Область изобретения
Изобретение касается рекомбинантных связывающих белков, специфичных к VEGF-A, а также нуклеиновых кислот, кодирующих такие VEGF-A связывающие белки, фармацевтических композиций, содержащих такие белки, и применения таких белков в лечении опухолей и заболеваний глаз.
Предпосылки создания изобретения
Ангиогенез, рост новых кровеносных сосудов из уже существующих сосудов, является ключевым процессом в ряде патологических состояний, включая рост опухоли и заболевания глаз, в частности глазные болезни, связанные с неоваскуляризацией, такие как возрастная макулярная дегенерации (AMD) или диабетический макулярный отек (DME) (Carmeliet, P., Nature 438, 932-936, 2005). Сосудистые эндотелиальные факторы роста (VEGF) стимулируют ангиогенез и лимфангиогенез путем активации VEGF рецепторных (VEGFR) тирозинкиназ в клетках эндотелия (Ferrara, N., Gerber, Н.P. and LeCouter, J., Nature Med. 9, 669-676, 2003).
Семейство VEGF млекопитающих состоит из пяти гликопротеинов, известных под названием VEGF-A, VEGF-B, VEGF-C, VEGF-D (также известный как FIGF), и фактора роста плаценты (P1GF, также известный как PGF). VEGF-A, как было показано, является эффективной мишенью для антиангиогенной терапии (Ellis, L. М. and Hicklin, D.J., Nature Rev. Cancer 8, 579-591, 2008). VEGF-A лиганды связываются и активируют три структурно подобные рецепторные тирозинкиназы типа III, названные VEGFR-1 (также известная как FLT1), VEGFR-2 (также известная как KDR) и VEGFR-3 (также известная как FLT4). VEGF лиганды обладают отличительными связывающими особенностями для каждой из этих рецепторных тирозинкиназ, которые способствуют разнообразию их функций.
В ответ на связывание лиганда, VEGFR тирозинкиназа активирует сеть различных сигнальных нисходящих путей. VEGFR-1 и VEGFR-2 обнаруживаются, главным образом, на сосудистом эндотелии, в то время как VEGFR-3, в основном, обнаруживается на лимфатическом эндотелии. У всех этих рецепторов есть внеклеточный домен, один трансмембранный участок и консенсусная тирозинкиназная последовательность, прерванная доменом киназной вставки. Совсем недавно нейропилин (NRP-1), первоначально определенный в качестве рецептора для медиаторов нейронов семейства семафорина/коллапсина, как показано, действует в качестве изоформ-специфического рецептора к VEGF-A.
Различные изоформы VEGF-A, как известно, получают путем альтернативного сплайсинга из восьми экзонов в VEGF-A гене. Все изоформы содержат экзоны 1-5 и терминальный экзон, экзон 8. Экзоны 6 и 7, которые кодируют гепарин-связывающие домены, могут быть включены или исключены. Это дает начало семейству белков, называемых в соответствии с их номером аминокислоты: VEGF-A165, VEGF-A121, VEGF-A189, и так далее. Однако экзон 8 состоит из двух 3' сайтов сращивания в нуклеотидных последовательностях, которые могут быть использованы клеткой для создания двух семейств изоформ с одинаковой длиной, но разными С-терминальными аминокислотными последовательностями (Varey, A.H.R. et al., British J. Cancer 98, 1366-1379, 2008). VEGF-Axxx ("xxx" обозначает номер аминокислоты зрелого белка), проангиогенное семейство изоформ получается при использовании наиболее проксимальной последовательности в экзоне 8 (что приводит к включению экзона 8а). Совсем недавно описанные антиангиогенные VEGF-Axxxb изоформы получаются при использовании дистального сайта сплайсинга, 66 bp, дальше вдоль гена от проксимального сайта сплайсинга. Это приводит к сплайсингу из экзона 8а и производству последовательностей мРНК, кодирующих семейство VEGF-Axxxb. VEGF-A 165 является преобладающей проангиогенной изоформой и обычно сверхэкспрессируется в различных человеческих солидных опухолях. VEGF-A165b было первой из установленных кодируемых экзоном 8b изоформ и, как показано, обладает антиангиогенными эффектами (Varey et al., loc. cit.; Konopatskaya, O. et al., Molecular Vision 12, 626-632, 2006). Это эндогенная ингибиторная форма VEGF-A, которая уменьшает VEGF-A индуцированную пролиферацию и миграцию клеток эндотелия. Хотя он может связываться с VEGFR-2, VEGF-A165b связывание не приводит к фосфорилированию рецептора или активации нисходящих сигнальных путей.
Существует несколько подходов к ингибированию VEGF-сигнализации, в том числе нейтрализация лиганда или рецептора антителами, и блокирование активации и сигнализации VEGF-A рецептора с помощью ингибиторов тирозинкиназы. VEGF-A-направленная терапия, как было показано, является эффективной в качестве монотерапии при AMD, DME, почечно-клеточной карциноме и гепатоцеллюлярной карциноме, а то время как она приносит пользу только в комбинации с химиотерапией у пациентов с метастатическим колоректальным раком, немелкоклеточным раком легких и метастатическим раком молочной железы (Narayanan, R. et al., Nat Rev. Drug Discov. 5, 815-816,2005; Ellis and Hicklin, loc. cit).
Помимо антител, для нейтрализации лиганда или рецептора могут быть использованы другие связывающие домены (Skerra, A., J. Mol. Recog. 13, 167-187, 2000; Binz, Н.К., Amstutz, P. and Pliickthun, A., Nat. Biotechnol. 23, 1257-1268, 2005). Один из таких новых классов связывающих доменов основан на спроектированных повторных доменах (WO 02/20565; Binz, Н.К., Amstutz, P., Kohl, A., Stumpp, М.Т., Briand, С., Forrer, P., Grütter, М.G., and Pliickthun, A., Nat. Biotechnol. 22, 575-582, 2004). WO 02/20565 описывает, насколько большие библиотеки повторных белков могут быть построены и их общее применение. Тем не менее, WО 02/20565 не раскрывает ни выбора повторных доменов со специфичностью связывания с VEGF-Axxx, ни конкретных повторных мотивов последовательности повторных доменов, которые специфически связываются с VEGF-Axxx.
Таргетинг VEGF-A с помощью имеющихся в настоящее время терапий не является эффективным у всех пациентов или от всех болезней (например, EGFR-экспрессирующие типы рака). Стало даже более очевидно, что терапевтический эффект, связанный с VEGF-A направленной терапией, является сложным и, вероятно, включает в себя несколько механизмов (Ellis and Hicklin, loc. cit.). Например, присутствующие на рынке анти-VEGF препараты, такие как бевацизумаб (Авастин®) или ранибизумаб (Lucentis®) (см. WO 96/030046, WO 98/045331 и WO 98/045332), или препараты в клинической разработке, такие как VEGF-Trap® (WO 00/075319) не делают различий между про- и антиангиогенными формами VEGF-A, так что они ингибируют обе. Как результат, они подавляют ангиогенез, а также лишают здоровые ткани существенного фактора выживания, а именно VEGF-Axxxb, что приводит к развитию цитотоксичности и ограничивающих дозу побочных эффектов, которые в свою очередь ограничивают эффективность. Побочные эффекты, которые присущи текущим анти-VEGF-A видам терапии, включают желудочно-кишечные перфорации, кровотечение, гипертензию, тромбоэмболии и протеинурию (Kamba, Т. and McDonald, D.M., Br. J. Cancer 96, 1788-95, 2007). Таким образом, существует необходимость улучшения антиангиогенных препаратов для лечения рака и других патологических состояний.
Технические проблемы, лежащие в основе изобретения, заключаются в выявлении новых антиангиогенных агентов, таких как повторные домены со специфичностью связывания с VEGF-Axxx, для улучшения лечения рака и других патологических состояний, например глазных болезней, таких как AMD или DME. Решение этой технической задачи достигается путем предоставления вариантов осуществления, которые характеризуются в формуле изобретения.
Краткое описание изобретения
Изобретение касается связывающего белка, включающего связывающий домен, в котором указанный связывающий домен ингибирует связывание VEGF-Axxx с VEGFR-2 и в котором температура денатрурации средней точки (Tm) указанного связывающего домена превышает 40°С при термальном развертывании, и он образует менее 5% (по весу) нерастворимых агрегатов при концентрации до 10 г/л в течение инкубации при температуре 37°С в течение 1 дня в PBS. Более конкретно, изобретение касается рекомбинантного связывающего белка, содержащего, по меньшей мере, один повторный домен, причем указанный повторный домен связывается с VEGF-Axxx с Kd ниже 10-7 М и ингибирует связывание VEGF-Axxx с VEGFR-2. В частности, такой связывающий белок ингибирует прорастание сфероидов HUVEC со значением IC50 ниже 10 нМ и такой связывающий белок имеет константу диссоциации для взаимодействия с VEGF-Axxxb, которая, по крайней мере, в 10 раз выше по сравнению с Kd для взаимодействие с VEGF-Axxx.
В частности, изобретение касается рекомбинантного связывающего белка, включающего связывающий домен со специфичностью к VEGF-A, который представляет собой повторный домен, например анкириновый повторный домен, в частности анкириновый повторный домен, включающий повторный модуль с анкириновым повторным мотивом последовательности

где 1, 2, 3, 4, 5, 6 и 7, представляют, независимо друг от друга, аминокислотный остаток, выбранный из группы, состоящей из A, D, Е, F, Н, I, К, L, М, N, Q, R, S, Т, V, W и Y.
Изобретение также касается рекомбинантного связывающего белка, включающего повторный домен со специфичностью связывания к VEGF-A, который имеет по крайней мере 70% идентичность аминокислотной последовательности с анкириновым повторным доменом настоящего изобретения или который включает в себя повторный модуль по крайней мере с 70% идентичностью аминокислотной последовательности с анкириновым повторным модулем настоящего изобретения, или в котором один или более аминокислотных остатков анкириновых повторных модулей обмениваются на аминокислотный остаток, обнаруженный в соответствующей позиции при выравнивании анкиринового повторного звена.
Изобретение также касается связывающих белков, включающих рекомбинантный связывающий белок изобретения, связанный с одним или несколькими дополнительными фрагментами, например фрагментом, который также связывается с VEGFR-2 или другой мишенью, фрагментом мечения, фрагментом, который облегчает очистку белка, или фрагментом, который обеспечивает улучшенную фармакокинетику, например полиэтиленгликолевый фрагмент. В некоторых вариантах изобретения дополнительный фрагмент представляет собой белковый фрагмент. В некоторых других вариантах изобретения дополнительный фрагмент представляет собой небелковый полимерный фрагмент.
Кроме того, изобретение касается молекул нуклеиновой кислоты, кодирующих рекомбинантные связывающие белки настоящего изобретения, а также фармацевтической композиции, содержащей один или несколько из вышеупомянутых связывающих белков или молекул нуклеиновых кислот.
Кроме того, изобретение касается способа лечения рака и других патологических состояний, например глазных болезней, таких как AMD или DME, с использованием белков изобретения.
Краткое описание фигур
Фигура 1. Специфическое связывание собачьего VEGF-Al64 выбранных спроектированных анкириновых повторных белков.
Взаимодействие выбранных клонов с собачьим VEGF-A164 (VEGF) и белком отрицательного контроля (МВР, мальтоза-связывающий белок Е. coli) показан с помощью ИФА неочищенного экстракта. Биотинилированный собачий VEGF-A164 и МВР были иммобилизированы над NeutrAvidin. Номера относятся к отдельным клонам DARPin, выбранных в рибосомном дисплее в сравнении с собачьим VEGF-A164 или соответствующим человеческим VEGF-A165. А = абсорбция. Белые колонки указывают на связывание с собачьим VEGF-A164, черные колонки показывают неспецифическое фоновое связывание с МВР.
Фигура 2. Ингибирование прорастания сфероидов выбранным DARPin.
Длина прорастаний в анализе ингибирования прорастания сфероидов показана в присутствии различных концентраций (a) DARPin №30 (SEQ ID NO: 29), DARPin со специфичностью к VEGF-Axxxx, или (b) DARPin NC, DARPin отрицательного контроля без специфичности к VEGF-Axxx.
Фигура 3. Специфичное распознавание VEGF-изоформ.
Анализ поверхностного плазмонного резонанса (SPR) связывающих белков на VEGF-A изоформах.
(а) и (b): SPR анализ Avastin®. 250 нМ Авастина® внесли в проточную ячейку с иммобилизованным собачьим VEGF-A164 (а) или собачьим VEGF-A164b (b) на 100 секунд, с последующей промывкой потоком буфера.
(с) и (d): SPR анализ DARPin №27 (SEQ ID NO: 16). 250 нМ DARPin №27 внесли в проточную ячейку с иммобилизованным собачьим VEGF-A164 (с) или собачьим VEGF-A164b (d) на 100 секунд, с последующей промывкой потоком буфера. RU = единицы резонанса.
Фигура 4. Эффективное ингибирование человеческого VEGF-A165 в глазу кролика. Кроличья модель сосудистой утечки для демонстрации эффективности DARPin в ингибировании человеческого VEGF-A165 в глазу по сравнению с Lucentis®. На 1-й день либо PBS, DARPin №30 или Lucentis® вводили путем интравитреальной инъекции в один глаз каждого кролика (обработанный глаз). На 4-й день или 30-й день в оба глаза каждого кролика ввели путем интравитреальной инъекции 500 нг человеческого VEGF-A165. Оба глаза оценивали через 48 часов после инъекции VEGF-А165 путем измерения содержания флуоресцеина в стекловидном теле и сетчатке глаза через один час после внутривенного введения натрия флуоресцеина. R = соотношение измерений флуоресцеина в обработанном глазу/необработанном глазу. Стандартные отклонения показаны планкой погрешностей. 4-PBS = соотношение через 4 дня после инъекции PBS (контроль); 4-D = соотношение через 4 дня после введения DARPin №30; 30-D = соотношение через 30 дней после введения DARPin №30; 4-L = соотношение через 4 дня после введения Lucentis®; 30-L = соотношение через 30 дней после введения Lucentis®.
Подробное описание изобретения
VEGF-A млекопитающих существует в виде двух семейств альтернативных сплайсинговых изоформ: (I) проангиогенные «VEGF-Аххх» изоформы, полученные путем проксимального сплайсинга экзона 8, и (II) антиангиогенные «VEGF-Axxxb» изоформы, полученные путем дистального сплайсинга экзона 8. Предпочтительно, связывающий домен в соответствии с изобретением является специфическим для проангиогенного VEGF-Axxx собаки, кроликов, обезьян или человеческого происхождения. Более предпочтительно, связывающий домен в соответствии с изобретением является специфическим для проангиогенного VEGF-Axxx человеческого происхождения. Наиболее предпочтительно, связывающий домен в соответствии с изобретением является специфическим для человеческого VEGF-A165.
Термин «белок» касается полипептида, в котором по крайней мере часть полипептида имеет или может приобрести определенное трехмерное расположение путем формирования вторичных, третичных и четвертичных структур внутри и/или между полипептидной(-ыми) цепью (-ями). Если белок состоит из двух или более полипептидов, отдельные полипептидные цепи могут быть связаны нековалентно или ковалентно, например, с помощью дисульфидных связей между двумя полипептидами. Часть белка, каждая из которых имеет или может приобрести определенное трехмерное расположение путем формирования вторичной или третичной структуры, называется «домен белка». Такие домены белка хорошо известны практикующим специалистам в данной области.
