Изобретение относится к биотехнологии, генной инженерии, микробиологической и медицинской промышленности и касается новых генетических конструкций и продуцентов, обеспечивающих в промышленных масштабах синтез рекомбинантного белка флагеллина Salmonella (FliC).
Флагеллин, основной компонент бактериальных жгутиков, является фактором вирулентности, который распознается врожденной иммунной системой у таких разнообразных организмов, как насекомые, растения и млекопитающие (Hayashi, F. et al,.Nature 2001;410, 1099-1103). Являясь естественным агонистом человеческого toll-подобного рецептора 5 (TLR5), флагеллин активирует врожденный иммунный ответ, который считается важным для инициирования и регулирования адаптивного иммунного ответа. TLR5, связывается со специфическим лигандом FliC, при этом происходит:
1) инициирование внутриклеточного сигнального пути, ведущего к выработке про-воспалительных цитокинов TNF, IL-6 и IL-12 (Pasare С, Medzhitov R, Science 2003; 299:1033-1036);
2) регуляция экспрессии генов, связанных с фагоцитозом;
3) усиление экспрессии основного комплекса гистосовместимости класса II (МНС II) в антигенпрезентирующих клетках (APCS) (Medzhitov R, et al, Nature 1997;388:394-397; Yamamoto M at al, Nature 2002;420:324-329; A. N. Honko, S. B. Mizel, Immunologic Research 2005;33/l:83-101).
TLR5 присутствует на поверхности эпителиальных и иммунных клеток, таких как дендритные клетки (DC), моноциты, естественные клетки-киллеры (NK) и Т-лимфоциты. FliC активирует эпителиальные клетки, хорошо известные своей способностью противостоять патогену и защищать организм от него. Т.е. FliC зависимая активация TLR5 прямо или косвенно изменяет активность клеток иммунной системы, которые участвуют как в гуморальных, так и в клеточно-опосредованных адаптивных иммунных реакциях. Таким образом, иммуномодулирующий белок флагеллина (FHC), полипептид или его модификация, действует как адъювант для стимуляции общего иммунного ответа.
Применение мономера флагеллина S. typhimurium типа 2 (FHC), в сочетании с протективным антигеном возбудителя (бактериального, протозойного или вирусного), повышает адаптивный иммунитет, снижает количество соответствующего антигена на дозу вакцины (Honko, and Mizel, 2004; Infect Immun;72:6676-6679; Honko et al., 2006; Infect Immun 74:1113-1120) и вызывает увеличение концентрации антител в сыворотке крови (Bargieri et al., 2008; Vaccine;26:6132-6142, Weimer et al., 2009; Vaccine; 27:6762-6769, Song et al., 2009; Vaccine; 27:5875-5884, Tussey et al., 2016; Open Forum Infect Dis;3. DOI: 10.1093/ofid/ofw015). Более того, его применение приводит к снижению аллергенности, гиперактивности вакцины и обеспечению долгосрочной защиты организма от соответствующего возбудителя инфекции. Минимальная доза мономера FliC, необходимая для воздействия на врожденную иммунную систему, была определена как 1-10 мкг в исследованиях на приматах (Weimer et al, 2009; Vaccine; 27:6762-6769).
За последние несколько лет возник большой интерес к флагеллину с целью использования его в качестве адъюванта в вакцинах для стимуляции гуморальных и клеточно-опосредованных иммунных реакций, вследствие этого появилась потребность в масштабном получение белка FliC.
Бактериальные жгутики Salmonella являются гомо-полимерами, состоят из 20000 - 100000 субъединиц флагеллина с молекулярной массой примерно 51,6 kD, каждая субъединица содержит 495 аминокислот без цистеиновых остатков. Мономер флагеллина состоит из четырех доменов: 1) N- и С-концевой D0, 2) высоко консервативный D1, 3) D2 и 4) гипервариабельный D3 (Yonekura et al, 2003; Nature; 424:643-650). Активации TLR5 в первую очередь способствует конфигурация N- и С-концевых доменов D0 и D1 (Yang et al., 2013; Hum Vaccines Immunother; 9:1084-1092). Т.к. D0 и D1 домены расположены внутри бактериальных жгутиков, активность полимерных форм FliC выражена слабо. Гипервариабельный домен D3, находится на поверхности жгутиков и его удаление или модификация, если сохраняется конформация доменов D1 и D0, не влияет на активацию TLR5 (Yang et al., 2013; Hum Vaccines Immunother; 9:1084-1092; Liu et al., 2011; PLOS ONE 201 l;6:e20928). Т.е., модификация структуры флагеллина путем делеции домена D3 и введение чужеродной антигенной последовательности, или присоединение к флагеллину с N- конца или С- конца белковой последовательности антигена возбудителя, позволяет создавать рекомбинантные структуры белков, обладающих новыми иммуногенными и протективными свойствами.
Были созданы и испытаны вакцины на основе следующих рекомбинантных белков: флагеллина-гемагглютинина вируса гриппа (Li Song, et al. 2015; BMC Biotechnology 15:79), флагеллина-F антигена Yersinia pestis, флагеллина-V антигена Yersinia pestis, флагеллина-DENV2 поверхностного белка вируса Денге (Dengue virus) (Bennett et al. 2015; BMC Biotechnology 15:71), флагеллина-иммуногенных эпитопов белков VP6 и VP8 ротавируса (RU 2649132, RU 2539913). Проведенные на животных испытания рекомбинантных вакцин на основе рекомбинантного флагеллина, продемонстрировали низкую реактогенность, высокую иммуногенность и высокую протективность по сравнению с использованием антигенов возбудителей без флагеллина (Tussey L, et.al., 2016; Open Forum Infect Dis; DOI: 10.1093/ofid/ofw015).
Известен способ получения флагеллина из Salmonella typhimurium (В. Н. Oliveira, et.al., 2011; Brazilian Journal of Chemical Engineering, Vol.28, No. 04, pp. 575 -584, October - December). Основными недостатками этого способа получения флагеллина являются: использование в технологии патогенного штамма Salmonella typhimurium, способного вызывать заболевание человека и низкий выход продукта, около 300 мг/л культуральной среды. Напротив, геном кишечной палочки хорошо охарактеризован, а производство белков в клетках Е. coli является быстрым, безопасным и экономичным (Lee, 1996; Trends Biotechnol; 14:98-105, Song L, et al, 2015, US9200042). Таким образом, производство флагеллина и рекомбинантных продуктов на его основе в клетках Е. coli является предпочтительным.
Все известные продуценты флагеллина сконструированы на основе векторов семейства рЕТ, в которых экспрессия находится под контролем промотора фага Т7 (рТ7), регулируется lac оператором, который индуцируется индуктором IPTG (RU 2649132, RU 2539913, RU 2524133, S. A. Mirhosseini, et al.; 2017, Brazilian journal of microbiology 48, 2017; 774-781;I. A. Hajam et al., 2013, Vet. Ital. Apr-Jun 2013;49(2):181-6, J.Renfroe; 2018, https://scholarworks.gsu.edu/cgi/viewcontent.cgi?article=1215&context=biology_diss). Недостатками этих продуцентов являются: использование богатых сред для ферментации, применение индуктора IPTG (изопропил-β-D-тиогалактозид), невозможность получения высоких значений оптической плотности (о. п. ) культуры при ферментации и, соответственно, низкий выход целевого продукта - не более 100 мг на литр ферментационной среды. Все это, в конечном итоге, приводит к удорожанию производства.
