Настоящее изобретение относится к области репродуктивной медицины (MAP) и, более конкретно, ко всем медицинским, ветеринарным или другим вариантам применения, в которых желательно стимулировать митохондриальную активность или, в более общем плане, энергетическую активность клеток, в частности, в протоколах, включающих стадию культивирования клеток или поддержание клеток ex vivo.
Более 15% семейных пар прибегают к медикаментозному продолжению рода в период своей половой жизни. При патологиях спермы обычно прибегают к методам микроинъекции для оплодотворения. Этот метод является высокотравматичным для ооцита. Кроме того, при оплодотворении in vitro (ЭКО) с использованием здоровой спермы регистрируется от 3 до 5% необъяснимых неудач оплодотворения.
В предварительной заявке на патент (WO 2005/051799 A2) группой изобретателей был описан циклический трипептид (Phe, AcGlu, AcGlu), имеющий формулу C-S-F-E-E-C (SEQ ID NO: 1) с циклической связью между двумя концевыми цистеинами (FEEc), а также его действие, увеличивающее возможности оплодотворения для человеческих гамет. Эквиваленты молекулы существуют у различных видов животных, и они имеют одинаковые свойства и также описаны в указанном документе.
При выполнении исследований изобретатели продемонстрировали, что молекула FEEc оказывает влияние на сперматозоиды (см. пример 1 ниже). В частности, она улучшает параметры подвижности сперматозоидов, анализируемые CASA (Компьютерным анализом спермы). Так, например, линейная скорость и амплитуда латерального смещения головки соответственно увеличиваются на 7% и 8% (P <0,05 и P <0,002) (см. пример 1). Это приводит к почти 30%-у увеличению числа гиперактивированных сперматозоидов (P <0,009). Именно эти гиперактивированные сперматозоиды являются оплодотворяющими сперматозоидами. Для того, чтобы сперматозоид мог увеличить свою скорость перемещения, логично предположить, что ему необходимо увеличить потребление АТФ или по меньшей мере повысить его энергетический метаболизм, поскольку его движение обуславливается путем связывания плечей динеина с трубочками вблизи аксонемы. Динеины представляют собой АТФазы. При исследовании метаболизма митохондрий сперматозоидов, подвергшихся воздействию FEEc, изобретатели выдвинули гипотезу о том, что FEEc вызывает увеличение потенциала митохондриальной мембраны, обеспечивая либо увеличение синтеза АТФ митохондриями, либо снижение потребления. Таким образом, FEEc улучшает энергетический метаболизм митохондрий или, в более общем случае, клеточный энергетический метаболизм сперматозоидов либо путем увеличения продукции АТФ, либо путем нормирования его использования клеткой.
Поэтому, настоящее изобретение, в общем, относится к применению циклического пептида, содержащего трипептид, воспроизводящий сайт связывания фертилина-бета с интегрином ооцита, или трипептида с формулой EEP (SEQ ID No. 2), для использования в качестве лекарственного средства для улучшения активности митохондрий или, в более общем плане, для улучшения клеточного энергетического метаболизма. Варианты применения такого пептида проиллюстрированы в данном документе на двух разных типах клеток, а именно, на сперматозоиде и ооците, а также на развитии многоклеточного организма, который является эмбрионом. Результаты, представленные в экспериментальном разделе, подтверждают возможность использования данного пептида в других вариантах применения, где желательно улучшить метаболизм клеток. В частности, этот пептид может быть полезен для улучшения выхода при культивировании клеток, в частности, лимфоцитов. В одном конкретном варианте осуществления пептид векторизован, то есть вводится в комбинации с агентом, молекулой, композицией или любым другим типом вектора, который облегчает его проникновение в клетки. В более общем смысле, векторизация служит для модуляции и контроля распределения действующего ингредиента в мишени путем объединения его с вектором.
Указанный выше «циклический пептид, содержащий трипептид, воспроизводящий сайт связывания фертилина-бета с интегрином ооцита», соответствует пептиду, описанному в заявке на патент WO 2005/051799 A2, указанной выше. Как указано в этой предшествующей заявке, в частности, в таблице 1, трипептид варьирует в зависимости от биологического вида. Он может циклизоваться любыми способами, известными специалистам в данной области, в частности, с помощью двух цистеиновых остатков, расположенных по обе стороны трипептида. В общем, все вариации, описанные в заявке WO 2005/051799 A2, считаются соответствующими определению «циклического пептида, содержащего трипептид, образующий сайт связывания фертилина-бета с интегрином ооцита» для целей настоящего изобретения. Чтобы облегчить прочтение настоящего текста, данный циклический пептид, а также лекарственное средство, содержащее его в качестве действующего ингредиента, будет обозначено в настоящем документе формулой «FEEc». Специалистам в данной области будет очевидно, что это обозначение также охватывает формы, которые могут быть использованы у других биологических видов, кроме человека, такие как, например, циклический трипептид TDE, который следует использовать у крупного рогатого скота.
При прочтении нижеследующего подробного описания изобретения, а также примеров, будет понятно, что объем настоящего изобретения не ограничивается применением в области репродуктивной медицины, а изобретение открывает реальные перспективы во многих других областях. Таким образом, изобретение относится, в общем, к применению FEEc в области медицины или в других областях, где желательна стимуляция клеточного энергетического метаболизма.
Действие на сперматозоиды при внутриутробном осеменении
Путем улучшения параметров подвижности сперматозоидов FEEc также способен улучшить частоту беременностей при внутриутробном осеменении (IUI). Согласно первому конкретному аспекту изобретения FEEc используется для увеличения скорости продвижения сперматозоидов в протоколе репродуктивной медицины (MAP). Еще, в контексте MAP, FEEc может использоваться для улучшения параметров подвижности сперматозоидов и повышения содержания гиперактивированных сперматозоидов. Поэтому, использование FEEc особенно выгодно в протоколе внутриутробного осеменения (IUI), как у людей, так и у других млекопитающих. Во время реализации этого аспекта изобретения сперматозоиды предпочтительно инкубировать в течение периода времени от одной минуты до 3 часов в присутствии от 10 до 100 мкМ пептида, затем промыть перед внутриутробным осеменением.
Действие на созревание ооцитов in vitro
Эта молекула также действует на ооциты. In vitro созревание ооцитов человека, блокированное на стадии зародышевого пузырька, увеличивается с 37,71% до 59,30% (P<5,7×105) в присутствии FEEc (пример 2). У пациентов в возрасте 37 лет и старше этот показатель увеличивается с 36,96% до 68,29% (P <0,003), что показывает, что эта молекула особенно эффективна в этом возрастном диапазоне. Ооциты женщин в возрасте от 37 до 40 лет являются анеуплоидными по меньшей мере в 50% случаев и, обычно, примерно в 80% случаев из-за снижения своей митохондриальной активности. Таким образом, молекула способна улучшить плоидность ооцитов и, тем самым, эмбрионов, потенциал развития которых и способность к имплантации также зависят от митохондриальной активности оплодотворенного ооцита. Значительное увеличение процента беременностей у женщин в возрасте до 37 лет в контексте «фертилинового» клинического исследования, описанного ниже, показывает, что этот благоприятный эффект наблюдается у любого ооцита и, в частности, при его оплодотворении и раннем эмбриональном развитии. Согласующаяся с этими результатами гипотеза заключается в том, что фертилин способен повысить плоидность ооцитов.
Поэтому, добавление молекулы в культуральную среду, а также в среду для оплодотворения in vitro и в инкубационную среду для эмбрионов, позволяет улучшить созревание ооцитов и эмбрионов (в частности, у женщин в возрасте до 30 лет, и у тех, кому 37 лет или больше), и частоту оплодотворений с помощью обычного ЭКО, а также с помощью ЭКО с ИКСИ (интрацитоплазматической инъекцией сперматозоидов).
Таким образом, добавление молекулы FEEc в культуральную среду при оплодотворении in vitro с микроманипуляцией или без нее с позволяет увеличить частоту беременностей и процент рожденных детей, в частности, у женщин в возрасте до 37 лет, при используемых экспериментальных условиях.
Действие на ооцит в протоколах созревания in vitro (IVM)
Молекула FEEc также способна быть эффективной в протоколах созревания ооцитов in vitro для сохранения фертильности (IVM).
В протоколе оплодотворения in vitro, созревание ооцита обычно завершается к моменту сбора ооцитов. Однако некоторые ооциты иногда все еще остаются незрелыми. Кроме того, у некоторых женщин наблюдаются аномалии функционирования яичников или клиническое состояние, затрудняющее стимуляцию. Поэтому, на добровольной основе проводили пункцию на стадии незрелых ооцитов для созревания in vitro. Пептид мог эффективно способствовать этому созреванию (созревание in vitro для сохранения фертильности).
Полностью незрелый ооцит имеет большое ядро, называемое зародышевым пузырьком (GV). Зрелый ооцит характеризуется наличием 1-го полярного тельца (PG) в перивителлиновом пространстве (между поверхностью ооцита и зоной пеллюцида). Только зрелые ооциты способны к оплодотворению.
Авторы изобретения продемонстрировали, что FEEc позволяет улучшить созревание ооцитов in vitro. Это объясняется влиянием трипептида на митохондриальную активность или на энергетический метаболизм ооцита. Действительно, активность митохондрий или, в общем, энергетический метаболизм снижаются с возрастом, а созревание ооцита включает в себя несколько стадий с большим потреблением энергии:
- разрыв зародышевого пузырька
- конденсация хромосом
- формирование метафазной пластинки
- формирование веретена
- синтез белков контрольных точек
- телофаза
- отделение полярного тельца.
На деле, данные проблемы созревания ооцитов, которые усиливаются с возрастом, корректируются микроинъекцией митохондрий, полученных из молодых клеток (ооцитов от молодых доноров или оогониальных стволовых клеток). Это подкрепляет вероятность гипотезы, согласно которой FEEc фактически исправляет дефект клетки, связанный с митохондриальной недостаточностью или, в более общем случае, с энергетической недостаточностью.
Согласно другому аспекту настоящее изобретение относится к применению FEEc для улучшения созревания ооцитов in vitro. Улучшение качества прохождения мейоза, вероятно, связано с уменьшением частоты выкидышей, которая также наблюдается у молодых женщин, так что этот аспект изобретения также полезен для улучшения созревания ооцитов in vitro у женщин в возрасте до 37 лет или даже в возрасте до 30 лет. При осуществлении этого аспекта изобретения, ооцит инкубируют в течение периода времени от 1 часа до 4 дней, в частности, до 3 дней или до 24 часов, в присутствии от 10 до 100 мкМ пептида.
Действие на активацию оплодотворенного ооцита
Авторы также продемонстрировали увеличение деконденсации головки сперматозоидов после оплодотворения. Это отражает улучшение активации ооцитов во время оплодотворения. Более того, это связано с митохондриальной активностью ооцита.
Действие на образование бластоцисты
Изобретатели также продемонстрировали (на мышах), что FEEc позволяет улучшить образование бластоцисты. Это также можно объяснить влиянием трипептида на активность митохондрий или, в более общем плане, на метаболизм клетки. Действительно, известно, что рекомбинация митохондриальной ДНК во время преимплантационного эмбриогенеза отсутствует. В течение первой недели развития, зигота (или яйцо) делится последовательными митозами, начиная с 2 и затем 4 клеток, и проходит стадию морулы до достижения стадии бластоцисты, преимущественно, используя митохондрии, первоначально присутствующие в ооците. Таким образом, митохондриальный дефицит (с точки зрения количества или функций) может отвечать за хромосомную нестабильность бластомеров во время мейоза и митозов и может привести к остановке развития зиготы, эмбриона или даже беременности. Это является частой причиной спонтанного выкидыша после естественного оплодотворения или после переноса эмбрионов во время ЭКО. Поэтому, настоящее изобретение относится также к применению FEEc для улучшения плоидности бластомеров в течение первой недели развития зиготы. В результате изобретение относится к использованию FEEc для уменьшения числа выкидышей. Изобретение также относится к применению FEEc для снижения риска анеуплоидии, в частности трисомии. При осуществлении этого аспекта изобретения эмбрион инкубируют в течение периода времени от 24 часов до 6 или 7 дней в присутствии от 10 до 100 мкМ пептида.
