ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
По настоящей заявке испрашивается приоритет Предварительных заявок на патент США с порядковыми номерами 60/933307, 60/933331 и 60/933312, поданных 6 июня 2007 года, включенных в настоящее описание в качестве ссылки в полном объеме.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Данное изобретение относится к способам конструирования белков для оптимизации их эффективности при определенных представляющих интерес условиях окружающей среды. В некоторых вариантах осуществления, данное изобретение относится к способам конструирования ферментов для оптимизации их каталитической активности при конкретных условиях окружающей среды. В некоторых предпочтительных вариантах осуществления, данное изобретение относится к способам изменения суммарного заряда поверхности и/или распределения заряда поверхности ферментов (например, металлопротеаз или сериновых протеаз) для получения вариантов ферментов, которые обладают улучшенной эффективностью в детергентных составах в сравнении с первоначальным или исходным ферментом.
УРОВЕНЬ ТЕХНИКИ
Свойства белков, функционирующих вне их природной среды, часто являются субоптимальными. Например, ферменты (например, протеазы, липазы, амилазы, целлюлазы и т.д.) часто используются для очистки от пятен в ткани в детергентах (моющих средствах) для стирки, которые обычно включают комплексную комбинацию активных ингредиентов. Фактически, большинство очищающих продуктов включают систему поверхностно-активных веществ, отбеливающие агенты, модифицирующие добавки, подавители мыльной пены, суспендирующие грязь агенты, высвобождающие грязь агенты, оптические отбеливатели, смягчающие агенты, диспергирующие агенты, ингибирующие перенос красителя агенты, абразивы, бактерициды и ароматизаторы, а также ферменты для очистки. Таким образом, несмотря на сложность существующих детергентов, имеются многочисленные пятна, которые трудно удалить полностью вследствие частично субоптимальной эффективности ферментов. Несмотря на многочисленные исследования в развитии ферментов, в данной области остается потребность в способах конструирования белков для конкретных применений и условий. В действительности, в данной области сохраняется потребность в способах быстрого и системного приспособления электростатических свойств для оптимизации их эффективности в коммерческих применениях. В частности, в данной области остается потребность в способах конструирования промышленно применимых ферментов, в том числе но не только, липаз, амилаз, кутиназ, маннаназ, оксидоредуктаз, целлюлаз, пектиназ, протеаз и других ферментов, для обеспечения улучшенной активности, стабильности и растворимости в очищающих растворах.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение относится к способам конструирования белков для оптимизации их эффективности при определенных представляющих интерес условиях окружающей среды. В некоторых вариантах осуществления, данное изобретение относится к способам конструирования ферментов для оптимизации их каталитической активности при конкретных условиях окружающей среды. В некоторых предпочтительных вариантах осуществления, данное изобретение относится к способам изменения суммарного заряда поверхности и/или распределения заряда поверхности ферментов (например, металлопротеаз или сериновых протеаз) для получения вариантов ферментов, которые демонстрируют улучшенную эффективность в детергентных составах в сравнении с первоначальным или исходным ферментом.
В некоторых вариантах осуществления, данное изобретение относится к способам замены заряда в белках, в частности ферментах. В некоторых предпочтительных вариантах осуществления, данное изобретение относится к способам получения ферментов с улучшенной моющей способностью. Данное изобретение находит применение в конструировании различных ферментов, а также других белков. В частности, данное изобретение находит применение в развитии улучшенных ферментов, которые находят применение в промышленности, в том числе, но не только, для очистки (например, белья, посуды, твердой поверхности и т.д.). Однако авторы изобретения не имеют в виду, что это изобретение ограничивается каким-либо конкретным ферментом или белком.
Данное изобретение относится к способам получения нейтрального варианта металлопротеазы с улучшенной моющей эффективностью в сравнении с исходной нейтральной металлопротеазой, предусматривающим: замену аминокислотного остатка в одном или нескольких положениях в исходной нейтральной металлопротеазе с получением варианта нейтральной металлопротеазы, имеющего более положительный заряд или более отрицательный заряд в сравнении с исходным ферментом. В некоторых особенно предпочтительных вариантах осуществления, эти способы дополнительно предусматривают тестирование моющей эффективности этого варианта сравнением способности исходного фермента и этого варианта удалять пятно, причем моющая эффективность исходного фермента принимается за величину 1,0, а вариант с улучшенной моющей эффективностью достигает величины более 1,0. В дополнительных вариантах осуществления, данное изобретение относится к способам получения варианта, имеющего улучшенную моющую эффективность. В некоторых вариантах осуществления, исходной нейтральной металлопротеазой является зрелая форма дикого типа нейтральной металлопротеазы. В некоторых других вариантах осуществления, этот вариант получен из нейтральной металлопротеазы Bacillaceae. В некоторых предпочтительных вариантах осуществления, этот вариант получен из нейтральной металлопротеазы Bacillus. В некоторых особенно предпочтительных вариантах осуществления, моющую эффективность тестируют в порошкообразной или жидкой детергентной композиции, имеющей рН от 6,5 до 12,0. В некоторых предпочтительных вариантах осуществления, моющую эффективность тестируют в жидком детергенте (моющем средстве) для стирки, имеющем основный рН. В некоторых альтернативных предпочтительных вариантах осуществления, одно или несколько положений в исходной нейтральной металлопротеазе являются положениями, имеющими доступную для растворителя поверхность (SAS), большую чем приблизительно 50%. В некоторых дополнительных предпочтительных вариантах осуществления, одно или несколько положений в исходной нейтральной металлопротеазе являются положениями, имеющими доступную для растворителя поверхность (SAS), большую чем приблизительно 65%.
Данное изобретение также относится к способам получения варианта нейтральной металлопротеазы с улучшенной моющей эффективностью в сравнении с исходной нейтральной металлопротеазой, предусматривающим: замену аминокислотного остатка в одном или нескольких положениях в исходной нейтральной металлопротеазе с получением варианта нейтральной металлопротеазы, имеющего более положительный заряд или менее отрицательный заряд в сравнении с исходным ферментом; и замену аминокислотного остатка в одном или нескольких положениях в исходной нейтральной металлопротеазе с получением варианта нейтральной металлопротеазы, имеющего более отрицательный заряд или менее положительный заряд в сравнении с исходным ферментом. В некоторых предпочтительных вариантах осуществления, эти способы дополнительно предусматривают тестирование моющей эффективности этого варианта сравнением способности исходного фермента и этого варианта удалять пятно, причем моющая эффективность исходного фермента принимается за величину 1,0, а вариант с улучшенной моющей эффективностью достигает величины более 1,0. В дополнительных вариантах осуществления, эти способы предусматривают получение варианта, имеющего улучшенную моющую эффективность. Предполагается, что эти стадии проводятся в любом удобном порядке. В некоторых вариантах осуществления, исходной нейтральной металлопротеазой является зрелая форма дикого типа нейтральной металлопротеазы. В некоторых других вариантах осуществления, этот вариант получен из нейтральной металлопротеазы Bacillaceae. В некоторых предпочтительных вариантах осуществления, этот вариант получен из нейтральной металлопротеазы Bacillus. В некоторых особенно предпочтительных вариантах осуществления, моющую эффективность тестируют в порошкообразной или жидкой детергентной композиции, имеющей рН от 6,5 до 12,0. В некоторых предпочтительных вариантах осуществления, моющую эффективность тестируют в жидком детергенте (моющем средстве) для стирки, имеющем основный рН. В некоторых альтернативных предпочтительных вариантах осуществления, одно или несколько положений в исходной нейтральной металлопротеазе являются положениями, имеющими доступную для растворителя поверхность (SAS), большую чем приблизительно 50%. В некоторых дополнительных предпочтительных вариантах осуществления, одно или несколько положений в исходной нейтральной металлопротеазе являются положениями, имеющими доступную для растворителя поверхность (SAS), большую чем приблизительно 65%. В некоторых предпочтительных вариантах осуществления по меньшей мере один кислый аминокислотный остаток заменен по меньшей мере одним основным аминокислотным остатком, тогда как в других вариантах осуществления, по меньшей мере один кислый аминокислотный остаток заменен по меньшей мере одним нейтральным аминокислотным остатком и, в некоторых дополнительных вариантах осуществления, по меньшей мере один нейтральный аминокислотный остаток заменен основным аминокислотным остатком. В некоторых вариантах осуществления, обеспечены различные комбинации замен. В дополнительных вариантах осуществления, по меньшей мере один основный аминокислотный остаток заменен по меньшей мере одним кислым аминокислотным остатком, тогда как в других вариантах осуществления по меньшей мере один основный аминокислотный остаток заменен по меньшей мере одним нейтральным аминокислотным остатком, и в других вариантах осуществления по меньшей мере один нейтральный аминокислотный остаток заменен по меньшей мере одной кислой аминокислотой. В дополнительных вариантах осуществления, по меньшей мере один нейтральный аминокислотный остаток в исходной нейтральной металлопротеазе заменен по меньшей мере одним нейтральным аминокислотным остатком с получением варианта нейтральной металлопротеазы, имеющего тот же самый заряд в сравнении с исходным ферментом. Авторы изобретения не предполагают ограничения данного изобретения какими-либо конкретными комбинациями замен. Они также не предполагают, что эти замены должны выполняться в каком-либо конкретном порядке.
Данное изобретение относится к способам получения варианта сериновой протеазы с улучшенной моющей эффективностью в сравнении с исходной сериновой протеазой, предусматривающим: замену аминокислотного остатка в одном или нескольких положениях в исходной сериновой протеазе с получением варианта сериновой протеазы, имеющего более положительный заряд или более отрицательный заряд в сравнении с исходным ферментом. В некоторых особенно предпочтительных вариантах осуществления, эти способы дополнительно предусматривают тестирование моющей эффективности этого варианта сравнением способности исходного фермента и этого варианта удалять пятно, причем моющая эффективность исходного фермента принимается за величину 1,0, а вариант с улучшенной моющей эффективностью достигает величины более 1,0. В дополнительных вариантах осуществления, данное изобретение относится к способам получения варианта, имеющего улучшенную моющую эффективность. В некоторых вариантах осуществления, исходной сериновой протеазой является зрелая форма дикого типа сериновой протеазы. В некоторых других вариантах осуществления, этот вариант получен из сериновой протеазы Bacillaceae. В некоторых предпочтительных вариантах осуществления, этот вариант получен из сериновой протеазы Bacillus. В некоторых особенно предпочтительных вариантах осуществления, моющую эффективность тестируют в порошкообразной или жидкой детергентной композиции, имеющей рН от 6,5 до 12,0. В некоторых предпочтительных вариантах осуществления, моющую эффективность тестируют в жидком детергенте (моющем средстве) для стирки, имеющем основный рН. В некоторых альтернативных предпочтительных вариантах осуществления, одно или несколько положений в исходной сериновой протеазе являются положениями, имеющими доступную для растворителя поверхность (SAS), большую чем приблизительно 50%. В некоторых дополнительных предпочтительных вариантах осуществления, одно или несколько положений в исходной сериновой протеазе являются положениями, имеющими доступную для растворителя поверхность (SAS), большую чем приблизительно 65%.
Данное изобретение также относится к способам получения варианта сериновой протеазы с улучшенной моющей эффективностью в сравнении с исходной сериновой протеазой, предусматривающим: замену аминокислотного остатка в одном или нескольких положениях в исходной сериновой протеазе с получением варианта сериновой протеазы, имеющего более положительный заряд или менее отрицательный заряд в сравнении с исходным ферментом; и замену аминокислотного остатка в одном или нескольких положениях в исходной сериновой протеазе с получением варианта сериновой протеазы, имеющего более отрицательный заряд или менее положительный заряд в сравнении с исходным ферментом. В некоторых предпочтительных вариантах осуществления, эти способы дополнительно предусматривают тестирование моющей эффективности этого варианта сравнением способности исходного фермента и этого варианта удалять пятно, причем моющая эффективность исходного фермента принимается за величину 1,0, а вариант с улучшенной моющей эффективностью достигает величины более 1,0. В дополнительных вариантах осуществления, эти способы предусматривают получение варианта, имеющего улучшенную моющую эффективность. Предполагается, что эти стадии проводятся в любом удобном порядке. В некоторых вариантах осуществления, исходной сериновой протеазой является зрелая форма дикого типа сериновой протеазы. В некоторых других вариантах осуществления, этот вариант получен из сериновой протеазы Micrococcineae. В некоторых предпочтительных вариантах осуществления, этот вариант получен из сериновой протеазы Cellulomonas. В некоторых особенно предпочтительных вариантах осуществления, моющую эффективность тестируют в порошкообразной или жидкой детергентной композиции, имеющей рН от 6,5 до 12,0. В некоторых предпочтительных вариантах осуществления, моющую эффективность тестируют в жидком детергенте (моющем средстве) для стирки, имеющем основный рН. В некоторых альтернативных предпочтительных вариантах осуществления, одно или несколько положений в исходной сериновой протеазе являются положениями, имеющими доступную для растворителя поверхность (SAS), большую чем приблизительно 50%. В некоторых дополнительных предпочтительных вариантах осуществления, одно или несколько положений в исходной сериновой протеазе являются положениями, имеющими доступную для растворителя поверхность (SAS), большую чем приблизительно 65%. В некоторых предпочтительных вариантах осуществления, по меньшей мере один кислый аминокислотный остаток заменен по меньшей мере одним основным аминокислотным остатком, тогда как в других вариантах осуществления по меньшей мере один кислый аминокислотный остаток заменен по меньшей мере одним нейтральным аминокислотным остатком и, в некоторых дополнительных вариантах осуществления, по меньшей мере один нейтральный аминокислотный остаток заменен основным аминокислотным остатком. В некоторых вариантах осуществления, обеспечены различные комбинации замен. В дополнительных вариантах осуществления, по меньшей мере один основный аминокислотный остаток заменен по меньшей мере одним кислым аминокислотным остатком, тогда как в других вариантах осуществления по меньшей мере один основный аминокислотный остаток заменен по меньшей мере одним нейтральным аминокислотным остатком и, в других вариантах осуществления, по меньшей мере один нейтральный аминокислотный остаток заменен по меньшей мере одной кислой аминокислотой. В дополнительных вариантах осуществления, по меньшей мере один нейтральный аминокислотный остаток в исходной сериновой протеазе заменен по меньшей мере одним нейтральным аминокислотным остатком с получением варианта нейтральной металлопротеазы, имеющего тот же самый заряд в сравнении с исходным ферментом. Авторы изобретения не предполагают ограничения данного изобретения какими-либо конкретными комбинациями замен. Они также не предполагают, что эти замены должны выполняться в каком-либо конкретном порядке.
Данное изобретение также относится к способам получения варианта сериновой протеазы с улучшенной моющей эффективностью в сравнении с исходной сериновой протеазой, предусматривающим: замену аминокислотного остатка в одном или нескольких положениях в исходной сериновой протеазе с получением варианта сериновой протеазы, имеющего более положительный заряд или менее отрицательный заряд в сравнении с исходным ферментом; замену аминокислотного остатка в одном или нескольких положениях в исходной сериновой протеазе с получением варианта сериновой протеазы, имеющего более отрицательный заряд или менее положительный заряд в сравнении с исходным ферментом, и получение варианта сериновой протеазы, полученного при помощи этих стадий. В дополнительных вариантах осуществления, эти способы предусматривают тестирование моющей эффективности этого варианта сравнением способности исходного фермента и этого варианта удалять пятно, причем моющая эффективность исходного фермента принимается за величину 1,0, а вариант с улучшенной моющей эффективностью достигает величины более 1,0. В дополнительных вариантах осуществления, эти способы предусматривают получение варианта, имеющего улучшенную моющую эффективность. Предполагается, что эти стадии проводятся в любом удобном порядке. В некоторых вариантах осуществления, исходной сериновой протеазой является зрелая форма дикого типа сериновой протеазы. В некоторых других вариантах осуществления, этот вариант получен из сериновой протеазы Micrococcineae. В некоторых предпочтительных вариантах осуществления, этот вариант получен из сериновой протеазы Cellulomonas. В некоторых особенно предпочтительных вариантах осуществления, моющую эффективность тестируют в порошкообразной или жидкой детергентной композиции, имеющей рН от 6,5 до 12,0. В некоторых предпочтительных вариантах осуществления, моющую эффективность тестируют в жидком детергенте (моющем средстве) для стирки, имеющем основный рН. В некоторых альтернативных предпочтительных вариантах осуществления, одно или несколько положений в исходной сериновой протеазе являются положениями, имеющими доступную для растворителя поверхность (SAS), большую чем приблизительно 50%. В некоторых дополнительных предпочтительных вариантах осуществления, одно или несколько положений в исходной сериновой протеазе являются положениями, имеющими доступную для растворителя поверхность (SAS), большую чем приблизительно 65%. В некоторых предпочтительных вариантах осуществления, по меньшей мере один кислый аминокислотный остаток заменен по меньшей мере одним основным аминокислотным остатком, тогда как в других вариантах осуществления по меньшей мере один кислый аминокислотный остаток заменен по меньшей мере одним нейтральным аминокислотным остатком и, в некоторых дополнительных вариантах осуществления, по меньшей мере один нейтральный аминокислотный остаток заменен основным аминокислотным остатком. В некоторых вариантах осуществления, обеспечены различные комбинации замен. В дополнительных вариантах осуществления, по меньшей мере один основный аминокислотный остаток заменен по меньшей мере одним кислым аминокислотным остатком, тогда как в других вариантах осуществления по меньшей мере один основный аминокислотный остаток заменен по меньшей мере одним нейтральным аминокислотным остатком и, в других вариантах осуществления, по меньшей мере один нейтральный аминокислотный остаток заменен по меньшей мере одной кислой аминокислотой. В дополнительных вариантах осуществления, по меньшей мере один нейтральный аминокислотный остаток в исходной нейтральной металлопротеазе заменен по меньшей мере одним нейтральным аминокислотным остатком с получением варианта нейтральной металлопротеазы, имеющего тот же самый заряд в сравнении с исходным ферментом. Авторы изобретения не предполагают ограничения данного изобретения какими-либо конкретными комбинациями замен. Они также не предполагают, что эти замены должны выполняться в каком-либо конкретном порядке.
Данное изобретение относится к способам получения по меньшей мере одного варианта белка с улучшенной эффективностью в сравнении с исходным белком, предусматривающим модификацию по меньшей мере одного аминокислотного остатка в одном или нескольких положениях в исходном белке с получением по меньшей мере одного варианта белка, имеющего более положительный, более отрицательный, менее положительный или менее отрицательный заряд в сравнении с исходным белком. В некоторых вариантах осуществления, эта модификация предусматривает замену, добавление и/или делецию, тогда как в других вариантах осуществления модификация включает химическую модификацию. В некоторых вариантах осуществления, этот белок является ферментом. В некоторых особенно предпочтительных вариантах осуществления, этим белком является протеаза, амилаза, целлюлаза, полиэстераза, эстераза, липаза, кутиназа, пектиназа, оксидаза, трансфераза, алкалаза или каталаза. В некоторых дополнительных особенно предпочтительных вариантах осуществления, эта протеаза является сериновой протеазой или нейтральной металлопротеазой. В некоторых дополнительных вариантах осуществления, эффективность по меньшей мере одного варианта белка оценивают с использованием по меньшей мере одного представляющего интерес теста. В некоторых дополнительных вариантах осуществления, этот по меньшей мере один представляющий интерес тест предусматривает измерение связывания субстрата, ингибирования фермента, уровней экспрессии, стабильности детергента, термостабильности, скорости реакции, степени реакции, термальной активности, разжижения крахмала, гидролиза сложных эфиров, ферментативного отбеливания, моющей эффективности, деградации биомассы, растворимости, стабильности к хелаторам и/или осахаривания. В некоторых дополнительных вариантах осуществления, по меньшей мере один вариант белка проявляет улучшенную эффективность по меньшей мере в одном представляющем интерес тесте в сравнении с исходным белком.
Данное изобретение также относится к способам получения по меньшей мере одного варианта фермента с улучшенной моющей эффективностью в сравнении с исходным ферментом, предусматривающим модификацию по меньшей мере одного аминокислотного остатка в одном или нескольких положениях исходного фермента для получения по меньшей мере одного варианта фермента, имеющего более положительный, более отрицательный, менее положительный или менее отрицательный заряд в сравнении с исходным ферментом. В некоторых вариантах осуществления, эта модификация включает замену, добавление и/или делецию, тогда как в альтернативных вариантах осуществления, модификация включает химическую модификацию. В некоторых дополнительных вариантах осуществления, эти способы дополнительно предусматривают тестирование моющей эффективности этого варианта фермента и исходного фермента для получения показателей эффективности для этого варианта фермента и исходного фермента. В некоторых вариантах осуществления, показатель эффективности этого варианта фермента имеет величину, большую чем 1,0, а моющая эффективность исходного фермента имеет показатель эффективности 1,0. В некоторых особенно предпочтительных вариантах осуществления, эти способы дополнительно предусматривают получение вариантного фермента, имеющего улучшенную моющую эффективность. В некоторых дополнительных вариантах осуществления, этим ферментом является протеаза, амилаза, целлюлаза, полиэстераза, эстераза, липаза, кутиназа, пектиназа, оксидаза, трансфераза, алкалаза или каталаза. В некоторых предпочтительных вариантах осуществления, эта протеаза является сериновой протеазой или нейтральной металлопротеазой. В некоторых особенно предпочтительных вариантах осуществления, исходным ферментом является протеаза Bacillus. В некоторых вариантах осуществления, моющую эффективность тестируют в порошкообразной или жидкой детергентной композиции, имеющей рН между 5 и 12,0. В некоторых вариантах осуществления, моющую эффективность тестируют в жидком детергенте (моющем средстве) для стирки, имеющем основный рН, тогда как в некоторых других вариантах осуществления моющую эффективность тестируют в жидком детергенте для холодной воды, имеющем основный рН. В некоторых вариантах осуществления, эти замены находятся в положениях в исходном ферменте, имеющих доступную для растворителя поверхность (SAS), большую чем приблизительно 25%. В некоторых дополнительных вариантах осуществления, эти замены находятся в положениях в исходном ферменте, имеющих доступную для растворителя поверхность (SAS), большую чем приблизительно 50%, или большую чем приблизительно 65%.
Данное изобретение также относится к способам получения вариантов ферментов с улучшенной моющей эффективностью в сравнении с исходным ферментом, предусматривающим: а) модификацию по меньшей мере одного аминокислотного остатка в одном или нескольких положениях в исходном ферменте для получения первого варианта фермента, имеющего более положительный, более отрицательный, менее положительный или менее отрицательный заряд в сравнении с исходным ферментом; и b) модификацию по меньшей мере одного аминокислотного остатка в одном или нескольких положениях в исходном ферменте для получения второго варианта фермента, имеющего более положительный, более отрицательный, менее положительный или менее отрицательный заряд в сравнении с исходным ферментом. В некоторых вариантах осуществления, эта модификация включает замену, добавление и/или делецию, тогда как в некоторых альтернативных вариантах осуществления, эта модификация включает химическую модификацию. В некоторых дополнительных вариантах осуществления, эти стадии повторяют для получения множества вариантов фермента. В некоторых дополнительных вариантах осуществления, этим исходным ферментом является протеаза, амилаза, целлюлаза, полиэстераза, эстераза, липаза, кутиназа, пектиназа, оксидаза, трансфераза, алкалаза или каталаза. В некоторых предпочтительных вариантах осуществления, эта протеаза является нейтральной металлопротеазой или сериновой протеазой. В некоторых особенно предпочтительных вариантах осуществления, исходным ферментом является протеаза Bacillus. В некоторых дополнительных вариантах осуществления, эти способы дополнительно предусматривают тестирование моющей эффективности вариантных ферментов и исходного фермента и сравнение способности исходного фермента и вариантных ферментов удалять пятно в тестировании моющей эффективности, причем моющая эффективность исходного фермента принимается за величину 1,0, а вариант с улучшенной моющей эффективностью достигает величины, большей чем 1,0. В некоторых вариантах осуществления, эти способы дополнительно предусматривают получение варианта, имеющего улучшенную моющую эффективность в сравнении с исходным ферментом. В некоторых предпочтительных вариантах осуществления, этим исходным ферментом является сериновая протеаза. В некоторых особенно предпочтительных вариантах осуществления, этой сериновой протеазой является сериновая протеаза Bacillus или сериновая протеаза Cellulomonas. В некоторых дополнительных вариантах осуществления, моющую эффективность тестируют в порошкообразной или жидкой детергентной композиции, имеющей рН между 5 и 12,0. В некоторых дополнительных вариантах осуществления, моющую эффективность тестируют в жидком детергенте (моющем средстве) для стирки, имеющем основный рН. В других дополнительных вариантах осуществления, моющую эффективность тестируют в жидком детергенте для холодной воды, имеющем основный рН. В некоторых альтернативных вариантах осуществления, эти замены находятся в положениях в исходном ферменте, имеющих доступную для растворителя поверхность (SAS), большую чем приблизительно 25%, тогда как в некоторых других вариантах осуществления эти замены находятся в положениях в исходном ферменте, имеющих доступную для растворителя поверхность (SAS), большую чем приблизительно 50%, или большую чем приблизительно 65%. В некоторых вариантах осуществления, по меньшей мере один кислый аминокислотный остаток заменен по меньшей мере одним основным аминокислотным остатком, тогда как в других вариантах осуществления по меньшей мере один кислый аминокислотный остаток заменен по меньшей мере одним нейтральным аминокислотным остатком, по меньшей мере один нейтральный аминокислотный остаток заменен по меньшей мере одним основным аминокислотным остатком, по меньшей мере один основный аминокислотный остаток заменен по меньшей мере одним кислым аминокислотным остатком, по меньшей мере один основный аминокислотный остаток заменен по меньшей мере одним нейтральным аминокислотным остатком, по меньшей мере один нейтральный аминокислотный остаток заменен по меньшей мере одной кислой аминокислотой и/или по меньшей мере один нейтральный аминокислотный остаток в исходном ферменте заменен по меньшей мере одним нейтральным аминокислотным остатком с получением варианта фермента, имеющего один и тот же заряд в сравнении с исходным ферментом. Предполагается, что любая подходящая комбинация замен найдет применение в данном изобретении, если желательно.
Данное изобретение также относится к способам получения по меньшей мере одного варианта белка с улучшенной эффективностью в сравнении с исходным белком, предусматривающим модификацию по меньшей мере одного аминокислотного остатка в одном или нескольких положениях в исходном белке для получения по меньшей мере одного варианта белка, имеющего более положительный, более отрицательный, менее положительный или менее отрицательный заряд в сравнении с исходным белком, причем эти одно или несколько положений имеют доступную для растворителя поверхность (SAS), большую чем приблизительно 25%. В некоторых вариантах осуществления, эти одно или несколько положений не являются консервативными в выравниваниях аминокислот гомологичных белковых последовательностей, содержащих исходный белок и по меньшей мере один дополнительный белок. В некоторых предпочтительных вариантах осуществления, этот исходный белок является ферментом. В некоторых особенно предпочтительных вариантах осуществления, этим ферментом является протеаза, амилаза, целлюлаза, полиэстераза, эстераза, липаза, кутиназа, пектиназа, оксидаза, трансфераза, алкалаза или каталаза. В некоторых дополнительных вариантах осуществления, эта улучшенная эффективность включает увеличение одного или нескольких свойств, выбранных из связывания субстрата, ингибирования фермента, экспрессии, стабильности в детергенте, термостабильности, скорости реакции, степени реакции, термальной активности, разжижения крахмала, деградации биомассы, осахаривания, гидролиза сложных эфиров, ферментативного отбеливания, моющей эффективности, растворимости, стабильности к хелаторам и/или модификации текстиля. В некоторых дополнительных вариантах осуществления, эта модификация включает замену, добавление и/или делецию, тогда как в других вариантах осуществления модификация предусматривает химическую модификацию. В некоторых вариантах осуществления, по меньшей мере одна замена включает изменение суммарного заряда 0, -1 или -2 относительно исходного белка, в то время как в других вариантах осуществления по меньшей мере одна замена включает изменение суммарного заряда +1 или +2 относительно исходного белка. В некоторых дополнительных вариантах осуществления, по меньшей мере одна из замен в исходном белке включает изменение суммарного заряда 0, -1 или -2 относительно исходного белка, причем по меньшей мере одна дополнительная замена в исходном белке включает изменение заряда +1 или +2 относительно исходного белка. В некоторых дополнительных вариантах осуществления, этот вариант белка имеет изменение суммарного заряда +1 или +2 относительно исходного белка, в то время как в других вариантах осуществления этот вариант белка имеет изменение суммарного заряда 0, -1 или -2 относительно исходного белка. В некоторых дополнительных вариантах осуществления, эти замены находятся в положениях в исходном белке, имеющих доступную для растворителя поверхность (SAS), большую чем приблизительно 50%, или большую чем приблизительно 65%.
КРАТКОЕ ОПИСАНИЕ чертежей
Фиг.1A изображает относительную активность в микролоскутках (маленьких образцах ткани) с кровью, молоком, чернилами (BMI) (нормализованную относительно наилучшего моющего агента) вариантов ASP как функцию изменения суммарного заряда относительно ASP дикого типа, измеренную в жидком детергенте AATCC (черные треугольники) и буфере (белые кружки) с одинаковыми рН и проводимостью (5 мМ HEPES pH 8,0, 2,5 мМ NaCl). Подобным образом, фиг.1B изображает относительную активность в микролоскутках BMI как функцию изменения заряда относительно дикого типа для комбинаторной библиотеки зарядов ASP (CCL).
Фиг.2 изображает относительную активность в микролоскутках (маленьких образцах ткани) BMI (нормализованную относительно наилучшего моющего агента) вариантов ASP как функцию изменения суммарного заряда относительно ASP дикого типа, измеренную в 5 мМ HEPES рН 8,0 с варьирующейся концентрацией NaCl: 2,5 мМ (белые кружки), 16 мМ (серые кружки) и 100 мМ (черные кружки).
Фиг.3A изображает очищающую BMI эффективность FNA CCL в Североамериканском детергенте (моющем средстве) для стирки как функцию изменения заряда. Подобным образом, фиг.3B изображает очищающую BMI эффективность GG36 CCL в Североамериканском детергенте (моющем средстве) для стирки как функцию изменения заряда.
Фиг.4A изображает очищающую BMI эффективность FNA CCL в Западноевропейском детергенте (моющем средстве) как функцию изменения заряда. Подобным образом, фиг.4В изображает очищающую BMI эффективность GG36 CCL в Западноевропейском жидком детергенте (моющем средстве) как функцию изменения заряда.
Фиг.5A изображает очищающую BMI эффективность FNA CCL в Японском порошкообразном детергенте (моющем средстве) как функцию изменения заряда. Подобным образом, фиг.5В изображает очищающую BMI эффективность GG36 CCL в Японском порошкообразном детергенте (моющем средстве) как функцию изменения заряда.
Фиг.6A изображает очищающую от сваренного яичного желтка эффективность FNA CCL в детергенте для автоматической мойки посуды как функцию изменения заряда. Подобным образом, фиг.6В изображает очищающую от сваренного яичного желтка эффективность GG36 CCL в детергенте для автоматической мойки посуды как функцию изменения заряда.
