Перекрестные ссылки на родственные заявки
Данная заявка на патент испрашивает приоритет на основе предварительной заявки на патент США №61/495, 085, поданной 9 июня 2011, и предварительной заявки на патент США №61/535, 668, поданной 16 сентября 2011, каждая из которых включена здесь посредством ссылки в полном объеме.
Включение посредством ссылки материалов, представленных в электронном виде
В настоящий документ включен посредством ссылки во всей своей полноте машиночитаемый список нуклеотидных/аминокислотных последовательностей, представленный с настоящей заявкой, и определенный следующим образом: один ASCII (текстовый) файл 53884 байтов с именем «710339_ST25.TXT» от 2 июня 2012.
Уровень техники
Экзотоксин A Pseudomonas (РЕ) является бактериальным токсином с цитотоксической активностью, который может быть эффективным для уничтожения или ингибирования роста нежелательных клеток, например, раковых клеток. Соответственно, РЕ может быть полезен для лечения или профилактики заболеваний, таких как, например, рак. Тем не менее, РЕ может иметь высокую иммуногенность. Соответственно, введение РЕ может стимулировать иммунный ответ на РЕ, включая, например, выработку антител к РЕ и/или Т-клеток против него, что нежелательно нейтрализует цитотоксическую активность РЕ. Такая иммуногенность может уменьшить количество РЕ, которое может быть дано пациенту, что может, в свою очередь, снизить эффективность РЕ для лечения заболевания, например, рака. Таким образом, существует потребность в улучшенном РЕ.
Краткое описание изобретения
Один вариант осуществления изобретения обеспечивает экзотоксин А Pseudomonas (РЕ), включающий аминокислотную последовательность, содержащую замены одного или более аминокислотных остатков из L294, L297, Y298, L299 и R302, при условии, что, когда аминокислотная последовательность содержит замену на аланин в аминокислотном остатке R302, по крайней мере один из аминокислотных остатков L294, L297, Y298 и L299 заменен, где аминокислотные остатки L294, L297, Y298, L299 и R302 определены относительно SEQ ID NO:1, необязательно с заменами одного или нескольких аминокислотных остатков в одном или нескольких В-клеточных эпитопах SEQ ID NO:1 и/или с заменами одного или нескольких аминокислотных остатков в одном или более Т-клеточных эпитопах в пределах аминокислотных остатков R421, L422, L423, А425, R427, L429, Y439, Н440, F443, L444, А446, А447, I450, 463-519, R551, L552, Т554, I555, L556 и W558 в SEQ ID NO:1.
Еще один вариант воплощения изобретения относится к РЕ, представленному аминокислотной последовательностью, имеющей формулу I:
где:
m, n, и р - независимо представляют собой 0 или 1;
FCS включает последовательность аминокислотных остатков, расщепляемую фурином, причем последовательность расщепляется фурином;
R1 включает один или более последовательных аминокислотных остатков из остатков 285-293 из SEQ ID NO:1;
R2 включает X1VAX2X3X4AAX5LSW (SEQ ID NO:2), где X1, X2, и Х4 независимо представляют собой лейцин, аланин, глицин, серин или глутамин; Х3 представляет собой тирозин, аланин, глицин, серин, или глутамин, и X5 представляет собой аргинин, аланин, глицин, серин или глутамин, при условии, что РЕ не содержит LVALYLAARLSW (SEQ ID NO:3) и, когда X5 представляет собой аланин, по крайней мере один из Х1, X2, Х3, и Х4 представляет собой аланин, глицин, серин или глутамин;
R3 включает 1 или более последовательных аминокислотных остатков из остатков 306-394 из SEQ ID NO:1; и
функциональный домен III РЕ включает остатки 395-613 из SEQ ID NO:1,
необязательно с заменами одного или нескольких аминокислотных остатков в одном или более В-клеточных эпитопах SEQ ID NO:1 и/или с заменами одного или нескольких аминокислотных остатков в одном или более Т-клеточных эпитопах в пределах аминокислотных остатков R421, L422, L423, А425, R427, L429, Y439, Н440, F443, L444, А446, А447, 1450, 463-519, R551, L552, Т554, 1555, L556 и W558 из SEQ ID NO:1.
Еще один вариант изобретения обеспечивает РЕ, имеющий аминокислотную последовательность, имеющую замену одного или нескольких аминокислотных остатков в положениях R421, L422, L423, А425, R427, L429, Y439, Н440, F443, L444, А446, А447, I450, 463-519, R551, L552, Т554, I555, L556 и W558 из SEQ ID NO:1; при условии, что
когда аминокислотный остаток в положении Q485 или L516 заменен на аланин, по крайней мере, один дополнительный аминокислотный остаток в позиции R421, L422, L423, А425, R427, L429, Y439, Н440, F443, L444, А446, А447, I450, 463-519, R551, L552, Т554, I555, L556 и W558 из SEQ ID NO:1 является замененным, и
когда аминокислотный остаток в положении R427, R467, R490, R505, R513 или R551 заменен на аланин, глицин, серин или глутамин, или когда аминокислотный остаток в положении R490 заменен на валин, лейцин, или изолейцин, по крайней мере, один аминокислотный остаток в позиции R421, L422, L423, А425, R427, L429, Y439, Н440, F443, L444, А446, А447, I450, 463-519, R551, L552, Т554, I555, L556 и W558 из SEQ ID NO:1 дополнительно заменен, что не включает замену на аланин, глицин, серин или глутамин для аминокислотных остатков в положениях R427, R467, R490, R505, R513 или R551 или замену на валин, лейцин или изолейцин для аминокислотного остатка в положении 490,
где аминокислотные остатки R421, L422, L423, А425, R427, L429, Y439, Н440, F443, L444, А446, А447, I450, 463-519, R551, L552, Т554, I555, L556 и W558 определены относительно SEQ ID NO:1.
Еще один вариант воплощения изобретения предусматривает экзотоксин A Pseudomonas (РЕ), включающий аминокислотную последовательность РЕ, имеющую замену одного или нескольких аминокислотных остатков из D463, Y481 и L516, определенных относительно SEQ ID NO:1, при условии, что, когда аминокислотный остаток в положении 516 заменен аланином, по крайней мере один из аминокислотных остатков D463 и Y481 также заменен, причем РЕ необязательно имеет дополнительные замены одного или нескольких аминокислотных остатков в одном или более В-клеточных эпитопах и/или дополнительные замены одного или нескольких аминокислотных остатков в одном или нескольких Т-клеточных эпитопах и/или делеции одного или более последовательных аминокислотных остатков из 1-273 и 285-394 в SEQ ID NO:1.
Дополнительные варианты воплощения изобретения обеспечивают родственные химерные молекулы, а также родственные нуклеиновые кислоты, рекомбинантные экспрессионные векторы, клетки-хозяева, популяции клеток и фармацевтические композиции.
Еще один вариант воплощения настоящего изобретения относится к способу лечения или профилактики рака у млекопитающего, включающему введение млекопитающему РЕ согласно изобретению, химерной молекулы согласно изобретению, нуклеиновой кислоты согласно изобретению, рекомбинантного экспрессионного вектора согласно изобретению, клетки-хозяина согласно изобретению, популяции клеток согласно изобретению или фармацевтической композиции согласно изобретению, в количестве, эффективном для лечения или профилактики рака у млекопитающего.
Еще один вариант воплощения настоящего изобретения относится к способу ингибирования роста клеток-мишеней, включающему контактирование клетки с РЕ согласно изобретению, химерной молекулой согласно изобретению, нуклеиновой кислотой согласно изобретению, рекомбинантным экспрессионным вектором согласно изобретению, клеткой-хозяином согласно изобретению, популяцией клеток согласно изобретению, или фармацевтической композицией согласно изобретению, в количестве, эффективном для ингибирования роста клетки-мишени.
Дополнительные варианты воплощения настоящего изобретения обеспечивают способы получения РЕ согласно изобретению и способы получения химерной молекулы согласно изобретению.
Краткое описание фигур
Фигура 1 представляет собой график, показывающий частоту аллелей (ось y) различных аллелей главного комплекса гистосовместимости II класса DR бета 1 (DRB1) (ось x) в мировой популяции (незакрашеные столбцы) и группе доноров (закрашенные столбцы).
Фигура 2А представляет собой график, показывающий количество образующих пятна клеток (SFC) на 1×106 клеток (ось y), указывающий на реакцию Т-клеток наивного донора 031810aph после размножения in vitro и инкубации со средой (М) (без пептида), пептидным пулом 3, пептидным пулом 16 или пептидным пулом 22 (ось х), измеренное методом ELISpot с интерлейкином (IL)-2.
Фигура 2 В представляет собой график, показывающий количество SFC на 1×106 клеток (ось y), указывающий на реакцию Т-клеток наивного донора 031810aph после инкубации без пептида, с пептидным пулом 3, пептидным пулом 16 или пептидным пулом 22 (ось х) без размножения in vitro, измеренную с помощью IL-2 ELISpot.
Фигура 3 представляет собой график, показывающий общее количество SFC на 1×106 клеток (ось y) для Т-клеток от каждого из доноров 1-50 при отсутствии пептида или для каждого из пептидных пулов 1-22 (ось х) после 14 дней размножения in vitro. Пунктирная линия показывает величину, в три раза превышающую фоновое значение.
На Фигуре 4 определены конкретные пептиды и области в пептидах (выделенные серым цветом участки) из пептидного пула 3, которые стимулируют ответ Т-клеток различной интенсивности у различных доноров.
Фигура 5 представляет собой график, показывающий цитотоксическую активность (% от контроля) (ось y) в зависимости от концентрации НА22 дикого типа (Fv фрагмент антитела к CD22, стабилизированный дисульфидными связями, конъюгированный с РЕ38) (кружки), L297A НА22 (квадраты), или R302A НА22 (ромбы) (нг/мл) (ось х) на СА46 клетках.
Фигура 6А представляет собой график, показывающий ответ Т-клеток донора 010710 (SFC на 1×106 клеток) (ось y) после повторной стимуляции без пептида, пептидом дикого типа (WT15) или R302A (ось y) после культивирования в течение 14 дней с НА22 дикого типа (закрашенные столбцы) или R302A НА22 (незакрашенные столбцы).
Фигуре 6 В представляет собой график, показывающий ответ Т-клеток донора 111909 (SFC на 1×106 клеток) (ось y) после повторной стимуляции без пептида, пептидом дикого типа (WT15) или R302A (ось y) после культивирования в течение 14 дней с НА22 дикого типа (закрашенные столбцы) или R302A НА22 (незакрашенные столбцы).
Фигура 7А представляет собой график, показывающий ответ Т-клеток донора 031510 (SFC на 1×106 клеток) (ось y) на стимуляцию НА22 (содержащим РЕЗВ) (закрашенные столбцы) или LR RIT (LR) (содержащим аминокислотные остатки 274-284 и 395-613 из SEQ ID NO:1) (незакрашеные столбцы) и повторную стимуляцию одним из пептидных пулов 1-22 (ось х). Контроль включает клетки, выращенные в присутствии цефтазидима (CEFT), клетки без стимуляции антигеном в день 0 и без стимуляции антигеном в день 14 («М линии»), и клетки со стимуляцией LMB9 в день 0 и без стимуляции антигеном в день 14 («без пептида»).
Фигура 7В представляет собой график, показывающий ответ Т-клеток донора 021610 (SFC на 1×106 клеток) (ось y) на стимуляцию НА22 (содержащим РЕ38) (закрашенные столбцы) или LR RIT (LR) (содержащим аминокислотные остатки 274-284 и 395-613 из SEQ ID NO:1) (незакрашенные столбцы) и повторную стимуляцию одним из пептидных пулов 1-22 (ось х). Контроль включает клетки, выращенные а присутствии цефтазидима (CEFT), клетки без стимуляции антигеном в день 0 и без стимуляции антигеном в день 14 («М линии»), и клетки со стимуляцией LMB9 в день 0 и без стимуляции антигеном в день 14 («без пептида»).
Фигура 7С представляет собой график, показывающий ответ Т-клеток донора 101509 (SFC на 1×106 клеток) (ось y) на стимуляцию НА22 (содержащим РЕ38) (закрашенные столбцы) или LR RIT (LR) (содержащим аминокислотные остатки 274-284 и 395-613 из SEQ ID NO:1) (незакрашенные столбцы) и повторную стимуляцию одним из пептидных пулов 1-22 (ось х). Контроль включает клетки, выращенные а присутствии цефтазидима (CEFT), клетки без стимуляции антигеном в день 0 и без стимуляции антигеном в день 14 («М линии»), и клетки со стимуляцией LMB9 в день 0 и без стимуляции антигеном в день 14 («без пептида»).
На Фигуре 8 определены конкретные пептиды (выделенные серым цветом участки) пептидов SEQ ID NO:102-111, которые стимулируют ответ Т-клеток, измеренный по образованию IL-2 для различных доноров.
Фигура 9 представляет собой график, показывающий суммарный процент общего ответа доноров для 50 доноров для каждой из SEQ ID NO:31-141.
Фигура 10 представляет собой график, показывающий реактивность фага анти-РЕ38 (домен III) против НА22 с точечными заменами. Черные ячейки означают реактивность менее 10%, пустые ячейки означают реактивность более 10%, и серые ячейки указывают на отсутствие проверки. Замены отсортированы по их расположению от М-конца (слева) к С-концу (справа).
Фигуры НА и 11 В представляют собой графики, показывающие результаты сравнительных экспериментов, определяющих концентрацию каждого из замещенных иммунотоксинов НА22 («НА», закрашенные кружки), HA22-LR («LR», не закрашенные кружки), HA22-L05 («L05», закрашенные треугольники), HA22-L06 («L06», не закрашенные треугольники), HA22-LR-8M («LR8M» закрашенные квадраты), и HA22-L010 («L010», не закрашенные квадраты), которая уменьшает уровень антител, реагирующих с НА22, на 50% (пунктирная линия) в сыворотке первого (фигура 11А) и второго (фигура 11В) пациентов, проходящих клинические испытания с НА22.
На Фигуре 12 приведен график, показывающий процент связывания антител с НА22, HA22-LR-8M, HA22-L010 (HA22-LRL010) или HA22-LRL010R в сыворотке пациентов, получающих лечение РЕ38.
Подробное описание изобретения
Экзотоксин A Pseudomonas («РЕ») является бактериальным токсином (молекулярная масса 66 кДа), секретируемым синегнойной палочкой Pseudomonas aeruginosa. Нативная последовательность РЕ дикого типа (SEQ ID NO:1) приведена в патенте США 5602095, который включен здесь посредством ссылки. Нативный РЕ дикого типа включает три структурных домена, которые опосредуют цитотоксичность. Домен la (аминокислоты 1-252) опосредует связывание с клеткой, домен II (аминокислоты 253-364) опосредует перемещение в цитозоль и домен III (аминокислоты 400-613) опосредует ADP-рибозилирование фактора элонгации 2. Хотя считается, что структурная граница домена III РЕ начинается с остатка 400, предполагается, что домену III может требоваться фрагмент домена Ib, чтобы сохранить ADP-рибозилирующую активность. Соответственно, функциональный домен III определяется как остатки 395-613 РЕ. Функция домена Ib (аминокислоты 365-399) остается неопределенной. Не желая быть связанными какой-либо конкретной теорией или механизмом, полагают, что цитотоксическая активность РЕ происходит через ингибирование синтеза белка в эукариотических клетках, например, путем инактивации ADP-рибозилированием фактора элонгации 2 (EF-2).
Замены в РЕ определены здесь по отношению к аминокислотной последовательности РЕ. Таким образом, замены РЕ описаны здесь в виде аминокислотного остатка, присутствующего в определенном положении, после которого указана аминокислота, на которую указанный остаток был заменен в конкретной обсуждаемом случае. В связи с этим позиции аминокислотной последовательности конкретного варианта воплощения РЕ упоминаются здесь как позиции аминокислотной последовательности конкретного варианта воплощения или как позиции, определяемые по SEQ ID NO:1. Когда позиции определены по SEQ ID NO:1, то фактические позиции аминокислотной последовательности конкретного варианта воплощения РЕ определяются относительно соответствующих положений в SEQ ID NO:1 и могут иметь номера позиций остатков, отличные от номеров позиций остатков из SEQ ID NO:1. Так, например, при замене, относящейся к замене аминокислотного остатка в аминокислотной последовательности конкретного варианта воплощения РЕ, соответствующей указанной позиции 613 аминокислотной последовательности SEQ ID NO:1, следует понимать, что фактические позиции в соответствующих аминокислотных последовательностях могут быть различными. Например, если позиции определены по SEQ ID NO:1, термин «R490» относится к аргинину, обычно присутствующему в положении 490 в SEQ ID NO:1, «R490A» означает, что обычно присутствующий аргинин в положении 490 в последовательности SEQ ID NO:1 заменен на аланин, a «K590Q» означает, что обычно присутствующий в положении 590 в SEQ ID NO:1 лизин заменен на глутамин. В случае множественных замен в двух или более позициях две или более замены могут быть одинаковыми или различными, т.е. каждый аминокислотный остаток из двух или нескольких замененных аминокислотных остатков может быть заменен одинаковыми или разными аминокислотными остатками, если явно не указано иное.
Термины «экзотоксин Pseudomonas» и «РЕ», используемые в данном изобретении, включают РЕ, который был изменен относительно нативного белка, чтобы уменьшить или устранить иммуногенность. Такие модификации могут включать устранение домена 1а, различные аминокислотные делеции в областях Ib, II, и III, отдельные аминокислотные замены и добавление одной или нескольких последовательностей на карбоксильный конец, таких как DEL и REDL (SEQ ID NO:7), но не ограничиваются ими. Смотри Siegall et al., J. Biol. Chem., 264: 14256-14261 (1989). Такие модифицированные РЕ могут быть дополнительно модифицированы, чтобы включать любую замену (замены) согласно изобретению одного или нескольких аминокислотных остатков в одном или нескольких Т-клеточных и/или В-клеточных эпитопах, описанных здесь. В одном из вариантов модифицированный РЕ может быть цитотоксическим фрагментом нативного РЕ дикого типа. Цитотоксические фрагменты РЕ могут включать те, которые являются цитотоксическими при последующем протеолитическом или другом процессинге в клетке-мишени или без него (например, как белок или белок-предшественник). В предпочтительном варианте воплощения цитотоксический фрагмент РЕ сохраняет по меньшей мере около 20%, предпочтительно по меньшей мере примерно 40%, более предпочтительно около 50%, еще более предпочтительно 75%, более предпочтительно по меньшей мере примерно 90%, иеще более предпочтительно 95% цитотоксичности нативного РЕ. В особенно предпочтительных вариантах воплощения цитотоксический фрагмент имеет по меньшей мере цитотоксичность нативного РЕ, и предпочтительно увеличенную цитотоксичность по сравнению с нативным РЕ.
Модифицированный РЕ с уменьшенной или устраненной иммуногенностью включает, например, РЕ4Е, РЕ40, РЕ38, РЕ25, PE38QQR, PE38KDEL и РЕ35. В одном варианте воплощения РЕ может быть любым из РЕ4Е, РЕ40, РЕ38, РЕ25, PE38QQR (в котором РЕ38 имеет последовательность QQR, добавленную на С-конец), PE38KDEL (в котором РЕ38 имеет KDEL последовательность (SEQ ID NO:5), добавленную на С-конец), PE-LR (устойчивый к деградации лизосомами) и РЕ35.
В одном варианте РЕ был изменен, чтобы уменьшить иммуногенность, путем удаления домена la, как описано в патенте США 4892827, который включен здесь посредством ссылки. РЕ могут быть также модифицированы путем замены определенных остатков домена Ia. В одном варианте воплощения изобретения РЕ может представлять собой РЕ4Е, который является РЕ с заменами, в котором домен 1а присутствует, но в котором основные остатки домена 1а в положениях 57, 246, 247, и 249 заменены кислотными остатками (например, глутаминовой кислотой), как описано в патенте США 5512658, который включен здесь посредством ссылки.
РЕ40 является усеченным производным РЕ (Pai et al., Proc. Nat'l Acad. Sci. USA, 88: 3358-62 (1991) и Kondo et al„ Biol. Chem., 263: 9470-9475 (1988)). PE35 является 35 кДа карбокси-концевым фрагментом РЕ, в котором аминокислотные остатки 1-279 удалены и молекула начинается с Met в положении 280 с последующими аминокислотами 281-364 и 381-613 нативного РЕ. PE35 и РЕ40 раскрыты, например, в патентах США 5602095 и 4892827, каждый из которых включен в данное описание посредством ссылки. РЕ25 содержит 11-аминокислотный фрагмент домена II и весь домен III. В некоторых вариантах осуществления РЕ содержит только домен III.
В предпочтительном варианте воплощения РЕ представляет собой РЕ38. РЕ38 содержит транслоцирующий и ADP-рибозилирующий домены РЕ, но не часть, связывающуюся с клеткой (Hwang J. et al., Cell, 48: 129-136 (1987)). РЕ38 представляет собой усеченный белок-предшественник РЕ, состоящий из аминокислот 253-364 и 381-613 (SEQ ID NO:144), который активируется в его цитотоксическую форму после процессинга в клетке (см., например, патент США 5608039, который включен здесь посредством ссылки, и Pastan et al., Biochim. Biophys. Acta, 1333: C1-C6 (1997)).
В другом предпочтительном варианте воплощения РЕ представляет собой PE-LR. PE-LR содержит делецию домена II за исключением последовательности, расщепляемой фурином (FCS), соответствующей аминокислотным остаткам 274-284 в SEQ ID NO:1 (RHRQPRGWEQL (SEQ ID NO:8)) и делецию аминокислотных остатков 365-394 домена Ib. Таким образом, PE-LR содержит аминокислотные остатки 274-284 и 395-613 из SEQ ID NO:1. PE-LR описан в международной патентной заявке WO 2009/032954, которая включена здесь посредством ссылки. PE-LR может, при необходимости, дополнительно содержать соединительный пептид GGS между FCS и аминокислотными остатками 395-613 в SEQ ID NO:1.
Как отмечалось выше, альтернативно или дополнительно, часть или весь домен Ib может быть удален, а остальные части соединены линкером или непосредственно пептидной связью. Альтернативно или дополнительно, некоторые части домена II могут быть удалены. Альтернативно или дополнительно, С-конец может содержать нативную последовательность остатков 609-613 (REDLK) (SEQ ID NO:6), или может содержать вариации, которые могут поддерживать способность РЕ к транслокации в цитозоль, такие как KDEL (SEQ ID NO:5) или REDL (SEQ ID NO:7), и повторы этих последовательностей. См., например, патенты США 5854044; 5821238; и 5602095 и международную патентной заявку WO 1999/051643, которые включены здесь посредством ссылки. Любая форма РЕ, в котором иммуногенность была устранена или уменьшена, может быть использована в сочетании с любой из замен согласно изобретению одного или нескольких аминокислотных остатков в одном или более Т-клеточном и/или В-клеточном эпитопах, описанных здесь, до тех пор, пока он остается цитотоксичным для клеток-мишеней, например, путем транслокации и рибозилирования EF-2 в клетках-мишенях.
Вариант воплощения изобретения обеспечивает экзотоксин А Pseudomonas (РЕ), включая любую из модификаций нативного белка РЕ, описанного здесь, представленный аминокислотной последовательностью, имеющей замену одного или нескольких аминокислотных остатков из L294, L297, Y298, L299 и R302, при условии, что когда аминокислотная последовательность содержит замену остатка R302 на аминокислоту аланин, по крайней мере, еще один дополнительный аминокислотный остаток заменен, где аминокислотные остатки L294, L297, Y298, L299 и R302 определены относительно SEQ ID NO:1, необязательно с замещением одного или нескольких аминокислотных остатков в одном или более В-клеточных эпитопах SEQ ID NO:1 и/или с замещением одного или нескольких аминокислотных остатков в одном или более Т-клеточных эпитопах в аминокислотных остатках R421, L422, L423, А425, R427, L429, Y439, Н440, F443, L444, А446, А447, I450, 463-519, R551, L552, Т554, I555, L556 и W558 из SEQ ID NO:1. Предпочтительно, замещение одного или нескольких аминокислотных остатков в одном или более Т-клеточных эпитопах представляет собой замену одного или нескольких аминокислотных остатков в положениях R421, L422, L423, А425, R427, L429, Y439, Н440, F443, L444, А446, А447, I450, Y470, 1471, А472, Р475, А476, L477, 1493, R494, N495, L498, L499, R500, V501, Y502, V503, R505, L508, Р509, R551, L552, Т554, I555, L556 и W558.
Еще один вариант воплощения изобретения относится к экзотоксину А Pseudomonas (РЕ), включая любую из модификаций нативного белка РЕ, описанного здесь, представленному аминокислотной последовательностью, имеющей замену одного или нескольких аминокислотных остатков из L294, L297, Y298, L299 и R302, при условии, что когда аминокислотная последовательность содержит замену остатка R302 на аминокислоту аланин, по меньшей мере один из аминокислотных остатков L294, L297, Y298, и L299 является замещенным, где аминокислотные остатки L294, L297, Y298, L299 и R302 определены относительно SEQ ID NO:1, необязательно с заменой одного или нескольких аминокислотных остатков в одном или нескольких В-клеточных эпитопах SEQ ID NO:1 и/или заменой одного или нескольких аминокислотных остатков в одном или нескольких Т-клеточных эпитопах в аминокислотных остатках R421, L422, L423, А425, R427, L429, Y439, Н440, F443, L444, А446, А447, I450, 463-519, R551, L552, Т554, I555, L556 и W558 из SEQ ID NO:1. Было обнаружено, что аминокислотные остатки L294, L297, Y298, L299 и R302 расположены в пределах одного или нескольких Т-клеточных эпитопов РЕ. Таким образом, замена одного или нескольких аминокислотных остатков из L294, L297, Y298, L299 и R302 может, предпочтительно, удалять один или несколько Т-клеточных эпитопов. Соответственно, РЕ согласно изобретению могут быть, предпочтительно, менее иммуногенными, чем незамещенные РЕ (например, дикого типа).
Заменой одного или более аминокислотных остатков из L294, L297, Y298, L299 и R302 может быть замена любого одного или более аминокислотных остатков из L294, L297, Y298, L299 и R302. В одном из вариантов воплощения изобретения, один или несколько аминокислотных остатков из L294, L297, Y298, L299 и R302 заменяют на аланин, глицин, серин или глутамин вместо одного или нескольких аминокислотных остатков из L294, L297, Y298, L299 и R302.
В одном из вариантов воплощения изобретения РЕ содержит X1VAX2X3X4AAX5LSW РЕ (SEQ ID NO:2), где X1, Х2, и Х4 независимо представляют собой лейцин, аланин, глицин, серин или глутамин; Х3 представляет собой тирозин, аланин, глицин, серин или глутамин; и Х5 представляет собой аргинин, аланин, глицин, серин или глутамин, при условии, что РЕ не содержит LVALYLAARLSW (SEQ ID NO:3) и, когда Х5 представляет собой аланин, по крайней мере один из Х1, Х2, Х3 и Х4 представляет собой аланин, глицин, серин или глутамин.
