ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Настоящая заявка испрашивает приоритет предварительной заявки на патент США 61/576,138, поданной 15 декабря 2011 года.
УРОВЕНЬ ТЕХНИКИ
Трансформация растений в целом охватывает методики, требуемые и применяемые для введения экспрессируемого растением чужеродного гена в растительные клетки, в результате чего может быть получено фертильное потомство растения, которое стабильно сохраняет и экспрессирует чужеродный ген. Были трансформированы многочисленные представители однодольных и двудольных растений. Трансгенные сельскохозяйственные культуры, а также плодовые и овощные культуры, представляют коммерческий интерес. Такие культуры включают, без ограничения, кукурузу, рис, сою, канолу, подсолнечник, люцерну, сорго, пшеницу, хлопок, арахис, томат, картофель и т.п.
Известно несколько методик введения чужеродного генетического материала в растительные клетки и получения растений, которые стабильно сохраняют и экспрессируют введенный ген. Такие методики включают ускорение генетического материала, нанесенного на микрочастицы, непосредственно в клетки (например, патент США 4945050 и патент США 5141131). Другая технология трансформации включает технологию WHISKERS™ (см., например, патент США 5302523 и патент США 5464765). Для трансформации растений также использовали методику электропорации. См., например, WO 87/06614, патент США 5472869, патент США 5384253, WO 92/09696 и WO 93/21335. Кроме того, может применяться слияние протопластов растения с липосомами, содержащими доставляемую ДНК, прямая инъекция ДНК, а также другие возможные методы.
После интеграции введенной ДНК в геном растения, она обычно остается относительно стабильной в последующих генерациях. Трансформированные клетки в растениях нормально растут. Они могут формировать зародышевые клетки и передавать трансформированный признак(и) потомству растения. Такие растения можно выращивать обычным способом и скрещивать с растениями, у которых есть такие же трансформированные наследственные факторы или другие наследственные факторы. Получаемые гибридные особи обладают соответствующими фенотипическими свойствами, например, способностью контролировать питание насекомых-вредителей растений.
Ряд альтернативных методик также может использоваться для введения ДНК в растительную клетку-хозяина. Такие методики включают, без ограничения, трансформацию T-ДНК, доставляемой Agrobacterium tumefaciens или Agrobacterium rhizogenes в качестве трансформирующего агента. Растения могут быть трансформированы с использованием агробактериальной методики, как описано, например, в патенте США 5177010, патенте США 5104310, заявке на европейский патент 0131624B1, заявке на европейский патент 120516, заявке на европейский патент 159418B1, заявке на европейский патент 176112, патенте США 5149645, патенте США 5469976, патенте США 5464763, патенте США 4940838, патенте США 4693976, заявке на европейский патент 116718, заявке на европейский патент 290799, заявке на европейский патент 320500, заявке на европейский патент 604662, заявке на европейский патент 627752, заявке на европейский патент 0267159, заявке на европейский патент 0292435, патенте США 5231019, патенте США 5463174, патенте США 4762785, патенте США 5004863 и патенте США 5159135. Применение T-DNA-содержащих векторов для трансформации растительных клеток было тщательно исследовано и в достаточной мере описано в заявке на европейский патент 120516; An et al., (1985, EMBO J. 4:277-284); Fraley et al., (1986, Crit. Rev. Plant Sci. 4: 1-46), а также в Lee and Gelvin (2008, Plant Physiol. 146:325-332), и хорошо известно в данной области.
Наиболее важный первый этап в трансформации растительных клеток посредством Agrobacterium spp. состоит в непосредственном контакте, прикреплении или адгезии бактериальных клеток к клеткам трансформируемого растения-хозяина. После связывания клеток, известна биология переноса T-ДНК из агробактерии в растительные клетки. См., например, Gelvin, 2003, Microbiol. Molec. Biol. Rev. 67: 16-37; и Gelvin, 2009, Plant Physiol. 150: 1665-1676. Как минимум, по меньшей мере, правый T-ДНК бордерный повтор, но часто и правый бордерный повтор, и левый бордерный повтор Ti или Ri плазмиды присоединены в качестве фланкирующей области генов, которые требуется ввести в растительную клетку. Левые и правые T-ДНК бордерные повторы являются ключевыми цис-действующими последовательностями, требуемыми для переноса T-ДНК. Различные транс-действующие компоненты кодируются полным геномом агробактерии. Наиболее важными из них являются белки, кодируемые vir генами, которые обычно присутствуют в виде ряда оперонов на Ti или Ri плазмидах. Различные Ti и Ri плазмиды несколько отличаются дополнительным элементам vir генов, например, не всегда присутствует virF. Белки, кодируемые vir генами, выполняют множество различных функций, в том числе распознавание и сигнализацию взаимодействия растительной клетки/бактерий, индукцию транскрипции vir генов, формирование канала секреции IV типа, распознавание бордерных повторов T-ДНК, формирование T-цепей, перенос T-цепей в растительную клетку, импорт T-цепей в ядро растительной клетки и интеграцию T-цепей в ядерную хромосому растения, и многие другие. См., например, Tzfira and Citovsky, 2006, Curr. Opin. Biotechnol. 17:147-154.
Если для трансформации используются штаммы Agrobacterium, ДНК, которую предстоит ввести в растительную клетку, можно клонировать в специальные плазмиды, например, либо в промежуточный (шаттл) вектор или в бинарный вектор. Промежуточные векторы не способны к независимой репликации в клетках Agrobacterium, но могут подвергаться манипуляциям и реплицироваться в обычных штаммах Escherichia coli для молекулярного клонирования. Как правило, такие промежуточные векторы включают последовательности, фланкированные правой и левой областями T-ДНК бордерных повторов, которые могут включать ген селективного маркера, функциональный для отбора трансформированных растительных клеток, клонирующий линкер, клонирующий полилинкер или другую последовательность, которая может функционировать как сайт введения генов, предназначенных для трансформации растительной клетки. Клонирование и манипуляции с генами, которые требуется перенести в растения, могут быть, таким образом, легко выполнены с помощью стандартных методов в E. coli при использовании шаттл-вектора в качестве клонирующего вектора. Полностью подготовленный шаттл-вектор может быть затем введен в штаммы Agrobacterium для трансформации растения в целях дальнейшей работы. Промежуточный вектор может быть перенесен в Agrobacterium с помощью хелперной плазмиды (посредством бактериальной конъюгации), электропорации, химически опосредованной прямой ДНК трансформации или с использованием других известных методик. Шаттл-векторы могут быть интегрированы в Ti или Ri плазмиду или их производные при гомологичной рекомбинации благодаря последовательностям, которые являются гомологичными между Ti или Ri плазмидой, или их производными и промежуточной плазмидой. Такое событие гомологичной рекомбинации (то есть интеграции плазмиды) обеспечивает, таким образом, способ стабильного сохранения измененного шаттл-вектора в агробактерии, с точкой начала репликации и другими функциями сохранения плазмиды, которые обеспечиваются частью Ti или Ri плазмиды в коинтегрантной плазмиде. Ti или Ri плазмида также включает vir области, включающие vir гены, необходимые для переноса T-ДНК. Как правило, плазмида, несущая vir область, представляет собой мутантную Ti или Ri плазмиду (хелперную плазмиду), из которой была удалена область T-ДНК, включающая правый и левый T-ДНК бордерные повторы. Такие pTi-производные плазмиды, содержащие функциональные vir гены и лишенные всей или почти всей T-области и связанных элементов, описательно именуются в настоящей заявке как хелперные плазмиды.
Супербинарная система является специальным примером системы шаттл-вектора/гомологичной рекомбинации (см. обзор Komari et al., 2006, Methods in Molecular Biology (K. Wang, ed.) No. 343: Agrobacterium Protocols, pp.15-41; и Komori et al., 2007, Plant Physiol. 145:1155-1160). Штамм LBA4404 (pSB1) несет две независимо реплицирующихся плазмиды, pAL4404 и pSB1. pAL4404 является хелперной плазмидой, полученной на основе Ti-плазмиды, и содержит интактный набор vir генов (из Ti плазмиды pTiACH5), но не имеет T-ДНК области (и, следовательно, не содержит левой и правой последовательностей T-ДНК бордерных повторов). Плазмида pSB1 предоставляет дополнительный частичный набор vir генов, полученных из pTiBo542; этот частичный набор vir генов включает оперон virB и оперон virC, а также гены virG и virD1. Одним из примеров шаттл-векторов, используемых в супербинарной системе является pSB11, который содержит клонирующий полилинкер, который служит сайтом введения генов, предназначенных для трансформации растительной клетки, и фланкирован правой и левой областями T-ДНК бордерных повторов. Шаттл-вектор pSB11 не способен к независимой репликации в агробактерии, но стабильно сохраняется как коинтегрантная плазмида при интеграции в pSB1 посредством гомологичной рекомбинации между обычными последовательностями, присутствующими в pSB1 и pSB11. Таким образом, на полностью модифицированную область T-ДНК, введенную в LBA4404 (pSB1) на модифицированном векторе pSB11, эффективно воздействуют и она переносится в растительные клетки Vir белками, полученными из двух различных агробактериальных Ti-плазмид (pTiACH5 и pTiBo542). Штаммом-хозяином Agrobacterium tumefaciens, используемым с супербинарной системой, является LBA4404 (pSB1). Супербинарная система оказалась особенно полезной при трансформации видов однодольных растений. См. Hiei et al., (1994) Plant J. 6:271-282; и Ishida et al., (1996) Nat. Biotechnol. 14:745-750.
В дополнение к vir генам, содержащимся в Ti плазмидах Agrobacterium, другие, хромосомные гены, регулирующие вирулентность (называемые chv генами), как известно, регулируют некоторые аспекты взаимодействия клеток Agrobacterium и растительных клеток, и таким образом влияют на общую частоту трансформации растения (Pan et al., 1995, Molec. Microbiol. 17:259-269). Несколько хромосомных генов, требуемых для вирулентности и прикрепления, сгруппированы в хромосомном локусе длиной 29 т.п.н. (Matthysse et al., 2000, Biochim. Biophys. Acta 1490:208-212).
