Область, к которой относится изобретение
Настоящее изобретение относится к улучшенным N-терминальным кэппинг модулям для сконструированных белков с анкириновым повтором (DARPin), которые придают улучшенную термальную стабильность DARPin, а также к нуклеиновым кислотам, кодирующим такие белки, фармацевтическим композициям, включающим такие белки и применению таких белков в лечении заболеваний.
Предпосылки создания изобретения
Кроме антител существуют новые связывающие белки или связывающие домены, которые могут использоваться для специфического связывания целевой молекулы (например, Binz, H.K., Amstutz, Р. и Plückthun, A., Nat. Biotechnol. 23, 1257-1268, 2005). Один такой новый класс связывающих белков или связывающих доменов основывается на сконструированных белках повторов или сконструированных повторяемых доменах (WO 2002/020565; Binz, H.K., Amstutz, P., Kohl, A., Stumpp, M.T., Briand, C., Forrer, P., Grütter, M.G., и Plückthun, A., Nat. Biotechnol. 22, 575-582, 2004; Stumpp, M.T., Binz, H.K и Amstutz, P., Drug Discov. Today 13, 695-701, 2008). WO 2002/020565 описывает, как могут быть сконструированы библиотеки белков повторов и их общее применение. Такие сконструированные повторяемые домены используют модулярную природу белков повторов и имеют N-терминальные и C-терминальные кэппинг модули для предотвращения агрегации сконструированных повторяемых доменов путем экранирования гидрофобного ядра домена (Forrer, P., Stumpp, М.Т., Binz, H.K. и Plückthun, A., FEBS letters 539, 2-6, 2003). Эти кэппинг модули основываются на кэппирующих повторах природного гуанин-аденин-связывающего белка (GA-связывающего белка). Было показано, что термальная и термодинамическая стабильность этих доменов сконструированных анкириновых повторов может быть дополнительно повышена путем усовершенствования С-терминального кэппирующего повтора, который имеет происхождение от GA-связывающего белка (Interlandi, G., Wetzel, S.K, Settanni, G., Plückthun, А. и Caflisch, A., J. Mol. Biol. 375, 837-854, 2008; Kramer, M.A, Wetzel, S.K., Plückthun, A., Mittl, P.R.E, и Grütter, M.G., J. Mol. Biol. 404, 381-391, 2010). Авторы вводили в общей сложности восемь мутаций в этот кэппинг модуль и удлиняли его C-терминальную спираль путем прибавления трех различных аминокислот. Тем не менее, введение этих модификаций в C-терминальный кэппинг модуль приводило к тенденции нежелательной димеризации сконструированного повторяемого домена, который несет этот мутированный C-терминальный кэппинг модуль. Таким образом, существует необходимость в получении дополнительных оптимизированных C-терминальных кэппинг модулей или C-терминальных кэппирующих повторов сконструированных доменов анкиринового повтора.
В общем случае, существует потребность в специфических для мишени улучшенных белках с анкириновым повтором с улучшенной стабильностью для лечения рака и других патологических состояний.
Техническая проблема, которая лежит в основе настоящего изобретения, заключается в идентификации новых белков с анкириновыми повторами с улучшенной стабильностью для улучшенного лечения рака и других патологических состояний. Решение этой технической проблемы достигается путем обеспечения воплощений, охарактеризованных в формуле изобретения.
Краткое изложение сущности изобретения
Настоящее изобретение относится к связывающему белку, включающему, по крайней мере, один домен анкиринового повтора, где указанный домен анкиринового повтора включает N-терминальный кэппирующий модуль, имеющий аминокислотную последовательность
GSDLGKKLLE AARAGQDDEV RILLKAGADV NA (SEQ ID NO: 14) или
GSDLGKKLLE AARAGQDDEV RELLKAGADV NA (SEQ ID NO: 15), в которой аминокислотный остаток L в положении 24 последовательности SEQ ID NO: 14 или SEQ ID NO: 15 является необязательно замененным V, I или A;
вплоть до 9 аминокислот SEQ ID NO: 14 или SEQ ID NO: 15 в других положениях, отличных от положения 24, необязательно являются замененными любыми аминокислотами; и
в котором G в положении 1 и/или S в положении 2 SEQ ID NO: 14 или SEQ ID NO: 15 являются необязательно отсутствующими.
В частности, изобретение относится к связывающему белку, где указанный N-терминальный кэппирующий модуль включает последовательность GSX1LX2KKLLE AARAGQDDEV X3X4LX5X6X7GADV NA (SEQ ID NO: 5), в которой G в положении 1 и/или S в положении 2 последовательности SEQ ID NO: 5 являются необязательно отсутствующими;
X1 представляет собой аминокислотный остаток G, A, или D;
X2 представляет собой аминокислотный остаток G или D;
X3 представляет собой аминокислотный остаток R или E;
X4 представляет собой аминокислотный остаток I, Е или V;
X5 представляет собой аминокислотный остаток L, V, I или A;
X6 представляет собой аминокислотный остаток A, K или E; и
X7 представляет собой аминокислотный остаток, выбранный из группы, состоящей из A, H, Y, K и R.
В другом воплощении изобретение относится к связывающему белку, включающему, по крайней мере, один домен анкиринового повтора, где указанный домен анкиринового повтора включает N-терминальный кэппирующий модуль, имеющий аминокислотную последовательность, по крайней мере, с 70%-ной идентичностью аминокислотной последовательности с последовательностью
GSDLGKKLLE AARAGQDDEV RILLKAGADV NA (SEQ ID NO: 14) или
GSDLGKKLLE AARAGQDDEV RELLKAGADV NA (SEQ ID NO: 15), и при условии, что аминокислотный остаток в положении 24 в аминокислотной последовательности указанного N-терминального кэппирующего модуля представляет собой L, V, I или A;
и такие N-терминальные кэппирующие молекулы, в которых аминокислоты в положении 1 и/или 2 являются отсутствующими.
Такие связывающие белки демонстрируют улучшенную термальную стабильность по сравнению с таким же связывающим белком, отличающимся только одним N-терминальным кэппирующим модулем, например, по сравнению со связывающим белком с N-терминальными кэппинг модулями, известными из уровня техники, такими как N-терминальный кэппирующий модуль, имеющий аминокислотную последовательность с аминокислотой М (метионин) в положении 24, например, SEQ ID NO: 14 или SEQ ID NO: 15, в которых L в положении 24 является замененной М.
Изобретение также относится к связывающему белку, включающему, по крайней мере, один домен анкиринового повтора, где указанный домен анкиринового повтора включает C-терминальный кэппирующий модуль, который имеет аминокислотную последовательность X1DKX2GKTX3X4DX5X6X7DX8GX9EDX10AEX11LQKAA (SEQ ID NO: 6).
Изобретение также относится к молекулам нуклеиновой кислоты, которые кодируют связывающие белки в соответствии с настоящим изобретением, и к фармацевтической композиции, включающей упомянутые выше связывающие белки или молекулы нуклеиновой кислоты.
Изобретение также относится к способу лечения патологического состояния при использовании связывающих белков в соответствии с изобретением.
Краткое описание фигур
Фигура 1. Термальная стабильность DARPin#17 и DARPin#18.
Представлены результаты термальной денатурации DARPin#17 и DARPin#18.
Термальная денатурация сопровождалась повышением интенсивности флуоресценции красителя SYPRO оранжевого, присутствующего в PBS при pH 7,4. Значения Tm для DARPin#17 и DARPin#18 были оценены как такие на уровне 64,5°C и 71,0°C, соответственно.
F, относительные единицы флуоресценции (RFUs), возбуждение при 515-535 нм, определение при 560-580 нм; T, температура в °C; определение DARPin смотри ниже.
Фигура 2. Термальная стабильность DARPin#19 и DARPin 20.
Представлены результаты термальной денатурации DARPin #19 и DARPin#20.
Термальная денатурация сопровождалась CD сигналом на уровне 222 нм в PBS при pH 7,4. Значения Tm для DARPin#19 и DARPin#20 были оценены как такие на уровне 72,3°C и 74,8°C, соответственно.
FU, фракция развернутого белка; T, температура в °C; определение DARPin смотри ниже.
Фигура 3. Термальная стабильность DARPin#21 и DARPin#23
Представлены результаты термальной денатурации DARPin#21 и DARPin#23.
Термальная денатурация сопровождалась CD сигналом на уровне 222 нм в PBS при pH 7,4. Значения Tm для DARPin#21 и DARPin#23 были оценены как такие на уровне 56.5°C и 63.5°C, соответственно.
FU, фракция развернутого белка; T, температура в °C; определение DARPin смотри ниже.
Фигура 4. Термальная стабильность DARPin#24 и DARPin#26
Представлены результаты термальной денатурации DARPin#24 и DARPin#26.
Термальная денатурация сопровождалась CD сигналом на уровне 222 нм в PBS при pH 7,4. Значения Tm для DARPin#24 и DARPin#26 были оценены как такие на уровне 83°C и 89°C, соответственно.
RE, относительный CD сигнал при 222 нм, нормализованный к сигналу, измеренному при 20°C; T, температура в °C; определение DARPin смотри ниже.
Подробное описание изобретения
Термин "белок" относится к полипептиду, в котором, по крайней мере, часть полипептида имеет, или обладает способностью приобретать определенное пространственное расположение путем формирования вторичных, третичных или четвертичных структур в пределах полипептидного(ых) цепи(ей) и/или между ними. В том случае, если белок включает два или более полипептидов, то индивидуальные полипептидные цепи могут быть связаны нековалентно или ковалентно, например, с помощью дисульфидной связи между двумя полипептидами. Часть белка, которая индивидуально имеет или обладает способностью приобретать определенное пространственное расположение путем формирования вторичных или третичных структур, называется "белковым доменом". Такие белковые домены являются хорошо известными квалифицированному специалисту в данной области техники.
Термин "рекомбинантный", как используется в словосочетании рекомбинантный белок, рекомбинантный белковый домен, рекомбинантный связывающий белок и тому подобное, означает, что указанные полипептиды получают с помощью методик рекомбинантной ДНК, хорошо известных квалифицированному специалисту в релевантной области техники. Например, рекомбинантная молекула ДНК (например, полученная с помощью синтеза генов), которая кодирует полипептид, может быть клонирована в бактериальную экспрессионную плазмиду (например, pQE30, Qiagen), дрожжевую экспрессионную плазмиду или экспрессионную плазмиду млекопитающих. Когда, например, такая сконструированная рекомбинантная бактериальная экспрессионная плазмида встраивается в приемлемую бактерию (например, Escherichia coli), то эта бактерия может продуцировать полипептид, который кодируется этой рекомбинантной ДНК. Соответствующим образом полученный полипептид называется рекомбинантным полипептидом.
В контексте настоящего изобретения термин "полипептид" относится к молекуле, которая состоит из одной или более цепей многочисленных аминокислот, то есть, из двух или более, аминокислот, связанных с помощью пептидных связей. Предпочтительно, полипептид состоит более чем из восьми аминокислот, связанных с помощью пептидных связей.
Термин "полипептидная метка" относится к аминокислотной последовательности, которая присоединяется к полипептиду/белку, где указанная аминокислотная последовательность является полезной для очистки, определения или мечения указанного полипептида/белка, или где указанная аминокислотная последовательность улучшает физико-химическое поведение полипептида/белка, или где указанная аминокислотная последовательность обладает эффекторной функцией. Индивидуальные полипептидные метки, остатки и/или домены связывающего белка могут быть связаны друг с другом непосредственно или с помощью полипептидных линкеров. Эти полипептидные метки являются хорошо известными в области техники и полностью доступны квалифицированному специалисту в данной области техники. Примеры полипептидных меток представляют собой малые полипептидные последовательности, например, His (например, His-метка последовательности SEQ ID NO: 16), myc, FLAG или Strep-метки или остатки, такие как ферменты (например, ферменты, подобные щелочной фосфатазе), которые позволяют осуществлять определение указанного полипептида/белка, или остатки, которые могут использоваться для мечения (такие как иммуноглобулины или их фрагменты) и/или в качестве эффекторных молекул.