Еще один вариант осуществления изобретения относится к экзотоксину А Pseudomonas (РЕ), представленному аминокислотной последовательностью РЕ, имеющей замену одного или более аминокислотных остатков из D463, Y481 и L516, определенных относительно SEQ ID NO:1, при условии, что, когда аминокислотный остаток в положении 516 заменен на аланин, по крайней мере один из аминокислотных остатков D463 и Y481 также заменен, где РЕ необязательно имеет дополнительные замены одного или нескольких аминокислотных остатков в одном или более В клеточных эпитопах и/или также замены одного или нескольких аминокислотных остатков в одном или более Т-клеточных эпитопах и/или делеции одного или более последовательных аминокислотных остатков, выбранных из остатков 1-273 и 285-394 в SEQ ID NO:1. Предпочтительно, замены одного или более аминокислотных остатков D463, Y481 и L516 являются, независимо, заменами на аланин, глицин, серин или глутамин одного или более аминокислотных остатков из D463, Y481 и L516. Было обнаружено, что аминокислотные остатки D463, Y481 и L516 расположены в пределах одного или нескольких В-клеточных эпитопов РЕ. Таким образом, замещение одного или более аминокислотных остатков из D463, Y481 и L516 может, предпочтительно, удалять один или несколько Т-клеточных и/или В-клеточных эпитопов. Соответственно, РЕ согласно изобретению могут, предпочтительно, быть менее иммуногенными, чем незамещенный РЕ (например, дикого типа).
В варианте воплощения изобретения дополнительным замещением аминокислот в одном или нескольких В-клеточных эпитопах является замена одного или более из аминокислотных остатков Е282, Е285, Р290, R313, N314, Р319, D324, Е327, Е331, Q332, D403, D406, R412, R427, Е431, R432, R458, D461, R467, R490, R505, R513, Е522, R538, Е548, R551, R576, Q592 и L597, определенных относительно последовательности SEQ ID NO:1. Предпочтительно, дальнейшее замещение аминокислот в одном или нескольких В-клеточных эпитопах представляет собой замену, независимо, на аланин, глицин или серин одного или нескольких аминокислотных остатков из R427, R458, R467, R490, R505 и R538. В особенно предпочтительном варианте заменами одного или нескольких аминокислотных остатков из D463, Y481 и L516 является замена на аланин аминокислотного остатка D463 и дальнейшее замещение аминокислот в одном или нескольких В-клеточных эпитопах: (а) замена аминокислотного остатка R427 на аланин, (б) замена аминокислотного остатка R458 на аланин, (в) замена аминокислотного остатка R467 на аланин, (г) замена аминокислотного остатка R490 на аланин; (д) замена аминокислотного остатка R505 на аланин и (е) замена аминокислотного остатка R538 на аланин относительно SEQ ID NO:1.
В дополнение к замене (заменам) одного или нескольких аминокислотных остатков в одном или более Т-клеточных и/или В-клеточных эпитопах РЕ, описанных здесь, РЕ согласно изобретению может, по желанию, также содержать дополнительную замену (замены) одного или нескольких аминокислотных остатков в одном или нескольких В-клеточных эпитопах SEQ ID NO:1. В связи с этим, в одном из вариантов воплощения изобретения РЕ имеет замену одной или нескольких аминокислот в одном или нескольких В-клеточных эпитопах SEQ ID NO:1. В предпочтительном варианте воплощения настоящего изобретения замещение одной или более аминокислот в одном или нескольких В-клеточных эпитопах SEQ ID NO:1 включает замену на аланин, глицин, серин или глутамин одной или нескольких аминокислот в одном или более В-клеточных эпитопах из SEQ ID NO:1. Замена (замены) в пределах одного или нескольких В-клеточных эпитопов может, предпочтительно, еще больше снизить иммуногенность путем удаления одного или более В-клеточных эпитопов. Замена (замены) могут быть расположены в любом подходящем В-клеточном эпитопе РЕ. Примеры В-клеточных эпитопов раскрыты, например, в международной патентной заявке WO 2007/016150, WO 2009/032954 и WO 2011/032022, каждая из которых включена в данное описание посредством ссылки. В предпочтительном варианте воплощения замена одной или нескольких аминокислот в одном или нескольких В-клеточных эпитопах SEQ ID NO:1 представляет собой замену на аланин, глицин, серин или глутамин, независимо, одного или нескольких из аминокислотных остатков из Е282, Е285, Р290, R313, N314, Р319, D324, Е327, Е331, Q332, D403, D406, R412, R427, Е431, R432, R458, D461, D463, R467, Y481, R490, R505, R513, L516, Е522, R538, Е548, R551, R576, К590, Q592 и L597, где аминокислотные остатки Е282, Е285, Р290, R313, N314, Р319, D324, Е327, Е331, Q332, D403, D406, R412, R427, Е431, R432, R458, D461, D463, R467, Y481, R490, R505, R513, L516, Е522, R538, Е548, R551, R576, К590, Q592 и L597 определены относительно SEQ ID NO:1. В особенно предпочтительном варианте замены аминокислот в одном или нескольких В-клеточных эпитопах SEQ ID NO:1 представляет собой замены на аланин, глицин или серин одного или нескольких аминокислотных остатков из D406, R432, R467, R490, R513, Е548, К590 и Q592. В особенно предпочтительном варианте замены аминокислот в одном или нескольких В-клеточных эпитопах SEQ ID NO представляют собой:1: (а) замену аминокислотного остатка D406 на аланин, (б) замену аминокислотного остатка R432 на глицин, (в) замену аминокислотного остатка R467 на аланин, (г) замену аминокислотного остатка R490 на аланин; (д) замену аминокислотного остатка R513 на аланин; (е) замену аминокислотного остатка Е548 на серин; (ж) замену аминокислотного остатка К590 на серин и (з) замену аминокислотного остатка Q592 на аланин.
В одном из вариантов воплощения изобретения РЕ представлен аминокислотной последовательностью, имеющей замену одного или нескольких аминокислотных остатков в положениях R421, L422, L423, А425, R427, L429, Y439, Н440, F443, L444, А446, А447, I450, 463-519, R551, L552, Т554, I555, L556 и W558 из SEQ ID NO:1, отдельно или в сочетании с любыми другими заменами, описанными здесь. В одном из вариантов воплощения изобретения замена одного или нескольких аминокислотных остатков в положениях R421, L422, L423, А425, R427, L429, Y439, Н440, F443, L444, А446, А447, I450, 463-519, R551, L552, Т554, I555, L556 и W558 из SEQ ID NO:1 представляет собой замену одного или нескольких аминокислотных остатков в положениях R421, L422, L423, А425, R427, L429, Y439, Н440, F443, L444, А446, А447, I450, Y470, 1471, А472, Р475, А476, L477, I493, R494, N495, L498, L499, R500, V501, Y502, V503, R505, L508, Р509, R551, L552, Т554, I555, L556 и W558.
Замена одного или нескольких аминокислотных остатков в положениях R421, L422, L423, А425, R427, L429, Y439, Н440, F443, L444, А446, А447, I450, 463-519, R551, L552, Т554, I555, L556 и W558 из SEQ ID NO:1 может быть заменой на любой аминокислотный остаток в одном или нескольких положениях из R421, L422, L423, А425, R427, L429, Y439, Н440, F443, L444, А446, А447, I450, 463-519, R551, L552, Т554, I555, L556 и W558 из SEQ ID NO: 1. Замена одного или нескольких аминокислотных остатков в положениях R421, L422, L423, А425, R427, L429, Y439, Н440, F443, L444, А446, А447, I450, 463-519, R551, L552, Т554, I555, L556 и W558 из SEQ ID NO: 1 может включать, например, замену на аланин, глицин, серин или глутамин одного или нескольких аминокислотных остатков в положениях 421, 422, 423, 425, 427, 429, 439, 440, 443, 444, 446, 447, 450, 463, 464, 465, 466, 467, 468, 469, 470, 471, 472, 473, 474, 475, 476, 477, 478, 479, 480, 481, 482, 483, 484, 485, 486, 487, 488, 489, 490, 491, 492, 493, 494, 495, 496, 497, 498, 499, 500, 501, 502, 503, 504, 505, 506, 507, 508, 509, 510, 511, 512, 513, 514, 515, 516, 517, 518, 519, 551, 552, 554, 555, 556 и 558 из SEQ ID NO: 1. В предпочтительном варианте воплощения изобретения замена одного или нескольких аминокислотных остатков в положениях R421, L422, L423, А425, R427, L429, Y439, Н440, F443, L444, А446, А447, I450, 463-519, R551, L552, Т554, I555, L556 и W558 из SEQ ID NO: 1 представляет собой замену на аланин, глицин, серин или глутамин одного или более аминокислотных остатков из R421, L422, L423, А425, R427, L429, Y439, Н440, F443, L444, А446, А447, I450, Y470, I471, А472, Р475, А476, L477, I493, R494, N495, L498, L499, R500, V501, Y502, V503, R505, L508, Р509, R551, L552, Т554, I555, L556 и W558. Одна или несколько замен в одном или нескольких Т-клеточных эпитопах, расположенных в положениях R421, L422, L423, А425, R427, L429, Y439, Н440, F443, L444, А446, А447, I450, 463-519, R551, L552, Т554, I555, L556 и W558 в РЕ, определенных относительно SEQ ID NO: 1, могут еще больше уменьшить иммуногенность РЕ. В одном варианте воплощения аминокислотная последовательность не имеет замен одного или нескольких аминокислотных остатков в положениях 427, 467, 485,490, 505, 513, 516 и 551.
В другом варианте воплощения изобретения РЕ представлен аминокислотной последовательностью, имеющей замену одного или нескольких аминокислотных остатков в положениях R421, L422, L423, А425, R427, L429, Y439, Н440, F443, L444, А446, А447, I450, 463-519, R551, L552, Т554, I555, L556 и W558 из SEQ ID NO: 1, при условии, что, когда аминокислотный остаток в положении Q485 или L516 заменен аланином, по крайней мере, один дополнительный аминокислотный остаток также заменен, и когда аминокислотный остаток в положении R427, R467, R490, R505, R513 или R551 заменен на аланин, глицин, серин или глутамин, или когда аминокислотный остаток в положении R490 заменен на валин, лейцин или изолейцин, по крайней мере, один дополнительный аминокислотный остаток также заменен, что не включает в себя замену на аланин, глицин, серин или глутамин аминокислотного остатка в положении 282, 285, 290, 313, 314, 319, 324, 327, 331, 332, 403, 406, 412, 427, 431, 432, 458, 461, 467, 490, 505, 513, 522, 538, 548, 551, 576, 590, 592 или 597, или замену на валин, лейцин или изолейцин аминокислотного остатка в положении 490, где аминокислотные остатки 282, 285, 290, 302, 313, 314, 319, 324, 327, 331, 332, 403, 406, 412, 427, 431, 432, 458, 461, 463-519, 522, 538, 548, 551, 576, 590, 592 и 597 определены относительно SEQIDNO:1.
В еще одном варианте воплощения настоящего изобретения РЕ представлен аминокислотной последовательностью, имеющей замену одного или нескольких аминокислотных остатков в положениях R421, L422, L423, А425, R427, L429, Y439, Н440, F443, L444, А446, А447, I450, 463-519, R551, L552, Т554, I555, L556 и W558 из SEQ ID NO:1, при условии, что, когда аминокислотный остаток в положении Q485 или L516 заменен на аланин, по крайней мере один дополнительный аминокислотный остаток в позиции R421, L422, L423, А425, R427, L429, Y439, Н440, F443, L444, А446, А447, I450, 463-519, R551, L552, Т554, I555, L556 и W558 из SEQ ID NO:1 также заменен, и, когда аминокислотный остаток в положении R427, R467, R490, R505, R513 или R551 заменен аланином, глицином, серином или глутамином, или когда аминокислотный остаток в положении R490 заменен на валин, лейцин или изолейцин, то, по крайней мере, один дополнительный аминокислотный остаток в позиции R421, L422, L423, А425, R427, L429, Y439, Н440, F443, L444, А446, А447, I450, 463-519, R551, L552, Т554, I555, L556 и W558 из SEQ ID NO:1 также заменен, что не включает в себя замену на аланин, глицин, серин или глутамин аминокислотного остатка в положении R427, R467, R490, R505, R513 или R551, или замену на валин, лейцин или изолейцин аминокислотного остатка в положении R490, где аминокислотные остатки R421, L422, L423, А425, R427, L429, Y439, Н440, F443, L444, А446, А447, 1450, 463-519, R551, L552, Т554, I555, L556 и W558 определены относительно SEQ ID NO:1.
Предпочтительно, РЕ содержит одну или несколько замен, которые повышают цитотоксичность, как раскрыто, например, в международной патентной заявке WO 2007/016150, которая включена здесь посредством ссылки. В этом отношении вариант воплощения настоящего изобретения обеспечивает РЕ с заменой аминокислоты в одном или нескольких В-клеточных эпитопах SEQ ID NO:1, где заменой аминокислоты в одном или нескольких В-клеточных эпитопах SEQ ID NO:1 является замена на валин, лейцин или изолейцин аминокислотного остатка R490, где аминокислотный остаток R490 определен относительно SEQ ID NO:1. В одном варианте воплощения изобретения замена одного или нескольких аминокислотных остатков в положениях 313, 327, 331, 332, 431, 432, 505, 516, 538 и 590, определенных относительно SEQ ID NO:1, на аланин или глутамин может обеспечить РЕ с повышенной цитотоксичностью, как раскрыто, например, в международной патентной заявке WO 2007/ 016150, которая включена здесь посредством ссылки. Повышение цитотоксической активности и снижение иммуногенности могут происходить одновременно, и не являются взаимоисключающими. Замены, которые и увеличивают цитотоксическую активность, и уменьшают иммуногенность, такие как замена R490 на глицин или, более предпочтительно, аланин, являются особенно предпочтительными.
В одном варианте воплощения изобретения РЕ представлен аминокислотной последовательностью, имеющей формулу I:
где:
m, n, и р независимо равны 0 или 1;
FCS представляет собой аминокислотную последовательность расщепления фурином, которая расщепляется фурином;
R1 включает один или более последовательных аминокислотных остатков из остатков 285-293 из SEQ ID NO:1;
R2 представляет собой X1VAX2X3X4X5LSW (SEQ ID NO:2), где X1, X2 и Х4 независимо друг от друга представляют собой лейцин, аланин, глицин, серин или глутамин; Х3 представляет собой тирозин, аланин, глицин, серин или глутамин; и X5 представляет собой аргинин, аланин, глицин, серин или глутамин, при условии, что РЕ не включает LVALYLAARLSW (SEQ ID NO:3), и что когда Х5 представляет собой аланин, по крайней мере один из X1, Х2, Х3 и Х4 представляет собой аланин, глицин, серин или глутамин;
R3 включает один или более последовательных аминокислотных остатков из остатков 306-394 из SEQ ID NO:1; и
функциональный домен III РЕ включает остатки 395-613 из SEQ ID NO:1, необязательно, с заменами одного или нескольких аминокислотных остатков в одном или нескольких В-клеточных эпитопах SEQ ID NO:1 и/или заменами одного или нескольких аминокислотных остатков в пределах одного или нескольких Т-клеточных эпитопов в пределах аминокислотных остатков R421, L422, L423, A425, R427, L429, Y439, Н440, F443, L444, А446, А447, I450, 463-519, R551, L552, Т554, I555, L556 и W558 из SEQ ID NO:1. В одном варианте воплощения замена одного или нескольких аминокислотных остатков из R421, L422, L423,A425, R427, L429, Y439, Н440, F443, L444, А446, А447, I450, 463-519, R551, L552, Т554, I555, L556 и W558 из SEQ ID NO:1 представляет собой замену одного или нескольких аминокислотных остатков из R421, L422, L423, А425, R427, L429, Y439, Н440, F443, L444, А446, А447, I450, Y470, 1471, А472, Р475, А476, L477, I493, R494, N495, L498, L499, R500, V501, Y502, V503, R505, L508, Р509, R551, L552, Т554, I555, L556 и W558.
В одном из вариантов воплощения изобретения m, n и/или р Формулы I представляют собой 0. В варианте воплощения изобретения, когда m, n и р каждый, представляют собой 0, РЕ Формулы I может дополнительно содержать соединяющий пептид GGS между FCS и функциональным доменом III РЕ.
Не желая быть связанными какой-либо конкретной теорией или механизмом, авторы полагают, что РЕ, содержащие последовательность, расщепляемую фурином (FCS), претерпевают протеолитический процессинг внутри клеток-мишеней, активирующий цитотоксическую активность токсина. FCS РЕ согласно изобретению может представлять собой любую подходящую аминокислотную последовательность, расщепляемую фурином, которая расщепляется фурином. Примеры последовательностей, расщепляемых фурином, описаны в работе Duckert et al., Protein Engineering, Design & Selection, 17(1): 107-112 (2004) и международной патентной заявке WO 2009/ 032954, каждая из которых включена в данное описание посредством ссылки. В одном варианте воплощения изобретения FCS включает остатки 274-284 из SEQ ID NO:1 (т.е. RHRQPRGWEQL (SEQ ID NO:8)), где заменой аминокислоты в одном или нескольких В-клеточных эпитопах SEQ ID NO:1 является замена на аланин, глицин, серин или глутамин аминокислотного остатка Е282 из SEQ ID NO:1. Другие подходящие аминокислотные последовательности FCS включают R-X1-X2-R, где X1 представляет собой любую встречающуюся в природе аминокислоту и Х2 представляет собой любую встречающуюся в природе аминокислоту (SEQ ID NO:9), RKKR (SEQ ID NO:10), RRRR (SEQ ID NO:11), RKAR (SEQ ID NO:12), SRVARS (SEQ ID NO:13), TSSRKRRFW (SEQ ID NO:14), ASRRKARSW (SEQ ID NO:15), RRVKKRFW (SEQ ID NO:16), RNWRRDW (SEQ ID NO:17), TRAVRRRSW (SEQ ID NO:18), RQPR (SEQ ID NO:19), RHRQPRGW (SEQ ID NO:20), RHRQPRGWE (SEQ ID NO:21), HRQPRGWEQ (SEQ ID NO:22), RQPRGWE (SEQ ID NO:23), RHRSKRGWEQL (SEQ ID NO:24), RSKR (SEQ ID NO:25), RHRSKRGW (SEQ ID NO:26), HRSKRGWE (SEQ ID NO:27), RSKRGWEQL (SEQ ID NO:28), HRSKRGWEQL (SEQ ID NO:29), RHRSKR (SEQ ID NO:30) и R-X1-X2-R, где X1 представляет собой любую встречающуюся в природе аминокислоту, и Х2 представляет собой аргинин или лизин (SEQ ID NO:4), но не ограничиваются ими.
В одном варианте воплощения изобретения m Формулы I представляет собой 1 и R1 из Формулы I включает остатки 285-293 из SEQ ID NO:1, где замена аминокислот в одном или нескольких В-клеточных эпитопах SEQ ID NO:1 включает замену на аланин, глицин, серин или глутамин аминокислотного остатка Е285 и/или Р290 из SEQ ID NO:1.
В другом варианте воплощения настоящего изобретения п Формулы I равно 1 и R3 Формулы I включает остатки 306-394 из SEQ ID NO:1, где замена аминокислот в одном или нескольких В-клеточных эпитопах SEQ ID NO:1 включает замену на аланин, глицин, серин или глутамин одного или более аминокислотных остатков из R313, N314, Р319, D324, Е327, Е331 и Q332 из SEQ ID NO:1.
В еще одном варианте воплощения настоящего изобретения функциональный домен III РЕ включает остатки 395-613 из SEQ ID NO:1, где замена аминокислоты в одном или нескольких В-клеточных эпитопах SEQ ID NO:1 включает замену на аланин, глицин, серин или глутамин одного или нескольких аминокислотных остатков из D403, D406, R412, R427, Е431, R432, R458, D461, D463, R467, Y481, R490, R505, R513, L516, Е522, R538, Е548, R551, R576, К590, Q592 и L597 из SEQ ID NO:1. В предпочтительном варианте воплощения изобретения функциональный домен III РЕ включает SEQ ID NO:142. В особенно предпочтительном варианте воплощения настоящего изобретения функциональный домен III РЕ включает SEQ ID NO:143.
РЕ согласно изобретению может быть менее иммуногенным, чем РЕ без замен в соответствии с изобретением, если иммунный ответ на РЕ согласно изобретению уменьшен, количественно или качественно, по сравнению с иммунным ответом на незамещенный РЕ. Количественное уменьшение иммуногенности включает уменьшение величины или степени иммунного ответа. Величина или степень иммуногенности может быть измерена на основе любого количества известных параметров, таких как снижение уровня выработки цитокинов (например, антиген-специфической выработки цитокина), уменьшение количества активированных лимфоцитов (например, пролиферации лимфоцитов (например, антиген-специфической пролиферации лимфоцитов)) или привлеченных лимфоцитов и/или уменьшение выработки антител (антиген-специфических антител), и т.д. Качественное снижение иммуногенности охватывает любое изменение в характере иммунного ответа, которое делает иммунный ответ менее эффективным в отношении снижения цитотоксической активности РЕ. Методы измерения иммуногенности известны в данной области. Например, можно измерить иммуногенность, измеряя типы и уровни продуцируемых цитокинов. Альтернативно или дополнительно, можно измерять иммуногенность, измеряя связывание РЕ антителами (например, антителами, которые ранее были выработаны на РЕ) и/или измеряя способность РЕ индуцировать выработку антител при введении млекопитающему (например, людям, мышам, и/или мышам, у которых мышиная иммунная система заменена на иммунную систему человека). Менее иммуногенный РЕ может характеризоваться снижением выработки цитокинов, таких как один или несколько из ИФН-у, ФНО-а и гранзим В, и/или сниженной стимуляцией клеточного иммунного ответа, например, уменьшением пролиферации и активации Т-клеток и/или макрофагов, специфичных для РЕ, по сравнению с результатами, полученными для РЕ без замен. Альтернативно или дополнительно, менее иммуногенный РЕ может характеризоваться увеличением выработки TGF-бета и/или IL-10 по сравнению с результатами, полученными для РЕ без замен. В предпочтительном варианте воплощения сниженную иммуногенность характеризуют одним или более параметров, включающих уменьшение стимуляции Т-клеток, снижение пролиферации Т-клеток, а также уменьшение секреции Т-клетками ИФН-у и/или секреции гранзима В. Альтернативно или дополнительно, менее иммуногенные РЕ могут быть охарактеризованы по уменьшению стимуляции и/или активации В-клеток, специфичных для РЕ, по сравнению с результатами, полученными с РЕ без замен. Например, менее иммуногенные РЕ могут быть охарактеризованы по уменьшению дифференцировки В-клеток в секретирующие антитела плазматические клетки и/или клетки памяти, по сравнению с данными, полученными с РЕ без замен. Снижение иммуногенности может характеризоваться одним или более из параметров, включающих уменьшение стимуляции В-клеток, уменьшение пролиферации В-клеток и уменьшение секреции антител к РЕ. Качественное и количественное уменьшение иммуногенности может происходить одновременно и не являются взаимоисключающими.
Специалист в данной области техники легко поймет, что РЕ согласно изобретению могут быть изменены любым количеством способов, таким образом, что терапевтическая или профилактическая эффективность предлагаемого в изобретении РЕ увеличится посредством модификации. Например, РЕ согласно изобретению могут быть конъюгированы или конденсированы либо напрямую, либо не напрямую через линкер с нацеливающим агентом. В этом отношении вариант воплощения настоящего изобретения обеспечивает химерную молекулу, содержащую (а) нацеливающий агент, конъюгированный или слитый с (б) любым из РЕ согласно изобретению, описанных здесь. Практика конъюгирования веществ, например, РЕ согласно изобретению, с нацеливающим агентом известна в данной области. См., например, Wadwa et al., J. Drug Targeting, 3: 111 (1995), патент США 5087616.
Термин «нацеливающий агент», используемый здесь, относится к любой молекуле или агенту, который специфически распознает и связывается с маркером клеточной поверхности, таким образом, что нацеливающий агент направляет доставку РЕ согласно изобретению к популяции клеток, на поверхности которых экспрессируется рецептор. Нацеливающие агенты включают антитела (например, моноклональные антитела) или их фрагменты, пептиды, гормоны, факторы роста, цитокины и любые другие природные или неприродные лиганды, но не ограничиваются ими.
Термин «антитело», используемый здесь, относится к целому (также известному как «интактное») антителу или его антигенсвязывающим участкам, которые сохраняют способность распознавания антигена и способность к связыванию. Антитело или его антигенсвязывающие участки могут быть природными антителами или их антигенсвязывающими участками, например, антителом или его антигенсвязывающим участком, выделенным и/или очищенным из млекопитающего, например, мыши, кролика, козы, лошади, курицы, хомяка, человека и т.д. Антитело или его антигенсвязывающий участок может быть в мономерной или полимерной форме. Кроме того, антитело или его антигенсвязывающий участок может быть любого уровня аффинности или авидности для маркера клеточной поверхности. Желательно, чтобы антитело или его антигенсвязывающий участок являлся специфическим для маркера клеточной поверхности, чтобы существовала минимальная кросс-реактивность с другими пептидами или белками.
Антитело может быть моноклональным или поликлональным и относиться к любому изотипу, например, IgM, IgG (например, IgG, IgG2, IgG3 или IgG4), IgD, IgA или IgE. Участки, определяющие комплементарность (CDR) антитела, или одноцепочечные вариабельные фрагменты (Fv) антитела к маркеру поверхности клеток-мишеней могут быть пересажены или вставлены в выбранное антитело для придания этому антителу специфичности к маркеру поверхности клеток-мишеней. Например, CDR из антитела против маркера поверхности клетки-мишени могут быть присоединены к каркасной области человеческого антитела с известной трехмерной структурой (см., например, международную патентную заявку WO 1998/ 045322 и WO 1987/ 002671; патенты США 5859205; 5585089; и 4816567, Европейскую патентную заявку 0173494; Jones et al., Nature, 321:522 (1986); Verhoeyen et al., Science, 239: 1534 (1988), Riechmann et al., Nature, 332: 323 (1988); and Winter & Milstein, Nature, 349: 293 (1991)) с образованием антител, которые могут незначительно повысить или вообще не вызывать иммунную реакцию при введении человеку. В предпочтительном варианте воплощения нацеливающий агент представляет собой моноклональное антитело.
