Область, к которой относится изобретение
Настоящее изобретение относится к сконструированным белкам повторов со связывающей специфичностью для сывороточного альбумина, а также к нуклеиновым кислотам, кодирующим такие белки, которые связывают сывороточный альбумин, фармацевтическим композициям, включающим такие белки, применению таких белков для модификации фармакокинетических характеристик биоактивных соединений и к применению таких белков в лечении заболеваний.
Предпосылки создания изобретения
Фармацевтическая промышленность является весьма заинтересованной в повышении эффективности биоактивных соединений, таких как белковые терапевтические агенты, путем модуляции или повышения их фармакокинетических (РК) in vivo свойств. Это, в частности, является правдой для биоактивных соединений, которые быстро выводятся из системы циркуляции путем почечного клиренса. В общем случае почки отфильтровывают из системы циркуляции молекулы, которые имеют средний молекулярный вес ниже 60 кДа. Одна стратегия для улучшения фармакокинетических свойств таких малых биоактивных соединений заключается в простом повышении их среднего молекулярного размера (то есть, повышении их гидродинамического радиуса), например, посредством прибавления небелковых полимерных остатков, таких как полимеры полиэтиленгликоля или остатки сахара, или прибавления белковых полимерных остатков, таких как глобулярные белки или неструктурированные полипептиды, такие как те, что описаны в WO 2007/103515 и WO 2008/155134.
Другие стратегии используют длительный период полувыведения из системы циркуляции сывороточных белков, таких как иммуноглобулины и сывороточный альбумин. Сывороточный альбумин, который имеет молекулярный вес 67 кДа, представляет собой белок, который находится в плазме крови в избыточном количестве и присутствует при концентрации 50 мг/мл (0,6 мМ), а также имеет период полувыведения из сыворотки крови 19 дней у людей. Сывороточный альбумин помогает поддерживать значение рН в плазме крови, осуществляет свой вклад в давление коллоидной крови, функционирует как носитель для многих метаболитов и жирных кислот и служит в качестве основного транспортного белка для лекарственных агентов в плазме крови. Существует несколько основных сайтов связывания малых молекул в альбумине, которые были описаны.
Было показано, что нековалентная ассоциация с сывороточным альбумином может удлинить период полувыведения малых молекул или полипептидов с коротким периодом полураспада (WO 1991/001743). Полипептиды, являющиеся такими, которые специфически связываются с сывороточным альбумином, и которые, таким образом, могут продлевать in vivo период полувыведения других молекул, слитых с ними, включают варианты альбуминсвязывающих доменов (например, WO 2005/097202 и WO 2009/016043), малые пептиды (например, Dennis, M.S., и др., J. Biol. Chem. 277(3), 35035-43, 2002 и WO 2001/045746) и фрагменты иммуноглобулинов (например, WO 2008/043822, WO 2004/003019; WO 2008/043821; WO 2006/040153; WO 2006/122787 и WO 2004/041865). WO 2008/043822 относится к связывающим белкам, отличным от фрагментов иммуноглобулинов, таких как молекулы на основе доменов белка А, тендамистат, фибронектин, липокалин, CTLA-4, рецепторы Т-клеток, сконструированные анкириновые повторы и PDZ домены, которые могут быть получены для специфического связывания с сывороточным альбумином. Тем не менее, WO 2008/043822 не раскрывает ни селекцию доменов сконструированного анкиринового повтора со связывающей специфичностью для сывороточного альбумина (SA), ни конкретных последовательностей мотивов повторяемых доменов, которые специфически связываются с SA. Кроме того, было описано, что in vivo период полувыведения полипептидов может быть удлинен путем их генетического слияния с сывороточным альбумином (например, WO 1991/001743). Такое изменение in vivo периода полураспада лекарственных средств может позитивно изменить их фармакокинетические (РК) и/или фармакодинамические (PD) свойства. В этом заключается ключевая проблема разработки новых и эффективных терапевтических агентов и способов лечения заболеваний. Таким образом, существует потребность в области техники в новых путях изменения РК и/или PD биоактивных соединений.
Кроме антител существуют новые связывающие белки или связывающие домены, которые могут использоваться для специфического связывания с целевой молекулой (например, Binz, H.K., Amstutz, P. и Plückthun, A., Nat. Biotechnol. 23, 1257-1268, 2005). Один такой новый класс связывающих белков или связывающих доменов основывается на сконструированных белковых повторах или сконструированных повторяемых доменах (WO 2002/020565; Binz, H.K., Amstutz, P., Kohl, A., Stumpp, M.T., Briand, С., Forrer, P., Grütter, M.G., и Plückthun, A., Nat. Biotechnol. 22, 575-582, 2004; Stumpp, M.T., Binz, H.K и Amstutz, P., Drug Discov. Today 13, 695-701, 2008). WO 2002/020565 описывает, как могут быть сконструированы большие библиотеки белковых повторов и их общее применение. Тем не менее, WO 2002/020565 не раскрывает ни селекцию повторяемых доменов со связывающей специфичностью для сывороточного альбумина SA, ни конкретные последовательности мотивов повторяемых доменов, которые специфически связываются с SA. Кроме того, WO 2002/020565 не предполагает, что повторяемые домены со связывающей специфичностью для SA могут использоваться для модуляции РК или PD других молекул. Такие сконструированные повторяемые домены используют модулярную природу белковых повторов и имеют N-терминальные и С-терминальные кэппинг модули для предотвращения агрегации сконструированных повторяемых доменов путем экранирования гидрофобного ядра домена (Forrer, P., Stumpp, M.T., Binz, Н.К. и Plückthun, A., FEBS letters 539, 2-6, 2003). Эти кэппинг модули основываются на кэппирующих повторах природного гуанин-аденин-связывающего белка (GA-связывающего белка). Было показано, что термальная и термодинамическая стабильность этих доменов сконструированных анкириновых повторов может быть дополнительно повышена путем усовершенствования С-терминального кэппирующего повтора, который имеет происхождение от GA-связывающего белка (Interlandi, G., Wetzel, S.K, Settanni, G., Plückthun, А. и Caflisch, A., J. Mol. Biol. 375, 837-854, 2008; Kramer, M.A, Wetzel, S.K., Plückthun, A., Mittl, P.R.E, и Grütter, M.G., J. Mol. Biol. 404, 381-391, 2010). Авторы вводили в общей сложности восемь мутаций в этот кэппинг модуль и удлиняли его С-терминальную спираль путем прибавления трех различных аминокислот. Тем не менее, введение этих модификаций в С-терминальный кэппинг модуль приводило к тенденции нежелательной димеризации сконструированного повторяемого домена, который несет этот мутированный С-терминальный кэппинг модуль. Таким образом, существует необходимость в получении дополнительных оптимизированных С-терминальных кэппинг модулей или С-терминальных кэппирующих повторов доменов анкиринового повтора.
Нацеливание SA для модуляции РК и/или PD с помощью доступных в настоящее время подходов не всегда является эффективным. При этом также стало более отчетливо видно, что модуляция РК и/или PD молекул путем захвата SA является сложной и все еще не совсем понятной.
В общем случае, существует потребность в улучшенных связывающих белках со специфичностью для SA, способных улучшать РК и/или PD терапевтически релевантных молекул или полипептидов для лечения рака и других патологических состояний.
Техническая проблема, которая лежит в основе настоящего изобретения, заключается в идентификации новых связывающих белков, таких как повторяемые домены, со связывающей специфичностью для SA, способных к модификации РК и/или PD терапевтически релевантных молекул для улучшенного лечения рака и других патологических состояний. Решение этой технической проблемы достигается путем обеспечения воплощений, охарактеризованных в формуле изобретения.
Краткое изложение сущности изобретения
Настоящее изобретение относится к связывающим белкам, включающим, по крайней мере один домен анкиринового повтора, где указанный домен анкиринового повтора имеет связывающую специфичность для сывороточного альбумина млекопитающих и где указанный домен анкиринового повтора включает модуль анкиринового повтора, имеющий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 49, 50, 51 и 52, а также последовательностей, в которых вплоть до 9 аминокислот SEQ ID NO: 49, 50, 51 и 52 являются замененными какой-либо аминокислотой.
В дополнительном воплощении изобретение относится к связывающим белкам, включающим, по крайней мере, один домен анкиринового повтора, где указанный повторяемый домен имеет связывающую специфичность для сывороточного альбумина млекопитающих, и где указанный домен анкиринового повтора включает аминокислотную последовательность, которая имеет, по крайней мере, 70%-ную идентичность аминокислотной последовательности с одним доменом анкиринового повтора, выбранным из группы, состоящей из SEQ ID NO: 17-31 и 43-48, где G в положении 1 и/или S в положении 2 указанного домена анкиринового повтора являются необязательно отсутствующими.
В частности, изобретение относится к связывающим белкам, как определяется в данной заявке выше, где домен анкиринового повтора конкурирует за связывание с сывороточным альбумином млекопитающих с доменом анкиринового повтора, выбранным из группы, состоящей из SEQ ID NO: 17-31 и 43-48.
Кроме того, изобретение относится к таким связывающим белкам, включающим биоактивное соединение, в частности, к связывающим белкам, включающим биоактивное соединение, которое обладает, по крайней мере, в два раза более высоким конечным периодом полувыведения у млекопитающего по сравнению с конечным периодом полувыведения указанного немодифицированного биоактивного соединения.
Изобретение также относится к молекулам нуклеиновой кислоты, которые кодируют связывающие белки в соответствии с настоящим изобретением, и к фармацевтической композиции, включающей один или или более упомянутых выше связывающих белков или молекул нуклеиновой кислоты.
Изобретение также относится к способу лечения патологического состояния при использовании связывающих белков в соответствии с изобретением.
Краткое описание чертежей
Фигура 1. Анализ стабильности выбранных DARPin с помощью SEC.
Профили элюирования прогонов эксклюзионной хроматографии размеров (SEC) DARPin со специфичностью для xSA перед инкубацией (Фиг.1а), после инкубации при концентрации 30 мг/мл (~2 мМ) в PBS в течение 28 дней при 40°С (Фиг.1b) или после хранения в течение 1 месяца при -80°С (Фиг.1с), проанализированные с помощью Superdex 200 колонки 5/150 (Фиг.1а или Фиг.1b), или с помощью колонки Superdex 200 10/300GL (Фиг.1с). Все образцы подвергали экспрессии и очищали так, как описано в Примере 1. Для проведения SEC анализа образцы разводили до концентрации 500 мкМ. Стандарты молекулярной массы Апротинин (АР) 6,5 кДа, Карбоангидраза (СА) 29 кДа и Кональбумин (СО) 75 кДа обозначены стрелками.
xSA, сывороточный альбумин млекопитающих, А, поглощение при 280 нм; t, время удержания в минутах;
DARPin #19 (SEQ ID NO: 19 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом);
DARPin #20 (SEQ ID NO: 20 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом);
DARPin #21 (SEQ ID NO: 21 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом);
DARPin #22 (SEQ ID NO: 22 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом);
DARPin #27 (SEQ ID NO: 27 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом);
DARPin #28 (SEQ ID NO: 28 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом);
DARPin #29 (SEQ ID NO: 29 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом);
DARPin #30 (SEQ ID NO: 30 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом).
DARPin #43 (SEQ ID NO: 43 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом).
DARPin #44 (SEQ ID NO: 44 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом).
DARPin #45 (SEQ ID NO: 45 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом).
DARPin #46 (SEQ ID NO: 46 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом).
DARPin #47 (SEQ ID NO: 47 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом).
DARPin #48 (SEQ ID NO: 48 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом).
Фигура 2. Термальная стабильность выбранных DARPin.
Запись для термальной денатурации DARPin со специфичностью для xSA (которая сопровождается повышением интенсивности флуоресценции красителя SYPRO оранжевого, присутствующего в буфере) в PBS при значении рН 7,4 (Фиг.2а) и в MES буфере при значении рН 5,8 (Фиг.2b) (250 мМ (2-N-морфолино)этансульфоновая кислота рН 5,5), 150 мМ NaCl, смешанный с PBS рН 7,4 1 к 4 (об./об.) и доведенный до значения рН 5,8).
F, относительные единицы флуоресценции (RFU), возбуждение при 515-535 нм, определение при 560-580 нм; Т, температура в °С; определение DARPin смотри выше.
Фигура 3. Выведение выбранных DARPin из плазмы крови у мышей.
Выведение из плазмы крови DARPin со специфичностью для MSA (мышиный сывороточный альбумин) и контрольных DARPin оценивали у мышей.
(Фиг.3а) DARPin, включающие только один повторяемый домен со связывающей специфичностью для MSA по сравнению с DARPin #32 (смотри ниже), не имеющим связывающей специфичности для MSA.
(Фиг.3b) DARPin, включающие два белковых домена (один из которых представляет собой повторяемый домен со связывающей специфичностью для MSA) по сравнению с DARPin #32, не имеющим связывающей специфичности для MSA.
DARPin метили с помощью His-Tag при использовании соединения 99mTc-карбонил и вводили внутривенно мышам. Радиоактивность у мышей, которым делали инъекцию, измеряли в различные моменты времени после инъекции и представляли в виде соотношения инъецируемой дозы, скорректированной для радиоактивного распада 99mTc (% ID). Апроксимированные кривые нелинейных регрессий радиоактивности, измеренной в различные моменты времени - двухфазный распад (Graphpad Prism). Каждая точка данных показывает среднее значение двух мышей на группу.
% ID, процент введенной дозы, скорректированный для радиоактивного распада 99mTc; t, время в часах;
DARPin #18 (SEQ ID NO: 18 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом);
DARPin #23 (SEQ ID NO: 23 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом);
DARPin #25 (SEQ ID NO: 25 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом);
DARPin #32 (негативный контроль DARPin без связывающей специфичности по отношению к xSA, SEQ ID NO: 32 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом);
DARPin #33 (DARPin, включающий два домена повтора, один со связывающей специфичностью для xSA, SEQ ID NO: 33 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом);
DARPin #35 (DARPin, включающий два домена повтора, один со связывающей специфичностью для xSA, SEQ ID NO: 35 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом);
DARPin #36 (DARPin, включающий два домена повтора, один со связывающей специфичностью для xSA, SEQ ID NO: 36 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом).
Фигура 4. Выведение выбранных DARPin из плазмы крови у обезьян циномолгус.
Выведение DARPin со специфичностью для CSA (сывороточный альбумин обезьян циномолгус) и контрольных DARPin из плазмы крови оценивали у обезьян циномолгус.
(Фиг.4а) DARPin #26 сравнивали с DARPin #32, не имеющим связывающей специфичности по отношению к CSA.
(Фиг.4b) DARPin #24, 34 и 17 сравнивали с DARPin #32, не имеющим связывающей специфичности по отношению к CSA. Следующие DARPin внутривенно вводили обезьянам циномолгус в момент времени t=0 часов в концентрации 0,5 мг/мл (DARPin #26, DARPin #24, DARPin #17 и DARPin #32) или 1 мг/мл (DARPin #34): Концентрацию DARPin в плазме крови обезьян измеряли с помощью ELISA в различные моменты времени после инъекции. Кривые показывают результат нелинейных регрессий концентраций, измеренных в различные моменты времени - двухфазный распад (Graphpad Prism). Из второй фазы может быть определен конечный период полувыведения DARPin. Каждая единичная точка данных указывает среднее значение двух независимых измерений ELISA того же образца сыворотки.
С, концентрация DARPin в нМ; t, время в часах;
DARPin #17 (SEQ ID NO: 17 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом);
DARPin #24 (SEQ ID NO: 24 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом);
DARPin #26 (SEQ ID NO: 26 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом);
DARPin #32 (негативный контроль DARPin без связывающей специфичности по отношению к xSA, SEQ ID NO: 32 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом);
DARPin #34 (DARPin, включающий два домена повтора, один со связывающей специфичностью для xSA, SEQ ID NO: 34 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом).
Подробное описание изобретения
Связывающий домен в соответствии с изобретением является специфическим для сывороточного альбумина млекопитающих (xSA). Предпочтительно, связывающий домен в соответствии с изобретением является специфическим для сывороточного альбумина, который имеет происхождение от мышей, крыс, собак, кролей, обезьян или человека. Более предпочтительно, связывающий домен в соответствии с изобретением является специфическим для сывороточного альбумина человеческого происхождения (HSA).
Термин "белок" относится к полипептиду, в котором, по крайней мере, часть полипептида имеет, или обладает способностью приобретать определенное пространственное расположение путем формирования вторичных, третичных или четвертичных структур в пределах полипептидного(ых) цепи(ей) и/или между ними. В том случае, если белок включает два или более полипептидов, то индивидуальные полипептидные цепи могут быть связаны нековалентно или ковалентно, например, с помощью дисульфидной связи между двумя полипептидами. Часть белка, которая индивидуально имеет или обладает способностью приобретать определенное пространственное расположение путем формирования вторичных или третичных структур, называется "белковым доменом". Такие белковые домены являются хорошо известными квалифицированному специалисту в данной области техники.
Термин "рекомбинантный", как используется в словосочетании рекомбинантный белок, рекомбинантный белковый домен, рекомбинантный связывающий белок и тому подобное, означает, что указанные полипептиды получают с помощью методик рекомбинантной ДНК, хорошо известных квалифицированному специалисту в релевантной области техники. Например, рекомбинантная молекула ДНК (например, полученная с помощью синтеза генов), которая кодирует полипептид, может быть клонирована в бактериальную экспрессионную плазмиду (например, pQE30, Qiagen), дрожжевую экспрессионную плазмиду или экспрессионную плазмиду млекопитающих. Когда, например, такая сконструированная рекомбинантная бактериальная экспрессионная плазмида встраивается в приемлемую бактерию (например, Escherichia coli), эта бактерия может продуцировать полипептид, который кодируется этой рекомбинантной ДНК. Соответствующим образом полученный полипептид называется рекомбинантным полипептидом.
В контексте настоящего изобретения термин "полипептид" относится к молекуле, которая состоит из одной или более цепей многочисленных, то есть, из двух или более, аминокислот, связанных с помощью пептидных связей. Предпочтительно, полипептид состоит из более, чем восемь аминокислот, связанных с помощью пептидных связей.
Термин "полипептидная метка" относится к аминокислотной последовательности, которая присоединяется к полипептиду/белку, где указанная аминокислотная последовательность является полезной для очистки, определения или мечения указанного полипептида/белка, или где указанная аминокислотная последовательность улучшает физико-химическое поведение полипептида/белка, или где указанная аминокислотная последовательность обладает эффекторной функцией. Индивидуальные полипептидные метки, остатки и/или домены связывающего белка могут быть связаны друг с другом непосредственно или с помощью полипептидных линкеров. Эти полипептидные метки являются хорошо известными в области техники и полностью доступны квалифицированному специалисту в данной области техники. Примеры полипептидных меток представляют собой малые полипептидные последовательности, например, His (например, His-метка последовательности SEQ ID NO: 15), myc, FLAG или Strep-метки или остатки, такие как ферменты (например, ферменты, подобные щелочной фосфатазе), которые позволяют осуществлять определение указанного полипептида/белка, или остатки, которые могут использоваться для мечения (такие как иммуноглобулины или их фрагменты) и/или в качестве эффекторных молекул.
Термин "полипептидный линкер" относится к аминокислотной последовательности, которая является способной к связыванию, например, двух белковых доменов, полипептидной метки и белкового домена, белкового домена и остатка, отличного от полипептида, такого как полиэтиленгликоль или двух последовательностей метки. Такие дополнительные домены, метки, остатки, отличные от полипептидов, и линкеры являются хорошо известными квалифицированному специалисту в релевантной области техники. Список примеров обеспечивается в описании к патентной заявке WO 2002/020565. Частные примеры таких линкеров представляют собой глицин-серин линкеры и пролин-треонин линкеры варьирующей длины; является предпочтительным, когда указанные линкеры имеют длину от 2 до 24 аминокислот; более предпочтительно, когда указанные линкеры имеют длину от 2 до 16 аминокислот. Пример глицин-серин линкера обеспечивается в SEQ ID NO: 16.