Фиг.7A изображает удельную ферментативную активность на крахмале BODIPY для AmyS-S242Q CCL как функцию изменения заряда. Подобным образом, фиг.7В изображает вязкость после разжижения кукурузного крахмала для вариантов поверхности заряда AmyS, охватывающих «лестницу» (лэддер) изменения заряда -12 - +4 относительно исходного фермента AmyS enzyme.
Фиг.8 изображает уровни экспрессии вариантов ASP в Bacillus subtilis как функцию изменения суммарного заряда относительно ASP дикого типа.
Фиг.9 изображает стабильность к LAS/EDTA вариантов FNA как функцию изменения суммарного заряда относительно исходного FNA.
Фиг.10 изображает термостабильность первой лестницы (лэддера) зарядов вариантов ASP как функцию изменения суммарного заряда относительно ASP дикого типа.
Фиг.11 изображает термостабильность первой лестницы (лэддера) зарядов вариантов AmyS как функцию изменения суммарного заряда относительно AmyS дикого типа.
Фиг.12 обеспечивает очищающую от рисового крахмала активность первой лестницы (лэддера) зарядов AmyS как функцию рН. pH 3,0-4,25 создается смесью 200 мМ Na-формиат + 0,01% Твин-80. pH 4,25-5,5 создается смесью 200 мМ Na-ацетат + 0,01% Твин-80. Эти данные построены в виде кривых титрования, каждая из которых имеет единственную величину pKa.
Фиг.13 обеспечивает величины pKa, определенные на фиг.13, построенные в зависимости от изменения заряда относительно AmyS дикого типа.
ОБЩЕЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Данное изобретение относится к способам конструирования белков для оптимизации их эффективности при определенных представляющих интерес условиях окружающей среды. В некоторых вариантах осуществления, данное изобретение относится к способам конструирования ферментов для оптимизации их каталитической активности при конкретных условиях окружающей среды. В некоторых предпочтительных вариантах осуществления, данное изобретение относится к способам изменения суммарного заряда поверхности и/или распределения заряда поверхности ферментов (например, металлопротеаз или сериновых протеаз) для получения вариантов ферментов, которые демонстрируют улучшенную эффективность в детергентных композициях в сравнении с первоначальным или исходным ферментом.
Протеаза субтилизин является главным ферментом, используемым в детергентах для стирки (моющих средствах) и, возможно, наиболее широко используемым ферментом в мире. Почти двадцать лет назад было замечено, что электростатические эффекты поверхности могут модулировать каталитическую активность субтилизина (см., например, Russell and Fersht, Nature 328:496-500 [1987]). Недавно наблюдали, что мутации, которые вовлекали изменение суммарного заряда субтилизина, имеют разительное действие на моющую эффективность в детергентах (см., например, патент EP № 0479870 B1, включенный здесь в качестве ссылки). Считается, что это полезное действие является результатом смещения pI (изоэлектрической точки) субтилизина к рН моющего раствора. Однако более позднее исследование продемонстрировало, что этот вывод не всегда применим (см., например, Патент США № 6673590 B1, включенный здесь в качестве ссылки). Как указывается в этом патенте, действие мутаций зарядов в субтилизине зависит разительно от концентраций детергентов, причем мутации, понижающие pI, обеспечивают фермент, который является более эффективным при высоких концентрациях детергента. Это имеет большую ценность, так как концентрация детергента в моющих растворах в значительной степени варьируется по земному шару. Таким образом, для специалистов в данной области становится понятно, что имеется оптимальная pI для моющей эффективности субтилизина, которая зависит от рН и концентрации детергента в моющем растворе. Были описаны дополнительные попытки улучшения активности субтилизина в детергентах (моющих средствах) для стирки (см. Публикацию патента США № 2005/0221461). Неожиданно было обнаружено, что варианты субтилизина, имеющие тот же самый суммарный электростатический заряд, что и исходный субтилизин, увеличивали моющую эффективность в условиях как высокой, так и низкой концентрации детергента.
Если нет других указаний, практика данного изобретения включает общепринятые способы, обычно используемые в получении белков инженерией (конструированием), молекулярной биологии, микробиологии, и способы рекомбинантных ДНК, которые находятся в пределах обычной квалификации в данной области. Такие способы известны квалифицированным в данной области специалистам и описаны в многочисленных руководствах и ссылочных исследованиях, хорошо известных в данной области. Все вышеуказанные патенты, заявки на патенты, статьи и публикации, как supra, так и infra, особо включены здесь в качестве ссылки.
Если нет других определений здесь, все технические и научные термины, используемые здесь, имеют значения, обычно понимаемые специалистом с обычной квалификацией в области, к которой относится это изобретение. Хотя в практике данного изобретения используются любые способы и материалы, сходные со способами, описанными здесь, или эквивалентными способами, описанными здесь, в этом изобретении описаны предпочтительные способы и материалы. Таким образом, термины, определенные непосредственно ниже, описаны более полно ссылкой на это описание в целом.
Кроме того, в данном контексте формы единственного числа включают формы множественного числа, если контекст не указывает явно другое. Если нет других указаний, нуклеиновые кислоты пишутся слева направо в ориентации 5'→3'; аминокислоты пишутся слева направо в ориентации амино → карбокси соответственно. Должно быть понятно, что это изобретение не ограничивается конкретной методологией, конкретными протоколами и описанными реагентами, так как они могут варьироваться в зависимости от контекста, в котором они используются специалистами с квалификацией в данной области.
Предполагается, что каждое максимальное численное ограничение, приводимое во всем этом описании, включает любое более низкое ограничение, как если бы такие более низкие численные ограничения были специально написаны здесь. Каждое минимальное численное ограничение, приводимое во всем этом описании, включает любое более высокое ограничение, как если бы такие более высокие численные ограничения были специально написаны здесь. Любой численный диапазон, приводимый во всем этом описании, будет включать любой более узкий численный диапазон, который попадает в такой более широкий численный диапазон, как если бы такой более узкий диапазон был специально написан здесь.
Кроме того, обеспеченные здесь заголовки не являются ограничениями различных аспектов или вариантов этого изобретения, которые могут иметься, при ссылке на это описание как на целое. Таким образом, термины, определенные непосредственно ниже, являются наиболее полно определенными со ссылкой на это описание как на целое. Тем не менее, для облегчения понимания этого изобретения, ряд терминов определен ниже.
Определения
В данном контексте, термины "протеаза" и "протеолитическая активность" относятся к белку или пептиду, проявляющему способность гидролизовать пептиды или субстраты, имеющие пептидные связи. Имеется много хорошо известных процедур для измерения протеолитической активности (см., например, Kalisz, "Microbial Proteinases," In: Fiechter (ed.), Advances in Biochemical Engineering/Biotechnology [1988]). Например, протеолитическая активность может быть определена сравнительными анализами, которые анализируют способность соответствующей протеазы гидролизовать коммерческий субстрат. Примеры субстратов, применимых в таких анализах протеазы или протеолитической активности, включают, но не ограничиваются ими, диметилказеин (Sigma C-9801), бычий коллаген (Sigma C-9879), бычий эластин (Sigma E-1625) и бычий кератин (ICN Biomedical 902111). Колориметрические анализы, использующие эти субстраты, хорошо известны в данной области (см., например, WO 99/34011 и Патент США № 6376450, оба из которых включены здесь в качестве ссылки). pNA-анализ (см., например, Del Mar et al., Anal Biochem, 99:316-320 [1979]) также используется в определении концентрации активного фермента для фракций, собранных во время градиентной элюции. Этот анализ измеряет скорость, при которой высвобождается п-нитроанилин при гидролизе этим ферментом растворимого синтетического субстрата, сукцинил-аланин-аланин-пролин-фенилаланин-п-нитроанилид (sAAPF-pNA). Скорость образования желтой окраски из реакции гидролиза измеряют при 410 нм на спектрофотометре, и эта скорость пропорциональна концентрации активного фермента. Кроме того, измерения оптической плотности при 280 нм могут быть использованы для определения общей концентрации белка. Отношение активный фермент-белок дает чистоту этого фермента.
В данном контексте, термины "протеаза ASP", "протеаза Asp" и "Asp" относятся к сериновым протеазам, описанным в настоящем документе и описанным в Заявке на патент США с порядковым номером 10/576331, включенной в настоящий документ в качестве ссылки. В некоторых предпочтительных вариантах осуществления, протеаза Asp является протеазой, обозначаемой здесь как протеаза 69B4, полученная из штамма Cellulomonas 69B4. Таким образом, в предпочтительных вариантах осуществления, термин "протеаза 69B4" относится к природной зрелой протеазе, полученной из штамма Cellulomonas 69B4 (DSM 16035), имеющей по существу идентичную аминокислотную последовательность с последовательностью, обеспеченной в SEQ ID NO:8. В альтернативных вариантах осуществления, данное изобретение обеспечивает части протеазы ASP.
Термин "гомологи протеазы Cellulomonas" относится к природным протеазам, имеющим по существу идентичные аминокислотные последовательности со зрелой протеазой, полученной из штамма Cellulomonas 69B4, или полинуклеотидные последовательности, которые кодируют такие природные протеазы, причем эти протеазы сохраняют функциональные характеристики сериновой протеазы, кодируемой такими нуклеиновыми кислотами. В некоторых вариантах осуществления, эти гомологи протеазы называют "целлюломонадинами".
В данном контексте, термины "вариант ASP", "вариант протеазы ASP" и "вариант протеазы 69B" относятся к протеазам, которые являются сходными с ASP дикого типа, в частности в их функции, но имеют мутации в их аминокислотной последовательности, которые делают их отличающимися по последовательности от протеазы дикого типа.
В данном контексте, "Cellulomonas ssp." обозначает все виды в пределах рода "Cellulomonas", которые являются грамположительными бактериями, классифицированными как члены семейства Cellulomonadaceae, подотряда Micrococcineae, отряда Actinomycetales, класса Actinobacteria. Известно, что род Cellulomonas продолжает подвергаться таксономической реорганизации. Так, предполагается, что этот род включает виды, которые были переклассифицированы.
В данном контексте, "Streptomyces ssp." обозначает все виды в пределах рода "Streptomyces," которые являются грамположительными бактериями, классифицированными как члены семейства Streptomycetaceae, подотряда Streptomycineae, отряда Actinomycetales, класса Actinobacteria. Известно, что род Streptomyces продолжает подвергаться таксономической реорганизации. Так, предполагается, что этот род включает виды, которые были переклассифицированы.
В данном контексте, "род Bacillus" включает все виды в пределах рода "Bacillus," известные квалифицированным в данной области специалистам, включающие, но не ограничивающиеся ими, B. subtilis, B. licheniformis, B. lentus, B. brevis, B. stearothermophilus, B. alkalophilus, B. amyloliquefaciens, B. clausii, B. halodurans, B. megaterium, B. coagulans, B. circulans, B. lautus и B. thuringiensis. Известно, что род Bacillus продолжает подвергаться таксономической реорганизации. Так, предполагается, что этот род включает виды, которые были переклассифицированы, включающие, но не ограничивающиеся им, такой организм, как B. stearothermophilus, который теперь называют "Geobacillus stearothermophilus". Образование устойчивых эндоспор в присутствии кислорода считается определяющим признаком рода Bacillus, хотя этот признак приложим также к недавно названным Alicyclobacillus, Amphibacillus, Aneurinibacillus, Anoxybacillus, Brevibacillus, Filobacillus, Gracilibacillus, Halobacillus, Paenibacillus, Salibacillus, Thermobacillus, Ureibacillus и Virgibacillus.
Термины "полинуклеотид" и "нуклеиновая кислота", используемые взаимозаменяемо здесь, относятся к полимерной форме нуклеотидов любой длины, либо рибонуклеотидов, либо дезоксирибонуклеотидов. Эти термины включают, но не ограничиваются ими, одно-, двух- или трехцепочечную ДНК, геномную ДНК, кДНК, РНК, гибрид ДНК-РНК или полимер, содержащий пуриновые и пиримидиновые основания, или другие природные, химически, биохимически модифицированные, неприродные или дериватизованные нуклеотидные основания. Следующие неограничивающие примеры полинуклеотидов включают: гены, фрагменты генов, фрагменты хромосом, EST, экзоны, интроны, мРНК, тРНК, рРНК, рибозимы, кДНК, рекомбинантные полинуклеотиды, разветвленные полинуклеотиды, плазмиды, векторы, выделенную ДНК любой последовательности, выделенную РНК любой последовательности, зонды и праймеры нуклеиновых кислот. В некоторых вариантах осуществления, полинуклеотиды содержат модифицированные нуклеотиды, такие как метилированные нуклеотиды и аналоги нуклеотидов, урацил, другие сахара и связывающие группы, такие как фторрибоза и тиоат, и нуклеотидные разветвления. В альтернативных вариантах осуществления, последовательность нуклеотидов прерывается ненуклеотидными компонентами.
В данном контексте, термины "конструкция ДНК" и "трансформирующая ДНК" используются взаимозаменяемо для обозначения ДНК, используемой для введения последовательностей в клетку-хозяина или организм-хозяин. Эта ДНК может быть генерирована in vitro при помощи ПЦР или любого другого подходящего способа (любых других подходящих способов), известных квалифицированным в данной области специалистам. В особенно предпочтительных вариантах осуществления, конструкция ДНК содержит представляющую интерес последовательность (например, входящую последовательность). В некоторых вариантах осуществления, эта последовательность функционально связана с дополнительными элементами, такими как регуляторные элементы (например, промоторы и т.д.). Эта конструкция ДНК может дополнительно содержать селектируемый маркер. Она может дополнительно содержать входящую последовательность, фланкированную боксами гомологии. В дополнительном варианте осуществления, эта трансформирующая ДНК содержит другие негомологичные последовательности, добавленные на концах (например, последовательности-наполнители, или «лишние» фрагменты, или «фланги»). В некоторых вариантах осуществления, концы входящей последовательности являются замкнутыми, так что эта трансформирующая ДНК образует замкнутое кольцо. Трансформирующие последовательности могут быть последовательностями дикого типа, мутантными или модифицированными. В некоторых вариантах осуществления, конструкция ДНК содержит последовательности, гомологичные хромосоме клетки-хозяина. В некоторых вариантах осуществления, эта конструкция ДНК содержит негомологичные последовательности. После сборки конструкции ДНК in vitro она может быть использована для: 1) инсерции гетерологичных последовательностей в желаемую последовательность-мишень клетки-хозяина; и/или 2) мутагенизации области хромосомы клетки-хозяина (т.е. замены эндогенной последовательности гетерологичной последовательностью); и/или 3) делеции генов-мишеней; и/или введения плазмиды репликации в этого хозяина.
В данном контексте, термины "экспрессионная кассета" и "экспрессирующий вектор" относятся к конструкциям нуклеиновых кислот, генерированным рекомбинантно или синтетически, с рядом указанных элементов нуклеиновых кислот, которые делают возможной транскрипцию конкретной нуклеиновой кислоты в клетке-мишени. Рекомбинантная экспрессионная кассета может быть включена в плазмиду, хромосому, митохондриальную ДНК, пластидную ДНК, вирус или фрагмент нуклеиновой кислоты. Обычно, участок рекомбинантной экспрессионной кассеты экспрессирующего вектора включает, среди других последовательностей, последовательность нуклеиновой кислоты, которая должна быть транскрибирована, и промотор. В предпочтительных вариантах осуществления, экспрессирующие векторы способны включать фрагменты гетерологичной ДНК в клетку-хозяина и экспрессировать их в этой клетке. Многие прокариотические и эукариотические экспрессирующие векторы являются коммерчески доступными. Выбор подходящих экспрессирующих векторов находится в пределах компетенции квалифицированных в данной области специалистов. Термин “экспрессионная кассета” используется здесь взаимозаменяемо с термином “конструкция ДНК” и их грамматическими эквивалентами. Выбор подходящих экспрессирующих векторов находится в пределах компетенции квалифицированных в данной области специалистов.
В данном контексте, термин "вектор" относится к полинуклеотидной конструкции, предназначенной для введения нуклеиновых кислот в один или несколько типов клеток. Векторы включают клонирующие векторы, экспрессирующие векторы, челночные векторы, плазмиды, кассеты и т.п. В некоторых вариантах осуществления, полинуклеотидная конструкция содержит ДНК-последовательность, кодирующую протеазу (например, предшественник протеазы или зрелую протеазу), которая функционально связана с подходящей пропоследовательностью (например, секреторной и т.д.), способной влиять на экспрессию этой ДНК в подходящем хозяине.
В данном контексте, термин "плазмида" относится к кольцевой двухцепочечной (ds) конструкции ДНК, используемой в качестве клонирующего вектора, которая образует внехромосомный самореплицирующийся генетический элемент в некоторых эукариотах или прокариотах или интегрируется в хромосому хозяина.
В настоящем документе в контексте введения последовательности нуклеиновой кислоты в клетку, термин "введенная" относится к любому способу, подходящему для переноса последовательности нуклеиновой кислоты в клетку. Такие способы введения включают, но не ограничиваются ими, слияние протопластов, трансфекцию, трансформацию, конъюгацию и трансдукцию (см., например, Ferrari et al., "Genetics", in Hardwood et al., (eds.), Bacillus, Plenum Publishing Corp., pages 57-72 [1989]).
В данном контексте, термины “трансформированная” и "стабильно трансформированная" относятся к клетке, которая имеет неприродную (гетерологичную) полинуклеотидную последовательность, интегрированную в ее геном или находящуюся в виде эписомной плазмиды, которая сохраняется по меньшей мере в течение двух генераций.
В данном контексте, термин “кодирующая селектируемый маркер нуклеотидная последовательность” относится к нуклеотидной последовательности, которая способна к экспрессии в клетках-хозяевах, и когда экспрессия селектируемого маркера придает клеткам, содержащим этот экспрессированный ген, способность расти в присутствии соответствующего селективного агента или в отсутствие незаменимого нутриента.
В данном контексте, термин "селектируемый маркер” и “селективный маркер” относятся к нуклеотидной последовательности (например, к гену), способной к экспрессии в клетке-хозяирне, которая позволяет выполнять легкий отбор хозяев, содержащих этот вектор. Примеры таких селектируемых маркеров включают, но не ограничиваются ими, антимикробные агенты. Таким образом, термин “селектируемый маркер” относится к генам, которые обеспечивают указание на то, что клетка-хозяин поглотила входящую представляющую интерес ДНК или произошла некая другая реакция. Обычно, селектируемыми маркерами являются гены, которые придают антимикробную устойчивость или метаболическое преимущество клетке-хозяину, что позволяет отличать клетки, содержащие эту экзогенную ДНК, от клеток, которые не получили никакой экзогенной ДНК во время трансформации. "Селектируемый маркер нахождения (локализации)” является маркером, который локализован на хромосоме трансформируемого микроорганизма. Этот селектируемый маркер нахождения (локализации) кодирует ген, который отличается от селектируемого маркера на трансформирующей ДНК-конструкции. Селективные маркеры хорошо известны квалифицированным в данной области специалистам. Как указано выше, предпочтительно этот маркер является устойчивым к антимикробным агентам маркером (например, ampR; phleoR; specR; kanR; eryR; tetR; cmpR и neoR (см., например, Guerot-Fleury, Gene, 167:335-337 [1995); Palmeros et al., Gene 247:255-264 [2000] и Trieu-Cuot et al, Gene, 23:331-341 [1983]). Другие маркеры, применимые в соответствии с этим изобретением, включают, но не ограничиваются ими, ауксотрофные маркеры, такие как триптофан, и детектирующие маркеры, такие как β-галактозидаза.
В данном контексте, термин "промотор" относится к последовательности нуклеиновой кислоты, которая функционирует для управления транскрипцией расположенного справа от нее гена. В предпочтительных вариантах осуществления, этот промотор является подходящим для клетки-хозяина, в которой экспрессируется этот ген-мишень. Промотор, вместе с другими транскрипционными и трансляционными регуляторными последовательностями (также называемыми “регуляторными последовательностями”), является необходимым для экспрессии конкретного гена. Обычно, эти транскрипционные и трансляционные регуляторные последовательности включают, но не ограничиваются ими, промоторные последовательности, сайты связывания рибосом, стартовые последовательности и стоп-последовательности транскрипции, стартовые последовательности и стоп-последовательности трансляции и энхансерные или активаторные последовательности.
Нуклеиновая кислота является "функционально связанной", когда она помещена в функциональную связь с другой последовательностью нуклеиновой кислоты. Например, ДНК, кодирующая секреторный лидер (т.е. сигнальный пептид), функционально связана с ДНК для полипептида, если он экспрессируется в виде пребелка, который участвует в секреции этого полипептида; промотор или энхансер функционально связаны с кодирующей последовательностью, если он влияет на транскрипцию этой последовательности; или сайт связывания рибосом функционально связан с кодирующей последовательностью, если он расположен таким образом, что он облегчает трансляцию. Обычно, термин "функционально связанные" означает, что эти являющиеся связанными ДНК-последовательности являются смежными и, в случае секреторного лидера, смежными и находящимися в рамке считывания. Однако энхансеры не должны быть обязательно смежными. Связывание осуществляется лигированием в подходящих сайтах рестрикции. Если такие сайты не существуют, используют синтетические олигонуклеотидные адапторы или линкеры в соответствии с общепринятой практикой.
В данном контексте, термин "ген" относится к полинуклеотиду (например, сегменту ДНК), который кодирует полипептид и включает области, предшествующие кодирующим областям или следующие за кодирующими областями, а также промежуточные последовательности (интроны) между индивидуальными кодирующими сегментами (экзонами).
В данном контексте, "гомологичными генами" называют пару генов из различных, но обычно родственных видов, которые соответствуют друг другу и которые являются идентичными или очень сходными друг с другом. Этот термин включает гены, которые разделены видообразованием (т.е. развитием новых видов) (например, ортологичные гены), а также гены, которые разделены генетической дупликацией (например, паралогичные гены).
В данном контексте, "ортолог" и "ортологичные гены" обозначают гены в разных видах, которые эволюционировали из общего предкового гена (т.е. гомологичного гена) в ходе эволюции. Обычно, ортологи сохраняют одну и ту же функцию в ходе эволюции. Идентификация ортологов находит применение в надежном предсказании функции гена в новых секвенированных геномах.
В данном контексте, термины "паралог" и "паралогичные гены" относятся к генам, которые являются родственными вследствие дупликации в пределах одного генома. В то время как ортологи сохраняют одну и ту же функцию в ходе эволюции, паралоги развивают новые функции, даже хотя некоторые функции часто являются родственными исходной функции. Примеры паралогичных генов включают, но не ограничиваются ими, гены, кодирующие трипсин, химотрипсин, эластазу и тромбин, все из которых являются сериновыми протеазами и встречаются вместе в одних и тех же видах.
В данном контексте, “гомологией” называют сходство или идентичность последовательности, причем термин “идентичность” является предпочтительным. Эту гомологию определяют с использованием стандартных способов, известных в данной области (см., например, Smith and Waterman, Adv. Appl. Math., 2:482 [1981]; Needleman and Wunsch, J. Mol. Biol., 48:443 [1970]; Pearson and Lipman, Proc. Natl. Acad. Sci. USA, 85:2444 [1988]; программы, такие как GAP, BESTFIT, FASTA и TFASTA в Wisconsin Genetics Software Package, Genetics Computer Group, Madison, WI; и Devereux et al., Nucl. Acid Res., 12:387-395 [1984]).
В данном контексте, "аналогичной последовательностью" является последовательность, в которой функция этого гена является по существу той же самой, что и функция этого гена на основе исходного гена (например, протеазы штамма Cellulomonas 69B4). Кроме того, аналогичные гены включают по меньшей мере 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99% или 100% идентичность последовательности с последовательностью исходного гена. Альтернативно, аналогичные последовательности имеют совпадение 70%-100% генов, обнаруженных в области исходного гена (например, протеазы штамма Cellulomonas 69B4) и/или имеют по меньшей мере 5-10 генов, обнаруженных в области, сопоставленной с этими генами в хромосоме, содержащей этот исходный ген (например, хромосоме штамма Cellulomonas 69B4). В дополнительных вариантах осуществления, более чем одно из указанных выше свойств могут быть отнесены к этой последовательности. Аналогичные последовательности определяют известными способами сопоставления последовательностей. Обычно используемым способом сопоставления является BLAST, хотя, как указано выше и ниже, имеются и другие способы, которые также находят применение в сопоставлении последовательностей.
Одним примером применимого алгоритма является PILEUP. PILEUP создает сопоставление множественных последовательностей из группы родственных последовательностей с использованием прогрессивных парных сопоставлений. Может быть также построено дерево, показывающее образующие кластеры взаимосвязи, используемые для создания этого сопоставления. PILEUP использует упрощение способа прогрессивного сопоставления Feng and Doolittle (Feng and Doolittle, J. Mol. Evol., 35:351-360 [1987]). Этот способ сходен со способом, описанным Higgins and Sharp (Higgins and Sharp, CABIOS 5:151-153 [1989]). Применимые параметры PILEUP включают вес гэпа по умолчанию 3,00, вес длины гэпа по умолчанию 0,10 и взвешенные конечные гэпы.
Другим примером применимого алгоритма является алгоритм BLAST, описанный Altschul et al, (Altschul et al., J. Mol. Biol., 215:403-410 [1990]; и Karlin et al, Proc. Natl. Acad. Sci., USA, 90:5873-5787 [1993]). Особенно применимой программой BLAST является программа WU-BLAST-2 (см. Altschul et al, Meth. Enzymol., 266:460-480 [1996]). WU-BLAST-2 использует несколько параметров поиска, большинство из которых устанавливают на величины по умолчанию. Эти регулируемые параметры установлены на следующие величины: протяженность перекрывания = 1, доля перекрывания = 0,125, предельная величина слова (T) = 11. Параметры HSP S и HSP S2 являются динамичными величинами и устанавливаются самой программой в зависимости от состава конкретной последовательности и состава конкретной базы данных, против которой проводится поиск представляющей интерес последовательности. Однако эти величины могут регулироваться для увеличения чувствительности. %-ную идентичность аминокислотной последовательности определяют по количеству совпадающих идентичных остатков, деленному на общее количество остатков «более длинной» последовательности в сопоставляемой области. Эта «более длинная» последовательность является последовательностью, имеющей наиболее актуальные остатки в сопоставляемой области (гэпы, вводимые WU-Blast-2 для максимизации балла сопоставления, игнорируются).
Таким образом, “процентная (%) идентичность последовательности нуклеиновой кислоты" определяется как процент нуклеотидных остатков в кандидатной последовательности, которые идентичны нуклеотидным остаткам исходной последовательности (т.е. представляющей интерес последовательности). Один предпочтительный способ использует модуль BLASTN WU-BLAST-2, установленный на параметры по умолчанию, причем протяженность перекрывания и доля перекрывания устанавливаются на 1 и 0,125 соответственно.
В данном контексте, термин "гибридизация" относится к процессу, при помощи которого цепь нуклеиновой кислоты соединяется с комплементарной цепью через спаривание оснований, известное в данной области.
Последовательность нуклеиновой кислоты считается "селективно гибридизуемой" со ссылочной последовательностью нуклеиновой кислоты, если эти две последовательности специфически гибридизуются друг с другом при условиях гибридизации и промывки средней или высокой строгости. Условия гибридизации основаны на температуре плавления (Tm) связывающего нуклеиновую кислоту комплекса или зонда. Например, "максимальная строгость" обычно имеет место при приблизительно Tm -5°C (5° ниже Tm этого зонда); "высокая строгость" при приблизительно 5-10°C ниже Tm; "средняя строгость" при приблизительно 10-20°C ниже Tm этого зонда; и "низкая строгость” при приблизительно 20-25°C ниже Tm. Функционально, условия максимальной строгости могут быть использованы для идентификации последовательностей, имеющих строгую идентичность или близкую к строгой идентичность с гибридизационным зондом; тогда как умеренная или низкая гибридизация может быть использована для идентификации или детектирования гомологов полинуклеотидных последовательностей.
Условия гибридизации умеренной и высокой строгости хорошо известны в данной области. Один пример условий высокой строгости включает гибридизацию при приблизительно 42°C в 50% формамиде, 5X SSC, 5X растворе Денхардта, 0,5% ДСН и 100 мкг/мл денатурированной ДНК-носителя с последующими промывками два раза в 2X SSC и 0,5% ДСН при комнатной температуре и еще два раза в 0,1X SSC и 0,5% ДСН при 42°C. Один пример условий умеренной (средней) строгости включает инкубирование в течение ночи при 37°C в растворе, содержащем 20% формамид, 5 x SSC (150 мМ NaCl, 15 мМ тринатрийцитрат), 50 мМ фосфат натрия (pH 7,6), 5 x раствор Денхардта, 10% декстрансульфат и 20 мг/мл денатурированной гидродинамически фрагментированной ДНК спермы лосося, с последующей промывкой фильтров в 1x SSC при приблизительно 37°С-50°C. Квалифицированным в данной области специалистам известно, как регулировать температуру, ионную силу и т.д. при необходимости приспособления к таким факторам, как длина зонда и т.п.
В данном контексте, термин "рекомбинантный" включает ссылку на клетку или вектор, которые были модифицированы введением последовательности гетерологичной нуклеиновой кислоты, или клетку, полученную из модифицированной таким образом клетки. Так, например, рекомбинантные клетки экспрессируют гены, которые не обнаруживаются в идентичной форме в природной (нерекомбинантной) форме этой клетки или экспрессируют природные гены, которые в других отношениях атипично экспрессируются, недостаточно экспрессируются или вообще не экспрессируются в результате преднамеренного вмешательства человека. "Рекомбинация", "рекомбинирование" и генерирование "рекомбинантной" нуклеиновой кислоты обозначают обычно сборку двух или более фрагментов нуклеиновых кислот, причем эта сборка приводит к возникновению химерного гена.
В одном предпочтительном варианте осуществления, мутантные ДНК-последовательности генерируют сайт-насыщающим мутагенезом по меньшей мере в одном кодоне. В другом предпочтительном варианте осуществления, сайт-насыщающий мутагенез выполняют для двух или более кодонов. В следующем варианте осуществления, мутантные ДНК-последовательности имеют более чем 50%, более чем 55%, более чем 60%, более чем 65%, более чем 70%, более чем 75%, более чем 80%, более чем 85%, более чем 90%, более чем 95% или более чем 98% гомологию с последовательностью дикого типа. В альтернативных вариантах осуществления, мутантную ДНК генерируют in vivo с использованием любой известной мутагенной процедуры, такой как, например, облучение, применение нитрозогуанидина и т.п. Затем эту желаемую ДНК-последовательность выделяют и используют в обеспеченных здесь способах.
В данном контексте, термин "последовательность-мишень" относится к ДНК-последовательности в клетке-хозяине, которая кодирует эту последовательность, когда желательно, чтобы входящая последовательность была инсертирована в геном клетки-хозяина. В некоторых вариантах осуществления, эта последовательность-мишень кодирует функциональный ген дикого типа или оперон, тогда как в других вариантах осуществления эта последовательность-мишень кодирует функциональный мутантный ген или оперон или нефункциональный ген или оперон.