Термин «рекомбинантный», используемый в терминах рекомбинантный белок, рекомбинантный домен белка и подобных означает, что указанные полипептиды производятся с использованием рекомбинантных ДНК-технологий, хорошо известных практикующим специалистам в соответствующих областях. Например, рекомбинантная молекула ДНК (например, полученная путем генного синтеза), кодирующая полипептид, может быть клонирована в бактериальную плазмиду экспрессии (например, pQE30, Qiagen). Когда такая сконструированная рекомбинантная плазмида экспрессии вставляется в бактерии (например, Е. coli), такие бактерии могут вырабатывать полипептид, кодируемый этой рекомбинантной ДНК. Соответственно, вырабатываемый полипептид называется рекомбинантным полипептидом.
Термин «полипептидный тег» касается аминокислотной последовательности, присоединенной к полипептиду/белку, отличающейся тем, что указанная аминокислотная последовательность используется для очистки, обнаружения или таргетирования указанного полипептида/белка, или в которой указанная аминокислотная последовательность улучшает физико-химическое поведение полипептида/белка, или отличающейся тем, что указанная аминокислотная последовательность обладает эффекторной функцией. Отдельные полипептидные теги, фрагменты и/или домены связывающего белка могут быть связаны друг с другом непосредственно или с помощью полипептидных линкеров. Эти полипептидные теги хорошо известны в данной области и в полной мере доступны для специалиста в данной области. Примеры полипептидных тегов включают малые полипептидные последовательности, например, His, myc, FLAG или Strep-теги или фрагменты, такие как ферменты (например, ферменты, такие как щелочная фосфатаза), которые позволяют обнаруживать указанный полипептид/белок, или фрагменты, которые могут быть использованы для таргетинга (например, иммуноглобулины или их фрагменты) и/или в качестве эффекторных молекул.
Термин «полипептидный линкер» касается аминокислотной последовательности, которая может связать, например, два белковых домена, полипептидный тег и домен белка, домен белка и не-полипептидный фрагмент, такой как полиэтиленгликоль или две тега последовательности. Такие дополнительные домены, теги, не полипептидные фрагменты и линкеры известны специалистам в соответствующей области. Список примеров приводится в описании патентной заявки WO 02/20565. Конкретные примеры таких линкеров включают глицин-сериновые линкеры разной длины, предпочтительно, длина указанных линкеров составляет от 2 до 16 аминокислот.
В контексте настоящего изобретения термин «полипептид» касается молекулы, состоящей из одной или нескольких цепей множества, то есть двух или более, аминокислот, связанных с помощью пептидных связей. Предпочтительно, полипептид состоит более чем из восьми аминокислот, связанных с помощью пептидных связей.
Термин «связывающий белок» касается белка, включающего один или несколько связывающих доменов, как далее описывается ниже. Предпочтительно, указанный связывающий белок составляет до четырех связывающих доменов. Более предпочтительно, указанный связывающий белок содержит до двух связывающих доменов. Наиболее предпочтительно, указанный связывающий белок содержит только один связывающий домен. Кроме того, любой такой связывающий белок может содержать дополнительные домены белка, которые не являются связывающими доменами, фрагментами мультимеризации, полипептидными тегами, полипептидными линкерами и/или не белковыми полимерными молекулами.
Примеры фрагментов мультимеризации включают иммуноглобулиновые тяжелоцепочечные константные участки, которые спариваются, чтобы обеспечить функциональные иммуноглобулиновые Fc домены, и лейциновые зипперы или полипептиды, содержащие свободный тиол, который образует межмолекулярную дисульфидную связь между двумя такими полипептидами. Примеры небелковых полимерных молекул включают гидроксиэтилкрахмал (ГЭК), полиэтиленгликоль (ПЭГ), полипропиленгликоль или полиоксиалкилен.
Термин «пегилированный» означает, что ПЭГ фрагмент ковалентно связанный, например, с полипептидом изобретения.
Термин «связывающий домен» означает домен белка, имеющий ту же «складку» (трехмерное расположение), что и белковая платформа и с заданным свойством, как это определено ниже. Такой связывающий домен может быть получен путем рациональных, или чаще всего, комбинаторных методов белковой инженерии, техник, которые известны в данной области (Skerra, 2000, loc. cit.; Binz et al., 2005, loc. cit.). Например, связывающий домен с заданным свойством можно получить с помощью метода, включающего стадии: (а) предоставление разнообразной коллекции белковых доменов, имеющих ту же «складку», что и белковая платформа, как это определено ниже, и (б) скрининг указанной разнообразной коллекции и/или выбор из указанной разнообразной коллекции для получения по крайней мере одного домена белка, обладающего указанным заданным свойством. Разнообразная коллекция белковых доменов может быть предоставлена с помощью нескольких методов в соответствии с используемой системой скрининга и/или выбора и может включать в себя использование методов, хорошо известных специалистам в данной области, таких как фаговый дисплей или рибосомный дисплей.
Термин «белковая платформа» означает белок с открытыми площадями поверхности, в которых хорошо переносятся аминокислотные вставки, замены или делеции. Примеры белковых платформ, которые могут быть использованы для создания связывающих доменов настоящего изобретения, представляют собой антитела или их фрагменты, такие как одноцепочечные Fv или Fab фрагменты, белок А из Staphylococcus aureus, билин-связывающий белок из Pieris brassicae или другие липокалины, анкириновые повторные белки или другие повторные белки и человеческий фибронектин. Белковые платформы известны специалистам в данной области (Binz et al., 2005, loc. cit.; Binz et al., 2004, loc. cit.).
Термин «заданное свойство» касается свойства, такого как связывание с мишенью, блокирование мишени, активация мишень-опосредованной реакции, ферментативная активность, и связанные с ними другие свойства. В зависимости от типа желаемого свойства, специалист с обычными навыками сможет определить формат и необходимые шаги для проведения скрининга и/или выбора связывающего домена с нужным свойством. Предпочтительно, указанное заданное свойство является связыванием с мишенью.
Предпочтительно, связывающий белок из изобретения не представляет собой антитело или его фрагмент, такой как Fab или ScFv фрагменты. Антитела и их фрагменты хорошо известны специалистам в данной области. Также желательно, чтобы связывающий домен изобретения не содержал иммуноглобулиновой складки, как в антителах и/или фибронектиновом домене типа III. Иммуноглобулиновая складка является характерной складкой для всех β-белков, которая состоит из 2-слойного сэндвича из примерно 7 антипараллельных β-нитей, расположенных в два β-листа. Иммуноглобулиновые складки хорошо известны специалистам в данной области. Например, такие связывающие домены, включающие иммуноглобулиновую складку, описаны в WO 07/080392 и WO 08/097497.
Далее предпочтительно, чтобы связывающий домен изобретения не содержал иммуноглобулин-подобный домен, который содержится в VEGFR-1 или VEGFR-2. Такие связывающие домены описаны в WO 00/075319.
Предпочтительный связывающий домен является связывающим доменом с антиангиогенным эффектом. Антиангиогенный эффект связывающего домена может быть определен анализами, которые хорошо известны специалисту в данной области, такими как анализ прорастания сфероидов HUVEC, описанный в Примере 2.
Далее предпочтительным является связывающий домен, содержащий от 70 до 300 аминокислот, в частности 100-200 аминокислот.
Далее предпочтительным является связывающий домен, лишенный свободного остатка Cys. Свободный остаток Cys не участвует в образовании дисульфидных связей. Еще более предпочтительным является связывающий домен без каких-либо остатков Cys.
Предпочтительный связывающий домен изобретения представляет собой повторный домен или спроектированный повторный домен, предпочтительно, как описано в WO 02/20565.
Особенно предпочтительным связывающим доменом является спроектированный анкириновый повторный домен (Binz, Н. К. et al., 2004, loc. cit.), желательно, как описано в WO 02/20565. Примеры спроектированных анкириновых повторных доменов показаны в примерах.
Определения в дальнейшем для повторных белков основаны на указанных в патентной заявке WO 02/20565. Патентная заявка WO 02/20565 дополнительно содержит общее описание характеристик, методов и применений повторных белков.
Термин «повторный белок» касается белка, включающего один или несколько повторных доменов. Предпочтительно, каждый из указанных повторных белков составляет до четырех повторных доменов. Более предпочтительно, каждый из указанных повторных белков содержит до двух повторных доменов. Наиболее предпочтительно, каждый из повторных белков включает в себя только один повторный домен. Кроме того, указанный повторный белок может содержать дополнительные неповторные белковые домены, полипептидные теги и/или полипептидные линкеры.
Термин «повторный домен» касается домена белка, состоящего из двух или более последовательных повторяющихся звеньев (модулей) в качестве структурных звеньев, причем указанные структурные звенья имеют ту же складку и компонуются плотно, для создания, например, сверхспиральной структуры, имеющей совместное гидрофобное ядро.
Термин «спроектированный повторный белок» и «спроектированный повторный домен» касается повторного белка или повторного домена, соответственно, полученного в результате изобретательской процедуры, объясненной в патентной заявке WO 02/20565. Спроектированные повторные белки и спроектированные повторные домены являются синтетическими, а не взятыми из природы. Они представляют собой полученные человеком белки или домены, соответственно, полученные путем экспрессии соответственно спроектированных нуклеиновых кислот. Предпочтительно, экспрессия осуществляется в эукариотических или прокариотических клетках, например бактериальных клетках, или с помощью бесклеточных систем экспрессии in vitro.
Термин «структурное звено» касается локально упорядоченной части полипептида, образованной путем трехмерных взаимодействий между двумя или более сегментами вторичной структуры, которые расположены вблизи друг друга вдоль полипептидной цепи. Такое структурное звено имеет структурный мотив. Термин «структурный мотив» касается трехмерного расположения элементов вторичной структуры, присутствующих хотя бы в одном структурном звене. Структурные мотивы хорошо известны специалистам в данной области. Структурные звенья сами по себе не в состоянии приобрести определенное трехмерное расположение, однако их последовательное расположение, например, в виде повторных модулей в повторном домене приводит к взаимной стабилизации соседних звеньев, что приводит к образованию сверхспиральной структуры.
Термин «повторное звено» касается аминокислотных последовательностей, включающих повторные мотивы последовательности из одного или нескольких природных повторных белков, причем указанные «повторные звенья» находятся в нескольких копиях, и которые обладают определенной топологией складывания, общей для всех указанных мотивов, определяющих складывание белка. Такие повторяющиеся звенья содержат каркасные остатки и остатки взаимодействия. Примерами таких повторяющихся единиц являются повторяющиеся звенья броненосца, лейцин-богатые повторяющиеся звенья, анкириновые повторяющиеся звенья, тетратрикопептидные повторяющиеся звенья, повторяющиеся звенья HEAT, и богатые лейцином вариантные повторяющиеся звенья. Природные белки, содержащие два или более таких повторяющихся звеньев, называют «природными повторными белками». Аминокислотные последовательности отдельных повторных звеньев повторного белка могут иметь значительное число мутаций, замен, добавлений и/или делеций при сравнении друг с другом, в то же время существенно сохраняя общий характер или мотив из повторяющихся звеньев.
Термин «каркасные остатки» касается аминокислотных остатков повторяющихся звеньев или соответствующих аминокислотных остатков повторных модулей, которые способствуют топологии складывания, т.е. которые вносят вклад в складывание указанного повторного звена (или модуля), или которые способствуют взаимодействию с соседним звеном (или модулем). Такой вклад может представлять собой взаимодействие с другими остатками в повторном звене (модуле), или влияние на конформацию полипептидной основы, которые содержатся в α-спирали или β-листах, или аминокислотных отрезков, образующих линейные полипептиды или петли.
Термин «остатки мишеневого взаимодействия» касается аминокислотных остатков повторяющихся звеньев или соответствующих аминокислотных остатков повторный модулей, которые способствуют взаимодействию с мишеневыми веществами. Такой вклад может представлять собой прямое взаимодействие с мишеневыми веществами, или влияние на другие, непосредственно взаимодействующие остатки, например, путем стабилизации конформации полипептида повторного звена (модуля), чтобы разрешить или повысить взаимодействие непосредственно взаимодействующих остатков с указанной мишенью. Такие остатки взаимодействия каркаса и мишени могут быть идентифицированы путем анализа структурных данных, полученных физико-химическими методами, такими как рентгеновская кристаллография, ЯМР и/или CD-спектроскопия, или путем сравнения с известной и связанной структурной информацией, хорошо известной практикующим специалистам в структурной биологии и/или биоинформатике.
Предпочтительно, чтобы повторные звенья, используемые для вывода повторного мотива последовательности, представляли собой гомологичные повторяющиеся звенья, в которых повторяющиеся звенья содержат тот же структурный мотив и в которых более 70% каркасных остатков указанных повторяющихся звеньев гомологичны друг другу. Предпочтительно, более 80% каркасных остатков указанных повторяющихся звеньев являются гомологичными. Наиболее предпочтительно, более 90% каркасных остатков указанных повторяющихся звеньев являются гомологичными. Компьютерные программы для определения процента гомологии между полипептидами, такие как Fasta, Blast или Gap, известны специалисту в данной области. Далее предпочтительно, чтобы повторные звенья, используемые для вывода повторного мотива последовательности, представляли собой гомологичные повторяющиеся звенья, полученные из повторных доменов, выбранных на мишени, например, как описано в Примере 1 и имеющих ту же мишень-специфичность.
Термин «повторный мотив последовательности» касается аминокислотной последовательности, которая выводится из одного или нескольких повторяющихся звеньев. Предпочтительно, указанные повторные звенья получают из повторных доменов, имеющих специфичность связывания для одной и той же мишени. Такие повторные мотивы последовательности содержат положения каркасных остатков и положения остатков мишеневых взаимодействий. Указанные положения каркасных остатков соответствуют положениям остатков мишеневого взаимодействия повторяющихся звеньев. Кроме того, указанные положения остатков мишеневого взаимодействия соответствуют положениям остатков мишеневого взаимодействия повторяющихся звеньев. Повторные мотивы последовательности содержат фиксированные положения и рандомизированные положения. Термин «фиксированное положение» касается положения аминокислоты в повторном мотиве последовательности, причем указанное положение установлено в конкретной аминокислоте. Чаще всего такие фиксированные положения соответствуют положениям каркасных остатков и/или положениям остатков мишеневых взаимодействий, которые являются специфическими для определенной мишени. Термин «рандомизированное положение» означает положение аминокислоты в повторном мотиве последовательности, в котором две или более аминокислот допускаются в указанном положении аминокислоты, например, в котором допускается любая из обычных двадцати природных аминокислот или в котором допускается большинство из двадцати природных аминокислот, таких как аминокислоты, отличные от цистеина, или аминокислоты, отличные от глицина, цистеина и пролина. Чаще всего такие рандомизированные положения соответствуют положениям остатков мишеневых взаимодействий. Однако могут быть также рандомизированы некоторые положения каркасных остатков.
Термин «топология складывания» касается третичной структуры указанных повторяющихся единиц. Топология складывания будет определяться участками аминокислот, образующими по крайней мере части α-спиралей или β-листов, или аминокислотных отрезков, образующих линейные полипептиды или петли, или любую комбинацию α-спиралей, β-листов и/или линейных полипептидов/петель.
Термин «последовательный» касается расположения, в котором повторяющиеся звенья или повторные модули расположены в тандеме. В спроектированных повторных белках существует по крайней мере 2, как правило, от 2 до 6, в частности по меньшей мере, 6, часто 20 или более повторных звеньев. В большинстве случаев повторяющиеся звенья будут обладать высокой степенью идентичности последовательности (те же самые аминокислотные остатки в соответствующих положениях) или сходством последовательности (аминокислотные остатки разные, но имеют сходные физико-химические свойства), а некоторые из аминокислотных остатков могут быть ключевыми остатками, которые сильно сохраняются в различных повторных звеньях, обнаруженных в природных белках. Однако высокая степень изменчивости последовательности аминокислотными вставками и/или делециями, и/или заменами между различными повторными звеньями, обнаруженными в природных белках, будет возможной до тех пор, пока поддерживается общая топология складывания.
