Область техники, к которой относится изобретение
Настоящая заявка относится к новому гену фосфатазы фосфатидной кислоты и его применению.
Уровень техники
Известно, что жирные кислоты, содержащие две или более ненасыщенные связи, в совокупности, именуемые полиненасыщенные жирные кислоты (ПНЖК), включают арахидоновую кислоту, дигомо-γ-линоленовую кислоту, эйкозапентаеновую кислоту и докозагексаеновую кислоту. Некоторые из полиненасыщенных жирных кислот не могут быть синтезированы в организме животных, и такие полиненасыщенные жирные кислоты должны поступать в организм с пищей, как незаменимые жирные кислоты. Полиненасыщенные жирные кислоты широко распространены. Например, арахидоновая кислота выделена из липидов, экстрагированных из надпочечников или печени животных. Однако, полиненасыщенные жирные кислоты содержатся в органах животных в небольших количествах, и экстрагирование и выделение полиненасыщенных жирных кислот только из органов животных является недостаточным для получения больших количеств полиненасыщенных жирных кислот. Поэтому были разработаны микробиологические методы для получения полиненасыщенных жирных кислот путем культивирования различных микроорганизмов. В частности, известно, что микроорганизмы рода Mortierella продуцируют липиды, содержащие полиненасыщенные жирные кислоты, такие как арахидоновая кислота.
Также были предприняты другие попытки получения полиненасыщенных жирных кислот в растениях. Известно, что полиненасыщенные жирные кислоты образуют запасные липиды, такие как триацилглицерин (также называемый триглицерид или ТГ), который накапливается в клетках микроорганизмов или семенах растений.
В качестве запасного липида триацилглицерин синтезируется в организме следующим образом. Ацильная группа вводится в глицерин-3-фосфат при помощи глицерин-3-фосфат-ацилтрансферазы с образованием лизофосфатидной кислоты, в которую ацильная группа вводится при помощи лизофосфатацилтрансферазы с образованием фосфатидной кислоты. Фосфатидная кислота затем дефосфорилируется фосфатазой фосфатидной кислоты с образованием диацилглицерина. Ацильная группа вводится в диацилглицерин при помощи диацилглицерин-ацилтрансферазы с образованием триацилглицерина.
В этом метаболическом пути фосфатидная кислота (далее именуемая также «ФК» или 1,2-диацил-sn-глицерин-3-фосфат) является предшественником триацилглицерина, а также биосинтетическим предшественником диацилглицерофосфолипида. В таких клетках как дрожжи, фосфатидная кислота-цитидилтрансфераза воздействует на ФК и цитидин-5'-трифосфат (ЦТФ), синтезируя ЦТФ диацилглицерин (ЦТФ-ДГ), который используется для биосинтеза различных фосфолипидов.
Как описано выше, известно, что дефосфорилирование ФК для биосинтеза диацилглицерина (далее именуемого также «ДГ») катализируется фосфатазой фосфатидной кислоты (К.Ф, 3.1.3.4, далее именуемая также «ФФК»). Известно, что ФФК присутствует во всех организмах от бактерий до позвоночных.
Известно, что дрожжи (Saccharomyces cerevisiae), которые относятся к низшим грибам, имеют два типа ФФК (Непатентные документы 1, 2 и 7). Одним из них является Mg2+-зависимая ФФК (ФФК1), а другим - Mg2+-независимая ФФК (ФФК2). Ген PAH1 известен как ген, кодирующий ФФК1 (Непатентные документы 3-5). Мутант pah1Δ также обладает ФФК1 активностью, что позволяет предположить существование других генов, проявляющих ФФК1 активность. У мутанта pah1Δ ядерная мембрана и мембрана ЭПС ненормально увеличены, и чрезмерно повышена экспрессия генов, играющих ключевую роль в биосинтезе фосфолипидов (Непатентный документ 6).
С другой стороны, известны гены DPP1 и LPP1, кодирующие ФФК2, которые обладают наибольшей ФФК2 активностью в дрожжах. Ферменты, кодируемые этими генами, обладают широкой субстратной специфичностью, и известно, что они дефосфорилируют, например, диацилглицеринпирофосфат (ДГПФ), лизофосфатидную кислоту, фосфаты сфингооснований и фосфаты изопреноидов.
Известно, что липид-продуцирующий грибок Mortierella alpina имеет два типа генов, т.е. MaPAH1.1 и MaPAH1.2, которые являются гомологами Mg2+-зависимой ФФК1 (Патентный документ 1), и ген MaPAP1, который является гомологом Mg2+-независимой ФФК2 (Патентный документ 2).
Список библиографических ссылок
Патентные документы
Патентный документ 1: Международная публикация No. WO 2011/081135
Патентный документ 2: Международная публикация No. WO 2009/008466
Непатентные документы
Непатентный документ 1: Biochem. Biophys. Acta, 1348, 45-55, 1997
Непатентный документ 2: Trends Biochem. Sci., 31(12), 694-699, 2006
Непатентный документ 3: EMBO J., 24, 1931-1941, 2005
Непатентный документ 4: J. Biol. Chem., 281(14), 9210-9218, 2006
Непатентный документ 5: J. Biol. Chem., 281(45), 34537-34548, 2006
Непатентный документ 6: J. Biol. Chem., 282(51), 37026-37035, 2007
Непатентный документ 7: J. Biol. Chem., 284(5), 2593-2597, 2009
Сущность изобретения
Техническая проблема
Большинство генов ФФК, о которых сообщалось ранее, не было исследовано относительно их способности изменять составное соотношение жирных кислот в композициях жирных кислот, продуцируемых клетками-хозяевами, в которых экспрессируются введенные гены ФФК. Существует потребность в ведении в практику нового гена, способного продуцировать жир с необходимым составным соотношением жирных кислот или повышать содержание необходимой жирной кислоты, при введении в клетку-хозяин или экспрессии в ней.
Целью настоящего изобретения является новый ген фосфатазы фосфатидной кислоты, кодируемый им белок и способы его применения.
Решение проблемы
Для решения вышеупомянутой проблемы авторами настоящего изобретения были проведены тщательные исследования. Был проанализирован геном липид-продуцирующего грибка Mortierella alpina с целью идентифицировать последовательности, обладающие гомологией с известными генами Mg2+-независимой фосфатазы фосфатидной кислоты (ФФК2). При помощи скрининга библиотеки кДНК или ПЦР была клонирована кДНК, чтобы получить полную открытую рамку считывания (ОРС), кодирующую ФФК. Этот ген был введен в высоко пролиферативные клетки-хозяева (например, клетки дрожжей), для подтверждения того, что белок, кодируемый клонированной кДНК, обладает активностью фосфатазы фосфатидной кислоты. Таким образом, ген новой фосфатазы фосфатидной кислоты (ФФК) был успешно клонирован, что привело к осуществлению настоящего изобретения. Соответственно, в одном варианте осуществления настоящее изобретение может быть следующим.
(1) Нуклеиновая кислота по любому из приведенных ниже пунктов (a)-(g):
(a) нуклеиновая кислота, содержащая нуклеотидную последовательность, кодирующую белок, который состоит из аминокислотной последовательности с делецией, заменой или добавлением одной или более аминокислот в аминокислотной последовательности, приведенной в SEQ ID NO: 2, и который обладает активностью фосфатазы фосфатидной кислоты;
(b) нуклеиновая кислота, содержащая нуклеотидную последовательность, которая гибридизуется в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, приведенной в SEQ ID NO: 1, и которая кодирует белок, обладающий активностью фосфатазы фосфатидной кислоты;
(c) нуклеиновая кислота, содержащая нуклеотидную последовательность, которая имеет идентичность 70% или более нуклеотидной последовательности, приведенной в SEQ ID NO: 1, и которая кодирует белок, обладающий активностью фосфатазы фосфатидной кислоты;
(d) нуклеиновая кислота, содержащая нуклеотидную последовательность, кодирующую белок, который состоит из аминокислотной последовательности, имеющей идентичность 70% или более аминокислотной последовательности, приведенной в SEQ ID NO: 2, и который обладает активностью фосфатазы фосфатидной кислоты;
(e) нуклеиновая кислота, содержащая нуклеотидную последовательность, которая гибридизуется в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, кодирующей белок, состоящий из аминокислотной последовательности, приведенной в SEQ ID NO: 2, и которая кодирует белок, обладающий активностью фосфатазы фосфатидной кислоты;
(f) нуклеиновая кислота, содержащая нуклеотидную последовательность, которая гибридизуется в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, приведенной в SEQ ID NO: 4, и которая включает экзон, кодирующий белок, обладающий активностью фосфатазы фосфатидной кислоты; и
(g) нуклеиновая кислота, содержащая нуклеотидную последовательность, которая имеет идентичность 70% или более нуклеотидной последовательности, приведенной в SEQ ID NO: 4, и которая включает экзон, кодирующий белок, обладающий активностью фосфатазы фосфатидной кислоты.
(2) Нуклеиновая кислота по любому из приведенных ниже пунктов (a)-(g):
(a) нуклеиновая кислота, содержащая нуклеотидную последовательность, кодирующую белок, который состоит из аминокислотной последовательности с делецией, заменой или добавлением от 1 до 110 аминокислот в аминокислотной последовательности, приведенной в SEQ ID NO: 2, и который обладает активностью фосфатазы фосфатидной кислоты;
(b) нуклеиновая кислота, содержащая нуклеотидную последовательность, которая гибридизуется с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, приведенной в SEQ ID NO: 1, в условиях 2x SSC буфер при температуре 50°C, и которая кодирует белок, обладающий активностью фосфатазы фосфатидной кислоты;
(c) нуклеиновая кислота, содержащая нуклеотидную последовательность, которая имеет идентичность 90% или более нуклеотидной последовательности, приведенной в SEQ ID NO: 1, и которая кодирует белок, обладающий активностью фосфатазы фосфатидной кислоты;
(d) нуклеиновая кислота, содержащая нуклеотидную последовательность, кодирующую белок, который состоит из аминокислотной последовательности, имеющей идентичность 90% или более аминокислотной последовательности, приведенной в SEQ ID NO: 2, и который обладает активностью фосфатазы фосфатидной кислоты;
(e) нуклеиновая кислота, содержащая нуклеотидную последовательность, которая гибридизуется с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, кодирующей белок, состоящий из аминокислотной последовательности, приведенной в SEQ ID NO: 2, в условиях 2x SSC буфер при температуре 50°C, и которая кодирует белок, обладающий активностью фосфатазы фосфатидной кислоты;
(f) нуклеиновая кислота, содержащая нуклеотидную последовательность, которая гибридизуется с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, приведенной в SEQ ID NO: 4, в условиях 2x SSC буфер при температуре 50°C, и которая включает экзон, кодирующий белок, обладающий активностью фосфатазы фосфатидной кислоты; и
(g) нуклеиновая кислота, содержащая нуклеотидную последовательность, которая имеет идентичность 90% или более нуклеотидной последовательности, приведенной в SEQ ID NO: 4, и которая включает экзон, кодирующий белок, обладающий активностью фосфатазы фосфатидной кислоты.
(3) Нуклеиновая кислота по любому из приведенных ниже пунктов (a)-(d):
(a) нуклеиновая кислота, содержащая нуклеотидную последовательность, приведенную в SEQ ID NO: 1, или ее фрагмент;
(b) нуклеиновая кислота, содержащая нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности, приведенной в SEQ ID NO: 2, или ее фрагмент;
(c) нуклеиновая кислота, содержащая нуклеотидную последовательность, приведенную в SEQ ID NO: 4, или ее фрагмент; и
(d) нуклеиновая кислота, содержащая нуклеотидную последовательность, приведенную в SEQ ID NO: 5, или ее фрагмент.
(4) Нуклеиновая кислота по пункту (1) или (2), отличающаяся тем, что активность фосфатазы фосфатидной кислоты имеет более высокую субстратную специфичность к фосфатидной кислоте, содержащей C18-ацильную группу, чем к фосфатидной кислоте, содержащей C17-ацильную группу.
(5) Белок по любому из приведенных ниже пунктов (a) или (b):
(a) белок, содержащий аминокислотную последовательность с делецией, заменой или добавлением одной или более аминокислот в аминокислотной последовательности, приведенной в SEQ ID NO: 2, и обладающий активностью фосфатазы фосфатидной кислоты, и
(b) белок, содержащий аминокислотную последовательность, которая имеет идентичность 70% или более аминокислотной последовательности, приведенной в SEQ ID NO: 2, и обладающий активностью фосфатазы фосфатидной кислоты.
(6) Белок по любому из приведенных ниже пунктов (a) или (b):
(a) белок, содержащий аминокислотную последовательность с делецией, заменой или добавлением от 1 до 110 аминокислот в аминокислотной последовательности, приведенной в SEQ ID NO: 2, и обладающий активностью фосфатазы фосфатидной кислоты; и
(b) белок, содержащий аминокислотную последовательность, которая имеет идентичность 90% или более аминокислотной последовательности, приведенной в SEQ ID NO: 2, и обладающий активностью фосфатазы фосфатидной кислоты.
(7) Белок по пункту (5) или (6), отличающийся тем, что активность фосфатазы фосфатидной кислоты имеет более высокую субстратную специфичность к фосфатидной кислоте, содержащей C18-ацильную группу, чем к фосфатидной кислоте, содержащей C17-ацильную группу.
(8) Белок, состоящий из аминокислотной последовательности, приведенной в SEQ ID NO: 2.
(9) Рекомбинантный вектор, содержащий нуклеиновую кислоту по любому из пунктов (1)-(4).
(10) Трансформант, трансформированный рекомбинантным вектором по пункту (9).
Полезный эффект изобретения
Настоящее изобретение относится к новому гену ФФК, кодируемому им белку и способам его применения. Ожидается, что ФФК настоящего изобретения будет продуцировать жирные кислоты в клетке-хозяине, эти жирные кислоты будут иметь составное соотношение, отличающееся от составного соотношения жирных кислот, продуцируемых в клетке-хозяине, в которую не вводилась ФФК. Это может обеспечить получение липидов, обладающих желаемыми характеристиками и действиями, и в связи с этим пригодных для применения, например, в пищевых продуктах, косметике, фармацевтических препаратах и мыле.
Краткое описание чертежей
На Фигуре 1-1 показана геномная последовательность и кодирующая нуклеотидная последовательность MaPAP2.2.
Фигура 1-2 является продолжением Фигуры 1-1.
На фигуре 2 показано выравнивание аминокислотной последовательности MaPAP2.2 и аминокислотных последовательностей предполагаемого белка, полученного из Laccaria bicolor, и ScDPP1 (YDR284C: регистрационный номер AAS56070), полученного из дрожжей.
На фигуре 3 показано выравнивание аминокислотных последовательностей MaPAP2.2 и MaPAP1, известного как Mg2+-независимая ФФК (ФФК2), полученная из Mortierella alpina (WO2009/008466). Три дважды подчеркнутых сегмента представляют собой консервативные области среди представителей семейства ферментов Mg2+-независимые фосфатазы фосфатидной кислоты типа 2 (ФФК2) (домены 1, 2 и 3, считая от N-конца), и знаком «*» обозначены аминокислотные остатки, существенные для активности ФФК.
