Настоящая заявка заявляет приоритет в отношении находящейся на рассмотрении временной патентной заявки США с регистрационным номером 60/831732.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение предлагает композиции протеаз, особенно хорошо подходящие для мытья посуды.
УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
Обычно традиционные композиции для мытья посуды коммунального и промышленного назначения основаны на комбинации высокощелочных детергентов и хлорсодержащего отбеливателя для мытья и санитарной обработки посуды. Системы такого рода как правило эффективны для удаления отбеливающихся пятен. Однако они могут оказаться недостаточными для удаления загрязнений, содержащих белки, зачастую присутствующих на домашней посуде, посуде в больницах, кафетериях, на предприятиях общественного питания и т.д. Кроме того, композиции, содержащие высокощелочные детергенты и хлор, не считаются безопасными для потребителей и окружающей среды.
Предпринимались различные попытки производства композиций для мытья посуды, эффективных для удаления белковых загрязнений. Обычно эти композиции включают протеазы, активные в щелочных условиях (например, при pH не менее 9,5). Однако подобные композиции имеют значительные недостатки, так как затруднительно разработать рецептуру их получения в виде жидкости или геля, которые потребители обычно предпочитают в качестве моющих средств. Кроме того, щелочные композиции для мытья посуды часто рассматриваются как вещества, вызывающие раздражение.
Было предпринято несколько попыток получения композиций для мытья посуды с пониженным pH (например, pH ниже 9,5). Эти композиции безопасны для потребителей и окружающей среды и могут быть разработаны в форме жидкости или геля. Однако современные композиции для мытья посуды с пониженным pH оказались очень неэффективными при удалении белковых загрязнений, даже при введении в композиции для мытья посуды высоких концентраций ферментов (например, протеаз).
Следовательно, в данной области сохраняется потребность в композициях для мытья посуды, удаляющих белковые загрязнения на посуде с высокой эффективностью. Кроме того, сохраняется потребность в композициях для мытья посуды, которые более безопасны для потребителей и окружающей среды и находятся в форме, экономичной и легкой в использовании.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение предоставляет мутантные протеазы, проявляющие улучшенные свойства для применения в составе средств для мытья посуды. В некоторых предпочтительных вариантах осуществления, мутантные протеазы имеют не менее 70% гомологии с аминокислотной последовательностью сериновой протеазы PB92, имеющей следующую аминокислотную последовательность: H2N-AQSVPWGISRVQAPAAHNRGLTGSGVKVAVLDTGISTHPDLNIRGGASFVPGEPSTQDGNGHGTHVAGTIAALNNSIGVLGVAPNAELYAVKVLGASGSGSVSSIAQGLEWAGNNGMHVANLSLGSPSPSATLEQAVNSATSRGVLVVAASGNSGAGSISYPARYANAMAVGATDQNNNRASFSQYGAGLDIVAPGVNVQSTYPGSTYASLNGTSMATPHVAGAAALVKQKNPSWSNVQIRNHLKNTATSLGSTNLYGSGLVNAEAATR-COOH (SEQ ID №2). В дальнейших вариантах осуществления, мутантные протеазы имеют, по меньшей мере, 75%, по меньшей мере, 80%, по меньшей мере, 85%, по меньшей мере, 90%, по меньшей мере, 95% или, по меньшей мере, 99% гомологию с SEQ ID №2. В каждом предпочтительном варианте осуществления, приведенном в данном тексте, мутантные протеазы снабжены улучшенными моющими характеристиками и/или повышенной стабильностью по сравнению с протеазой PB92 дикого типа.
В некоторых предпочтительных вариантах осуществления, настоящее изобретение предоставляет варианты протеаз с улучшенными моющими характеристиками, по сравнению с исходной протеазой. В некоторых особенно предпочтительных вариантах осуществления, ферменты включают таковые, обозначенные как 049, 045, 046, 047/048, 050, 051/052, 053, 054, 055/056, 057, 058, 059 и 060, имеющие замены, описанные в Таблице 1 данного текста. В дополнительных вариантах осуществления, настоящее изобретение предоставляет эти ферменты с дополнительными мутациями (например, заменами, вставками и/или делециями).
В некоторых особенно предпочтительных вариантах осуществления, настоящее изобретение предоставляет фермент, обозначенный как 049, имеющий нижеследующую аминокислотную последовательность:
AQSVPWGISR VQAPAAHNRG LTGSGVKVAV LDTGISTHPD LNIRGGASFV PGEPSTQDGN GHGTHVAGTI AALNNSIGVL GVAPNAELYA VKVLGASGSG SVSSIAQGLE WAGNNVMHVA NLSLGLQAPS ATLEQAVNSA TSRGVLVVAA SGNSGAGSIS YPARYANAMA VGATDQNNNR ASFSQYGAGL DIVAPGVNVQ STYPGSTYAS LNGTSMATPH VAGAAALVKQ KNPSWSNVQI RNHLKNTATS LGSTNLYGSG LVNAEAATR (SEQ ID №3).
Настоящее изобретение также предоставляет новые ферментативные моющие средства, содержащие протеолитические ферменты, среди которых содержится, по меньшей мере, одна мутантная протеаза, приведенная в данном тексте.
В дополнительных вариантах осуществления, настоящее изобретение предоставляет композиции средств для мытья посуды, включающие модифицированный субтилизин, где указанный субтилизин содержит, по меньшей мере, одну замену в последовательности, приведенную в SEQ ID №2, где каждое положение соответствует положению аминокислотной последовательности субтилизина BPN', и где замены выбраны из следующих положений: G118, S128, P129, S130 и S166.
В некоторых вариантах осуществления, модифицированный субтилизин содержит замены, сделанные в следующих положениях: G118, S128, P129 и S130. В некоторых предпочтительных вариантах осуществления, модифицированный субтилизин содержит мутацию G118V и, по меньшей мере, одну дополнительную мутацию. В некоторых особо предпочтительных вариантах осуществления, дополнительные мутации выбраны из группы, состоящей из S128F, S128L, S128N, S128R, S128V, P129E, P129L, P129M, P129N, P129L, P129Q, P129S, S130A, S130K, S130P, S130T, S130V и S166D. В еще более предпочтительных вариантах осуществления, модифицированный субтилизин содержит замены, сделанные в следующих положениях: S128, P129 и S130. В дополнительных предпочтительных вариантах осуществления, замены выбраны из группы, состоящей из S128C, S128R, P129Q, P129R, S130D и S130G. В некоторых особо предпочтительных вариантах осуществления, аминокислотная последовательность указанного модифицированного субтилизина приведенна в SEQ ID №3.
Настоящее изобретение также предоставляет варианты осуществления, включающие композиции для мытья посуды, содержащие модифицированный субтилизин, где указанный субтилизин содержит, по меньшей мере, одну замену в последовательности SEQ ID №2, где каждое положение соответствует положению аминокислотной последовательности субтилизина BPN', и где замена представляет собой S130T. Настоящее изобретение также предоставляет выделенную нуклеиновую кислоту, кодирующую модифицированный субтилизин, где указанный субтилизин содержит, по меньшей мере, две замены в последовательности, приведенной в SEQ ID №2, где каждое положение соответствует положению аминокислотной последовательности субтилизина BPN', и где замены выбраны из следующих положений: G118, S128, P129, S130 и S166. В некоторых вариантах осуществления, нуклеиновая кислота кодирует модифицированные субтилизины, содержащие замены, сделанные в следующих положениях: Gl18, S128, P129 и S130. В некоторых предпочтительных вариантах осуществления, нуклеиновая кислота кодирует модифицированные субтилизины, содержащие мутацию G118V и, по меньшей мере, одну дополнительную мутацию. В некоторых особо предпочтительных вариантах осуществления, нуклеиновая кислота далее содержит дополнительные мутации, выбранные из группы, состоящей из S128F, S128L, S128N, S128R, S128V, P129E, P129L, P129M, P129N, P129L, P129Q, P129S, S130A, S130K, S130P, S130T, S130V и S166D. В даже более предпочтительных вариантах осуществления, нуклеиновая кислота кодирует модифицированный субтилизин, содержащий замены, сделанные в следующих положениях: S128, P129 и S130. В некоторых дополнительных предпочтительных вариантах осуществления, нуклеиновая кислота содержит замены, выбранные из группы, состоящей из S128C, S128R, P129Q, P129R, S130D и S130G.
В некоторых особо предпочтительных вариантах осуществления, аминокислотная последовательность модифицированного субтилизина представляет собой последовательность, приведенную в SEQ ID №3. В некоторых дополнительных вариантах осуществления, настоящее изобретение предоставляет выделенную нуклеиновую кислоту, кодирующую аминокислотную последовательность, приведенную в SEQ ID №3.
В дополнительных вариантах осуществления, настоящее изобретение предоставляет векторы, содержащие, по меньшей мере, одну выделенную нуклеиновую кислоту, описанную выше. В дальнейших вариантах осуществления настоящее изобретение предоставляет векторы, содержащие, по меньшей мере, одну выделенную нуклеиновую кислоту, последовательность которой приведена выше. В дополнительных вариантах осуществления настоящее изобретение предоставляет клетки-хозяева, содержащие, по меньшей мере, один вектор, описанный выше.
Настоящее изобретение также предоставляет способы мытья посуды, включая этапы: обеспечение, по меньшей мере, одного модифицированного субтилизина, как описано выше, и посуды, нуждающейся в очистке; и приведение посуды в контакт, по меньшей мере, с одним модифицированным субтилизином в условиях, эффективных для обеспечения очистки посуды.
ОПИСАНИЕ ЧЕРТЕЖЕЙ
На Фигуре 1А показано конструирование мутантного вектора, содержащего ген протеазы PB92.
На Фигуре 1В представлено схематическое изображение процедуры введения мутации, применяемой в некоторых вариантах осуществления настоящего изобретения.
На Фигуре 1С показано конструирование экспрессионного вектора, содержащего мутантный ген протеазы PB92.
На фигуре 2 приведена нуклеотидная последовательность гена протеазы PB92 (SEQ ID № 1) и аминокислотная последовательность (SEQ ID №2) кодируемых пре-белка, про-белка и зрелого белка.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение предоставляет способы и композиции, содержащие, по меньшей мере, одну мутантную протеазу для применения при мытье посуды.
Если не указано иное, практическое применение настоящего изобретения подразумевает традиционные методики, обычно используемые в молекулярной биологии, микробиологии, очистке белков, белковой инженерии, секвенировании белков и ДНК, областях рекомбинантных ДНК и применении и разработке промышленных ферментов, которые все включены в круг компетентности в данной области техники. Все патенты, патентные заявки, статьи и публикации, упоминаемые в данном тексте как ранее, так и далее по тексту, настоящим безоговорочно введены в данный текст ссылкой.
Кроме того, приведенные в тексте рубрики не являются ограничением различных аспектов или вариантов осуществления изобретения, которые могут быть получены исходя из описания в целом. Следовательно, термины, чье определение приводится непосредственно ниже, полностью определены исходя из описания в целом. Тем не менее, с целью облегчения понимания изобретения, определения ряда терминов приводятся ниже.
Если в данном тексте не указано иное, все технические и научные термины, использованные здесь, имеют то же значение, что обычно подразумевается специалистами в области, к которой относится изобретение. Например, Singleton and Sainsbury, Dictionary of Microbiology and Molecular Biology, 2d Ed., John Wiley and Sons, NY (1994) и Hale and Markham, The Harper Collins Dictionary of Biology, Harper Perennial, NY (1991) обеспечили специалистов в данной области общими словарями многих терминов, применяемых в описании изобретения. Хотя любые методы и материалы, схожие либо эквивалентные описанным здесь, находят применение в практическом использовании настоящего изобретения, предпочтительными методами и материалами являются описанные в данном тексте. Соответственно, термины, определяемые непосредственно ниже, наиболее полно описаны исходя из определения в целом. Также, при использовании в данном тексте, формы единственного числа включают множественное число, если контекст в явном виде не указывает на иное. Если не указано иное, нуклеиновые кислоты пишутся слева направо в ориентации от 5' конца к 3' концу; аминокислотные последовательности пишутся слева направо в ориентации от амино- к карбокси-концу, соответственно. Должно быть понятно, что это изобретение не ограничено частными методиками, протоколами и описанными реагентами, так как они могут варьироваться в зависимости от контекста, при котором их применяет специалист в данной области.
Подразумевается, что каждое наибольшее численное ограничение, данное в этом описании, включает все меньшие численные пределы так же, как если бы данные меньшие численные ограничения были введены в текст в явном виде. Каждое наименьшее численное ограничение, данное в этом описании, включает все большие численные пределы так же, как если бы данные большие численные ограничения были введены в текст в явном виде. Каждый числовой интервал, данный в этом описании, будет включать все более узкие числовые интервалы, попадающие внутрь этого более широкого числового интервала так же, как если бы данные более узкие числовые интервалы были введены в текст в явном виде.
При использовании в данном тексте, термин "совместимый" обозначает, что моющие средства композиции не снижают ферментативную активность протеолитического(их) фермента(ов), предусмотренных данным текстом, до такой степени, что протеаза(ы) становится не достаточно эффективна(ы) при нормальных условиях использования. В качестве примеров далее в тексте приведены конкретные моющие вещества композиции.
При использовании в данном тексте, "эффективное количество фермента" относится к количеству фермента, необходимому для достижения ферментативной активности, требуемой при специализированном применении. Подобные эффективные количества легко устанавливаются специалистом в данной области и основаны на многих факторах, таких как применяемый конкретный вариант фермента, область применения в очистке, конкретный состав моющей композиции и требуется ли жидкая или твердая (например, гранулярная) композиция, и тому подобных факторов.
При использовании в данном тексте, термин "имеющий улучшенные свойства", применяемый в связи с термином "мутантный протеолитический фермент", относится к протеолитическим ферментам с улучшенным действием и/или повышенной стабильностью при той же силе действия, по сравнению с соответствующей протеазой дикого типа.
В некоторых особенно предпочтительных вариантах осуществления, улучшенные свойства выбраны из группы, состоящей из усиленного моющего действия и повышенной стабильности, равно как и комбинации усиленного моющего действия и повышенной стабильности.
При использовании в данном тексте, фраза "стабильность моющего средства" относится к стабильности композиции детергента. В некоторых вариантах осуществления, стабильность оценивается во время использования детергента, тогда как в других вариантах осуществления данный термин относится к стабильности композиции детергента во время хранения.
Термин "повышенная стабильность" применяется для обозначения лучшей стабильности мутантных(ой) протеаз(ы) в композиции во время хранения и/или лучшей стабильности в мыльной пене. В предпочтительных вариантах осуществления, мутантные(ая) протеазы(а) проявляют повышенную стабильность в мягких моющих средствах во время хранения и/или повышенную стабильность в мыльной пене, что включает устойчивость к окислительным агентам, комплексообразующим соединениям, саморазрушению, поверхностно-активным веществам и высокой щелочи, по сравнению с соответствующим ферментом дикого типа.
При использовании в данном тексте, фраза "устойчивость к протеолизу" относится к способности белка (например, фермента) противостоять протеолизу. Не подразумевается, что термин ограничен применением конкретной протеазы для оценки стабильности белка.
При использовании в данном тексте, "устойчивость к окислению" относится к способности белка функционировать в окислительных условиях. В частности, термин относится к способности белка функционировать в присутствии различных концентраций H2O2, перкислоты и других окислителей. Стабильность при различных окисляющих условиях может быть измерена либо традиционными методиками, известными специалистам в данной области, и/или способами, описанными в данном тексте. Индивидуальные изменения устойчивости к окислению подтверждаются, по меньшей мере, примерно 5% или более повышением или снижением (в большинстве вариантов осуществления, предпочтительно повышением) времени полужизни ферментативной активности, по сравнению с ферментативной активностью в отсутствие окислительных соединений.
