Изобретение относится к области молекулярной биологии, молекулярной генетики, биохимии и может быть использовано в медицине, в радиобиологии и экологии окружающей среды. Изобретение предназначено для определения скрытых хронических процессов в организме, которые сопровождаются увеличенной гибелью клеток организма, но не приводят к значительному увеличению общей концентрации циркулирующей внеклеточной ДНК.
Гибель клеток в организме в результате апоптоза, некроза и других причин сопровождается диффузией из погибших клеток структур, которые содержат ДНК погибших клеток. В результате в плазме крови человека возрастает концентрация циркулирующей внеклеточной ДНК (вкДНК). Известно, что концентрация вкДНК возрастает при острых и критических состояниях, таких как острый инфаркт миокарда, травма, сепсис [1-4]. Возрастание концентрации вкДНК имеет место у онкологических больных, благодаря тому, что в этом случае снижается активность ферментов, отвечающих за элиминацию вкДНК из кровотока [5]. В случае острого лучевого поражения организма ионизирующим излучением, например при лучевой терапии раковых больных [6] или при облучении экспериментальных животных [7], также происходит увеличение концентрации циркулирующей ДНК. Во всех перечисленных выше случаях концентрация вкДНК рассматривается в качестве интегрального показателя, который отражает общий высокий уровень гибели клеток организма на момент анализа [1-7].
Однако с течением времени или при развитии в организме хронических заболеваний, которые сопровождаются нерезким, но длительным увеличением уровня гибели клеток организма, происходит увеличение активности системы элиминации вкДНК из кровотока. При этом общая концентрация вкДНК может снижаться ниже контрольного уровня, который характерен для здорового организма [7]. В условиях относительно невысокого уровня гибели клеток больного органа и/или при значительно высоких значениях эндонуклеазной активности крови общая концентрация вкДНК более не является маркером гибели клеток, а ее сниженные значения могут создать ложное представление о благополучии в организме.
Основным результатом изобретения является нахождение конкретных последовательностей генома человека в составе вкДНК плазмы крови, которые обладает повышенной устойчивостью к возникновению двунитевых разрывов, возникающих в ДНК при действии клеточных и внеклеточных эндонуклеаз. Устойчивость к фрагментации приводит к тому, что фрагменты ДНК, включающие эти устойчивые последовательности, циркулируют в крови в виде высокомолекулярных соединений и медленно элиминируются из кровотока, в отличие от большинства фрагментов вкДНК. В результате в составе вкДНК происходит накопление этих последовательностей, по сравнению с их содержанием в составе клеточной (геномной ДНК). При скрытых хронических процессах гибели клеток в организме, когда активирована система элиминации вкДНК из кровотока, устойчивые фрагменты вкДНК являются маркером гибели клеток. Повышенное содержание этих фрагментов в пуле вкДНК может являться также маркером острого процесса, который имел место в организме в недавнем прошлом.
Стоит отметить, что в Российском патентном пространстве практически отсутствуют патенты и заявки на исследуемую тему. Те, что есть, принадлежал либо участникам выполняемого проекта, либо далеки по содержанию. Наиболее близки к тематике выполненной работы следующие патенты.
СПОСОБ ОЦЕНКИ ПОВРЕЖДЕНИЯ ДНК (ВАРИАНТЫ), автор ВАН ХОУТЕН Беннетт (US), патент RU №2213782 С2. Настоящее изобретение относится к области молекулярной биологии и биохимии и касается способа оценки повреждения ДНК. Представленный способ включает получение контрольной двухцепочечной ДНК-матрицы с первой и второй комплементарными цепями. Способ по настоящему изобретению позволяет производить анализ ДНК длиной более 5000 п.о., а также производить контроль генетических повреждений, которые произошли вследствие самоиндуцированного воздействия генотоксинов.
СПОСОБ ОТБОРА В ГРУППУ РИСКА ПО ФОРМИРОВАНИЮ ХРОНИЧЕСКИХ ВИДОВ ПАТОЛОГИИ У ДЕТЕЙ РАЗЛИЧНЫХ ВОЗРАСТНЫХ ГРУПП, авторы Крыночкина М.Ю., Бережанска С.Б., Амелина С.С., Спивак И.Г., патент RU №2172139 С2. Способ характеризуется тем, что у детей, рожденных в семьях мужчин, имевших контакт с источниками ионизирующего излучения, с целью всесторонней оценки состояния здоровья детей выделено 25 потенциальных неспецифических факторов риска, действовавших в различные периоды онтогенеза, начиная с антенатального. Изобретение относится к медицине, а именно к педиатрии, и может быть использовано для определения степени риска формирования хронических расстройств жизненно-важных систем организма в раннем возрасте и на период до поступления в школу у детей, родившихся в семьях мужчин, имевших контакт с источниками малых доз ионизирующего излучения.