Термин "полимерный остаток" относится либо к остатку белкового полимера, либо к остатку небелкового полимера. Термин "остаток белкового полимера" предпочтительно представляет собой полипептид, который не формирует стабильной третичной структуры не более чем в 10%, предпочтительно, не более чем в 5%; также предпочтительно не более чем в 2%; даже более предпочтительно, не более чем в 1%; и наиболее предпочтительно, в неспособных к определению количествах, как определяется с помощью эксклюзионной хроматографии размеров (SEC), олигомеров или агрегатов при хранении в концентрации приблизительно 0,1 мМ в забуференном фосфатом физиологическом растворе (PBS) при комнатной температуре (RT) в течение одного месяца. Такие белковые полимерные остатки разгоняют при среднем молекулярном весе в SEC, который является выше, чем их эффективный молекулярный вес при использовании глобулярных белков в качестве стандартов молекулярного веса для SEC. Предпочтительно, когда средний молекулярный вес указанных белковых полимерных остатков, определяемый с помощью SEC, является в 1,5 раза, 2 раза или 2,5 раза выше, чем их эффективный молекулярный вес, подсчитанный из их аминокислотной последовательности. Также является предпочтительным, когда средние молекулярные массы указанных небелковых полимерных остатков, определенные с помощью SEC, являются в 2 раза, 4 раза или 8 раз выше, чем их эффективный молекулярный вес, подсчитанный из их молекулярного состава. Предпочтительно, когда более чем 50%, 70% или даже 90% аминокислот указанных остаток белкового полимера не формируют стабильных вторичных структур при концентрации приблизительно 0,1 мМ в PBS при комнатной температуре, как определяется с помощью измерений циркулярного дихроизма (CD). Наиболее предпочтительно, когда указанный белковый полимер демонстрирует типичный спектр циркулярного дихроизма вблизи УФ статистической спиральной конформации. Такие CD являются хорошо известными квалифицированному специалисту в данной области техники. Также предпочтительными являются остатки белковых полимеров, которые состоят более чем из 50, предпочтительно более чем из 100, 200, 300, 400, 500, 600, 700, или наиболее предпочтительно более чем из 800 аминокислот. Примеры белковых полимерных остатков представляют собой XTEN® (зарегистрированный товарный знак Amunix; WO 2007/103515) полипептиды или полипептиды, включающие остатки пролина, аланина и серина, как описывается в WO 2008/155134. Такие белковые полимерные остатки могут быть ковалентно присоединены, например, к связывающему домену в соответствии с изобретением, путем получения полипептидов генетическим слиянием при использовании стандартных методик клонирования ДНК, после чего осуществляют их стандартную экспрессию и очистку.
Полимерный остаток в соответствии с изобретением может варьировать в широких пределах по своей молекулярной массе (то есть, от приблизительно 1 кДа до приблизительно 150 кДа). Предпочтительно, полимерный остаток имеет молекулярный вес, по крайней мере, 2, более предпочтительно, по крайней мере, 5, 10, 20, 30, 50, 70, или наиболее предпочтительно, по крайней мере, 100 кДа. Предпочтительно, когда указанный полимерный остаток соединяется с помощью полипептидного линкера со связывающим доменом.
Примеры небелковых полимерных остатков представляют собой гидроксиэтил крахмал (HES), полиэтиленгликоль (PEG), полипропиленгликоль или полиоксиалкилен. Термин "ПЭГилированный" означает, что остаток ПЭГ является ковалентно присоединенным, например, к полипептиду в соответствии с изобретением.
В специфическом воплощении остаток ПЭГ или любого другого небелкового полимера может, например, может быть слитым с тиолом цистеина с помощью малеимидного линкера, при этом цистеин является слитым с помощью пептидного линкера с N- или С-терминальным концом связывающего домена, как описывается в данной заявке.
Термин "связывающий белок" относится к белку, включающему один или более связывающих доменов, одно или более биоактивных соединений и один или более полимерных остатков, как дополнительно объясняется ниже. Предпочтительно, когда указанный связывающий белок включает вплоть до четырех связывающих доменов. Более предпочтительно, когда указанный связывающий белок включает вплоть до двух связывающих доменов. Наиболее предпочтительно, когда указанный связывающий белок включает только один связывающий домен. Кроме того, любой такой связывающий белок может включать дополнительные белковые домены, которые не являются связывающими доменами, остатки мультимеризации, полипептидные метки, полипептидные линкеры и/или единичный остаток Cys. Примеры остатков мультимеризации представляют собой константные участки тяжелой цепи иммуноглобулина, которые спариваются с обеспечением функциональных Fc доменов иммуноглобулина, и лейциновые "застежки" или полипептиды, включающие свободный тиол, который образует межмолекулярную дисульфидную связь между двумя такими полипептидами. Единичный остаток Cys может использоваться для конъюгации других остатков к полипептиду, например, путем использования малеимидной связи, которая является хорошо известной квалифицированному специалисту в данной области техники. Предпочтительно, когда указанный связывающий белок представляет собой рекомбинантный связывающий белок. Также является предпочтительным, когда связывающие домены связывающего белка обладают различными целевыми специфичностями.
Термин "связывающий домен" означает белковый домен, который демонстрирует такую же сборку (пространственное размещение), что и белковый каркас и обладает предварительно определенным свойством, как определяется ниже. Такой связывающий домен может быть получен с помощью целесообразных, или в наиболее общем случае, комбинаторных методик конструирования белка, методов, которые являются известными в области техники (Binz и др., 2005, в приведенном выше месте). Например, связывающий домен, имеющий предварительно определенное свойство, может быть получен с помощью способа, включающего этапы (а) обеспечения разнообразной коллекции белковых доменов, которые демонстрируют такую же сборку, что и белковый каркас, как определено дополнительно ниже; и (b) скрининга указанной разнообразной коллекции и/или селекции из указанной разнообразной коллекции для получения, по крайней мере, одного белкового домена, обладающего указанным предварительно определенным свойством. Разнообразная коллекция белковых доменов может обеспечиваться с помощью нескольких способов в соответствии с используемой системой скрининга и/или селекции и может включать применение способов, хорошо известных квалифицированному специалисту в данной области техники, таких как фаговый дисплей или рибосомальный дисплей. Предпочтительно, указанный связывающий домен представляет собой рекомбинантный связывающий домен.
Термин "белковый каркас" означает белок с внешними участками, в которых аминокислотные инсерции, замены или делеции являются в высокой степени допустимыми. Примеры белковых каркасов, которые могут использоваться для получения связывающих доменов настоящего изобретения, представляют собой антитела или их фрагменты, такие как одноцепочечные Fv или Fab фрагменты, белок А из Staphylococcus aureus, связывающий билин белок из Pieris brassicae или другие липокалины, белки анкириновых повторов или другие повторяемые белки, а также человеческий фибронектин. Белковые каркасы являются хорошо известными квалифицированному специалисту в данной области техники (Binz и др., 2005, в приведенном выше месте; Binz и др., 2004, в приведенном выше месте).
Термин "предварительно определенное свойство" относится к свойству, такому как, связывание с мишенью, блокирование мишени, активация опосредованной мишенью реакции, ферментативная активность и другие близкие свойства. В зависимости от типа желаемого свойства, средний специалист в данной области техники будет способен идентифицировать формат и необходимые этапы для осуществления скрининга и/или селекции связывающего домена с желаемым свойством. Предпочтительно, указанное предварительно определенное свойство представляет собой связывание с мишенью.
Определения, приведенные в данной заявке ниже для белков повторов, основываются на таких, которые описаны в патентной заявке WO 2002/020565. Патентная заявка WO 2002/020565 дополнительно содержит общее описание характеристик белковых повторов, методик и применений.
Термин "повторяемый белок" относится к белкам, включающим один или более повторяемых доменов. Предпочтительно, каждый указанный белок повторов включает вплоть до четырех повторяемых доменов. Более предпочтительно, когда каждый указанный белок повторов включает вплоть до двух повторяемых доменов. Наиболее предпочтительно, когда каждый указанный повторяемый белок включает только один повторяемый домен. Кроме того, указанный белок повторов может включать дополнительные небелковые повторяемые домены, полипептидные метки и/или полипептидные линкеры.
Термин "повторяемый домен" относится к белковым доменам, включающим две или более последовательных повторяемых единиц (модулей) в качестве структурных единиц, где указанные структурные единицы имеют такую же сборку и плотно упаковываются с образованием, например, сферической структуры, имеющей соединительное гидрофобное ядро. Предпочтительно, повторяемый домен дополнительно включает N-терминальную и/или С-терминальную кэппирующую единицу (или модуль). Даже более предпочтительно, когда указанные N-терминальные и/или С-терминальные кэппирующие единицы (или модули) представляют собой кэппирующие повторы.
Термин "сконструированные повторяемый белок" и "сконструированный повторяемый домен" относится к повторяемому белку или повторяемому домену, соответственно, полученному в результате изобретательской процедуры, которая объясняется в патентной заявке WO 2002/020565. Сконструированные повторяемые белки и сконструированные повторяемые домены являются синтетическими, а не природными. Они могут представлять собой созданные человеком белки или домены, соответственно, полученные путем экспрессии соответственным образом сконструированных нуклеиновых кислот. Предпочтительно, экспрессию осуществляют в прокариотических или эукариотических клетках, таких как бактериальные клетки, или путем использования бесклеточной экспрессионной системы in vitro. В соответствии с этим, сконструированный белок с анкириновым повтором (то есть, DARPin) соответствует связывающему белку в соответствии с изобретением, включающему, по крайней мере, один домен анкиринового повтора.
Термин "структурная единица" относится к локально упорядоченной части полипептида, образованной пространственными взаимодействиями между двумя или более сегментами вторичной структуры, которые находятся рядом друг с другом в полипептидной цепи. Такая структурная единица демонстрирует структурный мотив. Термин "структурный мотив" относится к пространственному расположению элементов вторичной структуры, присутствующих, по крайней мере, в одной структурной единице. Структурные мотивы являются хорошо известными квалифицированному специалисту в данной области техники. Структурные единицы, взятые отдельно, не являются способными приобретать определенное пространственное расположение; однако их последовательное расположение, например, в виде повторяемых модулей в повторяемом домене, приводит к взаимной стабилизации соседних единиц, что вызывает образование сферической структуры.
Термин "повторяемая единица" относится к аминокислотной последовательности, включающей мотивы повторяемой последовательности одного или более существующих в природе повторяемых белков, где указанные "повторяемые единицы" обнаруживаются во множественных копиях, и которые демонстрируют определенную топологию сборки, общую для всех указанных мотивов, определяющих сборку этого белка. Такие повторяемые единицы соответствуют "повторяемым структурным единицам (повторам)" повторяемых белков, как описывается Forrer и др., 2003, в приведенном выше месте, или "последовательным структурным единицам (повторам)" повторяемых белков, как описывается Binz и др., 2004, в приведенном выше месте. Такие повторяемые единицы включают каркасные остатки и остатки взаимодействия. Примеры таких повторяемых единиц представляют собой повторяемые единицы армадилло, обогащенные лейцином повторяемые единицы, единицы анкиринового повтора, повторяемые единицы тетратрикопептида, повторяемые единицы HEAT и вариантные повторяемые единицы, обогащенные лейцином. Существующие в природе белки, содержащие две или более таких повторяемых единиц, называются "существующими в природе повторяемыми белками". Аминокислотные последовательности индивидуальных повторяемых единиц повторяемых белков могут иметь значительное количество мутаций, замен, инсерций и/или делеций по сравнению с каждым другим, в то время как все еще существенно сохраняют общую модель, или мотив повторяемой единицы.
Термин "единица анкиринового повтора" будет означать повторяемую единицу, которая представляет собой анкириновый повтор, как описывается, например, Forrer и др., 2003, в приведенном выше месте. Анкириновые повторы являются хорошо известными квалифицированному специалисту в данной области техники.
Термин "каркасные участки" относится к аминокислотным остаткам повторяемой единицы, или соответствующим аминокислотным остаткам повторяемых модулей, которые осуществляют свой вклад в топологию сборки, то есть те, которые осуществляют свой вклад в сборку указанной повторяемой единицы (или модуля), или те, которые осуществляют свой вклад во взаимодействие с соседней единицей (или модулем). Такой вклад может представлять собой взаимодействие с другими остатками в повторяемой единице (или модуле), или влияние на конформацию полипептидного скелета, как обнаруживается в α-спиралях или β-складках, или аминокислотных цепочках, которые формируют линейные полипептиды или петли.
Термин "целевые остатки взаимодействия" относится к аминокислотным остаткам повторяемой единицы, или к соответствующим аминокислотным остаткам повторяемых модулей, которые осуществляют свой вклад во взаимодействие с целевыми веществами. Такой вклад может представлять собой непосредственное взаимодействие с целевыми веществами, или влияние на другие непосредственно взаимодействующие остатки, например, путем стабилизации конформации полипептида повторяемой единицы (или модуля) для того, чтобы позволить осуществить или улучшить взаимодействие непосредственно взаимодействующих остатков с указанной целью. Такие каркасные остатки и целевые остатки взаимодействия могут быть идентифицированы с помощью анализа структурных данных, полученных с помощью физико-химических методов, таких как рентгеноструктурная кристаллография, ЯМР и/или спектроскопия циркулярного дихроизма, или путем сравнения с известной и близкой структурной информацией, которая является хорошо известной практикующим специалистам в структурной биологии и/или биоинформатике.
Предпочтительно, когда повторяемые единицы, используемые для дедукции мотива повторяемой последовательности, являются гомологичными повторяемыми единицам, где повторяемые единицы включают тот же структурный мотив и в которых более чем 70% каркасных участков указанных повторяемых единиц являются гомологичными друг другу. Предпочтительно, более чем 80% каркасных участков указанных повторяемых единиц являются гомологичными. Наиболее предпочтительно, когда более чем 90% каркасных участков указанных повторяемых единиц являются гомологичными.
Компьютерные программы для определения процента гомологии между полипептидами, такие как Fasta, Blast или Gap, являются известными квалифицированному специалисту в данной области техники. Также является предпочтительным, когда повторяемые единицы, используемые для дедукции мотива повторяемой последовательности, являются гомологичными повторяемыми единицам, полученным из повторяемых доменов, отобранных на мишени, например, так, как описывается в Примере 1, и имеют такую же целевую специфичность.
Термин "мотив повторяемой последовательности" относится к аминокислотной последовательности, которая выводится из одной или более повторяемых единиц или повторяемых модулей. Предпочтительно, когда указанные повторяемые единицы или повторяемые модули являются такими из повторяемых доменов, имеющих связывающую специфичность для той же мишени. Такой мотив повторяемых последовательностей включает положения остатков каркасного участка и положения остатков целевого взаимодействия. Указанные положения остатков каркасного участка соответствуют положениям каркасных участков повторяемых единиц (или модулей). Подобно этому, указанные положения остатков целевого взаимодействия соответствуют положениям остатков целевого взаимодействия повторяемых единиц (или модулей). Мотив повторяемых последовательностей включает фиксированные положения и случайные положения. Термин "фиксированное положение" относится к аминокислотному положению в мотиве повторяемой последовательности, где указанное положение принимается за определенную аминокислоту. Наиболее часто, такие фиксированные положения соответствуют положениям каркасных участков и/или положениям остатков целевого взаимодействия, которые являются специфическими для определенной мишени. Термин "рандомизированное положение" относится к аминокислотному положению в мотиве повторяемой последовательности, в котором две или более аминокислот допускаются в указанном аминокислотном положении, например, в котором допускаются любые из обычных двадцати существующих в природе аминокислот, или в котором допускаются более двадцати существующих в природе аминокислот, таких, как аминокислоты, отличные от цистеина, или аминокислоты, отличные от глицина, цистеина и пролина. Наиболее часто, такие рандмизированные положения соответствуют положениям остатков целевого взаимодействия. Однако некоторые положения каркасных участков также могут быть рандомизированы.
Термин "топология сборки" относится к третичной структуре указанных повторяемых единиц или повторяемых модулей. Топология сборки будет определяться цепочками аминокислот, формирующими, по крайней мере, части α-спиралей или β-складок, или цепочками аминокислот, формирующими линейные полипептиды или петли, или любой комбинацией α-спиралей, β-складок и/или линейных полипептидов/петель.
Термин "последовательный" относится к размещению, в котором повторяемые единицы или повторяемые модули размещаются последовательно друг за другом. В сконструированных повторяемых белках, существует, по крайней мере, 2, обычно приблизительно от 2 до 6, в частности, по крайней мере, приблизительно 6, часто 20 или более повторяемых единиц (или модулей). В большинстве случаев повторяемые единицы (или модули) повторяемого домена будут демонстрировать высокую степень идентичности последовательности (одинаковые аминокислотные остатки в соответствующих положениях) или подобия последовательности (аминокислотные остатки являются различными, но имеют подобные физико-химические свойства), и некоторые из аминокислотных остатков могут представлять собой ключевые остатки, которые являются строго консервативными. Однако высокая степень вариабельности последовательности благодаря аминокислотным инсерциям и/или делециям, и/или заменам между различными повторяемыми единицами (или модулями) повторяемого домена может быть возможной при условии, что поддерживается общая топология сборки повторяемых единиц (или модулей).
Способы непосредственного определения топологии сборки белков повторов при использовании физико-химических средств, таких как рентгеновская кристаллография, ЯМР или спектроскопия циркулярного дихроизма, являются хорошо известными практикующему специалисту в данной области техники. Способы идентификации и определения повторяемых единиц или мотивов повторяемых последовательностей или идентификации семейств родственных белков, включающих такие повторяемые единицы или мотивы, такие, как поиски гомологии (BLAST и т.д.), являются хорошо установленными в области биоинформатики и хорошо известны практикующему специалисту в данной области техники. Этап уточнения исходного мотива повторяемой последовательности может включать итеративный процесс.
Термин "повторяемые модули" относится к повторяемой аминокислотной последовательности сконструированных повторяемых доменов, которые исходно имеют происхождение от повторяемой единицы существующих в природе повторяемых белков. Каждый повторяемый модуль, который содержится в повторяемом домене, имеет происхождение от одной или более повторяемых единиц семейства или подсемейства существующих в природе повторяемых белков, например, семейства белковых повторов армадилло или белков с анкириновым повтором.
"Повторяемые модули" могут включать положения с аминокислотными остатками, присутствующими во всех копиях соответствующих повторяемых модулей ("фиксированные положения"), и положения с отличающимися или "рандомизированными" аминокислотными остатками ("рандомизированные положения").
Связывающий белок в соответствии с изобретением включает, по крайней мере один домен анкиринового повтора, где указанный повторяемый домена имеет связывающую специфичность для сывороточного альбумина млекопитающих (xSA).
Термины "имеет связывающую специфичность для мишени", "которые специфически связываются с мишенью" или "целевая специфичность" и тому подобные означают, что связывающий белок или связывающий домен связываются в PBS с мишенью с более низкой константой диссоциации, чем с неродственным белком, например, таким как связывающий мальтозу белок Е.coli (МВР). Предпочтительно, когда константа диссоциации в PBS для мишени является, по крайней мере в 10, более предпочтительно в 102, даже более предпочтительно в 103, или наиболее предпочтительно в 104 раз ниже, чем соответствующая константа диссоциации для МВР.
Связывающий белок в соответствии с изобретением не является антителом или его фрагментом, таким как Fab или scFv фрагменты. Антитела и их фрагменты являются хорошо известными квалифицированному специалисту в данной области техники.
Кроме того, связывающий домен в соответствии с изобретением не включает специфической укладки цепи иммуноглобулинов, которая присутствует в антителах и/или домене фибронектина типа III. Специфическая укладка цепи иммуноглобулинов является общей для всех - это β-укладка белков, которая состоит из приблизительно 7 параллельных β-цепей, размещенных в двух β-складках. Укладки иммуноглобулинов являются хорошо известными квалифицированному специалисту в данной области техники. Например, такие связывающие домены, включающие иммуноглобулиновую укладку, описываются в WO 2007/080392 или WO 2008/097497.
В частности, изобретение относится к связывающему белку, включающему, по крайней мере, один домен анкиринового повтора, где указанный домен анкиринового повтора имеет связывающую специфичность для сывороточного альбумина млекопитающих и где указанный домен анкиринового повтора включает модуль анкиринового повтора, имеющий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 49, 50, 51 и 52 и последовательностей, в которых вплоть до 9 аминокислот в SEQ ID NO: 49, 50, 51 и 52 являются замененными какой-либо аминокислотой.