При использовании в данном тексте, "pH стабильность" относится к способности белка функционировать при конкретном pH. Как правило, большинство ферментов имеют определенный интервал pH, при котором они работают. В дополнение к ферментам, которые функционируют при средних значениях pH (то есть pH около 7), существуют ферменты, способные работать в условиях очень высоких либо очень низких значений pH. Стабильность при различных значениях pH может быть измерена либо традиционными методиками, известными специалистам в данной области, и/или способами, описанными в данном тексте. Индивидуальные изменения в pH стабильности подтверждаются, по меньшей мере, примерно 5% или большим повышением или снижением (в большинстве вариантов осуществления, предпочтительно повышение) времени полужизни ферментативной активности, по сравнению с ферментативной активностью при оптимальном для данного фермента pH. Однако не предполагается, что настоящее изобретение ограничено каким-либо уровнем pH стабильности или интервалом pH.
При использовании в данном тексте, "термостабильность" относится к способности белка функционировать при конкретной температуре. Как правило, большинство ферментов имеют определенный интервал температур, при которых они работают. В дополнение к ферментам, которые функционируют при средних значениях температур (например, комнатной температуре), существуют ферменты, способные работать при очень высоких либо очень низких температурах. Термостабильность может быть измерена с помощью любой известной методики либо способами, описанными в данном тексте. Индивидуальные изменения термостабильности подтверждаются, по меньшей мере, примерно 5% или большим повышением или снижением (в большинстве вариантов осуществления, предпочтительно повышением) времени полужизни каталитической активности мутантного белка при воздействии данной температуры. Однако не предполагается, что настоящее изобретение ограничено каким-либо уровнем термостабильности или интервалом температур.
При использовании в данном тексте, термин "химическая стойкость" относится к устойчивости белка (например, фермента) к химическим реагентам, которые могут отрицательно влиять на его активность. В некоторых вариантах осуществления, подобные химические реагенты включают, но не ограничены ими, перекись водорода, перкислоту, анионные детергенты, катионные детергенты, неионные детергенты, хелатирующие агенты и им подобные. Однако не предполагается, что настоящее изобретение ограничено каким-либо конкретным уровнем химической стойкости либо интервалом химической стойкости.
При использовании в данном тексте, термины "очищенный" и "выделенный" относятся к удалению загрязнений из образца. Например, представляющий интерес фермент очищается удалением загрязняющих белков и других соединений в растворе или препарате, которые не являются ферментом, представляющим интерес. В некоторых вариантах осуществления, представляющие интерес рекомбинантные ферменты экспрессируются в бактериальных или грибных клетках-хозяевах, и эти представляющие интерес рекомбинантные ферменты очищаются удалением других компонентов клетки-хозяина; таким образом, в образце повышается процентное содержание полипептида представляющего интерес рекомбинантного фермента.
При использовании в данном тексте, "представляющий интерес белок" относится к белку (например, к ферменту или "представляющему интерес ферменту"), который анализируется, идентифицируется и/или модифицируется. В настоящем изобретении находят применение как природные, так и рекомбинантные (например, мутантные) белки.
При использовании в данном тексте, термин "белок" относится к любому соединению, включающему в себя аминокислоты и определенному как белок специалистами в данной области. Термины "белок", "пептид" и "полипептид" в данном тексте используются как взаимозаменяемые. В случае, когда пептид является частью белка, специалисты в данной области поймут использование термина по контексту.
При использовании в данном тексте, функционально и/или структурно схожие белки рассматриваются как "родственные белки". В некоторых вариантах осуществления, эти белки происходят из разных родов и/или видов, включая различия между классами организмов (например, бактериальный белок и белок грибов). В некоторых вариантах осуществления, эти белки происходят из разных родов и/или видов, включая различия между классами организмов (например, бактериальный фермент и фермент грибов). В дополнительных вариантах осуществления, родственные белки получены из одного и того же вида. В действительности, не предполагается, что настоящее изобретение ограничено родственными белками из какого-либо конкретного источника(ов). Кроме того, термин "родственные белки" охватывает гомологи по третичной структуре и гомологи по первичной последовательности (например, ферменты настоящего изобретения). В дальнейших вариантах осуществления, термин охватывает белки, которые иммунологически являются перекрестно реагирующими антигенами. При использовании в данном тексте, термин "производное" относится к белку, который получен из белка путем добавления (то есть, вставки) одной или более аминокислот к какому-либо или обоим C- и N-концам, замены в одной или более аминокислот в одном или нескольких положениях аминокислотной последовательности, и/или делеции одной или более аминокислот с какого-либо или обоих C- и N-концов, либо в одном или нескольких положениях аминокислотной последовательности, и/или вставки одной или более аминокислот в одном или нескольких положениях аминокислотной последовательности. Получение производного белка предпочтительно осуществляется путем модификации последовательности ДНК, которая кодирует природный белок, трансформации данной последовательности ДНК в подходящий организм-хозяин и экспрессии модифицированной последовательности ДНК для образования производного белка.
Родственные (и производные) белки составляют "вариантные белки". В некоторых предпочтительных вариантах осуществления, вариантные белки отличаются от родительского белка и других вариантов на малое количество аминокислотных остатков. Число отличающихся аминокислотных остатков может составлять один или более, предпочтительно 1, 2, 3, 4, 5, 10, 15, 20, 30, 40, 50 или более аминокислотных остатков. В некоторых предпочтительных вариантах осуществления, число отличающихся аминокислотных остатков между вариантами составляет от 1 до 10. В некоторых особо предпочтительных вариантах осуществления, аминокислотные последовательности родственных белков и особенно вариантных белков идентичны, по меньшей мере, на примерно 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98% или 99%. Дополнительно, при использовании в данном тексте, родственный белок или вариантный белок относится к белку, который отличается от другого родственного белка или родительского белка по числу выдающихся областей. Например, в некоторых вариантах осуществления, вариантные белки имеют 1, 2, 3, 4, 5 или 10 соответствующих выдающихся областей, которые отличаются от родительского белка.
В данной области техники известно несколько способов, пригодных для получения вариантов протеолитических ферментов настоящего изобретения, включая, но не ограничиваясь ими, насыщающий мутагенез, сканирующий мутагенез, инсерционный мутагенез, неспецифический мутагенез, сайт-специфический мутагенез и направленную эволюцию, равно как и различные иные подходы из области рекомбинантных ДНК.
При использовании в данном тексте, термин "экспрессионный вектор" относится к конструкции ДНК, содержащей последовательность ДНК, которая функционально связана с подходящей регуляторной последовательностью, способной обеспечить экспрессию ДНК в подходящем организме-хозяине. Подобные регуляторные последовательности включают промотор для обеспечения транскрипции, дополнительную последовательность оператора для контроля такой транскрипции, последовательность, кодирующую соответствующие сайты связывания рибосомы на мРНК и последовательности, контролирующие окончание транскрипции и трансляции. Вектор может представлять собой плазмиду, фаговую частицу или просто потенциальную геномную вставку. Будучи трансформирован в подходящий организм-хозяин, вектор может реплицироваться и функционировать независимо от генома хозяина либо, в некоторых случаях, может интегрироваться в геном. В данном описании, термины "плазмида", "экспрессионная плазмида" и "вектор" часто используются как взаимозаменяемые, так как в настоящее время плазмида является наиболее распространенной формой вектора. Однако подразумевается, что изобретение включает иные формы экспрессионных векторов, которые несут те же функции и которые уже известны либо станут известны в данной области техники.
В некоторых предпочтительных вариантах осуществления, ген протеазы лигируется с соответствующей экспрессионной плазмидой. Затем клонированный ген протеазы используется для трансформации или трансфекции клеток-хозяев с целью экспрессии гена протеазы. Данная плазмида может реплицироваться в организме-хозяине в случае, если она содержит хорошо известные элементы, необходимые для репликации плазмиды, либо плазмида может быть спроектирована для интеграции в хромосому хозяина. Для эффективной экспрессии гена имеются необходимые элементы (например, промотор, функционально связанный с представляющим интерес геном). В некоторых вариантах осуществления, эти необходимые элементы снабжены промотором, гомологичным промотору гена, при условии, что он распознается (то есть транскрибируется клеткой-хозяином), и терминатором транскрипции, который является либо экзогенным, либо снабжен эндогенной терминаторной областью гена протеазы. В некоторых вариантах осуществления, также включен ген селекции, такой как ген устойчивости к антибиотику, который обеспечивает непрерывное поддержание роста в культуре клеток-хозяев, несущих плазмиду, в среде с антибиотиком.
Следующий кассетный метод мутагенеза может применяться для облегчения конструирования вариантов протеаз настоящего изобретения, хотя другие способы также могут использоваться.
Во-первых, как описано в данном тексте, природный ген, кодирующий протеазу, получен и секвенирован полностью либо частично. Затем последовательность сканируется в поиске точек, в которые требуется ввести мутацию (делецию, вставку или замену) одной или более аминокислот в кодируемой протеазе. Последовательности, примыкающие к этой точке, анализируются на предмет наличия сайтов рестрикции, чтобы провести замену короткого участка гена на последовательность из пула (набора) олигонуклеотидов, при экспрессии это приведет к образованию различных мутантов. Подобные рестрикционные сайты предпочтительно должны быть уникальными в гене протеазы, что облегчит процедуру замены сегмента. Однако может применяться и какой-либо удобный сайт рестрикции, который не слишком часто встречается на протяжении последовательности гена, что при рестрикции приведет к образованию нескольких фрагментов, которые затем могут быть собраны в правильной последовательности. Если на удобном расстоянии от выбранной точки (от 10 до 15 нуклеотидов) нет сайтов рестрикции, подобные сайты получаются заменой нуклеотидов в гене таким образом, что не нарушается рамка считывания и не заменяется кодируемая аминокислота в конечной конструкции. Мутация гена с целью изменения его последовательности до соответствия требуемой последовательности осуществляется методом удлинения затравки с помощью системы фага М13 согласно распространенным методикам. Задача определения местоположения подходящих примыкающих областей и анализа изменений, требуемых для получения последовательностей двух удобных сайтов рестрикции, выполняется рутинно с применением знаний о вырожденности генетического кода, рестрикционной карты гена и большого количества различных ферментов рестрикции. Обратите внимание, что если доступен удобный сайт рестрикции в примыкающей области, то вышеописанный метод может потребоваться только при работе с примыкающими областями, которые подобный сайт не содержат.
Как только клонирована природная и/или синтетическая ДНК, сайты рестрикции, примыкающие к положениям, которые будут мутированы, разрезаются узнающими эти сайты ферментами рестрикции, и в ген лигируется ряд олигонуклеотидных кассет с концами, комплементарными полученным в исходной ДНК гена. Процедура мутагенеза упрощается при использовании этого метода, так как все олигонуклеотиды могут быть синтезированы с одинаковыми сайтами рестрикции, и не требуется синтетических линкеров для создания сайтов рестрикции.
При использовании в данном описании, термин "соответствующий" относится к остатку на пронумерованном положении в белке или пептиде или остатку, который аналогичен, гомологичен или эквивалентен пронумерованному остатку в белке или пептиде.
При использовании в данном тексте, термин "соответствующая область", как правило, относится к аналогичному положению на протяжении последовательности родственных белков или родительского белка.
Термины "кодирующая молекула нуклеиновой кислоты", "кодирующая последовательность нуклеиновой кислоты", "кодирующая последовательность ДНК" и "кодирующая ДНК" относятся к порядку или последовательности дезоксирибонуклеотидов на протяжении цепи дезоксирибонуклеиновой кислоты. Порядок этих дезоксирибонуклеотидов определяет порядок аминокислот на протяжении полипептидной (белковой) цепи. Таким образом, последовательность ДНК кодирует аминокислотную последовательность.
При использовании в данном тексте, термин "аналогичная последовательность" относится к последовательности внутри белка, обеспечивающей схожую функцию, третичную структуру и/или содержащую консервативные остатки, что и представляющий интерес белок (то есть обычно исходный представляющий интерес белок). Например, область антигенной детерминанты, которая содержит структуры альфа-спирали или бета-слоя, при замене аминокислот в аналогичной последовательности предпочтительно сохранит ту же специфическую структуру. Термин также относится к нуклеотидным последовательностям, как и к аминокислотным последовательностям. В некоторых вариантах осуществления, аналогичные последовательности разработаны таким образом, что замена аминокислот приводит к образованию варианта фермента с той же или улучшенной функцией. В некоторых предпочтительных вариантах осуществления, третичная структура и/или консервативные остатки аминокислот в представляющем интерес белке расположены внутри или около представляющего интерес сегмента или фрагмента. Таким образом, в случае если сегмент или фрагмент содержит, например, структуры альфа-спирали или бета-слоя, при замене аминокислот предпочтительно сохранится данная специфическая структура.
При использовании в данном тексте, термин "гомологичный белок" относится к белку (например, протеазе), который обладает схожей активностью и/или структурой с представляющим интерес белком (например, протеазой из другого источника). Не подразумевается, что гомологи обязательно родственны в эволюционном смысле. Таким образом, предполагается, что термин охватывает одинаковые или схожие ферменты (то есть на основе структуры и функции), полученные из различных видов. В некоторых предпочтительных вариантах осуществления, целесообразно идентифицировать гомолог, имеющий четвертичную, третичную и/или первичную структуры, схожие с таковыми представляющего интерес белка, так как замена сегмента или фрагмента в представляющем интерес белке на аналогичный сегмент гомолога снизит разрушающее воздействие замены. В некоторых вариантах осуществления, гомологичные белки вызывали схожий иммунологический ответ с представляющим интерес белком.
При использовании в данном тексте, термин "гомологичные гены" относится, по меньшей мере, к двум генам из разных видов, чьи гены соответствуют друг другу и которые являются идентичными либо очень схожими. Термин охватывает гены, которые разделены видообразованием (то есть развитием нового вида) (например, ортологичные гены), равно как и гены, которые были разделены генетической дупликацией (например, паралогичные гены). Эти гены кодируют "гомологичные белки".
При использовании в данном тексте, термины "ортолог" и "ортологичные гены" относятся к генам разных видов, которые эволюционировали из общего предкового гена (то есть гомологичные гены) при видообразовании. Обычно ортологи сохраняют ту же функцию в ходе эволюции. Идентификация ортологов находит применение в достоверном прогнозировании функции гена в новых секвенированных геномах.
При использовании в данном тексте, термины "паралог" и "паралогичные гены" относятся к генам, которые образовались путем дупликации (удвоения) внутри генома. В то время как ортологи сохраняют свою функцию в ходе эволюции, паралоги приобретают новые функции, даже если некоторые функции часто взаимосвязаны с исходной. Примеры паралогичных генов включают, но не ограничены ими, гены, кодирующие трипсин, химотрипсин, эластазу и тромбин, которые все являются сериновыми протеазами и сосуществуют в одних и тех же видах организмов.
При использовании в данном тексте, белки "дикого типа" и "природные" представляют собой белки, встречающиеся в природе. Термины "последовательность дикого типа" и "ген дикого типа" в данном тексте употребляются как взаимозаменяемые для обозначения последовательности, встречающейся в природе или содержащейся в клетке-хозяине в природе. В некоторых вариантах осуществления, последовательность дикого типа обозначает представляющую интерес последовательность, представляющую собой исходный материал для проекта белковой инженерии. Гены, кодирующие природные белки, могут быть получены традиционными методами, известными специалистам в данной области. Данные методы, как правило, включают синтез меченых проб на основе предполагаемых последовательностей областей, кодирующих представляющий интерес белок, приготовление геномных библиотек из организмов, экспрессирующих данный белок, и скрининг библиотек в поисках представляющего интерес гена с помощью гибридизации с пробами. Затем положительные по результатам гибридизации клоны картируются и секвенируются.