В дополнение к многочисленным технологиям трансформации растений, тип ткани, которая контактирует с чужеродными генами, также может быть различным. Такая ткань может включать, без ограничения, эмбриогенную ткань, каллусную ткань I и II типов, гипокотиль и меристему. Почти все растительные ткани могут быть трансформированы во время дедифференцировки при использовании соответствующих методик, известных специалисту. Специалисту в области трансформации растений будет известно, что множество методик доступно для получения трансформированных растений, и что они могут быть изменены и специализированы в соответствии с биологическими различиями между различными видами растений-хозяев. Растительные экспланты (например, части листа, сегменты стебля, меристемы, корни, но также и протопласты или клетки, выращенные в суспензионной культуре) можно успешно культивировать с Agrobacterium tumefaciens или Agrobacterium rhizogenes для переноса ДНК в растительную клетку.
Каллусные культуры. Культуры растительных тканей можно успешно культивировать с Agrobacterium tumefaciens или Agrobacterium rhizogenes для переноса ДНК в растительную клетку, при этом их обычно получают из стерильных частей целого растения, которые могут состоять из частей органов, таких как листья или корни, или могут быть определенными типами клеток, такими как пыльца или эндосперм. Многие свойства экспланта, как известно, влияют на эффективность закладки культуры. Считается, что любая растительная ткань может использоваться в качестве экспланта, если подобраны правильные условия. Как правило, более молодая, более быстро растущая ткань (или ткань на ранней стадии развития) является наиболее эффективной. Экспланты, культивируемые на подходящей среде, могут давать начало неорганизованной, растущей и делящейся массе клеток (каллус). В культуре каллус можно поддерживать более или менее бессрочно, при условии, что его периодически переносят на свежую среду. Во время формирования каллуса присутствует некоторая степень дедифференцировки, как в морфологии (каллус обычно состоит из неспециализированных клеток паренхимы), так и в метаболизме.
Каллусные культуры чрезвычайно важны в биотехнологии растений. Манипуляция соотношениями растительных гормонов в питательной среде может приводить к развитию побегов, корней или соматических зародышей, из которых могут быть впоследствии получены целые растения (регенерация). Каллусные культуры могут также использоваться для инициирования суспензий клеток, которые используются различными способами в исследованиях трансформации растений.
Суспензионные культуры клеток. Каллусные культуры, вообще говоря, относятся к одной из двух категорий: плотные или рыхлые. В плотном каллусе клетки соединены плотно, тогда как в рыхлом каллусе клетки лишь слабо связаны друг с другом, при этом каллус становится мягким и легко разделяется. Рыхлый каллус дает инокулят для формирования суспензионных клеточных культур. Экспланты из некоторых видов растений или определенные типы клеток не склонны к формированию каллуса, что затрудняет закладку суспензии клеток. Рыхлость каллуса можно иногда повышать при манипуляции компонентами сред, путем повторного пересевания или его выращивания на полутвердой среде (среде с низкой концентрацией желирующего вещества). При помещении рыхлого каллуса в жидкую среду и последующем перемешивании, одиночные клетки и/или мелкие комки клеток переходят в среду. При правильных условиях такие высвобожденные клетки продолжают расти и делиться, с получением, в конечном счете, суспензионной клеточной культуры. Суспензии клеток можно поддерживать относительно просто в виде периодических культур в конических колбах и размножать при повторном пересеве в свежую среду. После пересева клетки делятся, и биомасса культуры растет характерным образом. Культуры суспензий клеток можно успешно выращивать с Agrobacterium tumefaciens или Agrobacterium rhizogenes для переноса ДНК в растительную клетку.
Культура апикальных клеток и меристемы. Верхушки побегов (которые содержат апикальные меристемы) можно культивировать in vitro с получением скоплений побегов либо из пазушных, либо из придаточных почек, и можно успешно культивировать с Agrobacterium tumefaciens или Agrobacterium rhizogenes для переноса ДНК в растительную клетку. Культуры апикальных меристем используются для регенерации зерновых культур (рассада может использоваться в качестве донорного материала).
Зародышальная культура Зародыши могут использоваться в качестве эксплантов для получения каллусных культур или соматических зародышей. В качестве эксплантов могут использоваться как незрелые, так и зрелые зародыши. Незрелый, полученный из зародышей, эмбриогенный каллус представляет собой ткань, используемую при регенерации однодольных растений, и может успешно культивироваться с Agrobacterium tumefaciens для переноса ДНК в растительную клетку. Незрелые зародыши представляют собой интактную ткань, которая способна к делению клеток, давая начало каллусным клеткам, которые могут дифференцироваться с развитием тканей и органов целого растения. Незрелые зародыши могут быть получены из опыленных початков зрелого растения кукурузы, например, из растений, опыленных с использованием методов, описанных в Neuffer et al. (1982, Growing maize for genetic purposes. In: Maize for Biological Research. W. F. Sheridan, Ed. UNIVERSITY PRESS, University of North Dakota, Grand Forks, ND). Примеры методов выделения незрелых зародышей из кукурузы описаны в Green and Phillips (Crop Sci. 15:417-421 (1976)). Незрелые зародыши предпочтительно выделяют из развивающегося початка при использовании асептических методов и выдерживают в стерильной среде до применения. Использование агробактерии в трансформации незрелых зародышей раскрыто в Sidorov & Duncan, (2009, Methods in Molecular Biology: Transgenic Maize, vol.526 Chapter 4, M. Paul Scott (Ed.)) и в патенте США 5981840.
Культура микроспор. Гаплоидную ткань можно культивировать in vitro при использовании пыльцы или пыльников в качестве экспланта и можно успешно культивировать с Agrobacterium tumefaciens для переноса ДНК в растительную клетку. Каллус и зародыши могут быть получены из пыльцы. Для получения культуры in vitro из гаплоидной ткани могут быть применены два подхода. В первом, пыльники (соматическая ткань, которая окружает и содержит пыльцу) культивируют на твердой среде. Полученные из пыльцы зародыши затем получают при раскрытии зрелых пыльников. Раскрытие пыльника зависит от его выделения на правильной стадии и от правильных условий культивирования. У некоторых видов уверенность в естественном раскрытии можно обеспечить, надрезав стенку пыльника. Во втором методе пыльники культивируют в жидкой среде, и пыльца, высвобождаемая из пыльников, может использоваться для формирования зародышей. Незрелую пыльцу также можно извлекать из развивающихся пыльников и культивировать непосредственно.
Многие зерновые культуры (рис, пшеница, ячмень и кукуруза) требуют среды с добавкой регуляторов роста растения для культуры пыльников или пыльцы. Регенерация из эксплантов микроспор может быть достигнута прямым эмбриогенезом или через стадию каллуса и последующего эмбриогенеза.
Культуры гаплоидных тканей также могут быть инициированы из женского гаметофита (яйцеклетки). В некоторых случаях, это более эффективный метод, чем использование пыльцы или пыльников.
Растения, полученные из гаплоидных культур, могут не быть гаплоидными. Это может быть следствием удвоения хромосом во время периода культивирования. Удвоение хромосом (которое может быть вызвано обработкой такими химическими веществами, как колхицин) может быть преимуществом, поскольку во многих случаях гаплоидные растения не являются желаемым результатом регенерации из гаплоидных тканей. Такие растения часто называются дигаплоидами, поскольку они содержат две копии одного и того же гаплоидного генома.
После трансформации любого из вышеуказанных растительных материалов при культивировании с Agrobacterium tumefaciens для переноса ДНК в растительную клетку, из зараженного растительного материала могут быть регенерированы целые растения после помещения в подходящие условиях роста и питательную среду, которая может содержать антибиотики или гербициды для селекции трансформированных растительных клеток. Растения, полученные таким образом, могут быть затем проверены на присутствие встроенной ДНК.
Трансформация клетки (в том числе трансформация растительной клетки) может включать конструирование вектора экспрессии, который будет функционировать в конкретной клетке. Такой вектор может содержать ДНК, которая включает ген, находящийся под контролем или функционально связанный с регуляторным элементом (например, промотором). Вектор экспрессии может содержать одну или более таких комбинаций функционально связанного гена/ регуляторного элемента. Вектор(ы) может быть в форме плазмиды и может использоваться один или в комбинации с другими плазмидами для получения трансформированных клеток с применением способов трансформации, описанных в настоящей заявке, в целях введения трансгена(ов) в генетический материал растительной клетки.
Векторы экспрессии для растительных клеток могут включать по меньшей мере один генетический маркер, функционально связанный с регуляторным элементом (например, промотором), который позволяет трансформированным клеткам, содержащим маркер, регенерироваться либо в условиях отрицательной селекции (то есть ингибирующей рост клеток, которые не содержат селективного маркерного гена), либо положительной селекции (то есть путем скрининга продукта, кодируемого генетическим маркером). В технологии трансформации известны множество селективных маркерных генов, подходящих для трансформации растений, которые включают, например, гены, кодирующие ферменты, которые метаболически детоксифицируют селективный химический агент, который может быть антибиотиком или гербицидом, или гены, которые кодируют измененную мишень, которая может быть нечувствительной к ингибитору. Несколько методов положительной селекции также известны в уровне техники. Индивидуально используемый селективный маркерный ген может соответственно обеспечить возможность отбора трансформированных клеток, тогда как рост клеток, которые не содержат встроенную ДНК, может быть подавлен селективным соединением. Предпочтение конкретному селективному маркерному гену отдается на усмотрение специалиста, но может использоваться любой из следующих селективных маркеров, а также любой другой ген, не перечисленный в настоящем описании, который мог бы функционировать как селективный маркер. Примеры селективных маркеров включают, без ограничения, устойчивость или резистентность к канамицину, G418, гигромицину, блеомицину, метотрексату, фосфинотрицину (Биалафосу), глифосату, имидазолинонам, сульфонилмочевинам и триазолопиримидиновым гербицидам, таким как хлорсульфурон, бромоксинил и далапон.
В дополнение к селективному маркеру может быть желательным использование репортерного гена. В некоторых случаях репортерный ген может использоваться без селективного маркера. Репортерные гены являются генами, которые обычно не обеспечивают преимуществ роста для реципиентного организма или ткани. Репортерный ген обычно кодирует белок, который предусматривает некоторое фенотипическое изменение или ферментативное свойство. Подходящие репортерные гены включают, без ограничения, гены, которые кодируют бету-глюкуронидазу (GUS), люциферазу светляка или флуоресцентные белки, такие как зеленый флуоресцентный белок (GFP) или желтый флуоресцентный белок (YFP, по существу как раскрыто в патенте США 7951923).
