ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к рекомбинантному микроорганизму, полезному для производства L-метионина и/или его производных и к способу получения L-метионина. Микроорганизм по изобретению модифицируется таким образом, что выход метионина/источник углерода увеличивается посредством сверхэкспрессии генов, гомологичных генам ygaZ и ygaH из Е. coli, в данном рекомбинантном микроорганизме.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Серосодержащие соединения, такие как цистеин, гомоцистеин, метионин или S-аденозилметионин, являются критически важными для клеточного метаболизма. В частности, L-метионин, незаменимая аминокислота, которая не может синтезироваться животными, играет важную роль во многих функциях организма. Большую часть промышленно произведенного метионина широко используют в качестве корма животных и пищевой добавки.
С уменьшением применения белков животного происхождения в результате BSE (губкообразная энцефалопатия крупного рогатого скота) и птичьего гриппа увеличилась потребность в чистом метионине. Обычно D,L-метионин производится химически из акролеина, метилмеркаптана и гидроцианида. Однако данная рацемическая смесь не работает так же хорошо, как чистый L-метионин (Saunderson, 1985). Кроме того, хотя чистый L-метионин и может быть получен из рацемического метионина, например, посредством ацилазной обработки N-ацетил-D,L-метионина, это кардинально увеличивает себестоимость. Соответственно, возрастающая потребность в чистом L-метионине в сочетании с заботой об окружающей среде делают микробную продукцию метионина привлекательной перспективой.
Другие важные аминокислоты, такие как лизин, треонин и триптофан, производятся посредством ферментации для применения в корме животных. Следовательно, данные аминокислоты можно получать с использованием глюкозы и других возобновляемых ресурсов в качестве исходных веществ. Промышленное производство L-метионина посредством фементации еще не было успешным, но разработка данной технологии продолжается.
Ранее были описаны разные подходы для оптимизации получения L-метионина в микроорганизмах (см., например, патенты или патентные заявки US 7790424, US 7611873, WO 2002/10209, WO 2005/059093 и WO 2006/008097); однако, для промышленного производства L-метионина из микроорганизмов требуются дальнейшие улучшения.
При синтезе L-метионина на определенном или более высоком уровне он ингибирует его собственную дальнейшую продукцию посредством петель обратной связи и нарушает физиологию клетки. Следовательно, одним из данных улучшений является уменьшение накопления L-метионина в микроорганизме для обеспечения эффективной продукции посредством усиления оттока L-метионина в рекомбинантном микроорганизме-сверхпродуценте L-метионина.
Экспорт метионина в Escherichia coli опосредован комплексом YgaZH, а в Corynebacterium glutamicum - гомологичным комплексом BrnFE ( et al., 2005). YgaZ представляет собой члена семейства экспортера аминокислот с разветвленной цепью (LIV-E), ответственного за экспорт L-валина и L-метионина. YgaZ образует комплекс с YgaH, прогнозируемым белком внутренней мембраны, для экспорта аминокислот при условиях, в которых их уровни были бы токсичными для клетки.
et al., 2005). YgaZ представляет собой члена семейства экспортера аминокислот с разветвленной цепью (LIV-E), ответственного за экспорт L-валина и L-метионина. YgaZ образует комплекс с YgaH, прогнозируемым белком внутренней мембраны, для экспорта аминокислот при условиях, в которых их уровни были бы токсичными для клетки.
В патентных заявках ЕР 1239041 и WO 2008/082211 описана сверхэкспрессия экспортера аминокислот с разветвленной цепью (YgaZH), ответственного за экспорт L-валина и L-метионина в Escherichia coli. Данная сверхэкспрессия приводит к улучшенной продукции метионина в Е. coli.
Авторы изобретения идентифицировали несколько генов, гомологичных генам ygaZ и ygaH из E. coli, которые неожиданно являются более эффективными для производства метионина, чем гены ygaZ и ygaH из Е. coli.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Изобретение относится к рекомбинантному микроорганизму и способу оптимизации продукции метионина и/или его производных, где усилен экспорт метионина. В данном рекомбинантном микроорганизме отток метионина усиливается посредством сверхэкспрессии генов, гомологичных генам ygaZ и ygaH из Escherichia coli.
В частности, данное изобретение относится к рекомбинантному микроорганизму и его применению в способе оптимизации производства метионина и/или его производных, где в указанном рекомбинантном микроорганизме усиливается отток метионина посредством сверхэкспрессии генов ygaZH, гомологичных генам ygaZ и ygaH из Escherichia coli, при условии, что данная сверхэкспрессия не объединяется ни со сверхэкспрессией metH и возможно fldA и fpr из Е. coli или гомологичных генов из С. glutamicum, ни с ослаблением экспрессии по меньшей мере одного гена, выбранного среди metN, metI или metQ, причем указанные гомологичные гены ygaZH выбраны среди группы генов видов Citrobacter, видов Shigella, видов Raoultella, видов Enterobacter, видов Yersinia и видов Photorhabdus.
Рекомбинантный микроорганизм по изобретению также может содержать другие генетические модификации, такие как:
- повышенная экспрессия по меньшей мере одного из следующих генов: ptsG, рус, pntAB, cysP, cysU, cysW, cysA, cysM, cysJ, cysI, cysH, gcvT, gcvH, gcvP, Ipd, serA, serB, serC, cysE, metF, metA, аллель metA*, кодирующий фермент с пониженной чувствительностью к S-аденозилметионину и/или метионину по механизму обратной связи, thrA или аллель thrA*, кодирующий фермент с ослабленным ингибированием треонином по механизму обратной связи и/или
- ослабленная экспрессия одного из следующих генов: metJ, pykA, pykF, purU, ybdL, udhA, dgsA, metE или yncA.
В конкретном воплощении настоящее изобретение относится к рекомбинатному микроорганизму, в котором: а) сверхэкспрессируются гены, гомологичные генам ygaZH из Escherichia coli, происходящие из Citrobacter koseri, Shigella flexneri, Raoultella ornithinolytica, Enterobacter sp., Yersinia enterocolitica, Photorhabdus luminescens, Citrobacter youngae или Citrobacter freundii; при условии или без условия того, что данная сверхэкспрессия не объединяется ни со сверхэкспрессией metH и возможно fldA и fpr из Е. coli или гомологичных генов из С. glutamicum, ни с ослаблением экспрессии по меньшей мере одного гена, выбранного среди metN, metI или metQ и б) усиливается экспрессия по меньшей мере одного из генов metA*, cysPUWAM, cysJIH, gcvTHP, metF, serA, serB, serC, cysE, thrA* и рус; и в) ослабляется экспрессия по меньшей мере одного из генов metJ, pykA, pykF, purU, ybdL, yncA, dgsA, metE и udhA.
Предпочтительно рекомбинантный микроорганизм представляет собой Escherichia coli или Corynebacterium glutamicum.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Перед подробным описанием настоящего изобретения следует понимать, что данное изобретение не ограничивается способами, конкретно проиллюстрированными примерами, и, естественно, они могут варьировать. Также следует понимать, что терминология, используемая в данном документе, служит лишь для цели описания конкретных воплощений изобретения, и они не предназначены для того, чтобы быть ограничивающими, причем изобретение будет ограничиваться только приложенной формулой изобретения.
Все публикации, патенты и патентные заявки, процитированные в данном документе, выше или ниже, являются тем самым включенными посредством ссылки во всей их полноте.
Кроме того, в воплощении настоящего изобретения на практике, если не указано иное, используются традиционные микробиологические и молекулярно-биологические методики, находящиеся в пределах квалификации в данной области. Такие методики хорошо известны квалифицированному сотруднику и полностью объясняются в литературе.
Следует отметить то, что формы единственного числа в том виде, в котором они используются в данном документе и в приложенной формуле изобретения, включают ссылку на множественное число, если контекст явно не диктует иное. Таким образом, например, ссылка на «микроорганизм» включает множество таких микроорганизмов, и ссылка на «эндогенный ген» представляет собой ссылку на один или более чем один эндогенный ген и так далее. Если не определено иначе, все использованные в данном документе технические и научные термины имеют такие значения, которые обычно понятны обычному специалисту в области, к которой принадлежит данное изобретение. Несмотря на то, что в воплощении на практике или в тестировании настоящего изобретения можно использовать любые материалы и методы, аналогичные или эквивалентные описанным в данном документе, сейчас описаны предпочтительные материалы и методы.
В формуле изобретения, которая следует далее, и в последующем описании изобретения, за исключением тех случаев, когда контекст требует иного из-за прямой формулировки или необходимого подтекста, слово «вмещать», «содержать» или «включать» или вариации, такие как «вмещает», «вмещающий», «содержащий», «включенный», «включает», «включающий», используются во включительном смысле, т.е. для определения присутствия утверждаемых характеристик, но не для исключения присутствия или добавления дополнительных характеристик в разных воплощениях изобретения.
Термин «метионин» и «L-метионин» обозначает незаменимую серосодержащую аминокислоту с химической формулой HO2CCH(NH2)CH2CH2SCH3 и номером CAS 59-51-8 или 63-68-3 для специфического L-изомера.
Фраза «производные метионина» относится к молекулам-аналогам метионина, которые имеют такой же химический остов, но отличаются от метионина по меньшей мере одной химической группой. В данном изобретении предпочтительными производными метионина являются N-ацетилметионин (NAM), S-аденозилметионин (SAM) и гидрокси-метионин (или гидрокси аналог метионина, или МНА).
Термин «микроорганизм» в том виде, как он используется в данном документе, относится к бактерии, дрожжам или грибку, которые не являются искусственно модифицированными. Предпочтительно микроорганизм выбран среди Enterobacteriaceae, Bacillaceae, Streptomycetaceae и Corynebacteriaceae. Более предпочтительно микроорганизм представляет собой вид Escherichia, Klebsiella, Pantoea, Salmonella или Corynebacterium. Даже более предпочтительно микроорганизм по изобретению представляет собой либо вид Escherichia coli, либо Corynebacterium glutamicum.
Термин «рекомбинантный микроорганизм» или «генетически модифицированный микроорганизм» в том виде, как он используется в данном документе, относится к бактерии, дрожжам или грибку, который не встречается в природе и является генетически отличным от его эквивалента, обнаруженного в природе. Это означает то, что он модифицирован либо путем введения, либо делеции, либо модификации генетических элементов. Он также может быть трансформирован посредством принуждения к развитию и эволюции новых метаболических путей посредством объединения направленного мутагенеза и эволюции под специфичным давлением отбора (см., например, WO 2004/076659 или WO 2007/011939).
Микроорганизм может быть модифицирован для экспрессии экзогенных генов, если данные гены вводятся в микроорганизм со всеми элементами, обеспечивающими их экспрессию в микроорганизме-хозяине. Модификация или «трансформация» микроорганизмов экзогенной ДНК представляет собой рутинную задачу для специалистов в данной области.
Микроорганизм может быть модифицирован для модуляции уровня экспрессии эндогенного гена.
Термин «эндогенный ген» означает то, что ген присутствовал в микроорганизме до любой генетической модификации. Эндогенные гены могут сверхэкспрессироваться посредством введения гетерологичных последовательностей, помимо или для того, чтобы заменять эндогенные регуляторные элементы, или посредством введения одной или более чем одной дополнительной копии гена в хромосому или плазмиду. Эндогенные гены также могут быть модифицированы для модуляции их экспрессии и активности соответствующего кодируемого белка. Например, в кодирующую последовательность могут быть введены мутации для модификации генного продукта, или могут быть введены гетерологичные последовательности, помимо или для замены эндогенных регуляторных элементов. Модуляция эндогенного гена может приводить к повышающей регуляции и/или усилению активности генного продукта, или, в качестве альтернативы, к понижающей регуляции и/или уменьшению активности эндогенного генного продукта.
Другим способом модуляции их экспрессии является замена эндогенного промотора гена (например, промотора дикого типа) на более сильный или более слабый промотор для повышающей или понижающей регуляции экспрессии эдогенного гена. Данные промоторы могут быть гомологичными или гетерологичными. Выбор подходящих промоторов находится целиком в пределах способностей специалиста в данной области.
Наоборот, термин «экзогенный ген» означает то, что ген был введен в микроорганизм способами, хорошо известными специалисту в данной области, в то время как данный ген не встречается в природе в данном микроорганизме. Экзогенные гены могут быть интегрированы в хромосому хозяина, или могут экспрессироваться внехромосомно плазмидами или векторами. В данной области хорошо известен целый ряд плазмид, которые отличаются в отношении их репликатора и числа их копий в клетке. Данные гены могут быть гомологичными.
В контексте данного изобретения термин «гомологичный ген» не ограничивается обозначенными генами, имеющими теоретического общего генетического предка, но и включает гены, которые могут не быть родственными генетически, которые, тем не менее, эволюционировали так, что кодируют белок, который выполняет аналогичные функции и/или имеет аналогичную структуру. Следовательно, термин «функциональный гомолог» для цели настоящего изобретения относится к тому факту, что определенная ферментативная активность может быть не только предоставлена специфическим белком с определенной аминокислотной последовательностью, но также и белками с аналогичной последовательностью из других (не)родственных микроорганизмов.
С использованием ссылок, приведенных в Genbank для известных генов, специалисты в данной области могут определять эквивалентные гены в других организмах, бактериальных штаммах, дрожжах, грибах, млекопитающих, растениях и т.д. Эта рутинная работа преимущественно осуществляется с использованием консенсусных последовательностей, которые могут быть определены посредством проведения выравниваний последовательностей с генами, происходящими из других микроорганизмов, и разработки вырожденных зондов для клонирования соответствующего гена в другом организме. Эти традиционные методы молекулярной биологии хорошо известны специалистам в данной области.
Термины «улучшенная продукция метионина», «улучшать продукцию метионина» и их грамматические эквиваленты в том виде, в котором они используются в данном документе, относятся к повышенному выходу метионина/источник углерода (отношение грамм/моль метионина, продуцированного на грамм/моль потребленного источника углерода, которое может быть выражено в процентах). Способы определения количества потребленного источника углерода и продуцированного метионина хорошо известны специалистам в данной области. Выход выше у рекомбинантного микроорганизма по сравнению с соответствующим немодифицированным микроорганизмом.
Фраза «микроорганизм, оптимизированный для ферментативной продукции метионина» относится к микроорганизмам, эволюционировавшим и/или генетически модифицированным для осуществления улучшенной продукции метионина по сравнению с эндогенной продукцией соответствующих микроорганизмов дикого типа. Такие микроорганизмы, «оптимизированные» для продукции метионина, хорошо известны в данной области и были раскрыты, в частности, в патентных заявках WO 2005/111202, WO 2007/077041, WO 2009/043803, WO 2010/020681, WO 2011/073738, WO 2011/080542, WO 2011/080301, WO 2012/055798, WO 2013/001055, WO 2013/190343, WO 2015/028675 и WO 2015/028674.
Согласно данному изобретению термины «ферментативная продукция», «культура» или «ферментация» используются для обозначения роста бактерий. Этот рост обычно проводится в ферментерах с подходящей культуральной средой, адаптированной к используемому микроорганизму и содержащей по меньшей мере один простой источник углерода и, при необходимости, косубстраты.
Фраза «подходящая культуральная среда» обозначает среду (например, стерильные жидкие среды), содержащую незаменимые или полезные питательные вещества для поддержания и/или роста клетки, такие как источники углерода или углеродные субстраты, источники азота, например, пептон, дрожжевые экстракты, мясные экстракты, экстракты солода, мочевина, сульфат аммония, хлорид аммония, нитрат аммония и фосфат аммония; источники фосфора, например, монокалия фосфат или дикалия фосфат; микроэлементы (например, соли металлов), например, соли магния, соли кобальта и/или соли марганца, а также факторы роста, такие как аминокислоты и витамины.
Термин «углеродный источник» или «углеродный субстрат», или «источник углерода» согласно настоящему изобретению обозначает любой источник углерода, который может использоваться специалистами в данной области для поддержки нормального роста микроорганизма, включая моносахариды (такие как глюкоза, галактоза, ксилоза, фруктоза или лактоза), олигосахариды, дисахариды (такие как сахароза, целлобиоза или мальтоза), меласса, крахмал или его производные, гемицеллюлозы и их комбинации. Особенно предпочтительным простым источником углерода является глюкоза. Другим предпочтительным простым источником углерода является сахароза. Источник углерода может происходить из возобновляемого сырья. Возобновляемое сырье определяется как сырье, требующееся для определенных промышленных процессов, которое может быть регенерировано в пределах короткого промежутка времени и в достаточном количестве для обеспечения его превращения в желательный продукт. Растительная биомасса, обработанная или необработанная, является интересным возобновляемым углеродным источником.
Термин «источник серы» согласно данному изобретению относится к сульфату, тиосульфату, гидросульфиду, дитионату, дитиониту, сульфиту, метилмеркаптану, диметилсульфиду и другим сульфидам с метильной группой или к комбинации разных источников. Более предпочтительно, источник серы в культуральной среде представляет собой сульфат или тиосульфат, или их смесь.
Термин «источник азота» соответствует либо соли аммония, либо газообразному аммиаку. Источник азота поставляется в форме аммония или аммиака.
Термины «ослабление» или с «ослабленной экспрессией» в данном контексте означают то, что экспрессия гена и/или продукция фермента уменьшается или подавляется по сравнению с немодифицированным микроорганизмом, приводя к уменьшению внутриклеточной концентрации рибонуклеиновой кислоты, белка или фермента, по сравнению с немодифицированным микроорганизмом. Специалист в данной области знает разные средства и способы для измерения концентрации рибонуклеиновой кислоты или концентрации белка в клетке, включая, например, применение полимеразной цепной реакции, сопряженной с обратной транскрипцией (ПЦР-ОТ), для определения концентрации рибонуклеиновой кислоты, и применение специфичного антитела для определения концентрации специфического белка.
Уменьшение или подавление продукции фермента получают посредством ослабления экспрессии гена, кодирующего указанный фермент.
Ослабление экспрессии генов может достигаться средствами и способами, известными специалисту в данной области. В общем, ослабление экспрессии гена может достигаться посредством:
- мутирования кодирующей области или промоторной области, или
- делеции всей или части промоторной области, необходимой для экспрессии гена, или
- делеции всей или части кодирующей области гена посредством гомологичной рекомбинации, или
- вставки в кодирующую область или в промоторную область внешнего элемента, или
- проведения экспрессии гена под контролем слабого промотора или индуцибельного промотора.
Специалист в данной области знает целый ряд промоторов, которые демонстрируют разную силу, и какой промотор использовать для слабой или индуцибельной генетической экспрессии.
Термин «активность» фермента используется взаимозаменяемо с термином «функция» и обозначает, в контексте изобретения, реакцию, которая катализируется ферментом. Специалист в данной области знает как измерять ферментативную активность указанного фермента.
Термины «ослабленная активность» или «пониженная активность» фермента означают либо пониженную удельную каталитическую активность белка, полученную мутацией в аминокислотной последовательности, и/или пониженные концентрации белка в клетке, полученные мутацией нуклеотидной последовательности или делецией кодирующей области гена.
Термины «повышенная активность» или «увеличенная активность» фермента обозначают или увеличенную удельную каталитическую активность фермента и/или увеличенное количество/доступность фермента в клетке, полученные, например, сверхэкспрессией гена, кодирующего фермент.
Термины «увеличенная экспрессия», «повышенная экспрессия» или «сверхэкспрессия» и их грамматические эквиваленты используются в тексте взаимозаменяемо и имеют аналогичное значение. Данные термины означают то, что экспрессия гена или продукция фермента увеличивается по сравнению с немодифицированным микроорганизмом, приводя к увеличению внутриклеточной концентрации рибонуклеиновой кислоты, белка или фермента по сравнению с немодифицированным микроорганизмом. Специалист в данной области знает разные средства и способы для измерения концентрации рибонуклеиновой кислоты или концентрации белка в клетке, включая, например, применение полимеразной цепной реакции, сопряженной с обратной транскрипцией (ПЦР-ОТ), для определения концентрации рибонуклеиновой кислоты, и применение специфичного антитела для определения концентрации специфического белка.
Увеличение продукции фермента получают посредством увеличения экспрессии гена, кодирующего указанный фермент.
Для увеличения экспрессии гена специалист в данной области знает разные методики, такие как:
- Увеличение числа копий гена в микроорганизме. Данный ген кодируется хромосомно или внехромосомно. При локализации гена на хромосоме, на хромосому можно вводить несколько копий гена способами рекомбинации, известными эксперту в данной области (включая замену гена). При внехромосомной локализации гена ее (экспрессию) можно проводить посредством разных типов плазмид, которые отличаются в отношении их репликатора и, таким образом, их числа копий в клетке. Данные плазмиды присутствуют в микроорганизме в 1-5 копиях или примерно в 20 копиях, или вплоть до 500 копий, в зависимости от природы плазмиды: плазмиды с малым числом копий с ограниченной репликацией (pSC101, RK2), плазмиды с малым числом копий (pACYC, pRSF1010) или плазмиды с большим числом копий (pSK bluescript II).
- Применение промотора, приводящего к высокому уровню экспрессии гена. Специалист в данной области знает, какие промоторы являются самыми удобными, например, широко используются промоторы Ptrc, Ptac, Plac или промоторы лямбда PR и PL. Данные промоторы могут быть «индуцибельными» посредством конкретного соединения или посредством конкретного внешнего условия, подобного температуре или свету. Данные промоторы могут быть гомологичными или гетерологичными.
- Ослабление активности или экспрессии репрессора транскрипции, специфичного или неспецифичного по отношению к гену.
- Применение элементов, стабилизирующих соответствующую матричную РНК (Carrier and Keasling, 1998), или элементов, стабилизирующих белок (например, GST (глутатион-S-трансфераза) метки, GE Healthcare).
Термины «кодирующий» или «кодирование» относятся к процессу, посредством которого полинуклеотид, посредством механизмов транскрипции и трансляции, продуцирует аминокислотную последовательность. Ген(ны), кодирующий(щие) фермент(ты), может быть экзогенным или эндогенным.
Термины «чувствительность по механизму обратной связи» или «ингибирование по механизму обратной связи» относятся к клеточному механизму контроля, в котором один или несколько ферментов, которые катализируют продукцию определенного вещества в клетке, ингибируются или становятся менее активными при накоплении данного вещества до определенного уровня. Таким образом, термины «пониженная чувствительность по механизму обратной связи» или «ослабленное ингибирование по механизму обратной связи» означают то, что активность такого механизма уменьшается или подавляется по сравнению с немодифицированным микроорганизмом. Специалист в данной области знает как модифицировать фермент для получения данного результата. Такие модификации были описаны в патентной заявке WO 2005/111202 или в патенте US 7611873.