Термин "рекомбинантная молекула ДНК", при использовании в данном тексте, относится к молекуле ДНК, которая содержит сегменты ДНК, соединенные вместе с помощью методик молекулярной биологии.
Термин "рекомбинантный олигонуклеотид" относится к олигонуклеотиду, созданному при помощи молекулярно-биологических манипуляций, включая, но не ограничиваясь этим, лигирование двух или более олигонуклеотидных последовательностей, полученных при расщеплении ферментами рестрикции полинуклеотидной последовательности, синтез олигонуклеотидов (например, синтез праймеров или олигонуклеотидов) и тому подобное.
Степень гомологии между последовательностями может быть определена с применением любого подходящего метода, известного в данной области (см., к примеру, Smith and Waterman, Adv. Appl. Math., 2:482 [1981]; Needleman and Wunsch, J. Mol. Biol., 48:443 [1970]; Pearson and Lipman, Proc. Natl. Acad. Sci. USA 85:2444 [1988]; такие программы, как GAP, BESTFIT, FASTA и TFASTA в Wisconsin Genetics Software Package (Genetics Computer Group, Madison, WI); и Devereux et al., Nucl. Acid Res., 12:387-395 [1984]).
Например, PILEUP является полезной программой для определения уровня гомологии последовательностей. PILEUP создает множественное выравнивание последовательностей из группы родственных последовательностей, используя прогрессивные парные выравнивания. Эта программа также может построить график направляющего дерева, показывающий группирование взаимосвязей, применяемое для получения выравнивания. PILEUP использует упрощенную версию прогрессивного способа выравнивания представленного авторами Feng и Doolittle (Feng and Doolittle, J. Mol. Evol., 35:351-360 [1987]). Этот способ похож на способ, описанный авторами Higgins и Sharp (Higgins and Sharp, CABIOS 5:151-153 [1989]). Удобные параметры PILEUP включают параметры по умолчанию: вес пропуска 3,00, вес длины пропуска 0,10 и взвешенные концевые пробелы. Другим примером удобного алгоритма является алгоритм BLAST, описанный Altschul и соавт. (Altschul et al, J. Mol. Biol., 215:403-410, [1990]; и Karlin et al.f Proc. Natl. Acad. Sci. USA 90:5873-5787 [1993]). Особенно полезной программой BLAST является программа WU-BLAST-2 (см. Altschul et al., Meth. Enzymol., 266:460-480 [1996]). Параметры "W", "T" и "X" определяют чувствительность (точность) и скорость выравнивания. Программа BLAST по умолчанию использует длину слова (W) 11, выравнивания по матрице сравнения BLOSUM62 (см., Henikoff and Henikoff, Proc. Natl. Acad. Sci. USA 89:10915 [1989]) (B) 50, ожидание (E) 10, M`5, N'-4 и сравнение обеих цепей.
При использовании в данном тексте, "процентная тождественность (%) последовательности нуклеиновой кислоты" определяется как процентное содержание нуклеотидных остатков кандидатной последовательности, идентичных нуклеотидным остаткам данной последовательности.
При использовании в данном тексте, термин "гибридизация" относится к процедуре, при которой цепь нуклеиновой кислоты соединяется с комплементарной цепью спариванием оснований, как известно в данной области техники.
При использовании в данном тексте, фраза "условия гибридизации" относится к условиям, при которых проводится реакция гибридизации. Эти условия обычно классифицируются по степени "жесткости" условий, при которых измеряется гибридизация. Степень жесткости может основываться на, например, значении температуры плавления (Tm) комплекса нуклеиновой кислоты или пробы. Например, "наибольшая жесткость" обычно составляет примерно Tm-5°C (5° ниже Tm пробы); "высокая жесткость" составляет примерно 5-10° ниже Tm; "средняя жесткость" составляет примерно 10-20° ниже Tm пробы; и "низкая жесткость" составляет примерно 20-25° ниже Tm. Альтернативно либо дополнительно, условия гибридизации могут быть подобраны на основе солености или ионной силы и/или одной или более жестких отмывок. Например, буфер 6xSSC = очень низкая жесткость; 3xSSC = от низкой до средней жесткости; 1xSSC = средняя жесткость; и 0,5xSSC = высокая жесткость. Функционально, условия наибольшей жесткости могут применяться для обнаружения последовательностей нуклеиновых кислот, имеющих строгую тождественность гибридизационной пробе либо почти строго тождественных ей последовательностей; тогда как условия высокой жесткости используются для обнаружения последовательностей нуклеиновых кислот, тождественных пробе на 80% и более.
Для областей применения, требующих высокой избирательности, обычно необходимо использовать относительно жесткие условия формирования гибридов (например, применяются условия относительно низкой соли и/или высокой температуры).
Фразы "существенно схожие" и "существенно тождественные" в контексте, по меньшей мере, двух нуклеиновых кислот или полипептидов обычно обозначают, что полинуклеотид или полипептид содержит последовательность, которая тождественна эталонной последовательности (то есть дикого типа), по меньшей мере, на примерно 50%, более предпочтительно, по меньшей мере, на примерно 60%, даже более предпочтительно, по меньшей мере, на примерно 75%, более предпочтительно, по меньшей мере, на примерно 80%, еще более предпочтительно, по меньшей мере, на примерно 90%, даже более предпочтительно примерно 95%, наиболее предпочтительно примерно 97%, особенно предпочтительно на 98% и на примерно 99%. Тождественность последовательности может быть определена с применением таких известных программ, как BLAST, ALIGN и CLUSTAL с использованием стандартных параметров (см., например, Altschul, et al., J. Mol. Biol. 215:403-410 [1990]; Henikoff et aL, Proc. Natl. Acad. Sci. USA 89:10915 [1989]; Karin et al., Proc. Natl. Acad. Sci USA 90:5873 [1993]; и Higgins et al, Gene 73:237-244 [1988]). Программное обеспечение для проведения анализа с помощью BLAST доступно публично через ресурс National Center for Biotechnology Information. Также поиск баз данных может осуществляться с помощью FASTA (Pearson et al, Proc. Natl. Acad. Sci. USA 85:2444-2448 [1988]). Одним из указаний на то, что два полипептида существенно тождественны, является то, что первый полипептид со вторым перекрестно реагируют иммунологически. Обычно полипептиды, которые отличаются заменами консервативных аминокислот, являются перекрестно реагирующими антигенами. Таким образом, полипептид является существенно тождественным второму полипептиду, например, в случае, если два полипептида отличаются только консервативной заменой. Другим указанием на то, что две нуклеотидные последовательности являются существенно тождественными, служит то, что две молекулы гибридизуются друг с другом в жестких условиях (например, в интервале от средней до высокой жесткости).
При использовании в данном тексте, термин "эквивалентные остатки" относится к белкам, имеющим общие особые аминокислотные остатки. Например, эквивалентные остатки могут быть идентифицированы путем определения гомологии на уровне третичной структуры для белка (например, протеазы), чья третичная структура установлена рентгеноструктурной кристаллографией. Эквивалентные остатки определяются как остатки, для которых атомные координаты двух или более главных атомов цепочки конкретного аминокислотного остатка белка с очевидными эквивалентными остатками относительно представляющего интерес белка находятся в пределах 0,13 нм и предпочтительно 0,1 нм, после выравнивания. Выравнивание осуществляется после того, как наилучшая модель была ориентирована и позиционирована таким образом, чтобы обеспечить максимальное перекрывание атомных координат неводородных атомов анализируемых белков. Предпочтительной моделью является кристаллографическая модель, дающая наиболее низкий R фактор для экспериментальных данных по дифракции при наибольшем доступном разрешении, определенная с применением методик, известных специалистам в области кристаллографии и белкового исследования/анализа.
Термин "регуляторный элемент" при использовании в данном тексте относится к генетическому элементу, который контролирует некоторые аспекты экспрессии последовательности нуклеиновой кислоты. Например, промотор представляет собой регуляторный элемент, который облегчает инициацию транскрипции функционально связанной с ним кодирующей области. Дополнительные регуляторные элементы включают сигналы сплайсинга, сигналы полиаденилирования и сигналы терминации.
При использовании в данном тексте, "клетки-хозяева" являются, как правило, прокариотическими или эукариотическими организмами-хозяевами, которые трансформированы или трансфецированы векторами, сконструированными с применением технологий рекомбинантных ДНК, известных в данной области техники. Трансформированные клетки-хозяева способны как к репликации векторов, кодирующих варианты белков, так и к экспрессии требуемого варианта белка. В случае векторов, которые кодируют пре- или препро-форму варианта белка, такие варианты при экспрессии обычно выделяются из клетки-хозяина в среду.
Термин "введенный" в контексте доставки последовательности нуклеиновой кислоты в клетку подразумевает трансформацию, трансдукцию или трансфекцию. Способы трансформации включают, но не ограничены ими, любые подходящие методики, известные в данной области, такие как трансформация протопласта, осаждение хлоридом кальция, электропорация, применение «оголенной» (депротеинизированной) ДНК и тому подобные, известные в данной области техники (см. Chang and Cohen, Mol. Gen. Genet, 168:111-115 [1979]; Smith et al., Appl. Env. Microbiol., 51:634 [1986]; и обзорная статья Ferrari и соавт. в Harwood, Bacillus, Plenum Publishing Corporation, pp. 57-72 [1989]).
Термин "промотор/энхансер" обозначает сегмент ДНК, содержащий последовательности, которые способны осуществлять функции и промотора, и энхансера. Энхансер/промотор может быть "эндогенным" либо "экзогенным", или "гетерологичным". Эндогенный энхансер/промотор представляет собой сегмент, природно связанный с данным геном в геноме. Экзогенный (гетерологичный) энхансер/промотор представляет собой сегмент, помещенный в смежное с геном положение при помощи генетических манипуляций (то есть молекулярно-биологических методик).
Наличие "сигналов сплайсинга" в экспрессионном векторе часто приводит к повышенному уровню экспрессии рекомбинантного транскрипта. Сигналы сплайсинга опосредуют удаление интронов из первичного РНК-транскрипта и состоят из донорного и акцепторного сайтов сплайсинга (Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd ed., Cold Spring Harbor Laboratory Press, New York [1989], pp. 16.7-16.8).
Термин "стабильная трансфекция" или "стабильно трансфецированный" относится к введению и интеграции инородной ДНК в геном трансфецированной клетки. Термин "стабильный трансфектант" относится к клетке, которая содержит инородную или экзогенную ДНК, стабильно интегрированную в геномную ДНК трансфецированной клетки.
Термины "селектируемый маркер" или "продукт гена селекции" при использовании в данном тексте относятся к применению гена, который кодирует ферментативную активность, обеспечивающую устойчивость к антибиотику или препарату клетки, в которой селектируемый маркер экспрессируется.
При использовании в данном тексте, термины "амплификация" и "амплификация гена" относятся к процессу, при котором специфические последовательности ДНК непропорционально реплицируются таким образом, что амплифицированный ген присутствует в виде большего количества копий, чем изначально присутствовало в геноме. В некоторых вариантах осуществления, отбор клеток при наращивании в присутствии препарата (например, ингибитора подавляемого фермента) приводит к амплификации эндогенного гена, кодирующего генный продукт, необходимый для роста в присутствии препарата, либо амплификации экзогенных (то есть введенных) последовательностей, кодирующих этот генный продукт, либо обоих. Отбор клеток путем наращивания в присутствии препарата (например, ингибитора подавляемого фермента) может привести к амплификации эндогенного гена, кодирующего генный продукт, необходимый для роста в присутствии препарата, либо амплификации экзогенных (то есть введенных) последовательностей, кодирующих этот генный продукт, либо обоих.
"Амплификация" является частным случаем репликации нуклеиновой кислоты, использующим специфичность матрицы. Этот случай противопоставлен репликации, неспецифической относительно матрицы (то есть репликации, которая зависит от матрицы, но не зависит от ее специфического типа). Специфичность по матрице здесь отличается от точности репликации (то есть синтеза правильной полинуклеотидной последовательности) и специфичности по нуклеотидам (рибо- или дезоксирибо-). Специфичность по матрице часто описывается в терминах "целевой" специфичности. Целевые последовательности являются "целями" в том смысле, что они выбираются из других нуклеиновых кислот. Методики амплификации в первую очередь разрабатывались для подобной сортировки.
При использовании в данном тексте, термин "совместная амплификация" относится к введению в одиночную клетку амплифицируемого маркера, соединенного с другими генными последовательностями (то есть содержащими один или более ген, не являющийся маркером селекции, такой как содержащийся в экспрессионном векторе), и амплификации в условиях подходящего селективного давления таким образом, что клетка амплифицирует и амплифицируемый маркер, и другие неселектируемые генные последовательности. Амплифицируемый маркер может быть физически соединен с другими генными последовательностями либо, альтернативно, система представляет собой две отдельных молекулы ДНК (одну, содержащую амплифицируемый маркер, и другую, содержащую неселектируемый маркер), которые вводятся в одну и ту же клетку.
При использовании в данном тексте, термины "амплифицируемый маркер", "амплифицируемый ген" и "вектор амплификации" относятся к маркеру, гену или вектору, кодирующему ген, который обеспечивает возможность амплификации этого гена при соответствующих условиях роста.
При использовании в данном тексте, термин "амплифицируемая нуклеиновая кислота" относится к нуклеиновым кислотам, которые могут быть амплифицированы каким-либо способом амплификации. Предполагается, что "амплифицируемая нуклеиновая кислота " обычно будет содержать "образец матрицы".
При использовании в данном тексте, термин " образец матрицы" относится к нуклеиновой кислоте, полученной из образца, анализируемого на предмет наличия "цели" (определено ниже). В противоположность, термин "фоновая матрица" используется для обозначения нуклеиновой кислоты, не являющейся образцом матрицы и которая может либо присутствовать, либо нет в образце. Фоновая матрица чаще всего является случайной. Наличие в образце фоновой матрицы может быть результатом примеси или присутствия загрязнения нуклеиновыми кислотами, от которых образец желательно очистить. Например, в качестве фона в тестовом образце помимо детектируемых могут присутствовать другие нуклеиновые кислоты из организмов.
В большинстве методик амплификации "специфичность по матрице" обеспечивается выбором фермента. Амплифицирующие ферменты представляют собой ферменты, которые, при условиях, в которых они работают, будут работать только со специфическими последовательностями нуклеиновых кислот в разнородной смеси нуклеиновых кислот. Например, в случае репликазы Qβ, РНК MDV-1 является специфической матрицей для репликазы (см., например, Kacian et al., Proc. Natl. Acad. Sci. USA 69:3038 [1972]). Другие нуклеиновые кислоты не реплицируются этим ферментом амплификации. Подобно этому, в случае РНК полимеразы фага T7, этот фермент амплификации строго специфичен для собственных промоторов (см. Chamberlin et al., Nature 228:227 [1970]). В случае ДНК лигазы T4, фермент не будет лигировать два олигонуклеотида или полинуклеотида, если есть несоответствие между субстратом (олигонуклеотидным или полинуклеотидным) и матрицей в месте соединения лигазой (см. Wu and Wallace, Genomics 4:560 [1989]). Наконец, полимеразы Taq и Pfu благодаря их свойству функционировать при высоких температурах демонстрируют высокую специфичность к последовательностям, связанным и таким образом определенным праймерами (затравками); высокая температура позволяет воспроизвести такие термодинамические условия, которые благоприятствуют гибридизации праймера и целевой последовательности и не допускают гибридизации с нецелевыми последовательностями.
