Настоящее изобретение относится к фармацевтическим композициям, содержащим смесь специфического антигена ВИЧ и непатогенной бактерии. Указанный специфический антиген ВИЧ содержит один или несколько эпитопов из белков Gag и/или Pol и, предпочтительно, находится в крупнодисперсной форме. Указанная бактерия предпочтительно является Lactobacillus plantarum. Эти фармацевтические композиции полезны для профилактики и(или) лечения заболевания ВИЧ у людей.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Спустя более двадцати пяти лет после открытия вируса иммунодефицита человека (ВИЧ) прогнозы, содержащиеся в программе Всемирной организации здравоохранения и Организации объединенных наций по ВИЧ/СПИД, свидетельствуют о том, что если развитие пандемии будет продолжаться с нынешней скоростью, к 2011 г. будет инфицировано более 30 млн. человек.
Однако, несмотря на существенные усилия в области научных исследований по выявлению эффективных способов терапии для профилактики инфекции ВИЧ, две недавно испытанные профилактические вакцины либо не дали результата (Me Elrath et al., 2008), либо дали скромные результаты (Rerks-Ngarm et al., 2009).
В публикации: Jae-Sung Yu et al. (Clinical and Vaccine Immunology, Nov. 2006, vol 13, No. 11, 1204-1211) описан рекомбинантный вектор для Mycobacterium smegmatis, сконструированный таким образом, чтобы он мог обеспечивать экспрессию консенсусного гена env из ВИЧ-1 группы М - CON6 - в виде либо поверхностного, либо секретируемого белка. Авторы смогли продемонстрировать, что при введении мышам рекомбинантная М. smegmatis была иммуногенна и индуцировала ВИЧ-1-специфические Т-клеточные ответы на слизистых оболочках.
В публикации Ke-Qin Xin et al. (Blood, 1 July 2003, vol 102, No. 1, 223-228) описан рекомбинантный вектор для Lactococcus lactis, обеспечивающий экспрессию петли V2-V4 белка Env из ВИЧ-1 на поверхности ее клетки. Пероральная иммунизация мышей этим вектором вызывала следующие эффекты:
- как мукозный, так и гуморальный иммунные ответы, что было показано путем выявления высоких уровней ВИЧ-специфических IgG в сыворотке и антител IgA в фекалиях; и
- клеточный иммунный ответ, что было показано по повышению количества ВИЧ-специфических клеток, секретирующих IFN-гамма.
Для того чтобы экспрессия на поверхности клеток L. lactis проходила правильно, можно применять сегменты генов размером 1 т.п.н. или менее.
Большинство ученых, участвующих в исследованиях патогенеза с участием ВИЧ и его профилактики, полагают, что перед тем как тестировать профилактические вакцины против ВИЧ или другие биологические композиции для профилактики или лечения ВИЧ-инфекции у человека, было бы полезнее протестировать их аналоги на приматах, отличных от человека (Morgan С, et al., 2008). Из приматов, отличных от человека, выбран макак-резус, а среди макаков, как было убедительно показано, макаки китайского происхождения, инфицированные вирусом иммунодефицита обезьян (SIV) 239, представляют собой самую лучшую модель, воспроизводящую большинство клинических, вирологических и иммунологических особенностей развития ВИЧ-инфекции у человека (Marcondes МС, et al. 2006; Stahl-Hennig С, et al. 2007; Chen S, et al. 2008).
Наконец, в настоящее время научное сообщество пришло к согласию о том, что как только будет открыта эффективная профилактическая биологическая композиция или вакцина против SIV239 для макака, ее, по всей вероятности, можно будет успешно адаптировать для человека с целью защиты его от СПИД.
Несмотря на постоянные исследовательские усилия, прилагаемые научным сообществом, все еще имеется потребность в эффективных стратегиях профилактики и терапии, необходимых для борьбы с всемирной пандемией СПИД.
Было описано, что различные бактерии обладают интересными свойствами адъювантности и иммуномодуляции при введении пациентам. В частности, сообщалось, что молочнокислые бактерии способствуют развитию эффекта толерантности иммунной системы.
Например, в WO 2006/123230, опубликованной 23 ноября 2006 г. от имени Stallergenes S.A., описано применение бактерии, выбранной из Bifidobacteria и молочнокислых бактерий, в качестве адъюванта в иммуногенной композиции, способной индуцировать антиген-специфическую толерантность при сублингвальном, перилингвальном или пероральном введении пациенту. Предполагается, что иммуногенная композиция полезна для лечения аллергии, аутоиммунных заболеваний или для профилактики отторжения трансплантата.
Например, в WO 2009/093900 опубликованной 30 июля 2009 г. от имени организации Stichting Top Institute Food and Nutrition, описана толерогенная композиция, содержащая существенное количество молочнокислых бактерий, находящихся в середине логарифмической фазы роста. Эта композиция при введении пациенту индуцирует не специфическую к антигену иммунологическую толерантность. Предполагается, что композиция будет применяться для профилактики, замедления и/или лечения состояний или заболеваний, связанных с воспалительными ответами, которые могут приводить к повреждению тканей, таких как аллергии, аутоиммунные заболевания и воспалительные заболевания кишечника.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Авторам изобретения удалось показать, что, неожиданно, исходные фармацевтические композиции, описанные в разделе примеров, индуцировали эффективную антиген-специфическую иммунную защиту против вируса SIV у макаков. Более того, авторы изобретения показали, что при индукции указанной SIV-специфической иммунной защиты она предотвращает репликацию/диссеминацию вируса SIV и последующее возникновение инфекции in vivo.
Действительно, авторы изобретения неожиданно продемонстрировали, что при введении фармацевтической композиции, раскрытой в этом документе, либо на слизистую оболочку, либо внутрикожно или интраэпителиально, репликация вируса была существенно подавлена или даже прекращена или предотвращена.
Фактически, авторы изобретения впервые могли наблюдать, что нецитотоксический CD8+ T-клеточный ответ супрессировал раннюю активацию CD4+ Т-клеток, презентирующих антиген вируса SIV, у макаков. Таким образом, не следуя какой-то определенной теории, фармацевтические композиции по настоящему изобретению индуцируют неожиданный новый тип вирусоспецифической иммунотолерантности при введении пациентам на слизистые, или внутрикожно, или интраэпителиально. Эта иммунотолерантность, по-видимому, является специфической к антигенам ВИЧ - Gag и/или Pol - иммунотолерантностью, индуцируемой супрессорными CD8+ T-клетками (также называемой в этом документе «Ts»-иммунотолерантностью, что означает «Т-супрессорная» иммунотолерантность), которая ограничена МНС («главный комплекс гистосовместимости»)-Ib/Е-и не является цитотоксической.
В свете сообщаемых здесь результатов в настоящем изобретении предлагается новая фармацевтическая композиция, способная обеспечивать достижение «Ts»-иммунотолерантности, как указано выше, для профилактики и/или лечения заболевания ВИЧ человека.
Целью настоящего изобретения является, таким образом, предложение фармацевтической композиции, содержащей смесь антигена и непатогенной живой бактерии, при этом, предпочтительно, указанный антиген является крупнодисперсным и/или содержит один или несколько эпитопов из белков ВИЧ - Gag и/или Pol, и при этом указанная бактерия предпочтительно является Lactobacillus plantarum.
Еще одной целью настоящего изобретения является предложение фармацевтической композиции, описанной выше, для применения в качестве вакцины.
Еще одной целью настоящего изобретения является предложение способа профилактики и/или лечения заболевания ВИЧ у человека, которому это необходимо, включающего по меньшей мере этап введения через слизистые (предпочтительно - перорально), или внутрикожно, или интраэпителиально эффективного количества фармацевтической композиции, упомянутой выше, указанному человеку.
Еще одной целью настоящего изобретения является предложение способа защиты человека от заболевания ВИЧ, включающего по меньшей мере этап введения через слизистые (предпочтительно - перорально), или внутрикожно, или интраэпителиально эффективного количества фармацевтической композиции, упомянутой выше, указанному человеку.
Еще одной целью настоящего изобретения является предложение способа защиты человека от сероконверсии под действием ВИЧ, включающего по меньшей мере этап введения через слизистые (предпочтительно - перорально), или внутрикожно, или интраэпителиально эффективного количества фармацевтической композиции, упомянутой выше, указанному человеку.
И еще одной целью настоящего изобретения является предложение фармацевтического набора для профилактики и/или лечения заболевания ВИЧ у человека, которому это необходимо, включающего:
- в первом контейнере - антиген; и
- во втором контейнере - непатогенную бактерию,
при этом указанный антиген и указанная бактерия находятся в фармацевтически приемлемых носителях для введения через слизистые, или внутрикожно, или интраэпителиально, при этом, предпочтительно, указанный антиген является крупнодисперсным и/или содержит один или несколько эпитопов из белков ВИЧ - Gag и/или Pol, и при этом указанная бактерия предпочтительно является Lactobacillus plantarum.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Настоящее изобретение проиллюстрировано следующими фигурами, на которые сделаны ссылки в неограничивающих примерах, приведенных ниже.
Фиг.1. Внутривенная (в/в) провокационная проба SIVmac239 макакам-резус, после предварительного интравагинального введения iSIV/BCG (БЦЖ).
Фиг.2. Интраректальная (и/р) провокационная проба SIVmac239 макакам-резус, после предварительного интравагинального введения iSIV/BCG (БЦЖ).
Фиг.3. Повторные провокационные пробы SIVmac239 (3 раза в/в и 2 раза и/р) макакам-резус, после предварительного интравагинального введения iSIV/BCG (БЦЖ).
Фиг.4. Внутривенная провокационная проба SIVmac239 макакам-резус, после предварительного интравагинального введения iSIV/BCG (БЦЖ) и внутрикожной бустер-инъекции.
Фиг.5. Интраректальная провокационная проба SIVmac239 макакам-резус, после предварительного интравагинального введения iSIV/BCG (БЦЖ) и внутрикожной бустер-инъекции..
Фиг.6. Интраректальная провокационная проба SIVmac239 макакам-резус, после предварительного перорального введения iSIV/BCG (БЦЖ).
Фиг.7. Антивирусная активность in vitro CD8+ Т-клеток, полученных от макак-резусов после предварительного интравагинального введения iSIV/BCG.
Фиг.8. Антивирусная активность in vitro CD8+ Т-клеток, полученных от 4 макак-резусов после предварительного перорального введения iSIV/BCG.
Фиг.9. SIV-специфическая супрессия активации CD4+ Т-клеток аутологичными CD8+ Т-клетками, полученными от 4 макаков-резусов после предварительного перорального введения iSIV/BCG.
Фиг.10а. Титры антител IgG против SIV в пробах плазмы, взятых у макаков-резусов, которым предварительно вводили iSIV/LP, iSIV или LP.
Фиг.10b. Пролиферация SIV-специфических Т-клеток в пробах МКПК (мононуклеарных клеток периферической крови), взятых у макаков-резусов, которым предварительно вводили iSIV/LP, iSIV или LP.
Фиг.10c. SIV-специфические Т-клетки, секретирующие IFN-гамма при стимуляции in vitro в присутствии или в отсутствие CD8 или CD25 Т-клеток.
Фиг.10d. SIV-специфическая супрессия активации CD4+ Т-клеток аутологичными CD8+ Т-клетками, полученными от 8 макаков-резусов, которым предварительно перорально вводили iSIV/LP, в сравнении с животными, которым предварительно перорально вводили LP (n=4) или iSIV (n=3).
Фиг.10е. SIV-специфические CD8+ Т-клетки через 60 дней после внутрижелудочного введения препарата iSIV/LP: цитотоксичность АТ-2 SIV-стимулированных CD4+ Т-клеток в присутствии CD8+ Т-клеток или клеток K562 в присутствии натуральных киллерных клеток человека (hNK) (в качестве контролей), со стимуляцией SEB и антителами против CD3/CD28 или без такой стимуляции.
Фиг.11a. Антивирусная активность in vitro (в CD4 клетках) аутологичных CD8+ Т-клеток, полученных от 8 макаков-резусов, которым предварительно перорально вводили iSIV/LP, в сравнении с животными, которым предварительно перорально вводили LP (n=4) или iSIV (n=3).
Фиг.11b. Антивирусная активность in vitro (в CD4 клетках) гетерологичных или аллогенных CD8+ Т-клеток, полученных от 4 из 8 макаков-резусов через 80 дней после перорального введения iSIV/LP.
Фиг.11c-g. Активность CD8+ Т-клеток против SIV через 60 дней после пероральной иммунизации в системе клеточной культуры с замедленной пролиферацией (с), культивируемой на вставках (insert) (d), аллогенной (е), в присутствии антител против MHC-Ia/ABC или антител против MHC-Ib/E (f), и в CD8+ Т-клетках, обедненных TCRγδ+ или Vβ8+ подгруппы (g).
Фиг.12а. Уровни вирусной нагрузки плазмы (количество копий SIV РНК на мл плазмы) после интраректальной и внутривенной провокационной пробы SIVmac239, проведенной макакам-резусам, которым предварительно перорально вводили iSIV/LP, в сравнении с животными, которым предварительно перорально вводили LP или iSIV.
Фиг.12b. Уровни вирусной нагрузки клеток (количество копий SIV ДНК на миллион мононуклеарных клеток периферической крови (МКПК)) после интраректальной и внутривенной провокационной пробы SIVmac239, проведенной макакам-резусам, которым предварительно перорально вводили iSIV/LP, в сравнении с животными, которым предварительно перорально вводили LP или iSIV.
Фиг.13. Истощение CD8+ Т-клеток периферической крови и лимфатических узлов у 8 предварительно обработанных iSIV/LP макаков путем инфузии антитела против CD8 - сМТ807. а. Количество CD8+ Т-клеток периферической крови до и после введения трех инъекций сМТ807; b. % от CD8+ Т-клеток лимфатических узлов до и после введения трех инъекций сМТ807; с. Вирусная нагрузка плазмы до и после введения трех инъекций сМТ807; d. Нагрузка SIV ДНК в клетках МКПК до и после введения трех инъекций сМТ807; е. Нагрузка SIV ДНК в лимфатических узлах до и после введения трех инъекций сМТ807.
Фиг.14. Вирусная нагрузка плазмы (а) и клеток МКПК (b) после третьей интраректальной провокационной пробы, введенной интраректально с применением SIVB670 восьми макакам-резусам, иммунизированным пероральным препаратом, приготовленным из iSIV и LP, и у 2 дополнительных неиммунизированных обезьян.
Фиг.15. Антивирусная активность in vitro и in vivo, опосредованная CD8+ Т-клетками, после внутрижелудочной иммунизации с применением iSIV и LP (иммунизация iSIV/LP №2). а. Активность против SIV (кратность супрессии вируса) CD8+ Т-клеток во время 60-420 дней после иммунизации у 8 макаков-резусов, которым будет проведена интраректальная провокационная проба; b и с. Вирусная нагрузка плазмы и клеток после интраректальной провокационной пробы SIVmac239, проведенной 8 макакам-резусам, иммунизированным с применением перорального препарата iSIV/LP, и у 8 контрольных обезьян, которым вводили только LP (n=4) или только iSIV (n=4).
Фиг.16. Нагрузка SIV ДНК и РНК в интраэпителиальных лимфоцитах слизистой оболочки прямой кишки (IPL) (a-b), клетках базальной мембраны (LPC) (c-d) и в тазовых лимфоузлах (PLN) (е) после интраэпителиальной провокационной пробы SIVmac239, введенной 8 макакам (иммунизация iSIV/LP №2).
ДЕТАЛЬНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к фармацевтической композиции, содержащей смесь антигена и непатогенной живой бактерии.
Антиген
Из-за высокой вариабельности генома ВИЧ, вызванной мутациями, рекомбинацией, вставками и/или делециями, классификацию ВИЧ подразделяют на группы, подгруппы, типы, подтипы и генотипы. Существует две основные группы ВИЧ (ВИЧ-1 и ВИЧ-2) и множество подгрупп, потому что геном ВИЧ постоянно мутирует. Основное различие между группами и подгруппами связано с оболочкой вируса. ВИЧ-1 классифицирую на основную подгруппу (М), при этом указанная подгруппа М разделяется на девять подтипов (кладов или подтипов), обозначаемых буквами с А по J (Hu et al., JAMA 275:210-216, 1996; Korber et al., Science 280:1868-1871, 1998), и на 10-ую резко отличную подгруппу (О). Также существует множество других подгрупп, возникающих в результате рекомбинаций in vivo предшествующих подгрупп (Papathanasopoulos MA, et al. Virus Genes 2003, 26:151-163). Предпочтительно, вирус ВИЧ является ВИЧ-1 или ВИЧ-2, включая все известные и еще не известные их клады. Еще более предпочтительно, он является ВИЧ-1.
В контексте настоящего изобретения «антиген» имеет происхождение от ВИЧ, что означает, что он относится к специфичной группе, подгруппе, типу, подтипу ВИЧ или к комбинации нескольких подтипов. Предпочтительно, указанный антиген ВИЧ является антигеном ВИЧ-1 или ВИЧ-2.
Указанный антиген не является инфекционным.
Долгое время научное сообщество подозревало, что активация CD4+ Т-клеток, основной мишени как для ВИЧ-1, так и для SIV, непосредственно участвует в репликации вирусов (Andrieu and Lu, 1995; Korin and Zack, 1999). Однако только недавно эта связь между активацией CD4+ Т-клеток и последующими этапами инфекционного процесса SIV или ВИЧ была выявлена. В покоящихся CD4+ Т-клетках вслед за проникновением вируса в течение 2 ч после входа происходила презентация на плазматической мембране эпитопов, происходящих от белков Gag и Pol входящих вирионов, тогда как для белков Env и Nef был необходим синтез de novo (Sacha et al., 2007). Однако следующие далее фазы инфекционного процесса, т.е., обратная транскрипция с последующей интеграцией вируса, в покоящихся клетках развивались очень неэффективно (Vatakis et al., 2009a and 2009b). В противоположность этому, если CD4+ T-клетки были активированы перед или в течение 48 ч после презентации эпитопов белков Gag и Pol на плазматической мембране, обратная транскрипция ВИЧ/SIV и интеграция его ДНК проходили крайне активно, что позволяло достичь очень высокого уровня репликации вируса и его высвобождения (Vatakis et al., 2009a and 2009b).
Следовательно, авторы изобретения постулируют, что специфическая блокировка in vivo этого раннего развития специфической, опосредуемой белками ВИЧ/SIV Gag или Pol, активации CD4+ Т-клеток после экспозиции ВИЧ/SIV приведет к предотвращению активной репликации вируса.
С учетом этого и с целью индуцировать супрессию активации CD4+ Т-клеток, презентирующих антигены белков ВИЧ - Gag и/или Pol, и, в свою очередь предотвратить репликацию и диссеминацию ВИЧ in vivo у людей, подвергшихся воздействию вируса, фармацевтическая композиция по настоящему изобретению содержит антиген ВИЧ, который предпочтительно имеет один или несколько эпитопов из белков ВИЧ - Gag и/или Pol. Такой антиген предпочтительно либо содержит белки ВИЧ - Gag и/или Pol, либо происходит от них.
Таким образом, термины «антиген, который содержит белки ВИЧ - Gag и/или Pol - или происходит от них» обозначают антиген ВИЧ:
- который содержит по меньшей мере Gag и/или Pol (и является «антигеном, который содержит белки Gag и/или Pol»); или
- который содержит один или несколько белков, кодируемых геном GAG, таких как белок капсида (p24) и белок матрикса (p17), и/или один или несколько белков, кодируемых геном POL, таких как интеграза, обратная транскриптаза и протеаза (и является «антигеном, который происходит от белков Gag и/или Pol»); или
- который содержит один или несколько эпитопов из этих белков (и тоже и является «антигеном, который происходит от белков Gag и/или Pol»).
В частности, любые другие вирусные белки или их эпитопы, выбранные из группы, состоящей из ENV, VIF, VPR, VPU для ВИЧ-1, VPX для ВИЧ-2, REV, NEF, TAT и т.п., не являются существенными компонентами антигена, содержащегося в фармацевтической композиции, раскрытой в этом документе. Любой из этих белков, если он присутствует, является всего лишь необязательным компонентом антигена, который будет применяться в фармацевтической композиции, раскрытой в этом документе.
Антиген предпочтительно является крупнодисперсным антигеном. Это означает, что он предпочтительно выбран из вирусных частиц, рекомбинантных вирусных частиц, вирусоподобных частиц, экспрессирующих Gag и/или Pol рекомбинантных бактерий или грибов, полимерных микрочастиц, презентирующих на своей поверхности один или несколько вирусных белков или пептидов или эпитопов (содержащих белки ВИЧ - Gag и/или Pol или происходящих от них). Предпочтительно, один или несколько эпитопов из Gag и/или Pol продуцируются или экспрессируются указанным антигеном или содержатся в нем. Когда применяют рекомбинантные вирусные частицы или вирусные частицы или экспрессирующие Gag и/или Pol рекомбинантные бактерии или грибы, они предпочтительно являются инактивированными микроорганизмами.
Антиген может являться вирусной частицей, рекомбинантной вирусной частицей, вирусоподобной частицей или экспрессирующими Gag и/или Pol рекомбинантными бактериями или грибами. Он также может являться одним или несколькими вирусными белками или пептидами (содержащими белки ВИЧ - Gag и/или Pol - или происходящими от них), рекомбинантными или нет, как в форме конъюгатов, так и в форме конкатемеров. Дополнительно, антиген не зависит от нуклеиновых кислот, то есть не зависит от вирусной ДНК или вирусной РНК.
Антиген может быть получен в результате экспрессии последовательности нуклеиновой кислоты вируса, предпочтительно, содержащейся в соответствующем рекомбинантном микроорганизме.
Если антиген, содержащийся в фармацевтической композиции по настоящему изобретению, является экспрессирующей Gag и/или Pol рекомбинантной бактерией, то указанная рекомбинантная бактерия предпочтительно отличается от непатогенной живой бактерии, которая также присутствует в композиции.
Когда антиген в фармацевтической композиции по настоящему изобретению является одним или несколькими вирусными белками или пептидами (содержащими белки ВИЧ - Gag и/или Pol - или происходящими от них) он, предпочтительно, находится в крупнодисперсной форме. Фактически, соответствующие крупнодисперсные антигены могут продуцироваться живыми микроорганизмами, такими как дрожжи, таким же способом, что и вакцины против вируса гепатита В, полученные с применением технологий рекомбинантных ДНК, где экспрессируемый полипептид HBsAg в результате самосборки образует иммуногенные сферические частицы, сильно напоминающие природные частицы размером 22 нм, которые обнаруживаются в сыворотке пациентов с хронической инфекцией HBV (Plotkin et al., 2008).
В соответствии с другим вариантом, когда антиген в фармацевтической композиции по настоящему изобретению является одним или несколькими вирусными белками или пептидами (содержащими белки ВИЧ - Gag и/или Pol - или происходящими от них) он, предпочтительно, находится в форме конъюгатов. В таком варианте воплощения, как это хорошо известны специалистам в данной области, целевые белки или пептиды ковалентно конъюгированы с соответствующим носителем. Стандартными носителями, которые доступны для приобретения, являются, среди прочего, белки, такие как белок KLH (гемоцианин фиссуреллы), белок БСА (бычий сывороточный альбумин), белок OVA (овальбумин) и т.п. (которые, предпочтительно, могут быть безопасно введены человеку перорально). Способы получения соответствующих конъюгатов известны специалистам в данной области.
