Изобретение относится к биотехнологии, в частности к генетической инженерии, и касается получения рекомбинантного штамма Escherichia coli - продуцента пенициллин G ацилазы путем введения в штамм-реципиент модифицированного гена рас в составе рекомбинантной плазмидной ДНК и разработки способа микробиологического синтеза продуцента пенициллин G ацилазы.
Фермент пенициллин G ацилазу (PGA) широко используют в промышленности в качестве биокатализаторов для гидролиза пенициллина G до 6-аминопенициллановой кислоты (6-АПК), которая является ключевым предшественником для синтеза β-лактамных антибиотиков [1-4], которые занимают доминирующее место на рынке системных антибиотиков [5, 6]. PGA также широко используют для ферментативного синтеза широко применяемых цефалоспориновых антибиотиков [7-9]. Несомненный практический интерес представляет усовершенствование известных способов получения ферментов, необходимых для синтеза этих антибиотиков, в частности с помощью технологий рекомбинантных ДНК.
Пенициллин G ацилазы представляют собой гетеродимеры, состоящие из 2-х альфа и 2-бета- субъединиц [9], которые формируются из одноцепочечного полипептидного предшественника.
Известны первичные структуры генов, кодирующих предшественники ферментов с активностью пенициллин G ацилазы из различных бактерий, в частности ген рас, кодирующий предшественник пенициллин G ацилазы из бактерии E. coli (ECOPGA) [7].
Общими недостатками известных методов гетерологичной экспрессии PGA являются невысокий уровень синтеза рекомбинантного продукта [10-12], сложности процессов масштабирования ферментации штаммов [13], накопление телец включения и непроцессированного предшественника, что в свою очередь приводит к замедлению роста штаммов, их лизису, высвобождению синтезированной PGA в среду культивирования [14-18].
Устранение указанных недостатков может быть достигнуто на пути разработки новых систем экспрессии пенициллин G-ацилазы, обеспечивающих повышение выхода функционально-активного фермента [19], в том числе за счет модификации сигнального пептида PGA, играющего важную роль в транслокации функционально-активного фермента из цитоплазмы в периплазму с участием Sec-системы E. coli [15, 20-23].
В качестве ближайшего аналога заявляемого штамма рассмотрим штамм-продуцент ECOPGA E. coli JM109(DE3) (pETKn29a, pARDegP), а в качестве ближайшего аналога заявляемого способа - способ получения ECOPGA с помощью указанного продуцента, обеспечивающий синтез целевого фермента с активностью 0,94 ME на мл культуры [10]. Удельная активность ECOPGA составляет 0,45 МЕ/мл на 1 ОД (оптических единиц). Эти штамм и способ его культивирования не позволяют достичь высокой плотности культуры.
Задача заявляемой группы изобретений - расширение арсенала способов микробиологического синтеза ECOPGA.
Задача решена путем:
- конструирования модифицированного гена рас бактерии Escherichia coli, кодирующего предшественник фермента с активностью пенициллин G ацилазы [(ans) ECOPGA] и соответствующего последовательности SEQ ID N17, часть которого, кодирующая природную сигнальную последовательность ECOPGA, заменена на фрагмент, кодирующий сигнальную последовательность секреции фермента L-аспарагиназы Erwinia carotovora;
- конструирования рекомбинантного штамма E. coli BL21(DE3)/pMD704 - продуцента ECOPGA, полученного путем введения заявляемого модифицированного гена рас в составе вектора pSVH0107 в штамм-реципиент Escherichia coli BL21 (DE3;
- разработки способа микробиологического синтеза ECOPGA путем культивирования заявляемого штамма-продуцента в подходящих условиях.
Для этого:
а) из штамма E. coli ВКПМ В-10182 выделен ген рас, кодирующий ECOPGA;
б) получен модифицированный ген рас, в котором последовательность, кодирующая природную сигнальную последовательность секреции ECOPGA, заменена на последовательность, кодирующую сигнальную последовательность секреции L-аспарагиназы Erwinia carotovora;
в) сконструирована рекомбинантная плазмида pMD704, полученная путем введения модифицированного гена рас в вектор экспрессии pSVH0107 [24];
г) в результате трансформации плазмидой pMD704 клеток реципиентного штамма Е. соli BL21(DE3) получен рекомбинантный штамм E. coli BL21(DE3)/pMD704, который при культивировании заявляемым способом позволяет получать целевой фермент с выходом более чем в 5 раз превышающим продукцию фермента, полученного с использованием способа - ближайшего аналога.
Выбор сигнальной последовательности секреции L-аспарагиназы Erwinia carotovora (SIGANS) [25] обусловлен тем, что эта последовательность является типичным secB-зависимым сигнальным пептидом для системы секреции 2 типа, и, в отличие от природной сигнальной последовательности ECOPGA, не содержит аминокислотных остатков с объемными боковыми группами, что облегчает процесс транспорта синтезированного (экспрессированного) белка в периплазму штаммов-продуцентов [7].
