ПРЕДПОСЫЛКИ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
Многочисленные заболевания связаны с дегенеративными изменениями в суставном хряще или процессами неправильной дифференцировки в костном мозге. Заболевания опорно-двигательного аппарата представляют третий из наиболее затратных факторов в лечении заболеваний в Германии после заболеваний сердечно-сосудистой системы и заболеваний пищеварительной системы. Одной из предпосылок для успешного лечения таких заболеваний является понимание клеточных процессов в костном мозге, которые лежат в основе этих заболеваний или связаны с ними. Таким образом, например, возможно лечение лейкоза путем замещения гемопоэтической системы пациента. В то же время, также аутоиммунные заболевания, такие как ревматоидный артрит или красная волчанка, регулируются долгоживущими плазматическими клетками и Т-клетками памяти, которые находятся в специализированных нишах костного мозга.
Костный мозг является многофункциональной многоклеточной тканью, которая, помимо прочего, расположена в полости трубчатых костей. Ткань развивается во время эмбрионального развития примерно на 4 месяце в условиях эндохондрального окостенения при развитии трубчатых костей. Костный мозг характеризуется пористой структурой, образованной губчатой костью и специализированной соединительной тканью, пронизанной многочисленными тонкостенными кровеносными сосудами, синусоидами костного мозга. Одной из главных функций костного мозга является обеспечение подходящего окружения для дифференцировки кроветворных клеток гемопоэтической системы. Гемопоэтическая стволовая клетка (HSC) представляет собой исходную точку для этих процессов дифференцировки, при этом гемопоэтическая стволовая клетка находится в недифференцированном состоянии в нишах гемопоэтических стволовых клеток в структурах губчатой кости в костном мозге. Ниши гемопоэтических стволовых клеток делятся на два функционально различных подтипа. В остеобластной нише гемопоэтические HSC взаимодействуют со специализированными остеобластами посредством различных молекул адгезии и сигнальных молекул, в результате чего HSC удерживаются в округленном непролиферирующем состоянии. После мобилизации HSC стволовые клетки мигрируют в сосудистые ниши стволовых клеток, расположенные в синусоидах, где HSC проявляют повышенную пролиферацию. В соответствии с сегодняшним пониманием ниш гемопоэтических стволовых клеток в костном мозге наименьшая единица в системе ниш стволовых клеток формируется комбинацией остеобластной ниши и сосудистой ниши, расположенной в синусоидном кровеносном сосуде губчатой кости. Помимо функциональных структур ниш стволовых клеток в современных исследованиях раскрывается, что костный мозг является вместилищем для иммунологической памяти. Мезенхимальные стромальные клетки (MSC) служат в качестве клеточных структурных элементов ниш, обеспечивающих выживание плазматических клеток и Т-клеток памяти. В заключение, биологические функциональные свойства костного мозга зависят от ряда клеток, таких как HSC, MSC, эндотелиальные клетки и остеобласты, и их надлежащего взаимодействия в пространственном компартменте.
Гиалиновый хрящ суставов, благодаря своим свойствам высокой эластичности при сжатии и очень гладкой поверхности, обеспечивает возможность движения суставов со слабым трением. Основная часть этой ткани образована множеством белков внеклеточного матрикса, тогда как клеточный компонент, хондроциты, представляет собой редкие вкрапления в этом матриксе, либо по отдельности, либо малыми группами (хондрон). Отсутствие нервных волокон и кровеносных сосудов и, как результат, низкая обеспеченность ткани кислородом и питательными веществами создает особое микроокружение для хондроцитов. Ткань хряща можно разделить на несколько слоев, при этом, вследствие горизонтальной ориентации молекул матрикса самый верхний слой может противодействовать трению, и вследствие вертикальной ориентации расположенные ниже средний и нижний слои могут амортизировать усилие, создаваемое движением. Хотя хондроциты появляются на ранних этапах эндохондрального окостенения при первоначальном процессе мезенхимальной конденсации, и эти клетки вносят существенный вклад в процесс роста костей в ходе формирования эпифизов, ткань гиалинового хряща в суставе развивается только на очень позднем этапе во время эмбриогенеза. Полностью дифференцированные, полностью функциональные слои хряща образуются только после рождения при начале механической стимуляции ткани. У взрослого организма гомеостаз хрящевой ткани сильно зависит от вида и силы механической нагрузки и микроокружения заключенных в ней хондроцитов.
Были предприняты многочисленные попытки разработки и получения in vitro моделей и тест-систем для костной и хрящевой ткани. Тем не менее, эти подходы были направлены на раздельное обеспечение либо костной, либо хрящевой ткани. Следовательно, эти модели все-таки только частично и недостаточно представляют естественное окружение, в котором происходит развитие и гомеостаз ткани кости, костного мозга и хряща.
Целью настоящего изобретения является преодоление одной или нескольких проблем известного уровня техники.
ОПИСАНИЕ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
В первом аспекте настоящее изобретение направлено на трехмерную (3D) in vitro модель, в которой учитывается тесное взаимодействие и совместное функционирование хрящевой и костной ткани. Настоящее изобретение предусматривает 3D in vitro двухфазный костно-хрящевой «органоид», содержащий:
a) слой искусственной хрящевой ткани и
b) слой искусственной костной ткани, где искусственная костная ткань содержит придающий структуру каркас и структуру костного мозга;
где слой искусственной хрящевой ткани контактирует по меньшей мере с одной поверхностью слоя искусственной костной ткани.
Искусственную хрящевую ткань в 3D in vitro двухфазном костно-хрящевом «органоиде» можно получить с использованием мезенхимальных клеток-предшественников, предпочтительно с использованием мезенхимальных клеток-предшественников, включающих по меньшей мере 50% клеток или состоящих из клеток, которые представляют собой CD105+, CD106+, CD44+, CD73+, CD90+ и CD13+, более предпочтительно из выделенных первичных хондроцитов. В частности, искусственную хрящевую ткань можно получить с помощью способа согласно настоящему изобретению, определенного ниже.
Структуру костного мозга в искусственной костной ткани в 3D in vitro двухфазном костно-хрящевом «органоиде» согласно настоящему изобретению получают путем посева мезенхимальных стволовых клеток на придающий структуру каркас. Структуру костного мозга в искусственной костной ткани получают предпочтительно путем посева мезенхимальных стволовых клеток на придающий структуру каркас и путем добавления гемопоэтических стволовых клеток после культивирования мезенхимальных стволовых клеток на придающем структуру каркасе в течение 2-10 дней, предпочтительно в течение 5-8 дней, более предпочтительно в течение 7 дней.
Придающий структуру каркас из искусственной костной ткани в 3D in vitro двухфазном костно-хрящевом «органоиде» согласно настоящему изобретению предпочтительно содержит биологический или небиологический материал или состоит из него, более предпочтительно 3D керамический каркас.
В соответствии с другим аспектом настоящее изобретение направлено на способ получения 3D in vitro двухфазного костно-хрящевого «органоида» согласно настоящему изобретению. Указанный способ включает следующие этапы или состоит из таких этапов:
посев мезенхимальных стволовых клеток на придающий структуру каркас;
культивирование указанных мезенхимальных стволовых клеток в течение 2-10 дней на указанном придающем структуру каркасе, предпочтительно в течение 5-8 дней, более предпочтительно в течение 7 дней;
добавление гемопоэтических стволовых клеток к мезенхимальным стволовым клеткам, культивируемым на придающем структуру каркасе, и культивирование полученной смеси в течение по меньшей мере 5 дней, предпочтительно в течение от 5 дней до 8 недель, более предпочтительно в течение от 1 недели до 4 недель для того, чтобы получить слой искусственной костной ткани, содержащей придающий структуру каркас, засеянный структурой костного мозга;
обеспечение искусственной хрящевой ткани, полученной с помощью способа по любому из пп. 13-15, и
размещение слоя указанной искусственной хрящевой ткани по меньшей мере на одной поверхности указанного слоя искусственной костной ткани.
В соответствии с другим аспектом настоящее изобретение направлено на трансплантат, содержащий 3D in vitro двухфазный костно-хрящевой «органоид» согласно настоящему изобретению.
Кроме того, настоящее изобретение предусматривает фармацевтическую композицию, содержащую 3D in vitro двухфазный костно-хрящевой «органоид» согласно настоящему изобретению или трансплантат согласно настоящему изобретению и по меньшей мере одно фармацевтически приемлемое вспомогательное вещество.
Настоящее изобретение также направлено на способ in vitro скрининга веществ, которые модулируют свойства хрящевой или костной ткани, включающий следующие этапы:
обеспечение образца 3D in vitro двухфазного костно-хрящевого «органоида» согласно настоящему изобретению;
разделение соответствующего образца на порции;
инкубирование по меньшей мере одной порции с веществом, подлежащим скринингу; и
сравнение параметров, измеренных у обработанной порции и другой порции, которую не инкубировали с веществом, подлежащим скринингу.
В соответствии с дополнительным аспектом настоящего изобретения предусматривается автономное устройство по типу 3D in vitro двухфазного костно-хрящевого «органоида»-на-чипе, содержащее по меньшей мере один резервуар для подачи среды, по меньшей мере одну секцию для роста органа, содержащую по меньшей мере одну полость для органа, вмещающую 3D in vitro двухфазный костно-хрящевой «органоид» согласно настоящему изобретению, где резервуар для подачи среды соединен по меньшей мере с одной секцией для роста органа микроструйным питающим каналом.
Настоящее изобретение также направлено на способ получения агрегатов хондрогенных клеток, включающий следующие этапы:
a) обеспечение мезенхимальных клеток-предшественников; и
b) культивирование мезенхимальных клеток-предшественников в не обеспечивающих адгезию условиях с образованием агрегатов хондрогенных клеток.
Мезенхимальные клетки-предшественники, применяемые в этом способе, предпочтительно включают по меньшей мере 50% клеток или состоят из клеток, которые представляют собой CD105+, CD106+, CD44+, CD73+, CD90+ и CD13+. Более предпочтительно, указанные мезенхимальные клетки-предшественники включают или состоят из выделенных первичных хондроцитов.
Настоящее изобретение основывается на данных, что на развитие и гомеостаз как костной ткани, так и хрящевой ткани оказывают влияние присутствие и взаимодействие обеих тканей. Давно известно, что на формирование и дифференцировку костной ткани оказывают влияние присутствие, длительность воздействия и направление физических усилий, прикладываемых к развивающейся кости. В естественном окружении эти усилия зачастую прикладываются к костной ткани через ее поверхность, контактирующую с хрящевой тканью. Одной функцией хрящевой ткани является надлежащая передача ударных усилий, которые создаются движением тела, нижележащей костной ткани. Это взаимодействие не только оказывает влияние на ориентацию и прочность костной ткани, но также воздействует на определение границ и организацию костного мозга и ниш стволовых клеток внутри кости. Не подлежит сомнению, что организация и дифференцировка костного мозга и ниши стволовых клеток оказывает значительное воздействие на иммунную систему индивида. Следовательно, обе ткани, костную ткань и хрящевую ткань, следует рассматривать как формирующие один общий орган. 3D in vitro двухфазный костно-хрящевой «органоид» согласно настоящему изобретению отражает это положение и обеспечивает возможность имитации влияния, которое оказывает присутствие обеих тканей на развитие и дифференцировку костной ткани, соответственно, на организацию и дифференцировку костного мозга и ниш стволовых клеток, и хрящевой ткани. Таким образом, 3D in vitro двухфазный костно-хрящевой «органоид» согласно настоящему изобретению впервые обеспечивает возможность изучения влияния тесного взаимного влияния обоих типов тканей друг на друга. В частности, когда 3D in vitro двухфазный костно-хрящевой «органоид» применяется в тест-системе, где слой искусственной хрящевой ткани может подвергаться воздействию определенных физических усилий посредством механической стимуляции, обеспечивается модельная система, которая точно имитирует естественные условия и, следовательно, обеспечивает возможность получения новых данных о биологии этих тканей, а также подходящая система для скрининга в отношении веществ, которые взаимодействуют с этими тканями или процессами.
Далее приведено более подробное описание различных аспектов настоящего изобретения. Каждый определенный таким образом аспект можно комбинировать с любым другим аспектом или аспектами, если только явным образом не указано иное. В частности, любой признак, указанный как предпочтительный или полезный, можно комбинировать с любым другим признаком или признаками, указанными как являющиеся частью настоящего изобретения или указанными как предпочтительные или преимущественные.
Настоящее изобретение направлено на 3D in vitro двухфазный костно-хрящевой «органоид». В контексте настоящего изобретения выражение «органоид» обозначает искусственный, созданный de novo, агрегат функциональных клеток из различных типов клеток in vitro, который проявляет по меньшей мере одну функцию, являющуюся типичной для соответствующего органа или ткани, предпочтительно, который проявляет большинство функций, являющиеся типичными для соответствующего органа или ткани.
3D in vitro двухфазный костно-хрящевой «органоид» согласно настоящему изобретению содержит слой искусственной хрящевой ткани. Искусственная хрящевая ткань представляет собой хрящевую ткань, которая была получена и собрана de novo и in vitro. Обычно искусственная хрящевая ткань содержит полностью или по меньшей мере частично дифференцированные хондроциты и компоненты внеклеточного матрикса, включающие коллаген II, IX и XI.
Существуют многочисленные методики получения искусственной хрящевой ткани in vitro, доступные специалисту в данной области техники. Поскольку хрящевая ткань представляет собой более чем простой 2D слой хондроцитов, предпочтительной является искусственная хрящевая ткань, созданная с помощью методик 3D культивирования. Подходящие методики кратко изложены в Tortelli and Cancedda («Three-dimensional cultures of osteogenic and chondrogenic cells: A tissue engineering approach to mimic bone and cartilage in vitro», European Cells and Materials (2009), 17, 1-14). Слой искусственной хрящевой ткани можно, например, получить с помощью методик, включающих культивирование и/или организацию клеток или клеточных агрегатов посредством инкапсулирования в гидрогеле, культивирования в микромассе, образования клеточных осадков высокой плотности и применения биосовместимых разлагаемых или неразлагаемых каркасов. Другой предпочтительный способ получения искусственной хрящевой ткани по настоящему изобретению, дополнительно описывается ниже как способ получения агрегатов хондрогенных клеток.
Чтобы обеспечить слой искусственной хрящевой ткани, искусственную хрящевую ткань получают с использованием мезенхимальных стволовых клеток (MCS), также называемых мезенхимальными клетками-предшественниками. Предпочтительно, применяют выделенные мезенхимальные клетки-предшественники. Выражение «выделенный» означает, что мезенхимальные клетки-предшественники представляют собой клетки, которые были выделены из естественного источника, или их соответствующее потомство, которое, например, было получено в результате пролиферации клеток. Применяемые мезенхимальные клетки-предшественники предпочтительно получают из хрящевой ткани донора, индивида или пациента. Предпочтительно, мезенхимальные клетки-предшественники представляют собой первичные клетки, которые не были трансформированы или иммортализированы. В частности, мезенхимальные клетки-предшественники могут включать или состоять из взрослых мезенхимальных клеток-предшественников. Такие взрослые мезенхимальные клетки-предшественники получают из не являющейся эмбриональной хрящевой ткани донора, индивида или пациента. Мезенхимальные клетки-предшественники можно получить из хрящевой ткани из любой донорской ткани, которая дифференцировалась так, что содержит хрящевую ткань. Предпочтительно, мезенхимальные клетки-предшественники получают из хрящевой ткани сустава, более предпочтительно из хряща колена или локтя донора. Мезенхимальные клетки-предшественники предпочтительно представляют собой человеческие клетки. Человеческие мезенхимальные клетки-предшественники получают из хрящевой ткани, взятой от человека. Мезенхимальные клетки-предшественники могут включать по меньшей мере 50% клеток, которые представляют собой CD105+, CD106+, CD44+, CD73+, CD90+ и CD13+, или могут состоять из клеток, которые представляют собой CD105+, CD106+, CD44+, CD73+, CD90+ и CD13+. Предпочтительно, мезенхимальные клетки-предшественники включают или состоят из выделенных первичных хондроцитов, таких как, например, человеческие первичные хондроциты.
3D in vitro двухфазный костно-хрящевой «органоид» согласно настоящему изобретению содержит слой искусственной костной ткани, где искусственная костная ткань содержит придающий структуру каркас и структуру костного мозга. Искусственная костная ткань представляет собой костную ткань, которая была получена и собрана de novo и in vitro. Обычно искусственная костная ткань содержит полностью или частично дифференцированные остеобласты, остеокласты и гемопоэтические стволовые клетки.
Существуют многочисленные методики получения искусственной костной ткани in vitro, доступные специалисту в данной области техники. Поскольку костная ткань обычно представляет собой более чем простой 2D слой клеток определенного типа, предпочтительной является искусственная костная ткань, созданная с помощью методик 3D культуры. Подходящие методики кратко изложены в Tortelli and Cancedda («Three-dimensional cultures of osteogenic and chondrogenic cells: A tissue engineering approach to mimic bone and cartilage in vitro», European Cells and Materials (2009), 17, 1-14). Слой искусственной костной ткани можно, например, получить с помощью методик, включающих культивирование и/или организацию клеток или клеточных агрегатов посредством культивирования в микромассе и применения биосовместимых разлагаемых или неразлагаемых каркасов. Подходящие методики культивирования и каркасы известны в уровне техники. Например, можно применять биоразлагаемые или неразлагаемые каркасы, содержащие или состоящие из биологического или небиологического материала. Было описано применение в качестве каркаса синтетических полимеров, а также применение каркасов на основе коллагена, каркасов на основе металлов, таких как, например, каркасы на основе титана, или каркасов на основе керамики.
Клетки, применяемые для получения искусственной костной ткани, могут включать мезенхимальные стволовые клетки (MSC) и гемопоэтические стволовые клетки (HSC). Предпочтительно, применяют выделенные MSC и/или выделенные HSC. Выражение «выделенный» означает, что MSC и/или HSC представляют собой клетки, которые были выделены из естественного источника или их соответствующее потомство, которое, например, было получено вследствие пролиферации клеток. Применяемые MSC и/или HSC предпочтительно получают из подходящей ткани донора, индивида или пациента. Предпочтительно, MSC и/или HSC представляют собой первичные клетки, которые не были трансформированы или иммортализированы. В частности, MSC и/или HSC могут включать или состоять из взрослых MSC и/или HSC. Такие взрослые MSC и/или HSC получают из ткани, не являющейся эмбриональной, предпочтительно из костной ткани или ткани костного мозга донора, индивида или пациента. MSC и/или HSC предпочтительно представляют собой человеческие клетки. Человеческие MSC и/или HSC предпочтительно получают из кости или ткани костного мозга человеческого происхождения.
Предпочтительный способ получения искусственной костной ткани по настоящему изобретению включает посев и культивирование MSC на керамическом 3D каркасе. Заселенный керамический 3D каркас культивируют в течение предварительно определенного периода времени, а затем могут добавлять HSC. В предпочтительном варианте осуществления HSC добавляют после культивирования MSC на керамическом каркасе в течение 2-10 дней. Затем заселенный керамический 3D каркас дополнительно культивируют либо в условиях статического культивирования, либо в перфузируемой системе для постоянной подачи питательных веществ. Методики создания искусственной костной ткань можно дополнительно изменить. Например, можно добавлять факторы роста или другие сигнальные молекулы для поддержки функций костного мозга. Можно применять условия гипоксии, которые подобны тем, с которыми сталкивается костный мозг в естественных условиях. Может потребоваться оптимизировать период культивирования, а также последовательность введения различных типов клеток в каркас. Помимо MSC и HSC для создания искусственной костной ткани можно применять дополнительные типы клеток. Эти дополнительные клетки могут включать типы клеток, которые в норме встречаются в структурах кости или костного мозга, например, иммунные клетки, эндотелиальные клетки и т.п.
3D in vitro двухфазный костно-хрящевой «органоид» согласно настоящему изобретению можно применять в качестве инструмента для исследований in vitro и/или in vivo. Его можно применять для изучения биологии одного или нескольких его компонентов. В качестве альтернативы, его можно применять в способе in vitro и/или in vivo скрининга в отношении веществ, которые модулируют свойства хрящевой или костной ткани.
