Настоящее изобретение относится к способу получения алкенов с помощью биологического процесса. Более специфически, изобретение относится к способу получения алкенов (например, пропилена, этилена, 1-бутилена, изобутилена или изоамилена) из молекул 3-гидроксиалканоатного типа.
В настоящее время много химических продуктов получают из нефтяного сырья. Алкены (такие, как этилен, пропилен, различные бутены, или, например, также пентены) используют в производстве пластмасс, например, для получения полипропилена или полиэтилена, и в других областях химической промышленности и топлив.
Этилен, самый простой алкен, занимает центральное место в промышленной органической химии: он является органическим соединением, которое вырабатывается в наибольшем количестве во всем мире. Он используется, в частности, для получения полиэтилена, основной пластмассы. Этилен также может быть превращен во многие промышленно используемые продукты путем реакции (окисления, галогенирования).
Пропилен занимает сходную важную роль: при его полимеризации получают пластмассовый материал, полипропилен. Технические свойства этого продукта в отношении устойчивости, плотности, твердости, деформируемости, и прозрачности несравненны. Во всем мире производство полипропилена непрерывно увеличивается с его изобретения в 1954 г.
Бутилен существует в четырех формах, один из которых, изобутилен, входит в состав метил-трет-бутилового эфира (МТВЕ), антидетонационной присадки для автомобильного топлива. Изобутилен также может использоваться для получения изооктена, который, в свою очередь, может быть восстановлен до изооктана (2,2,4-триметилпентан); очень высокое октановое число изооктана делает его наилучшим топливом для так называемых "бензиновых" двигателей.
Амилен, гексен и гептен существуют во многих формах в соответствии с положением и конфигурацией двойной связи. Эти продукты имеют реальное промышленное применение, но являются менее важными, чем этилен, пропилен или бутены.
Все эти алкены в настоящее время продуцируются путем каталитического крекинга нефтепродукты (или с помощью производного процесса Фишера-Тропша в случае гексена, из угля или газа). Их стоимость соответственно естественно индексирует цену нефти. Кроме того, каталитический крекинг иногда связан со значительными техническими сложностями, которые повышают сложность процесса и себестоимость производства.
Несмотря на представленный выше анализ, биополучение пластмасс ("биопластиков") является бурно развивающейся областью. Этот подъем обусловлен экономическим интересом, связанным с ценой на нефть, и экологическими соображениями, которые являются как глобальными (продукты с нулевым балансом выбросов углерода) и локальными (организация сбора и удаления отходов).
Основным семейством биопластиков является семейство полигидроксиалканоатов (РНА). Они представляют собой полимеры, получаемые путем конденсации молекул, содержащих как кислотную группу, так и спиртовую группу. Конденсация происходит путем эстерификации кислоты на спирте последующего мономера. Эта сложноэфирная связь не является настолько стабильной, как прямая углерод-углеродная связь, присутствующая в полимерах общепринятых пластмасс, что объясняет, почему РНА имеют способность к биологическому разложению от нескольких недель до нескольких месяцев.
РНА семейство включает, в частности, поли-3-гидроксибутират (РНВ), полимер 3-гидроксибутирата, и полигидроксибутират-валерата (PHBV), сополимер с регулярным чередованием мономерных единиц 3-гидроксибутирата и 3-гидроксивалерата.
РНВ естественно продуцируется некоторыми штаммами бактерий, таких как Alcaligenes eutrophus и Bacillus megaterium. Были сконструированы лабораторные бактерии, такие как Е.coli, имеющие интегрированные синтетические пути, приводящие к РНВ или к РНА в целом. Соединение или его полимер может, в определенных лабораторных условиях, обуславливать вплоть до 80% бактериальной массы (Wong MS и др., Biotech. Bioeng.99 (2008), 919-928). Попытки получения РНВ в промышленном масштабе осуществлялись в 1980х гг., но стоимость получения соединения путем ферментации считалась слишком высокой в то время. Разрабатываются проекты, направленные на прямое получение этих соединений в генетически модифицированных растениях (имеющие интегрированные ключевые ферменты пути синтеза РНВ, присутствующие в бактериях-продуцентах) и они должны быть менее затратными.
Биологическое получение алканов или молекул других углеводородов, которые можно использовать в качестве топлива или в качестве предшественников синтетических смол, рассматривается в контексте экологически безвредной промышленной эксплуатации в гармонии с геохимическим циклом. Первое поколение биотоплив включает продукты ферментации этанола, в качестве процессов ферментации и перегонки, которые уже существуют в пищевой промышленности. Получение биотполива второго поколения находится на стадии исследования, охватывая получение длинноцепочечных спиртов (бутанол и пентанол), терпенов, линейных алканов и жирных кислот. В двух недавних обзорах представлен обзор исследований в этой области: Ladygina N и др., Process Biochemistry, 2006, 41: 1001; и Wackett LP, Current Opinions in Chemical Biology, 2008, 21: 187.
В химическом семействе алкенов, изопрен (2-метил-1,3-бутадиент) представляет собой терпеновый мотив, который, посредством полимеризации, приводит к каучуку. Могут быть разработаны другие терпены, с помощью химических, биологических или смешанных путей, в качестве полезных продуктов, таких как биотопливо или для приготовления пластмасс. В недавно опубликованной литературе показано, что мевалонатный путь (ключевой промежуточный продукт при биосинтезе стероидов во многих организмах) можно использовать для эффективного получения продуктов терпенового семейства в промышленных выходах (Withers ST и др., Appl. Environ. Microbiol., 2007, 73: 6277).
Получение алкенов, в частности концевых алкенов, [этилен моно- или ди-замещенных в положении 2: H2C=C(R1)(R2)], по всей видимости, менее интенсивно исследуется. Было описано превращение изовалерата в изобутилен с помощью дрожжей Rhodotorula minuta (Fujii Т. и др., Appl. Environ. Microbiol., 1988, 54:583), но эффективность этой реакции, охарактеризована очень низким значением числа оборотов (kcat составляет 1×10-5 с-1), не позволяет реализовать промышленное применение. Механизм реакции был описан Fukuda H и др. (BBRC, 1994, 201(2): 516) и задействует фермент цитохром Р450, который декарбоксилирует изовалерат путем восстановления окоферрильной группы Fev=O. Ни при каких условиях реакция не включает гидроксилирование изовалерата. Изовалерат также представляет собой промежуточное соединение в катаболизме лейцина. Крупномасштабный биосинтез изобутилена с помощью такого пути представляется чрезвычайно неблагоприятным, поскольку для него будет необходим синтез и разложение одной молекулы лейцина с образованием одной молекулы изобутилена. Кроме того, фермент, катализирующий реакцию, использует гем в качестве кофактора, плохо поддается рекомбинантной экспрессии в бактерии и улучшению параметров фермента. Учитывая все эти факторы, кажется очень маловероятным, что этот путь из уровня техники может служить в качестве основы для промышленного использования. Были описаны другие микроорганизмы в качестве незначительно способных пригодно продуцировать изобутилен из изовалерата; полученные выходы были даже еще ниже, чем выходы, полученные с Rhodotoruia minuta (Fukuda Н. и др, Agric. Biol. Chem., 1984, 48: 1679).
В некоторых исследованиях также было описано природное получение пропилена: многие микроорганизмы способны продуцировать пропилен, однако также с чрезвычайно низким выходом. Продукция этилена растениями давно известна (Meigh и др., 1960, Nature, 186:902). В соответствии с описанным метаболическим путем, метионин является предшественником этилена (Adams и Yang, PNAS, 1979, 76:170). Также было описано превращение 2-окоглутарата (Ladygina N и др., Process Biochemistry 2006, 41:1001). Поскольку для получения двухуглеродной молекулы этилена расходуется четырех- или пяти-углеродная молекула предшественника, то эти пути являются материально и энергетически неблагоприятными для их промышленного применения.
Таким образом, существует потребность в эффективных методах получения алкенов, таких как этилен, пропилен, 1-бутилен, изобутилен, 1-амилен или изоамилен.
В WO 2010/001078 описан процесс получения алкенов путем ферментативного превращения 3-гидроксиалкановых кислот с помощью фермента, имеющего активность декарбоксилазы. Такой способ является благоприятным, поскольку он помогает избежать применения нефтепродуктов, снижает стоимость получения пластмасс и топлива и может иметь существенное глобальное влияние на окружающую среду вследствие предоставления возможности хранения углерода в твердой форме. Несмотря на то, что способ, описанный в WO 2010/001078, предоставляет возможность получения алкенов путем ферментативного превращения 3-гидроксиалканоатов, он все еще нуждается в улучшении, в частности относительно эффективности процесса таим образом, чтобы сделать его пригодным для промышленных целей. Настоящая заявка решает эту проблему.
В настоящей заявке описан способ получения алкеновых соединений, используя в качестве исходного соединения 3-гидроксиалканоат с помощью биологического процесса, в особенности ферментативный процесс, в котором два типа ферментов комбинированы для повышения эффективности производительности. Более специфически, настоящее изобретение относится к способу получения алкена, который отличается тем, что он включает превращение 3-гидроксиалканоата в указанный алкен с помощью
(i) первого фермента, который обладает активностью превращать 3-гидроксиалканоат в соответствующий 3-фосфоноксиалканоат; и
(ii) второго фермента, который отличается от первого фермента и, который обладает активностью превращать указанный 3-фосфоноксиалканоат в указанный алкен.
Настоящее изобретение также относится к применению по меньшей мере двух ферментов, где один фермент выбирают из (i), как указано выше, и другой фермент выбирают из (ii), как указано выше, или микроорганизма, продуцирующего указанную комбинацию ферментов, для получения алкенового соединения из 3-гидроксиатакажэата. Настоящее изобретение также относится к организмам, предпочтительно микроорганизмам, которые продуцируют по меньшей мере два фермента, где один фермент выбирают из (i), как указано выше, и другой фермент выбирают из (ii), как указано выше.
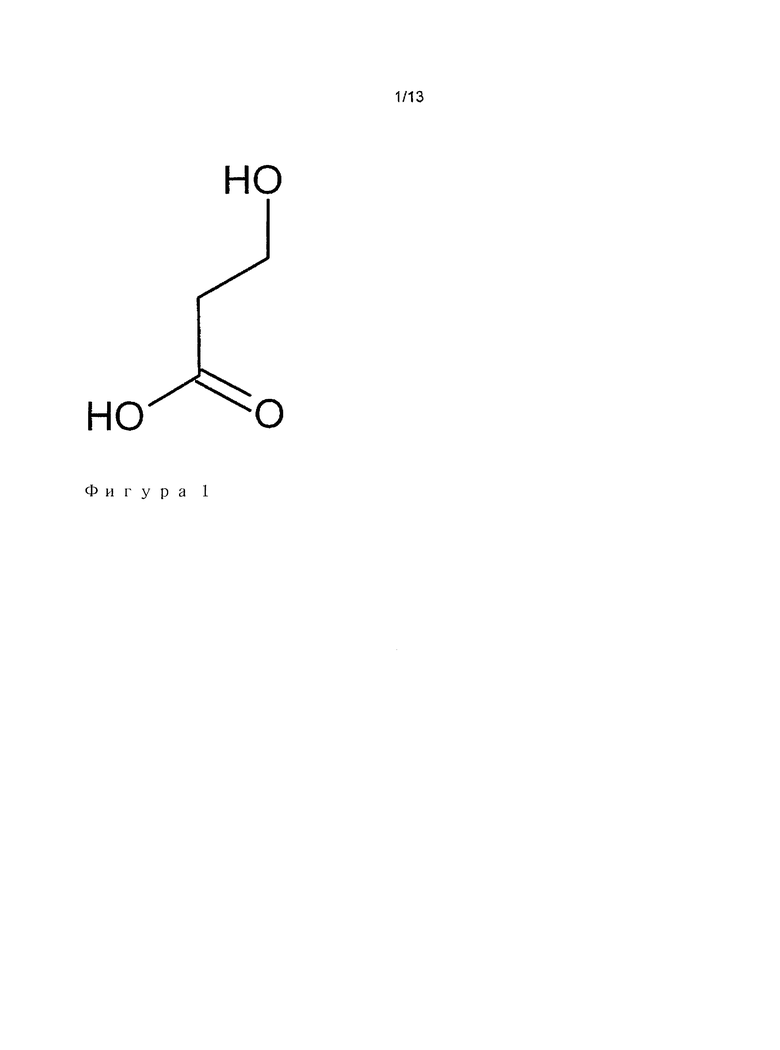
"3-гидроксиалканоат", как используется в настоящей заявке, обозначает молекулу в соответствии со следующей общей формулой:
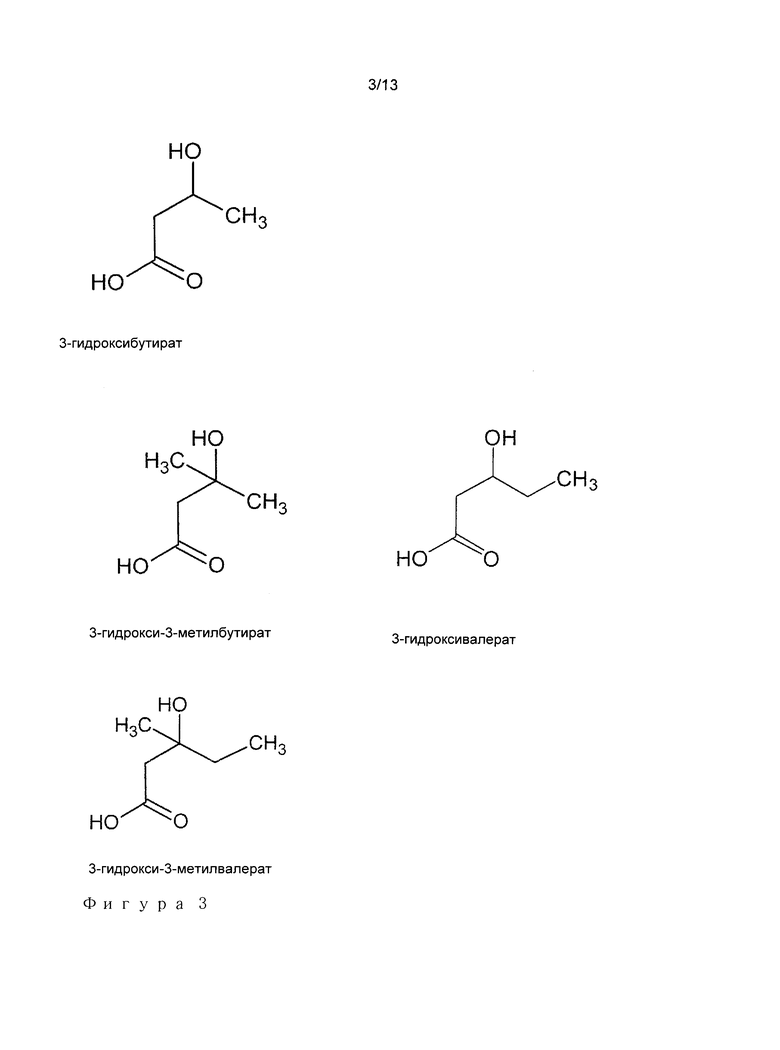
Cn+1H2n+2O3,
с 1<n<7, и содержащую 3-гидроксипропионат в качестве общего мотива (Фигура 1), и необязательно один или два алкильных заместителя на углероде 3. Указанные алкильные остатки или группы могут быть линейными или разветвленными. Как используется в настоящей заявке, термины "алкоил" и "алкил" имеют аналогичное значение и являются взаимозаменяемыми. Аналогичным образом, термины "остаток" и "группа" имеют аналогичное значение и являются взаимозаменяемыми. Метильная, этильная, пропильная, изопропильная, бутильная, изобутильная группы являются примерами указанных алкильных групп. Углерод 3 становится хиральным центром, если два алкильных заместителя являются различными. Настоящее определение охватывает две хиральные формы, даже если одна из двух форм, например, R форма, является основной формой, продуцируемой в естественных условиях. Примеры 3-гидроксиалканоатов представлены на Фигуре 3. Необязательно, алкильные заместители могут быть добавлены на углероде 2, который впоследствии также может становиться хиральным (если два заместителя являются разными). Равным образом, конфигурации 3-гидроксиалканоатных субстратов в настоящем изобретении охватывает все стереоизомеры. В предпочтительном варианте осуществления, 3-гидроксиалканоаты соответствуют либо 3-гидроксмпропионату или вариантам или производным 3-гидроксипропионата, в которых один из двух или два атома водорода, которые находятся на углероде 3, замещены мотивом, состоящим только из атомов углерода и водорода, количество атомов углерода указанных заместителей находится в диапазоне от 1 до 5, предпочтительно от 1 до 3, такие как метил, этил, пропил, изопропил, бутил или изобутил. Суффикс "оат", как используется в настоящей заявке, может взаимозаменяемо обозначать либо карбоксилатный ион (СОО-) или карбоновую кислоту (СООН). Он не используется для обозначения сложного эфира. В предпочтительном варианте осуществления, 3-гидроксиалканоаты представлены следующей формулой:
HO-CO-CH2-C(R1)(R2)-OH или O--CO-CH2-C(R1)(R2)-OH.