В данном контексте, "фланкирующей последовательностью" называют любую последовательность, которая находится выше или ниже по ходу транскрипции от обсуждаемой последовательности (например, для генов A-B-C, ген B фланкирован последовательностями генов A и C). В одном предпочтительном варианте осуществления, входящая последовательность фланкирована боксом гомологии на каждой стороне. В другом варианте осуществления, эта входящая последовательность и боксы гомологии содержат единицу, которая фланкирована последовательностью-наполнителем («лишним» фрагментом) на каждой стороне. В некоторых вариантах осуществления, фланкирующая последовательность присутствует только на одной стороне (либо 3', либо 5'), но в предпочтительных вариантах осуществления она находится на каждой стороне фланкируемой последовательности. В некоторых вариантах осуществления, фланкирующая последовательность присутствует только на одной стороне (либо 3', либо 5'), тогда как в предпочтительных вариантах осуществления она присутствует на каждой стороне фланкируемой последовательности.
В данном контексте термин "последовательность-наполнитель" («лишний» фрагмент) относится к любой лишней ДНК, которая фланкирует боксы гомологии (обычно векторные последовательности). Однако этот термин включает любую негомологичную ДНК-последовательность. Без связывания с какой-либо теорией авторы считают, что последовательность-наполнитель («лишний» фрагмент) обеспечивает некритическую мишень для клетки для инициации поглощения ДНК.
В данном контексте, термины "амплификация" и "амплификация гена" относятся к процессу, при помощи которого конкретные ДНК-последовательности непропорционально реплицируются, так что амплифицированный ген становится присутствующим в более высокой копийности, чем копийность, присутствующая в этом геноме. В некоторых вариантах осуществления, отбор клеток по росту в присутствии лекарственного средства (например, ингибитора подавляемого фермента) приводит к амплификации либо эндогенного гена, кодирующего генный продукт, необходимый для роста в присутствии этого лекарственного средства, либо к амплификации экзогенных (т.е. введенных) последовательностей, кодирующих этот генный продукт, или с использованием обоих способов.
"Амплификация" является особым случаем репликации нуклеиновых кислот, включающим матричную специфичность. Она должна быть противопоставлена неспецифической матричной репликации (т.е. репликации, которая является зависимой от матрицы, но не зависимой от специфической матрицы). Матричная специфичность отличается здесь от точности репликации (т.е. синтеза правильной полинуклеотидной последовательности) и нуклеотидной (рибо- или дезоксирибо-) специфичности. Матричная специфичность часто описывается в терминах специфичности в отношении "мишени". Последовательности-мишени является "мишенями" в том смысле, что они должны быть отобраны («отсортированы») от других нуклеиновых кислот. Способы амплификации были первично созданы для такого отбора («отсортировки»).
В данном контексте, термин "ко-амплификация" относится к введению в единственную клетку амплифицируемого маркера вместе с другими последовательностями генов (т.е. содержащими один или несколько неселектируемых генов, таких как гены, содержащиеся в экспрессирующем векторе) и применению подходящего давления отбора, так что эта клетка амплифицирует как амплифицируемый маркер, так и другие, неселектируемые последовательности генов. Этот амплифицируемый маркер может быть физически связан с другими последовательностями генов, или альтернативно два отдельных фрагмента ДНК, один, содержащий амплифицируемый маркер, и другой, содержащий неселектируемый маркер, могут быть введены в одну и ту же клетку.
В данном контексте, термины "амплифицируемый маркер", "амплифицируемый ген" и "вектор амплификации" относятся к гену или вектору, кодирующему ген, который делает возможной амплификацию этого гена при подходящих условиях выращивания.
"Матричная специфичность" достигается в большинстве способов амплификации посредством выбора фермента. Ферменты амплификации являются ферментами, которые, в условиях, в которых их используют, будут процессировать только специфические последовательности нуклеиновой кислоты в гетерогенной смеси нуклеиновых кислот. Например, в случае Qβ-репликазы, РНК MDV-1 является специфической матрицей для этой репликазы (см., например, Kacian et al., Proc. Natl. Acad. Sci. USA 69:3038 [1972]), а другие нуклеиновые кислоты не реплицируются этим ферментом амплификации. Подобным образом, в случае РНК-полимеразы T7, этот фермент амплификации имеет строгую специфичность в отношении его собственных промоторов (см. Chamberlin et al., Nature 228:227 [1970]). В случае ДНК-лигазы Т4 этот фермент не будет лигировать эти два олигонуклеотида или полинуклеотида, если имеется ошибочное спаривание между этим олигонуклеотидным или полинуклеотидным субстратом и матрицей в сайте лигирования (см. Wu and Wallace, Genomics 4:560 [1989]). Наконец, обнаружено, что полимеразы Taq и Pfu, вследствие их способности функционировать при высокой температуре, проявляют высокую специфичность в отношении последовательностей, связываемых и, следовательно, определяемых праймерами; высокая температура приводит к термодинамическим условиям, которые благоприятствуют гибридизации праймеров с последовательностями-мишенями и отсутствию гибридизации с последовательностями, которые не являются мишенями.
В данном контексте, термин "амплифицируемая нуклеиновая кислота" относится к нуклеиновым кислотам, которые могут быть амплифицированы любым способом амплификации. Предполагается, что "амплифицируемая нуклеиновая кислота" будет обычно содержать "матрицу пробы".
В данном контексте, термин "матрица пробы" относится к нуклеиновой кислоте, происходящей из пробы, которую анализируют на присутствие "мишени" (определенной ниже). В противоположность этому, термин "матрица фона" используют при ссылке на нуклеиновую кислоту, другую, чем матрица пробы, которая может присутствовать или может не присутствовать в пробе. Наиболее часто матрица фона является неумышленно внесенной. Она может быть результатом неумышленного выноса, или она может быть обусловлена присутствием загрязнений нуклеиновых кислот, которые должны быть удалены из этой пробы. Например, нуклеиновые кислоты из организмов, другие чем нуклеиновые кислоты, подлежащие детектированию, могут присутствовать в качестве фона в тест-пробе.
В данном контексте, термин "праймер" относится к олигонуклеотиду, независимо от того, встречается ли он природно в очищенном продукте расщепления рестриктазами или получен синтетически, который способен действовать в качестве точки инициации синтеза при помещении в условия, в которых индуцируется синтез продукта удлинения праймера, который является комплементарным цепи нуклеиновой кислоты (т.е. в присутствии нуклеотидов и индуцирующего агента, такого как ДНК-полимераза, и при подходящих температуре и рН). Этот праймер предпочтительно является одноцепочечным для максимальной эффективности в амплификации, но может быть, альтернативно, двухцепочечным. В случае двухцепочечного праймера его сначала обрабатывают для разделения его цепей перед использованием для получения продуктов удлинения. Предпочтительно, этот праймер является олигодезоксирибонуклеотидом. Праймер должен быть достаточно длинным для праймирования синтеза продуктов удлинения в присутствии индуцирующего агента. Точная длина этих праймеров будет зависеть от многих факторов, в том числе температуры, источника праймера и применения конкретного способа.
В данном контексте, термин "зонд" относится к олигонуклеотиду (т.е. последовательности нуклеотидов), независимо от того, встречается ли он природно в очищенном продукте расщепления рестриктазами или получен синтетически, рекомбинантно или ПЦР-амплификацией, который способен гибридизоваться с другим представляющим интерес олигонуклеотидом. Зонд может быть одноцепочечным или двухцепочечным. Зонды применимы в детектировании, идентификации и выделении конкретных последовательностей генов. Предполагается, что любой зонд, используемый в данном изобретении, будет помечен любой "репортерной молекулой", так что он является детектируемым в любой системе детектирования, в том числе, но не только, в ферментной (т.е. ELISA, а также гистохимических анализах на основе ферментов), флуоресцентной, радиоактивной и люминесцентной системах. Предполагается, что данное изобретение не ограничивается какой-либо конкретной системой детектирования или меткой.
В данном контексте, термин "мишень", при использовании в ссылке на полимеразную цепную реакцию, относится к области нуклеиновой кислоты, связываемой праймерами, используемыми для полимеразной цепной реакции. Таким образом, эта "мишень" должна быть найдена для отбора («отсортировки») от других последовательностей нуклеиновых кислот. "Сегмент" определяется как область нуклеиновой кислоты в пределах последовательности-мишени.
В данном контексте, термин "полимеразная цепная реакция" ("ПЦР") относится к способам Патентов США с номерами 4683195, 4683202 и 4965188, включенными здесь в качестве ссылки, которые включают способы увеличения концентрации сегмента последовательности-мишени в смеси геномной ДНК без клонирования или очистки, как известно квалифицированным в данной области специалистам. Поскольку желаемые амплифицируемые сегменты этой последовательности-мишени становятся преимущественными последовательностями (в отношении концентрации) в этой смеси, их называют "ПЦР-амплифицированными".
В данном контексте, термин "реагенты амплификации" относится к реагентам (дезоксирибонуклеотидтрифосфатам, буферу и т.д.), необходимым для амплификации, за исключением праймеров, нуклеиновой кислоты-матрицы и фермента амплификации. Обычно, реагенты амплификации вместе с другими компонентами реакции помещают в реакционный сосуд (тест-пробирку, микролунку и т.д.).
При помощи ПЦР можно амплифицировать единственную копию конкретной последовательности-мишени в геномной ДНК на уровне, детектируемом несколькими различными методологиями (например, гибридизацией с меченым зондом; включением биотинилированных праймеров с последующим детектированием конъюгата авидин-фермент; включением 32Р-меченых дезоксинуклеотидтрифосфатов, таких как dCTP или dATP, в амплифицируемый сегмент). Кроме геномной ДНК, любая олигонуклеотидная или полинуклеотидная последовательность может быть амплифицирована с подходящим набором праймерных молекул. В частности, амплифицированные сегменты, созданные при помощи самого процесса ПЦР, являются сами эффективными матрицами для последующих ПЦР-амплификаций.
В данном контексте, термины "ПЦР-продукт", "ПЦР-фрагмент" и "продукт амплификации" относятся к полученной смеси соединений после завершения двух или более циклов ПЦР-стадий денатурации, отжига и удлинения. Эти термины включает случай, в котором произошла амплификация одного или нескольких сегментов последовательностей-мишеней.
В данном контексте, термин "ОТ-ПЦР" относится к репликации и амплификации РНК-последовательностей. В этом способе, обратная транскрипция связана с ПЦР, наиболее часто с использованием одной ферментной процедуры, в которой используют термостабильную полимеразу, как описано в Патенте США № 5322770, включенном здесь в качестве ссылки. В ОТ-ПЦР, РНК-матрица превращается в кДНК вследствие активности обратной транскриптазы этой полимеразы и затем амплифицируется с использованием полимеразной активности этой полимеразы (т.е. как и в других ПЦР-способах).
В данном контексте, термины "рестрикционные эндонуклеазы" и "рестрикционные ферменты" относятся к бактериальным ферментам, каждый из которых разрезает двухцепочечную ДНК в конкретной нуклеотидной последовательности или вблизи конкретной нуклеотидной последовательности.
"Сайт рестрикции" обозначает нуклеотидную последовательность, узнаваемую и расщепляемую конкретной рестрикционной эндонуклеазой, и часто является сайтом для инсерции ДНК-фрагментов. В некоторых вариантах осуществления этого изобретения сайты рестрикции конструируют в селективном маркере и на 5'- и 3'-концах конструкции ДНК.
В данном контексте, термин "хромосомная интеграция" относится к процессу, при помощи которого входящую последовательность вводят в хромосому клетки-хозяина. Гомологичные области трансформирующей ДНК сопоставляют с гомологичными областями хромосомы. Затем, последовательность между боксами гомологии заменяют входящей последовательностью в двойном кроссинговере (т.е. гомологичной рекомбинацией). В некоторых вариантах осуществления данного изобретения, гомологичные участки инактивирующего хромосомного сегмента конструкции ДНК сопоставляют с фланкирующими гомологичными областями природной хромосомной области хромосомы Bacillus. Затем, природную хромосомную область удаляют конструкцией ДНК в двойном кроссинговере (т.е. гомологичной рекомбинацией).
"Гомологичная рекомбинация" обозначает обмен ДНК-фрагментами между двумя молекулами ДНК или парными хромосомами в сайте идентичных или почти идентичных нуклеотидных последовательностей. В предпочтительном варианте осуществления, хромосомная интеграция является гомологичной рекомбинацией.
“Гомологичные последовательности” обозначают в данном контексте последовательность нуклеиновой кислоты и полипептидную последовательность, имеющую 100%, 99%, 98%, 97%, 96%, 95%, 94%, 93%, 92%, 91%, 90%, 88%, 85%, 80%, 75% или 70% идентичность последовательности с другой последовательностью нуклеиновой кислоты или аминокислотной последовательностью при оптимальном сопоставлении для сравнения. В некоторых вариантах осуществления, гомологичные последовательности имеют 85%-100% идентичность последовательности, тогда как в других вариантах осуществления имеется 90%-100% идентичность последовательности и в более предпочтительных вариантах осуществления имеется 95%-100% идентичность последовательности.
В данном контексте, "аминокислотными" называют пептидные или белковые последовательности или их части. Термины "белок", "пептид" и "полипептид" используются взаимозаменяемо.
В данном контексте, "представляющий интерес белок" и "представляющий интерес полипептид" обозначают белок/полипептид, который является желаемым и/или который оценивают. В некоторых вариантах осуществления, "представляющий интерес белок" является "исходным белком" (т.е. первоначальным белком). В некоторых вариантах осуществления, исходным белком является фермент дикого типа, который используют в качестве начальной точки для конструирования/создания белка. В некоторых вариантах осуществления, представляющий интерес белок экспрессируется внутриклеточно, в то время как в других вариантах он является секретируемым полипептидом. В особенно предпочтительных вариантах осуществления, эти ферменты включают сериновые протеазы и металлопротеазы, описанные здесь. В некоторых вариантах осуществления, представляющий интерес белок является секретируемым полипептидом, слитым с сигнальным пептидом (т.е. амино-концевым удлинением на белке, который должен быть секретирован). Почти все секретируемые белки используют амино-концевое удлинение белка, которое играет решающую роль в нацеливании на мембрану и транслокации белков-предшественников через мембрану. Это удлинение протеолитически удаляется сигнальной пептидазой во время или сразу же после мембранного транспорта.
В данном контексте, термин "гетерологичный белок" относится к белку или полипептиду, который не встречается природно в клетке-хозяине. Примеры гетерологичных белков включают ферменты, такие как гидролазы, в том числе протеазы. В некоторых вариантах осуществления, гены, кодирующие эти белки, являются природными генами, в то время как в других вариантах осуществления используют мутированные и/или синтетические гены.
В данном контексте, "гомологичный белок" обозначает белок или полипептид, нативный или природный для клетки. В предпочтительных вариантах осуществления, эта клетка является грамположительной клеткой, хотя в особенно предпочтительных вариантах осуществления этой клеткой является клетка-хозяин Bacillus. В альтернативных вариантах осуществления, гомологичным белком является нативный белок, продуцируемый другими организмами, включающими, но не ограничивающимися ими, E. coli, Cellulomonas, Bacillus, Streptomyces, Trichoderma и Aspergillus. Это изобретение включает клетки-хозяева, продуцирующие гомологичный белок посредством технологии рекомбинантных ДНК.
В данном контексте, "область оперона" содержит группу смежных генов, которые транскрибируются в виде единой единицы транскрипции от общего промотора и являются посредством этого субъектом для ко-регуляции. В некоторых вариантах осуществления, оперон включает регуляторный ген. В наиболее предпочтительных вариантах осуществления, используют опероны, которые высоко экспрессируются, как измерено по уровням РНК, но имеют неизвестную или необязательную функцию.
В данном контексте, "антимикробная область" является областью, содержащей по меньшей мере один ген, который кодирует антимикробный белок.
Говорят, что полинуклеотид "кодирует” РНК или полипептид, если в его природном состоянии или при манипулировании способами, известными квалифицированным в данной области специалистам, он может быть транскрибирован и/или транслирован для получения РНК, полипептида или их фрагмента. Также говорят, что антисмысловая цепь такой нуклеиновой кислоты кодирует эти последовательности.
В данной области известно, что ДНК может быть транскрибирована РНК-полимеразой с получением РНК, но РНК может быть обратно транскрибирована обратной транскриптазой с получением ДНК. Таким образом, ДНК может кодировать РНК и наоборот.
Термин "регуляторный сегмент" или "регуляторная последовательность" или "последовательность регуляции экспрессии" относится к полинуклеотидной последовательности ДНК, которая функционально связана с полинуклеотидной последовательностью ДНК, которая кодирует аминокислотную последовательность полипептидной цепи, для выполнения экспрессии кодируемой аминокислотной последовательности. Эта регуляторная последовательность может ингибировать, репрессировать или стимулировать экспрессию функционально связанной полинуклеотидной последовательности, кодирующей аминокислоту.
"Штаммом-хозяином" или "клеткой-хозяином” называют подходящего хозяина для экспрессирующего вектора, содержащего ДНК в соответствии с данным изобретением.
Фермент является "сверхэкспрессируемым" в клетке-хозяине, если этот фермент экспрессируется в этой клетке при более высоком уровне, чем уровень, при котором он экспрессируется в соответствующей клетке дикого типа.
Термин "белок" и "полипептид" используются здесь взаимозаменяемо. Во всем этом описании используется 3-буквенный код для аминокислот, определенный в соответствии с IUPAC-IUB Joint Commission on Biochemical Nomenclature (JCBN). Понятно также, что полипептид может кодироваться более чем одной нуклеотидной последовательностью вследствие вырожденности генетического кода.
"Пропоследовательность" является аминокислотной последовательностью между сигнальной последовательностью и зрелой протеазой, которая необходима для секреции этой протеазы. Отщепление этой пропоследовательности будет приводить к зрелой активной протеазе.
Термин "сигнальная последовательность" или "сигнальный пептид" относится к любой последовательности нуклеотидов и/или аминокислот, которые участвуют в секреции зрелой формы или формы предшественника этого белка. Это определение сигнальной последовательности является функциональным определением, включающим все аминокислотные последовательности, кодируемые геном N-концевой части этого белка, которые участвуют в выполнении секреции белка. Они часто, но не универсально, связаны с N-концевой частью белка или с N-концевой частью белка-предшественника. Сигнальная последовательность может быть эндогенной или экзогенной. Сигнальная последовательность может быть сигнальной последовательностью, которая в норме ассоциирована с этим белком (например, протеазой), или она может происходить из гена, кодирующего другой секретируемый белок. Одна примерная экзогенная сигнальная последовательность содержит первые семь аминокислотных остатков сигнальной последовательности из субтилизина B. subtilis, слитые с остатком сигнальной последовательности субтилизина B. lentus (ATCC 21536).
Термин "гибридная сигнальная последовательность" относится к сигнальным последовательностям, в которых часть последовательности получена из экспрессии хозяина, слитого с сигнальной последовательностью экспрессируемого гена. В некоторых вариантах осуществления, используют синтетические последовательности.
Термин "по существу та же самая сигнальная активность" относится к сигнальной активности, как показано по существу той же самой секрецией этой протеазы в ферментационной среде, например, уровнем протеазы в ферментационной среде, составляющим по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% уровней секретируемой протеазы в ферментационной среде, обеспечиваемым сигнальной последовательностью SEQ ID NO:9.
Термин "зрелая" форма белка или пептида относится к конечной функциональной форме этого белка или пептида. В качестве примера, зрелая форма протеазы ASP данного изобретения включает по меньшей мере аминокислотную последовательность SEQ ID NO:8, тогда как зрелая форма протеазы NprE данного изобретения включает по меньшей мере аминокислотную последовательность SEQ ID NO:3.
Термин форма “предшественника" белка или пептида относится к зрелой форме этого белка, имеющей пропоследовательность, функционально связанную с амино- или карбонильным концом этого белка. Этот предшественник может также иметь "сигнальную" последовательность, функционально связанную с амино-концом этой пропоследовательности. Этот предшественник может также иметь дополнительные полинуклеотиды, которые участвуют в посттрансляционной активности (например, полинуклеотиды, отщепляемые от него с оставлением зрелой формы белка или пептида).
"Природный фермент" и "природный белок" обозначает фермент или белок, имеющий немодифицированную аминокислотную последовательность, идентичную последовательности, обнаруживаемой в природе. Природные ферменты включают нативные ферменты, ферменты, природно экспрессируемые или обнаруживаемые в конкретном микроорганизме.
Термины "произведенные из" и "полученные из" относятся не только к ферменту (например, протеазе, продуцированной или продуцируемой рассматриваемым штаммом организма), но также к ферменту, кодируемому ДНК-последовательностью, выделенной из такого штамма и продуцируемой в организме-хозяине, содержащем такую ДНК-последовательность. Кроме того, этот термин относится к ферменту, который кодируется ДНК-последовательностью синтетического и/или кДНК-происхождения и который имеет идентифицирующие характеристики рассматриваемого фермента.
"Производное" в объеме этого определения обычно сохраняет характерную протеолитическую активность, наблюдаемую в нативной или исходной форме дикого типа, до такой степени, что это производное применимо для тех же целей, для которых используют нативную или исходную форму дикого типа. Функциональные производные фермента включают природные, синтетически или рекомбинантно полученные пептиды или фрагменты пептидов, имеющие общие характеристики исходного фермента.
Термин "функциональное производное" относится к производному нуклеиновой кислоты, имеющему функциональные характеристики нуклеиновой кислоты, кодирующей фермент. Функциональные производные нуклеиновой кислоты, кодирующей фермент по данному изобретению, включают природные аллели и гомологи на основе вырожденности генетического кода, известные в данной области.
Термин "идентичные" в контексте двух нуклеиновых кислот или полипептидных последовательностей относится к остаткам в этих двух последовательностях, которые являются одинаковыми при сопоставлении для максимального соответствия, как измерено с использованием следующих алгоритмов сравнения или анализа последовательностей.
Термин "оптимальное выравнивание" относится к выравниванию, дающему наивысший балл процентной идентичности. "Процентная идентичность последовательности", "процентная идентичность аминокислотной последовательности", "процентная идентичность последовательности гена" и/или “процентная идентичность последовательности нуклеиновой кислоты/полинуклеотидной последовательности" в отношении двух аминокислотных, полинуклеотидных и/или генных последовательностей (соответственно) обозначают процент остатков, которые являются идентичными в этих двух последовательностях при оптимальном сопоставлении этих последовательностей. Таким образом, 80% идентичность аминокислотной последовательности означает, что 80% аминокислот в двух оптимально сопоставленных полипептидных последовательностях являются идентичными.
Так, фраза "по существу идентичные" в контексте двух нуклеиновых кислот или полипептидов относится к полинуклеотиду или полипептиду, которые имеют по меньшей мере 70% идентичность последовательности, предпочтительно по меньшей мере 80%, предпочтительно по меньшей мере 85%, предпочтительно по меньшей мере 90%, предпочтительно по меньшей мере 95%, предпочтительно по меньшей мере 97%, предпочтительно по меньшей мере 98% и предпочтительно по меньшей мере 99% идентичность последовательности в сравнении со ссылочной последовательностью, с использованием программ или алгоритмов (например, BLAST, ALIGN, CLUSTAL) при использовании стандартных параметров. Одним указанием на то, что два полипептида являются по существу идентичными, является то, что первый полипептид является иммунологически перекрестно-реактивным со вторым полипептидом. Обычно, полипептиды, которые различаются консервативными аминокислотными заменами, являются иммунологически перекрестно-реактивными. Таким образом, полипептид по существу идентичен второму полипептиду, например, когда эти два пептида различаются только одной консервативной заменой. Другим указанием на то, что две последовательности нуклеиновых кислот являются по существу идентичными, является то, что эти две молекулы гибридизуются друг с другом при строгих условиях в диапазоне от средней до высокой строгости.
Термин "выделенный" или "очищенный" относится к материалу, который удален из его исходного окружения (например, природного окружения, если он является природным). Например, говорят, что материал является "очищенным", когда он присутствует в конкретной композиции в более высокой или в более низкой концентрации, чем концентрация, которая существует в природном организме или организме дикого типа, или в комбинации с компонентами, в норме не присутствующими после экспрессии из природного организма или организма дикого типа. Например, природный полинуклеотид или полипептид, присутствующий в живом животном, не является выделенным, но тот же самый полинуклеотид или полипептид, отделенный от некоторых или всех из сосуществующих материалов в природной системе, является выделенным. В некоторых вариантах осуществления, такие полинуклеотиды являются частью вектора и/или такие полинуклеотиды или полипептиды являются частью композиции и все еще являются выделенными в том смысле, что такой вектор или такая композиция не являются частью его природного окружения. В некоторых предпочтительных вариантах осуществления, говорят, что нуклеиновая кислота или белок являются очищенными, если они приводят к появлению по существу одной полосы в электрофоретическом геле или блоте.
Термин "выделенная" при применении в ссылке на ДНК-последовательность относится к ДНК-последовательности, которая была удалена из ее природной генетической среды и, следовательно, не содержит других посторонних или нежелательных кодирующих последовательностей и находится в форме, подходящей для применения в полученных генной инженерией системах получения белков. Такие выделенные молекулы являются молекулами, которые отделены от их природного окружения и включают кДНК- и геномные клоны. Выделенные ДНК-молекулы данного изобретения не содержат других генов, с которыми они обычно ассоциированы, но могут включать природные 5'- и 3'-нетранслируемые области, такие как промоторы и терминаторы. Идентификация ассоциированных областей будет очевидна для специалиста с обычной квалификацией в данной области (см., например, Dynan and Tijan, Nature 316:774-78, 1985). Термин "выделенная ДНК-последовательность" альтернативно называют "клонированной ДНК-последовательностью".
Термин "выделенный", при ссылке на белок, относится к белку, который обнаружен в состоянии, другом, чем его природное окружение. В предпочтительной форме, этот выделенный белок не содержит других белков, в частности других гомологичных белков. Выделенный белок имеет чистоту более 10%, предпочтительно более 20% и даже более предпочтительно более 30%, как определено электрофорезом в ДСН-ПААГ. Дополнительные аспекты этого изобретения включают белок в высокоочищенной форме (т.е. с чистотой более 40%, более 60%, более 80%, более 90%, более 95%, более 97% и даже более 99%), как определено электрофорезом в ДСН-ПААГ.
В данном контексте, термин "комбинаторный мутагенез" относится к способам, в которых генерируют библиотеки вариантов исходной последовательности. В этих библиотеках, эти варианты содержат одну или несколько мутаций, выбранных из предварительно определенного набора мутаций. Кроме того, эти способы обеспечивают средства для введения случайных мутаций, которые не были членами предварительно определенного набора мутаций. В некоторых вариантах осуществления, эти способы включают способы, представленные в Заявке на патент США № 09/699250, поданной 26 октября 2000 года, включенной здесь в качестве ссылки. В альтернативных вариантах осуществления, способы комбинаторного мутагенеза включают коммерчески доступные наборы (например, QUIKCHANGE® Multisite, Stratagene, La Jolla, CA).
В данном контексте, термин "библиотека мутантов" относится к популяции клеток, которые являются идентичными в большей части их генома, но включают различные гомологи одного или нескольких генов. Такие библиотеки могут быть использованы, например, для идентификации генов или оперонов с улучшенными свойствами.
В данном контексте, термин "исходный ген" относится к представляющему интерес гену, который кодирует представляющий интерес белок, который должен быть улучшен и/или изменен с использованием данного изобретения.
В данном контексте, термин "вариант" относится к белку, который был получен из белка-предшественника (например, "исходного" белка) добавлением, заменой или делецией одной или нескольких аминокислот. В некоторых вариантах осуществления, вариант содержит по меньшей мере одну модификацию, которая включает изменение заряда по сравнению с белком-предшественником. В некоторых вариантах осуществления, этот белок-предшественник является исходным белком, который является белком дикого типа.
В данном контексте, термины "выравнивание множественных последовательностей" и "MSA" относятся к последовательностям множественных гомологов исходного гена, которые выравнивают при помощи алгоритма (например, Clustal W).
В данном контексте, термины "консенсусная последовательность" и "каноническая последовательность" относятся к архетипичной аминокислотной последовательности, с которой сравнивают все варианты конкретных представляющих интерес белков или последовательностей. Эти термины относятся также к последовательности, которая представляет нуклеотиды, которые наиболее часто представлены в представляющей интерес ДНК-последовательности. Для каждого положения гена, эта консенсусная последовательность дает аминокислоту, которая является наиболее часто представленной в этом положении в MSA.
В данном контексте, термин "консенсусная мутация" относится к различию в последовательности исходного гена и консенсусной последовательности. Консенсусные мутации идентифицируют сравнением последовательностей исходного гена и консенсусной последовательности, полученной из MSA. В некоторых вариантах осуществления, консенсусные мутации вводят в исходный ген, так что он становится более сходным с этой консенсусной последовательностью. Консенсусные мутации включают также изменения аминокислот, которые изменяют аминокислоту в исходном гене на аминокислоту, которая наиболее часто обнаруживается в MSA в этом положении относительно частоты этой аминокислоты в исходном гене. Таким образом, термин консенсусная мутация включает все изменения единственной аминокислоты, которые заменяют аминокислоту исходного гена аминокислотой, которая чаще представлена, чем эта аминокислота в MSA.
В данном контексте, термин "начальное попадание" (начальный хит) относится к варианту, который был идентифицирован скринингом комбинаторной библиотеки консенсусного мутагенеза. В предпочтительных вариантах осуществления, начальные попадания (хиты) имеют улучшенные характеристики эффективности в сравнении с исходным геном.
В данном контексте, термин "улучшенное попадание" (улучшенный хит) относится к варианту, который идентифицировали скринингом увеличенной комбинаторной библиотеки консенсусного мутагенеза.
В данном контексте, термины "улучшающая мутация" и "увеличивающая эффективность мутация" относятся к мутации, которая приводит к улучшенной эффективности при введении в исходный ген. В некоторых предпочтительных вариантах осуществления, эти мутации идентифицируют секвенированием попаданий (хитов), идентифицированных во время стадии скрининга этого способа. В большинстве вариантов осуществления, мутации, которые наиболее часто обнаруживаются в хитах, являются, по-видимому, улучшающими мутациями в сравнении с не подвергнутой скринингу комбинаторной библиотеки консенсусного мутагенеза.
В данном контексте, термин "увеличенная комбинаторная библиотека консенсусного мутазенеза" относится к CCM-библиотеке, которая спроектирована и сконструирована на основе результатов скрининга и/или секвенирования из более раннего раунда CCM-мутагенеза и скрининга. В некоторых вариантах осуществления, эта увеличенная CCM-библиотека основана на последовательности начального попадания (хита), происходящего из более раннего раунда CCM. В дополнительных вариантах осуществления, эта увеличенная CCM спроектирована так, что мутации, которые часто наблюдались в начальных попаданиях (хитах) из более ранних раундов мутагенеза и скрининга, являются наиболее предпочтительными. В некоторых предпочтительных вариантах осуществления, это осуществляется удалением из применения праймеров, которые кодируют уменьшающие эффективность мутации, или увеличением концентрации праймеров, которые кодируют увеличивающие эффективность мутации, относительно других праймеров, которые использовали в более ранних ССМ-библиотеках.
