Область техники, к которой относится изобретение
Данное изобретение относится к антителам к Р-селектину и их фрагментам, которые специфически связывают конформационные эпитопы Р-селектина, и к способам их применения и идентификации.
Уровень техники
При нормальном гемостазе и иммунологическом надзоре лейкоциты свободно циркулируют в потоке крови и отвечают на повреждение и инфекцию в процессе последовательной активации адгезии, опосредованной молекулами клеточной алгезии (1-3). В случае воспалительных и тромболических болезней этот процесс нарушается и может развиваться патология, когда лейкоциты атакуют ткани организма и могут вызывать серьезные, иногда смертельные осложнения. Хорошо известно, что адгезия лейкоцитов играет основную роль в развитии патологии многих воспалительных и тромботических расстройств, таких как вазоокклюзия при серповидноклеточном заболевании, реперфузионное повреждение, тромбоз, атеросклероз, астма, ревматоидный артрит, псориаз и метастазирование (4-15), глубокий венозный тромбоз (DVT). Р-селектин участвует также в развитии других болезней, таких как повреждение тканей, связанное с воспалением, например, ишемическое реперфузионное повреждение. Таким образом, Р-селектин является целевой мишенью при воспалительных и тромботических заболеваниях у людей.
Селектины относятся к семейству адгезивных белков, которые, как известно, играют ключевую роль в рекрутинге лейкоцитов в активированный эндотелий и в тромбоциты. Р-селектин (тромбоцитарный селектин) является членом селектинового семейства адгезивных гликопротеинов, которое также включает L-селектин и Е-селектин (16). Селектины опосредуют рекрутинг, первичную фиксацию, катание (роллинг) и адгезию лейкоцитов к сайтам воспаления (1). Р-селектин находится в тельцах Вейбеля-Паладе эндотелиальных клеток и в альфа-гранулах тромбоцитов и быстро мобилизуется в плазменной мембране после стимуляции вазоактивными веществами, такими как гистамин и тромбин (17).
Серповидноклеточное заболевание
Серповидноклеточное заболевание (SCD) представляет собой редкое наследственное заболевание крови, которое вызывает хроническую анемию и вазоокклюзию, поражая в основном людей афро-американского происхождения в США. Серповидноклеточное заболевание вызвано повреждением одного гена у афроамериканцев, которое бывает у примерно 1 из 375-600 людей афроамериканского происхождения (18-19). Серповидноклеточное заболевание встречается также среди людей в средиземноморских странах, у африканцев в странах Карибского бассейна и в некоторых частях Южной и Центральной Америки (18, 19).
SCD является аутосомной рецессивной болезнью, вызванной миссенс-мутацией (Val6Ala) в гене бета-глобина, которая делает мутантный гемоглобин менее растворимым и склонным к полимеризации после дезоксигенации. Полимеризация гемолобина вызывает деформацию эритроцитов, придавая клетке серповидную форму (20).
Существуют три варианта SCD: гомозиготая серповидноклеточная анемия (вызванная наличием гемоглобина SS), двойная гетерозиготная серповидноклеточная анемия, вызванная наличием гемоглобина С (гемоглобина SC) и серповидноклеточная β-талассемия. Наиболее распространенная и серьезная форма этой болезни встречается у людей, которые наследуют две копии варианта HbS (HbSS), исходный гемоглобин в их красных кровяных тельцах представляет собой серповидный гемоглобин. Другие люди с различной степенью развития этой болезни могут быть гетерозиготными носителями. У них есть одна копия варианта HbS, спаренная с копией другого варианта β-глобина. HbSC приводит к развитию умеренной формы этого заболевания.
Варианты Hb в случае бета-талассемии (приводящей к неспособности продуцировать цепь нормальныого βА-глобина (вариант болезни β0) или уменьшению этого продуцирования (вариант болезни β+)) приводят к ряду клинических осложнений. Форма HbS β0 является серьезным заболеванием, в то время как форма HbS β+ может быть выражена умеренно из-за вклада каждого варианта в состав общего гемоглобина у пациента. Другие более редкие варианты могут возникнуть, если в сочетании с геном S от другого родителя наследуется другой вид аномального гемоглобина, такой как гемоглобин D, G или О. Преобладающая серповидная форма клетки содержится у людей с одной копией HbS и одной копией нормального гена β-глобина (HbA). У этих людей наблюдаются признаки серповидноклеточного заболевания (18).
SCD поражает 50000-100000 человек в США (21-24). Все люди, которые являются гомозиготными или гетерозиготными носителями гена гемоглобина S, характеризуются некоторыми клиническими проявления SCD. Симптомы обычно проявляются в первые 6 мес жизни, но степень развития SCD может быть очень разной (25). Больные HbSS страдают от болезни больше всего. За ними следуют пациенты с HbS β0-талассемией (22, 26). Кроме генотипа, на степень серьезности заболевания влияют дополнительные факторы, такие как уровень фетального гемоглобина и гаплотип β-глобинового кластера, участка, который содержит 5 генов, которые кодируют β-часть гемоглобина. Несмотря на то, что можно определить факторы генетического риска, способность предсказать течение болезни от момента рождения ограничена (27).
В США во всех 50 штатах и в округе Колумбия при рождении требуется скрининг серповидных клеток, он дает возможность осуществить раннее вмешательство. Методика диагностического тестирования обычно включает полный анализ крови в сочетании с одним или более методами - электрофорезом гемоглобина, изоэлектрическим фокусированием, жидкостной хроматографией высокого разрешения и анализом ДНК (22).
Хроническая анемия и гемолиз
Серповидный эритроцит обладает более коротким временем полужизни по сравнению с нормальным эритроцитом и появляется вследствие нестабильности HbS и действия повторяющихся эпизодов полимеризации/деполимеризации гемоглобина в кровотоке. Это влияет на ионную проницаемость мембраны, вязкость клеточной мембраны и деформируемость (20) и ускоряет окислительное повреждение мембраны (29). Пациенты, больные SCD, страдают от анемии уже во время 2-3 мес жизни, у них развиваются симптомы и осложнения, связанные с хронической анемией и гемолизом (22, 30), такие как болезнь почек, офтальмологические расстройства, язвы на ногах, приапизм и легочная гипертензия (26, 31-37). Содержание гемоглобина у пациентов, страдающих от SCD, колеблется от 6 до 10 г/дл, а молекула гемоглобина S имеет низкое сродство к кислороду. Величина содержания гемоглобина равная 5 г/дл или менее или стремительное падение этого содержания до 2 г/дл и менее побуждают осуществлять проведение трансфузии пациентам. Трансфузии обычно проводят для восстановления содержания гемоглобина до нормального уровня, определяемого для каждого пациента, так как избыточный гематокрит может вызывать осаждение серповидноклеточных эритроцитов (38). Пациенты с SCD более восприимчивы к инфекции, вызываемой парвовирусом В19, который может остановить эритропоэз и привести к появлению апластической анемии (39).
Вазоокклюзивный болевой криз
Сосудистая окклюзия является центральным событием в клиническом течении SCD и, вероятно, затрагивает как микро-, так и макрокровоток. Закупорка, происходящая в мелких сосудах, может закончиться приступами острой боли или вазоокклюзивным болевым кризом. Вазоокклюзивный болевой криз является клиническим признаком микрососудистых окклюзии и причиной госпитализации более 90% взрослых пациентов, больных SCD. Хорошо известно, что полимеризация гемоглобина S во время дезоксигенации и образования серповидных клеток приводит к блокаде микроциркуляторной части сосудистого русла (40). Однако недавно выяснилось, что не только полимеризация гемоглобина S отвечает за вазоокклюзию. Было установлено, что такие события как лизис серповидных красных клеток, повреждение клеточной мембраны и окислительный стресс, повторяющиеся ишемические повреждения и повреждение микрососудов из-за адгезионных взаимодействий между серповидными красными клетками и эндотелием, заканчиваются появлением провоспалительной среды (41-43). В этой среде хронического воспаления сосудов прилипание лейкоцитов, тромбоцитов и серповидных красных клеток к активированному эндотелию кровеносных сосудов и друг к другу считается основной причиной микрососудистой блокады и вазоокклюзивного болевого криза (43-47). В процесс вазоокклюзии могут вносить свой вклад дополнительные факторы, такие как строгость серповидных красных клеток, повышенная вязкость крови и локальные сосудистые спазмы (вазоконстрикция).
Длительные повторяющиеся вазоокклюзивные события и окклюзии, происходящие в макроциркуляторной части сосудистого русла, могут вызвать осложнения, опасные для жизни, которые могут привести к повреждению органов и расстройствам, нарушению мозгового кровообращения и к смерти (40). У больных серповидноклеточным заболеванием, продолжительность жизни уменьшается примерно на 20-30 лет (48).
Хроническая боль при развитии SCD не является продолжением болевых ощущений от закупорки сосудов, она обычно является вторичной после васкулярного (лишенного сосудов) некроза костей в различных суставах (49). Серповидные красные клетки могут захватываться селезенкой, вызывая ее увеличение и кризис секвестрации селезенки, приводящие к внезапной и сильной анемии. Функциональная аспления делает больных восприимчивыми к развитию инфекций (18). Замедление роста костей, почечные (32), офтальмологические (33) и цереброваскулярные осложнения (от клинически очевидного нарушения мозгового кровообращении до временного бессимптомного (немого) инфаркта) (50) рассматриваются как основные клинические последствия SCD и вазоокклюзивного повреждения (22). Острый грудной синдром является еще одним серьезным осложнением (51) и важной причиной заболеваемости и смертности (52).
Приступы боли вызываются рядом факторов, включая охлаждение, стресс и физическое истощение (38, 53). Частота, серьезность, месторасположение и продолжительность болевого криза могут меняться даже в случае конкретного подтипа болезни. Пациенты с гомозиготной серповидноклеточной болезнью и серповидноклеточной β0-талассемией характеризуются большей частотой вазоокклюзивного болевого криза, чем пациенты с гемоглобином SC и генотипом β0-талассемии (54). Считается, что степень серьезности болезни зависит от сложного взаимодействия генетических, реологических и гематологических факторов. Криз обычно характеризуется болью в спине, ногах, коленях, руках, в груди и в брюшной полости (53). Частота появления криза и степень болевых ощущений различны у разных пациентов с серповидноклеточной болезнью от новорожденных до пациентов в возрасте 50 лет, 5,2% пациентов с этой болезнью имеют от 3 до 10 эпизодов сильной боли каждый год (54). Независимое исследование показало, что у более, чем 30% пациентов с SCD в США (около 27000) болевые кризы случались три и более раз в год (55). Более того, независимое исследование (PiSCES), которое оценивало качество жизни, связанное со здоровьем у пациентов с SCD, показало, что наличие болевого криза часто не сообщалось пациентами (56).
Существующие в настоящее время способы лечения сосудистых окклюзии
Сосудистые окклюзии у пациентов с SCD могут проявляться различным образом, включая вазоокклюзивный болевой криз, острый грудной синдром, сердечно-сосудистые расстройства и многочисленные виды поражения органов. Следовательно, способы лечения сосудистых окклюзии зависят от клинического течения заболевания и степени серьезности болезни, и, в общем, являются симптоматическими или паллиативными по своей природе. Санитарно-просветительная работа среди населения, направленная на исключение появления начальных факторов, которые провоцируют вазоокклюзивный болевой криз, дает свои профилактические преимущества. Два наиболее распространенных способа симптоматического лечения включают трансфузию крови и применение анальгетиков. У большей части пациентов с SCD содержание гемоглобина составляет обычно от 6 до 10 г/дл, во время вазоокклюзивного болевого криза эта величина падает до 1 г/дл. Сильную боль, появляющуюся в результате вазоокклюзивного болевого криза, можно лечить при помощи наркотиков, но их применение противоречит принципам борьбы со склонностью к наркотикам и их переносимости. Другие осложнения при применении наркотиков включают привыкание к наркотикам, сонливость и дыхательную депрессию. Для лечения вазоокклюзивного болевого криза использовали терапию с применением кислорода несмотря на отсутствие убедительных доказательств ее эффективности. Иногда во время вазоокклюзивного болевого криза применяют регидратацию, дающую некоторую пользу (22, 38).
Можно также использовать трансплантацию костного мозга, она может обеспечить лечебный эффект, но ее применение сдерживается ограниченным числом пациентов и высокой степенью риска заболеваемости и смертности (22).
Гидроксимочевина (Droxia) представляет собой единственное лекарство, одобренное FDA (Комиссией по контролю за лекарствами и питательными веществами) в качестве препарата для лечения болевых кризов при SCD. Механизмы ее благоприятного действия не известны, но они могут включать повышение уровня гемоглобина F в RBCs (эритроцитах) при снижении степени полимеризации гемоглобина S. Гидроксимочевина является цитотоксичным миелосупрессивным и тератогенным лекарством (57, 58), которое вызывает канцерогенный риск у пациентов с SCD. Однако влияние ее длительного применения на гематологическую токсичность, поражение органов и канцерогенность в настоящее время неизвестно (59, 60).
Подводя итог, следует указать, что большая часть способов лечения вазоокклюзивного болевого криза у пациентов с SCD приводит к симптоматическому облегчению и не учитывает причину, лежащую в основе этого изнуряющего состояния.
К настоящему времени FDA одобрила только один способ лечения болевого криза, таким образом, пациенты с SCD все еще нуждаются в лечении этого заболевания, угрожающего жизни с высоким процентом смертности.
Р-селектин как терапевтическая мишень при SCD
Как отмечено выше, при наличии SCD взаимодействия между серповидными красными клетками, тромбоцитами, лейкоцитами и системой микрососудов представляют собой процессы, зависимые от Р-селектина, и приводят к возникновению вазоокклюзивного болевого криза. Исследования, проведенные на трансгенных мышах, созданных для экспрессии человеческого β-гемоглобина S (βS), показали, что опосредованное антителами ингибирование функции Р-селектина может предотвратить и/или снизить вазоокклюзию, что свидетельствует о терапевтическом потенциале в отношении этой мишени. Кроме того, у мышей, экспрессирующих гемоглобин βS, у которых отсутствует Р-селектин (благодаря делеции гена) не бывает вазоокклюзии, что также подтверждает ключевую роль этой молекулы в развитии этого заболевания.
Гипервоспалительное состояние у пациентов с SCD характеризуется наличием активированных моноцитов и активированного сосудистого эндотелия 961-63). Похожий провоспалительный фенотип наблюдался в состоянии покоя у мышей с геном βS, которые имели повышенные уровни периферических лейкоцитов и нейтрофилов, повышенное количество роллинг-лейкоцитов и адгезивных лейкоцитов и пониженный объем кровообращения и пониженную скорость роста содержания красных кровяных клеток (64). Мыши с геном βS были гиперчувствительными к гипоксии/реоксигенации, что приводило к воспалительному ответу, характеризующемуся значительным увеличением количества адгезивных и эмигрировавших лейкоцитов. Этот воспалительный ответ был полностью остановлен при функциональном блокировании мышиного антитела к Р-селектину, но не путем функционального блокирования мышиного антитела к Е-селектину, что свидетельствует о критической роли Р-селектина в этом процессе.
Р-селектин играет центральную роль в рекрутинге лейкоцитов в воспалительном и тромботическом сайтах путем связывания его противорецептора, гликопротеинового лиганда-1 Р-селектина (PSGL-1) (или PSGL-1-подобного рецептора) в серповидных красных кровяных клетках, который представляет собой муциноподобный гликопротеин, постоянно экспрессируемый в клетках лейкоцитов, включая нейтрофилы, моноциты, тромбоциты, и в некоторых эндотелиальных клетках (68). Конечная физиологическая функция селектинов состоит в промотировании экстравазации лейкоцитов в воспаленные или поврежденные ткани. Первоначальное связывание Р-селектина на поверхности эндотелия с PSGL-1 на поверхности лейкоцитов является важным и центральным событием этого процесса. Преобладающий механизм роллинга и фиксации лейкоцитов на поверхности активированного эндотелия и тромбоцитов заключается в связывании PSGL-1 на поверхности лейкоцитов с Р-селектином на поверхности указанных клеток (68, 69). PSGL-1 связывается с Р-, L- и Е-селектинами (70). Р-селектин и SPG-3, гликосульфопептид, моделируемый из N-конца PSGL-1, подвергаются процессу сокристаллизации, при этом были идентифицированы контактные остатки для связывания лектина с лигандом (71).
Селектины содержат общие структурные фрагменты, включая домен лектина (или углевод-распознающий домен лектина), эпидермального фактора роста-подобный домен (EGF), различные виды повтора консенсуса, трансмембранный домен и цитоплазменный хвост (70). Как уже отмечалось, начальные фиксация и роллинг лейкоцитов опосредованы взаимодействием Р-селектина и PSGL-1, Таким образом, блокирование функции Р-селектина при помощи (1) антител к Р-селектину, (2) антител к PSGL-1, (3) фрагментов PSGL-1 или рекомбинантных форм PSGL-1, (4) малых молекул, которые имитируют связывающий домен PSGL-1 и (5) других молекул, которые разрушают связывание Р-селектина с PSGL-1, может блокировать роллинг и фиксацию лейкоцитов и, следовательно, предотвращать прочное прилипание к клеткам эндотелия и тромбоцитов. Мыши с дефицитом Р-селектина также не в состоянии поддерживать фиксацию и роллинг лейкоцитов на поверхности клеток активированного эндотелия (72, 74). L-селектин играет двойную роль, заключающуюся в том, что он постоянно экспрессируется на поверхности циркулирующих лейкоцитов и может инициировать "вторичное связывание" путем взаимодействия с PSGL-1 на поверхности других лейкоцитов (75). Это процесс приводит к дальнейшему рекрутингу новых лейкоцитов в воспаленную область. Связывание L-селектина с PSGL-1 также играет роль в хоминге (хоуминге) лейкоцитов в наружных эндотелиальных венулах (HEV) во вторичных органах лимфатической системы (76). Е-селектин регулируется транскрипционно и экспрессируется на поверхности активированных клеток эндотелия через определенное время после событий, опосредованных Р-селектином. Е-селектин может связывать PSGL-1 с низким сродством, но также может связывать и другие лиганды. На примере отдельных мышей с "выключенным" геном (нокаутированных мышей) было показано, что эти молекулы обладают компенсаторным механизмом хоминга и роллинга лейкоцитов (77).
С учетом вышесказанного следует указать, что до сих пор существует настоятельная необходимость в разработке новых способов лечения воспалительных и тромботических заболеваний, например, при помощи антител, направленных на Р-селектиновую мишень, путем нарушения связывания Р-селектина и PSGL-1. Следовательно, целью заявленного изобретения является блокирование связывания Р-селектина с PSGL-1 с одновременным блокированием адгезии кровяных клеток, которые участвуют в процессе вазоокклюзии при наличии SCD и других тромботических заболеваний, как описано далее в данной заявке.
РАСКРЫТИЕ ИЗОБРЕТЕНИЯ
Данное изобретение использует "двойную функцию"антител, которые специфически связываются с Р-селектином и которые не только блокируют связывание PSGL-1 с Р-селектином, но и приводят также к диссоциации ранее образовавшихся комплексов Р-селектин/PSGL-1. Данное изобретение содержит описание до сих пор нераспознанного домена связывания антитела (конформационного эпитопа) с доменом лектина (например, с углевод-распознающим доменом лектина, CRD) в Р-селектине, с которым связываются антитела с двойной функцией (которые могут быть химерными, человеческими или гуманизированными или, например, их фрагментами). Следовательно, настоящее изобретение относится к антителам к Р-селектину или к их фрагментам, которые связываются с конформационным эпитопом, описанным в данной заявке, и которые выполняют двойную функцию, участвуя в (1) блокировании связывания PSGL-1 с Р-селектином и (2) в процессе диссоциации ранее образовавшихся комплексов Р-селектин/PSGL-1. В частности, данное изобретение относится к применению указанной двойной функции антител к Р-селектину или их фрагментов, описанных в данной заявке. в процессе лечения воспалительных, тромботических или других заболеваний и расстройств у приматов (включая людей), в развитии которых принимают участие тромбоцит, серповидная красная кровяная клетка, лейкоцит, лимфоцит и/или адгезивность клеток эндотелия, при этом такие заболевание или расстройство представляют собой или связаны (но без ограничения) с по меньшей мере одним вазоокклюзивным болевым кризом, воспалением кишечника (например, болезнью Крона, язвенным колитоми энтеритом), артритом (например, ревматоидным артритом, остеоартритом или псориазным артритом), отторжением имплантатов, реакцией "трансплантат против хозяина", астмой, хронической обструктивной болезнью легких, псориазом, дерматитом, сепсисом, нефритом, эритематозной волчанкой, склеродермией, ринитом, анафилаксией, диабетом, рассеянным склерозом, атеросклерозом, тромбозом, метастазированием, аллергическими реакциями, тироидитом, ишемически-реперфузионным повреждением (например, в случае инфаркта миокарда, инсульта или трансплантации органов) и состояниями, связанными с обширной травмой, хроническим воспалением, таким как, например, повышенная чувствительность замедленного типа IV, ассоциируемая, например, с инфекцией, вызванной туберкулезной бациллой, или с синдромом системного воспалительного ответа или полиорганной недостаточностью. Важно то, что применение таких антител с двойной функцией, описанных в данной заявке, или их фрагментов при лечении указанных воспалительных заболеваний позволяет не только предотвращать воспаление, но также обеспечить механизм лечения происходящих в данное время воспалительных заболеваний, заключающийся в том, что эти антитела могут приводить к диссоциации ранее образовавшихся комплексов Р-селектин/PSGL-1. Например, в случае вазоокклюзивного болевого криза, вызванного наличием серповидных клеток, антитела с двойной функцией не только ингибируют или предотвращают появление будущих вазоокклюзивных событий, но также позволяют лечить происходящую в данное время вазоокклюзию. Другие варианты данного изобретения, описанного в этой заявке, станут понятны после ознакомления со следующим ниже подробным описанием.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
На Фигуре 1 показаны данные по сравнению гомологических аминокислотных последовательностей мышиного и человеческого Р-селектина, показано расположение доменов лектина, EGF, CR1 и CR2 (переход между доменами показан стрелкой). Нелинейные конформационные домены А, В, C1, D, Е1, С2, Е2, С3 и F показаны в прямоугольниках, обозначенных пунктиром. Различие в аминокислотах указано жирным шрифтом.
Фигура 2 отражает репрезентативные данные (полученные при помощи двухстадийной биосенсорной системы BIACORE), характеризующие химерное связывание Р-селектина в случае связывания антител G1, G3, G4 и G5 с последовательностями SEQ ID NO:1-4, 7-10, 18 и 19. Следует отметить, что связывание антитела G5 при осуществлении двухстадийного метода анализа в системе BIACORE осуществлялось только с SEQ ID NOs:1, 2, 18 и 19. Антитело G4 представляет собой новое мышиное моноклональное антитело к Р-селектину, которое было продуцировано клетками гибридомы, депонированной под номером РТА-12154 в Американской коллекции типовых культур, 10801 University Blvd., Manassas, VA, 20110-2209, которая является общепризнанным депозитарием.
На Фигуре 3 приведены сенсограммы BIACORE, демонстрирующие блокирование взаимодействия Р-селектина с PSGL-1 при применении антител G1, G3, G4 и hSeI001 (hSeI001 известно также как SeIG1, гуманизированная форма G1), описанными в данной заявке способами. Показано, что антитела G1, G3, G4 и hSeI001 блокируют взаимодействие Р-селектина с гликосульфопептидом GSP-6, являющимся миметиком PSGL-1. Антитело G5 связывает Р-селектин, но не блокирует связывание. Контрольный опыт показывает устойчивое состояние связывания Р-селектина с GSP-6.
На Фигуре 4 показаны сенсограммы BIACORE, демонстрирующие диссоциацию ранее образовавшегося комплекса Р-селектин/PSGL-1 после действия антител к Р-селектину, представляющих собой антитела G1, G4 и hSeI001 с двойной функцией. PSGL-1 представлен пептидом GSP-6, являющимся миметиком PSGL-1. Первоначальный рост RU (единицы ответа) показывает связывание Р-селектина с биотинилированным GSP-6, соединенным с чипом BIACORE, содержащим покрытие из стрептавидина. Как только достигается устойчивое состояние связывания Р-селектина с GSP-6, (то есть после того, как нормальная диссоциация указанного комплекса почти достигнет равновесия) впрыскивали испытуемые антитела и оценивали характеристики диссоциации. Антитело G5 связывало предварительно образовавшийся комплекс, но не вызывало его диссоциацию. Антитело G3 не связывало этот комплекс и не вызывало диссоциацию этого предварительно образовавшегося комплекса Р-селектин/PSGL-1. Антитела G1, G4 и hSeI001 и связывали этот комплекс, и вызывали его диссоциацию, что свидетельствовало об их новой двойной функции.
На Фигуре 5 показано 3D-изображение молекулы человеческого Р-селектина, связанного с GSP-6. Домены лектина и EGF обозначены пунктирными линиями. Участок связывания 1 определяет конформационный эпитоп кластера А, который отдален от домена связывания лектин/лиганд. Испытуемое антитело G1 связывало участок 1 в кластере А.
На Фигуре 6 показаны графики, отображающие результаты in vitro способа анализа роллинга клеток в потоке человеческих нейтрофилов при наличии Р-селектина низкой и высокой плотности. Эти результаты показывают, что происходит блокирование и/или диссоциация ранее образовавшегося комплекса Р-селектин/PSGL-1 и последующее высвобождение нейтрофилов после действия антител G1, G3, G4 и hSeI001. Во время исследования антитела вводили в эквивалентных концентрациях равных 20 мкг/мл. Существовал латентный период, составлявший около 1 мин, до достижения антителами камеры из-за мертвого объема системы. Затем с интервалами в 1 мин подсчитывали количество клеток, оставшихся связанными, и выражали его в процентах связанных клеток. Панели (А) и (С) на графике показывают роллинг нейтрофилов со средними скоростями 5 мкм/с и 6,5 мкм/с, соответственно, по мембране Р-селектина низкой плотности (50 сайтов/мкм2). Панели (В) и (D) на графике показывают роллинг нейтрофилов со средними скоростями 1 мкм/с по мембране Р-селектина высокой плотности (380 сайтов/мкм2).
На Фигуре 7 показана сенсограмма, иллюстрирующая кинетику связывания антител G1 и hSeI001 с Р-селектином при одной концентрации Р-селектина.
ОСУЩЕСТВЛЕНИЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Перед раскрытием в подробностях по меньшей мере одного варианта данного изобретения путем описания примеров рисунков, экспериментов, результатов и лабораторных методов, следует указать, что данное изобретение не ограничивается подробностями состава и применения компонентов, приведенными в следующем ниже описании или проиллюстрированными рисунками, примерами и/или результатами. Данное изобретение содержит и другие варианты и может быть осуществлено различным образом. Терминология, используемая в данном описании, предназначена для описания самого широкого возможного объема изобретения и объяснения его смысла, варианты настоящего изобретения являются примерными и не исчерпывающими. Следует также иметь в виду, что фразеология и терминология, используемые в данной заявке, служат для описания, но не для ограничения настоящего изобретения.
Если иное не указано, научные и технические термины, используемые в соответствии с изобретательским замыслом, имеют значения, общепринятые среди средних специалистов в данной области. Кроме того, если из контекста не следует иное, термины в единственном числе включают и множественное число, а термины во множественном числе подразумевают и единственное число. Для рекомбинантной ДНК, синтеза олигонуклеотидов, описания тканевых культур и процессов обработки (например, электропорации, липофекции) используются стандартные методики. Ферментативные реакции и методы очистки осуществляются в соответствии с инструкциями производителей или обычными способами, известными из уровня техники или описанными в данной заявке. Описываемые ниже методики и способы обычно осуществляют общепринятыми методами, хорошо известными в уровне техники и описанными в различных общих и более конкретных источниках, которые цитируются и обсуждаются в данной заявке. См., например, источники Sambrook et al. Molecular Cloning: A Laboratory Manual (2nd ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989), а также Coligan et al. Current Protocols in Immunology (Current Protocols, Wiley Interscience (1994)), которые включены в данную заявку посредством отсылок. Системы номенклатуры и лабораторные методики в области аналитической химии, синтетической органической химии и медицинской и фармакологической химии, описанные в данной заявке, хорошо известны и общеприняты в уровне техники. При описании химического синтеза, химических анализов, получения и доставки фармацевтических композиций и лечения пациентов приведены стандартные методы.
Все публикации и патентные документы, упомянутые в данном описании, отражают уровень знаний средних специалистов в той области техники, к которой относится данное изобретение. Все эти публикации и патентные документы включены в данную заявку посредством отсылок в той степени, в которой была бы включена посредством отсылки в отдельности любая конкретная публикация. В свете данного описания все раскрытые композиции и способы, описанные и заявленные в данной заявке, могут быть осуществлены без излишнего экспериментирования. Хотя эти композиции и способы согласно настоящему изобретению описаны на примере предпочтительных вариантов, среднему специалисту в данной области ясно, что могут быть сделаны и другие изменения этих композиций и/или способов, а также стадий или их последовательности в способах, описанных в данном описании без выхода за замысел, сущность и объем настоящего изобретения. Все такие похожие изменения и модификации, очевидные для специалиста в данной области, охвачены замыслом и сущностью изобретения и входят в объем данного изобретения, который определен прилагаемой формулой изобретения.
В соответствии с данным изобретением используемые термины, если не указано иное, следует понимать следующим образом.
Применение какого-либо термина в единственном числе в сочетании с другим термином, таким как "представляющий собой" или "содержащий" ("включающий") в формуле изобретения и/или в описании, означает не только "один", но и "один или более", "по меньшей мере один" и "один или более, чем один". Использование термина "или" в формуле изобретения означает также "и/или", если при этом четко не указано, что этот термин относится только к альтернативам или эти альтернативы являются взаимоисключающими, даже если в описании есть подтверждение определения, которое относится только к альтернативам и "и/или" альтернативам.
По всему тексту данной заявки термин "примерно" или "около" используется для указания того, что эта величина включает неотъемлемое отклонение, вызванное погрешностью прибора или способа, которые применялись для определения этой величины, или же того, что среди изучаемых предметов имеются отличия. Применение термина "по меньшей мере один" подразумевает наличие одного, а также любого количества, превышающего один, в том числе, но без ограничения, 2, 3, 4, 5, 10, 15, 20, 30, 40, 50, 100 и т.д. Термин "по меньшей мере один" может распространяться до 100 или 1000 или далее, в зависимости от понятия, к которому он относится; кроме того, количество 100/1000 не рассматривается как ограничительное, так как большие пределы также могут давать удовлетворительные результаты.
Термин "примерно" или "около" используется для указания того, что эта величина включает неотъемлемое отклонение, вызванное погрешностью прибора или способа, которые применялись для определения этой величины, или же того, что среди изучаемых предметов имеются отличия.
Применяемые в данном описании и в формуле изобретения термины "охватывающий" (и любая форма этого слова, такая как "охватывают" или "охватывает"), "имеющий" (и любая форма этого слова, такая как "имеют" и "имеет"), "включающий" (и любая форма этого слова, такая как "включают" и "включает") или "содержащий" (и любая форма этого слова, такая как "содержат" и "содержит") являются неограничительными и открытыми и не исключают наличия дополнительных, не упомянутых элементов или стадий способа.
Термин "или их комбинации", используемый в данной заявке, относится ко всем сочетаниям и комбинациям перечисленных предметов, указанных перед этим выражением. Например, выражение "А, В, С или их комбинации" подразумевает включение по меньшей мере одного предмета из группы А, В, С, АВ, АС, ВС или АВС, и, если в конкретном контексте важен порядок перечисления, также ВА, СА, СВ, СВА, ВСА, АСВ, ВАС или CAB. Продолжая этот пример, следует указать, что при этом определенно указаны комбинации, которые содержат один или более повторяющихся предметов или терминов, например, ВВ, ААА, MB, ВВС, АААВСССС, СВААА, САВАВВ и т.д. Специалисту в данной области понятно, что обычно количеству предметов или терминов в любой комбинации нет предела, если только из контекста очевидным образом не следует иное.
Термины "пептид", "полипептид" и "белок", используемые в данной заявке, относятся к полимеру, состоящему из остатков аминокислот. Термин "полипептид", применяемый в данной заявке, является родовым термином, который относится к нативному белку, фрагментам белка или аналогам полипептидных последовательностей. Следовательно, нативный белок, фрагменты белка и аналоги представляют собой виды из рода полипептидов. Термин "выделенный пептид/полипептид/белок", применяемый в данной заявке, относится к пептиду/полипептиду/белку на основе кДНК, рекомбинантной ДНК и пептиду/полипептиду/белку синтетического происхождения или к некоторым их комбинациям, когда в соответствии с их происхождением или источником получения "выделенный пептид/полипептид/белок" (1) не связан с пептидом/полипептидом/белком, обнаруженными в природе, (2) не содержит других пептида/полипептида/белка из того же источника, например, мышиных белков, (3) экспрессирован клеткой различных видов и/или (4) не существует в природе.
Применяемый в настоящей заявке термин "аминокислота" охватывает все молекулы, природные или синтетические, которые включают как аминные группы, так и кислотные группы, и способны входить в состав полимера на основе аминокислот природного происхождения. Примеры аминокислот включают аминокислоты природного происхождения, их аналоги, производные и родственные соединения, аналоги аминокислот, содержащие вариантные боковые цепи, и все стереоизомеры любых указанных выше соединений. Используемый в данной заявке термин "мышиная аминокислота" носится к остатку аминокислоты, которая обнаружена в мышином Р-селектине, но не найдена в соответствующем положении человеческого Р-селектина.
Применяемые в данной заявке названия двадцати стандартных аминокислот и их аббревиатуры являются общеупотребительными. См. публикацию Immunology, A Synthesis (2nd Edition, E.S. Golub and D.R. Gren, Eds., Sinauer Associates Sunderland Mass. (1991), которая включена в данную заявку посредством отсылки. Стереоизомеры (например, D-аминокислоты) двадцати стандартных аминокислот не природного происхождения, таких как α,α-дизамещенные аминокислоты, N-алкиламинокислоты, молочная кислота и другие нестандартные аминокислоты также могут быть подходящими компонентами полипептидов согласно настоящему изобретению. Примеры нестандартных аминокислот включают 4-гидроксипролин, α-карбоксиглутамат, ε- N-ацетиллизин, O-фосфосерин, N-ацетилсерин, N-формилметионин, 3-метилгистидин, 5-гидроксилизин, сигма-Н-метиларгинин и другие похожие аминокислоты и иминокислоты (например, 4-гидроксипролин). В соответствии со стандартными правилами при индексации полипептидов левосторонне направление означает направление цепи в сторону аминоконца и правостороннее направление означает направление цепи в сторону карбоксиконца полипептида.
Используемые в данной заявке термины "полинуклеотид" и "нуклеиновая кислота" являются взаимозаменяемыми. Они относятся к полимерным формам нуклеотидов любой длины или к дезоксирибонуклеотидам или к рибонуклеотидам или к их аналогам. Неограничивающие примеры полинуклеотидов включают кодирующие или не кодирующие участки гена или фрагмента гена, локус (локусы), определенные путем анализа сцепления генов, экзоны, интроны, матричную (информационную) РНК (мРНК), транспортную РНК (тРНК), рибосомальную РНК, рибозимы, кДНК, рекомбинантные полинуклеотиды, разветвленные полинуклеотиды, плазмиды, векторы, выделенную РНК с любыми последовательностями, выделенную ДНК с любыми последовательностями, зонды для нуклеиновых кислот и праймеры. Полинуклеотид может включать модифицированные нуклеотиды, такие как метилированные нуклеотиды и нуклеотидные аналоги. Если это требуется, может осуществляться модификация нуклеотидов до или после сборки полимера. Последовательность в нуклеотидах может прерываться ненуклеотидными компонентами. Полинуклеотид может быть дальше модифицирован, например, путем добавления метки. Термины "выделенная нуклеиновая кислота" и "выделенный полинуклеотид"используются как взаимозаменяемые. Нуклеиновая кислота или полинуклеотид считаются выделенными, если они: (1) не связаны с полинуклеотидом или его частью, когда "выделенный полинуклеотид" обнаружен в природе, (2) связаны с полинуклеотидом, который не обнаружен в природе, или (3) они не встречаются в природе как часть последовательности большей длины.
