Область техники, к которой относится изобретение
Настоящее изобретение относится к способу повышения частоты имплантации эмбриона в материнской матке у млекопитающих путем введения в матку млекопитающего эффективного количества бета-галактозид-связывающего лектина или его производных, а также продукта, включающего указанный лектин.
Описание предшествующего уровня техники
Сельскохозяйственная промышленность играет ключевую роль в экономике Бразилии благодаря значительному вкладу в валовой национальный продукт и позитивному вкладу в баланс внешней торговли.
Для уверенности в прибыльности сельскохозяйственного рынка, сельскохозяйственная собственность должна сохранять максимальную репродуктивную эффективность в животноводстве с целью соответствования запросам потребителей в высококачественных продуктах. В этом смысле сохранение адекватного уровня рождаемости является ключевым фактором в цепочке событий, которые приводят к прибыльности сельскохозяйственной собственности. Кроме того, во многих случаях сохранение оптимальных уровней репродуктивности является тяжелой задачей из-за высокого уровня потерь эмбрионов на ранних стадиях беременности.
Репродукцию определяют как зачатие живых существ от того же вида после последовательности физиологических событий, которые, в свою очередь, основываются на множестве факторов. Применение современных репродуктивных технологий в сельскохозяйственной области может играть ключевую роль в генетическом улучшении животноводства, повышению потенциала беременности (имплантации эмбрионов в материнскую матку) с целью обеспечения максимальной производительности в течение срока эксплуатации животного.
Тем не менее, повсюду в мире потеря эмбриона является одной из наиболее распространенных причин для экономических потерь разводчиков; хотя предотвращение ранней потери эмбриона до сих пор является проблемой ввиду комплексных механизмов, вовлеченных в диагностику и сохранение беременности.
Ранняя диагностика, которая определяет частоту имплантации эмбриона в материнскую матку у млекопитающих, является ключевым инструментов для сельскохозяйственных процедур, поскольку она позволяет разводчикам принимать профилактические меры и минимизировать конечные экономические потери. Так, новые технологии развиваются ввиду улучшения знаний репродуктивной физиологии яичников крупного рогатого скота для достижения лучших результатов в репродуктивности и для снижения затрат на лечение.
Сходным образом с in vitro оплодотворением крупного рогатого скота, вспомогательные репродуктивные технологии становятся все более популярными из-за нужд рынка. Эффективная репродуктивная способность крупного рогатого скота напрямую связана с рождением большого количества коров у разводчиков молочного скота и бычков у разводчиков мясного скота, таким образом обеспечивая экономические выгоды для сельскохозяйственной собственности.
Другие рыночные особенности, такие как мировой потребительский спрос на качество, взвешивание и стандартизация продуктов животного происхождения, вместе с борьбой за место для разведения, сельскохозяйственные и биоэнергетические области стимулируют поиск более высокой продуктивности, вызывающей улучшение репродуктивных технологий быстрыми темпами.
В ряду с производственными и потребительскими нуждами мирового основного рынка сельскохозяйственная промышленность Бразилии опирается на вспомогательные репродуктивные технологии для повышения средней продуктивности стада крупного рогатого скота, поскольку указанные технологии значительно повышают рождаемость генетически улучшенных животных.
Главные репродуктивные технологии, применяемые в сельскохозяйственных собственностях, включают: (i) спаривание или воспроизводство при помощи природных процессов; (ii) искусственное осеменение; (iii) in vivo производство с последующим перемещением эмбриона; (iv) in vitro производство с последующим перемещением эмбриона. Независимо от выбранной технологии главным объектом способа воспроизводства является обеспечение максимальной способности репродуцирующих видов для достижения положительных результатов.
Спаривание является широко применяемой технологией, поскольку она является менее затратной для заводчиков, но она позволяет ограниченно контролировать заболевания, передающиеся половым путем, и обеспечивает медленное генетическое улучшение стада. В этой процедуре эмбрионы состоят на 50% из генетического материала отца (таким образом, отличного от генетического материала матери), что может вызвать образование большого числа аллоантигенов и привести к потере эмбриона из-за недостаточной толерантности в системе мать-плод.
Аллоантиген представляет собой молекулу, содержащую генетический материал одного из организмов, принадлежащих к тому же виду. Поскольку указанная молекула отличается и вводится в другой организм, иммунная система последнего индуцирует производство серий ответов против «чужеродного» материала. Примером является только что оплодотворенный эмбрион, который содержит половину генетического материала от матери, и половину генетического материала от отца; последняя половина может быть чужеродной для материнской иммунной системы и, таким образом, может отторгаться материнским организмом. Отсутствие иммунной толерантности в системе мать-плод (процесс, при помощи которого материнский организм опознает/принимает плод без запуска иммунного ответа против него) является главной причиной раннего аборта.
Другой широко применяемой в сельскохозяйственной области репродуктивной технологией является искусственное осеменение (AI), суть которого заключается в искусственном помещении семенной жидкости (сперматозоида/мужских гамет и семенной плазмы) в репродуктивный тракт самки при помощи специальных технологий, подходящих для анатомических особенностей каждой самки. Успешность AI измеряют при помощи уровня беременности - или количества беременных самок по отношению к числу не осемененных самок. В этой процедуре развившиеся эмбрионы также состоят на 50% из генетического материала отца (таким образом, отличающегося от генетического материала матери), что может вызвать образование большого числа аллоантигенов и привести к потере эмбриона из-за отсутствия толерантности в системе мать-плод.
Дополнительно, in vivo производство с последующим переносом эмбриона (ET) является репродуктивной технологией, где несколько эмбрионов развиваются в репродуктивном тракте или самке-доноре (которая дает свой генетический материал), и переносятся в матку самки-реципиента того же вида, которая вынашивает беременность. Образование эмбрионов в матке доноров включает суперовуляцию, искусственное осеменение и эмбрион-стимулирующие технологии. После стимулирования эмбрионы оценивают, и жизнеспособные эмбрионы переносят или криосохраняют. Перенос (помещение) эмбрионов продукции in vivo в матку или рог матки реципиента зависит от процессов синхронизации эструса (то есть, адаптации матки реципиента к той же стадии развития эмбриона). В этой процедуре эмбрионы, переносимые в организм реципиента, состоят вплоть до 100% из аллоантигенов (другой генетический материал), поскольку они полностью состоят из генетического материала доноров и отца, что вызывает больший риск потери эмбриона из-за отсутствия толерантности в системе мать-плод.
С другой стороны, оплодотворение in vitro с последующим переносом эмбриона относится к образованию эмбрионов в лаборатории, и указанные эмбрионы переносят в матку самок-реципиентов тех же видов, которые вынашивают беременность до срока. Лабораторное образование эмбрионов зависит от следующего: (i) аспирации ооцитов из донорского яичника при помощи пункции фолликула под контролем ультразвука; (ii) IVM - in vitro созревания ооцитов с последующим in vitro индуцированием созревания цитоплазмы ооцита и ядра ооцита, таким образом, подготовки яйца к оплодотворению; (iii) IVF - in vitro оплодотворения или процесса сингамии зрелых ооцитов и функционирующих сперматозоидов; (iv) IVC - in vitro культивирования эмбрионов после оплодотворения до того, как указанные эмбрионы достигнут определенный стадии (морулы и/или бластоцисты) для переноса через 5-7 дней культивирования; (v) переноса эмбриона, когда - после периода культивирования - эмбрион следует оценить, а имеющих хорошие показатели можно переносить, криосохранять и/или витрифицировать. Перенос образованных in vitro эмбрионов в матку или рог матки реципиента также зависит от процессов синхронизации эструса (то есть, адаптирования матки реципиента к той же стадии эмбрионального развития). В этой процедуре овулированные в организме реципиента эмбрионы состоят вплоть до 100% из аллоантигенов (другой генетический материал), поскольку они полностью состоят из генетического материала доноров и отца, что вызывает больший риск потери эмбриона из-за отсутствия толерантности в системе мать-плод.
Как описано выше, толерантность в системе мать-плод представляет собой иммунный процесс, который регулирует ответ материнского организма против эмбриона или плода. Иммунная система организма несет ответственность за комплекс реакций на внешние факторы и/или агрессоров, которые могут нарушить ее обычное физиологическое состояние.