На фигуре 4 показаны диаграммы, иллюстрирующие результаты изучения зависимости активности MaPAP2.2, которая превращает 18:2-ФК в 18:2-ДГ (n = 1), от ионов Mg2+. На диаграмме A показаны результаты добавления Mg2+. На диаграмме B показаны результаты добавления ЭДТА (без Mg2+). По вертикальной оси показаны количества 18:2-ДГ (обнаруженные во фракциях ДГ) по отношению к белку (мкг/мг белка) в неочищенных ферментных растворах.
На фигуре 5 показаны диаграммы, иллюстрирующие результаты изучения активности MaPAP2.2 (A) и MaPAP1 (B), которая превращает 18:2-ФК в 18:2-ДГ (n=3), в реакционном растворе, не содержащем ионы Mg2+. По вертикальной оси показаны количества 18:2-ДГ (обнаруженные во фракциях ДГ) по отношению к белку (мкг/мг белка) в неочищенных ферментных растворах.
На фигуре 6A показаны диаграммы, иллюстрирующие результаты изучения количества 18:1-ДГ без добавления в качестве субстрата фосфатидной кислоты, в реакционных растворах, содержащих MaPAP2.2, и контролях (n=3). На Фигуре 6B показаны диаграммы, иллюстрирующие результаты изучения количества 18:1-ДГ при добавлении 18:1-ФК в качестве субстрата, в реакционных растворах, содержащих MaPAP2.2, и контролях (n=3). По каждой вертикальной оси показаны количества 18:1-ДГ (обнаруженные во фракциях ДГ) по отношению к белку (мкг/мг белка) в неочищенном ферментном растворе.
На фигуре 7 показаны диаграммы, иллюстрирующие результаты изучения количества 17:0-ДГ при добавлении 17:0-ФК в качестве субстрата, в реакционных растворах, содержащих MaPAP2.2, и контролях (n=3). По вертикальной оси показаны количества 17:0-ДГ (обнаруженные во фракциях ДГ) по отношению к белку (мкг/мг белка) в неочищенных ферментных растворах.
Описание вариантов осуществления изобретения
Настоящее изобретение относится к новому гену фосфатазы фосфатидной кислоты, полученному из рода Mortierella, где эта фосфатаза фосфатидной кислоты дефосфорилирует фосфатидную кислоту с образованием диацилглицерина, кодируемому им белку и способам его применения.
Фосфатаза фосфатидной кислоты представляет собой фермент, который катализирует реакцию образования диацилглицерина путем дефосфорилирования фосфатидной кислоты. Субстрат ФФК настоящего изобретения, как правило, представляет собой фосфатидную кислоту, но не ограничивается ею.
Нуклеиновая кислота, кодирующая фосфатазу фосфатидной кислоты
Фосфатаза фосфатидной кислоты (ФФК) настоящего изобретения включает MaPAP2.2 и ее мутанты. Соответствие между кДНК, кодирующей последовательностью, ОРС, которые представляют собой нуклеиновые кислоты, кодирующие MaPAP2.2, и аминокислотной последовательностью MaPAP2.2 обобщено в Таблице 1.
А именно, последовательности, относящиеся к MaPAP2.2, включают SEQ ID NO: 2, соответствующую аминокислотной последовательности MaPAP2.2; SEQ ID NO: 1, соответствующую последовательности области ОРС MaPAP2.2; SEQ ID NO: 3, соответствующую последовательности области кодирующей последовательности MaPAP2.2; и SEQ ID NO: 5, соответствующую последовательности кДНК MaPAP2.2. Среди этих последовательностей SEQ ID NO: 1, соответствует нуклеотидам 75-1163 в последовательности, приведенной в SEQ ID NO: 5; и SEQ ID NO: 3, соответствует нуклеотидам 75-1166 в последовательности, приведенной в SEQ ID NO: 5. SEQ ID NO: 4 соответствует геномной нуклеотидной последовательности, кодирующей MaPAP2.2. Геномная последовательность, приведенная в SEQ ID NO: 4, состоит из трех экзонов и двух интронов, и области экзонов соответствуют нуклеотидам 1-207, 445-582 и 889-1632 в SEQ ID NO: 4.
Нуклеиновые кислоты настоящего изобретения включают одноцепочечные и двухцепочечные ДНК и комплементарные им РНК, которые могут быть либо природного происхождения, либо искусственно синтезированными. Примеры ДНК включают без ограничений геномные ДНК, кДНК, соответствующие геномным ДНК химически синтезированные ДНК, ПЦР-амплифицированные ДНК, их комбинации и ДНК/РНК гибриды.
Предпочтительные варианты осуществления для нуклеиновых кислот настоящего изобретения включают нуклеиновые кислоты, содержащие (a) нуклеотидную последовательность, приведенную в SEQ ID NO: 1, (b) нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности, приведенной в SEQ ID NO: 2, и (c) нуклеотидную последовательность, приведенную в SEQ ID NO: 5.
Для получения этих нуклеотидных последовательностей данные о нуклеотидных последовательностях из баз данных маркерных экспрессируемых последовательностей (EST) или геномных ДНК организмов, обладающих активностью ФФК, могут быть использованы для поиска нуклеотидной последовательности, кодирующей белок, обладающий гомологией с известными белками, обладающими активностью ФФК. Предпочтительными организмами, обладающими активностью ФФК, являются липид-продуцирующие грибки, включая без ограничения M. alpina.
Для анализа EST сначала получают библиотеку кДНК. Библиотека кДНК может быть получена методами, описанными в «Molecular Cloning, A Laboratory Manual, 3rd ed.» (Cold Spring Harbor Press (2001)). В другом варианте может быть использован коммерчески доступный набор реагентов для получения библиотеки кДНК. Библиотека кДНК, пригодная для настоящего изобретения, может быть получена, например, по следующей методике. А именно, подходящим штаммом липид-продуцирующего грибка M. alpina инокулирут соответствующую питательную среду и прекультивируют в течение соответствующего периода. В число условий культивирования, подходящих для этого прекультивирования, входят, например, состав среды, включающий 1,8% глюкозы и 1% дрожжевого экстракта, значение pH 6,0, период культивирования от 3 до 4 суток и температуру культивирования 28°C. Продукт, полученный в результате прекультивирования, затем подвергают основному культивированию в соответствующих условиях. Состав среды, подходящей для основного культивирования, включает в себя, 1,8% глюкозы, 1% соевого порошка, 0,1% оливкового масла, 0,01% Адеканола, 0,3% KH2PO4, 0,1% Na2SO4, 0,05% CaCl2·2H2O и 0,05% MgCl2·6H2O и имеет значение pH 6,0. Условия культивирования, подходящие для основного культивирования, включают в себя, например, аэрацию и перемешивание культуры при 300 об./мин, аэрации 1 vvm и температуре 26°C в течение 8 суток. Необходимое количество глюкозы может быть добавлено в процессе культивирования. Во время основного культивирования в определенный момент времени из культуры отбирают образцы, из которых выделяют клетки для получения тотальной РНК. Тотальная РНК может быть получена любым известным способом, таким как методика с использованием гуанидин гидрохлорида и CsCl. Поли(A)+РНК может быть выделена из полученной тотальной РНК при помощи коммерчески доступного набора реагентов, и библиотека кДНК может быть сконструирована с использованием коммерчески доступного набора реагентов. Нуклеотидную последовательность любого клона из полученной библиотеки кДНК определяют с использованием праймеров, которые сконструированы для вектора и позволяют определить нуклеотидную последовательность вставки. В результате могут быть получены маркерные экспрессируемые последовательности. Например, получение библиотеки кДНК с использованием набора реагентов ZAP cDNA Gigapack III Gold Cloning Kit (STRATAGENE) позволяет проводить прямое клонирование.
При анализе геномной ДНК культивируют клетки организма, обладающего активностью ФФК, из которых затем получают геномную ДНК. Определяют нуклеотидную последовательность полученной геномной ДНК и объединяют определенную нуклеотидную последовательность. В полученной в конечном итоге последовательности суперконтига ищут последовательность, кодирующую аминокислотную последовательность, обладающую высокой гомологией аминокислотной последовательности известного белка, обладающего активностью ФФК. Из последовательностей суперконтига, которые были идентифицированы как кодирующие такую аминокислотную последовательность, получают праймеры. Затем проводят ПЦР, используя библиотеку кДНК в качестве матрицы, и полученные фрагменты ДНК встраивают в плазмиду для клонирования. Клонированную плазмиду в качестве матрицы и вышеупомянутые праймеры используют для ПЦР для получения зонда, который затем используют для скрининга библиотеки кДНК.
Поиск гомологии аминокислотных последовательностей MaPAP2.2 проводили среди аминокислотных последовательностей, зарегистрированных в GenBank, с помощью программы BLASTp. Полученная аминокислотная последовательность имела совпадение с самым высоким баллом с предсказанным белком Laccaria bicolor (SEQ ID NO: 10, регистрационный номер: XP_001878243), имея идентичность по нуклеотидной последовательности 36,7%. Идентичность между аминокислотной последовательностью MaPAP1, которая является известной ФФК2 (Mg2+-независимая ФФК), полученной из Mortierella alpina, и аминокислотной последовательностью MaPAP2.2 составила 20,5%.
Настоящее изобретение также включает нуклеиновые кислоты, функционально эквивалентные нуклеиновой кислоте, включающей вышеупомянутую нуклеотидную последовательность, приведенную в SEQ ID NO: 1, или нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности, приведенной в SEQ ID NO: 2. Предполагается, что термин «функционально эквивалентный» означает, что белок, кодируемый нуклеотидной последовательностью настоящего изобретения, и белок, состоящий из аминокислотной последовательности настоящего изобретения, обладают активностью фосфатазы фосфатидной кислоты (ФФК). Используемый здесь термин «активность ФФК» относится к активности катализирования реакции дефосфорилирования фосфатидной кислоты с образованием диацилглицерина. Активность ФФК может обладать, но не ограничивается, более высокой субстратной специфичностью по отношению к фосфатидной кислоте, содержащей C18-ацильную группу, чем к фосфатидной кислоте, содержащей C17-ацильную группу. Кроме того, активность ФФК может быть, но не ограничивается, независимой от ионов Mg2+.
Такие нуклеиновые кислоты, которые являются мутантами нуклеиновых кислот, содержащих нуклеотидную последовательность, приведенную в SEQ ID NO: 1, или нуклеиновой кислоты, содержащей нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности, приведенной в SEQ ID NO: 2, и которые являются функционально эквивалентными этих нуклеиновых кислот, включают нуклеиновые кислоты, содержащие нуклеотидные последовательности, указанные ниже в любом из пунктов (a)-(g).
(a) Нуклеиновые кислоты, содержащие нуклеотидную последовательность, кодирующую белок, который состоит из аминокислотной последовательности с делецией, заменой или добавлением одной или более аминокислот в аминокислотной последовательности, приведенной в SEQ ID NO: 2, и который обладает активностью ФФК
Примеры нуклеотидной последовательности, содержащейся в нуклеиновой кислоте настоящего изобретения, включают нуклеотидные последовательности, кодирующие белок, который состоит из аминокислотной последовательности с делецией, заменой или добавлением одной или более аминокислот в аминокислотной последовательности, приведенной в SEQ ID NO: 2, и который обладает активностью ФФК.
В частности, нуклеотидная последовательность, содержащаяся в нуклеиновой кислоте настоящего изобретения, представляет собой нуклеотидную последовательность, кодирующую белок, обладающий активностью ФФК, и состоящий из:
(i) аминокислотной последовательности с делецией одной или более (предпочтительно, одной или нескольких (например, от 1 до 110, от 1 до 100, от 1 до 75, от 1 до 50, от 1 до 30, от 1 до 25, от 1 до 20 или от 1 до 15, более предпочтительно, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1)) аминокислот в аминокислотной последовательности, приведенной в SEQ ID NO: 2;
(ii) аминокислотной последовательности, имеющей замену одной или более (предпочтительно, одной или нескольких (например, от 1 до 110, от 1 до 100, от 1 до 75, от 1 до 50, от 1 до 30, от 1 до 25, от 1 до 20 или от 1 до 15, более предпочтительно, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1)) аминокислот другими аминокислотами в аминокислотной последовательности, приведенной в SEQ ID NO: 2;
(iii) аминокислотной последовательности, имеющей добавление одной или более (предпочтительно, одной или нескольких (например, от 1 до 110, от 1 до 100, от 1 до 75, от 1 до 50, от 1 до 30, от 1 до 25, от 1 до 20 или от 1 до 15, более предпочтительно, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1)) других аминокислот в аминокислотной последовательности, приведенной в SEQ ID NO: 2; или
(iv) аминокислотную последовательность в любой комбинации (i)-(iii).
Из них замена является предпочтительно консервативной заменой, что означает замещение конкретного аминокислотного остатка другим остатком, обладающим аналогичными физико-химическими свойствами, и может быть любой заменой, которая не оказывает существенного влияния на структурные свойства исходной последовательности. Например, любая замена является допустимой, при условии, что замененные аминокислоты не нарушают спираль исходной последовательности или не нарушают любую другую вторичную структуру, характеризующую исходную последовательность.
Консервативную замену, как правило, вводят путем синтеза в биологической системе или химического синтеза пептидов, предпочтительно, путем химического синтеза пептидов. В этом случае, заместители могут включать не встречающийся в природе аминокислотный остаток, пептидомиметик или перевернутую или инвертированную форму, где незамещенная область является перевернутой или инвертированной по аминокислотной последовательности.
Неограниченные примеры взаимозаменяемых аминокислотных остатков систематизированы и перечислены ниже:
Группа A: лейцин, изолейцин, норлейцин, валин, норвалин, аланин, 2-аминобутановая кислота, метионин, O-метилсерин, трет-бутилглицин, трет-бутилаланин и циклогексилаланин;
Группа B: аспарагиновая кислота, глутаминовая кислота, изоаспарагиновая кислота, изоглутаминовая кислота, 2-аминоадипиновая кислота и 2-аминосубериновая кислота;
Группа C: аспарагин и глутамин;
Группа D: лизин, аргинин, орнитин, 2,4-диаминобутановая кислота и 2,3-диаминопропионовая кислота;
Группа E: пролин, 3-гидроксипролин и 4-гидроксипролин;
Группа F: серин, треонин и гомосерин; и
Группа G: фенилаланин и тирозин.
В случае неконсервативной замены, член одной из этих групп может быть заменен членом другой группы. В этом случае для поддержания биологических функций белка настоящего изобретения, предпочтительно учитываются гидропатические индексы аминокислот (индексы гидропатичности аминокислот) (Kyte, et al., J. Mol. Biol., 157: 105-131 (1982)).
В случае неконсервативной замены, замена аминокислоты может быть выполнена на основе гидрофильности.
В тексте всего описания и на чертежах, нуклеотиды, аминокислоты и аббревиатуры обозначаются в соответствии с номенклатурой, утвержденной Комиссией по Биохимической Номенклатуре ИЮПАК-ИЮБ, или с использованием названий, традиционно используемых в данной области техники, например, как описано в Immunology: A Synthesis (second edition, edited by E.S. Golub and D.R. Gren, Sinauer Associates, Sunderland, Massachusetts (1991)). Кроме того, подразумевается, что аминокислоты, которые могут иметь оптические изомеры, представлены своими L-изомерами, если не указано иное.
Стереоизомеры, такие как D-аминокислоты вышеупомянутых аминокислот, не встречающиеся в природе аминокислоты, такие как α,α-дизамещенные аминокислоты, N-алкиламинокислоты, молочная кислота и другие нетипичные аминокислоты также могут быть мономерами, образующими белки настоящего изобретения.