Термин "3-фосфоноксиалканоат" обозначает молекулу, которая соответствует следующей общей формуле:
Cn+1H2n+3О6Р,
с 1<n<7, и содержащей 3-фосфоноксипропионат в качестве общего мотива, и необязательно один или два алкильных заместителя на углероде 3.
Термин "алкен", как используется в настоящей заявке, обозначает молекулы, состоящие только из углерода и водорода, содержащие одну углерод-углеродную двойную связь и имеющие химическую формулу мононенасыщенного углеводорода, CnH2n, где n равно по меньшей мере двум. Предпочтительно, п равно по меньшей мере 3, 4, 5 или 6. Наиболее предпочтительно п представляет собой самое большее 6. Таким образом, в целом, термин "алкен" относится к молекуле, соответствующей формуле CnH2n, с 1<n<7.
В предпочтительном варианте осуществления алкены представлены структурной формулой H2C=C(R1)(R2), где R1 и R2 выбирают, независимо, из группы, включающей атом водорода и линейный или разветвленный алкильный радикал, таким образом, что общее количество атомов углерода в алкеновой молекуле составляет самое большее 6.
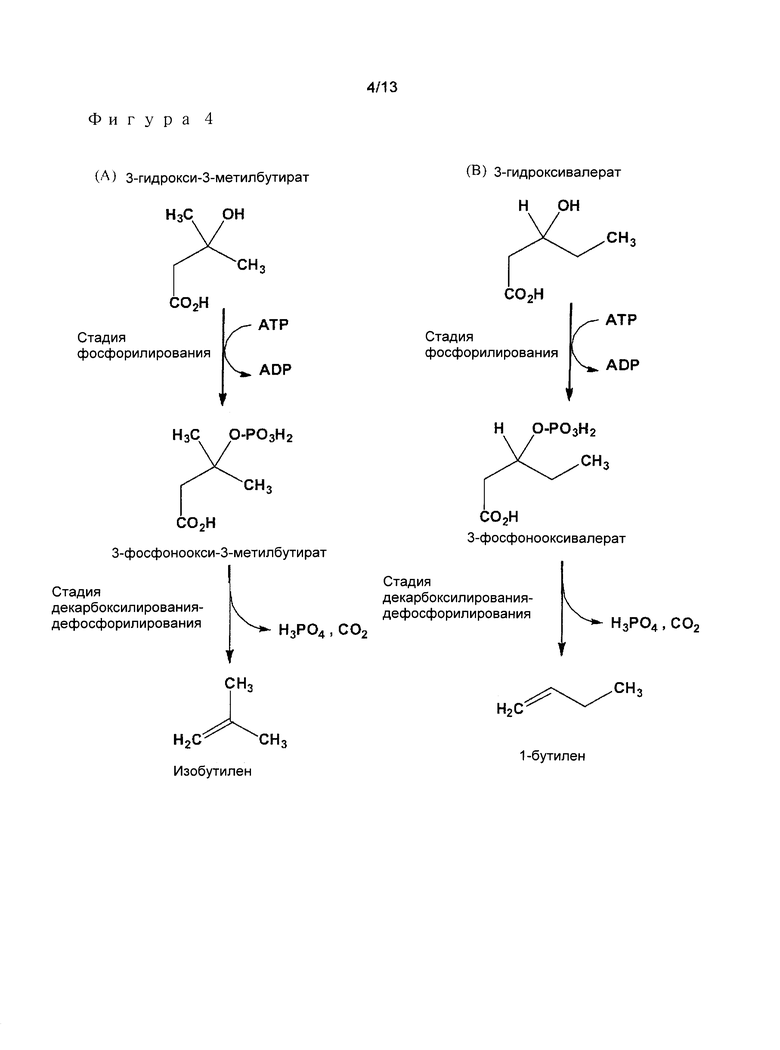
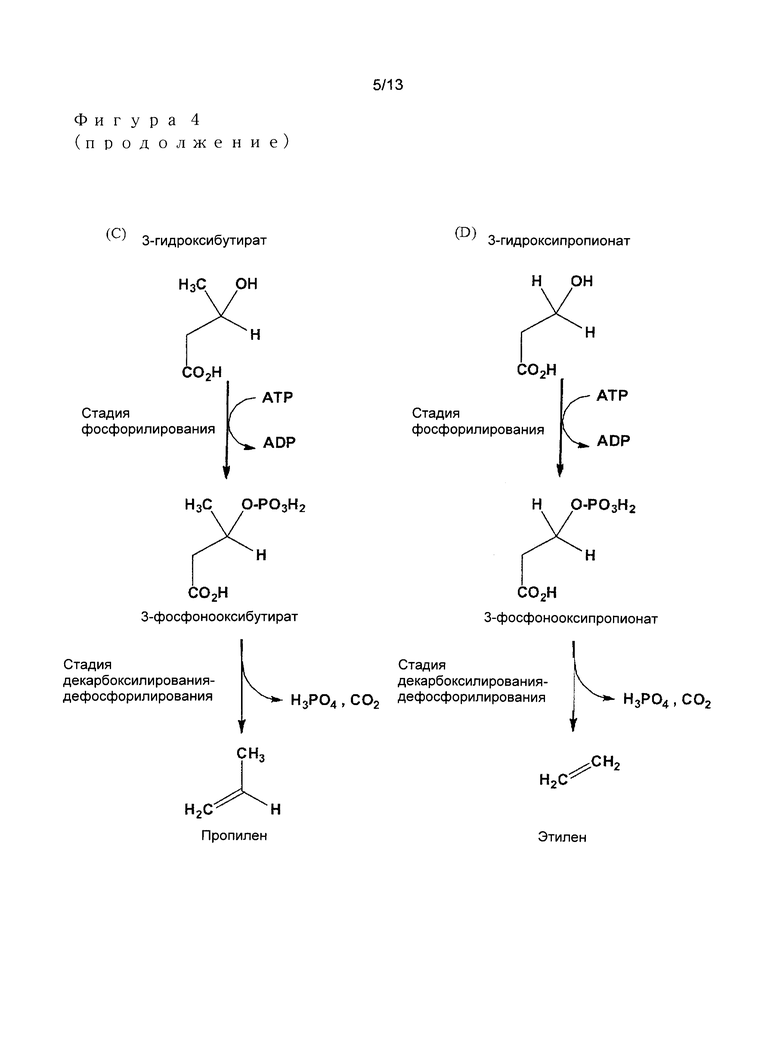
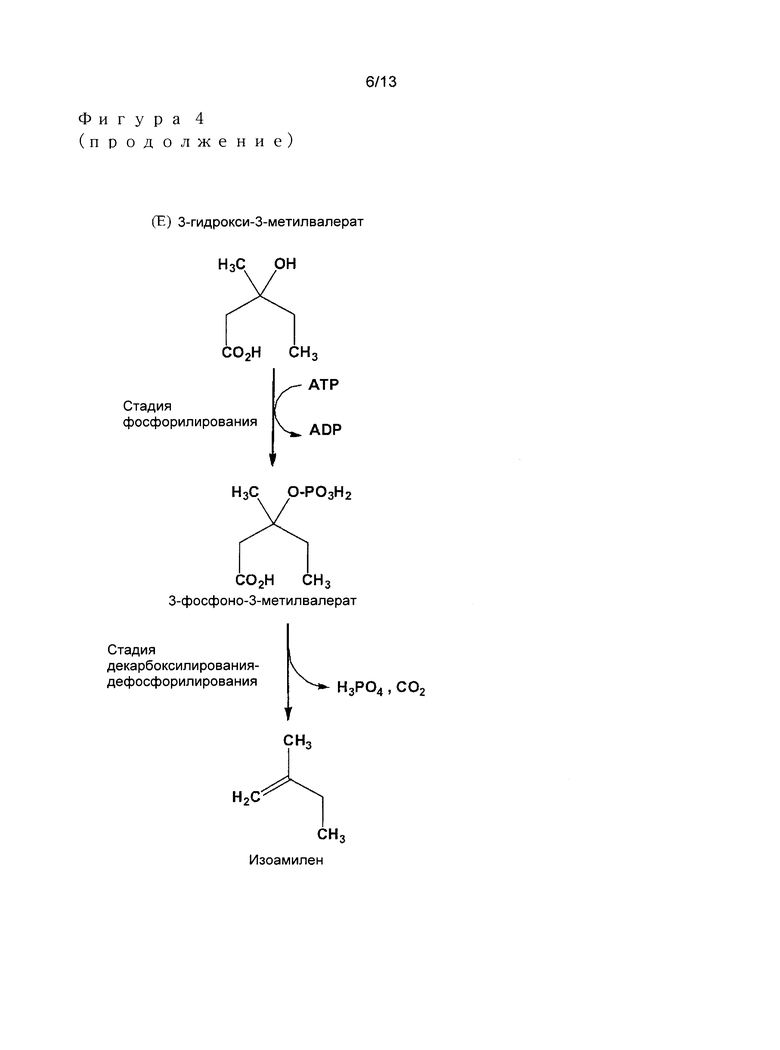
Предпочтительными примерами алкеновых соединений в соответствии с изобретением являются в особенности этилен, пропилен, изобутилен, и изоамилен (Фигура 4), или также 1-бутилен и 1-амилен.
"Источник углерода", как используется в настоящей заявке, обозначает любое углеродное соединение, которое можно использовать в качестве субстрата для организмов в соответствии с изобретением. Указанный термин включает глюкозу или любую другую гексозу, ксилозу или любую другую пентозу, полиолы, такие как глицерин, сорбит или маннит, или также полимеры, такие как крахмал, целлюлоза или гемицеллюлоза, или также поли-3-гидроксиалканоаты, такие как поли-3-гидроксибутират.Это может быть любой субстрат, предоставляющий возможность роста микроорганизмов, такой как, например, формиат. Это также может быть CO2 в том случае, если микроорганизмы способны осуществлять фотосинтез.
"Рекомбинантный", как используется в настоящей заявке, обозначает искусственную генетическую модификацию организма, либо путем добавления, удаления, или модификации хромосомного или экстра-хромосомного гена или регуляторного мотива, такого как промотор, или путем слияния организмов, или путем добавления вектора любого типа, например, плазмидного. Термин "рекомбинантная экспрессия" обозначает продукцию белка, включающего генетическую модификацию, предпочтительно для получения белка экзогенного или гетерологического происхождения по отношению к его хозяину, то есть, который в естественных условиях не встречается в продуцирующем хозяине, или для получения модифицированного или мутированного эндогенного белка.
"Сверхэкспрессия" или "сверхэкспрессирующий", как используется в настоящей заявке, обозначает рекомбинантную экспрессию белка в организме-хозяине, предпочтительно имеющего происхождение из организма, отличающегося от того, в котором он экспрессируется, повышенную по меньшей мере на 10% и предпочтительно на 20%, 50%, 100%, 500% и возможно больше по сравнению в естественной экспрессией указанного белка, встречающейся в указанном организме-хозяине. Это определение также охватывает случае, где отсутствует естественная экспрессия указанного белка.
"Ко-субстрат" представляет собой соединение или молекулу, добавляемую к ферментативной реакции, для того, чтобы улучшить их определенные параметры, и главным образом их активность, указанный продукт и основной субстрат расходуются в равных количествах. Таким образом, ко-субстрат следует добавлять к реакции в концентрации, сопоставимой с таковой основного субстрата. В зависимости от фермента, присутствие ко-субстрата может быть необходимым для ферментативной реакции.