Связывающая антиген часть может быть любой частью, которая имеет по крайней мере один сайт связывания антигена, например, вариабельными участками или CDR из интактного антитела. Примеры антигенсвязывающей части антител включают тяжелую цепь, легкую цепь, вариабельные или константные участки тяжелой или легкой цепей, одноцепочечные вариабельные фрагменты (scFv), или Fc, Fab, Fab', Fv, или F(ab)2' фрагменты; однодоменные антитела (см., например, Wesolowski, Med Microbiol Immunol., 198(3): 157-74 (2009); Saerens et al., Curr. Opin. Pharmacol., 8(5):6 00-8 (2008); Harmsen and de Haard, Appl. Microbiol. Biotechnol., 77(1): 13-22 (2007), антитела, стабилизированные спиралью (см., например, Arndt et al., J. Mol. В/о/., 312: 221-228 (2001); триатела; диатела (Европейская патентная заявка 0404097; международная патентная заявка WO 1993/011161, и Hollinger et al., Proc. Natl. Acad. Sci. USA, 90: 6444-6448 (1993)); одноцепочечные антитела («scFvs» см., например, патент США 5888773); антитела, стабилизированные дисульфидными связями («dsFvs», см., например, патенты США 5747654 и 6558672), а также доменные антитела («dAbs», см., например, Holt et al., Trends Biotech, 21(11):484-490 (2003), Ghahroudi et al., FEBS Lett., 414:521-526 (1997), Lauwereys et al., EMBO J 17:3512-3520 (1998), Reiter et al., J. Mol. Biol. 290:685-698 (1999); и Davies and Riechmann, Biotechnology, 13:475-479 (2001)), но не ограничиваются ими.
Способы оценки антитела или его антигенсвязывающей части на способность связываться с любым маркером клеточной поверхности известны в данной области и включают любой анализ связывания антитело-антиген, например, радиоиммуноанализ (RIA), ELISA, Вестерн блот, иммунопреципитацию и анализ конкурентного ингибирования (см., например, Janeway et al., ниже, и заявку США 2002/0197266 А1).
Подходящие способы получения антител известны в данной области. Например, стандартные методы с использованием гибридом описаны, например, Kohler and Milstein, Eur. J. Immunol., 5, 511-519 (1976), Harlow and Lane (eds.), Antibodies: A Laboratory Manual, CSH Press (1988), и С.А. Janeway et al. (eds.), Immunobiology, 5th Ed., Garland Publishing, New York, NY (2001)). С другой стороны, в данной области техники известны другие методы, такие как методы EBV-гибридом (Haskard and Archer, J. Immunol. Methods, 74(2), 361-67 (1984), и Roder et al., Methods Enzymol., 121, 140-67 (1986)), а также фаговые системы экспрессионных векторов (см., например, Huse et al., Science, 246, 1275-81 (1989)). Кроме того, способы получения антител в животных, отличных от человека, описаны, например, в патентах США 5545806, 5569825 и 5714352, и заявке США 2002/0197266 А1.
Фаговый дисплей также может быть использован для создания антител, которые могут быть использованы в химерных молекулах согласно изобретению. Фаговая библиотека, кодирующая антигенсвязывающие вариабельные (V) домены антител, может быть получена с использованием стандартных молекулярно-биологических методов и технологии рекомбинантных ДНК (см., например, Sambrook et al. (eds.), Molecular Cloning, A Laboratory Manual, 3rd Edition, Cold Spring Harbor Laboratory Press, New York (2001)). Фаг, кодирующий вариабельный домен с требуемой специфичностью, выбирают по специфическому связыванию желаемого антигена, и полное или частичное антитело восстанавливают, включая выбранный вариабельный домен. Последовательности нуклеиновых кислот, кодирующие восстановленное антитело, вводят в подходящую клеточную линию, такую как клетка миеломы, используемую для получения гибридом, так что антитела, имеющие характеристики моноклональных антител, секретируются клеткой (см., например, Janeway et al., выше, Huse et al., выше, и патент США 6265150).
В качестве альтернативы антитела могут быть получены с помощью трансгенных мышей, которые трансгенны по конкретным генам тяжелой и легкой цепей иммуноглобулинов. Такие способы известны в данной области и описаны, например, в патентах США 5545806 и 5569825, а также Janeway et al., см. выше.
Альтернативно, антитело, например, гуманизированное антитело или химерное антитело, может быть получено с использованием генной инженерии. Гуманизированные антитела преимущественно обеспечивают более низкий риск побочных эффектов, и могут дольше циркулировать в крови. Способы получения гуманизированных антител известны в данной области техники и подробно описаны в, например, Janeway et al., см. выше, патентах США 5225539, 5585089 и 5693761, европейском патенте 0239400 В1, и патенте Великобритании 2188638. Гуманизированные антитела также могут быть получены с использованием технологии изменения поверхности антитела, описанной, например, в патенте США 5639641 и Pedersen et al., J. Mol. Biol., 235, 959-973 (1994).
Нацеливающий агент может специфически связываться с любым подходящим маркером клеточной поверхности. Выбор конкретного нацеливающего агента и/или маркера клеточной поверхности может быть сделан в зависимости от конкретной клеточной популяции, являющейся мишенью. Маркеры клеточной поверхности известны в данной области (см., например, Mufson et al., Front. Biosci., 11:337-43 (2006); Frankel et al., Clin. Cancer Res., 6:326-334 (2000); and Kreitman et al., AAPS Journal, 8(3): E532-E551 (2006)), и могут быть, например, белками или углеводами. В одном из вариантов воплощения изобретения нацеливающий агент представляет собой лиганд, который специфически связывается с рецептором на поверхности клетки. Примеры лигандов включают фактор роста эндотелия сосудов (VEGF), Fas, родственный ФНО индуцирующий апоптоз лиганд (TRAIL), цитокины (например, IL-2, IL-15, IL-4, IL-13), лимфокины, гормоны и факторы роста (например, трансформирующий фактор роста (TGFa), фактор роста нейронов, эпидермальный фактор роста), но не ограничиваются ими.
Маркер клеточной поверхности может быть, например, раковым антигеном. Термин «раковый антиген», используемый здесь, относится к любой молекуле (например, белку, пептиду, липиду, углеводу и др.) исключительно или преимущественно экспрессирующейся или чрезмерно экспрессирующейся опухолевой клеткой или раковой клеткой, таким образом, что антиген ассоциирован с опухолью или раком. Раковый антиген может дополнительно экспрессироваться нормальной, неопухолевой или нераковой клеткой. Тем не менее, в таких случаях экспрессия ракового антигена на нормальной, неопухолевой или нераковой клетке не так выражена, как экспрессия опухолевыми или раковыми клетками. В связи с этим, опухолевые или раковые клетки могут чрезмерно экспрессировать антиген или экспрессируют антиген на значительно более высоком уровне по сравнению с экспрессией антигена на нормальной, неопухолевой или нераковой клетке. Кроме того, раковый антиген дополнительно может экспрессироваться клетками в другом состоянии развития или созревания. Например, раковый антиген может дополнительно экспрессироваться клетками эмбриональной или фетальной стадии, которые обычно не встречаются во взрослом хозяине. Альтернативно, раковый антиген может дополнительно экспрессироваться стволовыми клетками или клетками-предшественниками, которые обычно не встречаются во взрослом хозяине.
Раковый антиген может быть антигеном, экспрессируемым любой клеткой любого рака или опухоли, в том числе злокачественных новообразований и опухолей, описанных здесь. Раковый антиген может быть раковым антигеном только из одного типа рака или опухоли, таким, что раковый антиген ассоциирован или характерен только для одного типа рака или опухоли. Альтернативно, раковый антиген может быть раковым антигеном (например, может быть характерным) для более чем одного типа рака или опухоли. Например, раковый антиген может экспрессироваться как клетками рака груди, так и раковыми клетками рака предстательной железы, и не экспрессироваться совсем всеми нормальными, неопухолевыми или нераковыми клетками.
Примерны раковых антигенов, с которыми может специфически связываться нацеливающий агент, включают муцин 1 (MUC1), антиген меланомы (MAGE), преимущественно экспрессируемый меланомой антиген (PRAME), раково-эмбриональный антиген (СЕА), простатический специфический антиген (PSA), простатический специфический мембранный антиген (PSMA), рецептор гранулоцитарно-макрофагального колониестимулирующего фактора (GM-CSFR), CD56, человеческий рецептор эпидермального фактора роста 2 (HER2/neu) (также известный как ЕгЬВ-2), CD5, CD7, опухолевый антиген тирозиназа, белок, связанный с тирозиназой (TRP)1, TRP2, NY-ESO-1, теломеразу и р53, но не ограничиваются ими. В предпочтительном варианте воплощения поверхностный маркер, с которым специфически связывается нацеливающий агент, выбирают из группы, состоящей из белков кластера дифференцировки (CD) 19, CD21, CD22, CD25, CD30, CD33, CD79b, рецептора трансферрина, рецептора EGF, мезотелина, кадгерина и Lewis Y. Мезотелин экспрессируется, например, клетками рака яичников, мезотелиомы, немелкоклеточного рака легкого, аденокарциномы легких, рака фаллопиевых труб, рака головы и шеи, рака шейки матки, рака поджелудочной железы. CD22 экспрессируется, например, при волосатоклеточном лейкозе, хроническом лимфоцитарном лейкозе (CLL), пролимфоцитарной лейкемии (PLL), неходжкинской лимфоме, лимфоме малых лимфоцитов (SLL) и остром лимфолейкозе (ALL). CD25 экспрессируется, например, при лейкозах и лимфомах, включая волосатоклеточный лейкоз и лимфому Ходжкина. Lewis Y антиген экспрессируется, например, при раке мочевого пузыря, раке молочной железы, раке яичников, раке толстой кишки, раке пищевода, раке желудка, раке легкого и раке поджелудочной железы. CD33 экспрессируется, например, при остром миелоидном лейкозе (AML), хроническом миеломоноцитарном лейкозе (CML) и миелопролиферативных заболеваниях.
В одном варианте воплощения изобретения нацеливающий агент представляет собой антитело, которое специфически связывается с раковым антигеном. Примеры антител, которые специфически связываются с раковыми антигенами включают антитела к рецептору трансферрина (например, НВ21 и его варианты), антитела к CD22 (например, RFB4 и его варианты), антитела к CD25 (например, анти-Тас и их варианты), антитела к мезотелину (например, SS1, Morab-009, CC, HN1, HN2, MN, MB и их варианты) и антитела к антигену Lewis Y (например, В3 и его варианты), но не ограничиваются ими. В связи с этим нацеливающий агент может представлять собой антитело, выбранное из группы, состоящей из ВЗ, RFB4, SS, SS1, MN, MB, HN1, HN2, НВ21 и Morab-009,и их антигенсвязывающих частей. Дальнейшие примеры нацеливающих агентов, пригодных для использования в химерных молекулах согласно изобретению, раскрыты, например, в патентах США 5242824 (к рецептору трансферрина); 5846535 (анти-CD25); 5889157 (анти-Lewis Y); 5981726 (анти-Lewis Y); 5990296 (анти-Lewis Y); 7081518 (анти-мезотелин); 7355012 (анти-CD22 и анти-CD25); 7368110 (анти-мезотелин); 7470775 (анти-CD30); 7521054 (анти-С025) и 7541034 (анти-CD22); заявка на патент США 2007/0189962 (анти-CD22); Frankel et al., Clin. Cancer Res., 6: 326-334 (2000), и Kreitman et al., AAPS Journal, 8(3): E532-E551 (2006), каждый из которых включен в данное описание путем ссылки. В другом варианте воплощения изобретения нацеливающий агент может включать нацеливающий агентиммунотоксинов, известных в данной области. Примеры иммунотоксинов включают LMB-2 (Anti-Tac(Fv)-PE38), BL22 и НА22 (RFB4(dsFv)-PE38), SS1P (SS1 (dsFv)-PE38), HB21-PE40 и их варианты, но не ограничиваются ими. В предпочтительном варианте воплощения нацеливающий агент является антигенсвязывающим участком НА22. НА22 включает Fv фрагмент антитела к CD22, связанный дисульфидными связями, конъюгированный с РЕ38. НА22 и их варианты описаны в международных патентных заявках WO 2003/027135 и WO 2009/032954, которые включены здесь посредством ссылки.
В одном варианте воплощения изобретения химерная молекула включает линкер. Термин «линкер», используемый здесь, относится к любому агенту или молекуле, которая соединяет РЕ согласно изобретению с нацеливающим агентом. Специалисту в данной области техники очевидно, что участки РЕ согласно настоящему изобретению, которые не являются необходимыми для функций РЕ согласно изобретению, являются идеальными участками для прикрепления линкера и/или нацеливающего агента, при условии, что линкер и/или нацеливающий агент, будучи прикрепленными к РЕ согласно изобретению, (а) не мешает функционированию РЕ согласно изобретению, т.е. цитотоксической активности, ингибированию роста клеток-мишеней или лечению или профилактике рака. Линкер может быть способен образовывать ковалентные связи как с РЕ, так и с нацеливающим агентом. Подходящие линкеры известны в данной области и включают линкеры с простой или разветвленной углеродной цепью, гетероциклические углеродные линкеры и пептидные линкеры, но не ограничиваются ими. Если РЕ и нацеливающий агент представляют собой полипептиды, линкер может быть соединен с аминокислотами через боковые группы (например, через дисульфидную связь с цистеином). Предпочтительно присоединять линкер к альфа-углеродному атому амино-и карбоксильных групп концевых аминокислот.
В объем настоящего изобретения включены функциональные части РЕ согласно изобретению и химерные молекулы согласно изобретению, описанные здесь. Термин «функциональная часть» при использовании в отношении РЕ или химерной молекулы, относится к любой части или фрагменту РЕ или химерной молекулы согласно изобретению, где часть или фрагмент сохраняет биологическую активность РЕ или химерной молекулы, частью которого она является (исходных РЕ или химерной молекулы). Функциональные части охватывают, например, те части РЕ или химерной молекулы, которые сохраняют способность специфически связывать и уничтожать или подавлять рост клеток-мишеней или способность лечения или профилактики рака, примерно в той же степени, в той же степени, или более высокой степени, как исходные РЕ или химерная молекула. Относительно исходного РЕ или химерной молекулы функциональная часть может включать, например, около 10% или более, около 25% или более, около 30% или более, около 50% или более, около 68% или более, около 80% или более, около 90% или более, или около 95% или более исходного РЕ или химерной молекулы.
Функциональная часть может содержать дополнительные аминокислоты на амино-или карбоксильном конце, или на обоих концах, где такие дополнительные аминокислоты, которые не встречаются в аминокислотной последовательности исходных РЕ или химерной молекулы. Желательно, чтобы дополнительные аминокислоты не мешали биологической функции функциональной части, например, специфически связывать и уничтожать или ингибировать рост клеток-мишеней, способности лечить или предотвращать рак и т.д. Более желательно, дополнительные аминокислоты повышают биологическую активность по сравнению с биологической активностью исходных РЕ или химерной молекулы.
В объем настоящего изобретения включены функциональные варианты предлагаемых в изобретении РЕ и химерных молекул, описанных здесь. Термин «функциональный вариант», используемый здесь, относится к РЕ или химерной молекуле, имеющей существенную или значительную идентичность последовательности или близость к исходным РЕ или химерной молекуле, где функциональный вариант сохраняет биологическую активность РЕ или химерной молекулы, вариантом которых он является. Функциональные варианты включают, например, те варианты РЕ или химерной молекулы, описанные здесь (исходных РЕ или химерной молекулы), которые сохраняют способность специфически связывать и уничтожать или ингибировать рост клеток-мишеней примерно в той же степени, в той же степени, или более высокой степени, чем исходные РЕ или химерная молекула. Относительно исходного РЕ или химерной молекулы функциональный вариант может иметь, например, около 30% или более, около 50% или более, около 75% или более, около 80% или более, около 90% или более, около 95% или более, около 96% или более, около 97% или более, около 98% или более, или около 99% или более идентичности аминокислотной последовательности с исходными РЕ или химерной молекулой.
Функциональный вариант может, например, содержать аминокислотную последовательность исходных РЕ или химерной молекулы по меньшей мере с одной консервативной аминокислотной заменой. Консервативные аминокислотные замены известны в данной области и включают аминокислотные замены, в которых одну аминокислоту, имеющую определенные химические и/или физические свойства, заменяют другой аминокислотой, которая имеет те же химические или физические свойства. Например, консервативная замена на аминокислоту может быть заменой кислой аминокислоты на другую кислую аминокислоту (например, Asp или Glu), заменой аминокислоты с неполярной боковой цепью на другую аминокислоту с неполярной боковой цепью (например, Ala, Gly, Val, Ile, Leu, Met, Phe, Pro, Trp, Val и т.д.), основной аминокислоты на другой основную аминокислоту (Lys, Arg и т.д.), аминокислоты с полярной боковой цепью на другую аминокислоту с полярной боковой цепью (Asn, Cys, Gln, Ser, Thr, Туг и т.п.) и т.д.
Альтернативно или дополнительно, функциональные варианты могут включать аминокислотную последовательность исходных РЕ или химерной молекулы по меньшей мере с одной неконсервативной аминокислотной заменой. В этом случае предпочтительно, чтобы неконсервативная аминокислотная замена не мешала или не ингибировала биологическую активность функционального варианта. Предпочтительно, чтобы неконсервативная аминокислотная замена усиливала биологическую активность функционального варианта, так, чтобы биологическая активность функционального варианта увеличивалась по сравнению с исходными РЕ или химерной молекулой.
РЕ или химерные молекулы согласно изобретению могут состоять преимущественно из указанной последовательности аминокислотных остатков или последовательностей, описанных здесь, так что другие компоненты функционального варианта, например, другие аминокислоты, существенно не изменяют биологическую активность функционального варианта.
РЕ или химерные молекулы согласно изобретению (в том числе функциональные части и функциональные варианты) согласно изобретению могут содержать синтетические аминокислоты вместо одной или нескольких встречающихся в природе аминокислот. Такие синтетические аминокислоты хорошо известны в данной области и включают, например, аминоциклогексан карбоновую кислоту, норлейцин, α-амино-н-декановую кислоту, гомосерин, S-ацетиламинометил-цистеин, транс-3-и транс-4-гидроксипролин, 4-аминофенилаланин, 4-нитрофенилаланин, 4-хлорфенилаланин, 4-карбоксифенилаланин, β-фенилсерин, β-гидроксифенилаланин, фенилглицин, а-нафтилаланин, циклогексилаланин, циклогексилглицин, индолин-2-карбоновую кислоту, 1,2,3,4-тетрагидроизохинолин-3-карбоновую кислоту, аминомалоновую кислоту, моноамид аминомалоновой кислоты, N'-бензил-N'-метил-лизин, N',N'-дибензил-лизин, 6-гидроксилизин, орнитин, α-аминоциклопентан карбоновую кислоту, α-аминоциклогексан карбоновую кислоту, α-аминоциклогептан карбоновую кислоту, α-(2-амино-2-норборнан)-карбоновую кислоту, α,γ-диаминобутановую кислоту, α,β-диаминопропионовую кислоту, гомофенилаланин и α-трет-бутил глицин.
РЕ или химерные молекулы изобретения (в том числе функциональные части и функциональные варианты) могут быть гликозилированы, амидированы, карбоксилированы, фосфорилированы, этерифицированы, N-ацилированы, циклизованы, например, дисульфидной связью, или превращены в кислотно-аддитивную соль и/или необязательно димеризованы или полимеризованы, или конъюгированы.
Вариант воплощения изобретения обеспечивает способ получения РЕ согласно изобретению, включающий (а) рекомбинантную экспрессию РЕ и (б) очистку РЕ. РЕ и химерные молекулы согласно изобретению (в том числе функциональные части и функциональные варианты) могут быть получены с использованием способов получения белков и полипептидов, известных в данной области. Подходящие способы синтеза полипептидов и белков de novo описаны в справочниках, таких как Chan et al., Fmoc Solid Phase Peptide Synthesis, Oxford University Press, Oxford, United Kingdom, 2005; Peptide and Protein Drug Analysis, ed. Reid, R., Marcel Dekker, Inc., 2000; Epitope Mapping, ed. Westwood et al., Oxford University Press, Oxford, United Kingdom, 2000 и патенте США 5449752. Кроме того, РЕ и химерные молекулы согласно изобретению могут быть рекомбинантно экспрессированы с использованием нуклеиновых кислот, описанных здесь, используя стандартные рекомбинантные методы. См., например, Sambrook et al., Molecular Cloning: A Laboratory Manual, 3rd ed., Cold Spring Harbor Press, Cold Spring Harbor, NY 2001; и Ausubel et al., Current Protocols in Molecular Biology, Greene Publishing Associates and John Wiley & Sons, NY, 1994.
Способ дополнительно включает очистку РЕ. После экспрессии РЕ согласно изобретению могут быть очищены в соответствии с методиками очистки, известными в данной области. Примеры способов очистки включают осаждение сульфатом аммония, аффинную колоночную хроматография, колоночную хроматографию или способы, описанные, например, в R. Scopes, Protein Purification, Springer-Verlag, NY (1982), но не ограничиваются ими.
Еще один вариант воплощения настоящего изобретения обеспечивает способ получения химерной молекулы согласно изобретению, включающий (а) рекомбинантную экспрессию химерной молекулы и (б) очистку химерной молекулы. Химерная молекула может быть рекомбинантно экспрессирована и очищена, как описано здесь по отношению к другим аспектам изобретения. В одном варианте воплощения изобретения рекомбинантная экспрессия химерной молекулы включает вставку нуклеотидной последовательности, кодирующей нацеливающий агент, и нуклеотидной последовательности, кодирующей РЕ, в вектор. Способ может включать вставку нуклеотидной последовательности, кодирующей нацеливающий агент, и нуклеотидной последовательности, кодирующей РЕ, с сохранением рамки считывания таким образом, что она кодирует один непрерывный полипептид, включающий участок функционального нацеливающего агента и участок функционального РЕ. В одном варианте воплощения настоящего изобретения способ включает лигирование нуклеотидной последовательности, кодирующей РЕ, с нуклеотидной последовательностью, кодирующей нацеливающий агент, таким образом, чтобы при экспрессии РЕ находился на карбоксильном конце нацеливающего агента. В альтернативном варианте воплощения способ включает лигирование нуклеотидной последовательности, кодирующей РЕ, с нуклеотидной последовательностью, кодирующей нацеливающий агент, таким образом, чтобы при экспрессии РЕ располагался на амино-конце нацеливающего агента.
Еще один вариант воплощения изобретения предусматривает способ получения химерной молекулы согласно изобретению, включающий (а) рекомбинантную экспрессию РЕ согласно изобретению, (б) очистку РЕ, и (с) ковалентное связывание нацеливающего агента с очищенным РЕ. РЕ по изобретению может быть рекомбинантно экспрессирован, как описано здесь по отношению к другим аспектам изобретения. Способ дополнительно включает ковалентное связывание нацеливающего агента с очищенным РЕ. Способ присоединения РЕ к нацеливающему агенту может меняться в зависимости от химической структуры нацеливающего агента. Например, способ может включать взаимодействие любой одной или нескольких из различных функциональных групп, например, группы карбоновой кислоты (СООН), свободной аминогруппы (-NH2) или сульфгидрильной группы (-SH), присутствующих на РЕ, с соответствующей функциональной группой на нацеливающем агенте, тем самым образовывая ковалентную связь между РЕ и нацеливающим агентом. Альтернативно или дополнительно, способ может включать дериватизацию нацеливающего агента или РЕ, чтобы сделать доступными или прикрепить дополнительные реакционноспособные функциональные группы. Дериватизация может также включать присоединение одного или более линкеров к нацеливающему агенту или РЕ.
В другом варианте воплощения настоящего изобретения РЕ и химерные молекулы согласно изобретению могут быть получены с использованием не рекомбинантных способов. Например, РЕ и химерные молекулы согласно изобретению, описанные здесь (в том числе функциональные части и функциональные варианты), могут быть коммерчески синтезированы компаниями, такими как Synpep (Dublin, CA), Peptide Technologies Corp. (Gaithersburg, MD), и Multiple Peptide Systems (San Diego, CA). В этом отношении РЕ и химерные молекулы согласно изобретению могут быть синтетическими, рекомбинантными, выделенными и/или очищенными.
Может быть желательным при некоторых обстоятельствах освободить РЕ от нацеливающего агента, когда химерная молекула достигла одной или нескольких клеток-мишеней. В связи с этим химерные молекулы согласно изобретению могут содержать расщепляемый линкер. Линкер может расщепляться с помощью любых подходящих средств, например, ферментативно. Например, когда клетка-мишень представляет собой раковую (например, опухолевую) клетку, химерные молекулы могут включать линкер, расщепляемый в условиях, присутствующих в месте опухоли (например, под воздействием ассоциированных с опухолью ферментов или кислого рН).
Вариант воплощения изобретения обеспечивает нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую любой из РЕ или химерных молекул согласно изобретению, описанных здесь. Термин «нуклеиновая кислота», используемый здесь, включает «полинуклеотид», «олигонуклеотид» и «молекулу нуклеиновой кислоты» и обычно означает полимер ДНК или РНК, который может быть одноцепочечным или двухцепочечным, которые могут быть синтезированы или получены (например, выделены и/или очищены) из природных источников, которые могут содержать природные, неприродные или измененные нуклеотиды, и которые могут содержать природные, неприродные или измененные межнуклеотидные связи, такие как фосфороамидатная или фосфоротиоатная связи вместо фосфодиэфирной, находящейся между нуклеотидами немодифицированного олигонуклеотида. Как правило, предпочтительно, чтобы нуклеиновая кислота не содержала никаких вставок, делеций, инверсий и/или замен. Тем не менее, в некоторых случаях могут быть пригодны описанные здесь нуклеиновые кислоты, содержащие одну или более вставку, делецию, инверсию и/или замену.
Предпочтительно, нуклеиновые кислоты согласно данному изобретению являются рекомбинантными. Используемый здесь термин «рекомбинантный» относится к (I) молекулам, которые создаются вне живых клеток путем соединения природных или синтетических сегментов нуклеиновой кислоты, или (II) молекулам, которые являются результатом репликации молекул, описанных в (I) выше. Для целей данного изобретения, репликация может быть in vitro репликацией или in vivo репликацией.