В основе поиска устойчивых к двунитевой фрагментации последовательностей ДНК лежит информация, полученная из работ, в которых было впервые показано увеличение содержания GC-пар в составе вкДНК облученных крыс [8] и больных системной красной волчанкой (СКВ) [9]. Если в геноме грызунов содержание GC-пар составляет 40%, то в составе клонов, полученных из фрагментов вкДНК, оно варьировало от 42 до 60%. ДНК, выделенная из сыворотки больных СКВ, содержала 43-51% GC-пар, при среднем содержании в геноме здорового человека 38% GC-пар [8, 9]. Одной из причин накопления GC-ДНК в вкДНК может являться повышенная устойчивость GC-ДНК к фрагментации при накоплении в цепях однонитевых разрывов ДНК. Эта устойчивость обусловлена большей энергией связи оснований в GC-паре по сравнению с АТ-парой, в результате чего конкатемерные структуры в GC-ДНК (участки ДНК, содержащие однонитевые разрывы в цепях) имеют более высокую температуру плавления, чем в АТ-ДНК. Сформулированы следующие требования к маркерной последовательности:
(1) это должна быть GC-богатая последовательность с повышенной температурой плавления;
(2) последовательность должна быть представлена в геноме в виде повтора, который можно было бы легко определять количественно;
(3) для количественного определения этой последовательности должен быть разработан метод, который, в отличие от ПЦР, не обладает повышенной чувствительностью к повреждениям ДНК-мишени. В литературе приводятся доказательства того, что метод ПЦР дает очень заниженные результаты при определении концентрации вкДНК [10].
Всем перечисленным требованиям удовлетворяет последовательность транскрибируемой области рибосомного повтора человека (ТОрДНК). Участки ТОрДНК содержат от 60 до 85% GC-пар, что позволяет предположить высокую температуру плавления этой структуры, даже при наличии множественных однонитевых разрывов в двойной спирали ДНК.
В геноме человека рибосомный повтор представлен в количестве 300-700 копий на диплоидный геном [11].
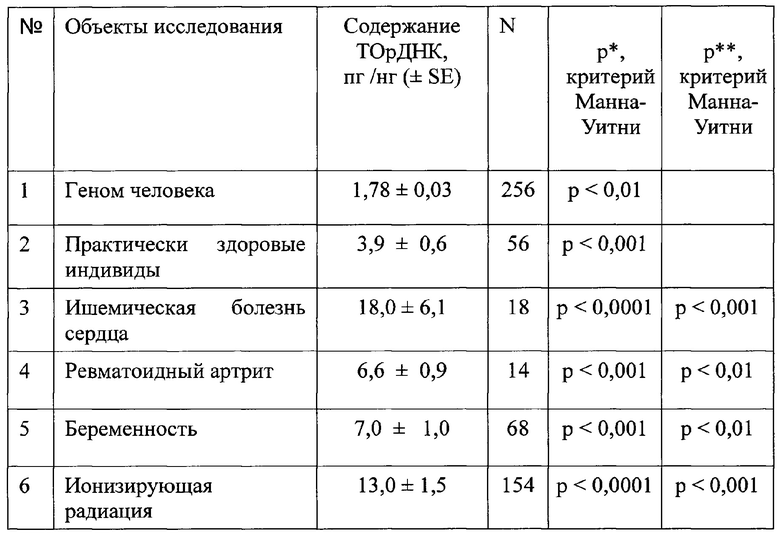
На фиг. 1 приводятся доказательства повышенной устойчивости ТОрДНК к действию эндонуклеаз. Образцы ДНК из различных источников подвергали электрофорезу в агарозном геле и далее анализировали содержание ТОрДНК методом нерадиоактивной блот-гибридизации по Саузерну.
(1) Геномную ДНК человека гидролизовали нуклеазой из ростков фасоли 0 мин (дорожка 1), 10 мин (дорожка 2) и 30 мин (дорожка 3) и анализировали размер фрагментов методом электрофореза в агарозном геле. Далее проводили блот-гибридизацию с зондом на фрагмент 28S рДНК (дорожки 1а-3а).
(2) В лимфоцитах человека индуцировали апоптоз хроматом калия. Далее выделяли клеточную ДНК и анализировали методом электрофореза (дорожка 1-3) и блот-гибридизации с зондом на 28S рДНК (дорожки 1а-3а).
(3) Из плазмы крови больного инсультом выделяли циркулирующую вкДНК и анализировали методом электрофореза (дорожка 1) и методом блот-гибридизации с зондом на 28S рДНК.
В составе ДНК ядер апоптотической клетки, в составе вкДНК больного инсультом, а также в составе депротеинизированной ДНК, обработанной эндонуклеазой, не смотря на высокий уровень фрагментации ДНК, ТОрДНК представлена длинными фрагментами (от 15 т.п.н.).