Методы прямого определения топологии складывания повторных белков физико-химическими средствами, такими как рентгеновская кристаллография, ЯМР-спектроскопия или CD-спектроскопия, хорошо известны практикующим специалистам в данной области. Методы для выявления и определения повторяющихся звеньев или повторных мотивов последовательности или для выявления семейств родственных белков, включающих такие повторяющиеся звенья или мотивы, как поиск гомологии (BLAST и др.), хорошо зарекомендовали себя в области биоинформатики и хорошо известны практикующим специалистам в данной области. Шаг очистки начального повторного мотива последовательность может включать итеративный процесс.
Термин «повторные модули» касается повторных аминокислотных последовательностей спроектированных повторных доменов, которые изначально получены из повторных звеньев природных повторных белков. Каждый модуль, содержащийся в повторном домене, происходит от одного или нескольких повторяющихся звеньев семейства или подсемейства природных повторных белков, например семейства повторных белков броненосца или анкириновых повторных белков.
«Повторные модули» могут включать положения с аминокислотными остатками, присутствующими во всех копиях соответствующих повторных модулей («фиксированные положения»), и положения с различными или «рандомизированными» аминокислотными остатками («рандомизированные положения»).
Термин «кеппинг-модуль» касается полипептида, слитого с N- или С-терминальным повторным модулем повторного домена, в котором указанный кеппинг-модуль образует жесткие третичные взаимодействия с указанным повторным модулем, тем самым обеспечивая покрытие, прикрывающее гидрофобное ядро указанного повторного модуля на стороне, не контактирующей с последовательным повторным модулем, от растворителя. Указанный N- и/или С-терминальный «кеппинг-модуль» может быть получен из кеппинг-единицы или другого домена, обнаруженного в природном повторном белке, соседствующем с повторным звеном. Термин «кеппинг-единица» касается природного сложенного полипептида, в котором указанный полипептид определяет особенности структурной единицы, которая N- или С-терминально слитая с повторным звеном, причем указанный полипептид формирует жесткие третичные взаимодействия с указанным повторным звеном, тем самым обеспечивая покрытие, прикрывающее гидрофобное ядро указанного повторного звена с одной стороны от растворителя. Такие кеппинг-единицы могут иметь сходство с последовательностью указанного повторного мотива последовательности. Кеппинг-модули и кеппинг-повторы описаны в WO 02/020565. Например, N-терминальный кеппинг-модуль SEQ ID NO: 21 кодируется аминокислотами из положения с 1 по 32.
Также предпочтительным является такой N-терминальный кеппинг-модуль, содержащий глициновый или аспартатный остаток в положении 5.
Термин «мишень» касается отдельной молекулы, такой как молекула нуклеиновой кислоты, полипептид или белок, углевод или любая другая природная молекула, включая любую часть такой отдельной молекулы или комплексы двух или более таких молекул. Мишень может представлять собой целую клетку или образец ткани, или может представлять собой любую не-природную молекулу или фрагмент. Предпочтительно, мишень представляет собой природный или неприродный полипептид или полипептид, содержащий химические модификации, например, модифицированный путем природного или неприродного фосфорилирования, ацетилирования или метилирования. В частном применении настоящего изобретения мишень представляет собой VEGF-Axxx или VEGFR-2.
Термин «консенсусная последовательность» касается аминокислотной последовательности, причем указанная консенсусная последовательность получается путем структурного выравнивания и/или выравнивания последовательности нескольких повторяющихся звеньев. Используя два или более структурных и/или выравненных по последовательности повторяющихся звеньев и учитывая пробелы в выравнивании, можно определить наиболее частые аминокислотные остатки в каждом положении. Консенсусная последовательность представляет собой последовательность, которая содержит аминокислоты, которые чаще всего представлены в каждом положении. В случае, если две или более аминокислот представлены выше среднего в одном положении, консенсусная последовательность может включать в себя подгруппу этих аминокислот. Указанные два или несколько повторяющихся звеньев могут быть взяты из повторяющихся звеньев, входящих в один повторный белок, или из двух или более различных повторных белков.
Консенсусные последовательности и методы их определения хорошо известны специалистам в данной области.
«Консенсусный аминокислотный остаток» представляет собой аминокислоту, обнаруженную в определенном положении в консенсусной последовательности. Если два или более, например три, четыре или пять, аминокислотных остатков обнаружены с аналогичными вероятностями в указанных двух или более повторяющихся звеньях, консенсусная аминокислота может представлять собой одну из наиболее часто встречающихся аминокислот или комбинацию указанных двух или более аминокислотных остатков.
Далее предпочтительными являются неприродные связывающие белки или связывающие домены.
Термин «неприродный» означает синтетический или не из природы, в частности, термин означает сделанный руками человека. Термин «неприродный связывающий белок» или «неприродный связывающий домен» означает, что указанный связывающий белок или указанный связывающий домен является синтетическим (т.е. производится путем химического синтеза из аминокислот) или рекомбинантным, а не из природы. «Неприродный связывающий белок» или «неприродный связывающий домен» представляет собой искусственный белок или домен, соответственно, полученный путем экспрессии соответственно спроектированных нуклеиновых кислот. Предпочтительно, экспрессия осуществляется в эукариотических или бактериальных клетках, или с помощью бесклеточных систем экспрессии in vitro. Кроме того, термин означает, что последовательность указанного связывающего белка или указанного связывающего домена не присутствует в виде ввода не-искусственной последовательности в базу данных последовательностей, например, в GenBank, EMBL-Bank или Swiss-Prot. Эти базы данных и другие аналогичные базы данных последовательностей хорошо известны специалистам в данной области.
Изобретение касается связывающего белка, включающего связывающий домен, в котором указанный связывающий домен ингибирует связывание VEGF-Axxx с VEGFR-2 и в котором указанный связывающий белок и/или связывающий домен имеет температуру денатурации (Tm) средней точки выше 40°С при термическом разворачивании и образует менее 5% (по весу) нерастворимых агрегатов при концентрации до 10 г/л при инкубации при температуре 37°С в течение 1 дня в фосфатном буферном растворе (PBS).
Связывающий домен может ингибировать связывание VEGF-Axxx с VEGFR-2 либо путем связывания с VEGF-Axxx или путем связывания с VEGFR-2 таким образом, что очевидная константа диссоциации (Kd) между VEGF-Axxx и VEGFR-2 увеличивается более 102 -кратно, желательно более чем 103-кратно, более предпочтительно, чем 104-кратно, более предпочтительно, чем 105-кратно, и наиболее предпочтительно более чем 106-кратно. Предпочтительно, Kd для взаимодействия связывающего домена либо с VEGF-Axxx, либо с VEGFR-2 ниже 10-7 М, предпочтительно ниже 10-8 М, более предпочтительно ниже 10-9 М, более предпочтительно ниже 10-10 М и наиболее предпочтительно ниже 10-11 М. Методы для определения константы диссоциации белок-белковых взаимодействий, такие как технологии на основе поверхностного плазмонного резонанса (SPR), хорошо известны специалистам в данной области.
Предпочтительный связывающий домен связывает VEGF-Axxx. Еще более предпочтительным является связывающий домен, который связывает человеческий VEGF-A165.
Термин «PBS» означает фосфатный буферный раствор, содержащий 137 мМ NaCl, 10 мМ фосфат и 2,7 мМ KCl и с рН 7,4.
Предпочтительно, связывающий белок и/или связывающий домен имеет температуру денатурации (Tm) средней точки выше 45°С, более предпочтительно выше 50°С, более предпочтительно выше 55°С, а наиболее предпочтительно выше 60°С при термическом разворачивании. Связывающий белок или связывающий домен изобретения обладает определенной вторичной и третичной структурой при физиологических условиях. Тепловое развертывание такого полипептида приводит к потере его третичной и вторичной структуры, которое может сопровождаться, например, путем измерений кругового дихроизма (CD). Температура денатурации средней точки связывающего белка или связывающего домена при тепловом разворачивании соответствует температуре в средней точке кооперативного перехода в физиологическом буфере при тепловой денатурации указанного белка или домена путем медленного повышения температуры от 10°С до 100°С. Определение температуры денатурации средней точки при термическом разворачивании хорошо известно специалистам в данной области. Эта температура денатурации средней точки связывающего белка или связывающего домена при тепловом разворачивании свидетельствует о термической стабильности указанного полипептида.
Также предпочтительным является связывающий белок и/или связывающий домен, формирующий менее чем 5% (по весу) нерастворимых агрегатов при концентрации до 20 г/л, предпочтительно до 40 г/л, более предпочтительно до 60 г/л, еще более предпочтительно до 80 г/л и наиболее предпочтительно до 100 г/л при инкубации в течение 5 дней, желательно в течение 10 дней, более предпочтительно в течение 20 дней, более предпочтительно в течение 40 дней и наиболее предпочтительно в течение 100 дней при 37°С в PBS. Образование нерастворимых агрегатов может быть обнаружено путем появления видимых осадков, гель-фильтрации или динамического рассеяния света, что сильно увеличивается при образовании нерастворимых агрегатов.
Нерастворимые агрегаты могут быть удалены из образца белка путем центрифугирования при 10'000xg в течение 10 минут. Предпочтительно, связывающий белок и/или связывающий домен образует менее 2%, 1%, 0,5%, 0,2%, 0,1%, или на 0,05% (по весу) нерастворимых агрегатов при указанных условиях инкубации при 37°С в PBS. Процент нерастворимых агрегатов может быть определен путем отделения нерастворимых агрегатов от растворимого белка, что сопровождается определением количества белка в растворимой и нерастворимой фракции путем стандартных методов количественной оценки.
Также предпочтительным является связывающий белок и/или связывающий домен, который не теряет своей родной трехмерной структуры при инкубации в PBS, содержащем 100 мМ дитиотреитола (DTT) в течение 1 или 10 часов при 37°С.
В одном конкретном варианте осуществления изобретение касается связывающего белка, включающего связывающий домен, ингибирующий связывание VEGF-Axxx с VEGFR-2, и имеющего указанную или предпочтительную температуру денатурации средней точки и не агрегирующие свойства, определенные выше, причем указанный связывающий белок подавляет прорастание HUVEC сфероидов со значением IC50 ниже 100 нМ.
Термин «HUVEC» означает эндотелиальные клетки пупочной вены человека (human umbilical vein endothelial cells - HUVEC), которые могут быть изолированы из нормальной человеческой пуповинной вены и которые отвечают за VEGF-A стимуляцию. Анализы для измерения прорастания сфероидов HUVEC, такие как описаны в Примере 2, хорошо известны специалистам в данной области.
Значение IC50 представляет собой концентрацию вещества, такого как связывающий белок или связывающий домен, которая требуется для 50% ингибирования in vitro экспериментального определенного параметра, такого как прорастание сфероидов HUVEC. Значения IC50 могут быть легко определены специалистом в данной области (KorffT. and Augustin H.G., J. Cell Biol. 143(5), 1341-52, 1998).
Предпочтительным является связывающий белок и/или связывающий домен, который ингибирует прорастание HUVEC сфероида со значением IC50 менее 10 нМ, предпочтительно менее 1 нМ, более предпочтительно менее 0,1 нМ, а наиболее предпочтительно ниже 0,05 нМ.
Далее предпочтительным является мономерный связывающий белок и/или связывающий домен, который ингибирует прорастание сфероидов HUVEC со значением IC50, ниже, чем соответствующее значение IC50 ранибизумаба (Lucentis®, зарегистрированная торговая марка компании Genentech), бевацизумаба (Авастин®, зарегистрированная торговая марка Genentech), афлиберцепт (VEGF Trap®, зарегистрированная торговая марка Regeneron) или пегаптаниб (Macugen®, зарегистрированная торговая марка компании Pfizer).
В частности, изобретение касается связывающего белка, включающего связывающий домен, ингибирующий связывание VEGF-Axxx с VEGFR-2 и имеющие указанную или предпочтительную температуру денатурации средней точки и не агрегирующие свойства, определенные выше, причем Kd для взаимодействия указанного связывающего домена с VEGF-Axxxb, по крайней мере, в 10 раз выше по сравнению с Kd для взаимодействия указанного связывающего домена с соответствующим VEGF-Axxx.
Предпочтительно, Kd для взаимодействия связывающего домена с VEGF-Axxxb, по крайней мере, в 102 раза выше, желательно в 103 раза выше, более предпочтительно в 104 раза выше, более предпочтительно в 105 раза выше, а еще лучше в 106 раза выше по сравнению с Kd для взаимодействия связывающего домена с соответствующим VEGF-Axxx.
Также желательно, Kd для взаимодействия связывающего домена с VEGF-Axxxb выше 103 нМ и Kd для взаимодействия связывающего домена с VEGF-Axxx ниже 10 или 1 нМ.
Kd для взаимодействия предпочтительного связывающего домена с VEGF-B, VEGF-C, VEGF-D, PIGF или PDGF превышает 1 нМ, предпочтительно выше 10 нМ, более предпочтительно выше 102 нМ, еще более предпочтительно выше 103 нМ, а наиболее предпочтительно выше 104 нМ.
Предпочтительно, VEGF-Axxx является либо собачьим VEGF-A164 или обезьяним VEGF-A165 или человеческим VEGF-A165, и VEGF-Axxxb является либо собачьим VEGF-A164b, или обезьяньим VEGF-A165b, или человеческим VEGF-A165b.
Другим предпочтительным вариантом осуществления является рекомбинантный связывающий белок, содержащий связывающий домен, причем указанный связывающий домен ингибирует связывание VEGF-Axxxx с VEGFR-2 и причем указанный связывающий домен является повторным доменом или спроектированным повторным доменом. Такой повторный домен может содержать один, два, три или более внутренних повторных модулей, которые будут принимать участие в связывании с VEGF-Axxx. Предпочтительно, такой повторный домен включает в себя N-терминальный кеппинг-модуль, от двух до четырех внутренних повторных модулей и С-терминальный кеппинг-модуль. Предпочтительно, указанный связывающий домен является анкириновым повторным доменом или спроектированным анкириновым повторным доменом.
Предпочтительным является рекомбинантный белок, причем анкириновый повторный домен или указанный спроектированный анкириновый повторный домен включает в себя повторный модуль с анкириновым повторным мотивом последовательности
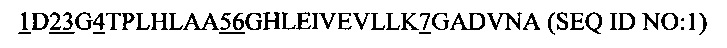
где 1, 2, 3, 4, 5, 6 и 7, представляют, независимо друг от друга, аминокислотный остаток, выбранный из группы A, D, Е, F, Н, I, К, L, М, N, Q, R, S, Т, V, W и Y.
Особенно предпочтительным является рекомбинантный связывающий белок, причем указанный анкириновый повторный домен или указанный спроектированный анкириновый повторный домен включает в себя повторный модуль с анкириновым повторным мотивом последовательности
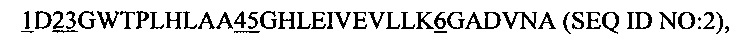
где
1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из F, Т, N, R, V, А, I, К, Q, S и Y, предпочтительно F, Т, N, R и V; более предпочтительно F и Т;
2 представляет аминокислотный остаток, выбранный из группы, состоящей из W, Y, Н и F, предпочтительно W, Y и Н;
3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из М, I, F и V; предпочтительно М и I;
4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Н, А, К, G, L, М, N, Т, V, W и Y, предпочтительно Н, А и К;
5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Е, Y, F, V, Н, I, L, N и R, предпочтительно Е, Y, F, V и Н, более предпочтительно Е, Y, F и V; и
6 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, N, Y, Н и R.