Нуклеиновые кислоты могут быть построены на основе химического синтеза и/или ферментативных реакций лигирования с использованием процедур, известных в данной области. См., например, Sambrook et al., выше, и Ausubel и др., выше. Например, нуклеиновая кислота может быть химически синтезирована с использованием природных нуклеотидов или различным образом модифицированных нуклеотидов, направленных на увеличение биологической стабильности молекулы или на увеличение физической стабильности дуплекса, образованного при гибридизации (например, фосфоротиоатные связи и замены нуклеотидов на акридин). Примеры модифицированных нуклеотидов, которые можно использовать для получения нуклеиновых кислот включают 5-фторурацил, 5-бромурацил, 5-хлорурацил, 5-йодурацил, гипоксантин, ксантин, 4-ацетилцитозин, 5-(карбоксигидроксиметил)урацил, 5-карбоксиметиламинометил-2-тиоуридин, 5-карбоксиметиламинометилурацил, дигидроурацил, бета-D-галактозилквеозин, инозин, N6-изопентениладенин, 1-метил гуанин, 1-метилинозин, 2,2-диметилгуанин, 2-метиладенин, 2-метилгуанин, 3-метилцитозин, 5-метилцитозин, N6-замещенный аденин, 7-метилгуанин, 5-метиламинометилурацил, 5-метоксиаминометил-2-тиоурацил, бета-D-маннозилквеуозин, 5'-метоксикарбоксиметилурацил, 5-метоксиурацил, 2-метилтио-N6-изопентиниладенин, урацил-5-оксиуксусную кислоту (V), вибутоксозин, псевдоурацил, квеозин, 2-тиоцитозин, 5-метил-2-тиоурацил, 2-тиоурацил, 4-тиоурацил, 5-метилурацил, метиловый эфир урацил-5-оксиуксусной кислоты, 3-(3-амино-3-N-2-карбоксипропил)урацил и 2,6-диаминопурин, но не ограничиваются ими. В качестве альтернативы одна или более нуклеиновых кислот согласно изобретению могут быть приобретены у компаний, таких как Macromolecular Resources (Fort Collins, CO) и Synthegen (Houston, TX).
Настоящее изобретение также относится к нуклеиновой кислоте, включающей нуклеотидную последовательность, комплементарную нуклеотидной последовательности любой из нуклеиновых кислот, описанных здесь, или нуклеотидную последовательность, которая гибридизуется в жестких условиях с нуклеотидной последовательностью любой из нуклеиновых кислот, описанных здесь.
Нуклеотидная последовательность, которая гибридизуется в жестких условиях, предпочтительно гибридизуется в условиях высокой жесткости. «Условия высокой жесткости» означают, что нуклеотидная последовательность специфически гибридизуется с последовательностью-мишенью (нуклеотидной последовательностью любой из нуклеиновых кислот, описанных здесь) в количестве, обнаруживаемом лучше, чем при неспецифической гибридизации. Условия высокой жесткости включают условия, которые отличают полинуклеотид с точной комплементарной последовательностью, или полинуклеотид, содержащий только несколько непоследовательных несоответствий, от случайной последовательности, которая случайным образом имеет только несколько небольших областей (например, 3-10 оснований), совпадающих с нуклеотидной последовательностью. Такие небольшие комплементарные области легче плавятся, чем участки, комплементарные полностью на протяжении 14-17 или большего количества оснований, и гибридизация в условиях высокой жесткости делает их легко отличимыми. Условия относительно высокой жесткости включают, например, низкое содержанием соли и/или высокие температуры, например, около 0,02-0,1 М NaCl или его эквивалента при температуре около 50-70°С. Такие условия высокой жесткости позволяют гибрид изоваться последовательностям с несколькими несоответствиями или вообще без несоответствий между нуклеотидной последовательностью и матричной цепью или цепью-мишенью, и особенно подходят для определения экспрессии любого из РЕ или химерных молекул согласно изобретению. Как правило, понятно, что условия могут быть сделаны более жесткими путем добавления возрастающих количеств формамида.
Изобретение также обеспечивает нуклеиновую кислоту, содержащую нуклеотидную последовательность, которая, на примерно 70% или более, например, примерно 80% или более, примерно 90% или более, примерно 91% или более, примерно 92% или более, примерно 93% или более, примерно 94% или более, примерно 95% или более, примерно 96% или более, примерно 97% или более, примерно 98% или более, или около 99% или более идентична любой из нуклеиновых кислот, описанных здесь.
Нуклеиновые кислоты согласно изобретению могут быть включены в рекомбинантный экспрессионный вектор. В связи с этим данное изобретение обеспечивает рекомбинантные экспрессионные векторы, содержащие любую из нуклеиновых кислот согласно изобретению. Для целей данного изобретения термин «рекомбинантный экспрессионный вектор» означает генетически модифицированный олигонуклеотид или полинуклеотидную конструкцию, которая делает возможной экспрессию мРНК, белка, полипептида или пептида в клетке-хозяине, где конструкция включает нуклеотидную последовательность, кодирующую мРНК, белок, полипептид или пептид, а вектор контактирует с клеткой в условиях, достаточных, чтобы получить мРНК, белок, полипептид или пептид, экспрессируемый в клетке. Векторы согласно изобретению в виде одного целого не встречаются в природе. Однако части векторов могут встречаться в природе. Предлагаемые в изобретении рекомбинантные экспрессионные векторы могут содержать любой тип нуклеотидов, включая, но не ограничиваясь ими, ДНК и РНК, которые могут быть одноцепочечными или двухцепочечными, которые могут быть синтезированы или получены частично из природных источников, и которые могут содержать природные, неприродные или измененные нуклеотиды. Рекомбинантные экспрессионные векторы могут содержать встречающиеся в природе, не встречающиеся в природе межнуклеотидные связи, или оба типа связей. Предпочтительно, не встречающиеся в природе или измененные нуклеотиды или межнуклеотидные связи не мешают транскрипции или репликации вектора.
Рекомбинантный экспрессионный вектор согласно настоящему изобретению может быть любым подходящим рекомбинантным экспрессионным вектором, и может быть использован для трансформации или трансфекции любой подходящей клетки-хозяина. Подходящие векторы включают векторы, которые предназначены для воспроизведения и размножения или для экспрессии, или и для того, и другого, такие как плазмиды и вирусы. Вектор может быть выбран из группы, состоящей из серии pUC (Fermentas Life Sciences), серии pBluescript (Stratagene, LaJolla, CA), серии рЕТ (Novagen, Madison, WI), серии pGEX (Pharmacia Biotech, Uppsala, Sweden) и серии pEX (Clontech, PaloAlto, CA). Бактериофаговые векторы, такие какЛСТЮ, AGT11, AZapll (Stratagene), AEMBL4 и ANM1149, также могут быть использованы. Примеры экспрессионных векторов для растений включают рВ101, рВ1101.2, рВ1101.3, рВ1121 и pBIN19 (Clontech). Примеры экспрессионных векторов для животных включают pEUK-CI, pMAM, и pMAMneo (Clontech). Предпочтительно, рекомбинантный экспрессионный вектор представляет собой вирусный вектор, например, ретровирусный вектор.
Рекомбинантные экспрессионные векторы согласно изобретению могут быть получены с использованием стандартных методик рекомбинантной ДНК, описанных в, например, Sambrook et al., см. выше, и Ausubel et al., см. выше. Конструкции векторов для экспрессии, которые могут быть кольцевыми или линейными, могут быть получены так, чтобы содержать функциональную систему репликации в прокариотической или эукариотической клетке-хозяине. Система репликации может быть получена, например, из ColEI, плазмиды 2μ, λ, SV40, вируса бычьей папилломы и тому подобное.
Желательно, чтобы рекомбинантный экспрессионный вектор включал регуляторные последовательности, такие как кодоны инициации и терминации транскрипции и трансляции, которые являются специфическими для типа хозяина (например, бактерии, грибов, растений или животных), в которого вектор должен быть введен, и с учетом того, выполнен ли вектор на основе ДНК или РНК.
Рекомбинантный экспрессионный вектор может включать один или несколько маркерных генов, которые позволяют отбор трансформированных или трансфицированных хозяев. Гены-маркеры включают устойчивость к биоцидам, например, устойчивость к антибиотикам, тяжелым металлам и т.д., комплементацию ауксотрофного хозяина с обеспечением прототрофии и тому подобное. Подходящие маркерные гены для экспрессионных векторов согласно изобретению включают, например, гены устойчивости к неомицину/0418, гены устойчивости к гигромицину, гены устойчивости к гистидинолу, гены устойчивости к тетрациклину, а также гены устойчивости к ампициллину.
Рекомбинантный экспрессионный вектор может содержать нативный или ненативный промотор, функционально связанный с нуклеотидной последовательностью, кодирующей РЕ или химерную молекулу согласно изобретению (в том числе функциональные части и функциональные варианты), или нуклеотидную последовательность, которая комплементарна или с которой гибридизуется нуклеотидная последовательность, кодирующая РЕ или химерную молекулу. Выбор промоторов, например, сильного, слабого, индуцибельного, тканеспецифичного и специфичного для определенной стадии, находится в пределах обычной квалификации специалиста в данной области. Кроме того, объединение нуклеотидной последовательности с промотором также находится в пределах обычной квалификации специалиста в данной области техники. Промотор может быть не вирусным промотором или вирусным промотором, например, цитомегаловирусным (CMV) промотором, промотором SV40, промотором RSV или промотором, обнаруживаемый в длинном концевом повторе мышиного вируса стволовых клеток.
Рекомбинантные экспрессионные векторы согласно изобретению могут быть разработаны для временной экспрессии, для стабильной экспрессии или для обоих видов экспрессии. Кроме того, рекомбинантные экспрессионные векторы могут быть разработаны для конститутивной экспрессии или индуцибельной экспрессии.
Еще один вариант воплощения настоящего изобретения дополнительно обеспечивает клетку-хозяина, содержащую любой из рекомбинантных экспрессионных векторов, описанных здесь. Используемый здесь термин «клетка-хозяин» относится к любому типу клеток, который может содержать рекомбинантный экспрессионный вектор согласно изобретению. Клетка-хозяин может быть эукариотической клеткой, например, растений, животных, грибов или водорослей, или может быть прокариотической клеткой, например, бактерией, или простейшим. Клетка-хозяин может быть культивируемой клеткой, прикрепленной или растущей в суспензии клеткой, то есть клеткой, которая растет во взвешенном состоянии. Для целей получения рекомбинантного РЕ или химерной молекулы согласно изобретению, клетка-хозяин предпочтительно является прокариотической клеткой, например, клеткой E. coli.
Изобретением, кроме того, обеспечивается популяция клеток, содержащих по меньшей мере одну клетку-хозяин, описанную здесь. Популяция клеток может быть гетерогенной популяцией, содержащей клетку-хозяина, содержащую любой из рекомбинантных экспрессионных векторов, описанных здесь, в дополнение к по меньшей мере одной другой клетке, например, клетке-хозяину, которая не включает какой-либо из рекомбинантных экспрессионных векторов. С другой стороны, популяция клеток может быть по существу гомогенной популяцией, в которой популяция включает в себя в основном клетки (например, состоит в основном из клеток-хозяев), содержащие рекомбинантный экспрессионный вектор. Популяция также может быть клональной популяцией клеток, в которой все клетки популяции являются клонами одной клетки-хозяина, содержащей рекомбинантный экспрессионный вектор, так что все клетки популяции содержат рекомбинантный экспрессионный вектор. В одном варианте воплощения изобретения популяция клеток представляет собой популяцию клонов клеток-хозяев, содержащих рекомбинантный экспрессионный вектор, как описано здесь.
РЕ, химерные молекулы (в том числе функциональные части и функциональные варианты), нуклеиновые кислоты, рекомбинантные экспрессионные векторы, клетки-хозяева (включая их популяции) и популяции клеток согласно изобретению могут быть выделены и/или очищены. Термин «выделенный» при использовании здесь означает выделение из его природного окружения. Термин «очищенный», используемый здесь, означает увеличенную чистоту, где «чистота» является относительным термином, и не должна быть обязательно истолкована как абсолютная чистота. Например, чистота может быть около 50% или более, около 60% или более, около 70% или более, около 80% или более, около 90% или более или приблизительно 100%. Предпочтительно чистота около 90% или более (например, от около 90% до около 95%) и более предпочтительно около 98% или более (например, от около 98% до около 99%).
РЕ по изобретению, химерные молекулы (в том числе функциональные части и функциональные варианты), нуклеиновые кислоты, рекомбинантные экспрессионные векторы, клетки-хозяева (в том числе их популяция) и популяции клеток согласно изобретению, которые все вместе называются далее «материал РЕ согласно изобретению», могут быть введены в композицию, такую как фармацевтическая композиция. В связи с этим настоящее изобретение относится к фармацевтической композиции, содержащей любые из РЕ, химерных молекул (в том числе функциональных частей и функциональных вариантов), нуклеиновых кислот, рекомбинантных экспрессионных векторов, клеток-хозяев (включая их популяции) и популяций клеток, и фармацевтически приемлемый носитель. Фармацевтическая композиция согласно изобретению, содержащая любой из предлагаемых материалов РЕ согласно изобретению, может содержать более чем один материал РЕ согласно изобретению, например, полипептид и нуклеиновую кислоту, или два или более различных РЕ. Кроме того, фармацевтическая композиция может содержать материалы РЕ в сочетании с одним или несколькими другими фармацевтически активными агентами или препаратами, такими как химиотерапевтические агенты, например аспарагиназа, бусульфан, карбоплатин, цисплатин, даунорубицин, доксорубицин, фторурацил, гемцитабин, гидроксимочевина, метотрексат, паклитаксел, ритуксимаб, винбластин, винкристин и т.п.
Предпочтительно носитель представляет собой фармацевтически приемлемый носитель. Что касается фармацевтических композиций, носитель может быть любым из обычно используемых носителей, и ограничивается только физико-химическими свойствами, такими как растворимость и отсутствие реактивности с активным соединением (соединениями), и путем введения. Описанные здесь фармацевтически приемлемые носители, например, формообразующие, добавки, наполнители и разбавители, хорошо известны специалистам в данной области и легко доступны. Предпочтительно, чтобы фармацевтически приемлемый носитель был химически инертен к активному агенту (агентам) и не имел вредных побочных эффектов или токсичности при условиях использования.
Выбор носителя будет частично определяться конкретным материалом РЕ согласно изобретению, а также конкретным способом, используемым для введения материала РЕ согласно изобретению. Соответственно, существует множество подходящих препаратов для фармацевтической композиции согласно изобретению. Следующие препараты для парентерального (например, подкожного, внутривенного, внутриартериального, внутримышечного, внутрикожного, внутрибрюшинного и интратекального), орального и аэрозольного введения являются иллюстративными и не должны быть истолкованы как ограничение изобретения. Для введения материалов РЕ согласно изобретению может быть использовано более одного пути введения, а в некоторых случаях конкретный путь может обеспечить более быструю и более эффективную реакцию, чем другой путь.
Композиции для местного применения хорошо известны специалистам в данной области. Такие композиции особенно пригодны в контексте настоящего изобретения для нанесения на кожу.
Композиции, пригодные для перорального введения, могут включать: (а) жидкие растворы, такие как эффективное количество материала РЕ согласно изобретению, растворенное в разбавителе, таком как вода, солевой раствор или апельсиновый сок, (б) капсулы, порошок для приготовления раствора, таблетки, таблетки для рассасывания и пастилки, каждая из которых включает заранее определенное количество активного ингредиента, в виде твердых веществ или гранул, (в) порошки, (г) суспензии в соответствующей жидкости, и (д) подходящие эмульсии. Жидкие композиции могут включать разбавители, такие как вода и спирты, например, этанол, бензиловый спирт и полиэтиленовые спирты, с добавлением фармацевтически приемлемых поверхностно-активных веществ или без них. Капсульные формы могут иметь обычную жесткую или мягкую желатиновую оболочку, содержать, например, поверхностно-активные вещества, смазывающие вещества и инертные наполнители, такие как лактоза, сахароза, фосфат кальция и кукурузный крахмал. Таблетки могут включать один или более наполнителей из лактозы, сахарозы, маннита, кукурузного крахмала, картофельного крахмала, альгиновой кислоты, микрокристаллической целлюлозы, камеди, желатина, гуаровой камеди, коллоидного диоксида кремния, кроскармеллозы натрия, талька, стеарата магния, стеарата кальция, стеарата цинка, стеариновой кислоты, и другие наполнители, красители, разбавители, буферные агенты, дезинтегрирующие агенты, увлажняющие агенты, консерванты, ароматизирующие агенты и другие фармакологически совместимые наполнители. Таблетки для рассасывания могут содержать материалы РЕ согласно изобретению в ароматизирующей добавке, обычно сахарозе и камеди или трагаканте, в то время как пастилки содержат материалы РЕ согласно изобретению в инертной основе, такой как желатин и глицерин, или сахароза и гуммиарабик, эмульсии, геле и тому подобное, дополнительно содержащей такие наполнители, которые известны в данной области техники.
Материалы РЕ по изобретению, отдельно или в комбинации с другими подходящими компонентами, могут быть превращены в аэрозольные препараты для введения посредством ингаляции. Эти аэрозольные композиции могут быть помещены в приемлемые для сжатия газы-вытеснители, такие как дихлордифторметан, пропан, азот и тому подобное. Аэрозольные композиции также могут быть приготовлены в виде фармацевтических препаратов не под давлением, например, для распылителей или пульверизаторов. Такие композиции для распыления также можно использовать для распыления на слизистую оболочку.
Препараты, пригодные для парентерального введения, включают водные и неводные изотонические стерильные растворы для инъекций, которые могут содержать антиоксиданты, буферы, бактериостатические и растворенные вещества, которые делают препарат изотоническим крови предполагаемого реципиента, а также водные и неводные стерильные суспензии, которые могут содержать суспендирующие агенты, солюбилизаторы, загустители, стабилизаторы и консерванты. Материалы РЕ согласно изобретению можно вводить в физиологически приемлемом разбавителе, в фармацевтическом носителе, например, стерильной жидкости или смеси жидкостей, включая воду, физиологический раствор, водный раствор декстрозы и растворы родственных Сахаров, спирт, такой как этанол или гексадециловый спирт, гликоли, такие как пропиленгликоль или полиэтиленгликоль, диметилсульфоксид, глицерин, кетали, такие как 2,2-диметил-1,3-диоксолан-4-метанол, простые эфиры, поли(этиленгликоль) 400, масла, жирные кислоты, сложные эфиры жирных кислот или глицериды, или ацетилированные глицериды жирных кислот, с добавлением или без добавления фармацевтически приемлемых поверхностно-активных веществ, таких как мыло или детергент, суспендирующего агента, такого как пектин, карбомер, метилцеллюлоза, гидроксипропилметилцеллюлоза, карбоксиметилцеллюлоза, или эмульгирующих агентов или других фармацевтических адъювантов.
Масла, которые могут быть использованы в парентеральных препаратах, включают выделенные из нефти, животные, растительные или синтетические масла. Конкретные примеры масел включают арахисовое, соевое, кунжутное, хлопковое, кукурузное, оливковое масло, вазелин и минеральное масло. Подходящие жирные кислоты для использования в парентеральных препаратах включают олеиновую кислоту, стеариновую кислоту и изостеариновую кислоту. Этилолеат и изопропилмиристат являются примерами подходящих сложных эфиров жирных кислот.
Подходящие мыла для использования в парентеральных препаратах включают соли жирных кислот и щелочных металлов, аммония и триэтаноламина, и подходящие детергенты включают: (а) катионные детергенты, такие как, например, диметил диалкиламмоний галогениды и алкилпиридиний галогениды, (б) анионные детергенты, такие как, например, алкил, арил и олефин сульфонаты, алкил, олефин, моноглицерид сульфаты и сульфосукцинаты и сульфаты и сульфосукцинаты простых эфиров, (в) неионные детергенты, такие как, например, оксиды жирных аминов, алканоламиды жирных кислот и сополимеры полиоксиэтиленполипропилена, (г) амфотерные детергенты, такие как, например, алкил-β-аминопропионаты и четвертичные аммониевые соли 2-алкил-имидазолина, и (д) их смеси.
Парентеральные препараты обычно содержат от около 0,5 масс.% до около 25 масс.% материала РЕ согласно изобретению в растворе. Могут быть использованы консерванты и буферы. Чтобы свести к минимуму или устранить раздражение в месте инъекции, такие композиции могут содержать один или несколько неионных поверхностно-активных веществ, имеющих гидрофильно-липофильный баланс (HLB) от около 12 до около 17. Количество поверхностно-активного вещества в таких препаратах, как правило, находится в интервале от около 5 масс.% до около 15 масс.%. Подходящие поверхностно-активные вещества включают полиэтиленгликоль, сложные эфиры сорбитана и жирных кислот, такие как моноолеат сорбитана, и высокомолекулярные аддукты этиленоксида и гидрофобного основания, образованные путем конденсации пропиленоксида с пропиленгликолем. Парентеральные препараты могут быть представлены в виде единичных дозированных форм или в многодозовых герметизированных контейнерах, например ампулах и флаконах, и могут храниться в высушенном замораживанием (лиофилизированном) состоянии, требующем только добавления стерильного жидкого вспомогательного вещества, например, воды для инъекций, непосредственно перед использованием. Растворы и суспензии для инъекций могут быть также получены из стерильных порошков, гранул и таблеток ранее описанного типа. Требования к эффективным фармацевтическим носителям для парентеральных композиций хорошо известны специалистам в данной области техники (см., например, Pharmaceutics and Pharmacy Practice, J.B. Lippincott Company, Philadelphia, PA, Banker and Chalmers, eds., pages 238-250 (1982), и ASHP Handbook on Injectable Drugs, Toissel, 4th ed., pages 622-630 (1986)).
Специалистам в данной области техники следует понимать, что, в дополнение к описанным выше фармацевтическим композициям, материалы РЕ согласно изобретению могут быть приготовлены в виде комплексов включения, таких как комплексы включения циклодекстрина или липосомы.
Для целей настоящего изобретения количество или доза вводимого материала РЕ согласно изобретению должно быть достаточным, чтобы осуществить желаемую реакцию, например, терапевтический или профилактический ответ, у млекопитающего в течение разумного периода времени. Например, доза материала РЕ согласно изобретению должна быть достаточной для ингибирования роста клетки-мишени или лечения или профилактики рака в течение периода времени от около 2 часов или дольше, например, от 12 до 24 или более часов, с момента введения. В некоторых вариантах воплощения период времени может быть даже больше. Доза будет зависеть от эффективности конкретного материала РЕ согласно изобретению и состояния млекопитающего (например, человека), а также от веса тела млекопитающего (например, человека), подлежащего лечению.
Многие анализы для определения введенной дозы известны в данной области. Вводимая доза может быть определена in vitro (например, на клеточной культуре) или in vivo (например, при исследованиях на животных). Например, вводимая доза может быть определена путем определения IC50 (дозы, которая достигает полумаксимального ингибирования симптомов), LD50 (дозы, летальной для 50% популяции), ED50 (дозы, терапевтически эффективной для 50% популяции) и терапевтического индекса в культуре клеток и/или исследованиях на животных. Терапевтический индекс представляет собой отношение LD50 к ED50 (т.е. LD50/ED50).
Доза материала РЕ согласно изобретению также будет определяться наличием, природой и степенью любых нежелательных побочных эффектов, которые могут сопровождать введение конкретного материала РЕ согласно изобретению. Как правило, лечащий врач определяет дозировку материала РЕ согласно изобретению, используемую для лечения каждого индивидуального пациента, принимая во внимание целый ряд факторов, таких как возраст, вес тела, общее состояние здоровья, диету, пол, вводимый материал РЕ согласно изобретению, путь введения и тяжесть состояния, которое лечат. В качестве примера и без намерения ограничивать изобретение, доза материала РЕ согласно изобретению может составлять от около 0,001 до около 1000 мг/кг массы тела субъекта, подлежащего лечению/день, от около 0,01 до около 10 мг/кг массы тела/день, от около 0,01 мг до около 1 мг/кг массы тела/день, от около 1 до около до 1000 мг/кг массы тела/день, от около 5 до около 500 мг/кг массы тела/день, около от 10 до около 250 мг/кг массы тела/день, от около 25 до около 150 мг/кг массы тела/день, или около 10 мг/кг массы тела/день.
Кроме того, материалы РЕ согласно изобретению могут быть изготовлены в форме депо, так что способ, которым материал РЕ согласно изобретению поступает в организм, в который его вводят, контролируется в отношении времени и места введения (см. например, патент США 4450150). Материалы РЕ согласно изобретению в форме депо могут быть, например, имплантируемыми композициями, содержащими материалы РЕ согласно изобретению и пористый или непористый материал, такой как полимер, где материалы РЕ согласно изобретению инкапсулируются или диффундируют по всему материалу и/или непористый материал деградирует. Депо затем имплантируют в нужном месте в тело, и материалы РЕ согласно изобретению освобождаются из имплантата с заданной скоростью.
Предлагаемые в изобретении материалы РЕ можно проанализировать на цитотоксичность с помощью анализов, известных в данной области техники. Примеры анализов на цитотоксичность включают WST-анализ, который измеряет пролиферацию клеток с использованием соли тетразолия WST-1 (реагенты и наборы, доступные от Roche Applied Sciences), как описано в международной патентной заявке WO 2011/032022.
Следует понимать, что фармацевтические композиции, РЕ, химерные молекулы, нуклеиновые кислоты, рекомбинантные экспрессионные векторы, клетки-хозяева или популяции клеток согласно изобретению могут быть использованы в способах лечения или профилактики рака. Не желая быть связанными какой-либо конкретной теорией или механизмом, авторы полагают, что предлагаемые в изобретении РЕ уничтожают клетки или ингибируют рост клеток посредством ингибирования синтеза белка в эукариотических клетках, например, путем инактивации ADP-рибозилированием фактора элонгации 2 (EF-2). Б Не желая быть связанными какой-либо конкретной теорией или механизмом, авторы полагают, что химерные молекулы согласно изобретению распознают и специфически связываются с маркерами клеточной поверхности, тем самым доставляя цитотоксический РЕ в популяцию клеток, экспрессирующих маркер клеточной поверхности с минимальной перекрестной реактивностью с клетками, которые не экспрессируют маркер клеточной поверхности, или вообще без такой реактивности. Таким образом, цитотоксичность РЕ может быть направлена на уничтожение или ингибирование роста конкретной популяции клеток, например, раковых клеток. В связи с этим, настоящее изобретение относится к способу лечения или профилактики рака у млекопитающего, включающему введение млекопитающему любых РЕ, химерных молекул, нуклеиновых кислот, рекомбинантных экспрессионных векторов, клеток-хозяев, популяций клеток или фармацевтических композиций, описанных здесь, в количестве, эффективном для лечения или профилактики рака у млекопитающего.
Термины «лечить» и «предотвращать», а также слова, являющиеся производными от них, используемые здесь, не обязательно подразумевают 100% или полное излечение или профилактику. Напротив, есть различные степени лечения или профилактики, которые любой специалист в данной области признает как имеющие потенциальное преимущество или терапевтический эффект.В этом отношении изобретенные способы могут обеспечить любую степень лечения или профилактики рака у млекопитающего. Кроме того, лечение или профилактика, обеспечиваемые способами согласно настоящему изобретению, могут включать лечение или профилактику одного или нескольких состояний или симптомов заболевания, например, рака, подвергаемого лечению или профилактике. Кроме того, для целей данного изобретения термин «предупреждение» может включать задержку начала болезни или симптомов, или состояния.