Термин "вектор", применяемый в данной заявке, относится к молекуле нуклеиновой кислоты, способной транспортировать другую нуклеиновую кислоту, с которой она была связана. Одним из видов векторов является "плазмида", этот термин относится к кольцевой двухцепочечной ДНК, где в вирусный геном могут быть замкнуты дополнительные сегменты ДНК. Некоторые векторы способны к автономной репликации в хозяйской клетке, в которую они вводятся (например, бактериальные векторы, имеющие бактериальное происхождение репликации и эписомальные векторы млекопитающих). Другие векторы (например, не эписомальные векторы млекопитающих) могут быть интегрированы в геном хозяйской клетки после введения в хозяйскую клетку и, следовательно, реплицируются вместе с хозяйским геномом. Более того, некоторые векторы способны управлять экспрессией генов. Такие векторы в данной заявке называются "рекомбинантными экспрессирующими векторами" (или просто "экспрессирующими векторами").
Термин "встречающийся в природе", применяемый в данной заявке в отношении какого-либо объекта, относится к тому факту, что этот объект может быть обнаружен в природе. Например, полинуклеотид или полипептидная последовательность, которые могут иметься в организме (включая вирусы), которые могут быть выделены из природного источника и которые не были намеренно модифицированы человеком в лаборатории или еще где-либо, являются встречающимися в природе. Термины "встречающийся в природе" и "нативный" могут применяться как взаимозаменяемые.
Термин "роллинг лейкоцитов", также применяемый в данной заявке, включает явление слабой адгезии лейкоцитов в клеткам эндотелия кровеносных сосудов и прокатывание лейкоцитов вдоль эндотелиальных клеток кровеносных сосудов до возникновения прочной адгезии и трансмиграции лейкоцитов в эндотелиальную ткань. Вслед за роллингом эти адгезивные лейкоциты могут мигрировать через эндотелий и разрушать ишемическую ткань во время реперфузии. Соответственно, снижение роллинга лейкоцитов приводит к уменьшению повреждения тканей и органов, вызванного острыми воспалительными ответами.
Применяемый в настоящей заявке термин "антагонист Р-селектина" относится к любому агенту, который способен оказывать антагонистическое действие по отношению к Р-селектину, например, путем ингибирования взаимодействия между Р-селектином и гликопротеиновым лигандом-1 Р-селектина, в том числе, путем ингибирования взаимодействия клеток эндотелия, экспрессирующих Р-селектин, и активированных тромбоцитов с лейкоцитами, экспрессирующими PSGL-1.
Термины "выделенная" или "очищенная" относятся к молекуле нуклеиновой кислоты, которая практически не содержит компонентов природной среды и содержится как преобладающий компонент (например, на молярной основе), например, в количестве более 50% в расчете на композицию. Например, выделенный белок практически не содержит клеточного материала или других белков из клетки или тканевого источника, из которых он был выделен. Этот термин относится также к препаратам, в которых выделенный белок составляет, по меньшей мере, 60% (вес/вес) в чистом виде, или по меньшей мере 70% % (вес/вес) в чистом виде, или по меньшей мере, 75% (вес/вес) в чистом виде, или по меньшей мере, 80% (вес/вес) в чистом виде, или по меньшей мере, 85% (вес/вес) в чистом виде, или по меньшей мере, 90% (вес/вес) в чистом виде, или по меньшей мере, 92% (вес/вес) в чистом виде, или по меньшей мере, 95% (вес/вес) в чистом виде, или по меньшей мере, 96% (вес/вес) в чистом виде, или по меньшей мере, 97% (вес/вес) в чистом виде, или по меньшей мере, 98% (вес/вес) в чистом виде, или по меньшей мере, 99% (вес/вес) в чистом виде, или по меньшей мере, 100% (вес/вес) в чистом виде. Согласно некоторым вариантам данного изобретения выделенная молекула обладает достаточной степенью чистоты, позволяющей использовать ее в фармацевтических композициях.
Термин "ингибирующая активность" относится к степени снижения активности Р-селектина ингибитором Р-селектина (таким как антитело к Р-селектину или фрагмент этого антитела) по сравнению с активностью Р-селектина в отсутствие этого ингибитора. Нейтрализующее антитело может уменьшать один или более видов активности Р-селектина. Например, степень снижения активности Р-селектина (например, связывания Р-селектина с PSGL-1) предпочтительно составляет по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95% или более. В другом примере степень диссоциирующей активности антитела или его фрагмента, обладающих двойной функцией (а именно, %-ное содержание предварительно образовавшегося комплекса Р-селектин/PSGL-1, который подвергается диссоциации), может быть равна по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95% или более.
Термин "ингибитор Р-селектина", применяемый в данной заявке, относится к любому агенту, такому как, например, нейтрализующее антитело, способному вызывать ингибирование активности, экспрессию, процессинг, связывание и локализацию Р-селектина на поверхности клеток. Такие ингибиторы "ингибируют", "нейтрализуют" или "снижают" биологическую активность Р-селектина.
Термин "эффективное количество" относится к количеству биологически активного вещества или его конъюгата или производного, которое является достаточным для создания заметного терапевтического эффекта, предпочтительно, без проявления неблагоприятных побочных эффектов (таких как токсичность, раздражение и аллергическая реакция) при разумном отношении польза/риск при использовании согласно данному изобретению. Термин "фармацевтически приемлемые" относится к соединениям и композициям, которые пригодны для введения людям и/или животным без проявления неблагоприятных побочных эффектов (таких как токсичность, раздражение и аллергическая реакция) при разумном отношении польза/риск при использовании согласно данному изобретению. Соединения по данному изобретению могут обеспечивать отсроченное, контролируемое и пролонгированное высвобождение при применении методик их получения, которые хорошо известны из уровня техники.
Термин "эпитоп" относится к антигенной детерминанте, которая взаимодействует с сайтом связывания специфичного антигена, известным как паратоп. Эпитопы могут быть линейными или конформационными. Конформационный эпитоп образуется пространственно расположенными бок о бок аминокислотами из разных сегментов линейной цепи полипептида.
Термин "моноклональное антитело", используемый в данной заявке, относится к антителу, которое получается из популяции практически гомогенных антител, то есть, отдельные антитела, содержащиеся в этой популяции, являются идентичными за исключением наличия возможных встречающихся в природе мутаций, которые могут содержаться в незначительном количестве. Моноклональные антитела являются высоко специфическими, обычно направленными против одного эпитопного сайта. Кроме того, в противоположность обычным поликлональным антителам, которые включают разные антитела, направленные против различных детерминант (эпитопов), каждое моноклональное антитело направлено против одной детерминанты. В дополнение к их специфичности моноклональные антитела являются предпочтительными, так как согласно одному из вариантов они получаются из гибридомной культуры, не загрязненной другими иммуноглобулинами. Прилагательное "моноклональное" указывает на характер антитела, полученного из практически гомогенной популяции антител, это антитело не должно рассматриваться как требующее получения антитела конкретным способом.
АНТИТЕЛА
Антитела относятся к семейству белков плазмы, называемых иммуноглобулинами, у которых основной структурный элемент, характерная для них укладка цепи или домен в различных формах используется во многих молекулах иммунной системы и других биологических систем распознавания. Типичный иммуноглобулин содержит четыре полипептидных цепи, содержащих антиген-связывающую область, известную как вариабельная область, и неизменный участок, известный как константная область.
Нативные антитела и иммуноглобулины обычно представляют собой гетеротетрамерные гликопротеины с молекулярным весом, составляющим примерно 150000 Да, состоящие из двух идентичных легких (L) цепей и двух идентичных тяжелых (Н) цепей. Каждая легкая цепь связана с тяжелой цепью одной ковалентной дисульфидной связью, количество дисульфидных связей в тяжелых цепях разных изотипов иммуноглобулинов различно. Каждая тяжелая и легкая цепь также содержат расположенные с регулярными интервалами межцепные дисульфидные мостики. Каждая тяжелая цепь содержит на одном конце вариабельный домен (VH) и затем ряд константных доменов. Каждая легкая цепь содержит на одном конце вариабельный домен (VL) и константный домен на другом своем конце. Константный домен легкой цепи соединен с первым константным доменом тяжелой цепи, а вариабельный домен легкой цепи - с вариабельным доменом тяжелой цепи.
В зависимости от вида аминокислотных последовательностей константного домена тяжелых цепей иммуноглобулинов они могут относиться к различным классам. Известны по меньшей мере пять основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, и несколько из них в свою очередь подразделяются на подклассы (изотипы), например, IgG1, IgG2, IgG3 и IgG4, а также IgA1 и IgA2. Константные домены тяжелых цепей, которые соответствуют различным классам иммуноглобулинов, называются альфа (α), дельта (δ), эпсилон (ε), гамма (γ) и мю (µ), соответственно. Легкие цепи антител могут быть отнесены к одному из двух различающихся типов, называемых каппа и лямбда, определяемых аминокислотными последовательностями их константного домена. Структуры подъединиц и трехмерные конфигурации различных классов иммуноглобулинов хорошо известны. Термин "вариабельный" в контексте вариабельного домена антител относится к тому факту, что некоторые части вариабельного домена сильно отличаются по своим последовательностям. Вариабельные домены предназначены для связывания и определяют специфичность каждого конкретного антитела по отношению к его конкретному антигену. Однако вариабельность распределена неравномерно среди вариабельных доменов антител. Она концентрируется в трех сегментах одной цепи, которые называются определяющими комплементарность областями (CDR), известными также как гипервариабельные участки, вариабельные домены как легкой, так и тяжелой цепи. В большинстве случаев в вариабельном участке легкой цепи содержатся три CDR (CDRL1, CDRL2 и CDRL3) и три CDR находятся в вариабельном участке тяжелой цепи (CDRH1, CDRH2 и CDRH3). CDR вносят свой вклад в функциональную активность молекулы антитела и разделены аминокислотными последовательностями, которые включают поддерживающие или каркасные области. Среди различных CDR последовательности CDR3 и, в частности, CDRH3, являются самыми разнообразными и, следовательно, больше всего влияют на специфичность антител. Существуют по меньшей мере два способа определения CDR3: (1) способ, основанный на межвидовой вариабельности последовательностей (см. публикацию Kabat et al., Sequences of Proteins of Immunological Interest (National Institute of Health, Bethesda, Md. (1987), включенную в данную заявку посредством отсылки); и (2) способ, основанный на кристаллографических исследованиях комплексов антиген-антитело (см. публикацию Chotia et al., Nature, 342:877 (1989), также включенную в данную заявку посредством отсылки).
Более высоко консервативные части вариабельных доменов называют остовом или каркасными областями (FR). Вариабельные домены нативных легких и тяжелых цепей каждый включают четыре FR области, в основном принимающие конфигурацию β-слоя, связанные тремя CDR, которые формируют петли, соединяющие и в некоторых случаях образующие часть структуры β-слоя. CDR в каждой легкой и тяжелой цепи удерживаются вместе в непосредственной близости от FR областей и участвуют в образовании антиген-связывающего сайта антитела. Константные домены не участвуют непосредственно в связывании антитела с антигеном, но проявляют различные эффекторные функции, такие как участие антитела в антитело-зависимой клеточной токсичности.
Таким образом, в соответствии с данным изобретением антитело может быть в любой из различных форм, включая целый иммуноглобулин, фрагмент антитела, такой как Fv, Fab и похожие фрагменты, единичную цепь антитела, которая содержит определяющие комплементарность области вариабельных доменов (CDR), и подобные формы, все из которых подпадают под общее понятие "антитело", применяемое в данной заявке. Согласно предпочтительным вариантам в контексте терапевтических способов и способов скрининга, описанных ниже, применяется антитело или его фрагмент, которые являются иммуноспецифичными по отношению к антигену или эпитопу по изобретению, описанным в данной заявке.
Термин "фрагмент антитела", используемый в данной заявке, относится к части антитела полной длины, обычно к антиген-связывающему или вариабельному участку. Примеры фрагментов антител включают Fab, Fab′, F(ab′)2 и Fv. Расщепление антител папаином приводит к получению двух идентичных антиген-связывающих фрагментов, называемых фрагментами Fab, каждый из которых содержит один антиген-связывающий сайт и остаточный "Fc"-фрагмент, называемый так из-за его способности к легкой кристаллизации. Обработка пепсином приводит к получению фрагмента F(ab′)2, который содержит два антиген-связывающих фрагмента, которые способны к сшиванию антигена, и остаточный фрагмент (который обозначается pFc′). Дополнительные фрагменты могут включать диатела, линейные антитела, молекулы одноцепочечных антител, а также мультиспецифичные антитела, образовавшиеся из фрагментов антител. Используемый в данной заявке термин "функциональный фрагмент" в случае антител относится к фрагментам Fv, Fab и F(ab′)2. Фрагменты антител по данному изобретению могут быть такими малыми, что они содержат, например, примерно 4-29 или от 30 до 35, до 40, до 45, до 50, до 75, до 100 или до 150-200, до 250 (включительно) или более аминокислот.
Некоторые типы фрагментов антитела определяются следующим образом:
Fab представляет собой фрагмент, который содержит одновалентный антиген-связывающий фрагмент антитела. Фрагмент Fab может быть получен путем расщепления целого антитела ферментом папаином с образованием интактной легкой цепи и части одной тяжелой цепи.
Fab′ является фрагментом молекулы антитела, который может быть получен путем обработки целого антитела пепсином с последующим восстановлением с образованием интактной легкой цепи и части тяжелой цепи. На одну молекулу антитела получаются два фрагмента Fab′.
Fab′-фрагменты отличаются от Fab-фрагментов наличием нескольких добавленных остатков на карбоксильном конце домена СН1 тяжелой цепи, включая один или более остатков цистеина из шарнирной области антитела.
F(ab′)2 представляет собой фрагмент антитела, который может быть получен путем обработки целого антитела ферментом пепсином без последующего восстановления. F(ab′)2 является димером двух фрагментов F(ab′), которые удерживаются вместе двумя дисульфидными связями.
Fv представляет собой фрагмент антитела минимального размера, который содержит полный центр распознавания и связывания антигена. Эта область состоит из димера вариабельных доменов одной тяжелой и одной легкой цепи в тесной нековалентной связи (димер VH-VL). именно в этой конфигурации три CDR каждого вариабельного домена взаимодействуют между собой, определяя антиген-связывающий сайт антитела на поверхности димера VH-VL. Шесть CDR вместе наделяют антитело специфичностью связывания. Однако даже один вариабельный домен (или половина Fv, содержащая только три CDR, специфичные к антигену) имеет способность распознавать и связывать антиген, хотя и с меньшим сродством, чем полный центр связывания.
Одноцепочечные антитела (SCA) в контексте данной заявке определяются как генетически сконструированные молекулы, содержащие вариабельную область легкой цепи и вариабельную область тяжелой цепи, соединенные подходящим полипептидным линкером с образованием генетически слитой одноцепочечной молекулы. Такие одноцепочечные антитела называются также фрагментами "одноцепочечный Fv" или "sFv" или "scFv". Обычно полипептид Fv содержит также полипептидный линкер между доменами VH и VL, который обеспечивает образование из sFv желательной для связывания структуры.
Согласно одному варианту данного изобретения оно относится к антителам, которые специфически связывают человеческий Р-селектин. CDR в таких антителах не ограничены конкретными последовательностями в областях VH и VL и могут включать варианты этих последовательностей, которые сохраняют способность вызывать блокирование и диссоциацию связывания Р-селектина с PSGL-1/ Такие варианты могут быть легко получены специалистом в данной области хорошо известными способами. Например, FR и/или CDR могут содержать замены, делеции и добавления аминокислот, как описано далее в данной заявке.
Хотя изменения в FR обычно осуществляют для повышения стабильности и снижения иммуногенности антитела, изменения CDR обычно нужны для увеличения сродства антитела к его мишени. Варианты FR включают также аллотипы иммуноглобулинов, встречающиеся в природе. Такие изменения, увеличивающие сродство антитела, могут быть определены эмпирически обычными методами, которые включают изменение CDR и определение сродства антитела к его мишени.
Например, консервативные замены аминокислот могут быть произведены в любой из описанных CDR. Различные изменения могут быть сделаны способами, которые хорошо известны специалистам в данной области (78). Они включают, но без ограничения, изменения в нуклеотидных последовательностях, которые осуществляются путем замены различных кодонов, которые кодируют идентичный или функционально эквивалентный остаток аминокислоты ("консервативные замены") в последовательности, при этом получается "молчащее" изменение. Например, неполярные аминокислоты могут быть консервативно заменены на любую другую кислоту, включают аланин, лейцин, изолейцин, валин, пролин, фенилаланин, триптофан и метионин. Полярные нейтральные аминокислоты, которые могут быть консервативно заменены на любую другую кислоту, включают глицин, серин, треонин, цистеин, тирозин, аспарагин и глутамин. Положительно заряженные (основные) аминокислоты, которые могут быть консервативно заменены на любую другую кислоту, включают аспартовую кислоту и глутаминовую кислоту. Заменяющие аминокислоты в последовательности могут быть также выбраны из других членов класса, к которому принадлежит аминокислота.
Производные и аналоги антител согласно данному изобретению могут быть получены способами, которые хорошо известны в уровне техники, включая рекомбинантный и синтетический способы (79, 80). Антитела, в которых последовательности CDR отличаются только несущественно и консервативно от последовательностей вариабельных областей антител к Р-селектину, таких как hSeI001, описанных подробно ниже, также входят в объем данного изобретения. Как отмечено выше, обычно аминокислоту заменяют родственной аминокислотой, имеющей подобные заряд, гидрофобные и стереохимические свойства. Такие замены являются очевидными для среднего специалиста в данной области. Кроме того, специалисту известно, что такие изменения могут быть сделаны в FR без неблагоприятного изменения связывающих свойств антитела. Изменения в FR включают, но без ограничения, гуманизацию некоторых не человеческих или сконструированных остатков в каркасных областях, которые важны для контакта с антигеном или для стабилизации сайта связывания, например, путем изменения класса или подкласса константной области, изменения конкретных остатков аминокислот, которые могут изменить эффекторную функцию, такую как связывание Fc-рецептора.
Согласно одному варианту данное изобретение относится к антителам, которые специфически связываются с Р-селектином, при этом CDR родительского антитела нечеловеческого происхождения прививаются к FR человеческих акцепторных антител, этот процесс называется "гуманизацией" антител. Процесс гуманизации антител предназначен для снижения иммуногенности антитела нечеловеческого происхождения при одновременном поддержании в максимально возможной степени, насколько это возможно, уровня исходного сродства. Согласно одному из вариантов человеческие FR тяжелой цепи и/или легкой цепи представляют собой немутированные аминокислотные последовательности зародышевых линий, из которых они получены. Ожидается, что такие FR будут неиммуногенными ввиду того, что эти последовательности имеются в организме всех людей до перестройки антител или созревания их сродства. Кроме того, аминокислотные остатки в таких FR антитела, особенно остатки смежные с CDR или расположенные вблизи CDR, могут потребовать замены аминокислот для лучшей сохранности сродства антитела к связыванию. Например, когда ключевые аминокислоты вариабельной FR родительского мышиного моноклонального антитела и человеческой акцепторной вариабельной FR отличаются друг от друга, аминокислоты человеческой FR в этих положениях могут быть заменены мышиными аминокислотными остатками, тем не менее, ожидается, что FR тяжелой и легкой цепи могут не содержать аминокислотных замен, и антитела, полученные при такой гуманизации, могут обладать большей частью, если не всей, величины сродства родительского антитела. Кроме того, в соответствии с данным изобретением антитела могут содержать человеческие FR VH и/или VL, которые представляют собой последовательности только зародышевой линии и не содержат замен в одной или обеих из этих последовательностей в FR.
Применяемый в данной заявке термин "сродство" антитела к Р-селектину или его конформационному эпитопу определяет его Kd или константу диссоциации. Более высокое сродство антитела характеризуется меньшей величиной Kd, в то время как более низкое сродство характеризуется большей величиной Kd. Как таковое, антитело в соответствии с настоящим изобретением предпочтительно имеет сродство к конформационному эпитопу Р-селектина, характеризующееся Kd≤1000 нМ или 500≤ нМ, или ≤100 нМ, или ≤50 нМ, или более предпочтительно Kd≤10 нМ, или еще более предпочтительно Kd≤5 нМ, или ≤1 нМ, или ≤0,1 нМ.
Термин "гомолог" антитела или его фрагмента согласно данному изобретения означает, что релевантная аминокислотная последовательность (предпочтительно, находящаяся, например, в CDR и/или вариабельных доменах VH и/или VL) белка или пептида идентична данной последовательности по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или на 100%. Например, такие последовательности могут быть вариантами, полученными из различных видов, или же гомологическая последовательность может быть получена рекомбинантным способом. Такая последовательность может быть получена путем процессинга, делеции, аминокислотной замены и добавления аминокислоты. Процент идентичности двух аминокислотных последовательностей определяется при помощи стандартных алгоритмов выравнивания, например, алгоритма Basic Local Alignment Tool (BLAST) и других алгоритмов выравнивания и способов, которые хорошо известны из уровня техники (81, 84).
Получение моноклональных антител хорошо известно среднему специалисту в данной области техники. Моноклональные антитела могут быть выделены из гибридомных культур с последующей очисткой различными хорошо известными методами. Такие методы выделения включают аффинную хроматографию с носителем белок А-сефароза, гель-хроматографию и ионообменную хроматографию. В случае альтернативного способа получения гибридом, секретирующих моноклональные антитела, моноклональное антитело к Р-селектину может быть идентифицировано и выделено путем комбинаторного скрининга библиотеки рекомбинантных иммуноглобулинов (например, библиотеки с фаговым дисплеем антител) с конформационными эпитопами, описанными в данной заявке в отношении выделенных при этом членов библиотеки иммуноглобулинов, которые связывают Р-селектин в соответствии с данным изобретением. Наборы для получения и скрининга библиотек с фаговым дисплеем являются коммерчески доступными (например, Pharmacia Recombinant Phage Antibody System, Catalog №27-9400-01 and the Stratagene SurJZAP™/Phage Display Kit, Catalog №240612).
In vitro и in vivo способы манипуляции с моноклональными антителами хорошо известны специалистам в данной области. Например, моноклональные антитела, которые можно применять в соответствии с данным изобретением, могут быть получены методом гибридомной технологии, впервые описанным Kohler and Milstein (85) или же рекомбинантным способом, например, описанным в патенте США №4816567.
Другой метод включает гуманизацию моноклонального антитела при помощи рекомбинантных средств для получения антител, содержащих, например, человеческие специфические и распознаваемые последовательности или такие последовательности из приматного источника.
Способы получения антител согласно данному изобретению, которые в высоким сродством связывают Р-селектин человеческого происхождения или его эпитопы, как описано в данной заявке, могут включать трансфекцию конструкта ДНК в клетку, при этом этот конструкт включает последовательность ДНК, кодирующую по меньшей мере часть специфических антител, нейтрализующих Р-селектин согласно изобретению, культивирование клетки в таких условиях, когда белок антитела экспрессируется клеткой, и выделение белка антитела.
Предпочтительно, когда константная область модифицирована для модулирования (а именно, ослабления или усиления) эффекторной функции по сравнению с эффекторной функцией Fc-области в тяжелой цепи иммуноглобулина дикого типа. Согласно различным вариантам константная область IgG обладает ослабленной эффекторной функцией или же она проявляет усиленную функцию. Эффекторная функция Fc включает, например, антитело-зависимую клеточную токсичность (ADCC), фагоцитоз, комплемент-зависимую клеточную токсичность и период полужизни и клиренс антитела. Аминокислотная последовательность Fc-домена в IgG может быть изменена с целью влияния на связывание с гамма-рецепторами Fc (и таким образом, на функцию ADCC или фагоцитоза) или изменения взаимодействия с системой комплемента (функция комплемент-зависимой клеточной токсичности).
Согласно одному из вариантов антитело по изобретению включает константную область или участок Fc, которые обладают низким сродством к по меньшей мере одному Fc-рецептору или не имеют такого сродства. По альтернативному варианту другой полипептид имеет низкое сродство к комплементному белку C1q или не имеет такого сродства. Сродство к связыванию обычно меняется при модификации эффекторного сайта связывания молекулы. Описание модификаций Ig, которые приводят к изменению взаимодействия с эффекторными молекулами, такими как Fc-рецептор, можно найти в патентах США №№5624821 и 5648260.
Согласно данному изобретению антитела-белки могут быть получены способами, которые хорошо известны из уровня техники. Например, эти белки могут быть получены по рекомбинантной технологии в клетках (79, 86). При рекомбинантном получении полинуклеотидная последовательность, кодирующая антитело, инсерцируется (включается) в соответствующий носитель экспрессивности, такой как вектор, который содержит необходимые элементы для транскрипции и трансляции инсерцированной кодирующей последовательности. Вектор экспрессивности затем трансфецируется в подходящую клетку-мишень, которая экспрессирует пептид. Методы трансфекции, известные из уровня техники, включают, но без ограничения, осаждение фосфатом кальция (84) и электропорацию (88). Для экспрессии белков-антител, описанных в данной заявке, могут быть использованы различные системы клетка-хозяин-экспрессирующий вектор, предпочтительно включающие эукариотные экспрессионные системы.
Данное изобретение предусматривает также выделенные нуклеиновые кислоты, кодирующие антитела, описанные в данной заявке. Эти нуклеиновые кислоты могут включать ДНК или РНК или могут быть полностью или частично синтетическими или рекомбинантными. Нуклеотидная последовательность в соответствии с данным изобретением является специфической последовательностью молекулы РНК, в которой U (урацил) заменен на Т (тимин), если только из контекста не следует иное.
Согласно другому варианту данного изобретения молекулы нуклеиновой кислоты, которые кодируют антитела по изобретению, включают также последовательности, которые идентичны описанным в данной заявке последовательностям по меньшей мере на 50%. Предусмотрены также варианты, согласно которым такие последовательности идентичны описанным в данной заявке последовательностям по меньшей мере на 60%, идентичны по меньшей мере на 70%, идентичны по меньшей мере на 75%, идентичны по меньшей мере на 80%, идентичны по меньшей мере на 85%, идентичны по меньшей мере на 90%, идентичны по меньшей мере на 91%, идентичны по меньшей мере на 91%, идентичны по меньшей мере на 92%, идентичны по меньшей мере на 93%, идентичны по меньшей мере на 94%, идентичны по меньшей мере на 95%, идентичны по меньшей мере на 96%, идентичны по меньшей мере на 97%, идентичны по меньшей мере на 98%, идентичны по меньшей мере на 99%, идентичны по меньшей мере на 99,5%, и/или которые гибридизуются с последовательностями, раскрытыми в данной заявке, в строгих условиях высокой или умеренной строгости. Процент идентичности последовательностей может быть определен путем визуального осмотра или математического расчета.
Термин "строгость", включая выражение "высокая строгость", используемое в данной заявке, обозначает условия гибридизации, которые легко определяются специалистом в данной области на основе, например, длины ДНК. Обычно эти условия определяются как условия гибридизации в присутствии 50%-ного раствора формамида, 6×SSC, при температуре 42°С (или похожего раствора для гибридизации, такого как, например, раствор Старка, содержащий 50%-ный раствор формамида при температуре 42°С) и с промывкой при температуре около 68°C с 0,2×SSC и 0,1% SDS. Специалисту в данной области известно, что температуру и концентрацию соли в промывочном растворе можно регулировать в зависимости от различных факторов, таких как длина зонда.
Термин "умеренная строгость", применяемый в данной заявке, включает условия гибридизации, которые легко определяются специалистом в данной области на основе, например, длины ДНК. Основные условия такой гибридизации были описаны Sambrook et al. (79) и включают применение раствора для предварительной промывки на нитроцеллюлозных фильтрах в присутствии 5×SSC, 0,5% SDS, 1,0 мМ ЭДТК (рН 8,0), гибридизацию в присутствии 50%-ного раствора формамида, 6×SSC, при температуре 42°С (или похожего раствора для гибридизации, такого как, например, раствор Старка, содержащий 50%-ный раствор формамида при температуре 42°С) и промывку при температуре 60°С в присутствии 0,5×SSC, 0,1% SDS.
Моноклональные антитела согласно данному изобретению включают "химерные" антитела, в которых часть тяжелой или легкой цепи идентична соответствующей последовательности в антителах, полученных из конкретных биологических видов или принадлежащих к конкретному классу или подклассу антител, или гомологичны им, а также фрагменты таких антител до тех пор, пока они проявляют желаемую биологическую активность (см. патент США №4816567).
Способы получения фрагментов антител также известны из уровня техники (89) (этот источник включен в данную заявку посредством отсылки). Фрагменты антител в соответствии с данным изобретением могут быть получены путем протеолитического гидролиза антитела или путем экспрессии ДНК, кодирующей эти фрагменты, в клетках Е.coli. Фрагменты антител, как отмечалось выше, могут быть также получены путем расщепления целых антител при помощи пептина или папаина обычными методами. Например, фрагменты антитела могут быть получены ферментативным расщеплением антител пепсином с образованием фрагмента 5S, обозначаемого как F(ab′)2. Этот фрагмент в свою очередь может быть расщеплен с помощью тиольного восстанавливающего агента и необязательной блокирующей группы для сульфгидрильных групп, образовавшихся при расщеплении дисульфидных связей с образованием одновалентных фрагментов Fab′-фрагментов 3.5S или же ферментативным расщеплением пепсином, непосредственно приводящим к получению двух одновалентных фрагментов Fab′. Эти методы описаны, например, в патенте США №4036945 и в патенте США №4331647 и в источниках, указанных в данной заявке и включенных в нее посредством отсылок.
Другие способы расщепления антител, такие как выделения тяжелых цепей с образованием одновалентных фрагментов или другие ферментативные, химические и генетические способы также могут быть использованы, если эти фрагменты связывают конформационный эпитоп, который распознается интактным антителом. Например. фрагменты Fv включают ассоциацию VH и VL, связанных пептидным линкером. Эти одноцепочечные белки, связывающие антигены (sFv), получают путем конструирования структурного гена, содержащего ДНК-последовательности, кодирующие домены VH и VL, связанные олигонуклеотидом. Структурный ген затем инсерцируется в экспрессирующий вектор, который вводится затем в хозяйскую клетку, такую как клетка E.coli. Рекомбинантные клетки хозяина синтезируют одну полипептидную цепь с пептидным линкером, связывающим два вариабельных домена. Другая форма получения фрагмента антитела представляет собой пептидное кодирование одной области CDR. CDRV пептидов ("минимальные единицы распознавания") часто участвуют в распознавании и связывании антигена. CDR пептидов могут быть получены путем клонирования или конструирования генов, кодирующих CDR нужного антитела. Такие гены получают, например, используя полимеразную цепную реакцию для синтеза вариабельной области из клеток РНК, продуцирующих антитела.
Данное изобретение относится также к сконструированным антителам, включающим полностью человеческие и гуманизированные формы нечеловеческих антител (например, приматного или мышиного происхождения). Такие гуманизированные антитела представляют собой химерные иммуноглобулины, цепи иммуноглобулинов или их фрагменты (такие как Fv, Fab, Fab′, F(ab′)2 или другие антиген-связывающие последовательности антител), которые содержат минимальные последовательности, происходящие из иммуноглобулинов нечеловеческого происхождения. Гуманизированные антитела большей частью представляют собой иммуноглобулины человеческого происхождения, в которых остатки в CDR человеческого акцепторного антитела заменены остатками CDR донорского антитела нечеловеческого происхождения, такого как мышиное, крысиное и кроличье, обладающего желаемыми специфичностью, сродством и антиген-связывающей способностью. Примером гуманизированного антитела по изобретению является гуманизированнон антитело, содержащее CDR антитела G4 и последовательности каркасной области антитела человеческого происхождения, которые гомологичны последовательностям каркасной области антитела G4.
При получении сконструированного антитела ДНК-последовательность, кодирующая молекулу антитела согласно данному изобретению, получается стандартными методами синтеза. Например, в соответствии с фосфоамидиновым способом синтезируются олигонуклеотиды, например, в автоматическом синтезаторе ДНК, затем они очищаются, ренатурируются, лигируются и клонируются в подходящих векторах. Вслед за этим ДНК-последовательность может быть введена в рекомбинантный экспрессирующий вектор, который может представлять собой любой вектор, который затем может быть подвергнут рекомбинации. Выбор вектора часто зависит от клетки-хозяина, куда он вводится, то есть он подгоняется к клетке-хозяину. Таким образом, вектор может быть автономно реплицирующимся вектором, то есть вектором, который существует как внехромосомная единица, репликация которой не зависит от хромосомной репликации плазмиды. Или же вектор может быть таким вектором, который, будучи введен в клетку-хозяина, интегрируется в геном хозяйской клетки и реплицируется вместе с хромосомой(-ами), в которую он встроился.
В векторе последовательность ДНК, кодирующая белок, должна быть функционально связана с последовательностью подходящего промотора. Промотор может быть любой ДНК-последовательностью, которая обладает транскрипционной активностью в выбранной хозяйской клетке и может быть получена из генов, кодирующих белки, либо гомологичные, либо гетерологичные хозяйской клетке. Примеры подходящих промоторов, подходящих для управления транскрипцией кодирующей ДНК-последовательности в клетках млекопитающих, включают, но без ограничения, промотор LTR, промотор SV40, промотор МТ-1 (металотиениновый промотор) или главный поздний промотор аденовируса 2. Подходящий промотор, используемый в клетках насекомых представляет собой полиэдриновый промотор. Промоторы, которые подходят для применения в дрожжевых клетках-хозяевах, представляют собой промоторы гликолитических генов дрожжей или генов алкогольдегидрогеназы, или промоторы TPI1 или ADH2-4c. Для использования в клетках-хозяевах нитчатых грибов подходит промотор ADH3 или промотор tpiA.
Кодирующая ДНК-последовательность может быть также функционально связана с подходящим терминатором, таким как терминатор гормона роста человека или (в случае грибных клеток-хозяев) промоторы TPI1 или ADH2-4c. Далее, вектор может включать такие элементы, как сигналы полиаденилирования (например, из энхансера SV40 или из области 5 ELB аденовируса) и трансляционные последовательности энхансера (например. последовательности, кодирующие РНК аденовируса VA).
Рекомбинантный экспрессирующий вектор может также включать ДНК-последовательность, способствующую репликации указанной клетки-хозяина. Примером такой последовательности (когда клеткой-хозяином является клетка млекопитающего) является источник репликации ДНК вируса SV 40. Такой вектор может также содержать селектируемый маркер, например, ген, продукт которого компенсирует дефект в клетке-хозяине такой как ген, кодирующий дигидрофолатредуктазу (DHFR) или ген, который придает резистентность к лекарству, например, неомицину, гидроксимицину или метотрексату. Методы, применяемые для лигирования последовательностей ДНК, кодирующих белки, промотор и терминатор, соответственно, и для инсерции их в подходящие векторы, содержащие информацию, необходимую для репликации, хорошо известны специалистам в данной области.
Для получения рекомбинантных белков в соответствии с данным изобретением кодирующие ДНК-последовательности могут быть эффективно слиты со второй последовательностью, кодирующей пептид, и кодирующей последовательностью сайта расщепления протеазы с образование конструкта ДНК, кодирующего слитый белок, при этом кодирующая последовательность сайта расщепления протеазы расположена между фрагментом НВР и второй ДНК, кодирующей пептид, инсерцированной в рекомбинантный экспрессирующий вектор и экспрессированной в рекомбинантных хозяйских клетках. Согласно одному из вариантов данного изобретения указанный второй пептид, выбран, но без ограничения, из группы, включающей глютатион-S-редуктазу, телячий тимозин, бактериальный тиоредоксин или природные или синтетические варианты убиквитина или их пептиды. Согласно другому варианту изобретения пептидная последовательность, содержащая сайт для расщепления протеазы, может представлять собой аминокислотную последовательность IEGR распознавания фактора свертывания крови Ха, аминокислотную последовательность DDDDK, представляющую собой сайт распознавания энтерокиназы, аминокислотную последовательность LVPR/GS сайта распознавания тромбина или аминокислотную последовательность XKX сайта распознавания Acharombacter lyticus.