Иммунорегуляторные ответы во время беременности представляют собой случаи, происходящие после овуляции, соития и оплодотворения, которые, главным образом, нацелены на рост и развитие оплодотворенного яйца (эмбриона или плода и связанных оболочек).
В этом смысле Lewis S. K. с соавт., 2007 (Galectin-15 [LGALS15]: A Gene, Uniquely Expressed in the Uteri of Sheep and Goats that Functions in Trophoblast Attachment) наблюдал, что у жвачных животных эмбрион на стадии MO (морула, между 4-6 днями) проникает в матку и продолжает развиваться до стадии BL (бластоциста, между 6-7 днями), которая содержит монослой клеток, называемых трофэктодермальными клетками. После отрыва блестящей оболочки вплоть до дня D12 (12-го дня) у овец или D15 (15-го дня) у коз эмбрион остается в стадии элонгации. Во время указанной стадии трофэктодерма продуцирует интерферон-тау (IFNT), который, в свою очередь, несет ответственность за ингибирование лютеолиза (инволюция желтого тела). Когда желтое тело (фолликул яичника) активно, поддерживается продукция прогестерона (P4), и эндометрий (слизистая оболочка, которая покрывает стенку матки) становится готовым для наступления беременности. Исследование также утверждает, что Р4 и IFNT регулируют транскрипцию Галектина-15 в эпителии эндометрия. Галектин-15 действует в маточной среде, поскольку указанный Галектин принимает участие в фиксации/адгезии трофэктодермы оплодотворенного яйца к эндометрию матки, таким образом, стимулируя биологические ответы, такие как адгезию и миграцию, которые являются принципиальными событиями во время стадии элонгации бластоцисты и, соответственно, в развитии беременности.
Другой момент, в соответствии с исследованием, проведенным Farmer J.L. с соавт., 2008 (Galectin-15 (LGALS15) Functions in Trophectoderm Migration and Attachment) Галектин-15 стимулирует клеточную пролиферацию и ингибирование апоптоза, которые являются важными событиями во время стадии имплантации. Авторы доказали, что, хотя ген Галектин-15 присутствует у овечьих, козьих и бычьих видов, мРНК (РНК-мессенджер) Галектина-15 экспрессируется в стадии элонгации только у козьих и овечьих, экспрессия при этом варьирует в соответствии с фазой цикла эструса. Дополнительно, отмечено, что введение экзогенного IFNT путем внутриматочной инфузии только повышает экспрессию гена Галектин-15, если женская особь получает лечение P4, таким образом, доказывая необходимость применения IFNT и P4 совместно в качестве индукторов транскрипции мРНК Галектина-15.
Satterfield M.C. с соавт., 2006 (Progesterone Ragulation of Preimplantation Conceptus Growth and Galectin-15 [LGALS15] in the Ovine Uterus) пришел к выводу, что Р4 индуцирует экспрессию генов некоторых протеинов, секретируемых эндометрием, таких как Галектин-15 и секретируемый фосфопротеин 1 (SPP1), которые считаются регуляторами выживаемости и роста оплодотворенного яйца, а также клеточной адгезии во время стадии имплантации. В этом смысле исследование Burghardt R.C. с соавт., 2009 (Enhanced Focal Adhesion Assembly Reflects Increased Mechanosensation and Mechanotransduction at Maternal - Conceptus Interface and Uterine Wall During Ovine Pregnancy) отмечает, что SPP1 и Галектин-15 являются механосенсорами на поверхности среды матки и оплодотворенного яйца.
Следуя другому стилю мышления, исследование, проведенное Than N.G. с соавт., 2008 (Emergence of Hormonal and Redox Regulation of Galectin-1 in Placental Mammals: Implication in Maternal-Fetal Immune Tolerance), привело к выводу, что Галектин-1 показывает высокие уровни структурной сохранности, димеризации и связывающих свойств с углеводами и интегринами (адгезивными протеинами), предполагая, что данные свойства сохранены у позвоночных животных и поддерживают стандартную экспрессию генов среди различных типов плаценты (как децидуальной, так и нет). Авторы также наблюдают, что Галектин-1 может передавать материнскую иммунотолерантность аллоантигенам плода, регулировать действие натуральных киллеров (NK) матки и действовать в качестве регулятора T-клеток и смягчающего средства (Т-клетки принимают участие в клеточном иммунитете). Наконец, авторы подтверждают синергическое действие Р4 в стимулировании продукции Галектина-1 эндометрием.
Благодаря исследованиям фертильности можно подтвердить, что роль Галектинов связана с модулированием иммунных ответов, также как элонгацией эмбриона и адгезией эмбриона к эндометрию.
Галектины известны как лиганды для бета-галактозид лектинов млекопитающих и могут быть экспрессированы большим числом тканей. Эти лектины обычно являются растворимыми и не содержат пептидный сигнал, будучи секретированными при помощи механизма, который не зависит от эндоплазматического ретикулума и комплекса Гольджи. На сегодняшний день существуют описания 15 Галектинов млекопитающих, у всех из которых имеется домен узнавания углеводов с приблизительно 130 аминокислотными остатками.
Известно, что взаимодействие Галектинов с гликанами с поверхности клеток иммунной системы во внеклеточном пространстве может модулировать продуцирование цитокинов и медиаторов, клеточную адгезию, апоптоз, хемотаксис и эндоцитоз. Во внутриклеточной среде Галектины могут принимать участие в маршрутах сигнализации и модулировать некоторые биологические ответы, такие как апоптоз, регулирование клеточного роста и сплайсинг пре-мРНК.
Farmer J.L. с соавт., 2008 (вышеупомянутая статья) также раскрывает тот факт, что кроме Галектина-15 и Галектина-1 другие Галектины могут быть экспрессированы эндометрием и плацентой млекопитающих и могут показывать важные свойства по отношению к дифференцировке эндометрия, имплантации бластоцисты и дифференцировки трофобласта; аналогичным образом, Poppovich с соавт., 2005 (Galectin-9: a New Endometrial Epithelial Marker for the Mid- and Late-Secretory and Decidual Phases in Human) раскрывает свойства Галектина-9, и Lee с соавт., 1998 (Spatio-Temporal Pattern for Expression of Galectin-3 in the Murine Utero-Placental Complex? Evidence for Differential Regulation) относится к экспрессии Галектина-3. Более того, существует несколько других статей в литературе, говорящих о потенциальном терапевтическом применении рекомбинантных Галектинов или специфических ингибиторов этих протеинов.
Галектин-1 является мультифункциональной молекулой, которая принимает участие в таких биологических процессах, как адгезия, пролиферация, дифференцировка и клеточные циклы; апоптоз; сплайсинг РНК, контролирование воспалительного процесса и адаптивный иммунный ответ. Экспрессия эндогенного Галектина-1 уже наблюдается в эпителиальных клетках тимуса, праймированных антигеном Т-клетках, активированных макрофагах, активированных В-клетках (принимающих участие в гуморальном иммунитете), эндотелиальных клетках, стромальных клетках и мышиных лимфоидных органах, таких как тимус и лимфатические узлы. В виду его иммунорегуляторных свойств Галектин-1 (как эндогенный, так и экзогенный) является важным медиатором для предотвращения потери плода и/или гибели эмбриона.
Галектин-1 можно получить от млекопитающих (человека, быков, овец, коз, лошадей и/или свиней) при помощи системы гетерогенной экспрессии в активной, стерильной, алкилированной и свободной от эндотоксинов форме. Способ получения рекомбинантного Галектина-1 широко известен в литературе и обычно включает следующие стадии: (i) получения сырого экстракта бактерий, содержащих Галектин-1; (ii) очищение Галектина-1; (iii) сохранение лектиновой активности Галектина-1 путем алкилирования с йодацетамидом; и (iv) устранение бактериального эндотоксина (LPS) йодацетамид-алкилированного лекарственного препарата Галектина-1. Среди раскрытых в литературе возможностей для проведенных в настоящем эксперименте тестов, производят Галектин-1 на основании следующих процедур.
