Область техники, к которой относится данное изобретение
Данное изобретение относится к способам лечения рака, агентам для ингибирования пролиферации раковых клеток и противораковым агентам.
Уровень техники
Рак является главной причиной смертности в индустриальных странах. Многие химиотерапевтические агенты были разработаны на протяжении последних 50 лет с целью лечения рака. Большинство химиотерапевтических агентов могут быть классифицированы как алкилирующие агенты, антиметаболиты, антрациклины, алкалоиды растений, ингибиторы топоизомеразы и противоопухолевые агенты. Все из этих фармацевтических агентов воздействуют на деление клеток или синтез ДНК и вызывают терапевтические эффекты посредством механизма, который функционирует некоторым определенным образом.
Эффективность конкретного химиотерапевтического агента является разной среди раков или пациентов или является разной в зависимости от временного хода в индивидуальных пациентах. Раковые клетки, подвергнутые химиотерапевтическим агентам, развивают устойчивость к этим химиотерапевтическим агентам и также часто развивают перекрестную устойчивость к множеству других противораковых агентов. Кроме того, для контроля побочных эффектов, происходящих из повреждения клеток этими химиотерапевтическими агентами на нормальных клетках посредством вышеупомянутого механизма этих агентов, доза или применение этих агентов часто являются ограниченными.
Вместо общепринятых химиотерапевтических агентов, недавно были разработаны молекулярно-нацеленные лекарственные средства, которые экспрессируются специфически на раковых клетках. С появлением этих молекулярно-нацеленных лекарственных средств побочные эффекты, присущие общепринятым химиотерапевтическим агентам, могли избегаться, и лечение рака, которое способствует QOL раковых пациентов, становится осуществимым. Такие молекулярно-нацеленные лекарственные средства включают в себя фармацевтические агенты с малыми молекулами, а также высокомолекулярные фармацевтические агенты, такие как антитела. Терапевтические антитела являются молекулами, которые врожденно присутствуют в организме и имеют преимущество низкой токсичности в отношении живых организмов, а также преимущество проявления терапевтических эффектов, специфически разрушающих клетки-мишени посредством механизма действия, другого, чем механизм фармацевтических агентов с малой молекулой, такого как цитотоксическая активность, опосредуемая эффекторными функциями. Таким образом, многие терапевтические антитела недавно были представлены на рынок.
Нацеливающий на терапевтические антитела эпирегулин, который высоко экспрессируется в раке ободочной кишки, аденокарциноме легкого, панкреатическом раке, раке желудка и раке почки, был описан в качестве антител, которые специфически повреждают клетки-мишени посредством механизма действия, другого, чем механизм действия низкомолекулярных агентов, такой как цитотоксическая активность, опосредуемая такими эффекторными функциями (Патентный Документ 1). Конкретно, измерение активности комплементзависимой цитотоксичности (CDC) и активности антителозависимой клеточноопосредованной цитотоксичности (ADCC) выявило, что анти-Эпирегулин-антитела имеют CDC-активность и ADCC-активность на Эпирегулин-экспрессирующих клетках. Кроме того, было обнаружено, что анти-Эпирегулин-антитела оказывают ингибирующее пролиферацию действия на линиях раковых клеток посредством нейтрализующего действия. Кроме того, из вышеупомянутых открытий было выявлено, что анти-Эпирегулин-антитела являются эффективными для диагностики, предотвращения и лечения различных первичных и метастатических раков.
Любой новый кандидатный фармацевтический агент, включающий в себя противораковые агенты, такие как описанные выше, должен проходить строгие испытания, чтобы стать коммерчески доступным. Например, эти испытания классифицируются как преклинические испытания и клинические испытания. Обычно последние классифицируются в испытаниях фазы I, фазы II и фазы III и выполняются на человеке, тогда как первые испытания выполняются с использованием животных. Обычно целью преклинических исследований является демонстрация того, что кандидатное лекарственное средство является сильным, а также эффективным и безопасным. Конкретно, целями этих исследований на животных является демонстрация того, что этот фармацевтический агент не является канцерогенным, мутагенным или тератогенным, а также понимание фармакокинетики этого фармацевтического агента. Клинические исследования с использованием введения тестируемого фармацевтического агента разрешаются только тогда, когда безопасность и эффективность тестируемого фармацевтического агента в отношении животных установлены в преклинических испытаниях.
Во многих случаях действие низкомолекулярного тестируемого фармацевтического агента (например, нового противоракового агента, произведенного из антрациклина) в животных может становиться индикатором для ожидаемых действий этого фармацевтического агента при введении человеку. Таким образом, обычно данные, полученные из таких преклинических испытаний, могут быть высокопредсказуемыми из действий, которые могут иметь место при введении людям. Однако такая предсказуемость не получается в любом типе тестируемого фармацевтического агента; и предсказуемость на основе результатов преклинических исследований, и вероятность того, что эти кандидатные фармацевтические агенты будут одобрены в клинических исследованиях, значительно уменьшается.
Обычно антитела могут функционировать посредством высокоспецифического узнавания молекул-мишеней, которые являются обычно белковыми. В большинстве случаев тестируемые фармацевтические агенты в виде антител являются моноклональными антителами и узнают только единственный сайт или единственный эпитоп на молекуле-мишени. Поскольку моноклональные антитела обычно имеют высокую мишень-идентифицирующую функцию, антитела становятся представляющими большой интерес кандидатами для развития фармацевтических агентов, но, с другой стороны, эта идентифицирующая функция в некоторых случаях делает преклинические исследования трудными. Это бывает потому, что имеются видоспецифические вариации в последовательностях молекул-мишеней, связанных этими антителами. Например, моноклональное антитело, которое специфически узнает молекулу Y via эпитоп Х в людях и связывается с этой молекулой, будет тестироваться на соответствующий эпитоп X' в соответствующей молекуле-мишени (ортологе) Y' в видах животных, используемых для преклинических исследований, но X' может отличаться от X, присутствующего в соответствующей молекуле-мишени в людях. Таким образом, часто это моноклональное антитело не может специфически узнавать ортолог Y' и связываться с этой молекулой. Даже среди групп моноклональных антител, которые имеют реактивность в отношении антигенов человека и приматов, имеется много примеров антител, которые реагируют только с гомологами антигена человека и шимпанзе. Например, такие случаи наблюдали для моноклональных анти-CD3-антител. Одним из наиболее широко используемых CD3-комплекс-специфических моноклональных антител, которые имеют большинство определяемых свойств, является OKT-3. OKT-3 реагирует с CD3 шимпанзе, но не реагирует с гомологами CD3 других приматов, таких как макак-резус или CD3 собаки (Непатентный Документ 2). С другой стороны, имеются примеры моноклональных антител, которые узнают антигены резуса, но не их ортологи человека. Одним примером в этой группе является FN-18, который является моноклональным антителом против полученного из макака-резус CD3 (Непатентный Документ 2).
Несколько стратегий были заимствованы для противостояния проблемам с преклиническими исследованиями на животных, вызываемыми высокой специфичностью таких моноклональных антител.
Первым известным подходом является выполнение преклинических исследований на тестируемых содержащих антитела фармацевтических препаратах с использованием модели шимпанзе. Шимпанзе являются ближайшим генетическим родственником людей, и поскольку их геном имеет 99% идентичность относительно генома человека, вариации молекулы-мишени, специфически связанные тестируемым содержащим антитело фармацевтическим препаратом в шимпанзе, являются с высокой вероятностью идентичными вариациям этой молекулы в людях. Действительно, Schereth et al. обнаружили, что эти вариации в CD3 являются общими между людьми и шимпанзе (Непатентный Документ 3). Таким образом, считается, что риск того, что эта молекула не будет распознаваться тестируемым содержащим антитело фармацевтическим препаратом в шимпанзе, является низким. Однако исследования с использованием шимпанзе являются очень дорогостоящими и также имеют этические проблемы. Кроме того, поскольку шимпанзе являются животными с риском вымирания, и количество животных, которые могут быть использованы в экспериментах, является в сильной степени ограниченным, такие преклинические исследования на шимпанзе исключены из развития большинства тестируемых содержащих антитела фармацевтических препаратов.
Вторым подходом является подход адаптирования молекулы, используемой в преклинических исследованиях к животному, используемому в этих исследованиях. В этом подходе информацию незаменимой безопасности получают в преклинических исследованиях конструированием так называемого ʺсуррогатногоʺ антитела для введения тест-животным. Обычно таким суррогатным антителом является антитело, которое специфически распознает ортолог тест-животного из молекулы-мишени, связанной несуррогатным антителом (подлинным тестируемым содержащим антитело фармацевтическим препаратом для человека), и является антителом, которое было модифицировано для связывания этого ортолога. Таким образом, в этом подходе с использованием такого ʺсуррогатногоʺ антитела, можно индивидуально сконструировать две разные молекулы: клинический тестируемый фармацевтический агент и преклинический тестируемый фармацевтический агент для использования в преклинических исследованиях на виде животного, которое имеет специфичность-мишень, соответствующую этому клиническому фармацевтическому агенту, и безопасность которого, как такового, должна быть испытана. Большим недостатком такого суррогатного подхода является то, что этим суррогатным антителом для преклинических исследований является модифицированный продукт этого клинического тестируемого содержащего антитело фармацевтического препарата. Таким образом, данные, полученные в преклинических исследованиях с использованием суррогатного антитела, часто не могут быть применимы непосредственно к людям. Таким образом, предсказуемость результатов клинического исследования, основанная на результатах преклинического исследования с использованием этих подходов, может уменьшаться.
Вышеупомянутый подход адаптирует тестируемый фармацевтический агент таким образом, что он является подходящим для животного, используемого в преклинических исследованиях. С другой стороны, другие известные подходы адаптируют животных, используемых в преклинических исследованиях, к кандидатному фармацевтическому агенту, подлежащему введению в людей.
Примером адаптации тест-животного к содержащему тест-антитело фармацевтическому препарату, предназначенному для введения людям, является продуцирование трансгенного животного, которое экспрессирует молекулу человека, с которой этот тестируемый содержащий антитело фармацевтический препарат специфически связывается, вместо молекулы не-человека, присущей этому виду тест-животного. Ожидается, что в этом способе, этот тестируемый содержащий антитело фармацевтический препарат связывается с антигеном человека в этом трансгенном тест-животном. Например, в исследовании, проведенном Bugelski et al., оценивание преклинической безопасности выполняли на моноклональном антителе keliximab с использованием CD4-трансгенных мышей человека для предсказания долгосрочного лечения ревматоидного артрита в пациентах-людях. Keliximab является моноклональным антителом, которое имеет специфичность в отношении CD4 человека и шимпанзе. Bugelski et al. делают вывод, что использование экспрессирующей белок человека трансгенной мыши обеспечивает полезный альтернативный способ для исследований, проводимых в шимпанзе, с использованием биологических фармацевтических препаратов, которые имеют ограниченную межвидовую специфичность. Однако получение трансгенных животных для тест-целей является трудоемким и дорогостоящим, так как оно требует большой работы.
Документы известного уровня техники
Патентные Документы
Патентный Документ 1 WO2008/047723.
Непатентные Документы]
Непатентный Документ 1 J. Med. Primatol. (1986 15, 441-451.
Непатентный Документ 2 J. Med. Primatol. (2001) 40, 141-147.
Непатентный Документ 3 Cancer Immunol. Immunother. Май 2006; 55 (5), 503-514.
Непатентный Документ 4 Hum. Exp. Toxicol. (2000) 19, 230-243.
Сущность изобретения
Проблемы, подлежащие решению этим изобретением
Данное изобретение относится к анти-Эпирегулин-антителам, обнаруживающим перекрестную межвидовую реактивность между животными, не являющимися человеком, и людьми. Дополнительно данное изобретение относится к анти-Эпирегулин-антителам с ослабленной химической деградацией. Данное изобретение относится также к анти-Эпирегулин-антителам с уменьшенной изоэлектрической точкой. Кроме того, данное изобретение относится к анти-Эпирегулин-антителам с уменьшенным количеством агрегата. Кроме того, данное изобретение относится к фармацевтическим композициям или терапевтическим агентам для рака, содержащим вышеупомянутые анти-антитела. Данное изобретение относится также к способам получения вышеупомянутых анти-Эпирегулин-антител.
Средства для решения этих проблем
Авторы этого изобретения обнаружили, что отношение активности связывания с Эпирегулином, выделенным из собакоподобной обезьяны, к активности связывания с Эпирегулином человека увеличивается заменой аминокислотного остатка в последовательности вариабельного района гуманизированного антитела EP27, которое ингибирует рост раковых клеток проявлением цитотоксической активности и нейтрализующей активности против раковых клеток, которые экспрессируют Эпирегулин, остатком аргинина. Более конкретно, авторы данного изобретения сконструировали анти-Эпирегулин-антитела, обнаруживающие перекрестную межвидовую реактивность между людьми и собакоподобными обезьянами, которые являются животными, не являющимися человеком. Кроме того, конструировали анти-Эпирегулин-антитела с уменьшенной химической деградацией заменой подходящим образом аминокислотных остатков в последовательности вариабельного района гуманизированного антитела EP27. Кроме того, конструировали анти-Эпирегулин-антитела с уменьшенной изоэлектрической точкой заменой подходящим образом аминокислотных остатков в последовательности вариабельного района гуманизированного антитела EP27. Конструировали анти-Эпирегулин-антитела с уменьшенным количеством агрегата заменой подходящим образом аминокислотных остатков в последовательности вариабельного района гуманизированного антитела EP27. Авторы данного изобретения выявили, что анти-Эпирегулин-антитела, имеющие эти свойства, обнаруживают эффекты ингибирования роста посредством нейтрализующей активности и цитотоксической активности на линиях раковых клеток. Кроме того, из вышеупомянутых открытий, авторы данного изобретения обнаружили, что анти-Эпирегулин-антитела являются эффективными для лечения различных первичных и метастатических раков, и тем самым завершили данное изобретение.
Более конкретно, данное изобретение относится к следующему:
[1] анти-Эпирегулин-антителу, которое является антителом, которое связывается с эпитопом, связанным анти-Эпирегулин-антителом, содержащим CDR вариабельного района тяжелой цепи SEQ ID NO: 9, 10 и 11 и CDR вариабельного района легкой цепи SEQ ID NO: 12, 13 и 14, где это антитело характеризуется наличием меньшего отношения величины KD для анти-Эпирегулин-антитела обезьяны SEQ ID NO: 170 (cEREG KD) к величине KD для Эпирегулина человека SEQ ID NO: 34 (hEREG KD) (cEREG KD/hEREG KD), чем это отношение (cEREG KD)/hEREG KD) анти-Эпирегулин-антитела, содержащего CDR вариабельного района тяжелой цепи SEQ ID NO: 9, 10 и 11 и CDR вариабельного района легкой цепи SEQ ID NO: 12, 13 и 14:
[2] антителу [1], где cEREG KD/hEREG KD меньше чем 40;
[3] антителу [1], где cEREG KD/hEREG KD меньше чем 10;
[4] антителу [1], где cEREG KD/hEREG KD меньше чем 6;
[5] антителу [1], где cEREG KD/hEREG KD меньше чем 4;
[6] антителу по любому [1]-[5], которое содержит вариабельный район тяжелой цепи, содержащий CDR1 тяжелой цепи SEQ ID NO: 9, CDR2 тяжелой цепи, выбранный из группы, состоящей из SEQ ID NO: 161, 160, 159, 157, 156, 155, 153, 108, 107, 106, 105, 104, 103, 102, 101 и 100, и CDR3 тяжелой цепи, выбранный из группы, состоящей из SEQ ID NO: 158, 154, 152, 151, 112, 111, 110 и 11; и вариабельный район легкой цепи, содержащий CDR1 легкой цепи, выбранный из группы, состоящей из SEQ ID NO: 163, 68, 67 и 12, CDR2 легкой цепи, выбранный из группы, состоящей из SEQ ID NO: 71, 69 и 13, и CDR3 легкой цепи, выбранный из группы, состоящей из SEQ ID NO: 164, 48, 47 и 14;
[7] антителу по любому из [1]-[5], которое содержит вариабельный район тяжелой цепи, выбранный из группы, состоящей из SEQ ID NO: 150, 149, 148, 147, 146, 145, 144, 143, 142, 140, 139, 138, 137, 135, 134, 133, 132, 131, 127, 126, 125, 124, 123, 122, 121, 120, 119, 118, 117, 116 и 115, и вариабельный район легкой цепи, выбранный из группы, состоящей из SEQ ID NO: 141, 136, 130, 129, 128, 99, 85, 84, 83, 82, 81, 80, 58, 57 и 29;
[8] анти-Эпирегулин-антителу, выбранному из любого ниже:
(1) анти-Эпирегулин-антитела, содержащего вариабельный район тяжелой цепи, выбранный из группы, состоящей из SEQ ID NO: 150, 149, 148, 147, 146, 145, 144, 143, 142, 140, 139, 138, 137, 135, 134, 133, 132, 131, 127, 126, 125, 124, 123, 122, 121, 120, 119, 118, 117, 116, 115, 98, 97, 96, 95, 94, 93, 92, 79, 78, 77, 76, 75, 74, 73, 72, 56, 55, 54, 53, 52, 51, 50, 49 и 38;
(2) анти-Эпирегулин-антитела, содержащего вариабельный район легкой цепи, выбранный из группы, состоящей из SEQ ID NO: 141, 136, 130, 129, 128, 99, 85, 84, 83, 82, 81, 80, 58, 57 и 29; и
(3) анти-Эпирегулин-антитела, содержащего вариабельный район тяжелой цепи, выбранный из группы, состоящей из SEQ ID NO: 150, 149, 148, 147, 146, 145, 144, 143, 142, 140, 139, 138, 137, 135, 134, 133, 132, 131, 127, 126, 125, 124, 123, 122, 121, 120, 119, 118, 117, 116, 115, 98, 97, 96, 95, 94, 93, 92, 79, 78, 77, 76, 75, 74, 73, 72, 56, 55, 54, 53, 52, 51, 50, 49 и 38; и вариабельный район легкой цепи, выбранный из группы, состоящей из SEQ ID NO: 141, 136, 130, 129, 128, 99, 85, 84, 83, 82, 81, 80, 58, 57 и 29;
[9] антителу по любому из [6]-[8], которое содержит константный район тяжелой цепи SEQ ID NO: 26;
[10] антителу по любому из [6]-[9], которое содержит константный район легкой цепи SEQ ID NO: 27;
[11] антителу по любому из [1]-[10], которое имеет нейтрализующую активность;
[12] антителу по любому из [1]-[11], которое имеет цитотоксичность;
[13] антителу по [12], где этой цитотоксичностью является CDC и/или ADCC;
[14] антителу по любому из [1]-[12], где ингибитор роста или цитотоксическое вещество связаны с этим антителом;
[15] антителу по [14], где это антитело является низкомолекулярным антителом;
[16] антителу по любому из [12]-[15], где константный район тяжелой цепи SEQ ID NO: 26 содержит по меньшей мере одну замену аминокислоты в положении, выбранном из группы, состоящей из 230, 240, 244, 245, 247, 262, 263, 266, 273, 275, 299, 302, 313, 323, 325, 328 и 332, как показано EU-нумерацией;
[17] вектору, содержащему полинуклеотид, кодирующий вариабельный район тяжелой цепи, который содержит CDR1 тяжелой цепи SEQ ID NO: 9, CDR2 тяжелой цепи, выбранный из группы, состоящей из SEQ ID NO: 161, 160, 159, 157, 156, 155, 153, 108, 107, 106, 105, 104, 103, 102, 101 и 100, и CDR3 тяжелой цепи, выбранный из группы, состоящей из SEQ ID NO: 158, 154, 152, 151, 112, 111, 110 и 11;
[18] вектору по [17], который содержит полинуклеотид, кодирующий константный район тяжелой цепи SEQ ID NO: 26;
[19] вектору, содержащему полинуклеотид, кодирующий вариабельный район легкой цепи, который содержит CDR1 легкой цепи, выбранный из группы, состоящей из SEQ ID NO: 163, 68, 67 и 12, CDR2 легкой цепи, выбранный из группы, состоящей из SEQ ID NO: 71, 69 и 13, и CDR3 легкой цепи, выбранный из группы, состоящей из SEQ ID NO: 164, 48, 47 и 14;
[20] вектору по [19], который содержит полинуклеотид, кодирующий константный район легкой цепи SEQ ID NO: 27;
[21] вектору, содержащему:
(1) полинуклеотид, кодирующий вариабельный район тяжелой цепи, содержащий CDR1 тяжелой цепи SEQ ID NO: 9, CDR2 тяжелой цепи, выбранный из группы, состоящей из SEQ ID NO: 161, 160, 159, 157, 156, 155, 153, 108, 107, 106, 105, 104, 103, 102, 101 и 100, и CDR3 тяжелой цепи, выбранный из группы, состоящей из SEQ ID NO: 158, 154, 152, 151, 112, 111, 110 и 11; и
(2) полинуклеотид, кодирующий вариабельный район легкой цепи, который содержит CDR1 легкой цепи, выбранный из группы, состоящей из SEQ ID NO: 163, 68, 67 и 12, CDR2 легкой цепи, выбранный из группы, состоящей из SEQ ID NO: 71, 69 и 13, и CDR3 легкой цепи, выбранный из группы, состоящей из SEQ ID NO: 164, 48, 47 и 14;
[22] вектору, содержащему;
(1) полинуклеотид, кодирующий вариабельный район тяжелой цепи, содержащий CDR1 тяжелой цепи SEQ ID NO: 9, CDR2 тяжелой цепи, выбранный из группы, состоящей из SEQ ID NO: 161, 160, 159, 157, 156, 155, 153, 108, 107, 106, 105, 104, 103, 102, 101 и 100, и CDR3 тяжелой цепи, выбранный из группы, состоящей из SEQ ID NO: 158, 154, 152, 151, 112, 111, 110 и 11; и полинуклеотид, кодирующий константный район легкой цепи SEQ ID NO: 26;
(2) полинуклеотид, кодирующий вариабельный район легкой цепи, содержащий CDR1 легкой цепи, выбранный из группы, состоящей из SEQ ID NO: 163, 68, 67 и 12, CDR2 легкой цепи, выбранный из группы, состоящей из SEQ ID NO: 71, 69 и 13, и CDR3 легкой цепи, выбранный из группы, состоящей из SEQ ID NO: 164, 48, 47 и 14; и полинуклеотид, кодирующий константный район легкой цепи SEQ ID NO: 27;
[23] вектору, содержащему полинуклеотид [18] или [22], который содержит мутированный нуклеотид, кодирующий константный район тяжелой цепи по меньшей мере с одной заменой аминокислоты в положении, выбранном из группы, состоящей из 221, 222, 223, 224, 225, 227, 228, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 243, 244, 245, 246, 247, 249, 250, 251, 254, 255, 256, 258, 260, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 278, 279, 280, 281, 282, 283, 284, 285, 286, 288, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 311, 313, 315, 317, 318, 320, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 339, 376, 377, 378, 379, 380, 382, 385, 392, 396, 421, 427, 428, 429, 434, 436 и 440, как показано EU-нумерацией в константном районе тяжелой цепи SEQ ID NO: 26;
[24] клетке-хозяину, содержащей векторы [17] и [19], векторы [18] и [20] или вектор [21] или [22].
[25] клетке-хозяину, содержащей вектор [23];
[26] клетке-хозяину по [25], где способность добавления фукозы к сахарной цепи в клетке-хозяине является низкой;
[27] клетке-хозяину по [26], где эта клетка-хозяин с низкой способностью добавления фукозы к сахарной цепи является клеткой-хозяином с недостаточностью в одном или нескольких функциональных белках, выбранных из группы, состоящей из фукозилтрансферазы, транспортера фукозы, GMD (GDP-маннозо-4,6-дегидратазы), Fx(GDP-кето-6-дезоксиманнозо-3,5-эпимеразы, 4-редуктазы) и GFPP (GDP-β-L-фукозопирофосфорилазы);
[28] клетке-хозяину [25], где эта клетка-хозяин способна образовывать двурассеченную структуру N-ацетилглюкозамина на сахарной цепи;
[29] клетке-хозяину [28], где эта клетка-хозяин, имеющая способность образовывать двурассеченную структуру N-ацетилглюкозамина на сахарной цепи, является клеткой-хозяином, которая имеет β(1,4)-галактозилтрансферазную активность и содержит вектор, содержащий полинуклеотид, кодирующий функциональный домен локализации Гольджи обитающего в аппарате
Гольджи полипептида;
[30] клетке-хозяину [29], содержащей вектор, который содержит полинуклеотид, кодирующий функциональный домен локализации Гольджи, выбранный из группы, состоящей из домена локализации маннозидазы II, домена локализации β(1,2)-N-ацетилглюкозаминилтрансферазы I, домена локализации β(1,2)-N-ацетилглюкозаминилтрансферазы II, домена локализации маннозидазы I и домена локализации α1-6-кора-фукозилтрансферазы; и полинуклеотид, кодирующий слитый полипептид, содержащий каталитический домен β(1,4)-галактозилтрансферазы;
[31] клетке-хозяину по любому из [24]-[30], где эта клетка-хозяин выбрана из группы, состоящей из СНО-клетки, ВНК-клетки, NS0-клетки, SP2/0-клетки, миеломной YO-клетки, P3X63-клетки мышиной миеломы, PER-клетки, PER.С6-клетки, HEK293-клетки и клетки гибридомы;
[32] способу получения антитела по любому из [1]-[16], который включает сбор клеток-хозяев по любому из [24]-[31] из культурального раствора;
[33] антителу, полученному по способу [32];
[34] антителу по [33], где агент ингибирования роста или цитотоксическое вещество связаны с этим антителом;
[35] фармацевтической композиции, содержащей антитело по любому из [1]-[16] или [33] или [34] в качестве активного ингредиента;
[36] терапевтическому агенту для рака или агенту для ослабления рецидива или метастазирования рака, который содержит антитело по любому из [1]-[16] или [33] или [34] в качестве активного ингредиента;
[37] терапевтическому агенту для рака или агенту для ослабления рецидива или метастазирования рака по [36], где этот рак является любым раком, выбранным из группы, состоящей из рака ободочной кишки, аденокарциномы легкого, рака поджелудочной железы, рака желудка и рака почки;
[38] терапевтическому агенту для рака или агенту для ослабления рецидива или метастазирования рака по [37], где этим раком является слабо дифференцированный рак ободочной кишки, умеренно дифференцированный рак ободочной кишки или хорошо дифференцированный рак ободочной кишки; и
[39] терапевтическому агенту для рака или агенту для ослабления рецидива или метастазирования рака по [36]-[38], где субъект, которому вводят этот терапевтический агент для рака, является субъектом, несущим экспрессирующие белок Эпирегулин раковые клетки, детектированные в выделенной пробе ткани.