Настоящее изобретение также направлено на трансплантат, содержащий 3D in vitro двухфазный костно-хрящевой «органоид» согласно настоящему изобретению, и на фармацевтическую композицию, содержащую 3D in vitro двухфазный костно-хрящевой «органоид» согласно настоящему изобретению или трансплантат согласно настоящему изобретению и по меньшей мере одно фармацевтически приемлемое вспомогательное вещество.
В дополнительном аспекте настоящее изобретение направлено на способ in vitro скрининга веществ, которые модулируют свойства хрящевой или костной ткани, включающий следующие этапы:
обеспечение образца 3D in vitro двухфазного костно-хрящевого «органоида» согласно настоящему изобретению;
разделение соответствующего образца на порции;
инкубирование по меньшей мере одной порции с веществом, подлежащим скринингу; и
сравнение параметров, измеренных у обработанной порции и другой порции, которую не инкубировали с веществом, подлежащим скринингу.
В предпочтительном варианте осуществления, порцию помещают в автономное устройство по типу орган-на-чипе перед инкубированием порции с веществом, подлежащим скринингу.
Вкратце, способ согласно настоящему изобретению делает возможным идентификацию и анализ веществ, которые оказывают влияние на хрящевую или костную ткань, при помощи 3D in vitro двухфазного костно-хрящевого «органоида» согласно настоящему изобретению. Образец, который, как следует понимать, содержит определенное количество продуктов-объектов настоящего изобретения, разделяют на несколько порций. Получают по меньшей мере две подгруппы; одну используют для скрининга, тогда как другая служит в качестве отрицательного контроля. Предпочтительно, количество частей для скрининга превышает количество частей для контроля. Обычно многочисленные части подвергают скринингу с высокой пропускной способностью. Вещества, подлежащие скринингу в способе согласно настоящему изобретению, не ограничены каким-либо образом. В варианте осуществления настоящего изобретения вещества выбраны из группы нуклеиновых кислот, в том числе интерферирующие РНК, рибозимов, аптамеров, антител, пептидов, углеводов, полимеров, малых молекул с молекулярным весом от 50 до 1000 Да и белков, предпочтительно антител, цитокинов и липокалинов. Эти вещества зачастую доступны в библиотеках. Предпочтительным является инкубирование отдельного соединения в отдельной порции образца. Тем не менее, также возможно изучать суммарный эффект веществ при помощи инкубирования по меньшей мере двух веществ в одной порции. Дополнительную подгруппу объектов одновременно инкубируют в отсутствие веществ. Процесс инкубирования зависит от различных параметров, например, типов клеток и чувствительности выявления, при оптимизации которых следуют обычным процедурам, известным специалистам в данной области техники. Идентификацию эффективных веществ в контексте настоящего изобретения предпочтительно осуществляют косвенно, например, путем определения паттернов экспрессии и/или жизнеспособности клеток, которые изменяются. Определение можно осуществлять в определенный момент и сопоставлять с силой сигнала вначале эксперимента и у отрицательного контроля. Подходящие тесты известны специалистам в данной области техники или могут быть легко разработаны в обычном порядке.
В настоящем изобретении также раскрывается автономное устройство по типу 3D in vitro двухфазный костно-хрящевой «органоид»-на-чипе, включающее автономное устройство по типу орган-на-чипе, которое описано в WO 2009/146911, и 3D in vitro двухфазный костно-хрящевой «органоид» согласно настоящему изобретению.
Автономное устройство по типу 3D in vitro двухфазного костно-хрящевого «органоида»-на-чипе согласно настоящему изобретению создано для обеспечения возможности получения или поддержания 3D in vitro двухфазного костно-хрящевого «органоида» согласно настоящему изобретению в формате миниатюризированного чипа, подходящем для наблюдения в реальном времени при помощи визуализации живых клеток и, например, двухфотонной микроскопии, и его применения, например, для тестирования активности, фармакодинамики и фармакокинетики соединений или для изучения самосборки, гомеостаза, повреждения, регенерации или взаимодействия органов или «органоидов» и ниш стволовых клеток, а также явлений созревания, старения, смерти и хронобиологии. Для этой цели автономное устройство по типу 3D in vitro двухфазного костно-хрящевого «органоида»-на-чипе согласно настоящему изобретению содержит по меньшей мере один резервуар для подачи среды, по меньшей мере одну секцию для роста органа, содержащую по меньшей мере одну полость для органа, вмещающую 3D in vitro двухфазный костно-хрящевой «органоид» согласно настоящему изобретению, где резервуар для подачи среды соединен по меньшей мере с одной секцией для роста органа микроструйным питающим каналом.
Объект настоящего изобретения обеспечивает возможность получения миниатюризированной in vitro культуры «органоида», содержащего хрящевую и костную ткань. Полученный 3D in vitro двухфазный костно-хрящевой «органоид» согласно настоящему изобретению подходит для долговременного культивирования вплоть до 3 месяцев, при культивировании, например, в условиях, обеспечивающих возможность постоянной подачи кислорода и питательных веществ, как, например, в условиях постоянной перфузии.
Объект настоящего изобретения обеспечивает возможность интеграции механических стимулов для имитации, например, физических усилий, создаваемых движением тела, и индукции сосудистых структур в «органоиде» и, следовательно, обеспечения окружения, которое приближается к естественным условиям. Исходя из этих свойств, объект настоящего изобретения обеспечивает возможность получения различных научных данных в области тканевой инженерии, молекулярной биологии клетки и дифференцировки тканей, а также в регенеративной медицине. Объект настоящего изобретения также может привести к замене дорогостоящих и трудоемких экспериментов на животных на тесты in vitro.
На данный момент не было попыток получить 3D in vitro «органоид», который объединяет функционально взаимосвязанные ткани кости, костного мозга и хряща в одной единой интегрированной системе, которая совместима с методиками длительного культивирования, такими как перфузионная культура, и которая позволяет осуществлять индукцию васкуляризации. Воздействуя на систему механическими силами или другими стимулами, естественное окружение будут имитировать в такой степени, которая не достигалась никогда ранее.
В другом аспекте настоящее изобретение направлено на in vitro способ получения агрегата хондрогенных клеток. Способ включает следующие этапы:
a) обеспечение выделенных мезенхимальных клеток-предшественников; и
b) культивирование мезенхимальных клеток-предшественников в не обеспечивающих адгезию условиях с образованием агрегата хондрогенных клеток.
Было обнаружено, что выделенные мезенхимальные клетки-предшественники способны формировать трехмерный клеточный агрегат, который проявляет хондрогенную дифференцировку без влияния эмбриональных клеток. Этот эффект достигается при помощи культивирования мезенхимальных клеток-предшественников в не обеспечивающих адгезию условиях. Можно продемонстрировать, что при таких не обеспечивающих адгезию условиях культивирования мезенхимальные клетки-предшественники располагаются свободно по отношению друг к другу и конденсируются в клеточный агрегат, который проявляет экспрессию маркеров, специфических для хондрогенной дифференцировки, и который, следовательно, обозначают как агрегат хондрогенных клеток.
Ниже предусматривается более подробное описание различных аспектов настоящего изобретения. Каждый определенный таким образом аспект можно комбинировать с любым другим аспектом или аспектами, если явно не утверждается иное. В частности, любой признак, указанный как предпочтительный или полезный, можно комбинировать с любым другим признаком или признаками, указанными как являющиеся частью настоящего изобретения или указанными как предпочтительные или преимущественные.
Способ согласно настоящему изобретению направлен на in vitro препарат из агрегата хондрогенных клеток. В контексте настоящего изобретения выражение «агрегат хондрогенных клеток» относится к агрегату функциональных клеток, проявляющему по меньшей мере одну функцию, которая является специфической для хондроцитов или хрящевой ткани. Предпочтительно, агрегат хондрогенный клеток согласно настоящему изобретению проявляет большую часть или, по сути, все функции органа или ткани для дифференцированных хондроцитов или хрящевой ткани. В частности, агрегат хондрогенных клеток согласно настоящему изобретению может вести себя подобно функциональной хрящевой ткани и/или способен индуцировать in vitro и/или in vivo развитие или дифференцировку хряща. Агрегат хондрогенных клеток согласно настоящему изобретению может характеризоваться повышенной экспрессией маркерных генов или белков, которые связаны с развитием или дифференцировкой хондроцитов или хрящевой ткани. Предпочтительно, агрегат хондрогенных клеток согласно настоящему изобретению может характеризоваться повышенной экспрессией коллагена II, IX и XI, а также пониженной экспрессией коллагена I и XII по сравнению с экспрессией в мезенхимальных клетках-предшественниках непосредственно перед культивированием в не обеспечивающих адгезию условий.
Агрегат хондрогенных клеток согласно настоящему изобретению формируют с помощью способа согласно настоящему изобретению. Агрегат хондрогенных клеток может представлять собой клеточный агрегат практически округлой или сферической формы. Он может иметь средний диаметр от 0,5 мм до 3 мм, предпочтительно средний диаметр от 0,5 мм до 2 мм. Агрегат хондрогенных клеток согласно настоящему изобретению не содержит какого-либо искусственного биологического или небиологического каркаса или носителя, который не происходит из клеток, применяемых в получении хондрогенного клеточного агрегата. Агрегат хондрогенных клеток согласно настоящему изобретению предпочтительно состоит из клеточного агрегата, сформированного при помощи способа согласно настоящему изобретению, где способ осуществляли без применения или добавления какого-либо биологического или небиологического каркаса или носителя, который не происходит непосредственно из клеток, применяемых в указанном способе.
В способе согласно настоящему изобретению применяются выделенные мезенхимальные клетки-предшественники. Выражение «выделенный» означает, что мезенхимальные клетки-предшественники представляют собой клетки, которые были выделены из естественного источника или их потомство, которое, например, было получено в результате пролиферации клеток. Применяемые мезенхимальные клетки-предшественники предпочтительно получают из хрящевой ткани донора, субъекта или пациента. Предпочтительно, мезенхимальные клетки-предшественники представляют собой первичные клетки, которые не были трансформированы или иммортализированы. В частности, мезенхимальные клетки-предшественники могут включать или состоять из взрослых мезенхимальных клеток-предшественников. Такие взрослые мезенхимальные клетки-предшественники получают из не являющейся к эмбриональной хрящевой ткани донора, субъекта или пациента. Мезенхимальные клетки-предшественники можно получить из хрящевой ткани из любой донорской ткани, которая дифференцировалась так, что содержит хрящевую ткань. Предпочтительно, мезенхимальные клетки-предшественники получают из хрящевой ткани сустава, более предпочтительно из хряща колена или локтя донора. Мезенхимальные клетки-предшественники, применяемые в способе согласно настоящему изобретению, предпочтительно представляют собой человеческие клетки. Человеческие мезенхимальные клетки-предшественники получают из хрящевой ткани, взятой от человека. Мезенхимальные клетки-предшественники могут включать по меньшей мере 50% клеток, которые представляют собой CD105+, CD106+, CD44+, CD73+, CD90+ и CD13+, или могут состоять из клеток, которые представляют собой CD105+, CD106+, CD44+, CD73+, CD90+ и CD13+. Предпочтительно, мезенхимальные клетки-предшественники включают или состоят из выделенных первичных хондроцитов, таких как, например, человеческие первичные хондроциты.
В способе согласно настоящему изобретению мезенхимальные клетки-предшественники можно культивировать в не обеспечивающих адгезию условиях на любой стадии после выделения. Однако формирование клеточных агрегатов дополнительно улучшается, если мезенхимальные клетки-предшественники перенесли культивирование, по меньшей мере в течение некоторого времени, в условиях прикрепленного 2D монослоя. Предпочтительно, мезенхимальные клетки-предшественники культивировались в 2D монослойной культуре в течение по меньшей мере 5 пассажей до культивирования в не обеспечивающих адгезию условиях. Для того чтобы гарантировать дедифференцированный фенотип, желательно культивировать мезенхимальные клетки-предшественники в условиях прикрепленного 2D монослоя в течение по меньшей мере 10 пассажей. Высокая эффективность формирования клеточных агрегатов сохраняется на протяжении широкого диапазона пассажей. Однако было обнаружено, что наилучшие результаты достигаются, если мезенхимальные клетки-предшественники культивировались в 2D монослойной культуре в течение по меньшей мере 5 пассажей и не более 20 пассажей. Предпочтительно, мезенхимальные клетки-предшественники культивируют в не обеспечивающих адгезию условиях после культивирования в 2D монослое в течение по меньшей мере 5 пассажей и не более 15 пассажей после выделения.
В способе согласно настоящему изобретению после обеспечения выделенных мезенхимальных клеток-предшественников мезенхимальные клетки-предшественники культивируют при не обеспечивающих адгезию условиях для формирования агрегатов хондрогенных клеток.
Формирование клеточных агрегатов является особенно эффективным, если мезенхимальные клетки-предшественники культивируют в не обеспечивающих адгезию условиях при определенной концентрации. Если концентрация является слишком низкой, клетки лишь изредка контактируют друг с другом, и конденсация в клеточные агрегаты является менее эффективной. С другой стороны, если концентрация мезенхимальных клеток-предшественников является слишком высокой, клетки являются менее гибкими или подвижными, и, следовательно, формирование клеточных агрегатов является менее эффективным. Предпочтительно, мезенхимальные клетки-предшественники культивируют в не обеспечивающих адгезию условиях при концентрации от 5×104 до 5×107 клеток на мл. Даже лучшие результаты достигаются, если мезенхимальные клетки-предшественники культивируют в не обеспечивающих адгезию условиях при концентрации от 1×105 до 1×107 клеток на мл, предпочтительно от 5×105 до 5×106 клеток на мл и наиболее предпочтительно от 9×105 до 1,1×106 клеток на мл.
В способе согласно настоящему изобретению выделенные мезенхимальные клетки-предшественники культивируют в не обеспечивающих адгезию условиях культивирования. Это означает, что мезенхимальные клетки-предшественники культивируют в условиях, при которых клетки не принимают уплощенную, распластанную форму, указывающую на сильное прикрепление и адгезию к поверхности для культивирования. Предпочтительно, мезенхимальные клетки-предшественники после посева сохраняют округлую форму и лишь слабо связываются с поверхностью для культивирования, если вообще связываются с ней. Подходящие средства для культивирования клеток без обеспечения адгезии хорошо известны в уровне техники. Не обеспечивающие адгезию условия культивирования могут включать культивирование мезенхимальных клеток-предшественников в культуральных сосудах с поверхностью для культивирования, которая не поддерживает адгезию мезенхимальных клеток-предшественников. Например, можно применять культуральные сосуды с поверхностями для культивирования со сверхнизким прикреплением клеток. С этой целью можно применять культуральные сосуды с нейтрально или положительно заряженной поверхностью для культивирования. Предпочтительно, поверхность для культивирования можно покрыть слоем материала, который дополнительно снижает взаимодействие мезенхимальных клеток-предшественников и поверхности для культивирования. Поверхность для культивирования можно покрыть гидрофильным гидрогелем.
Оказывается, что в не обеспечивающих адгезию условиях культивирования мезенхимальные клетки-предшественники связываются и конденсируются в клеточные агрегаты весьма быстро. Уже через 24 часов культивирования клеток без обеспечения адгезии мезенхимальные клетки-предшественники агрегируют в один большой комплекс. Однако для получения клеточных агрегатов, которые представляют агрегаты хондрогенных клеток с успешным развитием, дифференцировкой и/или функционированием, целесообразным является культивирование без обеспечения адгезии в течение периода времени, превышающего 24 часов. В способе согласно настоящему изобретению мезенхимальные клетки-предшественники можно культивировать в не обеспечивающих адгезию условиях в течение по меньшей мере 48 часов, предпочтительно в течение по меньшей мере 72 часов, более предпочтительно в течение по меньшей мере 1 недели, еще более предпочтительно в течение по меньшей мере 2 недель, наиболее предпочтительно в течение по меньшей мере 4 недель. Следовательно, мезенхимальные клетки-предшественники можно культивировать в не обеспечивающих адгезию условиях в течение от 48 часов до 8 недель, предпочтительно в течение от 60 часов до 6 недель, более предпочтительно в течение от 72 часов до 5 недель. При культивировании в обычных чашках для культивирования агрегаты прилипают к поверхности чашек, и клетки начинают мигрировать и пролиферировать, что приводит к распаду структуры агрегата. Таким образом, предпочтительно, агрегаты содержат в не обеспечивающих адгезию условиях в течение всего периода культивирования.
В способе согласно настоящему изобретению мезенхимальные клетки-предшественники предпочтительно культивируют в не обеспечивающих адгезию условиях по меньшей мере до тех пор, пока не сформируется клеточный агрегат округленной формы со средним диаметром от 0,5 мм до 3 мм, более предпочтительно со средним диаметром от 0,5 мм до 2 мм.
Пригодность агрегата хондрогенных клеток, созданного способом согласно настоящему изобретению, зависит от его способности индуцировать или обеспечивать дифференцированные хондроциты или хрящевую ткань. Следовательно, предпочтительно культивировать выделенные мезенхимальные клетки-предшественники в не обеспечивающих адгезию условиях до тех пор, пока полученный в результате клеточный агрегат не начнет проявлять свойства и/или функции частично или полностью дифференцированных хондроцитов или хрящевой ткани. Ход дифференцировки можно отслеживать по относительной экспрессии соответствующих маркерных генов, белков или структур.
Предпочтительно, выделенный мезенхимальные клетки-предшественники культивируют в не обеспечивающих адгезию условиях по меньшей мере до тех пор, пока сформированный клеточный агрегат не будет проявлять повышенную экспрессию коллагена II, IX и XI, а также пониженную экспрессию коллагена I и XII по сравнению с экспрессией в мезенхимальных клетках-предшественниках непосредственно культивированием в не обеспечивающих адгезию условиях. Поскольку при анализе экспрессии стадиеспецифических маркерных молекул обнаруживали поддерживаемую экспрессию N-кадгерина и повышенные уровни коллагена типа II, но не экспрессию коллагена типа X, предполагают дифференцированный фенотип у хондрогенного предшественника и столбчатого хондроцита, но без дальнейшей дифференцировки в прегипертрофическую стадию. Этот фенотип воспроизводит фенотип задержанного развития хондроцитов в хрящевых тканях сустава. Относительные уровни экспрессии генов можно легко определить с помощью хорошо известных способов, таких как, например, количественная или полуколичественная RT-PCR или нозерн-блоттинг. Относительные уровни экспрессии белков можно легко определить с помощью хорошо известных способов, таких как, например, методики вестерн-блоттинга и ELISA. В дополнение к анализу паттерна экспрессии на уровне мРНК и белков для определения качества агрегата будут включены дальнейшая подробная характеристика структуры агрегата с помощью иммуногистохимического окрашивания, а также определение сопротивления давлению.
Одним из недостатков, с которыми сталкивалось большинство способов из уровня техники, является то, что в этих способах, необходимо присутствие искусственного биологического или небиологического каркаса или носителя, на котором или внутри которого клетки культивируют для формирования хрящевой ткани. Биологический или небиологический каркас или носитель считается искусственным или добавленным, если указанный каркас или носитель обеспечивается извне и не формируется непосредственно клетками, применяемыми в способе согласно настоящему изобретению в ходе формирования клеточных агрегатов. В способе согласно настоящему изобретению применение или присутствие такого искусственного биологического или небиологического каркаса или носителя не требуется. Способ согласно настоящему изобретению дает агрегат хондрогенных клеток согласно настоящему изобретению без применения какого-либо такого искусственного биологического или небиологического каркаса или носителя, который не происходит из применяемых клеток. В предпочтительном варианте осуществления способ согласно настоящему изобретению осуществляют так, что не осуществляют добавления биологического или небиологического каркаса при формировании хондрогенных клеточных агрегатов.
В способе согласно настоящему изобретению мезенхимальные клетки-предшественники культивируют в обеспечивающих адгезию или не обеспечивающих адгезию условиях со стандартной средой. Не существует специализированной культуральной среды, необходимой для индукции надлежащей агрегации клеток в не обеспечивающих адгезию условиях. Специалист в данной области техники хорошо осведомлен о подходящей среде. Как правило, применяют стандартную DMEM с определенным содержанием фетальной телячьей сыворотки (FCS), предпочтительно FCS присутствует в концентрации от 5% до 15%, более предпочтительно в концентрации 10% FCS.
