Изобретение относится к микробиологической промышленности, медицинской биотехнологии, генной инженерии и представляет собой сконструированную in vitro рекомбинантную плазмидную ДНК, обеспечивающую гетерологичную экспрессию растворимого стрептавидина из Streptomyces avidinii, и содержащий рекомбинантную плазмиду штамм Escherichia coli - продуцент стрептавидина.
Стрептавидин (SAV) представляет собой четырехсубъединичный негликозилированный белок, специфически связывающий витамин H (d-биотин) [Biochemistry. 1992. V. 31. P. 9350-9354]. Биологическая функция этого белка в клетках S. avidinii окончательно не выяснена, но предполагается, что он, как и гликопротеин авидин (белок куриных яиц), играет защитную роль, подавляя рост биотинзависимых микроорганизмов. К настоящему времени аналоги стрептавидина, белки V1 и V2, обнаружены в S. venezuelae [Biochim. Biophys. Acta, Gene Struct. Expr. 1995. V. 1263. P. 60-66].
Тетрамерный белок стрептавидин связывает четыре молекулы биотина (Kd= 10-15). В современной медицине и биологии это свойство стрептавидина нашло широкое применение при создании спектра препаратов, предназначенных для высокочувствительной нерадиоактивной детекции биомолекул [Биоорган. химия. 1989. Т. 15. С. 354-357; Clin. Chem. 1991. V. 37. Р. 625-636; Proc. Natl. Acad. Sci. 1995. V. 92. P. 7590-7594]. В последнее время повышенное внимание вызывают исследования, направленные на создание терапевтических препаратов на основе стрептавидина, применяемых для лечения онкологических заболеваний человека. Известны работы по изучению распределения стрептавидина при его экзогенном введении в организм млекопитающих, по селективному накоплению SAV в почках человека [Kidney Intern. 1995. V. 47. P. 1327-1335], а также по использованию стрептавидина в химерных конструкциях в качестве белка-носителя, к которому присоединена адресующая часть и терапевтический агент химической или белковой природы [Proc. Natl. Acad. Sci. 1992. V.89. P.1534-1538; Bioconjugate Chem. 1995. V. 6. P. 139-144].
Промышленное получение стрептавидина в настоящее время базируется на культивировании S. avidinii АТСС 27419 и обладает рядом существенных недостатков: препарат часто загрязнен биотином, что приводит к уменьшению свободных валентностей стрептавидина при связывании биотина. Кроме того, длительное время культивирования продуцента S. avidinii (5 - 8 суток) и высокая природная антибиотикоустойчивость рода Streptomyces (устойчив к ряду стандартных доступных селективных агентов) делает процесс недостаточно эффективным [Proc. Natl. Acad. Sci. 1980. V. 77. Р. 4666-4668; Biochem. Biophys. Methods. 1986. V. 13. P. 103-112; J. lmmunol. Methods. 1988. V. 113. P. 83-91].
К настоящему времени проведены исследования, направленные на преодоление вышеперечисленных недостатков и создание более дешевого источника рекомбинантного стрептавидина, не содержащего примесей биотина. Нуклеотидная последовательность гена стрептавидина из S.avidinii известна [Nucleic Acids Res. 1986. V. 14. P. 1871-1882], в литературе описан ряд продуцентов рекомбинантного стрептавидина, его мутантных форм и химерных белков на его основе [Gene. 1993. V. 136. Р. 243-246; Proc. Natl. Acad. Sci. V. 87. Р. 142-146; BioTechnology. 1995. V. 13. P. 11]. Однако, оптимальной конструкции экспрессионной плазмиды для наработки стрептавидина не было найдено. Ближайшими аналогами к заявляемой группе изобретений являются рекомбинантная плазмидная ДНК и штамм-продуцент стрептавидина в работе Sano Т., Cantor C. [Proc.Natl. Acad.Sci. 1990. V. 87. Р. 142-146], в которой приведено получение продуцента рекомбинантного стрептавидина на основе клеток Е.coli. Первоначально, конструкция гибридной плазмиды включала полную нуклеотидную последовательность гена стрептавидина с лидерной частью, а также дополнительный фрагмент плазмидной ДНК, кодирующий 25 аминокислот. Полученный белок не обнаруживал основной биологической функции стрептавидина - связывания биотина. Авторы отказались от использования фрагмента лидерного пептида, предполагая, что либо лидерный пептид, либо пептид, кодируемый фрагментом плазмидной ДНК, приводят к инактивации белка. Дальнейшее конструирование привело к получению рекомбинантной плазмиды, где под регуляцией индуцибельного промотора Ф10 клонирована только структурная область гена стрептавидина от 15-й аминокислоты зрелого белка. Недостатками штамма Е. coli, продуцирующего стрептавидин на основе вышеприведенной экспрессионной системы, являются:
1) накопление стрептавидина в цитоплазме клеток в нерастворимом биологически неактивном состоянии в составе телец включения, в связи с чем дальнейшее выделение и очистка этого белка требуют проведения ряда стадий денатурации-ренатурации с использованием ионных детергентов типа гуанидинхлорида, что может приводить к необратимой утрате биологической активности белка, снижает выход конечного продукта и повышает его себестоимость.