"Кофактор" представляет собой продукт, добавляемый к ферментативной реакции, для того, чтобы улучшить их определенные параметры, и главным образом их активность, указанный продукт не израсходуется при осуществлении реакции, и поэтому его необходимо добавлять только в низких концентрациях, пропорционально количеству фермента, указанная концентрация, следовательно, обозначается как "каталитическая".
"Часть" аминокислотной последовательности обозначает фрагмент, содержащий по меньшей мере 10, предпочтительно по меньшей мере 20, 30, 40 или 50 последовательных аминокислотных остатков указанной последовательности.
"Гомология", как используется в настоящей заявке, обозначает существование сходства между двумя последовательностями, что измеряется и помощью процента идентичности между указанными двумя последовательностями. В предпочтительном варианте осуществления термин "гомология" обозначает идентичность последовательности.
Химические соединения часто известны под несколькими названиями, официальными или тривиальными. В настоящей заявке, предпочтительными являются тривиальные названия. Таким образом:
- "этилен" используется для обозначения этена
- "пропилен" используется для обозначения пропена
- "бутилен" используется для обозначения бутена
- "изобутилен" используется для обозначения 2-метилпропена или изобутена
- "амилен" используется для обозначения пентена
- "изоамилен" используется для обозначения 2-метил-бут-1-ена или изопентена
- "пропионат" используется для обозначения пропановой кислоты или пропаноатного иона
- "бутират" используется для обозначения бутановой кислоты или бутаноатного иона
- "валерат" используется для обозначения пентановой кислоты или пентаноатного иона.
В настоящей заявке описан способ получения алкеновых соединений, используя в качестве исходного соединения 3-гидроксиалканоат с помощью биологического процесса, в особенности ферментативный процесс, в котором два типа ферментов комбинированы для повышения эффективности производительности. Более специфически, настоящее изобретение относится к способу получения алкена, который отличается тем, что он включает превращение 3-гидроксиалканоата в указанный алкен с помощью
(i) первого фермента, который обладает активностью превращать 3-гидроксиалканоат в соответствующий 3-фосфоноксиалканоат; и
(ii) второго фермента, который отличается от первого фермента и, который обладает активностью превращать указанный 3-фосфоноксиалканоат в указанный алкен.
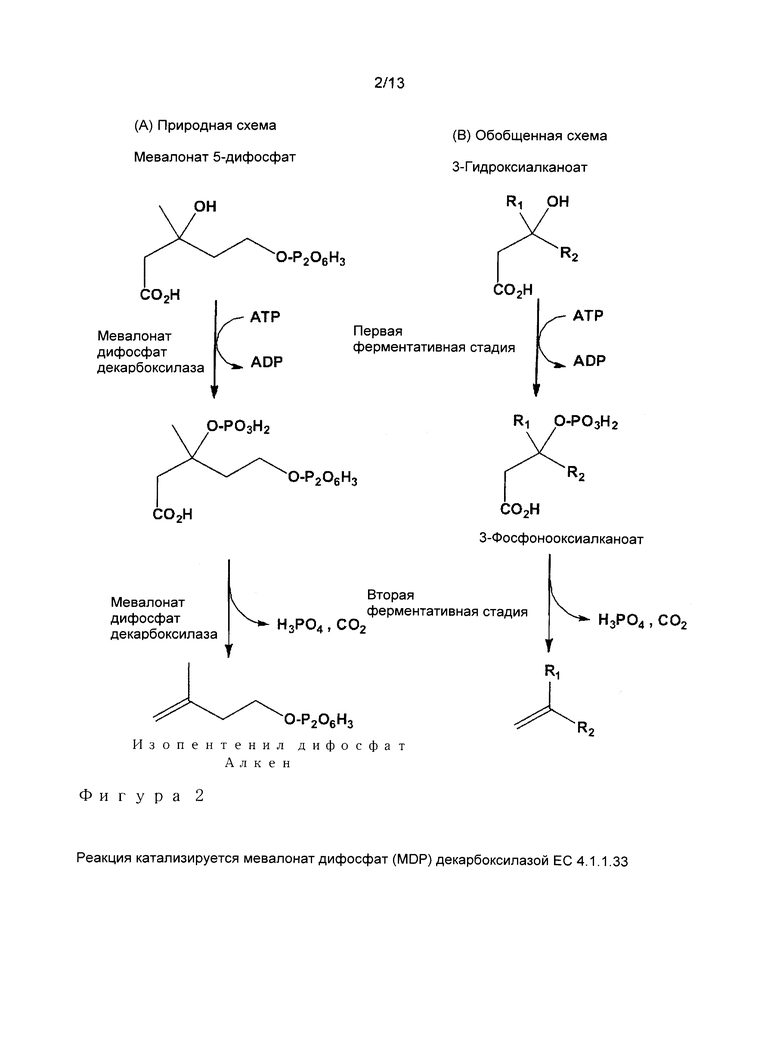
Как указано выше, в WO 2010/001078 описан способ получения алкенов путем ферментативного превращения 3-гидроксиалкановых кислот с помощью фермента, имеющего активность декарбоксилазы. В WO 2010/001078 было описано, что в целом превращение 3-гидроксиалканоата в алкен с помощью фермента, имеющего активность декарбоксилазы, например, мевалонат дифосфат (MDP) декарбоксилазы (Е.С.4.1.1.33), происходит путем превращения 3-гидроксиалканоата в соответствующий 3-фосфоноксиалканоат, который затем декарбоксилируют, что приводит к получению соответствующего алкена. Характерная реакция, осуществляемая MDP декарбоксилазой с использованием различных 3-гидроксиалканоатов, представлена на Фигуре 2В. Сейчас было обнаружено, что различные декарбоксилазы, в особенности мевалонат дифосфат декарбоксилазы, катализируют две вышеуказанные стадии с различными эффективностями, то есть что некоторые декарбоксилазы катализируют первую стадию с более высокой эффективностью по сравнению с другими декарбоксилазами и что некоторые декарбоксилазы проявляют предпочтение для второй стадии, то есть стадии декарбоксилирования, и что, следовательно, эффективность превращения 3-гидроксиалканоата в алкен, как описано в WO 2010/001078, может быть существенно повышена путем комбинирования соответствующих ферментов. Таким образом, настоящее изобретение в особенности относится к способу достижения более высокой эффективности при ферментативном получении алкенов из 3-гидроксиалканоатов, то есть способу улучшения эффективности такого ферментативного получения.
Термин "фермент, который обладает активностью превращать 3-гидроксиалканоат в соответствующий 3-фосфоноксиалканоат" обозначает фермент, который может фосфорилировать 3-гидроксиалканоат в соответствующий 3-фосфоноксиалканоат.Фосфатная группа предпочтительно поступает из молекулы АТФ.
Эта активность может быть измерена, например, как описано в приложенных примерах, в особенности Примере 5. Таким образом, одной возможностью является инкубирование соответствующего фермента с 3-гидроксиалканоатом и АТФ и измерение продукции АДФ (то отображает продукцию соответствующего 3-фосфоноксиалканоата). Исследования для измерения продукции АДФ известны специалисту в данной области техники. Одним из этих методом является исследование пируват киназа/лактат дегидрогеназа, описанное в примере 5. В этом случае, в исследовании измеряют скорость снижения абсорбции NADH при 340 нм, которая пропорциональна количеству АДФ. В предпочтительном варианте осуществления термин "фермент, который обладает активностью превращать 3-гидроксиалканоат в соответствующий 3-фосфоноксиалканоат" обозначает фермент, который может превращать 3-гидроксиизовалерат и АТФ в 3-фосфоноксиизовалерат и АДФ. Еще более предпочтительно, такой фермент может катализировать реакцию превращения 3-гидроксиалканоата в соответствующий 3-фосфоноксиалканоат, предпочтительно реакцию превращения 3-гидроксиизовалерата и АТФ в 3-фосфоноксиизовалерат и АДФ, с КМ 10 мМ или ниже, например, с КМ 5 мМ или ниже, предпочтительно 1 мМ или ниже и более предпочтительно 0,1 мМ или ниже. В особенно предпочтительном варианте осуществления, такой фермент может катализировать реакцию превращения 3-гидроксиалканоата в соответствующий 3-фосфоноксиалканоат, предпочтительно реакцию превращения 3-гидроксиизовалерата и АТФ в 3-фосфоноксиизовалерат и АДФ, с kcat по меньшей мере 0,2 с-1, предпочтительно kcat по меньшей мере 0,5 с-1, особенно предпочтительно kcat по меньшей мере 1,0 с-1, более предпочтительно по меньшей мере 2,0 с-1 и еще более предпочтительно kcat по меньшей мере 5,0 с-1.
В особенно предпочтительном варианте осуществления способность превращать 3-гидроксиизовалерат и АТФ в 3-фосфоноксиизовалерат и АДФ измеряют в исследовании, как описано в примере 5.
Термин "фермент, который обладает активностью превращать указанный 3-фосфоноксиалканоат в указанный алкен" обозначает фермент, который может катализировать реакцию, посредством которой происходит декарбоксилирование и дефосфорилирование 3-фосфоноксиалканоата, что впоследствии приводит к получению соответствующего алкена.
Эту активность можно измерить, например, как описано в приложенных примерах, в особенности в примере 8. Одной возможностью является инкубирование соответствующего фосфоноксиалканоата в условиях, которые в принципе предоставляют возможность декарбоксилирования и дефосфорилирования и обнаружение продукции соответствующего алкена, например, путем газовой хроматографии. В предпочтительном варианте осуществления термин "фермент, который обладает активностью превращать указанный 3-фосфоноксиалканоат в указанный алкен" обозначает фермент, который может превращать 3-фосфоноксиизовалерат в изобутен, предпочтительно в условиях, описанных в примере 8. Более предпочтительно такой фермент может катализировать реакцию превращения 3-фосфоноксиалканоата в соответствующий алкен (посредством декарбоксилирования и дефосфорилирования) с КМ 100 мМ или ниже, например, с КМ 75 мМ или ниже, или с КМ 50 мМ или ниже, предпочтительно 10 мМ или ниже или 5 мМ или ниже или 1 мМ или ниже, и более предпочтительно 0,1 мМ или ниже. В особенно предпочтительном варианте осуществления, такой фермент может катализировать реакцию превращения 3-фосфоноксиалканоата в соответствующий алкен, предпочтительно реакцию превращения 3-фосфоноксиизовалерата в изобутен, kcat по меньшей мере 10-6 с-1, предпочтительно kcat по меньшей мере 10-4 с-1, например, kcat по меньшей мере 10-3 с-1 или kcat по меньшей мере 10-2 с-1, как, например, kcat по меньшей мере 10-1 с-1, например kcat по меньшей мере 0,2 с-1, предпочтительно kcat по меньшей мере 0,5 с-1, особенно предпочтительно kcat по меньшей мере 1,0 с-1, более предпочтительно по меньшей мере 2,0 с-1 и еще более предпочтительно kcat по меньшей мере 5,0 с-1.
В особенно предпочтительном варианте осуществления способность превращать 3-фосфоноксиизовалерат в изобутен измеряют в исследовании, как описано в примере 8.
В одном предпочтительном варианте осуществления фермент, указанный в (i) и (ii), выше, представляет собой фермент, который можно найти в NCBI или эквивалентном источнике как имеющий COG3407 домен.
В предпочтительном варианте осуществления способа в соответствии с изобретением первый фермент (i), который обладает активностью превращать 3-гидроксиалканоат в соответствующий 3-фосфоноксиалканоат, выбирают из группы, включающей
(A) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO:1, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 15% идентична аминокислотной последовательности, представленной в SEQ ID NO:1, и проявляет активность превращать 3-гидроксиалканоат в соответствующий 3-фосфоноксиалканоат, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO:1;
(B) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO:2, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 15% идентична аминокислотной последовательности, представленной в SEQ ID NO:2, и проявляет активность превращать 3-гидроксиалканоат в соответствующий 3-фосфоноксиалканоат, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO:2;
(C) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO:3, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 15% идентична аминокислотной последовательности, представленной в SEQ ID NO:3, и проявляет активность превращать 3-гидроксиалканоат в соответствующий 3-фосфоноксиалканоат, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO:3; и
(D) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO:4, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 15% идентична аминокислотной последовательности, представленной в SEQ ID NO:4, и проявляет активность превращать 3-гидроксиалканоат в соответствующий 3-фосфоноксиалканоат, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO:4.
SEQ ID NO:1, представлена аминокислотная последовательность фермента из Picrophilus torridus DSM 9790 (GenBank номер доступа ААТ43941; Swissprot/TrEMBL номер доступа Q6KZB1).
SEQ ID NO:2, представлена аминокислотная последовательность фермента из Thermoplasma acidophilum (GenBank номер доступа САС12426; Swissprot/TrEMBL номер доступа Q9HIN1).
SEQ ID NO:3, представлена аминокислотная последовательность фермента из Thermoplasma volcanium (GenBank номер доступа ВАВ59465; Swissprot/TrEMBL номер доступа Q97BY2).
SEQ ID NO:4, представлена аминокислотная последовательность фермента из Ferroplasma acidarmanus fer1 (GenBank номер доступа ZP_05571615).
В дальнейшем предпочтительном варианте осуществления способа в соответствии с изобретением, второй фермент (ii), который обладает активностью превращать указанный 3-фосфоноксиалканоат в указанный алкен, выбирают из группы, включающей
(a) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO:5, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 15% идентична аминокислотной последовательности, представленной в SEQ ID NO:5, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO:5;
(b) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO:6, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 15% идентична аминокислотной последовательности, представленной в SEQ ID NO:6, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO:6;
(c) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO:7, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 15% идентична аминокислотной последовательности, представленной в SEQ ID NO:7, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO:7;
(d) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO:8, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 15% идентична аминокислотной последовательности, представленной в SEQ ID NO:8, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO:8;
(e) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO:9, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 15% идентична аминокислотной последовательности, представленной в SEQ ID NO:9, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO:9;
(f) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO:10, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 15% идентична аминокислотной последовательности, представленной в SEQ ID NO:10, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO:10;
(g) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO:11, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 15% идентична аминокислотной последовательности, представленной в SEQ ID NO:11, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO:11;
(h) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO:12, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 15% идентична аминокислотной последовательности, представленной в SEQ ID NO:12, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO:12;
(i) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO:13, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 15% идентична аминокислотной последовательности, представленной в SEQ ID NO:13, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO:13;
(j) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO:14, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 15% идентична аминокислотной последовательности, представленной в SEQ ID NO:14, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO:14; и
(k) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO:15, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 15% идентична аминокислотной последовательности, представленной в SEQ ID NO:15, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO:15.
