Изобретение относится к области биотехнологии, биоинженерии, биохимии, диагностической медицинской микробиологии и разработки диагностических тест-систем. Представляет собой одноцепочечные молекулы ДНК, полученные применением негативной селекции из пула олигонуклеотидов против бычьего сывороточного альбумина и α-казеина, как наиболее часто используемых компонентов блокирующих растворов, используемых в иммунохимии, биотинилированные по 5′-концу, примененимые в качестве зонда в составе диагностических медицинских тест-систем на основе иммунодетекции, сопряженной с полимеразной цепной реакцией, для снижения уровня неспецифической сорбции нуклеотидной матрицы и обеспечения максимального соотношения сигнал/фон при постановке анализа.
Основными областями применения неаффинных последовательностей ДНК являются медицинские и биологические исследования, клиническая диагностика заболеваний, в том числе создание высокочувствительных и высокоэффективных систем детекции мишеней белковой природы.
В основе метода иммуно-ПЦР лежит сочетание преимуществ двух широко распространенных диагностических стратегий: иммуноанализа и полимеразной цепной реакции. Сочетание этих стратегий позволяет достоверно выявлять малые и ультрамалые количества белковых мишеней в аналитах, что достигается за счет специфического связывания мишеней высокоаффинными антителами, комбинированного с экспоненциальной амплификацией сигнала от детектирующего комплекса, связанного с нуклеотидной матрицей, посредством ПЦР, что многократно увеличивает уровень чувствительности в сравнении с рутинным иммуноферментным анализом [Malou Ν., Raoult D. // Trends Microbiol. - 2011. - V. 19. - N. 6. - P. 295-302].
Технология иммуно-ПЦР востребована в тех случаях, когда диагностическое значение имеют ненуклеотидные структуры (протеины, липиды и др.), продуцируемые в количествах ниже порога детекции в иммуноанализе, а также когда выявление малых количеств мишеней (токсинов, онкомаркеров и т.п.) на ранних стадиях заболевания имеет значение для назначения своевременной специфической терапии [Zhang Η. et al. // Analyst. - 2007. - V. 132. - N. 8. - P. 724-737].
Одной из ключевых задач в разработке тест-систем с использованием высокочувствительного подхода иммуно-ПЦР является редукция фонового сигнала, обусловленного неспецифическим связыванием молекул ДНК с компонентами тест-системы [Adler Μ., Wacker R., Niemeyer С.Μ. // Analyst. - 2008. - V. 133. - Ν. 6. - P. 702-718]. Данная проблема может быть обусловлена неспецифическим взаимодействием ДНК-маркера с компонентами, входящими в состав тест-системы, а также возникновением ложноположительного сигнала при контаминации пробы нуклеиновыми кислотами, попавшими извне [Kakizaki Ε. et al. // Lett. Appl. Microbiol. - 1996. - V. 23. - N. 2. - P. 101-103].
Известен подход к решению описанной проблемы, заключающийся в конструировании адаптированных конъюгатов ДНК-антитело [Niemeyer С.М., Adler Μ., Wacker R. // Nature Protocols. - 2007. - V. 2. - N. 8. - P. 1918-1930; Lind K., Kubista M. // J. Immunol. Meth. - 2005. - V. 304. - N. 1. - P. 107-116]. Данный подход позволяет снизить количество манипуляций в процессе постановки анализа, а также уменьшить вероятность неспецифического взаимодействия молекулы ДНК с прочими компонентами вследствие более высокой вероятности аффинного связывания конъюгированного антитела с мишенью. Однако в этом случае невозможно полностью исключить вариант неспецифической сорбции ДНК прочими составляющими системы.
Известно решение проблемы возникновения ложноположительных результатов амплификации с использованием искусственно синтезированных последовательностей ДНК [Не X. et al. // Applied and environmental microbiology. - 2011. - V. 77. - N. 11. - P. 3558-3564], включение которых в состав тест-системы в качестве маркерной ДНК позволяет избежать ошибок амплификации, поскольку используемая ДНК уникальна и негомологична последовательностям, встречаемым в окружающей среде. Чаще всего такими последовательностями выступают фрагменты бактериальных векторов (pUC19, Bluescript и др.) [Kuczius Т. et al. // Analyt. Biochem. - 2012. - V. 431. - Ν. 1. - P. 4-10; Saito K. et al. // Clin. Chem. - 1999. - V. 45. - N. 5. - P. 665-669; Niemeyer С.M. et al. // Nucl. Acids Res. - 1999. - V. 27. - N. 23. - P. 4553-4561]. Известно также применение фаговых дисплеев как неспецифической амплифицируемой матрицы [Monjezi R. et al. // J. of Virol. Meth. - 2013. - V. 187. - Ν. 1. - P. 121-126; Guo Y. C. et al. // Nucl. Acids Res. - 2006. - V. 34. - N. 8. - P. е62-е62]. Однако невозможно полностью исключить вероятность возникновения ложноположительных результатов, особенно в условиях исследовательских лабораторий, поскольку данные конструкции широко используются в лабораторной практике. Также применение подобных последовательностей не отменяет вероятных неспецифических взаимодействий нуклеотидных матриц с прочими компонентами системы.
Известны способы негативной селекции ДНК в отношении структур различной природы. Негативный отбор обычно является одним из этапов отбора (преселекция) специфических аффинных последовательностей ДНК (аптамеров) по технологии SELEX ("Systematic Evolution of Ligands by Exponential Enrichment") [US Patent 5567588] и проводится для минимизации обогащения селектированного пула неспецифически связывающими олигонуклеотидами [Stoltenburg R., Reinemann С., Strehlitz В. // Biomol. Engineering. - 2007. - V. 24. - Ν. 4. - P. 381-403].
Метод направленной негативной селекции против заданных мишеней не применялся в отношении получения маркерных последовательностей, применимых в составе тест-систем на основе метода иммуно-ПЦР, позволяющих снизить или элиминировать фоновый сигнал амплификации.