2) недостаточно высокий уровень накопления SAV, который до очистки составляет 40-65 мг/л культуральной жидкости.
Задачей заявляемой группы изобретений является создание эффективного продуцента стрептавидина на основе штаммов Е.coli, обеспечивающего повышенный уровень накопления белка в биологически активном растворимом состоянии.
Для решения задачи получают рекомбинантную плазмидную ДНК pSAV27, состоящую из фрагмента хромосомной ДНК Streptomyces avidinii, включающего структурную часть гена стрептавидина (нуклеотидная последовательность лидерного пептида совместно с нуклеотидной последовательностью зрелого белка) и мультикопийного вектора, обеспечивающего высокоэффективную экспрессию гена стрептавидина в клетках Е.coli. Причем лидерный пептид прострептавидина обеспечивает секрецию прострептавидина в периплазматическое пространство клеток Е. coli, где происходит отщепление лидерного пептида, и стрептавидин локализуется в биологически активном растворимом состоянии. Штамм Е.coli JM110, трансформированный рекомбинантной плазмидой pSAV27 за 12-14 часов ферментации при 37oC на полусинтетической среде накапливает 180-240 мг растворимого стрептавидина на 1 л культуральной жидкости. Наблюдаемый эффект достигается, во-первых, за счет введения в реципиентную клетку Е.coli мультикопийного вектора (количество копий клонированного гена составляет 300-500 копий на геномный эквивалент), во-вторых, за счет использования сильного, конститутивного в применяемых условиях промотора, обеспечивающего эффективную транскрипцию клонированного гена, и, в-третьих, за счет использования лидерного пептида стрептавидина, приводящего к экскреции белкового продукта в периплазму и пост - либо коэкскреторному его отщеплению.
Заявляемый штамм E.coli JM110/pSAV27 депонирован во Всероссийской коллекции промышленных микроорганизмов и имеет регистрационный номер ВКПМ В- 7758.
Конструирование штамма состояло из нескольких этапов.
Этап 1. Выделение из Streptomyces avidinii M170791 фрагмента ДНК, кодирующего лидерный пептид стрептавидина совместно с зрелым белком (прострептавидин), и введение в данный фрагмент сайтов для эндонуклеаз рестрикции.
Этап 2. Конструирование рекомбинантной плазмиды рSAV27, основными характеристиками которой являются высокая копийность (300-500 копий на геномный эквивалент), удобный селективный маркер (ампициллинустойчивость), сильный промотор, обеспечивающий в штаммах Е.coli эффективную экспрессию клонированного гена стрептавидина.
Этап 3. Трансформация штамма E.coli JM110 сконструированной рекомбинантной плазмидой, подтверждение соответствия нуклеотидной последовательности клонированного гена стрептавидина вместе с лидерной последовательностью природному гену из S.avidinii.
Этап 4. Определение продуктивности штамма Е.coli ВКПМ В-7758 - продуцента растворимого стрептавидина из S.avidinii.