В первом аспекте изобретения рекомбинантный микроорганизм оптимизирован для ферментативной продукции метионина и/или его производных посредством усиления оттока метионина в указанном микроорганизме. Предпочтительно рекомбинантный микроорганизм выбран среди Enterobacteriaceae или Corynebacteriaceae. Более предпочтительно рекомбинантный микроорганизм выбран среди Escherichia coli или Corynebacterium glutamicum. Даже более предпочтительно микроорганизм по изобретению представляет собой рекомбинатный штамм Е. coli.
Как описано выше, в микроорганизмах-продуцентах аминокислот метионин выводится специфичным экспортирующим переносчиком. Примечательно то, что у Е. coli данный переносчик называется YgaZH и кодируется генами ygaZ и ygaH, тогда как у С. glutamicum он называется BrnFE и кодируется генами brnF и brnE. Функциональные гомологи данной системы оттока метионина были идентифицированы у нескольких других микроорганизмов. В качестве альтернативы, рекомбинантный микроорганизм по изобретению может сверхэкспрессировать функциональных гомологов систем YgaZH или BrnFE. Белки, гомологичные YgaZ и YgaH, представлены в Таблице 1 и Таблице 2 соответственно.
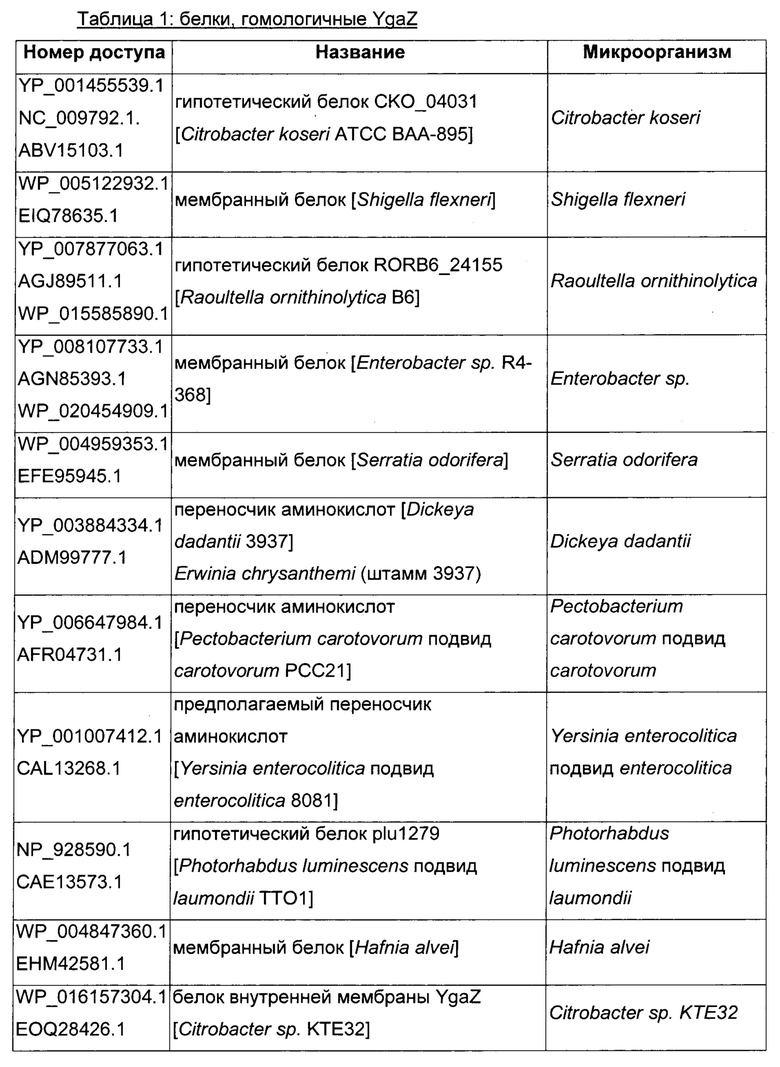
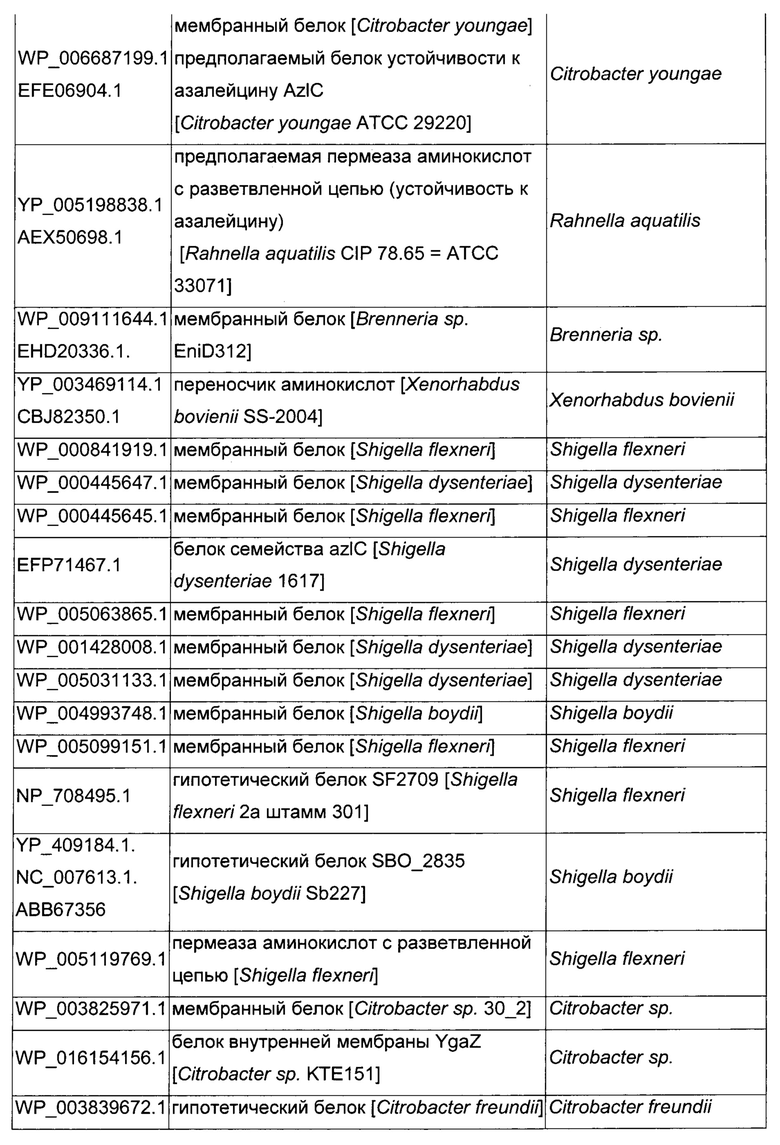
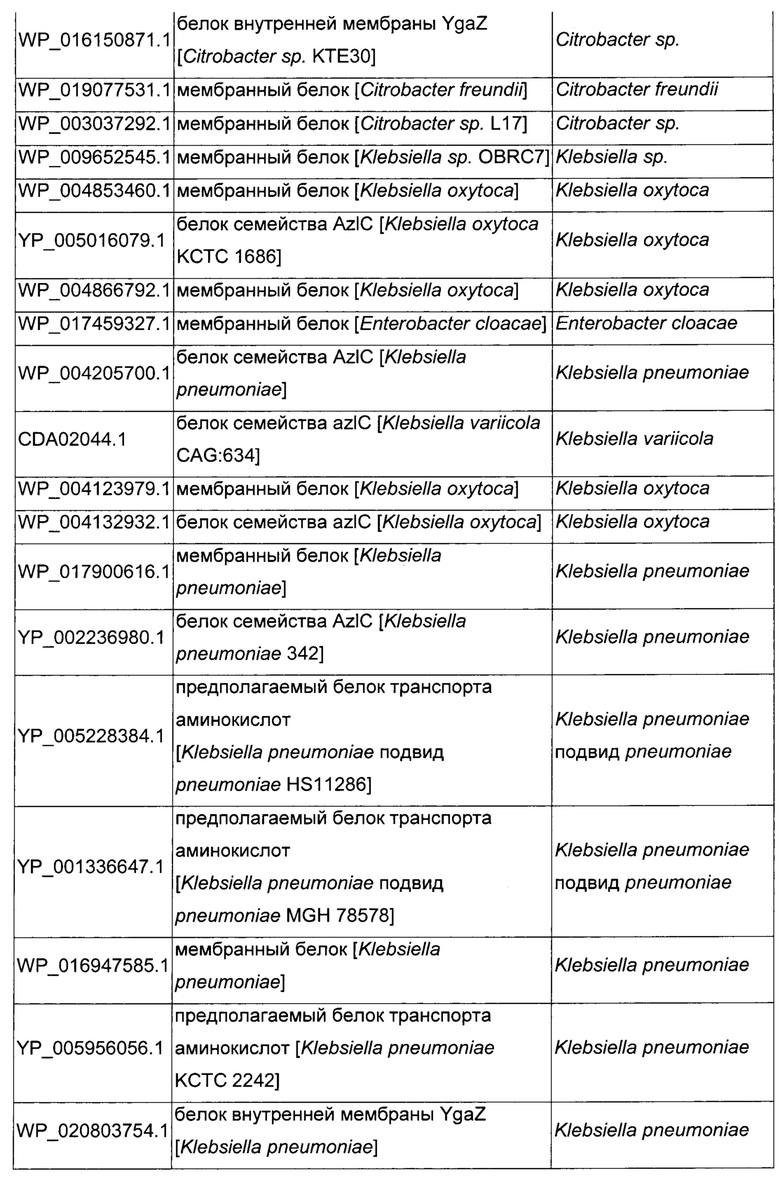
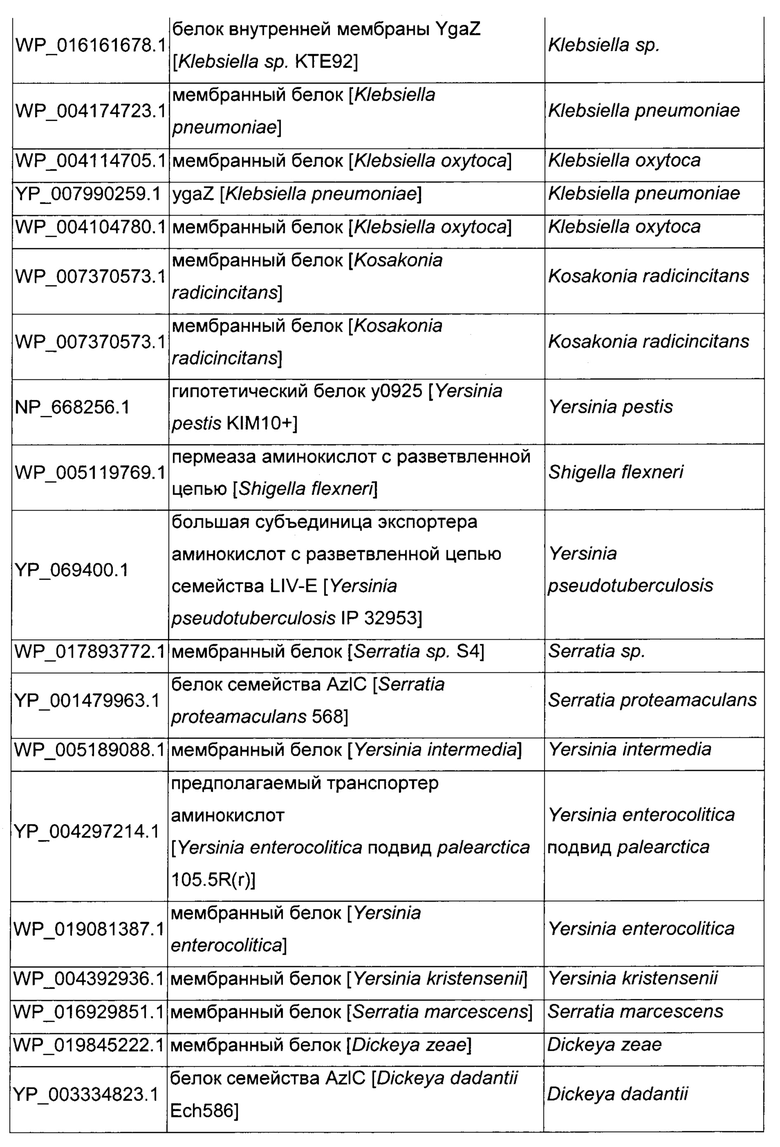
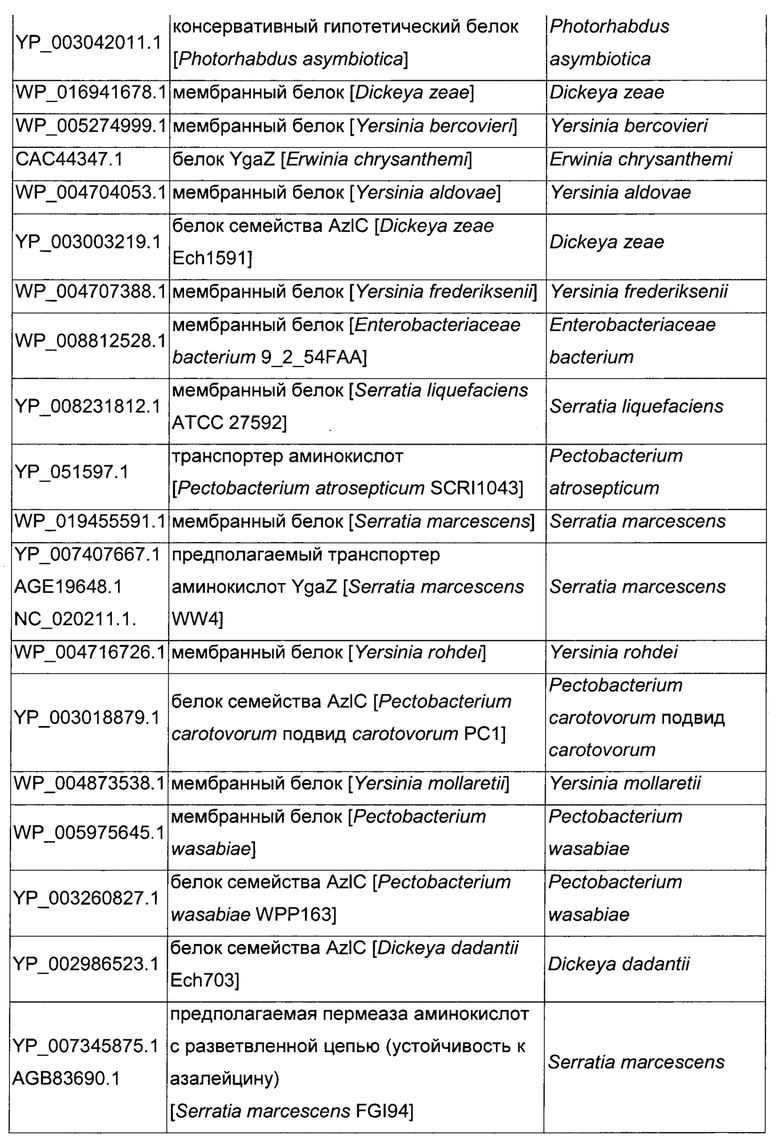
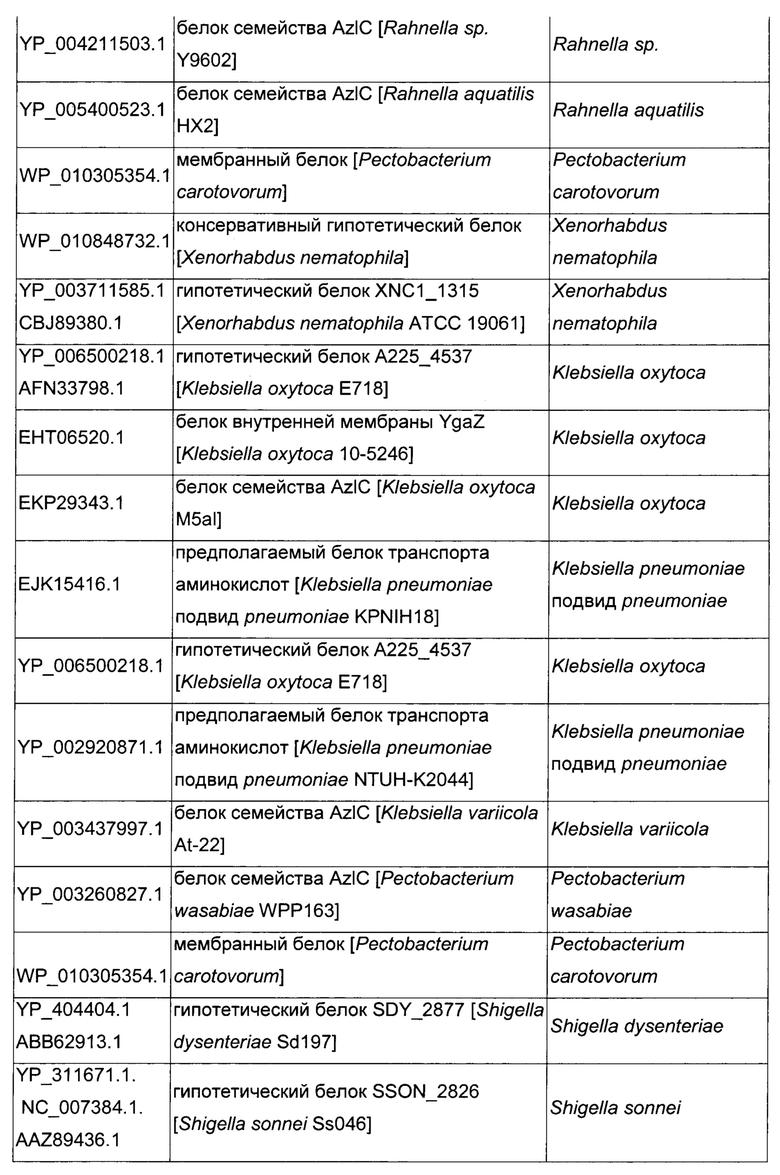
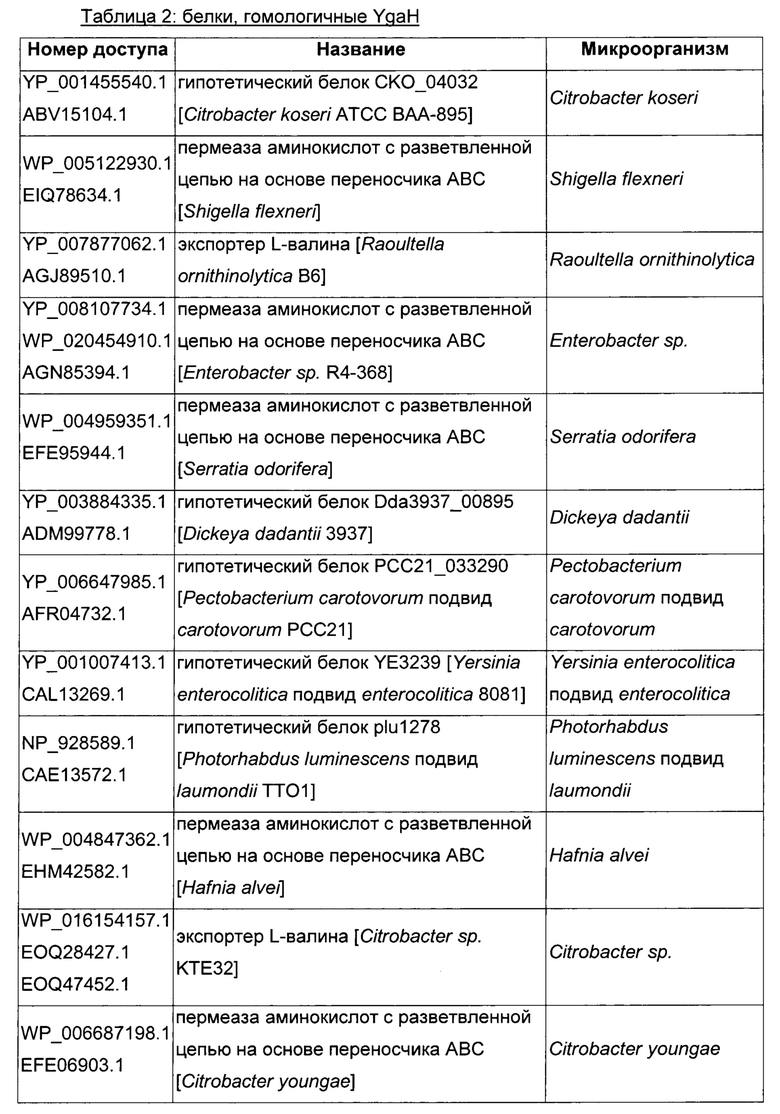
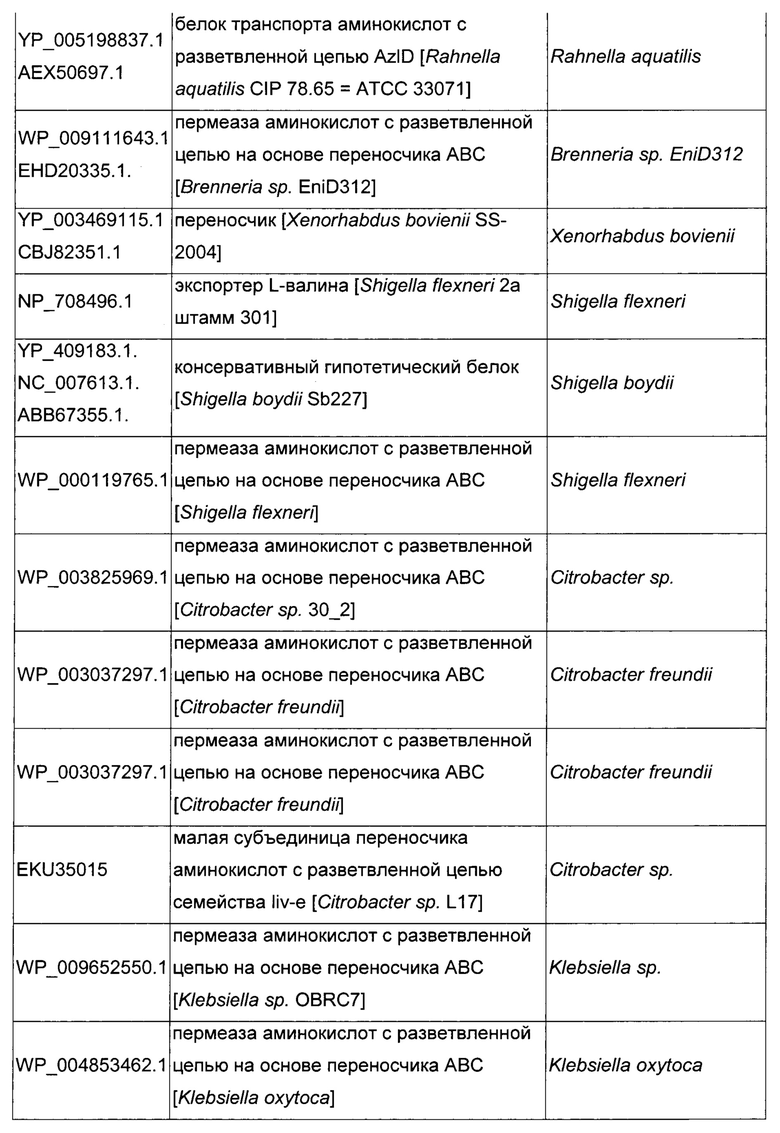
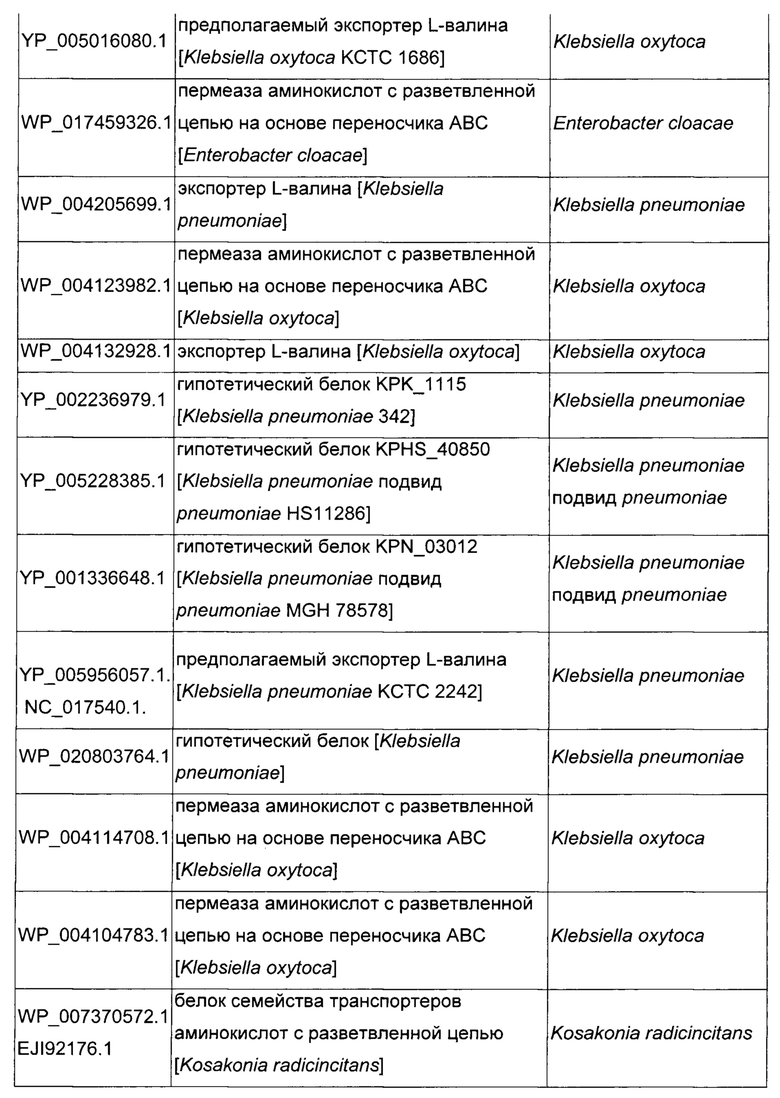
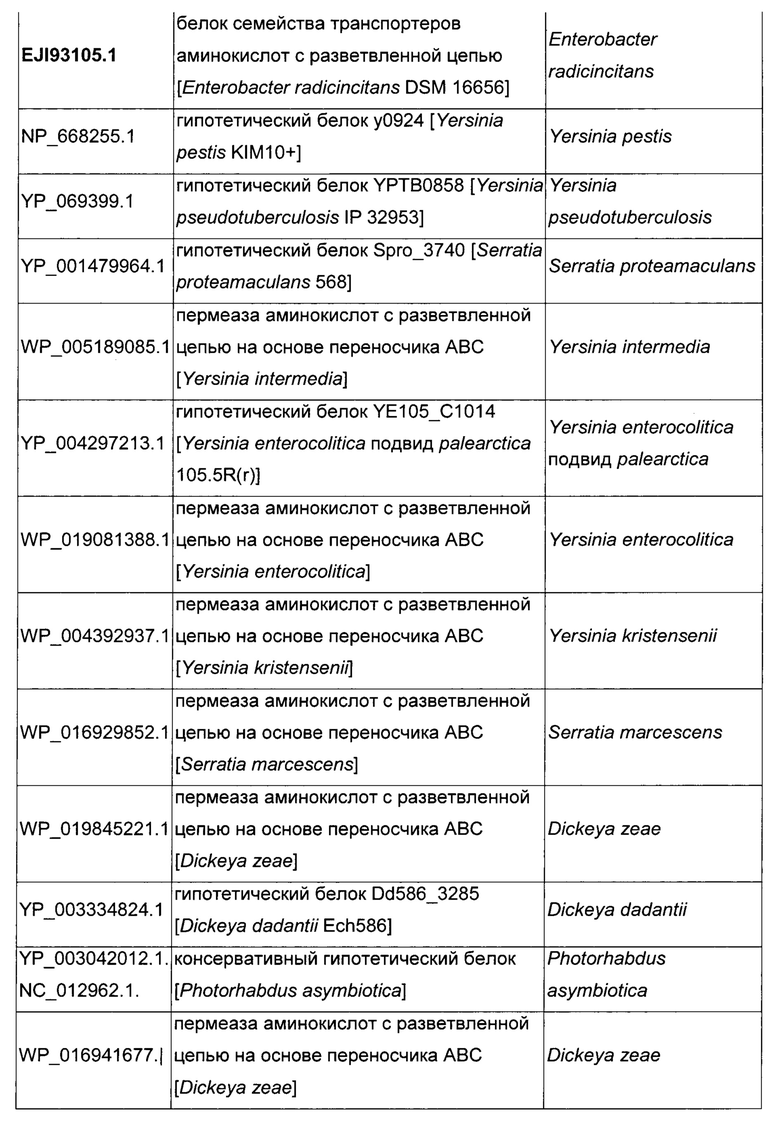
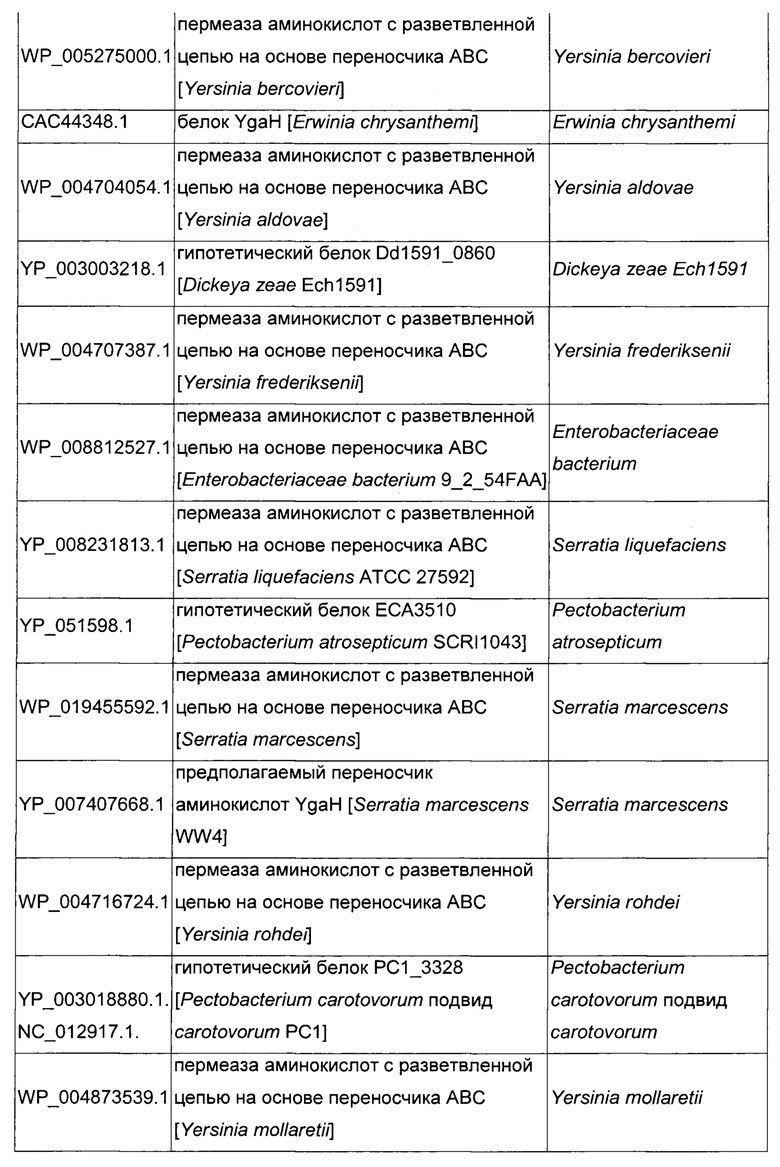
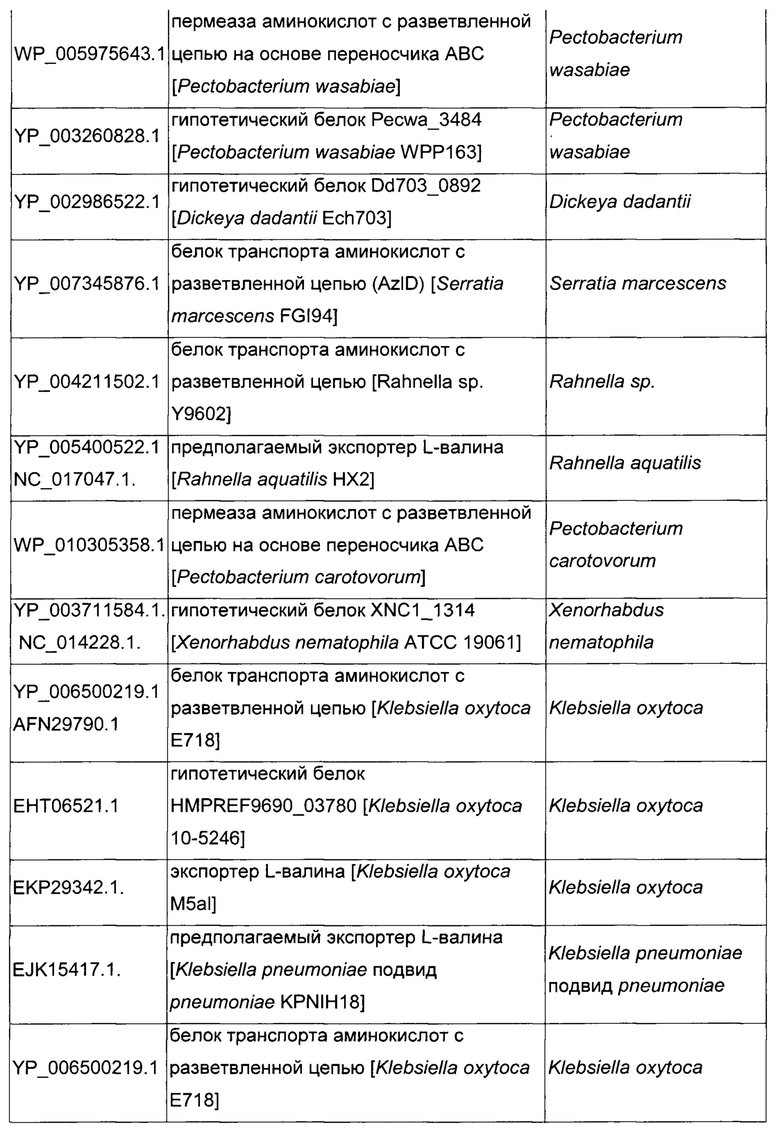
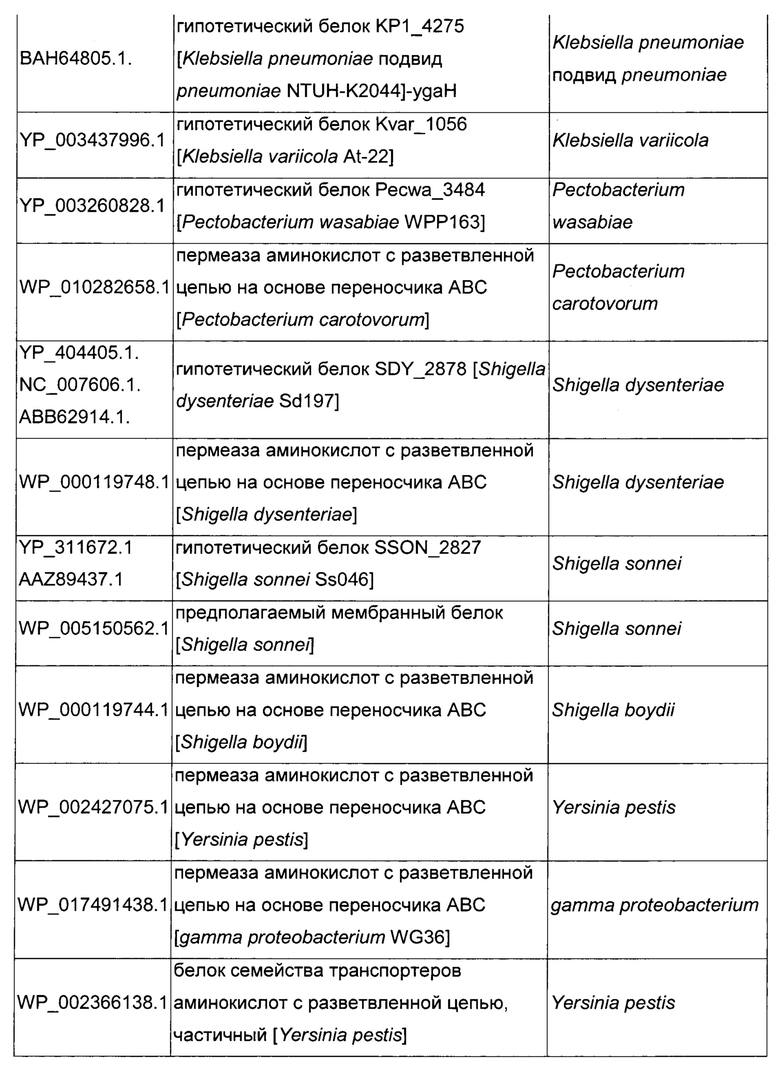
С номером доступа, раскрытым в таблицах для каждого гомолога, специалист в данной области способен получить аминокислотную последовательность и кодирующую ее нуклеотидную последовательность, например, в базах данных NCBI (Национальный центр биотехнологической информации).
Для специалиста в данной области получение генов, кодирующих данные гомологи, из аминокислотной последовательности или нуклеотидной последовательности является рутинной задачей. Это можно сделать либо искусственным синтезом гена, кодирующего интересующий белок, на основе его аминокислотной последовательности, либо посредством ПЦР (полимеразная цепная реакция) амплификации интересующей кодирующей области из соответствующей геномной ДНК. В контексте данного изобретения эти гены называются «генами, гомологичными ygaZ или ygaH». Последовательности данных гомологичных генов ygaZH можно корректировать в соответствии с отклонением использования кодонов у микроорганизма-хозяина.