Далее предпочтительным является рекомбинантный связывающий белок, причем указанный анкириновый повторный домен или указанный спроектированный анкириновый повторный домен включает в себя повторный модуль с анкириновым повторным мотивом последовательности

где
1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Т, E, A, D, F, К, N, Q, R, S, W и Y, предпочтительно Т и Е;
2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из V, F, Y, А, Н, I, К, R, Т и W; предпочтительно V, F и Y;
3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из S, A, N, F и М; предпочтительно S, А и N, более предпочтительно S и А;
4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Y, F, S и W;
5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, S, L и Y; предпочтительно А и S;
6 представляет собой аминокислотный остаток, выбранный из группы, состоящей из D, N, М, А, I, К и Y; предпочтительно D, N и М; более предпочтительно D и N;
7 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, Y, Н, N и D, а также
8 представляет собой аминокислотный остаток Т или А.
Далее предпочтительным является рекомбинантный связывающий белок, причем указанный анкириновый повторный домен или указанный спроектированный анкириновый повторный домен включает в себя повторный модуль с анкириновым повторным мотивом последовательности

где
1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из К, Т и Y;
2 представляет собой аминокислотный остаток N или М;
3 представляет собой аминокислотный остаток Т или F;
4 представляет собой аминокислотный остаток S или А;
5 представляет собой аминокислотный остаток Н или R;
6 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, Y, H и N, и
7 представляет собой аминокислотный остаток А или Т.
Еще более предпочтительным является рекомбинантный связывающий белок, причем указанный анкириновый повторный домен или указанный спроектированный анкириновый повторный домен включает в себя повторный модуль с анкириновым повторным мотивом последовательности SEQ ID NO: 3, причем указанному повторному модулю предшествует повторный модуль с анкириновым повторным мотивом последовательности SEQ ID NO: 2 и/или он сопровождается повторным модулем с анкириновым повторным мотивом последовательности SEQ ID NO: 4.
Далее предпочтительным является рекомбинантный связывающий белок, причем указанный анкириновый повторный домен или указанный спроектированный анкириновый повторный домен включает в себя повторный модуль с анкириновым повторным мотивом последовательности

где
1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, N, R, V, Y, Е, Н, I, К, L, Q, S и Т, предпочтительно A, N, R, V и Y; более предпочтительно А и R;
2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из S, A, N, R, D, F, L, Р, Т и Y, предпочтительно S, А, N и R;
3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Т, V, S, A, L и F; предпочтительно Т, V, S, А и L, более предпочтительно, Т, V и S;
4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из W, F и H;
5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Р, I, A, L, S, Т, V и Y, предпочтительно Р и I;
6 представляет собой аминокислотный остаток, выбранный из группы, состоящей из W, F, I, L, T и V;
7 представляет собой аминокислотный остаток L или Р, и
8 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, Н, N и Y.
Далее предпочтительным является рекомбинантный связывающий белок, причем указанный анкириновый повторный домен или указанный спроектированный анкириновый повторный домен включает в себя повторный модуль с анкириновым повторным мотивом последовательности

где
1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Н, Q, А, К, R, D, I, L, М, N, V и Y, предпочтительно Н, Q, А, К и R; более предпочтительно А и R;
2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Y, F и H;
3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Q, F и T;
4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из W, М, G, Н, N и Т, предпочтительно W и М;
5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Т, А, М, L и V, предпочтительно Т, А и М;
6 представляет собой аминокислотный остаток, выбранный из группы, состоящей из I, L, V, D и Т, предпочтительно I, L и V, и
7 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, Н, N и Y.
Еще более предпочтительным является рекомбинантный связывающий белок, причем указанный анкириновый повторный домен или указанный спроектированный анкириновый повторный домен включает в себя повторный модуль с анкириновым повторным мотивом последовательности SEQ ID NO: 6, причем повторному модулю предшествует повторный модуль с анкириновым повторным мотивом последовательности SEQ ID NO: 5.
Далее предпочтительным является рекомбинантный белок, причем указанный анкириновый повторный домен или указанный спроектированный анкириновый повторный домен включает в себя повторный модуль с анкириновым повторным мотивом последовательности
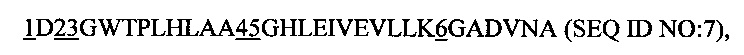
где
1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из К, М, N, R и V;
2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Y, Н, М и V;
3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из F, L, М и V;
4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из R, Н, V, А, К и N, предпочтительно R, Н, V и А;
5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из F, D, Н, Т, У, М и К; предпочтительно F, D, Н, Т и Y, а
6 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, H, N и Y.
Далее предпочтительным является рекомбинантный связывающий белок, причем указанный анкириновый повторный домен или указанный спроектированный анкириновый повторный домен включает в себя повторный модуль с анкириновым повторным мотивом последовательности

где
1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Т, А, Н, I, N и S;
2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из F, I, Q, R, V и N;
3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, G, N, Q и V;
4 представляет собой аминокислотный остаток W или Y;
5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, S, Т и М;
6 представляет собой аминокислотный остаток, выбранный из группы, состоящей из N, V, S, F, М и W;
7 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, H, N и Y, а
8 представляет собой аминокислотный остаток Т или А.
Далее предпочтительным является рекомбинантный связывающий белок, причем указанный анкириновый повторный домен или указанный спроектированный анкириновый повторный домен включает в себя повторный модуль с анкириновым повторным мотивом последовательности

где
1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из К, А, V и N;
2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из N, I и Y;
3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Т, F, Y и W;
4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из W, D и Y;
5 представляет собой аминокислотный остаток S или А;
6 представляет собой аминокислотный остаток, выбранный из группы, состоящей из D, I и М;
7 представляет собой аминокислотный остаток, выбранный из группы, состоящей из L, Т и Y, a
8 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, Н, Y и N.
Еще более предпочтительным является рекомбинантный связывающий белок, причем указанный анкириновый повторный домен или указанный спроектированный анкириновый повторный домен включает в себя повторный модуль с анкириновым повторным мотивом последовательности SEQ ID NO: 8, причем указанному повторному модулю предшествует повторный модуль с анкириновым повторным мотивом последовательности SEQ ID NO: 7 и/или он сопровождается повторным модулем с анкириновым повторным мотивом последовательности SEQ ID NO: 9.
Далее предпочтительным является рекомбинантный связывающий белок, причем указанный анкириновый повторный домен или указанный спроектированный анкириновый повторный домен включает в себя повторный модуль с анкириновым повторным мотивом последовательности
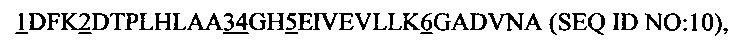
где
1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из L, S и T;
2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из G, S и С, предпочтительно G и S;
3 представляет собой аминокислотный остаток S или А;
4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Q, S, М и N, предпочтительно Q и S;
5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из L, М и Q, и
6 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, Н, N, Y и D.
Далее предпочтительным является рекомбинантный связывающий белок, причем указанный анкириновый повторный домен или указанный спроектированный анкириновый повторный домен включает в себя повторный модуль с анкириновым повторным мотивом последовательности

где
1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Y, Н, F, I, L и W, предпочтительно Y и Н;
2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из М, D, I, L, V, предпочтительно М и D;
3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из G, S и V;
4 представляет собой аминокислотный остаток W или F;
5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, G и T;
6 представляет собой аминокислотный остаток D или W;
7 представляет собой аминокислотный остаток L или F, а
8 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, Н, N и Y.
Еще более предпочтительным является рекомбинантный связующий белок, причем указанный анкириновый повторный домен или указанный спроектированный анкириновый повторный домен включает в себя повторный модуль с анкириновым повторным мотивом последовательности SEQ ID NO: 11, причем указанному повторному модулю предшествует повторный модуль с анкириновым повторным мотивом последовательности SEQ ID NO: 10.
Далее предпочтительным является рекомбинантный связывающий белок, причем указанный анкириновый повторный домен или указанный спроектированный анкириновый повторный домен включает в себя повторный модуль с анкириновым повторным мотивом последовательности
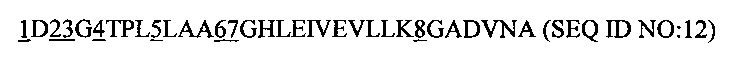
где
1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из К, S, I, N, Т и V, предпочтительно К и S;
2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из К, N, W, А, Н, М, Q и S; предпочтительно К и N;
3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из F, Q, L, Н и V, предпочтительно F, Q и L;
4 представляет собой аминокислотный остатка F или Т;
5 представляет собой аминокислотный остаток Q или Н;
6 представляет собой аминокислотный остаток Y или S;
7 представляет собой аминокислотный остаток, выбранный из группы, состоящей из N, Н, Y и М; предпочтительно N и Н, а
8 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, Н, N и Y.
Далее предпочтительным является рекомбинантный связывающий белок, причем указанный анкириновый повторный домен или указанный спроектированный анкириновый повторный домен включает в себя повторный модуль с анкириновым повторным мотивом последовательности

где
1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из F, Y, Н и W; предпочтительно F, Y и Н;
2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из I, М, D и V; предпочтительно I, М и D;
3 представляет собой аминоксилотный остаток F или L;
4 представляет собой аминокислотный остаток L или Р;
5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Н, L и Y, a
6 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, Н, N, С и Y.
Еще более предпочтительным является рекомбинантный связывающий белок, причем указанный анкириновый повторный домен или указанный спроектированный анкириновый повторный домен включает в себя повторный модуль с анкириновым повторным мотивом последовательности SEQ ID NO: 13, причем указанному повторному модулю предшествует повторный модуль с анкириновым повторным мотивом последовательности SEQ ID NO: 12.
Другим предпочтительным вариантом является рекомбинантный связывающий белок, содержащий по крайней мере один повторный домен со специфичностью связывания к VEGF-Axxx, причем указанный повторный домен конкурирует за связывание с VEGF-Axxx с повторным доменом, выбранным из группы, состоящей из SEQ ID NO: 16, 22, 23, 29, 30 и 33, или повторным доменом, выбранным из группы, состоящей из SEQ ID NO: 16,22,23,29, 30, 33, 34, 36, 39 и 40.
Термин «конкурируют за связывание» означает неспособность двух различных связывающих доменов изобретения связываться одновременно с той же мишенью, в то время как оба из них способны связываться с этой же мишенью индивидуально. Таким образом, два таких связывающих домена конкурируют за связывание с указанной мишенью. Методы, такие как конкурентный ИФА или конкурентные SPR измерения (например, с помощью инструмента Proteon от BioRad), для определения, конкурируют ли два связывающих домена за связывание с мишенью, хорошо известны практикующим специалистам в данной области.
Рекомбинантный белок, который конкурирует за связывание с VEGF-Axxx с выбранным повторным белком, может быть определен методами, хорошо известными специалисту с навыками в данной области, такими как конкурентный иммуноферментный анализ (ИФА).
Другим предпочтительным вариантом является рекомбинантный связывающий белок, содержащий повторный домен со специфичностью связывания к VEGF-Axxx, выбранный из группы, состоящей из SEQ ID NO: 14-33, или выбранный из группы, состоящей из SEQ ID NO: 14-40.
Далее предпочтительным является рекомбинантный связывающий белок, причем повторный домен со специфичностью связывания к VEGF-Axxx содержит аминокислотную последовательность, которая имеет по крайней мере 70% идентичность аминокислотной последовательности с повторным доменом указанной группы повторных доменов. Предпочтительно, указанная идентичность аминокислотной последовательности составляет, по крайней мере 75%, более предпочтительно 80%, более предпочтительно 85%, более предпочтительно 90%, а наиболее предпочтительно 95%.
Далее предпочтительным является рекомбинантный связывающий белок, причем указанный повторный домен со специфичностью связывания к VEGF-Axxx включает повторный модуль, который имеет по крайней мере 70% идентичность аминокислотной последовательности с повторным модулем повторного домена указанной группы повторных доменов. Предпочтительно, указанная идентичность аминокислотной последовательности составляет, по крайней мере 75%, более предпочтительно 80%, более предпочтительно 85%, более предпочтительно 90%, а наиболее предпочтительно 95%.
В предпочтительном варианте осуществления рекомбинантного связывающего белка, содержащего повторный домен в соответствии с настоящим изобретением, один или несколько аминокислотных остатков повторных модулей указанного повторного домена обмениваются на аминокислотный остаток, обнаруженный в соответствующем положении при выравнивании повторного звена. Предпочтительно, до 30% аминокислотных остатков обмениваются, более предпочтительно, до 20%, и даже более предпочтительно, до 10% аминокислотных остатков обмениваются. Наиболее предпочтительно, такое повторное звено является природным повторным звеном. Еще более предпочтительно, указанный повторный домен имеет специфичность связывания к VEGF-Axxx или VEGFR-2.
В еще одном частном варианте осуществления, до 30% аминокислотных остатков, более предпочтительно, до 20%, и даже более предпочтительно, до 10% аминокислотных остатков обмениваются на аминокислоты, которые не встречаются в соответствующем положении повторяющихся звеньев.
В других вариантах осуществления, любой из VEGF-Axxx связывающих белков или доменов, описанных авторами, может быть ковалентно связан с одним или несколькими дополнительными фрагментами, в том числе, например, фрагментом, который также связывается с VEGFR-2 (например, VEGFR-2 связывающий полипептид), фрагментом, который связывается с другой мишенью, такой как PIGF, человеческий сывороточный альбумин, клеточный рецептор (например, Her2), иммуноглобулин (например, IgG1), цитокин (например, TNF-альфа или интерлейкин) или фактор роста для создания связующего агента с двойной специфичностью, фрагмент для мечения (например, флуоресцентная метка, такая как флуоресцеин, или радиоактивный индикатор), фрагмент, который облегчает очистку белка (например, небольшой пептидный тег, такой, как His- или strep-тег), фрагмент, который обеспечивает эффекторные функции для улучшения терапевтической эффективности (например, Fc часть антител для обеспечения антител-зависимой клеточной цитотоксичности, токсичный фрагмент белка, такой как экзотоксин А (ЕТА) Pseudomonas aeruginosa или небольшой молекулярный токсичный агент, такой как майтанзиноиды или ДНК алкилирующие агенты) или фрагмент, который обеспечивает улучшенную фармакокинетику. Улучшенные фармакокинетические свойства могут быть оценены в соответствии с воспринимаемой терапевтической необходимостью. Часто желательно увеличить биодоступность и/или увеличить время между дозами, возможно, за счет увеличения времени, в течение которого белок остается доступным в сыворотке крови после приема препарата. В некоторых случаях желательно улучшить непрерывность сывороточной концентрации белка с течением времени (например, уменьшение разницы в сывороточной концентрации белка вскоре после приема и незадолго до следующего введения). Фрагменты, которые имеют тенденцию к медленному клиренсу белка из крови, включают гидроксиэтилкрахмал (ГЭК), полиэтиленгликоль (ПЭГ), сахара (например, сиаловые кислоты), хорошо переносимые белковые фрагменты (например, Fc фрагмент или сывороточный альбумин), а также связывающие домены или пептиды со специфичностью и сродством к доминирующим сывороточным белкам, таким как Fc фрагменты антитела или сывороточный альбумин. Рекомбинантный связывающий белок изобретения может быть присоединен к фрагменту, который уменьшает скорость клиренса полипептидов у млекопитающих (например, у мыши, крысы или человека) больше чем в три раза по сравнению с немодифицированными полипептидами.
