает иммуноглобулин А, или IgA. В некоторых вариантах реализации, модифицированная Fc-область, используемая в гетеромультимерных конструкциях, описанных в данной заявке, включает иммуноглобулин D, или IgD. В некоторых вариантах реализации, модифицированная Fc-область, используемая в гетеромультимерных конструкциях, описанных в данной заявке, включает иммуноглобулин Е, или IgE. В некоторых вариантах реализации, модифицированная Fc-область, используемая в гетеромультимерных конструкциях, описанных в данной заявке, включает все варианты изотипов иммуноглобулинов G, например, иммуноглобулины, которые называют иммуноглобулинами класса 1 (IgG1), класса 2 (IgG2), класса 3 (IgG3) или класса 4 (IgG4).
КРАТКОЕ ОПИСАНИЕ ФИГУР
На Фигуре 1 представлена трехмерная графическая структура антитела дикого типа, на которой показаны СН3 (вверху), СН2 (посередине) и участки рецептора. Пунктирный прямоугольник слева представлен в увеличенном виде справа, и на нем показаны два участка, Участок 1 и Участок 2, целевой области СН3.
На Фигуре 2 показано трехмерное графическое представление остатка дикого типа в положении 368.
На Фигуре 3 показано трехмерное графическое представление Участка 1, на котором показана мутация в положении 368.
На Фигуре 4 показано трехмерное графическое представление дополнительных мутаций на Участке 2.
На Фигуре 5 представлена таблица компьютерных расчетов показателя столкновения, различия в площади контакта, различия в укладке, различия в энергии электростатического взаимодействия и общего "показателя аффинности" для первых трех вариантов AZ1, AZ2 и AZ3.
На Фигуре 6 представлено трехмерное графическое изображение, на котором показаны варианты AZ2 и AZ3, которые "построены на" варианте AZ1.
На Фигуре 7 показаны трехмерные графические представления вариантов AZ2 и AZ3.
На Фигуре 8 показана такая же таблица, как и на Фигуре 5, но только для гетеродимеров AZ1, AZ2 и AZ3, и потенциальных гомодимеров. Показатель аффинности не показан для гомодимеров, так как он не применим.
На Фигуре 9 показано трехмерное графическое представление AZ4 дикого типа (слева) и мутированного AZ4 (справа).
На Фигуре 10 представлена такая же таблица, как и на Фигуре 5, на которой показаны компьютерные расчеты для гетеродимера AZ4 и потенциальных гомодимеров.
На Фигуре 11 показано графическое представление вариантов СН3 AZ5 (слева) и AZ6 (справа).
На Фигуре 12 представлена такая же таблица, как и на Фигуре 5, на которой показаны результаты компьютерных расчетов для AZ4, AZ5 и AZ6.
На Фигуре 13 показано трехмерное графическое представление антитела слева, с изображением вариантов особенностей связывания на участке рецептора с применением гетеродимерного подхода.
На Фигуре 14 показано схематическое представление молекулы IgG.
На Фигуре 15 показано выравнивание множества последовательностей Fcγ-рецепторов. Идентификационные номера последовательностей в Genebank/Uniprot: FcγRIIA (sp Ρ12318), FcγRIIB (sp P31994), FcγRIIC (gi 1261 16592), FcγRIIIA (sp P08637), FcγRIIIB (sp 075015).
На Фигуре 16 представлена схема кристаллической структуры комплекса Fc-FcγRIIIb [PDB ID: 1T83, Radaev & Sun]. Наблюдается комплекс 1:1 Fc и рецептора Fcγ с асимметричным контактом между двумя цепями Fc и FcγR.
На Фигуре 17 показана схема мультифункциональных молекул на основе асимметричного Fc-каркаса, образованного гетеродимерными вариантами, описанными в данной заявке: Асимметричный Fc-каркас и Асимметричный Fc-мономерное плечо IgG.
На Фигуре 18 показана схема мультифункциональных молекул на основе асимметричного Fc-каркаса, образованного гетеродимерными вариантами, описанными в данной заявке: Асимметричный Fc-моноспецифические плечи IgG и Асимметричный Fc-биспецифические плечи IgG (общая легкая цепь).
На Фигуре 19 показано изображение мультифункциональных молекул на основе асимметричного Fc-каркаса, образованного гетеродимерными вариантами, описанными в данной заявке. Асимметричный Fc-биспецифические плечи IgG и функциональная молекула, такая как токсин.
На Фигуре 20 проиллюстрированы мультифункциональные молекулы на основе асимметричного Fc-каркаса, образованного гетеродимерными вариантами, описанными в данной заявке: Асимметричный Fc-отдельное плечо scFv и Асимметричный Fc-биспецифические плечи scFv.
На Фигуре 21 проиллюстрированы альтернативные мультифункциональные молекулы на основе асимметричного Fc-каркаса, образованного гетеродимерными вариантами, описанными в данной заявке: Асимметричный Fc-триспецифические плечи scFv и Асимметричный Fc-тетраспецифические плечи scFv.
На Фигуре 22 показана асимметричная схема мутаций на одной стороне Fc для улучшения селективности к FcγR и представлена продуктивная сторона для взаимодействий с FcγR и непродуктивная сторона с взаимодействиями, свойственными дикому типу. Мутации на непродуктивной стороне Fc можно ввести, чтобы блокировать взаимодействия с FcR и сместить полярность Fc, для того чтобы взаимодействие происходило только на продуктивной стороне.
На Фигуре 23 показана последовательность аминокислот IgG1 человека дикого типа.
На Фигуре 24 показан итерационный процесс разработки гетеродимера Fc, сочетающий положительную и отрицательную стратегии разработки, подробно описанные ниже.
На Фигурах 25А-25С показан анализ in vitro, используемый для определения чистоты гетеродимера. Данный анализ осуществляют на основе полноразмерного каркаса моноспецифического антитела с двумя тяжелыми цепями Fc с различной молекулярной массой; тяжелая цепь А содержит C-концевую гистидиновую метку (His) и тяжелая цепь В содержит C-концевую отщепляемую метку - мономерный красный флуоресцентный белок (RFP). Две тяжелые цепи A (His) и В (RFP) экспрессируются в различных относительных соотношениях вместе с фиксированным количеством легкой цепи, позволяя получить 3 возможных вида димеров с различными молекулярными массами: а) гомодимер Цепь A (His)/Цепь A (His) (~150 кДа); b) гетеродимер Цепь A (His)Цепь В (RFP) (~175 кДа); с) гомодимер Цепь В (RFP)/Цепь В (RFP) (~200 кДа). После экспрессии, описанной в Примере 2, соотношение гетеродимера по сравнению с двумя гомодимерами определяют с помощью электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия (ПААГ/ДСН) в невосстановительных условиях, который обеспечивает возможность разделения 3 видов димера по молекулярной массе. Гели после ПААГ/ДСН окрашивают кумасси бриллиантовым голубым. На Фигуре 25А: исследованные варианты представляли собой только Цепь А дикого типа (His); только Цепь В дикого типа (RFP); Цепь А дикого типа (His) плюс цепь В (RFP); Контроль 1 цепь A (His) плюс цепь В (RFP), чистота гетеродимера которого была известна и составляла >95%. Состав полос димера проверяли с помощью вестерн-блоттинга с антителами, направленными против IgG-Fc (анти-Fc), против метки мономерного RFP (анти-mRFP) и против гистидиновой метки (анти-His), как проиллюстрировано выше. На геле ПААГ/ДСН была видна отдельная полоса гомодимера His/His, двойная полоса гетеродимера His/RFP и несколько полос гомодимера RFP. Указанные несколько полос представляли собой артефакт мечения мономерным RFP, и удостоверились, что это не влияет на физические свойства гетеродимера Fc. На Фигуре 25В: подтвердили достоверность анализа ПААГ/ДСН с помощью опубликованных Контролей 1-4 вариантов гетеродимера Fc в качестве контролей, смотри Таблицу А. Указанные варианты экспрессировали с различными относительными соотношениями цепи A (His) и цепи В (RFP): В частности, для LC,HC_His,HC_mRFP соотношение 1:3 эквивалентно соотношению 25%, 10%, 65%; соотношение 1:1 эквивалентно соотношению 25%, 20%, 55% и соотношение 3:1 эквивалентно соотношению 25%, 40%, 35%, соответственно (видимое соотношение 1:1 экспрессии цепи A (His) к цепи В (RFP) определили как близкое к 20%/55% (His/RFP) для Fc дикого типа (ДТ)). На Фигуре 25С показан анализ ПААГ/ДСН в невосстановительных условиях для определения чистоты гетеродимера вариантов Каркаса 1. Варианты Fc экспрессировали с различными относительными соотношениями цепи A (His) к цепи В (RFP) и анализировали с помощью ПААГ/ДСН в невосстановительных условиях, описанного на Фигуре 2. В частности, для LC,HC_His,HC_mRFP соотношение 1:3 эквивалентно соотношению 25%, 10%, 65%; соотношение 1:1 эквивалентно соотношению 25%, 20%, 55% и соотношение 3:1 эквивалентно соотношению 25%, 40%, 35%, соответственно (видимое соотношение 1:1 экспрессии цепи A (His) к цепи В (RFP) определили как близкое к 20%/55% (His/RFP) для Fc ДТ).
На Фигурах 26А-26В показаны варианты гетеродимера Fc, экспрессированные с определенным соотношением цепи A (His) к цепи В (RFP) (см. Таблицу 2), очищенные посредством аффинной хроматографии на белке А и проанализированные с помощью ПААГ/ДСН в невосстановительных условиях, описанного на Фигурах 25А-25С. На Фигуре 26А проиллюстрирована классификация гетеродимеров на основании чистоты, наблюдаемой путем визуального контроля результатов ПААГ/ДСН. Для сравнения, на гель загружали эквивалентное количество очищенного на белке А продукта. Такое определение чистоты на основании ПААГ/ДСН в невосстановительных условиях подтверждали для выбранных вариантов с помощью сочетания жидкостной хроматографии с масс-спектрометрией (ЖХ/МС) (см. Фигуру 28). На Фигуре 26В представлены примеры результатов ПААГ/ДСН для выбранных вариантов гетеродимера, очищенных на белке A (AZ94, AZ86, AZ70, AZ33 и AZ34).
На Фигурах 27А-27В проиллюстрированы анализы методом дифференциальной сканирующей калориметрии (ДСК) для определения температуры плавления гетеродимерного домена СН3-СН3, образованного вариантами гетеродимера, описанного в данной заявке. Для определения температур плавления использовали два независимых способа. На Фигуре 27А приведены термограммы, подогнанные по 4 независимым недвухфазным переходам и оптимизированные для получения значений для переходов СН2 и Fab, близких к опубликованным в литературе значениям для герцептина, составляющих ~72°С (СН2) и ~82°С (Fab). На Фигуре 27В показаны нормированные и скорректированные по фону термограммы для вариантов гетеродимера, которые вычли из ДТ для получения положительного и отрицательного разностного пика только для перехода СН3.
На Фигуре 28 проиллюстрирован анализ ЖХ/МС на примере варианта AZ70, как описано в примере 2. Указаны ожидаемые (средние расчетные) массы гликозилированного гетеродимера и гомодимеров. Участок, соответствующий массе гетеродимера, включает основные пики, соответствующие утрате глицина (-57 Да) и вставке 1 или 2 гексоз (+162 Да и +324 Да, соответственно). Чистоту гетеродимера классифицируют как >90%, если отсутствуют существенные пики, соответствующие любому из гомодимеров.
На Фигурах 29A-29D показана граница СН3: Fc ДТ на Фиг. 29А, AZ6 на Фиг. 29В, AZ33 на Фиг. 29С, AZ19 на Фиг. 29D. Подробный компьютерный анализ, описанный в разделе подробного описания, и сравнение вариантов с ДТ показали, что одной из причин уменьшения стабильности исходного гетеродимера AZ33 по сравнению с ДТ явилась утрата взаимодействия/укладки ядра Y407 и Т366. У исходного AZ33 наблюдается неоптимальная укладка данного гидрофобного ядра, проиллюстрированная на Фиг. 29В, позволяя предложить, что оптимизация данного участка, особенно в положении Т366, улучшит стабильность AZ33. Такая оптимизация показана на Фиг. 29С и Фиг. 29D с заменами T366I и T366L. Результаты данного эксперимента коррелируют с результатами структурного анализа и показывают, что замена T366L приводит к наиболее значительному улучшению Tm. См. Пример 5.
На Фигуре 30 проиллюстрирована польза и важность анализа конформационной динамики на примере исходного варианта AZ8 каркаса 1. Структуру после компьютерного мутагенеза (конформация каркаса близка к таковой у ДТ) наложили на типичную структуру, полученную с помощью моделирования методом молекулярной динамики (МД) в течение 50 нс. На фигуре выделено большое конформационное различие на участке петли D399-S400 варианта AZ8 по сравнению с ДТ, который, в свою очередь, экспонирует гидрофобное ядро в растворитель, что приводит к уменьшению стабильности гетеродимера AZ8.
На Фигурах 31А-31С проиллюстрировано, как информацию, полученную в результате подробного компьютерного анализа и моделирования методом МД, используют для описания стратегии положительной разработки. На Фигуре 30 проиллюстрировано, что одной из причин пониженной по сравнению с ДТ стабильности AZ8 является ослабленное взаимодействие петли 399-400 с 409, что преимущественно обусловлено утратой взаимодействий укладки F405 (см. сравнение Фиг. 31А (ДТ) с Фиг. 31В (AZ8)). Одной из стратегий положительной разработки являлась оптимизация гидрофобной укладки участка, чтобы стабилизировать конформацию петли 399-400. Этого добились путем введения мутации K392M, которая показана на Фигуре 31С. На Фиг. 31С представлен гетеродимер AZ33, который имеет Tm, составляющую 74°С, по сравнению с 68°С у исходного варианта AZ8 отрицательной разработки.
На Фигурах 32А-32В проиллюстрирована динамика молекулы Fc, наблюдаемая при применении анализа главных компонент молекулярно-динамической траектории. На Фиг. 32А показан след каркаса структуры Fc в качестве эталона. На Фиг. 32В и С представлено наложение динамики, наблюдаемой для 2 самых главных мод движения структуры Fc. У доменов СН2 цепи А и В выявили существенное движение открытия/закрытия по отношению друг к другу, тогда как домены СН3 были относительно неподвижны. Мутации на границе СН3 повлияли на относительную гибкость и динамику такого движения открытия/закрытия в доменах СН2.
На Фигурах 33А-33С проиллюстрирована укладка гидрофобного ядра двух вариантов каркаса 2 по сравнению с ДТ. На Фиг. 33А представлен Fc ДТ; на Фиг. 33В представлен AZ63; и на Фиг. 33С представлен AZ70. Подробный компьютерный анализ исходного варианта каркаса 2 позволил предположить, что утрата взаимодействия Y407-Т366 в ядре ДТ является одной из причин пониженной относительно ДТ стабильности исходных вариантов каркаса 2. Утрата Y407-T366 частично компенсируется мутациями K409F, но, как показано на Фигуре 33В, в частности, мутация Т366А оставляет полость в гидрофобном ядре, которая дестабилизирует данный вариант по сравнению с ДТ. Введение в данное гидрофобное ядро дополнительных мутаций T366V L351Y, как показано для варианта Fc AZ70 на Фиг. 33С, оказалось успешным; AZ70 имеет определенную путем эксперимента Tm, составляющую 75,5°С. См. Таблицу 4 и Пример 6.
На Фигурах 34А-34С проиллюстрированы взаимодействия петли 399-400 двух вариантов каркаса 2 по сравнению с ДТ: на Фиг. 34А представлен Fc ДТ; на Фиг. 34В представлен AZ63; и на Фиг. 34С представлен AZ94. Подробный компьютерный анализ исходного варианта каркаса 2 позволил предположить, что утрата солевого мостика K409-D399 ДТ (Фиг. 34А) в результате мутации K409F и, следовательно, ненасыщенность D399 (Фиг. 34В) приводит к более "открытой" конформации петли 399-400. Более того, это приводит к большему экспонированию гидрофобного ядра в растворитель и дополнительной дестабилизации данного варианта по сравнению с ДТ. Одной из стратегий, используемых для стабилизации петли 399-400 и компенсации утраты взаимодействия K409-D399, является создание дополнительных солевых мостиков D399R-T411E и S400R-K392E, показанных на Фиг. 34С для варианта AZ94. Результаты эксперимента показали чистоту >95% и Tm, составляющую 74°С. Смотри Таблицу 4 и Пример 6. Кроме того, хотя AZ94 имел значительно большую чистоту и стабильность по сравнению с исходным вариантом каркаса 2 (чистота <90%, Tm составляет 71°С), мутации в гидрофобном ядре в AZ94 являются менее предпочтительными, чем "лучшие" мутации в гидрофобном ядре, обнаруженные в варианте AZ70 (Фигура 33). Поскольку мутации в гидрофобном ядре в AZ70 (T366V_L351Y) отдалены от мутаций солевого мостика в петле 399-400 AZ94, ожидается, что комбинация мутаций аминокислот AZ70 и дополнительных мутаций AZ94 приводит к более высокой температуре плавления, чем у AZ70 или AZ94. Такую комбинацию можно исследовать, как описано в Примерах 1-4.
На Фигуре 35 проиллюстрирована константа ассоциации (Ka(М-1)) гомодимерного Fc IgG1, гетеродимерных вариантов гет1 (Контроль 1) A:Y349C_T366S_L368A_Y407V/B:S354C_T366W и гет2 (Контроль 4) A:K409D_K392D/B:D399K_D356K для связывания с шестью рецепторами Fc-гамма. Видна тенденция гетеродимерных вариантов Fc к небольшому изменению связывания с рецепторами Fc-гамма по сравнению с Fc IgG1 дикого типа. См. Пример 7.
На Фигуре 36А показана относительная сила связывания Fc IgG1 дикого типа и различных гомодимерных и асимметрично мутированных его форм с рецепторами IIbF, IIbY и IIaR, по сравнению с силой связывания дикого типа в качестве контроля. (ГомоFc + S267D) относится к силе связывания гомодимерного Fc с мутацией S267D на обеих цепях. (ГетFc + асиммS267D) относится к силе связывания гетеродимерного Fc с мутацией S267D, введенной в одну из двух цепей Fc. Приведено среднее значение силы связывания, полученное для введения мутации в каждую из двух цепей Fc. Введение данной мутации в одну цепь уменьшало силу связывания приблизительно до половины силы, наблюдаемой для той же мутации в гомодимере. (ГетFc + асиммS267D + асиммЕ269K) относится к силе связывания гетеродимерного Fc с обеими мутациями S267D и E269K, введенными асимметричным способом в одну из двух цепей Fc. Мутация E269K блокирует взаимодействие FcγR с одной из поверхностей Fc и способна снижать силу связывания приблизительно вдвое относительно наблюдаемой для варианта с асимметричной S267D (ГетFc + S267D) отдельно. Здесь ГетFc включает мутации СН3, указанные для варианта гет2 (Контроль 4) на Фигуре 35.
На Фигуре 36В показана константа ассоциации (Ka(М-1)) различных Fc и их вариантов с множеством аллотипов FcγRIIa, FcγRIIb и FcγRIIIa. Ka Fc IgG1 дикого типа с различными рецепторами Feγ представлена в виде столбцов с горизонтальной штриховкой. Столбцы с вертикальной штриховкой (гомодимерная основа2) представляют Ka гомодимерного Fc с мутациями S239D/D265S/I332E/S298A. Столбцы с наклонной штриховкой представляют Ka гетеродимерного Fc с асимметричными мутациями A:S239D/D265S/I332E/E269K и B:S239D/D265S/S298A в домене СН2. Введение асимметричных мутаций помогает добиться повышенной селективности в отношении рецепторов IIIa и IIa/IIb. Здесь гетеродимерный Fc включает мутации СН3, указанные для варианта гет2 (Контроль 4) на Фигуре 35.
На Фигуре 36С показана константа ассоциации ассоциации (Ka (М-1)) для IgG1 дикого типа и трех других вариантов, включающих гомодимерные или асимметричные мутации в домене СН2 Fc-области. Ka Fc дикого типа представлена в виде столбцов с сетчатой штриховкой. Ka варианта Fc с основной мутацией в основе S239D/K326E/A330L/I332E/S298A, введенной гомодимерным способом (гомодимерная основа 1) в обе цепи Fc, представлена в виде столбцов с наклонной штриховкой. Введение соответствующих мутаций асимметричным способом в цепи А и В гетеродимерного Fc (гетерооснова1) представлено в виде столбцов с горизонтальной штриховкой. Столбцы с вертикальной штриховкой представляют асимметричный вариант, включающий мутацию E269K (гетерооснова1 +PD). Здесь гетеродимерный Fc включает мутации СН3, указанные для варианта гет2 (Контроль 4) на Фигуре 35.
На Фигуре 37 показана Таблица 6, на которой представлен перечень вариантов доменов СН3 на основании третьей фазы разработки, описанной в Примере 5 для Каркаса 1.
На Фигуре 38 показана Таблица 7, на которой представлен перечень модифицированных доменов СН3 на основании третьей фазы разработки, описанной в Примере 6 для Каркаса 2.
На Фигуре 39А-39В проиллюстрировано определение чистоты вариантов без каких-либо C-концевых меток с помощью ЖХ/МС. На Фиг. 39А показаны спектры ЖХ/МС одного типичного варианта (AZ162: L351Y_F405A_Y407V/T366L_K392L_T394W). Указанный вариант экспрессировали путем временной совместной экспрессии, как описано в Примерах, применяя 3 различных соотношения тяжелой цепи А к тяжелой цепи В: 1:1,5 (AZ133-1), 1:1 (AZ133-2) и 1,5:1 (AZ133-3). Образцы очищали и дегликозилировали с помощью эндогликозадазы Endo S в течение 1 ч при 37°С. Перед анализом МС, образцы наносили на колонку Poros R2 и элюировали градиентом ацетонитрила 20-90%, 0,2% муравьиной кислоты (МК) в течение 3 минут. Пик с колонки ЖХ анализировали с помощью масс-спектрометра LTQ-Orbitrap XL (напряжение на конусе: 50 В, цилиндрическая линза: 215 В; разрешение при использовании преобразования Фурье (ПФ): 7500) и установленного программного обеспечения Promass с получением профилей молекулярных масс. На Фиг. 39В показаны спектры ЖХ/МС для образца Контроль 2, который представляет собой вариант с укладкой выступы-во-впадины. Указанный вариант экспрессировали путем временной совместной экспрессии, как описано в Примерах, применяя 3 различных соотношения тяжелой цепи А к тяжелой цепи В: 1:1,5 (Контроль 2-1), 1:1 (Контроль 2-2) и 1,5:1 (Контроль 2-3). Образцы очищали и дегликозилировали с помощью Endo S в течение 1 ч при 37°С. Перед анализом МС, образцы наносили на колонку Poros R2 и элюировали градиентом ацетонитрила 20-90%, 0,2% МК в течение 3 минут. Пик с колонки ЖХ анализировали с помощью масс-спектрометра LTQ-Orbitrap XL (напряжение на конусе: 50 В, цилиндрическая линза: 215 В; разрешение при использовании ПФ: 7500) и установленного программного обеспечения Promass с получением профилей молекулярных масс.
На Фигурах 40А-40В продемонстрировали биспецифическое связывание, применяя scFv против HER2 (анти-HER2) и HER3 (анти-HER3) гетеродимера Fc, слитые с N-концом цепи А и цепи В гетеродимера Fc. Полученные варианты - биспецифический вариант HER2/HER3 и два моновалентных-моноспецифических варианта HER2, HER3 - показаны на Фигуре 40-А (цепь А темно-серая; цепь В светло-серая). На Фигуре 40В продемонстрирован тест на биспецифическое связывание.
На Фигуре 41 проиллюстрирована вычислительная модель, сравнивающая IgG1 Fc дикого типа и AZ3003. Вычислительная модель для AZ3002 такая же, как и для AZ3003 в положении Т350. В таблице резюмированы выбранные варианты гетеродимера и стабилизирующее влияние мутации T350V на температуру плавления СН3. На фигуре показаны варианты гетеродимера, экспрессированные и очищенные, как описано в Примере 11. Дифференциальную сканирующую калориметрию (ДСК) осуществляли, как подробно описано в Примере 3, и количественный анализ методом ЖХ/МС осуществляли, как подробно описано в Примере 11.
На Фигуре 42 проиллюстрировано сравнение кристаллической структуры и предсказанной модели основного гетеродимера. Мутированные остатки на поверхности (указаны в таблице) выделены на графическом изображении.
На Фигуре 43 изображен анализ паттерна гликозилирования очищенного основного гетеродимера.
На Фигуре 44 проиллюстрированы результаты оценки форсированной деградации очищенного основного гетеродимера.
На Фигуре 45 изображена стандартизованная в промышленности схема процесса очистки антитела.
На Фигуре 46 резюмирована оценка последующей очистки варианта гетеродимера AZ3003 и показаны выходы на каждом этапе и общий выход (см. Пример 15 для получения более подробной информации). Указанный гетеродимер получили в 10 л временно трансфицированных клеток CHO, как подробно описано в Примере 11.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В данной заявке предложены модифицированные домены СН3, включающие определенные модификации аминокислот, чтобы способствовать образованию гетеромультимера. В одном варианте реализации, модифицированные домены СН3 включают определенные модификации аминокислот, чтобы способствовать образованию гетеродимера (см., например, Таблицы 1.1-1.3). В другом варианте реализации, модифицированные домены СН3 включают определенные модификации аминокислот, чтобы способствовать образованию гетеродимера с повышенной стабильностью (см., например, Таблицу 4, Таблицу 6 и Таблицу 7). Стабильность измеряли в виде температуры плавления (Tm) домена СН3, и повышенная стабильность относится к Tm, составляющей приблизительно 70°С или более. Домены СН3 образуют часть Fc-области гетеромультимерного мультиспецифического антитела. В одном варианте реализации данной заявки предложены гетеромультимеры, включающие гетеродимерную Fc-область, при этом гетеродимерная Fc-область содержит модифицированный домен СН3, включающий мутации аминокислот, способствующие образованию гетеродимера, при этом модифицированные домены СНЗ выбраны из вариантов, перечисленых в Таблице 1. Во втором варианте реализации предложены гетеромультимеры, включающие гетеродимерную Fc-область, при этом гетеродимерная Fc-область содержит модифицированный домен СН3, включающий мутации аминокислот, способствующие образованию гетеродимера с повышенной стабильностью, при этом модифицированный домен СН3 имеет температуру плавления (Tm), составляющую приблизительно 70°С или более.
Модификации аминокислот, используемые для получения модифицированного домена СН3, включают, но не ограничены перечисленными, вставки, делеции, замены и перестановки аминокислот. Модификации домена СН3 и модифицированные домены СН3 в данной заявке собирательно называют "модификациями СН3", "модифицированными доменами СН3" или "вариантами СН3". В некоторых вариантах реализации, модифицированные домены СН3 входят в состав выбранной молекулы. Соответственно, в одном варианте реализации предусмотрены молекулы, например полипептиды, такие как иммуноглобулины (например, антитела) и другие связывающие белки, содержащие Fc-область (в данной заявке в объем термина "Fc-область" и близких терминов входит любой домен константной области тяжелой цепи, содержащий по меньшей мере часть домена СН3), содержащую модифицированный домен СН3. Молекулы, содержащие Fc-области, содержащие модифицированный домен СН3 (например, домен СН3, включающий одну или более вставок, делеций, замен или перестановок аминокислот), в данной заявке относятся к "вариантам Fc", "гетеродимерам" или "гетеромультимерам". Настоящие варианты Fc включают домен СН3, который асимметрично модифицировали с получением вариантов или участков гетеродимера Fc. Fc-область состоит из двух полипептидов константного домена тяжелой цепи - цепи А и цепи В, и эти термины можно использовать взаимозаменяемо в том случае, если каждая Fc-область содержит один полипептид цепи А и один полипептид цепи В. Модификации аминокислот вводят в СН3 асимметричным путем с получением гетеродимера, в котором два модифицированных домена СН3 образуют вариант Fc (см., например, Таблицу 1). В данной заявке, асимметричные модификации аминокислот представляют собой любую модификацию, при которой аминокислота в определенном положении на одном полипептиде (например, на "цепи А") отлична от аминокислоты на втором полипептиде (например, на "цепи В") в том же положении гетеродимера или варианта Fc. Этого можно добиться путем модификации только одной из двух аминокислот или модификации обеих аминокислот на две отличные аминокислоты в цепи А и цепи В варианта Fc. Должно быть очевидно, что модифицированные домены СН3 включают одну или более асимметричных модификаций аминокислот.
Аминокислота, которая находится на границе между первым и вторым полипептидами домена СН3, представляет собой любую аминокислоту на первом или втором полипептиде домена СН3, которая взимодействует с аминокислотой на другом полипептиде домена СН3, что приводит к образованию димерного домена СН3. Аминокислота, которая не находится на границе между первым и вторым полипептидами домена СН3, представляет собой любую аминокислоту на первом или втором полипептиде домена СН3, которая не взаимодействует с аминокислотой на другом полипептиде домена СН3. В вариантах реализации, описанных в данной заявке, модифицированная аминокислота, которая не находится на границе между первым и вторым полипептидами домена СН3, представляет собой любую аминокислоту на первом или втором полипептиде домена СН3, которая, после того, как она была модифицирована, как описано в данной заявке, не взаимодействует с аминокислотой на другом полипептиде домена СН3. Например, в некоторых вариантах реализации, описанных в данной заявке, предложены модификации аминокислоты в положении Т350. Кристаллическая структура, представленная в Примере 12 и показанная на Фигуре 42, демонстрирует, что Т350 не участвует во взаимодействиях между двумя полипептидами домена СН3. Было показано, что любые модификации в положении Т350 оказывают незначительное воздействие на образование димеров СН3, как описано у Carter и др. Biochemistry 1998, 37, 9266. Обнаружили, что в гетеромультимерных конструкциях Fc, описанных в данной заявке, модификации в положении Т350 оказывают неожиданное стабилизирующее действие на варианты домена СН3, несмотря на то, что они непосредственно не участвуют в образовании самого димера СН3. Например, варианты, включающие по меньшей мере одну модификацию Т350Х, где X представляет собой природную или неприродную аминокислоту, выбранную из валина, изолейцина, лейцина, метионина и производных или вариантов указанных аминокислот, образуют очень стабильные варианты домена СН3. В некоторых вариантах реализации, описанных в данной заявке, предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, включающая по меньшей мере одну модификацию T350V. В некоторых вариантах реализации, первый и второй полипептиды варианта домена СН3 включают модификацию T350V, которая придает неожиданную стабильность варианту домена СН3 по сравнению с соответствующим доменом СН3, не включающим указанную модификацию.
В настоящем описании следует понимать, что любой диапазон концентраций, диапазон процентов, диапазон соотношений или диапазон целых чисел включает значение любого целого числа в рамках описанного диапазона и, когда это уместно, его части (такие как одна десятая и одна сотая целого числа), если не указано иначе. В данной заявке, "приблизительно" означает ± 10% от указанного диапазона, значения, последовательности или структуры, если не указано иначе. Должно быть очевидно, что термины единственного числа в данной заявке относятся к "одному или более" перечисленным компонентам, если не указано иначе или не обусловлено контекстом. Применение альтернативы (например, союза "или") следует понимать означающим любой один, оба или любую комбинацию альтернатив. В данной заявке, термины "содержат" и "включают" используют синонимично. Кроме того, должно быть очевидно, что отдельные одноцепочечные полипептиды или гетеродимеры, полученные из различных комбинаций структур и заместителей (например, модифицированных доменов СН3), описанных в данной заявке, описаны в настоящей заявке в равной мере, как если бы каждый одноцепочечный полипептид или гетеродимер был описан отдельно. Таким образом, выбор конкретных компонентов для получения отдельных одноцепочечных полипептидов или гетеродимеров входит в объем настоящего описания.
"Первый полипептид" представляет собой любой полипептид, который должен быть связан со вторым полипептидом, и его также называют в данной заявке "цепью А". Первый и второй полипептид "встречаются" на "границе". "Второй полипептид" представляет собой любой полипептид, который должен быть связан с первым полипептидом через "границу", и его также называют в данной заявке "цепью В". "Граница" включает такие "контактирующие" аминокислотные остатки в первом полипептиде, которые взаимодействуют с одним или более "контактирующими" аминокислотными остатками на границе второго полипептида. В данной заявке, граница включает домен СН3 Fc-области, который предпочтительно получен из антитела IgG и, наиболее предпочтительно, из антитела IgG1 человека.
В данной заявке, "изолированный" гетеромультимер означает гетеромультимер, который обнаружили и отделили и/или извлекли из компонентов природного окружения его культуры клеток. Контаминирующие компоненты его природного окружения представляют собой материалы, которые будут препятствовать применениям гетеромультимера в диагностике или терапии, и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества.
Аминокислота с объемом боковой цепи, "существенно не превышающим" таковой у первой аминокислоты, представляет собой любую аминокислоту, объем боковой цепи которой не более чем на 20Å3 превышает таковой у первой аминокислоты, на основании значений объемов боковых цепей из A.A. Zamyatnin, Prog. Biophys. Mol. Biol. 24: 107-123, 1972. В некоторых вариантах реализации, указанный объем не более чем на 10Å3 превышает таковой у первой аминокислоты. В некоторых вариантах реализации, указанный объем не более чем на 5Å3 превышает таковой у первой аминокислоты. Например, в некоторых вариантах реализации, описанных в данной заявке, предложены мутации лизина (K), такие как K392J, где J выбран из L, I или аминокислоты с объемом боковой цепи, значительно не превышающим объем боковой цепи K.
Варианты гетеродимера Fc, как правило, очищают до по существу гомогенного состояния. Формулировки "по существу гомогенный", "по существу гомогенная форма" и "по существу гомогенное состояние" используют для обозначения того, что продукт по существу свободен от побочных продуктов, возникающих в результате образования нежелательных комбинаций полипептидов (например, гомодимеров). Выражаясь в терминах чистоты, по существу гомогенное состояние означает, что количество побочных продуктов не превышает 10%, и оно предпочтительно ниже 5%, более предпочтительно ниже 1%, наиболее предпочтительно ниже 0,5%, где проценты указаны по массе.
Все термины, понимаемые специалистами в области технологии антител, имеют значения, приобретенные в данной области, если в данной заявке явно не дано другое определение. Известно, что антитела имеют вариабельные области, шарнирную область и константные домены. Обзор структуры и функции иммуноглобулинов приведен, например, в Harlow и др., ред., Antibodies: A Laboratory Manual, глава 14 (Cold Spring Harbor Laboratory, Колд-Спринг-Харбор, 1988 г.).
Разработка вариантов гетеродимера Fc из гомодимеров дикого типа проиллюстрирована на принципах положительной и отрицательной разработки в контексте белковой инженерии путем достижения баланса между стабильностью и специфичностью, при этом мутации вводят с такой целью, чтобы вызвать образование гетеродимера, но не гомодимера, когда полипептиды экспрессируются в условиях культуры клеток. Стратегии отрицательной разработки максимизируют неблагоприятные взаимодействия для образования гомодимеров, либо путем введения объемных боковых цепей на одной цепи и малых боковых цепей на противоположной, например, стратегия выступы-во-впадины, разработанная Genentech (Ridgway JB, Presta LG, Carter P. 'Knobs-into-holes' engineering of antibody СН3 domains for heavy chain heterodimerization. Protein Eng. 1996 Jul; 9(7):617-21; Atwell S, Ridgway JB, Wells JA, Carter P. Stable heterodimers from remodeling the domain interface of a homodimer using a phage display library. J Mol Biol. 270(1):26-35 (1997))), либо с помощью электростатической инженерии, что приводит к отталкиванию и препятствует образованию гомодимера, например, стратегия электростатического управления, разработанная Amgen (Gunaskekaran K, и др. Enhancing antibody Fc heterodimer formation through electrostatic steering effects: applications to bispecific molecules and monovalent IgG. JBC 285(25): 19637-19646 (2010)). В двух данных примерах отрицательной разработки, ввели асимметричные точечные мутации в домен СН3 дикого типа, чтобы вызвать образование гетеродимера. На сегодняшний день для разработки гетеродимеров Fc использовали только стратегии отрицательной разработки. Опубликованные результаты показывают, что разработка гетеродимеров с применением только подхода отрицательной разработки приводит к высокой специфичности с >95% гетеродимеров, но также к значительной дестабилизации комплекса (выше). Гетеродимеры, полученные применением подхода отрицательной разработки, имеют температуру плавления модифицированного домена СН3, составляющую 69°С или менее, и в них отсутствуют дополнительные дисульфидные связи по сравнению с диким типом. См. Таблицу А ниже.
Таблица А: Опубликованные антитела с гетеродимерным Fc
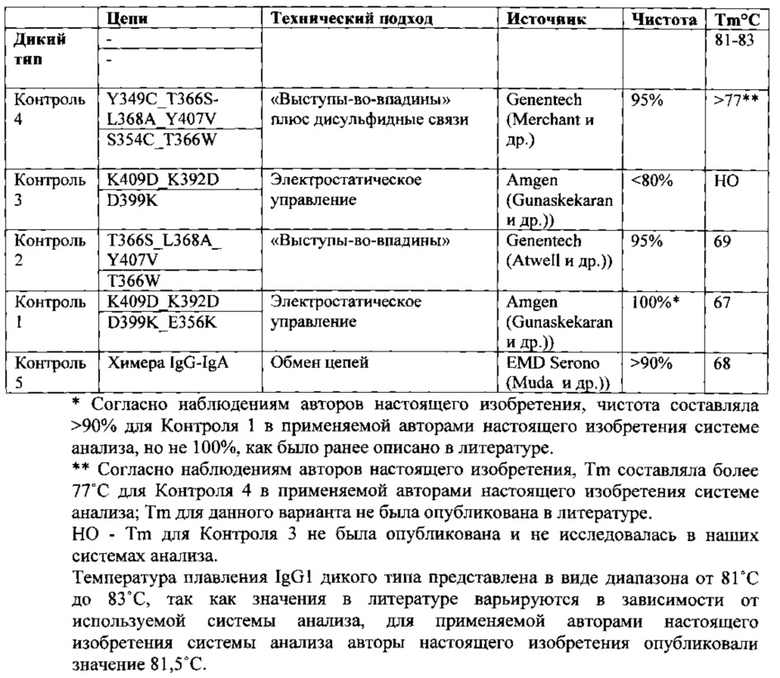
В противоположность отрицательной разработке, основным принципом, используемым для разработки белков, является положительная разработка. В этом случае модификации аминокислот вводят в полипептиды, чтобы максимизировать благоприятные взаимодействия внутри или между белками. В данной стратегии предполагают, что при введении нескольких мутаций, которые специфично стабилизируют желательный гетеродимер, при этом пренебрегая влиянием на образование гомодимеров, результирующим эффектом будет лучшая специфичность по отношению к желательным гетеродимерным, но не гомодимерным, взаимодействиям и, следовательно, большая специфичность по отношению к гетеродимерам. В контексте белковой инженерии очевидно, что стратегии положительной разработки оптимизируют стабильность желательных белковых взаимодействий, но редко добиваются специфичности >90% (Havranek JJ & Harbury РВ. Automated design of specificity in molecular recognition. Nat Struct Biol. 10(1):45-52 (2003); Bolon DN, Grant RA, Baker ТА, Sauer RT. Specificity versus stability in computational protein design. Proc Natl Acad Sci USA. 6; 102(36):12724-9 (2005); Huang PS, Love JJ, Mayo SL. A de novo designed protein-protein interface. Protein Sci. 16(12):2770-4 (2007)). До настоящего описания, стратегии положительной разработки не применялись для разработки гетеродимеров Fc для производства и разработки терапевтического антитела, так как большее внимание уделялось специфичности, но не стабильности. Кроме того, полезные мутации в стратегии положительной разработки может быть сложно предсказать. Другие методики улучшения стабильности, такие как введение дополнительных дисульфидных связей, пробовали использовать для повышения стабильности гетеродимеров Fc с ограниченным успехом улучшения молекулы (см. Таблицу А). Это может быть обусловлено тем, что все искусственные дисульфидные связи Fc доменов СН3 экспонированы в растворитель, что приводит к быстрому распаду дисульфидной связи и, следовательно, оказывает существенное влияние на долговременную стабильность гетеродимера, особенно если искусственный домен СН3 имеет Tm, меньшую чем 70°С, без дополнительной дисульфидной связи (как в Контроле 4, который имеет Tm, составляющую 69°С, без дисульфидной связи (см. Контроль 2)). Предполагается, что для настоящих вариантов Fc также можно применять другие методики повышения стабильности, отличные от образования дисульфидных связей, при условии, что внутренняя стабильность (измеренная в виде температуры плавления) домена СН3 составляет 70°С или более без дисульфидной связи, в частности, если внутренняя стабильность (измеренная в виде температуры плавления) домена СН3 составляет 72°С или более без дисульфидной связи.
Следовательно, в данной заявке авторами настоящего изобретения предложен новый способ разработки гетеродимеров Fc, который позволяет получить как стабильный, так и высокоспецифичный гетеродимер. В данном способе разработки объединены стратегии отрицательной и положительной разработки, наряду со структурным и компьютерным моделированием с использованием методик белковой инженерии. Этот эффективный способ позволил нам разработать новые комбинации мутаций в домене СН3 IgG1, причем при применении лишь стандартных условий культивирования клеток гетеродимеры образовывались с чистотой более чем 90% относительно гомодимеров, и полученные в результате этого гетеродимеры имели температуру плавления 70°С или более. В типичных вариантах реализации, варианты гетеродимера Fc имеют температуру плавления, составляющую 73°С или более, и чистоту более 98%. В других типичных вариантах реализации, варианты гетеродимера Fc имеют температуру плавления, составляющую 75°С или более, и чистоту более 90%. В некоторых вариантах реализации вариантов гетеродимера Fc, описанных в данной заявке, варианты гетеродимера Fc имеют температуру плавления, составляющую 77°С или более, и чистоту более 98%. В некоторых вариантах реализации вариантов гетеродимера Fc, описанных в данной заявке, варианты гетеродимера Fc имеют температуру плавления, составляющую 78°С или более, и чистоту более чем 98%. В некоторых вариантах реализации вариантов гетеродимера Fc, описанных в данной заявке, варианты гетеродимера Fc имеют температуру плавления, составляющую 79°С или более, и чистоту более чем 98%. В некоторых вариантах реализации вариантов гетеродимера Fc, описанных в данной заявке, варианты гетеродимера Fc имеют температуру плавления, составляющую 80°С или более, и чистоту более чем 98%. В некоторых вариантах реализации вариантов гетеродимера Fc, описанных в данной заявке, варианты гетеродимера Fc имеют температуру плавления, составляющую 81°С или более, и чистоту более чем 98%.
В некоторых вариантах реализации предложен изолированный гетеромультимер, включающий гетеродимерную Fc-область, при этом гетеродимерная Fc-область содержит модифицированный домен СН3, включающий мутации аминокислот, способствующие образованию гетеродимера с повышенной стабильностью, и при этом модифицированный домен СН3 имеет температуру плавления (Tm), составляющую 70°С или более. В данной заявке, "повышенная стабильность" или "стабильный гетеродимер" относится к модифицированному домену СН3, в составе гетеродимера, с температурой плавления, составляющей приблизительно 70°С или более. В некоторых вариантах реализации, "повышенная стабильность" или "стабильный гетеродимер" относится к модифицированному домену СН3, в составе гетеродимера, с температурой плавления, составляющей приблизительно 72°С или более. В некоторых вариантах реализации, "повышенная стабильность" или "стабильный гетеродимер" относится к модифицированному домену СН3, в составе гетеродимера, с температурой плавления, составляющей приблизительно 74°С или более. В некоторых вариантах реализации, "повышенная стабильность" или "стабильный гетеродимер" относится к модифицированному домену СН3, в составе гетеродимера, с температурой плавления, составляющей приблизительно 75°С или более. В некоторых вариантах реализации, "повышенная стабильность" или "стабильный гетеродимер" относится к модифицированному домену СН3, в составе гетеродимера, с температурой плавления, составляющей приблизительно 76°С или более. В некоторых вариантах реализации, "повышенная стабильность" или "стабильный гетеродимер" относится к модифицированному домену СН3, в составе гетеродимера, с температурой плавления, составляющей приблизительно 78°С или более. В некоторых вариантах реализации, "повышенная стабильность" или "стабильный гетеродимер" относится к модифицированному домену СН3, в составе гетеродимера, с температурой плавления, составляющей приблизительно 79°С или более. В некоторых вариантах реализации, "повышенная стабильность" или "стабильный гетеродимер" относится к модифицированному домену СН3, в составе гетеродимера, с температурой плавления, составляющей приблизительно 80°С или более. В некоторых вариантах реализации, "повышенная стабильность" или "стабильный гетеродимер" относится к модифицированному домену СН3, в составе гетеродимера, с температурой плавления, составляющей приблизительно 81°С или более. Кроме того, должно быть очевидно, что термин "способствующие образованию гетеродимера" в данной заявке относится к мутациям аминокислот в домене СН3, которые приводят к образованию гетеродимера более чем в 90% случаев по сравнению с образованием гомодимера.
В дополнительном варианте реализации, такая повышенная стабильность наблюдается в отсутствии дополнительной дисульфидной связи. В частности, повышенная стабильность наблюдается в отсутствии дополнительной дисульфидной связи в домене СН3. В одном варианте реализации, модифицированный домен СН3 не содержит дополнительную дисульфидную связь по сравнению с доменом СН3 дикого типа. В альтернативном варианте реализации, модифицированный домен СН3 содержит по меньшей мере одну дисульфидную связь по сравнению с доменом СН3 дикого типа, в том случае, если модифицированный домен СН3 имеет температуру плавления, составляющую 70°С или более, в отсутствии дисульфидной связи. В одном варианте реализации, модифицированный домен СН3 содержит по меньшей мере одну дисульфидную связь по сравнению с доменом СН3 дикого типа, и указанный модифицированный домен СН3 имеет температуру плавления (Tm), составляющую приблизительно 77,5°С или более. В одном варианте реализации, модифицированный домен СН3 содержит по меньшей мере одну дисульфидную связь по сравнению с доменом СН3 дикого типа, и указанный модифицированный домен СН3 имеет температуру плавления (Tm), составляющую приблизительно 78°С или более. В другом варианте реализации, модифицированный домен СН3 содержит по меньшей мере одну дисульфидную связь по сравнению с доменом СН3 дикого типа, и указанный модифицированный домен СН3 имеет температуру плавления (Tm) большую чем приблизительно 78°С, или большую чем приблизительно 78,5°С, или большую чем приблизительно 79°С, или большую чем приблизительно 79,5°С, или большую чем приблизительно 80°С, или большую чем приблизительно 80,5°С, или большую чем приблизительно 81°С, или большую чем приблизительно 81,5°С, или большую чем приблизительно 82°С, или большую чем приблизительно 82,5°С, или большую чем приблизительно 83°С.
В одном варианте реализации, модифицированный домен СН3 имеет температуру плавления большую чем приблизительно 70°С, или большую чем приблизительно 70,5°С, или большую чем приблизительно 71°С, или большую чем приблизительно 71,5°С, или большую чем приблизительно 72°С, или большую чем приблизительно 72,5°С, или большую чем приблизительно 73°С, или большую чем приблизительно 73,5°С, или большую чем приблизительно 74°С, или большую чем приблизительно 74,5°С, или большую чем приблизительно 75°С, или большую чем приблизительно 75,5°С, или большую чем приблизительно 76°С, или большую чем приблизительно 76,5°С, или большую чем приблизительно 77°С, или большую чем приблизительно 77,5°С, или большую чем приблизительно 78°С, или большую чем приблизительно 78,5°С, или большую чем приблизительно 79°С, или большую чем приблизительно 79,5°С, или большую чем приблизительно 80°С, или большую чем приблизительно 80,5°С, или большую чем приблизительно 81°С, или большую чем приблизительно 81,5°С, или большую чем приблизительно 82°С, или большую чем приблизительно 82,5°С, или большую чем приблизительно 83°С. В другом варианте реализации, модифицированный домен СН3 имеет температуру плавления, составляющую приблизительно 70°С, или приблизительно 70,5°С, или приблизительно 71°С, или приблизительно 71,5°С, или приблизительно 72°С, или приблизительно 72,5°С, или приблизительно 73°С, или приблизительно 73,5°С, или приблизительно 74°С, или приблизительно 74,5°С, или приблизительно 75°С, или приблизительно 75,5°С, или приблизительно 76°С, или приблизительно 76,5°С, или приблизительно 77°С, или приблизительно 77,5°С, или приблизительно 78°С, или приблизительно 78,5°С, или приблизительно 79°С, или приблизительно 79,5°С, или приблизительно 80°С, или приблизительно 80,5°С, или приблизительно 81°С. В еще одном варианте реализации, модифицированный домен СН3 имеет температуру плавления, составляющую от приблизительно 70°С до приблизительно 81°С, или от приблизительно 70,5°С до приблизительно 81°С, или от приблизительно 71°С до приблизительно 81°С, или от приблизительно 71,5°С до приблизительно 81°С, или от приблизительно 72°С до приблизительно 81°С, или от приблизительно 72,5°С до приблизительно 81°С, или от приблизительно 73°С до приблизительно 81°С, или от приблизительно 73,5°С до приблизительно 81°С, или от приблизительно 74°С до приблизительно 81°С, или от приблизительно 74,5°С до приблизительно 81°С, или от приблизительно 75°С до приблизительно 81°С, или от приблизительно 75,5°С до приблизительно 81°С, или от приблизительно 76°С до приблизительно 81°С, или от приблизительно 76,5°С до приблизительно 81°С, или от приблизительно 77°С до приблизительно 81°С, или от приблизительно 77,5°С до приблизительно 81°С, или от приблизительно 78°С до приблизительно 81°С, или от приблизительно 78,5°С до приблизительно 82°С, или от приблизительно 79°С до приблизительно 81°С. В еще одном варианте реализации, модифицированный домен СН3 имеет температуру плавления, составляющую от приблизительно 71°С до приблизительно 76°С, или от приблизительно 72°С до приблизительно 76°С, или от приблизительно 73°С до приблизительно 76°С, или от приблизительно 74°С до приблизительно 76°С.
Вдобавок к повышенной стабильности, гетеродимерная Fc-область содержит модифицированный домен СН3, включающий мутации аминокислот, способствующие образованию гетеродимера. Должно быть очевидно, что данные мутации аминокислот, способствуют образованию гетеродимера по сравнению с образованием гомодимера. Это образование гетеродимера по сравнению с образованием гомодимера совокупно называют в данной заявке "чистотой", или "специфичностью", или "чистотой гетеродимера", или "специфичностью гетеродимера". Должно быть очевидно, что чистота гетеродимера относится к проценту желательного гетеродимера по сравнению с видами гомодимера, образовавшимися в растворе при стандартных условиях культивирования клеток до селективной очистки видов гетеродимера. Например, чистота гетеродимера, составляющая 90%, указывает на то, что 90% видов димера в растворе представляют собой желательный гетеродимер. В одном варианте реализации, варианты гетеродимера Fc имеют чистоту, большую чем приблизительно 90%, или большую чем приблизительно 91%, или большую чем приблизительно 92%, или большую чем приблизительно 93%, или большую чем приблизительно 94%, или большую чем приблизительно 95%, или большую чем приблизительно 96%, или большую чем приблизительно 97%, или большую чем приблизительно 98%, или большую чем приблизительно 99%. В другом варианте реализации, варианты гетеродимера Fc имеют чистоту, составляющую приблизительно 90%, или приблизительно 91%, или приблизительно 92%, или приблизительно 93%, или приблизительно 94%, или приблизительно 95%, или приблизительно 96%, или приблизительно 97%, или приблизительно 98%, или приблизительно 99%, или приблизительно 100%.
В конкретном варианте реализации, изолированный гетеромультимер включает гетеродимерную Fc-область, при этом гетеродимерная Fc-область содержит модифицированный домен СН3, включающий мутации аминокислот, способствующие образованию гетеродимера с повышенной стабильностью, и при этом модифицированный домен СН3 имеет температуру плавления (Tm), составляющую 70°С или более, и полученный в результате этого гетеродимер имеет чистоту большую чем 90%. В одном аспекте, полученный в результате этого вариант гетеродимера Fc имеет чистоту большую чем 98%, и указанный модифицированный домен СН3 имеет температуру плавления большую чем приблизительно 70°С, или большую чем приблизительно 71°С, или большую чем приблизительно 72°С, или большую чем приблизительно 73°С, или большую чем приблизительно 74°С, или большую чем приблизительно 75°С, или большую чем приблизительно 76°С, или большую чем приблизительно 77°С, или большую чем приблизительно 78°С, или большую чем приблизительно 79°С, или большую чем приблизительно 80°С, или большую чем приблизительно 81°С. В дополнительном аспекте, модифицированный домен СН3 имеет температуру плавления, составляющую 70°С или более, и полученный в результате этого вариант гетеродимера Fc имеет чистоту большую чем приблизительно 90%, или большую чем приблизительно 91%, или большую чем приблизительно 92%, или большую чем приблизительно 93%, или большую чем приблизительно 94%, или большую чем приблизительно 95%, или большую чем приблизительно 96%, или большую чем приблизительно 97%, или большую чем приблизительно 98%, или большую чем приблизительно 99%.
Для разработки данных вариантов Fc с повышенной стабильностью и чистотой авторы настоящего изобретения итерационный процесс автоматизированного проектирования и экспериментальный скрининг для выбора наиболее удачных комбинаций стратегий положительной и отрицательной разработки (см. Фигуру 24).
В частности, на начальной фазе разработки путем отрицательной разработки получили различные варианты гетеродимера Fc и исследовали их экспрессию и стабильность, как описано в Примерах 1-3. Начальная фаза разработки включала варианты гетеродимера Fc AZI-AZ16 (см. Таблицу 1). Из этого исходного набора вариантов гетеродимера Fc, полученных путем отрицательной разработки, у которых ожидалась низкая стабильность (например, Tm меньшая чем 71°С), для дальнейшей разработки выбирали варианты гетеродимера Fc с чистотой более 90% и температурой плавления, составляющей приблизительно 68°С или более. Выбранные варианты включали варианты гетеродимера Fc AZ6, AZ8 и AZ15. На второй фазе разработки, эти выбранные варианты гетеродимера Fc дополнительно модифицировали, чтобы повысить как стабильность, так и чистоту, применяя стратегии положительной разработки, после подробного компьютерного и структурного анализа. Каждый из выбранных вариантов гетеродимера Fc (AZ6, AZ8 и AZ15) анализировали с помощью вычислительных способов и подробного структурно-функционального анализа, чтобы определить структурные причины того, почему данные варианты Fc имеют более низкую стабильность, чем гомодимер Fc дикого типа, у которого температура плавления составляет 81°С для IgG1. Перечень вариантов гетеродимера Fc и значений Tm смотри в Таблице 4.
В некоторых вариантах реализации, модифицированный домен СН3 выбирают из AZ1, или AZ2, или AZ3, или AZ4, или AZ5, или AZ6, или AZ7, или AZ8, или AZ9, или AZ10, или AZ11, или AZ12, или AZ13, или AZ14, или AZ15, или AZ16. В выбранных вариантах реализации, модифицированный домен СН3 представляет собой AZ6, или AZ8, или AZ15.
Вычислительные средства и структурно-функциональный анализ включали, но не ограничивались перечисленными: молекулярно-динамический анализ (МД), пересворачивание боковой цепи/каркаса, статистический потенциал (KBP), анализ полостей и (гидрофобной) укладки (LJ, CCSD, SASA, dSASA (углерода/всех атомов)), электростатические-GB расчеты и анализ связывания (на Фигуре 24 представлен обзор вычислительной стратегии).
Аспект подхода белковой инженерии основывался на объединении структурных сведений о Fc белка IgG, полученных с помощью рентгеноструктурного анализа, с компьютерным моделированием и моделированием домена СН3 дикого типа и его вариантных форм. Данный подход позволил нам получить новое структурное и физико-химическое представление о потенциальной роли отдельных аминокислот и их совместном действии. Данное структурное и физико-химическое представление, полученное для нескольких модифицированных доменов СН3, наряду с полученными эмпирическими данными относительно их стабильности и чистоты, помогло нам понять взаимосвязь между чистотой и стабильностью гетеродимера Fc, по сравнению с гомодимерами Fc и смоделированными структурными моделями. Для осуществления моделирования, авторы настоящего изобретения начали с построения полных и реалистичных моделей и улучшения качества структуры Fc дикого типа антитела IgG1. В белковых структурах, полученных с помощью рентгеноструктурного анализа, отсутствовала детализация, касающаяся некоторых свойств белка в водной среде при физиологических условиях, и наши процедуры детализации были направлены на устранение данных ограничений. Они включали построение отсутствующих участков в структуре белка, зачастую, подвижных участков белка, таких как петли и боковые цепи некоторых остатков, оценку и определение уровней протонирования нейтральных и заряженных остатков и внесение потенциально функционально значимых молекул воды, связанных с белком.
Алгоритмы молекулярной динамики (МД) представляют собой одно средство, которое авторы настоящего изобретения использовали при моделировании белковой структуры для оценки внутренней динамической природы гомодимера Fc и модифицированных доменов СН3 в водной среде. Молекулярно-динамическое моделирование позволяет отследить динамическую траекторию молекулы, возникшую в результате движений, порожденных взаимодействиями и силами, действующими между всеми атомными единицами в белке и локальной среде, окружающей его, в данном случае, между атомами, составляющими Fc, и окружающими его молекулами воды. После молекулярно-динамического моделирования, анализировали различные аспекты траекторий, чтобы разобраться в структурных и динамических свойствах гомодимера Fc и варианта гетеродимера Fc, которые использовали авторы настоящего изобретения, чтобы определить конкретные мутации аминокислот для улучшения как чистоты, так и стабильности молекулы.
Следовательно, полученные МД-траектории исследовали, применяя такие способы, как метод главных компонент, чтобы обнаружить внутренние низкочастотные колебания в движении структуры Fc. Это дало нам представление о потенциальных конформационных подсостояниях белка (см. Фигуру 32). Несмотря на то, что критические белок-белковые взаимодействия между цепями А и B в Fc-области возникали на границе доменов СН3, моделирование, выполненное авторами настоящего изобретения, показало, что эта граница действует как шарнир при движении, которое включает "открытие" и "закрытие" N-концов доменов СН2 относительно друг друга. Домен СН2 взимодействует этим концом с FcγR, что видно на Фигуре 16. Таким образом, не привязываясь к какой-либо конкретной теории, похоже, что введение мутаций аминокислот на границе СН3 оказывает влияние на амплитуду и природу движения открытия/закрытия N-конца Fc и, следовательно, на то, как Fc взимодействует с FcγR. См. Пример 4 и Таблицу 5.
Также исследовали полученные МД-траектории, чтобы определить возможность введения мутаций в определенные положения аминокислотных остатков в структуре Fc на основании определения профиля их гибкости и анализа их окружения. Данный алгоритм позволил нам определить остатки, которые могут влиять на структуру и функцию белка, и дал уникальное представление о свойствах остатков и возможности введения мутаций для последующих фаз разработки модифицированных доменов СН3. Данный анализ также позволил нам сравнить несколько моделей и оценить возможность введения мутаций на основании выпадающих значений после определения профиля.
Также исследовали полученные МД-траектории, чтобы определить согласованные движения остатков в белке и образование сетей остатков в результате образования связей между ними. Обнаружение динамических взаимосвязей и сетей остатков внутри структуры Fc является важным этапом для понимания белка как динамической молекулы и для получения представления о влиянии мутаций в периферических областях. См., например, Пример 6.
Таким образом, авторы настоящего изобретения подробно исследовали вклад мутаций в локальное окружение сайта мутации. Образование плотно упакованного ядра на границе СН3 между цепями А и В необходимо для самопроизвольного соединения двух цепей в стабильную структуру Fc. Плотная укладка достигается в результате сильной структурной комплементарности между взаимодействующими молекулярными партнерами, в сочетании с благоприятными взаимодействиями между контактирующими группами. Благоприятные взаимодействия образуются в результате спрятанных гидрофобных контактов, хорошо удаленных от взаимодействия с растворителем, и/или в результате образования комплементарных электростатических контактов между гидрофильными полярными группами. Данные гидрофобные и гидрофильные контакты вносят энтропийный и энтальпийный вклад в свободную энергию образования димера на границе СН3. Авторы настоящего изобретения используют множество алгоритмов для создания точной модели укладки на границе СН3 между цепью А и цепью В, а затем оцениваем термодинамические свойства границы путем оценки множества важных физико-химических свойств.
Авторы настоящего изобретения используют множество способов укладки белка, включая гибкие каркасы, для оптимизации и подготовки модельных структур для большого количества вариантов, которые авторы настоящего изобретения подвергли компьютерному скринингу. После укладки авторы настоящего изобретения оценили множество показателей, включая плотность контакта, показатель столкновений, водородные связи, гидрофобность и электростатику. Применение моделей сольватации позволило нам более тщательно изучить влияние окружения растворителя и сравнить различия в свободной энергии после мутации в определенных положениях белка на альтернативные типы остатков. Плотность контакта и показатель столкновений позволили измерить комплементарность, решающий аспект для эффективной укладки белка. Данные процедуры скрининга основаны на применении наукоемкого потенциала или схем анализа связывания, опирающихся на расчеты энергии попарного взаимодействия остатков и энтропии.
Такой подробный компьютерный анализ обеспечил углубленное понимание различий каждого варианта Fc по сравнению с Fc дикого типа в отношении горячих точек мутагенеза на границе, сайтов асимметрии, полостей и плохо упакованных участков, структурной динамики отдельных сайтов и сайтов с локальным разворачиванием. Данные объединенные результаты описанного компьютерного анализа позволили обнаружить конкретные остатки, мотивы последовательности/структуры и полости, которые не были оптимизированы, и, в комбинации, обуславливали более низкую стабильность (например, Tm, составляющую 68°С) и/или более низкую специфичность с чистотой <90%. На второй фазе разработки авторы настоящего изобретения использовали направленную положительную разработку для специально для проработки данной гипотезы с помощью введения дополнительных точечных мутаций, и исследовали их посредством вычислительной инженерии, применяя описанные выше методику и анализ (см. Фигуру 24). Экспрессию и стабильность вариантов гетеродимера Fc, разработанных таким образом, чтобы улучшить стабильность и чистоту для каждой направленной разработки во второй фазе (варианты гетеродимера Fc AZ17-AZ101), проверяли опытным путем, как описано в Примерах 1-4.
В некоторых вариантах реализации, в данной заявке предложены изолированные гетеромультимеры, включающие гетеродимерную Fc-область, при этом гетеродимерная Fc-область содержит модифицированный домен СН3, включающий мутации аминокислот, способствующие образованию гетеродимера с повышенной стабильностью, и при этом модифицированный домен СН3 представляет собой AZ17, или AZ18, или AZ19, или AZ20, или AZ21, или AZ22, или AZ23, или AZ24, или AZ25, или AZ26, или AZ27, или AZ28, или AZ29, или AZ30, или AZ31, или AZ32, или AZ33, или AZ34, или AZ35, или AZ36, или AZ37, или AZ38, или AZ39, или AZ40, или AZ41, или AZ42, или AZ43, или AZ44, или AZ45, или AZ46, или AZ47, или AZ48, или AZ49, или AZ50, или AZ51, или AZ52, или AZ53, или AZ54, или AZ55, или AZ56 или AZ57, или AZ58, или AZ59, или AZ60, или AZ61, или AZ62, или AZ63, или AZ64, или AZ65, или AZ66, или AZ67, или AZ68, или AZ69, или AZ70, или AZ71, или AZ72, или AZ73, или AZ74, или AZ75, или AZ76, или AZ77, или AZ78, или AZ79, или AZ80, или AZ81, или AZ82, или AZ83, или AZ84, или AZ85, или AZ86, или AZ87, или AZ88, или AZ89, или AZ90, или AZ91, или AZ92, или AZ93, или AZ94, или AZ95, или AZ96, или AZ97, или AZ98, или AZ99, или AZ100 или AZ101. В типичном варианте реализации, модифицированный домен СН3 представляет собой AZ17, или AZ18, или AZ19, или AZ20, или AZ21, или AZ22, или AZ23, или AZ24, или AZ25, или AZ26, или AZ27, или AZ28, или AZ29, или AZ30, или AZ31, или AZ32, или AZ33, или AZ34, или AZ38, или AZ42, или AZ43, или AZ 44, или AZ45, или AZ46, или AZ47, или AZ48, или AZ49, или AZ50, или AZ52, или AZ53, или AZ54, или AZ58, или AZ59, или AZ60, или AZ61, или AZ62, или AZ63, или AZ64, или AZ65, или AZ66, или AZ67, или AZ68, или AZ69, или AZ70, или AZ71, или AZ72, или AZ73, или AZ74, или AZ75, или AZ76, или AZ77, или AZ78, или AZ79, или AZ81, или AZ82, или AZ83, или AZ84, или AZ85, или AZ86, или AZ87, или AZ88, или AZ89, или AZ91, или AZ92, или AZ93, или AZ94, или AZ95, или AZ98, или AZ99, или AZ100 или AZ101. В конкретном варианте реализации, модифицированный домен СНЗ представляет собой AZ33 или AZ34. В другом варианте реализации, модифицированный домен СН3 представляет собой AZ70 или AZ90.
В типичном варианте реализации, домен СН3 содержит первый и второй полипептид (также называемые в данной заявке цепью А и цепью В), при этом первый полипептид включает модификации аминокислот L351Y, F405A и Y407V и при этом второй полипептид включает модификации аминокислот T366I, K392M и T394W. В другом варианте реализации, первый полипептид включает модификации аминокислот L351Y, S400E, F405A и Y407V и второй полипептид включает модификации аминокислот T366I, N390R, K392M и T394W.
Данный итерационный процесс компьютерного структурно-функционального анализа, направленной разработки и экспериментального подтверждения применяли для разработки остальных вариантов Fc, перечисленых в Таблице 1, в последующих фазах разработки, и что позволило получить варианты гетеродимера Fc с чистотой более 90%, и с повышенной стабильностью с температурой плавления домена СН3 более 70°С. В некоторых вариантах реализации, варианты Fc включают мутации аминокислот, выбранные из AZ1 - AZ136. В дополнительных вариантах реализации, варианты Fc включают мутации аминокислот, выбранные из вариантов Fc, перечисленых в Таблице 4.
В ходе первой и второй фаз разработки обнаружили два основных каркаса, Каркас 1 и Каркас 2, при этом в данные каркасы ввели дополнительные модификации аминокислот для точного подбора чистоты и стабильности вариантов гетеродимера Fc. В Примере 5 представлено подробное описание разработки Каркаса 1, включая AZ8, AZI 7-62 и варианты, перечисленные в Таблице 6. В Примере 6 представлено подробное описание разработки Каркаса 2, включая AZ15 и AZ63-101 и варианты, перечисленные в Таблице 7.
Мутации в ядре Каркаса 1 включают L351Y_F405A_Y407V/T394W. Каркас 1а включает мутации аминокислот T366I_K392M_T394W/F405A_Y407V и Каркас 1b включает мутации аминокислот T366L_K392M_T394W/F405A_Y407V. См. Пример 5.
В некоторых вариантах реализации, модифицированный домен СН3 содержит первый и второй полипептид (также называемые в данной заявке цепью А и цепью В), при этом первый полипептид включает модификации аминокислот L351Y, F405A и Y407V и второй полипептид включает модификацию аминокислоты T394W. В одном аспекте, модифицированный домен СН3 дополнительно включает точечные мутации в положениях F405 и/или K392. Данные мутации в положении K392 включают, но не ограничены перечисленными, K392V, K392M, K392R, K392L, K392F или K392E. Данные мутации в положении F405 включают, но не ограничены перечисленными, F405I, F405M, F405S, F405S, F405V или F405W. В другом аспекте, модифицированный домен СН3 дополнительно включает точечные мутации в положениях Т411 и/или S400. Данные мутации в положении Т411 включают, но не ограничены перечисленными, T411N, T411R, T411Q, T411K, T411D, Т411Е или T411W. Данные мутации в положении S400 включают, но не ограничены перечисленными, S400E, S400D, S400R или S400K. В еще одном варианте реализации, модифицированный домен СН3 включает первый и второй полипептид, при этом первый полипептид включает модификации аминокислот L351Y, F405A и Y407V и второй полипептид включает модификацию аминокислоты T394W, и при этом первый и/или второй полипептид включает дополнительные модификации аминокислот в положениях Т366 и/или L368. Данные мутации в положении Т366 включают, но не ограничены перечисленными, Т366А, T366I, T366L, Т366М, T366Y, T366S, Т366С, T366V или T366W. В типичном варианте реализации, мутация аминокислоты в положении Т366 представляет собой T366I. В другом типичном варианте реализации, мутация аминокислоты в положении Т366 представляет собой T366L. Мутации в положении L368 включают, но не ограничены перечисленными, L368D, L368R, L368T, L368M, L368V, L368F, L368S и L368A.
В некоторых вариантах реализации, модифицированный домен СН3 включает первый и второй полипептид (также называемые в данной заявке цепью А и цепью В), при этом первый полипептид включает модификации аминокислот L351Y, F405A и Y407V и второй полипептид включает модификации аминокислот T366L и T394W. В другом варианте реализации, модифицированный домен СН3 включает первый и второй полипептид, при этом первый полипептид включает модификации аминокислот L351Y, F405A и Y407V и второй полипептид включает модификации аминокислот T366I и T394W.
В других конкретных вариантах реализации, модифицированный домен СН3 включает первый и второй полипептид (также называемые в данной заявке цепью А и цепью В), при этом первый полипептид включает модификации аминокислот L351Y, F405A и Y407V и второй полипептид включает модификации аминокислот T366L, K392M и T394W. В другом варианте реализации, модифицированный домен СН3 включает первый и второй полипептид, при этом первый полипептид включает модификации аминокислот L351Y, F405A и Y407V и второй полипептид включает модификации аминокислот T366I, K392M и T394W.
В еще одном варианте реализации, модифицированный домен СН3 включает первый и второй полипептид (также называемые в данной заявке цепью А и цепью В), при этом первый полипептид включает модификации аминокислот F405A и Y407V и второй полипептид включает модификации аминокислот T366L, K392M и T394W. В другом варианте реализации, модифицированный домен СН3 включает первый и второй полипептид, при этом первый полипептид включает модификации аминокислот F405A и Y407V и второй полипептид включает модификации аминокислот T366I, K392M и T394W.
В некоторых вариантах реализации, модифицированный домен СН3 включает первый и второй полипептид (также называемые в данной заявке цепью А и цепью В), при этом первый полипептид включает модификации аминокислот F405A и Y407V и второй полипептид включает модификации аминокислот T366L и T394W. В другом варианте реализации, модифицированный домен СНЗ включает первый и второй полипептид, при этом первый полипептид включает модификации аминокислот F405A и Y407V и второй полипептид включает модификации аминокислот T366I и T394W.
В типичном варианте реализации, в данной заявке предложены изолированные гетеромультимеры, включающие гетеродимерную Fc-область, при этом гетеродимерная Fc-область содержит модифицированный домен СН3, включающий мутации аминокислот, способствующие образованию гетеродимера с повышенной стабильностью, при этом модифицированный домен СН3 имеет температуру плавления (Tm), составляющую приблизительно 74°С или более. В другом варианте реализации, в данной заявке предложены изолированные гетеромультимеры, включающие гетеродимерную Fc-область, при этом гетеродимерная Fc-область содержит модифицированный домен СН3, включающий мутации аминокислот, способствующие образованию гетеродимера с повышенной стабильностью, при этом модифицированный домен СН3 имеет температуру плавления (Tm), составляющую приблизительно 74°С или более, и гетеродимер имеет чистоту, составляющую приблизительно 98% или более.
В некоторых вариантах реализации предложен изолированный гетеромультимер, включающий гетеродимерную Fc-область, при этом гетеродимерная Fc-область содержит модифицированный домен СН3, включающий мутации аминокислот, способствующие образованию гетеродимера с повышенной стабильностью, и при этом модифицированный домен СН3 имеет температуру плавления (Tm) более чем 70°С, и указанные модифицированные домены СН3 выбраны из Таблицы 6.
Мутации в ядре Каркаса 2 включают L351Y_Y407A/T366A_K409F. Каркас 2а включает мутации аминокислот L351Y_Y407A/T366V_K409F и Каркас 2b включает мутации аминокислот Y407A/T366A_K409F. См. Пример 6.
В некоторых вариантах реализации, модифицированный домен СН3 включает первый и второй полипептид (также называемые в данной заявке цепью А и цепью В), при этом первый полипептид включает модификации аминокислот L351Y и Y407A и второй полипептид включает модификации аминокислот Т366А и K409F. В одном аспекте, модифицированный домен СН3 дополнительно включает точечные мутации в положениях Т366, L351, и Y407. Данные мутации в положении Т366 включают, но не ограничены перечисленными, T366I, T366L, Т366М, T366Y, T366S, Т366С, T366V или T366W. В конкретном варианте реализации, мутация в положении Т366 представляет собой T366V. Мутации в положении L351 включают, но не ограничены перечисленными, L351I, L351D, L351R или L351F. Мутации в положении Y407 включают, но не ограничены перечисленными, Y407V или Y407S. См. варианты СН3 AZ63-AZ70 в Таблице 1 и Таблице 4 и в Примере 6.
В типичном варианте реализации, модифицированный домен СН3 включает первый и второй полипептид (также называемые в данной заявке цепью А и цепью В), при этом первый полипептид включает модификации аминокислот L351Y и Y407A и второй полипептид включает модификации аминокислот T366V и K409F.
В типичном варианте реализации, в данной заявке предложены изолированные гетеромультимеры, включающие гетеродимерную Fc-область, при этом гетеродимерная Fc-область содержит модифицированный домен СН3, включающий мутации аминокислот, способствующие образованию гетеродимера с повышенной стабильностью, при этом модифицированный домен СН3 имеет температуру плавления (Tm), составляющую приблизительно 75,5°С или более. В другом варианте реализации, в данной заявке предложены изолированные гетеромультимеры, включающие гетеродимерную Fc-область, при этом гетеродимерная Fc-область содержит модифицированный домен СН3, включающий мутации аминокислот, способствующие образованию гетеродимера с повышенной стабильностью, при этом модифицированный домен СН3 имеет температуру плавления (Tm), составляющую приблизительно 75°С или более, и гетеродимер имеет чистоту, составляющую приблизительно 90% или более.
В других некоторых вариантах реализации, модифицированный домен СН3 включает первый и второй полипептид (также называемые в данной заявке цепью А и цепью В), при этом первый полипептид включает модификации аминокислот L351Y и Y407A и второй полипептид включает модификации аминокислот Т366А и K409F, и при этом модифицированный домен СН3 включает одну или более модификаций аминокислот в положениях Т411, D399, S400, F405, N390 и/или K392. Данные мутации в положении D399 включают, но не ограничены перечисленными, D399R, D399W, D399Y или D399K. Мутации в положении Т411 включают, но не ограничены перечисленными, T411N, T411R, T411Q, Т411К, T411D, Т411Е или T411W. Мутации в положении S400 включают, но не ограничены перечисленными, S400E, S400D, S400R или S400K. Мутации в положении F405 включают, но не ограничены перечисленными, F405I, F405M, F405S, F405S, F405V или F405W. Мутации в положении N390 включают, но не ограничены перечисленными, N390R, N390K или N390D. Мутации в положении K392 включают, но не ограничены перечисленными, K392V, K392M, K392R, K392L, K392F или K392E. См. варианты СН3 AZ71-101 в Таблице 1 и Таблице 4 и в Примере 6.
В типичном варианте реализации, модифицированный домен СН3 включает первый и второй полипептид (также называемые в данной заявке цепью А и цепью В), при этом первый полипептид включает модификацию аминокислоты Y407A и второй полипептид включает модификации аминокислот Т366А и K409F. В одном аспекте, данный модифицированный домен СН3 дополнительно включает модификации аминокислот K392E, Т411Е, D399R и S400R. В дополнительном варианте реализации, модифицированный домен СН3 включает первый и второй полипептид, при этом первый полипептид включает модификации аминокислот D399R, S400R и Y407A и второй полипептид включает модификации аминокислот Т366А, K409F, K392E и Т411Е.
В типичном варианте реализации, в данной заявке предложены изолированные гетеромультимеры, включающие гетеродимерную Fc-область, при этом гетеродимерная Fc-область содержит модифицированный домен СН3, включающий мутации аминокислот, способствующие образованию гетеродимера с повышенной стабильностью, при этом модифицированный домен СН3 имеет температуру плавления (Tm), составляющую приблизительно 74°С или более. В другом варианте реализации, в данной заявке предложены изолированные гетеромультимеры, включающие гетеродимерную Fc-область, при этом гетеродимерная Fc-область содержит модифицированный домен СН3, включающий мутации аминокислот, способствующие образованию гетеродимера с повышенной стабильностью, при этом модифицированный домен СН3 имеет температуру плавления (Tm), составляющую приблизительно 74°С или более, и гетеродимер имеет чистоту, составляющую приблизительно 95% или более.
В некоторых вариантах реализации, в данной заявке предложены изолированные гетеромультимеры, включающие гетеродимерную Fc-область, при этом гетеродимерная Fc-область содержит модифицированный домен СН3, включающий мутации аминокислот, способствующие образованию гетеродимера с повышенной стабильностью, и при этом модифицированный домен СН3 имеет температуру плавления (Tm) более 70°С, и модифицированные домены СН3 выбраны из Таблицы 7.
Более того, данный новый способ разработки вариантов гетеродимера Fc с повышенной стабильностью и чистотой можно применять и к другим классам и изотипам Fc-областей. В некоторых вариантах реализации, Fc-область представляет собой Fc-область IgG человека. В дополнительных вариантах реализации, Fc-область IgG человека представляет собой Fc-область IgG1, IgG2, IgG3 или IgG4 человека. В некоторых вариантах реализации, Fc-области принадлежат иммуноглобулинам, выбранным из группы, состоящей из IgG, IgA, IgD, IgE и IgM. В некоторых вариантах реализации, IgG принадлежит к подтипу, выбранному из группы, состоящей из IgG1, IgG2a, IgG2b, IgG3 и IgG4.
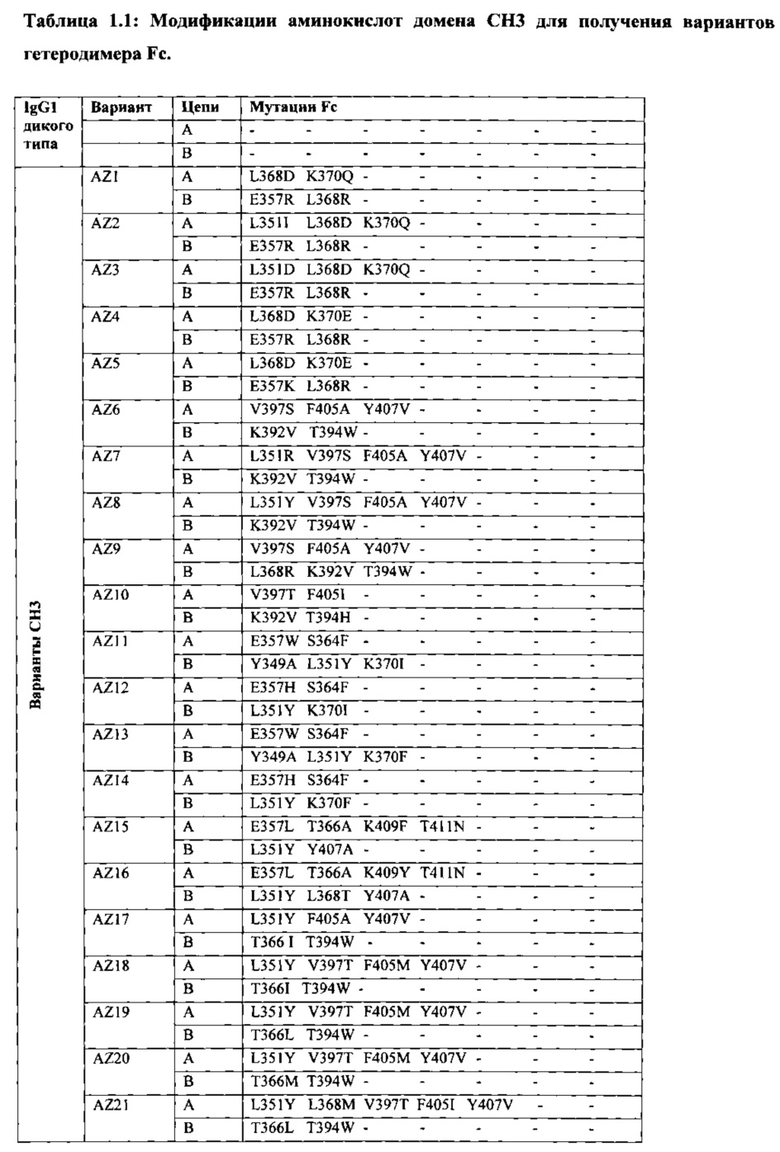
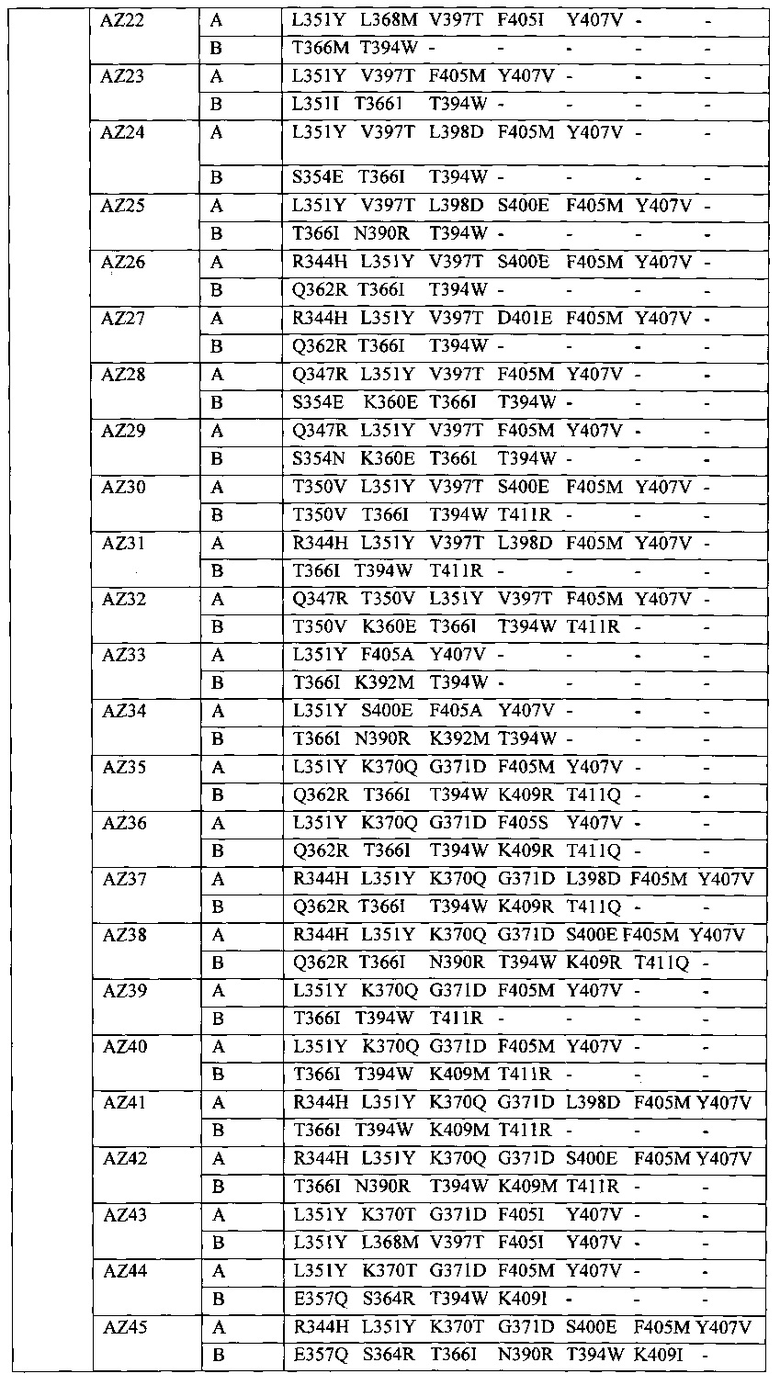
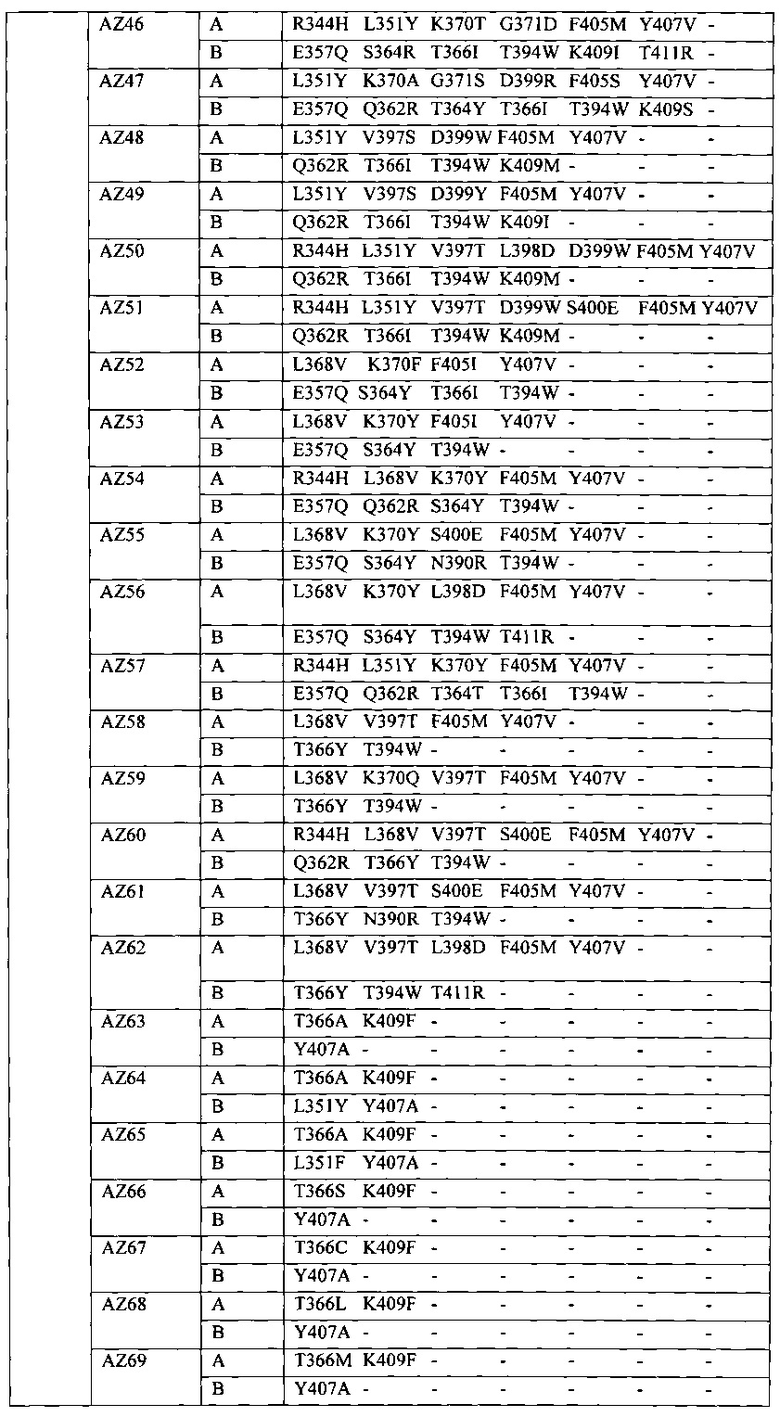
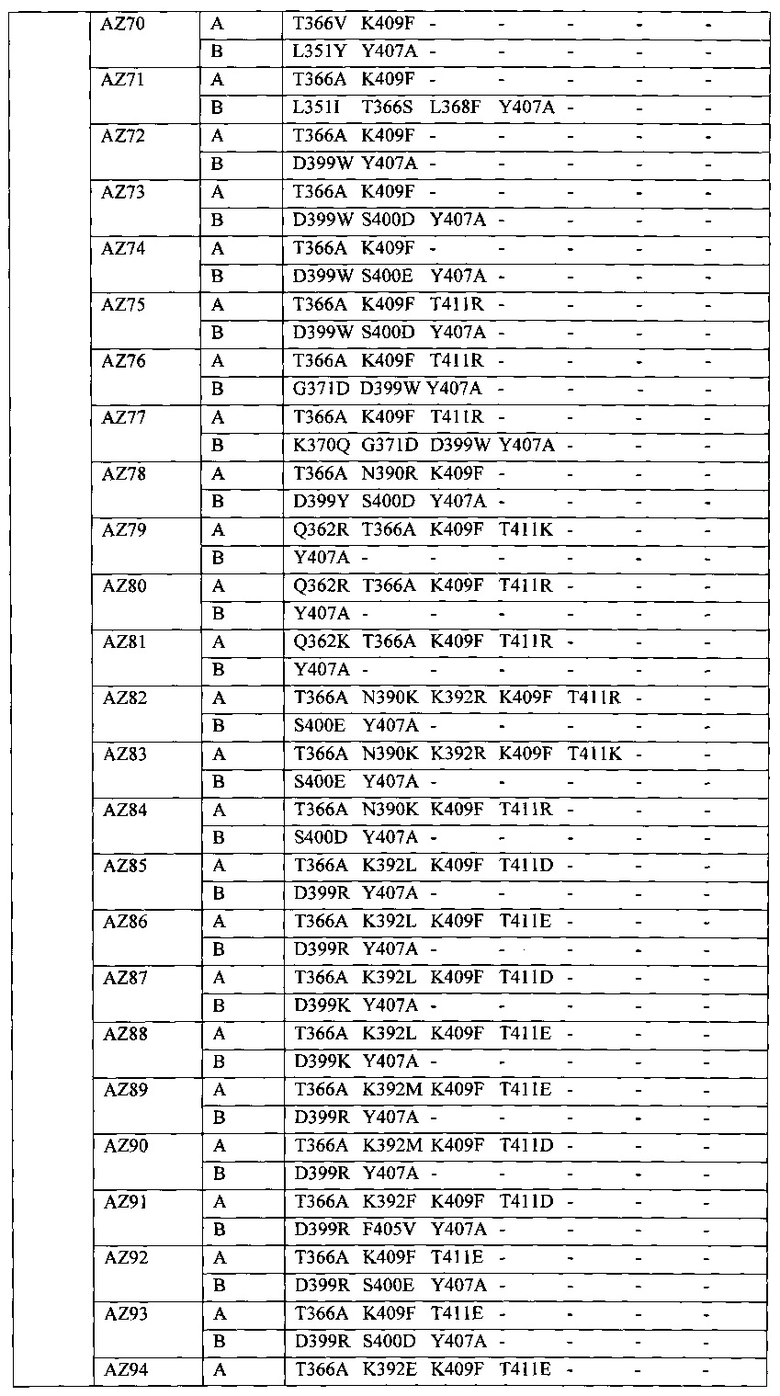
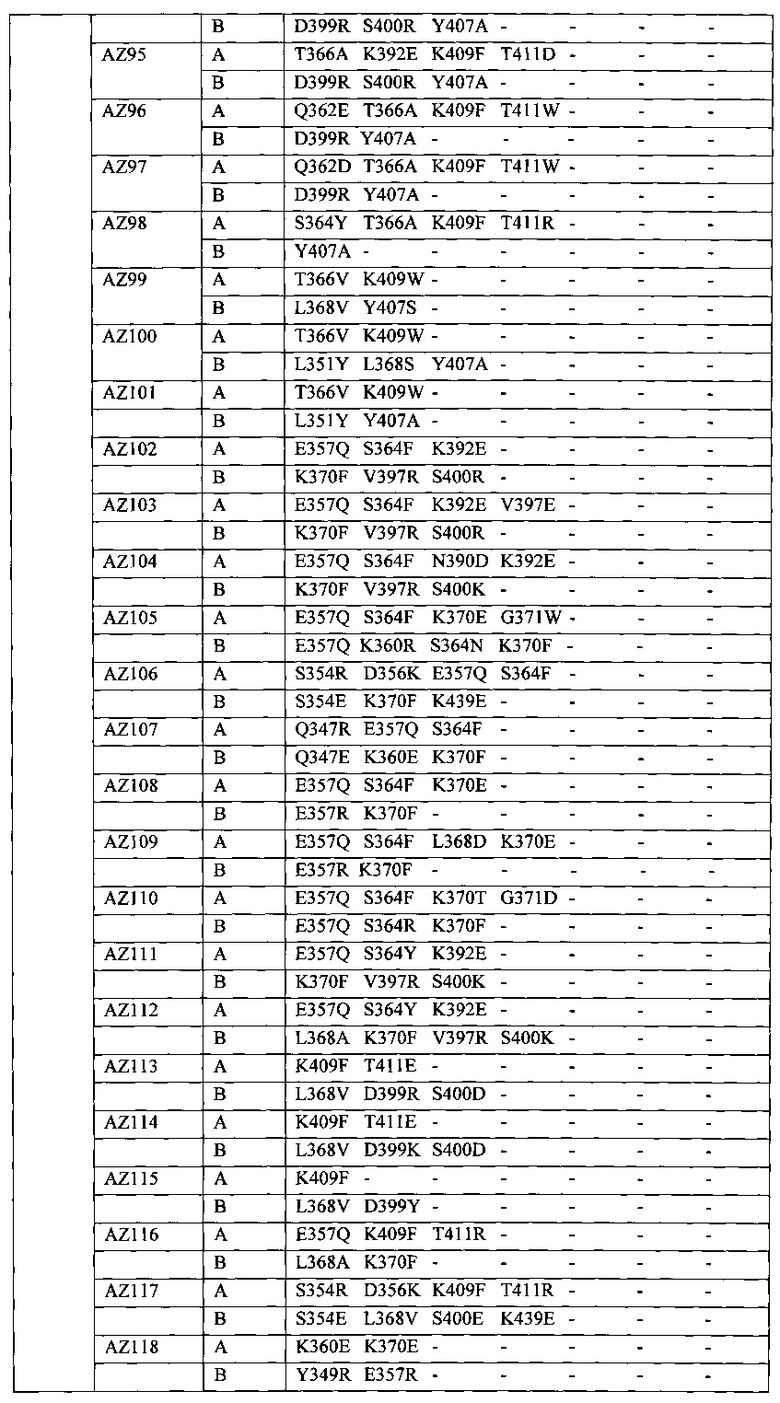
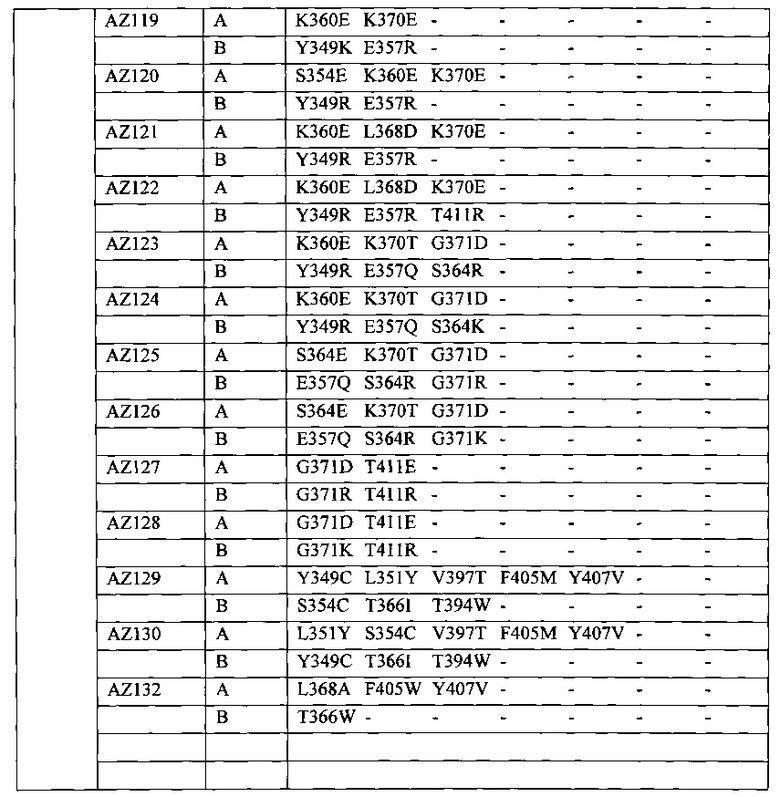
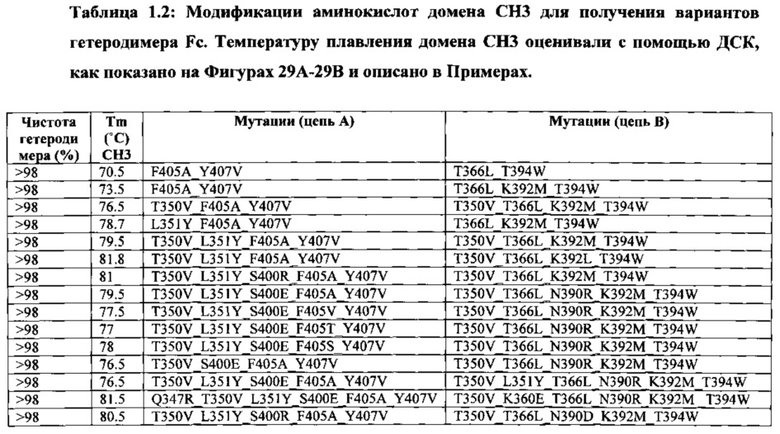