Варианты применения, описанные выше, особенно полезны в протоколе оплодотворения in vitro (ЭКО). У человека они позволяют женщинам всех возрастов иметь детей с помощью MAP с их собственными ооцитами, не прибегая к инъекциям митохондрий, описанным в литературе (основной эффект, наиболее широко документированный на сегодняшний день).
Снижение риска выкидышей
Настоящее изобретение также относится к использованию FEEc для снижения риска выкидышей, как было указано ранее и проиллюстрировано в экспериментальном разделе ниже.
Снижение риска трисомии
Настоящее изобретение также относится к использованию FEEc для снижения риска трисомии или, в более общем смысле, анеуплоидии во время ЭКО, у всех женщин, особенно, у женщин 35, 36, 37, 38, 39 или 40 лет и старше.
Улучшение кинетики развития эмбрионов in vitro
Настоящее изобретение также относится к применению FEEc для улучшения предимплантационного развития эмбриона in vitro. В одном конкретном варианте осуществления изобретения предимплантационное развитие получается в условиях длительного культивирования. Улучшение в кинетике развития эмбриона позволяет повысить процент рождений.
Действие на естественное размножение
Хотя эффекты FEEc на ооцит и зиготу были продемонстрированы изобретателями в контексте ЭКО, очевидно, что эти эффекты могут быть также получены при естественном оплодотворении, например, путем введения FEEc вагинально во время овуляции, причем трипептид соединен со средством, подходящим для направленного переноса («векторизации») его к ооциту.
Действие при криоконсервации гамет и эмбрионов
Было также показано, что выживаемость криоконсервированных ооцитов зависит, в частности, от их митохондриальной активности. Вероятно, то же самое является верным и для эмбрионов. Таким образом, пептид FEEc способен улучшить выживаемость и/или качество криоконсервированных гамет и эмбрионов во время их размораживания.
Действие на другие типы клеток
Циклический трипептид получали для связывания с интегрином α6β1 на ооците. Он способен связываться с цитоплазматической мембраной различных типов клеток, потому что этот интегрин экспрессируется на большинстве типов клеток. Поэтому, эта молекула, вероятно, способна увеличивать энергетическую активность множества типов клеток. Таким образом, пептид можно использовать в различных вариантах применения, кроме ЭКО.
Поскольку механизм действия молекулы, вероятно, опосредован интегрином α6β1, FEEc способен оказывать влияние на множество других типов клеток. Эту молекулу можно использовать для улучшения выхода любой клеточной культуры.
Как указано выше, FEEc, возможно, действует через интегрин α6β1 или другой рецептор, присутствующий на множестве типов клеток. Поэтому, его можно использовать для улучшения активности митохондрий или энергетического метаболизма любой клетки, несущей данный интегрин или этот другой рецептор. Прямым результатом этого свойства является улучшение выхода любой клеточной культуры. Таким образом, настоящее изобретение также относится к способу улучшения активности митохондрий или, в более общем случае, энергетической активности клеток in vitro, включающему в себя стадию приведения соответствующих клеток в контакт с FEEc. Этот способ можно, предпочтительно, проводить на первичных клетках, культивируемых ex vivo, в перспективе введения пациенту в контексте клеточной терапии. В качестве неограничивающих примеров клеточных культур, для которых был бы полезен этот способ, можно указать культуры клеток кожи для кожных трансплантатов, культуры лимфоцитов для клеточной иммунотерапии и т.д.
Изобретение относится к применению пептида FEEc для улучшения культивирования ex vivo всех типов клеток, экспрессирующих рецептор FEEc, для медицинских применений (таких как клеточная терапия) или немедицинских применений (таких как поддержание клеток в культура для экспериментальных целей или для производства белка).
Действие на млекопитающих
Молекула обладает видовой специфичностью. Ее изоформы могут быть адаптированы для использования у всех домашних или диких животных, включая сельскохозяйственных животных, репродукция некоторых видов которых затруднена (скаковые лошади, голштинские коровы).
Действие на митохондриальные патологии и антивозрастное действие
Свойства FEEc, как молекулы для стимуляции митохондриальной активности или, в более общем плане, энергетического метаболизма, также могут быть использованы in vivo при любом типе патологии, связанной с дефектом митохондриальной активности. В этом отношении можно упомянуть, в основном, старение. Например, взаимосвязь между митохондриальной дисфункцией и нейродегенеративными заболеваниями была установлена несколькими научными группами. Поэтому, FEEc можно использовать в качестве лекарственного средства для лечения нейродегенеративных заболеваний, таких как болезнь Альцгеймера или болезнь Паркинсона. Также была показана взаимосвязь между длиной теломер хромосом и митохондриальной активностью клетки. Действительно, сокращение теломер связано с процессами старения. Поэтому возможно, что, путем стимуляции митохондриальной или энергетической активности клетки можно отложить эффекты старения.
Митохондриальные заболевания имеют разнообразные проявления, но часто сочетают глазные проявления типа пигментного ретинита или офтальмоплегии. Для последней патологии местное введение FEEc в глаз, например, в лосьоне для глаз, может улучшить симптомы, связанные с митохондриальной или энергетической недостаточностью. Поэтому, другим объектом изобретения является лосьон для глаз, содержащий циклический пептид, включающий трипептид, способный образовывать сайт связывания фертилина-бета с интегрином ооцита. Такой лосьон для глаз может содержать, помимо FEEc, другой агент, такой как загуститель, антисептик, антибиотик или любое другое соединение, которое может быть использовано для этого типа продукта. Среда, описанная в заявке WO 2005/051799 A2, конечно, исключается из определения термина «лосьон для глаз» для целей настоящего изобретения.
Действие в области косметологии
Настоящее изобретение также относится к применению FEEc в косметической или терапевтической композиции, предназначенной для местного применения. В качестве примеров можно указать использование FEEc для стимуляции фибробластов для производства коллагена или для стимуляции волосяных фолликулов для усиления роста волос, например, для предотвращения или замедления алопеции. Поэтому, настоящее изобретение относится также к косметической или дерматологической композиции, содержащей FEEc в качестве действующего ингредиента. Термин «косметическая или дерматологическая композиция» означает в данном документе композицию, которая, помимо FEEc, содержит ингредиенты, обычно используемые в области косметологии. Согласно изобретению косметическая или дерматологическая композиция может иметь любую форму, известную специалистам в данной области. Она может представлять собой, например, эмульсию типа «масло-в-воде», «вода-в-масле» или «вода-в-силиконовой эмульсии», множественную эмульсию, микроэмульсию, наноэмульсию, твердую эмульсию, водный или водно-спиртовый гель, крем, молочко, лосьон, притирку, масло, бальзам, мазь, маску, порошок, пропитанный носитель, например, трансдермальный пластырь, водный или водно-спиртовый лосьон и/или воск, косметический продукт, например, основу для макияжа, шампунь, кондиционер, маску, сыворотку для местного применения или лосьон для волос. Среда, описанная в заявке WO 2005/051799 A2, конечно, исключается из определения косметических или дерматологических композиций для целей настоящего изобретения.
Согласно изобретению косметическая или дерматологическая композиция может представлять собой, например, композицию для ухода за лицом, телом или волосами, например, композицию для лица и/или тела, и/или волос.
Приведенные ниже примеры иллюстрируют изобретение, но не ограничивают его объем.
Описание фигур
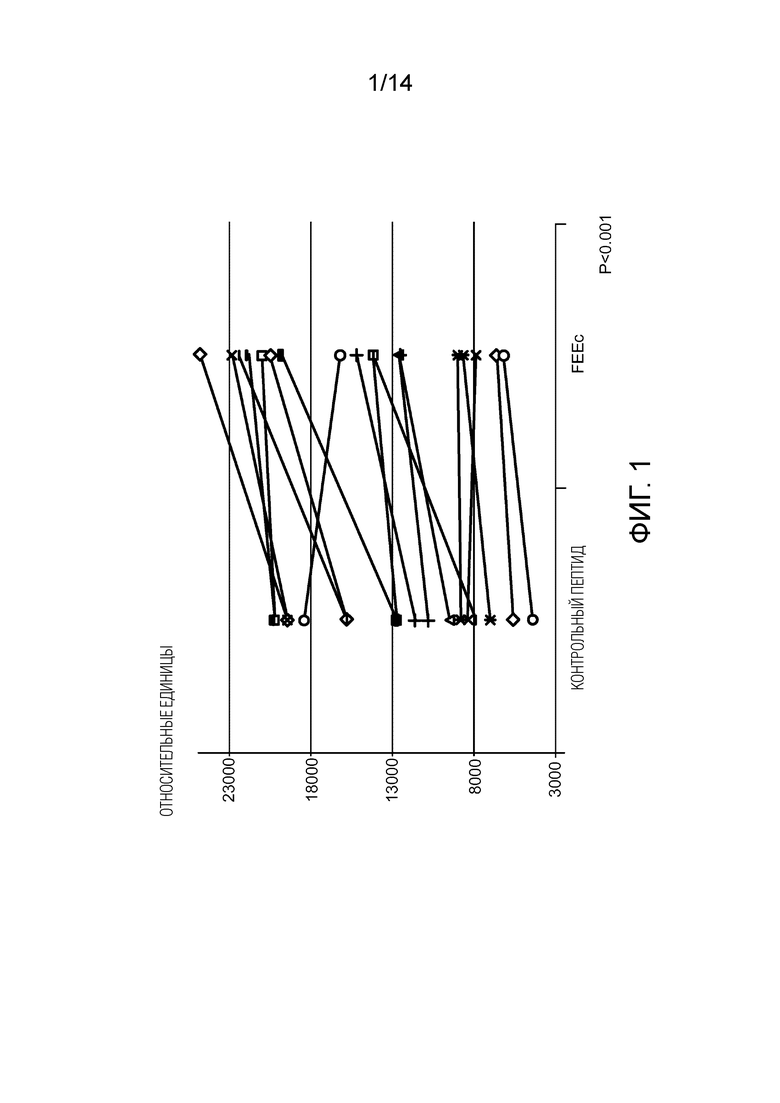
Фигура 1: изменение потенциала митохондриальной мембраны в присутствии FEEc (справа) по сравнению с «перетасованным» контрольным пептидом (слева). Следует отметить увеличение мембранного потенциала у обработанных сперматозоидов у большинства пациентов.