Один или несколько фрагментов полиэтиленгликоля могут быть присоединены в различных положениях в связывающем белке и такое присоединение может быть достигнуто в результате реакции с аминами, тиолами или другими подходящими реактивными группами. Присоединение фрагментов полиэтиленгликоля (пегилирование) может быть сайт-направленным, в котором подходящая реактивная группа вводится в белок, чтобы создать сайт, в котором преимущественно происходит пегилирование, или изначально присутствует в связывающем белке. Тиоловая группа может находиться на остатке цистеина, а аминогруппа может представлять собой, например, первичный амин, обнаруженный на N-конце полипептида, или аминогруппу, присутствующую в боковой цепи аминокислоты, такой как лизин или аргинин. В предпочтительном варианте осуществления, связывающий белок модифицируется так, чтобы содержать остаток цистеина в нужном положении, что позволяет осуществление сайт-направленного пегилирования на цистеине, например, путем реакции с производным полиэтиленгликоля, обладающим функцией малеимида. Фрагмент полиэтиленгликоля может сильно отличаться по молекулярной массе (т.е. примерно от 1 кДа до 100 кДа) и может быть разветвленным или линейным. Предпочтительно, молекулярная масса полиэтиленгликоля составляет от около 1 до около 50 кДа, предпочтительно от около 10 до около 40 кДа, еще более предпочтительно от около 15 до около 30 кДа и наиболее предпочтительно около 20 кДа.
В другом варианте осуществления, изобретение касается молекул нуклеиновой кислоты, кодирующих конкретные рекомбинантные связывающие белки. Кроме того, рассматривается вектор, содержащий указанную молекулу нуклеиновой кислоты.
Кроме того, рассматривается фармацевтическая композиция, содержащая один или несколько из вышеупомянутых связывающих белков, в частности рекомбинантные связывающие белки, содержащие повторные домены, или молекулы нуклеиновых кислот, кодирующие специфичные рекомбинантные связывающие белки, и, возможно, фармацевтический приемлемый носитель и/или разбавитель. Фармацевтические приемлемые носители и/или растворители известны специалисту с навыками в данной области и более подробно описаны ниже. Еще дополнительно, рассматривается диагностическая композиция, содержащая один или несколько из вышеупомянутых рекомбинантных связывающих белков, в частности связывающих белков, содержащих повторные домены.
Связывающий белок изобретения подавляет или предотвращает VEGF индуцированный патологический ангиогенез, сосудистые утечки (отек), легочную гипертензию, образование опухолей и/или воспалительные заболевания. Под «подавлением» следует понимать, что рекомбинантный белок препятствует упомянутым патологиям в некоторой степени, например, до 10% или 20%, более предпочтительно 50%, в частности 70%, 80% или 90% или даже 95%.
Термин «отек» означает состояние, которое вызвано сосудистой утечкой. Расширение сосудов и повышение проницаемости при воспалении могут представлять преобладающие патогенетические механизмы. Например, отек способствует расширению инфаркта после инсульта и может привести к опасной для жизни внутричерепной гипертензии у больных раком. Кроме того, экстравазация белков плазмы способствует метастазированию оккультных опухолей, а закупорка дыхательных путей может привести к летальным астматическим приступам. Увеличение сосудистой утечки, которое возникает при воспалении, может привести к развитию дыхательной недостаточности, асцита, перитонеального склероза (у пациентов на диализе), спаек (хирургия брюшной полости) и метастазирования.
Термин «ангиогенез» означает фундаментальный процесс, посредством которого образуются новые кровеносные сосуды. Первичный период ангиогенеза в организме человека происходит в течение первых трех месяцев эмбрионального развития, но также ангиогенез происходит как нормальный физиологический процесс в периоды роста тканей, таких как увеличение мышц или жира и во время менструального цикла и беременности.
Термин «патологический ангиогенез» касается образования и роста кровеносных сосудов во время поддерживания и прогрессирования нескольких болезненных состояний. Конкретные примеры патологического ангиогенеза имеют место в кровеносных сосудах (атеросклероз, гемангиомы, гемангиоэндотелиома), костях и суставах (ревматоидный артрит, синовит, разрушение костей и хряща, остеомиелит, рост паннуса, формирование остеофитов, новообразований и метастазов), коже (бородавки, гнойные гранулемы, рост волос, саркома Калоши, келоидные рубцы, аллергический отек, новообразования), печени, почках, легких, ухе и других эпителиях (воспалительные и инфекционные процессы, включая гепатит, гломерулонефрит, пневмонию и астму, полипы носа, отиты, расстройства при трансплантации, нарушения регенерации печени, новообразования и метастазы), матке, яичниках и плаценте (дисфункциональные маточные кровотечения в связи с внутриутробными противозачаточными средствами, образование фолликулярных кист, синдром гиперстимуляции яичников, эндометриоз, новообразования), головном мозге, нервах и глазах (ретинопатия недоношенных, диабетическая ретинопатия, нарушения сосудистой оболочки глаза и другие внутриглазные нарушения, лейкомаляция, новообразования и метастазы), сердце и скелетных мышцах в связи с перегрузками, жировой ткани (ожирение), эндокринных органах (тиреоидит, увеличение щитовидной железы, нарушения при трансплантации поджелудочной железы), гемопоезе (синдром Капоши при СПИДе), гематологических злокачественных новообразованиях (лейкозы), и лимфатических сосудах (опухолевые метастазы, лимфопролиферативные расстройства).
Термин «ишемическая болезнь сетчатки» означает, что питание сетчатки кровью и кислородом уменьшается, периферическая часть сетчатки теряет источник питания и прекращает надлежащее функционирование. Конкретный пример ишемической болезни сетчатки - ретинопатия. Общие заболевания, которые приводят к ретинопатии, включают диабетическую ретинопатию, окклюзию центральных вен сетчатки, стеноз сонной артерии, серповидно-клеточную ретинопатию. Диабетическая ретинопатия является основной причиной потери зрения у больных сахарным диабетом. В ишемической сетчатке происходит рост новых кровеносных сосудов (неоваскуляризация). Эти сосуды часто растут на поверхности сетчатки, в зрительном нерве или в передней части глаза на радужной оболочке. Новые сосуды не могут заменить поток необходимых питательных веществ и, наоборот, могут вызвать много проблем, таких как стекловидное кровотечение, отслойка сетчатки и неконтролируемая глаукома. Эти проблемы возникают из-за того, что новые сосуды хрупкие и подвержены кровотечению. В случае обнаружения на ранней стадии пролиферативная диабетическая ретинопатия иногда может быть остановлена путем панретинальной фотокоагуляции. Однако, в некоторых случаях хирургия по поводу эктомии стекловидного тела является единственным вариантом лечения.
Кроме этих видов ретинопатии, сосудистые заболевания глаз также включают глазные болезни, связанные с неоваскуляризацией, такие как макулярная дегенерация и диабетический макулярный отек (DME). Макулярная дегенерация образуется в результате неоваскулярного роста хороидного сосуда под макулой. Существует две формы макулярной дегенерации: сухая и влажная. Хотя влажная форма макулярной дегенерации составляет только 15% всех макулярных дегенераций, почти все влажные дегенерации приводят к слепоте. Кроме того, влажная форма макулярной дегенерации почти всегда образуется в результате сухой макулярной дегенерации. Как только один глаз поражается влажной макулярной дегенерацией, это состояние почти всегда оказывает влияние на второй глаз. Влажная макулярная дегенерация часто называется возрастной влажной макулярной дегенерацией (влажной AMD), как это встречается в основном у пожилых людей.
Диабетическая ретинопатия (DR) и DME являются основной причиной слепоты у трудоспособного населения большинства развитых стран. Увеличение числа людей с диабетом во всем мире свидетельствует, что DR и DME будут по-прежнему основными факторами для потери зрения и связанными с ними функциональными нарушениям на долгие годы. Несколько биохимических механизмов, в том числе активация протеинкиназы С-β, повышение выработки сосудистого эндотелиального фактора роста, окислительный стресс и накопление внутриклеточного сорбитола и повышенного количества конечных продуктов гликозилирования, может способствовать разрыву сосудов, которые характеризуют DR/DME. Ингибирование этих путей является многообещающим для борьбы с DR и DME.
Термин «легочная гипертензия» означает заболевание, при котором артериальное давление в легочных артериях является слишком высоким. В отсутствие других заболеваний сердца или легких она называется первичной легочной гипертензией. Диффузное сужение легочных артериол происходит в результате патологического артериогенеза, что сопровождается легочной гипертензией в ответ на повышение резистентности к кровотоку. Заболеваемость составляет 8 из 100'000 человек. Однако легочная гипертензия может также возникать как осложнение хронических обструктивных заболеваний легких (ХОЗЛ), таких как эмфизема, хронический бронхит или диффузный интерстициальный фиброз и у пациентов с астмообразным ХОЗЛ. Заболеваемость ХОЗЛ составляет примерно 5 из 10'000 человек.
Кроме того, связывающие белки изобретения могут быть использованы для лечения воспаления и, более конкретно, воспалительных заболеваний.
Термин «воспаление», используемый авторами, означает местную реакцию на повреждение живых тканей, особенно местную реакцию мелких кровеносных сосудов, их содержания и связанных с ними структур. Прохождение компонентов крови через стенки сосудов в ткани является признаком воспаления, и их накопление в ткани, образуемое таким образом, называется экссудатом или отеком. Любой вредный процесс, который повреждает живые ткани, например инфекции бактериями, чрезмерная жара, холод, механические повреждения, такие как дробление, кислоты, щелочи, облучение или заражение вирусами, может вызвать воспаление независимо от вовлеченного органа или ткани. Должно быть ясно, что болезни, классифицированные как «воспалительные заболевания», и тканевые реакции от ожогов до пневмоний, проказы, туберкулеза, ревматоидного артрита все представляют собой «воспаления».
Связывающие белки в соответствии с изобретением могут быть использованы для лечения опухолей. Термин «опухоль» означает массу патологичной ткани, которая возникает без видимой причины из уже существующих клеток организма, не имеет целенаправленной функции и характеризуется тенденцией к автономному и безудержному росту. Опухоли существенно отличаются от воспалительных и других отеков, поскольку клетки опухоли являются патологичными по их внешнему виду и других характеристикам. Патологичные клетки, т.е. вид клеток, которые обычно составляют опухоли, отличаются от нормальных клеток тем, что претерпели одно или несколько из следующих изменений: (1) гипертрофия или увеличение размера отдельных клеток; (2) гиперплазия или увеличение числа клеток в данной зоне; (3) анаплазия или регресс физических характеристик клетки до более примитивных или недифференцированного типа. Опухоли могут быть доброкачественными, например липомы, ангиомы, остеомы, хондромы и аденомы. Примеры злокачественных опухолей включают карциномы (например, опухоли молочной железы, карциномы дыхательных путей и желудочно-кишечного тракта, эндокринных желез и мочеполовой системы), саркомы (в соединительной ткани, в том числе фиброзных тканях, адипозных (жировых) тканях, мышцах, кровеносных сосудах, костях и хряще), карциносаркомы (в эпителиальной и соединительной ткани), лейкозы и лимфомы, опухоли нервной ткани (в том числе головного мозга), а также меланомы (рак пигментных клеток кожи). Использование связывающих белков настоящего изобретения против опухолей также может проводиться в сочетании с любой другой терапией опухолей, известной в данной области, такой как облучение, фотодинамическая терапия, химиотерапия или хирургическое вмешательство.
Фармацевтическая композиция содержит связывающие белки, как описано выше, и фармацевтически приемлемый носитель, наполнитель или стабилизатор (Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. [1980]). Подходящие носители, наполнители или стабилизаторы, известные квалифицированному специалисту, включают физиологический раствор, раствор Рингера, раствор декстрозы, раствор Хэнка, жирные масла, этилолеат, 5% раствор декстрозы в физиологическом растворе, вещества, которые повышают изотоничность и химическую стабильность, буфер и консерванты. Другие подходящие носители включают любой носитель, который сам по себе не вызывает выработку антител, вредных для пациента, получающего композицию, такие как белки, полисахариды, полимолочные кислоты, полигликолевые кислоты, полимерные аминокислоты и сополимеры аминокислот. Фармацевтическая композиция также может быть комбинированной формулировкой, включающей дополнительные активные вещества, такие как противораковый агент или антиангиогенный агент (например, человеческий VEGF-Axxxb; предпочтительно человеческий VEGF-A165b).
Предпочтительная фармацевтическая композиция для лечения глазных заболеваний включает в себя связывающие белки, как описано выше, и детергент, такой как полисорбат 20 (например, около 0,04%), буфер, такой как гистидин, фосфат или молочная кислота, и сахар, такой как сахароза или трегалоза. Предпочтительно, такая композиция содержит связывающие белки, как описано выше, и PBS. Указанные фармацевтические композиции могут вводиться местно либо местно на часть глаза или вводится в глаз, например, в субконъюктивальное, пери- или ретробульбарное пространство или прямо в глаз. Кроме того, указанные композиции могут быть введены системно путем парентерального введения. Предпочтительно, указанная фармацевтическая композиция применяется для глаз путем интравитреальной инъекции. Также желательно, указанная фармацевтическая композиция применяется для глаз местно и в качестве глазных капель. Глазные капли могут наноситься на роговицу (прозрачная часть в центре глаза), что позволяет молекулам проникать в глаз. Для лечения заболеваний, поражающих заднюю часть глаза, может быть наиболее желательным, чтобы связывающий белок проникал в склеру при введении под конъюнктиву или вокруг яблока. Введение белка может осуществляться после предварительного шага модуляции поверхности глаза, чтобы улучшить проникновение молекул. Предпочтительно, чтобы эпителиальный слой, такой как эпителий роговицы, модулировался с помощью усилителя проникновения для обеспечения достаточного и быстрого проникновения молекул, как, например, описанных выше. Использование связывающих белков настоящего изобретения для лечения глазных заболеваний может проводиться в комбинации с любым другим видом терапии, известным в данной области, таким как фотодинамическая терапия.
Композиции, которые будут использоваться для введения in vivo, должны быть асептическими или стерильными. Это легко осуществляется путем фильтрации через мембраны для стерильной фильтрации.
Могут быть получены препараты для пролонгированного высвобождения. В одном из вариантов осуществления изобретения может быть использован внутриглазной имплантат для обеспечения доставки связывающего белка изобретения. Подходящие примеры препаратов пролонгированного высвобождения включают полупроницаемые матрицы твердых гидрофобных полимеров, содержащих полипептид изобретения, матрицы которых находятся в виде изделий определенной формы, например, пленок или микрокапсул. Примеры матриц пролонгированного высвобождения включают сложные полиэфиры, гидрогели (например, поли(2-гидроксиэтил-метакрилат), или поли (виниловый спирт)), полилактиды, сополимеры L-глутаминовой кислоты и гамма-этил-L-глутамата, неразлагаемый этилен-винилацетат, разлагаемые сополимеры молочной кислоты и гликолевой кислоты, такие как ЛЮПРОН ДЕПО® (инъекционные микросферы, состоящие из сополимера молочной кислоты и гликолевой кислоты и лейпролида ацетата), и поли-D-(-)-3-оксимасляную кислоту.