В данном контексте, термин "уменьшающие эффективность мутации" относится к мутациям в комбинаторной библиотеке консенсусного мутагенеза, которые менее часто обнаруживаются в попаданиях (хитах), происходящих из скрининга, в сравнении с не подвергнутой скринингу комбинаторной библиотеки консенсусного мутагенеза. В предпочтительных вариантах осуществления, процесс скрининга удаляет и/или уменьшает обилие вариантов, которые содержат "уменьшающие эффективность мутации".
В данном контексте, термин "функциональный анализ" относится к анализу, который обеспечивает указание на активность белка. В особенно предпочтительных вариантах осуществления, этот термин относится к системам анализов, в которых белок анализируют на его способность функционировать в его обычной компетенции. Например, в случае ферментов, функциональный анализ включает определение эффективности фермента в катализе реакции.
В данном контексте, термин "свойство-мишень" относится к свойству исходного гена, которое должно быть изменено. Не предполагается, что данное изобретение ограничивается каким-либо конкретным свойством-мишенью. Однако, в некоторых предпочтительных вариантах осуществления, этим свойством-мишенью является стабильность продукта гена (например, устойчивость к денатурации, протеолизу или другим деградирующим факторам), в то время как в других вариантах осуществления, изменяется уровень получения в хозяине-продуценте. Фактически, предполагается, что любое свойство исходного гена найдет применение в данном изобретении.
Термин "свойство" или его грамматические эквиваленты в контексте нуклеиновой кислоты, в применении здесь, относится к любой характеристике или любому признаку нуклеиновой кислоты, которые могут быть выбраны или детектированы. Эти свойства включают, но не ограничиваются ими, свойство, влияющее на связывание с полипептидом, свойство, сообщаемое клетке, содержащей конкретную нуклеиновую кислоту, свойство, влияющее на транскрипцию гена (например, силу промотора, узнавание промотора, регуляцию промотора, энхансерную функцию), свойство, влияющее на процессинг РНК (например, сплайсинг РНК, стабильность РНК, конформацию РНК и посттрансляционную модификацию), свойство, влияющее на трансляцию (например, уровень, регуляцию, связывание мРНК с рибосомными белками, посттрансляционную модификацию). Например, сайт связывания фактора транскрипции, полимеразы, регуляторного фактора и т.д. нуклеиновой кислоты может быть изменен для получения желаемых характеристик или для идентификации нежелательных характеристик.
Термины "свойство", "представляющее интерес свойство" или их грамматические эквиваленты в контексте полипептида, в применении здесь, относятся к любой характеристике или любому признаку полипептида, которые могут быть выбраны или детектированы. Эти свойства включают, но не ограничиваются ими, оксилительную стабильность, субстратную специфичность, каталитическую активность, теплоустойчивость, устойчивость к щелочи, профиль активности рН, устойчивость к протеолитической деградации, KM, kcat, отношение kcat/kM, фолдинг белка, индукцию иммунной реакции, способность связываться с лигандом, способность связываться с рецептором, способность секретироваться, способность размещаться на поверхности клетки, способность к олигомеризации, способность передавать сигнал, способность стимулировать пролиферацию клеток, способность ингибировать пролиферацию клеток, способность индуцировать апоптоз, способность подвергаться модификации фосфорилированием или гликозилированием, способность лечить заболевание.
В данном контексте, термин "скрининг" имеет его обычное значение в данной области и является обычно многостадийным процессом. В первой стадии, обеспечивают мутантную нуклеиновую кислоту или вариантный полипептид, полученный из нее. Во второй стадии, определяют свойство мутантной нуклеиновой кислоты или вариантного полипептида. В третьей стадии, это определенное свойство сравнивают со свойством соответствующей исходной нуклеиновой кислоты, со свойством соответствующего природного полипептида или со свойством исходного материала (например, первоначальной последовательности) для генерирования мутантной нуклеиновой кислоты.
Квалифицированному в данной области специалисту будет понятно, что процедура скрининга для получения нуклеиновой кислоты или белка с измененным свойством зависит от конкретного свойства исходного материала, модификацию которого должно облегчить генерирование мутантной нуклеиновой кислоты. Таким образом, квалифицированному в данной области специалисту будет понятно, что это изобретение не ограничивается каким-либо конкретным свойством для скрининга и что следующее описание свойств перечисляет только иллюстративные примеры. Способы скрининга на любое конкретное свойство обычно описаны в данной области. Например, можно измерять связывание, рН, специфичность и т.д. до и после мутации, где перемена свидетельствует об изменении. Предпочтительно, эти скрининги выполняют высокопроизводительным способом, включающим многочисленные пробы, подвергаемые одновременному скринингу, в том числе, но не только, анализы, использующие чипы (микропроцессоры), фаговый дисплей и множественные субстраты и/или индикаторы.
В данном контексте, в некоторых вариантах осуществления, скрининги включают стадии отбора, в которых представляющие интерес варианты обогащаются из популяции вариантов. Примеры этих вариантов осуществления включают выбор вариантов, которые придают преимущество роста организму-хозяину, а также фаговый дисплей или любой другой способ дисплея, где варианты могут захватываться из популяции вариантов на основе их связывающих или каталитических свойств. В предпочтительном варианте осуществления, библиотеку вариантов подвергают действию стресса (тепла, протеазы, денатурации) и затем варианты, которые являются все еще интактными, идентифицируют в скрининге или обогащают посредством отбора. Предполагается, что этот термин включает любые подходящие средства для отбора. Фактически не предполагается, что данное изобретение ограничивается каким-либо конкретным способом скрининга.
В данном контексте, термин "нацеленная рандомизация" относится к процессу, который производит множество последовательностей, где одно или несколько положений были рандомизированы. В некоторых вариантах осуществления, рандомизация является полной (т.е. все четыре нуклеотида, A, T, G и C, могут встречаться в рандомизированном положении. В альтернативных вариантах осуществления, рандомизация нуклеотида ограничена субпопуляцией этих четырех нуклеотидов. Нацеленная рандомизация может быть применена к одному или нескольким кодонам последовательности, кодирующей один или несколько представляющих интерес белков. При экспрессии полученные библиотеки продуцируют популяции белков, в которых одно или несколько положений аминокислот могут содержать смесь из всех 20 аминокислот или субпопуляцию аминокислот, определяемую схемой рандомизации рандомизируемого кодона. В некоторых вариантах осуществления, индивидуальные члены популяции, происходящей из нацеленной рандомизации, различаются в количестве аминокислот вследствие нацеленной или случайной инсерции или делеции кодонов. В дополнительных вариантах осуществления в полученные популяции белков включают синтетические аминокислоты. В некоторых предпочтительных вариантах осуществления, большинство членов популяции, происходящей из нацеленной рандомизации, обнаруживают более высокую гомологию последовательности с консенсусной последовательностью, чем исходный ген. В некоторых вариантах осуществления, эта последовательность кодирует один или несколько представляющих интерес белков. В альтернативных вариантах осуществления, эти белки имеют различающиеся биологические функции. В некоторых предпочтительных вариантах осуществления, входящая последовательность содержит по меньшей мере один селектируемый маркер.
Термины "модифицированная последовательность" и "модифицированные гены" используются взаимозаменяемо здесь для обозначения последовательности, которая включает делецию, инсерцию или прерывание природной последовательности нуклеиновой кислоты. В некоторых предпочтительных вариантах осуществления, продуктом экспрессии этой модифицированной последовательности является укороченный белок (например, если модификация является делецией или прерыванием этой последовательности). В некоторых особенно предпочтительных вариантах осуществления, этот укороченный белок сохраняет биологическую активность. В альтернативных вариантах осуществления, продуктом экспрессии модифицированной последовательности является удлиненный белок (например, модификации, включающие инсерцию в последовательность нуклеиновой кислоты). В некоторых вариантах осуществления, инсерция приводит к укороченному белку (например, когда инсерция приводит к образованию стоп-кодона). Таким образом, инсерция может приводить либо к укороченному белку, либо к удлиненному белку в качестве продукта экспрессии.
В данном контексте, термины "мутантная последовательность" и "мутантный ген" являются взаимозаменяемыми и относятся к последовательности, которая имеет изменение по меньшей мере в одном кодоне, встречающемся в последовательности дикого типа клетки-хозяина. Продуктом экспрессии мутантной последовательности является белок с измененной аминокислотной последовательностью относительно дикого типа. Этот продукт экспрессии может иметь измененную функциональную способность (например, увеличенную ферментативную активность).
Термины "мутагенный праймер" или "мутагенный олигонуклеотид" (используемые здесь взаимозаменяемо) относятся к олигонуклеотидным композициям, которые соответствуют части матричной последовательности и которые способны гибридизоваться с этой матричной последовательностью. Что касается мутагенных праймеров, этот праймер будет точно соответствовать матричной нуклеиновой кислоте, причем ошибочное спаривание или ошибочные спаривания в этом праймере используют для введения желаемой мутации в библиотеку нуклеиновых кислот. В данном контексте, "немутагенный праймер" или "немутагенный олигонуклеотид" обозначают олигонуклеотидные композиции, которые точно соответствуют матричной нуклеиновой кислоте. В одном варианте этого изобретения, используют только мутагенные праймеры. В другом предпочтительном варианте осуществления этого изобретения, праймеры сконструированы таким образом, что по меньшей мере для одной области, в которую был включен мутагенный праймер, имеется также немутагенный праймер, включенный в олигонуклеотидную смесь. Посредством добавления смеси мутагенных праймеров и немутагенных праймеров, соответствующих по меньшей мере одному из мутагенных праймеров, можно производить полученную библиотеку нуклеиновых кислот, в которой представлено разнообразие комбинаторных мутационных паттернов. Например, если желательно, чтобы некоторые члены библиотеки мутантных нуклеиновых кислот сохранили их исходную последовательность в определенных положениях, тогда как другие члены являются мутантными в таких сайтах, эти немутагенные праймеры обеспечивают способность получения конкретного уровня немутантных членов в этой библиотеке нуклеиновых кислот для конкретного остатка. Способы этого изобретения используют мутагенные и немутагенные олигонуклеотиды, которые обычно имеют длину 10-50 оснований, более предпочтительно приблизительно 15-45 оснований. Однако может быть необходимо использовать праймеры, которые являются либо более короткими, чем 10 оснований, либо более длинными, чем 50 оснований, для получения желаемого результата мутагенеза. Что касается соответствующих мутагенных и немутагенных праймеров, необходимо, чтобы соответствующие олигонуклеотиды имели идентичную длину, но только чтобы имелось перекрывание в области, соответствующей добавляемой мутации.
В некоторых вариантах осуществления, праймеры добавляют в предварительно определенном соотношении. Например, если желательно, чтобы полученная библиотека имела значительный уровень определенной специфической мутации и меньшее количество другой мутации в том же самом или другом сайте, коррекцией количества добавляемого праймера можно получить желаемую смещенную библиотеку. Альтернативно, добавлением меньших или больших количеств немутагенных праймеров, можно корректировать частоту, с которой продуцируются соответствующие мутации в библиотеке мутантных нуклеиновых кислот.
В данном контексте, фраза "смежные мутации" относится к мутациям, которые представлены в пределах одного и того же олигонуклеотидного праймера. Например, смежные мутации могут быть смежными или находиться вблизи друг друга, однако они будут вводиться в полученные мутантные матричные нуклеиновые кислоты одним и тем же праймером.
В данном контексте, фраза "несмежные мутации" относится к мутациям, которые представлены на разных олигонуклеотидых праймерах. Например, несмежные мутации будут вводиться в полученные мутантные матричные нуклеиновые кислоты отдельно приготовленными олигонуклеотидными праймерами.
Термины "последовательность дикого типа," "последовательность нуклеиновой кислоты дикого типа" и "ген дикого типа" используются здесь взаимозаменяемо, в отношении последовательности, которая является нативной или природной в клетке-хозяине. В некоторых вариантах осуществления, последовательностью дикого типа называют представляющую интерес последовательность, которая является стартовой точкой конструирующего белок проекта. Последовательность дикого типа может кодировать либо гомологичный, либо гетерологичный белок. Гомологичным белком является белок, который клетка-хозяин могла бы продуцировать без вмешательства. Гетерологичным белком является белок, который клетка-хозяин могла бы производить, но вследствие вмешательства.
Термин "устойчивые к окислению" относится к протеазам данного изобретения, которые сохраняют указанное количество ферментативной активности на протяжении конкретного периода времени при условиях, преобладающих во время протеолитического, гидролизующего, очищающего или другого процесса этого изобретения, например при воздействии отбеливающих агентов или окисляющих агентов или контактировании с отбеливающими или окисляющими агентами. В некоторых вариантах осуществления, эти протеазы сохраняют по меньшей мере приблизительно 50%, приблизительно 60%, приблизительно 70%, приблизительно 75%, приблизительно 80%, приблизительно 85%, приблизительно 90%, приблизительно 92%, приблизительно 95%, приблизительно 96%, приблизительно 97%, приблизительно 98% или приблизительно 99% протеолитической активности после контакта с отбеливающим или окисляющим агентом на протяжении конкретного периода времени, например по меньшей мере 1 минуты, 3 минут, 5 минут, 8 минут, 12 минут, 16 минут, 20 минут и т.д.
Термин "устойчивые к хелатору" относится к протеазам данного изобретения, которые сохраняют указанное количество ферментативной активности на протяжении конкретного периода времени при условиях, преобладающих во время протеолитического, гидролизующего, очищающего или другого процесса этого изобретения, например при воздействии хелатообразующих агентов (хелаторов) или контактировании с хелатообразующими агентами (хелаторами). В некоторых вариантах осуществления, эти протеазы сохраняют по меньшей мере приблизительно 50%, приблизительно 60%, приблизительно 70%, приблизительно 75%, приблизительно 80%, приблизительно 85%, приблизительно 90%, приблизительно 92%, приблизительно 95%, приблизительно 96%, приблизительно 97%, приблизительно 98% или приблизительно 99% протеолитической активности после контакта с хелатообразующим агентом на протяжении конкретного периода времени, например по меньшей мере 10 минут, 20 минут, 40 минут, 60 минут, 100 минут и т.д.
Термины "термически стабильные" и "термостабильные" относится к протеазам данного изобретения, которые сохраняют указанное количество ферментативной активности после подвергания действию идентифицированных температур на протяжении конкретного времени при условиях, преобладающих во время протеолитического, гидролизующего, очищающего или другого процесса этого изобретения, например под воздействием измененных температур. Измененные температуры включают повышенные и пониженные температуры. В некоторых вариантах осуществления, эти протеазы сохраняют по меньшей мере приблизительно 50%, приблизительно 60%, приблизительно 70%, приблизительно 75%, приблизительно 80%, приблизительно 85%, приблизительно 90%, приблизительно 92%, приблизительно 95%, приблизительно 96%, приблизительно 97%, приблизительно 98% или приблизительно 99% протеолитической активности после подвергания действию измененных температур на протяжении конкретного периода времени, например по меньшей мере 60 минут, 120 минут, 180 минут, 240 минут, 300 минут и т.д.
Термин "увеличенная стабильность" в контексте устойчивой к окислению, хелатору, термостабильной и/или рН-стабильной протеазы относится к более высокой сохраненной протеолитической активности на протяжении времени в сравнении с другими сериновыми протеазами (например, протеазами типа субтилизина) и/или ферментами дикого типа.
Термин "уменьшенная стабильность" в контексте устойчивой к окислению, хелатору, термостойкой и/или рН-стабильной протеазы относится к меньшей сохраненной протеолитической активности на протяжении времени в сравнении с другими сериновыми протеазами (например, протеазами типа субтилизина) и/или ферментами дикого типа.
В данном контексте, термин "очищающая композиция" включает, если нет других указаний, гранулированные или порошкообразные моющие агенты для всех целей или имеющие большую мощность, в частности моющие детергенты; жидкие, гелеобразные или имеющие форму пасты моющие агенты для всех целей, в частности так называемые жидкие типы большой мощности; детергенты для тонких («деликатных») тканей; агенты для ручной мойки посуды или легкие (маломощные) агенты для мойки посуды, в частности средства с высоким пенообразованием; агенты для машинной мойки посуды, в том числе различные таблетки, гранулированные, жидкие и предназначенные для ополаскивания типы для домашнего применения и для применения в учреждениях; жидкие чистящие и дезинфицирующие агенты, в том числе антибактериальные типы для мытья рук, чистящие бруски, жидкости для промывания полости рта, моющие средства для зубных протезов, шампуни для машин и ковров, моющие средства для ванных комнат; шампуни для волос и ополаскиватели для волос; гели для душа, пены для ванн и чистящие средства для металлов; а также вспомогательные средства, такие как отбеливающие добавки и «пятновыводящий карандаш» или средства для предварительной обработки.
Если нет других указаний, уровни всех компонентов и композиций приводятся в виде ссылки на активный уровень компонентов или композиций и исключают примеси, например остаточные растворители или побочные продукты, которые могут присутствовать в коммерчески доступных источниках.
Массы ферментных компонентов основаны на общем активном белке. Все проценты и отношения рассчитаны на массу, если нет других указаний. Все проценты и отношения рассчитаны на основе всей композиции, если нет других указаний.
Термин "очищающая активность" относится к эффективности очистки, достигаемой протеазой при условиях, преобладающих во время протеолитического, гидролизующего, очищающего или другого процесса этого изобретения. В некоторых вариантах осуществления, эффективность очистки определяют с применением различных анализов очистки относительно чувствительных к ферментам пятен, например, травы, крови, молока или яичного белка, с использованием различных хроматографических, спектрофотометрических или других количественных методологий после подвергания этих пятен стандартным моющим условиям. Примеры анализов включают, но не ограничиваются ими, анализы, описанные в WO 99/34011 и Патент США № 6605458 (оба из которых включены здесь в качестве ссылки), а также способы, включенные в раздел примеров.
Термин "эффективное для очистки количество" протеазы относится к количеству описанной здесь выше протеазы, которое достигает желаемого уровня ферментативной активности в конкретной очищающей композиции. Такие эффективные количества могут быть легко получены специалистом с обычной квалификацией в данной области, и они основаны на многих факторах, таких как конкретная используемая протеаза, конкретное применение для очистки, конкретный состав чистящей композиции, и от того, требуется ли жидкая или сухая (например, гранулированная, в виде бруска) композиция, и т.д.
Термин "дополнительные чистящие материалы" означает, в данном контексте, любой жидкий, твердый или газообразный материал, выбранный для конкретного типа желаемой чистящей композиции и формы этого продукта (например, жидкой, гранулированной, порошкообразной, в виде бруска, пасты, спрея, таблетки, геля; или пенообразующей композиции), причем эти материалы предпочтительно также совместимы с протеазным ферментом, используемым в этой композиции. В некоторых вариантах осуществления, гранулированные композиции находятся в «компактной» форме, в то время как в других вариантах осуществления эти жидкие композиции находятся в «концентрированной» форме.
Термины "увеличенная эффективность" и "улучшенная моющая эффективность" в контексте очищающей активности относятся к увеличенной или более высокой очищающей активности в отношении некоторых чувствительных к ферментам пятен, таких как яйцо, молоко, трава или кровь, определяемой обычным оцениванием после одного стандартного моющего цикла и/или множественных моющих циклов.
Термин "уменьшенная эффективность" в контексте очищающей активности относится к уменьшенной или меньшей очищающей активности в отношении некоторых чувствительных к ферментам пятен, таких как яйцо, молоко, трава или кровь, определяемой обычным оцениванием после одного стандартного моющего цикла.
Термин "сравнительная эффективность" в контексте очищающей активности относится по меньшей мере к 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95% очищающей активности сравнительной протеазы (например, коммерчески доступных протеаз). Эффективность очистки может быть определена сравнением протеаз этого изобретения в различных анализах очистки относительно чувствительных к ферментам пятен, таких как кровь, молоко и/или чернила (BMI) с использованием определения обычными спектрофотометрическими или аналитическими методологиями после стандартных условий моющего цикла.
В данном контексте, система "с низкой концентрацией детергента" включает детергенты, в которых менее чем приблизительно 800 м.д. детергентных компонентов присутствуют в моющей воде. Японские детергенты обычно считаются системами с низкой концентрацией детергента, так как они обычно имеют приблизительно 667 м.д. детергентных компонентов в моющей воде.
В данном контексте, системы "со средней концентрацией детергента" включают детергенты, в которых приблизительно 800 м.д. - приблизительно 2000 м.д. детергентных компонентов присутствуют в моющей воде. Североамериканские детергенты обычно считаются системами со средней концентрацией детергента, так как они имеют обычно приблизительно 975 м.д. детергентных компонентов в моющей воде. Бразильские детергенты обычно имеют приблизительно 1500 м.д. детергентных компонентов, присутствующих в моющей воде.
В данном контексте, система "с высокой концентрацией детергента" включает детергенты, в которых менее чем приблизительно 2000 м.д. детергентных компонентов присутствуют в моющей воде. Европейские детергенты обычно считаются системами с высокой концентрацией детергента, так как они обычно имеют приблизительно 3000-8000 м.д. детергентных компонентов в моющей воде.
В данном контексте, "очищающие ткани композиции" включают детергентные композиции для ручной и машинной стирки, включающие дополнительные композиции для стирки и композиции, подходящие для использования в замачивании и/или предварительной обработке имеющих пятна тканей (например, скатертей, белья и других текстильных материалов).
В данном контексте, "очищающие композиции не для тканей" включают очищающие композиции не для текстиля (т.е. не для тканей), включающие, но не ограничивающиеся ими, детергентные композиции для мойки посуды, очищающие композиции для полости рта, очищающие композиции для зубных протезов и очищающие композиции для личной гигиены.
"Компактная" форма описанных здесь очищающих композиций лучше всего характеризуется плотностью и, в отношении состава, количеством неорганической соли в качестве наполнителя. Неорганические соли в качестве наполнителей являются общепринятыми ингредиентами детергентных композиций в порошкообразной форме. В общепринятых детергентных композициях, эти соли-наполнители присутствуют в существенных количествах, обычно 17-35% по массе общей композиции. В отличие от этого, в компактных композициях соль-наполнитель присутствует в количествах, не превышающих 15% общей композиции. В некоторых вариантах осуществления, соль-наполнитель присутствует в количествах, которые не превышают 10% или, более предпочтительно 5% по массе этой композиции. В некоторых вариантах осуществления, эти неорганические соли-наполнители выбраны из сульфатных и хлоридных солей щелочных или щелочно-земельных металлов. Предпочтительной солью-наполнителем является сульфат натрия.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Данное изобретение относится к способам конструирования белков для оптимизации их эффективности при определенных представляющих интерес условиях окружающей среды. В некоторых вариантах осуществления, данное изобретение относится к способам конструирования ферментов для оптимизации их каталитической активности при конкретных условиях окружающей среды. В некоторых предпочтительных вариантах осуществления, данное изобретение относится к способам изменения суммарного заряда поверхности и/или распределения заряда поверхности ферментов (например, металлопротеаз или сериновых протеаз) для получения вариантов ферментов, которые демонстрируют улучшенную эффективность в детергентных препаратах в сравнении с первоначальным или исходным ферментом.
В некоторых предпочтительных вариантах осуществления, данное изобретение обеспечивает способы и композиции, содержащие по меньшей мере одну вариантную нейтральную металлопротеазу и/или вариантную сериновую протеазу, которая улучшала моющую эффективность по меньшей мере в одном детергентном препарате. В некоторых особенно предпочтительных вариантах осуществления, данное изобретение обеспечивает варианты нейтральной металлопротеазы Bacillus amyloliquefaciens. В других особенно предпочтительных вариантах осуществления, данное изобретение обеспечивает варианты сериновой протеазы изолята Cellulomonas bogoriensis 69B4. Данное изобретение находит применение в приложениях, включающих, но не ограничивающихся ими, очистку, отбеливание и дезинфицирование. Кроме того, данное изобретение относится к способам конструирования фермента для оптимизации его каталитической активности при неблагоприятных условиях окружающей среды. В частности, данное изобретение относится к способам изменения суммарного заряда поверхности и/или распределения заряда поверхности металлопротеазы или сериновой протеазы для получения вариантов ферментов, которые демонстрируют улучшенную эффективность в детергентных препаратах.
Многие белки и ферменты являются высокочувствительными к денатурации и подвергаются необратимой денатурации при хранении в детергентах (моющих средствах) для стирки. Известно, что детергенты для стирки содержат анионогенные, катионогенные и неионогенные поверхностно-активные вещества, где это поверхностно-активное вещество классифицируется по его ионогенным свойствам (электрическому заряду) в воде. Эти ингредиенты могут взаимодействовать с зарядом поверхности молекулы белка, что приводит к денатурации белка (например, потере структуры и функции).
Было показано, что две протеазы, ASP (сериновая протеаза) и NprE (нейтральная металлопротеаза) являются в высокой степени нестабильными при хранении в детергентном препарате, включающем такое поверхностно-активное вещество, как LAS. LAS является анионогенным поверхностно-активным веществом, в котором общий отрицательный заряд усиливает взаимодействие с положительно заряженными боковыми цепями аминокислот, локализованных на поверхности белка. Такие электростатические взаимодействия влияют на внутреннюю стабильность белка ослаблением или разрушением стабилизирующих электростатических взаимодействий. Затем дестабилизированный белок разворачивается и становится неактивным.
Было обнаружено, что распределение заряженных остатков на поверхности протеазы в сильной степени влияет на моющую эффективность. Способы конструирования белков данного изобретения эффективно оптимизируют протеазы в отношении увеличенной эффективности в одном или нескольких свойствах в детергентных препаратах, оптимизацией суммарного заряда поверхности и/или распределения заряда поверхности этой протеазы. Хотя для иллюстрации способов, обеспечиваемых данным изобретением, используют металлопротеазу и сериновую протеазу, не предполагается, что данное изобретение ограничивается этими конкретными ферментами. Фактически, данное изобретение находит применение с различными ферментами и другими белками.
Вкратце, в некоторых вариантах осуществления данного изобретения, эти способы включают создание сайт-оценивающих (site-evaluation) библиотек в ряде аминокислотных остатков в представляющем интерес ферменте и анализ вариантных ферментов на представляющие интерес свойства. Это позволяет идентифицировать полезные, нейтральные и вредные мутации, а также оптимальное изменение заряда (относительно исходного фермента) для представляющего интерес свойства (представляющих интерес свойств). В некоторых альтернативных вариантах осуществления, получают сканы зарядов всех этих остатков для генерирования вариантов с мутациями, которые изменяют заряд в каждом сайте (например, мутируют нейтральные остатки в положительные и/или отрицательные остатки и мутируют заряженные остатки в противоположно заряженные и/или нейтральные остатки). В некоторых дополнительных предпочтительных вариантах осуществления, эти способы включают создание комбинаторных библиотек со “сбалансированным зарядом”, которые включают полезные мутации, которые изменяют заряд фермента в желаемом направлении, и полезные или нейтральные мутации, которые изменяют этот заряд в противоположном направлении, и затем анализ библиотеки со сбалансированным зарядом на представляющее интерес свойство (на представляющие интерес свойства). Таким образом, заряд поверхности и распределение заряда поверхности фермента одновременно оптимизируются, и можно идентифицировать варианты фермента, имеющие улучшения множественных свойств.
Способы данного изобретения находят применения в улучшении эффективности различных классов ферментов, а также протеаз (например, амилаз, целлюлаз, оксидаз, кутиназ, маннаназ, пектиназ, амилаз, липаз и т.д.). Фактически, не предполагается, что данное изобретение ограничивается каким-либо конкретным ферментом или классом ферментов. Кроме того, данное изобретение находит применение в оптимизации свойств неферментативных белков, которые требуют конкретного заряда поверхности и распределения заряда (например, экспрессии, связывания с клеточной поверхностью, пригодности для приготовления в виде препарата и т.д.).
I. Получение вариантов ASP с улучшенными свойствами
Конструировали сайт-оценивающие библиотеки (SEL), в которых каждую аминокислоту зрелого белка заменяли большинством других аминокислот (см., например, Заявку на патент США с порядковым номером 10/576331 и WO 2005/052146, которые включены здесь в качестве ссылки, так как они имеют отношение к SEL). Затем отдельные мутации, которые улучшают эффективность, объединяли и повторно тестировали на эффективность. Последующий анализ SEL и результатов объединения мутантов выявил существенное ослабление эффективности удаления пятен вследствие изменений в заряде поверхности этой молекулы.
После определения действия заряда поверхности на эффективность удаления пятен ASP в детергенте конструировали определенную библиотеку, в которой заряд поверхности молекулы систематически варьировали и определяли эффективность этих вариантов. Для улучшенной моющей эффективности в жидком TIDE изменение в заряде молекулы ASP относительно дикого типа было сжато (например, диапазон +2 - -2, с оптимальной эффективностью при приблизительно 0 - -1). Таким образом, объединение улучшенных вариантов вместе не является аддитивным, если это добавление нарушает границы изменения общего заряда для ASP при этих условиях (например, менее чем -2, или более чем +2). Определение этого ранее неизвестного изменения заряда позволяет конструировать библиотеки мутантов для получения молекул с оптимальным зарядом для улучшенной эффективности.
Проектировали, конструировали и подвергали скринингу библиотеку со "сбалансированным зарядом" (см. Заявку на патент США с порядковым номером 11/583334, включенную здесь в качестве ссылки, так как она относится к библиотекам со сбалансированным зарядом). Эта библиотека содержала четыре полезные мутации с отрицательным зарядом и четыре невредные мутации с положительным зарядом (для баланса мутаций с отрицательным зарядом) почти во всех возможных комбинациях (230/256 возможных вариантов). Эту библиотеку подвергали скринингу на ряд свойств и идентифицировали варианты фермента, имеющие увеличенную активность в одном или нескольких представляющих интерес свойствах.
В некоторых вариантах осуществления, после определения оптимального заряда для конкретного фермента, выполняли также скрининг природных изолятов для идентификации вариантов фермента с оптимальным зарядом/оптимальным распределением заряда.
II. Получение вариантов NprE с улучшенными свойствами
Этот подход, примененный с ASP, распространили затем на молекулярный скелет полностью отличающейся протеазы. Получали SEL NprE и подвергали их скринингу на эффективность удаления пятен в детергенте (см. Заявку на патент США с порядковым номером 11/581102, включенную здесь в качестве ссылки, так как она относится к SEL). Идентифицировали мутанты со значимо улучшенной эффективностью удаления пятен BMI. Все улучшенные мутанты добавляли положительный заряд молекуле NprE. Подход баланса заряда использовали для оптимизации суммарного заряда поверхности и распределения заряда поверхности NprE. В случае NprE, моющая эффективность значимо улучшалась, когда общий заряд этой молекулы был более положительным, чем заряд белка дикого типа. Оптимальную моющую эффективность на BMI получали, когда заряд на этом белке был +1 или +2, относительно белка дикого типа.