Независимо от используемой методики трансформации чужеродный ген может быть включен в вектор переноса гена, предназначенный для экспрессии чужеродного гена в растительной клетке при включении в вектор растительного промотора. В дополнение к растительным промоторам, в растительных клетках для экспрессии чужеродных генов могут эффективно использоваться промоторы из множества источников. Например, могут использоваться промоторы бактериального происхождения, такие как промотор октопинсинтазы, промотор нопалинсинтазы, промотор маннопинсинтазы; промоторы вирусного происхождения, такие как 35S и 19S промоторы вируса мозаики цветной капусты (CaMV), промотор из палочковидного вируса сахарного тростника и т.п. Полученные из растений промоторы включают, без ограничения, промотор малой субъединицы (ssu) рибулоза-1,6-бисфосфат (RUBP) карбоксилазы, промотор бета-конглицинина, промотор фазеолина, промотор ADH (алкогольдегидрогеназы), промоторы теплового шока, промотор ADF (фактора деполимеризации актина) и тканеспецифические промоторы. Промоторы могут также содержать некоторые энхансерные элементы последовательности, которые могут повышать эффективность транскрипции. Типичные энхансеры включают, без ограничения, интрон 1 алкогольдегидрогеназы 1 (ADH1) и ADH1-интрон 6. Могут использоваться конститутивные промоторы. Конститутивные промоторы направляют постоянную экспрессию гена практически во всех типах клеток и почти во всех случаях (например, актин, убиквитин, 35S CaMV). Тканеспецифические промоторы отвечают за экспрессию гена в определенных типах клеток или тканей, таких как листья или семена. Примеры других промоторов, которые могут использоваться, включает промоторы, активные во время определенной стадии развития растения, а также промоторы, активные в определенных растительных тканях и органах. Примеры таких промоторов включают, без ограничения, промоторы, которые являются корнеспецифическими, специфическими для пыльцы, эмбриоспецифическими, специфическими для кукурузных рыльцев, специфическими для хлопкового волокна, специфическими для эндосперма семян и флоэма-специфическими.
При некоторых обстоятельствах может быть желательным использование индуцируемого промотора. Индуцируемый промотор отвечает за экспрессию генов в ответ на специфический сигнал, такой как: физический стимул (например, гены теплового шока); свет (например, рибулозо-1,5-бисфосфаткарбоксилаза); гормон (например, глюкокортикоид); антибиотик (например, тетрациклин); метаболиты; и стресс (например, засуха). Также могут использоваться другие желательные элементы транскрипции и трансляции, которые функционируют в растениях, такие как, например, 5’-нетранслируемые лидерные последовательности, а также последовательности 3’ терминации транскрипции РНК и сигнальные последовательности присоединения полиаденилата. Может использоваться любой подходящий растительный вектор переноса гена, известный в уровне техники.
Трансгенные зерновые культуры, содержащие признаки устойчивости к насекомым (IR), широко распространены в растениях кукурузы и хлопка на территории Северной Америки, при этом использование таких признаков возрастает в мировом масштабе. Коммерческие трансгенные зерновые культуры, которые сочетают в себе признаки IR и устойчивости к гербицидам (HT), были разработаны многими компаниями-производителями семян. Они включают комбинации IR признаков, придаваемых инсектицидными белками Bacillus thuringiensis (B.t.), и HT признаков, таких как устойчивость к ингибиторам ацетолактатсинтазы (ALS), таким как сульфонилмочевины, имидазолиноны, триазолопиримидин, сульфонанилиды и т.п., ингибиторам глутаминсинтетазы (GS), таким как биалафос, глуфосинат и т.п., ингибиторам 4-гидроксифенилпируватдиоксигеназы (HPPD), таким как мезотрион, изоксафлютол и т.п., ингибиторам 5-енолпирувилшикимат-3-фосфатсинтазы (EPSPS), таким как глифосат и т.п., и ингибиторам ацетил-кофермент A карбоксилазы (ACCase), таким как галоксифоп, квизалофоп, диклофоп и т.п. Известны другие примеры, в которых трансгенно продуцируемые белки обеспечивают устойчивость растения к классам химических гербицидов, таким как гербициды - производные феноксикислот и пиридилоксиацетатные ауксиновые гербициды (см. WO 2007/053482 A2) или гербициды - производные феноксикислот и арилоксифеноксипропионатные гербициды (см. WO 2005/107437A1). Возможность контролировать несколько проблемных вредителей посредством IR признаков является ценной концепцией коммерческого продукта, причем удобство такой концепции продукта повышается, если признаки контроля насекомых и признаки контроля сорняков сочетаются в одном растении. Более того, более высокая ценность может быть достигнута посредством комбинации в одном растении IR признаков, придаваемых инсектицидным белком B.t., с одним или более дополнительными HT признаками, такими как указанные выше, плюс один или более дополнительных вводных признаков (например, устойчивость к другому насекомому, придаваемая B.t.-производными или другими инсектицидный белками, устойчивость против насекомого, придаваемая такими механизмами, как РНКи и т.п., устойчивость к болезням, устойчивость к стрессу, улучшенная утилизация азота и т.п.) или выходных признаков (например, высокое содержание масел, полезный состав масел, повышенная пищевая ценность и т.п.). Такие комбинации могут быть получены либо с помощью обычного скрещивания (например, селекционный блок) или совместно в качестве нового события трансформации, включающего одновременное введение нескольких генов (например, молекулярный блок). Благоприятные эффекты включают способность контролировать насекомых-вредителей и улучшенный контроль сорняков при выращивании сельскохозяйственного растения, что обеспечивает дополнительные выгоды для производителя и/или потребителя. Таким образом, способы в настоящем описании могут применяться для получения трансформированных растений с комбинациями признаков, которые включают полный агрономический набор для улучшения качества сельскохозяйственной культуры со способностью к гибкому и рентабельному решению любого количества сельскохозяйственных проблем.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Описаны способы трансформации растительной клетки. Указанные способы включают контакт растительных клеток с клетками агробактерии в жидкой среде, содержащей поверхностно-активное вещество. Клетки агробактерии могут быть соскоблены с твердой среды или выращены в жидкой питательной среде перед суспендированием в жидкой среде, содержащей поверхностно-активное вещество. Концентрация поверхностно-активного вещества может находиться в пределах от 0,001 процента по весу до 0,08 процента по весу. Поверхностно-активное вещество может быть неионным трисилоксановым поверхностно-активным веществом, при этом может использоваться больше одного поверхностно-активного вещества. Растительные клетки могут быть клетками кукурузы. Растительные клетки могут быть помещены в условия постоянного освещения после контакта с клетками агробактерии.
ОПИСАНИЕ ФИГУР
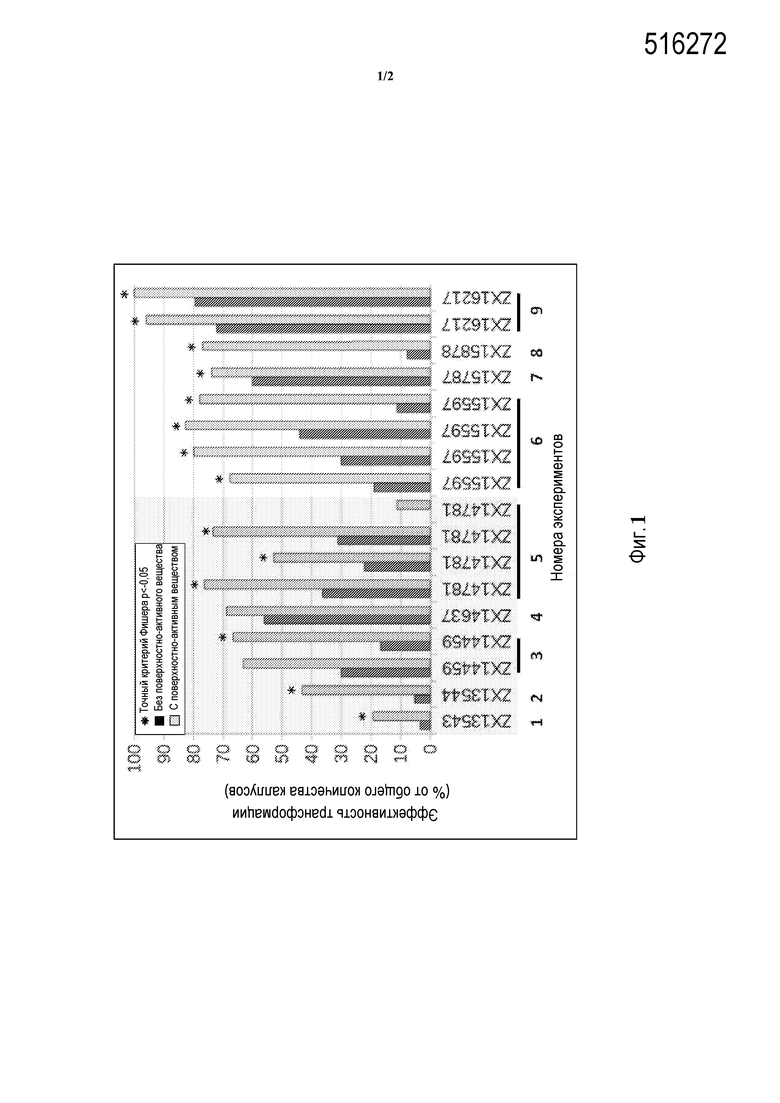
Фиг. 1 - гистограмма, на которой показано улучшение трансформации незрелых зародышей кукурузы при добавлении поверхностно-активного вещества BREAK-THRU® S 233 в среду инфицирования, используемую для приготовления суспензии клеток агробактерии (несущих плазмиду pEPS1083) перед совместным культивированием.