Для очистки рекомбинантного флагеллина в настоящее время используют следующие методы: водная двухфазная экстракция, осаждение сульфатом аммония или полиэтиленгликолем, аффинная металл-хелатная хроматография, ионообменные хроматографии, хроматография гидрофобного взаимодействия, гель-хроматография (Song et al., 2015; US 9200042)
Предлагаемое изобретение решает задачу конструирования бактериального штамма - продуцента, позволяющего получать чистый рекомбинантный флагеллин Salmonella typhimurium с высоким выходом (до 2 г субстанции фармакопейного качества с 1 л культуральной среды) при ферментации на синтетических средах, не содержащих продуктов животного происхождения.
Технический результат заключается в получении нового продукта-рекомбинантной плазмиды pFM-FliC_syn, обеспечивающей экспрессию рекомбинантного белка флагеллина Salmonella (FliC) и моноплазмидного штамма бактерий Escherichia coli FM-FliC_syn, а также в повышении выхода целевого продукта, упрощении технологии получения белка флагеллина Salmonella за счет конструирования рекомбинантной плазмидной ДНК pFM-FliC syn и создания на ее основе высокопродуктивного стабильного штамма Escherichia coli FM-FliC_syn, при культивировании которого на синтетических средах оптическая плотность культуры достигает высоких значений (до 50-60 о. е. и выше), а уровень экспрессии целевого продукта составляет до 3 г с 1 л культуральной среды.
Изобретение поясняется следующими рисунками.
На фиг. 1 приведена нуклеотидная последовательность синтетического гена kanR.
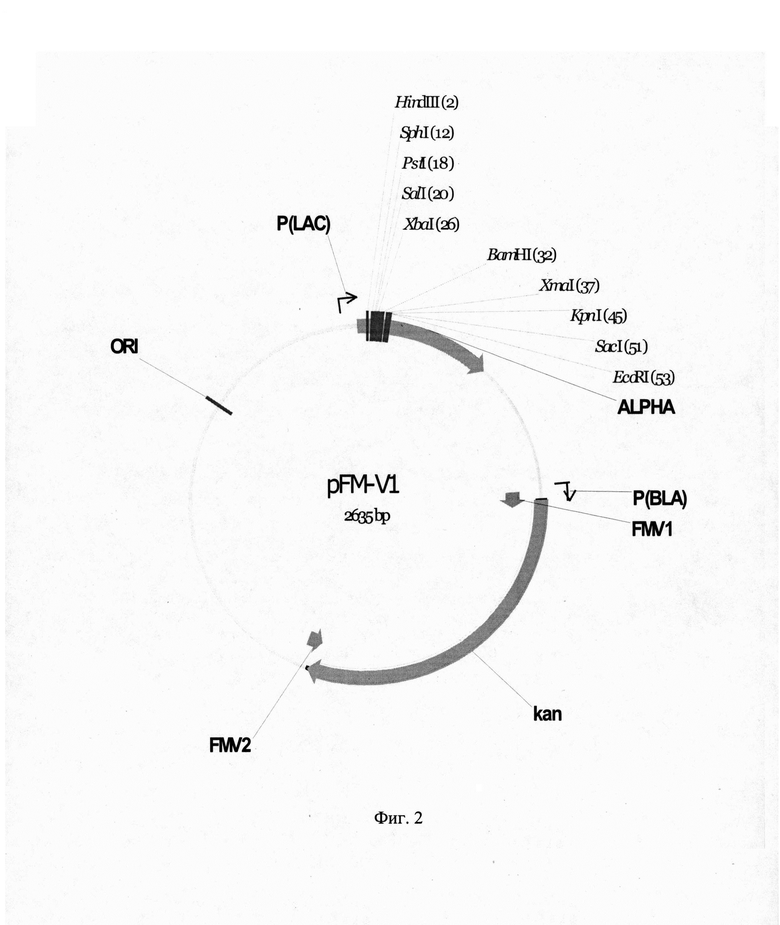
На фиг. 2 представлена рестрикционная карта векторной плазмиды pFM-V1.
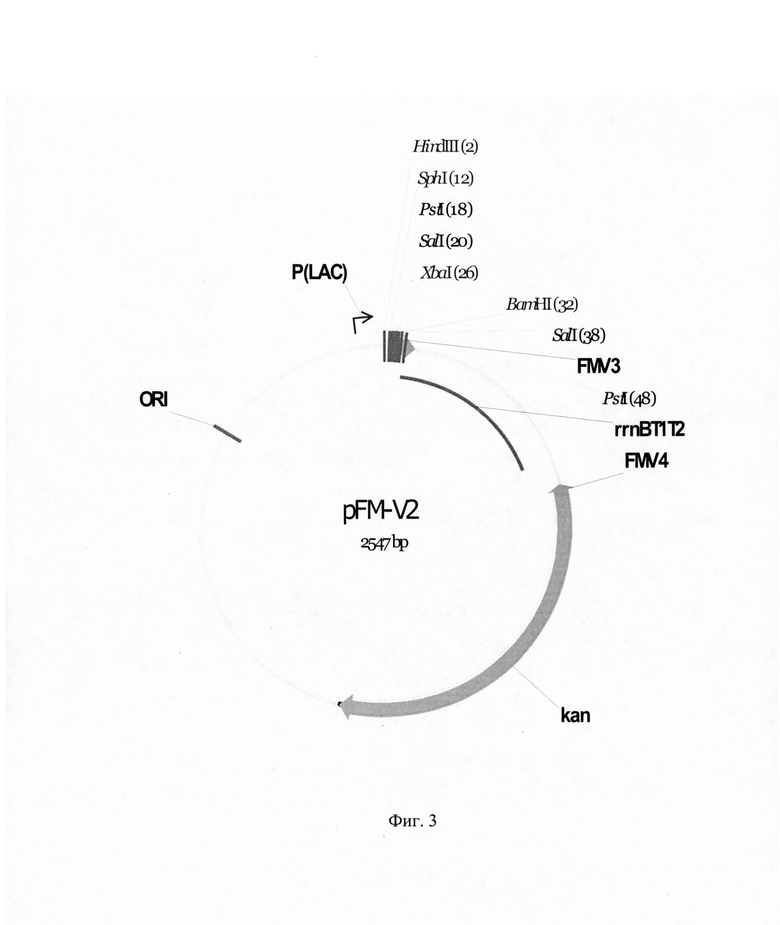
На фиг. 3 приведена рестрикционная карта векторной плазмиды pFM-
V-2.