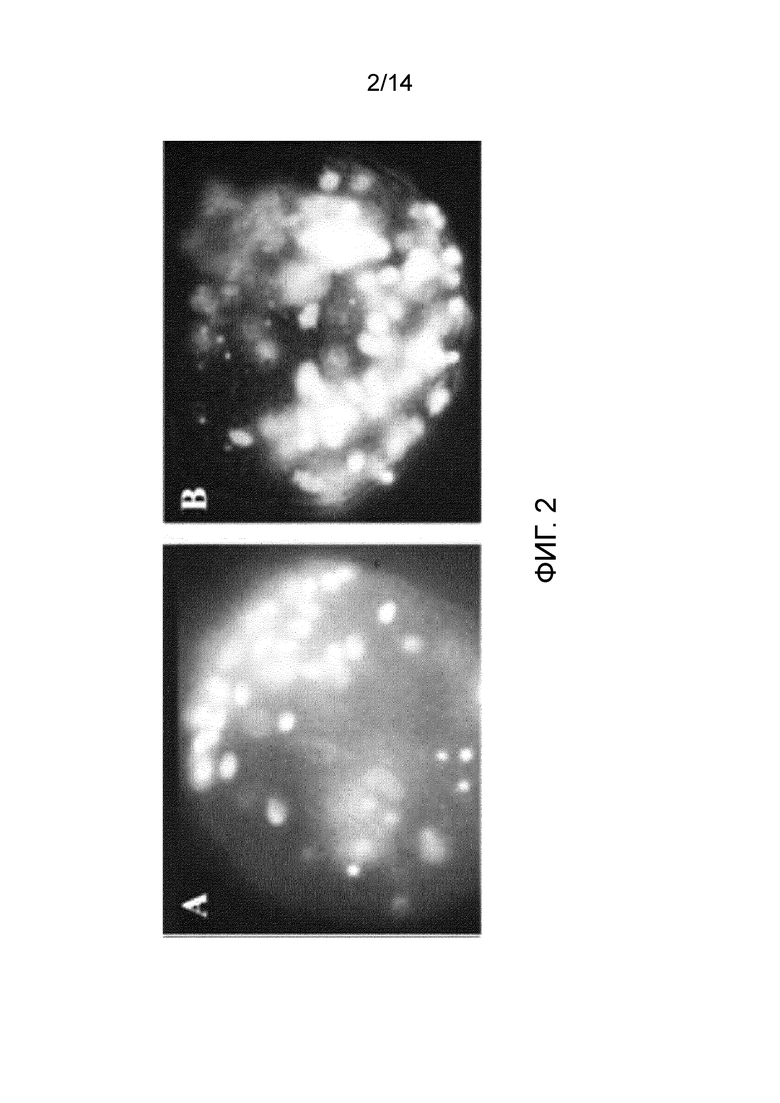
Фигура 2: подсчет после УФ-возбуждения сперматозоидов человека, слитых с ооцитами человека с удаленной зоной пеллюцида, инкубированных в отсутствие (А) или в присутствии FEEc в концентрации 100 мкМ (В). Следует отметить увеличение количества сперматозоидов и более быструю деконденсацию их головки.
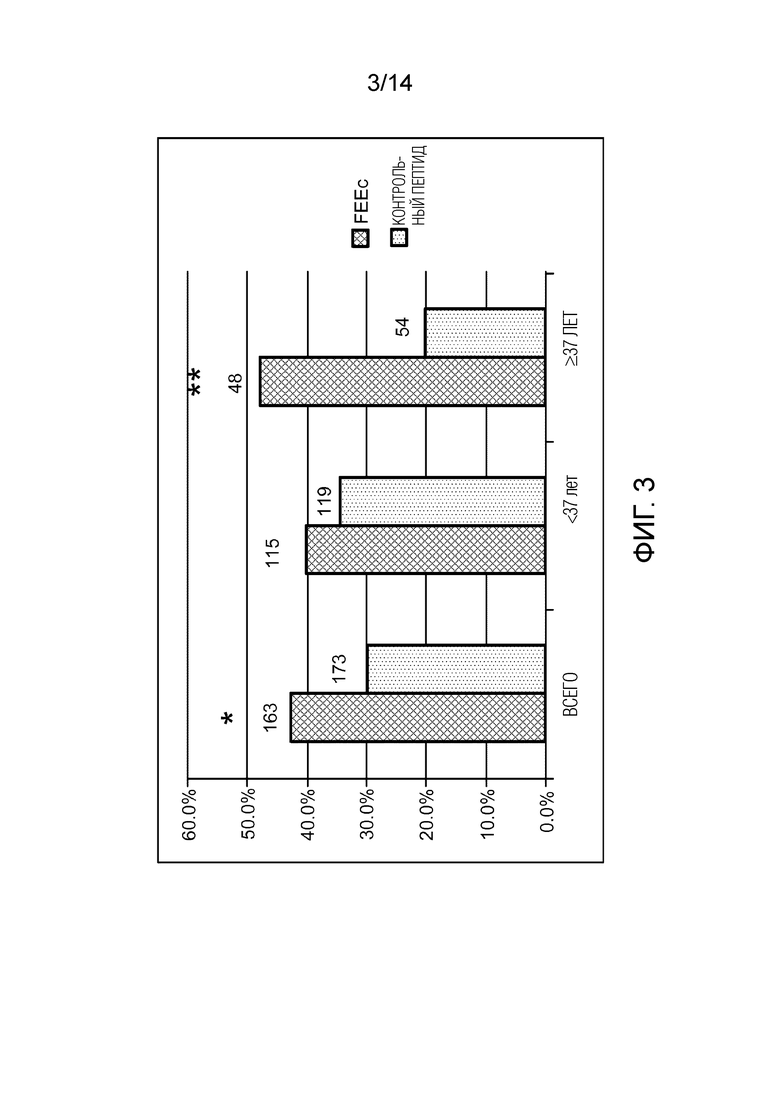
Фигура 3: доля зрелых ооцитов в D1 созревания in vitro (IVM). Графическое представление доли ооцитов человека в метафазе II (MII) из стадии GV после IVM в контрольной среде или среде, дополненной FEEc в концентрации 100 мкМ. *p=0,02. **p=0,003. D1=24 ч от IVM.
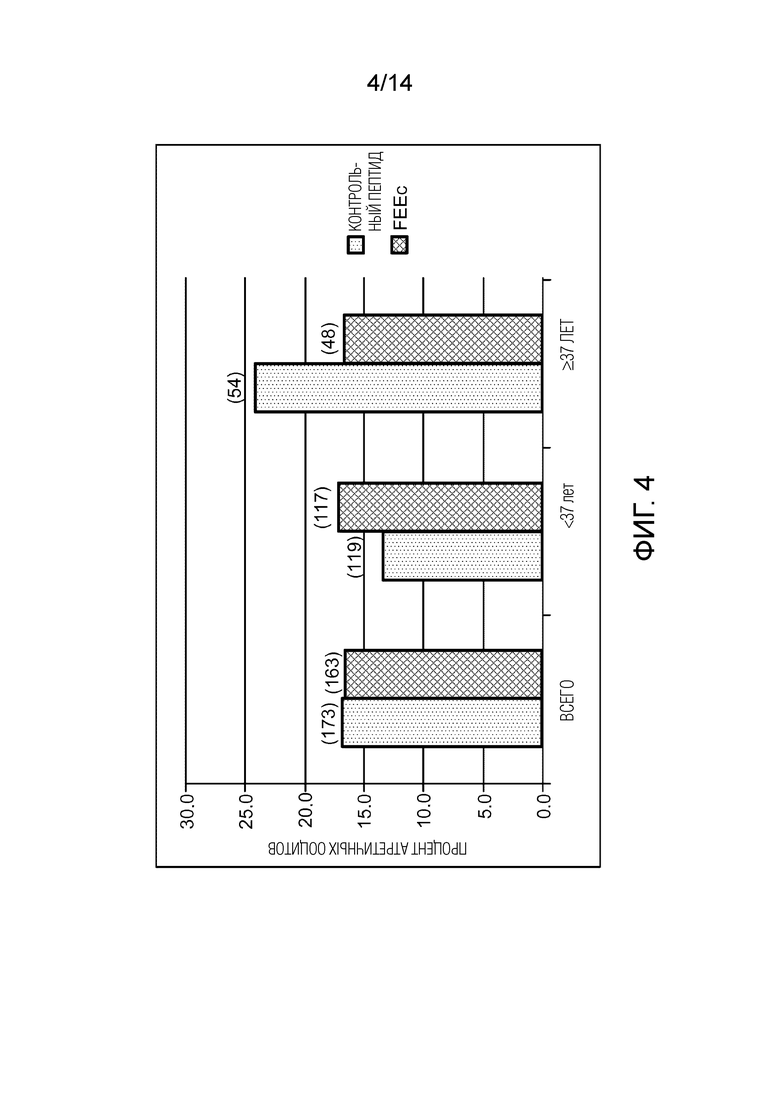
Фигура 4: доля атретических ооцитов в D1 созревания in vitro (IVM). Графическое представление доли атретических ооцитов человека. Результаты, наблюдаемые в D1, начиная со стадии GV после созревания in vitro (IVM) в контрольной среде или среде, дополненной FEEc в концентрации 100 мкМ. D1=24 ч IVM.
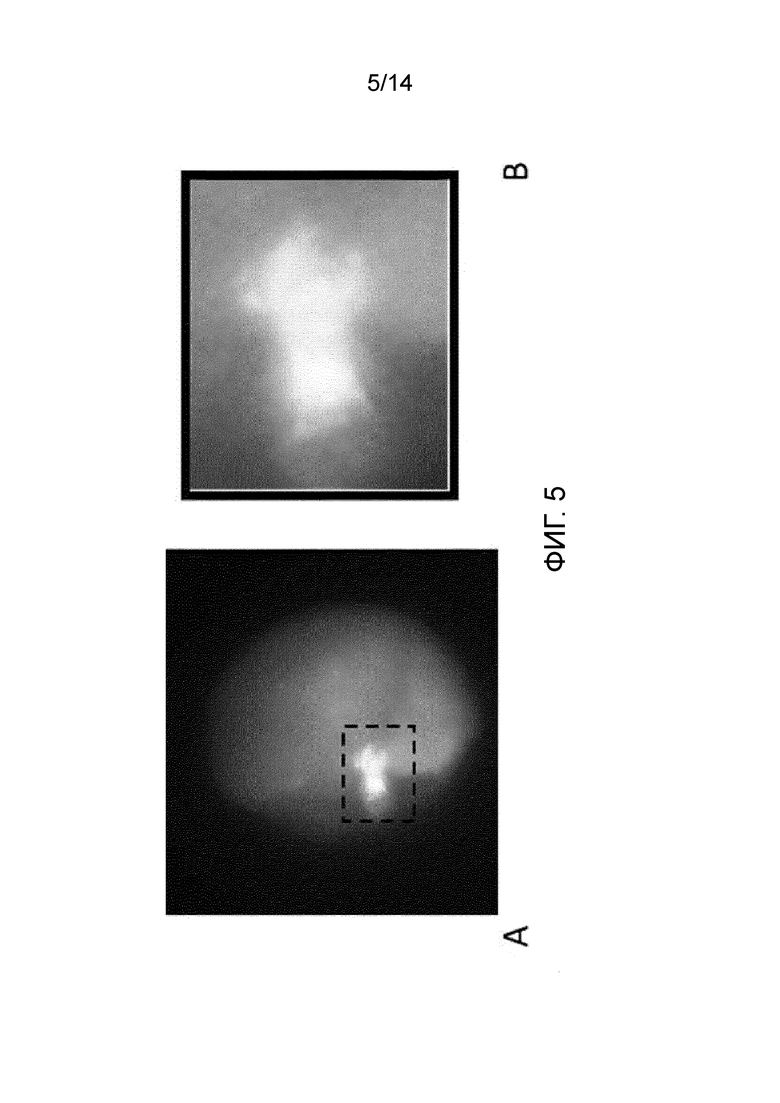
Фигура 5: мечение мейотического веретена, полученного на ооците человека в MII после IVM в стандартной среде (24 часа). Мечение веретена осуществляли анти-α-тубулиновым антителом и хромосомы окрашивали DAPI. Изображение получено на конфокальном микроскопе. A: весь ооцит. B: увеличение метафазной пластинки.