Для целей предлагаемых способов, в которых вводят клетки-хозяева или популяции клеток, клетки могут быть клетками, которые аллогенны или аутологичны хозяину. Предпочтительно, клетки аутологичны хозяину.
В предлагаемых способах рак может быть любым раком, в том числе любым видом рака, выбранным из рака надпочечников, саркомы (например, синовиальной саркомы, остеогенной саркомы, лейомиосаркомы матки, ангиосаркомы, фибросаркомы, рабдомиосаркомы, липосаркомы, миксомы, рабдомиомы, фибромы, липомы, и тератомы), лимфомы (например, лимфомы из малых лимфоцитов, лимфомы Ходжкина и неходжкинской лимфомы), гепатоцеллюлярной карциномы, глиомы, рака головы (например, плоскоклеточного рака), рака шеи (например, плоскоклеточного рака), острого лимфоцитарного рака, лейкозов (например, волосатоклеточного лейкоза, миелоидного лейкоза (острого и хронического), лимфатического лейкоза (острого и хронического), пролимфоцитарной лейкемия (PLL), миеломоноцитарного лейкоза (острого и хронического) и лимфоцитарного лейкоза (острого и хронического)), рака кости (остеогенной саркомы, фибросаркомы, злокачественной фиброзной гистиоцитомы, хондросаркомы, саркомы Юинга, злокачественной лимфомы (ретикулярно-клеточной саркомы), множественной миеломы, гигантоклеточной саркомы, хордомы, остеохондромы (костно-хрящевого экзостоза), доброкачественной хондромы, хондробластомы, хондромиксоидной фибромы, остеоид-остеомы и гигантоклеточных опухолей), рака мозга (астроцитомы, медуллобластомы, глиомы, эпендимомы, герминомы (пинеаломы), мультиформной глиобластомы, олигодендроглиомы, шванномы и ретинобластомы), рака фаллопиевых труб, рака молочной железы, рака ануса, анального канала или аноректального рака, рака глаза, рака внутрипеченочного желчного протока, рака суставов, рака шеи, рака желчного пузыря или плевры, рака носа, носовой полости или среднего уха, рака полости рта, рака вульвы (например, плоскоклеточного рака, интраэпителиальной карциномы, аденокарциномы и фибросаркомы), миелопролиферативных заболеваний (например, хронического миелоидного рака), рака толстой кишки (например, карциномы толстой кишки), рака пищевода (например, плоскоклеточного рака, аденокарциномы, лейомиосаркомы и лимфомы), рака шейки матки (рака шейки матки и преинвазивной дисплазии шейки матки), рака желудка, желудочно-кишечного карциноида, рака гортаноглотки, рака гортани, рака печени (например, гепатоцеллюлярной карциномы, холангиокарциномы, гепатобластомы, ангиосаркомы, гепатоцеллюлярной аденомы и гемангиомы), рака легких (например, бронхогенной карциномы (плоскоклеточного рака, недифференцированного мелкоклеточного рака, недифференцированного крупноклеточного рака и аденокарциномы), альвеолярной (бронхиолярной) карциномы, бронхиальной аденомы, хондроматозной гамартомы, немелкоклеточного рака легкого, мелкоклеточного рака легких и аденокарциномы легких), злокачественных мезотелиом, рака кожи (например, меланомы, базально-клеточной карциномы, плоскоклеточного рака, саркомы Капоши, невусов, диспластических невусов, липомы, ангиомы, дерматофибромы и келоидных рубцов), множественной миеломы, рака носоглотки, рака яичников (например, карциномы яичников (серозной, муцинозной цистаденокарциномы, эндометриоидной карциномы, светлоклеточной аденокарциномы), гранулезотекаклеточной опухоли, опухоли Сертоли-Лейдига, дисгерминомы и злокачественной тератомы), рака поджелудочной железы (например, аденокарциномы протоков, инсулиномы, глюкагономы, гастриномы, карциноидных опухолей и ВИПомы), рака брюшины, сальника, брыжейки, рака глотки, рака предстательной железы (например, аденокарциномы и саркомы), рака прямой кишки, рака почки (например, аденокарциномы, опухоли Вильмса (нефробластомы) и рака почки), мелкоклеточного рака кишки (аденокарциномы, лимфомы, карциноидных опухолей, саркомы Капоши, лейомиомы, гемангиомы, липомы, нейрофибромы и фибромы), опухолей мягких тканей, рака желудка (например, карциномы, лимфомы и лейомиосаркомы), рака яичка (например, семиномы, тератомы, эмбриональной карциномы, тератокарциномы, хориокарциномы, саркомы, опухоли клеток Лейдига, фибромы, фиброаденомы, аденоматоидной опухоли и липомы), рака матки (например, рака эндометрия), рака щитовидной железы и уротелиального рака (например, плоскоклеточной карциномы, переходно-клеточного рака, аденокарциномы, рака мочеточника и рака мочевого пузыря).
Используемый здесь термин «млекопитающее» относится к любому млекопитающему, в том числе млекопитающим отряда Rodentia, таким как мыши и хомяки, и млекопитающим отряда Logomorpha, таким как кролики, но не ограничиваясь ими. Предпочтительно, чтобы млекопитающие были из отряда Carnivora, в том числе семейства Felines (кошки) и Canines (собаки). Наиболее предпочтительно, чтобы млекопитающие были из отряда Artiodactyla, в том числе Bovines (коровы) и Swines (свиньи) или отряда Perssodactyla, в том числе Equines (лошади). Наиболее предпочтительно, чтобы млекопитающие были из отряда приматов, Ceboids или Simoids (обезьяны) или отряда Anthropoids (человек и человекообразнык обезьяны). Особенно предпочтительным млекопитающим является человек.
Настоящее изобретение также относится к способу ингибирования роста клетки-мишени, включающему контактирование клетки с РЕ, выбранным из любых РЕ, химерной молекулы, нуклеиновой кислоты, рекомбинантного экспрессионного вектора, клетки-хозяина, популяции клеток или фармацевтической композиции, описанных в данном описании, в количестве, эффективном для ингибирования роста клетки-мишени. Рост клетки-мишени может быть ингибирован в любой степени, например, на около 10% или более, около 15% или более, около 20% или более, около 25% или более, около 30% или более, около 35% или более, около 40% или более, около 45% или более, около 50% или более, около 55% или более, около 60% или более, около 65% или более, около 70% или более, около 75% или более, около 80% или более, около 85% или более, около 90% или более, около 95% или более или на около 100%. Клетка-мишень может быть предоставлена в биологическом образце. Биологический образец может быть получен из млекопитающего любым подходящим способом и из любого подходящего источника. Биологический образец может, например, быть получен путем сбора крови, лейкофереза и/или биопсии опухоли или аутопсии. Стадия контактирования может происходить in vitro или in vivo по отношению к млекопитающему. Предпочтительно, чтобы контактирование происходило in vitro.
В одном варианте воплощения изобретения клетка-мишень представляет собой раковую клетку. Клетка-мишень может быть раковой клеткой любого из описанных здесь видов рака. В одном из вариантов воплощения изобретения мишень может экспрессировать маркер клеточной поверхности. Маркер клеточной поверхности может быть любым маркером клеточной поверхности, описанным здесь по отношению к другим аспектам изобретения. Маркер клеточной поверхности может быть выбран, например, из группы, состоящей из CD19, CD21, CD22, CD25, CD30, CD33, CD79b, рецептора трансферрина, рецептора EGF, мезотелина, кадгерина и Lewis Y.
Следующие примеры дополнительно иллюстрируют изобретение, но не должны истолковываться как каким-либо образом ограничивающие его объем
Пример 1
Этот пример демонстрирует частоту аллелей HLA класса 2 в группе доноров, не получавших предварительного лечения, по сравнению с частотой мировой популяции.
Мононуклеарные клетки периферической крови (РВМС) выделяли из образцов 65 здоровых доноров, полученных из банка крови NIH, и образцов 50 пациентов, проходящих лечение иммунотоксином в NCI. РВМС выделяли из лейкоцитарных пленок при центрифугировании в плотности фиколла. Гаплотипы РВМС HLA класса I и класса II пациентов и здоровых доноров определяли с помощью набора для тканевого типирования на основе PCR-SSP/SSO. РВМС затем замораживали в инактивированной нагреванием сыворотке крови человека АВ и стандартной среде для замораживания и хранили в жидком азоте до использования.
Группу из 65 здоровых доноров изучали, чтобы предоставить информацию о количестве и частоте аллотипов HLA DR, экспрессируемых в мировой популяции. Анализ аллотипов, экспрессированных в группе доноров, не получавших лечение, сравнивали с мировой популяцией (Middleton, D. et al., NewAllele Frequency Database: www.allelefrequencies.net, Tissue Antigens, 61(5): 403-7(2003)), и анализ показал, что была разумная репрезентативность основных аллелей HLA класса II DRB1 в группе доноров, не получавших предварительного лечения. Статистический анализ показал корреляцию между мировой популяцией и группой доноров с R2=0,52. Фигура 1 сравнивает частоту различных типов аллелей HLA класса 2 у мировой популяции и когорты наивных доноров. В Таблице 1 приведены гаплотипы HLA класса II DRB1 популяции доноров.
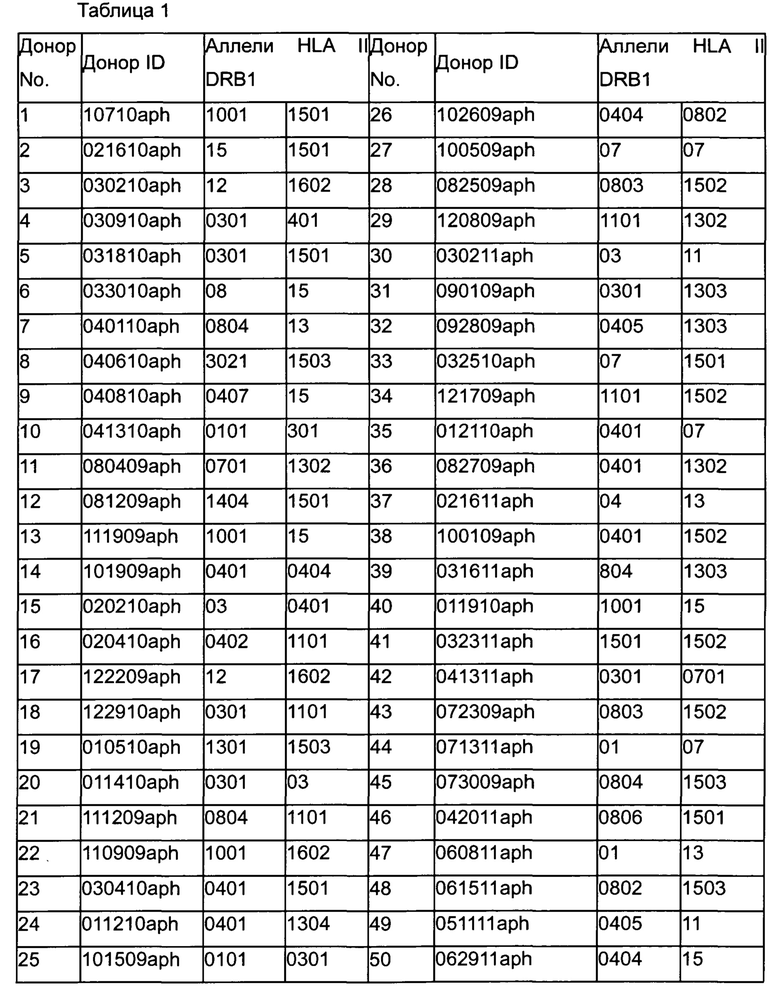
Пример 2
Этот пример демонстрирует получение библиотеки пептидов, которые будут использоваться, чтобы стимулировать Т-клетки.
Чтобы определить иммуногенность фрагмента токсина, разработали библиотеку из 111 пептидов, охватывающую всю последовательность РЕ38. Каждый пептид имел размер 15 аминокислот и перекрывался на 12 аминокислот, за исключением пептидов SEQ ID NO:31 и 32, которые перекрывались на 11 аминокислот. Пептиды синтезировали с >95% чистотой по данным высокопроизводительной жидкостной хроматографии (ВЭЖХ) (American Peptide А.Ш., Sunnyvale, CA). Лиофилизированные синтетические пептиды растворяли в диметилсульфоксиде (ДМСО) с получением 10 мкМ растворов.
Пептиды сгруппировали в 22 пула, как показано в Таблице 2. В Таблице 2 приведены последовательности, SEQ ID NO и номера пулов пептидов, используемых для картирования эпитопов.
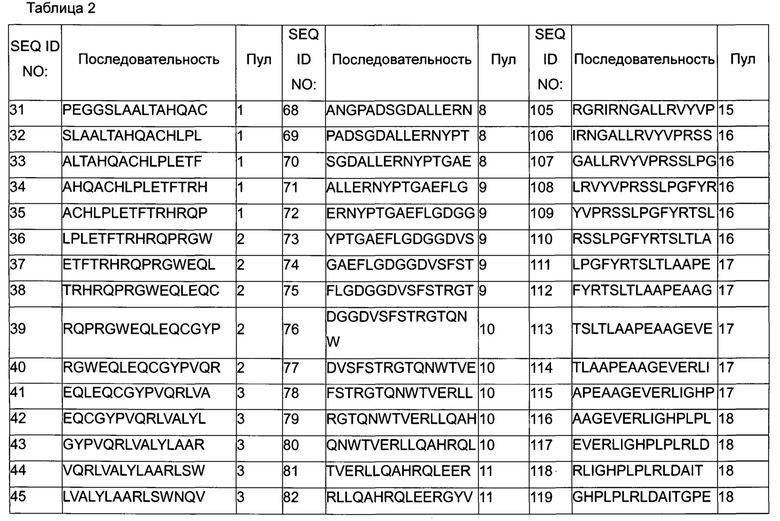

Пример 3
В этом примере показано, что размножение Т-клеток доноров, не получавших предварительного лечения, in vitro улучшает чувствительность и уровень ответной реакции Т-клеток на пептидные пулы.
Для того, чтобы имитировать иммунный ответ, который происходит у пациентов после лечения, и преодолеть низкую чувствительность и невосприимчивость образцов доноров, не получавших предварительного лечения, в краткосрочном анализе, выполнили стадию 17-дневного размножения in vitro, чтобы размножить конкретные популяции Т-клеток (Oseroff et al., J. Immunol., 185(2): 943-55 (2010)). Клетки инкубировали в течение 14-17 дней после стимуляции иммунотоксином. Для стимуляции использовали иммунотоксин LMB9 или ВЗ (dsFv)-PE38, направленный на LeY (Brinkmann et al., Proc. Natl. Acad. Sci. USA, 90(16): 7538-42 (1993)), антиген, отсутствующий на человеческих иммунных клетках. В день 14-17 клетки собирали и анализировали с помощью анализа ELISpot на интерлейкин 2 (IL)-2 с использованием 22 пептидных пулов из Таблицы 2. Добавление стадии размножения in vitro улучшало чувствительность и уровень ответа анализа ELISpot (например, наблюдали больше образующих пятна клеток (SFC)). Стадия размножения in vitro позволила обнаружить CD4-специфичный ответ доноров-добровольцев, а также уменьшить количество клеток, используемых для каждого анализа.
Типичные данные приведены на Фигурах 2А и 2В. На Фигуре 2А показан подсчет лунок IL-2 ELISpot, указывающих на ответ Т-клеток наивного донора 031810aph на исходную стимуляцию LMB9 (1,6 мкг/мл) с последующей рестимуляцией только средой (без пептидов) (М) или пептидными пулами 3, 16, или 22 после 17 дней размножения in vitro. На Фигуре 2 В показан подсчет лунок IL-2 ELISpot, указывающих на ответ Т-клеток наивного донора 031810aph на исходную стимуляцию LMB9 (1,6 мкг/мл) с последующей рестимуляцией только средой (без пептидов) (М) или пептидными пулами 3, 16, или 22 без размножения in vitro.
Не желая быть связанными какой-либо конкретной теорией или механизмом, авторы полагают, что этот подход имитирует иммунный ответ, потому что, как и in vivo, весь рекомбинантный иммунотоксин (RIT) был поглощен антигенпрезентирующими клетками (АРС), процессирован и представлен АРС в течение первых нескольких дней культивирования. Конкретные Т-клетки, которые ответили на представленные естественным образом пептиды, сохранились и размножились с помощью IL-2. Эпитопы, которые эти Т-клетки распознали после 17 дней размножения, были процессированными и представленными естественным образом пептидами.
Пример 4
Этот пример демонстрирует, что LVALYLAARLSW (SEQ ID NO:3) стимулирует Т-клеточный ответ для большинства доноров.
После первоначальной стимуляции иммунотоксином и после 14 дней размножения половину клеток собирали и подвергали воздействию пептидных пулов для первоначального скрининга пулов с IL-2 ELISpot. На 17-й день остальные клетки использовали для повторного скрининга пулов, и для наблюдаемых положительных пулов проводили более детальный скриниг. На 14 день исследования у 28 доноров наблюдали различные модели реагирования. Некоторые скрининги доноров показали ответ на единственный пул (например, доноры 1, 4, 7 и 8), а для других доноров (например, доноры 15, 16 и 23) выявили ответы на 3-4 пула. Так как пептиды перекрываются, было также перекрытие между пулами, как, например, в случаях, когда наблюдали ответы на последовательные пулы (например, реакция на пулы 15 и 16 для доноров 15 и 18). Порог был определен как 85 SFC/(1×106). Порог был определен на уровне этого значения, поскольку для значений ниже 85 SFC/(1×106) ни один из ответов в день 14 не был воспроизводимым в день 17 или при повторении размножение in vitro. Ответы более 85 SFC/(1×106) воспроизводились.
Четыре донора (доноры 25-28) не дали ответа (выше определенного порога) на любой пул и считались не отвечающими. Из начальных 28 доноров, которые были полностью проскринированы, 24 доноров дали ответ по меньшей мере на один пул. Различные доноры имели разные максимальные уровни реагирования. Некоторые доноры (например, доноры 2, 7 и 10) имели высокий максимальный отклик (более 1100 SFC для пула 3), в то время как другие доноры (например, доноры 3 и 9) имели ответ 250-320 SFC на один пул.
Число скринируемых доноров увеличили до 50. Количество образующих пятна клеток, наблюдаемых у всех 50 доноров, подсчитывали для каждого из 22 пептидных пулов. Результаты показаны в Таблице 3 и на Фигуре 3. Анализ всех 50 доноров показывает, что пул 3 (во II домене РЕ) вызвал большинство ответов (Таблица 3; Фигура 3). Результаты показывают, что пул 3 был иммунодоминантным пулом, стимулирующим ответы у многих доноров с различными HLA.
Подробный скрининг пула 3 для обнаружения иммунодоминантного участка (участков) показал, что для большинства доноров пептиды SEQ ID NO:44 и 45 были ответственны за ответ в пуле 3 (Фигура 4). Фигура 4 показывает, что пептиды SEQ ID NO:44 и 45 содержат общую область (LVALYLAARLSW) (SEQ ID NO:3) для стимуляции Т-клеток у большинства пациентов, несмотря на разный статус HLA.
Пример 5
Этот пример демонстрирует, что замены L294A, L297A, Y298A, L299A или R302A в LVALYLAARLSW (SEQ ID NO:3) снижают иммуногенность LVALYLAARLSW (SEQ ID NO:3).
Пептиды SEQ ID NO:44 и 45 (из пула 3) имеют 12 одинаковых аминокислот.Этот общий участок, LVALYLAARLSW (SEQ ID NO:3), соответствует положениям аминокислотных остатков 294-305 из SEQ ID NO:1 и включает как сайт связывания МНС, так и сайт связывания с Т-клеточным рецептором. Каждую аминокислоту из LVALYLAARLSW (SEQ ID NO:3) заменяли аланином. Образцы от 9 наивных доноров и 2 пациентов стимулировали LMB9 в течение 17 дней и анализировали с использованием IL-2 ELISpot, как описано в Примере 3. В Таблице 4 приведены данные из скрининга 11 образцов против замещенных пептидов. Для 10 из 11 образцов, одна или более замен снижали ответ до 7% от ответа на пептид дикого типа (WT). Для всех доноров, кроме одного, замещение L297A уменьшило реакцию до 7% или меньше. Y298A уменьшила ответ в 9 из 11 образцов, и R302A уменьшила ответ в 7 из 11 образцов. Таблица 4 показывает реакцию ответа в процентах от реакции, полученной с пептидом дикого типа (WT).
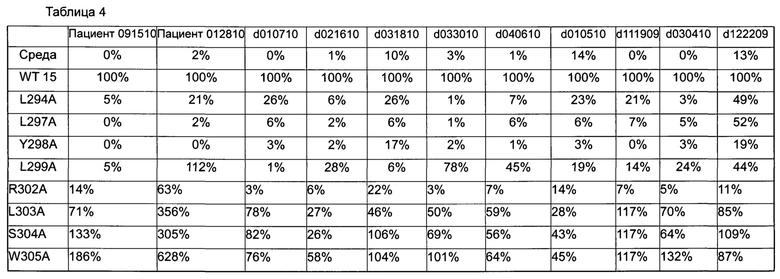
Не ограничиваясь конкретной теорией или механизмом, авторы полагают, что уменьшенный ответ при изменении одной аминокислоты может быть отнесен к нарушению связывания пептида с желобком (желобками) молекулы HLA или неспособности Т-клеточного рецептора узнать измененный пептид. В любом случае, замещенные пептиды L279A и Y298A вызвали сниженный ответ и, следовательно, можно считать, что они обеспечивают уменьшенную иммуногенность.
Пример 6
Этот пример демонстрирует, что замена R302A не создает никаких новых Т-клеточных эпитопов и обеспечивает пониженный ответ Т-клеток.
Для получения RIT R302 НА22 и L297A НА22 использовали сайт-направленный мутагенез, как описано ранее (Pastan et al., Methods Mol. Biol., 248: 503-18 (2004)). Цитотоксическую активность на клетках СА46 сравнивали с НА22 дикого типа (Фигура 5). На Фигуре 5 показано, что конструкции НА22 с заменами L297A или R302A были цитотоксическими. IC50 НА22, НА22 L297A и НА22 R302A составила 1,1, 1,8, и 1,8 нг/мл, соответственно.
РВМС от доноров 010710 и 111909 культивировали в течение 14 дней с любым из WT НА22 или с HA22-R302A и анализировали на Т-клеточный ответ при повторной стимуляции без пептида, пептидом дикого типа LVALYLAARLSWNQV (SEQ ID NO:45) (WT15) или LVALYLAAALSWNQV (SEQ ID NO:145) (R302A). Для обоих доноров 010710 (Фигура 6А) и 111909 (Фигура 6Б) никаких новых эпитопов не наблюдали, и Т-клеточный ответ на замещенные пептиды уменьшился.
Пример 7
В этом примере показано, что удаление части домена II, содержащей иммунодоминантный эпитоп, уменьшает ответ Т-клеток на пептиды РЕ38.
Как показано в Примере 4, домен II включает иммунодоминантный и смешанный эпитоп (включены в SEQ ID NO:44 и 45 из пула 3). PE-LR (устойчивый к деградации лизосомами) (также известный как LR или LR RIT) включает делецию домена II кроме последовательности, расщепляемой фурином RHRQPRGWEQL (SEQ ID NO:8), которая присутствует в SEQ ID NO:37 и 38. Таким образом, LR RIT включает аминокислотные остатки 274-284 и 395-613 из SEQ ID NO:1.
Ответ донорских Т-клеток, которые стимулировали НА22 (содержащим РЕ38), на 22 пептидных пула из Таблицы 2 сравнивали с ответом донорских Т-клеток, которые стимулировали PE-LR (у которого полностью отсутствует иммунодоминантный эпитоп).
В день 0 Т-клетки от трех доноров (донор 031510, донор 021610 или донор 101509) высевали в 6-луночные планшеты и стимулировали 10 мкг/мл либо НА22, либо PE-LR. В день 14 клетки собирали и помещали в планшет для IL-2 ELISpot и инкубировали с пептидами одного из 22 пулов из Таблицы 2 в четырех повторностях. Контроль включал клетки, выращенные с цефтазидимом (CEFT), клетки без стимуляции антигеном в день 0 и без стимуляции антигеном в день 14 («М линии»), и клетки со стимуляцией LMB9 в день 0 и без стимуляции антигеном вдень 14 («без пептида»).
Результаты приведены на Фигурах 7А (донор 031510), 7Б (донор 021610) и 7 В (донор 101509). Т-клетки от всех трех доноров продемонстрировали реакцию при стимуляции НА22 (содержащим РЕ38) и повторной стимуляции пептидами пула 3. Для клеток, которые стимулировали PE-LR, никакого ответа не наблюдали.
Пример 8
Этот пример демонстрирует идентификацию Т-клеточных эпитопов в домене III РЕ.
После первоначальной стимуляции иммунотоксином и 14 дней размножения Т-клетки от 20 доноров собирали и подвергали воздействию пептидных пулов для начального скрининга пептидных пулов с использованием IL-2 ELISpot, как описано в Примере 4. На 17-й день остальные клетки использовали для повторного скрининга и проводили подробный скрининг пептидов с SEQ ID NO:102-111. Результаты показаны на Фигуре 8. Пептиды, которые стимулировали образование IL-2 Т-клетками каждого из доноров 1-20, выделены серым цветом на Фигуре 8.
Пример 9
Этот пример показывает, что замена на аланин 1493, R494, N495, G496, L498, L499, R500, V501 или Y502 снижает Т-клеточный ответ, измеренный по уровню продукции IL-2, по сравнению с ответом на пептид дикого типа (WT).
Каждую аминокислоту, отличную от аланина, в пептиде дикого типа IRNGALLRVYVPRSS (SEQ ID NO:106), заменяли аланином. Образцы от доноров, не получавших предварительного лечения, стимулировали LMB9 в течение 17 дней и анализировали с использованием IL-2 ELISpot, как описано в Примере 3. В Таблице 5 приведены данные скрининга шести образцов против замещенных пептидов.
Как показано в Таблице 5, для всех шести образцов, одна или более замен уменьшали ответ до 10-30% относительно ответа на WT пептид. Для пяти из шести образцов одна или более замен уменьшали ответ до менее чем 10% от ответа на WT. Замены 1493А, R494A, N495A, G496A, L498A, L499A, R500A, V501A или Y502A снижали ответ Т-клеток на 70% и более по сравнению с ответом на WT по меньшей мере в трех из шести образцов. Таблица 5 показывает ответную реакцию в процентах от ответа на пептид WT.
Пример 10
В этом примере показано, что SEQ ID NO:38, 39, 44, 45, 81, 82, 86-88, 97, 98, 105-108 и 123-125 содержат Т-клеточные эпитопы.