Фармацевтическая композиция может быть введена с помощью любого подходящего метода в области знания квалифицированного специалиста. Предпочтительный путь введения - парентерально. При парентеральном введении лекарственное средство этого изобретения будет сформулировано в виде инъекционной формы единицы дозирования, такой как раствор, суспензия или эмульсия, в сочетании с фармацевтически приемлемыми наполнителями, как это определено выше. Дозировка и способ применения будут зависеть от конкретного пациента, получающего лечение, и конкретного заболевания. Как правило, фармацевтическую композицию вводят так, чтобы связывающий белок из настоящего изобретения вводился в дозе от 1 мкг/кг до 20 мг/кг, более предпочтительно от 10 мкг/г до 5 мг/кг, наиболее предпочтительно от 0,1 до 2 мг/кг. Предпочтительно, он вводится в виде болюса. Может также использоваться непрерывная инфузия, которая включает в себя непрерывную подкожную доставку через осмотический мининасос. Если это так, фармацевтическая композиция может вводится инфузией в дозе от 5 до 20 мкг/кг/ мин, более предпочтительно от 7 до 15 мкг/кг/ мин. В частности, фармацевтическую композицию вводят с помощью инъекций в глаз таким образом, чтобы связывающий белок изобретения вводится в дозе от 0,1 мг до 10 мг на инъекцию, более предпочтительно от 0,3 до 6 мг на инъекцию, наиболее предпочтительно от 1 мг до 4 мг на инъекцию. Кроме того, фармацевтическая композиция вводится в виде глазных капель в глаз таким образом, чтобы в глаз попадала одна капля раствора, содержащего концентрацию связывающего белка изобретения от 10 до 120 мг/мл, более предпочтительно от 20 до 100 мг/мл, наиболее предпочтительно от 40 до 80 мг/мл.
В другом варианте осуществления изобретения связывающий белок, ингибирующий активность VEGF-Axxx, как описано выше, может быть использован в комбинации со связывающим белком или малой молекулой, ингибирующей активность PIGF, с тем же уровнем ингибирования PIGF, как описано выше для VEGF-Axxx. Этот вариант осуществления основан на том, что PIGF ангиогенный в участках, где уровни VEGF-Axxx повышаются. Кроме того, связывающий белок, ингибирующий активность VEGF-Axxx, как описано выше, может быть использован в комбинации со связывающим белком или малой молекулой, ингибирующей активность тромбоцитарного фактора роста (PDGF), VEGF-C или других членов VEGF семейства белков, фактора некроза опухоли альфа (TNFalpha), дельта-лиганда подобного 4 (DU4), интерлейкина 6 (IL-6), нейропилина или ангиопоэтина 2 (Ang2).
Кроме того, изобретение предлагает новые методы лечения. С одной стороны, предлагается метод лечения ретинопатии, метод, включающий введение пациенту, нуждающемуся в этом, терапевтически эффективного количества связывающего белка изобретения, в частности связывающего белка, который ингибирует взаимодействие между человеческим VEGF- Аххх и человеческим VEGFR-2, но не взаимодействие между человеческим VEGF-Axxxb и человеческим VEGFR-2, и связывающий белок ингибирует VEGFR-2 опосредованный ангиогенез.
Кроме того, изобретение касается способов использования связывающего белка, как описано, для ингибирования VEGF-A биологической активности в клетке или ингибирования биологической активности, опосредованной VEGFR-2. Клетка может находиться in vivo или ex vivo, а может представлять, например, клетку живого организма, культурную клетку или клетку в образце ткани. Способ может включать контактирование указанной клетки с любым из ингибирующих VEGF-A/VEGFR-2 взаимодействие связывающих белков, описанных авторами, в количестве и в течение периода времени, достаточного для ингибирования такой биологической активности.
Изобретение предлагает способ лечения пациента, имеющего состояние, которое реагирует на ингибирование VEGF-Axxx или VEGFR-2. Такой способ включает введение указанному пациенту эффективного количества связывающего белка, описанного авторами. Состояние может представлять собой такое, которое характеризуется неправильным ангиогенезом. Состояние может представлять собой гиперпролиферативное состояние. Примеры состояний (или расстройств), подходящих для лечения, включают аутоиммунные расстройства, воспалительные заболевания, ретинопатии (в частности, пролиферативные ретинопатии) и рак, в частности, одно из заболеваний, описанных выше. Любой из связывающих белков, описанных авторами, может быть использован для получения лекарственного препарата для лечения таких расстройств, в частности расстройства, выбранного из группы, состоящей из: аутоиммунного расстройства, воспалительного заболевания, ретинопатии и рака. Предпочтительные состояния (или расстройства), подходящие для лечения, включают метастатическую почечно-клеточную карциному первой линии, рецидивирующую мультиформную глиобластому, адъювантный рак толстой кишки, адъювантный HER2-отрицательный рак молочной железы, адъювантный HER2-положительный рак молочной железы, адъювантный немелкоклеточный рак легких, диффузную крупную В-клеточную лимфому, распространенный рак желудка первой линии, HER2-отрицательный метастатический рак молочной железы первой линии, HER2-положительный метастатический рак молочной железы первой линии, метастатический рак яичников первой линии, желудочно-кишечные стромальные опухоли, карциноид высокого риска, гормон-рефрактерный рак простаты, недавно диагностированную мультиформную глиобластому, метастатический рак головы и шеи, рецидивирующий чувствительный к платине рак яичников, метастатический рак молочной железы второй линии, обширный мелкоклеточный рак легких, неплоскоклеточный, немелкоклеточный рак легких с ранее леченными ЦНС метастазами и рецидивирующей множественной миеломой, рак предстательной железы, немелкоклеточный рака легких (НМРЛ), колоректальный рак и рак поджелудочной железы, распространенный рак яичника (АОС), пациенты с АОС с симптоматическими злокачественными асцитами и неходжкинской лимфомой.
Рекомбинантный связывающий белок в соответствии с изобретением может быть получен и/или в дальнейшем разработан несколькими методами, такими как дисплей на поверхности бактериофагов (WO 90/02809, WO 07/006665) или бактериальной клетки (WO 93/10214), рибосомный дисплей (WO 98/48008), дисплей на плазмидах (WO 93/08278) или с помощью ковалентных РНК-повторных белковых гибридных конструкций (WO 00/32823), или внутриклеточная экспрессия и выбор/скрининг, такой как анализ дополнения белка (WO 98/341120). Такие методы известны специалисту в данной области.
Библиотека анкириновых повторных белков, используемых для выбора/скрининга рекомбинантного связывающего белка в соответствии с изобретением, может быть получена в соответствии с протоколами, известными специалистам в данной области (WO 02/020565, Binz, Н.К. et al., JMB, 332, 489-503, 2003, and Binz et al., 2004, loc. cit). Использование такой библиотеки для выбора VEGF-Axxx специфичных DARPins приведено в Примере 1. По аналогии, анкириновые повторные мотивы последовательности, представленные выше, могут использоваться для создания библиотек анкириновых повторных белков, которые могут быть использованы для выбора или скрининга VEGF-Axxx специфичных DARPins. Кроме того, повторные домены настоящего изобретения могут быть модульно собраны из повторных модулей в соответствии с текущими изобретениями и соответствующими кеппинг-модулями (Forrer, P., et al., FEBS letters 539, 2-6, 2003) с использованием стандартных технологий рекомбинантных ДНК (например, WO 02/020565, Binz et al., 2003, loc. cit. and Binz et al., 2004, loc. cit).
Изобретение не ограничивается конкретными вариантами осуществления, описанными в примерах. Другие источники могут быть использованы и обработаны, следуя общим рекомендациям, описанным ниже.
Примеры
Все исходные материалы и реагенты, указанные ниже, известны специалистам в данной области, и являются коммерчески доступными или могут быть получены с использованием известных методов.
Материалы
Химические вещества были приобретены у Fluka (Швейцария). Олигонуклеотиды от Microsynth (Швейцария). Если не указано иное, ДНК-полимеразы, ферменты рестрикции и буферы были получены от New England Biolabs (США) или Fermentas (Литва). Штаммом клонирования и выработки белка была Е. coli XL 1-blue (Stratagene, США). VEGF варианты были приобретены у R&D Systems (Миннеаполис, США), либо были произведены в клетках яичников китайского хомячка или в Pichia pastoris и очищены по стандартным протоколам (Rennel, Е.S. et al., European J. Cancer 44, 1883-94, 2008; Pichia expression system from Invitrogen). Биотинилированные VEGF варианты были получены химически через связь фрагмента биотина с первичными аминами из очищенных VEGF вариантов с использованием стандартных реактивов и методов биотинилирования (Pierce, США).
Молекулярная биология
Если не указано иное, методы выполняются в соответствии с описанными протоколами (Sambrook J., Fritsch Е. F. and Maniatis Т., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory 1989, New York).
Библиотеки спроектированных анкириновых повторных белков
Описаны библиотеки N2C и N3C спроектированных анкириновых повторных белков (WO 02/20565; Binz et al. 2003, loc. cit.; Binz et al. 2004, loc. cit.). Цифра в N2C и N3C описывает число рандомизированных повторных модулей, присутствующих между N-терминальным и С-терминальным кеппинг-модулями. Номенклатура, используемая для определения положений внутри повторяющихся звеньев и модулей, основана на Binz et al. 2004, loc. cit. с тем исключением, что границы повторных модулей и повторяющихся звеньев сдвинуты на одно положение аминокислоты. Например, положение 1 повторного модуля Binz et al. 2004 (loc. cit.) соответствует положению 2 повторного модуля текущего раскрытия и, следовательно, положение 33 повторного модуля Binz et al. 2004, loc. cit. соответствует положению 1 из следующего повторного модуля текущего раскрытия.
Все последовательности ДНК были подтверждены путем секвенирования, а расчетный молекулярный вес всех описанных белков был подтвержден методом масс-спектрометрии.
Пример 1: Выбор связывающих белков, содержащих повторный домен со специфичность связывания к VEGF-Axxx
С использованием рибосомного дисплея (Hanes, J. and Plückthun, A., PNAS 94, 4937-А2, 1997) из N2C или N3C DARPin библиотек, описанных Binz et al. 2004 (loc. cit.) были отобраны многие спроектированные анкириновые повторные белки (DARPins) со специфичностью связывания к VEGF-Axxx.
Связывание выбранных клонов со специфичными (VEGF-Axxx) и неспецифичными (МВР, мальтоза-связывающий белок Е. coli) мишенями оценивали с помощью ИФА неочищенного экстракта, указывающего на то, что VEGF-Axxx связывающие белки были успешно выбраны (Фиг. 1). SEQ ID NO: 14-40 представляют аминокислотные последовательности выбранных связывающих белков, содержащих повторный домен со специфичностью связывания к VEGF-Axxx. Анализ последовательности отдельных связующих веществ выявил специфичные анкириновые повторные мотивы последовательности, присущие определенным выбранным семействам связующих веществ. Такие анкириновые повторные мотивы последовательности, присутствующие в повторных доменах со специфичностью связывания к VEGF-Axxx, приводятся в SEQ ID NO: 1-13.
Выбор VEGF-Axxx специфичных анкириновых повторных белков путем рибосомного дисплея
Выбор VEGF-Axxx специфичных анкириновых повторных белков проводили путем рибосомного дисплея (Hanes and Pliickthun, loc. cit.)) с использованием собачьего VEGF-A 164 или человеческого VEGF-A 165 в качестве мишеневых белков, библиотеки спроектированных анкириновых повторных белков, как описано (WO 02/020565, Binz et al., 2003, loc. cit. и Binz et al., 2004, loc. cit) и установленных протоколов (ahnd, С, Amstutz, P. and Plückthun, A., Nat. Methods 4, 69-79, 2007). Раунды выбора рибосомного дисплея проводились на собачьих или человеческих VEGF вариантах (в том числе биотинилированных вариантах, иммобилизованных над нейтравидином или стрептавидином) с обеими N2C и N3C DARPin библиотеками с использованием установленных протоколов (Binz et al. 2004, loc. cit.). Количество циклов ГЩР обратной транскрипции (RT) после каждого отборочного раунда постоянно сокращалось с 40 до 30, приспосабливаясь к выходу за счет обогащения связующих веществ. Четыре начальных отборочных раунда на собачьем VEGF позволили получить пулы наномолярных-аффинных DARPins, как было показано с помощью ИФА и SPR измерений отдельных клонов. Для обнаружения DARPins с дальнейшим повышенным сродством, проводились дополнительные выборы «off-rate» на биотинилированном человеческом или собачьем VEGF, иммобилизованном над нейтравидином или стрептавидином, путем отбора пулов после второго и третьего первоначальных раундов отбора рибосомного дисплея, а затем раунда отбора «on-rate» на человеческом VEGF.
Выбранные клоны специфично связываются с VEGF-Axxx, как показано с помощью ИФА неочищенного экстракта
Индивидуальные выбранные DARPins, специфически связывающие VEGF-Axxx, были определены иммуноферментньгм анализом (ИФА) с использованием неочищенных экстрактов Escherichia coli клеток экспрессии DARPin с помощью стандартных протоколов. Выбранные клоны были клонированы в вектор экспрессии pQE30 (Qiagen), трансформированный в Е. coli XL 1-Blue (Stratagene), а затем их выращивали в течение ночи при 37°С в 96-глубоколуночном планшете (каждый клон в одной лунке), содержащем 1 мл среды роста (2YT, содержащая 1% глюкозы и 100 мкг/мл ампициллина). 1 мл свежей 2YT, содержащей 50 мкг/мл ампициллина, засевали с 100 мкл ночной культуры в свежий 96-глубоколуночный планшет. После инкубации в течение 2 ч. при 37°С, экспрессию вызывали с помощью IPTG (1 мМ конечная концентрация) и она продолжалась в течение 3 ч. Клетки были собраны, повторно суспендированы в 100 мкл B-PERII (Pierce) и инкубировались 15 мин при комнатной температуре при встряхивании. Затем добавили 900 мкл PBS-TB (PBS с добавкой 0,2% BSA, 0,1% Твин-20, рН 7,4) и остатки клеток были удалены с помощью центрифугирования. 100 мкл каждого лизированного клона внесли в лунку NeutrAvidin покрытого планшета MaxiSorp, содержащего либо VEGF-Axxx вариант или не связанный МВР иммобилизованный через их биотиновый фрагмент и инкубировали 1 ч при комнатной температуре. После интенсивной промывки с помощью PBS-T (PBS с добавкой 0,1% Твин-20, рН 7,4) планшет проявили с использованием стандартных процедур ИФА с использованием моноклонального анти-RGS(His)4 антитела (34650, Qiagen) в качестве первичного антитела и поликлонального коза антимыша антитела, конъюгированного со щелочной фосфатазой (А3562, Sigma) в качестве вторичного реагента. Затем связывание обнаруживали с помощью динатрия 4-нитрофенилфосфата (4NPP, Fluka) в качестве субстрата для щелочной фосфатазы. Цвет измеряли при 405 нм. Результаты с примера ИФА неочищенного экстракта, используемого для определения DARPins связывания с VEGF-Axxx, показаны на Фиг. 1. Скрининг нескольких сотен клонов такого ИФА неочищенного клеточного экстракта выявил более сотни различных DARPins со специфичностью к VEGF-Axxx. Эти связывающие белки были выбраны для дальнейшего анализа. Примеры аминокислотных последовательностей выбранных анкириновых повторных доменов, которые специфически связываются с VEGF-Axxx, приводятся в SEQ ID NO: 14-40.