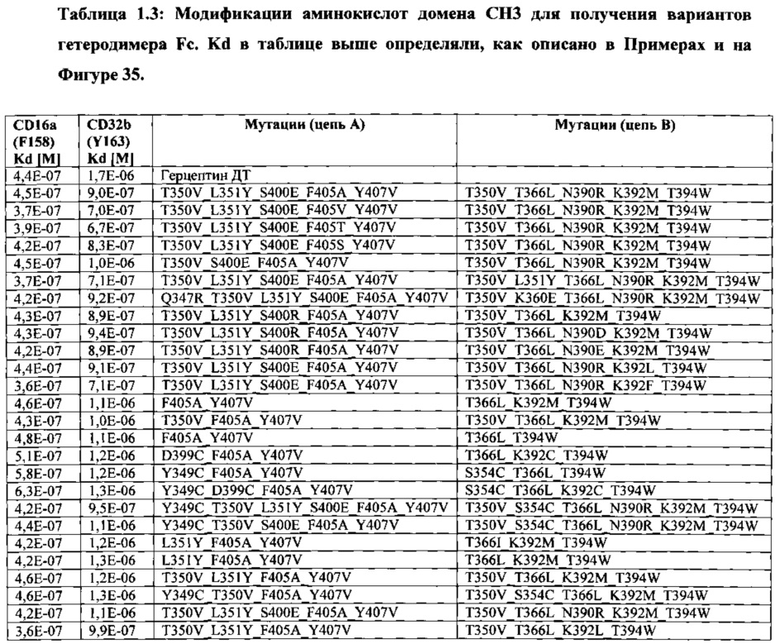
Fc-область, определенная в данной заявке, включает домен СН3 или его фрагмент, и может дополнительно включать один или более дополнительных доменов константной области, или их фрагментов, включая шарнирную область, СН1 или СН2. Очевидно, что нумерация аминокислотных остатков Fc соответствует индексу EU, описанному у Kabat и др., 1991, NIH Publication 91-3242, Национальная служба технической информации, Спрингфилд, Вирджиния. "Индекс EU, описанный у Kabat", относится к нумерации индекса EU по Kabat антитела IgG1 человека. Для удобства, в Таблице В представлены аминокислоты, пронумерованные согласно индексу EU, описанному у Kabat, домена СН2 и СН3 IgG1 человека.
Согласно одному аспекту настоящего изобретения предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный гетеродимерный домен СН3, где указанный модифицированный домен СН3 включает: первый модифицированный полипептид домена СН3, включающий по меньшей мере три модификации аминокислот по сравнению с полипептидом домена СН3 дикого типа, и второй модифицированный полипептид домена СН3, включающий по меньшей мере три модификации аминокислот по сравнению с полипептидом домена СН3 дикого типа; причем по меньшей мере один из указанных первого и второго полипептидов домена СН3 включает модификацию аминокислоты K392J, где J выбран из L, I или аминокислоты с объемом боковой цепи, значительно не превышающим объем боковой цепи K; при этом указанные первый и второй модифицированные полипептиды домена СН3 предпочтительно образуют гетеродимерный домен СН3 с температурой плавления (Tm), составляющей по меньшей мере приблизительно 74°С, и чистотой по меньшей мере 95%; и при этом по меньшей мере одна модификация аминокислоты не затрагивает аминокислоту, которая находится на границе между указанным первым и указанным вторым полипептидами домена СН3. В некоторых вариантах реализации предложена гетеромультимерная конструкция Fc, описанная в данной заявке, которая включает по меньшей мере одну модификацию Т350Х, где X представляет собой природную или неприродную аминокислоту, выбранную из валина, изолейцина, лейцина, метионина и производных или вариантов указанных аминокислот. В некоторых вариантах реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, которая включает по меньшей мере одну модификацию T350V. В одном варианте реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой модифицированный домен СН3 имеет температуру плавления (Tm), составляющую по меньшей мере приблизительно 75°С или более. В одном варианте реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой модифицированный домен СН3 имеет Tm, составляющую приблизительно 77°С или более. В некоторых вариантах реализации, модифицированный домен СН3 имеет Tm, составляющую приблизительно 80°С или более. В некоторых вариантах реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой по меньшей мере один полипептид домена СН3 представляет собой модифицированный полипептид домена СН3, включающий модификацию по меньшей мере одной из аминокислот L351, F405 и Y407. В некоторых вариантах реализации предложена изолированная гетеромультимерная конструкция Fc, в которой по меньшей мере один полипептид домена СН3 представляет собой модифицированный полипептид домена СН3, дополнительно включающий модификацию аминокислоты Т366. В некоторых вариантах реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой первый полипептид домена СН3 представляет собой модифицированный полипептид домена СН3, включающий модификации аминокислот в положениях L351, F405 и Y407, и второй полипептид домена СН3 представляет собой модифицированный полипептид домена СН3, включающий модификации аминокислот в положениях Т366, K392 и Т394. В одном варианте реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой указанный первый полипептид домена СН3 включает модификации аминокислот L351Y, F405A и Y407V, и указанный второй полипептид домена СН3 включает модификации аминокислот T366L, K392M и T394W. В некоторых вариантах реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой указанный первый полипептид домена СН3 включает модификации аминокислот L351Y, F405A и Y407V, и указанный второй полипептид домена СН3 включает модификации аминокислот T366L, K392L и T394W. В дополнительном варианте реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой указанный первый полипептид домена СН3 включает модификации аминокислот L351Y, F405A и Y407V, и указанный второй полипептид домена СН3 включает модификации аминокислот T366I, K392M и T394W. В некоторых вариантах реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой указанный первый полипептид домена СН3 включает модификации аминокислот L351Y, F405A и Y407V, и указанный второй полипептид домена СН3 включает модификации аминокислот T366I, K392L и T394W. В некоторых вариантах реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой по меньшей мере один из указанных первого и второго полипептидов домена СН3 представляет собой модифицированный полипептид домена СН3, включающий модификацию аминокислоты в положении S400. В дополнительном варианте реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, включающая модификацию S400Z, где Ζ выбран из положительно заряженной аминокислоты и отрицательно заряженной аминокислоты. В некоторых вариантах реализации, положительно заряженная аминокислота представляет собой лизин или аргинин и отрицательно заряженная аминокислота представляет собой аспарагиновую кислоту или глутаминовую кислоту. В некоторых вариантах реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой указанный первый полипептид домена СН3 включает модификацию аминокислоты, выбранную из S400E и S400R. В некоторых вариантах реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой по меньшей мере один из указанных первого и второго полипептидов домена СН3 представляет собой модифицированный полипептид домена СН3, включающий модификацию аминокислоты в положении N390. В некоторых вариантах реализации, модификация N390 представляет собой N390Z, где Ζ выбран из положительно заряженной аминокислоты и отрицательно заряженной аминокислоты. В одном варианте реализации, Ν390Ζ представляет собой N390R. В некоторых вариантах реализации изолированной гетеромультимерной конструкции Fc, описанной в данной заявке, указанный первый полипептид домена СН3 представляет собой модифицированный полипептид домена СН3, включающий модификацию аминокислоты S400E, и указанный второй полипептид домена СН3 представляет собой модифицированный полипептид домена СН3, включающий модификацию аминокислоты N390R. В некоторых вариантах реализации изолированной гетеромультимерной конструкции Fc, описанной в данной заявке, каждый из первого и второго полипептидов домена СН3 представляет собой модифицированный полипептид домена СН3, при этом один указанный модифицированный полипептид домена СН3 включает модификацию аминокислоты Q347R и другой модифицированный полипептид домена СН3 включает модификацию аминокислоты K360E.
В одном аспекте предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный гетеродимерный домен СН3, где указанный модифицированный домен СН3 включает: первый модифицированный полипептид домена СН3, включающий по меньшей мере три модификации аминокислот по сравнению с полипептидом домена СН3 дикого типа, и второй модифицированный полипептид домена СН3, включающий по меньшей мере три модификации аминокислот по сравнению с полипептидом домена СН3 дикого типа; причем по меньшей мере один из указанных первого и второго полипептидов домена СН3 включает модификацию аминокислоты K392J, где J выбран из L, I или аминокислоты с объемом боковой цепи, значительно не превышающим объем боковой цепи K; при этом указанные первый и второй модифицированные полипептиды домена СН3 предпочтительно образуют гетеродимерный домен СН3 с температурой плавления (Tm), составляющей по меньшей мере приблизительно 74°С, и чистотой по меньшей мере 95%; и при этом по меньшей мере одна модификация аминокислоты не затрагивает аминокислоту, которая находится на границе между указанным первым и указанным вторым полипептидами домена СН3. В некоторых вариантах реализации предложена гетеромультимерная конструкция Fc, описанная в данной заявке, которая включает по меньшей мере одну модификацию Т350Х, где X представляет собой природную или неприродную аминокислоту, выбранную из валина, изолейцина, лейцина, метионина и производных или вариантов указанных аминокислот. В некоторых вариантах реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, которая включает по меньшей мере одну модификацию T350V. В одном варианте реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой модифицированный домен СН3 имеет температуру плавления (Tm), составляющую по меньшей мере приблизительно 75°С или более. В одном варианте реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой модифицированный домен СН3 имеет Tm, составляющую приблизительно 77°С или более. В некоторых вариантах реализации, модифицированный домен СН3 имеет Tm, составляющую приблизительно 80°С или более. В одном варианте реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой по меньшей мере один полипептид домена СН3 представляет собой модифицированный полипептид домена СН3, включающий модификацию по меньшей мере одной из аминокислот K409 и Т411. В некоторых вариантах реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, включающая по меньшей мере одну из модификаций K409F, Т411Е и T411D. В некоторых вариантах реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой по меньшей мере один полипептид домена СН3 представляет собой модифицированный полипептид домена СН3, включающий модификацию аминокислоты D399. В некоторых вариантах реализации, модификация аминокислоты D399 представляет собой по меньшей мере одну из модификаций D399R и D399K.
В одном аспекте предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный гетеродимерный домен СН3, где указанный модифицированный домен СН3 включает: первый модифицированный полипептид домена СН3, включающий по меньшей мере три модификации аминокислот по сравнению с полипептидом домена СН3 дикого типа, и второй модифицированный полипептид домена СН3, включающий по меньшей мере три модификации аминокислот по сравнению с полипептидом домена СН3 дикого типа; причем по меньшей мере один из указанных первого и второго полипептидов домена СН3 включает модификацию аминокислоты K392J, где J выбран из L, I или аминокислоты с объемом боковой цепи, значительно не превышающим объем боковой цепи K; при этом указанные первый и второй модифицированные полипептиды домена СН3 предпочтительно образуют гетеродимерный домен СН3 с температурой плавления (Tm), составляющей по меньшей мере приблизительно 74°С, и чистотой по меньшей мере 95%; и при этом по меньшей мере одна модификация аминокислоты не затрагивает аминокислоту, которая находится на границе между указанным первым и указанным вторым полипептидами домена СН3. В некоторых вариантах реализации предложена гетеромультимерная конструкция Fc, описанная в данной заявке, которая включает по меньшей мере одну модификацию Т350Х, где X представляет собой природную или неприродную аминокислоту, выбранную из валина, изолейцина, лейцина, метионина и производных или вариантов указанных аминокислот. В некоторых вариантах реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, которая включает по меньшей мере одну модификацию T350V. В одном варианте реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой модифицированный домен СН3 имеет температуру плавления (Tm), составляющую по меньшей мере приблизительно 75°С или более. В одном варианте реализации предложена изолированная гетеромультимерная конструкция Fc, описанная в данной заявке, в которой модифицированный домен СН3 имеет Tm, составляющую приблизительно 77°С или более. В некоторых вариантах реализации, модифицированный домен СН3 имеет Tm, составляющую приблизительно 80°С или более. В некоторых вариантах реализации изолированной гетеромультимерной конструкции Fc, описанной в данной заявке, первый полипептид домена СН3 представляет собой модифицированный полипептид домена СН3, включающий по меньшей мере одну модификацию аминокислоты, выбранную из K409F, Т411Е и T411D, и второй полипептид домена СН3 представляет собой модифицированный полипептид домена СН3, включающий по меньшей мере одну модификацию аминокислоты, выбранную из Y407A, Y407I, Y407V, D399R и D399K. В некоторых вариантах реализации предложена любая из изолированных гетеромультимерных конструкций Fe, описанных в данной заявке, дополнительно содержащая первый модифицированный домен СН3, включающий одну из модификаций аминокислот T366V, T366I, Т366А, Т366М и T366L; и второй модифицированный домен СН3, включающий модификацию аминокислоты L351Y. В некоторых вариантах реализации предложена любая из изолированных гетеромультимерных конструкций Fc, описанных в данной заявке, содержащая первый модифицированный домен СН3, включающий одну из модификаций аминокислот K392L или K392E; и второй модифицированный домен СН3, включающий одну из модификаций аминокислот S400R или S400V.
В данной заявке предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный домен СН3, содержащий первый модифицированный полипептид домена СН3 и второй модифицированный полипептид домена СН3, при этом каждый модифицированный полипептид домена СН3 включает по меньшей мере четыре мутации аминокислот, при этом по меньшей мере один из указанного первого и указанного второго модифицированных полипептидов домена СН3 включает мутацию, выбранную из N390Z и S400Z, где Ζ выбран из положительно заряженной аминокислоты и отрицательно заряженной аминокислоты, и причем указанные первый и второй модифицированные полипептиды домена СН3 предпочтительно образуют гетеродимерный домен СН3 с температурой плавления (Tm), составляющей по меньшей мере приблизительно 70°С, и чистотой по меньшей мере 90%. В одном варианте реализации предложена изолированная гетеромультимерная конструкция Fc, в которой указанный первый модифицированный полипептид домена СН3 включает модификации аминокислот в положениях F405 и Y407 и указанный второй модифицированный полипептид домена СН3 включает модификацию аминокислоты в положении Т394. В одном варианте реализации предложена изолированная гетеромультимерная конструкция Fc, в которой первый модифицированный полипептид домена СН3 включает модификацию аминокислоты в положении L351. В некоторых вариантах реализации предложен изолированный гетеромультимер, описанный в данной заявке, в котором указанный второй модифицированный полипептид домена СН3 включает модификацию в по меньшей мере одном из положений Т366 и K392. В некоторых вариантах реализации предложен изолированный гетеромультимер, описанный в данной заявке, в котором модифицированный домен СН3 имеет температуру плавления (Tm), составляющую по меньшей мере приблизительно 75°С, и получен с чистотой по меньшей мере приблизительно 95%. В некоторых вариантах реализации предложен изолированный гетеромультимер, описанный в данной заявке, в котором по меньшей мере один модифицированный полипептид домена СН3 включает по меньшей мере одну из модификаций аминокислот N390R, S400E и S400R. В некоторых вариантах реализации предложен изолированный гетеромультимер, описанный в данной заявке, в котором один из указанных первого и второго модифицированных полипептидов домена СН3 включает модификацию аминокислоты в положении 347 и другой модифицированный полипептид домена СН3 включает модификацию аминокислоты в положении 360. В некоторых вариантах реализации предложен изолированный гетеромультимер, описанный в данной заявке, в котором по меньшей мере один из указанных первого и второго модифицированных полипептидов домена СН3 включает модификацию аминокислоты T350V. В конкретных вариантах реализации предложен изолированный гетеромультимер, описанный в данной заявке, в котором указанный первый модифицированный полипептид домена СН3 включает по меньшей мере одну модификацию аминокислоты, выбранную из L351Y, F405A и Y407V; и указанный второй модифицированный полипептид домена СН3 включает по меньшей мере одну модификацию аминокислоты, выбранную из T366L, T366I, K392L, K392M и T394W. В некоторых вариантах реализации, описанных в данной заявке, предложен изолированный гетеромультимер, в котором первый модифицированный полипептид домена СН3 включает модификации аминокислот в положениях D399 и Y407 и второй модифицированный полипептид домена СН3 включает модификации аминокислот в положениях K409 и Т411. В некоторых вариантах реализации предложен изолированный гетеромультимер, описанный в данной заявке, в котором первый полипептид домена СН3 включает модификацию аминокислоты в положении L351 и второй модифицированный полипептид домена СН3 включает модификации аминокислот в положениях Т366 и K392. В конкретных вариантах реализации предложены изолированные гетеромультимеры, описанные в данной заявке, в которых по меньшей мере один из указанных первого и второго полипептидов домена СН3 включает модификацию аминокислоты T350V. В некоторых вариантах реализации предложены изолированные гетеромультимеры, описанные в данной заявке, в которых модифицированный домен СН3 имеет температуру плавления (Tm), составляющую по меньшей мере приблизительно 750С или более, и получен с чистотой по меньшей мере приблизительно 95%.
В некоторых вариантах реализации предложены изолированные гетеромультимерные конструкции Fc, описанные в данной заявке, в которых указанный первый модифицированный полипептид домена СН3 включает модификации аминокислот, выбранные из L351Y, D399R, D399K, S400D, S400E, S400R, S400K, Y407A и Y407V; и указанный второй модифицированный полипептид домена СН3 включает модификации аминокислот, выбранные из T366V, T366I, T366L, Т366М, N390D, N390E, K392L, K392I, K392D, K392E, K409F, K409W, T411D и Т411Е.
В данной заявке предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный домен СН3, содержащий первый модифицированный полипептид домена СН3 и второй модифицированный полипептид домена СН3, причем каждый модифицированный полипептид домена СН3 включает мутации по меньшей мере трех аминокислот, при этом один из указанного первого и указанного второго модифицированных полипептидов домена СН3 включает мутацию, выбранную из Т411Е и T411D, и при этом указанные первый и второй модифицированные полипептиды домена СН3 предпочтительно образуют гетеродимерный домен СН3 с температурой плавления (Tm), составляющей по меньшей мере приблизительно 70°С, и чистотой по меньшей мере 90%. В одном варианте реализации предложена изолированная гетеромультимерная конструкция Fc, в которой указанный первый модифицированный полипептид домена СН3 включает модификации аминокислот в положениях F405 и Y407 и указанный второй модифицированный полипептид домена СН3 включает модификацию аминокислоты в положении Т394. В одном варианте реализации предложена изолированная гетеромультимерная конструкция Fc, в которой первый модифицированный полипептид домена СН3 включает модификацию аминокислоты в положении L351. В некоторых вариантах реализации предложен изолированный гетеромультимер, описанный в данной заявке, в котором указанный второй модифицированный полипептид домена СН3 включает модификацию в по меньшей мере одном из положений Т366 и K392. В некоторых вариантах реализации предложен изолированный гетеромультимер, описанный в данной заявке, в котором модифицированный домен СН3 имеет температуру плавления (Tm), составляющую по меньшей мере приблизительно 75°С, и получен с чистотой по меньшей мере приблизительно 95%. В некоторых вариантах реализации предложен изолированный гетеромультимер, описанный в данной заявке, в котором по меньшей мере один модифицированный полипептид домена СН3 включает по меньшей мере одну из модификаций аминокислот N390R, S400E и S400R. В некоторых вариантах реализации предложен изолированный гетеромультимер, описанный в данной заявке, в котором один из указанных первого и второго модифицированных полипептидов домена СН3 включает модификацию аминокислоты в положении 347 и другой модифицированный полипептид домена СН3 включает модификацию аминокислоты в положении 360. В некоторых вариантах реализации предложен изолированный гетеромультимер, описанный в данной заявке, в котором по меньшей мере один из указанных первого и второго модифицированных полипептидов домена СН3 включает модификацию аминокислоты T350V. В конкретных вариантах реализации предложен изолированный гетеромультимер, описанный в данной заявке, в котором указанный первый модифицированный полипептид домена СН3 включает по меньшей мере одну модификацию аминокислоты, выбранную из L351Y, F405A и Y407V; и указанный второй модифицированный полипептид домена СН3 включает по меньшей мере одну модификацию аминокислоты, выбранную из T366L, T366I, K392L, K392M и T394W. В некоторых вариантах реализации, описанных в данной заявке, предложен изолированный гетеромультимер, в котором первый модифицированный полипептид домена СН3 включает модификации аминокислот в положениях D399 и Y407 и второй модифицированный полипептид домена СН3 включает модификации аминокислот в положениях K409 и Т411. В некоторых вариантах реализации предложен изолированный гетеромультимер, описанный в данной заявке, в котором первый полипептид домена СН3 включает модификацию аминокислоты в положении L351 и второй модифицированный полипептид домена СН3 включает модификации аминокислот в положениях Т366 и K392. В конкретных вариантах реализации предложены изолированные гетеромультимеры, описанные в данной заявке, в которых по меньшей мере один из указанных первого и второго полипептидов домена СН3 включает модификацию аминокислоты T350V. В некоторых вариантах реализации предложены изолированные гетеромультимеры, описанные в данной заявке, в которых модифицированный домен СН3 имеет температуру плавления (Tm), составляющую по меньшей мере приблизительно 750С или более, и получен с чистотой по меньшей мере приблизительно 95%.
В некоторых вариантах реализации предложены изолированные гетеромультимерные конструкции Fc, описанные в данной заявке, в которых указанный первый модифицированный полипептид домена СН3 включает модификации аминокислот, выбранные из L351Y, D399R, D399K, S400D, S400E, S400R, S400K, Y407A и Y407V; и указанный второй модифицированный полипептид домена СН3 включает модификации аминокислот, выбранные из T366V, T366I, T366L, Т366М, N390D, N390E, K392L, K392I, K392D, K392E, K409F, K409W, T411D и Т411Е.
В данной заявке предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный домен СН3, содержащий первый модифицированный полипептид домена СН3, включающий модификации аминокислот L351Y, F405A и Y407V; и второй модифицированный полипептид домена СН3, включающий модификации аминокислот T366I, K392M и T394W.
В некотором аспекте предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный домен СН3, содержащий первый модифицированный полипептид домена СН3, включающий модификации аминокислот L351Y, F405A и Y407V; и второй модифицированный полипептид домена СН3, включающий модификации аминокислот T366I, K392L и T394W.
В некотором аспекте предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный домен СН3, содержащий первый модифицированный полипептид домена СН3, включающий модификации аминокислот L351Y, F405A и Y407V; и второй модифицированный полипептид домена СН3, включающий модификации аминокислот T366L, K392M и T394W.
В некоторых аспектах предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный домен СН3, содержащий первый модифицированный полипептид домена СН3, включающий модификации аминокислот L351Y, F405A и Y407V; и второй модифицированный полипептид домена СН3, включающий модификации аминокислот T366L, K392L и T394W.
В некотором аспекте предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный домен СН3, содержащий первый модифицированный полипептид домена СН3, включающий модификации аминокислот T350V, L351Y, F405A и Y407V; и второй модифицированный полипептид домена СН3, включающий модификации аминокислот T350V, T366L, K392L и T394W.
В некотором аспекте предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный домен СН3, содержащий первый модифицированный полипептид домена СН3, включающий модификации аминокислот T350V, L351Y, S400R, F405A, Y407V; и второй модифицированный полипептид домена СН3, включающий модификации аминокислот T350V, T366L, K392M и T394W.
В некотором аспекте предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный домен СН3, содержащий первый модифицированный полипептид домена СН3, включающий модификации аминокислот T350V, L351Y, S400E, F405A, Y407V; и второй модифицированный полипептид домена СН3, включающий модификации аминокислот T350V, T366L, N390R, K392M и T394W.
В некотором аспекте предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный домен СН3, содержащий первый модифицированный полипептид домена СН3, включающий модификации аминокислот T350V, L351Y, F405A, Y407V; и второй модифицированный полипептид домена СН3, включающий модификации аминокислот T350V, T366L, K392L и T394W.
В одном аспекте предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный домен СН3, содержащий первый модифицированный полипептид домена СН3, включающий модификации аминокислот T366V, K392L, K409F и Т411Е; и второй модифицированный полипептид домена СН3, включающий модификации аминокислот L351Y, D399R и Y407A.
В одном аспекте предложена изолированная гетеромультимерная конструкция Fc, содержащая модифицированный домен СН3, содержащий первый модифицированный полипептид домена СН3, включающий модификации аминокислот T366V, K392LE K409F и Т411Е; и второй модифицированный полипептид домена СН3, включающий модификации аминокислот L351Y, D399R, S400R и Y407A.
В некоторых вариантах реализации, вариант Fc содержит домен СН2. В некоторых вариантах реализации, домен СН2 представляет собой вариант домена СН2. В некоторых вариантах реализации, варианты домена СН2 включают асимметричные замены аминокислот в первой и/или второй полипептидной цепи. В некоторых вариантах реализации, гетеромультимер включает такие асимметричные замены аминокислот в домене СН2, что одна цепь указанного гетеромультимера селективно связывает рецептор Fc.
В некоторых вариантах реализации, гетеромультимер селективно связывает рецептор Fc. В некоторых вариантах реализации, рецептор Fc представляет собой член семейства рецепторов Feγ. В некоторых вариантах реализации, рецептор выбирают из FcγRI, FcγRIIa, FcγRIIb, FcγRIIc, FcγRIIIa и FcγRIIIb. В одном варианте реализации, домен СН2 включает асимметричные модификации аминокислот, которые способствуют селективному связыванию рецепторов Fc-гамма.
В некоторых вариантах реализации, гетеромультимер селективно связывается с FcγRIIIa. В некоторых вариантах реализации, гетеромультимер включает асимметричные замены аминокислот, выбранные из S267D, K392D и K409D. В некоторых вариантах реализации, гетеромультимер селективно связывается с FcγRIIa. В некоторых вариантах реализации, гетеромультимер включает асимметричные замены аминокислот, выбранные из S239D, K326E, A330L и I332E. В некоторых вариантах реализации, гетеромультимер селективно связывается с FcγRIIb. В некоторых вариантах реализации, гетеромультимер включает асимметричные замены аминокислот, выбранные из S239D, D265S, E269K и I332E. В некоторых вариантах реализации, гетеромультимер селективно связывается с FcγRIIIa и FcγRIIa. В некоторых вариантах реализации, гетеромультимер включает асимметричные замены аминокислот, выбранные из S239D, D265S и S298A. В некоторых вариантах реализации, гетеромультимер селективно связывается с FcγRIIIa и FcγRIIb. В некоторых вариантах реализации, гетеромультимер включает асимметричные замены аминокислот, выбранные из S239D, S298A, K326E, A330L и I332E. В некоторых вариантах реализации, гетеромультимер селективно связывается с FcγRIIa и FcγRIIb. В некоторых вариантах реализации, гетеромультимер включает асимметричные замены аминокислот, выбранные из S239D, D265S, S298A и I332E.
В некоторых вариантах реализации предложен способ разработки многофункционального лекарства, содержащего гетеромультимер, описанный в данной заявке. В некоторых вариантах реализации предложен способ разработки бифункционального лекарства, содержащего вариант гетеродимера Fc. В некоторых вариантах реализации предложен способ разработки асимметричных мутаций в домене СН2 варианта гетеродимера Fc, полученного путем введения мутаций в домен СН3. В некоторых вариантах реализации предложен способ разработки селективности к различным рецепторам Fc-гамма на основании мутаций в асимметричном Fc. В некоторых вариантах реализации предложен способ разработки мутаций, которые вызывают связывание рецепторов Fc-гамма с одной стороной молекулы Fc. В некоторых вариантах реализации предложен способ разработки факторов полярности, которые вызывают взаимодействие рецепторов Fcγ только с одной стороной асимметричного каркаса Fc гетеромультимера, описанного в данной заявке.
В некоторых вариантах реализации, предложен полипептид, включающий мутации в домене СН2 асимметричного Fc, которые приводят к предпочтительным профилям селективности в отношении рецептора Fc-гамма. В некоторых вариантах реализации, мутации в домене СН3 приводят к предпочтительному образованию гетеродимерного Fc. В некоторых вариантах реализации предложен способ разработки биспецифических терапевтических соединений на основе асимметричного Fc, описанного в данной заявке. В некоторых вариантах реализации предложен способ разработки мультиспецифичных терапевтических соединений на основе асимметричного Fc, описанного в данной заявке.
Моноклональные антитела, такие как IgG, представляют собой симметричные молекулы, состоящие из двух эквивалентных тяжелых и двух легких полипептидных цепей (Фигура 14), каждая из которых содержит несколько структурных доменов иммуноглобулина (Ig). Класс IgG МАТ существует в четырех изоформах: IgG1, IgG2, IgG3 или IgG4. Тяжелая цепь состоит из четырех (VH, CH1, СН2 и СН3), а легкая цепь состоит из двух (VL и CL) доменов Ig, соответственно. Домены VH и СН1 каждой из тяжелых цепей соединяются с доменами VL и CL легкой цепи с образованием двух Fab ("антигенсвязывающих фрагментов") плеч МАТ. Домены СН3 и СН2 двух тяжелых цепей взаимодействуют посредством белок-белковых контактов вдоль доменов СН3 и гликозилирования в доменах СН2 с образованием гомодимерной области Fc ("кристаллизуемый фрагмент"). Линкерная область между доменами СН1 и СН2 антитела образует шарнирную область молекулы антитела. Помимо соединения областей Fab и Fc МАТ, шарнир также сохраняет дисульфидные связи вдоль двух тяжелых цепей и скрепляет их. Количество аминокислот и дисульфидных связей в шарнирной области значительно отличается среди четырех изотипов IgG. Паттерн гликозилирования молекул IgG может значительно отличаться, на молекулах IgG можно наблюдать приблизительно 30 различных молекул углевода [Arnold J.N.; Wormald M.R.; Sim R.B.; Rudd P.M. и Dwek R.A. (2007) Annual Reviews of Immunology 25, 21-50].
Симметричная природа структуры моноклональньгх антител приводит к тому, что оба плеча Fab обладают способностью связывать антиген с аффинностью, созревшей для узнавания одного и того же эпитопа. С другой стороны, Fc-область молекулы антитела участвует во взаимодействиях с различными рецепторными молекулами на поверхности иммунных или "эффекторных" клеток, и некоторые из данных взаимодействий обуславливают опосредование эффекторных функций, таких как антителозависимая клеточная цитотоксичность (АЗКЦ), антителозависимый клеточный фагоцитоз (АЗКФ) и активация комплемента. Как правило, эффекторная функция вызывает иммунные ответы, что приводит к нейтрализации и элиминированию патогенов или токсинов, активации комплемента и фагоцитарному ответу гуморальной иммунной системы. Молекулы рецептора Fcγ (FcγR) на поверхности эффекторных клеток контактируют с Fc активированного антитела IgG в составе интегрального иммунного комплекса антитело-антиген, чтобы опосредовать и регулировать эффекторный ответ. Оптимизация взаимодействия белковых терапевтических агентов на основе моноклональных антител с данными рецепторами Fcγ может приводить к улучшению эффективности данных кандидатных лекарственных средств.
У людей существует три известных класса FcγR с дополнительными полиморфными типами в каждом классе. Известно, что Fc в молекуле IgG1 связывает FcγRI (CD64) с константами диссоциации в наномолярном диапазоне, тогда как связывание FcγRII (CD32) и FcγRIII (CD 16) происходит с константами диссоциации в микромолярном диапазоне [Bruhns P.; Iannascoli В.; England P.; Mancardi D.A.; Fernandez Ν.; Jorieux S. и Daeron M. (2009) Blood 113: 3716-25]. Высокоаффинные рецепторы FcγRI могут связывать IgG в мономерных формах, тогда как низкоаффинные рецепторы FcγRII и FcγRIII могут связывать только иммунные комплексы антиген-антитело или агрегаты IgG благодаря эффектам авидности. Различные формы IgG обладают различными аффинностями к различным FcγR; в частности, IgG1 и IgG3 проявляют более сильную активность. Рецепторы Fcγ представляют собой внеклеточные домены трансмембранных белков, которые также содержат цитоплазматические домены, участвующие в регуляции путей передачи сигналов в клетке. При кластеризации на поверхности иммунной клетки в результате ассоциации с образованными антителом иммунными комплексами, в зависимости от природы участвующих в передаче сигналов соединений, связанных с FcγR на цитоплазматической стороне данных рецепторов клеточной поверхности, эти молекулы регулируют эффекторный ответ [Nimmerjahn F. и Ravetch J.V. (2008) Nature Immu Rev 8(1):34-47].
На уровне хромосом человека, три гена кодируют FcγRI (FcγRIA, FcγRIB, FcγRIC) и FcγRII (FcγRIIA, FcγRIIB, FcγRIIC) и два гена кодируют FcγRIII (FcγRIIIA, FcγRIIIB). Было показано, что среди рецепторов Fcγ человека, связывающих IgG, типы рецепторов FcγRIA, FcγRIC и FcγRIIIA связаны с мембраной через общий сигнальный адаптерный белок с γ-цепью, который содержит цитоплазматический иммунорецепторный тирозиновый активирующий мотив (ITAM), который приводит к активации эффекторной функции. FcγRIIA и FcγRIIC также содержат цитоплазматический ITAM, но без общего сигнального адаптерного белка с γ-цепью. В то же время, FcγRIIB связан с иммунорецепторным тирозиновым ингибирующим мотивом (ITIM). Активация FcγRIIB, приводящая к фосфорилированию ITIM, приводит к ингибированию активации каскада передачи сигналов. FcγRIIIB, несмотря на то, что у него нет ни одного из тирозиновых иммуномодулирующих цитоплазматических хвостов, содержит гликозилфосфатидилинозитольный (GPI) якорь, и было показано, что он способствует активации некоторых гранулоцитов в присутствии FcγRIIA.
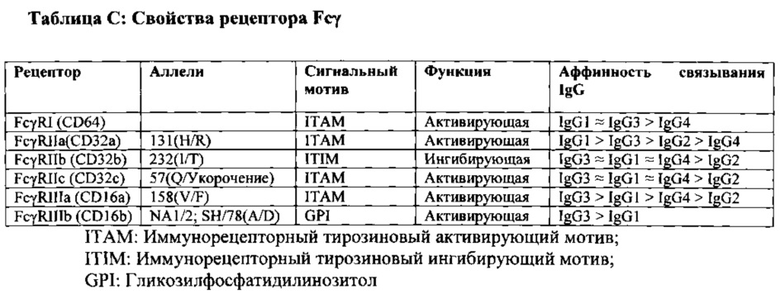
Несмотря на то, что функциональная роль мотивов ITAM и ITIM и связанных с ними молекул рецепторов известна, природа и механизмы модуляции передачи сигналов в комбинации не вполне понятны, особенно в комбинации с активностью других рецепторов на поверхности иммунных клеток и адаптерных молекул хозяина (например, BCR, CD22, CD45 и т.д.), вовлеченных в передачу сигнала. В данном контексте, разработка подобных Fc молекул, которые могут взаимодействовать с данными рецепторами Fcγ с совершенными профилями селективности, является полезной платформой для любой попытки развернуть и модулировать действие таких рецепторных молекул со слабовыраженными регуляторными активностями.
В контексте разработки молекул антител, которые могут различать рецепторы Fcγ, работа усложняется тем фактом, что внеклеточные связывающие Fc участки рецепторов типа FcγRII и FcγRIII обладают высоким подобием последовательностей (Фигура 15), что может объясняться по меньшей мере отчасти дупликациями фрагментов предкового гена. Два основных типа рецепторов FcγRII - А и В - обладают идентичностью последовательностей 69%, тогда как FcγRIIA и FcγRIIIA обладают идентичностью последовательностей приблизительно 44%. FcγRIIB и FcγRIIC отличаются только 2 остатками во внеклеточном участке, но при этом они значительно отличаются во внутриклеточном участке, стоит отметить, что в них присутствуют мотивы ITIM и ITAM, соответственно. В результате можно ожидать, что терапевтические молекулы антител, которые должны связывать один рецептор, также потенциально могут связываться с другими классами рецепторов, что, возможно, может приводить к неожиданным терапевтическим действиям.
Еще больше осложняет дело то, что в каждом из классов рецепторов присутствует множество однонуклеотидных полиморфизмов (SNP) и вариаций числа копий (CNV). Возникшее в результате этого разнообразие рецепторов различным образом влияет на их аффинность к IgG и механизм действия. Данные генетические вариации могут влиять на аффинность конкретных подклассов IgG к рецепторам Fcγ, изменять нижележащие эффекторные события или влиять на механизмы, которые изменяют уровни экспрессии рецептора, что приводит к образованию соответствующих по функциям фенотипов, нефункциональных вариантов рецептора или вариантов рецептора с неизвестной функцией (Bournazos S.; Woof J.M.; Hart S.P. и Dransfield I. (2009) Clinical and Experimental Immunology 157(2):244-54). Они потенциально могут приводить к сложным эффектам, изменяющим равновесие между активирующей и ингибирующей передачей сигналов от рецептора, что приводит к созданию восприимчивых к заболеванию фенотипов.
Некоторые из данных аллельных вариантов перечислены в Таблице С. Стоит отметить, что вариант FcγRIIa с R131 проявляет выраженный ответ на IgG1, в то время как альтернативные варианты H131 проявили более эффективные взаимодействия с IgG2 и IgG3. В случае FcγRIIIa, у доноров, гомозиготных по V в положении 158, наблюдалась повышенная активность клеток NK по сравнению с гомозиготными F/F158 индивидами, вследствие высокой аффинности первого аллотипа к IgG1, IgG3 и IgG4 человека. Аллельные варианты NA1 и NA2 FcγRIIIb образовались в результате четырех замен аминокислот, что, в свою очередь, привело к различиям в гликозилировании рецептора. Аллель NA1 вызывает повышенное связывание с нейтрофилами и фагоцитоз ими иммунного комплекса. У FcγRIIB есть два известных аллельных варианта - 232I и 232Т. Известно, что у варианта 232Т сильно нарушена отрицательная регуляторная активность. Были описаны частоты полиморфизмов FcγR и их связь с различной восприимчивостью к инфекциям или предрасположенностью к заболеваниям, таким как системная красная волчанка (СКВ), ревматоидный артрит (РА), васкулит, идиопатическая (аутоиммунная) тромбоцитопеническая пурпура (ИТП), миастения гравис, множественный склероз (МС) и иммунные нейропатии (синдром Гийена-Барре (СГБ)).
Была продемонстрирована вариация числа копий в локусе генов FcγR, в частности, для FcγRIIIB, FcγRIIC и FcγRIIIA, и была отмечена дополнительная корреляция этих различий с экспрессией на поверхности клетки данных рецепторов. Напротив, у FcγRIIa и FcγRIIb не было обаружено вариации числа копий генов. Известно, что малое число копий FcγRIIIb было ассоциировано с гломерулонефритом при аутоиммунном заболевании - системной красной волчанке (СКВ) [Aitman TJ и др. (2006) Nature 16; 439(7078):851-5]. Это представляет особенный интерес, учитывая, что несигнальный модуль GPI заякоревает рецептор FcγRIIIb. Можно предположить, что данные рецепторы FcγRIIIb потенциально могут действовать как конкурентные ингибиторы взаимодействия Fc с другими сигнальными FcγR. Также особый интерес представляет влияние вариаций числа копий в FcγRIIc. Однонуклеотидный полиморфизм С/Т в положении 202 в FcγRIIc превращает остаток глутамина в стоп-кодон, предотвращая образование функционального белка. Функциональная открытая рамка считывания FcγRIIc экспрессируется у 9% здоровых индивидов (в белой популяции), и наблюдается существенное доминирование (19%) данной аллели в популяции ИТП, что дает основания полагать, что данные фенотипы предрасполагают к ИТП [Breunis WB и др. (2008) Blood 111 (3): 1029-38]. Продемонстрировали, что у индивидов, экспрессирующих функциональный FcγRIIc на поверхности клеток NK, достигаемая АЗКЦ опосредована данными рецепторами в большей степени, чем FcγRIIIa. Такая сложность, связанная с данными полиморфизмами и генетическими вариациями, выдвигает на первый план необходимость персонифицированных стратегий лечения, требующих высокоспециализированных лекарств.
Различные эффекторные клетки отличаются по презентации данных рецепторов Fcγ, а также по их гуморальному и тканевому распределению, что способствует варьированию механизма их активации и действия [Таблица D]. Регулировка селективности терапевтических антител по отношению к узнаванию определенных типов FcγR и модуляция вклада некоторых классов эффекторных клеток приводит к оптимизации эффекторных механизмов для лечения конкретных заболеваний. Это означает избирательную активацию или ингибирование определенных эффекторных способов воздействия в зависимости от заболевания, от которого лечат.
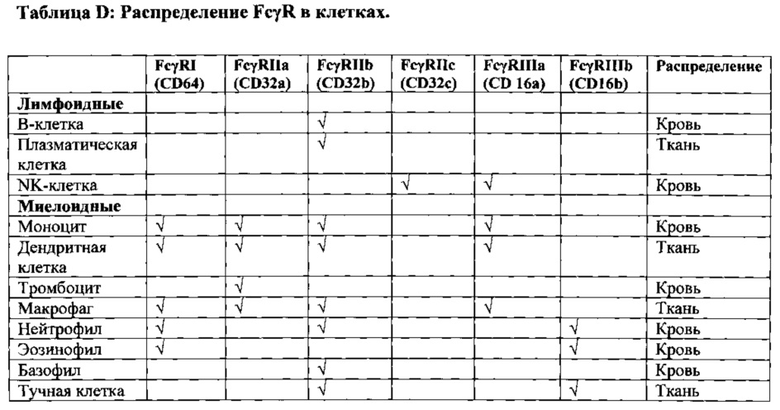
Кроме того, FcγR также экспрессируются фолликулярными дендритными клетками, эндотелиальными клетками, микроглиальными клетками, остеокластами и мезангиальными клетками. На сегодняшний день, функциональная значимость экспрессии FcγR на данных других клетках не известна.
Высокоаффинные FcγRI состоят из трех доменов типа С суперсемейства иммуноглобулинов (IgSF), тогда как каждый из низкоаффинных FcγRII и FcγRIII состоит из двух доменов типа С IgSF. Структуру рецепторных белков FcγRIIa, FcγRIIb, FcγRIIIa и FcγRIIIb выявили с помощью кристаллографии. Два домена IgSF в данных структурах расположены под углом 50-55 градусов по отношению друг к другу и соединены шарниром.
Общедоступная структура комплекса Fc-FcγR соответствует системе Fc-FcγRIIIb, и геометрия FcγR в данном комплексе сохраняется очень близкой к наблюдаемой в апо-состоянии белка (Sondermann P.; Huber R.; Oosthuizen V. и Jacob U. (2000) Nature 406, 267-273.; Radaev S.; Motyaka S.; Fridman W.; Sautes-Fridman С. и Sun P.D. (2001) J Biol Chem 276, 16469-16477; Sondermann P. и др. Biochem Soc Trans. 2002 Aug; 30(4):481-6; Sondermann P, Oosthuizen V. Immunol Lett. 2002 Jun 3; 82(1-2):51-6; Radaev S, Sun P. Mol Immunol. 2002 May; 38(14):1073-83). (Фигура 16). Сильное подобие последовательностей и структур между рецепторами служит основой для сравнительных моделей Fc, связанного с другими рецепторами. С другой стороны, подобие последовательностей и структур между данными рецепторными молекулами также затрудняет разработку Fc с точной селективностью между рецепторами и разнообразными их изотипами.
Перед структурной оценкой комплекса Fc-FcγR на основе кристаллографии, существовали вопросы, свидетельствует ли ось симметрии 2 порядка в молекуле Fc о наличии двух потенциальных сайтов связывания и эффективной стехиометрии 2:1 для ассоциации Fc-FcγR. Структурные исследования взаимодействий Fc - FcγR на основе ядерного магнитного резонанса (ЯМР) свидетельствуют о том, что связывание Fc с одним FcγR на одной стороне молекулы вызывает изменение конформации, которое исключает связывание второй молекулы FcγR с Fc той же молекулой антитела (Kato K. и др. (2000) J Mol Biol. 295(2):213-24). Геометрия доступного сокристаллизованного комплекса Fc-FcγRIIIb подтверждает ассоциацию FcγR с Fc в асимметричной ориентации со стехиометрией 1:1. На Фигуре 16 показано, что FcγR связывается с карманом на одной стороне молекулы Fc в форме подковы и контактирует с доменами СН2 на обеих цепях.
Сканирующий аланином мутагенез (Shields RL и др. (2001) JBC 276(9):6591-604) дает представление об остатках Fc, взаимодействующих с различными типами рецептора, и, следовательно, вовлеченных во взаимодействие и узнавание Fc-FcγR. По традиции, оптимизация терапевтических антител была сосредоточена вокруг мутаций, которые приводят к повышенному связыванию с активирующими рецепторами FcγRIII (патент США номер 6737056) или пониженной аффинности к FcγRIIb (US 2009/0010920A1). Во всех данных альтернативных вариантах, мутации вводили одновременно в обе цепи.
Моноклональные антитела часто проявляют терапевтическую активность путем пространственной локализации мишени и эффекторной иммунной клетки. Природное антитело осуществляет такую локализацию путем взаимодействия с мишенью посредством Fab доменов, а с эффекторной клеткой - посредством домена Fc. Они способны сближать иммунный комплекс с эффекторной клеткой таким образом, что может быть вызван клеточно-опосредованный ответ. Эффект авидности, необходимый для передачи сигналов FcγR, возникающих при образовании иммунных комплексов, что влечет за собой нацеливание на одну мишень множества молекул антител, представляет собой другой пример значимости пространственно-временной организации в работе иммунной системы.
Также существует пространственно-временной аспект передачи сигналов в клетке, который индуцируется как часть эффекторной активности молекул МАТ. Передача сигналов в клетке, например, возникающих при активации молекулы FcγR, включает локализацию соответствующих рецепторных молекул внутри участков доменов на мембране, называемых липидными рафтами. Липидные рафты обогащены гликосфинголипидом и холестерином и несколькими классами вышележащих сигнальных трансдукторов, включая киназы семейства Src. При стимуляции клетки, привлекаются различные сигнальные молекулы, адаптерные белки и сигнальные киназы, а также фосфатазы. Собрание молекул в липидных рафтах важно для передачи сигнала.
Стратегия синтетической разработки, сочетающая различные антигенные специфичности и повышенную авидность для получения лучших свойств связывания, представляет собой основу разработки биспецифического терапевтического средства. Биспецифические антитела или другие формы биспецифических или мультифункциональных белковых лекарственных средств разработаны таким образом, чтобы они опосредовали взаимодействия между мишенью и множеством эффекторных клеток [Müller & Kontermann (2010) BioDrugs 24(2):89-98]. Мультиспецифические терапевтические молекулы разработаны таким образом, чтобы они перенацеливали хелперные Т-клетки или другие иммунные эффекторные клетки на определенные целевые клетки.
В другом варианте реализации, настоящее изобретение относится к способу компьютерной идентификации полипептидов варианта Fc на основании рассчитанных аффинностей связывания с FcγRIIa, FcγRIIb и/или FcγRIIIa. В другом варианте реализации, способ дополнительно включает компьютерное вычисление электростатики, сольватации, укладки, плотности укладки, связывания водорода и энтропийных эффектов указанных полипептидов варианта Fc. В еще одном варианте реализации, способ согласно настоящему изобретению дополнительно включает конструирование полипептидов варианта Fc и экспрессию указанных полипептидов в виде терапевтического антитела, и дальнейшую экспрессию указанного антитела в клетках млекопитающих. В еще одном варианте реализации, способ согласно настоящему изобретению включает конструирование полипептидов варианта Fe, идентифицированных in silico с помощью сайт-направленного мутагенеза, мутагенеза на основе ПЦР, кассетного мутагенеза или синтеза de novo.
Факторы, принимаемые во внимание при разработке синтетического Fc-каркаса, включают компьютерные расчеты стерического отталкивания, изменения площади внутренней поверхности, относительной плотности контакта, относительной сольватации и электростатического эффекта. Все полученные матрицы применяли для получения показателя аффинности.
В одном аспекте настоящей заявки описана молекулярная разработка для достижения совершенных профилей селективности FcγR посредством разработки асимметричного каркаса, построенного на гетеродимерном Fc. Данный каркас обеспечивает возможность введения асимметричных мутаций в домен СН2 для достижения множества новых профилей селективности. Кроме того, каркас обладает внутренними свойствами для разработки на его основе мультифункциональных (би-, три-, тетра- или пентафункциональных) терапевтических молекул.
В некоторых вариантах реализации, асимметричный каркас оптимизируют по рН-зависимым свойствам связывания с неонатальным Fc-рецептором (FcRn), чтобы обеспечить лучшее возобновление молекулы и увеличить время ее полужизни и связанные с этим фармакокинетические свойства.
Асимметричный каркас можно оптимизировать для связывания с функционально значимыми аллотипами рецептора FcγRI. FcγRI представляет собой известный маркер на макрофагах, которые участвуют в хронических воспалительных расстройствах, таких как ревматоидный артрит, атопический дерматит, псориаз и множество заболеваний легких.
Асимметричный каркас можно оптимизировать для связывания белка А. Связывание белка А часто используют для разделения и очистки молекул антител. В асимметричный каркас можно ввести мутации, чтобы избежать агрегации терапевтического средства во время хранения.
Следовательно, особенно предполагается, что варианты Fc согласно настоящему изобретению могут включать, среди прочего, одну или более дополнительных замен, мутаций и/или модификаций аминокислотных остатков, которые придают антителу предпочтительные свойства, включая, но не ограничиваясь перечисленными: увеличенное время полужизни в сыворотке, повышенную аффинность связывания, пониженную иммуногенность, повышенную продукцию, повышенную или пониженную активность АЗКЦ или КЗЦ, изменение гликозилирования и/или дисульфидных связей и модифицированную специфичность связывания.
Предполагается, что у вариантов Fc согласно настоящему изобретению могут быть изменены другие свойства, включая увеличенное время полужизни in vivo (например, время полужизни в сыворотке) у млекопитающего, в частности, у человека; повышенную стабильность in vivo (например, время полужизни в сыворотке) и/или in vitro (например, время хранения) и/или повышенную температуру плавления (Tm), по сравнению со сравнимой молекулой. В одном варианте реализации, вариант Fc согласно настоящему изобретению имеет время полужизни in vivo более 15 дней, более 20 дней, более 25 дней, более 30 дней, более 35 дней, более 40 дней, более 45 дней, более 2 месяцев, более 3 месяцев, более 4 месяцев или более 5 месяцев. В другом варианте реализации, вариант Fc согласно настоящему изобретению имеет время полужизни in vitro (например, в виде жидкой или порошковой лекарственной формы) более 15 дней, более 30 дней, более 2 месяцев, более 3 месяцев, более 6 месяцев, или более 12 месяцев, или более 24 месяцев, или более 36 месяцев, или более 60 месяцев.
Для специалиста в данной области также должно быть очевидно, что варианты Fc согласно настоящему изобретению могут обладать измененной иммуногенностью при введении субъекту. Соответственно, предполагается, что модифицированный домен СН3, который минимизирует иммуногенность варианта Fc, как правило, более желателен для применения в терапии.
Варианты Fc согласно настоящему изобретению можно комбинировать с другими модификациями Fc, включая, но не ограничиваясь модификациями, которые изменяют эффекторную функцию. В объем настоящего изобретения входит сочетание варианта Fc согласно настоящему изобретению с другими модификациями Fc, чтобы обеспечить вспомогательные, синергичные или новые свойства антител или Fc-слитых белков.
Такие модификации могут присутствовать в шарнире, доменах СН1 или СН2 (или СН3, в том случае, если они не изменяют в худшую сторону стабильность и чистоту модифицированных доменов СН3 согласно настоящему изобретению) или в комбинация перечисленных доменов. Предполагается, что варианты Fc согласно настоящему изобретению улучшают свойства модификации, с которой они объединены. Например, если вариант Fc согласно настоящему изобретению объединен с мутантом, который, как известно, связывает FcγRIIIA с более высокой аффинностью, чем сравнимая молекула, содержащая Fc-область дикого типа; комбинация с мутантом согласно настоящему изобретению приводит к увеличению аффинности к FcγRIIIA в большее число раз.
В одном варианте реализации, варианты Fc согласно настоящему изобретению можно комбинировать с другими известными вариантами Fc, такими как описанные у Duncan и др., 1988, Nature 332:563-564; Lund и др., 1991, J Immunol 147:2657-2662; Lund и др., 1992, Mol Immunol 29:53-59; Alegre и др., 1994, Transplantation 57:1537-1543; Hutchins и др., 1995, Proc Natl. Acad Sci USA 92:1 1980-1 1984; Jefferis и др., 1995, Immunol Lett. 44:111-117; Lund и др., 1995, Faseb J 9:115-119; Jefferis и др., 1996, Immunol Lett 54:101-104; Lund и др., 1996, Immunol 157:4963-4969; Armour и др., 1999, Eur J Immunol 29:2613-2624; Idusogie и др., 2000, J Immunol 164:4178-4184; Reddy и др., 2000, J Immunol 164:1925-1933; Xu и др., 2000, Cell Immunol 200: 16-26; Idusogie и др., 2001, J Immunol 166:2571-2575; Shields и др., 2001, J Biol Chem 276:6591-6604; Jefferis и др., 2002, Immunol Lett 82:57-65; Presta и др., 2002, Biochem Soc Trans 30:487-490); в патентах США №№5624821; 5885573; 6194551; в заявках на патенты США №№60/601634 и 60/608852; в публикациях согласно РСТ №№ WO 00/42072 и WO 99/58572.
Для специалиста в данной области должно быть очевидно, что варианты Fc согласно настоящему изобретению могут обладать измененными свойствами связывания лиганда Fc (например, FcγR, C1q) (примеры свойств связывания включают, но не ограничены перечисленными: специфичность связывания, равновесную константу диссоциации (KD), скорости диссоциации и ассоциации (Koff и Kon, соответственно), аффинность связывания и/или авидность), и что некоторые изменения являются в большей или меньшей степени желательными. В данной области хорошо известно, что равновесная константа диссоциации (KD) рассчитывается как koff/kon. Как правило, очевидно, что связывающая молекула (например, антитело) с низкой KD предпочтительна по сравнению со связывающей молекулой (например, антителом) с высокой KD. Тем не менее в некоторых случаях значение kon или koff может быть более важным, чем значение KD. Специалист в данной области может определить, какой кинетический параметр наиболее важен для данного применения антитела. Например, модифицированные СН3 и/или СН2, которые повышают связывание Fc с одним или более положительными регуляторами (например, FcγRIIIA), при этом оставляя неизмененным или даже снижая связывание Fc с отрицательным регулятором FcγRIIB, будут наиболее полезными для усиления активности АЗКЦ. В качестве альтернативы, модифицированные СН3 и/или СН2, которые снижают связывание с одним или более положительными регуляторами и/или повышают связывание с FcγRIIB, будут наиболее полезными для снижения активности АЗКЦ. Соответственно, соотношение аффинностей связывания (например, равновесных констант диссоциации (KD)) может указывать на то, повышена или понижена активность АЗКЦ варианта Fc. Например, уменьшение соотношения равновесных констант диссоциации (KD) FcγRIIIA/FcγRIIB будет коррелировать с повышением активности АЗКЦ, тогда как увеличение данного соотношения будет коррелировать со снижением активности АЗКЦ.
В рамках описания вариантов Fc, исследовали их аффинность связывания с FcγRIIIA (CD16a) и FcγRIIB(CD32b), которую описали в виде соотношения по сравнению с IgG1 дикого типа (см. Пример 4 и Таблицу 5). В данном случае, оказалась возможной оценка влияния мутаций в домене СН3 на связывание с данными активирующими и ингибирующими рецепторами Fc. В одном варианте реализации, в данной заявке предложены изолированные гетеромультимеры, включающие гетеродимерную Fc-область, при этом гетеродимерная Fc-область содержит модифицированный домен СН3, включающий мутации аминокислот, способствующие образованию гетеродимера с повышенной стабильностью, и при этом модифицированный домен СН3 имеет температуру плавления (Tm) более 70°С, при этом связывание гетеродимера с CD16a приблизительно такое же, как и у гомодимера дикого типа. В некоторых вариантах реализации, связывание гетеродимера с CD16a повышено по сравнению с гомодимером дикого типа. В альтернативном варианте реализации, связывание гетеродимера с CD16a снижено по сравнению с гомодимером дикого типа.
В некоторых вариантах реализации, в данной заявке предложены изолированные гетеромультимеры, включающие гетеродимерную Fc-область, при этом гетеродимерная Fc-область содержит модифицированный домен СН3, включающий мутации аминокислот, способствующие образованию гетеродимера с повышенной стабильностью, и при этом модифицированный домен СН3 имеет температуру плавления (Tm) более 70°С, при этом связывание гетеродимера с CD32b приблизительно такое же, как и у гомодимера дикого типа. В некоторых вариантах реализации, связывание гетеродимера с CD32b повышено по сравнению с гомодимером дикого типа. В альтернативном варианте реализации, связывание гетеродимера с CD32b снижено по сравнению с гомодимером дикого типа.
Для специалиста в данной области должно быть очевидно, что вместо представления KD связывания CD16a и CD32b в виде сравнения варианта Fc с гомодимером дикого типа, KD можно представить в виде отношения связывания варианта Fc с CD16a к связыванию варианта Fc с CD32b (результаты не представлены). Данное отношение дает критерий влияния мутации в модифицированном домене СН3 на АЗКЦ (неизмененная, повышенная или пониженная по сравнению с диким типом), что подробнее описано ниже.
Аффинности и свойства связывания вариантов Fc согласно настоящему изобретению с FcγR изначально определяли, применяя тесты in vitro (биохимические или иммунологические анализы), известные в данной области для определения взаимодействия Fc-FcγR, т.е. специфичного связывания Fc-области с FcγR, включая, но не ограничиваясь перечисленными: твердофазный иммуноферментный анализ (анализ ELISA), анализ методом поверхностного плазмонного резонанса, анализы иммунопреципитации (см. ниже в разделе, названном "Характеристические и функциональные анализы") и другие способы, такие как непрямые анализы связывания, анализы конкурентного ингибирования, резонансный перенос энергии флуоресценции (FRET), электрофорез в геле и хроматография (например, гель-фильтрация). В данных и других способах можно использовать метку на одном или более исследуемых компонентах и/или применять множество способов детектирования, включая, но не ограничиваясь перечисленными: хромогенную, флуоресцентную, люминесцентную или изотопную метки. Подробное описание аффинностей и кинетики связывания можно найти в Paul, W.Ε., ред., Fundamental Immunology, 4ое изд., Lippincott-Raven, Филадельфия (1999), которая сфокусирована на взаимодействиях антитело-иммуноген.
Предполагается, что свойства связывания молекул согласно настоящему изобретению также можно описать с помощью функциональных анализов in vitro для определения одной или более опосредованных FcγR функций эффекторных клеток (см. выше в разделе, названном "Характеристические и функциональные анализы"). В некоторых вариантах реализации, молекулы согласно настоящему изобретению обладают свойствами связывания в моделях in vivo (такими как описанные и раскрытые в данной заявке), сходными с таковыми в анализах in vitro. Тем не менее настоящее изобретение не исключает молекулы согласно настоящему изобретению, которые не проявляют желательный фенотип в анализах in vitro, но проявляют желательный фенотип in vivo.
В объем настоящего изобретения входят варианты Fc, которые связывают FcγRIIIA (CD16a) с повышенной аффинностью по сравнению с аналогичной молекулой. В конкретном варианте реализации, варианты Fc согласно настоящему изобретению связывают FcγRIIIA с повышенной аффинностью и связывают FcγRIIB (CD32b) с аффинностью связывания, которая либо не изменена, либо снижена по сравнению с аналогичной молекулой. В еще одном варианте реализации, варианты Fc согласно настоящему изобретению имеют меньшее отношение равновесных констант диссоциации (KD) с FcγRIIIA/FcγRIIB, чем у аналогичной молекулы.
Также в объем настоящего изобретения входят варианты Fc, которые связывают FcγRIIIA (CD16a) с пониженной аффинностью по сравнению с аналогичной молекулой. В конкретном варианте реализации, варианты Fc согласно настоящему изобретению связывают FcγRIIIA с пониженной аффинностью по сравнению с аналогичной молекулой и связывают FcγRIIB с аффинностью связывания, которая либо не изменена, либо повышена по сравнению с аналогичной молекулой.
В одном варианте реализации, варианты Fc связываются с FcγRIIIA с повышенной аффинностью. В конкретном варианте реализации, указанные варианты Fc обладают аффинностью к FcγRIIIA, которая по меньшей мере в 2 раза, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 7 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 30 раз, или по меньшей мере в 40 раз, или по меньшей мере в 50 раз, или по меньшей мере в 60 раз, или по меньшей мере в 70 раз, или по меньшей мере в 80 раз, или по меньшей мере в 90 раз, или по меньшей мере в 100 раз, или по меньшей мере в 200 раз больше, чем таковая у сравнимой молекулы. В других вариантах реализации, варианты Fc обладают аффинностью к FcγRIIIA, которая повышена на по меньшей мере 10%, или на по меньшей мере 20%, или на по меньшей мере 30%, или на по меньшей мере 40%, или на по меньшей мере 50%, или на по меньшей мере 60%, или на по меньшей мере 70%, или на по меньшей мере 80%, или на по меньшей мере 90%, или на по меньшей мере 100%, или на по меньшей мере 150%, или на по меньшей мере 200% по сравнению с аналогичной молекулой.
В другом варианте реализации, вариант Fc имеет равновесную константу диссоциации (KD) с лигандом Fc (например, FcγR, C1q), которая снижена в диапазоне приблизительно от 2 до 10 раз, или в диапазоне приблизительно от 5 до 50 раз, или в диапазоне приблизительно от 25 до 250 раз, или в диапазоне приблизительно от 100 до 500 раз, или в диапазоне приблизительно от 250 до 1000 раз по сравнению с аналогичной молекулой.
В другом варианте реализации, указанные варианты Fc имеют равновесную константу диссоциации (KD) с FcγRIIIA, которая снижена по меньшей мере в 2 раза, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 7 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 30 раз, или по меньшей мере в 40 раз, или по меньшей мере в 50 раз, или по меньшей мере в 60 раз, или по меньшей мере в 70 раз, или по меньшей мере в 80 раз, или по меньшей мере в 90 раз, или по меньшей мере в 100 раз, или по меньшей мере в 200 раз, или по меньшей мере в 400 раз, или по меньшей мере в 600 раз по сравнению с аналогичной молекулой. В другом варианте реализации, варианты Fc имеют равновесную константу диссоциации (KD) с FcγRIIIA, которая снижена на по меньшей мере 10%, или на по меньшей мере 20%, или на по меньшей мере 30%, или на по меньшей мере 40%, или на по меньшей мере 50%, или на по меньшей мере 60%, или на по меньшей мере 70%, или на по меньшей мере 80%, или на по меньшей мере 90%, или на по меньшей мере 100%, или на по меньшей мере 150%, или на по меньшей мере 200% по сравнению с аналогичной молекулой.
В одном варианте реализации, вариант Fc связывается с FcγRIIB с аффинностью, которая не изменена или снижена. В конкретном варианте реализации, указанные варианты Fc обладают аффинностью к FcγRIIB, которая не изменена или снижена по меньшей мере в 1 раз, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 50 раз, или по меньшей мере в 100 раз по сравнению с аналогичной молекулой. В других вариантах реализации, варианты Fc обладают аффинностью к FcγRIIB, которая не изменена или снижена на по меньшей мере 10%, или на по меньшей мере 20%, или на по меньшей мере 30%, или на по меньшей мере 40%, или на по меньшей мере 50%, или на по меньшей мере 60%, или на по меньшей мере 70%, или на по меньшей мере 80%, или на по меньшей мере 90%, или на по меньшей мере 100%, или на по меньшей мере 150%, или на по меньшей мере 200% по сравнению с аналогичной молекулой.
В другом варианте реализации, варианты Fc имеют равновесную константу диссоциации (KD) с FcγRIIB, которая не изменена или повышена по меньшей мере в 2 раза, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 7 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 30 раз, или по меньшей мере в 40 раз, или по меньшей мере в 50 раз, или по меньшей мере в 60 раз, или по меньшей мере в 70 раз, или по меньшей мере в 80 раз, или по меньшей мере в 90 раз, или по меньшей мере в 100 раз, или по меньшей мере в 200 раз по сравнению с аналогичной молекулой. В другом конкретном варианте реализации, варианты Fc имеют равновесную константу диссоциации (KD) с FcγRIIB, которая не изменена или повышена на по меньшей мере 10%, или на по меньшей мере 20%, или на по меньшей мере 30%, или на по меньшей мере 40%, или на по меньшей мере 50%, или на по меньшей мере 60%, или на по меньшей мере 70%, или на по меньшей мере 80%, или на по меньшей мере 90%, или на по меньшей мере 100%, или на по меньшей мере 150%, или на по меньшей мере 200% по сравнению с аналогичной молекулой.
В другом дополнительном варианте реализации, варианты Fc связывают FcγRIIIA с повышенной аффинностью по сравнению с аналогичной молекулой и связывают FcγRIIB с аффинностью, которая не изменена или снижена по сравнению с аналогичной молекулой. В конкретном варианте реализации, варианты Fc обладают аффинностью к FcγRIIIA, которая повышена по меньшей мере в 1 раз, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 50 раз, или по меньшей мере в 100 раз по сравнению с аналогичной молекулой. В другом конкретном варианте реализации, варианты Fc обладают аффинностью к FcγRIIB, которая либо не изменена, либо снижена по меньшей мере в 2 раза, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 7 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 50 раз, или по меньшей мере в 100 раз по сравнению с аналогичной молекулой. В других вариантах реализации, варианты Fe обладают аффинностью к FcγRIIIA, которая повышена на по меньшей мере 10%, или на по меньшей мере 20%, или на по меньшей мере 30%, или на по меньшей мере 40%, или на по меньшей мере 50%, или на по меньшей мере 60%, или на по меньшей мере 70%, или на по меньшей мере 80%, или на по меньшей мере 90%, или на по меньшей мере 100%, или на по меньшей мере 150%, или на по меньшей мере 200% по сравнению с аналогичной молекулой, и указанные варианты Fc обладают аффинностью к FcγRIIB, которая либо не изменена, либо повышена на по меньшей мере 10%, или на по меньшей мере 20%, или на по меньшей мере 30%, или на по меньшей мере 40%, или на по меньшей мере 50%, или на по меньшей мере 60%, или на по меньшей мере 70%, или на по меньшей мере 80%, или на по меньшей мере 90%, или на по меньшей мере 100%, или на по меньшей мере 150%, или на по меньшей мере 200% по сравнению с аналогичной молекулой.
В еще одном варианте реализации, варианты Fc имеют отношение равновесных констант диссоциации (KD) с FcγRIIIA/FcγRIIB, которое снижено по сравнению с аналогичной молекулой. В конкретном варианте реализации, варианты Fe имеют отношение равновесных констант диссоциации (KD) с FcγRIIIA/FcγRIIB, которое снижено по меньшей мере в 1 раз, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 50 раз, или по меньшей мере в 100 раз по сравнению с аналогичной молекулой. В другом конкретном варианте реализации, варианты Fc имеют отношение равновесных констант диссоциации (KD) с FcγRIIIA/FcγRIIB, которое снижено на по меньшей мере 10%, или на по меньшей мере 20%, или на по меньшей мере 30%, или на по меньшей мере 40%, или на по меньшей мере 50%, или на по меньшей мере 60%, или на по меньшей мере 70%, или на по меньшей мере 80%, или на по меньшей мере 90%, или на по меньшей мере 100%, или на по меньшей мере 150%, или на по меньшей мере 200% по сравнению с аналогичной молекулой.
В другом варианте реализации, варианты Fc связывают FcγRIIIA с пониженной аффинностью по сравнению с аналогичной молекулой. В конкретном варианте реализации, указанные варианты Fc обладают аффинностью к FcγRIIIA, которая снижена по меньшей мере в 1 раз, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 50 раз, или по меньшей мере в 100 раз по сравнению с аналогичной молекулой. В других вариантах реализации, варианты Fc обладают аффинностью к FcγRIIIA, которая снижена на по меньшей мере 10%, или на по меньшей мере 20%, или на по меньшей мере 30%, или на по меньшей мере 40%, или на по меньшей мере 50%, или на по меньшей мере 60%, или на по меньшей мере 70%, или на по меньшей мере 80%, или на по меньшей мере 90%, или на по меньшей мере 100%, или на по меньшей мере 150%, или на по меньшей мере 200% по сравнению с аналогичной молекулой.
В другом дополнительном варианте реализации, варианты Fc связывают FcγRIIIA с пониженной аффинностью и связывают FcγRIIB с аффинностью, которая либо не изменена, либо повышена по сравнению с аналогичной молекулой. В конкретном варианте реализации, варианты Fe обладают аффинностью к FcγRIIIA, которая снижена по меньшей мере в 1 раз, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 50 раз, или по меньшей мере в 100 раз по сравнению с аналогичной молекулой. В другом конкретном варианте реализации, варианты Fc обладают аффинностью к FcγRIIB, которая по меньшей мере в 2 раза, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 7 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 50 раз, или по меньшей мере в 100 раз больше, чем таковая у сравнимой молекулы. В других вариантах реализации, варианты Fe обладают аффинностью к FcγRIIIA, которая снижена на по меньшей мере 10%, или на по меньшей мере 20%, или на по меньшей мере 30%, или на по меньшей мере 40%, или на по меньшей мере 50%, или на по меньшей мере 60%, или на по меньшей мере 70%, или на по меньшей мере 80%, или на по меньшей мере 90%, или на по меньшей мере 100%, или на по меньшей мере 150%, или на по меньшей мере 200% по сравнению с аналогичной молекулой и указанные варианты Fc обладают аффинностью к FcγRIIB, которая повышена на по меньшей мере 10%, или на по меньшей мере 20%, или на по меньшей мере 30%, или на по меньшей мере 40%, или на по меньшей мере 50%, или на по меньшей мере 60%, или на по меньшей мере 70%, или на по меньшей мере 80%, или на по меньшей мере 90%, или на по меньшей мере 100%, или на по меньшей мере 150%, или на по меньшей мере 200% по сравнению с аналогичной молекулой.
В другом дополнительном варианте реализации, варианты Fc имеют равновесную константу диссоциации (KD) с FcγRIIIA, которая повышена по меньшей мере в 1 раз, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 50 раз по сравнению с таковой у сравнимой молекулы. В конкретном варианте реализации, указанные варианты Fc имеют равновесную константу диссоциации (KD) с FcγRIIB, которая снижена по меньшей мере в 2 раза, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 7 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 50 раз или по меньшей мере в 100 раз по сравнению с аналогичной молекулой.
Варианты СН2 для селективности FcγR
Белок-белковое взаимодействие Fc-FcγR в данном комплексе указывает на то, что две цепи в молекуле Fc взаимодействуют с двумя различными сайтами на молекуле FcγR. Хотя в природных молекулах Fc две тяжелые цепи симметричны, локальное окружение FcγR вблизи остатков на одной цепи отличается от остатков FcγR, окружающих остаток в том же положении на противоположной цепи Fc. Два симметричных положения взаимодействуют с различными выборками остатков FcγR.
Принимая во внимание асимметрию при соединении Fc с FcγR, одновременные мутации цепей А и В молекулы Fc не влияют на взаимодействия с FcγR симметричным образом. При введении мутаций для оптимизации взаимодействий на одной цепи Fc с ее локальным окружением FcγR, в гомодимерной структуре Fc, соответствующая мутация во второй цепи может оказаться благоприятной, неблагоприятной или не способствующей необходимому профилю связывания и селективности FcγR.
С использованием структуры и компьютеризированного подхода, в двух цепях Fc разработали асимметричные мутации, чтобы преодолеть указанные ограничения традиционных стратегий разработки Fc, в которых одни и те же мутации вводили в обе цепи Fc. Если две цепи Fc оптимизированы независимо для повышенного связывания с соответствующей стороной рецепторной молекулы, можно добиться лучшей селективности связывания между рецепторами.
Например, мутации в конкретном положении на одной цепи Fc можно разработать таким образом, чтобы они повышали селективность к конкретному остатку, реализация положительной разработки, тогда как остаток в том же положении можно мутировать, чтобы он неблагоприятно взаимодействовал с локальным окружением в альтернативном типе рецептора Fcγ, реализация отрицательной разработки, таким образом достигается лучшая селективность между двумя рецепторами. В некоторых вариантах реализации, предложен способ разработки асимметричных модификаций аминокислот в домене СН2, которые избирательно связывают один рецептор Fc-гамма по сравнению с отличным рецептором Fc-гамма (например, избирательно связывают FcγRIIIa, но не FcγRIIb). В других некоторых вариантах реализации, предложен способ разработки асимметричных модификаций аминокислот в домене СН2 варианта гетеродимера Fc, включающего модификации аминокислот в домене СН3, чтобы вызвать образование гетеродимера. В другом варианте реализации, предложен способ достижения селективности к различным рецепторам Fc-гамма, основанный на варианте гетеродимера Fc, включающем асимметричные модификации аминокислот в домене СН2. В еще одном варианте реализации, предложен способ разработки асимметричных модификаций аминокислот, которые способствуют связыванию рецепторов Fc-гамма с одной стороной молекулы Fc. В других некоторых вариантах реализации, предложен способ разработки факторов полярности, которые вызывают взаимодействие рецепторов Fc-гамма только с одной стороной варианта гетеродимера Fc, включающего асимметричные модификации аминокислот в домене СН2.
Асимметричную разработку мутаций в домене СН2 можно приспособить для узнавания FcγR на одной стороне молекулы Fc. Эта сторона представляет собой продуктивную сторону асимметричного Fc-каркаса, тогда как противоположная сторона имеет склонность к взаимодействию, подобному молекуле дикого типа, без разработанного профиля селективности, и ее можно рассматривать как непродуктивную сторону. Стратегию отрицательной разработки можно применять для введения мутаций в непродуктивную сторону, чтобы блокировать взаимодействия FcγR с этой стороной асимметричного Fc-каркаса, тем самым усиливая стремление рецепторов Fcγ к желательному взаимодействию.