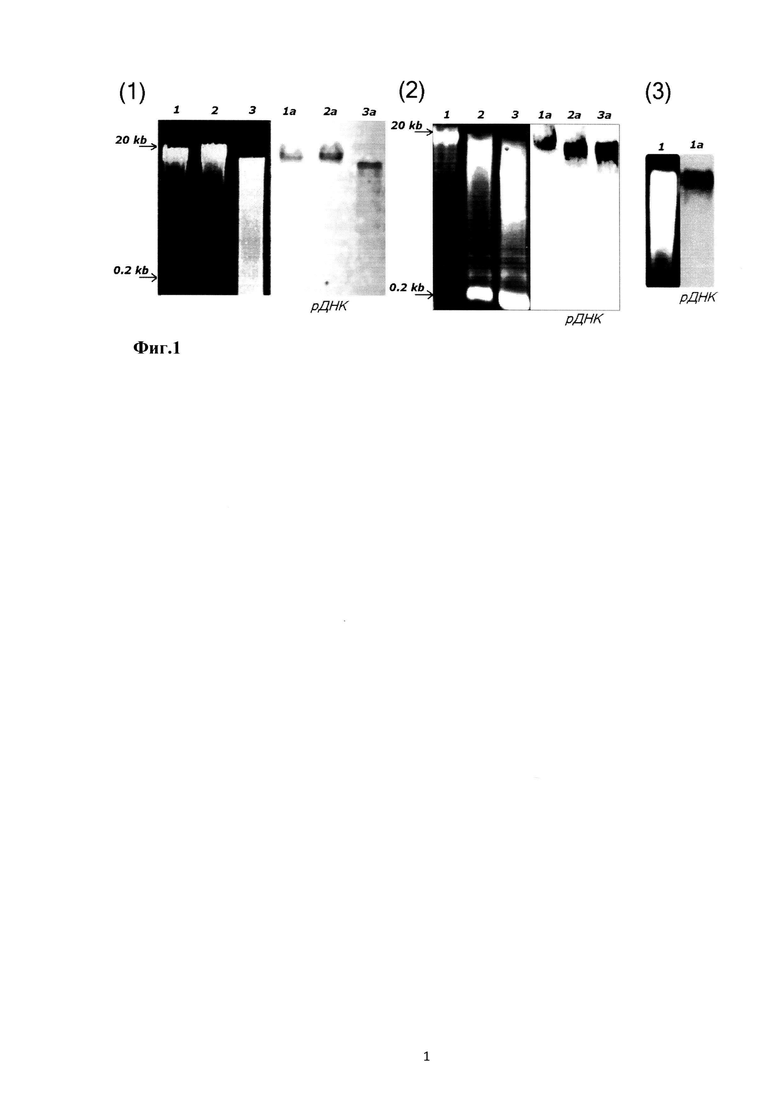
Ранее специально для ТОрДНК была разработана схема количественной гибридизации с использованием биотинированных ДНК-зондов и нерадиоактивной детекции с применением конъюгата стрептавидина с щелочной фосфатазой [11]. Принцип метода отображен на фиг. 2, где проиллюстрирована гибридизация ДНК с нерадиоактивным зондом.
(1) Общий вид фильтра с нанесенными и визуализированными образцами ДНК. На фильтр наносили по 2 мкл денатурированного образца гДНК. После иммобилизации нагреванием проводили гибридизацию с биотинированным зондом на 28S рДНК. Биотин выявляли с помощью конъюгата стрептавидина с щелочной фосфатазой. В качестве субстрата использовали стандартную смесь BCIP и NBT.
(2) Пример калибровочной зависимости сигнала от концентрации рДНК в стандартном растворе ДНК с известным содержанием ТОрДНК.
На мембрану наносятся денатурированные щелочью образцы вкДНК в количестве 8-10 точек для одного образца. На фильтр также наносятся различные количества образцов ДНК с известным содержанием ТОрДНК для построения калибровочной зависимости. Обычно наносили несколько калибровочных образцов разной степени фрагментации. На уровень сигнала степень фрагментации ДНК мишени в широком диапазоне длин фрагментов практически не влияет, в отличие от метода ПЦР. Проводится гибридизация с биотинированным зондом на фрагмент 28S рРНК и нерадиоактивная детекция. Для количественного анализа результата гибридизации использовали программу, которая автоматически находит пятно, определяет интегральную интенсивность сигнала, анализирует и учитывает возможный фон (Images 6.0, зарегистрирована в ЦИТИСе). Количество ТОрДНК в образце определяли с использованием калибровочной зависимости.
