Изобретение относится к области биохимии, в частности к гистохимии, для определения состава мышечных волокон у спортсменов, занимающихся различными видами спорта.
Большинство скелетных мышц человека представляют собой гетерогенные морфофункциональные системы, состоящие из мышечных волокон (MB), отличающихся по структуре, метаболизму и функциям. Многие авторы предлагают свои схемы классификации этих структур при характеристике свойств мышечных волокон различных типов, например [1, 4], в основе которых - различное количество ферментов, жиров, углеводов, площадь поперечного сечения мышечных волокон, скорость их сокращения, устойчивость к утомлению.
При использовании метода определения миофибриллярной АТФазы после преинкубации в щелочной среде в скелетных мышцах обнаружили два основных типа MB, которые по окрашиванию в светлые и темные структуры назвали волокнами I и II типа. При различной чувствительности активности миозина АТФазы к сдвигу рН в кислую сторону MB II типа были разделены на две подгруппы - IIА и IIВ.
MB I типа обозначают как красные, окислительные, медленные, устойчивые к утомлению, у них отмечается самое высокое содержание миоглобина, они характеризуются высокой степенью кровообращения. Каждое MB I типа контактирует с 5-6 кровеносными капиллярами. Содержание АТФ в MB I типа такое же, как в MB IIА, но более низкое, чем в MB IIВ. По активности ферментов обмена аминокислот MB типа I занимают промежуточное положение между MB IIА и MB IIВ.
MB IIВ обозначают как белые, быстрые, гликолитические, быстроутомляемые, они контактируют с меньшим количеством капилляров, чем MB других типов. В этих волокнах имеется очень мало миоглобина, именно этим объясняется их более светлая окраска, поэтому они и получили название «белых» MB.
MB IIА обозначают как промежуточные, окислительно-гликолитические, быстрые, устойчивые к утомлению. В связи с тем, что MB IIА способны длительное время производить работу достаточно большой мощности, они рассматриваются как универсальные MB в плане их функциональных возможностей. Поэтому для спортсменов, специализирующихся в видах спорта аэробной направленности, очень важно знать тип MB и их количество.
Преобладание в мышцах волокон I типа с их окислительным метаболизмом определенным образом влияет на функцию дыхательной системы, существует прямая зависимость между содержанием MB I в мышцах и показателем максимального потребления кислорода организмом.
До сих пор не могут прийти к единому мнению, какое же волокно является истинно гликолитическим, а какое - окислительным.
С одной стороны, это связано с обилием химических определений различных ферментов, с другой - невозможностью быстро и четко определить, какой фермент принадлежит тому или иному MB.
Прежде использовалась преинкубация только в щелочной среде, из-за чего происходила путаница как в определении типа MB, так и химических свойств активности АТФазы миозина мышечных структур.
Однако никто при определении различных типов MB не изучал одновременно их реакцию в кислой и щелочной среде. Поэтому-то и возникло так много разнообразных классификаций, каждый автор выдвигал свою собственную версию, исследуя множество различных параметров в MB: наличие гликогена, липидов, ферментов лактатдегидрогеназы, фумаратдегидрогеназы, сукцинатдегидрогеназы и т.д. Отсюда и разночтения в том, какое же MB является истинным - аэробным или анаэробным.
Считается, что в кислой среде активность АТФазы миозина наиболее высока в медленных (окислительных) и промежуточных (окислительно-гликолитических) MB, а в щелочной среде - наиболее высока в быстрых (гликолитических) MB.
Наиболее близкой к заявляемому способу является классификация, в основе которой - содержание в мышечных волокнах АТФазы миозина и СДГ (сукцинатдегидрогеназы); АТФаза миозина рассматривалась как показатель скорости гидролиза аденозинтрифосфорной кислоты (АТФ) и, следовательно, скорости мышечного сокращения, а сукцинатдегидрогеназа - как маркер активности окислительных процессов, причем использовали только кислую преинкубацию [5, 6].
Задача изобретения - разработать эффективный, объективный и доступный в использовании способ определения соотношения различных типов мышечных волокон.
Поставленная задача достигается тем, что для определения активности АТФазы миозина серийные гистологические срезы помещают в кислую (рН=4,6) или щелочную (рН=10,4) среду (этот этап называется «преинкубацией»). В кислой среде инактивируется «быстрый» миозин гликолитических волокон, в щелочной среде инактивируется «медленный» миозин окислительных волокон.
В кислой среде определяют три типа MB: окислительные - тип I, окислительно-гликолитические - тип IIА и гликолитические - тип IIВ, а в щелочной - два типа: окислительно-гликолитические (тип IIА) и гликолитические (тип IIВ); в связи с этим отпадает необходимость в применении большого количества ферментных и морфометрических показателей для определения соотношения различных типов MB.