Несмотря на тот факт, что надлежащая хондрогенная дифференцировка индуцируется при формировании агрегатов без дополнительных обработок, может быть полезна дополнительная оптимизация. Таким образом, в способе согласно настоящему изобретению после того, как процесс агрегации завершается, дополнительные этапы оптимизации, к которым относится введение фактора роста или культивирование в условиях гипоксии, будут дополнительно улучшать развитие и дифференцировку.
Настоящее изобретение также направлено на трансплантат, содержащий или состоящий из искусственного агрегата хондрогенных клеток согласно настоящему изобретению или ткани или структуры, полученных из него.
В другом аспекте настоящего изобретения предусматривается фармацевтическая композиция, содержащая искусственный агрегат хондрогенных клеток согласно настоящему изобретению, ткань или структуру, полученные из него, или трансплантат согласно настоящему изобретению и по меньшей мере одно фармацевтически приемлемое вспомогательное вещество.
Искусственный агрегат хондрогенных клеток согласно настоящему изобретению, трансплантат согласно настоящему изобретению или фармацевтическую композицию согласно настоящему изобретению можно применять при лечении повреждения хряща и/или разрушения или потери хряща.
Поскольку способ согласно настоящему изобретению работает с выделенными мезенхимальными клетками-предшественниками, полученными из не являющихся эмбриональными источников, способ обеспечивает возможность получения искусственного агрегата хондрогенных клеток, берущего начало из клеток, полученных от конкретного донора или пациента. Следовательно, возможно получать искусственные агрегаты хондрогенных клеток, которые были получены из клеток от лица, подлежащего лечению искусственными агрегатами хондрогенных клеток, фармацевтической композицией или трансплантатом согласно настоящему изобретению. Следовательно, по-видимому, можно получать трансплантат, который преимущественно, по существу или полностью получен из собственных клеток реципиента трансплантата так, чтобы реакции отторжения были сведены к минимуму или полностью отсутствовали. Искусственный агрегат хондрогенных клеток согласно настоящему изобретению можно применять для in vitro или in vivo получения дифференцированных хондроцитов или хрящевой ткани.
Искусственный агрегат хондрогенных клеток согласно настоящему изобретению, трансплантат согласно настоящему изобретению или фармацевтическую композицию согласно настоящему изобретению можно применять в качестве инструмента для исследований, который можно применять in vitro и in vivo.
Искусственный агрегат хондрогенных клеток согласно настоящему изобретению, трансплантат согласно настоящему изобретению или фармацевтическую композицию согласно настоящему изобретению можно применять в способе скрининга в отношении веществ in vitro или in vivo, которые модулируют свойства хондроцитов или хрящевой ткани.
В настоящем изобретении дополнительно представлен способ in vitro скрининга веществ, которые модулируют свойства хондроцитов или хрящевой ткани, включающий следующие этапы:
обеспечение образца искусственного агрегата хондрогенных клеток согласно настоящему изобретению или клеточного агрегата, полученного с помощью способа согласно настоящему изобретению, или ткани, полученной из них;
разделение соответствующего образца на порции;
инкубирование по меньшей мере одной порции с веществом, подлежащим скринингу; и
сравнение параметров, измеренных у обработанной порции и другой порции, которую не инкубировали с веществом, подлежащим скринингу.
В предпочтительном варианте осуществления, порцию помещают в автономное устройство по типу орган-на-чипе перед инкубированием порции с веществом, подлежащим скринингу.
Вкратце, способ согласно настоящему изобретению делает возможным идентификацию и анализ веществ, которые оказывают влияние на хондроциты или хрящевую ткань, при помощи искусственного агрегата хондрогенных клеток согласно настоящему изобретению. Образец, который, как следует понимать, содержит определенное количество продуктов-объектов настоящего изобретения, разделяют на несколько порций. Получают по меньшей мере две подгруппы; одну используют для скрининга, тогда как другая служит в качестве отрицательного контроля. Предпочтительно, количество частей для скрининга превышает количество частей для контроля. Обычно многочисленные части подвергают скринингу с высокой пропускной способностью. Вещества, подлежащие скринингу в способе согласно настоящему изобретению, не ограничены каким-либо образом. В варианте осуществления настоящего изобретения вещества выбраны из группы нуклеиновых кислот, в том числе интерферирующие РНК, рибозимов, аптамеров, антител, пептидов, углеводов, полимеров, малых молекул с молекулярным весом от 50 до 1000 Да и белков, предпочтительно антител, цитокинов и липокалинов. Эти вещества зачастую доступны в библиотеках. Предпочтительным является инкубирование отдельного соединения в отдельной порции образца. Тем не менее, также возможно изучать суммарный эффект веществ при помощи инкубирования по меньшей мере двух веществ в одной порции. Дополнительную подгруппу объектов одновременно инкубируют в отсутствие веществ. Процесс инкубирования зависит от различных параметров, например, типов клеток и чувствительности выявления, при оптимизации которых следуют обычным процедурам, известным специалистам в данной области техники. Идентификацию эффективных веществ в контексте настоящего изобретения предпочтительно осуществляют косвенно, например, путем определения паттернов экспрессии и/или жизнеспособности клеток, которые изменяются. Определение можно осуществлять в определенный момент и сопоставлять с силой сигнала вначале эксперимента и у отрицательного контроля. Подходящие тесты известны специалисту в данной области техники или могут быть легко разработаны в обычном порядке.
Поскольку искусственный агрегат хондрогенных клеток согласно настоящему изобретению можно рассматривать как орган, «органоид» или предшественник органа или их часть, особенно целесообразным может быть применение тест-системы, где искусственный агрегат хондрогенных клеток можно получить и/или культивировать в течение длительного времени в условиях, которые имитируют естественную перфузию. Особенно подходящим представляется комбинирование способа согласно настоящему изобретению и/или искусственного агрегата хондрогенных клеток согласно настоящему изобретению в аналитической системе на основе системы автономного устройства по типу орган-на-чипе, описанной в заявке на европейский патент с номером заявки ЕР 10008244 или в РСТ заявке с номером публикации WO 2009/146911.
В настоящем изобретении также раскрыто автономное устройство по типу «органоид»-на-чипе на основе искусственного агрегата хондрогенных клеток, содержащее автономное устройство по типу орган-на-чипе, как описано в WO 2009/146911, и искусственный агрегат хондрогенных клеток согласно настоящему изобретению.
Автономное устройство по типу «органоид»-на-чипе на основе искусственного агрегата хондрогенных клеток согласно настоящему изобретению создано для обеспечения возможности получения или поддержания искусственного агрегата хондрогенных клеток согласно настоящему изобретению в формате миниатюризированного чипа, подходящем для наблюдения в реальном времени при помощи визуализации живых клеток и, например, двухфотонной микроскопии, и его применения, например, для тестирования активности, фармакодинамики и фармакокинетики соединений или для изучения самосборки, гомеостаза, повреждения, регенерации или взаимодействия органов или «органоидов» и ниш стволовых клеток, а также явлений созревания, старения, смерти и хронобиологии. Для этой цели автономное устройство по типу «органоид»-на-чипе на основе искусственного агрегата хондрогенных клеток согласно настоящему изобретению содержит по меньшей мере один резервуар для подачи среды, по меньшей мере одну секцию для роста органа, содержащую по меньшей мере одну полость для органа, вмещающую искусственный агрегат хондрогенных клеток согласно настоящему изобретению, где резервуар для подачи среды соединен по меньшей мере с одной секцией для роста органа микроструйным питающим каналом.
В настоящем изобретении впервые предусматривается искусственный агрегат хондрогенных клеток и способ его получения, который характеризуется сборкой без воздействий внешних сил, отсутствием потребности в каком-либо искусственном биологическом или небиологическом каркасе или носителе, и который получают из взрослых клеток так, что не требуются эмбриональные клетки или ткани.
Искусственный агрегат хондрогенных клеток согласно настоящему изобретению представляет собой функциональный индукционный агрегат, который способен индуцировать или инициировать развитие хрящевой ткани in vitro и in vivo. Полученная хрящевая ткань характеризуется подтверждаемым присутствием структур, как правило, формирующих часть хрящевой ткани, которые расположены в физиологическом порядке.
ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На фиг. 1 показано, что дедифференцированные хондроциты проявляют характерные особенности мезенхимальных стволовых клеток. [А] Дедифференцированные хондроциты окрашивали в отношении панели мезенхимальных поверхностных маркеров. Клетки были положительными по CD105, CD106, CD44, CD73, CD90 и CD13 и отрицательными по гемопоэтическим маркерам CD34 и CD45 (показан один типичный эксперимент, n=6). Адипогенная дифференцировка. [B-D] Дедифференцированные хондроциты обрабатывали адипогенной средой в течение 28 дней. Дифференцировку в адипоциты оценивали с помощью световой микроскопии и окрашивания масляным красным О (Oil-Red-O). В) отрицательный контроль из клеток монослоя, неокрашенный; С) отрицательный контроль из клеток монослоя, окрашенный масляным красным О; D) дифференцировка в адипоциты после 28 дней подтверждается образованием заполненных липидами капель в цитоплазме; окрашивание масляным красным О заполненных липидами капель в дифференцированных адипоцитах (показан один типичный эксперимент, n=6); Дифференцировка в остеобласты. [F-G] Дедифференцированные хондроциты обрабатывали остеогенной средой в течение 28 дней. Успешную дифференцировку оценивали с помощью окрашивания по Коссу. Е) окрашенный отрицательный контроль; F) накопление кальцифицированного матрикса демонстрируется окрашиванием по Коссу (показан один типичный эксперимент, n=6); риска=50 мкм.
На фиг. 2 показаны кинетика конденсации на планшетах со слабым прикреплением. В этой системе для 3D культивирования используют специально обработанную поверхность чашки для культивирования, к которой не могут прикрепляться клетки. Суспензию клеток (106 клеток/мл) подавали в планшеты со слабым прикреплением. Клетки немедленно начинали взаимодействовать друг с другом после индукции 3D культуры (показан один типичный эксперимент, n=6).
На фиг. 3 показана кинетика экспрессии генов у связанных с хондрогенезом генов во время стадии конденсации. Анализ экспрессии генов с помощью ПЦР в реальном времени в пределах первых 90 или 72 часов после индукции 3D культуры в сравнении с экспрессией в монослое. Отношение к конститутивным генам представлено на логарифмической шкале, (n=6) статистический анализ с помощью U-критерия Манна-Уитни.
На фиг. 4 показана экспрессия генов у связанных с хондрогенезом генов через 28 дней культивирования со слабым прикреплением. Анализ экспрессии генов с помощью ПЦР в реальном времени через 28 дней после индукции 3D культуры сравнивали с экспрессией в монослое. Отношение к конститутивным генам представлено на логарифмической шкале, (n=6) статистический анализ с помощью U-критерия Манна-Уитни.
На фиг. 5 показана классификация дифференциально экспрессируемых генов. Оценка в двух независимых экспериментах с использованием микроматриц на [А] стадии конденсации (24 часа) и [В] стадии дифференцировки (28 дней), соответственно. Количества генов, подвергающихся понижающей и повышающей регуляции, в выбранных группах генной онтологии показаны в соответствии базой данных генной онтологии Panther.
На фиг. 6 показана клеточная конденсация клеток, обработанных РТХ (коклюшным токсином). Коклюшный токсин вводили в трех разных концентрациях к дедифференцированным хондроцитам во время процесса конденсации. Не наблюдали существенных изменений при обработке РТХ в том, что касается характера конденсации (показан один типичный эксперимент, n=3).
На фиг. 7 показано взаимодействие клеток и соединение суб-агрегатов во время процесса конденсации. Конденсацию индуцировали с помощью 3D культивирования со слабым прикреплением; сетчатые соединения различимы через 12 часов после индукции культуры (показан один типичный эксперимент, n=6).
На фиг. 8 показано схематическое изображение автономного устройства по типу 3D in vitro двухфазного костно-хрящевого «органоида»-на-чипе согласно настоящему изобретению.
На фиг. 9 показано поведение MSC в 3D керамической системе для культивирования. (А) Иммуногистохимический анализ и 2-фотонная микроскопия компонентов ниши внеклеточного матрикса (ЕСМ), (i) коллаген-1, (ii) фибронектин и (iii) интегрин 4а экспрессируются на высоком уровне от (а) дня 1 до (b) дня 7 культивирования (масштабная метка: 100 мкм). (В) Изображение, полученные с помощью сканирующей электронной микроскопии (SEM), иллюстрирующее структурное подобие между (i, iii) MSC после 2 недель 3D культивирования и (ii, iv) губчатой костью (масштабная метка: 10 мкм). (С) Спонтанная остеогенная дифференцировка MSC после 1 недели 3D культивирования, визуализированная с помощью положительного окрашивания по Коссу (i) и окрашивания ализариновым красным (iii), в сравнении с незасеянным керамическим материалом (ii, iv). (D) Анализ с помощью ПЦР в реальном времени известных молекул, характерных для ниш, в системе для совместного культивирования после 1 и 4 недель в сравнении с монослоем. (n=3, «усы»: SD среднего значения, ***р<0,001).
На фиг. 10 показаны выживаемость и фенотип гемопоэтических стволовых клеток/клеток-предшественников (HSPC). (A) HSPC выживают в системе для совместного 3D культивирования через (i) 1 неделю, (ii) 2 недели, (iii) 4 недели и (iv) 8 недель после посева, как визуализируется с помощью флуоресцентной микроскопии. Ядра MSC контрастно окрашивают синим с помощью DAPI (масштабная метка: 100 мкм). (В) Типичные диаграммы рассеяния FACS и (С, D) количественное определение примитивных (CD34+CD38-) и дифференцированных (CD34+CD38+) HSPC в системах для совместного 3D культивирования и традиционных системах для совместного культивирования. (n=8, «усы»: SD среднего значения, **р<0,01, ***р<0,001).
На фиг. 11 показана пролиферация HSPC. (А) Типичные гистограммы CFSE-FACS анализа, показывающего дифференциальную пролиферацию HSPC в различных системах для культивирования. Пунктирными линиями ограничены фракции медленно (справа) и быстро пролиферирующих (слева) клеток. (В) Количественная оценка медленно пролиферирующих клеток в четырех системах для культивирования. (С, D, Е) Количественная оценка примитивных (CD34+CD38-) HSPC во фракциях медленно и быстро пролиферирующих клеток в каждой системе для культивирования после 1, 2 и 4 недель культивирования. (n=6, «усы»: SD среднего значения, *р<0,05, ***р<0,001).
На фиг. 12 показана жизнеспособность и функциональность HSPC. (А) Типичные диаграммы рассеивания FACS и (В, С) количественная оценка некротических (положительных по PI) и апоптических (положительных по аннексину V) HSPC в системах для совместного 3D культивирования и традиционных системах для совместного культивирования. (n=8, «усы»: SD среднего значения, *р<0,05 **р<0,01, ***р<0,001). (D, Е) Потенциал дифференцировки в несколько линий у HSPC из системы для совместного 3D культивирования. Типичные колонии (i) GEMM, (ii) BFU-E, (iii) GM и (E) оценка колоний после CFU-GEMM анализов (n=3).
На фиг. 13 показано взаимодействие компонентов в системе для совместного 3D культивирования. (А) Иммуногистохимический анализ и 2-фотонная микроскопия системы для 3D совместного культивирования после 2 недель культивирования. Видно, что отслеживаемые по зеленому окрашиванию HSC и отслеживаемые по красному окрашиванию MSC присутствуют совместно с окрашенными Alexa 350 (синий) компонентами ЕСМ (ii) коллагеном I и (iii) фибронектином, а также с сигнальными молекулами, (iv) C-Kit, (v) интегрином 4а, (vi) N-кадгерином, по сравнению с неокрашенным контролем (i) (масштабная метка: 100 мкм). (В) SEM-изображение, иллюстрирующее физическое взаимодействие между более мелкими круглыми HSPC и более крупными плоскими прикрепленными MSC после 2 недель 3D культивирования (масштабная метка: 10 мкм).
ПРИМЕРЫ
Пример А
I. Формирование искусственной костной ткани Клетки
Мезенхимальные стволовые клетки (MSC) выделяли с помощью стандартных процедур. Вкратце, пунктаты костного мозга интенсивно промывали с помощью PBS и РВМС (мононуклеарные клетки периферической крови) выделяли путем центрифугирования в градиенте плотности фиколла. Мезенхимальные предшественники выделяли по адгезии к пластику с последующим периодом увеличения численности до 4 пассажей. Гемопоэтические стволовые клетки человека выделяли из образцов пуповинной крови по экспрессии CD34 с использованием MACS технологии (Miltenyi Biotec). HSC культивировали в дополненной среде (Stemspan®+ 100 нг/мл ТРО, FLT3, IL6 и SCF).
Платформа
Платформу для искусственного костного мозга обеспечивали с помощью керамического 3D каркаса (Sponceram® Zellwerk). Для того чтобы индуцировать согласованную пролиферацию, дифференцировку и реорганизацию микроокружения в соответствии с накоплением внеклеточного матрикс и экспрессией сигнальных молекул, мезенхимальные стволовые клетки высевали на данный керамический материал за одну неделю до добавления гемопоэтических стволовых клеток. Затем керамический материал дополнительно культивировали либо в статической культуре, либо в перфузируемой системе для постоянной подачи питательных веществ. Функциональность костного мозга оценивали с помощью анализа экспрессии генов, по составу различных клеточных популяций с помощью технологии FACS и путем определения типичных биологических функций (поддержание и пролиферация HSC, развитие отдельных ниш стволовых клеток, иммунологические аспекты).
II. Формирование агрегатов хондрогенных клеток или искусственной хрящевой ткани
Как остеоартрит, так и ревматоидный артрит представляют собой дегенеративные заболевания, которые поражают структуру хряща. Будучи спровоцированной различными факторами (например, механической перегрузкой, возрастом, повреждением и воспалением), потеря структуры ткани является необратимой, поскольку клетки внутри хряща характеризуются лишь очень слабой врожденной способностью к самовосстановлению. Таким образом, за последние несколько десятилетий было предпринято множество попыток поиска альтернативных стратегий индукции процесса регенерации в хряще. В последние годы применение аутологичных клеток в регенеративной медицине является объектом интенсивного изучения. Однако как идеальный источник мезенхимальных клеток-предшественников, так и оптимальные условия культивирования для in vitro дифференцировки все еще должны быть установлены. Мезенхимальные стромальные/стволовые клетки выделяются из общего ряда вследствие их способности дифференцироваться во все мезенхимальные ткани, хотя количество клеток, которые можно получать из костного мозга или других тканей-источников, ограничено. Таким образом, применение аутологичных клеток, выделенных из здоровой ткани с последующим увеличением численности, представляет одну перспективную стратегию для решения этой задачи.
К сожалению, клетки теряют свой дифференцированный фенотип после удаления из их естественного тканевого микроокружения. Следовательно, выделение хондроцитов из их естественного трехмерного матрикса в суставном хряще с помощью ферментативного расщепления и дальнейшее культивирование в двухмерном монослое приводит к изменению клеток, во время этого процесса клетки опять входят в клеточный цикл, теряют свой круглый фенотип и переключают свою продукцию молекул матрикса с коллагена типов II, IX и XI на типы I, III и V. Кроме того, было продемонстрировано, что 20-культивируемые хондроциты экспрессируют поверхностные маркеры мезенхимальных стволовых клеток и обладают потенциалом дифференцировки в несколько линий. Этот процесс называется дедифференцировкой и подчеркивает важность интактного 3D окружения для поддержания фенотипа хондроцитов. В связи с этим, в последние годы обнаружилось, что процессы развития контролируются не только растворимыми факторами, но, кроме того, пространственным расположением, обеспечиваемым молекулами адгезии, которые не только опосредуют межклеточные взаимодействия и взаимодействия клетка-матрикс, но также вносят вклад в регуляцию экспрессии генов путем передачи сигналов, связанных с адгезией. Таким образом, культивирование клеток-предшественников в 3D структуре оказывается одним из ключевых стимулирующих факторов во время процесса хондрогенной редифференцировки in vitro. Вследствие этого, для этой цели были созданы различные системы для 3D культивирования. Обычно используются четыре методики культивирования мезенхимальных клеток или клеток-предшественников в 3D системе: 1) инкапсулирование в гидрогель, 2) культивирование в микромассе, 3) образование клеточных осадков высокой плотности и 4) использование биосовместимых каркасов.