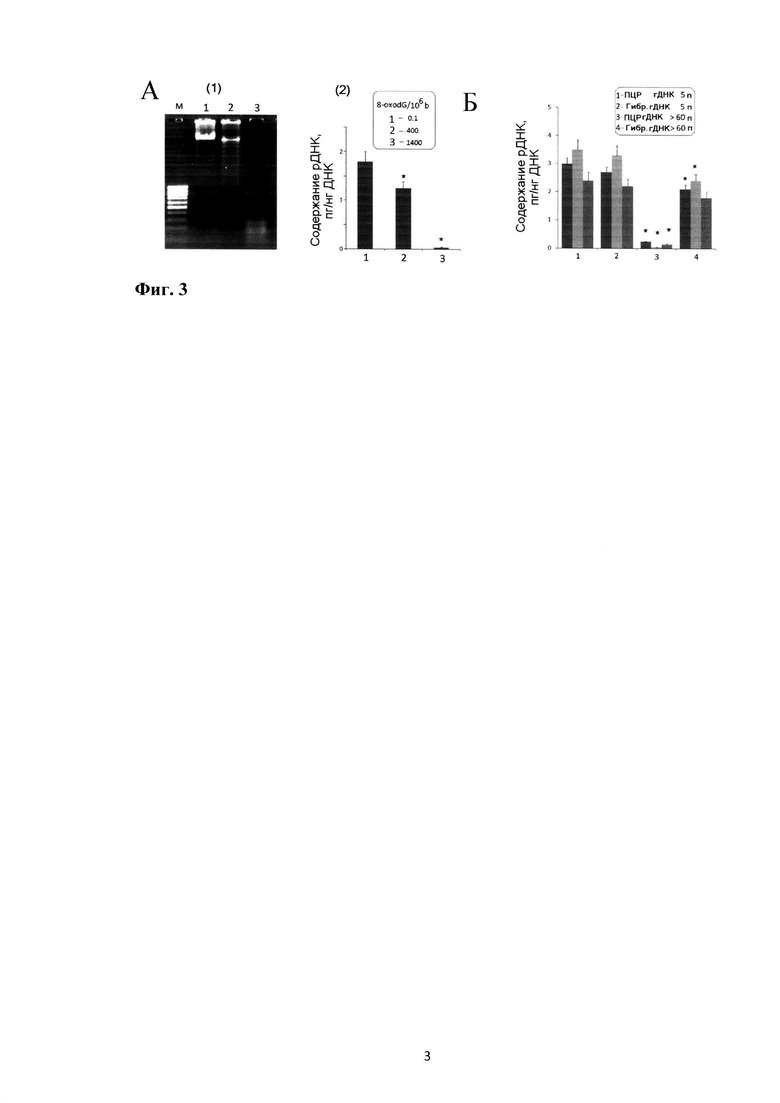
На фиг. 3 приводятся доказательства преимущества метода нерадиоактивной дот-гибридизации по сравнению с методом ПЦР при анализе матрицы ДНК, поврежденной активными формами кислорода (АФК).
А. (1) Длины фрагментов образцов гДНК. Интактный образец (дорожка 1), ДНК окислена метиленовым синим (дорожка 2) и пероксидом водорода при облучении 321 нм (дорожка 3).
(2) Определение содержания ТОрДНК методом ПЦР. В рамке приводится содержание в образцах маркера окисления 8-oxodG, определенное методом масс-спектрометрии.
Б. Определение содержания ТОрДНК в геномной ДНК, выделенной из молодых (5 пассаж) и старых (достигших предела Хейфлика) фибробластов кожи взрослых людей (три различных линии клеток). В рамке указан метод.
При патологии очень часто в организме повышается уровень АФК, которые окисляют клеточную ДНК. При действии ионизирующего излучения ДНК клеток также подвергается окислительной модификации. Поэтому вкДНК при патологии и внешнем воздействии сильно повреждена и является плохой матрицей для фермента Taq-полимеразы. Поскольку метод гибридизации не предполагает применения фермента, то он гораздо менее чувствителен к окислительным повреждениям ДНК и к однонитевым разрывам ДНК. Современные мембраны удерживают фрагменты ДНК длиной от 30 нуклеотидов, а использование длинного зонда позволяет нивелировать наличие окисленных оснований.
Чтобы доказать, что ТОрДНК действительно накапливается в вкДНК методом нерадиоактивной гибридизации были проанализированы 310 образцов вкДНК, выделенных из плазмы крови человека, и 256 образцов геномной ДНК, выделенной из лимфоцитов человека (таблица 1). Выборка образцов вкДНК включала образцы здоровых лиц (строка 2, таблица); образцы больных ишемической болезнью сердца и ревматоидным артритом с хроническим, неострым течением заболеваний (строки 3 и 4); образцы беременных женщин (1 триместр беременности, строка 5). Кроме того, проанализированы образцы вкДНК лиц, которые работают с источниками рентгеновского излучения, получая небольшие дозы радиации в течение продолжительного времени (строка 6).
В таблице указаны: p* - Достоверность различий содержания ТОрДНК в образцах вкДНК при хронических заболеваниях, при физиологических состояниях (беременность) и при хроническом действии неблагоприятных факторов окружающей среды на организм человека по сравнению с геномом.
p** - Достоверность различий содержания ТОрДНК в образцах вкДНК при хронических заболеваниях, при физиологических состояниях (беременность) и при хроническом действии неблагоприятных факторов окружающей среды на организм человека по сравнению со здоровым контролем.
Источники ДНК:
(1) Коллекция образцов ДНК здоровых людей (ФГБНУ «МГНЦ»).
(2) Коллекция образцов вкДНК плазмы здоровых людей (ФГБНУ «МГНЦ»).
(3) Образцы плазмы получены из клинической больницы РАН.
(4) Образцы плазмы получены из НИИ Ревматологии РАМН.
(5) Коллекция образцов вкДНК беременных женщин (ФГБНУ «МГНЦ»).
(6) Образцы плазмы получены из Медицинского радиологического научного центра РАМН, Обнинск.
Образцы плазмы для анализа подбирались таким образом, чтобы общая концентрация вкДНК в плазме крови не превышала контрольных значений (тестируемых в плазме здоровых людей), которые варьировали в пределах 1-1000 нг/мл (при использовании для тестирования концентрации вкДНК метода связывания ДНК с красителем RiboGreen [12]).