Необходимо отметить, что в используемой в настоящем изобретении номенклатуре белка направление влево является аминоконцевым направлением, а направление вправо является карбоксиконцевым направлением, в соответствии со стандартной практикой и сложившимися правилами в данной области техники.
Аналогично, как правило, если не указано иное, то левый конец одноцепочечной полинуклеотидной последовательности является 5’-концом, а направление влево двухцепочечной полинуклеотидной последовательности обозначается как 5’-направление.
Специалисты в данной области техники могут теоретически спланировать и получить соответствующие мутанты описанных здесь белков при помощи способов, известных из уровня техники. Так, например, они могут идентифицировать подходящие области в белковой молекуле, структура которых может быть изменена без ухудшения биологической активности белка настоящего изобретения, полагая, что выбранные области являются менее важными для биологической активности белка. Специалисты в данной области техники также могут идентифицировать остатки и области в молекулах, которые являются консервативными среди аналогичных белков, и также могут ввести консервативные аминокислотные замены в области, которые могут быть важными для биологической активности или структуры белка настоящего изобретения, без ухудшения биологической активности и без неблагоприятного воздействия на полипептидную структуру белка.
В частности, аминокислотная последовательность MaPAP2.2 (SEQ ID NO: 2) содержит три области, дважды подчеркнутые на Фигуре 3, которые являются консервативными среди представителей семейства ферментов Mg2+-независимые фосфатазы фосфатидной кислоты типа 2 (ФФК2), они соответствуют остаткам 115-123, 172-175 и 229-233. Известно, что в этих трех консервативных областях в семействе ферментов ФФК2 аргинин в домене 1 и гистидин в доменах 2 и 3 являются аминокислотами, существенными для активности, и эти аминокислоты также являются консервативными у MaPAP2.2, т.е. аргинин в положении остатка 122 и гистидин в положении остатков 175 и 229 в SEQ ID NO: 2. Вышеуказанные консервативные области являются существенными для семейства ферментов ФФК2, и они также являются важными для ФФК настоящего изобретения. Таким образом, в соответствии с настоящим изобретением мутанты могут содержать вышеуказанные консервативные области.
Специалисты в данной области техники могут провести так называемое структурно-функциональное исследование, которое выявляет остатки пептида, которые являются важными для биологической активности или структуры белка настоящего изобретения, и пептида, который является аналогичным пептиду этого белка, сравнивая аминокислотные остатки этих двух пептидов, и таким образом предсказывая, какой остаток в белке, аналогичном белку настоящего изобретения, является аминокислотным остатком, соответствующим аминокислотному остатку, важному для биологической активности или структуры. Они также могут выбрать мутант, который сохраняет биологическую активность белка настоящего изобретения, выбирая аминокислотный заместитель, химически аналогичный предсказанному аминокислотному остатку. Также специалисты в данной области техники могут проанализировать трехмерную структуру и аминокислотную последовательность этого белкового мутанта. Результаты анализа могут в дальнейшем быть использованы для предсказания расположения аминокислотных остатков, вовлеченных в трехмерную структуру белка. На основе вышеуказанных результатов анализа специалисты в данной области техники могут получить мутант, не имеющий изменений в тех аминокислотных остатках, которые теоретически могут находиться на поверхности белка, и которые могут быть вовлечены в важные взаимодействия с другими молекулами. Специалисты в данной области техники также могут получить мутант, имеющий одну аминокислотную замену любого из аминокислотных остатков, которые образуют белок настоящего изобретения. Эти мутанты могут быть подвергнуты скринингу любым известным способом анализа для сбора информации об отдельных мутантах, которая позволит оценить функциональность отдельных аминокислотных остатков, образующих белок настоящего изобретения, на основе сравнения биологической активности в следующем случае: мутант, имеющий замену конкретного аминокислотного остатка, обладает более низкой биологической активностью по сравнению с белком настоящего изобретения; такой мутант не обладает биологической активностью; или такой мутант обладает неприемлемой активностью, ингибирующей биологическую активность белка настоящего изобретения. Кроме того, специалисты в данной области техники смогут без труда определить аминокислотную замену, нежелательную для мутантов белка настоящего изобретения, на основе информации, собранной в результате только таких стандартных экспериментов или в сочетании с другими мутациями.
Как описано выше, белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением одной или более аминокислот в аминокислотной последовательности, приведенной в SEQ ID NO: 2, может быть получен таким способом как сайт-направленный мутагенез, как описано, например, в «Molecular Cloning, A Laboratory Manual 3rd ed.» (Cold Spring Harbor Press (2001)); «Current Protocols in Molecular Biology» (John Wiley & Sons (1987-1997); Kunkel, (1985), Proc. Natl. Acad. Sci. USA, 82: 488-92; или Kunkel, (1988), Method Enzymol., 85: 2763-6. Получение мутанта с такой мутацией, включающей аминокислотную делецию, замену или добавление, может быть выполнено, например, при помощи известных методик, таких как способ Кункеля или метод с разрывом дуплекса, с использованием набора реагентов для введения мутаций на основе сайт-направленного мутагенеза, такого как набор реагентов для сайт-направленного мутагенеза QuikChange™ (производства Stratagene), система для сайт-направленного мутагенеза GeneTailor™ (производства Invitrogen) или система для сайт-направленного мутагенеза TaKaRa (например, Mutan-K, Mutan-Super Express Km; производства Takara Bio Inc.).
Способы введения делеции, замены или добавления одной или более аминокислот в аминокислотную последовательность белка с сохранением его активности включают в себя, кроме вышеупомянутого сайт-направленного мутагенеза, способ обработки гена мутагеном и способ избирательного расщепления гена для делеции, замены или добавления выбранного нуклеотида и последующего лигирования сегментов.
Нуклеотидная последовательность, содержащаяся в нуклеиновой кислоте настоящего изобретения, предпочтительно представляет собой нуклеотидную последовательность, кодирующую белок, который состоит из аминокислотной последовательности с делецией, заменой или добавлением от 1 до 30, от 1 до 20 или от 1 до 10 аминокислот в аминокислотной последовательности, приведенной в SEQ ID NO: 2, и который обладает активностью ФФК.
Количество и места аминокислотных мутаций или модификаций в белке, который кодируется нуклеиновой кислотой настоящего изобретения, не ограничены, при условии сохранения активности ФФК.
Активность ФФК может быть определена известным способом, например, см. J. Biol. Chem., 273, 14331-14338 (1998).
Например, активность ФФК может быть измерена следующим образом: Неочищенный ферментный раствор получают путем разрушения трансформированных клеток, экспрессирующих ФФК настоящего изобретения, центрифугирования клеточного гомогената и отбора супернатанта. Полученный неочищенный ферментный раствор может быть далее подвергнут выделению ФФК настоящего изобретения. Неочищенный ферментный раствор, содержащий ФФК настоящего изобретения, или очищенную ФФК настоящего изобретения добавляют в реакционный раствор, содержащий 100 мкг/мл фосфатидной кислоты и 50 мМ Трис-HCl (pH 7,5), с последующей реакцией при температуре от 25°C до 28°C в течение соответствующего времени. Реакцию останавливают путем добавления смеси хлороформа и метанола, и затем экстрагируют липиды. Полученные липиды фракционируют при помощи тонкослойной хроматографии, чтобы определить количество образовавшегося диацилглицерина.
(b) Нуклеиновая кислота, содержащая нуклеотидную последовательность, которая гибридизуется в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, приведенной в SEQ ID NO: 1, и которая кодирует белок, обладающий активностью ФФК
Примеры нуклеотидной последовательности, содержащейся в нуклеиновой кислоте настоящего изобретения, включают нуклеотидные последовательности, которые гибридизуются в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, приведенной в SEQ ID NO: 1, и которые кодируют белок, обладающий активностью ФФК.
Такая нуклеотидная последовательность может быть получена способом, известным специалисту в данной области техники, например, из библиотеки кДНК или геномной библиотеки при помощи известного процесса гибридизации, такого как гибридизация колоний, гибридизация бляшек или Саузерн-блоттинг с использованием зонда, который получают способом, известным специалистам в данной области техники, используя соответствующий фрагмент.
Подробное описание методики гибридизации можно найти в «Molecular Cloning, A Laboratory Manual 3rd ed.» (Cold Spring Harbor Press (2001), в частности, Разделы 6 и 7), «Current Protocols in Molecular Biology» (John Wiley & Sons (1987-1997), в частности, Разделы 6.3 и 6.4), и «DNA Cloning 1: Core Techniques, A Practical Approach 2nd ed.» (Oxford University (1995), in particular, Раздел 2.10 для условий гибридизации).
Жесткость гибридизации определяется в первую очередь условиями гибридизации, более предпочтительно, условиями гибридизации и условиями отмывки. Предполагается, что используемый здесь термин «жесткие условия» включает умеренно или очень жесткие условия.
В частности, примеры умеренно жестких условий включают условия гибридизации в от 1x до 6x SSC буфере при температуре от 42°C до 55°C, более предпочтительно, в от 1x до 3x SSC буфере при температуре от 45°C до 50°C, наиболее предпочтительно, в 2x SSC буфере при температуре 50°C. В случае, когда раствор для гибридизации содержит, например, около 50% формамида, гибридизацию можно проводить при температуре ниже вышеуказанной на 5-15°C. Условия отмывки, например, являются следующими: от 0,5x до 6x SSC буфер при температуре от 40°C до 60°C. Как правило, от 0,05% до 0,2% ДСН, предпочтительно, примерно 0,1% ДСН может быть добавлено в раствор для гибридизации и отмывочный раствор.
Очень жесткие (высоко жесткие) условия включают гибридизацию и/или отмывку при более высокой температуре и/или более низкой концентрации соли, по сравнению с умеренно жесткими условиями. Примеры таких условий гибридизации включают гибридизацию в от 0,1x до 2x SSC буфере при температуре от 55°C до 65°C, более предпочтительно, в от 0,1x до 1x SSC буфере при температуре от 60°C до 65°C, наиболее предпочтительно, в 0,2x SSC буфере при температуре 63°C. Условия отмывки, например, являются следующими: от 0,2x до 2x SSC буфер при температуре от 50°C до 68°C и, более предпочтительно, 0,2x SSC буфер при температуре от 60°C до 65°C.
Примеры условий гибридизации, особенно применимых в настоящем изобретении, включают без ограничений предгибридизацию в 5x SSC буфере, содержащем 1% ДСН, 50 мМ Трис-Hcl (pH 7,5) и 50% формамида, при температуре 42°C с последующей гибридизацией с зондом в течение ночи при температуре 42°C, и затем три раза отмывки в 0,2x SSC буфере, содержащем 0,1% ДСН, при температуре 65°C, каждый раз в течение 20 минут.
Также могут быть использованы коммерчески доступные наборы реагентов для гибридизации, включающие нерадиоактивный зонд. В частности, для гибридизации используется, например, набор реагентов для DIG детекции нуклеиновых кислот (Roche Diagnostics) или система для прямого мечения и ECL детекции (производства Amersham).
Предпочтительные примеры нуклеотидной последовательности настоящего изобретения включают нуклеотидные последовательности, которые гибридизуются в условиях 2x SSC буфера при температуре 50°C с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, приведенной в SEQ ID NO: 1, и которые кодируют белок, обладающий активностью ФФК.
(c) Нуклеиновая кислота, содержащая нуклеотидную последовательность, которая имеет идентичность 70% или более нуклеотидной последовательности, приведенной в SEQ ID NO: 1, и которая кодирует белок, обладающий активностью ФФК
Нуклеотидная последовательность, содержащаяся в нуклеиновой кислоте настоящего изобретения, включает нуклеотидную последовательность, которая имеет, по меньшей мере, 70% идентичности нуклеотидной последовательности, приведенной в SEQ ID NO: 1, и которая кодирует белок, обладающий активностью ФФК.
Предпочтительно, например, нуклеиновая кислота содержит нуклеотидную последовательность, имеющую, по меньшей мере, 75%, более предпочтительно, 80% или более идентичности (например, 85% или более, более предпочтительно, 90% или более и, наиболее предпочтительно, 95%, 98% или 99% или более) нуклеотидной последовательности, приведенной в SEQ ID NO: 1, и кодирующую белок, обладающий активностью ФФК.
Процент идентичности между двумя нуклеотидными последовательностями может быть определен путем визуального исследования и математического вычисления, но предпочтительно определяется путем сравнения информации о последовательности двух нуклеиновых кислот с использованием компьютерной программы. Компьютерные программы для сравнения последовательностей включают в себя, например, программу BLAST (Altschul et al., (1990), J. Mol. Biol., 215: 403-10) в версии 2.2.7, доступную на веб-сайте Национальной медицинской библиотеки США: http://www.ncbi.nlm.nih.gov/blast/bl2seq/bls.html, или алгоритм WU-BLAST 2.0. Стандартные настройки параметров по умолчанию для WU-BLAST 2.0, которые описаны на следующем Интернет-сайте: http://blast.wustl.edu, могут быть использованы.
(d) Нуклеиновая кислота, содержащая нуклеотидную последовательность, кодирующую белок, который состоит из аминокислотной последовательности, имеющей идентичность 70% или более аминокислотной последовательности, приведенной в SEQ ID NO: 2, и который обладает активностью ФФК
Нуклеотидная последовательность, содержащаяся в нуклеиновой кислоте настоящего изобретения, включает нуклеотидную последовательность, кодирующую белок, который состоит из аминокислотной последовательности, имеющей идентичность 70% или более аминокислотной последовательности, приведенной в SEQ ID NO: 2, и который обладает активностью ФФК.
Белок, кодируемый нуклеиновой кислотой настоящего изобретения, может представлять собой любой белок, обладающий идентичностью аминокислотной последовательности MaPAP2.2, при условии, что указанный белок обладает активностью ФФК. Конкретные примеры аминокислотной последовательности белка, кодируемого нуклеиновой кислотой настоящего изобретения, включают аминокислотную последовательность, обладающую идентичностью 75% или более, более предпочтительно, 85% или более и наиболее предпочтительно, 90% или более (например, 95% или более, а также 98% или более) аминокислотной последовательности, приведенной в SEQ ID NO: 2.
Нуклеотидная последовательность, содержащаяся в нуклеиновой кислоте настоящего изобретения, предпочтительно, представляет собой нуклеотидную последовательность, состоящую из аминокислотной последовательности, имеющей, идентичность 90% или более аминокислотной последовательности, приведенной в SEQ ID NO: 2, и кодирующую белок, обладающий активностью ФФК, более предпочтительно, нуклеотидную последовательность, состоящую из аминокислотной последовательности, имеющей, идентичность 95% или более аминокислотной последовательности, приведенной в SEQ ID NO: 2, и кодирующую белок, обладающий активностью ФФК.
Процент идентичности между двумя аминокислотными последовательностями может быть определен путем визуального исследования и математического вычисления. В другом варианте, процент идентичности может быть определен с использованием компьютерной программы. Примеры такой компьютерной программы включают BLAST, FASTA (Altschul et al., J. Mol. Biol., 215: 403-410, (1990)) и ClustalW. В частности, различные условия (параметры) для поиска идентичности с использованием программы BLAST описаны в Altschul et al. (Nucl. Acids. Res., 25, pp. 3389-3402, 1997) и находятся в открытом доступе на веб-сайте Национального центра биотехнологической информации США (NCBI) или японской базы данных ДНК (DDBJ) (BLAST Manual, Altschul et al., NCB/NLM/NIH Bethesda, MD 20894; Altschul et al.). Процент идентичности также может быть определен с использованием программ обработки генетической информации, таких как GENETYX Ver.7 (Genetyx), DNASIS Pro (Hitachisoft) и Vector NTI (Infomax).