Данное изобретение относится также к способам ослабления пролиферации клеток, способам предотвращения рака и способам ослабления рецидива или метастазирования рака, которые включают стадию введения этому субъекту антитела по данному изобретению или антитела, полученного способом получения по данному изобретению. Кроме того, данное изобретение относится к применениям антитела по данному изобретению или антитела, полученного способом получения по данному изобретению, в получении агентов для ингибирования пролиферации клеток, агентов для предотвращения или лечения рака или агентов для ослабления рецидива или метастазирования рака. Данное изобретение относится также к антителам по данному изобретению или антителам, полученным способом получения по данному изобретению, для применения в ослаблении пролиферации клеток, предотвращении или лечении рака или ослабления рецидива или метастазировании рака. Кроме того, данное изобретение относится к способам получения агентов для ингибирования пролиферации клеток, агентов для предотвращения или лечения рака или агентов для ослабления рецидива или метастазирования рака, которые включают стадию применения антитела по данному изобретению или антитела, полученного способом получения по данному изобретению.
Краткое описание фигур
Фиг. 1 изображает диаграмму, показывающую отношение аффинности каждого из этих антител к Эпирегулину человека (величина KD каждого антитела/величина KD химерного антитела EP27). Хотя изображение (нотация) названия антитела описывается названием только вариабельного района, эта фигура изображает тест-результаты антител, содержащих константные районы тяжелой цепи G1d и легкой цепи каппа.
Фиг. 2 изображает диаграмму, показывающую отношение аффинности каждого из этих антител (величина KD для Эпирегулина обезьяны/величина KD для Эпирегулина человека). Хотя изображение (нотация) названия антитела описывается названием только вариабельного района, эта фигура изображает тест-результаты антител, содержащих константные районы тяжелой цепи G1d и легкой цепи каппа.
Фиг. 3 изображает диаграмму, показывающую отношение аффинности каждого из этих антител к Эпирегулину человека (величина KD каждого антитела/величина KD химерного антитела EP27).
Фиг. 4 изображает диаграмму, показывающую отношение аффинности каждого из этих антител к Эпирегулину человека (величина KD для Эпирегулина обезьяны/величина KD для Эпирегулина человека).
Фиг. 5 изображает график, показывающий активность ADCC каждого из этих антител (специфическая скорость выделения кальцеина АМ).
Фиг 6 изображает график, показывающий нейтрализующую активность каждого из этих антител в виде степени ингибирования зависимой от Эпирегулина пролиферации клеток человека BAF_EGFR.
Фиг. 7 изображает график, показывающий нейтрализующую активность каждого из этих антител в виде степени ингибирования зависимой от Эпирегулина пролиферации клеток обезьяны BAF_EGFR.
Фиг. 8 изображает активность каждого из этих антител в отношении ослабления in vivo роста опухоли человека в виде противоопухолевой активности в мышиных моделях, трансплантированных раковыми клеток человека.
Фиг. 9 является изображением, показывающим электрофоретические картины Эпирегулина человека (hsEREG-His) и Эпирегулина обезьяны (cysEREG-His).
Фиг. 10 показывает результаты флуоресцентного окрашивания клеток, каждые из которых экспрессировали различные количества белка Эпирегулина, и иммуногистологического окрашивания мышей, трансплантированных этими клетками.
Фиг. 11 изображает графики, показывающие активность ADCC против клеток, каждые из которых экспрессировали различные количества белка Эпирегулина.
Фиг. 12 изображает графики, показывающие in vivo активность ингибирования роста опухоли человека в виде противоопухолевой активности в мышиных моделях, трансплантированных клетками, каждые из которых экспрессировали различные количества белка Эпирегулина.
Фиг. 13 представляет фотографии, показывающие экспрессию Эпирегулина в клинических случаях слабо дифференцированного рака ободочной кишки.
Фиг. 14 является диаграммой, показывающей лекарственную эффективность гуманизированного EP27 антитела Glycomab против онкогенеза стволовых клеток PLR123 рака ободочной кишки, которые обнаруживают лекарственную эффективность против метастазирования рака ободочной кишки.
Фиг. 15 показывает лекарственную эффективность гуманизированного EP27 антитела Glycomab против легочного метастазирования стволовых клеток PLR123 рака ободочной кишки, которые обнаруживают лекарственную эффективность против метастазирования рака ободочной кишки.
Фиг. 16 является графиком, показывающим лекарственную эффективность гуманизированного EP27 антитела Glycomab против модели слабо дифференцированного рака ободочной кишки.
Фиг. 17 является графиком, показывающим лекарственную эффективность гуманизированного EP27 антитела Glycomab против умеренно дифференцированного рака ободочной кишки с использованием модели PLR379 умеренно дифференцированного рака ободочной кишки.
Фиг. 18 предоставляет фотографии, показывающие экспрессию Эпирегулина в клинических случаях аденокарциномы легкого.
Фиг. 19 является графиком, показывающим ADCC-активность (степень выделения специфического кальцеина АМ) гуманизированного EP27 антитела Glycomab против линии клеток Calu-3 аденокарциномы легких человека.
Фиг. 20 является графиком, показывающим лекарственную эффективность гуманизированного EP27 антитела Glycomab против аденокарциномы легких с использованием модели Calu-3 аденокарциномы легких.
Фиг. 21 показывает, что конъюгат антитело-лекарственное средство, содержащий гуманизированное EP27 антитело Glycomab, становится интернализованным в линии клеток DLD-1, которые экспрессируют Эпирегулин, и вызывают повреждение клеток против линии клеток DLD-1.
Способ проведения этого изобретения
Данное изобретение относится к анти-Эпирегулин-антителам, обнаруживающим перекрестную межвидовую реактивность между животными, не являющимися человеком, и людьми. Данное изобретение дополнительно относится к анти-Эпирегулин-антителам с ослабленной химической деградацией. Данное изобретение относится также к анти-Эпирегулин-антителам с уменьшенной изоэлектрической точкой. Кроме того, данное изобретение относится к анти-Эпирегулин-антителам с уменьшенным количеством агрегата. Кроме того, данное изобретение относится к фармацевтическим композициям или терапевтическим агентам для рака, содержащего вышеупомянутые анти-Эпирегулин-антитела. Данное изобретение относится также к способам получения вышеупомянутых анти-Эпирегулин-антител.
Определения и подробное описание ниже обеспечены для содействия пониманию данного изобретения, иллюстрированного здесь.
Определения
Аминокислоты
Здесь описаны аминокислоты в одно- или трехбуквенных кодах или обоих, например, Ala/A, Leu/L, Arg/R, Lys/K, Asn/N, Met/M, Asp/D, Phe/F, Cys/C, Pro/P, Gln/Q, Ser/S, Glu/E, Thr/T, Gly/G, Trp/W, His/H, Tyr/Y, Ile/I или Val/V. Аминокислоты, содержащиеся в аминокислотных последовательностях по данному изобретению, могут быть посттрансляционно модифицированы (например, модификация N-терминального глутамина в пироглутаминовую кислоту хорошо известна квалифицированным в данной области специалистам). Разумеется, такие посттрансляционно модифицированные аминокислоты включены в аминокислотные последовательности в данном изобретении.
Антигены
Антитела, обеспеченные данным изобретением, связываются с анти-Эпирегулином в качестве антигена. Эпирегулин является мембраносвязанным белком эпидермального фактора роста. Его аминокислотная последовательность описана в GenBank Acession Number NP_001423 (SEQ ID NO: 167). В данном изобретении определение Эпирегулин включает в себя как полноразмерный белок, так и его фрагменты. Термин ʺфрагментыʺ относится к полипептидам, содержащим любой район Эпирегулина, и они могут не иметь функции нативного Эпирегулина. Один пример этих фрагментов включает в себя фрагмент, содержащий внеклеточный район Эпирегулина. Положения 30-118 в аминокислотной последовательности SEQ ID NO: 167 соответствуют внеклеточному району Эпирегулина. Положения 119-140 в аминокислотной последовательности 167 соответствуют трансмембранному району. Здесь Эпирегулин может называться как EREG, и эти термины используют синонимически.
ʺЭпитопʺ обозначает антигенную детерминанту в антигене и относится к сайту антигена, с которым связывается антигенсвязывающий домен анти-Эпирегулин-антитела, описанного здесь. Так, например, этот эпитоп может быть определен в соответствии с его структурой. Альтернативно, этот эпитоп может быть определен в соответствии с антигенсвязывающей активностью анти-Эпирегулин-антитела, которое распознает этот эпитоп. Когда этот антиген является пептидом или полинуклеотидом, этот эпитоп может быть установлен по аминокислотным остатком, образующим этот эпитоп. Альтернативно, когда этот эпитоп является сахарной цепью, этот эпитоп может быть установлен по его специфической структуре сахарной цепи.
Линейным эпитопом является эпитоп, первичная аминокислотная последовательность которого распознается как эпитоп, состоящий из ряда последовательных аминокислот в этой первичной аминокислотной последовательности. Такой линейный эпитоп обычно содержит по меньшей мере три и наиболее часто по меньшей мере пять, например, приблизительно 8-10 или 6-20 аминокислот в этой специфической последовательности.
В противоположность линейному эпитопу, ʺконформационным эпитопомʺ является эпитоп, в котором первичная аминокислотная последовательность, содержащая этот эпитоп, не является единственной детерминантой этого распознаваемого эпитопа (например, первичная аминокислотная последовательность конформационного эпитопа необязательно распознается эпитоп-определяющим антителом). Конформационные эпитопы могут содержать большее количество аминокислот в сравнении с линейными эпитопами. Конформационный эпитоп-узнающее антитело распознает трехмерную структуру пептида или белка. Например, когда молекула белка складывается и образует трехмерную структуру, аминокислоты и/или основные цепи полипептида, которые образуют конформационный эпитоп, сопоставляются, и эпитоп становится распознаваемым этим антителом. Способы для определения конформаций эпитопа включают, например, рентгеновскую кристаллографию, двухмерный ядерный магнитный резонанс, сайт-специфическое спиновое мечение и электронный парамагнитный (спиновый) резонанс, но не ограничиваются ими. См., например, Epitope Mapping Protocols in Methods in Molecular Biology (1996), Vol. 66, Morris (ed.).
Связывающая активность
Примеры способа для подтверждении связывания антитела с Эпирегулином или, более конкретно, эпитопом, присутствующим в молекуле Эпирегулина, показаны ниже, но этот способ не ограничивается следующими способами, и квалифицированный в данной области специалист может подходящим образом использовать известные способы для измерения антигенсвязывающей активности антитела.
Например, узнает ли анти-Эпирегулин-антитело линейный эпитоп в молекуле Эпирегулина, может быть подтверждено, например, как описано ниже. Линейный пептид, содержащий аминокислотную последовательность, образующую внеклеточный домен Эпирегулина, синтезируют для вышеуказанной цели. Этот пептид может быть синтезирован химически или получен способами генетической инженерии с использованием района, кодирующего аминокислотную последовательность, соответствующую внеклеточному домену, в кДНК, кодирующей Эпирегулин (примеры включают в себя CR5419887 (SEQ ID NO: 169) и, например, кДНК-последовательность и NM_001432 и, например, мРНК-последовательность). Затем анти-Эпирегулин-антитело оценивают на его связывающую активность относительно линейного пептида, содержащего аминокислотную последовательность, образующую внеклеточный домен. Например, иммобилизованный линейный пептид может быть использован в качестве антигена при помощи ELISA для оценивания связывающей активности в отношении этого пептида. Альтернативно, связывающая активность относительно линейного пептида может оцениваться на основе уровня, до которого этот линейный пептид ингибирует связывание этого антитела с Эпирегулин-экспрессирующими клетками. Эти тесты могут демонстрировать связывающую активность этого антитела относительно линейного пептида.
Узнает ли анти-Эпирегулин-антитело конформационный эпитоп, может оцениваться следующим образом. Для вышеуказанной цели готовят Эпирегулин-экспрессирующие клетки. Анти-Эпирегулин-антитело может быть определено для распознавания конформационного эпитопа, когда он сильно связывается с Эпирегулин-экспрессирующими клетками после контакта, но по существу не связывается с иммобилизованным линейным пептидом, содержащим аминокислотную последовательность, образующую внеклеточный домен Эпирегулина. Здесь, ʺпо существу не связываетсяʺ означает, что связывающая активность равна 80% или менее, обычно 50% или менее, предпочтительно 30% или менее и, особенно предпочтительно, 15% или менее в сравнении со связывающей активностью относительно клеток, экспрессирующих Эпирегулин человека.
Способы для анализа связывающей активности анти-Эпирегулин-антитела относительно анти-Эпирегулин-экспрессирующих клеток включают, например, способы, описанные в Antibodies: A Laboratory Manual (Ed Harlow, David Lane, Cold Spring Harbor Laboratory (1988) 359-420). Конкретно, оценивание может проводиться на основе принципа ELISA или активируемого флуоресценцией сортинга клеток (FACS) с использованием Эпирегулин-экспрессирующих клеток в качестве антигена.
В формате ELISA связывающая активность анти-Эпирегулин-антитела в отношении Эпирегулин-экспрессирующих клеток может оцениваться количественно сравнением уровней сигнала, генерируемого ферментативной реакцией. Конкретно, тест-антитело добавляют к планшету ELISA, на котором иммобилизованы Эпирегулин-экспрессирующие клетки. Затем тест-антитело, связанное с этими клетками, детектируют с использованием меченного ферментом антитела, которое узнает это тест-антитело. Альтернативно, при использовании FACS, готовят серию разведений тест-антитела, и титр связывания антитела для Эпирегулин-экспрессирующих клеток может быть определен для сравнения связывающей активности этого тест-антитела относительно Эпирегулин-экспрессирующих клеток.
Связывание тест-антитела относительно антигена, экспрессируемого на поверхности клеток, суспендированных в буфере или т.п., может быть определено с использованием проточного цитометра. Известные проточные цитометры включают в себя, например, следующие устройства:
FACSCanto™ II;
FACSAria™;
FACSArray™;
FACSVantage™ SF;
FACSCalibur™ (все являются товарными названиями BD Biosciences);
EPICS ALTRA HyPerSort;
Cytomics FC 500;
EPICS XL-MCL ADC EPICS XL ADC;
Cell Lab Quanta/Cell Lab Quanta SC (все являются товарными названиями Beckman Coulter).
Предпочтительные способы для оценивания связывающей активности анти-Эпирегулин-антитела относительно Эпирегулина включают, например, следующий способ. Сначала Эпирегулин-экспрессирующие клетки реагируют с тест-антителом, и затем проводят окрашивание меченным FITC вторичным антителом, которое распознает это тест-антитело. Это тест-анти-Эпирегулин-антитело подходящим образом разводят подходящим буфером для получения этого антитела при желаемой концентрации. Например, это антитело может быть использовано при концентрации в диапазоне 10 мкг/мл-10 нг/мл. Затем определяют интенсивность флуоресценции и количество клеток с использованием FACSCalibur (BD). Эта интенсивность флуоресценции, полученная анализом с использованием CELL QUEST Software (BD), то есть величина геометрического среднего, отражает количество антитела, связанного с клетками. То есть связывающая активность тест-антитела, которая представлена количеством связанного тест-антитела, может быть определена измерением величины геометрического среднего.
Имеет ли анти-Эпирегулин-антитело общий эпитоп с другим антителом, может оцениваться на основе конкуренции между этими двумя молекулами за один и тот же эпитоп. Эта конкуренция между антителами может быть детектирована перекрестным блокирующим анализом или т.п. Например, предпочтительным перекрестным блокирующим анализом является конкурентный анализ ELISA.
Конкретно, в перекрестном блокирующем анализе белок Эпирегулин, иммобилизованный в лунках микротитрационного планшета, предварительно инкубируют в присутствии или в отсутствие кандидатного конкурентного антитела и затем к ним добавляют тест-антитело. Количество тест-антитела, связанного с белком Эпирегулином в этих лунках, непосредственно коррелирует со связывающей способностью кандидатного конкурентного антитела, которое конкурирует за это связывание с тем же самым эпитопом. То есть, чем больше аффинность этого конкурентного антитела в отношении того же самого эпитопа, тем ниже связывающая активность этого тест-антитела в отношении покрытых белком Эпирегулином лунок.
Количество тест-антитела, связанного с этими лунками через белок Эпирегулин, может быть легко определено мечением этого антитела заранее. Например, меченное биотином антитело измеряют с использованием конъюгата авидин/пероксидаза и подходящего субстрата. В частности, перекрестный блокирующий анализ, который использует ферментные метки, такие как пероксидаза, называют ʺконкурентным анализом ELISAʺ. Это антитело может быть также помечено другими метящими веществами, которые делают возможными детектирование или измерение. Конкретно, известны радиоактивные метки, флуоресцентные метки и т.п.
Когда кандидатное конкурентное антитело может блокировать связывание антителом относительно Эпирегулина по меньшей мере на 20%, предпочтительно по меньшей мере на 20-50% и более предпочтительно на по меньшей мере 50% в сравнении со связывающей активностью в контрольном эксперименте, проводимом в отсутствие этого конкурентного антитела, определяют, что это тест-антитело по существу связывается с тем же самым эпитопом, связанным конкурентным антителом, или конкурирует за связывание с тем же самым эпитопом.
Когда структура эпитопа, связанного анти-Эпирегулин-антителом, уже была идентифицирована, имеют ли тест-антитело и контрольное антитело общий эпитоп, может оцениваться сравнением связывающих активностей этих двух антител относительно пептида, полученного введением аминокислотных мутаций в пептид, образующий этот эпитоп.
Для измерения вышеуказанных связывающих активностей, например, эти связывающие активности тест- и контрольного антител относительно линейного пептида, в который введена мутация, сравнивают в вышеуказанном формате ELISA. Наряду со способами ELISA, связывающая активность относительно мутантного пептида, связанного с колонкой, может быть определена протеканием тест-антител и контрольных антител в колонке и затем определением количества антитела, элюированного в растворе для элюции. Известны способы адсорбирования мутантного пептида в колонку, например, в форме GST-слитого пептида.
Альтернативно, когда идентифицированный эпитоп является конформационным эпитопом, имеют ли тест-антитело и контрольное антитело общий эпитоп, может оцениваться следующим способом. Сначала готовят Эпирегулин-экспрессирующие клетки и клетки, экспрессирующие Эпирегулин с мутацией, введенной в эпитоп. Эти тест- и контрольные антитела добавляют к суспензии клеток, приготовленной суспендированием этих клеток в подходящем буфере, таком как PBS (ЗФР). Затем эти клеточные суспензии подходящим образом промывают буфером и добавляют к ним меченное FITC антитело, которое распознает тест- и контрольные антитела. Интенсивность флуоресценции и количество клеток, окрашенных меченым антителом, определяют с использованием FACSCalibur (BD). Эти тест- и контрольные антитела подходящим образом разводят с использованием подходящего буфера при желаемых концентрациях. Например, они могут быть использованы в концентрации в диапазоне 10 мкг/мл-10 нг/мл. Эта интенсивность флуоресценции, определенная анализом с использованием CELL QUEST Software (BD), то есть величина Геометрического Среднего, отражает количество меченого антитела, связанного с клетками. То есть связывающая активность тест- и контрольного антител, которая представлена количеством связанного тест-антитела, может быть определена измерением величины Геометрического Среднего.
В приведенном выше способе, действительно ли антитело ʺпо существу не связывается с клетками, экспрессирующими мутантный Эпирегулинʺ, может оцениваться, например, следующим способом. Сначала, тест- и контрольные антитела, связанные с клетками, экспрессирующими мутантный Эпирегулин, окрашивают меченым антителом. Затем определяют интенсивность флуоресценции этих клеток. При использовании FACSCalibur для детектирования флуоресценции проточной цитометрией, определяемая интенсивность флуоресценции может анализироваться с использованием CELL QUEST Software (BD). Из величин Геометрического Среднего в присутствии и в отсутствие антитела, величина сравнения (ΔGeo-Mean) может быть рассчитана в соответствии со следующей формулой (Формула 1) для определения отношения (степени) увеличения в интенсивности флуоресценции, как результата связывания этим антителом.
Формула 1
ΔGeo-Mean = Geo-Mean (в присутствии этого антитела)/Geo-Mean (в отсутствие этого антитела).
Сравнительную величину Геометрического Среднего (величину ΔGeo-Mean для мутантной молекулы Эпирегулина), определенную приведенным выше анализом, которая отражает количество тест-антитела, связанного с клетками, экспрессирующими мутантный Эпирегулин, сравнивают со сравнительной величиной ΔGeo-Mean, которая отражает количество тест-антитела, связанного с Эпирегулин-экспрессирующими клетками. В этом случае концентрации тест-антитела, используемые для определения сравнительных величин ΔGeo-Mean для Эпирегулин-экспрессирующих клеток и клеток, экспрессирующих мутантный Эпирегулин, особенно предпочтительно корректируют таким образом, чтобы они были равными или по существу равными. Антитело, которое, как было подтверждено, узнают эпитоп в Эпирегулине, используют в качестве контрольного антитела.
Если сравнительная величина ΔGeo-Mean тест-антитела для клеток, экспрессирующих мутантный Эпирегулин, является меньшей, чем сравнительная величина ΔGeo-Mean тест-антитела для Эпирегулин-экспрессирующих клеток, по меньшей мере на 80%, предпочтительно 50%, более предпочтительно 30% и особенно предпочтительно 15%, то тест-антитело действительно ʺпо существу не связывается с клетками, экспрессирующими мутантный Эпирегулинʺ. Формула для определения величины Geo-Mean (Геометрического Среднего) описана в CELL QUEST Software User's Guide (BD biosciences). Когда это сравнение показывает, что сравнительные величины являются по существу эквивалентными, может быть определено, что эпитоп для тест- и контрольного анти-Эпирегулин-антитела является одним и тем же.
Антитела
Здесь термин ʺантителоʺ относится к природному иммуноглобулину или иммуноглобулину, полученному частичным или полным синтезом. Антитела могут быть выделены из природных источников, таких как природно встречающиеся плазма и сыворотка, или культуральные супернатанты антитело-продуцирующих гибридом. Альтернативно, антитела могут быть частично или полностью синтезированы с использованием таких способов, как генетическая рекомбинация. Предпочтительные антитела включают в себя, например, антитела изотипа иммуноглобулина или подкласса, принадлежащего к ним. Известные иммуноглобулины человека включают в себя антитела следующих девяти классов (изотипов): IgG1, IgG2, IgG3, IgG4, IgА1, IgА2, IgD, IgЕ и IgМ. Из этих антител, антитела по данному изобретению включают в себя IgG1, IgG2, IgG3 и IgG4.
Способы получения антитела с желаемой связывающей активностью известны квалифицированным в данной области специалистам. Ниже приведен неограничивающий пример, который описывает способ получения антитела, которое связывается с Эпирегулином (анти-Эпирегулин-антитела).
Анти-Эпирегулин-антитела могут быть получены в виде поликлональных или моноклональных антител с использованием известных способов. Эти предпочтительно получаемые анти-Эпирегулин-антитела являются моноклональными антителами, полученными из млекопитающих. Такие полученные из млекопитающих моноклональные антитела включают в себя антитела, продуцируемые гибридомами, или клетками-хозяевами, трансформированными экспрессирующим вектором, несущим ген антитела, способами генетической инженерии. ʺГуманизированные антителаʺ или ʺхимерные антителаʺ включены в моноклональные антитела по данному изобретению.
Моноклональное антитело-продуцирующие гибридомы могут быть получены с использованием известных способов, например, таких, как описанные ниже. Конкретно, млекопитающих иммунизируют общепринятыми способами иммунизации с использованием белка Эпирегулина в качестве сенсибилизирующего агента. Полученные иммунные клетки сливают с известными родительскими клетками общепринятыми способами слияния клеток. Затем гибридомы, продуцирующие анти-Эпирегулин-антитело, могут быть отобраны скринингом на моноклональные клетки, продуцирующие антитело, которое связывается с эпитопом в молекуле Эпирегулина, с использованием общепринятых способов скрининга.
Конкретно, моноклональные антитела готовят, как описано ниже. Сначала ген Эпирегулина человека, нуклеотидная последовательность которого описана в SEQ ID NO: 169, может быть экспрессирована для получения белка Эпирегулина, показанного в SEQ ID NO: 167, который будет использован в качестве сенсибилизирующего антигена для получения антитела. То есть последовательность гена, кодирующую Эпирегулин человека, инсертируют в известный экспрессирующий вектор, и подходящие клетки-хозяева трансформируют этим вектором. Этот желаемый белок Эпирегулина человека очищают из клеток-хозяев известными способами. Для получения растворимого Эпирегулина человека из культуральных супернатантов, например, полипептид, содержащий аминокислоты в положении 30-118 в полипептидной последовательности SEQ ID NO: 167, или белок включали в эти аминокислоты в положениях 30-108, показанных как SEQ ID NO: 34. Очищенный природный белок Эпирегулин может быть также использован в качестве сенсибилизирующего антигена.