Клетка-хозяин, в которую вставляется экспрессирующий вектор, может быть любой клеткой, которая способна экспрессировать пептиды или белки полной длины и является, предпочтительно, эукариотной клеткой, такой как клетки беспозвоночных (насекомых) или клетки позвоночных, например, ооциты Xenopus laevis или клетки млекопитающих, в частности, клетки насекомых и млекопитающих. Примеры подходящих клеток млекопитающих включают, но без ограничения, клетки линий HEK293 (АТСС CRL-1573), COS (АТСС CRL-1650), BHK (АТСС CRL-1632, АТСС CCL-10) или СНО (АТСС CCL-61). Способы трансфецирования клеток млекопитающих и экспрессии ДНК-последовательностей, введенных в клетку, хорошо известны из уровня техники. Или же в качестве клеток-хозяев могут быть использованы клетки грибов (включая клетки дрожжей). Примеры подходящих клеток дрожжей включают клетки Saccharomyces spp. или Schizosaccharomyces spp., в частности, штаммы дрожжей Saccharomyces cerevisiae. Примерами других клеток грибов служат клетки нитчатых грибов, например, Aspergillus spp. или Neurospora spp., в частности, штаммы грибов Aspergillus oryzae или Aspergillus niger. Применение грибов Aspergillus spp. для осуществления экспрессии белков описано, например, в патенте ЕР №238023.
Среда, используемая для культивирования клеток, может быть любой стандартной средой, которая подходит для выращивания клеток млекопитающих, такая как среда, содержащая или не содержащая сыворотку, включающая соответствующие добавки, или среда, пригодная для выращивания клеток насекомых, дрожжей или грибов или любой клетки, применяемой для экспрессирования белков. Подходящие среды доступны у частных поставщиков или могут быть приготовлены в соответствии с описанными методиками.
Рекомбинантные белки, продуцируемые клетками, затем могут быть выделены из культуральной среды обычными способами, включая выделение клеток-хозяев из среды путем центрифугирования или фильтрования, осаждения белковоподобных компонентов из надосадочной жидкости или фильтрата при помощи соли, например, сульфата аммония, очистки различными хроматографическими методами, например, методами ВЭЖХ, ионообменной хроматографии, аффинной хроматографии или т.п. Например, как только антитела были получены, отдельные В-клетки были идентифицированы и/или моноклональные антитела были получены, получали последовательности, кодирующие вариабельные области этих антител. Последовательности вариабельных областей могут быть, например, получены путем секвенирования белка антитела, продуцированного гибридомой, В-клеткой или фагом и детерминации кодирующей последовательности нуклеиновой кислоты. Согласно одному из вариантов данного изобретения вместо этого может быть секвенирована вариабельная область (VH и VL) ДНК или кДНК иммуноглобулина. В том случае, когда антитело получено из клеточной линии гибридомы или выделенной В-клетки, кДНК, кодирующие вариабельные области, могут быть амплифицированы при помощи ПЦР способами, описанными, например, в публикации Babcook et al. (Proc. Natl. Acad. Sci. USA, 93:7843-7848 (1996)) и в опубликованной международной заявке № WO 92/02551. Содержание обоих этих источников полностью включено в данную заявку посредством отсылки.
Термин "химерное антитело" относится к антителу, полученному из компонентов из по меньшей мере двух источников. Согласно некоторым вариантам химерное антитело включает часть антитела, полученного из первого вида живых существ, слитую с другой молекулой, например, частью антитела, полученного из второго вида живых существ. В соответствии с некоторыми такими вариантами химерное антитело содержит часть антитела, полученного от животного, не являющегося человеком, слитую с частью антитела, полученного от людей. Согласно другим таким вариантам химерное антитело содержит всю вариабельную область антитела, полученного от одного животного или его часть, слитые с частью антитела от второго животного. Например, без всякого ограничения, химерное антитело может включать всю вариабельную область антитела, полученного от животного, не являющегося человеком, или ее часть, слитые с константной областью антитела, полученного от человека.
Применение моноклональных антител согласно настоящему изобретению может потребовать их введения субъекту, такому как, но без ограничения, человек. Однако, когда моноклональные тела продуцируются в организме животного, не являющегося человеком, такого как грызун, введение таких антител человеку обычно будет вызывать иммунный ответ, причем этот иммунный ответ направлен против последовательности введенных антител. Такие реакции ограничивают продолжительность и эффективность применяемой терапии. Для того, чтобы решить указанную проблему, моноклональные тела по данному изобретению "гуманизируют", то есть конструируют такие антитела, в которых одна или более антигенных частей удалены и вместо этого заменены подобными частями человеческого антитела, в то время как сродство антител к желательному эпитопу сохраняется. Это конструирование может затрагивать только несколько аминокислот или может включать все каркасные области антитела, оставляя только области, определяющие комплементарность интактного антитела. Несколько способов гуманизации антител известны из уровня техники и описаны в следующих патентах США: в патенте №6180370 на имя Queen et al., выданном 30 января 2001 г.; в патенте №6054927, выданном на имя Brickell 25 апереля 2000 г.; в патенте №5869619, выданном на имя Studnicka 9 февраля 1999 г.; в патенте №5861155, выданном на имя Lin 19 января 1999 г.; в патенте №5712120, выданном на имя Rodriquez et al. 27 января 1998 г.; и в патенте №4816567, выданном на имя Cabilly et al. 28 марта 1989 г., содержание которых полностью включено в данную заявку посредством отсылок.
Как упомянуто выше, термин "гуманизированное антитело" относится к антителу нечеловеческого происхождения, которое было модифицировано таким образом, что оно близко соответствует (по аминокислотной последовательности) антителу человеческого происхождения. Как описано выше, антитела взаимодействуют с целевыми антигенами, в основном по аминокислотным остаткам, которые расположены в определяющих комплементарность областях тяжелой и легкой цепи (CDR). По этой причине аминокислотные последовательности с CDR у отдельных антител могут отличаться от последовательностей, расположенных вне CDR. Ввиду того, что последовательности в CDR отвечают за большую часть взаимодействий антитело-антиген, можно экспрессировать рекомбинантные антитела, которые имитируют свойства специфических антител природного происхождения путем конструирования экспрессирующих векторов, в которых последовательности в CDR антитела природного происхождения привиты на последовательности каркасных областей другого антитела с другими свойствами, таких как каркасные области антитела человеческого происхождения. Такие последовательности каркасных областей могут быть получены из публичных баз ДНК-данных или опубликованных источников, которые включают последовательности гена антитела зародышевой линии. Например, последовательности ДНК зародышевой линии для генов вариабельных областей тяжелой и легкой цепи могут быть найдены в базе данных "VBase" последовательностей зародышевой линии человека (доступной в Интернете на сайте mrc-cpe.cam.ac.uk/vbase), а также в публикациях Kabat, E. A., et al. (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242; Tomlinson, I. M., et al. (1992) "The Repertoire of Human Germline VH Sequences Reveals about Fifty Groups of VH Segments with Different Hypervariable Loops" J. Mol. Biol. 227:776-798; и Сох, J. P. L. et al. (1994) "A Directory of Human Germline VH Segments Reveals a Strong Bias in their Usage" Eur. J. Immunol. 24:827-836; содержание которых полностью включено в данную заявку посредством отсылки.
Гуманизированные формы антител представляют собой иммуноглобулины, цепи иммуноглобулинов или их фрагменты (такие как Fv, Fab, Fab′, F(ab′)2 и другие антиген-связывающие последовательности антител), которые в основном образованы последовательностью человеческого иммуноглобулина и содержат минимальную последовательность, полученную из иммуноглобулина нечеловеческого происхождения. Гуманизация может быть проведена в соответствии со способом Winter и его сотрудников (Jones et al., 1986; Riechmann et al., 1988; Verhoeyen et al., 1988) путем замены последовательностей в CDR грызунов соответствующими последовательностями человеческого антитела. (См. также патент США №5225539). В некоторых случаях остатки в каркасной области Fv иммуноглобулина человеческого происхождения заменяются соответствующими остатками нечеловеческого происхождения. Гуманизированные антитела могут также содержать остатки, которые не встречаются ни в антителе реципиента, ни в последовательностях импортированных CDR или каркасных областей. В целом, Гуманизированные антитела будут включать практически все из по меньшей мере одной и обычно двух вариабельных областей, в которых все или практически все из областей CDR соответствуют таким областям в нечеловеческом иммуноглобулине и все или практически все из каркасных областей представляют собой каркасные области консенсусной последовательности человеческого иммуноглобулина.
Оптимально, когда гуманизированное антитело содержит также часть константной области иммуноглобулина (Fc), обычно часть константной области человеческого иммуноглобулина (Jones et al., 1986; Riechmann et al., 1988; and Presta, 1992).
Настоящее изобретение относится также к применению полностью человеческих моноклональных антител. Полностью человеческие моноклональные антитела в принципе представляют собой молекулы антител, в которых полная последовательность и легкой цепи, и тяжелой цепи, включая CDR, происходит из генов человека. Такие антитела называют в данной заявке "человеческими антителами" или "полностью человеческими антителами". "Человеческие антитела" содержат последовательности антитела человеческого происхождения и не содержат последовательностей антитела от животного, не являющегося человеком. В соответствии с некоторыми вариантами человеческое антитело может также включать синтетические последовательности, не обнаруживаемые в нативных антителах. Этот термин не ограничен способом, который применяется для получения антител.
Человеческие моноклональные антитела могут быть получены с применением триом, по гибридомной технологии из человеческих В-клеток (см. Kozbor et al., Hybridoma, 2:7 (1983)) и EBV-гибридомной технологии получения моноклональных антител (см. et al., PNAS, 82:859 (1985)).
Кроме того, человеческие антитела могут быть получены путем введения локуса человеческого иммуноглобулина в организм трансгенных животных, например, мышей, у которых эндогенные гены иммуноглобулина были частично или полностью инактивированы. За продуцированном человеческих антител осуществляется наблюдение, что очень напоминает наблюдение за людьми во всех отношениях, включая перегруппировку генов, сборку и репертуар антител. Этот подход описан, например, но без всякого ограничения, в следующих источниках: в патентах США №№5545807; 5545806; 5569825; 5625126; 5633425; 5661016, и в публикациях Marks et al., J Biol. Chem., 267:16007 (1992); Lonberg et al., Nature, 368:856 (1994); Morrison, 1994; Fishwild et al., Nature Biotechnol., 14:845 (1996); Neuberger, Nat. Biotechnol., 14:826 (1996); и Lonberg and Huszar, Int Rev Immunol., 13:65 (1995).
Человеческие антитела могут также продуцироваться с использованием трансгенных животных, не являющихся людьми, которые модифицированы таким образом, что они продуцируют полностью человеческие антитела, а не эндогенные антитела животных в ответ на антигенную стимуляцию. (См. опубликованную международную заявку № WO 94/02602). Эндогенные гены, кодирующие тяжелые и легкие цепи иммуноглобулинов в хозяине нечеловеческого происхождения, были вырезаны из структуры и в хозяйский геном были введены тяжелые и легкие цепи человеческих иммуноглобулинов. Гены человека вводятся, например, с помощью искусственных хромосом дрожжей, содержащих необходимые сегменты ДНК. Животное, которое обеспечивает все желательные модификации, получается в виде потомства путем кросс-бридинга промежуточных трансгенных животных, содержащих менее полного комплемента модификаций. Одним из вариантов такого животного нечеловеческого происхождения является мышь, она называется XENOMOUSE™, как указано в опубликованных международных заявках №№ WO 96/33735 и WO 96/34096. Это животное продуцирует В-клетки, которые секретируют полностью человеческие иммуноглобулины. Антитела могут быть получены непосредственно от животного после иммунизации нужным иммуногеном, таким как, например, препарат поликлонального антитела или, альтернативно, из иммортализованных В-клеток, полученных от этого животного, таких как клетки гибридомы, продуцирующие моноклональные антитела. Кроме того, гены, кодирующие иммуноглобулины с вариабельными областями человеческого происхождения могут быть выделены и экспрессированы для непосредственного получения антител или могут быть в дальнейшем модифицированы для получения аналогов антител, таких как, например. одноцепочечные молекулы Fv.
Пример способа получения животного-хозяина нечеловеческого происхождения, например, мышиного, без экспрессирования тяжелой цепи эндогенного иммуноглобулина описан в патенте США №5939598, выданном на имя Kucherlapati et al. 17 авуста 1999 г., который включен в данную заявку посредством отсылки. Оно может быть получено путем делеции J-генных сегментов из по меньшей мере одного эндогенного локуса тяжелой цепи в эмбриональной стволовой клетке для предотвращения перегруппировки локуса и для предотвращения образования транскрипта перегруппированного локуса тяжелой цепи иммуноглобулина, при этом делеция осуществляется целевым вектором, содержащим ген, кодирующий селектируемый маркер и продуцирующий из эмбриональной стволовой клетки трансгенную мышь, у которой соматические клетки и зародышевые клетки зародышевой линии содержат ген, кодирующий селектируемый маркер.
Способ получения нужного антитела, такого как антитело человеческого происхождения, описан в патенте США №5916771, выданном на имя Hori et al. 29 июня 1999 г. и включенном в данную заявку посредством отсылки. Этот способ включает введение экспрессирующего вектора, который содержит нуклеотидную последовательность, кодирующую тяжелую цепь, в одну хозяйскую клетку млекопитающего в культуральной среде, введение экспрессирующего вектора, который содержит нуклеотидную последовательность, кодирующую легкую цепь, в другую хозяйскую клетку млекопитающего и слияние этих двух клеток с образованием гибридной клетки. Гибридная клетка экспрессирует антитело, содержащее тяжелую цепь и легкую цепь. Термин "нейтрализующее антитело" или " антитело, которое нейтрализует" относится к антителу, которое снижает по меньшей мере один вид активности полипептида, содержащего эпитоп, с которым специфически связывается антитело. Согласно некоторым вариантам нейтрализующее антитело уменьшает активность in vitro и/или in vivo. Термин "антиген-связывающий сайт" относится к части антитела, способной к специфическому связыванию антигена. Согласно некоторым вариантам антиген-связывающий сайт обеспечивается одной или более вариабельными областями антитела. Антитела согласно данному изобретению предпочтительно включают одну или более модификаций, которые инактивируют комплемент. Термин "активность комплемента" в широком смысле охватывает виды биохимической и физиологической активности, связанные с активацией системы комплемента, молекул комплемента, связанных с каскадом реакций, а также генов, кодирующих эти молекулы. Таким образом, виды активности комплемента включают, например, структуру и экспрессию гена, кодирующего молекулы комплемента при протекании каскадных реакций, биохимическую активность (например, ферментативную или регуляторную) молекулы комплемента, клеточную активность, которая инициирует активацию системы комплемента или возникает вследствие этой активации и появление аутоантител сыворотки крови к молекулам каскадных реакций комплемента. В антителе hSe1001 предпочтительной модификацией при инактивации комплемента является замена остатка лизина остатком аланина в положении 342 в тяжелой цепи константной области СН2. Другие замены в том же самом положении могут включать, например, любой остаток из gly, leu, trp, tyr, pro, thr, ser, met, asp, asn, glu, gln, phe, ile, val, thr и cys, при условии, что эта замена является также эффективной в устранении способности константной области активировать комплемент.
Термины "молекулы, ассоциируемые с каскадными реакциями комплемента", "молекулы каскадных реакций комплемента" и "белки, ассоциируемые с каскадными реакциями комплемента" используются как взаимозаменяемые и относятся к различным молекулам, которые играют роль в процессе активации комплемента и клеточной активности в направлении 5′-3′, опосредованной чувствительной активированной системой комплемента или запускаемой этой системой. Они включают инициаторы каскадных реакций комплемента (то есть молекулы, которые непосредственно или косвенно запускают процесс активации системы комплемента), молекулы, которые продуцируются или играют роль во время активации комплемента (например, белки/ферменты, такие как С3, С5, C5b-9, Фактор В, MASP-1 и MASP-2), рецепторы или ингибиторы комплемента (например, кластерин, витронектин, CR1 или CD59) и молекулы, регулируемые или запускаемые активированной системой комплемента (например, мембрано-атакующий комплекс-ингибирующий фактор, MACIF). Таким образом, в добавление к белкам комплемента, упомянутым выше, молекулы системы комплемента, связанные с каскадом реакций, включают также, например, регуляторы С3/С5-конвертазы (RCA), такие как рецептор комплемента 1 типа (называемый также CR1 или CD35), рецептор комплемента 2 типа (называемый также CR2 или CD21), мембранный белок-кофактор (МСР или CD46) и C4bBP; регуляторы MAC, такие как вибронектин, кластерин (называемый также "SP40,40"), CRP, CD59 и фактор гомологичной рестрикции (HRF); цепи иммуноглобулинов, таких как Ig каппа, Ig лямбда или Ig гамма; ингибитор С1; и другие белки, такие как CR3, CR4 (CD11b/18) и DAF (CD55).
Двойная функция антител к P-селектину
Данное изобретение предусматривает использование двойной функции антител к Р-селектину и их фрагментов, которые специфически связываются с Р-селектином и не только блокируют связывание PSGL-1 с Р-селектином, но также вызывают диссоциацию ранее образовавшихся комплексов Р-селектин/PSGL-1. Данное изобретение раскрывает неизвестную до сих пор антиген-связывающую область антитела (конформационный эпитоп) в пектиновом домене (например, углевод-распознающем домене, CRD) Р-селектина, с которым связываются антитела с двойной функцией (которые могут представлять собой химерные, человеческие или гуманизированные антитела или их фрагменты). Настоящее изобретение относится также к антителам к Р-селектину, которые связываются с конформационным эпитопом, описанным в данной заявке, и которые обладают двойной функцией, проявляющейся в (1) блокировании связывания PSGL-1 с Р-селектином и (2) диссоциации ранее образовавшихся комплексов Р-селектин/PSGL-1. В частности, данное изобретение направлено на применение такой двойной функции антител к Р-селектину или их фрагментов при лечении воспалительных, тромботических или других патологических состояний или расстройств у приматов (включая людей), в которых играет роль адгезия тромбоцитов, серповидных красных кровяных клеток, лейкоцитов, лимфоцитов и/или эндотелиальных клеток, при этом такое состояние или расстройство охватывает (но без ограничения) по меньшей мере одно из заболеваний, включающих вазоокклюзивный болевой криз вследствие наличия серповидных клеток, воспалительные заболевания кишечника (например, болезнь Крона, язвенный колит и энтерит), артрит (например, ревматоидный артрит, остеоартрит и псориазный артрит), отторжение имплантатов, реакцию трансплантат против хозяина, астму, хроническую обструктивную болезнь легких, псориаз, дерматит, сепсис, нефрит, эритематозную волчанку, склеродермию, ринит, анафилаксию, диабет, рассеянный склероз, атеросклероз, тромбоз, метастазирование опухолей, аллергические реакции, тироидит, ишемически-реперфузионное повреждение, например, вызванное инфарктом миокарда, инсультом или трансплантаией органов) и состояния, связанные с тяжелой травмой или хроническим воспалением, такие как, например, замедленная гиперчувствительность TV типа, связанная, например, с инфекцией, вызванной Tubercle bacilli, или синдромом системного воспалительного ответа или множественным воспалением органов, или связано с перечисленными заболеваниями.
Важно отметить, что применение такой двойной функции антител в процессе лечения этих воспалительных болезней, позволит не только предотвратить, снизить или ингибировать воспаление, но также обеспечит механизм лечения продолжающейся болезни, состоящий в том, что антитела могут вызывать диссоциацию ранее образовавшихся комплексов Р-селектин/PSGL-1. Например, в случае вазоокклюзионного болевого криза вследствие наличия серповидных клеток двойная функция антител способна не только предотвратить возникновение будущих вазоокклюзивных событий, но может также использоваться при лечении происходящей в настоящее время вазоокклюзии. Данное изобретение относится также к способам скрининга для детекции антител к Р-селектину, которые связываются с конформационным эпитопом Р-селектина, который впервые описан в данной заявке, и которые не только блокируют связывание PSGL-1 с Р-селектином, но также вызывают изменение связывания PSGL-1 с Р-селектином (а именно, диссоциацию ранее образовавшихся комплексов Р-селектин/PSGL-1).
Как было отмечено в данной заявке, настоящее изобретение относится также к очищенным антителам (включая, но без ограничения, химерные, человеческие или гуманизированные антитела или их фрагменты), которые распознают Р-селектин (то есть связываются с ним) (SEQ ID NO:1) и которые блокируют связывание PSGL-1 (или PSGL-1-подобных рецепторов) с Р-селектином и вызывают также диссоциацию ранее образовавшихся комплексов Р-селектин/PSGL-1, которые были опосредованы взаимодействиями Р-селектин/PSGL-1, и к терапевтическим способам применения таких антител (или их связывающих фрагментов).
Более конкретно, настоящее изобретение относится к очищенным антителам (или их фрагментам), направленным против Р-селектина, хозяйским клеткам, которые продуцируют такие антитела к Р-селектину (или их фрагменты), способам скрининга для идентификации антител к Р-селектину (или их фрагментов), которые блокируют адгезию лейкоцитов, серповидных эритроцитов, лимфоцитов, тромбоцитов и эндотелиальных клеток, опосредованную Р-селектином, и дополнительно вызывают диссоциацию ранее образовавшихся адгезивных или клеточных комплексов, появление которых было обусловлено взаимодействиями Р-селектин/PSGL-1, и к терапевтическим способам применения таких антител (или их связывающих фрагментов). Данное изобретение охватывает новые антитела к Р-селектину у приматов (включая людей) и их связывающие фрагменты, в частности включающие, но без ограничения, G4, гуманизированные формы G4 и антитела hSe1001. В соответствии с данным изобретением предпочтительными антителами являются антитела, способные к специфическому связыванию Р-селектина у приматов (особенно у людей) и к ингибированию одного или более видов in vitro и/или in vivo активности Р-селектина. Применяемый в данной заявке термин "PSGL-1" включает также "PSGL-1-подобный рецептор" на поверхности серповидных красных кровяных клеток (эритроцитов).
В общем, антитело или фрагмент антитела согласно данному изобретению могут иметь верхний предел их размера, обеспечивающий похожие или иммунологические свойства в отношении антитела, которое специфически связывается с Р-селектин-связывающим сайтом, описанным в данной заявке и которое блокирует связывание PSGL-1 с Р-селектином и вызывает диссоциацию ранее образовавшихся комплексов Р-селектин/PSGL-1.
Применяемый в данной заявке термин "включительно" относится ко всем целым числам, находящимся между любыми двумя числами, указанными в данной заявке.
Как отмечается в данной заявке, фрагменты антител согласно данному изобретению сохраняют способность к селективному связыванию полного связывающего эпитопа Р-селектина или его части, описанных в данной заявке. Предпочтительно, когда антитело или связывающий фрагмент антитела по данному изобретению способны связывать эпитоп, включающий один или более остатков аминокислот в положении 1-35, более предпочтительно в положении 4-23, последовательности SEQ ID NO:1.
Как указывалось выше, антитела или фрагменты этих антител в соответствии с данным изобретением представляют собой химерные иммуноглобулины изотипов IgG1, IgG2, IgG3, IgG4 или IgG2/G4, которые предпочтительно связываются с Р-селектином с высоким сродством (например, когда Kd≤1000 нМ) и предпочтительно содержат константную область человеческого происхождения, и предпочтительно ингибируют связывание PSGL-1 с Р-селектином и, еще более предпочтительно, вызывают изменение связывания в ранее образовавшемся комплексе Р-селектин/PSGL-1. Кроме того, антитело к Р-селектину или его связывающий фрагмент предпочтительно не активируют комплемент по классическому пути за счет взаимодействия с C1q и предпочтительно не связывают Fc-рецепторы, что совместно образует эффекторную функцию антитела. Данное изобретение относится к применению указанной двойной функции антител к Р-селектину или их фрагментов, описанных и идентифицированных в данной заявке, при лечении воспалительных, тромботических или других патологических состояний или расстройств у приматов (включая людей), в которых играет роль адгезия тромбоцитов, серповидных красных кровяных клеток, лейкоцитов, лимфоцитов и/или эндотелиальных клеток, при этом такое состояние или расстройство охватывает (но без ограничения) по меньшей мере одно из заболеваний, включающих вазоокклюзивный болевой криз вследствие наличия серповидных клеток, воспалительные заболевания кишечника (например, болезнь Крона, язвенный колит и энтерит), артрит(например, ревматоидный артрит, остеоартрит и псориазный артрит), отторжение имплантатов, реакцию трансплантат против хозяина, астму, хроническую обструктивную болезнь легких, псориаз, дерматит, сепсис, нефрит, эритематозную волчанку, склеродермию, ринит, анафилаксию, диабет, рассеянный склероз, атеросклероз, тромбоз, метастазирование опухолей, аллергические реакции, тироидит, ишемически-реперфузионное повреждение, например, вызванное инфарктом миокарда, инсультом или трансплантаией органов) и состояния, связанные с тяжелой травмой или хроническим воспалением, такие как, например, замедленная гиперчувствительность IV типа, связанная, например, с инфекцией, вызванной туберкулезной бациллой, или синдромом системного воспалительного ответа или полиорганной недостаточностью, или связано с перечисленными заболеваниями.
Как указывалось выше, Р-селектин играет центральную роль в рекрутинге лейкоцитов и лимфоцитов в воспалительные и тромботические участки путем связывания с поверхностью лиганда (PSGL-1) на этих клетках и в связывании серповидных красных кровяных клеток с эндотелием, содержащим активированные эндотелиальные клетки. PSGL-1 постоянно экспрессируется в лейкоцитах, включая нейтрофилы и моноциты, и в некоторых эндотелиальных клетках. PSGL-1-подобный рецептор экспрессируется на серповидных красных кровяных клетках и дает им возможность связывать Р-селектин в активированных клетках эндотелия.
Не желая ограничиваться какой-либо конкретной теорией, полагают, что лечение вазоокклюзионного болевого криза, вызванного наличием серповидных клеток, например, при помощи антитела к Р-селектину по данному изобретению является эффективным за счет ингибирования любого одного или более следующих взаимодействий: (1) связывания PSGL-1 лейкоцитов с Р-селектином клеток активированного эндотелия, (2) связывания PSGL-1-подобного лиганда серповидных красных кровяных клеток с Р-селектином клеток активированного эндотелия, (3) связывания Р-селектина на поверхности активированных тромбоцитов с PSGL-1 в эндотелиальных клетках. (4) связывание лейкоцитов серповидными красными кровяными клетками по механизму взаимодействия неохарактеризованного лиганда с рецептором, (5) связывания Р-селектина активированных тромбоцитов с PSGL-1-подобным рецептором красных кровяных клеток и/или (6) связывания Р-селектина на поверхности активированных тромбоцитов с PSGL-1 на поверхности лейкоцитов.
Полагают, что двойная функция антитела к Р-селектину блокирует инициацию и распространение вазоокклюзии на многих уровнях взаимодействий клетка-клетка в микроциркуляторной части сосудистого русла. Кроме того, как отмечалось в данной заявке, двойная функция антитела к Р-селектину приводит к диссоциации комплексов Р-селектин/PSGL-1 и поэтому может быть использована в терапевтических целях для вмешательства в происходящую вазоокклюзию.
Р-селектин опосредует взаимодействие активированных тромбоцитов или клеток эндотелия с кровяными клетками, включая красные кровяные клетки (а именно, серповидные красные кровяные клетки) и лейкоциты, включая моноциты, нейтрофилы, эозинофилы, клетки CD+Т, клетки CD8+Т, В-клетки и клетки естественных киллеров (NK) клетки. Как уже отмечалось, известно, что Р-селектин принимает участие в формировании ряда клеточных ответов на воспаление, возникающее в результате травмы, инфекции или физико-химической агрессии. Атеросклероз, характеризующийся атеросклеротическими повреждениями на внутренних поверхностях кровеносных сосудов, является одним из примеров состояния, включающего связывание некоторых лейкоцитов эндотелиальными клетками, содержащими Р-селектин на внутренней выстилке стенок кровеносных сосудов.
Как указано выше, способы лечения, направленные на блокирование функции Р-селектина, например, путем применения антител к Р-селектину для предотвращения задержки и роллинга лейкоцитов и адгезии красных кровяных клеток (а именно серповидных красных кровяных клеток) могут оказывать глубокое влияние на протекание различных видов воспалительных и тромботических заболеваний. С учетом ключевой роли Р-селектина в инициации роллинга и задержки лейкоцитов на поверхности эндотелия и тромбоцитов он является имеющей первостепенное значение мишенью для терапевтического вмешательства при лечении воспалительных и тромботических заболеваний. Например, временная природа острой фазы серповидно-клеточной анемии, сочетающейся с повторяющимися хроническими явлениями повреждения органов и связанными с этим осложнениями и заболеваемостью, позволяет предположить, что терапевтическое вмешательство, которое вызывает и блокирование исходной адгезии вследствие связывания Р-селектина с PSGL-1, и индукцию разрушения ранее возникшей, имеющейся в настоящее время и установившейся заранее адгезии, найдет новое применение для лечения этих и других воспалительных и тромботических заболеваний. Таким образом, настоящее изобретение охватывает способ применения конформационного связывающего эпитопа антитела к Р-селектину для скрининга и идентификации "двойной функции" антител к Р-селектину, которые не только блокируют связывание Р-селектина с PSGL-1, но которые также вызывают диссоциацию ранее образовавшегося комплекса Р-селектин/PSGL-1 (и тем самым, комплекса клетка-клетка) и способ применения таких антител для терапевтического лечения болезней, таких как, но без ограничения, воспалительные и тромботические заболевания.
Как отмечалось выше, были получены новые конформацонные эпитопы Р-селектина, описанные в данной заявке. Обнаружение этих конформационных связывающих эпитопов затем привело к открытию двойной функции антител Р-селектину (поэтому эти антитела называются в данной заявке антителами с двойной функцией), которые связываются с конформационным эпитопом и которые не только блокируют связывание Р-селектина с PSGL-1, но которые также вызывают диссоциацию ранее образовавшихся комплексов Р-селектин/PSGL-1 (то есть индуцируют изменение связывания Р-селектина с PSGL-1), приводя при этом к диссоциации клеточных комплексов, таких как комплексы лейкоцит/клетка эндотелия, лейкоцит/тромбоцит, лимфоцит/клетка эндотелия, лимфоцит/тромбоцит, серповидная красная кровяная клетка/клетка эндотелия или серповидная красная кровяная клетка/тромбоцит.
Области связывания некоторых антител к Р-селектину ранее были картированы с помощью конструктов больших функциональных доменов, включающих лектин, эпидермальный фактор роста (EGF) и консенсусные повторяющиеся области (CR) нативного белка Р-селектина мышиного и человеческого происхождения (90, 70, 91-95).
Результаты картирования показали, что основные области связывания в некоторых функционально-блокирующих антителах к Р-селектину были в лектиновом связывающем домене, области, которая охватывает аминокислотные остатки 1-120 нативного Р-селектина, или в домене EGF, охватывающем аминокислоты 121-154.
Примеры по изобретению представлены ниже. Однако данное изобретение не ограничено этими конкретными примерами, описанными результатами и лабораторными методиками. Напротив, эти примеры представлены как различные варианты и не являются исчерпывающими для понимания данного изобретения.
Настоящее изобретение описывает новые нелинейные (например, конформационные) эпитопы, обнаруженные с помощью мышино-человеческих химерных антител, содержащих лектин, домены EGF, CR1 и CR2 Р-селектина, которые были зондированы функционально-блокирующими испытуемыми антителами к Р-селектину. Более ранние исследования показали, что для укладки (фолдинга) и обеспечения нужной конформации конструктов Р-селектина по меньшей мере требуется экспрессия доменов лектина и EGF (91). Сравнение аминокислотных последовательностей человеческого и мышиного Р-селектина показало, что в лектиновом домене имеется гомология с определенным числом различий аминокислотных остатков мышиного и человеческого происхождения. Поэтому для сравнения доменов лектина, EGF, CR1 и CR2 Р-селектина (см. Фигуру 1) с целью идентификации различий между аминокислотными остатками Р-селектинов мышиного и человеческого происхождения (последовательностей и номенклатуры в соответствии со зрелыми белками) была использованы трехмерная (3-D) разновидность гомологии. Этот метод заключался в 3-D-моделировании Р-селектина для сравнения положений различных аминокислот в Р-селектинах мышиного и человеческого происхождения на экспонированной части белка и для идентификации различий в аминокислотных кластерах в лектиновом домене и домене EGF мышиного и человеческого происхождения, которые расположены на поверхности свернутого белка. Это 3-D-моделирование отражает кластеры различных аминокислот, которые образуются при соприкосновении непрерывных аминокислот, которое возникло при свертывании белка. Например, некоторые аминокислоты будут образовывать конформционные эпитопы в силу нахождения на одной и той же поверхности, например, на наружной поверхности спиральных структур. Сравнение гомологии таких кластеров позволяет осуществить выбор аминокислотных замен с целью определения влияния таких изменений на связывание функционально блокирующего (PSGL-1-блокирующего) антитела с Р-селектином человеческого происхождения.
Указанный метод включал также картирование консервативных сайтов рестрикции в открытых рамках считывания кДНК для определения стратегии конструирования химерных белков, которые охватывают домены лектина, EGF, CR1 и CR2, которое позволит осуществить замену одной или многих аминокислот в специфических участках Р-селектина мышиного и человеческого происхождения для идентификации тех аминокислот, которые оптимизируют связывание антитела с человеческим Р-селектином. Были сконструированы химерные антитела, включающие ATG через домен CR2 с подходящим вектором, таким как pBluescript (pBS-hPsel и pBS-mPsel). Эти химеры были вставлены в другой подходящий вектор, такой как pIG1 (pIG-hPsel и pIG-mPsel), где конструкты были слиты с областью Fc человеческого IgG1, содержащего шарнирный участок, СН2 и СН3 области. Эти конструкты сохраняли структуры, которые соответствуют нативной конформации Р-селектина. Таким образом, домены, которые были экспонированы на поверхности нативного белка, также содержались в химерных конструктах и служили также в качестве предполагаемых эпитопов для связывания испытуемых антител. Такие конструкты могут быт трансфецированы и временно экспрессированы при помощи методов молекулярной и клеточной экспрессии, известных специалистам в данной области.
При помощи этого метода как такового испытуемые антитела можно оценить в отношении связывания с химерами мышиного и человеческого происхождения, при этом используют такой метод, но без всякого ограничения, как метод флуоресцентно-активированного клеточного сортинга (FACS) и метод поверхностного плазменного резонанса (BIACORE), которые известны специалистам в данной области. При этом оценивали влияние изменений в аминокислотах в различных положениях в химерных конструктах, осуществляемых путем замены аминокислот мышиного происхождения, например, последовательностью человеческого происхождения или, наоборот, человеческих аминокислот мышиными последовательностями, которые, например, исключили или позволили связывание антител, направленных против человеческого Р-селектина, и это дало возможность идентифицировать конкретные аминокислоты для оптимального связывания.
Характеристика химерных конструктов
Аминокислоты мышиного Р-селектина, которые были заменены последовательностями человеческого Р-селектина в химерах, описанных ниже, указаны полужирным шрифтом. Аминокислоты человеческого Р-селектина, которые были заменены мышиной последовательностью, указаны полужирным шрифтом курсивом. Замена глутамина гистидином указана подчеркнутым полужирным шрифтом.
Конструкты нативного белка
SEQ ID NO:1
Домены лектина, EGF, CR1, CR2 в Р-селектине человеческого происхождения