Термин "полипептидный линкер" относится к аминокислотной последовательности, которая является способной к связыванию, например, двух белковых доменов, полипептидной метки и белкового домена, белкового домена и остатка, отличного от полипептида, такого как полиэтиленгликоль или двух последовательностей метки. Такие дополнительные домены, метки, остатки, отличные от полипептидов, и линкеры являются хорошо известными квалифицированному специалисту в релевантной области техники. Список примеров обеспечивается в описании к патентной заявке WO 2002/020565. Частные примеры таких линкеров представляют собой глицин-серин линкеры и пролин-треонин линкеры варьирующей длины; является предпочтительным, когда указанные линкеры имеют длину от 2 до 24 аминокислот; более предпочтительно, когда указанные линкеры имеют длину от 2 до 16 аминокислот.
Термин "связывающий белок" относится к белку, включающему один или более связывающих доменов, одно или более биоактивных соединений и один или более полимерных остатков, как дополнительно объясняется ниже. Предпочтительно, когда указанный связывающий белок включает вплоть до четырех связывающих доменов. Более предпочтительно, когда указанный связывающий белок включает вплоть до двух связывающих доменов. Наиболее предпочтительно, когда указанный связывающий белок включает только один связывающий домен. Кроме того, любой такой связывающий белок может включать дополнительные белковые домены, которые не являются связывающими доменами, остатки мультимеризации, полипептидные метки, полипептидные линкеры и/или единичный остаток Cys. Примеры остатков мультимеризации представляют собой константные участки тяжелой цепи иммуноглобулина, которые спариваются с обеспечением функциональных Fc доменов иммуноглобулина, и лейциновые "застежки" или полипептиды, включающие свободный тиол, который образует межмолекулярную дисульфидную связь между двумя такими полипептидами. Единичный остаток Cys может использоваться для конъюгации других остатков к полипептиду, например, путем использования малеимидной связи, которая является хорошо известной квалифицированному специалисту в данной области техники. Предпочтительно, когда указанный связывающий белок представляет собой рекомбинантный связывающий белок. Также является предпочтительным, когда связывающие домены связывающего белка обладают различными целевыми специфичностями.
Термин "связывающий домен" означает белковый домен, который демонстрирует такую же сборку (пространственное размещение), что и белковый каркас и обладает предварительно определенным свойством, как определяется ниже. Такой связывающий домен может быть получен с помощью целесообразных, или в наиболее общем случае, комбинаторных методик конструирования белка, методов, которые являются известными в области техники (Binz и др., 2005, в приведенном выше месте). Например, связывающий домен, имеющий предварительно определенное свойство, может быть получен с помощью способа, включающего этапы (a) обеспечения разнообразной коллекции белковых доменов, которые демонстрируют такую же сборку, что и белковый каркас, как определено дополнительно ниже; и (b) скрининга указанной разнообразной коллекции и/или селекции из указанной разнообразной коллекции для получения, по крайней мере, одного белкового домена, обладающего указанным предварительно определенным свойством. Разнообразная коллекция белковых доменов может обеспечиваться с помощью нескольких способов в соответствии с используемой системой скрининга и/или селекции и может включать применение способов, хорошо известных квалифицированному специалисту в данной области техники, таких как фаговый дисплей или рибосомальный дисплей. Предпочтительно, указанный связывающий домен представляет собой рекомбинантный связывающий домен.
Термин "предварительно определенное свойство" относится к свойству, такому как, связывание с мишенью, блокирование мишени, активация опосредованной мишенью реакции, ферментативная активность и другие близкие свойства. В зависимости от типа желаемого свойства, средний специалист в данной области техники будет способен идентифицировать формат и необходимые этапы для осуществления скрининга и/или селекции связывающего домена с желаемым свойством. Предпочтительно, указанное предварительно определенное свойство представляет собой связывание с мишенью.
Предпочтительный связывающий белок включает, по крайней мере, один повторяемый домен.
Термины "имеет связывающую специфичность для мишени", "которые специфически связываются с мишенью" или "целевая специфичность" и тому подобные означают, что связывающий белок или связывающий домен связываются в PBS с мишенью с более низкой константой диссоциации, чем с неродственным белком, например, таким как связывающий мальтозу белок Е.coli (MBP). Предпочтительно, когда константа диссоциации в PBS для мишени является, по крайней мере в 10, более предпочтительно в 102, даже более предпочтительно в 103, или наиболее предпочтительно в 104 раз ниже, чем соответствующая константа диссоциации для MBP.
Способы определения констант диссоциации белок-белковых взаимодействий, такие как методики, которые основываются на поверхностном плазмонном резонансе (SPR) (например, SPR равновесный анализ) или изотермическая титрационная калориметрия (ITC), являются хорошо известными квалифицированному специалисту в данной области техники. Измеренные значения Kd конкретного белок-белкового взаимодействия могут варьировать, если измеряются при различных условиях (например, концентрации соли, pH). Таким образом, измерение значений Kd предпочтительно осуществляют при использовании стандартизированных растворов белка и стандартизированного буфера, такого как PBS.
Термин "мишень" относится к индивидуальной молекуле, такой как молекула нуклеиновой кислоты, полипептида или белка, углевода, или любой другой существующей в природе молекуле, включая любую часть такой индивидуальной молекулы, или к комплексам двух или более таких молекул. Мишень может представлять собой цельную клетку или образец ткани, или она может представлять собой любую не существующую в природе молекулу или остаток. Предпочтительно, когда мишень представляет собой существующий в природе или неприродный полипептид или полипептид, содержащий химические модификации, например, модифицированный с помощью природного или неприродного фосфорилирования, ацетилирования или метилирования.
Определения, приведенные в данной заявке ниже для белков повторов, основываются на таких, которые описаны в патентной заявке WO 2002/020565. Патентная заявка WO 2002/020565 дополнительно содержит общее описание характеристик белковых повторов, методик и применений.
Термин "повторяемые белки" относится к белкам, включающим один или более повторяемых доменов. Предпочтительно, каждый указанный белок повторов включает вплоть до четырех повторяемых доменов. Более предпочтительно, когда каждый указанный белок повторов включает вплоть до двух повторяемых доменов. Наиболее предпочтительно, когда каждый указанный повторяемый белок включает только один повторяемый домен. Кроме того, указанный белок повторов может включать дополнительные небелковые повторяемые домены, полипептидные метки и/или полипептидные линкеры.
Термин "повторяемый домен" относится к белковым доменам, включающим две или более последовательных повторяемых единиц (модулей) в качестве структурных единиц, где указанные структурные единицы имеют такую же сборку и плотно упаковываются с образованием, например, сферической структуры, имеющей соединительное гидрофобное ядро. Предпочтительно, повторяемый домен дополнительно включает N-терминальную и/или C-терминальную кэппирующую единицу (или модуль). Даже более предпочтительно, когда указанные N-терминальные и/или C-терминальные кэппирующие единицы (или модули) представляют собой кэппирующие повторы.
Термин "сконструированный повторяемый белок" и "сконструированный повторяемый домен" относится к повторяемому белку или повторяемому домену, соответственно, полученному в результате изобретательской процедуры, которая объясняется в патентной заявке WO 2002/020565. Сконструированные повторяемые белки и сконструированные повторяемые домены являются синтетическими, а не природными. Они могут представлять собой созданные человеком белки или домены, соответственно, полученные путем экспрессии соответственным образом сконструированных нуклеиновых кислот. Предпочтительно, экспрессию осуществляют в прокариотических или эукариотических клетках, таких как бактериальные клетки, или путем использования бесклеточной экспрессионной системы in vitro. В соответствии с этим, сконструированный белок с анкириновым повтором (то есть, DARPin) соответствует связывающему белку в соответствии с изобретением, включающему, по крайней мере, один домен анкиринового повтора.
Термин "структурная единица" относится к локально упорядоченной части полипептида, образованной пространственными взаимодействиями между двумя или более сегментами вторичной структуры, которые находятся рядом друг с другом в полипептидной цепи. Такая структурная единица демонстрирует структурный мотив. Термин "структурный мотив" относится к пространственному расположению элементов вторичной структуры, присутствующих, по крайней мере, в одной структурной единице. Структурные мотивы являются хорошо известными квалифицированному специалисту в данной области техники. Структурные единицы, взятые отдельно, не являются способными приобретать определенное пространственное расположение; однако их последовательное расположение, например, в виде повторяемых модулей в повторяемом домене, приводит к взаимной стабилизации соседних единиц, что приводит к образованию суперспиральной структуры.
Термин "повторяемая единица" относится к аминокислотной последовательности, включающей мотивы повторяемой последовательности одного или более существующих в природе повторяемых белков, где указанные "повторяемые единицы" обнаруживаются во множественных копиях, и которые демонстрируют определенную топологию сборки, общую для всех указанных мотивов, определяющих сборку этого белка. Такие повторяемые единицы соответствуют "повторяемым структурным единицам (повторам)" повторяемых белков, как описывается Forrer и др., 2003, в приведенном выше месте, или "последовательным структурным единицам (повторам)" повторяемых белков, как описывается Binz и др., 2004, в приведенном выше месте. Такие повторяемые единицы включают каркасные остатки и остатки взаимодействия. Примеры таких повторяемых единиц представляют собой повторяемые единицы армадилло, обогащенные лейцином повторяемые единицы, единицы анкиринового повтора, повторяемые единицы тетратрикопептида, повторяемые единицы HEAT и вариантные повторяемые единицы, обогащенные лейцином. Существующие в природе белки, содержащие две или более таких повторяемых единиц, называются "существующими в природе повторяемыми белками". Аминокислотные последовательности индивидуальных повторяемых единиц повторяемых белков могут иметь значительное количество мутаций, замен, инсерций и/или делеций по сравнению с каждым другим, в то время как все еще существенно сохраняют общую модель, или мотив повторяемой единицы.
Термин "единица анкиринового повтора" будет означать повторяемую единицу, которая представляет собой анкириновый повтор, как описывается, например, Forrer и др., 2003, в приведенном выше месте. Анкириновые повторы являются хорошо известными квалифицированному специалисту в данной области техники.
Термин "каркасные участки" относится к аминокислотным остаткам повторяемой единицы, или соответствующим аминокислотным остаткам повторяемых модулей, которые осуществляют свой вклад в топологию сборки, то есть те, которые осуществляют свой вклад в сборку указанной повторяемой единицы (или модуля), или те, которые осуществляют свой вклад во взаимодействие с соседней единицей (или модулем). Такой вклад может представлять собой взаимодействие с другими остатками в повторяемой единице (или модуле), или влияние на конформацию полипептидного скелета, как обнаруживается в α-спиралях или β-складках, или аминокислотных цепочках, которые формируют линейные полипептиды или петли.
Термин "остатки целевого взаимодействия" относится к аминокислотным остаткам повторяемой единицы, или к соответствующим аминокислотным остаткам повторяемых модулей, которые осуществляют свой вклад во взаимодействие с целевыми веществами. Такой вклад может представлять собой непосредственное взаимодействие с целевыми веществами, или влияние на другие непосредственно взаимодействующие остатки, например, путем стабилизации конформации полипептида повторяемой единицы (или модуля) для того, чтобы позволить осуществить или улучшить взаимодействие непосредственно взаимодействующих остатков с указанной целью. Такие каркасные остатки и целевые остатки взаимодействия могут быть идентифицированы с помощью анализа структурных данных, полученных с помощью физико-химических Методов, таких как рентгеноструктурная кристаллография, ЯМР и/или CD спектроскопия, или путем сравнения с известной и близкой структурной информацией, которая является хорошо известной практикующим специалистам в структурной биологии и/или биоинформатике.
Предпочтительно, когда повторяемые единицы, используемые для дедукции мотива повторяемой последовательности, являются гомологичными повторяемыми единицам, где повторяемые единицы включают тот же структурный мотив и в которых более чем 70% каркасных участков указанных повторяемых единиц являются гомологичными друг другу. Предпочтительно, более чем 80% каркасных участков указанных повторяемых единиц являются гомологичными. Наиболее предпочтительно, когда более чем 90% каркасных участков указанных повторяемых единиц являются гомологичными. Компьютерные программы для определения процента гомологии между полипептидами, такие как Fasta, Blast или Gap, являются известными квалифицированному специалисту в данной области техники. Также является предпочтительным, когда повторяемые единицы, используемые для дедукции мотива повторяемой последовательности, являются гомологичными повторяемыми единицам, полученным из повторяемых доменов, отобранных на мишени и имеющих такую же целевую специфичность.