Согласно изобретению рекомбинантный микроорганизм сверхэкспрессирует гены, гомологичные генам ygaZ и ygaH из Е. coli, кодирующие белки, последовательности которых, соответственно, раскрыты в SEQ ID NO: 1 и SEQ ID NO: 2, при условии или без условия того, что данная сверхэкспрессия не объединяется ни со сверхэкспрессией metH, возможно fldA и fpr из Е. coli, или гомологичных генов из С. glutamicum, ни с ослаблением экспрессии по меньшей мере одного гена, выбранного среди metN, metI или metQ. Предпочтительно гены, гомологичные ygaZ и ygaH, представлены парой генов, происходящих из того же самого организма, и представлены геном, гомологичным ygaZ, и геном, гомологичным ygaH. Однако можно было бы использовать несоответствующую пару гена, гомологичного ygaZ, из первого организма и гена, гомологичного ygaH, из второго организма.
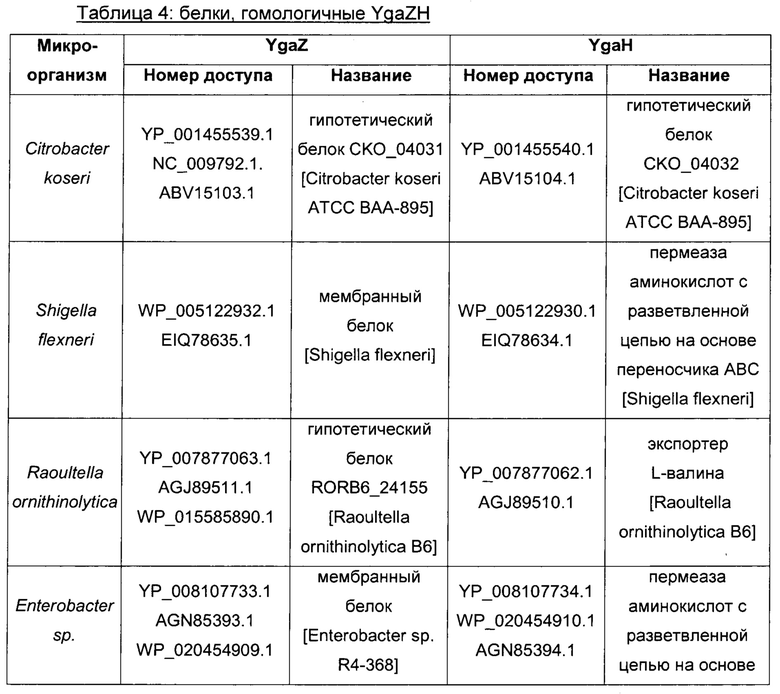
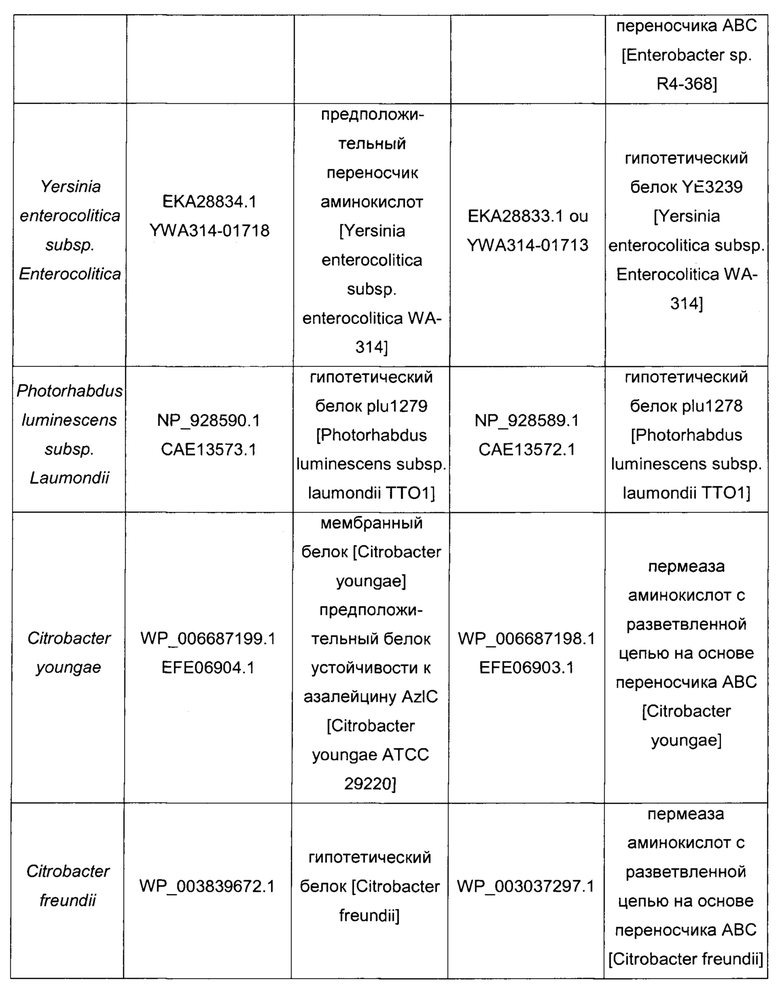
Гомологичные гены ygaZH выбраны среди генов, кодирующих гомологи YgaZ и YgaH, раскрытые в Таблице 1 и в Таблице 2 соответственно. Предпочтительно гомологичные гены ygaZH выбраны среди генов, кодирующих гомологи YgaZH из видов Citrobacter, видов Shigella, видов Raoultella, видов Enterobacter, видов Yersinia и видов Photorhabdus. Более предпочтительно гены, гомологичные ygaZH, происходят из Citrobacter koseri, Shigella flexneri, Raoultella ornithinolytica, Enterobacter sp., Yersinia enterocolitica, Photorhabdus luminescens, Citrobacter youngae или Citrobacter freundii. Наиболее предпочтительно гомологичные гены ygaZH происходят из Citrobacter koseri, Citrobacter youngae, Citrobacter freundii или Enterobacter sp.
Следовательно, гомологичные гены ygaZH выбраны среди генов, кодирующих пару гомолога YgaZH, определенные, соответственно, SEQ ID NO: 3 и SEQ ID NO: 4 из Citrobacter koseri, SEQ ID NO: 5 и SEQ ID NO: 6 из Shigella flexneri, SEQ ID NO: 7 и SEQ ID NO: 8 из Raoultella ornithinolytica, SEQ ID NO: 9 и SEQ ID NO: 10 из Enterobacter sp. (R4-368), SEQ ID NO: 11 или 12 и SEQ ID NO: 13 или 14 из Yersinia enterocolitica подвид enterocolitica, SEQ ID NO: 15 и SEQ ID NO: 16 из Photorhabdus luminescens подвид laumondii, SEQ ID NO: 17 и SEQ ID NO: 18 из Citrobacter youngae, SEQ ID NO: 19 и SEQ ID NO: 20 из Citrobacter freundii.