Очищенный белок Эпирегулин может быть использован в качестве сенсибилизирующего антигена для иммунизации млекопитающих. Частичный пептид Эпирегулина может быть также использован в качестве сенсибилизирующего антигена. В этом случае частичный пептид может быть приготовлен химическим синтезом на основе аминокислотной последовательности Эпирегулина человека, или инсертированием частичного гена Эпирегулина человека в экспрессирующий вектор для экспрессии. Альтернативно, частичный пептид может быть получен деградацией белка Эпирегулина человека протеазой. Длина и район частичного пептида Эпирегулин человека не ограничивается конкретными вариантами. Предпочтительный район может быть выбран произвольно из аминокислотной последовательности в положениях 30-118 или в положениях 30-108 в аминокислотной последовательности SEQ ID NO: 167. Количество аминокислот, образующих пептид, подлежащий применению в качестве сенсибилизирующего антигена, равно предпочтительно пяти или более, например, шести или более или семи или более. Более конкретно, пептид из 8-50 остатков, более предпочтительно 10-30 остатков, может быть использован в качестве сенсибилизирующего антигена.
Для сенсибилизирующего антигена можно использовать слитый белок, приготовленный слиянием желаемого частичного полипептида или пептида белка Эпирегулина с другим полипептидом. Например, Fc-фрагменты и пептидные метки антитела предпочтительно используют для получения слитых белков, подлежащих использованию в качестве сенсибилизирующих антигенов. Векторы для экспрессии таких слитых белков могут быть сконструированы слиянием в рамке считывания генов, кодирующих два или более желаемых полипептидных фрагментов и инсертированием этого слитого гена в экспрессирующий вектор, как описано выше. Способы получения слитых белков описаны в Molecular Cloning 2nd ed., (Sambrook, J. et al., Molecular Cloning 2nd ed., 9,47-9,58 (1989) Cold Spring Harbor Lab Press, Cold Spring Harbor (1989). Способы получения Эпирегулина в качестве сенсибилизирующего агента и способы иммунизации с использованием Эпирегулина описаны в WO 2008/047723.
Нет конкретного ограничения в отношении млекопитающих, подлежащих иммунизации сенсибилизирующим антигеном. Однако предпочтительно выбирать млекопитающих с учетом их совместимости с родительскими клетками, подлежащими использованию для слияния клеток. Обычно используются предпочтительно мыши, крысы и хомячки, кролики и обезьяны.
Вышеуказанных животных иммунизируют сенсибилизирующим антигеном известными способами. Обычно выполняемые способы иммунизации включают, например, внутриперитонеальную или подкожную инъекцию сенсибилизирующего антигена в млекопитающих. Конкретно, сенсибилизирующий антиген подходящим образом разводят PBS (забуференным фосфатом солевым раствором (ЗФР), физиологическим солевым раствором или т.п. Если желательно, общепринятый адъювант, такой как полный адъювант Фрейнда, смешивают с этим антигеном и эту смесь эмульгируют. Затем этот сенсибилизирующий антиген вводят млекопитающему несколько раз с интервалами 4-21 день. В иммунизации этим сенсибилизирующим антигеном могут быть использованы подходящие носители. В частности, при использовании низкомолекулярного частичного пептида в качестве сенсибилизирующего антигена, иногда желательно связывание этого сенсибилизирующего антигена с белком-носителем, таким как альбумин или гемоцианин фиссуреллы, для иммунизации.
Альтернативно, гибридомы, продуцирующие желаемое антитело, могут быть приготовлены с использованием ДНК-иммунизации, как указано ниже. ДНК-иммунизация является способом иммунизации, который придает иммуностимуляцию экспрессированием сенсибилизирующего антигена в иммунизированном животном в результате введения вектора ДНК, сконструированного для возможности экспрессии гена, кодирующего белок-антиген в этом животном. Ожидается, что в сравнении с общепринятыми способами иммунизации, в которых белковый антиген вводят животным, подлежащим иммунизации, ДНК-иммунизация будет превосходящей в том смысле, что:
- иммуностимуляция может быть обеспечена при сохранении структуры мембранного белка, такого как Эпирегулин, и
- нет необходимости очистки антигена для иммунизации.
Для получения моноклонального антитела по данному изобретению с использованием ДНК-иммунизации, сначала ДНК, экспрессирующую белок Эпирегулин, вводят животному, подлежащему иммунизации. Эта Эпирегулин-кодирующая ДНК может быть синтезирована известными способами, такими как ПЦР. Полученную ДНК инсертируют в подходящий экспрессирующий вектор и затем этот вектор вводят животному, подлежащему иммунизации. Предпочтительно используемые экспрессирующие векторы включают в себя, например, коммерчески доступные экспрессирующие векторы, такие как pcDNA3.1. Векторы могут вводиться в организм с использованием общепринятых способов. Например, иммунизацию ДНК выполняют с использованием генного пистолета для введения покрытого частицами золота экспрессирующего вектора в тело животного, подлежащего иммунизации. Антитела, которые распознают Эпирегулин, могут быть также получены способами, описанными в WO 2003/104453.
После иммунизации млекопитающего, как описано выше, увеличение в титре Эпирегулин-связывающего антитела подтверждается в сыворотке. Затем иммунные клетки собирают из этого млекопитающего и затем подвергают слиянию клетки. В частности, спленоциты предпочтительно используют в качестве иммунных клеток.
Миеломные клетки млекопитающих используют в качестве клеток, подлежащих слиянию с вышеуказанными иммунными клетками. Эти миеломные клетки предпочтительно содержат подходящий селектируемый маркер для скрининга. Селектируемый маркер придает клеткам характеристики для их выживания (или смерти) при конкретном условии культивирования. В качестве маркеров селекции известны дефицит гипоксантин-гуанинфосфорибозилтрансферазы (далее сокращаемый здесь как дефицит HGPRT) и дефицит тимидинкиназы (далее сокращаемый здесь как дефицит TK). Клетки с дефицитом HGPRT или TK имеют чувствительность к гипоксантин-аминоптерин-тимидину (далее сокращаемую здесь как HAT-чувствительность). HAT-чувствительные клетки не могут синтезировать ДНК в HAT-селекционной среде и таким образом убиваются. Однако, когда эти клетки сливают с нормальными клетками, они могут продолжать синтез ДНК с использованием пути регенерации нормальных клеток, и, следовательно, они могут расти даже в HAT-селекционной среде.
HGPRT-недостаточные и TK-недостаточные клетки могут быть отобраны в среде, содержащей 6-тиогуанин, 8-азагуанин (сокращаемый здесь далее как 8AG) или 5'-бромдезоксиуридин, соответственно. Нормальные клетки убиваются, так как они включает в себя эти пиримидиновые аналоги в их ДНК. Между тем, клетки, которые являются недостаточными в отношении этих ферментов, могут выживать в среде для селекции, так как они не могут включать в себя эти пиримидиновые аналоги. Кроме того, селектируемый маркер, называемый как G418-резистентность, обеспеченный неомицин-резистентным геном, придает резистентность к 2-дезоксистрептаминовым антибиотикам (аналогам гентамицина). Известны различные типы миеломных клеток, которые являются подходящими для клеточного слияния.
Например, предпочтительно могут быть использованы миеломные клетки, в том числе следующие клетки:
Р3(Р3х63Ag8.653) (J. Immunol. (1979 (4), 1548-1550);
P3x63AgU.1 (Current Topics in Microbiology and Immunology (1978) 81, 1-7);
NS-1 (C. Eur.J. Immunol. (1976) 6(7), 511-519);
MPС-11 (Cell (1976) 8(3), 405-415);
SP2/0 (Nature (1978) 276 (5685), 269-270);
FO (J. Immunol. Methods (1980) 34(1-2), 1-21);
S194/5.XX0.BU.1 (J. Exp. Med. (1978) 148(1), 313-323);
R210 (Nature (1979) 277 (5692), 131-133) и т.д.
Клеточные слияния между иммуноцитами и миеломными клетками важно проводить с использованием известных способов, например, способом Kohler and Milstein et al. (Methods Enzymol. (1981) 73-46).
Более конкретно, клеточное слияние может проводиться, например, в общепринятой культуральной среде в присутствии стимулирующего слияние клеток агента. Эти стимулирующие слияние клеток агенты включают в себя, например, полиэтиленгликоль (PEG) и вирус Сендай (HVJ). Если требуется, для улучшения эффективности слияния добавляют также вспомогательное вещество, такое как диметилсульфоксид.
Отношение иммунных клеток к миеломным клеткам может определяться по собственному усмотрению, предпочтительно, например, одна миеломная клетка на каждые один-десять иммуноцитов. Культуральная среда для использования для клеточных слияний, включает в себя, например, среды, которые являются подходящими для роста миеломных клеточных линий, такие как среда RPMI 1640 и среда МЕМ, и другие общепринятые культуральные среды, используемые для этого типа клеточной культуры. Кроме того, к этой культуральной среде могут быть предпочтительно добавлены сывороточные добавки, такие как фетальная телячья сыворотка (FCS).
Для клеточного слияния заранее определенные количества вышеуказанных иммунных клеток и миеломных клеток хорошо смешивают в вышеупомянутой культуральной среде. Затем раствор PEG (например, со средней молекулярной массой приблизительно 1000-6000), предварительно нагретый до приблизительно 37°С, добавляют к смеси обычно при концентрации 30%-60% (масс./об.). Этот раствор осторожно смешивают с получением желаемых слитых клеток (гибридом). Затем к этим клеткам постепенно добавляют подходящую культуральную среду, упомянутую выше, и эту смесь повторно центрифугируют для удаления супернатанта. Таким образом, могут быть удалены агенты клеточного слияния и т.п., которые являются неблагоприятными для роста гибридомы.
Полученные таким образом гибридомы могут быть отобраны культивированием с использованием общепринятой селективной среды, например, HAT-среды (культуральной среды, содержащей гипоксантин, аминоптерин и тимидин). Клетки, другие, чем желаемые гибридомы (неслитые клетки), могут убиваться продолжением культивирования в вышеуказанной HAT-среде в течение достаточного периода времени. Обычно этот период является периодом от нескольких дней до нескольких недель. Затем гибридомы, продуцирующие желаемое антитело, подвергают скринингу и по отдельности клонируют общепринятыми способами лимитирующих разведений.
Полученные таким образом гибридомы могут быть отобраны с использованием селективной (избирательной) среды на основе селекционного маркера, который имеет миелома, используемая для слияния клеток. Например, HGPRT- или TK-недостаточные клетки могут быть отобраны культивированием с использованием HAT-среды (культуральной среды, содержащей гипоксантин, аминоптерин и тимидин). Конкретно, когда HAT-чувствительные миеломные клетки используются для слияния клеток, клетки, успешно слитые с нормальными клетками, могут селективно пролиферировать в HAT-среду. Клетки, другие, чем желаемые гибридомы (неслитые клетки), могут убиваться продолжающимся культивированием в вышеуказанной HAT-среде в течение достаточного периода времени. Конкретно, желаемые гибридомы могут быть отобраны культивированием обычно в течение нескольких дней-нескольких недель. Затем гибридомы, продуцирующие желаемое антитело, подвергают скринингу и по отдельности клонируют общепринятыми способами лимитирующих разведений.
Желаемые антитела могут быть предпочтительно отобраны и по отдельности клонированы способами скрининга, основанными на известной реакции антиген/антитело. Например, Эпирегулин-связывающее моноклональное антитело может связываться с Эпирегулином на поверхности клетки. Такое моноклональное антитело может быть подвергнуто скринингу клеточным сортингом с возбуждением флуоресценции (FACS). FACS является системой, которая оценивает связывание антитела с клеточной поверхностью анализом клеток, контактированных с флуоресцентным антителом с использованием лазерного луча, и измерением флуоресценции, испускаемой из индивидуальных клеток.
Для скрининга на гибридомы, которые продуцируют моноклональное антитело по данному изобретению при помощи FACS, сначала готовят Эпирегулин-экспрессирующие клетки. Клетки, предпочтительно используемые для скрининга, являются клетками млекопитающих, в которых форсированно экспрессируется Эпирегулин. В качестве контроля, активность антитела в связывании Эпирегулина клеточной поверхности может быть селективно детектирована с использованием нетрансформированных клеток млекопитающих в качестве клеток-хозяев. Конкретно, гибридомы, продуцирующие моноклональное анти-Эпирегулин-антитело, могут быть выделены отбором гибридом, которые продуцируют антитело, которое связывается с клетками, принуждаемыми экспрессировать Эпирегулин, но не с клетками-хозяевами.
Альтернативно, активность антитела в связывании с иммобилизованными Эпирегулин-экспрессирующими клетками может оцениваться на основе принципа ELISA. Например, Эпирегулин-экспрессирующие клетки иммобилизуют в лунках планшета ELISA. Культуральные супернатанты гибридом контактируют с этими иммобилизованными клетками в этих лунках, и детектируют антитела, которые связываются с этими иммобилизованными клетками. Когда эти моноклональные антитела произведены из мыши, антитела, связанные с этими клетками, могут быть детектированы с использованием антитела против мышиного иммуноглобулина. Гибридомы, продуцирующие желаемое антитело, имеющее антигенсвязывающую способность, отбирают вышеуказанным скринингом, и они могут быть клонированы способом лимитирующих разведений или т.п.
Моноклональные антитело-продуцирующие гибридомы, приготовленные таким образом, могут быть пассированы в общепринятой культуральной среде и храниться в жидком азоте в течение продолжительного периода времени.
Вышеупомянутые гибридомы культивируют общепринятым способом, и желаемые моноклональные антитела могут быть приготовлены из культуральных супернатантов. Альтернативно, эти гибридомы вводят и выращивают в совместимых млекопитающих, и моноклональные антитела готовят из этих асцитов. Предшествующий способ является подходящим для получения антител с высокой чистотой.
Антитела, кодируемые генами антител, которые клонируют из антитело-продуцирующих клеток, таких как вышеуказанные гибридомы, могут быть также предпочтительно использованы. Ген клонированного антитела инсертируют в подходящий вектор, и вводят его в хозяина для экспрессии антитела, кодируемого этим геном. Способы выделения генов антител, инсертирования этих генов в векторы и трансформирования клеток-хозяев уже были установлены, например, Vandamme et al. (Eur. J. Biochem. (1990) 192(3), 767-775). Способы получения рекомбинантных антител также известны, как описано ниже.
Например, кДНК, кодирующую вариабельный район (V-район) анти-Эпирегулин-антитела, получают из гибридомных клеток, экспрессирующих анти-Эпирегулин-антитело. Для этой цели сначала экстрагируют общую РНК из гибридом. Способы, используемые для экстракции мРНК из клеток, включают, например:
- способ ультрацентрифугирования гуанидина (Biochemistry (1979) 18(24), 5294-5299), и
- AGPC-способ (Anal. Biochem. (1987) 162(1), 156-159).
Экстрагированные мРНК могут быть очищены с использованием набора для очистки мРНК (GE Healthcare Bioscience) или т.п. Альтернативно, наборы для экстракции общей мРНК непосредственно из клеток, такие как набор для очистки мРНК QuickPrep (GE Healthcare Bioscience), также являются доступными. мРНК могут быть получены из гибридом с использованием таких наборов. кДНК, кодирующие V-район антитела, могут быть синтезированы из полученных мРНК с использованием обратной транскриптазы. кДНК могут быть синтезированы с использованием набора AMV Reverse Transcriptase First-strand cDNA Synthesis (Seikagaku Co.) или т.п. Кроме того, набор амплификации кДНК SMART RACE (Clontech) и способ 5'-RACE на основе ПЦР (Proc. Natl. Acad. Sci. USA (1988) 85(23), 8998-9002; Nucleic Acids Res. (1989) 17(8), 2919-2932) могут быть подходящим образом использованы для синтеза и амплификации кДНК. В таком процессе синтеза кДНК, подходящие сайты рестрикционных ферментов, описанные ниже, могут быть введены в обоих концах кДНК.
Представляющий интерес фрагмент кДНК очищают из полученного продукта ПЦР и затем его лигируют в векторную ДНК. Таким образом конструируют рекомбинантный вектор и вводят его в E. coli или т.п. После отбора колоний, желаемый рекомбинантный вектор может быть получен из колониеобразующих E. coli. Затем тестируют, имеет ли рекомбинантный вектор представляющую интерес нуклеотидную последовательность кДНК, при помощи известного способа, такого как способ дидезокси-терминации нуклеотидной цепи.
Способ 5'-RACE, который использует праймеры для амплификации гена вариабельного района, удобным образом используется для выделения гена, кодирующего этот вариабельный район. Сначала конструируют библиотеку 5'-RACE кДНК (быстрой амплификации концов кДНК) синтезом кДНК с использованием РНК, экстрагированных из гибридомных клеток, в качестве матрицы. Коммерчески доступный набор, такой как набор амплификации кДНК SMART RACE, используют подходящим образом для синтеза библиотеки кДНК 5'-RACE.
Ген антитела амплифицируют при помощи ПЦР с использованием полученной библиотеки кДНК 5'-RACE в качестве матрицы. Праймеры для амплификации гена антитела мыши могут быть сконструированы на основе известных последовательностей генов антител. Эти нуклеотидные последовательности праймеров варьируются в зависимости от подкласса иммуноглобулина. Таким образом, предпочтительно этот подкласс определяют заранее с использованием коммерчески доступного набора, такого как набор для изотипирования моноклонального антитела мыши Iso Strip (Roche Diagnostics).
Конкретно, например, праймеры, которые позволяют амплификацию генов, кодирующих тяжелые цепи γ1, γ2a, γ2b и γ3 и легкие цепи κ и λ, используют для выделения мышиных IgG-кодирующих генов. В общем, праймер, который отжигается с сайтом константного района вблизи вариабельного района, используют в качестве праймера 3'-стороны для амплификации вариабельного района IgG. Тем временем, праймер, присоединенный к библиотеке кДНК 5'-RACE набора для конструирования, используют в качестве праймера 5'-стороны.
Амплифицированные таким образом продукты ПЦР используют для реконструирования иммуноглобулинов, состоящих из сочетания тяжелых и легких цепей. Желаемое антитело может быть выбрано с использованием Эпирегулин-связывающей активности реконструированного иммуноглобулина в качестве индикатора. Когда целью является выделение антитела против Эпирегулина, более предпочтительным является то, что связывание антитела с Эпирегулином является специфическим. Эпирегулин-связывающее антитело может быть подвергнуто скринингу, например, с использованием следующих стадий:
(1) контактирование Эпирегулин-экспрессирующей клетки с антителом, содержащим V-район, кодируемый кДНК, выделенной из гибридомы;
(2) детектирование связывания этого антитела с Эпирегулин-экспрессирующей клеткой; и
(3) отбор антитела, которое связывается с Эпирегулин-экспрессирующей клеткой.
Способы детектирования связывания антитела с Эпирегулин-экспрессирующими клетками известны. Конкретно, связывание антитела с Эпирегулин-экспрессирующими клетками может быть детектировано вышеописанными способами, такими как FACS. Иммобилизованные пробы Эпирегулин-экспрессирующих клеток подходящим образом используют для оценивания связывающей активности антитела.
Предпочтительные способы скрининга антител, которые используют связывающую активность в качестве индикатора, включают также способы пэннинга, использующие фаговые векторы. Способы скрининга, использующие фаговые векторы, являются предпочтительными, когда гены антитела выделены из библиотек подклассов тяжелой цепи и легкой цепи из экспрессирующей поликлональное антитело популяции клеток. Гены, кодирующие вариабельные районы тяжелой цепи и легкой цепи, могут быть соединены подходящей линкерной последовательностью с образованием одноцепочечного Fv (scFv). Фаги, презентирующие scFv на их поверхности, могут быть получены инсертированием гена, кодирующего scFv, в фаговый вектор. Эти фаги контактируют с представляющим интерес антигеном. Затем ДНК, кодирующая scFv, имеющий представляющий интерес связывающую активность, может быть выделена сбором фагов, связанных с антигеном. Этот процесс может повторяться при необходимости для обогащения scFv, имеющего представляющую интерес связывающую активность.
После выделения кДНК, кодирующей V-район представляющего интерес анти-Эпирегулин-антитела, эту кДНК расщепляют рестрикционными ферментами, которые узнают сайты рестрикции, введенные в оба конца этой кДНК. Предпочтительные рестрикционные ферменты узнают и расщепляют нуклеотидную последовательность, которая встречается в нуклеотидной последовательности гена этого антитела при низкой частоте. Кроме того, сайт рестрикции для фермента, который продуцирует липкий конец, предпочтительно вводят в вектор для инсертирования однокопийного расщепленного фрагмента в правильной ориентации. Эту кДНК, кодирующую V-район анти-Эпирегулин-антитела, расщепляют, как описано выше, и инсертируют в подходящий экспрессирующий вектор для конструирования экспрессирующего вектора антитела. В этом случае, если ген, кодирующий константный район антитела (С-район), и ген, кодирующий вышеупомянутый V-район, слиты в рамке считывания, получают химерное антитело. Здесь ʺхимерное антителоʺ обозначает, что происхождение константного района отличается от происхождения вариабельного района. Таким образом, кроме гетерохимерных антител мышь/человек, аллохимерные антитела человек/человек включены в химерные антитела по данному изобретению. Экспрессирующий вектор химерного антитела может быть сконструирован инсертированием вышеуказанного гена V-района в экспрессирующий вектор, который уже имеет константный район. Конкретно, например, последовательность узнавания для рестрикционного фермента, который вырезает вышеуказанный ген V-района, может быть подходящим образом помещена на 5'-стороне экспрессирующего вектора, несущего ДНК, кодирующую желаемый константный район антитела (С-район). Экспрессирующий вектор химерного антитела конструируют слиянием в рамке считывания этих двух генов, расщепленных одной и той же комбинацией рестрикционных ферментов.
Для получения моноклонального антитела, которое связывается с Эпирегулином, гены антитела инсертируют в экспрессирующий вектор таким образом, что эти гены экспрессируются под контролем регуляторного района экспрессии. Этот регуляторный район экспрессии для экспрессии антитела включает в себя, например, энхансеры и промоторы. Кроме того, подходящая сигнальная последовательность может быть присоединена к амино-концу таким образом, что это экспрессируемое антитело секретируется наружу из клеток. В описанных позже Примерах, в качестве сигнальной последовательности использовали пептид, имеющий аминокислотную последовательность MGWSCIILFLVATATGVHS (SEQ ID NO: 168). Между тем, могут быть присоединены другие подходящие сигнальные последовательности. Экспрессированный полипептид отщепляется при карбокси-конце выше описанной последовательности, и полученный полипептид секретируется наружу из клеток в виде зрелого полипептида. Затем подходящие клетки-хозяева трансформируют этим экспрессирующим вектором, и получают рекомбинантные клетки, экспрессирующие анти-Эпирегулин-антитело-кодирующую ДНК.
ДНК, кодирующие тяжелую цепь (Н-цепь) и легкую цепь (L-цепь) антитела, отдельно инсертируют в различные экспрессирующие векторы для экспрессии гена антитела. Молекула антитела, имеющая Н- и L-цепи, может быть экспрессирована котрансфекцией одной и той же клетки-хозяина векторами, в которые инсертированы гены Н-цепи и L-цепи, соответственно. Альтернативно, клетки-хозяева могут быть трансформированы единственным экспрессирующим вектором, в который инсертированы ДНК, кодирующие Н- и L-цепи (см. WO 1994/011523).
Известны различные комбинации клетка-хозяин/экспрессирующий вектор для получения антител введением выделенных генов антител в подходящих хозяев. Все из этих экспрессионных систем являются применимыми для выделения антигенсвязывающих доменов по данному изобретению. Подходящие эукариотические клетки, используемые в качестве клеток-хозяев, включают в себя клетки животных, клетки растений и клетки грибов. Конкретно, клетки животных включают в себя, например, следующие клетки:
(1) клетки млекопитающих: CHO, COS, миеломы, почки детеныша хомячка (BHK), HeLa, Vero, эмбриональной почки человека (HEK) 293 или т.п.;
(2) клетки земноводных: ооциты Xenopus или т.п.; и
(3) клетки насекомых: sf9, sf21, Tn5 или т.п.
Кроме того, в качестве клетки растения известна система экспрессии гена антитела, использующая клетки, полученные из рода Nicotiana, такая как Nicotiana tabacum. Культивируемые в виде каллюса клетки могут быть подходящим образом использованы для трансформации клеток растений.
Кроме того, следующие клетки могут быть использованы в качестве грибных клеток:
- дрожжи: род Saccharomyces, такой как Saccharomyces cerevisiae, и род Pichia, такой как Pichia pastoris; и
- мицелиальные грибы: род Aspergillus, такой как Aspergillus niger.
Кроме того, известны также системы экспрессии генов антител, которые используют прокариотические клетки. Например, при использовании бактериальных клеток, клетки E. coli, клетки Bacillus subtilis и т.п. могут быть подходящим образом использованы в данном изобретении. Экспрессирующие векторы, несущие представляющие интерес гены антител, вводят в эти клетки трансфекцией. Трансфицированные клетки культивируют in vitro, и желаемое антитело может быть получено из культуры трансформированных клеток.