SEQ ID NO:2
Домены лектина, EGF, CR1, CR2 в Р-селектине мышиного происхождения - различие в аминокислотах показано полужирным шрифтом.
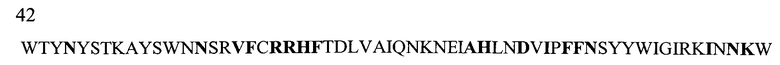

Челочеческие/мышиные химерные конструкты
SEQ ID NO:3
Химера-1
(замена мышиными остатками в человеческом кластере А - N4N14V17F18R20R21H22F23)

SEQ ID NO:4
Химера-2
(человеческий кластер от А до I35 - затем мышиная последовательность)
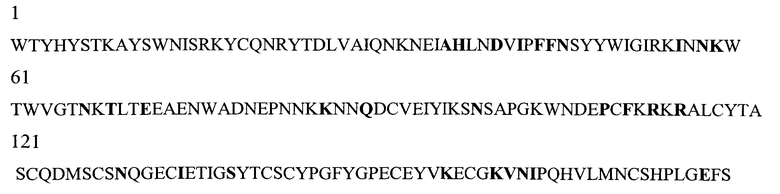
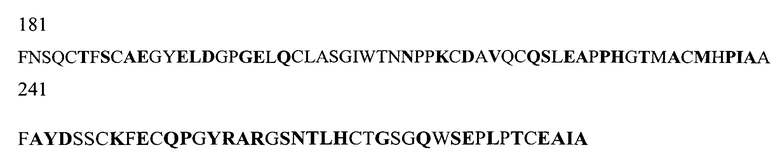
SEQ ID NO:5
Химера-3
(замена в человеческом кластере А - на мышиный N4N14V17F18)