В предпочтительном воплощении изобретения данные гены или, в общем, гомологичные гены ygaZH сверхэкспрессируются под контролем индуцибельного промотора. Специалист в данной области знает такие индуцибельные промоторы. Например, для сверхэкспрессии гомологичных генов ygaZH, в частности, гомологичных генов ygaZH, происходящих из Citrobacter koseri, Shigella flexneri, Raoultella ornithinolytica, Enterobacter sp., Yersinia enterocolitica, Photorhabdus luminescens, Citrobacter youngae или Citrobacter freundii, в рекомбинантном микроорганизме по изобретению могут быть использованы промоторы, подобные λPR или λPL.
Возможно эндогенные гены ygaZH или brnFE подвергаются делеции в рекомбинантном микроорганизме по изобретению.
Предпочтительным воплощением изобретения является именно сверхэкспрессия в микроорганизме-продуценте аминокислот гомологичных генов ygaZH, одних или в комбинации с другими генетическими модификациями, как раскрыто ниже, и, в частности, сверхэкспрессия гомологичных генов ygaZH, кодирующих гомологи YgaZ и YgaH, раскрытые, соответственно, в таблице 1 и в таблице 2, одних или в комбинации с другими генетическими модификациями, как раскрыто ниже.
Оптимизация пути биосинтеза метионина
Рекомбинантный микроорганизм согласно изобретению модифицируется для улучшения продукции метионина. Гены, участвующие в продукции метионина, хорошо известны в данной области и включают гены, участвующие в специфическом пути биосинтеза метионина, а также гены, участвующие в путях, дающих предшественники, и гены, участвующие в путях, потребляющих метионин.
Для эффективной продукции метионина требуется оптимизация специфического пути биосинтеза метионина и нескольких путей, дающих предшественники. Штаммы, продуцирующие метионин, уже были описаны, в частности, в патентных заявках WO 2005/111202, WO 2007/077041 и WO 2009/043803. Данные заявки включены в эту заявку в качестве ссылки.
За исключением случаев, когда утверждается иное, все гены, упомянутые ниже, имеющие отношение к оптимизации пути биосинтеза метионина, относятся к генам из Е. coli.
В конкретном воплощении изобретения рекомбинантный микроорганизм модифицирован, как описано ниже: увеличена экспрессия по меньшей мере одного гена, выбранного среди ptsG, рус, pntAB, cysP, cysU, cysW, cysA, cysM, cysJ, cysI, cysH, gcvT, gcvH, gcvP, Ipd, serA, serB, serC, cysE, metF, metA, аллеля metA*, кодирующего фермент с пониженной чувствительностью к S-аденозилметионину и/или метионину по механизму обратной связи, thrA и аллеля thrA*, кодирующего фермент с ослабленным ингибированием треонином по механизму обратной связи.
- ptsG кодирует фермент PTS IICBGlc, как описано в патентной заявке WO 2013/001055,
- рус кодирует пируваткарбоксилазу, как описано в патентной заявке WO 2013/001055. В предпочтительном воплощении ген рус является гетерологичным и выбран из генов рус из Rhizobium etli, Bacillus subtilis, Lactococcus lactis, Pseudomonas fluorescens или видов Corynebacterium,
- pntAB кодирует субъединицы мембраносвязанной трансгидрогеназы, такой как описанная в патентной заявке WO 2012/055798,
- cysP кодирует периплазматический сульфатсвязывающий белок, как описано в WO 2007/077041 и в WO 2009/043803,
- cysU кодирует компонент ABC переносчика сульфата, как описано в WO 2007/077041 и в WO 2009/043803,
- cysW кодирует мембраносвязанный белок, транспортирующий сульфат, как описано в WO 2007/077041 и в WO 2009/043803,
- cysA кодирует сульфатпермеазу, как описано в WO 2007/077041 и в WO 2009/043803,
- cysM кодирует О-ацетилсеринсульфгидралазу, как описано в WO 2007/077041 и в WO 2009/043803,
- cysI и cysJ кодируют, соответственно, альфа и бета субъединицы сульфитредуктазы, как описано в WO 2007/077041 и в WO 2009/043803. Предпочтительно cysI и cysJ сверхэкспрессируются совместно,
- cysH кодирует аденилилсульфатредуктазу, как описано WO 2007/077041 и в WO 2009/043803.
Усиление С1 метаболизма также представляет собой модификацию, которая приводит к улучшенной продукции метионина. Оно относится к увеличению активности по меньшей мере одного фермента, участвующего в С1 метаболизме, выбранного среди GcvTHP, Lpd или MetF. В предпочтительном воплощении изобретения одноуглеродный метаболизм усиливается путем увеличения экспрессии и/или активности по меньшей мере одного из следующих:
- gcvT, gcvH, gcvP и lpd, кодирующих комплекс расщепления глицина, как описано в патентной заявке WO 2007/077041. Комплекс расщепления глицина (GCV) представляет собой мультиферментный комплекс, который катализирует окисление глицина, давая диоксид углерода, аммиак, метилен-THF (тетрагидрофолат) и восстановленный пиридиновый нуклеотид. Комплекс GCV состоит из четырех белковых компонентов: указанного Р-белка - глициндегидрогеназы (GcvP), указанного Н-белка - липоил-CcvH-белка (GcvH), указанного Т-белка - аминометилтрансферазы (GcvT) и указанного L-белка - дигидролипоамиддегидрогеназы (GcvL или Lpd). Р-белок катализирует пиридоксальфосфат-зависимое высвобождение CO2 из глицина, оставляя метиламинную группировку. Метиламинная группировка переносится на группу липоевой кислоты Н-белка, который связан с Р-белком до декарбоксилирования глицина. Т-белок катализирует высвобождение NH3 из метиламинной группы и переносит остающееся С1 звено на THF, образуя метилен-THF. L-белок затем окисляет компонент в виде липоевой кислоты Н-белка и переносит электроны на НАД+, образуя НАДН;
- metF, кодирующей метилентетрагидрофолатредуктазу, как описано в патентной заявке WO 2007/077041;
Сверхэкспрессия по меньшей мере одного из следующих генов, участвующих в биосинтезе серина, также уменьшает продукцию побочного продукта - изолейцина:
- serA, который кодирует фосфоглицератдегидрогеназу, как описано в WO 2007/077041 и в WO 2009/043803,
- serB, который кодирует фосфосеринфосфатазу, как описано в WO 2007/077041 и в WO 2009/043803,
- serC, который кодирует фосфосеринаминотрансферазу, как описано в WO 2007/077041 и в WO 2009/043803.
Уже было показано, что сверхэкспрессия следующих генов улучшает продукцию метионина:
- cysE кодирует серинацилтрансферазу; его сверхэкспрессия обеспечивает усиление продукции метионина, как описано в WO 2007/077041;
- metA кодирует гомосеринсукцинилтрансферазу. Аллель metA* кодирует фермент с пониженной чувствительностью к S-аденозилметионину и/или метионину по механизму обратной связи. Предпочтительно используется аллель metA*, описанный в патентной заявке WO 2005/111202;
- thrA кодирует аспартокиназу/гомосериндегидрогеназу; аллель thrA* кодирует фермент с пониженным ингибированием треонином по механизму обратной связи, как описано в WO 2005/111202.
В конкретном воплощении изобретения гены могут находиться под контролем индуцибельного промотора. В предпочтительном воплощении изобретения по меньшей мере один из данных генов находится под контролем промотора, индуцируемого температурой. Предпочтительно экспрессия по меньшей мере одного из генов: thrA, cysE, metA прямо или опосредованно находится под контролем индуцибельного промотора. Более предпочтительно гены thrA, cysE и metA прямо или опосредованно находятся под контролем индуцибельного промотора. В предпочтительном воплощении изобретения экспрессия гена thrA находится под прямым контролем индуцибельного промотора, и экспрессия гена cysE подвергается полярному влиянию индуцибельной экспрессии гена thrA. В другом предпочтительном воплощении изобретения экспрессия гена thrA находится под прямым контролем индуцибельного промотора, и экспрессия генов cysE и metA подвергается полярному влиянию индуцибельной экспрессии гена thrA.
В наиболее предпочтительном воплощении промотор, индуцируемый температурой, принадлежит к семейству промоторов PR. Штамм, продуцирующий метионин, имеющий гены, находящиеся под контролем индуцибельных промоторов, описан в патентной заявке WO 2011/073122.
В другом конкретном воплощении изобретения микроорганизм был дополнительно модифицирован, и была ослаблена экспрессия по меньшей мере одного из следующих генов: metJ, pykA, pykF, purU, ybdL, yncA, metE, dgsA или udhA.
- Ген metJ кодирует репрессорный белок MetJ (GenBank 1790373), ответственный за понижающую регуляцию метионинового регулона, как было предложено в патентной заявке JP 2000/157267,
- Гены pykA и pykF кодируют ферменты «пируваткиназы». Ослабление экспрессии по меньшей мере одной или обеих пируваткиназ снижает потребление фосфоенолпирувата (PEP). Повышенная доступность PEP может увеличивать продукцию оксалоацетата, важного предшественника аспартата, который, в свою очередь, является предшественником метионина, как описано в WO 2007/077041 и в WO 2009/043803,
- purU кодирует формилтетрагидрофолатдеформилазу, фермент, который катализирует формил-THF-деформилазную реакцию. Ослабление деформилазной активности увеличивает продукцию метил-THF, который требуется для метилирования гомоцистеина. Потеря С1 метаболитов посредством деформилирования приводит к повышенной продукции гомоцистеина, который не может превращаться в метионин. Гомоцистеин затем может быть субстратом фермента цистатионин-гамма-синтазы (MetB), который может катализировать реакцию между О-сукцинилгомосерином и гомоцистеином, приводящую к образованию гомолантионина, как описано в WO 2007/077041 и в WO 2009/043803,
- ybdL кодирует аминотрансферазу, как описано в патентной заявке WO 2012/090021,
- yncA кодирует N-ацилтрансферазу, как описано в патентной заявке WO 2010/020681,
- metE кодирует кобаламин-независимую метионинсинтазу, как описано в патентной заявке WO 2013/190343,
- dgsA, лучше известный как MIc, кодирует двойной транскрипционный регулятор, который контролирует экспрессию генов, кодирующих ферменты систем фосфотрансферазы (PTS) и фосфоенолпирувата (PEP), как описано в патентной заявке WO 2013/001055,
- udhA кодирует растворимую трансгидрогеназу пиридиновых нуклеотидов, как описано в патентной заявке WO 2012/055798.
В более предпочтительном воплощении изобретения ферментативная продукция метионина и/или его производных рекомбинантным микроорганизмом, в котором усилен отток метионина, из глюкозы в качестве главного источника углерода может достигаться посредством комбинации обсуждавшихся выше модификаций в указанном микроорганизме, например:
- экспрессия гена metJ ослабляется, и экспрессия аллеля metA*, кодирующего фермент с пониженной чувствительностью к S-аденозилметионину и/или метионину по механизму обратной связи (MetA*), усиливается;
- экспрессия гена metJ ослабляется; экспрессия аллеля metA*, кодирующего фермент с пониженной чувствительностью к S-аденозилметионину и/или метионину по механизму обратной связи (MetA*), усиливается; и экспрессия аллеля thrA*, кодирующего фермент с ослабленным ингибированием треонином по механизму обратной связи (thrA*), усиливается;
- экспрессия гена metJ ослабляется; экспрессия аллеля metA*, кодирующего фермент с пониженной чувствительностью к S-аденозилметионину и/или метионину по механизму обратной связи (MetA*), усиливается; экспрессия аллеля thrA*, кодирующего фермент с ослабленным ингибированием треонином по механизму обратной связи (thrA*), усиливается; и экспрессия гена cysE усиливается;
- экспрессия гена metJ ослабляется; экспрессия аллеля metA*, кодирующего фермент с пониженной чувствительностью к S-аденозилметионину и/или метионину по механизму обратной связи (MetA*), усиливается; экспрессия аллеля thrA*, кодирующего фермент с ослабленным ингибированием треонином по механизму обратной связи (thrA*), усиливается; экспрессия гена cysE усиливается; и экспрессия гена metF усиливается.
В конкретном аспекте изобретения рекомбинантный микроорганизм содержит следующие генетические модификации:
- гомологичные гены ygaZH, происходящие из Citrobacter koseri, Shigella flexneri, Raoultella ornithinolytica, Enterobacter sp., Yersinia enterocolitica, Photorhabdus luminescens, Citrobacter youngae или Citrobacter freundii, сверхэкспрессируются; при условии, что данная сверхэкспрессия не объединяется ни со сверхэкспрессией metH и, возможно, fldA и fpr из Е. coli или гомологичных генов из С. glutamicum, ни с ослаблением экспрессии по меньшей мере одного гена, выбранного среди metN, metI или metQ,
- экспрессия генов metA*, cysPUWAM, cysJIH, gcvTHP, metF, serA, serB, serC, cysE, thrA*, ptsG и рус усиливается и
- экспрессия генов metJ, руkА, pykF, purU, metE, dgsA и yncA ослабляется.
В другом конкретном аспекте изобретения рекомбинантный микроорганизм содержит следующие генетические модификации:
- гомологичные гены ygaZH, происходящие из Citrobacter koseri, Shigella flexneri, Raoultella ornithinolytica, Enterobacter sp., Yersinia enterocolitica, Photorhabdus luminescens, Citrobacter youngae или Citrobacter freundii, сверхэкспрессируются;
- экспрессия генов metA* cysPUWAM, cysJIH, gcvTHP, metF, serA, serB, serC, cysE, thrA*, ptsG и рус усиливается и
- экспрессия генов metJ, руkА, pykF, purU, metE, dgsA и yncA ослабляется.
В другом конкретном аспекте изобретения рекомбинантный микроорганизм содержит следующие генетические модификации:
- гомологичные гены ygaZH, происходящие из Citrobacter koseri, Shigella flexneri, Raoultella ornithinolytica, Enterobacter sp., Yersinia enterocolitica, Photorhabdus luminescens, Citrobacter youngae или Citrobacter freundii, сверхэкспрессируются; при условии, что данная сверхэкспрессия не объединяется ни со сверхэкспрессией metH и, возможно fldA и fpr из Е. coli или гомологичных генов из С. glutamicum, ни с ослаблением экспрессии по меньшей мере одного гена, выбранного среди metN, metl или metQ,
- экспрессия генов metA*, cysPUWAM, cysJIH, gcvTHP, metF, serA, serB, serC, cysE и thrA* усиливается и
- экспрессия генов metJ, pykA, pykF и purU ослабляется.
В другом конкретном аспекте изобретения рекомбинантный микроорганизм содержит следующие генетические модификации:
- гомологичные гены ygaZH, происходящие из Citrobacter koseri, Shigella flexneri, Raoultella ornithinolytica, Enterobacter sp., Yersinia enterocolitica, Photorhabdus luminescens, Citrobacter youngae или Citrobacter freundii, сверхэкспрессируются;
- экспрессия генов metA*, cysPUWAM, cysJIH, gcvTHP, metF, serA, serB, serC, cysE и thrA* усиливается и
- экспрессия генов metJ, руkА, pykF и purU ослабляется.
В другом конкретном аспекте изобретения рекомбинантный микроорганизм содержит следующие генетические модификации:
- гомологичные гены ygaZH, происходящие из Citrobacter koseri, Shigella flexneri, Raoultella ornithinolytica, Enterobacter sp., Yersinia enterocolitica, Photorhabdus luminescens, Citrobacter youngae или Citrobacter freundii, сверхэкспрессируются; при условии, что данная сверхэкспрессия не объединяется ни со сверхэкспрессией metH и, возможно fldA и fpr из Е. coli или гомологичных генов из С. glutamicum, ни с ослаблением экспрессии по меньшей мере одного гена, выбранного среди metN, metI или metQ,
- экспрессия генов metA*, cysPUWAM, cysJIH, gcvTHP, metF, serA, serB, serC, cysE и thrA* усиливается и
- экспрессия генов metJ, pykA, pykF, metE, yncA и purU ослабляется.
В другом конкретном аспекте изобретения рекомбинантный микроорганизм содержит следующие генетические модификации:
- гомологичные гены ygaZH, происходящие из Citrobacter koseri, Shigella flexneri, Raoultella ornithinolytica, Enterobacter sp., Yersinia enterocolitica, Photorhabdus luminescens, Citrobacter youngae или Citrobacter freundii, сверхэкспрессируются;
- экспрессия генов metA*, cysPUWAM, cysJIH, gcvTHP, metF, serA, serB, serC, cysE и thrA* усиливается и
- экспрессия генов metJ, pykA, pyk, metE, yncA и purU ослабляется.
В другом конкретном аспекте изобретения рекомбинантный микроорганизм содержит следующие генетические модификации:
- гомологичные гены ygaZH, происходящие из Citrobacter koseri, Shigella flexneri, Raoultella ornithinolytica, Enterobacter sp., Yersinia enterocolitica, Photorhabdus luminescens, Citrobacter youngae или Citrobacter freundii, сверхэкспрессируются; при условии, что данная сверхэкспрессия не объединяется ни со сверхэкспрессией metH и, возможно fldA и fpr из Е. coli или гомологичных генов из С. glutamicum, ни с ослаблением экспрессии по меньшей мере одного гена, выбранного среди metN, metI или metQ,
- экспрессия генов metA*, cysPUWAM, cysJIH, gcvTHP, metF, serA, serB, serC, cysE, thrA* и рус усиливается и
- экспрессия генов metJ, pykA, pykF, metE, yncA и purU ослабляется.
В другом конкретном аспекте изобретения рекомбинантный микроорганизм содержит следующие генетические модификации:
- гомологичные гены ygaZH, происходящие из Citrobacter koseri, Shigella flexneri, Raoultella ornithinolytica, Enterobacter sp., Yersinia enterocolitica, Photorhabdus luminescens, Citrobacter youngae или Citrobacter freundii, сверхэкспрессируются;
- экспрессия генов metA*, cysPUWAM, cysJIH, gcvTHP, metF, serA, serB, serC, cysE, thrA* и рус усиливается и
- экспрессия генов metJ, pykA, pyk, metE, yncA и purU ослабляется.
Наиболее предпочтительно гомологичные гены ygaZH происходят из Citrobacter koseri, Citrobacter youngae, Citrobacter freundii или Enterobacter sp.
В конкретном воплощении изобретения рекомбинантный микроорганизм происходит из бактериального семейства Enterobacteriaceae или Corynebacteriaceae.
Предпочтительно рекомбинантный микроорганизм представляет собой Escherichia coli или Corynebacterium glutamicum. Более предпочтительно рекомбинантный микроорганизм по изобретению представляет собой Е. coli.
Культуральные условия
Во втором аспекте изобретения способ оптимизирован для ферментативного производства метионина и/или его производных. Он включает следующие стадии:
- культивирование рекомбинантного микроорганизма, в котором усилен отток метионина посредством сверхэкспрессии гомологичных генов ygaZH; при условии, что данная сверхэкспрессия не объединяется ни со сверхэкспрессией metH и, возможно, генов fldA и fpr из Е. coli или гомологичных им генов из С. glutamicum, ни с ослаблением экспрессии по меньшей мере одного гена среди metN, metI или metQ, причем указанные гомологичные гены ygaZH выбирают среди группы генов видов Citrobacter, видов Shigella, видов Raoultella, видов Enterobacter, видов Yersinia и видов Photorhabdus в подходящей культуральной среде, содержащей ферментируемый источник углерода и источник серы, и
- выделение метионина и/или его производных из культуральной среды.
В конкретном воплощении способ по изобретению представляет собой способ, оптимизированный для ферментативной продукции метионина и/или его производных, включающий следующие этапы:
- культивирование рекомбинантного микроорганизма, в котором усилен отток метионина посредством сверхэкспрессии генов ygaZH, гомологичных генам ygaZH из Escherichia coli, в подходящей культуральной среде, содержащей ферментируемый источник углерода и источник серы, причем указанные гомологичные гены ygaZH выбраны среди группы генов видов Citrobacter, видов Shigella, видов Raoultella, видов Enterobacter, видов Yersinia и видов Photorhabdus и
- выделение метионина и/или его производных из культуральной среды.
Специалисты в данной области способны определять культуральные условия для микроорганизмов согласно изобретению. В частности бактерии осуществляют ферментацию при температуре от 20°С до 55°С, предпочтительно от 25°С до 40°С и более конкретно - при примерно 30°С для С. glutamicum и примерно 37°С для Е. coli.
Для Е. coli культуральная среда может иметь состав, идентичный или аналогичный среде М9 (Anderson, 1946), среде М63 (Miller, 1992); или такой среде, как определено Schaefer et al., (1999).
Для С. glutamicum культуральная среда может иметь состав, идентичный или аналогичный среде BMCG (Liebl et al., 1989) или такой среде, как описано Riedel et al., (2001).
Согласно кокретному аспету изобретения сособ осуществляется с рекомбинантным микроорганизмом, и включает сверхэкспрессию генов ygaZH, гомологичных генам ygaZH из Е. coli, в частности генов, кодирующих белки YgaZH из Таблиц 1 и 2.
В способе по изобретению гомологичные гены ygaZH, которые сверхэкспрессируются в рекомбинантном микроорганизме, предпочительно выбраны среди группы, состоящей из гомологичных генов из видов Citrobacter, видов Shigella, видов Raoultella, видов Enterobacter, видов Yersinia и видов Photorhabdus, и более предпочтительно происходят из Citrobacter koseri, Shigella flexneri, Raoultella ornithinolytica, Enterobacter sp., Yersinia enterocolitica, Photorhabdus luminescens, Citrobacter youngae или Citrobacter freundii.
Согласно другому конкретному аспекту способа по изобретению указанные гомологичные гены ygaZH, которые сверхэкспрессируются в рекомбинантном микроорганизме, подлежащем культивированию в данном способе, выбраны среди группы, состоящей из гомологичных генов из Citrobacter koseri, Citrobacter youngae, Citrobacter freundii или Enterobacter sp.