Этап 5. Определение биологической активности растворимого стрептавидина, продуцируемого штаммом Е.coli ВКПМ В-7758.
Штамм E. coli ВКПМ В-7758 обладает следующими культурально-морфологическими и физиолого- биохимическими признаками.
1. Морфологические признаки. Клетки - прямые палочки, 1,1-1,5 • 2.0-6,0 мкм, подвижные за счет перитрихальных жгутиков, грамотрицательные, неспороносные.
2. Культуральные признаки. Клетки штамма хорошо растут на стандартных, описанных для Е. coli средах. При выращивании на полноценных агаризованных средах (LB-агар, агаризованная среда Хоттингера) колонии блестящие, гладкие, круглые, с ровными краями. При 37oC достаточно 12 - 14 часов роста. При выращивании в жидких средах в пробирках клетки образуют равномерную муть, достигая оптической плотности 2,4-2,6 при A600 за 10-12 часов роста при 37oC.
3. Физиолого-биохимические признаки. Температурный оптимум для роста клеток 37oC, факультативный анаэроб, оксидазоотрицательные, каталазоположительные, отрицательные по признакам образования H2S, гидролиза мочевины и активности липазы, катаболизируют D-глюкозу и другие углеводы с образованием кислоты и газа, источником азота могут служить минеральные соли в аммонийной форме, а также органические соединения в виде пептона, триптона, дрожжевого экстракта и аминокислот.
4. Генотипические признаки.
Основной генотипический признак заявляемого штамма Е.coli ВКПМ В-7758 (thi, thr, leu, lacY, supE44, galK, tonA, recA, dam, dcm, Δ(lac-proAB), [F', traD36, proAB, lacIqZΔM15] ) состоит в наличии под регуляцией промоторного фрагмента гена udp из Е.coli нуклеотидной последовательности кодирующей области гена стрептавидина с лидерной последовательностью, интегрированной в состав многокопийного экспрессионного вектора, содержащего в качестве селективного маркера ген β-лактамазы.
5. Устойчивость к антибиотикам. Устойчивость к ампициллину у штамма не ниже 150 мкг/мл на твердых агаризованных средах, при наращивании в жидких полноценных средах - не менее 100 мкг/мл.
6. Стабильность плазмиды pSAV27 в штамме Е.coli ВКПМ В-7758.
При хранении клеток на агаризованной среде (сроком до 1 месяца), при серии последовательных пересевов (в течение не менее 6 месяцев) и в процессе культивирования в жидкой среде с антибиотиком не происходит потери и перестройки плазмиды pSAV27.
Процесс биосинтеза стрептавидина в штаммах Е.coli включает стадии получения посевного материала, основную ферментацию, выделение биомассы и получение периплазматической фракции клеток. Выращивание посевного материала и основную ферментацию Е.coli ВКПМ В-7758 осуществляют в аэробных условиях на традиционно используемых для культивирования Е.coli питательных средах, содержащих ассимилируемые источники углерода, азота, минеральные соли, стимуляторы роста, например, в виде триптона, пептона, дрожжевых автолизатов или экстрактов. В питательные среды вносят ампициллин в концентрации 100 мг/л. Биомассу выращивают при pH 7-7.4 и температуре 37oC до концентрации, определяемой составом питательной среды и массообменными характеристиками ферментера.
После завершения процесса ферментации бактериальные клетки отделяют от культуральной среды центрифугированием и ресуспендируют в соответствующем буферном растворе, после чего центрифугированием отделяют периплазматическую фракцию растворимых белков клетки (в виде супернатанта), которую и используют для последующего выделения целевого продукта. Для этого применяют методы ионообменной хроматографии на колонке с Q-сефарозой, либо аффинную хроматографию на колонке с 2-иминобиотином. Данные методы очистки обеспечивают высокий выход целевого белка и его 95-97% чистоту. Степень очистки стрептавидина в процессе выделения анализируют с помощью SDS-полиакриламидного электрофореза. Выход конечного продукта составляет 180-240 мг/л культуральной жидкости. Определение N-концевой последовательности белка проводят методом деградации по Эдману на автоматическом секвенаторе "Beckman" 890В. Идентификацию Pth-производных аминокислот осуществляют методом ВЭЖХ на колонках "Zorbax РТН".