Настоящее изобретение также относится к слитым полипептидам, содержащим связывающий домен, слитый Fc-областью, при этом Fc-область содержит модифицированный домен СН3, включающий мутации аминокислот, способствующие образованию гетеродимера с повышенной стабильностью, и при этом модифицированный домен СН3 имеет температуру плавления (Tm) более 70°С. В частности, предполагается, что молекулы, включающие гетеродимер, содержащий модифицированный домен СН3, можно получить с помощью способов, хорошо известных специалисту в данной области. Вкратце, такие способы включают, но не ограничены перечисленными, объединение вариабельного участка или связывающего домена, обладающего желательной специфичностью (например, вариабельного участка, выделенного из библиотеки фагового дисплея или экспрессионной библиотеки, или полученного из человеческого или не принадлежащего человеку антитела или связывающего домена рецептора), с вариантами гетеродимера Fc. В качестве альтернативы, специалист в данной области может получить вариант гетеродимера Fc путем модификации домена СН3 в Fc-области молекулы, содержащей Fc-область (например, антитела).
В одном варианте реализации, варианты Fe представляют собой антитела или Fc-слитые белки. В конкретном варианте реализации, согласно настоящему изобретению предложены антитела, включающие Fc-область, содержащую модифицированный домен СН3, включающий мутации аминокислот, способствующие образованию гетеродимера с повышенной стабильностью, и при этом модифицированный домен СН3 имеет температуру плавления (Tm) более 70°С. Такие антитела включают молекулы IgG, которые по природе содержат Fc-область, содержащую домен СН3, который можно модифицировать с получением варианта Fc, или производные антител, в которые искусственно включили Fc-область, содержащую модифицированный домен СН3. Варианты Fc согласно настоящему изобретению включают любую молекулу антитела, которая связывается, предпочтительно, специфично (т.е., успешно конкурирует с неспецифичным связыванием, что определяют с помощью иммуноанализов, хорошо известных в данной области, для анализа специфичного связывания антиген-антитело), с антигеном, который включает Fc-область, содержащую модифицированный домен СН3. Такие антитела включают, но не ограничены перечисленными: поликлональные, моноклональные, моноспецифичные, биспецифические, мультиспецифичные, человеческие, гуманизированные, химерные антитела, одноцепочечные антитела, фрагменты Fab, фрагменты F(ab')2, связанные дисульфидными связями Fv и фрагменты, содержащие любой из доменов VL или VH, или даже определяющую комплементарность область (гипервариабельный участок, CDR), которая специфично связывает антиген, в некоторых случаях, в который искусственно включили вариант гетеродимера Fc или слили с ним.
"Антителозависимая клеточная цитотоксичность" или "АЗКЦ" относится к форме цитотоксичности, при которой секретируемое антитело, связанное с рецепторами Fc (FcR), присутствующими на некоторых цитотоксических клетках (например, на естественных клетах-киллерах (NK), нейтрофилах и макрофагах), позволяет данным цитотоксическим эффекторным клеткам специфично связываться с несущей антиген клеткой-мишенью и, впоследствии, убивать клетку-мишень цитотоксинами. Специфичные высокоаффинные антитела IgG, направленные на поверхность целевых клеток, нацеливают цитотоксические клетки и абсолютно необходимы для такого уничтожения. Лизис клетки-мишени является внеклеточным, требует непосредственного межклеточного контакта и происходит без участия комплемента.
Способность какого-либо конкретного антитела опосредовать лизис клетки-мишени путем АЗКЦ можно проанализировать. Чтобы оценить активность АЗКЦ, интересующее антитело добавляют к целевым клеткам в комбинации с иммунными эффекторными клетками, которые могут активироваться комплексами антиген-антитело, что приводит к цитолизу клетки-мишени. Цитолиз, как правило, обнаруживают по высвобождению метки (например, радиоактивных субстратов, флуоресцентных красителей или природных внутриклеточных белков) из лизированных клеток. Полезные эффекторные клетки для таких анализов включают мононуклеарные клетки периферической крови (РВМС) и естественные клети-киллеры (NK). Конкретные примеры анализов АЗКЦ in vitro описаны в Wisecarver и др., 1985, 79:277; Braggemarm и др., 1987, J Exp Med 166:1351; Wilkinson и др., 2001, J Immunol Methods 258:183; Patel и др., 1995 J Immunol Methods 184:29 и в данной заявке (см. ниже в разделе, названном "Характеристические и функциональные анализы"). В качестве альтернативы или дополнения, активность АЗКЦ интересующего антитела можно оценить in vivo, например, в модели на животном, такой как описанная у Clynes и др., 1998, PNAS USA 95:652.
Предполагается, что варианты Fc согласно настоящему изобретению можно описать с помощью функциональных анализов in vitro для определения одной или более опосредованных FcγR функций эффекторных клеток. В конкретных вариантах реализации, молекулы согласно настоящему изобретению обладают свойствами связывания и функциями эффекторных клеток в моделях in vivo (такими как описанные и раскрытые в данной заявке), сходными с таковыми в анализах in vitro. Тем не менее, настоящее изобретение не исключает молекулы согласно настоящему изобретению, которые не проявляют желательный фенотип в анализах in vitro, но проявляют желательный фенотип in vivo.
Согласно настоящему изобретению дополнительно предложены варианты Fc с усиленной функцией КЗЦ. В одном варианте реализации, варианты Fc обладают повышенной активностью КЗЦ. В одном варианте реализации, варианты Fe обладают активностью КЗЦ, которая по меньшей мере в 2 раза, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 10 раз, или по меньшей мере в 50 раз, или по меньшей мере в 100 раз больше, чем таковая у сравнимой молекулы. В другом варианте реализации, варианты Fc связывают C1q с аффинностью, которая по меньшей мере в 2 раза, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 7 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 50 раз, или по меньшей мере в 100 раз больше, чем таковая у сравнимой молекулы. В еще одном варианте реализации, варианты Fc обладают активностью КЗЦ, которая повышена на по меньшей мере 10%, или на по меньшей мере 20%, или на по меньшей мере 30%, или на по меньшей мере 40%, или на по меньшей мере 50%, или на по меньшей мере 60%, или на по меньшей мере 70%, или на по меньшей мере 80%, или на по меньшей мере 90%, или на по меньшей мере 100%, или на по меньшей мере 150%, или на по меньшей мере 200% по сравнению с аналогичной молекулой. В конкретном варианте реализации, варианты Fc согласно настоящему изобретению связывают C1q с повышенной аффинностью; обладают повышенной активностью КЗЦ и специфично связываются с по меньшей мере одним антигеном.
Согласно настоящему изобретению также предложены варианты Fc со сниженной функцией КЗЦ. В одном варианте реализации, варианты Fc обладают пониженной активностью КЗЦ. В одном варианте реализации, варианты Fc обладают активностью КЗЦ, которая по меньшей мере в 2 раза, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз или по меньшей мере в 10 раз или по меньшей мере в 50 раз или по меньшей мере в 100 раз меньше, чем таковая у сравнимой молекулы. В другом варианте реализации, вариант Fc связывает C1q с аффинностью, которая снижена по меньшей мере в 1 раз, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 50 раз, или по меньшей мере в 100 раз по сравнению с аналогичной молекулой. В другом варианте реализации, варианты Fc обладают активностью КЗЦ, которая снижена на по меньшей мере 10%, или на по меньшей мере 20%, или на по меньшей мере 30%, или на по меньшей мере 40%, или на по меньшей мере 50%, или на по меньшей мере 60%, или на по меньшей мере 70%, или на по меньшей мере 80%, или на по меньшей мере 90%, или на по меньшей мере 100%, или на по меньшей мере 150%, или на по меньшей мере 200% по сравнению с аналогичной молекулой. В конкретном варианте реализации, варианты Fc связываются с C1q с пониженной аффинностью, обладают пониженной активностью КЗЦ и специфично связываются с по меньшей мере одним антигеном.
В некоторых вариантах реализации, варианты Fc содержат одну или более сконструированных гликоформ, т.е., углеводную композицию, которая ковалентно присоединена к молекуле, содержащей Fc-область. Сконструированные гликоформы могут быть полезны для множества целей, включая, но не ограничиваясь перечисленными, усиление или снижение эффекторной функции. Сконструированные гликоформы можно получить с помощью любого способа, известного специалисту в данной области, например, путем применения сконструированных или измененных экспрессионных штаммов, путем совместного экспрессирования с одним или более ферментами, например, с β(1,4)-N-ацетилглюкозаминилтрансферазой III (GnT-IIII11), путем экспрессирования молекулы, включающей Fc-область, в различных организмах или линиях клеток из различных организмов, или путем модификации углевода(ов) после экспрессирования молекулы, включающей Fc-область. Способы получения сконструированных гликоформ известны в данной области и включают, но не ограничены перечисленными, гликоформы, описанные в Umana и др., 1999, Nat. Biotechnol 17: 176-180; Davies и др., 20017 Biotechnol Bioeng 74:288-294; Shields и др., 2002, J Biol Chem 277:26733-26740; Shinkawa и др., 2003, J Biol Chem 278:3466-3473) в патенте США номер 6602684; в заявке на патент США номер 10/277370; в заявке на патент США номер 10/113929; РСТ WO 00/61739 А1; РСТ WO 01/292246 А1; РСТ WO 02/311140 А1; РСТ WO 02/30954 А1; в технологии Potillegent™ (Biowa, Inc. Princeton, N.J.); в технологии разработки гликозилирования GlycoMAb™ (GLYCART biotechnology AG, Цюрих, Швейцария). См., например, WO 00061739; ЕА 01229125; US 20030115614; Okazaki и др., 2004, JMB, 336:1239-49.
Предполагается, что варианты Fe включают антитела, содержащие вариабельный участок и гетеродимерную Fc-область, при этом гетеродимерная Fc-область содержит модифицированный домен СН3, включающий мутации аминокислот, способствующие образованию гетеродимера с повышенной стабильностью, и при этом модифицированный домен СН3 имеет температуру плавления (Tm) более 70°С. Варианты Fc, которые представляют собой антитела, можно получить "de novo" путем соединения вариабельного домена, или его фрагмента, который специфично связывает по меньшей мере один антиген, с гетеродимерной Fc-областью, содержащей модифицированный домен СН3. В качестве альтернативы, варианты гетеродимера Fc можно получить путем модификации домена СН3 антитела, содержащего Fc-область, которое связывает антиген.
Антитела согласно настоящему изобретению могут включать, но не ограничены перечисленными: синтетические антитела, моноклональные антитела, рекомбинантные антитела, интраантитела, моноспецифические антитела, мультиспецифические антитела, биспецифические антитела, человеческие антитела, гуманизированные антитела, химерные антитела, синтетические антитела, одноцепочечные FvFc (scFvFc), одноцепочечные Fvs (scFv) и антиидиотипические (анти-Ид) антитела. В частности, антитела, используемые в способах согласно настоящему изобретению, включают молекулы иммуноглобулина и иммунологически активные области молекул иммуноглобулина. Молекулы иммуноглобулина согласно настоящему изобретению могут принадлежать к любому типу (например, IgG, IgE, IgM, IgD, IgA и IgY), классу (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2) или подклассу молекул иммуноглобулина.
Антитела согласно настоящему изобретению могут быть любого животного происхождения, включая птиц и млекопитающих (например, человека, мыши, осла, овцы, кролика, козы, морской свинки, верблюда, лошади или цыпленка). В конкретном варианте реализации, антитела представляют собой человеческие или гуманизированные моноклональные антитела, в частности, биспецифические моноклональные антитела. В данной заявке, "человеческие" антитела включают антитела, имеющие последовательность аминокислот иммуноглобулина человека, и включают антитела, выделенные из библиотек иммуноглобулинов человека или из мышей, которые экспрессируют антитела с человеческих генов.
Антитела, подобно всем полипептидам, имеют изоэлектрическую точку (pI), которую обычно определяют как pH, при котором полипептид несет нулевой суммарный заряд. В данной области известно, что растворимость белка обычно наиболее низкая, когда рН раствора равен изоэлектрической точке (pI) белка. Можно оптимизировать растворимость путем изменения количества и расположения ионизируемых остатков в молекуле антитела, чтобы скорректировать pI. Например, pI полипептида можно регулировать путем введения подходящих замен аминокислот (например, путем замены заряженной аминокислоты, такой как лизин, на незаряженный остаток, такой как аланин). Без привязки к какой-либо конкретной теории, замены аминокислот в антителе, которые приводят к изменениям pI указанного антитела, могут улучшать растворимость и/или стабильность антитела. Специалист в данной области поймет, какие замены аминокислот будут наиболее подходящими для конкретного антитела, чтобы добиться желательной pI. pI белка можно определить с помощью множества способов, включая, но не ограничиваясь перечисленными, изоэлектрическое фокусирование и различные компьютерные алгоритмы (см., например, Bjellqvist и др., 1993, Electrophoresis 14:1023). В одном варианте реализации, pI вариантов Fc согласно настоящему изобретению находится между рН 6,2 и рН 8,0. В другом варианте реализации, pI антител согласно настоящему изобретению находится между рН 6,8 и рН 7,4. В одном варианте реализации, замены, приводящие к изменениям pI варианта Fc согласно настоящему изобретению, не будут значительно уменьшать его аффинность связывания с антигеном. Предполагается, что модифицированный домен СН3 с повышенной стабильностью также можно получить путем изменения pI. В одном варианте реализации, варианты гетеродимера Fc специально выбирают таким образом, чтобы они обладали как повышенной стабильностью и чистотой, так и любым желательным изменением pI.
Антитела согласно настоящему изобретению могут быть моноспецифическими, биспецифическими, триспецифическими или обладать большей мультиспецифичностью. Мультиспецифические антитела могут специфично связываться с различными эпитопами желательной целевой молекулы или могут специфично связываться как с целевой молекулой, так и с гетерологичным эпитопом, таким как гетерологичный полипептид или материал твердой подложки. См., например, международные публикации с номерами WO 94/04690; WO 93/17715; WO 92/08802; WO 91/00360; и WO 92/05793; Tutt, и др., 1991, J. Immunol. 147:60-69; патенты США номер; 4474893; 4714681; 4925648; 5573920 и 5601819 и Kostelny и др., 1992, J. Immunol, 148:1547).
Различные варианты реализации мультифункциональных нацеливающих молекул можно разработать на основе данного асимметричного каркаса, как показано на Фигуре 20.
Мультиспецифические антитела обладают специфичностями связывания с по меньшей мере двумя различными антигенами. Хотя такие молекулы обычно будут связывать только два антигена (т.е. биспецифические антитела, БсАТ), антитела с дополнительными специфичностями, такие как триспецифические антитела, также входят в объем настоящего изобретения. Примеры БсАТ включают, без ограничения, такие антитела, у которых одно плечо направлено против антигена опухолевой клетки, а другое плечо направлено против цитотоксической молекулы, или оба плеча направлены против двух различных антигенов опухолевой клетки, или оба плеча направлены против двух различных растворимых лигандов, или одно плечо направлено против растворимого лиганда, а другое плечо направлено против рецептора клеточной поверхности, или оба плеча направлены против двух различных рецепторов клеточной поверхности. Способы получения биспецифических антител известны в данной области.
Согласно отличному подходу, вариабельные домены антитела с желательными специфичностями связывания (сайтами связывания антитело-антиген) сливают с последовательностями константного домена иммуноглобулина. Слияние можно осуществить с константным доменом тяжелой цепи иммуноглобулина, содержащим по меньшей мере часть шарнира, областей СН2 и СН3. Предполагается, что первая константная область тяжелой цепи (СН1), содержащая сайт, необходимый для связывания легкой цепи, присутствует в по меньшей мере одной из слитых молекул. ДНК, кодирующая слитую тяжелую цепь иммуноглобулина и, при необходимости, легкую цепь иммуноглобулина, встраивают в отдельные векторы экспрессии, которыми совместно трансфицируют подходящий организм хозяина. Это обеспечивает большую гибкость регулировки взаимного соотношения трех полипептидных фрагментов в вариантах реализации, когда неравные соотношения трех полипептидных цепей, используемых для конструирования, дают оптимальные выходы. См. Пример 1 и Таблицу 2. Тем не менее, можно вставить кодирующие последовательности для двух или всех трех полипептидных цепей в один вектор экспрессии, если экспрессия по меньшей мере двух полипептидных цепей в равных соотношениях приводит к высокому выходу или если соотношения не имеют особого значения.
Биспецифические антитела включают сшитые поперечными связями или "гетероконъюгированные" антитела. Например, одно из антител в гетероконъюгате может быть связано с авидином, а другое - с биотином. Такие антитела, например, были предложены для нацеливания клеток иммунной системы на нежелательные клетки (патент США номер 4676980) и для лечения ВИЧ-инфекции (WO 91/00360, WO 92/200373 и ЕР 03089). Гетероконъюгированные антитела можно получить, применяя любые удобные способы введения поперечных связей. Подходящие перекрестносшивающие агенты хорошо известны в данной области и описаны в патенте США номер 4676980, наряду с множеством методик введения поперечных связей.
Согласно настоящему изобретению предложены антитела с более чем двумя валентностями, включающие модифицированные домены СН3 и полученные гетеродимеры Fc. Например, можно получить триспецифические антитела. См., например, Tutt и др. J. Immunol. 147:60(1991).
Антитела согласно настоящему изобретению также включают такие антитела, которые имеют время полужизни (например, время полужизни в сыворотке) у млекопитающего (например, у человека) более 15 дней, более 20 дней, более 25 дней, более 30 дней, более 35 дней, более 40 дней, более 45 дней, более 2 месяцев, более 3 месяцев, более 4 месяцев или более 5 месяцев. Увеличенное время полужизни антител согласно настоящему изобретению у млекопитающего (например, у человека) приводит к более высокому титру указанных антител или фрагментов антител в сыворотке млекопитающего и, таким образом, позволяет уменьшить частоту введения указанных антител или фрагментов антител и/или уменьшить вводимую концентрацию указанных антител или фрагментов антител. Антитела, имеющие увеличенное время полужизни in vitro, можно получить с помощью способов, известных специалистам в данной области. Например, антитела с увеличенным временем полужизни in vivo можно получить путем модификации (например, замены, делеции или вставки) аминокислотных остатков, которые идентифицированы как участвующие во взаимодействии между доменом Fc и рецептором FcRn (см., например, международные публикации №№ WO 97/34631; WO 04/029207; патент США №6737056 и патентную публикацию США №2003/0190311).
В конкретном варианте реализации, вариант гетеродимера Fc, содержащий модифицированный домен СН3, представляет собой мультиспецифическое антитело (названное в данной заявке антителом согласно настоящему изобретению), указанное антитело согласно настоящему изобретению специфично связывает интересующий антиген. В частности, антитело согласно настоящему изобретению представляет собой биспецифическое антитело. В одном варианте реализации, антитело согласно настоящему изобретению специфично связывает полипептидный антиген. В другом варианте реализации, антитело согласно настоящему изобретению специфично связывает неполипептидный антиген. В еще одном варианте реализации, введение антитела согласно настоящему изобретению млекопитающему, страдающему от заболевания или расстройства, может привести к терапевтической пользе для данного млекопитающего.
Практически любая молекула может быть мишенью и/или может быть включена в конструкцию варианта гетеродимера Fc, предложенную в данной заявке (например, антитела, Fc-слитые белки), включая, но не ограничиваясь следующим перечнем белков, а также субъединиц, доменов, мотивов и эпитопов, принадлежащих следующему перечню белков: ренин; гормон роста, включая гормон роста человека и гормон роста быка; фактор, стимулирующий выделение гормона роста; паратиреоидный гормон; тиреостимулирующий гормон; липопротеины; альфа-1-антитрипсин; A-цепь инсулина; В-цепь инсулина; проинсулин; фолликулостимулирующий гормон; кальцитонин; лютеинизирующий гормон; глюкагон; факторы свертывания крови, такие как фактор VII, фактор VIIIC, фактор IX, тканевый фактор (TF), и фактор фон Виллебранда; антикоагулянты, такие как белок С; предсердный натрийуретический фактор; легочный сурфактант; активатор плазминогена, такой как урокиназный, или из человеческой мочи, или тканевой активатор плазминогена (t-PA); бомбезин; тромбин; гемопоэтический фактор роста; фактор некроза опухоли-альфа и -бета; энкефалиназа; RANTES (хемокин, экспрессируемый и секретируемый Т-клетками при активации); воспалительный белок макрофагов (MIP-1-альфа) человека; сывороточный альбумин, такой как сывороточный альбумин человека; мюллерова ингибирующая субстанция; A-цепь релаксина; В-цепь релаксина; прорелаксин; гонадотропин-ассоциированный пептид мыши; микробный белок, такой как бета-лактамаза; ДНКаза; IgE; антиген цитотоксических Τ лимфоцитов (CTLA), такой как CTLA-4; ингибин; активин; фактор роста эндотелия сосудов (VEGF); рецепторы гормонов или факторов роста, такие как, например, EGFR, VEGFR; интерфероны, такие как интерферон альфа (α-IFN), интерферон бета (β-IFN) и интерферон гамма (γ-IFN); белок А или D; ревматоидные факторы; нейротрофический фактор, такой как нейротрофический фактор головного мозга (BDNF), нейротрофин-3, -4, -5 или -6 (NT-3, NT-4, NT-5 или NT-6), или фактор роста нервов; фактор роста тромбоцитов (PDGF); фактор роста фибробластов, такой как AFGF и PFGF; эпидермальный фактор роста (EGF); трансформирующий фактор роста (TGF), такой как TGF-альфа и TGF-бета, включая TGF-1, TGF-2, TGF-3, TGF-4 или TGF-5; инсулинподобный фактор роста-I и -II (IGF-I и IGF-II); des(1-3)-IGF-I (IGF-I мозга), белки, связывающие инсулинподобный фактор роста; белки CD, такие как CD2, CD3, CD4, CD8, CD11a, CD14, CD18, CD19, CD20, CD22, CD23, CD25, CD33, CD34, CD40, CD40L, CD52, CD63, CD64, CD80 и CD147; эритропоэтин; остеоиндуктивные факторы; иммунотоксины; костный морфогенетический белок (BMP); интерферон, такой как интерферон альфа, бета и гамма; колониестимулирующие факторы (CSF), такие как M-CSF, GM-CSF и G-CSF; интерлейкины (IL), например, с IL-1 по IL-13; TNFα, супероксиддисмутаза; Т-клеточные рецепторы; поверхностные мембранные белки; стимулятор гемолиза; вирусный антиген, такой как, например, часть оболочки вируса СПИД, например, gp120; транспортные белки; хоминг-рецепторы; адрессины; регуляторные белки; молекулы клеточной адгезии, такие как LFA-1, Mac1, p 150.95, VLA-4, ICAM-1, ICAM-3 и VCAM, интегрин а4/р7, и (интегрин Xv/р3, включая любую из его субъединиц, субъединицы интегрина альфа, такие как CD49a, CD49b, CD49c, CD49d, CD49e, CD49f, альфа7, альфа8, альфа9, альфаD, CD11a, CD11b, CD51, CD11c, CD41, альфаIIb, альфаIELb; субъединицы интегрина бета, такие как CD29, CD18, CD61, CD104, бета5, бета6, бета7 и бета8; комбинации субъединиц интегрина, включая, но не ограничиваясь перечисленными, αVβ3, αVβ5 и α4β7; участник апоптозного пути; IgE; антигены групп крови; рецептор flk2/flt3; рецептор ожирения (ОВ); рецептор mp1; CTLA-4; белок С; рецептор Eph, такой как EphA2, EphA4, EphB2 и т.д.; лейкоцитарный антиген человека (HLA), такой как HLA-DR; белки комплемента, такие как рецептор комплемента CR1, C1Rq и другие факторы комплемента, такие как С3 и С5; рецептор гликопротеина, такой как GpIbα, GPIIb/IIIa и CD200; и фрагменты любого из перечисленных выше полипептидов.
Также предложены антитела согласно настоящему изобретению, которые специфично связывают раковые антигены, включая, но не ограничиваясь перечисленными: рецептор ALK (рецептор плеиотрофина), плеиотрофин, карциномный пан-антиген KS 1/4; антиген карциномы яичника (СА125); простатический кислотный фосфат; простатоспецифический антиген (PSA); ассоциированный с меланомой антиген р97; антиген меланомы gp75; высокомолекулярный антиген меланомы (HMW-MAA); простатоспецифичный мембранный антиген; карциноэмбриональный антиген (CEA); полиморфный эпителиальный муциновый антиген; антиген жировых шариков человеческого молока; ассоциированные с колоректальной опухолью антигены, такие как: CEA, TAG-72, СО17-1A, GICA 19-9, СТА-1 и LEA; антиген-38.13 лимфомы Беркитта; CD19; антиген CD20 человеческой В-лимфомы; CD33; специфические для меланомы антигены, такие как ганглиозид GD2, ганглиозид GD3, ганглиозид GM2 и ганглиозид GM3; опухолеспецифический трансплантационный антиген, находящийся на клеточной поверхности (TSTA); антигены индуцируемой вирусом опухоли, включая Т-антиген ДНКовых вирусов, вызывающих опухоли, и оболочечные антигены РНКовых вирусов, вызывающих опухоли; онкофетальный антиген - альфа-фетопротеин, такой как CEA толстой кишки, онкофетальный гликопротеин трофобластов 5Т4 и онкофетальный антиген опухоли мочевого пузыря; дифференцировочный антиген, такой как антигены L6 и L20 человеческой карциномы легкого; антигены фибросаркомы; антиген Gp37 человеческого Т-клеточного лейкоза; неогликопротеин; сфинголипиды; антигены рака молочной железы, такие как EGFR (рецептор эпидермального фактора роста); NY-BR-16; антиген NY-BR-16 и HER2 (p185HER2); полиморфный эпителиальный муцин (РЕМ); антиген АРО-1 злокачественных человеческих лимфоцитов; дифференцировочный антиген, такой как I-антиген, обнаруженный в фетальных эритроцитах; I-антиген первичной эндодермы, обнаруженный в эритроцитах взрослых; в зародышах до имплантации; I(Ма), обнаруженный при желудочных аденокарциномах; M18, М39, обнаруженный в эпителии молочной железы; SSEA-1, обнаруженный в миелоидных клетках; VEP8; VEP9; Myl; Va4-D5; D156-22, обнаруженный при колоректальном раке; TRA-1-85 (группа крови H); SCP-1, обнаруженный при раке яичка и яичников; С14, обнаруженный при аденокарциноме толстой кишки; F3, обнаруженный при аденокарциноме легкого; АН6, обнаруженный при раке желудка; Y-гаптен; Ley, обнаруженный в клетках эмбриональных карциномы; TL5 (группа крови А); рецептор EGF, обнаруженный в клетках А431; антиген серии Ε1 (группа крови В), обнаруженный при раке поджелудочной железы; FC10.2, обнаруженный в клетках эмбриональной карциномы; антиген желудочной аденокарциномы СО-514 (группа крови Lea), обнаруженный при аденокарциноме; NS-10, обнаруженный при аденокарциномах; СО-43 (группа крови Leb); G49, обнаруженный в рецепторе EGF клеток А431; МН2 (группа крови ALeb/Ley), обнаруженный при аденокарциноме толстой кишки; 19.9, обнаруженный при раке толстой кишки; муцины рака желудка; Т5А7, обнаруженный в миелоидных клетках; R24, обнаруженный при меланоме; 4.2, GD3, D1.1, OFA-1, GM2, OFA-2, GD2 и M1:22:25:8, обнаруженные в клетках эмбриональной карциномы, и SSEA-3 и SSEA-4, обнаруженные в зародышах на стадии 4-8 клеток; антиген кожной Т-клеточной лимфомы; антиген MART-1; антиген Sialy Τn (STn); антиген рака толстой кишки NY-CO-45; антиген рака легкого NY-LU-12, вариант А; антиген аденокарциномы ART1; паранеопластический антиген, ассоциированный с раком головного мозга-яичек (онконейрональный антиген МА2; паранеопластический нейрональный антиген); нейроонкологический вентральный антиген 2 (NOVA2); ген 520 антигена печеночно-клеточной карциномы; ассоциированный с опухолью антиген СО-029; ассоциированные с опухолью антигены MAGE-C1 (антиген рака яичек СТ7), MAGE-B1 (антиген MAGE-XP), MAGE-B2 (DAM6), MAGE-2, MAGE-4-a, MAGE-4-b и MAGE-X2; антиген рака яичек (NY-EOS-1) и фрагменты любого из перечисленных выше полипептидов.
В некоторых вариантах реализации, гетеромультимер, описанный в данной заявке, включает по меньшей мере одно терапевтическое антитело. В некоторых вариантах реализации, терапевтическое антитело связывает целевой раковый антиген. В одном варианте реализации, терапевтическое антитело может быть выбрано из группы, состоящей из абаговомаба, адалимумаба, алемтузумаба, аурограба, бапинейзумаба, базиликсимаба, белимумаба, бевацизумаба, бриакинумаба, канакинумаба, катумаксомаба, цертолизумаба пегола, цетуксимаба, даклизумаба, деносумаба, эфализумаба, галиксимаба, гемтузумаба озогамицина, голимумаба, ибритумомаба тиуксетана, инфликсимаба, ипилимумаба, лумиликсимаба, меполизумаба, мотавизумаба, муромонаба, микограба, натализумаба, нимотузумаба, окрелизумаба, офатумумаба, омализумаба, паливизумаба, панитумумаба, пертузумаба, ранибизумаба, реслизумаба, ритуксимаба, теплизумаба, тоцилизумаба/атлизумаба, тозитумомаба, трастузумаба, ПроксиниумаТМ, РенкарексаТМ, устекинумаба, залутумумаба и любых других антител.
Антитела согласно настоящему изобретению включают производные, которые были модифицированы (т.е., путем присоединения любого типа молекулы к антителу, например, путем ковалентного присоединения). Например, но без ограничения, производные антител включают антитела, которые были модифицированы, например, путем гликозилирования, ацетилирования, пегилирования, фосфорилирования, амидирования, дериватизации путем введения известных защитных/блокирующих групп, протеолитического расщепления, присоединения к клеточному лиганду или другому белку и т.д. Любую из множества химических модификаций можно осуществить с помощью известных методик, включая, но не ограничиваясь перечисленными, специфическое химическое расщепление, ацетилирование, формилирование, метаболический синтез туникамицина и т.д. Кроме того, производное может содержать одну или более неклассических аминокислот.
Антитела или фрагменты антител с увеличенным временем полужизни in vivo можно получить путем присоединения полимерных молекул, таких как высокомолекулярный полиэтиленгликоль (ПЭГ), к антителам или фрагментам антител. ПЭГ можно присоединить к антителам или фрагментам антител с помощью или без мультифункционального линкера, либо посредством сайт-специфичного конъюгирования ПЭГ с N- или С-концом указанных антител или фрагментов антител, либо через эпсилон-аминогруппы, присутствующие на остатках лизина. Следует использовать такую дериватизацию линейного или разветвленного полимера, которая приводит к минимальной потере биологической активности. Степень конъюгирования будут тщательно контролировать с помощью ЭФ в ПААГ/ДСН и масс-спектрометрии, чтобы обеспечить правильное конъюгирование молекул ПЭГ с антителами. Непрореагировавший ПЭГ можно отделить от конъюгатов антитело-ПЭГ, например, с помощью эксклюзионной или ионнообменной хроматографии.
Кроме того, антитела можно конъюгировать с альбумином, чтобы получить антитело или фрагмент антитела, более стабильный in vivo или имеющий большее время полужизни in vivo. Такие методики хорошо известны в данной области, см., например, международные публикации №№ WO 93/15199, WO 93/15200 и WO 01/77137; и европейский патент номер EP 413622. В объем настоящего изобретения входит применение антител или фрагментов антител, конъюгированных или слитых с одной или более молекулами, включая, но не ограничиваясь перечисленными, пептиды, полипептиды, белки, слитые белки, молекулы нуклеиновых кислот, малые молекулы, мимикрирующие агенты, синтетические лекарственные средства, неорганические молекулы и органические молекулы.
В объем настоящего изобретения входит применение антител или фрагментов антител, рекомбинантно слитых или химически конъюгированных (включая как ковалентное, так и нековалентное конъюгирование) с гетерологичным белком или полипептидом (или его фрагментом, например, с полипептидом, состоящим из по меньшей мере 10, по меньшей мере 20, по меньшей мере 30, по меньшей мере 40, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90 или по меньшей мере 100 аминокислот), с получением слитых белков. Слияние не обязательно должно быть непосредственным, оно может осуществляться через линкерные последовательности. Например, антитела можно применять для нацеливания гетерологичных полипептидов на конкретные типы клеток, либо in vitro, или in vivo, путем слияния или конъюгирования антител с антителами, специфичными к конкретным рецепторам клеточной поверхности. Антитела, слитые или конъюгированные с гетерологичными полипептидами, также можно использовать в иммуноанализах и способах очистки in vitro, с применением способов, известных в данной области. См., например, международную публикацию номер WO 93/21232; европейский патент номер ЕР 439095; Naramura и др., 1994, Immunol. Lett. 39:91-99; патент США номер 5474981; Gillies и др., 1992, PNAS 89:1428-1432; и Fell и др., 1991, J Immunol. 146:2446-2452.
Настоящее изобретение дополнительно включает композиции, содержащие гетерологичные белки, пептиды или полипептиды, слитые или конъюгированные с фрагментами антител. Например, гетерологичные полипептиды можно слить или конъюгировать с фрагментом Fab, фрагментом Fd, фрагментом Fv, фрагментом F(ab)2, доменом VH, доменом VL, гипервариабельным участком VH, гипервариабельным участком VL, или с фрагментом перечисленных молекул. Способы слияния или конъюгирования полипептидов с участками антитела хорошо известны в данной области. См., например, патенты США с номерами 5336603; 5622929; 5359046; 5349053; 5447851 и 5112946; европейские патенты с номерами ЕР 307434 и ЕР 367166; международные публикации с номерами WO 96/04388 и WO 91/06570; Ashkenazi и др., 1991, Proc. Natl. Acad. Sci. USA 88: 10535-10539; Zheng и др., 1995, J. Immunol. 154:5590-5600; и Vi1 и др., 1992, Proc. Natl. Acad Sci. USA 89:11337-11341.
Дополнительные слитые белки, например, с антителами, которые специфично связывают антиген (например, см. выше), можно получить с помощью методик перетасовки генов, перетасовки мотивов, перетасовки экзонов и/или перетасовки кодонов (совместно именуемые "перетасовками ДНК"). Перетасовки ДНК можно использовать для изменения активностей антител согласно настоящему изобретению или фрагментов указанных антител (например, для получения антител или фрагментов антител с более высокими аффинностями и более низкими скоростями диссоциации). См., в общем, патенты США с номерами 5605793; 5811238; 5830721; 5834252 и 5837458, и Patten и др., 1997, Curr. Opinion Biotechnol. 8:724-33; Harayama, 1998, Trends Biotechnol. 16(2):76-82; Hansson, и др., 1999, J. Mol. Biol. 287:265-76; и Lorenzo и Blasco, 1998, Biotechniques 24(2):308-313. Антитела или фрагменты антител, или кодированные антитела или фрагменты антител, можно изменить путем осуществления неспецифического мутагенеза с помощью ПЦР с использованием неточной полимеразы, произвольной вставки нуклеотидов или других способов, перед проведением рекомбинации. Один или более участков полинуклеотида, кодирующего антитело или фрагмент антитела, из участков, которые специфично связываются с антигеном, можно рекомбинировать с одним или более компонентами, мотивами, сегментами, частями, доменами, фрагментами и т.д. одной или более гетерологичных молекул.
В объем настоящего изобретения дополнительно входят применения вариантов гетеродимера Fe или его фрагментов, конъюгированных с терапевтическим агентом.
Антитело или фрагмент антитела можно конъюгировать с терапевтической молекулой, такой как цитотоксин, например, цитостатический или разрушающий клетки агент, терапевтический агент или ион радиоактивного металла, например, альфа-излучатели. Цитотоксин или цитотоксический агент включает любой агент, который неблагоприятен для клеток. Примеры включают рибонуклеазу, монометилауристатин Ε и F, паклитаксел, цитохалазин В, грамицидин D, бромистый этидий, эметин, митомицин, этопозид, тенопозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин, дигидроксиантрациндион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропранолол, пуромицин, эпирубицин и циклофосфамид и аналоги или гомологи перечисленных агентов. Терапевтические агенты включают, но не ограничены перечисленными, антиметаболиты (например, метотрексат, 6-меркаптопурин, 6-тиогуанин, цитарабин, 5-фторурацилдекарбазин), алкилирующие агенты (например, мехлоретамин, тиотепа, хлорамбуцил, мелфалан, кармустин (BCNU) и ломустин (CCNU), циклофосфамид, бусульфан, дибромоманнит, стрептозотоцин, митомицин С и диаминдихлорплатину (II) (DDP, цисплатин), антрациклины (например, даунорубицин (ранее называемый дауномицином) и доксорубицин), антибиотики (например, дактиномицин (ранее называемый актиномицином), блеомицин, митрамицин и антрамицин (АМС)), и антимитотические агенты (например, винкристин и винбластин). Более широкий перечень терапевтических молекул можно найти в публикации согласно РСТ WO 03/075957.
Кроме того, антитело или фрагмент антитела можно конъюгировать с терапевтическим агентом или лекарственной молекулой, которая изменяет исходный биологический ответ. Терапевтические агенты или лекарственные молекулы не должны рассматриваться как ограниченные классическими химическими терапевтическими агентами. Например, лекарственная молекула может представлять собой белок или полипептид, обладающий желательной биологической активностью. Такие белки могут включать, например, токсин, такой как абрин, рицин А, онконаза (или другая цитотоксическая РНКаза), экзотоксин синегнойной палочки, холерный токсин или дифтерийный токсин; белок, такой как фактор некроза опухоли, α-интерферон, β-интерферон, фактор роста нервов, тромбоцитарный фактор роста, тканевой активатор плазминогена, апоптотический агент, например, TNF-α, TNF-β, AIM I (см. международную публикацию номер WO 97/33899), AIM II (см., международную публикацию номер WO 97/34911), Fas-лиганд (Takahashi и др., 1994, J. Immunol., 6:1567), и VEGI (см. международную публикацию номер WO 99/23105), тромботический агент или антиангиогенный агент, например, ангиостатин или эндостатин; или модификатор биологического ответа, такой как, например, лимфокин (например, интерлейкин-1 ("IL-1"), интерлейкин-2 ("IL-2"), интерлейкин-6 ("IL-6"), гранулоцитарно-макрофагальный колониестимулирующий фактор ("GM-CSF"), гранулоцитарный колониестимулирующий фактор ("G-CSF") или фактор роста (например, гормон роста ("GH")).
Более того, антитело можно конъюгировать с терапевтическими молекулами, такими как радиоактивные материалы или макроциклические хелаторы, подходящие для конъюгирования ионов радиоактивных металлов (примеры радиоактивных материалов см. выше). В некоторых вариантах реализации, макроциклический хелатор представляет собой 1,4,7,10-тетраазациклододекан-N,N',N'',N''-тетрауксусную кислоту (DOTA), которую можно присоединить к антителу через линкерную молекулу. Такие линкерные молекулы широко известны в данной области и описаны в Denardo и др., 1998, Clin Cancer Res. 4:2483; Peterson и др., 1999, Bioconjug. Chem. 10:553; и Zimmerman и др., 1999, Nucl. Med. Biol. 26:943.
Способы слияния или конъюгирования антител с полипептидными молекулами известны в данной области. См., например, патенты США с номерами 5336603; 5622929; 5359046; 5349053; 5447851 и 5112946; ЕР 307434; ЕР 367166; публикации согласно РСТ WO 96/04388 и WO 91/06570; Ashkenazi и др., 1991, PNAS USA 88:10535; Zheng и др., 1995, J Immunol 154:5590; и Vil и др., 1992, PNAS USA 89:1 1337. Слияние антитела с молекулой не обязательно должно быть непосредственным, оно также может осуществляться через линкерные последовательности. Такие линкерные молекулы широко известны в данной области и описаны в Denardo и др., 1998, Clin Cancer Res 4:2483; Peterson и др., 1999, Bioconjug Chem 10:553; Zimmerman и др., 1999, Nucl Med Biol 26:943; Garnett, 2002, Adv Drug Deliv Rev 53:171.
Рекомбинантная экспрессия варианта Fc или его производного, аналога или фрагмента (например, антитела или слитого белка согласно настоящему изобретению) требует конструирования вектора экспрессии, содержащего полинуклеотид, который кодирует вариант Fc (например, антитело или слитый белок). Сразу после получения полинуклеотида, кодирующего вариант Fc (например, антитело или слитый белок), вектор для продукции варианта Fc (например, антитела или слитого белка) можно получить с помощью технологии рекомбинантной ДНК, применяя методики, хорошо известные в данной области. Таким образом, способы получения белка путем экспрессии полинуклеотида, содержащего последовательность нуклеотидов, кодирующую вариант Fc (например, антитело или слитый белок), описаны в данной заявке. Способы, которые хорошо известны специалистам в данной области, можно применять для конструирования векторов экспрессии, содержащих последовательности, кодирующие вариант Fe (например, антитело или слитый белок), и подходящие сигналы, контролирующие транскрипцию и трансляцию. Данные способы включают, например, методики рекомбинантной ДНК in vitro, методики синтеза и генетическую рекомбинацию in vivo. В настоящем изобретении, таким образом, предложены реплицируемые векторы, содержащие последовательность нуклеотидов, кодирующую вариант Fc согласно настоящему изобретению, функционально связанную с промотором. Такие векторы могут включать последовательность нуклеотидов, кодирующую константную область молекулы антитела (см., например, международную публикацию номер WO 86/05807; международную публикацию номер WO 89/01036; и патент США номер 5122464) и вариабельный домен антитела, или полипептид для получения варианта Fc можно клонировать в такой вектор для экспрессии полноразмерной цепи антитела (например, тяжелой или легкой цепи) или полного варианта Fc, включая слияние полипептида, не относящегося к антителу, и Fc-области, содержащей по меньшей мере модифицированный домен СН3.
Вектор экспрессии переносят в клетку-хозяина с помощью обычных методик, и трансфицированные клетки затем культивируют с помощью обычных методик для получения варианта Fc согласно настоящему изобретению. Таким образом, настоящее изобретение включает клетки-хозяева, содержащие полинуклеотид, кодирующий вариант Fc согласно настоящему изобретению, функционально связанный с гетерологичным промотором. В конкретных вариантах реализации, для экспрессии вариантов Fc, включающих двухцепочечные антитела, векторы, кодирующие как тяжелые, так и легкие цепи, можно совместно экспрессировать в клетке-хозяине для экспрессии целой молекулы иммуноглобулина, как подробно описано ниже.
Множество систем хозяин-вектор экспрессии можно использовать для экспрессии вариантов Fe согласно настоящему изобретению (например, молекул антитела или слитого белка) (см., например, патент США номер 5807715). Такие системы хозяин-вектор экспрессии представляют собой средства, с помощью которых интересующие кодирующие последовательности можно получить, а затем очистить, а также представляют собой клетки, которые после трансформирования или трансфицирования подходящими кодирующими последовательностями нуклеотидов могут экспрессировать in situ вариант Fc согласно настоящему изобретению. Такие системы включают, но не ограничены перечисленными: микроорганизмы, такие как бактерии (например, Е. coli и В. subtilis), трансформированные рекомбинантными векторами экспрессии ДНК бактериофага, плазмидной ДНК или космидной ДНК, содержащими последовательности, кодирующие вариант Fc; дрожжи (например, Saccharomyces Pichia), трансформированные рекомбинантными дрожжевыми векторами экспрессии, содержащими последовательности, кодирующие вариант Fc; системы клеток насекомого, зараженных рекомбинантными вирусными векторами экспрессии (например, бакуловирусными), содержащими последовательности, кодирующие вариант Fc; системы клеток растения, зараженных рекомбинантными вирусными векторами экспрессии (например, вирусом мозаики цветной капусты, CaMV; вирусом табачной мозаики, TMV) или трансформированных рекомбинантными плазмидными векторами экспрессии (например, плазмидой Ti), содержащими последовательности, кодирующие вариант Fc; или системы клеток млекопитающего (например, клеток COS, СНО, BHK, 293, NS0 и 3Т3), несущие рекомбинантные экспрессионные конструкции, включающие промоторы, полученные из генома клеток млекопитающих (например, промотор металлотионеинов) или из вирусов млекопитающих (например, из позднего промотора аденовируса; промотора вируса коровьей оспы 7.5K). В некоторых вариантах реализации, бактериальные клетки, такие как Escherichia coli, или эукариотические клетки используют для экспрессии варианта Fc, который представляет собой рекомбинантное антитело или молекулы слитого белка. Например, клетки млекопитающих, такие как клетки яичника китайского хомячка (СНО), в сочетании с вектором, таким как главный промоторный элемент промежуточно-ранних генов из цитомегаловируса человека, представляют собой эффективную систему экспрессии антител (Foecking и др., 1986, Gene 45:101; и Cockett и др., 1990, Bio/Technology 8:2). В конкретном варианте реализации, экспрессия последовательностей нуклеотидов, кодирующих вариант Fc согласно настоящему изобретению (например, антитело или слитый белок), регулируется конститутивным промотором, индуцируемым промотором или тканеспецифичным промотором.
В бактериальных системах можно успешно выбрать множество векторов экспрессии в зависимости от применения, которое предполагается для экспрессируемого варианта Fc (например, антитела или слитого белка). Например, если нужно получить большие количества такого белка для получения фармацевтических композиций варианта Fc, могут оказаться желательными векторы, которые направляют экспрессию высоких уровней продукта слитого белка, который легко очистить. Такие векторы включают, но не ограничены перечисленными, вектор экспрессии Е. coli pUR278 (Ruther и др., 1983, ЕМВО 12:1791), в котором последовательность, кодирующую вариант Fc, можно лигировать в вектор отдельно в одной рамке считывания с участком, кодирующим lac Ζ, таким образом, чтобы продуцировался lac Z-слитый белок; векторы pIN (Inouye и Inouye, 1985, Nucleic Acids Res. 13:3101-3109; Van Heeke и Schuster, 1989, J. Biol. Chem. 24:5503-5509); и тому подобные векторы. Векторы pGEX также можно применять для экспрессии чужеродных полипептидов в виде белков, слитых с глутатион-S-трансферазой (GST). Обычно, такие слитые белки оказываются растворимыми, и их легко можно очистить от лизированных клеток путем адсорбции и связывания с матриксом глутатион-агарозных гранул с последующим элюированием в присутствии свободного глутатиона. Векторы pGEX разработаны таким образом, чтобы они включали сайты расщепления протеазами тромбина или фактора Ха, таким образом, чтобы продукт клонированного целевого гена мог быть освобожден от молекул GST.
В системе насекомых, вирус ядерного полиэдроза Autographa califomica (AcNPV) используют в качестве вектора для экспрессии чужеродных генов. Вирус растет в клетках Spodoptera frugiperda. Последовательность, кодирующую вариант Fc (например, антитело или слитый белок), можно клонировать отдельно в несущественные области (например, в ген полиэдрина) вируса и поместить под контроль промотора AcNPV (например, промотора полиэдрина).
В клетках-хозяевах из млекопитающих можно применять множество систем экспрессии на основе вирусов. В случаях, когда в качестве вектора экспрессии используют аденовирус, представляющую интерес последовательность, кодирующую вариант Fc (например, антитело или слитый белок), можно лигировать с аденовирусным комплексом, контролирующим транскрипцию/трансляцию, например с поздним промотором и тройной лидерной последовательностью. Данный химерный ген можно затем встроить в геном аденовируса путем рекомбинации in vitro или in vivo. Вставка в несущественную область вирусного генома (например, в область E1 или Е3) позволит получить рекомбинантный вирус, который жизнеспособен и способен экспрессировать в вариант Fc (например, антитело или слитый белок) в инфицированных хозяевах (например, см. Logan и Shenk, 1984, Proc. Natl. Acad. Sci. USA 81:355-359). Для эффективной трансляции встроенных последовательностей, кодирующих антитело, могут также потребоваться определенные сигналы инициации. Данные сигналы включают инициирующий кодон ATG и прилегающие последовательности. Кроме того, инициирующий кодон должен быть в фазе с рамкой считывания желательной кодирующей последовательности для обеспечения трансляции всей вставки. Данные экзогенные сигналы контроля трансляции и инициирующие кодоны могут происходить из различных источников, как природных, так и синтетических. Эффективность экспрессии можно повысить путем включения подходящих энхансерных элементов транскрипции, терминаторов транскрипции и т.д. (см., например, Bittner и др., 1987, Methods in Enzymol. 153:516-544).
Экспрессию варианта Fc (например, антитела или слитого белка) можно контролировать с помощью любого промоторного или энхансерного элемента, известного в данной области. Промоторы, которые можно применять для контроля экспрессии гена, кодирующего вариант Fc (например, антитело или слитый белок), включают, но не ограничены перечисленными: область раннего промотора SV40 (Bernoist и Chambon, 1981, Nature 290:304-310), промотор, содержащийся в положении 3' длинного трминального повтора вируса саркомы Рауса (Yamamoto, и др., 1980, Cell 22:787-797), промотор тимидинкиназы герпеса (Wagner и др., 1981, Proc. Natl. Acad. Sci. U.S.A. 78:1441-1445), регуляторные последовательности гена металлотионеина (Brinster и др., 1982, Nature 296:39-42), промотор тетрациклина (Tet) (Gossen и др., 1995, Proc. Nat. Acad. Sci. USA 89:5547-5551); прокариотические векторы экспрессии, такие как промотор β-лактамазы (Villa-Kamaroff и др., 1978, Proc. Natl. Acad. Sci. U.S.A. 75:3727-3731), или tac-промотор (DeBoer и др., 1983, Proc. Natl. Acad. Sci. U.S.A. 80:21-25; см. также "Useful proteins from recombinant bacteria" в Scientific American, 1980, 242:74-94); векторы экспрессии в растениях, включая участок промотора нопалинсинтетазы (Herrera-Estrella и др., Nature 303:209-213) или промотор 35S РНК вируса мозаики цветной капусты (Gardner и др., 1981, Nucl. Acids Res. 9:2871), и промотор фотосинтетического фермента рибулозобисфосфаткарбоксилазы (Herrera-Estrella и др., 1984, Nature 310: 115-120); промоторные элементы из дрожжей или других грибов, такие как промотор Gal4, промотор ADC (алкогольдегидрогеназы), промотор PGK (фосфоглицеринкиназы), промотор щелочной фосфатазы, и следующие участки, контролирующие транскрипцию у животных, которые проявляют тканеспецифичность и используются у трансгенных животных: регуляторный участок гена эластазы I, который активен в ацинарных клетках поджелудочной железы (Swift и др., 1984, Cell 38:639-646; Ornitz и др., 1986, Cold Spring Harbor Symp. Quant. Biol. 50:399-409; MacDonald, 1987, Hepatology 7:425-515); регуляторный участок гена инсулина, который активен в бета-клетках поджелудочной железы (Hanahan, 1985, Nature 315:115-122), регуляторный участок гена иммуноглобулина, который активен в лимфоидных клетках (Grosschedl и др., 1984, Cell 38:647-658; Adames и др., 1985, Nature 318:533-538; Alexander и др., 1987, Mol. Cell. Biol. 7: 1436-1444), регуляторный участок вируса опухоли молочной железы мыши, который активен в тестикулярных, лимфоидных, тучных клетках и клетках молочной железы (Leder и др., 1986, Cell 45:485-495), регуляторный участок гена альбумина, который активен в печени (Pinkert и др., 1987, Genes and Devel. 1:268-276), регуляторный участок гена альфа-фетопротеина, который активен в печени (Krumlauf и др., 1985, Mol. Cell. Biol. 5: 1639-1648; Hammer и др., 1987, Science 235:53-58); регуляторный участок гена альфа-1-антитрипсина, который активен в печени (Kelsey и др., 1987, Genes and Devel. 1:161-171), регуляторный участок гена бета-глобина, активный в миелоидных клетках (Mogram и др., 1985, Nature 315:338-340; Kollias и др., 1986, Cell 46:89-94); регуляторный участок гена основного белка миелина, который активен в олигодендроцитах головного мозга (Readhead и др., 1987, Cell 48:703-712); регуляторный участок гена легкой цепи-2 миозина, который активен в скелетных мышцах (Sani, 1985, Nature 314:283-286); регуляторный участок гена нейрон-специфической енолазы (NSE), который активен в нервных клетках (Morelli и др., 1999, Gen. Virol. 80:571-83); регуляторный участок гена нейротрофического фактора головного мозга (BDNF), который активен в нервных клетках (Tabuchi и др., 1998, Biochem. Biophysic. Res. Com. 253:818-823); промотор глиального фибриллярного кислого белка (GFAP), который активен в астроцитах (Gomes и др., 1999, Braz J Med Biol Res 32(5): 619-631; Morelli и др., 1999, Gen. Virol. 80:571-83) и регуляторный участок гена гонадотропин-рилизинг гормона, который активен в гипоталамусе (Mason и др., 1986, Science 234:1372-1378).
Векторы экспрессии, содержащие вставки гена, кодирующего вариант Fc согласно настоящему изобретению (например, антитело или слитый белок), можно идентифицировать с помощью трех основных подходов: (а) гибридизация нуклеиновых кислот, (b) наличие или отсутствие функций "маркерного" гена и (с) экспрессия вставленных последовательностей. В первом подходе, присутствие гена, кодирующего пептид, полипептид, белок или слитый белок, в векторе экспрессии можно детектировать путем гибридизации нуклеиновых кислот, применяя пробы, включающие последовательности, которые гомологичны вставленному гену, кодирующему пептид, полипептид, белок или слитый белок, соответственно. Во втором подходе, систему рекомбинантный вектор/хозяин можно идентифицировать и выбрать на основании наличия или отсутствия некоторых функций "маркерного" гена (например, тимидинкиназной активности, устойчивости к антибиотикам, трансформированного фенотипа, образования телец включения в бакуловирусе и т.д.), обусловленных вставкой в вектор последовательности нуклеотидов, кодирующей антитело или слитый белок. Например, если последовательность нуклеотидов, кодирующую вариант Fc (например, антитело или слитый белок), вставить в последовательность маркерного гена вектора, рекомбинанты, содержащие встроенный ген, кодирующий антитело или слитый белок, можно идентифицировать по отсутствию функции маркерного гена. В третьем подходе, рекомбинантные векторы экспрессии можно идентифицировать, анализируя продукт гена (например, антитело или слитый белок), экспрессируемый рекомбинантом. Такие анализы могут основываться, например, на физических и функциональных свойствах слитого белка в системах анализа in vitro, например, на связывании с антителом против биологически активной молекулы.
Кроме того, можно выбрать штамм клеток-хозяев, который модулирует экспрессию встроенных последовательностей или модифицирует и процессирует продукт гена определенным необходимым образом. Экспрессию при использовании некоторых промоторов можно повысить в присутствии некоторых индукторов; таким образом, можно контролировать экспрессию полученного методом генетической инженерии слитого белка. Более того, у различных клеток-хозяев имеются характеристические и специфические механизмы трансляционного и посттрансляционного процессинга и модификации (например, гликозилирования, фосфорилирования) белков. Можно выбрать подходящие линии клеток или системы хозяев, чтобы обеспечить заданную модификацию и процессинг экспрессируемого чужеродного белка. Например, экспрессия в бактериальной системе позволит получить негликозилированный продукт и экспрессия в дрожжах позволит получить гликозилированный продукт.
Можно использовать эукариотические клетки-хозяева, которые обладают клеточным аппаратом для правильного процессинга первичного транскрипта (например, гликозилирования и фосфорилирования продукта гена). Такие клетки-хозяева млекопитающих включают, но не ограничены перечисленными: СНО, VERY, BHK, Hela, COS, MDCK, 293, 3Т3, WI38, NS0 и, в частности, линии нервных клеток, такие как, например, клетки нейробластом человека SK-N-AS, SK-N-FI, SK-N-DZ (Sugimoto и др., 1984, J. Natl. Cancer Inst. 73:51-57), нейробластомы человека SK-N-SH (Biochim. Biophys. Acta, 1982, 704:450-460), мозжечковой медуллобластомы человека Daoy (Не и др., 1992, Cancer Res. 52:1144-1148), клетки глиобластомы DBTRG-05MG (Kruse и др., 1992, In Vitro Cell. Dev. Biol. 28A: 609-614), нейробластомы человека IMR-32 (Cancer Res., 1970, 30:2110-2118), астроцитомы человека 1321N1 (Proc. Natl. Acad. Sci. USA, 1977, 74:4816), астроцитомы человека MOG-G-CCM (Br. J. Cancer, 1984, 49:269), глиобластомы-астроцитомы человека U87MG (Acta Pathol. Microbiol. Scand., 1968, 74:465-486), глиобластомы человека A172 (Olopade и др., 1992, Cancer Res. 52:2523-2529), клетки глиомы крысы С6 (Benda и др., 1968, Science 161:370-371), нейробластомы мыши Neuro-2а (Proc. Natl. Acad. Sci. USA, 1970, 65:129-136), нейробластомы мыши NB41A3 (Proc. Natl. Acad. Sci. USA, 1962, 48:1184-1190), хориоидного сплетения овцы SCP (Bolin и др., 1994, J. Virol. Methods 48:211-221), G355-5, нормальных астроцитов кошки PG-4 (Haapala и др., 1985, J. Virol. 53:827-833), головного мозга хорька Mpf (Trowbridge и др., 1982, In Vitro 18:952-960) и нормальные линии клеток, таких как, например, клетки нормальной мозговой коры крысы СТХ TNA2 (Radany и др., 1992, Proc. Natl. Acad. Sci. USA 89:6467-6471), такие как, например, CRL7030 и Hs578Bst. Более того, различные системы экспрессии вектор/хозяин могут в различной степени влиять на реакции процессинга.
Для длительного получения рекомбинантных белков с высоким выходом часто предпочтительна стабильная экспрессия. Например, можно разработать линии клеток, которые стабильно экспрессируют вариант Fc согласно настоящему изобретению (например, антитело или слитый белок). Вместо того, чтобы использовать векторы экспрессии, которые содержат вирусные точки начала репликации, клетки-хозяева можно трансформировать ДНК, контролируемой с помощью подходящих регулирующих экспрессию элементов (например, последовательностей промотора, энхансера, терминаторов транскрипции, сайтов полиаденилирования и т.д.) и селектируемого маркера. После введения чужеродной ДНК разработанным клеткам позволяют расти в течение 1-2 дней в обогащенной среде, а затем переводят их на селективную среду. Селектируемый маркер в рекомбинантной плазмиде придает устойчивость к селекции и дает клеткам возможность стабильно интегрировать плазмиду в хромосомы клеток и расти с образованием фокусов, которые, в свою очередь, можно клонировать и нарастить с получением линий клеток. Данный способ можно успешно применять для конструирования линий клеток, которые экспрессируют вариант Fc, который специфично связывается с антигеном. Такие сконструированные линии клеток могут быть особенно полезны для скрининга и оценки соединений, которые влияют на активность варианта Fc (например, полипептида или слитого белка), который специфично связывается с антигеном.
Можно применять множество систем селекции, включая, но не ограничиваясь перечисленными: гены тимидинкиназы вируса простого герпеса (Wigler и др., 1977, Cell 11:223), гипоксантингуанинфосфорибозилтрансферазы (Szybalska & Szybalski, 1962, Proc. Natl. Acad. Sci. USA 48:2026) и аденинфосфорибозилтрансферазы (Lowy и др., 1980, Cell 22:817) в клетках tk, hgprt или aprt, соответственно. Также можно использовать устойчивость к антиметаболитам в качестве основы для селекции следующих генов: dhfr, который придает устойчивость к метотрексату (Wigler и др., 1980, Natl. Acad. Sci. USA 77:3567; O'Hare и др., 1981, Proc. Natl. Acad. Sci. USA 78:1527); gpt, который придает устойчивость к микофеноловой кислоте (Mulligan & Berg, 1981, Proc. Natl. Acad. Sci. USA 78:2072); neo, который придает устойчивость к аминогликозиду G-418 (Colberre-Garapin и др., 1981, J. Mol. Biol. 150:1); и hygro, который придает устойчивость к гигромицину (Santerre и др., 1984, Gene 30:147).
Сразу после получения варианта Fc (например, антитела или слитого белка) согласно настоящему изобретению путем рекомбинантной экспрессии, его можно очистить с помощью любого способа очистки белка, известного в данной области, например, с помощью хроматографии (например, ионообменной, аффинной, в особенности аффинной к определенному антигену после белка А, и эксклюзионной колоночной хроматографии), центрифугирования, дифференциальной растворимости или с помощью любой другой стандартной методики очистки белков.
Вариант Fc, как правило, получают из культуральной среды в виде секретируемого полипептида, хотя его также можно получить из лизата клеток-хозяев, если он получен напрямую без секреторного сигнала. Если вариант Fc получен связанным с мембраной, его можно высвободить из мембраны, применяя подходящий раствор детергента (например, Triton -X100).
Если вариант Fc получен в рекомбинантной клетке, отличной от клетки человеческого происхождения, он полностью свободен от белков или полипептидов человеческого происхождения. Тем не менее необходимо очистить вариант Fc от белков или полипептидов рекомбинантной клетки, чтобы получить препараты, которые по существу гомогенны в отношении варианта Fc. На первом этапе культуральную среду или лизат, как правило, центрифугируют, чтобы удалить нерастворимые обломки клеток.
Гетеродимеры Fc, содержащие константные домены антитела, можно удобно очистить с помощью гидроксиапатитной хроматографии, электрофореза в геле, диализа или аффинной хроматографии, при этом аффинная хроматография представляет собой предпочтительный способ очистки. Также доступны другие методики очистки белка, такие как фракционирование на ионообменной колонке, преципитация этанолом, обращенно-фазовая ВЭЖХ, хроматография на силикагеле, хроматография на гепарин-сефарозе, хроматография на анион- или катионообменной смоле (например, на колонке с полиаспарагиновой кислотой), хроматофокусирование, ЭФ в ПААГ/ДСН и преципитация сульфатом аммония, в зависимости от очищаемого полипептида. Пригодность белка А в качестве аффинного лиганда зависит от используемых видов и изотипов домена Fc иммуноглобулина. Белок А можно применять для очистки Fc-областей иммуноглобулина, которые получены на основе тяжелых цепей γ1, γ2 или γ4 человека (Lindmark и др., J. Immunol. Meth. 62:1-13 (1983)). Белок G рекомендуют для всех изотипов мыши и для γ3 человека (Guss и др., ЕМВО J. 5:15671575 (1986)). Матрица, к которой присоединен аффинный лиганд, чаще всего представляет собой агарозу, но также доступны другие матрицы. Механически стабильные матрицы, такие как стекло с контролируемым размером пор или поли(стирендивинил)бензол, допускают более высокие скорости потока и более короткие времена обработки, чем те, которые могут быть достигнуты в случае агарозы. Условия для связывания иммуноадгезина с аффинной колонкой с белком А или G полностью обусловлены свойствами домена Fc; а именно, его видами и изотипами. Как правило, при правильном выборе лиганда происходит эффективное связывание непосредственно из неподготовленной культуральной жидкости. Связанные варианты гетеродимера Fc можно эффективно элюировать буфером либо с кислым pH (равным или более 3,0), либо с нейтральным pH, содержащим умеренно хаотропную соль. Данный этап аффинной хроматографии позволит получить препарат варианта гетеродимера Fc, чистый на >95%.
Уровни экспрессии варианта Fc (например, антитела или слитого белка) можно увеличить путем амплификации вектора (смотри обзор у Bebbington и Hentschel, The use of vectors based on gene amplification for the expression of cloned genes in mammalian cells in DNA cloning, том 3 (Academic Press, Нью-Йорк, 1987)). Например, если маркер в векторной системе, экспрессирующей антитело или слитый белок, амплифицируемый, повышение уровня ингибитора, присутствующего в культуре клеток-хозяев, повысит число копий маркерного гена. Поскольку амплифицированный участок связан с геном антитела, продукция антитела или слитого белка также возрастет (Crouse и др., 1983, Mol. Cell. Biol. 3:257).
Клетку-хозяина можно совместно трансфицировать двумя векторами экспрессии согласно настоящему изобретению. Например, первый вектор кодирует полипептид, разработанный на основе тяжелой цепи, и второй вектор кодирует полипептид, разработанный на основе легкой цепи. Указанные два вектора могут включать идентичные селектируемые маркеры, которые дают возможность экспрессировать полипептиды тяжелой и легкой цепи в равных количествах. В качестве альтернативы, можно использовать отдельный вектор, который кодирует и способен экспрессировать слитый белок или оба полипептида тяжелой и легкой цепей. Кодирующие последовательности для слитого белка или тяжелых и легких цепей могут включать кДНК или геномную ДНК.
Характеристические и функциональные анализы
Варианты Fc (например, антитела или слитые белки) согласно настоящему изобретению можно охарактеризовать множеством способов. В одном варианте реализации, чистоту вариантов гетеродимера Fc оценивают, применяя методики, хорошо известные в данной области, включая, но не ограничиваясь перечисленными: электрофорез в ПААГ/ДСН, вестерн-блоттинг, денситометрию или масс-спектрометрию. Стабильность белка можно охарактеризовать, применяя множество методик, не ограниченных перечисленными: эксклюзионную хроматографию, спектроскопию в УФ, видимом свете и спектроскопию кругового дихроизма, масс-спектроскопию, дифференциальное рассеяние света, анализ стабильности пробоподготовки, замораживание-оттаивание, в сочетании с другими методиками определения характеристик, дифференциальную сканирующую калориметрию, дифференциальную сканирующую флуориметрию, хроматографию гидрофобных взаимодействий, изоэлектрическое фокусирование, анализы на связывание рецептора или относительные уровни экспрессии белка. В типичном варианте реализации, стабильность вариантов гетеродимера Fc оценивают по температуре плавления модифицированного домена СН3, по сравнению с доменом СН3 дикого типа, применяя методики, хорошо известные в данной области, такие как дифференциальная сканирующая калориметрия или дифференциальная сканирующая флуориметрия.
Также можно проанализировать способность вариантов Fc согласно настоящему изобретению специфично связываться с лигандом (например, FcγRIIIA, FcγRIIB, C1q). Такой анализ можно осуществить в растворе (например, Houghten, Bio/Techniques, 13:412-421, 1992), на гранулах (Lam, Nature, 354:82-84, 1991), на микрочипах (Fodor, Nature, 364:555-556, 1993), на бактериях (патент США номер 5223409) на плазмидах (Cull и др., Proc. Natl. Acad. Sci. USA, 89: 1865-1869, 1992) или на фаге (Scott и Smith, Science, 249:386-390, 1990; Devlin, Science, 249:404-406, 1990; Cwirla и др., Proc. Natl. Acad. Sci. USA, 87:6378-6382, 1990; и Felici, J. Mol. Biol., 222:301-310, 1991). Молекулы, которые обнаружили специфично связанными с лигандом (например, с FcγRIIIA, FcγRIIB, C1q или антигеном), затем можно подвергнуть анализу на аффинность к лиганду.
Варианты Fc согласно настоящему изобретению можно подвергнуть анализу на специфичное связывание с молекулой, такой как антиген (например, раковый антиген и перекрестная реактивность с другими антигенами) или лиганд (например, FcγR), с помощью любого способа, известного в данной области. Иммуноанализы, которые можно применять для анализа специфичного связывания и перекрестной реактивности, включают, но не ограничены перечисленными: конкурентные и неконкурентные системы анализа с применением таких методик, как вестерн-блоттинг, радиоиммуноанализы, ELISA (твердофазный иммуноферментный анализ), "сэндвич"-иммуноанализы, анализы иммунопреципитации, реакции преципитации, реакции диффузной преципитации в геле, методы иммунодиффузии, агглютинационные анализы, анализы фиксации комплемента, иммунорадиометрические анализы, флуоресцентные иммуноанализы, иммуноанализы на белке А, среди прочих. Такие анализы стандартны и хорошо известны в данной области (см., например, Ausubel и др., ред., 1994, Current Protocols in Molecular Biology, том 1, John Wiley & Sons, Inc., Нью-Йорк).
Аффинность связывания вариантов Fc согласно настоящему изобретению с молекулой, такой как антиген или лиганд (например, FcγR), и скорость диссоциации взаимодействия можно определить с помощью конкурентных анализов на связывание. Один пример конкурентного анализа на связывание представляет собой радиоиммуноанализ, включающий инкубацию меченого лиганда, такого как FcγR (например, меченого 3Н или 125I), с интересующей молекулой (например, вариантами Fe согласно настоящему изобретению) в присутствии возрастающих количеств немеченого лиганда, такого как FcγR, и детектирование молекул, связанных с меченым лигандом. Аффинность молекул согласно настоящему изобретению к лиганду и скорости диссоциации связывания можно определить по результатам насыщения с помощью анализа Скэтчарда.
Кинетические параметры варианта Fc также можно определить, применяя анализы на основе поверхностного плазмонного резонанса (ППР), известные в данной области (например, кинетический анализ BIAcore). Обзор технологии на основе ППР смотри у Mullet и др., 2000, Methods 22:77-91; Dong и др., 2002, Review in Mol. Biotech., 82:303-23; Fivash и др., 1998, Current Opinion in Biotechnology 9:97-101; Rich и др., 2000, Current Opinion in Biotechnology 11:54-61. Кроме того, любое из устройств ППР и способов измерения белок-белковых взаимодействий на основе ППР, описанных в патентах СИТА с номерами 6373577; 6289286; 5322798; 5341215; 6268125, предусмотрены в способах согласно настоящему изобретению.
Сортировку клеток с возбуждением флуоресценции (FACS), с применением любой из методик, известных специалистам в данной области, можно применять для описания связывания вариантов Fc с молекулой, экспрессированной на поверхности клетки (например, FcγRIIIA, FcγRIIB). Проточные сортеры способны быстро проверить большое количество отдельных клеток, которые содержат вставки из библиотек (например, 10-100 миллионов клеток в час) (Shapiro и др., Practical Flow Cytometry, 1995). Проточные цитометры для сортировки и исследования биологических клеток хорошо известны в данной области. Известные проточные цитометры описаны, например, в патентах США с номерами 4347935; 5464581; 5483469; 5602039; 5643796; и 6211477. Другие известные проточные цитометры представляют собой систему FACS Vantage™ производства Becton Dickinson and Company и систему COPAS™ производства Union Biometrica.
Варианты Fc согласно настоящему изобретению можно охарактеризовать по их способности опосредовать FcγR-опосредованную функцию эффекторных клеток. Примеры функций эффекторных клеток, которые можно проанализировать, включают, но не ограничены перечисленными: антителозависимую клеточную цитотоксичность (АЗКЦ), фагоцитоз, опсонизацию, опсонофагоцитоз, связывание C1q и комплементзависимую клеточную цитотоксичность (КЗЦ). Для определения активации функции эффекторных клеток можно применять любой основанный на клеточной системе или бесклеточный анализ, известный специалистам в данной области (обзор анализов эффекторных клеток см. в Perussia и др., 2000, Methods Mol. Biol. 121: 179-92; Baggiolini и др., 1998 Experientia, 44(10):841-8; Lehmann и др., 2000 J. Immunol. Methods, 243(1-2):229-42; Brown Ε J. 1994, Methods Cell Biol., 45:147-64; Munn и др., 1990 J. Exp. Med, 172:231-237, Abdul-Majid и др., 2002 Scand. J. Immunol. 55:70-81; Ding и др., 1998, Immunity 8:403-411).
В частности, варианты Fc согласно настоящему изобретению можно подвергнуть анализу на опосредованную FcγR активность АЗКЦ в эффекторных клетках (например, естественных клетках-киллерах), применяя любой из стандартных способов, известных специалистам в данной области (см., например, Perussia и др., 2000, Methods Mol. Biol. 121: 179-92). Пример анализа для определения активности АЗКЦ молекул согласно настоящему изобретению основан на анализе высвобождения 51Cr, включающем: мечение целевых клеток [51Cr]Na2CrO4 (эту молекулу, проникающую через мембрану клетки, часто используют для мечения, поскольку она связывает цитоплазматические белки и, хотя она самопроизвольно высвобождается из клеток с замедленной кинетикой, она массово высвобождается после некроза клетки-мишени); опсонизацию целевых клеток вариантами Fc согласно настоящему изобретению; объединение опсонизированных радиоактивно меченых целевых клеток с эффекторными клетками в микротитрационном планшете при подходящем соотношении целевых клеток и эффекторных клеток; инкубацию смеси клеток в течение 16-18 часов при 37°С; сбор супернатантов; и анализ радиоактивности. Цитотоксичность молекул согласно настоящему изобретению затем можно определить, например, применяя следующую формулу: % лизиса = (экспериментальный имп/мин - имп/мин утечки из целевых клеток)/(имп/мин лизата после обработки детергентом - имп/мин утечки из целевых клеток) × 100%. В качестве альтернативы, % лизиса = (АЗКЦ-АНКЦ)/(максимальное высвобождение - самопроизвольное высвобождение). Специфичный лизис можно рассчитать, применяя формулу: специфичный лизис = % лизиса посредством молекул согласно настоящему изобретению - % лизиса в отсутствии молекул согласно настоящему изобретению. Можно построить график, изменяя либо соотношение мишень : эффекторная клетка, либо концентрацию антитела.
Способ описания способности вариантов Fc связывать C1q и опосредовать комплементзависимую цитотоксичность (КЗЦ) хорошо известны в данной области. Например, для определения связывания C1q, можно осуществить ELISA связывания C1q. Пример анализа может быть таким, как описано далее. Аналитические планшеты можно покрыть в течение ночи при 4С вариантом полипептида или исходным полипептидом (контролем) в буфере для покрытия. Планшеты затем можно промыть и блокировать. После промывки можно добавить в каждую лунку аликвоту C1q человека и инкубировать в течение 2 часов при комнатной температуре. После дополнительной промывки можно добавить в каждую лунку 100 мкл конъюгированного с пероксидазой антитела овцы против комплемента C1q и инкубировать в течение 1 часа при комнатной температуре. Планшет можно снова промыть буфером для промывки и можно добавить в каждую лунку 100 мкл субстратного буфера, содержащего OPD (дигидрохлорид орто-фенилендиамина (Sigma)). Реакции окисления, наблюдаемой по появлению желтого цвета, можно позволить протекать в течение 30 минут, а затем остановить путем добавления 100 мкл 4,5 н H2SO4. Затем можно считать поглощение на 492-405 нм.
Чтобы оценить активацию комплемента, можно провести анализ комплементзависимой цитотоксичности (КЗЦ) (например, как описано в Gazzano-Santoro и др., 1996, J. Immunol Methods 202:163). Вкратце, различные концентрации варианта Fc и комплемента человека можно разбавить в буфере. Клетки, которые экспрессируют антиген, с которым связан вариант Fc, можно разбавить до концентрации, составляющей приблизительно 1×106 клеток/мл. Смеси варианта Fc, разбавленного комплемента человека и клеток, экспрессирующих антиген, можно добавить в плоскодонный 96-луночный культуральный планшет и оставить для инкубации на 2 часа при 37С и 5% CO2, чтобы произошел опосредованный комплементом лизис клеток. Затем можно добавить в каждую лунку по 50 мкл аламарового синего (Accumed International) и инкубировать в течение ночи при 37°С. Поглощение измеряют, применяя 96-луночный фотометр с возбуждением при 530 нм и эмиссией при 590 нм. Результаты можно выразить в виде относительных единиц флуоресценции (RFU). Концентрации образцов можно вычислить по стандартной кривой, и можно привести процент активности для интересующего варианта Fc по сравнению с аналогичной молекулой (т.е. молекулой, содержащей Fc-область с немодифицированным доменом СН3 или доменом СН3 дикого типа).
Анализы на связывание комплемента можно осуществить в сыворотке морской свинки, кролика или человека. Лизис комплементом целевых клеток можно обнаружить, контролируя высвобождение внутриклеточных ферментов, таких как лактатдегидрогеназа (LDH), как описано в Korzeniewski и др., 1983, Immunol. Methods 64(3):313-20; и Decker и др., 1988, J. Immunol Methods 115(1):61-9; или высвобождение внутриклеточной метки, такой как европий, хром 51 или индий 111, которой помечены целевые клетки.
Способы
В объем настоящего изобретения входит введение одного или более вариантов Fc согласно настоящему изобретению (например, антител) животному, в частности млекопитающему, особенно, человеку, для предупреждения, лечения или облегчения одного или более симптомов, связанных с заболеванием, расстройством или инфекцией. Варианты Fc согласно настоящему изобретению особенно полезны для лечения или предупреждения заболевания или расстройства, когда желательно изменение эффективности функции эффекторных клеток (например, АЗКЦ, КЗЦ). Варианты Fc и содержащие их композиции особенно полезны для лечения или предупреждения первичного или метастатического неопластического заболевания (т.е. рака) и инфекционных заболеваний. Молекулы согласно настоящему изобретению можно предусмотреть в виде фармацевтически приемлемых композиций, известных в данной области или описанных в данной заявке. Ниже подробно описано, что молекулы согласно настоящему изобретению можно применять в способах лечения или предупреждения рака (особенно в пассивной иммунотерапии), аутоиммунного заболевания, воспалительных расстройств или инфекционных заболеваний.
Варианты Fc согласно настоящему изобретению также можно успешно применять в комбинации с другими терапевтическими агентами, известными в данной области для лечения или предупреждения рака, аутоиммунного заболевания, воспалительных расстройств или инфекционных заболеваний. В конкретном варианте реализации, варианты Fc согласно настоящему изобретению можно применять в комбинации с моноклональными или химерными антителами, лимфокинами или гематопоэтическими факторами роста (такими как, например, IL-2, IL-3 и IL-7), которые, например, используют для увеличения количества или активности эффекторных клеток, которые взаимодействуют с молекулами и повышают иммунный ответ. Варианты Fc согласно настоящему изобретению также можно успешно применять в комбинации с одним или более лекарственными средствами, применяемыми для лечения заболевания, расстройства или инфекции, такими как, например, противораковые агенты, противовоспалительные агенты или противовирусные агенты.
Соответственно, согласно настоящему изобретению предложены способы предупреждения, лечения или облегчения одного или более симптомов, связанных с раком и сопутствующими состояниями, путем введения одного или более вариантов Fc согласно настоящему изобретению. Без привязки к какому-либо конкретному механизму действия, вариант Fc согласно настоящему изобретению, который связывает FcγRIIIA и/или FcγRIIA с большей аффинностью, чем сравнимая молекула, и дополнительно связывает FcγRIIB с меньшей аффинностью, чем сравнимая молекула, и/или указанный вариант Fc, который обладает повышенной эффекторной функцией, например, АЗКЦ, КЗЦ, фагоцитоз, опсонизация и т.д., приведет к избирательному нацеливанию и эффективному разрушению раковых клеток.
В объем настоящего изобретения дополнительно входит введение одного или более вариантов Fc согласно настоящему изобретению в комбинации с другими методами лечения, известными специалистам в данной области для лечения или предупреждения рака, включая, но не ограничиваясь перечисленными: современную стандартную и экспериментальную химиотерапию, гормональные методы лечения, биологические методы лечения, иммунотерапию, лучевые методы лечения или хирургическое вмешательство. В некоторых вариантах реализации, молекулы согласно настоящему изобретению можно вводить в комбинации с терапевтически или профилактически эффективным количеством одного или более противораковых агентов, терапевтических антител или других агентов, известных специалистам в данной области, для лечения и/или предупреждения рака. Примеры схем приема и методов лечения, которые можно применять в комбинации с вариантами Fc согласно настоящему изобретению, хорошо известны в данной области и были подробно описаны в различных публикациях (см., например, публикации согласно РСТ WO 02/070007 и WO 03/075957).
Рак и сопутствующие расстройства, которые можно лечить или предупреждать с помощью способов и композиций согласно настоящему изобретению, включают, но не ограничены перечисленными далее: лейкемии, лимфомы, множественные миеломы, саркомы костей и соединительных тканей, опухоли головного мозга, рак молочной железы, рак надпочечников, рак щитовидной железы, рак поджелудочной железы, рак гипофиза, рак глаза, рак влагалища, рак вульвы, рак шейки матки, рак матки, рак яичников, рак пищевода, рак желудка, рак толстой кишки, рак прямой кишки, рак печени, рак желчного пузыря, холангиокарциномы, рак легких, рак яичка, рак предстательной железы, рак полового члена; рак ротовой полости, рак слюнной железы, рак глотки, рак кожи, рак почки, рак мочевого пузыря (обзор таких расстройств смотри в Fishman и др., 1985, Medicine, 2ое изд., J.B. Lippincott Co., Филадельфия, и Murphy и др., 1997, Informed Decisions: The Complete Book of Cancer Diagnosis, Treatment, and Recovery, Viking Penguin, Penguin Books U.S.A., Inc., Соединенные Штаты Америки).
В настоящем изобретении дополнительно предложена разработка любого из антител, известных в данной области, для лечения и/или предупреждения рака и сопутствующих расстройств, таким образом, чтобы антитела включали Fc-область, содержащую модифицированный домен СНЗ согласно настоящему изобретению.
В конкретном варианте реализации, молекула согласно настоящему изобретению (например, антитело, содержащее вариант гетеродимера Fc) ингибирует или уменьшает рост первичной опухоли или метастазирование раковых клеток на по меньшей мере 99%, по меньшей мере 95%, по меньшей мере 90%, по меньшей мере 85%, по меньшей мере 80%, по меньшей мере 75%, по меньшей мере 70%, по меньшей мере 60%, по меньшей мере 50%, по меньшей мере 45%, по меньшей мере 40%, по меньшей мере 45%, по меньшей мере 35%, по меньшей мере 30%, по меньшей мере 25%, по меньшей мере 20%, или на по меньшей мере 10% по сравнению с ростом первичной опухоли или метастазированием, наблюдаемым в отсутствии указанной молекулы согласно настоящему изобретению.
В объем настоящего изобретения входит применение одного или более вариантов Fc согласно настоящему изобретению для предупреждения, лечения или контролирования одного или более симптомов, связанных с воспалительным расстройством у субъекта. Без привязки к какому-либо конкретному механизму действия, варианты Fc с повышенной аффинностью к FcγRIIB приведут к угнетению активации рецепторов и, таким образом, к угнетению иммунного ответа, и обладают терапевтической эффективностью для лечения и/или предупреждения аутоиммунного расстройства. Более того, антитела, связывающие более чем одну мишень, связанную с воспалительным расстройством, такие как биспецифические антитела, содержащие вариант гетеродимера Fc, могут оказать синергичное действие при моновалентной терапии.
В объем настоящего изобретения дополнительно входит введение вариантов Fc согласно настоящему изобретению в комбинации с терапевтически или профилактически эффективным количеством одного или более противовоспалительных агентов. Согласно настоящему изобретению также предложены способы предупреждения, лечения или контролирования одного или более симптомов, связанных с аутоиммунным заболеванием, дополнительно включающие введение указанному субъекту варианта Fc согласно настоящему изобретению в комбинации с терапевтически или профилактически эффективным количеством одного или более иммуномодулирующих агентов. Примеры аутоиммунных расстройств, которые можно лечить путем введения вариантов Fc согласно настоящему изобретению, включают, но не ограничены перечисленными: очаговую алопецию, анкилозирующий спондилит, антифосфолипидный синдром, аутоиммунную болезнь Аддисона, аутоиммунные заболевания надпочечников, аутоиммунную гемолитическую анемию, аутоиммунный гепатит, аутоиммунный оофорит и орхит, аутоиммунную тромбоцитопению, болезнь Бехчета, буллезный пемфигоид, кардиомиопатию, дерматит при глютенчувствительной целиакии, синдром хронической усталости и иммунной дисфункции (СХУИД), хроническую воспалительную демиелинизирующую полинейропатию, синдром Черджа-Стросс, рубцовый пемфигоид, КРЕСТ-синдром, синдром холодовой агглютинации, болезнь Крона, дискоидную волчанку, идиопатическую смешанную криоглобулинемию, фибромиалгию-фибромиозит, гломерулонефрит, болезнь Грейвса, синдром Гийена-Барре, тиреоидит Хашимото, идиопатический фиброз легких, идиопатическую тромбоцитопеническую пурпуру (ИТП), IgA-нейропатию, ювенильный артрит, красный плоский лишай, красную волчанку, болезнь Меньера, смешанное заболевание соединительной ткани, множественный склероз, сахарный диабет 1 типа или иммуноопосредованный сахарный диабет, миастению гравис, обыкновенную пузырчатку, пернициозную анемию, нодозный полиартериит, полихондрит, полигландулярные синдромы, ревматическую полимиалгию, полимиозит и дерматомиозит, первичную агаммаглобулинемию, первичный билиарный цирроз печени, псориаз, псориатический артрит, феномен Рейно, синдром Рейтера, ревматоидный артрит, саркоидоз, склеродермию, синдром Шегрена, синдром мышечной скованности, системную красную волчанку, красную волчанку, артериит Такаясу, височный артериит/гигантоклеточный артериит, язвенный колит, увеит, васкулит, такой как герпетиформный дерматит, витилиго и гранулематоз Вегенера. Примеры воспалительных расстройств включают, но не ограничены перечисленными, астму, энцефалит, воспалительное заболевание кишечника, хроническую обструктивную болезнь легких (ХОБЛ), аллергические расстройства, септический шок, фиброз легких, недифференцированную спондилоартропатию, недифференцированную артропатию, артрит, воспалительный остеолиз и хроническое воспаление, возникшее в результате хронических вирусных или бактериальных инфекций. Некоторые аутоиммунные расстройства связаны с воспалительным состоянием, таким образом, понятия аутоиммунного расстройства и воспалительного расстройства перекрываются. Следовательно, некоторые аутоиммунные расстройства также можно описать как воспалительные расстройства. Примеры воспалительных расстройств, которые можно предупреждать, лечить или контролировать в соответствии со способами согласно настоящему изобретению, включают, но не ограничены перечисленными, астму, энцефалит, воспалительное заболевание кишечника, хроническую обструктивную болезнь легких (ХОБЛ), аллергические расстройства, септический шок, фиброз легких, недифференцированную спондилоартропатию, недифференцированную артропатию, артрит, воспалительный остеолиз и хроническое воспаление, возникшее в результате хронических вирусных или бактериальных инфекций.
Варианты Fc согласно настоящему изобретению также можно применять для уменьшения воспаления, проявляющегося у животных, особенно у млекопитающих, страдающих воспалительными расстройствами. В конкретном варианте реализации, вариант Fc согласно настоящему изобретению уменьшает воспаление у животного на по меньшей мере 99%, по меньшей мере 95%, по меньшей мере 90%, по меньшей мере 85%, по меньшей мере 80%, по меньшей мере 75%, по меньшей мере 70%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 50%, по меньшей мере 45%, по меньшей мере 40%, по меньшей мере 35%, по меньшей мере 30%, по меньшей мере 25%, по меньшей мере 20%, или на по меньшей мере 10% по сравнению с воспалением у животного, которому не вводили указанную молекулу.
В настоящем изобретении дополнительно предложена разработка любых антител, известных в данной области, для лечения и/или предупреждения аутоиммунного заболевания или воспалительного заболевания, таким образом, чтобы антитела содержали вариант гетеродимера Fc согласно настоящему изобретению.
В объем настоящего изобретения также входят способы лечения или предупреждения инфекционного заболевания у субъекта, включающие введение терапевтически или профилактически эффективного количества одного или более вариантов Fc согласно настоящему изобретению. Инфекционные заболевания, которые можно лечить или предупреждать с помощью вариантов Fc согласно настоящему изобретению, вызваны инфекционными агентами, включая, но не ограничиваясь перечисленными, вирусы, бактерии, грибы, простейшие и вирусы.
Вирусные заболевания, которые можно лечить или предупреждать, применяя варианты Fc согласно настоящему изобретению в сочетании со способами согласно настоящему изобретению, включают, но не ограничены заболеваниями, вызванными вирусом гепатита типа А, гепатита типа В, гепатита типа С, гриппа, ветряной оспы, аденовирусом, вирусом простого герпеса I типа (ΒΠΓ-Ι), вирусом простого герпеса II типа (ВПГ-II), вирусом чумы крупного рогатого скота, риновирусом, эховирусом, ротавирусом, респираторно-синцитиальным вирусом, вирусом папилломы, паповавирусом, цитомегаловирусом, эхиновирусом, арбовирусом, хантавирусом, вирусом Коксаки, вирусом свинки, вирусом кори, вирусом краснухи, полиовирусом, вирусом натуральной оспы, вирусом Эпштейна-Барр, вирусом иммунодефицита человека I типа (ВИЧ-I), вирусом иммунодефицита человека II типа (ВИЧ-II) и агентами вирусных заболеваний, таких как вирусный менингит, энцефалит, лихорадка денге или натуральная оспа.
Бактериальные заболевания, которые можно лечить или предупреждать, применяя варианты Fc согласно настоящему изобретению в сочетании со способами согласно настоящему изобретению, вызваны бактериями, включающими, но не ограниченными перечисленными: микобактерии, риккетсии, микоплазмы, нейссерии, S. pneumonia, Borrelia burgdorferi (болезнь Лайма), Bacillus antracis (сибирская язва), столбняк, стрептококк, стафилококк, микобактерии, столбняк, коклюш, холеру, чуму, дифтерию, хламидии, S. aureus и легионеллы. Протозойные болезни, которые можно лечить или предупреждать, применяя молекулы согласно настоящему изобретению в сочетании со способами согласно настоящему изобретению, вызваны простейшими, включающими, но не ограниченными перечисленными: лейшманию, кокцидии, трипаносому или малярию. Паразитарные болезни, которые можно лечить или предупреждать, применяя молекулы согласно настоящему изобретению в сочетании со способами согласно настоящему изобретению, вызваны паразитами, включающими, но не ограниченными перечисленными: хламидии и риккетсии.
В некоторых вариантах реализации, варианты Fc согласно настоящему изобретению можно вводить в комбинации с терапевтически или профилактически эффективным количеством одного или более дополнительных терапевтических агентов, известных специалистам в данной области для лечения и/или предупреждения инфекционного заболевания. В настоящем изобретении предложено применение молекул согласно настоящему изобретению в комбинации с другими молекулами, известными специалистам в данной области для лечения и/или предупреждения инфекционного заболевания, включая, но не ограничиваясь перечисленными, антибиотики, противогрибковые агенты и противовирусные агенты.
Согласно настоящему изобретению предложены способы и фармацевтические композиции, содержащие варианты Fc согласно настоящему изобретению (например, антитела, полипептиды). Согласно настоящему изобретению также предложены способы лечения, профилактики и уменьшения выраженности одного или более симптомов, связанных с заболеванием, расстройством или инфекцией, путем введения субъекту эффективного количества по меньшей мере одного варианта Fc согласно настоящему изобретению или фармацевтической композиции, содержащей по меньшей мере один вариант Fc согласно настоящему изобретению. В одном аспекте, вариант Fc является по существу очищенным (т.е. по существу свободным от веществ, которые ограничивают его действие или вызывают нежелательные побочные эффекты, которые включают гомодимеры и другой клеточный материал). В конкретном варианте реализации, субъект представляет собой животное, такое как млекопитающее, включая не относящихся к приматам животных (например, коров, свиней, лошадей, котов, собак, крыс и т.д.) и приматов (например, обезьян, таких как яванский макак, и человека). В конкретном варианте реализации, субъект представляет собой человека. В еще одном конкретном варианте реализации, антитело согласно настоящему изобретению получено из того же вида, к которому относится субъект.
Путь введения композиции зависит состояния, от которого лечат. Например, внутривенная инъекция может оказаться предпочтительной для лечения системного расстройства, такого как рак лимфатической системы или опухоль, которая метастазировала. Дозировку композиций, которую нужно вводить, может определить квалифицированный специалист без лишних экспериментов в соответствии со стандартными исследованиями дозозависимого эффекта. Обстоятельства, которые стоит рассматривать для принятия такого решения, включают состояние или состояния, от которых лечат, выбранную для введения композицию, возраст, массу тела и ответ конкретного пациента, и тяжесть симптомов пациента. В зависимости от состояния, композицию можно вводить пациенту перорально, парентерально, интраназально, вагинально, ректально, лингвально, сублингвально, буккально, интрабуккально и/или трансдермально.
Соответственно, композиции, разработанные для перорального, лингвального, сублингвального, буккального и интрабуккального введения, можно получить без проведения лишних экспериментов с помощью средств, хорошо известных в данной области, например, с добавлением инертного разбавителя или съедобного носителя. Композицию можно заключить в желатиновые капсулы или спрессовать в таблетки. Для целей перорального введения лекарственного средства, в фармацевтические композиции согласно настоящему изобретению можно включить вспомогательные вещества и применять их в виде таблеток, пастилок, капсул, эликсиров, суспензий, сиропов, облаток, жевательных резинок и тому подобных лекарственных форм.
Таблетки, пилюли, капсулы, пастилки и тому подобные лекарственные формы также могут содержать связующие вещества, вспомогательные вещества, дезинтегрирующие агенты, лубриканты, подслащивающие агенты и/или ароматизирующие агенты. Некоторые примеры связующих веществ включают микрокристаллическую целлюлозу, трагакантовую камедь и желатин. Примеры вспомогательных веществ включают крахмал и лактозу. Некоторые примеры дезинтегрирующих агентов включают альгиновую кислоту, кукурузный крахмал и тому подобные агенты. Примеры лубрикантов включают стеарат магния и стеарат калия. Пример скользящего вещества представляет собой коллоидный диоксид кремния. Некоторые примеры подслащивающих агентов включают сахарозу, сахарин и тому подобные агенты. Примеры ароматизирующих агентов включают перечную мяту, метилсалицилат, апельсиновый ароматизатор и тому подобные агенты. Материалы, используемые для получения данных различных композиций, должны быть фармацевтически чистыми и нетоксичными в применяемых количествах.
Фармацевтические композиции согласно настоящему изобретению можно вводить парентерально, например, путем внутривенной, внутримышечной, интратекальной и/или подкожной инъекции. Парентеральное введение можно осуществить путем включения композиций согласно настоящему изобретению в состав раствора или суспензии. Такие растворы или суспензии также могут включать стерильные разбавители, такие как вода для инъекций, солевой раствор, нелетучие масла, полиэтиленгликоли, глицерин, пропиленгликоль и/или другие синтетические растворители. Парентеральные лекарственные формы также могут включать антибактериальные агенты, такие как, например, бензиловый спирт и/или метилпарабены, антиоксиданты, такие как, например, аскорбиновая кислота и/или бисульфит натрия, и хелирующие агенты, такие как ЭДТА. Также можно добавить буферы, такие как ацетаты, цитраты и фосфаты, и агенты, регулирующие тоничность, такие как хлорид натрия и декстроза. Композиции для парентерального введения можно заключить в ампулы, одноразовые шприцы и/или флаконы на несколько доз, сделанные из стекла или пластика. Ректальное введение включает введение композиции в прямую кишку и/или толстую кишку. Это можно осуществить, применяя суппозитории и/или клизмы. Суппозиторные лекарственные формы можно получить с помощью способов, известных в данной области. Трансдермальное введение включает всасывание композиции через кожу. Трансдермальные лекарственные формы включают пластыри, мази, кремы, гели, бальзамы и тому подобные лекарственные формы. Композиции согласно настоящему изобретению можно вводить пациенту назально. В данной заявке, введение назально или назальное введение включает введение композиций в слизистые оболочки носового хода и/или назальной полости пациента.
Фармацевтические композиции согласно настоящему изобретению можно применять в соответствии со способами согласно настоящему изобретению для предупреждения, лечения или облегчения одного или более симптомов, связанных с заболеванием, расстройством или инфекцией. Предполагается, что фармацевтические композиции согласно настоящему изобретению стерильны и находятся в подходящей лекарственной форме для введения субъекту.
В одном варианте реализации, композиции согласно настоящему изобретению представляют собой апирогенные лекарственные формы, которые по существу свободны от эндотоксинов и/или аналогичных пирогенных веществ.
Эндотоксины включают токсины, которые содержатся внутри микроорганизма и высвобождаются, когда микроорганизмы разрушаются или погибают. Пирогенные вещества также включают вызывающие жар, термостабильные вещества (гликопротеины) из наружной мембраны бактерий и других микроорганизмов. Данные вещества могут вызывать жар, гипотонию и шок при введении людям. Вследствии потенциального неблагоприятного воздействия, предпочтительно удалить даже небольшие количества эндотоксинов из растворов для внутривенного введения фармацевтических лекарственных средств. Управление по контролю за пищевыми продуктами и лекарственными средствами ("FDA") установило верхний предел, составляющий 5 единиц эндотоксина (EU) на дозу на килограмм массы тела за одночасовой период внутривенного введения лекарственного средства (The United States Pharmacopeial Convention, Pharmacopeial Forum 26 (1):223 (2000)). Когда терапевтические белки вводят в количествах нескольких сотен или тысяч миллиграмм на килограмм массы тела, что может происходить в случае моноклональных антител, предпочтительно, чтобы были удалены даже следовые количества эндотоксина. В конкретном варианте реализации, уровни эндотоксина и пирогена в композиции составляют меньше чем 10 EU/мг, или меньше чем 5 EU/мг, или меньше чем 1 EU/мг, или меньше чем 0,1 EU/мг, или меньше чем 0,01 EU/мг, или меньше чем 0,001 EU/мг.
Согласно настоящему изобретению предложены способы предупреждения, лечения или облегчения одного или более симптомов, связанных с заболеванием, расстройством или инфекцией, указанные способы включают: (а) введение субъекту, нуждающемуся в этом, дозы профилактически или терапевтически эффективного количества композиции, содержащей один или более вариантов Fc, и (b) введение одной или более последующих доз указанных вариантов Fc для поддержания на желательном уровне концентрации в плазме (например, от приблизительно 0,1 до приблизительно 100 мкг/мл) варианта Fc, который непрерывно связывается с антигеном. В конкретном варианте реализации, концентрация в плазме варианта Fc поддерживается на уровне 10 г/мл, 15 г/мл, 20 г/мл, 25 мкг/мл, 30 мкг/мл, 35 мкг/мл, 40 мкг/мл, 45 мкг/мл или 50 мкг/мл. В конкретном варианте реализации, указанное эффективное количество варианта Fc, которое нужно вводить, находится в диапазоне от по меньшей мере 1 мг/кг до 8 мг/кг на дозу. В другом конкретном варианте реализации, указанное эффективное количество варианта Fc, которое нужно вводить, находится в диапазоне от по меньшей мере 4 мг/кг до 5 мг/кг на дозу. В еще одном конкретном варианте реализации, указанное эффективное количество варианта Fc, которое нужно вводить, находится в диапазоне от 50 мг до 250 мг на дозу. В еще одном конкретном варианте реализации, указанное эффективное количество варианта Fc, которое нужно вводить, находится в диапазоне от 100 мг до 200 мг на дозу.
В объем настоящего изобретения также входят протоколы для предупреждения, лечения или облегчения одного или более симптомов, связанных с заболеванием, расстройством или инфекцией, в которых вариант Fc применяют в комбинации с терапией (например, профилактическим или терапевтическим агентом), отличной от варианта Fc и/или варианта слитого белка. Настоящее изобретение, отчасти, основано на признании того, что варианты Fc согласно настоящему изобретению усиливают действие и оказывают синергичное действие, повышают эффективность, улучшают переносимость и/или уменьшают побочные действия, вызванные другими способами лечения рака, включая современную стандартную и экспериментальную химиотерапию. Комбинированная терапия согласно настоящему изобретению обладает аддитивной эффективностью, оказывает аддитивное терапевтическое действие или синергичное действие. Комбинированная терапия согласно настоящему изобретению позволяет применять более низкие дозировки лекарственного средства (например, профилактических или терапевтических агентов), применяемого в сочетании с вариантами Fc, для предупреждения, лечения или облегчения одного или более симптомов, связанных с заболеванием, расстройством или инфекцией, и/или позволяет менее частое введение таких профилактических или терапевтических агентов субъекту, страдающему заболеванием, расстройством или инфекцией, чтобы улучшить качество жизни указанного субъекта и/или достигнуть профилактического или терапевтического эффекта. Кроме того, комбинированная терапия согласно настоящему изобретению позволяет уменьшить или избежать нежелательных или вредных побочных действий, связанных с применением современных монотерапевтических методов лечения и/или существующей комбинированной терапии, что, в свою очередь, улучшает соблюдение пациентом протокола лечения. Множество молекул, которые можно применять в комбинации с вариантами Fc согласно настоящему изобретению, хорошо известны в данной области. См., например, публикации согласно РСТ WO 02/070007; WO 03/075957 и публикацию патента США 2005/064514.
Согласно настоящему изобретению предложены наборы, содержащие один или более вариантов Fc с измененной аффинностью связывания с рецепторами FcγR и/или с C1q и измененной активностью АЗКЦ и/или КЗЦ, которые специфично связываются с антигеном, конъюгированным или слитым с детектируемым агентом, терапевтическим агентом или лекарственным средством, в одном или более контейнерах, для применения для контролирования, диагностики, предупреждения, лечения или облегчения одного или более симптомов, связанных с заболеванием, расстройством или инфекцией.
ПРИМЕРЫ
Примеры приведены ниже для того, чтобы проиллюстрировать осуществление настоящего изобретения. Не предполагается, что они ограничивают или определяют весь объем настоящего изобретения.
Пример 1: Получение бивалентных моноспецифических антител с гетеродимерными доменами Fc.
Гены, кодирующие тяжелые и легкие цепи антитела, сконструировали путем генного синтеза, применяя кодоны, оптимизированные для экспрессии у человека/млекопитающего. Последовательности Fab получили из известного AT, связывающего Her2/neu (Carter Р. и др. (1992) Humanization of an anti Ρ185 Her2 antibody for human cancer therapy. Proc Natl Acad Sci 89, 4285), a Fc принадлежал изотипу IgG1 (SEQ ID NO: 1). Конечные продукты генов субклонировали в вектор экспрессии млекопитающих рТТ5 (NRC-BRJ, Канада) (Durocher, Y., Perret, S. и Kamen, A. High-level and high-throughput recombinant protein production by transient transfection of suspension-growing human HEK293-EBNA1 cells. Nucleic acids research 30, E9 (2002)). Мутации в домене СНЗ вводили путем сайт-направленного мутагенеза исходных векторов рТТ5. См. перечень мутаций, введенных в модифицированный домен СН3, в Таблице 1, Таблице 6 и Таблице 7.
Для оценки образования гетеродимеров и определения отношения гомодимеров к гетеродимерам, были разработаны две тяжелые цепи гетеродимера с C-концевыми удлинениями различного размера (в частности, цепь А с C-концевой гистидиновой меткой и цепь В с C-концевым mRFP плюс StrepTagII). Такое различие в молекулярной массе позволяет отличить гомодимеры от гетеродимеров с помощью электрофореза в ПААГ/ДСН в невосстановительных условиях, показанного на Фигуре 25А.
Клетки HEK293 трансфицировали в экспоненциальной фазе роста (от 1,5 до 2 миллионов клеток/мл) с помощью водного 1 мг/мл 25 кДа полиэтиленимина (ПЭИ, Polysciences) при соотношении ПЭИ:ДНК, составляющем 2,5:1 (Raymond С. и др. А simplified polyethylenimine-mediated transfection process for large-scale and high-throughput applications. Methods. 55(1):44-51 (2011)). Для определения оптимального диапазона концентраций для образования гетеродимеров, ДНК трансфицировали в трех отдельных соотношениях двух тяжелых цепей. Например, трансфекцию осуществляли в объеме культуры 2 мл, и трансфицированная ДНК состояла из 5% GFP, 45% ДНК из молок лососевых, 25% легкой цепи и 25% всех тяжелых цепей, при этом плазмиду с тяжелой цепью А (с C-концевой гистидиновой меткой) и плазмиду с тяжелой цепью В (с C-концевой меткой StrepTagll плюс RFP) в количестве 65%/55%/35% или 10%/20%/40%) использовали в 3 различных относительных соотношениях (цепь_A(His)/цепь_B(mRFP)), составляющих 10%/65%; 20%/55%; 40%/35% (наблюдаемое соотношение 1:1 экспрессии гетеродимера ДТ_His/ДТ_mRFP оказалось близким к соотношению ДНК 20%/55%). Через 4-48 часов после трансфекции в бессывороточной среде F17 (Gibco), добавляли пептон TN1 до конечной концентрации 0,5%. Экспрессированное антитело проанализировали с помощью электрофореза в ПААГ/ДСН, чтобы определить наилучшее соотношение тяжелой и легкой цепей для оптимального образования гетеродимера (см. Фигуру 25В и С).
Выбранное соотношение ДНК, например, 50% плазмиды с легкой цепью, 25% плазмиды с тяжелой цепью А, 25% плазмиды с тяжелой цепью В AZ33 и AZ34, с 5% GFP и 45% ДНК из молок лососевых использовали для трансфекции 150 мл культуры клеток, как описано выше. Трансфицированные клетки собирали через 5-6 дней, культуральную среду собирали после центрифугирования при 4000 об/мин и осветляли с применением фильтра с диаметром пор 0,45 мкм. См. Таблицу 2 ниже, в которой представлен перечень процентов плазмид с легкой и тяжелой А и В цепями, используемых для анализов трансфекции в увеличенном масштабе для каждого из антител с мутациями СН3, полученных для дальнейшего анализа, включая определение чистоты и температуры плавления.
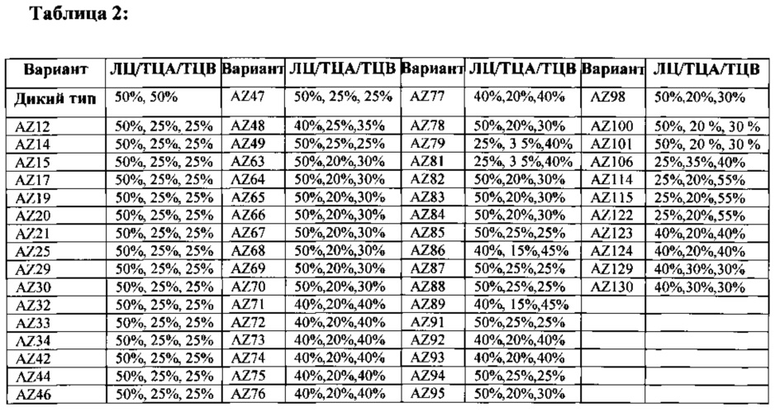
Пример 2: Очистка бивалентных моноспецифических антител с гетеродимерными доменами Fc.
Осветленную культуральную среду загружали на колонку с белком A MabSelect SuRe (GE Healthcare) и промывали 10 объемами колонки буфера ФБР при pH 7,2. Антитело элюировали 10 объемами колонки цитратного буфера при pH 3,6, и полученные объединенные фракции, содержащие антитело, нейтрализовали с помощью Трис при pH 11. Наконец, белок обессоливали, применяя колонку 10DG Econo-Pac (Bio-Rad). C-концевую метку mRFP удаляли с тяжелой цепи В путем инкубации антитела с энтерокиназой (NEB) при соотношении 1:10000 в ФБР в течение ночи при 25°С. Антитело очищали из смеси посредством гель-фильтрации. Для гель-фильтрации, 3,5 мг смеси с антителом концентрировали до 1,5 мл и загружали на колонку Sephadex 200 HiLoad 16/600 200 pg (GE Healthcare) с помощью системы АКТА Express FPLC при скорости потока 1 мл/мин. Буфер ФБР при pH 7,4 использовали при скорости потока 1 мл/мин. Фракции, соответствующие очищенному антителу, собирали, концентрировали до ~1 мг/мл и хранили при -80°С.
Образование гетеродимеров, по сравнению с гомодимерами, анализировали, применяя ЭФ в ПААГ/ДСН в невосстановительных условиях и масс-спектрометрию. Очищенное на белке А антитело разгоняли в градиентном 4-12% ПААГ/ДСН геле в невосстановительных условиях, чтобы определить процент гетеродимеров, образовавшихся до обработки энтерокиназой (EK) (см. Фигуру 26). Для масс-спектрометрии, все эксперименты времяпролетной ЖХ/МС с ионизацией электрораспылением и ионной ловушкой (ИЭР-ВП) осуществляли на системе Agilent 1100 HPLC, соединенной с масс-спектрометром Q-TOF2 Waters. Пять мкг очищенного гель-фильтрацией антитела впрыскивали в картридж Protein MicroTrap (1,0 на 8,0 мм), промывали 1% ацетонитрила в течение 8 минут, градиентом от 1 до 20% ацетонитрила/0,1% муравьиной кислоты в течение 2 минут, а затем элюировали градиентом от 20 до 60% ацетонитрила/0,1% муравьиной кислоты в течение 20 минут. Элюат (30-50 мкл/мин) направляли в спектрометр, который регистрировал спектр каждую секунду (отношению массы к заряду (m/z) составляло от 800 до 4000) (см. Фигуру 28). Варианты, образующие более чем 90% гетеродимеров, выбирали для дальнейшего анализа, за исключением AZ12 и AZ14, каждый из которых образовывал более чем 85% гетеродимеров.
Пример 3: Определение стабильности бивалентных моноспецифических антител с гетеродимерными доменами Fc с помощью дифференциальной сканирующей калориметрии (ДСК).
Все эксперименты ДСК осуществляли, применяя устройство VP-Capillary GE. У белков заменили буфер на ФБР (pH 7,4), и разбавили их до 0,4-0,5 мг/мл, из которых 0,137 мл загружали в кювету для образца и проводили измерения при скорости сканирования 1°С/мин от 20 до 100°С. Результаты анализировали, применяя программное обеспечение Origin (GE Healthcare), фон буфера ФБР вычитали (см. Фигуру 27). См. Таблицу 3, в которой представлен перечень исследованных вариантов и определенных температур плавления. См. Таблицу 4, в которой представлен перечень вариантов с температурой плавления 70°С и выше и Tm, определенных для каждого варианта.