Кроме вышеописанных клеток-хозяев, для получения рекомбинантного антитела может быть также использовано рекомбинантное антитело. То есть это антитело может быть получено из животного, в которое введен ген, кодирующий представляющее интерес антитело. Например, этот ген антитела может быть сконструирован в виде слитого гена инсертированием в рамке считывания, который кодирует белок, продуцируемый специфически в молоке. Например, может быть использован козий β-казеин и т.п. в качестве белка, секретируемого в молоке. ДНК-фрагменты, содержащие слитый ген, инсертированный с геном антитела, инъецируют в козий эмбрион, и затем этот эмбрион вводят в самку козы. Желаемые антитела могут быть получены в виде белка, слитого с белком молока, из молока, продуцируемого трансгенной козой, рожденной от козы-реципиента эмбриона (или ее потомства). Кроме того, для увеличения объема молока, содержащего желаемое антитело, продуцируемого трансгенной козой, в эту трансгенную козу, если необходимо, могут вводиться гормоны (Ebert, K. M. et al., Bio/Technology (1994) 12(7), 699-702).
При введении описанного здесь анти-Эпирегулин-антитела человеку, антигенсвязывающий домен, полученный из генетически рекомбинантного антитела, которое было искусственно модифицировано для уменьшения гетерологичной антигенности против человека и т.п., может быть подходящим образом использован в качестве антигенсвязывающего домена этого антитела. Такие генетически рекомбинантные антитела включают в себя, например, гуманизированные антитела. Эти модифицированные антитела подходящим образом продуцируются известными способами.
Вариабельный район антитела, используемый для получения антигенсвязывающего домена описанного здесь анти-Эпирегулин-антитела, обычно образуется тремя определяющими комплементарность районами (CDR), которые разделены четырьмя каркасными районами (FR). CDR является районом, который по существу определяет специфичность связывания антитела. Аминокислотные последовательности CDR являются крайне разнообразными. С другой стороны, FR-образующие аминокислотные последовательности часто имеют высокую идентичность даже среди антител с различными специфичностями связывания. Таким образом, обычно специфичность связывания определенного антитела может быть введена в другое антитело трансплантацией CDR.
Гуманизированное антитело называют также реконструированным антителом человека. Конкретно, известны гуманизированные антитела, полученные трансплантацией CDR антитела не-человека, такого как антитело мыши, на антитело человека и т.п. Общие способы генетической инженерии для получения гуманизированных антител также известны. Конкретно, например, ПЦР с перекрывающимся удлинением известна, как способ для трансплантации CDR мышиного антитела на FR человека. В ПЦР с перекрывающимся удлинением нуклеотидную последовательность, кодирующую CDR мышиного антитела, предназначающуюся для трансплантации, добавляют к праймерам для синтеза FR антитела человека. Праймеры готовят для каждого из четырех FR. Обычно считается, что при трансплантации мышиного CDR к FR человека отбор FR человека, который имеет высокую идентичность относительно FR мыши, является выгодным для сохранения функции CDR. То есть обычно является предпочтительным применение FR человека, содержащего аминокислотную последовательность, которая имеет высокую идентичность относительно аминокислотной последовательности FR, соседней с мышиным CDR, предназначенным для трансплантации.
Нуклеотидные последовательности, подлежащие лигированию, конструируют таким образом, что они будут соединены одна с другой в рамке считывания. FR человека индивидуально синтезируют с использованием соответствующих праймеров. В результате, получают продукты, в которых ДНК, кодирующая CDR мыши, присоединена к индивидуальным ДНК, кодирующим FR. Нуклеотидные последовательности, кодирующие CDR мыши каждого продукта, сконструированы таким образом, что они перекрываются друг с другом. Затем реакцию синтеза комплементарных цепей проводят для отжига перекрывающихся CDR-районов этих продуктов, синтезированных с использованием гена антитела человека в качестве матрицы. FR человека лигируют через последовательности CDR мыши посредством этой реакции.
Полноразмерный ген V-района, в котором в конечном счете лигированы три CDR и четыре FR, амплифицируют с использованием праймеров, которые отожжены на их 5’- или 3’-конце, которые дополняют подходящими последовательностями узнавания рестрикционных ферментов. Один экспрессирующий вектор для гуманизированного антитела может быть получен инсертированием ДНК, полученной, как описано выше, и ДНК, которая кодирует С-район антитела человека, так что они будут лигированы в рамке считывания. После трансфицирования этого рекомбинантного вектора в хозяина для установления рекомбинантных клеток, эти рекомбинантные клетки культивируют, и ДНК, кодирующая гуманизированное антитело, экспрессируется для получения гуманизированного антитела в культуре клеток (см. Europian Patent Publication № EP 239400 и International Patent Publication № WO 1996/002576).
Посредством качественного или количественного измерения и оценивания антигенсвязывающей активности гуманизированного антитела, полученного, как описано выше, специалист может удобным образом отобрать FR антитела человека, которые позволяют CDR образовать благоприятный антигенсвязывающий сайт при лигировании посредством этих CDR. Аминокислотные остатки в FR могут быть заменены в случае необходимости, так что CDR реконструированного антитела человека образуют подходящий антигенсвязывающий сайт. Например, аминокислотная последовательность может быть введена в FR с использованием способа ПЦР, используемого для трансплантации СDR мыши в FR человека. Более конкретно, частичные мутации нуклеотидной последовательности могут быть введены в праймеры, которые отжигаются с этим FR. Мутации нуклеотидных последовательностей вводят в FR, синтезированные с использованием таких праймеров. Последовательности мутантных FR, имеющие желаемые характеристики, могут быть отобраны измерением и оцениванием активности мутантного антитела с заменами аминокислот для связывания с антигеном при помощи вышеуказанного способа (Cancer Res. (1993) 53: 851-856).
Альтернативно, желаемые антитела человека могут быть получены иммунизацией трансгенных животных, имеющих весь репертуар генов антитела человека (см. WO 1993/012237; WO 1992/003918; WO 1994/002602; WO 1994/025585; WO 1996/034096; WO 1996/033735), иммунизацией ДНК.
Кроме того, известны также способы для получения антител человека при помощи пэннинга с использованием библиотек антител человека. Например, V-район антитела человека экспрессируют в виде одноцепочечного антитела (scFv) на поверхности фага с использованием способа фагового дисплея. Фаги, экспрессирующие scFv, который связывается с этим антигеном, могут быть отобраны. ДНК-последовательность, кодирующая V-район антитела человека, которая связывается с этим антигеном, может быть определена анализом генов отобранных фагов. Определяют ДНК-последовательность scFv, которая связывается с этим антигеном. Экспрессирующий вектор готовят слиянием V-района в рамке считывания с С-районом желаемого антитела человека и инсертированием продукта в подходящий экспрессирующий вектор. Этот экспрессирующий вектор вводят в клетки, подходящие для экспрессии, такие, как описанные выше. Это антитело человека может быть получено экспрессией гена, кодирующего антитело человека, в этих клетках. Эти способы уже известны (см. WO 1992/001047; WO 1992/020791; WO 1993/006213; WO 1993/011236; WO 1993/019172; WO 1995/001438; WO 1995/015388).
Кроме способов, описанных здеcь, способы клонирования B-клеток (идентификация каждой антитело-кодирующей последовательности, клонирование и ее выделение; применение в конструировании экспрессирующего вектора для приготовления каждого антитела (в частности, IgG1, IgG3 или IgG4); и т.п.) такие как описанные Bernasconi et al. (Science (2002) 298: 2199-2202) или в WO 2008/081008), могут быть подходящим образом использованы для выделения генов антител.
EU-нумерация и нумерация Кабата
Согласно способам, используемым в данном изобретении, положения аминокислот, приписанные CDR и FR антитела, указаны в соответствии с нумерацией Кабата (Sequences of Proteins of Immunological Interest (National Institute of Health, Bethesda, Md., 1987 и 1991)). Здесь аминокислоты вариабельного района анти-Эпирегулин-антитела указаны в соответствии с нумерацией Кабата, тогда как аминокислоты константного района указаны в соответствии с EU-нумерацией на основе положений аминокислот Кабата.
Изменение аминокислот
Известные способы, такие как сайт-направленный мутагенез (Kunkel et al., (Proc. Natl. Acad. Sci. USA (1985) 82, 488-492)) и ПЦР с перекрывающимся удлинением, могут быть подходящим образом использованы для модификации аминокислот в последовательностях аминокислот антител. Кроме того, различные известные способы могут быть также использованы в качестве альтернативного способа для замены аминокислот аминокислотами, другими, чем природные аминокислоты (Annu. Rev. Biophis. Biomol. Struct. (2006) 35, 225-249; Proc. Natl. Acad. Sci. U.S.A. (2003) 100(11), 6353-6357). Например, можно подходящим образом использовать систему бесклеточной трансляции (Clover Direct (Protein Express)), содержащую тРНК, соединенные с неприродной аминокислотой на тРНК амбер-супрессора, которые комплементарны кодону UAG (амбер-кодону), который является стоп-кодоном.
В данном контексте, при описании положения изменения аминокислоты, значение термина ʺи/илиʺ включает в себя любую комбинацию, где ʺиʺ и ʺилиʺ объединены подходящим образом. Конкретно, например, “аминокислота (аминокислоты) в положении (положениях) 33, 55 и/или 96 являются замененными” включает в себя следующие изменения аминокислот: аминокислота (аминокислоты) в (а) положении 33, (b) положении 55, (с) положении 96, (d) положениях 33 и 55, (e) положениях 33 и 96, (f) положениях 55 и 96 и (g) положениях 33, 55 и 96.
Уменьшение иммуногенности
Предпочтительно, предсказанная иммуногенность антитела по данному изобретению в хозяине-человеке уменьшается.
ʺНизкая иммуногенностьʺ обозначает, что в течение достаточного времени для достижения терапевтической эффективности антитело не индуцирует реакцию антитела по меньшей мере в большинстве индивидуумов, получающих достаточное количество антитела для уменьшения эффективности продолжающегося введения этого антитела.
Уровень иммуногенности в людях может быть предсказан с использованием программы предсказания связывания с использованием МНС класса II Propred (http://www.imtech.res.in/raghava/propred) с использованием анализа 1% пороговой величины всех аллелей. Другие программы, которые могут быть использованы, включают:
- Rankpep (http://bio.dfci.harvard.edu/Tools/rankpep.html); и
-Epibase (Algonomics proprietary software: algonomics.com).
В сравнении с инициирующей донорской молекулой, молекулы с уменьшенной иммуногенностью не содержат пептидов или содержат уменьшенное количество пептидов, для которых предсказано, что они связываются с аллелями МНС класса II, которые высоко экспрессируются в популяции-мишени (Flower et al., Frug Discov. Today (2004) 9(2), 82-90).
Функциональный анализ связывания МНС класса II может выполняться генерированием перекрывающихся пептидов, соответствующих представляющему интерес белку, и тестированием их способности вызывать Т-клеточную активацию (анализ пролиферации Т-клеток) или заменой известного МНС класса II-связывающего пептида, который является репортерным пептидом (Hammer J. et al., Exp. Med. (1994) 180, 2353-2358).
Несколько способов могут быть использованы для уменьшения иммуногенности анти-Эпирегулин-антител по данному изобретению. Первый способ является способом гуманизации вышеуказанных антител. Более конкретно, CDR анти-Эпирегулин-антитела, выделенные из животного (не-человека), такого как мышь, трансплантировали в антитело человека. Для поддержания или усиления связывания с Эпирегулином, аминокислотный остаток (аминокислотные остатки) FR может быть дополнительно заменен таким образом, что эти CDR гуманизированного анти-Эпирегулин-антитела будут образовывать подходящие антигенсвязывающие сайты.
Один неограничивающий вариант CDR анти-Эпирегулин-антитела, выделенного из мышей по данному изобретению, включает в себя CDR1 тяжелой цепи (HCDR1) SEQ ID NO: 9, CDR2 тяжелой цепи (НСDR2) SEQ ID NO: 10, и CDR3 тяжелой цепи (HCDR3) SEQ ID NO: 11. Кроме того, один неограничивающий вариант CDR анти-Эпирегулин-антитела, выделенного из мышей по данному изобретению, включает в себя CDR1 легкой цепи (LCDR1) SEQ ID NO: 12, CDR2 легкой цепи (LСDR2) SEQ ID NO: 13, и CDR3 легкой цепи (LCDR3) SEQ ID NO: 14.
Один неограничивающий вариант FR антитела человека по данному изобретению включает в себя FR1 тяжелой цепи (HFR1) SEQ ID NO: 1, FR2 тяжелой цепи (HFR2) SEQ ID NO: 2, FR3 тяжелой цепи (HFR3) тяжелой цепи SEQ ID NO: 3, и FR4 тяжелой цепи (HFR4) SEQ ID NO: 4. Кроме того, один неограничивающий вариант FR антитела человека по данному изобретению включает в себя FR1 легкой цепи (LFR1) SEQ ID NO: 5, FR2 легкой цепи (LFR2) SEQ ID NO: 6, FR3 легкой цепи (LFR3) SEQ ID NO: 7, и FR4 легкой цепи (LFR4) SEQ ID NO: 8.
Один неограничивающий вариант вариабельного района гуманизированного анти-Эпирегулин-антитела по данному изобретению включает в себя вариабельный район тяжелой цепи SEQ ID NO: 15. Кроме того, один неограничивающий вариант вариабельного района гуманизированного анти-Эпирегулин-антитела по данному изобретению включает в себя вариабельный район легкой цепи SEQ ID NO: 16.
Кроме того, один неограничивающий вариант FR антитела человека по данному изобретению включает в себя FR1 тяжелой цепи (HRF1) SEQ ID NO: 17 и FR3 тяжелой цепи (HFR3) SEQ ID NO:18. Кроме того, один неограничивающий вариант FR антитела человека по данному изобретению включает в себя FR2 легкой цепи (LFR2) SEQ ID NO: 20, FR3 легкой цепи (LFR3) SEQ ID NO: 21, и FR3 легкой цепи (LFR3) SEQ ID NO: 23.
Один неограничивающий вариант вариабельного района гуманизированного анти-Эпирегулин-антитела по данному изобретению включает в себя вариабельный район тяжелой цепи SEQ ID NO: 19. Кроме того, один неограничивающий вариант вариабельного района гуманизированного анти-Эпирегулин-антитела по данному изобретению включает в себя вариабельный район легкой цепи SEQ ID NO: 22 или 24.
Кроме того, один неограничивающий вариант FR гуманизированного антитела по данному изобретению включает в себя FR3 тяжелой цепи (HFR3) SEQ ID NO: 35 и FR3 тяжелой цепи (HFR3) SEQ ID NO: 36. Один неограничивающий вариант вариабельного района гуманизированного анти-Эпирегулин-антитела по данному изобретению включает в себя вариабельный район тяжелой цепи SEQ ID NO: 37 или 38.
Вторым способом является способ конструирования измененной последовательности с уменьшенной иммуногенностью анализом измененной последовательности с изменениями аминокислот в аминокислотной последовательности анти-Эпирегулин-антитела с использованием вышеупомянутой программы предсказания связывания МНС класса II. Подходящие неограничивающие примеры сайта (сайтов) изменений аминокислот для уменьшения иммуногенности анти-Эпирегулин-антител по данному изобретению включают в себя аминокислоту (аминокислоты) в положении (положениях) 87 и/или 101, как показано нумерацией Кабата, в последовательности тяжелой цепи анти-Эпирегулин-антитела SEQ ID NO: 38. Кроме того, предпочтительные неограничивающие примеры этого сайта изменения аминокислоты для уменьшения иммуногенности анти-Эпирегулин-антител включают в себя аминокислоту в положении 24, как показано нумерацией Кабата в последовательности легкой цепи анти-Эпирегулин-антитела SEQ ID NO: 29.
Один предпочтительный пример неограничивающего варианта вышеупомянутой аминокислотной замены (вышеупомянутых аминокислотных замен) включает в себя замену Ser(S) аминокислоты в положении 87 и/или заменой Tyr(Y) или Phe(F) аминокислоты в положении 101, как показано нумерацией Кабата, в последовательности тяжелой цепи анти-Эпирегулин-антитела SEQ ID NO: 38. Кроме того, предпочтительный пример неограничивающего варианта вышеупомянутой аминокислотной замены включает в себя замену Arg(R) аминокислоты в положении 24, как показано нумерацией Кабата, в последовательности легкой цепи анти-Эпирегулин-антитела SEQ ID NO: 29.
Третьим способом является способ подходящего отбора константного района IgG1-аллотипа, выбранного среди G1m3-типа (последовательности, полученной добавлением GK к С-концу последовательности SEQ ID NO: 31), G1m17,1-типа (SEQ ID NO: 26) и G1m17-типа (последовательности, полученной добавлением GK к С-концу последовательности SEQ ID NO: 30), при конструировании анти-Эпирегулин-антитела, содержащего константный район IgG1-антитела. Известно, что совместимость между аллотипом вида животного (не человека), которому водят фармацевтический препарат антитела, и аллотипом этого содержащего фармацевтический препарат антитела влияет на иммунные реакции в этих видах животных (Genes and Immunity (2011) 12, 213-221).
Один неограничивающий вариант тяжелой цепи гуманизированного анти-Эпирегулин-антитела по данному изобретению с уменьшенной иммуногенностью включает в себя тяжелые цепи, выбранные из группы, состоящей из SEQ ID NO: 37, 38, 49-56, 72-79, 92-98, 115-127, 131-135, 137-140 и 142-150. Кроме того, один неограничивающий вариант легкой цепи гуманизированного анти-Эпирегулин-антитела по данному изобретению включает в себя легкие цепи, выбранные из группы, состоящей из SEQ ID NO: 29, 57, 58, 80-85, 99, 128-130, 136 и 141.
Один неограничивающий вариант гуманизированного анти-Эпирегулин-антитела по данному изобретению с уменьшенной иммуногенностью включает в себя гуманизированные анти-Эпирегулин-антитела, содержащие тяжелые цепи, выбранные из группы, состоящей из SEQ ID NO: 37, 38, 49-56, 72-79, 92-98, 115-127, 131-135, 137-140 и 142-150, и легкие цепи, выбранные из группы, состоящей из SEQ ID NO: 29, 57, 58, 80-85, 99, 128-130 и 141.
Супрессия деамидирования, изомеризации и гидролиза
Предпочтительно то, что деамидирование, изомеризация и гидролиз ослабляются в анти-Эпирегулин-антителах по данному изобретению.
Очень важно контролировать количество агрегата в белковом фармацевтическом препарате с учетом контроля качества и влияний на эффективность лекарственного средства и иммуногенности (Curr. Opin. Biotechnol. (2009) 20(6), 708-714). Обычно образование агрегата подвергается действию как коллоидальной стабильности, происходящей из среды раствора белка, так и конформационной стабильности, происходящей из структуры белка (J.Pharm. Sci. (2010) 100(4), 1306-1315). Условия, эффективные для коллоидальной стабильности, могут быть получены скринингом концентрации антитела или рН, типа буферного раствора, ионной силы, добавки и т.п. испытанием предписания композиции антитела. С другой стороны, конформационная стабильность частично зависит от аминокислотной последовательности, и в случае антител важным считается сохранение характерных структур, таких как каноническая структура CDR, консенсусные последовательности FR и взаимосвязь VH/VL и т.п. (Jung et al., J. Mol. Biol. (2001) 309(3), 701-716; Xiang et al., J. Mol. Biol. (1995) 253(3), 385-390; Ewert et al., Methols. (2004) 34(2), Vargas-Madrazo et al., J. Mol. Recognit. (2003) 16(3) 113-120: и Morea et al., J. Mol. Biol. (1998) 275, 269-294).
Реакцией деамидирования называют реакции, которые имеют место неферментативно на боковых цепях аспарагина и глутамина, и изменяют амиды на боковых цепях аспарагина и глутамина в карбоновые кислоты. Изомеризация происходит из образования нестабильного циклического имидного промежуточного продукта деамидированием аспарагина или дегидратацией аспарагиновой кислоты, в результате атаки карбонильной группы, присутствующей на боковой цепи аспарагина или аспарагиновой кислоты, электронной парой азота на остатке, расположенном на С-конце. Этот промежуточный продукт изменяется большей частью в изоаспарагиновую кислоту расщеплением, и остальное становится аспарагиновой кислотой. В случае глутамина скорость деамидирования равна обычно одной десятой скорости деамидирования аспарагина, но этот механизм является по существу тем же самым и только требует молекул воды для прогрессирования реакции. Поскольку эти реакции деамидирования и изомеризации, которые имеют место во время хранения белков, таких как антитела, становятся причинами вышеописанной гетерогенности, они желательно ослабляются до возможного предела. Кроме того, сообщалось, что реакции деамидирования легко имеют место, особенно в сайтах, в которых аспарагин и глицин находятся рядом друг с другом (Asn-Gly) (Geiger et al., J. Biol. Chem. (1987) 262, 785-794). Кроме того, сообщалось, что расщепление пептидной цепи реакцией гидролиза имеет место в аспарагиновой кислоте, и сообщается, что гидролиз легко имеет место при кислотных условиях, в частности, в последовательности, в которой пролин присутствует на С-концевой стороне (Asp-Pro) (Segalas et al., FEBS Letters (1995) 371, 171-175).
При необходимости, для супрессии деамидирования, иммунизации и гидролиза, могут проводиться замены аминокислот и т.д. для удаления глутамильных и аспарагинильных остатков, которые являются сайтами, которые подвергаются деамидированию. Предпочтительным неограничивающим вариантом сайта деамидирования, удаление которого является особенно эффективным, включает в себя сайты, в которых реакция деамидирования является усиленной, или, более конкретно, остаток глицина, остаток аспарагина или остаток глутамина в мотивах, указанных как последовательности NG и GC. Замена любого из этих аминокислотных остатков (N, Q или G) может заметно ослаблять реакции деамидирования (WO 2003/057881, WO 2005/067620 или т.п.). Кроме того, способы получения антител с ослабленной реакцией деамидирования посредством контролирования этого способа для культивирования антитело-продуцирующих клеток могут быть также использованы при необходимости. Анти-Эпирегулин-антитела, обеспечиваемые данным изобретением, также включают в себя анти-Эпирегулин-антитела, к которым применялся вышеуказанный способ ослабления (супрессии) реакции деамидирования.
Неограничивающие примеры сайтов, которые подвергаются деамидированию, включают в себя аминокислоту (аминокислоты) в положении (положениях) 31, 52, 54 и/или 101, как указано нумерацией Кабата, в последовательности тяжелой цепи анти-Эпирегулин-антитела SEQ ID NO: 38. Кроме того, неограничивающие примеры сайтов, которые подвергаются деамидированию, предпочтительно включают в себя аминокислоту (аминокислоты) в положении (положениях) 28, 92 и/или 93, как указано нумерацией Кабата, в последовательности легкой цепи анти-Эпирегулин-антитела SEQ ID NO: 29.
Один неограничивающий вариант вышеупомянутых замены (замен) аминокислот предпочтительно включает в себя замену (замены) Ala(A) аминокислоты в положении 31 и/или замены Thr(T), Lys(K), Phe(F), Val(V), Arg(R) или Leu(L) аминокислоты в положении 55, как указано нумерацией Кабата, в последовательности тяжелой цепи анти-Эпирегулин-антитела SEQ ID NO: 38. Кроме того, один неограничивающий вариант вышеупомянутых замены (замен) аминокислот предпочтительно включает в себя замену (замены) Glu(E) аминокислоты в положении 92 и/или Arg(R) или Gln(Q) аминокислоты в положении 93, как указано нумерацией Кабата, в последовательности легкой цепи анти-Эпирегулин-антитела SEQ ID NO: 29.
Неограничивающие варианты тяжелой цепи гуманизированного анти-Эпирегулин-антитела по данному изобретению с ослабленными деамидированием, изомеризацией и гидролизом включают в себя тяжелые цепи, выбранные из группы, состоящей из SEQ ID NO: 49-56, 98, 115-127, 131-135, 137-140 и 142-150. Кроме того, неограничивающие варианты легкой цепи гуманизированного анти-Эпирегулин-антитела по данному изобретению с ослабленными деамидированием, изомеризацией и гидролизом включают в себя легкие цепи, выбранные из группы, состоящей из SEQ ID NO: 57, 58, 128 и 141.
Неограничивающие варианты тяжелой цепи гуманизированного анти-Эпирегулин-антитела по данному изобретению с ослабленными деамидированием, изомеризацией и гидролизом включают в себя гуманизированные анти-Эпирегулин-антитела, содержащие тяжелые цепи, выбранные из группы, состоящей из SEQ ID NO: 49-56, 98, 115-127, 131-135, 137-140 и 142-150, и легкие цепи, выбранные из группы, состоящей из SEQ ID NO: 57, 58, 128 и 141.