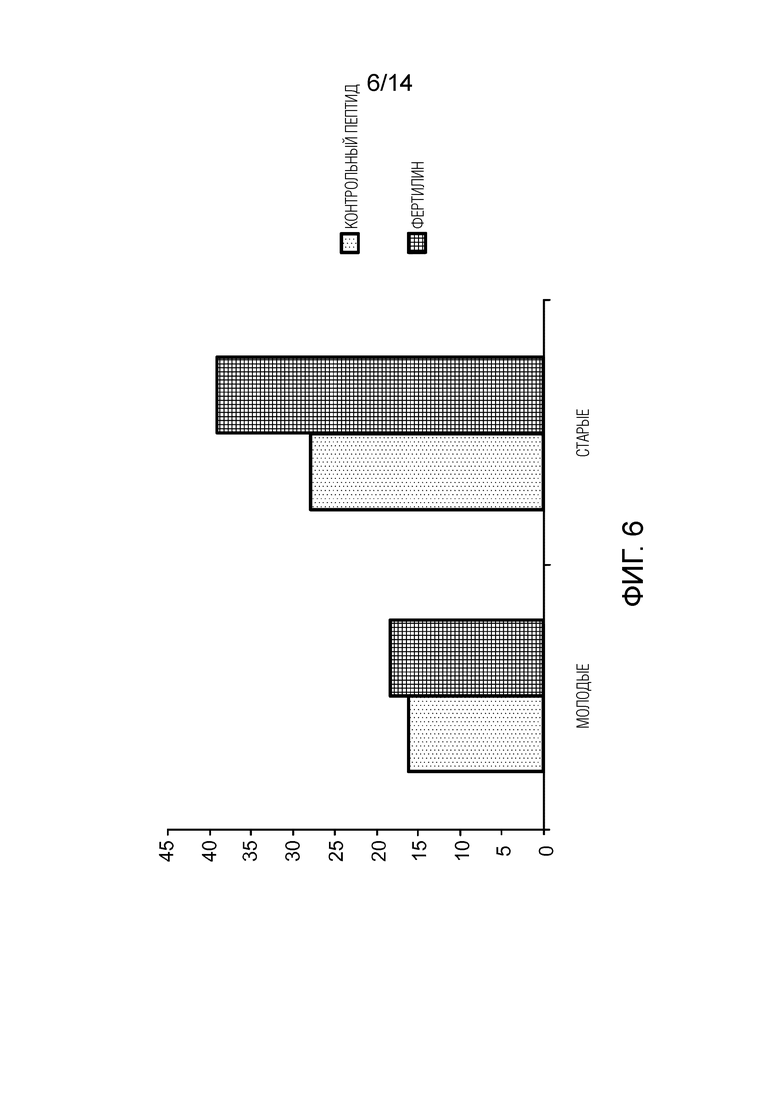
Фигура 6: сравнение скорости оплодотворения в D1 между молодыми и старыми мышами в присутствии или в отсутствие фертилина, который соответствует пептиду QDEc.
Молодые мыши: 7-недельные мыши B6CBAF1; старые мыши: 7-месячные мыши B6CBAF1.
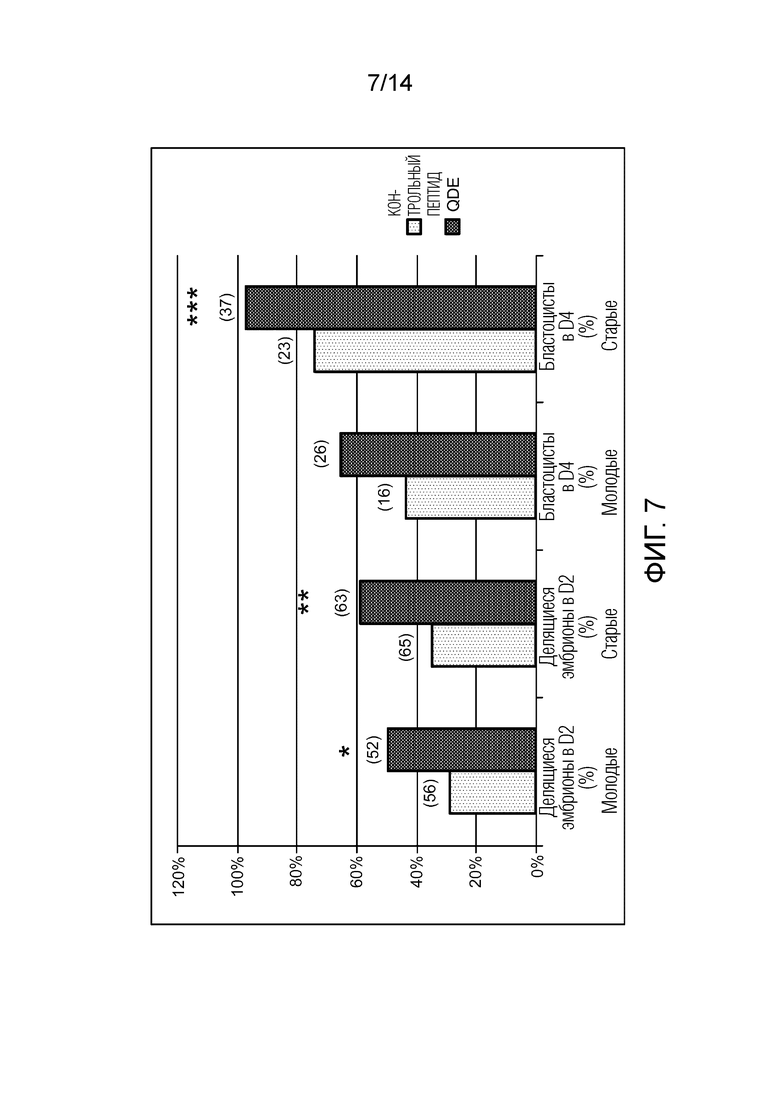
Фигура 7: Сравнение среднего процента делящихся эмбрионов на стадиях D2 и D4 между молодыми и старыми мышами в присутствии или в отсутствие пептида QDEc.
Молодые мыши: 7-недельные мыши B6CBAF1 (n=108 ооцитов, 56 - контрольных, 52 - QDEc); старые мыши: 7-месячные мыши B6CBAF1 (n=128 ооцитов, 65 - контрольных, 63 - QDEc); *p=0,02, **p=0,008, ***p=0,01.
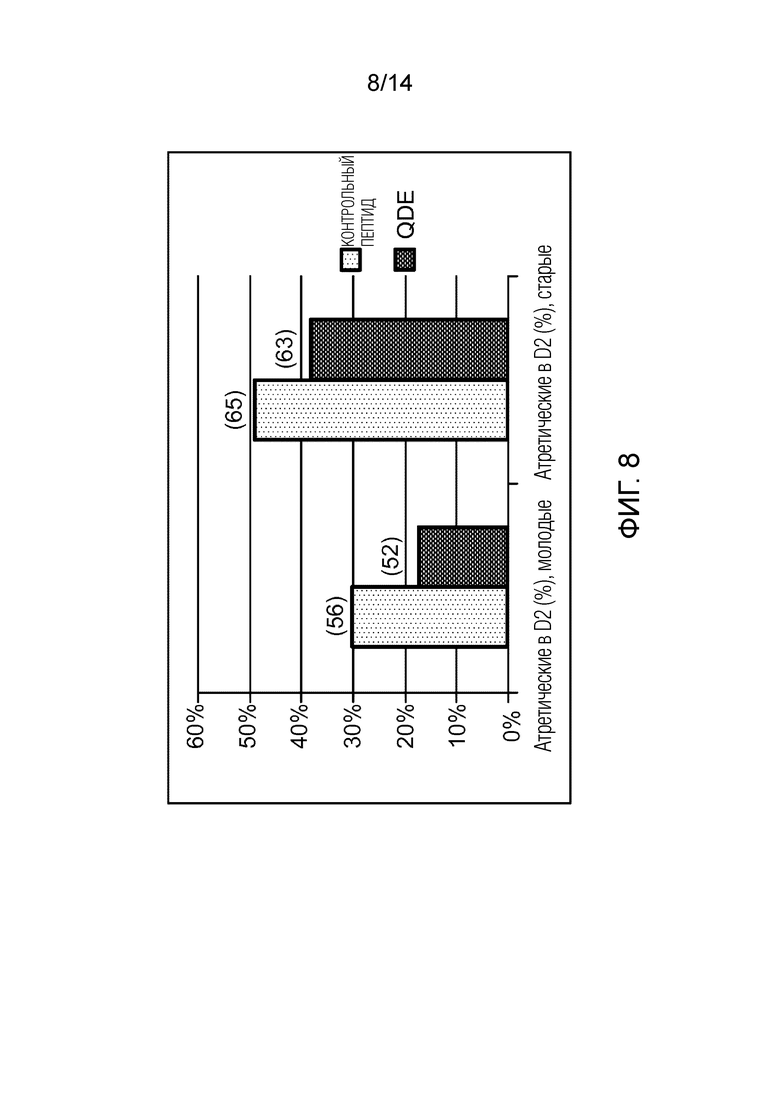
Фигура 8: сравнение среднего процента атрезии (ATR) в D2 между молодыми и старыми мышами в присутствии или в отсутствие пептида QDEc.
Молодые мыши: 7-недельные мыши B6CBAF1 (n=108 ооцитов, 56 - контрольных, 52 - QDEc); старые мыши: 7-месячные мыши B6CBAF1 (n=128 ооцитов, 65 - контрольных, 63 - QDEc).
Фигура 9: Графическое представление методологии клинического исследования, проведенного в примере 5.
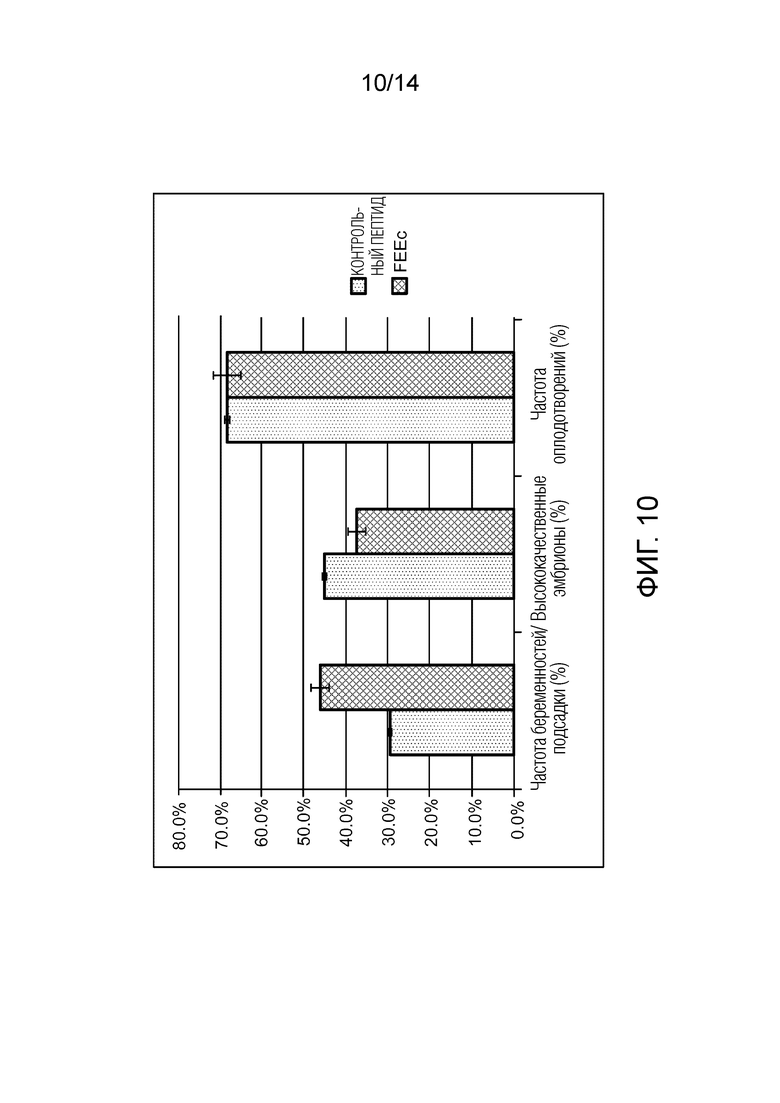
Фигура 10: предварительные результаты критериев основной и вторичной оценки в контексте клинического исследования. Частота беременностей при подсадке свежего или замороженного эмбриона (контрольная группа, n=17 подсадок/группа FEEc, n=13 подсадок). Процент «высококачественных эмбрионов» (контрольная группа, n=75 делящихся эмбрионов/группа FEEc, n=72). Частота оплодотворений (контрольная группа, n=259 MII/ группа FEEc, n=246 MII).
