ПЕРЕКРЕСТНЫЕ ССЫЛКИ НА РОДСТВЕННЫЕ ЗАЯВКИ
Настоящая заявка испрашивает приоритет временной заявки на патент США, серийный № 61/630345, поданной 9 декабря 2011 года, полное описание которой включается в настоящий документ посредством ссылки.
ИНФОРМАЦИЯ ОТНОСИТЕЛЬНО ИССЛЕДОВАНИЙ, СПОНСИРУЕМЫХ
ФЕДЕРАЛЬНЫМ ПРАВИТЕЛЬСТВОМ
Настоящее изобретение осуществлено при правительственной поддержке согласно грантам №№ 2007-35203-18274 и 2011-67015-20025 от USDA National Institute of Food and Agriculture (USDA-NIFA). Правительство имеет определенные права на настоящее изобретение.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к композиции и к способу ее применения для искусственного осеменения. Более конкретно, настоящее изобретение относится к добавке для стимуляции спермы для использования при искусственном осеменении сельскохозяйственных животных и при оплодотворении in vitro и в эмбриональной культуре в клиниках для лечения бесплодия у людей.
УРОВЕНЬ ТЕХНИКИ
Искусственное осеменение (AI) представляет собой распространенную технологию при разведении свиней и крупного рогатого скота. Семя борова непосредственно после эйакуляции должно храниться в растворе разбавителя для консервации при 15- 18°C или 4-5°C, а семя быка должно разбавляться перед криоконсервированием и хранением в жидком азоте. Разработаны разнообразные типы растворов разбавителей и соединений для уменьшения метаболической активности спермы и для создания возможности для консервации в разбавленном виде. Однако необходимы новые и улучшенные среды для культивирования и/или разбавители спермы для улучшения искусственного осеменения у животных и для оплодотворения in vitro и эмбриональной культуры у людей.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В одном из аспектов, настоящее изобретение предлагает среды для консервации спермы, содержащие неорганический пирофосфат (PPi). В одном из вариантов осуществления, концентрация PPi находится в пределах примерно между 1 мкМ и примерно 200 мкМ. В другом варианте осуществления, концентрация PPi находится в пределах примерно между 1 мкМ и примерно 20 мкМ. В другом варианте осуществления, концентрация PPi составляет примерно 10 мкМ. Еще в одном варианте осуществления, среда для консервации используется для консервации спермы свиней.
В другом аспекте, настоящее изобретение предлагает среды для переноса спермы, содержащей неорганический пирофосфат (PPi). В одном из вариантов осуществления, концентрация PPi находится в пределах примерно между 1 мкМ и примерно 200 мкМ. В другом варианте осуществления, концентрация PPi находится в пределах примерно между 1 мкМ и примерно 20 мкМ.
Другой аспект настоящего изобретения предлагает среды для оплодотворения in vitro (IVF) или искусственного осеменения (AI), содержащие неорганический пирофосфат (PPi). В одном из вариантов осуществления, концентрация PPi находится в пределах примерно между 1 мкМ и примерно 200 мкМ. В другом варианте осуществления, концентрация PPi находится в пределах примерно между 1 мкМ и примерно 20 мкМ.
В другом аспекте, настоящее изобретение предлагает способ сексинга спермы, включающий: (a) разделение смешанной суспензии спермы в первой среде для культивирования на популяцию спермы, несущей x-хромосому или несущей y-хромосому, с помощью элюирующей среды; (b) консервацию спермы, несущей x-хромосому или несущей y-хромосому, во второй среде для культивирования, где неорганический пирофосфат (PPi) добавляют в первую среду для культивирования, элюирующую среду или вторую среду для культивирования.
Еще в одном аспекте, настоящее изобретение предлагает способ консервации спермы, включающий хранение сперма в средах, содержащих неорганический пирофосфат (PPi). В одном из вариантов осуществления, концентрация PPi находится в пределах примерно между 1 мкМ и примерно 200 мкМ. В других вариантах осуществления, концентрация PPi находится в пределах примерно между 1 мкМ и примерно 20 мкМ или концентрация PPi составляет примерно 10 мкМ. Еще в одном варианте осуществления, сперму хранят в средах, содержащих PPi, в течение до 10 дней.
Другой аспект настоящего изобретения предлагает способ оплодотворения in vitro (IVF), включающий приведение в контакт ооцита со спермой в присутствии неорганического пирофосфата (PPi). В одном из вариантов осуществления, концентрация PPi находится в пределах примерно между 1 мкМ и примерно 200 мкМ. В другом варианте осуществления, концентрация PPi находится в пределах примерно между 1 мкМ и примерно 20 мкМ. В другом варианте осуществления, сперму хранят в присутствии PPi.
В другом аспекте, настоящее изобретение предлагает способ культивирования эмбрионов, включающий культивирование эмбриона в средах, содержащих неорганический пирофосфат.
Еще в одном аспекте, настоящее изобретение предлагает способ искусственного осеменения, включающий доставку спермы и неорганического пирофосфата (PPi) в половые пути самки. В вариант осуществления, PPi постепенно высвобождается в половых путях самки.
Другой аспект настоящего изобретения предлагает способ созревания ооцита in vitro, включающий культивирование ооцитов в средах, содержащих неорганический пирофосфат.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг. 1: Показывает измерение содержания пирофосфата (PPi) с помощью флуорометрического анализа. (A) Интенсивность флуоресценции стандартов PPi (конечная концентрация 0-200 мкМ PPi). (B) Анализ PPi с помощью семенной плазмы борова (SP), овидуктальных жидкостей свиней (pOVF), сыворотки кролика, сыворотки мыши (конечная концентрация 10 мкг/мл), сперматозоидов борова (1×106 сперматозоидов/мл) и 10 мМ рабочего раствора Η2O2 (отрицательный контроль). Интенсивности флуоресценции измеряют во множестве временных точек для отслеживания кинетики реакции (возбуждение на 530 нм; испускание на 590 нм). Эксперименты повторяют три раза. Значения выражают как среднее значение интенсивности флуоресценции.
Фиг. 2: Показывает генерирование PPi. PPi получается посредством гидролиза ATP до AMP в клетках. Неорганическая пирофосфатаза (PPA1) катализирует гидролиз PPi с образованием 2 ортофосфатов (2Pi), с получением в результате высвобождения энергии.
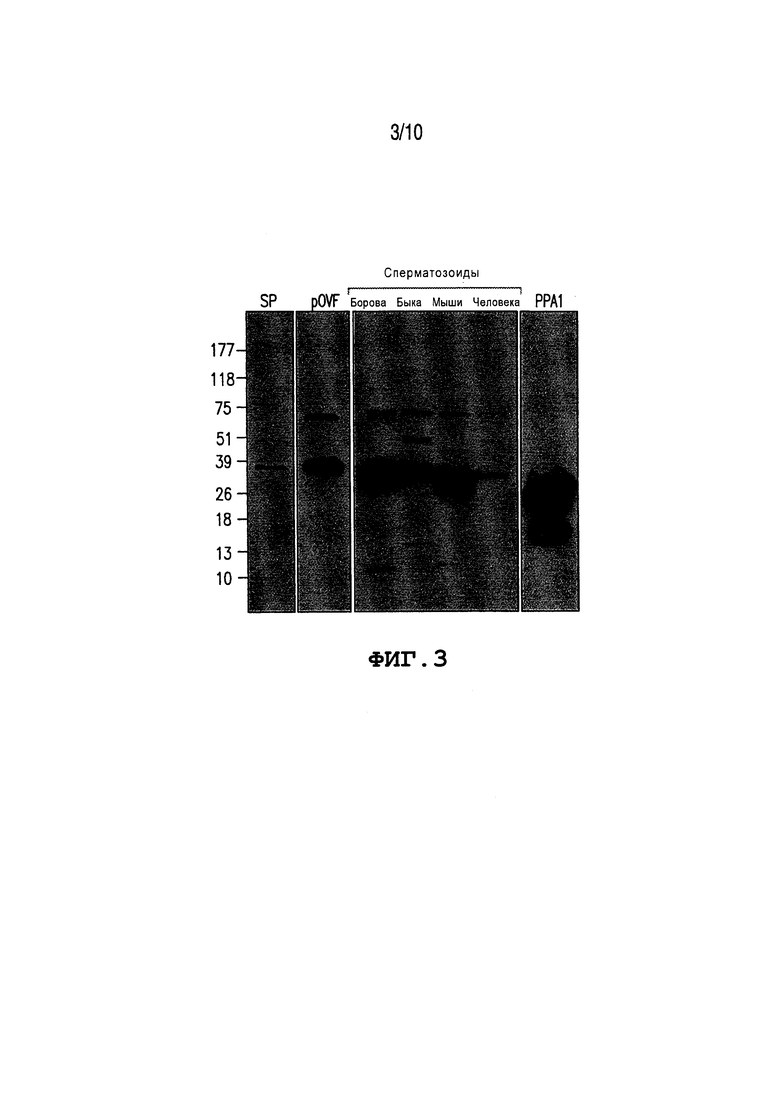
Фиг. 3: Показывает детектирование неорганической пирофосфатаазы (PPA1) с помощью вестерн-блоттинга. Семенную плазму борова (SP; 20 мкг/мл), овидуктальную жидкость свиней (pOVF; 100 мкг/мл) и сперматозоиды борова, быка, мыши и человека (все при 1×106 сперматозоидов/мл) экстрагируют для осуществления анализа белка. Используют одинаковые нагрузки белка. Отдельную полосу при ~32 кДа детектируют с помощью поликлонального антитела кролика анти-PPA1. Очищенную PPA1 (крайняя правая полоса; 1 мкг/мл; Sigma 11643) от S. cerevisiae используют в качестве контрольного белка.