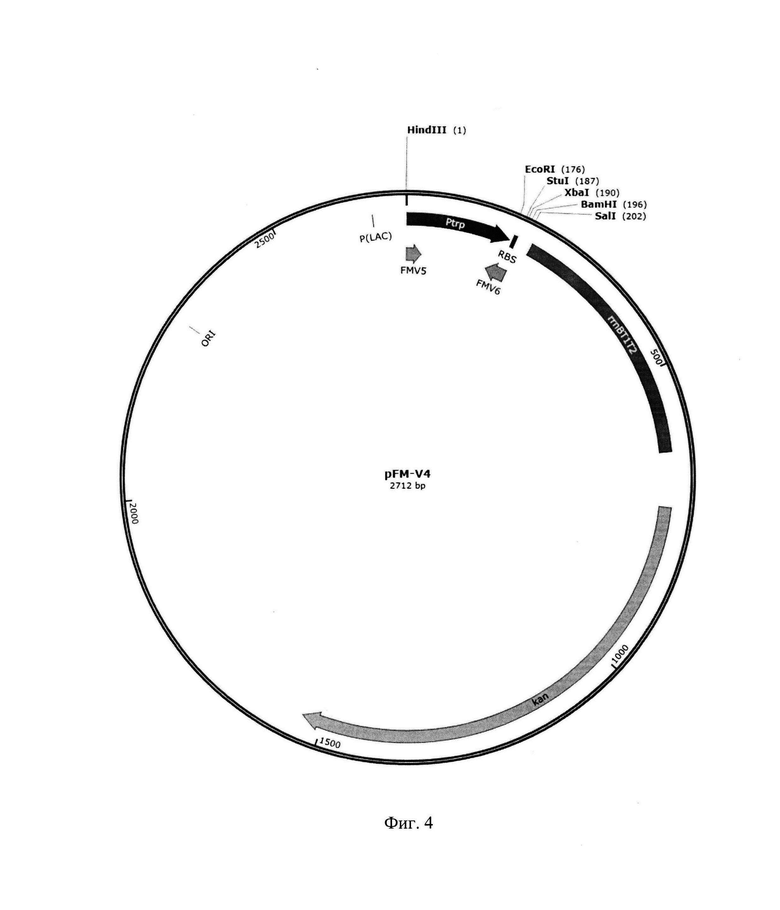
На фиг. 4 приведена рестрикционная карта рекомбинантной плазмиды pFM-V-4.
На фиг. 5 представлена нуклеотидная последовательность синтетического гена флагеллина Salmonella (FliC), встроенного в плазмиду pFM-FliC_syn.
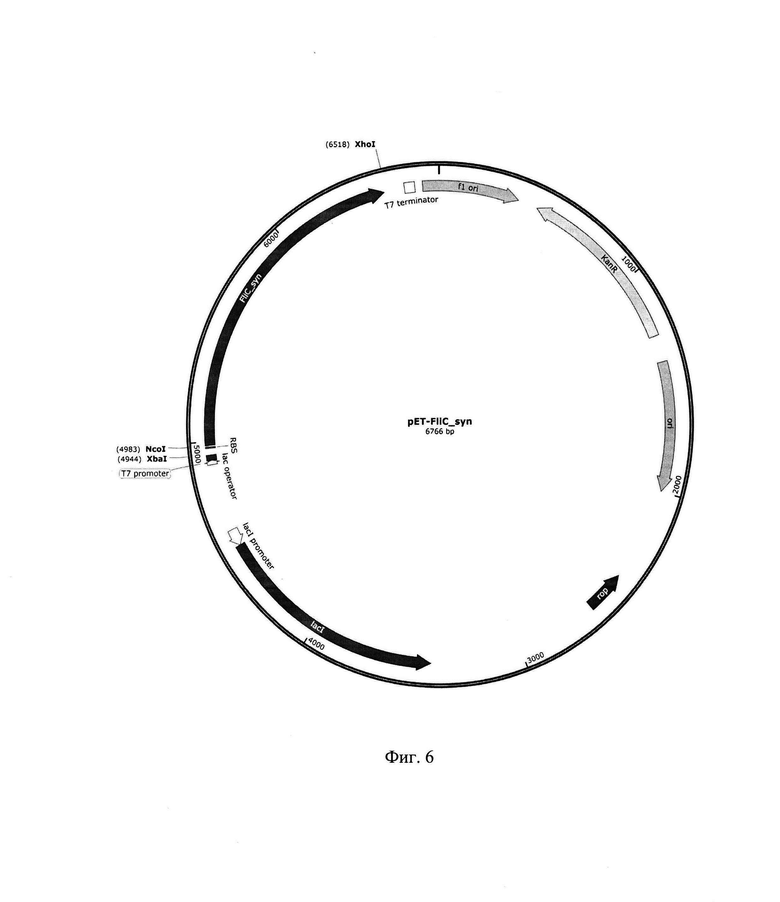
На фиг. 6 приведена рестрикционная карта рекомбинантной плазмиды pET-FliC_syn.
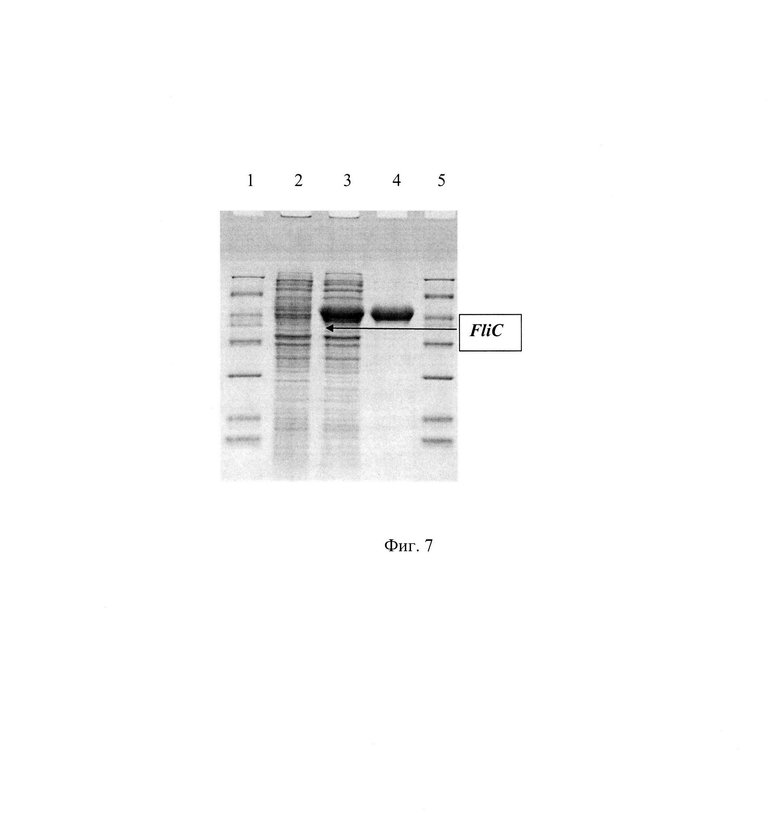
На фиг. 7 изображено накопление флагеллина в клетках штамма E.coli BL21(DE3)_pET-FliC_syn в процессе ферментации, где: лунка 1 - смесь маркерных белков; лунка 2 - ночная культура, посевной материал культуральной жидкости (КЖ); лунка 3 - восьмой час ферментации КЖ; лунка 4 - стандарт флагеллина FliC, 2,5 мкг; лунка 5 - смесь маркерных белков.