В заявляемом способе используют только два фермента: сукцинатдегидрогеназу - для оценки напряженности окислительных процессов в митохондриях и лактатдегидрогеназу - для определения интенсивности гликолиза в цитоплазме клетки, а по площади поперечного сечения рассчитывают тип MB, что позволяет достоверно подтвердить определяемый тип волокна.
Примеры конкретного выполнения. АТФаза миозина относится к классу гидролаз, к которому относятся ферменты, осуществляющие расщепление и синтез многих веществ при участии воды:
R1R2+НОН<→R1H+R2OH
АТФаза миозина катализирует течение реакции:
АТФ+Н2O→АДФ + ортофосфат
АТФаза миозина обладает функцией «переходного вентиля», доставляющего АТФ для продолжения протекающих процессов обмена веществ.
Вначале происходит гидролитическое расщепление соли АТФ под влиянием фермента. Далее полученный гидрофосфат натрия вступает в реакцию с САСI2:
Na2HPO4+СаСl2→СаНРO4+2NaCI
затем гистологические срезы обрабатывают солью кобальта:
CoCI2+СаНРO4→СoНРО4+СаСI2
и в завершение их обрабатывают раствором сульфида аммония:
СoНРО4+(NH4)2S→CoS+(NH4)2HPO4
Затем свежеприготовленные препараты помещают в ацетатный буфер - на 30 минут при комнатной температуре; рН=4,6, тип IIА - растворяют в 0,1 М растворе уксусной кислоты; тип IIВ - растворяют в 0,1 М растворе ацетата натрия. В завершение препараты споласкивают в дистиллированной воде.
По второму варианту преинкубации гистологические срезы смешивают с 20 мл 0,1 м раствора мединала (2,062%), 10 мл 0,18 м раствора СаСI2 (1,998%), 30 мл дистиллированной воды, 304 мг двунатриевой соли АТФ. Все компоненты растворяют, далее, после растворения, добавляют 0,1 м раствор NaOH до рН=9,4, а общий объем водой доводят до 100 мл.
Затем последовательно перемещают препараты сначала в 1% растворе СаСI2, затем в 2% растворе СоСI2 и 1% растворе сульфида аммония.
Свежеприготовленные гистологические срезы помещают в кислую среду - рН=4,6, время = 30 минут, температура = 22°С; в щелочную среду - рН=10,4, время = 30 минут, температура = 22°С.
Преинкубация в кислой среде (рН=4,6) инактивирует «быстрый» миозин гликолитических волокон. На срезе черным цветом окрашиваются окислительные волокна, серым цветом окислительно-гликолитические волокна и белым цветом - гликолитические волокна.
Преинкубация в щелочной среде (рН=10,4) инактивирует «медленный» миозин окислительных волокон и активирует «быстрый» миозин гликолитических волокон. На срезе черным цветом окрашиваются гликолитические волокна, а не окрашиваются окислительные и окислительно-гликолитические.
Последовательность химических реакций, происходящих в мышечных волокнах, при определении типа MB заключается в том, что структуры, обладающие ферментативной активностью, окрашиваются в черный цвет, то есть в местах размещения фермента выпадают осадки сульфида кобальта черного или серого цвета.
Все вышеприведенные операции выполняют не на одном срезе, а производят серийные срезы.
Последовательность выполнения серийных срезов:
- выделяют кусочек мышцы, ставят его на блок и замораживают в жидком азоте (-150°С);
- блок с замороженным материалом помещают в криостат и при температуре минус 20°С делают серийные срезы микротомным ножом. Размеры среза составляют 12 мкм;
- кончиком волоска беличьей кисточки сажают один микросрез на одно предметное стекло и прилепляют его к стеклу. На другое стекло прилепляют другой срез и.т.д.;
- таким способом можно сделать до 10 срезов на разных стеклах, но с одним и тем же рисунком. Одно и то же волокно с одним и тем же «лицом» будет на разных стеклах;
- на этих разных стеклах с одним и тем же рисунком можно ставить разные реакции, с разными ферментами. Каждое волокно будет вести себя по-разному в зависимости от того, какой тип энергообеспечения в нем присутствует.
Сукцинатдегидрогеназа (СДГ) относится к числу наиболее распространенных ферментов и является одним из центральных ферментов цикла трикарбоновых кислот Кребса. Относится к оксиредуктазам, по химической природе - флавинопротеид, молекула содержит железо, в составе активного центра имеются сульфгедрильные группы.
Является активным переносчиком протонов водорода по дыхательной цепи митохондрий, так как цикл Кребса происходит только в них. Митохондрий очень много именно в окислительных медленных волокнах типа I. При выявлении СДГ в MB гранулы моно- и диформазана откладываются в большом количестве только в окислительных волокнах, несколько меньше в окислительно-гликолитических и совсем мало в белых гликолитических быстрых волокнах.