Фиг. 4: Показывает локализацию неорганической пирофосфатаазы (PPA1; красный) в сперматозоидах с помощью иммунофлуоресценции. (A, B) Иммунофлуоресценция от цельного препарата сперматозоидов борова. Наиболее заметное присутствие метки наблюдают в соединительной части хвоста сперматозоида и в постакросомальной оболочке головки сперматозоида. (C) Идентичное присутствие метки наблюдают в сперматозоиде, прикрепившемся к вителлиновому слою ооцита, через 30 мин после гаметического смешивания во время IVF. (D) Отрицательный контроль с помощью антитела анти-PPA1, иммунологически насыщенного с помощью полноразмерного белка PPA1. ДНК контрастно окрашивают с помощью DAPI (голубой). Фотографии эпифлуоресценции перекрываются с фотографиями в парфокальном проходящем свете, полученными с помощью оптики с DIC.
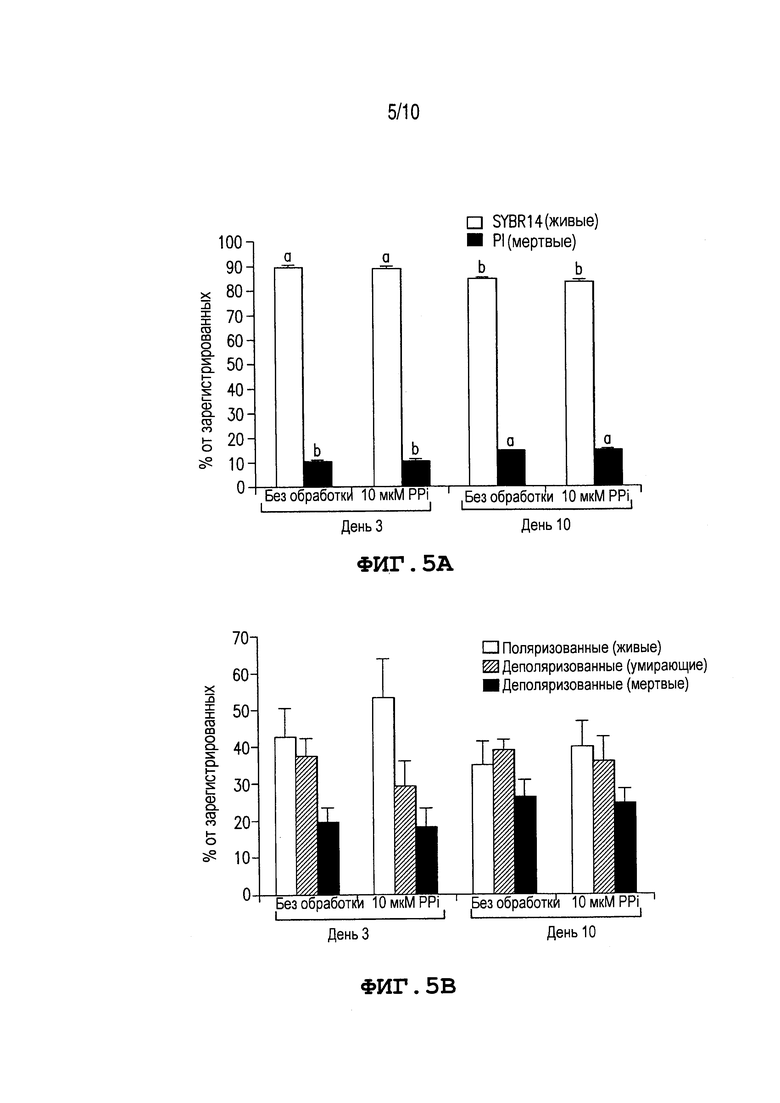
Фиг. 5: Показывает жизнеспособность спермы и митохондриальный мембранный потенциал во время хранения спермы с/без PPi. (A) Проценты жизнеспособных сперматозоидов на основе мечения SYBR14 (живая сперма) и PI (мертвая сперма). (B) Проценты сперматозоидов с поляризованными (живые), деполяризующимися (умирающие) и деполяризованными (мертвые) митохондриальными мембранами. Эксперименты повторяют три раза. Значения выражают как средний процент ± SEM. Различные верхние индексы a и b в каждой группе столбцов означают значимое различие при p≤0,05.
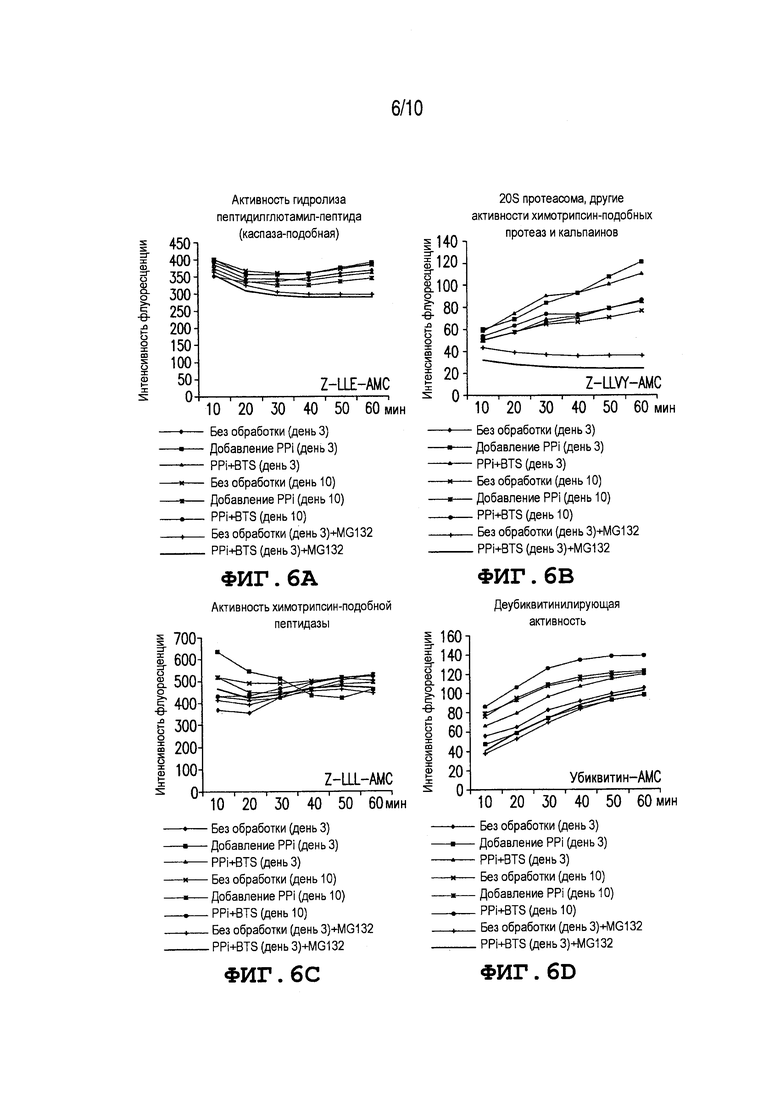
Фиг. 6: Показывает воздействие PPi на протеасомальные ферментные активности хранящихся сперматозоидов борова. Свежие сперматозоиды борова хранят в BTS с 10 мкМ PPi и без него в течение 3 или 10 дней (не обработанные/PPi+BTS). Протеасомальные протеолитические и деубиквитинирующие активности измеряют с использованием специфичных флуорометрических субстратов Z-LLE-AMC (A), Z-LLVY-AMC (B), Z-LLL-AMC (C) и убиквитин-AMC (D). При отдельной обработке, добавляют PPi перед измерением к сперматозоидам, консервируемым без PPi (Добавление PPi). В качестве отрицательного контроля, добавляют 10 мкМ MG132 (протеасомальный ингибитор) к сперматозоидам ʺбез обработкиʺ и ʺPPi+BTSʺ в день 3. Эксперименты повторяют три раза. Значения выражают как среднее значение интенсивности флуоресценции.