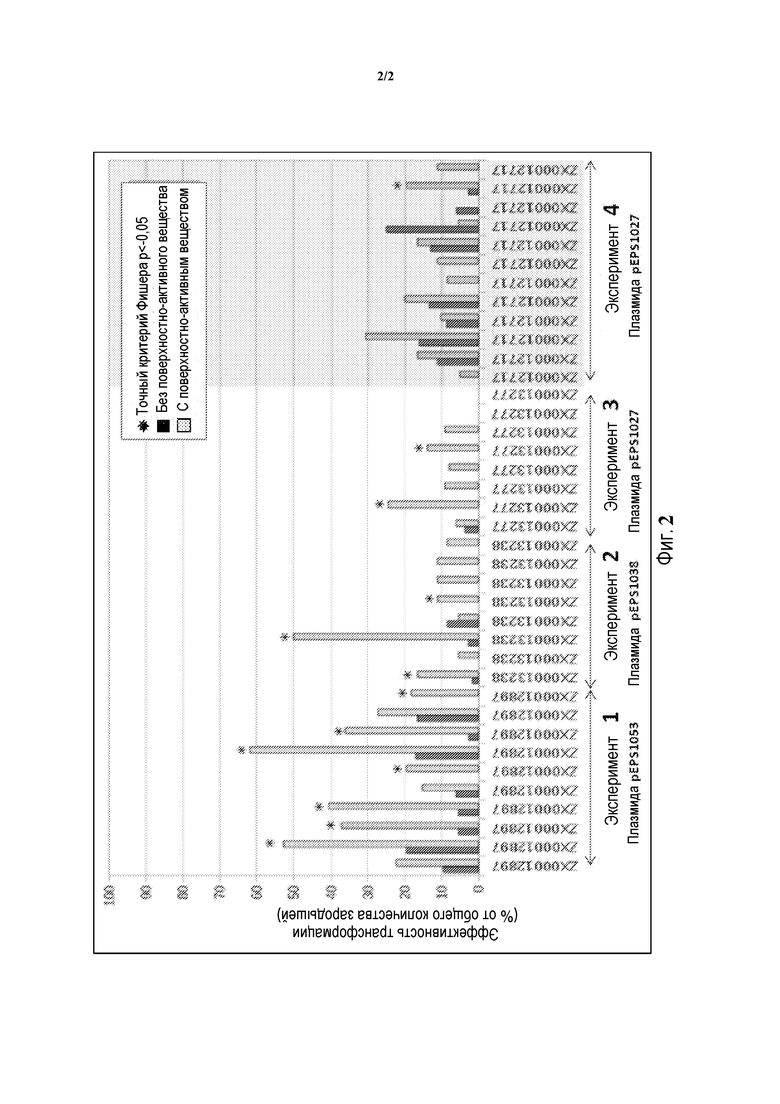
Фиг. 2 - гистограмма, на которой показано улучшение трансформации незрелых зародышей кукурузы при добавлении поверхностно-активного вещества BREAK-THRU® S 233 в среду инфицирования, используемую для приготовления суспензии клеток агробактерии перед совместным культивированием. Плазмиды, используемые для каждого эксперимента, показанного на Фигуре 2, включают: Эксперимент 1 = pEPS1053; GOI = IPT, селективный маркер = aad1. Эксперимент 2 = pEPS1038; GOI = GF14, селективный маркер = aad1. Эксперимент 3 и Эксперимент 4 = pEPS1027; без GOI, селективный маркер = aad1.
ПОДРОБНОЕ ОПИСАНИЕ
Описаны способы повышения частоты трансформации в растениях при использовании агробактерии. Способы включают контакт растительных клеток с клетками агробактерии в жидкой среде, содержащей поверхностно-активное вещество. Некоторые способы включают пребывание растительных клеток в условиях постоянного освещения после контакта с клетками агробактерии. Примеры растений, которые могут применяться с такими способами, включают растения кукурузы и незрелые зародыши кукурузы.
Штаммы агробактерии отличаются друг от друга по своей способности трансформировать клетки различных видов растений. Независимо от конкретной рассматриваемой комбинации штамма Agrobacterium/растения-хозяина, агробактерия действует посредством прикрепления к клетке-хозяину во время трансформации. См. McCullen and Binns, 2006, Ann. Rev. Cell. Dev. Biol. 22: 101-127; и Citovsky et al., 2007, Cell. Microbiol. 9:9-20. Поэтому способы, которые усиливают связывание клеток Agrobacterium с растительными клетками, такие как способы, раскрытые в настоящем описании, в которых применяются поверхностно-активные вещества, могут обеспечить повышение эффективности трансформации. Усиление связывания клеток агробактерии с растительными клетками отличается для различных видов и типов ткани, поскольку различные виды растений, а также различные ткани растения одного вида, могут иметь различный химический и биохимический состав клеточных стенок. Кроме того, такие различия могут также изменяться во время различных стадий развития одной растительной ткани.
Дополнительно, бактерии различных родов и видов, и фактически различные штаммы бактерий разных видов, часто отличаются по химическому и биохимическому составу своих клеточных стенок, причем эти различия могут изменяться во время цикла роста бактерий. Увеличение эффективности трансформации растений способами, раскрытыми в настоящем описании, таким образом, может являться результатом способности поверхностно-активных веществ уменьшать отталкивающие гидрофобные взаимодействия между клеточными стенками агробактерии и стенками растительной клетки, и допускать, таким образом, плотные межклеточные взаимодействия.
Поэтому химические различия между различными поверхностно-активными веществами можно использовать, чтобы способствовать клеточным взаимодействиям между клетками различных штаммов Agrobacterium (и различными фазами роста таких клеток) и клетками и тканями различных растений-хозяев во время различных фаз культивирования растительных тканей, в результате чего можно наблюдать повышение эффективности трансформации.
Поверхностно-активные вещества относятся к нескольким классам химических соединений, при этом специалисту в области трансформации растений будет известно, что для повышения эффективности трансформации растений с различными растениями-хозяевами могут использоваться различные химические классы поверхностно-активных веществ. Примеры поверхностно-активных веществ из этих химических классов, которые могут применяться в способах, раскрытых в настоящем описании, включают вспомогательные вещества, неионные поверхностно-активные вещества, анионные поверхностно-активные вещества, поверхностно-активные вещества на масляной основе, амфотерные поверхностно-активные вещества и полимерные поверхностно-активные вещества. Примером предпочтительного поверхностно-активного вещества, которое может применяться в способах, описанных в настоящей заявке, является неионное поверхностно-активное вещество, производное трисилоксана, такое как BREAK-THRU® S233 производства Evonik Industries (Essen, Germany). Примеры других предпочтительных поверхностно-активных веществ, которые могут применяться в способах, описанных в настоящей заявке, включают трисилоксан алкоксилаты, этоксилированные соевые масла, этоксилаты спиртов C-13, C12-C14-алкилдиметилбетаины, а также блок-сополимеры ди-втор-бутилфенол этиленоксида - пропиленоксида. В Таблице 1 представлен неограничивающий список поверхностно-активных веществ различных химических типов, которые могут применяться для практического осуществления способов, описанных в настоящей заявке.
S240
INDUSTRIES AG
(Essen, Germany)
S243
INDUSTRIES AG
HS 429
(Friendly, WV)
HS 312
(Aukland, NZ)
(Midland, MI)
PG 2069
(Berkeley, CA)
COMPANIES, INC
(Leominster, MA)
5993-A
MN)
пропиленоксида
(Cincinnati, OH)
(The Woodlands, TX)
EMGARD®
поверхностно-активные вещества
пропиленоксида
(Indianapolis, IN)
PRODUCTS INC.
(Greeley, CO)
(C-12)
SURFACE
CHEMISTRY LLC
(Chicago, IL)
G 850
(Cranberry, NJ)
(Vashi, India)
(Wayne, NJ)
SURFACE
CHEMISTRY LLC
**POE = моли пропиленоксида, прореагировавшие с конкретным гидрофобным веществом
В способах, раскрытых в настоящем описании, применяются усиливающие трансформацию свойства поверхностно-активных веществ для резкого повышения эффективности агробактериальной (например, Agrobacterium tumefaciens) трансформации в растениях, таких как незрелые зародыши кукурузы. Поверхностно-активные вещества, применяемые в способах, описанных в настоящей заявке, выбирают, как предложено выше, по способности усиливать межклеточные взаимодействия, которые повышают эффективность трансформации. Концентрация поверхностно-активного вещества в жидкой среде может составлять от 0,001 процента по весу до 0,08 процента по весу, от 0.001 процента по весу до 0,07 процента по весу, от 0,001 процента по весу до 0,06 процента по весу, от 0,001 процента по весу до 0,05 процента по весу, от 0,001 процента по весу до 0,04 процента по весу, от 0,001 процента по весу до 0,035 процента по весу, от 0,001 процента по весу до 0,03 процента по весу, от 0,001 процента по весу до 0,025 процента по весу, от 0,001 процента по весу до 0,02 процента по весу, от 0,001 процента по весу до 0,015 процента по весу, от 0,001 процента по весу до 0,01 процента по весу или от 0,005 процента по весу до 0,01 процента по весу.
Одно или более дополнительных поверхностно-активных веществ также могут применяться в способах, описанных в настоящей заявке. Как указано, эффективность трансформации зависит от множества факторов, в том числе от видов растений и штамма агробактерии, и типа ткани. Принимая во внимание тип включенных взаимодействий, система из двух или более поверхностно-активных веществ может обеспечивать повышенную эффективность трансформации. Дополнительные поверхностно-активные вещества, применяемые в системе двух или более поверхностно-активных веществ, могут быть выбраны, например, из Таблицы 1.
Способы, описанные в настоящей заявке, могут широко применяться к множеству видов и сортов растений, включая однодольные и двудольные. Целевые сельскохозяйственные культуры включают, без ограничения, кукурузу, рис, сою, канолу, подсолнечник, люцерну, сорго, пшеницу, хлопок, арахис, томаты, картофель и т.п. Способы в настоящей заявке могут применяться с клетками на различных стадиях развития, например, незрелыми зародышами. Таким образом, способы, описанные в настоящей заявке, могут применяться для трансформации незрелых зародышей кукурузы. Размер незрелых зародышей, применяемых в способах, описанных в настоящей заявке, может изменяться. Например, незрелые зародыши могут иметь длину больше или равную 1,5 мм и меньше или равную 2,5 мм.
Условия, в которых выдерживают клетки после контакта с Agrobacterium согласно способам, описанным в настоящей заявке, могут быть регулируемыми. Например, температура, pH и другие компоненты питательной среды, в которую помещают клетки после трансформации согласно способам, описанным в настоящей заявке, могут быть различными и обычно известны специалистам в данной области. Одной из таких переменных является освещение. Способы, описанные в настоящей заявке, могут включать освещение растительных клеток согласно стандартным режимам 18 часов света/6 часов темноты или, в альтернативе, постоянное освещение после контакта с клетками Agrobacterium. Например, клетки, обработанные согласно способам, описанным в настоящей заявке, могут выдерживаться в условиях 24 часового освещения лампами дневного света в течение нескольких недель после обработки, например, до регенерации и стадий выделения всходов при получении растения.