SEQ ID NO:6
Химера-4
(замена в человеческом кластере А - на мышиный R20R21H22F23)

SEQ ID NO:7
Химера-5
(единственная замена аминокислоты - человеческая Н4 на мышиную N4)

SEQ ID NO:8
Химера-5Q
(замена Q на Н4 в кластере А - удаление предполагаемого сайта гликозилирования)

SEQ ID NO:9
Химера-6
(человеческая последовательность EGF - S121 - мышиные EGF, CR1 и CR2)
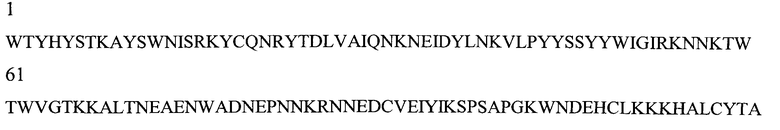

SEQ ID NO:10
Химера-7
(человеческая последовательность в G177 - затем мышиная последовательность)

SEQ ID NO:11
Химера-7В
(человеческая последовательность в конце EGF - V156 - затем мышиная последовательность)


SEQ ID NO:12
Химера-8
(единственное изменение аминокислоты - человеческая I14 на мышиную N14)

SEQ ID NO:13
Химера-9
(единственной изменение аминокислоты - человеческая K17 на мышиную V17)

SEQ ID NO:14
Химера-10
(единственное изменение аминокислоты - человеческая Y18 на мышиную F18)
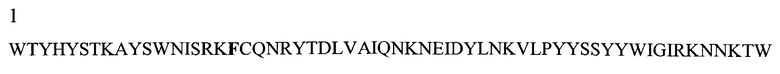

SEQ ID NO:15
Химера-11
(единственное изменение аминокислоты - человеческая Q20 на мышиную R20)

SEQ ID NO:1
Химера-12
(единственное изменение аминокислоты - человеческая N21 на мышиную R21)

SEQ ID NO:17
Химера-13
(единственное изменение аминокислоты - человеческая R22 на мышиную Н22)

SEQ ID NO:18
Химера-14
(единственное изменение аминокислоты - человеческая Y23 на мышиную F23)

SEQ ID NO:19
Химера-15
(единственное изменение аминокислоты - человеческая в кластере С2 - S97 - мышиная последовательность)


SEQ ID NO:20
Химера-16
(замена человеческой H4I14K17N21R22 на мышиный кластер А)

SEQ ID NO:21
Химера-17
(человеческая последовательность в кластере B в I35 - мышиный кластер B в I42 - человеческая последовательность CR1 в E154 - мышиные CR1, CR2)

SEQ ID NO:22
Химера-17B
(человеческая последовательность в кластере B в I35 - мышиный кластер B в I42 - затем человеческая последовательность)

SEQ ID NO:23
Химера-18
(человеческая последовательность в мышином кластере С (C1, C2, С3) и мышиные CR1, CR2)

SEQ ID NO:24
Химера-18B
(человеческая последовательность в мышином кластере С (C1, C2, С3)

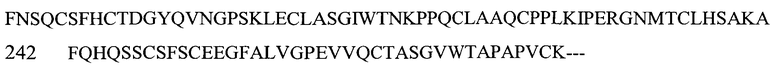
SEQ ID NO:25
Химера-19
(человеческая последовательность в мышином кластере D и мышиные CR1, CR2)

SEQ ID NO:26
Химера-19B
(человеческая последовательность в мышином кластере D)

SEQ ID NO:27
Химера-20
(человеческая последовательность - мышиный кластер Е и мышиные CR1, CR2)
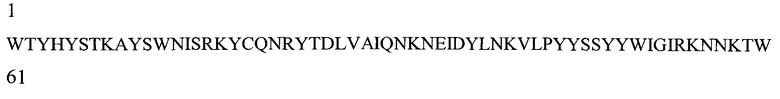

SEQ ID NO:28
Химера-20B
(человеческая последовательность - мышиный кластер Е)

SEQ ID NO:29
Химера-21
(человеческая последовательность - мышиным кластером F)