Специальная схема выравнивания для сравнения множества аминокислотных последовательностей также может показать совпадение последовательностей в конкретной короткой области, и поэтому может определить область, имеющую очень высокую идентичность по последовательности в такой короткой области, даже если полноразмерные последовательности не имеют между собой значительного сходства. Кроме того, алгоритм BLAST может использовать матрицу замен аминокислот BLOSUM62, и следующие параметры выбора могут быть использованы: (A) включение фильтров для маскирования сегмента в последовательности запроса, обладающего низкой структурной сложностью (как определено программой SEG, авторов Wootton and Federhen (Computers and Chemistry, 1993); см. также Wootton and Federhen, 1996, “Analysis of compositionally biased regions in sequence databases”, Methods Enzymol., 266: 554-71), или для маскирования сегментов, состоящих из часто повторяющихся внутренних повторов, (как определено программой XNU, авторов Claverie and States (Computers and Chemistry, 1993), и (B) порог статистической значимости для представления информации о соответствиях, обнаруженных в последовательностях из базы данных, или для ожидаемого правдоподобия соответствий, обнаруженных случайно, согласно статистической модели E-индекса (Karlin and Altschul, 1990); если статистическая значимость, приписанная соответствию, больше, чем этот порог E-индекса, то об этом соответствии не будет сообщено.
(e) Нуклеиновая кислота, содержащая нуклеотидную последовательность, которая гибридизуется в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, кодирующей белок, состоящий из аминокислотной последовательности, приведенной в SEQ ID NO: 2, и которая кодирует белок, обладающий активностью ФФК
Нуклеотидная последовательность, содержащаяся в нуклеиновой кислоте настоящего изобретения, включает нуклеотидную последовательность, которая гибридизуется в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, кодирующей белок, состоящий из аминокислотной последовательности, приведенной в SEQ ID NO: 2, и которая кодирует белок, обладающий активностью ФФК.
Белок, состоящий из аминокислотной последовательности, приведенной в SEQ ID NO: 2, и условия гибридизации являются такими, как описано выше.
(f) Нуклеиновая кислота, содержащая нуклеотидную последовательность, которая гибридизуется в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, приведенной в SEQ ID NO: 4, и которая включает экзон, кодирующий белок, обладающий активностью ФФК
Нуклеотидные последовательности, приведенные в SEQ ID NO: 4, являются последовательностями геномной ДНК, кодирующими MaPAP2.2.
Нуклеотидная последовательность, содержащаяся в нуклеиновой кислоте настоящего изобретения, включает нуклеотидную последовательность, которая гибридизуется в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, приведенной в SEQ ID NO: 4, и которая включает экзон, кодирующий белок, обладающий активностью ФФК.
Такая нуклеотидная последовательность может быть получена способом, известным специалистам в данной области техники, например, из геномной библиотеки при помощи известного процесса гибридизации, такого как гибридизация колоний, гибридизация бляшек или Саузерн-блоттинг с использованием зонда, который получают способом, известным специалистам в данной области техники, используя соответствующий фрагмент.
(g) Нуклеиновая кислота, содержащая нуклеотидную последовательность, которая имеет идентичность 70% или более нуклеотидной последовательности, приведенной в SEQ ID NO: 4, и которая включает экзон, кодирующий белок, обладающий активностью ФФК
Нуклеотидная последовательность, содержащаяся в нуклеиновой кислоте настоящего изобретения, включает нуклеотидную последовательность, которая имеет, по меньшей мере, 70% идентичности нуклеотидной последовательности, приведенной в SEQ ID NO: 4, и которая кодирует белок, обладающий активностью ФФК.
Предпочтительные примеры нуклеотидной последовательности включают нуклеотидные последовательности, имеющие, по меньшей мере, 75%, более предпочтительно, 80% или более идентичности (например, 85% или более, более предпочтительно, 90% или более и, наиболее предпочтительно, 95%, 98% или 99% или более), нуклеотидной последовательности, приведенной в SEQ ID NO: 4, и имеющие экзон, кодирующий белок, обладающий активностью ФФК. Процент идентичности между двумя нуклеотидными последовательностями может быть определен так, как описано выше.
Последовательность геномной ДНК SEQ ID NO: 4 состоит из 3 экзонов и 2 интронов. Области экзонов соответствуют нуклеотидам 1-207, 445-582 и 889-1632 в SEQ ID NO: 4.
В другом варианте осуществления изобретения примеры нуклеотидной последовательности, содержащейся в нуклеиновой кислоте настоящего изобретения, включают нуклеотидные последовательности, включающие области интронов, имеющие идентичность нуклеотидной последовательности 100% с последовательностью геномной ДНК, приведенной в SEQ ID NO: 4, и области экзонов, имеющие идентичность нуклеотидной последовательности, по меньшей мере, 70% или более, более предпочтительно, 75% или более и более предпочтительно 80% или более (например, 85% или более, более предпочтительно, 90% или более и, наиболее предпочтительно, 95%, 98% или 99% или более) последовательности, приведенной в SEQ ID NO: 4, где экзон кодирует белок, обладающий активностью ФФК.
В другом варианте осуществления изобретения примеры нуклеотидной последовательности, содержащейся в нуклеиновой кислоте настоящего изобретения, включают нуклеотидные последовательности, включающие области экзонов, имеющие идентичность нуклеотидной последовательности 100% с последовательностью геномной ДНК, приведенной в SEQ ID NO: 4, и области интронов, имеющие идентичность нуклеотидной последовательности, по меньшей мере, 70% или более, более предпочтительно, 75% или более и более предпочтительно 80% или более (например, 85% или более, более предпочтительно, 90% или более и, наиболее предпочтительно, 95%, 98% или 99% или более) последовательности, приведенной в SEQ ID NO: 4, где области интронов могут быть удалены путем сплайсинга, и таким образом области экзонов соединяются и кодируют белок, обладающий активностью ФФК.
В другом варианте осуществления изобретения примеры нуклеотидной последовательности, содержащейся в нуклеиновой кислоте настоящего изобретения, включают нуклеотидные последовательности, включающие области интронов, имеющие идентичность нуклеотидной последовательности, по меньшей мере, 70% или более, более предпочтительно, 75% или более и более предпочтительно 80% или более (например, 85% или более, более предпочтительно, 90% или более и, наиболее предпочтительно, 95%, 98% или 99% или более) последовательности геномной ДНК, приведенной в SEQ ID NO: 4, и области экзонов, имеющие идентичность нуклеотидной последовательности, по меньшей мере, 70% или более, более предпочтительно, 75% или более и более предпочтительно 80% или более (например, 85% или более, более предпочтительно, 90% или более и, наиболее предпочтительно, 95%, 98% или 99% или более) последовательности, приведенной в SEQ ID NO: 5 или SEQ ID NO: 10, где области интронов могут быть удалены путем сплайсинга, и таким образом области экзонов соединяются и кодируют белок, обладающий активностью ФФК.
Процент идентичности между двумя нуклеотидными последовательностями может быть определен вышеописанным способом.
Кроме того, нуклеиновые кислоты настоящего изобретения включают нуклеиновые кислоты, каждая из которых состоит из нуклеотидной последовательности, имеющей делецию, замену или добавление одного или более нуклеотидов в нуклеотидной последовательности, приведенной в SEQ ID NO: 1, и кодирующей белок, обладающий активностью ФФК. В частности, применимая нуклеиновая кислота включает любую нуклеиновую кислоту из следующих нуклеотидных последовательностей:
(i) нуклеотидная последовательность, имеющая делецию одного или более (предпочтительно, одного или нескольких (например, от 1 до 330, от 1 до 300, от 1 до 250, от 1 до 200, от 1 до 150, от 1 до 100, от 1 до 50, от 1 до 30, от 1 до 25, от 1 до 20 или от 1 до 15, более предпочтительно, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1)) нуклеотидов в нуклеотидной последовательности, приведенной в SEQ ID NO: 1;
(ii) нуклеотидная последовательность, имеющая замену одного или более (предпочтительно, одного или нескольких (например, от 1 до 330, от 1 до 300, от 1 до 250, от 1 до 200, от 1 до 150, от 1 до 100, от 1 до 50, от 1 до 30, от 1 до 25, от 1 до 20 или от 1 до 15, более предпочтительно, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1)) в нуклеотидной последовательности, приведенной в SEQ ID NO: 1;
(iii) нуклеотидная последовательность, имеющая добавление одного или более (предпочтительно, одного или нескольких (например, от 1 до 330, от 1 до 300, от 1 до 250, от 1 до 200, от 1 до 150, от 1 до 100, от 1 до 50, от 1 до 30, от 1 до 25, от 1 до 20 или от 1 до 15, более предпочтительно, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1)) нуклеотидов в нуклеотидной последовательности, приведенной в SEQ ID NO: 1; или
(iv) нуклеотидная последовательность в любой комбинации (i)-(iii), где эта нуклеотидная последовательность кодирует белок, обладающий активностью ФФК.
В другом варианте осуществления нуклеиновая кислота настоящего изобретения также включает нуклеиновую кислоту, содержащую фрагмент нуклеотидной последовательности, указанной в любом из нижеприведенных пунктов (a)-(d):
(a) нуклеотидная последовательность, приведенная в SEQ ID NO: 1;
(b) нуклеотидная последовательность, кодирующая белок, состоящий из аминокислотной последовательности, приведенной в SEQ ID NO: 2;
(c) нуклеотидная последовательность, приведенная в SEQ ID NO: 4; и
(d) нуклеотидная последовательность, приведенная в SEQ ID NO: 5.
Нуклеотидная последовательность (a), приведенная в SEQ ID NO: 1, нуклеотидная последовательность (b), кодирующая белок, состоящий из аминокислотной последовательности, приведенной в SEQ ID NO: 2, и нуклеотидная последовательность(d), приведенная в SEQ ID NO: 5, показаны в Таблице 1. Нуклеотидная последовательность, приведенная в SEQ ID NO: 4, также описана выше. Фрагментами этих последовательностей являются ОРС, кодирующая последовательность, биологически активная область, область, используемая в качестве праймера, как описано далее, и область, которая может служить в качестве зонда, которые содержатся в этих нуклеотидных последовательностях, и могут быть либо природного происхождения, либо получены искусственно.
Нуклеиновая кислота настоящего изобретения включает следующие нуклеиновые кислоты.
(1) Нуклеиновая кислота по любому из приведенных ниже пунктов (a)-(g):
(a) нуклеиновая кислота, содержащая нуклеотидную последовательность, кодирующую белок, который состоит из аминокислотной последовательности с делецией, заменой или добавлением одной или более аминокислот в аминокислотной последовательности, приведенной в SEQ ID NO: 2;
(b) нуклеиновая кислота, которая гибридизуется в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, приведенной в SEQ ID NO: 1;
(c) нуклеиновая кислота, содержащая нуклеотидную последовательность, кодирующую белок, состоящая из нуклеотидной последовательности, которая имеет идентичность 70% или более нуклеотидной последовательности, приведенной в SEQ ID NO: 1; и
(d) нуклеиновая кислота, содержащая нуклеотидную последовательность, кодирующую белок, который состоит из аминокислотной последовательности, имеющей идентичность 70% или более аминокислотной последовательности, приведенной в SEQ ID NO: 2;
(e) нуклеиновая кислота, которая гибридизуется в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, кодирующей белок, состоящий из аминокислотной последовательности, приведенной в SEQ ID NO: 2;
(f) нуклеиновая кислота, которая гибридизуется в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, приведенной в SEQ ID NO: 4; и
(g) нуклеиновая кислота, содержащая нуклеотидную последовательность, кодирующую белок, состоящая из нуклеотидной последовательности, которая имеет идентичность 70% или более нуклеотидной последовательности, приведенной в SEQ ID NO: 4.
(2) Нуклеиновая кислота, описанная в (1), по любому из приведенных ниже пунктов (a)-(g):
(a) нуклеиновая кислота, содержащая нуклеотидную последовательность, кодирующую белок, который состоит из аминокислотной последовательности с делецией, заменой или добавлением от 1 до 130 аминокислот в аминокислотной последовательности, приведенной в SEQ ID NO: 2;
(b) нуклеиновая кислота, которая гибридизуется в условиях 2x SSC буфер при температуре 50°C с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, приведенной в SEQ ID NO: 1;
(c) нуклеиновая кислота, содержащая нуклеотидную последовательность, которая имеет идентичность 90% или более нуклеотидной последовательности, приведенной в SEQ ID NO: 1;
(d) нуклеиновая кислота, содержащая нуклеотидную последовательность, кодирующую белок, который состоит из аминокислотной последовательности, имеющей идентичность 90% или более аминокислотной последовательности, приведенной в SEQ ID NO: 2;
(e) нуклеиновая кислота, которая гибридизуется в условиях 2x SSC буфер при температуре 50°C с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, кодирующей белок, состоящий из аминокислотной последовательности, приведенной в SEQ ID NO: 2;
(f) нуклеиновая кислота, которая гибридизуется в условиях 2x SSC буфер при температуре 50°C с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, приведенной в SEQ ID NO: 4; и
(g) нуклеиновая кислота, содержащая нуклеотидную последовательность, которая имеет идентичность 90% или более нуклеотидной последовательности, приведенной в SEQ ID NO: 4.
Белок фосфатазы фосфатидной кислоты
Белок настоящего изобретения, который может быть либо природного происхождения, либо получен искусственно, включает белок, состоящий из аминокислотной последовательности, приведенной в SEQ ID NO: 2, и его мутантный белок, функционально эквивалентный этому белку. Такой белок, состоящий из аминокислотной последовательности, приведенной в SEQ ID NO: 2, описан выше. Термин «функционально эквивалентный белок» относится к белкам, обладающим активностью ФФК, описанной выше в разделе «Нуклеиновая кислота, кодирующая фосфатазу фосфатидной кислоты».
В настоящем изобретении примеры белков, функционально эквивалентных белку, состоящему из аминокислотной последовательности, приведенной в SEQ ID NO: 2, приведены ниже в пунктах (a) и (b):
(a) белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением одной или более аминокислот в аминокислотной последовательности, приведенной в SEQ ID NO: 2, и обладающий активностью ФФК; и
(b) белок, состоящий из аминокислотной последовательности, которая имеет идентичность 70% или более аминокислотной последовательности, приведенной в SEQ ID NO: 2, и обладающий активностью ФФК.
Вышеуказанная аминокислотная последовательность с делецией, заменой или добавлением одной или более аминокислот в аминокислотной последовательности, приведенной в SEQ ID NO: 2, или аминокислотная последовательность, которая имеет идентичность 70% или более аминокислотной последовательности, приведенной в SEQ ID NO: 2, является такой, как описано выше в разделе «Нуклеиновая кислота, кодирующая фосфатазу фосфатидной кислоты». «Белок, обладающий активностью ФФК» включает мутанты белка, кодируемого нуклеиновой кислотой, содержащей нуклеотидную последовательность, приведенную в SEQ ID NO: 1, или белки, мутированные многими типами модификаций, такими как делеция, замена и добавление одной или более аминокислот в аминокислотной последовательности, приведенной в SEQ ID NO: 2, или модифицированные белки, имеющие, например, модифицированные боковые цепи аминокислот, или белки, слитые с другими белками, где эти белки обладают активностью ФФК.