Первая стадия для получения рекомбинантного Галектина-1 начинается с бактериальной культуры (предпочтителен штамм E. Coli в виде розетки), трансформируемой при помощи вектора экспрессии (предпочтительно pET29a плазмид), содержащего ген Галектина-1 в 200 мл основной жидкой среды LB (Invitrogen, Gibco, Карлсбад, Калифорния, США), содержащей 50 мкг/мл ампициллина (USB Corporation, США), что происходит в орбитальном встряхивателе при 200 об/мин в течение 16-18 часов при 37°С. После этого периода 25 мл этой культуры переносят к 1 л предварительно автоклавированной половины LB, содержащей 500 мкл ампициллина (50 мкг/мл). Далее эту бактериальную суспензию инкубируют снова в течение еще 2 часов при 37°С в орбитальном встряхивателе при 200 об/мин. Оптимум уровня бактериального роста показывает оптическую плотность (OD) в диапазоне от 0,5 до 0,5 на 600 нм. Далее, 0,36 г изопропил-D-тиогалактопиранозида (IPTG, Promega, Висконсин, США), разведенного в 1 мл половины LB, добавляют к культуре для индуцирования экспрессии Галектина-1 трансформированными бактериями. Культуру еще раз помещают во встряхиватель (37°С - от 250 до 300 об./мин) на 4 часа. По прошествии этого времени бактериальную суспензию центрифугируют при 5000 g в течение 15-20 минут при 4°С, и осадок культуры еще раз центрифугируют при 3000 g в течение 15-20 минут при 4°С. Наконец, надосадочную жидкость удаляют и осадок хранят при -80°С до момента применения.
Вторая стадия получения рекомбинантного Галектина-1 из бактериального осадка, который помещают в ледяную баню и далее ресуспендируют в буфер для лизиса, содержащий 7 мл PBS (солевой фосфатный буфер-NaCl (136,8 ммоль); KCl (2,7 ммоль); Na2HPO4 (6,4 ммоль); KH2PO4 (0,9 ммоль, pH 7,4); 14 ммоль меркаптоэтанола (2-МЕ) (Merck-Schuchardt, Германия), 1 таблетку свободных от EDTA ингибиторов протеаз (Roche Diagnostics GmbH, M, Германия), 1 мл лизоцима - 1мг/мл (Roche Diagnostics GmbH, M, Германия), 10 мкл РНКазы А типа 3А - 10 мг/мл (Sigma-Aldrich) и 10 мкл ДНКазы I типа IV - 10 мг/мл (Sigma-Aldrich), и далее инкубируют в течение 30 минут с буфером для лизиса в ледяной бане. Далее образец разрушают ультразвуком в течение 5 циклов по 20 секунд каждый при 40 Вт (Sonics Vibra cell; SONICS & MATERIALS INC.); между циклами суспензия находится в спокойном состоянии в течение 15 секунд. Лизат бактерий далее центрифугируют при 10000 g в течение 45 минут при 4°С. Далее надосадочную взвесь собирают и подвергают аффинной хроматографии в агароза-лактозной колонке (Sigma-Aldrich) с объемом слоя 5 мл. Нелиганд вымывается при помощи балансирующего буфера (PBS, добавляемые от 2-МЕ до 14 ммоль, pH 7,4), и собирают 20 фракций по 2 мл. Лиганд колонки для аффинной хроматографии вымывают при помощи буфера вымывания (содержащего 14 ммоль лактозы, pH 7,4) и собирают 10 фракций по 0,5 мл. Процедуру хроматографии мониторируют при помощи показателя оптической плотности, равного 280 нм (UV Mini 1240, Shimadzu), и электрофореза с полиакриламидным гелем. Концентрации белка в растворах Галектина-1 определяют при помощи спектрометрии, применяя показатели оптической плотности, равные 280 нм, или колориметрические исследования, имеющиеся на рынке, и выражаются в миллиграммах протеина на миллилитры (мг/мл). Хроматографические фракции, полученные при этом процессе очищения Галектина-1 на агарозе-лактозе, анализируют при помощи электрофореза (SDS-PAGE - «Электрофорез с натрий додецил сульфат полиакриламидным гелем»). Образцы, содержащиеся в восстанавливающем и диссоциирующем буфере (конечный объем 20 мкл), используют с полиакриламидным гелем (15%) и подвергают электрофоретическому движению при постоянном напряжении (150 В). Хроматографические фракции сырого лизата бактерий и нелигандного материала применяют в качестве образцов. Для контроля применяют известный стандарт молекулярного веса (LMWH - низкомолекулярный весовой калибровочный набор для электрофореза - GE, Амстердам - Biosciences, Упсала, Швеция). Гель окрашивают «Кумасси бриллиантовым голубым». Растворы Галектина-1, полученные после хроматографии на агарозе-лактозе, держат в вымывающем буфере с целью предотвращения активности лектина по отношению к этому протеину, и хранят при -80°С до момента применения. Для анализов эти растворы подвергают деминерализации в PD10 колонке путем молекулярно-вытеснительной хроматографии в соответствии с инструкциями производителя (Sephadex-G25M; Pharmacia LKB, Упсала, Швеция). Концентрации деминерализованных растворов Галектина-1 определяют при помощи спектрометрии или колориметрических реакций, как описано выше. С целью оценивания уровня активности сохранения лектинов (способность Галектина-1 распознавать сахара) в Галектине-1, очищенном в агарозно-лактозных смолах, образцы этого протеина деминерализуют в PD-10 колонке и немедленно рехроматографируют в агароза-лактозной колонке. Собирают 20 фракций по 1,0 мл каждая. Процесс вымывания контролируют по средней величине белковой концентрации (мг/мл). В последней процедуре вымывание Галектина-1 промывным буфером считается индикатором потери лектиновых свойств этого протеина, поскольку в таких условиях Галектин-1 не способен распознать лактозу и, таким образом, не удерживается в агарозно-лактозной смоле. Окисление сульфгидрильных групп Галектина-1 и его денатурация могут обеспечить снижение лектиновой активности этого протеина, свойств, связанных с некоторыми функциями этой молекулы. Учитывая то, что очищенные растворы Галектина-1 хранятся при -80°С и должны быть разморожены перед применением, следует оценить эффект указанных процедур на гемагглютинирующую активность этого лектина. Гемагглютинация развивается в присутствии или в отсутствие Галектин-1-специфического гаптенового сахара, то есть лактозы (20 ммоль). Только соединения Галектина-1, имеющие гемагглютинирующую активность при концентрациях, не превышающих 2 мкмоль, применяют в различных анализах.
Третья стадия приготовления рекомбинантного Галектина-1 включает контролирования лектиновой активности алкилированных препаратов этого лектина при помощи гемагглютинации и/или других способов, которые предоставляют возможность определять сохранность свойств лектина этих рекомбинантных препаратов Галектина-1. Окисление сульфгидрильных групп Галектина-1 обеспечивает денатурацию и потерю лектиновой активности этого протеина. Таким образом, с целью получения более стабильных образцов Галектина-1 препараты этого лектина подвергают алкилированию с применением йодацетамида, восстановительное составляющее которого взаимодействует ковалентным способом с сульфгидрильными группами, таким образом, образуя карбоксиамидометил-Галектин-1 (алкилированный Галектин-1). Кратко, 0,037 г йодацетамида (протеин-йодацетамид, Sigma-Aldrich; конечная концентрация 20 мкмоль) разводят в 1,0 мл очищенного раствора Галектина-1 в присутствии 100 ммоль лактозы. Далее этот раствор инкубируют в ледяной бане, защищая от света в течение 16-18 часов. После инкубирования раствор подвергают молекулярной вытеснительной хроматографии в PD10 для удаления свободного йодацетамида и лактозы. Концентрацию соединений алкилированного Галектина-1 определяют при помощи спектрометрии или выражают в мг/мл, как упомянуто выше.
Четвертая стадия приготовления рекомбинантного Галектина-1 относится к удалению бактериальных эндотоксинов (LPS). Учитывая то, что Галектин-1 получают из грамотрицательных бактерий, которые обладают LPS (липополисахаридом) в составе их клеточной стенки, после стадии алкилирования с йодацетамидом соединения Галектина-1 подвергают аффинной хроматографии на Полимиксин B-агарозной колонке (Detoxi-Gel Endotoxin Removing Gel, Pierce, Иллинойс, США). Эффективность процедуры удаления LPS из алкилированных соединений Галектина-1 оценивают, измеряя количество эндотоксина LPS при помощи набора QCL-1000 (Chromogenic Limulus Amebocyte Lysate Assay, Cambrex Company, Мэриленд, США). Активные и свободные от эндотоксина алкилированные соединения Галектина-1 далее подвергают стерилизации путем фильтрации (мембраны 0,22 мкм).
