Применение комбинаторных способов для идентификации, описания и модификации белков весьма успешно как в академических, так и в коммерческих исследованиях и разработках. В этом отношении технология дисплея с использованием нитчатого бактериофага, или технология фагового дисплея, была первой платформой, в которой были использованы библиотеки, и она до сих пор остается доминирующей технологией. Таким образом, фаговый дисплей широко используют как в фундаментальных, так и в прикладных исследованиях белков, а также в разработке как новых диагностических, так и новых терапевтических средств на основе белков, которые во всем мире являются наиболее быстро растущим классом соединений.
Принцип комбинаторной технологии фагового дисплея основан на связи генотипа и фенотипа, обеспеченной тем свойством, что на поверхности каждого вириона будут представлены только те белки, которые кодирует геном, заключенный в белковую оболочку вириона. Фаговая частица сама по себе высоко устойчива к множеству физико-химических условий; поэтому фаговый дисплей обеспечивает большую по сравнению с конкурирующими комбинаторными технологиями универсальность во многих схемах селекции.
Фаговый дисплей гетерологичных полипептидов был осуществлен с использованием всех пяти структурных белков оболочки нитчатого фага, но широкое распространение получило только применение дисплея на белке pIII (pIII-дисплея) и, в некоторой степени, дисплея на белке pVIII (pVIII-дисплея) (Фиг.1).
Когда гетерологичный слитый белок представляет собой лишь короткий пептид, предпочтительны мультивалентные дисплейные системы с использованием векторов на основе генома фага, в то время как для более крупных слитых белков с прошедшими фолдинг доменами чаще всего предпочтительны фагмидные системы. В последнем случае, в данной области техники значительно преобладает фаговый pIII-дисплей антител, но постепенно появляются альтернативные каркасы, обуславливающие необходимость расширения способов конструирования белков для применения в будущем. Особенно желательным применением является эффективное получение высокоаффинных специфичных пептидных или белковых связывающих агентов из фаговой дисплейной библиотеки посредством простого инфицирования бактерий вирионами фага, когда они все еще связаны с их мишенью.
Endemann и Model, 1995 (PMID: 7616570) сообщали, что минорный белок оболочки pIX не был функциональным при слиянии с его N-концом другого белка. Таким образом, в этом сообщении был сделан вывод о том, что pIX не может быть использован для фагового дисплея.
Позднее Gao et al. (PMID: 10339535, 12239343 и WO 0071694) и Khalil et al. (PMID: 17360403) продемонстрировали, что дисплей с использованием слитого белка pIX при слиянии с N-концевой областью возможен при его экспрессии с фагмид и использовании в комбинации с зависимой от сигнальной последовательности периплазматической локализацией. В этих системах происходит комплементация, поскольку геном фага-помощника обеспечивает белок pIX дикого типа (wt pIX) при спасении фагмиды.
Фагмида, раскрытая на Фиг.2А WO 0071694 (без вставки слитого белка) и Фиг.2В WO 0071694 (со вставкой слитого белка), очевидно содержит сигнальную последовательность peIB (описание графических материалов на странице 7, строки 2-14).
Как указано выше, ранее было предложено, что pIX не был функциональным при слиянии с его N-концами другого белка, и Gao et al. предложили две возможные причины их успеха, либо по отдельности, либо в комбинации.
Одна возможная причина состояла в том, что к N-концам слитых белков была присоединена прокариотическая лидерная последовательность (сигнальная последовательность), что обеспечивает, таким образом, направление рекомбинантного белка в периплазматическое пространство и предотвращение посредством этого его накопления в цитоплазме.
Другая возможная причина состояла в том, что рекомбинантные белки экспрессировали с фагмиды, а не с генома фага, как у Endemann и Model, поэтому для спасения фагмиды возникала неизбежная потребность в комплементации белка pIX дикого типа от фага-помощника с рекомбинантными слитыми белками pIX, сохраняя таким образом функциональность дикого типа, которая, в противном случае, могла бы быть потеряна в виду рекомбинантной модификации. То есть фаги будут содержать смесь белков дикого типа и слитых белков.
В Khalil et al. (PMID: 17360403) описана прикладная система с использованием свойства вириона биспецифичного нитчатого фага, где экзогенный пептид представлен на каждом дистальном конце одного и того же вириона. Это было осуществлено с использованием комбинации обычного pIII-вектора из фагового генома, обеспечивающего комплементацию с фагмидой для зависимого от прокариотической сигнальной последовательности pIX-дисплея. В этом случае вектор из генома фага выполнял функции фага-помощника при спасении фагмиды.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Задачей настоящего изобретения является предложение альтернативного каркаса для пептидов, представленных на нитчатых фагах.
Согласно первому аспекту изобретения предложены слитые белки pIX, имеющие происхождение из нитчатого фага, не содержащие прокариотической N-концевой сигнальной последовательности, и, таким образом, представляющие собой белки, непосредственно слитые с экзогенным пептидом.
Другой аспект изобретения относится к нуклеиновым кислотам, кодирующим слитые белки по изобретению.
Один аспект изобретения относится к нитчатым фагам, содержащим слитые белки по изобретению.
Другой аспект изобретения относится к библиотеке нитчатых фагов.
Другой аспект изобретения относится к системе фагового дисплея, включающей фагмиду и фаг-помощник, где фагмида содержит нуклеиновую кислоту, кодирующую слитые белки pIX по изобретению.
Один аспект относится к набору, включающему систему фагового дисплея, включающую фагмиду и фаг-помощник, где фагмида содержит нуклеиновую кислоту, кодирующую слитые белки pIX по изобретению.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг.1
Схематическое изображение структуры нитчатого фага. Вирион образуют пять структурных белков, расположенных вокруг молекулы одноцепочечной ДНК (ssDNA). В фаге дикого типа (wt) присутствуют приблизительно 2700 копий белка pVIII и приблизительно 3-5 копий каждого из четырех белков pIII, pVI, pVII и pIX, обнаруженных на каждом конце вириона. Размер вириона зависит от размера генома (приблизительно 2,3 нуклеотида на один белок оболочки pVIII), и, таким образом, длину частицы корректируют, увеличивая или уменьшая число включенных копий белка pVIII. Следует отметить, что структуры белков pIII и pVIII были описаны с использованием волоконной дифракции рентгеновских лучей (x-ray fiber diffraction), кристаллографии и ядерного магнитного резонанса (NMR). Минорный белок оболочки pIII содержит три отдельных домена, разделенных областями, богатыми глицином: N1 (связывается с TolA), N2 (связывается с F-пилями) и СТ (интегрирован в вирион и важен для нормальной сборки вириона).
Фиг.2
Схематическое изображение новых фагмид для pIX-дисплея pGALD9 (A) и pGALD97ΔL (Б). Векторный каркас обеих фагмид был основан на фагмиде для pIII-дисплея pSEX81 (SEQ ID NO:2), последовательность которой может быть получена из GenBank, регистрационный номер Y14584, и подробности их конструирования описаны в разделе «Материалы и методы». Обе фагмиды могут включать сегменты экзогенных внутрирамочных последовательностей (называемые E1 и Е2) посредством легкой кассетной замены частей NcoI/HindIII и MluI/NotI, соответственно. Кассеты соединены синтетической линкерной последовательностью, варьирующей у различных конструкций, описанных здесь. Аббревиатуры: lacPO, lac-промотор; sd, последовательность Шайна-Далгарно; pelB, сигнальная последовательность бактериальной пектатлиазы; ТР, сайт трипсиновой протеазы; t, терминатор транскрипции Т7.
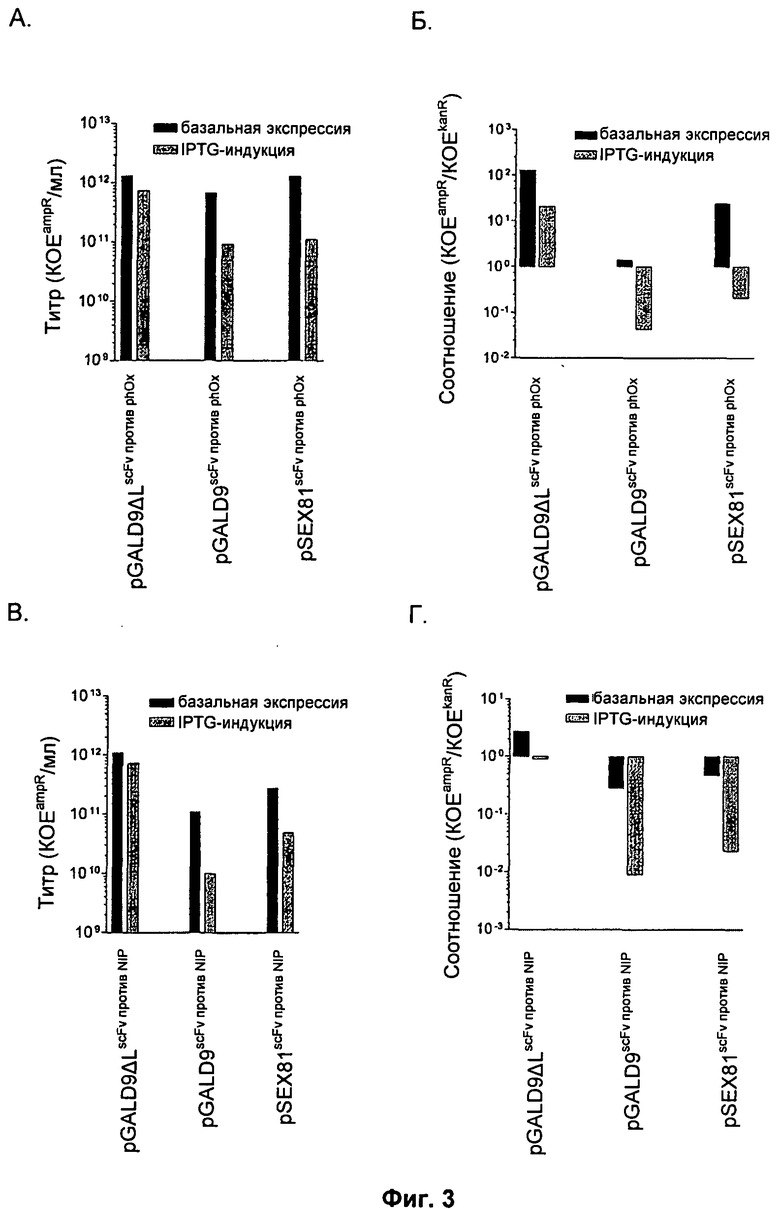
Фиг.3
Титры фагмид (A) scFv против phOx (SEQ ID NO:11) и (В) scFv против NIP, дисплей которых проводили с использованием pGALD9ΔL, pGALD9, pSEX81.