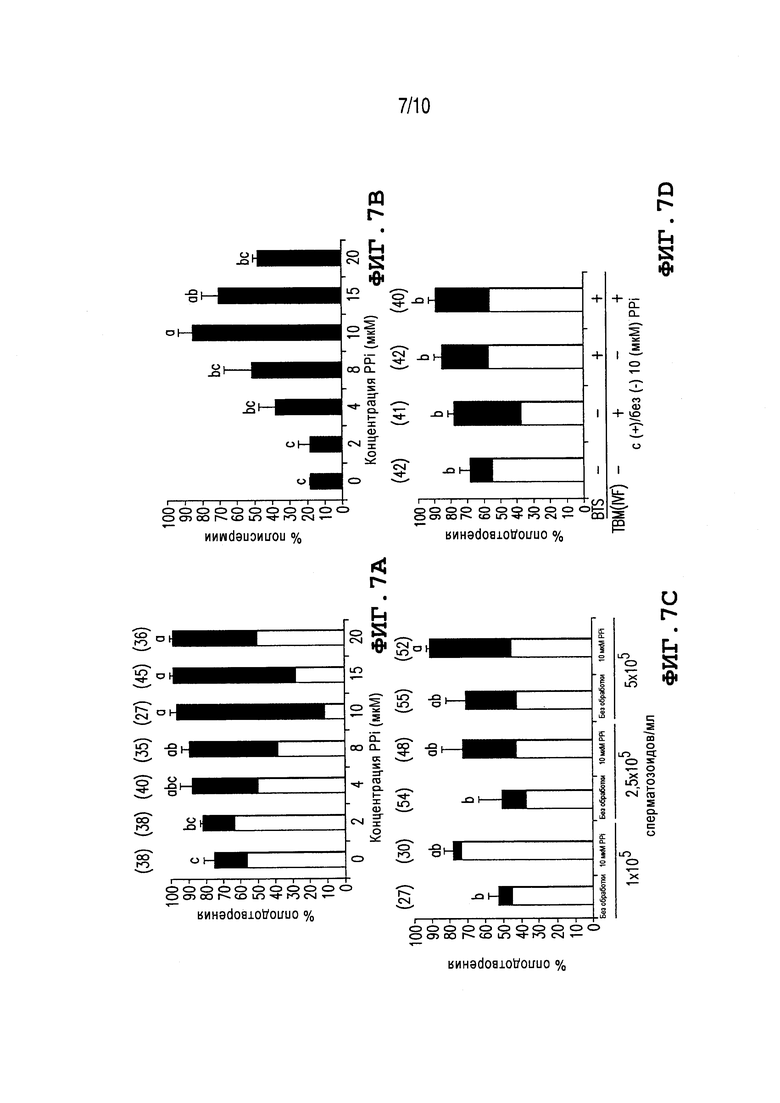
Фиг. 7: Показывает воздействие PPi на полное и полиспермное оплодотворение во время IVF свиней. Значения выражают как средние проценты ± SEM.  % моноспермные и
% моноспермные и  % полиспермные ооциты. Различные верхние индексы a-c в каждой группе столбцов обозначают значимое различие при p<0,05. Количества не осемененных яйцеклеток указаны в круглых скобках. (A) Ооциты свиней, созревающие in vitro, осеменяют при стандартной концентрации 1×106 сперматозоидов/мл, в присутствии увеличивающихся концентраций PPi. Эксперименты повторяют пять раз. (B) Доли полиспермии, отражающие способность спермы к оплодотворению in vitro (также панель A), кардинально повышаются в присутствии PPi. (C) Доли оплодотворения ооцитов свиньи, осемененных с помощью различных концентраций сперматозоидов в присутствии/отсутствии 10 мкМ PPi. Эксперименты повторяют три раза. (D) Доли оплодотворения ооцитов, осемененных сперматозоидами борова, консервируемыми в течение 3 дней в BTS с/без 10 мкМ PPi. Другие ооциты свиньи осеменяют (концентрация сперматозоидов 5×105 сперматозоидов/мл) с присутствием 10 мкМ PPi и без него, сперматозоидами, хранящимися с PPi и без него. Эксперименты повторяют три раза. (E) Воздействие постороннего фермента PPA1 на IVF свиньи. Ооциты осеменяют при различных концентрациях очищенного белка PPA1. Эксперименты повторяют дважды. (F) Ооциты свиньи осеменяют в присутствии поликлонального антитела кролика анти-PPA1 или неиммунной сыворотки кролика (контроль для антитела PPA1). Эксперименты повторяют дважды.
% полиспермные ооциты. Различные верхние индексы a-c в каждой группе столбцов обозначают значимое различие при p<0,05. Количества не осемененных яйцеклеток указаны в круглых скобках. (A) Ооциты свиней, созревающие in vitro, осеменяют при стандартной концентрации 1×106 сперматозоидов/мл, в присутствии увеличивающихся концентраций PPi. Эксперименты повторяют пять раз. (B) Доли полиспермии, отражающие способность спермы к оплодотворению in vitro (также панель A), кардинально повышаются в присутствии PPi. (C) Доли оплодотворения ооцитов свиньи, осемененных с помощью различных концентраций сперматозоидов в присутствии/отсутствии 10 мкМ PPi. Эксперименты повторяют три раза. (D) Доли оплодотворения ооцитов, осемененных сперматозоидами борова, консервируемыми в течение 3 дней в BTS с/без 10 мкМ PPi. Другие ооциты свиньи осеменяют (концентрация сперматозоидов 5×105 сперматозоидов/мл) с присутствием 10 мкМ PPi и без него, сперматозоидами, хранящимися с PPi и без него. Эксперименты повторяют три раза. (E) Воздействие постороннего фермента PPA1 на IVF свиньи. Ооциты осеменяют при различных концентрациях очищенного белка PPA1. Эксперименты повторяют дважды. (F) Ооциты свиньи осеменяют в присутствии поликлонального антитела кролика анти-PPA1 или неиммунной сыворотки кролика (контроль для антитела PPA1). Эксперименты повторяют дважды.