Заявляемая группа изобретений иллюстрируется следующими фигурами графических изображений.
Фиг. 1. Физико-генетическая карта рекомбинантной плазмиды pSAV27.
Рекомбинантная плазмида сконструирована с использованием следующих фрагментов ДНК: 1). Sac1-Sa1G1 фрагмент (размером 2684 н.п.) плазмиды pUC18, который содержит репликон pMB1, ген rop, участвующий в регуляции копийности плазмиды, ген β-лактамазы, определяющий устойчивость к антибиотику ампициллину. Фрагмент получен рестрикцией плазмиды pUC18. 2). Sacl-BamHI фрагмент (размером 169 н.п.) промоторной области гена уридинфосфорилазы из Е.coli, содержащий все элементы, необходимые для эффективной инициации транскрипции и трансляции. Сайты Sac1 и BamH1 введены в промоторный фрагмент методом ПЦР. Фрагмент получен методом рестрикции сконструированной нами плазмиды pUU18 [Биоорган. химия. 1995. Т. 21. С. 354-358]. 3). BamH1-Sa1G1 фрагмент (размером 571 н.п.) гена стрептавидина, включающий нуклеотидную последовательность лидерного пептида. Сайты для BamH1 и Sa1G1 введены при получении фрагмента методом ПЦР.
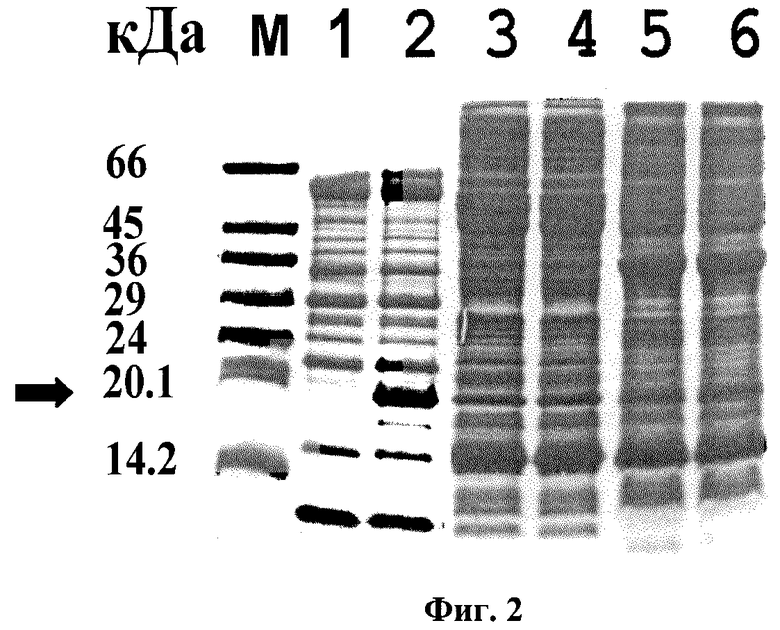
Фиг. 2. Электрофоретическое разделение в денатурирующем 12% ПААГ фракций клеточных белков Escherichia coli.
Белки периплазматических (дорожки 1 и 2), цитоплазматических (дорожки 3 и 4) и мембранных (дорожки 5 и 6) фракций штамма-продуцента (дорожки 2, 4, 6) и штамма-реципиента (дорожки 1, 3, 5). М - белковые маркеры с молекулярными массами 66, 45, 36, 29, 24, 20.1, 14.2 кДа. Стрелкой показана полоса, соответствующая стрептавидину (дорожка 2).