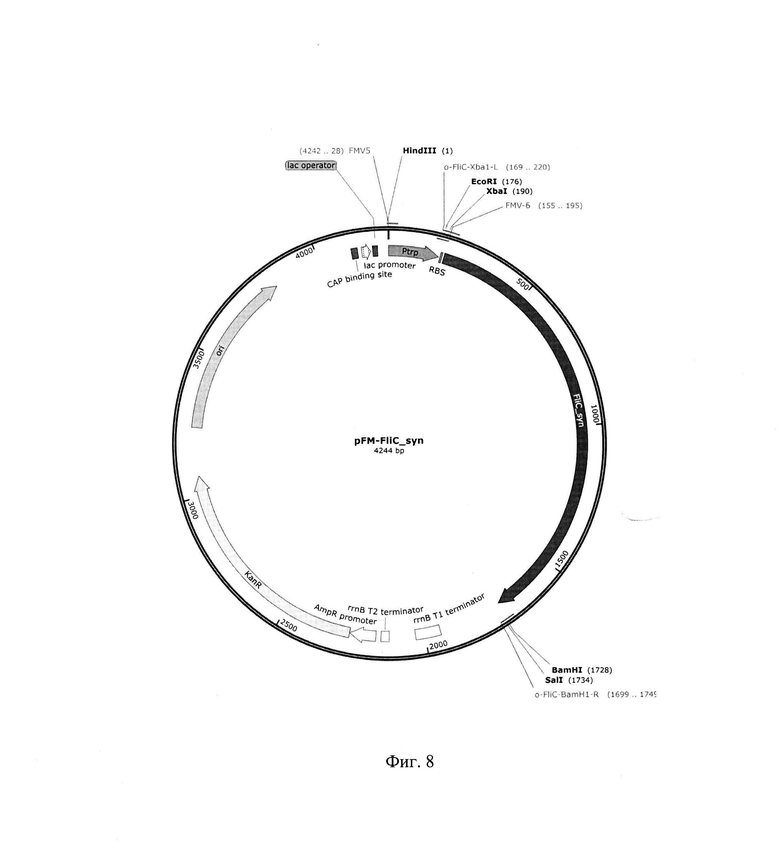
На фиг. 8 представлена рестрикционная карта рекомбинантной плазмиды pFM-FliC_syn.
Сущность изобретения
Рекомбинантная плазмидная ДНК pFM-FliC_syn кодирует синтез рекомбинантного белка флагеллина Salmonella (FliC) под контролем триптофанового и лактозного промоторов и терминатора транскрипции rrnBT1T2, а также содержит в качестве генетического маркера синтетический ген kanR, детерминирующий устойчивость к антибиотику канамицину.
Плазмида pFM-FliC_syn имеет 4244 пар оснований (п.о.), и характеризуется наличием следующих фрагментов:
- фрагмент ДНК размером 194 п. о., включающий триптофановый промотор E.coli и последовательность Шайн - Дельгарно (SD), ответственную за инициацию трансляции;
- синтетический фрагмент ДНК размером 1533 п. о., включающий оптимизированную кодирующую часть гена fliC (GenBank: AAL20871.1); при оптимизации гена флагеллина из последовательности удалены внутренние TATA-боксы, chi-сайты, RBS последовательности, АТ-богатые и GC-богатые последовательности, повторяющиеся последовательности и потенциальные вторичные структуры мРНК; частота использования кодонов была адаптирована для Е. coli, при этом параметр CAI (индекс адаптации кодонов)=0,81 (https://www.genscript.com/tools/rare-codon-analysis);
- фрагмент ДНК плазмиды рКК223-3 размером 534 п. о., включающий последовательность терминатора транскрипции rrnBT1T2, последовательности промотора и SD последовательность гена бета-лактомазы;
- синтетический фрагмент ДНК размером 810 п. о., включающий синтетический ген kanR, обеспечивающий устойчивость к канамицину - аналог гена kanR из транспозона Tn903, в котором удалены сайты рестрикции AvaI, ClaI, SmaI, Hindlll;
- фрагмент ДНК размером 1173 п. о. плазмиды pUC19, включающий последовательности: ответственную за репликацию плазмиды (ori) и лактозный промотор.
Моноплазмидный штамм E.coli BL21 FM-FliC_syn содержит рекомбинантную плазмиду pFM-FliC_syn, обладает высоким уровнем экспрессии белка флагеллина Salmonella (FliC). Рекомбинантный штамм бактерий E.coli FM-FliC_syn получен путем трансформации клеток штамма Е. coli BL21 рекомбинантной плазмидной ДНК pFM-FliC_syn.
Преимущества настоящего изобретения заключается в использовании стабильно наследуемой мультикопийной рекомбинантной плазмиды pFM-FliC_syn, в которой под контролем индуцибельного триптофанового и лактозного промоторов и rrnT1T2 терминатора транскрипции расположен синтетический ген флагеллина Salmonella (FliC) с оптимизированными для клеток бактерий E.coli кодонами, кодирующими соответствующие аминокислоты и детерминирующей высокий уровень экспрессии целевого продукта на минимальных синтетических средах.
Штаммы E.coli FM-FliC_syn характеризуется следующими признаками. Культурально-морфологические признаки
Штамм обладает свойствами типичного представителя вида Escherichia coli. Клетки мелкие, прямые, утолщенной палочковидной формы, грамотрицательные, неспороносные.
Штамм хорошо растет на простых питательных средах. Колонии при росте штамма на L-агаре (LA) - круглые, гладкие, выпуклые, мутные, блестящие, серые, с ровными краями. В жидких средах (в минимальной среде с глюкозой или в L- бульоне (LB)) штамм образует интенсивную ровную муть.
Физиолого-биохимические признаки
Штамм растет в аэробных условиях, при температуре в пределах от 4 до 42°С. Оптимум рН для роста составляет 6,5-7,5.
В качестве источника азота клетки штамма используют минеральные соли в аммонийной или нитратной формах; органические соединения, в частности, аминокислоты, пептон, триптон, дрожжевой экстракт и т.д.
В качестве источника углерода клетки штамма могут использовать аминокислоты, глицерин, углеводы.
Устойчивость к антибиотикам
Штамм проявляет устойчивость к канамицину (100 мкг/мл). Особенность штамма Продуцирует белок флагеллина. Условия хранения штамма
Штамм хранится: под маслом в L-агаре с добавлением канамицина до концентрации 100 мкг/мл; криоконсервированным в L-бульоне, содержащем 15% глицерина и антибиотик канамицин 100 мкг/мл, в ампулах при температуре минус 40°-70°С; или в лиофилизированном состоянии в ампулах при температуре плюс 4°С.