Полная природная кодирующая последовательность гена рас (SEQ ID №1) получена методом полимеразной цепной реакции (ПЦР) с использованием в качестве матрицы хромосомной ДНК, выделенной из штамма E. coli ВКПМ В-10182, а в качестве праймеров - синтетических олигонуклеотидов с нуклеотидными последовательностями SEQ ID №3 (праймер PGA_F) и SEQ ID №4 (праймер PGA_R). Полученная последовательность немодифицированного гена рас введена в вектор pSVH0107 [24]. В результате получена рекомбинантная плазмида pMD0107 (Пример 2).
Нуклеотидную последовательность, кодирующую SIGANS (SEQ ID №5), получили сборкой из двух синтетических олигонуклеотидов ansU (SEQ ID №7) и ansD (SEQ ID №8).
Положение участка, кодирующего SIGANS в составе модифицированного гена рас, определено на основании данных о структуре предшественника и зрелой ECOPGA (номера доступа номера доступа в PDB 1Е3А и 1GM7, соответственно), предшественника и зрелой ECAR-LANS (номера доступа в Genbank №ААР92666.3 и PDB 2GVN соответственно).
Объединение последовательностей, кодирующих SIGANS (SEQ ID №5) и зрелую ECOPGA проведено в составе вектора pMD0107, обеспечивающего экспрессию не модифицированного гена рас.
Для создания вектора экспрессии модифицированного гена рас, направляющего в клетках Е. coli синтез белка (ans)ECOPGA на основе плазмиды pMD0107, сконструирован промежуточный вектор pMD0107-1, полученный путем удаления сайта SphI в положении 123 SEQ ID №1 и введения дополнительного сайта рестрикции SacI в положении 84 SEQ ID №1, который использован для последующего встраивания последовательности S1GANS между сайтами NdeI и SacI гена рас. Полученная при этом экспрессирующая рекомбинантная плазмида обозначена как pMD704 (Фиг. 2).
Путем трансформации клеток реципиентного штамма E. coli BL21(DE3) [26] сконструированной плазмидой pMD704, отбора и культивирования клонов трансформантов с высоким уровнем синтеза функционально-активной ECOPGA получен рекомбинантный штамм E. coli BL21(DE3)/pMD704 - продуцент ECOPGA.
Для осуществления микробиологического синтеза ECOPGA заявляемый рекомбинантный штамм культивируют на среде с добавлением лактозы, которая служит как источником углерода, так и индуктором синтеза ECOPGA.
Таким образом, сконструированы:
A) модифицированный ген рас, который кодирует предшественник ECOPGA, состоящий из аминокислотной последовательности сигнального пептида L-аспарагиназы Erwinia carotovora и структурной части ECOPGA штамма E. coli ВКПМ В 10182 и характеризуется нуклеотидной последовательностью SEQ ID №17.
B) рекомбинантная плазмида pMD704, обеспечивающая синтез (ans)ECOPGA в клетках E. coli , которая образована вектором pSVH0107, содержащим промотор и терминатор РНК-полимеразы фага Т7, разделенные участком полилинкера, ген устойчивости к канамицину и ген lас-репрессора, и модифицированным геном рас, встроенным в полилинкерную область указанного вектора.
C) рекомбинантный штамм Е. coli BL21 (DE3)/pMD704 - продуцент ECOPGA.
Заявляемый рекомбинантный штамм Е.соli BL21(DE3)/pMD704 характеризуется следующими признаками.
Морфологические признаки: Клетки имеют продолговатую палочковидную форму, при делении не почкуются.
Культуральные признаки:
Клетки хорошо растут на обычно используемых питательных средах. Время генерации около 30 мин в жидкой LB-среде. На 1,5-2% питательном агаре "Difco" образуются круглые, гладкие, желтоватые колонии с ровными краями. При выращивании на жидких LB- и YT-средах образуется интенсивная ровная мутность.
Физиолого-биохимические признаки:
Оптимальная температура культивирования - 20°С, оптимум рН - 7,6. Источником азота служат органические соединения (в виде триптона, дрожжевого экстракта). Уровень активности ECOPGA при культивировании сконструированного штамма заявляемым способом составляет не менее 5 МЕ/мл при выходе влажной биомассы порядка 18 г/л.