При использовании в данном тексте, термин "праймер" относится к олигонуклеотиду, либо природному очищенному после расщепления рестриктазами, либо полученному синтетически, который, будучи позиционирован, способен выступить в роли точки инициации синтеза в условиях, вызывающих синтез продукта удлинения праймера, комплементарного цепи нуклеиновой кислоты (то есть в присутствии нуклеотидов и стимулирующего агента, такого как ДНК полимераза, и при подходящих температуре и pH). Праймер предпочтительно является одноцепочечным, что обеспечит наибольшую эффективность амплификации, но, альтернативно, может быть двуцепочечным. В последнем случае, праймер сначала обрабатывается для разделения его цепей перед применением в получении продуктов удлинения. Предпочтительно, праймер является олигодезоксирибонуклеотидом. Праймер должен быть достаточно длинным, чтобы начинать синтез продукта удлинения в присутствии стимулирующего агента. Точные длины праймеров зависят от многих факторов, включая температуру, источник праймера и применяемую методику.
При использовании в данном тексте, термин "проба" относится к олигонуклеотиду (то есть последовательности нуклеотидов), либо природному очищенному после расщепления рестриктазами, либо полученному синтетически с помощью рекомбинантных технологий или амплификацией с помощью ПЦР, который способен гибридизоваться с другим представляющим интерес олигонуклеотидом. Проба может быть одноцепочечной или двуцепочечной. Пробы полезны при детекции, идентификации и выделении конкретных генных последовательностей. Предполагается, что любая проба, применяемая в настоящем изобретении, будет мечена какой-либо "репортерной молекулой" таким образом, что может быть детектирована какой-либо детектирующей системой, включая, но не ограничиваясь ими, системы на основе действия ферментов (например, твердофазный иммуноферментный анализ, равно как и основанный на работе ферментов гистохимический анализ), флуоресцентные, радиоактивные и люминесцентные системы. Не подразумевается, что настоящее изобретение ограничено применением какой-либо конкретной системой детекции или типом мечения.
При использовании в данном тексте, термин "цель" в контексте описания способа амплификации (например, полимеразной цепной реакции) относится к области нуклеиновой кислоты, связанной с праймером, используемым для полимеразной цепной реакции. Следовательно, "цель" должна быть выбрана среди прочих последовательностей нуклеиновых кислот. "Сегмент" определяется как область нуклеиновой кислоты внутри целевой последовательности. При использовании в данном тексте, термин "полимеразная цепная реакция" ("ПЦР") относится к способам, описанным в патентах США №№ 4683195, 4683202 и 4965188, настоящим введенным в текст ссылкой, которые содержат способы увеличения концентрации сегмента целевой последовательности в смеси геномной ДНК без клонирования и очистки. Этот процесс амплификации целевой последовательности включает введение большого избытка двух олигонуклеотидных праймеров в смесь ДНК, содержащую требуемую целевую последовательность, и затем соблюдение определенной последовательности температурных циклов в присутствии ДНК полимеразы. Два праймера комплементарны соответствующим цепям двойной цепи целевой последовательности. Для осуществления амплификации смесь денатурируется, и праймеры сплавляются с комплементарными последовательностями на целевой молекуле. После сплавления ("отжиг" праймеров) праймеры удлиняются полимеразой до формирования новых пар комплементарных цепей. Этапы денатурации, отжига праймеров и удлинения полимеразой могут быть повторены много раз (то есть денатурация, отжиг и удлинение составляют один "цикл"; возможно осуществить много "циклов") для получения высокой концентрации амплифицированного сегмента требуемой целевой последовательности. Длина амплифицированного сегмента требуемой целевой последовательности определяется относительным расположением праймеров по отношению друг к другу, и, следовательно, эта длина является контролируемым параметром. Благодаря элементу повторения в ходе процесса данный способ назван "полимеразная цепная реакция" (здесь и далее "ПЦР"). Так как требуемые амплифицируемые сегменты целевой последовательности становятся преобладающими последовательностями (в смысле концентрации) в смеси, они называются "амплифицированными ПЦР". При использовании в данном тексте, термин "реагенты амплификации" относится к тем веществам (дезоксирибонуклеотид трифосфаты, буфер и т.д.), которые, помимо праймеров, матрицы нуклеиновой кислоты и фермента амплификации, необходимы для амплификации. Обычно сами реагенты амплификации с другими компонентами реакции помещаются и содержатся в реакционных емкостях (пробирках, микроплашках и т.д.).
С помощью ПЦР возможно амплифицировать единичную копию специфической целевой последовательности в геномной ДНК до уровня, детектируемого несколькими различными методиками (например, гибридизацией с меченой пробой; введением биотинилированных праймеров с последующей детекцией ферментом, слитым с авидином; введением меченых 32P (радиоактивный изотоп фосфора) дезоксинуклеотид трифосфатов, таких как дЦТФ или дАТФ, в амплифицированный сегмент). Помимо геномной ДНК, любая последовательность олигонуклеотидов или полинуклеотидов может быть амплифицирована с помощью соответствующего набора праймеров. В частности, амплифицированные сегменты, полученные ПЦР, сами по себе являются эффективной матрицей для последующих амплификаций ПЦР.
При использовании в данном тексте, термины "продукт ПЦР", "ПЦР фрагмент" и "продукт амплификации" относятся к полученной смеси веществ после двух или более завершенных циклов ПЦР ( этапов денатурации, отжига и удлинения). Эти термины охватывают случай, когда была проведена амплификация одного или более сегментов одной или более целевых последовательностей.
При использовании в данном тексте, термины "эндонуклеазы рестрикции" и "ферменты рестрикции" относятся к бактериальным ферментам, каждый из которых разрезает двуцепочечную ДНК рядом или внутри специфической нуклеотидной последовательности.
При использовании в данном тексте, "поверхностное свойство" применяется в отношении электростатического заряда, равно как и таких свойств, как гидрофобность и/или гидрофильность, проявляемые поверхностью белка.
При использовании в данном тексте, термины "моющая композиция" и "моющий состав" применяются в отношении смесей, которые предполагаются для использования в моющей среде для очистки загрязненных предметов. В предпочтительных вариантах осуществления, термин применяется по отношению к детергентам, используемым для очистки посуды, столовых приборов и т.д. (например, "детергенты для мытья посуды"). Не подразумевается, что настоящее изобретение ограничено какими-либо конкретными моющими составами или композициями. В действительности, предполагается, что в дополнение к детергентам, которые содержат, по меньшей мере, одну протеазу настоящего изобретения, термин охватывает детергенты, которые содержат поверхностно-активные вещества, трансферазу(ы), гидролитические ферменты, оксидоредуктазы, моющие компоненты, отбеливающие вещества, активаторы отбеливания, вещества, придающие синий цвет, и флуоресцентные красители, ингибиторы образования комков, маскирующие средства, активаторы ферментов, антиоксиданты и растворители.
При использовании в данном тексте, "композиция для мытья посуды" относится ко всем формам композиции для мытья посуды, в том числе столовых приборов, включая, но ими не ограничиваясь, гранулярные и жидкие формы. Не подразумевается, что настоящее изобретение ограничено каким-либо конкретным типом моющей композиции.
В действительности, настоящее изобретение находит свое применение в очистке посуды (например, столовой посуды, включая, но не ограничиваясь ими, тарелки, чашки, стаканы, миски и т.д.) и столовых приборов (например, кухонные принадлежности, включая, но не ограничиваясь ими, ложки, ножи, вилки, вспомогательные кухонные инструменты и т.д.) из любого материала, включая, но не ограничиваясь ими, керамические, пластмассовые, металлические, фарфоровые, стеклянные, эмалированные и т.д. Термин "посуда" используется в данном тексте по отношению и к посуде, и к столовым приборам.
При использовании в данном тексте, термин "моющее действие" мутантной протеазы относится к вкладу мутантного протеолитического фермента в мытье посуды, который обеспечивает дополнительное очищающее действие к действию детергента без добавления мутантной протеазы в композицию. Моющее действие сравнивается в соответствующих условиях мытья (посуды).
Термин "соответствующие условия мытья (посуды)" используется в данном тексте для обозначения условий, в частности температуры мытья, времени, механического воздействия при мытье, концентрации моющего вещества, типа детергента и жесткости воды, обычно применяемых в домашнем хозяйстве целевого потребителя на рынке моющих средств.
Термин "усиленное моющее действие" используется для обозначения того, что получен лучший конечный результат в смысле удаления загрязнений с посуды и/или столовых приборов при соответствующих условиях мытья, либо что для получения того же конечного результата требуется меньше мутантной протеазы (по весу), чем соответствующего фермента дикого типа.
Термин "сохраненное моющее действие" применяется для обозначения того, что моющее действие мутантного протеолитического фермента (количества по весу) составляет, по меньшей мере, 80% от действия соответствующего фермента дикого типа при соответствующих условиях мытья.
Моющее действие протеаз обычно измеряется по их способности удалять определенные характерные загрязнения при надлежащих тестовых условиях. В этих тестовых системах другие соответствующие факторы, такие как состав моющего средства, концентрация моющего средства, жесткость воды, механическое воздействие при мытье, время, pH и/или температура, могут контролироваться таким образом, чтобы воспроизвести условия, обычные для применения в домашнем хозяйстве целевого потребителя на рынке моющих средств. Лабораторная система тестирования при применении, описанная в данном описании, является представительной для применения в домашнем хозяйстве, когда используется для анализа действия протеолитических ферментов, модифицированных ДНК-мутагенезом. Таким образом, способы, предоставленные в данном тексте, облегчают тестирование больших количеств различных ферментов и отбор тех из них, которые особенно хорошо пригодны для применения моющего средства конкретного типа. Таким способом легко отбираются ферменты, "изготовленные на заказ" для конкретных условий применения.
При использовании в данном тексте, термин "обеззараживающий" относится к удалению загрязнений с поверхности, равно как и к подавлению роста или уничтожению микробов на поверхностях предметов. Не предполагается, что настоящее изобретение ограничено конкретной поверхностью, предметом или типом удаляемых загрязнения(ий) или микробов.
Некоторые сериновые протеазы бактерий называются "субтилизинами". Субтилизины включают сериновые протеазы Bacillus subtilis, Bacillus amyloliquefaciens ("субтилизин BPN`") и Bacillus licheniformis ("субтилизин Карлсберг (Carlsberg)") (см., например, Markland and Smith, в Boyer (ed.), Enzymes. The (Boyer, ed.) vol. 3, pp.561-608, Academic Press, New York, [1971]). Такие штаммы Bacillus, как щелочелюбивые штаммы Bacillus, производят другие протеазы. Примерами таковых из последней категории являются такие сериновые протеазы, как протеаза MAXACAL® (также относится к называемому в данном тексте "протеаза PB92", выделенная из Bacillus nov. spec. PB92) и протеаза SAVINASE®. Дополнительные протеазы включают, но не ограничены этим, протеазу PROPERASE®.
Аминокислотная (SEQ ID №2) и ДНК последовательности (SEQ ID №1) протеазы PB92 показаны на Фигуре 2. Зрелая протеаза содержит 269 аминокислот, имеет молекулярный вес около 27000 Дальтон и изоэлектрическую точку в высоком щелочном интервале. Активность протеазы PB92 в отношении белкового субстрата выражается в ADU (Щелочных Единицах Дельфта). Активность в ADU определяется согласно методике, описанной в Описании Изобретения Патента Великобритании № 1353317, за исключением того, что pH заменен с 8,5 на 10,0. Очищенная протеаза PB92 имеет активность 21000 ADU на мг. Число оборотов (kcat), измеренное относительно казеина в качестве субстрата, составляет 90 сек-1моль-1.
Специфическая активность очищенных препаратов субтилизина Карлсберг (Carlsberg) (см., Delange and Smith, J. Biol. Chem., 243:2184 [1968]) достигает 10000 ADU/мг и субтилизина BPN` (Matsubara et al., J. Biol. Chem., 240:1125 [1965]) до 7000 ADU/мг. Помимо вышеупомянутых параметров, таких как специфическая активность и число оборотов (kcat), протеаза PB92 отличается от таких протеаз, как субтилизин Карлсберг (Carlsberg), субтилизин BPN' и других протеаз, вводимых в состав моющих средств (например, MAXATASE® и ALCALASE®), тем, что имеет высокий положительный заряд, что можно увидеть при электрофорезе нативного белка в геле.
Так как протеаза PB92 активна при удалении загрязнений при щелочных значениях pH, то обычно используется как добавка в моющих средствах совместно с такими составляющими моющих средств, как поверхностно-активные вещества, моющие компоненты и окисляющие агенты. Последние чаще всего используются в форме порошков. Протеаза PB92 имеет высокую эффективность в удалении загрязнений по сравнению с другими протеазами, такими как вышеуказанные субтилизины. Это означает, что для достижения того же моющего действия требуется меньшее количество протеазы PB92. Чувствительность к окислению является существенным недостатком протеазы PB92 и всех других известных сериновых протеаз, применяемых в моющих средствах (см., например, Stauffer et al., J. Biol. Chem., 244:5333-5338 [1969]; and Estell et al, J. Biol. Chem., 263:6518-6521 [1985]). Окисление протеазы PB92 H2O2 или перкислотой, производимыми активаторной системой, содержащей тетрагидрат соли перборной кислоты и ТАЕД (тетраацетил этиленамид), приводит к образованию фермента с активностью 50% и 10%, соответственно, в ADU/мг, по сравнению с неокисленной протеазой PB92.
Настоящее изобретение предоставляет способы и композиции для производства, скрининга и отбора мутантных протеолитических ферментов, производных природных сериновых протеаз бактерий. Подобными мутантами являются, например, те, что кодируются геном, производным из гена дикого типа щелочелюбивого штамма Bacillus. В наиболее предпочтительных вариантах осуществления, штамм представляет собой штамм PB92.
Однако пригодны и мутанты, полученные на основе сериновой протеазы щелочелюбивых Bacillus SAVINASE®. Настоящее изобретение также находит применение в отборе модифицированных протеаз, происходящих из других протеаз, кроме сериновых протеаз щелочелюбивых штаммов Bacillus PB92. Например, гены Bacillus subtills, Bacillus amyloliquefaciens и Bacillus licheniformis, кодирующие сериновые протеазы, известны и могут использоваться как цели мутагенеза. Однако не подразумевается, что настоящее изобретение ограничено какими-либо конкретными способами, так как любой подходящий способ мутагенеза находит свое применение в настоящем изобретении, включая, но не ограничиваясь этим, опосредованный олигонуклеотидами сайт-направленный мутагенез либо неспецифический мутагенез генной области.
В некоторых предпочтительных вариантах осуществления, способы отбора мутантных протеолитических ферментов, предоставленные настоящим изобретением, включая производство и скрининг, состоят из следующих этапов: мутирование клонированного гена, кодирующего представляющий интерес протеолитический фермент или его фрагмент; выделение полученного(ых) гена или генов мутантной протеазы; введение указанных гена или генов мутантной протеазы, предпочтительно в составе подходящего вектора, в подходящий штамм организма-хозяина для экспрессии и производства; восстановление (выделение) полученной мутантной протеазы и идентификация тех мутантных протеаз, что имеют улучшенные свойства для применения в составе моющих средств.
Подходящие для производства мутантных протеаз штаммы организмов-хозяев включают трансформируемые микроорганизмы, в которых может быть осуществлена экспрессия протеазы. Особенно целесообразно применение в качестве хозяев штаммов тех же видов или родов, из которых происходят пртеазы, такие как штамм Bacillus, предпочтительно щелочелюбивый штамм Bacillus и наиболее предпочтительно Bacillus nov. spec. PB92 или его мутант, в значительной мере обладающий теми же свойствами. Также к предпочтительным штаммам относятся штаммы B. subtilis, B. licheniformis и B. amyloliquefaciens. Другие подходящие и предпочтительные штаммы организмов-хозяев включают те штаммы, которые в основном неспособны к производству внеклеточных протеолитических ферментов до трансформации мутантным геном. Особый интерес представляют штаммы организмов-хозяев Bacillus, дефицитные по протеазе, такие как дефицитный по протеазе производный штамм Bacillus nov. spec. PB92. Экспрессия протеаз достигается с помощью экспрессионных сигналов, которые функционируют в выбранном организме-хозяине. Сигналы экспрессии включают последовательности ДНК, регулирующие транскрипцию и трансляцию генов протеаз. Надлежащие векторы способны реплицироваться для получения в выбранном организме-хозяине достаточно высокого числа копий на клетку либо обеспечивают стабильное содержание гена протеазы в штамме хозяина путем встраивания в хромосому.