Предпочтительно, когда вплоть до 8 аминокислот в SEQ ID NO: 49, 50, 51 и 52 заменены другой аминокислотой, более предпочтительно вплоть до 7 аминокислот, более предпочтительно вплоть до 6 аминокислот, более предпочтительно вплоть до 5 аминокислот, даже более предпочтительно вплоть до 4 аминокислот, более предпочтительно вплоть до 3 аминокислот, более предпочтительно вплоть до 2 аминокислот, более предпочтительно вплоть до 1 аминокислоты, и наиболее предпочтительно ни одна аминокислота в SEQ ID NO: 49, 50, 51 и 52 не является замененной.
Предпочтительно, когда аминокислоты заменяются в SEQ ID NO: 49, 50, 51 и 52, эти аминокислоты являются выбранными из группы, состоящей из A, D, Е, F, Н, I, К, L, M, N, Q, R, S, Т, V, W и Y; более предпочтительно из группы, состоящей из А, D, Е, Н, I, К, L, Q, R, S, Т, V и Y. Также предпочтительно, когда замещение аминокислоты гомологичной аминокислотой; то есть, аминокислота заменяется аминокислотой, которая имеет боковую цепь с подобными биофизическими свойствами. Например, негативно заряженная аминокислота D может быть заменена негативно заряженной аминокислотой Е, или гидрофобная аминокислота, такая как L, может быть заменена А, I или V. Замещение аминокислоты гомологичной аминокислотой является хорошо известным квалифицированному специалисту в данной области техники.
Предпочтительный связывающий белок включает, по крайней мере, один домен анкиринового повтора, где указанный повторяемый домен связывает xSA с константой диссоциации (Kd) в PBS ниже 10-4 М. Предпочтительно, указанный повторяемый домен связывает xSA с Kd в PBS ниже 10-4 М, более предпочтительно ниже 10-5 М, 10-6 М, 10-7 М, или наиболее предпочтительно 10-8 М.
Способы определения констант диссоциации белок-белковых взаимодействий, такие как методики, которые основываются на поверхностном плазменном резонансе (SPR) (например, SPR равновесный анализ) или изотермическая титрационная калориметрия (ITC), являются хорошо известными квалифицированному специалисту в данной области техники. Измеренные значения Kd конкретного белок-белкового взаимодействия могут варьировать, если измеряются при различных условиях (например, концентрации соли, рН). Таким образом, измерение значений Kd предпочтительно осуществляют при использовании стандартизированных растворов белка и стандартизированного буфера, такого как PBS.
Связывающие белки, включающие домен анкиринового повтора, которые связывают xSA со значением Kd PBS ниже 10-4 М, являются показанными в разделе Примеры.
Домен анкиринового повтора связывающего белка в соответствии с изобретением связывает xSA. Предпочтительным является связывающий белок, включающий домен анкиринового повтора, который связывает человеческий сывороточный альбумин (HSA).
Также предпочтительным является связывающий домен, включающий от 70 до 300 аминокислот, в частности, от 100 до 200 аминокислот.
Также предпочтительным является связывающий белок или связывающий домен, в котором отсутствуют свободный Cys остаток. "Свободный Cys остаток" не вовлекается в образование дисульфидной связи. Даже более предпочтительным является связывающий белок или связывающий домен, который не содержит никакого Cys остатка.
Связывающий домен в соответствии с изобретением представляет собой домен анкиринового повтора или сконструированный домен анкиринового повтора (Binz и др., 2004, в приведенном выше месте), предпочтительно такой, как описывается в WO 2002/020565. Примеры сконструированных доменов анкиринового повтора являются показанными в разделе Примеры.
В дополнительном воплощении изобретение относится к связывающему белку, включающему, по крайней мере, один домен анкиринового повтора, где указанный повторяемый домен имеет связывающую специфичность для сывороточного альбумина млекопитающих и где указанный домен анкиринового повтора включает аминокислотную последовательность, которая имеет, по крайней мере, 70%-ную идентичность аминокислотной последовательности с одним доменом анкиринового повтора, выбранным из группы, состоящей из SEQ ID NOs: 17-31 и 43-48, в котором G в положении 1 и/или S в положении 2 указанного домена анкиринового повтора являются необязательно отсутствующими.
Предпочтительно, когда такой домен анкиринового повтора в связывающем белке в соответствии с изобретением включает аминокислотную последовательность, которая имеет, по крайней мере, 70% идентичности с аминокислотной последовательностью с одним доменом анкиринового повтора, выбранным из группы, состоящей из SEQ ID NO: 21, 27 и 46; предпочтительно 27 и 46. Как определяется выше, указанный домен анкиринового повтора связывает xSA с константой диссоциации (Kd) в PBS ниже 10-4 М. Предпочтительно, указанный повторяемый домен связывает xSA с Kd в PBS ниже 10-4 М, более предпочтительно ниже 10-5 М, 10-6 М, 10-7 М, или наиболее предпочтительно 10-8 М.
Предпочтительно, такой домен анкиринового повтора в связывающием белке в соответствии с изобретением включает аминокислотную последовательность, которая имеет, по крайней мере, 70% идентичности с аминокислотной последовательностью с "рандомизированной повторяемой единицей" рандомизированными положениями" в домене анкиринового повтора, выбранном из группы, состоящей из SEQ ID NOs: 17-31 и 43-48.
Предпочтительно, вместо 70%-ной идентичности аминокислотной последовательности такой домен анкиринового повтора в связывающем белке в соответствии с изобретением включает аминокислотную последовательность, которая имеет, по крайней мере, 75%, более предпочтительно, по крайней мере, 80%, более предпочтительно, по крайней мере, 85%, более предпочтительно, по крайней мере, 90%, или наиболее предпочтительно, по крайней мере, 95% идентичности аминокислотной последовательности.
В частном воплощении связывающий белок со связывающей специфичностью для сывороточного альбумина млекопитающих, который характеризуется замещением вплоть до 9 аминокислот в модулях анкиринового повтора последовательности SEQ ID NO: 49, 50, 51 и 52, или характеризуется, по крайней мере, 70% идентичности аминокислотной последовательности с одним доменом анкиринового повтора, выбранным из группы, состоящей из SEQ ID NO: 17-31 и 43-48, имеет, по крайней мере, в 5 раз более высокий конечный период полувыведения у млекопитающего по сравнению с соответствующими связывающими белками, которые не связываются с сывороточным альбумином млекопитающих, например, доменом анкиринового повтора последовательности SEQ ID NO: 32. В таком предпочтительном связывающем белке минимальный конечный период полувыведения у человека составляет, по крайней мере, 1 день, более предпочтительно, по крайней мере, 3 дня, даже более предпочтительно, по крайней мере, 5 дней.
В дополнительном воплощении изобретение относится к связывающему белку, где указанный домен анкиринового повтора включает повторяемый модуль с мотивом последовательности анкиринового повтора
X1DX2X3X4X5TPLHLAAX6X7GHLX8IVEVLLKX9GADVNA (SEQ ID NO: 53)
в котором X1, X2, Х3, Х4, X5, Х6, Х7, Х8 и Х9, представляют собой независимо друг от друга, аминокислотные остатки, выбранные из группы, состоящей из A, D, Е, F, Н, I, К, L, M, N, Q, R, S, Т, V, W и Y;
в котором предпочтительно
X1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, D, M, F, S, I, Т, N, Y и К; более предпочтительно К и А;
X2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Е, К, D, F, M, N, I и Y; более предпочтительно I, E и Y;
Х3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из W, R, N, Т, Н, К, А и F; более предпочтительно W, R и F;
Х4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из G и S;
Х5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из N, Т и Н;
Х6 представляет собой аминокислотный остаток, выбранный из группы, состоящей из N, V и R;
Х7 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Е, Y, N, D, Н, S, А, Q, Т и G; более предпочтительно Е, Y и N;
Х8 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Е и К;
Х9 представляет собой аминокислотный остаток, выбранный из группы, состоящей из S, A, Y, Н и N; более предпочтительно Y и Н; и
в котором необязательно вплоть до 5 аминокислот в другом положении, отличном от положений, обозначенных как Х в SEQ ID NO: 53, заменены какой-либо аминокислотой.
В частности, изобретение относится к связывающему белку, где домен анкиринового повтора включает повторяемый модуль с мотивом последовательности анкиринового повтора
X1DX2X3GX4TPLHLAAX5X6GHLEIVEVLLKX7GADVNA (SEQ ID NO: 10),
в котором
X1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из A, D, M, F, S, I, Т, N, Y и К; предпочтительно К и А;
Х2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Е, К, D, F, M, N, I и Y; предпочтительно I, E и Y;
Х3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из W, R, N, Т, Н, К, А и F; предпочтительно W, R и F;
Х4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из N, Т и Н;
Х5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из N, V и R;
Х6 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Е, Y, N, D, Н, S, A, Q, Т и G; предпочтительно Е, Y и N;
Х7 представляет собой аминокислотный остаток, выбранный из группы, состоящей из S, А, Y, Н и N; предпочтительно Y и Н; и
в котором необязательно вплоть до 5 аминокислот в положениях, отличных от тех, которые обозначены как Х в SEQ ID NO: 10, заменены какой-либо аминокислотой.
В дополнительном воплощении изобретение относится к связывающему белку, в котором домен анкиринового повтора включает повторяемый модуль с мотивом последовательности анкиринового повтора
X1DYFX2HTPLHLAARX3X4HLX5IVEVLLKX6GADVNA (SEQ ID NO: 11),
в котором
X1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из D, К и А; предпочтительно К и А;
Х2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из D, G и S; предпочтительно G и S;
Х3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Е, N, D, H, S, A, Q, Т и G; предпочтительно Q, D и N; более предпочтительно Q и N;
Х4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из G и D;
Х5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Е, К и G; предпочтительно Е и К;
Х6 представляет собой аминокислотный остаток, выбранный из группы, состоящей из H, Y, А и N; предпочтительно H, А и Y; более предпочтительно А и Y; и
в котором необязательно вплоть до 5 аминокислот в положениях, отличных от тех, которые обозначены как Х в SEQ ID NO: 11, заменены какой-либо аминокислотой.
Еще в одном воплощении изобретение относится к связывающему белку, в котором домен анкиринового повтора включает повторяемый модуль с мотивом последовательности анкиринового повтора
X1DFX2GX3TPLHLAAX4X5GHLEIVEVLLKX6GADVNA (SEQ ID NO: 54),
в котором
X1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из F, S, L и К; предпочтительно S и К;
Х2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из V и А;
Х3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из R и К;
Х4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из S и N;
Х5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из N, D, Q, S, А, Т и Е; предпочтительно D и Q;
Х6 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, Н, Y, S и N; предпочтительно А и Н; и
в котором необязательно вплоть до 5 аминокислот в положениях, отличных от тех, которые обозначены как Х в SEQ ID NO: 54, заменены какой-либо аминокислотой.
В частности, изобретение относится к связывающему белку, где домен анкиринового повтора включает повторяемый модуль с мотивом последовательности анкиринового повтора
X1DFX2GX3TPLHLAAX4DGHLEIVEVLLKX5GADVNA (SEQ ID NO: 12),
в котором
X1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из F, S, L и К; предпочтительно S и К;
Х2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из V и А;
Х3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из R и К;
Х4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из S и N;
Х5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, Н, Y, S и N; предпочтительно А и Н; и
в котором необязательно вплоть до 5 аминокислот в положениях, отличных от тех, которые обозначены как Х в SEQ ID NO: 12, заменены какой-либо аминокислотой.
Предпочтительным является связывающий белок, где указанный домен анкиринового повтора включает повторяемый модуль с мотивом последовательности анкиринового повтора SEQ ID NO: 12, с предшествующим повторяемым модулем с мотивом последовательности анкиринового повтора SEQ ID NO: 11.
Еще в одном воплощении изобретение относится к связывающему белку, где домен анкиринового повтора включает повторяемый модуль с мотивом последовательности анкиринового повтора
X1DX2X3GTTPLHLAAVYGHLEX4VEVLLKX5GADVNA (SEQ ID NO: 13),
в котором
X1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из К, A, D, M, F, S, I, Т, N, и Y; предпочтительно К и А;
Х2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Е, К, D, F, M, N и Y; предпочтительно Е, D и Y;
Х3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из R, N, Т, Н, К, А и F; предпочтительно R и F;
Х4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из I и M;
X5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Н, Y, К, А и N; предпочтительно К и А; и
в котором необязательно вплоть до 5 аминокислот в положениях, отличных от тех, которые обозначены как Х в SEQ ID NO: 13, заменены какой-либо аминокислотой.
Еще в одном воплощении изобретение относится к связывающему белку, где домен анкиринового повтора включает повторяемый модуль с мотивом последовательности анкиринового повтора
X1NETGYTPLHLADSSGHX2EIVEVLLKX3X4X5DX6NA (SEQ ID NO: 14),
в котором
X1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Q и К;
Х2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из L и Р;
Х3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Н, N, Y и А; предпочтительно Н и А;
Х4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из G и S;
Х5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, V, Т и S; предпочтительно S и А;
Х6 представляет собой аминокислотный остаток, выбранный из группы, состоящей из V и F; и в котором необязательно вплоть до 5 аминокислот в положениях, отличных от тех, которые обозначены как Х в SEQ ID NO: 14, заменены какой-либо аминокислотой.
Предпочтительным является связывающий белок, где указанный домен анкиринового повтора включает повторяемый модуль с мотивом последовательности анкиринового повтора SEQ ID NO: 14 с предшествующим повторяемым модулем с мотивом последовательности анкиринового повтора SEQ ID NO: 13.
Термин "кэппирующий модуль" относится к полипептиду, слитому с N- или С-терминальным повторяемым модулем повторяемого домена, где указанный кэппирующий модуль формирует плотные третичные взаимодействия (то есть, взаимодействия третичной структуры) с указанным повторяемым модулем, обеспечивая, таким образом, колпачок, который защищает гидрофобное ядро указанного повторяемого модуля со стороны, которая не находится в контакте с последовательным повторяемым модулем, от растворителя. Указанный N- и/или С-терминальный кэппирующий модуль может представлять собой, или может иметь происхождение от, кэппирующей единицы или другой структурной единицы, которая обнаруживается в существующих в природе белковых повторах, соседствующих с повторяемой единицей. Термин "кэппирующая единица" относится к существующему в природе собранному полипептиду, где указанный полипептид определяет конкретную структурную единицу, которая является N- или С-терминально слитой с повторяемой единицей, где указанный полипептид формирует плотные взаимодействия третичной структуры с указанной повторяемой единицей, обеспечивая, таким образом, колпачок, который защищает гидрофобное ядро указанной повторяемой единицы с одной стороны, от растворителя. Предпочтительно, кэппинг модули или кэппирующие единицы представляют собой кэппирующие повторы. Термин "кэппирующий повтор" относится к кэппирующему модулю или кэппирующей единице, которая имеет подобную или такую же сборку, что и указанная соседняя повторяемая единица (или модуль), и/или подобность последовательности указанной соседней повторяемой единицы (или модуля). Кэппинг модули и кэппирующие повторы описываются в WO 2002/020565 и Interlandi и др., 2008 (в приведенном выше месте). Например, WO 2002/020565 описывает N-терминальный кэппирующий модуль (то есть, кэппирующий повтор), который имеет аминокислотную последовательность GSDLGKKLLEAARAGQDDEVRILMANGADVNA (SEQ ID NO: 1) и С-терминальный кэппирующий модуль (то есть, кэппирующий повтор), имеющий аминокислотную последовательность QDKFGKTAFDISIDNGNEDLAEILQKLN (SEQ ID NO: 2).
Interlandi и др., 2008 (в приведенном выше месте) описывают С-терминальные кэппинг модули, имеющие аминокислотные последовательности QDKFGKTPFDLAIREGHEDIAEVLQKAA (SEQ ID NO: 3) и QDKFGKTPFDLAIDNGNEDIAEVLQKAA (SEQ ID NO: 4).
Например, N-терминальный кэппирующий модуль последовательности SEQ ID NO: 17 кодируется аминокислотами от положения 1 до 32, а С-терминальный кэппирующий модуль последовательности SEQ ID NO: 17 кодируется аминокислотами от положения 99 до 126.
Предпочтительный N-терминальный кэппирующий модуль включает мотив последовательности
X1LX2KKLLEAARAGQDDEVRILX3AX4GADVNA (SEQ ID NO: 5),
в котором X1 представляет собой аминокислотный остаток G, А или D;
в котором Х2 представляет собой аминокислотный остаток G или D;
в котором Х3 представляет собой аминокислотный остаток L, V, I, А или М; предпочтительно, L или М; и
в котором Х4 представляет собой аминокислотный остаток А, Н, Y, К, R или N; предпочтительно, А или N.
Дополнительно предпочтительным является любой такой N-терминальный кэппирующий модуль, включающий N-терминальный кэппирующий повтор, в котором одна или более аминокислотных остатков в указанном кэппирующем повторе заменены аминокислотным остатком, который обнаруживается в соответствующем положении на выравнивании соответствующей кэппирующей единицы или повторяемой единицы.
Предпочтительный С-терминальный кэппирующий модуль включает мотив последовательности
X1DKX2GKTX3X4DX5X6X7DX8GX9EDX10AEX11LQKAA (SEQ ID NO: 6),
в котором X1 представляет собой аминокислотный остаток Q или К;
в котором Х2 представляет собой аминокислотный остаток A, S или F; предпочтительно, S или F;
в котором Х3 представляет собой аминокислотный остаток А или Р;
в котором Х4 представляет собой аминокислотный остаток А или F;
в котором Х5 представляет собой аминокислотный остаток I или L;
в котором Х6 представляет собой аминокислотный остаток S или А;
в котором Х7 представляет собой аминокислотный остаток I или А;
в котором Х8 представляет собой аминокислотный остаток А, Е или N; предпочтительно, А или N;
в котором Х9 представляет собой аминокислотный остаток N или Н;
в котором Х10 представляет собой аминокислотный остаток L или I;
в котором Х11 представляет собой аминокислотный остаток I или V; и
в котором Х2 не является F, если Х4 представляет собой F и Х7 представляет собой I, и Х8 представляет собой N или Е.
Еще один предпочтительный С-терминальный кэппирующий модуль включает мотив последовательности
X1DKX2GKTX3ADX4X5X6DX7GX8EDX9AEX10LQKAA (SEQ ID NO: 7),
в котором X1 представляет собой аминокислотный остаток Q или К;
в котором Х2 представляет собой аминокислотный остаток А, S или F; предпочтительно, S или F;
в котором Х3 представляет собой аминокислотный остаток А или Р;
в котором Х4 представляет собой аминокислотный остаток I или L;
в котором Х5 представляет собой аминокислотный остаток S или А;
в котором Х6 представляет собой аминокислотный остаток I или А;
в котором Х7 представляет собой аминокислотный остаток А, Е или N; предпочтительно, А или N;
в котором Х8 представляет собой аминокислотный остаток N или Н;
в котором Х9 представляет собой аминокислотный остаток L или I; и
в котором Х10 представляет собой аминокислотный остаток I или V.
Еще один предпочтительный С-терминальный кэппирующий модуль включает мотив последовательности
X1DKX2GKTX3ADX4X5ADX6GX7EDX8AEX9LQKAA (SEQ ID NO: 8),
в котором X1 представляет собой аминокислотный остаток Q или К;
в котором Х2 представляет собой аминокислотный остаток A, S или F; предпочтительно, S или F;
в котором Х3 представляет собой аминокислотный остаток А или Р;
в котором Х4 представляет собой аминокислотный остаток I или L;
в котором X5 представляет собой аминокислотный остаток S или А;
в котором Х6 представляет собой аминокислотный остаток А, Е или N; предпочтительно, А или N;
в котором Х7 представляет собой аминокислотный остаток N или Н;
в котором Х8 представляет собой аминокислотный остаток L или I; и
в котором Х9 представляет собой аминокислотный остаток I или V.
Предпочтительно, такой С-терминальный кэппирующий модуль, включающий мотив последовательности SEQ ID NO: 6, 7 или 8, имеет аминокислотный остаток А, I или К; предпочтительно, I или К; в положении, которое соответствует положению 3 указанного мотива последовательности.