SEQ ID NO:5, представлена аминокислотная последовательность фермента, клонированного из Streptococcus gordonii. SEQ ID NO:6, представлена аминокислотная последовательность фермента из Streptococcus gordonii str. Challis substr. CH1 (GenBank номер доступа ААТ43941; Swissprot/TrEMBL номер доступа A8UU9). SEQ ID NO:7, представлена аминокислотная последовательность фермента из Streptococcus infantarius subsp infarrtarius ATCC BAA-102 (GenBank номер доступа EDT48420.1; Swissprot/TrEMBL номер доступа B1SCGO). SEQ ID NO:8, представлена аминокислотная последовательность фермента из Homo sapiens (GenBank номер доступа ААС50440.1; Swissprot/TrEMBL номер доступа Р53602.1). SEQ ID NO:9, представлена аминокислотная последовательность фермента из Lactobacilius delbrueckii (GenBank номер доступа САI97800.1; Swissprot/TrEMBL номер доступа Q1GAB2). SEQ ID NO:10, представлена аминокислотная последовательность фермента из Streptococcus mitis (штамм В6) (GenBank номер доступа CBJ22986.1). SEQ ID NO:11, представлена аминокислотная последовательность фермента из Streptococcus gallolyticus UCN34 (GenBank номер доступа СВ113757.1). SEQ ID NO:12, представлена аминокислотная последовательность фермента из Streptococcus sanguinis SK36 (GenBank номер доступа ABN43791.1). SEQ ID NO:13, представлена аминокислотная последовательность фермента из Streptococcus sp.M143 (GenBank номер доступа EFA24040.1). SEQ ID NO:14, представлена аминокислотная последовательность фермента из Streptococcus suis 89/1591 (GenBank номер доступа EEF63672.1). SEQ ID NO:15, представлена аминокислотная последовательность фермента из Streptococcus salivarius SK126 (GenBank номер доступа ЕЕК09252).
В предпочтительном варианте осуществления способа в соответствии с изобретением первый фермент (i) имеет значение, как определено в (А) выше, и второй фермент (ii) имеет значение, как определено в (а) или (b), указанных выше, более предпочтительно второй фермент имеет значение, как определено в (f), (g), (h), (i), (j) или (k), указанных выше. Как проиллюстрировано в примерах, комбинация этих ферментов особенно эффективна для получения алкеновык соединений в соответствии с настоящим изобретением.
В другом предпочтительном варианте осуществления способа в соответствии с изобретением второй фермент (ii), который обладает активностью превращать указанный 3-фосфоноксиалканоат в указанный алкен, выбирают из любого из белков, перечисленных в последующей таблице, или из белка, содержащего аминокислотную последовательность, которая по меньшей мере на 15% идентична аминокислотной последовательности такого белка и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность указанного белка.
Как указано выше, можно использовать не только белки, имеющие специфически указанные аминокислотные последовательности, представленные в соответствующих SEQ ID NO или в таблице 1, но также и белки, которые можно найти в NCBI или эквивалентом источнике в качестве имеющего COG3407 домен и, более предпочтительно, белки, аминокислотная последовательность которых имеет гомологию по меньшей мере 15% со специфической указанной аминокислотной последовательностью и которые имеют соответствующую ферментативную активность по меньшей мере, такую же высокую, как и активность белка, имеющего специфически указанную аминокислотную последовательность. Предпочтительные ферменты благоприятно имеют по меньшей мере х% гомологию, где х, выбирают из группы, включающей 20, 25, 20, 35, 40, 45, 50, 55 и 60. В дальнейшем предпочтительном варианте осуществления фермент имеет по меньшей мере 65% последовательности, гомологичную, предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 75%, еще более предпочтительно, по меньшей мере 80, 85, 90, 95, 96, 97, 98 или 99% гомологии к одной из последовательностей, представленных в SEQ ID NO:1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 или к одной из последовательностей, представленных в таблице 1. Процент гомологии последовательностей можно определить с помощью различных методов и с помощью программного обеспечения, известного специалисту в данной области техники, такого, как, например, CLUSTAL метод или BLAST и производных программ, или с помощью алгоритма сравнения последовательности, такого как, например, описанный Needleman и Wunsch (J. Mol. Biol., 1970, 48:443) или Smith и Waterman (J. Mol. Biol., 1981, 147:195).
Такие белки, проявляющие указанную степень гомологии, могут представлять собой, например, другие ферменты, которые встречаются в природе или которые были получены синтетически. Они включают, в особенности, ферменты, которые могут выбраны по их способности продуцировать алкены в соответствии с изобретением. Таким образом, тест для отбора включает контактирование очищенного фермента, или микроорганизма, продуцирующего фермент, с субстратом реакции и измерение продукции соответствующего соединения, то есть 3-фосфоноксиалканоата или алкена. Такие тесты описаны в экспериментальном разделе. Такие тесты для отбора также можно использовать для скрининга ферментов с оптимизированной ферментативной активностью к субстрату, которые превращается в 3-фосфоноксиалканоат или алкен, то есть имеющих оптимизированную активность по отношению к одному или нескольким 3-гидроксиалканоатам или 3-фосфоноксиалканоатам.
Такие методы скрининга хорошо известны в данной области техники и включают, например, технологии белковой инженерии, такие как, например, неспецифический мутагенез, массивный мутагенез, сайт-направленный мутагенез, перестановка в ДНК, синтетическое «перемешивание», in vivo эволюция, или полный синтез генов и последующий скрининг относительно желательной ферментативной активности.
Таким образом, фермент, используемый в изобретении, может быть природным или синтетическим, и продуцироваться с помощью химических, биологических или генетических средств. Он также может быть химически модифицирован, например, для улучшения его активности, резистентности, специфичности, очистки, или для его иммобилизации на подложке.
Было обнаружено, что ферменты, которые способны катализировать вышеописанные реакции для превращения 3-гидроксиалканоата в алкен посредством 3-фосфо-гидроксиалканоата, часто представляют собой ферменты, которые могут быть классифицированы в филогенетическом суперсемействе мевалонат дифосфат (МОР) декарбоксилазы (номенклатура ферментов ЕС 4.1.1.33). MDP декарбоксилаза представляет собой фермент, задействованный в биосинтез холестерина. Указанный фермент быть выделен из различных организмов, включая животных, грибов, дрожжей и некоторых бактерий. Также он может экспрессироваться некоторыми растениями (Lalitha и др., Phytochemistry 24 (11), (1985), 2569-2571). Были клонированы и секвенированы многие гены, кодирующие этот фермент. Эти ферменты, как правило, состоят из 300-400 аминокислот и используют АТФ в качестве субстратного кофактора, который превращается в процессе реакции в АДФ и неорганический фосфат.Фосфатная группа переносится с молекулы АТФ на третичный спирт мевалонат дифосфата, высвобождая АДФ. В промежуточной реакции происходит фосфорилирование на 3-гидроксильной группе, после этого происходит элиминирование фосфатной группы, в физиологическом случае с высвобождением изопентенил дифосфата (Фигура 2).
Таким образом, в предпочтительном варианте осуществления, фермент, определенный в (i) или (ii) выше, представляет собой MDP декарбоксилазу. В контексте настоящего изобретения MDP декарбоксилаза определяется как фермент, который может по меньшей мере катализировать превращение 5-дифосфо-3-фосфомевалоната в изопентенил-5-дифосфат и CO2 или который может по меньшей мере катализировать реакцию превращения мевалонат дифосфата и АТФ в 5-дифосфо-3-фосфомевалонат и АДФ. Предпочтительно, такой фермент может катализировать обе реакции.
В другом предпочтительном варианте осуществления фермент, определенный в (i) выше, представляет собой фермент, как определено в (i) (В). Последовательность, указанная в SEQ ID NO:2, представляет собой фермент, идентифицированный в Thermoplasma acidophilum. В Genbank этот фермент классифицирован как мевалонат дифосфат декарбоксилаза. Тем не менее, он известен из Chen и Poulter (Biochemistry 49 (2010), 207-217), который в Th. acidophilum существует в альтернативном мевалонатном пути, который вовлекает действие мевалонат-5-монофосфат декарбоксилазы. Таким образом, представляется возможным, что фермент, охарактеризованный в SEQ ID NO:2, действительно представляет собой мевалонат-5-монофосфат декарбоксилазу.
Аналогичное может быть верным и в отношении другие архебактерий. Следовательно, в другом предпочтительном варианте осуществления фермент, определенный в (i) или (ii) выше, представляет собой мевалонат-5-монофосфат декарбоксилазу. Такой фермент способен превращать мевалонат-5-монофосфат в изопентенилпирофосфат.
В предпочтительных вариантах осуществления изобретения:
- 3-гидроксипропионат превращают посредством 3-фосфоноксипропионата в этилен; или
- 3-гидроксибутират превращают посредством 3-фосфоноксибутирата в пропилен; или
- 3-гидроксивалерат превращают посредством 3-фосфоноксивалерата в 1-бутилен; или
- 3-гидрокси-3-метилбутират (или 3-гидроксиизовалерат) превращают посредством 3-фосфонокси-3-метилбутирата (3-фосфоноксиизовалерат) в изобутилен; или
- 3-гидрокси-3-метилвалерат превращают посредством 3-фосфонокси-3-метилвалерата в изоамилен.
Способ в соответствии с изобретением может быть осуществлен in vitro, в присутствии выделенных ферментов (или ферментных систем, дополнительно содержащих один или несколько кофакторов). In vitro предпочтительно обозначает в бесклеточной системе.
В одном варианте осуществления, ферменты, задействованные в способ, используют в очищенной форме, для превращения 3-гидроксиалканоатов в алкены. Тем не менее, такой способ может быть дорогостоящим, поскольку стоимость получения и очистки фермента и субстрата являются высокими.
Таким образом, в другом предпочтительном варианте осуществления, ферменты, вовлеченные в способ, присутствуют в реакции в виде неочищенного экстракта, или также в форме нелиофилизированной бактерии, для того, чтобы сэкономить стоимость очистки белка. Тем не менее, затраты, связанные с таким способом, все еще могут быть достаточно высокими вследствие стоимости получения и очистки субстратов.
Таким образом, в одном предпочтительном варианте осуществления, ферменты, нативные или рекомбинантные, очищенные или нет, используют для превращения 3-гидроксиалканоата в алкен. Для осуществления этого, ферменты инкубируют в присутствии субстрата в физико-химических условиях, предоставляющих возможность ферментам быть активными, и предоставляющих возможность осуществляться инкубированию в течение достаточного периода времени. После окончания инкубирования, необязательно измеряют присутствие алкена с помощью любой системы для обнаружения, известной специалисту в даммой области техники, такой как, например, газовая хроматография или колориметрические тесты для измерения образования алкенового продукта, или свободного фосфата, или также для измерения исчезновения 3-гаодроксиалканоатного субстрата или АТФ.
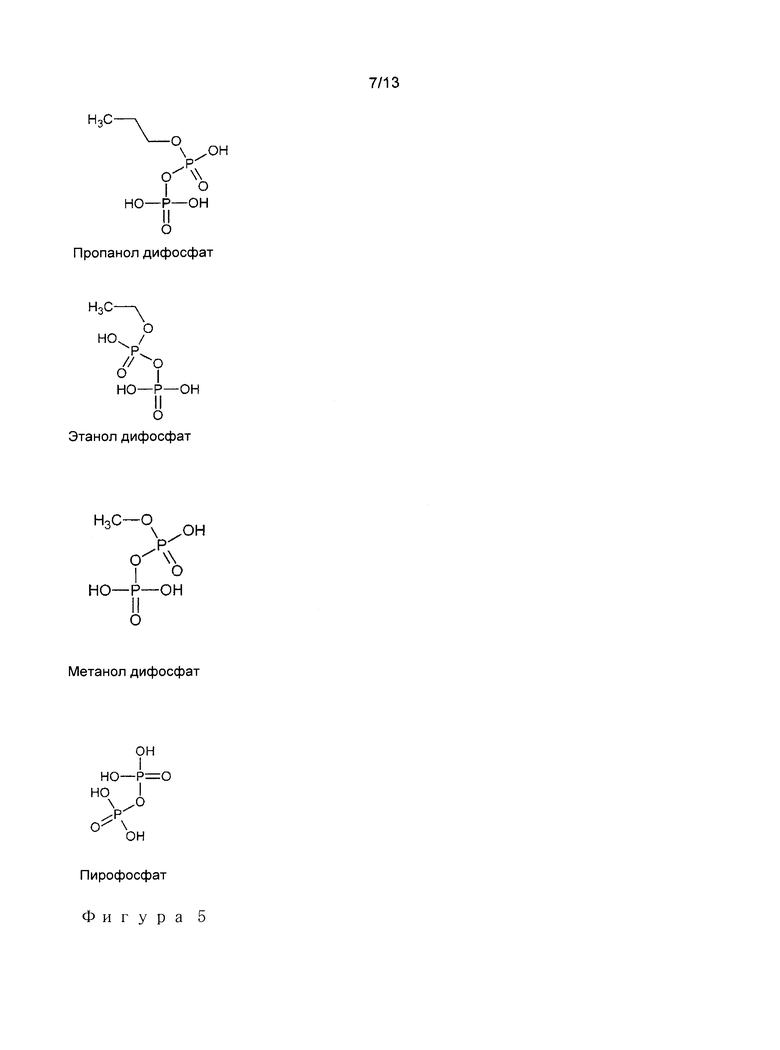
В предпочтительном варианте осуществления, добавляют кофакторы для того, чтобы наилучшим образом имитировать естественную реакцию или для того, чтобы обеспечить стерическую или электронную комлементацию в каталитической щели. Например, если один из ферментов, используемых в способе в соответствии с изобретением, представляет собой фермент, который в естественных условиях использует мевалонат дисфосфат (MDP) в качестве субстрата, то структура 3-гидроксиалканоатов оставляет большое пространство пустым в каталитической щели при осуществлении связывания фермент-субстрат, поскольку в целом 3-гидроксиалканоат соответствует фрагменту MDP. Заполнение это пространство с помощью кофактора для замены пропущенной части субстрата направлено на наиболее точное имитирование молекулы MDP. Поскольку кофактор не модифицируется при осуществлении реакции, то, следовательно, его необходимо добавлять в каталитических количествах. В том случае, если субстрат реакции представляет собой 3-гидроксипропионат, то комплементарный кофактор будет представлять собой пропил дифосфат. В том случае, если субстрат представляет собой 3-гидроксибутират или 3-гидрокси-3-метилбутират, то комплементарный кофактор будет представлять собой этил дифосфат. В том случае, если субстрат представляет собой 3-гидроксивалерат или 3-гидрокси-3-метилвалерат, то комплементарный кофактор будет представлять собой метил дифосфат. Эти различные молекулы представлены на Фигуре 5. Случайно, может случиться, что комплементарный кофактор реакции обладает положительным вливание на реакцию другого субстрата. В целом, кофактор может представлять собой любую молекулу, содержащую фосфоангидрид, и поэтому имеющий общую глобальную формулу R-PO2H-O-РО3Н2, в которой R представляет собой в особенности Н, линейную, разветвленную или циклическую алкильную группу, предпочтительно имеющую от 1 до 10 или от 1 до 5 атомов углерода, или любую другую моновалентную органическую группу. Аналогичные мотивы, соответствующие метилен дифосфонат моноэфирам, имеющим общую формулу R-O-PO2H-СН2-РО3Н2, в который фосфиангидрид заменен метиленовым мостиком, имеющим преимущество не подвергаться гидролизу, также являются частью изобретения. В целом, кофакторы могут представлять собой монофосфат, или даже безфосфатные, аналоги предшествующих молекул, или также любую другую молекулу, которая может улучшить выход реакции путем обеспечения стерической или электронной комплементации в каталическом сайте фермента. Кофактор благоприятно выбирают из группы, включающей пирофосфатный ион, метил дифосфат, этил дифосфат, или пропил дифосфат.