Способ микробиологического синтеза в общем виде
Посевной материал, представляющий собой клетки заявляемого штамма Е.соli BL21(DE3)/pMD704, подготавливают путем инкубации в течение 20-24 часов при температуре 20-22°С на среде LB, содержащей дополнительно 17 мг/л канамицина. Выросшую культуру переносят в соотношении 1:400 (по объему) в среду ZY5052 [27] содержащую мас. %: энзиматический гидролизат казеина - 1, дрожжевой экстракт - 0,5, (NH4)2SO4 -0,33, КН2РO4 - 0,68, Na2HPO4 - 0,71, глицерин - 0,5, глюкоза - 0,05, лактоза-0,2, MgSO4 - 0,012, вода - остальное. Дополнительно в среду добавляют 50 мг/л канамицина. Процесс культивирования ведут при температуре 20-22°С с аэрацией на качалке (250-300 об/мин). Синтез целевого белка происходит под действием содержащейся в составе среды лактозы по мере исчерпания в процессе роста культуры, входящей в состав среды глюкозы. Биомассу отделяют центрифугированием, промывают фосфатным буфером и повторно осаждают.
Уровень синтетазной активности в биомассе клеток составляет не менее чем 1500 МЕ/г влажн., 6000 МЕ/г сух.
Заявляемая группа изобретений проиллюстрирована следующими фигурами.
Фиг. 1 - Сравнение частичных нуклеотидных последовательностей гена PGA штамма ВКПМ В-10182 и ПЦР-фрагмента, выделенного с помощью праймеров PGA_F и PGA_R.
Фиг. 2 - физическая и генетическая карты плазмиды pMD0107. Обозначены положения гена PGA (заполнение черным), сигнального пептида L-аспарагиназы (SIGANS), индикаторных сайтов рестрикции, участков промотора и терминатора РНК-полимеразы фага Т7 (Т7 prom и Т7 term), область начала репликации pBR322 (pBR322 origin), область начала репликации f1 (f1 origin)ген устойчивости к канамицину (обозначен KanR, заполнение черным), гена lас-репрессора.
Фиг. 3 - физическая и генетическая карты плазмиды pMD704. Все обозначения - как на Фиг. 2.
Фиг. 4 - Выравнивание N-концевых аминокислотных последовательностей предшественника PGA дикого типа (ECOPGA) и варианта предшественника ECOPGA с сигнальным пептидом L-аспарагиназы (ans)ECOPGA.
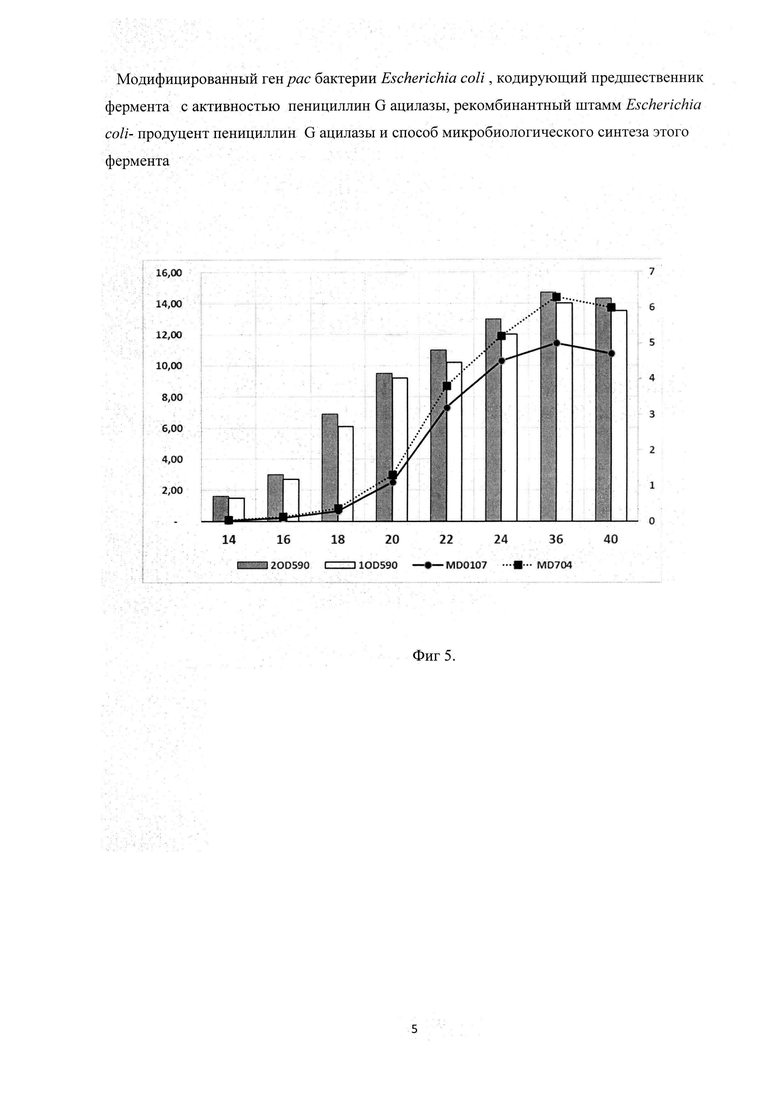
Фиг. 5 - Динамика накопления активности ECOPGA (столбцы, основная ось Y) и прироста биомассы (график, дополнительная ось Y) штаммами BL21(DE3)/pMD0107 и BL21(DE3)/pMD704.