В соответствии с еще одним вариантом, когда антиген в фармацевтической композиции по настоящему изобретению является одним или несколькими вирусными белками или пептидами (содержащими белки ВИЧ - Gag и/или Pol - или происходящими от них) он, предпочтительно, находится в форме конкатемеров. Как хорошо известно специалистам в данной области, конкатемеры состоят из множества копий целевых белков или пептидов, которые физически соединены друг с другом в одну макромолекулу. В конкатемерах копия целевого белка или пептида может быть соединена с еще одной копией либо напрямую, либо через разделяющий их синтетический спейсер. Таким образом, конкатемер содержит по меньшей мере две копии, предпочтительно - до 10 копий или более целевого белка или пептида. Способы получения соответствующих конкатемеров известны специалистам в данной области.
При использовании в этом документе, термин «вирусоподобная частица» (VLP) означает частицу, которая очень похожа на зрелые вирионы, но не содержит вирусный геномный материал указанного вируса. Более конкретно, VLP, которые также называют псевдо-вирионами, представляют собой субъединичные структуры, состоящие из множества копий белков вирусного капсида и/или других вирусных белков. Эти вирусные белки способны путем самосборки образовывать VLP с заданной сферической симметрией in vivo. Эти VLP не содержат каких-либо молекул нуклеиновой кислоты, кодирующих вирусные белки, и, более конкретно, не содержат каких-либо молекул нуклеиновой кислоты. Следовательно, VLP не способны к репликации и неинфекционны по своей природе, что делает их безопасными для введения в форме фармацевтической композиции. Способы получения частиц VLP хорошо известны специалистам в данной области (см., например, Liew et al., 2010; Plummer and Manchester, 2010). Неограничивающие примеры пригодных способов получения частиц VLP описаны в US 5919458, EP 386882, WO 91/07425, US 5861282 и WO 91/05864, где раскрыты частицы VLP ВИЧ (псевдо-вирионы), которые не содержат генома ВИЧ и вообще не содержат молекул нуклеиновой кислоты.
При использовании в этом документе термин «рекомбинантная вирусная частица» означает вирусную частицу, которая содержит или которая экспонирует на своей поверхности белки из различных вирусов. Кроме того, рекомбинантная вирусная частица также может означать бактерию или другую клетку-хозяина, которая содержит, которая продуцирует или которая экспонирует на своей поверхности один или несколько вирусных белков или пептидов или эпитопов, содержащих белки ВИЧ - Gag и/или Pol - или происходящих от них.
Фактически, большинство рекомбинантных вирусных частиц представляют собой вирусные частицы, в которых часть исходных структурных белков (т.е., в основном белки оболочки и белки кора) замещены аналогичными белками из другого вируса. В качестве примера, может быть произведен обмен белков оболочки. В таком случае рекомбинантные вирусные частицы содержат «химерный» геном, представляющий собой геном вируса, в котором последовательность, кодирующая белки оболочки, заменена последовательностью, кодирующей белки оболочки другого вируса. Большинство рекомбинантных вирусных частиц способны к репликации и являются инфекционными.
При использовании в этом документе рекомбинантный вирус, содержащий белки из другого вируса, означает, что рекомбинантная вирусная частица содержит один или несколько вирусных белков или пептидов или эпитопов, содержащих белки ВИЧ - Gag и/или Pol - или происходящие от них, которые либо находятся внутри, либо экспонированы на их поверхности. Неограничивающие примеры способов получения рекомбинантных вирусных частиц описаны для следующих объектов.
* Альфа-вирус: в WO 02/053757, где описан рекомбинантный альфа-вирус, экспрессирующий белок ВИЧ (белок ENV).
* Ретровирус: в ЕР 1499736, в котором раскрыты лентивирусные векторы, экспрессирующие химерные гликопротеины.
* Аденовирусы (например, типов 5, 7 или 35): в US 2007/077257, US 2007/054395, JP 2007037402, WO 2006/120034, US 2004/253210, US 2004/170647, US 2005/070017, US 2003/228329, US 2004/101957, US 2003/219458, US 2004/009936, US 2004/028652, WO 03/050238, WO 03/038057, WO 03/020893, WO 02/31168, WO 02/22080, WO 01/02607 и US6716823, в которых раскрыты рекомбинантные аденовирусы, экспрессирующие белки ВИЧ.
* Поксвирус (канарипокс, вирус коровьей оспы, вирус коровьей оспы Анкара и вирус оспы кур): в US 5766598, ЕР 0592546, US 2007/048861, US 2006/188961, US 2006/134133, ЕР 1789438, WO 2005/017208, WO 2004/035006, US 2004/146528, JP 2003321391, ЕР 1378516, WO 95/07099, JP 7170982, DE 4141741, ЕР 0449116, JP 1148183, JP 1085072, ЕР 0592546, ЕР 0243029, US 2005/287162, JP 2004105187, JP 2004089185, WO 03/095656, ЕР 0592546, WO 96/40880, US 6,136,318, US 5,670,367, в которых раскрыты рекомбинантные поксвирусы, экспрессирующие вирусные белки, в том числе белки ВИЧ.
* Бактерии, которые содержат, которые продуцируют или которые экспонируют на своей поверхности по меньшей мере один белок вируса: в US 7189402 и WO 96/11708, в которых раскрыты Salmonella или Е.coli, экспрессирующие гликопротеины ВИЧ (т.е., белки оболочки).
Предпочтительно, рекомбинантная вирусная частица соответствует поксвирусу, при этом поксвирус предпочтительно выбран из группы, содержащей канарипокс (например, вирусные векторов ALVAC, такие как раскрытый в патентах US 5766598 и ЕР 0592546), вирус коровьей оспы (например, вирус коровьей оспы, раскрытый в международной публикации патента WO 95/07099), вирус коровьей оспы Анкара (например, вирусные векторы NYVAC, такие как описанный в заявке на патент ЕР 1789438) и, вирус оспы кур (например, вирусные векторы TROVAC, такие как раскрыт в международной заявке на патент WO 03/095656).
Более предпочтительно, указанный поксвирус является канарипокс вирусом. В качестве примера рекомбинантной вирусной частицы, соответствующей канарипокс вирусу и экспрессирующей пептид/белок ВИЧ, можно дать ссылку на вирусные векторы ALVAC, раскрытые в патенте US 5766598 (содержание которого включено сюда начиная с колонки 6, строки 18 и до колонки 82, строки 36), при этом векторы ALVAC экспрессируют, например, белок gp120 из ВИЧ-1, gp160 из ВИЧ-1, нерасщепляемую секретируемую форму env из ВИЧ-1, gp120 из ВИЧ-1, заякоренный с помощью трансмембранной последовательности, gag/pol из ВИЧ-1, gag/pol и env (gp120) из ВИЧ-1, gag/pol и env (gp160) из ВИЧ-1 и gag/pol и env (gp120 с трансмембранной заякоривающей последовательностью) из ВИЧ-1. Предпочтительно, указанный вектор ALVAC экспрессирует gag/pol и env (gp120) из ВИЧ-1, и наиболее предпочтительно - указанный вектор ALVAC является вектором ALVAC VCP1521.
«Вирусная частица» предпочтительно представляет собой частицу SIV или ВИЧ, такую как вирусная частица SIV или ВИЧ, которая может содержать мутированный вирусный геном (например, путем внесения мутации, замены или вставки в нуклеиновую кислоту), что приводит к выработке неинфекционных вирусных частиц.
Вирусные частицы, содержащие мутированный вирусный геном, раскрыты в US 7229625, US 6121021, US 6923970, US 6544527, US 6451322 и US 6080408.
Предпочтительно, и для того, чтобы получить вирусные частицы или рекомбинантные вирусные частицы, которые можно безопасно вводить человеку, указанные вирусные частицы или рекомбинантные вирусные частицы перед введением инактивируют. Такая инактивация может быть необходима для рекомбинантных вирусных частиц, даже для тех из них, которые не способны к репликации.
При использовании в этом документе «инактивированная вирусная частица» (причем указанная вирусная частица может быть как рекомбинантной, так и не быть рекомбинантной) означает вирусную частицу, которая больше не является инфекционной и, предпочтительно, больше не способна к репликации.
Способы инактивации вирусных частиц или рекомбинантных вирусных частиц хорошо известны специалистам в данной области. Неограничивающие примеры инактивации вирусов включают химическую инактивацию, например, обработкой с помощью формалина, хлорамина таурина, формальдегида, параформальдегида, пропиолактена, бета-пропиолактона (REMUNE) или алдритиола-2 (aldrithiol-2, АТ-2, см. US 6001155), термическую инактивацию, физическую инактивацию, например, с помощью УФ- или гамма-облучения или обработки микроволновым излучением, и их сочетания. Информацию по инактивации ВИЧ см. в публикации: RAVIV et al. (J. Virol., vol.79(19), р: 12394-12400, 2005).
В соответствии с одним из вариантов воплощения указанная инактивация является химической инактивацией, выбранной из группы, состоящей из инактивации помощью формалина, хлорамина таурина, формальдегида, параформальдегида, пропиолактена, бета-пропиолактона (REMUNE) или алдритиола-2.
В соответствии с другим вариантом или дополнительно, указанная инактивация является термической инактивацией. Такая инактивация хорошо известна специалистам в данной области и, в качестве примера такого способа, можно дать ссылку на способ, раскрытый в разделе примеров. Действительно, авторы изобретения неожиданно установили, что у макаков химически (т.е. с помощью АТ-2) и/или термически инактивированный вирус индуцирует защитную иммунотолерантность при введении вместе с непатогенной живой бактерией.
Предпочтительно, для целей введения людям, вирусные частицы инактивируют по меньшей мере дважды, в типичном случае с применением по меньшей мере двух способов инактивации, упомянутых выше.
Предпочтительно, как указано выше, вирусные частицы (рекомбинантные или нет, VLP или нет), которые применяют в качестве антигенов в фармацевтических композициях по настоящему изобретению, не зависят от нуклеиновой кислоты (т.е., ДНК или РНК), что означает, что вирусные частицы не содержат какой-либо вирусной ДНК или РНК или, если они содержат ДНК или РНК, она не играет роли в иммуногенности.
В соответствии с другим вариантом, полимерные микрочастицы (в форме микрокапсул, микросфер и т.п.) с различными структурами и презентирующие на своей поверхности один или несколько вирусных белков или пептидов или эпитопов, содержащих белки ВИЧ - Gag и/или Pol - или происходящих от них, могут применяться в качестве антигенов в фармацевтических композициях по настоящему изобретению. Такие микрочастицы могут быть изготовлены из соответствующих биологических или химических полимеров, таких как метакрилированный декстран, метакрилированный полиэтиленгликоль и/или желатин, на которые можно прикрепить вирус ВИЧ или вирусные белки или пептиды или эпитопы, содержащие белки ВИЧ - Gag и/или Pol - или происходящие от них. Примеры таких полимерных микрочастиц можно найти в литературных источниках (например, см. публикации: Wei Li Lee et al. (2010), Sandri et al. (2007), Goldberg et al. (2003), Delie F. (1998), Ponchel et al. (1998), Mathiowitz et al. (1997), Fasano et al. (1997), Chickering et al. (1997)).
В предпочтительном варианте воплощения антиген в фармацевтической композиции ВИЧ-1 по настоящему изобретению является одной или несколькими вирусными частицами, способными экспрессировать один или несколько вирусных белков или пептидов или эпитопов, содержащих белки ВИЧ - Gag и/или Pol - или происходящих от них. В соответствии с другим вариантом воплощения антиген в фармацевтической композиции ВИЧ-1 по настоящему изобретению является одной или несколькими полимерными микрочастицами, презентирующими на своей поверхности один или несколько вирусных белков или пептидов или эпитопов, содержащих белки ВИЧ - Gag и/или Pol - или происходящих от них.
Предпочтительно, антиген, который будет применяться в фармацевтической композиции по настоящему изобретению, имеет размер не менее чем примерно 110 кДа. Предпочтительно, он имеет размер по меньшей мере примерно 120, 130, 140, 150, 160, 170, 180, 190, 200 кДа или даже больше.
Эффективное количество вирусного антигена, которое будет применяться в контексте изобретения, может легко определить специалист в данной области, применяя общеизвестные знания и с учетом примеров, раскрытых в этом документе далее, в связи с вирусами SIV и ВИЧ.
В качестве примера, когда указанный антиген является крупнодисперсным антигеном и, в частности, вирусной частицей, количество вирусных частиц составляет от примерно 106 до примерно 1012 на миллилитр указанной смеси.
Непатогенная бактерия
Как показали авторы изобретения в случае вируса SIV у макаков, при введении через слизистые, или внутрикожным, или интраэпителиальным путем вместе с соответствующим антигеном, указанным выше, непатогенная живая бактерия в составе фармацевтической композиции способна индуцировать и, предпочтительно, поддерживать состояние иммунотолерантности к вышеуказанному антигену. У человека это дает возможность предотвращать и/или лечить заболевание ВИЧ.
Указанная бактерия может, таким образом, рассматриваться как состоящий из частиц адъювант, который может обозначаться в этом документе как «толерогенный адъювант» или «толерогенный носитель» или «носитель толерантности» или «носитель толеризации», при этом данные термины являются синонимичными.
Предпочтительно, все эти эквивалентные термины означают непатогенную живую бактерию, которую применяют в сочетании с антигеном ВИЧ, указанным выше, для достижения специфической иммунной защиты (предпочтительно, иммунотолерантности) к антигену, тем самым предотвращая и/или обеспечивая лечение заболевания ВИЧ у человека.
Более предпочтительно, «толерогенный носитель» - это непатогенная живая бактерия, которую вводят в смеси с антигеном ВИЧ, указанным выше, с целью достижения одного или нескольких, предпочтительно - 2 или нескольких, еще более предпочтительно - 3 или нескольких из следующих иммунопротекторных эффектов:
1) «Толерогенный носитель» не индуцирует существенный уровень выработки системных антител, специфичных к антигену ВИЧ.
В частности, не наблюдается существенного уровня выработки системных антител против ВИЧ классов IgM и/или IgG. Например, не наблюдается существенного уровня системного гуморального ответа, то есть, либо отсутствует специфический детектируемый системный гуморальный иммунный ответ, который можно определить с применением классических клинических лабораторных способов, таких как анализ ELISA, либо, в случае, если детектируются системные антитела, они не могут служить защитой от инфекции вируса ВИЧ.
2) «Толерогенный носитель» не индуцирует существенный уровень специфической пролиферации CD4+ Т-клеток, вызванной антигеном ВИЧ.
В частности, не наблюдается существенного уровня пролиферации CD4 Т-клеток, специфических к антигену ВИЧ, при стимуляции in vitro антигеном ВИЧ, согласно измерению с применением стандартных аналитических способов, таких как описаны в сопутствующих примерах.
3) «Толерогенный носитель» не индуцирует существенный уровень выработки CD8+ Т-клетками гамма-интерферона при стимуляции in vitro антигеном ВИЧ.
В частности, уровень секреции гамма-интерферона CD8+ T-клетками, который наблюдается при стимуляции in vitro антигеном ВИЧ, находится ниже порогового уровня для иммуноферментного спот-анализа.
4) «Толерогенный носитель» индуцирует существенный уровень CD8+ Т-клеточного ответа, супрессирующего активацию презентирующих антиген ВИЧ CD4+ Т-клеток.
В частности, этот ответ может быть определен путем измерения in vitro уровня ингибирования CD8+ Т-клетками репликации вируса (который служит показателем «существенного уровня» CD8+ Т-клеточного ответа), как показано в сопутствующих примерах. Эти CD8+ Т-клетки также называют CD8+ «регуляторными» Т-клетками. Также в частности, этот ответ не является цитотоксичным при условии, что, например, он не индуцирует существенный уровень выработки гамма-интерферона. Также в частности, этот ответ ограничивается комплексом MHC-Ib/E. Также в частности, по-видимому, в CD8+ T-клеточный ответ, супрессирующий репликацию вируса, вовлечен TCR-αβ. Также в частности, этот ответ супрессирует активацию презентирующих антиген ВИЧ CD4+ T-клеток, по сравнению с той же популяцией клеток, обедненной CD8+ T-клетками. Предпочтительно, указанный ответ супрессирует раннюю активацию презентирующих антиген ВИЧ CD4+ T-клеток, при этом указанная «ранняя» активация измеряется по маркеру Ki67+ (Scholzen and Gerdes. J. Cell Physiol. 182, 311-322 (March 2000)).
Под терминами «не индуцирует», применяемыми выше в пп.1), 2) и 3), подразумевается, что результат ниже порогового уровня для соответствующего способа количественного определения, при этом указанный «пороговый уровень» представляет собой значение, определенное данным способом количественного определения на основании результатов отрицательных контрольных проб: при этом значении результат является отрицательным результатом. Это значение может варьировать от одного количественного определения к другому и от одного способа детекции к другому.
Предпочтительно, толерогенный носитель выбран из живых:
- непатогенных бактерий, в частности, пробиотических и симбиотических бактерий;
- ослабленных патогенных бактерий;
- инактивированных (необязательно, также предварительно ослабленных) патогенных бактерий.
Толерогенный носитель может быть рекомбинантным или нет.
«Непатогенные бактерии», которые будут применяться в качестве толерогенных носителей в контексте настоящего изобретения, в общем случае не вызывают какую-либо патологию у человека. По этой причине их относят к общепризнанным безопасным (GRAS). Безусловно, такие бактерии должны быть пригодными для введения человеку.
Предпочтительными непатогенными бактериями, которые можно применять в качестве толерогенных носителей, являются симбиотические бактерии. Такие бактерии хорошо известны специалистам в данной области. Неограничивающие примеры включают Bacillus sp. (например, В. coagulans), Bifidobacterium animalis, Bifidobacterium breve, Bifidobacterium infantis, Bifidobacterium longum, Bifidobacterium bifidum, Bifidobacterium lactis, Escherichia coli, Lactobacillus acidophilus, Lactobacillus bulgaricus, Lactobacillus casei, Lactobacillus paracasei, Lactobacillus johnsonii, Lactobacillus plantarum, Lactobacillus reuteri, Lactobacillus rhamnosus, Lactobacillus brevis, Lactobacillus gasseri, Lactobacillus salivarius, Lactococcus lactis, Streptococcus thermophilus и т.п.
«Симбиотические бактерии» для применения в качестве толерогенных носителей в контексте настоящего изобретения предпочтительно являются молочнокислыми бактериями или бифидобактериями, которые, в частности, выбраны из списка, приведенного выше, включая также их комбинации. Предпочтительной симбиотической бактерией является Lactobacillus sp., и более предпочтительно - Lactobacillus plantarum. В приведенных ниже примерах впервые показано, что Lactobacillus plantarum является толерогенным носителем, вызывающим иммунотолерантность к вирусу при введении вместе с антигеном, описанным выше.
Предпочтительно, комбинацию непатогенных бактерий, таких как два или несколько видов симбиотических бактерий, можно применять в качестве толерогенного носителя.
При использовании в этом документе термины «патогенные бактерии» означают бактерии, вызывающие патологии у человека. Такие бактерии хорошо известны специалистам в данной области и включают, среди прочего, Listeria разные виды (например, Listeria monocytogenes), Corynebacterium, разные виды, Mycobacterium, разные виды, Rhococcus, разные виды, Eubacteria, разные виды, Bortadella, разные виды и Nocardia, разные виды. Предпочтительно, патогенная бактерия выбрана из Mycobacterium, разные виды, и более предпочтительно - Mycobacterium bovis.
При использовании в этом документе «ослабленные патогенные бактерии» - это патогенные бактерии, которые в меньшей степени вирулентны, по сравнению со своими аналогами дикого типа, из-за одной или нескольких мутаций или одного или нескольких способов обработки для ослабления (например, химической обработки и/или последовательных пассажей на специфической среде). Такие ослабленные патогенные бактерии хорошо известны специалистам в данной области. Неограничивающие примеры ослабленных патогенных бактерий включают ослабленные Salmonella typhimurium и Mycobacteria, предпочтительно - ослабленную Mycobacteria. В качестве примера ослабленной Mycobacteria, можно назвать «бациллу Кальмета-Герена», также известную как «BCG (БЧЖ)», и, в частности, среди прочих, шесть широко применяемых штаммов BCG - эволюционно ранний штамм BCG Japanese, два эволюционно поздних штамма в DU2 группа III (BCG Danish и Glaxo) и три эволюционно поздних штамма в DU2 группа IV (BCG Connaught, Pasteur и Tice). В качестве еще одного примера ослабленной Mycobacteria можно также сослаться на рекомбинантную BCG, такую как штамм rBCG30, описанный в публикации: HOFT et al. (2008), рекомбинантную BCG, описанную в публикации: WANG et al (2008), а также рекомбинантную BCG, раскрытую в международных публикациях патентов WO 2005/111205 и WO 02/102409 и раскрытую в патентах US 7122195 и US 6261568.
Предпочтительно, вместо ослабления или в дополнение к ослаблению патогенные бактерии можно инактивировать для того, чтобы применять в качестве толерогенных носителей в контексте настоящего изобретения, но ослабление патогенных бактерий также можно применять после того, как они были инактивированы.
«Инактивированные патогенные бактерии» хорошо известны специалистам в данной области. Способы получения таких инактивированных патогенных бактерий хорошо известный специалистам в данной области. В качестве примеров таких способов можно сослаться на опосредованный фагом лизис, химическую инактивацию, например, обработку формалином (см. US 7393541), термическую инактивацию, физическую инактивацию, например, лиофилизацию (например, длительное замораживание-высушивание) или облучение УФ- или гамма-излучением (см. WO 2008/128065) или воздействие микроволнового излучения и их комбинации.
Предпочтительно, указанный толерогенный носитель является ослабленным производным патогенной бактерии типа BCG. В приведенных ниже примерах впервые показано, что BCG является толерогенным носителем, вызывающим иммунотолерантность к вирусу при введении вместе с антигеном, описанным выше.
Если он является рекомбинантным, то толерогенный носитель по настоящему изобретению не экспрессирует какие-либо белки или пептиды или эпитопы ВИЧ.
Предпочтительно, по меньшей мере существенное количество живых бактерий, применяемых в качестве толерогенного носителя, находятся в середине фазы логарифмического роста. Более предпочтительно, по меньшей мере примерно 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или даже примерно 100% от суммарного количества бактериальных клеток находятся в середине фазы логарифмического роста.
Эффективное количество толерогенного носителя может легко определить специалист в данной области, и примеры такого эффективного количества раскрыты в этом документе далее.
В качестве примера, количество указанной бактерии в фармацевтической композиции по настоящему изобретению составляет от примерно 104 до примерно 1014 КОЕ на 1 мл указанной смеси.
Фармацевтическая композиция
Указанная композиция также называется в этом документе «толерогенной композицией» или «толеро-иммуногенной композицией», при этом данные термины являются эквивалентными.
Толерогенный носитель и антиген, содержащий белки ВИЧ - Gag и/или Pol - или происходящий от них, - это два отдельных и различных компонента, которые содержатся в виде смеси в фармацевтической композиции по настоящему изобретению. Это означает, что указанный толерогенный носитель и указанный антиген присутствуют в виде отдельных компонентов в указанной композиции.
Предпочтительно, фармацевтическая композиция по изобретению не содержит какого-либо олигонуклеотида (например, CpG или дцРНК) в качестве адъюванта.