ВкДНК выделяли из плазмы периферической крови или из лейкоцитарной массы, обрабатывая образцы РНКазой А, протеиназой К и далее применяя стандартный метод фенольной экстракции и осаждения ДНК этанолос в присутствии 2 М ацетата аммония. Для определения ТОрДНК использовали зонд pHRGRR-28S (фрагмент гена 28S рРНК длиной 2,4 т.п.н.), клонированный в вектор pBR3222 [11]. Биотинирование ДНК-зондов осуществляли с использованием фотореагента, как подробно описано ранее [13, 14]. Методика количественной нерадиоактивной гибридизации подробно описана ранее [12-14]. Коротко, мембрану (Extra С) смачивали раствором 10×SSC и высушивали. К 10 мкл раствора ДНК в ТЕ-буфере добавляли 1 мкл 1 М гидроокиси натрия при 0°C. Раствор перемешивали и инкубировали 10 мин при 0°C. Далее проводили нейтрализацию раствора ДНК, добавляя 11 мкл 20×SSC (рН=5,0). Денатурированные пробы ДНК наносили на подготовленный фильтр в количестве 1.5-2 мкл на одно пятно. Для каждой пробы наносили от 5-10 параллельных точек. На этот же фильтр наносили стандартные образцы геномной ДНК с известным содержанием повторов для построения калибровочной зависимости сигнала от количества повтора в пробе. Концентрация ДНК в стандартной калибровочной пробе составляла от 0,5 до 50 нг/мкл. Фильтр прогревали при 80°C под вакуумом в течение 2 ч. Для проведения гибридизации использовали специальное оборудование - печь для гибридизации, которая позволяет изменять температуру и имеет качающийся столик для перемешивания раствора над мембраной. Мембрану блокировали в растворе (10× раствор Денхарда, 0,05 М фосфатный буфер, pH 7,0; 0,7 М хлорид натрия, 50% формамид, 100 мкг/мл тРНК E. coli) 5 ч при 42°C. Далее добавляли денатурированный ДНК-зонд в концентрации 20 нг/мл. Гибридизацию проводили 16 ч при 42°C. Фильтр отмывали раствором 2×SSC, 0,1% SDS (2×15 мин, 25°C), 0,01×SSC, 0,1% SDS (20 мин, 65°C) и 2×SSC (10 мин, 25°C). Мембрану после проведения гибридизации блокировали (30 мин, 37°C) раствором (0,1% обезжиренное молоко, 0,1% желатин, трис-HCL буфер, pH 7,5, 0,1 М хлорид натрия). Далее обрабатывали 20 мин (25°C) конъюгатом стрептавидина с щелочной фосфатазой (1 мкг/мл, Sigma) в растворе (0,1 М трис-HCL буфер, pH 7,5, 0,1 М хлорид натрия, 0,005 М хлорид магния). Фильтр отмывали (3×10 мин.) раствором (трис-HCL буфер, pH 7,5, 0,1 М хлорид натрия, 0,005 М хлорид магния). Далее фильтр помещали в раствор субстратов для щелочной фосфатазы (трис-HCL буфер, pH 9,5, 0,1 М хлорид натрия, 0,005 М хлорид магния, 4,4 мкл/мл NBT и 3,3 мкл/мл BCIP). Реакцию проводили в темноте при 37°C, контролируя визуально появление окрашенных фиолетовых пятен. После окончания реакции фильтр промывали водой и высушивали в темноте. Высушенный фильтр сканировали. Для количественного анализа результата гибридизации использовали программу (Images6, ФГБУ МГНЦ РАМН). Программа определяет местонахождение пятна, определяет близлежащий фоновый сигнал и определяет площадь и интегральную интенсивность пятна. Сигналы для одного образца усредняются, приводится значение среднего и стандартная ошибка. Концентрация повтора в конкретном образце рассчитывается по уравнению калибровочной прямой. Все концентрации для ТОрДНК приводятся в расчете на всю транскрибируемую область рибосомного повтора длиной 13334 п.н.
Таким образом, с помощью описанных методов можем количественно определить содержание ТОрДНК (пг/нг) в составе геномной ДНК и внеклеточной ДНК. Применяемый метод количественной нерадиоактивной гибридизации позволяет выявить присутствие ТОрДНК даже при сильном повреждении матрицы - вкДНК. Поскольку ТОрДНК устойчива к действию нуклеаз и накапливается в составе вкДНК с течением времени, содержание ТОрДНК во вкДНК отражает уровень гибели клеток организма даже при эффективных системах элиминации вкДНК. Очевидно, что, даже у здоровых лиц содержание ТОрДНК в вкДНК в среднем в 2 раза выше, чем в геноме (p<0,001, критерий Манна-Уитни).