Все фагмиды содержат маркер устойчивости к ампициллину; таким образом, титры показаны как количество устойчивых к ампициллину колониеобразующих единиц на миллилитр раствора (KOEampR/мл). Соотношения фагмиды и фага-помощника для (Б) scFv против phOx (SEQ ID NO:11) и (Г) scFv против NIP, дисплей которых проводили с использованием pGALD9ΔL, pGALD9, pSEX81, показаны как отношение титра фагмиды (KOEampR/мл) к титру фага-помощника (КОЕkanR/мл). Упаковку вириона проводили стандартным спасением фагмиды, как описано в разделе «Материалы и методы», без изопропил-бета-D-тиогалактопиранозида (IPTG) (базальная экспрессия) или при его конечной концентрации 0,1 мМ (IPTG-индукция) после суперинфекции.
Фиг.4
Антиген-специфичный твердофазный иммуноферментный анализ (ELISA), где проведено сравнение функционального pIX- и pIII-дисплея (А) scFv против phOx (SEQ ID NO:11) и (Б) scFv против NIP (SEQ ID NO:3) с сигнальной последовательностью (ΔL) и без нее. ELISA проводили, как описано в разделе «Материалы и методы», с использованием 100 мкл очищенной надосадочной жидкости, содержащей вирионы, на лунку. Антитело против М13, конъюгированное с пероксидазой хрена (HRP), является отрицательным контролем неспецифической адсорбции моноклонального антитела (mAb) для выявления вирионов на антигене и блокирующих реагентах. (В и Г) Повторяли ELISA А и Б, но проявку останавливали до насыщения сигнала и относительные уровни дисплея определяли с использованием реактивности в отношении антигена как функции титра.
Фиг.5
Кажущееся обогащение специфичности в отношении мишени при аффинной селекции зависит от каркаса для дисплея на капсиде, экспрессии рассматриваемого белка (POI) и условий элюирования. Равные объемы нетитрованной надосадочной жидкости, содержащей вирионы, от каждой из 8 библиотек, содержащих scFv против phOx-/NIP, после 1 и 2 раундов селекции оценивали на предмет реактивности в отношении антигена посредством ELISA. Раунд 0 соответствует внесению образцов с концентрацией 1×1010 KOEampR. Для оценки максимально возможного ответа включали супернатанты клонов, из которых имеют происхождение концентраты, использованные в 0 раунде, и показанные результаты получены на основании сигналов с вычтенным фоном как доля максимально возможного ответа, на который указывает коническая форма.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Следует отметить, что воплощения и признаки, описанные в контексте одного из аспектов настоящего изобретения, также применимы к другим аспектам изобретения.
Все ссылки на патентную и непатентную литературу, приведенные в настоящей заявке, полностью включены сюда посредством ссылки.
В данной заявке авторами настоящего изобретения предложена новая концепция, согласно которой проводят генетическую модификацию структурного белка оболочки pIX вириона нитчатого фага, таким образом, что модифицированный вариант кодирует N-концевой пептид или белковый домен.
Слитый белок pIX
В одном аспекте согласно настоящему изобретению предложен слитый белок pIX, имеющий происхождение из нитчатого фага, включающий слияние экзогенного пептида с N-концом белка pIX. Такой слитый белок полезен, например, в контексте фагового дисплея.
При ссылке на экзогенный пептид подразумевают пептид, не являющийся изначально частью белка pIX с любыми аминокислотами, связывающими его с N-концевой областью аминокислотной части pIX слитого белка, или без них.
В предпочтительном воплощении нуклеиновая кислота, кодирующая слитый белок, не содержит прокариотическую N-концевую сигнальную последовательность.
При использовании здесь термин «пептид» охватывает короткие пептиды, полипептиды, белки и их фрагменты.
Термин «белок pIX» относится к аминокислотной последовательности, раскрытой в SEQ ID NO:1 (MSVLVYSFASFVLGWCLRSGITYFTRLMETSS).
В одном воплощении белок pIX содержит аминокислотную последовательность с идентичностью последовательности в отношении последовательности SEQ ID NO:1 по меньшей мере 70%, такой как 75% идентичность, такой как 80% идентичность, такой как 81% идентичность, такой как 82% идентичность, такой как 83% идентичность, такой как 84% идентичность, такой как 85% идентичность, такой как 86% идентичность, такой как 87% идентичность, такой как 88% идентичность, такой как 89% идентичность, такой как 90% идентичность, такой как 91% идентичность, такой как 92% идентичность, такой как 93% идентичность, такой как 94% идентичность, такой как 95% идентичность, такой как 96% идентичность, такой как 97% идентичность, такой как 98% идентичность, такой как 99% идентичность.
Идентичность последовательности
Согласно обычному определению «идентичность» определяют здесь как идентичность последовательности генов или белков на нуклеотидном или аминокислотном уровне, соответственно.
Таким образом, в контексте настоящего изобретения «идентичность последовательности» является мерой идентичности белков на аминокислотном уровне и мерой идентичности нуклеиновых кислот на нуклеотидном уровне. Идентичность последовательности белков может быть определена сравнением аминокислотной последовательности в данном положении в каждой последовательности при выравнивании последовательностей. Сходным образом, идентичность последовательности нуклеиновых кислот может быть определена сравнением нуклеотидной последовательности в данном положении в каждой последовательности при выравнивании последовательностей.
Для определения процента идентичности двух аминокислотных последовательностей или двух последовательностей нуклеиновых кислот проводят выравнивание последовательностей в целях оптимального сравнения (например, могут быть введены разрывы в последовательность первой аминокислотной последовательности или последовательности нуклеиновой кислоты для оптимального выравнивания со второй аминокислотной последовательностью или последовательностью нуклеиновой кислоты). Затем сравнивают аминокислотные остатки или нуклеотиды в соответствующих аминокислотных положениях или нуклеотидных положениях. Когда положение в первой последовательности занято таким же аминокислотным остатком или нуклеотидом, как в соответствующем положении второй последовательности, молекулы являются идентичными в этом положении. Процент идентичности двух последовательностей является функцией числа идентичных положений в этих последовательностях (то есть, % идентичности = число идентичных положений / общее число положений (например, перекрывающихся положений) × 100). В одном воплощении две последовательности имеют одинаковую длину.
Выравнивание последовательностей и подсчет числа идентичных аминокислот могут быть проведены вручную. Альтернативно, выравнивание двух последовательностей для определения процента идентичности может быть проведено с использованием математического алгоритма. Такой алгоритм включен в программы NBLAST и XBLAST (Altschul et al. 1990). Поиск нуклеотидных последовательностей с использованием BLAST может быть проведен программой NBLAST, показатель (score) = 100, разрядность (wordlength) = 12, с получением нуклеотидных последовательностей, гомологичных молекулам нуклеиновых кислот по изобретению. Поиск белков с использованием BLAST может быть проведен программой XBLAST показатель = 50, разрядность = 3 с получением аминокислотных последовательностей, гомологичных молекуле белка по изобретению. Для получения выравниваний с разрывами в целях сравнения может быть использован Gapped BLAST. Альтернативно, для осуществления повторного поиска может быть использован PSI-Blast, выявляющий отдаленное родство молекул. При использовании программ NBLAST, XBLAST и Gapped BLAST могут быть использованы параметры по умолчанию для указанных программ. См. http://www.ncbi.nlm.nih.gov. Альтернативно, идентичность последовательностей может быть вычислена после выравнивания последовательностей, например, программой BLAST в базе данных EMBL (www.ncbi.nlm.gov/cgi-bin/BLAST). Обычно для выравнивания могут быть использованы настройки по умолчанию в отношении, например, «матрицы замен» ("scoring matrix") и «штрафа за разрыв в последовательности» ("gap penalty"). В контексте настоящего изобретения могут быть предпочтительны настройки BLASTN и PSI BLAST по умолчанию.
Процент идентичности двух последовательностей может быть определен с использованием методик, сходных с описанными выше, с использованием разрывов или без него. При вычислении процента идентичности учитывают только точные соответствия.