Изобретение решает задачу создания биотинилированных последовательностей ДНК, не имеющих сродства к компонентам блокирующих растворов - бычьему сывороточному альбумину (БСА) и α-казеину, а значит, не связывающихся в этими белками при использовании в составе тест-систем на основе иммунодетекции, сопряженной с ПЦР. Полученные инертные по отношению к данным белкам последовательности ДНК позволяют существенно снизить фоновый сигнал амплификации, получаемый при неспецифической сорбции ДНК-матрицы на компонентах блокирующих растворов, входящих в состав тест-системы.
Поставленная задача решается за счет получения методом негативной селекции из обширной комбинаторной библиотеки олигонуклеотидных одноцепочечных последовательностей ДНК, не имеющих сродства к белковым компонентам блокирующих растворов - бычьему сывороточному альбумину (БСА) и казеину.
Также поставленная задача решается за счет введения молекулы биотина в 5′-положение селектированных неаффинных к БСА и α-казеину последовательностей ДНК. Биотин позволяет эффективно связывать биотинилированное антитело и биотинилированную ДНК посредством их одновременного взаимодействия с тетравалентными белками авидинового ряда.
Техническим результатом изобретения являются последовательности одноцепочечной ДНК, несущие биотин в 5′-положении последовательности, длиной 81 нуклеотид, с молекулярной массой 28,2 кДа, содержащие области для связывания с праймерами длиной 18 нуклеотидов в крайних положениях последовательности, не образующие связей с БСА и α-казеином, пригодные для использования в системах, основанных на иммунодетекции, сопряженной с ПЦР, в качестве матрицы, апмлифицируемой для усиления сигнала при детекции мишени.
В предлагаемом техническом решении получение инертных к БСА и α-казеину коротких одноцепочечных биотинилированных последовательностей ДНК осуществляется за счет проведения последовательных раундов негативной селекции комбинаторной библиотеки последовательностей ДНК против заданных белков. Раунды отбора представляют собой чередование инкубации олигонуклеотидной библиотеки с заданными белками-мишенями, отделение несвязавшихся молекул ДНК от комплексов, образованных белками и ДНК, с применением гель-фильтрационной хроматографии на колонке Superdex 200™ (GE Healthcare, Великобритания), и последующей амплификацией олигонуклеотидов, не образовавших связей с БСА и α-казеином, для обогащения пула одноцепочечных ДНК последовательностями, не обладающими способностью связывать вышеперечисленные белки, чаще всего используемые в составе блокирующих буферов при проведении иммуноанализа.
Предлагаемое техническое решение предусматривает получение одноцепочечных фрагментов ДНК после проведения каждого раунда селекции, что позволяет использовать в каждом раунде селекции олигонуклеотиды сходной структуры. Также предусмотрено получение индивидуальных последовательностей одноцепочечных ДНК, которые в дальнейшем подвергаются скринингу на предмет аффинности к БСА и α-казеину, и возможности их успешного использования в дальнейшем как матричных молекул в составе иммуно-ПЦР тест-систем.
Также техническое решение предлагает введение молекулы биотина в крайнее 5′-положение последовательностей, показавших положительные результаты при предварительном скрининге и отвечающих заданным параметрам, которое позволяет использовать высокоаффинную (константа аффинности 1015 М-1) связь биотин-нейтравидин для связывания ДНК-матрицы с иммунным комплексом, содержащим биотинилированное детектирующее антитело.
Важным достоинством предлагаемого изобретения является уникальность полученных последовательностей, которые не встречаются в окружающей среде и не применяются в общелабораторной и исследовательской практике, таким образом снижается вероятность контаминации исследуемых образцов, а значит, и получения ложноположительных результатов.
Также преимуществом изобретения является снижение неспецифического фонового сигнала амплификации в связи с низкой вероятностью связывания маркерной ДНК с элементами блокирующего раствора.
Изобретение осуществляют следующим образом:
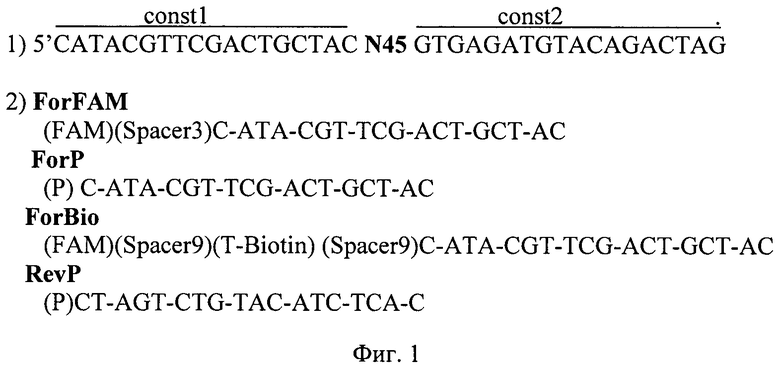
Синтезируют (ЗАО «Евроген», Россия; по заказу ФБУН ГНЦ ПМБ) комбинаторную библиотеку олигонуклеотидов вида 5′-const1-N45-const2, где N45 - вариабельная область, состоящая из 45 нуклеотидов, a const1 и const2 - константные фланкирующие области длиной по 18 нуклеотидов, которые служат для связывания со специфическими праймерами, позволяющими амплифицировать пул олигонуклеотидов после этапов селекции. Таким образом, олигонуклеотидная библиотека содержит 445 вариантов последовательности, содержащей 81 нуклеотид (Фиг. 1).