Принцип выявления СДГ:
- вначале СДГ отщепляет от сукцината протоны и электроны, которые акцептируются нитро-синим тетразолием, при этом нитро-синий тетразолий восстанавливается до моно- и диформазанов;
- в 10 мл забуференного раствора сукцината натрия добавляют 10 мл водного раствора нитро-СТ с концентрацией 1 мг/мл, затем срезы помещают в инкубационную среду на 5-20 минут при 37°С, затем промывают в солевом растворе и переносят в формалин-солевой раствор на 10 минут, снова промывают в 15% растворе этанола 5 мин.
В результате осадки моно- и диформазана, тетраформазана свидетельствуют о наличии и локализации СДГ. Высокие концентрации фермента выявляются в окислительных мышечных волокнах, низкие - в гликолитических.
Для определения активности СДГ в мышечных волокнах использовали метод L.S. Kaplov (Kaplov L.S. Polgar J. Cytochemical demonstration of acid phosphatase in hematological dicorders using azo dye technigues // J. Histochem., Cytochem. - 1964. - V. 12, N 6. - P. 805-811). С помощью прибора спектрофотометра определяли степень интенсивности реакции в волокнах различного типа. Были выделены 5 вариантов интенсивности окисления СДГ: 0 - крайне низкая активность реакции; 1 - слабая активность реакция; 2 - средняя активность реакции; 3 - высокая активность реакции; 4 - крайне высокая активность реакции. По интенсивности реакции, согласно этому методу, судят об активности сукцинатдегидрогеназы MB. В волокнах окислительного типа, которые имеют очень много митохондрий, откладываются гранулы диформазана и тетраформазана, что свидетельствует о высокой активности данного фермента в митохондриях. В гликолитических волокнах при выявлении СДГ гранулы формазана не откладываются, а в окислительно-гликолитических волокнах откладываются только гранулы моноформазана.
Определение активности ферментов энергетического метаболизма существенно дополняет оценку типов мышечных волокон по АТФазной активности. Метод выявления СДГ-активности, в отличие от АТФазного, позволяет не только выявить три типа волокон, но и, за счет различной интенсивности реакции, уловить градации активности СДГ во всех типах MB. При этом возникает возможность сопоставления вариантов развития окислительной энергетики в различных мышцах.
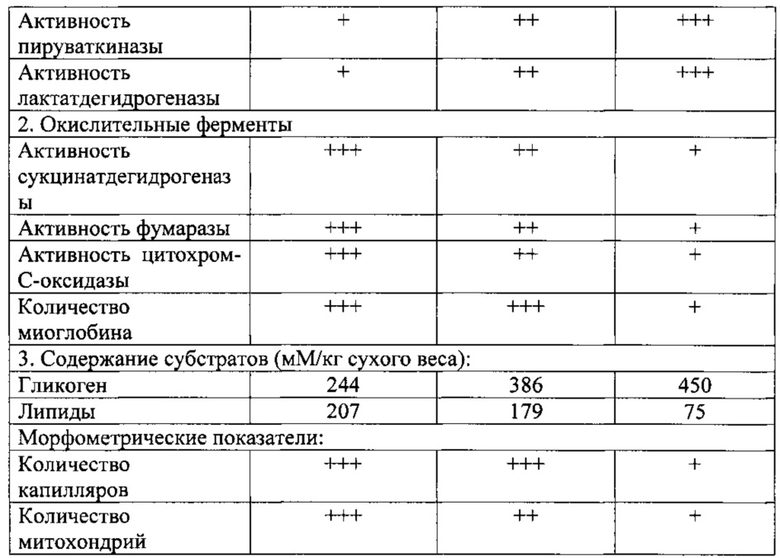
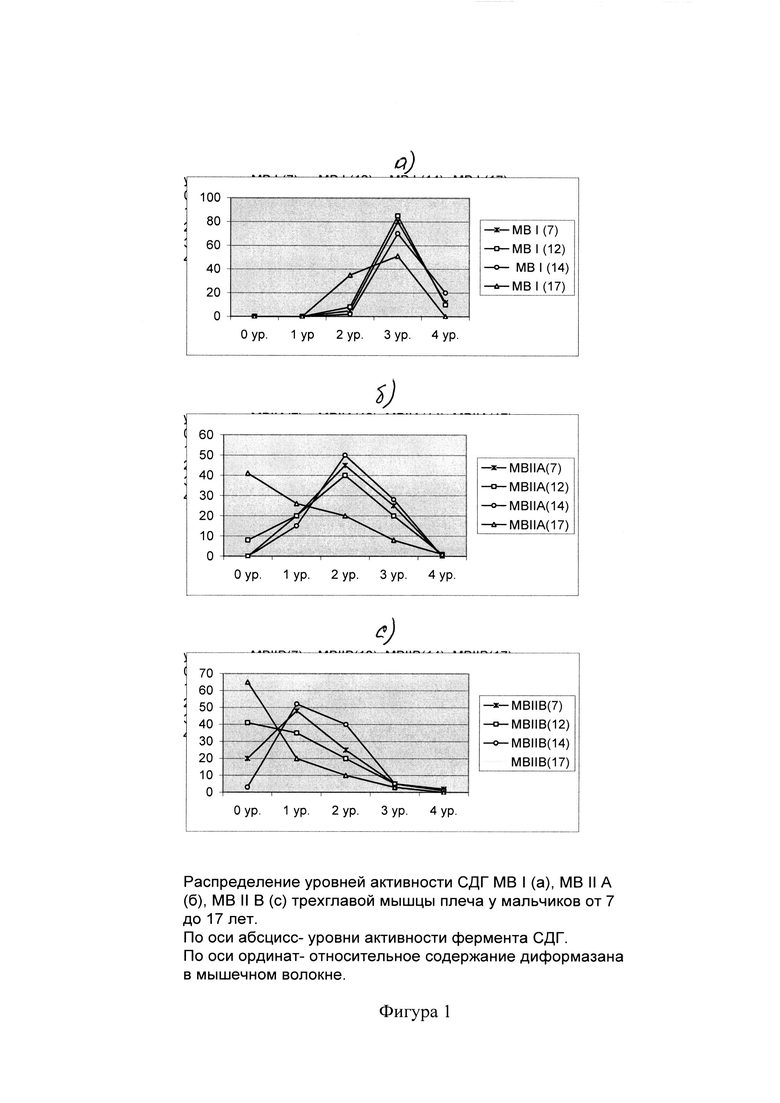
Были проведены исследования, где с помощью использованного метода выявлена динамика изменения окислительного потенциала MB у мальчиков, подростков и юношей в период от 7 до 17 лет (Тамбовцева Р.В. Изменение активности СДГ в мышечных волокнах четырехглавой мышцы бедра у мальчиков 7-17 лет // Ж. Морфология. 1996, №1, с. 68-70).