В предпочтительном варианте осуществления, превращение происходит в присутствии ко-субстрата, указанный ко-субстрат предпочтительно является соединением, содержащим фосфоангидрид, и предпочтительно является АТФ, rNTP, dNTP или смесью нескольких этих молекул, полифосфат, или пирофосфат. Ко-субстрат, как правило, присутствует в хозяине. Тем не менее, в другом особенно предпочтительном варианте осуществления, ко-субстрат можно добавлять в реакцию, предпочтительно, выбирают из группы, включающей АТФ, rNTP, dNTP, смесь нескольких rNTP или dNTP, полифосфат, и предпочтительно пирофосфат, или соединение, содержащее фосфоангидрид (представленное общей формулой Х-РО3Н2 на Фигуре 2).
Несмотря на то, что для стадии декарбоксилирования, то есть реакцию, определенная как (ii) в настоящей заявке выше, не требуется потребление АТФ, может быть показано, что присутствие АТФ в реакции может быть благоприятным. Это продемонстрировано в примере 7, используя 3-фосфоноксиизовалерат в качестве субстрата. Полагают, что АТФ может оказывать влияние на укладку белка путем связывания АТФ с АТФ-связывающим сайтом дифосфомевалонат декарбоксилазы. В действительности, это можно наблюдать наглядно: очищенный фермент имеет склонность к осаждению, и добавление АТФ предотвращает этот эффект. Полагают, что не только АТФ, но также и другие сходные соединения, такие как dATP, АДФ, AMP или другие NTP или dNTP имеют этот эффект. Таким образом, в предпочтительном варианте осуществления, способ в соответствии с настоящим изобретением осуществляют с АТФ, dATP, АДФ, AMP или NTP, отличающимся от АТФ или dNTP в качестве субстратного кофактора.
В другом предпочтительном варианте осуществления, способ в соответствии с изобретением осуществляют в культуре, в присутствии организма, предпочтительно микроорганизма, продуцирующего ферменты. Следовательно, в таком варианте осуществления изобретения, используют организм, предпочтительно микроорганизм, который продуцирует ферменты, указанные в (i) и (ii) выше. В предпочтительном варианте осуществления, (микро)организм является рекомбинантным, в том отношении, что ферменты, указанные в (i) и (ii), продуцируемые хозяином, являются гетерологичными по отношению к продуцирующему хозяину. Таким образом, способ можно осуществлять непосредственно в культуральной среде, без необходимости разделения или очистки ферментов. В особенно благоприятном методе, используют (микро)организм, имеющий естественное или искусственное свойство эндогенно продуцировать один или несколько 3-гидроксиалканоатов, и также экспрессировать или свырхэкспрессировать ферменты, указанные в (i) и (ii) выше, природные или модифицированные, для того, чтобы продуцировать алкены непосредственно из источника углерода, присутствующего в растворе.
Например, способ в соответствии с изобретением можно осуществлять путем применения микроорганизмов, которые продуцируют один или несколько 3-гидроксиалканоатов [например, Alcaligenes eutrophus или Bacillus megaterium, или также штамм Е.Coli, генетически модифицированный таким образом, чтобы продуцировать указанный(е) продукт(ы)] и которые были генетически сконструированы таким образом, чтобы они сверхэкспрессировали ферменты, как определено в (i) и (ii) выше, указанные ферменты предпочтительно имеют происхождение из организма, отличающегося от микроорганизма-хозяина. Генетическая модификация может состоять, например, в интеграции соответствующих генов, кодирующих ферменты, в хромосому, экспрессирование ферментов из плазмиды, содержащей промотор, расположенный против хода транскрипции относительно последовательности, кодирующей фермент, промотор и кодирующая последовательность предпочтительно имеют происхождения из разных организмов, или любой другой метод, известный специалисту в данной области техники. Альтернативно, другие бактерии или дрожжи могут иметь специфические преимущества и могут быть выбраны. Например, можно использовать дрожжи, такие, как, например, Saccharomyces cerevisiae, экстремофильную бактерию, такую, как, например, Thermus thermophilus, или анаэробную бактерию из семейства Clostridiae, микроводоросли, или фотосинтезирующую бактерию.
Организм, используемый в изобретении, может представлять собой прокариотический или эукариотический, предпочтительно, они представляют собой микроорганизмы, такие, как, например, бактерии, дрожжи, грибы или плесневые грибы, или клетки растений или клетки животных. В предпочтительном варианте осуществления, микроорганизмы представляют собой бактерии, предпочтительно родов Escherichia, Alcaligenes или Bacillus и более предпочтительно видов Escherichia coli, Alcaligenes eutrophus или Bacillus megaterium.
В другом предпочтительном варианте осуществления, микроорганизмы представляют собой рекомбинантные бактерии родов Escherichia, предпочтительно видов Escherichia coli, которые были модифицированы таким образом, чтобы эндогенно продуцировать один или несколько 3-гидроксиалканоатов, и превращать их в алкены.
В дальнейшем предпочтительном варианте осуществления микроорганизм представляет собой гриб, более предпочтительно гриб родов Saccharomyces, Schizosaccharomyces, Aspergillus или Trichoderma и более предпочтительно видов Saccharomyces cerevisiae, Schizosaccharomyces pombe, Aspergillus niger или видов Trichoderma reesei. В особенно предпочтительном варианте осуществления микроорганизм представляет собой рекомбинантные дрожжи, продуцирующие 3-гидроксиалканоаты и превращающие их в алкены благодаря экспрессии ферментов, указанных в (i) и (ii) выше.
В другом предпочтительном варианте осуществления, в способе в соответствии с изобретением используют фотосинтетический микроорганизм, экспрессирующий ферменты, как указано в (i) и (ii) выше. Предпочтительно, микроорганизм представляет собой фотосинтетическую бактерию, или микроводоросль. Более предпочтительно такой микроорганизм имеет естественное или искусственное свойство эндогенно продуцировать один или несколько 3-гидроксиалканоатов. В этом случае, микроорганизм будет способен продуцировать непосредственно из CO2, присутствующего в растворе.
Также представляется возможным использовать в способе в соответствии с изобретением один микроорганизм, который продуцирует фермент, как определено в (i) выше, и другой микроорганизм, который продуцирует фермент, как определено в (ii) выше. Кроме того, в дальнейшем варианте осуществления по меньшей мере один из микроорганизмов способе продуцировать один или несколько 3-гидроксиалканоатов или, в альтернативном варианте осуществления, другой микроорганизм используют в способе, который способен продуцировать один или несколько 3-гидроксиалканоатов.
В другом предпочтительном варианте осуществления в способе в соответствии с изобретением используют многоклеточный организм, экспрессирующий ферменты, как определено в (i) и (ii) выше. Примерами таких организмов являются растения или животные.
В предпочтительном варианте осуществления, способ вовлекает культивируемые микроорганизмы в стандартных культуральных условиях (30-37°С при 1 атм, в ферментере, предоставляющем возможность аэробного роста бактерий) или нестандартных условиях (более высокая температура, например, для соответствия культуральных условий термофильным организмам).
В дальнейшем предпочтительном варианте осуществления способ согласно изобретению осуществляют в микроаэрофильных условиях. Это обозначает, что количество инъецируемого воздуха ограничено для того, чтобы минимизировать остаточную концентрацию кислорода в выходящих газовых потоках, содержащих алкеновые углеводороды.
В другом предпочтительном варианте осуществления способ в соответствии с изобретением также включает стадию сбора газообразных алкенов, дегазирования реакции, то есть восстановление продуктов, которые дегазируют, например, из культуры. Таким образом, в предпочтительном варианте осуществления, способ осуществляют в присутствии системы для сбора алкена в газообразной форме при осуществлении реакции. По существу, короткие алкены, и в особенности этилен, пропилен и бутен изомеры, принимают газообразное состояние при комнатной температуре и атмосферном давлении. Следовательно, для способа в соответствии с изобретением не требуется экстрагирование продукта из жидкой культуральной среды, стадия, которая всегда очень дорогая при осуществлении в промышленном масштабе. Откачка и хранение газообразных углеводородов и их возможное последующее физическое разделение и химическое превращение можно осуществлять в соответствии с любым методом, известным специалисту в данной области техники. В предпочтительном варианте осуществления, способ также включает обнаружение алкена (например, пропилена, этилена или изобутилена), который присутствует в газообразной фазе. Присутствие соединения, которое продуцируется в окружающий воздух или другой газ, даже в небольших количествах, можно обнаружить с помощью различных технологий и в особенности путем применения систем газовой хроматографии с инфракрасным обнаружением или обнаружением путем плазменной ионизации, или путем спаривания с масс-спектрометрией
В предпочтительном варианте осуществления, алкены, продуцируемые с помощью способа в соответствии с изобретением, конденсируют, затем необязательно восстанавливают, путем применения техник, известных специалисту в данной области техники, для того, чтобы продуцировать длинноцепочечные алкены, или длинноцепочечные алканы. Например, изобутилен можно использовать для синтеза изооктана: каталитические методы для успешного осуществления этой реакции уже были полностью описаны.
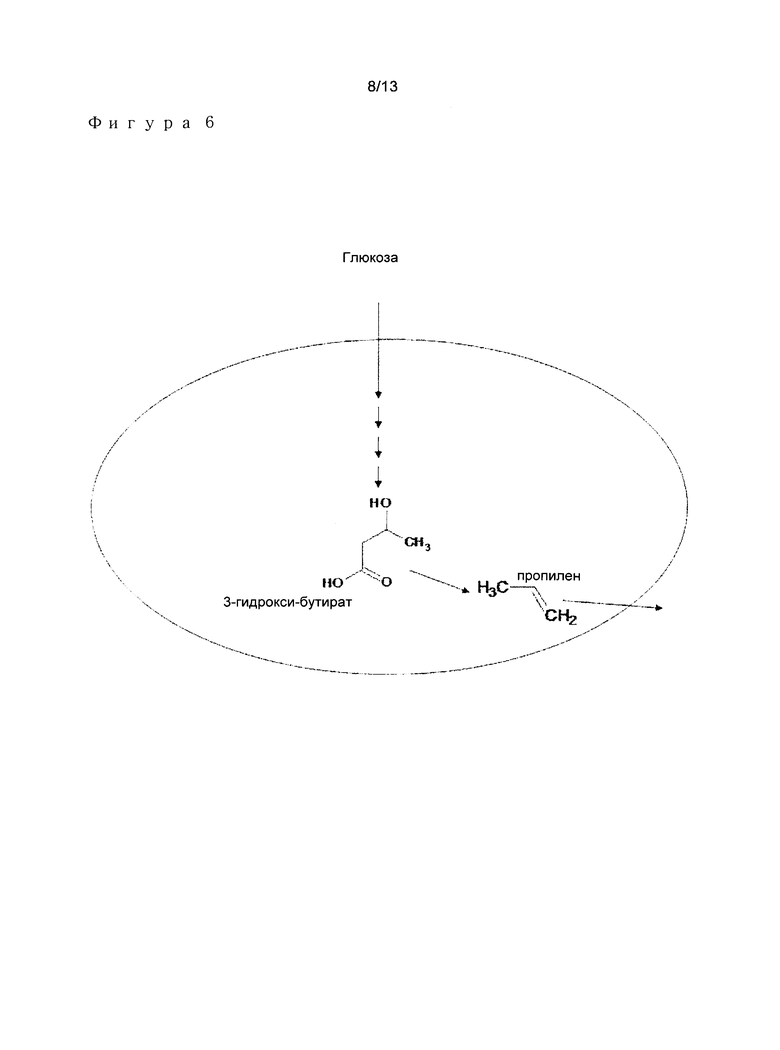
В другом варианте осуществления, способ в соответствии с изобретением характеризуется путем превращения источника углерода, такого как, например, глюкоза, в 3-гидроксиалканоат, с последующим превращением указанного 3-гидроксиалканоата в соответствующий алкен. Различные стадии указанного способа представлены на Фигуре 6.
В предпочтительном варианте осуществления, способ характеризуется путем превращения полигидроксиалканоатов в 3-гидроксиалканоат с использованием фермента или подходящего физико-химического метода, с последующим превращением указанного 3-гидроксиалканоата в указанный алкен. Необязательно, полигидроксиалканоат продуцируют с помощью микроорганизма или растения, чьи метаболические пути были модифицированы для того, чтобы продуцировать высокие выходы полигидроксиалканоата.
В другом варианте осуществления, способ в соответствии с изобретением включает продукцию алкенов из атмосферного СО2 или из СО2, искусственно добавленного в культуральную среду. В этом случае, способ осуществляют в организме, который способен осуществлять фотосинтез, таком, например, микроводоросли.