Поскольку раскрыт способ приготовления рекомбинантного Галектина-1, следует отметить, что любой другой бета-галактозид-связывающий лектин или его производные можно получить путем очень похожей процедуры.
Таким образом, зная, что Галектин-1 играет важную роль в модулировании иммунных расстройств, ингибировании воспалительного ответа и модулировании функции Т-клеток как в системе in vivo, так и in vitro, патентная заявка US 12/175227 раскрывает способ модулирования иммунного ответа указанного лектина в целью предотвращения и лечения иммунных расстройств, включая лимфому Ходжкина.
Что касается потенциала взаимодействия с другими молекулами и/или клетками, известно, что в присутствии Галектина-1 дендритные клетки становятся толерогенными и способными модулировать иммунный ответ у млекопитающих, предотвращая развитие аутоиммунных заболевания, а также противостоя отторжению трансплантатов. В этом смысле патентная заявка US 12/137004 раскрывает способ приготовления соединения, включающего дендритные клетки в присутствии указанного лектина, имея объектом лечения новообразования, а также аутоиммунные и инфекционные заболевания.
Исследование Blois S.M. с соавт., 2007 (A Pivotal Role for Galectin-1 in Fetomaternal Tolerance) основано на результатах экспериментальной лечебной модели, в которой Галектин-1 внутрибрюшинно вводят изогенным мышам с целью оценки индуцированной стрессом потери плода, вызванной усилением шума. Отмечают, что экспрессия Галектина-1 в матке мышей высокочувствительна к изменениям окружающей среды, таким образом, подвергая риску беременность самок и приводя к стрессу. С другой стороны, когда мыши получают рекомбинантный Галектин-1, частота потерь плода значительно снижается.
Известно, что для сохранения беременности материнский организм претерпевает значительные изменения, включая гормональные и иммунные. Имеются данные, что подготовка эндометрия к имплантации эмбриона не является только следствием гормональной стимуляции, она зависит от взаимодействия между бластоцистой и эндометрием, что также регулируют цитокины, факторы роста и адгезивные молекулы, которые продуцирует и секретирует эндометрий и бластоциста. Даже таким образом взаимодействие между различными физиологическими системами может привести к потере беременности, которую нельзя объяснить действием какого-либо изолированного фактора.
С точки зрения сельскохозяйственного рынка механизмы иммунной толерантности в системе мать-плод во время беременности, получившейся в результате спаривания или искусственного осеменения, по-прежнему являются проблемой для репродуктивной медицинской ветеринарии с точки зрения иммунного ответа против аллоантигенов плода.
С этой точки зрения следует отметить, что потеря беременности влечет за собой экономические потери с тяжелыми последствиями для разводчиков, которые, в свою очередь, должны поддерживать удовлетворительные уровни репродуктивной эффективности с целью удовлетворения потребительских нужд и сохранения прибыльности их собственности.
В дополнение, разводчики как молочного, так и мясного крупного рогатого скота часто применяют репродуктивные технологии для определения пола эмбриона после оплодотворения. Тем не менее, выращенные in vitro эмбрионы, которые дальше переносят в матку реципиентов, не показывают удовлетворительного уровня выживаемости из-за морфологических и структурных изменений, в основном развивающихся из-за поддерживающей среды, что вызывает раннюю потерю эмбриона.
Более того, одним из ограничений рыночного применения репродукционных биотехнологий является расстояние для покрытия между сторонами сбора и помещения донорских гамет самца (семенной жидкости) или самки (ооцита), или эмбриона, выращенного как in vivo, так и in vitro. Известно, что сохранение функции клеток напрямую зависит от вариаций pH и температуры, среди других свойств, и, таким образом, является проблемой сохранить ее при оптимальных условиях вплоть до момента переноса эмбриона.
C учетом изложенного выше, настоящее изобретение раскрывает решение настоящих репродуктивных трудностей, обычно обнаруживаемых на сельскохозяйственном рынке, главной целью которого является обеспечение повышения частоты имплантации эмбриона в материнской матке путем применения к материнской матке млекопитающих эффективного количества бета-галактозид-связывающего лектина или его производных.
Цели изобретения
Целью настоящего изобретения является применение эффективного количества бета-галактозид-связывающего лектина или его производных, предпочтительно члена группы, состоящей из Галектина-1, Галектина-3, Галектина-9, Галектина-13 или Галектина-15 или их производных, с целью улучшить частоту имплантации эмбриона в материнской матке коров, свиней, овец, коз, лошадей, буйволов, собак, кошек и человека, среди других видов, таким образом, предотвращая элиминацию эмбриона материнской иммунной системой.
Другой целью настоящего изобретения является повышение частоты имплантации эмбриона в матке млекопитающих путем переноса бета-галактозид-связывающего лектина и его производных в репродуктивный тракт млекопитающих при помощи искусственного осеменения, переноса in vitro или in vivo эмбриона или спаривания.
Более того, целью настоящего исследования является применение эффективного количества бета-галактозид-связывающего лектина или его производных с целью обеспечения оплодотворения: (i) свежей, охлажденной или криоконсервированной семенной жидкости; (ii) свежего, охлажденного или криоконсервированного ооцита; и (iii) свежего, охлажденного или витрифицированного эмбриона при переносе эмбриона или оплодотворении in vitro, или клонированного или трансгенного эмбриона.
Другой целью настоящего изобретения является предоставление бета-галактозид-связывающего лектина или его производных для повышение частоты имплантации эмбриона в матке млекопитающих.
Наконец, целью настоящего изобретения является применение продукта, включающего эффективное количества бета-галактозид-связывающего лектина или его производных для повышения частоты имплантации эмбриона в матке млекопитающих.
Краткое описание изобретения
Цели настоящего изобретения достигаются при помощи:
(а) способа повышения частоты имплантации эмбриона в матке млекопитающих путем введения в матку млекопитающих эффективного количества бета-галактозид-связывающего лектина или его производных;
(b) применения эффективного количества бета-галактозид-связывающего лектина или его производных для повышения частоты имплантации эмбриона в матке млекопитающих;
(c) бета-галактозид-связывающего лектина или его производных, применяемых для повышения частоты имплантации эмбриона в матке млекопитающих; и
(d) продукта, включающего эффективное количество бета-галактозид-связывающего лектина или его производных для повышения частоты имплантации эмбриона в матке млекопитающих.
Подробное описание изобретения
В настоящем изобретении частота имплантации относится к числу эмбрионов, которые прикрепляются к эндометрию млекопитающих после оплодотворения (объединения мужской и женской гамет), независимо от того, производится ли это при помощи вспомогательной репродуктивной технологии или нет.
Цели настоящего изобретения достигают путем помещения в материнскую матку млекопитающего, как через мочеточник, так и чрезвагинально, эффективного количества активной форму бета-галактозид-связывающего лектина или его производных в стандартном буферном растворе в одиночку или в смеси со спермой, ооцитом или эмбрионом в поддерживающей среде.
Буферный раствор служит для сохранения, обработки и перемещения семенной жидкости, ооцита или эмбриона, и им предпочтительно является носитель, состоящий из солевого буферного раствора (Phosphate Buffer Saline - PBS) или стерильной, стабильной, свободной от эндотоксинов, изотонической физиологической сыворотки с pH между 6,8 и 7,4.
Поддерживающая среда представляет собой комплекс и свободную от сыворотки среду для сохранения эмбрионов в атмосферном воздухе в течение различного времени в соответствии с температурой. Поддерживающая среда обычно состоит из изотонического буферного раствора, который содержит незаменимые аминокислоты, факторы роста, энзимы, энергетические субстраты, клеточные нутриенты и антибиотики.
Примером поддерживающей среды служит разбавитель, то есть, разбавляющая жидкость, добавляемая или смешиваемая с семенной жидкостью для сохранения способности к оплодотворению последней. Специальные разбавители для замораживания также обладают криогенными свойствами, таким образом, обеспечивая перенос, замораживание и разморозку. Разбавители состоят из изотонического буфера, энергетических субстратов и клеточных нутриентов, антибиотиков, антиоксидантов и криопротекторов.