На фиг. 1, 2 представлено распределение уровней СДГ-активности в мышечных волокнах различного типа. Видно, что у мальчиков 7-8 лет в двуглавой, трехглавой мышцах плеча и четырехглавой мышце бедра (группа MB I) наибольшее относительное количество составляют волокна с высокой активностью (3й уровень активности СДГ). Причем, в двуглавой мышцах плеча число данных структур достоверно выше (83,3±1,52; 81,9±1,41), чем в четырехглавой мышце бедра (54,2±1,72) (Р<0,001). В то время как относительное количество волокон со средней активностью (2й уровень активности СДГ) в этой же группе MB I в четырехглавой мышце бедра (35,4±1,73) значительно превышает (Р<0,001) таковые в трехглавой и двуглавой мышцах плеча (7,9±0,92; 1,3±0,52).
В группе MB IIА в этот же возрастной период наибольшее относительное количество во всех исследованных мышцах занимают волокна со средней активностью реакции (2й уровень) - около 50%.
В группе MB IIВ наибольшее содержание во всех исследованных мышцах приходится на волокна со слабой активностью реакции (1й уровень активности СДГ), причем их количество в трехглавой мышце бедра (50,5±0,90; 50,0±0,34) достоверно выше (Р<0,001), чем в двуглавой мышце плеча (40,9±1,32).
У мальчиков 7-8 лет MB всех трех типов в основной своей массе имеют более высокие окислительные возможности. Распределение уровней активности СДГ сдвинуто вправо. Уже с 7-летнего возраста варианты активности СДГ связаны с типом волокон.
В 11-12-летнем возрасте в различных типах MB мышц верхних и нижних конечностей наблюдается заметное снижение активности СДГ.
В 14-летнем возрасте интенсивность СДГ-активности в различных типах MB мышц верхних и нижних конечностей смещается в сторону увеличения волокон с окислительным типом энергообеспечения.
В 17-летнем возрасте, когда появляются признаки дефинитивной организации MB, происходят противоположные, в сравнении с предыдущей возрастной группой, изменения. Этот возраст характеризуется резким снижением окислительных возможностей всех трех типов MB четырехглавой мышцы бедра и трехглавой мышцы плеча. Поэтому кривые распределения интенсивности реакции в этих мышцах сдвинуты влево. В группе МВIIВ происходит значительное увеличение относительного количества волокон с крайне низкой активностью фермента (нулевой уровень).
Возрастные изменения окислительных возможностей выявляются во всех типах MB. При этом каждый тип MB характеризуется своеобразным спектром СДГ-активности и с возрастом положение этого спектра по отношению к шкале смещается в сторону уменьшения активности фермента, особенно в гликолитических волокнах.
Уровень окислительной активности СДГ в ходе постнатального онтогенеза постоянно меняется и устанавливается только к завершению пубертатного периода, когда наблюдается положительная корреляционная связь между уровнем активности СДГ и АТФазой миозина.