Термин "мотив повторяемой последовательности" относится к аминокислотной последовательности, которая выводится из одной или более повторяемых единиц или повторяемых модулей. Предпочтительно, когда указанные повторяемые единицы или повторяемые модули являются такими из повторяемых доменов, имеющих связывающую специфичность для той же мишени. Такой мотив повторяемых последовательностей включает положения остатков каркасного участка и положения остатков целевого взаимодействия. Указанные положения остатков каркасного участка соответствуют положениям каркасных участков повторяемых единиц (или модулей). Подобно этому, указанные положения остатков целевого взаимодействия соответствуют положениям остатков целевого взаимодействия повторяемых единиц (или модулей). Мотив повторяемых последовательностей включает фиксированные положения и случайные положения. Термин "фиксированное положение" относится к аминокислотному положению в мотиве повторяемой последовательности, где указанное положение принимается за определенную аминокислоту. Наиболее часто, такие фиксированные положения соответствуют положениям каркасных участков и/или положениям остатков целевого взаимодействия, которые являются специфическими для определенной мишени. Термин "рандомизированное положение" относится к аминокислотному положению в мотиве повторяемой последовательности, в котором две или более аминокислот допускаются в указанном аминокислотном положении, например, в котором допускаются любые из обычных двадцати существующих в природе аминокислот, или в котором допускаются более двадцати существующих в природе аминокислот, таких, как аминокислоты, отличные от цистеина, или аминокислоты, отличные от глицина, цистеина и пролина. Наиболее часто, такие рандомизированные положения соответствуют положениям остатков целевого взаимодействия. Однако некоторые положения каркасных участков также могут быть рандомизированы.
Термин "топология сборки" относится к третичной структуре указанных повторяемых единиц или повторяемых модулей. Топология сборки будет определяться цепочками аминокислот, формирующими, по крайней мере, части α-спиралей или β-складок, или цепочками аминокислот, формирующими линейные полипептиды или петли, или любой комбинацией α-спиралей, β-складок и/или линейных полипептидов/петель.
Термин "последовательный" относится к размещению, в котором повторяемые единицы или повторяемые модули размещаются последовательно друг за другом. В сконструированных повторяемых белках, существует, по крайней мере, 2, обычно приблизительно от 2 до 6, в частности, по крайней мере, приблизительно 6, часто 20 или более повторяемых единиц (или модулей). В большинстве случаев повторяемые единицы (или модули) повторяемого домена будут демонстрировать высокую степень идентичности последовательности (одинаковые аминокислотные остатки в соответствующих положениях) или подобия последовательности (аминокислотные остатки являются различными, но имеют подобные физико-химические свойства), и некоторые из аминокислотных остатков могут представлять собой ключевые остатки, которые являются строго консервативными. Однако высокая степень вариабельности последовательности благодаря аминокислотным инсерциям и/или делециям, и/или заменам между различными повторяемыми единицами (или модулями) повторяемого домена может быть возможной при условии, что поддерживается общая топология сборки повторяемых единиц (или модулей).
Способы для непосредственного определения топологии сборки белков повторов при использовании физико-химических средств, таких как рентгеновская кристаллография, ЯМР или спектроскопия циркулярного дихроизма, являются хорошо известными практикующему специалисту в данной области техники. Способы идентификации и определения повторяемых единиц или мотивов повторяемых последовательностей или идентификации семейств родственных белков, включающих такие повторяемые единицы или мотивы, такие, как поиски гомологии (BLAST и т.д.), являются хорошо установленными в области биоинформатики и хорошо известны практикующему специалисту в данной области техники. Этап уточнения исходного мотива повторяемой последовательности может включать итеративный процесс.
Термин "повторяемые модули" относится к повторяемой аминокислотной последовательности сконструированных повторяемых доменов, которые исходно имеют происхождение от повторяемой единицы существующих в природе повторяемых белков. Каждый повторяемый модуль, который содержится в повторяемом домене, имеет происхождение от одной или более повторяемых единиц семейства или подсемейства существующих в природе повторяемых белков, например, семейства белковых повторов армадилло или белков с анкириновым повтором.
"Повторяемые модули" могут включать положения с аминокислотными остатками, присутствующими во всех копиях соответствующих повторяемых модулей ("фиксированные положения"), и положения с отличающимися или "рандомизированными" аминокислотными остатками ("рандомизированные положения").
Термин "кэппирующий модуль" относится к полипептиду, слитому с N- или C-терминальным повторяемым модулем повторяемого домена, где указанный кэппирующий модуль формирует плотные третичные взаимодействия (то есть, взаимодействия третичной структуры) с указанным повторяемым модулем, обеспечивая, таким образом, колпачок, который защищает гидрофобное ядро указанного повторяемого модуля со стороны, которая не находится в контакте с последовательным повторяемым модулем, от растворителя. Указанный N- и/или C-терминальный кэппирующий модуль может представлять собой, или может иметь происхождение от кэппирующей единицы или другой структурной единицы, которая обнаруживается в существующих в природе белковых повторах, соседствующих с повторяемой единицей. Термин "кэппирующая единица" относится к существующему в природе собранному полипептиду, где указанный полипептид определяет конкретную структурную единицу, которая является N- или C-терминально слитой с повторяемой единицей, где указанный полипептид формирует плотные взаимодействия третичной структуры с указанной повторяемой единицей, обеспечивая, таким образом, колпачок, который защищает гидрофобное ядро указанной повторяемой единицы с одной стороны, от растворителя. Предпочтительно, кэппинг модули или кэппирующие единицы представляют собой кэппирующие повторы. Термин "кэппирующий повтор" относится к кэппирующему модулю или кэппирующей единице, которая имеет подобную или такую же сборку, что и указанная соседняя повторяемая единица (или модуль), и/или подобность последовательности указанной соседней повторяемой единицы (или модуля). Кэппинг модули и кэппирующие повторы описываются в WO 2002/020565 и Interlandi и др., 2008 (в приведенном выше месте). Например, WO 2002/020565 описывает N-терминальный кэппирующий модуль (то есть, кэппирующий повтор), который имеет аминокислотную последовательность
GSDLGKKLLEAARAGQDDEVRILMANGADVNA (SEQ ID NO: 1) и
C-терминальный кэппирующий модуль (то есть кэппирующий повтор), имеющий аминокислотную последовательность
QDKFGKTAFDISIDNGNEDLAEILQKLN (SEQ ID NO: 2).
Interlandi и др., 2008 (в приведенном выше месте) описывают C-терминальные кэппинг модули, имеющие аминокислотные последовательности
QDKFGKTPFDLAIREGHEDIAEVLQKAA (SEQ ID NO: 3) и
QDKFGKTPFDLAIDNGNEDIAEVLQKAA (SEQ ID NO: 4).
Например, N-терминальный кэппирующий модуль последовательности SEQ ID NO: 17 кодируется аминокислотами от положения 1 до положения 32, а C-терминальный кэппирующий модуль последовательности SEQ ID NO: 17 кодируется аминокислотами от положения 99 до положения 126.
Настоящее изобретение относится к связывающему белку, включающему, по крайней мере, один домен анкиринового повтора, где указанный домен анкиринового повтора включает N-терминальный кэппирующий модуль, имеющий аминокислотную последовательность
GSDLGKKLLE AARAGQDDEV RILLKAGADV NA (SEQ ID NO: 14) или
GSDLGKKLLE AARAGQDDEV RELLKAGADV NA (SEQ ID NO: 15), в которой аминокислотный остаток L в положении 24 SEQ ID NO: 14 или SEQ ID NO: 15 является необязательно замененным V, I или A;
вплоть до 9 аминокислот SEQ ID NO: 14 или SEQ ID NO: 15 в других положениях, отличных от положения 24, необязательно являются замененными любыми аминокислотами; и
в которой G в положении 1 и/или S в положении 2 SEQ ID NO: 14 или SEQ ID NO: 15 являются необязательно отсутствующими.
Было обнаружено, что положение 24 в N-терминальном кэппирующем модуле не должно представлять собой метионин (M). В последовательности SEQ ID NO: 14 и SEQ ID NO: 15, положение 24 представляет собой лейцин (L). Аминокислота в этом положении может представлять собой V, I или A. Предпочтительной является L в положении 24, или замещение с помощью А. Наиболее предпочтительным является L в положении 24.
Принцип замещения метионина в положении 24 может быть применен к разнообразию других N-терминальных кэппирующих модулей. Как следствие этого, предмет в соответствии с изобретением также включает все те N-терминальные кэппинг модули, которые отличаются от аминокислотной последовательности SEQ ID NO: 14 и SEQ ID NO: 15 путем замены вплоть до 9 аминокислот в других положениях, отличных от положения 24. Более предпочтительно такие N-терминальные кэппинг модули отличаются замещением 8 аминокислот, более предпочтительно 7 аминокислот, более предпочтительно 6 аминокислот, более предпочтительно 5 аминокислот, даже более предпочтительно 4 аминокислот, более предпочтительно 3 аминокислот, более предпочтительно 2 аминокислот, и наиболее предпочтительно 1 аминокислоты.
Замещение аминокислот может осуществляться при использовании любой из 20 наиболее часто встречающих в природе аминокислот, предпочтительно с помощью аминокислот, выбранных из группы, состоящей из A, D, E, F, H, I, K, L, M, N, Q, R, S, T, V, W и Y; и более предпочтительно из группы, состоящей из A, D, E, H, I, K, L, Q, R, S, T, V, и Y. Также предпочтительно, замена аминокислот осуществляется с гомологичной аминокислоты; аминокислота заменяется аминокислотой, которая имеет боковую цепь с подобными биофизическими свойствами. Например, негативно заряженная аминокислота D может быть заменена негативно заряженной аминокислотой E, или гидрофобная аминокислота, такая как L, может быть заменена A, I или V. Замещение аминокислоты гомологичной аминокислотой является хорошо известным квалифицированному специалисту в данной области техники.
Аминокислота G в положении 1 и/или S в положении 2 последовательности SEQ ID NO: 14 или SEQ ID NO: 15 может быть удалена из N-терминальных кэппинг модулей при отсутствии видимого влияния на свойства. Эти две аминокислоты служат в качестве линкеров для связывания домена анкиринового повтора с дополнительными аминокислотами и белками. Изобретение также включает такие N-терминальные кэппинг модули, в которых G в положении 1 и/или S в положении 2 удалены. При этом является понятным, что "положение 24", как определяется в данной заявке, соответствующим образом адаптируется, приводя к положению 23, если одна аминокислота отсутствует, или положению 22, если 2 аминокислоты отсутствуют, соответственно.
Замена метионина в положении 24 в N-терминальном кэппирующем модуле обеспечивает более высокую термальную стабильность, то есть более высокое значение Tm в PBS, домену анкиринового повтора по сравнению с доменом анкиринового повтора, который имеет идентичную аминокислотную последовательность, включая N-терминальный кэппирующий модуль за исключением того, что аминокислотный остаток его N-терминального кэппирующего модуля, соответствующий положению 24 SEQ ID NO: 14 или SEQ ID NO: 15 представляет собой M вместо L, V, I или A. Примеры таких пар доменов анкиринового повтора и связывающих белков (M в положении 24 против L, V, I или A в положении 24) и их значение Tm описываются в разделе Примеры и являются представленными на Фигурах. Предпочтительными являются N-терминальные кэппинг модули, в которых замена M в положении 24 другой аминокислотой приводит к повышению значения Tm, по крайней мере, на 1°C, предпочтительно, по крайней мере, на 2°C, более предпочтительно, по крайней мере, на 3°C, или наиболее предпочтительно, по крайней мере, на 4°C, в домене анкиринового повтора, который несет такие N-терминальные кэппирующие модули.