III. Общие способы получения полезных вариантов фермента
Как описано здесь, определяли взаимосвязь между моющей эффективностью в анализе с микролоскутками BMI и общим зарядом на поверхности фермента. Способы данного изобретения находят применение в улучшении эффективности различных ферментов и белков (например, амилаз, целлюлаз, оксидаз, кутиназ, маннаназ, пектиназ, липаз, протеаз и других ферментов). Кроме того, эти способы находят применение в улучшении других желательных свойств белков, в том числе, но не только, экспрессии, термостабильности, стабильности в поверхностно-активных веществах и/или хелаторах и взаимосвязях рН-активность. Вкратце, идентифицируют аминокислотные остатки, локализованные на поверхности фермента дикого типа, которые более чем на приблизительно 35% экспонированы растворителю, предпочтительно более чем на приблизительно 50% экспонированы растворителю и наиболее предпочтительно более чем на приблизительно 65% экспонированы растворителю, и создают сайт-оценивающие библиотеки, где каждый остаток дикого типа заменен множеством других природных аминокислот. Кроме того, отмечают изменение суммарного заряда вариантных ферментов, которые обнаруживают улучшенную эффективность в одном или нескольких свойствах для определения взаимосвязи структура-функция. В дополнительных вариантах осуществления, после определения оптимального заряда для конкретного фермента, подвергают скринингу природные изоляты для идентификации вариантов фермента с оптимальным зарядом и распределением заряда.
ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
Следующие примеры обеспечены для демонстрации и дополнительной иллюстрации некоторых предпочтительных вариантов осуществления и аспектов данного изобретения и не должны пониматься как ограничение объема изобретения.
В следующем экспериментальном описании используются следующие аббревиатуры: °С (градусы стоградусной шкалы); об/мин (обороты в минуты); Н2О (вода); HCl (хлористоводородная кислота); аа и АА (аминокислота); п.н. (пара оснований); т.п.н. (тысячи пар оснований); кД (килодальтон); г (граммы); мкг (микрограммы); мг (миллиграммы); нг (нанограммы); мкл (микролитры); мл (миллилитры); мм (миллиметры); нм (нанометры); мкм (микрометр); M (молярный); мМ (миллимолярный); мкМ (микромолярный); Е (единицы); V (вольт); MW (молекулярная масса); сек (секунды); мин (минута/минуты); ч (час/часы); MgCl2 (хлорид магния); NaCl (хлорид натрия); OD280 (оптическая плотность при 280 нм); OD405 (оптическая плотность при 405 нм); OD600 (оптическая плотность при 600 нм); ПААГ (электрофорез в полиакриламидном геле); EtOH (этанол); ЗФР (забуференный фосфатом солевой раствор [150 мМ NaCl, 10 мМ натрийфосфатный буфер, pH 7,2]); LAS (лаурилсульфонат натрия); ДСН (додецилсульфат натрия); Трис (трис(гидроксиметил)аминометан); TAED (N,N,N'N'-тетраацетилэтилендиамин); BES (полиэфирсульфон); MES (2-морфолиноэтансульфоновая кислота, моногидрат; f.w. 195,24; Sigma # M-3671); CaCl2 (хлорид кальция, безводный; f.w. 110,99; Sigma # C-4901); ДМФ (N,N-диметилформамид, f.w. 73,09, d = 0,95); Abz-AGLA-Nba (2-аминобензоил-L-аланилглицил-L-лейцил-L-аланино-4-нитробензиламид, f.w. 583,65; Bachem # H-6675, VWR catalog # 100040-598); SBG1% ("Супербульон с глюкозой"; 6 г Soytone [Difco], 3 г дрожжевого экстракта, 6 г NaCl, 6 г глюкозы); pH доводили до 7,1 при помощи NaOH перед стерилизацией с использованием способов, известных в данной области; м/о (масса/объем); об./об. (объем/объем); Npr и npr (нейтральная металлопротеаза); SEQUEST® (программа поиска базы данных SEQUEST, University of Washington); Npr и npr (ген нейтральной металлопротеазы); nprE и NprE (нейтральная металлопротеаза B. amyloliquefaciens); PMN (очищенная металлопротеаза MULTIFECT®); MTP (микротитрационный планшет); MS (масс-спектрометрия); SRI (Индекс Удаления Пятен); TIGR (Institute for Genomic Research, Rockville, MD); AATCC (Американская ассоциация химиков и колористов текстильной промышленности); Procter & Gamble (Procter & Gamble, Inc., Cincinnati, OH); Amersham (Amersham Life Science, Inc. Arlington Heights, IL); ICN (ICN Pharmaceuticals, Inc., Costa Mesa, CA); Pierce (Pierce Biotechnology, Rockford, IL); EMPA (Eidgenossische Material Prufungs und Versuch Anstalt, St. Gallen, Switzerland); CFT (Center for Test Materials, Vlaardingen, The Netherlands); Amicon (Amicon, Inc., Beverly, MA); ATCC (American Type Culture Collection, Manassas, VA); Becton Dickinson (Becton Dickinson Labware, Lincoln Park, NJ); Perkin-Elmer (Perkin-Elmer, Wellesley, MA); Rainin (Rainin Instrument, LLC, Woburn, MA); Eppendorf (Eppendorf AG, Hamburg, Germany); Waters (Waters, Inc., Milford, MA); Geneart (Geneart GmbH, Regensburg, Germany); Perseptive Biosystems (Perseptive Biosystems, Ramsey, MN); Molecular Probes (Molecular Probes, Eugene, OR); BioRad (BioRad, Richmond, CA); Clontech (CLONTECH Laboratories, Palo Alto, CA); Difco (Difco Laboratories, Detroit, MI); GIBCO BRL или Gibco BRL (Life Technologies, Inc., Gaithersburg, MD); Epicentre (Epicentre Biotechnologies, Madison, WI); Zymo Research (Zymo Research Corp., Orange, CA); Integrated DNA Technologies (Integrated DNA Technologies, Inc., Coralville, IA): New Brunswick (New Brunswick Scientific Company, Inc., Edison, NJ); Thermoelectron (Thermoelectron Corp., Waltham, MA); BMG (BMG Labtech, GmbH, Offenburg, Germany); Novex (Novex, San Diego, CA); Finnzymes (Finnzymes OY, Finland) Qiagen (Qiagen, Inc., Valencia, CA); Invitrogen (Invitrogen Corp., Carlsbad, CA); Sigma (Sigma Chemical Co., St. Louis, MO); DuPont Instruments (Asheville, NY); Global Medical Instrumentation или GMI (Global Medical Instrumentation; Ramsey, MN); MJ Research (MJ Research, Waltham, MA); Infors (Infors AG, Bottmingen, Switzerland); Stratagene (Stratagene Cloning Systems, La Jolla, CA); Roche (Hoffmann La Roche, Inc., Nutley, NJ); Ion Beam Analysis Laboratory (Ion Bean Analysis Laboratory, The University of Surrey Ion Beam Centre (Guildford, UK); TOM (Terg-o-Meter); BMI (кровь, молоко, чернила); BaChem (BaChem AG, Bubendorf, Switzerland); Molecular Devices (Molecular Devices, Inc., Sunnyvale, CA); MicroCal (Microcal, Inc., Northhampton, MA); Chemical Computing (Chemical Computing Corp., Montreal, Canada); NCBI (National Center for Biotechnology Information); GE Healthcare (GE Healthcare, UK).
ПРИМЕР 1
Анализы
Следующие анализы использовали в описанных ниже примерах. Любые отклонения от обеспеченных ниже протоколов указаны в примерах. В этих примерах использовали спектрофотометр для измерения оптической плотности продуктов, образованных после завершения реакций. Рефлектометр использовали для измерения отражательной способности (коэффициента отражения) маленьких образцов тканей (лоскутков).
А. Анализ ВСА для определения содержания белка в 96-луночных микротитрационных планшетах (MTP)
В этих анализах, анализ ВСА (бицинхониновой кислоты; Pierce) использовали для определения концентрации белка в пробах протеазы в формате МТР. В этой системе анализа, химическими и реагентными растворами были: реагент BCA для анализа белка и буфер для разведения Pierce (50 мМ MES, pH 6,5, 2 мМ CaCl2, 0,005% TWEEN®-80). Используемым оборудованием был SpectraMAX (типа 340) MTP-ридер. МТР получали из Costar (типа 9017). В этом тесте, 200 мкл реагента ВСА пипетировали в каждую лунку с последующим добавлением 20 мкл разведенного белка. После тщательного смешивания эти МТР инкубировали в течение 30 минут при 37°С. Пузырьки воздуха удаляли и оптическую плотность раствора (OD) в этих лунках считывали при 562 нм. Для определения концентрации белка показатель фона вычитали из показателей проб. Величины OD562 строили для стандартов белков (очищенной протеазы) с получением калибровочной кривой. Концентрации белка этих проб экстраполировали из этой калибровочной кривой.
В. Анализ микролоскутков для тестирования эффективности протеазы
Детергенты, используемые в этом анализе, не содержали ферментов. Используемым оборудованием был Термомиксер Эппендорфа и МТР-ридер SpectraMAX (типа 340; Molecular Devices). MTP получали из Costar (типа 9017).
Приготовление детергента (жидкого детергента для стирки белья TIDE® 2X Ultra, Clean Breeze (Procter & Gamble); условия стирки US)
Воду Milli-Q доводили до жесткости воды 6 gpg (Ca/Mg=3/1) и добавляли 0,78 г/л детергента TIDE® 2X Ultra Clean Breeze". Этот детергент нагревали предварительно при 95°C в течение одного часа для инактивации ферментов, присутствующих в этом препарате. Раствор детергента перемешивали в течение 15 минут. Затем добавляли 5 мМ HEPES (свободную кислоту) и рН доводили до 8,2.
Микролоскутки
Микролоскутки с диаметром круга 0,25 дюйма получали из CFT Vlaardingen. Перед нарезанием этих лоскутков ткань (EMPA 116) промывали водой. Один микролоскуток помещали в каждую лунку 96-микротитрационного планшета.
Тест-способ
Раствор желаемого детергента готовили, как описано выше. После уравновешивания Термомиксера при 25°С в каждую содержащую микролоскуток лунку МТР добавляли 190 мкл раствора детергента. К этой смеси добавляли 10 мкл разведенного раствора фермента, так что конечная концентрация была 1 мкг/мл (как определено анализом ВСА). Этот МТР герметизировали с использованием клейкой ленты и помещали в термостат на 30 минут с перемешиванием при 1400 об/мин. После инкубирования при подходящих условиях 100 мкл раствора из каждой лунки переносили в свежий МТР. Новый МТР, содержащий 100 мкл раствора на лунку, считывали при 405 нм с использованием МТР-ридера SpectraMax. Включали также холостые контроли, а также контроли, содержащие микролоскуток и детергент, но не фермент.
Анализ микролоскутков с использованием рисового крахмала
Анализ с использованием рисового крахмала является тестом на эффективность амилазы. Детергенты готовили, как описано в другом месте в этом документе. Используемое оборудование включало шейкер/термостат New Brunswick Innova 4230 и МТР-ридер SpectraMAX (типа 340). МТР получали из Corning (типа 3641). Выдержанный крахмал с лоскутками с оранжевым пигментом (CS-28) получали из Center for Test Materials (Vlaardingen, Netherlands). Перед нарезанием микролоскутков с диаметром круга 0,25 дюйма ткань промывали водой. В каждую лунку 96-луночного микротитрационного планшета помещали два микролоскутка. Тестируемый детергент уравновешивали при 20°C (Северная Америка) или 40°C (Западная Европа). 190 мкл раствора детергента добавляли в каждую лунку MTP, содержащую микролоскутки. К этой смеси добавляли 10 мкл разбавленного раствора фермента. МТР герметизировали клейкой фольгой и помещали в термостат на 1 час с перемешиванием при 750 об/мин при желаемой температуре тестирования (обычно 20°C или 40°C). После инкубирования 150 мкл раствора из каждой лунки переносили в свежий МТР. Этот МТР считывали при 488 нм с использованием МТР-ридера SpectraMax MTP для количественного определения очистки. Включали также холостые контроли, а также контроли, содержащие микролоскуток и детергент, но не фермент.
Расчет эффективности фермента
Полученную величину оптической плотности корректировали с использованием величины холостого контроля (т.е. величины, полученной после инкубирования в отсутствие фермента). Полученная оптическая плотность обеспечивала меру гидролитической активности тестируемого фермента.
Н. Инактивация детергента нагреванием
Инактивация нагреванием коммерческих смесей детергентов служит для разрушения ферментативной активности любого белкового компонента при сохранении свойств неферментных компонентов. Таким образом, этот способ пригоден для приготовления коммерчески продаваемых детергентов для применения в тестировании вариантов фермента данного изобретения. Для североамериканских (NA) и западноевропейских (WE) жидких детергентов для стирки в интенсивном режиме (HDL) инактивацию нагреванием выполняли помещением предварительно взвешенного жидкого детергента (в стеклянной бутыли) в водяную баню при 95°C на 2 часа. Время инкубации для инактивации нагреванием Североамериканского (NA) и Японского (JPN) гранулированного детергента (HDG) для стирки в интенсивном режиме было равно 8 часам, а время инкубации для Западноевропейского (WE) детергента HDG было равно 5 часам. Время инкубации для инактивации нагреванием детергентов NA и WE для автоматической мойки посуды (ADW) было равно 8 часам. Эти детергенты покупали в магазинах местного супермаркета. Как ненагреваемые, так и нагреваемые детергенты анализировали в течение 5 минут растворения детергента для точного определения процентной инактивации. Активность фермента анализировали при помощи suc-AAPF-pNA-анализа.
Для тестирования активности фермента в инактивированных нагреванием детергентах рабочие растворы детергентов готовили из инактивированных нагреванием исходных растворов. Подходящие количества жесткости воды (6 gpg или 12 gpg) и буфера добавляли к растворам детергентов для создания желаемых условий (таблица 1-1). Растворы перемешивали с использованием вортекса или переворачиванием бутылок.
Bodipy-крахмал-анализ для определения амилазной активности
Bodipy-крахмал-анализ выполняли с использованием набора для анализа амилазы (E33651, Invitrogen). Готовили исходный раствор 1 мг/мл субстрата крахмала DQ растворением содержимого флакона, содержащего лиофилизированный субстрат, в 100 мкл 50 мМ натрий-ацетатного буфера при pH 4,0. Флакон встряхивали на вортексе в течение приблизительно 20 секунд и оставляли при комнатной температуре, в темноте, с периодическим смешиванием до растворения. Добавляли 900 мкг буфера для анализа (50 мМ ацетат натрия с 2,6 мМ CaCl2 рН 5,8) и флакон встряхивали на вортексе в течение приблизительно 20 секунд. Этот раствор субстрата хранили при комнатной температуре, в темноте, до использования или при 4оС. Для анализа готовили 100 мкг/мл рабочего раствора субстрата DQ из раствора субстрата 1 мг/мл в буфере для анализа. 190 мкл раствора субстрата 100 мкг/мл добавляли в каждую лунку в 96-луночном плоскодонном планшете. 10 мкл проб фермента добавляли в эти лунки, смешивали в течение 30 секунд при помощи термомиксера при 800 об/мин. В анализ включали холостую пробу, которая содержит только буфер и субстрат (холостая проба без фермента). Измеряли скорость изменения интенсивности флуоресценции (возбуждение: 485 нм, испускание: 520 нм) в планшет-ридере для измерения флуоресценции в микротитрационном планшете при 25°C в течение 5 минут.
К. Определение уменьшения вязкости крахмала амилазой
В этом анализе измеряли уменьшение вязкости субстратного раствора кукурузного крахмала в вискозиметре. Субстратный кукурузный крахмал готовили в виде свежей взвеси в периодическом режиме с 30% сухого вещества кукурузной муки в дистиллированной воде и доводили до рН 5,8 серной кислотой. Для каждого опыта 50 г взвеси (15 г сухого вещества) взвешивали и предынкубировали в течение 10 минут для нагревания до 70°C. После добавления амилазы температура немедленно поднималась с 70°C до 85°C при скорости вращения 75 об/мин. После достижения смесью взвеси и амилазы 85°C эта температура держалась на этом уровне и вязкость наблюдали в течение дополнительных 30 минут.
ПРИМЕР 2
Получение протеазы NprE в B. subtilis
В этом примере описаны эксперименты, проводимые для получения протеазы NprE в B. subtilis. В частности, обеспечены способы, используемые в трансформации плазмиды pUBnprE в B. subtilis. Трансформацию выполняли, как известно в данной области (см., например, WO 02/14490 и Заявку на патент США с порядковым номером 11/581102, включенные здесь в качестве ссылки). ДНК-последовательность (nprE-лидер, nprE-про и зрелая nprE-последовательность ДНК из B. amyloliquefaciens), представленная ниже, кодирует белок-предшественник NprE:
GTGGGTTTAGGTAAGAAATTGTCTGTTGCTGTCGCCGCTTCCTTTATGAGTTTAACC
ATCAGTCTGCCGGGTGTTCAGGCCGCTGAGAATCCTCAGCTTAAAGAAAACCTGAC
GAATTTTGTACCGAAGCATTCTTTGGTGCAATCAGAATTGCCTTCTGTCAGTGACAA
AGCTATCAAGCAATACTTGAAACAAAACGGCAAAGTCTTTAAAGGCAATCCTTCTG
AAAGATTGAAGCTGATTGACCAAACGACCGATGATCTCGGCTACAAGCACTTCCGT
TATGTGCCTGTCGTAAACGGTGTGCCTGTGAAAGACTCTCAAGTCATTATTCACGTC
GATAAATCCAACAACGTCTATGCGATTAACGGTGAATTAAACAACGATGTTTCCGC
CAAAACGGCAAACAGCAAAAAATTATCTGCAAATCAGGCGCTGGATCATGCTTATA
AAGCGATCGGCAAATCACCTGAAGCCGTTTCTAACGGAACCGTTGCAAACAAAAAC
AAGCCGAGCTGAAAGCAGCAGCCACAAAAGACGGCAAATACCGCCTCGCCTATG
ATGTAACCATCCGCTACATCGAACCGGAACCTGCAAACTGGGAAGTAACCGTTGAT
GCGGAAACAGGAAAAATCCTGAAAAAGCAAAACAAAGTGGAGCAT GCCGCCACA
ACCGGAACAGGTACGACTCTTAAAGGAAAAACGGTCTCATTAAATATTTCTTCT
GAAAGCGGCAAATATGTGCTGCGCGATCTTTCTAAACCTACCGGAACACAAAT
TATTACGTACGATCTGCAAAACCGCGAGTATAACCTGCCGGGCACACTCGTAT
CCAGCACCACAAACCAGTTTACAACTTCTTCTCAGCGCGCTGCCGTTGATGCG
CATTACAACCTCGGCAAAGTGTATGATTATTTCTATCAGAAGTTTAATCGCAAC
AGCTACGACAATAAAGGCGGCAAGATCGTATCCTCCGTTCATTACGGCAGCAG
ATACAATAACGCAGCCTGGATCGGCGACCAAATGATTTACGGTGACGGCGACG
GTTCATTCTTCTCACCTCTTTCCGGTTCAATGGACGTAACCGCTCATGAAATGA
CACATGGCGTTACACAGGAAACAGCCAACCTGAACTACGAAAATCAGCCGGGC
GCTTTAAACGAATCCTTCTCTGATGTATTCGGGTACTTCAACGATACTGAGGAC
TGGGATATCGGTGAAGATATTACGGTCAGCCAGCCGGCTCTCCGCAGCTTATC
CAATCCGACAAAATACGGACAGCCTGATAATTTCAAAAATTACAAAAACCTTCC
GAACACTGATGCCGGCGACTACGGCGGCGTGCATACAAACAGCGGAATCCCG
AACAAAGCCGCTTACAATACGATTACAAAAATCGGCGTGAACAAAGCGGAGCA
GATTTACTATCGTGCTCTGACGGTATACCTCACTCCGTCATCAACTTTTAAAGA
TGCAAAAGCCGCTTTGATTCAATCTGCGCGGGACCTTTACGGCTCTCAAGATG
CTGCAAGCGTAGAAGCTGCCTGGAATGCAGTCGGATTGTAA (SEQ ID NO: 1)
В приведенной выше последовательности жирный шрифт указывает ДНК, которая кодирует зрелую протеазу NprE, стандартный шрифт указывает лидерную последовательность (nprE-лидер), а подчеркнутый шрифт указывает про-последовательности (nprE-pro). Аминокислотная последовательность (NprE-лидер, NprE-про и зрелая последовательность белка NprE), приведенная ниже (SEQ ID NO:2), соответствует полноразмерному белку NprE. В этой последовательности подчеркнутое указывает пропоследовательность, а жирный шрифт указывает зрелую протеазу NprE.
MGLGKKLSVAVAASFMSLTISLPGVQAAENPQLKENLTNFVPKHSLVQSELPSVSDKAI
KQYLKQNGKVFKGNPSERLKLIDQTTDDLGYKHFRYVPVVNGVPVKDSQVHHVDKSN
NVYAINGELNNDVSAKTANSKKLSANOALDHAYKAIGKSPEAVSNGTVANKNKAELK
AAATKDGKYRLAYDVTIRYIEPEPANWEVTVDAETGKILKKQNKVEH AATTGTGTTL
KGKTVSLNISSESGKYVLRDLSKPTGTQIITYDLQNREYNLPGTLVSSTTNQFTTSSQ
RAAVDAHYNLGKVYDYFYQKFNRNSYDNKGGKIVSSVHYGSRYNNAAWIGDQMI
YGDGDGSFFSPLSGSMDVTAHEMTHGVTQETANLNYENQPGALNESFSDVFGYFN
DTEDWDIGEDITVSQPALRSLSNPTKYGQPDNFKNYKNLPNTDAGDYGGVHTNSGI
PNKAAYNTITKIGVNKAEQIYYRALTVYLTPSSTFKDAKAALIQSARDLYGSQDAAS
VEAAWNAVGL (SEQ ID NO:2)
Зрелая последовательность NprE представлена в виде SEQ ID NO:3. Эту последовательность использовали в качестве основы для приготовления библиотек вариантов, описанных здесь.
AATTGTGTTLKGKTVSLNISSESGKYVLRDLSKPTGTQIITYDLQNREYNLPGTLVSSTT
NQFTTSSQRAAVDAHYNLGKVYDYFYQKFNRNSYDNKGGKIVSSVHYGSRYNNAAWI
GDQMIYGDGDGSFFSPLSGSMDVTAHEMTHGVTQETANLNYENQPGALNESFSDVFGY
FNDTEDWDIGEDITVSQPALRSLSNPTKYGQPDNFKNYKNLPNTDAGDYGGVHTNSGIP
NKAAYNTITKIGVNKAEQIYYRALTVYLTPSSTFKDAKAALIQSARDLYGSQDAASVEA
AWNAVGL (SEQ ID NO:3)
Экспрессирующий вектор pUBnprE конструировали амплификацией гена nprE из хромосомной ДНК B. amyloliquefaciens при помощи ПЦР с использованием двух специфических праймеров:
Олиго AB1740: CTGCAGGAATTCAGATCTTAACATTTTTCCCCTATCATTTTTCCCG (SEQ ID NO:4)
Олиго AB1741: GGATCCAAGCTTCCCGGGAAAAGACATATATGATCATGGTGAAGCC (SEQ ID NO:5)
ПЦР выполняли в термоциклере с ДНК-полимеразой Phusion High Fidelity (FINNZYMES). Смесь ПЦР содержала 10 мкл 5x буфера (Finnzymes Phusion), 1 мкл 10 мМ dNTP, 1,5 мкл ДМСО, 1 мкл каждого праймера, 1 мкл ДНК-полимеразы Finnzymes Phusion, 1 мкл раствора хромосомной ДНК 50 нг/мкл, 34,5 мкл воды MilliQ. Использовали следующий протокол:
Протокол ПЦР:
1) 30 сек при 98°C;
2) 10 сек при 98°C;
3) 20 сек при 55°C;
4) 1 мин при 72°C;
5) 25 циклов стадий 2-4; и
6) 5 мин при 72°C.
Эта ПЦР приводила к ДНК-фрагменту 1,9 т.п.н., который расщепляли рестрикционными ферментами ДНК BglII и BclI. Многокопийный вектор pUB110 Bacillus (см., например, Gryczan, J Bacteriol, 134:318-329 [1978]) расщепляли BamHI. Затем ПЦР-фрагмент x BglII x BclI лигировали в вектор pUB110 x BamHI с получением экспрессирующего вектора pUBnprE.
pUBnprE трансформировали в штамм B. subtilis (ΔaprE, ΔnprE, oppA, ΔspoIIE, degUHy32, ΔamyE::xylR,pxylA-comK). Трансформацию в B. subtilis выполняли, как описано в WO 02/14490, включенном здесь в качестве ссылки. Селективный рост трансформантов B. subtilis, несущих вектор pUBnprE, получали во встряхиваемых колбах, содержащих 25 мл среды MBD (среды определенного состава на основе MOPS) с 20 мг/л неомицина. Среду MBD готовили, по существу как известно в данной области (см., например, Neidhardt et al., J Bacteriol, 119: 736-747 [1974]), за исключением того, что NH4Cl2, FeSO4 и CaCl2 удаляли из базальной среды, использовали 3 мМ K2HPO4, и эту базальную среду дополняли 60 мМ мочевиной, 75 г/л глюкозы и 1% soytone. Готовили также микронутриенты в виде 100 Х исходного раствора, содержащего в одном литре 400 мг FeSO4.7H2O, 100 мг MnSO4.H2O, 100 мг ZnSO4.7H2O, 50 мг CuCl2.2H20, 100 мг CoCl2.6H2O, 100 мг NaMoO4.2H2O, 100 мг Na2B4O7.10H2O, 10 мл 1М CaCl2 и 10 мл 0,5M цитрата натрия. Эту культуру инкубировали в течение трех дней при 37°С в термостате/шейкере (Infors). Эта культура приводила к получению секретируемой протеазы NprE с протеолитической активностью, как показано анализами протеазы. Гель-анализ выполняли с использованием 10% Бис-Трис-гелей NuPage Novex (Invitrogen, Catalog No. NP0301BOX). Для приготовления проб для анализа 2 объема супернатанта смешивали с 1 объемом 1М HCl, 1 объемом 4xLDS-буфера для проб (Invitrogen, Catalog No. NP0007) и 1% PMSF (20 мг/мл) и затем нагревали в течение 10 минут при 70°C. Затем 25 мкл каждой пробы наносили на гель, вместе с 10 мкл предварительно окрашенных SeeBlue plus 2 белковых стандартов (Invitrogen, Catalog No.LC5925). Результаты ясно показали, что стратегия клонирования nprE, описанная в этом примере, пригодна для получения активного NprE в B. subtilis.
ПРИМЕР 3
Получение протеазы ASP в B. subtilis
В этом примере описаны эксперименты, проводимые для получения протеазы 69B4 (также называемой здесь "ASP", "Asp" и "протеазой ASP" и "протеазой Asp") в B. subtilis. В частности, обеспечены способы, используемые для трансформации плазмиды pHPLT-ASP-C1-2 в B. subtilis. Трансформацию выполняют, как известно в данной области (см., например, WO 02/14490 и Заявку на патент США с порядковым номером 11/583334, включенные здесь в качестве ссылки). Для оптимизации экспрессии ASP в B. subtilis получали синтетическую последовательность ДНК с использованием DNA2.0 и использовали в этих экспериментах экспрессии. ДНК-последовательность (синтетическая ДНК-последовательность ASP), обеспеченная ниже, с использованием кодонов, адаптированных в отношении видов Bacillus, кодирует белок-предшественник ASP дикого типа:
ATGACACCACGAACTGTCACAAGAGCTCTGGCTGTGGCAACAGCAGCTGCTACACT
CTTGGCTGGGGGTATGGCAGCACAAGCTAACGAACCGGCTCCTCCAGGATCTGCAT
CAGCCCCTCCACGATTAGCTGAAAAACTTGACCCTGACTTACTTGAAGCAATGGAA
CGCGATCTGGGGTTAGATGCAGAGGAAGCAGCTGCAACGTTAGCTTTTCAGCATGA
CGCAGCTGAAACGGGAGAGGCTCTTGCTGAGGAACTCGACGAAGATTTCGCGGGCA
CGTGGGTTGAAGATGATGTGCTGTATGTTGCAACCACTGATGAAGATGCTGTTGAA
GAAGTCGAAGGCGAAGGAGCAACTGCTGTGACTGTTGAGCATTCTCTTGCTGATTT
AGAGGCGTGGAAGACGGTTTTGGATGCTGCGCTGGAGGGTCATGATGATGTGCCTA
CGTGGTACGTCGACGTGCCTACGAATTCGGTAGTCGTTGCTGTAAAGGCAGGAGCG
CAGGATGTAGCTGCAGGACTTGTGGAAGGCGCTGATGTGCCATCAGATGCGGTCAC
TTTTGTAGAAACGGACGAAACGCCTAGAACGATG TTCGACGTAATTGGAGGCAAC
GCATATACTATTGGCGGCCGGTCTAGATGTTCTATCGGATTCGCAGTAAACGG
TGGCTTCATTACTGCCGGTCACTGCGGAAGAACAGGAGCCACTACTGCCAATC
CGACTGGCACATTTGCAGGTAGCTCGTTTCCGGGAAATGATTATGCATTCGTC
CGAACAGGGGCAGGAGTAAATTTGCTTGCCCAAGTCAATAACTACTCGGGCGG
CAGAGTCCAAGTAGCAGGACATACGGCCGCACCAGTTGGATCTGCTGTATGCC
GCTCAGGTAGCACTACAGGTTGGCATTGCGGAACTATCACGGCGCTGAATTCG
TCTGTCACGTATCCAGAGGGAACAGTCCGAGGACTTATCCGCACGACGGTTTG
TGCCGAACCAGGTGATAGCGGAGGTAGCCTTTTAGCGGGAAATCAAGCCCAAG
GTGTCACGTCAGGTGGTTCTGGAAATTGTCGGACGGGGGGAACAACATTCTTT
CAACCAGTCAACCCGATTTTGCAGGCTTACGGCCTGAGAATGATTACGACTGA
CTCTGGAAGTTCCCCT GCTCCAGCACCTACATCATGTACAGGCTACGCAAGAACG
TTCACAGGAACCCTCGCAGCAGGAAGAGCAGCAGCTCAACCGAACGGTAGCTATGT
TCAGGTCAACCGGAGCGGTACACATTCCGTCTGTCTCAATGGACCTAGCGGTGCGG
ACTTTGATTTGTATGTGCAGCGATGGAATGGCAGTAGCTGGGTAACCGTCGCTCAAT
CGACATCGCCGGGAAGCAATGAAACCATTACGTACCGCGGAAATGCTGGATATTAT
CGCTACGTGGTTAACGCTGCGTCAGGATCAGGAGCTTACACAATGGGACTCACCCT
CCCCTGA (SEO ID NO:6)
В приведенной выше последовательности жирный шрифт указывает ДНК, которая кодирует зрелую протеазу ASP, стандартный шрифт указывает лидерную последовательность (лидер ASP) и подчеркивание указывает N-концевую и С-концевую пропоследовательности. Аминокислотная последовательность, приведенная ниже (SEQ ID NO:7), соответствует полноразмерному белку ASP. В этой последовательности подчеркивания указывают пропоследовательности и жирный шрифт указывает зрелую протеазу ASP.