Выведение повторных мотивов последовательности из выбранных повторных доменов со специфичностью связывания к VEGF-Axxx
Аминокислотные последовательности выбранных повторных доменов с специфичностью связывания к VEGF-Axxx были дополнительно проанализированы с помощью инструментов анализа последовательности, известных практикующим специалистам в данной области (WO 02/020565; Forrer et al., 2003, loc. cit.; Forrer, P., Binz, H.K., Stumpp, M.T. and Plückthun, A., ChemBioChem, 5(2), 183-189, 2004). Тем не менее, в отличие от WO 02/020565, где для выведения повторных мотивов последовательности были использованы природные повторные мотивы, здесь повторные мотивы последовательности были выведены из повторяющихся звеньев выбранных повторных доменов со специфичностью связывания к VEGF-Axxx. Таким образом были определены семейства выбранных повторных доменов, содержащих общий повторный мотив последовательности. Такие повторные мотивы последовательности, присутствующие в повторных доменах со специфичность связывания к VEGF-Axxx, приводятся в SEQ ID NO: 1-13.
Высокий уровень и растворимая экспрессия DARPins
Для дальнейшего анализа выбранные клоны, показывающие специфичное VEGF-Axxx связывание в ИФА неочищенного клеточного экстракта, как описано выше, были экспрессированы в клетках Е. coli XL1-blue и очищены с использованием их His-тега с помощью стандартных протоколов. 25 мл стационарных ночных культур (LB, 1% глюкозы, 100 мг/л ампициллина; 37°С) были использованы для инокуляции 1 л культуры (та же среда). При А(600)=0,7 культуры были индуцированы с 0,5 мМ IPTG и инкубировали при 37°С в течение 4 ч. Культуры центрифугировали и полученные гранулы ресуспендировали в 40 мл TBS500 (50 мМ Трис-HCl, 500 мМ NaCl, рН 8) и обрабатывали ультразвуком. Лизат рецентрифугировали, в полученный супернатант были добавлены глицерин (10% (по объему) конечной концентрации) и имидазол (20 мМ конечная концентрация). Белки были очищены над колонкой с Ni-нитрилотриуксусной кислотой (2,5 мл объема колонки) в соответствии с инструкцией завода-изготовителя (Qiagen, Германия). До 200 мг хорошо растворимых DARPins со специфичностью связывания с VEGF-Axxx можно очистить из одного литра культуры Е. coli с чистотой выше 95% по оценкам SDS-15% PAGE. Такие очищенные DARPins используются для дальнейшей характеризации.
Пример 2: Определение значений ICsn выбранных DARPins со специфичностью связывания с VEGF-Axxx в анализе прорастания сфероидов
Добавление VEGF-Axxx к HUVEC сфероидам, встроенных в коллагеновые матрицы, приводит к прорастанию сфероидов. Добавление ингибитора VEGF-Axxx будет блокировать формирование прорастания, которые могут быть количественно определены статистически по количеству и длине прорастаний. При добавлении различных концентраций ингибитора и постоянного количества VEGF можно определить IC50.
Ингибирование прорастания сфероида с помощью VEGF-Axxx специфичных DARPins Анализы прорастания сфероидов были сделаны в соответствии со стандартными протоколами (Korff et al., loc. cit.). DARPins со специфичностью к VEGF-Axxx были отобраны и очищены до >96% чистоты, как описано в примере 1. Человеческие клетки пуповинной вены были выращены до слияния в монослойной культуре. После трипсинизации клеточную суспензию поместили в висячую каплю для образования сфероидов, т.е. около 500 организованных агрегированных HUVEC. Сфероиды были вложены в коллагеновую матрицу и их стимулировали с помощью VEGF-A165 для инициации прорастания. Ингибиторы проращивания были добавлены дополнительно для наблюдения их влияния на ингибирование прорастания. Количество прорастаний в сфероиде и длина прорастания были количественно определены с помощью графического программного обеспечения.
Результаты двух примеров анализов прорастания сфероидов приведены на Фиг. 2а (DARPin # 30 со специфичностью связывания для VEGF-Axxx) и Фиг. 2b (DARPin NC, DARPin отрицательного контроля без специфичности связывания для VEGF-Axxx; например, DARPin Е3_5 (Binz et al., 2005, loc. cit.). Самые эффективные DARPins в этом анализе показали значения IC50 в диапазоне от 10 до 50 пМ, в то время как Avastin®, Lucentis ® и Macugen® показали значения IC50 в параллельных опытах в диапазоне от 150 до 500 пМ.
Пример 3: Определение мишеневой специфичности DARPin №27 по сравнению с Avastin® с помощью анализа поверхностного плазменного резонанса
Собачий VEGF-A164 или собачий VEGF-A164b были иммобилизированы в проточной ячейке и анализировали взаимодействие DARPin №27 (SEQ ID NO: 16) и Avastin® с иммобилизованными мишенями.
Анализ поверхностного плазменного резонанса (SPR)
SPR измеряли с помощью инструмента Proteon (BioRad). Буфер представлял собой 20 мМ HEPES, рН 7,4, 150 мМ NaCl и 0,005% Твин 20. Около 1200 RU собачьего VEGF-А164 или собачьего VEGF-A164b были иммобилизованы на чипе GLC (BioRad). Взаимодействий были измерены при потоке 60 мкл/мин с 5 мин потоком буфера, 100 секундной инъекцией Авастина® или DARPin №27 в концентрации 250 нМ и измерении «off-rate» в течение нескольких минут с потоком буфера. Сигнал непокрытой ячейки сравнения был вычтен из измерений.
Результаты приведены на Фиг. 3а (взаимодействие Авастина с собачьим VEGF-A164), Фиг. 3b (взаимодействие Авастина с собачьим VEGF-A164b), Фиг. 3с (взаимодействие DARPin №27 с собачьим VEGF-A164) и Фиг. 3d (взаимодействие DARPin №27 с собачьим VEGF-A164b). В то время как Авастин четко взаимодействует с обеими иммобилизованными изоформами VEGF, DARPin №27 показывает только взаимодействие с VEGF-A164, но не с VEGF-A164b.
Пример 4: Эффективность in vivo DARPin №30 в ингибировании VEGF-A165 на кроличьей модели сосудистой утечки.
Пегилированный DARPin №30 (SEQ ID NO: 29) или Lucentis® вводили путем интравитреальной инъекции в глаз кролика для испытания их эффективности для ингибирования сосудистого утечки, индуцированной последующей интравитреальной инъекцией человеческого VEGF-A165.
Измерение ингибирования сосудистой утечки у кроликов
В 1-й день либо PBS, пегилированный DARPin №30 (125 мкг), или эквимолярное количество Lucentis® (162 мкг) вводили путем интравитреальной инъекции в один глаз каждого кролика (обработанный глаз). На 4-й день или 30-й день в обработанный глаз каждого кролика вводили интравитреальной инъекцией 500 нг человеческого VEGF-А165. Оба глаза всех животных оценивали через 48 часов после инъекций VEGF-A165 путем измерения содержания флуоресцеина в каждом глазу через 1 ч после внутривенного введения натрия флуоресцеина (50 мг/кг массы тела животного, 10% (м/об) в 0,9% (м/об) физиологического раствора). Соотношение количества флуоресценции в обработанном и необработанном глазу были рассчитаны для каждого животного. Соотношение, равное единице, соответствует отсутствию дополнительной утечки флуоресценции в обработанном глазу, соотношение больше единицы указывает на большую утечку флуоресценции в обработанном глазу, чем в необработанном контрольном глазу.
Подготовка пегилированного DARPin
Пегилирование белка за счет использования одного остатка Cys и химии малеимида хорошо известно специалистам в данной области и может быть выполнено в соответствии с установленными протоколами (например, от Pierce). DARPin №30, включающий дополнительный С-терминальный линкер (GGGSGGGSC, SEQ ID NO: 41), был очищен до близкой гомогенности с использованием стандартных хроматографических методов. Белок был полностью восстановлен с помощью DTT и очищен с помощью гель-фильтрации для удаления DTT и обмена буфера на PBS. PEG-малеимид (метокси-поли(этиленгликоль)-оксопропиламино-пропил-малеимид; NOF, № Sunbright МЕ-200МА), растворенный в PBS, смешивали с DARPin в PBS при приблизительно 15% молярном избытке PEG-малеимида в течение 2-4 часов при комнатной температуре. Пегилированный DARPin затем отделяли от не-реактивного DARPin и неактивных PEG фрагментов с помощью стандартной анионообменной хроматографии.
Результаты приведены на Фиг. 4. Оба и пегилированный DARPin №30 и Lucentis® были в состоянии защитить глаза кролика от индуцированной VEGF-A165 сосудистой утечки через 4 дня после того, как они были введены путем интравитреальной инъекции. Тем не менее, только пегилированный DARPin №30, а не Lucentis® был в состоянии защитить глаза кролика от индуцированной VEGF-A165 сосудистой утечки до 30 дней после интравитреальной инъекции.
| название | год | авторы | номер документа |
|---|---|---|---|
| СВЯЗЫВАЮЩИЕ БЕЛКИ, ИНГИБИРУЮЩИЕ ВЗАИМОДЕЙСТВИЯ VEGF-A РЕЦЕПТОРА | 2009 |
|
RU2550258C2 |
| МОДИФИЦИРОВАННЫЕ СВЯЗЫВАЮЩИЕ БЕЛКИ, ИНГИБИРУЮЩИЕ ВЗАИМОДЕЙСТВИЕ VEGF-A РЕЦЕПТОРА | 2011 |
|
RU2605309C2 |
| СКОНСТРУИРОВАННЫЕ БЕЛКИ ПОВТОРОВ, КОТОРЫЕ СВЯЗЫВАЮТСЯ С СЫВОРОТОЧНЫМ АЛЬБУМИНОМ | 2011 |
|
RU2625882C2 |
| УЛУЧШЕННЫЕ N-ТЕРМИНАЛЬНЫЕ КЭППИНГ МОДУЛИ ДЛЯ СКОНСТРУИРОВАННЫХ БЕЛКОВ С АНКИРИНОВЫМ ПОВТОРОМ | 2011 |
|
RU2636552C2 |
| Связывающие белки, содержащие по меньшей мере две повторяющиеся области-антагонисты HER2 | 2013 |
|
RU2664464C9 |
| РЕКОМБИНАНТНЫЕ СВЯЗЫВАЮЩИЕ БЕЛКИ И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2778346C2 |
| ИНГИБИТОРЫ РЕЦЕПТОРОВ ФАКТОРА РОСТА ЭНДОТЕЛИЯ СОСУДОВ ТИПА 2 | 2004 |