Настоящее изобретение также относится к способу получения алкена, который включает стадию ферментативного превращения 3-фосфоноксиалканоата в соответствующий алкен путем применения фермента, который может катализировать превращение посредством декарбоксилирования и дефосфорилирования.
Как указано по отношению к предпочтительному ферменту, используемому в таком способе, аналогичное применятся, как было указано выше в связи с (ii) способа в соответствии с изобретением, как описано в данной заявке выше. Кроме того, также по отношению к другим предпочтительным вариантам осуществления, описанным выше для способа в соответствии с изобретением, аналогичное применяется к способу получения алкена из 3-фосфоноксиалканоата.
Настоящее изобретение также относится к организмам, предпочтительно микроорганизмам, которые продуцируют по меньшей мере два фермента, где один фермент выбирают из (i), как указано выше, и другой фермент выбирают из (ii), как указано выше. В предпочтительном варианте осуществления, такой организм представляет собой рекомбинантный организм в том смысле, что он генетически модифицирован благодаря интродукции по меньшей мере одной молекулы нуклеиновой кислоты, кодирующей по меньшей мере один из вышеуказанных ферментов. Предпочтительно, такая молекула нуклеиновой кислоты является гетерологичной по отношению к организму, что обозначает, что она в естественных условиях не встречается в указанном хозяине. Таким, настоящее изобретение также относится к организму, предпочтительно микроорганизму, содержащему молекулу нуклеиновой кислоты, кодирующей фермент, как определено в (i) выше, и содержащему молекулу нуклеиновой кислоты, кодирующей фермент, как определено в (ii) выше. В предпочтительном варианте осуществления, по меньшей мере одна из молекул нуклеиновой кислоты является гетерологичной по отношению к организму, что обозначает, что она не встречается в естественных условиях в указанном организме. Микроорганизм предпочтительно представляет собой бактерию, дрожжи или грибы. В другом предпочтительном варианте осуществления организм представляет собой растение или животное, отличающееся от человека. Что касается других предпочтительных вариантов осуществления, аналогичное применяется, как и было указано выше в связи со способом в соответствии с изобретением.
Кроме того, настоящее изобретение также относится к композиции, содержащей микроорганизм в соответствии с настоящим изобретением, подходящую культуральную среду и 3-гидроксиалканоатное соединение или источник углерода, которое может быть превращено микроорганизмом в 3-гидроксиалканоатное соединение.
Настоящее изобретение также относится к применению комбинации по меньшей мере двух ферментов, где один фермент выбирают из следующих (i) и другой фермент выбирают из следующих (ii) или организма, предпочтительно микроорганизма, в соответствии с изобретением или композиции в соответствии с изобретением, для получения алкеновых соединений из 3-гидроксиалканоатов, где (i) и (ii) являются следующими:
(i) первый фермент, который обладает активностью превращать 3-гидроксиалканоат в соответствующий 3-фосфоноксиалканоат; и
(ii) второй фермент, который отличается от первого фермента и, который обладает активностью превращать указанный 3-фосфоноксиалканоат в указанный алкен.
По отношению к предпочтительным вариантам осуществления указанных различных соединений, аналогичное применяется, как и было указано выше в связи со способом в соответствии с изобретением.
Другие аспекты и преимущества изобретения будут описаны в последующих примерах, которые представлены с целью иллюстрации и не для ограничения.
Анализы:
1. Без фермента, 0 мМ АТФ
2. 2 мг/мл фермента, 0 мМ АТФ
3. Без фермента, 10 мМ АТФ
4. 2 мг/мл фермента, 10 мМ АТФ
Последующие примеры служат для иллюстрации изобретения.
Примеры
Пример 1: Клонирование, экспрессия и очистка библиотеки MDP декарбоксилазы.
Библиотеку 55 генов, кодирующих представителей семейства дифосфомевалонат декарбоксилазы (MDP декарбоксилазы) из эукариотических, прокариотических и простейших организмов конструировали и тестировали для идентификации наиболее активных кандидатов для улучшения продукции изобутена (IBN).
Клонирование, бактериальные культуры и экспрессия белков.
Гены, кодирующие мевалонат дифосфат (MDP) декарбоксилазу ЕС 4.1.1.33. клонировали в рЕТ 25b векторе (Novagen) для эукариотических генов и в рЕТ 22b (Novagen)для прокариотических генов. Фрагмент секвенирования из 6 гистидиновых кодонов вставляли после индицирующего метионинового кодона для обеспечения аффинной метки для очистки. Компетентные клетки Е.coli BL21(DE3) (Novagen) трансформировали с помощью этих векторов в соответствии с процедурой теплового шока. Трансформированные клетки выращивали при встряхивании (160 об. мин.) на ZYM-5052 авто-индукционной среде (Studier FW, Prot. Exp. Pur. 41, (2005), 207-234) в течение 6 часов при 37°С и экспрессия белка продолжалась при 28°С в течение ночи (приблизительно 16 часов). Клетки собирали путем центрифугирования при 4°С, 10 тыс.об/мин, в течение 20 минут и осадок после центрифугирования замораживали при -80°С.
Очистка и концентрация белка.
Осадок после центрифугирования из 200 мл культуральных клеток размораживали на льду и ресуспендировали в 5 мл Na2HPO4 pH 8, содержащем 300 мМ NaCl, 5 мМ MgCl2 и 1 мМ DTT. Добавляли двадцать микролитров лизоназы (Novagen). Клетки инкубировали 10 минут при комнатной температуре и после этого возвращали на лед на 20 минут. Лизис клеток завершали путем обработки ультразвуком в течение 3×15 секунд. После этого бактериальные экстракты осветляли путем центрифугирования при 4°С, 10 тыс.об./мин. в течение 20 минут. Осветленные бактериальные лизаты загружали на колонку PROTINO-1000 Ni-TED (Macherey-Nagel), предоставляя возможность абсорбции 6-His меченных белков. Колонки промывали и представляющие интерес ферменты элюировали в 4 мл 50 мМ Na2HPO4 pH 8, содержащем 300 мМ NaCl, 5 мМ MgCl2, 1 мМ DTT, 250 мМ имидазола. После этого элюаты концентрировали и обессоливали на фильтровальной установке Amicon Ultra-4 10 кДа (Millipore) и ресуспендировали на 0,25 мл 50 мМ Tris-HCl pH 7,4, содержащем 0,5 мМ DTT и 5 мМ MgCl2. Концентрации белков определяли количественно с помощью метода Бредфорда. Чистота белков, очищенных таким способом, колебалась от 40% до 90%.
Пример 2: Скрининг библиотеки МОР декарбоксилазы.
MDP декарбоксилазы оценивали с использованием анализа комплементации. MDP декарбоксилазу Р. torridus инкубировали совместно с каждым тестируемым ферментом из библиотеки. Ферментативный анализ осуществляли в следующих условиях:
50 мМ Tris HCl pH 7,0
10 мМ MgCl2
20 мМ KCl
40 мМ АТФ
50 мМ 3-гидроксиизовалерат (HIV)
Значение pH доводили до 7,0
100 мкг МОР декарбоксилазы из Р. torridus и 1 мг тестируемой МОР декарбоксилазы добавляли к 1 мл реакционной смеси. Реакционную смесь, содержащую только 100 мкг Р. torridus MOP декарбоксилазы, использовали в качестве сравнения. После этого смесь инкубировали без встряхивания при 45°С в течение 90 часов в запечатанном сосуде (Interchim).
Один мл газообразной фазы собирали и инъецировали на НР5890 газовый хроматограф (HP), оборудованный FIO детектором и СР SilicaPlot колонкой (Varian). Коммерчески доступный изобутен использовали в качестве сравнения.
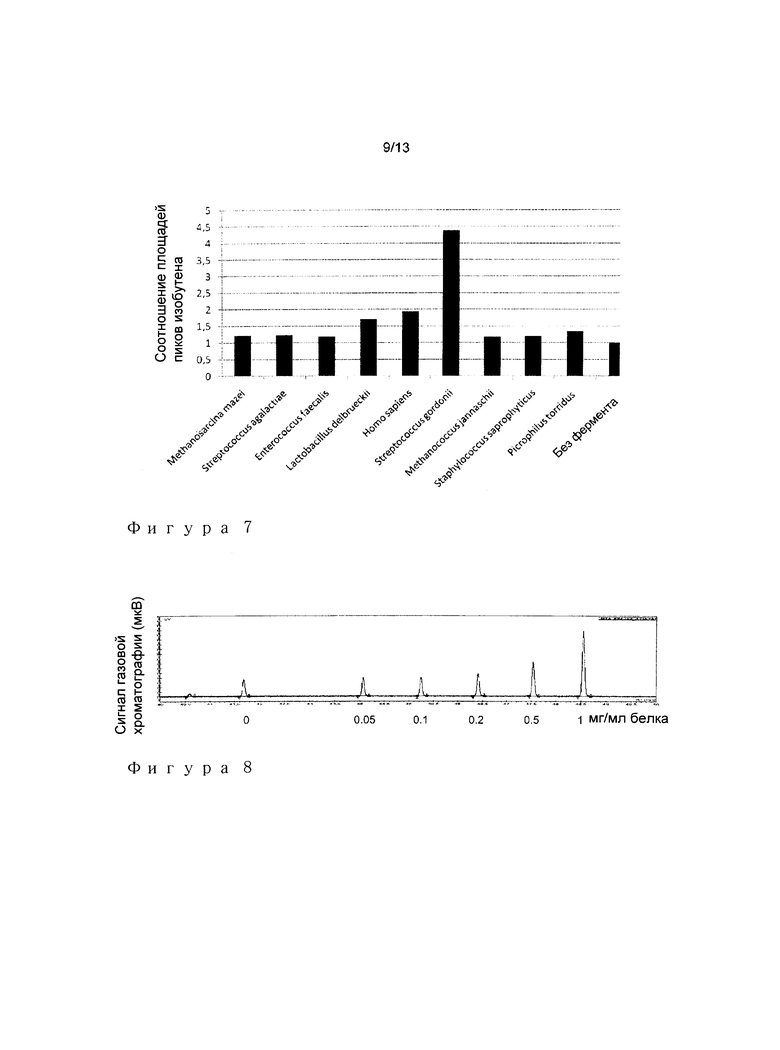
Эта процедура скрининга приводила к идентификации нескольких ферментов МОР декарбоксилазы, повышающих производительность изобутена. Как представлено на Фигуре 7, более высокую продукцию изобутена наблюдали для следующих МОР декарбоксилаз.
Кандидат 1:
Номер доступа Genbank: CAI97800
Номер доступа SwissProt/TrEMBL: Q1GAB2
Организм: Lactobacillus delbrueckii subsp. bulgaricus ATCC 11842
Кандидат 2:
Номер доступа Genbank: ААС50440.1
Номер доступа SwissProt/TrEMBL: P53602.1
Организм: Homo sapiens
Кандидат 3:
Номер доступа Genbank: ABV09606 Номер доступа SwissProt/TrEMBL: A8AUU9 Организм: Streptococcus gordonii str. Challis substr. CH1
Наиболее высокую продукцию изобутена наблюдали с очищенной МОР декарбоксилазой из Streptococcus gordonii.
Это указывало на то, что два фермента, представленных в анализе (один из Р. torridus и другой из S. gordonii) действовали взаимодополнительно на двух стадиях реакции, продуцируя IBN из HIV: перенос концевой фосфорильной группы из АТФ на С3-кислород 3-гидроксиизовалерата с последующим комбинированием дефосфорилированием-декарбоксилированием промежуточного 3-фосфоноксиизовалерата.
Пример 3: Влияние концентрации фермента на выход продукции изобутена.
Влияние концентрации MDP декарбоксилаза Streptococcus gordonii оценивали в следующих усвояли:
50 мМ Tris-HCl pH 7,0
10 мМ MgCl2
20 мМ KCl
40 мМ АТФ
50 мМ 3-гидроксиизовалерат (H1V)
Значение pH доводили до 7,0
100 мкг MDP декарбоксилазы из Р. torridus и различные количества (от 0 до 1 мг) очищенной MDP декарбоксилазы из Streptococcus gordonii добавляли к 1 мл реакционной смеси. После этого смесь инкубировали без встряхивания при 45°С в течение 90 часов в запечатанном сосуде (Interchim).
Один мл фазы равновесного пара собирали и инъецировали в НР5890 газовый хроматограф (HP), оборудованный FID детектором и СР SilicaPlot колонкой (Varian). Коммерчески доступный изобутен использовали в качестве сравнения.
Повышение концентрации фермента S. gordonii приводит к повышению количества продуцируемого изобутена (Фигура 8).
Пример 4: Скрининг библиотеки гомологов MDP декарбоксилазы Streptococcus gordonii.
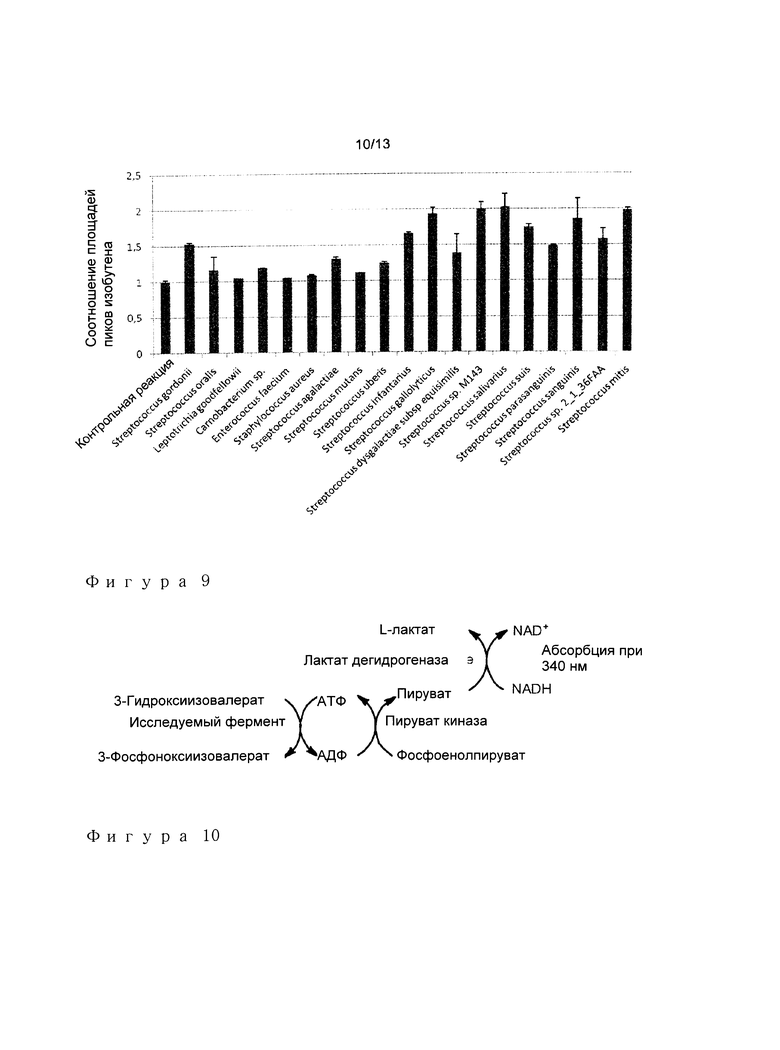
Используя он-лайн программу BLAST, представленную на NCBI, просматривали последовательности относительно базы данных не-избыточных белковых последовательностей для создания перечня ферментов с высоким сходством последовательности (>40% идентичности) к ферменту Streptococcus gordonii. Полученный перечень включал 18 кандидатов.