Белки, прошедшие фолдинг
В предпочтительном воплощении термин "пептид" относится исключительно к белкам, прошедшим фолдинг, таким как домены, имеющие происхождение из антител. Специалисту в данной области техники будет ясно, что белки, прошедшие фолдинг, могут представлять собой антитела или их фрагменты, включая Fv, scFv, Fab, отдельные домены, домен Z белка А или его фрагменты (аффитело), анкирин или его фрагменты, сконструированные белки с анкириновыми повторами (DARPin) или их фрагменты, Т-клеточный рецептор или его фрагменты, молекулы главного комплекса гистосовместимости (МНС) I или II класса или их фрагменты, фибронектин или его фрагменты, антикалины или их фрагменты, PDZ-домены или их фрагменты, новый иммуноглобулиновый антигенный рецептор (IgNAR) или его фрагменты, антиген 4, ассоциированный с цитотоксическими Т-лимфоцитами (CTLA4), или его фрагменты, иммунный белок колицина Е7 (ImmE7) или его фрагменты, ноттины или их фрагменты, авимер или его фрагменты, зеленый флуоресцентный белок (GFP) или его фрагменты и другие кодируемые генами биологические флуорофоры.
В принципе, можно создать библиотеку любых белков, если возможен их дисплей, таким образом, в самой простой форме, возможно только разделение между чем-либо, имеющим не структурированную конфигурацию, по сравнению с упорядоченной структурой, то есть, структурой, прошедшей фолдинг.
В другом предпочтительном воплощении термин «пептид» относится исключительно к коротким пептидам, содержащим от 2 до 50 аминокислот. При определенной длине короткий пептид, образующий случайный клубок, будет достаточно длинным для формирования определенной вторичной или третичной структуры и, таким образом, будет соответствовать определению домена, прошедшего фолдинг. Очевидно, что это будет зависеть от химического состава, следовательно, один пептид из 20 аминокислот все еще будет образовывать случайный клубок, в то время как другой пептид из 20 аминокислот будет способен проходить фолдинг и соответствовать определению домена, прошедшего фолдинг.
В другом предпочтительном воплощении слитый белок pIX по изобретению содержит последовательность, выбранную из группы, состоящей из положения 1-32, 2-32, 3-32, 4-32, 5-32, 6-32, 7-32, 8-32, 9-32, 10-32, 11-32 и 12-32 SEQ ID NO:1. В принципе, в контексте фагмиды можно предполагать любую N-концевую модификацию белка pIX, при условии сохранения трансмембранной части и возможности нормального включения белка в состав вириона и нормальной сборки вириона.
SEQ ID NO:1 (MSVLVYSFASFVLGWCLRSGITYFTRLMETSS) представляет собой аминокислотную последовательность структурного белка оболочки pIX нитчатого фага (белка pIX дикого типа). Наиболее предпочтительно, слитый белок pIX включает положения 1-32 SEQ ID NO:1.
SEQ ID NO:1 не следует путать с сигнальной/лидерной последовательностью, описанной ниже.
Сигнальная последовательность
Предпочтительно, экзогенный пептид слит непосредственно, с какими-либо связывающими аминокислотами или без них, с N-концевой областью последовательности pIX слитого белка.
В другом предпочтительном воплощении слитый белок pIX не содержит прокариотическую N-концевую лидерную последовательность.
Термин «лидерная последовательность» использован взаимозаменяемо с терминами «сигнальный пептид» и «сигнальная последовательность» и относится к аминокислотной последовательности, направляющей белок (частью которого является лидерная последовательность) в периплазматическое межмембранное пространство грамотрицательных бактерий. Примерами часто используемых лидерных последовательностей являются pelBss, OmpAss, TorAss, malEss, phoAss, lamBss, Blass и DspAss, mglBss, sfmCss, tolBss и TorTss. Известно, что такие сигнальные последовательности направляют нативный белок в секреторный аппарат Е.coli, который, как известно, включает по меньшей мере SRP-зависимое, SEC-зависимое, TatABC-зависимое или YidC-зависимое перемещение из цитозоля в периплазматическое пространство (Baneyx et al. PMID: 15529165). Таким образом, термин «N-концевая сигнальная последовательность» относится к сигнальной последовательности, расположенной в N-концевой части белка.
Сигнальные последовательности, свойством которых является направление белка (частью которого они являются) в секреторный аппарат Е.coli и, посредством этого, перемещение его из цитозоля в периплазматический компартмент, могут отчасти быть идентифицированы посредством характерных черт, или мотивов, определяемых химическими свойствами входящих в их состав аминокислот.
Однако существующее на данный момент множество функциональных сигнальных последовательностей превосходит имеющиеся знания по их идентификации, поэтому, при современном состоянии данной области техники определение пептидов как когнатных сигнальных последовательностей обычно проводят путем получения данных с использованием данных баз знаний в качестве матрицы, например, нейронной сетью или эвристическими методами. На данный момент к некоторым таким инструментам открыт свободный доступ через открытые каналы доступа, как например SignalP, PPSEARCH of PROSITE (EMBL-EBI), SecretomeP, TatP.
Проблема даже серьезнее при рассмотрении класса секреторных белков, в том смысле, что происходит их экспорт из цитозольного компартмента, при котором имеет место отклонение от правил, заключающееся в том, что невозможно определить мотив сигнальной последовательности, но посредством получения данных здесь можно также определить признаки сигнальных последовательностей или получить вероятность, с которой рассматриваемый эукариотический белок будет обладать секреторной способностью. На данный момент для прокариотических таксонов такого инструмента не существует.
Таким образом, единственным доступным в настоящее время способом, позволяющим однозначно определить пептид как сигнальную последовательность, является определение экспериментальными средствами с подтверждением свойства пептида для установления того, является ли он настоящей сигнальной последовательностью. Также очевидно, что может быть проведено конструирование в таких пептидах, таким образом, что данные аминокислотные положения в сигнальной последовательности могут быть изменены при сохранении ее функции в качестве сигнального пептида, либо с естественными функциональными свойствами, либо с измененными функциональными свойствами, как например с повышенной транспортной способностью. Также может быть использовано удаление или добавление аминокислот. Действительно, такой анализ и конструирование были осуществлены с сигнальной последовательностью Ff pVIII, g8pss в метаболическом пути Sec, и TorAss в метаболическом пути Tat. В особенности, результаты Shen et al. могут служить обоснованными руководствами для конструирования функциональных, но измененных мутантов сигнальной последовательности pIII и сигнальной последовательности бактериальной пектатлиазы.
Функциональные свойства сигнальной последовательности могут затем быть разделены на два следующих свойства.
1. Направление белка (частью которого она являются) в секреторный аппарат Е.coli и, посредством этого, перемещение его из цитозоля в периплазматический компартмент, в процессе чего происходит ее протеолитическое отщепление от остальной части белка специфическими протеазами, такими как липопротеиновая сигнальная пептидаза или лидерные пептидазы.
2. Направление белка (частью которого она являются) в секреторный аппарат Е.coli и, посредством этого, перемещение его из цитозоля в периплазматический компартмент, после чего она все еще остается частью белка.
Хотя для подавляющего большинства сигнальных последовательностей характерно свойство (1), приведенное выше, очевидно, что эти белки могут легко быть сконструированы с приданием им свойства (2). Таким образом, любые известные в настоящее время сигнальные последовательности, например мутантная pelBss и другие, для которых изначально характерно свойство (1), но измененные с приданием им свойства (2), все еще рассматривают как когнатные сигнальные последовательности.
Более того, можно либо изменить сигнальную последовательность, для которой характерно свойство (1), с приданием ей свойства (2), либо непосредственно выбрать сигнальную последовательность, для которой характерно свойство (2), и затем, после перемещения, удалить сигнальную последовательность. Это может быть осуществлено эндогенными протеазами хозяина и/или, в случае, например, фагового дисплея, при слиянии белка с белком капсида. Затем в нужную область сигнальной последовательности или белка, частью которого она является, будет включен искусственный протеазный сайт, таким образом, что можно будет осуществить определенное расщепление. Здесь можно предположить два разных типа выбранных протеазных сайтов:
А. Протеазный сайт не приводит к расщеплению рассматриваемого белка, а приводит к расщеплению только предсказанного сайта, как например, сайт карбоксипептидазы А или сайт протеазы 3С риновирусов в комбинации с антителами или другими интересующими каркасами, такими как молекулы главного комплекса гистосовместимости или Т-клеточные рецепторы. При использовании такого подхода можно предполагать, например, фаговый дисплей рассматриваемого белка с использованием сигнальной последовательности, для которой характерно свойство (2), описанное выше, и, перед использованием в селекции и тому подобном, искусственное удаление сигнального пептида с получением функциональности и гомогенности слитого белка капсида.
Б. Протеазный сайт приводит к расщеплению рассматриваемого белка в дополнение к сконструированному сайту, как например, сайт расщепления трипсином.
Тем не менее, оба варианта будут рассматривать как зависимый от сигнальной последовательности фаговый дисплей.
Экзогенный пептид
В предпочтительном воплощении экзогенный пептид слитого белка pIX выбран из группы, состоящей из антител или их фрагментов, включая Fv, scFv, Fab, отдельных доменов, домена Z белка А или его фрагментов (аффитела), анкирина или его фрагментов, сконструированных белков с анкириновыми повторами (DARPin) или их фрагментов, Т-клеточного рецептора или его фрагментов, молекул главного комплекса гистосовместимости (МНС) I или II класса или их фрагментов, фибронектина или его фрагментов, антикалинов или их фрагментов, PDZ-доменов или их фрагментов, нового иммуноглобулинового антигенного рецептора (IgNAR) или его фрагментов, антигена 4, ассоциированного с цитотоксическими Т-лимфоцитами (CTLA4), или его фрагментов, иммунного белка колицина Е7 (ImmE7) или его фрагментов, ноттинов или их фрагментов, авимера или его фрагментов, зеленого флуоресцентного белка (GFP) или его фрагментов и других кодируемых генами биологических флуорофоров.