Так как толерогенный носитель является бактерией, бактерию того же рода и/или вида можно отдельно применять в рекомбинантной форме как источник антигена. Например, рекомбинантная бактерия будет содержать нуклеиновую кислоту, кодирующую антиген, которая помещена под контроль соответствующих регуляторных последовательностей (включая промоторы, индуцибельные или конститутивные), либо в виде состоящего из нуклеиновой кислоты вектора, содержащегося внутри клетки, либо в виде последовательности нуклеиновой кислоты, интегрированной в бактериальную хромосому. Благодаря этому рекомбинантная бактерия будет способна экспрессировать или продуцировать указанный антиген. Таким образом, в соответствии с конкретным вариантом воплощения, фармацевтическая композиция по настоящему изобретению включает толерогенный носитель, который является бактерией, и антиген, который является одним или несколькими вирусными белками или пептидами или эпитопами, содержащими белки ВИЧ - Gag и/или Pol - или происходящими от них, и который был отдельно продуцирован рекомбинантной бактерией, принадлежащей к тому же роду и/или виду, что и толерогенный носитель.
В фармацевтической композиции по настоящему изобретению, если указанный антиген является крупнодисперсным антигеном и, более предпочтительно, вирусной частицей, отношение в указанной смеси количества указанной вирусной частицы (выраженного в частицах на мл указанной смеси) к количеству указанной бактерии (выраженному в КОЕ на мл указанной смеси) составляет от примерно 1:10 до примерно 1:1000, предпочтительно - от примерно 1:25 до примерно 1:750, еще предпочтительнее - от примерно 1:50 до примерно 1:500, даже еще предпочтительнее - от примерно 1:75 до примерно 1:250 и еще более предпочтительно - примерно 1:100.
Введение фармацевтической композиции по изобретению
Может быть возможно введение толерогенного носителя и антигена либо одновременно, либо по отдельности, либо последовательно.
Таким образом, целью настоящего изобретения является создание фармацевтического набора для профилактики и/или лечения заболевания ВИЧ у человека, которому это необходимо, при этом набор содержит:
- в первом контейнере - антиген, указанный выше; и
- во втором контейнере - непатогенную живую бактерию, указанную выше, при этом указанный антиген и указанная бактерия находятся в фармацевтически приемлемых носителях для ведения через слизистую или внутрикожно или интраэпителиально.
Также целью настоящего изобретения является создание продуктов, содержащих:
- непатогенную живую бактерию в качестве толерогенного носителя, указанную выше; и
- крупнодисперсный антиген или антиген, содержащий один или несколько эпитопов из белков ВИЧ - Gag и/или Pol, указанный выше,
в виде комбинированной фармацевтической композиции для одновременного, раздельного или последовательного применения для профилактики и/или лечения заболевания ВИЧ у человека, которому это необходимо. Указанная профилактика и/или лечение достигаются путем ведения через слизистую или внутрикожно или интраэпителиально указанной комбинированной фармацевтической композиции указанному человеку. Для достижения этого может быть возможно введение толерогенного носителя и антигена либо одновременно, либо по отдельности, либо последовательно.
В качестве примера, непатогенная живая бактерия может быть введена перорально (например, в виде лекарственного средства для перорального приема или пищевой добавки), тогда как антиген вводят через слизистую, или внутрикожно, или интраэпителиально.
Конечно, могут применяться соответствующие фармацевтические носители для обеспечения правильной доставки к каждому предполагаемому месту введения (например, к поверхности слизистой). Время введения и вводимую дозу каждого толерогенного носителя и антигена сможет легко скорректировать опытный специалист.
Предпочтительно, фармацевтическая композиция по настоящему изобретению является фармацевтической композицией для введения через слизистые, или внутрикожно, или интраэпителиально. Еще более предпочтительно, она является фармацевтической композицией для перорального приема.
При использовании в этом документе «фармацевтическая композиция для введения через слизистые, или внутрикожно, или интраэпителиально» - это фармацевтическая композиция, которая предназначена для введения через слизистые, или внутрикожно, или интраэпителиально, т.е., приготовлена в виде готовой лекарственной формы для такого введения.
В частности, фармацевтическая композиция может дополнительно включать один или несколько соответствующих фармацевтических носителей (или основ) для доставки через слизистые, или внутрикожно, или интраэпителиально указанного антигена и указанной бактерии.
Предпочтительно, «доставка через слизистые» в этом документе выбрана из интраназальной, пероральной, подъязычной, трахеальной, глоточной, бронхиальной, пищеводной, желудочной, дуоденальной, кишечной, ректальной, препуциальной и вагинальной доставки. «Доставка через слизистые» - это доставка к поверхности слизистой, такой как слизистая поверхность носа, ротовой полости, подъязычная, трахеальная, бронхиальная, глоточная, пищеводная, желудочная и слизистая двенадцатиперстной кишки, тонкого и толстого кишечника, включая прямую кишку, а также слизистая крайней плоти и влагалища. В данном контексте поверхность слизистой также включает наружную поверхность глаза, т.е., слизистую, окружающую глаз. Еще предпочтительнее, поверхность слизистой относится к слизистой влагалища или пищеварительного тракта, и более предпочтительно - к слизистой пищеварительного тракта. Еще предпочтительнее, доставка через слизистую - это пероральная доставка.
Таким образом, фармацевтическая композиция также может включать один или несколько фармацевтических носителей, в зависимости от пути введения. Специалистам в области фармацевтики известны, или легко могут быть установлены, носители для доставки лекарств к поверхности слизистой или для внутрикожной или интраэпителиальной доставки. Полезные ссылки для этого: Chien (Novel Drug delivery system, Chapters 3 through 6 and 9, Marcel Dekker, 1992), Ullmann's Encyclopedia of Industrial Chemistry, 6th Ed. (various editors, 1989-1998, Marcel Dekker); and Pharmaceutical Dosage Forms and Drug Delivery Systems (ANSEL et al., 1994, WILLIAMS & WILKINS).
Примеры способов и путей доставки лекарства, полезных применительно к изобретению, кратко описаны ниже.
Введение через слизистую бронхов, бронхиол, трахеи, носовой полости, ротовой полости, крайней плоти или глотки может быть достигнуто путем приготовления фармацевтической композиции в виде ингаляционной лекарственной формы, в виде лекарственной формы спрея и т.п. (например, назального спрея, аэрозольного спрея или спрея с насосом и т.п.), раствора, геля и т.д. Устройства типа небулайзера, пригодные для доставки фармацевтических композиций на слизистую носовой полости, трахеи или бронхиол, хорошо известны специалистам в данной области, и поэтому не будут описываться здесь подробно. Фармацевтическая композиция может дополнительно содержать носитель, выбранный из группы, содержащей растворы, эмульсии, микроэмульсии, эмульсии масло-в-воде, безводные липиды и эмульсии масло-в-воде, другие типы эмульсий.
Введение через слизистую влагалища может быть достигнуто путем приготовления фармацевтической композиции в виде лекарственной формы раствора, клизмы, пены, суппозитория, вагинальной таблетки или геля для местного применения. Предпочтительные носители для вагинальной доставки включают гидрофильные и гидрофобные носители, такие как носители, стандартно применяемые при приготовлении лекарственных форм эмульсии или геля (например, эмульсии или геля из масла и воды).
Введение через слизистую пищеварительного тракта может быть достигнуто путем приготовления фармацевтической композиции в виде лекарственной формы капсулы, микрокапсулы. Предпочтительные носители для доставки через слизистую пищеварительного тракта соответствуют капсулам и микрокапсулам (например, капсулам и микрокапсулам из пектина и/или альгината), обычно принимаемым перорально, таким как стандартно применяемые для приготовления лекарственных форм для доставки через пищеварительный тракт (например, микрокапсулы, раскрытые в международной заявке на патент WO 2007/140613). В соответствии с другим вариантом доставка через пищеварительный тракт может быть достигнута путем приема внутрь или введения соответствующих жидкостей и/или продуктов питания, таких как напитки, йогурты и т.п.
Внутрикожное или интраэпителиальное введение хорошо известны специалистам в данной области. Внутрикожное введение (например, инъекция) может, например, быть произведено с применением устройств с иглой, таких как раскрыты в патенте US 6933319 и в международной заявке на патент WO 2004/101025, или с применением безыгольных устройств.
Фармацевтическая композиция может дополнительно включать по меньшей мере одно вещество для абсорбции. «Вещества для абсорбции» хорошо известны специалистам в данной области. В качестве примеров, можно сослаться на поверхностно-активные вещества, такие как полиоксиэтиленовые производные неполных эфиров жирных кислот и ангидридов сорбитола (например, Твин® 80, полиоксил 40 стеарат, полиоксиэтилен 50 стеарат, полиоксиэтилен-9-лауриловый эфир и октоксинол), желчные кислоты, такие как гликохолат натрия, смешанные мицеллы, енамины, доноры оксида азота (например, S-нитрозо-N-ацетил-DL-пеницилламин, NOR1, NOR4, которые преимущественно вводят совместно с акцепторами NO, такими как карбокси-PITO или доклофенак натрия), салицилат натрия, сложные эфиры глицерина и ацетоуксусной кислоты (например, глицерил-1,3-диацетоацетат или 1,2-изопропилиденглицерин-3-ацетоацетат), циклодекстрина или бета-производные циклодекстрина (например, 2-гидроксипропил-бета-циклодекстрин и гептакис(2,6-ди-O-метил-бета-циклодекстрин)), жирные кислоты со средней длиной цепочки, такие как моно- и диглицериды (например, экстракты капрата натрия из кокосового масла, Капмул), или триглицериды (например, амилодекстрин, Эстарам 299, Миглиол 810), полимеры, такие как карбоксиметилцеллюлоза, карбопол, поликарбофил, трагакант и альгинат натрия, а также другие вещества для абсорбции, адаптированные для доставки через слизистые или внутрикожно или интраэпителиально. Информацию об общих принципах применения веществ для абсорбции, которые успешно применялись для доставки через слизистые или внутрикожно или интраэпителиально лекарственных препаратов, см. публикацию: Chien, Novel Drug Delivery Systems, Ch. 4 (Marcel Dekker, 1992).
Фармацевтическая композиция может дополнительно включать одну или несколько добавок (например, разбавителей, вспомогательных веществ, стабилизаторов, консервантов и т.п.) Общую информацию см. в публикации: Ullmann's Encyclopedia of Industrial Chemistry, 6th Ed. (various editors, 1989-1998, Marcel Dekker); and Pharmaceutical Dosage Forms and Drug Delivery Systems (ANSEL et al., 1994, WILLIAMS & WILKINS).
Как описано ниже, соответствующие дозировки фармацевтической композиции по настоящему изобретению, которые будут вводить пациенту, который является человеком, могут быть определены по одной или нескольким характеристикам указанного пациента, таким как пол, возраст, масса тела, состояние здоровья и т.д.
В качестве примера, когда антиген находится в крупнодисперсной форме и, в частности, когда он является вирусной частицей, то указанному человеку может быть введена доза от примерно 108 до примерно 1014 вирусных частиц в сутки. В качестве еще одного примера, указанному человеку может быть введена доза непатогенной живой бактерии от примерно 106 до примерно 1016 КОЕ в сутки.
Применение фармацевтической композиции по изобретению
Еще одной целью настоящего изобретения является предложение фармацевтической композиции, описанной выше, для применения в качестве лекарственного средства, предпочтительно - в качестве вакцины.
Настоящее изобретение также относится к способу профилактики и/или лечения заболевания ВИЧ у человека, которому это необходимо, включающему по меньшей мере этап введения через слизистые, или внутрикожно, или интраэпителиально эффективного количества фармацевтической композиции, описанной выше, указанному человеку.
В соответствии с настоящим изобретением для профилактических целей «человек, которому это необходимо» может являться любым человеком, предпочтительно - в возрасте не менее чем примерно 2 лет. Для терапевтических целей «человек, которому это необходимо» - это человек, которому необходимо лечение, так как он(а) страдает заболеванием ВИЧ.
«Заболевание ВИЧ» означает любое связанное с ВИЧ иммунное нарушение, включая СПИД, а также более ранние стадии прогрессирования заболевания, в том числе сероконверсию (развитие хронической инфекции).
Настоящее изобретение дополнительно относится к способу защиты человека от заболевания ВИЧ, включающему по меньшей мере этап введения через слизистые, или внутрикожно, или интраэпителиально эффективного количества фармацевтической композиции, описанной выше, указанному человеку.
В такой способ позволяет защитить человека от инфекции ВИЧ при воздействии на ВИЧ при введении через слизистые, и/или от репликации ВИЧ, при воздействии на ВИЧ при внутривенном введении.
Настоящее изобретение также дополнительно относится к способу защиты человека от сероконверсии под действием ВИЧ, включающему по меньшей мере этап введения через слизистые, или внутрикожно, или интраэпителиально эффективного количества фармацевтической композиции, описанной выше, указанному человеку. Таким образом, указанный человек не станет сероположительным и у него не будет наблюдаться существенный уровень антител против ВИЧ.
Термин «вакцинация» означает действие (или действия) (главным образом - введение фармацевтической композиции по настоящему изобретению), которое предпринимается для профилактики и/или лечения заболевания ВИЧ у человека. Предпочтительно, фармацевтическая композиция по изобретению полезна для индуцирования и, предпочтительно, поддержания иммунотолерантности к антигену, который содержит белки ВИЧ - Gag и/или Pol - или происходит от них, у человека, то есть, иными словами, для вакцинации (или «толеризации») указанного человека. Таким образом, вакцинирование человека с применением лекарственного средства по настоящему изобретению называют «толерогенной вакцинацией» (или «толеризацией»).
Если после ведения через слизистую или внутрикожно или интраэпителиально фармацевтической композиции по изобретению (т.е., после толерогенной вакцинации) у человека успешно была индуцирована иммунотолерантность, то указанный человек считается «вакцинированным» (или «толеризированным», или «толерантным»). Ответ, т.е., репликация вируса согласно оценке по нагрузке вирусной РНК в плазме, у «вакцинированного» человека при провокационной пробе с инфекционным вирусом in vivo снижен по меньшей мене примерно на 50%, более предпочтительно - по меньшей мене примерно на 70%, еще более предпочтительно - по меньшей мене примерно на 75% или 80% или 85% или 90% или 95% или 98% или 99% или даже больше (99,5%, 99,8%, 99,9%, 100%), относительно нагрузки вирусной РНК в плазме служащего контролем человека, которому ввели либо только антиген, либо антиген, ассоциированный со стандартным адъювантом (как указано выше), либо не вводили фармацевтическую композицию или плацебо.
В соответствии с настоящим изобретением толерогенная вакцинация может включать одно или несколько последовательных введений фармацевтической композиции. Предпочтительно, толерогенная вакцинация может включать по меньшей мере два или несколько последовательных введений (т.е., вакцинаций) и более предпочтительно - более чем два последовательных введения указанной композиции.
Предпочтительно, интервал между последовательными толерогенными вакцинациями составляет от 1 мин до 3 мес., предпочтительно - от 15 мин до 2 мес.
Также предпочтительно, толерогенные вакцинации по изобретению могут включать повторные толерогенные вакцинации через один год или несколько лет после первой толерогенной вакцинации через слизистую, или внутрикожно, или интраэпителиально (например, через срок от 1 до 10 лет).
Новые толерогенные вакцинации после первой толерогенной вакцинации через слизистую, или внутрикожно, или интраэпителиально могут быть выбраны из толерогенных вакцинаций через слизистую, или внутрикожно, или интраэпителиально. Следует отметить, что если новые толерогенные вакцинации являются внутрикожными или интраэпителиальными инъекциями, то может детектироваться специфический системный гуморальный и/или цитотоксический (с выработкой гамма-интерферона) ответ, но он не играет роли в профилактике или лечении заболевания.
В соответствии с настоящим изобретением эффективное количество фармацевтической композиции вводят человеку, которому это необходимо. Термин «эффективное количество» означает количество, достаточное для достижения требуемого биологического эффекта, который в этом документе является лечебным или профилактическим эффектом (иными словами, иммунопротекторным эффектом), через индукцию иммунотолерантности, предпочтительно - «Ts»-иммунотолерантности. При этом понимается, что эффективная дозировка будет зависеть от возраста, пола, состояния здоровья и массы тела пациента, который будет получать лечение, типа сопутствующего лечения, если оно имеется, частоты лечения и природы ожидаемого эффекта. Диапазоны эффективных доз, указанные ниже, не предназначены для ограничения изобретения и представляют собой предпочтительные диапазону доз. Однако предпочтительная дозировка может быть скорректирована для пациента по усмотрению и на основании оценки специалиста в данной области, без проведения чрезмерной экспериментальной работы. См. например, публикацию: Ebadi, Pharmacology, Little, Brown and Co., Boston, Mass. (1985).
Например, в отношении ВИЧ, типичная дозировка для взрослого человека будет составлять от примерно 106 до 1012 вирусных частиц ВИЧ (т.е., VLP, рекомбинантных или нерекомбинантных вирусных частиц) на дозу, при этом предпочтительно - 108-1010. Безусловно, какая бы доза ни применялась, это должно быть безопасное и эффективное количество, определенное известными способами, а также как описано в этом документе.
Более того, специалист в данной области также может определить с учетом своих общих знаний эффективное количество толерогенного носителя, которое необходимо ввести человеку для достижения требуемого биологического эффекта.
В качестве примера, указанное эффективное количество для ослабленной производной формы патогенных бактерий (например, BCG) находится в диапазоне от 104 до 1012, предпочтительно - от 105 до 1010 КОЕ (колониестимулирующих единиц) и более предпочтительно - от 106 до 108 КОЕ на дозу. В качестве еще одного примера, указанное эффективное количество ослабленной производной формы патогенных бактерий или инактивированных патогенных бактерий (например, BCG) находится в диапазоне от 0,001 мг до 1 г, предпочтительно - от 0,01 до 100 мг и более предпочтительно - от 0,1 до 10 мг на дозу.
В качестве еще одного примера, указанное эффективное количество непатогенных бактерий (например, Lactobacillus sp.) находится в диапазоне примерно от 106 до 10-10 КОЕ, и более предпочтительно - примерно от 1010 до 10-10 КОЕ на дозу.
Как описано выше, фармацевтические композиции по настоящему изобретению пригодны для профилактики заболевания ВИЧ в будущем у человека или для лечения человека, уже страдающего заболеванием ВИЧ.
Для терапевтических целей «антиген, который содержит белки ВИЧ - Gag и/или Pol - или происходит от них», описанный выше, может быть аутологичным, то есть, он может происходить от вируса ВИЧ, инфицирующего человека, которого будут лечить. В таком случае, например, вирус ВИЧ может быть изолирован из организма человека, затем его могут культивировать и инактивировать (предпочтительно - инактивировать по меньшей мере дважды) и наконец - соединить с толерогенным носителем таким образом, чтобы получить фармацевтическую композицию, описанную выше.
Например, фармацевтическая композиция, содержащая аутологичные или не аутологичный антиген, который содержит белки ВИЧ - Gag и/или Pol - или происходит от них, могут вводить человеку в течение периода стандартного противовирусного лечения, что в первую очередь, привело бы к недетектируемому уровню вирусной нагрузки. Затем стандартная противовирусная терапия может быть прекращена после одной или нескольких толерогенных вакцинаций с применением фармацевтической композиции, при условии, что достигается соответствующая супрессия репликации вируса ex vivo в не аутологичных CD4 Т-клетках с острой инфекцией под влиянием аутологичных вирусоспецифических CD8 Т-клеток, или при условии, что достигается соответствующая супрессия активации CD4 Т-клеток, индуцированная CD8 Т-клетками.
В частности, для терапевтических целей, фармацевтическая композиция может быть введена только один раз в течение жизни человека, которого будут лечить. В соответствии с другим вариантом, она может быть введена два или несколько раз в течение жизни человека, которого будут лечить, в тот же день или в разные дни, разделенные интервалом в диапазоне, например, от примерно 1 дня до примерно 1 года или более. В частности, она может вводиться каждый день или периодически, в течение периодов в диапазоне, например, от примерно 1 дня до примерно 1 года или более. При необходимости, фармацевтическая композиция может вводиться в течение всей жизни человека, которого будут лечить.
Настоящее изобретение дополнительно направлено на создание способа определения in vitro того, имеет ли человек защиту от ВИЧ, при этом способ включает:
a) выделение CD8 Т-клеток периферической крови из образца крови указанного вакцинированного человека;
b) культивирование в соответствующих условиях:
(i) указанных выделенных CD8 Т-клеток с аллогенными или аутологичными CD4+ T-клетками, которые были подвергнуты острой инфекции in vitro штаммом вируса, эквивалентным указанному ВИЧ; и
(ii) указанных аллогенных или аутологичных CD4+ T-клеток, которые были подвергнуты острой инфекции in vitro;
с) извлечение супернатанта культур клеток;
d) измерение вирусной нагрузки в указанных супернатантах; и
e) определение, имеется ли у указанного человека защита против указанного ВИЧ или нет.
Под «штаммом вируса, эквивалентным вирусу ВИЧ, тестирование которого будет проводиться», подразумевается, что указанный штамм вируса происходит от вируса дикого типа и обладает существенными характеристиками, схожими с таковыми вируса ВИЧ, тестирование которого будет проводиться (например, можно сослаться на штамм вируса HTLVIIIB, который происходит от отдельного ВИЧ-1: HTLVIIIB можно рассматривать как «штамм вируса, эквивалентный» ВИЧ-1). Предпочтительно, указанный штамм вируса будет происходить от вируса дикого типа, который является вирусом ВИЧ, тестирование которого будет проводиться. Таким образом, указанный штамм вируса представляет собой подходящую модель для изучения, включающего вирус ВИЧ, особенно вирус ВИЧ дикого типа. Штамм вируса, конечно же, хорошо приспособлен для таких исследований, особенно с точки зрения безопасности.
Все указанные выше этапы могут быть проведены с применением стандартных способов, хорошо известных специалистам в данной области. В частности, соответствующие условия культивирования для этапа b) составляют часть знаний в области, к которой относится изобретений (таких как стандартные способы, описанные в примерах ниже).
Вирусная нагрузка может быть измерена на этапе d) с применением стандартных способов, таких как описаны в примерах ниже.
Вирусная нагрузка в супернатанте, полученном из культуры указанных аллогенных или аутологичных CD4+ Т-клеток, которые были подвергнуты острой инфекции in vitro в соответствии с под-этапом b)(ii), будет применяться в качестве значения сравнения при проведении определения на этапе е). Можно преимущественно вычислить «процент супрессии (%)» или «коэффициент супрессии» или «противовирусный эффект», например, сравнив геометрические средние значения концентрации вируса в супернатанте для двух повторов (или трех повторов, или четырех повторов, или более) лунок, содержащих только клетки из аллогенных или аутологичных CD4+ Т-клеток, которые были подвергнуты острой инфекции in vitro, с геометрическими средними значениями концентрации вируса в супернатанте для двух повторов (или трех повторов, или четырех повторов, или более) лунок, содержащих CD8 Т-клетки и клетки из аллогенных или аутологичных CD4+ Т-клеток, которые были подвергнуты острой инфекции in vitro.