Все случаи, приведенные в строках 3-6 таблицы, характеризуются сравнительно небольшим, но хроническим повышением уровня гибели клеток в организме, определяемым по достоверному увеличению содержания ТОрДНК в составе вкДНК. Общая концентрация вкДНК плазмы крови не является эффективным показателем высокого уровня гибели клеток при высокой нуклеазной активности плазмы крови [16]. При хронических неострых процессах содержание ТОрДНК может возрастать в несколько раз по сравнению с содержанием в вкДНК здоровых людей (p<0,01, критерий Манна-Уитни). Так, при ишемической болезни сердца содержание ТОрДНК в составе вкДНК выше в 4,6 раз по сравнению с контрольной группой здоровых доноров (p<0,001, критерий Манна-Уитни). При ревматоидном артрите с хроническим, неострым течением заболевания, содержание ТОрДНК во вкДНК выше в 1,7 раз по сравнению с контрольной группой здоровых доноров (p<0,01, критерий Манна-Уитни). Известно, что нормальная беременность сопровождается увеличением уровня апоптоза клеток организма матери и плода (в основном, клеток плаценты) [15]. Апоптоз усиливается по мере увеличения срока беременности. ДНК погибших клеток поступает частично в циркуляцию, увеличивая пул вкДНК. Одним из важных звеньев механизма элиминации вкДНК является активность ДНКазы 1 крови [16]. В ранние сроки беременности нуклеазы эффективно элиминируют фрагменты вкДНК из кровотока, и содержание ТОрДНК, устойчивой к действию нуклеаз, в составе вкДНК, отражает реальный уровень гибели клеток организма. При нормально протекающей беременности в первом триместре у женщин уровень ТОрДНК в составе вкДНК выше в 1,8 раз по сравнению с контрольной группой небеременных женщин соответствующего возраста (p<0,01, критерий Манна-Уитни). Кроме того, с составе вкДНК лиц, которые работают с источниками рентгеновского излучения, получая небольшие дозы радиации в течение продолжительного времени, содержание ТОрДНК возрастает в 3,3 раз (p<0,001, критерий Манна-Уитни). Это также является свидетельством процесса повышенной гибели клеток в результате хронического воздействия малых доз радиации.
Таким образом, определение количества ТОрДНК в составе вкДНК плазмы крови с помощью описанного изобретения можно рекомендовать в качестве стандартного анализа, позволяющего оценить наличие скрытой хронической патологии, которая приводит к повышению уровня гибели клеток в организме. Патология может быть вызвана и внешним воздействием (курение, облучение, вредное производство и т.д.).
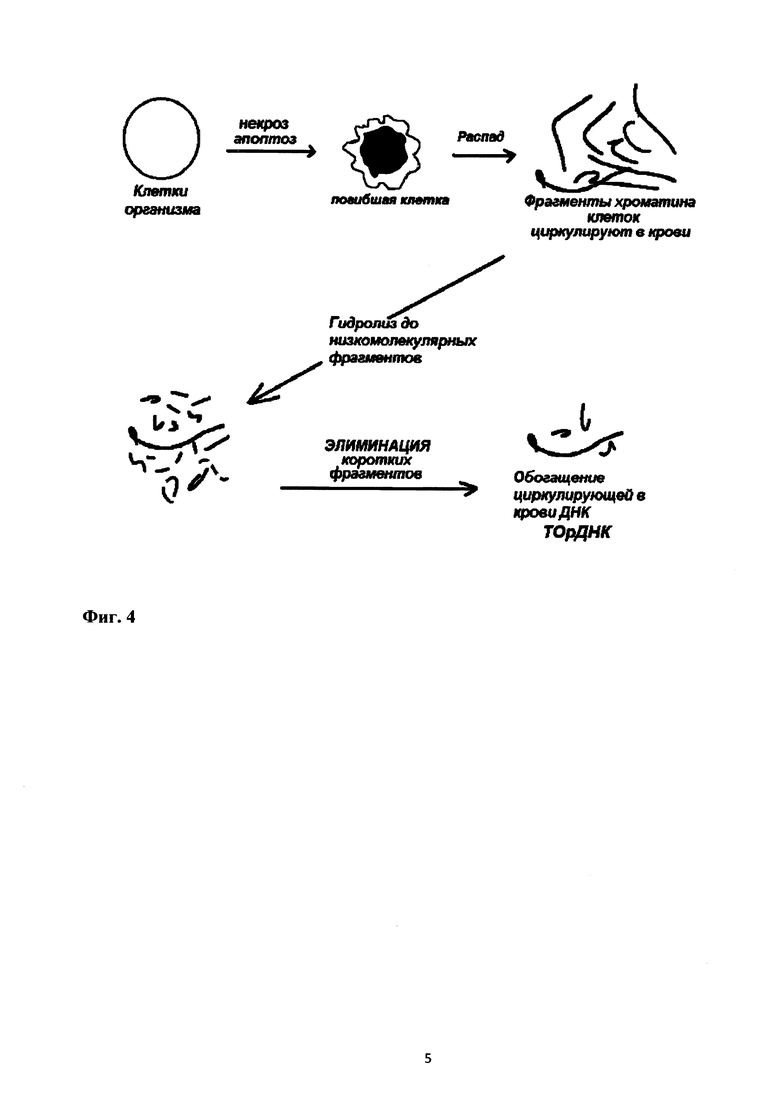
Процесс накопления ТОрДНК или другой GC-богатой последовательности генома в составе циркулирующей вкДНК при хроническом процессе в организме, который характеризуется повышением уровня гибели клеток, описан на схеме (фиг. 4). Погибшие клетки организма являются источником дополнительных количеств вкДНК в циркуляции. В плазме крови циркулируют ферменты, которые гидролизуют вкДНК до низкомолекулярных структур, внося однонитевые разрывы. Накопление однонитевых разрывов приводит к двунитевым разрывам ДНК. В составе конкатемерных GC-богатых структур ДНК двойные разрывы образуются медленнее, поскольку температура плавления этих структур выше и требуется большее количество однонитевых разрывов, чтобы реализовался двунитевой разрыв ДНК. Низкомолекулярные фрагменты элиминируются путем почечной фильтрации, высокомолекулярные GC-богатые структуры ДНК остаются в циркуляции.