Штамм E.coli FM-FliC_syn стабильно синтезирует белок флагеллина с высоким уровнем экспрессии (2-3 г с 1 л культуральной жидкости). Штамм технологичен и для достижения высокой продуктивности не требуют особых условий для культивирования, специальных индукторов экспрессии целевого продукта, а также применения сложных питательных сред, включающих компоненты животного происхождения.
Целевой продукт накапливается в количестве более 15-20% от суммарного белка клетки в растворимой форме. Использование нового продуцента, содержащий плазмиду pFM-FliC_syn, позволит значительно повысить выход целевого продукта, а также упростить технологический процесс получения белка флагеллина за счет упрощения стадии культивирования штаммов.
Изобретение иллюстрируется, но не ограничивается следующими примерами.
Пример 1
Конструирование рекомбинантной плазмиды pFM-FliC_syn, обеспечивающей экспрессию синтетического гена флагеллина Salmonella typhimurium (FliC) (GenBank: AAL20871.1) и конструирование рекомбинантного штамма Escherichia coli FM-FliC_syn
Способ получения рекомбинантной плазмиды pFM-FliC_syn включает несколько последовательных этапов:
- конструирование векторной плазмиды pFM-V-1 (2635 п. о.);
- конструирование векторной плазмиды pFM-V-2 (2547 п. о.);
- конструирование векторной плазмиды pFM-V-4 (2712 п. о.);
- конструирование рекомбинантной плазмиды pET-FliC_syn (6766
п. о.);
- конструирование рекомбинантной плазмиды pFM- FliC_syn (4244
п. о.);
- конструирование рекомбинантного штамма Е. coli FM-FliC_syn. 1.1. Конструирование векторной плазмиды pFM-V-1
Исходным вектором при получении векторной плазмиды pFM-V-1 служит вектор pUC19, в котором кодирующая последовательность гена бета-лактомазы, обеспечивающего устойчивость к ампициллину, заменена на кодирующую последовательность синтетического гена kanR, обеспечивающего устойчивость к канамицину. Для получения векторной плазмиды pFM-V-1 провели 2 раунда амплификации ДНК. Для первого раунда реакции амплификации использовали два синтетических олигонуклеотида FMV1 hFMV2: FMV1-5'-
ATAATATTGAAAAAGGAAGAGTATGAGCCATATTCAACGGGAAAC-
3'
FMV2-5'-
GAGTAAACTTGGTCTGACAGTTAGAAAAACTCATCGAGCATC-3'
и в качестве матрицы - ДНК синтетического гена kanR - аналога гена kanR из транспозона Тп903, в котором удалены сайты рестрикции AvaI, ClaI, SmaI, HindIII.
Данную амплификацию и все последующие манипуляции проводили в ДНК-амплификаторе в буферном растворе, содержащем: 20 тМ Tis- НС1, рН 8.8, 10 тМ (NH4)2S04, 10 тМ КС1, 2 mM MgCl2, 0.1% Triton Х100, 0.1 мг/мл BSA, 0.2 тМ каждого dNTP, 1.25 ед. Pfu ДНК полимеразы, 100 нг ДНК матрицы при следующих режимах: прогревание - 5 мин при 96°С, далее 35 циклов ПЦР (режимы одного цикла: 30 с при 96°С, 30 с при 58°С, 1 мин при 72°С) и затем финальная элонгация - 10 мин при 72°С. В результате полимеразной цепнаой реакции (ПЦР) получили фрагмент размером 852 п. о., который электрофоретически очистили.
Для второго раунда амплификации использовали векторную ДНК плазмиды pUC19 и фрагмент ДНК, полученный в первом раунде амплификации в эквимолярных количествах при следующих режимах: прогревание - 5 мин при 96°С, далее 6 циклов ПЦР (режимы одного цикла: 30 с при 96°С, 30 с при 58°С, 3 мин при 72°С) и затем финальная элонгация - 10 мин при 72°С.Полученную ДНК во втором раунде ПЦР трансформировали в клетки штамма E.coli DH5a и клетки высеяли на среду LA, содержащую 100 мкг/мл канамицина. После инкубирования в течение 12 ч при 37°С, клоны отсеяли, выделили плазмидную ДНК, провели рестрикционный анализ и определили первичную структуру ДНК. В результате получили плазмиду pFM-V-1 размером 2635 п. о. Рестрикционная карта рекомбинантной плазмиды pFM-V-1 приведена на рис. 2. 1.2. Конструирование векторной плазмиды pFM-V-2 Векторная плазмида pFM-V-2 представляет собой плазмиду pFM-V-1, в которой после сайта BamHI последовательность ДНК, кодирующуюя альфа LacZ пептид и промотор бета-лактомазы заменена на последовательность ДНК, кодирующую терминатор транскрипции rrnBT1T2 и промотор бета-лактомазы из ДНК вектора pKK223-3. Генетическое конструирование плазмиды pFV-V-2 осуществили в два раунда амплификации ДНК с помощью полимеразной цепной реакции (ПЦР). Для первого раунда реакции амплификации использовали два синтетических олигонуклеотида FMV3 и FMV4:
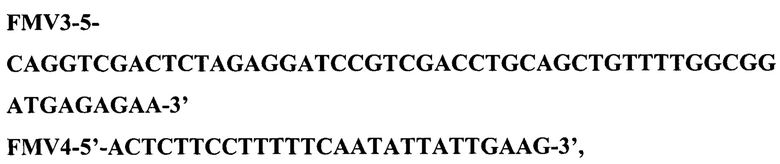
а в качестве матрицы - ДНК плазмиды рКК223-3. ПЦР провели при следующих режимах: прогревание - 5 мин при 96°С, далее 35 циклов ПЦР (режимы одного цикла: 30 с при 96°С, 30 с при 58°С, 1 мин при 72°С) и затем финальная элонгация - 10 мин при 72°С. В результате ПЦР получили фрагмент размером 549 п. о., который электрофоретически очистили.
Для второго раунда амплификации использовали в эквимолярных количествах векторную ДНК плазмиды pFM-V-1 и фрагмент ДНК, полученный в первом раунде амплификации. Амплификацию осуществляли при следующих режимах: прогревание - 5 мин при 96°С, далее 6 циклов ПЦР (режимы одного цикла: 30 с при 96°С, 30 с при 58°С, 3 мин при 72°С) и затем финальная элонгация - 10 мин при 72°С.Полученную ДНК во втором раунде ПЦР трансформировали в клетки штамма E.coli DH5α и высеяли на среду LA, содержащую 100 мкг/мл канамицина, IPTG, X-gal. После инкубирования в течение 12 ч при 37°С, белые колонии пересеяли, выделили плазмидную ДНК, провели рестрикционный анализ и определили первичную структуру ДНК. В результате получили плазмиду pFM-V-2 размером 2547 п. о.