Лактатдегидрогеназа (ЛДГ) - один из важнейших ферментов анаэробного и аэробного расщепления углеводов. Систематическое название L-лактат: НАД-оксиредуктаза, кофактором является цинк, фермент катализирует реакцию:
L-лактат + НАД → Пируват + НАД.Н2.
Фермент с помощью НАД отщепляет от субстрата протоны и электроны, которые также акцептируются нитро-синим тетразолием.
Реактивы: 1.0,1 мл 1 М раствора L-лактата натрия, 0,1 мл 0,1-1 М раствора НАД, 0,1 мл 0,1 М раствора цианида натрия, 0,25 мл 0,06 М фосфатного буферного раствора, 0,25 мл раствора нитро-СТ, 1 мл дистиллированной воды, 10%-1 раствор хлорида кальция в 10% растворе формалина.
В том случае, если при нагрузке организм не испытывает нужды в кислороде и происходит равномерное потребление кислорода, при использовании в качестве энергетического субстрата глюкозы или гликогена пировиноградная кислота последовательно расщепляется до ацетил-коА (активная форма уксусной кислоты) и через мембрану митохондрии уходит в цикл Кребса.
Но если в процессе физической нагрузки организм испытывает недостаток кислорода, то пировиноградная кислота не превращается в ацетил-коА, а переходит из-за нехватки кислорода в молочную кислоту. При умеренной физической нагрузке в работу вовлекаются только медленные двигательные единицы и работа идет с участием O2 с энергообеспечением по циклу Кребса. При работе в субмаксимальной зоне медленные двигательные единицы в работу не включаются, а работают только быстрые двигательные единицы и соответственно - белые MB, которые имеют очень мало митохондрий и работают только в режиме гликолиза в анаэробных условиях (когда ПВК переходит в молочную кислоту). Гликолиз протекает в клетке только в цитоплазме, в отличие от цикла Кребса (только в митохондриях).