Модуляция изоэлектрической точки
Предпочтительно, изоэлектрические точки анти-Эпирегулин-антител по данному изобретению являются модулированными. Способы контролирования заряда поверхности молекулы антитела изменением аминокислотных остатков, экспонированных на поверхности молекулы антитела, являются известными, как один из способов для контролирования периода полужизни антитела в плазме (WO 2007/114319 и WO 2009/041543). Конкретно, было известно, что уменьшение величины изоэлектрической точки (pI) антитела способно пролонгировать период полужизни этого антитела в плазме. Напротив, было известно, что увеличение изоэлектрической точки антитела укорачивает его период полужизни в плазме и улучшает свойства миграции в ткани этого антитела (Vaisitti et al., J. Biol. Homeost. Agents. (2005) 18(3-4), 195-112; Pardridge et al., J, Pharmacol. Exp. Ther. (1998) 286(1), 548-554). С другой стороны, известно, что противоопухолевое действие антитела, изоэлектрическая точка которого уменьшалась, увеличивается в сравнении с этим действием антитела перед изменением (WO 2009/041062).
pI белка, такого как антитело, определяется как рН, при котором полипептид имеет эффективный заряд. В этой области известно, что обычно растворимость белка является самой низкой, когда рН этого раствора является эквивалентным изоэлектрической точке (pI) этого белка. Таким образом, на основе pI может быть оценена растворимость белка при конкретном рН, например, рН 6. pI белка является также хорошим индикатором в отношении вязкости белка в жидком препарате. Высокая pI показывает высокую растворимость и низкую вязкость (которая особенно важна для высококонцентрированного препарата). В одном неограничивающем варианте осуществления данного изобретения, отбирали кандидатные домены, имеющие более высокую pI, чем предварительно определенный порог. pI антитела также играет роль в эффективности лекарственных средств и биологическом распределении этого антитела. Например, было известно, что когда pI антитела становится высоким, локализация в клетках и/или снаружи сосуда увеличивается. Наиболее желательно, должны быть определены свойства pI для конкретного антитела для достижения желаемой цели. В некоторых вариантах осуществления могут быть получены антитела с величиной pI приблизительно 5,0, 5,5, 6,0, 6,5, 7,0, 7,5, 8,0, 8,5, 9,0 или более. В другом неограничивающем варианте осуществления могут быть выбраны антитела с величиной pI приблизительно 9,0, 8,5, 7,5, 7,0, 6,5, 6,0, 5,5, 5,0 или менее. Квалифицированному в данной области специалисту будет понятно, что единственный белок может иметь множество заряженных форм. Без связывания себя с конкретной теорией, авторы изобретения считают, что заряд белка может модулироваться рядом различных механизмов действия; и, без ограничения, примеры таких механизмов включают в себя аминокислотную замену (аминокислотные замены), образование катионов, деаминирование, гетерогенность карбокси-концевой аминокислоты, фосфорилирование, гликозилирование и т.п. Используемая здесь pI определяется как pI форма главного заряда.
pI белка может быть определена различными способами и, без ограничения, один неограничивающий вариант таких способов включает в себя изоэлектрическое фокусирование и различные компьютерные алгоритмы (см., например, Bjelqvist et al., Electrophoresis (1993) 14, 1023). В одном неограничивающем варианте осуществления pI определяют с использованием системы электрофореза Pharmacia Biotech Multiphor 2, оборудованной узлом рециркуляции охлаждающей бани Multi-temp III и источником питания EPS 3501 XL. Гель Pre-cast Ampholin (Amersham Biosciences, диапазон pI: 2,5-10) загружают вместе с 5 мкг этого белка. Относительную pI антитела определяют с использованием широкого диапазона стандартного маркера pI (Amersham, диапазон рI: 3-10, 8 мкл). Электрофорез выполняют при условиях 1500 В и 50 мА в течение 105 минут. Затем гель фиксируют с использованием фиксирующего раствора Sigma (5х), разведенного до 1х очищенной водой. Затем этот гель окрашивают с использованием красителя Simple Blue Stain (Invitrogen) в течение ночи при комнатной температуре. Затем этот гель обеспечивают с использованием раствора, состоящего из 2,5% этанола, 8% уксусной кислоты и 67% очищенной воды. Изоэлектрическую точку определяют на основе стандартной калибровочной кривой с использованием денситометра Bio-Rad.
Для модуляции изоэлектрической точки анти-Эпирегулин-антитела по данному изобретению, изменения аминокислот и т.п., в том числе удаления заряженных аминокислот в аминокислотной последовательности анти-Эпирегулин-антитела, или добавления или инсертирования заряженных аминокислот в аминокислотную последовательность анти-Эпирегулин-антитела, может проводиться при необходимости. Известно, что заряженные аминокислоты присутствуют среди этих аминокислот. Обычно лизин (К), аргинин (R) и гистидин (H) известны в качестве аминокислот, имеющих положительный заряд (положительно-заряженные аминокислоты). Аспарагиновая аминокислота (D), глутаминовая аминокислота (E) и т.п. известны в качестве аминокислот, имеющих отрицательный заряд (отрицательно заряженные аминокислоты). Другие аминокислоты известны как незаряженные аминокислоты.
Вышеупомянутым ʺизмененным аминокислотным остаткомʺ является предпочтительно аминокислотный остаток, выбранный подходящим образом из аминокислот в любой группе (а) или группы (b), подвергнутых добавлению, инсертированию или удалению, но не ограничиваемый особо этими аминокислотами.
(a) Глутаминовая кислота (Е), аспарагиновая кислота (D);
(b) лизин (K), аргинин (R), гистидин (H).
Когда первоначальный аминокислотный остаток (перед изменением) уже является заряженным, изменение его в незаряженный аминокислотный остаток является также предпочтительным вариантом данного изобретения. Более конкретно, изменения в данном изобретении включают в себя (1) замену заряженной аминокислоты незаряженной аминокислотой, (2) замену заряженной аминокислоты противоположно заряженной кислотой в сравнении с первоначальной кислотой, и (3) замену незаряженной аминокислоты заряженной кислотой.
Неограничивающие примеры положений, где аминокислоты изменены для модуляции изоэлектрической точки анти-Эпирегулин-антитела по данному изобретению, предпочтительно включают в себя аминокислоту (аминокислоты) в положении (положениях) 13, 61, 62, 64, 65, 97 и/или 98, как указано нумерацией Кабата, в последовательности тяжелой цепи анти-Эпирегулин-антитела SEQ ID NO: 38. Кроме того, неограничивающие примеры положений, где аминокислоты изменены для модуляции изоэлектрической точки анти-Эпирегулин-антитела по данному изобретению, предпочтительно включают в себя аминокислоту (аминокислоты) в положении (положениях) 24, 55, 56 и/или 107, как указано нумерацией Кабата, в последовательности легкой цепи анти-Эпирегулин-антитела SEQ ID NO: 29.
Неограничивающие примеры вышеупомянутой аминокислотной замены (аминокислотных замен) предпочтительно включает в себя замену (замены) Asn(N) или Lys(K) аминокислоты в положении 14, замены Asp(D) или Glu(E) аминокислоты в положении 61, замену Ser(S) или Glu(Q) аминокислоты в положении 62, замену Asp(D) или Glu(E) аминокислоты в положении 64, замену Asp(D) или Glu(E) аминокислоты в положении 65, замену Arg(R) аминокислоты в положении 97 и/или замену Glu(E) аминокислоты в положении 98, как указано нумерацией Кабата, в последовательности тяжелой цепи анти-Эпирегулин-антитела SEQ ID NO: 38. Кроме того, неограничивающие примеры вышеупомянутых аминокислотной замены (аминокислотных замен) предпочтительно включает в себя замену (замены) Gln(Q) или Ser(S) аминокислоты в положении 24, замену Glu(E) в положении 55, замену Glu(E) аминокислоты в положении 56 или замену Glu(E) аминокислоты в положении 106(а), как указано нумерацией Кабата, в последовательности легкой цепи анти-Эпирегулин-антитела SEQ ID NO: 29.
Один неограничивающий вариант анти-Эпирегулин-антитела по данному изобретению с модулированной изоэлектрической точкой включает в себя тяжелые цепи, выбранные из группы, состоящей из SEQ ID NO: 72-79, 98, 115-127, 131-135, 137-140 и 142-150. Кроме того, один неограничивающий вариант легкой цепи гуманизированного анти-Эпирегулин-антитела по данному изобретению с модулированной изоэлектрической точкой включает в себя легкие цепи, выбранные из группы, состоящей из SEQ ID NO: 80-85 и 99.
Один неограничивающий вариант анти-Эпирегулин-антитела по данному изобретению с модулированной изоэлектрической точкой включает в себя тяжелые цепи, выбранные из группы, состоящей из 72-79, 98, 115-127, 131-135, 137-140 и 142-150, и легкие цепи выбранные из группы, состоящей из SEQ ID NO: 80-85 и 99.
Улучшение стабильности
Предпочтительно, стабильность анти-Эпирегулин-антител по данному изобретению является улучшенной. Один или несколько параметров, которые описывают стабильность антитела, включают в себя величину температуры теплового плавления (Tm) домена. Величина Tm антитела является отличным индикатором термической стабильности антитела и также может быть индикатором периода хранения для этого антитела. Более низкая величина Tm указывает на образование агрегата/более низкую стабильность, тогда как более высокая величина Tm указывает на необразование агрегатов/высокую стабильность. Таким образом, предпочтительно, чтобы были обеспечены антитела с высокими величинами Tm. В одном неограничивающем варианте осуществления данного изобретения, отбирают антитела с величинами Tm, более высокими, чем предварительно заданный порог. В некоторых вариантах осуществления отбирают антитела, имеющие величину Tm по меньшей мере 50°C, 55°C, 60°C, 65°C, 70°C, 75°C, 80°C, 85°C, 90°C, 95°C или 100°C или большую.
Величина температуры теплового плавления (Тm) антитела может быть измерена различными стандартными способами, известными в данной области. Например, Vermeer et al. исследовали развертывание и денатурацию моноклонального мышиного антитела против крысиного IgG изотипа 2b дифференциальной сканирующей калориметрией (DSC) и спектроскопией с круговым дихроизмом (CD) (Biophys. J. (2000) 78, 394-404; Colloid Surfaces A: Physicochem. Eng. Aspects (2000), 161, 139-150; J. Colloid Interface Sci. (2000) 225, 394-397, и Biophys. J. (2000) 79, 2150-2154). В результате, считается, что укладка и развертывание этого IgG может характеризоваться двумя главными транзициями, которые сами перекрывают различные стадии. Бимодальное распределение, наблюдаемое как в DSC-, так и в СD-экспериментах, не зависело от скорости сканирования в этих экспериментах. По-видимому, эти две транзиции являются независимыми, и развертывание было необратимым. Вторичную структуру, а также термодинамическую стабильность Fab и Fc, которые отщеплялись из IgG, сравнивали с этими параметрами интактного иммуноглобулина (Vermeer et al., Biophys. J. (2000) 79, 2150-2154). Было показано, что эти два пика, наблюдаемые для интактного IgG, могут быть отнесены к Fab-фрагменту и Fc-фрагменту, соответственно. Vermeer et al. показали также, что, кроме индукции нагреванием, структурное искажение IgG обычно могло также запускаться изменением рН (Biophys. J. (2000) 78, 394-404) или взаимодействием с гидрофобной средой, например, адсорбцией на тефлоновые поверхности или взаимодействием с поверхностно-активными веществами. (Vermeer et al., 1998, Biochim. Biophym. Acta. (1988) 1425, 1-12; Colloid Surfaces A: Physicochem. Eng. Aspects (2000) 161, 139-150; J. Colloid Interface Sci. (2000) 225, 394-397).
В одном неограничивающем варианте осуществления данного изобретения Tm антитела измеряют с использованием пробы, содержащей выделенные антитела. В одном варианте Tm антитела измеряют с использованием VP-DSC (MicroCal, LLC) при условиях скорости сканирования 1,0°C/мин и диапазона температур 25°C-120°C. Может быть использован период фильтрования восемь секунд вместе с пятиминутным термостатом предварительного сканирования. В одном конкретном примере пробы готовят диализом в 25 мМ гистидин-HCl с использованием диализных чашек Pierce (3,5 кДа). Средняя концентрация антител равна 50 мкг/мл, как определено абсорбцией при 280 нм. Температуры плавления определяют согласно процедурам изготовителя с использованием Origin-software, поставляемого с этой системой. Вкратце, множество фонов (базовых линий) загружают буфером как в пробе, так и в ссылочных клетках для установления термического равновесия. После вычитания фона из термограммы пробы, эти данные нормализовали относительно концентрации и подгоняли с использованием функции деконволюции. В другом варианте стабильность этих антител оценивают с использованием способа, описанного ниже. Одна или несколько метрик могут дополнительно включать в себя метрики, характеризующие стабильность этого антитела при одном или нескольких условиях, выбранных из группы, состоящей из различных величин рН, различных температур, различных срезающих стрессов и различных циклов замораживание/оттаивание.
В одном неограничивающем варианте осуществления данного изобретения, DSC-измерения могут выполняться с использованием Setaram Micro-DSC III (Setaram, Caluire, France). Эти пробы помещали в калориметр в 1-мл ячейке для пробы против 1-мл ссылочной ячейки, содержащей подходящее количество пустого раствора. Эти клетки стабилизировали в течение четырех часов при 25°C внутри калориметра перед нагреванием этих клеток до конечной температуры при выбранной скорости нагревания. Температура переходного состояния и энтальпия могут быть определены с использованием Setaram, Version 1.3.
В одном неограничивающем варианте осуществления данного изобретения, кривая термической денатурации/ренатурации может быть получена с использованием спектроскопии с круговым дихроизмом (СD). Изменения во вторичной структуре IgG в виде функции температуры и/или, например, рН, могут быть исследованы CD-спектроскопией (Fasman et al., Circular Dichroism and Confonnational Analysis of Biomolecules. (1996 Plenum Press)). Согласно de Jongh et al., преимуществом этого способа является то, что на спектроскопический сигнал не влияет присутствие окружающего раствора, и что хорошо установленные процедуры являются доступными для выяснения вторичной структуры на основе ссылочных спектров элементов с различной структурой (Biochemistry (1994) 33, 14521-14528). Фракции вторичных структурных элементов могут быть получены из CD-спектров.
В одном неограничивающем варианте осуществления данного изобретения могут быть сделаны измерения на спектрополяриметре JASCO, модели О-715 (Jasco International Co.). Может быть использована кварцевая кювета с длиной пути света 0,1 см. Регуляция температуры может проводиться с использованием термопары JASCO PTC-348WI (JASCO International). Температурные сканы регистрировали с использованием термопары Peltier c разрешением 0,2°C и временной константой 16 секунд. Сканы длин волн в дальней ультрафиолетовой области (разрешение 0,2 нм) могут быть получены накапливанием множества сканов с подходящей скоростью сканирования.
Кривая термическая денатурация/ренатурация может быть также измерена спетроскопическим способом. Когда белок в растворе денатурируется в реакции на нагревание, молекулы агрегируются и этот раствор рассеивает свет более сильно. Агрегация приводит к изменениям в оптической прозрачности этой пробы и может быть измерена мониторингом этого изменения в оптической плотности видимого или ультрафиолетового света определенной длины волны.
В одном неограничивающем варианте осуществления данного изобретения, для получения кривой термическая денатурация/ренатурация используют флуоресцентную спектроскопию. В одном варианте осуществления подвергают мониторингу внутреннюю флуоресценцию белка, например, внутреннюю флуоресценцию триптофана. В другом варианте осуществления мониторингу подвергают флуоресцентные молекулы пробы. Способы для проведения экспериментов с флуоресцентной спектроскопией хорошо известны квалифицированным в данной области специалистам. См., например, Bashford et al. (Spectrophotometry and Spectrofluorometry: A Practical Approach (1987) 91-114, IRL Press Ltd.), Bell, J.E. (Spectroscopy in Biochemistry (1981) Vol. 1, 155-194, CRC Press) или Brand et al. (Ann. Rev. Biochem. (1972) 41, 843).
Что касается биологических активностей композиций антител, различные способы могут быть использованы для оценивания их стабильности на основе физических и химических структур этих антител. Например, для исследования денатурации антител доступны такие способы, как абсорбция с переносом заряда, флуоресцентная спектроскопия, ЯМР, редуцирующий капиллярный гель-электрофорез (rCGE) и/или высокоэффективная эксклюзионная (ситовая) хроматография (HPSEC) (например, Wang et al. (J. Parental Science & Technology 42 (Suppl.), S4-S26)), наряду с вышеупомянутыми аналитическими способами.
Редуцирующий капиллярный гель-электрофорез и высокоэффективная эксклюзионная хроматография являются наиболее обычными и простейшими способами для оценивания образования агрегатов белков, продуктов деградации белков и белковых фрагментов. Таким образом, стабильность композиции, содержащей анти-Эпирегулин-антитело, обеспеченная этим изобретением, может также оцениваться этими способами.
Для модификации стабильности анти-Эпирегулин-антител по данному изобретению, могут проводиться подходящие изменения аминокислот в аминокислотной последовательности анти-Эпирегулин-антител и т.д.
В качестве неограничивающего варианта осуществления для модификации стабильности анти-Эпирегулин-антитела по данному изобретению, когда анти-Эпирегулин-антитело является IgG1-антителом, амидирование С-концевой аминогруппы вследствие делетирования С-концевого аминокислотного остатка лизина и делетирования двух С-концевых аминокислот глицина и лизина сообщается как гетерогенность, произведенная из С-концевой последовательности тяжелой цепи IgG-антитела человека (G1, SEQ ID NO: 25) (Anal. Biochem. (2007) 360(1), 75-83). Один предпочтительный способ для уменьшения такой гетерогенности включает изменение делеции двух С-концевых аминокислот тяжелой цепи, а именно делетирование глицина в положении 446 и лизина в положении 447, как показано EU-нумерацией (WO 2009/041613). Поскольку желательно, чтобы гетерогенность, произведенная из С-концевой последовательности тяжелой цепи в анти-Эпирегулин-антителах по данному изобретению, не присутствовала, IgG1-последовательность, в которой глицин в положении 446 и лизин в положении 447, указанные EU-нумерацией в IgG1-человека, делетированы (G1d, SEQ ID NO: 26), может быть использована в качестве последовательности константного района.
Один неограничивающий вариант тяжелой цепи анти-Эпирегулин-антитела по данному изобретению с улучшенной стабильностью включает в себя тяжелые цепи, выбранные из группы, состоящей из SEQ ID NO: 37, 38, 49-56, 72-79, 92-98, 115-127, 131-135, 137-140 и 142-150. Кроме того, один неограничивающий вариант легкой цепи гуманизированного анти-Эпирегулин-антитела по данному изобретению с улучшенной стабильностью включает в себя легкие цепи, выбранные из группы, состоящей из SEQ ID NO: 99, 128-130, 136 и 141.
Один неограничивающий вариант анти-Эпирегулин-антитела по данному изобретению с улучшенной стабильностью включает в себя анти-Эпирегулин-антитела, содержащие тяжелые цепи, выбранные из группы, состоящей из SEQ ID NO: 37, 38, 49-56, 72-79, 92-98, 115-127, 131-135, 137-140 и 142-150, и легкие цепи, выбранные из группы, состоящей из SEQ ID NO: 99, 128-130, 136 и 141.
Уменьшение количества агрегата
Количество агрегата анти-Эпирегулин-антител по данному изобретению предпочтительно уменьшается. Контролирование количества агрегата в фармацевтическом препарате белка является очень важным с учетом контроля качества и влияний на лекарственную эффективность и иммуногенность (Curr. Opin. Biotechnol. (2009) 20(6), 708-714). Обычно на образование агрегата влияет как коллоидальная стабильность, происходящая из окружающего белок раствора, так и конформационная стабильность, происходящая из структуры этого белка (J. Pharm. Sci. (2010) 100(4), 1306-1315). Желаемые условия, эффективные для коллоидальной стабильности, могут быть достигнуты скринингом концентрации антитела, типа буфера, ионной силы, добавок и т.п. испытанием предписанного рецепта фармацевтического препарата антитела. С другой стороны, конформационная стабильность, считающаяся важной для сохранения характерных структур, таких как каноническая структура CDR, и консенсусные последовательности FR и граница VH/VL и т.д. (Yung et al., J. Mol. Biol. (2001) 309(3), 701-716, Xiang et al., J. Mol. Biol. (1995) 253(3), 385-390, Ewert et al., (Methods. (2004) 34(2), 184-199, Vargas-Madrazo et al., (J. Mol. Recognit. (2003) 16(3), 113-120; Morea et al., J. Mol. Biol. (1998) 275, 269-294, и Vargas-Madrazo et al., J. Mol. Recognit. (2003) 16(3), 113-120).
Различные способы могут быть использованы для оценивания стабильности препаратов белка, содержащих препараты антител, на основе физических и химических структур этих белков, а также их биологических активностей. Например, для исследования денатурации белка, доступны такие способы, как абсорбция с переносом заряда, термический анализ, флуоресцентная спектроскопия, круговой дихроизм (CD), ЯМР, а также НPSEC, фильтрование с тангенциальным потоком (TFF), рассеяние статического света (SLS), инфракрасная спектроскопия с Фурье-преобразованием, технология индуцируемого мочевиной развертывания белка, внутренняя флуоресценция триптофана, дифференциальная сканирующая калориметрия и технология связывания белка 1-анилино-8-нафталинсульфонатом (ANS). Например, может быть подходящим образом использован способ, выбранный из способов, описанных в Wang et al., (J. Parental Science & Technology (1988) 42 (Suppl), S4-S26).
rCGE и HPSEC являются наиболее обычными и проcтейшими способами для оценивания образования белковых агрегатов, деградации белка и фрагментации белка. Таким образом, стабильность жидких композиций по данному изобретению может оцениваться этими способами.
Например, стабильность анти-Эпирегулин-антител по данному изобретению может быть оценена HPSEC или rCGE, где процентная площадь пика представляет недеградированное антитело или фрагмент недеградированного антитела. В одном неограничивающем варианте осуществления, приблизительно 250 мкг антитела (приблизительно 25 мкл жидкого препарата, содержащего 10 мг/мл вышеупомянутого антитела) инжектируют в колонку TosoH Biosep TSK G3000SWXL (7,8 мм × 30 см), снабженную защитной колонкой TSK SW x1 (6,0 мм × 4,0 см). Это антитело (включающее в себя фрагменты антитела) элюируют изократически 0,1 М динатрийфосфатом, содержащим 0,1 М сульфат натрия и 0,05% азид натрия, при скорости потока 0,8-1,0 мл/мин. Этот элюированный белок детектируют с использованием UF оптической плотности при 280 нм. Ссылочный стандарт присутствует в этом анализе в качестве контроля, и результаты сообщаются в виде площади в процентах пика мономера продукта для всех других пиков, за исключением пика объема, наблюдаемого при приблизительно 12-14 минутах. Пики, элюирующиеся раньше, чем пик мономера, регистрируются как агрегат в процентах.
Для уменьшения агрегата анти-Эпирегулин-антитела по данному изобретению, уменьшали гидрофобное взаимодействие между молекулами, содержащими тяжелую цепь и легкую цепь анти-Эпирегулин-антитела. Конкретно, изменение (изменения) аминокислот, которые заменяют гидрофильным остатком (гидрофильными остатками) гидрофобный остаток (гидрофобные остатки) в аминокислотной последовательности тяжелой цепи или легкой цепи анти-Эпирегулин-антитела и т.п., выполняют подходящим образом. В предпочтительном неограничивающем варианте осуществления, изменение (изменения) аминокислоты, которая заменяет гидрофильный остаток (гидрофильные остатки) гидрофобным остатком(ами) в CDR анти-Эпирегулин-антитела и т.д., выполняют подходящим образом. Известно, что аминокислоты включают в себя гидрофильные аминокислоты и гидрофобные аминокислоты. Обычно Asp(D), Glu(E), Arg(R), Lys(K), His(H), Gly(G), Ser(S), Thr(T), Cys(C), Asn(N), Gln(Q) и Tyr(Y) известны как гидрофильные аминокислоты. Ala(A), Val(V), Leu(L), Ile(I), Met(M), Trp(W), Phe(F) и Pro(P) известны как гидрофобные аминокислоты. Предпочтительно, вышеупомянутое изменение аминокислот подходящим образом выбирают из замены (замен) одной или нескольких аминокислот, выбранных среди вышеупомянутых гидрофобных остатков, аминокислотами, выбранными среди вышеупомянутых гидрофильных остатков, но они не ограничиваются конкретно специфическими аминокислотами.
Неограничивающие примеры положений, которые подвергаются изменениям аминокислот для уменьшения количества агрегата анти-Эпирегулин-антитела по данному изобретению, предпочтительно включают в себя аминокислоту (аминокислоты) в положении (положениях) 60 и/или 98, как указано нумерацией Кабата, в последовательности тяжелой цепи анти-Эпирегулин-антитела SEQ ID NO: 38.
Один неограничивающий вариант вышеупомянутой замены предпочтительно включает в себя замену His(H), Lys(K), Thr(T), Ser(S) или Arg(R) аминокислоты в положении 60 и/или замену Ser(S) или Glu E) аминокислоты в положении 98, как указано нумерацией Кабата, в тяжелой цепи анти-Эпирегулин-антитела SEQ ID NO: 38.
Один неограничивающий вариант тяжелой цепи анти-Эпирегулин-антитела по данному изобретению с уменьшенной величиной агрегата включает в себя тяжелые цепи, выбранные из группы, состоящей из SEQ ID NO: 92-98, 115-127, 131-135, 137-140 и 142-150.
Один неограничивающий вариант анти-Эпирегулин-антитела по данному изобретению с уменьшенной величиной агрегата включает в себя тяжелые цепи, выбранные из группы, состоящей из SEQ ID NO: 92-98, 115-127, 131-135, 137-140 и 142-150, и легкие цепи, выбранные из группы, состоящей из SEQ ID NO: 99, 128-130, 136 и 141.
Придание межвидовой реактивности в отношении связывания Эпирегулина животного (не-человека) и Эпирегулина человека
Анти-Эпирегулин-антитела по данному изобретению являются предпочтительно анти-Эпирегулин-антителами, обнаруживающими перекрестную межвидовую реактивность между животными (не-человеком) и человеком. Более конкретно, данное изобретение обеспечивает анти-Эпирегулин-антитело, которое связывается с эпитопом, связанным с анти-Эпирегулин-антителом, содержащим CDR вариабельного района тяжелой цепи SEQ ID NO: 9, 10 и 11, и CDR вариабельного района легкой цепи SEQ ID NO: 12, 13 и 14, где это антитело характеризуется тем, что его отношение величины KD для Эпирегулина обезьяны SEQ ID NO: 170 (cEREG KD) к величине KD для Эпирегулина человека SEQ ID NO: 34 (hERG KD) (cEREG KD/hEREG KD) является меньшим, чем отношение cEREG KD/hEREG KD анти-Эпирегулин-антитела, содержащего CDR вариабельного района тяжелой цепи SEQ ID NO: 9, 10 и 11 и CDR вариабельного района легкой цепи SEQ ID NO: 12, 13 и 14.