Дополнительный способ включает приготовление жидкой среды, содержащей поверхностно-активное вещество, суспендирование клеток агробактерии в жидкой среде и контакт растительных клеток с клетками агробактерии в жидкой среде, содержащей поверхностно-активное вещество. Клетки агробактерии могут быть соскоблены с твердой среды перед суспендированием в жидкой среде, содержащей поверхностно-активное вещество. Кроме того, клетки агробактерии могут быть выращены в жидкой питательной среде перед суспендированием в жидкой среде, содержащей поверхностно-активное вещество.
Методики и способы трансформации растений с применением агробактерий известны специалистам в области молекулярной биологии. Любые типы известных методов с использованием агробактерий в трансформации растений могут применяться в способах, описанных в настоящей заявке. В Примерах ниже представлены варианты осуществления способов, демонстрирующих эффективность способов, описанных в настоящей заявке, однако настоящие Примеры не следует рассматривать как ограничения объема изобретения.
Все патенты, заявки на патенты, предварительные заявки и публикации, указанные или процитированные в настоящее описании, полностью включены посредством отсылки в той степени, в которой они не противоречат принципам настоящего описания.
ПРИМЕРЫ
Следующие примеры иллюстрируют процедуры для практического осуществления изобретения. Примеры и варианты осуществления, описанные в настоящей заявке, служат исключительно в иллюстративных целях, при этом различные модификации или вариации с учетом изложенного в настоящем описании будут предложены специалистами, квалифицированными в данной области техники, и должны быть включены в сущность и объем заявленного изобретения. Все проценты приведены по весу, а все соотношения смеси растворителей приведены по объему, если не указано иное. Все температуры приведены в градусах Цельсия.
ПРИМЕР 1. Трансформация агробактерии с целью создания супербинарных векторов.
Агробактериальная супербинарная система успешно используется для трансформации однодольных растений-хозяев. Методики конструирования и проверки супербинарных векторов подробно описаны и включены в настоящее описание посредством отсылки (Operating Manual for Plasmid pSBl, Version 3.1, available from Japan Tobacco, Inc., Tokyo, Japan). При получении супербинарных плазмид использовали стандартные методы молекулярной биологии и микробиологии. Проверку правильной структуры супербинарной плазмиды выполняли при использовании методики, предложенной в Руководстве для плазмиды pSB1.
В данной работе использовали штаммы Agrobacterium, несущие различные супербинарные плазмиды. Все эти плазмиды содержали, в качестве селективного маркера/гена устойчивости к гербициду, кодирующую последовательность (CDS) белка AAD1 (патент США 7838733), экспрессия которого находилась под транскрипционным контролем промотора актина1 риса и связанного интрона 1, по существу как раскрыто в патенте США 5641876 и в GENBANK™ EU155408.1. Терминация транскрипции и полиаденилирование мРНК aad1 были определены 3'UTR липазы кукурузы, по существу как раскрыто в качестве оснований 921-1277 из GENBANK™ gb|L35913.1|MZELIPASE и в патенте США 7179902. Кроме того, супербинарные плазмиды несли ген, экспрессия которого, как ожидали, не должна была влиять на частоту трансформации. В частности, в плазмиде pEPS1083, CDS, кодирующая белок YFP (по существу как раскрыто в патенте США 7951923) (транскрипцию которого регулировал промотор убиквитина 1 кукурузы со связанным интроном 1; патент США 5510474), и мРНК которой терминированы 3'UTR Per5 кукурузы (патент США 6384207)), успешно использовали в качестве визуального маркера для контроля трансформации и определения относительной эффективности трансформации. Другие супербинарные плазмиды, используемые для иллюстрирования способов, раскрытых в настоящей заявке (плазмиды pEPS1013, pEPS1018, pEPS1028, pEPS1036, pEPS1038, pEPS1059, pEPS1064, pEPS1066, pEPS1068, pEPS6004 и pEPS6008), несли CDS, кодирующую белок, представляющий собственность Dow AgroSciences, экспрессию которого регулировали теми же элементами транскрипции/терминации, которые использовали для CDS YFP.
Экспрессию YFP использовали для измерения эффективности трансформации в некоторых экспериментах. Проценты эффективности трансформации вычисляли как количество каллусов, которые показали экспрессию YFP, деленное на количество обработанных зародышей и умноженное на 100. Экспрессию YFP измеряли посредством визуального наблюдения при использовании флуоресцентного микроскопа Olympus SZX12 (Olympus America Inc.; Center Valley, PA) или Leica M165FC (Leica Microsystems Inc.; Buffalo Grove, IL) с фильтрами YFP, охватывающими диапазоны возбуждения при 514 нм и эмиссии, измеряемой при 527 нм.
В других экспериментах, в которых использовали штаммы Agrobacterium, несущие супербинарные плазмиды без гена YFP, эффективность трансформации вычисляли согласно анализу TAQMAN® (Life Technologies; Carlsbad, CA) у дочерних растений, полученных из зародышей, которые подвергали селекции на устойчивость к галоксифопу. Используемые компоненты TAQMAN® были специфичными к кодирующей области aad1. Эффективность трансформации вычисляли из числа определенных TAQMAN®-положительных событий, деленного на число обработанных зародышей и умноженного на 100. "Событием" в этих целях считался зародыш, который давал одно или более TAQMAN®-проверенных растений. Отдельный зародыш, как полагали, являлся одним событием, независимо от того, сколько растений он мог бы дать.
ПРИМЕР 2. Трансформация кукурузы штаммами Agrobacterium (Методика трансформации 1).
Основной ход работы приведен ниже. Зародыши выделяли из незрелых початков кукурузы в стадии развития, на которой молодые зародыши имели длину приблизительно 1,4-1,9 мм. При необходимости сравнения различных условий трансформации, приблизительно равные количества зародышей, выделенных из одного початка, распределяли на все обработки. Зародыши инкубировали с суспензией, содержащей клетки агробактерии и поверхностно-активное вещество (или нет, для сравнения), затем переносили на чашки с твердой средой и проводили совместное культивирование в течение 3-5 дней. Обработанные зародыши переносили в среду, содержащую антибиотики (для подавления и лизиса клеток агробактерии) и соединения для селективного выделения генетически трансформированных тканей и растений кукурузы. Ткань кукурузы (обычно каллус, но без ограничения этим) выращивали на селективной среде до восстановления растений. Эти растения тестировали для подтверждения их генетической трансформации, и те растения, которые имели требуемую модификацию, выращивали для получения семян.
Получение незрелых зародышей. Семена из инбредного гибрида B104 высевали в горшки объемом 4 галлона (~15 л), содержащие смесь SUNSHINE CUSTOM BLEND 160; Bellevue, WA). Растения выращивали в теплице, используя комбинацию натриевых ламп высокого давления и металлогалогеновых ламп при световом периоде 16:8 ч света:темноты. Для получения незрелых зародышей для трансформации, проводили контролируемое опыление сибсов. Незрелых зародышей выделяли через 10-13 дней после опыления, когда зародыши имели размер приблизительно 1,4-1,9 мм. Початки кукурузы стерилизовали на поверхности после удаления листовой обертки и пестиков путем погружения в 50% коммерческий отбеливатель (CLOROX®, 5,25% гипохлорита натрия) с TWEEN®-20 (1 или 2 капли на 500 мл) на 10 минут и тройной промывки стерильной водой.
Незрелые зародыши в стерильных условиях выделяли непосредственно в микроцентрифужную пробирку, содержащую 2 мл инфицирующей среды с суспендированными клетками агробактерии и поверхностно-активное вещество, при необходимости. Зародыши инкубировали с суспензией клеток агробактерии, содержащей поверхностно-активное вещество (или нет, для контрольных экспериментов), в течение 5-30 минут.
Суспензию клеток агробактерии, содержащих супербинарный вектор, подготавливали, выращивая сначала клетки газоном в течение 4 дней при 25° или 3 дней при 28°, на чашках с твердым агаром, содержащих YEP (г/л: дрожжевой экстракт, 5; пептон, 10; NaCl, 5; агар 15) с 50 мг/л спектиномицина; 10 мг/л рифампицина и 50 мг/л стрептомицина. (В некоторых экспериментах клетки агробактерии выращивали на твердой среде LB (SIGMA ALDRICH; St. Louis, MO) 20 г/л, с антибиотиками, как указано выше.) Эту культуру сеяли штрихом из изолята одиночной колонии, культивируемого в таких же условиях. Одну или две петли клеток соскабливали с газона, затем однородно ресуспендировали (мягким пипетированием) в Инфицирующей среде (IfM) до оптической плотности при 600 нм (OD600) 0,35-0,45. Инфицирующая среда содержала: 4,33 г/л солей MS; 1X ISU модифицированные витамины MS; 68,4 г/л сахарозы; 36 г/л глюкозы; 700 мг/л L-пролина; 3,3 мг/л Dicamba-KOH и 100 мкΜ ацетосирингона (приготовленного в ДМСО); при pH 5,2. В зависимости от эксперимента соответствующее количество раствора поверхностно-активного вещества (например, BREAK-THRU® S 233 в конечной концентрации 0,01) добавляли в Инфицирующую среду после суспендирования клеток.
Раствор агробактерий и зародышей инкубировали в течение 5-30 минут при комнатной температуре, и затем зародыши переносили в Среду для совместного культивирования, которая содержала 4,33 г/л солей MS; 1X ISU модифицированные витамины MS; 30 г/л сахарозы; 700 мг/л L-пролина; 100 мг/л миоинозита; 3,3 мг/л Dicamba-KOH; 100 мг/л ферментативного гидролизата казеина; 15 мг/л AgNO3; 100 мкΜ ацетосирингона и 3 г/л GELZAN™; при pH 5,8. Инкубация совместной культуры проводили в течение 3-4 дней при 25° при 24-часовом освещении лампами дневного света (приблизительно 50 мкЭм-2с-1).