Целей настоящего изобретения также можно достичь путем последовательного или одновременного применения бета-галактозид-связывающего лектина или его производных совместно с семенной жидкостью, ооцитами или эмбрионами в матке млекопитающего (это стадии дополнительно обозначены).
Некоторые преимущества, обеспечиваемые настоящим изобретением, представляют собой следующие: повышение репродуктивной эффективности скота; качественные и количественные улучшения в потомстве; снижение времени между рождением и забоем; снижение затрат на кормление; повышение возможности развития гетерозигот; улучшения в обработке и снижение технических затрат, затрат на здоровье и репродуктивных затрат; среди других аспектов, описываемых здесь далее.
Настоящее изобретение включает применение бета-галактозид-связывающего лектина или его производного, предпочтительно Галектина-1, Галектина-3, Галектина-9, Галектина-13 и Галектина-15, для повышения частоты имплантации эмбриона в матке млекопитающих.
Повышения частоты имплантации эмбриона в матке млекопитающих при помощи репродуктивных технологий напрямую влияет на возможность применения таких процедур в качестве инструмента генетического прогресса, учитывая то, что это улучшает результаты и/или успешность технологий до экономически приемлемых уровней.
Дополнительно, лучше применять генетический материал от генетически более совершенных особей при репродуктивных биотехнологиях, поскольку это максимально увеличивает распространенность этих животных и обеспечивает альтернативу дефициту превосходящих особей. Репродуктивные биотехнологии также позволяют снизить интервалы между поколениями.
В дополнение, одной из главных проблем на сегодняшний день является победа над голодом в Мире, где культивируемые области земли сокращаются с постоянным 50% ростом популяции в следующие 25 лет. Таким образом, повышение частот имплантации эмбриона в материнской матке коров, свиней, овец и других видов означает повышение пищевой продукции в целом, что включает мясную продукцию, потребность в которой увеличится вдвое за следующие 25 лет, достигнув более 127 миллионов тонн в год.
Влияние повышенных частот имплантации эмбриона в матке млекопитающих у продуктивного крупного рогатого скота представляю собой следующее:
- Качественные и количественные улучшения поколений - у крупного рогатого скота, как в отношении веса, так и качества - повышение ежедневного набора веса, улучшение качества окраса, слабость здоровья, добавки и также улучшенная продукция высших сортов, что добавляет большую ценность к конечному продукту. У молочного скота это обеспечивает повышенную продуктивность и улучшение качества молока.
- Это также предоставляет снижение времени до забоя, то есть, повышение ежедневного набора веса, получаемые из-за генетических улучшений, снижает время от рождения до забоя вплоть до 30%. Таким образом, ранний рост животных значительно повышается.
- Снижение затрат на кормление: несмотря на растущие вложения, необходимые для достижения наивысшего генетического качества, существует более удовлетворительная связь со снижением затрат на производство животных, в основном из-за повышения акселерации, которая снижает время до забоя. В виду вышесказанного затраты на производство снижаются по меньшей мене на 10%.
- Повышение возможности появления гетерозигот: они становятся более жизнеспособными для гибридного скота, который не адаптирован к тропическому климату Бразилии, но который обеспечивает улучшения тела, акселерации и плодовитости стада.
- Улучшения в обработке и снижению технических затрат, затрат на здоровье и репродуктивных затрат: применение репродуктивных биотехнологий обеспечивает лучшие состояния здоровья, поскольку они предотвращают распространение заболеваний крупного рогатого скота, передающихся половым путем, снижает затраты на лекарственные средства для лечения заболеваний и, таким образом, снижает затраты на производство.
Более конкретно, настоящее изобретение является результатом тестов по переносу произведенного in vivo эмбриона коровам с целью улучшения и повышения частот имплантации эмбриона в матке млекопитающих в присутствии Галектина-1.
Примеры
Единичную дозу 100 мкг в 22 мкл стерильного, активного, алкилированного и свободного от эндотоксинов рекомбинантного человеческого Галектина-1 вводят реципиентам эмбриона чрезматочным путем. Следует отметить, что вышеобозначенная доза является критически низкой при сравнении с весом тела коровы, и способствует биосохранным аспектам лечащихся самок, которые, в отличие от мышей (обычно используемых в экспериментах над животными) имеют тело, готовое к одной беременности в данный момент времени.
С целью оценки эффектов Галектина-1 в качестве регулирующего средства для оплодотворения млекопитающих используют неизогенных, содержащихся в стойлах животных с нормальными рабочими условиями.
Самок-доноров крупного рогатого скота подвергают суперовуляции и осеменяют в D0 (день 0) оплодотворения. Параллельно реципиентов подвергают протоколам эструсной синхронизации с целью достижения овуляции на D0.
На D7 (7-ой день) доноры входят в стадию сбора и оценки эмбрионов в стадии морулы и/или бластоцисты. Слабые (степень 3), хорошие (степень 2) и отличные (степень 1) эмбрионы помещают в бутылки для последующего переноса в этот же день реципиентам, которых предварительно отбирают в соответствии с развитием желтого тела.
В целом классификацию эмбриона производят в свете морфологических параметров, и она охватывает три главные стадии, определяемые в зависимости от их стадии развития. В лучшем случае эмбрион не должен иметь видимых фрагментов, клетки должны быть гомогенными по размеру. Следует также отметить, что не все эмбрионы с отличной морфологией являются внутренне здоровыми, поскольку они являются ограниченными условиями, связанными с яйцеклеткой, сперматозоидом и самим процессом оплодотворения.
Эмбрионы в колонке с TQC® поддерживающей средой и Галектином-1, разведенном в стерильной физиологической сыворотке в другой колонке (эмбрион не контактирует с раствором Галектина во время хранения в бутылках), помещают в одинаковые пробирки и переносят в матку реципиентов, синхронизированных с донорами, при помощи пипетки для осеменения. Все содержимое пробирки переносят единовременно и, в этом варианте осуществления, эмбрионы и Галектин-1 достигают матки реципиента раздельно и одновременно.
Целью тестов является анализ числа подтвержденных имплантаций эмбриона в матку млекопитающих на 30 и 60 дни после даты переноса эмбриона (i) реципиентам, которые получали местное лечение Галектином-1, (ii) реципиентам перенесенного эмбриона без присутствия Галектина-1. Наконец, реципиентов перенесенного эмбриона разделяют на два отдельные группы для каждого спаривания (донор х семенная жидкость) следующим образом: контрольная группа (эмбрионы без присутствия Галектина-1) и лечебная группа (эмбрионы в присутствии Галектина-1).
Диагностику беременности производят как в лечебной группе, так и в контрольной группе при помощи ультразвукового исследования в два конкретных момента: первое измерение производят через 30 дней после переноса эмбриона (представлено в таблице ниже как «Р30»), и вторую оценку частоты имплантации эмбриона в матку млекопитающего производят через 60 дней после переноса эмбриона (представлено в таблице ниже как «Р60»).
Методы
Реципиентами являются самки крупного рогатого скота с хорошей выработкой молока и известными темпами размножения. Реципиентами являются самки Гольстейна с хорошей выработкой молока и известными темпами размножения.
Процедура отбора для спаривания относится к выбору производителя (источника семенной жидкости) для каждого донора. После сбора и оценки эмбрионов около половины созданных эмбрионов переносят в матку реципиентов, принадлежащих к контрольной группе, другую половину эмбрионов переносят в матку реципиентов, принадлежащих к группе, подвергнутой лечению. Имеет место попытка равного разделения эмбрионов в соответствии со степенью качества между контрольной группой и группой, подвергнутой лечению, с целью отображения сравнения групп.
Методы: КОНТРОЛЬНАЯ ГРУППА
Процедуру переноса эмбриона в контрольной группе осуществляют в соответствии с традиционным подходом к подобным событиям, то есть, перенос эмбриона происходит в рог матки, ипсилатерально от желтого тела (где овуляция происходит во время эструса) реципиентов. Применяемая пробирка состоит из трех главных колонок, наполненная первая колонка с TQC® поддерживающей средой, с последующей колонкой с воздухом с последующей колонкой с TQC® поддерживающей средой, содержащей эмбрион, с последующей другой колонкой с воздухом с последующей другой колонкой с TQC® поддерживающей средой. Все содержимое пробирки переносят единомоментно.