Белок настоящего изобретения может быть получен искусственно. В этом случае белок может быть получен путем химического синтеза, такого как синтез с использованием флуоренилметилоксикарбонила (Fmoc) или синтез с использованием трет-бутилоксикарбонила (tBoc). Кроме того, синтезаторы пептидов, производства компаний Advanced ChemTech, Perkin Elmer, Pharmacia, Protein Technology Instrument, Synthecell-Vega, PerSeptive, Shimadzu Corporation или других производителей также могут быть использованы для химического синтеза.
Белок настоящего изобретения также включает следующие белки:
(1) (a) белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением одной или более аминокислот в аминокислотной последовательности, приведенной в SEQ ID NO: 2; и
(b) белок, состоящий из аминокислотной последовательности, которая имеет идентичность 80% или более аминокислотной последовательности, приведенной в SEQ ID NO: 2; и
(2) белок по любому из приведенных ниже пунктов (a) и (b):
(a) белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением от 1 до 110 аминокислот в аминокислотной последовательности, приведенной в SEQ ID NO: 2; и
(b) белок, состоящий из аминокислотной последовательности, которая имеет идентичность 90% или более аминокислотной последовательности, приведенной в SEQ ID NO: 2.
Клонирование нуклеиновых кислот
Нуклеиновая кислота ФФК может быть клонирована, например, путем скрининга из библиотеки кДНК с использованием соответствующего зонда. Клонирование может быть выполнено путем ПЦР амплификации с использованием соответствующих праймеров и последующего лигирования в подходящий вектор. Клонированная нуклеиновая кислота может быть в дальнейшем клонирована в другой вектор.
Могут быть использованы коммерчески доступные плазмидные векторы, такие как pBlue-ScriptTM SK(+) (Stratagene), pGEM-T (Promega), pAmp (TM: Gibco-BRL), p-Direct (Clontech), or pCR2.1-TOPO (Invitrogen). При ПЦР амплификации праймерами могут быть любые области нуклеотидной последовательности, приведенной в SEQ ID NO: 5, Например, праймер PAP2.2-1f: 5’-TTCCGTCAGGACACTCCTCCAGT-3’ (SEQ ID NO: 6) может быть использован в качестве прямого праймера, а праймер PAP2.2-4r: 5’-GACAATGCCGAGGATCGAGCC-3’ (SEQ ID NO: 7) может быть использован в качестве обратного праймера. Например, ПЦР затем проводят с использованием этих праймеров и ДНК-полимеразой с кДНК, полученной из клеток M. alpina. Эта методика может быть без труда выполнена специалистами в данной области техники, например, в соответствии с «Molecular Cloning, A Laboratory Manual 3rd ed.» (Cold Spring Harbor Press (2001)). Например, ПЦР в настоящем изобретении может быть проведена в следующих условиях:
Температура денатурации: от 90°C до 95°C
Температура отжига: от 40°C до 60°C
Температура элонгации: от 60°C до 75°C, и
Количество циклов: 10 или более циклов.
ПЦР-продукт может быть очищен известным способом, например, способом с использованием набора реагентов, такого как GENECLEAN (Funakoshi Co., Ltd.), набора реагентов для очистки ПЦР-продуктов QIAquick (QIAGEN) или ExoSAP-IT (GE Healthcare Bio-Sciences); способом с использованием фильтра из ДЭАЭ-целлюлозы; или способом с использованием диализной трубки. В случае использования агарозного геля ПЦР-продукт подвергается электрофорезу в агарозном геле и выделяется из него, например, при помощи набора реагентов GENECLEAN (Funakoshi Co., Ltd.) или набора реагентов для экстракции из геля QIAquick (QIAGEN) или при помощи метода замораживание-сдавливание.
Нуклеотидную последовательность клонированной нуклеиновой кислоты можно определить при помощи секвенатора нуклеиновых кислот.
Векторная конструкция для экспрессии ФФК и получения трансформанта
Настоящее изобретение также относится к рекомбинантному вектору, содержащему нуклеиновую кислоту, кодирующую ФФК. Кроме того, настоящее изобретение относится к трансформанту, трансформированному таким рекомбинантным вектором.
Рекомбинантный вектор и трансформант могут быть получены следующим образом. Плазмиду, имеющую нуклеиновую кислоту, кодирующую MaPaP2.2 или его мутант, обрабатывают ферментом рестрикции. Примеры фермента рестрикции включают без ограничений EcoRI, KpnI, BamHI и SalI. Конец может быть сделан тупым при помощи T4 полимеразы. Вырезанный ферментом фрагмент ДНК выделяют при помощи электрофореза в агарозном геле. Этот фрагмент ДНК встраивают в экспрессионный вектор при помощи известного способа, чтобы получить вектор для экспрессии ФФК. Этот экспрессионный вектор вводят в клетку хозяин, чтобы получить трансформант, который предназначен для получения желаемого белка.
В этом случае экспрессионный вектор и хозяин могут быть любого типа, который обеспечивает экспрессию желаемого белка. Примеры хозяина включают низшие грибы, бактерии, растения, животные и их клетки. Примеры низших грибов включают мицелиальные грибы, такие как липид-продуцирующий грибок M. alpina и штаммы дрожжей, таких как Saccharomyces cerevisiae. Примеры бактерий включают Escherichia coli и Bacillus subtilis; и примеры растений включают масличные растения, такие как рапс, соя, хлопчатник, сафлор и лен.
Подходящими для использования липид-продуцирующими микроорганизмами являются, например, штаммы, описанные в MYCOTAXON, Vol. XLIV, NO. 2, pp. 257-265 (1992), и их конкретные примеры включают микроорганизмы, принадлежащие к роду Mortierella, например микроорганизмы, принадлежащие к подроду Mortierella, например, Mortierella elongata IFO8570, Mortierella exigua IFO8571, Mortierella hygrophila IFO5941, Mortierella alpina IFO8568, ATCC16266, ATCC32221, ATCC42430, CBS219.35, CBS224.37, CBS250.53, CBS343.66, CBS527.72, CBS528.72, CBS529.72, CBS608.70, и CBS754.68; и микроорганизмы, принадлежащие к подроду Micromucor, например, Mortierella isabellina CBS194.28, IFO6336, IFO7824, IFO7873, IFO7874, IFO8286, IFO8308, IFO7884, Mortierella nana IFO8190, Mortierella ramanniana IFO5426, IFO8186, CBS112.08, CBS212.72, IFO7825, IFO8184, IFO8185, IFO8287, и Mortierella vinacea CBS236.82. В частности, Mortierella alpina является предпочтительным.
В случае использования в качестве хозяина низшего гриба, нуклеиновая кислота настоящего изобретения предпочтительно является самореплицирующейся в хозяине или предпочтительно имеет структуру, обеспечивающую ее вставку в грибковую хромосому. Предпочтительно, нуклеиновая кислота также включает промотор и терминатор. В случае использования в качестве хозяина M. alpina, в качестве экспрессионного вектора может использоваться, например, pD4, pDuraSC, или pDura5. Любой промотор, который обеспечивает экспрессию в хозяине, может быть использован, и его примеры включают промоторы, полученные из M. alpina, такие как промотор гена гистона H4.1, промотор гена ГАФД (глицеральдегид-3-фосфатдегидрогеназы) и промотор гена ФЭТ (фактора элонгации трансляции).
Примеры способа введения рекомбинантного вектора в мицелиальные грибы, такие как M. alpina, включают электропорацию, метод сферопластов, метод доставки частиц и прямую микроинъекцию ДНК в ядра. В случае использования ауксотрофного штамма-хозяина трансформированный штамм может быть получен путем отбора штамма, который растет на селективной среде, в которой отсутствуют определенные питательные вещества. В другом случае, при трансформации с использованием маркерного гена устойчивости к лекарственному препарату, колонии устойчивых к лекарственному препарату клеток могут быть получены путем культивирования клеток-хозяев в селективной питательной среде, содержащей этот лекарственный препарат.
В случае использования в качестве хозяина дрожжей, в качестве экспрессионного вектора может быть использован, например, pYE22m. В другом варианте могут быть использованы коммерчески доступные экспрессионные векторы дрожжей, такие как pYES (Invitrogen) и pESC (STRATAGENE). Примеры хозяев, пригодных для использования в настоящем изобретении, включают без ограничений Saccharomyces cerevisiae штамм EH13-15 (trp1, MATα). Примеры промотора, который может быть использован, включают промоторы, полученные из дрожжей, такие как ГАФД промотор, ga11 промотор и ga110 промотор.
Примеры способов введения рекомбинантного вектора в дрожжи включают способ с использованием ацетата лития, электропорацию, метод сферопластов, декстран-опосредованную трансфекцию, осаждение фосфатом кальция, полибрен-опосредованную трансфекцию, слияние протопластов, инкапсуляцию полинуклеотида(ов) в липосомы и прямую микроинъекцию ДНК в ядра.
В случае использования в качестве хозяина бактерии, такой как E. coli, в качестве экспрессионного вектора может быть использован pGEX и pUC18 производства Pharmacia. Примеры промотора включают промоторы, полученные, например, из E. coli или фага, такие как trp промотор, lac промотор, PL промотор и PR промотор. Примеры способа введения рекомбинантного вектора в бактерии включают электропорацию и метод с использованием хлорида кальция.
Способ получения композиции жирных кислот
Настоящее изобретение относится к способу получения композиции жирных кислот из вышеописанных трансформантов, т.е. способу получения композиции жирных кислот из продукта культивации, полученного путем культивирования трансформанта. Композиция жирных кислот включает комбинацию в ней одной или нескольких жирных кислот. Эти жирные кислоты могут находиться в виде свободных жирных кислот или в виде их липидов, таких как триглицерид или фосфолипид. В частности, композиция жирных кислот настоящего изобретения может быть получена следующими способами. Однако эти способы не ограничиваются способами, перечисленными ниже, и композиция жирных кислот также может быть получена любым другим известным способом.
Среда, используемая для культивирования организма, экспрессирующего ФФК, может представлять собой любой культуральный раствор (среду), который имеет необходимое значение pH и осмотическое давление, и содержит биоматериалы, такие как питательные вещества, микроэлементы, сыворотки и антибиотики, необходимые для роста каждого хозяина. Например, в случае экспрессии ФФК трансформированными дрожжами, неограниченные примеры используемой питательной среды включают среду SC-Trp, среду YPD и среду YPD5. Типичным примером состава конкретной среды, т.е. среды SC-Trp, является следующий: один литр среды включает 6,7 г основы азотного агара для дрожжей без аминокислот (DIFCO), 20 г глюкозы и 1,3 г порошка аминокислот (смесь из 1,25 г аденин сульфата, 0,6 г аргинина, 3 г аспарагиновой кислоты, 3 г глутаминовой кислоты, 0,6 г гистидина, 1,8 г лейцина, 0,9 г лизина, 0,6 г метионина, 1,5 г фенилаланина, 11,25 г серина, 0,9 г тирозина, 4,5 г валина, 6 г треонина и 0,6 г урацила).
Любые условия культивирования, приемлемые для роста хозяина, и подходящие для стабильного сохранения продуцируемого фермента, могут быть использованы. В частности, отдельные условия, такие как степень анаэробности, период культивирования, температура, влажность и статическое культивирование или культивирование с использованием встряхивания-перемешивания, могут регулироваться. Культивирование может проводиться в одних и тех же условиях (одноэтапное культивирование) или путем, так называемого двухэтапного или трехэтапного культивирования, включающего два или более различных условий культивирования. Для крупномасштабного культивирования двух- или более этапное культивирование является предпочтительным, по причине высокой эффективности такого культивирования.
При двухэтапном культивировании с использованием дрожжей в качестве хозяина композиция жирных кислот настоящего изобретения может быть получена следующим образом: На этапе прекультивирования колонией трансформанта инокулируют, например, среду SC-Trp и культивируют при встряхивании-перемешивании при температуре 30°C в течение двух суток. Затем, на этапе основного культивирования 500 мкл раствора прекультивирования добавляют в 10 мл среды YPD5 (2% дрожжевого экстракта, 1% полипептона и 5% глюкозы) с последующим культивированием при встряхивании-перемешивании при температуре 30°C в течение двух суток.
Композиция жирных кислот
Настоящее изобретение также относится к композиции жирных кислот, содержащей агрегаты одной или нескольких жирных кислот в клетках экспрессирующих MaPAP2.2 и его мутанты, предпочтительно, к композиции жирных кислот, полученной путем культивирования трансформанта, экспрессирующего MaPAP2.2 и его мутанты. Жирные кислоты могут находиться в виде свободных жирных кислот или в виде их липидов, таких как триглицерид или фосфолипид.
Жирные кислоты, содержащиеся в композиции жирных кислот настоящего изобретения, представляют собой монокарбоновые кислоты линейных или разветвленных углеводов с длинной цепью, и их примеры включают без ограничений миристиновую кислоту (тетрадекановую кислоту) (14:0), миристолеиновую кислоту (тетрадеценовую кислоту) (14:1), пальмитиновую кислоту (гексадекановую кислоту) (16:0), пальмитолеиновую кислоту (9-гексадеценовую кислоту) (16:1), маргариновую кислоту (гептадекановую кислоту) (17:0), стеариновую кислоту (октадекановую кислоту) (18:0), олеиновую кислоту (цис-9-октадеценовую кислоту) (18:1(9)), вакценовую кислоту (11-октадеценовую кислоту) (18:1(11)), линолевую кислоту(цис,цис-9,12 октадекадиеновую кислоту) (18:2(9,12)), α-линоленовую кислоту (9,12,15-октадекатриеновую кислоту) (18:3(9,12,15)), γ-линоленовую кислоту (6,9,12-октадекатриеновую кислоту) (18:3(6,9,12)), стеаридоновую кислоту (6,9,12,15-октадекатетраеновую кислоту) (18:4(6,9,12,15)), арахидиновую кислоту (эйкозановую кислоту) (20:0), (8,11-эйкозадиеновую кислоту) (20:2(8,11)), мидовую кислоту (5,8,11-эйкозатриеновую кислоту) (20:3(5,8,11)), дигомо-γ-линоленовую кислоту (8,11,14- эйкозатриеновую кислоту) (20:3(8,11,14)), арахидоновую кислоту (5,8,11,14-эйкозатетраеновую кислоту) (20:4(5,8,11,14)), эйкозатетраеновую кислоту (8,11,14,17-эйкозатетраеновую кислоту) (20:4 (8,11,14,17)), эйкозапентаеновую кислоту (5,8,11,14,17- эйкозапентаеновую кислоту) (20:5(5,8,11,14,17)), бегеновую кислоту (докозановую кислоту) (22:0), (7,10,13,16- докозатетраеновую кислоту) (22:4(7,10,13,16)), (7,10,13,16,19- докозапентаеновую кислоту) (22:5(7,10,13,16,19)), (4,7,10,13,16-докозапентаеновую кислоту) (22:5(4,7,10,13,16)), (4,7,10,13,16,19-докозагексаеновую кислоту) (22:6(4,7,10,13,16,19)), лигноцериновую кислоту (тетракозановую кислоту) (24:0), нервоновую кислоту (цис-15-тетракозеновую кислоту) (24:1) и церотиновую кислоту (гексакозановую кислоту) (26:0). Необходимо отметить, что наименования веществ являются тривиальными названиями, определенными Биохимической Номенклатурой ИЮПАК, и их систематические наименования приведены в скобках вместе с числами, обозначающими количество атомов углерода и положения двойных связей.