MTPRTVTRALAVATAAATLLAGGMAAOANEPAPPGSASAPPRLAEKLDPDLLEAMER
DLGLDAEEAAATLAFOHDAAETGEALAEELDEDFAGTWVEDDVLYVATTDEDAVEEV
EGEGATAVTVEHSLADLEAWKTVLDAALEGHDDVPTWYVDVPTNSVVVAVKAGAOD
VAAGLVEGADVPSDAVTFVETDETPRTM FDVIGGNAYTIGGRSRCSIGFAVNGGFITA
GHCGRTGATTANPTGTFAGSSFPGNDYAFVRTGAGVNLLAQVNNYSGGRVQVAG
HTAAPVGSAVCRSGSTTGWHCGTITALNSSVTYPEGTVRGLIRTTVCAEPGDSGGS
LLAGNQAOGVTSGGSGNCRTGGTTFFOPVNPILQAYGLRMITTDSGSSP APAPTSCT
GYARTFTGTLAAGRAAAOPNGSYVOVNRSGTHSVCLNGPSGADFDLYVORWNGSSW
VTVAOSTSPGSNETITYRGNAGYYRYVVNAASGSGAYTMGLTLP (SEQ ID N0:7)
Зрелая последовательность ASP представлена в виде SEQ ID NO:8. Эту последовательность использовали в качестве основы для получения библиотек вариантов, описанных здесь.
FDVIGGNAYTIGGRSRCSIGFAVNGGFITAGHCGRTGATTANPTGTFAGSSFPGNDYAFV
RTGAGVNLLAQVNNYSGGRVQVAGHTAAPVGSAVCRSGSTTGWHCGTITALNSSVTY
PEGTVRGLIRTTVCAEPGDSGGSLLAGNQAQGVTSGGSGNCRTGGTTFFQPVNPILQAY
GLRMITTDSGSSP (SEQ ID NO:8)
Экспрессионные кассеты Asp конструировали в векторе pXX-KpnI и затем клонировали в вектор pHPLT для экспрессии ASP в B. subtilis. pXX-KpnI является вектором на основе pUC с промотором aprE (B. subtilis), запускающим экспрессию, геном cat и двойным промотором aprE для амплификации определенного числа копий в B. subtilis. Ген bla делает возможным селективный рост в E. coli. KpnI, введенная в сайт связывания рибосом, справа от области промотора aprE, вместе с сайтом HindIII позволяет клонировать экспрессионные кассеты Asp в pXX-KpnI. pHPLT-EBS2c2, производное pHPLT (Solingen et al., Extremophiles 5:333-341 [2001]), содержит промотор термостабильной амилазы LAT (PLAT) Bacillus licheniformis, за которым следуют сайты рестрикции XbaI и HpaI для клонирования экспрессионных конструкций ASP. Экспрессионную кассету Asp клонировали в вектор pXX-KpnI, содержащий ДНК, кодирующую гибридный сигнальный пептид (SEQ ID NO:9), сконструированный из 5 аминокислот N-концевого сигнального пептида AprE субтилизина, слитых с 25 аминокислотами С-концевого сигнального пептида Asp:
MRSKKRTVTRALAVATAAATLLAGGMAAQA (SEQ ID NO:9).
Этот гибридный сигнальный пептид ASP кодируется следующей последовательностью ДНК:
ATGAGAAGCAAGAAGCGAACTGTCACAAGAGCTCTGGCTGTGGCAACAGCAGCTG
CTACACTCTTGGCTGGGGGTATGGCAGCACAAGCT (SEQ ID NO:10)
Экспрессионную кассету Asp, клонированную в векторе pXX-KpnI, трансформировали в E. coli (Electromax DH10B, Invitrogen, Cat.No. 12033-015). Используемые праймеры и стратегия клонирования обеспечены ниже. Затем, эти экспрессионные кассеты клонировали из этих векторов и вводили в экспрессирующий вектор pHPLT для трансформации в штамм B. subtilis (ΔaprE, ΔnprE, oppA, ΔspoIIE, degUHy32, ΔamyE::xylR,pxylA-comK). Праймеры и стратегия клонирования для клонирования экспрессионных кассет ASP в pHPLT также приведены ниже.
Праймеры получали из MWG и Invitrogen. ДНК-полимеразу высокой точности Invitrogen Platinum Tag (Catalog No. 11304-029) использовали для ПЦР-амплификации (0,2 мкМ праймеры, 25-30 циклов) согласно протоколу Invitrogen. Лигазные реакции экспрессионных кассет Asp и векторов хозяев выполняли с использованием ДНК-лигазы Т4 Invitrogen (Cat. No. 15224-025) с использованием протокола, рекомендуемого для общего клонирования липких концов.
Экспрессию гена asp исследовали в штамме B. subtilis (ΔaprE, ΔnprE, oppA, ΔspoIIE, degUHy32, ΔamyE::xylR,pxylA-comK). Плазмиду pHPLT-ASP-C1-2 трансформировали в B. subtilis (ΔaprE, ΔnprE, oppA, ΔspoIIE, degUHy32, ΔamyE::xylR,pxylA-comK). Трансформацию выполняли, как известно в данной области (см., например, WO 02/14490, включенный здесь в качестве ссылки).
Селективный рост трансформантов B. subtilis (ΔaprE, ΔnprE, oppA, ΔspoIIE, degUHy32, ΔamyE::xylR,pxylA-comK), несущих вектор pHPLT-ASP-C1-2, выполняли во встряхиваемых колбах, содержащих 25 мл синтетической максатазной среды (SMM), с 0,97 г/л CaCl2.6H2O вместо 0,5 г/л CaCl2 (см. Патент США № 5324653, включенный здесь в качестве ссылки) с 20 мг/л неомицина. Этот рост приводил к получению секретируемой ASP, имеющей протеолитическую активность. Гель-анализ выполняли с использованием 10% Бис-Трис-гелей NuPage Novex (Invitrogen, Catalog No. NP0301BOX). Для приготовления проб для анализа 2 объема супернатанта смешивали с 1 объемом 1М HCl, 1 объемом 4xLDS-буфера для проб (Invitrogen, Catalog No. NP0007) и 1% PMSF (20 мг/мл) и затем нагревали в течение 10 минут при 70°C. Затем 25 мкл каждой пробы наносили на гель, вместе с 10 мкл предварительно окрашенных SeeBlue plus 2 белковых стандартов (Invitrogen, Catalog No.LC5925). Результаты ясно показали, что стратегия клонирования asp, описанная в этом примере, пригодна для получения активной Asp в B. subtilis.
TCATGCAGGGTACCATGAGAAGCA AGAAGCGAACTGTCACAAGAGCTC TGGCT (SEQ ID NO:11)
ASP-syntc-mature-RV
GTGTGCAAGCTTTCAAGGGGAACT TCCAGAGTCAGTC (SEQ ID NO:12)
TGAGCTGCTAGCAAAAGGAGAGGG TAAAGAATGAGAAGCAAGAAG
(SEQ ID NO:13)
pHPLT-ASPmat-RV
CATGCATCCCGGGTTAAGGGGAAC TTCCAGAGTCAGTC
(SEQ ID NO:14)
ПРИМЕР 4
Генерирование сайт-оценивающих библиотек (SEL) и библиотек сайт-насыщенного мутагенеза (SSML)
В этом примере описаны способы, используемые в конструировании SEL nprE и asp.
А. Генерирование SEL nprE
Вектор pUBnprE, содержащий описанную выше экспрессионную кассету nprE, служил в качестве матричной ДНК. Этот вектор содержит уникальный сайт рестрикции BglII, который использовали в конструировании сайт-оценивающей библиотеки. Вкратце, для конструирования сайт-оценивающей библиотеки nprE выполняли три ПЦР-реакции, включающие две ПЦР мутагенеза для введения мутированного представляющего интерес кодона в ДНК-последовательность зрелой nprE и третью ПЦР использовали для слияния этих двух ПЦР мутагенеза для конструирования экспрессирующего вектора pUBnprE, включающего желаемый мутированный кодон в зрелой последовательности nprE.
Способ мутагенеза основывался на подходе кодон-специфической мутации, в котором создание всех возможных мутаций одновременно выполняли с использованием прямого и обратного олигонуклеотидного праймера с длиной 25-45 нуклеотидов, включающего специфическую сконструированную тройную ДНК-последовательность NNS (N = A, C, T или G; и S = C или G), которая соответствовала последовательности мутируемого кодона и гарантировала случайное включение нуклеотидов в этом конкретном кодоне зрелого nprE. Цифра, перечисленная в названиях праймеров, соответствует конкретному положению кодона зрелого nprE. Оцененные сайты включали: 4, 12, 13, 14, 23, 24, 33, 45, 46, 47, 49, 50, 54, 58, 59, 60, 65, 66, 87, 90, 96, 97, 100, 186, 196, 211, 214, 228 и 280. Примерный перечень праймерных последовательностей описан в Заявке на патент США с порядковым номером 11/581102, включенной здесь в качестве ссылки.
Два дополнительных праймера, используемых для конструирования сайт-оценивающих библиотек, содержали сайт рестрикции BglII вместе с частью ДНК-последовательности pUBnprE, фланкирующей сайт рестрикции BglII. Эти праймеры получали из Invitrogen (шкала 50 нмоль, обессоленные):
pUB-BglII-FW GTCAGTCAGATCTTCCTTCAGGTTATGACC (SEQ ID NO:15); и pUB-BglII-RV GTCTCGAAGATCTGATTGCTTAACTGCTTC (SEQ ID NO:16).
Конструирование каждой библиотеки SEL начинали с двух первичных ПЦР-амплификаций с использованием праймера pUB-BglII-FW и специфического обратного праймера мутагенеза nprE. Для второй ПЦР использовали праймер pUB-BglII-RV и специфический прямой праймер мутагенеза nprE (одинаковые положения зрелого nprE для прямого и обратного праймеров мутагенеза).
Введение этих мутаций в зрелую последовательность nprE выполняли с использованием ДНК-полимеразы высокой точности (Phusion High-Fidelity DNA Polymerase) (Finnzymes; Catalog No. F-530L). Все ПЦР выполняли согласно протоколу Finnzymes, дополненному этой полимеразой. Условия ПЦР для первичных ПЦР были следующими:
Для первичной ПЦР 1:
Праймер pUB-BglII-FW и специфический обратный праймер мутагенеза NPRE - оба в количестве 1 мкл (10 мкМ);
Для первичной ПЦР 2:
Праймер pUB-BglII-RV и специфический обратный праймер мутагенеза NPRE - оба в количестве 1 мкл (10 мкМ); вместе с
автоклавированной водой
ПЦР-программа была следующей: 30 секунд при 98°C, 30х (10 секунд при 98°C, 20 секунд при 55°C, 1,5 минуты при 72°C) и 5 минут при 72°C, выполняемой в термоциклере PTC-200 Peltier (MJ Research). Эти ПЦР-эксперименты приводили к двум фрагментам приблизительно 2-3 т.п.н., которые имели приблизительно 30 нуклеотидных оснований, перекрывающихся около представляющего интерес зрелого кодона NprE. Фрагменты сливали в третьей ПЦР-реакции с использованием двух вышеупомянутых фрагментов и прямого и обратного праймеров BglII. Эту реакцию ПЦР-слияния проводили в следующем растворе:
Праймер pUB-BglII-FW и праймер pUB-BglII-RV - оба 1 мкл (10 мкМ) вместе с
автоклавированной водой
Программа ПЦР-слияния была следующей: 30 секунд при 98°C, 30x (10 секунд при 98°C, 20 секунд при 55°C, 2:40 минут при 72°C) и 5 мин при 72°C, в термоциклере PTC-200 Peltier (MJ Research).
Амплифицированный линейный фрагмент 6,5 т.п.н. очищали с использованием набора для очистки ПЦР QIAQUICK® (Qiagen, Catalog No. 28106) и расщепляли рестрикционным ферментом BglII для создания липких концов на обеих сторонах этого слитого фрагмента:
- 35 мкл очищенного линейного ДНК-фрагмента
- 4 мкл буфера REACT® 3 (Invitrogen)
- 1 мкл BglII, 10 единиц/мл (Invitrogen)
Условия реакции: 1 час, 30°C.
Лигирование расщепленного BglII и очищенного с использованием набора для очистки ПЦР QIAQUICK® (Qiagen, Catalog No. 28106) фрагмента приводило к кольцевой и мультимерной ДНК, содержащей желаемую мутацию:
- 30 мкл очищенного расщепленного BglII ДНК-фрагмента
- 8 мкл буфера для ДНК-лигазы Т4 (Invitrogen Catalog No. 46300-018)
- 1 мкл ДНК-лигазы Т4, 1 единицы/мкл (Invitrogen Catalog No. 15224-017)
Условия реакции: 16-20 часов при 16°C.
Затем эту смесь для лигирования трансформировали в штамм B. subtilis (ΔaprE, ΔnprE, oppA, ΔspoIIE, degUHy32, ΔamyE::xylR,pxylA-comK). Трансформацию в B. subtilis выполняли, как описано в WO 02/14490, включенном здесь в качестве ссылки. Для каждой библиотеки выскребали 96-отдельных колоний и выращивали на среде MOPS с неомицином и 1,25 г/л дрожжевым экстрактом для анализа последовательности (BaseClear) и для целей скрининга. Каждая библиотека включала максимально 19 сайт-специфических вариантов nprE.
Эти варианты получали выращиванием трансформантов B. subtilis SEL в 96-луночном MTP при 37°C в течение 68 часов в среде MBD с 20 мг/л неомицина и 1,25 г/л дрожжевого экстракта.
В. Генерирование SSML Asp
В этом примере описаны эксперименты, проводимые для развития библиотек сайт-насыщающего мутагенеза Asp (SSML). Каждая сайт-насышающая библиотека Asp содержала 96 клонов B. subtilis (ΔaprE, ΔnprE, oppA, ΔspoIIE, degUHy32, ΔamyE::xylR,pxylA-comK), несущих экспрессирующий вектор pHPLT-ASP-С1-2. Было обнаружено, что этот вектор, содержащий экспрессионную кассету, состоящую из синтетической последовательности ДНК, кодирующей гибридный сигнальный пептид Asp и N-концевой про- и зрелый белок Asp, делает возможными экспрессию белка, указанного ниже (сигнальный пептид и предшественник протеазы), и секрецию зрелой протеазы Asp.
MTPRTVTRALAVATAAATLLAGGMAAQANEPAPPGSASAPPRLAEKLDPDLLEAMER
DLGLDAEEAAATLAFQHDAAETGEALAEELDEDFAGTWVEDDVLYVATTDEDAVEEV
EGEGATAVTVEHSLADLEAWKTVLDAALEGHDDVPTWYVDVPTNSVVVAVKAGAQD
VAAGLVEGADVPSDAVTFVETDETPRTMFDVIGGNAYTIGGRSRCSIGFAVNGGFITAG
HCGRTGATTANPTGTFAGSSFPGNDYAFVRTGAGVNLLAQVNNYSGGRVQVAGHTAA
PVGSAVCRSGSTTGWHCGTITALNSSVTYPEGTVRGLIRTTVCAEPGDSGGSLLAGNQA
QGVTSGGSGNCRTGGTTFFQPVNPILQAYGLRMITTDSGSSP (SEQ ID NO:17)
Конструирование 189 библиотек сайт-насыщающего мутагенеза Asp (SSML) завершали с использованием экспрессирующего вектора pHPLT-ASP-C1-2 в качестве матрицы. Все праймеры мутагенеза, используемые в этих экспериментах, содержали тройную ДНК-последовательность, кодирующую NNS (N = A, C, T или G; и S = C или G), в положении, которое соответствует кодону зрелой последовательности Asp, который должен быть мутирован, и гарантировали случайное включение нуклеотидов в этом положении. Конструирование каждой библиотеки начинали с двух ПЦР-амплификаций с использованием праймера pHPLT-BglII-FW и специфического обратного праймера мутагенеза, и праймера pHPLT-BglII-RV и специфического обратного праймера мутагенеза (одинаковые положения для праймеров мутагенеза). Примерный перечень конкретных последовательностей прямого и обратного праймеров описан в WO 2005/052146, включенном здесь в качестве ссылки, так как он относится к последовательностям праймеров. Последовательность праймера pHPLT-BglII-FW представлена в SEQ ID NO:18 (GCAATCAGATCTTCCTTCAGGTTATGACC); а последовательность праймера pHPLT-BglII-RV представлена в SEQ ID NO:19 (GCATCGAAGATCTGATTGCTTAACTGCTTC).
ДНК-полимеразу высокой точности Platinum Taq (Invitrogen. Catalog No. 11304-029) использовали для ПЦР-амплификации (0,2 мкМ праймеры, 20-30 циклов) согласно протоколу, обеспечиваемому изготовителем. Вкратце, 1 мкл амплифицированного ДНК-фрагмента обеих специфических ПЦР-смесей, обе из которых нацелены на один и тот же кодон, добавляли к 48 мкл свежеприготовленного раствора для ПЦР-реакции вместе с праймерами pHPLT-BglII-FW и pHPLT-BglII-RV. Эта ПЦР-амплификация слияния (22 цикла) приводила к линейному ДНК-фрагменту pHPLT-ASP-c1-2 с конкретным зрелым кодоном Asp, мутированным случайным образом, и уникальным сайтом рестрикции BglII на обоих концах. Очистка этого ДНК-фрагмента (набор для очистки ПЦР Qiagen, Catalog No. 28106), расщепление его BglII, выполнение дополнительной стадии очистки и реакции лигирования (ДНК-лигаза Т4 Invitrogen (Catalog No. 15224-025) генерировали кольцевую и мультимерную ДНК, которую затем трансформировали в B. subtilis (ΔaprE, ΔnprE, oppA, ΔspoIIE, degUHy32, ΔamyE::xylR,pxylA-comK). Для каждой библиотеки, после ночного инкубирования при 37°С, 96 отдельных колоний выскребали из чашек с агаром с инфузионным раствором для сердца с 20 мг/л неомицина и выращивали в течение 4 дней при 37°С в среде MOPS с 20 мг/мл неомицина и 1,25 г/л дрожжевого экстракта (см. WO 03/062380, включенный здесь в качестве ссылки в отношении точного приготовления среды, используемой здесь). Проводили анализ последовательности (BaseClear) и экспрессию протеазы определяли для каждой колонии для целей скрининга. Номера библиотек находились в диапазоне 1-189, причем каждый номер обозначает кодон зрелой последовательности, который был мутирован случайным образом. После отбора каждая библиотека включала максимально 19 вариантов протеазы Asp.
ПРИМЕР 5
Генерирование вариантных протеаз посредством сайт-направленного мутагенеза
В этом примере описаны способы генерирования SEL nprE с использованием набора QUIKCHANGE® для направленного на множественные сайты мутагенеза (Stratagene). Однако способы, обеспеченные здесь, пригодны для получения SEL и других представляющих интерес ферментов (например, Asp). Как описано выше в примере 4, вектор pUBnprE, содержащий экспрессионную кассету nprE, служил в качестве источника матричной ДНК для генерирования SEL nprE и вариантов NprE. Основное различие между этими двумя способами заключается в том, что этот способ требует амплификации всего вектора с использованием комплементарных сайт-направленных мутагенных праймеров.
Материалы
Штамм Bacillus, содержащий вектор pUBnprE
Набор Qiagen Plasmid Midi Kit (Qiagen Catalog No. 12143)
Готовый для лизиса лизоцим (Epicentre Catalog No. R1802M)
Набор метилазы dam (New England Biolabs Catalog No. M0222L)
Набор для извлечения ДНК Zymoclean Gel (Zymo Research Catalog No. D4001)
Сайт-направленные мутагенные праймеры nprE, в пределах шкалы 100 нмоль, 5'-фосфорилированные, очищенные электрофорезом в ПААГ (Integrated DNA Technologies)
Набор для направленного на множественные сайты мутагенеза QUIKCHANGE® (Stratagene Catalog No. 200514)
Термоциклер MJ Research PTC-200 Peltier (Bio-Rad Laboratories)
1,2% агарозные гели для электрофореза (Invitrogen Catalog No. G5018-01)
Набор для амплификации TempliPhi (GE Healthcare Catalog No. 25-6400-10)
Компетентные клетки B. subtilis (ΔaprE, ΔnprE, oppA, ΔspoIIE, degUHy32, ΔamyE::xylR,pxylA-comK).
Способы
Для получения плазмид pUBnprE, содержащих одну мутацию (идентифицированную посредством скрининга на SEL nprE, как описано выше в примере 4 и в Заявке на патент США с порядковым номером 11/581102, включенном здесь в качестве ссылки), отдельную колонию каждого представляющего интерес штамма Bacillus использовали для инокуляции пробирки на 5 мл с LB+10 м.д. неомицина (например, культуры-закваски). Эту культуру выращивали при 37°C со встряхиванием при 225 об/мин в течение 6 часов. Затем 100 мл свежего LB+10ppm неомицина инокулировали с 1 мл исходной культуры. Эту культуру выращивали в течение ночи при 37оС со встряхиванием при 225 об/мин. После этого инкубирования осадок клеток собирали достаточным центрифугированием для обеспечения осадка клеток. Этот осадок клеток ресуспендировали в 10 мл буфере Р1 (Qiagen Plasmid Midi Kit). Затем, к этому ресуспендированному осадку клеток добавляли 10 мкл готового для лизиса лизоцима и инкубировали при 37оС в течение 30 мин. Протокол набора Qiagen Plasmid Midi Kit продолжали с использованием 10 мл буфера Р2 и Р3 для соответствия увеличенному объему культуры клеток. После выделения из Bacillus каждой плазмиды pUBnprE, содержащей отдельную мутацию nprE, определяли концентрацию каждой плазмиды. Затем эти плазмиды dam метилировали с использованием набора для dam метилазы (New England Biolabs) согласно инструкциям изготовителя для метилирования приблизительно 2 мкг каждой плазмиды pUBnprE на пробирку. Набор для извлечения ДНК в геле Zymoclean Gel использовали для очистки и концентрирования dam-метилированных плазмид pUBnprE. Затем эти dam-метилированные плазмиды pUBnprE определяли количественно и разбавляли до рабочей концентрации 50 нг/мкл для каждой. Смешанные сайт-направленные мутагенные праймеры готовили раздельно для каждой реакции. Например, при использовании плазмиды pUBnprE T14R в качестве источника матрицы, пробирка со смешанными сайт-направленными мутагенными праймерами будет содержать 10 мкл nprE-S23R, 10 мкл nprE-G24R, 10 мкл nprE-N46K и 10 мкл nprE-T54R (все праймеры при 10 мкМ, каждый). ПЦР-реакцию с использованием набора для направленного на множественные сайты мутагенеза QuikChange (Stratagene) выполняли согласно инструкциям изготовителя (например, 1 мкл dam-метилированной плазмиды pUBnprE, содержащей одну мутацию (50 нг/мкл), 2 мкл сайт-направленных мутагенных праймеров nprE (10 мкМ), 2,5 мкл 10x буфера QuikChange Multi Reaction, 1 мкл dNTP-смеси, 1 мкл смеси ферментов QuikChange Multi (2,5 Е/мкл) и 17,5 мкл дистиллированной, автоклавированной воды с получением 25 мкл общей реакционной смеси). Библиотеки вариантов nprE амплифицировали с использованием следующих условий: 95°C, в течение 1 мин (только в 1-м цикле), затем 95°C в течение 1 мин, 55°C в течение 1 мин, 65°C в течение 13,5 мин, и повторение циклов 29 раз. Продукт реакции хранили при 4°С в течение ночи. Затем эту реакционную смесь подвергали обработке рестриктазой DpnI (поставляемой с набором QUIKCHANGE® для направленного на множественные сайты мутагенеза) для расщепления исходной плазмиды pUB-nprE с использованием протокола изготовителя (т.е. 1,5 мкл рестриктазы DpnI добавляли в каждую пробирку и инкубировали при 37°С в течение 3 часов; затем 2 мкл расщепленной DpnI реакционной смеси ПЦР анализировали на 1,2% геле для электрофореза для гарантии того, что ПЦР-реакция работала и что исходная матрица была деградирована). Затем использовали амплификацию по типу катящегося кольца (разматывающегося рулона) для генерирования больших количеств ДНК для увеличения размера библиотеки мультивариантов nprE с использованием протокола изготовителя (т.е. 1 мкл DpnI-обработанная ПЦР мутагенеза QuikChange, направленная на множественные сайты, 5 мкл буфера для проб TempliPhi, 5 мкл реакционного буфера TempliPhi и 0,2 мкл смеси ферментов TempliPhi, для общей реакции ~11 мкл; инкубированная при 30°C в течение 3 часов; реакционную смесь TempliPhi разбавляли добавлением 200 мкл дистиллированной, автоклавированной воды и кратковременно перемешивали с использованием вортекса). Затем 1,5 мкл разбавленного материала TempliPhi трансформировали в компетентные клетки B. subtilis и мультиварианты nprE отбирали для использования чашек с LA+10 м.д. неомицина + 1,6% сепарированное молоко. Колонии выскребали и затем секвенировали для идентификации различных комбинаций библиотек вариантов nprE.
Таблица 5-1 обеспечивает название праймера и последовательность, используемую в этих экспериментах. Интегрированные DNA Technologies синтезировали все эти праймеры (на шкале 100 нмоль, 5'-фосфорилированные и очищенные электрофорезом в ПААГ). Дополнительные праймеры мутагенеза описаны в Заявке на патент с порядковым номером 11/581102, включенной здесь в качестве ссылки.

Этот пример описывает также получение лэддеров (лестниц) зарядов ферментов и комбинаторных библиотек зарядов как для протеаз, так и для амилаз.
Лэддеры зарядов ферментов
Множественные варианты белков, охватывающие диапазон представляющих интерес физических свойств, отбирают из существующих библиотек или генерируют способами сайт-направленного мутагенеза, как известно в данной области (см., например, Заявки на патент США с порядковыми номерами 10/576331, 11/581102 и 11/583334). Затем этот определенный набор белков-зондов анализируют в представляющем интерес тесте.
Примеры вариантов лэддеров зарядов протеаз показаны в следующих таблицах и анализированы, как описано здесь. В этих таблицах изменение заряда выражено относительно фермента дикого типа.



Аминокислотную последовательность зрелой протеазы FNA использовали в качестве основы для получения описанных здесь библиотек вариантов:
AQSVPYGVSQIKAPALHSQGYTGSNVKVAVIDSGIDSSHPDLKVAGGASMVPSETN
PFQDNNSHGTHVAGTVAALNNSIGVLGVAPSASLYAVKVLGADGSGQYSWIINGIE
WAIANNMDVINMSLGGPSGSAALKAAVDKAVASGWWAAAGNEGTSGSSSTVGY
PGKYPSVIAVGAVDSSNQRASFSSVGPELDVMAPGVSIQSTLPGNKYGALNGTSMA
SPHVAGAAALILSKHPNWTNTQVRSSLENTTTKLGDSFYYGKGLINVQAAAQ (SEQ ID NO:25).

Аминокислотную последовательность зрелой протеазы GG36 использовали в качестве основы для получения описанных здесь библиотек вариантов:
AQSVPWGISRVQAPAAHNRGLTGSGVKVAVLDTGISTHPDLNIRGGASFVPGEPSTQDG
NGHGTHVAGTIAALNNSIGVLGVAPSAELYAVKVLGASGSGSVSSIAQGLEWAGNNGM
HVANLSLGSPSPSATLEQAVNSATSRGVLVVAASGNSGAGSISYPARYANAMAVGATD
QNNNRASFSQYGAGLDIVAPGVNVQSTYPGSTYASLNGTSMATPHVAGAAALVKQKN
PSWSNVQIRNHLKNTATSLGSTNLYGSGLVNAEAATR (SEQ ID NO:26).
Примеры вариантов лэддеров зарядов амилаз показаны в следующих таблицах и анализированы, как описано здесь. В этих таблицах изменение заряда выражено относительно фермента дикого типа.
Последовательность гена AmyS была предоставлена Gene Oracle (Mountain View, CA) для синтеза 25 вариантов лэддеров заряда, показанных в таблице 5-5. Gene Oracle синтезировал и клонировал варианты AmyS в вектор pGov4 и трансформировал их в E. coli. Для каждого варианта были предоставлены ДНК, выделенные из минипрепаратов, а также агаровый столбик.
Эти варианты ПЦР-амплифицировали и клонировали в экспрессирующий вектор pHPLT B. subtilis.

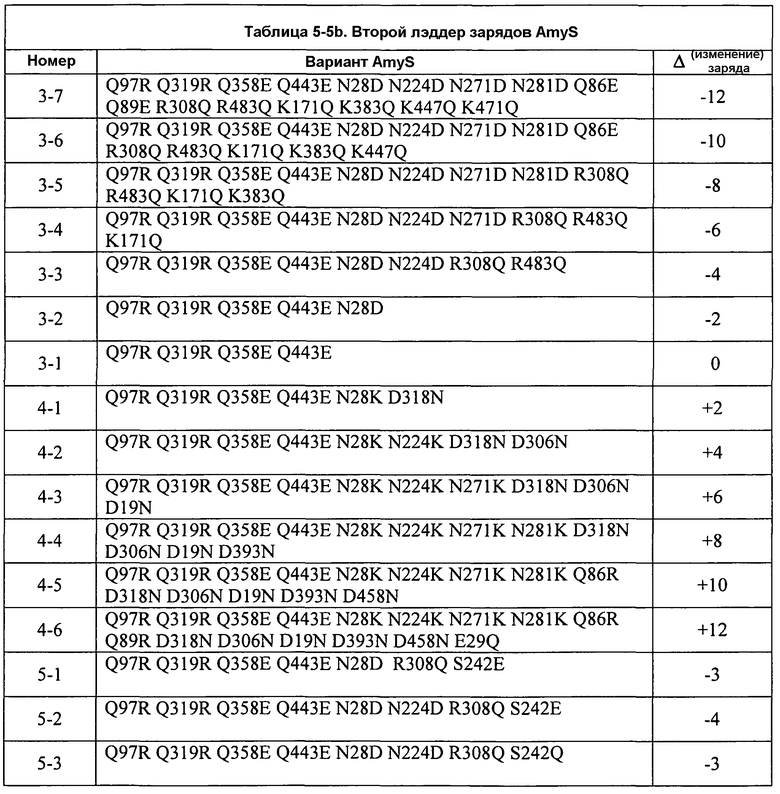
Аминокислотную последовательность зрелой амилазы AmyS использовали в качестве основы для получения описанных здесь библиотек вариантов:
AAPFNGTMMQYFEWYLPDDGTLWTKVANEANNLSSLGITALWLPPAYKGTSRSD
VGYGVYDLYDLGEFNQKGTVRTKYGTKAQYLQAIQAAHAAGMQVYADVVFDHK
GGADGTEWVDAVEVNPSDRNQEISGTYQIQAWTKFDFPGRGNTYSSFKWRWYHF
DGVDWDESRKLSRIYKFRGIGKAWDWEVDTENGNYDYLMYADLDMDHPEVVTE
LKNWGKWYVNTTNIDGFRLDAVKHIKFSFFPDWLSYVRSQTGKPLFTVGEYWSY
DINKLHNYITKTNGTMSLFDAPLHNKFYTASKSGGAFDMRTLMTNTLMKDQPTLA
VTFVDNHDTEPGQALQSWVDPWFKPLAY AFILTRQEGYPCVFYGDYYGIPQYNIPS
LKSKIDPLLIARRDYAYGTQHDYLDHSDIIGWTREGVTEKPGSGLAALITDGPGGS
KWMYVGKQHAGKVFYDLTGNRSDTVTINSDGWGEFKVNGGSVSVWVPRKTTVS
TIARPITTRPWTGEFVRWTEPRLVAWP (SEQ ID NO:27).