При осуществлении изобретения, помимо методов, подробно раскрытых в нижеследующих примерах, используют хорошо известные специалистам методики, описанные в руководствах по молекулярной биологии и генетической инженерии [28].
Пример 1. Выделение немодифицированного гена рас
Выделение гена рас, кодирующего предшественник ECOPGA размером 846 а.к.о., проводят на основании известных данных о его кодирующей последовательности, идентифицированной в ходе расшифровки и аннотации геномной последовательности штамма E. coli ВКПМ В- 10182 [29].
В качестве матрицы используют хромосомную ДНК штамма E. coli ВКПМ В-10182, а в качестве праймеров - олигонуклеотиды PGA_F (SEQ ID №3) и PGA_R (SEQ ID №4) специфичные к N- и С-концевым кодирующим последовательностям гена рас.
1 мкг геномной ДНК штамма E. coli ВКПМ В-10182 денатуририруют путем нагревания при 100° в течение 5 минут, помещают в ледяную баню, и подвергают 30 циклам ПЦР с использованием набора Expand Long Template PCR system («Roche Diagnostics GmbH”, Mannheim, Германия) в соответствии с инструкцией изготовителя.
Смесь для ПЦР (50 мкл):
5 мкл 10-кратного ПЦР-буфера («Roche Diagnostics GmbH");
5 мкл геномной ДНК (200 нг/мкл);
5 мкл 3 мкМ праймера PGA_F
5 мкл 3 мкМ праймера PGA_R
5 мкл 2 мМ dNTP каждого вида;
25 мкл деионизованной воды; 1 мкл ДНК полимеразы («Roche Diagnostics GmbH").
Условия проведения ПЦР: 94°, 5' (денатурация), 94°, 30''; 50°, 30''; 72°,2' (амплификация).
После амплификации 5 мкл ПЦР смеси анализируют электрофорезом в 1% агарозном геле и выявляют гомогенный фрагмент размером около 2,2 тпн. Фрагмент выделяют из геля с помощью набора Wizard PCR Preps Kit (Promega, США) в соответствии с инструкцией производителя и подвергают автоматическому секвенированию на приборе ABIPrizm 3100 DNA Sequencer с использованием праймеров PGA_F (SEQ ID №3) и PGA_R (SEQ ID №4).
Результат сравнения полученной нуклеотидной последовательности с последовательностью геномной копии гена рас штамма E. coli ВКПМ В-10182 с помощью программы ClustalX демонстрирует их идентичность (Фиг1).
Пример 2. Конструирование плазмиды рМР0107 для экспрессии немодифицированного гена рас
Для создания плазмиды, способной направлять синтез ECOPGA в клетках E. coli , проводят встраивание выделенного гена рас под контроль промотора фага Т7 вектора PSVH0107 [30].
Полученный методом ПЦР фрагмент геномной ДНК штамма E. coli ВКПМ Β-10182 содержит полную кодирующую последовательность ECOPGA и фланкирован для его последующего направленного встраивания под контроль промотора Т7 экспрессионных векторов уникальными сайтами рестрикции NdeI и XhoI, отсутствующими в кодирующей последовательности гена рас.
Выделенный из агарозного геля фрагмент гидролизуют рестриктазами NdeI/XhoI и клонируют в вектор pSVH0107. Затем отбирают канамицин-устойчивые трансформанты штамма E. coli JM109 путем «ПЦР скрининга» с помощью праймеров PGA3 (SEQ ID №9) и PGA4 (SEQ ID №10), специфичных к центральной части кодирующей последовательности гена PGA по критерию образования фрагментов размером 0,5 тпн в полученных образцах.
С целью выявления клона, несущего вставку с интактным геном рас без возможных мутаций, привнесенных за счет ПЦР-амплификации, из полученных «положительных» клонов (канамицин-устойчивых трансформантов) выделяют плазмидную ДНК и секвенируют с использованием праймеров PGA_F, PGA_R, PGA3, PGA4 и идентифицируют клон, несущий вставку с интактным геном рас, кодирующая последовательность которого идентична последовательности фрагмента ДНК SEQ ID №1. Плазмиду, выделенную из отобранного клона, обозначают как pMVD0107. Схема плазмиды приведена на Фиг 2.
Пример 3. Конструирование плазмиды рМD0107-1
А) Конструирование промежуточной плазмиды pMD007-a.