Метод: ПОДВЕРГНУТАЯ ЛЕЧЕНИЮ ГРУППА
Процедуру переноса эмбриона в лечебной группе осуществляют в соответствии с традиционным подходом к подобным событиям, то есть, перенос эмбриона происходит в рог матки, ипсилатерально от желтого тела (где овуляция происходит во время эструса) реципиентов. Применяемая пробирка состоит из трех главных колонок, наполненная первая колонка с TQC® поддерживающей средой, с последующей колонкой с воздухом с последующей колонкой с TQC® поддерживающей средой, содержащей эмбрион, с последующей другой колонкой с воздухом с последующей другой колонкой с поддерживающей средой, содержащей Галектин-1. Все содержимое пробирки переносят единомоментно, и в этом варианте осуществления эмбрионы и Галектин-1 достигают матки реципиента раздельно и одновременно.
Результаты: В двух иллюстрированных таблицах ниже первый столбец слева (Повторения) отображает повторения переносов эмбрионов, выполняемых каждой самкой крупного рогатого скота, в соответствии с общим числом в 12 повторений. Тем временем два других столбца разделены для разделения результатов, полученных в контрольной группе и лечебной группе, последующие два столбца показывают отношения между частотами дополнительной имплантации эмбриона в матку реципиентов, полученные при применении в процедуре Галектина-1.
Столбец, соответствующий «Контрольному количеству», отображает число эмбрионов, произведенных от переносов эмбрионов без присутствия Галектина-1. Следующий столбец справа (Контрольное Р30) разделен для демонстрации числа имплантаций через 30 дней после переноса эмбриона (Количество 30) с учетом соответствующих процентов, соответствующих частоте имплантации в течение того же периода (T30). Аналогичное объяснение применяют для следующего столбца справа (Контрольное Р60), разделенного для демонстрации как числа имплантаций через 60 дней после переноса эмбриона (Количество 60), так и соответствующих процентов, соответствующих частоте имплантации в течение того же периода (T60).
Столбец, соответствующий «Лечебному количеству» отображает число эмбрионов, произведенных от переносов эмбрионов в присутствии Галектина-1. Следующий столбец справа (Лечебное Р30) разделен для демонстрации числа имплантаций через 30 дней после переноса эмбриона (Количество 30) с учетом соответствующих процентов, соответствующих частоте имплантации в течение того же периода (T30). Аналогичное объяснение применяют для следующего столбца справа (Лечебное Р60), разделенного для демонстрации как числа имплантаций через 60 дней после переноса эмбриона (Количество 60), так и соответствующих процентов, соответствующих частоте имплантации в течение того же периода (T60).
Столбец, соответствующий «∆ частоты имплантации», разделен для отображения разницы в частоте имплантации, полученной в контрольной группе и лечебной группе Р30 (∆30), а также в контрольной группе и лечебной группе Р60 (∆60).
Таблицы:
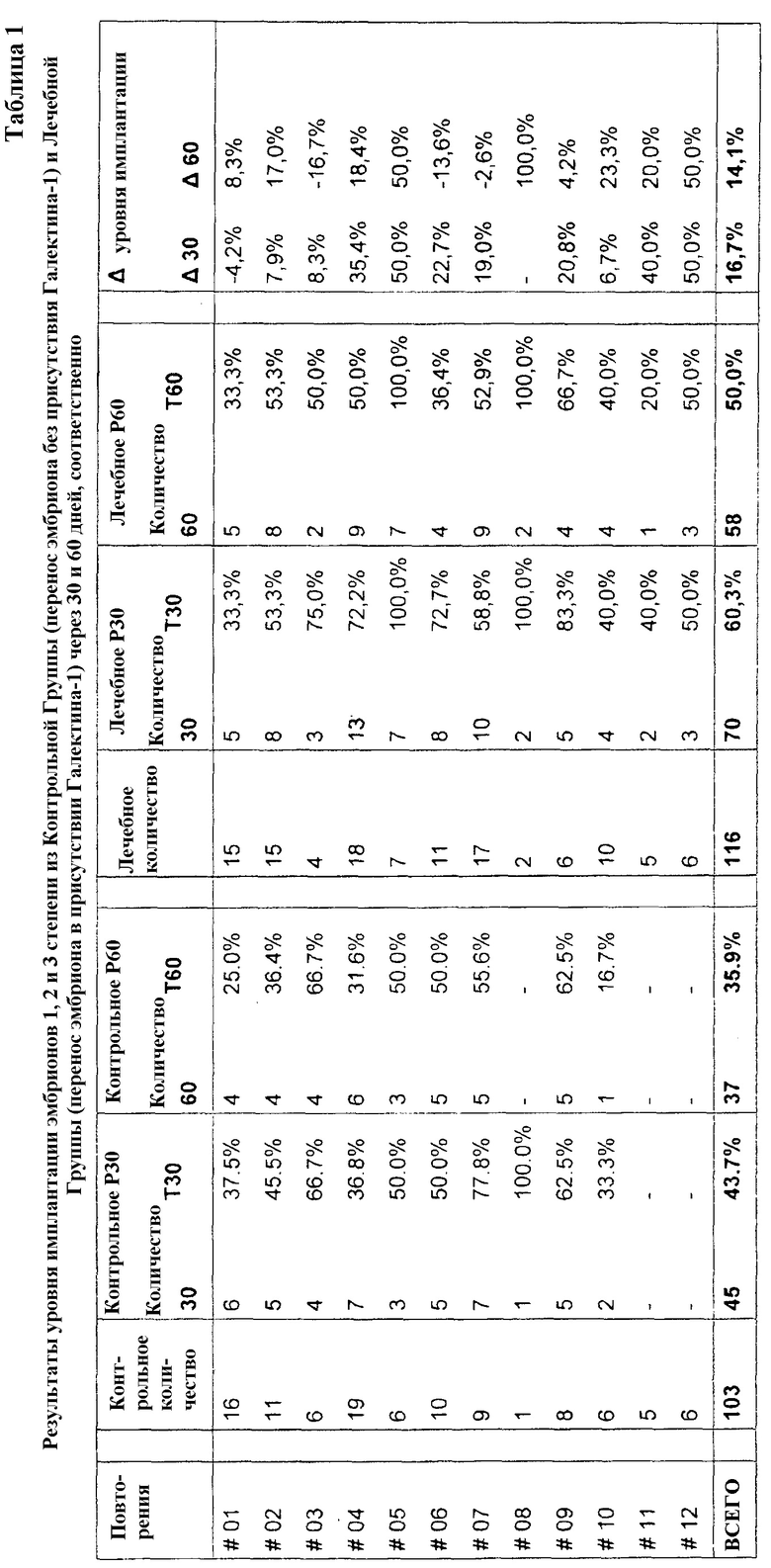
Как видно из таблицы 1, анализ конечного результата показывает, что через 30 дней после переноса эмбриона в лечебной группе наблюдают более высокую частоту имплантации в материнской матке, равную 60,3% по сравнению с 43,7% в контрольной группе. Анализ конечных результатов через 60 дней после переноса эмбриона показывает, что в лечебной группе наблюдается 50% имплантация в материнской матке, в то время как в контрольной группе - 35,9%.
В свете результатов таблицы 1 возможно сделать вывод о том, что применение Галектина-1 увеличивает число имплантаций эмбрионов в материнской матке на 16,7 процентов через 30 дней и 14,1 процентов через 60 дней.