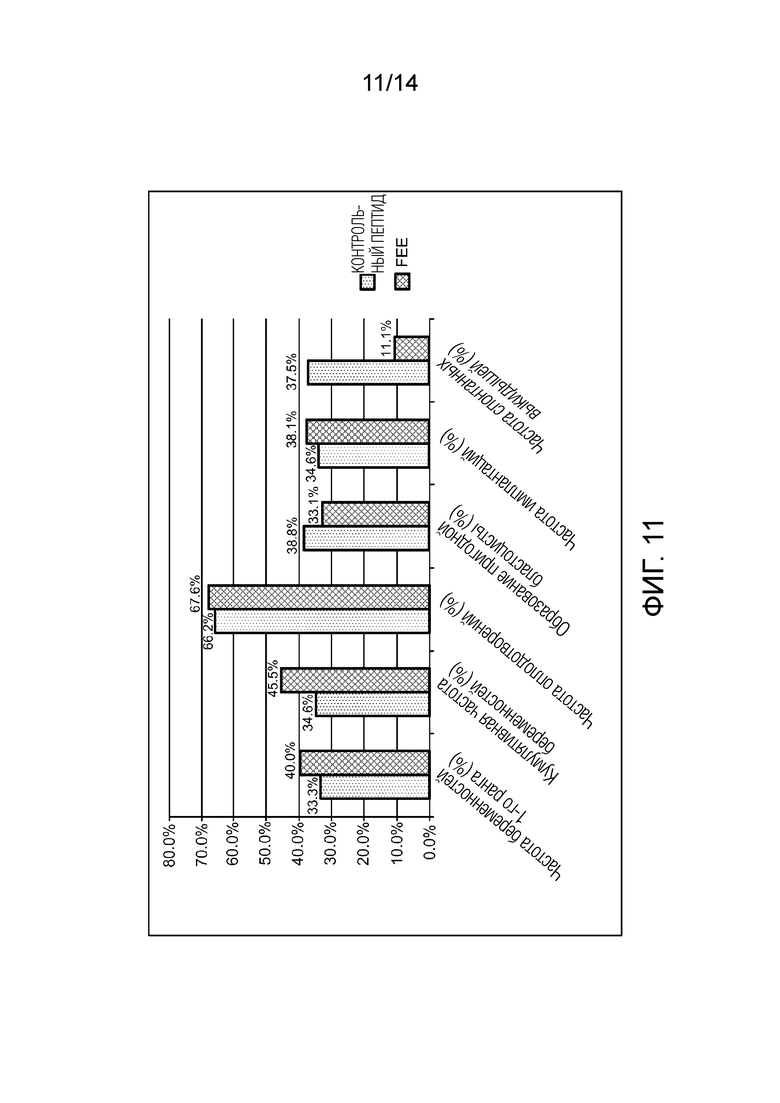
Фигура 11: частота беременностей, полученных в контексте клинического исследования.
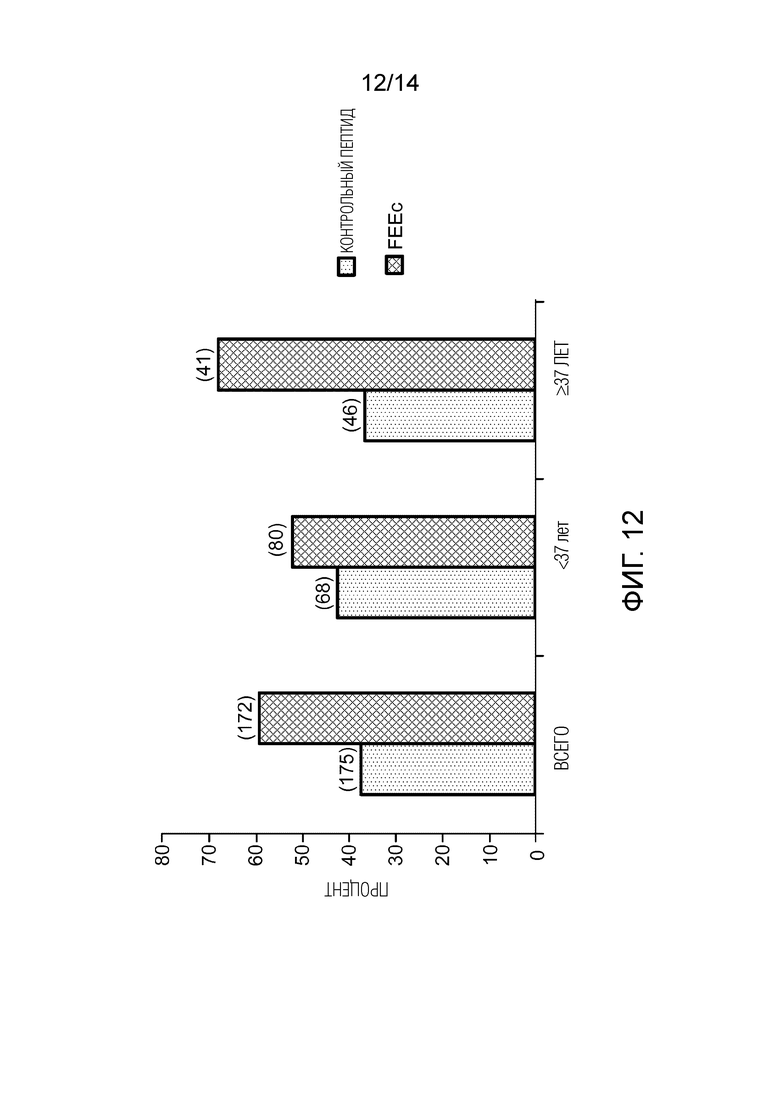
Фигура 12: увеличение степени созревания ооцитов, блокированных в GV, после инкубации в присутствии FEEc. Это исследование проводилось с учетом одного ооцита на женщину и путем инкубации указанного ооцита в присутствии FEEc или контрольного пептида.
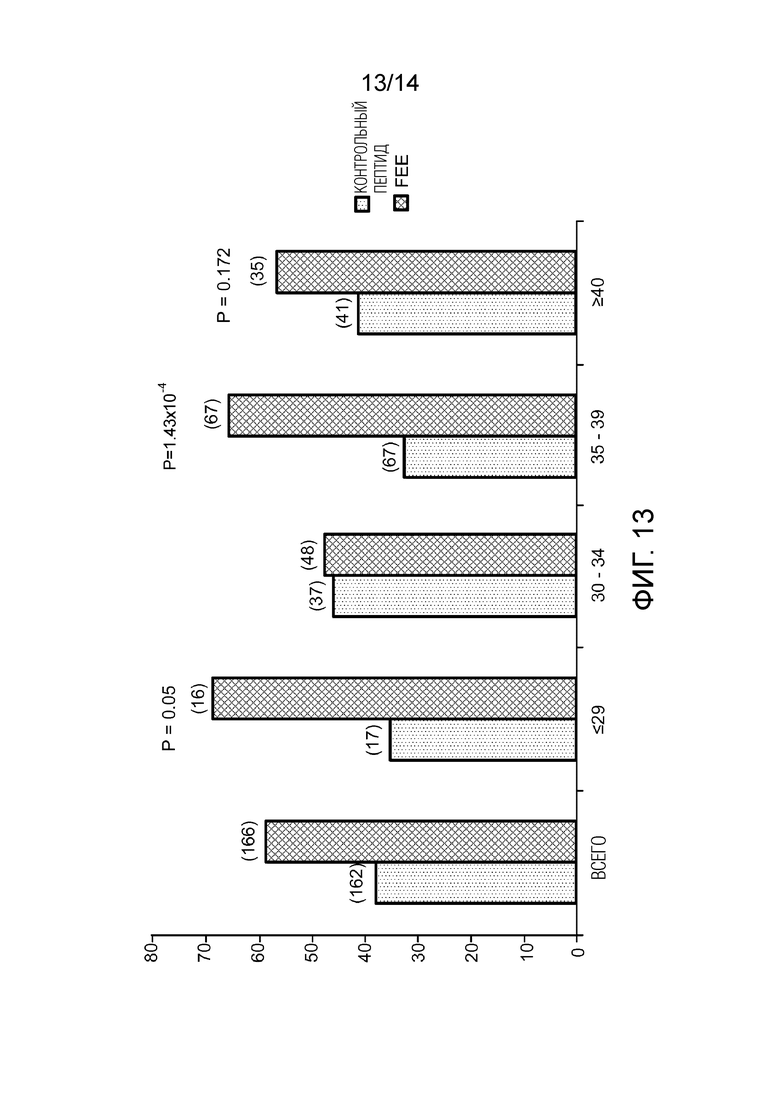
Фигура 13: эффекты FEEc на созревание ооцитов человека, блокированных в GV, с учетом возрастного диапазона.
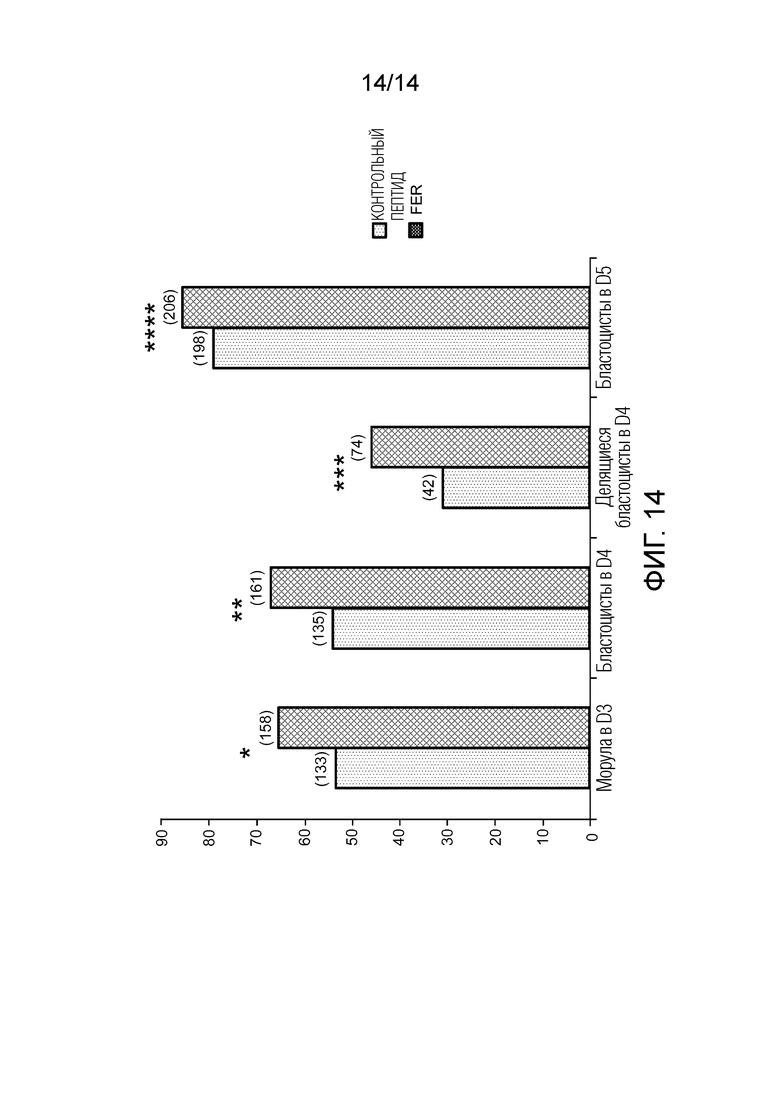
Фигура 14: стимуляция предимплантационного эмбрионального развития у молодых мышей с помощью QDEc (*P<0,00532; **P<0,00374; ***P<0,00913; ****P<0,068; () число эмбрионов).