В композиции жирных кислот настоящего изобретения эти жирные кислоты могут использоваться отдельно или в комбинации с двумя или более различными жирными кислотами.
Пищевые или другие продукты, содержащие композицию жирных кислот
Настоящее изобретение также относится к пищевому продукту, содержащему вышеописанную композицию жирных кислот. Композиция жирных кислот настоящего изобретения может быть использована для получения пищевых продуктов, содержащих жиры и масла, и получения промышленного сырья (например, сырья для косметических изделий, фармацевтических препаратов (например, для наружного применения на коже) и мыла) общепринятыми способами. Косметические изделия (косметические композиции) или фармацевтические препараты (фармацевтические композиции) могут быть приготовлены в любой лекарственной форме, включая без ограничений, раствор, пасту, гель, твердое вещество и порошок. Примеры форм пищевых продуктов включают фармацевтические препараты, такие как капсула, натуральная жидкая пища, рацион питания с полупереваренными питательными веществами и элементный рацион питания, где композиция аминокислот настоящего изобретения смешана с белками, сахарами, жирами, микроэлементами, витаминами, эмульгаторами и вкусоароматическими добавками; и переработанные формы, такие как питьевые препараты и питательные вещества для энтерального питания.
Кроме того, примеры пищевого продукта настоящего изобретения включают без ограничений пищевые добавки, оздоровительные пищевые продукты, функциональные пищевые продукты, продукты для детского питания, модифицированное молоко для питания грудных детей, модифицированное молоко для питания недоношенных детей и пищевые продукты для лиц пожилого возраста. Термин «пищевой продукт» используется здесь в качестве собирательного термина для обозначения съедобных материалов в виде твердого вещества, жидкости или их смеси.
Термин «пищевые добавки» относится к пищевым продуктам, обогащенным определенными питательными ингредиентами. Термин «оздоровительные пищевые продукты» относится к пищевым продуктам, которые являются оздоравливающими или полезными для здоровья, и включает пищевые добавки, натуральные пищевые продукты и диетические пищевые продукты. Термин «функциональные пищевые продукты» относится к пищевым продуктам, содержащим питательные ингредиенты, которые помогают организму контролировать свои функции, и является синонимом термина «пищевые продукты для специального медицинского применения». Термин «продукты для детского питания» относится к пищевым продуктам, предназначенным для питания детей в возрасте до примерно 6 лет. Термин «пищевые продукты для лиц пожилого возраста» относится к пищевым продуктам, обработанным для более легкого их переваривания и всасывания, по сравнению с необработанными пищевыми продуктами. Термин «модифицированное молоко для питания грудных детей» относится к модифицированному молоку, предназначенному для питания детей в возрасте до примерно одного года. Термин «модифицированное молоко для питания недоношенных детей» относится к модифицированному молоку, предназначенному для питания недоношенных детей до примерно 6 месяцев после рождения.
Примеры этих пищевых продуктов включают натуральные пищевые продукты (обработанные жиром или маслом), такие как мясо, рыба и орехи; пищевые продукты с добавлением жира или масла во время приготовления, такие как блюда китайской кухни, китайская лапша и супы; пищевые продукты, приготовленные с использованием жира или масла в качестве нагревающей среды, такие как темпура (жареные во фритюре рыба и овощи), жареные во фритюре пищевые продукты, жареный тофу, китайский жареный рис, пончики, японские жареные лепешки (каринто); пищевые продукты на основе жира и масла или переработанные пищевые продукты, в которые жир и масло были добавлены во время технологической обработки, такие как масло, маргарин, майонез, соус, шоколад, лапша быстрого приготовления, карамель, песочное печенье, печенье, пирожные и мороженное; и пищевые продукты, обрызганные или покрытые жиром и маслом после приготовления, такие как рисовое печенье, хрустящее печенье и булочки с начинкой из сладких бобов адзуки (анпан). Пищевые продукты настоящего изобретения, однако, не ограничиваются пищевыми продуктами, содержащими жир и масло, и другие примеры пищевых продуктов включают сельскохозяйственные пищевые продукты, такие как хлебобулочные изделия, лапша, подвергнутый тепловой обработке рис, сладости (например, конфеты, жевательная резинка, мармелад, пастилки, японские сладости), тофу и продукты их переработки; ферментированные пищевые продукты, такие как очищенное саке, лечебный алкогольный напиток, алкогольный напиток в качестве приправы (мирин), уксус, соевый соус, и мисо; пищевые продукты животного происхождения, такие как йогурт, ветчина, бекон и колбаса; морепродукты, такие как рыбная паста (камабоко), рыбная паста жареная во фритюре (агетен) и рыбные котлеты (ханпен); и фруктовые напитки, безалкогольные напитки, спортивные напитки, алкогольные напитки и чай.
Способ оценки или отбора штаммов с использованием нуклеиновой кислоты, кодирующей ФФК, или белка ФФК
Настоящее изобретение также относится к способу оценки или отбора липид-продуцирующего грибка с использованием нуклеиновой кислоты, кодирующей ФФК, или белка ФФК настоящего изобретения. Подробное описание приведено ниже.
(1) Способ оценки
Одним вариантом осуществления настоящего изобретения является способ оценки липид-продуцирующего грибка с использованием нуклеиновой кислоты, кодирующей ФФК, или белка ФФК настоящего изобретения. В этом способе оценки в соответствии с настоящим изобретением, например, штамм липид-продуцирующего грибка, в качестве проверяемого штамма, оценивают на наличие активности ФФК с использованием праймеров или зондов, разработанных на основе нуклеотидной последовательности нуклеиновой кислоты, кодирующей ФФК, настоящего изобретения. Такая оценка может быть проведена по известным методикам, описанным, например, в Публикации Международной заявки No. WO01/040514 и в заявке на Патент Японии JP-A-8-205900. Способ оценки будет кратко описан ниже.
Первым этапом является получение генома проверяемого штамма. Геном может быть получен любым известным способом, таким как метод Херефорда или метод с использованием ацетата калия (см., например, Methods in Yeast Genetics, Cold Spring Harbor Laboratory Press, p. 130 (1990)).
Праймеры или зонды разрабатывают на основе нуклеотидной последовательности нуклеиновой кислоты, кодирующей ФФК, настоящего изобретения, предпочтительно, последовательности, приведенной в SEQ ID NO: 1. Эти праймеры или зонды могут представлять собой любые области нуклеотидной последовательности нуклеиновой кислоты, кодирующей ФФК, настоящего изобретения, и могут быть разработаны по известной методике. Число нуклеотидов в полинуклеотиде, используемом в качестве праймера, как правило, составляет 10 или более, и предпочтительно, от 15 до 25. Число нуклеотидов, подходящее для области между праймерами, как правило, составляет от 300 до 2000.
Праймеры или зонды, полученные выше, используются для выяснения того, содержит ли геном проверяемого штамма последовательность, характерную для нуклеотидной последовательности нуклеиновой кислоты, кодирующей ФФК, настоящего изобретения. Последовательность, характерная для нуклеотидной последовательности настоящего изобретения может быть обнаружена любым известным способом. Например, полинуклеотид, содержащий часть нуклеотидной последовательности нуклеиновой кислоты, кодирующей ФФК, настоящего изобретения или полинуклеотид, содержащий нуклеотидную последовательность, комплиментарную этой нуклеотидной последовательности, используется в качестве одного праймера, в то время как полинуклеотид, содержащий часть последовательности, расположенной левее или правее этой последовательности, или полинуклеотид, содержащий нуклеотидную последовательность, комплиментарную этой нуклеотидной последовательности, используется в качестве другого праймера, и нуклеиновая кислота из проверяемого штамма амплифицируется при помощи ПЦР или любого другого метода. При помощи этой процедуры можно подтвердить наличие продукта амплификации и определить молекулярную массу продукта амплификации.
Условия ПЦР, подходящие для способа настоящего изобретения, которые могут быть использованы без любых ограничений, являются, например, следующими:
Температура денатурации: от 90°C до 95°C
Температура отжига: от 40°C до 60°C
Температура элонгации: от 60°C до 75°C, и
Количество циклов: 10 или более циклов.
Продукты реакции могут быть разделены при помощи электорфореза в агарозном геле или любым другим способом, для определения молекулярной массы продукта амплификации. Таким образом, активность ФФК проверяемого штамма может быть спрогнозирована или оценена путем подтверждения того, является ли молекулярная масса продукта амплификации достаточной для того, чтобы его можно было охарактеризовать как молекулу нуклеиновой кислоты, соответствующую области, специфичной для нуклеотидной последовательности настоящего изобретения. Кроме того, анализ нуклеотидной последовательности продукта амплификации при помощи вышеописанного способа позволяет спрогнозировать или оценить активность ФФК более точно. Способ оценки активности ФФК представляет собой способ, который описан выше.
В другом варианте, при проведении оценки в соответствии с настоящим изобретением, активность ФФК проверяемого штамма может быть оценена путем культивирования проверяемого штамма и измерения уровня экспрессии ФФК, кодируемой нуклеотидной последовательностью настоящего изобретения, например, последовательностью, приведенной в SEQ ID NO: 1. Уровень экспрессии ФФК может быть измерен путем культивирования проверяемого штамма в соответствующих условиях и количественного определения мРНК или белка ФФК. мРНК и белок могут быть количественно определены любым известным способом. Например, мРНК может быть количественно определена при помощи Нозерн-гибридизации или количественной ОТ-ПЦР, а белок может быть количественно определен при помощи Вестерн-блоттинга (Current Protocols in Molecular Biology, John Wiley & Sons, 1994-2003).
(2) Способ отбора
Другим вариантом осуществления настоящего изобретения является способ отбора липид-продуцирующего грибка с использованием нуклеиновой кислоты, кодирующей ФФК, или белка ФФК настоящего изобретения. При отборе в соответствии с настоящим изобретением, штамм, обладающий желаемой активностью, может быть отобран путем культивирования проверяемого штамма, измерения уровня экспрессии ФФК, кодируемой нуклеотидной последовательностью настоящего изобретения, например, последовательностью, приведенной в SEQ ID NO: 1, и отбора штамма с желаемым уровнем экспрессии. В другом варианте, желаемый штамм может быть отобран путем определения стандартного штамма, раздельного культивирования стандартного штамма и проверяемого штамма, измерения уровня экспрессии каждого штамма и сравнения уровня экспрессии стандартного штамма с уровнем экспрессии проверяемого штамма. В частности, например, стандартный штамм и проверяемые штаммы культивируют в соответствующих условиях и измеряют уровень экспрессии каждого штамма. Штамм, проявляющий желаемую активность, может быть отобран путем отбора проверяемого штамма, обладающего более высокой или более низкой экспрессией, чем у стандартного штамма. Желаемая активность может быть определена, например, путем измерения уровня экспрессии ФФК и составного соотношения композиции жирных кислот, продуцируемой ФФК, как описано выше.
При отборе в соответствии с настоящим изобретением, проверяемый штамм, обладающий желаемой активностью, может быть отобран путем культивирования проверяемых штаммов и отбора штамма, имеющего высокую или низкую активность в соответствии с настоящим изобретением. Желаемая активность может быть определена, например, путем измерения уровня экспрессии ФФК и составного соотношения композиции жирных кислот, продуцируемой ФФК, как описано выше.
Примеры проверяемого штамма и стандартного штамма включают без ограничений штаммы, трансформированные вектором настоящего изобретения, штаммы, модифицированные таким образом, чтобы в них подавлялась экспрессия нуклеиновой кислоты настоящего изобретения, мутагенизированные штаммы и штаммы, мутировавшие естественным путем. Активность ФФК может быть измерена, например, способом, описанным здесь в разделе «Нуклеиновая кислота, кодирующая фосфатазу фосфатидной кислоты настоящего изобретения». Примеры мутагенеза включают без ограничений физические методы, такие как УФ или радиоактивное облучение, и химические методы, такие как химическая обработка, например, ЭМС (этилметансульфонат) или N-метил-N-нитрозогуанидином (см., например, Yasuji Oshima ed., Biochemistry Experiments vol. 39, Experimental Protocols for Yeast Molecular Genetics, pp. 67-75, Japan Scientific Societies Press).
Примеры штаммов, используемых в качестве стандартного штамма и проверяемого штамма настоящего изобретения, включают без ограничений липид-продуцирующий грибок и дрожжи, описанные выше. В частности, стандартный штамм и проверяемый штамм могут представлять собой любую комбинацию штаммов, принадлежащих к разным родам или видам, а также один или более проверяемых штаммов могут использоваться одновременно.
Следующие примеры дополнительно иллюстрируют настоящее изобретение, но не предназначены для ограничения объема настоящего изобретения.
Примеры
Пример 1: Геномный анализ Mortierella alpina
Штаммом 1S-4 M. alpina инокулировали 100 мл среды GY2:1 (2% глюкозы, 1% дрожжевого экстракта, pH 6,0) и культивировали при встряхивании при температуре 28°C в течение 2 суток. Клетки собирали путем фильтрации для получения геномной ДНК с использованием набора реагентов DNeasy (QIAGEN).
Нуклеотидную последовательность геномной ДНК определяли на геномном секвенаторе Roche 454 GS FLX Standard. Этот процесс включал два прочтения нуклеотидной последовательности библиотеки фрагментов и три прочтения нуклеотидной последовательности библиотеки mate-pair. Полученные нуклеотидные последовательности были объединены в 300 суперконтигов.
Пример 2: Синтез кДНК и получение библиотеки кДНК
Штаммом 1S-4 M. alpina инокулировали 100 мл среды (1,8% глюкозы, 1% дрожжевого экстракта, pH 6,0) и прекультивировали при температуре 28°C в течение 3 суток. В сосуд для культивирования емкостью 10 л (Able Co., Tokyo) загружали 5 л среды (1,8% глюкозы, 1% соевого порошка, 0,1% оливкового масла, 0,01% Адеканола, 0,3% KH2PO4, 0,1% Na2SO4, 0,05% CaCl2·2H2O, 0,05% MgCl2·6H2O, pH 6,0), и которую инокулировали полностью продуктом прекультивирования, с последующим аэробным культивированием с постоянным перемешиванием в условиях 300 об./мин, 1 vvm и температуре 26°C в течение 8 суток. В дни 1, 2 и 3 культивирования добавляли глюкозу в количестве, соответствующем 2%, 2% и 1,5%, соответственно. Клетки собирали на каждом этапе культивирования (дни 1, 2, 3, 6 или 8) для получения тотальной РНК при помощи метода с использованием гуанидин гидрохлорида и CsCl. Поли(A)+ РНК очищали от тотальной РНК с использованием набора реагентов для выделения мРНК Oligotex-dT30<Super>mRNA Purification Kit (Takara Bio Inc.). Библиотеку кДНК для каждого этапа получали с использованием набора реагентов ZAP-cDNA Gigapack III Gold Cloning Kit (STRATAGENE).