При выявлении ЛДГ, гранулы формазана в большой концентрации откладываются только в белых MB непосредственно в цитоплазме.
Концентрация моно-диформазана определяется в MB при помощи спектрофотометра.
Для облегчения распознавания в полевых условиях, в частности в работе со спортивными командами, достаточно использовать выявление активности АТФазы миозина, но в разных преинкубациях: кислой и щелочной и использовать доступные ферменты: сукцинатдегидрогеназу и лактатдегидрогеназу, которые в волокнах с разным энергетическим профилем ведут себя по-разному.
В табл.1 представлены морфометрические, биохимические и гистохимические характеристики различных типов MB спортсменов различных типов конституции, а в табл.2 - их существующие номенклатуры.
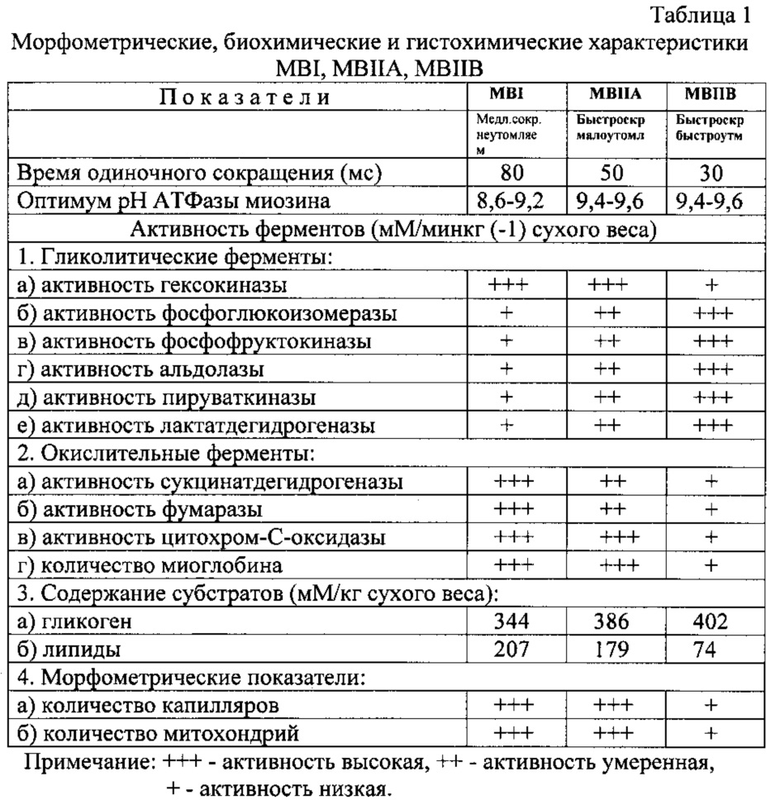
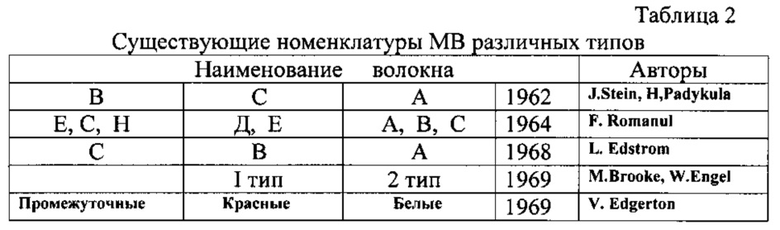
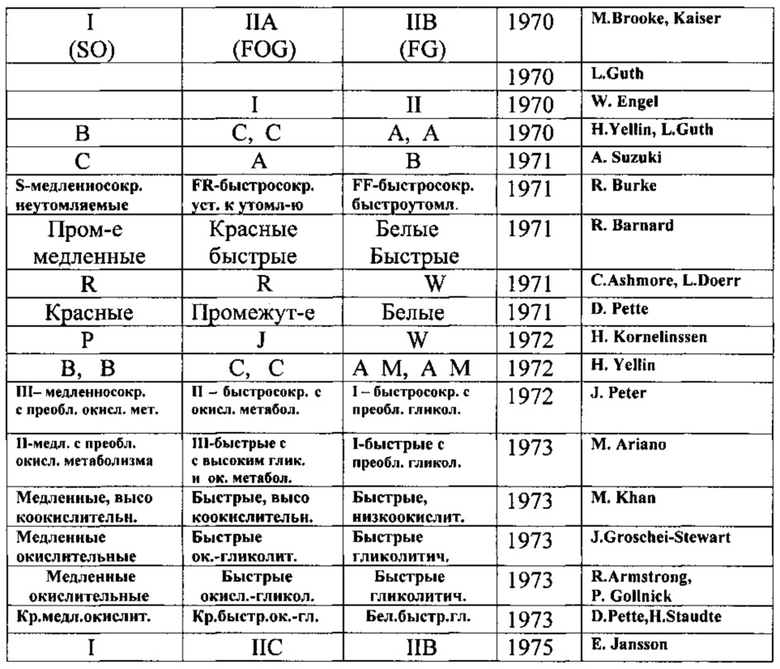
Признаки, по которым определяют тип MB, приведены в табл.3
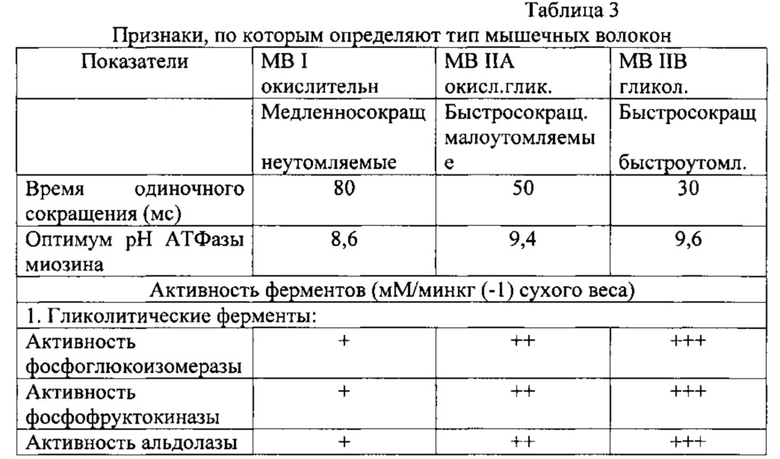
Площадь поперечного сечения MB определяют по одному из двух вариантов:
1. при помощи окуляр-микрометра (№ МОВ-1-15) измеряют большой (а) и малый (в) диаметры клетки и вычисляют абсолютную площадь поперечного сечения MB (по 100 волокон каждого типа с препарата) по формуле эллипса - S=а×b×пи. Далее рассчитывают суммарную относительную площадь каждого из типов MB: S=Sn/SI,IIA,IIB×100%, где Sn - сумма площадей поперечного сечения всех MB данного типа на поперченном срезе, SI,IIA,IIB на поперечном срезе. На поперечном срезе самые крупные волокна с большой площадью поперечного сечения - белые гликолитические быстро сокращающиеся волокна, средняя площадь поперечного сечения - окислительно-гликолитические, с малой площадью поперечного сечения - окислительные медленно сокращающиеся MB;
2. при помощи специальной программы в пакете Statgraf.