Далее, указанный этап определения е) предпочтительно выполняют следующим образом:
если коэффициент супрессии выше, чем примерно 100, можно сделать вывод о том, что указанный человек обладает защитой. В типичном случае, это будет иметь место тогда, когда не инфицированному БИЧ человеку провели эффективную профилактической противовирусной лечение, или когда инфицированному ВИЧ человеку провели эффективное терапевтическое лечение, при этом указанное эффективное профилактическое или терапевтическое лечение включает предпочтительно фармацевтическую композицию по настоящему изобретению, и, таким образом, не будет необходимости дополнительно проводить какое-либо профилактическое или терапевтическое лечение у человека, у которого сохраняется защита.
Если коэффициент супрессии ниже, чем примерно 100, можно сделать вывод о том, что указанный человек не обладает защитой от указанного вируса. Тогда человеку, либо не инфицированному ВИЧ человеку, либо инфицированному ВИЧ человеку, будет предпочтительно проведено профилактическое или терапевтическое лечение, включающее фармацевтическую композицию по настоящему изобретению, соответственно, и указанный выше способ in vitro будет осуществлен один раз или несколько раз, с соответствующими интервалами времени, для того, чтобы убедиться, что человек приобрел защиту.
Дополнительно, настоящее изобретение направлено на создание набора для определения in vitro, обладает ли человек защитой от вируса ВИЧ, при этом набор включает аллогенные или аутологичные CD4+ Т-клетки, которые могут быть инфицированы штаммом вируса, при этом указанный штамм вируса, как указано выше, эквивалентен указанному вирусу ВИЧ, тестирование которого будет проводиться. Набор также может включать соответствующий штамм вируса в подходящей концентрации для инфицирования указанных выше аллогенных или аутологичных CD4+ Т-клеток и/или соответствующие реактивы и/или контроли и/или среду (например, среду для суспензии клеток, культуры клеток, хранения клеток и т.д.) Набор по настоящему изобретению может быть специфическим для отдельного типа вируса ВИЧ, или же он может быть адаптирован к различным типам вируса, при этом указанные типы вируса являются близкими (в частности, филогенетически близкими).
Настоящее изобретение можно легко адаптировать с целью его применения для профилактики и/или лечения любых хронических инфекционных заболеваний. Неограничивающие примеры таких заболеваний включают: гепатит В и С, папилломавирус человека (HPV), EBV и другие вирусы герпеса, туберкулез, проказу, лейшманиоз и т.д.
В целом, каждый раз, когда один или несколько патогенных антигенов, ассоциированных с упомянутыми выше инфекциями или заболеваниями, вовлечены в специфическую активацию CD4+ Т-клеток, которые презентируют эпитопы, происходящие от упомянутых выше патогенных белков или пептидов, может быть вызвана специфическая супрессия/предотвращение активации CD4+ Т-клетки нецитотоксичными CD8+ T-клетками в результате воздействия вводимых через слизистую или внутрикожно или интраэпителиально фармацевтических композиций, объединяющих указанный(е) выше антиген(ы) и толерогенный носитель, описанные в этом документе.
В этом документе показано, что вирусные инфекции и связанные с ними заболевания можно предотвращать и/или лечить у млекопитающих/человека с применением фармацевтических композиций по настоящему изобретению. На основании этой идеи можно предложить другие фармацевтические композиции, содержащие (i) толерогенные носители, описанные в этом документе, и (ii) любой антиген вирусного, бактериального, грибкового, протозойного или паразитарного происхождения. Такие фармацевтические композиции созданы в виде лекарственной формы для соответствующего способа доставки (предпочтительно - через слизистую или внутрикожно или интраэпителиально) указанных толерогенных носителей и указанных антигенов. Они полезны для профилактики и/или лечения хронических инфекций у млекопитающих, вызванных вирусами, бактериями, грибками, простейшими или паразитическими организмами, от которых происходят антигены. Примером является бактериальная фармацевтическая композиция, содержащая (i) толерогенный носитель, описанный в этом документе; и (ii) антиген, происходящий от Mycobacterium tuberculosis. Эта бактериальная фармацевтическая композиция создана в виде лекарственной формы для соответствующего способа доставки (предпочтительно - через слизистую или внутрикожно или интраэпителиально) указанного антигена и указанного толерогенного носителя. Особый интерес с точки зрения профилактики и/или лечения туберкулеза у человека представляет такая бактериальная фармацевтическая композиция, в которой микобактериальный антиген происходит из палочки Коха.
Иммунная защита, достигаемая с помощью фармацевтической композиции по изобретению
Толерантность - это физиологическая способность иммунной системы распознавать антигены, проникшие через слизистую оболочку, и развивать анэргию, обычно ассоциированную с другими иммунологическими модификациями, к последующей встрече с теми же антигенами. Неоднократно было показано, что толерантность вызывает удержание находящихся на слизистой антигенов секретируемой формой иммуноглобулина A (sIgA), которая регулирует проникновение через слизистую оболочку, и при этом не происходит стимуляции системной части иммунной системы. TGF-бета, регуляторный цитокин также в некоторых случаях был вовлечен в развитие толерантности. Часто в качестве возможного механизма толерантности, реализуемой на уровне слизистой, рассматривают активную супрессию регуляторными CD25+ Т-клетками (Faria and Weiner, 2005; Mestecky et al., 2007). Однако ни одна из этих иммунологических модификаций не наблюдалась при индуцируемой фармацевтической композицией иммунологической толерантности, описанной в настоящем изобретении, для которой в основном характерна активность CD8+ T-клеток, супрессирующих активацию презентирующих антиген (вирусный эпитоп) CD4+ Т-клеток; такой тип иммунной реакции до настоящего времени не выявляли и, в частности, он представляет собой новый тип иммунологической толерантности.
Выражения «толерантность», «иммунологическая толерантность», «иммунотолерантность», «иммунотолерантность к вирусу», «новый тип вирусоспецифической толерантности», «иммунотолерантность к вирусным антигенам», «иммунотолерантность к вирусным иммуногенам» и «Ts-иммунотолерантность» являются синонимичными. Авторы изобретения показали, что это соответствует у макаков активно индуцируемому, сильному, нецитотоксическому, ограниченному комплексом MHC-Ib/E ответу с участием CD8+ T-клеток, который супрессирует раннюю активацию презентирующих антигены ВИЧ - Gag и/или Pol - CD4+ Т-клеток, и который ассоциирован с отсутствием пролиферации CD4+ Т-клеток, а также отсутствием секреции гамма-интерферона CD8+ T-клетками при стимуляции активированным антигеном SIV и отсутствием выработки системных антител против SIV: IgM и IgG. Кроме того, авторам изобретения удалось показать, что TCRγδ и Vβ8 не участвовали в супрессии CD8+ T-клетками репликации вируса, что свидетельствует о том, что TCRαβ должен играть центральную роль в узнавании презентации MHC-Ib/E-пептида инфицированными CD4+ Т-клетками.
«Обычный иммунный ответ на антиген, происходящий от вируса,» можно наблюдать, среди прочего, при вакцинации с применением стандартной профилактической вакцины, в состав которой входят антиген, включающий белки вируса ВИЧ - Gag и/или Pol, или происходящий от них, и стандартный обычно применяемый адъювант (т.е., любая форма физического, химического или биологического адъюванта, целью которого является стимуляция и/или облегчение развития и/или усиление иммунного ответа, ассоциированного с антигеном, такая как описаны в главах «Адъюванты» и «Вакцины» в публикации: S. Plotkin et al). Такой «обычный иммунный ответ на антиген, происходящий от вируса,» включает гуморальный, клеточный или одновременно гуморальный и клеточный иммунные ответы, и стандартно характеризуется по:
(i) пролиферации вирусоспецифических CD4 клеток при специфической стимуляции in vitro; и/или
(ii) индукции специфического системного гуморального ответа посредством выработки системных антител против вирусных антигенных белков и/или пептидов; и/или
(iii) индукции специфического клеточного ответа, ассоциированного с выработкой гамма-интерферона CD8 Т-клетками, и/или
(iv) отсутствию нецитотоксического, CD8+ T-клеточного ответа, супрессирующего активацию CD4+ Т-клеток, презентирующих антигены белков ВИЧ - Gag и/или Pol.
В противоположность этому, фармацевтическая композиция, содержащая антиген, который содержит белки ВИЧ - Gag и/или Pol - или происходит от них, и толерогенный носитель, предложенная по настоящему изобретению, вызывает «иммунотолерантность к вирусу» и, в частности, «Ts-иммунотолерантность», которая в примерах, приведенных ниже для вируса SIV у макаков, характеризуется следующим:
(i) отсутствием пролиферации вирусоспецифических CD4 клеток при специфической стимуляции in vitro; и
(ii) отсутствием существенного уровня системного гуморального ответа, то есть, либо отсутствует специфический детектируемый системный гуморальный иммунный ответ, который можно определить с применением классических клинических лабораторных способов, таких как анализ ELISA, либо, в случае, если детектируются системные антитела, они не могут служить защитой от инфекции вируса SIV; и
(iii) отсутствием какого-либо цитотоксического CD8+ Т-клеточного ответа, ассоциированного с выработкой гамма-интерферона (например, доступного для определения методом иммуноферментных пятен (ELIspot)) при соответствующей стимуляции in vitro, или, если детектируется цитотоксический CD8 Т-клеточный ответ, то он неспособен служить защитой против инфекции вируса SIV; и
(iv) являющемуся существенной особенностью активно индуцируемому, сильному, нецитотоксическому, не зависящему от CD25, ограниченному комплексом MHC-Ib/E ответу с участием CD8+ T-клеток, который супрессирует раннюю активацию CD4+ Т-клеток, презентирующих антигены SIV.
Как упомянуто выше, фармацевтическая композиция по настоящему изобретению содержит толерогенный носитель и антиген, который содержит белки ВИЧ - Gag и/или Pol - или происходит от них. Фактически, толерогенный носитель, объединенный с вирусным антигеном, индуцирует состояние вирусоспецифической иммунотолерантности у человека, вместо того, чтобы вызывать обычный иммунный ответ, определение которого дано выше. Таким образом, антиген, введенный через слизистые, или внутрикожным, или интраэпителиальным путем, сам по себе или со стандартным адъювантом, в общем случае способен вызывать обычный иммунный ответ, за исключением случаев, когда он ассоциирован с толерогенным носителем, как в фармацевтической композиции по настоящему изобретению. В этих специфических условиях ассоциация толерогенного носителя/антигена в фармацевтической композиции по настоящему изобретению индуцирует иммунотолерантность, как описано выше. Это означает, что иммунотолерантность может быть достигнута только при введении (через слизистые, или внутрикожным, или интраэпителиальным путем) соответствующей смеси толерогенного носителя и антигена, который содержит белки ВИЧ - Gag и/или Pol - или происходит от них. Если один компонент ввести человеку в отсутствие другого, то человек не будет «вакцинирован», или у него не будет «вызвана толерантность».
Как показано в примерах ниже (с применением вируса SIV в модели на макаках), иммунотолерантность (также называемая «Ts»-иммунотолерантностью), индуцированная или достигнутая при введении фармацевтической композиции по настоящему изобретению, характеризуется CD8+ T-клеточным ответом (в частности, нецитотоксическим и/или ограниченным комплексом MHC-Ib/E), который супрессирует активацию (в частности, раннюю активацию) CD4+ Т-клеток, презентирующих антигены белков SIV - Gag и/или Pol; а также, предпочтительно, одной или нескольких из следующих особенностей: 1) отсутствием пролиферации CD4+ Т-клеток; 2) отсутствием существенного уровня секреции гамма-интерферона CD8+ T-клетками при стимуляции антигеном SIV; и 3) отсутствием существенного уровня выработки системных антител против SIV: IgM и IgG.
Под терминами «отсутствие существенного уровня секреции гамма-интерферона CD8+ T-клетками при стимуляции антигеном ВИЧ или SIV» в этом документе подразумевается, что уровень секреции гамма-интерферона CD8+ T-клетками, который наблюдается при стимуляции антигеном ВИЧ или SIV, равен нулю или низок. «Низкий уровень» секреции гамма-интерферона CD8+ Т-клетками при стимуляции антигеном ВИЧ или SIV в типичном случае составляет менее чем примерно 80 пятнообразующих клеток на 2×105 МКПК (мононуклеарных клеток периферической крови).
Под терминами «отсутствием существенного уровня выработки системных антител против ВИЧ и против SIV: IgM и IgG» в этом документе подразумевается, что наблюдаемый уровень выработки системных антител против ВИЧ и против SIV - IgM и IgG - равен нулю или низок. «Низкий уровень» выработки системных антител против ВИЧ и против SIV в типичном случае составляет титр антител против ВИЧ и против SIV на уровне примерно 300 или ниже.
Таким образом, фармацевтическая композиция по настоящему изобретению индуцирует и предпочтительно поддерживает антиген-специфическую иммунную защиту против ВИЧ у человека, при этом указанная иммунная защита предпочтительно характеризуется следующими особенностями:
- снижением и, предпочтительно, супрессией вирусной нагрузки ВИЧ у указанного человека, по сравнению с соответствующими экспериментальными контролями; и/или
- CD8+ Т-клетками, которые супрессируют активацию CD4+ Т-клеток, презентирующих антигены белков ВИЧ - Gag и/или Pol - у указанного человека, по сравнению с соответствующими экспериментальными контролями.
Предпочтительно, указанная иммунная защита определяется у указанного человека путем детекции in vitro присутствия или активности CD8+ регуляторных Т-клеток указанного человека. Такую детекцию можно провести с применением стандартных способов in vitro, таких как способы, описанные в примерах ниже.
Все патенты, заявки на патенты и публикации, на которые даны ссылки выше, включены сюда в качестве ссылок.
ПРИМЕРЫ
Часть А. Общие материалы и способы
А-I. Животные. Воспитанные в колонии китайские макаки-резусы (Масаса mulatta) были помещены в клетки в соответствии с правилами «Руководства по уходу и использованию лабораторных животных» Национального института здоровья США. Все животные были полностью здоровы, их возраст составлял 2-4 года, вес - 4-6 кг, и все они были сероотрицательными в отношении SIV, SRV, Т-лимфотропного вируса обезьян 1, вируса гепатита В и вируса В. В начале эксперимента всем животным проводили рентгенологическое исследование и аллергическую кожную пробу (с применением PPD, очищенного от белков среды сухого туберкулина), чтобы исключить возможных носителей туберкулеза.
A-II. Типирование МНС класса I. Было проведено генотипирование классических для макаков-резусов аллелей МНС класса I в пробах мононуклеарных клеток периферической крови (МКПК) с применением ПЦР-анализа с сиквенс-специфическими праймерами (SSP) на присутствие последовательностей Mamu-A и Mamu-B, описанных ранее (Muhl et al., 2002; Loffredo et al., 2007).
A-III. Получение препаратов антигенных вирусов.
III-1. Выработку вируса SIV осуществляли в клетках СЕМ174, инокулированных штаммом SIVmac239 предоставленным Р.А. Marx). Собирали клеточные супернатанты при максимальном уровне выработки.
III-2. АТ-2-инактивированный вирус SIVmac239: SIVmac239 инактивировали с помощью 250 мкМ Алдритиола (AT-) (Sigma) в течение 2 ч и промывали три раза с помощью ультрацентрифугирования. АТ2-инактивированный вирус применяли в конечной дозе 109 вирусных частиц на каждое введение (т.е., вакцинацию).
III-3. Инактивированный нагреванием вирус SIVmac239: вирус SIVmac239 инкубировали при 56°С в течение 30 мин. Инактивированный нагреванием вирус применяли в конечной дозе 109 вирусных частиц на каждое введение.
III-4. Инактивированный АТ-2 и нагреванием вирус SIVmac239: SIVmac239 инактивировали с помощью 250 мкМ Алдритиола (АТ-2) (Sigma) в течение 2 ч и промывали три раза с помощью ультрацентрифугирования. Затем вирус подвергали обработке температурой 56°С в течение 30 мин. Инактивированный вирус применяли в конечной дозе 109 вирусных частиц на каждое введение.
III-5. Препаратами инактивированного вируса инокулировали клетки СЕМ174, чтобы убедиться в 100%-ом ингибировании инфекционности вируса.
A-IV. Количественное определение гуморальных иммунных ответов на вирус SIV. Проводили титрование антител против SIV - IgG, IgM и IgA - в сыворотке с применением количественного анализа с иммунофлуоресцентно меченными антителами (IFA) (Mederle et al., 2003). Коротко, пробы последовательных двукратных разведении испытуемой плазмы инкубировали с предметными стеклами, к которым были прикреплены инфицированные вирусом SIV клетки СЕМ174, при 37°С в течение 30 мин. После промывки реактивом Хэнкса добавляли конъюгированные с FITC (флуоресцеинизотиоцианатом) козьего антитела против IgG (Sigma), IgM (ADI, Сан-Антонио, штат Техас, США) или IgA (ADI) макака и дополнительно инкубировали в течение 30 мин (при 37°С). Титры антител определяли как обратную величину наибольшего разведения, при котором достигается положительное иммунофлуоресцентное окрашивание. Чувствительность анализа IFA составила титр 20 для IgG и титр 5 для IgM и IgA. Если пробы плазмы были негативными при анализе IFA (содержание ниже уровня чувствительности), для облегчения анализа данных пробе приписывали значение 1.
Также собирали пробы секрета слизистой, промывая прямую кишку фосфатно-солевым буферным раствором (PBS) с применением катетера для желудочной инстилляции, как описано ранее (Tsai et al., 1993). Кратко, добавили в пробы ингибитор трипсина (10 мкг/мл) и ЭДТА (5×10-4 М) (Sigma), затем пробы центрифугировали в течение 10 мин при 10000×g и температуре 4°С. Супернатанты собирали и добавляли фенилметилсульфонилфторид (10-3 М) и азид натрия (0,01%) (Sigma). Пробы хранили до применения при -80°С. Определение титров IgA против SIV в прямой кишке проводили с применением описанного выше анализа IFA.
A-V. Проточная цитометрия. Анализ проточной цитометрией проводили с применением системы FACScalibur (BD Biosciences, Сан-Хосе, штат Калифорния, США) и флуоресцентно меченных моноклональных антител против следующих молекул: CD3-PE-Cy7 (клон SP34-2), CD4-PE (клон МТ477), CD8-PerCP (клон RPA-Т8), а также второе антитело кролика против антител мыши, конъюгированных с АРС (аллофикоцианином) (BD Biosciences). Ki-67-PE (фикоэритрин) (BD Biosciences) и конъюгированное с FITC моноклональное антитело против Р27 (Fitzgerald, Конкорд, штат Массачусетс, США) или конъюгированное с биотином моноклональное антитело против Р27 (Fitzgerald) совместно с APC-SAv (стрептавидин-аллофикоцианин, BD Biosciences), применяли для внутриклеточного окрашивания после повышения проницаемости.
Конъюгированные с фикоэритрином (РЕ) моноклональные антитела против TCRγδ (клон В1), Vβ8 и CD-антигенов (CD7, CD16, CD28, CD62L, CD95, CD122, CD137, CD150, CD183, CD184, CD195, CD196, CD197, CD226, CD272 и CD305) приобретали в компании BD Biosciences; конъюгированные с фикоэритрином моноклональные антитела против CD-антигенов (CD11a, CD25, CD27, CD39, CD101, CD129, CD215, CD277 и CD357) приобретали в компании BioLegend (Сан-Диего, штат Калифорния, США); и конъюгированные с фикоэритрином моноклональные антитела против CD-антигенов (CD127, CD247 и CD279) приобретали в компании eBioscience (Сан-Диего, штат Калифорния, США).
A-VI. Пролиферация клеток. МКПК (мононуклеарные клетки периферической крови) получали, как описано ранее (Lu et al. 2003). 2003). Пролиферацию SIV-специфических CD4+ или CD8+ Т-клеток оценивали с применением количественного определения с мечением диацетатом сложного сукцинимидильного эфира карбоксифлуоресцеина (CFSE) (Molecular Probes, Юджин, штат Орегон, США) согласно инструкциям производителя. МКПК окрашивали 3 мкМ CFSE в течение 15 мин при 37°С. После промывки меченные CFSE клетки стимулировали в течение 5 дней с помощью 10 мкг/мл рекомбинантного коревого белка вируса SIV - Р27 (ImmunoDiagnostics, Вобун, штат Массачусетс, США), 2 мкг/мл 15-мерных пептидов из белка вируса SIV gag (GLS, Шанхай, Китай), 109/мл АТ-2-инактивированных частиц вируса SIV или только средой. После мечения антителами против CD3 и против CD4 или против CD8 клетки МКПК фиксировали в 1% параформальдегиде для проведения проточной цитометрии.
A-VII. Активация клеток. Свежевыделенные МКПК, обедненные (или не обедненные) CD8 или CD25 с помощью магнитных шариков, инфицировали в одном раунде АТ-2-обработанным вирусом SIVmac239 в течение 2 ч при концентрации вирусных частиц 1010/мл. Инфицированные клетки стимулировали в течение ночи эндотоксином стафилококка В (2,5 мкг/мл) и антителами против CD3 (2,5 мкг/мл)/против CD28 (2,5 мкг/мл). Внутриклеточное окрашивание белков SIV Р27 и Ki-67 проводили через 48 ч после стимуляции для определения процентной доли активации (Ki-67+) в инфицированных (Р27+) CD4+ Т-клетках.
A-VIII. Иммуноферментный слот-анализ (ELISPOT). Иммуноферментный слот-анализ IFN-γ макаков-резусов и IL-10 проводили в некультивируемых МКПК в присутствии или в отсутствие Р27 или АТ-2-инактивированного SIV, применяя коммерческий набор (Cell Sciences, Кантон, штат Массачусетс, США). Набор для иммуноферментного спот-анализа TGF-b1 покупали в компании R&D Systems (Миннеаполис, штат Миннесота, США). Данные считывали в автоматизированном ридере для иммуноферментного слот-анализа - ELISPOT reader (AID, GmbH, Штрасберг, Германия). Количество SIV-специфических пятнообразующих клеток (SFC) вычисляли вычитанием количества неспецифических SPC в присутствии только среды.
А-IX. Анализ антивирусной активности. Аутологичные CD4+ Т-клетки, полученные от каждого животного, очищенные с применением магнитного мечения с положительным отбором (MicroBeads, Miltenyi Biotec), подвергали острой инфекции вирусом SIVmac239 (10-3 MOI) в присутствии или в отсутствие очищенных с применением магнитного поля CD8+ Т-клеток при соотношении CD4/CD8, равном 1:2, а затем стимулировали с помощью SEB (стафилококкового энтеротоксина В, Sigma) в течение 16 ч. После промывки клетки культивировали в четырех повторах в конечном объеме 200 мкл на лунку среды RPMI 1640 (Invitrogen, Шанхай, Китай), содержащей 100 ME rlL2 человека, в 96-луночных планшетах в течение 5 дней при 37°С в присутствии 5% CO2. В клеточных культурах один раз заменили половину среды на свежую на 3-й день. Супернатанты клеточных культур, собранные на 5-й день, применяли для измерения вирусной нагрузки с помощью ОТ-ПЦР в реальном времени (см. ниже). Процент супрессии (%) вычисляли путем сравнения геометрического среднего концентрации вируса в супернатантах клеточных культур для двух повторных лунок, содержащих только инфицированные CD4+ Т-клетки, с геометрическим средним концентрации вируса в супернатантах из четырех повторных лунок, содержащих смесь CD8+ и CD4+ клеток. CD4+ Т-клетки также культивировали совместно с аллогенными CD8+ Т-клетками с целью определения корреляции между супрессией вируса и HLA-рестрикцией.