Плазмида pMD0107 содержит два сайта рестриктазы SphI в положениях 4929 и 5351 на физической карте плазмиды (Фиг. 2). Присутствие сайта SphI в положении 5351 в кодирующей последовательности гена рас затрудняет получение различных вариантов ECOPGA методами сайт-направленного мутагенеза.
Для удаления этого сайта проводят конструирование промежуточной плазмиды pMD007-a, содержащей в положении 123 последовательности SEQ ID №1 гена рас замену С>Т, не приводящую к изменению аминокислотной последовательности ECOPGA.
Удаление сайта SphI проводят с использованием метода «рекомбинантной» ПЦР [31]. Для этого на матрице плазмиды pMD007 получают сначала ПЦР фрагменты №1 и №2.
Фрагмент №1, размером 440 пн, получают с использованием прайм еров PromF (SEQ ID №11)/SphD (SEQ ID №12). Праймер PromF расположен в положении 4940 плазмиды pMD007 рядом с сайтом SphI (4929). Праймер SphD расположен в положении 5343-5383 последовательности SEQ ID №1 (по комплементарной цепи) и содержит замену C>Т в положении 5362, приводящую к удалению сайта SphI, без изменения аминокислотной последовательности ECOPGA.
Фрагмент №2, размером 445 пн, получают с использованием праймеров NcoR (SEQ ID №13) и SphU (SEQ ID №14). Праймер NcoR расположен в положении 5717-5747 плазмиды pMD007 (по комплементарной цепи) и перекрывает сайт NcoI 5738. Праймер SphU комплементарен праймеру SphD.
Фрагменты №1 и №2 выделяют из агарозного геля как описано в примере 1 и используют в качестве матрицы в ПЦР реакции, которую проводят, как описано ниже:
Смесь для ПЦР (50 мкл):
5 мкл 10-кратного ПЦР-буфера («RocheDiagnosticsGmbH");
1 мкл Фрагмента 1 (20 нг/мкл);
1 мкл Фрагмента 2 (20 нг/мкл)
4 мкл 5 мкМ праймера PromF
4 мкл 5 мкМ праймера NcoR;
5 мкл 2 мМ dNTP каждого вида;
30 мкл деионизованной воды;
1 мкл ДНК полимеразы («RocheDiagnosticsGmbH”).
Условия проведения ПЦР (30 циклов): 94°, 5' (денатурация), 94°, 30''; 55°, 30''; 72°,1' (амплификация).
После амплификации 5 мкл ПЦР смеси анализируют электрофорезом в 1% агарозном геле и выявляют гомогенный фрагмент размером около 0,9 тпн. Фрагмент выделяют из геля, гидролизуют рестриктазами SphI/NcoI и клонируют в SphI/NcoI вектор pMD007. Из полученных канамицин-устойчивых трансформантов выделяют плазмидную ДНК и анализируют путем совместного гидролиза рестриктазами SphI/NcoI. Отбирают клоны, образующие SphI/NcoI фрагменты размером 7000 и 800 пн в отличие от фрагментов размером 7000 пн, 420 пн и 370 пн, характерных для плазмиды pMD007. Полученные положительные клоны секвенируют и отбирают плазмиду с нужной мутацией, не содержащую других неспецифических мутаций, обусловленных ошибками ПЦР. Один из отобранных клонов обозначают pMD007-a и используют в дальнейшей работе.
Б) Конструирование плазмиды pMD007-l.
Для удобства получения вариантов ECOPGA с заменами и делециями в области сигнальной последовательности секреции в положение 83-88 кодирующей последовательности гена рас (SEQ ID №1) вводят уникальный сайт рестриктазы SacI без изменения аминокислотной последовательности ECOPGA.
Введение сайта проводят с использованием метода «рекомбинантной» ПЦР. Для этого на матрице плазмиды pMD007-1 получают сначала ПЦР фрагменты №3 и №4.
Фрагмент №3 размером 400 пн получают с использованием праймеров PromF (SEQ ID №11)/SacD (SEQ ID №15). Праймер SacD расположен в положении 63-108 последовательности SEQ ID №1 и содержит замену TCG>AGC в положении 85-87 приводящую к введению сайта SacI без изменения аминокислотной последовательности ECOPGA.
Фрагмент №4 размером 500 пн получают с использованием праймеров NcoR (SEQ ID №13) и SacU (SEQ ID №16). Праймер SacU комплементарен праймеру SacD.