1.3. Конструирование векторной плазмиды pFM-V-4
Плазмида pFM-V-4 представляет собой плазмиду pFM-V-2, в которую клонирован фрагмент ДНК, кодирующий бактериальный триптофановый промотор Е. coli и SD последовательность по сайтам рестрикции HindIII-XbaI, размером 194 п. о.
HindIII-XbaI фрагмент ДНК размером 194 п. о., кодирующий триптофановый промотор Е. coli и SD последовательность, получили методом ПЦР, используя в качестве матрицы тотальную ДНК E.coli и праймеры FMV5 и FMV6 с последующей обработкой амплифицированного фрагмента ферментами рестрикции Hindlll и Xbal.
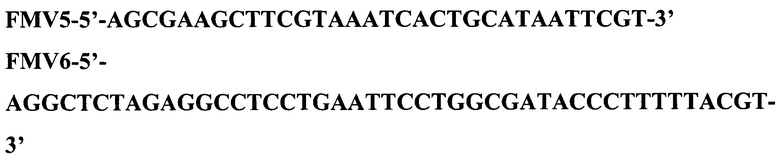
Далее, предварительно очищенный фрагмент HindIII-XbaI и плазмиду pFM-V-2, обработанную ферментами рестрикции HinIII и XbaI объединили, лигировали ферментом лигазой фага Т4, ДНК трансформировали в клетки штамма E.coli DH5α и высеяли на среду LA, содержащую 100 мкг/мл канамицина. После инкубирования в течение 12 ч при 37°С, клоны отсеяли, выделили плазмидную ДНК, провели рестрикционный анализ и определили первичную структуру ДНК.
В результате получили плазмиду pFM-V-4 размером 2712 п. о.
1.4. Конструирование рекомбинантной плазмиды pET-FliC_syn
Плазмида pET-FliC_syn представляет собой векторную плазмиду рЕТ28, в которую по сайтам рестрикции NcoI и XhoI клонирован фрагмент ДНК размером 1533 п. о., кодирующий синтетический ген FliC_syn, с оптимизированными для клеток бактерий E.coli кодонами, кодирующими соответствующие аминокислоты и детерминирующими высокий уровень экспрессии целевого продукта. При оптимизации гена FliCsyn из последовательности были удалены внутренние TATA-boxes, chi-sites, RBS последовательности, АТ-богатые и GC-богатые последовательности, повторяющиеся последовательности и потенциальные вторичные структуры мРНК; codon usage был адаптирован для Е. coli, при этом параметр CAI (codon adaptation index)=0,81.
Синтетический ген FliC_syn был получен стандартным методом химико-ферментативного синтеза. Рекомбинантную плазмиду pET-FliC_syn трансформировали в компетентные клетки штаммов E.coli DH5a и E.coli BL21(DE3). Клетки высеяли на среду LA, содержащую 100 мкг/мл канамицина. После инкубирования в течение 12 ч при 30°С, клоны отсеяли, выделили плазмидную ДНК, провели рестрикционный анализ и определили первичную структуру ДНК. В результате получили штаммы E.coli DH5a_pET-FliC_syn и E.coli BL21(DE3)_pET-FliC_syn. Так как в рекомбинантной плазмиде pET-FliC_syn ген флагеллина находится под контролем Т7 промотора и 1ас оператора, клетки штамма E.coli ЕТ-FliC_syn_BL21(DE3) проверили на способность к экспрессии рекомбинантного флагеллина по стандартной методике для штаммов, содержащих плазмиды серии рЕТ.
1.5. Конструирование рекомбинантной плазмиды pFM- FliC_syn Плазмида pFM-FliC_syn представляет собой векторную плазмиду pFM V-4, в которой ген, кодирующий синтез флагеллина Salmonella (FliC), клонирован по сайтам XbaI-BamHI. Для конструирования плазмиды pFM-FliC_syn осуществили амплификацию ДНК с помощью полимеразной цепной реакции (ПЦР) с использованием двух синтетических олигонуклеотидов: o-FliC-Xbal-L и o-FliC-BamH1-R: FliC-Xba1-L-5'-
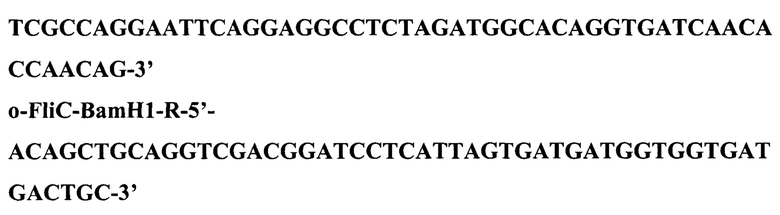
и в качестве матрицы - ДНК плазмиды pET-FliC_syn. ПЦР провели при следующих режимах: прогревание - 5 мин при 96°С, далее 35 циклов ПЦР (режимы одного цикла: 30 с при 96°С, 30 с при 58°С, 1 мин при 72°С) и затем финальная элонгация - 10 мин при 72°С. В результате ПЦР получили фрагмент размером 1533 п. о., который электрофоретически очистили.
ПЦР фрагмент ДНК размером 1533 п. о. и ДНК плазмиды pFM-V-4 обработали ферментами рестрикции Xbal и BamHI, ДНК слигировали ферментом лигазой фага Т4 и трансформировали в компетентные клетки штамма E.coli DH5α. Клетки высеяли на среду LA, содержащую 100 мкг/мл канамицина. После инкубирования в течение 12 ч при 30°С, клоны отсеяли, выделили плазмидную ДНК, провели рестрикционный анализ и определили первичную структуру ДНК. В результате получили плазмиду pFM-FliC_syn. размером 4244 п. о. Рестрикционная карта рекомбинантной плазмиды pFM-FliC_syn приведена на рис. 8.
1.6. Конструирование рекомбинантного штамма Escherichia coli FM-FliC_syn
Путем трансформации клеток штамма Е. coli BL21 рекомбинантной плазмидой pFM-FM-FliHC_syn получили трансформанты, которые высеяли на среду LA, содержащую 100 мкг/мл канамицина.
После инкубирования в течение 12 ч при 30°С, клоны отсеяли, и из одного клона с подтвержденной структурой ДНК плазмиды pFM-FliHC_syn получили штамм Escherichia coli FM-FliC_syn.
Пример 2
Культивирование штамма Е. coli FM-FliC_syn
Посевной материал штамма Е. coli FM-FliC_syn культивировали в течение 12 ч при 30°С в 3 л LB среды, содержащей 1хМ9, 0,4% глюкозы, 100 мг/мл канамицина и асептически внесли в ферментер, содержащий 27 л синтетической минерально-солевой среды, содержащей 1хМ9, 1% глюкозы, следовые количества металлов и 100 мг/мл канамицина.