А-Х. Измерение вирусной нагрузки. Содержание SIV РНК в плазме или ассоциированной с клетками SIV ДНК количественно определяли с помощью ОТ-ПЦР в реальном времени или ПЦР в реальном времени с применением праймеров (прямой, SEQ ID No. 1: 5'-GAGGAAAAGAAATTTGGAGCAGAA-3'; обратный, SEQ ID No. 2: 5'-GCTTGATGGTCTCCCACACAA-3') и зонда (SEQ ID No. 3: 5'-FAM-AAAGTTGCACCCCCTATGACATTAATCAGATGTTA-TAMRA-3'), специфически оптимизированных для штаммов SIVmac239 и SIVmac251.
A-XI. Анализ SIV-специфических супрессирующих Т-клеток. Свежевыделенные МКПК, обедненные (или не обедненные) либо CD8, либо CD25 с помощью конъюгированных с магнитными шариками антител против CD8 или против CD25 в соответствии с протоколом, предоставленным производителем (Miltenyi Biotec), инфицировали вирусом SIVmac239 в течение 2 ч при 0,5 множественности заражения (MOI). Инфицированные клетки обрабатывали в течение ночи эндотоксином стафилококка В (SEB) (2,5 мкг/мл; Sigma) и антителами против CD3 (2,5 мкг/мл)/против CD28 (2,5 мкг/мл; BD Biosciences). Одновременное внутриклеточное окрашивание белков SIV P27 и Ki-67 проводили через 48 ч после стимуляции in vitro для определения процентной доли (%) активации Т-клеток (Ki-67+) в инфицированных (Р27+) клеточных популяциях.
А-XII. Вирусные провокационные пробы.
XII-1. Выработку вируса SIV осуществляли в клетках МКПК макаков, инокулированных штаммом SIVmac239 предоставленным Р.А. Marx). Собирали клеточные супернатанты при максимальном уровне выработки.
XII-2. Интраректальная провокационная проба (IRC): После вакцинации животных инокулировали (с повторами) интраректально с помощью 5000 MID100 (100% минимальной инфицирующей дозы), т.е, 5×105 TCID50 (50% инфицирующей дозы для культуры клеток) патогенного штамма SIVmac239. Эта инфицирующая доза в общем случае вызывает системную инфекцию у 100% китайских макаков-резусов с максимальным значением вирусной нагрузки в плазме (1066-10 копий/мл) в период между 10-м днем и 14-м днем. У всех животных, которым делали провокационную пробу с применением вируса SIV, проводили клиническую и биологическую оценку каждые 2 недели в течение 1 месяца, а затем - каждый месяц. XII-3. Внутривенная провокационная проба (IVC): После вакцинации животных инокулировали (с повторами) внутривенно с помощью 5 MID100, т.е., 500 TCID50 (согласно титрованию на клеточной линии СЕМ174) патогенного вируса SIVmac239 (предоставленного д-ром Марксом (Р.А. Marx) из научно-исследовательского, центра Aaron Diamond по изучению СПИД, Нью-Йорк, США). Эта инфицирующая доза в общем случае вызывает системную инфекцию у 100% китайских макаков-резусов с максимальным значением вирусной нагрузки в плазме (1066-10 копий/мл) в период между 10-м днем и 14-м днем. У всех животных, которым делали провокационную пробу с применением вируса SIV, проводили клиническую и биологическую оценку каждые 2 недели в течение 1 месяца, а затем - каждый месяц.
A-XIII. Статистический анализ. Непарные данные для сравнения между животными из разных групп или парные данные для сравнения животных в одной группе до и после иммунизации анализировали с применением критерия Манна-Уитни и критерия Уилкоксона, соответственно.
Часть В. Специфические материалы и способы
В-I. ПРИМЕНЕНИЕ BCG В КАЧЕСТВЕ ТОЛЕРОГЕННОГО НОСИТЕЛЯ
В-I-I. Получение BCG
I-1. Живая BCG: живую BCG, полученную в Копенгагене, в Государственном институте сыворотки (Statens Serum Institut) (штамм SSI 1331), приобрели в компании Laboratories Sanofi-Pasteur Merck, Sharp and Dome (SPMSD) и применяли в конечной концентрации 5×106 КОЕ для кишечного или интравагинального введения или в конечной концентрации 5×105 КОЕ для каждой внутрикожной бустер-инъекции.
I-2. BCG, инактивированная длительной лиофилизацией: штамм SSI 13 живой3 BCG инактивировали посредством 5-дневной длительной лиофилизации в условиях вакуума (давления менее 20 мкм рт.ст) и применяли в конечной дозе, соответствующей 5×106 КОЕ для каждого кишечного и интравагинального введения или 5×105 КОЕ для каждого внутрикожного введения.
I-3. BCG, инактивированная нагреванием: штамм SSI 13 живой3 BCG автоклавировали в течение 15 мин. при 115°С в боратном буферном растворе и применяли в конечной дозе, соответствующей 5×106 КОЕ для каждого кишечного и интравагинального введения или 5×105 КОЕ для каждого внутрикожного введения.
B-I-II. Фармацевтические композиции
Композиции были свежеприготовленными, с применением среды RPMI 1640 (Invitrogen, Шанхай, Китай), содержащей один из антигенов вируса SIV и толерогенный носитель.
B-I-III. Иммунизация животных
Во время иммунизации животным делали анестезию с применением гидрохлорида тилетамина и золазепана (0,7 мг/кг), вводимых внутримышечной инъекцией.
III-1. Интравагинальная иммунизация (IVI): самок животных иммунизировали под анестезией путем интравагинальной инъекции в течение 4 ч с применением одного миллилитра фармацевтической композиции или одного толерогенного носителя, раскрытого ранее, в качестве контроля. Бустер-инъекцию с применением фармацевтической композиции или толерогенного носителя проводили в той же дозе в том же месте на 8-й неделе. После первой иммунизации проводили клиническую и биологическую оценку всех животных каждые две недели.
III-2. Пероральная (внутрижелудочная) иммунизация (IGI): самцам или самкам животных под анестезией вводили внутрь желудка 15 мл 0,1 М бикарбоната натрия за 15 мин до введения фармацевтической композиции или одного толерогенного носителя, раскрытого выше, в качестве контроля. Сразу же после введения давали дополнительные 15 мл раствора бикарбоната натрия. Такую же толерогенную вакцинацию, что и начальная, повторяли два раза с интервалом в 1 мес. у каждого животного. После первой иммунизации проводили клиническую и биологическую оценку всех животных каждые две недели.
III-3. Внутрикожная бустер-инъекция (IDI): самкам животных после интравагинальной иммунизации (см. выше в разделе «IVI») через 90 дней после первой иммунизации делали под анестезией внутрикожную бустер-инъекцию с применением 0,1 мл фармацевтической композиции, содержащей 109 копий АТ-2-инактивированного вируса SIV и 5×105 КОЕ живой BCG. После первой иммунизации проводили клиническую и биологическую оценку всех животных каждые две недели.
B-I-IV. Анализ антивирусной активности. Пороговое значение, соответствующее стерильному иммунитету после интраректальной провокационной пробы, составляет по меньшей мере 20.
B-II. ПРИМЕНЕНИЕ Lactobacillus plantarum В КАЧЕСТВЕ ТОЛЕРОГЕННОГО НОСИТЕЛЯ
B-II-I. Получение бактерий (получение толерогенного носителя). Lactobacillus plantarum (LP) (ATCC8014) культивировали при 37°С в среде MRS при скорости вращения 200 об./мин. Для получения LP в логарифмической (срединной логарифмической) фазе роста бактериальной культуры бактерии культивировали до достижения оптической плотности 1,0 при 600 нм, при этом конечная концентрация LP составляла примерно 1010 КОЕ/мл (по прошествии примерно 3,5 ч).
B-II-II. Иммунизация животных с пероральной (внутрижелудочной) доставкой. Животным не давали принимать пищу в течение ночи и не давали завтрак. Во время перорального введения животным делали анестезию с применением гидрохлорида тилетамина и золазепана (0,7 мг/кг), вводимых внутримышечной инъекцией.
Иммунизация №1: восьми животным внутрижелудочно вводили 30 мл смеси вирусно-бактериального препарата, содержащей 4×107 копий/мл дважды инактивированный (DI)-SIV и 3×109 КОЕ/мл живой LP в растворе мальтодекстрина (20%). После первой иммунизации обезьянам внутрижелудочно вводили 25 мл того же вирусно-бактериального препарата (т.е., фармацевтической композиции) каждые 30 мин в течение 3 ч. Этот протокол перорального введения выполняли 5 раз в течение 5 последовательных рабочих дней. В качестве контролей параллельно 4 животным вводили живую LP отдельно и еще 3 животным - только дважды инактивированный SIV.
Иммунизация №2: двенадцати животным (iSIV/LP №№9-20) внутрижелудочно вводили 30 мл препарата, содержащего 4×107 копий/мл iSIV (инактивированного АТ-2 и нагреванием SIVmac239) и 3×109 КОЕ/мл живой LP в растворе мальтодекстрина (20%). После этого животным вводили 25 мл того же препарата каждые 30 мин в течение 3 ч (6 раз) в течение 5 последовательных дней. Шести животным (LP №№5-10) внутрижелудочно вводили 30 мл раствора 3×109 КОЕ/мл живой LP в мальтодекстрине (20%). После этого животным вводили 25 мл того же препарата каждые 30 мин в течение 3 ч (6 раз) в течение 5 последовательных дней. Наконец, еще 6 животным (iSIV №№5-10) внутрижелудочно вводили 30 мл препарата, содержащего только 4×107 копий/мл iSIV. После этого животным вводили 25 мл того же препарата каждые 30 мин в течение 3 ч (6 раз) в течение 5 последовательных дней.
B-II-III. Истощение CD8+ Т-клеток in vivo. Сначала макакам делали анестезию, а потом делали внутривенную инъекцию химерного моноклонального антитела против CD8 (сМТ-807, Centocor Research & Development, Inc., Молверн, штат Пенсильвания, США) в дозе 5 мг/кг в 0-й, 4-й и 7-й день), как описано ранее (Schmitz et al., 1999). Пробы периферической крови (5 мл) отбирали от каждого животного на 0-й день и в различных временных точках после инъекции антитела.
B-II-IV. Анализ антивирусной активности. Пороговое значение, соответствующее стерильному иммунитету после интраректальной провокационной пробы, составляет по меньшей мере 100.
B-II-V. Анализ CD8+ Т-клеточной супрессии вируса SIV. Аутологичные CD4+ Т-клетки, полученные от каждого животного, очищенные с применением магнитного мечения с положительным отбором (MicroBeads, Miltenyi Biotec), подвергали острой инфекции вирусом SIVmac239 (множественность заражения: 10-3) в присутствии или в отсутствие очищенных с применением магнитного поля CD8+ Т-клеток при соотношении CD4/CD8, равном 1:3, а затем стимулировали с помощью SEB и антител против CD3/против CD28 в течение 16 ч. После промывки клетки культивировали в четырех повторах в 96-луночных планшетах. Культуры поддерживали в конечном объеме 200 мкл на лунку среды RPM11640, содержащей 100 ME rIL2 человека (Roche Diagnostics GmbH, Мангейм, Германия) в течение 5 дней. Супернатанты клеточных культур, собранные на 5-й день, применяли для измерения вирусной нагрузки с помощью ОТ-ПЦР в реальном времени (см. ниже). Кратность супрессии вычисляли как отношение геометрического среднего концентрации вируса в супернатантах клеточных культур из лунок, содержащих только инфицированные CD4+ целевые клетки, и геометрического среднего концентрации вируса в супернатантах из лунок, содержащих смесь CD8+ и CD4+ клеток.
В некоторых экспериментах CD8+ и CD4+ Т-клетки культивировали в отсутствие межклеточного контакта, применяя систему Multiwell Insert (BD Biosciences) (CD8 - во вставленной лунке, a CD4 - в нижней лунке); CD4+ Т-клетки культивировали совместно с аллогенными CD8+ Т-клетками для того, чтобы определить корреляцию между супрессией вируса и МНС-рестрикцией; и CD8+ и CD4+ Т-клетки также совместно культивировали в присутствии антител против МНС-АВС (BioLegend) или против МНС-Е (Cell Science) для определения типов МНС-рестрикции. Для выявления подгруппы CD8+ T-клеток, ассоциированных с антивирусной активностью, CD8+ T-клеток очищали из МКПК сразу же после их обеднения с применением конъюгированных с фикоэритрином антител против TCRγδ, против Vβ8 или против другого CD-антигена, применяя микрошарики с антителами против фикоэритрина и пропуская через колонку LD (Miltenyi Biotec).
B-II-VI. Анализ цитотоксичности SIV-специфических CD8+ Т-клеток. Очищенные CD8+ Т-клетки (эффекторные клетки) и очищенные CD4+ Т-клетки, стимулированные 1010 АТ-2-инактивированным SIVmac239 (целевые клетки) метили с применением 40 нМ 3,3'-дигексилоксакарбоцианина (DiOC6) (Marchetti et al., 1996) (Molecular Probes) в течение 10 мин при 37°С. Целевые клетки метили с применением PerCP-Су5-конъюгированного антитела против CD4 (BD Bioscience) в течение 20 мин на льду. После промывки (3 раза) эффекторные клетки смешали с целевыми клетками в 96-луночном планшете с лунками с U-образным дном при различных соотношениях эффекторные клетки/целевые клетки (Е/Т) (3:1, 1:1, 0.3:1) в трех повторах. Клетки K562 (целевые) с АРС-конъюгированным антителом против CD32 (BD Bioscience) и очищенные CD56+ клетки (натуральные киллерные клетки, эффекторные) от 4 здоровых доноров были включены в анализ в качестве контроля. После 4 ч инкубации при 37°С в присутствии SEB и антитела против CD3/против CD28 клетки собирали и анализировали проточной цитометрией. Цитотоксичность (в %) вычисляли следующим образом: 100 × (% суммы апоптотических целевых клеток - % спонтанных апоптотических целевых клеток)/(100 - % спонтанных апоптотических целевых клеток).
B-II-VII. Вирусные провокационные пробы.
Первое исследование: Через четыре месяца после перорального введения вакцины или контролей в первой серии экспериментов (иммунизация №1) 8 иммунизированных животных и 7 контрольным животным инокулировали интраректально 2500 MID100 (100000 TCID50) патогенного вируса SIVmac239. Через два месяца 4 вакцинированным и уже защищенным обезьянам сделали повторную провокационную пробу интраректально (100000 TCID50), тогда как 4 другим защищенным обезьянам сделали провокационную пробу внутривенным введением 5 MID100 (200 TCID50) вируса SIVmac239. В качестве контролей 2 обезьянам сделали провокационную пробу интраректально, а другим 2 - провокационную пробу внутривенно. Эти инфицирующие дозы в общем случае вызывают системную инфекцию у 100% китайских макаков-резусов с максимальным значением вирусной нагрузки в плазме (107-109 вирусных частиц/мл) в период между 10-м днем и 14-м днем.
Второе исследование (иммунизация №2): На 420-й день после иммунизации в ходе второй серии экспериментов 16 животным (8 обезьянам, иммунизированным вирусом iSIV и LP) и 8 контрольным животным (по 4 для iSIV и 4 LP) сделали провокационную пробу интраректально с применением 100000 TCID50 вируса SIVmac239.
Часть С. Результаты.
С-I. BCG ЯВЛЯЕТСЯ ТОЛЕРОГЕННЫМ НОСИТЕЛЕМ
С-I-I. Защита от внутривенной провокационной пробы SIVmac239 после интравагинального введения:
шести животным (композиции 1, 2, 3, 4, 5 и 6) ввели интравагинально один миллилитр толерогенной композиции, содержащей АТ2-инактивированный вирус в качестве антигена и живую BCG. Была сделана бустер-инъекция той же толерогенной композиции в ту же область через 8 недель.
Параллельно 5 другим животным (контроли 1, 2, 3, 4 и 5) вводили интравагинально один миллилитр композиции, содержащей только живую BCG. Была сделана бустер-инъекция той же композиции в ту же область через 8 недель.
Через четыре месяца после первого введения всем 11 животным (композиция 1-6 и контроли 1-5) сделали провокационную пробу путем внутривенной инокуляции вирусом.
Регулярно проводили определение вирусной нагрузки в плазме обработанных животных.
На фиг.1 показана вирусная нагрузка (количество копий SIV РНК на миллилитр плазмы) как функция от времени (в днях) у животных, которым вводили композицию (композиции 1-6), и контрольных животных (контроли 1-5) после разовой внутривенной провокационной пробы вирусом.
Результаты показывают, что после внутривенной провокационной пробы вирусом у 5 контрольных животных (контроли 1-5) наблюдалась типичная первичная инфекция с максимальным значением вирусной нагрузки в плазме (106-106 копий/мл) в период между 10 и 14 днями после введения провокационной пробы, как и ожидалось. Вирусная нагрузка в плазме у этой группы контрольных животных все еще оставалась высокой (>105 вирусных частиц/мл) в течение 60 дней после провокационной пробы вирусом и далее.
В противоположность этому, 4/6 животных, которым интравагинально вводили толерогенную композицию, приготовленную из АТ-2-инактивированного SIVmac239 и BCG, демонстрировали очень низкую максимальную вирусную нагрузку в плазме (<1000 вирусных частиц/мл; между 10 и 14 днями), которая быстро стала недетектируемой (<10 вирусных частиц/мл) (через один месяц после провокационной пробы вирусом). У 2 животных с высокой максимальной вирусной нагрузкой в плазме (>106 копий/мл) был более низкий уровень вирусной нагрузки в бессимптомный период инфекции (<1000 копий/мл), чем у контрольной группы (>105 копий/мл) на 60-й день.
C-I-II. Защита от интраректальной провокационной пробы SIVmac239 после интравагинального введения композиции:
семи животным (композиции 7, 8, 9, 10, 11, 12 и 13) ввели интравагинально один миллилитр толерогенной композиции, содержащей АТ2-инактивированный вирус в качестве антигена и живую BCG в качестве толерогенного носителя. Была сделана бустер-инъекция той же композиции в ту же область через 8 недель.
Параллельно пяти другим животным (контроли 6, 7, 8, 9 и 10) вводили интравагинально один миллилитр композиции, содержащей только живую BCG. Была сделана бустер-инъекция той же композиции в ту же область через 8 недель.
Через четыре месяца после первого введения всем 12 животным (композиции 7-13 и контроли 6-10) сделали провокационную пробу путем интраректальной инокуляции вирусом SIVmac239.
Регулярно проводили определение вирусной нагрузки в плазме обработанных и контрольных животных.
На фиг.2 показана вирусная нагрузка (количество копий SIV РНК на миллилитр плазмы) как функция от времени (в днях) у животных, которым вводили композицию (композиции 7-13), и контрольных животных (контроли 6-10) после интраректальных провокационных проб вирусом.
Результаты показывают, что после интраректальных провокационных проб вирусом у 5 животных, которым интравагинально вводили только живую BCG (контроли 1-5), наблюдалась типичная первичная инфекция с максимальным значением вирусной нагрузки в плазме (106-107 вирусных частиц/мл) в период между 10 и 14 днями после введения провокационной пробы, как и ожидалось. Вирусная нагрузка в плазме в бессимптомный период инфекции у этой группы контрольных животных все еще оставалась высокой (>105 копий/мл) в течение 60 дней после провокационной пробы вирусом.
В противоположность этому, 4/7 животных, которым интравагинально вводили АТ-2-инактивированный SIVmac239 с живой BCG, неожиданно продемонстрировали недетектируемый уровень вирусной нагрузки (<10 копий/мл) в течение периода 60 дней после провокационной пробы. Трое других животных демонстрировали типичную первичную инфекцию с максимальной вирусной нагрузкой в плазме между 10 и 14 днями после провокационной пробы. Однако их вирусная нагрузка в бессимптомный период инфекции (103-105 копий/мл) была существенно ниже, чем уровень у контрольных животных (>105).
C-I-III. Защита от повторных внутривенных или интраректальных провокационных проб SIVmac239 после интравагинального введения фармацевтической композиции:
через два и восемь месяцев 3 животным с недетектируемым уровнем вирусной нагрузки после внутривенной провокационной пробы (композиции 1, 2 и 3) ввели вторую и третью внутривенную провокационную пробу с той же дозой инокулята вируса.
После второй и третьей внутривенных провокационных проб у этой группы обезьян схожую низкую максимальную вирусную нагрузку в плазме наблюдали на 10-й день. Однако к 30-му дню после вирусной провокационной пробы вирусная нагрузка опять стала недетектируемой (фиг.3).
Через шестнадцать и двадцать три месяца после начального введения композиции 3 животным, которым уже вводили в сумме 3 внутривенные провокационные пробы (композиции 1, 2 и 3), сделали дополнительную интраректальную инокуляцию провокационной пробой.
Как и ожидалось, эти 3 животных (которым первоначально интравагинально ввели АТ-2-инактивированный SIVmac239 с BCG) опять продемонстрировали недетектируемую (<10 копий/мл) максимальную вирусную нагрузку в плазме после 2 последовательных интраректальных вирусных провокационных проб (фиг.3).
Эти результаты показали, что эффективность ингибирования вирусной репликации стабильно сохраняется, так как это ингибирование все еще наблюдается более чем через 20 месяцев после начального введения композиции.
C-I-IV. Защита от внутривенной или интраректальной провокационной пробы SIVmac239 после интравагинального введения фармацевтической композиции и внутрикожной бустер-инъекции:
как и ожидалось, после последующих внутривенных (контроли 17 и 18, фиг.4) или интраректальных (контроли 19 и 20, фиг.5) вирусных провокационных проб 4 животных, которым интравагинально вводили только живую BCG, демонстрировали типичную первичную инфекцию с максимальной вирусной нагрузкой в плазме (106-107 вирусных частиц/мл) между 10 и 14 днями после провокационной пробы, как и ожидалось. Вирусная нагрузка в плазме в бессимптомный период инфекции у этой группы контрольных животных все еще оставалась высокой (>105 копий/мл) к 60-му дню после вирусной провокационной пробы.
В противоположность этому, 3/4 (75%) животных (композиции 14, 15 и 17), которым интравагинально вводили композицию, приготовленную из АТ-2-инактивированного SIVmac239 и живой BCG, а также делали внутрикожную бустер-инъекцию той же самой композиции, продемонстрировали недетектируемый уровень вирусной нагрузки (<10 копий/мл) в течение периода 60 дней после внутривенной провокационной пробы (см. фиг.4). Оставшееся одно животное (композиция 16) демонстрировало первичную инфекцию с максимальной вирусной нагрузкой в плазме (>105 копий/мл) между 10 и 14 днями после провокационной пробы (см. фиг.4). Однако, его вирусная нагрузка в бессимптомный период инфекции достигала относительно низкого уровня (104 копий/мл) на 60-й день.
Более того, 4/4 (100%) животных (композиции 18-21), которым интравагинально вводили композицию, приготовленную из АТ-2-инактивированного SIVmac239 и живой BCG, а также делали внутрикожную бустер-инъекцию той же самой композиции, продемонстрировали недетектируемый уровень вирусной нагрузки (<10 копий/мл) в течение периода 60 дней после интраректальной провокационной пробы (см. фиг.5).