В предпочтительном воплощении экзогенный пептид слитого белка pIX является членом библиотеки.
Библиотека при использовании в контексте настоящего изобретения относится к совокупности разных пептидов. Пептиды могут представлять собой домены, прошедшие фолдинг, или короткие пептиды, например, состоящие из 2-50 аминокислот. Такие библиотеки представляют интерес, поскольку они могут быть использованы для идентификации новых лигандов, связывающихся сданной мишенью.
Есть несколько преимуществ использования белка pIX для дисплея библиотеки по сравнению с библиотеками, дисплей которых проводят с использованием белков pIII или pVIII. plX-дисплей включает те же средства, что и pIII-дисплей относительно направленности и валентности, но не оказывает влияния на инфекционную способность, явление, о котором известно при pIII-дисплее, привносящее в систему неконтролируемую и нежелательную гетерогенность, например, при спасении фагмиды после аффинной селекции.
Более того, pIX-дисплей может быть проведен без необходимости в прокариотической N-концевой сигнальной последовательности, что является необходимым условием как pIII-дисплея, так и pVIII-дисплея. В заключение, для любых разновидностей, иммобилизованных на мишени, при pIII-дисплее обычно необходим разрыв (обычно конкурентным элюированием или элюированием с высоким или низким рН) этой связи мишень-фаг. Например, известно, что это существенно препятствует получению высокоаффинных или стабильных связывающих агентов при pIII-дисплее. Поскольку при pIX-дисплее белок pIII, необходимый для инфекции, не изменен и легко доступен для альтернативных взаимодействий даже после взаимодействия фага и мишени, это полностью устраняет необходимость разрыва связи, например, элюированием в кислых условиях, поскольку иммобилизованные фаги сохраняют полную инфекционную способность и поэтому могут быть получены просто инфекцией при сохранении связи с мишенью.
Нуклеиновая кислота
Во втором аспекте изобретения предложена нуклеиновая кислота, кодирующая слитый белок по изобретению. Нуклеиновая кислота по изобретению может быть частью плазмиды, вектора, генома фага, фагмиды или фазмиды.
Термин «нуклеиновая кислота» относится к макромолекуле, состоящей из цепей мономерных нуклеотидов. В биохимии эти молекулы переносят генетическую информацию или образуют структуры внутри клеток. Наиболее распространенными нуклеиновыми кислотами являются дезоксирибонуклеиновая кислота (ДНК) и рибонуклеиновая кислота (РНК). В дополнение, термин «нуклеиновые кислоты» включает искусственные нуклеиновые кислоты, такие как пептид-нуклеиновая кислота (PNA), нуклеиновая кислота с морфолиновыми олигонуклеотидами и закрытая нуклеиновая кислота (LNA), а также гликоль-нуклеиновая кислота (glycol nucleic acid, GNA) и треозо-нуклеиновая кислота (TNA). Каждая из них отличается от встречающейся в природе ДНК или РНК изменениями в каркасе молекулы.
Фагмида или фазмида представляет собой тип клонирующего вектора, разработанный как гибрид нитчатого фага Ft и плазмид с получением вектора, способного к размножению, как плазмида, и также к упаковке в вирусные частицы в форме одноцепочечной ДНК. Подобно плазмиде, фагмида может быть использована для клонирования фрагментов ДНК и может быть введена в бактериального хозяина несколькими методиками (трансформацией, электропорацией). Однако для обеспечения нужных вирусных компонентов для репликации одноцепочечной ДНК и упаковки фагмидной ДНК в фаговые частицы необходимо инфицирование бактериального хозяина, содержащего фагмиду, фагом-помощником, например VCSM13 или M13K07.
Таким образом, один аспект настоящего изобретения относится к геному фага или фагмиде, содержащим нуклеиновую кислоту, кодирующую слитый белок pIX, имеющий происхождение из нитчатого фага, где слитый белок не содержит прокариотическую N-концевую сигнальную последовательность.
В одном воплощении настоящего изобретения геном фага или фагмида содержат нуклеиновую кислоту, кодирующую слитый белок pIX, имеющий происхождение из нитчатого фага, где слитый белок не содержит прокариотическую N-концевую сигнальную последовательность, и где слитый белок pIX содержит последовательность, выбранную из группы, состоящей из положения 1-32, 2-32, 3-32, 4-32 и 5-32 SEQ ID NO:1 (MSVLVYSFASFVLGWCLRSGITYFTRLMETSS).
В другом воплощении настоящего изобретения предложен экзогенный пептид, непосредственно слитый с N-концевой областью последовательности pIX.
Одно воплощение настоящего изобретения относится к геному фага или фагмиде по настоящему изобретению, где экзогенный пептид, слитый с белком pIX, выбран из группы, состоящей из антител или их фрагментов, включая Fv, scFv, Fab, отдельных доменов, домена Z белка А или его фрагментов (аффитела), анкирина или его фрагментов, сконструированных белков с анкириновыми повторами (DARPin) или их фрагментов, Т-клеточного рецептора или его фрагментов, молекул главного комплекса гистосовместимости (МНС) I или II класса или их фрагментов, фибронектина или его фрагментов, антикалинов или их фрагментов, PDZ-доменов или их фрагментов, нового иммуноглобулинового антигенного рецептора (IgNAR) или его фрагментов, антигена 4, ассоциированного с цитотоксическими Т-лимфоцитами (CTLA4), или его фрагментов, иммунного белка колицина Е7 (ImmE7) или его фрагментов, ноттинов или их фрагментов, авимера или его фрагментов, зеленого флуоресцентного белка (GFP) или его фрагментов и других кодируемых генами биологических флуорофоров.
В другом воплощении настоящего изобретения экзогенный пептид, слитый с белком pIX, является членом библиотеки.
Нитчатый фаг
Согласно третьему аспекту изобретения предложен нитчатый фаг, содержащий слитый белок по изобретению. Вирион нитчатого фага может содержать фагмиду.
Под фагом, часто называемым бактериофагом, здесь подразумевают вирус, инфицирующий бактерии, проходящий репликацию в бактериях и секрецию из них. Нитчатый бактериофаг, или нитчатый фаг, представляет собой фаг с одноцепочечной молекулой ДНК (ssDNA), упакованной в белки оболочки фага. Секретируемая частица нитчатого фага имеет фенотипически нитчатую структуру.
При использовании здесь термин «нитчатый фаг» охватывает как вирионы, имеющие происхождение из генома фага, так и вирионы, имеющие происхождение из фагмиды.
Термин «фаг-помощник» относится к вирусу, способствующему воспроизведению отдельного и неродственного дефектного вируса, определяемого как, например, фагмида, которая сама по себе не является ни геномом фага, ни функциональным вирусом, а является лишь плазмидой, содержащей один или несколько элементов, имеющих происхождение из генома фага, инфицированием той же самой клетки-хозяина, в которой уже присутствует дефектный вирус, и обеспечением белков, отсутствующих у дефектного вируса и необходимых для образования вирионов, содержащих фагмиду.
В одном воплощении нитчатый фаг содержит нуклеиновую кислоту, кодирующую слитый белок по изобретению. Особенно предпочтительным является фаг, содержащий фагмиду, содержащую нуклеиновую кислоту, кодирующую слитый белок по изобретению.
Фаговая библиотека представляет собой совокупность нитчатых фагов, на которых представлены пептиды или белки как часть одного или более чем одного белка оболочки нитчатого фага. Такие библиотеки могут включать два или более чем два фага, на которых представлены различные пептиды или белки.
Таким образом, в одном воплощении настоящего изобретения на нитчатых фагах представлены пептиды или белки как часть одного или более чем одного белка оболочки нитчатого фага.
В одном воплощении нитчатый фаг дополнительно содержит слитый белок pIII, слитый белок pVII или слитый белок pVIII.
В одном аспекте изобретения предложена библиотека нитчатых фагов по изобретению, на которых представлены экзогенные пептиды или белки как конструкции, слитые с одним или более чем одним из pIII, pVII, pVIII или pIX.
Библиотека представляет собой совокупность нитчатых фагов, на которых представлены пептиды или белки как часть одного или более чем одного белка оболочки нитчатого фага.
Такие библиотеки могут содержать два или более чем два фага, на которых представлены различные пептиды или белки.
Один аспект настоящего изобретения относится к фаговой библиотеке, включающей два или более чем два нитчатых фага, на которых представлены различные белки, по меньшей мере один из которых представляет собой слитый белок pIX, экспрессированный с генома фага или фагмиды по настоящему изобретению.
Одно воплощение настоящего изобретения относится к фаговой библиотеке, включающей два или более чем два нитчатых фага, на которых представлены различные пептиды или белки.
В определенном воплощении по меньшей мере один из этих пептидов или белков представляет собой слитый белок pIX по изобретению.
В одном воплощении пептиды представлены одновременно на pIX и pIII, pVII или pVIII.
В другом воплощении пептиды представлены одновременно на pIX и двух или трех белках, выбранных из группы, состоящей из pIII, pVII или pVIII.