Фрагменты №3 и №4 выделяют из агарозного геля как описано в примере 1 и используют в качестве матрицы в ПЦР реакции с праймерами PromF и NcoR. Полученный ПЦР фрагмент размером 900 пн выделяют из геля, гидролизуют рестриктазами SphI/NcoI и клонируют в SphI/NcoI вектор pMD007-a. Из полученных канамицин-устойчивых трансформантов выделяют плазмидную ДНК и анализируют путем совместного гидролиза рестриктазами SacI/NcoI. Отбирают клоны, образующие Sac/NcoI фрагменты размером 7374 и 412 пн в отличие от фрагмента размером 7777 пн, характерного для плазмиды pMD007a. Полученные положительные клоны секвенируют и отбирают плазмиду с нужной мутацией, несодержащую других неспецифических мутаций, обусловленных ошибками ПЦР. Один из отобранных клонов обозначают pMD007-1 и используют в дальнейшей работе.
Пример 3. Конструирование плазмиды рМР704 для экспрессии модифицированного гена рас
Для получения вектора, содержащего ген, кодирующий (ans)ECOPGA под контролем промотора и терминатора гена 10 фага Т7, проводят замену собственной сигнальной последовательности ECOPGA в составе гена рас плазмиды pMD007-1 на SIGANS.
В качестве источника SIGANS используют два синтетических олигонуклеотида ansU (SEQ ID №7) и ansD (SEQ ID №8). По 10 мкл 10 мкМ раствора каждого олигонуклеотида в буфере TEN (0,1 MNaCl, 10mMTris, 1 mMEDTA, pH 8.0) смешивают, денатурируют 5 мин при 95°С, плавно охлаждают до комнатной температуры и клонируют в NdeI/SacI вектор pMD007-1.
Из полученных канамицин-устойчивых трансформантов выделяют плазмидную ДНК, анализируют путем секвенирования и отбирают плазмиду со вставкой SIGANS, содержащую последовательность модифицированного гена рас, кодирующего (ans)ECOPGA. Плазмиду обозначают pMD704 и используют для конструирования заявляемого штамма (Фиг. 3).
Пример 4. Получение заявляемого штамма - продуцента ECOPGA
Полученными рекомбинантными плазмидными ДНК pMD0107 (контроль) и pMD704 с использованием стандартных методов проводят трансформацию реципиентного штамма E. coli BL21(DE3) [F-, ompT, hsdSB (rB-, mB-), dcm, gal, λ (DE3) [26]. Выбор этого штамма в качестве реципиента для продукции ECOPGA обусловлен тем, что он синтезирует РНК-полимеразу фага Т7, а также обладает сниженной протеазной активностью, что способствует повышению выхода целевых белков.
Для индукции синтеза ECOPGA штаммы BL21(DE3)/pMD0107 и BL21(DE3)/pMD704 выращивают на среде ZY5052 [27] содержащую мас. %: энзиматический гидролизат казеина - 1, дрожжевой экстракт - 0,5, (NH4)2SO4 -0,33, КН2РО4 - 0,68, Na2HPO4 - 0,71, глицерин - 0,5, глюкоза - 0,05, лактоза- 0,2, MgSO4 - 0,012, вода - остальное. Дополнительно в среду добавляют 50 мг/л канамицина и 30 мг/л хлорамфеникола. Культивирование проводят в объеме 25 мл среды ZY5052 в колбах объемом 250 мл на качалке при 210 об/мин в течение 40 часов при температуре 21°С. В процессе культиврования параметры роста культуры и активности фермента в бактериальных клетках.
Удельную пенициллинамидазную активность выражают в единицах фермента (мкмолей субстрата в мин при 37°С) на мл культуры штамма-продуцента при помощи метода, описанного в работе [32].
Как видно из представленных данных (Фиг. 5), общая активность фермента достигала максимума через 36 часов инкубации; при этом в штамме E. coli BL21(DE3)/ pMVD704 она выше, чем в контрольном штамме E. coli BL21(DE3)/ pMVD0107.
Таким образом, получен модифицированный ген рас бактерии Escherichia coli, кодирующий предшественник фермента с активностью пенициллин G ацилазы, с использованием которого сконструирован штамм-продуцент ECOPGA и на его основе разработан способ микробиологического синтеза ECOPGA, позволяющий получить этот фермент с активностью до 15 ME на мл культуры, превосходящий способ - ближайший-аналог - в 15 раз.
Список использованной информации:
1. Курочкина В.Б., Скляренко А.В. Ферментативный синтез бета-лактамных антибиотиков: аналитический обзор. // Антибиот Химиотер. 2005. Vol. 50, №№5-6. Р. 39-58.
2. Sklyarenko A.V., Kurochkina V.B. Е.А.М. Enzymatic transformation and synthesis of beta-lactam antibiotics. // Egorov A.M. Zaikov G. New Res. Biotechnol. Biol. Med. - Nov. Sci. Publ. 2006. № Chapter 8. P. 73-86.
3. Kurochkina V.B. S.A.V. Enzymatic synthesis of beta-lactam antibiotics. Analytical review. // Zaikov G.E. Biotechnol. state art Prospect. Dev. Nov. Sci. Publ. 2008. Vol. chapter 20. P. 175-204.