Для определения иммунодоминантного участка (участков) выполняли подробный скрининг всех положительных пулов для 50 доноров. Для каждого донора считали количество SFC и рассчитывали относительную долю каждого пептида от общего количества SFC, которую нормализовали в соответствии с формулой (I) ниже, где D означает донора, и x представляет собой одного из 50 доноров, Р означает пептид, у представляет собой один из пептидов SEQ ID NO:31-141 и Sy представляет собой количество SFC для пептида у:
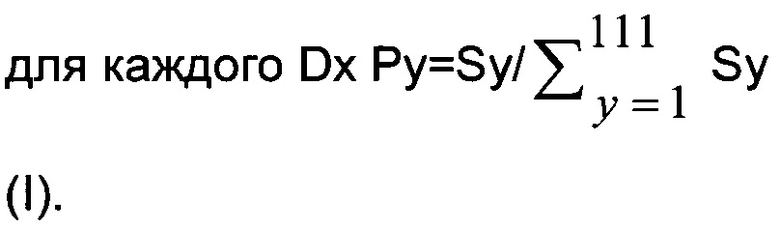
Нормализованное значение означает уровень иммуногенности каждого пептида, и подсчет этих относительных значений для 50 доноров показан на Фигуре 9. Фигура 9 показывает, что следующие пептиды содержат Т-клеточные эпитопы: SEQ ID NO:38, 39, 44, 45, 81, 82, 86, 87, 88, 97, 98, 105, 106, 107, 108, 123,124 и 125.
На основе изложенных на Фигуре 9 данных выбрали наиболее иммуногенные пептиды из домена III. Пептиды, число доноров и пациентов, которые отвечали на пептид, и последовательности, общие для пептидов, приведены в Таблице 6. Пациенты в Таблице 6 ранее получали РЕ38, и ответ, показанный в Таблице 6, является вторичным иммунным ответом. В Таблице 6 ответ определяли, как 5% SFC донора или пациента в ответ на пептид.
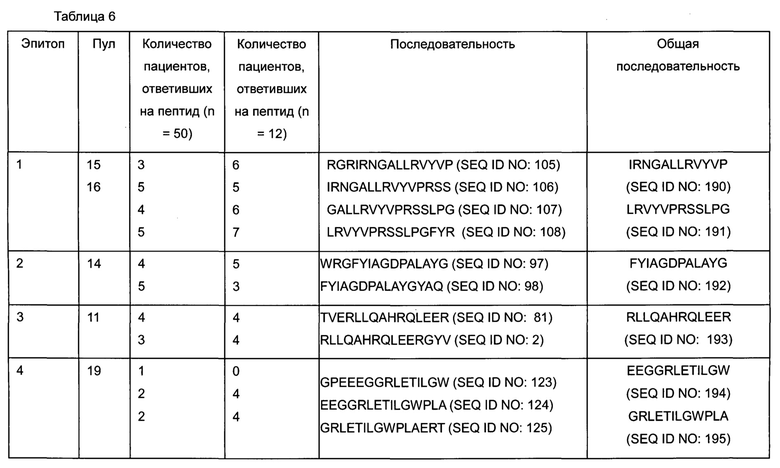
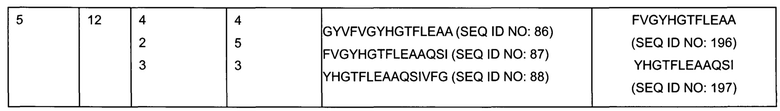
Пример 11
Этот пример демонстрирует, что замены R421A, L422A, L423A, A425G, R427A, L429A, Y470A, 1471 A, A472G, Р475А, A476G, L477A, I493A, N495A, R494A, L498A, L499A, R500A, V501A, Y502A, V503A, R505A, L508A или Р509А, определенные относительно SEQ ID NO:1, уменьшают иммуногенность РЕ.
SEQ ID NO:106 соответствует положениям аминокислотных остатков 493-507 из SEQ ID NO:1, SEQ ID NO:107 соответствует положениям аминокислотных остатков 496-510 из SEQ ID NO:1, SEQ ID NO:97 соответствует положениям аминокислотных остатков 466-480 из SEQ ID NO:1, и SEQ ID NO:81 соответствует положениям аминокислотных остатков 418-432 из SEQ ID NO:1. Каждую аминокислоту в SEQ ID NO:106 и SEQ ID NO:107 заменили аланином. Образцы от доноров и пациентов стимулировали LMB9 в течение 17 дней и анализировали с использованием IL-2 ELISpot, как описано в Примере 3. В Таблице 7 (SEQ ID NO:106), Таблице 8 (SEQ ID NO:107), Таблице 9 (SEQ ID NO:97) и Таблице 10 (SEQ ID NO:81) суммированы данные (% от ответа на пептид дикого типа (WT)) скрининга образцов против замещенных пептидов.
Как показано в Таблице 7, замены I493A, R494A, N495A, L498A, L499A, R500A Y502A, V501A или Y502A снижают ответ Т-клеток на 70% и более по сравнению с ответом на WT пептид.
Как показано в Таблице 8, замены L498A, L499A, R500A, V501A, Y502A, V503A, R505A, L508A или Р509А снижают ответ Т-клеток на 70% и более по сравнению с ответом на WT пептид.
Как показано в Таблице 9, замены Y470A, 1471А, A472G, Р475А, A476G или L477A снижают ответ Т-клеток на 70% и более по сравнению с ответом на WT пептид.
Как показано в Таблице 10, замены R421A, L422A L423A, A425G, R427A или L429A снижают ответ Т-клеток на 70% и более по сравнению с ответом на WT пептид.
Пример 12
Этот пример демонстрирует, что замены Y439A, Н440А, F443A, L444A, A446G, А447А, I450A, R551A, L552A, Т554А, I555A, L556A или W558A, определенные относительно SEQ ID NO:1, снижают иммуногенность РЕ.
Синтезировали 18-аминокислотный пептид (GPEEEGGRLETILGWPLA) (SEQ ID NO:198), включающий аминокислотные остатки из обоих SEQ ID NO:123 и 124. Синтезировали 18-аминокислотный пептид (FVGYHGTFLEAAQSIVFG) (SEQ ID NO:199), включающий аминокислотные остатки из обоих SEQ ID NO:87 и 88. SEQ ID NO:198 соответствует положениям аминокислотных остатков 544-561 из SEQ ID NO:1, и SEQ ID NO:199 соответствует положениям аминокислотных остатков 436-453 из SEQ ID NO:1. Каждую аминокислоту в SEQ ID NO:198 и 199 заменили аланином. Образцы от доноров и пациентов стимулировали LMB9 в течение 17 дней и анализировали с использованием IL-2 ELISpot, как описано в Примере 3. В Таблице 11 (SEQ ID NO:198) и Таблице 12 (SEQ ID NO:199) суммированы данные (% от ответа на пептид дикого типа (WT)) скрининга образцов против замещенных пептидов.
Как показано в Таблице 11, замены R551A, L552A, Т554А, I555A, L556A и W558A снижают ответ Т-клеток на 70% и более по сравнению с ответом на WT пептид.
Как показано в Таблице 12, замены Y439A, Н440А, F443A, L444A, A446G, А447А или I450A снижают Т-клеточный ответ на 70% и более по сравнению с ответом на WT пептид.
Пример 13
Этот пример демонстрирует идентификацию Т-клеточных эпитопов в РЕ.
На основании результатов, полученных в Примерах 4-12, аминокислотные последовательности, представленные в Таблице 13, идентифицировали как Т-клеточные эпитопы РЕ.
Примеры 14-16
Следующие материалы и способы использовали для примеров 14-16:
Поучение образцов цельной крови пациентов, хранение и выделение РНК. Образцы крови брали у 6 пациентов, которых лечили рекомбинантным иммунотоксином (RIT). 2,5 мл образца крови отбирали в пробирки, содержащие катионный детергент PAXGENE и добавочные соли (PreAnalytiX GmbH, Hombrechtikon, Switzerland), тщательно перемешивали, осторожно переворачивая пробирку 4-6 раз, и инкубировали при комнатной температуре 10 часов, а затем хранили при -80°С. Внутриклеточную РНК из образцов цельной крови пациента очищали с помощью набора PAXGENE Blood RNA (PreAnalytiX) в соответствии с инструкциями производителя и хранили при -80°С.
Синтез кДНК тяжелой цепи и легкой цепи, ПЦР-амплификация и сборка гена scFv. Сайт фермента рестрикции или линкер вектора присоединяли (Таблица 15) к праймерам. Репертуар тяжелых цепей и репертуар легких цепей готовили отдельно и соединяли линкером, чтобы обеспечить образование scFv. Репертуар тяжелых цепей получали из IgG из зрелых В-лимфоцитов. Синтез первой цепи кДНК проводили с использованием набора для синтеза первой цепи кДНК (GE Healthcare, NJ) с праймером для константной области IgG: HulgG1-4CH1FOR (Таблица 15). Репертуар легких цепей получали из Vκ генов с использованием праймера к константной области к: HuGKFOR (Таблица 15). 40 пмоль праймеров добавляли в 15 мкл реакционной смеси для синтеза кДНК.
VH и Vκ гены амплифицировали отдельно в три этапа с использованием синтеза первой цепи кДНК. Для извлечения гена VH из внутриклеточной РНК из цельной крови пациентов на первом этапе ПЦР использовали праймер к константной области IgG HulgG1-4CH1FOR и эквимолярную смесь соответствующих обратных праймеров для человеческих генов VH с учетом семейства (Таблица 15). Для гена Vκ использовали праймер к константной области к HuGKFOR, и соответствующие обратные праймеры для человеческих генов Vκ с учетом семейства (Таблица 15). Первый этап ПЦР проводили с использованием высококачественной полимеразы Phusion (New England Biolabs, Ipswich, MA) в конечном объеме 50 мкл реакционной смеси с 10 пмоль каждого праймера в соответствии с рекомендациями производителя.
Для второго этапа ПЦР, соединения с помощью удлинения перекрывающихся фрагментов (ЗОЕ), и последнего этапа, для подготовки вставки, использовали высокоточную полимеразу PrimeStar (Takara, Kyoto, Japan) с 10 пмоль каждого праймера в соответствии с рекомендациями производителя. Последовательность 5'-GCC CAG CCG GCC ATG GCC-3' (SEQ ID NO:185), включая сайт NcoI (подчеркнут), присоединяли к обратному праймеру для человеческого VH с получением обратного праймера для человеческого VH с псо (Таблица 15). Для получения фаговой библиотеки использовали вектор pCANTAB. Последовательность 5'-АСС ТСС AGA TCC GCC АСС АСС GGATCC GCC ТСС GCC-3' (SEQ ID NO:186), включая линкер pCANTAB, присоединяли к прямому праймеру для человеческого JH с получением прямых праймеров с линкером для человеческого JH. Обратный праймер для человеческого VH с nco и прямой праймер для человеческого JH с линкером использовали во втором ПЦР, чтобы добавить сайт NcoI на конец гена VH и линкер pCANTAB перед геном VH.
На втором этапе для амплификации гена Vκ последовательность 5'-GGA ТСС GGT GGC GGT TCT GGA GGA GGT GGC GGA AGC-3' (SEQ ID NO:187), включая линкер pCANTAB, присоединяли к обратному праймеру для человеческого Vκ с получением обратного праймера для человеческого Vκ с линкером. Последовательность 5'-GAG TCA TTC TCG ACT TGC GGC CGC-3' (SEQ ID NO:184), включающую сайт NotI (подчеркнут дважды), соединили с прямым праймером для человеческого Jκ с получением прямых праймеров для человеческого Jκ с Not. Обратные праймеры для человеческого Vκ и прямые праймеры для человеческого Jκ использовали на втором этапе ПЦР для добавления сайта NotI перед геном и линкера pCANTAB в конце гена Vκ.
VH и Vκ гены получали на третьем этапе по отдельности с помощью (а) пары праймеров - обратный праймер к человеческому VH с Nco и праймер с линкером pCANTAB с линкером R' (Таблица 15) для гена VH, и (б) пары праймеров - прямого праймера к человеческому Jκ с Not и праймера с линкером pCANTAB с линкером F' (Таблица 15) для гена Vκ.
Праймеры с линкерами F' и R', которые использовали на третьем этапе, были комплементарными праймерами. VH и Vκ гены объединяли, чтобы обеспечить формирование scFv, используя SOE-ПЦР. Наконец, библиотеку фрагментов scFv амплифицировали с использованием праймеров VHIgGFOR и VLREV (Таблица 15) для приготовления вставки.
Получение фазовой библиотеки. Амплифицированный scFv фрагмент расщепляли с помощью NcoI и NotI и клонировали в pCANTAB 5Е, расщепленный теми же ферментами, чтобы получить библиотеку scFv с использованием лигазы Т4. Лигазную смесь очищали экстракцией на колонке QIAQUICK spin column (Qiagen, Valencia, CA), и ресуспендировали в воде. Конечная концентрация составляла около 50 нг/мл. 4 мкл образца электропорировали в 50 мкл электрокомпетентных клеток TG1 (Lucigen, WI) с помощью кюветного электропоратора и импульсного контроллера (Bio-Rad Laboratories) и повторяли процедуру 6 раз для большого размера библиотеки. Клетки инкубировали в 6 мл SOC (Invitrogen, Carlsbad, CA) в течение 1 часа при 37°С при перемешивании со скоростью около 250 оборотов в минуту. 20 мкл образца отбирали, разбавляли и высевали на чашки с триптиказо-соевым агаром (TYE) с ампициллином для расчета размера библиотеки. Добавляли среду 2YT в количестве 6 мл с 200 мкг/мл ампициллина и 4% глюкозой и инкубировали еще 1 час. Среду доводили до объема 200 мл средой 2YT с 100 мг/мл ампициллина и 2% глюкозой. Клетки выращивали до OD600=0,4 и заражали 1011 БОЕ фага-помощника М13К07 (New England Biolaboratories) при перемешивании со скоростью 250 оборотов в минуту в течение 30 мин после 30 мин инкубации. Клетки собирали центрифугированием в течение 5 мин при 5000 оборотах в минуту в роторе GSA и ресуспендировали в 100 мл среды 2YT с 100 мкг/мл ампициллина и 50 мкг/мл канамицина в течение ночи при 30°С при перемешивании со скоростью 250 оборотов в минуту.
Фаги осаждали из супернатанта с 1/5 объема ПЭГ/NaCl (20% полиэтиленгликоль 6000, 2,5 М NaCl) и ресуспендировали в среде 2YT. Титр фаговой библиотеки определяли путем последовательных разведении 10 мкл фага, к которым добавляли 90 мкл клеток TG1, OD600=0,4, которые затем высевали на LB-агаре с добавлением 100 мкг/мл ампицилина и 1% глюкозы. Определяли количество колоний после ночного роста и рассчитывали титр.
Панорамирование фазовой библиотеки. Для панорамирования фаговой библиотеки использовали в качестве антигена LMB-Q (B3(dsFv)-PE38, специфичный для LewisY антигена). LMB9 биотинилировали с использованием EZ-Link сульфо-NHS-биотина (Thermo Scientific, Rockford, IL) в молярном соотношении 50:1, количество биотиновых групп на каждом LMBQ-биотине определяли с использованием набора для количественного определения биотина (Biotin quantitation kit, Thermo Scientific, Rockford, IL) в соответствии с инструкциями изготовителя. 350 мл фага и модифицированные стрептавидином магнитные шарики (Dynabeads MYONE Streptavidin T1, диаметр 1 мкм, связывающая способность биотинилированного Ig 40-50 мкг*мг-1, гидрофобные, активированные тозилом шарики (Invitrogen)) предварительно заблокировали в 3% BSA/ PBST (0,1% твин-20). Фаг наносили для де-селекции шариками.
Магнитный штатив использовали, чтобы отделить шарики от жидкой фазы, в результате чего шарики стали иммобилизованными вдоль стенок пробирки. Блокирующий буфер удалили и шарики ресуспендировали в растворе фага и инкубировали при комнатной температуре на качалке в течение 30 мин. Раствор фага переносили в другую пробирку с предварительно заблокированными шариками для дополнительной де-селекции. Де-селекцию повторяли с 1 мг шариков два раза и с 2 мг шариков 1 раз. Фаг переносили в предварительно заблокированную пробирку, и добавляли биотинилированный антиген LMBQ-биотин, чтобы позволить образоваться комплексам фаг-антиген-биотин с LMBQ-биотином в количестве 10 мкг для первого раунда и 5 мкг для последующего раунда. Реакционный раствор инкубировали при комнатной температуре на качалке в течение 2 часов и переносили в пробирку с 2 мг шариков для инкубации на роторе в течение дополнительных 45 мин. Надосадочную жидкость удаляли, а шарики промывали 12 раз PBST. Фаг освобождали от шариков путем добавления холодной 0,1 М HCl в количестве 1 мкл и рН нейтрализовали 200 мкл раствора Трис-HCl (рН 8,0). Получали результат панорамирования, и его сохраняли для дополнительных раундов панорамирования, и рассчитывали титр. Выход фага в количестве 0,6 мкл использовали для инфицирования 5 мл TG1 (OD600=0,4) для выделения фага.
Фазовый ИФА (ELISA) u секвенирование клонов фага. После трех или четырех раундов панорамирования и выделения фага 198 отдельных клонов из финального раунда панорамирования отбирали для дальнейшего анализа. Клоны переносили в 96-луночный планшет с круглодонными лунками и инкубировали с 150 мкл среды 2YT (100 мкг/мл ампициллина, 2% глюкоза) в течение 4 ч при 37°С при перемешивании со скоростью 250 оборотов в минуту, и в лунку после 30 мин инкубации при перемешивании со скоростью 250 оборотов в минуту в течение 30 минут добавляли 108 БОЕ фага-помощника М13К07 в 50 мкл среды 2YT (100 мкг/мл ампициллина, 2% глюкоза). Клетки собирали центрифугированием при 2700 оборотах в минуту в течение 10 мин с держателями для 96-луночных планшетов и ресуспендировали в среде 2YT в количестве 200 мкл со 100 мкг/мл ампициллина и 50 мк/ мл канамицина течение ночи при 30°С с перемешиванием со скоростью 250 оборотов в минуту. Осадок ресуспендировали в 100 мкл среды 2YT с 100 мкг/мл ампициллина, 2% глюкозой и 30% глицерином и хранили при -80°С как стоковый раствор. Фаги осаждали из супернатанта для фагового ELISA при 2700 оборотах в минуту в течение 10 мин. 96-луночные плоскодонные плашки NUNC MaxiSorp (Nunc США, Rochester, NY) покрывали LMB9 (5 мкг/мл в PBS) в течение ночи при 4°С. Планшет промывали и блокировали 2% обезжиренным молоком (Cell Signalling). Добавляли супернатант с фагом (50 мкл) и 2% молоко (50 мкл) и инкубировали в течение 1 часа при комнатной температуре. Планшет промывали 3 раза PBST, добавляли конъюгированные с пероксидазой антитела к М13 (1:1000, GE Healthcare, Waukesha, WI) и инкубировали в течение 1 часа при комнатной температуре. Планшет промывали три раза фосфатным солевым буфером и Твин 20 (PBST), и добавляли субстрат 3,3',5,5'-тетраметилбензидин (ТМВ) (Thermo Scientific, Rockford, IL) на 15 мин. Результаты фиксировали на спектрофотометре при 450 нм, чтобы определить положительные и отрицательные клоны. Положительный клон отбирали для выделения фага из малого объема из соответствующей лунки исходного планшета и проводили секвенирование с использованием набора BIGDYE Terminator v1.1 Cycle Sequencing (Applied Biosystems, Foster City, CA). Клоны с той же последовательностью удаляли, и полученные последовательности выравнивали с помощью IMGT/V-Quest (www.imgt.org/IMGT_vquest/vquest).
Конкурентный ICC-ИФА. Библиотеку фагов-антител получали вышеупомянутым методом из 20 мл культуры. Разведение фагов-антител определяли с помощью ИФА. К планшетам для ИФА, покрытым в течение ночи при 4°С rFc-мезотелином в количестве 50 мкл/лунку при концентрации 4 мкг/мл в PBS добавляли антитело SS1P, 50 мкл/лунку, 1 мкг/мл в 2% обезжиренного молока. Планшет 3 раза промывали PBST; добавляли фаг-антитело в различных разведениях и детектировали с использованием HRP-конъюгированного антитела к М13 и субстрата ТМВ. Разведение фагов-антител определяли по кривой разбавления, и желаемую А450 установили на уровне около 1,0. Конкурентный анализ ICC-ELISA проводили, чтобы определить эпитоп связывания антигена РЕ38 фагом-антителом с помощью сыворотки пациента, РЕ38 без Fv, или РЕ38 с единичной мутацией. Фаг-антитело смешивали с серийными разведениями мутанта в течение ночи при 4°С и добавляли к планшету ИФА с комбинацией SSIP-rFc-мезотелин. Конкуренцию мутанта за связывание фага-антитела с SS1P определяли путем измерения связывания оставшегося фага-антитела с использованием HRP-конъюгированного антитела к М13. Эффект конкуренции нормировали на связывание с HA22-LR, в котором в РЕ38 отсутствовали замены.
Антигенность сыворотки. Связывание НА22 или НА22 с заменами с антителами в сыворотке крови человека анализировали методом вытеснения. Человеческую сыворотку получили по протоколу 10С0066. Мезотелин-rFc добавляли в планшеты для ELISA (100 нг в 50 мкл PBS / лунку) и инкубировали в течение ночи при 4°С. После промывания добавляли антимезотелин/SSIP (100 нг в 50 мкл блокирующего буфера / лунку) и инкубировали в течение 1 ч, чтобы связать оставшиеся свободными человеческие антитела к РЕ38. В отдельных пробирках смешивали сыворотку (разведения от 97 до 30 658) с 2 мкг/мл НА22 или НА22 с заменами и инкубировали в течение ночи при 4°С. После промывания в каждую лунку планшета добавляли 50 мкл смеси иммунотоксин-антитело. Человеческие антитела, не связавшиеся с НА22 или НА22 с заменами, связались с SS1P, их детектировали с использованием конъюгированного с HRP кроличьих антител против человеческого Fc IgG (Jackson ImmunoResearch Laboratories, West Grove, PA), а затем набора с субстратом ТМВ (Thermo Scientific Inc, Waltham, MA). Кривые апроксимировали с использованием четырехпараметрической модели логистической кривой с помощью SoftMaxPro 4.0 (Molecular Devices). Значения IC50 показывают концентрацию RIT, которая ингибирует 50% реактивности антител с SS1P.
Статистика. Использовали непараметрический метод Манна-Уитни, р<0,05 считали статистически значимым.
Пример 14
Этот пример демонстрирует выделение и секвенирование человеческих scFv, специфичных для РЕ38.
Образцы крови получили у 6 пациентов, которых лечили различными рекомбинантными иммунотоксинами (RIT), содержащими РЕ38 (Таблица 14). РНК выделяли из образцов крови с использованием наборов PAXGENE Blood RNA (PreAnalytiX GmbH, Hombrechtikon, Switzerland). Первую цепь кДНК синтезировали из РНК с использованием праймеров к соответствующим константным областям (Таблица 15). Отдельные полосы нужного размера для кДНК VH и Vκ получили с использованием первой цепи кДНК в качестве матрицы. VH и VL фрагменты амплифицировали индивидуально в три этапа. Во фрагмент добавили сайт фермента рестрикции и линкер. 100 нг фрагментов VH и VL объединяли путем ПЦР с соединением путем удлинения перекрывающихся фрагментов (SOE-ПЦР) для формирования scFv. Фрагмент scFv расщепляли NcoI и NotI и клонировали в pCANTAB 5E, расщепленный теми же ферментами, для получения библиотеки scFv.
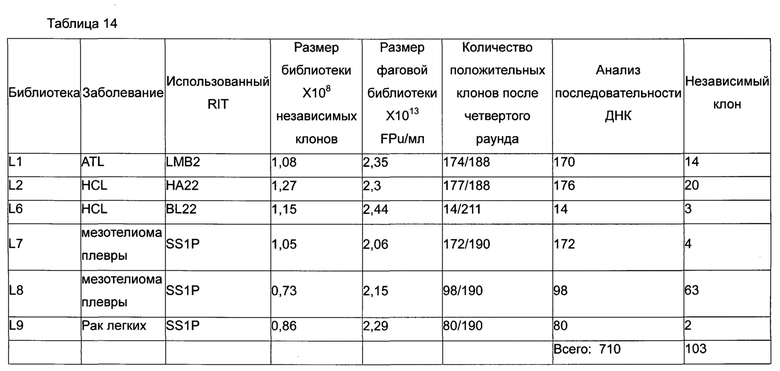


Биотинилированный иммунотоксин LMB-9 (B3-Fv-PE38) использовали в качестве антигена для отбора фагов, экспрессирующих Fv, связывающий РЕ38. Каждая молекула LMB-9 содержала 6 биотинов. 6 библиотек человеческих антител получили путем электропорации в клетки Escherichia coli (E.coli) TG1, содержащие 7,3×107-1,27×108 VH-VL scFv клонов (Таблица 15). Фаговую библиотеку воспроизводили благодаря суперинфекции фагом-помощником (Таблица 15), и 350 мл каждой библиотеки содержали около 7×1012 scFv фрагментов, экспрессируемых на поверхности фага.
710 Fv-содержащих клонов фагов получали и секвенировали. Секвенирование показало наличие 103 уникальных последовательностей человеческих тяжелой цепи и легкой цепи каппа, кроме 2 клонов, которые имели одинаковую последовательность легкой цепи. Чтобы показать, что Fv были получены из В-клеток, производивших антитела к иммунотоксину, провели конкурентный анализ и показали, что иммунная анти-сыворотка заблокировала связывание фага с частью LMB-9, соответствующей РЕ38, и ни один из клонов не связался с Fv частью иммунотоксина. Прочность связывания измеряли с использованием ICC-ELISA. 47 клонов продемонстрировали слабое связывание и не были изучены дополнительно. Другие 56 клонов использовали для определения конкретных человеческих эпитопов в РЕ38.
Пример 15
В этом примере показано расположение человеческих В-клеточных эпитопов.
LMB-9 включает II и III домены РЕ. Чтобы определить фаг, который связывается только с доменом III, определили связывание каждого клона с HA22-LR, который включает только домен III и лишен домена II. Пятнадцать из 56 клонов фагов не могли связываться с HA22-LR, что указывает, что эпитопы, распознаваемые этими 15 клонами фагов расположены в домене II. Остальные 41 клонов фагов использовали для определения остатков, которые образуют В-клеточные эпитопы в домене III, путем измерения их связывания с белками с заменами, в которых отдельные большие гидрофобные аминокислоты на поверхности домена III заменили на аланин и глицин. Эти замены устраняли большие гидрофобные боковые цепи, которые участвуют в узнавании и связывании антителом. Данные приведены на Фигуре 10, где клоны со слабым связыванием (<10%) показаны черными клетками, а белки с заменами с нормальной реактивностью показаны в виде пустых клеток. Результаты показывают, что единственная замена уменьшает связывание многих клонов, тем самым указывая, что они относятся к одной и той же группе эпитопов.