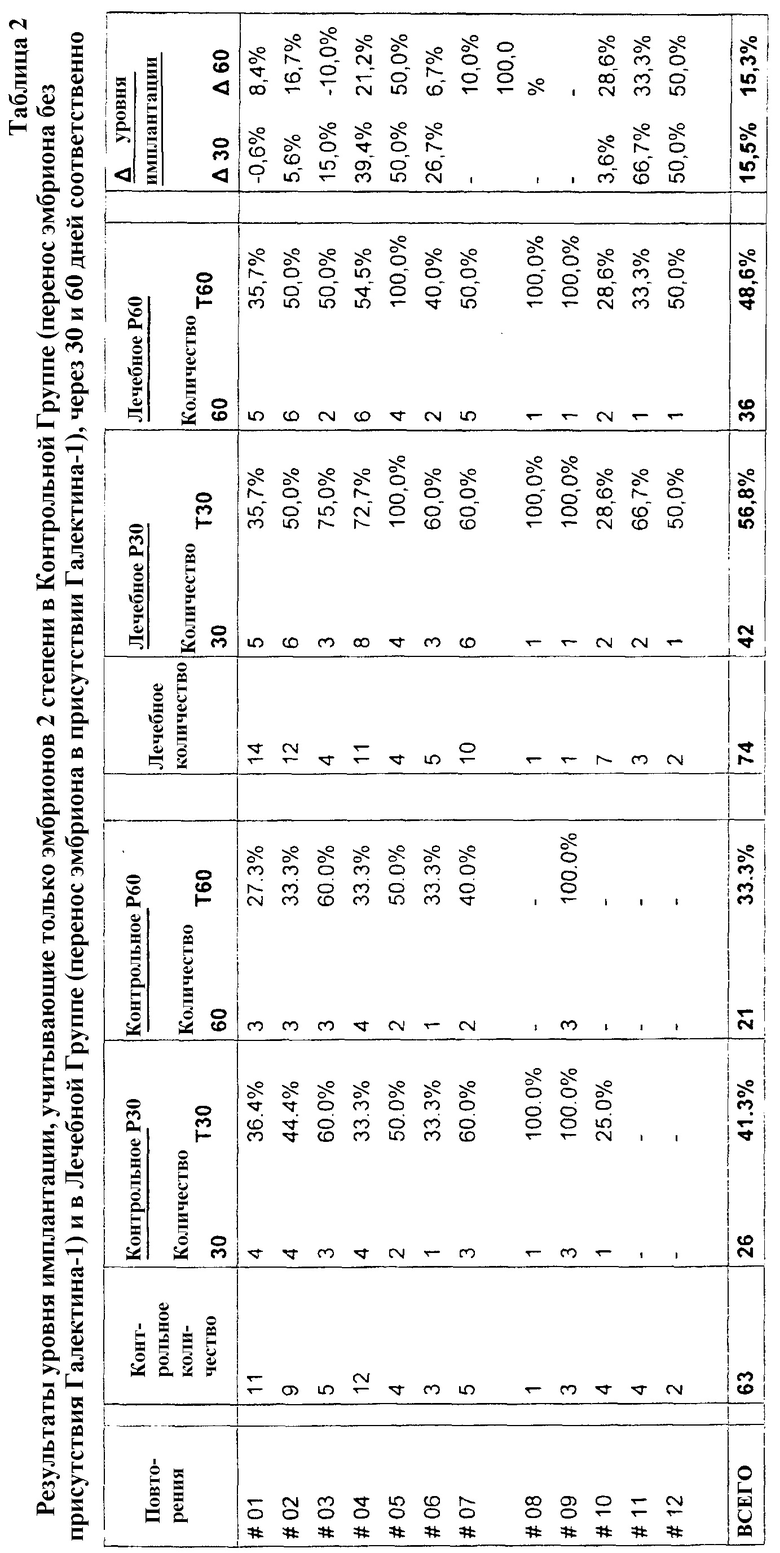
Результаты, отображенные в таблице 2, показывают, что в лечебной группе наблюдается большее число имплантаций в материнской матке и составляет 56,8% частоты имплантации по сравнению с 41,3% имплантации, достигнутой в контрольной группе через 30 дней после переноса эмбрионов на стадии 2. Анализ конечных результатов через 60 дней после переноса эмбрионов на стадии 2 показывает, что в лечебной группе наблюдается частота имплантации в материнской матке в 48,6%, в то время как в контрольной группе частота составляет 33,9%.
В виду результатов, показанных в таблице 2, возможно сделать вывод, что Галектин-1, вводимый в матку крупного рогатого скота-реципиента в лечебной группе, повышает число имплантаций на 15,5 процентов через 30 дней, и на 15,3 процента через 60 дней, улучшая частоту имплантации эмбрионов 2 степени в матке коров.
Учитывая то, что в этом эксперименте рекомбинантный человеческий Галектин-1 действует как антиген в материнской матке крупного рогатого скота, в основном потому, что он происходит от других видов, исследования проводятся с целью доказать (i) большее взаимодействие между Галектинами и гликанами одних и тех же видов, и, следовательно, (ii) лучшее влияние Галектина-1 на повышение частоты имплантации эмбриона в материнской матке млекопитающих.
Физиологические механизмы, запускаемые Галектином-1, который позволяет улучшить частоту имплантации эмбриона в матке млекопитающих, все еще находятся в процессе изучения, поскольку беременности реципиентов крупного рогатого скота, которым осуществлялся перенос эмбрионов, еще не подошли к своему сроку.
У крупного рогатого скота плацента имеет множество единиц различных размеров и форм, которые, в свою очередь, состоят из возвышений со сплетенными фетальными ворсинами, которые формируют инвагинации, соединяющиеся в матке. Для сохранения беременности эмбрион должен участвовать, посылая признаки своего существования в маточной среде путем продукции IFNT. Таким образом, лютеолиз остается ингибированным и, соответственно, уровни Р4 поддерживаются на высоком уровне. Таким образом, и IFNT, и P4 стимулируют мРНК к повышению уровня Галектинов в матке. После того как материнская среда распознает и принимает эмбрион, клетки трофобласта дифференцируются и объединяются с клетками маточного эпителия, таким образом, вступая в прямой контакт с материнскими тканями.
Считается, таким образом, что повышение частоты имплантации эмбриона в материнской матке млекопитающих объясняется участием Галектина-1 в регуляции механизмов, связанных с иммунной толерантностью и/или в запуске процесса элонгации бластоцисты и адгезии эмбриона к эндометрию.
Настоящее изобретение дополнительно раскрывает, что бета-галактозид-связывающий лектин или его производные, предпочтительно выбираемые из Галектина-1, Галектина-3, Галектина-9, Галектина-13 или Галектина-15 или его производных, можно применять в качестве средств регуляции фертильности семенной жидкости, ооцитов или эмбрионов, таким образом, повышая частоту имплантации эмбриона в матке млекопитающих.
Количество бета-галактозид-связывающего лектина или его производного, применяемого к материнской матке млекопитающего, может варьировать в соответствии с весом тела вида, и уровень его концентрации должен предпочтительно варьировать от 0,0000001 до 1,0 мг на кг веса тела млекопитающего.
В одном варианте осуществления бета-галактозид-связывающий лектин или его производные разводят в забуферированном растворе, предпочтительно в забуферированном фосфатом физиологическом растворе (PBS) или физиологической сыворотке, и применяют в матке при помощи стандартной пробирки, смешанной с семенной жидкостью, ооцитом и эмбрионом, содержащихся в поддерживающей среде (способной поддерживать выживание клеток и биологическую целостность во время обработки, переноса, замораживания и размораживания), таким образом, что все содержимое пробирки применяют к матке млекопитающего единовременно.
В предпочтительном варианте бета-галактозид-связывающий лектин или его производные запаковывают в две стадии, раздельно и после семенной жидкости, ооцита или эмбриона, и в этой процедуре используют две пробирки. В первой заявке семенную жидкость, ооцит или эмбрион помещают в поддерживающую среду и запаковывают в обычную пробирку, таким образом, чтобы все содержимое применить в матке млекопитающего. Во второй заявке бета-галактозид-связывающий лектин или его производные, разведенные в буферированном растворе, запаковывают в разные пробирки и далее применяют в матке млекопитающего. Промежуток времени между применением бета-галактозид-связывающего лектина или его производного и применением спермы, ооцита и эмбриона преимущественно зависит от вида млекопитающего и применяемого способа лечения, и может длиться до 17 дней в соответствии с применяемым лечением.
Также раскрыто, что бета-галактозид-связывающий лектин или его производные, разведенные в буферированном растворе, запаковывают в колонку данной обычной пробирки, вставленной между другими колонками, содержащими сперму, ооцит или эмбрион в поддерживающей среде, так что все содержимое пробирки применяют в матке млекопитающего.
Во всех предоставленных вариантах бета-галактозид-связывающий лектин или его производные вместе с семенной жидкостью, ооцитом или эмбрионом вводят через матку или через влагалище. Семенная жидкость может быть свежей, охлажденной или криоконсервированной; ооцит может быть свежим, криоконсервированным или витрифицированным: и эмбрион может быть свежим, криоконсервированным или витрифицированным и может существовать после переноса эмбриона, оплодотворения in vitro, а также из клона или трансгенного эмбриона.