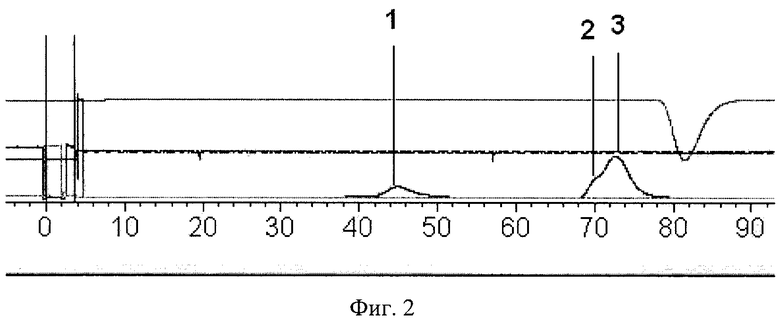
Проводят раунд негативной селекции последовательностей ДНК комбинаторной библиотеки против белковых компонентов блокирующих растворов - бычьего сывороточного альбумина и α-казеина. Для этого инкубируют пул олигонуклеотидов последовательно с БСА и α-казеином при избытке белка. Разделение свободных белка и ДНК, а также комплексов белок-ДНК проводят эксклюзионной хроматографией на колонке с Superdex 200™ (Фиг. 2). После выделения несвязавшихся молекул ДНК в отдельный пул суспензию доочищают фенол-хлороформом и концентрируют, используя метод преципитации этанолом. Полученный пул обогащают селектированными последовательностями. Для этого проводят амплификацию с использованием специфических к константным областям праймеров: прямого, меченного по 5'-концу карбоксифлуоресцеином (FAM метка), и обратного, несущего на 5′-конце остаток фосфорной кислоты. Фосфорилированную цепь расщепляют обработкой двуцепочечных фрагментов 5′-экзонуклеазой фага лямбда. Наличие FAM метки позволяет визуально контролировать полноту расщепления обратной цепи при проведении электрофореза в 10 % полиакриламидном геле, содержащем мочевину (Фиг. 3). Обогащенный пул одноцепочечных последовательностей ДНК применяют для проведения следующего раунда селекции. Всего проводят 5 раундов негативного отбора по вышеописанной методике.
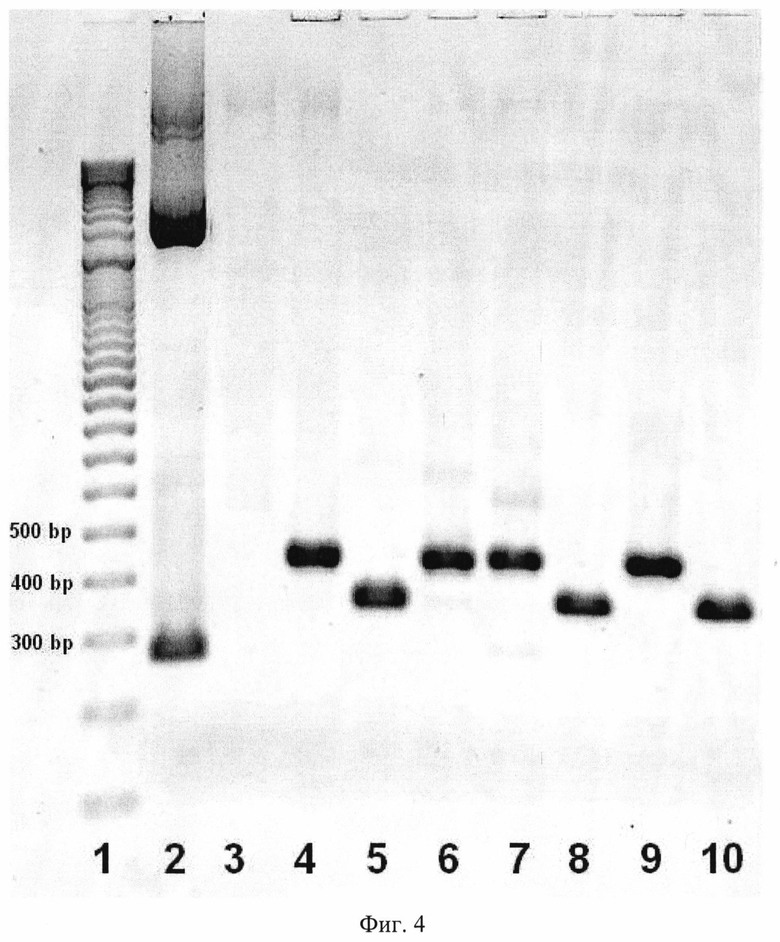
После максимального обогащения пула олигонуклеотидов неаффинными к БСА и казеину молекулами ДНК получают индивидуальные последовательности неаффинных ДНК. Для этого пул селектированных олигонуклеотидов клонируют в стандартный вектор pBluescriptll SK(-) (Agilent Technologies, США). Проводят скрининг единичных колоний на наличие в них плазмиды со встроенной последовательностью, соответствующей последовательности из библиотеки (Фиг. 4). Индивидуальные клонированные последовательности ДНК размножают и используют в дальнейшем скрининге.
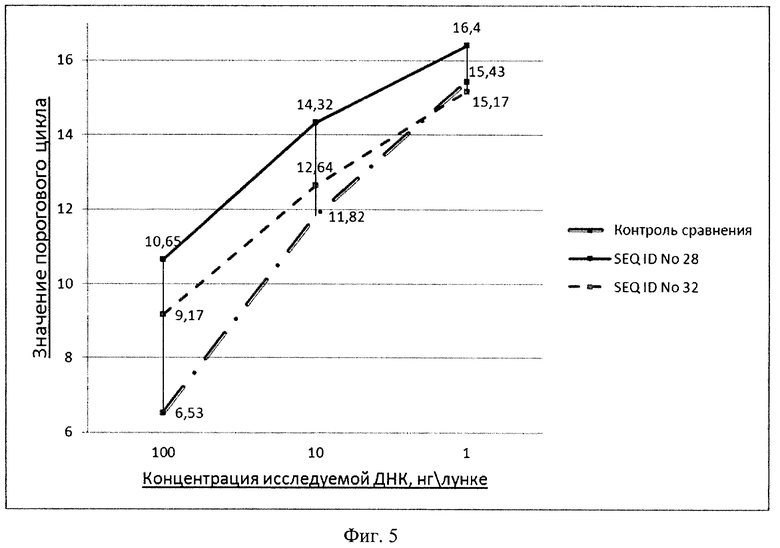
Проводят скрининг аффинности выделенных клонированных последовательностей в отношении БСА и казеина в формате ПЦР. Для этого на ПЦР-планшете иммобилизируют целевой белок, после чего инкубируют с клонами неаффинной ДНК. Результат визуализируют в ПЦР в режиме реального времени с применением интеркалирующего красителя SYBR-Green I (ЗАО «Синтол», Россия). В качестве контроля сравнения используют исходный пул олигонуклеотидов. Количество анализируемой ДНК в лунках с контролем и исследуемыми клонами уравнивают (Фиг. 5).