Последовательности ферментов MDP декарбоксилазы, выведенные из геномов вышеуказанных видов, а также из генома S. gordonii, генерировали путем олигонуклеотидной конкатенации для подгонки частоты использования кодона E.coli. Фрагмент секвенирования из 6 гистидиновых кодонов вставляли после инициирующего кодона метионина для обеспечения аффинной метки для очистки Таким образом синтезированные гены клонировали pET25b экспрессионном векторе (векторы конструировали путем GENEART AG). После трансформации штамма E.coli BL21(DE3), продуцировали белки в соответствии с протоколом, описанным в примере 1. Затем ферменты анализировали, используя способ, описанный в примере 2, используя MDP декарбоксилазу ТП. acidophilum вместо фермента Р. torridus. Эта процедура скрининга приводила к идентификации ферментов, которые более эффективны для продукции изобутена, чем фермент S. gordonii (Фигура 9), в особенности MDP декарбоксилазы из S. infantarius, S. gallolyticus, S. sp.M143, S. salivarius, S. suis, S. sanguinis и S. mitis.
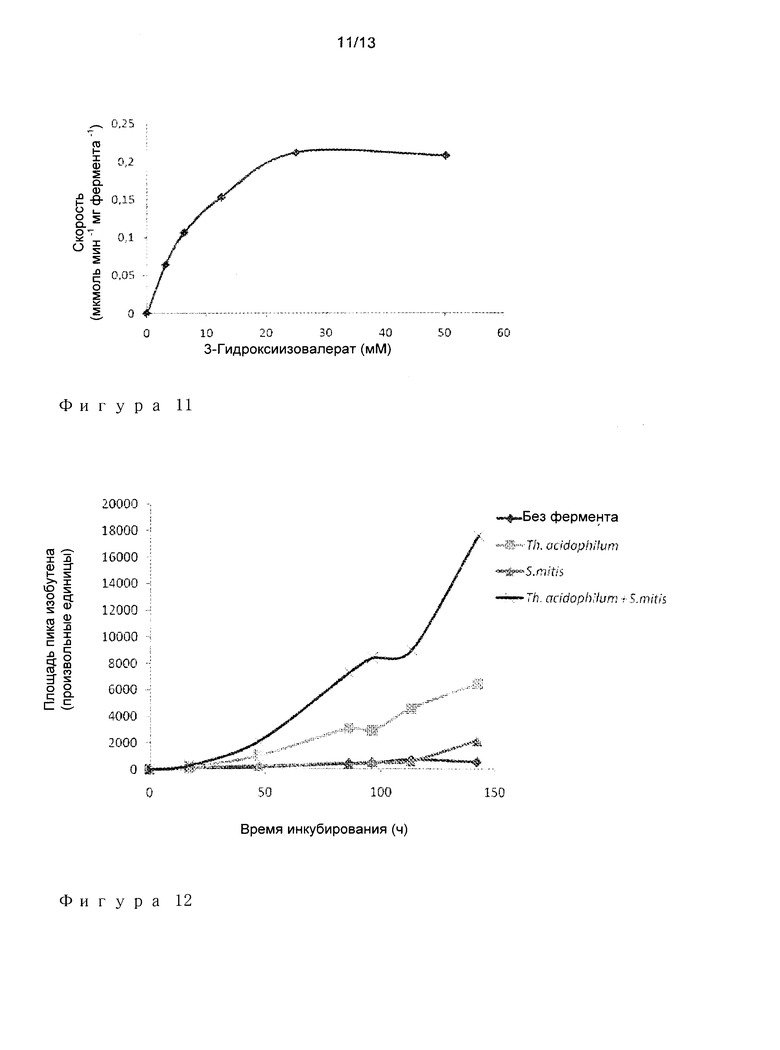
Пример 5: Характеристика фосфотрансферазной активности.
Высвобождение АДФ, которое ассоциировано с продукцией IBN из HIV, определяли количественно с использованием сопряженного анализа пируват киназа/лактат дегидрогеназа (Фигура 10). MDP декарбоксилазы из Р. torridus, Th. acidophilum, S. infantarius, S. mitis оценила относительно их способности фосфорилировать HIV, высвобождая АДФ.
Исследуемую ферментативную реакцию осуществляли в следующих условиях при 40°С:
50 мМ Tris-HCl pH 7,0
10 мМ MgCl2
100 мМ KCl
5 мМ АТФ
0,2 мМ NADH
0,5 мМ Фосфоенолпируват
3 Ед./мл Лактат дегидрогеназа
1,5 Ед./мл Пируват киназа
0-50 мМ 3-Гидроксиизовалерат (HIV)
Значение pH доводили до 7,0
Каждый анализ запускали путем добавления конкретного фермента (при концентрации от 0,05 до 1 мг/мл) и за израсходованием NADH наблюдали, отслеживая абсорбцию при 340 нМ.
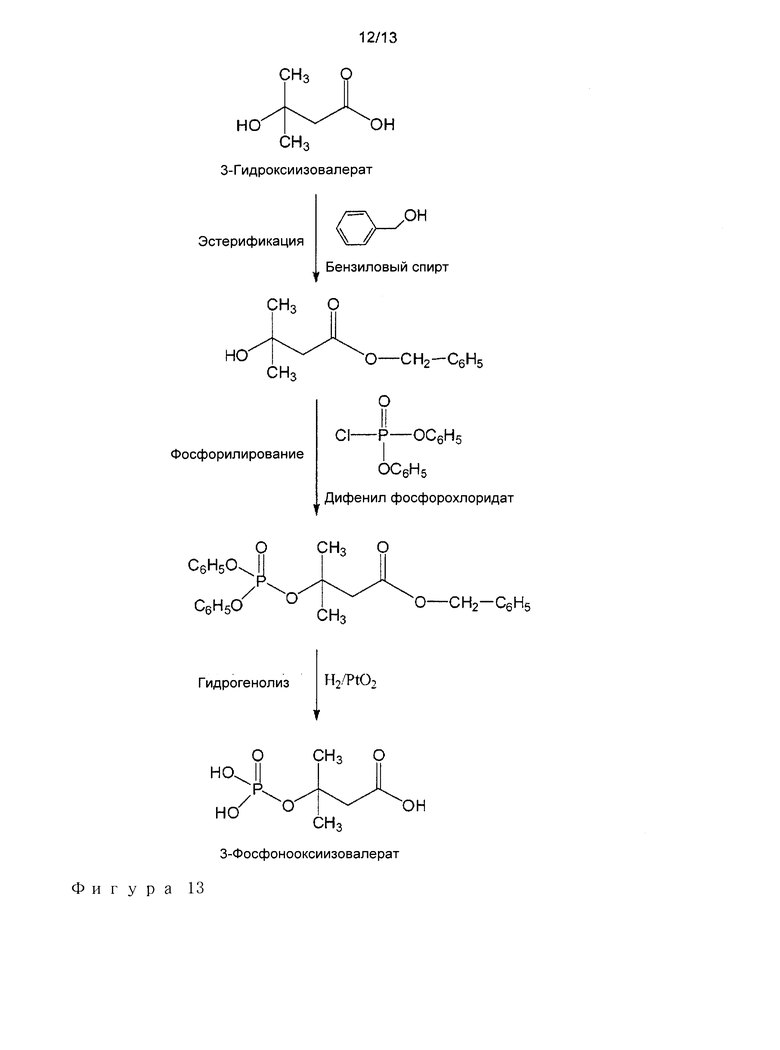
Исследования с MDP декарбоксилазами из Р. torridus phylum, а также из Streptococcus genus, приводили к воспроизводимому повышению продукции АДФ в присутствии HIV. На фигуре 11 показан пример графика Михаэлиса-Ментен, соответствующий данным, собранным для фермента Р. torridus. Кинетические параметры представлены в следующей таблице.
Ферменты из Р. torridus phylum проявляют более высокие фосфотрансферазные активности по сравнению с таковыми для ферментов из родов Streptococcus.
Пример 6: Продукция изобутена из 3-гидроксиизовалерата путем комбинирования двух ферментов.
Желательную ферментативную реакцию осуществляли в следующих условиях:
50 мМ Tris HCl рН 7,5
10 мМ MgCl2
20 мМ KCl
40 мМ АТФ
50 мМ HIV
Значение рН доводили до 7,5
100 мкг MDP декарбоксилазы из Th. acidophilum и 500 мкг MDP декарбоксилазы из S. mitis добавляли к 1 мл реакционной смеси. Контрольные реакции только с одним из двух ферментов выполняли параллельно. Пробы инкубировали без встряхивания при 37°С в запечатанном сосуде (Interchim).
Продукцию IBN измеряли путем анализа аликвот, отобранных в течение 142 часового периода инкубирования.
Один мл газообразной фазы собрали и инъецировали на НР5890 газовый хроматограф (HP), оборудованный FID детектором и СР SilicaPlot колонкой (Varian). Коммерчески доступный изобутен использовали в качестве сравнения.
Кинетики продукции изобутена представлены на фигуре 12. MDP декарбоксилаза из Th. acidophilum катализирует продукцию изобутена из HIV. Добавление MDP декарбоксилазы из S. mitis приводит к 3х кратному повышению продукции изобутена после инкубирования в течение 142 часов. MDP декарбоксилаза из S. mitis самостоятельно продуцирует только небольшие количества изобутена после инкубирования в течение 6 дней, указывая на низкую фосфотрансферазную активность.
Таким образом, продукция изобутена может быть увеличена путем комбинирования двух типов ферментов, действующих комплементарно на двух стадиях реакции.
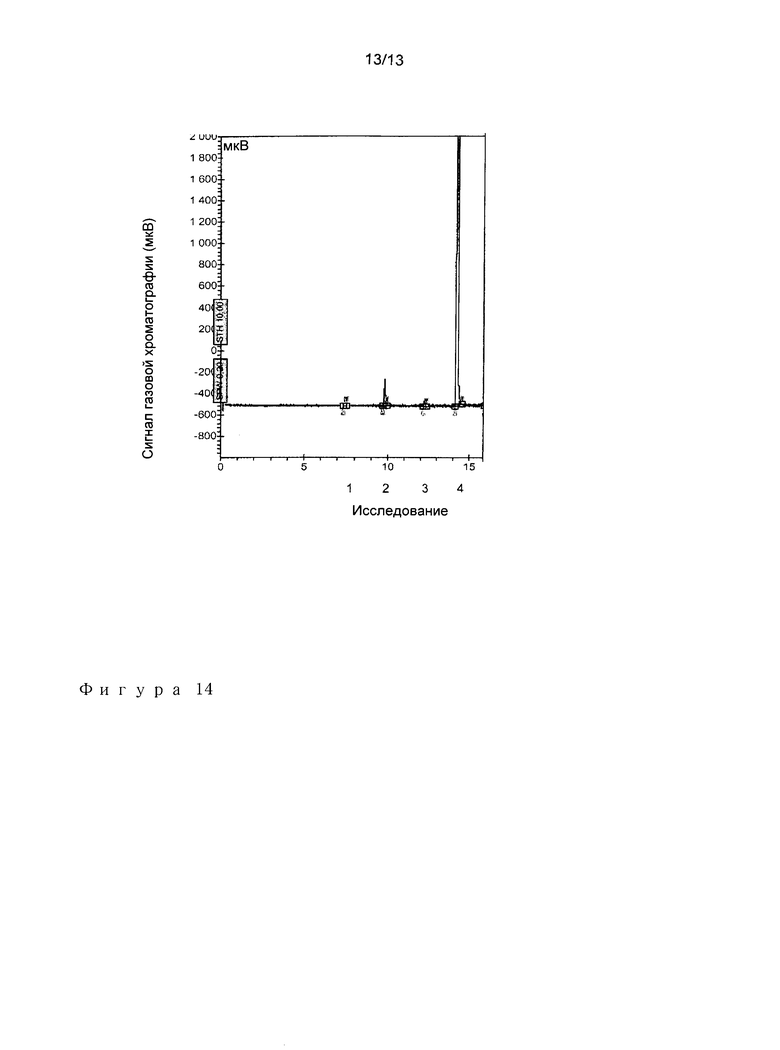
Пример 7: Влияние АТФ на продукцию изобутена из 3-фосфоноксиизовалерата (PIV).
Соединение 3-фосфоноксиизовалерат (PIV) химически синтезировали из 3-гидроксиизовалерата в соответствии со схемой, представленной на фигуре 13 с помощью SYNTHEVAL (France). Анализы продукции изобутена осуществляли в следующих условиях:
50 мМ Tris-HCl pH 7,5
10 мМ MgCl2
20 мМ KCl
О мМ АТФ (анализ №1 и №2)
10 мМ АТФ (анализ №3 и №4)
25 мМ 3-фосфоноксиизовалерат
Значение pH доводили до 7,5
Реакцию инициировали путем добавления 2 мг очищенной МОР декарбоксилазы из S. mitis к 0,5 мл реакционной смеси. Контрольные реакции осуществляли при отсутствии фермента (анализы №1 и №3). Смесь инкубировали без встряхивания при 37°С в течение 26 часов в запечатанном сосуде 2 мл (Interchim).