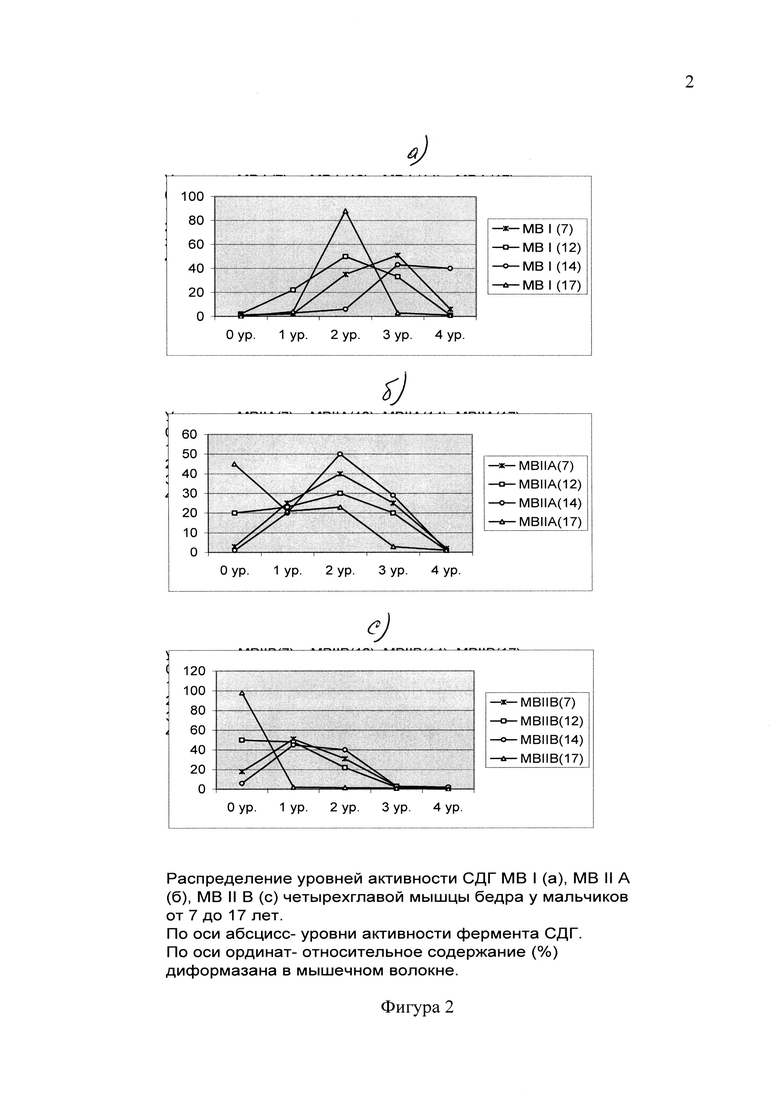
На фиг. 3 представлено распределение различных типов мышечных волокон - MB I, MB IIА, MB IIВ (а) и средней площади их поперечного сечения (в) четырехглавой мышцы бедра мужчин 20-летнего возраста литосомного и эурисомного типов сложения: по оси абцисс - MB I, MB IIА, MB IIВ, по оси ординат - %, S (мкм ∧ 2).
На фиг. 4 представлен поперечный срез латеральной головки четырехглавой мышцы бедра мужчины 20 лет. Обработка по Падикула-Герман при выявлении активности АТФазы миозина (рН=4,6).
На фиг. 5 изображены поперечные серийные срезы камбаловидной мышцы крысы самца 60 дней жизни. Обработка проводилась по Падикула-Герман для выявления активности АТФазы миозина и активности СДГ. Об. 20, ок. 10.
Заявляемый способ позволяет достаточно быстро и достоверно определить состав мышечных волокон в разных мышцах у спортсменов, занимающихся различными видами спорта, четко разделить их на три варианта: красные медленно-сокращающиеся тонические окислительные MB, красные быстро-сокращающиеся окислительно-гликолитические MB и белые быстро-сокращающиеся гликолитические MB. Этим можно предопределить правильное использование упражнений в различных зонах мощности.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОВЫШЕНИЯ УСТОЙЧИВОСТИ К УТОМЛЕНИЮ МЫШЦ ЧЕЛОВЕКА ПРИ СОХРАНЕНИИ МАКСИМАЛЬНОЙ ПРОИЗВОЛЬНОЙ СИЛЫ | 2005 |
|
RU2306960C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ГЕНЕТИЧЕСКОЙ ОСНОВЫ ОБЩИХ ТИПОВ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ | 2008 |
|
RU2386399C9 |
| Способ предотвращения развития утомления скелетной мышцы | 2023 |
|
RU2814077C1 |
| Способ предотвращения атрофии, повышенной утомляемости медленных мышечных волокон и их трансформации в быстрые мышечные волокна в камбаловидной мышце | 2024 |
|
RU2834246C1 |
| Способ определения дисфункции ферментов энергетического обмена при респираторной патологии у детей и его применение | 2020 |
|
RU2759652C1 |
| СПОСОБ ПРОГНОЗИРОВАНИЯ МИТОХОНДРИАЛЬНЫХ ЗАБОЛЕВАНИЙ У ДЕТЕЙ С НЕДИФФЕРЕНЦИРОВАННЫМИ ФОРМАМИ ЗАДЕРЖКИ НЕРВНО-ПСИХИЧЕСКОГО РАЗВИТИЯ (ВАРИАНТЫ) | 2008 |
|
RU2366959C1 |
| СПОСОБ ОЦЕНКИ СТЕПЕНИ ТКАНЕВОЙ АДАПТАЦИИ К ФУНКЦИОНАЛЬНОЙ НЕДОСТАТОЧНОСТИ ОРГАНОВ И СИСТЕМ С ПОМОЩЬЮ ОПРЕДЕЛЕНИЯ ФЕНОМЕНА КОМПЕНСАТОРНОЙ ПРОЛИФЕРАЦИИ МИТОХОНДРИЙ В КЛЕТКАХ СООТВЕТСТВУЮЩИХ ТКАНЕЙ | 2007 |
|
RU2357247C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ГЕНЕТИЧЕСКОЙ ОСНОВЫ ОБЩИХ ТИПОВ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ | 2009 |
|
RU2401071C1 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ПРОФИЛАКТИКИ ИЛИ ЛЕЧЕНИЯ МИОЗИТА, СОДЕРЖАЩАЯ ВЫДЕЛЕННЫЕ МИТОХОНДРИИ В КАЧЕСТВЕ АКТИВНОГО ИНГРЕДИЕНТА | 2020 |
|
RU2810508C2 |
| СРЕДСТВО ДЛЯ ЛЕЧЕНИЯ АЛКОГОЛЬНЫХ ПОРАЖЕНИЙ ВНУТРЕННИХ ОРГАНОВ | 1989 |
|
RU2013091C1 |