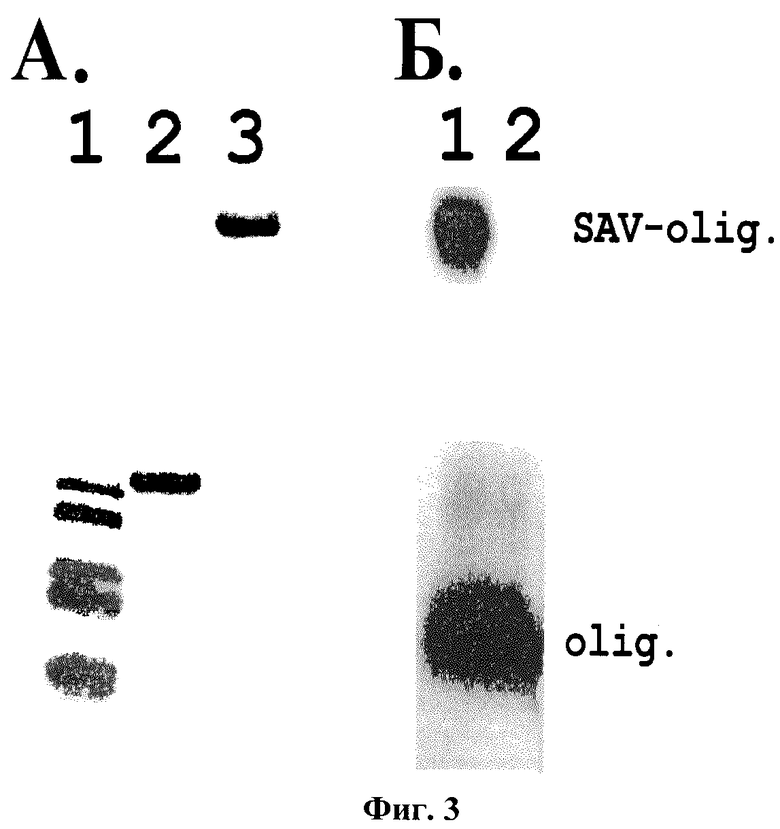
Фиг. 3. Электрофоретическое разделение в 12% ПААГ выделенного очищенного стрептавидина.
Электрофоретическая подвижность очищенного стрептавидина в денатурирующих (A2) и неденатурирующих (A3) условиях. Окрашивание геля Кумасси синим. Радиоавтограф электрофоретического разделения неденатурированного комплекса очищенного стрептавидина с небиотинилированным (Б2) и биотинилированным (Б1) радиоактивно меченным олигонуклеотидом. A1 - маркерные белки 16.4, 14.4, 10.6, 8.1, 6.2 кДа. SAV-olig - комплекс стрептавидин-радиоактивномеченный биотинилированный олигонуклеотид. Olig - радиоактивномеченный небиотинилированный олигонуклеотид в отсутствие связывания со стрептавидином.
Заявляемая группа изобретений иллюстрируется примерами.
Пример 1. Выделение фрагмента ДНК, содержащего структурную часть гена стрептавидина и введение в него участков для узнавания эндонуклеаз рестрикции.
Фрагмент гена стрептавидина, включающий нуклеотидную последовательность лидерного пептида и кодирующую область стрептавидина получают ПЦР-амплификацией соответствующего участка хромосомной ДНК из S.avidinii М 170791 с использованием синтетических олигонуклеотидов (1) 5'CGTGGGATC CATGCGCAAAATCGTCGTTG3' и (2) 5'CGGGGTCGACTTACTGCTGAACTGC GTC3'. Амплификационная смесь (50 мкл) содержит 10 мМ Трис-HCl (pH 8.4), 0.5 мМ MgCl2, 50 мМ KCl2, 0,2 мМ dNTP, 10 нг хромосомной ДНК S.avidinii, по 50 пмоль праймеров (1) и (2), 2.5 ед. акт. Taq-полимеразы. Режим амплификации (oC/с): 1 цикл -95/120 - денатурация хромосомной ДНК; 7 циклов - денатурация 95/10, отжиг 48/10, элонгация 72/30; 25 циклов - денатурация 95/10, отжиг 60/10, элонгация 72/30. Полученный фрагмент ДНК размером 571 н.п., кодирующий структурную часть гена прострептавидина, выделяют препаративным электрофорезом в 1.2% агарозном геле. В олигонуклеотиды при синтезе вводят нуклеотидные последовательности сайтов эндонуклеаз рестрикции BamH1 и Sa1G1, необходимые для последующего клонирования гена в составе экспрессионного вектора.