Покой и селекция. После совместного культивирования, зародыши (36 зародышей/чашка) осторожно переносили в свежую неселективную Среду для покоя, содержащую 4,33 г/л солей MS; 1X ISU модифицированные витамины MS; 30 г/л сахарозы; 700 мг/л L-пролина; 3,3 мг/л Dicamba в KOH; 100 мг/л миоинозита; 100 мг/л ферментативного гидролизата казеина; 15 мг/л AgNO3; 0,5 г/л MES; 250 мг/л карбенициллина и 2,3 г/л GELZAN™; при pH 5,8. Инкубирование продолжали в течение 7 дней при 28° в условиях 24-часового освещения лампами дневного света (приблизительно 50 мкЭм-2с-1).
После 7-дневного периода покоя, зародыши переносили в Селективную среду. Для отбора тканей кукурузы, трансформированных супербинарной плазмидой, содержащей экспрессируемый в растении селективный маркерный ген aad1, зародыши (36/чашка) сначала переносили в Селективную среду I, которая содержала Среду для покоя (выше), содержащую 100 нМ R-галоксифоп кислоты (0,0362 мг/л). Зародыши инкубировали в течение 1 недели (28°; постоянное освещение), и затем переносили на Селективную среду II, которая содержала Среду для покоя с 500 нМ R-галоксифоп кислоты (0,1810 мг/л), на которой их инкубировали при постоянном освещении в течение еще 7 дней. В это время их переносили в свежую Селективную среду II и продолжали инкубирование, как указано выше, в течение дополнительной недели.
Специалисту в области трансформации кукурузы будет известно, что существуют другие доступные методы отбора трансформированных растений, когда используется другой экспрессируемый в растении селективные маркерные гены (например, гены устойчивости к гербицидам).
Пререгенерация После процесса отбора, культуры переносили в Среду для пререгенерации, содержащую 4,33 г/л солей MS; 1X ISU модифицированные витамины MS; 45 г/л сахарозы; 350 мг/л L-пролина; 100 мг/л миоинозита; 50 мг/л ферментативного гидролизата казеина; 1,0 мг/л AgNO3; 0,25 г/л MES; 0,5 мг/л нафтилуксусной кислоты в NaOH; 2,5 мг/л абсцизовой кислоты в этаноле; 1 мг/л6-бензиламинопурина; 250 мг/л карбенициллина; 2,5 г/л GELZAN™ и 500 нМ R-галоксифоп кислоты; при pH 5,8. Инкубирование продолжали в течение 7 дней при 28° в условиях постоянного освещения лампами дневного света, как указано выше.
Регенерация и выделение всходов Для регенерации, культуры переносили в Среду для регенерации I, содержащую 4,33 г/л солей MS; 1X ISU модифицированные витамины MS; 60 г/л сахарозы; 100 мг/л миоинозит; 125 мг/л карбенициллина; 2,5 г/л GELZAN™ и 500 нМ R-галоксифоп кислоты; при pH 5,8, и всходам позволяли сформироваться, и расти при 28° в условиях постоянного освещения лампами дневного света в течение 3 недель.
Когда всходы достигали подходящей стадии роста, их вырезали с помощью пинцета и скальпеля и переносили в Среду для регенерации II, содержащую 4,33 г/л солей MS; 1X ISU модифицированные витамины MS; 30 г/л сахарозы; 100 мг/л миоинозита; 3,0 г/л GELZAN™; при pH 5,8; и инкубировали при 28° в условиях постоянного освещения лампами дневного света, как указано выше, чтобы обеспечить дальнейший рост и развитие стебля и корней.
Получение семян Растения пересаживали в беспочвенный субстрат METRO-MIX 360 (SUN GRO HORTICULTURE; BELLEVUE, WA) и оставляли укрепляться в климакамере. Затем растения пересаживали в почвенную смесь SUNSHINE CUSTOM BLEND 160 и выращивали до цветения в теплице. Проводили контролируемое опыление для получения семян.
ПРИМЕР 3. Трансформация кукурузы штаммами Agrobacterium (Методика трансформации 2).
Основной ход работы приведен ниже. Зародыши выделяли из незрелых початков кукурузы на стадии развития, в которой молодые зародыши имели длину приблизительно 1,8-2,4 мм. При необходимости сравнения различных условий трансформации, приблизительно равные количества зародышей, выделенных из одного початка, распределяли по всем обработкам. Зародыши инкубировали с суспензией, содержащей клетки Agrobacterium и поверхностно-активное вещество (или нет, для сравнения), затем переносили на чашки с твердой средой и проводили совместное культивирование в течение 1-4 дней. Обработанные зародыши переносили на среду, содержащую антибиотик (для подавления и лизиса клеток Agrobacterium) и соединения для селективного выделения генетически трансформированных тканей и растений кукурузы. Ткань кукурузы (обычно каллус, но не ограничивается этим) выращивали на селективной среде до регенерации растений. Эти растения тестировали для подтверждения их генетической трансформации, и те растения, которые имели требуемую модификацию, выращивали до зрелого состояния с целью получения семян.
Получение незрелых зародышей Семена имбредной линии кукурузы B104 (сорт штата Айова, выпущенный на рынок в начале 1980-ых), выращивали в горшках объемом 4 галлона (~15 л), содержащих SUNSHINE CUSTOM BLEND 160 (SUN GRO HORTICULTURE; Bellevue, WA). Растения выращивали в теплице, используя для освещения комбинацию натриевых ламп высокого давления и металлогалогеновых ламп со световым периодом 16:8 ч света:темноты. С целью получения незрелых зародышей для трансформации, проводили контролируемое опыление сибсов. Незрелые зародыши выделяли через 10-13 дней после опыления, когда размер зародышей составлял приблизительно 1,8-2,4 мм. Початки кукурузы стерилизовали на поверхности после удаления листовой обвертки и пестиков при погружении в 50% коммерческий отбеливатель (CLOROX®, 6,15% гипохлорита натрия) с TWEEN®-20 (1 или 2 капли на 500 мл) в течение 10 минут и три раза промывали стерильной водой.
В альтернативе, поверхность початков кукурузы можно стерилизовать путем тщательного напыления недавно приготовленного 70% раствора этанола до полной пропитки початка. Перед использованием початок оставляют высыхать на воздухе в течение получаса в стерильном ламинаре, чтобы раствор этанола мог полностью испариться.
Незрелые зародыши в стерильных условиях отделяли непосредственно в микроцентрифужную пробирку, содержащую 2 мл Инокуляционную среду с суспендированными клетками агробактерии и поверхностно-активным веществом, при необходимости. Зародыши инкубировали с суспензией клеток агробактерии, содержащей поверхностно-активное вещество (или нет, для контрольных экспериментов), в течение 5-30 минут.
Суспензию клеток агробактерии, содержащих супербинарный вектор, получали, выращивая сначала клетки в 125 мл (в колбе с отбойниками на 500 мл) среды LB (SIGMA ALDRICH; St. Louis, MO) 20 г/л, содержащей 50 мг/л спектиномицина; 10 мг/л рифампицина и на 50 мг/л стрептомицина с встряхиванием (250 об/мин в темноте) при 26° в течение 6 часов. Эту культуру закладывали при разведении 1:5 25 мл ночной культуры (выращенной в той же среде) в свежей среде. Клетки осаждали центрифугированием в течение 15 минут при 3500 об/мин при 4°, затем однородно ресуспендировали (мягким пипетированием) в Инокуляционной среде (InM) до оптической плотности приблизительно 1,0 при 600 нм (OD600). Инокуляционная среда содержала: 2,2 г/л солей MS (Frame et al., 2011, Genetic Transformation Using Maize Immature Zygotic Embryos, In Plant Embryo Culture Methods and Protocols: Methods in Molecular Biology. T. A. Thorpe and E. C. Yeung, (Eds), Springer Science and Business Media, LLC. pp 327-341); 1X ISU модифицированные витамины MS (Frame et al., 2011, выше); 68,4 г/л сахарозы; 36 г/л глюкозы; 115 мг/л L-пролина; 100 мг/л миоинозита и 200 мкΜ ацетосирингона (приготовленного в ДМСО); при pH 5,4. В зависимости от эксперимента, соответствующее количество раствора поверхностно-активного вещества (например, С BREAK-THRU® S 233 в конечной концентрации 0,01%) добавляли в Инокуляционную среду после суспендирования клеток.
Раствор агробактерий и зародышей инкубировали в течение 5-15 минут при комнатной температуре, и затем зародышей переносили в Среду для совместного культивирования, которая содержала 4,33 г/л солей MS; 1X ISU модифицированные витамины MS; 30 г/л сахарозы; 700 мг/л L-пролина; 3,3 мг/л Dicamba в KOH (3,6-дихлор-o-анисовая кислота или 3,6-дихлор-2-метоксибензойная кислота); 100 мг/л миоинозита; 100 мг/л ферментативного гидролизата казеина; 15 мг/л AgNO3; 100 мкΜ ацетосирингона в ДМСО; и 3 г/л GELZAN™ (SIGMA- ALDRICH); при pH 5,8. Инкубирование совместной культуры проводили в течение 3-4 дней при 25° в условиях постоянного освещения лампами дневного света (приблизительно 50 мкЭм-2с-1).
Покой и селекция После совместного культивирования, зародыши (36 зародышей/чашка) осторожно переносили в неселективную Среду для покоя, содержащую 4,33 г/л солей MS; 1X ISU модифицированные витамины MS; 30 г/л сахарозы; 700 мг/л L-пролина; 3,3 мг/л Dicamba в KOH; 100 мг/л миоинозита; 100 мг/л ферментативного гидролизата казеина; 15 мг/л AgNO3; 0,5 г/л MES (2-(N-морфолино)этансульфоновой кислоты моногидрат (PHYTOTECHNOLOGIES LABR.; Lenexa, KS)); 250 мг/л карбенициллина и 2,3 г/л GELZAN™; при pH 5,8. Инкубирование продолжали в течение 7 дней при 28° в условиях постоянного освещения лампами дневного света, как указано выше.
После 7-дневного периода покоя, зародышей переносили в Селективную среду. Для отбора тканей кукурузы, трансформированных супербинарной плазмидой, содержащей экспрессируемый в растении селективный маркерный ген aad1, зародыши (18 зародышей/чашка) сначала переносили в Селективную среду I, которая состояла из Среды для покоя (выше) и содержала 100 нМ R-галоксифоп кислоты (0,0362 мг/л). Зародыши инкубировали в течение 1 недели, а затем переносили (12 зародышей/чашка) на Селективную среду II, которая состояла из Среды для покоя (выше) с 500 нМ R-галоксифоп кислоты (0,1810 мг/л), на которой их инкубировали в течение еще 2 недель. Трансформированные изоляты получали в течение приблизительно 4-6 недель при 28° в условиях 24-часового освещения лампами дневного света (приблизительно 50 мкЭм-2с-1). Выделенные изоляты переносили на свежую Пререгенерационную среду для инициирования регенерации и дальнейшего анализа.