Последовательности, показавшие наилучший результат, соответствующий наименьшему сигналу относительно контроля сравнения, модифицируют путем амплификации с прямым праймером For Bio (ЗАО «Синтол», Россия), несущим молекулу биотина на 5′-конце, и обратным фосфорилированным праймером RevP (Фиг. 1). Для получения биотинилированной одноцепочечной последовательности обрабатывают ПЦР продукт 5′-экзонуклеазой фага лямбда (Фиг. 3).
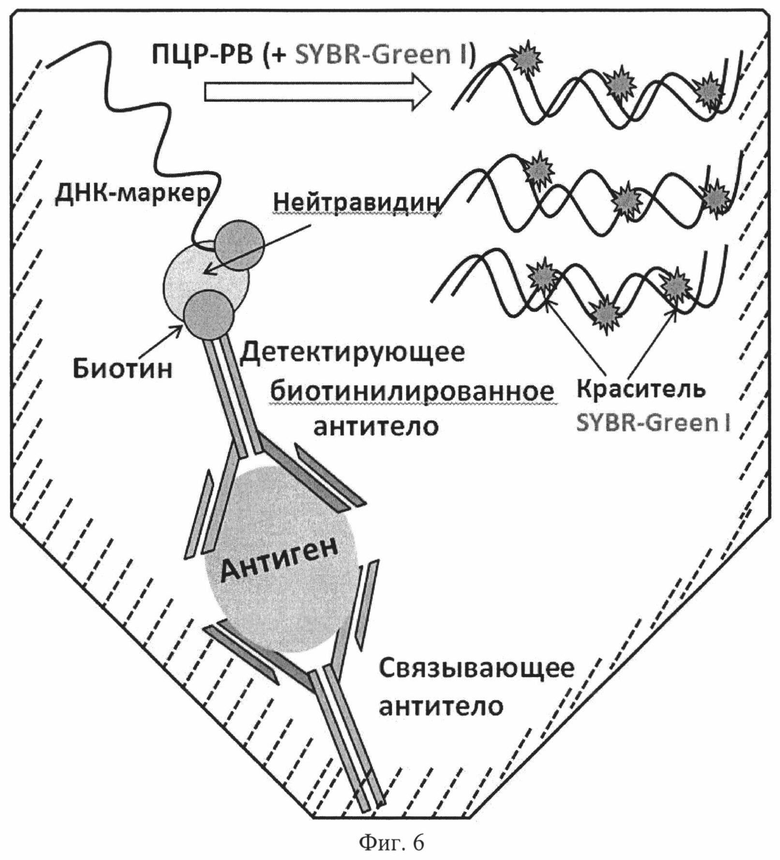
Проводят скрининг эффективности использования биотинилированных последовательностей в сэндвич-варианте иммуно-ПЦР. Для этого на поверхности планшета иммобилизируют связывающие антитела, после чего свободные валентности полученного днтекционного комплекса блокируют 3 % раствором БСА с добавлением 0,5 % казеина в фосфатно-солевом буфере. В лунки наносят антиген, аффинностью в которому обладают связывающие и детектирующие антитела. Далее инкубируют с детектирующими биотинилированными антителами, затем проводят цикл инкубации с раствором нейтравидина. На завершающем этапе инкубируют в лунках биотинилированную матричную ДНК, которая связывается с образовавшимся иммунным комплексом (Фиг. 6). Сигнал считывают в ПЦР в режиме реального времени с использованием интеркалирующего флуоресцентного красителя SYBR-Green I (Фиг. 7).
Изобретение иллюстрируют следующие графические материалы:
Фиг.1. Последовательности ДНК, используемые для селекции неаффинных молекул ДНК, для модификации полученных фрагментов и для проведения амплификации селектированных последовательностей в составе тест-систем.
1) Последовательность олигонуклеотидной библиотеки, используемой в селекции, где N45 - вариабельная область, const1 и const2 - константные области для отжига специфических праймеров;
2) Последовательности праймеров, используемых в реакциях обогащения библиотеки, для модификации клонированных последовательностей и в реакциях амплификации в процессе постановки анализа по методу иммуно-ПЦР.
Фиг. 2. Разделение свободных и связанных последовательностей из пула комбинаторной библиотеки после инкубации с α-казеином с применением эксклюзионной хроматографии на колонке с Superdex 200™.
1. Комплексы казеин-ДНК;
2. Казеин;
3. Свободные молекулы ДНК.