Пример 2. Конструирование рекомбинантной плазмиды pSAV27 и получение штамма Е.coli ВКПМ В-7758 - продуцента растворимого стрептавидина.
Для подготовки экспрессионного вектора используют мультикопийную плазмиду pUC18 [Маниатис Т. и др. Молекулярное клонирование. М.: Мир, 1984], репликон которой в благоприятных условиях обеспечивает накопление до 300-500 копий плазмидной ДНК на клетку. Для этого плазмидную ДНК pUC18 расщепляют рестриктазами Sac1 и BamH1 в условиях, рекомендуемых фирмой-изготовителем рестриктаз ("Fermentas") и лигируют с промоторным фрагментом гена udp, размером 169 п. н., полученным рестрикцией по Sac1 и BamH1 из ранее полученной нами [ДАН. 1994. Т. 339. N. 4. С. 1-3; Биоорганическая химия. 1995. Т. 21. N 5. С. 354-358] плазмиды pUU18, так как данный фрагмент ДНК промотора udp обеспечивает эффективную транскрипцию как гомологичных, так и гетерологичных генов в Е.coli.
После трансформации и отбора целевого клона, содержащего плазмидную ДНК с промотором udp, полученный вектор расщепляют рестриктазами BamH1 и Sa1G1 и лигируют с описанным в примере 1 и расщепленным по сайтам BamH1 и Sa1G1 фрагментом ДНК, включающим структурную часть гена стрептавидина с лидерной последовательностью. Трансформацию проводят в компетентную культуру штамма Е.coli JM110 (ВКПМ В-6527). Эффективность трансформации составляет около 105 клонов на 1 мкг плазмидной ДНК. Трансформанты отбирают на среде с ампициллином, изменение генотипа определяют методом ПЦР с помощью праймеров (1) и (2), структура которых приведена в описании примера 1.
Выделенную из целевых клонов плазмиду обозначили pSAV27 (фиг. 1). Нуклеотидную последовательность клонированного фрагмента подтверждают секвенированием по Сэнгеру [J.Mol.Biol. 1975. V. 94. P. 441-446]. Определение уровня накопления стрептавидина и его локализацию в клетках штамма-продуцента исследуют электрофорезом в 12.5% ПААГ в денатурирующих условиях. Для этого клетки из 1 мл ночной культуры Е.coli ВКПМ В-7758 осаждают центрифугированием (12000 g, 1 мин). К осадку добавляют 100 мкл буфера, содержащего 30 мМ Трис HCl pH 8.0, 5 мМ ЭДТА pH 8.0, 20% сахарозу, 1 мг/мл лизоцима. После выдерживания клеток в течение 10 мин в ледяной бане суспензию центрифугируют (12000 g, 1 мин), отобранный супернатант представляет собой периплазматическую фракцию клеток. К осадку добавляют 100 мкл 100 мМ Трис HCl pH 8.0 и три раза проводят быстрое замораживание при - 70oC, и медленное оттаивание при 37oC. После центрифугирования супернатант содержит суммарные белки цитоплазматической фракции клеток, а осадок - белки мембранных фракций. Для солюбилизации мембранных белков к осадку добавляют 100 мкл 1% Тритона X-100. До проведения анализа в ПААГ фракции хранят во льду. Анализ со става фракций в денатурирующем 12.5% ПААГ показал (фиг. 2), что описанная экспрессионная система приводит к накоплению стрептавидина (белок размером 18 кДа) именно в периплазматическом пространстве клеток штамма-продуцента. При этом белок остается активным, что подтверждается экспериментами по связыванию биотинсодержащего олигонуклеотида (см. пример 4) суммарными белками соответствующих фракций клеток штамма-продуцента.
В результате вышеописанных процедур получают штамм-продуцент растворимого стрептавидина coli ВКПМ В-7758.
Пример 3. Определение продуктивности штамма Е.coli ВКПМ В-7758 - продуцента растворимого стрептавидина из S.avidinii.