Также предпочтительно, когда такой С-терминальный кэппирующий модуль, включающий мотив последовательности SEQ ID NO: 6, 7 или 8, имеет аминокислотный остаток R или D в положении, которое соответствует положению 14 указанного мотива последовательности.
Предпочтительный С-терминальный кэппирующий модуль представляет собой С-терминальный кэппирующий модуль, имеющий аминокислотную последовательность QDKSGKTPADLAADAGHEDIAEVLQKAA (SEQ ID NO: 9).
Также предпочтительным является С-терминальный кэппирующий модуль, имеющий аминокислотную последовательность SEQ ID NO: 9, в которой
аминокислотный остаток в положении 1 представляет собой Q или К;
аминокислотный остаток в положении 4 представляет собой S или F;
аминокислотный остаток в положении 9 представляет собой А или F;
аминокислотный остаток в положении 13 представляет собой А или I;
аминокислотный остаток в положении 15 представляет собой А, Е или N; и
где указанный С-терминальный кэппирующий модуль не имеет аминокислотной последовательности SEQ ID NO: 2, 3 или 4.
Дополнительный предпочтительный С-терминальный кэппирующий модуль, имеет аминокислотную последовательность, которая имеет, по крайней мере, 70%, предпочтительно, по крайней мере, 75%, 80%, 85%, 90%, или наиболее предпочтительно, по крайней мере, 95% идентичности аминокислотной последовательности на выравнивании с SEQ ID NO: 9 или 2. Предпочтительно, аминокислотный остаток указанного С-терминального кэппирующего модуля в положении, которое соответствует положению 4 SEQ ID: 9 на выравнивании, представляет собой S, аминокислотный остаток указанного С-терминального кэппирующего модуля в положении, которое соответствует положению 9 SEQ ID: 9 на выравнивании, представляет собой А, аминокислотный остаток указанного С-терминального кэппирующего модуля в положении, которое соответствует положению 13 SEQ ID: 9 на выравнивании, представляет собой А, и/или аминокислотный остаток указанного С-терминального кэппирующего модуля в положении, которое соответствует положению 15 SEQ ID: 9 на выравнивании, представляет собой А. Также предпочтительно, когда аминокислотный остаток указанного С-терминального кэппирующего модуля в положении, которое соответствует положению 9 SEQ ID: 9 на выравнивании, представляет собой А и/или аминокислотный остаток указанного С-терминального кэппирующего модуля в положении, которое соответствует положению 13 SEQ ID: 9 на выравнивании, представляет собой А. Также предпочтительно, когда указанный С-терминальный кэппирующий модуль включает 28 аминокислот.
Предпочтительным также является С-терминальный кэппирующий модуль, имеющий аминокислотную последовательность SEQ ID NO: 2 или 9, в котором один или более аминокислотных остатков, указанного С-терминального кэппирующего модуля являются замененными с помощью аминокислоты, которая обнаружена в соответствующем положении выравнивания соответствующего С-терминального кэппирующего повтора или кэппирующей единицы и в котором
аминокислотный остаток в положении 4 представляет собой S;
аминокислотный остаток в положении 9 представляет собой А;
аминокислотный остаток в положении 13 представляет собой А; и/или
аминокислотный остаток в положении 15 представляет собой А.
Предпочтительно, вплоть до 30% аминокислотных остатков указанного С-терминального кэппирующего модуля являются замененными, более предпочтительно, вплоть до 20% и даже более предпочтительно, вплоть до 10% аминокислотных остатков являются замененными. Также является предпочтительным, когда такой С-терминальный кэппирующий модуль представляет собой существующий в природе С-терминальный кэппирующий повтор.
Также предпочтительным является С-терминальный кэппирующий модуль, включающий аминокислоты от положения 1 до 25 или от положения 1 до 26 любого из упомянутых выше С-терминальных кэппинг модулей на основе последовательности SEQ ID NO: 9.
Также предпочтительным является такой С-терминальный кэппирующий модуль, имеющий аминокислотную последовательность, которая не включает аминокислоты N, после которой располагается G.
Также предпочтительным является С-терминальный кэппирующий модуль, который имеет, по крайней мере, 70%, предпочтительно, по крайней мере, 75%, 80%, 85%, 90%, или наиболее предпочтительно, по крайней мере, 95% идентичности аминокислотной последовательности с каким-либо из упомянутых выше С-терминальных кэппинг модулей на основе последовательности SEQ ID NO: 9 или с самой SEQ ID NO: 9.
Дополнительно предпочтительным является С-терминальный кэппирующий модуль, который имеет, по крайней мере, 70%, предпочтительно, по крайней мере, 75%, 80%, 85%, 90%, или наиболее предпочтительно, по крайней мере, 95% идентичности аминокислотной последовательности с последовательностью SEQ ID NO: 2 или 9 и где указанный С-терминальный кэппирующий модуль имеет аминокислоту А в положении 9; предпочтительно, указанный С-терминальный кэппирующий модуль имеет аминокислоту А в положениях 9 и 13; более предпочтительно, указанный С-терминальный кэппирующий модуль имеет аминокислоту А в положениях 9, 13 и 15; и наиболее предпочтительно, указанный С-терминальный кэппирующий модуль имеет аминокислоту А в положениях 9, 13 и 15 и S в положении 4.
Дополнительно предпочтительным является такой С-терминальный кэппирующий модуль, который не имеет аминокислоты R в положении 14, и/или который не имеет аминокислоты Е в положении 15.
Также предпочтительным является такой С-терминальный кэппирующий модуль, который не имеет аминокислотной последовательности, идентичной таковой SEQ ID NO: 2, 3 или 4.
Дополнительно предпочтительным является такой С-терминальный кэппирующий модуль, который имеет аминокислотную последовательность на основе SEQ ID NO: 9, где указанный С-терминальный кэппирующий модуль имеет аминокислоты в положениях 26, 27 и 28, выбранные из группы, состоящей из A, L, R, М, К и N; более предпочтительно, A, L, R и К; и наиболее предпочтительно, К, А и L.
Кэппирующий модуль повторяемого домена может быть заменен на кэппирующий модуль в соответствии с изобретением путем комбинирования методик, таких как выравнивание аминокислотной последовательности, мутагенез и синтез генов, известных квалифицированному специалисту в данной области техники. Например, С-терминальный кэппирующий повтор SEQ ID NO: 17 может быть заменен на С-терминальный кэппирующий повтор SEQ ID NO: 9 с помощью (i) определения С-терминального кэппирующего повтора последовательности SEQ ID NO: 17 (то есть, положения последовательности от положения 99 до 126) путем выравнивания последовательности с SEQ ID NO: 9, (ii) замещения последовательности определенного С-терминального кэппирующего повтора последовательности SEQ ID NO: 17 последовательностью SEQ ID NO: 9, (iii) получения гена, который кодирует повторяемый домен, кодирующего замененный С-терминальный кэппирующий модуль, (iv) экспрессии модифицированного повторяемого домена в цитоплазме Е.coli и (v) очистки модифицированного повторяемого домена с помощью стандартных способов.
Кроме того, повторяемый домен в соответствии с изобретением может быть сконструирован генетически путем объединения N-терминального кэппирующего модуля (то есть, N-терминального кэппирующего повтора последовательности SEQ ID NO: 1), после которого идут один или более повторяемых модулей (то есть, повторяемых модулей, включающих аминокислотные остатки от положения 33 до 98 последовательности SEQ ID NO: 17), и С-терминального кэппирующего модуля (то есть, С-терминального кэппирующего повтора последовательности SEQ ID NO: 9) с помощью методов генетического синтеза. Генетически сконструированный ген повторяемого домена может потом экспрессироваться в Е.coli так, как описано выше.
Также предпочтительным является связывающий белок, где домен анкиринового повтора или сконструированный домен анкиринового повтора включает С-терминальный кэппирующий модуль с мотивом последовательности SEQ ID NO: 6, 7 или 8, где указанный кэппирующих модуль имеет аминоислоту I в положении 3 и где указанному повторяемому модулю предшествует повторяемый модуль с мотивом последовательности анкиринового повтора последовательности SEQ ID NO: 12.
Дополнительно предпочтительным является связывающий белок, повторяемый домен, N-терминальный кэппирующий модуль или С-терминальный кэппирующий модуль, имеющий аминокислотную последовательность, в которой отсутствуют аминокислоты С, М или N.
Дополнительно предпочтительным является связывающий белок, повторяемый домен, N-терминальный кэппирующий модуль или С-терминальный кэппирующий модуль, имеющий аминокислотную последовательность, в которой отсутствует аминокислота N, после которой следует G.
Дополнительно предпочтительным является любой такой С-терминальный кэппирующий модуль, включающий С-терминальный кэппирующий повтор, в котором один или более аминокислотных остатков в указанном кэппирующем повторе заменены аминокислотным остатком, который обнаруживается в соответствующем положении в выравнивании соответствующей кэппирующей единицы или повторяемой единицы.
Дополнительно предпочтительным является связывающий белок, включающий любой такой N-терминальный или С-терминальный кэппирующий модуль.
Примеры аминокислотных последовательностей таких С-терминальных кэппинг модулей представляют собой аминокислотные последовательности от положения 99 до 126 в SEQ ID NO: 19, 21, 27, 28, 38, 40 и 42. Пример 6 демонстрирует, что термальная стабильность повторяемого домена может быть повышена путем замещения его С-терминальных кэппинг модулей кэппирующим модулем в соответствии с изобретением.
Термин "мишень" относится к индивидуальной молекуле, такой как молекула нуклеиновой кислоты, полипептида или белка, углевода, или любой другой существующей в природе молекуле, включая любую часть такой индивидуальной молекулы, или к комплексам двух или более таких молекул. Мишень может представлять собой цельную клетку или образец ткани, или она может представлять собой любую не существующую в природе молекулу или остаток. Предпочтительно, когда мишень представляет собой существующий в природе или неприродный полипептид или полипептид, содержащий химические модификации, например, модифицированный с помощью природного или неприродного фосфорилирования, ацетилирования или метилирования. В конкретном применении в соответствии с настоящим изобретением мишень представляет собой xSA.
Термин "xSA" относится к сывороточному альбумину млекопитающих, такому как сывороточный альбумин, полученный из мыши, крысы, кролика, собаки, свиньи, обезьяны или человека. Термин "MSA" относится к мышиному сывороточному альбумину (UniProtKB/Swiss-Prot первичной номер депонирования Р07724), термин "CSA" относится к сывороточному альбумину обезьяны циномолгус (то есть, Масаса fascicularis) (UniProtKB/Swiss-Prot первичной номер депонирования A2V9Z4) и термин "HSA" относится к человеческому сывороточному альбумину (UniProtKB/Swiss-Prot первичной номер депонирования Р02768).
Термин "консенсусная последовательность" относится к аминокислотной последовательности, где указанную консенсусную последовательность получают с помощью структурного выравнивания и/или выравнивания последовательности множественных повторяемых единиц. При использовании двух или более структурно выровненных и/или выровненных по последовательности повторяемых единицы, и разрешая пробелы в выравнивании, является возможным определить наиболее часто встречаемый аминокислотный остаток в этом положении. Консенсусная последовательность представляет собой последовательность, которая включает аминокислоты, которые являются наиболее часто представленными в каждом положении. В случае, когда две или более аминокислоты являются представленными на уровне показателей выше среднего значения в одном положении, то консенсусная последовательность может включать набор таких аминокислот. Указанные две или более повторяемые единицы могут быть взяты из повторяемой единицы, которая входят в состав единичного повторяемого белка, или из двух или более различных повторяемых белков.
Коненсусные последовательности и способы их определения являются хорошо известными квалифицированному специалисту в данной области техники.
"Консенсусный аминокислотный остаток" представляет собой аминокислоту, которая обнаруживается в определенном положении в консенсусной последовательности. Если два или более, например, три, четыре или пять, аминокислотных остатка обнаруживаются с одинаковой вероятность в указанных двух или более повторяемых единицах, то консенсусная аминокислота может представлять собой одну из наиболее часто встречаемых аминокислот или комбинацию указанных двух или более аминокислотных остатков.
Также предпочтительными являются несуществующие в природе кэппирующие модули, повторяемые модули, связывающие белки или связывающие домены.
Термин "несуществующий в природе" означает синтетический или, в частности, неприродный, термин означает такой, который создан человеком. Термин "несуществующий в природе связывающий белок" или "несуществующий в природе связывающий домен" означает, что указанный связывающий белок или указанный связывающий домен является синтетическим (то есть, полученным с помощью химического синтеза из аминокислот) или рекомбинантным, а не природным. "Несуществующий в природе связывающий белок" или "несуществующий в природе связывающие домен" представляет собой созданный человеком белок или домен, соответственно, полученный путем экспрессии соответственно сконструированных нуклеиновых кислот. Предпочтительно, экспрессию осуществляют в эукариотических или бактериальных клетках, или путем использования бесклеточной in vitro экспрессионной системы. Кроме того, термин означает, что последовательность указанного связывающего белка или указанного связывающего домена не является присутствующей в виде неискусственной последовательности в базе данных последовательностей, например, в GenBank, EMBL-Bank или Swiss-Prot. Эти базы данных и другие базы данных последовательностей являются хорошо известными специалисту в данной области техники.
Изобретение относится к связывающему белку, включающему связывающий домен, где указанный связывающий домен представляет собой домен анкиринового повтора и специфично связывается с xSA и где указанный связывающий белок и/или связывающий домен имеет среднюю температуру денатурации (Tm) выше 40°С при термальном развертывании в PBS и формирует менее 5% (вес./вес.) нерастворимых агрегатов при концентрациях вплоть до 10 г/л при инкубации при температуре 37°С в течение 1 дня в PBS.
Термин "PBS" означает забуференный фосфатом водный раствор, содержащий 137 мМ NaCl, 10 мМ фосфата и 2,7 мМ KCl и имеющий значение рН 7,4.
Предпочтительно, когда связывающий белок и/или связывающий домен имеет среднюю температуру денатурации (Tm) выше 45°С, более предпочтительно выше 50°С, более предпочтительно выше 55°С, и наиболее предпочтительно выше 60°С при термальном развертывании в PBS при рН 7,4 или в MES буфере при рН 5,8. Связывающий белок или связывающий домен в соответствии с изобретением обладает определенной вторичной и третичной структурой при физиологических условиях. Термальное развертывание такого полипептида приводит к потере его третичной и вторичной структуры, что может быть продемонстрировано, например, с помощью измерений циркулярного дихроизма (CD). Среднее значение температуры денатурации связывающего белка или связывающего домена при термальном развертывании соответствует температуре в средней точке согласованного перехода в физиологическом буфере при тепловой денатурации указанного белка или домена путем медленного повышения температуры от 10°С до приблизительно 100°С. Определение средней точки температуры денатурации при термальном развертывании является хорошо известным квалифицированном специалисту в данной области техники. Эта средняя точка температуры денатурации связывающего белка или связывающего домена при термальном развертывании является показательной для термальной стабильности указанного полипептида.
Также предпочтительным является связывающий белок и/или связывающий домен, формирующий менее чем 5% (вес./вес.) нерастворимых агрегатов при концентрациях вплоть до 20 г/л, предпочтительно вплоть до 40 г/л, более предпочтительно вплоть до 60 г/л, даже более предпочтительно вплоть до 80 г/л, и наиболее предпочтительно вплоть до 100 г/л при инкубации в течение 5 дней, предпочтительно в течение 10 дней, более предпочтительно в течение 20 дней, более предпочтительно в течение 40 дней, и наиболее предпочтительно в течение 100 дней при 37°С в PBS. Образование нерастворимых агрегатов может определяться с помощью возникновения визуальной преципитации, гель-фильтрации или динамического рассеяния света, которое сильно повышается при образовании нерастворимых агрегатов. Нерастворимые агрегаты могут быть удалены из образца белка путем центрифугирования при 10'000×g в течение 10 минут. Предпочтительно, связывающий белок и/или связывающий домен образует менее чем 2%, более предпочтительно менее чем 1%, 0,5%, 0,2%, 0,1%, или наиболее предпочтительно менее чем 0,05% (вес./вес.) нерастворимых агрегатов при упомянутых выше условиях инкубации при 37°С в PBS. Процентное содержание нерастворимых агрегатов может определяться с помощью отделения нерастворимых агрегатов от растворимого белка, после чего осуществляют определение количеств белка в растворимой и нерастворимой фракции с помощью стандартных способов количественной оценки.
Также предпочтительным является связывающий белок и/или связывающий домен, который не утрачивает своей нативной пространственной структуры при инкубации в PBS, содержащем 100 мМ дитиотреитол (DTT) в течение 1 или 10 часов при 37°С.
В одном частном воплощении изобретение относится к связывающему белку, включающему связывающий домен, который является доменом анкиринового повтора, в частности, который связывается с xSA и имеет указанное или предпочтительное среднее значение температуры денатурации неагрегирующие свойства, как определяется выше, где указанный связывающий белок обладает, по крайней мере, в 5 раз более высоким конечным периодом полувыведения у млекопитающего по сравнению со связывающим доменом, который не связывается с сывороточным белком, таким как xSA.
Предпочтительно, указанный связывающий домен имеет, по крайней мере, в 10 раз, более предпочтительно, по крайней мере, в 20 раз, в 40 раз, в 100 раз, в 300 раз, или наиболее предпочтительно, по крайней мере, в 103 раз более высокий конечный период полувыведения у млекопитающего по сравнению со связывающим доменом, который не связывается с сывороточным белком, таким как xSA.
Также предпочтительно, когда указанный связывающий домен не связывает xSA при Kd выше 10-4 М, более предпочтительно выше 10-3 М или наиболее предпочтительно выше 10-2 М для связывания xSA. Пример связывающего домена, который не связывает xSA, представляет собой повторяемый домена последовательности SEQ ID NO: 32.
Также предпочтительным является, когда указанный связывающий домен представляет собой повторяемый домен и имеет, по крайней мере, в 5 раз, более предпочтительно, по крайней мере, в 10 раз, более предпочтительно, по крайней мере, в 20 раз, в 40 раз, в 100 раз, в 300 раз, или наиболее предпочтительно, по крайней мере, в 103 раз более высокий (то есть, более длительный) конечный период полувыведения у млекопитающего по сравнению повторяемым доменом последовательности SEQ ID NO: 32 или по сравнению с DARPin #32, DARPin #41 или DARPin #42.
Предпочтительный связывающий белок включает связывающий домен со связывающей специфичностью для HSA, который обладает конечным период полувыведения выше 1, более предпочтительно выше 3, 5, 7, 10, 15, или наиболее предпочтительно выше 20 дней у людей.
Конечный период полувыведения связывающего домена может быть определен с помощью анализов, хорошо известных квалифицированному специалисту в данной области техники (Toutain, P.L., и Bousquet-Mélou, A., J. Vet. Pharmacol. Ther. 27(5), 427-439, 2004). Примеры определения конечного периода полувыведения являются приведенными в разделе Примеры.
Термин "конечный период полувыведения из плазмы крови" лекарственного агента, такого как связывающий белок или связывающий домен в соответствии с изобретением, относится ко времени, которое требуется для достижения половины концентрации в плазме крови лекарственного средства, которое применяется к млекопитающему после достижения псевдоравновесия. Этот период полувыведения не определяется как время, необходимое для выведения половины дозы лекарственного средства, которое было введено млекопитающему.
В одном частном воплощении изобретение относится к связывающему белку, включающему связывающий домен, которой представляет собой домен анкиринового повтора, в частности, такому, который связывается с xSA и включает биоактивное соединение.
Термин "биоактивное соединение" относится к соединению, которое изменяет заболевание, когда применяется к млекопитающему, которое имеет указанное заболевание. Биоактивное соединение может иметь агонистические или антагонистические свойства и может представлять собой белковое биоактивное соединение или небелковое биоактивное соединение.
Такие белковые биоактивные соединения могут быть ковалентно присоединены, например, к связывающему домену в соответствии с изобретением путем получения генетически слитых полипептидов при использовании стандартных методик клонирования ДНК, после чего осуществляют их стандартную экспрессию и очистку. Например, DARPin #36 включает повторяемый домен со связывающей специфичностью для человеческого фактора роста (то есть, биоактивное соединение), после которого следует повторяемый домен со связывающей специфичностью для HSA.