Специалистам в области трансформации кукурузы будет известно, что существуют другие доступные способы отбора трансформированных растений, когда используются другие экспрессируемые в растениях селективные маркерные гены (например, гены устойчивости к гербицидам).
Пререгенерация. После процесса селекции, культуры, подвергнутые 24-часовому световому режиму, переносили (6-8 каллусов/чашка) в Среду для пререгенерации, содержащую 4,33 г/л солей MS; 1X ISU модифицированные витамины MS; 45 г/л сахарозы; 350 мг/л L-пролина; 100 мг/л миоинозита; 50 мг/л ферментативного гидролизата казеина; 1,0 мг/л AgNO3; 0,25 г/л MES; 0,5 мг/л нафтилуксусной кислоты в NaOH; 2,5 мг/л абсцизовой кислоты в этаноле; 1 мг/л 6-бензиламинопурина; 250 мг/л карбенициллина; 2,5 г/л GELZAN™ и 500 нМ R-галоксифоп кислоты; при pH 5,8. Инкубирование продолжали в течение 7-14 дней при 28° в условиях постоянного освещения лампами дневного света (приблизительно 50 мкЭм-2с-1).
Регенерация и выделение всходов Для регенерации, культуры переносили (до 12 каллусов на PHYTATRAY™ (PHYTOTECHNOLOGIES LABR.)) на первичную Среду для регенерации, содержащую 4,33 г/л солей MS; 1X ISU модифицированные витамины MS; 60 г/л сахарозы; 100 мг/л миоинозита; 125 мг/л карбенициллина; 3,5 г/л GELZAN GUM G434 (PHYTOTECHNOLOGIES LABR.); и 500 нМ R-галоксифоп кислоты; при pH 5,8, и всходам позволяли образоваться и расти в течение 3 недель.
Когда всходы достигли длины 3-5 см, их переносили (6 растений на PHYTATRAY™) на Среду для выращивания растений, содержащую 4,33 г/л солей MS; 1X ISU модифицированные витамины MS; 30 г/л сахарозы; 100 мг/л миоинозита; 3,5 г/л GELLAN GUM G434 и 0,5 мг/л индолуксусной кислоты в NaOH; при pH 5,8, и инкубировали при 25° в условиях 16-часового освещения лампами дневного света (приблизительно 50 мкЭм-2с-1) для обеспечения дальнейшего роста и развития стебля и корней.
Получение семян Растения пересаживали в беспочвенный субстрат METRO-MIX 360 (SUN GRO HORTICULTURE; BELLEVUE, WA) и оставляли укрепляться в климакамере. Затем растения пересаживали в почвенную смесь SUNSHINE CUSTOM BLEND 160 и выращивали до цветения в теплице. Проводили контролируемое опыление для получения семян.
ПРИМЕР 4. Эффективность трансформации при использовании клеток агробактерии, выращенных в жидкой среде.
Супербинарный штамм Agrobacterium, LBA4404 (pEPS1083), использовали для трансформации незрелых зародышей кукурузы способом, раскрытым в Примере 2 (Методика трансформации 1). Проводили сравнение эффективности трансформации, полученной при соскабливании клеток агробактерии с чашек с YEP агаром и ресуспендировании в Инфицирующей среде (IfM), и в экспериментах, проведенных в то же время при использовании клеток агробактерии, выращенных в жидкой среде LB, собранных центрифугированием и ресуспендированых в IfM. Сравнительную эффективность трансформации определяли на различных стадиях процесса путем подсчета количества желтых флуоресцентных бляшек (YFP+) на участках обработанной ткани через одну - пять недель после начала экспериментов трансформации. В Таблице 2 приведены полученные результаты.
Результаты, приведенные в Таблице 2, демонстрируют, что заражение зародышей кукурузы при использовании клеток агробактерии, только что собранных из жидкой культуры, обеспечивает значительно более высокую эффективность трансформации, чем при использовании клеток, соскабливаемых с агаровых чашек.
ПРИМЕР 5. Улучшение эффективности трансформации при включении поверхностно-активного вещества в Методику трансформации 1.
Супербинарный штамм Agrobacterium, LBA4404 (pDAB 108652), использовали для трансформации незрелых зародышей кукурузы способами, раскрытыми в Примере 2. Плазмида pDAB 108652 содержала область, кодирующую YFP, экспрессию которой направлял промотор ZmUbi1, а также несла область, кодирующую aad1 (устойчивость к гербициду) под экспрессионным контролем промотор актина1 риса. Проводили сравнение эффективности трансформации, полученной при суспендировании клеток агробактерии в IfM без поверхностно-активного вещества, и экспериментов, проводимых в это же время с IfM, содержащей в качестве добавки поверхностно-активное вещество BREAK-THRU® S 233 в различных концентрациях. Эффективность трансформации вычисляли путем подсчета каллусов с флуоресцентными секторами (каждый каллус развивается из одного зародыша) через 4 недели селекции с галоксифопом. В этот момент флуоресцентные сектора были большими, и поэтому ткани представляли собой стабильно трансформированные сектора. Результаты, приведенные в Таблице 3, демонстрируют, что использование поверхностно-активного вещества увеличивает эффективность трансформации, и что степень повышения эффективности зависит от концентрации используемого поверхностно-активного вещества.
Супербинарный штамм Agrobacterium, LBA4404 (pEPS1083), использовали для трансформации незрелых зародышей способом, раскрытым в Примере 2. Проводили сравнение эффективности трансформации, полученной при суспендировании клеток агробактерии в IfM без поверхностно-активного вещества, и в экспериментах, проводимых в это же время в присутствии поверхностно-активного вещества, добавленного в IfM. Сравнительную эффективность трансформации определяли на различных стадиях процесса путем подсчета количества желтых флуоресцентных бляшек (YFP+) на участках обработанной ткани через одну - пять недель после начала экспериментов трансформации. В Таблице 4 представлены полученные результаты.
В некоторых экспериментах клетки агробактерии перед этапом совместного культивирования промывали средой IfM (с или без поверхностно-активного вещества) путем суспендирования и мягкого центрифугирования ("промывка" в Таблице 4). Далее, в Эксперименте 5 (Таблица 4), 200 мкмМ ацетосирингона (а не 100 мкМ, как определено в Примере 2) использовали для индукции экспрессии vir генов, и клетки агробактерии выращивали на чашке со средой LB и соответствующими антибиотиками, а не со средой YEP.
танных зародышей
YFP+ зародышей
** IfM - Инфицирующая среда, используемая для суспендирования и/или промывки клеток агробактерии.
*** Клетки агробактерии выращивали на чашке со средой LB и антибиотиками, и экспрессию гена индуцировали 200 мкМ ацетосирингона.
Эксперименты, представленные в Таблице 4, четко показывают, что присутствие поверхностно-активного вещества BREAK-THRU® S 233 в Инфицирующей среде, используемой для ресуспендирования клеток агробактерии, соскобленных с чашек с твердой средой, резко увеличивает эффективность трансформации незрелых зародышей. Кроме того, поверхностно-активное вещество TACTIC™ оказывает положительное, но менее сильное влияние на повышение эффективности трансформации.
В дальнейшей иллюстрации способов настоящего описания незрелые зародыши кукурузы трансформировали клетками агробактерии штамма LBA4404 (pEPS1083) согласно способам Примера 2. Эффективность трансформации контролировали по появлению YFP+ бляшек или секторов при развитии каллусов из незрелых зародышей. На левой стороне Фиг.1 показаны пять экспериментов (Эксперименты 1-5), в которых использовали клетки агробактерии, соскобленные с чашек с твердым агаром, а на правой стороне Фигуры 1 показаны результаты трех экспериментов (Эксперименты 6-9), в которых клетки агробактерии собирали из культур, выращенных в жидкой среде. В комбинированных Экспериментах 1-5 эффективность трансформации повышалась в зародышах из всех девяти собранных початков (100%), причем увеличение эффективности трансформации было статистически значимым (точный критерий Фишера, p<-0,05) в зародышах шести из девяти початков (67%). В комбинированных Экспериментах 6-9 (агробактерия, выращенная в жидкой среде), зародыши из всех восьми собранных початков (100%) показали статистически значимое увеличение эффективности трансформации. Таким образом, из результатов, представленных на Фигуре 1, четко видно, что добавление BREAK-THRU® S233 в Инфицирующую среду резко увеличивает эффективность трансформации незрелых зародышей кукурузы, в некоторых случаях с достижением эффективности трансформации более 90%.
В другом примере способов настоящего описания незрелые зародыши кукурузы трансформировали клетками агробактерии штамма LBA4404, несущего различные плазмиды (все плазмиды содержали селективный маркерный ген aad1), согласно способам Примера 2. Как и прежде, в экспериментальных обработках сравнивали эффективность трансформации с применением 0,01% поверхностно-активного вещества BREAK-THRU® S 233, или без него. После регенерации зародыши подвергали селекции с галоксифопом для получения растений. Таким образом, данные собирали на существенно более поздней стадии, нежели данные, представленные на Фиг. 1. Процент эффективности трансформации вычисляли путем деления количества зародышей, которые давали трансгенное растение ("событие"), на количество обработанных незрелых зародышей и умножения на 100. С этой целью зародыша подсчитывали как одно событие, даже если он давал несколько трансгенных растений. Результаты трех экспериментов, в которых использовали клетки агробактерии, соскобленные с чашек с агаром, показаны на Фиг. 2 (Эксперименты 1, 2 и 3). Кроме того, на Фиг. 2 представлены результаты эксперимента (Эксперимент 4), в котором клетки агробактерии выращивали в жидкой среде, собирали центрифугированием и ресуспендировали в IM (с или без BREAK- THRU® S 233). Парные столбики на Фиг. 2 указывают ответы зародышей из отдельных початков.