Культивирование в 3D микроокружении и предотвращение образования монослоя является основой всех упомянутых систем для культивирования. Несмотря на тот факт, что все модели культивирования способны индуцировать хондрогенную дифференцировку, подтвержденную повышающей регуляцией коллагена типа II и протеогликана, все еще остаются существенные недостатки. Клетки заставляют формировать клеточные 3D агрегаты либо с помощью центрифугирования (культивирование осадков), либо под воздействием гравитации (в микромассе). С другой стороны, возможность клеточного взаимодействия ограничена или даже невозможна после инкапсулирования в гидрогель или культивирования на биосовместимых каркасах. Однако, по-видимому, оправдано имитировать полный in vivo процесс дифференцировки хондроцитов in vitro как для получения оптимальных предпосылок для применений в регенеративной медицине, так и для обеспечения модельной системы для культивирования, чтобы расширить соответствующие знания об этом многоэтапном процессе.
Мезенхимальная конденсация представляет собой начальный этап в развитии элементов скелета, на котором предшественники мигрируют в участок образования будущего скелета и начинают накапливаться и конденсироваться, формируя в результате клеточные агрегаты высокой плотности. Во время процесса in vitro редифференцировки главный интерес представляет осуществление этого начального этапа формирования агрегатов. К сожалению, ни одна из обычных система для 3D культивирования не подходит для такой цели, поскольку все они не соответствуют предварительному условию подвижности независимых отдельных клеток перед процессом конденсации.
Авторами настоящего изобретения была создана система для культивирования без использования каркаса, предусматривающая начальную конденсацию клеток и последующую хондрогенную редифференцировку выделенных дедифференцированных хондроцитов. Процесс конденсации, а также редифференцированные клетки были тщательно охарактеризованы с помощью всестороннего анализа экспрессии генов. Поскольку при хондрогенной дифференцировке демонстрируется сильная повышающая регуляция экспрессии коллагена типа II и массовая секреция и накопление протеогликанов, данная новая система для 3D культивирования впервые обеспечивает возможность изучения стимулирования процесса мезенхимальной конденсации in vitro. Ингибирование передачи сигнала с участием хемокинов в ходе агрегации клеток демонстрирует, что процесс конденсации не зависит от миграции клеток, а скорее опосредуется молекулами адгезии и усиливается межклеточными контактами.
Материалы и методы
Выделение и культивирование первичных хондроцитов
Суставной хрящ коленного сустава от доноров-добровольцев получали в Берлинском центре исследования скелетно-мышечных заболеваний при клинике Шарите (Musculoskeletal Research Center Berlin, Charité) (Университетская клиника Берлина, Германия), а также в Институте экспериментальной патологии при клинике Шарите (Institute of Experimental Pathology, Charité) в кампусе им. Вирхова. Исследование было санкционировано местным комитетом по этике (Charité Campus Mitte, Университетская клиника Берлина). Хрящевую ткань измельчали на маленькие кусочки и хондроциты выделяли с помощью ферментативного расщепления в течение ночи при 37°С с использованием коллагеназы (1 мг/мл, Sigma- Aldrich) в DMEM. Клетки отделяли от расщепляемой ткани путем фильтрации через 70 мкм нейлоновый фильтр для клеток (BD Falcon™, Бельгия), отмывали с помощью PBS и культивировали в DMEM, содержащей 10% FCS. Первичные хондроциты культивировали в термостате с водяной рубашкой в увлажняемой атмосфере (5% CO2/воздух) при 37°С в среде DMEM (Gibco), дополненной 10% FCS и пенициллином/стрептомицином (Gibco). Клетки пассировали по меньшей мере 10 раз для получения полностью дедифференцированного фенотипа.
Характеристика дедифференцированных хондроцитов
Анализ экспрессии на поверхности клеток с помощью FACS.
Дедифференцированные хондроциты анализировали в отношении экспрессии ими различных поверхностных маркерных молекул для мезенхимальных стволовых клеток (MSC) с помощью прямого иммунофлуоресцентного окрашивания. Клетки собирали при помощи трипсинизации и центрифугирования и клеточный осадок ресуспендировали в PBS/BSA (0,5%). Клетки инкубировали с антителами с флуоресцентной меткой в течение 15 минут при RT (комнатная температура). Для изотипических контролей клетки окрашивали маркерными молекулами для Т-клеток того же изотипа, что и антитела к MSC. Окрашенные анализируемые образцы затем отмывали с помощью PBS/BSA и центрифугировали для удаления несвязавшихся антител. Клеточные осадки ресуспендировали в 400 мкл PBS/BSA и анализировали на FACSCalibur™ (BD Biosciences). Для выявления мертвых клеток непосредственно перед цитометрическим анализом добавляли пропидиум йодид. 30000 событий обнаруживали и анализировали с использованием программного обеспечения CellQuest™.
Потенциал дифференцировки. Адипогенную дифференцировку индуцировали в 2D культурах с использованием общепринятых добавок в среду (DMEM с 10% FCS, 10 мкг/мл инсулина, 0,2 мМ индометацина, 1 мкМ дексаметазона, 0,5 мМ 3-изобутил-1-метил-ксантина (Sigma)). Дедифференцированные хондроциты высевали в 6-луночные планшеты (Corning - 100000 клеток на лунку) и культивировали в DMEM + 10% FCS до тех пор, пока клетки не достигали конфлуэнтности. Клетки стимулировали в течение 28 дней с заменой среды каждые 4 дня. Окрашивание масляным красным О на липиды. После 28 дней клетки фиксировали в 4% параформальдегиде в течение 10 минут, отмывали с помощью PBS и затем инкубировали со свежепрофильтрованным окрашивающим раствором масляного красного О (0,7% в пропиленглицерине) в течение 1 часа (Sigma). После двукратного промывания клеток дистиллированной водой окрашивание оценивали с помощью световой микроскопии. Остеогенную дифференцировку индуцировали в монослойных культурах при помощи подходящей среды для дифференцировки (DMEM + 10% FCS, 10 мМ β-глицерофосфата (Sigma), 10 нМ дексаметазона (Sigma), 0,1 мМ 2-фосфата L-аскорбиновой кислоты (Sigma). Вкратце, клетки высевали в 6-луночные планшеты (Corning - 100000 клеток на лунку) и культивировали в DMEM + 10% FCS до тех пор, пока клетки не достигали конфлуэнтности. Среду заменяли каждые 4 дня и после 21 дня остеогенной стимуляции клетки окрашивали красителем по Коссу на секретируемый минерализованный матрикс на основе Са2+ в качестве маркера дифференцировки в остеобласты. Клетки фиксировали 4% параформальдегидом (Sigma). К зафиксированным клеткам добавляли раствор нитрата серебра (5%, Sigma) и культуральный планшет помещали под УФ-излучение на 20 минут. Наконец, клетки промывали водой и оценивали при помощи световой микроскопии.
Системы для 3D культивирования
Для 3D культивирования со слабым прикреплением дедифференцированные хондроциты собирали и ресуспендировали в DMEM + 10% FCS с получением суспензии отдельных клеток с концентрацией 106 клеток на мл. Клеточную суспензию (1 мл на лунку) добавляли в 24 луночный планшет со слабым прикреплением (Ultra Low Cluster Plate, Corning, Германия). В отличие от отрицательно заряженной гидрофильной поверхности стандартных чашек для культивирования тканей планшеты со сверхслабым прикреплением обладают нейтральной, покрытой гидрофильным гидрогелем поверхностью, которая существенно уменьшает связывание белков, участвующих в прикреплении. При использовании этих специализированных чашек для культивирования клетки не оседают в виде олигослойной структуры вследствие адгезии клеток, как при культивировании в микромассе. Формирование 2D монослойной культуры предотвращалось и клетки сохраняли круглую форму в этих условиях поддержания суспензионной культуры. Кроме того, в отличие от системы для культивирования осадков, в которых клетки заставляют осаждаться при помощи центрифугирования, система для культивирования со слабым прикреплением обеспечивает возможность свободного движения клеток и межклеточного взаимодействия в ходе начального процесса конденсации. Чтобы гарантировать постоянные условия культивирования среду заменяли регулярно каждые 3 дня. Для того чтобы в культуре со слабым прикреплением воспрепятствовать передаче хемотаксических сигналов, вызываемых рецепторами, связанными с G-белками, в экспериментах по конденсации при слабом прикреплении в культуральную среду вводили коклюшный токсин (Sigma, Германия) в различных концентрациях (10, 100 и 1000 нг/мл).
Гистология
Срезы собранных агрегатов получали на криостате при толщине 5 мкм и монтировали на предметные стекла Superfrost Plus (Menzel). Срезы высушивали и фиксировали.
Окрашивание гематоксилином Срезы инкубировали в окрашивающем растворе гематоксилина (Sigma) в течение 3 минут, а затем промывали водопроводной водой. Срезы обезвоживали и заключали. Окрашенные срезы анализировали с помощью световой микроскопии.
Окрашивание альциановым синим. Микроскопические препараты предварительно инкубировали в 3% уксусной кислоте в течение 3 минут перед обработкой раствором альцианового синего (Sigma) (3% в 3% уксусной кислоте) в течение 30 минут при комнатной температуре. Срезы отмывали водой, обезвоживали и заключали. Окрашенные срезы анализировали с помощью световой микроскопии.
Окрашивание коллагена типа II. Микроскопические препараты обрабатывали блокирующим раствором для белков (Dako Cytomation) в течение 10 минут при RT, а затем инкубировали в течение ночи при 4°С с первичным моноклональным антителом к коллагену типа 2А (BD Biosciences). На следующий день срезы инкубировали в течение 1 часа с конъюгированным с Су3 вторичным антителом (Dako Cytomation) и ядра подвергали контрастному окрашиванию с помощью DAPI. Окрашенные срезы анализировали с помощью флуоресцентной микроскопии.
ПЦР в реальном времени. В указанные моменты времени клетки лизировали, РНК экстрагировали с использованием набора NucleoSpin® RNA II (Machery and Nagel). кДНК синтезировали с помощью обратной транскрипции 400 нг общей РНК (Taqman, Roche). Эксперименты с ПЦР в реальном времени осуществляли с использованием следующих условий: 1 мкл кДНК, 0,5 мкМ каждого праймера, 6 мМ MgCl2, 200 мМ dNTP, 50 мМ Tris (рН 8,8), 500 нг/мл BSA, 0,05 ед./мл Immolase (Bioline), lx Sybrgreen (Molecular Probes). Относительный уровень экспрессии приводится как отношение к экспрессии GAPDH.
Анализ ДНК на микроматрицах Микроматрицу CGH 44К на основе генома человека (Agilent) использовали в соответствии с протоколом производителя. Эксперимент на микроматрице основывался на гибридизации с использованием соотношения двух цветовых меток и набора Low RNA Input Fluorescent Linear Amplification (Agilent) для мечения РНК. Вкратце, 500 нг общей РНК подвергали обратной транскрипции с использованием праймера олиго(dT)-Т7 промотора и обратной транскриптазы вируса лейкоза мышей Молони (MMLV-RT) для синтеза первой и второй нитей кДНК. Флуоресцентную антисмысловую кРНК синтезировали с помощью РНК-полимеразы Т7, которая одновременно включает либо цианин-3-цитидин-5'-трифосфат (3-СТР), либо цианин-5-CTP. Очищенные продукты количественно оценивали по поглощению при А552 нм на цианин-3-СТР и А650 нм на цианин-5-CTP и эффективность мечения подтверждали с помощью фотометра Nanodrop (Kisker). Перед гибридизацией 2 мкг каждого меченного кРНК продукта фрагментировали и смешивали с контрольными мишенями и буфером для гибридизации в соответствии с протоколом поставщика (Agilent). Гибридизации проводили в течение ночи при 60°С. Затем препараты отмывали и сканирование микроматриц осуществляли с разрешением 5 мкм с использованием лазерного сканера для ДНК микроматриц (Agilent). Элементы изображения выделяли с помощью инструмента для анализа изображения версии А 6.1.1 (Agilent) с использованием настроек по умолчанию. Анализ данных проводили в системе Rosetta Informatics Platform Resolver, сборка 4.0. Изменения в паттернах экспрессии идентифицировали при строгой оценке данных и пороге отсечения с 2-кратным уровнем экспрессии в сочетании с требованиями, чтобы выбирались только экспериментальные точки с низким значением р (р<0,05). При использовании данной стратегии оценка данных была независима от моделей погрешностей, реализованных в системе Rosetta Resolver (Agilent).
Результаты
Для того чтобы выполнить полную фенотипическую дедифференцировку, выделенные хондроциты культивировали в течение по меньшей мере десяти пассажей в монослое. Как описано ранее, клетки принимали четко выраженный фенотип предшественника, характеризующийся экспрессией поверхностных маркеров MSC (CD105, CD106, CD44, CD73, CD90 и CD13) и потенциалом к дифференцировке в несколько типов клеток (адипоциты и остеоциты) (фиг. 1) (Rosowski et al., 2006). В соответствии с экспрессией на поверхности клеток наблюдали однородную популяцию дедифференцированных хондроцитов.
Клеточная конденсация при культивировании со слабым прикреплением
Основная идея данной методики культивирования заключается в препятствовании прикреплению клеток к поверхности чашки для культивирования и сохранении подвижности клеток. Для того чтобы предотвратить формирование прикрепленной монослойной культуры, клетки высевали в лунки планшетов со слабым прикреплением со специально обработанной поверхностью. Спустя несколько часов клетки начинали прилипать друг к другу, с формированием в результате сетчатой структуры. В течение следующих 72 часов переплетенная структура конденсировалась и, наконец, формировала в результате один клеточный агрегат высокой плотности на лунку со средним диаметром агрегата 1-2 миллиметров (фиг. 2). Клеточные агрегаты культивировали в течение дополнительных 4 недель без каких-либо явных изменений размера или формы. Процесс культивирования разделяли на раннюю и позднюю стадию и оценивали отдельно.
Мезенхимальная конденсация in vitro - ранние события
На первом этапе анализировали экспрессию генов, характерных для хондрогенной дифференцировки (фиг. 3). Уровни мРНК транскрипционного фактора SOX-5 были слегка повышены после 12 часов культивирования. В более поздние моменты времени экспрессия возвращалась к уровням, характерным для монослоя. Неожиданно оказалось, что экспрессия мРНК транскрипционного регулятора SOX-9 была снижена в 3 раза по сравнению с монослойными культурами. Этот пониженный уровень мРНК сохранялся в течение первых трех дней при формировании конденсации. Тем не менее, оба транскрипционных фактора экспрессировались на высоком уровне как в монослойных культурах, так и в культурах со слабым прикреплением, на что указывало отношение к GAPDH. Дедифференцированные хондроциты проявляли выраженную экспрессию TGF-B1; однако этот фактор роста не менялся при клеточной конденсации. Как ожидалось, экспрессия N-кадгерина была значительно повышена при конденсациях со слабым прикреплением в течение первых часов конденсации. В более поздние моменты времени, когда конденсат был сформирован, для молекулы адгезии выявляли экспрессию гена, ослабленную в 4 раза. Что неожиданно, значительные изменения уровней мРНК для выбранных молекул матрикса определяли даже на начальной фазе 3D культивирования. Обнаружено, что коллаген типа 1А1 в большом количестве экспрессируется в монослойной культуре, но экспрессия данной молекулы была понижена в 10 раз через 4 часа после индукции 3D культуры и проявляла дальнейшее постепенное снижение в течение следующих 90 часов. В отличие от этого, обнаружено, что экспрессия молекулы хрящевого матрикса, коллагена типа 2А1, индуцировалась в 12 раз после 90 часов культивирования со слабым прикреплением. Однако уровень мРНК для протеогликанового компонента аггрекан снижался практически в 10 раз в самые ранние моменты времени (4 часа) и далее снижался в течение следующих 4 дней 3D культивирования (фиг. 3).
Хондрогенная дифференцировка - поздние события
Для того чтобы проанализировать хондрогенный потенциал системы для 3D культивирования со слабым прикреплением, определяли накопление специфических компонентов внеклеточного матрикса (фиг. 4). Кроме того, экспрессию связанных с хондрогенезом генов количественно оценивали с помощью ПЦР в реальном времени (фиг. 4). После 28 дней из агрегатов получали срезы для гистологического анализа. Окрашивание гематоксилином срезов агрегатов демонстрировало высокую плотность клеток. Более того, при окрашивании альциановым синим можно наблюдать массивное накопление протеогликанов внеклеточного матрикса. Повышение экспрессии коллагена типа 2А1 на 3 порядка величины после 4 недель 3D культивирования было наиболее заметным результатом для определяемых молекул матрикса. Кинетический иммуногистологический анализ в отношении экспрессии коллагена типа 2А1 демонстрировал накопление коллагена типа 2А1 даже во время начальной фазы 3D культивирования. В течение первых двух дней после индукции культуры сигналы от коллагена 2А1 не выявлялись. Однако через 8 дней после индукции клеточной конденсации явно присутствовал значительный сигнал, а после четырех недель культивирования даже еще более масштабное накопление коллагена типа 2А. Кроме того, снижение экспрессии коллагена типа 1А1 доказывалось снижением уровня мРНК в 25 раз по сравнению с монослойными культурами. Что неожиданно, уровни мРНК аггрекана снижались в 5 раз по сравнению с пролиферирующими 2D культурами (фиг. 4). Неожиданно, транскрипционный фактор Sox-9 проявлял исключительно высокий уровень экспрессии генов в пролиферирующих клетках монослоя, на что указывали отношения к GAPDH. После временного снижения экспрессии во время формирования агрегата уровень мРНК возвращался к базовым значениям для монослоя. С другой стороны, можно выявить дополнительный прирост экспрессии Sox-5 в 14 раз. Следовательно, относительная экспрессия Sox-5 превышала относительную экспрессию Sox-9 через 4 недели культивирования со слабым прикреплением. Кроме того, в 3D культуре можно определить дополнительное повышение уровней мРНК TGF-βl и молекулы адгезии N-кадгерина (фиг. 4).
Сравнительный анализ процесса конденсации и дифференцировки
Для того чтобы воспользоваться системой для дифференцировки без использования каркаса, для дальнейшего изучения клеточной дифференцировки, а также для характеристики изменений паттерна экспрессии в ходе процесса конденсации проводили полногеномный анализ на микроматрицах. Осуществляли два независимых эксперимента с отдельными первичными клеточными культурами. Зонды с флуоресцентными метками гибридизировались на ДНК-чипах Agilent 44К. Полученные в результате данные от обоих чипов объединяли для оценки. Гены определялись как подвергающиеся регуляции гены, если обе матрицы демонстрировали по меньшей мере 2-кратное изменение экспрессии гена. Значение р порога отсечения было предварительно установлено как р<0,05. Для того чтобы получить общее представление об этом конкретном изучаемом биологическом процессе, подвергающиеся регуляции гены группировали в соответствии с классификацией Panther GO (таблицы 1-5). Группами с наибольшим количеством подвергающихся регуляции генов как в ходе ранней, так и поздней фазы являются группы межклеточной коммуникации, процесса развития и сигнальной трансдукции (фиг. 5), что указывает на массовую модуляцию экспрессии генов в функциональных группах, которые требуются для индукции процессов дифференцировки. Однако задачей системы для 3D культивирования является редифференцировка дедифференцированных хондроцитов. Соответственно, раздел «развитие скелетной системы» в классификации Panther исследовали более подробно.
Фаза конденсации
Как указано в таблицах 1 и 2, факторы роста TGF-β3 и GDF-15, а также транскрипционный фактор Fos представляют собой гены, подвергшиеся повышающей регуляции в наибольшей степени в этой классифицируемой группе. Как можно заметить, экспрессию нескольких представителей кластера Нох-А определяли как повышенную во время стадии конденсации. Представитель группы TGF-β, ингибин В, представляет собой ген, подвергающийся наибольшей понижающей регуляции, и его репрессия также была очевидной после 28 дней культивирования.