В некотором воплощении изобретения рост рекомбинантного микроорганизма подвергают ограничению или голоданию в отношении одного или нескольких неорганических субстратов, в частности, фосфата и/или калия в культуральной среде. Оно относится к условию, при котором рост микроорганизмов управляется количеством поставляемого неорганического химического соединения, которое все еще обеспечивает слабый рост. Такое ограничение роста микроорганизма было описано в патентной заявке WO 2009/043372. В предпочтительном воплощении изобретения культуру подвергают ограничению в отношении количества фосфата.
В конкретном воплощении способа по изобретению рекомбинантный микроорганизм происходит из бактериального семейства Enterobacteriaceae или Corynebacteriaceae. Предпочтительно рекомбинантный микроорганизм представляет собой Escherichia coli или Corynebacterium glutamicum, и более предпочтительно рекомбинантный микроорганизм по изобретению представляет собой Е. coli.
Действие «выделение метионина и/или его производных из культуральной среды» обозначает действие по выделению L-метионина и/или одного из его производных, в частности N-ацетилметионина (NAM) и S-аденозилметионина (SAM) и всех других производных, которые могут быть полезными, таких как гидросиметионин (или аналог гидросиметионина, или МНА). Действие «выделение метионина из культуральной среды» обозначает действие по выделению метионина из ферментационной среды, какой бы ни была степень его числоты. Термин «выделение» означает выделение первого продукта, непосредственно полученного из процесса ферментации (ферментационного сусла), которая содержит интересующий продукт (в данном случае - метионин) и другие сопродукты ферментации, таким образом, с более или менее приемлемой степенью чистоты.
Стадия «очистки» состоит из специфичной очистки интересующего продукта (в данном случае - метионина) для того, чтобы получать указанный интересующий продукт с улучшенной степенью чистоты.
Метионин мог бы быть выделен и очищен методиками и способами, хорошо известными специалисту в данной области, подбными перегонке, способам ионообменной хроматографии, осаждению, кристаллизации или комплексообразованию с солями и, в частности, с солями кальция или солями аммония. Такие способы выделения и очистки полученных соединений, в частности, раскрыты в WO 2005/007862 и в WO 2005/059155. Предпочтительно стадия выделения метионина и/или его производных включает стадию концентрирования метионина и/или его производных в ферментационном бульоне.
Количество продукта в ферментационной среде можно определять с использованием целого ряда способов, известных в данной области, например, высокоэффективной жидкостной хроматографии (ВЭЖХ) или газовой хроматографии (ГХ). Например, количество метионина, полученного в среде, измеряют ВЭЖХ после дериватизации OPA/Fmoc с использованием L-метионина (Fluka, Ref 64319) в качестве стандарта. Количество NAM определяется с использованием рефрактометрической ВЭЖХ с использованием NAM (Sigma, Ref 01310) в качестве стандарта.
ПРИМЕРЫ
Следующие эксперименты демонстрируют пользу сверхэкспрессии генов, кодирующих систему выведения L-метионина, из разных микроорганизмов в разных рекомбинантных штаммах Е. coli - продуцентах L-метионина в качестве фона.
В примерах, приведенных ниже, способы, хорошо известные в данной области, используют для конструирования штаммов E. coli, содержащих реплицируемые векторы и/или разные хромосомные вставки, делеции и замены, с использованием гомологичной рекомбинации, хорошо описанной Datsenko & Wanner, (2000).
Таким же образом, применение плазмид или векторов для экспрессии или сверхэкспрессии одного или нескольких генов в рекомбинантных микроорганизмах является хорошо известным для специалиста в данной области.
Примеры подходящих экспрессионных векторов Е. coli включают pTrc, pACYC184n, pBR322, pUC18, pUC19, рKС30, pRep4, pHS1, pHS2, pPLc236 и т.д.
ПРОТОКОЛЫ
Для конструирования штаммов, продуцирующих метионин, описанных в следующих примерах, использовали несколько протоколов.
Протокол 1 (хромосомные модификации посредством гомологичной рекомбинации, отбор рекомбинантов и вырезание кассеты устойчивости к антибиотику) и протокол 2 (трансдукция фага Р1), использованные в данном изобретении, были полностью описаны в патентной заявке WO 2013/001055.
Протокол 3: конструирование рекомбинантных плазмид
Генная инженерия хорошо описана и известна специалисту в данной области. Вкратце, фрагменты ДНК ПЦР-амплифицировали с использованием олигонуклеотидов (которые может сконструировать специалист в данной области) и геномной ДНК MG1655 в качестве матрицы. Фрагменты ДНК и отобранная плазмида расщепляются совместимыми рестрикционными ферментами, лигируются и затем трансформируются в компетентные клетки. Трансформанты анализируются, и интересующие рекомбинантные плазмиды подтверждаются посредством секвенирования ДНК.

ПРИМЕР 1: сверхпродукция эндогенной системы секреции L-метионина в рекомбинантном штамме Е. coli - сверхпродуценте L-метионина - описание штамма 1 и конструирование штаммов 2-7
Штамм 1 - контрольный штамм
Штамм 17, продуцирующий метионин, описанный в патентной заявке WO 2013/001055 (которая включена в данную заявку в виде ссылки), был переименован в штамм 1 в данной настоящей заявке. В том, что касается оставшейся части документа, данный штамм сверхэкспрессировал metH, благодаря искусственному промотору и сайту связывания рибосомы, интегрированным перед геном metH в его эндогенном локусе (относительно подробностей, см. патентную заявку WO 2007/077041. Данный штамм также содержит мутацию в гене metE, раскрытую в патентной заявке WO 2013/190343.
Конструирование штамма 6
Все из гена metH, кодирующего кобаламин-зависимую метионинсинтазу, и генов fldA и fpr, кодирующих систему реактивации MetH, сверхэкспрессировали на генетическом фоне штамма 1.
Перед использованием штамма 1 удаляли кассету устойчивости к антибиотику из модификации ΔdgsA с использованием рекомбиназы Flp, как описано Datsenko & Wanner, 2000 (согласно Протоколу 1). Отбирали чувствительных к канамицину трансформантов, и отсутствие кассеты устойчивости к антибиотику в локусе ΔdgsA подтверждали анализом ПЦР с подходящими олигонуклеотидами. Сохранившийся штамм называли штамм 2.
Для достижения сверхэкспрессии metH использовали стратегию гомологичной рекомбинации, описанную Datsenko & Wanner, 2000 (согласно Протоколу 1). Ген metH, связанный функциональным образом с таким же промотором и сайтом связывания рибосомы, как описано в патентной заявке WO 2007/077041, интегрировали на хромосоме в двух разных локусах ybeM и ypjC (выбранных из списка, раскрытого в патентной заявке WO 2011/073122, и делеция которых не имеет влияния на продукцию метионина).
Для обеих хромосомных интеграций фрагмент, несущий ген metH, связанный с его искусственным промотором и маркером устойчивости, которые обе фланкированы последовательностями ДНК, гомологичными целевому локусу интеграции ybeM или ypjC, ПЦР-амплифицировали посредством методики перекрывающейся ПЦР (с перекрывающимися олигонуклеотидами). Последовательности для рекомбинации в ybeM или ypjC называются SEQ ID NO: 21 и SEQ ID NO: 22, и SEQ ID NO: 23 и SEQ ID NO: 24 (перечислены в таблице 3) для ybeM или ypjC соответственно. Полученные ПЦР-продукты «ΔybeM::metH::Km» и «ΔypjC::metH::Cm» затем по отдельности вводили посредством электропорации в штамм MG1655 metA*11 (pKD46). Отбирали трансформантов с устойчивостью к антибиотику, и вставку гена metH с кассетой устойчивости в целевом локусе подтверждали анализом ПЦР с подходящими олигонуклеотидами. Сохранившиеся штаммы обозначали MG1655 metA*11 ΔybeM::metH::Km и MG1655 metA*11 ΔypjC::metH::Cm. Наконец, обе хромосомные интеграции ΔybeM::metH::Km и ΔypjC::metH::Cm последовательно переносили посредством трансдукции фагом Р1 (согласно Протоколу 2) из MG1655 metA*11 ΔybeM::metH::Km и MG1655 metA*11 ΔypjC::metH в штамм 2. Отбирали трансдуктантов, устойчивых к хлорамфениколу или канамицину, и присутствие хромосомных интеграций ΔybeM::metH::Km и ΔypjC::metH::Cm подтверждали посредством анализа ПЦР с подходящими олигонуклеотидами. Сохранившийся штамм называли штамм 3.
Кассеты устойчивости к антибиотику удаляли из хромосомных интеграций, сделанных в локусах ybeM или ypjC, в штамм 3 с использовнием рекомбиназы Flp, как описано Datsenko & Wanner, 2000 (согласно Протоколу 1). Отбирали трансформантов, чувствительных к канамицину и хлорамфениколу, и отсутствие кассеты устойчивости к антибиотику в обоих локусах подтверждали анализом ПЦР с подходящими олигонуклеотидами. Сохранившийся штамм называли штамм 4.
Для сверхэкспресии fldA и fpr данные гены связывали функциональным образом с искусственными промоторами и с искусственным сайтом связывания рибосомы и интегрировали на хромосому в локусе ytfA (такие же критерии отбора, что и с локусами ybeM или ypjC, см. выше). Для fldA конструировали искусственный промотор с SEQ ID NO: 25. Что касается fpr, используемый в штаммах искусственный промотор был описан для сверхэкспрессии оперона cysPUWAM в патентной заявке WO 2009/043803. Для обоих генов искусственные сайты связывания рибосомы являются такими же, как описано для сверхэкспрессии гена ptsG в штамме 17, раскрытом в патентной заявке WO 2013/001055.
Для введения копий fldA и fpr на хромосому для сверхэкспрессии использовали стратегию гомологичной рекомбинации, описанную Datsenko & Wanner, 2000 (согласно Протоколу 1). Фрагмент, несущий гены fldA и fpr с их соответствующими промоторами и маркером устойчивости, оба фланкированные последовательностью ДНК, гомологичной локусу интеграции ytfA, ПЦР-амплифицировали посредством методики перекрывающейся ПЦР (с перекрывающимися олигонуклеотидами). Последовательности для рекомбинации в локус ytfA называются SEQ ID NO: 26 и SEQ ID NO: 27 (перечислены в Таблице 3). Полученый ПЦР-продукт «ΔytfA::fldA-fpr::Km» затем вводили посредством электропорации в штамм MG1655 metA*11 (pKD46). Затем отбирали трансформантов с устойчивостью к антибиотику, и вставку генов fldA-fpr с кассетой устойчивости в локусе ytfA проверяли посредством ПЦР-анализа с подходящими олигонуклеотидами. Сохранившийся штамм обозначали MG1655 metA*11 ΔytfA::fldA-fpr::Km. Наконец, хромосомную интеграцию ΔytfA::fldA-fpr::Km переносили посредством трансдукции фагом Р1 (согласно Протоколу 2) из MG1655 metA*11 ΔytfA::fldA-fpr::Km в штамм 4. Отбирали трансдуктантов, устойчивых к канамицину, и присутствие хромосомной интеграции ΔytfA::fldA-fpr::Km подтверждали ПЦР-анализом с подходящими олигонуклеоидами. Сохранившийся штамм называли штамм 5.
Затем кассету устойчивости к антибиотику удаляли из хромосомной интеграции, сделанной в локусе ytfA, в штамм 5 с использованием рекомбиназы Flp, как описано Datsenko & Wanner, 2000 (согласно Протоколу 1). Отбирали трансформантов, чувствительных к канамицину, и отсутствие кассеты устойчивости к антибиотику в локусе ytfA подтверждали ПЦР-анализом с подходящими олигонуклеотидами. Сохранившйся штамм называли штамм 6, и он будет соответствовать генетическому фону А для следующих примеров.
Конструирование штамма 7 - сверхэкспрессия эндогенных генов ygaZH
Гены ygaZH из Е. coli, кодирующие экспортер метионина, клонировали на плазмиду с умеренным числом копий pCL1920 (Lerner & Inouye, 1990) с использованием природного промотора ygaZ. Данную плазмиду называли рМЕ1247. Наконец, плазмиду рМЕ1247 трансформировали в штамм 6 с получением штамма 7.
ПРИМЕР 2: сверхпродукция разных систем секреции L-метионина из разных микроорганизмов в штамме Е. coli - продуценте L-метионина, сверхпродуцирующем кобаламин-зависимую метионинсинтазу (MetH) - конструирование штаммов 8-15
Гомологичные гены ygaZH из видов Citrobacter, видов Raoultella, видов Klebsiella, видов Enterobacter, видов Yersinia и видов Photorhabdus сверхэкспрессировали на генетическом фоне А для сравнения со штаммом 7, несущим гены ygaZH Е. coli.
Конструирование штаммов 8-15 - сверхэкспрессия гомологичных генов ygaZH из рода и видов, перечисленных в Таблице 4
Для сверхэкспрессии гомологичных генов ygaZH, кодирующих белки, перечисленные в Таблице 4, в штамме-продуценте L-метионина клонировали каждую пару генов, как уже было описано выше для генов ygaZH Е. coli, на плазмиде с умеренным числом копий pCL1920 (Lerner & Inouye, 1990) с использованием природного промотора и сайта связывания рибосомы гена ygaZ Е. coli. Как определено в Таблице 5, гомологичные гены ygaZH либо амплифицировали из геномной ДНК соответствующего штамма, либо синтезировали химически с оптимизацией использования кодонов для Е. coli (как предложено службой GeneArt® Gene Synthesis с использованием программы GeneOptimizer® - Lifetechnologies) или без нее. Амплифицированные фрагменты ДНК, содержащие гомологичные гены ygaZH, раскрыты в SEQ ID, указанных в Таблице 5. Образующиеся в результате плазмиды были названы, как упомянуто в Таблице 5. Наконец, каждую плазмиду трансформировали в штамм 6 с получением штаммов 8-15, как упомянуто в Таблице 5.
ПРИМЕР 3: сверхпродукция разных систем секреции L-метионина из разных микроорганизмов в нескольких штаммах - сверхпродуцентах L-метионина - Штаммах 16 и 17, и конструирование штаммов 18-53
Полезное влияние сверхэкспрессии гомологичных генов ygaZH на продукцию L-метионина оценивали на разных фонах, ранее описанных в патентах. Они являются следующими:
- Генетический Фон В: MG1655 metA*11 ΔmetJ Ptrc-metH Ptrc36-ARNmst17-metF PtrcF-cysPUWAM PtrcF-cysJIH Ptrc09-gcvTHP ΔpykA ΔpykF ΔpurU::Km (pME101-thrA*1-cysE-PgapA-metA*11) (pCC1BAC-serB-serA-serC), названный в данной заявке штаммом 17.
Его материнский штамм без вектора pME101-thrA*1-cysE-PgapA-metA*11, названный в данной заявке штаммом 16, и MG1655 metA*11 ΔmetJ Ptrc-metH Ptrc36-ARNmst17-metF PtrcF-cysPUWAM PtrcF-cysJIH Ptrc09-gcvTHP ΔpykA ΔpykF ΔpurU::Km (pME101-thrA*1-cysE) (pCC1BAC-serB-serA-serC) описаны в патентной заявке WO 2009/043803. Для получения штамма 17 и, более конкретно, плазмиды pME101-thrA*1-cysE-PgapA-metA*11 ген metA*11 вместе с промотором гена gapA (Thouvenot, et al. 2004) клонировали ниже гена cysE в плазмиду pME101-thrA*1-cysE с использованием методологии, аналогичной методологии, описанной в патентной заявке WO 2011/073122, для конструирования плазмиды pCL1920-TTadc-CI857-PlambdaR*(-35)-thrA*1-cysE-PgapA-metA*11. Затем вектор рМЕ101-thrA*1-cysE-PgapA-metA*11 вводили в штамм 16 с получением штамма 17.
- Генетический фон С: штамм 10, описанный в патентной заявке WO 2012/055798.
- Генетический фон D: штамм 3, описанный в патентной заявке WO 2012/090021.
- Генетический фон Е: штамм 17, описанный в патентной заявке WO 2013/001055 и названный в данной настоящей заявке штаммом 1.
Некоторые из штаммов-сверхпродуцентов L-метионина, используемые в качестве реципиентов для сверхэкспрессируемых гомологичных генов ygaZH, уже несли плазмиду типа pCL1920 (например, фон В и D). Следовательно, стратегии для клонирования либо эндогенных, либо гетерологичных оперонов ygaZH, связанных с промотором PygaZ Е. coli, были адаптированы согласно генетическому фону реципиента. Точнее, (i) для генетических фонов С и Е эндогенные или гетерологичные опероны ygaZH клонировали в пустую плазмиду pCL1920, (ii) для генетического фона В эндогенные или гетерологичные опероны ygaZH клонировали в плазмиду pME101-thrA*1-cysE-PgapA-metA*11, и (iii) для генетического фона D эндогенные или гетерологичные опероны ygaZH клонировали в плазмиду pCL1920-PgapA-pycRe-TT07. Полученные в результате плазмиды называли, как упомянуто в Таблице 6.
Как видно в Таблице 6, для плазмид, несущих оперон ygaZH из того же самого вида, числовой индекс плазмиды является одинаковым, и единственным изменением в номенклатуре является буквенный суффикс (b или с) согласно типу pCL1920, использованному для клонирования оперона ygaZH.
Для разных штаммов Е. coli - сверхпродуцентов L-метионина, протестированных в данном примере, гомологичные гены ygaZH либо амплифицировали из геномной ДНК соответствующего штамма, либо синтезировали химически с оптимизацией использования кодонов для Е. coli (как предложено службой GeneArt® Gene Synthesis с использованием программы GeneOptimizer® - Lifetechnologies) или без нее, как было описано для штамма 6 генетического фона и определено в Таблице 5.
Наконец, штаммы 18-53 были получены введением разных плазмид, перечисленных в Таблице 6, в следующие фоны:
- для генетического фона В: типы плазмид pME101-thrA*1-cysE-PgapA-metA*11-PygaZ-ygaZH вводили в штамм 16 с получением штаммов 18-26,
- для генетического фона С: типы плазмид pCL1920-PygaZ-ygaZH вводили в штамм 10 заявки WO 2012/055798 с получением штаммов 27-35,
- для генетического фона D: типы плазмид pCL1920-PgapA-pycre-TT07-PygaZ-ygaZH вводили в штамм 3 заявки WO 2012/090021 с получением штаммов 36-44 и
- для генетического фона Е: типы плазмид pCL1920-PygaZ-ygaZH вводили в штамм 1 с получением штаммов 45-53.
Полученные в результате штаммы, объединяющие конкретный генетический фон для продукции L-метионина с конкретной плазмидой, несущей сверхэкспрессию конкретной пары гомологичных генов ygaZH, перечислены в Таблице 7 ниже.
ПРИМЕР 4: сверхпродукция разных систем секреции L-метионина из разных микроорганизмов в нескольких штаммах - сверхпродуцентах L-метионина, которые не имеют сверхэкспрессии генов metH, fldA и fpr - конструирование штаммов 54-73
Полезное влияние сверхэкспрессии гомологичных генов ygaZH на продукцию L-метионина также оценивали независимо от сверхэкспрессии генов metH, fldA и fpr. Нативный низкий уровень экспрессии metH восстанавливали на фоне В и Е и, посредством распространения, в штаммах 17-26, в штамме 1 и в штаммах 45-53.
Для восстановления природного уровня экспрессии metH осуществляли замену на его природный промотор перед геном metH, и, следовательно, в то же самое время удаляли искусственный промотор с использованием стратегии гомологичной рекомбинации, описанной Datsenko & Wanner, 2000, описанной в Протоколе 1. Прежде всего, в штамме дикого типа, обладающем немодифицированным metH под контролем его собственного промотора, добавляли ген устойчивости к антибиотику ниже гена metH. Точнее, получали ПЦР-продукт, несущий ген устойчивости к антибиотику (Тс для тетрациклина) вместе с сайтами FRT, окруженный последовательностями, гомологичными области, расположенной ниже гена metH и выше гена yjbB (SEQ ID NO: 28 и SEQ ID NO: 29), и вставляли в MG1655 metA*11, в который ранее был трансформирован вектор pKD46. Трансформантов, устойчивых к антибиотику, подтверждали с использованием подходящих олигонуклеотидов, и сохранившийся штамм представлял собой MG1655 metA*11 metH::Tc. Наконец, хромосомную модификацию metH::Tc переносили посредством трансдукции фагом Р1 (согласно Протоколу 2) из MG1655 metA*11 metH::Tc в штаммы 17-26 и в штамм 1, и в штаммы 45-53. Отбирали трансдуктантов, устойчивых к тетрациклину, и присутствие хромосомной интеграции metH::Tc, а также целостность хромосомных модификаций штаммов-реципиентов, которые закрыты в отношении локуса metH, подтверждали анализом ПЦР с подходящими олигонуклетидами. Сохранившиеся штаммы называли штаммы 54-73, как перечислено в Таблице 8 ниже.
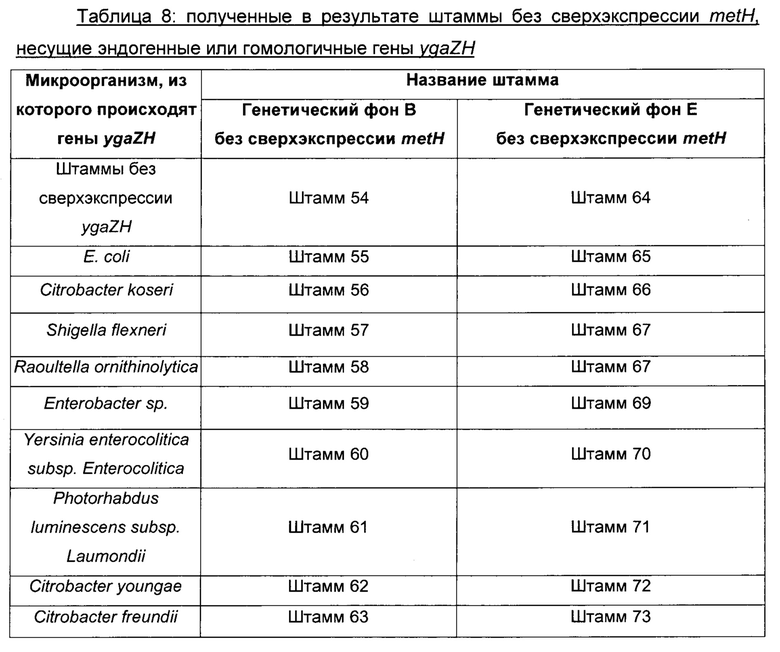
ПРИМЕР 5: продукция L-метионина
Продукционные штаммы оценивали в маленьких колбах Эрленмейера. Для штамма 17 и его производных штаммов, названных 18-26 и 54-63, культуральные условия описаны в патенте ЕР 2573189А1, для штамма 10 из заявки WO 2012/055798 и его производных штаммов, названных 27-35, и для штамма 3 из заявки WO 2012/090021 и его производных штаммов, названных 36-44, культуральные условия описаны в каждом соответствующем патенте.