Термин «дикий тип», иногда обозначаемый «wt», представляет собой типичную форму организма, штамма, гена или признака, как они встречаются в природе. Дикий тип относится к наиболее распространенному фенотипу в природной популяции. Дикий тип также относится к аллелю в каждом локусе, необходимому для получения фенотипа дикого типа. Дикий тип представляет собой контрольный эталон генотипа и фенотипа. В частности, в биологии он относится к различию между встречающимся в природе организмом и организмом, который был намеренно мутирован. Сайт-направленный мутагенез представляет собой исследовательскую методику, позволяющую осуществить мутацию определенных нуклеотидов в генной последовательности гена дикого типа. Белки дикого типа обозначают как «wt-(название белка)», например, белок pIX дикого типа обозначают wt pIX, wt-pIX или pIX дикого типа.
Таким образом, один аспект изобретения относится к нитчатому фагу по настоящему изобретению, не содержащему ген, кодирующий wt pIX, и/или белок wt pIX.
Другой аспект настоящего изобретения относится к нитчатому фагу по изобретению, дополнительно содержащему ген, кодирующий wt pIX, и/или белок wt pIX.
Другой аспект настоящего изобретения относится к нитчатому фагу, содержащему геном фага или фагмиду по настоящему изобретению.
В одном воплощении настоящего изобретения нитчатый фаг, содержащий геном фага или фагмиду по настоящему изобретению, дополнительно содержит ген, кодирующий wt pIX, и/или белок wt pIX.
В другом воплощении настоящего изобретения нитчатый фаг, содержащий геном фага или фагмиду по настоящему изобретению, не содержит ген, кодирующий wt pIX, и/или белок wt pIX.
Другой аспект настоящего изобретения относится к слитым белкам pIX, являющимся функциональными в фаговом дисплее без комплементации белком wt pIX. Один аспект настоящего изобретения относится к нитчатому фагу, содержащему геном фага или фагмиду по настоящему изобретению, дополнительно содержащему одну или более чем одну группу, выбранную из слитого белка pIII, слитого белка pVII и слитого белка pVIII.
Система фагового дисплея
Согласно пятому аспекту изобретения предложена система фагового дисплея, включающая фагмиду и фаг-помощник, где фаг-помощник содержит нуклеиновую кислоту, кодирующую слитый белок pIX по изобретению.
Система фагового дисплея, методика фагового дисплея, технология фагового дисплея или просто фаговый дисплей относятся к способу обнаружения и изучения взаимодействий белок-белок, белок-пептид и белок-ДНК, в котором используют бактериофаг для связывания белков с кодирующей их генетической информацией.
Представление белка или представленный белок относится к белку, слитому с белком оболочки фага, доступному для выявления или иммобилизации лигандом.
Согласно шестому аспекту изобретения предложена система фагового дисплея, включающая фагмиду, содержащую нуклеиновую кислоту, кодирующую слитый белок pIX по изобретению, и фаг-помощник.
Один аспект настоящего изобретения относится к системе фагового дисплея, включающей геном фага или фагмиду по настоящему изобретению.
Другой аспект настоящего изобретения относится к системе фагового дисплея, включающей геном фага или фагмиду по настоящему изобретению и фаг-помощник.
Один аспект настоящего изобретения относится к фаговой библиотеке, включающей два или более чем два нитчатых фага, на которых представлены различные белки, где по меньшей мере один из этих белков представляет собой слитый белок pIX, экспрессированный с генома фага или фагмиды по настоящему изобретению.
В одном воплощении настоящего изобретения фаговая библиотека по настоящему изобретению включает один или более чем один дополнительный слитый белок, выбранный из слитого белка pIII, слитого белка pVII и слитого белка pVIII.
Наборы
Согласно седьмому аспекту изобретения предложен набор, включающий систему фагового дисплея, состоящую из фагмиды, содержащей нуклеиновую кислоту, кодирующую слитый белок pIX по изобретению, и фага-помощника. Набор должен включать фагмиду с геном, кодирующим белок pIX, с сайтом множественного клонирования, расположенным ближе к N-концу кодирующей области, и фаг-помощник (например, M13K07, VCSM13 или другой). Набор должен включать протоколы инфицирования, экспрессии, иммобилизации, селекции и выявления фаговых клонов. Наборы должны также быть укомплектованы необходимыми прописями буферов и сред для проведения определенных анализов.
Набор здесь относится к совокупности реагентов для получения фаговых частиц с моно- или биспецифичными слитыми белками, либо в форме библиотеки для фагового дисплея, либо в форме отдельной фаговой частицы. Набор может включать фагмиды, фаги-помощники, бактериальные штаммы и протокол с прописями реагентов и описанием анализов. Набор может быть использован для разработки исследовательских, диагностических и терапевтических реагентов.
Один аспект настоящего изобретения относится к набору, включающему фаговый дисплей, включающий геном фага или фагмиду по настоящему изобретению и фаг-помощник.
Другой аспект настоящего изобретения относится к набору, включающему геном фага или фагмиду по настоящему изобретению.
Еще один аспект настоящего изобретения относится к набору, включающему нитчатый фаг, содержащий геном фага или фагмиду по настоящему изобретению.
Теперь изобретение будет описано более подробно в следующих неограничивающих примерах.
ПРИМЕРЫ
Пример 1: Фагмидный дисплей на белке pIX
Реагенты
По существу, все среды и буферы приготавливали, как описано в Sambrook et al. (2001) Molecular Cloning: A Laboratory Manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press. Антитело против М13, конъюгированное с HRP, приобретали у GE Healthcare Bio-Sciences AB (Uppsala, Sweden). Рестриктазы (RE) приобретали у New England Biolabs (Ipswich, MA, USA), за исключением DpnI, которую получали от Stratagene (LaJolla, CA, USA). Олигонуклеотиды ДНК приобретали у MWG Biotech AG (Ebersberg, Germany). Бычий сывороточный альбумин (BSA) и Tween 20 приобретали у Sigma-Aldrich (Oslo, Norway). ДНК-полимеразу Pfu Turbo приобретали у Stratagene (LaJolla, CA, USA). Гаптены 2-фенилоксазол-5-он (phOx) и 5-нитрофенилацетил (NIP), конъюгированные с BSA, по существу получали, как описано в других источниках (Nakela et al, PMID; 722243; и Michaelsen et al, PMID: 2125362). Изопропил-бета-D-тиогалактопиранозид (IPTG) приобретали у Fermentas (Burlington, Canada). Триэтиламин (TEA) и трипсин/этилендиаминтетрауксусную кислоту (EDTA) приобретали у Sigma-Aldrich (Oslo, Norway) и BioWhittaker (Lonza Group Ltd., Visp, Switzerland), соответственно. Штамм E.coli XL1-Blue приобретали у Stratagene (LaJolla, CA, USA). Фаг-помощник М13К07 приобретали у GE Healthcare Bio-Sciences AB (Uppsala, Sweden). Фагмида pSEX81 (SEQ ID NO:2) (дисплей с использованием pIII) с одноцепочечным Fv (scFv), специфичным против phOx-BSA, была любезно предоставлена Affitech AS (Oslo, Norway). Прокариотический вектор экспрессии pSG1 с scFv против NIP (SEQ ID NO:3) (не опубликовано) основан на pHOG21 (Kiprianov et al, PMID: 9005945) и был изготовлен собственными силами из вариабельных генов антител, имеющих происхождение из pLNOH2 и pLNOK (Norderhaug et al, PMID: 9202712).
Конструирование новых фагмидных векторов pGALD9 и pGALD9ΔL для pIX-дисплея
В качестве начальной матрицы для каркаса вектора была выбрана фагмида pSEX81 (SEQ ID NO:2), описанная выше (GenBank, регистрационный номер: Y14584). Во-первых, для удаления прокариотической сигнальной последовательности pelB (N-MKYLLPTAAAGLLLLAAQPAMA-C) (SEQ ID NO:4), кодирующей участок в данном векторе, сайт рестрикции NcoI вводили в крайнюю N-концевую область (extreme N-terminus) посредством in vitro мутагенеза QuikChange™ с использованием пары праймеров a41g-frwd/a41g-rev (5'-AGAGGAGAAATTAACCATGGAATACCTATTGCCTACGGC-3'/5'-GCCGTAGGCAATAGGTATTCCATGGTTAATTTCTCCTCT-3') (SEQ ID NO:5 и SEQ ID NO:6, соответственно), изменяя посредством этого первый нуклеотид второго кодона открытой рамки считывания (ORF) pelB с А на G. После мутагенеза вектор расщепляли с использованием NcoI, проводили повторное лигирование и использовали его в качестве матрицы во второй полимеразной цепной реакции (ПЦР) с получением подходящей части вектора с использованием пары праймеров pHOG_EcoRI_frwd/scTCR_rev (5'-TAGCTCACTCATTAGGCACCC-3'/5'-TTTGGATCCAGCGGCCGC-3') (SEQ ID NO:7 и SEQ ID NO:8, соответственно). Этот ПЦР-фрагмент затем перемещали в исходную pSEX81 (SEQ ID NO:2) на совместимые сайты рестрикции EcoRI/HindIII с использованием стандартных методик и результат подтверждали секвенированием ДНК. На этой стадии полностью удаляли кодирующую часть сигнальной последовательности pelB, но сохраняли стартовый кодон и его относительное расположение по отношению к lacPO и последовательности Шайна-Далгарно (SD), важной для нормальной транскрипции и трансляции, а также добавляли только один остаток Ala перед экзогенной последовательностью, определенной сайтами рестрикции NcoI/NotI, обнаруженными в исходной pSEX81 (SEQ ID NO:2). Новая конструкция была обозначена pSEX81ΔL.