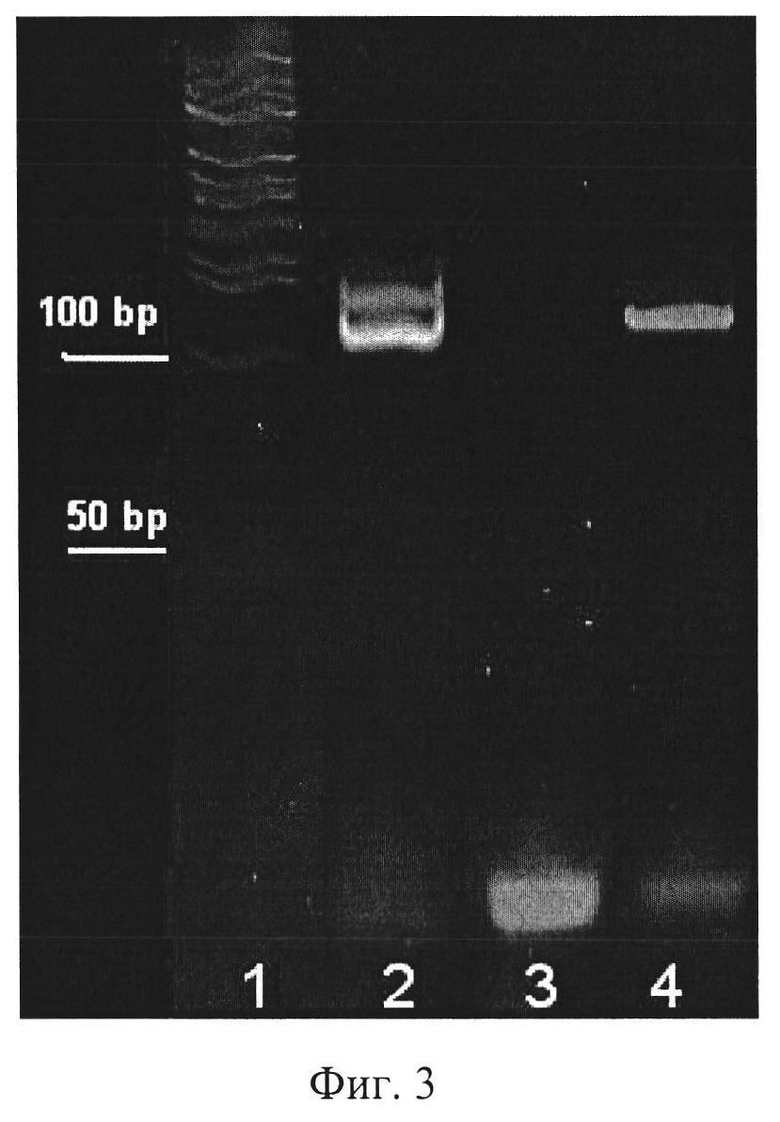
Фиг. 3. Расщепление обратной цепи ДНК при обработке двуцепочечного фрагмента 5'-экзонуклеазой фага лямбда.
1. Маркер молекулярной массы (50 bp DNA Ladder, New England BioLabs, США)
2. Двуцепочечный фрагмент ДНК
3. Отрицательный контроль (амплификация без внесения целевой ДНК)
4. Одноцепочечный фрагмент ДНК, полученный после обработки ПЦР-продукта 5′-экзонуклеазой фага лямбда.
Изображение получено с использованием лазерного сканера Typhoon FLA 9500.
Фиг. 4. Типичные результаты ПЦР со стандартными праймерами М13 отдельных выделенных колоний после клонирования селектированного пула ДНК в вектор pBluescriptll SK(-).
1. Маркер молекулярной массы (O'RangeRuler 100+500 bp DNA Ladder, Thermo Scientific, США)
2. Контроль (апмлификация исходной плазмиды pBluescriptll SK(-) со стандартными праймерами М13.
3. Отрицательный контроль (амплификация без внесения целевой ДНК)
4. 6, 7, 9. Результаты амплификации клонов, содержащих две копии вставки.
5. 8, 10. Результаты амплификации клонов, содержащих одну копию вставки, соответствующей 81 нуклеотиду.
Изображение получено с использованием лазерного сканера Typhoon FLA 9500.
Фиг. 5. Типичные результаты скрининга индивидуальных клонов селектированных последовательностей в отношении БСА с применением ПЦР в режиме реального времени для определения эффективности связывания ДНК с белком.
Фиг. 6. Принципиальная схема метода, используемого для скрининга биотинилированных неаффинных индивидуальных последовательностей ДНК в сэндвич-варианте иммуно-ПЦР.
Фиг. 7. Типичные результаты скрининга биотинилированных неаффинных индивидуальных последовательностей ДНК в сэндвич-варианте иммуно-ПЦР.
Фиг. 8. Последовательности неаффинных к БСА и казеину ДНК.
Для лучшего понимания сущности изобретения ниже следуют примеры его конкретного выполнения.
Пример 1. Проведение раунда негативной селекции последовательностей ДНК из пула комбинаторной библиотеки олигонуклеотидов против БСА и казеина
Бычий сывороточный альбумин, имеющий массу 69 кДа, в количестве 500 мкг растворяют в 1 мл буферного раствора, содержащего 20 мМ Tris-HCl, 50 мМ NaCl и 5 мМ EDTA (pH 7,4). К полученному раствору добавляют 20 мкг (4×1014 молекул) исходной комбинаторной библиотеки олигонуклеотидов. Таким образом, в смеси представлен молярный избыток белка по отношению к ДНК. Смесь инкубируют в течение 1 часа при температуре 37°C со встряхиванием на орбитальном шейкере. По окончании инкубации смесь разделяют при помощи эксклюзионной хроматографии на колонке, содержащей сорбент низкого давления Superdex 200™ (GE Healthcare, Великобритания), в буферном растворе состава 20 мМ Tris-HCl, 300 мМ NaCl, 5 мМ EDTA (pH 7,4), при скорости потока 0,5 мл/мин на фракции белка, несвязавшихся молекул ДНК и молекул ДНК, ассоциировавших с белком (Фиг. 2).
Пул отделенных несвязавшихся олигонуклеотидов доочищают с использованием метода фенол-хлороформной экстракции нуклеиновых кислот, затем концентрируют преципитацией ДНК этанолом в присутствии копреципитанта Pellet Paint® Co-Precipitant (Novagen, Германия).