Список цитируемой литературы.
1. V. Swarup, M.R. Rajeswari. Circulating (cell-free) nucleic acids-a promising, non-invasive tool for early detection of several human diseases. FEBS Lett. 581 (2007) 795-799.
2. M. Cui, M. Fan, R. Jing, H. Wang, J. Qin, H. Sheng, Y. Wang, X. Wu, L. Zhang, J. Zhu, S. Ju. Cell-Free circulating DNA: a new biomarker for the acute coronary syndrome. Cardiology, 124 (2013) 76-84.
3. B. Ren, F. Liu, F. Xu, J. He, H. Zhu, G. Zou. Is plasma cell-free DNA really a useful marker for diagnosis and treatment of trauma patients? Clin. Chim. Acta. 424 (2013) 109-113.
4. Saukkonen K., Lakkisto P.,  V., Varpula M., Karlsson S., Ruokonen E., Pulkki K. Finnsepsis Study Group. Cell-free plasma DNA as a predictor of outcome in severe sepsis and septic shock. Clin Chem. 2008 Jun; 54(6):1000-7.
V., Varpula M., Karlsson S., Ruokonen E., Pulkki K. Finnsepsis Study Group. Cell-free plasma DNA as a predictor of outcome in severe sepsis and septic shock. Clin Chem. 2008 Jun; 54(6):1000-7.
5. E. Gormally, E. Caboux, P. Vineis, P. Hainaut. Circulating free DNA in plasma or serum as biomarker of carcinogenesis: practical aspects and biological significance, Mutat. Res. 635 (2007) 105-117.
6. C. Cheng, M. Omura-Minamisawa, Y. Kang, T. Hara, I. Koike, T. Inoue. Quantification of circulating cell-free DNA in the plasma of cancer patients during radiation therapy. Cancer Sci. 100 (2009) 303-309.
7. V.G. Vladimirov, A.S. Belokhvostov, S.S. Sherlina, I.N. Vasilyeva, A.M. Voskresensky. Extracellular DNA level in the blood of irradiated rats, Int. J. Radiat. Biol. 62 (1992) 667-71.
8. I.N. Vasilyeva. Low-molecular-weight DNA in blood plasma as an index of the influence of ionizing radiation, Ann. N Y Acad. Sci. 945 (2001) 221-228.
9. Van Helden PD. Potential Z-DNA-forming elements in serum DNA from human systemic lupus erythematosus. J Immunol. 1985 Jan; 134(l):177-9.
10. M. Fleischhacker, B. Schmidt, Circulating nucleic acids (CNAs) and cancer - a survey, Biochim. Biophys. Acta 1775 (2007) 181-232.
11. Н.Н. Вейко, Н.А. Еголина. Г.Г. Радзивил. С.Д. Нурбаев, Н.В. Косякова, Н.О. Шубаева, Н.А. Ляпунова. Количественное определение повторяющихся последовательностей в геномной ДНК человека. Обнаружение увеличенного количества рибосомных повторов в геномах больных шизофренией (результаты молекулярного и цитогенетического анализа). // Молекулярная биология. 2003. Т. 37. №3. С. 409-419.
12. Вейко Н.Н., Конорова И.Л., Неверова М.Е., Фиделина О.В., Мкртумова Н.А., Ершова Е.С., Конькова М.С., Постнов А.Ю. Влияние CpG-богатых фрагментов ДНК на формирование гипертензии у спонтанно-гипертензивных крыс (SHR). Биомедицинская химия. 2010. Т. 56. №6. С. 686-699.
13. Вейко Н.Н., Карпухин А.В., Салимов А.Г., Немцова М.В., Спитковский Д.М. Биотинирование ДНК с использованием фотоактивируемого реагента N-(4-азидо-2-нитробензоил)-1,7-диаминогептана // Биотехнология. 1989. Т. 5. С. 414-418.
14. Карпухин А.В., Вейко Н.Н., Салимов А.Г., Спитковский Д.М. (1988) Способ биотинирования нуклеиновых кислот. Авторское свидетельство на изобретение №1443404.
15. Al Nakib М.,  R., Bonello N., Bretelle F., Boubli L., Gabert J., Levy-Mozziconacci A. Total and fetal cell-free DNA analysis in maternal blood as markers of placental insufficiency in intrauterine growth restriction. Fetal DiagnTher. (2009) P. 24-28. DOI:10.1159/000236355.