Агрегаты после 28 дней (таблицы 3 и 4).
Генами, подвергающимися наиболее выраженной повышающей регуляции после 28 дней, являются гены связанных с хрящом матриксных молекул, коллагена II, IX и XI. У матриксного Gla-белка и кислого белка хряща также проявлялись повышенные уровни мРНК, тогда как, неожиданно, но в соответствии с данными ПЦР в реальном времени оказалось, что аггрекан и ферменты синтеза аггрекана подвергались понижающей модуляции. В числе генов, подвергавшихся наиболее сильной повышающей регуляции в этой классифицируемой группе, транскрипционные факторы Fos и Runx2, а также фактор роста GDF-15, тогда как уровень не характерных для хряща матриксных молекул коллагена I и XII сильно снижался.
Процесс конденсации опосредован межклеточной адгезией
Процесс мезенхимальной конденсации характеризуется агрегацией рыхлой мезенхимы. Тем не менее, немного известно о том, как опосредуется это повышение плотности клеток-предшественников. Вследствие многофункциональности нескольких семейств белков в классификации Panther молекулы адгезии и хемотаксические молекулы были выделены в подгруппу в более узком смысле в соответствии с базой данных KEGG. Поскольку данные анализа ДНК на микроматрицах демонстрировали дифференциальную экспрессию хемокинов и соответствующих рецепторов (таблица 5), в качестве первой попытки использования данной 3D системы для культивирования в ходе процесса конденсации следует проанализировать потенциальное вовлечение хемокинов и миграционную активность клеток. Подавляющее большинство хемокиновых рецепторов представляют собой часть семейства из семи трансмембранных рецепторов. Данный класс белков вызывает передачу ими сигналов с помощью связанной с G-белком сигнальной трансдукции. Коклюшный токсин (РТХ, Bordetella pertussis) нарушает этот путь посредством АДФ-рибозилирования G-субъединицы комплекса гетеротримерного G белка. Для того чтобы исключить хемокиновую активность в ходе конденсации, эксперименты по культивированию со слабым прикреплением проводили при добавлении РТХ в различных концентрациях. Что неожиданно, продукты конденсации клеток в обработанных РТХ культурах, по-видимому, не отличаются от контрольного эксперимента в течение первых 72 часов вне зависимости от применяемой концентрации (фиг. 6). Непосредственно после индукции культуры, клетки формировали небольшие клеточные кластеры, в дальнейшем уплотняющиеся через сетеподобную промежуточную стадию до тех пор, пока, наконец, не оформлялся один клеточный агрегат высокой плотности, независимо от введения РТХ и с аналогичной кинетикой. Данные наблюдения указывают на то, что опосредованная хемокинами миграция клеток не вовлечена в процесс агрегации клеток. Интересно, что результаты оценки с помощью микроматриц демонстрировали, что 75% дифференциально экспрессируемых молекул адгезии, по-видимому, подвергались повышающей регуляции, что предполагает повышенное межклеточное взаимодействие во время этапа конденсации (таблица 5). Данное предположение подтверждалось более подробными микроскопическими наблюдениями. На отдельной промежуточной стадии существования многочисленных субагрегатов становились видны длинные дендритоподобные клеточные отростки. Соединения формировались либо непосредственно от одного клеточного суб-центра к следующему (фиг. 7А-С), либо создавались одной клеткой с двумя удлинениями в противоположных направлениях (фиг. 7D). На более поздних стадиях эти тонкие нити использовались для соединения всех субагрегатов в конечный агрегат. Следовательно, процесс конденсации запускается скорее повышенной экспрессией молекул адгезии, приводящей в результате к увеличенному взаимодействию клеток, нежели опосредованной хемокинами миграцией клеток.
Краткое изложение
Применение аутологичных клеток-предшественников представляет перспективную стратегию восстановления поврежденной хрящевой ткани при остеоартрите. В этих клетках можно запускать дифференцировку в функциональные активные хондроциты с получением в результате вновь синтезированного хряща. Для этой цели на сегодняшний день было создано несколько систем для культивирования. Поскольку хондрогенная дифференцировка in vivo инициируется этапом мезенхимальной конденсации, большой интерес представляет характеристика первого линиеспецифического этапа in vitro. По этой причине разработали систему для 3D культивирования со слабым прикреплением без использования каркаса, имитирующую полный процесс хондрогенеза, включающий стадию начальной конденсации. Система для культивирования способна индуцировать хондрогенную дифференцировку, на которую указывает повышенная экспрессия генов ключевого транскрипционного фактора Sox-5 и коллагена типа II и IX. По сравнению с другими известными системами для 3D культивирования применение культивирования в планшетах со слабым прикреплением обеспечивает преимущество при анализе различных фаз агрегации клеток, начиная с суспензии отдельных клеток из ранее выделенных и размноженных человеческих первичных клеток мезенхимального происхождения. Анализ ранних стадий демонстрировал, что процесс конденсации скорее опосредован повышенной межклеточной адгезией, нежели передачей сигнала с участием хемокинов. Следовательно, система для 3D культивирования со слабым прикреплением, наконец, может помочь идентифицировать ключевые факторы, которые являются незаменимыми для индукции хондрогенной дифференцировки in vivo или в случае in vitro созданных трансплантатов для клинических применений.
III. Автономное устройство по типу 3D in vitro двухфазного костно-хрящевого «органоида»-на-чипе
На фигуре 8 изображено автономное устройство по типу 3D in vitro двухфазного костно-хрящевого «органоида»-на-чипе согласно настоящему изобретению.
Автономное устройство 1 по типу 3D in vitro двухфазного костно-хрящевого «органоида»-на-чипе содержит подложку 2, в которой сформированы два резервуара 5 для подачи среды, которые соединены друг с другом через канал 6 для подачи среды. Автономное устройство 1 по типу 3D in vitro двухфазного костно-хрящевого «органоида»-на-чипе дополнительно содержит полость 7 для органа, которая вмещает 3D in vitro двухфазный костно-хрящевой «органоид», содержащий слой искусственной хрящевой ткани 4 и слой искусственной костной ткани 3, где искусственная хрящевая ткань 4 непосредственно контактирует с поверхностью искусственной костной ткани 3. Для того чтобы обеспечить приложение механических сил к 3D in vitro двухфазному костно-хрящевому «органоиду», может присутствовать исполнительный механизм 8.
Пример В
Материалы и методы
1. Выделение и размножение MSC
Человеческие MSC выделяли из костного мозга головки бедренной кости, полученной после операции по протезированию сустава, при наличии письменного согласия в соответствии с руководством Совета по этике при клинике Шарите и университетской клинике Берлина (Charité Universitatsmedizin Berlin), как описывалось ранее. Затем клетки размножали в среде Игла в модификации Дульбекко (DMEM) с 4,5 г/л глюкозы (РАА Laboratories, Австрия), содержащей 10%FBS (РАА Laboratories, Австрия) и пенициллин-стрептомицин (РАА laboratories). MSC от пассажа 4 до 7 использовали для экспериментов по совместному культивированию.
2. Выделение HSPC
Человеческие HSPC выделяли из пуповинной крови, при наличии письменного согласия в соответствии с руководством Совета по этике при клинике Шарите и университетской клинике Берлина (Charité - Universitatsmedizin Berlin). Пуповинную кровь собирали в раствор PBS-BSA-EDTA и мононуклеарные клетки выделяли путем центрифугирования в градиенте плотности. Затем HSPC отделяли при помощи иммуномагнитного разделения с использованием набора для выделения CD34+ клеток с помощью MACS (Miltenyi Biotec, Германия) и исходя из инструкций производителя. Затем свежевыделенные клетки помещали в различные системы для культивирования при плотности 2×104 клеток/культура.
3. Керамический материал
Покрытые гидроксиапатитом керамические диски на основе оксида циркония Sponceram НА® толщиной 1 мм и диаметром 1 см приобретали у Zellwerk GmbH, Германия. Диски автоклавировали перед применением.
4. Системы для культивирования клеток
Керамические диски засевали MSC с плотностью примерно 106 клеток/диск за 7 дней до посева HSC. Керамические диски погружали в DMEM с высоким содержанием глюкозы (РАА Laboratories, Австрия), содержащую 10% FBS (РАА Laboratories, Австрия) и пенициллин-стрептомицин, в 24-луночных планшетах со сверхслабым прикреплением (Corning Inc., США). Планшеты выдерживали при 37°С, 5% CO2. Среду заменяли каждые 48 часов. Одновременно в 6-луночные планшеты также высевали MSC так, что они достигали конфлуэнтности за 24 часа. Один набор этих планшетов культивировали с той же средой, что и керамический материал, тогда как другой обрабатывали остеиндуктивной средой, содержащей дексаметазон, аскорбиновую кислоту и β-глицерофосфат (Sigma, США). Через 7 дней после посева MSC свежевыделенные HSPC вводили в 4 варианта условий культивирования: 1. совместное 3D культивирование с MSC, высеянными на керамический материал, 2. совместное 2D культивирование с монослоем MSC, высеянных в 6-луночные планшеты, 3. совместное 2D культивирование с монослоем остеогенно индуцированных MSC, высеянных в 6-луночные планшеты, и 4. суспензионное культивирование в среде Stemspan (Stemcell Technologies Inc., Канада), дополненной 100 нг/мл IL-6, SCF, ТРО и FLT-3L (Peprotech, Великобритания). В данном исследовании использовали 9 независимых образцов MSC и HSC. Клетки из каждой системы для культивирования анализировали с помощью проточной цитометрии и иммуногистохимического анализа через 1, 2 и 4 недели после начала культивирования. 3D культуру анализировали в дополнительный момент времени 8 недель для подтверждения потенциала длительного культивирования.
5. Выделение РНК, получение кДНК и количественная ПЦР
Выделение РНК осуществляли с использованием набора NucleoSpin® RNA II (Macherey-Nagel, Германия), следуя предоставленным инструкциям. Клетки, культивируемые в керамическом материале, лизировали непосредственно на керамическом материале. Обратную транскрипцию мРНК осуществляли с использованием набора реактивов для обратной транскрипции кДНК TaqMan® (Applied Biosystems, США) в соответствии с инструкциями производителя. ПЦР в реальном времени осуществляли с использованием 1 мкл кДНК с 1 мкл смеси праймеров и набора SensiFAST™ Sybr No-ROX (Bioline, Германия) в 96-луночных планшетах для ПЦР (Biozym Scientific, Германия) и сканировали с помощью многоканальной системы для количественной ПЦР Stratagene MX 3005Р™ (Agilent Technologies, США). Праймеры заказывали в TIBMolBiol (Германия).
6. Отслеживание клеток
Для долговременного отслеживания HSPC и MSC с помощью флуоресцентной микроскопии клетки перед культивированием метили с использованием наборов для мечения клеток Qtracker® 525 и CellTracker™ Red СМТРХ (Invitrogen, США), соответственно, согласно инструкциям производителя. Для отслеживания деления клеток HSPC непосредственно после выделения метили сложным диацетат-сукцинимидильным эфиром карбоксифлуоресцеина (CFSE, Invitrogen, США) в концентрации 2,5 мкМ в соответствии с инструкциями производителя. Затем выделяли сигнал от меченных зеленым клеток и отслеживали клеточное деление после 1, 2 и 4 недель при помощи проточной цитометрии.
7. Иммуногистохимический анализ
Для того чтобы визуализировать ЕСМ и сигнальные молекулы в керамических каркасах, керамический материал с клетками фиксировали в ацетоне (Sigma, США) при -20°С через 2 недели после посева отслеживаемых HSPC. Затем при помощи скальпеля вырезали диски и окрашивали в 96-луночных планшетах в общем объеме 150 мкл первичными антителами к коллагену I (мышиное антитело к человеческому белку, Sigma, США), C-Kit (мышиное антитело к человеческому белку, Santa Cruz Biotechnology Inc, США), фибронектину (мышиное антитело к человеческому белку, Millipore, США), интегрину 4а (мышиное антитело к человеческому белку, Abeam, Великобритания) и N-кадгерину (мышиное антитело к человеческому белку, Santa Cruz Biotechnology Inc, США). Затем образцы окрашивали вторичным козьим антителом к мышиному белку, конъюгированным с Alexa 350 или Alexa 594 (Invitrogen, США), отмывали и визуализировали с помощью микроскопии.
8. Флуоресцентная и 2-фотонная микроскопия
Присутствие отслеживаемых по зеленому сигналу Qtracker® HSPC в системе для совместного культивирования после 1, 2, 4 и 8 недель подтверждали путем их визуализации с помощью цифрового флуоресцентного микроскопа (BZ 9000, Keyence, Германия) после контрастного окрашивания ядер с помощью DAPI (Sigma, США). ЕСМ и сигнальные молекулы визуализировали в качестве 3D пачек срезов с использованием 2-фотонного микроскопа (Trimscope II, LaVision BioTec, Германия) и преобразовывали для просмотра с использованием Imaris версии 7.5 (Bitplane Scientific Software, Швейцария).
9. Сканирующая электронная микроскопия (SEM)
SEM использовали для визуализации структурного подобия между засеянной клетками керамической системы для культивирования и костным мозгом, а также физического взаимодействия между HSPC и MSC. Керамические диски с MSC и HSPC после 2 недель совместного культивирования и кусочки костного мозга размером 1 см2, вырезанные из головок бедренных костей, фиксировали и обезвоживали с использованием ацетона и получали путем сушки при критической температуре. Затем эти образцы покрывали золотом и серебром и визуализировали с использованием SEM Hitachi S-520 (Hitachi, Япония).
10. Проточная цитометрия
Фенотип HSPC из всех систем для культивирования сравнивали на основании экспрессии поверхностных маркеров при анализе с помощью проточной цитометрии. Клетки собирали из всех прикрепленных систем для культивирования путем инкубирования с 1х раствором трипсина-EDTA (РАА Laboratories, Австрия) в течение 30 минут при 37°С в термостате. Клетки в суспензии просто переносили пипеткой в пробирки для сбора. Затем клетки окрашивали следующими антителами в соответствии с инструкциями производителя: CD34-APC (Miltenyi Biotec, Германия), CD38-PE (eBioscience®, США), аннексии V- Pacific Blue (BioLegend, США), пропидиум йодид (Sigma, США). Анализ с помощью проточной цитометрии выполняли с использованием проточного цитометра с анализатором MACSQuant® (Miltenyi, Германия) и данные анализировали с использованием программного обеспечения FlowJo версия 7.6.5 (Tree Star Inc., США).
11. Анализ CFU-GEMM
Потенциал к дифференцировке в миелоидные клетки у HSPC, полученных из керамической системы для совместного культивирования через 1, 2, 4 и 8 недель после посева тестировали с использованием анализа колониеобразующих единиц - гранулоцитов, эритроцитов, макрофагов, мегакариоцитов. 1000 клеток высевали в среду для CFU-GEMM (Miltenyi Biotec, Германия) и колонии визуализировали и подсчитывали через 2 недели.
12. Статистический анализ
2-факторный дисперсионный анализ (ANOVA) анализ с последующим использованием поправок Бонферони применяли к наборам данных при помощи программного обеспечения GraphPad Prism® версии 5.0 (GraphPad Software Inc., США). Значения р больше или равные 0,05 рассматривали как значимые. Данные представлены как средние значения +/- стандартное отклонение.
Результаты
1. MSC образуют микроокружение, характеризующееся близким структурным подобием с костным мозгом, за 7 дней культивирования в керамическом материале
Имеется множество документальных подтверждений влияния условий 3D культивирования на дифференцировку MSC костного мозга и их продукцию ЕСМ. Было показано, что жесткие каркасы, такие как керамический материал, используемый в данном исследовании, предрасполагают MSC к остеогенной дифференцировке. Таким образом, перед введением HSPC определяли эффект 3D культивирования в керамических каркасах на MSC.
Иммуногистохимический анализ показал, что MSC образуют паутиноподобные сети из ЕСМ, состоящего из коллагена I и фибронектина, через 7 дней после посева (фиг. 9А (i, ii)), внутри которых находятся MSC. Также продуцируется интегрин 4а (фиг. 9А (iii)). Известно, что коллаген I и фибронектин вовлечены в поддержание ниши HSC в костном мозге, и, как полагают, они опосредуют прикрепление HSPC к костному мозгу. Известно, что интегрин 4а играет роль во взаимодействии MSC-HSC в пределах ниши. Сканирующая электронная микроскопия (фиг. 9В) выявила поразительное подобие структуры засеянного MSC керамического материала (фиг. 9В (i, iii)) с таковой матрикса человеческого костного мозга (фиг. 9В (ii, iv)).
Положительное окрашивание ализариновым красным (фиг. 9С (i, ii)), окрашивание по Коссу (фиг. 9С (iii, iv)) указывали на то, что MSC, высеянные в керамический материал, подвергаются спонтанной остеогенной дифференцировке после 7 дней культивирования.
Анализ экспрессии раннего остеогенного маркера, остеопонтина, с помощью количественной ПЦР (фиг. 9D) в сравнении с монослойными культурами MSC подтверждал по меньшей мере частичную спонтанную остеогенную дифференцировку MSC. Экспрессия других молекул с предполагаемой ролью в эндостальной нише, а именно Jagged 1, N-кадгерина, BMPR1a, ICAM1 и CXCR4, также подвергалась повышающей регуляции в 3D культуре в сравнении с монослоем, что позволяет предположить, что при культивировании в керамическом каркасе MSC спонтанно образуют микроокружение, создающее условия для поддержания HSC. Было определено, что 7 дней является подходящим моментом времени для введения в систему гемопоэтических стволовых клеток и клеток-предшественников.
2. Примитивные CD34+CD38- HSPC поддерживаются в системе для совместного 3D культивирования в течение периода до 8 недель
Первое соображение для проверки системы для совместного 3D культивирования в качестве средства, поддерживающего HSPC, заключалось в том, попадают ли HSPC, высеянные в керамический материал, внутрь этого материала и остаются ли они там в течение значительных периодов времени. С помощью флуоресцентной микроскопии HSPC, меченные зеленым красителем для отслеживания клеток, выявляли в системе для совместного культивирования через 1, 2, 4 и 8 недель (фиг. 10А) после посева, что указывает на то, что HSPC инфильтрируют в засеянный MSC керамический материал и стабильно поддерживаются в течение периода до 8 недель. Дальнейшие моменты времени не тестировали.
Для определения того сохраняют ли HSPC свой примитивный (CD34+CD38-) фенотип и изучения преимуществ системы для совместного 3D культивирования над уже применяемыми стратегиями культивирования HSPC, проводилось сравнение процентного соотношения примитивных HSPC, поддерживаемых в указанной керамической системе для совместного культивирования, с таковой в традиционной суспензионной культуре и совместной культуре с монослоями индуцированных и неиндуцированных к остеогенной дифференцировке MSC в течение периода до 4 недель путем выявления с помощью цитометрии клеток, экспрессирующих CD34 и CD38 (фиг. 10В, С, D).
Обнаружено, что процентное соотношение CD34+CD38- клеток, поддерживаемых в керамической системе для совместного культивирования, остается стабильной на уровне около 50% вплоть до 8 недель после посева. При этом во всех других условиях культивирования доля CD34+CD38- клеток непрерывно уменьшалась (фиг. 10С) до тех пор, пока не оставалось менее 5% примитивных HSPC. Обнаружено, что большой процент HSPC (приблизительно 50%) в монослойных совместных культурах составляли дифференцирующиеся (CD34+CD38+) клетки, которые не выявлялись через 2 недели. Малый процент этих клеток (<5%) также систематически наблюдали в 3D системе (фиг. 10D).
3. HSPC, поддерживаемые в системе для совместного культивирования на основе керамического материала, являются медленно делящимися и неактивными
Далее сравнивали пролиферацию HSPC в системе для совместного 3D культивирования и при традиционных способах культивирования, упомянутых ранее, при помощи анализа с помощью проточной цитометрии HPSC, меченных CFSE, в 1, 2 и 4 неделю. Клетки, которые претерпевали 1 или менее делений (первые 2 пика справа на гистограмме CFSE) считали медленно пролиферирующими, тогда как все остальные клетки считали быстро пролиферирующими.