Такие небелковые биоактивные соединения могут быть ковалентно присоединены, например, к связывающему домену в соответствии с изобретением, химическим путем, например, путем слияния тиола цистеина через малеимидный линкер с цистеином, слитым с помощью пептидного линкера с N- или С-терминальным концом связывающего домена, как описывается в данной заявке.
Примеры белковых биоактивных соединений представляют собой связывающие домены, которые обладают отличной специфичностью по отношению к мишени (например, нейтрализация фактора роста с помощью его связывания), цитокинам (например, интерлейкинам), фактору роста (например, человеческому гормону роста), антителам и их фрагментам, гормонам (например, GLP-1) и любому возможному белковому лекарственному средству.
Примеры небелковых биоактивных соединений представляют собой токсины (например, DM1 от ImmunoGen), малые молекулы, нацеливающие GPCR, антибиотики и любое возможное белковое лекарственное средство.
В одном частном воплощении изобретение относится к связывающему белку, включающему домен анкиринового повтора, в частности, такой, который связываются с xSA, и дополнительно включающий биоактивное соединение, где указанный связывающий белок имеет, по крайней мере, в 2 раза более высокий период полувыведения у млекопитающего по сравнению с конечным периодом полувыведения указанного немодифицированного биоактивного соединения, где указанный более высокий конечный период полувыведения указанного связывающего белка обеспечивается с помощью указанного повторяемого домена.
Предпочтительно, когда указанный связывающий белок имеет, по крайней мере, в 10 раз, более предпочтительно, по крайней мере, в 20 раз, в 40 раз, в 100 раз, в 300 раз, или наиболее предпочтительно, по крайней мере, в 103 раз более высокий конечный период полувыведения у млекопитающего по сравнению по сравнению с указанным немодифицированным биоактивным соединением.
Другое предпочтительное воплощение представляет собой рекомбинантный связывающий белок, включающий связывающий домен, который специфически связывается с xSA, и где указанный связывающий домен представляет собой домен анкиринового повтора или сконструированный домен анкиринового повтора. Такой домен анкиринового повтора может включать один, два, три или более внутренних повторяемых модулей, которые будут участвовать в связывании с xSA. Предпочтительно, такой домен анкиринового повтора включает N-терминальный кэппирующий модуль, от двух до четырех повторяемых модулей и С-терминальный кэппирующий модуль. Предпочтительно, указанный связывающий домен представляет собой домен анкиринового повтора или сконструированные домен анкиринового повтора. Также предпочтительно, когда указанные кэппинг модули представляют собой кэппирующие повторы.
В частности, изобретение относится к связывающему белку, как определяется в данной заявке выше, где домен анкиринового повтора конкурирует за связывание с сывороточным альбумином млекопитающих с доменом анкиринового повтора, выбранным из группы, состоящей из SEQ ID NO: 17-31 и 43-48; предпочтительно SEQ ID NO: 17-31; более предпочтительно SEQ ID NO: 19, 21, 27 и 28, в частности SEQ ID NO: 19 и 27.
Наиболее предпочтительным является связывающий белок, в котором домен анкиринового повтора является выбранным из группы, состоящей из SEQ ID NO: 17-31 и 43-48, в котором G в положении 1 и/или S в положении 2 указанного домена анкиринового повтора являются необязательно отсутствующими.
Также предпочтительно, когда указанный повторяемый домен конкурирует за связывание с xSA со связывающим белком, выбранным из группы DARPin #17-31 и 43-48. Предпочтительно, указанный повторяемый домен конкурирует за связывание с xSA со связывающим белком из группы DARPin #19, 21, 27, 28, 45, 46, 47 и 48. Более предпочтительно, указанный повторяемый домен конкурирует за связывание с xSA со связывающим белком DARPin #19, 45, 46, 48 или 27; даже более предпочтительно, указанный повторяемый домена конкурирует за связывание с xSA со связывающим белком DARPin #46 или 27.
Термин "конкурирует за связывание" означает неспособность двух различных связывающих доменов в соответствии с изобретением связываться одновременно с той же мишенью, в то время как они оба являются способными связываться с той же мишенью индивидуально. Таким образом, такие два связывающих домена конкурируют за связывание с указанной мишенью. Предпочтительно, когда указанные два конкурирующих связывающих домена связываются с перекрывающимся эпитопом или тем же связывающим эпитопом на указанной мишени. Способы, такие как конкурентный твердофазный иммуноферментный анализ (ELISA) или конкурентные SPR измерения (например, при использовании аппарата Proteon от BioRad), для определения, будут ли связывающие домены конкурировать за связывание с мишенью, являются хорошо известными практикующему специалисту в данной области техники.
Другое предпочтительное воплощение представляет собой связывающий белок, включающий повторяемый домен со связывающей специфичностью для xSA, выбранный из группы, состоящей из повторяемых доменов SEQ ID NO: 17-31. Предпочтительно, указанный повторяемый домен представляет собой повторяемый домен последовательности SEQ ID NO: 19, 21, 27 или 28. Более предпочтительно, указанный повторяемый домен представляет собой повторяемый домен SEQ ID NO: 19. Также более предпочтительно, когда указанный повторяемый домен представляет собой повторяемый домен SEQ ID NO: 21. Также более предпочтительно, когда указанный повторяемый домен представляет собой повторяемый домен SEQ ID NO: 27. Также более предпочтительно, когда указанный повторяемый домен представляет собой повторяемый домен SEQ ID NO: 28.
Дополнительно предпочтительным является связывающий белок, в котором указанный повторяемый домен со связывающей специфичностью для xSA включает аминокислотную последовательность, которая имеет, по крайней мере, 70% идентичности аминокислотной последовательности с повторяемым доменом указанной группы повторяемых доменов. Предпочтительно, указанная идентичность аминокислотной последовательности составляет, по крайней мере, 75%, более предпочтительно, по крайней мере, 80%, более предпочтительно, по крайней мере, 85%, более предпочтительно, по крайней мере, 90%, и наиболее предпочтительно, по крайней мере, 95%.
Дополнительно предпочтительным является связывающий белок, где указанный повторяемый домен со связывающей специфичностью для xSA включает повторяемый модуль, который имеет, по крайней мере, 70% идентичности аминокислотной последовательности с повторяемым модулем повторяемого домена указанной группы повторяемых доменов. Указанная идентичность аминокислотной последовательности составляет, по крайней мере, 75%, более предпочтительно, по крайней мере, 80%, более предпочтительно, по крайней мере, 85%, более предпочтительно, по крайней мере, 90%, и наиболее предпочтительно, по крайней мере, 95%. Дополнительно предпочтительным является связывающий белок, где указанный связывающий белок включает два или более указанных повторяемых доменов со связывающей специфичностью для xSA. Предпочтительно, указанный связывающий белок включает 2 или 3 указанных повторяемых домена. Указанные два или более повторяемых доменов имеют ту же или отличную аминокислотную последовательность.
В дополнительном предпочтительном воплощении связывающий белок включает повторяемый домен в соответствии с настоящим изобретением, а один или более аминокислотных остатков повторяемых модулей указанного повторяемого домена являются замененными аминокислотными остатками, которые обнаруживаются в соответствующих положениях на выравнивании повторяемой единицы. Предпочтительно, вплоть до 30% аминокислотных остатков являются измененными, более предпочтительно, вплоть до 20%, и даже более предпочтительно, вплоть до 10% аминокислотных остатков являются измененными. Наиболее предпочтительно, когда такая повторяемая единица представляет собой существующую в природе повторяемую единицу.
В дополнительном предпочтительном воплощении связывающий белок включает повторяемый домен в соответствии с настоящим изобретением, а один или более аминокислотных остатков N-терминального кэппирующего модуля указанного повторяемого домена являются замененными аминокислотными остатками, которые обнаруживаются в соответствующих положениях на выравнивании N-терминальной кэппирующей единицы. Предпочтительно, вплоть до 30% аминокислотных остатков являются измененными, более предпочтительно, вплоть до 20%, и даже более предпочтительно, вплоть до 10% аминокислотных остатков являются измененными. Наиболее предпочтительно, когда такая N-терминальная кэппирующая единица представляет собой существующую в природе N-терминальную кэппирующую единицу.
В дополнительном предпочтительном воплощении связывающий белок включает повторяемый домен в соответствии с настоящим изобретением, а один или более аминокислотных остатков С-терминального кэппирующего модуля указанного повторяемого домена являются замененными аминокислотными остаткками, которые обнаруживаются в соответствующих положениях на выравнивании С-терминальной кэппирующей единицы. Предпочтительно, когда вплоть до 30% аминокислотных остатков являются измененными, более предпочтительно, вплоть до 20%, и даже более предпочтительно, вплоть до 10% аминокислотных остатков являются измененными. Наиболее предпочтительно, такая С-терминальная кэппирующая единица представляет собой существующую в природе С-терминальную кэппирующую единицу.
Еще в одном частном воплощении вплоть до 30% аминокислотных остатков, более предпочтительно, вплоть до 20%, и даже более предпочтительно, вплоть до 10% аминокислотных остатков заменяются аминокислотами, которые не обнаруживаются в соответствующих положениях в повторяемых единицах, N-терминальных кэппирующих единицах или С-терминальных кэппирующих единицах.
В дополнительных воплощениях любой из белков, связывающих xSA, или доменов, описанных в данной заявке, может быть ковалентно связан одним или более дополнительных остатков, включая, например, остаток, который связывается с различными мишенями для создания связывающего агента с двойной специфичностью, биоактивного соединения, остатка для мечения (например, флуоресцентной метки, такой как флуоресцеин, или радиоактивной метки), остатка, который способствует очистке белка (например, небольшая пептидная метка, такая как His- или strep-метка), остаток, который обеспечивает эффекторные функции для улучшенной терапевтической эффективности (например, Fc часть антитела для обеспечения зависимой от антитела клеточной цитотоксичности, токсический белковый остаток, такой как экзотоксин A Pseudomonas aeruginosa (ETA) или небольшой молекулярный токсический агент, такой как майтансиноиды или ДНК алкилирующие агенты) или остатка, который обеспечивает улучшенные фармакокинетические характеристики. Улучшенные фармакокинетические характеристики могут быть оценены в соответствии с имеющейся терапевтической необходимостью. Часто является желательным повысить биодоступность и/или увеличить время между дозами, возможно, посредством увеличения времени, в течение которого белок остается доступным в сыворотке крови после введения дозы. В некоторых случаях является желательным улучшить непрерывность концентрации белка в сыворотке крови в течение периода времени (например, снизить различия в сывороточной концентрации белка между концентрацией непосредственно после введения и концентрацией непосредственно перед последующим введением).
В дополнительном воплощении изобретение относится к молекулам нуклеиновой кислоты, которые кодируют конкретные связывающие белки, конкретные N-терминальные кэппинг модули или конкретные С-терминальные кэппирующие модули. Кроме того, рассматривается вектор, включающий указанные молекулы нуклеиновой кислоты.
Кроме того, рассматривается также фармацевтическая композиция, включающая один или более из упомянутых выше связывающих белков, в частности, связывающих белков, включающих повторяемые домены, или молекулы нуклеиновой кислоты, которые кодируют конкретные связывающие белки, и необязательно, фармацевтически приемлемый носитель и/или разбавитель. Фармацевтически приемлемые носители и/или разбавители являются известными квалифицированному специалисту в данной области техники и рассматриваются более подробно ниже. Кроме того, рассматривается также диагностическая композиция, включающая один или более из упомянутых выше связывающих белков, в частности, связывающие белки, включающие повторяемые домены.
Фармацевтическая композиция включает связывающие белки так, как описано выше, и фармацевтически приемлемый носитель, наполнитель или стабилизатор, например, такой, как описывается в Remington's Pharmaceutical Sciences 16ое издание, Osol, А. ред. [1980]. Приемлемые носители, наполнители или стабилизаторы, известные специалисту, представляют собой физиологический раствор, раствор Рингера, раствор декстрозы, раствор Хэнкса, жирные растительные масла, этилолеат, 5% декстрозу в физиологическом растворе, вещества, которые улучшают изотоничность и химическую стабильность, буферы и консервирующие агенты. Другие приемлемые носители включают любой носитель, который сам по себе не индуцирует выработку антител, которые являются вредными для индивидуума, получающего композицию, такой, как белки, полисахариды, полимеры молочной кислоты, полигликолевые кислоты, полимерные аминокислоты и сополимеры аминокислот. Фармацевтические композиции могут также представлять собой комбинационные композиции, включающие дополнительный активный агент, такой как противораковый агент или антиангиогенный агент.
Композиции, используемые для введения in vivo, должны быть асептическими или стерильными. Это может легко осуществляться с помощью фильтрации через стерильные фильтрационные мембраны.
Фармацевтические композиции могут вводиться с помощью любого приемлемого способа, который является известным специалисту в данной области техники. Предпочтительный путь введения представляет собой парентеральный. При парентеральном введении лекарственное средство в соответствии с изобретением будет рецептироваться в виде единичной дозированной формы, которая вводится путем инъекции, такой как раствор, суспензия или эмульсия, в сочетании с фармацевтически приемлемыми наполнителями, как определяется выше. Дозированная форма и способ введения будут зависеть от индивидуума, которого подвергают лечению, и частного заболевания. В общем случае фармацевтические композиции вводятся так, что связывающий белок в соответствии с настоящим изобретением вводится при дозе от 1 мкг/кг до 20 мг/кг, более предпочтительно от 10 мкг/кг и 5 мг/кг, наиболее предпочтительно от 0,1 до 2 мг/кг. Предпочтительно, композицию вводят в виде болюсной дозы. Беспрерывная инфузия может также использоваться и включает беспрерывную подкожную доставку с помощью осмотического мининасоса. В этом случае фармацевтические композиции могут вводиться путем инфузии при дозе от 5 до 20 мкг/кг/минута, более предпочтительно от 7 до 15 мкг/кг/минута.
Кроме того, любая из упомянутых выше фармацевтических композиций предполагается для лечения заболевания.
Изобретение также обеспечивает способы лечения. Такой способ включает введение пациенту, который в этом нуждается, терапевтически эффективного количества связывающего белка в соответствии с изобретением.
Также предполагается способ лечения патологического состояния у млекопитающего, включая человека, включающий введение пациенту, который в этом нуждается, эффективного количества упомянутых выше фармацевтических композиций.
Связывающий белок в соответствии с изобретением может также быть получен и/или выделен с помощью нескольких способов, таких как дисплей на поверхности бактериофагов (WO 1990/002809, WO 2007/006665) или бактериальных клеток (WO 1993/010214), рибосомальный дисплей (WO 1998/048008), дисплей на плазмидах (WO 1993/008278) или путем использования гибридных конструкций на основе ковалентных РНК-белковых повторов (WO 2000/032823), или внутриклеточной экспрессии и селекции/скрининга, например, комплементационного анализа белка (WO 1998/341120). Такие способы являются известными квалифицированному специалисту в данной области техники.
Библиотека белков анкириновых повторов, которая используется для селекции/скрининга связывающих белков в соответствии с изобретением, может быть получена в соответствии с прописями, известными квалифицированному специалисту в области техники (WO 2002/020565, Binz, Н.К., и др., J. Mol. Biol., 332, 489-503, 2003, и Binz и др., 2004, в упомянутом выше месте). Применение такой библиотеки для селекции специфических для xSA DARPin является приведенным в Примере 1. Аналогично, последовательность мотивов анкиринового повтора, как представлено выше, может использоваться для построения библиотек белков с анкириновым повтором, которые могут использоваться для селекции или скрининга специфических для xSA DARPin. Кроме того, повторяемые домены в соответствии с настоящим изобретением могут быть модульно собраны из повторяемых модулей в соответствии с настоящим изобретением и приемлемых кэппинг модулей или кэппирующих повторов (Forrer, P., и др., FEBS letters 539, 2-6, 2003) при использовании стандартных методик рекомбинантной ДНК (например, WO 2002/020565, Binz и др., 2003, в приведенном выше месте и Binz и др., 2004, в приведенном выше месте).
Изобретение не является ограниченным частными воплощениями, описанными в Примерах. Все источники могут использоваться и подвергаться развитию в соответствии с общей процедурой, описанной выше.
Примеры
Все исходные материалы и реагенты, раскрытые ниже, являются известными квалифицированному специалисту в данной области техники и являются коммерчески доступными или могут быть получены при использовании хорошо известных методик.
Материалы
Реактивы закупали от Fluka (Швейцария). Олигонуклеотиды были такими от Microsynth (Швейцария). Если не указано иное, то ДНК полимеразы, рестрикционные ферменты и буферы были такими от New England Biolabs (USA) или E.coli XL1-blue (Stratagene, USA) или BL21 (Novagen, USA). Очищенный сывороточный альбумин и сыворотку, полученную от различных видов, закупали (например, у Sigma-Aldrich, Швейцария или Innovative Research, США). Биотинилированный сывороточный альбумин различных видов получали химическим путем с помощью слияния остатка биотина с первичными аминами очищенных сывороточных альбуминов при использовании стандартных реагентов и способов для биотинилирования (Pierce, USA).
Молекулярная биология
Если не указано иное, то способы осуществляли в соответствии с описанными прописями (Sambrook J., Fritsch E.F. и Maniatis Т., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory 1989, New York).
Сконструированные библиотеки белков с анкириновым повтором
Описываются библиотеки N2C и N3C сконструированных белков с анкириновым повтором (WO 2002/020565; Binz и др. 2003, в приведенном выше месте; Binz и др. 2004, в приведенном выше месте). Символ в N2C и N3C описывает количество рандомизированных повторяемых модулей, которые присутствуют между N-терминальным и С-терминальным кэппирующими модулями. Номенклатура, которая используется для определения положений внутри повторяемых единиц и модулей, основывается на Binz и др. 2004, в приведенном выше месте, с модификацией, которая граничит с модулями анкиринового повтора, а единицы анкиринового повтора смещаются на одно положение аминокислоты. Например, положение 1 модуля анкиринового повтора в соответствии с Binz и др. 2004 (в приведенном выше месте) соответствует положению 2 модуля анкиринового повтора в соответствии с настоящей заявкой и, следовательно, положению 33 модуля анкиринового повтора в соответствии Binz и др. 2004, в приведенном выше месте, соответствует положению 1 следующего модуля анкиринового повтора в соответствии с настоящей заявкой.
Все последовательности ДНК были подтверждены секвенированием, а подсчитанный молекулярный вес для всех описанных белков был подтвержден с помощью масс-спектометрии.
Пример 1: Селекция связывающих белков, включающих повторяемый домен со связывающей специфичностью для xSA
При использовании рибосомального дисплея (Hanes, J. и Plückthun, A., PNAS 94, 4937-42, 1997) были подвергнуты селекции многочисленные сконструированные белки с анкириновыми повторами (DARPin) со связывающей специфичностью для xSA из библиотек N2C или N3C DARPin, описанных Binz и др. 2004 (в приведенном выше месте). Оценивали связывание отобранных клонов со специфическими (xSA; то есть MSA, HSA или CSA) и неспецифическими (МВР, белок, связывающий мальтозу Е.coli) мишенями при использовании сырьевого экстракта ELISA, что свидетельствовало о том, что специфические для xSA связывающие белки были успешно отобраны. Повторяемые домены SEQ ID NO: 17-31 составляли аминокислотные последовательности отобранных связывающих белков, включающих повторяемый домен со связывающей специфичностью для xSA. Анализ последовательностей отобранных связывающих агентов выявил специфическую последовательность мотивов анкиринового повтора, присущую некоторым отобранным семействам связывающих агентов. Такие мотивы последовательностей анкиринового повтора, которые присутствовали в повторяемых доменах со связывающей специфичностью для xSA, обеспечиваются в SEQ ID NO: 11-14.