Материалы и способы
Приведенные ниже экспериментальные примеры были получены с использованием следующих материалов и способов:
Наблюдение параметров подвижности сперматозоидов
Исследуемые сперматозоиды разделяли на 2 аликвоты, одну из которых инкубировали с FEEc, а другую инкубировали с «перетасованным» пептидом, содержащим такие же аминокислоты, но в случайном порядке. Авторы изобретения инкубировали в течение 3 ч при 37°С сперматозоиды человека в присутствии 100 мкМ пептида FEEc или «перетасованного» пептида, а затем наблюдали параметры подвижности сперматозоидов с помощью автоматизированного анализа (Компьютерного анализа спермы, CASA).
Проверенными параметрами спермы были следующие: сглаженные VAP, VSL, VCL и ALH. Они соответствуют, соответственно, средней скорости пути, линейной скорости, криволинейной скорости и амплитуде латерального смещения головки. Исследование показало значительное увеличение процентного содержания гиперактивированных сперматозоидов (в соответствии с критериями Mortimer et al.). Это может объяснить увеличение процента оплодотворений, наблюдаемое в присутствии пептида.
Измерение потенциала митохондриальной мембраны
Авторы изобретения инкубировали в течение 3 ч при 37°С сперматозоиды человека в присутствии либо пептида FEEc, либо «перетасованного» пептида, который содержит такие же аминокислоты в случайном порядке и, следовательно, представляет собой контрольную группу. После промывки сперматозоиды маркируют с использованием липофильного флуоресцентного красителя, DIOC6.
Митохондриальный мембранный потенциал (протонный градиент на уровне митохондриальной внутренней мембраны) затем измеряли проточной цитометрией. Обнаружено, что он увеличивается у сперматозоидов после воздействия FEEc.
Измерение индекса фертильности
Ооциты человека с удаленной зоной пеллюцида инкубировали с сперматозоидами человека в отсутствие или в присутствии FEEc в концентрации 100 мкМ. Слитые сперматозоиды подсчитывали после УФ-возбуждения. Сперматозоиды считались слитыми, когда ядро было помечено Hoechst 33342. Кроме того, головки сперматозоидов, проникшие в ооцит в присутствии пептида FEEc, были не только больше по численности, но также имели размытый вид, свидетельствующий о деконденсации головок сперматозоидов. Эта деконденсация является одним из первых этапов активации ооцита после проникновения сперматозоида. Из этого можно сделать вывод, что FEEc не только улучшает удобрения способность сперматозоидов к оплодотворению, но также активирует оплодотворенный ооцит.
Сбор ооцитов человека для созревания in vitro (IVM)
Незрелые ооциты человека, полученные для исследования, были собраны в лаборатории оплодотворения in vitro (ЭКО) Центра репродукции (MAP) больницы Коши (Париж, Франция). Через два часа после забора ооцитов пункцией, у ооцитов, предназначенных для микроинъекции, удаляли корону с помощью гиалуронидазы (Origio, Limonest, Франция). После наблюдения под инвертированным микроскопом (Hoffman) незрелые ооциты на стадии зародышевого пузырька (GV) сохраняли для остальных экспериментов.
Созревание in vitro (IVM) незрелых ооцитов человека
Незрелые ооциты человека на стадии GV были рандомизированы либо в контрольной культуральной среде (Global, JCD, La Mulatière, Франция) (n=203), либо в той же среде, дополненной 100 мкМ FEEc (n=193). Незрелые ооциты человека были разделены на две группы: ооциты от женщин в возрасте до 37 лет, и ооциты от женщин в возрасте 37 лет и старше. В день фолликулярной пункции две группы ооцитов на стадии GV инкубировали в 20-микролитровых каплях, покрытых маслом, и оставляли при 37°С в 5% CO2, чтобы наблюдать их под инвертированным микроскопом (Hoffman) в D1 (24 ч инкубации) и D2 (48 ч инкубации). Ооциты классифицировали как находящиеся в метафазе II (1-е полярное тельце в перивителлинном пространстве), в стадии зародышевого пузырька (GV), метафазе I (разрыв зародышевого пузырька без вытеснения полярного тельца) или атретические.
Сбор ооцитов мыши для созревания in vitro (IVM)
Самок мышей B6CBAF1 (в возрасте от 5 до 8 недель), поставленных лабораторией Чарльз-Ривер (L'Arbresle, Франция), стимулировали инъекцией PMSG (гонадотропина из сыворотки жеребой кобылы) в количестве 10 МЕ (Sigma-Aldrich, Saint-Quentin Fallavier, Франция) без запуска овуляции. Незрелые ооциты собирали из яичников через 48 ч после последней инъекции, затем удаляли их кумулюс с помощью гиалуронидазы и три раза промывали в культуральной среде М2. Только ооциты, классифицированные как GV, сохраняли для остальных экспериментов.
Созревание in vitro (IVM) незрелых ооцитов мыши
Ооциты мышей инкубировали, рандомизируя между стандартной средой, и средой, дополненной 100 мкМ QDEc. Культуральные чашки приготавливали накануне и инкубировали при 37°С и 5% СО2. Ооциты наблюдали в D0 (через 8 ч после забоя) и D1 (24 ч).
Иммунофлуоресценция
Ооциты человека, полученные в результате IVM, фиксировали в 2%-м параформальдегиде (PFA) в течение 1 часа при температуре окружающей среды и затем промывали в PBS, содержащем 0,5% BSA. Пермеабилизацию проводили путем инкубации ооцитов в растворе, содержащем 0,5% БСА, 0,1% Тритона-Х100, 0,05% Твина-20 и 5% нормальной козьей сыворотки. Ооциты затем промывали в PBS с 0,5%-м БСА перед инкубацией в течение ночи в разбавлении 1/200 антитела против α-тубулина человека (Sigma-Aldrich) в PBS, содержащем 0,5% БСА. Ооциты затем инкубировали в течение 1 часа в присутствии вторичного IgG-антитела, конъюгированного с Alexa Fluor (Life Technologies, Alfortville, France). После стадии промывания ооциты инкубировали в течение 10 мин в DAPI (разбавленном до 1/1000) перед монтированием на слайдах и наблюдали с помощью конфокальной микроскопии в темноте. Для анализа веретена ооциты с четкими и хорошо организованными волокнами микротрубочек, ассоциированные с идеальным выравниванием хромосом на метафазной пластинке, определяли, как нормальные.
Стимуляция и спаривание мышей
7-недельных «молодых» и 7-месячных «старых» самок мышей B6CBAF1 спаривали с самцами C57N после суперовуляции, причем последняя заключалась в инъекции PMSG в количестве 10 МЕ (Sigma-Aldrich) с последующим запуском овуляции путем введения hCG (хорионического гонадотропина человека) в количестве 10 МЕ (Sigma-Aldrich) через 46-48 ч.
На следующий день после спаривания мышей, имеющих вагинальную пробку, умерщвляли. Ооциты собирали из яйцеводов через 15-16 ч после инъекции hCG, а процент оплодотворения оценивали по наличию второго полярного тельца в перивителлиновом пространстве.
Инкубация оплодотворенных ооцитов мыши
Ооциты мыши, оплодотворенные после спаривания и собранные, случайным образом распределяли на 4 группы (молодые, обработанные циклическим пептидом QDE (QDEc); молодые контрольные; старые, обработанные QDEc; старые контрольные) и помещали в капли культуральной среды (KSOM) по 20 мкл для контрольных мышей и в капли культуральной среды (KSOM), дополненной 100 мкМ QDEc, для «обработанных» групп. Воздействие пептида длилась от D1 до D4 после спаривания in vivo (D0). QDEc эквивалентен человеческому FEEc. Культурные чашки инкубируют при 37°С в 5% СО2 и покрывают минеральным маслом.
Ооциты наблюдали каждый день в бинокуляре, чтобы оценить признаки эмбрионального развития. Нормальная кинетика развития соответствует, как минимум, в D2, эмбриону, разделившемуся на 2 клетки, и в D5 - эмбриону на стадии морулы или бластоцисты.
Рандомизированное проспективное исследование в ЭКО человека
Клиническое исследование было начато в лаборатории ЭКО Центра репродукции Коши 09/08/2014 на 66 парах, где средний возраст для женщин составлял 34,3±4,2 года, а для партнеров - 37,0±5,2. Это было рандомизированное одноцентровое проспективное исследование оплодотворения in vitro (ЭКО), проведенного в присутствии или отсутствии FEEc. Ооциты, освобожденные от кумулюса, были разделены на две группы поочередно в одну, а затем в другую, в соответствии с порядком их выделения. Когда все ооциты были освобождены, лаборант, который не участвовал в удалении кумулюса, определял случайным образом, какую из двух групп оплодотворяли в присутствии FEEc, а какая служила контролем. Одну часть ооцитов инкубируют в стандартной культуральной среде (Global, JCD), другую часть в этой же среде, дополненной 100 мкМ FEEc.
Ниже представлена методология этого исследования, представленная схематически на фигуре 9. Ооциты человека, полученные от женщин в возрасте от 18 до 43 лет, были выделены гинекологами пункцией яичников после гормональной стимуляции. Их разделяли случайным образом на 2 группы: ооциты, инкубированные в присутствии стандартной культуральной среды, дополненной 100 мкМ FEEc, или стандартной культуральной среды (Global, JCD), затем помещали в инкубатор при 37°C в атмосфере 5%-го CO2. Сперматозоиды партнера выделяли в лаборатории, и наиболее подвижные отбирали в соответствии со стандартной методикой получения центров репродукции. IVF состоит в приведении сперматозоидов в контакт с ооцитами в среде для оплодотворения при концентрации 105 отобранных сперматозоидов/мл в каплях по 20 мкл под маслом. Оплодотворение проводят в инкубаторе при 37°С в 5%-м СО2 в течение 18 часов. Через 18 ч после оплодотворения (D1) у ооцитов удаляют корону. Оплодотворенные ооциты промывают и переносят в другую каплю среды, и помещают культивировать в течение еще 24 часов. Для подсадки в матку их промывают три раза, затем помещают в среду для переноса и помещают в полость матки. Эмбрионы подсаживают в соответствии с их кажущимся качеством, не обращая внимания на группу происхождения. Подсаживают один или несколько эмбрионов в зависимости от возраста, показаний к ЭКО, степени важности попытки и качества полученных эмбрионов, а также с согласия пар. Некоторые эмбрионы могут культивироваться в течение длительного времени (в течение 5 дней, на стадии бластоцисты) либо сразу, либо после подсадки эмбрионов в D2.