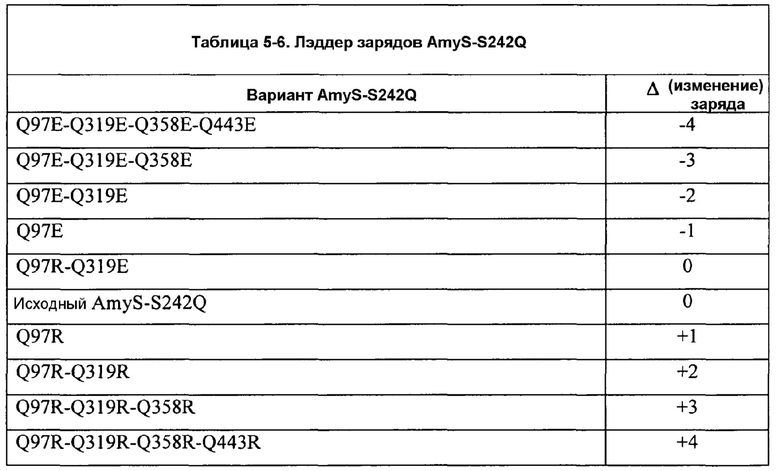
Аминокислотную последовательность зрелой укороченной амилазы S242Q с замененными аминокислотами, показанными курсивом, использовали в качестве основы для получения описанных здесь библиотек вариантов:
AAPFNGTMMQYFEWYLPDDGTLWTKVANEANNLSSLGITALWLPPAYKGTSRSD
VGYGVYDLYDLGEFNQKGTVRTKYGTKAQYLQAIQAAHAAGMQVYADVVFDHK
GGADGTEWVDAVEVNPSDRNQEISGTYQIQAWTKFDFPGRGNTYSSFKWRWYHF
DGVDWDESRKLSRIYKFRGIGKAWDWEVDTENGNYDYLMYADLDMDHPEVVTE
LKNWGKWYVNTTNIDGFRLDAVKHIKFGFFPDWLSYVRSQTGKPLFTVGEYWSY
DINKLHNYITKTNGTMSLFDAPLHNKFYTASKSGGAFDMRTLMTNTLMKDQPTLA
VTFVDNHDTEPGQALQSWVDPWFKPLAYAFILTRQEGYPCVFYGDYYGIPQYNIPS
LKSKIDPLLIARRDYAYGTQHDYLDHSDHGWTREGVTEKPGSGLAALITDGPGGS
KWMYVGKQHAGKVFYDLTGNRSDTVTINSDGWGEFKVNGGSVSVWVPRKTT
(SEQ ID NO:28).
Комбинаторные библиотеки зарядов ферментов
Генерирование комбинаторных библиотек зарядов субтилизина B. lentus (=GG36)
Плазмиду pAC-GG36ci, содержащую ген GG36 с улучшенным кодоном, отсылали в DNA 2.0 Inc. (Menlo Park, CA) для генерирования комбинаторных библиотек зарядов (CCL). Их обеспечивали также штаммом Bacillus subtilis (генотип: ΔaprE, ΔnprE, ΔspoIIE, amyE::xylRPxylAcomK-phleo) для трансформации. Кроме того, был сделан запрос в DNA2.0 Inc. в отношении генерирования позиционных библиотек в каждом из четырех сайтов в протеазе GG36, которые показаны в таблице 5-7. Варианты поставлялись в виде глицериновых исходных растворов в 96-луночных планшетах.
Эти CCL GG36 конструировали идентификацией четырех хорошо распределенных, экспонированных на поверхности, незаряженных полярных аминокислотных остатков вне активного центра. Этими остатками являются Ser-85, Gln-107, Ser-182 и Asn-242 (остатки 87, 109, 188 и 248 в нумерации BPN). Создавали 81-членную комбинаторную библиотеку (G-1 - G-81) выполнением всех комбинаций трех возможностей в каждом сайте: аминокислоты дикого типа, аргинина или аспарагиновой кислоты.

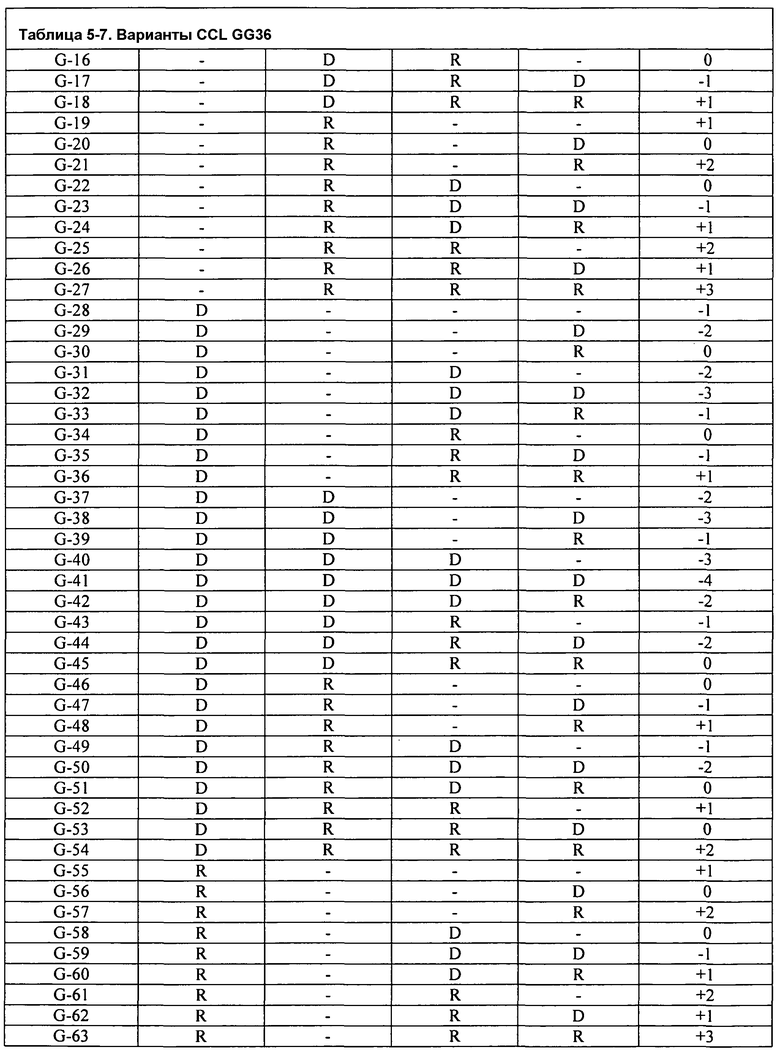
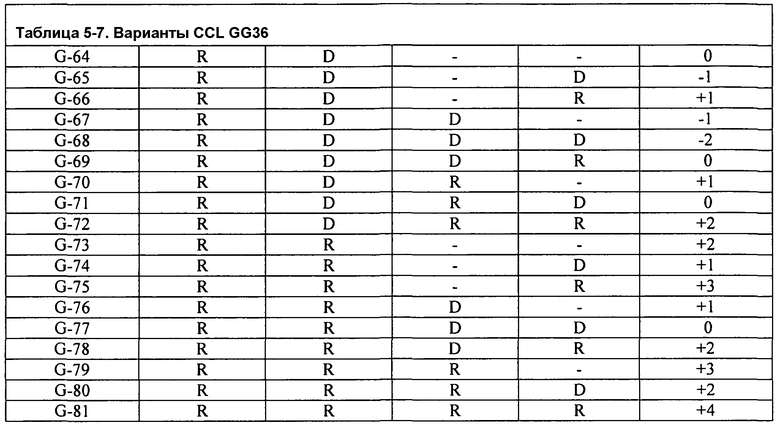
Генерирование CCL BPN'-Y217L (=FNA) субтилизина B. amyloliquefaciens
Плазмиду pAC-FNAre, содержащую ген FNA, отсылали в DNA 2.0 Inc. (Menlo Park, CA) для генерирования комбинаторных библиотек зарядов (CCL). Их обеспечивали также штаммом Bacillus subtilis (генотип: ΔaprE, ΔnprE, ΔspoIIE, amyE::xylRPxylAcomK-phleo) для трансформации. Был сделан запрос в DNA 2.0 Inc. в отношении генерирования позиционных библиотек в каждом из четырех сайтов протеазы FNA, которые показаны в таблице 5-8. Варианты поставлялись в виде глицериновых исходных растворов в 96-луночных планшетах.
Комбинаторную библиотеку зарядов субтилизина BPN'-Y217L конструировали идентификацией четырех хорошо распределенных, экспонированных на поверхности, незаряженных полярных аминокислотных остатков вне активного центра. Этими остатками являются Ser-87, Asn-109, Ser-188 и Ser-248. Создавали 81-членную комбинаторную библиотеку (F-1 - F-81) выполнением всех комбинаций трех возможностей в каждом сайте: аминокислоты дикого типа, аргинина или аспарагиновой кислоты.
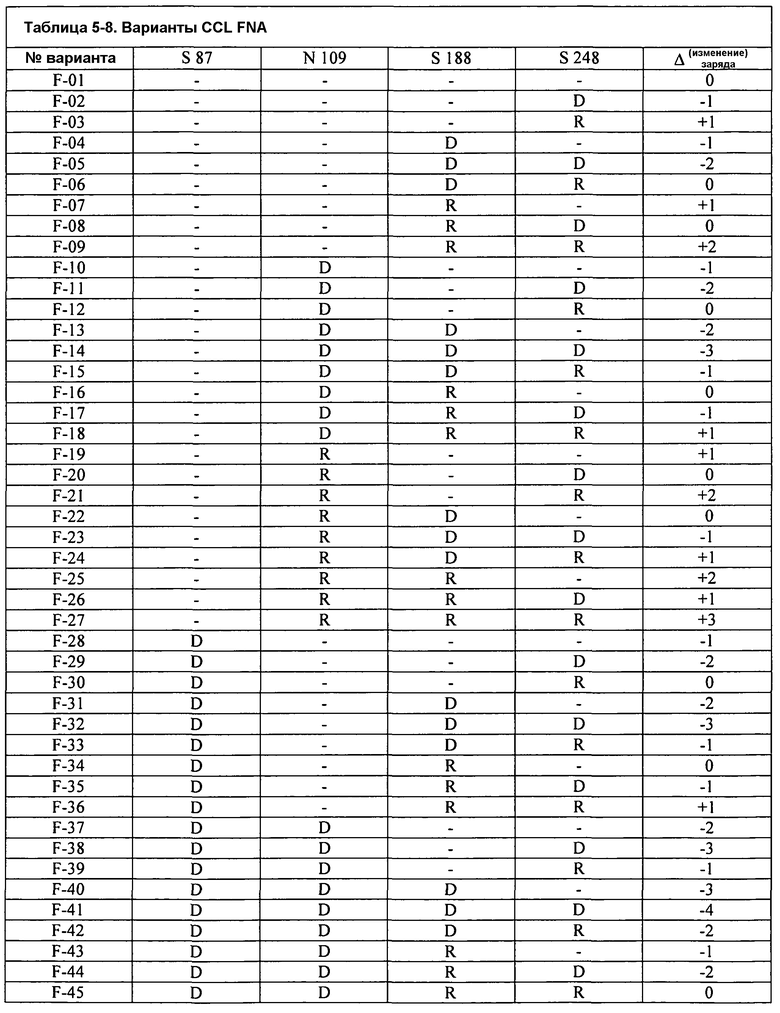
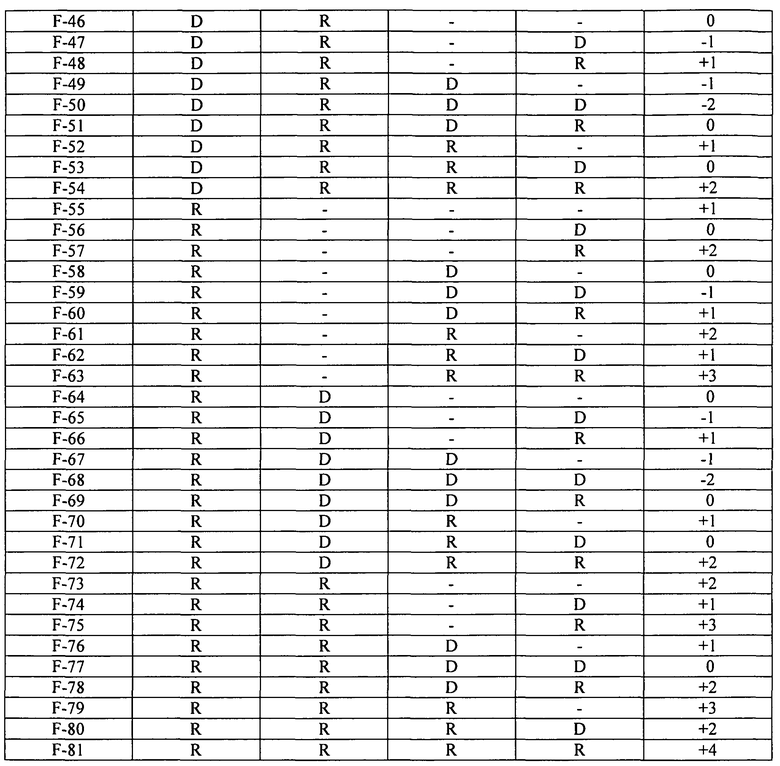
Генерирование CCL AmyS-S242Q B. stearothermophilus
ДНК плазмиды AmyS-S242Q выделяли из трансформированного штамма B. subtilis (генотип: ΔaprE, ΔnprE, amyE::xylRPxylAcomK-phleo) и отсылали в DNA2.0 Inc. в качестве шаблона для конструирования CCL. Был сделан запрос в DNA2.0 Inc. (Mountain View, CA) в отношении генерирования позиционных библиотек в каждом из четырех сайтов в амилазе AmyS-S242Q, которые показаны в таблице 5-9. Варианты поставлялись в виде глицериновых исходных растворов в 96-луночных планшетах.
Комбинаторную библиотеку зарядов AmyS S242Q конструировали идентификацией следующих четырех остатков: Gln-97, Gln-319, Gln-358 и Gln-443. Четырехсайтную, 81-членную CCL создавали выполнением всех комбинаций трех возможностей в каждом сайте: аминокислоты дикого типа, аргинина или аспарагиновой кислоты.


ПРИМЕР 6
Очистка и характеристика вариантных протеаз
Этот пример описывает способы, используемые для очистки протеаз, экспрессируемых трансформированным B. subtilis предыдущих примеров.
После 36 часов инкубации при 37°C ферментационный бульон извлекали и центрифугировали при 12000 об/мин (центрифуга SORVALL® модели RC5B). Секретируемые нейтральные металлопротеазы выделяли из культуральной жидкости и концентрировали приблизительно в 10 раз с использованием фильтрующей системы Amicon 8400 с пределом отсечения BES (полиэфирсульфоном) 10 кДа.
Этот концентрированный супернатант диализовали в течение ночи при 4°C против 25 мМ MES-буфера, рН 5,4, содержащего 10 мМ NaCl. Затем диализат наносили на катионообменную колонку Poros HS20 (общий объем ~83 мл; связывающая способность ~4,5 г белка/мл колонки; Waters), как описано ниже. Эту колонку предварительно уравновешивали 25 мМ MES-буфером, pH 5,4, содержащим 10 мМ NaCl. Затем на эту колонку наносили приблизительно 200-300 мл пробы. Связанный белок элюировали с использованием градиента рН 5,4-6,2 в объеме 10 колонок MES-буфера. Элюцию белка осуществляли при рН 5,8-6,0 и оценивали с использованием протеолитической активности, как описано здесь, и электрофорезом в 10% (м/о) ДСН NUPAGE®-РААГ (Novex). Затем содержащие нейтральную металлопротеазу объединяли. Хлоридные соли кальция и цинка в соотношении 3:1 добавляли перед доведением рН до 5,8. Для очистки белка использовали Perceptive Biosystems BIOCAD® Vision (GMI).
Было определено, что очищенный белок, оцениваемый с использованием электрофореза в 10% (м/о) NUPAGE® ДСН-ПААГ, является гомогенным, имеющим чистоту более 95%. Обычно, очищенные препараты обнаруживали незначительную активность сериновой протеазы при оценке с использованием стандартного анализа сериновых протеаз с субстратом N-сукцинил-L-Ala-L-Ala-L-Pro-L-Phe-п-нитроанилидом (Bachem). Этот белок готовили для хранения с использованием 25 мМ MES-буфера, pH 5,8, содержащего 1 мМ хлорид цинка, 4 мМ хлорид кальция и 40% пропиленгликоль.
ПРИМЕР 7
Моющая эффективность
Этот пример описывает тестирование вариантов NprE и ASP в анализе микролоскутков (маленьких образцов ткани) с BMI (кровь, молоко, чернила) при 0,25 мкг/мл в жидком детергенте (анализе эффективности BMI-TIDE® 2X Ultra Clean Breeze").
Таблица 7-1а суммирует данные, полученные для NprE дикого типа (WT) и различных вариантов NprE. Эта таблица дает перечни положений и замен аминокислот, показывает эффективность очистки BMI и изменение суммарного заряда относительно NprE WT.
Таблица 7-1b дает перечни мутаций, содержащихся в вариантах NprE, которые имеют обозначения "AA" в таблице 7-1а.

Таблицы 7-2a и 7-2b суммируют данные, полученные для ASP дикого типа (WT) и различных вариантов ASP. Эти таблицы дают перечень положений и замен аминокислот, эффективность очистки BMI и изменение суммарного заряда относительно ASP WT.


Таким образом, мутации заряда поверхности в протеазе влияют на ее моющую эффективность на BMI, и оптимизация заряда поверхности белка увеличивает его моющую эффективность.
ПРИМЕР 8
Стабильность в отношении LAS
В этом примере измеряли стабильность в отношении LAS после инкубации тест-протеазы в присутствии 0,06% LAS (додецилбензолсульфоната натрия), и остаточную активность определяли с использованием AAPF-анализа.
Реагенты:
Додецилбензолсульфонат, натриевая соль (=LAS): Sigma D-2525
TWEEN®-80: Sigma P-8074
Трис-буфер (свободная кислота): Sigma T-1378); 6,35 г растворяют в приблизительно 960 мл воды; рН доводят до 8,2 4н HCl. Конечная концентрация Триса равна 52,5 мМ.
Исходный раствор LAS: готовят 10,5% раствор LAS в воде MQ (=10,5 г на 100 мл MQ)
Трис-буфер-100 мМ/pH 8,6 (100 мМ Трис/0,005% Tween80)
Трис-Са-буфер, pH 8,6 (100 мМ Трис/10 мМ CaCl2/0,005% Tween80)
Оборудование:
Плоскодонные MTP: Costar (#9017)
Biomek FX
Многоканальная пипетка ASYS
МТР-ридер Spectramax
Инкубатор/Шейкер iEMS
Инкубатор/Шейкер Innova 4330
Многоканальная пипетка Biohit
Термостат/Шейкер BMG
Способ:
10 мкл 0,063% раствора LAS готовили в 52,5 мМ Трис-буфере рН 8,2. Рабочий раствор AAPF готовили добавлением 1 мл 100 мг/мл исходного раствора AAPF (в ДМСО) к 100 мл (100 мМ) Трис-буфера, рН 8,6. Для разведения супернатантов плоскодонные планшеты наполняли буфером для разведения и аликвоту супернатанта добавляли и хорошо перемешивали. Коэффициент разведения зависел от концентрации ASP-контролей в планшетах для выращивания (AAPF-активности). Желаемая концентрация белка была равна 80 м.д.
Десять мкл разведенного супернатанта добавляли к 190 мкл 0,063% LAS-буфера на лунку. Планшет МТР закрывали с использованием клейкой ленты, встряхивали в течение нескольких секунд и помещали в термостат (Innova 4230) при 25°C на 60 минут при встряхивании при 200 об/мин. Начальную активность (t=10 минут) определяли после 10 минут инкубирования перенесением 10 мкл этой смеси в каждой лунке в свежий MTP, содержащий 190 мкл рабочего раствора AAPF. Эти растворы хорошо смешивали и активность AAPF измеряли с использованием МТР-ридера (20 считываний в течение 5 минут и при 25°C).
Конечную активность (t=60 минут) определяли взятием другой порции 10 мкг раствора из инкубируемого планшета после 60 минут инкубирования. Затем активность AAPF определяли, как описано выше. Расчеты выполняли следующим образом: %-ная остаточная активность была равна [величина t-60] * 100/[величина t-10].
В некоторых вариантах осуществления, предпочтительным путем анализа вариантов является анализ с использованием различия свободной энергии для варианта против исходного белка в представляющем интерес процессе. Для конкретного процесса, изменение свободной энергии Гиббса относительно исходного фермента (ΔΔ G) получают следующим образом:
ΔΔG = -RT ln (kвариант/kисходный)
где kвариант является константой скорости для вариантного фермента, а kисходный является константой скорости исходного фермента, R обозначает константу уравнения состояния газа и Т обозначает абсолютную температуру. Большинство анализов не позволяли определить истинные свободные энергии, так что изменение средней (кажущейся) свободной энергии (ΔΔGapp) определяют следующим образом:
ΔΔGapp = -RT ln (Pвариант/Pисходный)
где Pвариант обозначает величину эффективности для варианта, а Pисходный обозначает величину эффективности для исходного фермента при тех же самых условиях. Для расчета величин ΔΔGapp LAS-стабильности остаточную активность дикого типа определяют как меру эффективности молекулы дикого типа (Pисходный), а остаточную активность варианта определяют как эффективность вариантной молекулы (Pвариант). Отрицательная величина ΔΔGapp указывает на улучшение LAS-стабильности варианта, тогда как положительная величина ΔΔGapp является показателем варианта с уменьшенной LAS-стабильностью.
Затем рассчитывали среднюю величину ΔΔGapp для накопителей изменения заряда относительно фермента дикого типа, для диапазона +2 - -7. Из этого анализа видно, что увеличение общего отрицательного заряда фермента ASP увеличивает стабильность этого фермента в отношении LAS.
ПРИМЕР 9
Эффективность фермента
Этот пример описывает тестирование вариантов ASP в анализе BMI (кровь, молоко, чернила) микролоскутков (маленьких образцов ткани) при 1,0 мкг/мл в детергенте AATCC HDL или 5 мМ HEPES-буфере при варьирующейся ионной силе. Описано также тестирование вариантов FNA и GG36 в анализах BMI на микролоскутках и анализах с вареным яйцом в детергентах, представляющих различную географию рынка (например, различные рН, Т и/или жесткость воды), в применениях как для стирки, так и для автоматической мойки посуды. Этот пример описывает также тестирование вариантов альфа-амилазы в очищающих приложениях, а также в ожижении крахмала. Использовали способы, обеспеченные в примере 1 (см. "Enzyme Performance Assays" и "Corn Four Hydrolysis").
Как показано на фиг.1А, имеется оптимальное изменение суммарного заряда для очищающей эффективности для ASP в детергенте AATCC HDL. Эффективность измеряют по относительной очищающей эффективности, наблюдаемой в BMI-микроанализах с микролоскутками. Величина около 1,0 указывает наилучшую очищающую эффективность в этом анализе. Как видно из этого чертежа, накапливание очень отрицательных (-5) или положительных (+3) зарядов относительно дикого типа приводит к слабой очищающей эффективности. Имеется явный оптимум заряда для очищающей эффективности, сосредоточенный при -2 относительно ASP дикого типа. Это является примером оптимизации физического свойства белка (например, суммарного заряда) для улучшения конкретного выхода или пользы (например, очищающей эффективности в жидком детергенте для стирки). Оптимум заряда, идентифицированный с этим ограниченным набором белков-зондов, согласуется с оптимальным зарядом, наблюдаемым при измерении всей комбинаторной библиотеки зарядов ASP, как показано на фиг.1В. Таким образом, применение белков-зондов предсказывает поведение всей библиотеки.
Согласно теории Debye-Hϋckel (Israelachivili, Intermolecular and Surface Forces, Second Edition: With Applications to Colloidal and Biological Systems, Academic Press 2nd Ed. [1992]), электростатические взаимодействия управляются прежде всего прочностью двухслойных сил между взаимодействующими молекулярными частицами при константном потенциале или константном заряде (ферментов, субстратов, ткани и детергента), их размером и диэлектрической константой окружающей среды. Для характеристики электростатического поведения частиц в комплексной среде, такой как препарат детергента, их взаимодействие в восстановленной среде, имеющей ту же самую длину скринига Debye, является достаточным. Это выполняется выбором буфера для соответствия рН и проводимости рН и проводимости детергента при условиях мойки. Как показано на фиг.1А, скрининг лэддера зарядов ASP в этом буфере точно предсказал оптимум заряда при -2, наблюдаемый с детергентом AATCC (черные кружки). Фиг.2 изображает относительное удаление пятна BMI как функцию изменения заряда относительно ASP дикого типа, в 5 мМ HEPES-буфере при рН 8,0 с варьирующимися количествами постороннего электролита, в этом случае NaCl. Добавление 2,5 мМ NaCl к этому буферу соответствует pH и проводимости типичных условий мойки в Северной Америке. Добавление более высоких концентраций NaCl является характерным для Японских и Европейских условий мойки, обычно более высоких по ионной силе вследствие как увеличенной жесткости воды, так и увеличенных концентраций детергента. Таким образом, оптимум заряда ASP является функцией окружающего раствора (например, препарата детергента).
Имеются два признака, которые становятся сразу же очевидными. Во-первых, использование модельной системы, состоящей из ограниченного количества белков-зондов для конкретного физического свойства (например, вариантов ASP с лэддером зарядов) в восстановленной буферной среде соответствующих рН и проводимости, является предсказывающим поведение большой библиотеки ASP, подвергаемой скринингу при детергентных условиях. Действительно, оптимум заряда, показанный на фиг.1А, измеренный в буфере, содержащем 2,5 мМ NaCl (белые кружки), является идентичным оптимуму, наблюдаемому для этого лэддера зарядов ASP, подвергнутого скринингу в детергенте AATCC в условиях мойки в Северной Америке. Во-вторых, местоположение оптимума заряда в сильной степени зависит от ионной силы. С дальнейшим добавлением NaCl оптимум заряда смещается в сторону вариантов с положительным зарядом относительно ASP дикого типа. Вкратце, использование белков-зондов с лэддером зарядов делает возможным быстрое предсказание эффективности различных вариантов фермента в препаратах, репрезентативных для различных географических рынков.
Приведенные выше наблюдения относятся также и к другим сериновым протеазам, таким как субтилизины FNA и GG36. Например, фиг.3А и 3В показывают оптимальный заряд для FNA и GG36 соответственно, в очищающей эффективности в условиях североамериканской стирки с использованием детергента TIDE 2X. Левые Y-оси показывают очищающую микролоскутки эффективность, где более высокое число указывает лучшее удаление пятна BMI. Правые Y-оси показывает показатель эффективности, определяемый как очищающая эффективность вариантов (черные символы) относительно исходной молекулы (белые символы). Горизонтальные линии указывают показатель эффективности при отклонении либо 2, либо 3 выше фона этого анализа. Комбинаторная библиотека (CCL) зарядов FNA проявляет оптимум заряда при изменениях заряда, равных нулю, в отношении исходного FNA, тогда как CCL GG36 проявляет оптимум при отрицательных двух зарядах относительно исходного GG36.
Фиг.4A, 4B, 5A и 5B демонстрируют, что местоположение оптимума заряда зависит от окружающего раствора, определяемого составом детергента, рН, температурой и ионной силой, обусловленной жесткостью воды и концентрацией детергента. Например, оптимум заряда для CCL FNA смещается разительно от нуля при североамериканских условиях стирки к более положительным зарядам при западноевропейских и японских условиях. Кроме того, оптимум заряда наблюдается как для жидких, так и для гранулированных (порошкообразных) препаратов детергентов для стирки. Подобным образом, оптимум заряда наблюдали как для CCL FNA, так и для CCL GG36 в детергенте для автоматической мойки посуды (ADW) в отношении (например, Reckitt Benckiser Calgonit 40°C, 12 gpg, pH 10) сваренного яйца в качестве субстрата фермента, как показано на фиг.6А и 6В.
Как показано во время разработки данного изобретения, очищающая эффективность вариантов зарядов протеазы (например, ASP, GG36, FNA и т.д.) в различных детергентах в значительной степени доминируется рН и проводимостью рабочего раствора. Конечная проводимость является мерой ионной силы и обусловлена жесткостью воды, концентрацией и составом детергентов. Например, имеется корреляция между очищающей эффективностью вариантов GG36 и FNA в отношении пятен сваренного яйца при европейском и североамериканском детергенте при проведении стирки при рН 10,6 и проводимости 3,0 мС/см. В частности, очищающая эффективность вариантов зарядов хорошо коррелирует при условии, что рН и проводимость являются одинаковыми. Это открытие делает возможным скрининг эффективности фермента с использованием конкретного детергента для экстраполяции этих результатов на другой детергент соответствующих рН и проводимости. Подобным образом, может подвергать скринингу эффективность фермента в буфере совпадающих рН и проводимости для экстраполяции этих результатов на детергент, проявляющий сходные рабочие рН и проводимость.
Имеется оптимум заряда для очищающей эффективности вариантов зарядов амилаз (например, AmyS-S242Q и AmyTS23t и т.д.) в очищающих приложениях, который в сильной степени зависит от рН и проводимости рабочего раствора. Конкретно, как определено во время разработки данного изобретения, варианты изменения положительного заряда S242Q являются лучшими для очистки микролоскутков от рисового крахмала при североамериканских условиях стирки (например, TIDE 2X), в то время как варианты изменения отрицательного заряда AmyTS23t являются лучшими для очистки микролоскутков от рисового крахмала при западноевропейских условиях стирки. Кроме того, эти наблюдения являются верными для амилазы, используемой в реакциях гидролиза крахмала. Как показано на фиг.7А, положительные варианты S242Q проявляют более высокую удельную активность в отношении гидролиза крахмала BODIPY в качестве субстратов.
Разжижение крахмала вариантами AmyS с лэддером зарядов определяли мониторингом конечной вязкости после разжижения кукурузного крахмала. Низкая величина вязкости указывает на разрушение полисахаридов крахмала. Как показано на фиг.7В, наблюдали оптимум заряда (например, -4 - -2) для разжижения. Варианты AmyS, которые были слишком отрицательными (например, -12 - -10), проявляли очень высокие конечные вязкости, а варианты, которые были слишком положительными (например, +6 или более), проявляли даже более высокие конечные вязкости (например, за пределами границ лабораторного инструментария вследствие перегрузки вращающего момента).
ПРИМЕР 10
Экспрессия белка
Этот пример описывает определение взаимосвязи между зарядом белка и экспрессией белка.