Мутантные протеолитические ферменты, согласно изобретению, получаются путем культивирования трансформированного штамма хозяина, содержащего ген или гены требуемого протеолитического фермента, при целесообразных условиях ферментации и выделения произведенных ферментов.
Предпочтительно, экспрессируемые протеазы секретируются в культуральную среду, что облегчает их выделение, либо, в случае грамотрицательных бактерий в качестве штамма хозяина, в периплазматическое пространство. Для обеспечения секреции используется подходящая аминоконцевая сигнальная последовательность, предпочтительно сигнальная последовательность, кодируемая исходным геном, если она функциональна в выбранном штамме организма-хозяина.
В некоторых вариантах осуществления, свойства природных или мутированных естественным путем протеаз в моющем средстве усиливаются путем введения в фермент ряда мутаций. В большинстве случаев, мутации представляют собой замены как консервативные, так и неконсервативные, хотя делеции и вставки также находят свое применение в некоторых вариантах осуществления.
В случае консервативных замен применяется следующая таблица:
где любая аминокислота может быть заменена любой аминокислотой в той же категории, особенно в той же линии. Кроме того, полярные аминокислоты N, Q могут замещать сами или быть замещенными на заряженные аминокислоты. Для целей настоящего изобретения особый интерес представляют замены, приводящие к повышению анионных свойств протеазы, особенно в сайтах, не связанных напрямую с активным центром.
Области особого интереса для введения мутации являются теми аминокислотами, которые находятся на расстоянии в пределах 4 Å от молекулы ингибитора Eglin C, когда Eglin C связан с активным центром.
Следующая нумерация основана на последовательности протеазы PB92, но расчеты верны и для других сериновых протеаз, имеющих в значительной степени гомологичную структуру, особенно для тех протеаз, которые имеют степень гомологии более чем примерно 70% и предпочтительнее имеющих степень гомологии более чем примерно 90%. Особо представляющие интерес положения включают 32, 33, 48-54, 58-62, 94-107, 116, 123-133, 150, 152-156, 158-161, 164, 169, 175-186, 197, 198, 203-216 (нумерация по PB92), так как большинство этих положений доступны для прямого контакта с субстратом белковой природы. Обычно аминокислоты в положениях 32, 62, 153 и 215 не заменяются, так как мутации этих сайтов имеют тенденцию приводить к снижению моющего действия. В некоторых наиболее предпочтительных вариантах осуществления, мутации введены по положениям 116, 126, 127 и 128 (нумерация по PB92). В альтернативных вариантах осуществления, вводится дополнительная мутация по положению 160.
В дальнейших вариантах осуществления, особенно представляющие интерес положения для замен включают 60, 94, 97-102, 105, 116, 123-128, 150, 152, 160, 183, 203, 211, 212, 213, 214 и 216 (нумерация по PB92). В некоторых положениях происходит замена нестабильной аминокислоты (например, метионина) на более устойчивую к окислению аминокислоту (например, треонин), при этом сохраняется основная конформация и объем остатка аминокислоты в этом сайте. В некоторых иных вариантах осуществления, замена природной аминокислоты на почти любую другую аминокислоту приводит к лучшему результату, особенно в случае замен аминокислот с гидроксильными группами, S и/или T, на полярную или неполярную аминокислоту, или даже на ароматическую аминокислоту. В некоторых наиболее предпочтительных вариантах осуществления, замены включают (нумерация по PB92):
Gl16 I, V, L
S126 любая аминокислота
P127 любая аминокислота
S128 любая аминокислота
S160 анионная или нейтральная алифатическая, либо R
A166 заряженная, особенно анионная
M169 нейтральная алифатическая, предпочтительно неполярная
N212 анионная
M216 алифатическая полярная, особенно S, T, N, Q
Как ни удивительно, в то время как многие мутации приводили к снижению специфической активности протеазы в отношении обычных субстратов, моющее действие было сравнимо либо повышено по сравнению с природным ферментом, и во многих случаях улучшалась стабильность при хранении. В дополнение, моющее действие некоторых мутантных протеаз на основе PB92 по сравнению с природной протеазой PB92 составило от 120 до примерно 180 процентов. Следовательно, настоящее изобретение предоставляет варианты протеаз со значительно усиленным действием по сравнению с природной протеазой.
В некоторых вариантах осуществления скомбинированы несколько мутаций с целью повышения стабильности протеазы в составе моющих средств. Несколько мутаций, положительно влияющих на моющее действие той же протеазы, могут быть скомбинированы в одном гене мутантной протеазы, обеспечивая возможность производства, возможно, еще более эффективных протеаз (например, S126M, P127A, S128G, S160D и G116V, S126N, P127S, S128A, S160D; нумерация по PB92). Дополнительные мутанты протеазы получаются путем комбинирования хороших свойств в смысле моющего действия, например G116V и S16OD, и свойств стабильности, привносимых другими мутациями (нумерация по PB92).
Полезные мутанты также получаются путем комбинирования каких-либо мутаций или набора мутаций, описанных в данном тексте. В дополнительных вариантах осуществления, полезные мутации, особенно предоставленные в данном тексте, комбинируются с мутациями по другим сайтам. В некоторых вариантах осуществления, эти комбинации приводят к существенным изменениям свойств ферментов, тогда как в других вариантах осуществления изменения менее существенны.
Данное изобретение также охватывает применение одного или более мутантного протеолитического фермента, как определено в данном тексте, в композиции(ях) моющих средств и/или процедуре мытья. Наконец, будет понятно, что при делециях или вставках аминокислот в полипептидную цепь протеазы как искусственно введенных мутагенезом, так и присутствующих в природе у протеаз, гомологичных протеазе PB92, нумерация аминокислотных остатков в цепи может измениться. Однако должно быть понятно, что положения, гомологичные положениям аминокислот в протеазе PB92, включены в пределы формулы изобретения.
ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
Следующие примеры приведены с целью демонстрации и дальнейшей иллюстрации некоторых предпочтительных вариантов осуществления и аспектов настоящего изобретения и не должны толковаться как ограничивающие его рамки.
В следующем раскрытии эксперимента применяются аббревиатуры: 0C (градусы Цельсия); об/мин (обороты в минуту); H2O (вода); HCl (соляная кислота); аа (аминокислота); п.н. (пары нуклеотидов); т.п.н. (тысяча пар нуклеотилов); кДа (тысяча Дальтон); г (граммы); мкг (микрограммы); мг (миллиграммы); нг (нанограммы); мкл (микролитры); мл (миллилитры); мм (миллиметры); нм (нанометры); мкм (микрометры); M (молярный); мМ (миллимолярный); мкМ (микромолярный); Ед (Единицы); В (Вольты); МВ (молекулярный вес); сек (секунды); мин (минуты); час; MgCl2 (хлорид магния); NaCl (хлорид натрия); OD280 (оптическая плотность при 280 нм); OD600 (оптическая плотность при 600 нм); PAGE (электрофорез в полиакриламидном геле); EtOH (этанол); PBS (фосфатно-солевой буфер [150 мМ NaCl, 10 мМ натриево-фосфатный буфер, pH 7,2]); SDS (додецилсульфат натрия); Tris(трис(гидроксиметиламинометан)); TAED (N,N,N'N'-тетраацетилэтилендиамин); в/о (вес к объему); в/в (вес к весу); МС (масс-спектроскопия); TIGR (Институт Геномных Исследований, Rockville, MD); AATCC (Американская Ассоциация Специалистов по Красителям для Тканей); SR (удаление загрязнений или пятен); WFK (wfk Testgewebe GmbH, Bruggen-Bracht, Germany); Amersham (Amersham Life Science, Inc. Arlington Heights, IL); ICN (ICN pharmaceuticals, Inc., Costa Mesa, CA); Pierce (Pierce Biotechnology, Rockford, IL); Amicon (Amicon, Inc., Beverly, MA); ATCC (Американская Коллекция Типовых Культур, Manassas, VA); Amersham (Amersham Biosciences, Inc., Piscataway, NJ); Becton Dickinson (Becton Dickinson Labware, Lincoln Park, NJ); BioRad (BioRad, Richmond, CA); Clontech (CLONTECH Laboratories, Palo Alto, CA); Difco (Difco Laboratories, Detroit, MI); GIBCO BRL или Gibco BRL (Life Technologies, Inc., Gaithersburg, MD); Novagen (Novagen, Inc., Madison, WI); Qiagen (Qiagen, Inc., Valencia, CA); Invitrogen (Invitrogen Corp., Carlsbad, CA); Finnzymes (Finnzymes Oy, Espoo, Finland); Macherey-Nagel (Macherey-Nagel, Easton, PA); Merieux (Instirut Merieux, Codex, FR); Kelco (CP Kelco, Atlanta, GA); Genaissance (Genaissance Pharmaceuticals, Inc., New Haven, CT); DNA 2.0 (DNA 2.0, Menlo Park, CA); MIDI (MIDI Labs, Newark, DE) InvivoGen (InvivoGen, San Diego, CA); Sigma (Sigma Chemical Co., St. Louis, MO); Sorvall (Sorvall Instruments, a subsidiary of DuPont Co., Biotechnology Systems, Wilmington, DE); Stratagene (Stratagene Cloning Systems, La Jolla, CA); Roche (Hoffmann La Roche, Inc., Nutley, NJ); Agilent (Agilent Technologies, Palo Alto, CA); Minolta (Konica Minolta, Ramsey, NJ); Zeiss (Carl Zeiss, Inc., Thornwood, NY); Henkel (Henkel, GmbH, Dusseldorf, Germany); Cognis (Cognis Corp, USA, Cincinnati, OH); Finnzymes (Finnzymes Oy, Espoo, Finland); Reckitt Benckiser, Berks, United Kingdom); BASF (BASF Corp., Florham Park, NJ); IKW (Industrieverband Korperflege und Waschmittel, Frankfurt, Germany); и WFK (Testgewebe GmbH, Bruggen-Bracht, Germany).
Синтетическая вода, содержащая 3,00 ммоль Ca+Mg (16,8°d), применяемая в некоторых экспериментальных процедурах мытья посуды, готовилась следующим образом. Сначала были приготовлены три исходных раствора. Раствор 1 представлял собой 800 ммоль/л NaHCO3 (67,2 г/л); раствор 2 представлял собой 154,2 ммоль/л MgSO4 * 7 H2O (38,0 г/л); и раствор 3 представлял собой 446,1 ммоль/л CaCl2 * 2 H2O (65,6 г/л). После того как растворы были приготовлены, в сосуд с 7 литрами деминерализованной воды добавлялось по 50 мл каждого из растворов 1, 2 и 3, и затем в сосуд добавлялась деминерализованная вода до общего объема 10 литров. Перед использованием значение pH синтетической воды доводилось до 7,5 с помощью HCl или NaOH.
В следующей Таблице (Таблица 1) приведены описания мутантов, произведенных и проанализированных в процессе разработки настоящего изобретения. В этой Таблице для удобства приведена нумерация по последовательностям как BPN', так и PB92.
Мутанты PB92
В таблице 1 все штаммы были получены и охарактеризованы с применением методик, описанных в Примерах. Штаммы, обозначенные звездочкой (*), были получены, как описано в EP 0 571 049 B1 (см. Пример 1A-C). Все остальные варианты были получены, как описано в Примере 1D.
ПРИМЕР 1
Создание мутантов протеазы PB92
В этом примере описаны способы, применяемые для получения некоторых мутантов протеазы PB92, приведенных в данном тексте. Базовый (исходный) конструкт, с которого начиналась работа по мутагенезу, обозначается как "pM58", описан в EP 0283075 и в EP 571049. Соблюдаемая стратегия содержала три фазы:
A. Создание вектора для мутагенеза 13M1
B. Процедура мутации
C. Создание pM58Eco и субклонирование мутированного фрагмента ДНК в вектор
Дополнительно, часть D ("Получение вариантов PB92") включает описание конструирования различных вариантов PB92, которые были показаны как полезные в настоящем изобретении.
A. Создание вектора для мутагенеза M13M1
Исходный конструкт pM58 был расщеплен ферментами рестрикции HpaI и BalI. Фрагмент длиной 1400 п.н., содержащий ген протеазы PB92, был очищен с применением низкоплавкой агарозы по методике, известной в данной области техники. Вектор M13MP11 (см. Messing et al., Nucl. Acids Res., 9:303-321 [1981]) был расщеплен SmaI. Представляющий интерес фрагмент ДНК длиной 1400 п.н. лигировался в этот вектор с последующей трансформацией в E. coli JM101 с применением методик, известных в данной области (см. Cohen et al., Proc. Natl. Acad. Sci. USA 69:2110-2114 [1972]).
После размножения бактериофага в E. coli JM101 была выделена одноцепочечная ДНК фага с применением методик, известных в данной области (см. Heidecker et al., Gene 10:69-73 [1980], и путем секвенирования ДНК по методу Сэнгера анализировались наличие вставки и ее ориентация (см. Sanger, Proc. Natl. Acad. Sci. USA 74:6463 [1977]). Был получен вектор, подходящий для мутагенеза, и назван "M13M1". Вышеописанная процедура схематически изображена на Фигуре 1А.
B. Процедуры мутации
Мутагенез конструкции M13M1 осуществлялся с применением одноцепочечной ДНК этого вектора и двуцепочечной ДНК M13mp19 (Messing et al. Nucl. Acids Res., 9:303-321 [1988]), где последний был расщеплен рестриктазами EcoRI и HindIII с последующей очисткой большего фрагмента с применением низкоплавкой агарозы.
Мутагенез осуществлялся по методикам, известным в данной области техники (см. Kramer et al., Nucl. Acids Res., 12:9441-9456 [1984]), с той модификацией, что для отбора мутантов использовался штамм E. coli JM105, а не E. coli WK30-3.
Длина олигонуклеотидов, использованных для введения специфических мутаций, составляла 22 нуклеотида. Мутагенез специфической области применялся для одновременного получения нескольких мутаций в конкретной последовательности ДНК и осуществлялся с помощью олигонуклеотидов длиной в 40 нуклеотидов со всеми четырьмя нуклеотидами, случайно введенными в положения, соответствующие аминокислотам, которые требовалось заменить.
После мутагенеза потенциальные мутанты анализировались на наличие надлежащей мутации путем дидезокси-секвенирования по методу Сэнгера (см. Sanger, supra). Одноцепочечный интервал (см. Фиг. 1B) секвенировался полностью для подтверждения отсутствия вторичных мутаций. Процедура схематически изображена на Фигуре 1B.
Описанная процедура была полезна при получении фрагментов ДНК с мутациями, расположенных в 3'-концевой области гена протеазы (аминокислоты 154-269).
Однако не подразумевается, что настоящее изобретение ограничено указанными конкретными методами, так как применяться может любой подходящий метод, известный в данной области техники. В действительности, специалисты в данной области признают, что с целью получения фрагментов ДНК с мутациями из 5'-концевой области гена протеазы в векторе Bacillus, для конструирования методами, аналогичными методу, показанному на Фигуре 1А, могут применяться альтернативные ферменты рестрикции и модифицированные гены протеазы.
C. Конструирование pM58Eco и субклонирование фрагментов ДНК, содержащих мутации, в вектор
Для создания pM58Eco, pM58 был расщеплен ферментом рестрикции EcoR1 и лигирован с помощью лигазы фага T4 в условиях (сильного) разведения, как известно в данной области техники. Лигазная смесь применялась для трансформации B. subtilis 1-A40 (Bacillus Genetic Stock Centre, Ohio) с применением методик, известных в данной области (см. Spizizen et al., J. Bacteriol., 81:741-746 [1961]).