Полученный пул одноцепочечных коротких фрагментов ДНК амплифицируют для обогащения его молекулами ДНК, имеющими низкий показатель аффинности в отношении БСА. Амплификацию проводят с использованием праймеров: прямого ForFAM, несущего в положении 5′ флуоресцентную метку карбоксифлуоресцеин, и обратного RevP, фосфорилированного по 5′-концу последовательности (температура отжига праймеров 54°C, время элонгации 15 секунд) (Фиг. 1). В результате проведенной ПЦР получают двуцепочечный фрагмент, одна из цепей которого мечена FAM.
Для получения обогащенного пула одноцепочечных фрагментов ПЦР-продукт обрабатывают 5′-экзонуклеазой фага лямбда, которая расщепляет обратную фосфорилированную цепь фрагмента [Higuchi R. G., Ochman Η. // Nucleic Acids Research. - 1989. - V. 17. - N. 14. - P. 5865]. Эффективность расщепления цепи контролируют проведением электрофореза в 10%-ном полиакриламидном геле, содержащем мочевину, с последующим окрашиванием бромистым этидием, при этом наличие в пробе только полосы, флуоресцирующей зеленым, свидетельствует об успешном расщеплении обратной цепи фрагмента, а присутствие в пробе второй полосы с красным цветом флуоресценции свидетельствует о неуспешном или неполном расщеплении фосфорилированной цепи ДНК (Фиг. 3)
Полученные одноцепочечные ДНК очищают и концентрируют, используя фенол-хлороформную экстракцию и преципитацию этанолом. Пул обогащенных олигонуклеотидов в количестве 20 мкг примешивают к 1 мл буферного раствора (20 мМ Tris-HCl, 50 мМ NaCl и 5 мМ EDTA, pH 7,4), содержащего 250 мкг α-казеина (молекулярная масса 20 кДа), что также составляет молярный избыток белка по отношению к ДНК. Инкубируют смесь 1 час при 37°C, после чего разделяют гель-фильтрационной хроматографией и обогащают амплификацией, как описано выше.
Проводят еще 4 аналогичных раунда селекции.
Пример 2. Получение индивидуальных последовательностей олигонуклеотидов, подвергшихся негативной селекции
Фрагменты, пригодные для клонирования в коммерческий вектор pBluescriptll SK(-) (Agilent Technologies, США), синтезируют из пула селектированных олигонуклеотидов при помощи ПЦР с использованием прямого и обратного фосфорилированных праймеров ForP и RevP (отжиг праймеров 54°C, элонгация 15 секунд). Плазмиду pBluescriptII SK(-) обрабатывают эндонуклеазой рестрикции SmaI в буфере Tango (Thermo Scientific, США), после чего обрабатывают щелочной фосфатазой (Invitrogen, США) для предотвращения лигирования внутри вектора. Фосфорилированные двуцепочечные фрагменты ДНК лигируют в подготовленную плазмиду.
Электрокомпетентные клетки Е. coli DH12S трансформируют подготовленной ранее плазмидой pBluescriptII SK(-), содержащей вставки селектированных последовательностей, применив электротрансформацию (прибор Eppendorf Electroporator 2510 (Eppendorf, Германия), режим 1,7 KB). Трансформированные клетки высевают на плотную среду с 2хYТ-агаром, содержащим 50 мкг/мл ампициллина, 80 мкг/мл X-Gal и 20 мМ IPTG, и выращивают в течение ночи при 37°C.
Отсевают единичные колонии, потенциально содержащие вектор со вставкой (по принципу бело-голубой селекции [Ullmann A, Jacob F, Monod J. // J Mol Biol. - 1967. - V. 24. - N. 2. - P. 339-343]), и анализируют наличие встроенного фрагмента ДНК искомой длины в плазмиде с помощью ПЦР со стандартными прямым и обратным праймерами M13/pUC (Fermentas, США) (Фиг. 4). Увеличивают количество копий индивидуальных последовательностей клонированных фрагментов ДНК амплификацией с использованием праймеров ForFAM и RevP (Фиг. 1). Получают одноцепочечный продукт для каждого из клонов обработкой 5′-экзонуклеазой фага лямбда с визуальным контролем в электрофорезе (Фиг. 3).
Пример 3. Скрининг аффинности индивидуальных селектированных последовательностей в отношении белков, входящих в состав блокирующих растворов
На поликарбонатном ПЦР-планшете (Corning, США) иммобилизируют белок - БСА или казеин. Для этого в лунки планшета вносят 150 мкл раствора белка с концентрацией 100 мкг/мл в фосфатно-солевом буфере (pH 7,4) (ФСБ). Инкубируют в течение часа при 37°C. Трижды отмывают ФСБ с добавлением 0,05% Tween-20. Затем в лунки планшета вносят одноцепочечную ДНК в объеме 50 мкл. Количество ДНК во всех пробах уравнено и раститровано следующим образом: 0,5 мкг/лун - 0,1 мкг/лун - 0,05 мкг/лун - 0,01 мкг/лун - 0,005 мкг/лун - 0,001 мкг/лун. В качестве контроля сравнения используют соответствующие разведения исходной комбинаторной библиотеки. Все пробы исследуют в трех повторностях. Буфер для разведения ДНК содержит 20 мМ Tris-HCl, 50 мМ NaCl, 5 мМ EDTA (pH 7,4). Инкубируют планшет с ДНК в течение часа при 37°C. Отмывку от несвязавщихся молекул ДНК производят раствором, содержащим 20 мМ Tris-HCl, 300 мМ NaCl, 5 мМ EDTA (pH 7,4) пятикратно.
Контролируют реакцию связывания ДНК с белком в ПЦР в режиме реального времени. Для этого в лунки планшета вносят амплификационную смесь, содержащую флуоресцентный интеркалирующий краситель SYBR-Green I (ЗАО "Синтол", Россия) с добавлением праймеров ForP и RevP (Фиг. 1). Амплификацию проводят в режиме: температура отжига праймеров 54°C, время элонгации 10 секунд, детекцию сигнала проводят по каналу SYBR 1(FAM) (прибор IQ™5, Bio-Rad, США). За положительный результат принимают показания прибора для пороговых циклов, которые превышают пороговые циклы контроля сравнения не менее чем на один цикл (Фиг. 5).