Анализ с помощью проточной цитометрии и последующее построение гистограмм (фиг. 11А) указывали на то, что клетки в системе для совместного 3D культивирования являются наиболее медленно пролиферирующими из всех систем для культивирования. Обнаружено, что большую долю (>70%) HSPC в совместных 3D культурах составляют медленно пролиферирующие клетки. Обнаружено, что этот процент незначительно снижается (<10%) между 2 и 4 неделями (фиг. 11В). Обнаружено, что во всех традиционных культурах доля медленно пролиферирующих клеток была относительно ниже, чем в системе для совместного 3D культивирования, и к 4 неделе культивирования она снижалась до менее 10% (фиг. 11В).
При изучении фенотипа медленно и быстро пролиферирующих HSPC после 1, 2 и 4 недель культивирования при помощи анализа с помощью проточной цитометрии на CD34 и CD38 было обнаружено, что HSPC из системы для совместного 3D культивирования сохраняли наибольшую долю клеток с примитивным CD34+CD38- фенотипом (фиг. 11С, D, Е) после 1, 2 и 4 недель культивирования. Более 50% медленно и быстро пролиферирующих HSPC из совместной 3D культуры сохраняли примитивный фенотип в течение периода до 4 недель.
Как и ожидалось, доля CD34+CD38- клеток среди быстропролиферирующих клеток была меньше, нежели среди медленно пролиферирующих клеток. Процент примитивных клеток в медленно и быстро пролиферирующих фракциях из традиционных совместных культур неизменно снижался со временем (фиг. 11С, D, Е).
4. HSPC, поддерживаемые в системе для совместного культивирования на основе керамического материала, являются жизнеспособными и неапоптическими.
Установив, что примитивный фенотип HSPC наилучшим образом сохраняется в 3D культуре, во всех системах для культивирования определяли жизнеспособность клеток путем измерения экспрессии аннексина V, широко используемого индикатора раннего апоптоза, и измерения пропидиум йодида (PI), который поглощается апоптическими и некротическими клетками.
Очень малая доля (<0,2%) HSPC в системе для совместного 3D культивирования экспрессировала аннексии V (фиг. 12С) и поглощала PI (фиг. 12В) вплоть до 2 недели, после чего эти клетки больше не присутствовали.
Обнаружено, что в традиционных системах, в частности в суспензионной культуре и совместной культуре с индуцированными к остеогенной дифференцировке MSC, немного большая доля HSPC (0,5-1,5%) были апоптическими или некротическими (фиг. 12В, С). Исходя из этих данных, был сделан вывод, что при всех условиях культивирования большинство HSPC остаются жизнеспособными и неапоптическими в течение периода до 8 недель.
5. HSPC из совместных культур на керамическом материале функциональны и способны к дифференцировке в несколько линий клеток.
После подтверждения того, что HSPC, поддерживаемые в системе для совместного 3D культивирования сохраняют свой примитивный фенотип и являются жизнеспособными, при помощи анализа CFU-GEMM тестировалось, сохраняют ли эти клетки потенциал к дифференцировке в несколько линий клеток, характерный для HSPC.
CD34+ клетки, выделенные из системы для совместного 3D культивирования через 1, 2 и 4 недели после посева, давали колонии CFU-GEMM, BFU-E и CFU-GM (фиг. 12D). При подсчете этих колоний не было обнаружено существенного отличия в разные моменты времени (фиг. 12Е). Следовательно, CD34+клетки, поддерживаемые в системе для совместного 3D культивирования, являются не только жизнеспособными, но также сохраняют характерный для них потенциал к дифференцировке в несколько линий клеток.
6. HSPC в системе для совместного 3D культивирования взаимодействуют с ЕСМ и клеточными компонентами своего микроокружении.
Наконец, для того чтобы проиллюстрировать, что в керамической системе для совместного культивирования в значительной степени были сымитированы взаимодействия в нише HSPC в костном мозге, изучали физическое взаимодействие HSPC и MSC в пределах керамической системы. Хотя ранее наблюдали совместное расположение HSPC и MSC (фиг.10А), не было получено доказательств физического контакта или какого-либо указания на механизм взаимодействия.
Сканирующая электронная микроскопия (SEM) выявила, что в керамической системе для совместного культивирования мелкие округлые HSPC, которые имели диаметр приблизительно 10 мкм, всегда оказывались в непосредственной близости от одной из более плоских крупных MSC (фиг. 13В). Для дальнейшего выяснения факторов, вовлеченных в эти взаимодействия, проводили иммуногистологические анализы.
2-фотонная микроскопия выявила, что меченные флуоресцентной меткой MSC (красный) и HSPC (зеленый) были заключены в сеть ЕСМ, состоящую преимущественно из фибронектина и коллагена I (фиг. 13А ii, iii). Иммуноокрашивание на сигнальные молекулы C-k.it, интегрин 4а и N-кадгерин показало, что эти молекулы расположены в участках, где HSPC контактируют с MSC (фиг. 13А iv, v, vi), что указывает на то, что HSPC и MSC в разработанной системе для совместного 3D культивирования взаимодействуют через эти молекулы, что очень напоминает взаимодействия, которые, как полагают, существуют в эндостальной гемопоэтической нише в костном мозге.
Обсуждение
Эндостальная гемопоэтическая ниша, в которой гемопоэтические стволовые клетки и клетки-предшественники (HSPC) поддерживаются в виде медленно пролиферирующих примитивных клеток, как известно, является очень сложной системой. Было выявлено, что несколько различных типов клеток, в том числе остеобласты, остеогенные клетки-предшественники и мезенхимальные стволовые клетки, вносят вклад в регуляцию ниши посредством ряда механизмов, таких как продукция молекул ЕСМ, прямой межклеточный контакт и продукция сигнальных молекул. Хотя отдельные типы клеток, пути передачи сигнала и даже физические критерии были вовлечены в различные аспекты поддержания ниши, взаимосвязь этих факторов и их точная роль в нише остается неясной. Разработка in vitro модели, которая эффективно имитирует эндостальную нишу, следовательно, найдет отличное применение для изучения взаимодействий в нише.
Недавние исследования указали на то, что для успешной имитации взаимодействий в эндостальной нише, необходимо присутствие стромальных поддерживающих клеток и 3D структуры. В нескольких независимых исследованиях было выявлено, что мезенхимальные стволовые клетки (MSC) костного мозга являются особенно эффективными в поддержании и обеспечении пролиферации примитивных покоящихся HSC in vitro. Полагают, что MSC опосредуют поддержание ниши путем продукции сигнальных молекул, таких как интегрины, компонентов ЕСМ, в том числе коллагена I и фибронектина, а также с помощью клеточного взаимодействия. Известно, что коллаген I и фибронектин опосредуют хоминг HSC, связываясь с поверхностными рецепторами и улавливая секретируемые факторы. Также известно, что MSC являются предшественниками для остеобластных клеток, которые являются известными партнерами HSPC в пределах ниши. Было показано, что остеобластные клетки, участвующие в обеспечении ниши HSPC в костном мозге, являются смешанной популяцией, включающей клетки с различной степенью дифференцировки.