Во-вторых, амплифицировали кодирующую последовательность pXI из M13K07 с использованием пары праймеров с рестрикционными метками на 5'-концах pIX_EcoRV/pIX_NheI (5'-ATATGATATCAGAATGAGTGTTTTAGTGTATTCTTTCGCC-3'/5'-ATATGCTAGCTTATCATGAGGAAGTTTCCATTAAACGGG-3') (SEQ ID NO:9 и SEQ ID NO:10). Этот ПЦР-фрагмент затем перемещали как в фагмиду pSEX81 (SEQ ID NO:2), так и в фагмиду pSEX81ΔL на совместимые сайты рестрикции, заменяя посредством этого область, кодирующую белок pIII, в обеих фагмидах и получая N-концевое слияние кассеты, определенной сайтами NcoI/NotI, с pIX с сохранением рамки считывания в исходной pSEX81 (SEQ ID NO:2). Получение новых конструкций подтверждали секвенированием ДНК и обозначали их pGALD9 и pGALD9ΔL, соответственно. Замену единицы scFv против phOx (SEQ ID NO:11) в различных фагмидах, описанных выше, единицей scFv против NIP (SEQ ID NO:3) из pSG1 проводили заменой кассеты, определенной сайтами рестрикции NcoI/NotI, с применением стандартных методик. Все фагмиды, описанные здесь, вводили в Е.coli XL1-Blue электропорацией с применением стандартных методик.
Получение фаговых частиц
Спасение фагмиды из Е.coli XL1-Blue с использованием фагов-помощников M13K07 и сборку вирионов контролировали точечным титрованием (spot titration) согласно описаниям (Welschof et al, PMID: 9050877 и Koch et al, PMID: 11126120).
Твердофазные иммуноферментные анализы (ELISA) иммобилизации фагов
phOx-BSA или NIP-BSA сорбировали на лунках титрационного микропланшета MaxiSorp™ (Nunc, Roskilde, Denmark) в концентрации 5 мкг/мл в забуференном фосфатом физиологическом растворе (PBS), рН 7,4, в течение ночи при 4°С. Лунки блокировали 2% BSA в PBS (масс./об.) в течение 1 часа при комнатной температуре (к.т.), затем добавляли препараты вирионов и оставляли их взаимодействовать от 1 до 2 часов при к.т.перед выявлением иммобилизованных вирионов антителом против М13, конъюгированным с HRP (1:5000), в течение 1 часа при к.т. После каждой стадии лунки промывали 3 раза PBST (PBS/0,05% Tween 20). Окрашивание лунок проводили растворимым тетраметилбензидиновым (ТМВ) субстратом, останавливали 1М HCl через 30 минут и считывали оптическую плотность при 450 нм (А450нм). Для определения реактивности в отношении антигена как функции представленных слитых белков из расчета на вирион окрашивание при ELISA останавливали добавлением 1М HCl до того, как в каком-либо образце наблюдали насыщение сигнала, показатели оптической плотности преобразовывали согласно следующей формуле: относительный уровень дисплея = (А450нм/титр фагмиды) × 1012.
phOx-/NIP-BSA селекция с внесением scFv
Получали свежие образцы вирионов, с индукцией 1 мМ IPTG или без нее, проводили осаждение с использованием полиэтиленгликоля (PEG) и титрование согласно описаниям.
Затем вещество, специфичное в отношении мишени, вносили в постороннюю среду в отношении 1:107 с получением известного разнообразия 107, что соответствует комбинаторной библиотеке среднего размера.
Для NIP-BSA-селекции scFv против NIP вносили в аналог scFv против phOx и наоборот. Изначальное вносимое количество составляло 1×1010 КОЕampR, приводя к степени сложности 103 в 1 раунде пэннинга для всех 12 модельных библиотек.
Кратко, мишень иммобилизовали на лунках титрационного микропланшета MaxiSorp™ (Nunc, Roskilde, Denmark) в трех экземплярах на одном и том же планшете с использованием объемов 100 мкл с концентрациями 1 мкг/мл и 0,1 мкг/мл для 1 и 2 раунда пэннинга, соответственно.
Перед пэннингом лунки блокировали PBSTM в течение 1-2 часов при к.т. до того, как добавляли 100 мкл соответствующих предварительно блокированных (в PBSTM) препаратов вирионов и оставляли их взаимодействовать в течение 1,5 часов при к.т. при перемешивании.
Лунки отмывали 9 раз в PBST с последующей пятикратной отмывкой в дистиллированной воде с использованием устройства для мойки микропланшетов перед элюированием вирионов, связанных с мишенью (в представленных в трех экземплярах лунках), одним из следующих способов: 1) добавлением 100 мкл 100 мМ TEA (pH 12) на лунку на 5 минут при к. т. с последующей нейтрализацией переносом в новую лунку, содержащую 100 мкл трис-HCl на лунку, pH 6,8; 2) добавлением 100 мкл трипсина/EDTA на лунку на 10 минут при к.т. с последующим переносом в новые лунки; 3) добавлением 200 мкл Е.coli XL1-Blue в логарифмической фазе роста (оптическая плотность при 600 нм (А600нм) приблизительно 0,5, соответствует ≥5×107 клеток) на лунку на 30 минут при 37°С при перемешивании с последующим переносом в 10 мл предварительно нагретой среды YT-TAG (среды 2х YT, содержащей 30 мкг/мл тетрациклина, 100 мкг/мл ампициллина и 0,1 М глюкозы), дополненной фагом-помощником M13K07 при множественности заражения (MOI) 10.
Инкубацию продолжали 15 минут при 37°С при неинтенсивном перемешивании и затем 30 минут при 37°С при интенсивном перемешивании. Параллельно образцы, элюированные TEA и трипсином, использовали для инфицирования культур Е.coli XL1-Blue в логарифмической фазе роста в 9 мл среды YT-TAG, инкубировали при неинтенсивном перемешивании в течение 15 минут при 37°С перед добавлением 1 мл среды YT-TAG, дополненной фагом-помощником M13K07 при MOI 10.
Инкубацию продолжали 15 минут при 37°С при неинтенсивном перемешивании и затем 30 минут при 37°С при интенсивном перемешивании. Все образцы затем центрифугировали при 3000 g в течение 10 минут при к.т., надосадочные жидкости удаляли и осадки осторожно ресуспендировали в 10 мл предварительно нагретой среды 2х YT, содержащей 100 мкг/мл ампициллина и 50 мкг/мл канамицина.
В соответствующие образцы добавляли 1 мМ IPTG и все образцы инкубировали в течение ночи при 30°С при интенсивном перемешивании. На следующий день культуры центрифугировали при 4000 g в течение 10 минут при к.т., и надосадочную жидкость фильтровали в новые пробирки объемом 15 мл через стерилизующие 0,2 мкм фильтры.
Эти надосадочные жидкости затем использовали в следующем раунде пэннинга согласно описаниям с использованием объемов 50 мкл на образец, что соответствует вносимому количеству по меньшей мере 109 KOEampR на образец. После 2 раунда селекции надосадочные жидкости, содержащие вирионы, использовали в антиген-специфичном ELISA, как описано выше.
Результаты
Известно, что все пять структурных белков оболочки нитчатого фага (fd, М13 и f1) представляют собой интегральные мембранные белки, которые обнаруживают во внутренней мембране грамотрицательного хозяина перед их включением в образуемый вирион (Endeman and Model, PMID: 7616570). В этом сообщении также был сделан вывод о том, что хотя pIX и не позволял проводить какие-либо модификации N-концевым слиянием, в интактном вирионе белок капсида сам по себе был гидрофильным. Позднее Gao et al. (PMID: 10339535 и 12239343) и Khalil et al. (PMID: 17360403) продемонстрировали, что дисплей слитых белков pIX при N-концевом слиянии возможен при их экспрессии с фагмид и использовании в комбинации с зависимой от сигнальной последовательности периплазматической локализацией. В этих системах происходит комплементация, поскольку геном фага-помощника является источником белка wt pIX при спасении фагмиды. Для определения возможности такого дисплея без какой-либо зависимой от сигнальной последовательности периплазматической локализации авторы изобретения сконструировали две новые фагмиды, названные pGALD9 и pGALD9AL, позволяющие проводить pIX-дисплей при N-концевом слиянии с такой сигнальной последовательностью или без нее, соответственно (Фиг.2).
Для анализа с использованием этих новых фагмид были выбраны два разных слитых белка pIX, и их сравнивали с их аналогами с использованием стандартного pIII-дисплея. Оба слитых белка представляли собой scFv-фрагменты антител, основанные либо на сегментах вариабельных генов человеческих антител, специфичных в отношении конъюгата гаптена phOx-BSA (scFv против phOx), либо на сегментах вариабельных генов мышиных антител, специфичных в отношении конъюгата гаптена NIP-BSA (scFv против NIP). To есть, scFv против phOx (SEQ ID NO:11) был выбран из библиотеки scFv человеческих антител, и известно, что он довольно хорошо экспрессируется в E.coli (Marks et al., PMID: 1748994). В отличие от этого, хорошо известна плохая экспрессия многих вариабельных генов мышиных гибридом в Е.coli и также при фаговом дисплее (Krebber et al, PMID: 9032408).
pIX-дисплей scFv не должен мешать нормальной сборке вириона. Поэтому авторы изобретения сравнили эффективность этих фагмид для дисплея scFv с сигнальной последовательностью и без нее и также стандартным pIII-дисплеем (для которого абсолютно необходима зависимая от сигнальной последовательности периплазматическая локализация) с использованием стандартного спасения фагмиды и титрования, как описано в разделе «Материалы и методы» (Фиг.3).