Более того, известно, что системы продуцирования эмбриона in vitro позволяют определить пол эмбриона после оплодотворения и, таким образом, указанные системы являются важным инструментом для разводчиков молочных и мясных стад. Тем не менее, произведенные in vitro эмбрионы не показывают удовлетворительных уровней выживаемости в матке млекопитающих. Следовательно, изобретение обеспечивает решение регулирования фертильной спермы, ооцитов и эмбрионов, тем самым увеличивая частоту имплантации эмбриона в матку млекопитающих.
С другой стороны, одним из ограничений промышленного применения репродуктивных биотехнологий является проходимое расстояние между местом сбора семенной жидкости, ооцита и донорского эмбриона и местом нахождения вида-реципиента; а также расстояние, разделяющее видов-доноров от видов-реципиентов. В этом смысле криосохранение (процесс замораживания) и витрифицирование (ультрабыстрый метод замораживания) стали распространенной практикой в производстве животных. Тем не менее, хотя технологии криосохранения и витрифицирования имеют промышленные преимущества, сперматозоиды могут подвергаться изменениям в мембране, ранней активности, изменениям ДНК и оксидативному стрессу во время замораживания, что негативно влияет на их фертильность.
В предпочтительном варианте осуществления настоящее изобретение раскрывает применение продукта, включающего эффективное количество бета-галактозид-связывающего лектина или его производных, для повышения частоты имплантации эмбриона в матке млекопитающих.
Как раскрыто в нескольких примерах предпочтительных вариантов осуществления, следует понимать, что объем настоящего изобретения охватывает другие возможные варианты осуществления и ограничен только содержимым прилагаемой формулой изобретения, включающим возможные эквиваленты.
Изобретение относится к способу повышения частоты имплантации эмбриона в материнской матке млекопитающих. Способ осуществляют путем введения в матку млекопитающего эффективного количества бета-галактозид-связывающего лектина или его алкилированной формы перед предимплантационным периодом. Изобретение касается применения эффективного количества бета-галактозид-связывающего лектина или его алкилированной формы для повышения частоты имплантации эмбриона в матке млекопитающих путем осуществления способа, а также продукта для повышения частоты имплантации эмбриона в матке здоровых млекопитающих, включающего эффективное количество бета-галактозид-связывающего лектина или его алкилированной формы. Применение продукта увеличивает число имплантаций эмбрионов в материнской матке. 3 н. и 14 з.п. ф-лы, 2 табл.
1. Способ повышения частоты имплантации эмбриона в матке здоровых млекопитающих, отличающийся тем, что он включает введение в матку млекопитающего эффективного количества бета-галактозид-связывающего лектина или его алкилированной формы перед предимплантационным периодом.
2. Способ по п. 1, отличающийся тем, что количество, варьируемое от 0,0000001 до 1,0 мг активной формы бета-галактозид-связывающего лектина или его алкилированной формы, вводят на килограмм веса тела млекопитающего.
3. Способ по п. 2, отличающийся тем, что активная форма бета-галактозид-связывающего лектина или его алкилированной формы находится в форме стерильного, стабильного, свободного от эндотоксина, изотонического носителя и имеет рН между 6,8 и 7,4.
4. Способ по п. 3, отличающийся тем, что носитель представляет собой буферированный раствор.
5. Способ по п. 4, отличающийся тем, что буферированный раствор выбирают из солевого фосфатного буфера (PBS) или физиологической сыворотки.
6. Способ по п. 1, отличающийся тем, что бета-галактозид-связывающий лектин или его алкилированную форму выбирают из Галектина-1, Галектина-3, Галектина-9, Галектина-13, Галектина-15 или их алкилированной формы.
7. Способ по п. 1, отличающийся тем, что бета-галактозид-связывающий лектин или его алкилированную форму применяют в матке млекопитающего смешанным со семенной жидкостью, ооцитом или эмбрионом.
8. Способ по п. 1, отличающийся тем, что бета-галактозид-связывающий лектин или его алкилированную форму применяют в матке млекопитающего вместе с семенной жидкостью, ооцитом или эмбрионом, как раздельно, так и одновременно.
9. Способ по п. 1, отличающийся тем, что бета-галактозид-связывающий лектин или его алкилированную форму применяют в матке млекопитающего вместе с семенной жидкостью, ооцитом или эмбрионом, как раздельно, так и последовательно.
10. Способ по п. 9, отличающийся тем, что бета-галактозид-связывающий лектин или его алкилированную форму применяют в матке млекопитающего вместе с семенной жидкостью, ооцитом или эмбрионом последовательно, и временной промежуток между применением бета-галактозид-связывающего лектина или его алкилированной формы и применением семенной жидкости, ооцита или эмбриона продолжается до 17 дней.
11. Способ по п. 7, отличающийся тем, что семенную жидкость, ооцит или эмбрион предоставляют в поддерживающейся среде.
12. Способ по п. 11, отличающийся тем, что семенная жидкость является свежей, охлажденной или замороженной.
13. Способ по п. 11, отличающийся тем, что эмбрион является свежим, криоконсервированным или витрифицированным и получен путем переноса эмбриона (ЕТ), оплодотворения in vitro (IVF) или из клона или трансгенного эмбриона.
14. Способ по п. 11, отличающийся тем, что ооцит является свежим, охлажденным или витрифицированным.
15. Способ по п. 7, отличающийся тем, что бета-галактозид-связывающий лектин или его алкилированную форму вместе с семенной жидкостью, ооцитом или эмбрионом вводят через матку или влагалище.
16. Применение эффективного количества бета-галактозид-связывающего лектина или его алкилированной формы для повышения частоты имплантации эмбриона в матке млекопитающих путем осуществления способа по пп. 1-15.
17. Продукт для повышения частоты имплантации эмбриона в матке здоровых млекопитающих, отличающийся тем, что он включает от 0,0000001 до 1,0 мг бета-галактозид-связывающего лектина или его алкилированной формы и поддерживающую среду, где бета-галактозид-связывающий лектин или его алкилированную форму выбирают из Галектина-1, Галектина-3, Галектина-9, Галектина-13, Галектина-15 или их алкилированной формы, и поддерживающую среду выбирают из солевого фосфатного буфера (PBS) или физиологической сыворотки.
| US 2007185014 A1, 09.08.2007 | |||
| СПОСОБ ПОВЫШЕНИЯ ВЫЖИВАЕМОСТИ ЭМБРИОНОВ У КОРОВ | 2004 |
|
RU2272630C2 |
| Shaye K | |||
| Lewis et al | |||
| Прибор для нагревания перетягиваемых бандажей подвижного состава | 1917 |
|
SU15A1 |
| Спускная труба при плотине | 0 |
|
SU77A1 |
| ПРИВОД ДЛЯ ПЛОСКОЙ ВЯЗАЛЬНОЙ МАШИНЫ | 1923 |
|
SU1027A1 |
| Shaye K | |||
| Lewis et al | |||
| Прибор для нагревания перетягиваемых бандажей подвижного состава | 1917 |
|
SU15A1 |
| Спускная труба при плотине | 0 |
|
SU77A1 |
| ПРИВОД ДЛЯ ПЛОСКОЙ ВЯЗАЛЬНОЙ МАШИНЫ | 1923 |
|
SU1027A1 |
| US 5869081 A, 09.02.1999 | |||
| DAVID G | |||
| PORTER | |||
| The local effect of intra-uterine progesterone treatment on myometrial activity in rabbits | |||
| J | |||
| Reprod | |||
| Fert | |||
| Приспособление для контроля движения | 1921 |
|
SU1968A1 |
| Способ образования окрасок на волокнах | 1925 |
|
SU437A1 |
| Choe YS et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Mol Reprod Dev | |||
| Электрическое сопротивление для нагревательных приборов и нагревательный элемент для этих приборов | 1922 |
|
SU1997A1 |
| Найдено в PubMed, PMID: 9291476 | |||
| Junko Nio and Toshihiko Iwanaga | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Journal of Histochemistry & Cytochemistry | |||
| Устройство двукратного усилителя с катодными лампами | 1920 |
|
SU55A1 |