Один мл газообразной фазы собрали и инъецировали на гаховый хроматограф Varian 430-GC, оборудованный FID детектором и колонкой СР SilicaPlot (Varian). Коммерчески доступный изобутен использовали в качестве сравнения.
Добавление 10 мМ АТФ к реакционной смеси повышает в 120 раз продукцию изобутена из 3-фосфоноксиизовалерата (P1V) (Фигура 14).
Пример 8: Кинетические параметры продукции изобутена из 3-фосфоноксиизовалерата (PIV).
Кинетические параметры продукции изобутена измеряли в следующих условиях:
50 мМ Tris-HCl рН 7,5
10 мМ MgCl2
50 мМ KCl
40 мМ АТФ
0-100 мМ 3-фосфоноксиизовалерат
Значение рН доводили до 7,5
Реакцию инициировали путем добавления 1 мг очищенной MDP декарбоксилазы из S. mitis к 0,5 мл реакционной смеси. После этого смесь инкубировали без встряхивания при 37°С в течение 44 часов в запечатанном сосуде 2 мл (Interchim).
Один мл газообразной фазы собрали и инъецировали на газовый хроматограф Varian 430-GC, оборудованный FID детектором и СР SilicaPlot колонкой (Varian). Коммерчески доступный изобутен использовали в качестве сравнения.
Анализы с MDP декарбоксилазой из S. mitis показали 160-400-кратное увеличение продукции IBN по сравнению с фоновым уровнем (спонтанное разложение 3-фосфоноксиизовалерата) в присутствии АТФ в качестве кофактора (см. последующую таблицу).
Было обнаружено, что MDP декарбоксилаза из S. mitis имеет КМ выше чем 60 мМ и kcat по меньшей мере 1,3×10-3 с-1.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОЛУЧЕНИЕ ТЕРМИНАЛЬНЫХ АЛКЕНОВ С ПОМОЩЬЮ ФЕРМЕНТАТИВНОГО ДЕКАРБОКСИЛИРОВАНИЯ 3-ГИДРОКСИАЛКАНОЕВЫХ КИСЛОТ | 2009 |
|
RU2571933C2 |
| НИТРИЛГИДРАТАЗА ИЗ Rhodococcus | 2005 |
|
RU2385932C2 |
| ПОЛУЧЕНИЕ 1,4 ДИАМИНОБУТАНА | 2010 |
|
RU2577967C2 |
| УСТОЙЧИВЫЕ К ЦИАНИДАМ НИТРИЛГИДРАТАЗЫ | 2005 |
|
RU2385876C2 |
| МНОГОКОМПОНЕНТНАЯ ИММУНОГЕННАЯ КОМПОЗИЦИЯ ДЛЯ ПРЕДУПРЕЖДЕНИЯ ЗАБОЛЕВАНИЯ, ВЫЗВАННОГО β-ГЕМОЛИТИЧЕСКИМИ СТРЕПТОКОККАМИ (БГС) | 2009 |
|
RU2478396C2 |
| МОДИФИЦИРОВАННЫЕ МИКРООРГАНИЗМЫ И СПОСОБЫ ПОЛУЧЕНИЯ БУТАДИЕНА С ИХ ПРИМЕНЕНИЕМ | 2012 |
|
RU2639564C2 |
| СПОСОБ ФЕРМЕНТАТИВНОГО ПОЛУЧЕНИЯ L-МЕТИОНИНА ИЗ О-ФОСФО-L-ГОМОСЕРИНА И МЕТАНТИОЛА | 2013 |
|
RU2650859C2 |
| ГЕН И БЕЛОК ВИРУЛЕНТНОСТИ И ИХ ИСПОЛЬЗОВАНИЕ | 2000 |
|
RU2240327C2 |
| МИКРООРГАНИЗМ, ЭКСПРЕССИРУЮЩИЙ КСИЛОЗОИЗОМЕРАЗУ | 2009 |
|
RU2553537C2 |
| СПОСОБ ПОЛУЧЕНИЯ 3-ГИДРОКСИ-3-МЕТИЛМАСЛЯНОЙ КИСЛОТЫ ИЗ АЦЕТОНА И АЦЕТИЛ-СОА | 2010 |
|
RU2573389C2 |





















Группа изобретений относится к области биотехнологии. Представлен способ получения алкена, включающий превращение 3-гидроксиалканоата в указанный алкен с помощью первого фермента, который обладает активностью превращать 3-гидроксиалканоат в соответствующий 3-фосфоноксиалканоат, и второго фермента, который структурно отличается от первого фермента и который обладает активностью превращать указанный 3-фосфоноксиалканоат в указанный алкен, при этом первый и второй ферменты представляют собой ДФМ (дифосфат мевалонат) декарбоксилазу. Представлен рекомбинантный микроорганизм для получения алкена, который был трансформирован гетерологичными молекулами нуклеиновой кислоты, кодирующими по меньшей мере два вышеуказанных фермента. Представлена реакционная смесь для ферментативного производства алкена, содержащая указанный микроорганизм, культуральную среду и 3-гидроксиалканоатное соединение. Группа изобретений позволяет улучшить общую эффективность способа получения алкена из 3-гидроксиалканоата и увеличить продукцию получаемого алкена. 4 н. и 14 з.п. ф-лы, 14 ил., 4 табл., 8 пр.
1. Способ получения алкена, который характеризуется тем, что он включает превращение 3-гидроксиалканоата в указанный алкен с помощью
(i) первого фермента, который обладает активностью превращать 3-гидроксиалканоат в соответствующий 3-фосфоноксиалканоат; и
(ii) второго фермента, который структурно отличается от первого фермента и который обладает активностью превращать указанный 3-фосфоноксиалканоат в указанный алкен, где первый и второй ферменты представляют собой ДФМ (дифосфат мевалонат) декарбоксилазу.
2. Способ по п. 1, где
(i) первый фермент, который обладает активностью превращать 3-гидроксиалканоат в соответствующий 3-фосфоноксиалканоат, выбирают из группы, включающей
(A) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 1, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 17% идентична аминокислотной последовательности, представленной в SEQ ID NO: 1, и проявляет активность превращать 3-гидроксиалканоат в соответствующий 3-фосфоноксиалканоат, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 1;
(B) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 2, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 17% идентична аминокислотной последовательности, представленной в SEQ ID NO: 2, и проявляет активность превращать 3-гидроксиалканоат в соответствующий 3-фосфоноксиалканоат, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 2;
(C) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 3, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 17% идентична аминокислотной последовательности, представленной в SEQ ID NO: 3, и проявляет активность превращать 3-гидроксиалканоат в соответствующий 3-фосфоноксиалканоат, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 3; и
(D) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 4, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 17% идентична аминокислотной последовательности, представленной в SEQ ID NO: 4, и проявляет активность превращать 3-гидроксиалканоат в соответствующий 3-фосфоноксиалканоат, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 4.
3. Способ по п. 1, где
(ii) второй фермент, который обладает активностью превращать указанный 3-фосфоноксиалканоат в указанный алкен, выбирают из группы, включающей
(a) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 5, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 22% идентична аминокислотной последовательности, представленной в SEQ ID NO: 5, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 5;
(b) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 6, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 22% идентична аминокислотной последовательности, представленной в SEQ ID NO: 6, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 6;
(c) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 7, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 22% идентична аминокислотной последовательности, представленной в SEQ ID NO: 7, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 7;
(d) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 8, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 22% идентична аминокислотной последовательности, представленной в SEQ ID NO: 8, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 8;
(e) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 9, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 22% идентична аминокислотной последовательности, представленной в SEQ ID NO: 9, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 9;
(f) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 10, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 22% идентична аминокислотной последовательности, представленной в SEQ ID NO: 10, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 10;
(g) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 11, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 22% идентична аминокислотной последовательности, представленной в SEQ ID NO: 11, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 11;
(h) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 12, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 22% идентична аминокислотной последовательности, представленной в SEQ ID NO: 12, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 12;
(i) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 13, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 22% идентична аминокислотной последовательности, представленной в SEQ ID NO: 13, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 13;
(j) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 14, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 22% идентична аминокислотной последовательности, представленной в SEQ ID NO: 14, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 14; и
(k) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 15, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 22% идентична аминокислотной последовательности, представленной в SEQ ID NO: 15, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 15.
4. Способ по любому из пп. 1-3, который включает стадию превращения 3-гидроксипроприоната в этилен.
5. Способ по любому из пп. 1-3, который включает стадию превращения 3-гидроксибутирата в пропилен.
6. Способ по любому из пп. 1-3, который включает стадию превращения 3-гидроксивалерата в 1-бутилен.
7. Способ по любому из пп. 1-3, который включает стадию превращения 3-гидрокси-3-метилбутирата в изобутилен.
8. Способ по любому из пп. 1-3, который включает стадию превращения 3-гидрокси-3-метилвалерата в изоамилен.
9. Способ по любому из пп. 1-3, который отличается тем, что стадию превращения осуществляют in vitro, в бесклеточной системе.
10. Способ по любому из пп. 1-3, который отличается тем, что способ осуществляют в присутствии микроорганизма, продуцирующего указанные ферменты, как определено в (i) и (ii) пункта 1.
11. Способ по любому из пп. 1-3, который отличается тем, что используют микроорганизм, продуцирующий указанные ферменты, как определено в (i) и (ii) п. 1, причем указанный микроорганизм представляет собой Е.coli.
12. Способ по п. 1, который отличается тем, что способ осуществляют с АТФ, dATP, АДФ, AMP, NTP, отличающимся от АТФ, dNTP, или пирофосфатом в качестве субстратного кофактора.
13. Способ по любому из пп. 1-3, который включает стадию сбора газообразных алкенов дегазирования реакции.
14. Рекомбинантный микроорганизм, предназначенный для получения алкена, который трансформирован гетерологичными молекулами нуклеиновой кислоты, кодирующими по меньшей мере два фермента, где один фермент выбирают из (i) и другой фермент выбирают из (ii), где (i) и (ii) являются следующими:
(i) первый фермент, который обладает активностью превращать 3-гидроксиалканоат в соответствующий 3-фосфоноксиалканоат; и
(ii) второй фермент, который структурно отличается от первого фермента и который обладает активностью превращать указанный 3-фосфоноксиалканоат в указанный алкен,
где первый и второй ферменты представляют собой ДФМ (дифосфат мевалонат) декарбоксилазу, а молекулы нуклеиновой кислоты, которые кодируют эти ферменты, являются гетерологичными для этого организма, причем он был трансформирован такими молекулами нуклеиновых кислот.
15. Реакционная смесь для ферментативного производства алкена, которая содержит микроорганизм по п. 14, подходящую культуральную среду и 3-гидроксиалканоатное соединение или источник углерода, который может быть превращен микроорганизмом в 3-гидроксиалканоатное соединение.
16. Применение комбинации двух ферментов, где один фермент выбирают из следующих (i) и другой фермент выбирают из следующих (ii), или микроорганизма по п. 14, или реакционной смеси по п. 15 для получения алкеновых соединений из 3-гидроксиалканоатов,
где (i) и (ii) являются следующими:
(i) первый фермент, который обладает активностью превращать 3-гидроксиалканоат в соответствующий 3-фосфоноксиалканоат; и
(ii) второй фермент, который структурно отличается от первого фермента и который обладает активностью превращать указанный 3-фосфоноксиалканоат в указанный алкен.
17. Применение по п. 16, где
(i) первый фермент, который обладает активностью превращать 3-гидроксиалканоат в соответствующий 3-фосфоноксиалканоат, выбирают из группы, включающей
(А) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 1, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 17% идентична аминокислотной последовательности, представленной в SEQ ID NO: 1, и проявляет активность превращать 3-гидроксиалканоат в соответствующий 3-фосфоноксиалканоат, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 1;
(B) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 2, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 17% идентична аминокислотной последовательности, представленной в SEQ ID NO: 2, и проявляет активность превращать 3-гидроксиалканоат в соответствующий 3-фосфоноксиалканоат, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 2;
(C) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 3, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 17% идентична аминокислотной последовательности, представленной в SEQ ID NO: 3, и проявляет активность превращать 3-гидроксиалканоат в соответствующий 3-фосфоноксиалканоат, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 3; и
(D) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 4, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 17% идентична аминокислотной последовательности, представленной в SEQ ID NO: 4, и проявляет активность превращать 3-гидроксиалканоат в соответствующий 3-фосфоноксиалканоат, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 4.
18. Применение по п. 16 или 17, где
(ii) второй фермент, который обладает активностью превращать указанный 3-фосфоноксиалканоат в указанный алкен, выбирают из группы, включающей
(а) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 5, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 22% идентична аминокислотной последовательности, представленной в SEQ ID NO: 5, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 5;
(b) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 6, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 22% идентична аминокислотной последовательности, представленной в SEQ ID NO: 6, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 6;
(c) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 7, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 22% идентична аминокислотной последовательности, представленной в SEQ ID NO: 7, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 7;
(d) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 8, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 22% идентична аминокислотной последовательности, представленной в SEQ ID NO: 8, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 8;
(e) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 9, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 22% идентична аминокислотной последовательности, представленной в SEQ ID NO: 9, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 9;
(f) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 10, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 22% идентична аминокислотной последовательности, представленной в SEQ ID NO: 10, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 10;
(g) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 11, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 22% идентична аминокислотной последовательности, представленной в SEQ ID NO: 11, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 11;
(h) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 12, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 22% идентична аминокислотной последовательности, представленной в SEQ ID NO: 12, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 12;
(i) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 13, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 22% идентична аминокислотной последовательности, представленной в SEQ ID NO: 13, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 13;
(j) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 14, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 22% идентична аминокислотной последовательности, представленной в SEQ ID NO: 14, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 14; и
(k) белок, содержащий аминокислотную последовательность, как представлено в SEQ ID NO: 15, или белок, содержащий аминокислотную последовательность, которая по меньшей мере на 22% идентична аминокислотной последовательности, представленной в SEQ ID NO: 15, и проявляет активность превращать указанный 3-фосфоноксиалканоат в указанный алкен, которая по меньшей мере такая же высокая, как и соответствующая активность белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 15.
| WO2010001078 A2, 07.01.2010 | |||
| EP0205303 А2, 17.12.1986 | |||
| СПОСОБ ПОЛУЧЕНИЯ ЭТИЛЕНА | 2005 |
|
RU2281316C1 |