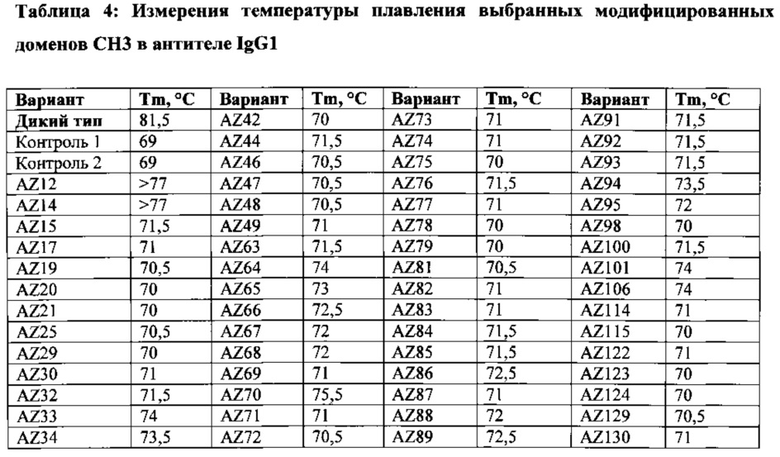
Пример 4: Оценка связывания рецептора Fc-гамма с применением поверхностного плазменного резонанса
Все эксперименты на связывание осуществляли, применяя устройство ProteOn XPR36 BioRad при 25°C с добавлением 10 мМ HEPES, 150 мМ NaCl, 3,4 мМ ЭДТА и 0,05% Tween 20 при pH 7,4. Рекомбинантный белок HER-2/neu (р185, ErbB-2 (eBiosciences, Inc.)) закрепляли на активированном сенсорном чипе GLM путем впрыскивания 4,0 мкг/мл в 10 мМ NaOAc (pH 4,5) при скорости потока 25 мкл/мин до тех пор, пока не иммобилизовалось приблизительно 3000 резонансных единиц (RU), оставшиеся активные группы гасили, 40 мкг/мл очищенных антител к HER-2/neu, содержащих модифицированные домены СН3, захватывались сенсорным чипом посредством связывания с белком Her-2/neu, когда их впрыскивали при скорости потока 25 мкл/мин в течение 240 с (всего приблизительно 500 RU), вслед за впрыскиванием буфера для получения стабильного фона. Рецептор Fc-гамма (CD16a (аллотип f) и CD32b) в концентрациях (6000, 2000, 667, 222 и 74,0 нМ) впрыскивали при скорости потока 60 мкл/мин в течение 120 сек с фазой диссоциации 180 сек. с получением набора сенсограмм связывания. Результирующие значения KD определяли по изотермам связывания, применяя модель равновесной подгонки, при этом значения представляли в виде среднего по трем независимым экспериментам. Сравнения осуществляли с доменом Fc IgG1 дикого типа и связывание выражали в виде отношения KD ДТ к KD варианта (см. Таблицу 5).
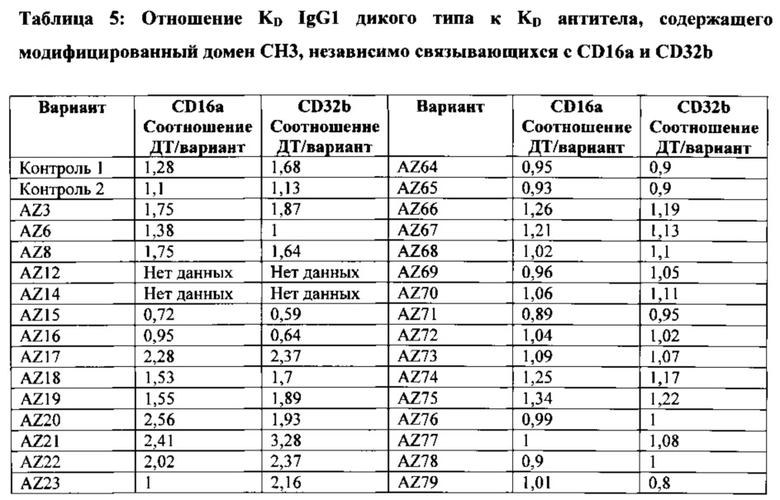
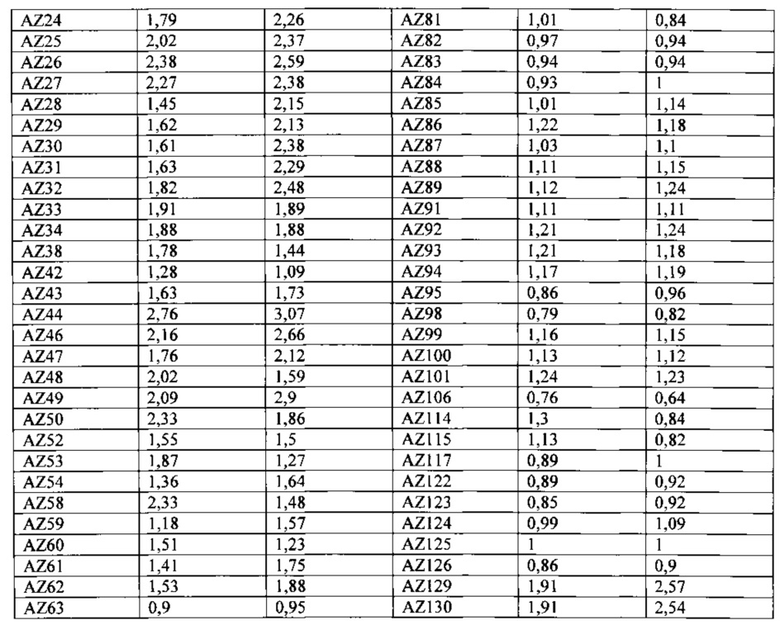
Пример 5: Рациональный дизайн вариантов Fc с применением разработки Fc_CH3 - каркас 1 (1а и 1b) и разработка AZ17-62 и AZ133-AZ2438
Для получения вариантов AZ с высокой стабильностью и чистотой, использовали структурные и вычислительные стратегии, описанные выше (см. Фигуру 24). Например, углубленный структурно-функциональный анализ AZ8 обеспечил углубленное понимание значения каждой из введенных в AZ8 мутаций L351Y_V397S_F405A_Y407V/K392V_T394W, по сравнению с IgG1 человека дикого типа, и показал, что важными основными мутациями для гетеродимера были L351Y_F405A_Y407V/T394W, тогда как V397S, K392V не влияли на образование гетеродимера. Мутации в ядре (L351Y_F405A_Y407V/T394W) в данной заявке называют мутациями "каркаса 1". Анализ дополнительно выявил, что важные горячие точки мутагенеза на границе, которые были утрачены по сравнению с образованием гомодимера дикого типа (ДТ), представляли собой взаимодействия ДТ F405-K409, Y407-T366 и укладку Y407-Y407 и -F405 (см. Фигуру 29). Это отразили в анализе укладки, полостей и МД, который выявил большое различие конформации участка петли D399-S400-D401 (см. Фигуру 30) и ассоциированного складчатого бета-слоя в положении КЗ 70. Это привело к утрате межцепочечных взаимодействий K409-D399 (см. Фигуру 30) и ослаблению сильной водородной связи K370 с Е357 (K370 больше непосредственно не контактировал с S364 и Е357, и был полностью экспонирован в растворитель). В домене СН3 IgG1 ДТ эти участки связывают границу, и край защищает взаимодействия в ядре от большей части контактов с растворителем и повышает динамическое возникновение благоприятных гидрофобных ван-дер-ваальсовских взаимодействий. Следствием были меньшая площадь заглубленной поверхности AZ8 по сравнению с ДТ и более высокая доступность для растворителя гидрофобного ядра. Это позволило указать наиболее важные факторы пониженной стабильности AZ8 по сравнению со стабильностью ДТ, представляющие собой а) утрату взаимодействия ДТ F405-K409 и укладки F405 и b) утрату сильного взаимодействия укладки Y407-Y407 и Y407-T366. Смотри Фигуру 29.
Следовательно, авторы настоящего изобретения обнаружили ключевые остатки/мотивы последовательности, ответственные за низкую стабильность AZ8 по сравнению с ДТ. Для улучшения стабильности и специфичности гетеродимера AZ8, последующие усилия положительной разработки, по этой причине, были направлены конкретно на стабилизацию конформации петли в положениях 399-401 путем получения более "закрытой" конформации, подобно ДТ (см. Фигуру 30), и на корректирование в целом слегка ослабленной (неплотной) укладки гидрофобного ядра в положениях Т366 и L368 (см. Фигуру 29). Для достижения такой стабилизации конформации петли в положениях 399-401 применяли описанный вычислительный подход, чтобы оценить наши различные идеи направленной разработки. В частности, проанализировали три различные независимые возможности для варианта Fc AZ8, чтобы оптимизировать обнаруженные ключевые участки для улучшения стабильности. Во-первых, оценили связывающий карман, расположенный рядом с положениями K409 и F405A, на предмет лучшей гидрофобной укладки, чтобы защитить гидрофобное ядро, а также стабилизировать конформацию петли в положениях 399-400 (см. Фигуру 30). Для этого ввели, не ограничиваясь перечисленными, дополнительные точечные мутации в положения F405 и K392. Во-вторых, оценили возможности улучшения электростатических взаимодействий положений 399-409, чтобы стабилизировать конформацию петли 399-400 и защитить гидрофобное ядро. Для этого ввели, не ограничиваясь перечисленными, дополнительные точечные мутации в положения Т411 и S400. В-третьих, оценили связывающий карман в положениях укладки ядра Т366, T394W и L368, чтобы улучшить гидрофобную укладку ядра (см. Фигуру 29). Для этого ввели, не ограничиваясь перечисленными, дополнительные точечные мутации в положения Т366 и L368. Различные независимые идеи положительной разработки исследовали компьютерными способами, и экспрессию и стабильность некоторых хороших вариантов, выявленных с применением вычислительных средств (AZ17-AZ62), проверяли опытным путем, как описано в Примерах 1-4. См. Таблицу 4, в которой приведен перечень некоторых гетеродимерных конструкций на основе Fc, разработанных с помощью данной стратегии разработки, с температурой плавления 70°С или выше.
Вариант Fc AZ33 представляет собой пример разработки варианта Fc, в котором каркас 1 модифицировали с получением каркаса 1а, содержащего мутации для повышения стабильности и чистоты. Данный вариант Fc был разработан на основе AZ8 с целью улучшения гидрофобной укладки в положениях 392-394-409 и 366, чтобы защитить гидрофобное ядро, а также стабилизировать конформацию петли 399-400. Данный вариант Fc, гетеродимер AZ33, содержал две дополнительные точечные мутации, отличные от мутаций в ядре AZ8, - K392M и T366I. Мутации T366I_K392M_T394W/F405A_Y407V в данной заявке называют мутациями "каркаса 1а". Мутацию K392M разработали таким образом, чтобы она улучшила укладку в полости, расположенной рядом с положениями K409 и F405A, чтобы защитить гидрофобное ядро и стабилизировать конформацию петли 399-400 (см. Фигуру 31). Мутацию T366I разработали таким образом, чтобы она улучшила гидрофобную укладку ядра и исключила образование гомодимеров цепи с T394W (см. Фигуру 29). Результаты эксперимента с AZ33 показали значительное улучшение стабильности по сравнению с другими вариантами Fc отрицательной разработки, такими как AZ8 (Tm 68°С), при этом AZ33 имел Tm, составляющую 74°С, и содержание гетеродимера составляло >98%. (см. Фигуру 25С).
Разработка вариантов Fc с применением мутаций каркаса 1 в третьей фазе разработки вариантов гетеродимера Fc
Хотя AZ33 обладает существенно улучшенной стабильностью и специфичностью (или чистотой) по сравнению с исходным вариантом AZ8, наш анализ показал, что можно добиться дополнительного улучшения стабильности варианта гетеродимера Fc путем дополнительных модификаций аминокислот, применяя результаты эксперимента для AZ33 и описанные выше способы разработки. Экспрессию и стабильность вариантов, разработанных с применением различных идей разработки, исследовали независимо, но независимые идеи разработки можно комбинировать, и наиболее удачный гетеродимер будет получен с применением комбинации различных способов разработки. В частности, для оптимизации укладки AZ8, мутации в полости, расположенной рядом с положениями K409-F405A-K392, оценивали независимо от мутаций, которые оптимизируют укладку ядра, в остатках L366T-L368. Данные два участка 366-368 и 409-405-392 отдалены друг от друга и рассматриваются как независимые. Вариант Fc AZ33, например, оптимизировали в отношении укладки в положениях 409-405-392, но не в положениях 366-368, так как данные оптимизирующие мутации оценивали отдельно. Сравнение мутаций в положениях 366-368 позволяет предположить, что T366L придавала улучшенную стабильность по сравнению с Т366, а также с T366I, последнюю точечную мутацию использовали при разработке варианта Fc AZ33. Следовательно, представленные результаты эксперимента прямо предполагают дополнительную оптимизацию AZ33 путем введения T366L, например, вместо T366I. Следовательно, мутации аминокислот в домене СН3 T366L_K392M_T394W/F405A_Y407V в данной заявке называют мутациями "каркаса 1b".
Аналогичным образом были проанализированы все результаты эксперимента, чтобы определить точечные мутации, которые можно вводить для дополнительного улучшения настоящего варианта гетеродимера Fc AZ33. Данные обнаруженные мутации анализировали с помощью описанного выше вычислительного подхода и упорядочили с получением перечня дополнительных вариантов гетеродимера Fc, основанных на AZ33, показанных в Таблице 6.
Пример 6: Рациональный дизайн вариантов Fc с помощью разработки Fc_CH3: каркас 2 (a и b) и разработка AZ63-101 и AZ2199-AZ2524
Для улучшения стабильности и чистоты исходного варианта Fc фазы отрицательной разработки - AZ15, использовали структурные и вычислительные стратегии, описанные выше (см. Фигуру 24). Например, углубленный структурно-функциональный анализ варианта Fc AZ15 дал углубленное понимание каждой из введенных в AZ15 мутаций, L351Y_Y407A/E357L_T366A_K409F_T411N, по сравнению с IgG1 человека дикого типа (ДТ), и показал, что важными мутациями в ядре гетеродимера были L351Y_Y407A/T366A_K409F, тогда как E357L и T411N не оказывали непосредственного влияния на образование и стабильность гетеродимера. Мутации ядра (L351Y_Y407A/T366A_K409F) в данной заявке называют мутациями "каркаса 2". Проведенный анализ, кроме того, показал, что важные горячие точки мутагенеза на границе, которые утрачиваются по сравнению с образованным гомодимером дикого типа (ДТ), представляют собой солевой мостик D399-K409, водородную связь Y407-T366 и укладку Y407-Y407. В подробном анализе, приведенном ниже, описано, как авторы настоящего изобретения улучшили стабильность исходного варианта Fc AZ15 и какие модификации аминокислот ввели в каких положениях, чтобы получить данные варианты Fc с повышенной стабильностью.
Разработка вариантов Fc с применением мутаций каркаса 2 и дополнительная разработка мутаций каркаса 2а.
Компьютерный анализ показал неоптимальную укладку разработанных ранее вариантов Fc, например, мутаций K409F_T366A_Y407A AZ15, и ослабленную укладку гидрофобного ядра в целом вследствие утраты взаимодействий ДТ Y407-Y407. Гетеромультимеры, описанные в данной заявке, разработаны с более оптимальной укладкой. Некоторые из исследований положительной разработки, описанных в данной заявке, были сфокусированы на точечных мутациях для компенсации недостаточности укладки исходного варианта Fc AZ15. Целевые остатки включали положения Т366, L351 и Y407. Различные комбинации мутаций в данных положениях исследовали компьютерным способом, и экспрессию и стабильность лучших вариантов Fc, выявленных с помощью вычислительных средств (AZ63-AZ70), проверяли опытным путем, как описано в Примерах 1-4.
Вариант Fc AZ70 представляет собой пример разработки варианта Fc, в котором каркас 2 модифицировали путем введения мутаций каркаса 2а для повышения стабильности и чистоты. Данный вариант Fc был разработан на основе AZ15 с целью достижения лучшей укладки гидрофобного ядра, как описано выше. Вариант Fc AZ70 содержал те же мутации в ядре каркаса 2 (L351Y_Y407A/T366A_K409F), описанные выше, за исключением того, что Т366 мутировали с получением T366V вместо Т366А (Фигура 33). Мутация L351Y улучшала температуру плавления варианта 366A_409F/407A от 71,5°С до 74°С, и дополнительная замена 366А на 366V улучшала Tm до 75,5°С (см. AZ63, AZ64 и AZ70 в Таблице 4, с Tm, составляющими 71,5°С, 74°С и 75,5°С, соответственно). Мутации в ядре (L351Y_Y407A/T366V_K409F) в данной заявке называют мутациями "каркаса 2а". Результаты эксперимента для варианта Fc AZ70 показали значительное улучшение стабильности по сравнению с исходным вариантом Fc отрицательной разработки - AZ15 (Tm 71°С), при этом AZ70 имел Tm, составляющую 75,5°С, и содержание гетеродимера составляло >90% (Фигура 33 и 27).
Разработка вариантов Fc с применением мутаций каркаса 2 и дополнительная разработка мутаций каркаса 2b.
Моделирование методом молекулярной динамики (МД) и анализ укладки выявили предпочтительную более "открытую" конформацию петли 399-400, которая, вероятно, обусловлена утратой солевого мостика K409-D399 в ДТ. Это также приводило к ненасыщенности D399, который, в свою очередь, предпочтитал компенсирующее взаимодействие с K392, что приводило к более "открытой" конформации петли. Такая более "открытая" конформация петли приводит к ослабленной укладке в целом и к повышенной доступности воздействию растворителя остатков границы ядра домена СН3, что, в свою очередь, значительно дестабилизирует гетеродимерный комплекс. Следовательно, одним из целевых исследований положительной разработки было придание более "закрытой", подобной ДТ, конформации петли путем введения дополнительных точечных мутаций, которые компенсируют утрату солевого мостика D399-K409 и взаимодействий укладки K409. Целевые остатки включали положения Т411, D399, S400, F405, N390, K392 и комбинации перечисленных положений. Различные укладки, гидрофобные и электростатические стратегии положительной разработки исследовали компьютерным способом в отношении указанных выше положений, и экспрессию и стабильность лучших вариантов Fc, выявленных с помощью вычислительных средств (AZ71-AZ101), проверяли опытным путем, как описано в Примерах 1-4.
Вариант Fc AZ94 представляет собой пример разработки варианта Fc, в котором каркас 2 модифицировали путем введения мутаций каркаса 2b, наряду с введением дополнительных точечных мутаций для повышения стабильности и чистоты. Данный вариант Fc был разработан с целью придания петле 399-400 более "закрытой", подобной ДТ, конформации и компенсации утраты солевого мостика D399-K409, как описано выше. Вариант Fc AZ94 содержал четыре дополнительные точечные мутации в каркасе 2 (L351Y Y407A/T366A K409F), и в нем заменили L351Y на L351 дикого типа, оставив (Y407A/T366A K409F) в качестве мутаций в ядре для данного варианта Fe. Мутации в ядре Y407A/T366A K409F в данной заявке называют мутациями "каркаса 2b". Четыре дополнительные точечные мутации AZ94 представляли собой K392E_T411E/D399R_S400R. Мутации T411E/D399R введены для образования дополнительного солевого мостика и компенсации утраты взаимодействия K409/D399 (Фигура 34). Кроме того, данный солевой мостик был разработан для предотвращения образования гомодимера, препятствуя взаимодействиям заряд-заряд в обоих потенциальных гомодимерах. Дополнительные мутации K392E/S400R предназначались для образования другого солевого мостика и, следовательно, для дополнительного придания петле 399 400 более "закрытой", подобной ДТ конформации (Фигура 34). Результаты экспериментов для AZ94 показали улучшенную стабильность и чистоту по сравнению с исходным вариантом Fc отрицательной разработки - AZ15 (Tm 71°С, чистота >90%), при этом вариант Fc AZ94 имел Tm, составляющую 74°С, и содержание гетеродимеров или чистоту >95%.
Разработка вариантов Fc с применением мутаций каркаса 2 в третьей фазе разработки вариантов гетеродимера Fc
Варианты Fc AZ70 и AZ94 обладают существенными улучшениями в стабильности и чистоте по сравнению с исходными вариантами Fc отрицательной разработки, такими как AZ15, но наш анализ и непосредственное сравнение AZ70 и AZ94 свидетельствуют о том, что можно добиться неожиданных улучшений в стабильности данного варианта гетеродимера Fc с помощью дополнительных модификаций аминокислот. Например, варианты Fc AZ70 и AZ94 были разработаны с целью изменения двух различных неоптимизированных участков в исходном варианте AZ15, что осуществили путем улучшения укладки гидрофобного ядра и введения мутаций за пределами остатков границы ядра, что позволило получить дополнительные солевые мостики и водородные связи для стабилизации конформации петли в положениях 399-401. Дополнительные точечные мутации вариантов Fc AZ70 и AZ94 отдалены друг от друга и, следовательно, они независимы и их можно перенести на другие варианты Fc, разработанные на основе тех же мутаций в ядре каркаса 2, включая мутации 2а и 2b. В частности, AZ70 несет только оптимизированные мутации в ядре L351Y_Y407A/T366A_K409F, без дополнительных солевых мостиков, тогда как AZ94 включает четыре дополнительных электростатических мутации (K392E_T411E/D399R_S400R), но содержит на одну мутацию меньше на границе гидрофобного ядра (Y407A/T366A_K409F). Данные мутации каркаса 2b менее стабильны, чем в AZ70 (см., например, AZ63, который содержит мутации в ядре, эквивалентные AZ94, и имеет Tm, составляющую 72°С), что компенсируется путем введения дополнительных мутаций K392E_T411E/D399R_S400R. Представленные экспериментальные результаты относительно стабильности и чистоты свидетельствуют о том, что сочетание мутаций AZ70, которые оптимизируют гидрофобное ядро, и электростатических мутаций AZ94 должно дополнительно улучшать стабильность и чистоту вариантов гетеродимера Fc. Аналогичным образом были проанализированы все результаты эксперимента для вариантов каркаса 2 Fc (AZ63-101), чтобы определить точечные мутации, которые можно вводить для дополнительного улучшения вариантов гетеродимера Fc AZ70 и AZ94. Данные обнаруженные мутации дополнительно анализировали с помощью описанного выше вычислительного подхода и упорядочили с получением перечня дополнительных вариантов гетеродимера Fc, основанных на AZ70 и AZ94, показанных в Таблице 7.
Пример 7: Влияние гетеродимерного СН3 на связывание FcγR
В качестве примера прототипа активности гетеродимерного Fc по отношению к FcγR, исследовали два варианта антитела с гетеродимерной Fc-областью A:K409D_K392D/B:D399K_D356K (контроль 1 (гет1 на Фигуре 35)) и A:Y349C_T366S_L368A_Y407V/B:S354C_T366W (контроль 4 (гет2 на Фигуре 35)) со связывающими Her2 Fab-плечами с помощью анализа ППР, описанного в Примере 4 для связывания FcγR. Как показано на Фигуре 35, авторы настоящего изобретения обнаружили, что обе гетеродимерные Fc-области связывали различные рецепторы Fc-гамма с такой же относительной силой, как и Fc-область IgG1 дикого типа, но в целом гетеродимерная Fc-область связывала каждый из FcγR несколько лучше, чем антитело дикого типа. Это свидетельствует о том, что мутации на границе СН3 Fc могут влиять на силу связывания Fc-области с рецепторами Fc-гамма по всем доменам СН2, что наблюдалось в молекулярно-динамическом моделировании и анализе, выполненном авторами настоящего изобретения.
Пример 8: Влияние асимметричных мутаций в СН2 гетеродимерного Fc на связывание FcγR
Известно, что замена серина в положении 267 в домене СН2 Fc-области на аспарагиновую кислоту (S267D) увеличивает связывание с рецепторами Fc-гамма IIbF, IIbY и IIaR, если ее ввести гомодимерным способом в обе цепи домена СН2. Данную мутацию можно ввести только в один из доменов СН2 в гетеродимерной молекуле Fc, чтобы добиться увеличения силы связывания приблизительно вдвое по сравнению со случаем, когда данная мутация введена в гомодимерный СН2 Fc, о чем свидетельствуют результаты, представленные на Фигуре 36А. С другой стороны, мутация E269K в гомодимерном домене СН2 Fc предотвращает связывание Fc-области с FcγR. Авторами настоящего изобретения представлена схема улучшенного управления силой связывания Fc-области с рецепторами Fcγ путем асимметричного введения данных благоприятных и неблагоприятных мутаций в одну из двух цепей в домене СН2 Fc. Введение мутации E269K асимметричным способом в одну цепь СН2 гетеродимерного Fc приводит к возникновению полярности путем блокирования связывания FcγR со стороной, где присутствует данная мутация, при этом позволяя другой стороне Fc взаимодействовать с FcγR обычным путем. Результаты данного эксперимента представлены на Фигуре 36А. Возможность избирательно изменять силу связывания обеими цепями Fc независимым образом дает расширенную возможность контролировать силу связывания и селективность между Fc и рецепторами Fcγ. Таким образом, такая асимметричная разработка мутаций в домене СН2 дает нам возможность внедрить стратегии положительной и отрицательной разработки для того, чтобы способствовать или препятствовать некоторым моделям связывания, предоставляя расширенную возможность привнесения селективности.
В последующем эксперименте, авторы настоящего изобретения изменили профиль селективности основного мутанта Fc S239D_D265S_I332E_S298A, который проявил повышенную силу связывания с рецепторами Fc-гамма IIIaF и IIIaV, при этом продолжая проявлять более слабое связывание с рецепторами Fc-гамма IIaR, IIbFn IIbY. Это показано в профиле связывания, представленном на Фигуре 36В. Путем введения асимметричных мутаций E269K в цепь А и исключения мутации I332E из цепи В, авторы настоящего изобретения смогли получить новый профиль связывания FcγR, который дополнительно ослабляет связывание с рецепторами IIa и IIb и обеспечивает более специфичное связывание Fc с рецептором IIIa.
В другом примере, показанном на Фигуре 36С, асимметричные мутации выделены по сравнению с гомодимерным Fc, включая мутации S239D/K326E/A330L/I332E/S298A в домене СН2. По сравнению с Fe IgG1 дикого типа, данный вариант проявил повышенное связывание с рецептором IIIa, но он также связывает рецепторы IIa и IIb слегка сильнее, чем Fc дикого типа. Введение данных мутаций асимметричным способом A:S239D/K326E/A330L/I332E и B:S298A не только уменьшает связывание IIIa, но также повышает связывание с рецептором IIa/IIb, что приводит к утрате селективности данного процесса. При введении асимметричной мутации E269K в данный гетеродимерный вариант, т.е. A:S239D/K326E/A330L/I332E/E269K и B:S298A, связывание IIa/IIb снижается обратно до уровней дикого типа. Это подчеркивает тот факт, что использование асимметричных мутаций в домене СН2 Fc может предоставить существенную возможность разработки улучшенной селективности к рецепторам Fc-гамма.
Реагенты, используемые в примерах, доступны для приобретения, или их можно получить, применяя доступные для приобретения оборудование, способы или реагенты, известные в данной области. Предшествующие примеры проиллюстрировали различные аспекты настоящего изобретения и осуществления способов согласно настоящему изобретению. Примеры не предназначены для предоставления исчерпывающего описания множества различных вариантов реализации настоящего изобретения. Таким образом, хотя описанное выше изобретение было описано довольно подробно с помощью иллюстрирования и приведения примеров с целью ясности понимания, для средних специалистов в данной области очевидно, что можно внести в него множество изменений и модификаций, не отклоняясь от сущности или объема прилагаемой формулы изобретения.
Пример 9: Связывание FcRn, определенное с помощью ППР.
Связывание с FcRn определяли с помощью ППР в двух различных ориентациях.
1. Пропускание потока варианта гетеродимера через иммобилизованный FcRn. В данном эксперименте получали поверхности с высокой плотностью иммобилизации, составляющей приблизительно 5000 RU, применяя стандартное связывание комбинацией N-гидроксисукцинимида/1-этил-3-(3-диметиламинопропил)карбодиимида (NHS/EDC). 100 нМ ДТ и каждого варианта впрыскивали в трех повторах при скорости потока 50 мкл/мин в течение 120 сек с диссоциацией в течение 600 сек в подвижном буфере MES, pH 6.
2. Пропускание потока FcRn через опосредованно захваченные варианты гетеродимера. В данном эксперименте ППР, использовали поверхность с иммобилизованными антителами козы к IgG человека, чтобы опосредованно захватить антитела (приблизительно по 400 RU каждого антитела), а затем впрыскивали FcRn в 3-кратных разведениях (начиная с концентрации 6000 нМ и ниже). Подвижный буфер представлял собой 10 мМ MES/150 мМ NaCl/3,4 мМ ЭДТА/0,05 Tween 20 при pH 6. Не наблюдалось существенного связывания FcRn с поверхностью с иммобилизованными поликлональными антителами козы. Сенсограммы для всех вариантов оказались анлогичны ДТ. В Таблице 8 ниже показаны Kd, определенные путем опосредованной иммобилизации с пропусканием потока FcRn (2).
Таблица 8: Kd, определенные путем опосредованной иммобилизации с пропусканием потока FcRn
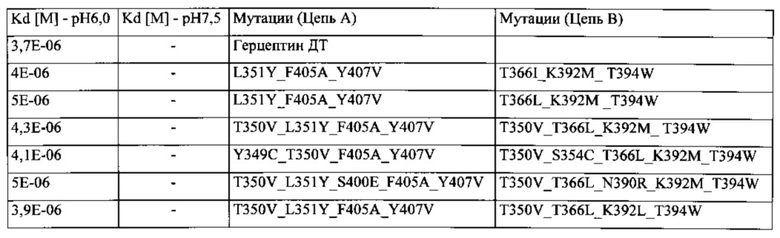
Пример 10: Биспецифическое связывание гетеродимера Fc, описанного в данной заявке
Биспецифическое связывание продемонстрировали на примере гетеродимера Fc с мутациями в цепи A: L351Y_F405A_Y407V, цепи В: T366L_K392M_T394W, слитого с scFv против HER2 (анти-HER2) и HER3 (анти-HER3) на N-конце цепи А и цепи В гетеродимера Fc. Полученные варианты: биспецифический вариант HER2/HER3 и два моновалентных-моноспецифических варианта с HER2 или HER3, - показаны на Фиг. 40А. Для анализа биспецифического связывания, диапазон доз двух моновалентных вариантов (моновалентный анти-HER2 и моновалентный anra-HER2, показаны на Фиг. 40А) и биспецифического гетеродимера анти-HER2/HER3 инкубировали с клетками меланомы MALME-M3, а затем с помощью анализа FACS определяли видимую аффинность связывания каждой молекулы (показано на Фиг. 40В). Систему анализа настраивали согласно протоколам, описанным в: "Antitumor activity of a novel bispecific antibody that targets the ErbB2/ErbB3 oncogenic unit and inhibits heregulin-induced activation of ErbB3", McDonagh CF и др., Mol Cancer Ther. 11 (3):582-93 (2012).
Пример 11: Экспрессия и очистка бивалентных моноспецифических антител с гетеродимерными доменами Fc и количественный анализ чистоты с помощью ЖХ/МС
Гетеродимерные варианты AZ133 (A: L351Y/F405A/Y407V, В: T366L/K392M/T394W), AZ138 (A: F405A/Y407V, В: T366L/K392M/T394W), AZ3002 (А: T350V/L351Y/F405A/Y407V, В: T350V/T366L/K392M/T394W), AZ3003 (А: T350V/L351Y/F405A/Y407V, В: T350V/T366L/K392L/T394W), и другие конструкции AZ (AZ3000-AZ3021) получали и очищали, как описано в Примерах 1 и 2. Для того, чтобы оценить устойчивость образования гетеродимера и влияние избытка одной из цепей гетеродимера на чистоту гетеродимера, выбранные гетеродимеры временно экспрессировали с использованием 3 различных соотношений ДНК двух тяжелых цепей А и В (например, соотношения А:В=1:1,5; 1:1; 1,5:1).
Гены, кодирующие тяжелые и легкие цепи гетеродимера, сконструировали путем генного синтеза, используя кодоны, оптимизированные для экспрессии в человеке/млекопитающем, как подробно описано в Примере 1. Последовательности Fab получали из известного AT, связывающего Her2/neu (Carter Р. и др. (1992) Humanization of an anti P185 Her2 antibody for human cancer therapy. Proc Natl Acad Sci 89, 4285), и Fc принадлежал изотипу IgG1 (SEQ ID NO: 1). Указанный вариант экспрессировали путем временной совместной экспрессии, как описано в Примерах 1-2, применяя 3 различных соотношения тяжелой цепи А к тяжелой цепи В: 1:1,5, 1:1 и 1,5:1. Образцы очищали с помощью аффинной хроматографии на белке А и препаративной гель-фильтрации (см. Пример 2 для получения более подробной информации). Очищенные образцы дегликозилировали с помощью PNGaзы F в течение ночи при 37°С. Перед анализом МС, образцы наносили на колонку Poros R2 и элюировали градиентом ацетонитрила 20-90%, 0,2% МК в течение 3 минут. Пик с колонки ЖХ анализировали с помощью масс-спектрометра LTQ-Orbitrap XL (напряжение на конусе: 50 В, цилиндрическая линза: 215 В; разрешение при использовании ПФ: 7500) и установленного программного обеспечения Promass с получением профилей молекулярных масс.
Относительные высоты пиков гетеродимера и гомодимеров использовали для оценки чистоты гетеродимера (см. Фигуру 39).
Пример 12: Кристаллическая структура гетеромультимеров AZ3002 и AZ3003
Гетеродимерные конструкции Fc AZ3002 и AZ3003 временно экспрессировали в клетках СНО и очищали до гомогенного состояния с помощью хроматографии на белке А и эксклюзионной хроматографии. Очищенные гетеродимеры Fc кристаллизовали при 18°С после ~24 часов инкубации с применением способа висячей капли посредством диффузии в парах при соотношении 2:1 выше маточного раствора, состоящего из 5% (об./об.) этиленгликоля, 18% (вес/об.) полиэтиленгликоля 3350 и 0,15 M йодида аммония, с помощью микрозатравок. Кристаллы защищали от воздействия низкой температуры путем повышения концентрации этиленгликоля до 30% (об./об.), а затем мгновенно охлаждали жидким азотом. Данные о дифракции обоих кристаллов собирали при 100 K, используя колебания в 0,5 градуса для общих 200 градусов, и обрабатывали с помощью XDS1. Структуру AZ3002 разрешали путем молекулярного замещения с использованием программы Phaser, используя структуру из Международного банка белковых структур PDB ID: 2J6E в качестве запрашиваемого белка2. Структуру AZ3002 затем использовали для разрешения AZ3003 аналогичным способом. Для того, чтобы обеспечить идеальную обратную взаимообусловленность гетеродимера Azymetric, присутствующего в кристаллографической асимметричной единице (например, степень заполнения молекулой А можно в равной степени описать молекулой В, и наоборот), смоделировали две возможные пары гетеродимера, каждая со степенью заполнения атомами 0,5, с помощью программы Coot, и уточнили структуры в программе Refmac3,4. Обработка результатов дифракции и статистика уточнения структуры представлены в Таблице 9.
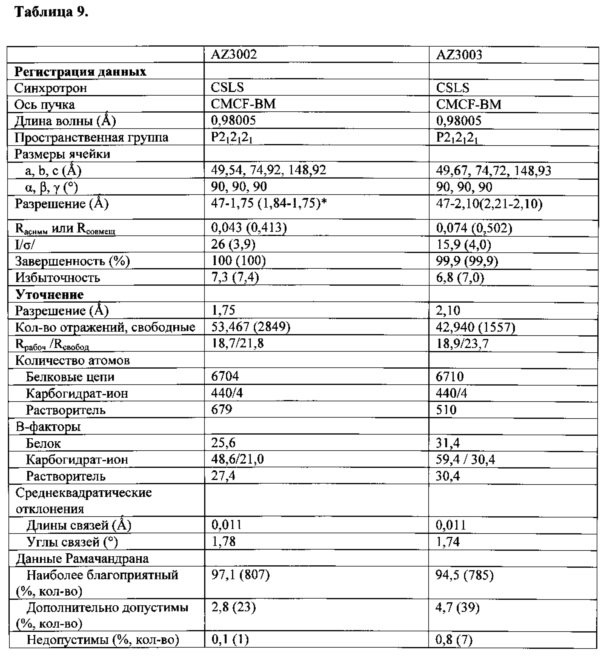
1. Kabsch, W. XDS. Acta Crystallogr D Diol Crystallogr 66, 125-132 (2010).
2. McCoy, A.J. Solving structures of protein complexes by molecular replacement with Phaser. Acta Crystallogr. D. Biol. Crystallogr. 63, 32-41 (2007).
3. Emsley, P. & Cowtan, K. Coot: model-building tools for molecular graphics. Acta Crystallogr. D. Biol. Crystallogr. 60, 2126-2132 (2004).
4. Murshudov, G.N., Vagin, A.A. & Dodson, E.J. Refinement of macromolecular structures by the maximum-likelihood method. Acta Crystallogr. D. Biol. Crystallogr. 53, 240-255 (1997).
Наложение кристаллических структур показано на Фигуре 42. Кристаллическая структура гетеродимеров AZ3002 и AZ3003 очень хорошо согласуется с компьютерными моделями (среднеквадратическое отклонение для всех атомов = 0,706 