R., Bonello N., Bretelle F., Boubli L., Gabert J., Levy-Mozziconacci A. Total and fetal cell-free DNA analysis in maternal blood as markers of placental insufficiency in intrauterine growth restriction. Fetal DiagnTher. (2009) P. 24-28. DOI:10.1159/000236355.
16. Черепанова A.B., Тамкович C.H., Власов B.B., Лактионов П.П. Активность дезоксирибонуклеаз крови в норме и при патологии. Биомедицинская химия (2007). Т. 53, С. 488-496.
Таблица 1.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ диагностики тяжести ишемически-гипоксических поражений ЦНС на основе установления целостности геномных ДНК | 2023 |
|
RU2827183C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ АНЕУПЛОИДИИ ПЛОДА В ОБРАЗЦЕ КРОВИ БЕРЕМЕННОЙ ЖЕНЩИНЫ | 2021 |
|
RU2777072C1 |
| СПОСОБ НЕИНВАЗИВНОЙ ДИАГНОСТИКИ АНЕУПЛОИДИЙ ПЛОДА МЕТОДОМ СЕКВЕНИРОВАНИЯ | 2014 |
|
RU2543155C1 |
| СПОСОБ НЕИНВАЗИВНОЙ ПРЕНАТАЛЬНОЙ ДИАГНОСТИКИ АНЕУПЛОИДИЙ ПЛОДА | 2014 |
|
RU2583830C2 |
| ТЕХНОЛОГИЯ ОПРЕДЕЛЕНИЯ АНЕУПЛОИДИИ МЕТОДОМ СЕКВЕНИРОВАНИЯ | 2012 |
|
RU2529784C2 |
| Способ получения панели ДНК-зондов для определения хромосомных транслокаций, делеций и амплификаций | 2023 |
|
RU2828103C1 |
| СПОСОБЫ И СИСТЕМЫ МОНИТОРИНГА СОСТОЯНИЯ ЗДОРОВЬЯ И ПАТОЛОГИИ ОРГАНОВ | 2020 |
|
RU2818052C2 |
| Технология полноэкзомного обогащения на основе различия в кинетике реассоциации фрагментов ДНК | 2023 |
|
RU2839144C1 |
| СПОСОБ НЕИНВАЗИВНОЙ ПРЕНАТАЛЬНОЙ ДИАГНОСТИКИ АНЕУПЛОИДИЙ ПЛОДА | 2015 |
|
RU2627673C2 |
| НАБОР ЗОНДОВ ДЛЯ АНАЛИЗА ОБРАЗЦОВ ДНК И СПОСОБЫ ИХ ИСПОЛЬЗОВАНИЯ | 2016 |
|
RU2753883C2 |

Изобретение относится к области медицины и предназначено для определения наличия в организме беременных женщин хронического процесса, который сопровождается увеличенной гибелью клеток. В составе циркулирующей внеклеточной ДНК плазмы периферической крови определяют содержание GC-богатой последовательности генома (GC-ДНК) - фрагмента транскрибируемой области рибосомного повтора человека методом нерадиоактивной количественной гибридизации. Если содержание повтора повышено более чем в 2 раза по сравнению с его содержанием в геномной ДНК, то делают вывод о наличии в организме хронического процесса, который сопровождается увеличенной гибелью клеток. Изобретение позволяет оценить наличие скрытой хронической патологии, которая может быть вызвана внешним воздействием (курение, облучение, вредное производство и т.д.). 4 ил., 1 табл., 1 пр.
Способ определения наличия в организме беременных женщин хронического процесса, который сопровождается увеличенной гибелью клеток, заключающийся в определении содержания GC-богатой последовательности генома (GC-ДНК) в составе циркулирующей внеклеточной ДНК плазмы периферической крови, отличающийся тем, что содержание GC-богатой последовательности генома - фрагмента транскрибируемой области рибосомного повтора человека - в составе выделенной внеклеточной ДНК определяют методом нерадиоактивной количественной гибридизации и, если определенное содержание повтора повышено более чем в 2 раза по сравнению с его содержанием в геномной ДНК, делают вывод о наличии в организме хронического процесса, который сопровождается увеличенной гибелью клеток.
| ВЕЙКО Н.Н | |||
| и др | |||
| Фрагменты транскрибируемой области рибосомного повтора в составе внеклеточной ДНК - маркер гибели клеток организма | |||
| Биомедицинская химия | |||
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| VEIKO N.N | |||
| et al | |||
| Stimulatory effect of fragments from transcribed region of ribosomal repeat on human peripheral blood lymphocytes | |||
| Bull Exp Biol Med | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| АЛЕКСЕЕВА А.Ю | |||
| Влияние внеклеточной ДНК на функциональную активность клеток эндотелия | |||
| Автореф | |||
| дисс | |||
| канд | |||
| биол | |||
| наук | |||
| Москва, 2013, 27 c | |||
| СПОСОБ ОЦЕНКИ ПОВРЕЖДЕНИЯ ДНК (ВАРИАНТЫ) | 1996 |
|
RU2213782C2 |