Для того чтобы имитировать все эти факторы настолько точно, насколько это возможно, использовали покрытый гидроксиапатитом керамический 3D каркас Sponceram®, засеянный MSC костного мозга, в качестве системы для культивирования для HSPC, полученных из пуповинной крови. Керамические каркасы оптимизированы для адгезии клеток и, как сообщалось, индуцируют спонтанную остеогенную дифференцировку MSC.
Однако было показано, что степень дифференцировки была меньшей, чем в культурах, которые индуцировали с помощью BMP или других факторов, это позволило предположить, что культивирование MSC исключительно на керамическом каркасе будет приводить к образованию популяции клеток с различными степенями остеогенной дифференцировки, как в нише костного мозга.
При иммуногистохимическом анализе керамического материала, засеянного MSC, наблюдали экспрессию молекул ЕСМ, коллагена I и фибронектина, которые составляют главные компоненты плотных сетеподобных структур, которые наблюдались в керамическом материале.
Также выявляли повышенные уровни интегрина 4а. Анализ засеянного MSC керамического материала и губчатой кости из головки бедренной кости с помощью сканирующей электронной микроскопии выявил высокую степень структурного подобия между ними. Анализ этой системы для культивирования с помощью ПЦР в реальном времени показал, что экспрессия раннего остеогенного маркера, остеопонтина, а также молекул с известной ролью в эндостальной нише: Jagged-1, С-Х-С хемокинового рецептора типа 4 (CXCL12), BMP рецептора 1А (BMPR1A), N-кадгерина и молекулы межклеточной адгезии-1 (ICAM-1), была повышена в 3D культурах. Известно, что остеопонтин способствует поддержанию ниши и состоянию покоя HSPC путем ограничения пролиферации и мобилизации HSPC. Полагают, что Jagged-1 опосредует самообновление HSPC через передачу сигнала с участием Notch. Все из CXCL12, BMPR1A и N-кадгерина были вовлечены в хоминг HSC, a ICAM-1, как известно, является медиатором межклеточного связывания. Следовательно, в течение 1 недели после посева MSC в керамические каркасы наблюдали, что образовывалось микроокружение, характеризующееся структурным и молекулярным подобием с нишей костного мозга. Для того чтобы заполнить эту предполагаемую искусственную нишу, в эту систему затем вводили CD34+HSPC.
При помощи флуоресцентной микроскопии смогли выявлять меченые флуоресцентной меткой HSPC в системе для совместного 3D культивирования вплоть до 8 недель после их введения в керамический материал, что указывает на то, что HSPC не только проникают в керамический материал, но также сохраняются в нем в течение длительного времени. Путем анализа с помощью проточной цитометрии продемонстрировали, что большая доля (более 50%) HSPC в совместной культуре в керамическом материале стабильно сохраняет примитивный CD34+CD38- фенотип с первой недели вплоть до 8 недель культивирования. В отличие от этого было обнаружено, что HSPC, культивируемые в традиционных монослойных совместных культурах или в виде суспензионных культур в определенной дополненной цитокинами среде постоянно теряли свой примитивный фенотип. Также подтвердилось, что клетки во всех системах для культивирования не были апоптическими или некротическими.
Поскольку примитивные HSPC в эндостальной нише также характеризуются низкой скоростью пролиферации, изучали пролиферацию HSPC в каждой из систем для культивирования с использованием исследований разведения CFSE. Было продемонстрировано, что HSPC в системе для совместного 3D культивирования претерпевали очень малое число делений (<2) за 4 недели по сравнению с таковыми из традиционных культур. В частности, высокую скорость пролиферации проявляли клетки в суспензионной культуре. Также обнаружили, что больший процент медленно пролиферирующих клеток в совместной 3D культуре сохранял CD34+CD38- фенотип.
Установив таким образом, что разработанная система для совместного 3D культивирования способна поддерживать жизнеспособные медленно пролиферирующие CD34+CD38- HSPC в течение длительных периодов времени in vitro; были использованы анализы CFU-GEMM, чтобы продемонстрировать, что эти клетки являются функциональными и способны образовывать эритроидные и миелоидные колонии. Колониеобразующая способность у HSPC из керамического материала не менялась со временем в культуре, что указывает на стабильное поддержание функциональных HSPC.
При оценке молекулярных взаимодействий в пределах системы для совместного культивирования обнаружили, что MSC и HSPC располагались в непосредственной в близости внутри описанной ранее сети, состоящей из фибронектина и коллагена I. Наблюдали, что молекулы-посредники в нише: рецептор фактора роста стволовых клеток (C-kit), N-кадгерин и интегрин 4а также находились совместно с HSPC и MSC, что указывает на то, что эти два типа клеток могут взаимодействовать посредством этих молекул. C-kit представляет собой рецептор для фактора роста стволовых клеток и, как полагают, способствует поддержанию HSC в нише, опосредуя адгезию к матриксу ниши. Также полагают, что N-кадгерин и интегрин 4а выполняют аналогичные роли. Дополнительная визуализация при помощи SEM выявила физическое взаимодействие между примыкающими MSC и HSPC, что явно указывает на клеточные взаимодействия в пределах системы совместной 3D культуры, подобные таковым в нише.
Недавние исследования достигли успеха в поддержании HSPC в 3D матрицах на основе геля, состоящих из коллагена I и фибрина, соответственно, в совместной культуре с MSC. Эти исследования фактически продемонстрировали потребность в комбинации 3D каркаса, подходящего ЕСМ и клеток-партнеров для успешного поддержания HSPC in vitro. В другой работе было продемонстрировано, что остеогенно дифференцированные клетки представляют собой эффективные опорные клетки для HSPC в 3D. Исследования монослойных совместных культур и трансплантации показали, что MSC сильно улучшают поддержку HSPC как in vitro, так и in vivo. Разработанная система для совместного 3D культивирования комбинирует структурированный жесткий каркас, MSC костного мозга, которые спонтанно продуцируют соответствующие компоненты ЕСМ и подвергаются спонтанной дифференцировке, и примитивные HSPC для получения искусственной системы, которая точно имитирует нишу HSPC костного мозга в том, что касается клеточного, молекулярного и структурного состава. Данная система способна поддерживать примитивные HSPC в течение более длительного времени, чем ранее описанные системы.
Вывод
Разработанная система для совместного 3D культивирования на основе керамического материала точно имитирует наиболее важные аспекты эндостальной ниши HSPC и будет обеспечивать полезную in vitro модель для изучения взаимодействий в нише как в здоровом, так и в пораженном болезнью состоянии. Она также имеет большой потенциал в качестве платформы тестирования веществ для лекарственных средств, нацеленных на любую часть ниши. Данная система также может создавать основу для эффективной стратегии размножения HSPC для трансплантации.