Термальная стабильность белка и домена анкиринового повтора, в частности, может подвергаться анализу с помощью анализа термальной стабильности на основе флуоресценции (Niesen, F.H., Nature Protocols 2(9): 2212-2221, 2007). Таким образом, температуру, при которой белок развертывается, измеряется путем повышения флуоресценции красителя с аффинностью для гидрофобных частей белка, которые подвергаются развертыванию белка. Полученное таким образом среднее значение перехода флуоресценции (от более низкой интенсивности флуоресценции до более высокой интенсивности флуоресценции) соответствует среднему значению температуры денатурации (Tm) анализируемого белка. Альтернативно, термальную стабильность белка можно анализировать с помощью CD спектроскопии; то есть путем измерения его тепловой денатурации в соответствии с его сигналом циркулярного дихроизма (CD) при 222 нм с помощью методик, известных квалифицированному специалисту в данной области техники.
В одном воплощении тогда, когда вплоть до 9 аминокислот SEQ ID NO: 14 или SEQ ID NO: 15 в других положениях, отличных от положения 24, являются необязательно замененными другими аминокислотами, предпочтительно аминокислотный остаток A в положении 26 SEQ ID NO: 14 или SEQ ID NO: 15 заменяется H, Y, K или R. Более предпочтительно, однако, когда аминокислотный остаток А в положении 26 не замещается.
В дополнительном воплощении тогда, когда вплоть до 9 аминокислот SEQ ID NO: 14 или SEQ ID NO: 15 в других положениях, отличных от положения 24, являются необязательно замененными другими аминокислотами, предпочтительно аминокислотный остаток R в положении 21 SEQ ID NO: 14 или SEQ ID NO: 15 заменяется Е. Более предпочтительно, однако, когда аминокислотный остаток R в положении 26 не замещается.
В дополнительном воплощении тогда, когда вплоть до 9 аминокислот SEQ ID NO: 14 или SEQ ID NO: 15 в других положениях, отличных от положения 24, являются необязательно замененными другими аминокислотами, предпочтительно аминокислотный остаток I в положении 22 SEQ ID NO: 14 или аминокислотный остаток E в положении 22 последовательности SEQ ID NO: 15 заменяется V. Более предпочтительно, однако, когда аминокислотный остаток I или Е, соответственно, в положении 22 не замещается; смотри, например, пару соединений, показанных на Фигуре 2.
В дополнительном воплощении тогда, когда вплоть до 9 аминокислот SEQ ID NO: 14 или SEQ ID NO: 15 в других положениях, отличных от положения 24, являются необязательно замененными другими аминокислотами, предпочтительно аминокислотный остаток K в положении 25 SEQ ID NO: 14 или SEQ ID NO: 15 заменяется A или E. Более предпочтительно, однако, когда аминокислотный остаток K в положении 25 не замещается или замещается A, как продемонстрировано парой соединений на Фигуре 1.
В дополнительном воплощении тогда, когда вплоть до 9 аминокислот SEQ ID NO: 14 или SEQ ID NO: 15 в других положениях, отличных от положения 24, являются необязательно замененными другими аминокислотами, предпочтительно аминокислотные остатки RILLKA от положения 21 до положения 26 последовательности SEQ ID NO: 14 или аминокислотные остатки RELLKA от положения 21 до положения 26 SEQ ID NO: 15 не замещаются.
Дополнительный предпочтительный N-терминальный кэппирующий модуль включает мотив последовательности
GSX1LX2KKLLE AARAGQDDEV X3X4LX5X6X7GADV NA (SEQ ID NO: 5), в которой G в положении 1 и/или S в положении 2 SEQ ID NO: 5 являются необязательно отсутствующими;
X1 представляет собой аминокислотный остаток G, A, или D; предпочтительно, A или D;
X2 представляет собой аминокислотный остаток G или D;
X3 представляет собой аминокислотный остаток R или E;
X4 представляет собой аминокислотный остаток I, E или V; предпочтительно, I или E;
X5 представляет собой аминокислотный остаток L, V, I или A; предпочтительно, L или А;
X6 представляет собой аминокислотный остаток A, K или E; предпочтительно, A или K; и
X7 представляет собой аминокислотный остаток, выбранный из группы, состоящей из A, H, Y, K и R; предпочтительно A или H.
В другом воплощении N-терминальный кэппирующий модуль включает последовательность
X1LX2KKLLEAARAGQDDEVRILX3AX4GADVNA (SEQ ID NO: 13),
в которой X1 представляет собой аминокислотный остаток G, A или D;
в которой X2 представляет собой аминокислотный остаток G или D;
в которой X3 представляет собой аминокислотный остаток L, V, I или A; предпочтительно L; и
в которой X4 представляет собой аминокислотный остаток A, H, Y, K, R или N; предпочтительно, A или N.
Наиболее предпочтительными являются связывающие белки, включающие, по крайней мере, один домен анкиринового повтора, где указанный домен анкиринового повтора включает N-терминальный кэппирующий модуль, имеющий аминокислотную последовательность
GSDLGKKLLE AARAGQDDEV RILLKAGADV NA (SEQ ID NO: 14) или
GSDLGKKLLE AARAGQDDEV RELLKAGADV NA (SEQ ID NO: 15), в которой G в положении 1 и/или S в положении 2 SEQ ID NO: 14 и SEQ ID NO: 15 являются необязательно отсутствующими.
В другом воплощении изобретение относится к связывающему белку, включающему, по крайней мере, один домен анкиринового повтора, где указанный домен анкиринового повтора включает N-терминальный кэппирующий модуль, имеющий аминокислотную последовательность, характеризующуюся, по крайней мере, 70%-ной идентичностью аминокислотной последовательности с
GSDLGKKLLE AARAGQDDEV RILLKAGADV NA (SEQ ID NO: 14) или
GSDLGKKLLE AARAGQDDEV RELLKAGADV NA (SEQ ID NO: 15), и при условии, что аминокислотный остаток в положении 24 в аминокислотной последовательности указанного N-терминального кэппирующего модуля представляет собой L, V, I или A;
и такие N-терминальные кэппирующие молекулы, где аминокислоты в положении 1 и/или 2 являются отсутствующими.
Предпочтительно, когда соответствующие N-терминальные кэппинг модули имеют аминокислотные последовательности, характеризующиеся, по крайней мере, 75%-ной идентичностью аминокислотной последовательности с SEQ ID NO: 14 или SEQ ID NO: 15, более предпочтительно 80%-ной идентичностью аминокислотной последовательности, даже более предпочтительно 85%-ной идентичностью аминокислотной последовательности, более предпочтительно 90%-ной идентичностью аминокислотной последовательности, и наиболее предпочтительно 95%-ной идентичностью аминокислотной последовательности, всегда при условии, что аминокислотный остаток в положении 24 в аминокислотной последовательности указанного N-терминального кэппирующего модуля, представляет собой L, V, I или A, более предпочтительно L или A, и наиболее предпочтительно L.
В частных воплощениях N-терминальные кэппинг модули имеют указанный процент идентичности аминокислотной последовательности с SEQ ID NO: 14 или SEQ ID NO: 15, и аминокислотный остаток A, H, Y, K или R в положении 26, и/или аминокислотный остаток R или E в положении 21, всегда при условии, что аминокислотный остаток в положении 24 в аминокислотной последовательности указанного N-терминального кэппирующего модуля, представляет собой L, V, I или A.
Дополнительно предпочтительным является любой такой N-терминальный кэппирующий модуль, который включает N-терминальный кэппирующий повтор, в котором один или более аминокислотных остатков в указанном кэппирующем повторе заменены аминокислотным остатком, который обнаруживается в соответствующем положении в выравнивании соответствующей кэппирующей единицы или повторяемой единицы.
Связывающий белок в соответствии с изобретением, включающий домен анкиринового повтора, включает N-терминальный кэппирующий модуль, как определяется в данной заявке, при этом указанный домен анкиринового повтора может дополнительно содержать один из следующих C-терминальных кэппирующих модулей. Предпочтительный C-терминальный кэппирующий модуль включает мотив последовательности
X1DKX2GKTX3X4D X5X6X7DX8GX9EDX10 AEX11LQKAA (SEQ ID NO: 6), в которой X1 представляет собой аминокислотный остаток Q или K;
X2 представляет собой аминокислотный остаток A, S или F; предпочтительно, S или F;
X3 представляет собой аминокислотный остаток A или P;
X4 представляет собой аминокислотный остаток A или F;
X5 представляет собой аминокислотный остаток I или L;
X6 представляет собой аминокислотный остаток S или A;
X7 представляет собой аминокислотный остаток I или A;
X8 представляет собой аминокислотный остаток A, E или N; предпочтительно, A или N;
X9 представляет собой аминокислотный остаток N или H;
X10 представляет собой аминокислотный остаток L или I;
X11 представляет собой аминокислотный остаток I или V; и
X2 не является F, если X4 представляет собой F, и X7 представляет собой I, а X8 представляет собой N или E.
Дополнительный предпочтительный C-терминальный кэппирующий модуль включает мотив последовательности
X1DKX2GKTX3AD X4X5X6DX7GX8EDX9 AEX10LQKAA (SEQ ID NO: 7), в которой
X1 представляет собой аминокислотный остаток Q или K;
X2 представляет собой аминокислотный остаток A, S или F; предпочтительно, S или F;
X3 представляет собой аминокислотный остаток A или P;
X4 представляет собой аминокислотный остаток I или L;
X5 представляет собой аминокислотный остаток S или A;
X6 представляет собой аминокислотный остаток I или А;
X7 представляет собой аминокислотный остаток A, E или N; предпочтительно, A или N;
X8 представляет собой аминокислотный остаток N или H;
X9 представляет собой аминокислотный остаток L или I; и
X10 представляет собой аминокислотный остаток I или V.
Дополнительный предпочтительный C-терминальный кэппирующий модуль включает мотив последовательности
X1DKX2GKTX3AD X4X5ADX6GX7EDX8 AEX9LQKAA (SEQ ID NO: 8), в которой
X1 представляет собой аминокислотный остаток Q или K;
X2 представляет собой аминокислотный остаток A, S или F; предпочтительно, S или F;
X3 представляет собой аминокислотный остаток A или P;
X4 представляет собой аминокислотный остаток I или L;
X5 представляет собой аминокислотный остаток S или A;
X6 представляет собой аминокислотный остаток A, E или N; предпочтительно, A или N;
X7 представляет собой аминокислотный остаток N или H;
X8 представляет собой аминокислотный остаток L или I; и
X9 представляет собой аминокислотный остаток I или V.
Предпочтительно, когда такой C-терминальный кэппирующий модуль, включающий мотив последовательности SEQ ID NO: 6, 7 или 8, имеет аминокислотный остаток A, I или K; предпочтительно, I или K; в положении, которое соответствует положению 3 указанного мотива последовательности.
Также предпочтительно, когда такой C-терминальный кэппирующий модуль, включающий мотив последовательности SEQ ID NO: 6, 7 или 8, имеет аминокислотный остаток R или D в положении, которое соответствует положению 14 указанного мотива последовательности.
Предпочтительный C-терминальный кэппирующий модуль представляет собой С-терминальный кэппирующий модуль, имеющий аминокислотную последовательность QDKSGKTPADLAADAGHEDIAEVLQKAA (SEQ ID NO: 9).
Изобретение также относится к связывающему белку, включающему, по крайней мере, один домен анкиринового повтора, где указанный домен анкиринового повтора включает C-терминальный кэппирующий модуль, имеющий аминокислотную последовательность SEQ ID NO: 6, 7 или 8, как определяется в данной заявке выше.
Домен анкиринового повтора в соответствии с изобретением может быть сконструирован генетически путем объединения N-терминального кэппирующего модуля (то есть, N-терминального кэппирующего повтора последовательности SEQ ID NO: 14), после которого идут один или более повторяемых модулей (то есть, повторяемых модулей, включающих аминокислотные остатки от положения 33 до положения 98 последовательности SEQ ID NO: 17), и C-терминального кэппирующего модуля (то есть, C-терминального кэппирующего повтора последовательности SEQ ID NO: 9) с помощью методов генетического синтеза. Генетически сконструированный ген повторяемого домена может потом экспрессироваться в Е.coli так, как описано выше.
Дополнительно предпочтительным является связывающий белок, повторяемый домен, N-терминальный кэппирующий модуль или C-терминальный кэппирующий модуль, имеющий аминокислотную последовательность, в которой отсутствуют аминокислоты C, M или N.