Клетки из трансформационной смеси были высажены на чашки с минимальной питательной средой, содержащей 20 г/мл неомицина, согласно известным в данной области методикам (см. Пример 1 в EP 0283075).
Плазмидная ДНК трансформированных организмов была выделена с применением известных в данной области методик (см. Birnboim and Doly, Nucl. Acids Res., 7:1513-1523 [1979]) и охарактеризована с помощью рестрикционного анализа. Следовательно, таким способом был выделен pM58Eco (см. Фиг. 1С).
Для получения мутантного фермента фрагменты ДНК M13M1, содержащие требуемые мутации, полученные как описано в части B выше, были субклонированы в pM58Eco. Затем двуцепочечная ДНК (дцДНК) M13Ml (описана выше) была расщеплена ферментом EcoRI и лигирована по сайту EcoRI в pM58Eco. Лигазная смесь использовалась для трансформации B. subtilis DB104 (см. Doi, J. Bacteriol., 160:442-444 [1984]) с применением методик, известных в данной области (см. Spizizen et al., supra).
Клетки из трансформационной смеси были высажены на чашки с минимальной питательной средой, содержащей 20 г/мл неомицина и 0,4% казеин, согласно известным в данной области методикам (см. EP 0283075). Из трансформированных организмов, продуцирующих протеазы, была выделена ДНК согласно известным в данной области методикам (см. Birnboim and Doly, supra) и охарактеризована с помощью рестрикционного анализа.
D. Получение вариантов PB982
Варианты PB92, обозначенные с 053 по 060, были получены путем слияния фрагментов при прохождении реакции ПЦР, методика известна в данной области (см., например, Патентная заявка США № 10/541737, введен в данный текст ссылкой во всей полноте). В следующей Таблице приведены последовательности праймеров, применяемые для слияния фрагментов в процессе ПЦР («фьюжн ПЦР»), описанном в данном тексте.
Для осуществления этих реакций ПЦР применялась полимераза Phusion™ (Finnzymes). В данных экспериментах, 2 мкл 10 мМ прямого и обратного праймера, 1 мкл 10 мМ dNTP (смесь дезоксинуклеотид трифосфатов), 5 мкл 10x HF буфера для полимеразы Phusion, 1,5 мкл DMSO и l мкл матрицы добавлялись в реакцию с суммарным объемом 50 мкл. Использовалась следующая программа: 3 минуты денатурации при 95°C, отжиг праймеров 1 минуту при 65ºC и элонгация (удлинение праймера) 1 минуту 15 секунд при 72ºC, всего 30 циклов, с последующим выдерживанием 7 минут при 72ºC.
По завершении продукты реакции хранились при комнатной температуре.
Мутант, обозначенный как "047/048", применялся в качестве матрицы для получения мутантов с 053 по 058. Праймер BglII-Fw комбинировался в реакции с праймером 118V-Rv, а второй фрагмент был получен комбинированием праймера BglII-Rv с праймерами 128-130-Fw. В случае мутанта 057, комбинировались праймеры BglII-Fw /S166D-Rv и BglII-Rv/S166D-Fw.
Кроме того, мутант "051/052" применялся в качестве матрицы для получения мутантов 059 и 060.
Праймер 126-up-Rv комбинировался в реакции с праймером BglII-Fw, тогда как BglII-Rv комбинировался с праймерами 128-130Fw.
Фрагменты ожидаемых размеров были очищены из агарозных гелей с помощью колонок для очистки продуктов ПЦР от производителя Macherey-Nagel. Правильные фрагменты были слиты и амплифицированы с применением праймеров BglII и используя в реакции ПЦР полимеразу Phusion™, по следующей программе: время денатурации 3 минуты при 95°C, отжиг праймеров 1 минуту при 65ºC и элонгация в течение 2 минут при 72°C, всего 25 циклов, с последующими 7 минутами при 72°C. По завершении продукты реакции хранились при комнатной температуре.
Фрагменты расщеплялись BglII, очищались из агарозных гелей и лигировались в течение ночи при 14°C с 1 мкл T4 ДНК лигазой, 8 мкл 5x T4 буфера для лигирования в объеме 40 мкл, согласно методикам, известным в данной области техники.
Высокотрансформируемый штамм B. subtilis BG3594 comK использовался для получения положительных по протеазе трансформантов. Экспрессионные векторы, полученные как описано выше, были трансформированы в составе 10 мкл продукта лигирования. Компетентные клетки Bacillus subtilis, BG3594comK были трансформированы экспрессионными плазмидами, описанными в Примере 1. Компетентность бактерий достигалась при индукции гена comK под контролем промотора, индуцируемого ксилозой (см., например, Hahn et al., Mol. Microbiol., 21:763-775 [1996]). Клоны, положительные по протеазе, были отобраны, выделены, секвенированы и трансформированы в B. clausii PBT125, как описано в Примере 2.
ПРИМЕР 2
Мутанты, экспрессированные в B. clausii
В этом Примере описаны способы, использованные для разработки мутантных протеаз, экспрессированных в B. clausii PBT125, трансформированных экспрессионными векторами, описанными в Примере 1. Данный штамм является дефицитным по протеазе производным штамма PBT110, который был получен из штамма PB92 с применением классических методов усовершенствования штаммов, с последующим скринингом по аспорогенности и повышенной производительности протеазы.
Процедура трансформации дефицитного по протеазе производного PB92: штамма PBT125
Для трансформации B. clausii PBT125 экспрессионными векторами, описанными в Примере 1, применялся способ трансформации с индукцией протопласта полиэтиленгликолем (авторы Chang и Cohen), известный в данной области техники (см., например, Chang and Cohen, Mol. Gen. Genet., 168:111-115 [1979]), со следующими модификациями. Во-первых, протопласты были приготовлены в щелочесодержащей среде с 0,5 M сахарозы, 0,02 M MgCl2 и 0,02 M трис-малеатного буфера (pH 8,0), к которой добавлялся лизоцим до концентрации 0,4 мг на мл. Затем протопласты осаждались и суспендировались в 5 мл щелочесодержащей среды, к которой были добавлены 3,5% (вес/объем) среды Bacto-Penassay (Difco) и 0,04% альбумин (Merieux). Трансформированные протопласты восстанавливались на модифицированных плашках DM3 (5), содержащих 8,0 г GELRITE® gellam Gum (Kelco), 0,3 г CaCl2•2H2O, 4,06 г MgCl2•H2O, 5,0 г буфера N-трис(гидроксиметил)метил-2-аминоэтансульфоновой кислоты (Sigma), 5,0 г казаминовых кислот, 5 г дрожжевого экстракта, 1,5 мл 4 M NaOH, растворенных в 750 мл H2O и смешанных после стерилизации с 250 мл 2 M сахарозы и 10 мл 50% (вес/объем) глюкозы, 1 мл альбумина (Merieux), 10 мг тиамина, 5 мг биотина и 50 мг неомицина. Среда в плашках застывала приблизительно при 70ºC. Трансформация компетентных клеток B. clausii осуществлялась согласно методикам, известным в данной области техники (см., например, Tanaka, "Construction of Bacillus subtilis Plasmid and Molecular Cloning in Bacillus subtilis", в D. Schlesinger (ed.), Microbiology. American Society for Microbiology, Washington, D.C., pp.15-18, [1982]).
Условия ферментации и производство протеазы
B. clausii PBT125 наращивались в ферментерах на 6 или 3 литра в среде, содержащей 22 г (сухого вещества) дрожжей на литр, 5 г K2HPO4•3H2O на литр, 0,05 г MgSO4•7H2O на литр, 0,05 г CaCI2 на литр, 0,005 г FeSO4•7H2O на литр и 0,05 г MnSO4•4H2O на литр. Компоненты среды растворялись в 90% конечного объема и стерилизовались при pH 7,0 при температуре 120ºC в течение 1 часа. Высеянная культура была получена путем высевания B. clausii PBT125 в 100 мл TSB; после стерилизации из наклонной трубки было добавлено 4 мл 1 M раствора карбоната натрия. Высеянная культура инкубировалась при 37°C в течение 24 часов на аппарате для встряхивания. В среду при 37°C и pH 8,0 высевался 1 объем высеянной культуры на 100 объемов среды. Основная ферментация проводилась при 37°C в перемешивающих ферментерах, оснащенных приборами для контроля pH, температуры и вспенивания и приспособлением для непрерывного отслеживания концентрации растворенного кислорода и скорости потребления кислорода. Через 17 часов после высева добавлялся 30% раствор глюкозы, стерилизованный при 120ºC в течение 1 часа, до конечной концентрации 30 г глюкозы на литр среды.
Среды откручивались 30 минут при 11800xg. Надосадочная фракция фильтровалась через воронку Бюхнера с стекловолоконным фильтром Whatman и 0,8 мкм фильтром с последующей фильтрацией с применением целлюлозных подкладок. Полученный материал хранился при 4°C до непосредственного использования. Затем материал был сконцентрирован с применением фильтровальной установки Pall UF с ограничением пропускания 10 кДа. Полученный ультрафильтрованный концентрат был дополнен пропиленгликолем и формиатом натрия. Для доведения pH до 6,0 использовалась муравьиная кислота.
ПРИМЕР 3
Аналитические методики для определения чистоты очищенных протеаз
В этом Примере описаны способы, применяемые для определения чистоты очищенных протеаз. Препараты протеаз считались чистыми, если при электрофорезе и ВЭЖХ (высокоэффективная жидкостная хроматография) была получена уникальная полоса или пик, соответственно.
Электрофорез в полиакриламидном геле (PAGE) в присутствии додецилсульфата натрия (SDS) проводился согласно методике, известной в данной области (см. Laemmli, Nature, 227:680-685 [1970]). Однако перед денатурацией образцов белка в присутствии SDS при 100ºC требовалась инактивация протеолитической активности с целью предотвращения саморасщепления. Это осуществлялось путем инкубации с фенилметилсульфонил флуоридом (PMSF) (1 мМ, 30 мин, комнатная температура) или осаждения трихлоруксусной кислотой (TCA, 8%, 30 мин, на льду). Нативный (не денатурирующий) PAGE проводился при pH 7,45 (буфер для геля, содержащий 20 мМ гистидин (His) и 50 мМ 3-[N-морфолинпропансульфониевую] кислоту (MOPS)) в 5% полиакриламидных гелях (соотношение акриламид:бисакриламид составляло 20:1). Образцы белка вносились в верхнюю часть гелевого блока, электрофорез осуществлялся в направлении к катоду. Тот же 5 His/MOPS буфер использовался в качестве буфера для электрофореза, но его pH составил 6,3. После электрофореза (1-2 часа при 350 В) гель пропитывался 8% уксусной кислотой с целью фиксации белков в геле и затем окрашивался кумасси бриллиантовым синим R250 по методике, известной в данной области.
Для анализа чистоты методом ВЭЖХ применялись катионообменные колонки (MonoS; Pharmacia) и колонки для гель-фильтрации (TSK 2000; SW-LKB). Внесение в колонку проводилось в 10 мМ натрий-фосфатном буфере с pH 5,5. Элюция связанной с носителем протеазы осуществлялась с применением линейного градиента 10-300 мМ фосфата натрия pH 5,5. Гель-фильтрация на колонках осуществлялась в 0,25 M натрий-ацетатном буфере с pH 5,5.
ПРИМЕР 4
Определение концентрации протеазы
В этом Примере описаны способы определения концентраций протеаз. В некоторых экспериментах измерения затухания проводились при 280 нм с использованием подсчитанного коэффициента затухания (M), и проводилось титрование активного центра для определения концентрации белка в растворе очищенной протеазы (как описано ниже). В дополнительных экспериментах применялись методики, приведенные в Патентной заявке США № 11/011666, введенной ссылкой во всей полноте.
Коэффициент затухания при 280 нм был подсчитан по числу триптофанов (M =5600 M-1·см-1) и тирозинов (M = 1330 M-1·см-1) на молекулу фермента. Для протеазы PB92 использовалось значение M, которое составило 26100 M-1·см-1 (3 Trp, 7 Tyr остатков), эквивалентное E1 l%см, измеренному при 280 нм = 9,7 (Mr = 26729 Да). В случае мутантов с измененным числом остатков Trp и Tyr были внесены соответствующие поправки.
Оценка количества активных молекул фермента производилась титрованием активных центров. Так как широко применяемая методика с использованием N-трансциннамоилимидазола (см. Bender et al., J. Am. Chem. Soc, 88:5890-5931 [1966]), как было установлено, не работает удовлетворительно в отношении протеазы PB92, был разработан альтернативный метод с использованием PMSF.
В этой методике раствор протеазы с оцененной концентрацией фермента (по поглощению при 280 нм) смешивался с 0,25, 0,50, 0,75, 1,00 и 1,25 эквивалентами PMSF, соответственно, и проводилась реакция в течение одного часа при комнатной температуре в 10 мМ натрий-фосфатном буфере pH 6,5. Концентрация фермента должна быть, по меньшей мере, 50 M.
Остаточная активность измерялась спектрофотометрически с применением сукцинил-L-аланил-L-аланил-L-пролил-L-фенилаланил-пара-нитроанилида (sAAPFpNA) в качестве субстрата. Чистота (и, следовательно, концентрация) PMSF определялась ЯМР-спектроскопией, исходный раствор готовился в изопропаноле. Результаты титрования активных центров оказались соответствующими результатам анализа чистоты методом ВЭЖХ.
ПРИМЕР 5
Определение кинетических параметров мутантных протеаз и протеаз дикого типа
В этом Примере описаны способы определения кинетических параметров протеаз дикого типа и мутантных протеаз.
Активность в отношении белковых субстратов (казеин) измерялась при pH 10,0 как описано в Описании Патента Великобритании 1353317 (выражены в ADU = щелочных единицах Дельфта).
Число оборотов в отношении казеина как субстрата измерялось с применением pH-стата. Реакционная камера pH-стат (Radiometer, Copenhagen) содержала 10 мл 0,1 M KCl с 50 мг казеина (Hammerstein, Merck). Протоны, высвобождаемые при гидролизе казеина протеазой PB92, титровались 10 мМ NaOH, в то время как pH поддерживался на величине 10,0 (при 40ºC и при протекании газообразного азота).
Активность синтетических пептидов измерялась с использованием sAAPFpNA. Образующийся паранитроанилид (желтый) (pNA) определялся спектрофотометрически при 410 нм: M = 8480 M-1·см-1 согласно методикам, известным в данной области (см. Delmar et al., Anal. Biochem., 94:316-320 [1979]), с помощью спектрофотометра UVIKON 860 (KONTRON), оснащенного шестилуночной термостатируемой камерой. Значения кинетических параметров kcat и Km были получены при измерениях начальной скорости при различных концентрациях субстрата (для протеазы PB92 0,1-6,0 мМ) и нанесении данных на график гиперболической функции способом нелинейной регрессии, используя многомерный вариант итерационного метода секущих. Затем подсчитывалось значение константы специфичности kcat/Km. Измерения проводились при 25°C, в конечном объеме 1 мл, содержащем 0,1 M TRIS-HCl + 0,1 M NaCl pH 8,6. Присутствие хлорида натрия являлось необходимым, так как в его отсутствие протеаза PB92 демонстрировала нелинейные координаты Лайнуивера-Берка, что могло быть вызвано субстратным ингибированием. Сначала субстрат растворялся в DMSO до концентрации 200 мМ и затем был разведен 0,1 M TRIS-HCl pH 8,6 для получения исходного раствора с концентрацией 20 мМ (определено спектрофотометрически при 315 нм; M - 14000 M-1·см -1). Для различных концентраций DMSO (0,05-3,0% вес/вес) не вводилось никаких поправок.