Последовательности с наименьшей аффинностью к БСА и казеину биотинилируют по 5′-концу с применением праймеров ForBio и RevP (Фиг. 1) (температура отжига 54°C, время элонгации 15 секунд). Полученный двуцепочечный биотинилированный продукт обрабатывают 5′-экзонуклеазой фага лямбда для получения одноцепочечного фрагмента ДНК, биотинилированного по 5′-концу. Полученные последовательности используют для дальнейшего скрининга в отношении применения их в качестве компонента тест-системы. (Фиг. 8).
Пример 4. Скрининг эффективности использования модифицированных неаффинных последовательностей в диагностической системе на основе метода иммуно-ПЦР
Для определения эффективности использования модифицированных неаффинных последовательностей в качестве маркерной ДНК в тест-системах на базе иммуно-ПЦР проводят модельный эксперимент по внедрению полученных биотинилированных одноцепочечных ДНК в тест-систему для определения ботулинического нейротоксина типа А по методу иммуно-ПЦР.
Поликлональные антитела, специфичные в отношении тяжелой цепи ботулинического токсина, иммобилизируют на поверхности поликарбонатного ПЦР-планшета (Corning, США). Для этого в лунки планшета вносят по 50 мкл суспензии, содержащей 0,5 мкг антител в ФСБ (pH 7,4). Инкубируют планшет в течение часа при температуре 37°C. После этого трижды производят отмывку лунок планшета, используя ФСБ с добавление 0,05% Tween-20. Блокируют свободные центры связывания планшета комплексным раствором, состоящим из обезжиренного (массовое содержание жиров 0,5%) молока с добавлением 3% БСА, внося в лунки по 200 мкл блокирующего буфера, и выдерживают 40 минут при 37°C. Отмывку проводят ФСБ с 0,05% Tween-20 трижды.
Вносят в лунки разведения в ФСБ образца тяжелой цепи ботулотоксина А в объеме 50 мкл и следующих концентрациях: 10 нг/мл - 1 нг/мл - 0,1 нг/мл - 10 пг/мл - 1 пг/мл - 0,1 пг/мл - 10 фг/мл - 1 фг/мл. В качестве отрицательного контроля используют ФСБ, не содержащий целевой антиген. Каждую пробу исследуют в двух повторностях. Инкубируют планшет при 37°C один час, после чего отмывают ФСБ с 0,05% Tween-20 трижды.
Инкубируют планшет в течение часа при 37°C с суспензией биотинилированных антител, которые вносят в каждую лунку в объеме 50 мкл с содержанием антител 0,025 мкг. Отмывку от несвязавшихся антител проводят аналогично предыдущим этапам.
Образованный иммунный комплекс инкубируют с нейтравидином. Для этого вносят в лунки 50 мкл ФСБ с содержанием нейтравидина 0,2 мкг. Инкубацию проводят в течение 30 минут при 37°C, отмывку проводят аналогично предыдущим этапам.
Иммунный комплекс через нейтравидин связывают с биотинилированной маркерной ДНК. Сравнивают селектированные инертные по отношению к БСА и α-казеину биотинилированные ДНК с биотинилированной ДНК, входящей в состав тест-системы «IPCR-ΒοΝΤ/Α», которая представляет собой двуцепочечную ДНК длиной 400 п.н., каждая цепь которой биотинилирована по 5′-концу. При этом молекулярная масса такого маркера составляет 280 кДа, что в десять раз больше массы любого из селектированных одноцепочечных биотинилированных маркеров длиной 81 нуклеотид (28 кДа). Количество вносимых в лунки маркерных ДНК уравнивают эквимолярно по количеству копий одноцепочечной ДНК. Таким образом, биотинилированные одноцепочечные неаффинные маркерные ДНК длиной 81 нуклеотид вносят в лунки планшета по 50 мкл из концентрации 1×105 мкг/мл, а ДНК-маркер, входящий в состав тест-системы, вносят в том же объеме из концентрации 5×105 мкг/мл. Разведения ДНК готовят в буфере следующего состава: 20 мМ Tris-HCl, 50 мМ NaCl, 5 мМ EDTA (pH 7,4). Выдерживают час при температуре 37°C, отмывают лунки планшета от свободных молекул ДНК буфером 20 мМ Tris-HCl, 300 мМ NaCl, 5 мМ EDTA (pH 7,4), содержащим 0,05 % Tween-20, шестикратно.
Для регистрации результатов детекции молекулы ботулотоксина и образования иммунного «сэндвича» в каждую лунку ПЦР-планшета вносят амплификационную смесь, содержащую интеркалирующий краситель SYBR-Green I (ЗАО "Синтол", Россия), а также специфические праймеры: для селектированных неаффинных последовательностей такими праймерами являются ForP и RevP, для ДНК-маркера тест-системы «IPCR-ΒοΝΤ/Α» существуют собственные праймеры, входящие в состав набора реагентов. Сигнал детектируют на протяжении 35 циклов амплификации в ПЦР в режиме реального времени по каналу SYBR-1 в условиях: температура отжига праймеров 54°C, время элогнации 30 секунд. Предел детекции тест-системы на основе метода иммуно-ПЦР для определения ботулинического нейротоксина типа А с исходной матричной ДНК составляет 1 пг/мл (в реакционном объеме 50 мкл) для тяжелой цепи белка, при использовании селектированных неаффинных олигонуклеотидов в составе системы на той же иммунологической основе предел детекции достигает 10 фг/мл, что на два порядка больше.