Дополнительно предпочтительным является связывающий белок, повторяемый домен, N-терминальный кэппирующий модуль или C-терминальный кэппирующий модуль, имеющий аминокислотную последовательность, в которой отсутствует аминокислота N, после которой следует G.
Также предпочтительными являются несуществующие в природе кэппирующие модули, повторяемые модули, связывающие белки или связывающие домены.
Термин "несуществующий в природе" означает синтетический или, в частности, неприродный, термин означает такой, который создан человеком. Термин "несуществующий в природе связывающий белок" или "несуществующий в природе связывающий домен" означает, что указанный связывающий белок или указанный связывающий домен является синтетическим (то есть, полученным с помощью химического синтеза из аминокислот) или рекомбинантным, а не природным. "Несуществующий в природе связывающий белок" или "несуществующий в природе связывающие домен" представляет собой созданный человеком белок или домен, соответственно, полученный путем экспрессии соответственно сконструированных нуклеиновых кислот. Предпочтительно, экспрессию осуществляют в эукариотических или бактериальных клетках, или путем использования бесклеточной in vitro экспрессионной системы. Кроме того, термин означает, что последовательность указанного связывающего белка или указанного связывающего домена не является присутствующей в виде неискусственной последовательности в базе данных последовательностей, например, в GenBank, EMBL-Bank или Swiss-Prot. Эти базы данных и другие базы данных последовательностей являются хорошо известными специалисту в данной области техники.
Термин "PBS" означает забуференный фосфатом водный раствор, содержащий 137 мМ NaCl, 10 мМ фосфата и 2,7 мМ KCl и имеющий значение pH 7,4.
В одном частном воплощении изобретение относится к связывающему белку, включающему домен анкиринового повтора, включающий N-терминальный кэппирующихй модуль в соответствии с изобретением и включающий биоактивное соединение.
Термин "биоактивное соединение" относится к соединению, которое изменяет заболевание, когда применяется к млекопитающему, которое имеет указанное заболевание. Биоактивное соединение может иметь агонистические или антагонистические свойства и может представлять собой белковое биоактивное соединение или небелковое биоактивное соединение.
Такие белковые биоактивные соединения могут быть ковалентно присоединены, например, к домену анкиринового повтора в соответствии с изобретением путем получения генетически слитых полипептидов при использовании стандартных методик клонирования ДНК, после чего осуществляют их стандартную экспрессию и очистку.
Такие небелковые биоактивные соединения могут быть ковалентно присоединены, например, к связывающему домену в соответствии с изобретением, химическим путем, например, путем слияния тиола цистеина через малеимидный линкер с цистеином, слитым с помощью пептидного линкера с N- или C-терминальным концом связывающего домена, как описывается в данной заявке.
Примеры белковых биоактивных соединений представляют собой связывающие домены, которые обладают отличной специфичностью по отношению к мишени (например, нейтрализация фактора роста с помощью его связывания), цитокинам (например, интерлейкинам), фактору роста (например, человеческому гормону роста), антителам и их фрагментам, гормонам (например, GLP-1) и любому возможному белковому лекарственному средству.
Примеры небелковых биоактивных соединений представляют собой токсины (например, DM1 от ImmunoGen), малые молекулы, нацеливающие GPCR, антибиотики и любое возможное белковое лекарственное средство.
Другое предпочтительное воплощение представляет собой рекомбинантный связывающий белок, включающий связывающий домен, где указанный связывающий домен представляет собой домен анкиринового повтора или сконструированный домен анкиринового повтора. Такой домен анкиринового повтора может включать один, два, три или более внутренних повторяемых модулей, которые будут участвовать в связывании с мишенью. Предпочтительно, такой домен анкиринового повтора включает нетерминальный кэппирующий модуль, от двух до четырех повторяемых модулей и C-терминальный кэппирующий модуль. Предпочтительно, указанный связывающий домен представляет собой домен анкиринового повтора или сконструированный домен анкиринового повтора.
Предпочтительным является связывающий белок, как определяется выше, где указанный домен анкиринового повтора или указанный сконструированные домен анкиринового повтора включает повторяемый модуль с мотивом последовательности анкиринового повтора
X1DX2X3GX4TPLHLAAX5X6GHLEIVEVLLKX7GADVNA (SEQ ID NO: 10),
в которой X1, X2, X3, X4, X5, X6 и X7, независимо друг от друга представляют собой аминокислотный остаток, выбранный из группы, состоящей из A, D, E, F, H, I, K, L, M, N, Q, R, S, T, V, W и Y; предпочтительно,
X1 представляет собой аминокислотный остаток, выбранный из группы, состоящей из A, D, М, F, S, I, Т, N, Y и K; более предпочтительно K и A; и
X7 представляет собой аминокислотный остаток, выбранный из группы, состоящей из S, A, Y, H и N; более предпочтительно, Y или H.
В дополнительных воплощениях любой из связывающих белков или доменов, описанных в данной заявке, может быть ковалентно связан с одним или более дополнительных остатков, включая, например, остаток, который связывается с различными мишенями для создания связывающего агента с двойной специфичностью, биоактивного соединения, остатка для мечения (например, флуоресцентной метки, такой как флуоресцеин, или радиоактивной метки), остатка, который способствует очистке белка (например, небольшая пептидная метка, такая как His- или strep-метка), остаток, который обеспечивает эффекторные функции для улучшенной терапевтической эффективности (например, Fc часть антитела для обеспечения зависимой от антитела клеточной цитотоксичности, токсический белковый остаток, такой как экзотоксин A Pseudomonas aeruginosa (ETA) или небольшой молекулярный токсический агент, такой как майтансиноиды или ДНК алкилирующие агенты) или остатка, который обеспечивает улучшенные фармакокинетические характеристики. Улучшенные фармакокинетические характеристики могут быть оценены в соответствии с имеющейся терапевтической необходимостью. Часто является желательным повысить биодоступность и/или увеличить время между дозами, возможно, посредством увеличения времени, в течение которого белок остается доступным в сыворотке крови после введения дозы. В некоторых случаях является желательным улучшить непрерывность концентрации белка в сыворотке крови в течение периода времени (например, снизить различия в сывороточной концентрации белка между концентрацией непосредственно после введения и концентрацией непосредственно перед последующим введением).
В дополнительном воплощении изобретение относится к молекулам нуклеиновой кислоты, которые кодируют частные связывающие белки, частные домены анкиринового повтора и частные N-терминальные кэппирующие модули. Также предполагается вектор, включающий указанные молекулы нуклеиновой кислоты.
Кроме того, рассматривается также фармацевтическая композиция, включающая один или более из упомянутых выше связывающих белков, включающих домены анкиринового повтора, или молекулы нуклеиновой кислоты, которые кодируют конкретные связывающие белки, и необязательно, фармацевтически приемлемый носитель и/или разбавитель. Фармацевтически приемлемые носители и/или разбавители являются известными квалифицированному специалисту в данной области техники и рассматриваются более подробно ниже. Кроме того, предполагаются также диагностическая композиция, включающая один или более из упомянутых выше связывающих белков, в частности, связывающие белки, включающие домены анкиринового повтора.
Фармацевтические композиции включают связывающие белки так, как описано выше, и фармацевтически приемлемый носитель, наполнитель или стабилизатор, например, такой, как описывается в Remington’s Pharmaceutical Sciences 16ое издание, Osol, А. ред. [1980]. Приемлемые носители, наполнители или стабилизаторы, известные квалифицированному специалисту, представляют собой физиологический раствор, раствор Рингера, раствор декстрозы, раствор Хэнкса, жирные растительные масла, этилолеат, 5% декстрозу в физиологическом растворе, вещества, которые улучшают изотоничность и химическую стабильность, буферы и консервирующие агенты. Другие приемлемые носители включают любой носитель, который сам по себе не индуцирует выработку антител, которые являются вредными для индивидуума, получающего композицию, такой, как белки, полисахариды, полимеры молочной кислоты, полигликолевые кислоты, полимерные аминокислоты и сополимеры аминокислот. Фармацевтические композиции могут также представлять собой комбинационные композиции, включающие дополнительный активный агент, такой как противораковый агент или антиангиогенный агент.
Композиции, используемые для введения in vivo, должны быть асептическими или стерильными. Это может легко осуществляться с помощью фильтрации через стерильные фильтрационные мембраны.
Фармацевтические композиции могут вводиться с помощью любого приемлемого способа, который является известным специалисту в данной области техники. Предпочтительный путь введения представляет собой парентеральный. При парентеральном введении лекарственное средство в соответствии с изобретением будет рецептироваться в виде единичной дозированной формы, которая вводится путем инъекции, такой как раствор, суспензия или эмульсия, в сочетании с фармацевтически приемлемыми наполнителями, как определяется выше. Дозированная форма и способ введения будут зависеть от индивидуума, которого подвергают лечению, и частного заболевания. В общем случае фармацевтические композиции вводятся так, что связывающий белок в соответствии с настоящим изобретением вводится при дозе от 1 мкг/кг до 20 мг/кг, более предпочтительно от 10 мкг/кг до 5 мг/кг, наиболее предпочтительно от 0,1 до 2 мг/кг. Предпочтительно, композицию вводят в виде болюсной дозы. Беспрерывная инфузия может также использоваться и включает беспрерывную подкожную доставку с помощью осмотического мини-насоса. В этом случае фармацевтические композиции могут вводиться путем инфузии при дозе от 5 до 20 мкг/кг/минута, более предпочтительно от 7 до 15 мкг/кг/минута.
Кроме того, любая из упомянутых выше фармацевтических композиций предполагается для лечения заболевания. Изобретение также обеспечивает способы лечения. Такой способ включает введение пациенту, который в этом нуждается, терапевтически эффективного количества связывающего белка в соответствии с изобретением.
Также предполагается способ лечения патологического состояния у млекопитающего, включая человека, включающий введение пациенту, который в этом нуждается, эффективного количества упомянутых выше фармацевтических композиций.
Связывающий белок в соответствии с изобретением может также быть получен и/или выделен с помощью нескольких способов, таких как дисплей на поверхности бактериофагов (WO 1990/002809, WO 2007/006665) или бактериальных клеток (WO 1993/010214), рибосомальный дисплей (WO 1998/048008), дисплей на плазмидах (WO 1993/008278) или путем использования гибридных конструкций на основе ковалентных РНК-белковых повторов (WO 2000/032823), или внутриклеточной экспрессии и селекции / скрининга, например, комплементационного анализа белка (WO 1998/341120). Такие способы являются известными квалифицированному специалисту в данной области техники.
Библиотека белков анкириновых повторов, которая используется для селекции/скрининга связывающих белков в соответствии с изобретением, может быть получена в соответствии с прописями, известными квалифицированному специалисту в области техники (WO 2002/020565, Binz, Н.K., и др., J. Mol. Biol., 332, 489-503, 2003, и Binz и др., 2004, в упомянутом выше месте). Повторяемые домены в соответствии с настоящим изобретением могут быть модульно собраны из повторяемых модулей в соответствии с настоящим изобретением и приемлемых кэппинг модулей или кэппирующих повторов (Forrer, Р., и др., FEBS letters 539, 2-6, 2003) при использовании стандартных методик рекомбинантной ДНК (например, WO 2002/020565, Binz и др., 2003, в приведенном выше месте и Binz и др., 2004, в приведенном выше месте).
Изобретение не является ограниченным частными воплощениями, описанными в Примерах. Все источники могут использоваться и подвергаться развитию в соответствии с общей процедурой, описанной ниже.
Примеры
Все исходные материалы и реагенты, раскрытые ниже, являются известными квалифицированному специалисту в данной области техники и являются коммерчески доступными или могут быть получены при использовании хорошо известных методик.
Материалы
Реактивы закупали от Fluka (Швейцария). Олигонуклеотиды были такими от Microsynth (Швейцария). Если не указано иное, то ДНК полимеразы, рестрикционные ферменты и буферы были такими от New England Biolabs (USA) или Fermentas (Литва). Штамм для клонирования и продукции белка представлял собой Е.coli XL1-blue (Stratagene, USA) или BL21 (Novagen, USA).