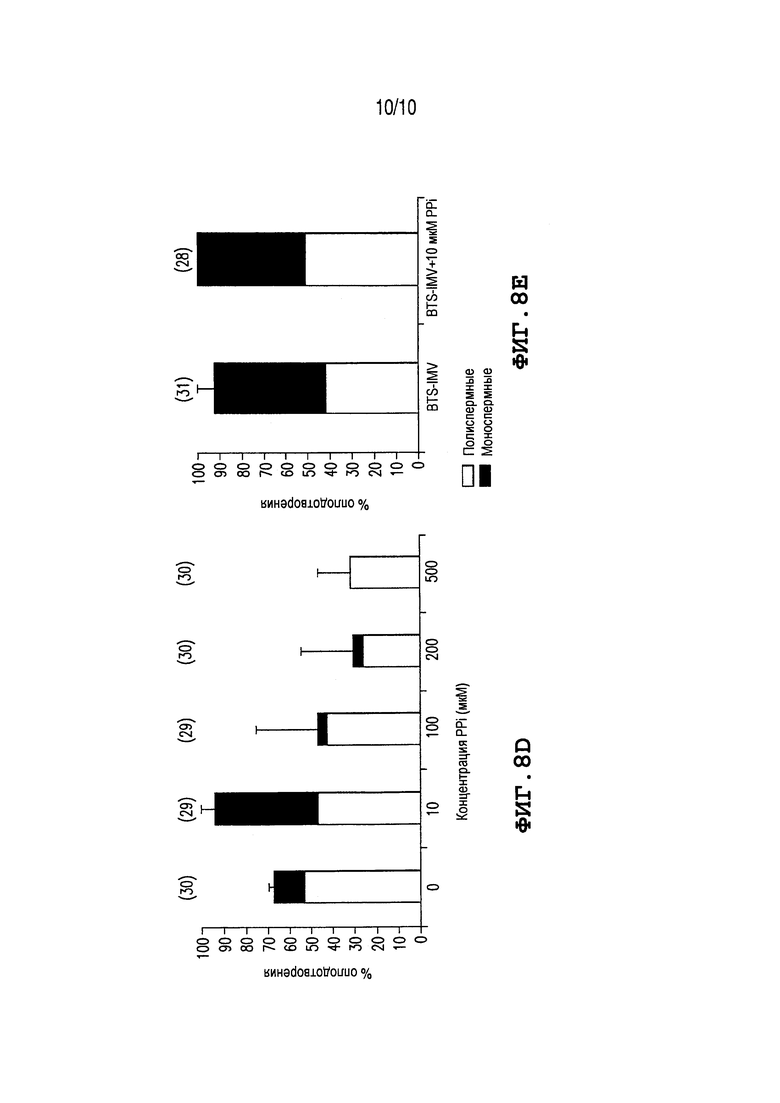
Фиг. 8: (A) Показывает воздействие PPi на связывание сперматозоид - вителлиновый слой. Ооциты свиньи осеменяют (концентрация сперматозоидов 5×105 сперматозоидов/мл) при различных концентрациях PPi в течение 30 мин, фиксируют и окрашивают с помощью красителя DAPI для ДНК. Количества сперматозоидов, связанных с вителлиновым слоем (ZP), считают под эпифлуоресцентным микроскопом. Значения выражают как среднее значение × SEM. Различные верхние индексы a-c в каждой группе столбцов обозначают значимое различие при p×0,05. Количества не осемененных яйцеклеток показаны в круглых скобках. (B) Процент сперматозоидов панели A с прореагировавшей акросомой (PNA с окрашиванием FITC). Значения выражают как средние проценты × SEM. Различные верхние индексы a и b в каждой группе столбцов обозначают значимое различие при p×0,05. (C) Воздействие дополнения PPi на жизнеспособность сперматозоидов, хранящихся в коммерческих и приготовленных в лаборатории разбавителях на основе BTS. Сперматозоиды борова консервируют в BTS-IMV (Technologies IMV, France) или BTS-HM (приготовлен в лаборатории) с 10 мкМ PPi и без него в течение 7 дней при комнатной температуре. Подвижности сперматозоидов оценивают с помощью наблюдения под световым микроскопом при 37,5°C. Наблюдается подвижность сперматозоидов в BTS-IMV с PPi в день 6, более высокая, чем в любой другой группе. Эксперименты повторяют дважды. Различные верхние индексы a, b в каждой группе столбцов обозначают значимое различие при p<0,05. (D) Добавление избыточных концентраций PPi в среду для IVF. Доли оплодотворения уменьшаются при высоких концентрациях PPi. Эксперименты повторяют дважды. Значения выражают как средние проценты ± SEM. % моноспермные и % полиспермные ооциты. Количества не осемененных яйцеклеток указаны в круглых скобках. (E) Воздействие PPi на оплодотворение с помощью сперматозоидов, хранящихся в коммерческом разбавителе BTS-IMV. Достигается почти 100% оплодотворение с использованием сперматозоидов, консервируемых в BTS-IMV с 10 мкМ PPi (день 3). Эксперименты повторяют дважды. Значения выражают как средние проценты ± SEM. % моноспермные и % полиспермные ооциты. Количества неосемененных яйцеклеток указаны в круглых скобках.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение предлагает новые среды и способы консервации спермы, эмбриональной культуры, оплодотворения in vitro (IVF), искусственного осеменения (AI). В частности, настоящее изобретение представляет собой прогресс в данной области в том, что оно сообщает и подтверждает, что неорганический пирофосфат (PPi) существует в сперматозоидах, семенной плазме (SP) и овидуктальных жидкостях (OVF) некоторых видов млекопитающих, хотя предыдущие исследования показали, что концентрация цитозольных PPi прецизионно регулируется в клетках млекопитающих (Baykov et al., Prog Mol Subcell Biol 23: 127-150, 1999; Sivula et al, FEBS Lett 454:75-80, 1999). В одном из аспектов настоящего изобретения, PPi мог бы, следовательно, использоваться в качестве источника энергии для жизнеспособности сперматозоидов.
В одном из вариантов осуществления, настоящее изобретение предлагает новые и улучшенные среды для консервации спермы, также упоминаемые как разбавители спермы, которые могут продлить период хранения семени и поддержать жизнеспособность сперматозоидов и таким образом улучшить AI у животных. Настоящее изобретение также предлагает новые и улучшенные среды для культивирования, для переноса эмбрионов у животных.
В другом варианте осуществления настоящее изобретение предлагает новый и улучшенный способ IVF и AI, а также среды для культивирования для эмбрионов в клинике животных и человека.
Еще в одном аспекте настоящего изобретения описывается новый и улучшенный способ сексинга спермы, использующий PPi. Настоящий способ сексинга спермы включают стадию добавления определенного количества PPi в среды в течение процедуры сексинга спермы для повышения продолжительности жизни и жизнеспособности спермы. Например, в соответствии с определенными вариантами осуществления настоящего изобретения, настоящий способ сексинга спермы может включать стадию добавления PPi в среды для начальной обработки спермы (со сперматозоидами, несущими как x-, так и y-хромосому; перед обычной стадией разделения/сортировки), в элюирующие среды или в среды со спермой, разделенной с помощью сексинга.
Как правило, оттаивающий раствор Белтсвилля (BTS) добавляют к замороженной оттаянной сперме в качестве оттаивающего раствора, и его используют также для хранения в жидком виде в течение 3-5 дней (Johnson et al., Zuchthygiene 23:49-55, 1988). Жидкое семя, разбавленное BTS, обычно используют для AI благодаря его простой композиции и разработанной системе транспортировки. Однако подвижность спермы, консервируемой в разбавителе, последовательно уменьшается во время хранения из-за естественного старения, потери ATP и cAMP, а также из-за пониженного потребления кальция (Johnson et al., Anim Reprod Sci 62 143-172, 2000). Разбавленное семя, хранимое в течение 5 дней после сбора, показывает уменьшение долей опороса приблизительно 50% по сравнению с семенем, консервируемым в течение 2 дней после сбора, которое показывает уменьшение долей опороса приблизительно 65-70% (Johnson et al., Anim Reprod Sci 62 143-172; Johnson et al., Zuchthygiene 23:49-55, 1988; Johnson and Rath, (Eds), Proc. 2nd Int. Conf. Deep Freezing Boar Semen. Reprod. Domest. Anim., Suppl. 1, p. 402, 1991; Rath et al., (Eds) Proc. Int. Conf. Deep Freezing of Boar Semen. Reprod. Domest. Anim. Suppl. 1, p. 342, 1996; Johnson, Proc. 15th Int. Pig Vet. Sci. Congress 1, 225-229, 1998). В последнее время, Yeste et al. (Anim Reprod Sci 108: 180-195, 2008) сообщили, что добавление простагландина F2α (PGF2α) к сперме, разбавленной в BTS, поддерживает лучшую жизнеспособность и подвижность спермы после 6 дней охлаждения.
Неорганический пирофосфат (PPi) представляет собой сильнодействующий, связывающий минеральные вещества низкомолекулярный ингибитор нуклеации и роста кристаллов (Fleisch et al, Nature 212:901-903, 1966) и присутствует во внеклеточном матриксе большинства тканей и телесных жидкостей, включая плазму (Fleisch et al, Am J Physiol 203:671-675, 1962; Russell et al, J Clin Invest 50:961-969, 1971). Метаболизм PPi наблюдают в культивируемых гепатоцитах и хондроцитах (Davidson et al., Biochem J 254:379-384, 1988; Johnson et al., 1999; Rosen et al., Arthritis Rheum 40: 1275-1281, 1997; Rosenthal et al., Calcif Tissue Int 59: 128-133, 1996; Rosenthal et al., J Rheumatol 26:395-401, 1999; Ryan et al., Arthritis Rheum 42:555-560, 1999). Внутриклеточный PPi генерируется в митохондриях, и внутри- и внеклеточные концентрации PPi регулируются с помощью митохондриального метаболизма энергии (Davidson et al., Biochem J 254:379-384, 1988; Johnson et al., Arthritis Rheum 43: 1560-1570, 2000). В прокариотах PPi обеспечивает "высокоэнергетическое" соединение, и он способен замещать ATP в реакциях, родственных гликолизу, при ослабленном дыхании (Chi et al., J Biol Chem 275:35677-35679, 2000). Кроме того, PPi создает митохондриальный мембранный потенциал с помощью PPA (Pereira-da-Silva et al., Arch Biochem Biophys 304:310-313, 1993), и PPi, полученный из ATP, служит в качестве донора фосфатов при фосфорилировании белков в митохондриях дрожжей, а также в клетках млекопитающих (da Silva et al., Biochem Biophys Res Commun 178: 1359-1364, 1991; Terkeltaub et al., Am J Physiol Cell Physiol 281:C1-C11, 2001). Как следствие, PPi может использоваться в качестве источника энергии для жизнеспособности.