Пример 13: Анализ гликозилирования AZ3003
Гетеродимер AZ3003 экспрессировали и очищали, как описано в Примере 11. Гликаны анализировали с помощью платформы GlykoPrep™ для быстрого получения N-гликанов с InstantAB™ (Prozyme), используя стандартный протокол производителя.
Результаты показаны на Фигуре 43, и проиллюстрировано, что у AZ3003 наблюдается обычный паттерн гликозилирования.
Пример 14: Оценка стабильности AZ3003 в условиях форсированной деградации
Стабильность гетеродимера AZ3003 оценивали путем инкубации в условиях форсированной деградации. Стабильность МАТ в условиях форсированной деградации может представлять собой хорошую оценку долгосрочной стабильности и стабильности при получении лекарственной формы.
Очищенный образец гетеродимера (который экспрессировали и очищали, как описано в Примере 11) концентрировали до концентрации 100 мг/мл без признаков агрегации. Образец разбавляли в подходящем буфере и оценивали в условиях форсированной деградации, как описано в Таблице 10, ниже. Обработанные образцы анализировали с помощью ЭФ в ПААГ/ДСН и ВЭЖХ-ЭХ.
ЭФ в ПААГ/ДСН осуществляли в восстановительных (В) и в невосстановительных (НВ) условиях на готовых градиентных гелях, приобретенных у LONZA. Белковые полосы визуализировали путем окрашивания кумасси бриллиантовым голубым G-250.
Аналитическую ЭХ-ВЭЖХ осуществляли, применяя колонку для ВЭЖХ либо BIOSEP-SEC-S4000, Phenomenex, либо Bio-Sil TSK 4000, BioRad, при скорости потока 0,8 мл/мин с 10 мМ фосфатом натрия, 0,14 М NaCl, 10% изопропанолом в качестве подвижного буфера. Это позволило осуществить количественный анализ потенциальных высоко- и низкомолекулярных молекул.
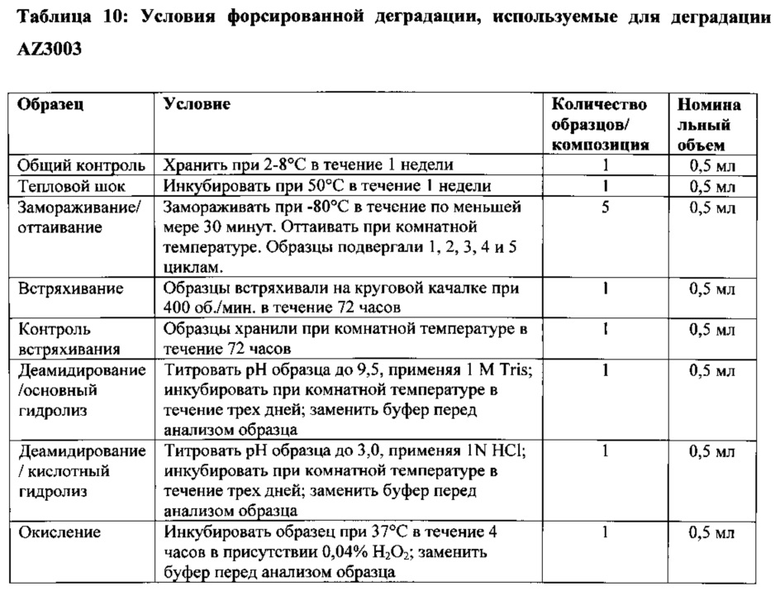
Результаты показаны на Фигуре 44 и демонстрируют, что гетеродимер AZ3003 стабилен и обладает профилем стабильности, который соответствует таковому для стандартных промышленных МАТ.
Пример 15: Последующая оценка очистки AZ3003
Проводили оценку возможности производства AZ3003, чтобы оценить свойства AZ3003, применяя стандартизованную в промышленности схему процесса очистки антитела, показанную на Фигуре 45. Данный процесс включал три этапа очистки на колонках, включая аффинную хроматографию на белке А для захвата продукта, после которого следовала катионообменная хроматография (КОХ) для удаления агрегатов, выщелоченного белка А и белков клетки-хозяина (БКХ), и, наконец, анионообменная хроматография (АОХ) в режиме непрерывного потока для захвата вирусов, ДНК и отрицательно заряженных контаминант.Данную оценку использовали, чтобы определить потенциальные проблемы при производстве (например, стабильность процесса, стабильность и качество продукта) с кандидатным(и) лекарственным(и) средством(ами) на ранней стадии исследования/разработки.
В ходе оценки возможности производства, авторы настоящего изобретения оценивали хроматографические свойства, стабильность белка и качество продукта, применяя стандартизованный в промышленности процесс очистки, показанный на Фигуре 46. В Таблице 11 (ниже) перечислены основные критерии, используемые для оценки, т.е. выход на разных этапах, содержание высокомолекулярных агрегатов (ВМА) и объем элюирования. Высокий выход на этапах и малые объемы элюирования в процессе очистки свидетельствовали о стабильности и хороших свойствах белка. Контроль содержания ВМА и их удаление в процессе очистки является крайне необходимым, так как присутствие белковых агрегатов (видов ВМА) в конечном продукте может приводить к пониженной активности, иммуногенным реакциям у пациента и/или образованию дисперсных загрязнений в течение срока хранения фармацевтического продукта. Экспрессия МАТ с минимальным исходным содержанием ВМА (<4%) необходима, так как высокие уровни агрегата потребуют дополнительных этапов очистки для их удаления, следовательно, повышая время и стоимость производства.
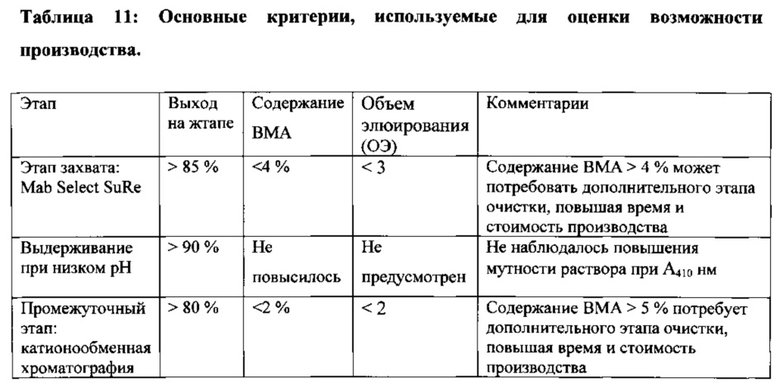
Стандартные процессы промышленной очистки применяли, чтобы проверить стабильность, хроматографические свойства и качество продукта AZ3003.
1.1 Захват белком А
Культуральную среду с экспрессированным AZ3003 фильтровали через фильтр с диаметром пор 0,22 мкм, применяя фильтр на горловину флакона (PES) от Millipore, и наносили на колонку Mab Select SuRe (1,6×25 см), уравновешенную 5 объемами колонки (ОК) 20 мМ Трис-HCl, 0,14 M NaCl, pH 7,5. После загрузки, колонку тщательно промывали уравновешивающим буфером до тех пор, пока поглощение на А280 не достигло стабильного исходного уровня. AZ3003 элюировали 0,1 M ацетатным буфером, рН 3,6, и немедленно титровали до рН 5,2 путем добавления 1/10 объема 1 M Трис-основания.
После этапа элюирования, колонку промывали 0,1 M ацетатом, pH 3,0. Анализ путем ЭФ в ПААГ/ДСН показал, что все МАТ были связаны с колонкой, так как МАТ не были обнаружены в потоке с колонки. Хорошо очищенные МАТ были обнаружены в элюирующем буфере при рН 3,6. Исходный этап захвата и очистки с применением аффинной хроматографии на белке А позволил получить продукт с чистотой >90%.
1.2 Исследование с выдерживанием при низком pH
Следующий этап в процессе выделения и очистки продукта представлял собой выдерживание при низком pH, которое осуществляли, чтобы инактивировать вирусы. После элюирования с колонки с белком А, МАТ (~10 мг/мл, pH 4,0) титровали до pH 3,6 с помощью 10% уксусной кислоты и инкубировали при комнатной температуре при перемешивании в течение 90 мин. Стабильность AZ3003 к воздействию низкого pH оценивали путем ЭФ в ПААГ/ДСН, ЭХ-ВЭЖХ и путем измерения мутности при A410 нм. AZ3003 хорошо переносил этап выдерживания при низком рН, не проявляя изменений при ЭФ в ПААГ/ДСН или ЭХ-ВЭЖХ. Кроме того, не было обнаружено повышения мутности после 90 мин инкубации, что указывает на отсутствие образования нерастворимых агрегатов, которые могут представлять собой проблему в процессе очистки (т.е. засорение фильтров и колонок в процессе очистки, утрату продукта). Полученные результаты показали, что AZ30003 оказался стабилен на этапе выдерживания при низком pH.
1.3 Катионообменная хроматография (КОХ)
КОХ исследовали в качестве второго этапа в процессе очистки. Оценивали две смолы: Fractogel EMD SO3 (M) от Merk/Millipore и SP HP от GE Lifesciences.
Fractogel EMD SO3 (M), pH 5,2: Объединенные фракции с колонки Mab Select SuRe (35 мг) титровали до pH 5,2 путем добавления 10% (об./об.) 1 M Трис-основания, а затем разбавляли в 2 раза уравновешивающим буфером, 20 мМ ацетатом, рН 5,2. Эти объединенные фракции наносили на колонку Fractogel EMD SO3 (M), уравновешенную 5 OK 20 мМ ацетата, рН 5,2. Колонку промывали уравновешивающим буфером до тех пор, пока поглощение на А280 не достигло стабильного исходного уровня. МАТ элюировали с колонки 10 ОК линейного градиента соли от 0 до 600 мМ NaCl, рН 5,2. Оставшиеся контаминаты снимали с колонки 20 мМ ацетатом, 1 M NaCl, рН 5,2, с последующей обработкой 1 N NaOH. Осуществляли анализ путем ЭФ в ПААГ/ДСН и ЭХ-ВЭЖХ для отслеживания уровней ВМА и их удаления из основной фракции МАТ с данной колонки. Выход на данном этапе (на основании считывания на А280 нм) составлял 73%.
SP HP, pH 5,2: Объединенные фракции с колонки Mab Select SuRe (50 мг) титровали до pH 5,2 путем добавления 10% (об./об.) 1 M Трис-основания, а затем в равной степени разбавляли уравновешивающим буфером, 20 мМ ацетатом, рН 5,2. Эти объединенные фракции наносили на колонку SP HP (1,6×2,5 см/5 мл), уравновешенную 5 ОК 20 мМ ацетата, pH 5,2 (Фиг. 21). Колонку промывали уравновешивающим буфером до тех пор, пока поглощение на А280 не достигло стабильного исходного уровня. МАТ элюировали с колонки 10 ОК линейного градиента соли от 0 до 600 мМ NaCl, рН 5,2. Оставшиеся контаминаты снимали с колонки 20 мМ ацетатом, 1 M NaCl, pH 5,2, с последующей обработкой 1 N NaOH. Осуществляли анализ путем ЭФ в ПААГ/ДСН и ЭХ-ВЭЖХ для отслеживания уровней агрегации и их разделения на данной колонке. Выход на данном этапе (на основании считывания на А280 нм) составлял 87%.
1.4 Анионообменная хроматография (АОХ)
Анионообменники (например, четвертичный амин, Q) широко применяют для очистки моноклональных антител. Среда АОХ используется в режиме непрерывного потока, и МАТ обнаруживаются в потоке с колонки, тогда как БКХ, ДНК, вирусы и эндотоксины задерживаются.
Объединенные фракции с колонки Fractogel SO3 M при рН 5,2 (25 мг) титровали до рН 7,0 путем добавления 1 M Трис-основания и наносили на 1 мл колонку HiTrap Q FF, уравновешенную 5 ОК 10 мМ фосфата, рН 7,0. Колонку промывали уравновешивающим буфером. Тем не менее, в данном случае, стабильный исходный уровень не был достигнут. Следовательно, колонку промывали ФБР, чтобы элюировать все остаточные связанные МАТ. Затем колонку промывали 10 мМ фосфатом, 1 M NaCl, рН 7,0, чтобы удалить все связанные контаминаты. Данный этап требует дополнительной оптимизации, чтобы все фракции МАТ присутствовали в потоке с колонки. Выход на данном этапе (на основании считывания на А280 нм) составлял 82%, с чистотой, оцененной с помощью ЭХ-ВЭЖХ, составляющей >98%.
1.5 Анализ путем ЭФ в ПААГ/ДСН очистки на этапах
ЭФ в ПААГ/ДСН (Фиг. 35) осуществляли с элюатами из каждого этапа очистки, чтобы отследить удаление контаминантов на всем протяжении процесса и чтобы оценить качество и чистоту конечного продукта. Анализ гелей показал ожидаемый паттерн движения МАТ в невосстановительных условиях и в восстановительных условиях. Сравнение гелей с нанесенными объединенными фракциями с двух смол КОХ не выявило существенных различий в профилях продуктов. Все конечные объединенные фракции после КОХ, хроматографии на керамическом гидроксиапатите (СНТ) и хроматографии гидрофобных взаимодействий (ХГВ), рН 5,0 и рН 7,0, были аналогичны в отношении чистоты и контаминирующих полос.
1.7 Оценка чистоты с помощью ЭХ-ВЭЖХ
Очищенный AZ3003 после трех этапов колоночной хроматографии: на белке А; КОХ; АОХ в режиме непрерывного потока, - оценивали с помощью ЭХ-ВЭЖХ при нативных условиях (Фигура 46).
AZ3003 элюировался одиночным пиком в рамках ожидаемой области 150 кДа для нативного IgG1. Чистоту оценили >98%.
1.8 Выходы процесса для очистки AZ3003
Выходы процесса рассчитали для дальнейшего процесса очистки и перечислили на Фигуре 46. Выходы AZ3003 на этапах были типичными для IgG, очищенных с применением стандартизованного в промышленности трехколоночного процесса очистки.
AZ3003 успешно очищали из культуральной среды, применяя стандартизованный в промышленности процесс очистки (аффинную смолу с белком А, КОХ, а затем АОХ).
Полученные результаты показали, что AZ3003 сравним со стандартными МАТ по общему выходу продукта (см. Kelley В. Biotechnol. Prog. 2007, 23, 995-1008) и наблюдаемой на минимальном уровне агрегации. AZ3003 также оценивали в отношении выдерживания при низком рН и с помощью хроматографии на СНТ (керамическом гидроксиапатите) и ХГВ (хроматографии гидрофобных взаимодействий) (Phenyl HP pH 5 и рН 7), с получением хорошего выхода в отсутствии агрегатов. На Фигуре 46 проиллюстрировано, что основной гетеродимер успешно очистили из культуральной среды, применяя стандартизованный в промышленности процесс очистки (аффинную смолу с белком А, КОХ, а затем АОХ).
Все публикации, патенты и заявки на патент, упомянутые в данном описании, включены в данную заявку посредством ссылки в равной мере, как если бы каждая отдельная публикация, патент или заявка на патент были конкретно и отдельно указаны, как включенные в данную заявку посредством ссылок.
| название | год | авторы | номер документа |
|---|---|---|---|
| РАЗРАБОТКА СТАБИЛЬНОГО ГЕТЕРОДИМЕРНОГО АНТИТЕЛА С МУТАЦИЯМИ В ДОМЕНЕ Fc | 2012 |
|
RU2810540C2 |
| БИСПЕЦИФИЧЕСКИЕ АСИММЕТРИЧНЫЕ ГЕТЕРОДИМЕРЫ, СОДЕРЖАЩИЕ АНТИ-CD3 КОНСТРУКЦИИ | 2013 |
|
RU2650868C2 |
| ДИЗАЙН УСТОЙЧИВОГО ГЕТЕРОДИМЕРНОГО АНТИТЕЛА С МУТАЦИЯМИ В Fc ДОМЕНЕ | 2011 |
|
RU2604490C2 |
| ГЕТЕРОМУЛЬТИМЕРЫ С УМЕНЬШЕННОЙ ИЛИ ПОДАВЛЕННОЙ ЭФФЕКТОРНОЙ ФУНКЦИЕЙ | 2014 |
|
RU2655439C2 |
| МОДИФИЦИРОВАННЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ ПОЛИПЕПТИДНЫЕ КОНСТРУКЦИИ И ИХ ПРИМЕНЕНИЕ | 2015 |
|
RU2832921C1 |
| ПОЛИВАЛЕНТНЫЕ МОДУЛЯТОРЫ РЕГУЛЯТОРНЫХ T-КЛЕТОК | 2017 |
|
RU2769871C2 |
| ПРЕДПОЧТИТЕЛЬНОЕ СПАРИВАНИЕ ДОМЕНОВ АНТИТЕЛ | 2018 |
|
RU2792440C2 |
| БЕЛКИ, СВЯЗЫВАЮЩИЕ СПЕЦИФИЧЕСКИЙ МЕМБРАННЫЙ АНТИГЕН ПРОСТАТЫ, И СООТВЕТСТВУЮЩИЕ КОМПОЗИЦИИ И СПОСОБЫ | 2012 |
|
RU2632647C2 |
| МОДИФИЦИРОВАННЫЕ АСИММЕТРИЧНЫЕ АНТИТЕЛА, СВЯЗЫВАЮЩИЕ Fc-РЕЦЕПТОР, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2687043C2 |
| ВАРИАНТЫ FC-ОБЛАСТИ С УЛУЧШЕННОЙ СПОСОБНОСТЬЮ СВЯЗЫВАТЬСЯ С БЕЛКОМ А | 2015 |
|
RU2698969C2 |