4. Volpato G., Rodrigues R.C., Fernandez-Lafuente R. Use of enzymes in the production of semi-synthetic penicillins and cephalosporins: drawbacks and perspectives. // Curr. Med. Chem. 2010. Vol.17, №32. P. 3855-3873.
5. К.B. The evolution of β-lactamases. Antibiotic resistance: origins, evolution, selection and spread. // John Wiley Sons, Ltd. 2007. P. 152-166.
6. Parmar A. et al. Advances in enzymatic transformation of penicillins to 6-aminopenicillanic acid (6-APA) // Biotechnol. Adv. 2000. Vol.18, №4. P. 289-301.
7. Srirangan K. et al. Biotechnological advances on penicillin G acylase: pharmaceutical implications, unique expression mechanism and production strategies. // Biotechnol. Adv. 2013. Vol.31, №8. P. 1319-1332.
8. Schmidt F.-R. The beta-lactam antibiotics: current situation and future prospects in manufacture and therapy. // Hofrichter M. Ind. Appl. Mycota X. 2nd Edn Springer-Verlag Berlin Heidelb. 2010. Vol.chapter 5. P. 101-121.
9. Sudhakaran V.K. et al. Molecular aspects of penicillin and cephalosporin acylases // Process Biochem. 1992. Vol.27, №3. P. 131-143.
10. Xu Y. et al. Characterization of the T7 promoter system for expressing penicillin acylase in Escherichia coli. // Appl. Microbiol. Biotechnol. 2006. Vol.72, №3. P. 529-536.
11. Narayanan N. et al. Arabinose-induction of lac-derived promoter systems for penicillin acylase production in Escherichia coli. // Biotechnol. Prog. 2006. Vol.22, №3. P. 617-625.
12. Narayanan N., Xu Y., Chou CP. High-level gene expression for recombinant penicillin acylase production using the araB promoter system in Escherichia coli. // Biotechnol. Prog. 2006. Vol.22, №6. P. 1518-1523.
13. Vélez A.M. et al. High-throughput strategies for penicillin G acylase production in E. coli fed-batch cultivations // BMC Biotechnol. 2014. Vol.14, №1. P. 6.
14. Ignatova Z. et al. The relative importance of intracellular proteolysis and transport on the yield of the periplasmic enzyme penicillin amidase in Escherichia coli* // Enzyme Microb. Technol. 2000. Vol. 26, №2-4. P. 165-170.
15. Ignatova Z. et al. Improvement of posttranslational bottlenecks in the production of penicillin amidase in recombinant Escherichia coli strains. // Appl. Environ. Microbiol. 2003. Vol.69, №2. P. 1237-1245.
16. Scherrer S. et al. Periplasmic aggregation limits the proteolytic maturation of the Escherichia coli penicillin G amidase precursor polypeptide. // Appl. Microbiol. Biotechnol. 1994. Vol.42, №1. P. 85-91.
17. Sriubolmas N. et al. Localization and characterization of inclusion bodies in recombinant Escherichia coli cells overproducing penicillin G acylase. // Appl. Microbiol. Biotechnol. 1997. Vol.47, №4. P. 373-378.
18. Pan K.-L. et al. Roles of DegP in prevention of protein misfolding in the periplasm upon overexpression of penicillin acylase in Escherichia coli. // J. Bacteriol. 2003. Vol.185, №10. P. 3020-3030.
19. Jana S., Deb J.K. Strategies for efficient production of heterologous proteins in Escherichia coli. // Appl. Microbiol. Biotechnol. 2005. Vol.67, №3. P. 289-298.
20. Berks B.C., Sargent F., Palmer T. The Tat protein export pathway. // Mol. Microbiol. 2000. Vol.35, №2. P. 260-274.
21. Akkaya  . et al. Mutations in the translation initiation region of the pac gene resulting in increased levels of activity of penicillin G acylase. // World J. Microbiol. Biotechnol. 2012. Vol.28, №5. P. 2159-2164.
. et al. Mutations in the translation initiation region of the pac gene resulting in increased levels of activity of penicillin G acylase. // World J. Microbiol. Biotechnol. 2012. Vol.28, №5. P. 2159-2164.
22. Topping T.B. et al. Direct demonstration that homotetrameric chaperone SecB undergoes a dynamic dimer-tetramer equilibrium. // J. Biol. Chem. 2001. Vol.276, №10. P. 7437-7441.
23. Chou CP. et al. Effect of SecB chaperone on production of periplasmic penicillin acylase in Escherichia coli. // Biotechnol. Prog. 1999. Vol.15, №3. P. 439-445.