Молекулярная биология
Если не указано иное, то способы осуществляли в соответствии с описанными прописями (Sambrook J., Fritsch E.F. и Maniatis Т., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory 1989, New York).
DARPin, используемые в Примерах
DARPin#17 (SEQ ID NO: 17 с гистидиновой меткой (SEQ ID NO: 16), слитой с его N-терминальным концом);
DARPin#18 (SEQ ID NO: 18 с гистидиновой меткой (SEQ ID NO: 16), слитой с его N-терминальным концом);
DARPin#19 (SEQ ID NO: 19 с гистидиновой меткой (SEQ ID NO: 16), слитой с его N-терминальным концом);
DARPin#20 (SEQ ID NO: 20 с гистидиновой меткой (SEQ ID NO: 16), слитой с его N-терминальным концом);
DARPin#21 (SEQ ID NO: 21 с гистидиновой меткой (SEQ ID NO: 16), слитой с его N-терминальным концом);
DARPin#22 (SEQ ID NO: 22 с гистидиновой меткой (SEQ ID NO: 16), слитой с его N-терминальным концом);
DARPin#23 (SEQ ID NO: 23 с гистидиновой меткой (SEQ ID NO: 16), слитой с его N-терминальным концом);
DARPin#24 (SEQ ID NO: 24 с гистидиновой меткой (SEQ ID NO: 16), слитой с его N-терминальным концом);
DARPin#25 (SEQ ID NO: 25 с гистидиновой меткой (SEQ ID NO: 16), слитой с его N-терминальным концом);
DARPin#26 (SEQ ID NO:26 с гистидиновой меткой (SEQ ID NO: 16), слитой с его N-терминальным концом);
DARPin#27 (SEQ ID NO: 27 с гистидиновой меткой (SEQ ID NO: 16), слитой с его N-терминальным концом);
DARPin#28 (SEQ ID NO: 28 с гистидиновой меткой (SEQ ID NO: 16), слитой с его N-терминальным концом);
DARPin#29 (SEQ ID NO: 29 с гистидиновой меткой (SEQ ID NO: 16), слитой с его N-терминальным концом);
DARPin#30 (SEQ ID NO: 30 с гистидиновой меткой (SEQ ID NO: 16), слитой с его N-терминальным концом);
DARPin#31 (SEQ ID NO: 31 с гистидиновой меткой (SEQ ID NO: 16), слитой с его N-терминальным концом);
DARPin#32 (SEQ ID NO: 32 с гистидиновой меткой (SEQ ID NO: 16), слитой с его N-терминальным концом).
Сконструированные библиотеки белков с анкириновым повтором
Описываются библиотеки N2C и N3C сконструированных белков с анкириновым повтором (WO 2002/020565; Binz и др. 2003, в приведенном выше месте; Binz и др. 2004, в приведенном выше месте). Символ в N2C и N3C описывает количество рандомизированных повторяемых модулей, которые присутствуют между N-терминальным и C-терминальным кэппирующими модулями. Номенклатура, которая используется для определения положений внутри повторяемых единиц и модулей, основывается на Binz и др., 2004, в приведенном выше месте, с модификацией, которая граничит с модулями анкиринового повтора, а единицы анкиринового повтора смещаются на одно положение аминокислоты. Например, положение 1 модуля анкиринового повтора в соответствии с Binz и др. 2004 (в приведенном выше месте) соответствует положению 2 модуля анкиринового повтора в соответствии с настоящей заявкой и, следовательно, положению 33 модуля анкиринового повтора в соответствии Binz и др. 2004, в приведенном выше месте, соответствует положению 1 следующего модуля анкиринового повтора в соответствии с настоящей заявкой.
Все последовательности ДНК были подтверждены секвенированием, а подсчитанный молекулярный вес для всех описанных белков был подтвержден с помощью масс-спектометрии.
Пример 1: Конструирование, экспрессия и очистка DARPin
DARPin, которые имеютопределенные аминокислотные последовательности, могут быть получены с помощью генного синтеза соответствующей обратно транслированной последовательности нуклеиновой кислоты, субклонирования в приемлемый экспрессионный вектор экспрессионной системы (например экспрессионной системы Е.coli), экспрессии и очистки белка. Такие способы являются известными квалифицированному специалисту в данной области техники.
Замена кэппирующих модулей/повторов
N- или C-терминальные кэппирующие повторы домена анкиринового повтора могут быть заменены N- или C-терминальными кэппирующими повторами в соответствии с изобретением, соответственно, путем комбинирования методик, таких как выравнивание аминокислотной последовательности, мутагенез и синтез генов, известных квалифицированному специалисту в данной области техники.
Например, N-терминальный кэппирующий повтор SEQ ID NO: 17 может быть заменен на N-терминальный кэппирующий повтор SEQ ID NO: 14 путем (i) определения N-терминального кэппирующего повтора последовательности SEQ ID NO: 17 (то есть последовательности от положения 1 до 32) путем выравнивания последовательности с SEQ ID NO: 14, (ii) замещения последовательности определенного C-терминального кэппирующего повтора последовательности SEQ ID NO: 17 последовательностью SEQ ID NO: 14, что приводит к последовательности SEQ ID NO: 18, (iii) получения гена, который кодирует повторяемый домен, кодирующего замененный C-терминальный кэппирующий повтор (то есть, последовательности SEQ ID NO: 18), (iv) экспрессии модифицированного повторяемого домена в цитоплазме Е.coli и (v) очистки модифицированного повторяемого домена с помощью стандартных способов.
В качестве дополнительного примера, C-терминальный кэппирующий повтор SEQ ID NO: 17 может быть заменен на C-терминальный кэппирующий повтор SEQ ID NO: 9 путем (i) определения C-терминального кэппирующего повтора последовательности SEQ ID NO: 17 (то есть, последовательности от положения 99 до положения 126) путем выравнивания последовательности с SEQ ID NO: 9, (ii) замещения последовательности определенного C-терминального кэппирующего повтора последовательности SEQ ID NO: 17 последовательностью SEQ ID NO: 9, (iii) получения гена, который кодирует повторяемый домен, кодирующий замененный С-терминальный кэппирующий модуль, (iv) экспрессии модифицированного повторяемого домена в цитоплазме Е.coli и (v) очистки модифицированного повторяемого домена с помощью стандартных способов.
Высокий уровень и экспрессия растворимых DARPin
DARPin экспрессировали в клетках Е.coli BL21 или XL1-Blue и очищали с помощью их His-метки при использовании стандартных прописей. 25 мл стационарных ночных культур (LB, 1% глюкозы, 100 мг/л ампициллина; 37°C) использовали для инокуляции 1 л культур (та же среда). При значении поглощения 1 при 600 нм культуры индуцировали с помощью 0,5 мМ IPTG и инкубировали при 37°C в течение 4 часов. Центрифугировали культуры и полученные осадки ресуспендировали в 40 мл TBS500 (50 мМ Трис-HCl, 500 мМ NaCl, pH 8) и подвергали обработке ультразвуком. Лизаты повторно центрифугировали и прибавляли глицерин (заключительная концентрация 10% (об./об.)) и имидазол (заключительная концентрация 20 мМ) к полученному супернатанту. Белки очищали при использовании колонки с Ni-нитрилтрехуксусной кислотой (2,5 мл объема колонки) в соответствии с инструкциями производителя (QIAgen, Germany). Альтернативно, DARPin или домены анкиринового повтора, не содержащие 6xHis-MeTKH, очищали с помощью анионобменной хроматографии, после чего проводили эксклюзионную хроматографию размеров при использовании стандартной смолы и прописей, известных квалифицированному специалисту в данной области техники. Вплоть до 200 мг высоко растворимых DARPin может быть очищено из 1 л культуры Е.coli со степенью чистоты >95%, как оценивается с помощью SDS-15% ПАГЭ. Такие очищенные DARPin использовались для дальнейшей характеристики.
Пример 2: Более высокая термальная стабильность DARPin с улучшенным N-терминальным кэппирующим модулем.
Термальную стабильность очищенного DARPin (в соответствии с Примером 1) анализировали с помощью анализа термальной стабильности на основе флуоресценции. (Niesen, F.H., Nature Protocols 2(9): 2212-2221, 2007). Таким образом, температура, при которой белок (то есть, DARPin) разворачивается, измеряется с помощью повышения флуоресценции красителя (например, SYPRO оранжевого; Invitrogen, кат. номер S6650)) с аффинностью для гидрофобных частей белка, которые подвергаются воздействию при развертывании белка. Среднее значение температуры перехода флуоресценции, полученное таким образом (от более низкой интенсивности флуоресценции до более высокой интенсивности флуоресценции) соответствует среднему значению температуры денатурации (Tm) анализируемого белка. Альтернативно, термальную стабильность такого очищенного DARPin анализировали с помощью CD спектроскопии; то есть, путем измерения его тепловой денатурации в соответствии с его сигналом циркулярного дихроизма (CD) при 222 нм при использовании методик, хорошо известных квалифицированному специалисту в данной области техники.
Анализ термальной стабильности на основе флуоресценции
Термальную денатурацию DARPin при использовании SYPRO оранжевого как флуоресцентного красителя измеряли при использовании способа ПЦР в реальном времени (то есть, C1000 термоблока для проведения реакций (BioRad) в комбинациии с CFX96 оптической системой (BioRad)). DARPin получали при концентрации 50 мкМ либо в PBS при pH 7,4, либо в MES буфере при pH 5,8 (250 мМ (2-N-морфолино)этансульфоновая кислота pH 5,5, 150 мМ NaCl, смешанного с PBS pH от 7,4 1 до 4 (об./об.) и при доведении значения pH до 5,8), содержащем 1x SYPRO оранжевого (разведенного из маточного раствора 5'000x SYPRO оранжевого, Invitrogen), и 50 мкл таких белковых растворов или только буфер прибавляли к содержимому ячеек белого планшета на 96 ячеек для проведения ПЦР (Bio-Rad). Планшеты закрывали при использовании Microseal 'В' Adhesive изоляционной прокладки (Bio-Rad) и нагревали в устройстве для проведения ПЦР в реальном времени от 20°C до 95°C при приращениях 0,5°C включая этап выдерживания в течение 25 секунд после каждого приращения температуры, и после проведения термальной денатурации DARPin осуществляли измерение относительных единиц флуоресценции образцов при каждом приращении температуры. Относительные единицы флуоресценции в ячейках планшета измеряли при использовании канала 2 аппарата для проведения ПЦР в реальном времени (то есть, возбуждение осуществляли при 515-535 нм, а определение проводили при длине волны 560-580 нм), и вычитали соответствующие значения, полученные при использовании только буфера. Из полученных таким образом средних значений перехода термальной денатурации, могут быть определены Tm значения для анализируемых DARPin.
Анализ термальной стабильности на основе CD спектроскопии
CD сигнал DARPin регистрировали при 222 нм в устройстве Jasco J-715 (Jasco, Japan) при медленном нагревании белка при концентрации 0,02 мМ в PBS pH 7,4 от 20C до 95°C при использовании скорости изменения 1°C за минуту. Этот способ представляет собой эффективное средство длчя отслеживания денатурации DARPin, поскольку они в основном состоят из альфа-спиралей, которые демонстрируют сильное изменение своего CD сигнала при 222 нм при развертывании. Срединная точка наблюдаемого перехода такого измеренного CD сигнала для DARPin соответствует их значению Tm.
Результаты термальной денатурации DARPin в PBS при pH 7,4 с последующим измерением повышения интенсивности флуоресценции SYPRO оранжевого или с последующей CD спектроскопией являются показанными в Фигурах и Таблице 1.
Термальную стабильность DARPin#17 сравнивали с термальной стабильностью DARPin#18 при использовании анализа термальной стабильности на основе флуоресценции (Таблица 1, Фигура 1). Эти два DARPin обладают идентичными аминокислотными последовательностями за исключением одной аминокислоты в N-терминальном кэппирующем модуле их повторяемых доменов. Повторяемый домен DARPin#18, но не DARPin#17, включает улучшенный N-терминальный кэппирующий модуль, как описывается в данной заявке; то есть N-терминальный кэппирующий модуль DARPin#18 содержит остаток лейцина (L) в положении 24 его N-терминального кэппирующего модуля, в то время как DARPin#17 содержит метионин (М) в этом положении. Неожиданно было обнаружено, что это изменение одной аминокислоты приводило к повышению значения Tm приблизительно на 6,5°C.