Клеточный PPi получается с помощью различных процессов биосинтеза и гидролизуется до двух неорганических фосфатов (Pi) с помощью неорганической пирофосфатазы (PPA1). PPA1 представляет собой распространенный металлзависимый фермент, обеспечивающий термодинамический пул для многих реакций биологического синтеза, таких как синтез ДНК, РНК, белков, полисахаридов, и для жизни клетки (Chen et al., 1990, Lundin et al., 1991, Sonnewald 1992, Lahti 1983, Peller 1976). PPA1 детектируется в бактериях (Chen et al., 1990) и дрожжах (Lundin et al., 1991), и растворимая PPA1 идентифицирована и характеризована в Mycoplasma suis, которые принадлежат к хемотропным бактериям, которые прикрепляются к поверхности эритроцитов-хозяев (Hoelzle et al.). Однако PPi не использовался в каких-либо средах, относящихся к консервации спермы или к процедурам AI или IVF.
Настоящее изобретение идентифицирует путь PPi как важный компонент физиологии сперматозоидов млекопитающих. Обращаясь к Фиг. 2, здесь PPi (P2O74-) образуется посредством гидролиза ATP до AMP в клетках, затем гидролизуется с помощью неорганической пирофосфатазы (PPA1) до двух молекул неорганического ортофосфата (Pi). PPA1, важный фермент для метаболизма энергии (Chen et al., J Bacteriol 172:5686-5689, 1990; Lundin et al., J Biol Chem 266: 12168-12172, 1991), участвует в регуляции метаболизма, роста и развития в растениях (Sonnewald, Plant J 2:571-581, 1992), и даже в развитии и линьке у паразитных круглых червей Ascaris (Islam et al., Infect Immun 73: 1995-2004, 2005). Во время деления клеток S. cerevisiae, PPA1 является самым важным веществом для репликации генома митохондрии (Lundin et al., Biochim Biophys Acta 1098; 217-223, 1992).
В то время как PPA1 детектируется в соединительной части хвоста сперматозоида, в области удерживания центриоли сперматозоида и при фиксации флагеллярных внешних плотных волокон и дублетов микротрубочек, настоящее изобретение говорит, что из этих областей, путь метаболизма PPi может переносить энергию для флагеллярного движения и для акросомальной функции во время проникновения сперматозоида в вителлиновый слой. В добавление к этому, настоящее изобретение также говорит, что путь PPi в головке и жгутике сперматозоида могут поддерживать фосфорилирование белков во время капацитации сперматозоидов, что наблюдается как in vitro, так и in vivo, в овидуктальном резервуаре спермы.
Кроме того, настоящее изобретение описывает способность сперматозоидов млекопитающих к использованию PPi в качестве источника энергии во время переноса сперматозоидов и взаимодействий сперматозоид - яйцеклетка, когда сперматозоиды подвергаются воздействию капацитации, реакции акросомы и проникновения сперматозоида в вителлиновый слой. В настоящее время описано, что PPi можно использовать в качестве стабильного, недорогого источника энергии для улучшения жизнеспособности сперматозоидов во время хранения и переноса семени для биотехнологии больших животных и для повышения доли проникновения сперматозоидов и повышения долей оплодотворения для вспомогательной репродуктивной терапии у некоторых видов млекопитающих (включая людей). Кроме того, настоящее изобретение дополнительно предлагает добавление PPi в среды для культивирования, в разбавитель спермы, в среды для IVF или в среды, используемые при сексинге спермы, с целью обеспечения полезных воздействий при консервации спермы и оплодотворении, таких как увеличение продолжительности жизни и жизнеспособность сперматозоидов, во время консервации и переноса спермы и поддержания и улучшения жизнеспособности, проникновения сперматозоидов и долей оплодотворения во время процедур оплодотворения.
Следовательно, среды по настоящему изобретению могут содержать PPi, концентрация которого может изменяться в зависимости от видов животных. В определенных вариантах осуществления, среда по настоящему изобретению может содержать примерно 1-200 мкМ PPi. В другом варианте осуществления, среда по настоящему изобретению может содержать примерно 1-20 мкМ PPi. Настоящее изобретение можно использовать для различных млекопитающих, включая сельскохозяйственных животных, таких как боров и бык.
Для IVF или AI, PPi можно непосредственно добавлять или постепенно высвобождать в среды. При необходимости, высвобождение PPi может точно контролироваться/модулироваться во время AI, особенно, когда гель для замедленного высвобождения, содержащий PPi, используют в качестве части катетера для AI, для постепенного высвобождения PPi в половых путях самки.
Если не определено иного, все технические и научные термины, используемые в настоящем документе, имеют такое же значение, как обычно понимается специалистами в области, к которой принадлежит настоящее изобретение. Все публикации, заявки на патенты, патенты и другие ссылки, рассмотренные в настоящем документе, включаются в качестве ссылок во всей их полноте.
ПРИМЕРЫ
Следующие далее описанные варианты осуществления являются всего лишь иллюстрациями настоящего изобретения, которое может быть воплощено в различных формах. Таким образом, конкретные структурные, функциональные и процедурные детали, описанные в следующих далее примерах, не должны интерпретироваться в качестве ограничивающих.
Пример 1
Сбор и обработки семени
Семя собирают от проверенных способных к зачатию взрослых боровов Дьюрок в возрасте 15-22 месяцев согласно инструкциям одобренных протоколов Animal Care and Use Committee (ACUC) of University of Missouri-Columbia (UM-C). Для боровов осуществляют рутинную процедуру сбора, по одному сбору в неделю. Обогащенная сперматозоидами фракция эйакулята собирается в изолированную вакуумную емкость. Используют обогащенную сперматозоидами фракцию эйакулятов с более чем 85% подвижных сперматозоидов. Объемы семени определяют с помощью градуированного цилиндра. Концентрации сперматозоидов оценивают с помощью гемоцитометра (Fisher Scientific, Houston, TX). Процент подвижных сперматозоидов оценивают при 38,5° с помощью световой спектроскопии при увеличении 250°. Семя медленно охлаждают до комнатной температуры (20-23°C) за 2 час после сбора и разбавляют разбавителем на основе оттаивающего раствора Белтсвилля (BTS; 3,71 г глюкозы, 0,60 г тринатрия цитрата, 1,25 г этилендиаминтетрауксусной кислоты, 1,25 г бикарбоната натрия, 0,75 г хлорида калия, 0,06 г пенициллина G и 0,10 г стрептомицина в 100,0 мл дистиллированной воды) (Pursel and Johnson, 1975) до конечной концентрации 35×106 сперматозоидов/мл в 100 мл разбавителя BTS. Разбавленное семя хранят в боксах Styrofoam™ при комнатной температуре в течение 10 дней. Если не указано иного, все химикалии, используемые в этом исследовании, покупают от Sigma Chemical Co. (St. Louis, MO).
Пример 2
Сбор и созревание in vitro (IVM) ооцитов свиней
Яичники собирают у препуберантных подсвинков на местной бойне и переносят в лабораторию в теплом боксе (25-30°C). Комплексы кумулюс-ооцит (COCs) освобождают с отсосом от антральных фолликул (диаметром 3-6 мм), промывают три раза в среде HEPES-лактатный буфер Тироде (TL-HEPES-PVA), содержащей 0,01% (масс/объем) поливинилового спирта (PVA), и затем промывают три раза средой для созревания (Abeydeera et al., Biol Reprod 58: 1316-1320, 1998). Каждый раз, 50 в целом COC переносят в 4-луночный планшет (Nunc, Roskilde, Denmark), содержащий 500 мкл среды для созревания, который покрывают сверху минеральным маслом и уравновешивают при 38,5°C с 5% CO2 на воздухе. Среда, используемая для созревания ооцитов, представляет собой среду для культивирования тканей (TCM) 199 (Gibco, Grand Island, NY), дополненную 0,1% PVA, 3,05 мМ D-глюкозы, 0,91 мМ пирувата натрия, 0,57 мМ цистеина, 0,5 мкг/мл LH (L5269, Sigma), 0,5 мкг/мл FSH (F2293, Sigma), 10 нг/мл эпидермального фактора роста (E4127, Sigma), 10% фоликуллярной жидкости свиней, 75 мкг/мл пенициллина G и 50 мкг/мл стрептомицина. После 22 час культивирования ооциты промывают дважды и культивируют в TCM 199 без LH и FSH в течение 22 час при 38,5°C, 5% CO2.