Расположение остатков, которые при замене снижали связывание фага с различными эпитопами >90% показаны в Таблице 16. Аминокислоты, ассоциированные с каждым человеческим (Н1, Н2, Н3, Н4, Н5 и Н6) и мышиным (2с, 4а, 4b, 5, 6а, 6b и 7) эпитопом, показаны в Таблице 16. Человеческий эпитоп Н1 включает D403, R427 и Е431. R427 и Е431 принадлежат к мышиному эпитопу 4а, и эти остатки были вовлечены в связывание и человеческих, и мышиных антител. Человеческий эпитоп Н2 включает остатки R467 и D463, которые входят и в мышиный эпитоп 2 с, и Е548, который входит в мышиный эпитоп 6а. Y481, L516, Е522 и R551 были специфичными для человеческих эпитопов. Человеческий эпитоп НЗ содержал только R458, который принадлежал мышиному эпитопу 4b. Человеческий эпитоп Н4 включает R432 и R505. R432 принадлежал мышиному эпитопу 4а, a R505 был специфическим для человека остатком. Человеческий эпитоп Н5 состоял из R490 и R576, которые принадлежали мышиному эпитопу 5. Человеческий эпитоп Н6 включал R538. R538 принадлежит мышиному эпитопу 2 с.D406, R412, R513, L597, Q592 и К590 были специфичными для мышиных эпитопов и не участвовали в связывании человеческих антител.
На фаговые клоны, реагирующие с эпитопом Н1, сильно воздействовали замены в остатках D403, R427 и Е431. Замена любого из этих остатков на аланин сильно влияла на связывание многих фагов, которые узнавали эпитоп (Фигура 10). Как и ожидалось для замен, которые составляют эпитоп, эти остатки были пространственно смежными на домене III. Эпитоп Н2 был сложным. На фаги, реагирующие с эпитопом Н2, влияли замены 8 остатков. Замена R467 на аланин уничтожала связывание шести из восьми фагов, которые определяли эпитоп Н2. Замена остатка D463 предотвращала связывание четырех фагов, замена Y481 предотвращала связывание трех фагов, замена R551 предотвращала связывание двух фагов и замена остатков D461, L516, Е522 или Е548 предотвращала связывание одного фага. Структурно эти остатки находились в ограниченной зоне и составляли кластер. Эпитоп НЗ узнавали 2 фага, которые связывались с R458. Эпитоп Н4 узнавали 11 фагов, и связывание нарушалось заменой R505A. Замена R432, который находится близко к R505, нарушила связывание 1 из 11 фагов. Эпитоп Н5 узнавали 4 фага. На связывание со всеми четырьмя повлияла замена R490, а замена R576 нарушила связывание трех из четырех фагов. Эти остатки находятся пространственно близко на домене III, хотя они разделены 86 аминокислотами в последовательности. Замена R538 нарушала связывание одного из двух фагов. Таким образом, замена сильно экспонированных поверхностных остатков на аланин позволила определить остатки, которые связываются фагами, которые связывают домен III, показывая, что эпитопы располагались на различных участках на поверхности домена III.
Пример 16
Этот пример демонстрирует получение рекомбинантного иммунотоксина (RIT) с низкой антигенностью для человека.
Идентификацию отдельных остатков, которые были вовлечены в связывание с человеческой антисывороткой, использовали для проектирования и создания иммунотоксинов с заменами, которые устраняют реактивность с человеческой антисывороткой, сохраняя цитотоксическую активность, и могут быть получены в достаточном количестве, чтобы быть полезными. В большинстве случаев остатки заменяли на аланин, потому что его маленькая боковая цепь плохо реагирует с антителами и это, как правило, не влияет на сворачивание белков. Также использовали серин, чтобы избежать сильно гидрофобной поверхности.
На основании информации, полученной в исследованиях по картированию эпитопов, замены, выбранные из различных аминокислот, которые нарушали связывание человеческих Fv с доменом III из HA22-LR, объединили. Замены показаны в Таблице 17 ниже. LR05 имел все замены, присутствующие в HA22-LR-8M, и 4 новые замены, LR06 имел только 2 замены из HA22-LR-8M и 4 новые замены, и L010 был похож на LR06, но имел дополнительную замену 463А (Таблица 17).
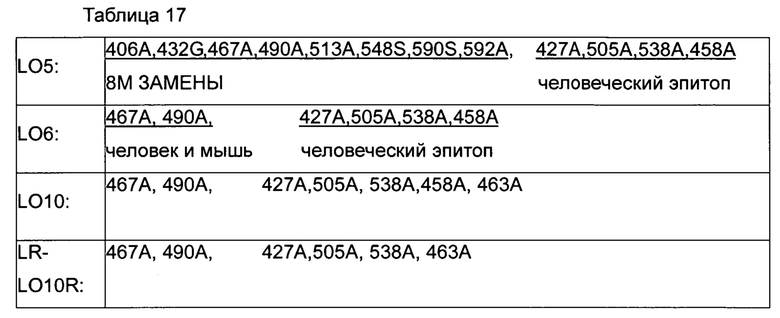
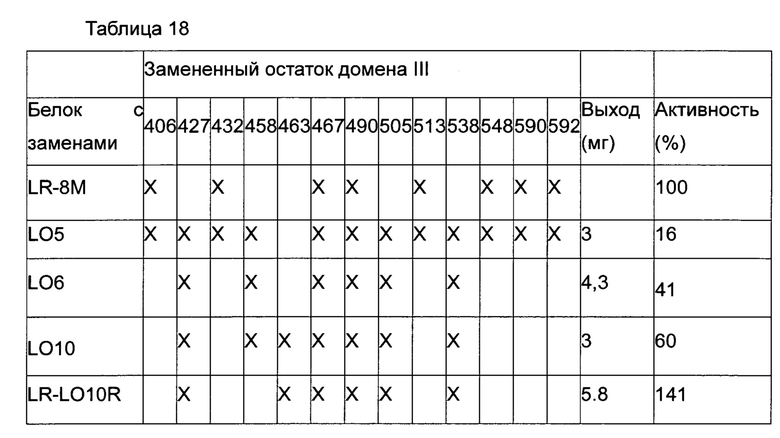
Белки с заменами экспрессировали и очищали. SDS-электрофорез показал, что замещенные белки были более чем на 95% гомогенными. Очищенные белки затем анализировали на цитотоксическую активность на нескольких CD22-положительных клеточных линиях и на антигенность с точки зрения их способности связываться с антителами, присутствующими в сыворотке пациентов, у которых образовались нейтрализующие антитела к иммунотоксинам, содержащим РЕ38. Анализировали 25 сывороток от пациентов, которые получали несколько различных иммунотоксинов (LMB-9, SS1P и НА22).
Данные в Таблице 19 показывают, что все три новых иммунотоксина были активны на С022-положительных линиях лимфомы с IC50 около 1 нг/мл, но менее активны, чем HA22-LR. Наиболее активным был HA22-L010, который имел 60% активности HA22-LR на клетках Дауди, 27% активности на клетках Raji и 29% активности на клетках СА46. Эти новые иммунотоксины были CD22-специфичными и не показали никакой активности на клетках А431, которые не экспрессируют CD22 (Таблица 19).
Антигенность определяли как связывание иммуногенов с уже существующими антителами. Для оценки антигенности HA22-LO с заменами с человеческой сывороткой пациента проводили конкурентный анализ, в котором измеряли концентрацию каждого из иммунотоксинов с заменами, которые снижали уровень антител, реагирующих с НА22, на 50%. Типичные результаты конкуренции с двумя сыворотками пациентов показаны на Фигурах 11А и 11Б. 11А показывает, что концентрация НА22, HA22-LR, HA22-L05, HA22-L06, НА22-LR-8M и HA22-L010, при которой связывание с РЕ38 ингибируется на 50% (IC50), составляет 84,8, 38,1, 4580, 1440, 3610,>396 ООО нМ, соответственно. Отношение связывания (IC50) НА22 к HA22-LR, HA22-L05, HA22-L06, HA22-LR-8М и HA22-L010 составило 223, 1,85, 5,89, 2,35, и 0,0214%, соответственно. Фигура 11 В показывает, что концентрация НА22, HA22-LR, HA22-L05, HA22-L06, HA22-LR-8M и HA22-L010, при которой связывание с РЕ38 ингибируется на 50% (IC50), составила 50,9, 67700, >396000, >396000, >396000, >396000 нМ, соответственно. Отношение связывания (IC50) НА22 к HA22-LR, HA22-L05, HA22-L06, HA22-LR-8M и HA22-L010 составило 0,752, <0,0129, <0,0129, <0,0129 и <0,0129%.
Анализировали сыворотку 32 пациентов, которых лечили в течение более чем 10 лет иммунотоксинами SS1P, НА22 и LMB9, содержащими РЕ38. Отношение связывания для иммунотоксинов с заменами показано в Таблице 20. Обнаружили, что антигенность HA22-LR-L010 с человеческой сывороткой была существенно снижена по сравнению с НА22, HA22-LR и HA22-LR8M. На Фигуре 12 приведен график, показывающий процентное связывание антител с НА22, HA22-LR-8M, HA22-LRL010 или HA22-LR-L010R в сыворотке пациентов, которых лечили с использованием РЕ38. HA22-LR-L010R похож на HA22-L010 за исключением того, что HA22-LR-L010R не имеет замену R458A, которая присутствует в HA22-L010 (Таблицы 17 и 18). На Фигуре 12 показано, что двадцать три из тридцати двух пациентов продемонстрировали связывание (антигенность), которая была снижена более чем в 100 раз (100-10000-раз). Только у четырех из тридцати двух пациентов не смогли обнаружить снижение антигенности.
Дополнительные сведения, подтверждающие цитотоксическую активность РЕ с заменами, представлены в статье авторов настоящего изобретения, опубликованной в Proceedings of the National Academy of Sciences, 2014, vol. 111, no. 23, p. 8571-76, и дополняющих ее таблицах S1 и S2. В этой статье показано, что РЕ, содержащий замены в положениях L477, R427 и F443, является цитотоксичным для СР22-экспрессирующих клеточных линий (фиг. 4А и 4В публикации) и клеток пациентов с волосистоклеточным лейкозом (HCL) и хронической лимфоцитарной лейкемией (CLL) (фиг. 4D и 4Е публикации) (см. также стр. 8573 публикации, правая колонка, 2-й абзац), а также приведены данные о том, что лечение опухоленесущих мышей с помощью РЕ, содержащих замены в положениях L477, R427 и F443, приводит к полной регрессии опухоли (фиг. 4F публикации, стр. 8573, правая колонка, 3-й абзац). Кроме того, в дополняющей таблице S2 публикации показано, что РЕ, имеющий мутацию в положении L423, R427, F443, I471 или L477, был цитотоксичным в отношении одной или нескольких клеточных линий CD22+ СА46, Raji и лимфомы Дауди Беркитта.
Все ссылки, включая публикации, патентные заявки и патенты, цитируемые здесь, включены посредством ссылок в той же степени, как если бы о каждой ссылке было индивидуально и конкретно указано, что она включена посредством ссылки во всей своей полноте.