Для других штаммов (штамм 1 и его производные штаммы, названные 45-53 и 64-73) 5,5 мл предкультуры выращивали при 30°С в течение 21 часа в смешанной среде (10%-ная среда LB (Sigma 25%) с 2,5 г×л-1 глюкозы и 90%-ная минимальная среда РС1, Таблица 8). Ее использовали для инокуляции 50 мл культуры до ОП600 (оптическая плотность при 600 нм) 0,2 в среде РС1. Когда это было необходимо, добавляли спектиномицин и канамицин в концентрации 50 г×л-1, гентамицин в концентрации 10 г×л-1 и тетрациклин в концентрации 5 мг×л-1. Когда это было необходимо, добавляли IPTG (изопропилтиогалактозид) в концентрации 0,0048 г×л-1. Температура культур составляла 37°С и скорость встряхтвания составляла 200 об./мин. При достижении культурой ОП600 от 5 до 7 внеклеточные аминокислоты количественно измеряли посредством ВЭЖХ (высокоэффективная жидкостная хроматография) после дериватизации OPA/Fmoc, и другие релевантные метаболиты анализировали с использованием ВЭЖХ с рефрактометрическим выявлением (органические кислоты и глюкоза) и ГХ-МС (газовая хроматография-масс-спектрометрия) после силилирования.
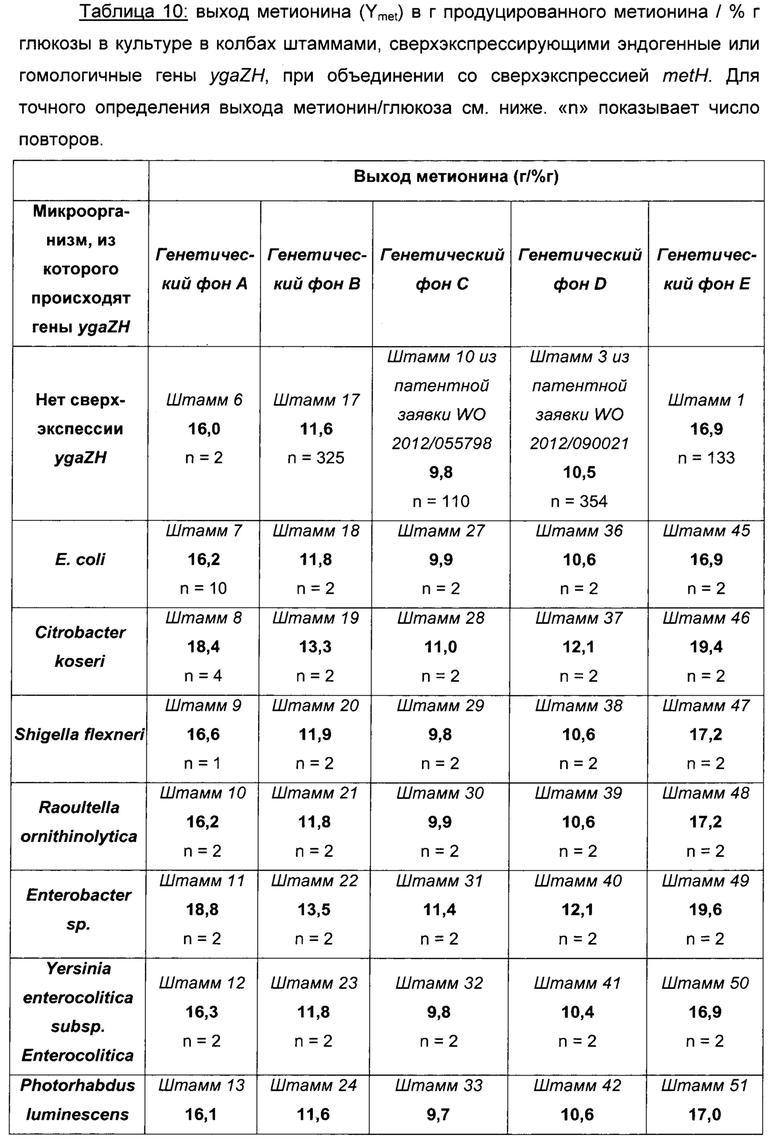
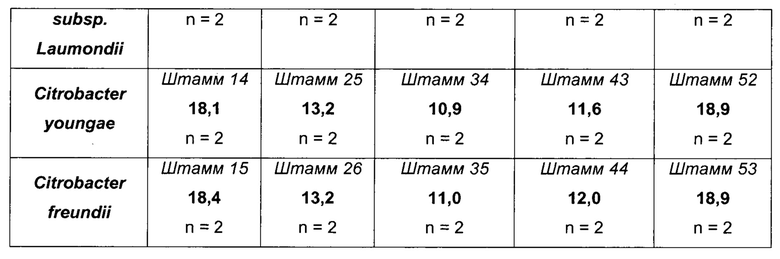
Как можно видеть в Таблице 10 выше, эффективности продукции L-метионина протестированных штаммов, несущих гомологичную пару YgaZH, происходящую из разных микроорганизмов, являются либо равными, либо даже большими, чем у эквивалентного им штамма, несущего эндогенный YgaZH из Е. coli (штаммы 8-15 по сравнению со штаммом 7). Кроме того, данные результаты получаются, какой бы генетический фон не использовался для сверхпродукции гомологичной системы секреции L-метионина, как, например, в штаммах 15, 26, 35, 44 и 53 по сравнению со штаммами 7, 18, 27, 36 и 45 соответственно. Наилучшие результаты достигаются для продукции L-метионина с использованием генов ygaZH, происходящих из Citrobacter koseri, Enterobacter sp, Citrobacter youngae и Citrobacter freundii и сверхэкспрессированных в штаммах-сверхпродуцентах L-метионина.
Выход метионина выражали следующим образом:


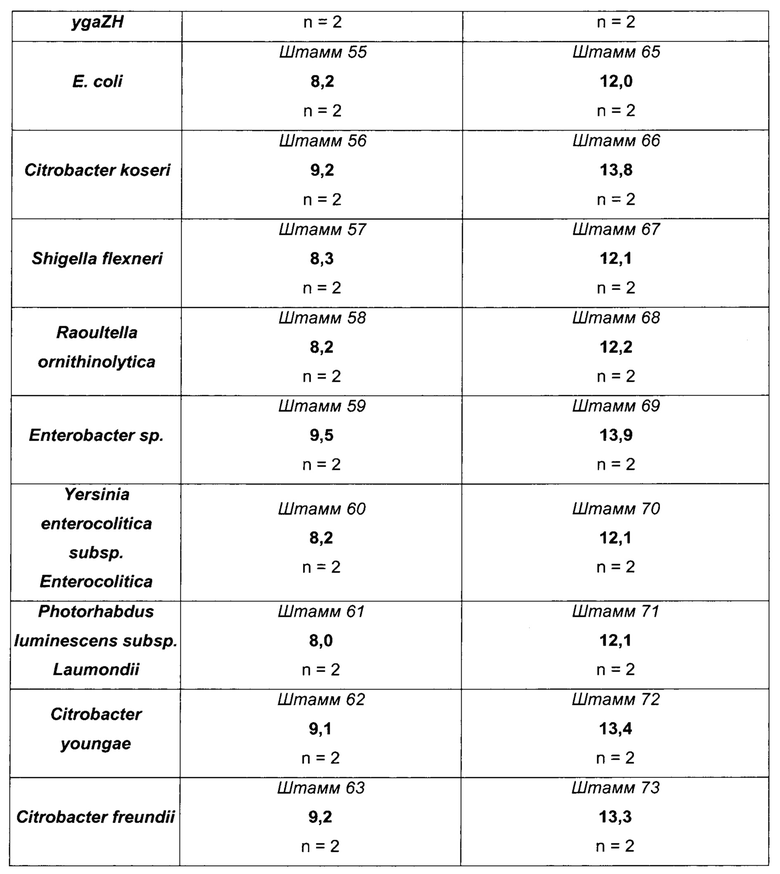
Как можно видеть в таблице 11 выше, эффективности продукции L-метионина протестированных штаммов, несущих гомологичную пару YgaZH, происходящую из разных микроорганизмов, являются либо равными, либо даже большими, чем у эквивалентного им штамма, несущего эндогенный YgaZH из Е. coli (см., например, штаммы 56-63 по сравнению со штаммом 55). Кроме того, данные результаты получаются независимо от генетического фона, как, например, для штаммов 63 и 73, сравниваемых, соответственно, со штаммами 55 и 65, и, более конкретно, без сверхэкспрессии metH. Наилучшие результаты в показателях продукции L-метионина достигаются с генами ygaZH, происходящими из Citrobacter koseri, Enterobacter sp, Citrobacter youngae и Citrobacter freundii, и сверхэкспрессируемыми в штаммах-сверхпродуцентах L-метионина.
В заключение, сверхпродукция гомологичных белков YgaZH в рекомбинантных штаммах-продуцентах L-метионина является полезной для продукции данной аминокислоты, независимо от сверхэкспрессий генов metH и fldA-frp. Увеличение продукции при использовании сверхэспрессии генов ygaZH, происходящих из Citrobacter koseri, Enterobacter sp, Citrobacter youngae и Citrobacter freundii, больше, чем при использовании сверхэкспрессии эндогенного гена ygaZH из Е. coli.
ССЫЛКИ
- Anderson, 1946, Proc. Natl. Acad. Sci. USA 32:120-128.
- Carrier Т., Keasling J.D., 1999, Biotechnology Progress, 15:58-64
- Datsenko K.A., Wanner B.L., 2000, Proceedings of the National Academy of Sciences of the USA, 97:6640-6645
- Lerner C.G. and Inouye M., 1990, Nucleic Acids Research, 18(15):4631
- Liebl et al., 1989, Appl. Microbiol. Biotechnol. 32: 205-210.
- Miller, 1992; "A Short Course in Bacterial Genetics: A Laboratory Manual and Handbook for Escherichia coli and Related Bacteria", Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
- Riedel et al., 2001, J. Mol. Microbiol. Biotechnol. 3: 573-583.
- Saunderson C.L., 1985, British Journal of Nutrition, 54:621-633
- Schaefer et al. 1999, Anal. Biochem. 270: 88-96.
-  C., Deutenberg D., Bathe В., Burkovski A.,
C., Deutenberg D., Bathe В., Burkovski A.,  R., 2005, Journal of Bacteriology. 187(11):3786-3794
R., 2005, Journal of Bacteriology. 187(11):3786-3794
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> METABOLIC EXPLORER
<120> СПОСОБ И МИКРООРГАНИЗМ ДЛЯ ПРОИЗВОДСТВА МЕТИОНИНА ПОСРЕДСТВОМ
ФЕРМЕНТАЦИИ С УЛУЧШЕННЫМ ОТТОКОМ МЕТИОНИНА
<130> B368292D34164
<150> EP14306346.9
<151> 2014-09-01
<160> 37
<170> PatentIn version 3.5
<210> 1
<211> 245
<212> ПРТ
<213> Escherichia coli
<400> 1
Met Glu Ser Pro Thr Pro Gln Pro Ala Pro Gly Ser Ala Thr Phe Met
1 5 10 15
Glu Gly Cys Lys Asp Ser Leu Pro Ile Val Ile Ser Tyr Ile Pro Val
20 25 30
Ala Phe Ala Phe Gly Leu Asn Ala Thr Arg Leu Gly Phe Ser Pro Leu
35 40 45
Glu Ser Val Phe Phe Ser Cys Ile Ile Tyr Ala Gly Ala Ser Gln Phe
50 55 60
Val Ile Thr Ala Met Leu Ala Ala Gly Ser Ser Leu Trp Ile Ala Ala
65 70 75 80
Leu Thr Val Met Ala Met Asp Val Arg His Val Leu Tyr Gly Pro Ser
85 90 95
Leu Arg Ser Arg Ile Ile Gln Arg Leu Gln Lys Ser Lys Thr Ala Leu
100 105 110
Trp Ala Phe Gly Leu Thr Asp Glu Val Phe Ala Ala Ala Thr Ala Lys
115 120 125
Leu Val Arg Asn Asn Arg Arg Trp Ser Glu Asn Trp Met Ile Gly Ile
130 135 140
Ala Phe Ser Ser Trp Ser Ser Trp Val Phe Gly Thr Val Ile Gly Ala
145 150 155 160
Phe Ser Gly Ser Gly Leu Leu Gln Gly Tyr Pro Ala Val Glu Ala Ala
165 170 175
Leu Gly Phe Met Leu Pro Ala Leu Phe Met Ser Phe Leu Leu Ala Ser
180 185 190
Phe Gln Arg Lys Gln Ser Leu Cys Val Thr Ala Ala Leu Val Gly Ala
195 200 205
Leu Ala Gly Val Thr Leu Phe Ser Ile Pro Val Ala Ile Leu Ala Gly
210 215 220
Ile Val Cys Gly Cys Leu Thr Ala Leu Ile Gln Ala Phe Trp Gln Gly
225 230 235 240
Ala Pro Asp Glu Leu
245
<210> 2
<211> 111
<212> ПРТ
<213> Escherichia coli
<400> 2
Met Ser Tyr Glu Val Leu Leu Leu Gly Leu Leu Val Gly Val Ala Asn
1 5 10 15
Tyr Cys Phe Arg Tyr Leu Pro Leu Arg Leu Arg Val Gly Asn Ala Arg
20 25 30
Pro Thr Lys Arg Gly Ala Val Gly Ile Leu Leu Asp Thr Ile Gly Ile
35 40 45
Ala Ser Ile Cys Ala Leu Leu Val Val Ser Thr Ala Pro Glu Val Met
50 55 60
His Asp Thr Arg Arg Phe Val Pro Thr Leu Val Gly Phe Ala Val Leu
65 70 75 80
Gly Ala Ser Phe Tyr Lys Thr Arg Ser Ile Ile Ile Pro Thr Leu Leu
85 90 95
Ser Ala Leu Ala Tyr Gly Leu Ala Trp Lys Val Met Ala Ile Ile
100 105 110
<210> 3
<211> 244
<212> ПРТ
<213> Citrobacter koseri
<400> 3
Met Glu Ser Pro Ala Pro Gln Ser Glu Pro Arg Pro Ala Thr Leu Thr
1 5 10 15
Glu Gly Phe Lys Asp Ser Leu Pro Ile Val Ile Ser Tyr Ile Pro Val
20 25 30
Ala Phe Ala Phe Gly Leu Asn Ala Thr Arg Leu Gly Phe Thr Pro Leu
35 40 45
Glu Ser Val Phe Phe Ser Cys Ile Ile Tyr Ala Gly Ala Ser Gln Phe
50 55 60
Val Ile Thr Thr Met Leu Ala Ala Gly Ser Thr Leu Trp Val Ala Ala
65 70 75 80
Leu Thr Val Met Ala Met Asp Val Arg His Val Leu Tyr Gly Pro Ser
85 90 95
Leu Arg Ser Arg Ile Ser Gln Arg Leu Ser Lys Pro Lys Thr Ala Leu
100 105 110
Trp Ala Phe Gly Leu Thr Asp Glu Val Phe Ala Ala Ala Thr Ala Lys
115 120 125
Leu Val Arg Asp Asn Arg Arg Trp Ser Glu Trp Met Ile Gly Ile Ala
130 135 140
Phe Cys Ser Trp Ala Ser Trp Val Leu Gly Thr Val Ile Gly Ala Phe
145 150 155 160
Ser Gly Ser Gly Leu Leu Lys Gly Phe Pro Ala Val Glu Ala Ala Leu
165 170 175
Gly Phe Met Leu Pro Ala Leu Phe Met Ser Phe Leu Leu Ala Ser Phe
180 185 190
Gln Arg Lys Gln Thr Leu Cys Val Thr Ala Ala Leu Ile Gly Ala Leu
195 200 205
Ala Gly Val Thr Leu Phe Ser Ile Pro Ala Ala Ile Leu Ala Gly Ile
210 215 220
Ala Ser Gly Cys Leu Thr Ala Leu Ile Gln Ser Phe Trp Gln Gly Ala
225 230 235 240
Pro Asp Glu Leu
<210> 4
<211> 111
<212> ПРТ
<213> Citrobacter koseri
<400> 4
Met Ser Tyr Glu Val Leu Leu Leu Gly Leu Leu Val Gly Cys Ala Asn
1 5 10 15
Tyr Cys Phe Arg Tyr Leu Pro Leu Arg Leu Arg Met Gly Asn Thr Arg
20 25 30
Pro Ala Arg Arg Gly Ala Thr Gly Val Leu Leu Asp Thr Ile Gly Ile
35 40 45
Ala Ser Ile Cys Ala Leu Leu Val Val Ser Thr Ala Pro Glu Val Met
50 55 60
His Asp Ala Ser Arg Phe Ile Pro Thr Leu Val Gly Phe Ala Val Leu
65 70 75 80
Gly Val Ser Phe Tyr Lys Thr Arg Ser Ile Ile Ile Pro Thr Leu Leu
85 90 95
Ser Ala Leu Gly Tyr Gly Leu Ala Trp Lys Ile Glu Val Ile Leu
100 105 110
<210> 5
<211> 245
<212> ПРТ
<213> Shigella flexneri
<400> 5
Met Glu Ser Pro Val Pro Gln Ser Glu Ser Arg Ser Ala Thr Leu Thr
1 5 10 15
Glu Gly Phe Lys Asp Ser Leu Pro Ile Val Ile Ser Tyr Ile Pro Val
20 25 30
Ala Phe Ala Phe Gly Met Asn Ala Thr Arg Leu Gly Phe Thr Pro Val
35 40 45
Glu Ser Val Phe Phe Ser Cys Ile Ile Tyr Ala Gly Ala Ser Gln Phe
50 55 60
Val Ile Thr Thr Met Leu Ala Ala Gly Ser Ser Leu Trp Val Ala Ala
65 70 75 80
Leu Thr Val Met Ala Met Asp Val Arg His Val Leu Tyr Gly Pro Ser
85 90 95
Leu Arg Ser Arg Ile Ala Arg Gln Leu Ser Lys Pro Lys Ser Ala Leu
100 105 110
Trp Ala Phe Gly Leu Thr Asp Glu Val Phe Ala Ala Ala Thr Ala Lys
115 120 125
Leu Val Arg Asp Asn Arg Arg Trp Ser Glu Asn Trp Met Ile Gly Ile
130 135 140
Ala Leu Cys Ser Trp Ala Ser Trp Val Leu Gly Thr Val Ile Gly Ala
145 150 155 160
Phe Ser Gly Thr Gly Leu Leu Lys Gly Phe Pro Ala Val Glu Ala Ala
165 170 175
Leu Gly Phe Met Leu Pro Ala Leu Phe Met Ser Phe Leu Leu Ala Ser
180 185 190
Phe Gln Arg Asn Gln Thr Leu Cys Val Thr Ala Ala Leu Ala Gly Ala
195 200 205
Leu Ala Gly Val Thr Leu Phe Ser Ile Pro Ala Ala Ile Leu Ala Gly
210 215 220
Ile Val Cys Gly Cys Leu Thr Ala Leu Ile Gln Ser Phe Trp Gln Gly
225 230 235 240
Gly Pro Asp Glu Leu
245
<210> 6
<211> 111
<212> ПРТ
<213> Shigella flexneri
<400> 6
Met Ser Tyr Glu Val Leu Leu Leu Gly Leu Leu Val Gly Cys Val Asn
1 5 10 15
Tyr Cys Phe Arg Tyr Leu Pro Leu Arg Leu Arg Met Gly Asn Val Arg
20 25 30
Pro Thr Lys Arg Gly Ala Thr Gly Ile Leu Leu Asp Thr Ile Gly Ile
35 40 45
Ala Ser Ile Cys Ala Leu Leu Val Val Ser Thr Ala Pro Glu Val Met
50 55 60
His Asp Ala Arg Arg Phe Val Pro Thr Leu Val Gly Phe Val Val Leu
65 70 75 80
Gly Ala Ser Phe Tyr Lys Thr Arg Ser Ile Ile Ile Pro Thr Leu Leu
85 90 95
Ser Ala Leu Gly Tyr Gly Leu Ala Trp Lys Met Leu Phe Val Leu
100 105 110
<210> 7
<211> 242
<212> ПРТ
<213> Raoultella ornithinolytica
<400> 7
Met Glu Lys Pro Ala Pro Ala Ser Glu Ala Thr Leu Pro Glu Gly Ile
1 5 10 15
Lys Asp Ser Leu Pro Ile Val Ile Ser Tyr Ile Pro Val Ala Phe Ala
20 25 30
Phe Gly Leu Asn Ala Thr Arg Leu Gly Phe Thr Pro Leu Glu Ser Leu
35 40 45
Phe Phe Ser Cys Ile Ile Tyr Ala Gly Ala Ser Gln Phe Val Ile Thr
50 55 60
Ala Met Leu Ala Ala Gly Ser Ser Leu Trp Val Ala Ala Leu Thr Val
65 70 75 80
Met Ala Met Asp Val Arg His Val Leu Tyr Gly Pro Ser Leu Arg Ser
85 90 95
Arg Ile His Arg Ala Leu Asp Lys Arg Lys Thr Ala Leu Trp Ala Phe
100 105 110
Gly Leu Thr Asp Glu Val Phe Ala Ala Ala Thr Ala Lys Leu Val Arg
115 120 125
Asp Asn Arg Arg Trp Ser Glu Ser Trp Met Leu Gly Ile Ala Phe Thr
130 135 140
Ser Trp Ile Ser Trp Val Phe Gly Thr Leu Ile Gly Ala Tyr Ser Gly
145 150 155 160
Ser Gly Leu Leu Val Gly Phe Pro Ala Val Glu Ala Ala Leu Ser Phe
165 170 175
Met Leu Pro Ala Leu Phe Met Ser Phe Leu Leu Ala Ser Phe Gln Arg
180 185 190
Lys Gln Ser Leu Ser Val Thr Ala Ala Leu Ala Gly Ala Leu Gly Gly
195 200 205
Ile Ile Leu Phe Ser Ile Pro Ala Ala Ile Leu Ala Gly Ile Val Cys
210 215 220
Gly Cys Leu Ala Ala Leu Ile Gln Ala Ser Ile Gln Gly Met Pro Asp
225 230 235 240
Glu Gln
<210> 8
<211> 113
<212> ПРТ
<213> Raoultella ornithinolytica
<400> 8
Met Asn Asn Asn Val Leu Ile Ile Gly Ile Val Val Gly Cys Val Asn
1 5 10 15
Tyr Leu Phe Arg Tyr Leu Pro Leu Arg Leu Arg Ala Gly Asn Ala Arg
20 25 30
Pro Thr Arg Arg Gly Pro Leu Ser Val Leu Leu Asp Thr Ile Gly Ile
35 40 45
Ala Ser Ile Cys Ala Leu Leu Ile Val Ser Ser Val Pro Glu Ile Leu
50 55 60
Ser Asp Ser Arg Arg Leu Leu Pro Thr Leu Val Gly Phe Thr Val Leu
65 70 75 80
Gly Leu Ala Phe Trp Lys Thr Arg Ser Ile Ile Met Pro Thr Leu Leu
85 90 95
Ser Ala Leu Ala Tyr Gly Ile Ala Trp Lys Ile Thr Thr Phe Leu Tyr
100 105 110
Phe
<210> 9
<211> 250
<212> ПРТ
<213> Enterobacter sp. (R4-368)
<400> 9
Met Asp Met Asp Ser Ser Val Thr Ala Thr Lys Ser Thr Ser Asp Gln
1 5 10 15
Ser Ala Thr Phe Leu Glu Gly Ile Lys Asp Ser Leu Pro Ile Val Leu
20 25 30
Ser Tyr Val Pro Val Ala Phe Ala Phe Gly Met Asn Ala Thr Lys Leu
35 40 45
Gly Phe Thr Pro Leu Glu Ser Val Phe Phe Ser Cys Ile Ile Tyr Ala
50 55 60
Gly Ala Ser Gln Phe Val Ile Thr Thr Met Leu Ala Ala Gly Ser Ala
65 70 75 80
Leu Trp Val Ala Ala Leu Thr Val Met Ala Met Asp Val Arg His Val
85 90 95
Leu Tyr Gly Pro Ser Leu Arg Ser Arg Ile Leu Gln Pro Leu Lys Asn
100 105 110
Arg Lys Thr Ala Val Trp Ala Phe Gly Leu Thr Asp Glu Val Phe Ala
115 120 125
Ala Ala Thr Ala Lys Leu Val Arg Asp Asn Arg Arg Trp Ser Glu Asn
130 135 140
Trp Met Ile Gly Ile Ala Leu Phe Ser Trp Leu Ser Trp Val Ala Gly
145 150 155 160
Thr Val Leu Gly Ala Phe Ser Gly Asp Gly Leu Leu Asp Gly Tyr Pro
165 170 175
Ala Val Glu Ser Ala Leu Gly Phe Met Leu Pro Ala Leu Phe Met Ser
180 185 190
Phe Leu Leu Ala Ser Phe Gln Arg Arg Gln Ile Ser Ala Val Thr Ala
195 200 205
Ala Leu Leu Gly Ala Leu Ala Gly Val Thr Leu Phe Ser Ile Pro Ala
210 215 220
Ala Ile Leu Ala Gly Ile Phe Ala Gly Cys Leu Ala Ala Leu Val Gln
225 230 235 240
Ala Phe Tyr Gln Gly Ala Ser Asp Ala Gln
245 250
<210> 10
<211> 111
<212> ПРТ
<213> Enterobacter sp. (R4-368)
<400> 10
Met Arg Asn Glu Val Leu Leu Leu Gly Leu Leu Val Gly Cys Val Asn
1 5 10 15
Phe Leu Phe Arg Tyr Leu Pro Leu Arg Ile Arg Ala Gly Gln Ser Arg
20 25 30
Pro Ala Lys Arg Gly Val Ser Gly Val Phe Leu Asp Thr Ile Gly Ile