Изобретение относится к диагностике, в частности к гистохимии, и может быть использовано для оценки типа мышечных волокон. Для этого гистологические срезы для определения окислительного, окислительно-гликолитического и гликолитического типов мышечных волокон последовательно помещают вначале в 1% раствор CaCI2, затем в 2% раствор CoCI2 и далее в 1% раствор сульфида аммония, в последующем их помещают в кислую среду, в которой инактивируется «быстрый» миозин гликолитических волокон, и в щелочную среду для определения окислительно-гликолитического и гликолитического типов, в которой инактивируется «медленный» миозин окислительных волокон, при этом окислительные волокна на срезе окрашиваются черным цветом, окислительно-гликолитические волокна - серым цветом, а гликолитические волокна - белым цветом, тип же мышечных волокон определяют по площади их поперечного сечения, а в качестве маркеров используют ферменты: сукцинатдегидрогеназу - для оценки активности окислительных процессов в митохондриях и лактатдегидрогеназу - для определения интенсивности гликолиза в цитоплазме клетки. Способ позволяет достоверно определить тип мышечного волокна при его доступности. 3 табл., 5 ил.
Cпособ оценки типов мышечных волокон, заключающийся в использовании ферментов при изучении окислительных процессов, отличающийся тем, что гистологические срезы для определения окислительного, окислительно-гликолитического и гликолитического типов мышечных волокон последовательно помещают вначале в 1% раствор CaCI2, затем в 2% раствор CoCI2 и далее в 1% раствор сульфида аммония, в последующем их помещают в кислую среду, в которой инактивируется «быстрый» миозин гликолитических волокон, и в щелочную среду для определения окислительно-гликолитического и гликолитического типов, в которой инактивируется «медленный» миозин окислительных волокон, при этом окислительные волокна на срезе окрашиваются черным цветом, окислительно-гликолитические волокна - серым цветом, а гликолитические волокна - белым цветом, тип же мышечных волокон определяют по площади их поперечного сечения, а в качестве маркеров используют ферменты: сукцинатдегидрогеназу - для оценки активности окислительных процессов в митохондриях и лактатдегидрогеназу - для определения интенсивности гликолиза в цитоплазме клетки.
| BROOKE M.H | |||
| et al | |||
| Muscle Fiber Types: How Many and What Kind?// Arch Neurol | |||
| Кинематографический аппарат | 1923 |
|
SU1970A1 |
| СПОСОБ ДИФФЕРЕНЦИРОВАННОЙ ОЦЕНКИ СТРУКТУРЫ МЫШЕЧНОЙ ТКАНИ НИЖНИХ КОНЕЧНОСТЕЙ | 2001 |
|
RU2204945C2 |
| Устройство для запирания составного триода | 1960 |
|
SU133493A1 |
| BROOKE M.H | |||
| et al | |||
| The histographic analysis human muscle biopsies with regard to types// neurology, 1969, vol | |||
| Способ изготовления электрических сопротивлений посредством осаждения слоя проводника на поверхности изолятора | 1921 |
|
SU19A1 |
| САМСОНОВА А.В | |||
| и др | |||
| Методы оценки композиции мышечных волокон в скелетных мышцах человека// Труды кафедры биомеханики НГУ им | |||
| П.Ф | |||
| Лесгафта, СПб, 2012, вып.6, с.18-27. | |||