Более конкретно, в данном изобретении аминокислотные последовательности CDR вариабельного района тяжелой цепи SEQ ID NO: 9, 10 и 11 и CDR вариабельного района легкой цепи SEQ ID NO: 12, 13 и 14 изменяли для модификации связывающей активности в отношении Эпирегулина обезьяны анти-Эпирегулин-антитела, содержащего вышеупомянутые CDR. Поскольку структурные различия между Эпирегулином обезьяны и Эпирегулином человека не были выяснены, не имелось абсолютно никакого руководства в отношении того, в каких аминокислотных остатках должно быть изменено анти-Эпирегулин-антитело для увеличения связывающей активности в отношении Эпирегулина обезьяны. В данном изобретении были сконструированы множественные последовательности антител с заменой (заменами) аминокислотных остатков в произвольном сайте (в произвольных сайтах) в этих CDR. Конкретно, были получены анти-Эпирегулин-антитела с заменой остатком Arg(R) аминокислоты в вышеупомянутых CDR-последовательностях.
Эти анти-Эпирегулин-антитела с заменой остатком Arg(R) аминокислоты в этих CDR-последовательностях неожиданно имели увеличенную связывающую активность в отношении Эпирегулина обезьяны. То есть замена остатком Arg(R) аминокислоты в CDR-последовательностях анти-Эпирегулин-антитела способна придавать перекрестную межвидовую реактивность связывания Эпирегулину животного (не-человека) и Эпирегулину человека.
Величина KD для Эпирегулина обезьяны SEQ ID NO: 170 (cEREG KD) может быть рассчитана способом, описанным в вышеупомянутом разделе по ʺсвязывающей активностиʺ. Эпирегулин-связывающая активность анти-Эпирегулин-антитела может быть измерена способами, известными квалифицированным в данной области специалистам, и условия измерения могут быть определены подходящим образом квалифицированными в данной области специалистами. Эпирегулин-связывающая активность анти-Эпирегулин-антитела может быть оценена как KD (константа диссоциации), видимая KD (видимая константа диссоциации), kd, которая является скоростью диссоциации (константой скорости диссоциации), видимая kd (видимая константа диссоциации) или т.п. Они могут измеряться способами, известными квалифицированным в данной области специалистам, и, например, могут быть использованы Biacore (GE-healthcare), Scatchard plot, FACS и т.п. Отношение величины KD для Эпирегулина обезьяны (cEREG KD) к величине KD для Эпирегулина человека (hEREG KD) (cEREG KD/hEREG KD) может быть определено делением этой величины KD на величину KD для Эпирегулина человека SEQ ID NO: 34.
В данном изобретении, cEREG KD/hEREG KD равно менее 40. Кроме того, в одном неограничивающем варианте осуществления cEREG KD/hEREG KD равно предпочтительно менее 10. В другом не ограничивающем варианте осуществления cEREG KD/hEREG KD равно предпочтительно менее 6, и в другом неограничивающем варианте осуществления cEREG KD/hEREG KD равно предпочтительно менее 4.
Неограничивающие примеры положения (положений), где аминокислота (аминокислоты) изменена для придания перекрестной межвидовой реактивности в отношении связывания с Эпирегулином животного (не-человека) и Эпирегулином человека, обеспеченным данным изобретением, предпочтительно включают в себя аминокислоту (аминокислоты) в положении (положениях) 33, 51, 54, 55, 56, 57, 58, 59, 60, 62, 65, 96, 97 и/или 98, как указано нумерацией Кабата, в последовательности тяжелой цепи анти-Эпирегулин-антитела SEQ ID NO: 38. Кроме того, неограничивающие примеры положения (положений), где аминокислота (аминокислоты) изменена для придания перекрестной межвидовой реактивности в отношении связывания с Эпирегулином животного (не-человека) и Эпирегулином человека, обеспеченным данным изобретением, предпочтительно включают в себя аминокислоту (аминокислоты) в положениях 24, 93 и/или 94, как указано нумерацией Кабата, в последовательности легкой цепи анти-Эпирегулин-антитела SEQ ID NO: 29. Один предпочтительный пример изменения аминокислот включает в себя замену остатком Arg(R) одной или нескольких вышеупомянутых аминокислот.
Примеры неограничивающего варианта тяжелой цепи анти-Эпирегулин-антитела по данному изобретению, наделенного перекрестной межвидовой реактивностью, включают в себя тяжелые цепи, выбранные из группы, состоящей из SEQ ID NO: 115-127, 131-135, 137-140 и 142-150. Примеры неограничивающего варианта легкой цепи гуманизированного анти-Эпирегулин-антитела по данному изобретению, наделенного перекрестной межвидовой реактивностью, включают в себя легкие цепи, выбранные из группы, состоящей из SEQ ID NO: 128-130, 136 и 141.
Примеры неограничивающего варианта анти-Эпирегулин-антитела по данному изобретению, наделенного перекрестной межвидовой реактивностью, включают в себя анти-Эпирегулин-антитела, содержащие тяжелые цепи, выбранные из группы, состоящей из SEQ ID NO: 98, 115-127, 131-135, 137-140 и 142-150, и легкие цепи, выбранные из группы, состоящей из SEQ ID NO: 29, 128-130, 136 и 141.
Нейтрализующая активность
Анти-Эпирегулин-антитело по данному изобретению является предпочтительно антителом, которое имеет нейтрализующую активность. Обычно ʺнейтрализующая активностьʺ относится к активности ингибирования биологической активности лиганда, такого как вирусы или токсины, в отношении клеток. Более конкретно, ʺвещества, имеющие нейтрализующую активностьʺ относится к веществам, которые связываются с этим лигандом или с рецептором, связывающимся с этим лигандом, и ингибируют связывание между этим лигандом и этим рецептором. Когда связывание этого лиганда с рецептором блокируется нейтрализующей активностью, опосредованная рецептором биологическая активность не может вызываться. Антитела, которые имеют такую нейтрализующую активность, обычно называют нейтрализующими антителами. Эта нейтрализующая активность может быть измерена сравнением биологических активностей в присутствии и в отсутствие тест-вещества, нейтрализующая активность которых должна быть оценена, в присутствии представляющего интерес лиганда.
В данном изобретении связывание лиганда рецептора EGF, который считается главным рецептором Эпирегулина, приводит к димеризации этого рецептора и активации домена внутриклеточной тирозинкиназы этого рецептора. Активированная тирозинкиназа образует фосфорилированные тирозинсодержащие пептиды аутофосфорилированием, и эти пептиды ассоциируются с различными вспомогательными молекулами трансдукции сигнала. Ими являются в основном PLCγ (фосфолипаза γ), Shc, Grb2 и т.п. Из этих вспомогательных молекул, первые две дополнительно фосфорилируются тирозинкиназой рецептора EGF. Основным путем трансдукции сигнала от рецептора EGF является путь, в котором фосфорилирование трансдуцируется в порядке Shc, Grb2, Sos, Ras, Raf/MAPK-киназа/MAP-киназа. Кроме того, считается, что существует альтернативный путь от PLCγ к PRC.
Поскольку такие каскады внутриклеточных сигналов варьируются в зависимости от типа клеток, подходящие молекулы могут быть нацелены на представляющую интерес клетку-мишень, и эти молекулы-мишени не ограничиваются вышеупомянутыми факторами. Коммерчески доступные наборы для измерения in vivo сигнала активации могут быть использованы подходящим образом (например, система анализа активности протеинкиназы С (GE Healthcare Bio-Sciences)).
Кроме того, активация передачи сигнала in vivo может быть детектирована с использованием в качестве индекса индуцирующего транскрипцию эффекта на гене-мишени, представленном ниже в каскаде передачи сигнала in vivo. Изменения в транскрипционной активности могут быть детектированы на основе принципа анализа репортера. Более конкретно, репортерный ген, такой как зеленый флуоресцентный белок (GFP) или люцифераза, помещают справа от района транскрипционного фактора или промотора гена мишени и измеряют репортерную активность. Изменение транскрипционной активности может быть измерена на основе этой репортерной активности.
Кроме того, поскольку рецептор EGF обычно функционирует для увеличения пролиферации клеток, активация передачи сигнала in vivo может оцениваться измерением активности пролиферации клеток-мишеней. В настоящем изобретении нейтрализующая активность нейтрализующего антитела по настоящему изобретению оценивается оценкой активности пролиферации клетки. Однако данное изобретение не ограничивается этим способом, и нейтрализующая активность может быть оценена подходящим применением вышеупомянутых способов для отбора клеток-мишеней.
Конкретно, например, измерением упомянутой ниже активности пролиферации клеток, нейтрализующая активность анти-Эпирегулин-антитела может быть оценена и измерена. Например, используют способ, который измеряет включение [3Н]-меченого тимидина, добавленного к среде, живыми клетками в качестве индекса способности репликации ДНК. В качестве более удобного способа, используют способ исключения красителя, который измеряет под микроскопом способность клетки высвобождать краситель, такой как трипановый синий, наружу из клеток, или MTT-способ. Последний использует способность живых клеток превращать 3-(4,5-диметилтиазол-2-ил)-2,5-дифенилтетразолийбромид (MTT), который является солью тетразолия, в синий продукт формазан. Более конкретно, тест-антитело добавляют к культуральному раствору тест-клетки, и после определенного периода времени этот MTT-раствор добавляют к культуральному раствору и оставляют его стоять в течение определенного периода времени, чтобы MTT включился в эту клетку. В результате, MTT, который является желтым соединением, превращается в синее соединение действием сукцинатдегидрогеназы в митохондриях этой клетки. После растворения этого синего продукта для окрашивания, оптическую плотность измеряют и используют в качестве индикатора для ряда жизнеспособных клеток.
Кроме МТТ, такие реагенты, как MTS, XTT, WST-1 и WST-8, являются коммерчески доступными (Nacalai Tesque и т.п.) и могут быть подходящим образом использованы. Кроме того, известны способы, которые оценивают активность пролиферации клеток с использованием клеточного АТФ или импеданса культуры клеток в качестве индикатора. Для измерений активности связывающее антитело, которое имеет тот же самый изотип, что и анти-Эпирегулин-антитело, но не имеет нейтрализующей активности, может быть использовано в качестве контрольного антитела таким же образом, как анти-Эпирегулин-антитело, и может быть определено, что эта активность присутствует, когда это анти-Эпирегулин-антитело имеет более сильную нейтрализующую активность, чем это контрольное антитело.
Цитотоксичность
Антитело, используемое в данном изобретении, является предпочтительно антителом, имеющим цитотоксичность.
В данном изобретении цитотоксичность включает в себя, например, активность антитело-зависимой клеточноопосредованной цитотоксичности (ADCC) и активность комплемент-зависимой цитотоксичности (CDC). В данном изобретении CDC-активность относится к опосредуемой системой комплемента цитотоксической активности. Между тем, ADCC-активность относится к активности повреждения клетки-мишени при присоединении специфического антитела к его антигену поверхности клеток. Удерживающая Fcγ-рецептор клетка (иммуноцит или т.п.) связывается с Fc-частью этого антитела через Fcγ-рецептор, и клетка-мишень разрушается. Антителом, используемым в данном изобретении, может быть антитело, имеющее CDC-активность или ADCC-активность, или может быть антителом, имеющим как CDC-активность, так и ADCC-активность.
Имеет или не имеет анти-Эпирегулин-антитело ADCC-активность или CDC-активность, может быть определено известными способами (например, Current protocols in Immunology, Chapter 7. Immunologic studies in humans, Editor, John E, Coligan et al., John Wiley & Sons, Inc., (1993) и т.п.).
Сначала, конкретно, готовят эффекторные клетки, раствор комплемента и клетки-мишени.
(1) Приготовление эффекторных клеток
Селезенку удаляют из мыши CBA/N или т.п., и клетки селезенки выделяют в RPMI1640 (Invitrogen). После промывания этих клеток той же самой средой, содержащей 10% фетальную телячью сыворотку (FBS, HyClone), концентрация промытых клеток селезенки может быть доведена до 5×106/мл для получения эффекторных клеток.
(2) Приготовление раствора комплемента
Комплемент детеныша кролика (CEDARLANE) разводят в 10 раз в культуральной среде (Invitrogen), содержащей 10% FBS для приготовления раствора комплемента.
(3) Приготовление клеток-мишеней
Клетки-мишени могут быть радиоактивно помечены культивированием клеток, экспрессирующих белок Эпирегулин с 0,2 мКи (милликюри) хроматом-51Cr натрия (GE Healthcare Bio-Sciences) в среде DMEM, содержащей 10% FBS (ФТС), в течение одного часа при 37°С. Для экспрессирующих белок Эпирегулин клеток можно использовать клетки, трансформированные геном, кодирующим белок Эпирегулин, первичные клетки рака ободочной кишки, метастатические клетки рака ободочной кишки, клетки аденокарциномы легких, клетки рака панкреатической железы, клетки рака желудка, клетки рака почки, клетки рака ободочной кишки, клетки рака пищевода или т.п. После радиоактивного мечения, клетки промывают три раза средой RPMI1640, содержащей 10% ФТС, и эти клетки-мишени могут быть приготовлены доведением концентрации клеток до 2×105 клеток/мл.
Активность ADCC и активность CDC могут быть измерены способом, описанным ниже. В случае измерения активности ADCC, 50 мкл клеток-мишеней и 50 мкл анти-Эпирегулин-антитела добавляют в каждую лунку 96-луночного планшета с U-дном ((Becton Dickinson), и этот материал оставляли стоять на льду в течение 15 минут. Затем 100 мкл этих эффекторных клеток добавляют в каждую лунку планшета, и этот планшет инкубируют в термостате с диоксидом углерода в течение четырех часов. Конечную концентрацию антитела можно доводить до 0 или 10 мкг/мл. После инкубирования 100 мкл этого супернатанта собирают, и радиоактивность измеряют в гамма-счетчике (COBRAII AUTO-GAMMA, MODEL D5005, Packard Instrument Company). Измеренную величину используют для расчета цитотоксической активности (%) в соответствии с Формулой 2:
Формула 2
(A-C)/(B-C)×100.
А представляет радиоактивность (импульсов в минуту, cpm) в каждой пробе, В представляет радиоактивность (cpm) в каждой пробе, к которой добавляют 1% NP-40 (Nacalai Tesque), и С представляет радиоактивность (cpm) пробы, содержащей только клетки-мишени.
Тем временем, в случае измерения CDC-активности, 50 мкл клеток-мишеней и 50 мкл анти-Эпирегулин-антитела добавляют в каждую лунку 96-луночного плоскодонного планшета (Becton Dickinson), и этот материал оставляют стоять на льду в течение 15 минут. Затем 100 мкл раствора комплемента добавляют в каждую лунку, и планшет инкубируют в инкубаторе с диоксидом углерода в течение четырех часов. Конечную концентрацию этого антитела доводят до 0 или 3 мкг/мл. После инкубации 100 мкл супернатанта собирают, и радиоактивность измеряют с использованием гамма-счетчика. Цитотоксичность может быть рассчитана таким же образом, что и в определении ADCC-активности.
Анти-Эпирегулин-антитела с увеличенной ADCC-активностью
Анти-Эпирегулин-антитела с увеличенной ADCC-активностью могут быть также использованы предпочтительно в качестве анти-Эпирегулин-антител по данному изобретению. Как описано выше, ADCC-активностью называют активность повреждения клетки-мишени, когда специфическое антитело прикрепляется к антигену ее клеточной поверхности, и Fcγ-рецептор-экспрессирующая клетка (иммунная клетка или т.п.) связывается с Fc-частью этого антитела через этот Fcγ-рецептор. Таким образом, опосредованная анти-Эпирегулин-антителом ADCC-активность может быть увеличена увеличением Fcγ-рецептор-связывающей активности Fcγ-рецептор-экспрессирующих клеток, таких как иммунные клетки. В качестве способов для увеличения Fcγ-рецептор-связывающей активности Fcγ-рецептор-экспрессирующих клеток, таких как иммунные клетки, могут быть использованы по меньшей мере три известных способа.
(1) Анти-Эпирегулин-антитела с изменением (изменениями) аминокислот Fc-района
Анти-Эпирегулин-антитела с увеличенной Fcγ-рецептор-связывающей активностью могут быть получены изменением аминокислоты (аминокислот) Fc-района, содержащихся в анти-Эпирегулин-антителах по данному изобретению. Примеры предпочтительных Fc-районов IgG-иммуноглобулинов для этого изменения включают в себя Fc-районы IgG человека (IgG1, IgG2, IgG3 или IgG4 и их варианты).
Аминокислоты в любых положениях могут быть изменены в другие аминокислоты, пока Fcγ-связывающая активность антитела может увеличиваться. Когда анти-Эпирегулин-антитело по данному изобретению содержит Fc-район IgG1 человека в качестве Fc-района, предпочтительно включать изменения, которые приводят к увеличению Fcγ-рецептор-связывания в сравнении со связывающей активностью Fc-района, произведенного из IgG1 человека. Аминокислотные изменения для увеличения Fcγ-рецептор-связывающей активности сообщались, например, в WO 2007/024249, WO 2007/021841, WO 2006/031370, WO 2000/042072, WO 2004/029207, WO 2004/099249, WO 2006/105338, WO 2007/041635, WO 2008/092117, WO 2005/070963, WO 2006/020114, WO 2006/116260 и WO 2006/023403.
Примеры таких аминокислот, которые могут быть изменены, включают в себя по меньшей мере одну или несколько аминокислот, выбранных из группы, состоящей из аминокислот в положениях 221, 222, 223, 224, 225, 227, 228, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 243, 244, 245, 246, 247, 249, 250, 251, 254, 255, 256, 258, 260, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 278, 279, 280, 281, 282, 283, 284, 285, 286, 288, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 311, 313, 315, 317, 318, 320, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 339, 376, 378, 379, 380, 382, 385, 392, 396, 421, 427, 428, 429, 434, 436 и 440, как указано EU-нумерацией. Изменение этих аминокислот может увеличивать Fcγ-рецептор-связывание Fc-района анти-Эпирегулин-антитела. Эти замены аминокислот могут увеличивать Fcγ-рецептор-связывание Fc-района анти-Эпирегулин-антитела по данному изобретению. Анти-Эпирегулин-антитело по данному изобретению может содержать по меньшей мере одну замену аминокислоты, например, в положении, выбранном из группы, состоящей из 230, 240, 244, 245, 247, 262, 263, 266, 273, 275, 299, 302, 313, 323, 325, 328 и 332, как указано EU-нумерацией, в константном районе тяжелой цепи SEQ ID NO: 26.
Особенно предпочтительные примеры изменений для использования в данном изобретении включают в себя по меньшей мере одно или несколько изменений аминокислот, выбранных из группы, состоящей из:
аминокислоты в положении 221 или в Lys, или в Tyr;
аминокислоты в положении 222 в любую из Phe, Trp, Glu и Tyr;
аминокислоты в положении 223 в любую из Phe, Trp, Glu и Lys;
аминокислоты в положении 224 в любую из Phe, Trp, Glu и Tyr;
аминокислоты в положении 225 в любую из Glu, Lys и Trp;
аминокислоты в положении 227 в любую из Glu, Gly, Lys и Tyr;
аминокислоты в положении 228 в любую из Glu, Gly, Lys и Tyr;
аминокислоты в положении 230 в любую из Ala, Glu, Gly и Tyr;
аминокислоты в положении 231 в любую из Glu, Gly, Lys, Pro и Tyr;
аминокислоты в положении 232 в любую из Glu, Gly, Lys и Tyr;
аминокислоты в положении 233 в любую из Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислоты в положении 234 в любую из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислоты в положении 235 в любую из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислоты в положении 236 в любую из Ala, Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислоты в положении 237 в любую из Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислоты в положении 238 в любую из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислоты в положении 239 в любую из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp и Tyr;
аминокислоты в положении 240 в любую из Ala, Ile, Met и Thr;
аминокислоты в положении 241 в любую из Asp, Glu, Leu, Arg, Trp и Tyr;
аминокислоты в положении 243 в любую из Leu, Glu, Leu, Gln, Arg, Trp и Tyr;
аминокислоты в положении 244 в His;
аминокислоты в положении 245 в Ala;
аминокислоты в положении 246 в любую из Asp, Glu, His и Tyr;
аминокислоты в положении 247 в любую из Ala, Phe, Gly, His, Ile, Leu, Met, Thr, Val и Tyr;
аминокислоты в положении 249 в любую из Glu, His, Gln и Tyr;
аминокислоты в положении 250 или в Glu, или в Gln;
аминокислоты в положении 251 в Phe;
аминокислоты в положении 254 в любую из Phe, Met и Tyr;
аминокислоты в положении 255 в любую из Glu, Leu и Tyr;
аминокислоты в положении 256 в любую из Ala, Met и Pro;
аминокислоты в положении 258 в любую из Asp, Glu, His, Ser и Tyr;
аминокислоты в положении 260 в любую из Asp, Glu, His и Tyr;
аминокислоты в положении 262 в любую из Ala, Glu, Phe, Ile и Thr;
аминокислоты в положении 263 в любую из Ala, Ile, Met и Thr;
аминокислоты в положении 264 в любую из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Trp и Tyr;
аминокислоты в положении 265 в любую из Ala, Leu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислоты в положении 266 в любую из Ala, Ile, Met и Thr;
аминокислоты в положении 267 в любую из Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp и Tyr;
аминокислоты в положении 268 в любую из Asp, Glu, Phe, Gly, Ile, Lys, Leu, Met, Pro, Gln, Arg, Thr, Val и Trp;
аминокислоты в положении 269 в любую из Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислоты в положении 270 в любую из Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Gln, Arg, Ser, Thr, Trp и Tyr;
аминокислоты в положении 271 в любую из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислоты в положении 272 в любую из Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Arg, Ser, Thr, Trp, Val и Tyr;
аминокислоты в положении 273 или в Phe, или в Ile;
аминокислоты в положении 274 в любую из Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислоты в положении 275 или в Leu, или в Trp;
аминокислоты в положении 276 в любую из Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислоты в положении 278 в любую из Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val и Trp;
аминокислоты в положении 279 в Ala;
аминокислоты в положении 280 в любую из Ala, Gly, His, Lys, Leu, Pro, Gln, Trp и Tyr;
аминокислоты в положении 281 в любую из Asp, Lys, Pro и Tyr;
аминокислоты в положении 282 в любую из Glu, Gly, Lys, Pro и Tyr;
аминокислоты в положении 283 в любую из Ala, Gly, His, Ile, Lys, Leu, Met, Pro, Arg и Tyr;
аминокислоты в положении 284 в любую из Asp, Glu, Leu, Asn, Thr и Tyr;
аминокислоты в положении 285 в любую из Asp, Glu, Lys, Gln, Trp и Tyr;
аминокислоты в положении 286 в любую из Glu, Gly, Pro и Tyr;
аминокислоты в положении 288 в любую из Asn, Asp, Glu и Tyr;
аминокислоты в положении 290 в любую из Asp, Gly, His, Leu, Asn, Ser, Thr, Trp и Tyr;
аминокислоты в положении 291 в любую из Asp, Glu, Gly, His, Ile, Gln и Thr;
аминокислоты в положении 292 в любую из Ala, Asp, Glu, Pro, Thr и Tyr;
аминокислоты в положении 293 в любую из Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислоты в положении 294 в любую из Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислоты в положении 295 в любую из Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислоты в положении 296 в любую из Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr и Val;
аминокислоты в положении 297 в любую из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислоты в положении 298 в любую из Ala, Asp, Glu, Phe, His, Ile, Lys, Met, Asn, Gln, Arg, Thr, Val, Trp и Tyr;
аминокислоты в положении 299 в любую из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp и Tyr;
аминокислоты в положении 300 в любую из Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val и Trp;
аминокислоты в положении 301 в любую из Asp, Glu, His и Tyr;
аминокислоты в положении 302 в Ile;
аминокислоты в положении 303 в любую из Asp, Gly и Tyr;
аминокислоты в положении 304 в любую из Asp, His, Leu, Asn и Thr;
аминокислоты в положении 305 в любую из Glu, Ile, Thr и Tyr;
аминокислоты в положении 311 в любую из Ala, Asp, Asn, Thr, Val и Tyr;
аминокислоты в положении 313 в Phe;
аминокислоты в положении 315 в Leu;
аминокислоты в положении 317 или в Glu, или в Gln;
аминокислоты в положении 318 в любую из His, Leu, Asn, Pro, Gln, Arg, Thr, Val и Tyr;
аминокислоты в положении 320 в любую из Asp, Phe, Gly, His, Ile, Leu, Asn, Pro, Ser, Thr, Val, Trp и Tyr;
аминокислоты в положении 322 в любую из Ala, Asp, Phe, Gly, His, Ile, Pro, Ser, Thr, Val, Trp и Tyr;
аминокислоты в положении 323 в Ile;
аминокислоты в положении 324 в любую из Asp, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Thr, Val, Trp и Tyr;
аминокислоты в положении 325 в любую из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислоты в положении 326 в любую из Ala, Asp, Glu, Gly, Ile, Leu, Met, Asn, Pro, Gln, Ser, Thr, Val, Trp и Tyr;
аминокислоты в положении 327 в любую из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Thr, Val, Trp и Tyr;
аминокислоты в положении 328 в любую из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислоты в положении 329 в любую из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислоты в положении 330 в любую из Cys, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислоты в положении 331 в любую из Asp, Phe, His, Ile, Leu, Met, Gln, Arg, Thr, Val, Trp и Tyr;
аминокислоты в положении 332 в любую из Ala, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислоты в положении 333 в любую из Ala, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Ser, Thr, Val и Tyr;
аминокислоты в положении 334 в любую из Ala, Glu, Phe, Ile, Leu, Pro и Thr;
аминокислоты в положении 335 в любую из Asp, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Val, Trp и Tyr;
аминокислоты в положении 336 в любую из Glu, Lys и Tyr;
аминокислоты в положении 337 в любую из Glu, His и Asn;
аминокислоты в положении 339 в любую из Asp, Phe, Gly, Ile, Lys, Met, Asn, Gln, Arg, Ser и Tyr;
аминокислоты в положении 376 или в Ala, или в Val;
аминокислоты в положении 377 или в Gly, или в Lys;
аминокислоты в положении 378 в Asp;
аминокислоты в положении 379 в Asn;
аминокислоты в положении 380 в любую из Ala, Asn и Ser;
аминокислоты в положении 382 или в Ala, или в Ile;
аминокислоты в положении 385 в Glu;
аминокислоты в положении 392 в Thr;
аминокислоты в положении 396 в Leu;
аминокислоты в положении 421 в Lys;
аминокислоты в положении 427 в Asn;
аминокислоты в положении 428 или в Phe, или в Leu;
аминокислоты в положении 429 в Met;
аминокислоты в положении 434 в Trp;
аминокислоты в положении 436 в Ile; и
аминокислоты в положении 440 в любую из Gly, His, Ile, Leu и Tyr,
в соответствии с EU-нумерацией в Fc-районе анти-Эпирегулин-антитела по данному изобретению. Количество аминокислот, которые являются измененными, особо не ограничивается. Может быть изменена аминокислота только в одном положении, или могут быть изменены аминокислоты в двух или нескольких положениях. Примеры комбинаций изменений аминокислот в двух или более положениях включают в себя комбинации, показанные в Таблицах 1-1 и 1-2.