35 40 45
Ala Ser Ile Cys Ala Leu Leu Val Val Ser Cys Val Pro Glu Ile Ala
50 55 60
Ala Asp Ser Arg Arg Leu Leu Pro Thr Leu Ala Gly Phe Ala Val Leu
65 70 75 80
Gly Val Ser Phe Trp Lys Thr Arg Ser Ile Ile Leu Pro Thr Leu Leu
85 90 95
Ser Ala Phe Ala Tyr Gly Ile Val Trp Lys Leu Leu Ala Asp Ala
100 105 110
<210> 11
<211> 257
<212> ПРТ
<213> Yersinia enterocolitica подвид Enterocolitica
<400> 11
Met Gln Ser Gln Thr Thr Asp Ser Pro Ser Thr Ala Gln Pro Thr Ala
1 5 10 15
Thr Phe Ile Glu Gly Ile Thr Asp Ser Leu Pro Ile Val Ile Gly Tyr
20 25 30
Leu Pro Val Ala Phe Ala Phe Gly Leu Ser Ser Val Lys Leu Gly Phe
35 40 45
Thr Pro Trp Glu Ala Ile Phe Phe Ser Cys Ile Ile Tyr Ala Gly Ala
50 55 60
Ser Gln Phe Val Ile Thr Ala Leu Leu Ser Ala Gly Met Ser Leu Trp
65 70 75 80
Val Ser Ala Leu Thr Val Met Ala Met Asp Val Arg His Ile Leu Tyr
85 90 95
Gly Pro Ala Leu Lys His Arg Ile Val Thr Arg Leu Ser Gly Lys Lys
100 105 110
Thr Ala Leu Trp Ala Phe Gly Leu Thr Asp Glu Val Phe Ala Ala Ala
115 120 125
Thr Thr Lys Leu Met Lys Asp Gln Arg Arg Trp Ser Glu Asn Trp Met
130 135 140
Leu Gly Ile Ala Phe Thr Ser Trp Leu Ser Trp Val Ala Gly Thr Ala
145 150 155 160
Ile Gly Ala Met Phe Gly His Gly Pro Leu Glu Asn Tyr Pro Ala Ile
165 170 175
Glu Ala Ser Leu Ser Phe Met Leu Pro Ala Leu Phe Leu Ser Phe Leu
180 185 190
Leu Ala Ser Phe Lys Arg Gln Tyr Ser Leu Thr Val Ile Ala Ser Leu
195 200 205
Thr Gly Ala Leu Leu Gly Val Leu Leu Phe Ser Ile Pro Val Ala Ile
210 215 220
Leu Ala Gly Ile Gly Gly Gly Cys Leu Ala Ala Leu Leu Gln Pro Val
225 230 235 240
Pro Glu Thr Val Ile Glu Asn Asn Glu Ser Asp Lys Glu Glu Pro Lys
245 250 255
Pro
<210> 12
<211> 257
<212> ПРТ
<213> Yersinia enterocolitica подвид Enterocolitica
<400> 12
Met Gln Ser Gln Thr Thr Asp Ser Pro Ser Thr Ala Gln Pro Thr Ala
1 5 10 15
Thr Phe Ile Glu Gly Ile Thr Asp Ser Leu Pro Ile Val Ile Gly Tyr
20 25 30
Leu Pro Val Ala Phe Ala Phe Gly Leu Ser Ser Val Lys Leu Gly Phe
35 40 45
Thr Pro Trp Glu Ala Ile Phe Phe Ser Cys Ile Ile Tyr Ala Gly Ala
50 55 60
Ser Gln Phe Val Ile Thr Ala Leu Leu Ser Ala Gly Met Ser Leu Trp
65 70 75 80
Val Ser Ala Leu Thr Val Met Ala Met Asp Val Arg His Ile Leu Tyr
85 90 95
Gly Pro Ala Leu Lys His Arg Ile Val Thr Arg Leu Ser Gly Lys Lys
100 105 110
Thr Ala Leu Trp Ala Phe Gly Leu Thr Asp Glu Val Phe Ala Ala Ala
115 120 125
Thr Thr Lys Leu Met Lys Asp Gln Arg Arg Trp Ser Glu Asn Trp Met
130 135 140
Leu Gly Ile Ala Phe Thr Ser Trp Leu Ser Trp Val Ala Gly Thr Ala
145 150 155 160
Ile Gly Ala Met Phe Gly His Gly Pro Leu Glu Asn Tyr Pro Ala Ile
165 170 175
Glu Ala Ser Leu Ser Phe Met Leu Pro Ala Leu Phe Leu Ser Phe Leu
180 185 190
Leu Ala Ser Phe Lys Arg Gln Tyr Ser Leu Thr Val Ile Ala Ser Leu
195 200 205
Thr Gly Ala Leu Leu Gly Val Leu Leu Phe Ser Ile Pro Val Ala Ile
210 215 220
Leu Ala Gly Ile Gly Gly Gly Cys Leu Ala Ala Leu Leu Gln Pro Val
225 230 235 240
Pro Glu Thr Val Ile Glu Asn Asn Glu Ser Asp Lys Glu Glu Pro Lys
245 250 255
Pro
<210> 13
<211> 113
<212> ПРТ
<213> Yersinia enterocolitica подвид Enterocolitica
<400> 13
Met Asn Met Asp Val Val Ile Ile Gly Leu Val Val Gly Thr Val Asn
1 5 10 15
Tyr Leu Phe Arg Tyr Leu Pro Leu Arg Leu Gly Pro Ala Arg Lys Gln
20 25 30
Ala Gly Leu Gln Arg Gly Lys Val Ser Leu Leu Leu Asp Ser Ile Gly
35 40 45
Ile Ala Ser Ile Cys Ala Leu Leu Val Val Ser Ser Thr Pro Glu Ile
50 55 60
Val His Thr Pro Gln Lys Leu Ile Pro Thr Leu Ile Gly Phe Leu Val
65 70 75 80
Ile Cys Gly Cys Phe Tyr Lys Thr Asn Ser Ile Ile Phe Ala Thr Leu
85 90 95
Leu Gly Ala Leu Ser Tyr Gly Leu Thr Phe Lys Leu Leu Met Ile Leu
100 105 110
Ala
<210> 14
<211> 113
<212> ПРТ
<213> Yersinia enterocolitica подвид Enterocolitica
<400> 14
Met Asn Met Asp Val Val Ile Ile Gly Leu Val Val Gly Thr Val Asn
1 5 10 15
Tyr Leu Phe Arg Tyr Leu Pro Leu Arg Leu Gly Pro Ala Arg Lys Gln
20 25 30
Ala Gly Leu Gln Arg Gly Lys Val Ser Leu Leu Leu Asp Ser Ile Gly
35 40 45
Ile Ala Ser Ile Cys Ala Leu Leu Val Val Ser Ser Thr Pro Glu Ile
50 55 60
Val His Asn Pro Gln Lys Leu Ile Pro Thr Leu Ile Gly Phe Leu Val
65 70 75 80
Ile Cys Gly Cys Phe Tyr Lys Thr Asn Ser Ile Ile Phe Ala Thr Leu
85 90 95
Leu Gly Ala Leu Ser Tyr Gly Leu Thr Phe Lys Leu Leu Met Ile Leu
100 105 110
Ala
<210> 15
<211> 252
<212> ПРТ
<213> Photorhabdus luminescens подвид laumondii
<400> 15
Met Pro Val Ser Asp Thr Ser Ser Pro Leu Thr Ser Lys Lys Ser Ser
1 5 10 15
Phe Thr Glu Gly Ile Ile Asp Ser Leu Pro Ile Val Ile Gly Tyr Ile
20 25 30
Pro Val Ala Phe Ala Phe Gly Leu Asn Ala Val Lys Leu Gly Phe Asn
35 40 45
Pro Met Glu Ala Ile Phe Phe Ser Cys Ile Ile Tyr Ala Gly Ala Ser
50 55 60
Gln Phe Val Ile Thr Ala Leu Leu Ser Ala Gly Thr Ser Leu Trp Ile
65 70 75 80
Ser Ala Leu Thr Ile Met Ala Met Asp Val Arg His Ile Leu Tyr Gly
85 90 95
Pro Ser Leu Arg His Arg Ile Lys Asp Lys Leu Thr Glu Lys Lys Thr
100 105 110
Val Ile Trp Ala Phe Gly Leu Thr Asp Glu Val Phe Ala Ala Ala Thr
115 120 125
Ala Lys Leu Ile Lys Asn His Arg Ser Trp Ser Glu Asn Trp Met Val
130 135 140
Ala Ile Ala Ile Cys Ser Trp Leu Ala Trp Gly Ala Gly Thr Ala Ala
145 150 155 160
Gly Ala Phe Leu Gly Asn Gly Tyr Leu Glu Ser Tyr Pro Ala Ile Glu
165 170 175
Ala Ala Met Ile Phe Met Leu Pro Ala Leu Phe Leu Ser Phe Leu Leu
180 185 190
Ala Ser Cys Arg Lys Gln Asn Ser Tyr Cys Val Ala Thr Ala Leu Thr
195 200 205
Gly Ala Leu Leu Gly Ile Thr Phe Phe Ser Ile Pro Val Ala Ile Leu
210 215 220
Ala Gly Ile Val Gly Gly Cys Ile Ala Ala Leu Leu Gln Pro Gln Asn
225 230 235 240
Asn Cys Asn Asp Ser Ser Glu Gln Lys Glu Thr Pro
245 250
<210> 16
<211> 112
<212> ПРТ
<213> Photorhabdus luminescens подвид laumondii
<400> 16
Met Ile Asp Ser Lys Ile Leu Leu Ile Gly Leu Phe Val Gly Leu Ala
1 5 10 15
Asn Phe Ser Phe Arg Tyr Leu Pro Leu Arg Phe Gly Lys Ala Arg Gln
20 25 30
Ser Ala Gly Arg Lys Ala Gly Lys Thr Ser Ile Ile Leu Asp Ser Ile
35 40 45
Gly Ile Ala Ser Ile Cys Ser Leu Leu Ile Val Ser Gly Val Pro Asp
50 55 60
Val Met Arg Glu Ser Gln Lys Leu Leu Pro Thr Leu Ile Gly Cys Leu
65 70 75 80
Thr Ile Cys Leu Val Phe Tyr Lys Thr Lys Gln Ile Ile Leu Ala Thr
85 90 95
Leu Phe Gly Ala Leu Leu Phe Gly Leu Thr Phe Lys Ile Phe Met Asn
100 105 110
<210> 17
<211> 245
<212> ПРТ
<213> Citrobacter youngae
<400> 17
Met Asp Ser Pro Ile Pro Gln Ser Gly Ser Arg Ser Ala Thr Leu Thr
1 5 10 15
Glu Gly Phe Lys Asp Ser Leu Pro Ile Val Ile Ser Tyr Ile Pro Val
20 25 30
Ala Phe Ala Phe Gly Leu Asn Ala Thr Arg Leu Gly Phe Thr Pro Val
35 40 45
Glu Ser Val Phe Leu Ser Cys Ile Ile Tyr Ala Gly Ala Ser Gln Phe
50 55 60
Val Ile Thr Thr Met Leu Ala Ala Gly Ser Ser Leu Trp Val Ala Ala
65 70 75 80
Leu Thr Val Met Ala Met Asp Val Arg His Val Leu Tyr Gly Pro Ser
85 90 95
Leu Arg Ser Arg Ile Ala Gln Arg Leu Ser Lys Pro Lys Ser Ala Leu
100 105 110
Trp Ala Phe Gly Leu Thr Asp Glu Val Phe Ala Ala Ala Thr Ala Lys
115 120 125
Leu Val Arg Asp Asn Arg Arg Trp Ser Glu Asn Trp Met Ile Gly Ile
130 135 140
Ala Leu Cys Ser Trp Ala Ser Trp Val Phe Gly Thr Ala Ile Gly Ala
145 150 155 160
Phe Ser Gly Ser Gly Leu Leu Lys Asp Tyr Pro Ala Val Glu Ala Ala
165 170 175
Leu Gly Phe Met Leu Pro Ala Leu Phe Met Ser Phe Leu Leu Ala Ser
180 185 190
Phe Gln Arg Lys Gln Ala Leu Cys Val Thr Val Ala Leu Thr Gly Ala
195 200 205
Leu Ala Gly Val Ile Leu Phe Ser Ile Pro Ala Ala Ile Leu Leu Gly
210 215 220
Ile Val Cys Gly Cys Leu Thr Ala Leu Leu Gln Ser Phe Trp Gln Gly
225 230 235 240
Gly Pro Asp Glu Leu
245
<210> 18
<211> 111
<212> ПРТ
<213> Citrobacter youngae
<400> 18
Met Ser Tyr Glu Val Leu Leu Leu Gly Leu Leu Val Gly Cys Val Asn
1 5 10 15
Tyr Cys Phe Arg Tyr Leu Pro Leu Arg Leu Gly Ala Gly Asn Val Arg
20 25 30
Pro Ala Arg Arg Gly Ala Thr Gly Ile Leu Leu Asp Thr Ile Gly Ile
35 40 45
Ala Ser Ile Cys Ala Leu Leu Val Val Ser Thr Ala Pro Glu Val Met
50 55 60
His Asp Ala Arg Arg Phe Val Pro Thr Leu Val Gly Phe Val Ile Leu
65 70 75 80
Gly Ala Ser Phe Tyr Lys Thr Arg Ser Ile Ile Ile Pro Thr Leu Leu
85 90 95
Ser Ala Leu Gly Tyr Gly Leu Ala Trp Lys Met Leu Val Gly Leu
100 105 110
<210> 19
<211> 245
<212> ПРТ
<213> Citrobacter freundii
<400> 19
Met Glu Ser Pro Val Pro Gln Ser Glu Ser Ser Ser Ala Thr Leu Thr
1 5 10 15
Glu Gly Phe Lys Asp Ser Leu Pro Ile Val Ile Ser Tyr Ile Pro Val
20 25 30
Ala Phe Ala Phe Gly Leu Asn Ala Thr Arg Leu Gly Phe Thr Pro Val
35 40 45
Glu Ser Val Phe Phe Ser Cys Ile Ile Tyr Ala Gly Ala Ser Gln Phe
50 55 60
Val Ile Thr Thr Met Leu Ala Ala Gly Ser Ser Leu Trp Val Ala Ala
65 70 75 80
Leu Thr Val Met Ala Met Asp Val Arg His Val Leu Tyr Gly Pro Ser
85 90 95
Leu Arg Ser Arg Ile Ala Gln Arg Leu Ser Lys Pro Lys Ser Ala Leu
100 105 110
Trp Ala Phe Gly Leu Thr Asp Glu Val Phe Ala Ala Ala Thr Ala Lys
115 120 125
Leu Val Arg Asp Asn Arg Arg Trp Ser Glu Asn Trp Met Ile Gly Ile
130 135 140
Ala Leu Cys Ser Trp Ala Ser Trp Val Phe Gly Thr Val Leu Gly Ala
145 150 155 160
Phe Ser Gly Ser Gly Leu Leu Lys Asp Tyr Pro Ala Val Glu Ala Ala
165 170 175
Leu Gly Phe Met Leu Pro Ala Leu Phe Met Ser Phe Leu Leu Ala Ser
180 185 190
Phe Gln Arg Lys Gln Ala Leu Cys Val Thr Ala Ala Leu Ala Gly Ala
195 200 205
Leu Ala Gly Val Met Leu Phe Ser Ile Pro Ala Ala Ile Leu Ala Gly
210 215 220
Ile Val Cys Gly Cys Leu Thr Ala Leu Ile Gln Ser Phe Trp Gln Gly
225 230 235 240
Gly Pro Asp Glu Leu
245
<210> 20
<211> 111
<212> ПРТ
<213> Citrobacter freundii
<400> 20
Met Ser Tyr Glu Val Leu Leu Leu Gly Leu Leu Val Gly Cys Val Asn
1 5 10 15
Tyr Cys Phe Arg Tyr Leu Pro Leu Arg Leu Gly Val Gly Asn Val Arg
20 25 30
Pro Thr Lys Arg Gly Ala Thr Gly Ile Leu Leu Asp Thr Ile Gly Ile
35 40 45
Thr Ser Ile Cys Ala Leu Leu Val Val Ser Thr Ala Pro Glu Val Met
50 55 60
His Asp Ala Arg Arg Phe Val Pro Thr Leu Val Gly Phe Val Ile Leu
65 70 75 80
Gly Ala Ser Phe Tyr Lys Thr Arg Ser Ile Ile Ile Pro Thr Leu Leu
85 90 95
Ser Ala Leu Gly Tyr Gly Leu Ala Trp Lys Met Leu Val Val Leu
100 105 110
<210> 21
<211> 100
<212> ДНК
<213> искусственная
<220>
<223> Олигонуклеотид YbeM
<400> 21
aacactgcaa aatcctgcta tttgatttgt atgagtgata agtgtaacgc cgaataatcg 60
tcgttggcga attttacgac tctgacagga ggtggcaatg 100
<210> 22
<211> 100
<212> ДНК
<213> искусственная
<220>
<223> Олигонуклеотид YbeM
<400> 22
gagaaagtaa acgtaacatg atgacgacaa ttctgacgat tcatgttcct tcaacgccgg 60
ggcgcgcatg gaatatgctg gtggcacttc aggcaggaaa 100
<210> 23
<211> 100
<212> ДНК
<213> искусственная
<220>
<223> Олигонуклеотид YpjC
<400> 23
tgaggaatag acaatgttag ttagtaaaag caacggattt aacgctagcg cagttttggg 60
tagtggaagt tataatgaaa ataaatcttc taaacacatg 100
<210> 24
<211> 100
<212> ДНК
<213> искусственная
<220>
<223> Олигонуклеотид YpjC
<400> 24
tgcgctaaaa gaaatgaata gaaccttttc gataatataa gaaaaagtga ttttcatgtt 60
ggtttactta agccaagtag tacgcgtagt gttattttag 100
<210> 25
<211> 45
<212> ДНК
<213> искусственная
<220>
<223> Олигонуклеотид FldA
<400> 25
aaattattct tgtatctttg ttataatatg ggaaagtgca accat 45
<210> 26
<211> 100
<212> ДНК
<213> искусственная
<220>
<223> Олигонуклеотид YtfA
<400> 26
cgttaatcag caggttagcc agccacaaaa agccattgag aaaattattg attttacatg 60
ggattattat attgctaatc cttggttttt aaaaattgtg 100
<210> 27
<211> 100
<212> ДНК
<213> искусственная
<220>
<223> Олигонуклеотид YtfA
<400> 27
tcatctaccg cgcacgaata aaactgccat ccggctggcg ggtgaacagg acctgttgat 60
tattccccgt atcaatggtt aagcccgtca ccacgccgct 100
<210> 28
<211> 50
<212> ДНК
<213> искусственная
<220>
<223> Олигонуклеотид YjbB
<400> 28
ttgagcgctg gctggcaccg aatctggggt atgacgcgga ctgattcaca 50
<210> 29
<211> 50
<212> ДНК
<213> искусственная
<220>
<223> Олигонуклеотид YjbB
<400> 29
aatctgtcac ttttccttac aacaaacagg gcgctcaatg agtgccctgt 50
<210> 30
<211> 1063
<212> ДНК
<213> Citrobacter koseri
<400> 30
atggaaagcc ctgcacccca gtctgagccc cgtccggcaa cattaacgga aggattcaaa 60
gacagtttac cgatagtcat aagttatatt ccggtggcgt ttgcgtttgg ccttaacgcc 120
acccgtctgg gctttactcc cctcgaaagc gtttttttct cctgcattat ttacgcaggc 180
gccagccagt tcgtcatcac caccatgctc gcggcgggca gcacattatg ggtcgccgcg 240
ctgaccgtga tggcgatgga cgtgcgtcat gtgctgtacg gcccttccct gcgtagtcgc 300
atcagccaac ggctcagtaa acctaaaacc gccctgtggg catttggcct caccgatgaa 360
gtgtttgctg ccgccacggc caaactggtg cgggataacc gccgctggag tgaaaactgg 420
atgatcggca tcgcgttctg ctcctgggcc tcctgggtgc tcggcacggt cattggcgca 480
ttttccggga gcggattgct gaaaggcttc cccgccgttg aggcggcatt aggttttatg 540
ctgccagccc tgtttatgag ctttttgctc gcttcttttc aacgcaaaca aacgctgtgc 600
gtcacggcgg cgttaatcgg cgcgctggca ggcgtcacgc tgttttccat tcctgcggct 660
atcctggcgg gtatcgccag cgggtgtctg accgccttga tccagtcgtt ctggcaagga 720
gcgcccgatg agttatgagg ttctgctgct gggactgctg gtcggctgcg ccaattattg 780
ttttcgttat ttaccgcttc gtctgcgaat gggaaacacc cgccccgcca ggcgcggcgc 840
aacgggcgtg ttgctcgaca ccattggcat cgcgtccatc tgcgccctgc tggtggtgtc 900
tacggctccc gaagtgatgc acgacgccag ccggttcatt ccgacgctgg tcgggtttgc 960
cgtcctgggc gtcagtttct acaagacgcg cagcatcatc atcccaacgc tactgagcgc 1020
tctgggctat ggactcgcct ggaagataga ggtcatttta taa 1063
<210> 31
<211> 1063
<212> ДНК
<213> Shigella flexneri
<400> 31
atggaaagcc ctgttcccca gtctgaatcc cgttctgcaa cgttaactga aggattcaaa 60
gacagcctac cgatagttat cagttatatt ccggtcgcat ttgcatttgg tatgaatgcg 120
actcgcctgg gctttactcc cgttgaaagc gtttttttct cctgcatcat ttacgctggc 180
gccagccagt ttgtcatcac aaccatgctc gccgcaggca gctcactgtg ggtcgcggct 240
ctgaccgtca tggcgatgga tgttcgccat gttttgtacg gcccttctct gcgcagccgt 300
atcgcccgac agctgagcaa acctaaaagc gcgctatggg cctttggcct caccgacgaa 360
gtctttgccg cggcaacggc caagctggtg cgggataacc ggcgctggag tgaaaactgg 420
atgatcggca tcgcgctatg ctcctgggct tcctgggtac ttggtacggt tatcggcgca 480