Основным критерием оценки является процент клинических беременностей при подсадке свежих или замороженных эмбрионов и процент выкидышей с учетом 3 групп: гомогенных подсадок (контрольный и исследуемый пептиды) и смешанных подсадок (смесь этих двух).
Вторичными критериями являются:
- частота оплодотворений, то есть соотношение числа зигот, имеющих две пронуклеуса в цитоплазме, через 18 часов после оплодотворения, относительно количества ооцитов в метафазе 2 в когорте.
- процент эмбрионов хорошего качества, т.е. эмбрионов, из которых последовательность дробления соответствует идеальной последовательности, то есть: от 4 до 5 клеток в D2, и от 8 до 9 клеток в D3, и у которых фрагментация бластомера является A-типа (когда объем, занимаемый фрагментами, составляет менее 10% объема эмбриона) или B-типа (когда объем, занимаемый фрагментами, составляет от 10% до 30% от общего объема эмбриона). Таким образом, эти «высококачественные» эмбрионы соответствуют соотношению числа эмбрионов каждого типа относительно общего числа эмбрионов для каждой группы.
Этот протокол был одобрен Комитетом по этике West VI 12/13/2012. Исследование получило разрешение от Агентства Франции по биомедицине 07/08/2013. ЭКО проводится со строгим соблюдением хорошей клинической практики. Каждая пара, соглашаясь участвовать в исследовании, подписала свободный и информированный документ согласия.
Статистический анализ
Количественные переменные изучались с использованием их численных значений, среднего и стандартного отклонения. Данные сравнивались между получавшей воздействие и не получавший воздействие группами с использованием соответствующего теста (теста Стьюдента или теста Уилкоксона) для количественных переменных. Сравнение процентов проводилось с использованием критерия Хи-квадрата (χ2) или точного теста Фишера. Различия между сравниваемыми данными считались статистически значимыми, когда значение p (порог значимости) было меньше 0,05.
Пример 1: улучшение параметров подвижности сперматозоидов и процента гиперактивированных сперматозоидов у мужчин
Предварительные эксперименты показали, что в 18-часовом тесте на выживаемость сперматозоидов, инкубированных в присутствии пептида FEE, выживаемость значительно улучшается в группе, обработанной FEE в концентрации 100 мкМ, по сравнению с контролем.
Пример 1а: автоматизированный анализ параметров подвижности сперматозоидов после инкубации в присутствии FEEc и «перетасованного» пептида в качестве контроля
Результаты, представленные в таблице 1 ниже, показывают увеличение в сглаженной VAP (p=0,008), в VSL (p=0,048), в VCL (p <0,0001) и в ALH (p=0,002), что дает 29%-е увеличение содержания гиперактивированных сперматозоидов (p=0,009) по сравнению с контрольной группой. Это улучшение в процентном содержании гиперактивированных сперматозоидов объясняет улучшение в их фузиогенной способности и увеличение степени оплодотворения, зарегистрированное у мышей на интактных комплексах ооцит-кумулюс.
Таблица 1: автоматизированный анализ параметров подвижности сперматозоидов в сравнении контрольной группы и группы после инкубации с FEEc.
Пример 1b: измерение потенциала митохондриальной мембраны
Установлено, что потенциал митохондриальной мембраны увеличивается на 21% в присутствии пептида FEEc по сравнению с «перетасованным» пептидом (p<0,001) (фигура 1).
Таким образом, FEEc улучшает параметры подвижности сперматозоидов, увеличивая потенциал митохондриальной мембраны спермы.
Пример 1c: исследование индекса фертильности
Результаты, представленные на фигуре 2, показывают, что не только большее число сперматозоидов сливается с ооцитами с удаленной зоной пеллюцида в присутствии пептида FEEc, но указанные сперматозоиды также деконденсируются в отличие от таковых в контрольной группе.
Среднее значение 19,0±4,6 сперматозоидов насчитывается в цитоплазме в контрольных ооцитах, тогда как увеличение до 36,9±11,7 сперматозоидов, слитых с ооцитами, было зарегистрировано после инкубации с FEEc в концентрации 100 мкМ (р<0,001). Это явление предполагает увеличение способности сперматозоидов к оплодотворению и активации ооцитов, опосредованное пептидом FEEc.
Параметры подвижности сперматозоидов у 37 пациентов были проанализированы в присутствии или отсутствии FEEc (инкубация в течение 3 часов). Наблюдается значительное увеличение процента гиперактивированных сперматозоидов в соответствии с критериями Мортимера, что объясняет увеличение их способности к оплодотворению (см. таблицу 1 выше).
Пример 2: улучшение в процентах in vitro созревания незрелых ооцитов человека
Достоверное улучшение созревания ооцитов с помощью FEEc было продемонстрировано в D1 для всех тестируемых ооцитах человека. Полученные результаты были следующими: 42,3% (69/163) ооцитов в метафазе II (MII) с FEEc относительно 30,0% (52/173) в контрольной группе, p=0,02 (фигура 3). Улучшение созревания еще более заметно для ооцитов от женщин в возрасте 37 лет и старше, сразу после D1. Полученные результаты были следующими: 47,9% (23/48) ооцитов в MII с использованием FEEc относительно 20,4% (11/54) в контрольной группе (p=0,003).
Несмотря на небольшую разницу для ооцитов, полученных от женщин моложе 37 лет, улучшение IVF для ооцитов у человека очень важно для ооцитов от пожилых женщин, так как была достигнута та же частота созревания, что и для ооцитов от молодых женщин (47,9% ооцитов в метафазе II в присутствии FEEc относительно 20,4% в контрольной группе, p=0,003).
Результаты, полученные с 336 ооцитами человека на стадии GV, инкубированными рандомизированным образом в присутствии или в отсутствие пептида FEEc, показывают, что присутствие FEEc увеличивает долю созревших ооцитов человека.
Не было обнаружено существенной разницы между показателями атрезии ооцитов среди 2 групп (16,5% в присутствии пептида против 16,8% для контрольной группы) (фигура 4). В образцах от женщин в возрасте 37 лет и старше наблюдалась незначительная тенденция к снижению частоты атрезии в присутствии пептида FEEc (16,7%, 8/48) по сравнению с контрольной средой (24,1%, 13/54) p>0,05.
Исследование было продолжено, и были получены дополнительные результаты. Эти результаты подтверждают описанные выше результаты и демонстрируют другие эффекты пептида FEEc, которые описано ниже.
В общей сложности, 600 ооцитов прошли созревание in vitro. Для анализа в исследование включали только один ооцит на женщину, чтобы все события были независимыми. В присутствии фертилина в среде процент созревания повышался с 38,3% до 59,0% (P<1,6×10-4) (фигура 12). Когда анализ проводится в зависимости от возраста женщины, от которой были получены ооциты в GV, видно, что улучшение созревания относительно скромно для женщин в возрасте до 37 лет (с 42,6% до 51,8%, P<0,2), но оно намного выше для ооцитов, полученных от женщин в возрасте 37 лет и старше (с 35,1% до 65,9%, P<3,91×10-5). Поэтому, фертилин способен стимулировать созревание ооцита in vivo с удалением короны и отделением первого полярного тельца. По-видимому, чем больше возрастной дефицит энергии в ооците, тем лучше фертилин выполняет эту стимуляцию. Это исследование также позволило продемонстрировать увеличение процента созревания, в особенности, ооцитов у женщин в возрасте от 37 и до 30 лет (фигура 13).
Пример 3: организация мейотического веретена ооцитов человека, созревших in vitro
На фигуре 5 показана неполная и аберрантная организация выравнивания хромосом на метафазной пластинке с невыравненными хромосомами. Изображение получено для ооцита человека в MII после 24 ч IVM в присутствии контрольной среды.
Пример 4: улучшение степени оплодотворения и раннего эмбрионального развития у мышей
В D1 степень оплодотворения остается неизменной у молодых мышей, тогда как у старых мышей она возрастает с 39% до 51% (р<0,03) (фигура 6).
В D2 у молодых мышей, 50,0% (26/52) ооцитов деляться в группе QDEc по сравнению с 32,4% (18/56) в контрольной группе (P=0,02) (фигура 7). Что касается старых мышей, то в D2 наблюдается значительно большее число делящихся эмбрионов в присутствии QDEc (58,7%, 37/63) по сравнению с контролем (35,4%, 23/65), p=0,008. В D4, процент делящихся эмбрионов у молодых мышей, которые достигают стадии морулы или бластоцисты, составляет 34,6% (7/16) для контрольной группы по сравнению с 63,0% (17/26) для среды, дополненной QDEc, p <0,03. Этот процент достигает 86,3% (63/73) для старых мышей в присутствии пептида по сравнению с 28,3% (15/53) в контрольной среде, p= 0,001.
Предпочтительно, образование бластоцист у мышей улучшается в присутствии пептида QDEc.
Процент атретических эмбрионов существенно не отличается между двумя группами у молодых мышей: 17,3% (9/52) в присутствии QDEc и 30,4% (17/56) для контроля (p=0,1). По сравнению с молодыми мышами процент атрезии выше у группы старых мышей со значениями, сходными между группами: QDEc (38,1%, 24/63) и контролем (49,2%, 32/65) (p=0,2) (фигура 8).
Для более тщательного изучения предимплантационного развития эмбрионов использовали следующий протокол: мышей спаривали в D0. В D1 ооциты извлекали путем разрезания яичников. Затем их распределяли на две группы рандомизированным способом и помещали в культуру с или без QDEc перед оплодотворением.
Результаты показывают увеличение в числе морул в D3, в числе бластоцист в D4, и среди них увеличение делящихся бластоцист при инкубации ооцита в присутствии QDEc (31,1% относительно 45,9%, P<0,009) (фигура 14). Кроме того, после подсадки этих бластоцист псевдогестационным самкам наблюдается значительное увеличение количества потомства, полученного подсадкой эмбрионов, в особенности, для эмбрионов, полученных от молодых мышей (таблица 2 ниже).
Таблица 2: рождаемость у мышей после подсадки контрольных эмбрионов и эмбрионов, обработанных QDEc.
Пример 5: увеличение процента клинических беременностей после оплодотворения in vitro
Результаты, представленные ниже, были получены в контексте исследования «фертилина».
На первом этапе было включено 56 пар. Средний возраст составляет 33,9±4,1 года среди женщин и 36,7±5,3 для партнеров. Не было отмечено существенной разницы между процентом оплодотворения или между процентом высококачественных эмбрионов среди 2 групп - FEEc и контролем. Незначительная тенденция к увеличению процента беременностей при пересадке отмечена в присутствии пептида по сравнению с контролем и со смешанной группой - 46,1% (6/13) по сравнению с 29,4% (5/17) и 40% (2/5), соответственно (фигура 10 и таблица 2 ниже). Сообщалось о выкидыше после подсадки одного эмбриона из контрольной группы. На сегодняшний день никаких побочных эффектов не обнаружено.
На сегодняшний день в исследование включено 66 пар. Было проведено 54 подсадки: 26 с контрольными эмбрионами, 22 с эмбрионами из группы FEE, 6 с эмбрионами из двух групп (смешанные подсадки). Кумулятивная частота беременностей составляла, соответственно, 34,6%, 45,5% и 33,3% в 3 группах. Частота самопроизвольных выкидышей составляла, соответственно, 33,3%, 10% и 0%. Таким образом, частота клинических беременностей составляет, соответственно, 23%, 41% и 33,3%. Существенным фактом является то, что когда частоту клинической беременности анализируют относительно возраста пациента, у молодых пациентов видно, что частота развивающейся клинической беременности (то есть беременности, развивающейся до родов) значительно увеличивается с 20% до 57,1% (P<0,03).