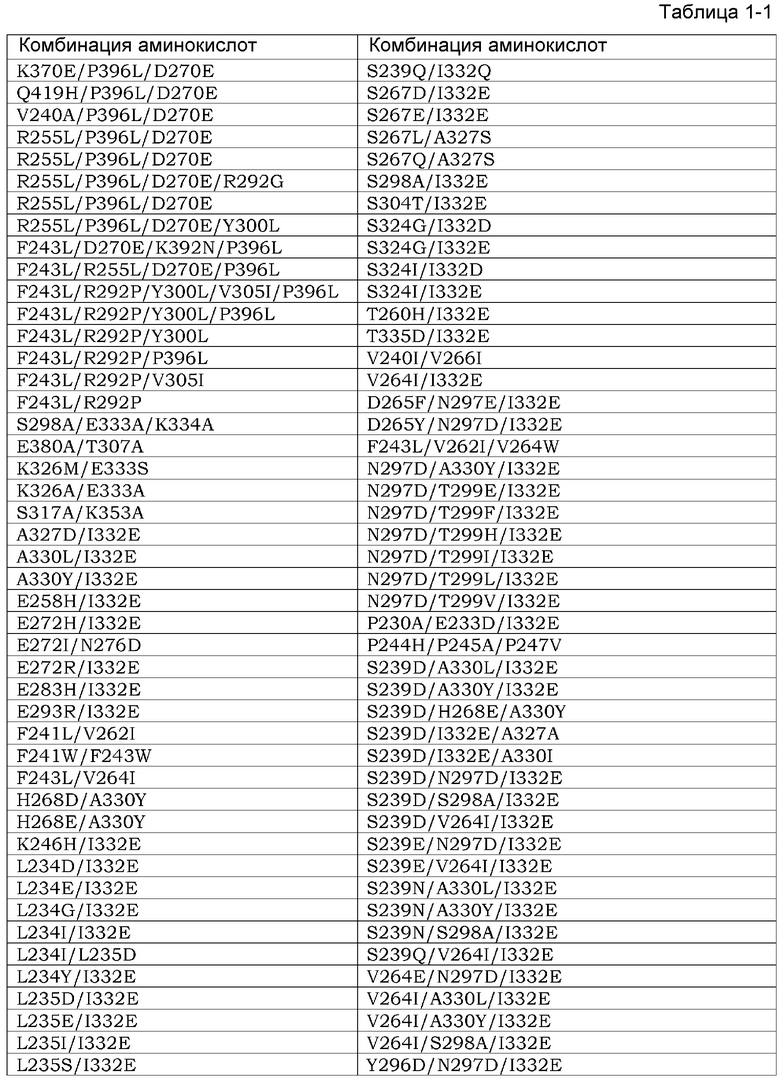
Таблица 1-2 является продолжением Таблицы 1-1.
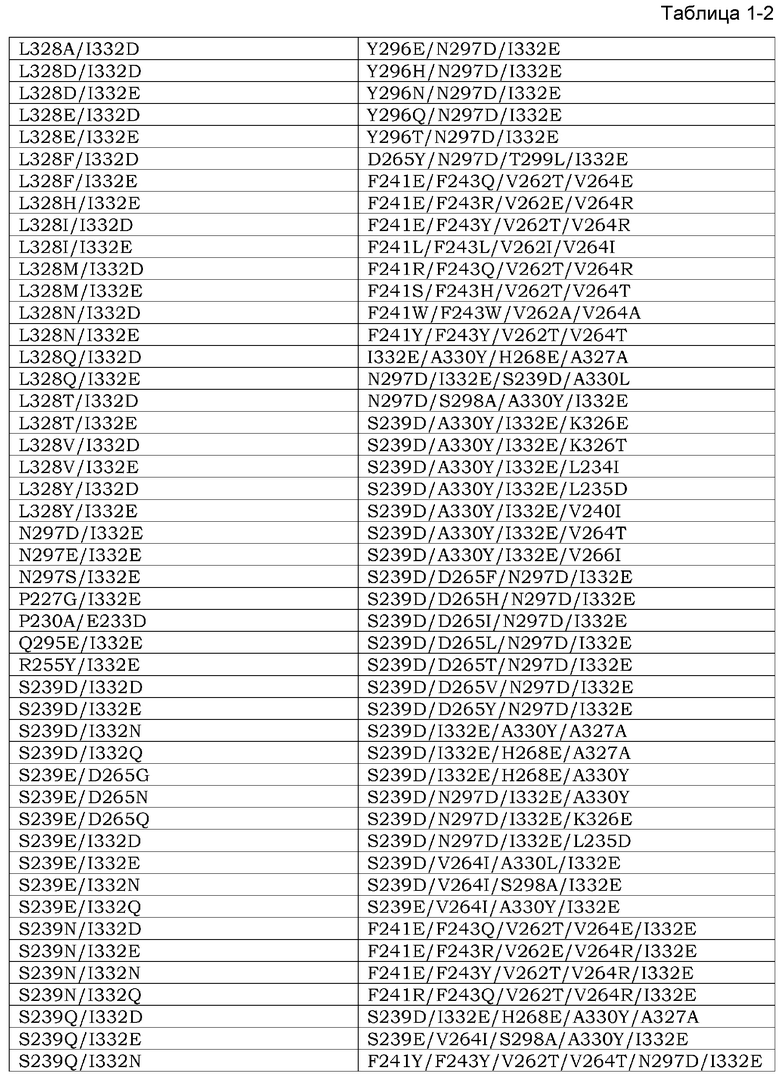
Здесь, фраза “Fcγ-рецептор-связывающая активность Fc-района анти-Эпирегулин-антитела является увеличенной” означает, что активность Fc-района анти-Эпирегулин-антитела с вышеупомянутыми изменениями аминокислот для связывания любых из Fcγ-рецепторов, FcγRI, FcγRIIa, FcγRIIb, FcγRIIIa и/или FcγRIIIb, является большей, чем активность Fc-района анти-Эпирегулин-антитела перед изменением аминокислот для связывания этих Fcγ-рецепторов человека. Например, на основе вышеупомянутого способа анализа, это означает, что связывающая активность анти-Эпирегулин-антитела после изменения равна 105% или более, предпочтительно 110% или более, 115% или более, 120% или более, 125% или более, особенно предпочтительно 130% или более, 135% или более, 140% или более, 145% или более, 150% или более, 155% или более, 160% или более, 165% или более, 170% или более, 175% или более, 180% или более, 185% или более, 190% или более, 195% или более, в два раза или более, в 2,5 раза или более, в 3 раза или более, в 3,5 раза или более, в 4 раза или более, в 4,5 раза или более, в 5 раз или более, в 7,5 раз или более, в 10 раз или более, в 20 раз или более, в 30 раз или более, в 40 раз или более, в 50 раз или более, в 60 раз или более, в 70 раз или более, в 80 раз или более, в 90 раз или более или в 100 раз или более в сравнении со связывающей активностью анти-Эпирегулин-антитела перед изменением, используемым в качестве контроля.
(2) Анти-Эпирегулин-антитела с уменьшенным содержанием фукозы в сахарных цепях, присоединенных к Fc-району
Один неограничивающий вариант анти-Эпирегулин-антитело по данному изобретению предпочтительно включает в себя анти-Эпирегулин-антитела, которые были модифицированы таким образом, что эта композиция сахарных цепей, присоединенных к Fc-районам анти-Эпирегулин-антитела, будет высокой в фукоза-недостаточных сахарных цепях. Квалифицированные в данной области специалисты могут подходящим образом отбирать анти-Эпирегулин-антитела с высокой долей Fc-района, связанной с фукоза-недостаточными сахарными цепями, посредством способа для анализа структур сахарных цепей, описанных ниже. Доля таких Fс-районов, связанных с фукоза-недостаточными сахарными цепями, может быть выбрана квалифицированными в данной области специалистами подходящим образом из 10%-100%. В одном неограничивающем варианте осуществления, эта доля может быть выбрана из 10%, 20%, 30%, 40%, 60%, 70%, 80%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% и 100%. Известно, что при удалении остатка фукозы из N-ацетилглюкозамина на редуцирующем конце N-гликозид-связанной в виде комплекса сахарной цепи, присоединенной к Fc-району, аффинность в отношении FcγRIIIa увеличивается (J. Biol. Chem. (2003) 278, 3466-3473). Поскольку известно, что IgG1-антитела, содержащие такие Fc-районы, имеют увеличенную ADCC-активность, как описано ниже, эти Fc-район-содержащие анти-Эпирегулин-антитела применимы также в качестве анти-Эпирегулин-антител, подлежащих включению в фармацевтические композиции по данному изобретению. Один неограничивающий вариант антител, в котором остаток фукозы удален из N-ацетилглюкозамина на редуцирующем конце N-гликозид-связанной в виде комплекса сахарной цепи, присоединенной к Fc-району, предпочтительно включает в себя модифицированное гликозилированием антитело (WO 1999/054342).
Другой неограничивающий вариант антитела, в котором остаток фукозы удален из N-ацетилглюкозамина на редуцирующем конце N-гликозид-связанной в виде комплекса сахарной цепи, присоединенной к Fc-району антитела, предпочтительно включает в себя антитела, которые являются недостаточными по фукозе, присоединенной к сахарным цепям (например, WO 2000/061739, WO 2002/031140 и WO 2006/067913). Клетки-хозяева, которые имеют низкую способность добавлять фукозу к сахарным цепям вследствие модификаций их активности в образовании структур сахарных цепей для полипептидов, подлежащих гликозилированию, используют для получения антител, недостаточных по фукозе, присоединенной к сахарным цепям. Посредством экспрессии желаемого гена антитела в этих клетках-хозяевах, антитело, которое является недостаточным по фукозе, может быть собрано из культуры клеток-хозяев. Неограничивающие предпочтительные примеры активности для образования структуры сахарных цепей полипептида включают в себя активность фермента или транспортера, выбранного из группы, состоящей из фукозилтрансферазы (EC 2.4.1.152), транспортера фукозы (SLC35C1), GMD (GDP-маннозо-4,6-дегидратазы) (ЕС 4.2.1.47), Fx (GDP-кето-6-дезоксиманнозо-3,5-эпимеразы) (ЕС-1.1.1.271) и GFPP (GDP-β-L-фукозо-пирофосфорилазы (EC-2.7.7.30). Структуры этих ферментов и транспортеров указываются необязательно, пока их активности не проявляются. Здесь, белки, которые могут проявлять эти активности, называют функциональными белками. Один неограничивающий вариант способа для модификации этих активностей включает потерю этих активностей. Для получения клеток-хозяев, недостаточных в этих активностях, известные способы, такие как способы разрушения генов этих функциональных белков, чтобы сделать их дисфункциональными, могут быть использованы подходящим образом (например, WO 2000/061739, WO 2002/031140 и WO 2006/067913). Клетки-хозяева, недостаточные по таким активностям, могут быть получены, например, способом, который разрушает эндогенные гены этих функциональных белков в клетках СНО, клетках BHK, клетках NS0, клетках SP2/0, YO миеломных клетках, Р3Х63 мышиных миеломных клетках, клетках PER, клетках RER.С6, клетках HEK293, гибридомных клетках и т.п., так что они неспособны функционировать.
Сбором антител из культуральных супернатантов вышеупомянутых клеток-хозяев, трансформированных экспрессирующим вектором, содержащим ген анти-Эпирегулин-антитела, собирают анти-Эпирегулин-антитела, в которых остаток фукозы удален из N-ацетилглюкозамина на редуцирующем конце N-гликозид-связанной в виде комплекса сахарной цепи, присоединенной к Fc-району антитела.
(3) Анти-Эпирегулин-антитела, содержащие Fc-районы, соединенные с делящим пополам (двурассеченным) N-ацетилглюкозамином
Один неограничивающий вариант анти-Эпирегулин-антител по данному изобретению предпочтительно включает в себя анти-Эпирегулин-антитела, содержащие Fc-районы, соединенные с делящим пополам N-ацетилглюкозамином. Антитела, имеющие сахарную цепь, содержащую делящую пополам структуру GlcNAc (WO 2002/079255 и т.п.), являются известными. Клетки-хозяева, имеющие активность в образовании сахарной цепи, содержащей делящую пополам структуру, используют для получения антител, к которым присоединены сахарные цепи, содержащие GlcNAc-структуру. В одном предпочтительном неограничивающем варианте, получали клетки-хозяева, экспрессирующие ген, кодирующий функциональный белок, имеющий активность GnTIII (β-1,4-маннозил-гликопротеина, 4-β-N-ацетилглюкозаминилтрансферазы) (EC.4.1.144) или активность GalT (β-1,4-галактозилтрансферазы) (EC 2.4.1.38). В другом предпочтительном неограничивающем варианте, продуцируются клетки-хозяева, которые коэкспрессируют ген, кодирующий функциональный белок, имеющий активность ManII (манозидазы II) (3.2.1.114), ген, кодирующий функциональный белок, имеющий активность GnTI (β-1,2-ацетилглюкозаминилтрансферазы I) (EC 2.4.1.94), ген, кодирующий функциональный белок, имеющий активность GnTII (β-1,2-ацетилглюкозаминилтрансферазы II) (EC 2.4.1.143), ген, кодирующий функциональный белок, имеющий активность ManI (маннозидазы) (EC 3.2.1.113), и α-1,6-фукозилтрансферазу (ЕС 2.4.1.68), в дополнение к вышеупомянутым функциональным белкам (WO 2004/065540). Вышеупомянутые клетки-хозяева, имеющие активность образования сахарной цепи, содержащей делящую пополам структуру GlcNAc, могут быть получены трансфекцией экспрессирующего вектора, содержащего ген для вышеупомянутого функционального белка, в клетки СНО, клетки BHK, клетки NS0, клетки SP2/0, YO миеломные клетки, Р3Х63 мышиные миеломные клетки, клетки PER, клетки RER.С6, клетки HEK293, гибридомные клетки и т.п.
Анти-Эпирегулин-антитела, содержащие Fc-районы, соединенные с делящим пополам N-ацетилглюкозамином, собирают сбором антител из супернатанта культуры вышеупомянутых клеток-хозяев, трансфицированных экспрессирующим вектором, содержащим ген анти-Эпирегулин-антитела. Известные способы могут быть использованы для анализа структуры сахарной цепи, присоединенной к этим антителам. В одном неограничивающем варианте, позволение N-гликозидазе F (Roche diagnostics) действовать на анти-Эпирегулин-антитело по данному изобретению, заставляет сахарную цепь присоединяться к этому анти-Эпирегулин-антителу для диссоциации от этого белка (J. Pharm. Sci. (1994) 83(12), 1670-1675). После удаления белка с использованием этанола (J. Clin. Invest. (2001), 108(11), 1687-95), свободную сахарную цепь концентрировали и сушили и затем проводили флуоресцентное мечение с использованием 2-аминопиридина или 2-аминобензамида (Anal. Biochem. (1995) 230(2), 229-238). Полученную 2-АР-меченую сахарную цепь или 2-АB-меченую сахарную цепь освобождают от реагентов экстракцией твердой фазы с использованием целлюлозного картриджа и затем концентрируют центрифугированием, и полученный материал подвергают последующим анализам в виде очищенной 2-АB-меченой сахарной цепи. Затем позволением β-галаксидазе (Seikagaku Corporation) действовать на очищенной 2-АВ-меченой сахарной цепи, готовят агалактозил-2-АВ-меченую сахарную цепь. Эту агалактозил-2-АВ-меченую сахарную цепь, полученную с использованием сахарных цепей, диссоциированных из анти-Эпирегулин-антител по данному изобретению, в качестве исходных материалов, анализируют посредством нормальной фазы высокоэффективной жидкостной хроматографии (ВЭЖХ) с использованием амидной колонки TSKgel Amide-80 (Tosoh) и их хроматограммы сравнивают.
Кроме того, сообщалось, что сила взаимодействия между Fc-районом и FсgR зависит от концентрации Zn2+ (Immunology Letters 143 (2012) 60-69). Это антитело обнаруживает более сильное взаимодействие между Fc-районом и FcgR, когда концентрация иона Zn2+ Fc-района является более высокой. Хелатообразование Zn2+ с использованием His310 и His435, присутствующих в СН3 Fc-района этого антитела, открывает каждый домен СН2 Fc-района в дистальном положении. Это облегчает взаимодействие между доменом СН2 и FcgR и это взаимодействие между FcgR увеличивается. Один неограничивающий вариант антитела по данному изобретению включает в себя антитело, в котором His в положении 310, His в положении 435, His в положении 433 и/или Asn в положении 434 (EU-нумерация) хелатируется Zn2+.
Конъюгат антитело-лекарственное средство
Один неограничивающий вариант анти-Эпирегулин-антител по данному изобретению включает в себя конъюгаты анти-Эпирегулин-антитело-лекарственное средство, получаемые связыванием анти-Эпирегулин-антитела с лекарственным средством, имеющим цитотоксичность. Здесь, далее, конъюгат антитело-лекарственное средство может называться как ADC. Более конкретно, конъюгаты анти-Эпирегулин-антитело-лекарственное средство, используемые в данном изобретении, могут быть связаны подходящим образом с ингибиторами роста или цитотоксическими веществами, такими как токсичные пептиды или радиоактивные химические вещества. Термин ʺрадиоактивные химические веществаʺ в данном изобретении относится к веществам, содержащим радиоизотоп. Этот радиоизотоп не ограничивается конкретно и, хотя может быть использован любой изотоп, могут быть использованы 32Р, 14С, 125I, 3Н, 131I, 186Re, 188Re и т.п. Такие конъюгаты анти-Эпирегулин-антитело-лекарственное средство могут быть получены химическим модифицированием полученных анти-Эпирегулин-антител. Более конкретно, для позволения ингибитору роста или цитотоксическому веществу химически конъюгировать (например, ковалентно связываться) с анти-Эпирегулин-антителом, используют линкерную молекулу для связывания ингибитора роста или цитотоксического вещества с антителом через химические связи (такие как описанные выше).
Предпочтительно, этот связывающий агент (линкер) является расщепляемым линкером. Более предпочтительно, этот линкер расщепляется при мягких условиях (т.е. условиях в клетке, при которых не затрагивается лекарственное средство). Примеры подходящих расщепляемых линкеров включают в себя дисульфидные линкеры, кислотолабильные линкеры, фотолабильные линкеры, пептидаза-лабильные линкеры и эстераза-лабильные линкеры. Дисульфидсодержащие линкеры являются линкерами, которые могут расщепляться через дисульфидный обмен, который может осуществляться при физиологических условиях. Кислотолабильные линкеры являются линкерами, расщепляемыми при кислотном рН. Например, определенные внутриклеточные компартменты, такие как эндосомы и лизосомы, имеют кислотный рН (рН 4-5) и обеспечивают условия, подходящие для расщепления кислотолабильных линкеров. Фотолабильные линкеры применимы на поверхности тела и во многих полостях тела, которые могут экспонироваться свету. Кроме того, инфракрасный свет может проникать в ткани. Пептидаза-лабильные линкеры могут быть использованы для расщепления некоторых пептидов внутри или снаружи клеток (см., например, Trouet et al., Proc. Natl. Acad. Sci. USA (1982) 79, 626-629; и Umemoto et al., Int. J. Cancer (1989) 43, 677-684).
Кроме вышеупомянутых химических модификаций, такие ADC могут быть получены в форме молекул биспецифического антитела, конструированных с использованием способов генетической инженерии таким образом, что это антитело может распознавать ингибиторы роста или цитотоксические вещества, такие как токсичные пептиды или радиоактивные химические вещества. Термин ʺанти-Эпирегулин-антителаʺ по данному изобретению включает в себя также такие антитела.
Примеры неограничивающего варианта конъюгата анти-Эпирегулин-антитело-лекарственное средство, обеспеченного данным изобретением, включают в себя анти-Эпирегулин-антитела, которые были модифицированы с использованием токсичного пептида, такого как рицин, абрин, рибонуклеаза, онконаза, ДНКказа I, энтеротоксин А Staphylococcus, антивирусный белок фитолакки американской, гелонин, дифтерийный токсин, экзотоксин Pseudomonas, эндотоксин Pseudomonas, L-аспарагиназа или PEG L-аспарагиназа. В другом варианте осуществления один, два или более из ингибиторов роста и цитотоксических веществ, таких как токсичный пептид, могут быть комбинированы и использованы для модификации анти-Эпирегулин-антитела. Как описано выше, анти-Эпирегулин-антитело может быть связано с вышеупомянутыми ингибитором роста или цитотоксичным веществом, таким как токсичный пептид или радиоактивное химическое вещество, через ковалентную связь или нековалентную связь. Способы для получения ADC, связанных с таким фактором роста или цитотоксическим веществом, таким как токсичный пептид или радиоактивное вещество, известны в данной области. Примеры связывающих групп, когда анти-Эпирегулин-антитело и ингибитор роста или цитотоксическое вещество, такое как токсичный пептид или радиоактивное вещество, непосредственно связаны вместе, включают в себя дисульфидную связь, использующую SH-группы. Конкретно, внутримолекулярная дисульфидная связь в Fc-районе анти-Эпирегулин-антитела восстанавливается восстанавливающим агентом, таким как дитиотреитол, и дисульфидная связь в молекуле ингибитора роста или цитотоксическом веществе восстанавливается, так что эти два компонента связываются этой дисульфидной связью. Перед связыванием либо это антитело, либо один из ингибитора роста или цитотоксического вещества может быть активирован стимулирующим активацию агентом, таким как реагент Эллмана, так что образование этой дисульфидной связи между этим двумя молекулами стимулируется. Предпочтительные неограничивающие примеры других способов для прямого связывания анти-Эпирегулин-антитела и ингибитора роста или цитотоксического вещества, такого как токсичный пептид, включают способ, использующий основание Шиффа, карбодиимидный способ, способ активного сложного эфира (N-гидроксисукцинимидный способ), способ с использованием смешанного ангидрида и способ, использующий диазореакцию.
Следующее может быть примером токсичных пептидов, используемых в данном изобретении.
- А-цепь дифтерийного токсина (Langone et al. (Methods in Enzymology (1993) 93, 307-308))
- Экзотоксин Pseudomonas (Pai et al. (Nat. Med (1996) 2(3) 350-353))
- Цепь рицина (А-цепь рицина)(Fulton et al. (J. Biol. Chem. (1986) 261, 5314-5319)); Sivam et al. (Cancer Res. (1987) 47, 3169-3173); Cumber et al. (J. Immunol. Methods (1990) 135, 15-24); Wawrzynczak et al. (Cancer Res. (1990) 50, 7519-7562); Gheeite et al. (J. Immunol. Methods (1991) 142, 223-230))
- Цепь А дегликозилированного рицина (Thrope et al., (Cancer Res. (1987) 47, 5924-5931))
- Цепь А Абрина (Wawrzynczak et al. (Br. J. Cancer (1992) 66, 361-366); Wawrzynczak et al. (Cancer Res (1990) 50, 7519-7562); Sivam et al. (Cancer Res. (1987) 47, 3169-3173); Thrope et al. (Cancer Res. (1987) 47, 5924-5931))
- Гелонин (Sivam et al. (Cancer Res. (1987) 47, 3169-3173); Cumber et al. (J. Immunol. Methods (1990) 135, 15-24); Wawrzynczak et al. (Cancer Res. (1990) 50, 7519-7562); Bolognesi et al. (Clin. Exp. Immunol. (1992) 89, 341-346))
- Антивирусный белок Pap-s или фитолакки американской из семян (Bolognesi et al. (Clin. Exp. Immunol. (1992) 89, 341-346))
- Бриодин (Bolognesi et al. (Clin. Exp. Immunol. (1992) 89, 341-346))
- Сапорин (Bolognesi et al. (Clin. Exp. Immunol. (1992) 89, 341-346))
- Момордин (Cumber et al. (J. Immunol. Methods (1990) 135, 15-24); Wawrzynczak et al. (Cancer Res. (1990) 50, 7519-7562); Bolognesi et al. (Clin. Exp. Immunol. (1992) 89, 341-346))
- Моморкохин (Bolognesi et al. (Clin. Exp. Immunol. (1992) 89, 341-346))
- Диантин 32 (Bolognesi et al. (Clin. Exp. Immunol. (1992) 89, 341-346))
- Диантин 30 (Stirpe et al. (FEBS Let. (1986) 195, 1-8))
- Модекцин (Stirpe et al. (FEBS Let. (1986) 195, 1-8))
- Вискумин (Stirpe et al. (FEBS Let. (1986) 195, 1-8))
- Волкесин (Stirpe et al. (FEBS Let. (1986) 195, 1-8))
- Додекандрин (Stirpe et al. (FEBS Let. (1986) 195, 1-8))
- Тритин (Stirpe et al. (FEBS Let. (1986) 195, 1-8))
- Луффин (Stirpe et al. (FEBS Let. (1986) 195, 1-8))
- Трихокирин (Casellas et al. (Eur. J. Biochem. (1988) 176, 581-588); Bolognesi et al. (Clin. Exp. Immunol. (1992) 89, 341-346))
Белковые или пептидные фармацевтические агенты или токсины могут быть также связаны с анти-Эпирегулин-антителом способами генетической инженерии. Конкретно, ДНК, кодирующая вышеупомянутый токсичный пептид, и ДНК, кодирующая анти-Эпирегулин-антитело по данному изобретению, могут быть слиты в рамке считывания для получения рекомбинантной ДНК и затем инерсертированы в экспрессирующий вектор для конструирования рекомбинантного вектора. Затем этот вектор вводят в подходящие клетки-хозяева для генерирования трансформированных клеток, которые экспрессируют инсертированную ДНК в этих клетках. Конъюгат анти-Эпирегулин-антитело-лекарственное средство, с которым связан токсичный пептид, может быть получен выделением и очисткой из культурального раствора. При получении слитых белков этого антитела обычно соответствующие ДНК связывают таким образом, что белковые лекарственные средства и токсины размещаются на С-конце этого антитела, но не ограничиваются этим. Можно размещать пептидный линкер между этим антителом и белковым фармацевтическим агентом или токсином.