Из данных, представленные на Фиг. 2, ясно, что добавление поверхностно-активного вещества BREAK-THRU® S 233 приводит к значительному увеличению эффективности агробактериальной трансформации незрелых зародышей кукурузы, независимо от предыдущей конфигурации роста клеток агробактерии и независимо от генного состава трансформирующей плазмиды. В комбинированных Экспериментах 1, 2 и 3 эффективность трансформации была увеличена в зародышах из 23 из 26 собранных початков (88%), причем увеличение эффективности трансформации было статистически значимым (точный критерий Фишера p<-0,05) в зародышах из 12 из 26 початков (46%). В Эксперименте 4 (агробактерия выращена в жидкой культуре) зародыши из 10 из 12 собранных початков (83%) показали увеличение эффективности трансформации, причем увеличение было статистически значимым в одном из 12 початков (8%).
ПРИМЕР 6. Сравнение усиливающего эффективность трансформации действия поверхностно-активных веществ различных химических классов.
В Таблице 1 приведен неограничивающий список поверхностно-активных веществ нескольких химических классов. Эксперименты трансформации незрелых зародышей проводили при использовании Методики трансформации 2, представленной в Примере 3. Клетки агробактерии, несущие различные плазмиды, суспендировали в Инокуляционной среде (InM), содержащей BREAK-THRU® S 233 или другие различные поверхностно-активные вещества (все в концентрации 0,01%), и сравнивали процент трансформации (измеренный с помощью анализа Taqman® гена aad1) через 7-10 недель после начала эксперимента.
Процент эффективности трансформации вычисляли путем деления количества зародышей, которые давали трансгенное растение ("событие"), на число обработанных незрелых зародышей и умножения на 100. С этой целью, зародышей подсчитывали как одно событие, даже если он давал несколько трансгенных растений. В Таблице 5 приведены полученные эффективности трансформации.
танных зародышей
Результаты, приведенные в Таблице 5, демонстрируют, что применение BREAK-THRU® S 233, при его включении в Инокуляционную среду, используемую для ресуспендирования инокулята клеток агробактерии, выращенных и собранных из жидкой среды, обеспечивало более высокую эффективность трансформации, чем полученную с большинством других протестированных поверхностно-активных веществ. В трех экспериментах (Эксперимент 2, Эксперимент 9 и Эксперимент 11) наблюдаемая эффективность трансформации была почти одинаковой для двух поверхностно-активных веществ.
ПРИМЕР 7. Результаты трансформации, полученные различными операторами.
Специалисту в области трансформации кукурузы будет известно, что методики трансформации растений часто требуют значительного объема экспертизы, на которую уходят месяцы или годы экспериментирования. Эффективность трансформации может изменяться в широких диапазонах вследствие несогласованности методов, в которых процедуры осуществляются различными операторами. Таким образом, выгодно предоставить такие методики трансформации кукурузы, которые повысят предсказуемость эффективности трансформации, проводимой различными операторами в разное время. Трансформацию незрелых зародышей кукурузы проводили в течение нескольких месяцев с применением способов Примера 3 (Agrobacterium, штамм LBA4404, несущий различные плазмиды) и с присутствием BREAK-THRU® S233 в Инокуляционной среде. Эффективность трансформации оценивали по количеству полученных каллусных тканей, устойчивых к галоксифопу. В Таблице 6 приведены полученные результаты.
зованных початков
дышей
**Средняя эффективность трансформации для всех операторов.
Результаты, приведенные в Таблице 6, показывают, что методика трансформации, раскрытая в Примере 3, при осуществлении с включением BREAK-THRU® S233 в Инокуляционную среду, обеспечивает надежный и предсказуемый метод, который позволяет снизить расхождение между операторами, влияющее на эффективность трансформации. Более того, повышение предсказуемости способов позволяет более точно определять размер эксперимента (например, количество зародышей, которые требуется обработать) с целью получения необходимого результата (например, количества полученных трансформированных объектов).
Объем настоящего изобретения не ограничивается вариантами осуществлениями, раскрытыми в настоящей заявке, которые предназначены как иллюстрация нескольких аспектов изобретения и любых вариантов осуществления, которые являются функционально эквивалентными в рамках настоящего изобретения. Различные модификации способов, в дополнение к показанным и описанным в настоящей заявке, будут очевидны специалистам в данной области техники и должны быть включены в объем прилагаемой формулы изобретения. Кроме того, хотя только некоторые репрезентативные комбинации этапов способов, раскрытых в настоящем описании, конкретно обсуждаются в вариантах осуществления выше, другие комбинации этапов способов будут очевидны специалистам в данной области техники и также должны быть включены в объем прилагаемой формулы изобретения. Таким образом, комбинация этапов может быть прямо указана в настоящей заявке, и, тем не менее, другие комбинации этапов также включены, хотя они и не представлены прямо. В том случае, если значение не указано прямо, следует понимать, что значения, которые имеют приблизительно ту же величину, что и указанное значение, также включены в объем изобретения. В том случае, если указан диапазон значений, каждое промежуточное целочисленное значение, и каждая его дробная часть, между указанными верхним и нижним пределами данного диапазона, также прямо раскрыта, наряду с каждым поддиапазоном между такими значениями. Термин "включающий" и его вариации, при использовании в настоящем описании, используются синонимично с термином "содержащий" и его вариациями и представляют собой открытые, неограничивающие термины. При использовании в настоящем описании, термины "модифицирует" или "изменяет", или любые их формы, означают модифицировать, изменить, заменить, удалить, заместить, варьировать или трансформировать.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ КУКУРУЗЫ | 2007 |
|
RU2351120C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ ОДНОДОЛЬНЫХ РАСТЕНИЙ | 2007 |
|
RU2351121C1 |
| ШТАММЫ AGROBACTERIUM, МОДИФИЦИРОВАННЫЕ ДЛЯ УВЕЛИЧЕНИЯ ЧАСТОТЫ ТРАНСФОРМАЦИИ РАСТЕНИЙ | 2011 |
|
RU2611188C2 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСФОРМИРОВАННЫХ РАСТЕНИЙ ЗЕРНОВЫХ КУЛЬТУР | 2004 |
|
RU2279211C2 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННОГО РАСТЕНИЯ КУКУРУЗЫ | 1991 |
|
RU2114911C1 |
| ЭНХАНСЕР ПАЛОЧКОВИДНОГО ВИРУСА САХАРНОГО ТРОСТНИКА (SCBV) И ЕГО ПРИМЕНЕНИЕ В ФУНКЦИОНАЛЬНОЙ ГЕНОМИКЕ РАСТЕНИЙ | 2013 |
|
RU2639517C2 |
| Белок устойчивости к гербицидам, кодирующий его ген и их применение | 2016 |
|
RU2692553C2 |
| Белок устойчивости к гербицидам, кодирующие его гены и их применение | 2016 |
|
RU2681162C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ СОРГО | 2002 |
|
RU2229793C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ МОРКОВИ, ПРОДУЦИРУЮЩИХ ИНТЕРЛЕЙКИН-10 ЧЕЛОВЕКА | 2007 |
|
RU2374321C2 |
Изобретение относится к области биохимии, в частности к способу трансформации растительной клетки, где способ включает контакт растительных клеток незрелых зародышей кукурузы с клетками Agrobacterium в жидкой среде, содержащей неионное трисилоксановое поверхностно-активное вещество. Способ включает контакт растительных клеток с клетками агробактерии в жидкой среде, содержащей неионное трисилоксановое поверхностно-активное вещество, где поверхностно-активное вещество имеет концентрацию от примерно 0,005 процента по весу до примерно 0,04 процента по весу в жидкой среде, совместное культивирование растительных клеток и Agrobacterium в течение 1-4 дней на свету, и где растительные клетки получены из незрелых зародышей кукурузы и имеют длину, большую или равную 1,5 мм и меньшую или равную 2,5 мм. Изобретение позволяет улучшить результаты трансформации незрелых зародышей кукурузы. 2 н. и 4 з.п. ф-лы, 2 ил., 6 табл., 7 пр.
1. Способ трансформации растительной клетки, где способ включает контакт растительных клеток незрелых зародышей кукурузы с клетками Agrobacterium в жидкой среде, содержащей неионное трисилоксановое поверхностно-активное вещество, где поверхностно-активное вещество имеет концентрацию от примерно 0,005 процента по весу до примерно 0,04 процента по весу в жидкой среде; и
совместное культивирование растительных клеток и Agrobacterium в течение 1-4 дней на свету,
где растительные клетки получены из незрелых зародышей кукурузы и имеют длину, большую или равную 1,5 мм и меньшую или равную 2,5 мм.
2. Способ трансформации растительной клетки по п. 1, дополнительно включающий дополнительное поверхностно-активное вещество.
3. Способ трансформации растительной клетки, включающий:
приготовление жидкой среды, содержащей неионное трисилоксановое поверхностно-активное вещество, имеющее концентрацию от примерно 0,005 процента по весу до 0,04 процента по весу в жидкой среде;
суспендирование клеток Agrobacterium в жидкой среде, содержащей неионное трисилоксановое поверхностно-активное вещество, имеющее концентрацию от примерно 0,005 процента по весу до 0,04 процента по весу; и
контакт растительных клеток незрелых зародышей кукурузы с клетками Agrobacterium в жидкой среде, содержащей поверхностно-активное вещество и
совместное культивирование растительных клеток и Agrobacterium в течение 1-4 дней на свету,
где растительные клетки получены из незрелых зародышей кукурузы и имеют длину, большую или равную 1,5 мм и меньшую или равную 2,5 мм.
4. Способ трансформации растительной клетки по п. 3, где клетки Agrobacterium соскабливают с твердой среды перед суспендированием в жидкой среде, содержащей поверхностно-активное вещество.
5. Способ трансформации растительной клетки по п. 3, где клетки Agrobacterium выращивают в жидкой питательной среде перед суспендированием в жидкой среде, содержащей поверхностно-активное вещество.
6. Способ трансформации растительной клетки по п. 3, дополнительно включающий дополнительное поверхностно-активное вещество.
| YANG et al., Improvement of Agrobacterium-mediated transformation of embryogenic calluses from maize elite inbred lines, In Vitro Cellular & Developmental Biology-Plant, 2006 | |||
| US2004148656 A1, 29.07.2004 | |||
| CN101451149 A, 10.06.2009 | |||
| CN102102109 A, 22.06.2011 | |||
| ВИКТОРЭК-СМАГУР и др., Сравнение двух методов трансформации Arabidopsis thaliana: погружение цветочных почек и вакуумная инфильтрация, Физиология растений, 2009. |