Пример 3: Поиск гомологов полученного из дрожжей DPP1
В геномной базе данных проводили поиск гомологов ScDPP1 (YDR284C: регистрационный номер AAS56070) и ScLPP1 (YDR503C: регистрационный номер AAT93210). В результате было идентифицировано два суперконтига. Один из них является суперконтигом, включающим геномную последовательность MaPAP1, а другой - суперконтигом, включающим SEQ ID NO: 4. Ген, представленный SEQ ID NO: 4, был назван MaPAP2.2.
Пример 4: Клонирование и анализ последовательности MaPAP2.2
(1) Клонирование
Для клонирования кДНК MaPAP2.2 были синтезированы следующие праймеры.
Праймер PAP2.2-1f: TTCCGTCAGGACACTCCTCCAGT (SEQ ID NO: 6)
Праймер PAP2.2-4r: GACAATGCCGAGGATCGAGCC (SEQ ID NO: 7)
Библиотеку, полученную как описано выше, использовали в качестве матрицы для постановки ПЦР с полимеразой ExTaq (Takara Bio Inc.) в следующих условиях: 94°C в течение 2 минут и 30 циклов, состоящих из 94°C в течение 1 минуты, 55°C в течение 1 минуты и 72°C в течение 1 минуты, используя комбинацию праймеров PAP2.2-1F и PAP2.2-4R. Полученный фрагмент ДНК размером примерно 0,4 т.п.н. клонировали при помощи набора реагентов для клонирования TOPO-TA Cloning (Invitrogen), чтобы определить нуклеотидную последовательность каждой вставки. Плазмида, содержащая нуклеотиды 534-904 последовательности, приведенной в SEQ ID NO: 3, была названа pCR-MaPAP2.2-P.
Затем плазмиду pCR-MaPAP2.2-P использовали в качестве матрицы для постановки ПЦР с вышеуказанными праймерами для получения зонда. Для реакции использовали полимеразу ExTaq (Takara Bio Inc.), но вместо смеси дНТФ использовали смесь для ПЦР-мечения (Roche Diagnostics), входящую в состав набора реагентов для получения зонда (MaPAP2.2 зонда), меченного дигоксигенином (DIG), из амплифицируемой ДНК. Этот зонд использовали для скрининга библиотеки кДНК.
Условия гибридизации были следующими.
Буфер: 5x SSC, 1% ДСН, 50 мМ Трис-Hcl (pH 7,5), 50% формамида;
Температура: 42°C (в течение ночи);
Условия отмывки: три раза в растворе 0,2x SSC, 0,1% ДСН, при температуре 65°C, в течение 20 минут.
Детекцию проводили с помощью набора реагентов для DIG детекции нуклеиновых кислот (Roche Diagnostics). Плазмиду отделяли путем вырезания in vivo из фаговых клонов, полученных в результате скринига, для получения плазмидной ДНК каждого клона. Плазмида, имевшая самую длинную вставку, содержала нуклеотидную последовательность, приведенную в SEQ ID NO: 5, и получила название плазмида pB-MaPAP2.2.
Нуклеотиды 75-1166 в последовательности SEQ ID NO: 5 (те же, что в SEQ ID NO: 3) были идентифицированы как кодирующая последовательность MaPAP2.2, по наличию старт и стоп кодонов и других сопоставлений с гомологами ФФК2.
(2) Анализ последовательности
Поиск гомологий нуклеотидной последовательности гена MaPAP2.2 и кодируемой им предсказанной аминокислотной последовательности проводили среди известных нуклеотидных и аминокислотных последовательностей при помощи программ BLAST и ClustalW. Среди последовательностей, обнаруженных при поиске гомологий среди аминокислотных последовательностей зарегистрированных в GenBank, при помощи программы BLASTX, последовательностью, имеющей самое низкое значение E-индекса, оказался предполагаемый белок, полученный из Laccaria bicolor (регистрационный номер: XP_001878243), который имел идентичность по аминокислотной последовательности 36,7%. На Фигуре 2 показано выравнивание аминокислотной последовательности MaPAP2.2 с предполагаемым белком, полученным из Laccaria bicolor, и ScDPP1 (YDR284C: регистрационный номер AAS56070), полученным из дрожжей.
Идентичность аминокислотной последовательности между MaPAP1, известным как Mg2+-независимая ФФК (ФФК2), полученная из Mortierella alpina (WO2009/008466), и MaPAP2.2 составила примерно 20,5%. Выравнивание аминокислотных последовательностей MaPAP2.2 и MaPAP1 показано на Фигуре 3. Ферменты семейства ФФК2 имеют три консервативные области, и также известны аминокислоты, существенные для их активности. Как показано на Фигуре 3, MaPAP2.2 также содержит эти три консервативные области (дважды подчеркнуты на Фигуре 3) и остатки, существенные для активности, т.е. аргинин в домене 1 и гистидины в доменах 2 и 3 (остатки, обозначенные знаком «*» на Фигуре 3).
Пример 5: Функциональный анализ MaPAP2.2
(1) Конструирование дрожжевого экспрессионного вектора
Для того чтобы экспрессировать MaPAP2.2 в клетках дрожжей, дрожжевые экспрессионные векторы конструировали следующим образом. Синтезировали праймеры Eco-PAP2-2-F и Kpn-PAP2-2-R для последующего проведения ПЦР с полимеразой ExTaq (Takara Bio Inc.), используя pB-MaPAP2.2 в качестве матрицы.
Праймер Eco-PAP2-2-F: GAATTCATGTTCTCGTCCATGCGCTTCAAG (SEQ ID NO: 8).
Праймер Kpn-PAP2-2-R: TGGTACCTCATGGTCCCAAGTATACATCGTTCC (SEQ ID NO: 9).
Полученный фрагмент ДНК размером 1,1 т.п.н. клонировали по TA при помощи набора реагентов для клонирования TOPO-TA Cloning (Invitrogen) для подтверждения его нуклеотидной последовательности. Плазмида, несущая правильную нуклеотидную последовательность кодирующей последовательности (SEQ ID NO: 3) MaPAP2.2 была названа pCR-MaPAP2.2. Фрагмент ДНК размером примерно 1,1 т.п.н., образующийся в результате расщепления плазмиды pCR-MaPAP2.2 ферментами рестрикции EcoRI и KpnI, и фрагмент ДНК размером примерно 8,3 т.п.н., образующийся в результате расщепления дрожжевого экспрессионного вектора pYE22m (Biosci. Biotech. Biochem., 59, 1221-1228, 1995) ферментами рестрикции EcoRI и KpnI, лигировали при помощи набора реагентов Ligation High (TOYOBO) для получения плазмиды pYE-MaPAP2.2.
(2) Получение трансформированных дрожжей
Каждую из плазмид pYE22m, pYE-MaPAP2.2 и pYE-MaPAP1 (WO2009/008466) использовали для трансформации дрожжей S. cerevisiae штамм EH13-15 (trp1, MATα) (Appl. Microbiol. Biotechnol., 30, 515-520, 1989) способом с использованием ацетата лития. Трансформированные штаммы скринировали на наличие способности расти на агаризованной среде SC-Trp (2% агара), содержащей 6,7 г основы азотного агара для дрожжей без аминокислот (DIFCO), 20 г глюкозы и 1,3 г порошка аминокислот (смесь из 1,25 г аденин сульфата, 0,6 г аргинина, 3 г аспарагиновой кислоты, 3 г глутаминовой кислоты, 0,6 г гистидина, 1,8 г лейцина, 0,9 г лизина, 0,6 г метионина, 1,5 г фенилаланина, 11,25 г серина, 0,9 г тирозина, 4,5 г валина, 6 г треонина и 0,6 г урацила) на один литр среды.
(3) Культура дрожжей
Трансформированные штаммы, полученные с каждым вектором, культивировали в следующих условиях. На этапе прекультивирования дрожжами инокулировали 10 мл среды SC-Trp и культивировали при встряхивании при температуре 30°C в течение 1 суток. На этапе основного культивирования раствор прекультивирования (1 мл) добавляли в 100 мл среды SC-Trp и эту смесь культивировали при встряхивании при температуре 30°C в течение 1 суток.
(4) Получение неочищенного ферментного раствора
Клетки собирали путем центрифугирования, промывали водой и временно хранили при температуре -80°C. К клеткам добавляли Буфер A (5 мл: 50 мМ Трис-HCl (pH 7,5), 0,3 М сахарозы 10 мМ ДТТ, 1 М ПМСФ), чтобы суспендировать в нем клетки. Затем клетки разрушали при помощи френч-пресса (16 кПа, три раза). Суспензию разрушенных клеток центрифугировали при 1500x g при температуре 4°C в течение 10 минут, и полученный супернатант использовали в качестве неочищенного ферментного раствора.
(5) Определение активности ФФК
Методика
Активность ФФК измеряли следующим образом. Готовили реакционный раствор общим объемом 500 мкл, содержащий 50 мМ Трис-HCl (рН 7,5), 50 мкг линолевой кислоты (18:2)-ФК, олеиновой кислоты (18:1)-ФК или маргариновой кислоты (17:0)-ФК (фосфатидная кислота в качестве субстрата), 0,5 мМ MgCl2 или 0,5 мМ ЭДТА, 10 мМ ДТТ и 100 мкл неочищенного ферментного раствора, и оставляли эту смесь реагировать при температуре 28°C в течение 30 минут. В реакционную смесь добавляли смесь хлороформа и метанола (1:2), чтобы остановить реакцию. Реакционный раствор, содержащий вместо неочищенного ферментного раствора супернатант клеточного гомогената дрожжей (контрольного штамма), трансформированных плазмидой pYE22m, не содержащей ген MaPAP2.2, использовали в качестве контроля. Липиды экстрагировали методом Блая-Дайера, центрифугировали на концентрирующей центрифуге до сухого состояния, растворяли в хлороформе и затем фракционировали при помощи ТСХ (пластина с силикагелем 60, гексан:диэтиловый эфир:уксусная кислота = 70:30:1). Пластину опрыскивали раствором примулина для визуализации липидов под УФ излучением, и затем счищали фракции фосфолипидов и диглицерида (ДГ). Все содержащиеся во фракциях жирные кислоты переводили в сложные метиловые эфиры, чтобы анализировать при помощи газовой хроматографии.
Активность ФФК может быть определена путем измерения превращения диглицерида (ДГ) из фосфатидной кислоты (ФК), добавленной в качестве субстрата.
Результаты
Результаты показаны на Фигурах с 4 по 7.
Как показано на Фигуре 4, количества 18:2-ДГ увеличились в реакционных растворах с добавлением неочищенного ферментного раствора MaPAP2.2, по сравнению с контролями. Это свидетельствует о том, что превращение 18:2-ФК в 18:2-ДГ увеличилось, благодаря наличию MaPAP2.2, что означает, что MaPAP2.2 обладает активностью ФФК. Кроме того, как видно из результатов, приведенных на Фигуре 4, степень превращения 18:2-ФК в 18:2-ДГ была сопоставима между контролем и штаммом, активно экспрессирующим MaPAP2.2, (обозначенным на Фигуре 4 как MaPAP2.2), независимо от наличия или отсутствия ионов Mg2+, что ясно указывает на то, что MaPAP2.2 обладает одинаковой активностью ФФК независимо от наличия или отсутствия ионов Mg2+, то есть активность ФФК MaPAP2.2 является независимой от ионов Mg2+.
На Фигуре 5 показаны диаграммы, иллюстрирующие результаты изучения количества 18:2-ДГ в реакционном растворе, не содержащем ионы Mg2+ и содержащем неочищенный ферментный раствор MaPAP2.2 или MaPAP1. MaPAP2.2 значительно увеличивал превращение 18:2-ФК в 18:2-ДГ, в то время как MaPAP1 давал только такое же количество 18:2-ДГ, что и в контроле. Эти результаты показывают, что MaPAP2.2 и MaPAP1 являются различными по субстратной специфичности.
На Фигуре 6 показаны результаты изучения количества 18:1-ДГ после реакции при добавлении или без добавления 18:1-ФК в качестве субстрата, в реакционных растворах, содержащих неочищенный ферментный раствор MaPAP2.2, и в контролях. Количества 18:1-ФК (фоновые) измеряли в условиях без добавления 18:1-ФК, так как 18:1-ФК является фосфатидной кислотой, изначально присутствующей в дрожжах, и никаких различий не было обнаружено между штаммами, экспрессирующими MaPAP2.2, и контрольными штаммами (Фигура 6A). Затем были измерены количества 18:1-ДГ после реакции с добавлением 18:1-ФК. Результаты показали, что большее количество 18:1-ФК образовывалось в случае использования неочищенного ферментного раствора контроля, чем в случае использования неочищенного ферментного раствора MaPAP2.2 (Фигура 6B). Этот результат свидетельствует о том, что MaPAP2.2 обладает активностью ФФК, даже когда 18:1-ФК используется в качестве субстрата.
На Фигуре 7 показаны результаты сравнения количеств 17:0-ДГ после реакции при добавлении 17:0-ФК в качестве субстрата, в реакционный раствор, содержащий неочищенный ферментный раствор MaPAP2.2, и ее добавлении в контроль. Результат, приведенный на Фигуре 7, показывает, что MaPAP2.2 обладает активностью ФФК, даже в случае использования в качестве субстрата 17:0-ФК.
Сравнение результатов, приведенных на Фигурах 5-7, показывает, что активность превращения 18:2-ФК в 18:2-ДГ была сопоставима с активностью превращения 18:1-ФК в 18:1-ДГ, в то время как активность превращения 17:02-ФК в 17:0-ДГ составляла примерно одну пятую от активности превращения 18:2-ФК в 18:2-ДГ или 18:1-ФК в 18:1-ДГ. Это свидетельствует о том, что активность фосфатазы фосфатидной кислоты MaPAP2.2 имеет более высокую субстратную специфичность к фосфатидной кислоте, содержащей C18-ацильную группу, чем к фосфатидной кислоте, содержащей C17-ацильную группу.