C-I-V. Защита от интраректальной провокационной пробы SIVmac239 после перорального введения фармацевтической композиции:
четырем животным (композиции 22, 23, 24 и 25) внутрижелудочно вводили один миллилитр композиции, содержащей АТ2-инактивированный вирус и живую BCG.
Параллельно четырем другим животным (контроли 21-24) внутрижелудочно вводили один миллилитр только живой BCG.
Такое же введение, что и первоначально, повторили каждому животному три раза на 15-й, 30-й и 60-й день после первого этапа введения.
Результаты демонстрируют, что после интраректальной вирусной провокационной пробы (проведенной на 90-й день) 4 животных, которым вводили только живую BCG (контроли 21-24) демонстрировали типичную первичную инфекцию с максимальной вирусной нагрузкой в плазме (106-107 копий/мл) между 10 и 14 днями после провокационной пробы, тогда как 4 животных (композиции 22-25), которым вводили АТ-2-инактивированный SIV с живой BCG в составе композиции, неожиданно продемонстрировали недетектируемый уровень вирусной нагрузки (<10 копий/мл; между 10 и 14 днями) (фиг.6).
C-I-VI. Корреляция иммунного ответа и защиты против провокационной пробы SIVmac239 после введения композиции, приготовленной из АТ2-инактивированного вируса и живой BCG:
в крови обработанных животных не было обнаружено системного антитела, выработанного против SIV. Однако был детектирован некоторый уровень специфического системного гуморального ответа при введении композиции в виде внутрикожной бустер-инъекции. Следовательно, наблюдаемая защита против инфекции SIV у обработанных животных не является результатом системного гуморального ответа.
Более того, иммуноферментный слот-анализ не выявил стандартных специфических к SIV цитотоксических Т-лимфоцитов, продуцирующих гамма-интерферон (данные не приведены). С целью оценки того, имеется ли SIV-специфический нестандартный клеточный ответ, были взяты пробы крови у каждого обработанного и контрольного животного и из каждой пробы очищены CD4+ и CD8+ клетки с применением стандартных способов. Полученные ранее CD4+ клетки культивировали, а затем инфицировали SIVmac239, применяя стандартные способы. После этого инфицированные SIV клетки CD4+ культивировали в присутствии или в отсутствие полученных ранее аутологичных CD8+ клеток в течение 5 дней. Концентрацию SIV в супернатантах анализировали с применением количественной ПЦР в реальном времени.
На фиг.7 показана кратность супрессии репликации вируса в инфицированных SIV клетках CD4+, полученная в присутствии или в отсутствие аутологичных клеток CD8+, полученных в ходе экспериментов, представленных на фиг.2. Испытываемые клетки CD8 были получены от животных, которым интравагинально вводили композицию АТ2-инактивированного вируса + BCG (композиции 7-13), или от контрольных животных (контроли 6-10).
Результаты показывают, что CD8 Т-клетки от животных, защищенных от вирусной инфекции (композиция 7, композиция 8, композиция 10 и композиция 11), обеспечивают уровень вирусной супрессии в SIV-инфицированных CD4 клетках более чем в 20 раз, тогда как CD8 Т-клетки от животных, не имеющих защиты от вирусной инфекции (композиция 9, композиция 12 и композиция 13) обеспечивают уровень вирусной супрессии, равный 10-кратному или менее (фиг.7). Более того, более чем 20-кратная вирусная супрессия также наблюдалась у 4 животных, у которых была защита от внутривенных вирусных провокационных проб, представленных на фиг.1 (данные не приведены).
На фиг.8 показаны уровни вирусной супрессии в инфицированных SIV клетках CD4+, полученной в присутствии или в отсутствие аутологичных клеток CD8+, полученных в ходе экспериментов, представленных на фиг.6. Испытываемые клетки CD8 были получены от 4 животных, которым перорально вводили композицию (композиции 22-25) АТ2-инактивированного вируса + BCG, или от 4 контрольных животных (контроли 21-24).
На фиг.9 показаны уровни активации Т-клеток (Ki-67+) в инфицированных SIV (P27+) клетках CD4, полученной в присутствии или в отсутствие аутологичных клеток CD8+, полученных в ходе экспериментов, представленных на фиг.6. Испытываемые клетки CD8 были получены от 4 животных, которым перорально вводили композицию (композиции 22-25) АТ2-инактивированного вируса + BCG, или от 4 контрольных животных (контроли 21-24). SIV-специфическая супрессия активации CD4+ Т-клеток аутологичными CD8+ Т-клетками наблюдалась у 4 животных, которым вводили композицию.
Результаты подтверждают, что предотвращение системной или мукозной инфекции SIV, достигнутое при интравагинальном или пероральном введении АТ2-инактивированного вируса вместе с BCG, вызывало состояние иммунотолерантности, характеризующееся нецитотоксическим CD8+ Т-клеточным ответом, ассоциированным с анергией SIV-инфицированных CD4 клеток. Принимая во внимание эти результаты, можно считать BCG толерогенным адъювантом.
Обобщая, можно сказать, что эти результаты продемонстрировали, что равновесное состояние иммунотолерантности к антигенам SIV впервые достигнуто путем интравагинального или перорального (или внутрижелудочного) введения композиции, приготовленной из инактивированного вируса SIV и живой BCG. В то же время, впервые было показано, что интравагинальное или пероральное введение фармацевтической композиции, содержащей АТ2-инактивированный вирус SIV и живую BCG, по изобретению эффективно (>50%) для профилактики хронической вирусной инфекции после интраректальной или внутривенной провокационной пробы.
С-II. ПРИМЕНЕНИЕ Lactobacillus plantarum В КАЧЕСТВЕ ТОЛЕРОГЕННОГО НОСИТЕЛЯ
С-II-I. Индукция SIV-специфической иммунотолерантности путем перорального совместного введения дважды инактивированного SIV и Lactobacillus plantarum (iSIV/LP)
С одной стороны, у животных, которым давали пероральную дозу iSIV/LP, наблюдались SIV-специфические антитела (IgG, IgM и IgA) (фиг.10а). С другой стороны, у животных, обработанных iSIV/LP, не наблюдалось существенной пролиферации SIV Р27-специфических CD4+ Т-клеток периферической крови, тогда как обработанные iSIV животные демонстрировали существенный уровень пролиферации Р27-специфических CD4+ Т-клеток периферической крови.
C-II-II. Противоактивирующая и противовирусная активности нецитотоксических CD8+ клеток
Пролиферация SIV Р27-специфических CD8+ Т-клеток периферической крови наблюдалась как у iSIV/LP-обработанных, так и у iSIV-обработанных животных (фиг.10b). Однако у iSIV/LP-обработанных животных не были детектированы секретирующие гамма-интерферон Т-клетки (после стимуляции in vitro), и обеднение либо CD8+, либо CD25+ клетками не отменяло отсутствие ответа у Р27-специфических эффекторных Т-клеток (фиг.10c). Более того, сильная супрессия активации (Ki-67+) инфицированных (Р27+) CD4+ Т-клеток нецитотоксическими CD8+ Т-клетками также наблюдалась при острой инфекции in vitro МКПК, взятых от iSIV/LP-обработанных животных, и обеднение CD25+ клетками не отменяло сильную супрессию, вызванную CD25-CD8+ T-клетками в отношении активации инфицированных CD4+ Т-клеток (фиг.10d).
Важно отметить тот факт, что при применении высокочувствительного способа анализа цитотоксичности не было выявлено лизиса клеток (Marchetti et al., 1996) после совместной инкубации CD8+ Т-клеток и CD4+ Т-клеток, которые стимулировали нерепликативной формой SIVmac239 в присутствии или в отсутствие SEB и антител против CDS/против CD28 (фиг.10е).
Наконец, CD8+ Т-клетки периферической крови, взятые от животных спустя не менее чем 2 мес. после обработки iSIV/LP, продемонстрировали сильную ингибирующую активность против репликации вируса в остро инфицированных in vitro аутологичных CD4+ Т-клетках (фиг.11а). Более того, такая сильная противовирусная активность CD8+ Т-клеток также наблюдалась в равной степени в остро инфицированных in vitro гетерологичных CD4+ Т-клетках (фиг.11b), свидетельствуя, что в супрессорную/ингибирующую активность CD8+ Т-клеток вовлечен неклассический механизм HLA1-рестрикции. Очищенные CD8+ Т-клетки периферической крови, взятые у макаков, иммунизированных LP/iSIV не менее чем за 2 мес. до этого, обладали сильной противовирусной активностью в отношении аутологичных остро инфицированных вирусом SIVmac239 CD4+ Т-клеток, которые стимулировали в течение ночи с помощью SEB и антител против CD3/против CD28, а затем совместно культивировали в течение 5 дней. Как только достигалась активация SIV-специфических CD4+ Т-клеток (через 48 ч после иммунизации), добавление CD8+ T-клеток уже не вызывало ингибирования репликации вирусов (фиг.11c). Это наблюдение свидетельствует против возможного лизиса целевых (продуктивно инфицированных) CD4+ Т-клеток CD8+ Т-клетками при длительном культивировании, как предполагалось в предшествующем исследовании аутоиммунного диабета 1-го типа у человека (Jiang et al., 2010). Эта опосредованная CD8+ Т-клетками противовирусная активность требует наличия межклеточных контактов (фиг.11d), а также не подвержена классической МНС1а-рестрикции, что видно по сильному ингибированию репликации вируса под действием CD8+ Т-клеток на остро инфицированные CD4+ Т-клетки других иммунизированных животных или контрольных животных (фиг.11е). Наконец, опосредованная CD8 клетками противовирусная активность блокировалась антителом против MHC-Ib/E, но не антителом против MHC-Ia/ABC, что указывает на неклассическую, ограниченную комплексом MHC-Ib/E, активность CD8+ Т-клеток (фиг.11f).
Установлено, что экспрессия TCR в CD8+ Т-клетках необходима для распознавания комплексов MHC-Ib/E-пептид, находящихся на целевых CD4+ Т-клетках (Sarantopoulos et al., 2004; Van Kaer, 2010). Применяя обеднение in vitro с помощью конъюгированных с антителами магнитных микрошариков, показали, что TCRγδ и Vβ8 не участвовали в супрессии CD8+ T-клетками репликации вируса (фиг.11g). Таким образом, по-видимому, TCRαβ играет центральную роль в узнавании презентирующего комплекса MHC-Ib/E-пептид на инфицированных CD4+ Т-клетках. Более того, путем обеднения CD8+ Т-клетками с помощью имеющихся антител против белков человека, которые дают перекрестную реакцию с мембранными антигенами CD (сокр. от «кластер дифференцировки») приматов, отличных от человека (CD7, CD11a, CD16, CD25 (IL-2RA), CD27, CD28, CD39, CD62L, CD95, CD101, CD122 (IL-2RB), CD127 (IL-7R), CD129 (IL-9R), CD137, CD150, CD183 (CXCR3), CD184 (CXCR4), CD195 (CCR5), CD196 (CCR6), CD197 (CCR7), CD215 (IL-15Ra), CD218 (IL-18Ra), CD223 (LAG3), CD226, CD247, CD272, CD277, CD279 (PD-1), CD305 (LAIR1), и CD357), не удалось выявить CD-антиген, ассоциированный с ограниченной комплексом MHC-Ib/E CD8+ Т-клеточной активностью (табл.1). В табл.1 ниже представлена противовирусная активность (кратность супрессии, геометрическое среднее ± станд. ошибка) CD8+ Т-клеток, взятых от 8 иммунизированных iSIV/LP животных до и после обеднения указанных подгрупп CD-антигенов* в первом исследовании по иммунизации (иммунизация №1).
* В каждой серии экспериментов проверяли противовирусную активность (кратность супрессии, геометрическое среднее ± станд. ошибка) CD8+ Т-клеток, взятых от 8 иммунизированных iSIV/LP животных после обеднения (или без обеднения), проведенного с применением антител против 2-х CD-антигенов.
С-II-III. Защита животных от интраректальных провокационных проб с помощью перорально вызванной иммунотолерантности
Через три месяца после введения iSIV/LP или контрольных препаратов 8 иммунизированным iSIV/LP и 7 контрольным животным сделали интраректальные провокационные пробы разовой высокой дозой (100000 TCID50) вируса SIVmac239. Восемь из 8 обработанных iSIV/LP животных имели защиту от интраректальной провокационной пробы патогенного SIVmac239, тогда как 4 животных, обработанных iSIV, и 4 животных, обработанных LP, были инфицированы такой же интраректальной вирусной провокационной пробой (фиг.12a и b, слева).
C-II-IV. Защита животных от внутривенной провокационной пробы с помощью перорально вызванной иммунотолерантности
Через два месяца после этой первой провокационной пробы, 4 из 8 обезьян сделали вторую провокационную пробу внутривенным путем (200 TCID50). У всех у них был отмечен небольшой пик репликации (<200 копий SIV ДНК/млн, клеток МКПК и 200 копий SIV РНК/мл плазмы) на 10-й день после провокационной пробы; однако к 30-у дню уровень SIV ДНК в МКПК снизился до 10 копий/млн, клеток, а уровень SIV РНК в плазме был недетектируемым (т.е., менее 10 копий/мл), свидетельствуя об отсутствии активной репликации вируса in vivo (фиг.12а и b, справа). В противоположность этому, 2 неиммунизированных животных, которым вводили ту же внутривенную провокационную пробу с SIVmac239 (200 TCID50), были успешно инфицированы. Оставшимся 4 обезьянам сделали повторную интраректальную провокационную пробу (100000 TCID50), и все они остались полностью защищенными (фиг.12а и b, справа).
C-II-V. Подтверждение роли CD8+ Т-клеток in vivo
Через пять месяцев после этой второй провокационной пробы, с целью подтверждения in vivo роли CD8+ Т-клеток, сделали 3 внутривенные инъекции химерного моноклонального антитела мышь-человек против CD8 (сМТ-807, Centocor) в течение периода в одну неделю (на 300-й, 304-й и 307-й день после иммунизации) 8 обезьянам, которые уже получали провокационные пробы, чтобы временно снизить у них количество CD8+ Т-клеток в периферической крови и лимфоидных органах (фиг.13а и и). У 4 макаков, которым делали повторную провокационную пробу интраректальный путем, не было выявлено вирусных РНК или ДНК, что снова свидетельствует об их полной антивирусной защите; в противоположность этому, сильная репликация вируса сопровождала обеднение лимфоидных органов CD8+ Т-клетками у 4 животных, которым сделали внутривенную провокационную пробу, что было видно по вирусной нагрузке у них в плазме, которая достигала уровня 106 копий РНК/мл, а также по провирусной нагрузке у них в МКПК и лимфатических узлах, которая достигала уровня 104 копий ДНК/106 клеток на 15-й день (в низшей точке обеднения CD8+ Т-клетками); к 60-90-у дню, когда у 4-х обезьян восстановился исходный уровень концентрации CD8+ Т-клеток, содержание SIV РНК в плазме и SIV ДНК в клетках МКПК и лимфатических узлах также вернулось к исходным уровням (фиг.13с, d и е). Это подтверждает уникальную роль индуцированных iSIV/LP CD8+ Т-клеток в контроле репликации вируса in vivo у животных, которым делают провокационную пробу с SIV, у которых способный к репликации вирус остается латентным, предположительно, в покоящихся CD4+ Т-клетках памяти.
Через восемь месяцев после второй провокационной пробы 4 обезьяны, которым делали интраректальную повторную провокационную пробу, а также 4 обезьяны, которым делали внутривенную повторную провокационную пробу, получили третью провокационную пробу, на этот раз интраректальным путем, с применением SIVB670 (100000 TCID50), другого инфекционного штамма SIV. Восемь животных остались полностью защищены в течение следующих12 месяцев, что было видно по недетектируемым уровням SIVB670 ДНК и РНК, тогда как 2 неиммунизированных животных были успешно инфицированы с помощью той же провокационной пробы с SIVB670, демонстрируя, что выработанные под действием LP/iSIVmac239 ограниченные MHC-Ib/E CD8+ Т-клетки обеспечивали перекрестную защиту, предотвращая активацию CD4+ Т-клеток, инфицированных другими штаммами SIV (фиг.14a и b).
Для определения длительности эффективного предотвращения заболевания SIV у обработанных iSIV/LP животных провели вторую иммунизацию с применением iSIV/LP у 8 новых макаков китайского происхождения и провели проверку противовирусной активности in vitro их CD8+ Т-клеток с течением времени, не производя ввода провокационной пробы с SIV. Такая противовирусная активность in vitro была детектирована, начиная с 60 дней после иммунизации, по сравнению с контрольными животными, которым вводили либо только LP (n=4), либо только iSIV (n=4).
Уровни активности ex vivo против SIV сохранялись вплоть до 420-го дня у 7 из 8 обезьян, тогда как у одной обезьяны противовирусная активность прогрессивно снижалась, начиная с 360-го дня, достигнув показателей на исходном уровне как у контрольных обезьян к 420-му дню (фиг.15а). На 420-й день после иммунизации 16 животным были сделаны интраректальные провокационные пробы с дозой 100000 TCID50 вируса SIVmac239. Семь из 8 иммунизированных iSIV/LP животных приобрели стерильный иммунитет, без появления SIV РНК и ДНК в плазме и МКПК (фиг.15b и c), а также в лимфоцитах слизистой прямой кишки (где измерение проводили, начиная с 1-го дня после провокационной пробы) и тазовых лимфатических узлах (фиг.16а-е), тогда как одна иммунизированная обезьяна была полностью инфицирована. Важно отметить, что развитие противовирусной активности ех vivo у 8 вакцинированных обезьян позволило предсказать, начиная с 360-го дня после иммунизации (т.е., за 60 дней до того, как им сделали провокационную пробу), 7 защищенных обезьян и одну незащищенную (фиг.15а-с).
C-II-VI. Выводы
В этом документе раскрыт, на модели макаков, тот факт, что введение инактивированного вируса SIVmac239 (iSIV) и симбиотической бактерии Lactobacillus plantarum (LP) (называемой толерогенным адъювантом) вырабатывает ограниченные комплексом MHC-Ib/E CD8+ Т-клетки, которые индуцируют супрессию активации презентирующих антигены SIV CD4+ Т-клеток и тем самым - супрессию репликации SIV и защиту макаков от провокационных проб с SIV.
Смесь, приготовленную из инактивированного iSIV и LP, внутрижелудочно вводили в сумме 16 животным, и 15 животных служили контролем. Через 4-14 месяцев всем животным делали интраректальные провокационные пробы с патогенным вирусом SIVmac239.
Полная защита от инфекции SIV наблюдалась у 15 из 16 животных, которым вводили iSIV/LP; в противоположность этому, инфицированными оказались все контрольные животные и одна вакцинированная обезьяна. Незащищенную обезьяну можно было выявить с помощью анализа противовирусной активности ех vivo за 60 дней до проведения интраректальной провокационной пробы. Восемь защищенных животных сохраняли защиту после второй провокационной пробы с SIVmac239, вводимой внутривенно 4 животным и интраректально другим 4 животным.
У 8 иммунизированных iSIV/LP животных наблюдалось полное отсутствие пролиферации SIV-специфических CD4+ Т-клеток периферической крови, и у них не вырабатывались системные SIV-специфические антитела (IgG, IgM или IgA).
Более того, их SIV-специфические CD8+ Т-клетки периферической крови имели несколько особенностей:
1) они активно пролиферировали, но не секретировали гамма-интерферон при стимуляции in vitro;
2) они сильно супрессировали активацию остро инфицированных аутологичных CD4+ Т-клеток;
3) обе эти функции не менялись после удаления CD25+ клеток;
4) они также ингибировали репликацию остро инфицированных аллогенных CD4+ Т-клеток; и
5) их супрессорное/ингибирующее действие было ограничено MHC-Ib/E.
Эти результаты показывают, что внутрижелудочное совместное введение iSIV и LP способствует развитию у макаков вирусоспецифических нецитотоксических, ограниченных MHC-Ib/E регуляторных CD8+ Т-клеток, которые обеспечивают выработку SIV-специфической иммунотолерантности, и, что было неожиданно, такая вирусоспецифическая иммунотолерантность ассоциирована с обеспечиваемой вакциной защитой животных против установления инфекции SIV.
Выше показано, что фармацевтическая композиция по настоящему изобретению предотвращает инфекцию ВИЧ и SIV у человека/млекопитающих. Это профилактическое действие достигается у макаков путем индукции «Ts»-иммунотолерантности у толерогенно-вакцинированных особей (т.е., млекопитающих, которым ввели фармацевтическую композицию). В этом документе продемонстрировано, что указанная «Ts»-иммунотолерантность включает в себя вирусоспецифические нецитотоксические, ограниченные MHC-Ib/E, супрессивные регуляторные CD8 Т-клетки, присутствие и активность которых, как было показано, позволяют достичь:
- ингибирования репликации SIV в остро инфицированных CD4+ T-клетках макаков, которым ввели фармацевтическую композицию по настоящему изобретению (in vitro); и/или
- предотвращения репликации SIV у толерогенно-вакцинированных макаков, которым делают провокационную пробу с инфекционным SIV (in vivo).
СПИСОК ЛИТЕРАТУРЫ
Morgan С, et al. Plos Medicine 2008, 5:1200-1204
Marcondes MC, et al. Viral Immunol 2006, 19:679-689
Stahl-Hennig C, et al. J Med Primatol 2007, 36:195-205
Chen S, et al. Zhongguo Yi Xue Ke Xue Yuan Xue Bao 2008, 30:156-160
Hu et al., JAMA 275:210-216, 1996
Korberet al., Science 280:1868-1871, 1998
Papathanasopoulos MA, et al. Virus Genes 2003, 26:151-163
RAVIV et al. J. Virol., vol.79(19), p: 12394-12400, 2005
Chien, Novel Drug Delivery Systems, Ch. 4 (Marcel Dekker, 1992)
Ullmann's Encyclopedia of Industrial Chemistry, 6th Ed. (various editors, 1989-1998, Marcel Dekker)
Pharmaceutical Dosage Forms and Drug Delivery Systems (ANSEL et al., 1994, WILLIAMS & WILKINS)
Ebadi, Pharmacology, Little, Brown and Co., Boston, Mass. (1985) WONG et al. Proc. Natl. Acad. Sci. U.S.A., vol.104(8), p:2591-2595, 2007 Fuller. J. Appl. Bacteriol. 66:365-378 (1989)
I. Mederle, R. Le Grand, B. Vaslin et al., Vaccine 21 (27-30), 4153 (2003)
W. Lu, X. Wu, Y. Lu et al., Nature medicine 9 (1), 27 (2003)
A. S. Fauci, Nature 384 (6609), 529 (1996)
Faria and Weiner, Immunol Rev 206, 232 (2005) Mestecky et al., J Immunol 179 (9), 5633 (2007) HOFT et al. (J Infect Dis., vol. 198(10), p: 1491-1501, 2008)
WANG et al (Med Microbiol Immunol., 2008 May 20)
Rerks-Ngarm et al. N. Engl. J. Med. Vaccination with ALVAC and AIDSVAX to prevent HIV-1 infection in Thailand, 2009 Oct. 20 (epub)
McElrath et al. The Lancet, Volume 372, Issue 9653, Pages 1894-1905, 29 November 2008
Tsai et al., Laboratory animal science 43 (5), 411 (1993)
Wei Li Lee et al. Small, (p 1003-1011) Published Online: Mar 31 2010 12:16PM DOI: 10.1002/smll. 200901985
Delie F. Adv Drug Deliv Rev. 1998; 34:221-233
Mathiowitz et al. Nature. 1997; 386:410-414 Goldberg et al. Nat Rev Drug Discov. 2003; 2:289-295 Fasanoetal. J Clin Invest. 1997; 99:1158-1164 Sandri et al. Eur J Pharm Biopharm. 2007; 65:68-77 Chickering et al. J Control Release. 1997; 48:35^*6 Ponchel et al. Adv Drug Deliv Rev. 1998; 34:191-219
Plotkin et al. Vaccines, Saunders Elsevier Fifth edition, 2008, pages 215 & 216
Korin and Zack, Journal of virology 73 (8), 6526 (1999) Vatakis et al., Journal of virology S3 (12), 6222 (2009a) Vatakis et al., Journal of virology 83 (7), 3374 (2009b) Andrieu and Lu, Immunol Today 16 (1), 5 (1995) Sacha et al., J Immunol 178 (5), 2746 (2007)
Schmitz et al., The American journal of pathology 154 (6), 1923 (1999) Liew et al., 2010.