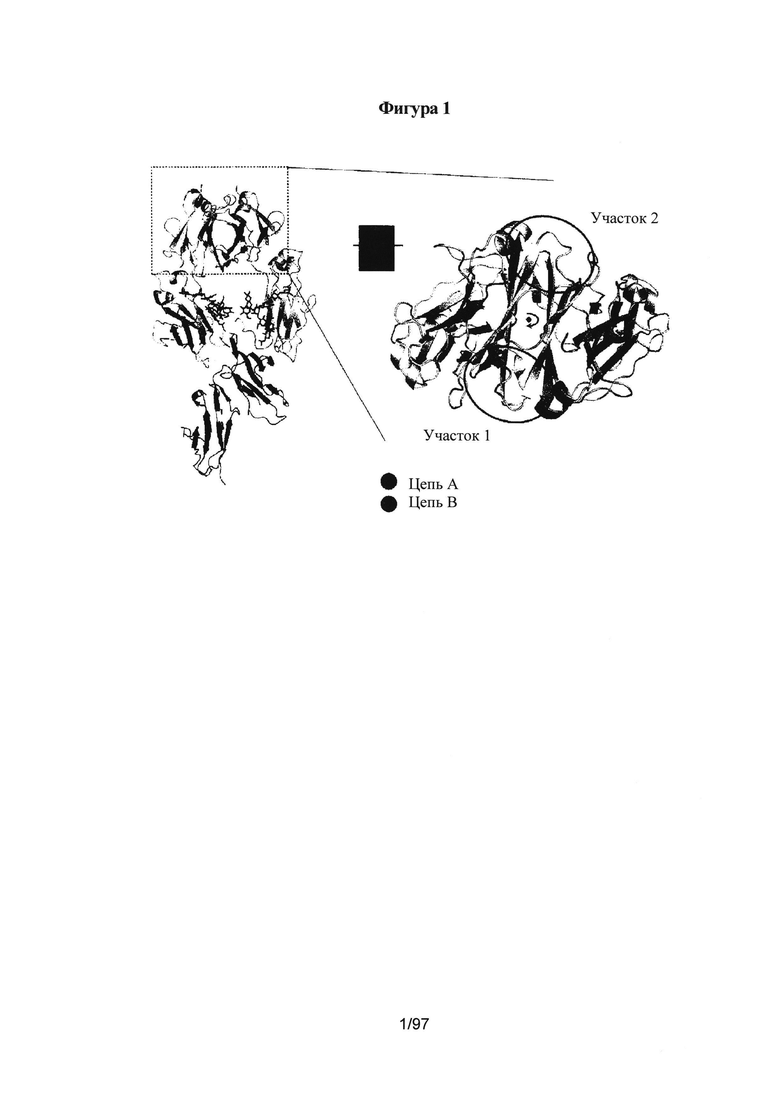



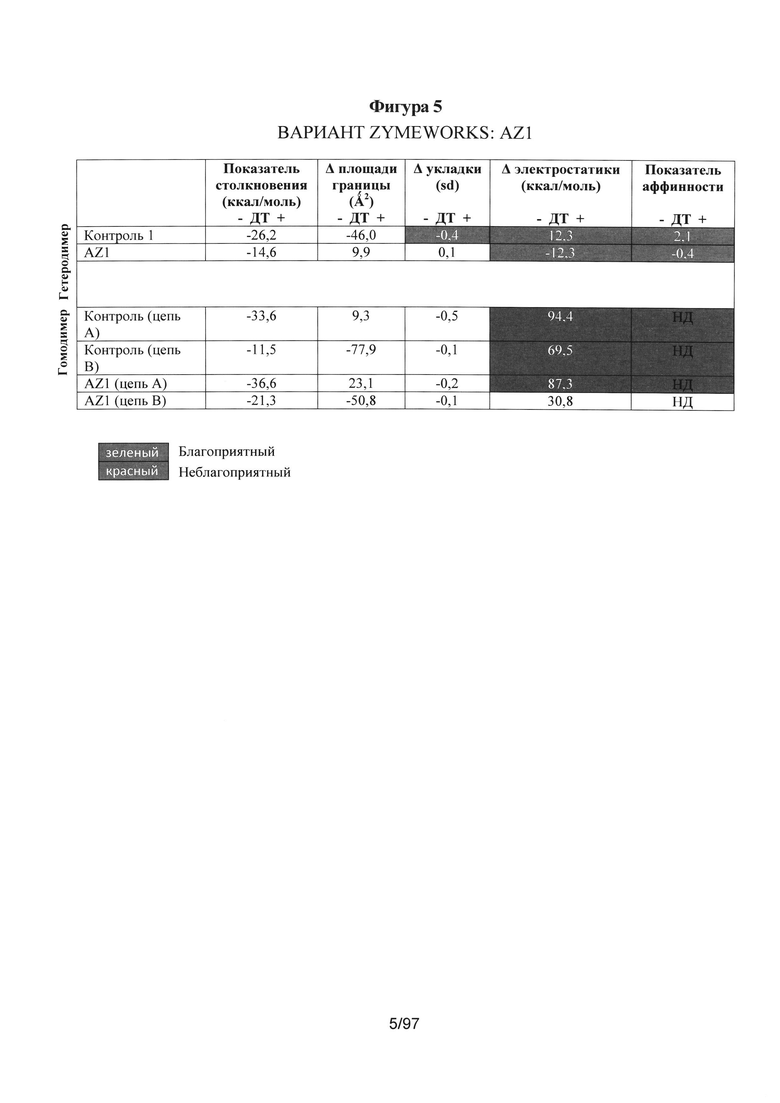


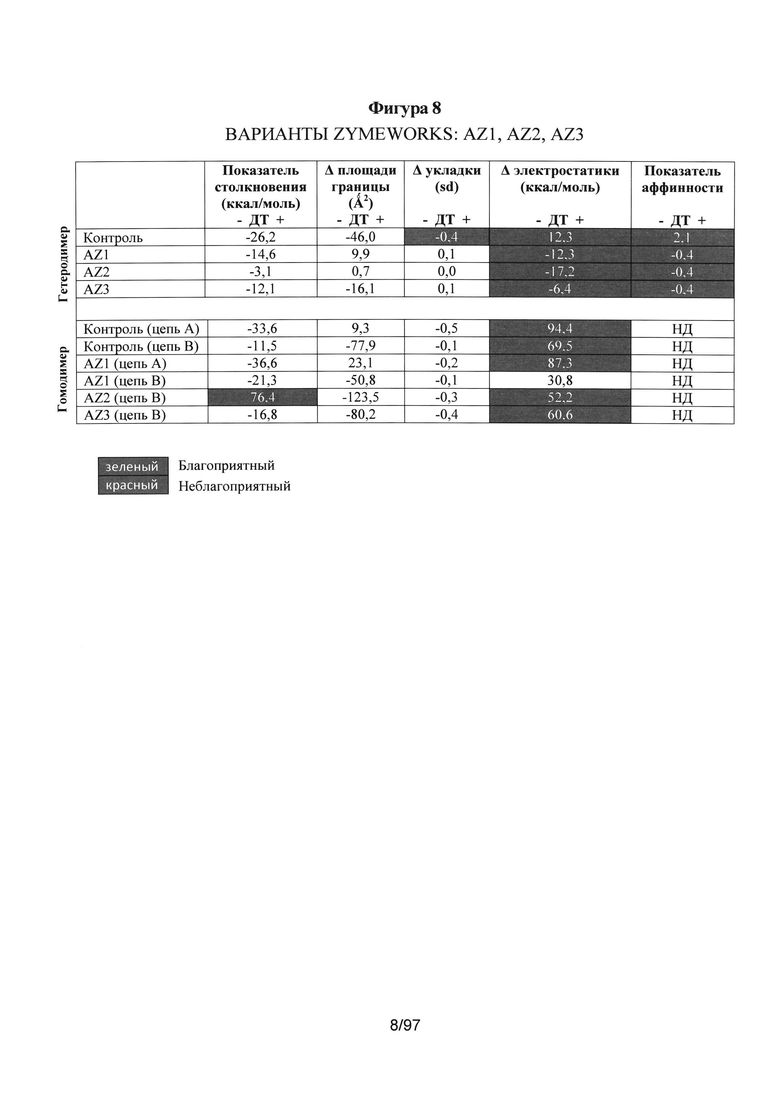

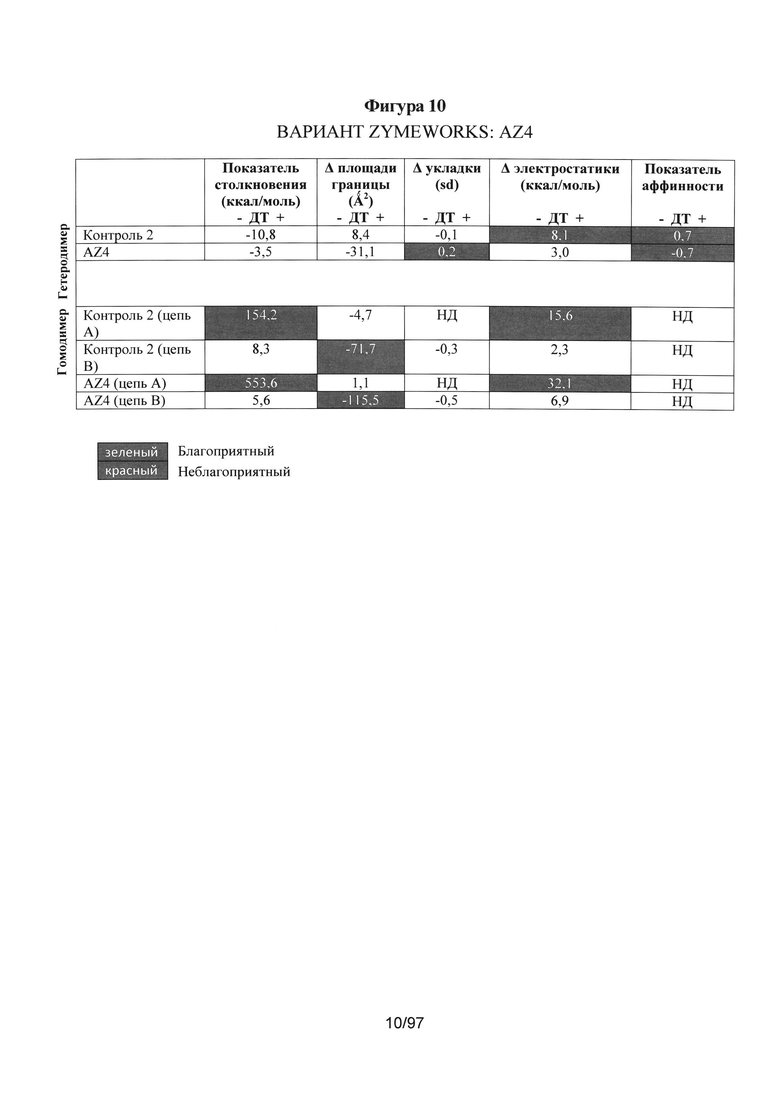

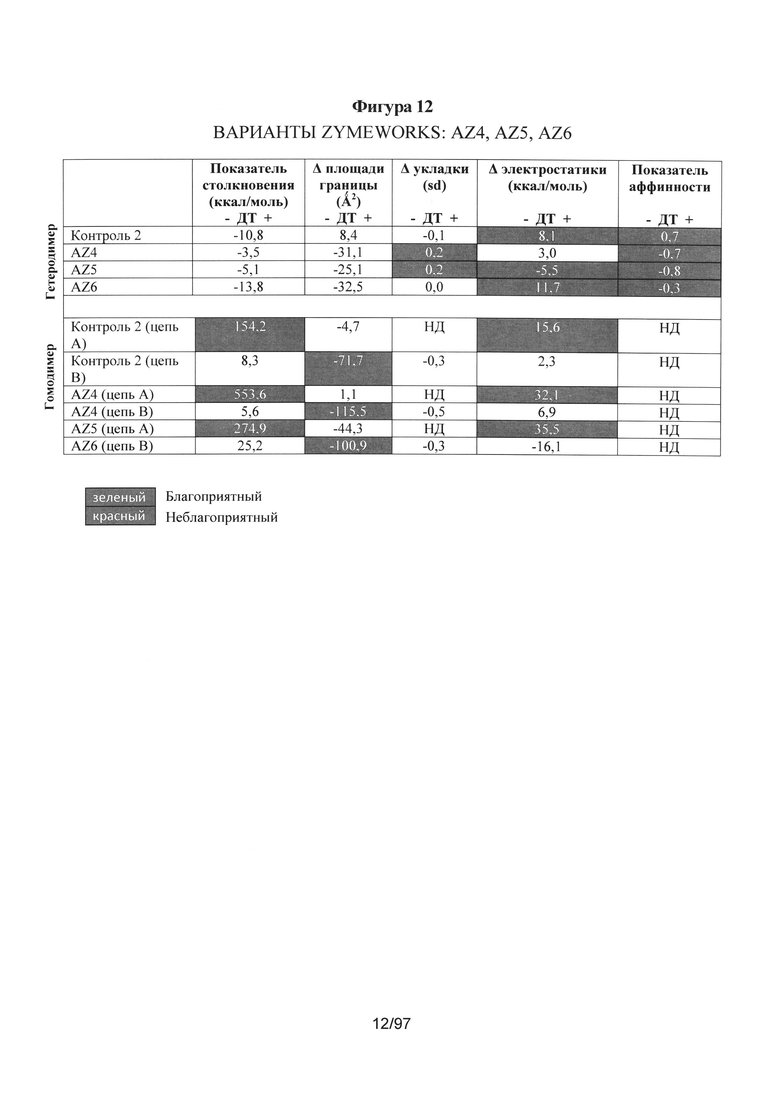

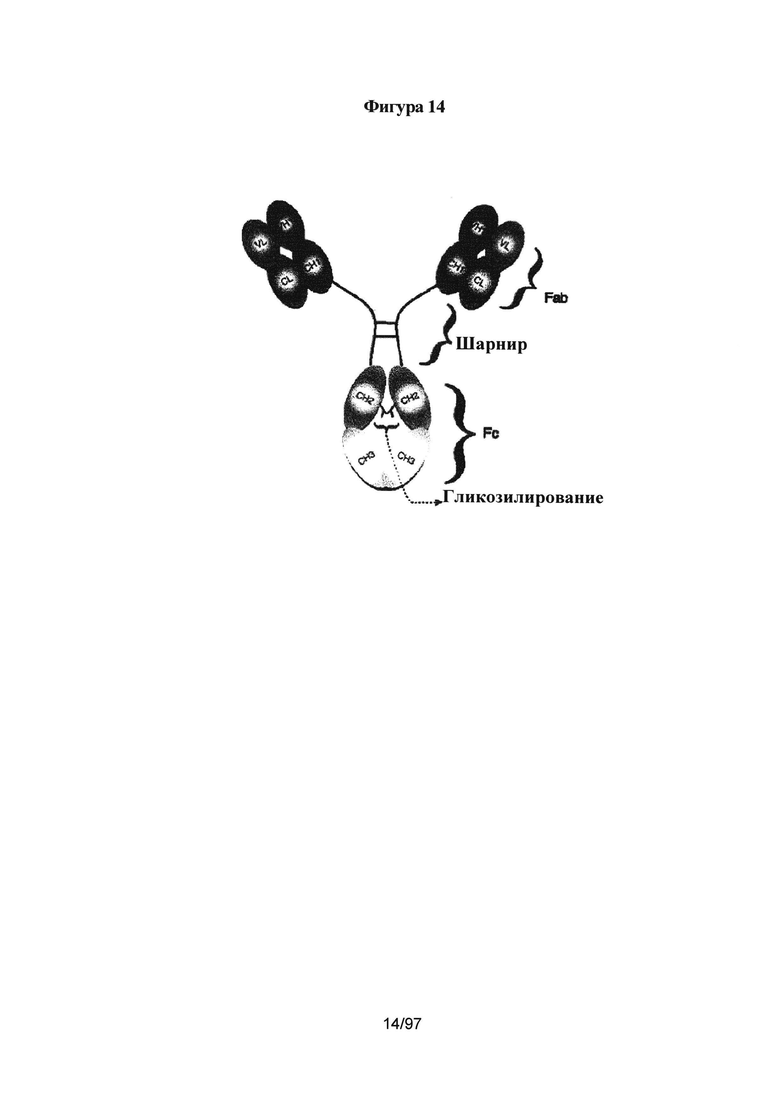










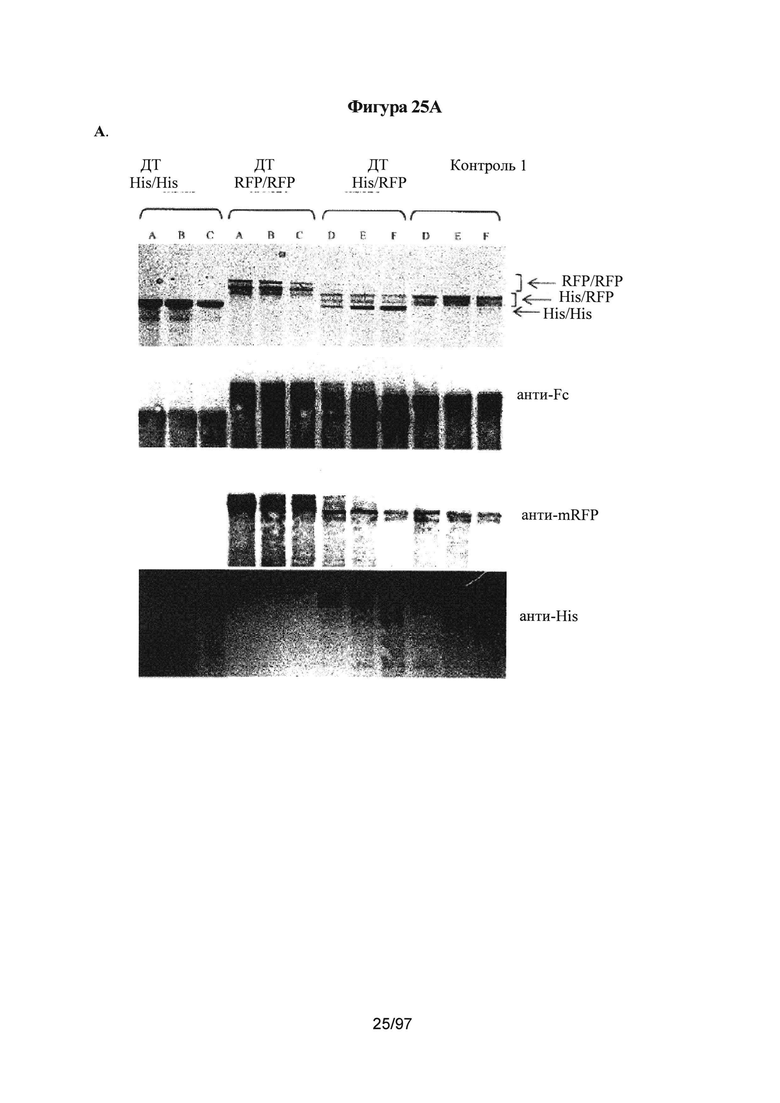
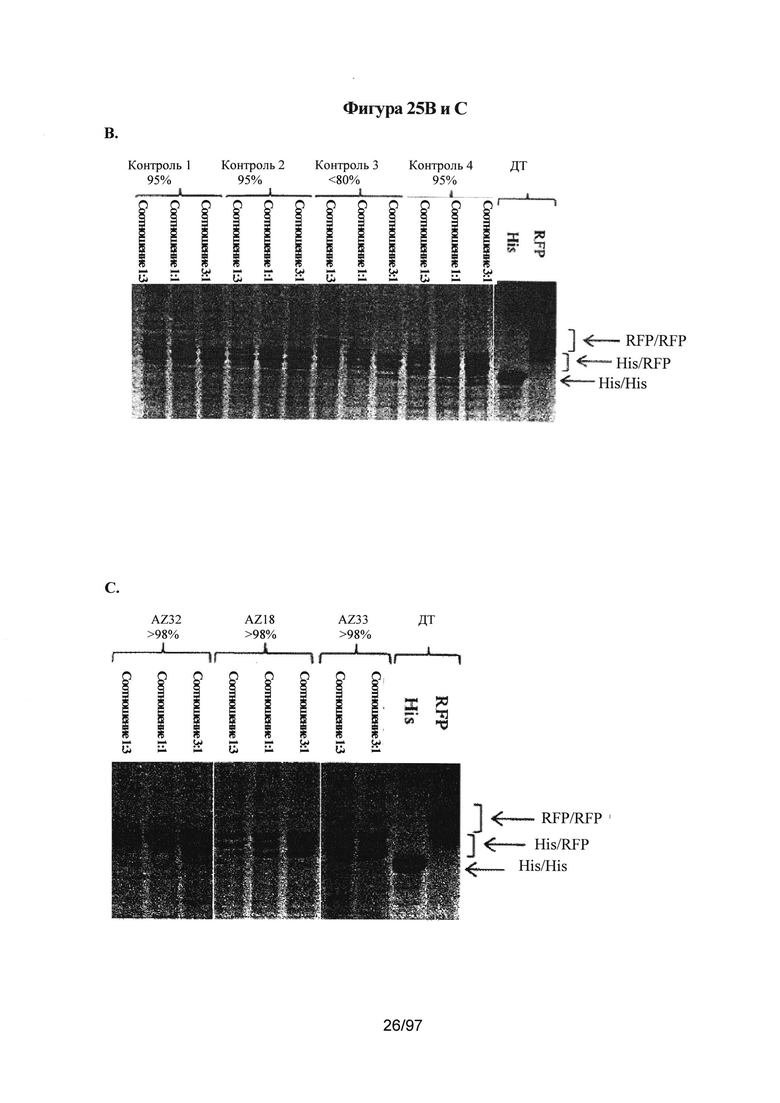
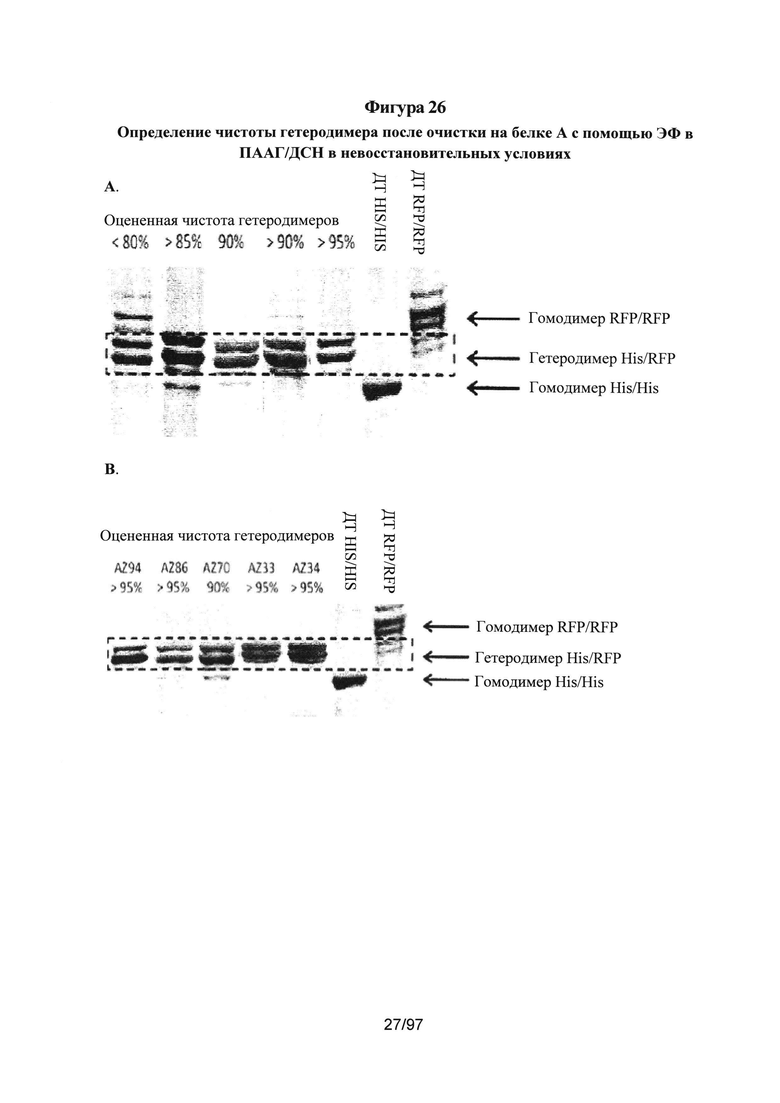
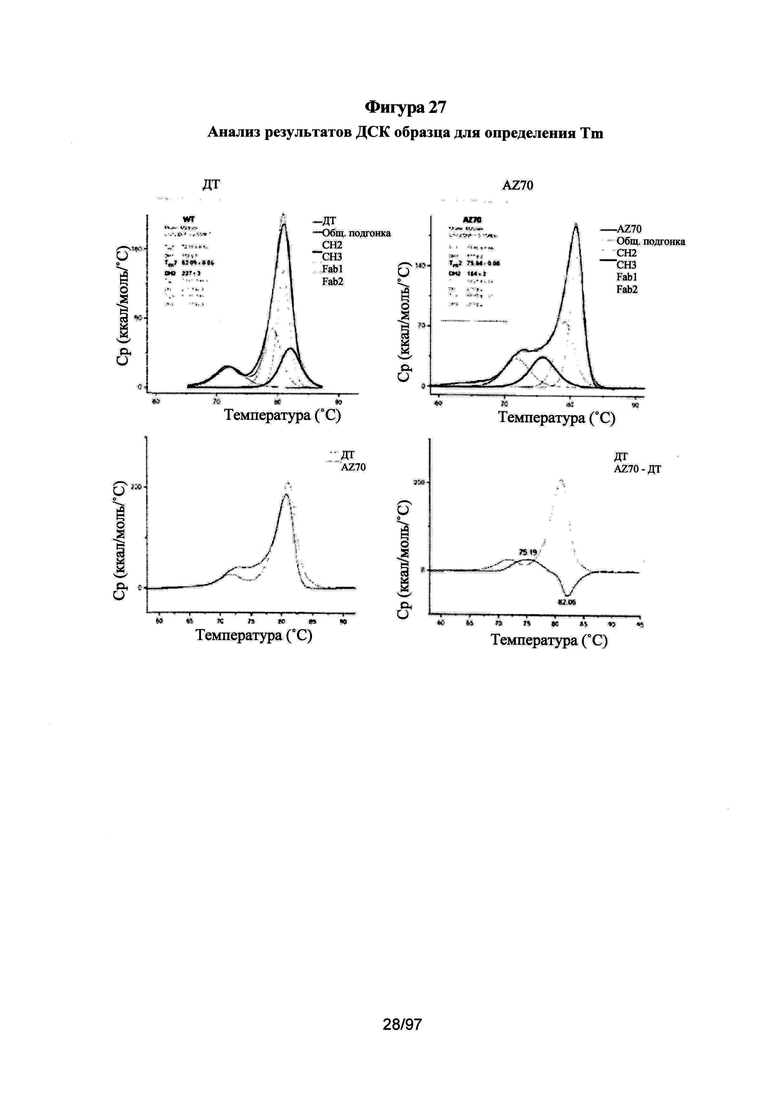
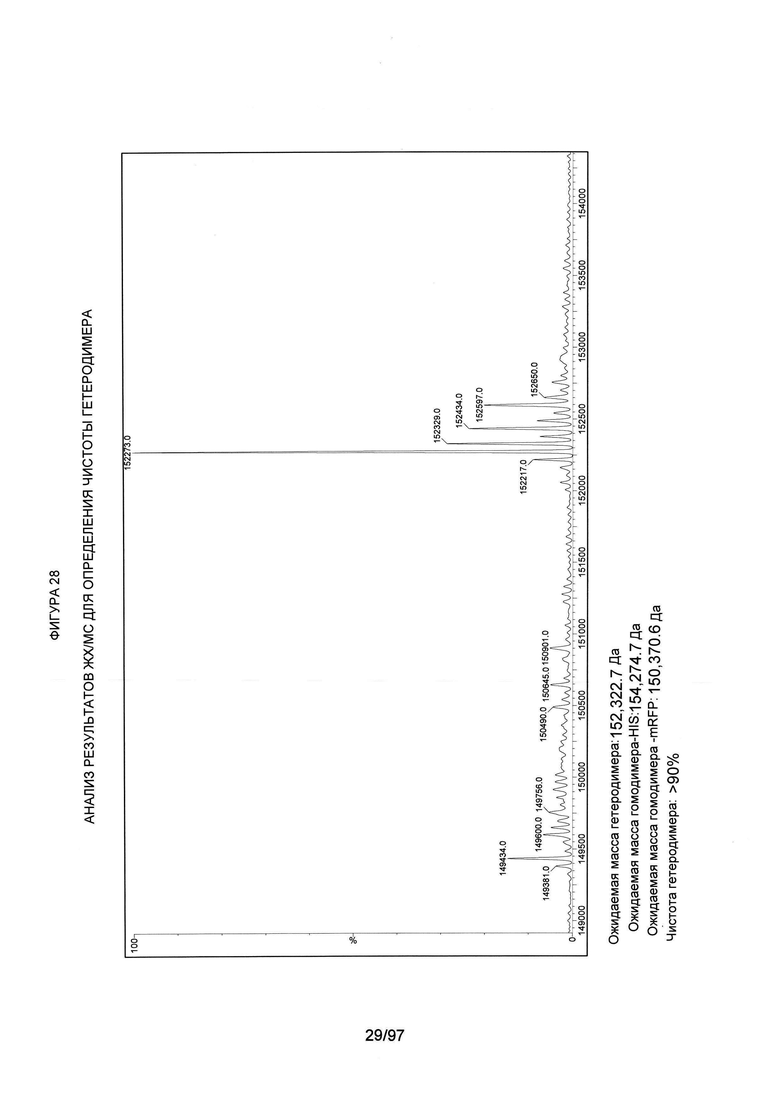
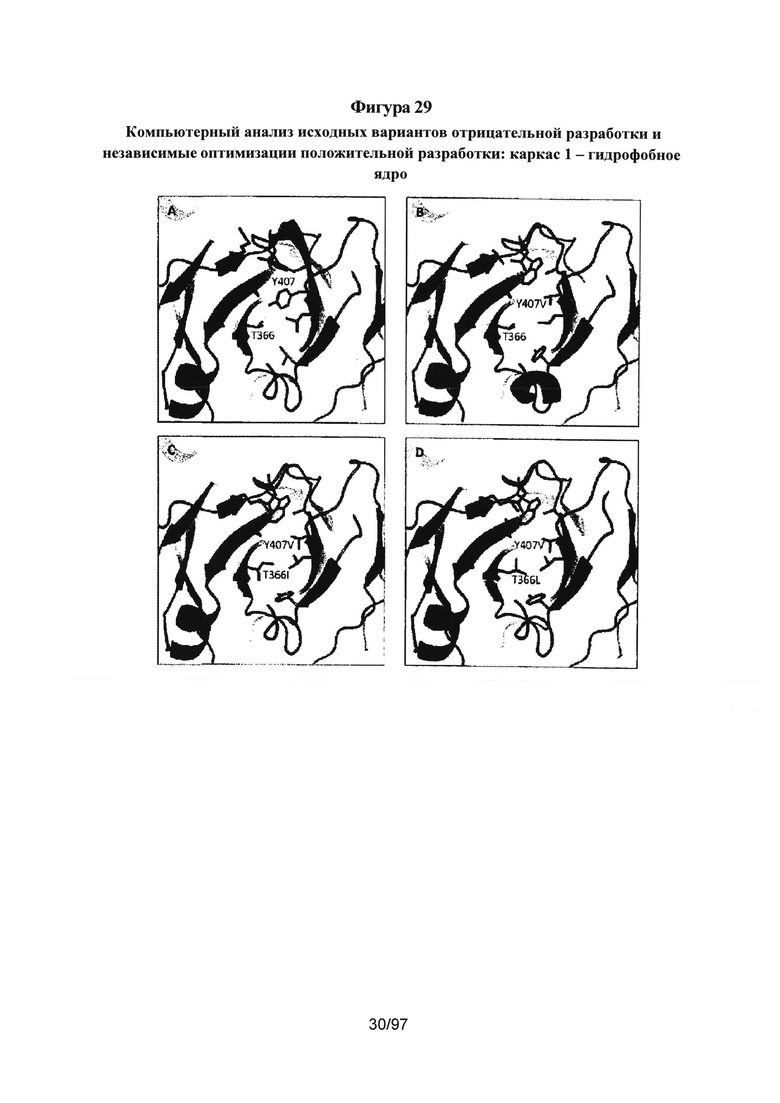

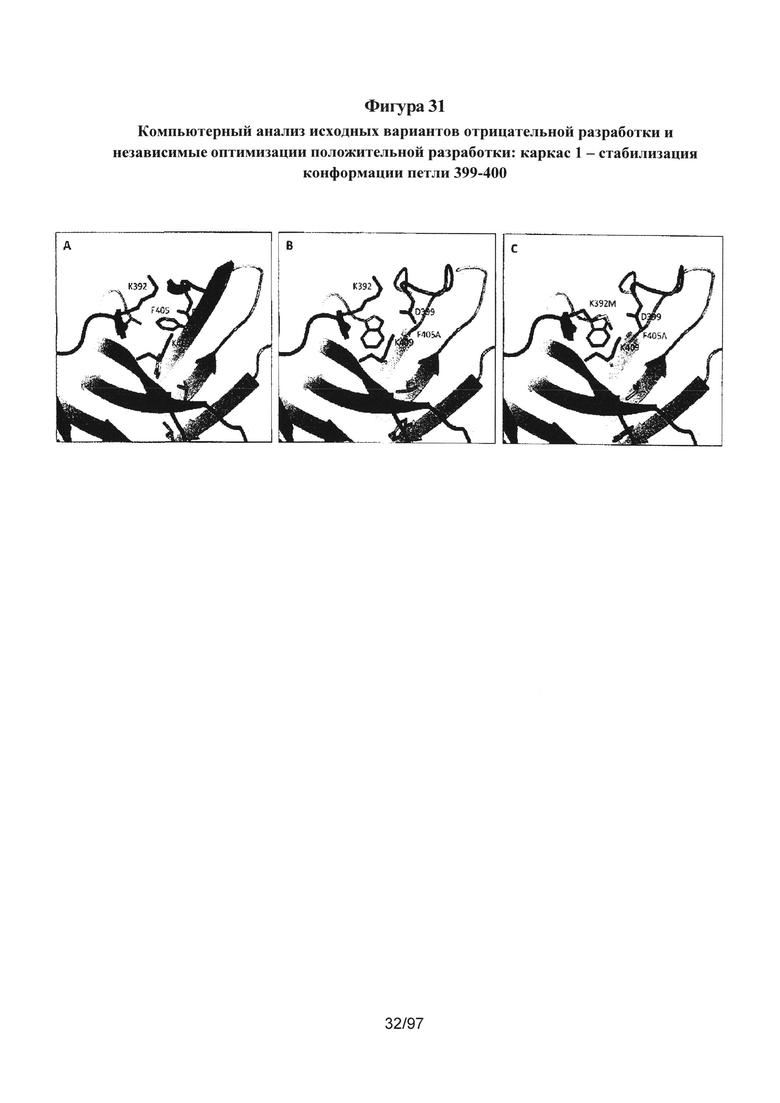

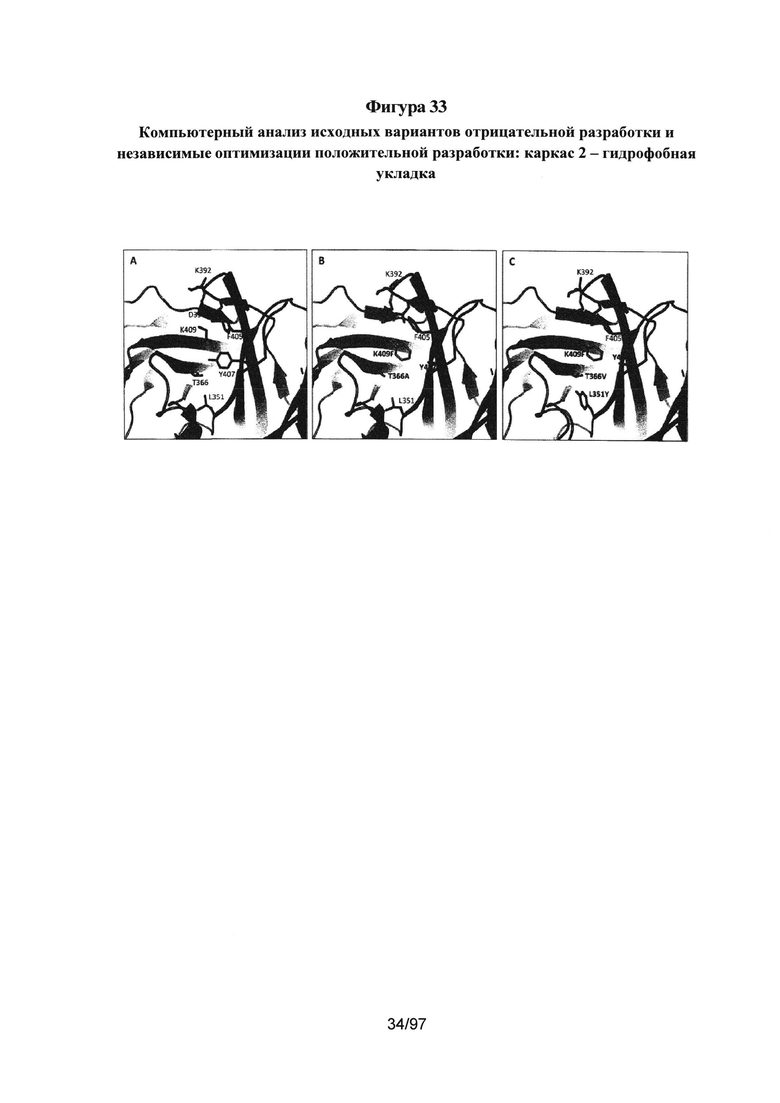

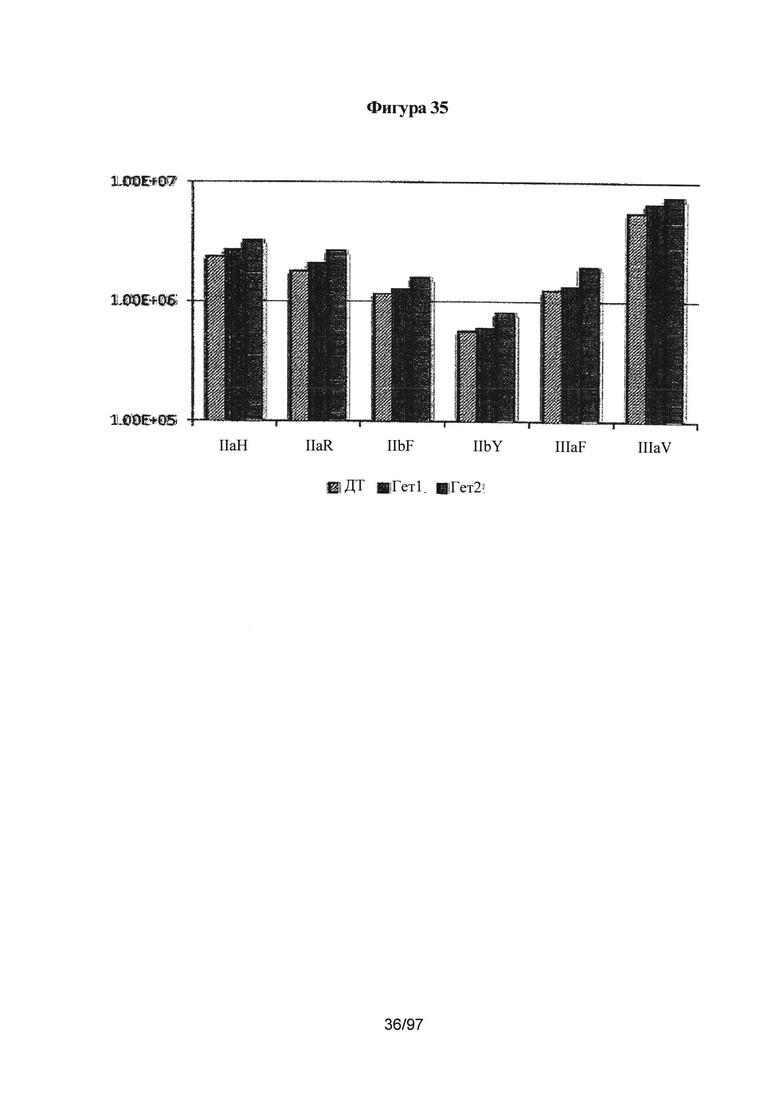
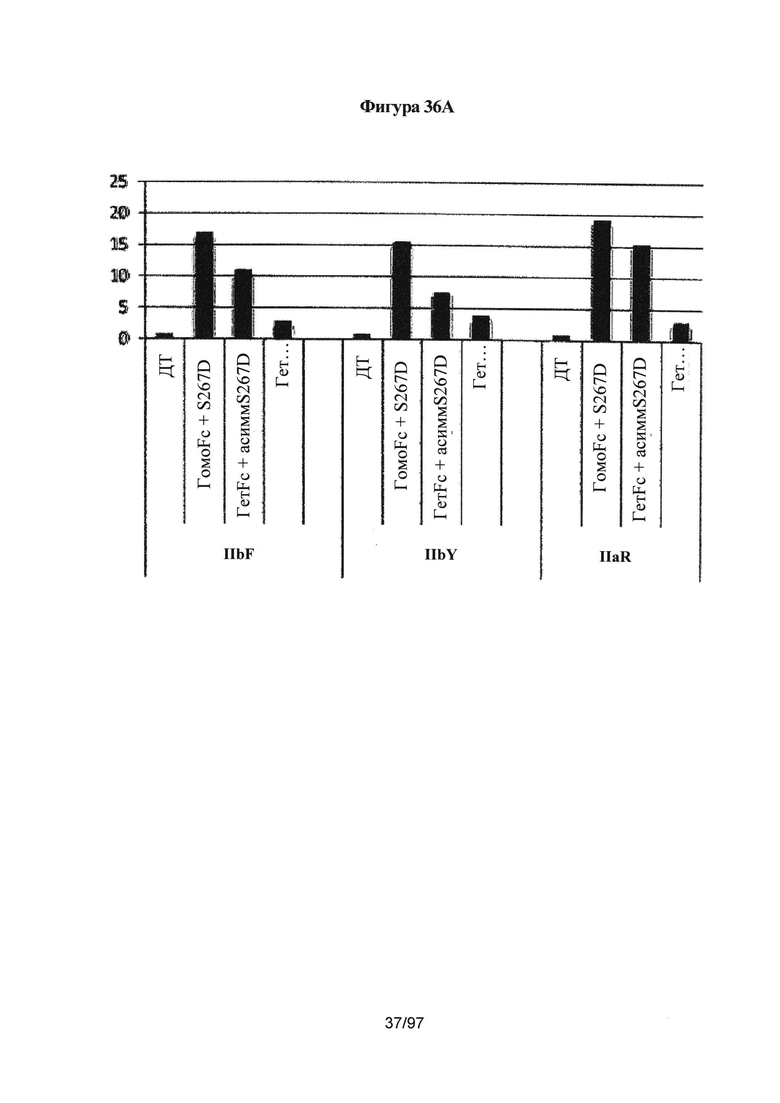
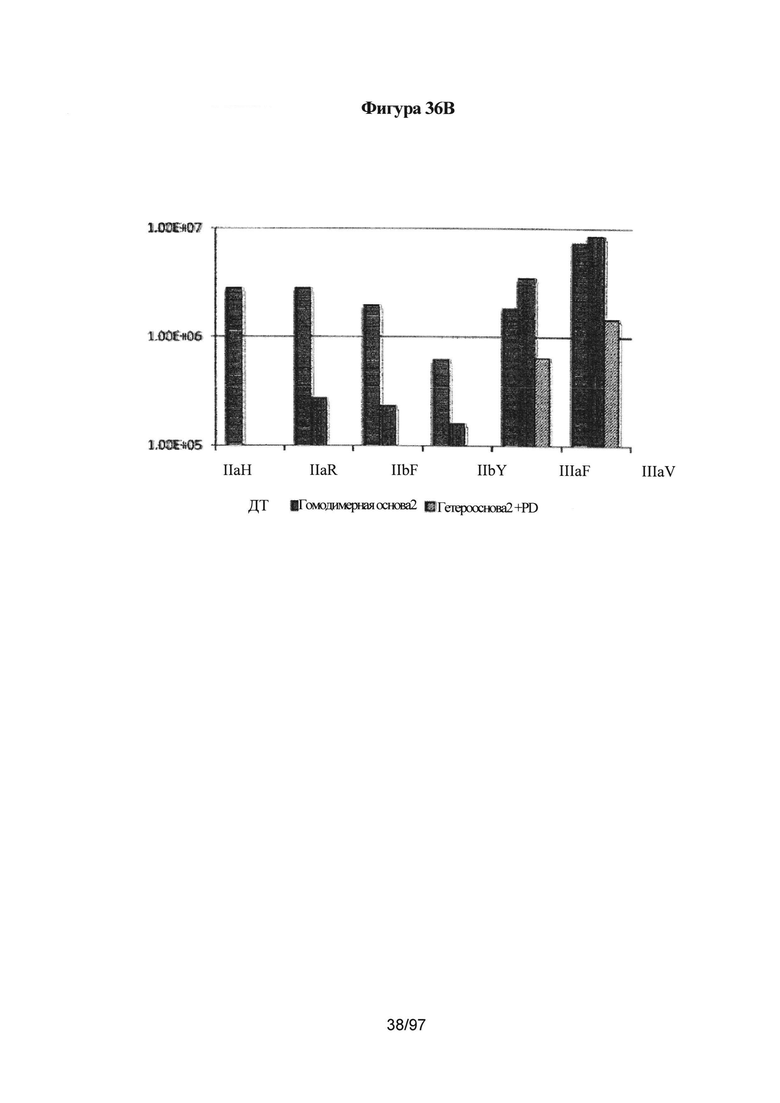
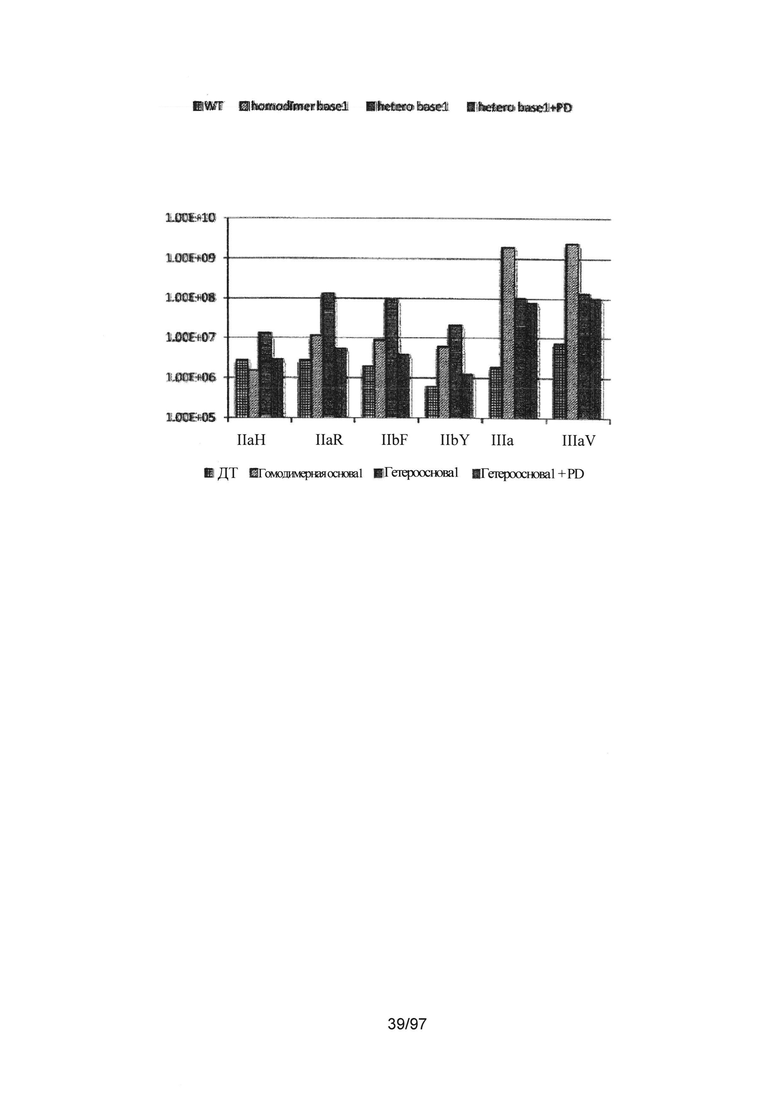
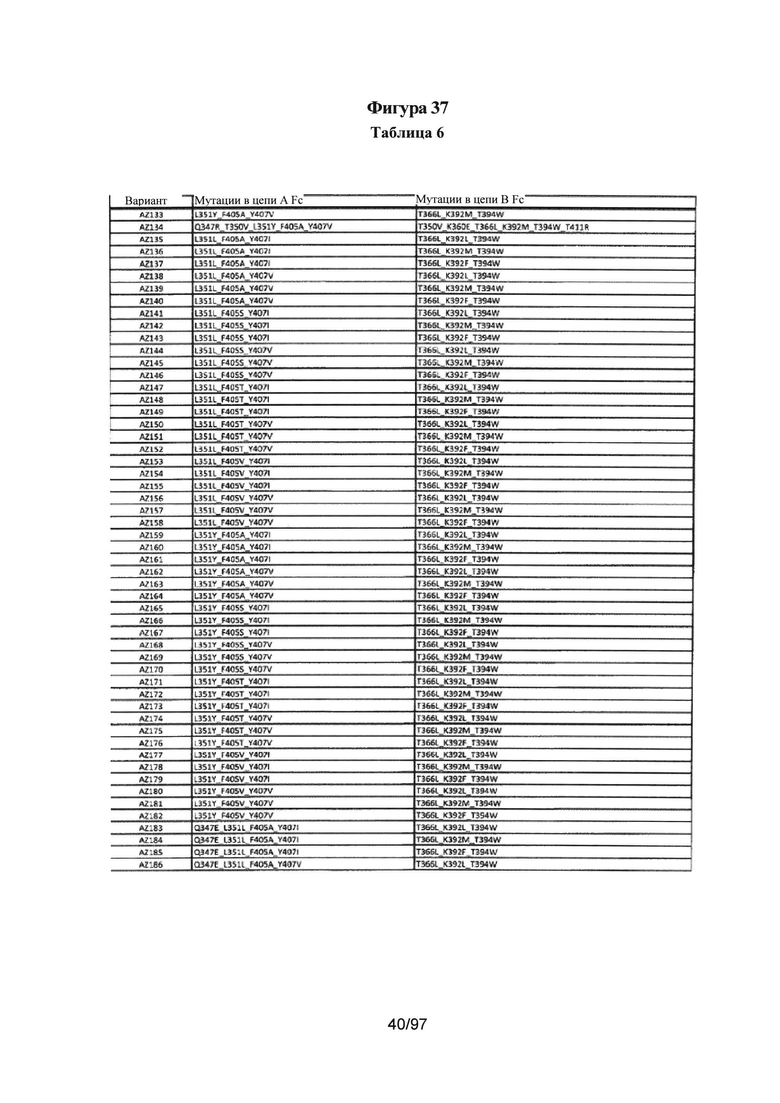

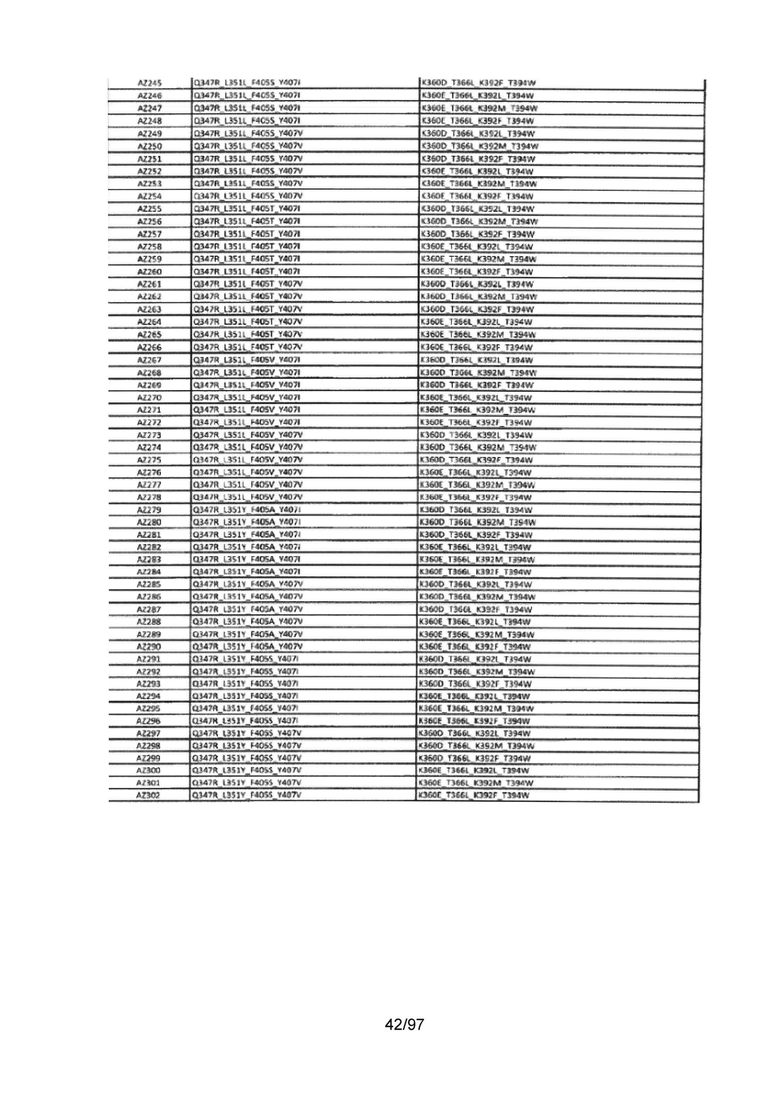
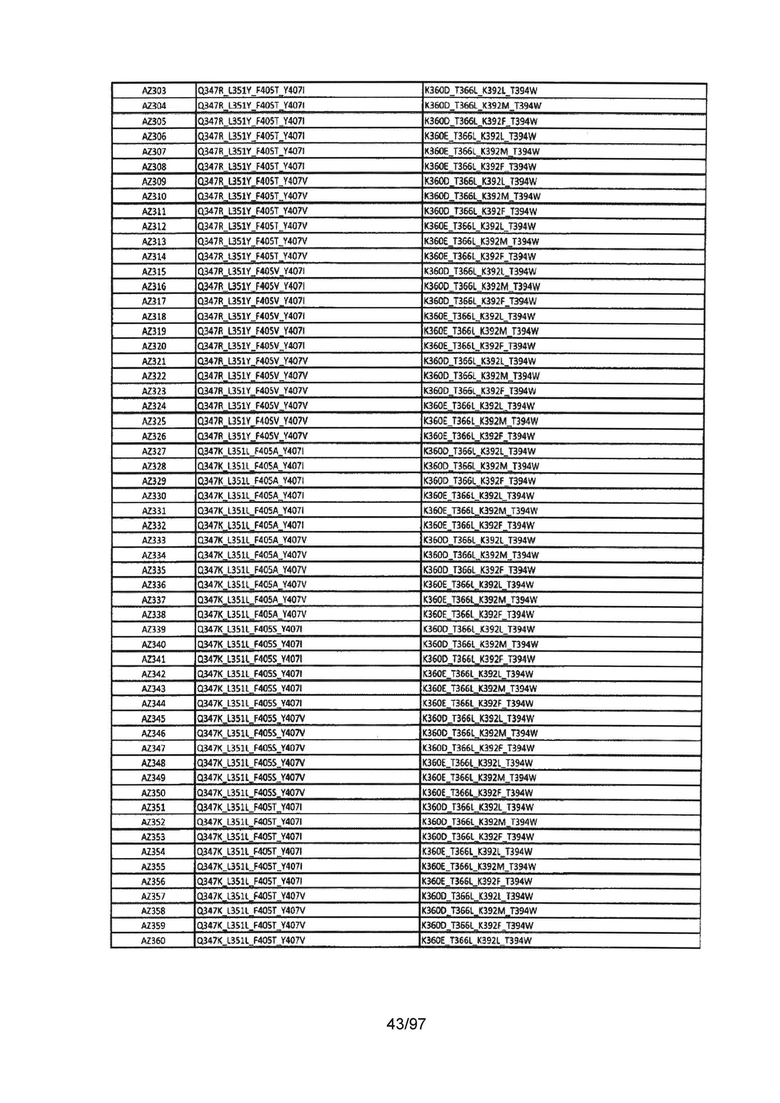
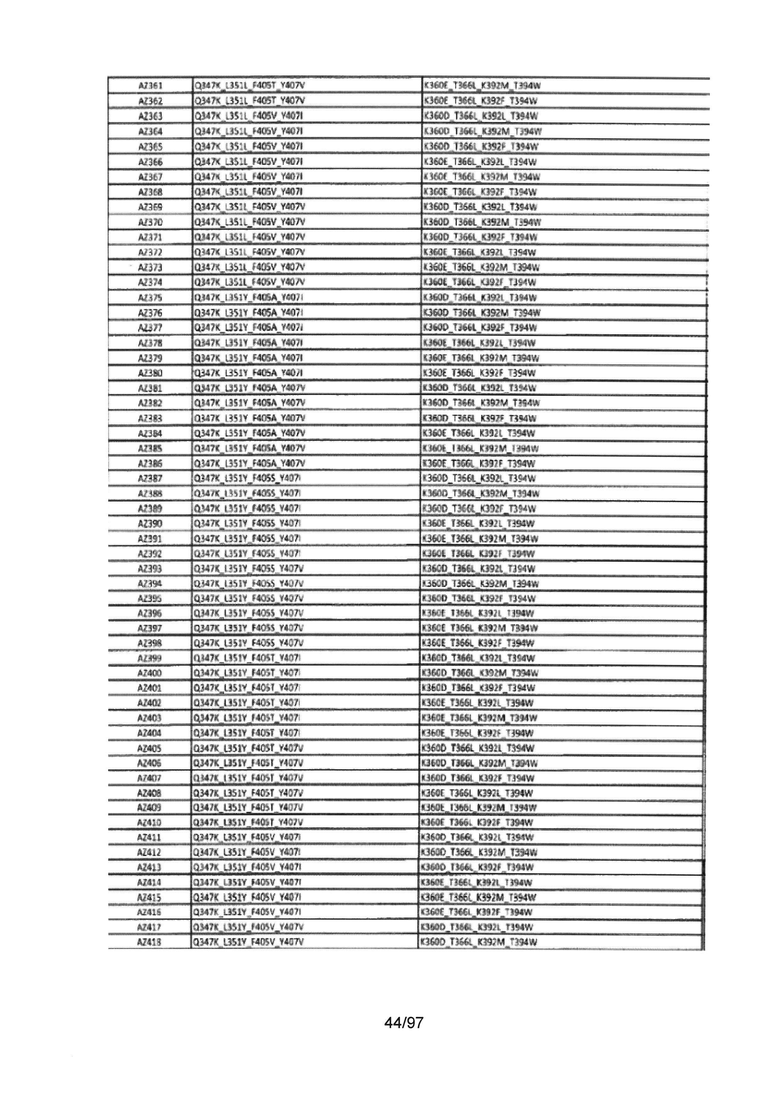
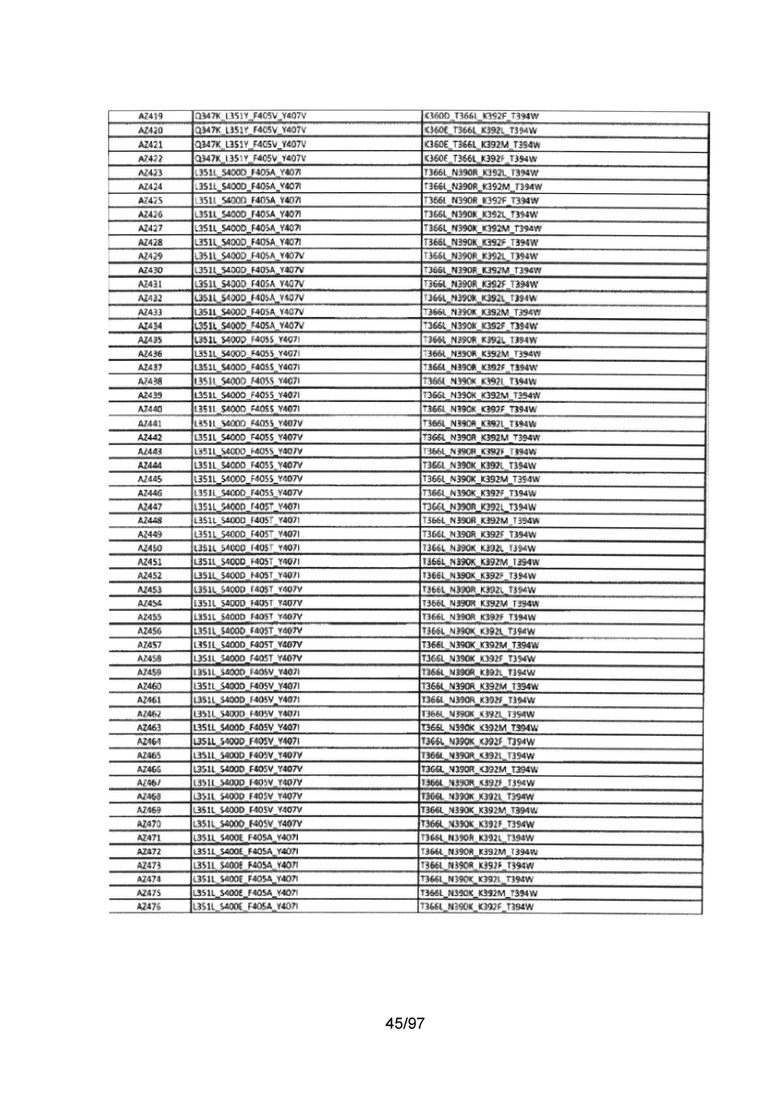
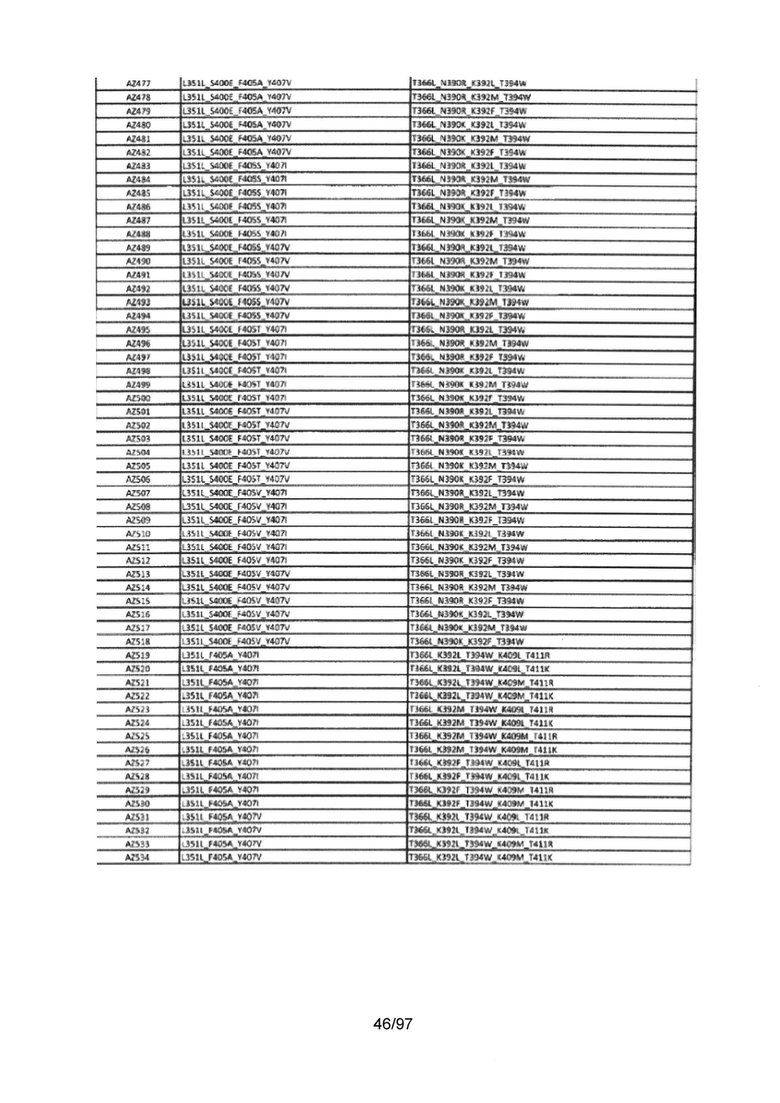
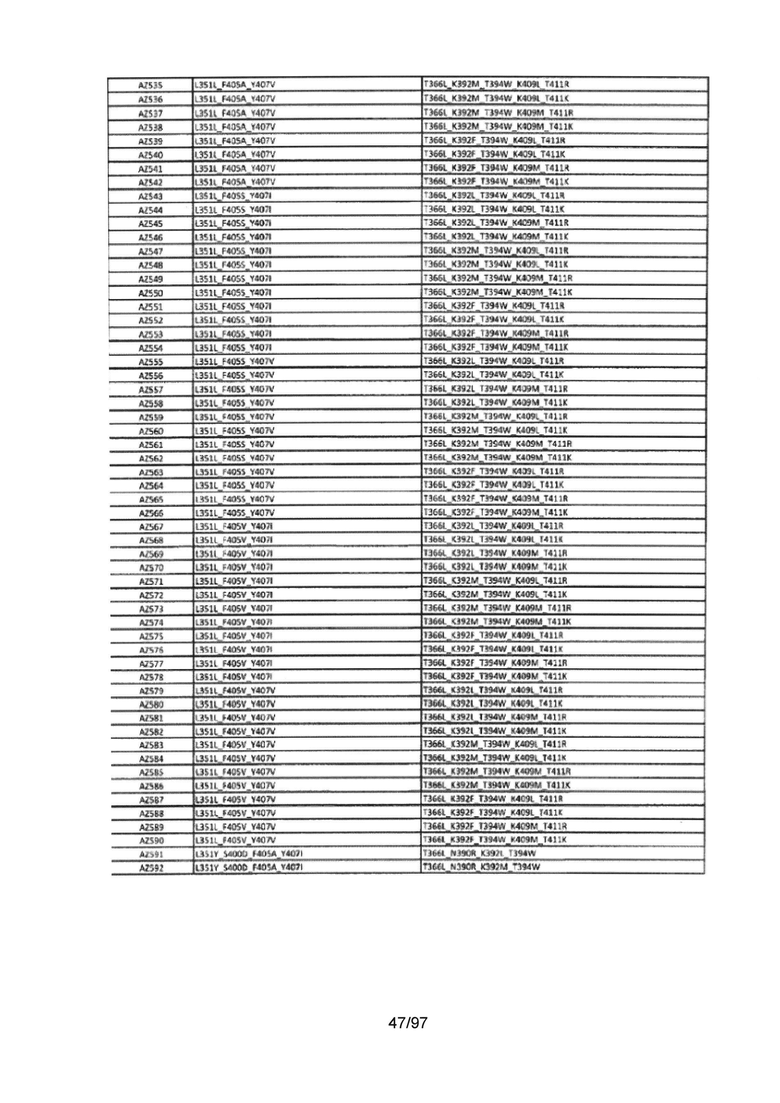
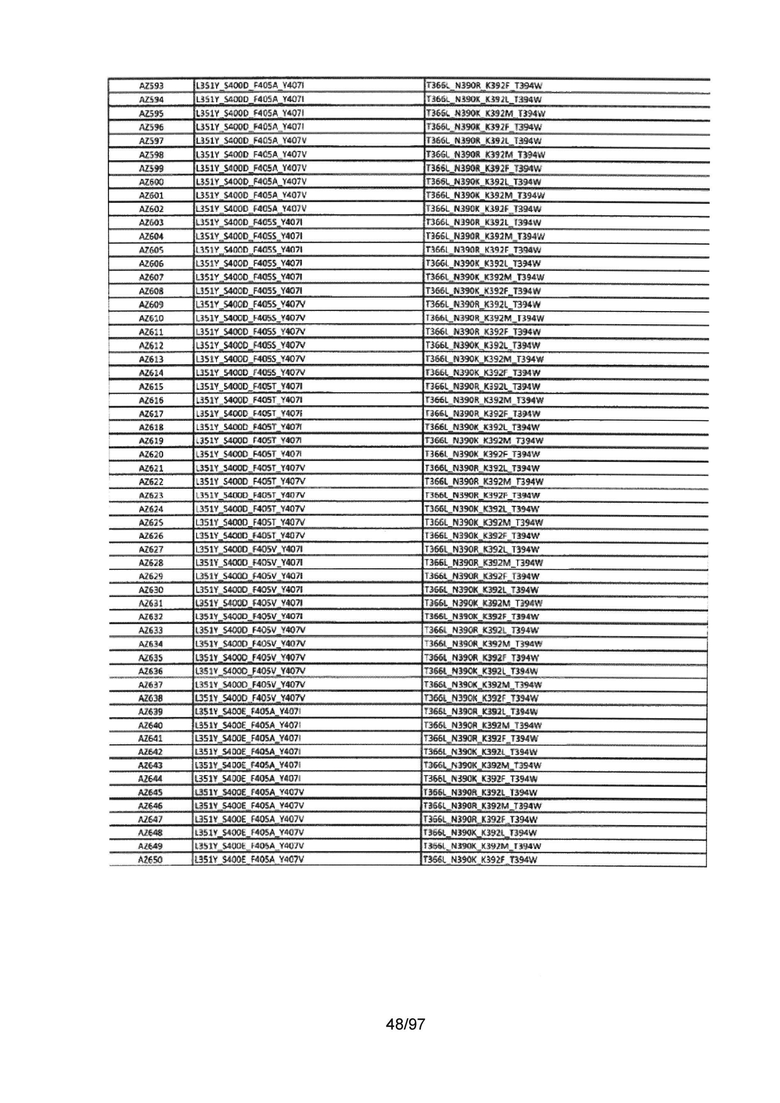
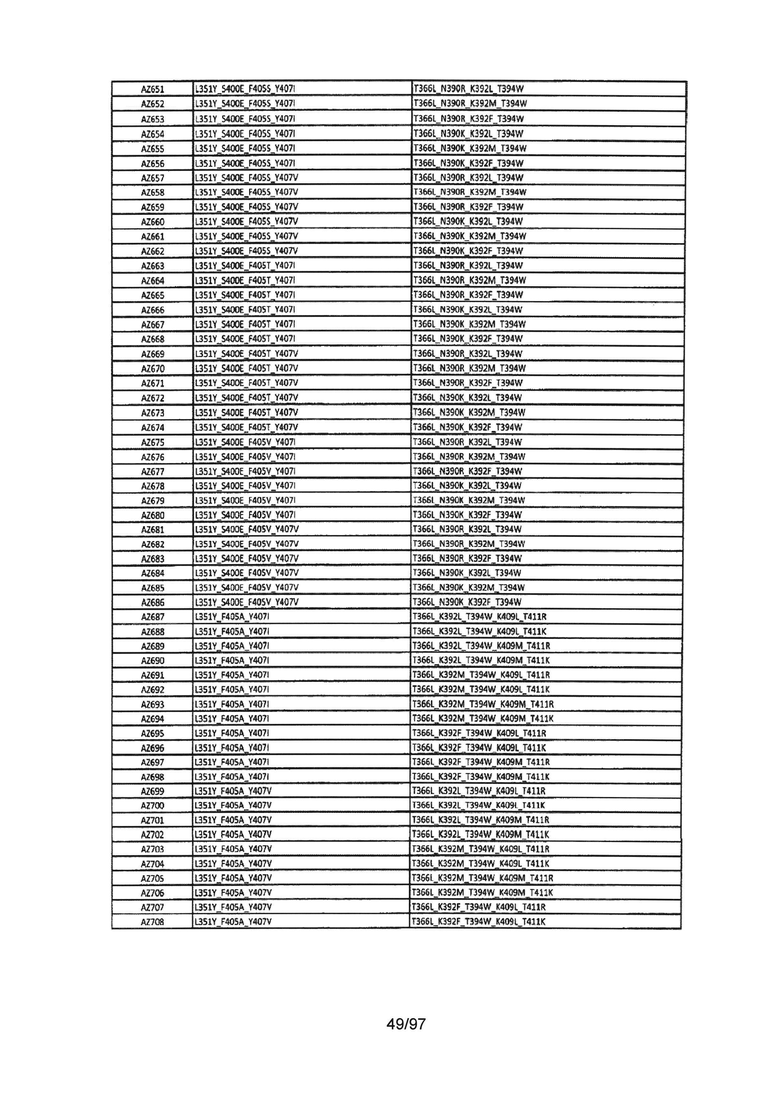
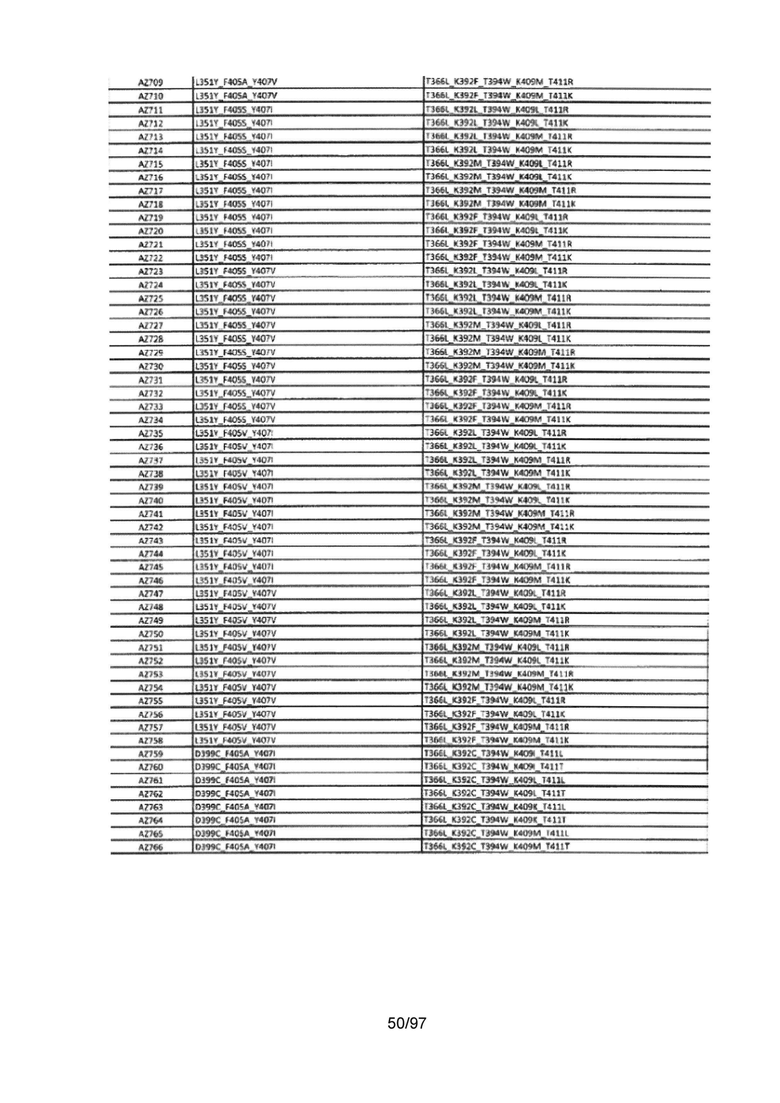