ttttccggca ctggcttact gaagggattc ccggcggtag aagcggcgct ggggtttatg 540
ctcccggcgc tgtttatgag ttttctgctg gcctctttcc agcgtaatca aacgctatgc 600
gtcacggcgg ctttagccgg tgcgctggct ggcgtgacgc tgttttctat cccggcagcc 660
atcctcgcag gcatagtctg cggatgcctg accgcgctca ttcagtcgtt ctggcaggga 720
ggtcctgatg agttatgagg ttctgctgct cggcctgctg gtcggctgcg tcaattactg 780
ttttcgctat ttaccactgc gtctgcgaat ggggaatgtg cgcccgacaa aacgcggagc 840
cactggaata ctactcgaca ccatcggtat tgcatcaatt tgcgccctgc tagtggtgtc 900
tactgcgcca gaagtgatgc acgatgcccg tcgttttgtg cctacgctgg tggggtttgt 960
ggtactgggt gcaagcttct ataagacccg cagcatcatc attccgacct tactgagtgc 1020
cctgggctat ggattagcct ggaaaatgct gtttgtctta tag 1063
<210> 32
<211> 1060
<212> ДНК
<213> Raoultella ornithinolytica
<400> 32
atggaaaaac cggcaccggc aagcgaagca accctgccgg aaggtattaa agatagcctg 60
ccgattgtga ttagctatat tccggttgca tttgcctttg gtctgaatgc aacccgtctg 120
ggttttacac cgctggaaag cctgtttttt agctgtatta tctatgccgg tgcaagccag 180
tttgttatta ccgcaatgct ggcagcaggt agcagcctgt gggttgcagc actgaccgtt 240
atggcaatgg atgttcgtca tgttctgtat ggtccgagcc tgcgtagccg tattcatcgt 300
gcactggata aacgtaaaac cgcactgtgg gcatttggcc tgaccgatga agtttttgca 360
gcagcaaccg caaaactggt tcgtgataat cgtcgttgga gcgaaagctg gatgctgggt 420
attgcattta ccagctggat tagctgggtt tttggcaccc tgattggtgc atatagcggt 480
agcggtctgc tggttggttt tccggcagtt gaagcagccc tgagctttat gctgcctgca 540
ctgtttatga gttttctgct ggcaagcttt cagcgtaaac agagcctgag cgttaccgca 600
gcactggcag gcgcactggg tggtattatt ctgtttagca ttccggcagc aattctggca 660
ggtattgttt gtggttgtct ggcagcgctg attcaggcaa gcattcaggg tatgccggat 720
gaacaataac gttctgatta ttggtattgt ggtgggctgt gtgaattacc tgtttcgtta 780
tctgccgctg cgtctgcgtg caggtaatgc acgtccgacc cgtcgtggtc cgctgagcgt 840
tctgctggat accattggca ttgcaagcat ttgtgcactg ctgattgtta gcagcgttcc 900
ggaaattctg agcgatagcc gtcgtctgct gccgaccctg gttggtttta ccgttctggg 960
tctggcattt tggaaaaccc gtagcattat tatgccgaca ctgctgagcg cactggccta 1020
tggtattgca tggaaaatta ccacctttct gtatttttga 1060
<210> 33
<211> 1078
<212> ДНК
<213> Enterobacter sp. (R4-368)
<400> 33
atggatatgg atagcagcgt taccgcaacc aaaagcacca gcgatcagag cgcaaccttt 60
ctggaaggta ttaaagatag cctgccgatt gttctgagct atgttccggt tgcatttgcc 120
tttggtatga atgcaaccaa actgggtttt acaccgctgg aaagcgtgtt ttttagctgt 180
attatctatg ccggtgcaag ccagtttgtt attaccacca tgctggcagc aggtagcgca 240
ctgtgggttg cagcactgac cgttatggca atggatgttc gtcatgttct gtatggtccg 300
agcctgcgta gccgtattct gcagccgctg aaaaatcgta aaaccgcagt gtgggcattt 360
ggtctgaccg atgaagtttt tgcagcagca accgcaaaac tggttcgtga taatcgtcgt 420
tggagcgaaa attggatgat tggtattgca ctgtttagct ggctgagctg ggttgccggt 480
acagttctgg gtgcatttag cggtgatggt ctgctggatg gttatccggc agttgaaagt 540
gcactgggct ttatgctgcc tgccctgttt atgagctttc tgctggcaag ctttcagcgt 600
cgtcagatta gcgcagttac cgcagcactg ctgggtgcac tggcaggcgt taccctgttt 660
agcattccgg cagcaattct ggcaggcatt tttgcaggtt gtctggcagc actggttcag 720
gccttttatc agggtgcaag tgatgcgcaa tgaggttctg ctgctgggcc tgctggttgg 780
ttgtgttaat tttctgtttc gttatctgcc gctgcgtatt cgtgcaggtc agagccgtcc 840
ggcaaaacgt ggtgttagcg gtgtttttct ggataccatt ggcattgcaa gcatttgtgc 900
cctgctggta gttagctgtg ttccggaaat tgcagcagat agccgtcgtc tgctgccgac 960
cctggcaggt tttgcagtgc tgggtgttag cttttggaaa acccgtagca ttattctgcc 1020
gacactgctg agcgcatttg cgtatggtat tgtttggaaa ctgctggcag atgcctaa 1078
<210> 34
<211> 1112
<212> ДНК
<213> Yersinia enterocolitica подвид enterocolitica
<400> 34
atgcaaagcc aaaccaccga ctccccctcg acggcccagc cgaccgccac ctttattgaa 60
ggaataaccg atagcctacc gattgttatc ggttatctac ccgttgcttt tgcctttggt 120
ttgagttcgg taaaacttgg ctttactccg tgggaagcta ttttcttttc ttgcattatt 180
tatgccggag ccagccaatt cgttattacc gccctgctca gcgcggggat gtcattgtgg 240
gtttccgcct tgaccgtgat ggctatggat gtccgccata tcttgtacgg gccagcactg 300
aaacaccgca ttgtaaccag gttatctggc aaaaaaacgg cgctgtgggc ctttggtctt 360
actgatgaag tgtttgccgc cgcaacaacc aagctaatga aagatcaacg gcgctggagt 420
gaaaactgga tgcttggcat cgcgttcacc tcttggttgt cttgggtagc tggcaccgct 480
atcggcgcga tgtttggtca tgggccgctg gaaaattacc cggcgattga agcatcactc 540
tcctttatgc tcccggcgct attcctcagc ttcttattgg cctcgttcaa acgccaatac 600
agccttaccg ttattgcttc actgaccgga gccttgctgg gcgtgctgct gttctctatt 660
ccggtggcta ttttagccgg tattggcggc ggatgcctgg cagccctgct ccaacccgtc 720
cccgagaccg ttatagaaaa taacgagagt gataaagagg agccgaagcc atgaatatgg 780
atgttgtgat cattggtttg gtggtgggaa cggtcaatta cctgtttcgt tatctgccgc 840
tgcgcctggg gcctgcccgt aaacaagcag gcctgcaacg agggaaagtc tccctgttgc 900
tagacagcat cgggatcgcc tctatctgtg cgttgttggt ggtttccagt accccggaga 960
tagtgcataa cccacagaaa ttaattccta cactaattgg ttttttagtt atctgtggat 1020
gcttttataa aaccaacagt attatcttcg ccaccttact gggagcactc agttacggtc 1080
tgacattcaa attactgatg attttggcat aa 1112
<210> 35
<211> 1094
<212> ДНК
<213> Photorhabdus luminescens подвид laumondii
<400> 35
atgcctgttt ctgatacatc atccccctta acgagtaaaa aatcttcttt tactgaagga 60
ataatagata gtttacccat tgttatcggt tatattcccg tcgcctttgc ttttggtctc 120
aatgccgtca aacttggctt caacccaatg gaagccattt tcttttcatg catcatctac 180
gccggtgcaa gccagttcgt catcacagct ttactgagtg cggggacatc attatggatt 240
tctgccctaa caattatggc aatggatgtc cgccatattc tttatggtcc atctttaagg 300
caccgtatca aagataagct aacggagaaa aaaaccgtta tctgggcttt cggcctgaca 360
gatgaagttt ttgccgccgc gactgcaaaa ctcattaaaa accaccggag ctggagtgaa 420
aactggatgg ttgctattgc aatctgttct tggctggcct ggggcgcagg taccgcagcc 480
ggtgcatttc ttggtaacgg ttatttggaa tcctatcccg ctatagaagc tgccatgatt 540
ttcatgttac cagcactatt tctcagtttt cttcttgctt cttgtagaaa acaaaatagt 600
tattgtgttg caaccgcact aaccggagca cttttaggga ttacattttt ctcaattcca 660
gttgctattc tggcaggtat tgtcggtggt tgtatcgcgg cactgttaca accgcaaaac 720
aattgcaatg actcttcaga acaaaaggaa acaccatgat tgatagcaag attttgctga 780
ttggactatt tgttgggtta gctaactttt catttcgcta tctgccacta cgatttggga 840
aagcacgcca atctgccggc agaaaagctg gaaaaacaag cattatcctt gacagtattg 900
gtattgcatc catttgttct ttactcatcg tatcaggtgt acctgatgtg atgagagaaa 960
gtcaaaaact acttcctacc ctcataggtt gtctgaccat ctgtttagtc ttttacaaaa 1020
caaagcaaat tatactcgca acactatttg gcgcactgct ttttggacta acattcaaaa 1080
tatttatgaa ttag 1094
<210> 36
<211> 1063
<212> ДНК
<213> Citrobacter youngae
<400> 36
atggatagcc cgattccgca gagcggtagc cgtagcgcaa ccctgaccga aggttttaaa 60
gatagcctgc cgattgtgat tagctatatt ccggttgcat ttgcctttgg tctgaatgca 120
acccgtctgg gttttacacc ggttgaaagc gtttttctga gctgtattat ctatgccggt 180
gcaagccagt ttgttattac caccatgctg gcagcaggta gcagcctgtg ggttgcagca 240
ctgaccgtta tggcaatgga tgttcgtcat gttctgtatg gtccgagcct gcgtagccgt 300
attgcacagc gtctgagcaa accgaaaagc gcactgtggg catttggcct gaccgatgaa 360
gtttttgcag cagcaaccgc aaaactggtt cgtgataatc gtcgttggag cgaaaattgg 420
atgattggta ttgcactgtg tagctgggca agctgggttt ttggcaccgc aattggtgca 480
tttagcggta gcggtctgct gaaagattat ccggcagttg aagcagcact gggctttatg 540
ctgcctgcac tgtttatgag ctttctgctg gcgagctttc agcgtaaaca ggcactgtgt 600
gttaccgttg ccctgaccgg tgcactggca ggcgttattc tgtttagcat tccggcagca 660
attctgctgg gtattgtttg tggttgtctg accgcactgc tgcagagctt ttggcagggt 720
ggtccggatg agctatgagg ttctgctgct gggtctgctg gttggttgtg tgaattattg 780
ttttcgttat ctgccgctgc gtctgggtgc aggtaatgtt cgtccggcac gtcgtggtgc 840
aaccggtatc ctgctggata caattggcat tgcaagcatt tgtgcactgc tggtagttag 900
caccgcaccg gaagttatgc atgatgcacg tcgttttgtt ccgaccctgg tgggttttgt 960
tattctgggt gccagcttct ataaaacccg tagcattatt atcccgaccc tgctgagcgc 1020
actgggttat ggtctggcat ggaaaatgct ggtaggtctg taa 1063
<210> 37
<211> 1063
<212> ДНК
<213> Citrobacter freundii
<400> 37
atggaaagtc cggttccgca gagcgaaagc agcagcgcaa ccctgaccga aggttttaaa 60
gatagcctgc cgattgtgat tagctatatt ccggttgcat ttgcctttgg tctgaatgca 120
acccgtctgg gttttacacc ggttgaaagc gtgtttttta gctgcattat ctatgccggt 180
gcaagccagt ttgttattac caccatgctg gcagcaggta gcagcctgtg ggttgcagca 240
ctgaccgtta tggcaatgga tgttcgtcat gttctgtatg gtccgagcct gcgtagccgt 300
attgcacagc gtctgagcaa accgaaaagc gcactgtggg catttggcct gaccgatgaa 360
gtttttgcag cagcaaccgc aaaactggtt cgtgataatc gtcgttggag cgaaaattgg 420
atgattggta ttgcactgtg tagctgggca agctgggttt ttggcaccgt tctgggtgca 480
tttagcggta gcggtctgct gaaagattat ccggcagttg aagcagcact gggctttatg 540
ctgcctgcac tgtttatgag ctttctgctg gcgagctttc agcgtaaaca ggcactgtgt 600
gttaccgcag ccctggcagg cgcactggct ggtgttatgc tgtttagcat tccggcagca 660
attctggcag gtattgtttg tggttgtctg accgcactga ttcagagctt ttggcagggt 720
ggtccggatg agctatgaag ttctgctgct gggtctgctg gttggttgtg tgaattattg 780
ttttcgttat ctgccgctgc gtctgggtgt tggtaatgtt cgtccgacca aacgtggtgc 840
aaccggtatt ctgctggata ccattggtat taccagcatt tgtgcactgc tggtagttag 900
caccgcaccg gaagttatgc atgatgcacg tcgttttgtt ccgaccctgg tgggttttgt 960
tatcctgggt gccagcttct ataaaacccg tagcattatt atcccgaccc tgctgagcgc 1020
actgggttat ggtctggcat ggaaaatgct ggttgttctg taa 1063
<---





























Изобретение относится к биотехнологии и представляет собой рекомбинантный штамм E.coli для ферментативной продукции метионина, где в указанном рекомбинантном штамме экспрессия генов ygaZH, гомологичных генам ygaZ и ygaH из Escherichia coli, увеличена по сравнению с немодифицированным штаммом E.coli, при условии, что этот рекомбинантный штамм не обладает сверхэкспрессией генов metH из E. coli и ослаблением экспрессии по меньшей мере одного гена среди metN, metI или metQ, причем указанные гены, гомологичные ygaZH, выбирают среди группы генов, кодирующих пару генов, гомологичных YgaZH, определенных соответственно в SEQ ID NO: 3 и SEQ ID NO: 4 из Citrobacter koseri, SEQ ID NO: 9 и SEQ ID NO: 10 из Enterobacter sp. (R4-368), SEQ ID NO: 17 и SEQ ID NO: 18 из Citrobacter youngae, SEQ ID NO: 19 и SEQ ID NO: 20 из Citrobacter freundii. Изобретение позволяет получать метионин с высокой степенью эффективности. 2 н. и 7 з.п. ф-лы, 11 табл., 5 пр.
1. Рекомбинантный штамм E.coli для ферментативной продукции метионина, где в указанном рекомбинантном штамме экспрессия генов ygaZH, гомологичных генам ygaZ и ygaH из Escherichia coli, увеличена по сравнению с немодифицированным штаммом E.coli, при условии, что этот рекомбинантный штамм не обладает сверхэкспрессией генов metH из E. coli и ослаблением экспрессии по меньшей мере одного гена среди metN, metI или metQ, причем указанные гены, гомологичные ygaZH, выбирают среди группы генов, кодирующих пару генов, гомологичных YgaZH, определенных соответственно в SEQ ID NO: 3 и SEQ ID NO: 4 из Citrobacter koseri, SEQ ID NO: 9 и SEQ ID NO: 10 из Enterobacter sp. (R4-368), SEQ ID NO: 17 и SEQ ID NO: 18 из Citrobacter youngae, SEQ ID NO: 19 и SEQ ID NO: 20 из Citrobacter freundii.
2. Рекомбинантный штамм E. coli по п. 1, в котором гомологичные гены ygaZH экспрессируются под контролем индуцибельного промотора.
3. Рекомбинантный штамм E. coli по любому из пп. 1, 2, в котором по сравнению с немодифицированным штаммом E. coli также увеличивается экспрессия по меньшей мере одного из следующих генов: ptsG, pyc, pntAB, cysP, cysU, cysW, cysA, cysM, cysJ, cysI, cysH, gcvT, gcvH, gcvP, lpd, serA, serB, serC, cysE, metF, metA, аллеля metA*, кодирующего фермент с пониженной чувствительностью к S-аденозилметионину и/или метионину по механизму обратной связи, thrA и аллеля thrA*, кодирующего фермент с ослабленным ингибированием треонином по механизму обратной связи.
4. Рекомбинантный штамм E. coli по п. 3, в котором по меньшей мере один из указанных генов находится под контролем индуцибельного промотора.
5. Рекомбинантный штамм E. coli по любому из пп. 1-4, в котором также ослабляется экспрессия по меньшей мере одного из следующих генов: metJ, pykA, pykF, purU, ybdL, yncA, metE, dgsA или udhA.
6. Рекомбинантный штамм E. coli по любому из пп. 1-5, в котором:
а. экспрессия генов, гомологичных ygaZ и ygaН, увеличена по сравнению с немодифицированным штаммом E. coli;
б. экспрессия генов metA*, cysPUWAM, cysJIH, gcvTHP, metF, serA, serB, serC, cysE, thrA*, ptsG и pyc усиливается; и
в. экспрессия генов metJ, pykA, pykF, purU, dgsA, metE и yncA ослабляется.
7. Способ ферментативного производства метионина, включающий следующие этапы:
а. культивирование рекомбинантного штамма E. coli, определенного в любом из пп. 1-6, в подходящей культуральной среде, содержащей ферментируемый источник углерода и источник серы, и
б. выделение метионина из культуральной среды.
8. Способ по п. 7, в котором рост рекомбинантного штамма E. coli подвергается ограничению или недостаточности в отношении фосфата и/или калия в культуральной среде.
9. Способ по любому из пп. 7, 8, в котором стадия выделения метионина включает стадию концентрирования метионина в ферментационном бульоне.
| Устройство для перемещения и подачи на рабочую позицию поддонов с изделиями | 1984 |
|
SU1239041A1 |
| WO 2008082211 A1, 10.07.2008 | |||
| WO 2012055798 A1, 03.05.2012 | |||
| WO 2012090021 A1, 05.07.2012 | |||
| WO 2013001055 A1, 03.01.2013 | |||
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ, ШТАММ Escherichia coli - ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ (ВАРИАНТЫ) | 2001 |
|
RU2215784C2 |
| TROTSCHEL CHRISTIAN et al., Characterization of Methionine Export in Corynebacterium glutamicum, JOURNAL OF BACTERIOLOGY Vol | |||
| Индукционная катушка | 1920 |
|
SU187A1 |
| Походная разборная печь для варки пищи и печения хлеба | 1920 |
|
SU11A1 |
| ПРИБОР ДЛЯ НАБЛЮДЕНИЯ И ЗАПИСИ МОРСКОГО ВОЛНЕНИЯ | 1926 |
|
SU3786A1 |