Культивирование в ферментере осуществляли при температуре 30°С, поддерживая рН 6,9+0,15 до оптической плотности 10-12 о. е., после чего температуру поднимали до 38°С и культивирование продолжали до достижения стационарной фазы. Концентрацию растворенного кислорода в диапазоне (30±0,5) % от насыщения поддерживали путем изменения скорости оборотов мешалки от 100 до 800 об/мин и подачи воздуха от 1 до 15 л/мин. Концентрацию субстратов, в частности глюкозы, поддерживали в течение ферментации путем подачи концентрированного 40% раствора глюкозы перистальтическим насосом.
Накопление флагеллина в клетках бактерий происходит, в основном, в виде растворимой формы и незначительное количество - в нерастворимой форме в виде "телец включений", которые контролировали с помощью фазово-контрастной микроскопии. Ферментацию останавливали по достижении максимальной оптической плотности (~ 50-60 о.е.). Содержание флагеллина в биомассе клеток, получаемой с 1 л культуры, составляло 2-3 г флагеллина.
По окончании ферментации культуральную жидкость сепарировали центрифугированием, бактериальную биомассу фасовали в полиэтиленовые пакеты и замораживали при температуре минус 70°С.
Пример 3
Способ получения рекомбинантного флагеллина Salmonella (FliC) из биомассы штамма Е. coli FM-FliC_syn
Получение флагеллина проводили в 5 этапов:
1 этап.Дезинтеграция биомассы и грубая очистка растворимой формы флагеллина.
2 этап.Хроматографическая очистка флагеллина на металл-хелатной смоле.
3 этап.Хроматографическая очистка флагеллина на катионообменной смоле.
4 этап.Хроматографическая очистка флагеллина на анионообменной смоле.
5 этап.Хроматографическая очистка флагеллина методом колоночной гель-фильтрации.
На первом этапе биомассу рекомбинантного штамма Е. coli FM-FliC syn дезинтегрировали в проточном дезинтеграторе типа Френч-пресс при давлении 800-900 атм. в высоко солевом фосфатном буфере 1 (50 mM Na2HPO4, 300 mM NaCl, 10 mM imidazole, рН 7-8). Клеточный дебрис удалили центрифугированием, а растворимую форму флагеллина осаждали 1,5-2,0 М сульфатом аммония и растворили в высоко солевом фосфатном буфере 1.
На втором этапе раствор белка флагеллина очищали методом аффинной хроматографии на Ni хелатных смолах типа Ni-NTA Agarose (Qiagen), Ni Sepharose Fast Flow (Cytiva), Profinity IMAC Resin, Ni-charged (Bio Rad). Флагеллин элюировали градиентом имидазола от 10mM до 300 mM в высоко солевом фосфатном буфере 1.
На третьем этапе полученный раствор флагеллина, разбавляли буфером 50 мМ уксусной кислоты рН 3,0 и очищали на катионообменной смолах топа Toyopearl SP-650M (Tosoh Bioscience), SP Sepharose Fast Flow (Cytiva). Флагеллин элюировали градиентом создаваемый буфером 50 mM уксусной кислоты, рН 3.0, и буфером 20 мМ Tris НС1, рН 8.0.
На четвертом этапе флагеллин очищали на анионообменниках типа Toyopearl Q-650M (Tosoh Bioscience) или Macro-Prep Q (Bio-Rad). Флагеллин элюировали градиентом создаваемый буферами 20 мМ Tris/HCl, рН 8.0 и 20 мМ Tris/HCl, рН 8.0, 1.0 М NaCl.
На пятом этапе проводили окончательную очистку флагеллина методом колоночной гель-фильтрации на смоле Sephacryl S75, S200 (GE Healthcare). Хроматографию проводили в буфере PBS (10 mM Na2HPO4, 100 mM NaCl, рН 8.0).
Описанный способ выделения и очистки рекомбинантного флагеллина дал возможность получить 3 грамма субстанции флагеллина фармакопейного качества за один цикл с 1,0 л ферментационной среды. Полученный в результате использования указанной технологии конечный продукт флагеллина, сохраняет свои свойства более 2 лет при температуре минус 15-20°С.
Характеристики препарата флагеллина, выделенного из штамма Е. coli FM-FliC_syn, определены методами электрофореза в 12% полиакриламидном геле (ПААГ), эксклюзионной гель-хроматографии (SE HPLC), хромогенного ЛАЛ-теста и биологической активности на культуре клеток. Электрофореграмму образцов выделенного флагеллина из штамма pFM-FliC_syn и стандартного образца флагеллина из S. typhimurium проводили на приборе Mini-Protean Tetra Cell, Bio-Rad. Концентрирующий гель - 5% ПААГ, разделяющий гель- 12% ПААГ. Параметры: напряжение -100 V. Нагрузка на лунку 10 мкл. Электрофорез проведен в восстанавливающих условиях. Окрашивание геля - раствором Кумасси.
Результаты исследования показали, что положение основной полосы на электрофореграмме образца выделенного флагеллина «RecFlic-FM» соответствует положению основной полосы на электрофореграмме раствора стандартного образца «FLA-ST Ultrapure», являющегося очищенным флагеллином из S. typhimurium (InvivoGen, кат.ном. tlrl-epstfla). Чистота выделенного флагеллина не уступает чистоте стандартного образца. В исследуемом образце «RecFlic-FM» с концентрацией 0,5 мг/мл нет полос с более высоким молекулярным весом, чем основная полоса. В исследуемом образце с концентрацией 0,5 мг/мл отсутствуют дополнительные полосы, более интенсивные, чем полоса, полученная для исследуемого раствора с концентрацией 0,01 мг/мл.
Хроматограмму образца флагеллина, выделенного из штамма pFM-FliC syn получали с использованием хроматографа Ultimate 3000, Dionex.
Хроматографическая колонка: - TSKGel (сорбент G2000SWXL, 300x7,8 мм, размер частиц 5 мкм). Температура колонки (25±0,1)°С. Детектирование при длине волны 210 нм. Мобильная фаза: фосфатно-солевой буфер 10 мМ Na2HPO4, 137 мМ NaCl, рН 7.5. Скорость потока мобильной фазы 0,5 мл/мин. Объем вводимой пробы: 0,005 мл из раствора с концентрацией 1,8 мг/мл. В результате на хроматограмме, полученной для образца, выделенного флагеллина, площадь основного пика составила 99,6% от общей площади всех пиков.
Определение эндотоксинов в образце выделенного флагеллина из штамма pFM-FliC syn проводили с использованием хромогенного теста согласно прописи производителя набора Chromogenic Endotoxin Quant Kit (Pierce, кат.ном. A39552).
Образец выделенного флагеллина разводили водой для ЛАЛ-теста до концентрации белка 0,005 мг/мл. Для построения калибровочной кривой использовали растворы стандартных образцов, входящие в состав набора.
В результате, измеренное содержание бактериальных эндотоксинов в исследуемом образце рекомбинантного флагеллина составило 2,5 ЕЭ на 1 мг белка.