Селекция белков с анкириновыми повторами, специфичных для сывороточного альбумина, с помощью рибосомального дисплея
Селекцию белков с анкириновыми повторами, специфичных для сывороточного альбумина, осуществляли с помощью рибосомального дисплея (Hanes и Plückthun, в приведенном выше месте) при использовании HSA, CSA или MSA в качестве белков-мишеней, библиотеки сконструированных белков с анкириновыми повторами, как описывается в литературе (WO 2002/020565, Binz и др., 2003, в приведенном выше месте и Binz и др., 2004, в приведенном выше месте) и установленных прописей (Zahnd, С., Amstutz, P. и Plückthun, A., Nat. Methods 4, 69-79, 2007). Селекцию с использованием циклов рибосомального дисплея осуществляли на HSA, CSA или MSA (включая биотинилированные варианты HSA или MSA, иммобилизованные на нейтравидине и стрептавидине) для обоих библиотек N2C и N3C DARPin при использовании установленных прописей (Binz и др. 2004, в приведенном выше месте). Количество циклов обратной транскрипции (ОТ)-ПЦР после каждого круга селекции уменьшали с 40 до 30, осуществляя подгонку выхода благодаря повышению количества связывающих агентов. Четыре цикла селекции на HSA, CSA или MSA обеспечивали получение пулов DARPin с микромолярной - наномолярной аффинностью, как было выявлено с помощью измерений ELISA и SPR единичных клонов. Аффинность определенных DARPin дополнительно повышали при использовании аффинного созревания с помощью способов, хорошо известных квалифицированному специалисту в данной области техники (например, путем диверсификации клонов DARPin при использовании склонной к ошибкам ПЦР, а также селекции и скрининга на усовершенствованные связывающие агенты так, как описано выше).
Отобранные клоны специфически связываются с сывороточным альбумином, как показано при использовании сырьевого экстракта ELISA
Индивидуальные отобранные DARPin, специфически связывающиеся с xSA, были идентифицированы с помощью с помощью твердофазного иммуно-ферментного анализа (ELISA) при использовании сырьевых экстрактов клеток Escherichia coli, экспрессирующих DARPin, при использовании стандартных прописей. С помощью рибосомального дисплея отобранные клоны клонировали в pQE30 (Qiagen) экспрессионный вектор, трансформировали в Е.coli XL1-Blue (Stratagene) и потом выращивали в течение ночи при 37°С в планшете, содержащем 96 глубоких ячеек (каждый клон в одной ячейке), при этом каждая ячейка содержала 1 мл ростовой среды (2YT, содержащая 1% глюкозы и 100 мкг/мл ампициллина). 1 мл свежей среды 2YT, содержащей 50 мкг/мл ампициллина, инокулировали в 100 мкл ночной культуры в новый планшет, содержащий 96 глубоких ячеек. После инкубации в течение 2 часов при 37°С, экспрессию индуцировали с помощью IPTG (заключительная концентрация 1 мМ) и продолжали инкубацию в течение 3 часов. Собирали клетки, ресуспендировали их в 100 мкл B-PERII (Pierce) и инкубировали в течение 15 минут при комнатной температуре при встряхивании. Потом прибавляли 900 мкл PBS-TC (PBS с добавкой 0,25% гидролизата казеина, 0,1% Твин 20®, рН 7,4), и удаляли клеточные дебрисы путем центрифугирования. 100 мкл каждого лизированного клона применяли к ячейке MaxiSorp планшета, покрытого нейтравидином, содержащей либо xSA или неродственный МВР, иммобилизованный с помощью остатка биотина, и инкубировали в течение 1 часа при комнатной температуре. После экстенсивного промывания с использованием PBS-T (PBS с добавкой 0,1% Твин 20®, рН 7,4) планшет подвергали обработке при использовании стандартной процедуры ELISA с использованием моноклонального анти-RGS(His)4 антитела (34650, Qiagen) в качестве первичного антитела и поликлонального козьего анти-мышиного антитела, конъюгированного со щелочной фосфатазой (А3562, Sigma), в качестве вторичного реагента. Потом определяли связывание при использовании динатрий 4-нитрофенил фосфата (4NPP, Fluka) в качестве субстрата для щелочной фосфатазы. Развитие окраски измеряли при 405 нм. Скрининг нескольких сотен клонов с помощью такого анализа сырьевого экстракта ELISA выявил более сотни различных DARPin со специфичностью для xSA. Эти связывающие белки были выбраны для дальнейшего анализа. Примеры аминокислотных последовательностей выбранных повторяемых доменов, которые специфически связываются с xSA, обеспечиваются в SEQ ID NO: 17-31, 37-40 и 43-48.
Дедукция мотивов повторяемой последовательности из отобранных повторяемых доменов со связывающей специфичностью для xSA
Аминокислотные последовательности выбранных повторяемых доменов со связывающей специфичностью для xSA подвергали дальнейшему анализу с помощью инструментов для анализа последовательности, которые являются известными практикующему специалисту в данной области техники (WO 2002/020565; Forrer и др., 2003, в приведенном выше месте; Forrer, P., Binz, H.K., Stumpp, M.T. и Plückthun, A., ChemBioChem, 5(2), 183-189, 2004). Тем не менее, в противовес WO 2002/020565, где существующие в природе мотивы повторов использовали для выведения мотивов повторяемых последовательностей, в данном случае мотивы повторяемых последовательностей были выведены из повторяемой единицы отобранных повторяемых доменов со связывающей специфичностью для xSA. Таким образом, определяли семейства отобранных повторяемых доменов, включающих общий мотив повторяемой последовательности. Такие мотивы повторяемых последовательностей, присутствующих в повторяемых доменах со связывающей специфичностью для xSA, обеспечиваются в SEQ ID NO: 11-14.
Высокий уровень и экспрессия растворимых DARPin
Для дальнейшего анализа отобранные клоны, демонстрирующие xSA связывание в сырьевом клеточном экстракте ELISA так, как описано выше, экспрессировали в клетках Е.coli BL21 или XL1-Blue и очищали с помощью их His-метки при использовании стандартных прописей. 25 мл стационарных ночных культур (LB, 1% глюкозы, 100 мг/л ампициллина; 37°С) использовали для инокуляции 1 л культур (та же среда). При значении поглощения 0,7 при 600 нм культуры индуцировали с помощью 0,5 мМ IPTG и инкубирвоали при 37°С в течение 4 часов. Центрифугировали культуры и полученные осадки ресуспендировали в 40 мл TBS500 (50 мМ Трис-HCl, 500 мМ NaCl, pH 8) и подвергали обработке ультразвуком. Лизаты повторно центрифугировали и прибавляли глицерин (заключительная концентрация 10% (об./об.)) и имидазол (заключительная концентрация 20 мМ) к полученному супернатанту. Белки очищали при использовании колонки с Ni-нитрилтрехуксусной кислотой (2,5 мл объема колонки) в соответствии с инструкциями производителя (QIAgen, Germany). Альтернативно, DARPin или отобранные повторяемые домены, не содержащие 6×His-метки, очищали с помощью анионобменной хроматографии, после чего проводили эксклюзионную хроматографию размеров при использовании стандартной смолы и прописей, известных квалифицированному специалисту в данной области техники. Вплоть до 200 мг высоко растворимых DARPin со связывающей специфичностью по отношению к сывороточному альбумину могут быть очищены из 1 л культуры Е.coli со степенью чистоты >95%, как оценивается с помощью SDS - 15% ПАГЭ. Такие очищенные DARPin использовались для дальнейшей характеристики.
Пример 2: Анализ стабильности и эксклюзионная хроматография размеров DARPin со связывающей специфичностью для xSA
DARPin #19-22 и DARPin #27-30 со связывающей специфичностью для xSA очищали почти до полной гомогенности при использовании их His-метки так, как описано выше, и хранили в PBS в течение 28 дней при концентрации 30 мг/мл (~2 мМ) при 40°С (изучение стабильности). В день 0 (Фигура 1а) и день 28 (Фигура 1b) брали образцы, разводили до концентрации 500 мкМ и анализировали при использовании эксклюзионной хроматографии размеров (SEC) для оценки их среднего молекулярного веса и стабильности (то есть, их агрегации, мультимеризации или тенденции к деградации) в течение периода времени.
В дополнительном эксперименте (Фиг.1с), DARPin #19 и 43-48 со связывающей специфичностью для xSA очищали почти до полной гомогенности при использовании их His-метки так, как описано выше, и хранили в PBS в течение 28 дней при концентрации приблизительно 100 мг/мл при -80°С, разводили до концентрации 500 мкМ и анализировали при использовании эксклюзионной хроматографии размеров (SEC) для их характеристики (то есть, агрегации, мультимеризации или тенденции к деградации). В частности, использовали большую колонку по сравнению с сериями первого анализа (смотри ниже).
Эксклюзионная хроматография размеров (SEC)
Аналитическую SEC осуществляли при использовании HPLC системы (Agilent 1200 series) при использовании либо колонки Superdex 200 5/150 (Фиг.1а и Фиг.1b), либо колонки Superdex 200 10/300GL (Фиг.1с) (GE Healthcare) при 20°С. Колонка Superdex 200 5/150 имела объем поглощающего слоя 3,0 мл, и свободного объема 1,08 мл (определен экспериментально при использовании голубого декстрана). Колонка Superdex 200 10/300GL имела объем поглощающего слоя 24 мл, и свободный объем приблизительно 8 мл. Измерения осуществляли в соответствии со стандартными процедурами, известными квалифицированному специалисту в данной области техники. Прогоны осуществляли при скорости истечения 0,2 мл/мин. и максимальном давлении 15 бар (Superdex 200 5/150) или 0,6 мл/мин. и максимальном давлении 18 бар (Superdex 200 10/300GL) в PBS. Образцы белков разводили в PBS до концентрации приблизительно 20-500 мкМ, фильтровали (0,22 мкМ), и 20-100 мкл разведенных образцов наносили на колонку для разделения. Профили элюирования образцов белка регистрировали путем считывания поглощения при 280 нм. Апротинин (АР) с молекулярным весом 6,5 кДа, карбоангидразу (СА) с молекулярным весом 29 кДа и кональбумин (СО) с молекулярным весом 75 кДа использовали в качестве стандартных белков для получения калибровочной кривой, из которой могут быть определены средние молекулярные массы белков образцов.
Эти результаты являются представленными на Фигуре 1. DARPin #19-22 и 27-30 продемонстрировали неразличимые SEC хроматограммы (то есть, неразличимые профили элюирования) в день 0 и день 28 исследования стабильности. Следовательно, все DARPin элюируются как мономеры при условиях анализа и DARPin #19-23 и 27-30 являются стабильными в течение, по крайней мере, 1 месяца при 40°С в PBS (то есть, их профили элюирования не выявляли какой-либо агрегации, мультимеризации или тенденции к деградации).
Пример 3: Термальная стабильность DARPin со связывающей специфичностью для xSA
Термальную стабильность DARPin со специфичностью для xSA анализировали с помощью анализа стабильности на основе флуоресценции (Niesen, F.H., Nature Protocols 2(9): 2212-2221, 2007). Таким образом, температуру, при которой белок (то есть, белок, такой как DARPin) развертывается, измеряли путем повышения флуоресценции красителя (например, SYPRO оранжевый; Invitrogen, кат. номер S6650) с аффинностью для гидрофобных частей белка, которые подвергаются развертыванию белка. Полученное таким образом среднее значение перехода флуоресценции (от более низкой интенсивности флуоресценции до более высокой интенсивности флуоресценции) соответствует среднему значению температуры денатурации (Tm) анализируемого белка.
Анализ термальной стабильности на основе флуоресценции
Термальную денатурацию DARPin при использовании SYPRO оранжевого как флуоресцентного красителя измеряли при использовании способа ПЦР в реальном времени (то есть, С1000 термоблока для проведения реакций (BioRad) в комбинациии с CFX96 оптической системой (BioRad)). DARPin получали при концентрации 80 мкМ либо в PBS при рН 7,4, либо в MES буфере при рН 5,8, содержащем 1× SYPR оранжевого (разведенного из маточного раствора 5'000× SYPRO оранжевого, Invitrogen), и 50 мкл таких белковых растворов или только буфер прибавляли к содержимому ячеек белого планшета на 96 ячеек для проведения ПЦР (Bio-Rad). Планшеты закрывали при использовании Microseal ’В’ Adhesive изоляционной прокладки (Bio-Rad) и нагревали в устройстве для проведения ПЦР в реальном времени от 20°С до 95°С при приращениях 0,5°С включая этап выдерживания в течение 25 секунд после каждого приращения температуры, и после проведения термальной денатурации DARPin осуществляли измерение относительных единиц флуоресценции образцов при каждом приращении температуры. Относительные единицы флуоресценции в ячейках планшета измеряли при использовании канала 2 аппарата для проведения ПЦР в реальном времени (то есть, возбуждение осуществляли при 535 нм, а определение проводили при длине волны 560-580 нм), и вычитали соответствующие значения, полученные при использовании только буфера. Из полученных таким образом срединных значений перехода термальной денатурации, могут быть определены Tm значения для анализируемых DARPin.
Результаты термальной денатурации DARPin либо в PBS при рН 7,4, либо в MES буфере при рН 5,8, после которой осуществляли повышение я интенсивности флуоресценции SYPRO оранжевого, являются представленными на Фигуре 2 и Фигуре 3. Измеренные переходы термальной денатурации демонстрируют, что все проанализированные DARPin со связывающей специфичностью для xSA имеют Tm значения выше 40°С (как при рН 7,4, так и при рН 5,8).
Пример 4: Характеристика DARPin со связывающей специфичностью для xSA с помощью поверхностного плазмонного резонанса
DARPin со связывающей специфичностью для xSA при использовании их His-метки иммобилизировали в проточной кювете, покрытой α-RGS-His антителом (Qiagen, кат. номер 34650), и анализировали взаимодействие сывороточного альбумина человека, обезьяны циномолгус (cyno), мыши, крысы, кролика и собаки с иммобилизованными DARPin.
Анализ с помощью поверхностного плазмонного резонанса (SPR)
SPR измеряли при использовании ProteOn устройства (BioRad), и измерения осуществляли в соответствии со стандартными процедурами, известными квалифицированному специалисту в данной области техники. Буфер для разгона представлял собой PBS, рН 7,4, содержащий 0,01% Твин 20®. Анти-RGS-His антитело ковалентно имммобилизовали на GLC чипе (BioRad) до уровня приблизительно 2000 резонансных единиц (RU). Потом проводили иммобилизацию DARPin на чипе, покрытом антителом, путем введения 150 мкл 1 мкМ раствора DARPin в течение 300 секунд (скорость истечения = 30 мкл/мин.). Взаимодействие сывороточного альбумина различных видов потом измеряли путем введение в течение 60 секунд буфера для разгона объемом 100 мкл (PBS, содержащий 0,01% Твин®), который содержал отличный сывороточный альбумин при концентрации 400, 200, 100, 50 нМ (измерение скорости ассоциации), после чего пропускали буфер для разгона в течение от 10 до 30 минут (скорость истечения = 100 мкл/мин.) (измерение скорости диссоциации). Сигналы (то есть, количественное значение резонансных единиц (RU)) стандартной ячейки без покрытия и эталонного введения (то есть, введение только буфера для разгона) вычитали из RU значений, полученных после введения сывороточных альбуминов (то есть, при использовании двух эталонов). Из значений SRP, полученных из измерений скорости диссоциации и скорости ассоциации, могут быть определены скорости диссоциации и скорости ассоциации взаимодействия соответствующих DARPin сывороточного альбумина.
Результаты подытожены в Таблице 1 и Таблице 2. Константы диссоциации (Kd) подсчитывали из оцененных скоростей диссоциации и ассоциации при использовании стандартных процедур, известных квалифицированному специалисту в данной области техники, и они оказывались в интервале от приблизительно 3 до приблизительно 300 нМ. В то время как сывороточный альбумин человека и обезьяны циномолгус связывался со всеми проанализированными DARPin, сывороточный альбумин кролика, мыши, крысы и собаки связывался только с поднабором этих DARPin.
Пример 5: Конечный период полувыведения DARPin со связывающей специфичностью для xSA
Конечный период полувыведения DARPin у мышей и обезьян циномолгус (Масаса fascicularis, также имеет аббревиатуру "cyno") определяли в соответствии со стандартными процедурами, известными квалифицированному специалисту в данной области техники (Toutain, и др., в приведенном выше месте). Определенное количество DARPin внутривенно вводили млекопитающим, и выведение DARPin из плазмы крови оценивали в течение периода времени в соответствии с его концентрацией в плазме крови. Концентрация DARPin сначала снижается до достижения псевдоравновесия (альфа-фаза), после чего происходит экспоненциальное дальнейшее снижение концентрации DARPin в плазме крови (бета-фаза). Из этой бета-фазы может быть подсчитан конечный период полувыведения DARPin.
Определение выведения DARPin из плазмы крови у мышей
Для того чтобы оценить выведение DARPin со связывающей специфичностью для xS из плазмы крови А, исследуемые белки метили радиоактивной меткой и вводили в хвостовую вену несенсибилизированных Balb/c мышей. При этом вводили следующие DARPin: DARPin #19, DARPin #21, DARPin #23, DARPin #25, DARPin #18, DARPin #32, DARPin #35, DARPin #36, DARPin #33, DARPin #34, DARPin #37, DARPin #38, DARPin #43, DARPin #44, DARPin #45, DARPin #46, DARPin #47 и DARPin #48. DARPin радиоактивно метили с помощью комплекса 99mTc-карбонил, как описано ранее (Waibel, R., и др., Nature Biotechnol. 17(9), 897-901, 1999). DARPin (40 мкг) инкубировали с 99mTc-карбонилом (0,8-1,6 мКи) в течение 1 часа перед разведением до 400 мкл в PBS (рН 7,4). Каждой мыши внутривенно вводили 100 мкл меченного таким образом раствора DARPin (эквивалентный 10 мкг белка и 0,2-0,4 мКи). Образцы крови мышей собирали через 1 час, 4 часа, 24 часа и 48 часов после исходной инъекции и измеряли радиоактивность образцов. Уровень радиоактивности, измеренный в определенные моменты времени, является непосредственной мерой для количества DARPin, которые все еще присутствуют в плазме крови в этот момент времени. % введенной путем инъекции дозы представляет собой процент общей радиоактивности крови у мыши (1,6 мл для 18 г мыши), измеренной в определенный момент времени, по отношению к общей радиоактивности введенного образца, скорректированной для радиоактивного распада 99mTc.
DARPin со связывающей специфичностью для MSA обладают значительно повышенным конечным периодом полувыведения у мышей по сравнению с DARPin #32, не имеющим связывающей специфичности для xSA (Фигура 4). DARPin #19, DARPin #21, DARPin #23, DARPin #33, DARPin #37, DARPin #43, DARPin #44, DARPin #45, DARPin #46, DARPin #47 и DARPin #48 имели конечный период полувыведения у мышей приблизительно 2-2,5 дня.
Определение выведения DARPin из плазмы крови у обезьян циномолгус
DARPin, разведенные в PBS, вводили в виде болюсной инъекции в подкожную латеральную вену конечности. Вводили следующие DARPin: DARPin #26 (0,5 мг/кг), DARPin #24 (0,5 мг/кг), DARPin #17 (0,5 мг/кг), DARPin #34 (1 мг/кг) и DARPin #32 (0,5 мг/кг). В различные моменты времени после инъекции получали плазму из крови, собранной из бедренной вены животных. Концентрацию DARPin в образцах плазмы крови потом определяли с помощью сэндвич ELISA при использовании стандартных прописей, известных квалифицированному специалисту в данной области техники, и приемлемой стандартной кривой DARPin с известными концентрациями DARPin.
Образцы плазмы крови обезьян циномолгус серийно разводили в PBS-C (PBS, содержащий 0,25% казеина, рН 7,4) на планшетах MaxiSorp ELISA, которые были покрыты специфическим для DARPin кроличьим моноклональным антителом. После экстенсивного промывания с помощью PBS-T (PBS с добавкой 0,1% Твин 20®, рН 7,4) планшеты инкубировали с моноклональным анти-RGS(His)4 антителом, меченным пероксидазой хрена HRP (Qiagen). Связывание определяли при использовании 100 мкл BM-Blue POD субстрата (Roche Diagnostics). Реакцию останавливали путем прибавления 50 мкл 1 М H2SO4 и измеряли поглощение при 450 нм (вычитая при этом поглощение при 620 нм). Концентрацию DARPin в образцах плазмы крови подсчитывали путем осуществления моноэкспоненциальной регрессии на стандартной кривой DARPin, разведенного в сыворотке крови обезьян (GraphPad Prism). Конечный период полувыведения из плазмы крови DARPin подсчитывали путем проведения нелинейных регрессий (двухфазный распад) на определенных значениях концентрации вплоть до 240 часов после инъекции. Период полувыведения второй (бета) фазы соответствует конечному периоду полувыведения из плазмы крови.