Получение вариантов ASP в масштабе ферментера на 14 литров
Проводили ряд периодических ферментаций с подпиткой в масштабе 14 литров для сравнения продукционных уровней вариантов комбинаторных библиотек зарядов протеазы ASP (R14I-N112E-T116E-R123F-R159F, R14I-N112E-T116E-R123F, R14I-N112E-T116E, R14I-N112E, R14I, R14I-D184T, R14I-D184T-T86K, R14I-T86K-D184T-A64K и R14I-T86K-D184T-A64K-Q81K), которые различаются по заряду от -5 до +3. Культуры посевного материала выращивали инокуляцией встряхиваемых колб без перегородок на 2 л, содержащих 600 мл культуральной среды (LB-бульон + 1% глюкоза + 20 мг/л неомицина) 1 мл исходного глицеринового раствора Bacillus subtilis, соответствующего каждому варианту. Эти культуры инкубировали при 37°С с перемешиванием при 175 об/мин во встряхиваемом термостате, пока OD550 не достигала 0,8-1,5. В этот момент, все культуры посевного материала переносили асептически в ферментеры на 14 литров, оборудованные интегрированным контроллером (регулятором) для мониторинга: температуры, процента растворенного кислорода (% DO), рН и перемешивания. Отходящие газы подвергали мониторингу при помощи подключенного масс-спектрофотометра. Используемая ферментационная среда (7 л) состояла из 10% соевой муки в буфере на основе фосфата, содержащем сульфат магния, следовые элементы и дополнительный неомицин при 20 мг/л. Начальные параметры ферментации устанавливали на: температуру 37°С, pH 6,8 (корректируемый гидроксидом аммония во время эксперимента), перемешивание при 750 об/мин, 40% DO (поддерживаемый во время эксперимента коррекцией воздуха и перемешивания), поток воздуха 11 slpm и давление 1 бар. Добавляли пеногаситель (Mazu DF204) при необходимости для контроля пенообразования. Периодический процесс с подпиткой с линейной подачей 0,5-2,1 г/мин глюкозы на протяжении 10 часов программировали (с использованием 60% глюкозы для подпитки) повышением рН в качестве пускового механизма. Взятие проб ферментации имело место каждые 4 часа, с отбором 15 мл полного бульона для выполнения следующих измерений: плотности клеток (измерением оптической плотности при 550 нм) на спектрофотометре, продуцирования варианта ASP, глюкозы, азота, фосфата и общего белка. Общее время эксперимента по ферментации находилось между 40 и 45 часами.
Измерение титра варианта ASP с использованием анализа Ааа-Pna
Пробы культур B. subtilis, полученные во время ферментации, анализировали на продуцирование вариантных протеаз ASP. Продуцируемые ферменты анализировали на активность против субстрата, N-сукцинил-Ala-Ala-Ala-п-нитроанилида (AAA-pNA). Этот анализ измерял продуцирование модифицированной протеазы в виде увеличения оптической плотности при 405 нм, происходящего из гидролиза и высвобождения п-нитроанилина (Estell et al, J Biol Chem, 260: 6518-6521 [1985]). Аликвоты осветленных супернатантов B. subtilis из ферментера анализировали в буфере, содержащем: 100 мМ Трис, 0,01 мМ CaC12, 0,005% Тритон Х-100, при рН 8,6. Стандарт протеазы ASP дикого типа служил для генерирования калибровочной кривой для расчета белка, продуцированного в г/л ферментационного бульона.
Фиг.8 изображает уровни экспрессии белков-зондов лэддеров зарядов ASP в Bacillus subtilis как функцию суммарного заряда относительно ASP дикого типа. Как видно из этого чертежа, накапливание крайне отрицательных (-5) или положительных (+3) зарядов относительно ASP дикого типа приводит к плохим уровням экспрессии. Применение белков-зондов с лэддером зарядов ASP позволяет быстро идентифицировать оптимальный суммарный заряд для улучшения экспрессии в конкретном организме-хозяине. В этом случае диапазон суммарного заряда между -2 и +1 относительно ASP дикого типа соответствует оптимальным уровням экспрессии. При самом оптимуме заряда наблюдали почти 4-кратное улучшение экспрессии для ASP (-2) в сравнении с вариантами, имеющими крайние изменения заряда. Эти наблюдения на уровне встряхиваемых колб подтверждали в масштабе ферментера на 14 литров. Таблица 8-1 показывает два измерения экспрессии в ферментерах на 14 литров приблизительный титр ASP при 40 часах, а также продуцирование ASP, рассчитанное из линейной части кривых экспрессии. Титры встряхиваемых колб обеспечены для ссылки в последнем столбце. Все титры нормализовали относительно уровней ASP-R14I. Диапазон изменения суммарного заряда -2 - +1 относительно ASP дикого типа соответствует оптимальным уровням экспрессии в масштабе ферментера. Это является другим примером оптимизации физического свойства белка, в этом случае суммарного заряда, для модуляции полностью отличающего преимущества, в этом случае экспрессии рекомбинантного белка.

Экспрессия и секреция белка в клетке-хозяине включает взаимодействие экспрессируемого белка с рядом белков хозяина. Оптимальное взаимодействие экспрессируемого белка с белками клетки-хозяина, в частности с ограничивающим скорость взаимодействием, является существенным для продуцирования белка. Это взаимодействие может быть оптимизировано модификацией заряда поверхности/гидрофобности этого экспрессируемого белка (или белка клетки-хозяина). Тем не менее, знание участвующего в этом механизма (участвующих в этом механизмов) не является обязательным для выполнения и применения данного изобретения.
ПРИМЕР 11
Стабильность в отношении LAS и хелатора
Этот пример описывает определение взаимосвязи между зарядом и стабильностью белка в реакционной среде, содержащей как анионогенное поверхностно-активное вещество, так и хелатор. Для определения протеазной активности подвергнутых и не подвергнутых стрессу проб использовали анализ suc-AAPF-pNA.
Использованные реагенты включали: контрольный буфер: 50 мМ HEPES, 0,005% Твин-80, pH 8,0; и стрессовый буфер: 50 мМ HEPES, 0,1% (м/о) LAS (натриевая соль додецилбензолсульфоната, Sigma D-2525), 10 мМ ЭДТА, pH 8,0. Варианты фермента (20 м.д.) разводили 1:20 в 96-луночный несвязывающий плоскодонный планшет, содержащий либо контрольный, либо стрессовый буфер, и перемешивали. Контрольный планшет инкубировали при комнатной температуре, тогда как стрессовый планшет немедленно помещали при 37°С на 30-60 минут (в зависимости от стабильности тестируемого фермента). После инкубации активность фермента измеряли с использованием анализа suc-AAPF-pNA. Фракция оставшейся или остаточной активности равна скорости реакции стрессовой пробы, деленной на скорость реакции контрольной пробы. Исходные ферменты и варианты являются стабильными в течение 60 минут в контрольном буфере.
Фиг.9 изображает стабильность в отношении LAS/ЭДТА в зависимости от изменения суммарного заряда относительно исходного FNA, для библиотеки, содержащей 80 вариантов. Эту библиотеку проектировали и конструировали согласно способам, описанным в примере 5, так что она включает несколько суммарных зарядов относительно исходной молекулы FNA. Как видно из этого чертежа, накапливание отрицательных зарядов (до -4) относительно исходной молекулы FNA является полезным для комбинированной стабильности в отношении LAS/хелатора. Это является примером оптимизации физического свойства белка, в этом случае суммарного заряда, для улучшения стабильности белка в среде комплексного жидкого детергента для стирки.
Для ASP и FNA существует зависимость LAS/ЭДТА-стабильности от заряда. Добавление отрицательного заряда увеличивает стабильность. Но, даже когда один или два заряда являются более положительными, чем в исходном ферменте, можно найти, по способу авторов изобретения, расположение мутаций зарядов, которые придают равную стабильность или стабильность, большую, чем стабильность исходной молекулы.
ПРИМЕР 12
Термостабильность
Этот пример описывает определение взаимосвязи между зарядом белка и термостабильностью. Анализы протеазы были основаны на гидролизе диметилказеина (DMC) до и после нагревания забуференного культурального супернатанта. Анализы амилазы основаны на гидролизе крахмала BODIPY до и после нагревания культурального супернатанта. Для этих анализов использовали растворы химических соединений и реагентов, описанные в примере 1.
Анализ термостабильности для протеаз
Отфильтрованные культуральные супернатанты разбавляли до 20 м.д. в PIPES-буфере (на основе концентрации контролей в чашках для выращивания). Сначала брали 10 мкл каждой разбавленной пробы фермента для определения начальной активности в анализе с диметилказеином и обрабатывали, как описано ниже. Затем 50 мкл каждого разбавленного супернатанта помещали в пустые лунки МТР. Планшет МТР инкубировали в iEMS-термостате/шейкере HT (Thermo Labsystems) в течение 90 минут при 60°C и 400 об/мин. Эти планшеты охлаждали на льду в течение 5 минут. Затем 10 мкл этого раствора добавляли в свежий МТР, содержащий 200 мкл субстрата диметилказеина на лунку, для определения конечной активности после инкубирования. Этот МТР покрывали с использованием липкой ленты, встряхивали в течение нескольких секунд и помещали в печь при 37°С на 2 часа без перемешивания.
Остаточную активность пробы выражали в виде отношения конечной оптической плотности и начальной оптической плотности, обе из которых корректировали относительно контролей. Фиг.10 показывает индекс термостабильности как функцию изменения суммарного заряда относительно ASP дикого типа для библиотеки SEL. Более высокий коэффициент указывает на более термостабильный вариант. Как видно из этого чертежа, накапливание экстремального отрицательного (-2) или положительного (+2) зарядов относительно фермента дикого типа является вредным для термостабильности. Имеется отчетливый оптимум заряда для термостабильности, сосредоточенный при изменениях суммарного заряда при нуле, относительно ASP дикого типа. Это является примером оптимизации физического свойства белка, в этом случае суммарного заряда, для улучшения термостабильности фермента для применения в жидком детергенте для стирки.
Анализ термостабильности для альфа-амилаз
Отфильтрованные культуральные супернатанты серийно разводили в 50 мМ ацетате натрия + 2 мМ CaCl2 pH 5,8 с 0,002% Твином. 10 мкл каждого разведенного культурального супернатанта анализировали для определения начальной амилазной активности при помощи анализа с использованием крахмала BODIPY. 50 мкл каждого разведенного культурального супернатанта помещали в 96-луночный планшет для ПЦР с низким профилем VWR. В каждую лунку добавляли 30 мкл минерального масла в качестве герметика. Этот планшет инкубировали в термоциклере BioRad DNA engine Peltier при 95°C в течение 30 или 60 минут в зависимости от стабильности исходного фермента. После инкубирования этот планшет охлаждали до 4°С в течение 5 минут и затем выдерживали при комнатной температуре. 10 мкл каждой пробы добавляли в свежий планшет и анализировали для определения конечной амилазной активности анализом с использованием крахмала BODIPY, как описано в примере 1.
Расчет термостабильности
Остаточную активность пробы выражали в виде отношения конечной оптической плотности и начальной оптической плотности, обе из которых корректировали относительно контролей. Эти наблюдения выполняли также с вариантами зарядов амилазы. Фиг.11 показывает остаточную активность первого леддера зарядов AmyS как функцию изменения заряда относительно дикого типа. Опять, накапливание экстремального отрицательного (-12) или положительного (+10) зарядов относительно фермента дикого типа является вредным для термостабильности. Это является примером оптимизации физического свойства белка, в этом случае суммарного заряда, для улучшения термостабильности фермента для применения в жидком детергенте для стирки.
ПРИМЕР 13
Модуляция профиля рН-активности фермента
Этот пример описывает применение мутаций зарядов поверхности для оптимизации профиля рН-активности для конкретной реакции.
Фиг.12 показывает активность очистки микролоскутков от рисового крахмала как функцию рН для первого лэддера зарядов AmyS примера 5. Диапазон рН от 3,0 до 4,25 находился в 200 мМ формиате Na, содержащем 0,01% Твин-80, тогда как диапазон рН от 4,25 до 5,5 находился в 200 мМ ацетате Na, содержащем 0,01% Твин-80. Эти данные построены в виде кривых титрования, каждая из которых имеет единственную величину pKa.
Фиг.13 показывает среднюю (кажущуюся) pKa для катализа AmyS как функцию изменения заряда для первого лэддера зарядов AmyS примера 5. Эти данные демонстрируют, что профили рН-активности для альфа-амилазы могут существенно смещаться мутациями заряда поверхности, даже в 200 мМ буфере. Хотя это сообщалось при очень низкой ионной силе для субтилизина (Russell et al., J Mol Biol, 193: 803-13 [1987]) и для D-ксилозоизомеразы (Cha et al., Mol Cell, 8: 374-82 [1998]), авторы считают, что это было впервые выполнено с альфа-амилазой и, неожиданно, даже при высокой ионной силе.
Хотя были иллюстрированы и описаны конкретные варианты осуществления данного изобретения, квалифицированным в данной области специалистам будет понятно, что различные другие изменения и модификации могут быть произведены без отклонения от идеи и объема этого изобретения. Таким образом, предполагается, что все такие изменения и модификации, которые находятся в объеме этого изобретения, включены в прилагаемую формулу изобретения.
Все патенты и публикации, упоминаемые в этом описании, являются показателями уровней специалистов, квалифицированных в области, к которой относится это изобретение. Все патенты и публикации включены здесь в качестве ссылки до той же степени, как если бы каждая отдельная публикация была особо и отдельно указана как включенная в качестве ссылки.
После описания предпочтительных вариантов данного изобретения специалистам с обычной квалификацией в данной области будет ясно, что различные модификации могут быть произведены в отношении описанных вариантов и что предполагается, что такие модификации находятся в объеме данного изобретения.
Специалистам с квалификацией в данной области будет легко понять, что данное изобретение хорошо приспособлено для достижения целей и получения результатов и преимуществ, упомянутых в нем и присущих ему. Композиции и способы, описанные здесь, являются типичными примерами предпочтительных вариантов осуществления и не предназначены для ограничений объема этого изобретения. Квалифицированному в данной области специалисту легко понятно, что варьирующие замены и модификации могут быть произведены в отношении описанного здесь изобретения без отклонения от объема и идеи этого изобретения.
Описанное здесь иллюстративно изобретение может использоваться на практике в отсутствие любого элемента или любых элементов, ограничения или ограничений, которые не описаны конкретно здесь. Термины и выражения, которые использовались, применяются в качестве терминов описания, а не ограничения, и не предполагается, что в применении таких терминов и выражений исключаются любые эквиваленты показанных и описанных здесь признаков или их частей, но признается, что различные модификации возможны в пределах объема заявленного изобретения. Таким образом, должно быть понятно, что, хотя данное изобретение было конкретно описано с использованием предпочтительных вариантов осуществления и необязательных признаков, модификация и варьирование описанных здесь принципов могут быть видоизменены квалифицированными в данной области специалистами и что предполагается, что такие модификации и вариации находятся в объеме этого изобретения, определенном здесь.
Данное изобретение было описано здесь широко и в общем виде. Каждый из более узких видовых признаков изобретения и каждая из подродовых групп, попадающих в пределы этого общего описания, также образуют часть этого изобретения. Это включает общее описание данного изобретения, с условием или отрицательным ограничением удаления любого объекта из этого класса объектов, независимо от того, описывается или не описывается здесь конкретно удаленный материал.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ИДЕНТИФИКАЦИИ УЛУЧШЕННЫХ ВАРИАНТОВ БЕЛКА | 2008 |
|
RU2520808C2 |
| СПОСОБЫ УЛУЧШЕНИЯ МНОЖЕСТВЕННЫХ СВОЙСТВ БЕЛКА | 2008 |
|
RU2553550C2 |
| ПРОДУЦИРОВАНИЕ ТЕРМОЛИЗИНА И ЕГО ВАРИАНТОВ И ЕГО ИСПОЛЬЗОВАНИЕ В ЖИДКИХ ДЕТЕРГЕНТАХ | 2008 |
|
RU2536255C2 |
| ПРОДУЦИРОВАНИЕ ТЕРМОЛИЗИНА И ЕГО ВАРИАНТОВ И ЕГО ИСПОЛЬЗОВАНИЕ В ЖИДКИХ ДЕТЕРГЕНТАХ | 2014 |
|
RU2733541C2 |
| ВАРИАНТЫ СЕРИНОВОЙ ПРОТЕАЗЫ С МНОЖЕСТВЕННЫМИ МУТАЦИЯМИ | 2007 |
|
RU2558261C2 |
| КОМПОЗИЦИИ И СПОСОБЫ, ВКЛЮЧАЮЩИЕ В СЕБЯ ПРИМЕНЕНИЕ ВАРИАНТНЫХ МИКРОБНЫХ ПРОТЕАЗ | 2009 |
|
RU2575073C2 |
| ПРИМЕНЕНИЕ И ПОЛУЧЕНИЕ СТАБИЛЬНОЙ ПРИ ХРАНЕНИИ НЕЙТРАЛЬНОЙ МЕТАЛЛОПРОТЕИНАЗЫ | 2006 |
|
RU2433182C2 |
| ПРОТЕАЗА Streptomyces | 2008 |
|
RU2486244C2 |
| CПОСОБ МЫТЬЯ ПОСУДЫ | 2007 |
|
RU2509152C2 |
| ПРИМЕНЕНИЕ БЕЛКОВЫХ ГИДРОЛИЗАТОВ ДЛЯ СТАБИЛИЗАЦИИ ДЕТЕРГЕНТНЫХ СОСТАВОВ МЕТАЛЛОПРОТЕАЗЫ | 2008 |
|
RU2484138C2 |


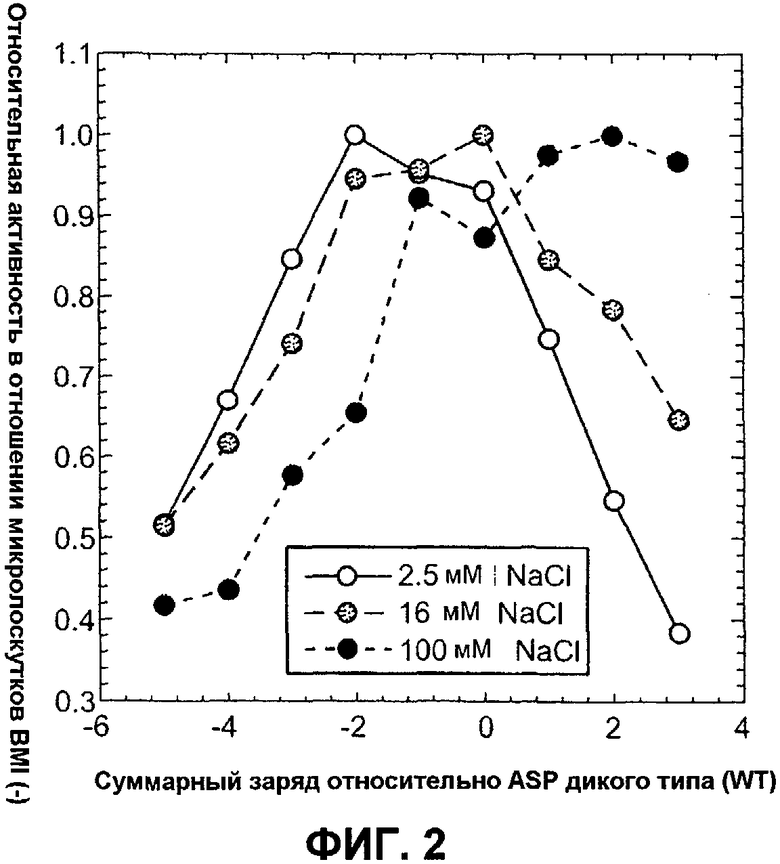





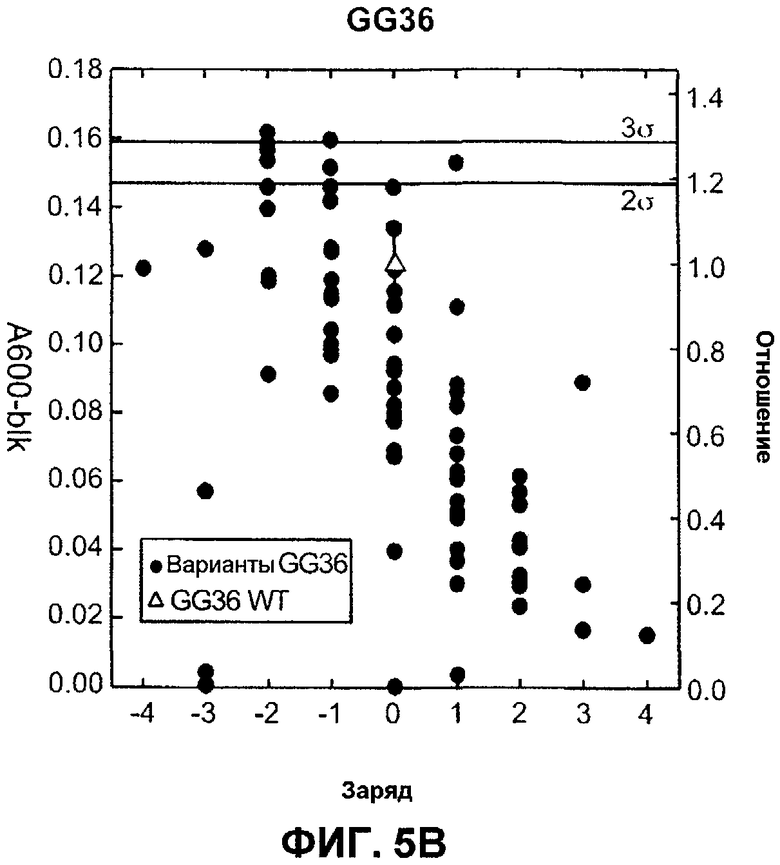







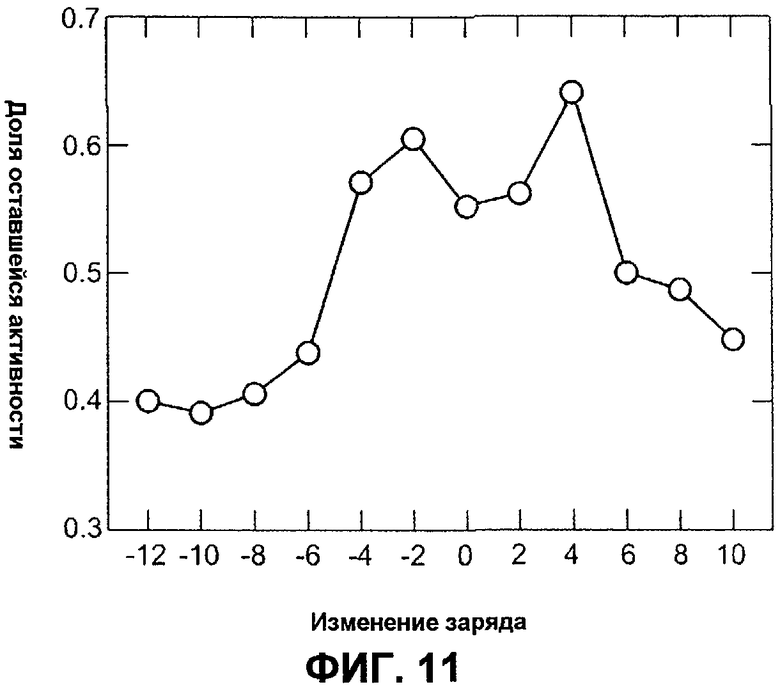

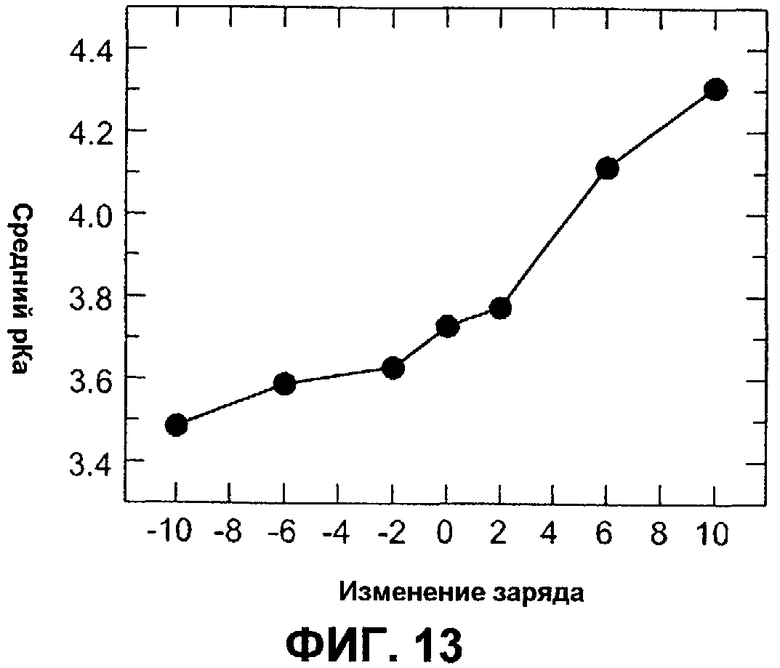
Изобретение относится к области биотехнологии и представляет собой способ получения по меньшей мере одного варианта протеазы с улучшенной моющей эффективностью в порошкообразной или жидкой детергентной композиции, имеющей pH между 5 и 12,0, в сравнении с исходной протеазой. Указанный способ предусматривает: a) определение оптимальных суммарных зарядов поверхности протеазы для моющей активности в порошкообразной или жидкой детергентной композиции, имеющей pH между 5 и 12,0, тестированием в указанной среде множества вариантов замещенных протеаз, имеющих изменение суммарного заряда поверхности молекулы, при этом оптимальные суммарные заряды поверхности протеаз представляют собой заряды -2, или -1, или +1, или +2 относительно исходной протеазы; b) модификацию по меньшей мере одного аминокислотного остатка в одном или нескольких положениях, имеющих доступную для растворителя поверхность, большую чем 50%, на поверхности указанной исходной протеазы для получения по меньшей мере одного варианта протеазы, имеющего более положительный или более отрицательный заряд поверхности в сравнении с исходной протеазой для того, чтобы варианты протеазы имели оптимальный суммарный заряд поверхности, как определено на стадии а); и c) оценку моющей эффективности по меньшей мере одного варианта белка со стадии b) для идентификации варианта белка, который проявляет улучшенную моющую эффективность в указанном представляющем интерес тесте по сравнению с исходным белком. Изобретение позволяет получать протеазы с улучшенной моющей эффективностью в порошкообразной или жидкой детергентной композиции, имеющей pH между 5 и 12,0, в сравнении с исходной протеазой. 18 з.п. ф-лы, 13 ил., 20 табл., 13 пр.
1. Способ получения по меньшей мере одного варианта протеазы с улучшенной моющей эффективностью в порошкообразной или жидкой детергентной композиции, имеющей pH между 5 и 12,0, в сравнении с исходной протеазой, предусматривающий:
a) определение оптимальных суммарных зарядов поверхности протеазы для моющей активности в порошкообразной или жидкой детергентной композиции, имеющей pH между 5 и 12,0, тестированием в указанной среде множества вариантов замещенных протеаз, имеющих изменение суммарного заряда поверхности молекулы, при этом оптимальные суммарные заряды поверхности протеаз представляют собой заряды -2, или -1, или +1, или +2 относительно исходной протеазы;
b) модификацию по меньшей мере одного аминокислотного остатка в одном или нескольких положениях, имеющих доступную для растворителя поверхность, большую чем 50%, на поверхности указанной исходной протеазы для получения по меньшей мере одного варианта протеазы, имеющего более положительный или более отрицательный заряд поверхности в сравнении с исходной протеазой для того, чтобы варианты протеазы имели оптимальный суммарный заряд поверхности, как определено на стадии а), и
c) оценку моющей эффективности по меньшей мере одного варианта белка со стадии b) для идентификации варианта белка, который проявляет улучшенную моющую эффективность в указанном представляющем интерес тесте по сравнению с исходным белком.
2. Способ по п.1, где указанная модификация включает замену, добавление и/или делецию по меньшей мере одной аминокислоты в указанной исходной протеазе.
3. Способ по п.1, где указанная модификация включает химическую модификацию.
4. Способ по п.1, где показатель эффективности варианта протеазы имеет величину, большую чем 1,0, а моющая эффективность исходной протеазы имеет показатель эффективности 1,0.
5. Способ по п.1, дополнительно предусматривающий: получение варианта протеазы, имеющего улучшенную моющую эффективность.
6. Способ по п.1, где указанная протеаза является сериновой протеазой или нейтральной металлопротеазой.
7. Способ по п. 6, где указанная протеаза является протеазой Bacillus.
8. Способ по п.1, где указанную моющую эффективность тестируют в жидком детергенте для стирки, имеющем основный рН.
9. Способ по п.1, где указанную моющую эффективность тестируют в жидком детергенте для холодной воды, имеющем основный рН.
10. Способ по п.1, где замены находятся в положениях в исходном ферменте, имеющих доступную для растворителя поверхность (SAS), большую чем приблизительно 65%.
11. Способ по п.1, где указанной протеазой является сериновая протеаза.
12. Способ по п.11, где указанной сериновой протеазой является сериновая протеаза Bacillus или сериновая протеаза Cellulomonas.
13. Способ по п.1, где указанная стадия модификации включает замену по меньшей мере одного кислого аминокислотного остатка в указанной исходной протеазе по меньшей мере одним основным аминокислотным остатком.
14. Способ по п.1, где указанная стадия модификации включает замену по меньшей мере одного кислого аминокислотного остатка в указанной исходной протеазе по меньшей мере одним нейтральным аминокислотным остатком.
15. Способ по п.1, где указанная стадия модификации включает замену по меньшей мере одного нейтрального аминокислотного остатка в указанной исходной протеазе по меньшей мере одним основным аминокислотным остатком.
16. Способ по п.1, где указанная стадия модификации включает замену по меньшей мере одного основного аминокислотного остатка в указанной исходной протеазе по меньшей мере одним кислым аминокислотным остатком.
17. Способ по п.1, где указанная стадия модификации включает замену по меньшей мере одного основного аминокислотного остатка в указанной протеазе по меньшей мере одним нейтральным аминокислотным остатком.
18. Способ по п.1, где указанная стадия модификации включает замену по меньшей мере одного нейтрального аминокислотного остатка в указанной протеазе по меньшей мере одним кислым аминокислотным остатком.
19. Способ по п.2, где указанные одно или несколько положений не являются консервативными в выравниваниях аминокислот гомологичных белковых последовательностей, содержащих указанный исходный белок и по меньшей мере один дополнительный белок.
| ВАРИАНТ СУБТИЛИЗИНА (ВАРИАНТЫ), КОДИРУЮЩАЯ ЕГО ДНК, ЭКСПРЕССИРУЮЩИЙ ВЕКТОР, ОЧИЩАЮЩАЯ КОМПОЗИЦИЯ, КОРМ ДЛЯ ЖИВОТНЫХ, КОМПОЗИЦИЯ ДЛЯ ОБРАБОТКИ ТКАНИ | 1998 |
|
RU2252958C2 |
| Неразряжающийся сифон | 1980 |
|
SU945502A1 |
| KATHERINE L., et al., "Increasing the Net Charge and Decreasing the Hydrophobicity of Bovine Carbonic Anhydrase Decreases the Rate of Denaturation with Sodium Dodecyl Sulfate", Biophysical Journal Volume 91 July 2006 298-310 | |||
| JIAN-ZHONG LIU and MIN WANG "Improvement of activity and stability of | |||