Флуоресцентно-активированный клеточный сортинг связывания антител к Р-селектину с химерами человеческого/мышиного происхождения
Связывание антитела к Р-селектину с человеческими/мышиными химерами изучали с помощью флуоресцентно-активированного клеточного сортинга в системе, такой как система BD BIOSCIENCES FACS ARIA CELL SORTER для определения связывания антител к Р-селектину с человеческими/мышиными химерами, которые иммобилизованы на гранулах, покрытых антителом козы к человеческму Fc. Такие гранулы, покрытые химерами, инкубировали вместе с испытуемыми антителами к Р-селектину и затем эти антитела детектировали с применением Fc мышиного антитела или изотип-специфичных антител, меченых репортерами, такими как FITC, подходящими для детекции в системе FACS.
Одностадийный метод поверхностного плазменного резонанса (BIACORE)
Согласно одному из аспектов данного изобретения для улавливания испытуемого антитела к Р-селектину использовали чипы BIACORE. Человеческие-мышиные химерные антитела к Р-селектину, описанные в данной заявке, прикреплялись к поверхности сенсорного чипа и затем добавлялись испытуемые антитела. Связывание химер с испытуемыми антителами измерялось в единицах резонансного ответа.
Двухстадийный метод поверхностного плазменного резонанса (BIACORE)
Согласно другому аспекту настоящего изобретения на биосенсорном чипе, таком как чип BIACORE, осуществляли иммобилизацию поликлонального антитела козы к человеческому Fc изотипа IgG путем ковалентного связывания его с поверхностью чипа. Человеческие-мышиные химерные конструкты к Р-селектину, содержащие домены CR1 лектина, EGF и CR2, впрыскивали на указанный чип и улавливали при концентрациях, которые обеспечивали стандартный уровень покрытия поверхности при осуществлении метода поверхностного плазменного резонанса. Желаемый уровень резонансного отклика, наблюдаемый (достигаемый) после введения каждой химерной конструкции Р-селектина принимали за (обозначали как) новый "вторичный базовый" уровень. Испытуемые антитела к Р-селектину (например, мышиные или гуманизированные моноклональные антитела к Р-селектину, включая G1, G3, G4, G5 и hSe1001) затем впрыскивали на поверхность чипа и инкубировали для осуществления связывания химерного конструкта с Р-селектином, уже уловленного поверхностью чипа. Этот метод может быть модифицирован для испытуемых гуманизированных антител путем создания конструктов селектина с мышиным Fc изотипа IgG и улавливания поликлональным антителом козы против Fc IgG с последующим зондированием при помощи испытуемых гуманизированных антител к Р-селектину. Антитела, которые связывают конструкты Р-селектина, вызывают увеличение степени резонансного ответа по сравнению со вторичным базовым уровнем. Полученное увеличение уровня резонансного ответа может быть измерено как "дополнительные резонансные единицы (RU)" и свидетельствует о степени связывания с конструктом Р-селектина, предварительно нанесенным на поверхность биосенсорного чипа, с испытуемым антителом. При помощи описанных методов были точно картированы оптимальные требования к связыванию антител к химерам Р-селектина с конкретными конформационнми эпитопами.
Идентификация двойной функции антител, направленных против Р-селектина (антител, которые и блокируют связывание, и вызывают диссоциацию комплекса Р-селектина с PSGL-1)
Для выявления двойной функциональности специфических антител к Р-селектину, которые, как уже обсуждалось выше, могут как блокировать, так и вызывать диссоциацию (изменение) продуктов реакций связывания между Р-селектином и PSGL-1, также использовали метод анализа в биосенсорной системе BIACORE.
При осуществлении этого метода PSGL-1 или малые молекулы миметиков связывающего эпитопа PSGL-1, такого как биотинилированный глюкосульфопептидный миметик (например, GSP-6), или химерные белки, содержащие N-конец PSGL-1, улавливались на чипе BIACORE, таком как чип со стрептавидином, способами, известными средним специалистам в данной области (GSP-6 представляет собой гликозилированный сульфированный пептидный миметик, содержащий аминокислоты 42-60 в экспонированном N-конце PSGL-1, например, подробно описанный в патенте США №6593459) Согласно одному из вариантов данного изобретения вначале для определения функционально блокирующей способности антитело к Р-селектину предварительно смешивали с растворимым Р-селектином и инкубировали в течение промежутка времени, достаточного для образования комплекса Р-селектин/антитело. Полученный комплекс антитело к Р-селектину/Р-селектин помещали на поверхность чипа иди другого субстрата, содержащего PSGL-1 (или миметик PSGL-1) и измеряли степень связывания PSGL-1 или его миметика. Антитела к Р-селектину, которые предотвращают связывание Р-селектина с PSGL-1 или миметиком PSGL-1 на поверхности чипа, представляют собой функционально блокирующие антитела.
Затем антитела, которые, как было показано (с помощью данного метода или другого похожего метода), являются функционально блокирующими антителами (то есть, которые блокируют связывание PSGL-1 с Р-селектином, могут быть испытаны с целью определения дополнительной функции, то есть наличия способности к диссоциации (изменению) ранее образовавшегося комплекса PSGL-1 с Р-селектином. Указанная двойная функция антител по изобретению может быть определена при помощи биосенсорного метода BIACORE, описанного в данной заявке. Согласно одному из вариантов для определения указанной двойной функции PSGL-1 или его миметик, такой как GSP-6, иммобилизуется на поверхности чипа BIACORE. Затем Р-селектин наносится на поверхность указанного чипа и осуществляется его связывание с PSGL-1 или его миметиком. После того как на сенсограмме будет показано достижение равновесного связывания Р-селектина с PSGL-1 или его миметиком, вводят функционально блокирующие антитела к Р-селектину и с помощью соответствующего метода измеряется диссоциация комплеса Р-селектина с PSGL-1 или его миметиком. Антитела, которые по данным этого метода, вызывают диссоциацию (обратную реакцию) комплекса Р-селектина с PSGL-1, называются "диссоциирующими антителами" и характеризуются как антитела с двойной функцией, то есть они обладают функционально блокирующими свойствами и диссоциирующими свойствами при разрушении связывания Р-селектина с PSGL-1. Такие антитела, обладающие двойной функцией, являются особенно предпочтительным вариантом согласно данному изобретению, так как они особенно пригодны для терапевтического применения при лечении острых и хронических воспалительных и тромботических заболеваний, описанных в данной заявке.
Выявление конформационных эпитопов
Проводили анализ трехмерной (3-D) структуры зрелых белов Р-селектина человеческого и мышиного происхождения и сравнивали полученные результаты по характеристике аминокислот с аминокислотами в доменах пектина и EGF. На экспонированной поверхности белков были идентифицированы шесть кластеров с различными конформационными аминокислотами. Они были обозначены как кластеры А, В, С, D, Е и F (см. Фигуру 1). N-концы человеческих и мышиных Р-селектинов, включающие остатки 1-35, содержат различия в 8 аминокислотах. Кластер А произвольно был определен границей между первой отличающейся аминокислотой (Н4) и последней отличающейся аминокислотой (Y23). Кластер А включал 20 аминокислот и образовывал жесткую структуру, называемую альфа-спиральной структурой, с цистеиновым остатком около N-конца белка (см. область "1" на Фигуре 5). Кластер В (см. Фигуру 1) представляет собой конформационный эпитоп, включающий аминокислотные остатки 36-42 и содержит 4 аминокислоты, которые отличаются друг от друга в человеческом и мышином Р-селектине. Применяемый в данной заявке термин "конформационный эпитоп" относится к эпитопу, который не распознается в условиях восстановления.
Кластеры С и Е (см. Фигуру 1) являются конформационными и дискретными и приводятся в соприкосновение друг с другом при фолдинге нативного белка Р-селектина. Кластер С содержит три конформационных области (C1, C2, С3), включающие 5 аминокислот, которые в человеческом Р-селектине отличаются от аминокислот в мышином Р-селектине. Кластер C1 отделен от C2 51 аминокислотой и кластер C2 отделен от кластера С3 15 аминокислотами. Точно так же кластер Е имеет два конформационных эпитопа (Е1 и Е2), в составе которых пять аминокислот, содержащихся в человеческом и мышином селектине, отличаются друг от друга, при этом кластер Е1 отделен от кластера Е2 19 аминокислотами. Кластеры А, В, С, D и Е находятся внутри консенсусного домена пектина в Р-селектине (см. Фигуру 1). Кластер F содержится в домене EGF и в нем находятся три отличающиеся друг от друга аминокислоты. Кластеры C1, Е1, C2, Е2 и С3 охватывают ключевые контактные остатки, которые были ранее идентифицированы как подходящие для взаимодействий Р-селектина с его физиологическим лигандом PSGL-1 (Somers et al). Кластеры А и В наиболее удалены от этих контактных участков (в 3′-5′ (обратном) направлении).
Были исследованы открытые рамки считывания кДНК для человеческого и мышиного Р-селектина с целью идентификации общих сайтов рестрикции, которые могут быть использованы для сборки химер, включающих указанные выше кластеры. Для проведения кодирования кДНК для специфических белковых и химерных конструктов, таких как SEQ ID NO:1-29 (описанные выше и в перечне последовательностей) были использованы ПЦР и химический синтез ДНК. Для конструктов-плазмид, кодирующих человеческие/мышиные химеры, применяли метод рестрикционного клонирования. Химеры временно экспрессировались в клетках COS-7 и применялись для осуществления методов FACS BIACORE. Химеры Р-селектина применяли для определения связывающей функции, используя метод в системе BIACORE путем анализа их связывания с GSP-6, иммобилизованным на поверхности чипа BIACORE. Как уже отмечалось выше, GSP-6 представляет собой малую молекулу, которая имитирует N-конец человеческого PSGL-1. Все испытуемые химеры связывались с GSP-6 на поверхности чипа, хотя и в разной степени, так как мышиный Р-селектин имеет более низкое сродство к связыванию с человеческим PSGL-1, чем человеческий Р-селектин. Это показывало, что химеры проявляли указанную функцию после экспрессии и очистки.
Метод FACS
Результаты флуоресцентно-активированного клеточного сортинга (FACS) антител к Р-селектину с применением конструктов или химер, соответствующих SEQ ID NOs.:1-29, показаны в Таблице 1. Четыре антитела к Р-селектину (G1, G3, G4 и G5) были выделены из гибридом, полученных путем иммунизации мышей с человеческим рекомбинантным Р-селектином, содержащим домены пектина и EGF (90 и неопубликованные работы). Эти исследования показали, что антитела G1, G3 и G4 представляют собой функционально блокирующие антитела, в то время как G5 не является блокирующим (90 и неопубликованные работы). Антитела G1, G3 и G5 анализировали методом FACS. Этот метод анализа показал, что G1, G3 и G5 все связывались с человеческим Р-селектином (SEQ ID NO:1), но не связывали мышиный Р-селектин (SEQ ID NO:2). Когда соответствующие восемь мышиных аминокислот заняли место в кластере А человеческой последовательности (химера 1, SEQ ID NO:3), антитело G1, но не антитела G3 или G5, утратили способность к связыванию, что свидетельствовало о том, что по меньшей мере одно или более соответствующих положений аминокислот в кластере А является существенным для связывания G1 с Р-селектином и что замена "мышиными" аминокислотами в этих одном или более положениях вызвало потерю способности к связыванию Р-селектина. Для того, чтобы подтвердить важное значение этих остатков, восемь различных человеческих аминокислот в положениях 1-23 были заменены в кластере А мышиной последовательности (SEQ ID NO:4, химера-2). Эти замены придали G1 способность к связыванию с мышиным белком. Химеры, содержавшие пектиновый домен человеческого Р-селектина с заменами мышиными аминокислотами в доменах EGF, CR1 и CR2 (SEQ ID NO:9, химера-6) и человеческую последовательность в домене EGF и в домене CR1 с мышиной последовательностью в остальной части домена CR1 и целый домен CR2 (SEQ ID NO:10, химера-7), были связаны с G1 и G3, что свидетельствовало о том, что основные связывающие эпитопы оставались интактными после замены этих мышиных аминокислот и что конформация связывающих эпитопов антитела при этом не нарушилась. Эти данные подтвердили, что эпитопы для связывания G1 и G3 находились в домене лектина.
При помощи метода FACS было показано, что испытуемое антитело G3 связывает человеческий Р-селектин, но не связывает Р-селектин мышиного происхождения и в противоположность антителу G1 связывалось с последовательностью SEQ ID NO:3 (химера-1), показывая, что антитело G3 связывается с эпитопом, который отличается от эпитопа, который связан с G1. Подробности картирования эпитопа G3 приведены в Таблице 1 ниже.
Испытуемое антитело G5, которое, как было указано ранее, является блокирующим, также изучалось при помощи этого метода. Было показано также, что G5 связывает человеческий селектин, но не мышиный Р-селектин. G5 связано с последовательностями SEQ ID NO:3 и SEQ ID NO:10, которые включают домен EGF и содержат также первую часть CR1 до N178, но антитело G5 не связано с последовательностью SEQ ID NO:9, которая включает S128. Эти результаты показывают, что антитело G5 связывается с первой частью CR1 и содержит по меньшей мере аминокислоты с R128 по N178.
Одностадийный метод поверхностного плазменного резонанса (BIACORE)
Для дальнейшего изучения значения домена кластера А (аминокислоты в положении 4-23) для связывания антитела G1 с Р-селектином были получены несколько конструктов, в которых одна или многие аминокислоты были инсерцированы в последовательность человеческого Р-селектина и затем анализировали связывание с помощью метода поверхностного плазменного резонанса ("одностадийный" метод в системе BIACORE), описанного в данной заявке. Результаты одностадийного метода, проводимого в системе BIACORE, показаны в Таблице 1. Испытуемое антитело G1 улавливалось поверхностью чипа BIACORE и результаты связывания измерялись как единицы ответа. Характерная сенсограмма, показывающая результаты связывания антитела G1 с конструктами человеческого Р-селектина (SEQ ID NO:1), мышиного Р-селектина с кластером А человеческого происхождения (SEQ ID NO:4) и мышиного Р-селектина (SEQ ID NO:2), показана на Фигуре 2.
При помощи этого метода было показано, что антитело G1 связывалось с человеческим Р-селектином и с последовательностью SEQ ID NO:4 (где человеческие аминокислоты были введены в кластер А мышиного Р-селектина), но не связывалось с мышиным Р-селектином (SEQ ID NO:2). Эти результаты свидетельствовали о том, что результаты, полученные указанным методом, согласовались с результатами метода FACS.
Р-селектин мышиного происхождения (SEQ ID NO:2) содержит предполагаемый сайт гликозилирования (N) в положении 4, в то время как Р-селектин человеческого происхождения (SEQ ID NO:1) не содержит этого сайта. Для определения значения этого различия аминокислота в положении 4 в человеческой последовательности SEQ ID NO:1 Р-селектина была заменена N (образуется SEQ ID NO:7) или Q (образуется SEQ ID NO:8) и с последующим определением влияния таких замен на связывание антитела G1. Инсерция остатка аспарагина (N) в молекулу Р-селектина человеческого происхождения в положении 4 уменьшало степень связывания антитела G1, что позволяет предположить, что гликозилирование в этом сайте человеческого Р-селектина будет мешать связыванию этого антитела. Замена остатком глутамина (Q) в этом положении не препятствовала связыванию антитела G1. Для дальнейшей идентификации положений аминокислот в кластере А, которые являются оптимальными или существенными для связывания антитела G1, осуществляли вставки одной аминокислоты в аминокислотной последовательности мышиного происхождения в последовательность (SEQ ID NO:1) человеческого Р-селектина и оценивали связывание полученных химер с антителом G1, используя одностадийный метод в системе BIACORE, описанный в дано заявке (Таблиц 1). Испытуемые химеры (с конкретными заменами, указанными в скобках) представляли собой SEQ ID NO:7 (H4N); SEQ ID NO:12 (I14N); SEQ ID NO:13 (K17V); SEQ ID NO:14 (Y18F); SEQ ID NO:15 (Q20R); SEQ ID NO:16 (N21R); SEQ ID NO:17 (R22H) и SEQ ID NO:18 (Y23F). Антитело G1 связывалось с SEQ ID NO:14, 15 и 18, но не связывалось с SEQ ID NO:12, 13 и 17 и только слабо связывалось с SEQ ID NO:7 и SEQ ID NO:16. Этот результат показал, что каждое из аминокислотных положений 4, 14, 17, 21 и 22 является положением, которое требуются для связывания антитела G1. Согласно предпочтительному варианту данного изобретения такие остатки аминокислот представляют собой Н4, I14, K17, N21 и R22, соответственно. Для подтверждения того, что гуманизированная форма антитела G1 (hSe1001) сохраняла идентичную специфичность эпитопа родительского антитела, оценивали связывание hSe1001 с последовательностями SEQ ID:1, 2 и 20 также с помощью указанного метода. Паттерн связывания hSe1001 был идентичен паттерну связывания G1, подтверждая тот факт, что специфичность эпитопа сохранялась в процессе гуманизации.
Двухстадийный метод поверхностного плазменного резонанса (BIACORE)
Для оценки связывания антител G1, G3, G4 и G5 (G5 является не блокирующим антителом) с дополнительными химерами применяли двухстадийный метод поверхностного плазмонного резонанса ("двухстадийный метод" BIACORE), описанный в данной заявке. Результаты "двухстадийного" метода анализа испытуемых антител приведены в Таблице 1 и на Фигуре 2. Этот метод показал, что ни одно из указанных выше антител не связывалось с Р-селектином мышиного происхождения, но все эти антитела связывались с человеческим Р-селектином. Испытуемые антитела G1, G3, G4 и G5 все связывались с последовательностью SEQ ID NO:10, свидетельствуя о том, что они все связываются с областью, включающей N-конец до доменов лектина и EGF Р-селектина человеческого происхождения. Не блокирующее связывание антитело G5 не связывалось с последовательностью SEQ ID NO:10, подтверждая тот факт, что G5 связывается с доменом CR1.
Дальнейшее исследование с помощью этого метода показало, что G3 не связывало ни последовательность SEQ ID NO:23, которая включала остатки мышиных аминокислот, инсерцированные в кластер C1, C2, С3, CR1 и CR2, ни последовательность SEQ ID NO:24, которая содержала мышиные аминокислоты в C1, C2, и С3. Антитело G3 также не связывалось с другими химерами, которые содержали мышиную последовательность в кластере С, то есть последовательности SEQ ID NO:19 и SEQ ID NO:20. Эти результаты показывают, что в блокирующем антителе G3 для связывания требуется наличие кластера С и показывают неизвестный ранее факт, заключающийся в том, что конформационные кластеры аминокислот, находящихся вблизи друг от друга в результате фолдинга белков, могут служить в качестве доменов связывания (конформационных эпитопов) для антител к Р-селектину. Этот метод также подтвердил, что антитело G3 может блокировать связывание PSGL-1 и Р-селектина и, таким образом, имеет функционально-блокирующие свойства (см. Фигуру 3). Однако этот метод также показал, что G3 не связывает Р-селектин с PSGL-1 и не вызывает диссоциацию (изменение) комплекса Р-селектин/PSGL-1 (см. Фигуру 4) и поэтому не обладает двойной функцией антител, являющихся предпочтительными согласно данному изобретению. Положения аминокислот, которые вносят свой вклад в связывание G1 и G4 с Р-селектином, были определены путем генерирования человеческого/мышиного гибрида в кластере А. Химера гибридного кластера A (SEQ ID NO:20) содержит аминокислоты человеческого Р-селектина в положениях 4, 14, 17, 21 и 22 (Н, I, K, N и R, соответственно) и аминокислоты Р-селектина мышиного происхождения в положениях 18, 20 и 23 (Y, Q, и Y, соответственно). Как показано в Таблице 1, и антитело G1, и антитело G4 связывались с последовательностью SEQ ID NO:20. Этот результат в совокупности с ранее полученными данными свидетельствует о том, что положения аминокислот 4, 14, 17, 21 и 22 представляют собой положения, из которых каждое требуется для оптимального связывания G1, G4 и гуманизированной формы G1 (hSe1001) с Р-селектином. Эти результаты представляют собой новое открытие того факта, что группа антител, включающая G1, G4 и hSe1001, связывает один и тот же или похожий эпитоп и что этот эпитоп находится в спиральной структуре кластера А, который удален от контактных остатков в домене, связывающем лектин с лигандом, которые были ранее идентифицированы для Р-селектина (71). В соответствии с предпочтительным вариантом данного изобретения аминокислоты человеческого в положениях 4, 14, 17, 21 и 22, дистальные по отношению к константным остаткам в домене, связывающем лектин с лигандом, ранее были идентифицированы как Р, I, K, N, и R, соответственно. Трехмерный анализ этого эпитопа выявил наличие жесткой спиральной структуры с требующимися аминокислотами, занимающими положения на той же поверхности спирали, поэтому кластер А обозначен как включающий конформационный эпитоп (см. Фигуру 5). Анализ в системе BIACORE, результаты которого показаны на Фигурах 3 и 4, подтвердил, что антитела G1, G4 и hSe1001 могут блокировать связывание Р-селектина и PSGL-1 и диссоциировать образовавшийся комплекс Р-селектин/PSGL-1, поэтому связывание описанного в данной заявке эпитопа этой группой антител имеет характер двойной функции согласно предпочтительным вариантам данного изобретения. Эти результаты показывают, что антитела, которые связываются с конформационным эпитопом, расположенным в аминокислотах 1-35 и, более конкретно, в положениях 4-23 последовательности SEQ ID NO:1, которая отдалена от домена связывания лектин-лиганд в Р-селектине человеческого происхождения, будут обладать уникальной двойной функцией. Не ограничиваясь какой-либо определенной теорией, полагают, что антитела, которые связываются с этим эпитопом, действуют за счет участия аллостерических сил, которые оказывают свое действие на межфазную поверхность связывания лектин-лиганд с индуцированном конформационного изменения, которое приводит к диссоциации комплекса Р-селектин/PSGL-1. Таким образом, антитело G1 способно связывать дистальный эпитоп в кластере А и блокировать и диссоциировать комплекс путем связывания и разрушения молекулярных взаимодействий в сайте связывания лектин-лиганд в Р-селектине. В противоположность этому, такие антитела, как G3, которое связывается с эпитопом в домене связывания лектин-лиганд Р-селектина, могут блокировать связывание Р-селектина с PSGL-1 за счет аллостерического затруднения, но могут вызывать диссоциацию комплекса Р-селектин/PSGL-1, так как эти антитела не могут связываться с конформационным эпитопом кластера С, когда он занят лигандом. Испытуемое антитело G5 связывалось с кластером CR1 в Р-селектине и, как было показано, является не блокирующим (см. Фигуру 3).
Суммируя сказанное выше, следует отметить, что антитела, которые связываются с Р-селектином, характеризовались как имеющие три возможных вида активности в отношении взаимодействия Р-селектина и его лиганда, PSGL-1.
Во-первых, антитела к Р-селектину могут связываться с Р-селектином, но не препятствовать связыванию PSGL-1 с Р-селектином ("не блокирующие антитела"). Например, как показано в данной заявке, антитело G5 связывает аминокислоты в положениях 157-164 и требует наличия R157, E161, L162, E163 и L164 в CR1 для связывания, но это связывание не препятствует связыванию Р-селектина с PSGL-1.
Во-вторых, антитела к Р-селектину могут связываться с Р-селектином и препятствовать связыванию PSGL-1 с Р-селектином ("блокирующие функцию антитела"), но не связываются с ранее образовавшимся комплексом Р-селектин/PSGL-1 и не диссоциируют этот комплекс. Например, результаты, описанные в данной заявке, показывают. Что антитело G3 связывается с конформационными кластерами в различных частях молекулы Р-селектина, которые включают домен связывания лектин-лиганд. G3 связывает кластер С и для этого требуются аминокислоты Y44, Y45, S46 and C2 в C1 и аминокислота P98 в С2 и аминокислота H114 в С3 и, таким образом, требуется конформационный эпитоп. Это антитело блокирует взаимодействие между Р-селектином и PSGL-1, но не может связывать или диссоциировать ранее образовавшийся комплекс Р-селектин/PSGL-1.
В-третьих, мы выявили антитела к Р-селектину с конкретной специфичностью, которые могут связываться с Р-селектином и не только блокируют связывание PSGL-1 с Р-селектином (блокирующее функцию антитело), но могут также вызывать изменение ранее образовавшегося комплекса Р-селектин/PSGL-1 (диссоциирующее связывание).
Такие антитела в данной заявке называют антителами с "двойной функцией". Результаты, описанные в данной заявке, показывают, что, например, антитело G1 связывается с конформационным эпитопом в кластере А и что это связывание предъявляет абсолютное требование к конформационному эпитопу, содержащему аминокислоты в положениях 4, 14, 17, 21 и 22, при этом предпочтительно, когда эти аминокислоты представляют собой Н, I, K, N и R, соответственно. Как описано в данной заявке, замена "человеческой аминокислоты (Н, I, K, N и R) на соответствующие "мышиные" аминокислоты (N, N, V, R и Н) приведет к отмене связывания антителами с двойной функцией, описанными в данной заявке.
Согласно другому варианту данного изобретения ранее не охарактеризованное мышиное моноклональное антитело к Р-селектину, обозначенное как G4 и полученное стандартным методом гибридомной технологии, было изучено на предмет связывания с конформационным эпитопом кластера А, определялась также двойная функция с помощью методов, описанных в данной заявке. Определяли связывание G4 с последовательностями SEQ ID NOs.:1-4, 7-10, 19 и 20 человеческих/мышиных химер. Было показано, что G4 связывается с SEQ ID NO:20 и имеет специфичность связывания, похожую на специфичность связывания антитела G1 (см. Table 1). Затем было показано, что G4 блокирует связывание Р-селектина с PSGL-1 (Фигура 3 3), а также вызывает диссоциацию ранее образовавшихся комплексов Р-селектин/PSGL-1 (см. Фигуру 4), что характеризует G4 как антитело к Р-селектину с двойной функцией, которое связывается с эпитопом (в кластере А), который наиболее удален от домена связывания лектин/лиганд Р-селектина и блокирует связывание Р-селектина с PSGL-1 и вызывает диссоциацию комплекса Р-селектин/PSGL-1. Таким образом, оба антитела G1 и G4 связывались с эпитопом в кластере А и оба проявляли двойную функцию. Этот результат показывает также возможность применения кластера А или конкретных положений его аминокислот в качестве эпитопа, который можно использовать для скрининга антител к Р-селектину с двойной функцией. Эта двойная функция антител имеет уникальные свойства, позволяющие применять их для лечения острых или хронических болезней, при которых инициирование адгезии, опосредованной Р-селектином, и/или уже имеющаяся адгезия, опосредованная Р-селектином, снимаются. С помощью способов, описанных в данной заявке, методом скрининга с использованием эпитопа кластера А или конкретных положений его аминокислот можно выявить другие антитела, обладающие двойной функцией.
Гуманизированное антитело к Р-селектину изотипа IgG2, у которого отсутствует эффекторная функция, обозначаемое как hSe1001 (гуманизированное антитело к Р-селектину, включающее CDR мышиного антитела G1, привитые на каркасные области антитела человеческого происхождения, как описано в международной заявке No. WO 2008/069999) также было подвергнуто скринингу методом, описанным в данной заявке.
Результаты, приведенные в Таблице 1, показывают, что антитело hSe1001 было связано с теми же самыми химерами (SEQ ID NO:1 и 20), что и антитело G1. Связывание hSe1001 было специфическим для конформационного эпитопа, описанного в данной заявке и расположенного внутри кластера А. Результат показали, что антитело hSe1001 обладает двойной функцией, заключающейся и в блокировании связывания Р-селектина с PSGL-1 (см. Фигуру 3), и в диссоциации комплексов Р-селектин/PSGL-1 (см. Фигуру 4). Таким образом, hSe1001 представляет собой другое антитело, охваченное данным изобретением, и может быть использовано для терапевтического лечения воспалительных и тромботических заболеваний, описанных в данной заявке, при котором блокируется связывание Р-селектина с PSGL-1 и ускоряется диссоциация ранее образовавшегося комплекса Р-селектин/PSGL-1.
Клеточный in vitro анализ роллинга в потоке человеческих нейтрофилов
Для дальнейшей оценки блокирующих и диссоциирующих свойств антител G1, G4 и hSe1001 был проведен клеточный in vitro анализ роллинга свежевыделенных человеческих нейтрофилов, которые вводились в виде потока с плотностью 1,0 дин/см2 в проточную камеру, стенки которой были покрыты мембранным Р-селектином в низкой и высокой концентрации. Р-селектин низкой плотности был нанесен с концентрацией 0,25 мкг/мл, а Р-селектин высокой плотности - с концентрацией 2,0 мкг/мл.
Величина плотности сайтов определялась при помощи I125-меченого mAb G1, она была равна 50 сайтов/мм2 (низкая плотность) и 380 сайтов/мм2 (высокая плотность). В случае Р-селектина низкой плотности нейтрофилы "перекатывались" вдоль сосудистой стенки со средней скоростью 5 мкм/мм2 или 6,5 мкм/мм2. В случае Р-селектина высокой плотности нейтрофилы "перекатывались" со средней скоростью 1 мкм/мм2. Нейтрофилы вводили в буферной среде в виде потока, затем начинались роллинг и прикрепление нейтрофилов. После достижения равновесия вводили испытуемые антитела (G1, G3, G4 и hSe1001) в потоке буфера, не содержащего клеток. Существует латентный период, равный примерно 1 мин до достижения антителами камеры для испытаний. Затем с интервалами в 1 мин подсчитывали клетки, оставшиеся связанными, и выражали это количество в процентах связанных клеток. Результаты записывали с помощью видеомикроскопа в течение примерно 0-20 мин и затем их анализировали.
Результаты, показанные на панелях (А) и (С) на Фигуре 6, показали, что нейтрофилы перекатывались с большей скоростью по стенке сосуда в случае Р-селектина низкой плотности. Так, по мере высвобождения комплекса Р-селектин/PSGL-1 вследствие нормальной кинетики, работающей по принципу "включено-выключено", нейтрофилы проходили до следующего сайта связывания в Р-селектине большее расстояние с более высокой скоростью. По мере того, как комплекс высвобождается, ранее занятый Р-селектин становится доступным для связывания с антителами к Р-селектину. Таким образом, все четыре антитела, G1, G3, G4 и hSe1001, через 1-4 мин показали эквивалентную блокирующую функциональность.
Результаты, показанные на панелях (В) и (D) на Фигуре 6, показали, что в случае Р-селектина высокой плотности нейтрофилы перекатываются вдоль стенок сосудов гораздо медленнее (скорость 1 мкм/с), так как доступно большее число связывающих сайтов Р-селектина. Многие нейтрофилы останавливаются на Р-селектине высокой плотности. При этих условиях мышиные антитела G1 и G4 и гуманизированное антитело hSe1001 были способны высвобождать перекатывающиеся и прикрепляющиеся нейтрофилы при диссоциации комплекса Р-селектин/PSGL-1 сразу же или через 1-8 мин. В противоположность этим антителам антителу G3, направленному против Р-селектина, потребовались 14-16 мин для блокирования роллинга нейтрофилов. Это свидетельствовало о том, что антитело G3 было способно только соединяться с незанятыми сайтами Р-селектина и таким образом блокировать комплексы Р-селектин/PSGL-1, но было неспособно связывать и диссоциировать эти ранее образовавшиеся комплексы. Эти анализы, проведенные в потоке клеток, подтверждают результаты метода BIACORE, описанные ранее, которые показывают, что мышиные антитела G1 и G4 и гуманизированное антитело hSe1001 все обладали двойной функцией, состоящей в блокировании и диссоциации ранее образовавшихся комплексов Р-селектин/PSGL-1.
hSe1001 обладает усиленным связыванием по сравнению с мышиным антителом G1
Сродство к связыванию антителами G1 и hSe1001 Р-селектина анализировали при помощи метода поверхностного плазменного резонанса (Biacore). Растворимый человеческий Р-селектин был ковалентно связан с чипом Biacore CM5, и мышиное антитело G1 и гуманизированное антитело hSe1001 испытывали независимо при их впрыскивании на поверхность чипа.
Для того, чтобы далее оценить взаимодействие антител к Р-селектину с Р-селектином (являющимся антигеном), проводили анализ кинетических данных с помощью метода поверхностного плазменного резонанса (SensiQ). Анализ свойств мышиного моноклонального антитела к Р-селектину G1 и гуманизированного антитела hSe1001 проводили методом поверхностного плазмонного резонанса (SensiQ). Для проведения анализа кинетики связывания этих антител поверхность сенсорного чипа была функционализирована путем ковалентного присоединения рекомбинантного белка G. Мышиное антитело G1 вводили и улавливали на поверхности чипа путем впрыскивания раствора с концентрацией равной 20 нМ в течение 1 мин. Гуманизированное антитело hSe1001 также вводили и улавливали на функционализированной таким же образом поверхности отдельного сенсорного канала путем впрыскивания раствора с концентрацией равной 10 нМ в течение 1 мин. Вводили растворимый Р-селектин человеческого происхождения с концентрацией 100 нМ - 1,56 нМ и измеряли степень связывания в единицах ответа (RU). Данные, полученные для каждого антитела, анализировали с помощью компьютерной программы Qdat.
На Фигуре 7 показаны результаты определения связывания обоих антител при одной концентрации Р-селектина.
На основании этих результатов рассчитывали Kd (константу диссоциации) для антитела hSe1001, она была равна 5,89 нМ, а для мышиного антитела G1 эта константа была равна 8,94 нМ (см. Таблицу 2). Таким образом, в случае гуманизированного антитела к Р-селектину hSe1001 наблюдалось увеличение сродства к связыванию на 34% по сравнению с G1, мышиным моноклональным антителом к Р-селектину. Кроме того, Ka (константа ассоциации для for hSe1001) была на 64,7% больше, чем для G1, означая, что гуманизированное антитело к Р-селектину hSe1001 связывается с Р-селектином быстрее. Оба этих результата показывают, что гуманизированное антитело к Р-селектину hSe1001 имеет повышенную Kd (сродство) и уменьшенную Ka (скорость начального связывания) по сравнению с G1.
Антитела согласно данному изобретению, полученные любым из описанных в данной заявке способов, предпочтительно применяются при получении фармацевтических композиций, применяемых для терапевтического лечения патологического состояния, при этом такое лечение введение фармацевтической композиции для ослабления, изменения или ингибирования воспалительного ответа или тромбоза, или другого состояния.
Важной целью данного изобретения является применение антител или функционально активных фрагментов или вариантов этих антител для приготовления фармацевтических композиций и применение этих композиций для предотвращения и/или лечения воспалительных ответов или тромбоза, таких как, но без исключения, описанные в данной заявке.
Например, данное изобретение в частности относится, но без ограничения, к применению двойной функции антител к Р-селектину или фрагментов этих антител, описанных и идентифицированных в данной заявке, для лечения воспалительных, тромботических или других состояний или расстройств у приматов (включая людей), которые включают адгезию тромбоцитов, серповидных красных кровяных клеток, лейкоцитов, лимфоцитов и/или клеток эндотелия, причем такое состояние или расстройство включает (но без ограничения) по меньшей мере одно заболевание, включающее вазоокклюзионный болевой криз, вызванный наличием серповидных клеток, воспалительные заболевания кишечника (например, болезнь Крона, язвенный колит и энтерит), артрит (например, ревматоидный артрит, остеоартрит и псориазный артрит), отторжение имплантатов, реакцию трансплантат против хозяина, астму, хроническую обструктивную болезнь легких, псориаз, дерматит, сепсис, нефрит, эритематозную волчанку, склеродермию, ринит, анафилаксию, диабет, рассеянный склероз, атеросклероз, тромбоз, метастазирование опухолей, аллергические реакции, тироидит, ишемическое реперфузионное повреждение, например, вызванное инфарктом миокарда, инсультом или трансплантаией органов) и состояния, связанные с тяжелой травмой или хроническим воспалением, такие как, например, замедленная гиперчувствительность IV типа, связанная, например, с инфекцией, вызванной туберкулезной бациллой, или синдромом системного воспалительного ответа или полиорганной недостаточностью, или связано с перечисленными заболеваниями. Термин "примат", применяемый в данной заявке, относится к людям, обезьянам, включая, например, бабуинов, яванских макак, и шимпанзе, горилл, гиббонов и орангутангов. Другие патологические состояния, не перечисленные в данной заявке, но которые относятся к воспалительным или тромботическим состояниям или расстройствам, также можно лечить с помощью антител и композиций, описанных в данной заявке.