Перечень последовательностей к заявке на получение патента «Последовательность ДНК, применимая в качестве зонда для обеспечения максимального соотношения «сигнал/фон» в тест-системах на основе иммуно-ПЦР»:
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК-АПТАМЕРОВ, СВЯЗЫВАЮЩАЯСЯ С ПРОТЕОЛИТИЧЕСКОЙ СУБЪЕДИНИЦЕЙ НЕЙРОТОКСИНА ТИПА A CLOSTRIDIUM BOTULINUM | 2014 |
|
RU2571210C1 |
| Последовательность ДНК-аптамера, связывающаяся с пептидогликан-ассоциированным липопротеином Legionella pneumophila | 2019 |
|
RU2737829C1 |
| ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК-АПТАМЕРОВ, СВЯЗЫВАЮЩАЯСЯ С БАКТЕРИЯМИ Escherichia coli O157:H7 | 2014 |
|
RU2566551C1 |
| ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК-АПТАМЕРОВ, СВЯЗЫВАЮЩАЯ ШИГА-ТОКСИН ТИПА 2 | 2014 |
|
RU2566552C1 |
| Способ направленного истощения олигонуклеотидных библиотек для снижения неспецифической адсорбции при твердофазной селекции аптамеров на основе нуклеиновых кислот | 2015 |
|
RU2618872C1 |
| СПОСОБ СПЕЦИФИЧЕСКОГО ОТБОРА ВЫСОКОАФФИННЫХ МОЛЕКУЛ ДНК (ДНК-АПТАМЕРОВ) К РЕКОМБИНАНТНОМУ БЕЛКУ-МИШЕНИ | 2012 |
|
RU2513700C1 |
| Способ получения аттенуированного бесплазмидного штамма F.tularensis 15 CMSA, синтезирующего микобактериальный антиген супероксиддисмутазу А | 2019 |
|
RU2745161C1 |
| АНТИТЕЛО ПРОТИВ С5 И СПОСОБ ПРЕДУПРЕЖДЕНИЯ И ЛЕЧЕНИЯ ОБУСЛОВЛЕННЫХ КОМПЛЕМЕНТОМ ЗАБОЛЕВАНИЙ | 2014 |
|
RU2663349C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ИЗМЕРЕНИЯ И КАЛИБРОВКИ СИСТЕМАТИЧЕСКОЙ ОШИБКИ АМПЛИФИКАЦИИ В МУЛЬТИПЛЕКСНЫХ ПЦР-РЕАКЦИЯХ | 2013 |
|
RU2631797C2 |
| Способ получения линии гуманизированных мышей, содержащих инсерцию 3974insT в гене mGrin2a (mice glutamate [NMDA] receptor subunit epsilon-1), приводящую к преждевременному прекращению трансляции белка grin2a | 2021 |
|
RU2764650C1 |
Изобретение относится к биотехнологии, а именно к последовательности ДНК, которую используют в качестве зонда для обеспечения максимального соотношения «сигнал/фон» в тест-системах на основе иммуно-ПЦР. Указанная последовательность ДНК имеет длину 81 нуклеотид и представляет собой последовательность CA-TAC-GTT-CGA-CTG-CTA-CTG-GTG-CAA-CGG-GTT-CGA-TCA-GCT-CCG-CCG-GGC-CTC-CCA-ATC-CCC-CGT-GAG-ATG-TAC-AGA-CTA-G и CA-TAC-GTT-CGA-CTG-CTA-CCG-AAT-GGA-TGT-GTC-CGA-GCT-CCA-TTT-CCG-TCC-AAT-CCT-CTT-CGT-CGT-GAG-ATG-TAC-AGA-CTA-G. Данная последовательность ДНК инертна по связыванию с бычьим сывороточным альбумином и α-казеином, являющимися наиболее часто используемыми инертными белками в составе блокирующих растворов, применяемых в иммуноанализе, несет молекулу биотина в крайнем 5'- положении для образования комплекса с биотинилированным детектирующим антителом посредством одновременного взаимодействия с белком авидинового ряда. Предложенное изобретение обеспечивает высокое отношение «сигнал/фон» при детекции белка-мишени по методу иммуно-ПЦР. 8 ил., 4 пр.
Последовательность ДНК, применимая в качестве зонда для обеспечения максимального соотношения «сигнал/фон» в тест-системах на основе иммуно-ПЦР длиной 81 нуклеотид, имеет последовательность CA-TAC-GTT-CGA-CTG-CTA-CTG-GTG-CAA-CGG-GTT-CGA-TCA-GCT-CCG-CCG-GGC-CTC-CCA-ATC-CCC-CGT-GAG-ATG-TAC-AGA-CTA-G и CA-TAC-GTT-CGA-CTG-CTA-CCG-AAT-GGA-TGT-GTC-CGA-GCT-CCA-TTT-CCG-TCC-AAT-CCT-CTT-CGT-CGT-GAG-ATG-TAC-AGA-CTA-G, инертна по связыванию с бычьим сывороточным альбумином и α-казеином, являющимися наиболее часто используемыми инертными белками в составе блокирующих растворов, применяемых в иммуноанализе, несет молекулу биотина в крайнем 5'- положении для образования комплекса с биотинилированным детектирующим антителом посредством одновременного взаимодействия с белком авидинового ряда, обеспечивает высокое отношение «сигнал/фон» при детекции белка-мишени по методу иммуно-ПЦР в связи с сокращением уровня неспецифического связывания олигонуклеотидного апмлифицируемого зонда с инертными белками в составе блокирующего раствора.
| US 0005567588 A1, 22.10.1996 | |||
| US 20130171652 A1, 04.07.2013 | |||
| SAITO K | |||
| ET AL., Detection of human serum tumor necrosis factor-alpha in healthy donors, using a highly sensitive immuno-PCR assay, Clin Chem, 1999, v.45, no.5, p | |||
| Приспособление для разгонки рельсов ударами | 1923 |
|
SU665A1 |
| СПОСОБ ДИАГНОСТИКИ ИНФЕКЦИОННОЙ ГУБЧАТОЙ ЭНЦЕФАЛОПАТИИ ЖИВОТНЫХ МЕТОДОМ ИММУНО-ПЦР | 2012 |
|
RU2486517C1 |