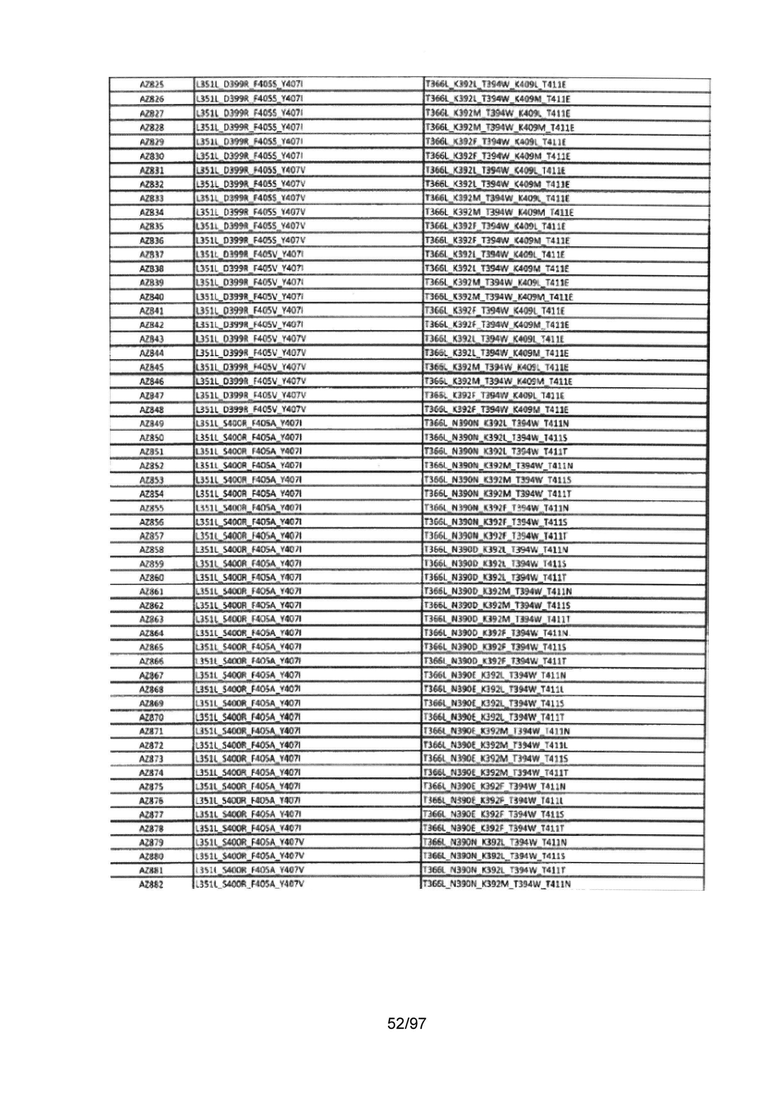
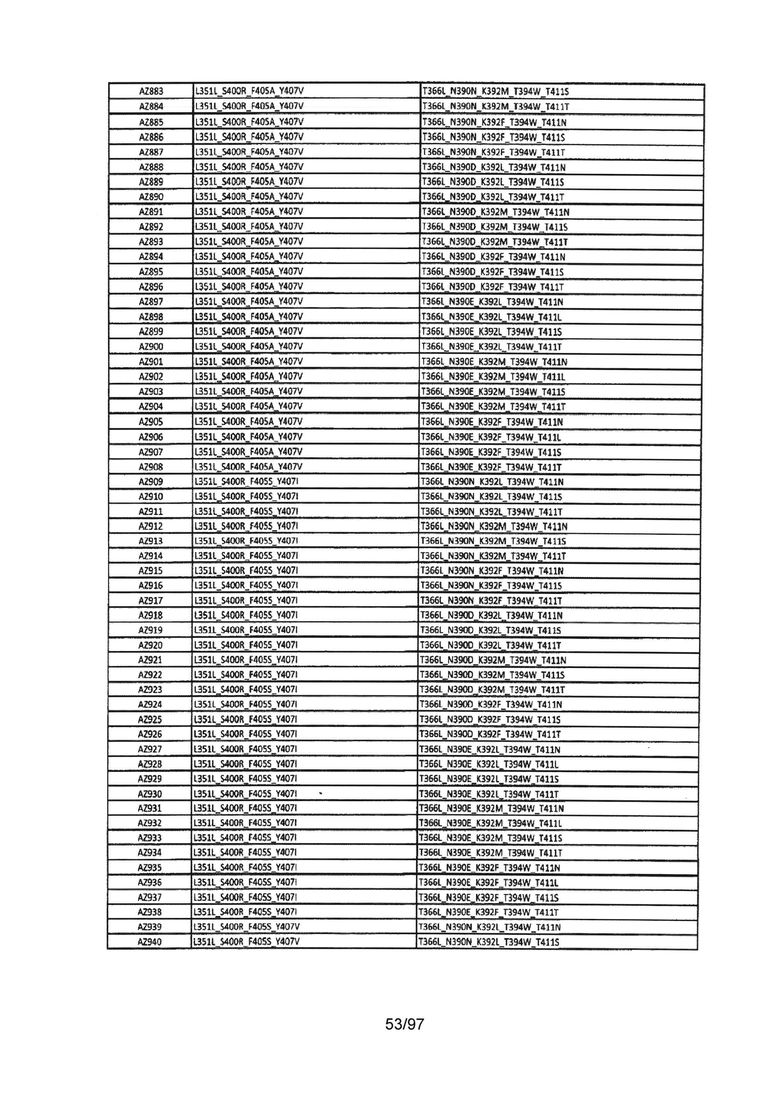
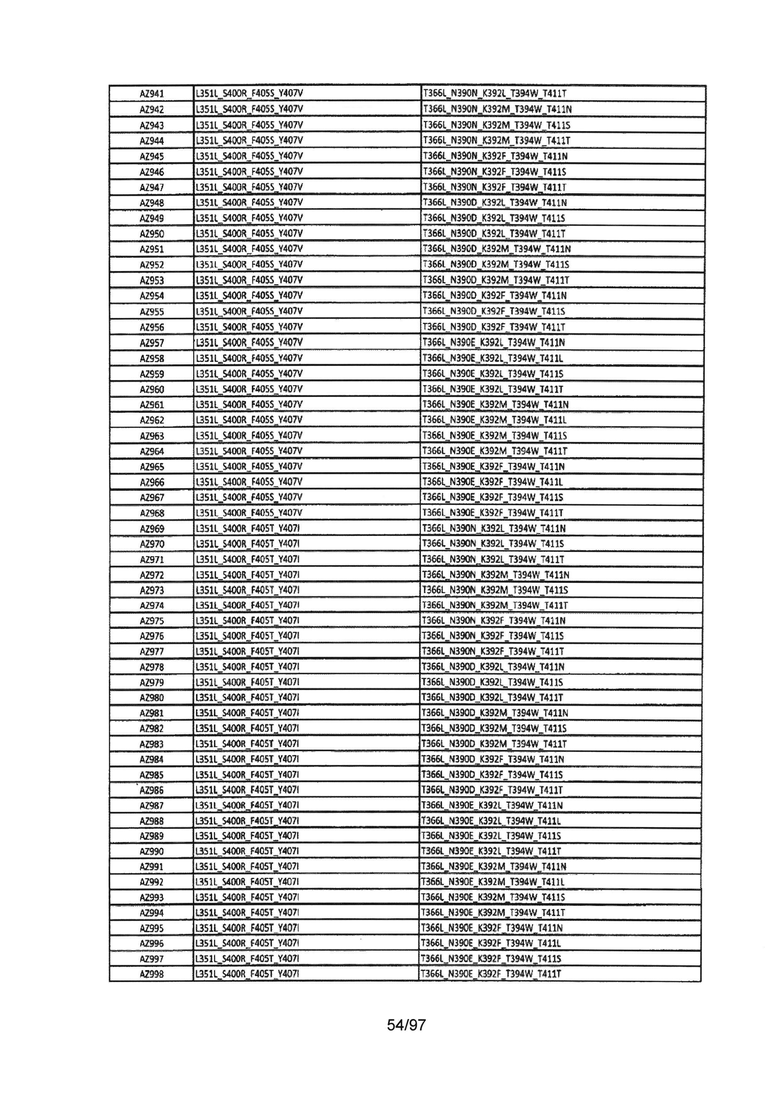
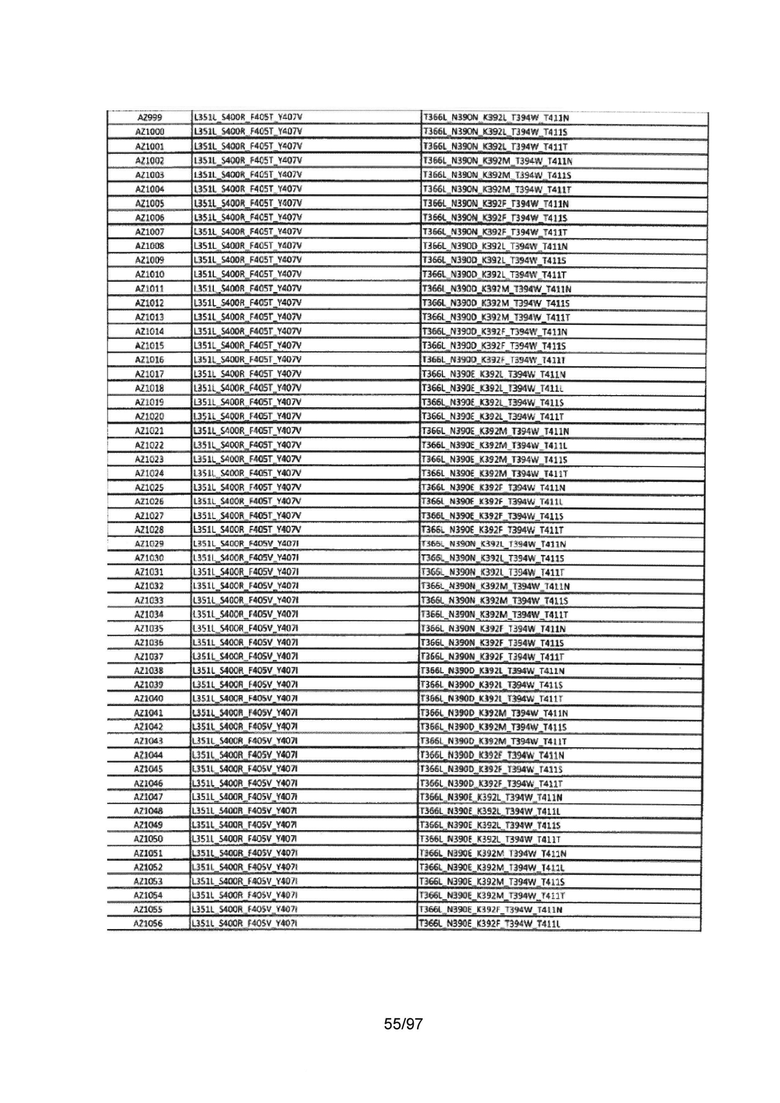
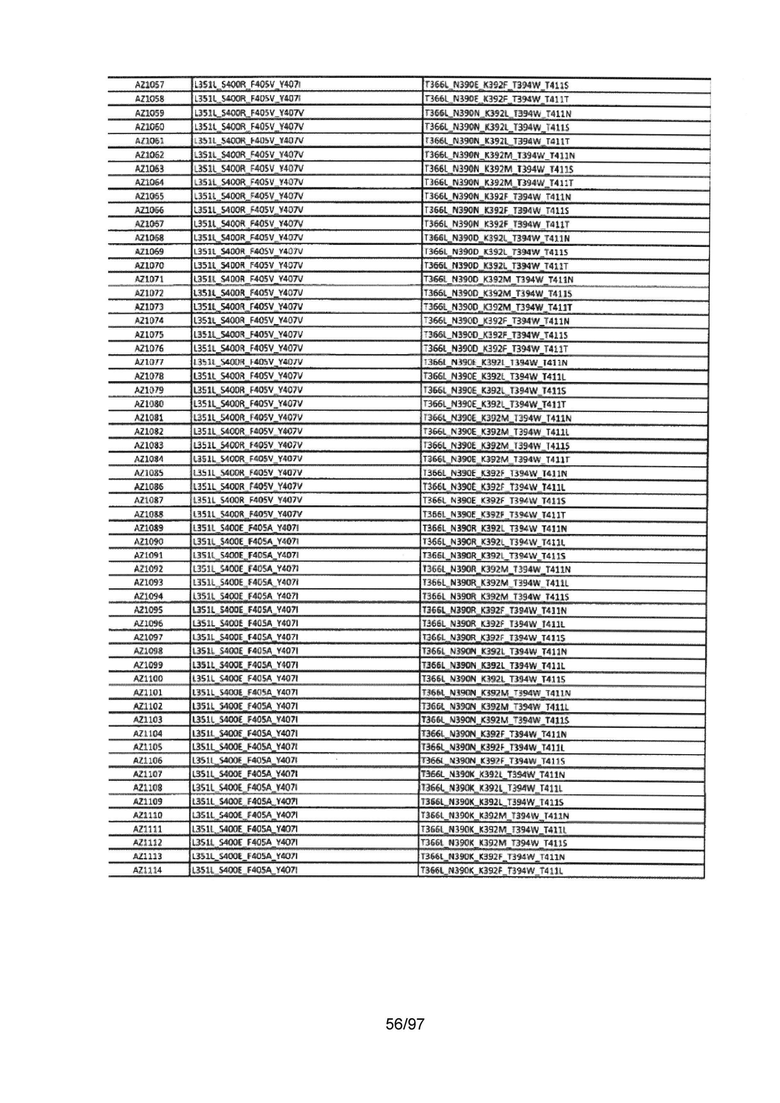
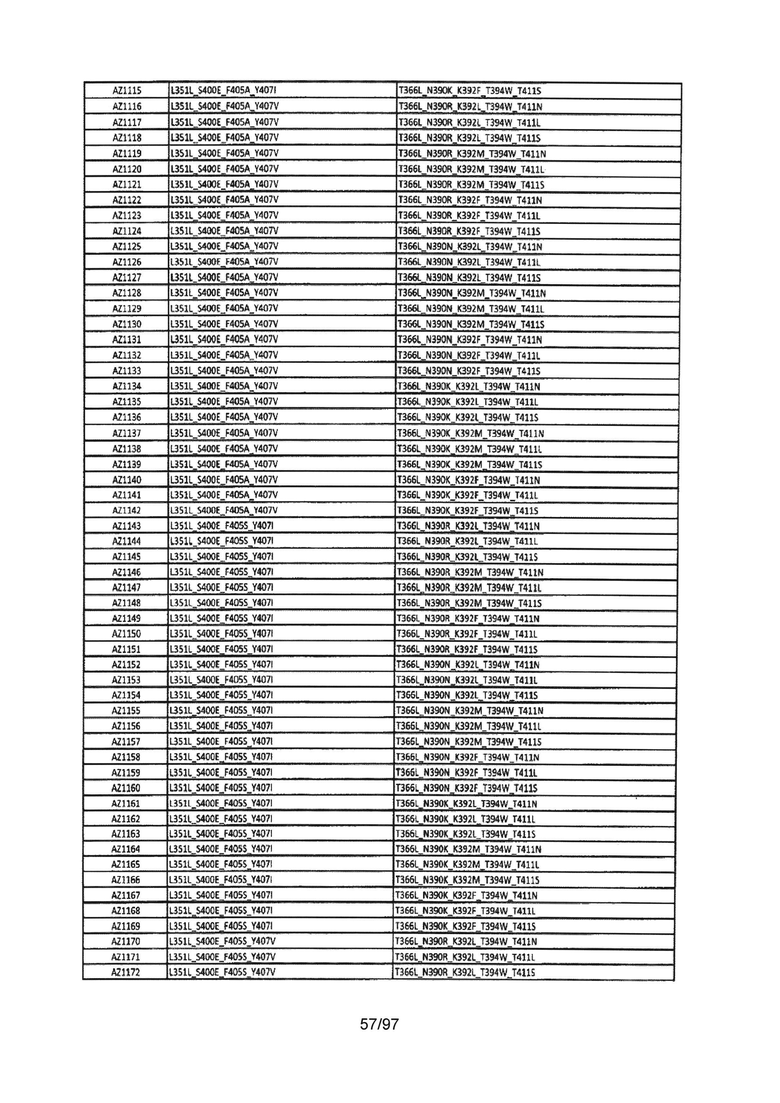
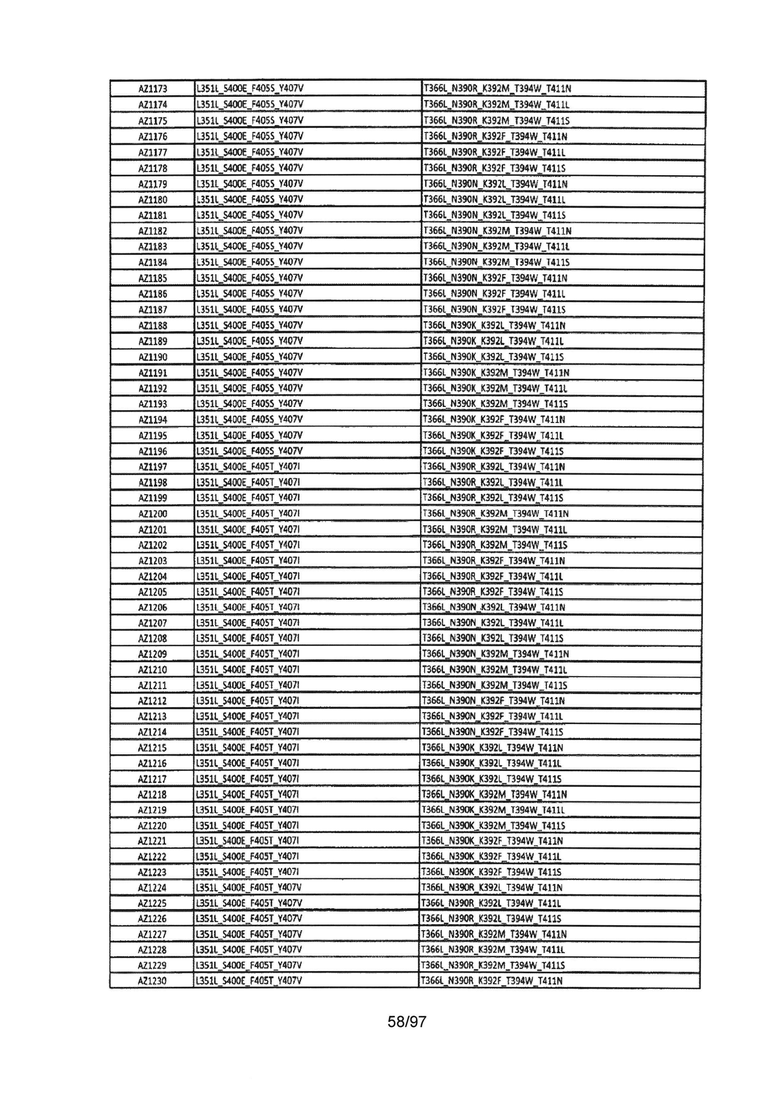
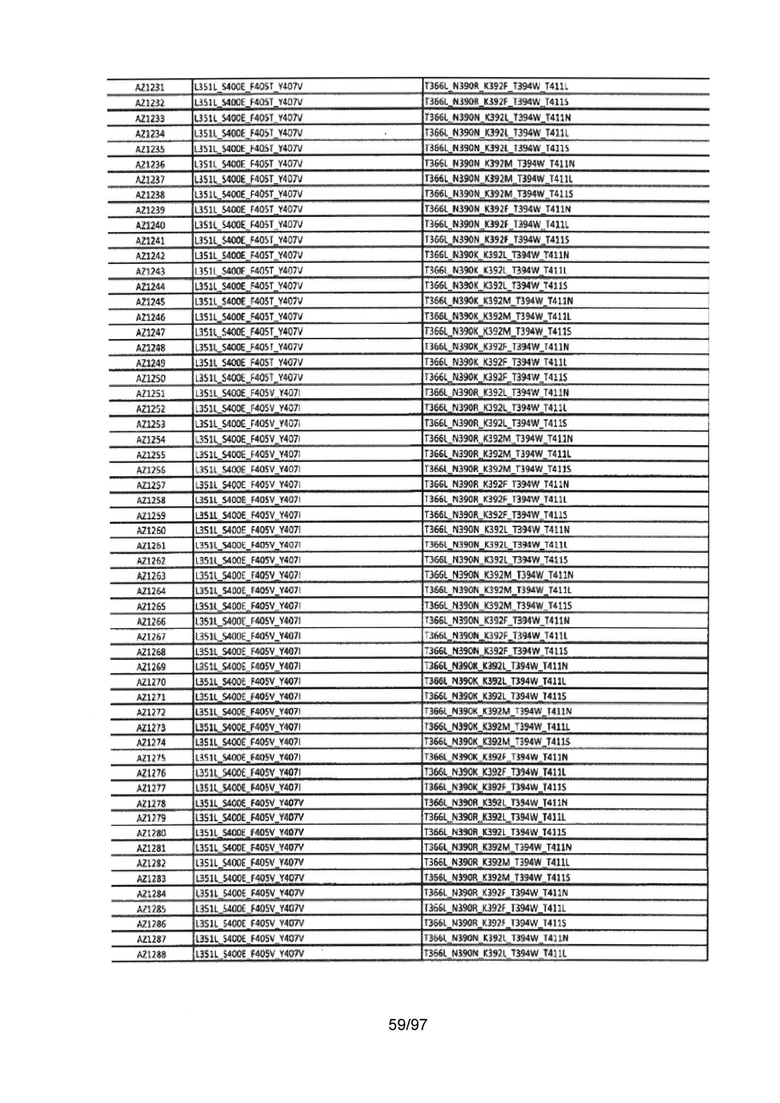
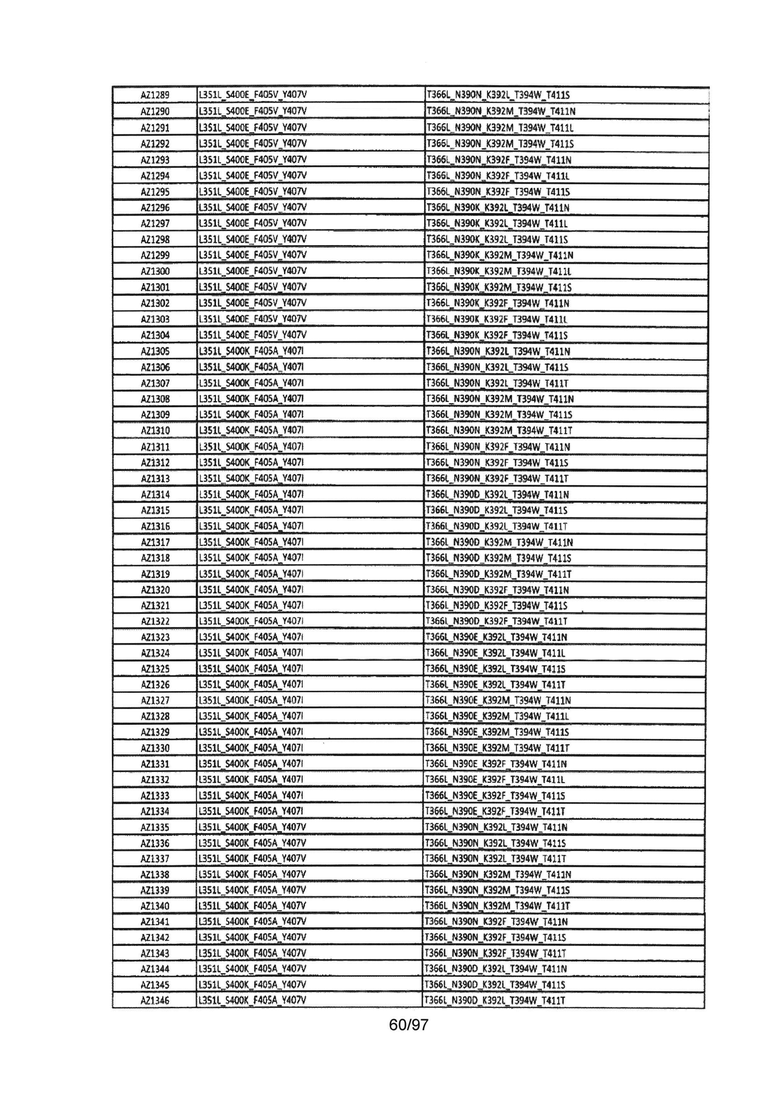
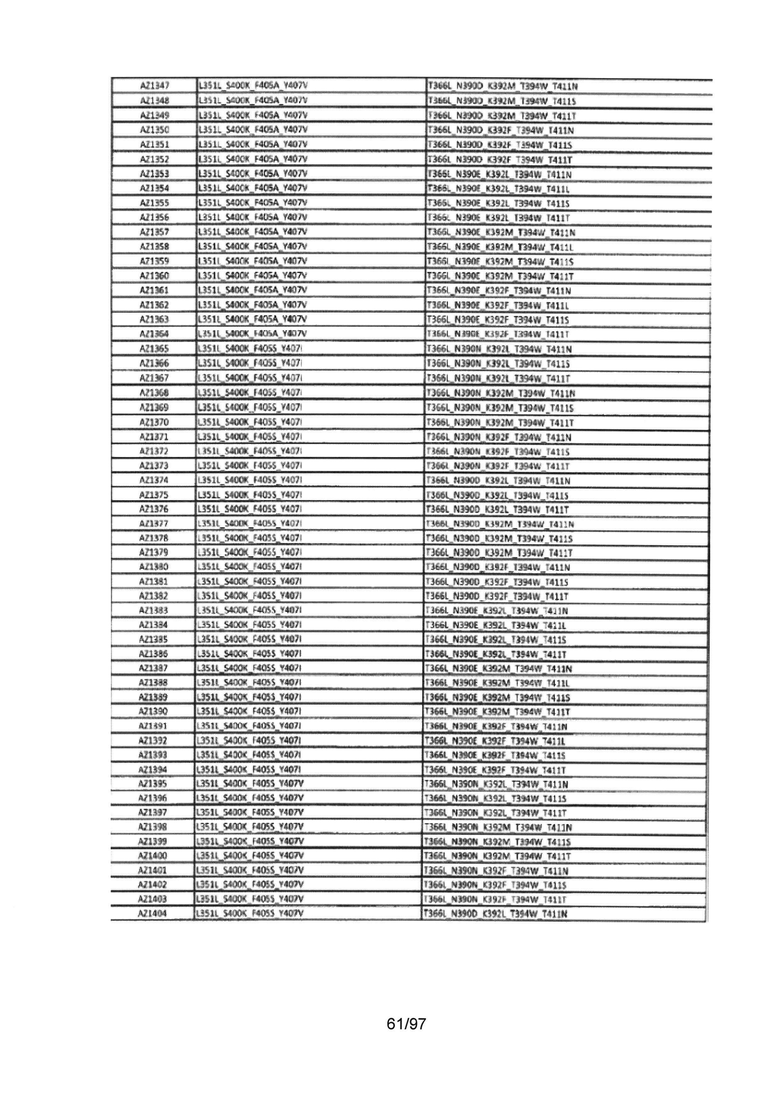
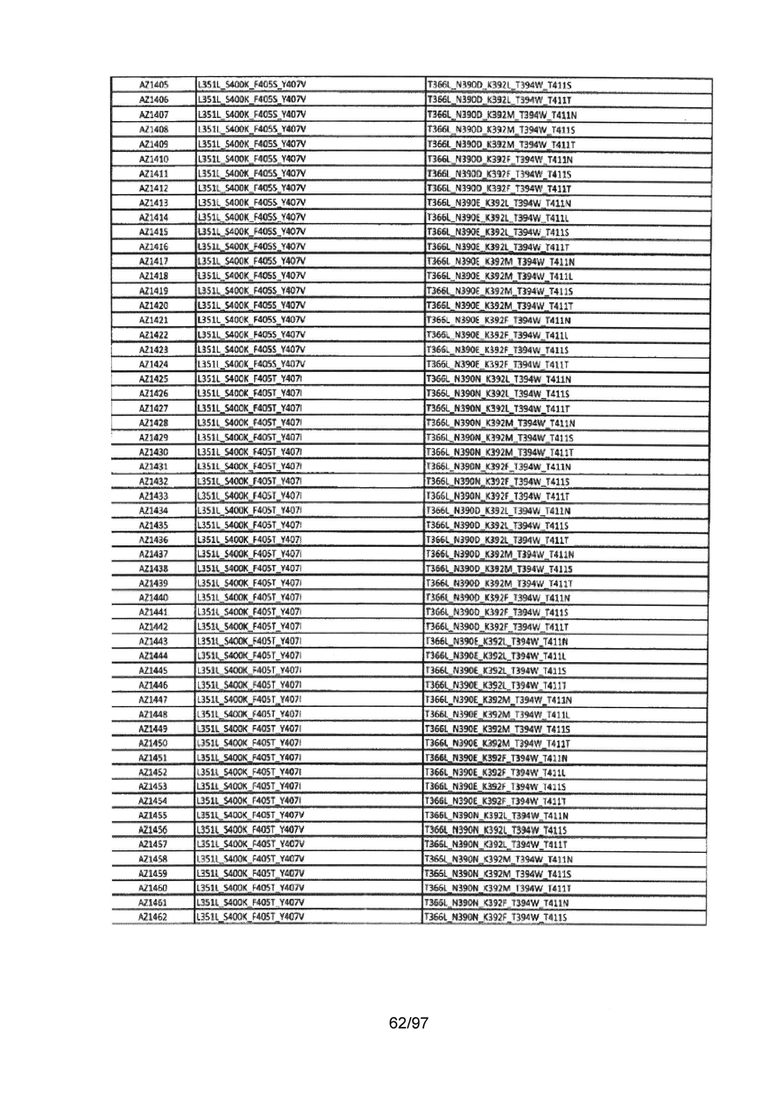
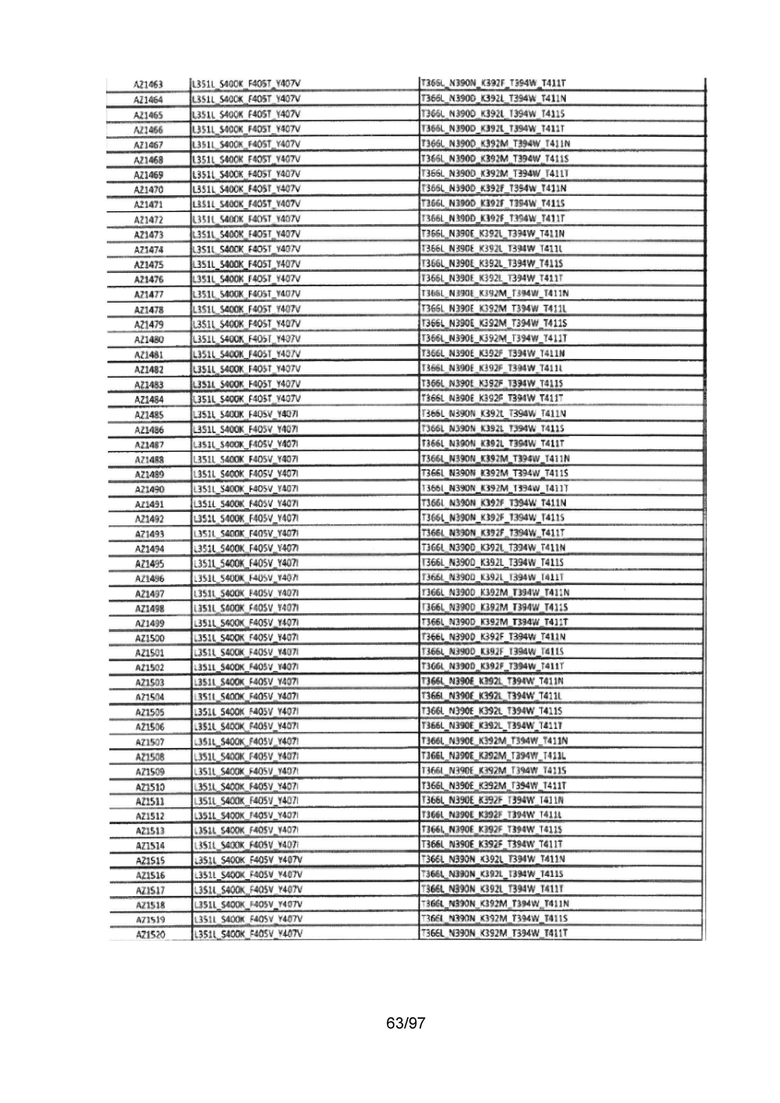
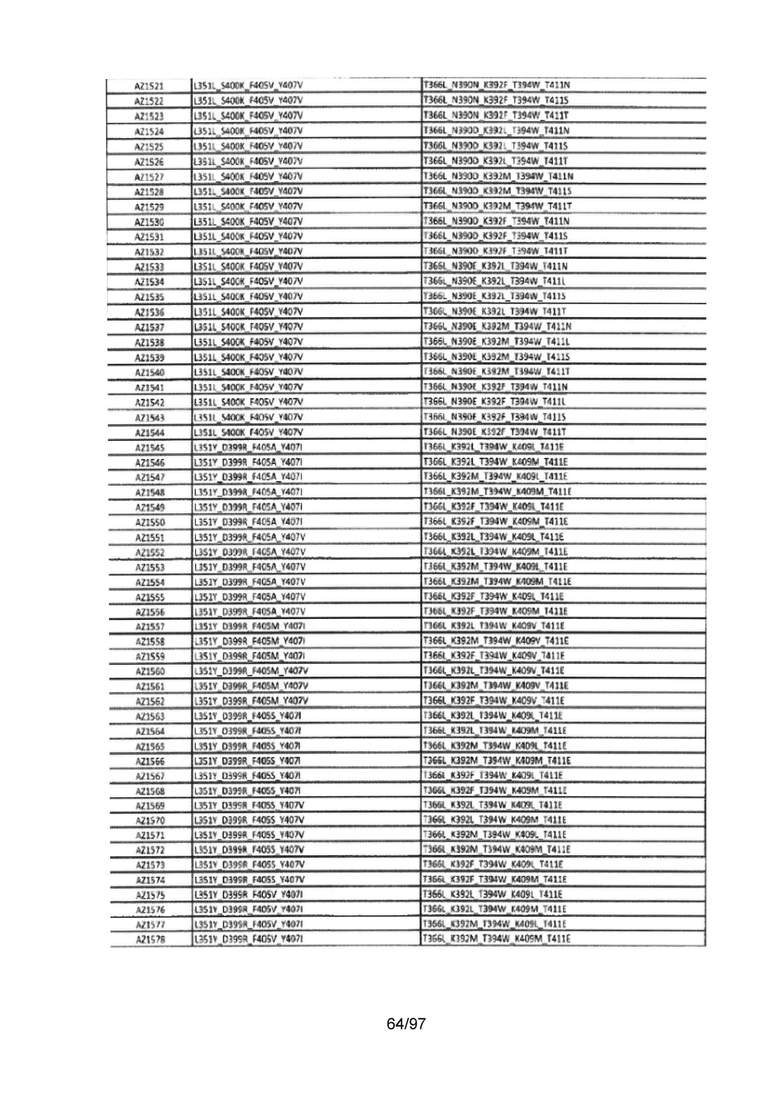
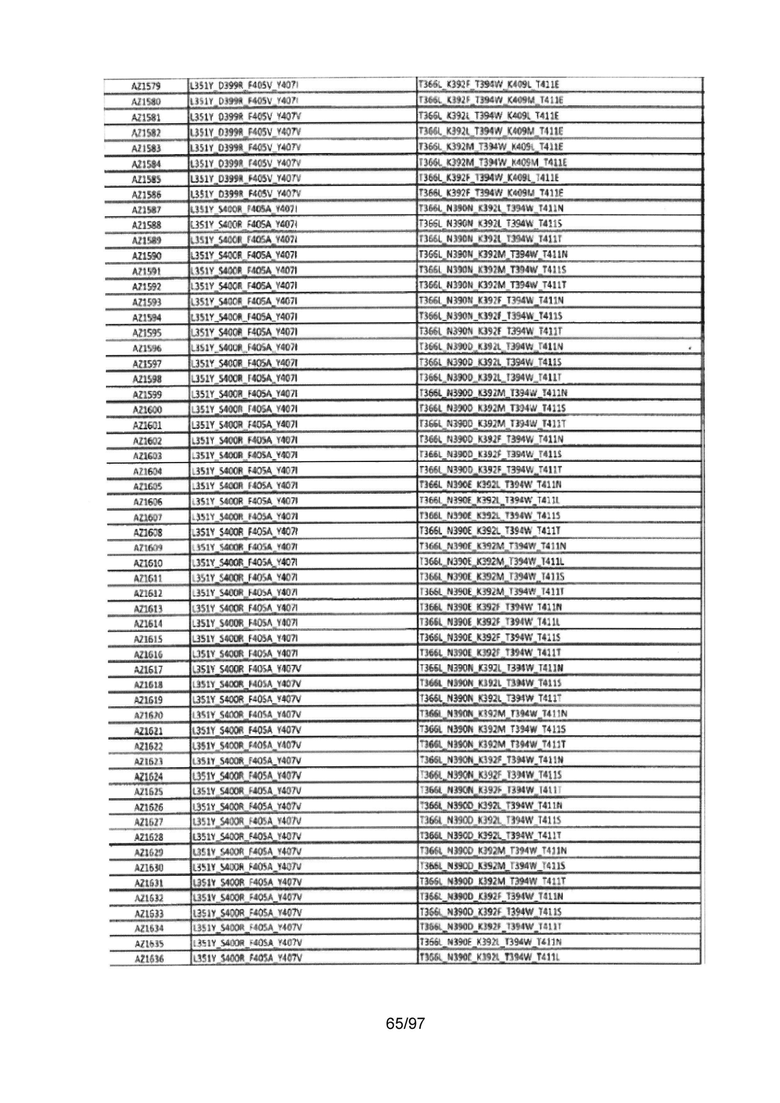
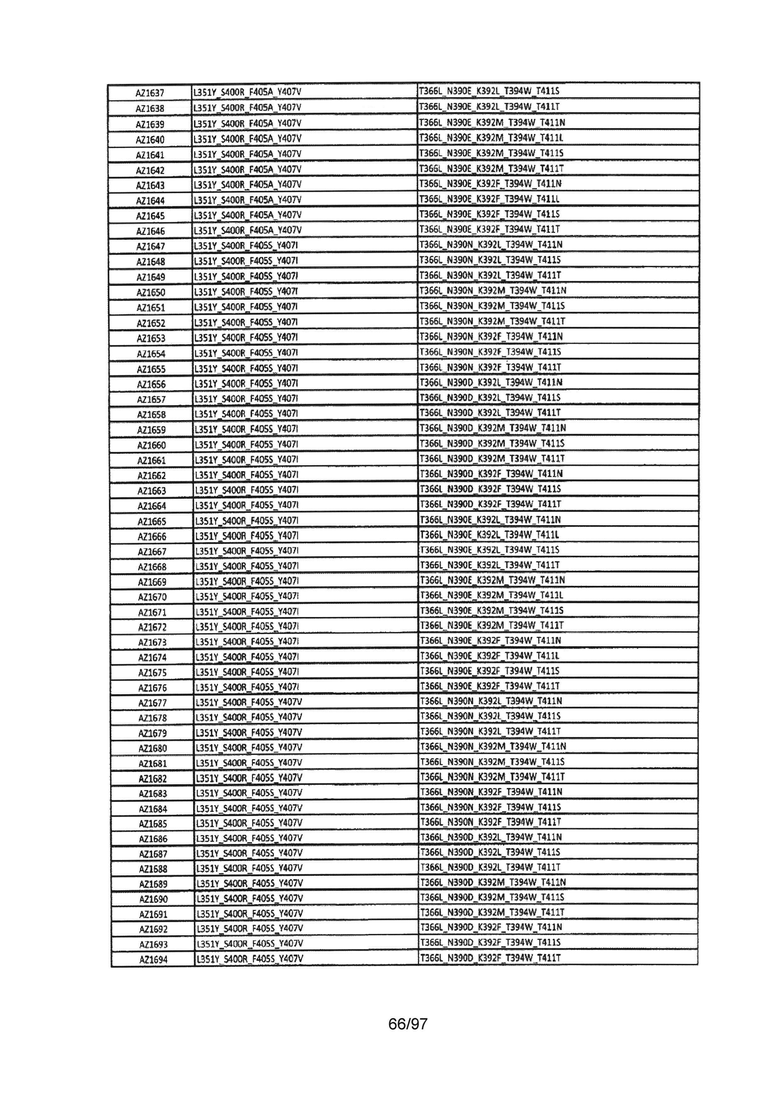
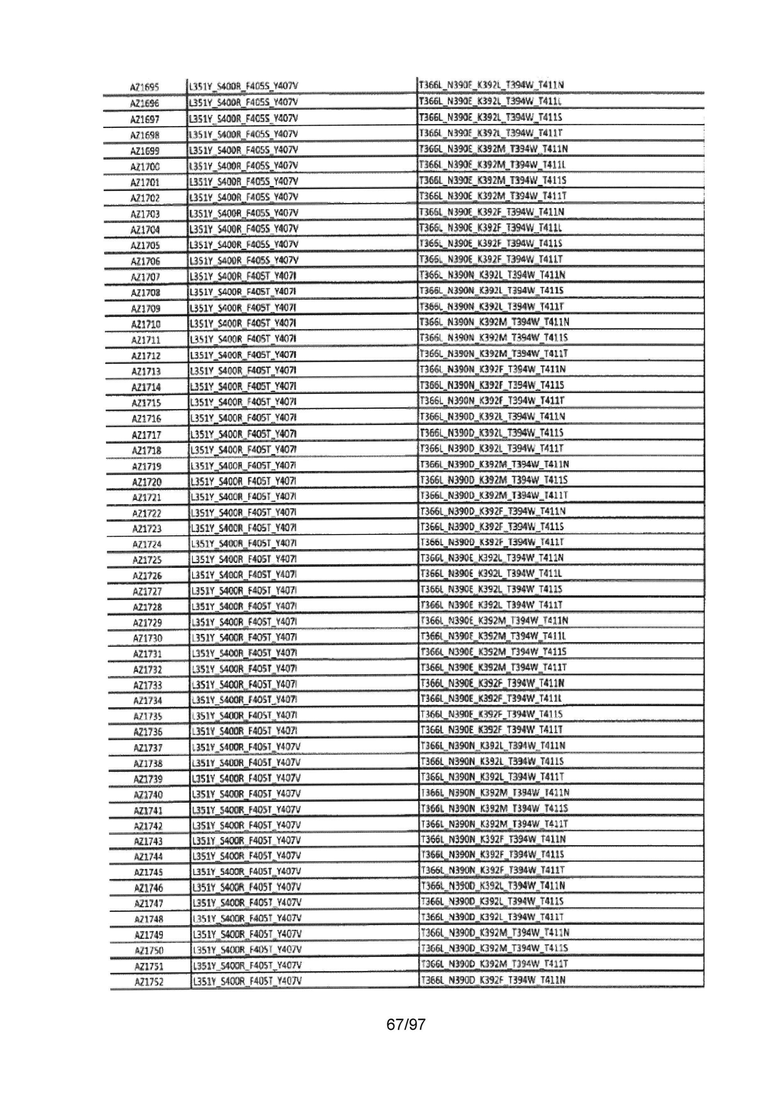
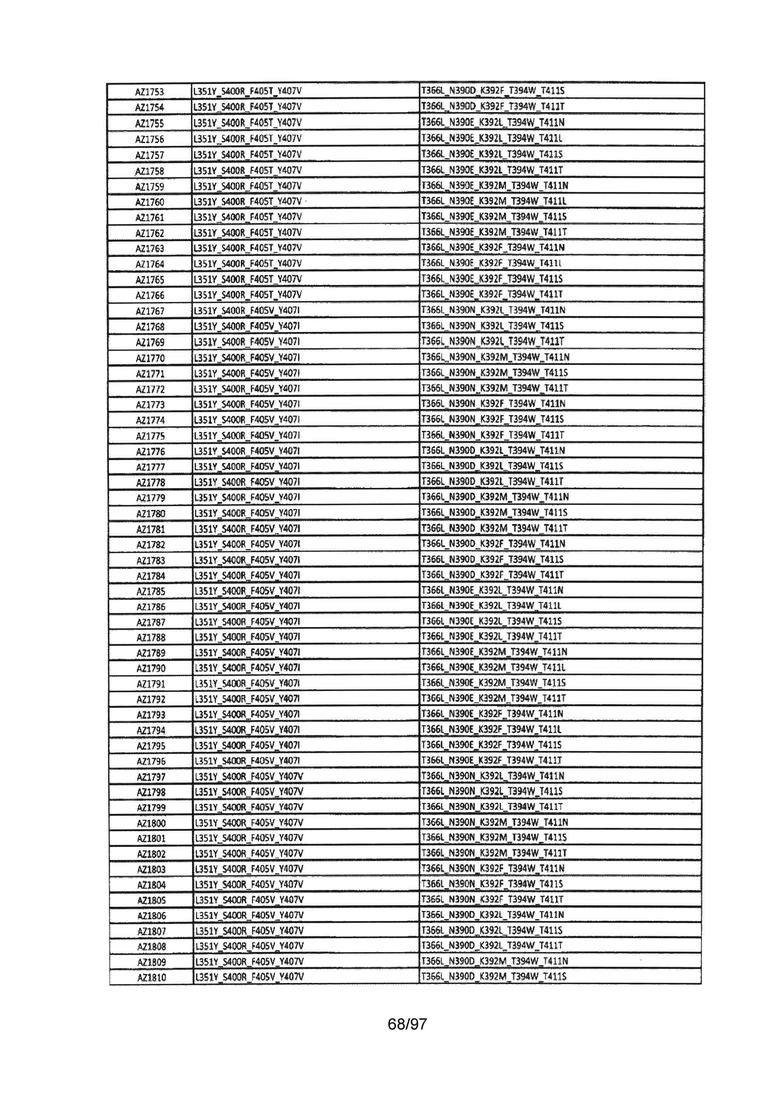
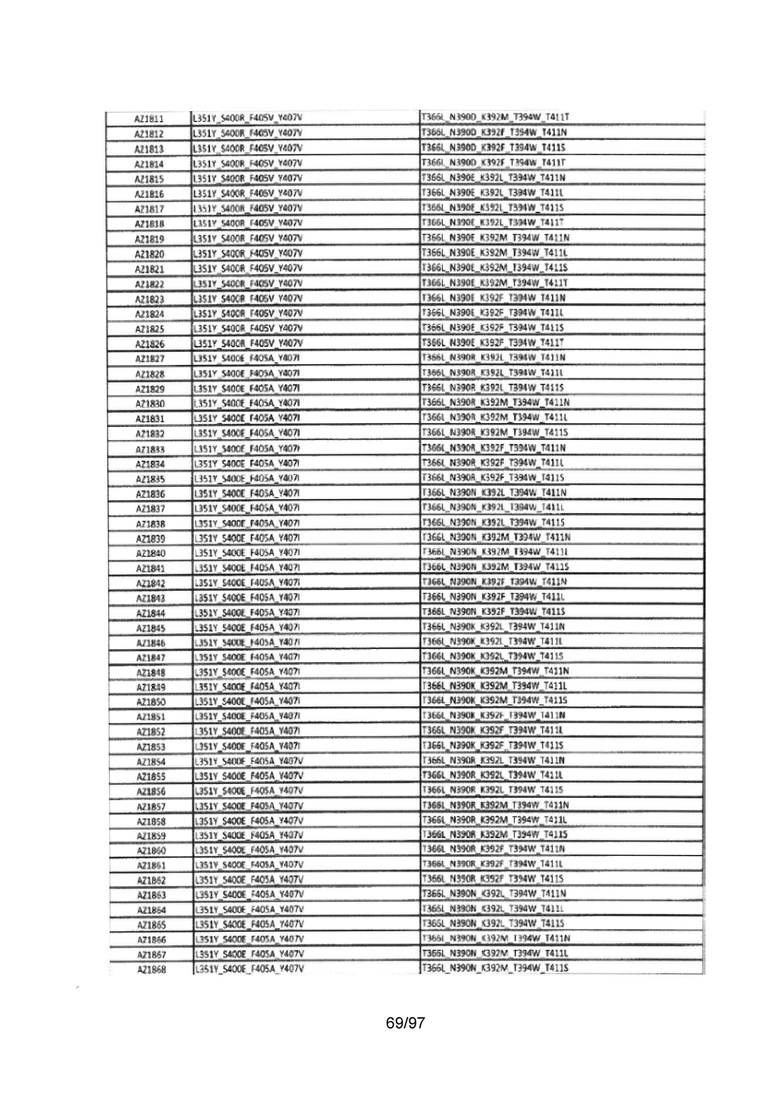
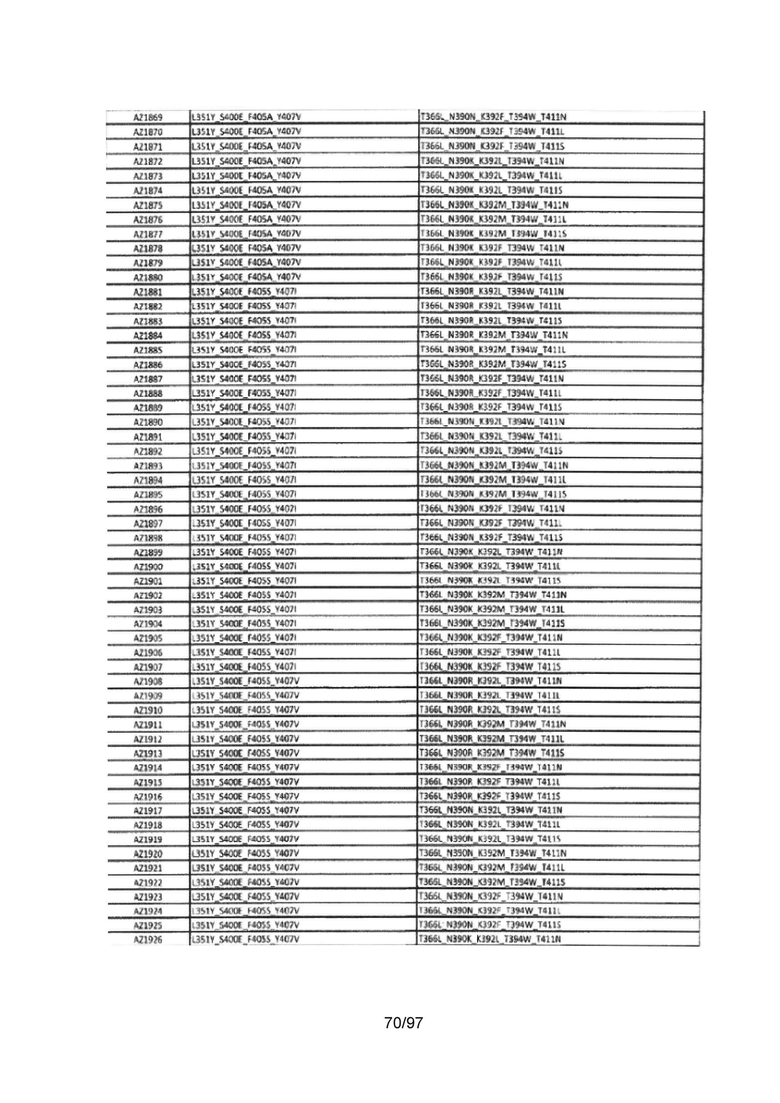
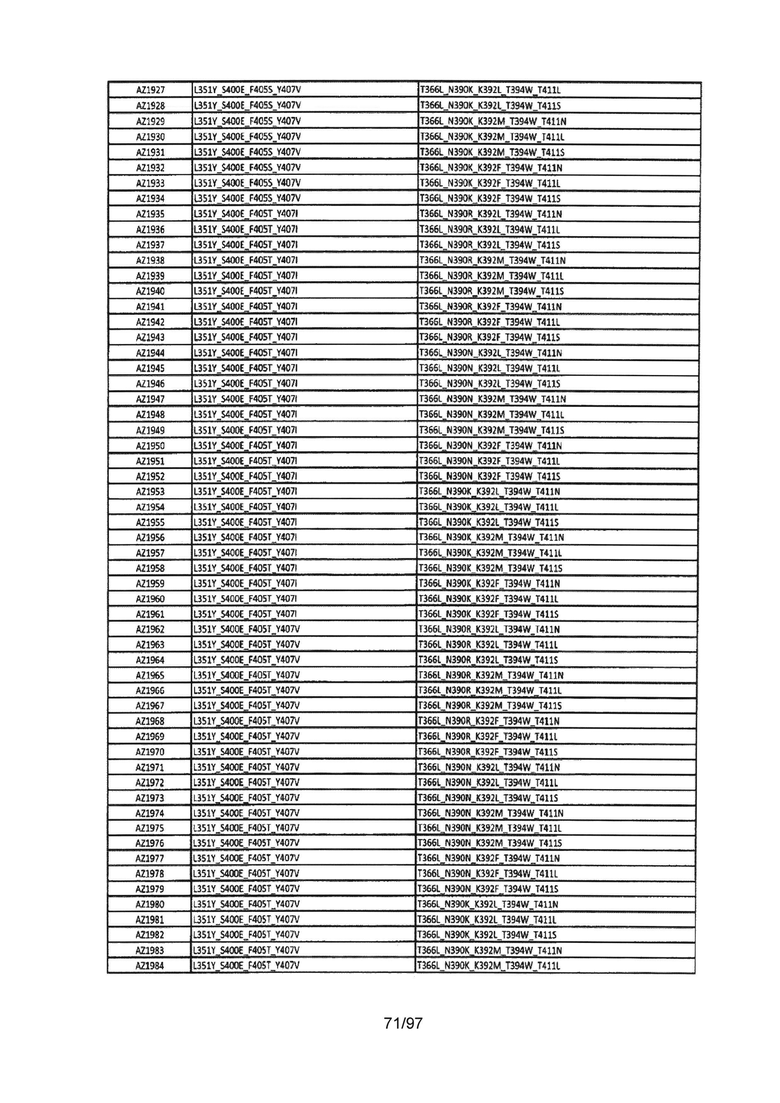
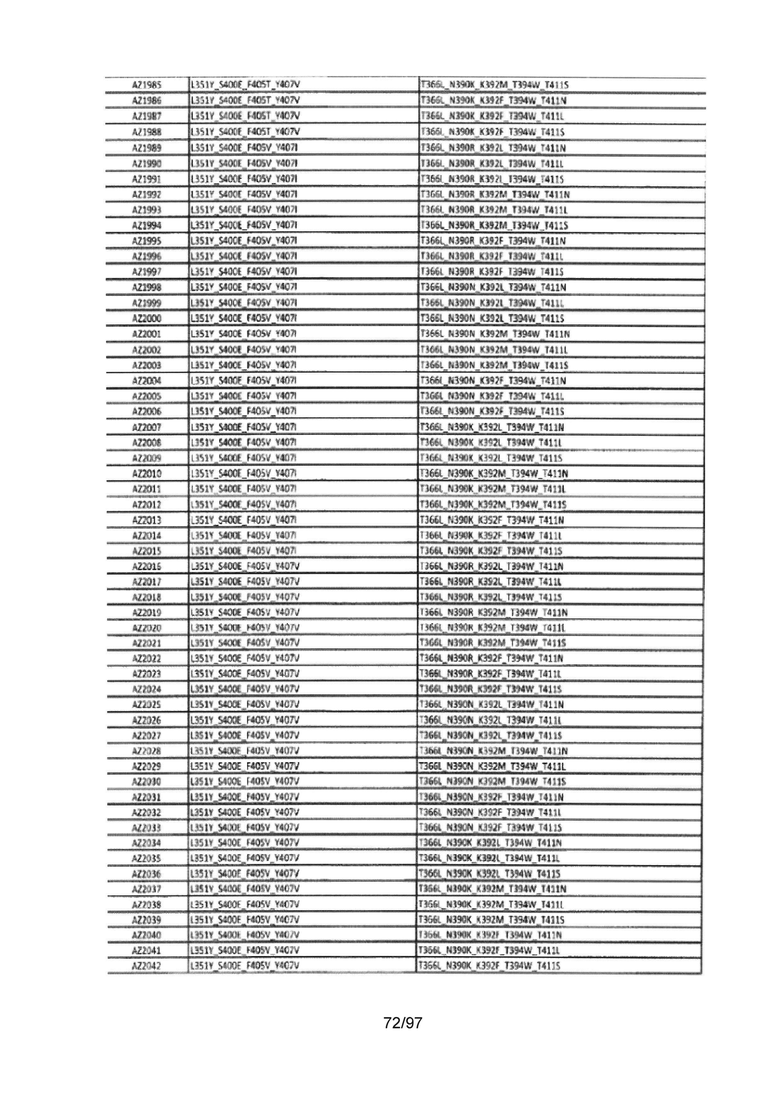