Антитела могут быть введены в фармацевтическую композицию или препарат согласно данному изобретению любым из обычных методов получения фармацевтических композиций, например, методами, описанными в последнем издании Remington′s Pharmaceutical Sciences или в других источниках. Композиция может быть в виде, подходящем для локальной или системной инъекции или инфузии и, как таковая, может быть получена в стерильной воде или в изотоничном физиологическом растворе или в растворе глюкозы. Эти композиции могут быть подвергнуты стерилизации обычными методами стерилизации, которые хорошо известны из уровня техники. Полученные водные растворы могут быть упакованы для последующего применения или отфильтрованы в асептических условиях и лиофилизированы, причем лиофилизированный препарат соединяются со стерильным водным раствором перед применением. Такая композиция может содержать фармацевтически приемлемые вспомогательные вещества в соответствии с физиологическими состояниями, такие как буферные вещества, агенты, регулирующие тоничность, и т.п., например, ацетат натрия, лактат натрия, хлорид натрия, хлорид калия и т.д. Концентрация белков может меняться в широких пределах, например, от менее примерно 0,01% до 15-20% или более по весу. Стандартная доза композиции может содержать, например, от примерно 1 мкг до примерно 1000 мг антитела или фрагмента антитела.
Антитела или фрагменты антител по данному изобретению могут вводиться топически или путем инъекции. Дозы прописываются лечащим врачом в соответствии с конкретным состоянием конкретного пациента, который подвергаются лечению. Дозы и частота введения тщательно подбираются и регулируются в соответствии с параметрами, определенными лечащим врачом. Предпочтительные методы введения могут представлять собой пероральный метод, введение путем ингаляции, подкожный, внутривенный, внутримышечный, интратрахеальный, внутрипузырный методы или интраперитонеальную инъекцию, введение может проводиться раз в 24-48 ч, или в неделю, раз каждые 14 дн, каждые 4 нед, например, в дозе равной от 0,01 до 1000 мг, особенно от 0.1 мг до 100 мг, в частности. 1-10 мг на кг веса. Доза может вводиться непрерывно при помощи катетера или в виде отдельных болюсов. Антитело по данному изобретению может быть введено в эффективном количестве, таком как, но без ограничения, количество между 1 нг/кг и 1 мкг/кг, от 0,01 мкг/кг до 50 мкг/кг, от 0,1 мкг/кг до 1 мкг/кг, от 1 мкг/кг до 5 мкг/кг, от 5 мкг/кг до 10 мкг/кг, от 10 мкг/кг до 50 мкг/кг, от 50 мкг/кг до 100 мкг/кг, от 100 мг/кг до 1 мг/кг, от 1 мг/кг до 10 мг/кг или от 10 мг/кг до 100 мг/кг веса.
Фармацевтические композиции, применяемые по данному изобретению, содержащие антитела, описанные в данной заявке, могут дополнительно включать другие терапевтические соединения, которые обычно прописываются лечащим врачом согласно конкретному состоянию и конкретному пациенту, который подвергается лечению, такие как противовоспалительные средства, причем такие соединения прописываются лечащим врачом в соответствии с конкретным состоянием конкретного пациента.
Как уже отмечалось в данной заявке, наличие связывания Р-селектина с PSGL-1 имеет функциональное значение в процессах взаимодействия серповидных красных кровяных клеток, лейкоцитов эндотелиальных клеток и тромбоцитов и/или адгезии микропузырьков, роллинга лейкоцитов, рекрутинга, агрегации, выделения лейкоцитами цитокинов, ускорения коагуляции и в других аспектах воспаления, тромбоза, коагуляции, иммунного ответа и сигнальной трансдукции, включая, но без ограничения, вазоокклюзивный болевой криз, вызванный наличием серповидных клеток, воспалительные заболевания кишечника (например, болезнь Крона, язвенный колит и энтерит), артрит (например, ревматоидный артрит, остеоартрит и псориазный артрит), отторжение имплантатов, реакцию трансплантат против хозяина, астму, хроническую обструктивную болезнь легких, псориаз, дерматит, сепсис, нефрит, эритематозную волчанку, склеродермию, ринит, анафилаксию, диабет, рассеянный склероз, атеросклероз, тромбоз, метастазирование опухолей, аллергические реакции, тироидит, ишемическое реперфузионное повреждение, например, вызванное инфарктом миокарда, инсультом или трансплантацией органов) и состояния, связанные с тяжелой травмой или хроническим воспалением, такие как, например, замедленная гиперчувствительность IV типа, связанная, например, с инфекцией, вызванной туберкулезной бациллой, или синдромом системного воспалительного ответа или полиорганной недостаточностью. Нейтрализующее антитело (или его фрагмент) к Р-селектину, описанное в данной заявке. будет ингибировать один или более видов указанной активности у пациента, опосредованных связыванием Р-селектина с PSGL-1 (или в случае наличия серповидных красных кровяных клеток связыванием Р-селектина с PSGL-1-подобным рецептором), например, in vivo or in vitro. Таким образом, ингибирование связывания Р-селектина с PSGL-1 нейтрализующим антителом (или его фрагментом), описанным в данной заявке, полезно при лечении различных состояний и расстройств у пациента, включая, но без ограничения, заболевания, описанные в данной заявке.
Антитела, специфические к Р-селектину, или их фрагменты связывания, описанные в данной заявке, могут быть связаны с другой молекулой. Например, эти антитела могут быть связаны с другим пептидом или белком, токсином, радиоизотопом, цитотоксичным или цитостатическим агентом.
Антитела могут быть связаны ковалентно за счет химического сшивания или рекомбинантными методами. Антитела могут быть также связаны с одним из многих небелковых полимеров, например, с полиэтиленгликолем, полипропиленгликолем или полиоксиалкиленами, способами, описанными в патентах США №№4,640,835; 4,496,689; 4,301,144; 4,670,417; 4,791,192 или 4,179,337. Антитела могут быть химически модифицированы путем ковалентного соединения с полимером, например, для увеличения их стабильности или периода полужизни. Примеры таких полимеров и методов их присоединения описаны также в патентах США №№4,766,106; 4,179,337; 4,495,285 и 4,609,546.
Антитела (или их фрагменты) могут быть также мечены детектируемой меткой. Детектируемая метка представляет собой молекулу, которая по своей химической природе обеспечивает появление идентифицируемого сигнала, который позволяет обнаружить молекулярное взаимодействие. Белок, включающий антитело, имеет детектируемую метку, если она ковалентно или не ковалентно связана с молекулой, которая распознается непосредственно (например, с помощью хромофора, флуорофора или радиоизотопа) или косвенно (например, путем катализа реакции, в результате которой образуется окрашенное, люминесцентное или флуоресцентное вещество).
Детектируемые метки включают радиометку, такую как 131I or 99Tc, тяжелый металл или флуоресцентный субстрат, такой как, например, европий, который может быть также присоединен к антителам обычными химическими методами. Детектируемые метки включают также ферментные метки, такие как пероксидазу хрена или щелочную фосфатазу. Они могут также включать химические вещества, такие как биотин, который может быть обнаружен при связывании с конкретной родственной обнаруживаемой молекулой, например, меченым авидином.
Данное изобретение относится также к способам скрининга антител к Р-селектину и их фрагментов, которые блокируют и связывание Р-селектина с PSGL-1 и/или вызывают диссоциацию ранее образовавшихся комплексов РН-селектин/PSGL-1. Как уже отмечалось выше, настоящее изобретение относится к антителам к Р-селектину, к хозяйским клеткам, которые продуцируют такие антитела к Р-селектину, к векторам, которые содержат ДНК, кодирующие такие антитела к Р-селектину, к методам экспрессии и скрининга с целью идентификации антител к Р-селектину, которые блокируют и связывание Р-селектина с PSGL-1 и согласно другому варианту имеют "двойную функцию", вызывая диссоциацию ранее образовавшихся комплексов РН-селектин/PSGL-1. Так, согласно одному варианту данного изобретения оно относится к способам идентификации антител к Р-селектину, которые специфически связываются с конформационным эпитопом в последовательности, содержащей аминокислоты 1-35 и, более предпочтительно, аминокислоты 4-23, человеческого Р-селектина (SEQ ID NO:1), таким как конформационный эпитоп, описанный в данной заявке), и которые удерживают PSGL-1 или его миметики от связывания с Р-селектином и которые могут менять направление такого связывания, тем самым проявляя двойную функцию, заключавшуюся при блокировании адгезии, опосредованной Р-селектином вследствие связывания Р-селектина с PSGL-1 и диссоциации ранее образовавшихся комплексов РН-селектин/PSGL-1.
Согласно предпочтительным вариантам способ скрининга включает in vitro анализы, основанные на использовании жидкой среды или субстратов, которые могут быть использованы для идентификации антител к Р-селектину или их фрагментов, которые ингибируют или разрушают связывание Р-селектина с PSGL-1 и, предпочтительно, также вызывают диссоциацию ранее образовавшихся комплексов Р-селектин/PSGL-1. Скрининг испытуемых антител проводят для выявления двойной функции с помощью ряда методов анализа, таких как, но без ограничения, методы, описанные в данной заявке, которые дают возможность идентифицировать такие антитела, которые связываются с конформационным эпитопом аминокислот 1-35 и более предпочтительно, аминокислот 4-23, в Р-селектине и которые блокируют связывание лиганда PSGL-1 с Р-селектином и которые, предпочтительно, вызывают диссоциацию ранее образовавшихся комплексов Р-селектин/PSGL-1. До сих пор не были выявлены антитела к Р-селектину, которые имели бы способность как блокировать связывание лиганда PSGL-1 с Р-селектином, так и вызывать диссоциацию ранее образовавшихся комплексов Р-селектин/PSGL-1. Используемый в данной заявке термин "испытуемое антитело" относится к целым антителам или к их фрагментам. Испытуемое антитело может быть членом библиотеки (например, фагов, дрожжей или бактерий) или фрагментом антитела в этой библиотеке. Библиотеку можно сузить, исключив из нее все члены, которые связывали нежелательные эпитопы.
Например, на первой стадии способа скрининга испытуемые антитела, направленные против Р-селектина, анализировались на способность к блокированию связывания PSGL-1 с Р-селектином. Испытуемые антитела, которые блокируют связывание PSGL-1 с Р-селектином, подвергали скринингу для определения их способности вызывать диссоциацию ранее образовавшихся комплексов Р-селектин/PSGL-1. Те антитела, которые были идентифицированы как обладающие двойной функцией, состоящей в блокировании связывания PSGL-1 с Р-селектином, а также способности вызывать диссоциацию комплекса Р-селектин/PSGL-1, являются особенно предпочтительным вариантом данного изобретения и могут быть использованы при осуществлении способов согласно настоящему изобретению. Примеры вариантов этого вида анализа с применением жидкой среды включают, но без ограничения, (1) метод FACS, осуществляемый с лейкоцитами или HL60/активированными тромбоцитами, которые анализируют клеточные агрегаты, которые были или не были подвергнуты действию антитела, способного связывать Р-селектин (например, антитела hSe1001) или антитела, вызывающего диссоциацию комплекса Р-селектин/PSGL-1, (2) метод анализа с применением жидкой среды, основанный на использовании комплекса Р-селектин/GSP-6-стрептавидин-биотин, который был или не был подвергнут действию антитела, способного связывать Р-селектин (например, антитела hSe1001) или испытуемого антитела и проанализирован при помощи SEC (эксклюзионной хроматографии, гель-хроматографии) и 93) метод с использованием гранул AlphaLisa в качестве субстрата, применяемого при проведении анализа с жидкой средой.
Согласно одному из вариантов такого способа испытуемые антитела, которые блокируют связывание PSGL-1 с Р-селектином, вначале идентифицируют способом скрининга. Например, согласно предпочтительному варианту этого метода скрининга способом, известным специалисту в данной области, получают PSGL-1 или синтетический миметик PSGL-1, такой как GSP-6, или концевой эпитоп, способный связывать Р-селектин (или, возможно, связываться с подложкой субстрата, такого как чип BIACORE). Затем PSGL-1 (или миметик PSGL-1) (который может быть связан с подложкой) подвергается воздействию комплекса испытуемое антитело к Р-селектину/Р-селектин. Затем оценивается степень связывания комплекса с подложкой для PSGL-1. Если комплекс испытуемое антитело/Р-селектин не связывается с PSGL-1, это испытуемое антитело рассматривается как " блокирующее функцию" антитело. Согласно одному из вариантов GSP-6 или миметик PSGL-1 связан с биотином. Комплекс GSP-6/миметик-биотин сам может быть, например, связан со стрептавидином в покрытии на носителе.
Согласно другому варианту метода скрининга берут Р-селектин или его часть, которая сохраняет целостность конформационного эпитопа, возможно, связанные с подложкой субстрата. Например, часть Р-селектина, которая сохраняет конформационный эпитоп, включает, но без ограничения, последовательность, содержащую связывающие домены лектина и EGF Р-селектина (например, аминокислоты 1-153 в SEQ ID NO:1). В соответствии с этим вариантом Р-селектин или его часть, включающая конформационный эпитоп, подвергаются действию испытуемого антитела, которое связывается с ними с образованием комплекса Р-селектин-антитело. Затем на PSGL-1 или его высокомолекулярный миметик, такой как комплекс GSP-6/биотин/авидин, действуют комплексом Р-селектин/испытуемое антитело и определяют степень его связывания с PSGL-1 (или его миметиком). Антитело, которое предотвращает или ингибирует связывание PSGL-1 с комплексом Р-селектин/испытуемое антитело, рассматривается как "блокирующее функцию" антитело.
Согласно еще одному варианту метода скрининга или PSGL-1, или его миметик является связанным, как описано выше, с подложкой субстрата (такой как гранула или чип BIACORE). Затем Р-селектин наносится на комплекс PSGL-1/подложка с образованием комплекса Р-селектин/PSGL-1 (или его миметик). Затем вносят испытуемое антитело и оценивают диссоциацию комплекса как уменьшение массы или как единицы ответа (RU) в результате диссоциации. Функционально блокирующее антитело к Р-селектину, которое индуцирует диссоциацию комплекса Р-селектин/PSGL-1, считается антителом к Р-селектину, обладающим двойной функцией.
В соответствии с альтернативным вариантом этого метода скрининга Р-селектин или часть молекулы Р-селектина, включающая конформационный эпитоп, может быть связана скорее с подложкой носителя, чем с PSGL-1. Затем PSGL-1 или его высокомолекулярный миметик, такой как комплекс GSP-6/биотин/авидин, подвергается действию Р-селектина на подложке носителя и образуется комплекс PSGL-1/Р-селектин. Затем на комплекс PSGL-1/Р-селектин действует блокирующее функцию антитело к Р-селектину и оценивается степень диссоциации комплекса, например, с применением биосенсорной системы BIACORE, что описано в данной заявке.
Согласно еще одному варианту этого метода само антитело к Р-селектину связано с подложкой субстрата, и комплекс PSGL-1/Р-селектин подвергается его действию, после чего оценивается степень диссоциации, как описано выше.
Хотя настоящее изобретение и его преимущества были подробно описаны, следует иметь в виду, что, не выходя за рамки и замысел изобретения, можно осуществить различные изменения и замены. Более того, объем данного изобретения не ограничивается конкретными вариантами способа, элементами, композициями, средствами, методами и стадиями, описанными в данной заявке. Как очевидно среднему специалисту в данной области из приведенного выше описания, в соответствии с данным изобретением могут быть использованы способы, элементы, композиции, средства, методы и стадии, уже существующие или те, которые будут позже разработаны, обладающие теми же функциями или приводящие к достижению такого же результата, что и описанные в данной заявке. Соответственно, изобретение, описанное в данной заявке, призвано охватывать такие способы, элементы, композиции, средства, методы или стадии.
Каждый из источников, патентов или публикаций, упомянутых в данной заявке, включая, но без ограничения, заявки США на патент №№12/974,539; 12/974,739; 12/516,987 и международную заявку WO 2008/069999, включены в данную заявку посредством отсылок.
Ссылки, приведенные в данной заявке
1. Springer ТА. Traffic signals for lymphocyte recirculation and leukocyte emigration: the multistep paradigm. Cell, 1994. 76:301-314. PubMed: 7507411.
2. McEver RP, Moore KL, Cummings RD. Leukocyte trafficking mediated by selectin-carbohydrate interactions. J Biol Chem., 1995. 12; 270(19):11025-8. Review. PMID: 7538108.
3. Zimmerman GA, McIntyre TM, Prescott SM. Adhesion and signaling in vascular cell-cell interactions. J Clin Invest., 1996. 15; 98(8):1699-702. No abstract available. PMID: 8878418.
4. Frenette PS. Sickle cell vasoocclusion: heterotypic, multicellular aggregations driven by leukocyte adhesion. Microcirculation, 2004. 11(2):167-77. Review. PMID: 15280090.
5. Weyrich AS, Ma XY, Lefer DJ, Albertine KH, Lefer AM. In vivo neutralization of P-selectin protects feline heart and endothelium in myocardial ischemia and reperfusion injury. J Clin Invest., 1993. 91(6):2620-9. PMID: 7685773.
6. Lefer DJ. Pharmacology of selectin inhibitors in ischemia/reperfusion states. Annu Rev Pharmacol Toxicol., 2000. 40:283-94. Review. PMID: 10836137.
7. Singbartl K, Green SA, Ley K. Blocking P-selectin protects from ischemia/reperfusion-induced acute renal failure. FASEB J., 2000. 14(1):48-54. PMID: 10627279.
8. Palabrica Т, Lobb R, Furie ВС, Aronovitz M, Benjamin C, Hsu YM, Sajer SA, Furie B. Leukocyte accumulation promoting fibrin deposition is mediated in vivo by P-selectin on adherent platelets. Nature, 1992. 29; 359(6398):848-51. PMID: 1279433.
9. Burger PC, Wagner DD. Platelet P-selectin facilitates atherosclerotic lesion development. Blood, 2003. 1; 101(7):2661-6. Epub 2002 Dec 12. PMID: 12480714.
10. Theoret JF, Bienvenu JG, Kumar A, Merhi Y. P-selectin antagonism with recombinant p-selectin glycoprotein ligand-1 (rPSGL-Ig) inhibits circulating activated platelet binding to neutrophils induced by damaged arterial surfaces. J Pharmacol Exp Ther., 2001. 298(2):658-64. PMID: 11454928.
11. Kumar A, Villani MP, Patel UK, Keith JC Jr, Schaub RG. Recombinant soluble form of PSGL-1 accelerates thrombolysis and prevents reocclusion in a porcine model. Circulation, 1999. 16; 99(10):1363-9. PMID: 10077522.
12. Romano SJ. Selectin antagonists: therapeutic potential in asthma and COPD. Treat Respir Med., 2005. 4(2):85-94. Review. PMID: 15813660.
13. Grober JS, Bowen BL, Ebling H, Athey B, Thompson CB, Fox DA, Stoolman LM. Monocyte-endothelial adhesion in chronic rheumatoid arthritis. In situ detection of selectin and integrin-dependent interactions. J Clin Invest., 1993. 91(6):2609-19. PMID: 7685772.
14. Friedrich M, Bock D, Philipp S, Ludwig N, Sabat R, Wolk K, Schroeter-Maas S, Aydt E, Kang S, Dam TN, Zahlten R, Sterry W, Wolff G. Pan-selectin antagonism improves psoriasis manifestation in mice and man. Arch Dermatol Res., 2006. 297(8):345-51. Epub 2005 Dec 16. PMID: 16362415.
15. Johnson BA, Haines GK, Harlow LA, Koch AE. Adhesion molecule expression in human synovial tissue. Arthritis Rheum, 1993. 36(2):137-46. PMID: 7679271.
16. Bevilacqua MP, Nelson RM. Selectins. J Clin Invest., 1993. 91(2):379-87. Review. PMID: 7679406.
17. McEver RP, Beckstead JH, Moore KL, Marshall-Carlson L, Bainton DF. GMP-140, a platelet alpha-granule membrane protein, is also synthesized by vascular endothelial cells and is localized in Weibel-Palade bodies. J Clin Invest., 1989. 84(1):92-9. PMID: 2472431.
18. Sickle cell disease: screening, diagnosis, management, and counseling in newboms and infants. Clinical Practice Guideline No. 6. AHCPR Pub. No. 93-0562. Rockville, MD: Agency for Health Care Policy and Research, Public Health Service, U.S. Department of Health and Human Services. April 1993 [cited; Available from: www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=hstat6.chapter.16946.
19. Nietert, PJ, Silverstein MD, Abboud MR. Sickle cell anaemia: epidemiology and cost of illness. Pharmacoeconomics, 2002. 20(6): p.357-66. PMID: 12052095.
20. Bookchin, RM and Lew VL. Pathophysiology of sickle cell anemia. Hematol Oncol Clin North Am, 1996. 10(6): p.1241-53. PMID: 8956013.
21. NHBLI. [cited; Available from: www.nhlbi.nih.gov/health/dci/Diseases/Sca/SCA_WhatIs.html.
22. NHLBI, The Management of Sickle Cell Disease. NIH Publication No.022117.
23. FDA Approves Droxia for Sickle Cell Anemia, in FDA Talk Paper. 1998. p.T98-11.
24. NIH, Hydroxurea Treatment for Sickle Cell Disease February 25-27 2008. National Institutes of Health Consensus Development Conference Statement, February 27 2008.
25. Gill FM, Sleeper LA, Weiner SJ, Brown AK, Bellevue R, Grover R, Pegelow CH, Vichinsky E. Clinical events in the first decade in a cohort of infants with sickle cell disease. Cooperative Study of Sickle Cell Disease. Blood, 1995. 86(2): p.776-83. PMID: 7921116.
26. Ashley-Koch A, Yang Q, and Olney RS. Sickle hemoglobin (HbS) allele and sickle cell disease: a HuGE review. Am J Epidemiol, 2000. 151(9): p.839-45. PMID: 10797557.
27. Thomas PW, Higgs DR, and Serjeant GR. Benign clinical course in homozygous sickle cell disease: a search for predictors. J Clin Epidemiol, 1997. Feb; 50(2): p.121-6. PMID: 9120504.
28. National Newborn Screening Status Report Updated 03/04/08.
29. Rice-Evans С, Omorphos SC, and Baysal E. Sickle cell membranes and oxidative damage. Biochem J, 1986. 237(1): p.265-9. PMID: 3800879.
30. Wethers, D.L., Sickle cell disease in childhood: Part II. Diagnosis and treatment of major complications and recent advances in treatment. Am Fam Physician. 2000 Sep 15; 62(6):1309-14. PMID: 11011859.
31. Gladwin MT, Sachev V, Jison ML, Shizukuda Y, Plehn JF, Minter K, Brown B, Coles WA, Nichols JS, Ernst I, Hunter LA, Blackwelder WC, Schechter AN, Rodgers GP, Castro O, Ognibene FP, Pulmonary hypertension as a risk factor for death in patients with sickle cell disease. N Engl J Med, 2004. 350(9): p.886-95. PMID: 14985486.
32. Saborio P and Scheinman JI, Sickle cell nephropathy. J Am Soc Nephrol, 1999. 10(1): p.187-92. PMID: 9890326.
33. Charache S, Eye disease in sickling disorders. Hematol Oncol Clin North Am, 1996. 10(6): p.1357-62. PMID: 8956022.
34. Kaul, DK, Liu XD, Fabry ME, Nagel RL. Impaired nitric oxide-mediated vasodilation in transgenic sickle mouse. Am J Physiol Heart Circ Physiol, 2000. 278(6): p. H1799-806. PMID: 10843875.
35. Nolan, VG, Adewoye A, Baldwin C, Wang L, Ma Q, Wyszynski DF, Farrell JJ, Sebastiani P, Parrer LA, Steinberg MH. Sickle cell leg ulcers: associations with haemolysis and SNPs in Klotho, TEK and genes of the TGF-beta/BMP pathway. Br J Haematol, 2006. 133(5): p.570-8. PMID: 1661647.
36. Nolan, VG, Wyszynski DF, Farrer LA, Steinberg MH. Hemolysis-associated priapism in sickle cell disease. Blood, 2005. 106(9): p.3264-7. PMID: 15985542.
37. Gladwin MT and Kato GJ. Cardiopulmonary complications of sickle cell disease: role of nitric oxide and hemolytic anemia. Hematology Am Soc Hematol Educ Program, 2005: p.51-7. PMID: 16304359.
38. Yale, SH, Nagib N, and Guthrie T. Approach to the vaso-occlusive crisis in adults with sickle cell disease. Am Fam Physician, 2000. 61(5): p.1349-56, 1363-4. PMID: 10735342.
39. Heegaard, ED and Brown KE. Human parvovirus B19. Clin Microbiol Rev, 2002. 15(3): p.485-505. PMID: 12097253.
40. Ballas, SK and Mohandas N. Pathophysiology of vaso-occlusion. Hematol Oncol Clin North Am, 1996. 10(6): p.1221-39. PMID: 8956012.
41. Embury, SH. The not-so-simple process of sickle cell vasoocclusion. Microcirculation, 2004. 11(2): p.101-13. PMID: 15280086.
42. Conran N and Costa FF. Hemoglobin disorders and endothelial cell interactions. Clin Biochem, 2009. 42(18): p.1824-38. PMID: 19580799.
43. Chiang EY and Frenette PS. Sickle cell vaso-occlusion. Hematol Oncol Clin North Am, 2005. 19(5): p.771-84, Review. PMID: 16214643.
44. Turhan A, Weiss LA, Mohandas N, Collier BS, Freette PS. Primary role for adherent leukocytes in sickle cell vascular occlusion: a new paradigm. Proc Natl Acad Sci USA, 2002. 99(5): p.3047-51. PMID: 1180644.
45. Hebbel RP, Osarogiagbon R, and Kaul D. The endothelial biology of sickle cell disease: inflammation and a chronic vasculopathy. Microcirculation, 2004. 11(2): p.129-51. Review. PMID: 15280088.
46. Okpala I, Leukocyte adhesion and the pathophysiology of sickle cell disease. Curr Opin Hematol, 2006. 13(1): p.40-4. Review. PMID: 16319686.
47. Matsui NM, Borsig L, Rosen SD, Yaghmai M, Varki A, Embury SH P-selectin mediates the adhesion of sickle erythrocytes to the endothelium. Blood, 2001. 98(6): p.1955-62. PMID: 11535535.
48. Platt OS, Brambilla DJ, Rosse WF, Milner PF, Castro O, Steinberg MH, Klug PP. Mortality in sickle cell disease. Life expectancy and risk factors for early death. N Engl J Med, 1994. 330(23): p.1639-44. PMID: 7993409.
49. Smith JA. Bone disorders in sickle cell disease. Hematol Oncol Clin North Am, 1996. 10(6): p.1345-56. Review. PMID: 8956021.
50. Platt OS. Prevention and management of stroke in sickle cell anemia. Hematology Am Soc Hematol Educ Program, 2006: p.54-7. Review. PMID: 17124040.
51. Maitre В, Habibi A, Roudot-Thoraval F, Bachir D, Belghiti DD, Galacteros F, Godeau B. Acute chest syndrome in adults with sickle cell disease. Chest, 2000. 117(5): p.1386-92. PMID: 1087826.
52. Vichinsky EP, Neumayr LD, Earles AN, Williams R, Lennette ET, Dean D, Nickerson B, Orringer E, McKie V, Bellevue R, Daeschner C, Manci EA. Causes and outcomes of the acute chest syndrome in sickle cell disease. National Acute Chest Syndrome Study Group. N Engl J Med, 2000. 342(25): p.1855-65. PMID: 10861320.
53. Serjeant GR, Ceulaer CD, Lethbridge R, Morris J, Singhal A, Thomas PW. The painful crisis of homozygous sickle cell disease: clinical features. Br J Haematol, 1994. 87(3): p.586-91. PMID: 7993 801.
54. Platt OS, Thorington BD, Brambilla DJ, Milner PF, Rosse WF, Vichinsky E, Kinney TR. Pain in sickle cell disease. Rates and risk factors. N Engl J Med, 1991. 325(1): p.11-6. PMID: 1710777.
55. Health Advances, LLC. Application of Selexys Technologies to Inflammatory and Thrombotic Diseases. In Private Study. 2004.
56. McClish DK, Health related quality of life in sickle cell patients: the PiSCES project. Health Qual Life Outcomes, 2005. 3: p.50. PMID: 16129027.
57. Charache S, Terrin ML, Moore RD, Dover GJ, Barton FB, Eckert SV, McMahon RP, Bonds DR. Effect of hydroxyurea on the frequency of painful crises in sickle cell anemia. Investigators of the Multicenter Study of Hydroxyurea in Sickle Cell Anemia. N Engl J Med, 1995. 332(20): p.1317-22. PMID: 7715639.
58. Patient Information Leaflet: Droxia. Bristol-Myers Squibb Company, 2006.
59. NIH. NIH State-of-the-Science Conference Statement on Hydroxyurea Treatment for Sickle Cell Disease. NIH Consensus and State-of-the Science Statements; February 27-29, 2008. 25(1):1-30. PMID: 18309362.
60. Hydroxyurea for the Treatment of Sickle Cell Disease, Agency for Healthcare Research and Quality; AHRQ Publication No. 08-E007. Feb. 2008.
61. Shet AS, Aras O, Gupta K, Hass MJ, Rausch DJ, Saba N, Koopmeiners L, Key NS, Hebbel RP. Sickle blood contains tissue factor-positive microparticles derived from endothelial cells and monocytes. Blood, 2003. 102(7): p.2678-83. PMID: 12805058.
62. Solovey A, Gui L, Key NS, Hebbel RP. Tissue factor expression by endothelial cells in sickle cell anemia. J Clin Invest, 1998. 101(9): p.1899-904. PMID: 9576754.
63. Solovey A, Lin Y, Browne P, Choong S, Wayner E, Hebbel RP. Circulating activated endothelial cells in sickle cell anemia. N Engl J Med, 1997. 337(22): p.1584-90. PMID: 9371854.
64. Kaul DK and Hebbel RP. Hypoxia/reoxygenation causes inflammatory response in transgenic sickle mice but not in normal mice. J Clin Invest, 2000. 106(3): p.411-20. PMID: 10930444.
65. Matsumoto S, Okabe Y, Setoyama H, Takayama K, Ohtsuka J, Funahashi H, Imaoka A, Okada Y, and Umesaki Y. Inflamatory bowel disease-like enteritis and caecitis in a senescence accelerated mouse P1/Yit strain. Gut, 1998. 43:71-78. PMID: 9771408.
66. Kosiewicz MM, Nast CC, Krishnan A, Rivera-Nieves J, Moskaluk CA, Matsumoto S, Kozaiwa K, and Cominelli F. Th1-type responses mediate spontaneous ileitis in a novel murine model of Crohn′s disease. J. Clin. Invest., 2001 107:695-702. PMID: 11254669.
67. Rivera-Nieves J, Burcin TL, Olson TS, Morris MA, McDuffie M, Cominelli R and Ley K. Critical role of endothelial P-selectin glycoprotein ligand 1 in chronic murine ileitis. JEM, 2006. 203(4):907-917. PMID: 16567389.
68. Moore KL, Stults NL, Diaz S, Smith DF, Cummings RD, Varki A, McEver RP. Identification of a specific glycoprotein ligand for P-selectin (CD62) on myeloid cells. J Cell Biol, 1992; 118:445-456. PMID: 1378449.
69. McEver RP, Cummings RD. Role of PSGL-1 binding to selectins in leukocyte recruitment. J Clin Invest, 1997; 100:S97-103. PMID: 9413410.
70. Kansas GS. Selectins and their ligands: current concepts and controversies. Blood, 1996; 88:3259-3287. PMID: 8896391.
71. Somers WS, Tang J, Shaw GD, Camphausen RT. Insights into the molecular basis of leukocyte tethering and rolling revealed by structures of P- and E-selectin bound to SLe(X) and PSGL-1. Cell, 2000. Oct 27; 103(3):467-79. Erratum in: Cell, 2001. Jun 29; 105(7):971. PMID: 11081633.
72. Mayadas TN, Johnson RC, Rayburn H, Hynes RO, Wagner DD. Leukocyte rolling and extravasation are severely compromised in P selectin-deficient mice. Cell, 1993. Aug 13; 74(3):541-54. PMID: 7688665.
73. Xia L, Sperandio M, Yago T, McDaniel JM, Cummings RD, Pearson-White S, Ley K, McEver RP. P-selectin glycoprotein ligand-1-deficient mice have impaired leukocyte tethering to E-selectin under flow. J Clin Invest, 2002. 109:939-950. PMID: 11927621.
74. Yang J, Hirata T, Croce K, Merrill-Skoloff G, Tchernychev B, Williams E, Flaumenhaft R, Furie ВС, Furie B. Targeted gene disruption demonstrates that P-selectin glycoprotein ligand 1 (PSGL-1) is required for P-selectin-mediated but not E-selectin-mediated neutrophil rolling and migration. J Exp Med, 1999. Dec 20; 190(12):1769-82. PMID: 10601352
75. Walcheck B, Moore KL, McEver RP, and Kishimoto TK. Neutrophil-neutrophil interactions under hydrodynamic shear stress involve L-selectin and PSGL-1. A mechanism that amplifies initial leukocyte accumulation on P-selectin in vitro. J. Clin. Invest., 1996. 98:1081-1087. PMID: 8787668.
76. Bargatze RF, Kurk S, Butcher EC, Jutila MA. Neutrophils roll on adherent neutrophils bound to cytokine-induced endothelial cells via L-selectin on the rolling cells. J Exp Med., 1994. Nov 1; 180(5):1785-92. PMID: 7525838.
77. Jung U and Ley K. Mice Lacking Two or All Three Selectins Demonstrate Overlapping and Distinct Functions for Each Selectin. J. Immunol., 1999. 162:6755-6762. PMID: 10352295.
78. Antibody Engineering, 2nd edition., Borrebaeck CA, Ed., Oxford University Press (1995).
79. Sambrook J and Russell D. Molecular Cloning: A Laboratory Manual, 2nd edition. Cold Spring Harbor Laboratory Press (1989).
80. Bodansky M and Bodansky A. The Practice of Peptide Synthesis, 2nd edition., Springer-Verlag, Berlin. 1995.
81. Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alaignment search tool. J. Mol. Biol., 1990. 215:403-410 PMID: 2231712.
82. Needleman SB, Wunsch CD. A general method applicable to the search for similarities in the amino acid sequence of two proteins. J. Mol. Biol., 1970. 48(3):444-453 PMID: 5420325.
83. Meyers EW and Miller W. Optimal alignments in linear space. Comput. Appl. Biosci., 1988. 4(1):11-17. PMID: 3382986.
84. Tatusova ТА and Madden TL. BLAST 2 Sequences, a new tool for comparing protein and nucleotide sequences. FEMS Microbiol. Lett., 1999. 174(2):247-250. PMID: 10339815.
85. Kohler G and Milstein C. Continuous cultures of fused cells secreting antibody of predefined specificity. Nature, 1975. 256(5517): 495-7. PMID: 1172191.
86. Ausubel FM. Current Protocols in Molecular Biology, Greene Publishing Associates and Wiley Interscience, N.Y., 1989).
87. Wigler M, Pellicer A, Silverstein S, Axel R. Biochemical transfer of single-copy eukaryotic genes using total cellular DNA as donor. Cell, 1978. 14(3):725-31. PMID: 210957.
88. Neumann E, Schaefer-Ridder M, Wang Y, Hofschneider PH. EMBO, 1982. J. 1(7):841-5. PMID: 6329708.
89. Harlow E and Lane D. Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, New York, 1988.
90. Geng JG, Moore KL, Johnson AE, McEver RP. Neutrophil recognition requires a Ca2+-induced conformational change in the lectin domain of GMP-140. J. Biol. Chem., 1991. 266(33):22313-22318. PMID: 1718992.
91. Mehta P, Patel KD, Laue TM, Erickson HP, Mcever RP. Souluble monomeric P-selectin containing only the lectin and epidermal growth factor domains binds to P-selectin glycoprotein ligand-1 on leukocytes. Blood, 1997. Sep.15; 90(6):2381-9. PMID: 9310489.
92. Hirose M, Kawashima H, Miyasaka M. A functional epitope on P-selectin that supports binding of P-selectin to P-selectin glycoprotein ligand-1 but not to sialyl Lewis X oligosaccharides. Int. Immunol., 1998. May; 10(5):639-49. PMID: 9645612.
93. Ruchaud-Sparangano MH, Malaud E, Gayet O, Chignier E, Buckland R, McGregor JL. Mapping the epitope of a functional P-selectin monoclonal antibody (LYP20) to a short complement-like repeat (SCR 4) domain: use of human-mouse chimera and homologue-replacement mutagenesis. Biochem. J., 1998. 1; 332(Pt2):309-14. PMID: 9601057.
94. Chestnut, RW, Polley, MJ, Paulson, JC, Hones, T, Saldanha, JW, Bendig, MM, Kriegler, M. Perez, C, Bayer, R Nunn, M. Antibodies to P-selectin and Their Uses. US Patent No. 5,800,815. Sep.1, 1998.
95. Berg EL, Fromm C, Melrose J, Tsurushita N. Antibodies cross-reactive with E- and P-selectin block both E- and P-selectin functions. Blood, 1995. Jan 1; 85(1):31-7. PMID: 7528571.
96. Leppanen A, Mehta P, Ouyang YB, Ju T, Helin J, Moore KL, van Die I, Canfield WM, McEver RP, Cummings RD. 1999. A novel glycosulfopeptide binds to P-selectin and inhibits leukocyte adhesion to P-selectin. J Biol Chem Aug 27; 274(35):24838-48. PMID: 10455156.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА | 2005 |
|
RU2482131C2 |
| АНТИ-PSGL-1-АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2650817C2 |
| ЧЕТЫРЕХВАЛЕНТНЫЕ АНТИТЕЛА К PSGL-1 И ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2766000C2 |
| МОДУЛЯТОРЫ Р-СЕЛЕКТИН ГЛИКОПРОТЕИН ЛИГАНДА 1 | 2004 |
|
RU2407539C2 |
| АНТИТЕЛО ПРОТИВ IGF-1R И ЕГО ПРИМЕНЕНИЕ В КАЧЕСТВЕ АДРЕСУЮЩЕГО ПЕРЕНОСЧИКА ДЛЯ ЛЕЧЕНИЯ РАКА | 2015 |
|
RU2698977C2 |
| СПОСОБЫ ЛЕЧЕНИЯ ВОСПАЛИТЕЛЬНОЙ БОЛИ | 2009 |
|
RU2467765C2 |
| АНТИТЕЛА К ГЕПСИДИНУ И ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2668391C2 |
| СПОСОБЫ ЛЕЧЕНИЯ ХРОНИЧЕСКОЙ БОЛИ | 2009 |
|
RU2522493C2 |
| АНТИТЕЛА ПРОТИВ SEZ6 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2691698C2 |
| РЕКОМБИНАНТНЫЕ IL4-АНТИТЕЛА, ИСПОЛЬЗУЕМЫЕ ДЛЯ ЛЕЧЕНИЯ НАРУШЕНИЙ, СВЯЗАННЫХ С ДЕЙСТВИЕМ IL4 | 1994 |
|
RU2162711C2 |