Действительно, результат титрования продемонстрировал, что вирионы, содержащие фагмиды, были получены во всех случаях (Фиг.3А и В). Когда слитые белки scFv экспрессируют с lacPO без индукции промотора (базальная экспрессия), оба варианта pIX-дисплея и контрольный pIII-дисплей приводят к сопоставимым титрам scFv против phOx (Фиг.2А). В отличие от этого, титр при независимом от сигнальной последовательности pIX-дисплее scFv против NIP в 5-10 раз выше, чем при pIII-дисплее и зависимом от сигнальной последовательности pIX-дисплее, соответственно (Фиг.3В). При стимуляции повышенной экспрессии слитого белка scFv IPTG-индукцией lacPO происходило снижение титров приблизительно в 10 раз, как при зависимом от сигнальной последовательности pIX-дисплее, так и при pIII-дисплее. В отличие от этого, эффекты на оба варианта независимого от сигнальной последовательности pIX-дисплея были лишь незначительными (Фиг.3А и В). Поскольку в данной системе происходит wt-комплементация белка pIX посредством фага-помощника, это обнаружение было и неожиданным, и важным, поскольку оно показывает, что зависимый от сигнальной последовательности pIX-дисплей и pIII-дисплей препятствуют процессу сборки вирионов, в то время как этот эффект был лишь незначительным в случае независимого от сигнальной последовательности pIX-дисплея, даже при IPTG-индукции. Этот эффект наиболее значителен при сравнении результата, полученного с использованием scFv против NIP, дисплей которого проводили на белке pIX с сигнальной последовательностью или без нее, где в первом случае показано снижение титра в 100 раз. Также заслуживает внимания то, что для обоих scFv сборка вирионов также эффективна или эффективнее, чем при дисплее с использованием pIII, когда scFv получают независимым от сигнальной последовательности вариантом pIX-дисплея.
Основой любой платформы для комбинаторной селекции является физическая связь фенотипа и генотипа, что делает возможным получение генотипа фенотипической селекцией. При нарушении или потере этой физической связи система становится нефункциональной. Относительно фагмидного дисплея это означает, что для любой селекции необходимо, чтобы при спасении фагмиды с использованием фага-помощника в вирионах была заключена именно фагмида, а не геном фага-помощника. Поскольку фагмида и фаг-помощник содержат различные селектируемые маркеры устойчивости к антибиотикам, это может быть легко оценено при определении инфекционного титра вычислением соотношения фагмиды и фага-помощника на основании их соответствующего числа колониеобразующих единиц (КОЕ), полученного при выращивании на подходящей селективной среде (с ампициллином (ampR) или канамицином (kanR)). В таком случае, для того, чтобы любая эффективная дальнейшая селекция была возможной, это соотношение должно быть больше 1.
При оценке соотношений фагмиды и фага-помощника в препаратах вирионов, описанных выше (Фиг.3Б и Г), были выявлены существенные различия этих трех способов дисплея. При использовании стандартной упаковки вириона без индукции промотора (базальная экспрессия) все три способа возможны для scFv против phOx (Фиг.2Б), в то время как для scFv против NIP (Фиг.3Г) был возможен только независимый от сигнальной последовательности вариант pIX-дисплея. При индукции промотора (IPTG-индукции), как для pIII-дисплея, так и для зависимых от сигнальной последовательности вариантов pIX-дисплея была продемонстрирована существенная потеря связи фенотипа и генотипа, и эффект был наиболее значительным для слитого белка scFv против NIP. В отличие от этого, данный эффект отсутствовал (scFv против phOx) или он был незначительным (scFv против NIP) для независимого от сигнальной последовательности варианта pIX-дисплея. Таким образом, результаты ясно показывают, что независимый от сигнальной последовательности pIX-дисплей обеспечивает лучшую связь фенотипа и генотипа по сравнению как со стандартным pIII-дисплеем, так и с зависимым от сигнальной последовательности pIX-дисплеем.
На основании образцов, описанных выше, авторы изобретения затем оценивали дисплей функциональных scFv на этих вирионах в антиген-специфичном ELISA (Фиг.4). Результат ясно показал дисплей функциональных scFv при всех трех способах дисплея (Фиг.4А и Б). Поскольку образцы не нормализовали по титру фагмид, интенсивности сигналов не являются непосредственно сопоставимыми. Более того, в нескольких образцах наблюдали насыщение сигнала. Для адекватной оценки функциональных различий трех способов дисплея в новом антиген-специфичном ELISA определяли относительные уровни дисплея (число функциональных дисплейных единиц на вирион), как описано в разделе «Материалы и методы» (Фиг.4В и Г). В то время как уровни дисплея при pIII-дисплее и при зависимом от сигнальной последовательности варианте pIX-дисплея были сопоставимы, уровень при независимом от сигнальной последовательности pIX-дисплее был существенно ниже. Это было справедливо для обоих scFv, как при базальной экспрессии, так и при IPTG-индукции. Как и следовало ожидать, все три способа дисплея с использованием обоих scFv демонстрировали более эффективный дисплей при IPTG-индукции. Однако крайне важно рассмотреть результаты, показанные на Фиг.4В и Г, в свете соотношений, приведенных на Фиг.3Б и Г. Например, ясно, что как pIII-дисплей, так и зависимый от сигнальной последовательности pIX-дисплей единицы scFv против NIP демонстрируют в 10-20 раз (базальная экспрессия) и приблизительно в 25 раз (при IPTG-индукции) более эффективный дисплей по сравнению с независимым от сигнальной последовательности pIX-аналогом. Тем не менее, оба два первых способа имеют очень слабую или утраченную связь фенотипа и генотипа, вследствие чего они будут нефункциональными в схеме комбинаторной селекции, поскольку их генотип будет потерян при фенотипической селекции. Этот эффект также имеет место при дисплее единицы scFv против phOx, но только при IPTG-индукции. Более того, уровень дисплея единицы scFv против phOx при независимом от сигнальной последовательности варианте pIX-дисплея с IPTG-индукцией сопоставим с вариантами pIII-дисплея и зависимого от сигнальной последовательности pIX-дисплея при базальной экспрессии. В свете этих данных интересно обнаружить, что в единственном сообщении, где используют белок pIX для (зависимого от сигнальной последовательности) дисплея scFv (Gao, PMID: 12239343), неизменно используют IPTG-индукцию при упаковке вирионов, но не сообщают соотношения фагмиды и фага-помощника.
Несмотря на частое использование формата scFv ввиду его благоприятных профилей экспрессии в Е.coli (Bradbury and Marks, PMID: 15261570), в некоторых сообщениях выявлено преимущество форматов дисплея более низкого уровня, таких как, например, Fab-фрагменты антител, при получении высокоаффинных связывающих агентов при аффинной селекции (de Haard, et al, PMID: 10373423, Hoogenboom, PMID: 16151404, и Rothe et al (PMID: 18191144)).
Таким образом, несмотря на то, что независимый от сигнальной последовательности pIX-дисплей scFv, по-видимому, обеспечивает низкоуровневый дисплей, на самом деле, это может стать значительным преимуществом при использовании для высокоаффинной селекции.
Поэтому для оценки эффективности независимого от сигнальной последовательности pIX-дисплея при аффинной селекции авторы изобретения проводили сравнение с обычным pIII-дисплеем.
Вирионы получали в присутствии или в отсутствие IPTG-индукции, и рассматриваемые белки представляли собой scFv либо против phOx, либо против NIP.
Таким образом, в общей сложности 4 популяции фагов оценивали в отношении двух мишеней, phOx- и NIP-BSA, соответственно. В каждом случае специфичный в отношении мишени scFv смешивали с scFv с посторонней специфичностью в соотношении 1:107.
Важно, что два scFv не являются перекрестно реактивными. Затем проводили два раунда аффинной селекции.
Использовали три разных способа элюирования: элюирование при высоком рН, протеолитическое элюирование или прямое инфицирование. Обогащение верифицировали антиген-специфичным поликлональным фаговым ELISA после второго раунда селекции (Фиг.5).
При дисплее рассматриваемого белка на pIX селекция была приблизительно одинаково эффективной и в большинстве случаев более эффективной, чем при дисплее POI на pIII. В частности, для стандартного способа pIII-дисплея с использованием элюирования при высоком рН (TEA) или протеолитического элюирования (трипсин) было продемонстрировано незначительное по сравнению с pIX-дисплеем обогащение.
Селекция без IPTG-индукции была более эффективной, чем с IPTG-индукцией, независимо от способа дисплея и условий элюирования, и отрицательный эффект, по-видимому, был наиболее тяжелым для способа pIII-дисплея.
Таким образом, склонность независимого от сигнальной последовательности pIX-дисплея к низкоуровневому дисплею не приводит к неэффективной селекции. Если белок pIII оставлен неизмененным и полностью гидрофильным, спасение вириона после стадии селекции из библиотеки может быть эффективно проведено без разрыва связи вириона и мишени, что делает стадию элюирования избыточной и ускоряет протоколы с высокой пропускной способностью.
Последнее может также способствовать выделению связывающих агентов с высокой аффинностью, в отношении которых неприменимы многие способы элюирования (Balass et al, PMID: 8954559).
ССЫЛКИ
1. Endemann, H. & Model, P. Location of Filamentous Phage Minor Coat Proteins in Phage and in Infected Cells. Journal of Molecular Biology 250, 496-506 (1995) (PMID: 7616570).
2. Gao, С. et al. Making artificial antibodies: A format for phage display of combinatorial heterodimeric arrays. PNAS 96, 6025-6030 (1999) (PMID: 10339535).