|
RU2402567C2 |
| СЛИТНЫЙ БЕЛОК АНТИАНГИОГЕННОГО ИНДУЦИРУЮЩЕГО ФАКТОРА И ЕГО ПРИМЕНЕНИЕ | 2012 |
|
RU2560589C2 |
| СЛИТЫЕ БЕЛКИ, СОДЕРЖАЩИЕ СВЯЗЫВАЮЩИЕ PDGF И VEGF ЧАСТИ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2692652C2 |
| РЕКОМБИНАНТНАЯ ЭКСПРЕССИЯ БЕЛКОВ В ДВУХЦЕПОЧЕЧНОЙ ФОРМЕ С ДИСУЛЬФИДНЫМ МОСТИКОМ | 2006 |
|
RU2412253C2 |
































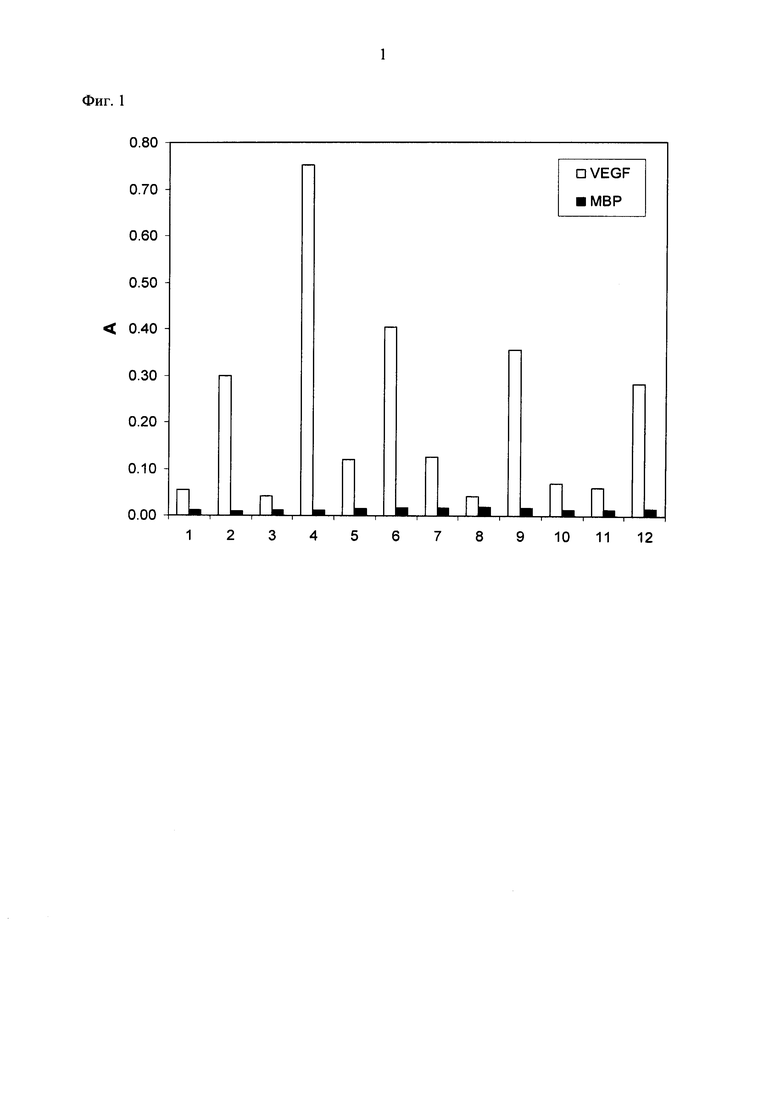

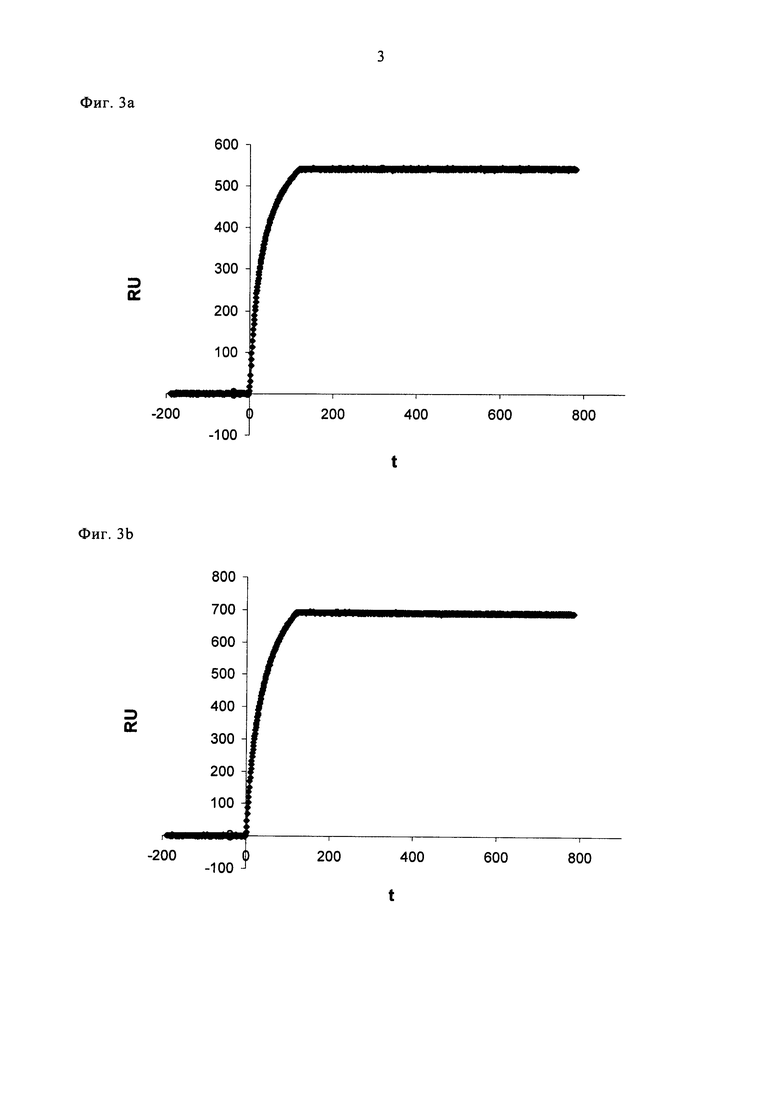
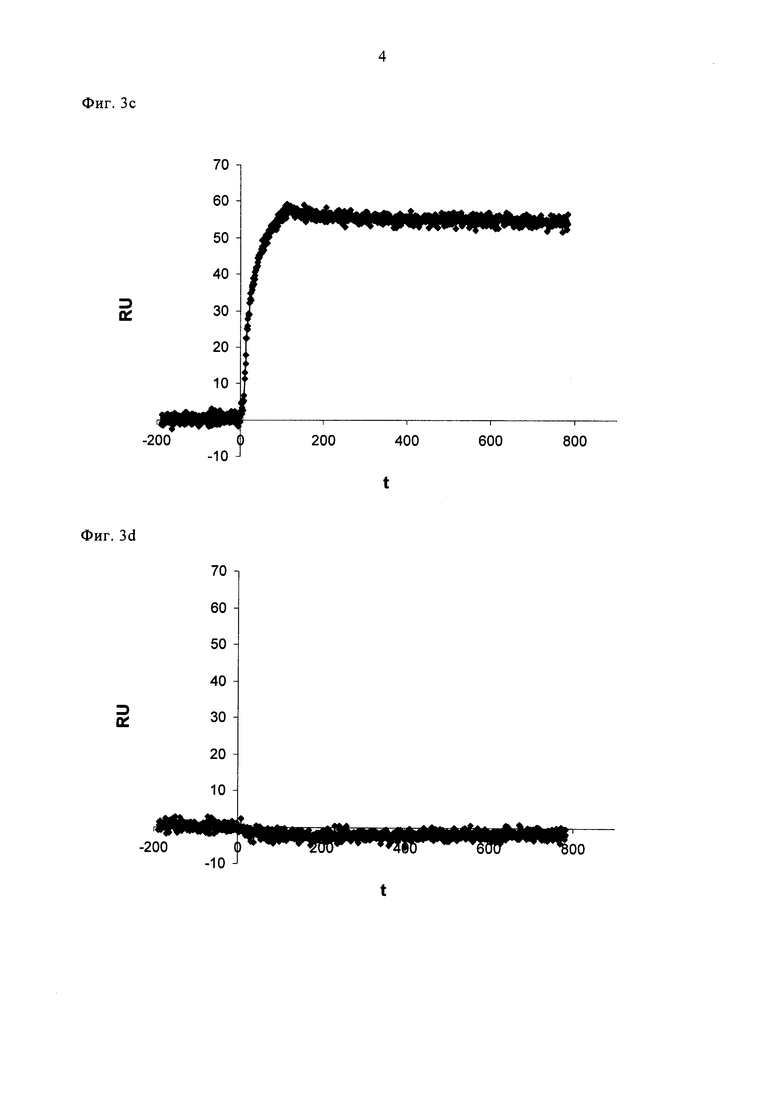
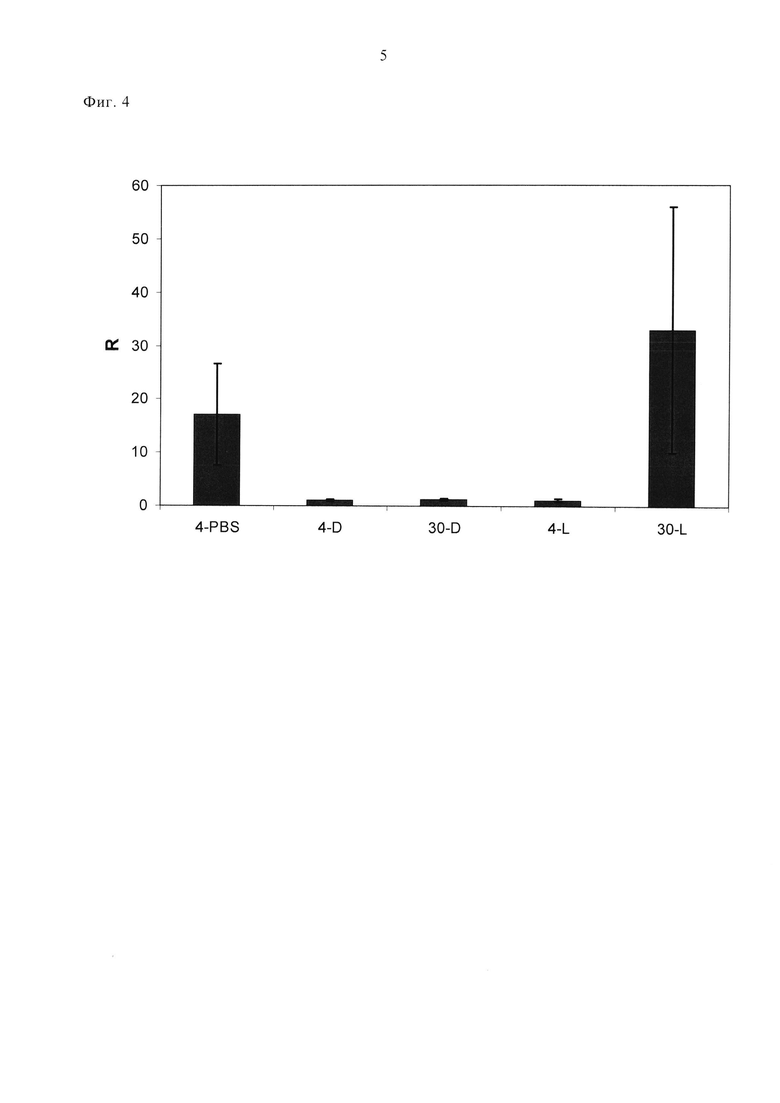
Настоящее изобретение относится к области биотехнологии, конкретно к связывающим белкам, специфичным к VEGF-A, и может быть использовано в медицине. Рекомбинантным путем получают связывающий белок, содержащий до четырех анкириновых повторных домена, которые способны ингибировать связывание VEGF-A165 с VEGFR-2. Изобретение позволяет получить эффективный ингибитор VEGF-A165, который не обладает рядом побочных эффектов, например способностью подавлять ангиогенез в клетках, в сравнении с известными пептидными и непептидными аналогами. 3 н. и 13 з.п. ф-лы, 4 ил., 4 пр.
1. Антиангиогенный рекомбинантный связывающий белок, включающий не более четырех доменов с анкириновым повтором, где указанные не более четырех доменов с анкириновым повтором связаны друг с другом через полипептидные линкеры, и причем по меньшей мере один домен с анкириновым повтором связывает VEGF-A165 с Kd ниже 10-7 М и ингибирует связывание VEGF-A165 с VEGFR-2 и где, дополнительно, указанный домен с анкириновым повтором состоит из
N-концевого кэппинг-модуля;
первого повторного модуля, который представляет собой мотив последовательности анкиринового повтора:
1D23G4TPLHLAA56GH7EIVEVLLK8GADVNA (SEQ ID NO: 5), где
1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, N, R, V, Y, Е, Н, I, К, L, Q, S и Т;
2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из S, A, N, R, D, F, L, Р, Т и Y;
3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Т, V, S, A, L и F;
4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из W, F и H;
5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Р, I, A, L, S, Т, V и Y;
6 представляет собой аминокислотный остаток, выбранный из группы, состоящей из W, F, I, L, T и V;
7 представляет собой аминокислотный остаток Р; и
8 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, Н, N и Y;
второго повторного модуля, который представляет собой мотив последовательности анкиринового повтора
1D23G4TPLHLAA56GHLEIVEVLLK7GADVNA (SEQ ID NO: 1),
где 1, 2, 3, 4, 5, 6 и 7 независимо друг от друга представляют собой аминокислотный остаток, выбранный из группы, состоящей из A, D, Е, F, Н, I, К, L, М, N, О, R, S, Т, V, W и Y, и
С-концевого кэппинг-модуля.
2. Связывающий белок по п. 1, где указанный по меньшей мере один домен с анкириновым повтором ингибирует прорастание HUVEC сфероидов со значением IC50 менее 10 нМ.
3. Связывающий белок по п. 1, причем Kd для взаимодействия указанного по меньшей мере одного домена с анкириновым повтором с VEGF-A165b по меньшей мере в 10 раз выше, чем Kd для взаимодействия указанного по меньшей мере одного домена с анкириновым повтором с соответствующим VEGF-A165.
4. Связывающий белок по п. 1, где указанный по меньшей мере один домен с анкириновым повтором конкурирует за связывание с VEGF-A165 с доменом с анкириновым повтором, выбранным из группы, которая состоит из последовательностей SEQ ID NO: 16, 22, 23, 29, 30, 33, 34, 36, 39 и 40.
5. Связывающий белок по п. 1, в котором указанный по меньшей мере один домен с анкириновым повтором включает аминокислотную последовательность, обладающую по меньшей мере 75% идентичностью с доменом с анкириновым повтором, выбранным из группы, состоящей из последовательностей SEQ NO 24-33 и 36-38.
6. Связывающий белок по п. 1, где указанный по меньшей мере один домен с анкириновым повтором выбран из группы, состоящей из последовательностей SEQ NO 24-26, 30, 32 и 36-38 и каждой из этих последовательностей, в которой глицин в положении 5 заменен на аспартат.
7. Связывающий белок по п. 1, в котором указанный по меньшей мере один домен с анкириновым повтором выбран из группы, состоящей из последовательностей: 24-26, 30, 32 и 36-38.
8. Связывающий белок по п. 1, где указанный первый повторный модуль представляет собой мотив последовательности анкиринового повтора
1D23G4TPLHLAA56GH7EIVEVLLK8GADVNA (SEQ ID NO: 5), в котором:
1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, N, R, V, Y, Е, Н, I, K, L, Q, S и Т;
2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из S, A, N и R;
3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Т, V и S;
4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из W, F и H;
5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Р и I;
6 представляет собой аминокислотный остаток, выбранный из группы, состоящей из W, F, I, L, T и V;
7 представляет собой аминокислотный остаток Р; и
8 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, Н, N и Y.
9. Нуклеиновая кислота, кодирующая домен с анкириновым повтором по любому из пп. 5-8.
10. Фармацевтическая композиция для лечения заболеваний, ассоциированных с VEGF-A165, включающая связывающий белок по любому из пп. 1-8 в терапевтически эффективном количестве и терапевтически приемлемый носитель и/или разбавитель.
11. Фармацевтическая композиция по п. 10 для лечения глазных заболеваний, ассоциированных с VEGF-A165.
12. Фармацевтическая композиция по п. 10 для ингибирования VEGF-A165 в сосудистой системе.
13. Способ лечения заболеваний, ассоциированных с VEGF-A165, у млекопитающих, включая человека, который включает введение нуждающемуся в таком лечении пациенту эффективного количества фармацевтической композиции по п. 10.
14. Способ по п. 13, при котором фармацевтическая композиция вводится парентерально.
15. Способ по п. 13, при котором фармацевтическая композиция вводится таким образом, что указанный связывающий белок вводится в дозе от 1 мкг/кг до 20 мг/кг.
16. Антиангиогенный рекомбинантный связывающий белковый конъюгат, включающий до четырех доменов с анкириновым повтором и полиэтиленгликолевую группу, где полиэтиленгликоль имеет молекулярную массу около 1-100 кДа, где полиэтиленгликоль является разветвленным или линейным, где полиэтиленгликоль ковалентно связан с тиольной группой остатка цистеина или с аминогруппой остатка лизина или аргинина, где указанные не более четырех доменов с анкириновым повтором связаны друг с другом через полипептидные линкеры, и причем по меньшей мере один домен с анкириновым повтором связывает VEGF-A165 с Kd ниже 10-7 М и ингибирует связывание VEGF-A165 с VEGFR-2 и где, дополнительно, указанный домен с анкириновым повтором состоит из
N-концевого кэппинг-модуля;
первого повторного модуля, который представляет собой мотив последовательности анкиринового повтора:
1D23G4TPLHLAA56GH7EIVEVLLK8GADVNA (SEQ ID NO: 5), где
1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, N, R, V, Y, Е, Н, I, К, L, Q, S и T;
2 представляет собой аминокислотный остаток, выбранный из группы, состоящей из S, A, N, R, D, F, L, Р, Т и Y;
3 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Т, V, S, A, L и F;
4 представляет собой аминокислотный остаток, выбранный из группы, состоящей из W, F и H;
5 представляет собой аминокислотный остаток, выбранный из группы, состоящей из Р, I, A, L, S, Т, V и Y;
6 представляет собой аминокислотный остаток, выбранный из группы, состоящей из W, F, I, L, T и V;
7 представляет собой аминокислотный осадок Р; и
8 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, Н, N и Y;
второго повторного модуля, который представляет собой мотив последовательности анкиринового повтора 1D23G4TPLHLAA56GHLEIVEVLLK7GADVNA (SEQ ID NO: 1),
где 1, 2, 3, 4, 5, 6 и 7 независимо друга представляют собой аминокислотный остаток, выбранный из группы, которая состоит из A, D, Е, F, Н, I, K, L, М, N, О, R, S, Т, V, W и Y, и
С-концевого кэппинг-модуля.
| BRADLEY C.M | |||
| et al | |||
| Limits of cooperativity in a structurally modular protein: response of the Notch ankyrin domain to analogous alanine substitutions in each repeat, J | |||
| Mol | |||
| Biol., 2002, v | |||
| Телефонный аппарат, отзывающийся только на входящие токи | 1921 |
|
SU324A1 |
| Устройство для одновременного приема и передачи по радиотелефону | 1921 |
|
SU373A1 |
| WO 2005087797 A1, 22.09.2005 | |||
| ЦИФРОВОЙ ТЕРМОМЕТР | 0 |
|
SU220565A1 |
| INOKI I | |||
| et al | |||
| Connective tissue growth factor binds vascular endothelial growth factor (VEGF) and inhibits VEGF-induced angiogenesis, The FASEB Journal, 2002, v | |||
| Устройство для электрической сигнализации | 1918 |
|
SU16A1 |
| Прибор для записи звуковых волн | 1920 |
|
SU219A1 |
| STUMPP M.T | |||
| et al | |||
| DARPins: A new generation of protein therapeutics, Drug Discovery Today, 2008, v | |||
| Насос | 1917 |
|
SU13A1 |
| Прибор для нагревания перетягиваемых бандажей подвижного состава | 1917 |
|
SU15A1 |
| АНТРАНИЛАМИДПИРИДИНАМИДЫ ИЗБИРАТЕЛЬНОГО ДЕЙСТВИЯ В КАЧЕСТВЕ ИНГИБИТОРОВ VEGFR-2 И VEGFR-3 | 2002 |
|
RU2299208C2 |