Термальную стабильность DARPin#19 сравнивали с термальной стабильностью DARPin #20 при использовании анализа термальной стабильности на основе флуоресценции и на основе CD (Таблица 1, Фигура 2). Эти два DARPin обладают идентичными аминокислотными последовательностями за исключением одной аминокислоты в N-терминальном кэппирующем модуле их повторяемых доменов. Повторяемый домен DARPin#20, но не DARPin#19, включает улучшенный N-терминальный кэппирующий модуль, как описывается в данной заявке; то есть N-терминальный кэппирующий модуль DARPin#20 содержит остаток L в положении 24 его N-терминального кэппирующего модуля, в то время как DARPin#19 содержит метионин (M) в этом положении. Неожиданно было обнаружено, что это изменение одной аминокислоты приводило к повышению значения Tm приблизительно на 2,5°C. Таким образом, термальная стабильность уже весьма стабильных DARPin может быть дополнительно повышена путем использования улучшенного N-терминального кэппирующего модуля в соответствии с изобретением.
Термальную стабильность DARPin#21 сравнивали с термальной стабильностью DARPin#22 и DARPin#23 при использовании анализа термальной стабильности на основе CD (Таблица 1, Фигура 3). Эти три DARPin обладают идентичными аминокислотными последовательностями за исключением одной аминокислоты в N-терминальном кэппирующем модуле их повторяемых доменов. Повторяемые домены DARPin#22 и DARPin#23, но не DARPin#21, включают улучшенный N-терминальный кэппирующий модуль, как описывается в данной заявке; то есть N-терминальный кэппирующий модуль DARPin#22 и DARPin#23 содержит остаток L в положении 24 (в то время как DARPin#21 содержит М в этом положении) и A остаток в положении 26 (в то время как DARPin#21 содержит N в этом положении); кроме того, N-терминальный кэппирующий модуль DARPin#23 содержит остаток K в положении 25 (в то время как DARPin#21 и DARPin#22 содержат A в этом положении). Таким образом, DARPin #23 включает улучшенный N-терминальный кэппирующий модуль, включающий аминокислотную последовательность RILLKA (SEQ ID NO: 11) от положения 21 до положения 26, как описывается в данной заявке. Неожиданно было обнаружено, что это эти небольшие изменения в N-терминальном кэппирующем модуле DARPins#22 и DARPin#23 приводили к повышению значения Tm приблизительно на 8,5°C или 7°C, соответственно, по сравнению с DARPin#21. Кроме того, DARPin#22 и DARPin#23 обладают почти идентичной термальной стабильностью, в то время как их аминокислотная последовательность отличается одной аминокислотой в N-терминальном кэппирующем модуле их повторяемых доменов; то есть, N-терминальный кэппирующий модуль DARPin#22 содержит остаток A в положении 25 своего N-терминального кэппирующего модуля, в то время как DARPin#23 содержит K в этом положении. Таким образом, это изменение одной аминокислоты в положении 25 такого N-терминального кэппирующего модуля, как оказалось, является хорошо переносимым и не оказывает влияния на термальную стабильность.
Термальную стабильность DARPin#24 сравнивали с термальной стабильностью DARPin#25 и DARPin#26 при использовании анализа термальной стабильности на основе флуоресценции и на основе CD (Таблица 1, Фигура 4). Эти три DARPin обладают идентичными аминокислотными последовательностями за исключением трех или четырех аминокислот в N-терминальном кэппирующем модуле их повторяемых доменов. Повторяемые домены DARPin#25 и DARPin#26, но не DARPin#24, включают улучшенный N-терминальный кэппирующий модуль, как описывается в данной заявке; то есть N-терминальный кэппирующий модуль DARPin#25 и DARPin#26 содержит остаток L в положении 24 (в то время как DARPin #4 содержит M в этом положении), содержит остаток K в положении 25 (в то время как DARPin#24 содержит A в этом положение) и остаток A в положении 26 (в то время как DARPin #24 содержит остаток N в этом положении); кроме того, N-терминальный кэппирующий модуль DARPin#26 содержит остаток E в положении 22 (в то время как DARPin#24 и DARPin#25 содержит I в этом положении). Таким образом, DARPin#25 и DARPin#26 включают улучшенный N-терминальный кэппирующий модуль, включающий аминокислотные последовательности RILLKA (SEQ ID NO: 11) и RELLKA (SEQ ID NO: 12), соответственно, от положения 21 до положения 26, как описывается в данной заявке. Неожиданно было обнаружено, что это небольшие изменения в N-терминальном кэппирующем модуле DARPins#25 и DARPin#26 приводили к повышению значения Tm приблизительно на 5°C или 6°C, соответственно, по сравнению с DARPin#24. Кроме того, DARPin#25 и DARPin#26 обладают почти идентичной термальной стабильностью, в то время как их аминокислотные последовательности отличаются одной аминокислотой в N-терминальном кэппирующем модуле их повторяемых доменов; то есть, N-терминальный кэппирующий модуль DARPin#25 содержит остаток I в положении 252 своего N-терминального кэппирующего модуля, в то время как DARPin#26 содержит E в этом положении. Таким образом, это изменение одной аминокислоты в положении 22 такого N-терминального кэппирующего модуля, как оказалось, является хорошо переносимым и не оказывает никакого существенного влияния на термальную стабильность.
В общем случае, термальная стабильность различных DARPin может быть существенно улучшена с помощью небольших изменений аминокислотной последовательности их N-терминальных кэппинг модулей, как описывается в данной заявке.
Пример 3: Более высокая термальная стабильность DARPin с улучшенными C-терминальными кэппируюшими модулями
Термальную стабильность DARPin подвергали анализу с помощью анализа стабильности на основе флуоресценции или с помощью CD спектроскопии так, как описывается в Примере 2.
Термальную стабильность DARPin#27 (SEQ ID NO: 27 с гистидиновой меткой (SEQ ID NO: 16), слитой с его N-терминальным концом) сравнивали с термальной стабильностью DARPin#28 (SEQ ID NO: 28 с гистидиновой меткой (SEQ ID NO: 16), слитой сего N-терминальным концом) при использовании анализа термальной стабильности на основе флуоресценции. Эти два DARPin обладают идентичной аминокислотной последовательностью за исключением C-терминальных кэппирующих модулей своих повторяемых доменов. Повторяемый домен DARPin#28, но не DARPin#27, включает улучшенный C-кэппирующий модуль, как описывается в данной заявке. Значения Tm в PBS pH 7,4, определенные для DARPin#27 и DARPin#28, составляли приблизительно 63°C и приблизительно 73°C, соответственно. Значения Tm в MES буфере pH 5,8, определенные для DARPin#27 и DARPin#28, составляли приблизительно 54,5°C и приблизительно 66°C, соответственно.
Термальную стабильность DARPin#29 (SEQ ID NO: 29 с гистидиновой меткой (SEQ ID NO: 16), слитой с его N-терминальным концом) сравнивали с термальной стабильностью DARPin#30 (SEQ ID NO: 30 с гистидиновой меткой (SEQ ID NO: 16), слитой сего N-терминальным концом) при использовании анализа термальной стабильности на основе флуоресценции. Эти два DARPin обладают идентичной аминокислотной последовательностью за исключением C-терминальных кэппирующих модулей своих повторяемых доменов. Повторяемый домен DARPin DARPin#30, но не DARPin#29, включает улучшенный C-кэппирующий модуль, как описывается в данной заявке.
Значения Tm в MES буфере pH 5,8, определенные для DARPin#39 и DARPin#40, составляли приблизительно 51°C и приблизительно 55°C, соответственно.
Термальную стабильность DARPin#31 (SEQ ID NO: 31) сравнивали с термальной стабильностью DARPin#32 (SEQ ID NO: 32) при использовании CD спектроскопии. Эти два DARPin обладают идентичной аминокислотной последовательностью за исключением C-терминальных кэппирующих модулей своих повторяемых доменов. Повторяемый домен DARPin#32, но не DARPin#31, включает улучшенный C-кэппирующий модуль, как описывается в данной заявке. Значения Tm в PBS pH 7,4, определенные для DARPin#31 и DARPin#32, составляли приблизительно 59,5°C и приблизительно 73°C, соответственно.




















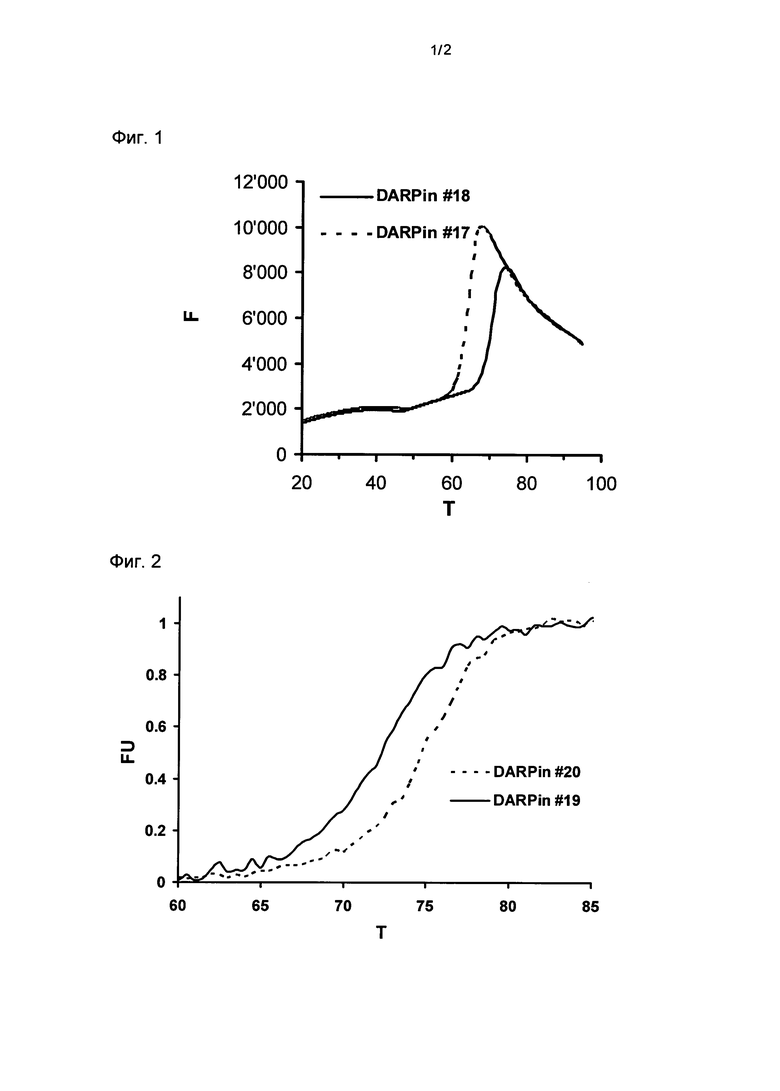
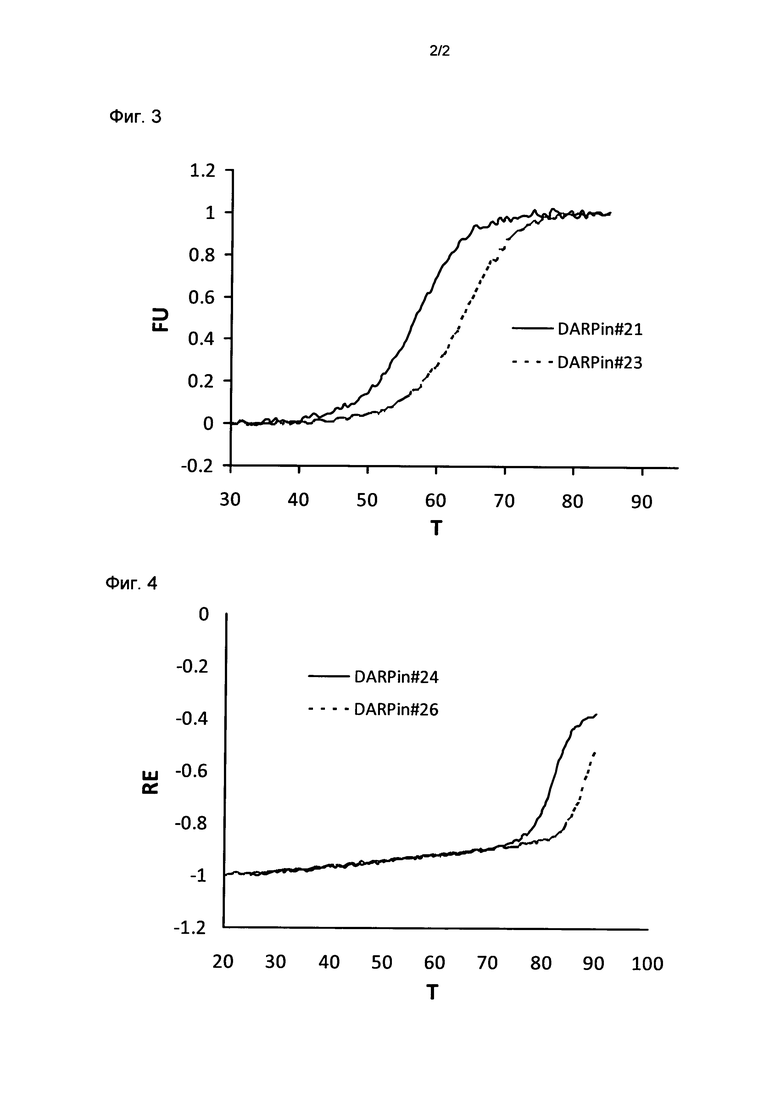
Настоящее изобретение относится к области биотехнологии, конкретно к N-терминальным кэппирующим модулям для белков. Полученные N-терминальные кэппинг-модули для сконструированных белков с анкириновым повтором (DARPin) обеспечивают их большую термальную стабильность DARPin по сравнению с немодифицированным белком. 5 н. и 16 з.п. ф-лы, 3 пр., 1 табл., 4 ил.