DARPin со связывающей специфичностью для xSA имеют повышенный конечный период полувыведения у обезьяны циномолгус по сравнению с DARPin #32, который не имеет связывающей специфичности для xSA (Фигура 5, Таблица 3). DARPin #19, DARPin #21, DARPin #43, DARPin #44, DARPin #45, DARPin #46, DARPin #47 и DARPin #48 имели конечный период полувыведения у обезьян циномолгус приблизительно от 10 до 15 дней.
Пример 6: Более высокая термальная стабильность DARPin с улучшенными С-терминальными кэппирующими модулями
Термальную стабильность DARPin подвергали анализу с помощью анализа стабильности на основе флуоресценции так, как описывается в Примере 3. Альтернативно, термальную стабильность DARPin анализировали при использовании CD спектрометрии; то есть путем измерения их тепловой денатурации в соответствии с их сигналом циркулярного дихроизма (CD) при 222 нм с помощью методик, хорошо известных квалифицированному специалисту в данной области техники. CD сигнал образца регистрировали при 222 нм в устройстве Jasco J-715 (Jasco, Japan) при медленном нагревании белка при концентрации 0,02 мМ в PBS рН 7,4 от 20°С до 95°С при использовании скорости изменения 1°С за минуту. Этот способ представляет собой эффективное средство отслеживать денатурацию DARPin, поскольку они в основном состоят из альфа-спиралей, которые демонстрируют сильное изменение своего CD сигнала при 222 нм при развертывании. Срединная точка наблюдаемого перехода такого измеренного CD сигнала для DARPin соответствует их значению Tm.
Термальная стабильность DARPin #37 (SEQ ID NO: 37 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом) сравнивали с термальной стабильностью DARPin #38 (SEQ ID NO: 38 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом) при использовании анализа термальной стабильности на основе флуоресценции. Эти два DARPin обладают идентичной аминокислотной последовательностью за исключением С-терминальных кэппирующих модулей своих повторяемых доменов. Повторяемый домен DARPin #38, но не DARPin #37, включает улучшенный С-кэппирующий модуль, как описывается в данной заявке. Значения Tm в PBS рН 7, определенные для DARPin #37 и DARPin #38, составляли приблизительно 63°С и приблизительно 73°С, соответственно. Значения Tm в MES буфере рН 5,8, определенные для DARPin #37 и DARPin #38, составляли приблизительно 54,5°С и приблизительно 66°С, соответственно.
Термальную стабильность DARPin #39 (SEQ ID NO: 39 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом) сравнивали с термальной стабильностью DARPin #40 (SEQ ID NO: 40 с гистидиновой меткой (SEQ ID NO: 15), слитой с его N-терминальным концом) при использовании анализа термальной стабильности на основе флуоресценции. Эти два DARPin обладают идентичной аминокислотной последовательностью за исключением С-терминальных кэппирующих модулей своих повторяемых доменов. Повторяемый домен DARPin DARPin #40, но не DARPin #39, включает улучшенный С-кэппирующий модуль, как описывается в данной заявке. Значения Tm в MES буфере рН 5,8, определенные для DARPin #39 и DARPin #40, составляли приблизительно 51°С и приблизительно 55°С, соответственно.
Термальную стабильность DARPin #41 (SEQ ID NO: 41) сравнивали с термальной стабильностью DARPin #42 (SEQ ID NO: 42) при использовании CD спектроскопии. Эти два DARPin обладают идентичной аминокислотной последовательностью за исключением С-терминальных кэппирующих модулей своих повторяемых доменов. Повторяемый домен DARPin #42, но не DARPin #41, включает улучшенный С-кэппирующий модуль, как описывается в данной заявке. Значения Tm в PBS рН 7,4, определенные для DARPin #41 и DARPin #42, составляли приблизительно 59,5°С и приблизительно 73°С соответственно.







































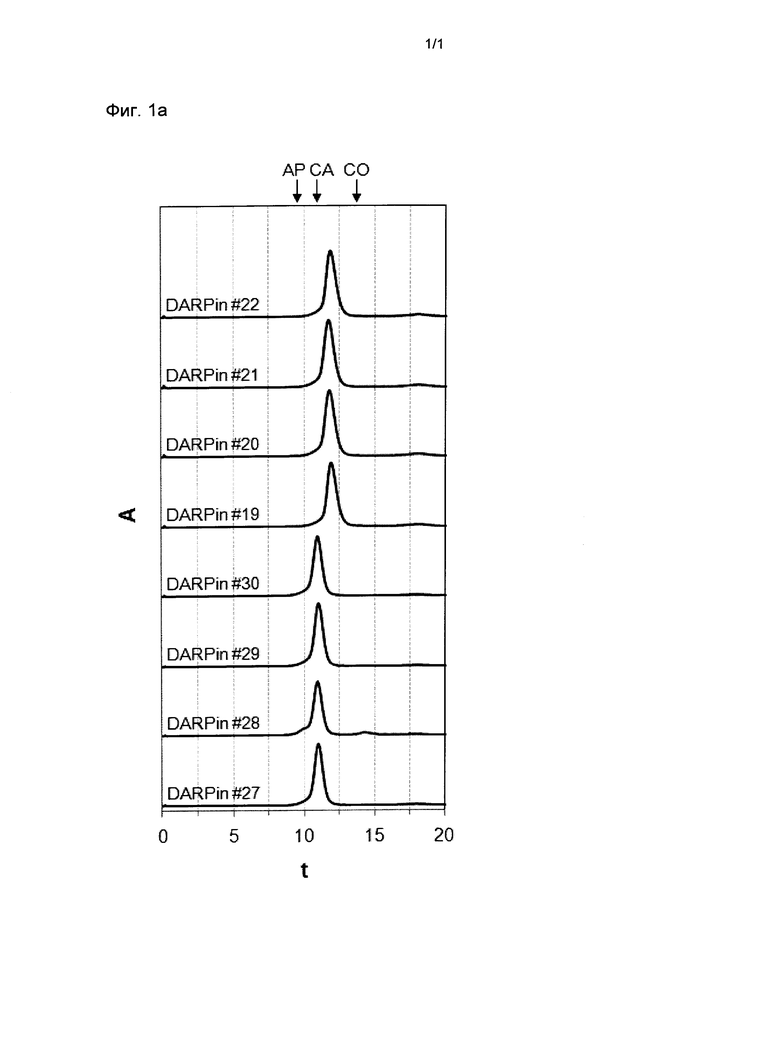
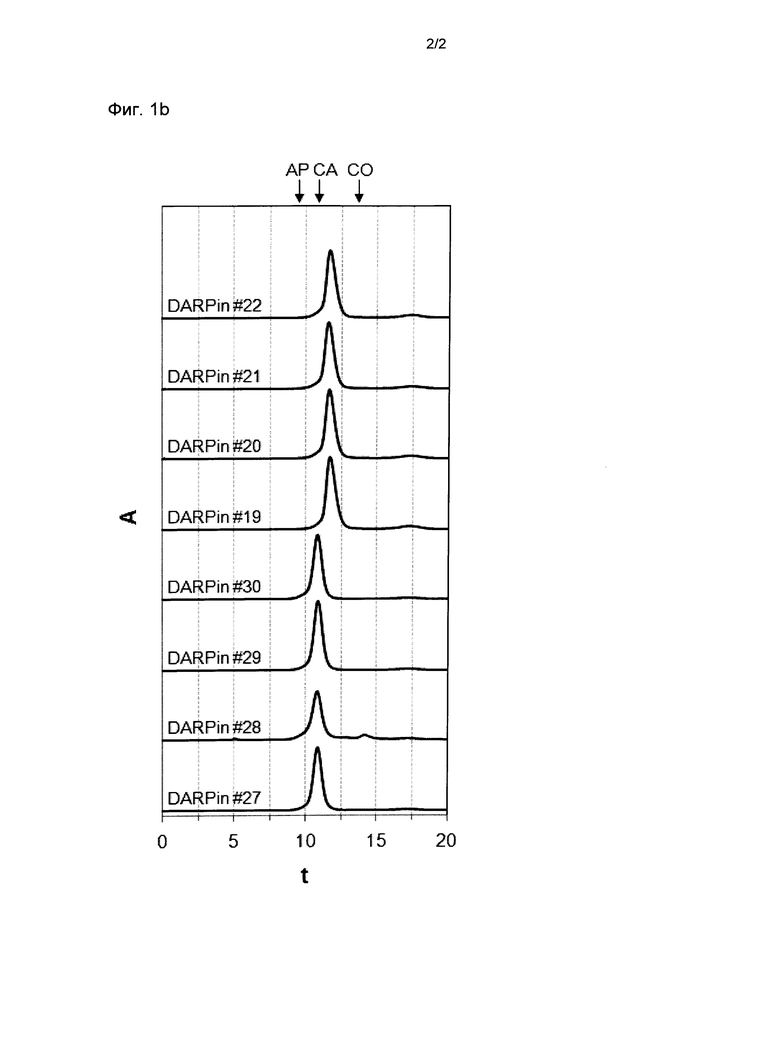
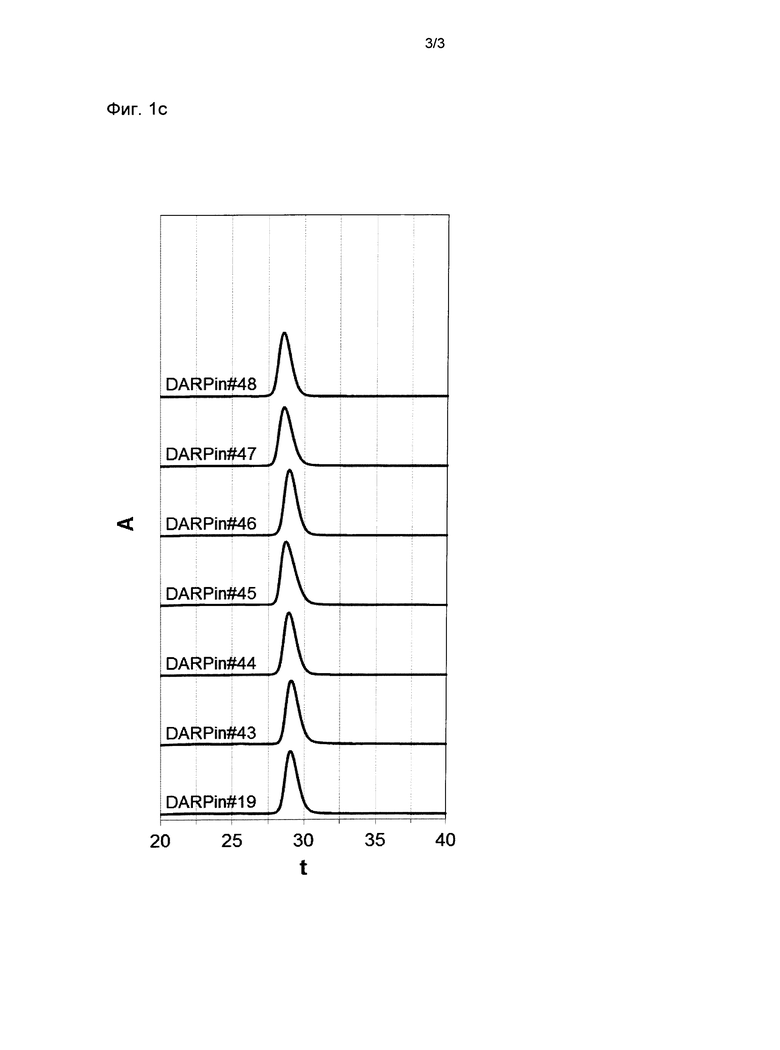
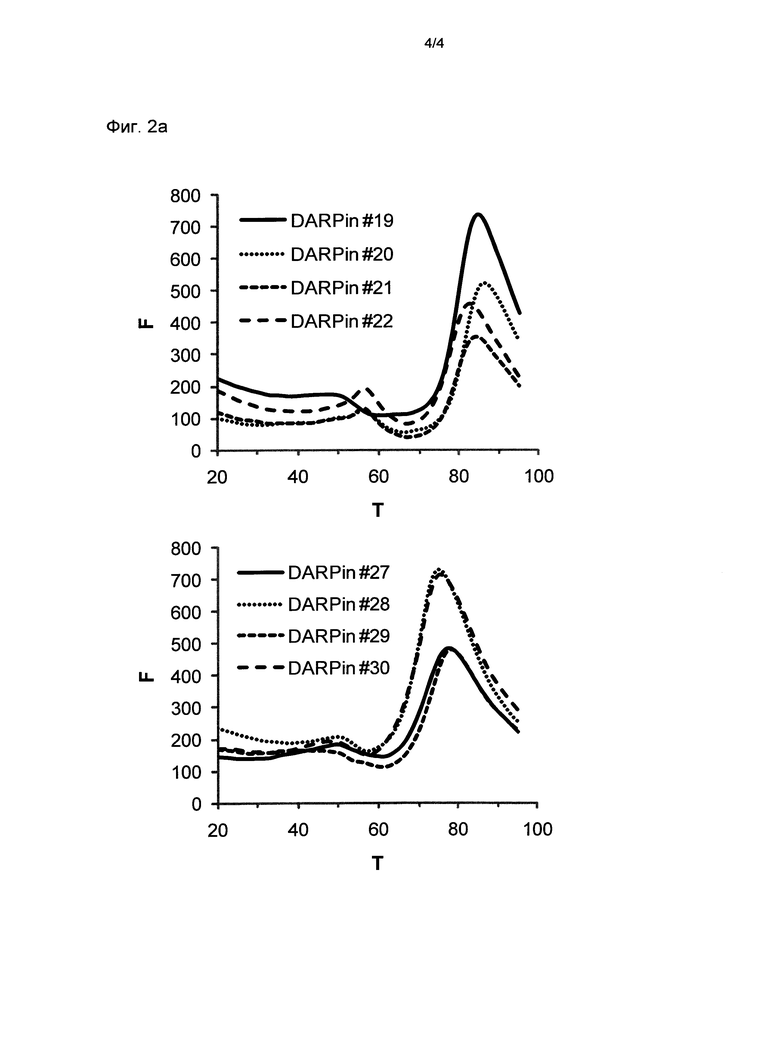
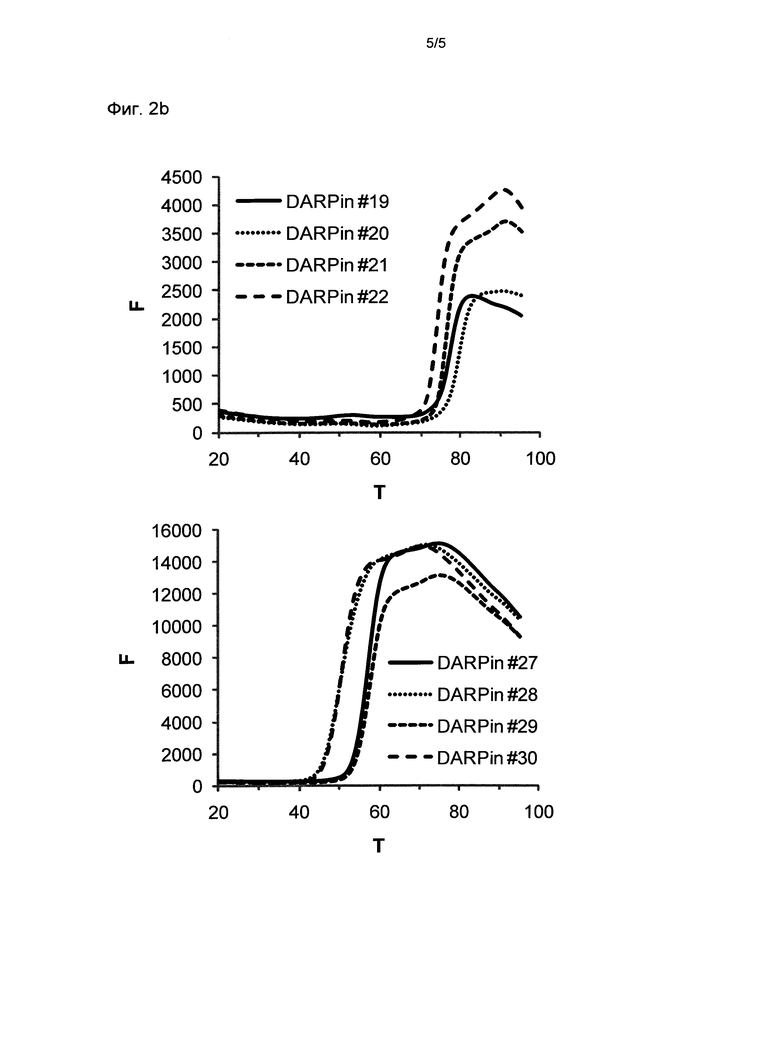
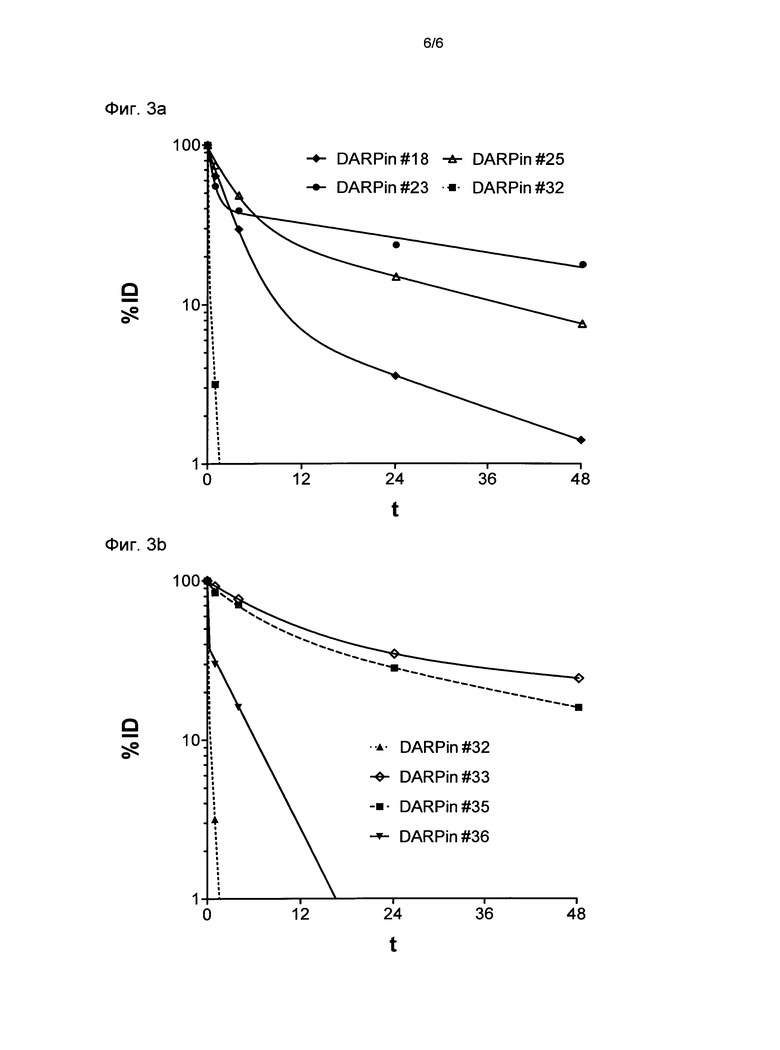
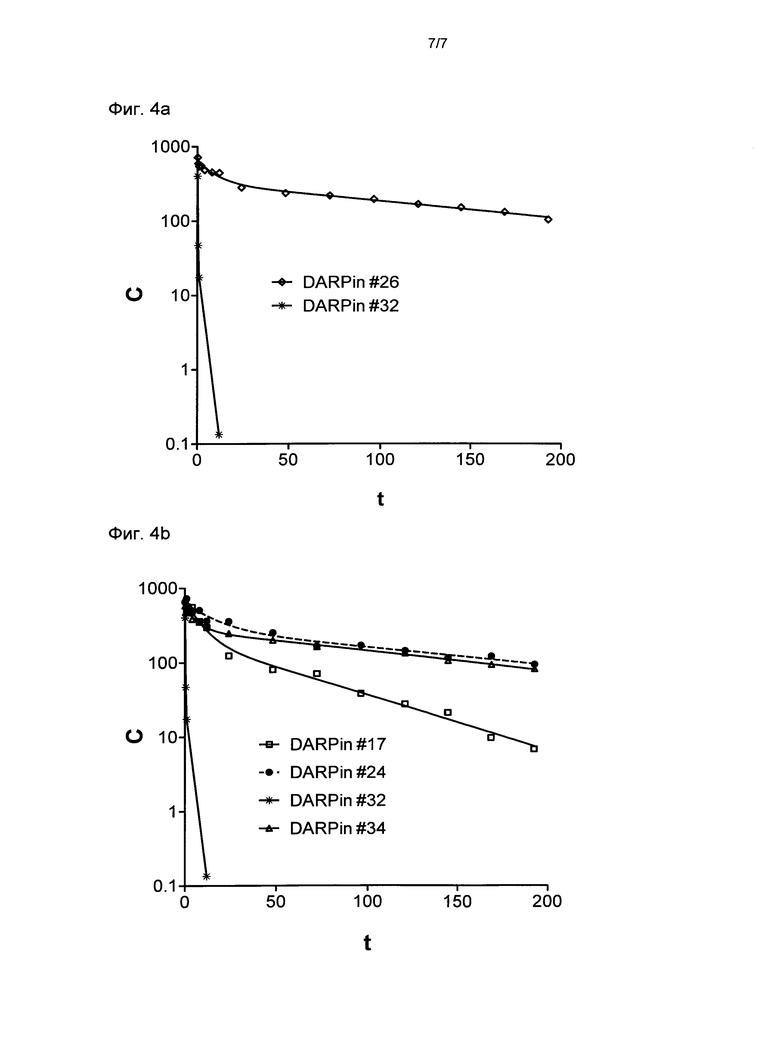
Изобретение относится к области биотехнологии, конкретно к получению новых сконструированных белков со связывающей специфичностью для сывороточного альбумина, что может быть использовано в медицине. Рекомбинантным путем получают белки, включающие домен анкиринового повтора, а также нуклеиновые кислоты, которые кодируют такие белки и фармацевтические композиции, включающие такие белки для лечения заболеваний. Изобретение позволяет существенно увеличить период полувыведения в плазме крови по сравнению с белками, которые не связывают сывороточный альбумин. 6 н. и 12 з.п. ф-лы, 4 ил., 3 табл., 6 пр.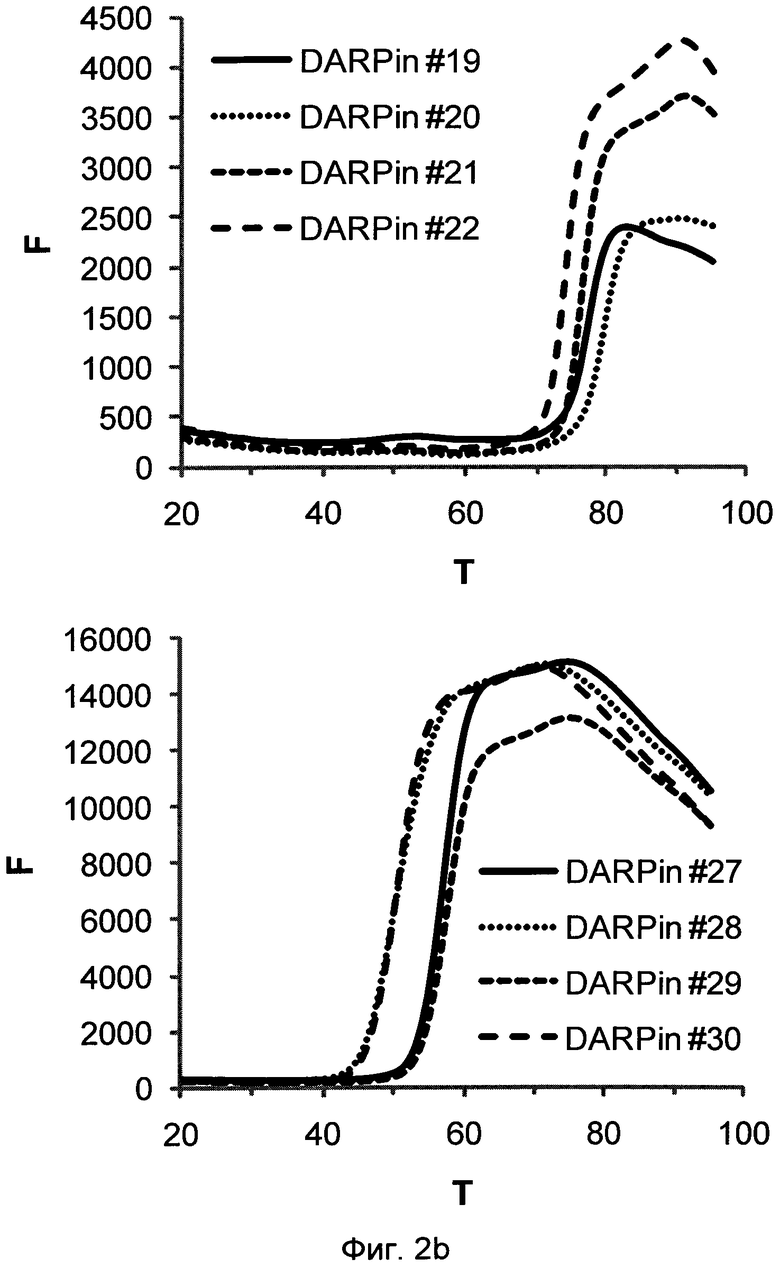
1. Связывающий белок, включающий один домен анкиринового повтора, где указанный домен анкиринового повтора имеет связывающую специфичность для сывороточного альбумина млекопитающих и где указанный домен анкиринового повтора включает
(а) N-терминальный кэппинг модуль, (б) С-терминальный кэппинг модуль и (в) модуль анкиринового повтора, имеющий аминокислотную последовательность, выбранную из группы, состоящей из
1) SEQ ID NO: 49, 50, 51 и 52,
(2) последовательностей, где вплоть до 3 аминокислот в SEQ ID NO: 49, 50, 51 и 52 заменены другими аминокислотами,
(3) аминокислотную последовательность
X1DX2X3X4X5TPLHLAAX6X7GHLX8IVEVLLKX9GADVNA (SEQ ID NO: 53),
где X1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из A, D, М, F, S, I, Т, N, Y и K;
Х2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Е, K, D, F, М, N, I и Y;
Х3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из W, R, N, Т, Н, K, А и F;
Х4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из G и S;
Х5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из N, Т и Н;
Х6 представляет собой аминокислотный остаток, выбранный из группы, состоящей из N, V и R;
Х7 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Е, Y, N, D, Н, S, A, Q, Т и G;
Х8 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Е и K;
Х9 представляет собой аминокислотный остаток, выбранный из группы, состоящей из S, A, Y, Н и N; и необязательно вплоть до 2 аминокислот в положениях, отличных от тех, которые обозначены как X в SEQ ID NO: 53, заменены какой-либо аминокислотой;
(4) аминокислотную последовательность
X1DX2X3GX4TPLHLAAX5X6GHLEIVEVLLKX7GADVNA (SEQ ID NO: 10),
где X1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из A, D, М, F, S, I, Т, N, Y и K;
Х2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Е, K, D, F, М, N, I и Y;
Х3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из W, R, N, Т, Н, K, А и F;
Х4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из N, Т и Н;
Х5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из N, V и R;
Х6 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Е, Y, N, D, Н, S, A, Q, Т и G;
Х7 представляет собой аминокислотный остаток, выбранный из группы, состоящей из S, A, Y, Н и N; и необязательно вплоть до 2 аминокислот в положениях, отличных от тех, которые обозначены как Х в SEQ ID NO: 10, заменены какой-либо аминокислотой;
(5) аминокислотную последовательность
X1DYFX2HTPLHLAARX3X4HLX5IVEVLLKX6GADVNA (SEQ ID NO: 11),
где X1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из D, K и А;
Х2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из D, G и S;
Х3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Е, N, D, Н, S, A, Q, Т и G;
Х4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из G и D;
Х5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Е, K и G;
Х6 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Н, Y, А и N; и необязательно вплоть до 2 аминокислот в положениях, отличных от тех, которые обозначены как X в SEQ ID NO: 11, заменены какой-либо аминокислотой.
(6) аминокислотную последовательность
X1DFX2GX3TPLHLAAX4X5GHLEIVEVLLKX6GADVNA (SEQ ID NO: 54),
где Х1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из F, S, L и K;
Х2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из V и А;
Х3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из R и K;
Х4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из S и N;
Х5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из N, D, Q, S, А, Т и Е;
Х6 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, Н, Y, S и N; и необязательно вплоть до 2 аминокислот в положениях, отличных от тех, которые обозначены как X в SEQ ID NO: 54, заменены какой-либо аминокислотой;
(7) аминокислотную последовательность
X1DFX2GX3TPLHLAAX4DGHLEIVEVLLKX5GADVNA (SEQ ID NO: 12),
где X1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из F, S, L и K;
Х2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из V и А;
Х3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из R и K;
Х4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из S и N;
Х5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, Н, Y, S и N; и необязательно вплоть до 2 аминокислот в положениях, отличных от тех, которые обозначены как X в SEQ ID NO: 12, заменены какой-либо аминокислотой;
(8) аминокислотную последовательность
X1DX2X3GTTPLHLAAVYGHLEX4VEVLLKX5GADVNA (SEQ ID NO: 13),
где X1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из K, A, D, М, F, S, I, Т, N, и Y;
Х2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Е, K, D, F, М, N и Y;
Х3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из R, N, Т, Н, K, А и F;
Х4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из I и М;
Х5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Н, Y, K, А и N; и необязательно вплоть до 2 аминокислот в положениях, отличных от тех, которые обозначены как X в SEQ ID NO: 13, заменены какой-либо аминокислотой;
(9) аминокислотную последовательность
X1NETGYTPLHLADSSGHX2EIVEVLLKX3X4X5DX6NA (SEQ ID NO: 14),
где X1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Q и K;
Х2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из L и Р;
Х3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Н, N, Y и А;
Х4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из G и S;
Х5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, V, Т и S;
Х6 представляет собой аминокислотный остаток, выбранный из группы, состоящей из V и F; и необязательно вплоть до 2 аминокислот в положениях, отличных от тех, которые обозначены как X в SEQ ID NO: 14, заменены какой-либо аминокислотой.