24. Патент RU 2388826. C12N 15/62, C12N 15/55, C12N 15/70, C12N 1/22. Рекомбинантная ДНК, кодирующая функционально активный гибридный белок g17аса-ацилазы с хитин-связывающим доменом (brdgl7aca-cbd), рекомбин. плазмида psvh0108, обеспечивающая его синтез в клетках. 2010.
25. RU 2221868. C12N 015/55, C12N 009/82, C12N 001/21, C12N 001/21, C12R 001/19. Ген L-аспарагиназы Erwinia carotovora и штамм Escherichia coli ВКПМ №В - 8174 - продуцент L-аспарагиназы Erwinia carotovora. 2004.
26. рЕТ System Manual, 11th edition. Novagen Inc. 2006.
27. Studier F.W. Protein production by auto-induction in high-density shaking cultures // Protein Expr. Purif. 2005. Vol.41, №1. P. 207-234.
28. J. Vennison. Laboratory Manual for Genetic Engineering. PHI Learning (January 30, 2010), 2010. P. 124.
29. Mardanov Α. V et al. Draft Genome Sequence of Escherichia coli Strain VKPM B-10182, Producing the Enzyme for Synthesis of Cephalosporin Acids. // Genome Announc. 2014. Vol.2, №6.
30. Eldarov M. et al. Probing the Substrate Specificity and Intersubunit Interactions of Brevundimonas Diminuta Glutaryl Acylase with Site-Directed Mutagenesis // Am. J. Biochem. Biotechnol. Science Publications, 2014. Vol.10, №3. P. 169-179.
31. Higuchi R., Krummel В., Saiki R.K. A general method of in vitro preparation and specific mutagenesis of DNA fragments: study of protein and DNA interactions. // Nucleic Acids Res. 1988. Vol.16, №15. P. 7351-7367.
32. Ныс П.С., Савицкая E.M. K.T.C. Метод определения активности пенициллинамидазы. //Антибиотики. 1973. Vol.17. P. 270-276.





















Изобретение относится к области биохимии, генетической инженерии и биотехнологии, в частности к модифицированному гену pac бактерии Escherichia coli. Нуклеотидная последовательность указанного гена представлена в SEQ ID NO: 17. Указанный ген кодирует предшественник фермента пенициллин G ацилазы, состоящего из аминокислотной последовательности сигнального пептида L-аспарагиназы Erwinia carotovora и структурной части пенициллин G ацилазы E.coli. Настоящее изобретение относится также к рекомбинантному штамму E.coli BL21(DE3)/pMD704. Настоящий штамм является продуцентом пенициллин G ацилазы. Штамм получен путем введения указанного модифицированного гена pac в составе вектора pSVH0107 в штамм-реципиент E.coli BL21(DE3). Изобретение также относится к способу микробиологического синтеза пенициллин G ацилазы. Указанный способ предусматривает культивирование штамма E.coli BL21(DE3)/pMD704 в подходящих условиях. Настоящее изобретение позволяет получать фермент пенициллин G ацилазу E.coli с высокой активностью. 3 н.п. ф-лы, 5 ил., 4 пр.
1. Модифицированный ген рас бактерии Escherichia coli, кодирующий предшественник фермента с активностью пенициллин G ацилазы и соответствующий последовательности SEQ ID NO: 17.
2. Рекомбинантный штамм Escherichia coli BL21(DE3)/pMD704 - продуцент пенициллин G ацилазы, полученный путем введения модифицированного гена рас бактерии Escherichia coli по п. 1 в составе вектора pSVH0107 в штамм-реципиент Escherichia coli BL21(DE3).
3. Способ микробиологического синтеза пенициллин G ацилазы путем культивирования штамма-продуцента по п. 2 в подходящих условиях.
| Xu Y et al., Characterization of the T7 promoter system for expressing penicillin acylase in Escherichia coli, Applied microbiology and biotechnology, 2006 | |||
| Choi JH et al., Secretory and extracellular production of recombinant proteins using Escherichia coli, Applied microbiology and biotechnology, 2004 | |||
| РЕКОМБИНАНТНАЯ ДНК, КОДИРУЮЩАЯ ФУНКЦИОНАЛЬНО АКТИВНЫЙ ГИБРИДНЫЙ БЕЛОК G17ACA-АЦИЛАЗЫ С ХИТИН-СВЯЗЫВАЮЩИМ ДОМЕНОМ (BrdG17ACA-cbd), РЕКОМБИНАНТНАЯ ПЛАЗМИДА pSVH0108, ОБЕСПЕЧИВАЮЩАЯ ЕГО СИНТЕЗ В КЛЕТКАХ Escherichia coli, И РЕКОМБИНАНТНЫЙ ШТАММ Escherichia coli BL21(DE3)/pSVH0108-ПРОДУЦЕНТ BrdG17ACA-cbd | 2006 |
|
RU2388826C2 |