Пример 3
Оплодотворение in vitro (IVF) и культивирование ооцитов свиней
После созревания ооцитов, кумулюсные клетки удаляют с помощью 0,1% гиалуронидазы в среде TL-HEPES-PVA и промывают три раза средой TL-HEPES-PVA и средой с Tris буфером (mTBM) (Abeydeera et al., Biol Reprod 58: 1316-1320, 1998), содержащей 0,2% BSA (A7888, Sigma), соответственно. Затем, 25-30 ооцитов помещают в каждую из четырех 50-мкл капель среды mTBM, которые покрывают минеральным маслом в 35-мм полистироловой чашке для культивирования. Чашкам позволяют уравновеситься в инкубаторе в течение 30 мин, пока не добавляют сперматозоиды для оплодотворения. Один мл жидкого семени, консервируемого в разбавителе BTS, промывают дважды в PBS, содержащем 0,1% PVA (PBS-PVA), при 800 г в течение 5 мин. В конце процедуры промывки, сперматозоиды повторно суспендируют в среде mTBM. После соответствующего разбавления, 50 мкл этой суспензии сперматозоидов добавляют в 50 мкл среды, которая содержит ооциты, с получением конечной концентрации сперматозоидов 1-10×105 сперматозоидов/мл. К каплям для оплодотворения (конечные концентрации: 0-20 мкМ) добавляют различные концентрации неорганического пирофосфата (PPi; S6422, Sigma) во время добавления сперматозоидов. Ооциты инкубируют вместе со сперматозоидами в течение 6 час при 38,5°C, 5% CO2. Через 6 час после IVF, ооциты переносят в 100 мкл NCSU23, содержащего 0,4% BSA (A6003, Sigma), для дальнейшего культивирования в течение 16-20 час.
Пример 4
Иммунофлуоресценция и оценка долей оплодотворения
Сперматозоиды/ооциты фиксируют в 2% формальдегида в течение 40 мин при комнатной температуре, промывают, пермеабилизируют в PBS с помощью 0,1% Triton-X-100 (PBS-TX) и блокируют в течение 25 мин в PBS-TX, содержащем 5% нормальную сыворотку козы. Сперматозоиды/ооциты инкубируют с помощью поликлонального антитела кролика анти-пирофосфатаза 1 (PPA1) (разбавление 1:200; #ab96099, Abeam, San Francisco, CA) или с поликлональным антителом кролика анти-ANKH (разбавление 1:200; #SAB1102581, Sigma) в течение 40 мин, затем инкубируют вместе с антителом козы-анти-кролик (GAR)-IgG-TRITC (разбавление 1/80; Zymed Inc., San Francisco, CA). Для оценки оплодотворения, ооциты/зиготы фиксируют в 2% формальдегиде в течение 40 мин при комнатной температуре, промывают три раза PBS, пермеабилизируют с помощью PBS-TX в течение 40 мин при комнатной температуре и окрашивают с помощью 2,5 мкг/мл DAPI (Molecular Probes, Eugene, OR) в течение 40 мин. Ооциты с двумя или более пронуклеусами и, по меньшей мере, с одним хвостом сперматозоидов в ооплазме регистрируются как оплодотворенные. Для подсчета количества сперматозоидов, связанных с вителлиновым слоем, или сперматозоидов с прореагировавшей акросомой, ооциты фиксируют и окрашивают с помощью DAPI и PNA-FITC с лектином, связывающимся с акросомой (Molecular Probes), через 30 мин после IVF (5×105 сперматозоидов/мл). Регистрацию изображений осуществляют на микроскопе Nikon Eclipse 800 (Nikon Instruments Inc., Melville, NY) с камерой Cool Snap (Roper Scientific, Tucson, AZ) и с программным обеспечением MetaMorph (Universal Imaging Corp., Downington, PA).
Как показано на Фиг. 4, иммунофлуоресценция детектирует четкое мечение PPA1 в соединительной части хвоста сперматозоида и в постакросомальной оболочке сперматозоидов борова. Идентичное мечение обнаруживается в сперматозоидах, прикрепившихся к вителлиновому слою ооцита, через 30 мин оплодотворения in vitro, в то время как отрицательный контроль с антителом анти-PPA1, иммунонасыщенным полноразмерным рекомбинантным белком PPA1, показывает отсутствие такой флуоресценции и никакого мечения непермеабилизированных сперматозоидов.
Пример 5
Вестерн-блоттинг и ииммунофлуоресценция
Для вестерн-блоттинга, экстракты 1×106 сперматозоидов/мл нагружают на полосу. Сперматозоиды промывают в PBS и кипятят вместе с буфером для загрузки (50 мМ Tris [pH 6,8], 150 мМ NaCl, 2% SDS, 20% глицерола, 5% β-меркаптоэтанола, 0,02% бромфенола голубого). Гель-электрофорез осуществляют на гелях с градиентом 4-20% (PAGEr® Precast gels, Lonza Rockland Inc., Rockland, ME), с последующим переносом на PVDF мембраны (Millipore) с использованием системы мокрого переноса Owl (Fisher Scientific) при постоянном напряжении 50 В в течение 4 час. Мембраны последовательно инкубируют вместе с 10% нежирным молоком в течение 1 час, затем с антителами анти-PPA1 или анти-ANKH (разбавление 1:2000) в течение ночи. Затем мембраны инкубируют вместе с HRP-конъюгированным IgG козы анти-кролик (GAR-IgG-HRP; разбавление 1:10000) в течение 1 час. Мембраны взаимодействуют с хемилюминесцентным субстратом (SuperSignal, Pierce, Rockford, IL) и визуализируются посредством экспонирования для пленки Kodak BioMax Light (Kodak, Rochester, NY).
PPA1 детектируют в семенной плазме млекопитающих, в овидуктальной жидкости и в сперме. Как показано на Фиг. 3, полоса белка, соответствующая вычисленной массе PPA1 (32 кДа), детектируется в семенной плазме борова, в овидуктальной жидкости свиней и в сперматозоидах борова, быка, мыши и человека с помощью вестерн-блоттинга с использованием поликлонального антитела кролика анти-PPA1. Малые полосы для более высокой (~51 и 75 кДа) или более низкой массы (~13 кДа у борова и ~18 кДа у быка) наблюдают в каждом образце спермы, вероятно, они соответствуют продуктам посттрансляционной модификации и деградации белков продуктов PPA1. Очищенная PPA1 из пекарских дрожжей (S. cerevisiae), используемая в качестве белка положительного контроля, также показывает дополнительные полосы при 32 и 13 кДа.
Пример 6
Анализ пирофосфата
Измерение пирофосфата (PPi) осуществляют с использованием PiPerТМ Pyrophosphate Assay Kit (Cat. No. P22062, Molecular Probes), следуя протоколу производителя. Образцы приготавливают с использованием реакционного буфера IX (Kit) с семенной плазмой борова (SP), овидуктальными жидкостями свиней (OVF), с сывороткой кролика, сывороткой мыши (конечная концентрация 10 мкг/мл), сперматозоидами борова (1×106 сперматозоидов/мл) и 10 мМ рабочего раствора H2O2 (отрицательный контроль). Стандарт PPi приготавливают посредством разбавления 50 мМ стандартного раствора PPi (конечная концентрация 0-200 мкМ PPi). Рабочий раствор 100 мкМ реагента Amplex® Red содержит 0,02 Ед/мл неорганической пирофосфатазы (PPA1), 4 Ед/мл мальтоза фосфорилазы, 0,4 мМ мальтозы, 2 Ед/мл глюкоза оксидазы и 0,4 Ед/мл HRP. В этой реакции, PPA1 гидролизирует PPi на два неорганических фосфата (Pi). В присутствии Pi, мальтоза фосфорилаза преобразует мальтозу в глюкозу 1-фосфат и глюкозу. Затем глюкоза оксидаза преобразует глюкозу в глюконолактон и H2O2. В присутствии пероксидазы хрена (HRP), H2O2 взаимодействует с реагентом Amplex®Red (10-ацетил-3,7-дигидроксифеноксазин) с генерированием резоруфина, который детектируется с помощью флуоресценции. Пятьдесятимикролитровые образцы загружают в черный 96-луночный планшет (Coster-Corning, Corning, NY), а затем к каждому образцу добавляют 50 мкл рабочих растворов, соответственно. 96-луночный планшет инкубируют при 37,5°C в течение 30 мин, и измеряют флуоресценцию во множестве временных точек для отслеживания кинетики реакции. Интенсивность флуоресценции измеряют с помощью Thermo Fluoroskan Ascent (ThermoFisher Scientific) с использованием длин волн 530 нм для возбуждения и 590 нм для испускания.
Результаты измерения содержания PPi в SP борова, pOVF и сперме борова с помощью флуорометрического анализа показаны на Фиг. 1A и 1B. В качестве стандартов измеряют различные концентрации PPi (0-200 мкМ PPi) и интенсивность флуоресценции повышается в соответствии с повышением концентрации PPi (Фиг. 1A). Интенсивность флуоресценции также повышается в pOVF, SP, сперме, сыворотке мыши и сыворотке кролика. Как показано на Фиг. 1B, сперма борова, сыворотка мыши и сыворотка кролика показывают более высокие интенсивности флуоресценции, чем SP или pOVF, через 40 мин регистрации (98,2-101,1 по сравнению с 83,8 и 85,5). Однако, интенсивности для pOVF, сыворотки мыши и сыворотки кролика постепенно уменьшаются. Только SP и сперма показывают непрерывное увеличение интенсивности флуоресценции во время измерения (интенсивности флуоресценции: 111,5 и 117,7 при 60 мин, p<0,05). Отрицательный контроль, 10 мМ H2O2, показывает уменьшающуюся картину, вероятнее всего, из-за гашения флуоресценции.