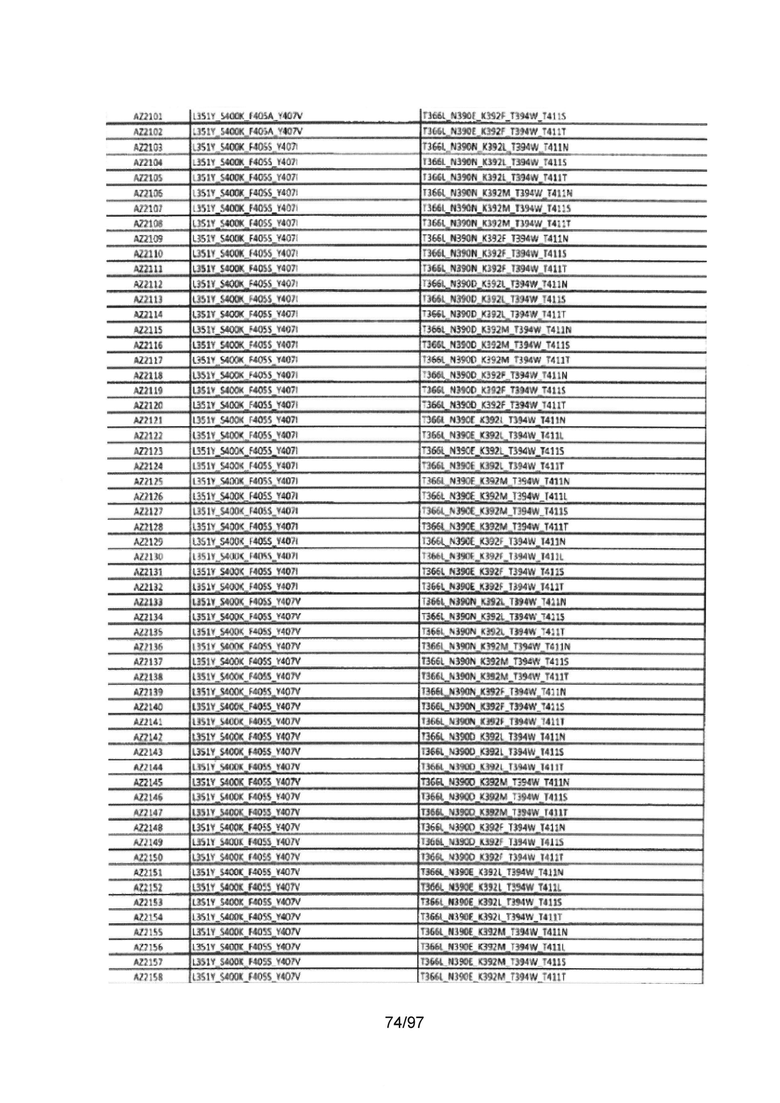
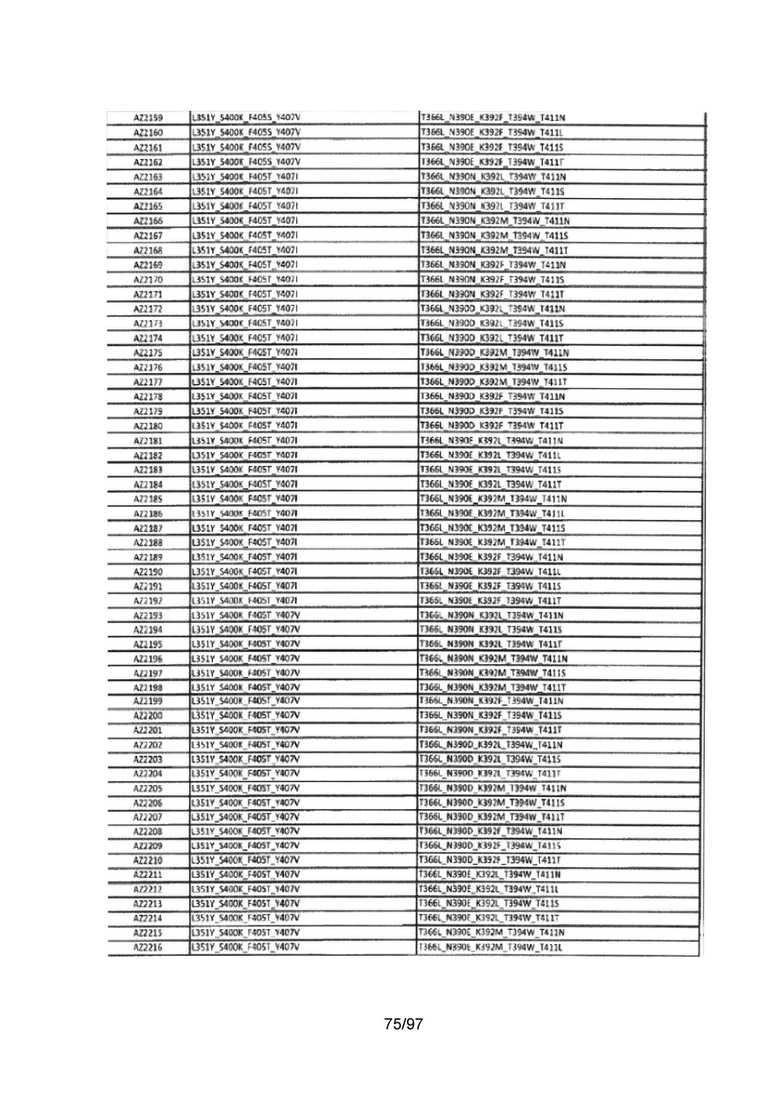
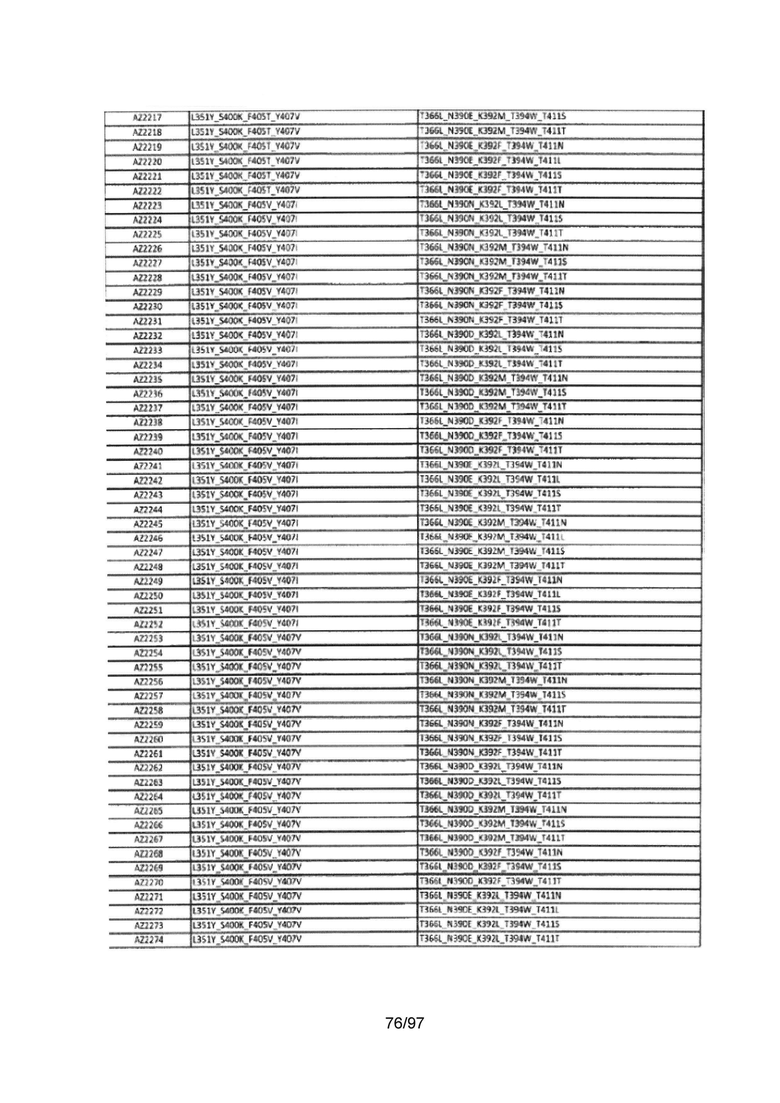
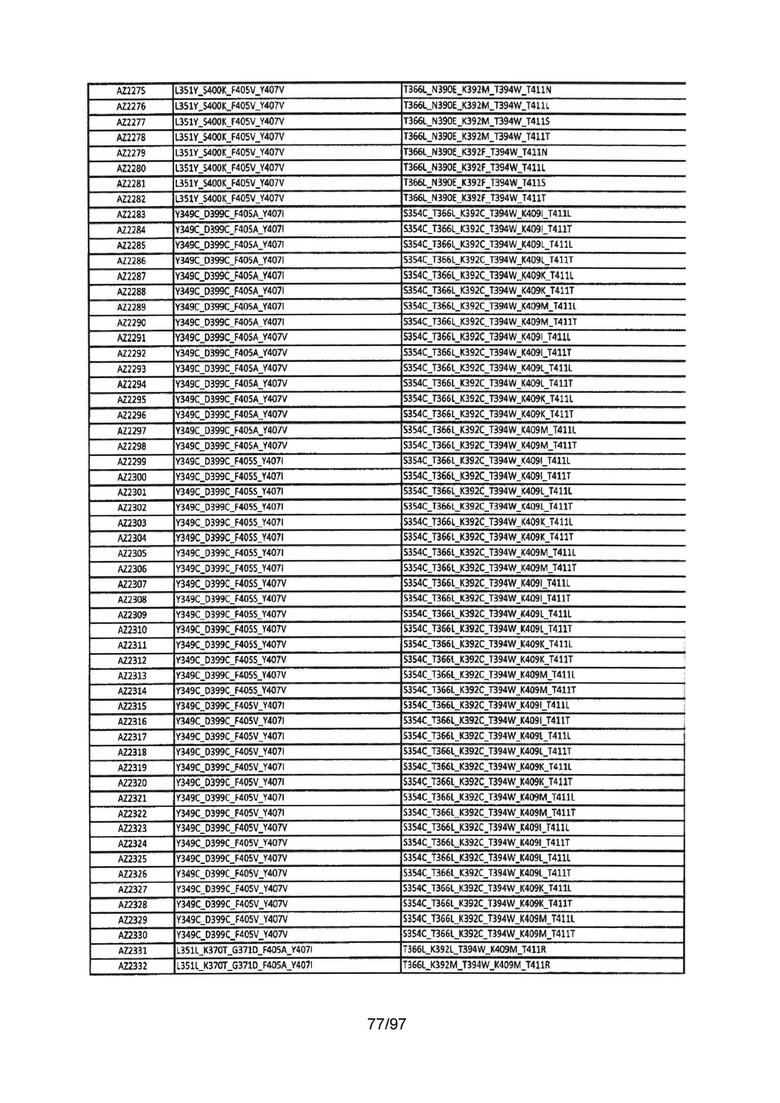
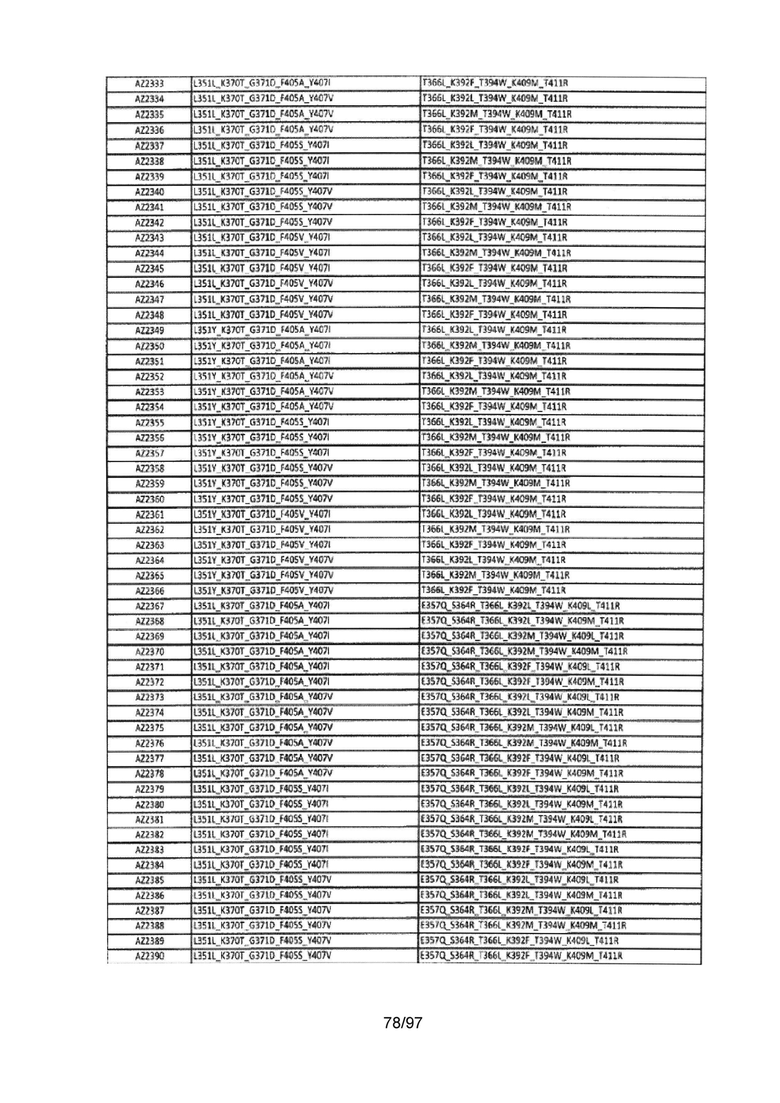
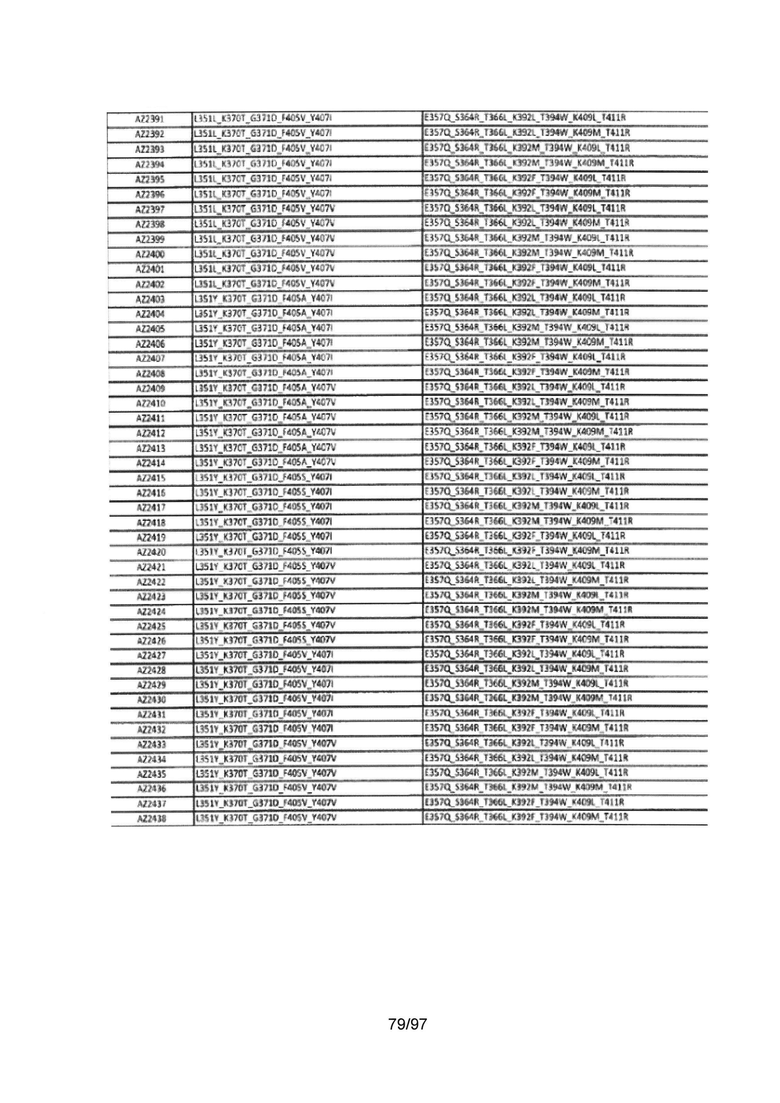
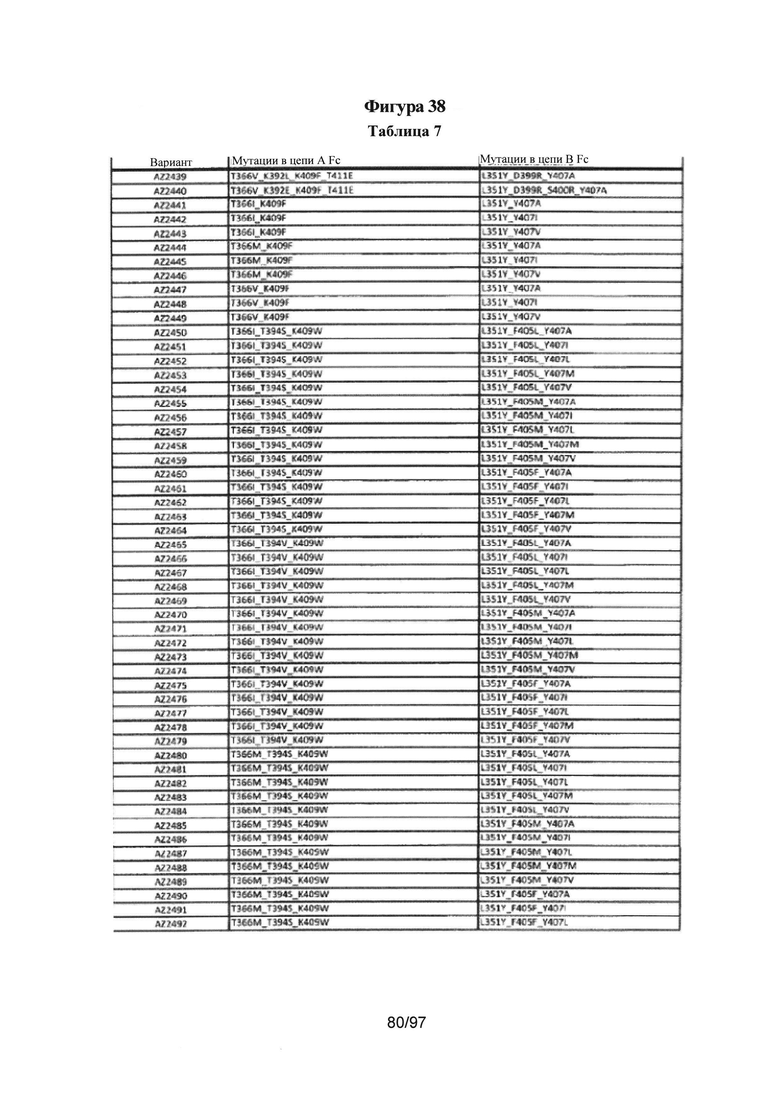
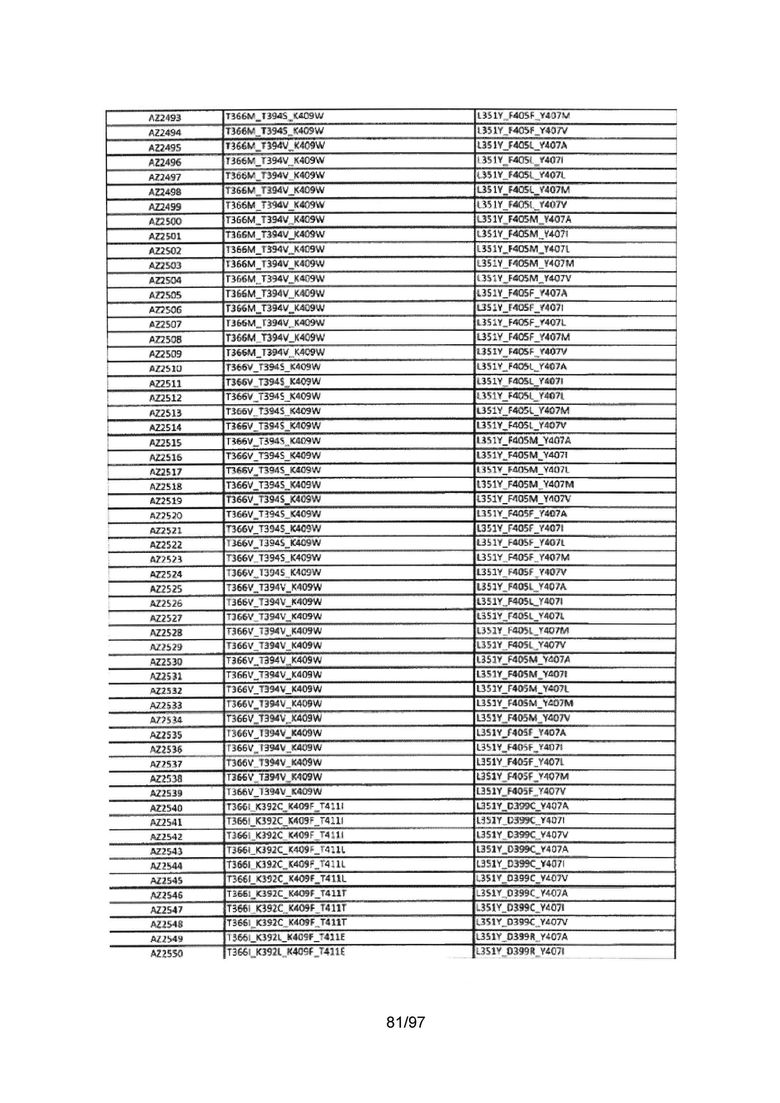
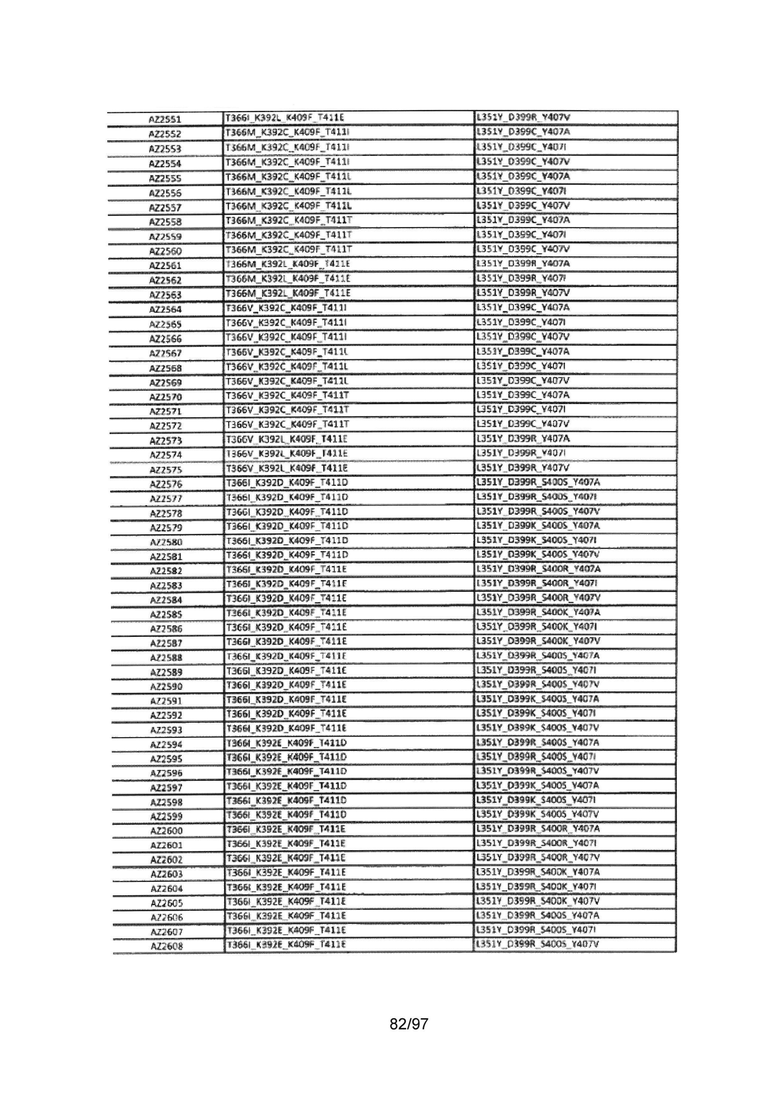
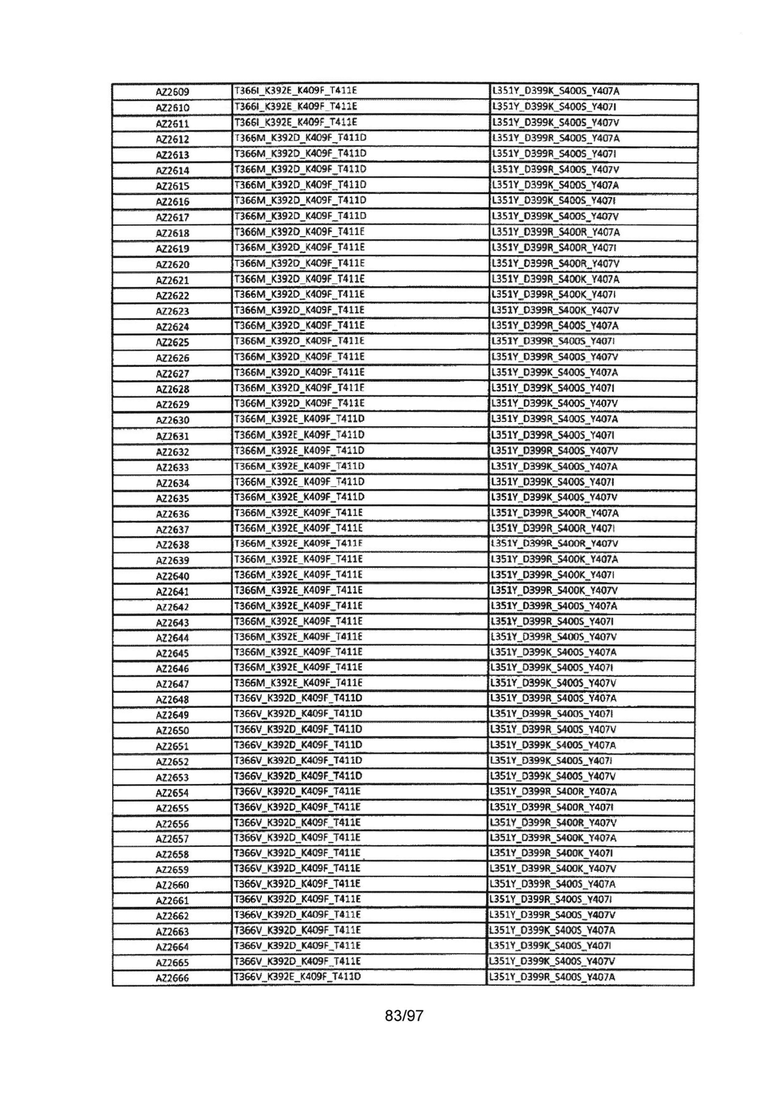
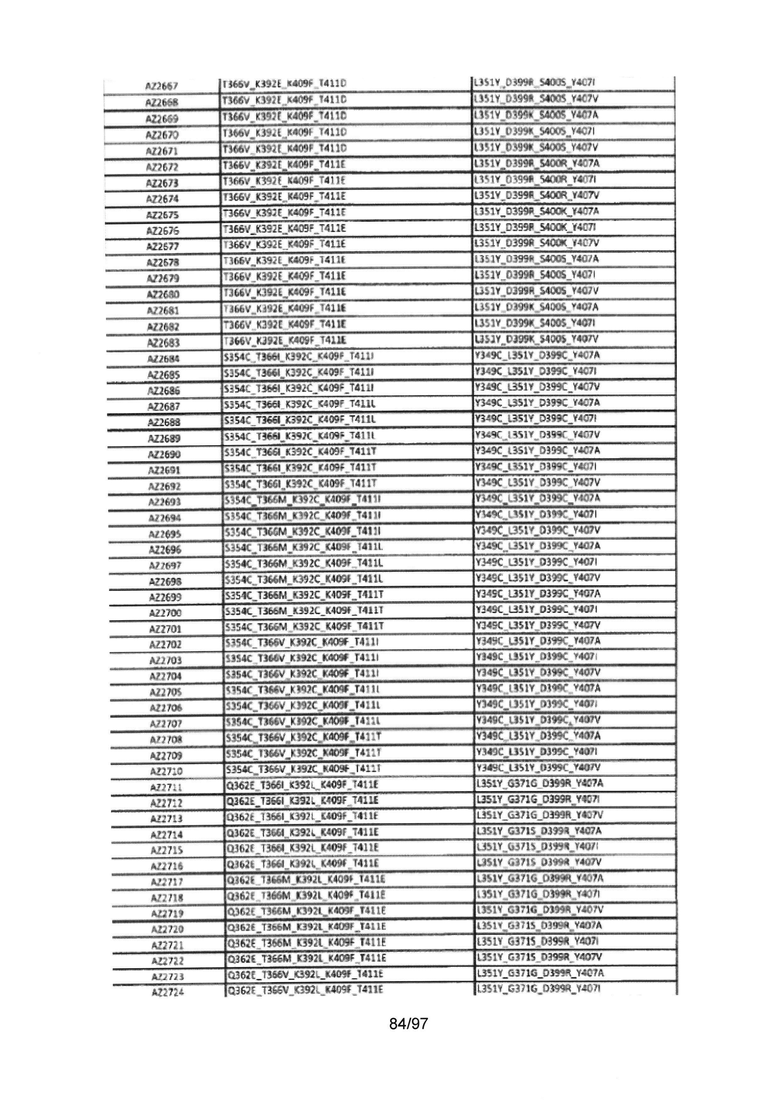
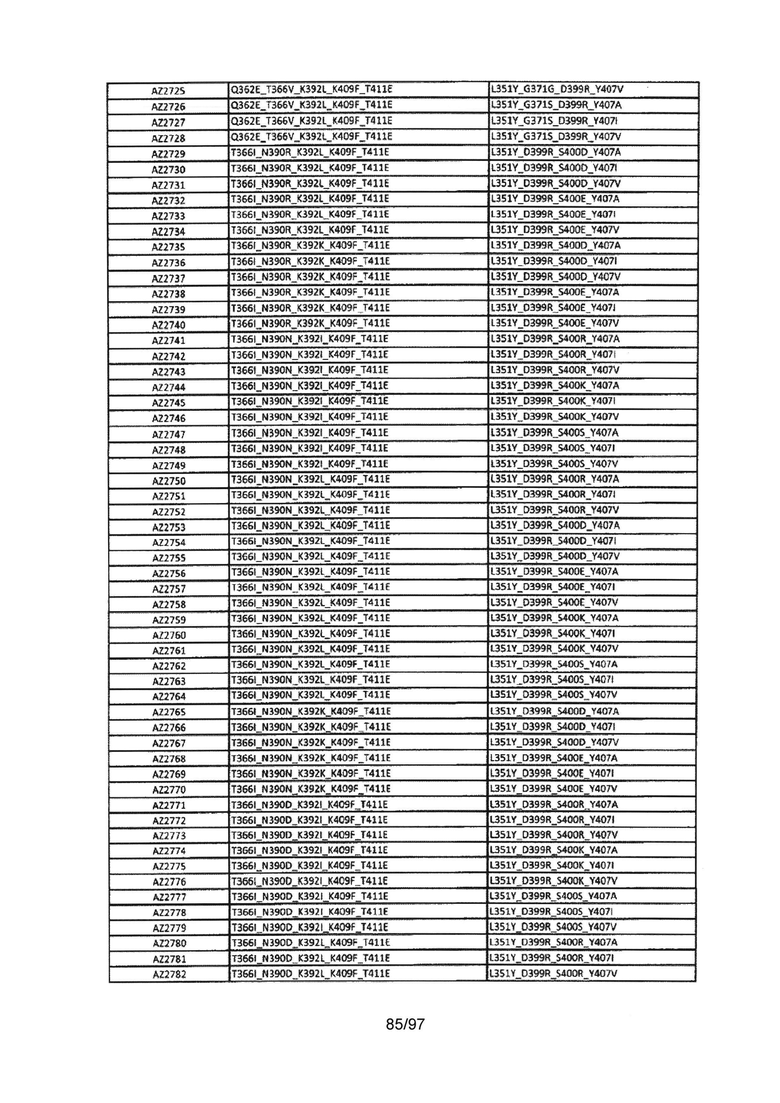
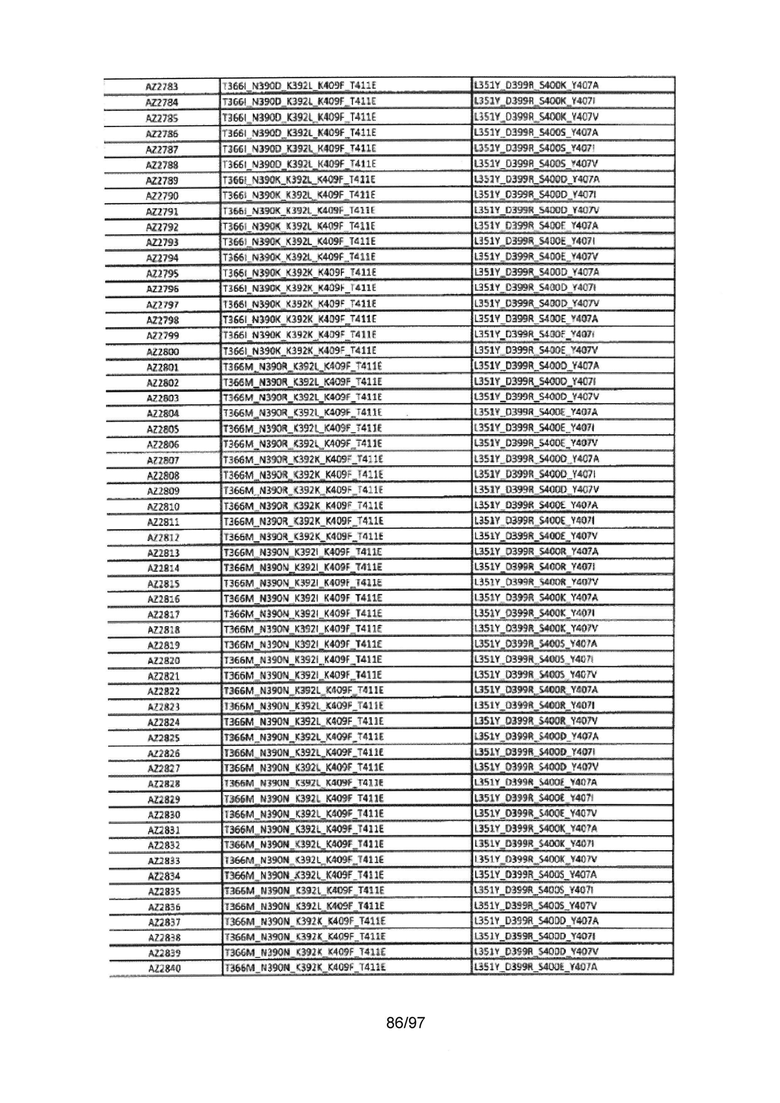
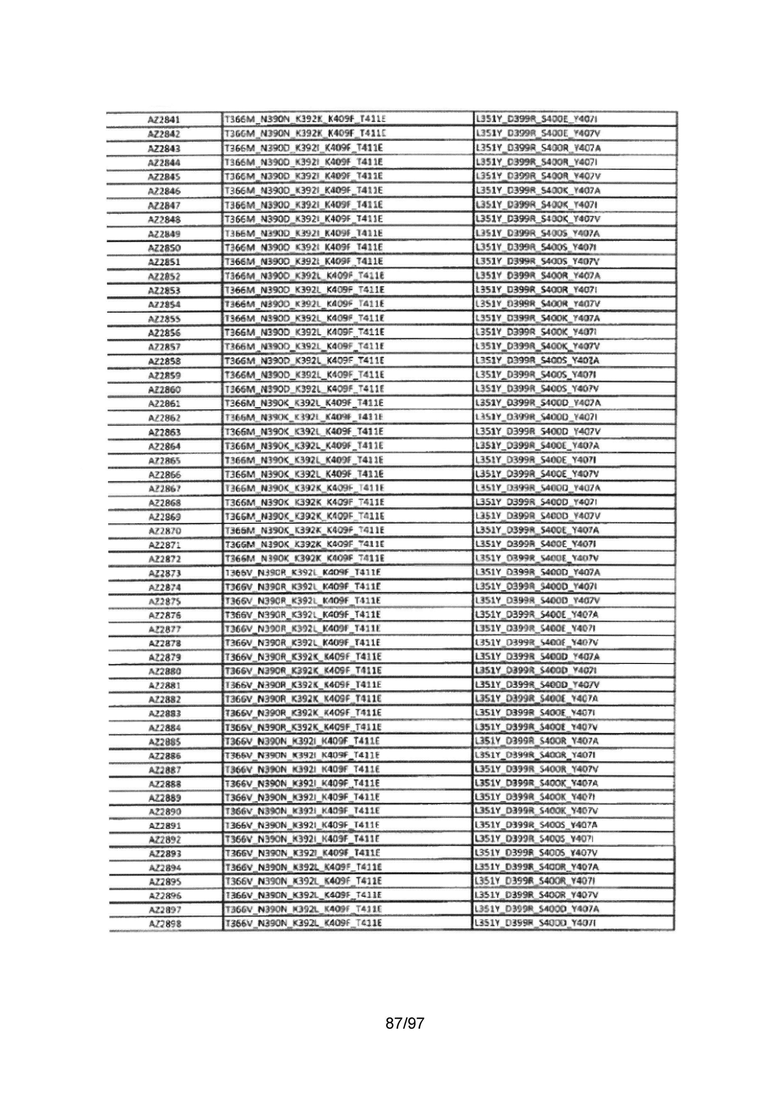
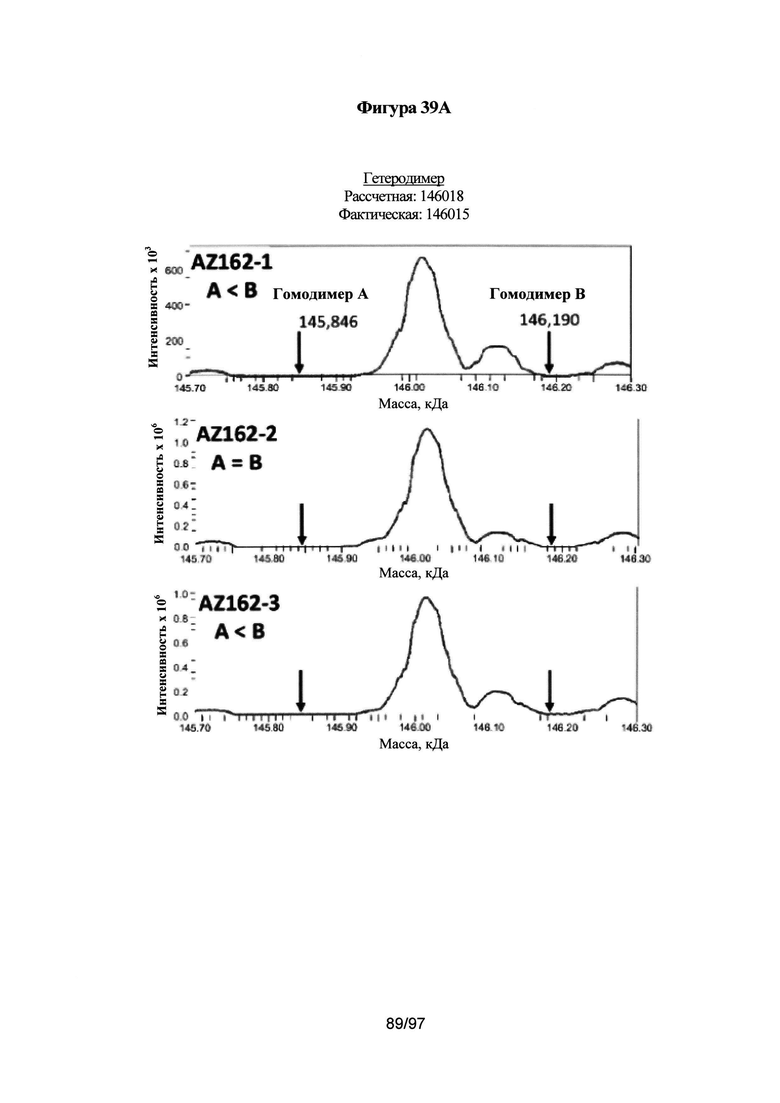
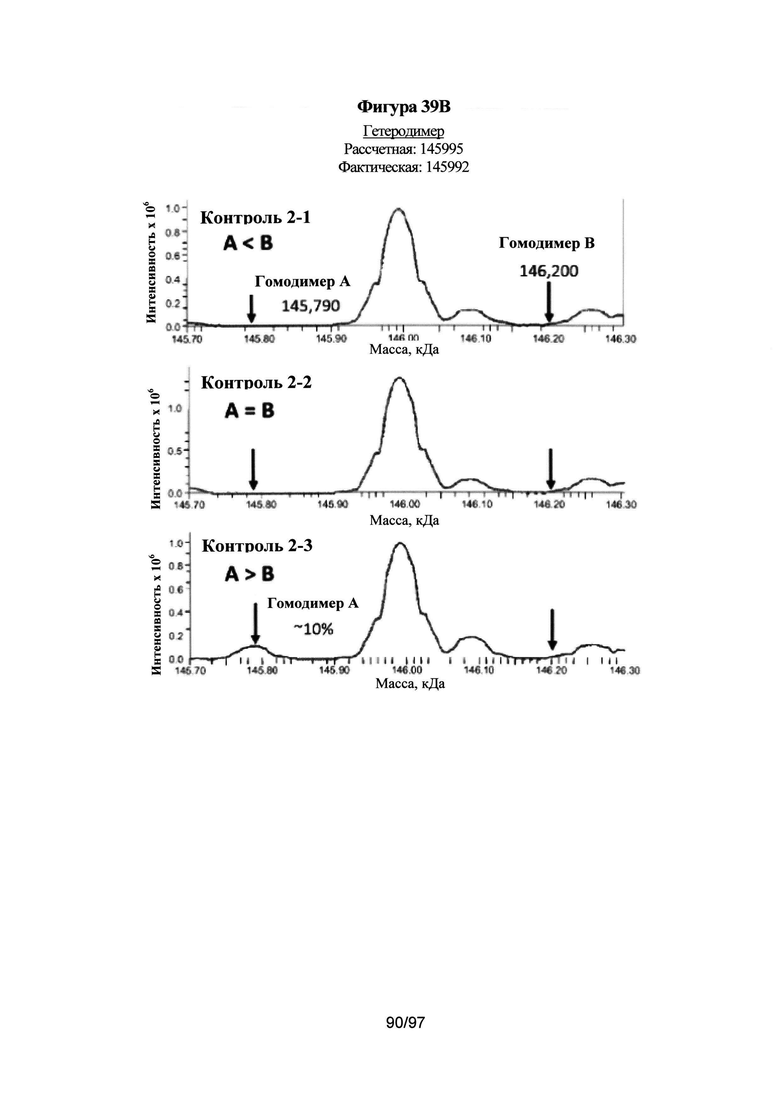

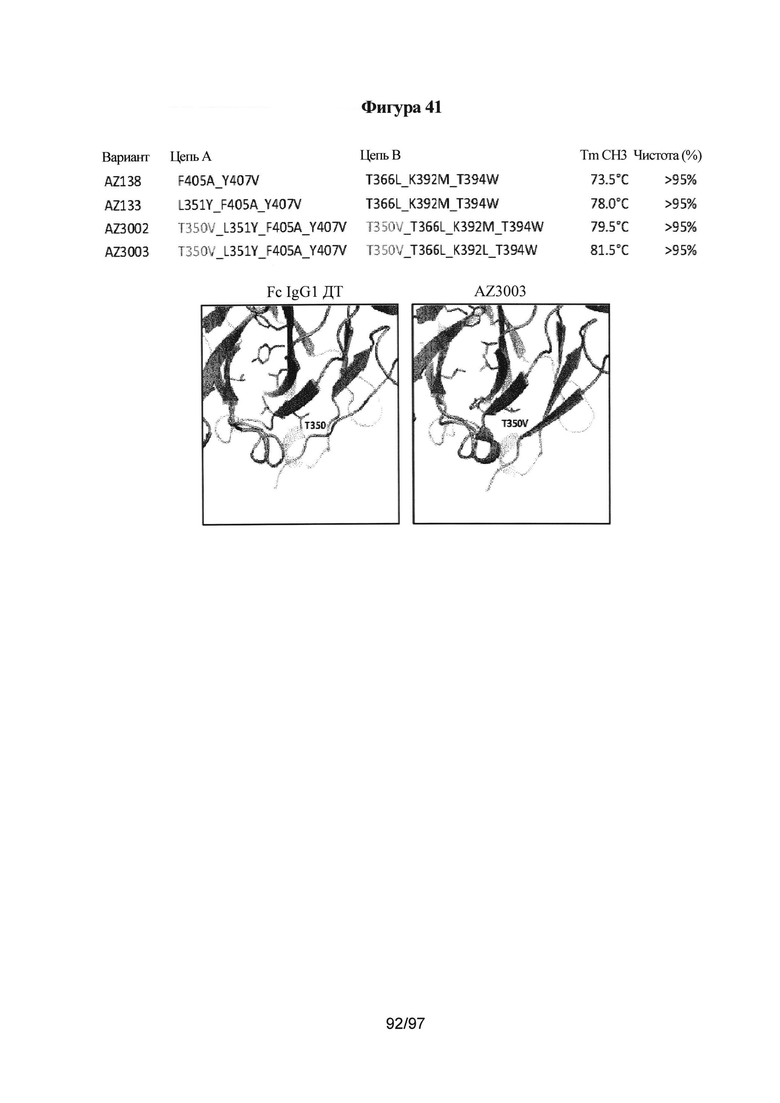
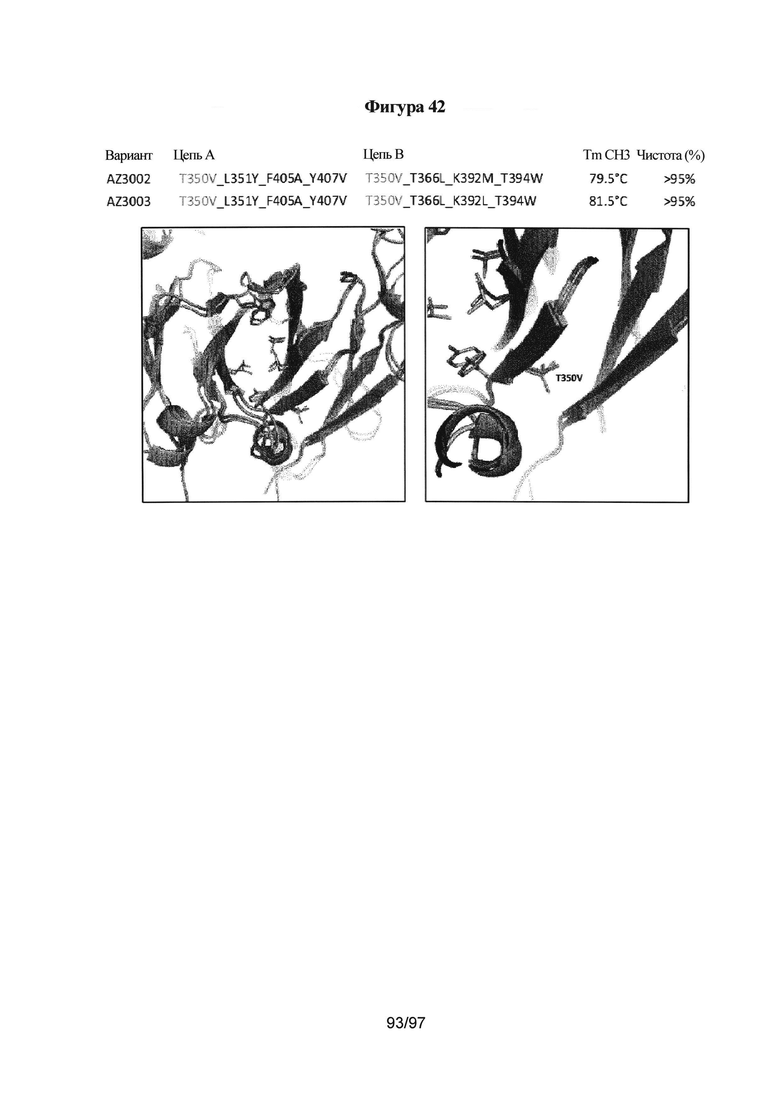
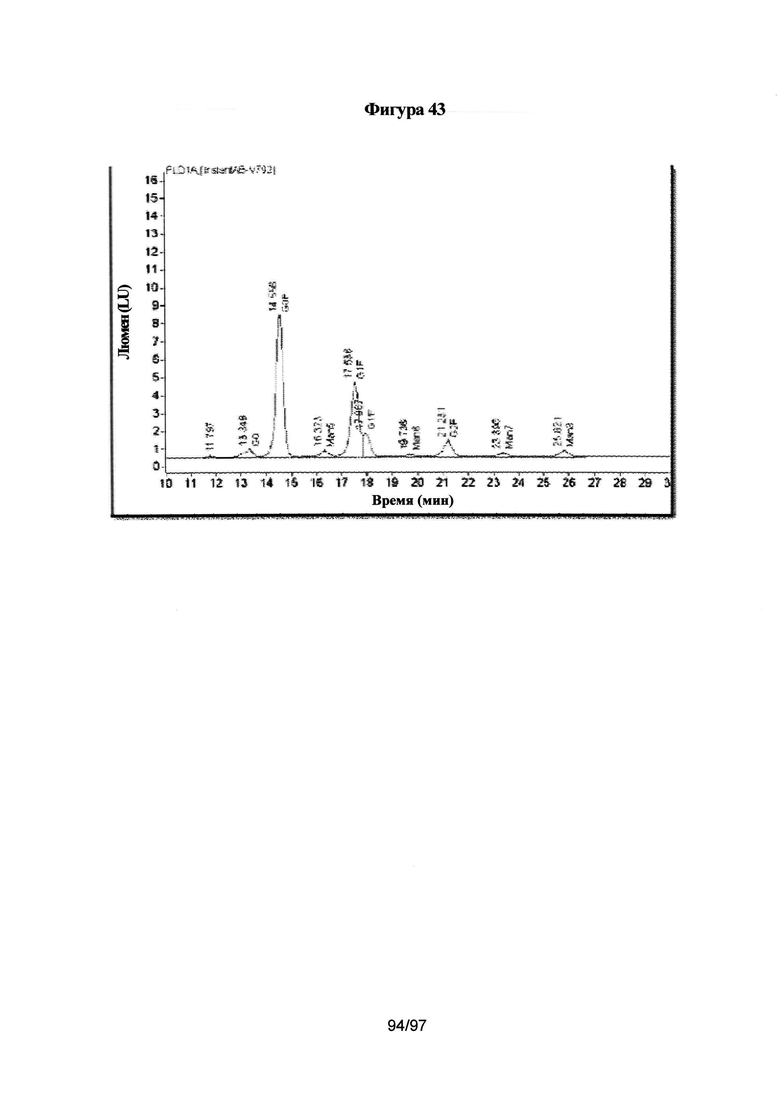
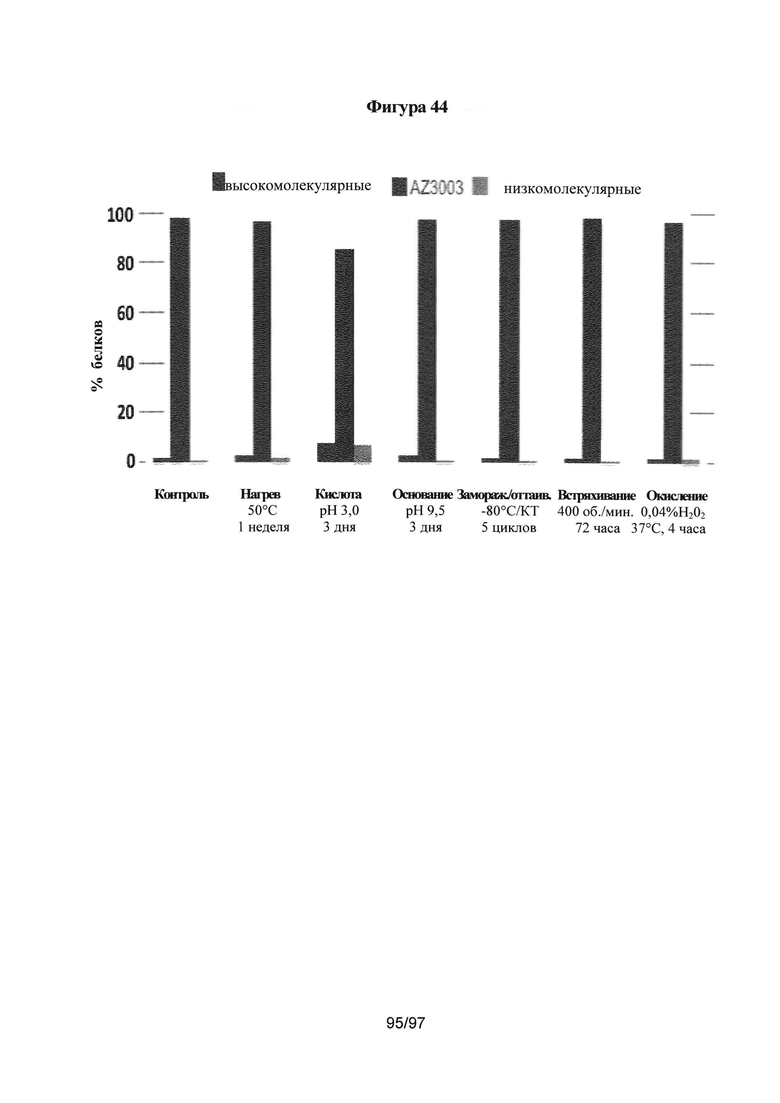
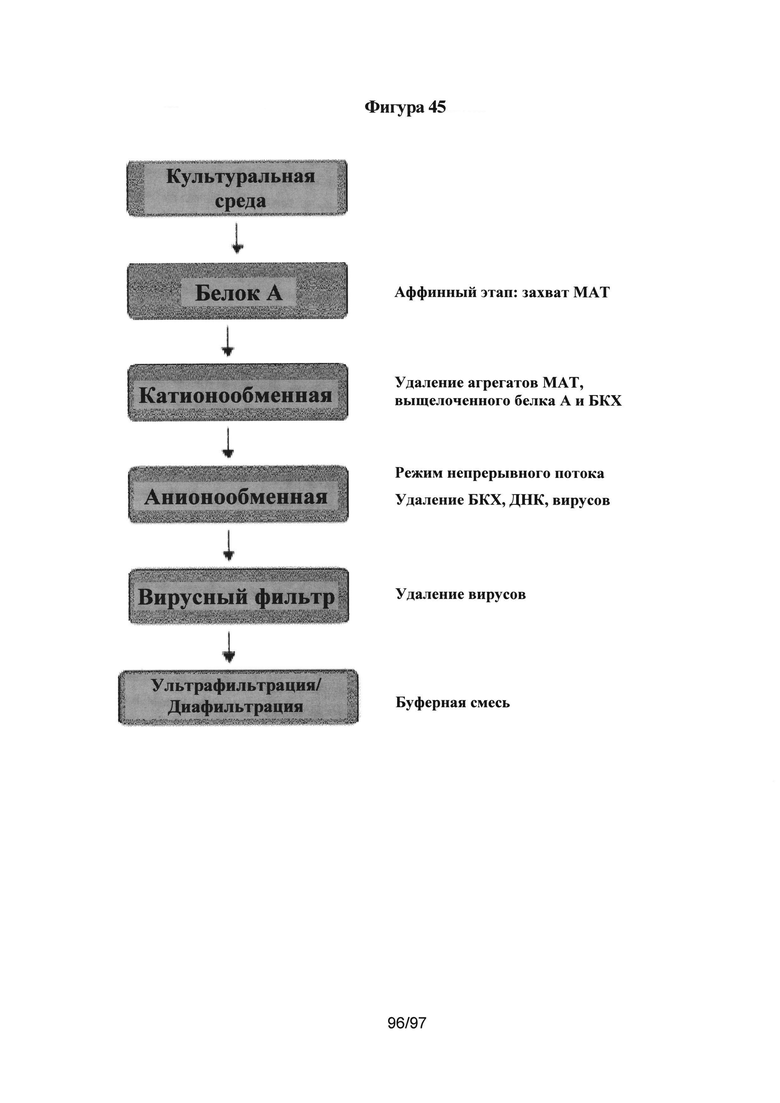
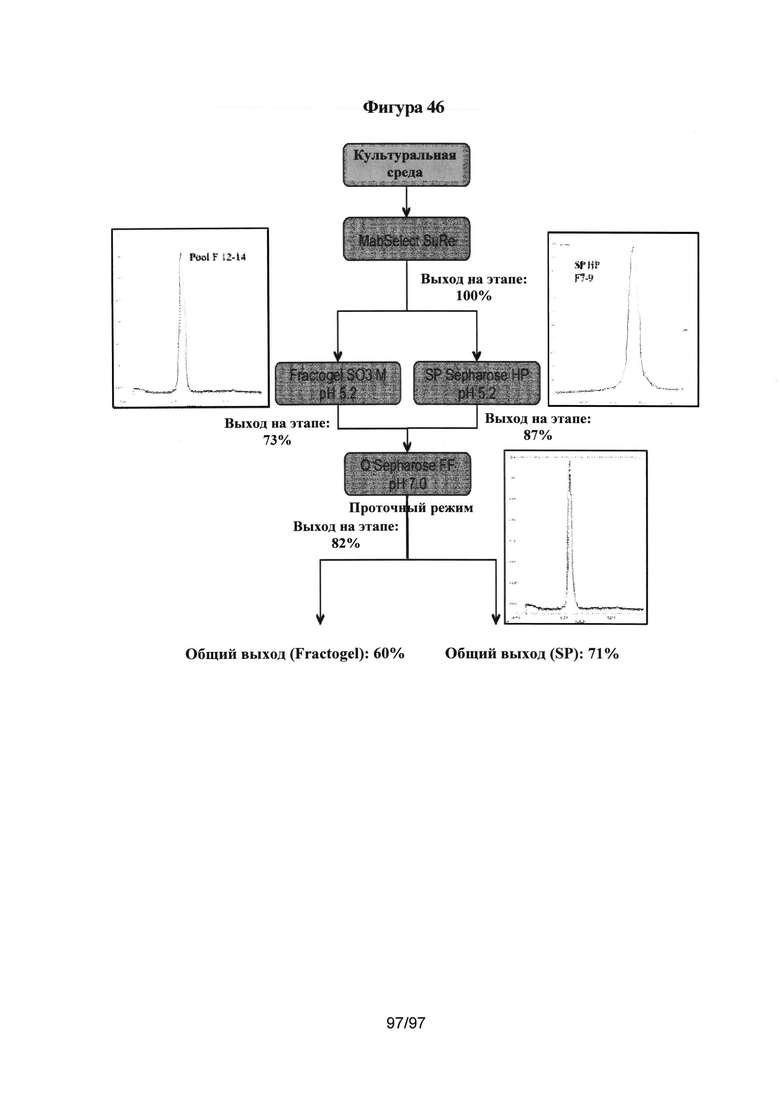
Данное изобретение относится к области биотехнологии и иммунологии. Предложена нуклеотидная композиция для получения гетеродимерной конструкции Fc, имеющей замены в CH3 доменах, обеспечивающие повышение температуры плавления. Также предложена клетка-хозяин и способ экспрессии гетеродимерной конструкции Fc. Настоящее изобретение может найти применение в разработке терапевтических антител. 3 н. и 24 з.п. ф-лы, 46 ил., 16 табл., 15 пр.
1. Нуклеотидная композиция для получения гетеродимерной конструкции Fc, содержащая один или более полинуклеотидов, кодирующих гетеродимерную конструкцию Fc, содержащую модифицированный домен СН3, содержащий:
первый полипептид домена СН3 и второй полипептид домена СН3, каждый из которых включает аминокислотные модификации по сравнению с полипептидом домена СН3 дикого типа;
причем первый полипептид домена СН3 содержит аминокислотные модификации в положениях F405 и Y407, и второй полипептид домена СН3 содержит аминокислотные модификации в положениях Т366 и Т394,
причем аминокислотная модификации в положении F405 представляет собой F405A, F405S, F405T или F405V; аминокислотная модификации в положении Y407 представляет собой Y407V; аминокислотная модификация в положении Т366 представляет собой T366I или T366L, и аминокислотная модификации в положении Т394 представляет собой T394W,
причем второй полипептид домена СН3 дополнительно содержит аминокислотную модификацию K392J, где J выбран из L, М или F;
причем по меньшей мере один из первого и второго полипептидов домена СН3 дополнительно содержит аминокислотную модификацию T350V; причем Fc конструкция основана на IgG1 человека;
при этом модифицированный домен СН3 обладает температурой плавления (Tm), составляющей по меньшей мере приблизительно 74°С; и
при этом нумерация аминокислотных остатков соответствует индексу EU, описанному у Kabat.
2. Нуклеотидная композиция по п. 1, причем каждый из первого и второго полипептидов домена СН3 содержит аминокислотную модификацию T350V.
3. Нуклеотидная композиция по п. 1 или 2, причем модифицированный домен СН3 имеет температуру плавления (Tm), составляющую приблизительно 75°С или более, приблизительно 77°С или более, или приблизительно 80°С или более.
4. Нуклеотидная композиция по п. 1 или 2, причем первый полипептид домена СН3 дополнительно содержит аминокислотную модификацию L351Y.
5. Нуклеотидная композиция по п. 4, причем второй полипептид домена СН3 содержит аминокислотную модификацию K392L или K392M.
6. Нуклеотидная композиция по п. 1 или 2, причем второй полипептид домена СН3 содержит аминокислотную модификацию K392L или K392M.
7. Нуклеотидная композиция по п. 1 или 2, причем аминокислотная модификация в положении F405 представляет собой F405A.
8. Нуклеотидная композиция по п. 1 или 2, причем аминокислотная модификация в положении F405 представляет собой F405A, и при этом J представляет собой L или М.
9. Нуклеотидная композиция по п. 8, причем первый полипептид домена СН3 дополнительно содержит аминокислотную модификацию L351Y.
10. Нуклеотидная композиция по п. 1 или 2, причем:
(a) указанный первый полипептид домена СН3 содержит аминокислотные модификации L351Y, F405A и Y407V, и указанный второй полипептид домена СН3 содержит аминокислотные модификации T366L, K392M и T394W;
(b) указанный первый полипептид домена СН3 содержит аминокислотные модификации L351Y, F405A и Y407V, и указанный второй полипептид домена СН3 содержит аминокислотные модификации T366L, K392L и T394W;
(c) указанный первый полипептид домена СН3 содержит аминокислотные модификации L351Y, F405A и Y407V, и указанный второй полипептид домена СН3 содержит аминокислотные модификации T366I, K392M и T394W, или
(d) указанный первый полипептид домена СН3 содержит аминокислотные модификации L351Y, F405A и Y407V, и указанный второй полипептид домена СН3 содержит аминокислотные модификации T366I, K392L и T394W.
11. Нуклеотидная композиция по п. 1 или 2, причем по меньшей мере один из первого и второго полипептидов домена СН3 дополнительно содержит модификацию S400Z, где Z выбран из положительно заряженной аминокислоты и отрицательно заряженной аминокислоты.
12. Нуклеотидная композиция по п. 11, причем первый полипептид домена СН3 содержит аминокислотную модификацию S400E или S400R.
13. Нуклеотидная композиция по п. 1 или 2, причем по меньшей мере один из первого и второго полипептидов домена СН3 дополнительно содержит модификацию N390Z, в которой Z выбран из положительно заряженной аминокислоты и отрицательно заряженной аминокислоты.
14. Нуклеотидная композиция по п. 13, причем второй домен СН3 содержит аминокислотную модификацию N390R.
15. Нуклеотидная композиция по п. 1 или 2, причем первый полипептид домена СН3 содержит аминокислотную модификацию S400E и второй полипептид домена СН3 содержит аминокислотную модификацию N390R.
16. Нуклеотидная композиция по п. 9, причем первый полипептид домена СН3 содержит аминокислотную модификацию S400E и второй полипептид домена СН3 содержит аминокислотную модификацию N390R.
17. Нуклеотидная композиция по п. 1 или 2, причем один из первого и второго полипептидов домена СН3 содержит аминокислотную модификацию Q347R, а другой полипептид домена СН3 содержит аминокислотную модификацию K360E.
18. Нуклеотидная композиция по п. 1, причем:
(a) первый полипептид домена СН3 содержит аминокислотные модификации T350V, F405A и Y407V, и второй полипептид домена СН3 содержит аминокислотные модификации T350V, T366L, K392M и T394W;
(b) первый полипептид домена СН3 содержит аминокислотные модификации T350V, L351Y, F405A и Y407V, и второй полипептид домена СН3 содержит аминокислотные модификации T350V, T366L, K392M и T394W;
(c) первый полипептид домена СН3 содержит аминокислотные модификации T350V, L351Y, F405A и Y407V, и второй полипептид домена СН3 содержит аминокислотные модификации T350V, T366L, K392L и T394W;
(d) первый полипептид домена СН3 содержит аминокислотные модификации T350V, L351Y, S400R, F405A и Y407V, и второй полипептид домена СН3 содержит аминокислотные модификации T350V, T366L, K392M и T394W;
(e) первый полипептид домена СН3 содержит аминокислотные модификации T350V, L351Y, S400E, F405A и Y407V, и второй полипептид домена СН3 содержит аминокислотные модификации T350V, T366L, N390R, K392M и T394W;
(f) первый полипептид домена СН3 содержит аминокислотные модификации T350V, L351Y, S400E, F405V и Y407V, и второй полипептид домена СН3 содержит аминокислотные модификации T350V, T366L, N390R, K392M и T394W;
(g) первый полипептид домена СН3 содержит аминокислотные модификации T350V, L351Y, S400E, F405T и Y407V, и второй полипептид домена СН3 содержит аминокислотные модификации T350V, T366L, N390R, K392M и T394W;
(h) первый полипептид домена СН3 содержит аминокислотные модификации T350V, L351Y, S400E, F405S и Y407V, и второй полипептид домена СН3 содержит аминокислотные модификации T350V, T366L, N390R, K392M и T394W;
(i) первый полипептид домена СН3 содержит аминокислотные модификации T350V, S400E, F405A и Y407V, и второй полипептид домена СН3 содержит аминокислотные модификации T350V, T366L, N390R, K392M и T394W;
(j) первый полипептид домена СН3 содержит аминокислотные модификации T350V, L351Y, S400E, F405A и Y407V, и второй полипептид домена СН3 содержит аминокислотные модификации T350V, L351Y, T366L, N390R, K392M и T394W;
(k) первый полипептид домена СН3 содержит аминокислотные модификации Q347R, T350V, L351Y, S400E, F405A и Y407V, и второй полипептид домена СН3 содержит аминокислотные модификации T350V, K360E, T366L, N390R, K392M и T394W;
(l) первый полипептид домена СН3 содержит аминокислотные модификации T350V, L351Y, S400R, F405A и Y407V, и второй полипептид домена СН3 содержит аминокислотные модификации T350V, T366L, N390D, K392M и T394W;
(m) первый полипептид домена СН3 содержит аминокислотные модификации T350V, L351Y, S400R, F405A и Y407V, и второй полипептид домена СН3 содержит аминокислотные модификации T350V, T366L, N390E, K392M и T394W;
(n) первый полипептид домена СН3 содержит аминокислотные модификации T350V, L351Y, S400E, F405A и Y407V, и второй полипептид домена СН3 содержит аминокислотные модификации T350V, T366L, N390R, K392L и T394W,
(о) первый полипептид домена СН3 содержит аминокислотные модификации T350V, L351Y, S400E, F405A и Y407V, и второй полипептид домена СН3 содержит аминокислотные модификации T350V, T366L, N390R, K392F и T394W;
(p) первый полипептид домена СН3 содержит аминокислотные модификации Y349C, T350V, L351Y, S400E, F405A и Y407V, и второй полипептид домена СН3 содержит аминокислотные модификации T350V, S354C, T366L, N390R, K392M и T394W;
(q) первый полипептид домена СН3 содержит аминокислотные модификации Y349C, T350V, S400E, F405A и Y407V, и второй полипептид домена СН3 содержит аминокислотные модификации T350V, S354C, T366L, N390R, K392M и T394W, или
(r) первый полипептид домена СН3 содержит аминокислотные модификации Y349C, T350V, F405A и Y407V, и второй полипептид домена СН3 содержит аминокислотные модификации T350V, S354C, T366L, K392M и T394W.
19. Нуклеотидная композиция по любому из пп. 1, 2 или 18, причем гетеродимерная конструкция Fc представляет собой антитело.
20. Нуклеотидная композиция по п. 19, причем антитело представляет собой биспецифическое антитело или мультиспецифическое антитело.
21. Нуклеотидная композиция по любому из пп. 1, 2 или 18, причем гетеродимерная конструкция Fc дополнительно содержит по меньшей мере один антигенсвязывающий домен, который связывает антиген рака.
22. Нуклеотидная композиция по п. 19, причем антитело содержит по меньшей мере антигенсвязывающий домен, который связывает антиген рака.
23. Нуклеотидная композиция по любому из п.п. 1, 2 или 18, причем гетеродимерная конструкция Fc дополнительно содержит по меньшей мере один антигенсвязывающий домен терапевтического антитела.
24. Нуклеотидная композиция по п. 23, причем терапевтическое антитело выбрано из группы, состоящей из абаговомаба, адалимумаба, алемтузумаба, аурограба, бапинейзумаба, базиликсимаба, белимумаба, бевацизумаба, бриакинумаба, канакинумаба, катумаксомаба, цертолизумаба пегола, цетуксимаба, даклизумаба, деносумаба, эфализумаба, галиксимаба, гемтузумаба озогамицина, голимумаба, ибритумомаба тиуксетана, инфликсимаба, ипилимумаба, лумиликсимаба, меполизумаба, мотавизумаба, муромонаба, микограба, натализумаба, нимотузумаба, окрелизумаба, офатумумаба, омализумаба, паливизумаба, панитумумаба, пертузумаба, ранибизумаба, реслизумаба, ритуксимаба, теплизумаба, тоцилизумаба/атлизумаба, тозитумомаба, трастузумаба, Проксиниума™ (Proxinium™), Ренкарекса™ (Rencarex™), устекинумаба и залутумумаба.
25. Нуклеотидная композиция по любому из пп. 1, 2 или 18, в которой один или более полинуклеотидов входят в состав одного или более экспрессионных векторов или мультицистронного экспрессионного вектора.
26. Клетка-хозяин для получения гетеродимерной конструкции Fc, содержащая нуклеотидную композицию по любому из пп. 1-25.
27. Способ экспрессии гетеродимерной конструкции Fc в клетках млекопитающего, включающий:
(a) трансфицирование по меньшей мере одной клетки млекопитающего нуклеотидной композицией по любому из пп. 1-25, для получения по меньшей мере одной временно или постоянно трансфицированной клетки млекопитающего, и
(b) культурирование по меньшей мере одной временно или постоянно трансфицированной клетки млекопитающего в условиях, подходящих для экспрессии гетеродимерной конструкции Fc.
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| KABAT E.A | |||
| et al., "Sequences of proteins of immunological interest", DIANE publishing, 5th ed., Vol.1, 1991, NIH Publication 91-3242; см | |||
| Колосниковая решетка с охлажденными водой колосниками | 1925 |
|
SU688A1 |
| ATWELL S | |||
| et al | |||
| "Stable heterodimers from remodeling the domain interface of a homodimer using a phage display library", Journal of molecular biology, 1997; 270(1): 26-35 | |||
| GUNASEKARAN K | |||
| et al., "Enhancing Antibody Fc Heterodimer Formation through Electrostatic Steering Effects | |||
| APPLICATIONS TO BISPECIFIC MOLECULES AND MONOVALENT IgG", Journal of Biological Chemistry, 2010; 285(25): 19637-19646 | |||
| DEMAREST S.J | |||
| et al., "Optimization of the antibody CH3 domain by residue frequency analysis of IgG sequences", Journal of molecular biology, 2004; 335(1): 41-48. | |||