ПРИМЕР 6
Тестирование моющего действия
В этом примере описаны способы определения моющего действия мутантных вариантов протеазы PB92 и коммерчески доступной сериновой протеазы PROPERASE® при применении для мытья посуды с использованием коммерчески доступных моющих средств.
В этом примере варианты PB92 (049: G116V, S126I, P127Q, S128A и 046: G116V, S126I, P127Q, S128A, S160D; нумерация по PB92) тестировались при различных условиях. Составы моющих детергентов приведены ниже. Эти моющие средства являются коммерчески доступными от WFK (Институт Технологий Химической Чистки) и соотносятся с обозначениями, приведенными ниже. Протоколы для каждого типа загрязнений (мясной фарш, яичный желток и яичный желток с молоком) приведены ниже. Перед тем как отдельные типы загрязнений будут нанесены на тестовую посуду, посуда должна быть полностью вымыта. Это условие необходимо, так как остатки некоторых устойчивых пятен все еще могут присутствовать на посуде после предыдущих тестов. Новая посуда также подвергалась троекратной промывке перед первым использованием в тесте.
Пятна яичного желтка на нержавеющей стали
Листы нержавеющей стали (10×15 см; покрытие одной стороны), применяемые в эксперименте, были тщательно вымыты при 95°C в лабораторной посудомоечной машине с высокощелочным коммерческим детергентом (например, моющее средство ECOLAB®; Henkel) для получения листов, которые были бы чистыми и обезжиренными. Перед первым использованием листы были сглажены (сняты заусеницы). Листы высушивались в течение 30 минут при 80ºC в тепловом шкафу перед загрязнением пятнами яичного желтка. К поверхностям, которые должны быть вычищены, не прикасались перед нанесением загрязнения. Также не допускалось наличие водяных пятен или ворсинок. Охлажденные листы взвешивались перед нанесением загрязнения.
Яичные желтки были приготовлены путем отделения желтков приблизительно 10-11 яиц (200 г яичного желтка) от белков. Желтки были перемешаны вилкой в мензурке для гомогенизации суспензии желтков. Затем желтки фильтровались (через сито приблизительно 0,5 мм) для удаления крупных частиц и каких-либо фрагментов яичной скорлупы.
Суспензия яичного желтка (1,0±0,1) г наносилась плоской щеткой (2,5") настолько равномерно, насколько возможно на площадь 140 см2 вычищенных поверхностей каждого из листов из нержавеющей стали, при этом края оставлялись незагрязненными на ширину примерно 1 см (при необходимости использовалась клейкая лента). Загрязненные листы высушивались в горизонтальном положении (для предотвращения образования капель на краях листа) при комнатной температуре в течение 4 часов (не более 24 часов).
Для денатурации листы на 30 секунд погружались в кипящую деминерализованную воду (при необходимости с использованием удерживающего приспособления). Затем листы вновь высушивались в течение 30 мин при 80ºC. После высушивания и охлаждения листы взвешивались. После взвешивания листы оставлялись на не менее чем 24 часа (20ºC, относительная влажность 40-60%) перед использованием в тесте моющего действия. Для соответствия требованиям теста в нем использовались только листы с 500 ± 100 мг/140 см2 (яичного желтка после денатурации). По завершении теста моющего действия листы высушивались в течение 30 мин при 80ºC в тепловом шкафу и вновь взвешивались после охлаждения. Процентное отношение чистящего действия определялось делением (мг яичного желтка, высвобожденного при мытье, x 100) на (мг нанесенного денатурированного желтка).
Мясной фарш на фарфоровых тарелках
Для этих экспериментов использовались десертные тарелки (Arzberg, белые, глазированный фарфор), соответствующие EN 50242, форма 1495, № 0219, диаметром 19 см. Всего 225 г постной свинины и говядины (равное соотношение) были измельчены и остужены после удаления видимого жира. Смесь была дважды пропущена через мясорубку. Не допускалось повышение температуры более 35ºC. Затем 225 г мясного фарша смешивались с 75 г яйца (перемешанные белок и желток). Затем препараты замораживались при -18°C на срок до 3 месяцев перед использованием. При отсутствии свинины использовалась говядина, так как они взаимозаменяемы.
Смесь мясного фарша и яйца (300 г) нагревалась до комнатной температуры и смешивалась с 80 мл синтетической воды. Затем смесь гомогенизировалась при помощи ручного кухонного блендера в течение 2 мин. Затем с помощью вилки 3 г смеси мясной фарш/яйцо/вода были распределены по каждой белой фарфоровой тарелке, при этом примерно 2 см по краю были оставлены не загрязненными. Наносимое количество смеси составило 11,8 ± 0,5 мг/см2. Тарелки высушивались в течение 2 часов при 120ºC в предварительно нагретом тепловом шкафу. Как только тарелки остывали, они были готовы к использованию. Тарелки были уложены в стопку, между тарелками помещены бумажные салфетки.
После мытья тарелки опрыскивались раствором нингидрина (1% этанол) для лучшего выявления остатков мясного фарша. Для ускорения цветной реакции тарелки нагревались в течение 10 мин при 80ºC в тепловом шкафу. Оценка моющего действия производилась путем визуального наблюдения цветных реакций остатков мясного фарша в соответствии с каталогом фотографий (IKW).
Яично-молочные пятна на нержавеющей стали
Листы нержавеющей стали (10×15 см; покрытие одной стороны), применяемые в эксперименте, были тщательно вымыты при 95°C в лабораторной посудомоечной машине с высокощелочным коммерческим детергентом для получения листов, которые были бы чистыми и обезжиренными. Сухие листы полировались целлюлозной тканью. К поверхностям, которые должны быть вычищены, не прикасались перед нанесением загрязнения. Также не допускалось наличие водяных пятен или ворсинок. Перед нанесением загрязнений листы были помещены в тепловой шкаф на 30 мин при 80°C. Охлажденные листы взвешивались перед нанесением загрязнения.
Желтки и белки целых сырых яиц (3-4 яйца; 160 г/яйцо) помещались в миску и взбивались венчиком для яиц. Затем к смеси добавлялось 50 мл полуобезжиренного молока UHT (1,5% жирности, обработка высокой температурой, гомогенизированное). Молоко и яйца перемешивались без образования пены. Для равномерного распределения 1,0±0,1 г смеси яйцо/молоко по вычищенной поверхности листов нержавеющей стали (распределение отслеживалось с помощью балансира) применялась плоская щетка. Полоса приблизительно 1,0 см шириной была оставлена чистой около коротких сторон листа. Загрязненные листы высушивались в горизонтальном положении (для предотвращения образования капель на краях листа) при комнатной температуре в течение 4 часов (не более 24 часов).
Затем листы на 30 секунд погружались в кипящую деминерализованную воду (при необходимости с использованием удерживающего приспособления). Затем листы вновь высушивались в течение 30 мин при 80ºC. После высушивания и охлаждения листы взвешивались. После взвешивания листы оставлялись на не менее чем 24 часа (20ºC, относительная влажность 40-60%) перед использованием в тесте моющего действия. Для соответствия требованиям теста в нем использовались только листы с 190 ± 10 мг яичного желтка.
По завершении теста моющего действия листы высушивались в течение 30 мин при 80ºC в тепловом шкафу и вновь взвешивались после охлаждения. Процентное отношение чистящего действия определялось делением (мг смеси яйцо/молоко, высвобожденной при мытье, x 100) на (мг нанесенной смеси яйцо/молоко).
Моечное оборудование и условия
Тестирование моющего действия проводилось в автоматических посудомоечных машинах (Miele: G690SC), которые загружались загрязненной посудой и листами нержавеющей стали, подготовленными как описано выше. Использовалось определенное количество детергента, которое указано ниже в таблицах результатов. Тесты проводились при температурах 45°C, 55°C и 65°C. Жесткость воды составила 9° или 21°GH (немецкие градусы жесткости) (374 миллионных долей Ca).
Как сказано выше, тарелки, загрязненные мясным фаршем, после мытья оценивались визуально, используя фотографическую шкалу оценок от 0 до 10, где "0" обозначало полностью грязную тарелку, а "10" обозначало чистую тарелку. Эти величины соответствуют способности детергента, содержащего фермент, удалять пятна или загрязнения (SR).
Вымытые листы из нержавеющей стали, загрязненные яичным желтком и/или смесью яиц и молока, анализировались взвешиванием для определения количества остаточных после мытья загрязнений. Мутантная протеаза PB92 и протеаза PROPERASE® и другие мутанты анализировались при количестве от 0 до 20,57 мг активного белка на одну процедуру мытья.
Моющие средства, применяемые в этих экспериментах, описаны ниже. Эти детергенты были получены из источников, не содержащих ферментов для тестирования ферментов в этих экспериментах.
IEC-60436 WFK тип B (pH=10,4 в 3 г/л)
IEC-60436 WFK тип С (pH=10,5 в 3 г/л)
В следующих таблицах приведены результаты различных экспериментов. В каждом из этих экспериментов использовалось 20,57 мг активного белка на одну процедуру мытья. В этих результатах за индекс было принято значение 100. Следовательно, результатам измерения моющего действия для фермента PROPERASE® было присвоено значение "100", а результаты для мутантов сравнивались с этой величиной. Например, если результат для PROPERASE® составил 45% SR (100 в виде индекса), а результат для мутанта составил 52% SR, то результат для мутанта будет равен 52/45 × 100 = 116 (индекс).
Все патенты и публикации, указанные в описании, являются показательными для специалистов в данной области, к которой относится изобретение. Все патенты и публикации настоящим введены в данный текст ссылкой в той же степени, как если бы каждая индивидуальная публикация была особым образом и индивидуально указана как введенная ссылкой.
На основе описанных предпочтительных вариантов осуществления настоящего изобретения для специалистов в данной области будет ясно, что в раскрытые варианты осуществления могут быть внесены различные модификации, и что подобные модификации подразумеваются как входящие в пределы настоящего изобретения.
Специалисты в данной области, равно как и те, кто связан с данной областью, охотно согласятся, что настоящее изобретение хорошо приспособлено для осуществления целей и получения указанных результатов и преимуществ. Композиции и способы, описанные в данном тексте, являются представительными в отношении предпочтительных вариантов осуществления, приведены в качестве примеров и не подразумеваются как ограничивающие пределы изобретения. Для специалиста в данной области очевидно, что в раскрытое в данном тексте изобретение могут быть внесены различные изменения и модификации без отклонения от рамок и смысла настоящего изобретения.
Изобретение, наглядно описанное в данном тексте, может надлежащим образом применяться на практике в отсутствие какого-либо элемента или элементов, ограничения или ограничений, которые не описаны в данном тексте особым образом. Использованные термины и выражения применены как термины описания, а не ограничения, и не подразумевается, что использование подобных терминов и выражений исключает какие-либо эквиваленты показанных и описанных свойств, в том числе частичных, но подтверждается, что в рамках формулы изобретения возможны различные модификации. Следовательно, должно быть понятно, что хотя настоящее изобретение было особым образом раскрыто предпочтительными вариантами осуществления и выборочными характеристиками, специалисты в данной области могут обратиться к модификациям и вариантам концепции, раскрытой в данном тексте, и что подобные модификации и варианты рассматриваются как входящие в пределы настоящего изобретения, как определено прилагаемой формулой изобретения.