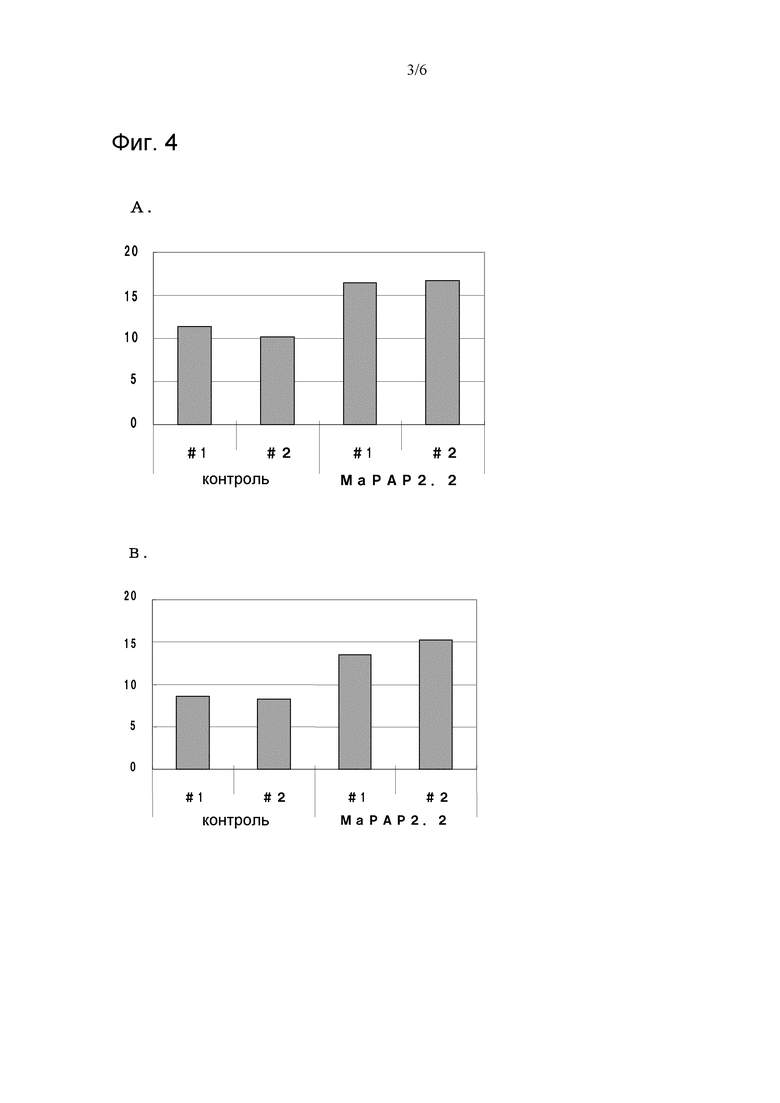
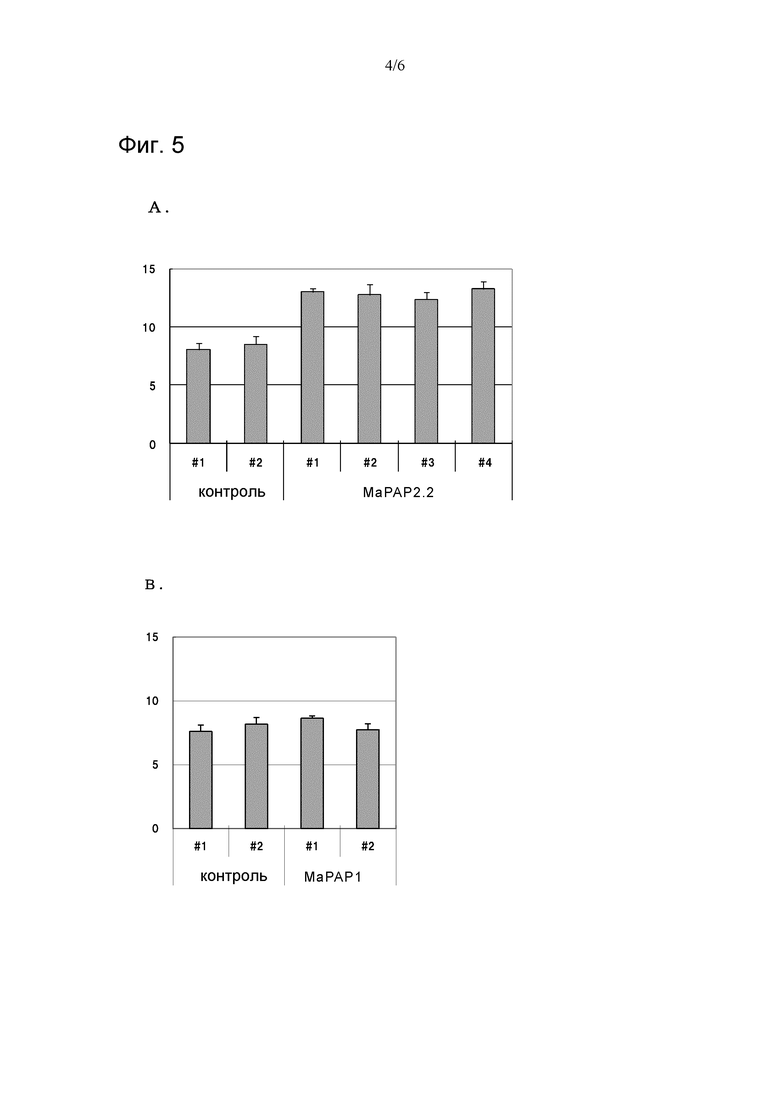
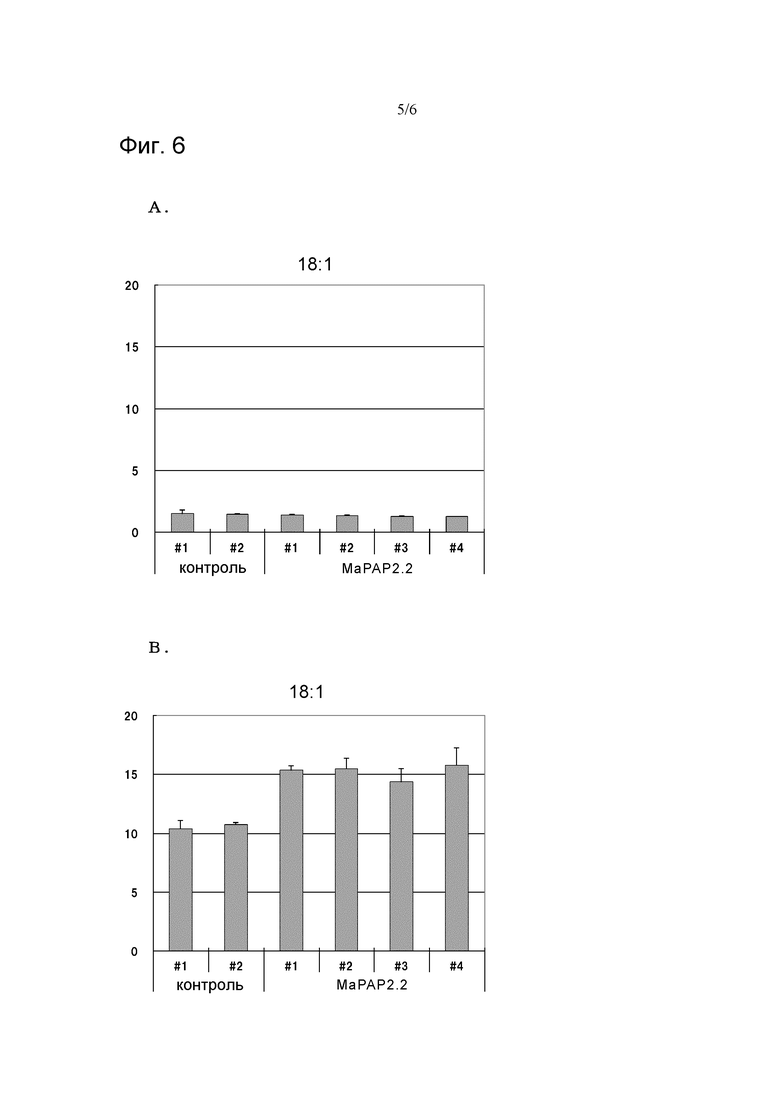
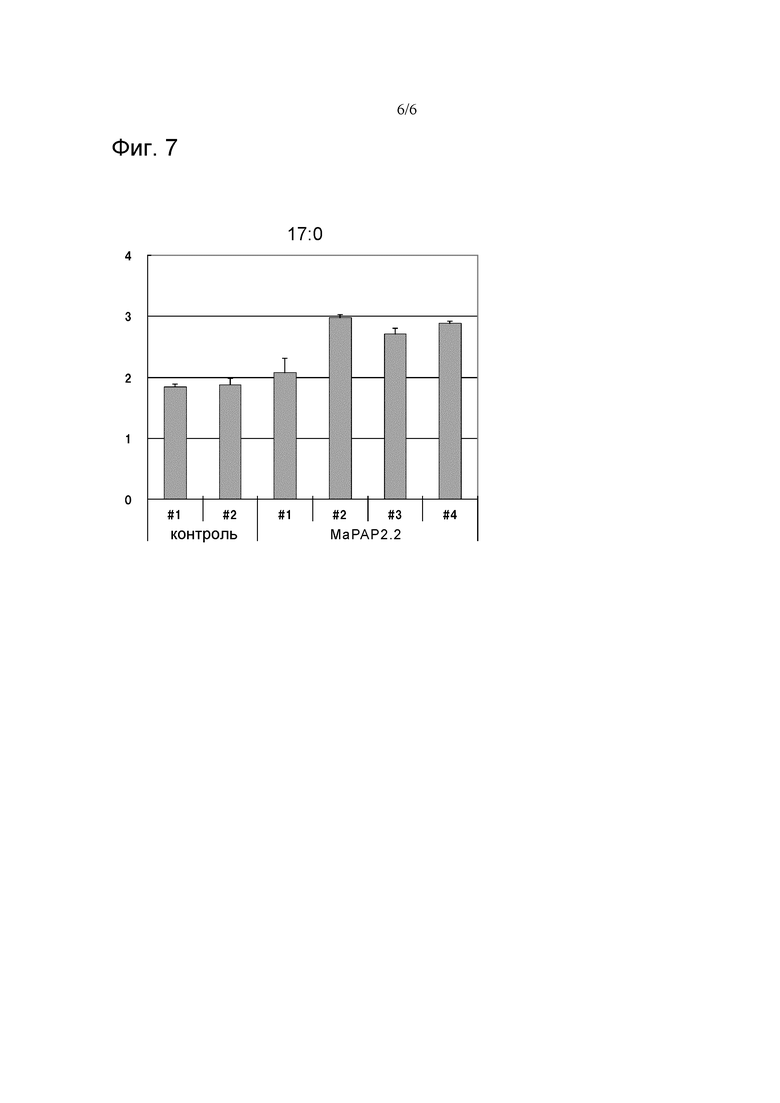
Изобретение относится к области биохимии, генной инженерии и биотехнологии, в частности к нуклеиновым кислотам, кодирующим белки, которые обладают активностью фосфатазы фосфатидной кислоты. Нуклеотидные последовательности указанных нуклеиновых кислот представлены в SEQ ID NO: 1, 4, 5. Изобретение также относится к мутантным вариантам данных нуклеотидных последовательностей. Изобретение относится к белку, обладающему активностью фосфатазы фосфатидной кислоты, и к его мутантным вариантам. Аминокислотная последовательность указанного белка представлена в SEQ ID NO: 2. Настоящее изобретение относится к экспрессионному рекомбинантному вектору, содержащему одну из указанных нуклеиновых кислот. Изобретение также относится к клетке-хозяину, трансформированной данным рекомбинантным вектором. Указанная клетка-хозяин предназначена для получения фосфатазы фосфатидной кислоты. Настоящее изобретение позволяет получать новые белки с активностью фосфатазы фосфатидной кислоты. 8 н. и 2 з.п. ф-лы, 7 ил., 1 табл., 5 пр.
1. Нуклеиновая кислота, которая кодирует белок, обладающий активностью фосфатазы фосфатидной кислоты, по приведенному ниже пункту (а), (с), (d) или (g):
(а) нуклеиновая кислота, состоящая из нуклеотидной последовательности, кодирующей белок, который состоит из аминокислотной последовательности с делецией, заменой или добавлением от 1 до 30 аминокислот в аминокислотной последовательности, приведенной в SEQ ID NO: 2;
(с) нуклеиновая кислота, состоящая из нуклеотидной последовательности, которая имеет идентичность 90% или более нуклеотидной последовательности, приведенной в SEQ ID NO: 1;
(d) нуклеиновая кислота, состоящая из нуклеотидной последовательности, кодирующей белок, который состоит из аминокислотной последовательности, имеющей идентичность 90% или более аминокислотной последовательности, приведенной в SEQ ID NO: 2; и
(g) нуклеиновая кислота, состоящая из нуклеотидной последовательности, которая имеет идентичность 90% или более нуклеотидной последовательности, приведенной в SEQ ID NO: 4.
2. Нуклеиновая кислота, которая кодирует белок, обладающий активностью фосфатазы фосфатидной кислоты, по приведенному ниже пункту (а), (с), (d) или (g):
(а) нуклеиновая кислота, состоящая из нуклеотидной последовательности, кодирующей белок, который состоит из аминокислотной последовательности с делецией, заменой или добавлением от 1 до 15 аминокислот в аминокислотной последовательности, приведенной в SEQ ID NO: 2;
(с) нуклеиновая кислота, состоящая из нуклеотидной последовательности, которая имеет идентичность 95% или более нуклеотидной последовательности, приведенной в SEQ ID NO: 1;
(d) нуклеиновая кислота, состоящая из нуклеотидной последовательности, кодирующей белок, который состоит из аминокислотной последовательности, имеющей идентичность 95% или более аминокислотной последовательности, приведенной в SEQ ID NO: 2; и
(g) нуклеиновая кислота, состоящая из нуклеотидной последовательности, которая имеет идентичность 95% или более нуклеотидной последовательности, приведенной в SEQ ID NO: 4.
3. Нуклеиновая кислота, которая кодирует белок, обладающий активностью фосфатазы фосфатидной кислоты, по любому из приведенных ниже пунктов (а)-(d):
(a) нуклеиновая кислота, состоящая из нуклеотидной последовательности, приведенной в SEQ ID NO: 1;
(b) нуклеиновая кислота, состоящая из нуклеотидной последовательности, кодирующей белок, состоящий из аминокислотной последовательности, приведенной в SEQ ID NO: 2;
(c) нуклеиновая кислота, состоящая из нуклеотидной последовательности, приведенной в SEQ ID NO: 4; и
(d) нуклеиновая кислота, состоящая из нуклеотидной последовательности, приведенной в SEQ ID NO: 5.
4. Нуклеиновая кислота по п. 1 или 2, где активность фосфатазы фосфатидной кислоты имеет более высокую субстратную специфичность к фосфатидной кислоте, содержащей C18-ацильную группу, чем к фосфатидной кислоте, содержащей C17-ацильную группу.
5. Белок, обладающий активностью фосфатазы фосфатидной кислоты, по любому из приведенных ниже пунктов (а) или (b):
(a) белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением от 1 до 30 аминокислот в аминокислотной последовательности, приведенной в SEQ ID NO: 2, и
(b) белок, состоящий из аминокислотной последовательности, которая имеет идентичность 90% или более аминокислотной последовательности, приведенной в SEQ ID NO: 2.
6. Белок, обладающий активностью фосфатазы фосфатидной кислоты, по любому из приведенных ниже пунктов (а) или (b):
(a) белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением от 1 до 15 аминокислот в аминокислотной последовательности, приведенной в SEQ ID NO: 2; и
(b) белок, состоящий из аминокислотной последовательности, которая имеет идентичность 95% или более аминокислотной последовательности, приведенной в SEQ ID NO: 2.
7. Белок по п. 5 или 6, где активность фосфатазы фосфатидной кислоты имеет более высокую субстратную специфичность к фосфатидной кислоте, содержащей C18-ацильную группу, чем к фосфатидной кислоте, содержащей C17-ацильную группу.
8. Белок, обладающий активностью фосфатазы фосфатидной кислоты, состоящий из аминокислотной последовательности, приведенной в SEQ ID NO: 2.
9. Экспрессионный рекомбинантный вектор, содержащий нуклеиновую кислоту по любому из пп.1-4.
10. Клетка-хозяин для получения фосфатазы фосфатидной кислоты, трансформированная рекомбинантным вектором по п.9.
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| RU 2005114507 A, 27.10.2005. | |||