2. Связывающий белок, включающий один домен анкиринового повтора, где указанный домен анкиринового повтора содержит N-терминальный кэппинг модуль, С-терминальный кэппинг модуль и модуль анкиринового повтора, где указанный домен анкиринового повтора имеет связывающую специфичность для сывороточного альбумина млекопитающих и где указанный домен анкиринового повтора включает аминокислотную последовательность, которая имеет по крайней мере 90%-ную идентичность аминокислотной последовательности с одним доменом анкиринового повтора, выбранным из группы, состоящей из SEQ ID NO: 17-31 и 43-48, в котором G в положении 1 и/или S в положении 2 указанного домена анкиринового повтора необязательно отсутствует.
3. Связывающий белок по п. 1 или 2, где аминокислотная последовательность указанного модуля анкиринового повтора выбрана из группы, состоящей из (1) SEQ ID NO: 49, 50, 51 и 52 и (2) последовательностей, где вплоть до 2 аминокислот в SEQ ID NO: 49, 50, 51 и 52 заменены какой-либо аминокислотой.
4. Связывающий белок по п. 1 или 2, где указанный модуль анкиринового повтора содержит аминокислотную последовательность SEQ ID NO: 49.
5. Связывающий белок, содержащий домен анкиринового повтора, где указанный домен анкиринового повтора имеет связывающую специфичность к сывороточному альбумину млекопитающих и где указанный домен анкиринового повтора содержит (а) N-терминальный кэппинг модуль, (б) С-терминальный кэппинг модуль и (в) модуль анкиринового повтора, имеющий аминокислотную последовательность, выбранную из группы, состоящей из (1) SEQ ID NO: 49, 50, 51 и 52 и (2) последовательностей, где вплоть до 3 аминокислот в SEQ ID NO: 49, 50, 51 и 52 заменены какой-либо аминокислотой и где указанный связывающий белок дополнительно включает биоактивное соединение и где указанный связывающий белок обладает по крайней мере в 2 раза более высоким конечным периодом полувыведения из плазмы у млекопитающего по сравнению с конечным периодом полувыведения указанного немодифицированного биоактивного соединения, где указанный более высокий конечный период полувыведения придается указанному связывающему белку с помощью указанного домена анкиринового повтора.
6. Связывающий белок п. 1 или 2, где указанный модуль анкиринового повтора имеет аминокислотную последовательность, выбранную из группы, состоящей из (1) SEQ ID NO: 50 и (2) последовательностей, где вплоть до 2 аминокислот в SEQ ID NO: 50 заменены другими аминокислотами.
7. Связывающий белок по п. 1 или 2, где указанный домен анкиринового повтора включает аминокислотную последовательность, которая имеет по меньшей мере 95%-ную идентичность аминокислотной последовательности с доменом анкиринового повтора SEQ ID NO: 46, где G в положении 1 и/или S в положении 2 указанного домена анкиринового повтора необязательно пропущены.
8. Связывающий белок по п. 1 или 2, где указанный модуль анкиринового повтора содержит аминокислотную последовательность
X1DYFX2HTPLHLAARX3X4HLX5IVEVLLKX6GADVNA (SEQ ID NO: 11), в которой
X1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из D, K и А;
Х2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из D, G и S;
Х3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Е, N, D, Н, S, A, Q, Т и G;
Х4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из G и D;
Х5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Е, K и G;
Х6 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Н, Y, А и N; и
необязательно вплоть до 2 аминокислот в положениях, отличных от тех, которые обозначены X в SEQ ID NO: 11, заменены какой-либо аминокислотой.
9. Связывающий белок по п. 1 или 2, где указанный модуль анкиринового повтора содержит аминокислотную последовательность
X1DFX2GX3TPLHLAAX4X5GHLEIVEVLLKX6GADVNA (SEQ ID NO: 54), в которой
X1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из F, S, L и K;
Х2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из V и А;
Х3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из R и K;
Х4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из S и N;
Х5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из N, D, Q, S, А, Т и Е;
Х6 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, Н, Y, S и N; и
необязательно вплоть до 2 аминокислот в положениях, отличных от тех, которые обозначены X в SEQ ID NO: 54, заменены какой-либо аминокислотой.
10. Связывающий белок по п. 1 или 2, где указанный модуль анкиринового повтора содержит аминокислотную последовательность
X1DYFX2HTPLHLAARX3X4HLX5IVEVLLKX6GADVNA (SEQ ID NO: 11),
где X1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из K и А;
Х2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из G и S;
Х3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Q и N;
Х4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из G и D;
Х5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Е и K;
Х6 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А и Y; и
необязательно вплоть до 2 аминокислот в положениях, отличных от тех, которые обозначены как Х в SEQ ID NO: 11, заменены какой-либо аминокислотой.
11. Связывающий белок по п. 1 или 2, где указанный модуль анкиринового повтора имеет аминокислотную последовательность
X1DYFX2HTPLHLAARX3X4HLX5IVEVLLKX6GADVNA (SEQ ID NO: 11),
где X1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из D, K и А;
Х2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из D, G и S;
Х3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Е, N, D, Н, S, A, Q, Т и G;
Х4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из G и D;
Х5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Е, K и G;
Х6 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Н, Y, А и N; и необязательно вплоть до 2 аминокислот в положениях, отличных от тех, которые обозначены как X в SEQ ID NO: 11, заменены какой-либо аминокислотой;
и где указанный домен анкиринового домена дополнительно содержит модуль анкиринового повтора, имеющий аминокислотную последовательность
X1DFX2GX3TPLHLAAX4DGHLEIVEVLLKX5GADVNA (SEQ ID NO: 12), в котором
X1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из F, S, L и K;
Х2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из V и А;
Х3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из R и K;
Х4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из S и N;
Х5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, Н, Y, S и N; и необязательно вплоть до 2 аминокислот в положениях, отличных от тех, которые обозначены как Х в SEQ ID NO: 12, заменены какой-либо аминокислотой,
и где модулю анкиринового повтора, содержащему аминокислотную последовательность SEQ ID NO: 12, предшествует модуль анкиринового повтора, содержащий аминокислотную последовательность SEQ ID NO: 11.
12. Связывающий белок, содержащий домен анкиринового повтора, где указанный домен анкиринового повтора содержит N-терминальный кэппинг модуль, С-терминальный кэппинг модуль и модуль анкиринового повтора, где указанный домен анкиринового повтора имеет связывающую специфичность к сывороточному альбумину млекопитающих и где указанный домен анкиринового повтора содержит аминокислотную последовательность, которая по меньшей мере на 90% идентична аминокислотной последовательности с одним доменом анкиринового повтора, выбранного из группы, состоящей из SEQ ID NOs - 17 до 31 и 43 до 48, где G в положении 1 и/или S в положении 2 указанного домена анкиринового повтора необязательно отсутствует и где указанный связывающий белок дополнительно включает биоактивное соединение и где указанный связывающий белок обладает по меньшей мере в два раза более высоким конечным периодом полувыведения из плазмы у млекопитающего по сравнению с конечным периодом полувыведения указанного немодифицированного биоактивного соединения, где указанный высокий конечный период полувыведения придается указанном связывающему белку с помощью указанного домена анкиринового повтора.
13. Связывающий белок в по любому из пп. 1, 2, 5 и 12, где указанный домен анкиринового повтора конкурирует за связывание с сывороточным альбумином млекопитающих с доменом анкиринового повтора, выбранным из группы, состоящей из SEQ ID NO: 17-31 и 43-48.
14. Связывающий белок по п. 1 или 2, где указанный домен анкиринового повтора включает аминокислотную последовательность, которая имеет по меньшей мере 95% идентичность с аминокислотной последовательностью домена анкиринового повтора выбранного из группы, состоящей из SEQ ID NO: 17-31 и 43-48, в котором G в положении 1 и/или S в положении 2 указанного домена анкиринового повтора являются необязательно отсутствующими.
15. Связывающий белок по любому из пп. 1-14, где указанный домен анкиринового повтора обладает по меньшей мере в пять раз более высоким конечным периодом полувыведения из плазмы у млекопитающего по сравнению с доменом анкиринового повтора SEQ ID NO: 32.
16. Связывающий белок по любому из пп. 1-14, где указанный домен анкиринового повтора обладает конечным периодом полувыведения из плазмы у человека по меньшей мере 1 день.
17. Нуклеиновая кислота, которая кодирует молекулу анкиринового повтора по любому из пп. 1, 3, 4, 6 и 8-10, два модуля анкиринового повтора по п. 11 или домен анкиринового повтора по любому из пп. 2, 7 и 14.
18. Фармацевтическая композиция для лечения рака и других патологических состояний, включающая эффективное количество связывающего белка по любому из пп. 1-16 или нуклеиновую кислоту по п. 17 и фармацевтически приемлемый носитель и/или разбавитель.
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| BRADLEY C.M | |||
| et al, Limits of cooperativity in a structurally modular protein: response of the Notch ankyrin domain to analogous alanine substitutions in each repeat, J | |||
| Mol | |||
| Biol., 2002, v.324, p.373-386 | |||
| INTERLANDI G | |||
| et al., Characterization and Further Stabilization of Designed Ankyrin Repeat Proteins by Combining Molecular Dynamics Simulations and Experiments J | |||
| Mol | |||
| Biol., 2008, v.375, p.837-854 | |||
| WO2005087797 A1, 22.09.2005 | |||
| WO 2009115919 A2, 24.09.2009 | |||
| АНТРАНИЛАМИДПИРИДИНАМИДЫ ИЗБИРАТЕЛЬНОГО ДЕЙСТВИЯ В КАЧЕСТВЕ ИНГИБИТОРОВ VEGFR-2 И VEGFR-3 | 2002 |
|
RU2299208C2 |