Journal of Biotechnology 150:224-231
Plummerand Manchester, 2010. Viral nanoparticles and virus-like particles: platforms for contemporary vaccine design. John Wiley & Sons, Inc. WIREs Nanomedicine and Nanobiotechnology. DOI: 10.1002/wnan.119
Ke-Qin Xin et al. Blood 102 (1), 223-228 (1 July 2003)
Jae-Sung Yu et al. Clinical and Vaccine Immunology 13(11), 1204-1211 (Nov. 2006)
Scholzen and Gerdes. J. Cell Physiol. 182, 311-322 (March 2000)
Muhl, et al. MHC class I alleles influence set-point viral load and survival time in simian immunodeficiency virus-infected rhesus monkeys. J Immunol 169, 3438-3446 (2002)
Loffredo et al. Mamu-B*08-positive macaques control simian immunodeficiency virus replication. Journal of virology 81, 8827-8832 (2007)
Sarantopoulos et al. Qa-1 restriction of CD8+ suppressor Т cells. The Journal of clinical investigation 114, 1218-1221 (2004)
Jiang et al. HLA-E-restricted regulatory CD8(+) Т cells are involved in development and control of human autoimmune type 1 diabetes. The Journal of clinical investigation 120, 3641-3650 (2010)
Van Kaer, L. Comeback kids: CD8(+) suppressor Т cells are back in the game. The Journal of clinical investigation 120, 3432-3434 (2010)
Marchetti, P., et al. Mitochondrial permeability transition is a central coordinating event of apoptosis. The Journal of experimental medicine 184, 1155-1160 (1996)



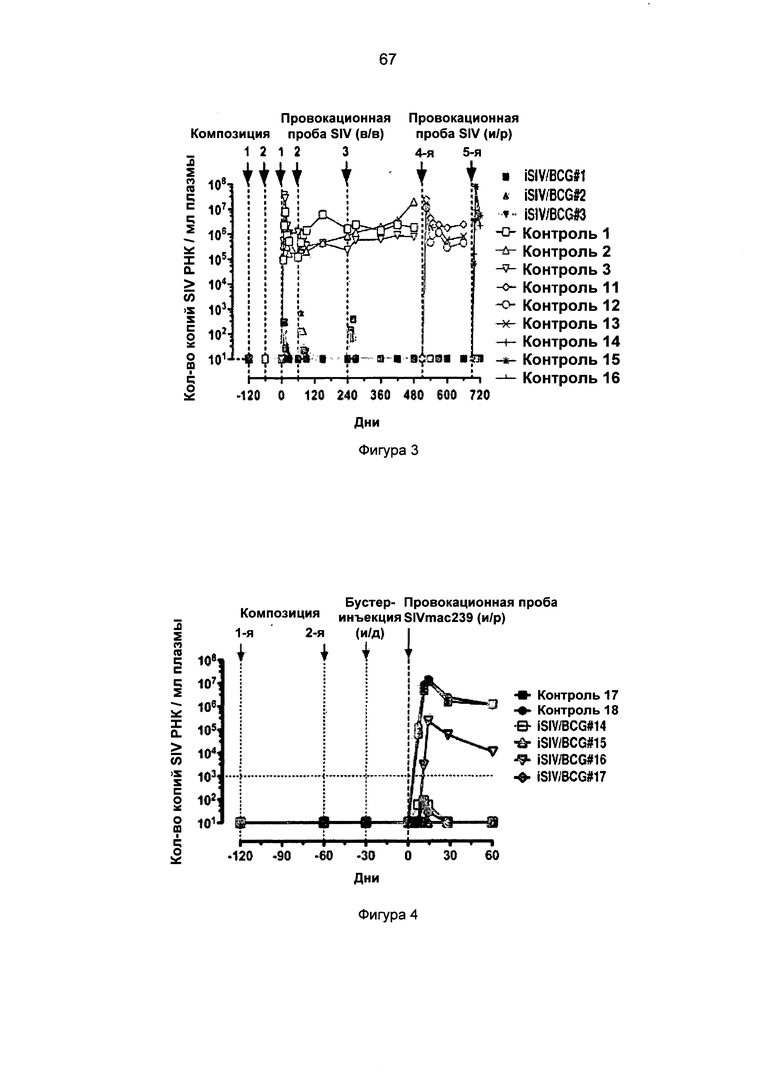
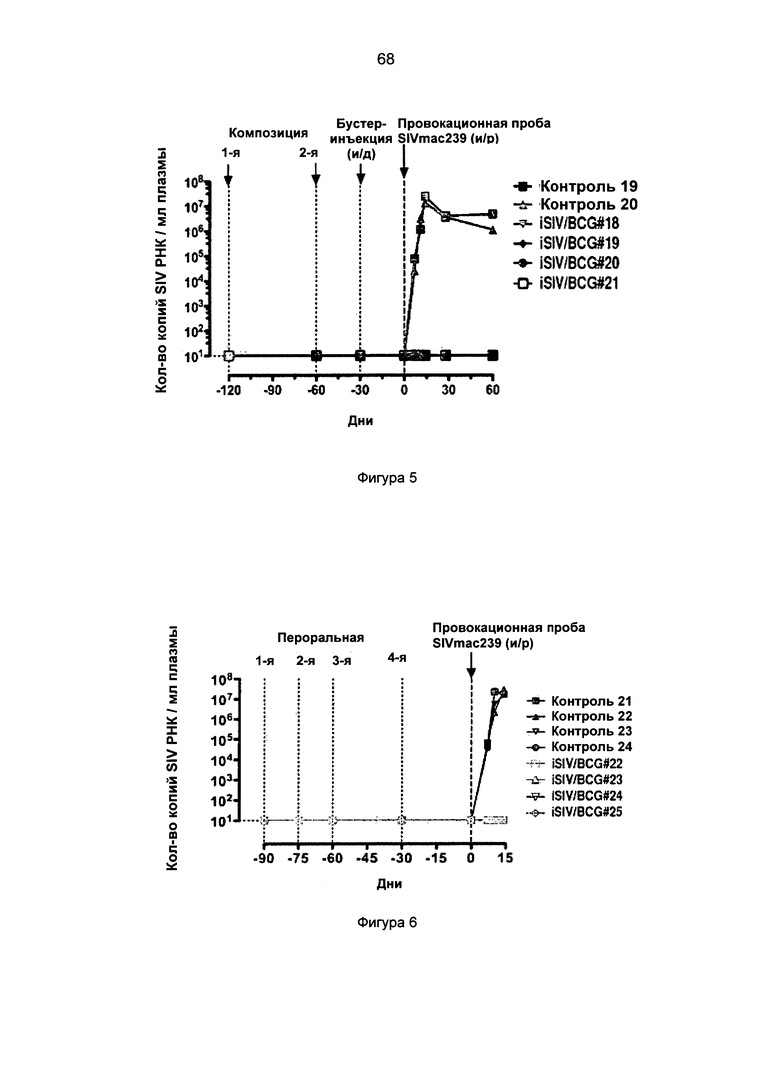
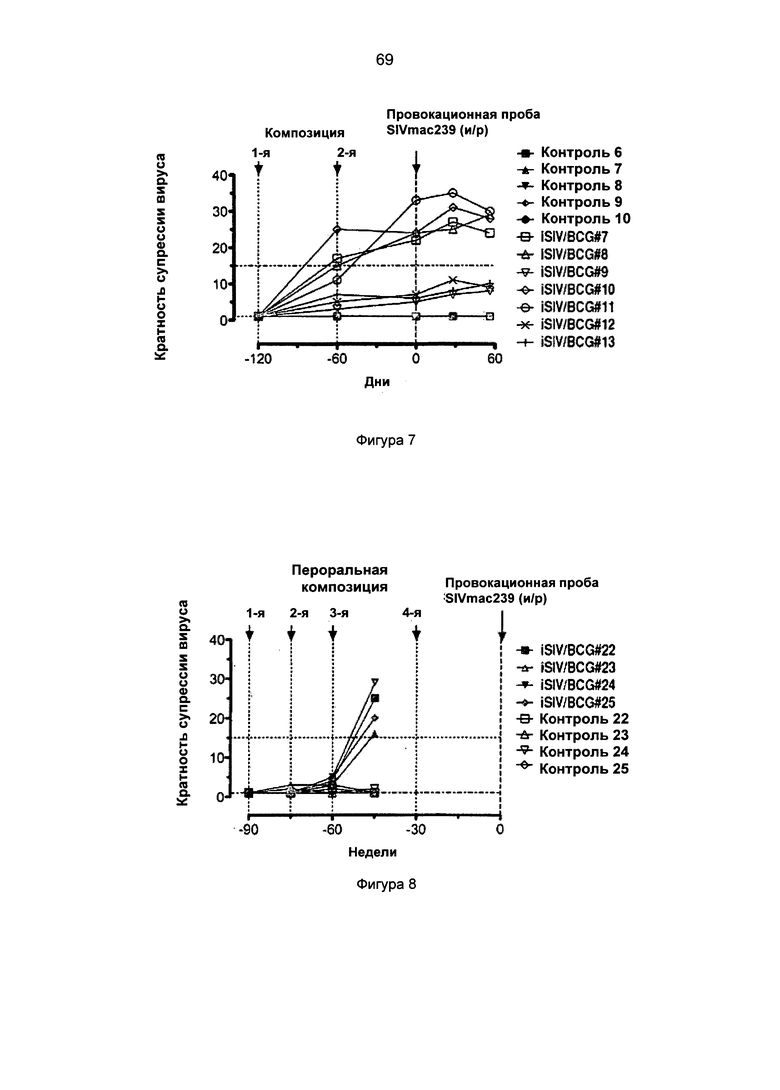
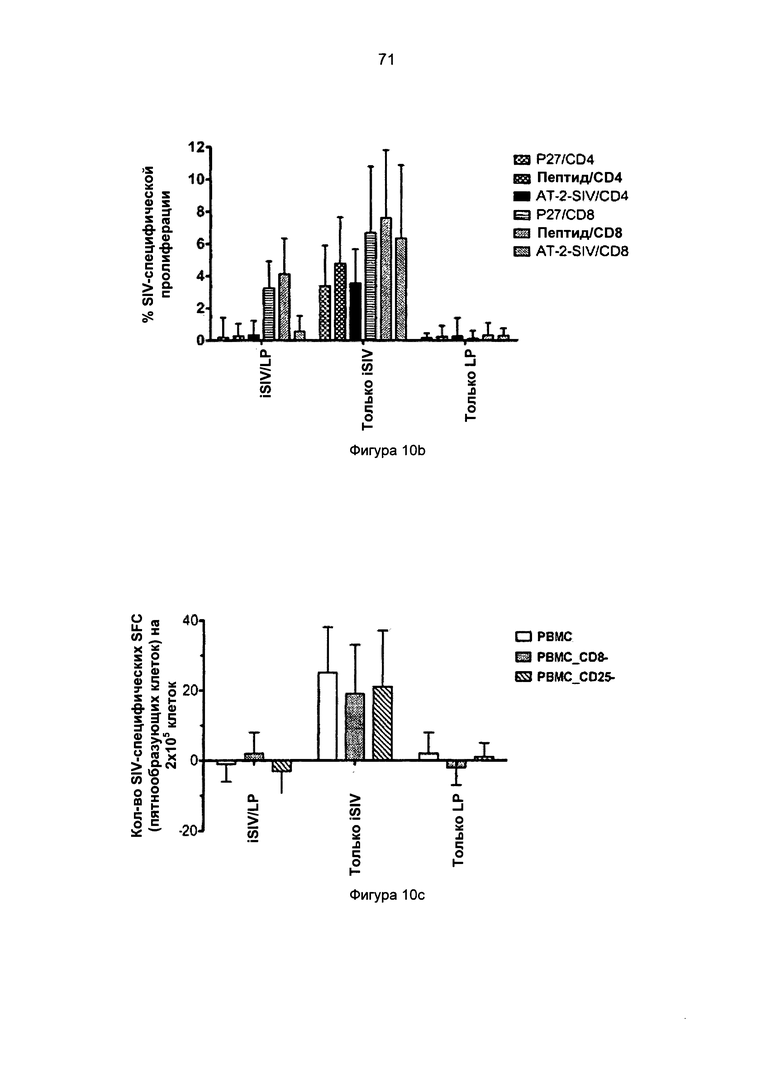


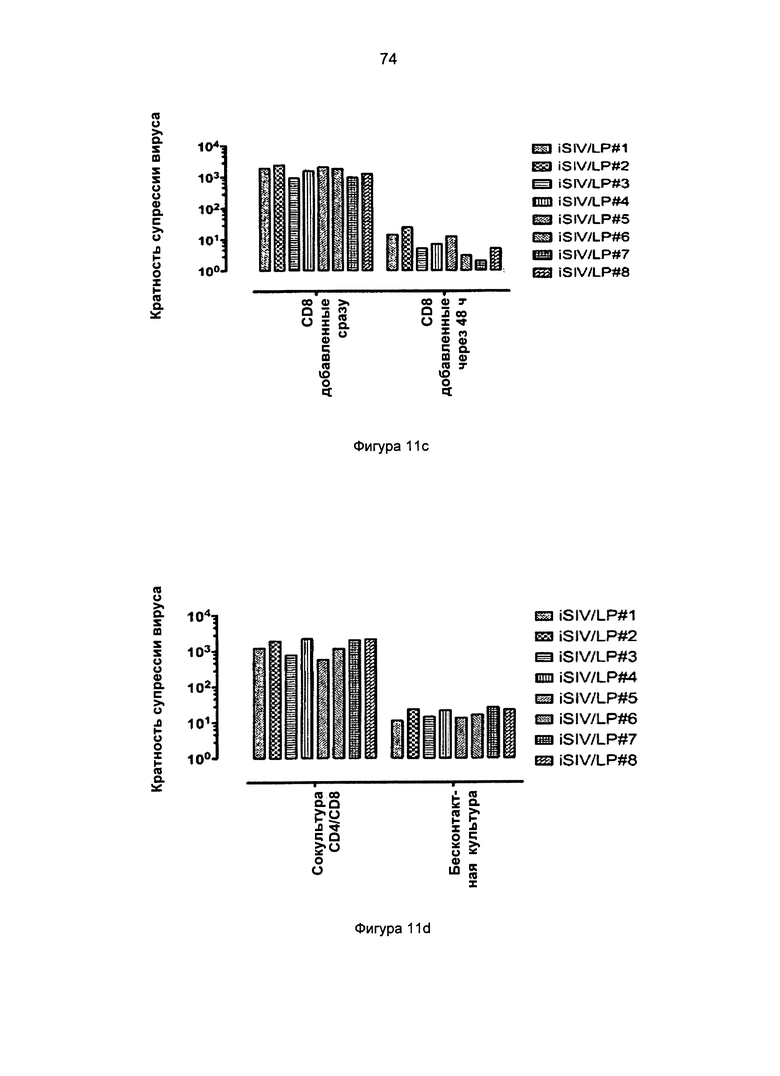
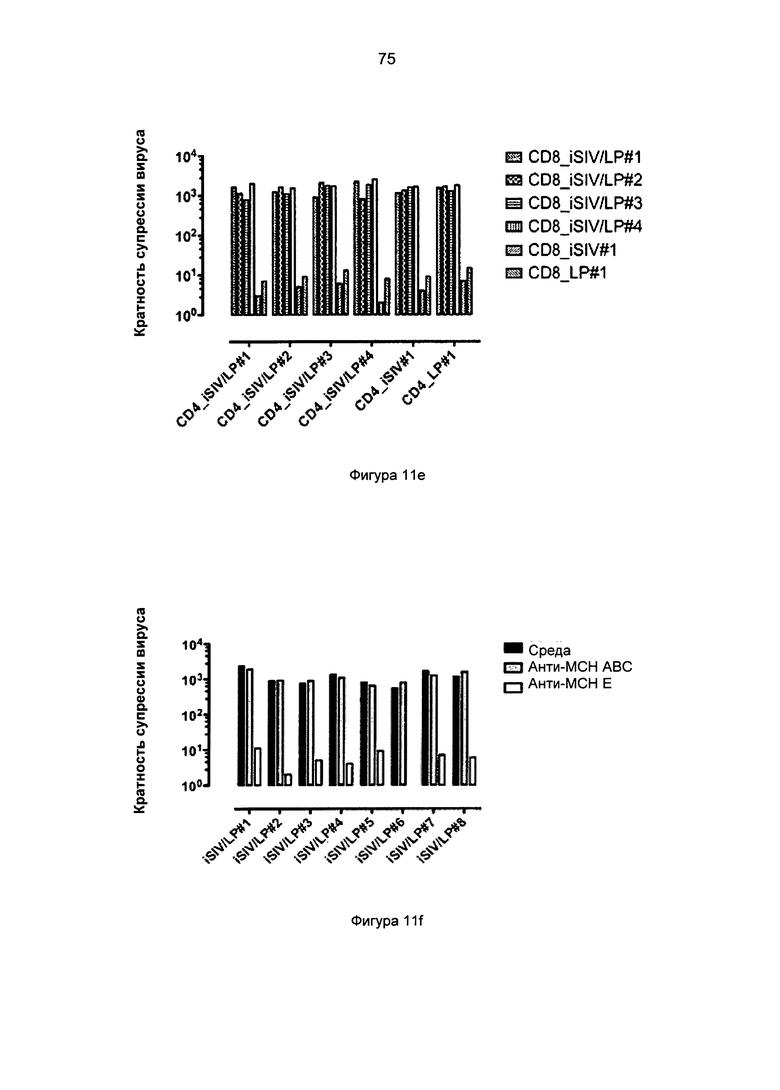
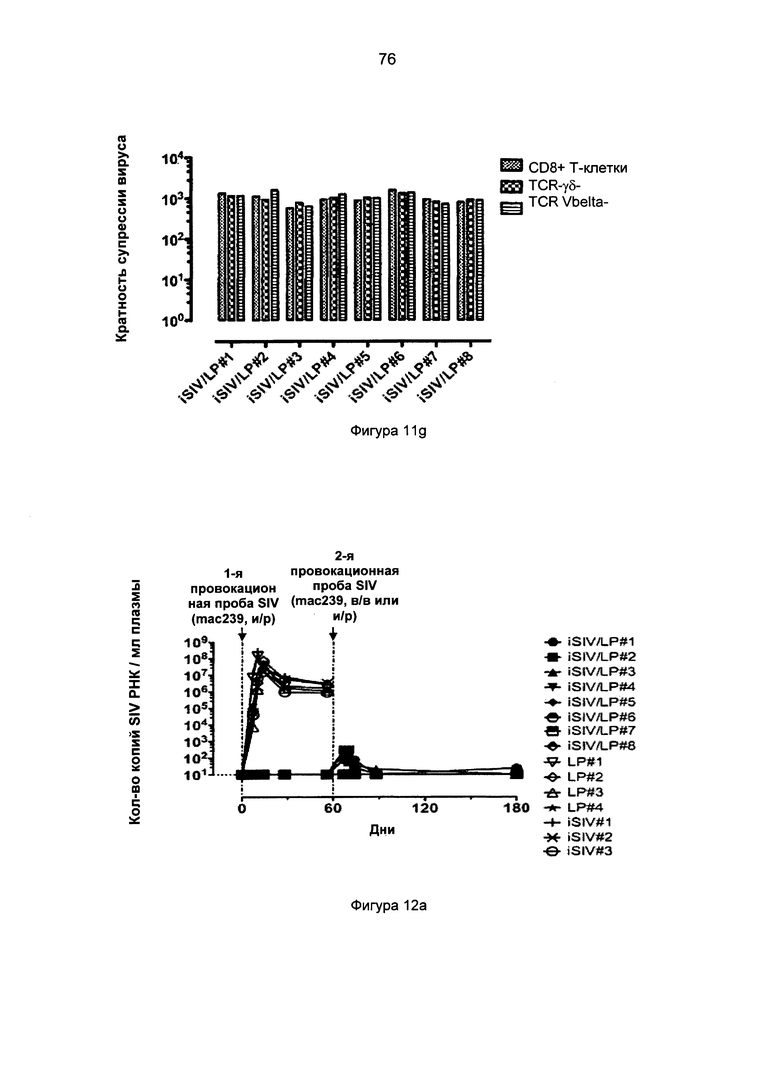
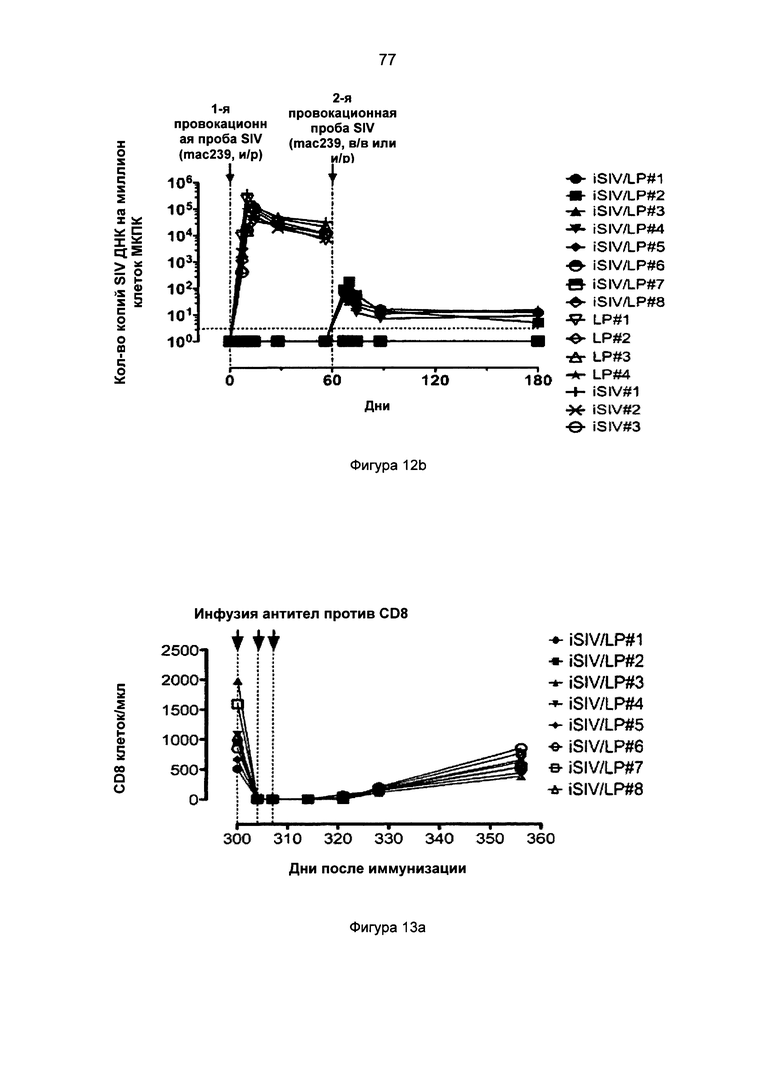
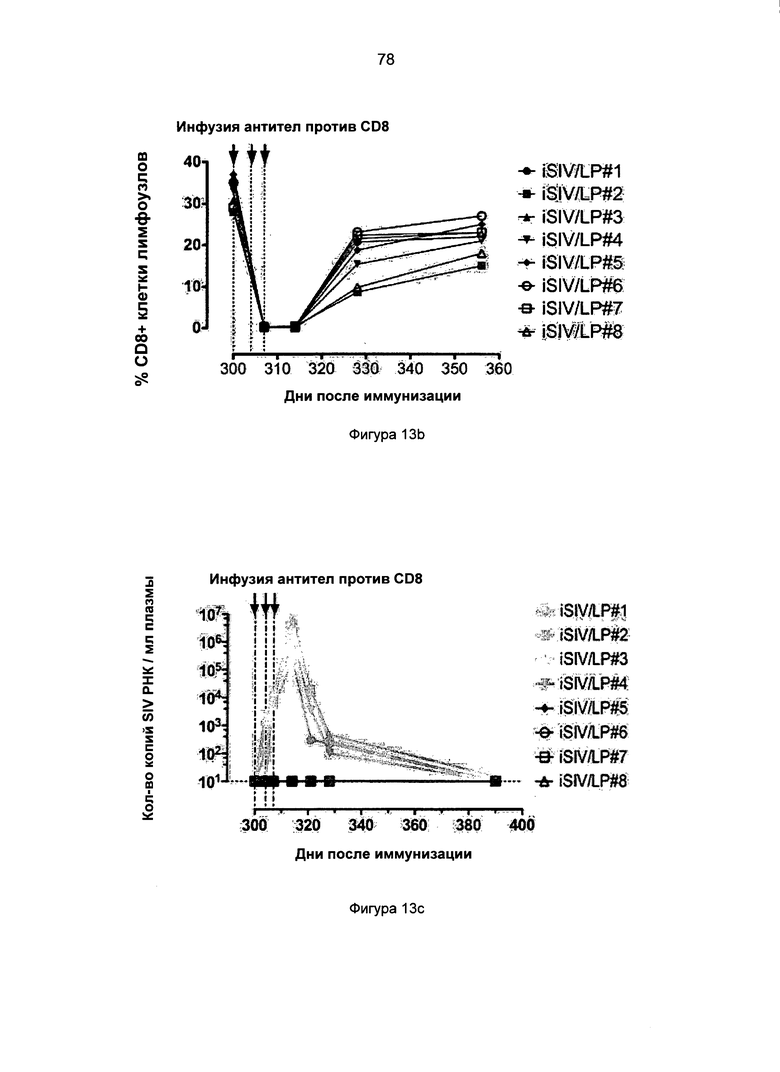
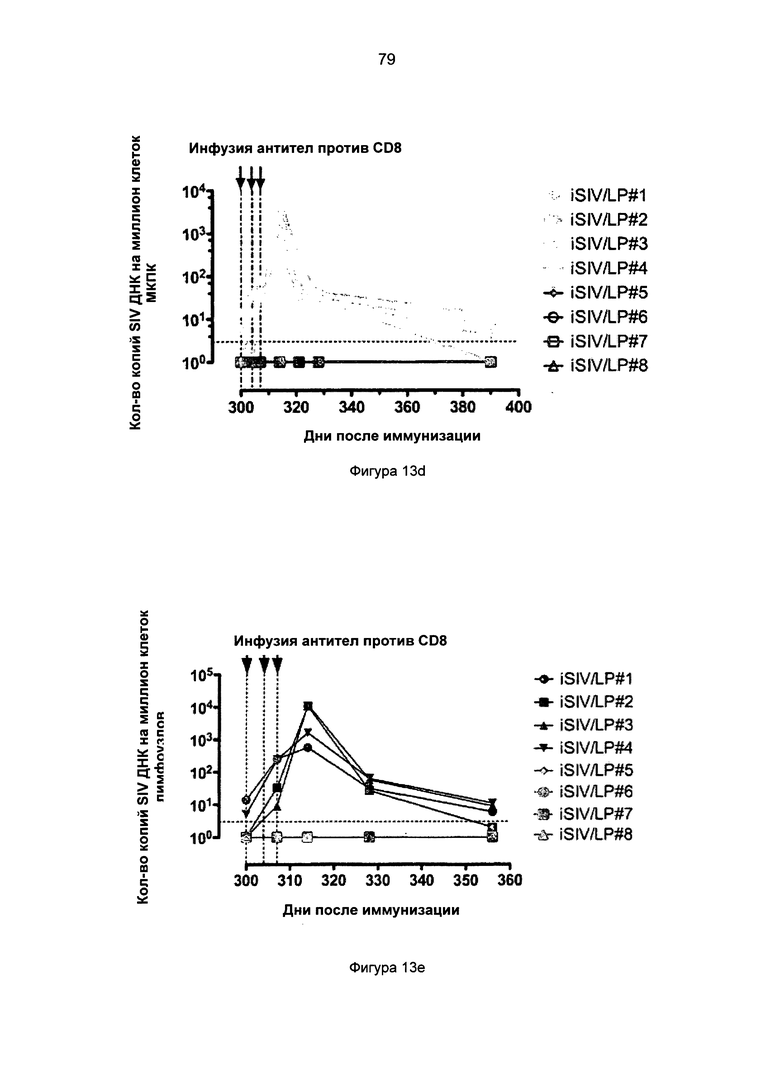
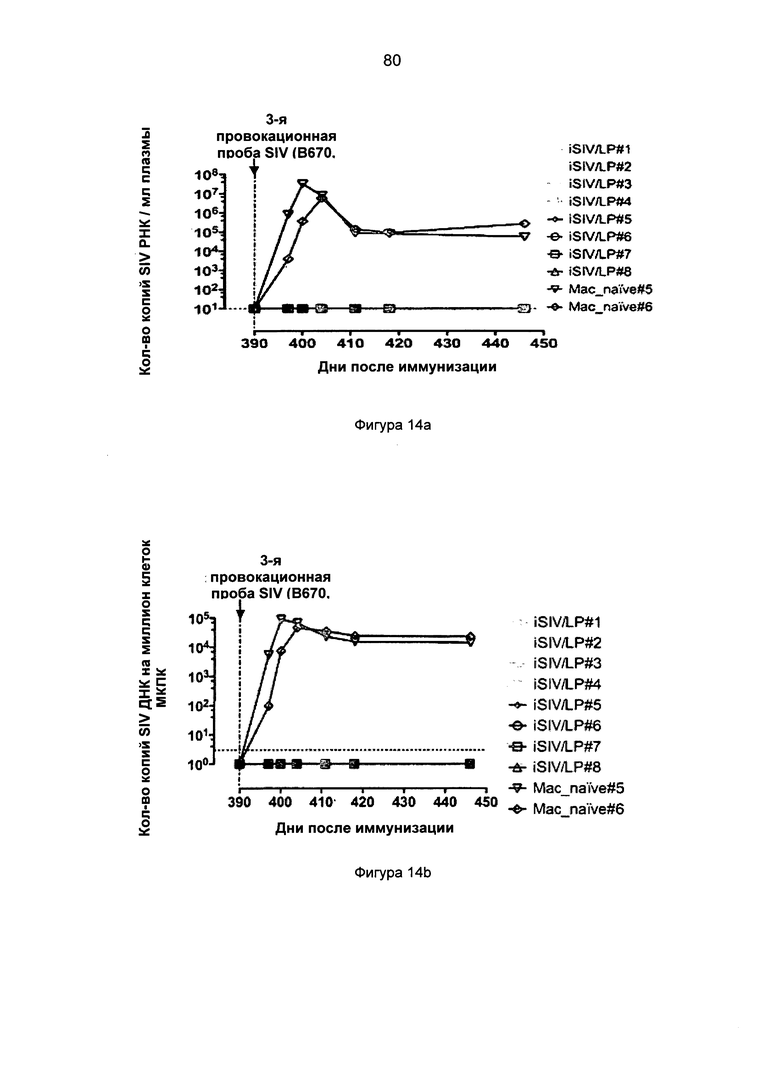
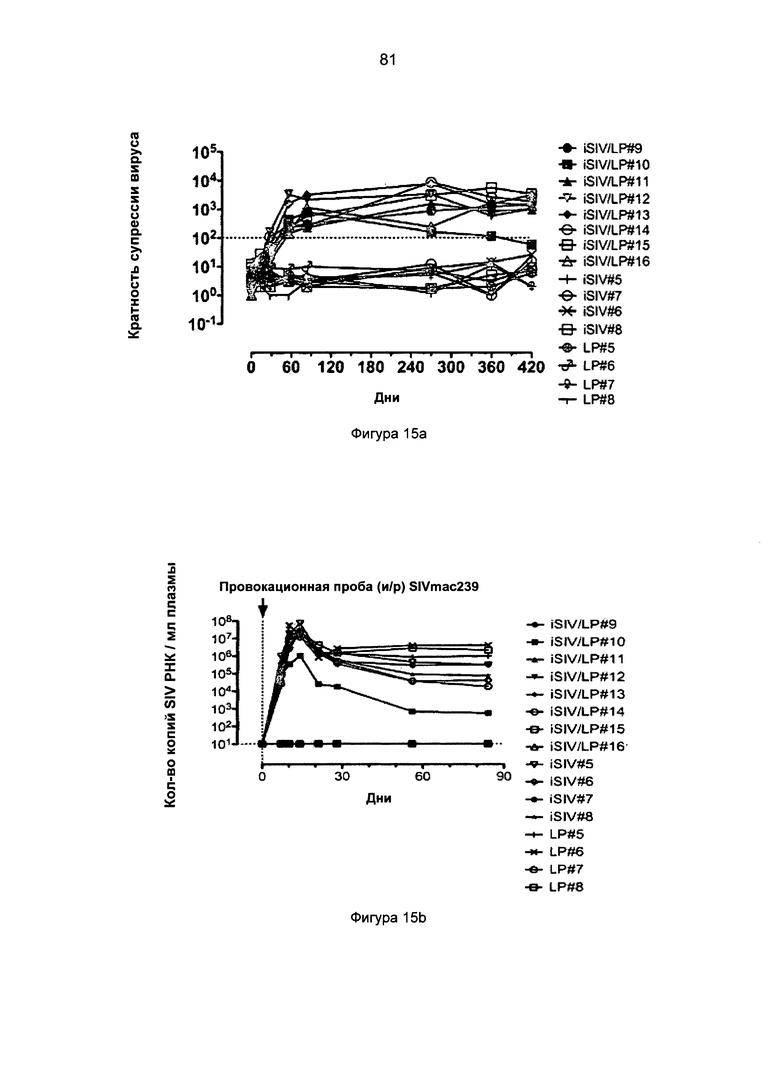
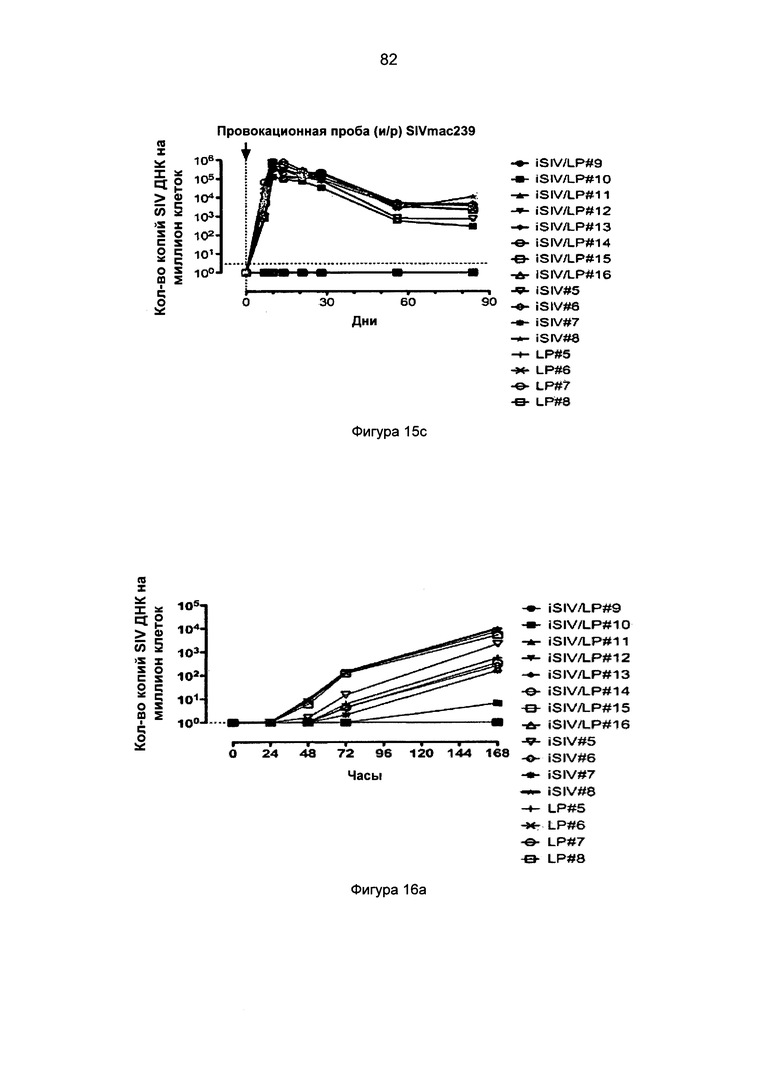
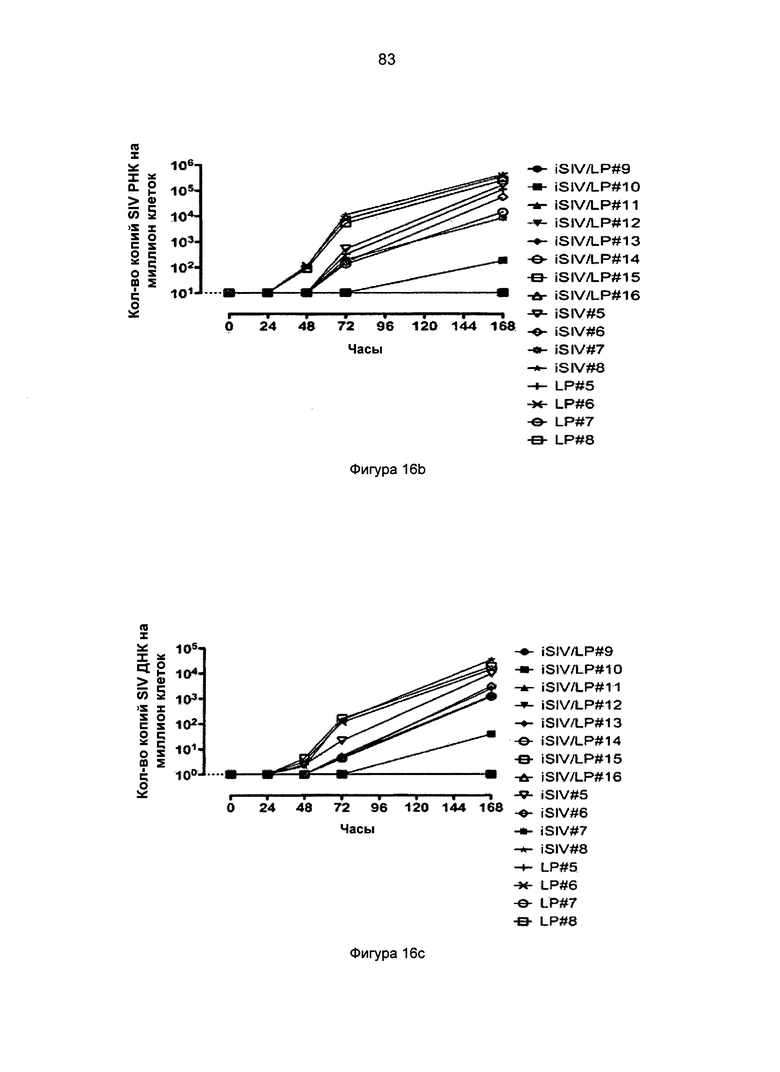
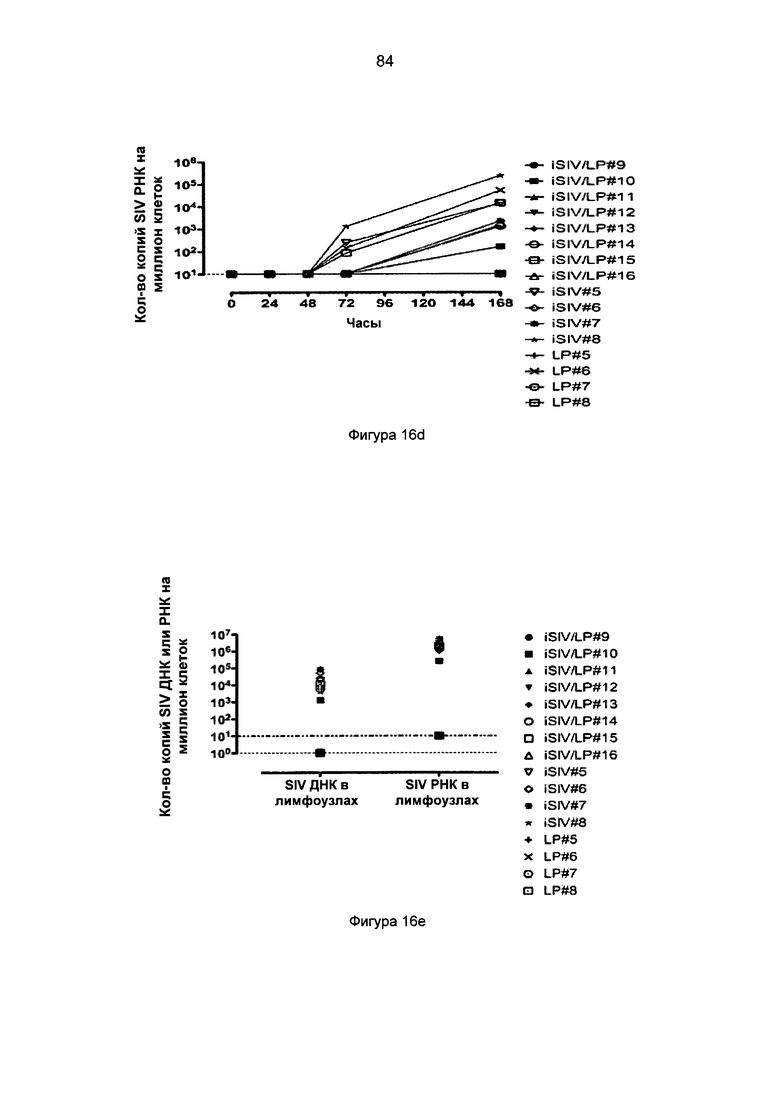
Группа изобретений относится к медицине и может быть использована для профилактики и/или лечения заболевания ВИЧ у человека, которому это необходимо. Пероральная фармацевтическая композиция включает смесь крупнодисперсного антигена, который содержит один или несколько эпитопов из белков ВИЧ - Gag и/или Pol, и непатогенную не являющуюся рекомбинантной бактерию, выбранную из группы, состоящей из бактерии рода Lactobacillus и BCG. Также предложен набор для профилактики и/или лечения заболевания ВИЧ у человека. Группа изобретений обеспечивает профилактику и/или лечение заболевания ВИЧ у человека за счет способности, указанной композиции, обеспечивать достижение «Ts»-иммунотолерантности против ВИЧ, в результате чего указанный вирус не способен реплицироваться в организме-хозяине. 2 н. и 9 з.п. ф-лы, 16 ил., 1 табл., 1 пр.