Пример 7
Проточный цитометрический анализ жизнеспособности сперматозоидов и митохондриального мембранного потенциала
Сперму борова промывают дважды PBS-PVA, и концентрацию сперматозоидов доводят до 1×106 сперматозоидов/мл в PBS-PVA. Жизнеспособность сперматозоидов оценивают с помощью LIVE/DEAD® Sperm Viability Kit (L-7011, Molecular Probes), который содержит красители ДНК SYBR14 и пропидий йодид (PI), следуя протоколу производителя. Образцы спермы (198 мкл) загружают в 96-луночный планшет. SYBR14 (1 мкл; конечная концентрация 100 нм) и PI (1 мкл; конечная концентрация 12 мкМ) добавляют в образцы спермы и инкубируют в течение 10 мин при 37,5°C в темноте. Проточный цитометрический анализ осуществляют с использованием проточного цитометра Guava EasyCyteTM Plus (Guava Technologies, IMV Technologies, L’Aigle, France). Для каждого образца анализируют 5000 событий с помощью программы Guava ExpressPro Assay с использованием стандартных настроек производителя. Для оценки митопотенциала сперматозоидов, сперматозоиды борова окрашивают JC-1 (Cat. No. 4500-0250, MitoPotential Kit, IMV) и измеряют с использованием настроек производителя. Для отрицательных контролей, добавляют в образцы спермы DMSO или окрашивающий раствор.
Следуя промышленной практике для хранения семени борова, свежее семя борова разбавляют в разбавителе BTS и хранят при комнатной температуре (15-17°C) в течение 10 дней. Основной разбавитель подбирается для кратковременного хранения (3-5 дней); однако период хранения пролонгируется до 10 дней для сравнения жизнеспособности сперматозоидов и митохондриального мембранного потенциала между днями хранения 3 и 10 в присутствии/в отсутствие 10 мкМ PPi. Как описано выше, жизнеспособность сперматозоидов оценивается с помощью проточной цитометрии с использованием набора для оценки жизнеспособности SYBR14/PI, и митопотенциал измеряют с использованием красителя JC-1. Дополнение PPi изменяет гистограммы и вносит разброс в диаграммы флуоресценции, получаемые с помощью указанных выше зондов, и контрольный носитель, DMSO не дает флуоресценции.
Фиг. 5A и 5B сравнивают жизнеспособность сперматозоидов и митохондриальные мембранные потенциалы во время хранения спермы с PPi и без него. Как показано на Фиг. 5A, процент живых сперматозоидов выше в день 3, чем в день 10 (p<0,05), но нет значимого различия между контрольными сперматозоидами и теми, которые дополнены 10 мкМ PPi. В противоположность жизнеспособности, дополнение PPi уменьшает содержание метаболически активных сперматозоидов с поляризованными митохондриальными мембранами в день 3 (Фиг. 5B). Сходная тенденция наблюдается для сперматозоидов, консервируемых с помощью 10 мкМ PPi в течение 10 дней (Фиг. 5B).
Пример 8
Измерение протеасомальной-протеолитической активности
Протеасомальные-протеолитические и деубиквитинирующие активности, которые являются главными для оплодотворения, анализируют с использованием специфичных флуорометрических субстратов Z-LLL-AMC, Z-LLVY-AMC, Z-LLE-AMC и убиквитин-AMC в сперматозоидах, хранящихся в течение 3 и 10 дней, с PPi или без него. Альтернативно, 10 мкМ PPi+BTS добавляют в семя, консервируемое без PPi, во время анализа (обработка ʺдобавление PPiʺ). В качестве отрицательного контроля, к образцам спермы перед анализом добавляют 10 мкМ MG132 (протеасомального ингибитора).
Сперматозоиды, консервируемые в BTS с 10 мкМ PPi и без него, загружают в 96-луночный черный планшет (конечная концентрация сперматозоидов 1×106 сперматозоидов/мл), и инкубируют при 37,5°C вместе с Z-LLE-AMC (специфичный субстрат для активности гидролиза 20S химотрипсин-подобного пептидил-глютамилпептида [PGPH] нечувствительного к MG132; конечная концентрация 100 мкМ; Enzo Life Sciences, Plymouth, PA), Z-LLVY-AMC (специфичный субстрат для 20S протеасомы и других химотрипсин-подобных протеаз, а также для кальпаинов; конечная концентрация 100 мкМ; Enzo), Z-LLL-AMC (специфичный субстрат для 20S химотрипсин-подобной активности, чувствительный к протеасомальному ингибитору MG132; конечная концентрация 100 мкМ; BostonBiochem, Cambridge, MA) или вместе с убиквитин-AMC (специфичный субстрат для активности убиквитин-гидролазы с C-окончанием; конечная концентрация 1 мкМ; Enzo) в течение 1 часа. Флуорогенные протеасомальные сердцевинные субстраты состоят из малого пептида (LLL/LLE/LLVY), соединенного с флуоресцентным зондом, аминометилкумарином (AMC). Интактный AMC-связанный субстрат не испускает флуоресценции. В присутствии соответствующей 20S сердцевинной активности, молекула AMC отщепляется и становится флуоресцентной. Это испускаемая флуоресценция измеряется каждые 10 мин в течение периода 1 час, с получением кривой относительной флуоресценции (без единиц). Интенсивность флуоресценции измеряют с помощью Thermo Fluoroskan Ascent (Thermo Scientific), с использованием возбуждения на 380 нм и испускания на 460 нм.
Фиг. 6A-6D показывают воздействия PPi на протеасомальные ферментные активности хранящихся сперматозоидов борова. Как показано на Фиг. 6A, более высокая химотрипсин-подобная активность PGPH (субстрат Z-LLE-AMC) измеряется при обработке Добавления PPi в день 3 (относительная флуоресценция 392,1; без единиц) и PPi+BTS в день 10 (относительная флуоресценция 388), по сравнению с другими видами обработки (363,1-386,1; p<0,05). Как показано на Фиг. 6B, химотрипсин-подобная протеасомальная сердцевинная активность (субстрат Z-LLVY-AMC) постепенно увеличивается во время измерения во всех группах и виды обработки PPi+BTS и Добавление PPi показывают более высокие интенсивности флуоресценции для этого субстрата, по сравнению с контролями (110,8-121,5 по сравнению с 85,7; p<0,05). Самая высокая интенсивность флуоресценции наблюдается при Добавлении PPi через 10 мин, но интенсивность уменьшается соответствующим образом во время измерения и химотрипсин-подобная активность не показывает различий между различными видами обработки (Фиг. 6C). Наоборот более высокая деубиквитинирующая активность (убиквитин-AMC) наблюдается при обработке PPi+BTS в день 10, по сравнению с другими видами обработки (относительная флуоресценция 138,9 по сравнению с 98,4-122,7; Фиг. 6D). Как и ожидалось, низкая химотрипсин-подобная активность детектируется у сперматозоидов, обработанных протеасомальным ингибитором MG132. В целом, дополнение с помощью PPi увеличивает протеасомальную-протеолитическую и деубиквитинирующую активность у сперматозоидов и показывает преимущественное воздействие во время консервации спермы.
Пример 9
PPi улучшает проникновение сперматозоида в вителлиновый слой во время оплодотворения и способность к оплодотворению после продолжительного хранения.