Использованные в контексте описания изобретения термины (особенно в контексте следующей формулы) должны быть истолкованы, как охватывающие и единственное, и множественное число, если прямо не указано иное или нет явного противоречия контексту. Термины «содержащий», «имеющий», «включающий» должны быть истолкованы как неограничивающие термины (т.е. означающие «включающий, но не ограниченный этим»), если не указано иное. Указание диапазонов значений здесь предназначено только в качестве краткого способа указания каждого отдельного значения, находящегося в пределах диапазона, если не указано иное, и каждое отдельное значение включено в описание, как если бы оно было индивидуально приведено в настоящем документе. Все описанные здесь способы могут быть выполнены в любом подходящем порядке, если не указано иное или иным образом явно не противоречит контексту. Использование любых и всех примеров или оборотов речи, обозначающих пример, (например, «такой как»), представленных в настоящем документе, предназначено только для лучшего освещения изобретения и не накладывает ограничений на объем изобретения, если не заявлено иное. Ни одну фразу в описании не следует толковать как указывающую на то, что какой-либо признак, не включенный в формулу изобретения, является существенным для практического осуществления изобретения.
Предпочтительные варианты воплощения настоящего изобретения описаны в данном документе, включая наилучший способ, известный изобретателям, для выполнения данного изобретения. Вариации этих предпочтительных вариантов воплощения могут быть очевидны специалистам в данной области техники при чтении вышеприведенного описания. Изобретатели ожидают, что специалисты используют такие вариации в зависимости от обстоятельств, и изобретатели предполагают, что изобретение может быть осуществлено на практике иначе, чем конкретно описано здесь. Соответственно, данное изобретение включает все модификации и эквиваленты объекта, изложенного в формуле изобретения, прилагаемой к настоящему документу, в соответствии с действующим законодательством. Кроме того, любая комбинация вышеописанных элементов во всех их возможных вариантах охватывается изобретением, если иное не указано здесь или явно не противоречит контексту.
| название | год | авторы | номер документа |
|---|---|---|---|
| МОДИФИЦИРОВАННЫЙ ЭКЗОТОКСИН А ПСЕВДОМОНАД | 2014 |
|
RU2687143C2 |
| РЕКОМБИНАНТНЫЙ ИММУНОТОКСИН, НАЦЕЛЕННЫЙ НА МЕЗОТЕЛИН | 2012 |
|
RU2600067C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ К CD22 | 2012 |
|
RU2644243C2 |
| ГЛИКАНЗАВИСИМЫЕ ИММУНОТЕРАПЕВТИЧЕСКИЕ МОЛЕКУЛЫ | 2016 |
|
RU2754661C2 |
| ИММУНОТЕРАПЕВТИЧЕСКИЕ КОМПОЗИЦИИ НА ОСНОВЕ ДРОЖЖЕЙ С BRACHYURY | 2012 |
|
RU2690180C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ М971 | 2013 |
|
RU2658485C2 |
| АНТИТЕЛА ПРОТИВ ФАКТОРА D И ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2799044C2 |
| СПОСОБЫ ЛЕЧЕНИЯ АУТОИММУННОГО ЗАБОЛЕВАНИЯ С ИСПОЛЬЗОВАНИЕМ АЛЛОГЕННЫХ Т-КЛЕТОК | 2017 |
|
RU2773831C2 |
| АНТИ-C5a-АНТИТЕЛА И ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2773779C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ M971 | 2018 |
|
RU2770411C1 |























































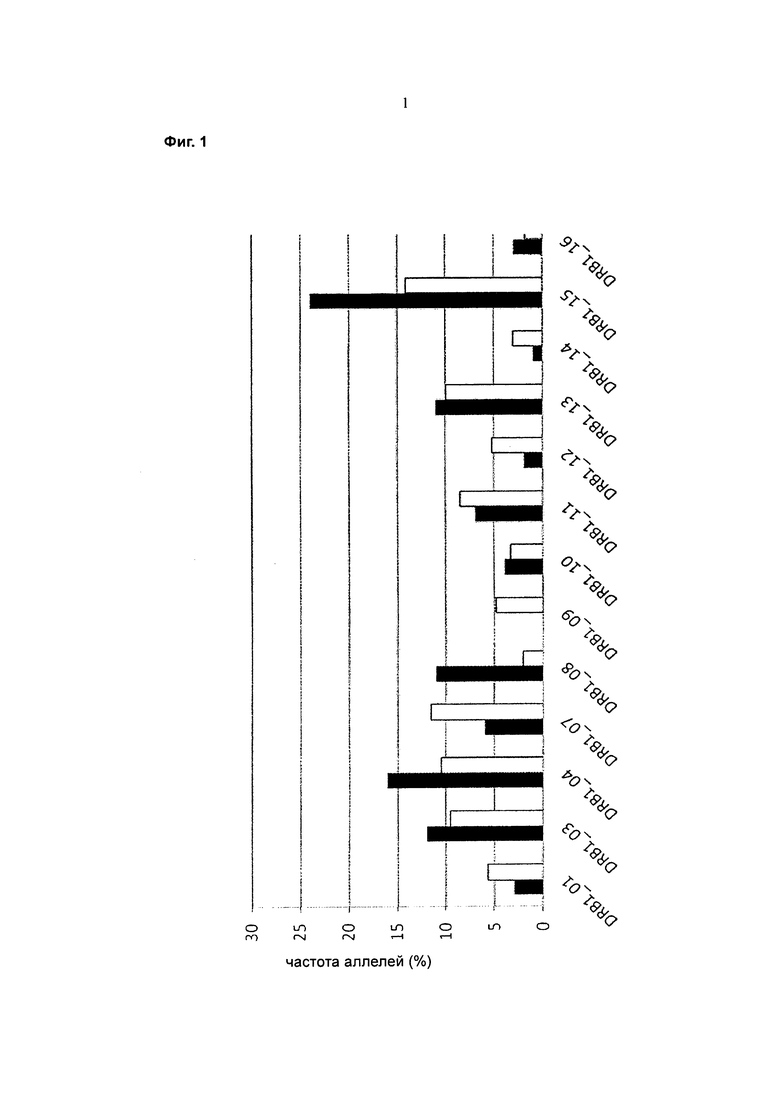
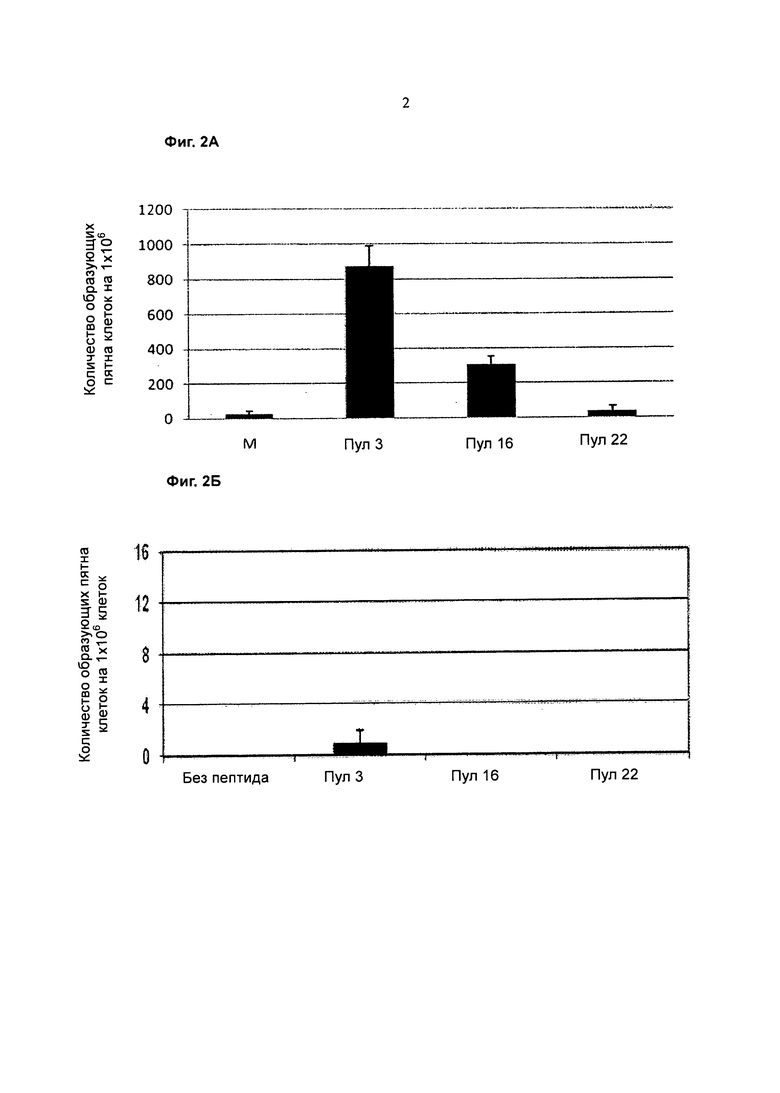
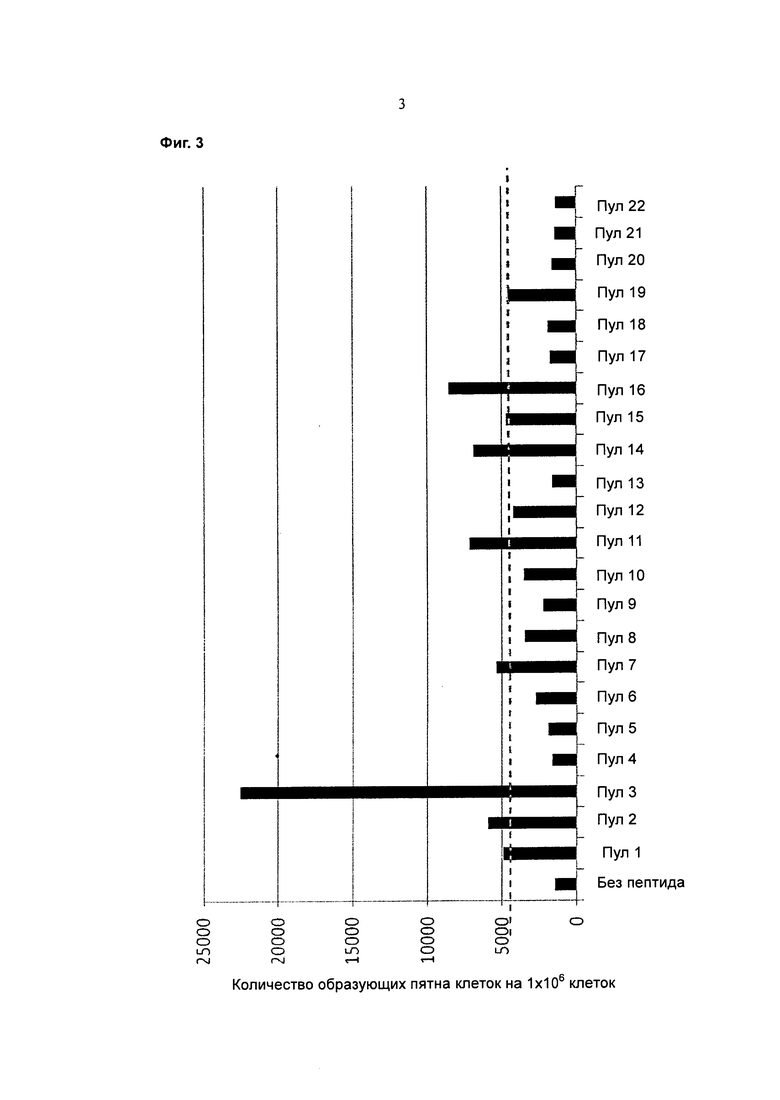

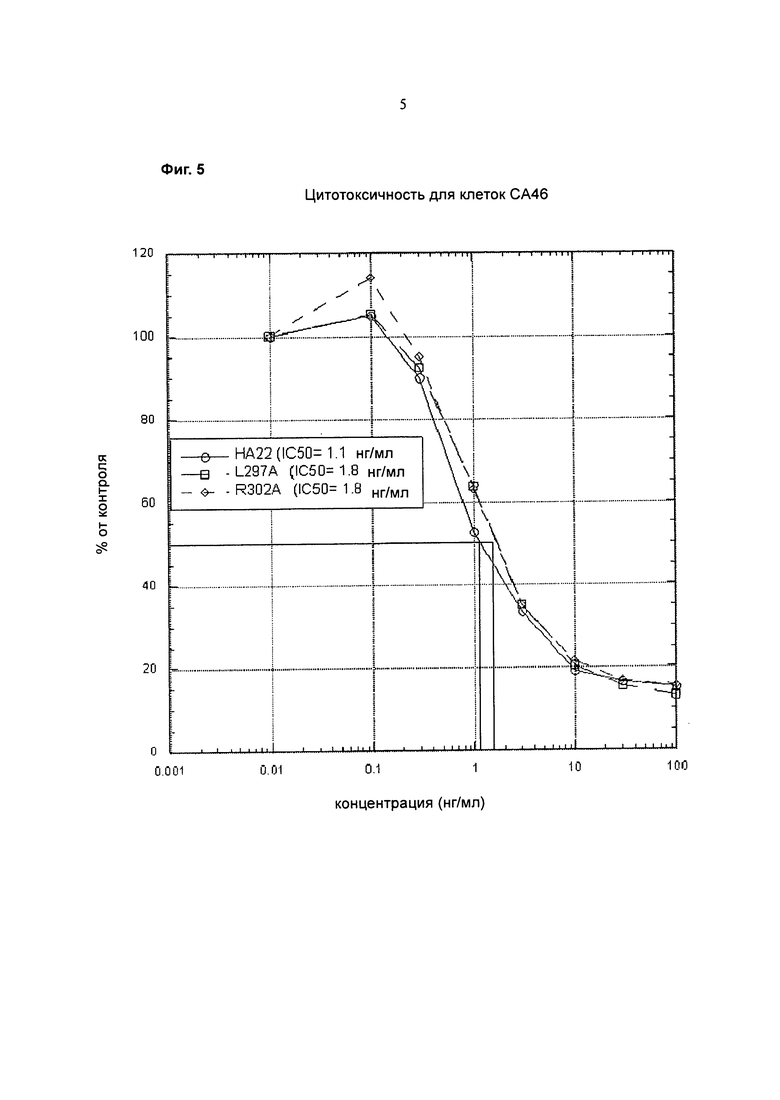
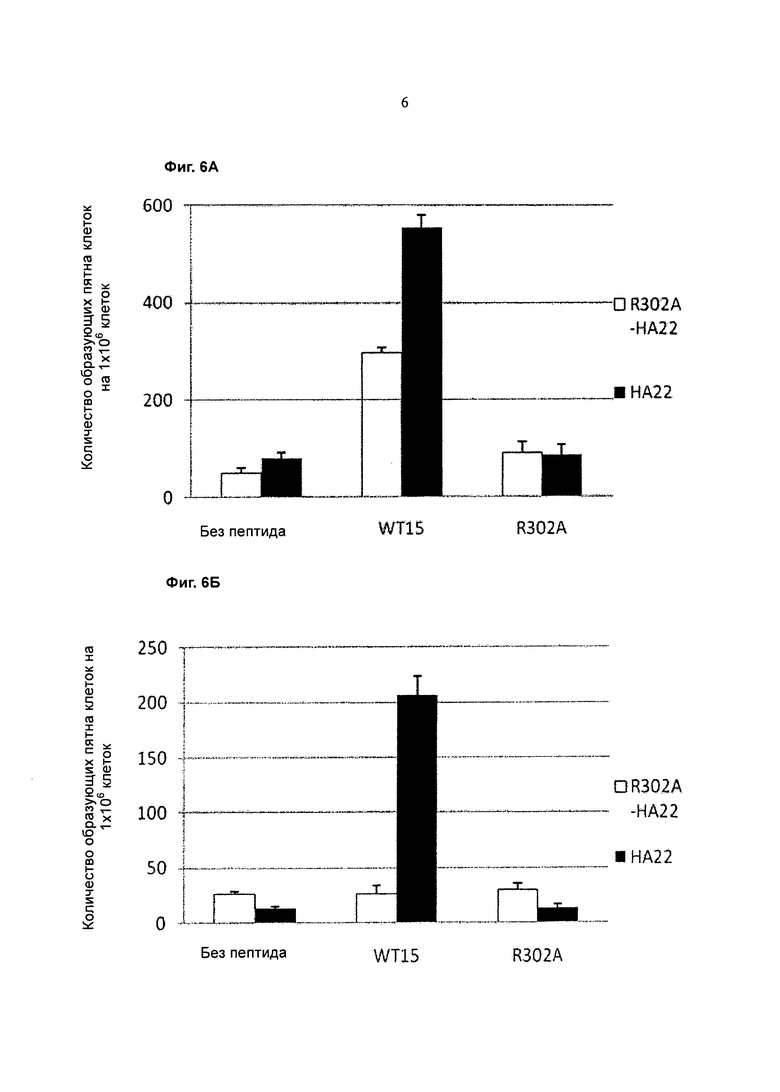
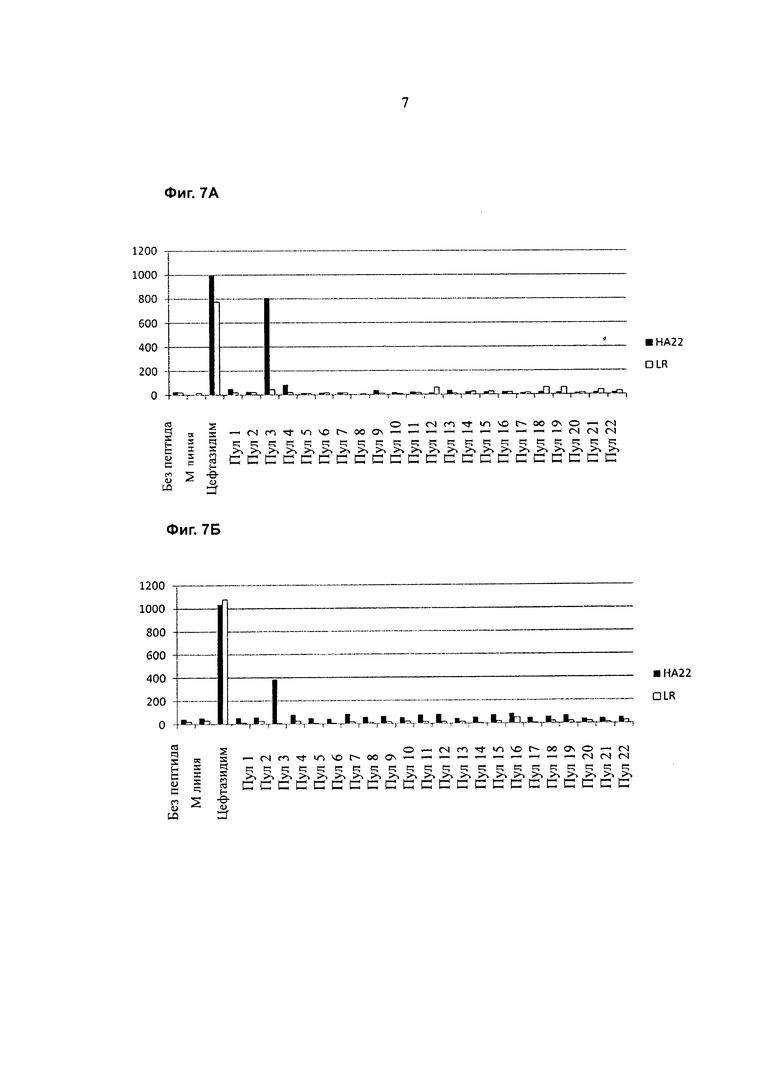
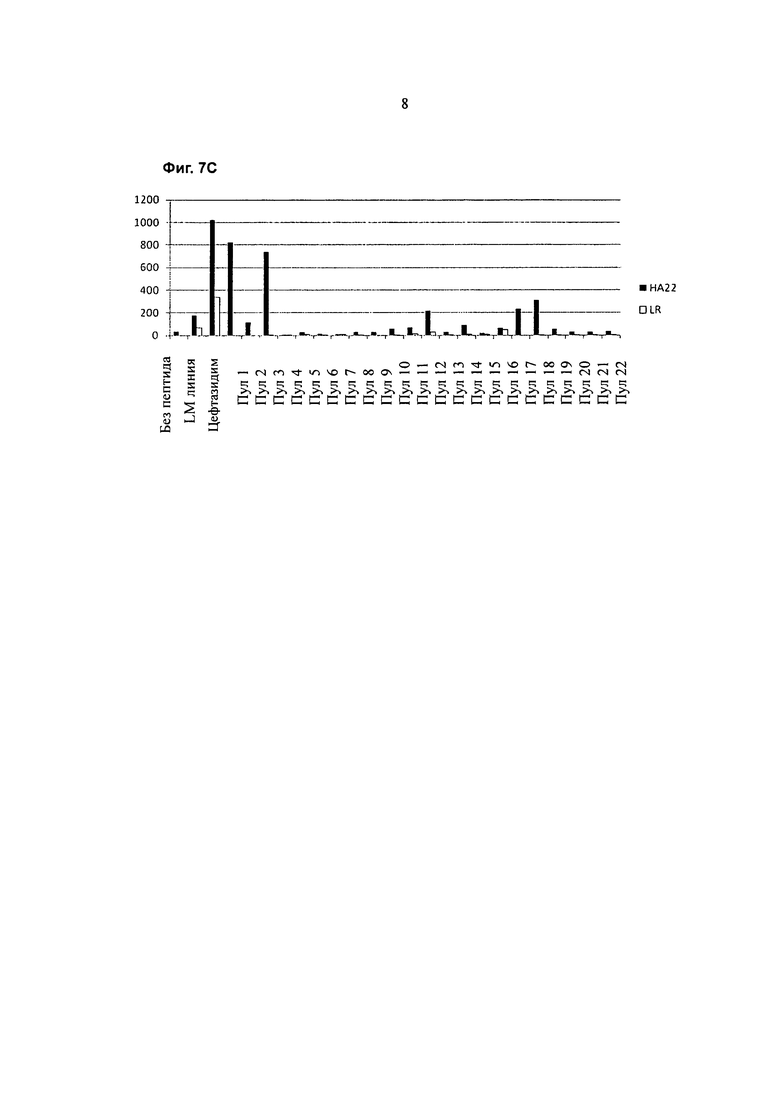
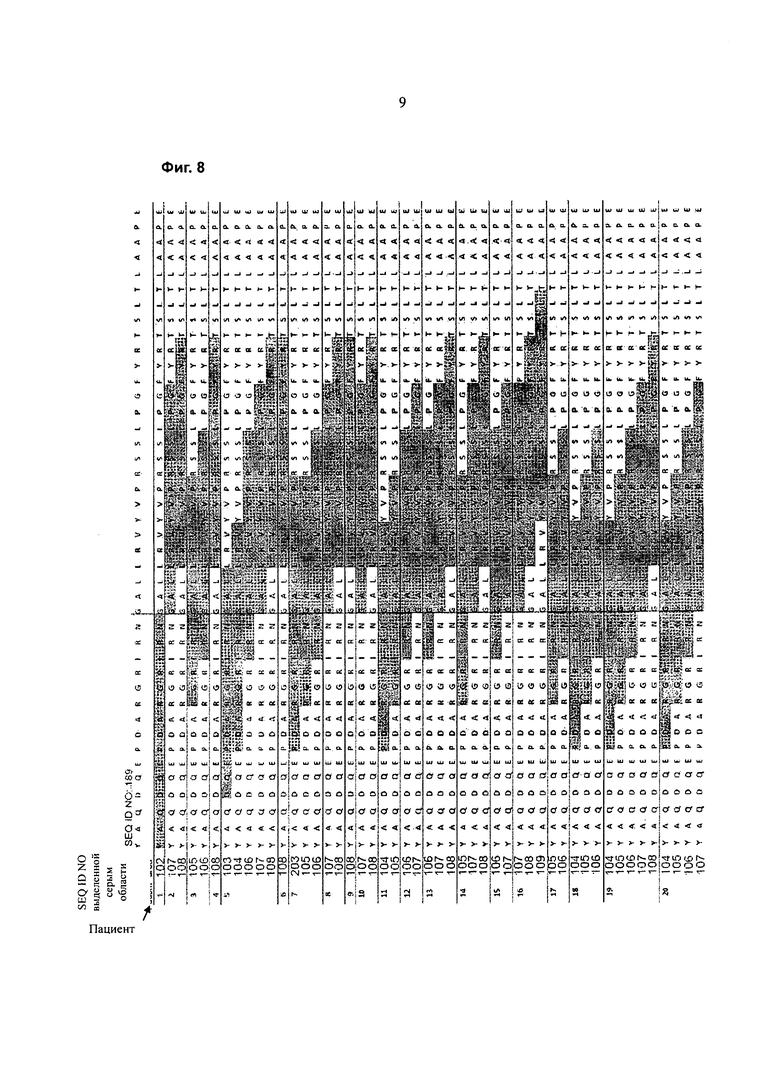
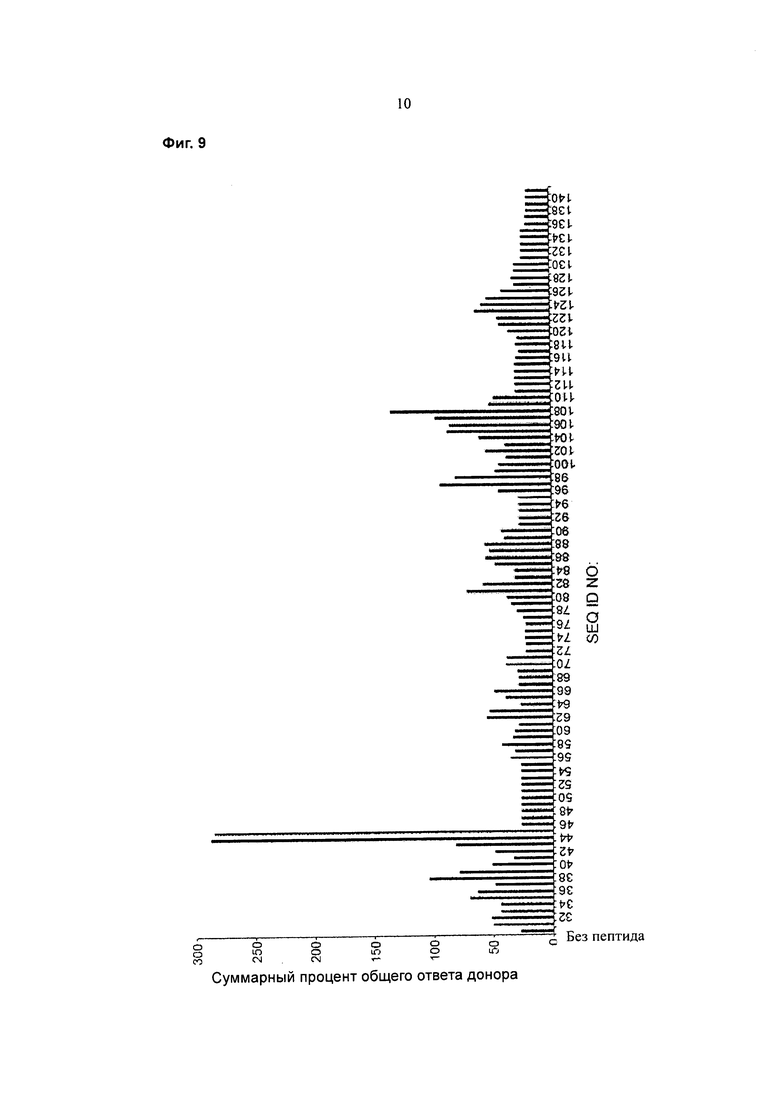
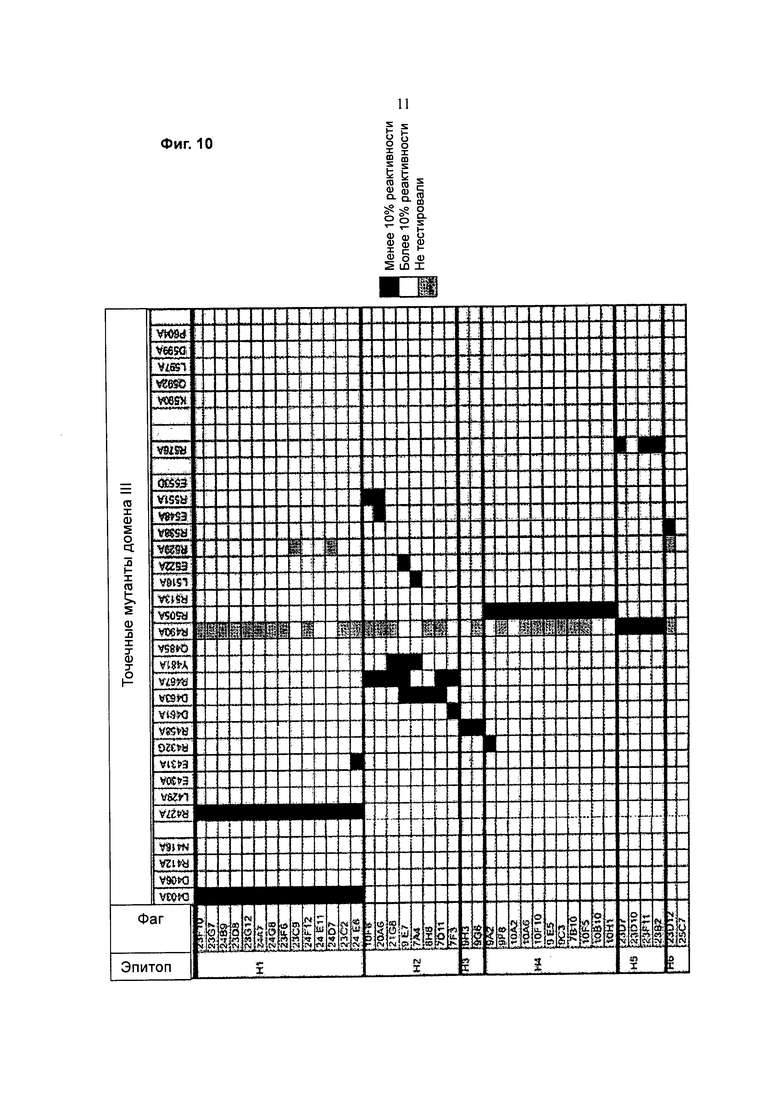
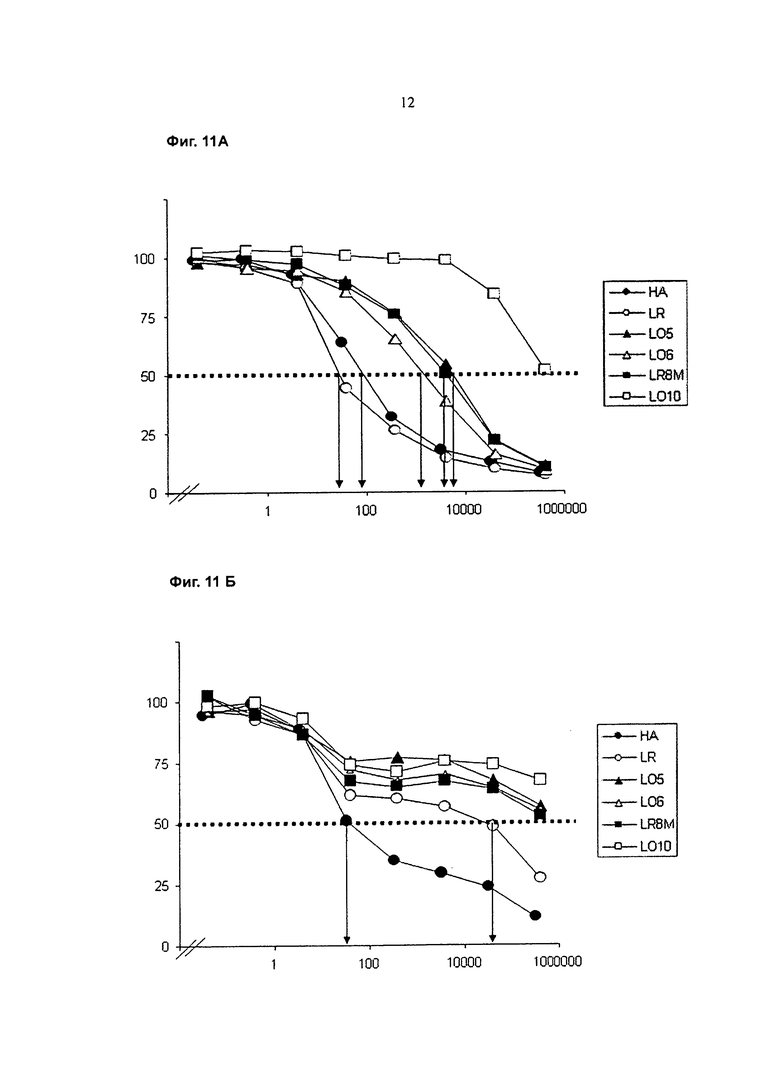
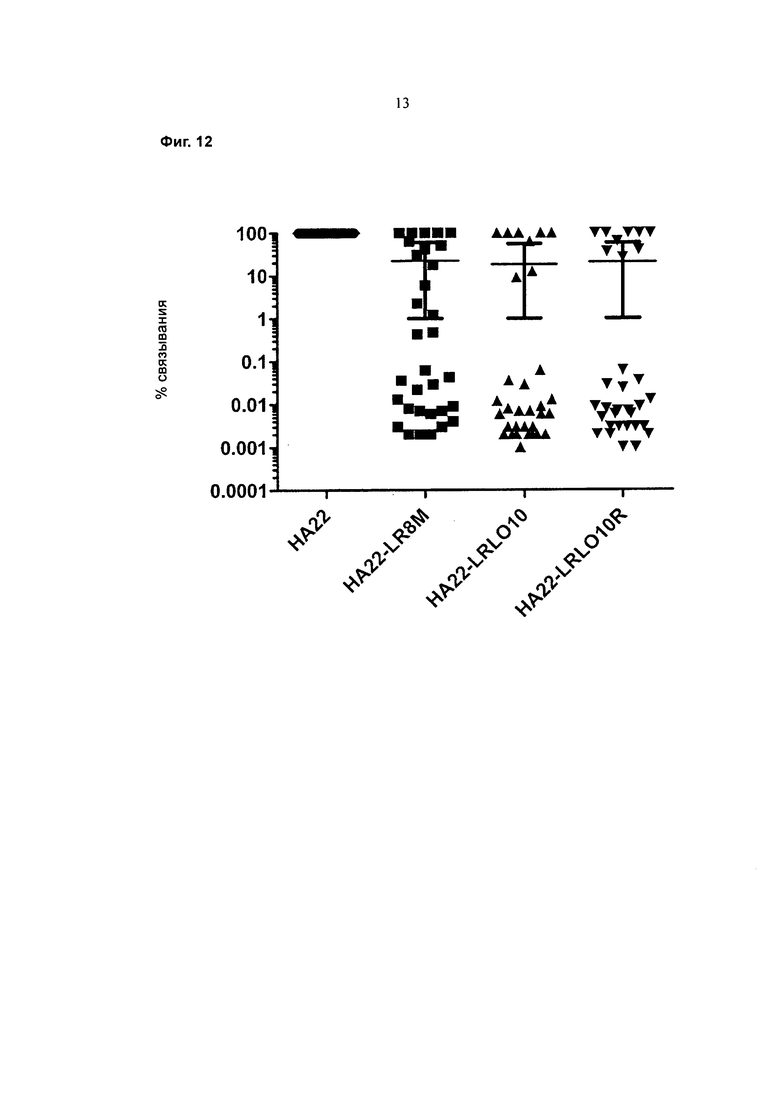
Изобретение относится к биотехнологии. Предложен экзотоксин A Pseudomonas (РЕ), имеющий замену одного или нескольких аминокислотных остатков в положениях L423, F443 и L477. Также рассмотрена химерная молекула, содержащая нацеливающий агент, слитый с РЕ, и молекула, содержащая нацеливающий агент, конъюгированный с РЕ. Кроме того, описаны нуклеиновые кислоты, кодирующие РЕ и химерную молекулу; рекомбинантный экспрессионный вектор; клетка-хозяин для получения РЕ; фармацевтическая композиция для ингибирования роста клеток; способ лечения или предупреждения рака у млекопитающего; способ ингибирования роста клетки-мишени; способы получения РЕ, химерной молекулы, содержащей нацеливающий агент, слитый с РЕ, и молекулы, где РЕ ковалентно связан с нацеливающим агентом. Экзотоксин A Pseudomonas по настоящему изобретению проявляет уменьшенную иммуногенность при сохранении цитотоксичности. В этой связи данное изобретение может найти дальнейшее применение в терапии. 13 н. и 10 з.п. ф-лы, 12 ил., 20 табл., 16 пр.
1. Экзотоксин A Pseudomonas (РЕ), включающий аминокислотную последовательность, имеющую замену одного или нескольких аминокислотных остатков в положениях L423, F443 и L477 из SEQ ID NO: 1, где аминокислотные остатки L423, F443 и L477 определены относительно SEQ ID NO: 1, где
замена аминокислотного остатка L423 представляет собой замену на аланин или серин в положении аминокислотного остатка L423,
замена аминокислотного остатка F443 представляет собой замену на аланин в положении аминокислотного остатка F443, и
замена аминокислотного остатка L477 представляет собой замену на аланин, глицин или гистидин в положении аминокислотного остатка L477.
2. Химерная молекула для ингибирования роста клеток-мишеней, содержащая (а) нацеливающий агент, слитый с (б) РЕ по п.1.
3. Химерная молекула по п.2, в которой нацеливающий агент представляет собой моноклональное антитело.
4. Химерная молекула по п.3, в которой моноклональное антитело специфически связывается с маркером клеточной поверхности, выбранным из группы, состоящей из CD19, CD21, CD22, CD25, CD30, CD33, CD79b, рецептора трансферрина, рецептора эпидермального фактора роста (EGF), мезотелина, кадгерина и Lewis Y.
5. Химерная молекула по п.2, в которой нацеливающий агент представляет собой антитело, выбранное из группы, состоящей из В3, RFB4, SS, SS1, MN, MB, HN1, HN2, НВ21, MORAb-009, НА22 и их антигенсвязывающих частей.
6. Химерная молекула по п.2, в которой нацеливающий агент является антигенсвязывающей частью НА22.
7. Нуклеиновая кислота, кодирующая РЕ по п.1.
8. Нуклеиновая кислота, кодирующая химерную молекулу по любому из пп.2-6.
9. Рекомбинантный экспрессионный вектор, содержащий нуклеиновую кислоту по п.7 или 8.
10. Клетка-хозяин для получения РЕ по п.1, содержащая рекомбинантный экспрессионный вектор по п.9.
11. Молекула для ингибирования роста клеток-мишеней, содержащая (а) нацеливающий агент, конъюгированный с (б) РЕ по п.1.
12. Молекула по п.11, в которой нацеливающий агент представляет собой моноклональное антитело.
13. Молекула по п.12, в которой моноклональное антитело специфически связывается с маркером клеточной поверхности, выбранным из группы, состоящей из CD19, CD21, CD22, CD25, CD30, CD33, CD79b, рецептора трансферрина, рецептора EGF, мезотелина, кадгерина и Lewis Y.
14. Молекула по п.11, в которой нацеливающий агент представляет собой антитело, выбранное из группы, состоящей из В3, RFB4, SS, SS1, MN, MB, HN1, HN2, НВ21, MORAb-009, НА22 и их антигенсвязывающих частей.
15. Молекула по п.11, в которой нацеливающий агент является антигенсвязывающей частью НА22.
16. Фармацевтическая композиция для ингибирования роста клеток, содержащая (а) эффективное количество РЕ по п.1 или химерной молекулы по любому из пп.2-6 и 11-15 и (б) фармацевтически приемлемый носитель.
17. Способ лечения или предупреждения рака у млекопитающего, включающий введение млекопитающему РЕ по п.1, химерной молекулы по любому из пп.2-6 и 11-15 или фармацевтической композиции по п.16 в количестве, эффективном для лечения или профилактики рака у млекопитающего.
18. Способ ингибирования роста клетки-мишени, включающий контактирование клетки с РЕ по п.1, химерной молекулой по любому из пп.2-6 и 11-15 или фармацевтической композицией по п.16 в количестве, эффективном для ингибирования роста клетки-мишени.
19. Способ по п.18, где клетка-мишень представляет собой раковую клетку.
20. Способ по п.18 или 19, где клетка-мишень экспрессирует маркер клеточной поверхности, выбранный из группы, состоящей из CD19, CD21, CD22, CD25, CD30, CD33, CD79b, рецептора трансферрина, рецептора EGF, мезотелина, кадгерина и Lewis Y.
21. Способ получения РЕ по п.1, включающий (а) рекомбинантную экспрессию РЕ по п.1 и (б) очистку РЕ.
22. Способ получения химерной молекулы по любому из пп.2-6, включающий (а) рекомбинантную экспрессию химерной молекулы по любому из пп.2-6 и (б) очистку химерной молекулы.
23. Способ получения молекулы по любому из пп.11-15, включающий (а) рекомбинантную экспрессию РЕ по п.1, (б) очистку РЕ и (с) ковалентное связывание нацеливающего агента с очищенным РЕ.
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| ONDA M | |||
| et al., "Characterization of the B cell epitopes associated with a truncated form of Pseudomonas exotoxin (PE38) used to make immunotoxins for the treatment of cancer patients", The Journal of Immunology, 2006, 177(12), 8822-8834 | |||
| Колосоуборка | 1923 |
|
SU2009A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| ROSCOE D | |||
| M | |||
| et al., "Identification of epitopes on a mutant form of Pseudomonas exotoxin using serum from humans treated with Pseudomonas exotoxin containing immunotoxins", European journal of immunology, 1997, 27(6), 1459-1468 | |||
| BECERRA J.C | |||
| et al | |||
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| ЛЯМИН А.В | |||
| "Особенности биологических свойств и факторов патогенности штаммов Pseudomonas aeruginosa, выделенных из разных источников." Аспирант | |||
| вестн | |||
| Поволжья, 2010, 3-4: 220-223. | |||