Изобретение в данном тексте было описано широко, в общем. Каждая из более узких категорий и субгенерических групп, охватываемая общим раскрытием, также образует часть изобретения. Это включает общее описание изобретения с условием или отрицательным ограничением удаления какого-либо объекта из заявляемого класса объектов вне зависимости от того, перечислены особо или нет исключенные материалы.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИИ И СПОСОБЫ, ВКЛЮЧАЮЩИЕ ВАРИАНТЫ СУБТИЛИЗИНА | 2009 |
|
RU2575602C2 |
| КОМПОЗИЦИИ И СПОСОБЫ, ВКЛЮЧАЮЩИЕ В СЕБЯ ПРИМЕНЕНИЕ ВАРИАНТНЫХ МИКРОБНЫХ ПРОТЕАЗ | 2009 |
|
RU2575073C2 |
| ПРИМЕНЕНИЕ ВАРИАНТОВ ПРОТЕАЗЫ | 2010 |
|
RU2639534C2 |
| КОМПОЗИЦИИ И СПОСОБЫ, ВКЛЮЧАЮЩИЕ ВАРИАНТЫ МИКРОБНЫХ ПРОТЕАЗ | 2009 |
|
RU2541786C2 |
| СПОСОБ ИДЕНТИФИКАЦИИ УЛУЧШЕННЫХ ВАРИАНТОВ БЕЛКА | 2008 |
|
RU2520808C2 |
| СПОСОБЫ УЛУЧШЕНИЯ ЭФФЕКТИВНОСТИ БЕЛКОВ | 2008 |
|
RU2569106C2 |
| СПОСОБЫ УЛУЧШЕНИЯ МНОЖЕСТВЕННЫХ СВОЙСТВ БЕЛКА | 2008 |
|
RU2553550C2 |
| ВАРИАНТЫ СЕРИНОВОЙ ПРОТЕАЗЫ С МНОЖЕСТВЕННЫМИ МУТАЦИЯМИ | 2007 |
|
RU2558261C2 |
| ПРОДУЦИРОВАНИЕ ТЕРМОЛИЗИНА И ЕГО ВАРИАНТОВ И ЕГО ИСПОЛЬЗОВАНИЕ В ЖИДКИХ ДЕТЕРГЕНТАХ | 2008 |
|
RU2536255C2 |
| ВАРИАНТЫ СУБТИЛАЗ | 2010 |
|
RU2651525C2 |
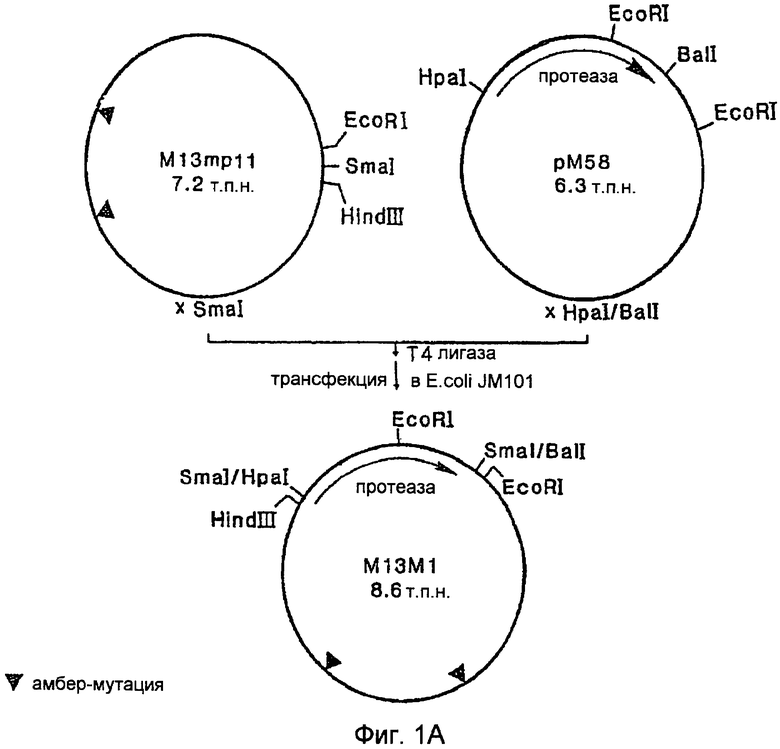
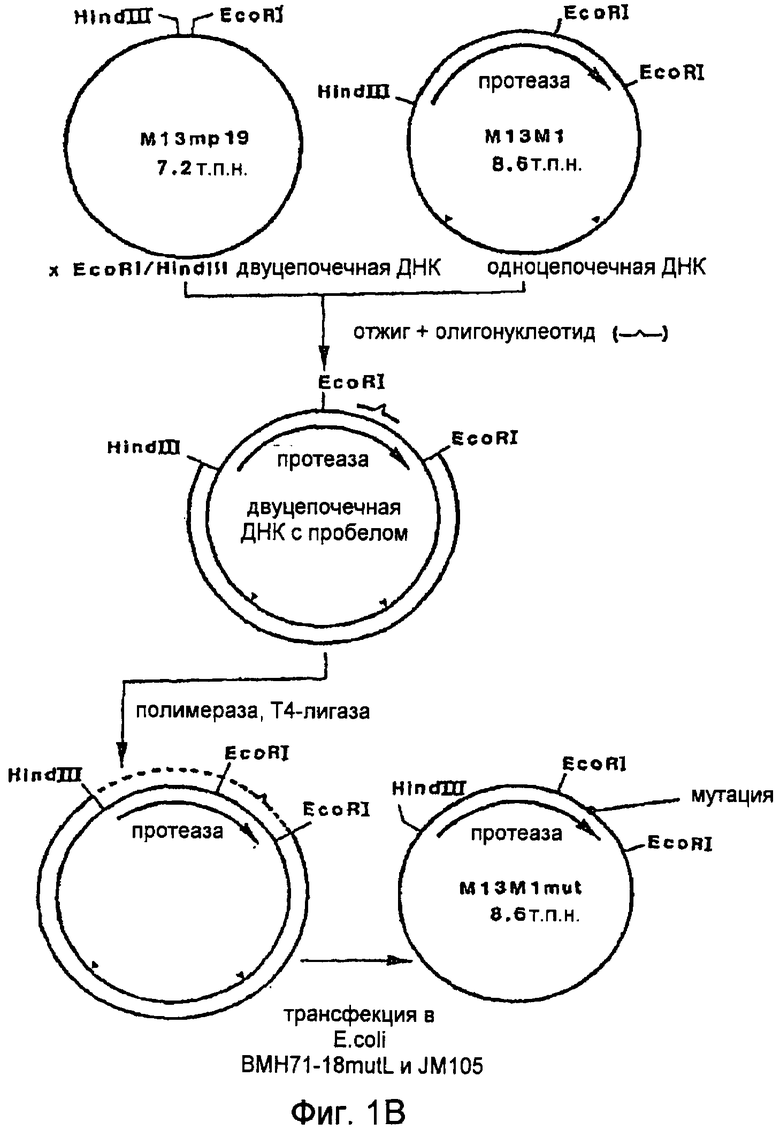
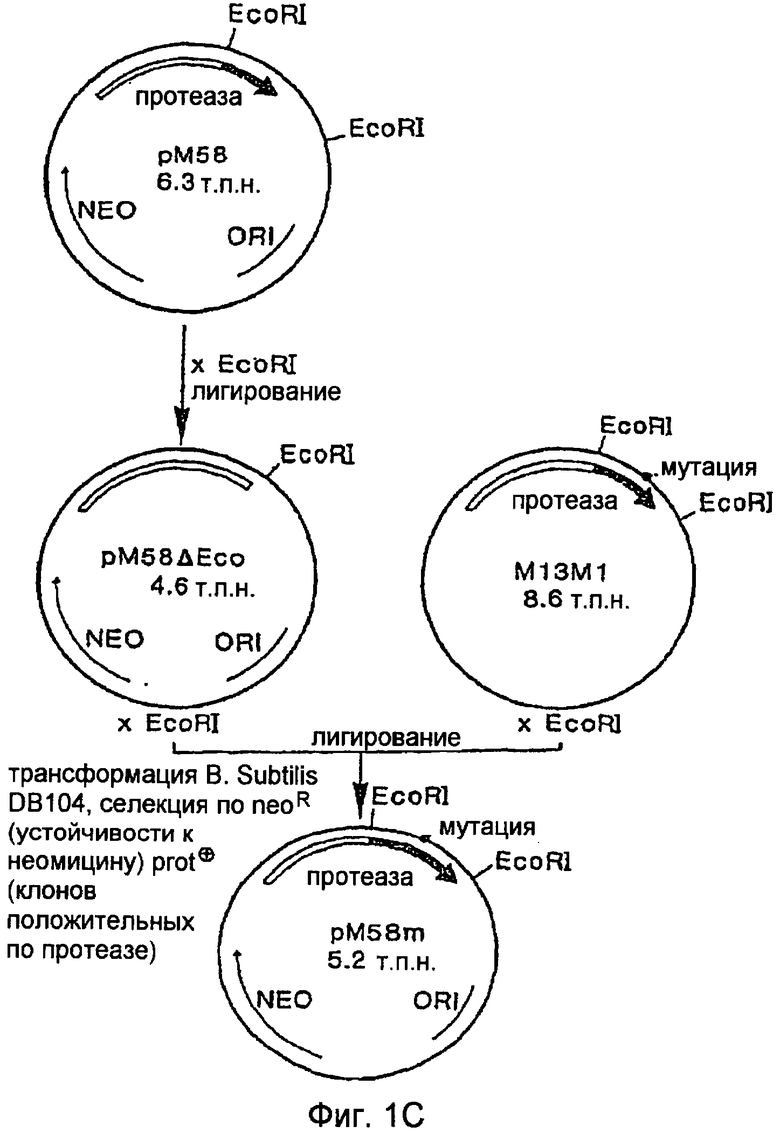
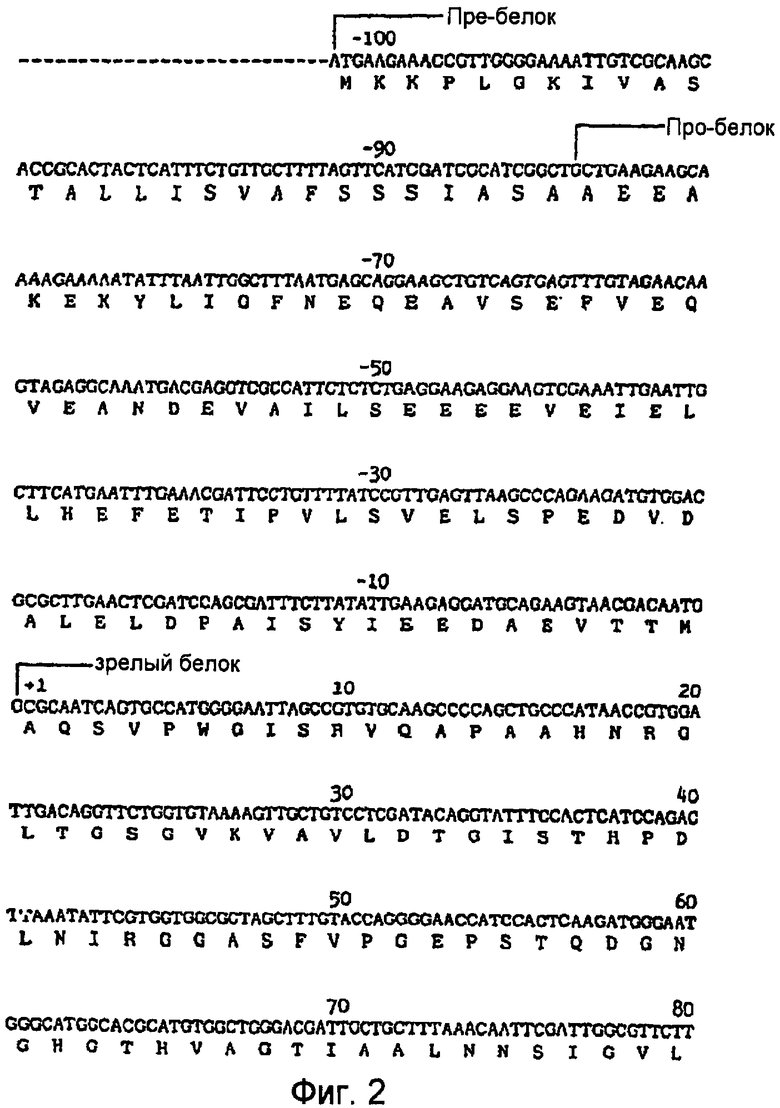
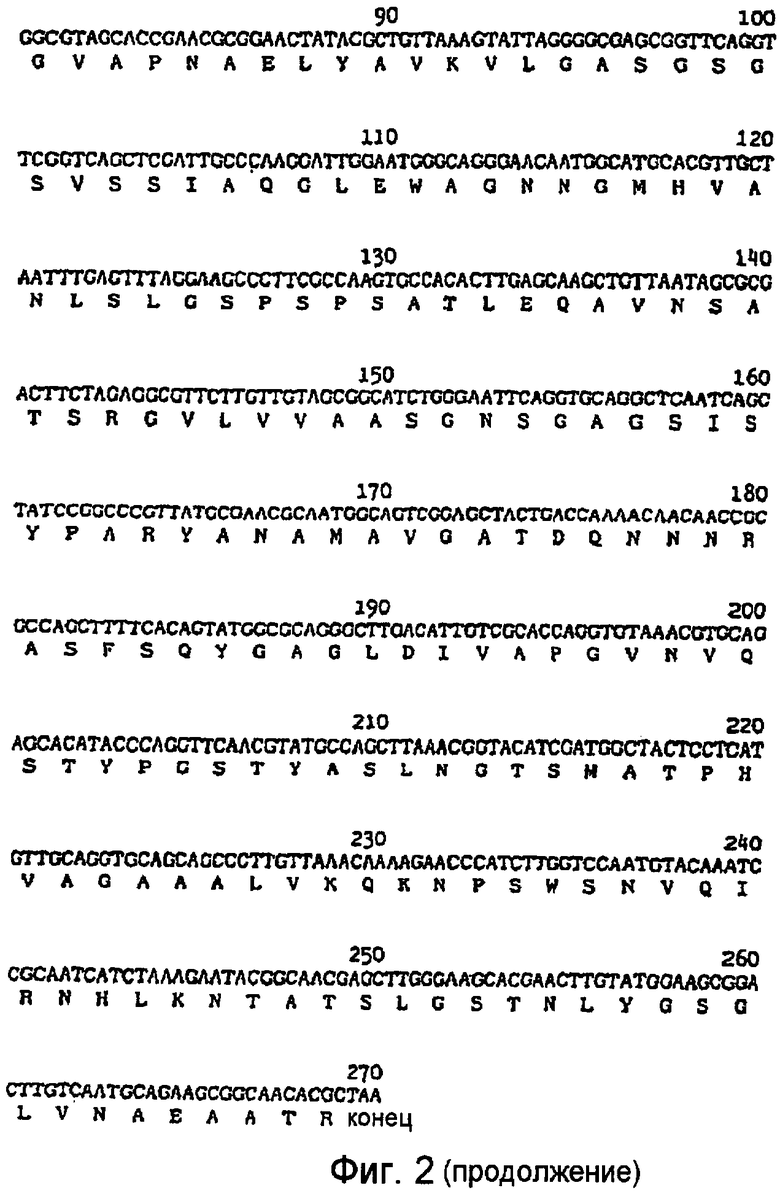
Изобретение относится к области биотехнологии и касается способа мытья посуды. Охарактеризованный способ заключается в обеспечении композиции для мытья посуды и посуды, нуждающейся в очистке, приведении указанной посуды в контакт с указанной композицией для мытья посуды при условиях, эффективно обеспечивающих очистку указанной посуды, где указанная композиция для мытья посуды содержит модифицированный субтилизин, где указанный модифицированный субтилизин, по меньшей мере, на 70% гомологичен последовательности AQSVPWGISRVQAPAAHNRGLTGSGVKVAVLDTGISTHPDLNIRGGASFVPGEPSTQDGNGHGTHVAGTIAALNNSIGVLGVAPNAELYAVKVLGASGSGSVSSIAQGLEWAGNNGMHVANLSLGSPSPSATLEQAVNSATSRGVLVVAASGNSGAGSISYPARYANAMAVGATDQNNNRASFSQYGAGLDIVAPGVNVQSTYPGSTYASLNGTSMATPHVAGAAALVKQKNPSWSNVQIRNHLKNTATSLGSTNLYGSGLVNAEAATR и содержит (i) замену G116V и, по меньшей мере, одну дополнительную мутацию или (ii) замену S126R. Представленное изобретение позволяет удалять белковые загрязнения на посуде с высокой эффективностью, является безопасным для потребителей и окружающей среды. 5 з.п. ф-лы, 2 табл., 2 ил., 6 пр.
1. Способ мытья посуды, включающий этапы обеспечения композиции для мытья посуды и посуды, нуждающейся в очистке, и приведения указанной посуды в контакт с указанной композицией для мытья посуды при условиях, эффективно обеспечивающих очистку указанной посуды, где указанная композиция для мытья посуды содержит модифицированный субтилизин, где указанный модифицированный субтилизин, по меньшей мере, на 70% гомологичен последовательности
AQSVPWGISRVQAPAAHNRGLTGSGVKVAVLDTGISTHPDLNIRGGASFVPGEPSTQDGNGHGTHVAGTIAALNNSIGVLGVAPNAELYAVKVLGASGSGSVSSIAQGLEWAGNNGMHVANLSLGSPSPSATLEQAVNSATSRGVLVVAASGNSGAGSISYPARYANAMAVGATDQNNNRASFSQYGAGLDIVAPGVNVQSTYPGSTYASLNGTSMATPHVAGAAALVKQKNPSWSNVQIRNHLKNTATSLGSTNLYGSGLVNAEAATR
и содержит
(i) замену G116V и, по меньшей мере, одну дополнительную мутацию или
(ii) замену S126R.
2. Способ мытья посуды по п.1, где указанный модифицированный субтилизин содержит замены, сделанные в положениях G116, S126, P127 и S128.
3. Способ мытья посуды по п.1, где указанный субтилизин содержит замену G116V, и дополнительная замена выбрана из группы, состоящей из S126F, S126L, S126N, S126R, S126V, P127E, P127L, P127M, P127N, P127L, P127Q, P127S, S128A, S128K, S128P, S128T,S128V и S160D.
4. Способ мытья посуды по п.1, где указанный модифицированный субтилизин содержит замены, сделанные в положениях S126, P127 и S128.
5. Способ мытья посуды по п.4, где указанные замены выбраны из группы, состоящей из S126C, S126R, P127Q, P127R, S128D и S128G.
6. Способ мытья посуды по п.1, где аминокислотная последовательность указанного модифицированного субтилизина представляет собой:
AQSVPWGISRVQAPAAHNRGLTGSGVKVAVLDTGISTHPDLNIRGGASFVPGEPSTQDGNGHGTHVAGTIAALNNSIGVLGVAPNAELYAVKVLGASGSGSVSSIAQGLEWAGNNVMHVANLSLGLQAPSATLEQAVNSATSRGVLVVAASGNSGAGSISYPARYANAMAVGATDQNNNRASFSQYGAGLDIVAPGVNVQSTYPGSTYASLNGTSMATPHVAGAAALVKQKNPSWSNVQIRNHLKNTATSLGSTNLYGSGLVNAEAATR
или
AQSVPWGISRVQAPAAHNRGLTGSGVKVAVLDTGISTHPDLNIRGGASFVPGEPSTQDGNGHGTHVAGTIAALNNSIGVLGVAPNAELYAVKVLGASGSGSVSSIAQGLEWAGNNVMHVANLSLGLQAPSATLEQAVNSATSRGVLVVAASGNSGAGSIDYPARYANAMAVGATDQNNNRASFSQYGAGLDIVAPGVNVQSTYPGSTYASLNGTSMATPHVAGAAALVKQKNPSWSNVQIRNHLKNTATSLGSTNLYGSGLVNAEAATR.
| WO 94002618 A1, 03.02.1994 | |||
| EP 0571049 A1, 24.11.1993 | |||
| СПОСОБ ПОЛУЧЕНИЯ МУТАНТНОЙ ПРОТЕАЗЫ | 1989 |
|
RU2069695C1 |