Для препаративной наработки стрептавидина и определения продуктивности штамма плазмидосодержащие клетки выращивают на агаризованной LB-среде с ампициллином (150 мкг/мл) при 37oC в течение 12-14 часов, затем используют выросшую биомассу для получения посевного материала. Для этого клетки переносят в колбу Эрленмейера на 750 мл со 100 мл LB-среды, содержащей 2 г/л глюкозы и 100 мкг/мл ампициллина. Культуру выращивают на качалке при интенсивном перемешивании при 37oC до плотности 2.4 - 2.6 при A600. Основное культивирование проводят в ферментерах "Anglicon" объемом 750 мл на среде следующего состава, г/л: бактопептон - 20, дрожжевой экстракт - 5, NaCl - 6, (NH4)2SO4 - 4, К2HPO4 - 2, MgSO4 - 0.4, FeSO4 - 0.02, MnSO4 - 0.02. Ферментацию проводят в условиях термо- и pH-статирования, доза засева 2%, ампициллин - 100 мкг/мл, содержание растворенного кислорода в среде поддерживают на уровне 5%, в качестве источника углерода используют глюкозу в дробной подаче в количестве 1-2 г/л/час, поддержание pH 7.0-7.2 достигают автоматической подтитровкой аммиачной водой, при 37oC культуру наращивают в течение 12-14 часов.
После завершения ферментации биомассу отделяют центрифугированием из предварительно охлажденной до 12oC культуральной жидкости и используют для получения периплазматической фракции клеток. Для этого клетки суспендируют в буфере, содержащем 30 мМ Трис-HCl pH 8.0, 5 мМ ЭДТА pH 8.0, 20% сахарозу, 1 мг/мл лизоцима, выдерживают 10 мин в ледяной бане и центрифугируют при 6-10 тыс. об. Супернатант используют для получения очищенного стрептавидина методом аффинной хроматографии на колонках с 2-иминобиотинагарозой ("Sigma") и для определения продуктивности штамма методом SDS-ПААГ, которая составляет 180 мг стрептавидина, нарабатываемого в 1 л культуральной жидкости.
Очистку стрептавидина от белков периплазматической фракции проводят с использованием 2-иминобиотинагарозы. Для этого полученную периплазматическую фракцию переводят с помощью ультрафильтрации в ячейке "Amicon" с мембраной YM30 в буфер следующего состава: 0.5 М NaCl, 50 mM Na2CO3, pH 11.0. Колонку, содержащую 10 мл геля 2-иминобиотинагарозы уравновешивают буфером 0.5 М NaCl, 50 mM Na2CO3, pH 11.0, вносят периплазматическую фракцию и колонку промывают 50 мл 1 М NaCl (pH 7.0). Элюцию стрептавидина ведут буфером, содержащим 50 mM CH3COOH, pH 4.0. Элюированный стрептавидин переводят в буфер 5 mM Tris-HCl, pH 7.5, содержащий 0.01% NaN3 в ячейке для ультрафильтрации на мембране YM30, и высушивают на установке для лиофилизации "Hetosicc" (США). Уровень очистки стрептавидина составляет 95-97%.
Пример 4. Определение биологической активности растворимого стрептавидина, продуцируемого штаммом Е.coli ВКПМ В-7758.
Для исследования связывания полученного стрептавидина с d-биотином используют синтетический биотинилированный олигонуклеотид, полученный по ранее предложенной нами схеме [Биоорган. химия. 1991. Т. 17. С. 625-629].
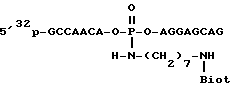
Данный олигонуклеотид содержит остаток биотина, введенный по межнуклеотидной фосфатной группе, что позволяет осуществлять его дополнительное радиоактивное мечение фосфорилированием с помощью полинуклеотидкиназы бактериофага T4 и [γ32P]АТР. После проведения комплексообразования (выдерживают смесь олигонуклеотида со стрептавидином при комнатной температуре 10 мин) смесь разделяют в ПААГ и образовавшийся комплекс выявляют радиоавтографией. Данный подход позволяет с помощью измерения радиоактивности соответствующих зон ПААГ провести сравнительную количественную оценку эффективности связывания. Полученные данные позволяют утверждать, что выделенный рекомбинантный стрептавидин связывает 3.7 - 3.8 ммоль остатка биотина на 1 ммоль тетрамера стрептавидина, что соответствует практически полной биологической активности.