1. Пероральная фармацевтическая композиция для профилактики и/или лечения заболевания ВИЧ у человека, которому это необходимо, включающая смесь крупнодисперсного антигена, который содержит один или несколько эпитопов из белков ВИЧ - Gag и/или Pol, и непатогенную не являющуюся рекомбинантной бактерию, выбранную из группы, состоящей из бактерии рода Lactobacillus и BCG.
2. Фармацевтическая композиция по п. 1, при этом указанный крупнодисперсный антиген выбран из вирусных частиц, вирусоподобных частиц, рекомбинантных вирусных частиц, конъюгата вирусных белков и конкатемера вирусных белков.
3. Фармацевтическая композиция по п. 2, при этом указанные рекомбинантные вирусные частицы или указанные вирусные частицы инактивированы.
4. Фармацевтическая композиция по п. 1 для защиты человека от ВИЧ.
5. Фармацевтическая композиция по п. 1 для защиты человека от сероконверсии, вызванной ВИЧ.
6. Фармацевтическая композиция по любому из пп. 1-5 для применения в качестве лекарственного средства, предпочтительно в качестве вакцины.
7. Фармацевтическая композиция по п. 1, которая индуцирует и, предпочтительно, поддерживает антиген-специфическую иммунную защиту против ВИЧ у человека.
8. Фармацевтическая композиция по п. 7, при этом указанная иммунная защита характеризуется снижением и, предпочтительно, супрессией вирусной нагрузки ВИЧ у указанного человека.
9. Фармацевтическая композиция по любому из пп. 7 или 8, при этом указанная иммунная защита характеризуется присутствием регуляторных CD8+ Т-клеток, супрессирующих активацию CD4+ Т-клеток, презентирующих антигены белков ВИЧ - Gag и/или Pol - у указанного человека.
10. Фармацевтическая композиция по п. 9, при этом указанная иммунная защита определяется у указанного человека путем детекции in vitro присутствия или активности регуляторных CD8+ Т-клеток указанного человека, и при этом указанные регуляторные CD8+ Т-клетки являются ограниченными MHC-Ib/E регуляторными CD8+ Т-клетками.
11. Фармацевтической набор для профилактики и/или лечения заболевания ВИЧ у человека, которому это необходимо, содержащий:
- в первом контейнере - крупнодисперсный антиген, содержащий один или несколько эпитопов из белков ВИЧ - Gag и/или Pol; и
- во втором контейнере - непатогенную не являющуюся рекомбинантной живую бактерию, выбранную из группы, состоящей из бактерии рода Lactobacillus и BCG.
при этом указанный антиген и указанная бактерия находятся в фармацевтически приемлемых носителях, пригодных для перорального введения.
| KARPENKO LI et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| Циркуль-угломер | 1920 |
|
SU1991A1 |
| XIN KQ et al | |||
| Immunogenicity and protective efficacy of orally administered recombinant Lactococcus lactis expressing surface-bound HIV Env.// Blood | |||
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |
| GERALD K | |||
| CHEGE et al | |||
| Устройство двукратного усилителя с катодными лампами | 1920 |
|
SU55A1 |