Таблица 3: частота беременностей после подсадки (для 66 пациентов)
Результаты также показывают увеличение на 21% в клинических беременностях после подсадки эмбриона из группы FEEc - 40,0% (8/20) по сравнению с 33,3% (8/24) в контрольной группе (p>0,05). Частота оплодотворений составляет 66,2% среди контролей по сравнению с 67,6% в группе, подвергшейся воздействию FEEc (p> 0,05). Частота ранних спонтанных выкидышей достигает 37,5% (3/8) в контрольной группе по сравнению с 11,1% (1/9), когда эмбрион подвергался воздействию FEEc (p> 0,05), (фигура 11).
Для женщин в возрасте до 37 лет, после воздействия FEEc частота оплодотворений составляет 70,9% по сравнению с 68,3% в контрольной группе. В случае, когда ооцит подвергался воздействию FEEc, частота беременностей достигает 57,1% по сравнению с 20,0% в контрольной группе (таблица 4 ниже).
Таблица 4: частота оплодотворений и частота беременностей в зависимости от возраста партнера (≥37 или <37 лет), *P<0,03.
У первых 66 пар, включенных в исследование до настоящего времени, была проведена 51 подсадка, остальные были отложены, и результаты показывают, что из этих 51 подсадок:
- 21 была проведена с использованием эмбрионов контрольной группы;
- 18 были проведены с использованием группы в присутствии FEEc;
- 12 были проведены с эмбрионами из двух групп.
Частота беременностей была выше и было меньше случаев выкидышей в группе FEEc в сравнении с другими группами (33% выкидышей по сравнению с 9% для эмбрионов в присутствии FEEc) (таблицы 5 и 6 ниже).
Частота беременностей для молодых женщин поднимается от 20% до 57,1% (р <0,03) в присутствии FEEc по сравнению с контролем.
Таблица 5: частота беременностей после подсадки (для 66 пациентов)
Таблица 6: частота беременностей при подсадке в зависимости от возраста женщин.
Продолжение исследования фертилина позволило получить дополнительные результаты. Всего в исследование было включено 66 пар. Результаты представлены в таблицах ниже.
Общие данные о попытках представлены в таблице 7. Общие показатели оплодотворения практически не изменялись. Однако процент попыток, при которых наблюдалось плохое оплодотворение (менее 20%), упал с 37,9% до 27,3% в присутствии FEEc, что говорит о лучшей способности гамет к оплодотворению в присутствии фертилина. Аналогичным образом, полиспермия в группе фертилина была того же порядка, что и в контрольной группе (4,0% относительно 5,2%), что показывает, что нормальный механизм блокирования полиспермии не был изменен.
Таблица 7: общие результаты исследования фертилина на 66 пациентах.
Результаты всех пар, которым пересадили эмбрионы, приведены в таблице 8. В этой таблице исключены пары, у которых была необъяснимая неудача оплодотворения в 2 группах ооцитов с или без фертилина (n=6).
(не достоверно)
Таблица 8: результаты, полученные на 66 парах в исследовании фетилина.
Как показано в таблице 8, у пациентов, перенесших подсадку эмбрионов, частота оплодотворения повышалась с 69,5% до 78,6%, что составляет улучшение на 13%, но которое не достигает значимости в данной когорте. Эмбриональное развитие до стадии бластоцисты не изменяется, равно как и частота имплантации эмбрионов в матку. С другой стороны, частота выкидышей снижается почти на 50% в группе фертилина (15,4% против 30,5%).
Результаты для пар, перенесших подсадку эмбрионов, и в которых женщина имеет возраст до 37 лет, приводятся в таблице 9.
Если учесть только пары, в которых женщина моложе 37 лет (n=47), и которые соответствуют более чем 70% пациентов, получавших лечение, видно, что частота оплодотворений достоверно повышается с 70,9% до 83,3% (Р<0,05). Частота выкидышей снижается от 36,3% для пациентов, получивших эмбрионы контрольной группы, до 9,1%, то есть в четыре раза относительно тех, кто получил эмбрион из группы фертилина. Фактически, частота развивающихся беременностей, приводящих к рождению ребенка, повышается с 28,6% в группе контрольных эмбрионов до 41,6% у эмбрионов из группы фертилина.
P<0,05
Таблица 9: результаты, полученные в парах, в которых женщина моложе 37 лет в контексте исследования фертилина.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПРИМЕНЕНИЕ РАСТВОРИМОГО CD146 В КАЧЕСТВЕ БИОМАРКЕРА ДЛЯ СЕЛЕКЦИИ ОПЛОДОТВОРЕННОГО IN VITRO ЭМБРИОНА ДЛЯ ИМПЛАНТАЦИИ МЛЕКОПИТАЮЩЕМУ | 2016 |
|
RU2735542C2 |
| СПОСОБ РЕГУЛИРОВАНИЯ ФОЛЛИКУЛЯРНОГО РЕЗЕРВА ЯИЧНИКА, СПОСОБ ЛЕЧЕНИЯ ОТКЛОНЕНИЙ В РОСТЕ ПОКОЯЩИХСЯ ФОЛЛИКУЛОВ У ЖЕНЩИН, СРЕДСТВО СТИМУЛЯЦИИ РАЗВИТИЯ ФОЛЛИКУЛОВ И СРЕДСТВО ОПРЕДЕЛЕНИЯ ВЛИЯНИЯ СОЕДИНЕНИЙ НА УСКОРЕНИЕ ИЛИ ЗАМЕДЛЕНИЕ РОСТА ФОЛЛИКУЛОВ ПРИ ПРОВЕДЕНИИ ТОКСИКОЛОГИЧЕСКИХ ИСПЫТАНИЙ (ВАРИАНТЫ) | 2004 |
|
RU2418604C9 |
| ДОНЕКОПРИД КАК НЕЙРОПРОТЕКТОРНЫЙ АГЕНТ ПРИ ЛЕЧЕНИИ НЕЙРОДЕГЕНЕРАТИВНЫХ ЗАБОЛЕВАНИЙ | 2019 |
|
RU2800802C2 |
| СПОСОБ ПРОГНОЗИРОВАНИЯ НАСТУПЛЕНИЯ БЕРЕМЕННОСТИ В ПРОГРАММЕ ЭКСТРАКОРПОРАЛЬНОГО ОПЛОДОТВОРЕНИЯ ПРИ СЕЛЕКТИВНОМ ПЕРЕНОСЕ ЭМБРИОНОВ ПУТЕМ ОЦЕНКИ МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКОГО ПРОФИЛЯ ГАМЕТ С ПОМОЩЬЮ ПЦР-РВ | 2014 |
|
RU2550965C1 |
| ТЕХНОЛОГИЯ ДОЗРЕВАНИЯ ООЦИТОВ ЧЕЛОВЕКА НА СТАДИИ GV С ПОМОЩЬЮ ВНЕКЛЕТОЧНЫХ ВЕЗИКУЛ ФОЛЛИКУЛЯРНОЙ ЖИДКОСТИ В ПРОГРАММАХ ЭКСТРАКОРПОРАЛЬНОГО ОПЛОДОТВОРЕНИЯ: EV-IVM (EXTRACELLULAR VESICLES IN VITRO MATURATION) | 2023 |
|
RU2807492C1 |
| Способ проведения экстракорпорального оплодотворения (ЭКО) в естественном цикле при отсутствии гиперпролактинемии | 2024 |
|
RU2834540C1 |
| НЕОРГАНИЧЕСКИЙ ПИРОФОСФАТ И ЕГО ПРИМЕНЕНИЕ | 2012 |
|
RU2626932C2 |
| СПОСОБ УПРАВЛЕНИЯ КАЧЕСТВОМ ООЦИТОВ И КОМПОЗИЦИЯ ДЛЯ ДОБАВЛЕНИЯ В СРЕДУ КУЛЬТИВИРОВАНИЯ ООЦИТОВ | 2005 |
|
RU2281777C1 |
| Способ получения in vitro эмбрионов крупного рогатого скота | 2023 |
|
RU2823596C1 |
| БЕЗБЕЛКОВАЯ КЛЕТОЧНАЯ КУЛЬТУРАЛЬНАЯ СРЕДА (ВАРИАНТЫ) И ЕЕ ПРИМЕНЕНИЕ | 2008 |
|
RU2462510C2 |

Настоящее изобретение относится области биотехнологии, конкретно к применению циклического пептида, воспроизводящего сайт связывания фертилина-бета с интегрином ооцита, для повышения способности гамет человека к оплодотворению in vitro и может быть использовано в медицинской практике. Изобретение позволяет стимулировать энергетический метаболизм половых клеток человека в контексте протоколов репродуктивной медицины (MAP), в частности, для улучшения созревания незрелого ооцита in vitro, гиперактивации сперматозоидов, а также частоты оплодотворений и частоты рождаемости. 3 н. и 7 з.п. ф-лы, 14 ил., 9 табл., 5 пр.
1. Применение in vitro циклического пептида, содержащего циклический пептид, имеющий формулу C-S-F-E-E-C с циклической связью между двумя концевыми цистеинами, воспроизводящий сайт связывания фертилина-бета с интегрином ооцита, для созревания in vitro незрелых ооцитов.
2. Применение in vitro по п. 1, где ооциты представляют собой ооциты человека.
3. Применение in vitro по п. 1 или 2, где ооциты инкубируют в течение периода от 1 минуты до 4 дней в присутствии от 1 до 100 мкМ пептида.
4. Применение in vitro по любому из пп. 1-3 для улучшения плоидности ооцитов.
5. Применение in vitro по любому из пп. 1-4 для увеличения вероятности беременности и/или снижения риска выкидыша.
6. Применение in vitro циклического пептида, содержащего циклический пептид, имеющий формулу C-S-F-E-E-C с циклической связью между двумя концевыми цистеинами, воспроизводящий сайт связывания фертилина-бета с интегрином ооцита, для гиперактивации сперматозоидов в протоколе репродуктивной медицины (MAP).
7. Применение in vitro по п.6 для увеличения скорости движения сперматозоидов.
8. Применение in vitro по п. 6 или 7, где сперматозоиды инкубируют в течение от 1 минуты до 3 часов в присутствии от 1 до 100 мкМ пептида до внутриутробного осеменения.
9. Применение in vitro циклического пептида, содержащего циклический пептид, имеющий формулу C-S-F-E-E-C с циклической связью между двумя концевыми цистеинами, воспроизводящий сайт связывания фертилина-бета с интегрином ооцита, для криоконсервации гамет и эмбрионов.
10. Применение in vitro по любому из предшествующих пунктов в протоколе экстракорпорального оплодотворения (IVF).
| WO 2005051799 A2, 09.06.2005 | |||
| ZIYYAT A | |||
| et al.: Cyclic FEE peptide increases human gamete fusion and potentiates its RGD-induced inhibition, HUMAN REPRODUCTION, 2005, v | |||
| Прибор для промывания газов | 1922 |
|
SU20A1 |
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| ПРИБОР ДЛЯ СТЕРИЛИЗАЦИИ И АСЕПТИЧЕСКОГО ХРАНЕНИЯ И РАСХОДОВАНИЯ ЖИДКОСТЕЙ | 1925 |
|
SU3452A1 |
| ROTIVAL R | |||
| et al.: Comprehensive determination of the cyclic FEE peptide chemical stability in solution, J.Pharm.Biochem.An., 2013, v | |||
| Способ размножения копий рисунков, текста и т.п. | 1921 |
|
SU89A1 |
| Устройство для выпрямления многофазного тока | 1923 |
|
SU50A1 |