Как показал анализ N-концевой последовательности выделенного белка, отщепление лидерного пептида в Е.coli происходит по связи -S-A22/23 S-A-D-P-S-K-, что отличается от указанного ранее места расщепления -S-A-S-A24/25D-P-S-K- для S.avidinii [Nucl. Acids Res. 1986. V. 14. P. 1871-1882].
Таким образом, заявляемая группа изобретений позволяет получать стрептавидин в бактериальных клетках Е.coli в виде растворимого белка с сохранением полной биологической активности и в количествах, в три и более раза превышающих таковые у ближайшего аналога.
Изобретение относится к микробиологической промышленности, медицинской биотехнологии и генной инженерии. Предложена рекомбинантная плазмидная ДНК pSAV27, кодирующая синтез растворимого, экскретируемого в периплазму стрептавидина, на основе которой сконструирован штамм Escherichia coli ВКПМ В-7758, являющийся продуцентом стрептавидина. За 1 - 14 ч ферментации данный штамм накапливает в культуральной жидкости 180 - 240 мг/л стрептавидина. 2 с.п. ф-лы, 3 ил.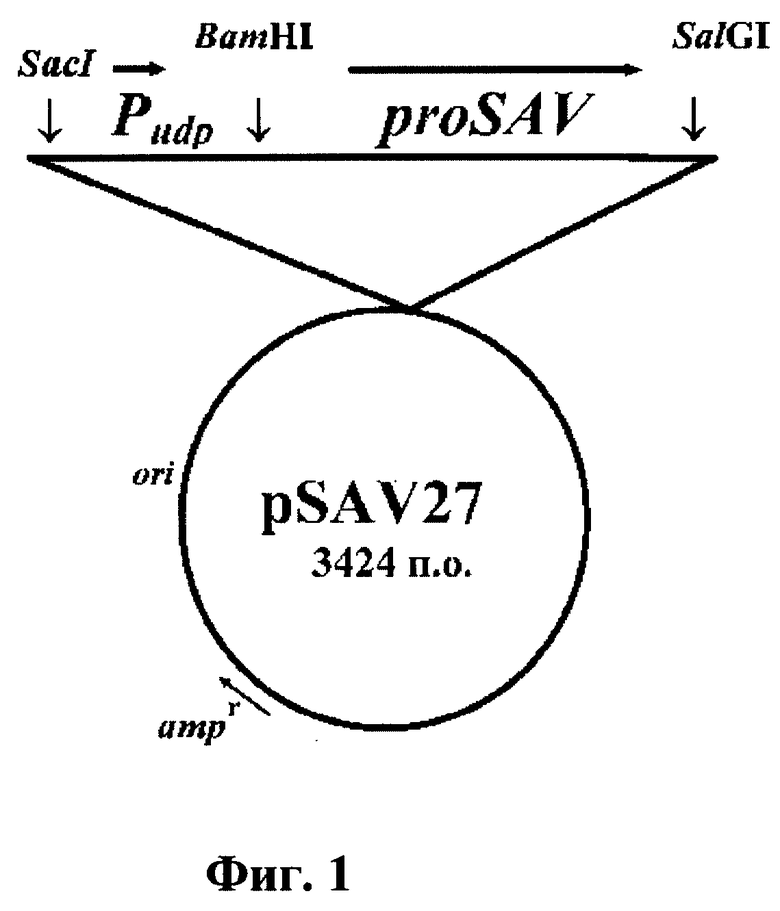
| Штамм стрептомицета SтRертомYсеS снRомоFUSсUS - продуцент биотинсвязывающего белка | 1988 |
|
SU1643610A1 |
| WO 9324631 A1, 09.12.1993 | |||
| US 5489528 A, 06.02.1996. | |||