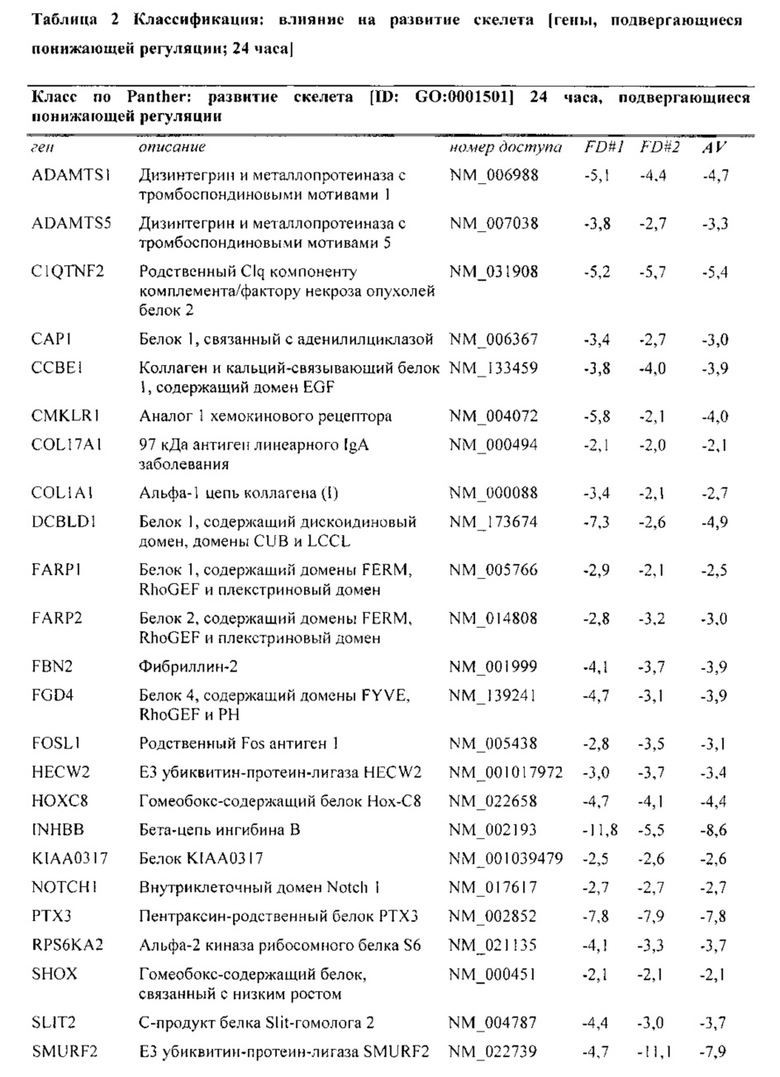
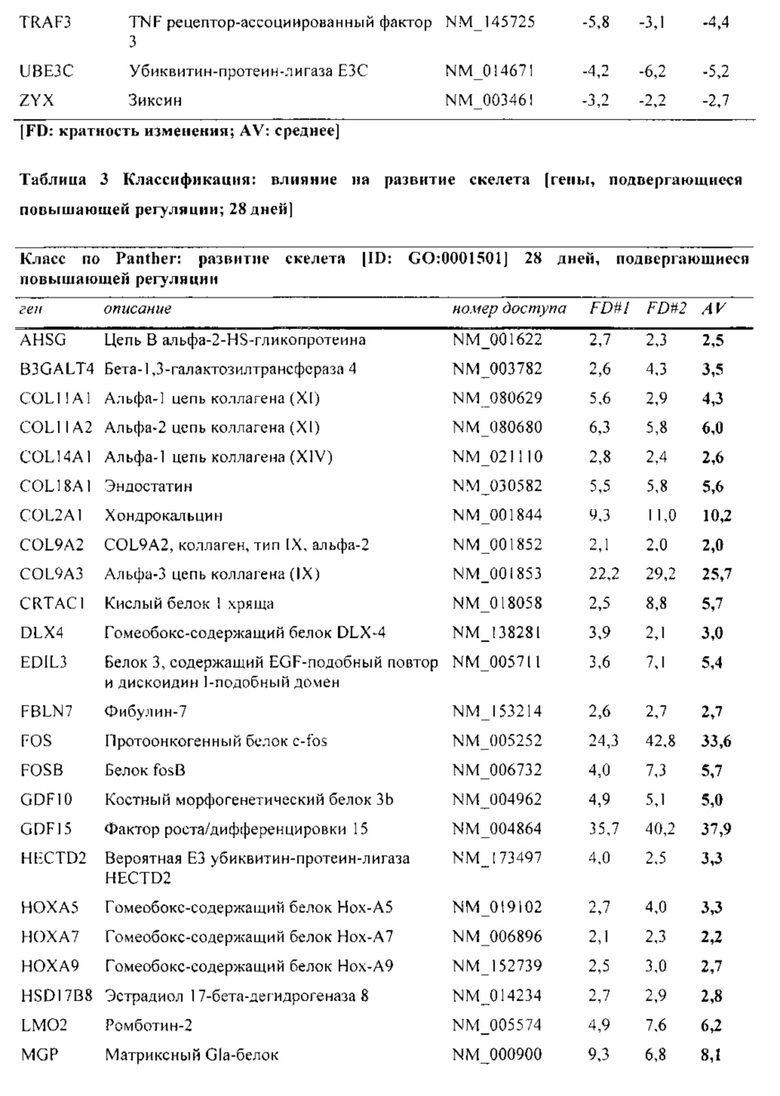
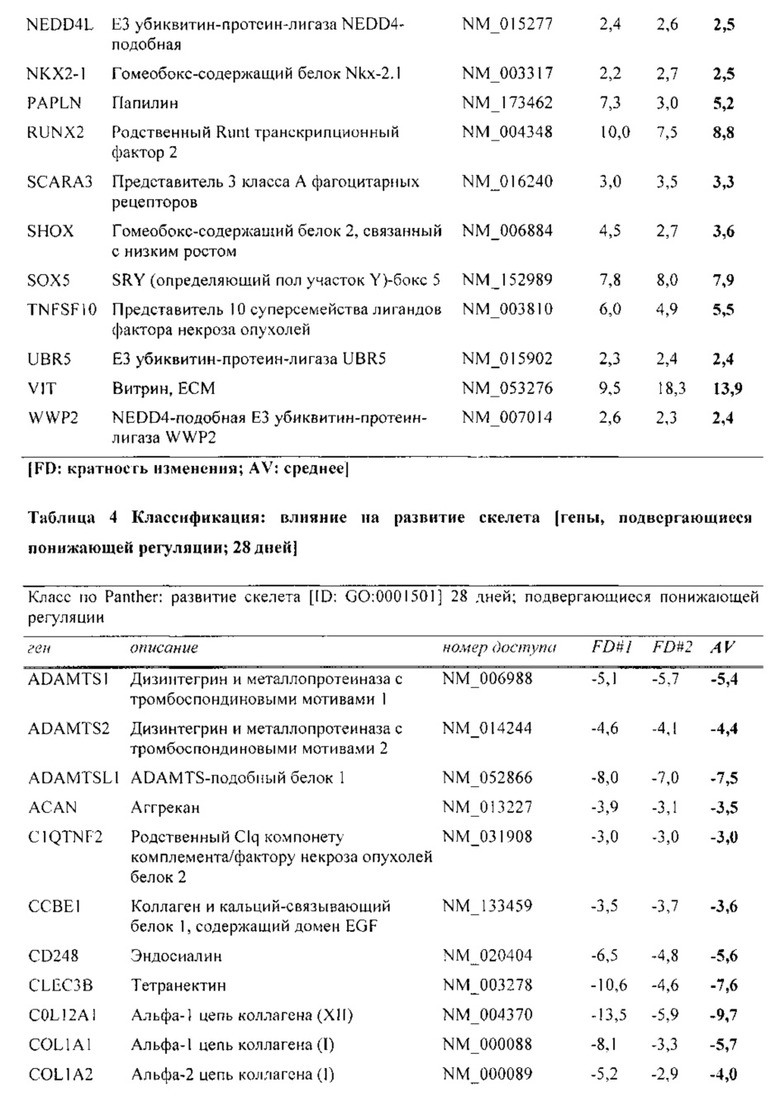
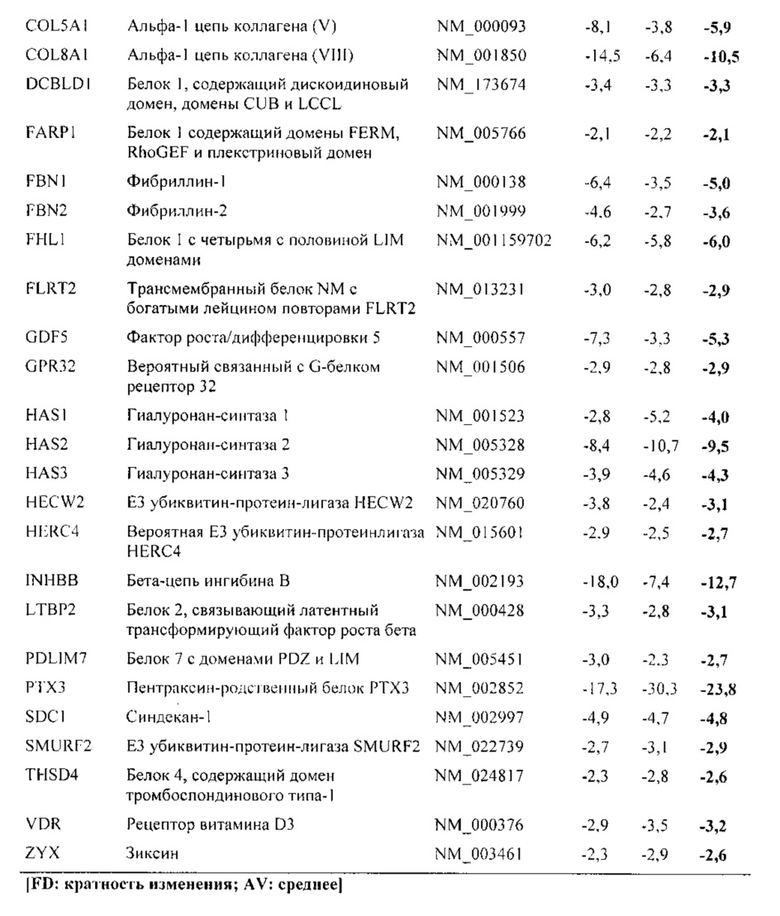

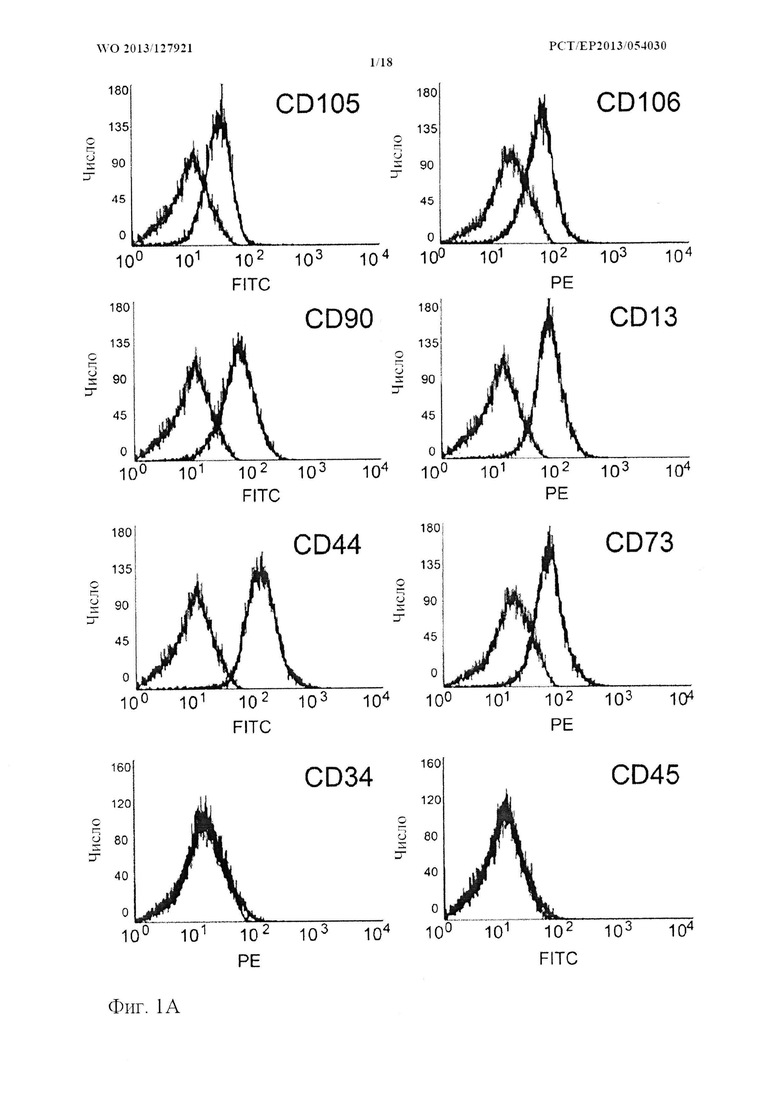


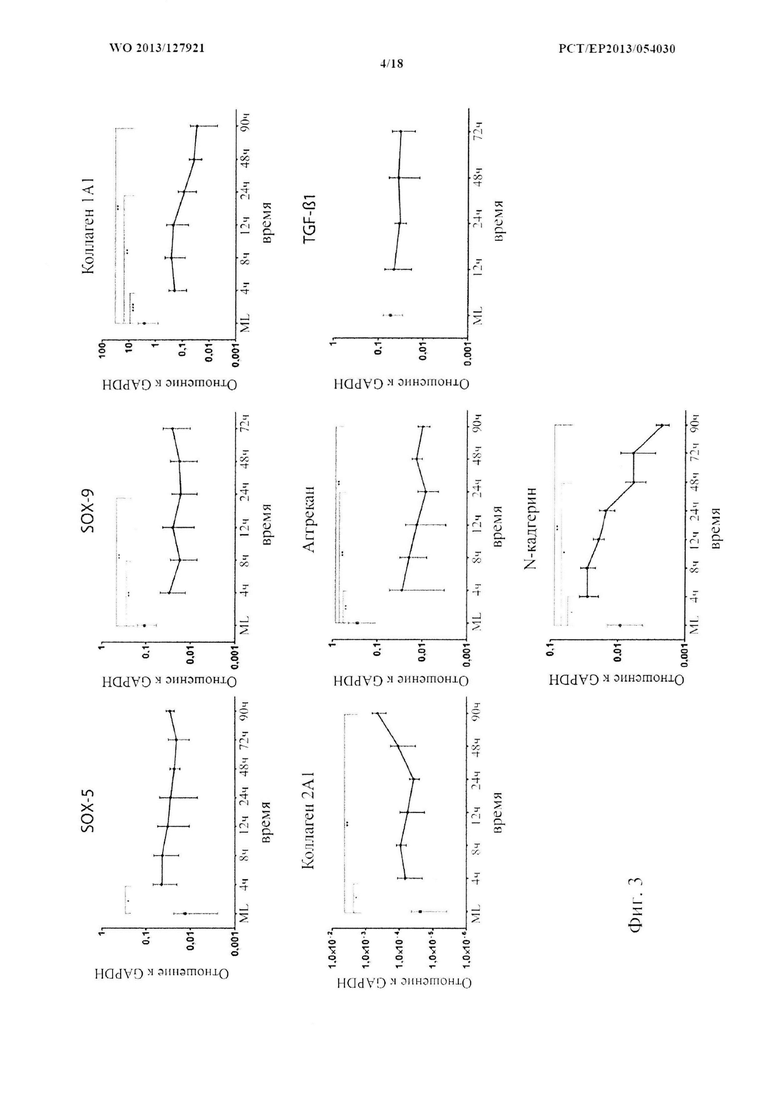
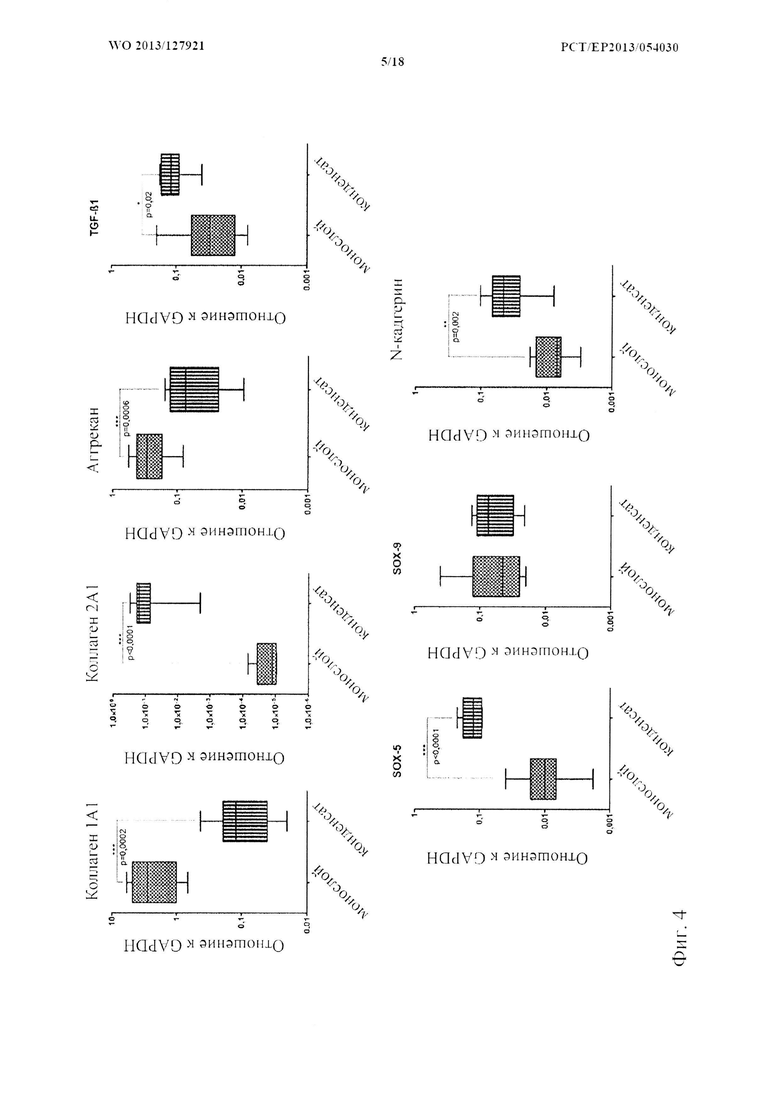
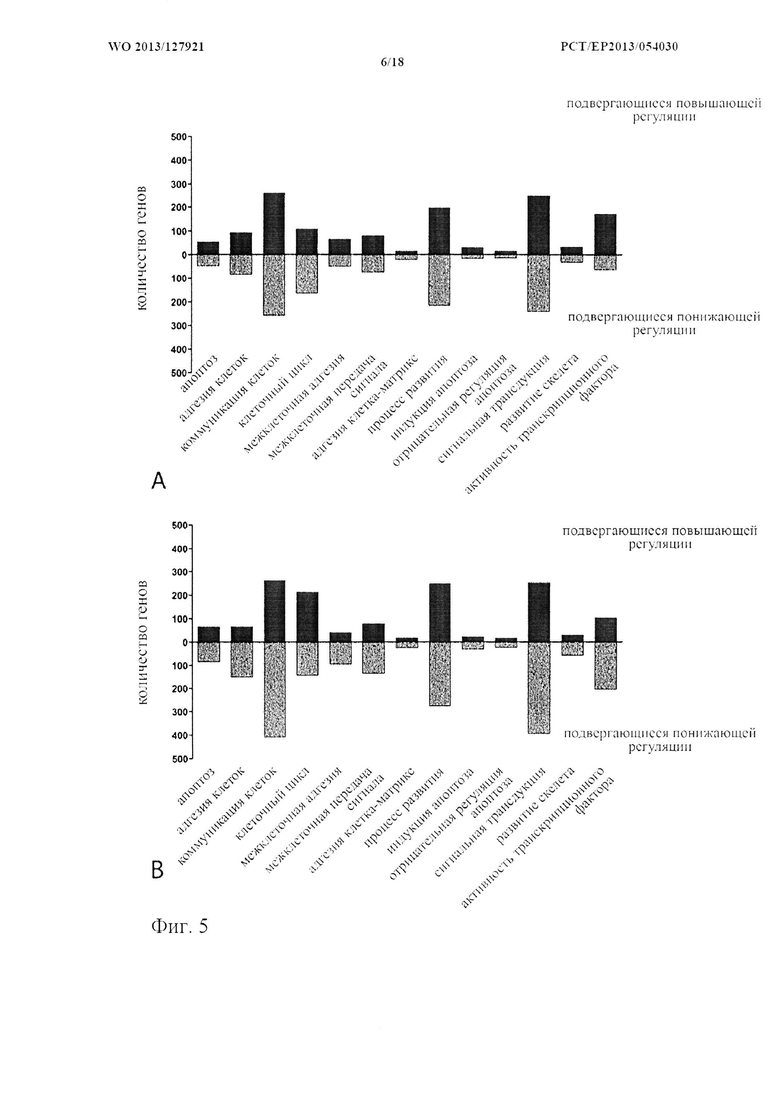
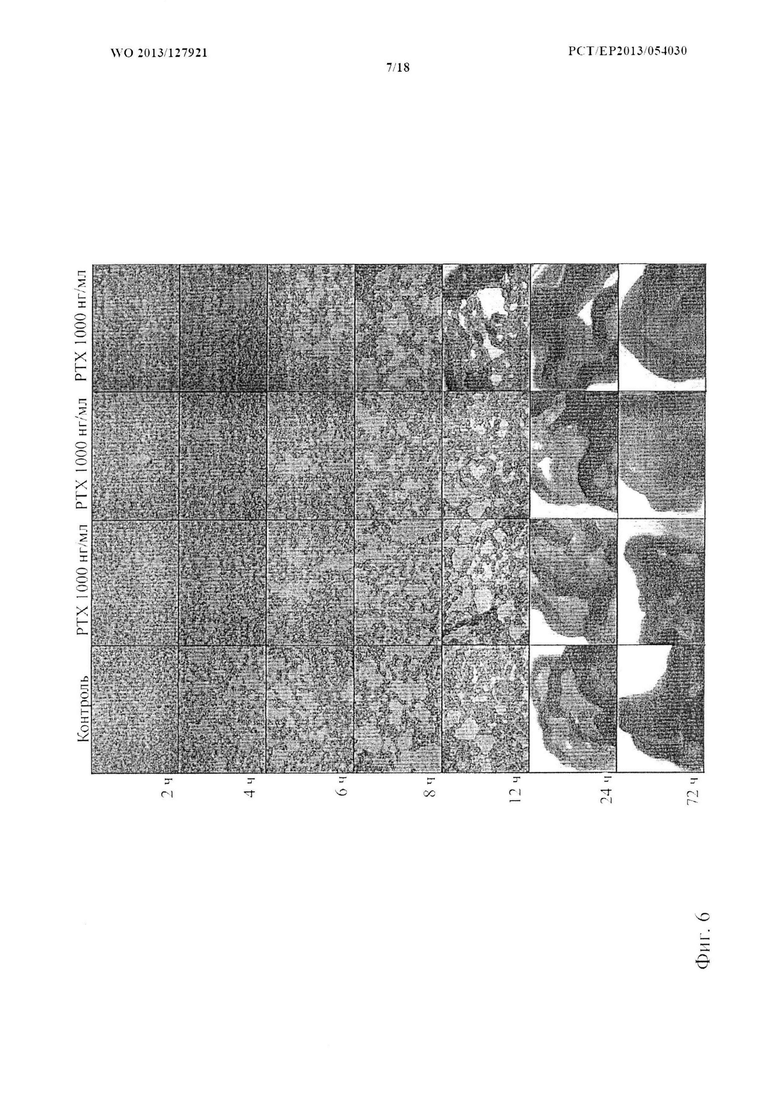

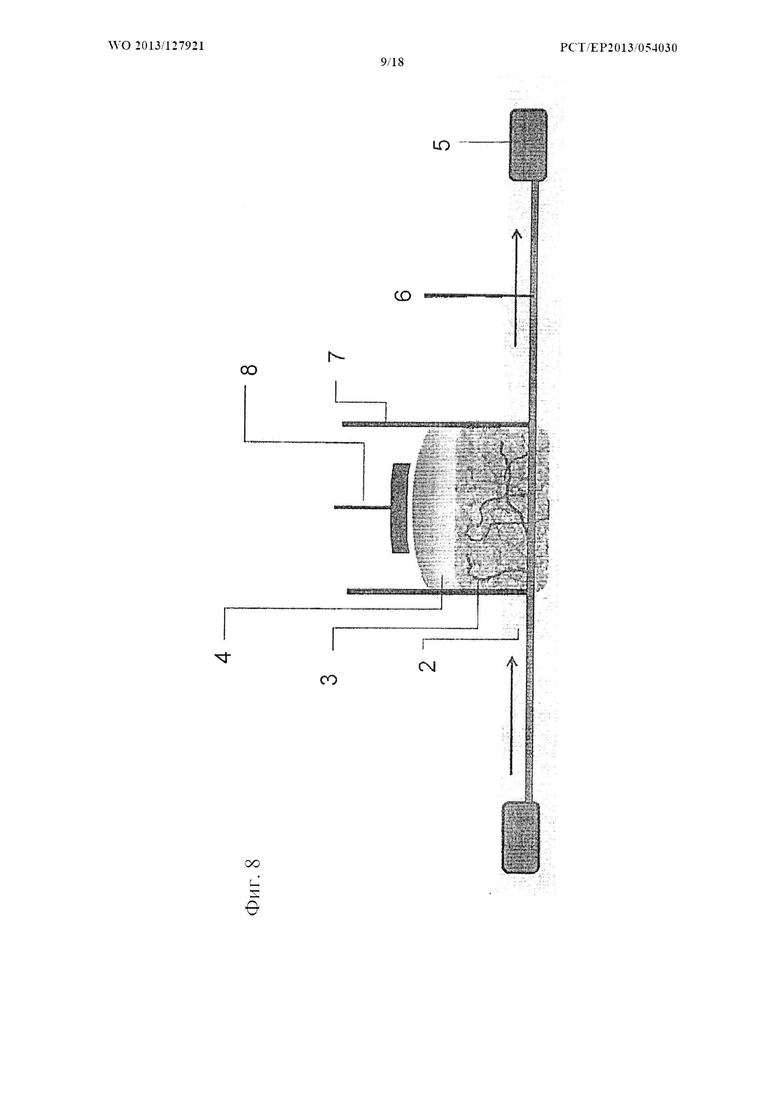



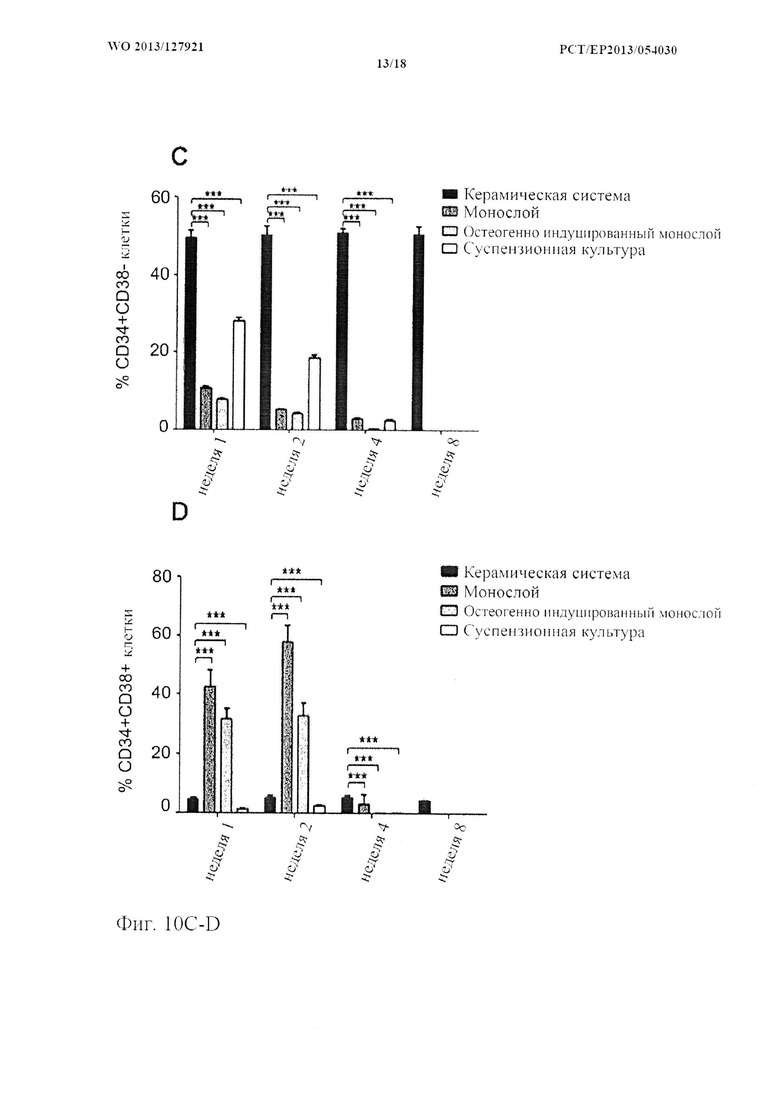
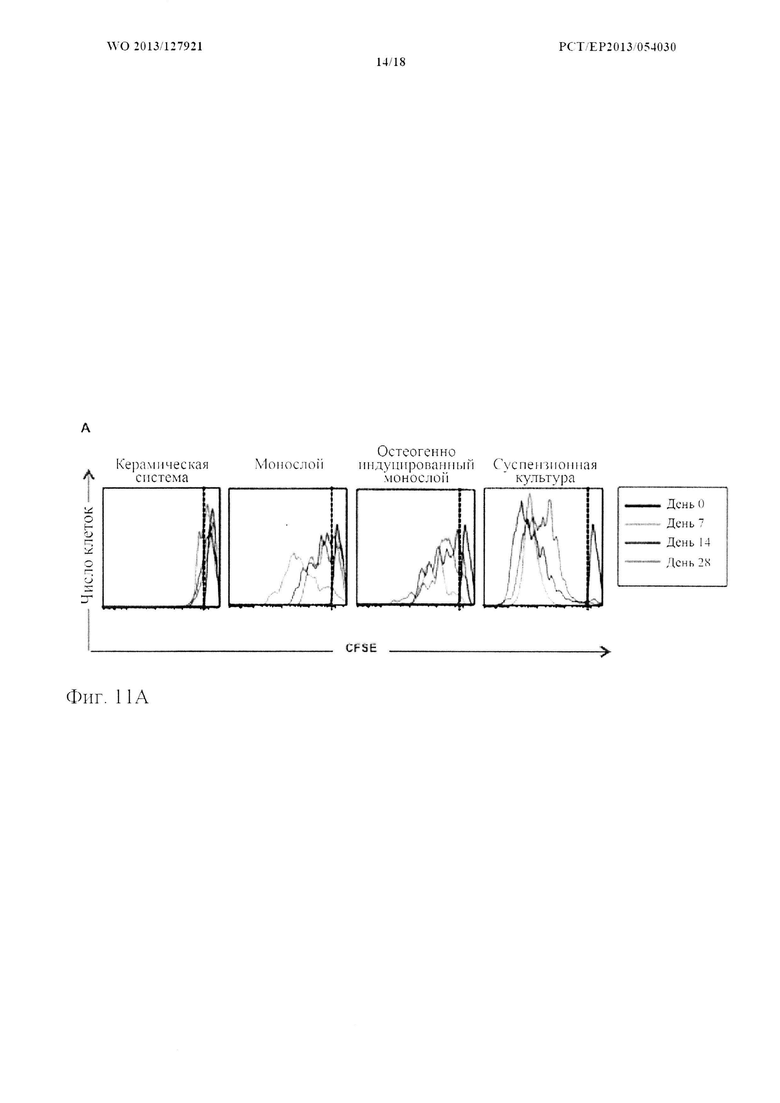
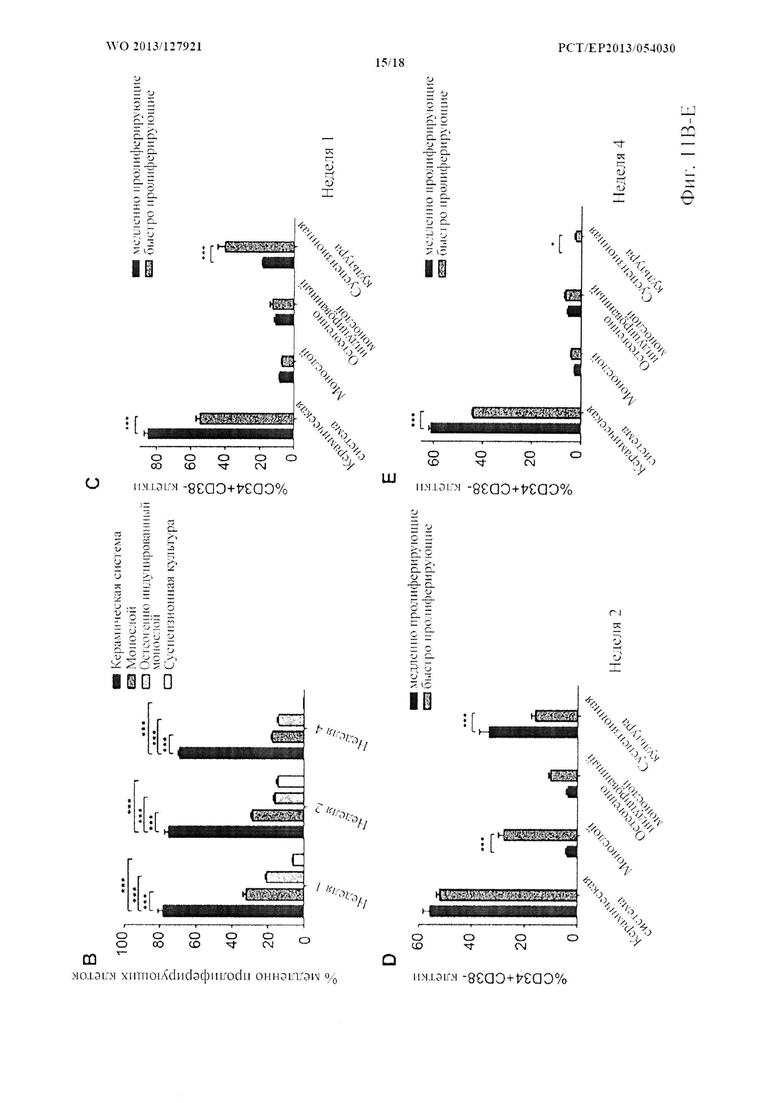
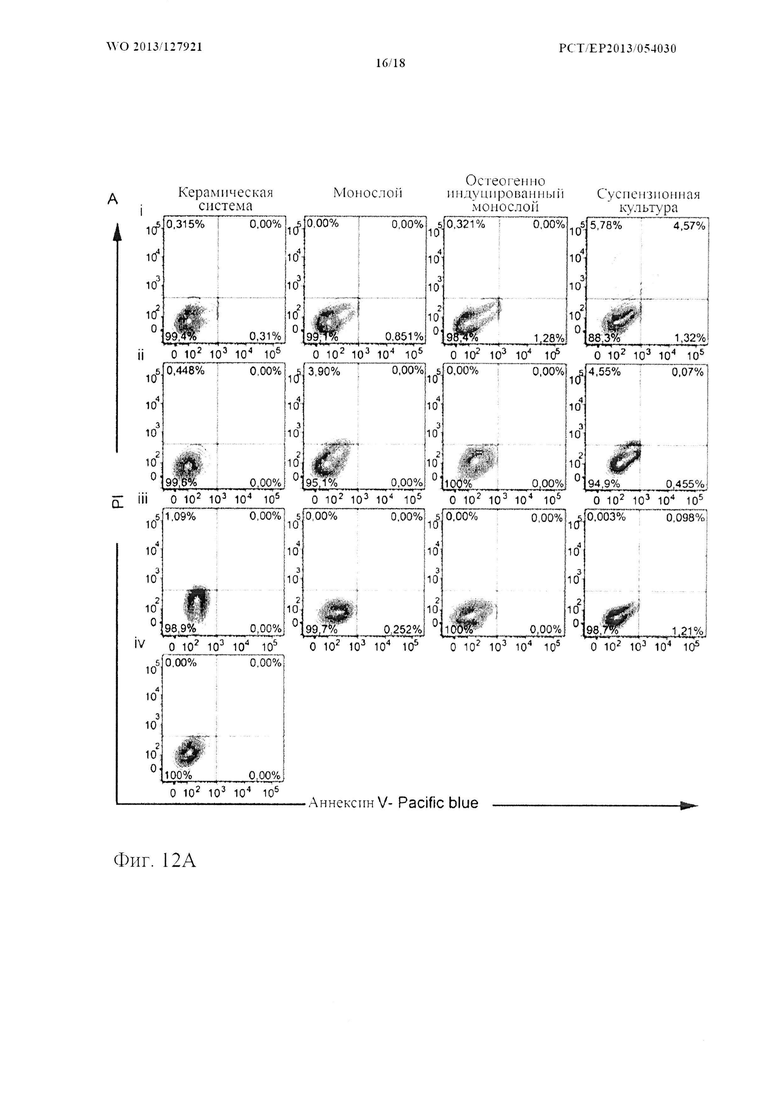
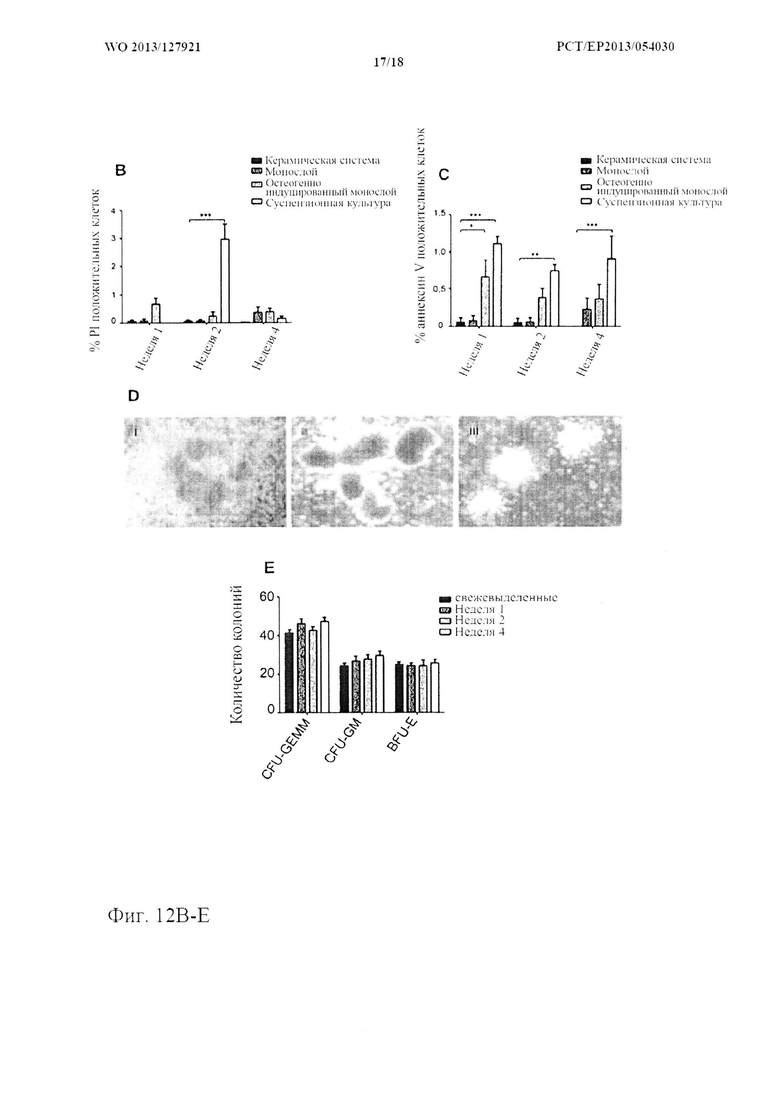

Изобретение относится к биотехнологии. Описан 3D in vitro двухфазный костно-хрящевой «органоид», содержащий слой искусственной хрящевой ткани/агрегатов хондрогенных клеток и слой искусственной костной ткани, где искусственная костная ткань содержит придающий структуру каркас и структуру костного мозга; где структура костного мозга в искусственной костной ткани получена путем посева мезенхимальных стволовых клеток на придающий структуру каркас и где слой искусственной хрящевой ткани/агрегатов хондрогенных клеток контактирует по меньшей мере с одной поверхностью слоя искусственной костной ткани, и способы его получения и применения. Изобретение расширяет арсенал средств для медицинской диагностики. 7 н. и 7 з.п. ф-лы, 13 ил., 5 табл., 2 пр.
1. Способ получения искусственной хрящевой ткани/агрегатов хондрогенных клеток, включающий следующие этапы:
a) обеспечение мезенхимальных клеток-предшественников; и
b) культивирование мезенхимальных клеток-предшественников в не обеспечивающих адгезию условиях в течение по меньшей мере двух недель с образованием искусственной хрящевой ткани/агрегатов хондрогенных клеток.
2. Способ по п. 1, где мезенхимальные клетки-предшественники включают по меньшей мере 50% клеток или состоят из клеток, которые представляют собой CD105+, CD106+, CD44+, CD73+, CD90+ и CD13+.
3. Способ по любому из пп. 1 или 2, где мезенхимальные клетки-предшественники включают выделенные дедифференцированные первичные хондроциты или состоят из них.
4. 3D in vitro двухфазный костно-хрящевой «органоид», содержащий:
a) слой искусственной хрящевой ткани/агрегатов хондрогенных клеток; и
b) слой искусственной костной ткани, где искусственная костная ткань содержит придающий структуру каркас и структуру костного мозга;
где структура костного мозга в искусственной костной ткани получена путем посева мезенхимальных стволовых клеток на придающий структуру каркас; и
где слой искусственной хрящевой ткани/агрегатов хондрогенных клеток контактирует по меньшей мере с одной поверхностью слоя искусственной костной ткани.
5. 3D in vitro двухфазный костно-хрящевой «органоид» по п. 4, где искусственная хрящевая ткань/агрегаты хондрогенных клеток получены с использованием мезенхимальных клеток-предшественников, предпочтительно с использованием мезенхимальных клеток-предшественников, включающих по меньшей мере 50% клеток или состоящих из клеток, которые представляют собой CD105+, CD106+, CD44+, CD73+, CD90+ и CD13+, более предпочтительно из выделенных первичных хондроцитов.
6. 3D in vitro двухфазный костно-хрящевой «органоид» по п. 4, где искусственная хрящевая ткань/агрегаты хондрогенных клеток получены с помощью способа по п. 1.
7. 3D in vitro двухфазный костно-хрящевой «органоид» по п. 4, где структура костного мозга в искусственной костной ткани получена путем посева мезенхимальных стволовых клеток на придающий структуру каркас и путем добавления гемопоэтических стволовых клеток после культивирования мезенхимальных стволовых клеток на придающем структуру каркасе в течение 2-10 дней.
8. 3D in vitro двухфазный костно-хрящевой «органоид» по п. 4, где придающий структуру каркас в искусственной костной ткани содержит биологический или небиологический материал или состоит из него.
9. 3D in vitro двухфазный костно-хрящевой «органоид» по одному из предыдущих пунктов, где придающий структуру каркас в искусственной костной ткани содержит 3D керамический каркас или состоит из него.
10. Способ получения 3D in vitro двухфазного костно-хрящевого «органоида» по одному из пп. 4-9, при этом способ включает следующие этапы:
посев мезенхимальных стволовых клеток на придающий структуру каркас;
культивирование указанных мезенхимальных стволовых клеток в течение 2-10 дней на указанном придающем структуру каркасе;
добавление гемопоэтических стволовых клеток к мезенхимальным стволовым клеткам, культивируемым на придающем структуру каркасе, и культивирование полученной смеси в течение по меньшей мере 5 дней для того, чтобы получить слой искусственной костной ткани, содержащий придающий структуру каркас, засеянный структурой костного мозга;
обеспечение искусственной хрящевой ткани/агрегатов хондрогенных клеток, полученных с помощью способа по одному из пп. 1-3, и
размещение слоя указанной искусственной хрящевой ткани/агрегатов хондрогенных клеток по меньшей мере на одной поверхности указанного слоя искусственной костной ткани.
11. Трансплантат, содержащий 3D in vitro двухфазный костно-хрящевой «органоид» по одному из пп. 4-9.
12. Фармацевтическая композиция для лечения повреждения хряща и/или разрушения или потери хряща, содержащая 3D in vitro двухфазный костно-хрящевой «органоид» по одному из пп. 4-9 или трансплантат по п. 11 и по меньшей мере одно фармацевтически приемлемое вспомогательное вещество.
13. Способ in vitro скрининга веществ, которые модулируют свойства хрящевой или костной ткани, включающий следующие этапы:
обеспечение образца 3D in vitro двухфазного костно-хрящевого «органоида» по одному из пп. 4-9;
разделение соответствующего образца на порции;
инкубирование по меньшей мере одной порции с веществом, подлежащим скринингу; и
сравнение параметров, измеренных у обработанной порции и другой порции, которую не инкубировали с веществом, подлежащим скринингу.
14. Автономное устройство по типу 3D in vitro двухфазного костно-хрящевого «органоида»-на-чипе, содержащее по меньшей мере один резервуар для подачи среды, по меньшей мере одну секцию для роста органа, содержащую по меньшей мере одну полость для органа, вмещающую 3D in vitro двухфазный костно-хрящевой «органоид» по одному из пп. 4-9, где резервуар для подачи среды соединен по меньшей мере с одной секцией для роста органа микроструйным питающим каналом.
| WIEGANDT KATHARINA, Improving in vitro generated cartilage-carrier-constructs by optimizing growth factor combination, Open Biomed Eng J | |||
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| WO 2009146911 A2, 10.12.2009 | |||
| PIERA-VELAZQUEZ SONSOLES, Increased life span of human osteoarthritic chondrocytes by exogenous expression of telomerase, Arthritis Rheum | |||
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| EA 200901126 A1, 30.04.2010. | |||