Пример 4
Биологическая активность рекомбинантного флагеллина Salmonella (FliC) получаемого из биомассы штамма Е. coli FM-FliC_syn
Определение биологической активности флагеллина проводили на перевиваемой культуре клеток HEK-Blue hTLR5 (InvivoGen, кат.ном. hkb-htlr5). Клетки HEK-Blue hTLR5 получены путем трансфекции гена TLR5 человека (hTLR5) и репортерного гена секретируемой эмбриональной щелочной фосфатазы (SEAP) в клетки НЕК293. Ген SEAP находится под контролем промотора, индуцируемого NF-Kb. Стимуляция флагеллином рецептора hTLR5 приводит к активации NF-Kb и АР-1, которые в свою очередь индуцируют выработку репортерного белка SEAP. Активность SEAP оценивается с помощью реагента для определения щелочной фосфатазы QUANTI-Blue (InvivoGen, кат.ном. rep-qbs).
Образец выделенного флагеллина «RecFlic-FM» сравнивали со стандартным образцом «FLA-ST Ultrapure», являющимся очищенным флагеллином из S. typhimurium (InvivoGen, кат.ном. tlrl-epstfla). Оба образца предварительно разводили до примерно одинаковой концентрации в диапазоне от 9 до 10 нг/мл. Коэффициенты предварительных разведений учитывали при расчете активностей. Растворы стандартного и испытуемого образцов для нанесения на планшет готовили из предварительно разведенных образцов путем одиннадцати последовательных двукратных разведений. Разведенные образцы вносили на планшет по 20 мкл на лунку по 4 лунки из каждого разведения. В четыре лунки вносили по 20 мкл ростовой среды без флагеллина (контроль клеток, КК).
На планшет с внесенными образцами высевали индикаторные клетки HEK-Blue hTLR5 по 18 тыс. на лунку в 180 мкл ростовой среды. В четыре лунки вносили по 200 мкл ростовой среды без клеток (холостая проба, Blank). Проводили инкубирование клеток с препаратом в течение ночи при 37°С и 5% CO2.
После инкубации клеток с образцами флагеллина добавляли во все лунки планшета по 50 мкл предварительно разведенного раствора QUANTI-Blue. Инкубировали с реагентом в течение 2,5 ч и измеряли оптическую плотность (ОП) растворов в лунках планшета при 620 нм с помощью планшетного фотометра.
Средняя оптическая плотность растворов составила по результатам измерений: для холостой пробы (контрольный образец) ОП Blanc=0,087 (требование ≤0,2); для среды без флагеллина (КК) с вычетом холостой пробы ОП кк - ОП Blanc=0,042 (требование ≤0,1); для полученного образца флагеллина с вычетом холостой пробы ОП CO1 - ОП Blanc=1,329 (требование ≥1,0).
Результаты анализа убедительно продемонстрировали, что выделенный образец рекомбинантного флагеллина обладает высокой активностью. Его удельная активность составляет более 250% от удельной активности стандартного образца очищенного флагеллина из S. typhimurium «FLA-ST Ultrapure».


Изобретение относится к биотехнологии, конкретно к рекомбинантной плазмиде pFM-FliC_syn, кодирующей экспрессию белка флагеллина Salmonella (FliC), к рекомбинантному штамму Escherichia coli FM-FliC_syn-продуценту рекомбинантного FliC и к способу получения рекомбинантного белка флагеллина и может быть использовано в качестве адъюванта для вакцин. Рекомбинантный штамм бактерий E.coli FM-FliC_syn получают путем трансформации клеток штамма Е. coli BL21 рекомбинантной плазмидной ДНК pFM-FliC_syn. Рекомбинантная плазмидная ДНК pFM-FliC_syn, размером 4244 п. о., кодирует синтез белка флагеллина Salmonella (FliC) под контролем лактозного и триптофанового промотора и терминатора транскрипции rrnBT1T2 и содержит в качестве генетического маркера синтетический ген kanR, детерминирующий устойчивость к канамицину. Изобретение позволяет получать рекомбинантный белок флагеллина Salmonella (FliC) с высоким выходом и по упрощенной технологии. 3 н.п. ф-лы, 8 ил., 4 пр.
1. Рекомбинантная плазмида pFM-FliC_syn, обеспечивающая экспрессию рекомбинантного белка флагеллина Salmonella, имеющая размер 4244 п. о. и состоящая из следующих элементов: фрагмент ДНК размером 194 п. о., включающий триптофановый промотор E.coli и последовательность Шайн-Дельгарно (SD), ответственную за инициацию трансляции; синтетический фрагмент ДНК размером 1533 п. о., включающий кодирующую часть гена флагеллина Salmonella (FliC), из которой удалены внутренние TATA-боксы, chi-сайты, RBS последовательности, АТ-богатые и GC-богатые последовательности, повторяющиеся последовательности и потенциальные вторичные структуры мРНК; частота использования кодонов, адаптированная для Е. coli, причем индекс адаптации кодонов равен 0,81; фрагмент ДНК плазмиды pKK223-3 размером 534 п. о, включающий последовательность строгого терминатора транскрипции rrnBT1T2, последовательности промотора и SD последовательность гена бета-лактомазы; синтетический фрагмент ДНК размером 810 п. о., включающий синтетический ген kanR, обеспечивающий устойчивость к канамицину - аналогу гена kanR из транспозона Tn903, в котором удалены сайты рестрикции AvaI, ClaI, SmaI, HindIII; фрагмент ДНК размером 1173 п. о. плазмиды pUC19, включающий последовательность, ответственную за репликацию плазмиды (ori) и лактозный промотор.
2. Моноплазмидный штамм бактерий Escherichia coli FM-FliC_syn, полученный путем трансформации клеток штамма Е. coli BL21 рекомбинантной плазмидной ДНК pFM-FliC_syn по п. 1 и содержащий рекомбинатную плазмиду pFM-FliC_syn - продуцент рекомбинантного флагеллина Salmonella (FliC).
3. Способ получения рекомбинантного белка флагеллина Salmonella (FliC), характеризующийся тем, что исходную биомассу моноплазмидного штамма Escherichia coli FM-FliC_syn по п. 2 подвергают дезинтеграции в фосфатно-солевом буфере и удаляют клеточный дебрис центрифугированием, а оставшийся раствор флагеллина в фосфатно-солевом буфере подвергают последовательной четырехстадийной хроматографии, на первой из которых осуществляют очистку флагеллина на металл-хелатной смоле, на второй - на катионообменной смоле, на третьей - на анионообменной смоле и на последней выделяют очищенный флагеллин методом колоночной гель-фильтрации.
| ШТАММ БАКТЕРИЙ Escherichia coli - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО ФЛАГЕЛЛИНА | 2012 |
|
RU2524133C2 |
| Рекомбинантная плазмида pHis6-flagG-protE, обеспечивающая синтез рекомбинантного химерного белка, включающего эпитопы гликопротеина Е вируса клещевого энцефалита и флагеллин G S.typhii и используемого в качестве основы для вакцины против вируса клещевого энцефалита | 2018 |
|
RU2702716C2 |
| Способ получения химерного рекомбинантного белка fliC:pagN | 2015 |
|
RU2627602C2 |
| WO 2009087508 A1, 16.07.2009. | |||