Фиг. 7A-7F и Фиг. 8A-8E иллюстрируют воздействия PPi на общее и полиспермное оплодотворение во время IVF свиней (Фиг. 7A-7F) и воздействие PPi на связывание сперматозоидов с вителлиновым слоем (Фиг. 8A-8E). Ооциты свиней оплодотворяются в присутствии PPi при различных концентрациях (Фиг. 7A). Доли общего и полиспермного оплодотворения увеличиваются значительно и соответствующим образом (до 10 мкМ PPi) при увеличении концентраций PPi (p<0,05). Самую высокую полиспермию наблюдают после добавления 10 мкМ PPi (полиспермия 84,9%; Фиг. 7B). Среднее количество сперматозоидов, связанных с ZP, слегка, но незначительно, уменьшается при увеличении концентраций PPi (Фиг. 8A). Однако процент сперматозоидов с прореагировавшей акросомой значительно выше в присутствии 20 мкМ PPi, чем при 0-15 мкМ PPi (p<0,05, Фиг. 8B). Поскольку желательным является уменьшение дозы осеменения в настройках AI, ооциты свиней осеменяются также при пониженных концентрациях сперматозоидов с 10 мкМ PPi и без него. Консистентно, процент общего и полиспермного оплодотворения увеличивается с помощью PPi при 1, 2, и 5×105 сперматозоидов/мл; увеличение, индуцируемое PPi, является статистически значимым при концентрации 5×105 сперматозоидов/мл (Фиг. 7C). Для определения того, имеет ли хранение спермы в разбавителе BTS, дополненном PPi, полезное воздействие на способность спермы к оплодотворению, сперматозоиды борова непосредственно после эйакуляции хранятся в BTS с 10 мкМ PPi или без него в течение 3-4 дней и используются для IVF в присутствии или в отсутствие 10 мкМ PPi. Доли оплодотворения выше и полиспермия выше для всех видов обработки с добавлением PPi во время IVF, в отсутствие PPi в TBM (Фиг. 7D; второй столбец). Однако, самая высокая доля комбинированного (одно + полиспермного) оплодотворения наблюдается для сперматозоидов, консервируемых вместе с 10 мкМ PPi в BTS, когда их используют для IVF без добавления PPi (Фиг. 7D; третий столбец), или с PPi в среде для IVF (Фиг. 7D; четвертый столбец). Вместе с тем, PPi показывает статистически значимые (p<0,05), полезные воздействия на консервацию спермы и на способность сперматозоидов к оплодотворению.
Осуществляют контрольные эксперименты для обеднения спермы PPi с помощью посторонней неорганической пирофосфатазы в форме очищенной PPA1. Для нейтрализации собственной PPA1 спермы, ооциты свиней оплодотворяют в присутствии антитела анти-PPA1. Специфичность обоих реагентов устанавливают с помощью вестерн-блоттинга (смотри Фиг. 3). Антитело, как PPA, так и анти-PPA1 уменьшает долю оплодотворения зависимым от дозы образом (Фиг. 7E и F). Не наблюдают значительных различий в долях оплодотворения, когда антитело анти-PPA1 заменяют нормальной сывороткой во время оплодотворения (Фиг. 7F).
Для оценки возможного разброса между средами для хранения спермы, загрузки спермы борова консервируют в коммерческом BTS (BTS-IMV, IMV Technologies, L’Aigle, France) или в BTS, приготовленном в лаборатории BTS (BTS-HM) (Pursel et al, J Anim Sci 40:99-102, 1975), в присутствии 10 мкМ PPi, в течение 7 дней. Обнаруживается более высокая подвижность сперматозоидов в BTS-IMV + 10 мкМ PPi в день 6, чем во всех других группах (Фиг. 8C). Избыток PPi, добавляемый в среду IVF (100-500 мкМ PPi), уменьшает доли оплодотворения образом, зависимым от дозы (Фиг. 8D). В отдельном опыте сперматозоиды борова хранят в BTS-IMV вместе с 10 мкМ PPi в течение 3 дней и используют для IVF. Наблюдают почти 100% оплодотворение с помощью сперматозоидов, консервируемых с 10 мкМ PPi, по сравнению с оплодотворением меньшим чем 70%, без PPi в BTS-IMV (Фиг. 8E).
Пример 10
Статический анализ
Анализы разброса (ANOVA) осуществляют с использованием пакета программ SAS в полностью рандомизированном дизайне. Критерий множественного ранжирования Данкона используют для сравнения значений для индивидуальной обработки, когда F-значение является значимым (p<0,05).
Хотя настоящее изобретение описывается в связи с конкретными вариантами его осуществления, будет понятно, что устройство по настоящему изобретению может быть подвергнуто дополнительным модификациям. Настоящая заявка на патент, как предполагается, перекрывает любые варианты, применения или адаптации настоящего изобретения, следуя в целом принципам настоящего изобретения и включая такие отклонения от настоящего описания как попадающие в пределы известной или обычной практики в области, к которой относится настоящее изобретение, и как можно применить к основным признакам, которые изложены ранее в настоящем документе.





Изобретение относится к области медицины и ветеринарии. Добавка для стимулирования спермы содержит неорганический пирофосфат (PPi) в количестве примерно между 1 мкМ и примерно 200 мкМ. Добавление PPi в среды для оплодотворения in vitro людей/животных (IVF) улучшает долю оплодотворения; добавление PPi в разбавитель семени для искусственного осеменения (AI) сельскохозяйственных животных может улучшить долю забеременевших; кроме того, ооциты млекопитающих, созревающие in vitro в среде, содержащей PPi, приобретают улучшенный потенциал оплодотворения и развития, в то время как эмбрионы, культивируемые в среде, дополненной PPi, имеют улучшенное развитие до бластоцистов. 9 н. и 11 з.п. ф-лы, 25 ил., 10 пр.
1. Среда для консервации спермы, содержащая неорганический пирофосфат (PPi), где концентрация PPi находится в пределах примерно между 1 мкМ и примерно 200 мкМ.
2. Среда для консервации спермы по п. 1, где концентрация PPi находится в пределах примерно между 1 мкМ и примерно 20 мкМ.
3. Среда для консервации спермы по п. 1, где концентрация PPi составляет примерно 10 мкМ.
4. Среда для консервации спермы по п. 1, где указанная среда для консервации используется для консервации спермы свиней.
5. Среда для обработки спермы, содержащая неорганический пирофосфат (PPi), причем концентрация PPi находится в пределах примерно между 1 мкМ и примерно 200 мкМ.
6. Среда по п. 5, где концентрация PPi находится в пределах примерно между 1 мкМ и примерно 20 мкМ.
7. Среда для оплодотворения in vitro (IVF) или искусственного осеменения (AI), содержащая неорганический пирофосфат (PPi), где концентрация PPi находится в пределах примерно между 1 мкМ и примерно 200 мкМ.
8. Среда по п. 7, где концентрация PPi находится в пределах примерно между 1 мкМ и примерно 20 мкМ.
9. Способ сексинга спермы, включающий:
(a) разделение смешанной суспензии сперматозоидов в первой среде для культивирования на популяции сперматозоидов, несущих х-хромосому или у-хромосому, с помощью элюирующей среды;
(b) консервацию сперматозоидов, несущих х-хромосому или у-хромосому, во второй среде для культивирования, где неорганический пирофосфат (PPi) добавляют в первую среду для культивирования, в элюирующую среду или вторую среду для культивирования и где концентрация PPi находится в пределах примерно между 1 мкМ и примерно 200 мкМ.
10. Способ консервации спермы, включающий хранение спермы в средах, содержащих неорганический пирофосфат (PPi), где концентрация PPi находится в пределах примерно между 1 мкМ и примерно 200 мкМ.
11. Способ по п. 10, где концентрация PPi находится в пределах примерно между 1 мкМ и примерно 20 мкМ.
12. Способ по п. 10, где концентрация PPi составляет примерно 10 мкМ.
13. Способ по п. 10, где сперму хранят в средах, содержащих PPi, в течение до 10 дней.
14. Способ оплодотворения in vitro (IVF), включающий приведение в контакт ооцита со сперматозоидами в присутствии неорганического пирофосфата (PPi), где концентрация PPi находится в пределах примерно между 1 мкМ и примерно 200 мкМ.
15. Способ по п. 14, где концентрация PPi находится в пределах примерно между 1 мкМ и примерно 20 мкМ.
16. Способ по п. 14, где сперму хранят в присутствии PPi.
17. Способ культивирования эмбрионов, включающий культивирование эмбрионов в средах, содержащих неорганический пирофосфат, причем концентрация неорганического пирофосфата находится в пределах примерно между 1 мкМ и примерно 200 мкМ.
18. Способ искусственного осеменения, включающий доставку спермы и неорганического пирофосфата (PPi) в половые пути самки, причем концентрация неорганического пирофосфата (PPi) находится в пределах примерно между 1 мкМ и примерно 200 мкМ.
19. Способ по п. 18, где PPi постепенно высвобождается в половых путях самки.
20. Способ созревания ооцита in vitro, включающий культивирование ооцитов в средах, содержащих неорганический пирофосфат, причем концентрация неорганического пирофосфата находится в пределах примерно между 1 мкМ и примерно 200 мкМ.
| PATE ET AL | |||
| "Resolution of the competitive inhibitory effects of lithium and AMPPNP on the beat frequency of ATP-reactivated, demembranated, sea urchin sperm flagella", J MUSCLE RES CELL MOTIL., (198508), vol | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| YOO ET AL | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Способ сужения чугунных изделий | 1922 |
|
SU38A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| US 6309815 B1, 30.10.2001 | |||
| US 20020115055 A1, 22.08.2002 | |||
| СИНТЕТИЧЕСКАЯ СРЕДА ДЛЯ РАЗБАВЛЕНИЯ СПЕРМЫ СЕЛЬСКОХОЗЯЙСТВЕННОЙ ПТИЦЫ | 1995 |
|
RU2099025C1 |