1. N-терминальный кэппирующий модуль синтезированного домена анкиринового повтора, где указанный N-терминальный кэппирующий модуль содержит аминокислотную последовательность
GSDLGKKLLE AARAGQDDEV RILLKAGADV NA (SEQ ID NO: 14) или
GSDLGKKLLE AARAGQDDEV RELLKAGADV NA (SEQ ID NO: 15), в которой
1) аминокислотный остаток L в положении 24 SEQ ID NO: 14 или SEQ ID NO: 15 необязательно заменен V, I или А;
2) вплоть до одной аминокислоты SEQ ID NO: 14 в положении, отличном от положения 24, или вплоть до двух аминокислот SEQ ID NO: 15 в других положениях, отличных от положения 24, необязательно заменено другими аминокислотами; и
3) G в положении 1 и/или S в положении 2 SEQ ID NO: 14 или SEQ ID NO: 15 необязательно отсутствуют;
и где указанный N-терминальный кэппирующий модуль обеспечивает более высокое значение Tm в PBS для указанного домена анкиринового повтора по сравнению с доменом анкиринового повтора, который имеет идентичную аминокислотную последовательность за исключением наличия аминокислотного остатка М в положении, соответствующем положению 24 SEQ ID NO: 14 или SEQ ID NO: 15.
2. N-терминальный кэппирующий модуль по п. 1, в котором вплоть до 1 аминокислоты SEQ ID NO: 14 или SEQ ID NO: 15 в другом положении, отличном от положения 24, необязательно заменено другой аминокислотой.
3. N-терминальный кэппирующий модуль по п. 1, где аминокислотный остаток R в положении 21 SEQ ID NO: 14 или SEQ ID NO: 15 не заменен или заменен Е.
4. N-терминальный кэппирующий модуль по п. 1, где аминокислотный остаток I в положении 22 SEQ ID NO: 14 или аминокислотный остаток Е в положении 22 SEQ ID NO: 15 не заменен или заменен V.
5. N-терминальный кэппирующий модуль по п. 1, где аминокислотный остаток K в положении 25 SEQ ID NO: 14 или SEQ ID NO: 15 не заменен или заменен А или Е.
6. N-терминальный кэппирующий модуль по п. 1, где аминокислотный остаток А в положении 26 SEQ ID NO: 14 или SEQ ID NO: 15 не заменен или заменен Н, Y, K или R.
7. N-терминальный кэппирующий модуль по п. 1, где аминокислотные остатки
RILLKA от положения 21 до положения 26 SEQ ID NO: 14 или аминокислотные остатки
RELLKA от положения 21 до положения 26 SEQ ID NO: 15 не заменены.
8. N-терминальный кэппирующий модуль по п. 1, где аминокислотная
последовательность указанного N-терминального кэппирующего модуля представляет собой
GSDLGKKLLEAARAGQDDEVRILLKAGADVNA (SEQ ID NO: 14) или
GSDLGKKLLEAARAGQDDEVRELLKAGADVNA (SEQ ID NO: 15), в которой
G в положении 1 и/или S в положении 2 SEQ ID NO: 14 и SEQ ID NO: 15 необязательно отсутствуют.
9. N-терминальный кэппирующий модуль по п. 1, где аминокислотная последовательность указанного N-терминального кэппирующего модуля содержит
GSDLGKKLLEAARAGQDDEVRILLKAGADVNA (SEQ ID NO: 14) или
GSDLGKKLLEAARAGQDDEVRELLKAGADVNA (SEQ ID NO: 15).
10. N-терминальный кэппирующий модуль по п. 1, где аминокислотная
последовательность указанного N-терминального кэппирующего модуля содержит GSDLGKKLLEAARAGQDDEVRILLKAGADVNA (SEQ ID NO: 14).
11. N-терминальный кэппирующий модуль по п. 1, где аминокислотная
последовательность указанного N-терминального кэппирующего модуля содержит GSDLGKKLLEAARAGQDDEVRELLKAGADVNA (SEQ ID NO: 15).
12. N-терминальный кэппирующий модуль по п. 1, где аминокислотная
последовательность указанного N-терминального кэппирующего модуля содержит GSDLGKKLLEAARAGQDDEVRILLKAGADVNA (SEQ ID NO: 14), в которой остаток аминокислоты в положении 25 заменен А.
13. N-терминальный кэппирующий модуль синтезированного домена анкиринового повтора, где указанный N-терминальный кэппирующий модуль включает аминокислотную последовательность
GSDLGKKLLEAARAGQDDEVRILLKAGADVNA (SEQ ID NO: 14) или
GSDLGKKLLEAARAGQDDEVRELLKAGADVNA (SEQ ID NO: 15), где
1) аминокислотный остаток L в положении 24 SEQ ID NO: 14 или SEQ ID NO: 15 необязательно заменен V, I или A;
2) вплоть до шести аминокислот SEQ ID NO: 14 или SEQ ID NO: 15 в положениях, отличных от положения 24, необязательно заменены другими аминокислотами;
3) G в положении 1 и/или S в положении 2 SEQ ID NO: 14 или SEQ ID NO: 15 необязательно отсутствуют;
4) аминокислотный остаток А в положении 26 SEQ ID NO: 14 или SEQ ID NO: 15 не заменен или заменен Н, Y, K или R.
14. N-терминальный кэппирующий модуль по п. 13, где вплоть до трех аминокислот SEQ ID NO: 14 или SEQ ID NO: 15 в положениях, отличных от положения 24, необязательно заменено другими аминокислотами.
15. N-терминальный кэппирующий модуль по п. 13, где аминокислотные остатки RILLKA от положения 21 до положения 26 SEQ ID NO: 14 или аминокислотные остатки RELLKA от положения 21 до положения 26 SEQ ID NO: 15 не заменены.
16. N-терминальный кэппирующий модуль по п. 13, где аминокислотный остаток L в положении 24 SEQ ID NO: 14 или SEQ ID NO: 15 не заменен.
17. N-терминальный кэппирующий модуль по п. 13, где аминокислотный остаток А в положении 26 SEQ ID NO: 14 или SEQ ID NO: 15 не заменен.
18. N-терминальный кэппирующий модуль синтезированного домена анкиринового повтора, где указанный N-терминальный кэппирующий модуль содержит последовательность
GSX1LX2KKLLE AARAGQDDEV X3X4LX5X6X7GADV NA (SEQ ID NO: 5), в которой
G в положении 1 и/или S в положении 2 SEQ ID NO: 5 необязательно отсутствуют;
X1 представляет собой аминокислотный остаток G, А, или D;
Х2 представляет собой аминокислотный остаток G или D;
Х3 представляет собой аминокислотный остаток R или Е;
Х4 представляет собой аминокислотный остаток I, Е или V;
Х5 представляет собой аминокислотный остаток L, V, I или А;
Х6 представляет собой аминокислотный остаток А, K или Е; и
Х7 представляет собой аминокислотный остаток, выбранный из группы, состоящей из А, Н, Y, K и R;
и где указанный N-терминальный кэппирующий модуль обеспечивает более высокое значение Tm в PBS для указанного домена анкиринового повтора по сравнению с доменом анкиринового повтора, который имеет идентичную аминокислотную последовательность N-терминального кэппирующего модуля за исключением наличия аминокислотного остатка М в положении, соответствующем положению 24 SEQ ID NO: 14 или SEQ ID NO: 15.
19. N-терминальный кэппирующий модуль синтезированного домена анкиринового повтора, где указанный N-терминальный кэппирующий модуль содержит аминокислотную последовательность, характеризующуюся по меньшей мере 95%-ной идентичностью с аминокислотной последовательностью
GSDLGKKLLE AARAGQDDEV RILLKAGADV NA (SEQ ID NO: 14) или
GSDLGKKLLE AARAGQDDEV RELLKAGADV NA (SEQ ID NO: 15), при
условии, что аминокислотный остаток в положении 24 в аминокислотной последовательности указанного N-терминального кэппирующего модуля представляет собой L, V, I или А и аминокислотный остаток в положении 26 указанного N-терминального кэппирующего модуля представляет собой А, Н, Y K или R;
и где аминокислоты в положениях 1 и/или 2 SEQ ID NO: 14 или SEQ ID NO: 15 необязательно отсутствуют;
и где указанный N-терминальный кэппирующий модуль обеспечивает более высокое значение Tm в PBS для указанного домена анкиринового повтора по сравнению с доменом анкиринового повтора, который имеет идентичную аминокислотную последовательность N-терминального кэппирующего модуля за исключением наличия аминокислотного остатка М в положении, соответствующем положению 24 SEQ ID NO: 14 или SEQ ID NO: 15.
20. N-терминальный кэппирующий модуль по п. 19, где аминокислотный остаток в положении 21 указанного N-терминального кэппирующего модуля представляет собой R или Е.
21. Нуклеиновая кислота, кодирующая N-терминальный кэппирующий модуль по любому из пп. 1-20.
| WO 2012060748 A1, 03.06.2010 | |||
| WO 2009115919 A2, 24.09.2009 | |||
| BINZ H | |||
| K | |||
| et al., Designing Repeat Proteins: Well-expressed, Soluble and Stable Proteins from Combinatorial Libraries of Consensus Ankyrin Repeat Proteins, J | |||
| Mol | |||
| Biol., 2003, v | |||
| Приспособление, обнаруживающее покушение открыть замок | 1910 |
|
SU332A1 |
| Подставка для настольных электрических ламп | 1923 |
|
SU489A1 |
| KRAMER M.A | |||
| et al., Structural determinants for improved stability of designed ankyrin repeat proteins with a redesigned C-capping module, 2010, v | |||
| Катодный усилитель с промежуточными контурами и батарейным коммутатором для цепей сетки | 1923 |
|
SU404A1 |
| Подвесная канатная дорога | 1920 |
|
SU381A1 |
| АНТРАНИЛАМИДПИРИДИНАМИДЫ ИЗБИРАТЕЛЬНОГО ДЕЙСТВИЯ В КАЧЕСТВЕ ИНГИБИТОРОВ VEGFR-2 И VEGFR-3 | 2002 |
|
RU2299208C2 |