3. Gao, С. et al. A method for the generation of combinatorial antibody libraries using pIX phage display. PNAS 2002 Oct 1; 99(20):12612-6. Epub 2002 Sep 18 (PMID: 12239343).
4. WO/2000/071694. Janda, Kim, D.
5. Khalil AS, Ferrer JM, Brau RR, Kottmann ST, Noren CJ, Lang MJ, Belcher AM. Single M13 bacteriophage tethering and stretching. Proc Natl Acad Sci USA. 2007 Mar20; 104(12):4892-7. Epub 2007 Mar 13 (PMID: 17360403).
6. Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990 Oct 5; 215(3):403-10 (PMID: 2231712).
7. Baneyx F, Mujacic M. Recombinant protein folding and misfolding in Escherichia coli. Nat Biotechnol. 2004 Nov; 22(11):1399-408 (PMID: 15529165).
8. Sambrook et al., Molecular cloning: a laboratory manual. Cold Spring Harbor Laboratory Press.
9. Michaelsen ТЕ, Aase A, Westby C, Sandlie I. Enhancement of complement activation and cytolysis of human IgG3 by deletion of hinge exons. Scand J Immunol. 1990 Nov; 32(5):517-28 (PMID: 2125362).
10. Nakela O, Kaartinen M, Pelkonen JL, Karjalainen K. Inheritance of antibody specificity V. Anti-2-phenyloxazolone in the mouse. J Exp Med. 1978 Dec 1; 148(6):1644-60 (PMID: 722243).
11. Kipriyanov SM, Moldenhauer G, Little M. High level production of soluble single chain antibodies in small-scale Escherichia coli cultures. J Immunol Methods. 1997 Jan 15:200(1-2):69-77 (PMID: 9005945).
12. Norderhaug L, Olafsen T, Michaelsen ТЕ, Sandlie I. Versatile vectors for transient and stable expression of recombinant antibody molecules in mammalian cells. J Immunol Methods, 1997 May 12; 204(1):77-87 (PMID: 9202712).
13. Welschof M, Terness P, Kipriyanov SM, Stanescu D, Breitling F, Dorsam H, Dubel S, Little M, Opelz G. The antigen-binding domain of a human IgG-anti-F(ab')2 autoantibody. Proc Natl Acad Sci USA. 1997 Mar 4; 94(5):1902-7 (PMID: 9050877).
14. Koch J, Breitling F, Dubel S. Rapid titration of multiple samples of filamentous bacteriophage (M13) on nitrocellulose filters. Biotechniques. 2000 Dec; 29(6):1196-8, 2002 (PMID: 11126120).
15. Marks JD, Hoogenboom HR, Bonnert TP, McCafferty J, Griffiths AD, Winter G. By-passing immunization. Human antibodies from V-gene libraries displayed on phage. J Mol Biol. 1991 Dec 5; 222(3):581-97 (PMID: 1748994).
16. Krebber A, Bornhauser S, Burmester J, Honegger A, Willuda J, Bosshard HR, Pluckthun A. Reliable cloning of functional antibody variable domains from hybridomas and spleen cell repertoires employing a reengineered phage display system. J Immunol Methods. 1997 Feb 14; 201(1):35-55 (PMID: 9032408).
17. Bradbury AR, Marks JD. Antibodies from phage antibody libraries. J Immunol Methods. 2004 Jul; 290(1-2):29-49 (PMID: 15261570).
18. de Haard HJ, van Neer N, Reurs A, Hufton SE, Roovers RC, Henderikx P, de 20 Bruine AP, Arends JW, Hoogenboom HR. A large non-immunized human Fab fragment phage library that permits rapid isolation and kinetic analysis of high affinity antibodies. A large non-immunized human Fab fragment phage library that permits rapid isolation and kinetic analysis of high affinity antibodies (PMID: 10373423).
19. Hoogenboom HR. Selecting and screening recombinant antibody libraries. Nat Biotechnol. 2005 Sep; 23(9):1105-16 (PMID: 16151404).
20. Rothe С, Urlinger S, Lohning С, Prassler J, Stark Y, Jager U, Hubner B, Bardroff M, Pradel I, Boss M, Bittlingmaier R, Bataa T, Frisch C, Brocks B, Honegger A, Urban M. The human combinatorial antibody library HuCAL GOLD combines diversification of all six CDRs according to the natural immune system with a novel display method for efficient selection of high-affinity antibodies. J Mol Biol. 2008 Feb 29; 376(4): 1182-200. Epub 2007 Dec 15 (PMID: 18191144).











Изобретение относится к области молекулярной биологии, биохимии и генетической инженерии. Предложен геном фага, фагмида и нитчатый фаг для использования в фаговом дисплее, система фагового дисплея, набор и фаговая библиотека для определения продукта экзогенного гена. Изобретение может быть использовано в исследованиях белков и при разработке диагностических и терапевтических средств на основе белков. 6 н. и 5 з.п. ф-лы, 5 ил., 1 пр.
1. Геном фага для использования в фаговом дисплее, содержащий нуклеиновую кислоту, кодирующую слитый белок pIX, имеющий происхождение из нитчатого фага, где белок pIX слит с экзогенным пептидом непосредственно N-концевой областью последовательности pIX, где слитый белок не содержит прокариотическую N-концевую сигнальную последовательность, где слитый белок pIX содержит последовательность, выбранную из группы, состоящей из положения 1-32, 2-32, 3-32, 4-32 и 5-32 SEQ ID NO:1 (MSVLVYSFASFVLGWCLRSGITYFTRLMETSS), и где экзогенный пептид выбран из группы, состоящей из антител или их фрагментов, включая Fv, scFv, Fab, отдельных доменов, домена Z белка A или его фрагментов (аффитела), анкирина или его фрагментов, сконструированных белков с анкириновыми повторами (DARPin) или их фрагментов, T-клеточного рецептора или его фрагментов, молекул главного комплекса гистосовместимости (МНС) I или II класса или их фрагментов, фибронектина или его фрагментов, антикалинов или их фрагментов, PDZ-доменов или их фрагментов, нового иммуноглобулинового антигенного рецептора (IgNAR) или его фрагментов, антигена 4, ассоциированного с цитотоксическими Т-лимфоцитами (CTLA4), или его фрагментов, иммунного белка колицина Е7 (ImmE7) или его фрагментов, ноттинов или их фрагментов, авимера или его фрагментов, зеленого флуоресцентного белка (GFP) или его фрагментов и других кодируемых генами биологических флуорофоров.
2. Геном фага по п.1, где экзогенный пептид является членом библиотеки.
3. Фагмида для использования в фаговом дисплее, содержащая нуклеиновую кислоту, кодирующую слитый белок pIX, имеющий происхождение из нитчатого фага, где белок pIX слит с экзогенным пептидом непосредственно N-концевой областью последовательности pIX, где слитый белок не содержит прокариотическую N-концевую сигнальную последовательность, где слитый белок pIX содержит последовательность, выбранную из группы, состоящей из положения 1-32, 2-32, 3-32, 4-32 и 5-32 SEQ ID NO:1 (MSVLVYSFASFVLGWCLRSGITYFTRLMETSS), и где экзогенный пептид выбран из группы, состоящей из антител или их фрагментов, включая Fv, scFv, Fab, отдельных доменов, домена Z белка A или его фрагментов (аффитела), анкирина или его фрагментов, сконструированных белков с анкириновыми повторами (DARPin) или их фрагментов, T-клеточного рецептора или его фрагментов, молекул главного комплекса гистосовместимости (МНС) I или II класса или их фрагментов, фибронектина или его фрагментов, антикалинов или их фрагментов, PDZ-доменов или их фрагментов, нового иммуноглобулинового антигенного рецептора (IgNAR) или его фрагментов, антигена 4, ассоциированного с цитотоксическими Т-лимфоцитами (CTLA4), или его фрагментов, иммунного белка колицина Е7 (ImmE7) или его фрагментов, ноттинов или их фрагментов, авимера или его фрагментов, зеленого флуоресцентного белка (GFP) или его фрагментов и других кодируемых генами биологических флуорофоров.
4. Фагмида по п.3, где экзогенный пептид является членом библиотеки.
5. Нитчатый фаг для использования в фаговом дисплее, содержащий геном фага по любому из п.п.1-2 или фагмиду по любому из п.п.3-4.
6. Нитчатый фаг по п.5, дополнительно содержащий ген, кодирующий белок р1Х дикого типа (wt pIX), и/или белок wt pIX.
7. Нитчатый фаг по п.5, не содержащий ген, кодирующий белок wt pIX, и/или белок wt pIX.
8. Нитчатый фаг по любому из п.п.5-7, дополнительно содержащий одну или более чем одну группу, выбранную из слитого белка pill, слитого белка pVII и слитого белка pVIII.
9. Система фагового дисплея для определения продукта экзогенного гена, включающая геном фага по любому из п.п.1-2 или фагмиду по любому из п.п.3-4 и фаг-помощник.
10. Набор для определения продукта экзогенного гена, включающий систему фагового дисплея по п.9 и протокол с прописями реагентов и описанием анализов.
11. Фаговая библиотека для определения продукта экзогенного гена, включающая два или более чем два нитчатых фага, на которых представлены различные белки, по меньшей мере один из которых представляет собой слитый белок pIX, экспрессированный с генома фага по п.1 или фагмиды по п.3.
| WO0071694 A1, 30.11.2000 | |||
| US 0006472147 B1, 29.10.2002 | |||
| US 20060252028 A1, 09.11.2006 | |||
| Контактный термометр | 1928 |
|
SU10828A1 |