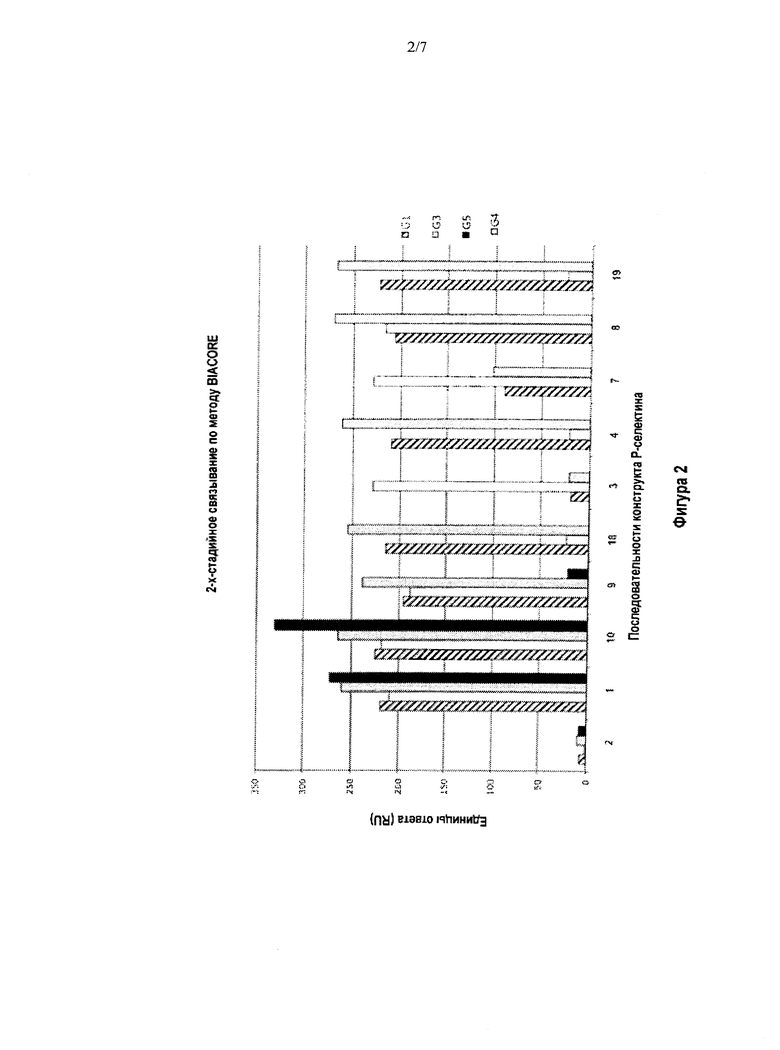
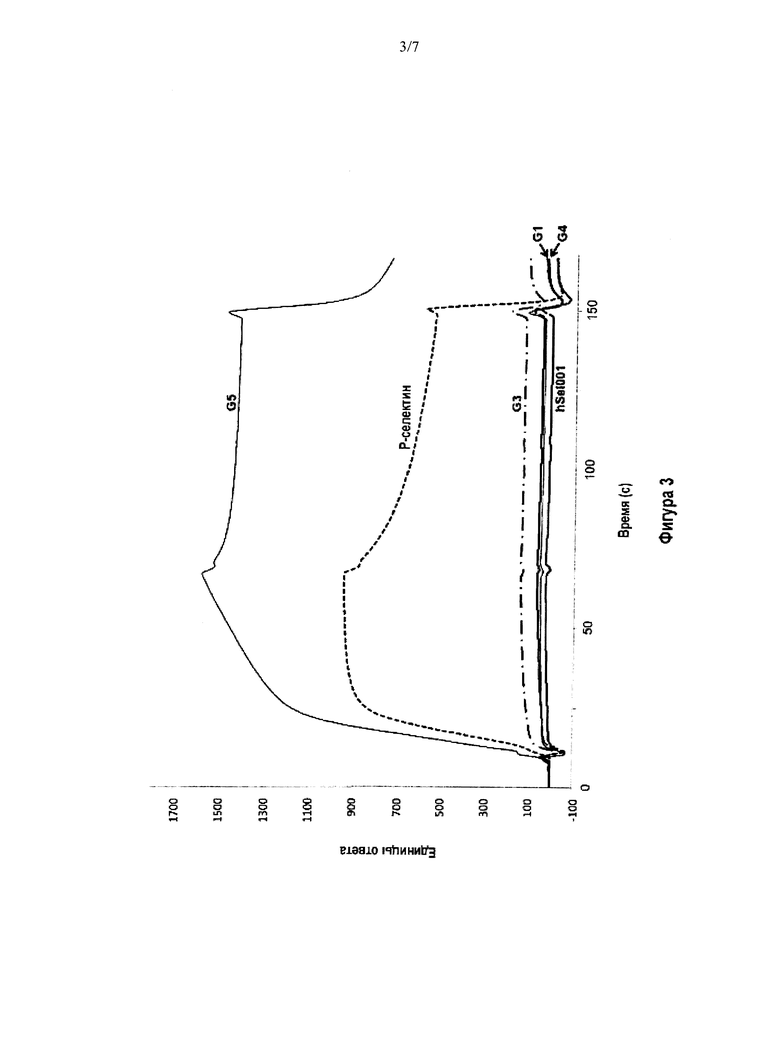
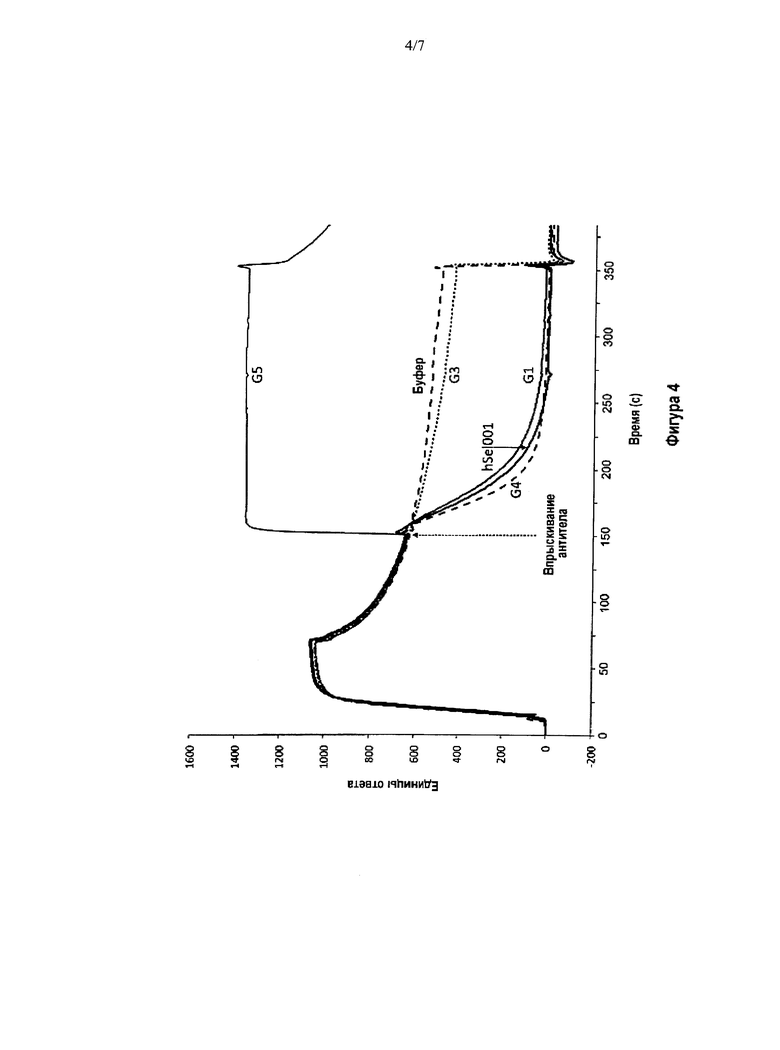
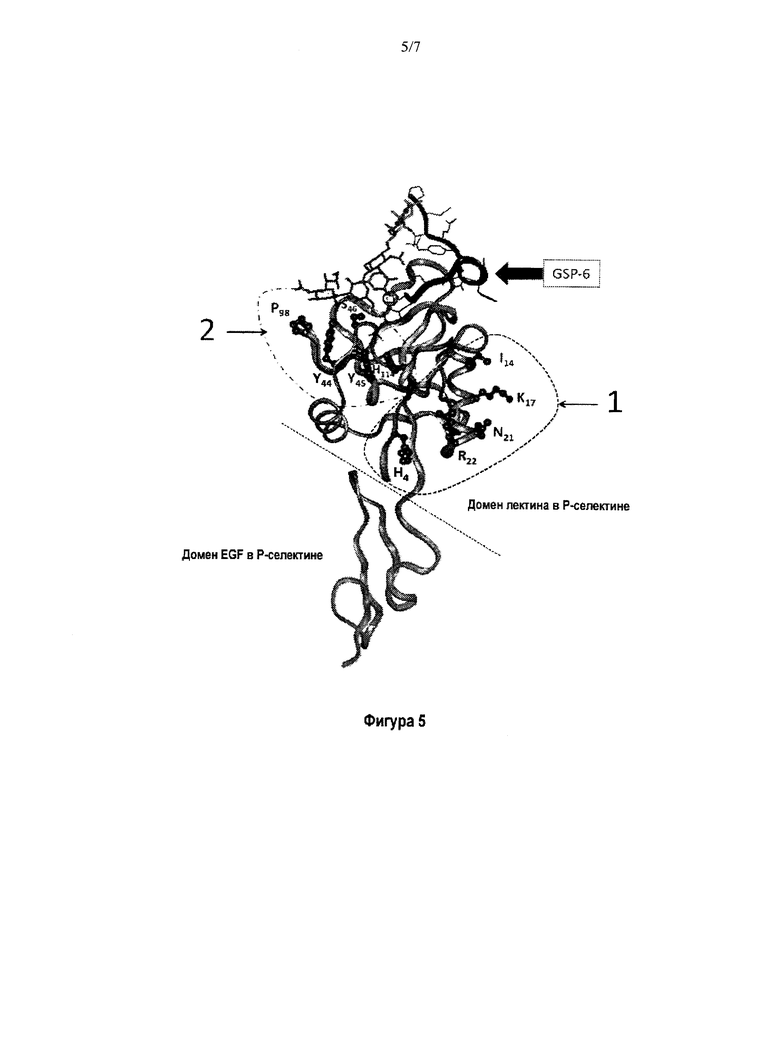
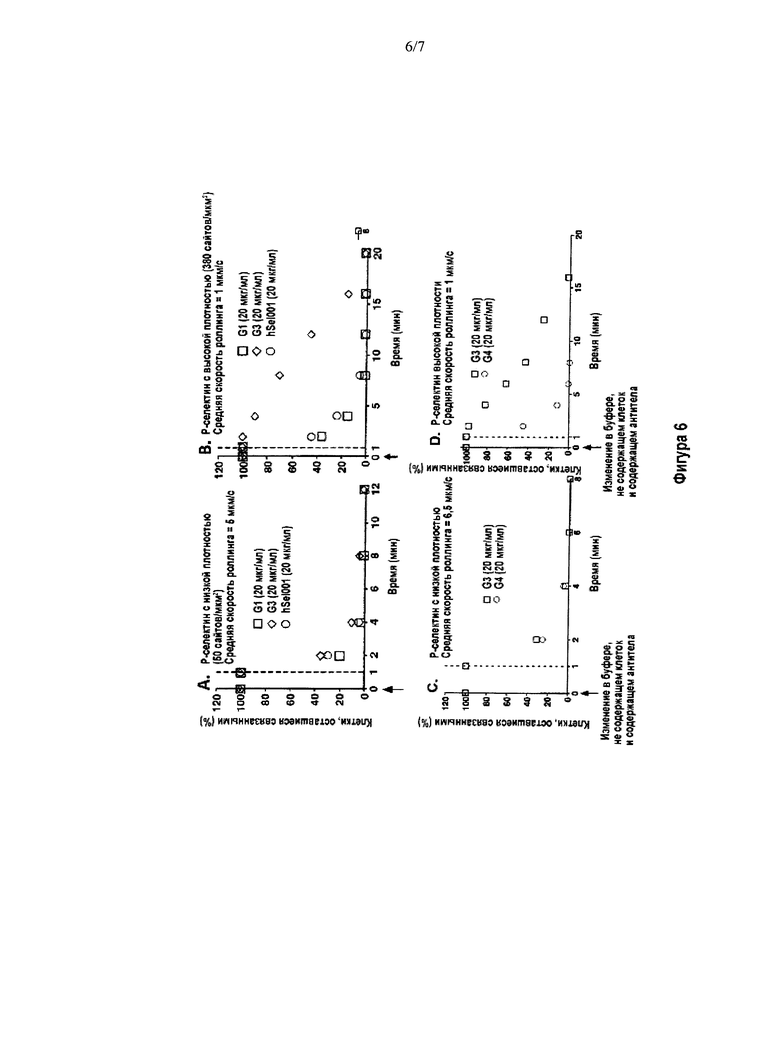
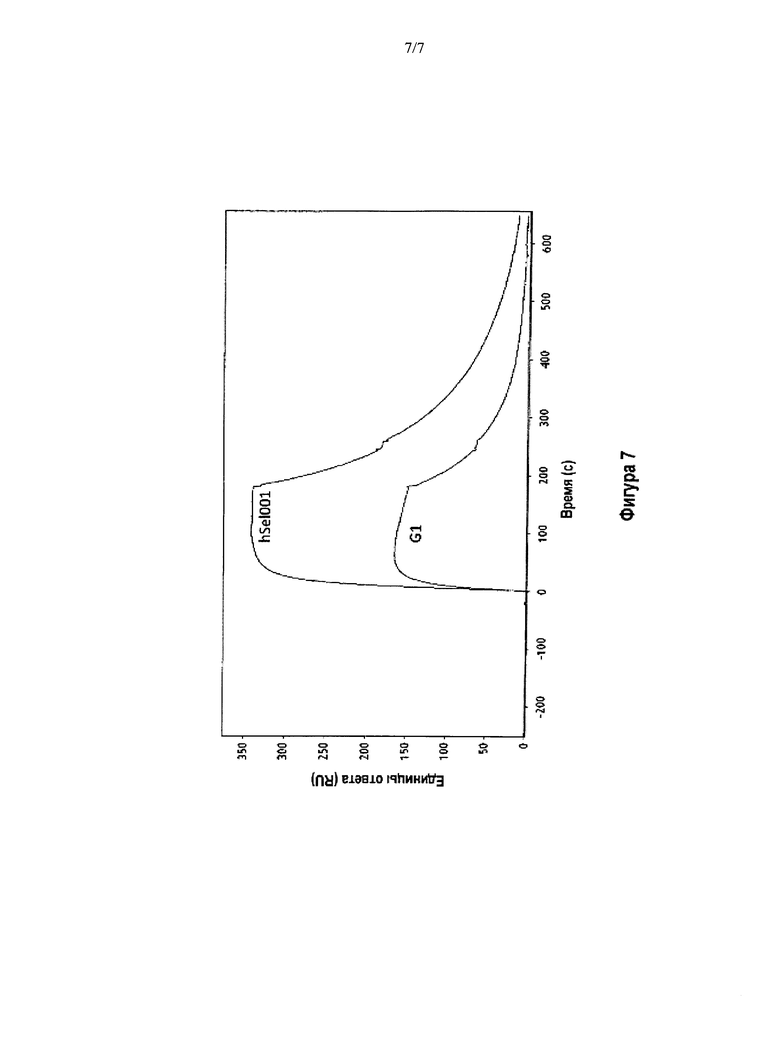
Изобретение относится к области биотехнологии и иммунологии. Описаны антитела, которые специфически связываются с Р-селектином, блокируют связывание PSGL-1 с Р-селектином и вызывают диссоциацию ранее образовавшихся комплексов Р-селектин/PSGL-1. Идентифицирован ранее неизвестный связывающий домен антитела вблизи N-конца (конформационный эпитоп) Р-селектина, с которым связываются антитела. Раскрытые антитела, связываются с конформационным эпитопом Р-селектина и которые обладают двойной функцией блокирования, заключающейся в связывании PSGL-1 с Р-селектином и диссоциации ранее образовавшихся комплексов Р-селектин/PSGL-1. Эта двойная функция антител к Р-селектину и их связывающих фрагментов может быть использована при лечении различных острых и хронических воспалительных и тромботических заболеваний и состояний. Предложенное изобретение может быть использовано в медицине. 13 з.п. ф-лы, 7 ил., 2 табл.
1. Антитело или его связывающий фрагмент, которые специфически связываются с конформационным эпитопом Р-селектина, находящимся в пределах аминокислотных остатков в положениях 1-35 в SEQ ID NO: 1, где указанное антитело или его связывающий фрагмент блокируют связывание PSGL-1 к Р-селектину и диссоциирует ранее образовавшиеся комплексы Р-селектин/PSGL-1, для применения для лечения или ингибирования происходящего в данное время воспалительного или тромботического состояния или расстройства у субъекта, который нуждается в диссоциации ранее образовавшегося комплекса Р-селектин/PSGL-1 для указанного лечения или ингибирования, и где антитело или его связывающий фрагмент характеризуются KD к конформационному эпитопу Р-селектина ≤1000 нМ.
2. Антитело или его связывающий фрагмент для применения по п. 1, где воспалительное или тромботическое состояние или расстройство или процессы воспалительных заболеваний, связанные с таким состоянием или расстройством, относятся по меньшей мере к одному или более выбранных из группы, состоящей из адгезии тромбоцитов к серповидным красным кровяным клеткам, лейкоцитам, лимфоцитам или клеткам эндотелия, адгезии клеток эндотелия к серповидным красным кровяным клеткам, лейкоцитам, лимфоцитам или тромбоцитам, вазоокклюзионного серповидно-клеточного болевого криза, тромбоза, атеросклероза, метастазирования опухолей, аллергических реакций, тироидита, псориаза, дерматита, нефрита, системной красной волчанки, склеродермии, сепсиса, ринита, анафилакции, диабета, рассеянного склероза, отторжения трансплантатов, реакции трансплантат против хозяина, астмы, хронической обструктивной болезни легких, воспалительного заболевания кишечника, артрита и ишемического реперфузионного повреждения, состояний, связанных с тяжелой травмой или хроническим воспалением, синдрома системного воспалительного ответа и полиорганной недостаточности.
3. Антитело или его связывающий фрагмент для применения по п. 2, где состояние или расстройство представляет собой серповидно-клеточный болевой криз и антитело или его связывающий фрагмент направлен на лечение происходящей в настоящее время вазоокклюзии.
4. Антитело или его связывающий фрагмент для применения по любому предыдущему пункту, где антитело предназначено для использования в количестве, эффективном для диссоциации ранее образовавшихся комплексов Р-селектин/PSGL-1.
5. Антитело или его связывающий фрагмент для применения по любому предыдущему пункту, где антитело обладает способностью вызывать диссоциацию связи клетка-клетка между активированными клетками эндотелия и лейкоцитами, лимфоцитами, серповидными красными кровяными клетками и/или тромбоцитами.
6. Антитело или его связывающий фрагмент для применения по любому предыдущему пункту, где антитело обладает способностью вызывать диссоциацию связи клетка-клетка между активированными тромбоцитами и лейкоцитами, лимфоцитами, серповидными красными кровяными клетками и/или клетками эндотелия.
7. Антитело или его связывающий фрагмент для применения по любому предыдущему пункту, где антитело является моноклональным.
8. Антитело или его связывающий фрагмент для применения по любому предыдущему пункту, где антитело является химерным, человеческим или гуманизированным.
9. Антитело или его связывающий фрагмент для применения по любому предыдущему пункту, где антитело содержит иммуноглобулин, выбранный из класса, состоящего из IgA, IgD, IgE, IgG и IgM.
10. Антитело или его связывающий фрагмент для применения по любому предыдущему пункту, где антитело или его связывающий фрагмент представляют собой IgG1, IgG2, IgG3, IgG4 или химеру IgG2/G4.
11. Антитело или его связывающий фрагмент для применения по любому предыдущему пункту, где связывающий фрагмент включает по меньшей мере один из фрагментов Fab, Fab′, F(ab)2 или scFv.
12. Антитело или его связывающий фрагмент для применения по любому предыдущему пункту, где указанное антитело предназначено для введения субъекту парентерально, внутримышечно, интраперитонеально, эпидурально или перорально, внутривенно, подкожно или в распыленном виде.
13. Антитело или его связывающий фрагмент для применения по любому предыдущему пункту, где указанное антитело или связывающий фрагмент предназначено для введения субъекту в количестве, составляющем от 0,1 мг/кг до 100 мг/кг.
14. Антитело или его связывающий фрагмент для применения по любому предыдущему пункту, где антитело представляет собой моноклональное антитело, обозначаемое в данной заявке как G1, hSel001, G4 или гуманизированное антитело, содержащее CDR указанного моноклонального антитела.
| US 2010209423 A1, 19.08.2009 | |||
| US 5593882 A, 14.01.1997 | |||
| US 2009285812 A1, 19.11.2009 | |||
| ГУМАНИЗИРОВАННЫЙ ИММУНОГЛОБУЛИН, СПЕЦИФИЧНЫЙ ДЛЯ БЕЛКА L-СЕЛЕКТИНА ЧЕЛОВЕКА | 1993 |
|
RU2151612C1 |