Ингибиторы роста
В одном неограничивающем варианте осуществления ингибиторы роста могут предпочтительно включать в себя разрушающие ДНК агенты, антимитотические агенты и/или антиметаболиты. Разрушающими ДНК агентами могут быть алкилирующие агенты, ингибиторы топоизомеразы и/или интеркаляторы ДНК. Предпочтительные неограничивающие примеры ингибиторов роста могут включать в себя карбоплатин (ДНК-алкилирующий агент), этопозид (ингибитор топизомеразы II), доксорубицин (интеркалятор ДНК), доцетаксел (антимитотический агент) и Gemzar (гемцитабин, антиметаболит).
По меньшей мере один из приведенных ниже может быть выбран алкилирующим агентом. То есть по меньшей мере один алкилирующий агент, выбранный из хлорамбуцила, циклофосфамида, ифосфамида, мехлорэтамина, мелфалана, урацилпроизводного горчичного газа (иприта), тиотепа, бусульфана, кармустина, ломустина, стрептозоцина, карбоплатина, цисплатина, сатраплатина, оксалиплатина, алтретамина, ET-743, XL119 (бекатекарина), дакарбазина, хлорметина, бендамустина, трофосфамида, урамустина, фотемустина, нимустина, преднимустина, ранимустина, семустина, недаплатина, тетранитрата триплатина, манносульфана, треосульфана, темозоломида, карбоквона, триазиквона, триэтиленмеламина и прокарбазина, может быть использован в качестве алкилирующего агента.
По меньшей мере один из приведенных ниже может быть выбран в качестве ингибитора топоизомеразы. То есть по меньшей мере один ингибитор топоизомеразы, выбранный из доксорубицина, (Doxil), даунорубицина, эпирубицина, идарубицина, антрацендиона (Novantrone), митоксантрона, митомицина С, блеомицина, дактиномицина, пликатомицина, илинотекана (Camptosar), камптотецина, рубитекана, белотекана, этопозида, тенипозида и топотекана (Hycamptin), может быть использован в качестве ингибитора топоизомеразы.
По меньшей мере один инкалятор ДНК, выбранный из профлавина, доксорубицина (адриамицина), даунорубицина, дактиномицина и талидомида, может быть использован в качестве интеркалятора ДНК.
По меньшей мере один из приведенных ниже может быть выбран в качестве антимитотического агента. По меньшей мере один антимитотический агент, выбранный из паклитаксела (Abraxane)/Таксола, доцетаксела (Taxotere), BMS-275183, Xyotax, Tocosal, винорлебина, винкристина, винбластина, виндесина, (VM-26, винзолидина, этопозида (VP-16), тенипозида, иксабепилона, ларотаксела, ортатаксела, тезетаксела и испинесиба, может быть использован в качестве антимитотического агента.
По меньшей мере один из приведенных ниже может быть выбран в качестве антиметаболита. По меньшей мере один метаболит, выбранный из фторурацила (5-FU), флоксуридина (5-FUdR), метотрексата, Xeloda, Arranon, лейковорина, гидроксимочевины, тиогуанина (6-TG), меркаптопурина (6-MP), цитарабина, пентостатина, флударабинфосфата, кладрибина (2-CDA), аспарагиназы, гемцитабина, пеметрекседа, бортезомиба, аминоптерина, ралтитрекседа, клофарабина, эноцитабина, сапацитабина, азацитидина и т.п., может быть использован в качестве антиметаболита.
При включении конъюгата анти-Эпирегулин-антитело-лекарственное средство в клетку, конъюгат-связанный ингибитор роста или цитотоксическое вещество, такое как токсичный пептид, может индуцировать смерть клетки, которая включает это антитело. Таким образом, антитело, с которым связаны предпочтительно ингибитор роста или цитотоксическое вещество, такое как токсичный пептид, также имеют активность интернализации. В данном изобретении термин ʺантитело, имеющее активность интернализацииʺ относится к антителу, которое транспортируется в клетку (например, в цитопазму, везикулы, другие органеллы и т.п.) после связывания с Эпирегулином на клеточной поверхности. Имеет или не имеет антитело активность интернализации, может быть подтверждено с использованием способов, известных квалифицированным в данной области специалистам. Например, активность интернализации может быть подтверждена способом контактирования анти-Эпирегулин-антитела, связанного с меченым веществом, с Эпирегулин-экспрессирующими клетками, и определения, включается ли это меченое вещество в эти клетки, или способом контактирования анти-Эпирегулин-антитела, связанного с ингибитором роста или цитотоксическим веществом, таким как токсичный пептид, с Эпирегулин-экспрессирующими клетками, и определения, индуцируется ли смерть клеток в этих Эпирегулин-экспрессирующих клетках. Более конкретно, имеет или не имеет антитело активность интернализации, может быть подтверждено, например, способом, описанным в описанных далее в Примерах.
После того как антитело, имеющее цитотоксичность, такую как ADCC, связывается с антигеном-мишенью, экспрессируемым на клеточной поверхности, это антитело остается на этой клеточной поверхности посредством связывания с этим антигеном, и эффекторные клетки, такие как NK-клетки, связываются с Fc-районами этого антитела, и это заставляет эти эффекторные клетки индуцировать цитотоксичность против клеток, экспрессирующих этот антиген-мишень. В противоположность этому, антитело, которое используется в качестве ADC, предпочтительно интернализуется в клетки после связывания с этим антигеном. Как можно определить из вышеупомянутого механизма действия, обычно считается, что, предпочтительно, антитело, которое повреждает клетки-мишени посредством цитотоксичности, такой как ADCC, имеет низкую активность интернализации, а антитело, которое повреждает клетки-мишени в форме ADC, имеет высокую активность интернализации (Anticancer Research (1993) 13, 2207-2212). Неожиданно, в противоположность этому, анти-Эпирегулин-антитела по данному изобретению не только ослабляют пролиферацию Эпирегулин-экспрессирующих клеток их нейтрализующей активностью, но также индуцируют повреждение Эпирегулин-экспрессирующих клеток цитотоксичностями, такими как ADCC; и, между тем, было обнаружено, что конъюгаты, содержащие анти-Эпирегулин-антитела по данной заявке, индуцируют цитотоксичность против Эпирегулин-экспрессирующих клеток.
Низкомолекулярное антитело
В одном неограничивающим варианте анти-Эпирегулин-антитела, включенные в конъюгат анти-Эпирегулин-антитело-лекарственное средство, могут быть низкомолекулярными антителами. Низкомолекулярное антитело содержит фрагмент антитела, лишенный части целого антитела (например, цельного IgG). Пока оно имеет активность связывания антигена Эпирегулина, частичные делеции молекулы этого антитела являются пермиссивными. Фрагменты антител по данному изобретению предпочтительно содержат вариабельный домен тяжелой цепи (VH) и/или вариабельный район легкой цепи (VL). Аминокислотная последовательность VH или VL может иметь замены, делеции, добавления и/или инсерции. Кроме того, пока она имеет активность связывания антигена Эпирегулина, VH и VL могут быть частично делетированными. Вариабельный район может быть химеризованным или гуманизированным. Специфические примеры этих фрагментов антител включают в себя Fab, Fab', F(ab')2 и Fv. Специфические примеры низкомолекулярных антител включают в себя Fab, Fab', F(ab')2, Fv, scFv (одноцепочечный Fv), диатело и sc(Fv)2 (одноцепочечный (Fv)2). Мультимеры этих антител (например, димеры, тримеры, тетрамеры и полимеры) также включены в антитела с низкомолекулярной массой по этому изобретению.
Низкомолекулярная масса антител может быть получена обработкой антитела ферментом для получения фрагментов антител. Известными ферментами, которые продуцируют фрагменты антител, являются, например, папаин, пепсин и плазмин. Альтернативно, гены, кодирующие эти фрагменты антител, могут быть инсертированы в экспрессирующие векторы; подходящие клетки-хозяева с введенными экспрессирующими векторами культивируют для экспрессии фрагмента антитела; и затем фрагмент антитела может быть получен выделением из культуральной среды и их очисткой (см., например, Co et al., (J. Immunol. (1994) 152, 2968-2976); Better et al., (Methods in Enzymology (1989) 178, 476-496); Plueckthun et al., (Methods in Enzymology (1989) 178, 476-496); Lamoyi ((Methods in Enzymology (1989) 121, 652-663); Rousseaux et al., (Methods in Enzymology (1989) 121, 663-669); Bird et al., (TIBTECH (1991) 9, 132-137)).
Термин диатело относится к бивалентному фрагменту антитела, сконструированному слиянием генов (Hollinger et al., Proc. Natl. Acad. Sci. USA (1993) 90: 6444-6448; EP 404097; WO 1993/11161; и т.п.). Диатело является димером, составленным из двух полипептидных цепей. Обычно, в каждой полипептидной цепи, составляющей димер, VL и VH связаны линкером в одной и той же цепи. Линкер в диателе является обычно достаточно коротким для предотвращения связывания между VL и VH. Конкретно, аминокислотные остатки, составляющие линкер, имеют, например, пять остатков или близкое количество остатков. Таким образом, VL и VH, которые кодируются одной и той же полипептидной цепью, не могут образовывать одноцепочечный фрагмент вариабельного района и образуют димер с другим фрагментом одноцепочечного вариабельного района. В результате, диатела имеют два антигенсвязывающего сайта. scFv может быть получен лигированием V-района Н-цепи и V-района L-цепи антитела. Конкретно, scFv может быть приготовлен лигированием V-района Н-цепи и V-района L-цепи через линкер, предпочтительно пептидный линкер (Huston et al., Proc. Natl. Acad. Sci. U.S.A., (1988) 85, 5879-5883). V-район Н-цепи и V-район L-цепи scFv могут быть произведены из любого из антител, описанных здесь в качестве анти-Эпирегулин-антител. Структура и свойства пептидного линкера для лигирования V-районов особо не ограничивается. Например, любой одноцепочечный пептид, состоящий из 3-25 остатков или т.п., может быть использован в качестве линкера.
sc(Fv)2 является низкомолекулярным антителом, получаемым лигированием двух VH и двух VL линкерами или т.п. для образования одноцепочечной цепи (Hudson et al., J. Immunol Methods (1999) 231, 177-189). sc(Fv)2 может быть получен, например, связыванием scFv с линкером.
Фамацевтические композиции
В другом аспекте данное изобретение обеспечивает фармацевтические композиции, содержащие анти-Эпирегулин-антитело по данному изобретению в качестве активного ингредиента. Кроме того, данное изобретение относится к ингибиторам роста клеток, в частности, противораковым агентам для супрессии рецидива и метастазирования рака, которые содержат анти-Эпирегулин-антитело по данному изобретению в качестве активного ингредиента. Предпочтительно, ингибиторы роста клеток или агенты для ослабления рецидива или метастазирования рака вводят субъекту, который страдает от рака или может страдать от рака в будущем.
В данном изобретении ингибиторы роста клеток, содержащие анти-Эпирегулин-антитело в качестве активного ингредиента, могут быть также экспрессированы в качестве способов для ослабления роста клеток, которые включают введение анти-Эпирегулин-антитела субъекту; применения анти-Эпирегулин-антитела в получении ингибиторов роста клеток; или анти-Эпирегулин-антител для использования в ослаблении роста клеток.
Кроме того, в данном изобретении, противораковые агенты или агенты для ослабления рецидива или метастазирования рака, содержащие анти-Эпирегулин-антитело в качестве активного ингредиента, могут также экспрессироваться в качестве способов для предотвращения или лечения рака или ослабления рецидива или метастазирования рака, которые включают стадию введения анти-Эпирегулин-антитела субъекту; применения анти-Эпирегулин-антитела в получении противораковых агентов или агентов для ослабления рецидива или метастазирования рака; или анти-Эпирегулин-антител для применения в предотвращении или лечении рака или ослаблении рецидива или метастазирования рака.
В данном изобретении термин ʺсодержащее анти-Эпирегулин-антитело в качестве одного активного ингредиентаʺ обозначает содержание анти-Эпирегулин-антитела в качестве главного активного ингредиента, и отсутствие ограничения в отношении содержания анти-Эпирегулин-антитела. Эти антитела, включенные в фармацевтические композиции по данному изобретению (например, ингибиторы роста клеток и противораковые агенты по данному изобретению; и те же самые здесь далее), не являются особо ограничивающими, пока они связываются с белком Эпирегулином, и могут быть иллюстрированы антителами, описанными здесь.
Фармацевтические композиции по данному изобретению могут вводиться субъектам (пациентам) или перорально, или парентерально. Предпочтительным является парентеральное введение. Такие способы введения специфически включают введение инъекцией, трансназальное введение, легочное введение и чрескожное введение. Для введения инъекцией, фармацевтическая композиция по данному изобретению может вводиться системно или локально, например, внутривенной инъекцией, внутримышечной инъекцией, внутрибрюшинной инъекцией и подкожной инъекцией. Способ введения может быть выбран подходящим образом в соответствии с возрастом и симптомами пациента. Доза может быть выбрана, например, в диапазоне от 0,0001 мг до 1000 мг на килограмм массы тела на одно введение. Альтернативно, эта доза может быть выбрана, например, в пределах диапазона от 0,001 мг массы тела до 100000 мг массы тела пациента. Однако фармацевтические композиции по данному изобретению не ограничиваются этими дозам.
Фармацевтические композиции по данному изобретению могут быть образованы в соответствии с общепринятыми способами (например, Remington’s Pharmaceutical Sciences, latest edition, Mark Publishing Company, Easton, U.S.A.) и могут также содержать фармацевтически приемлемые носители и добавки. Примеры включают в себя поверхностно-активные вещества, эксципиенты, красящие агенты, ароматизирующие агенты, консерванты, стабилизаторы, буферы, суспензидирующие агенты, изотонизирующие агенты, связывающие агенты, дезинтеграторы, смазывающие вещества, усиливающие текучесть агенты и корригенты. Без ограничения ими, могут быть подходящим образом использованы другие обычно используемые носители. Конкретные примеры носителей включают в себя слабую безводную кремниевую кислоту, лактозу, кристаллическую целлюлозу, маннит, крахмал, кальций-кармеллозу, натрий-кармеллозу, гидроксипропилцеллюлозу, гидроксипропилметилцеллюлозу, диэтиламиноацетат поливинилацеталя, поливинилпирролидон, желатин, триглицериды жирной кислоты со средней цепью, отверждаемое полиоксиэтиленом касторовое масло 60, сахарозу, карбоксиметилцеллюлозу, кукурузный крахмал, неорганические соли и т.п.
Анти-Эпирегулин-антитела описаны выше как Эпирегулин-белок-связывающие антитела, включенные в фармацевтические композиции по данному изобретению. Клетки, которые связаны анти-Эпирегулин-антителами, конкретно не ограничиваются, пока эти клетки являются Эпирегулин-экспрессирующими клетками. Предпочтительные Эпирегулин-экспрессирующие клетки по данному изобретению являются раковыми клетками. Более конкретно, этими клетками являются раковые клетки ободочной кишки, клетки аденокарциномы легкого, клетки панкреатического рака, клетки рака желудка, эзофагеальные раковые клетки и клетки рака почки. Способы по данному изобретению могут применяться как к первичным, так и к метастатическим очагам этих типов рака. Более предпочтительные раковые клетки являются первичными клетками ободочной кишки, метастатическими клетками рака ободочной кишки и панкреатическими раковыми клетки.
В данном изобретении ʺконтактированиеʺ выполняется, например, добавлением антитела в культуральный раствор Эпирегулин-экспрессирующих клеток, культивируемых в тест-пробирке. В этом случае антитело может быть добавлено в форме, например, раствора или твердого вещества, полученного лиофилизацией или т.п. При добавлении этого антитела в виде водного раствора, этот используемый водный раствор может содержать исключительно только это антитело, или этот раствор может включать в себя, например, вышеупомянутые поверхностно-активные вещества, эксципиенты, красящие агенты, отдушки, консерванты, стабилизаторы, буферы, суспендирующие агенты, изотонизирующие агенты, связывающие агенты, дезинтеграторы, смазывающие вещества, усиливающие текучесть агенты или вкусовые добавки. Концентрация для добавления не ограничивается конкретно, но конечная концентрация в культуре, которая может быть подходящим образом использована, находится предпочтительно в диапазоне 1 пг/мл-1 г/мл, более предпочтительно 1 нг/мл-1 мг/мл и, даже более предпочтительно, 1 мкг/мл-1 мг/мл.
Кроме того, в другом варианте осуществления, ʺконтактированиеʺ в данном изобретении проводится введением антитела животному (не-человеку), которому Эпирегулин-экспрессирующая клетка была трансплантирована в тело, или животному, несущему раковые клетки, эндогенно экспрессирующие Эпирегулин. Способ введения может быть пероральным или парентеральным введением. Этот способ введения является особенно предпочтительно парентеральным введением, и, конкретно, этот способ введения является, например, введением инъекцией, трансназальным введением или трансдермальным введением. Примеры введения инъекцией включают в себя системное и локальное введения фармацевтических композиций, ингибиторов пролиферации клеток и противораковых агентов по данному изобретению внутривенной инъекцией, внутримышечной инъекцией, внутрибрюшинной инъекцией, подкожной инъекцией и т.п. Подходящий способ введения может быть выбран в соответствии с возрастом тест-животного и симптомами. При введении в виде водного раствора, этот используемый водный раствор может исключительно содержать только это антитело, или этот раствор может включать в себя, например, вышеупомянутые поверхностно-активные вещества, эксципиенты, красящие агенты, отдушки, консерванты, стабилизаторы, буферы, суспендирующие агенты, изотонизирующие агенты, связывающие агенты, дезинтеграторы, смазывающие вещества, усиливающие текучесть агенты или вкусовые добавки. Доза может быть выбрана, например, в диапазоне 0,0001 мг-1000 мг на кг массы тела в каждом введении. Альтернативно, например, для каждого пациента может быть выбрана доза в диапазоне 0,001-100000 мг/антитело. Однако доза антитела по данному изобретению не ограничивается этими дозами.
Терапевтические агенты для рака или агенты для ослабления рецидива или метастазирования рака, которые вводят субъектам, несущим Эпирегулин-белок-экспрессирующие раковые клетки
Данное изобретение обеспечивает терапевтические агенты для рака или агенты для ослабления рецидива или метастазирования рака, где эти субъекты, которым вводят терапевтические агенты для рака или агенты для ослабления рецидива или метастазирования рака, являются субъектами, несущими экспрессирующие белок Эпирегулин раковые клетки, которые детектируют с использованием выделенной пробы ткани. Наряду со способами, описанными здесь, уровень экспрессии белка Эпирегулина может быть определен способами, известными квалифицированным в данной области специалистам. Для определения, следует или не следует вводить субъекту терапевтические агенты для рака или агенты для ослабления рецидива или метастазирования рака по данному изобретению, уровень экспрессии белка Эпирегулина может быть определен в пробе ткани, выделенной из кандидатного субъекта. Если в этой пробе детектируется белок Эпирегулин, субъет, из которого получена эта проба, может быть субъектом для введения терапевтических агентов для рака или агентов для ослабления рецидива или метастазирования рака по данному изобретению. Уровень экспрессии белка Эпирегулина не ограничивается конкретной численной величиной, но в одном неограничивающем варианте он может быть установлен подходящим образом из диапазона численных величин порядка 103, 104, 105, 106, 107 и 108. В качестве неограничивающего варианта, уровень экспрессии белка Эпирегулина, выбранный из любого из 1×103, 2×103, 3×103, 4×103, 5×103, 6×103, 7×103, 8×103, 9×103, 10×103, 1×104, 2×104, 3×104, 4×104, 5×104, 6×104, 7×104, 8×104, 9×104, 1×105, 2×105, 3×105, 4×105, 5×105, 6×105, 7×105, 8×105, 9×105, 1×106, 2×106, 3×106, 4×106, 5×106, 6×106, 7×106, 8×106, 9×106, 1×107, 2×107, 3×107, 4×107, 5×107, 6×107, 7×107, 8×107, 9×107, 1×108, 2×108, 3×108, 4×108, 5×108, 6×108, 7×108, 8×108 и 9×108, может быть показан как предпочтительный пример численной величины (пороговой величины), которая может быть установлена в качестве стандарта для выбора субъектов, которые будут получать это введение. Численные величины, упомянутые выше, могут быть величинами, рассчитанными для одной значащей цифры; и в этом случае, например, уровень экспрессии EREG 1×103, рассчитанный для одной значащей цифры, может быть уровнем экспрессии EREG, имеющем численную величину, любую из 0,5×103, 0,6×103, 0,7×103, 0,8×103, 0,9×103, 1,0×103, 1,1×103, 1,2×103, 1,3×103, 1,4×103, при расчете на две значащие цифры.
Пробы ткани
Термин ʺпроба тканиʺ, используемый в данном изобретении, относится к любой биологической пробе, которая может быть получена из индивидуума, жидкости тела (например, крови, сыворотки, плазмы и спинальной жидкости), культуры ткани, срезов ткани или т.п. Пробы из субъектов могут быть использованы в качестве предпочтительных примеров биологических проб. Предпочтительными пробами из субъектов являются ткани, полученные из субъектов, и более предпочтительно ткани ободочной кишки из субъектов. Биопсию, известный способ, предпочтительно используют в качестве способа для сбора тканей ободочной кишки. В качестве способов для биопсии могут быть использованы подходящим образом известные способы. Такие примеры включают в себя получение тканей с использованием биопсии, такой как аспирационная биопсия, зажимная биопсия, скарифицированная биопсия, биопсия для цитодиагностики, эндоскопическая биопсия, тонкоигольная (аспирационная) биопсия, пункционная биопсия, чрезбронхиальная биопсия, или получение тканей во время хирургии.
В данном изобретении, поскольку пробы ткани наблюдают с проходящим светом под микроскопом, их срезают таким образом, что свет, используемый на микроскопе, достаточно проходил через пробы тканей. Перед получением срезов пробы тканей фиксируют. Конкретно, пробы ткани отверждают дегидратацией или денатурацией белков в тканях/клетках для быстрого убивания клеток, составляющих эти ткани. Полученные ткани имеют стабилизированную и нерастворимую структуру. Сначала пробы ткани, подлежащие фиксации, нарезают на фрагменты, имеющие размер и форму, подходящую для приготовления залитых в парафине срезов с использованием режущего ножа, такого как скальпель. Затем эти фрагменты погружают в фиксирующий раствор, реагент, используемый для проведения фиксации. Этим фиксированным используемым раствором является предпочтительно формалин или более предпочтительно нейтральный забуференный формалин. Концентрацию этого нейтрального забуференного формалина подходящим образом выбирают в соответствии с характеристиками или физическими свойствами проб тканей. Эта концентрация может быть изменена подходящим образом и использована между 1% и 50%, предпочтительно между 5% и 25% и более предпочтительно между 10% и 15%. Фиксирующий раствор, в который погружали препараты ткани, подходящим образом деаэрируют с использованием вакуумного насоса. Фиксацию проводят оставлением проб ткани в фиксирующем растворе в течение нескольких часов при условиях нормального давления и комнатной температуры. Время, требуемое для фиксации, может быть выбрано подходящим образом в диапазоне от 1 часа до 7 дней, предпочтительно от 2 часов до 3 дней или предпочтительно от 3 часов до 24 часов, и более предпочтительно от 4 часов до 16 часов. Фиксированные пробы дополнительно погружают подходящим образом в фосфатный буфер или т.п. на несколько часов (это время может быть выбрано подходящим образом в диапазоне от 2 часов до 48 часов, предпочтительно от 3 часов до 24 часов и более предпочтительно от 4 часов до 16 часов).
Затем из этих фиксированных проб тканей могут быть приготовлены срезы предпочтительно с использованием способа получения лиофилизированных срезов или способа изготовления срезов тканей, погруженных (залитых) в парафин. Предпочтительные примеры получения лиофилизированных срезов включают в себя способ, который включает лиофилизацию этих тканей помещением их в O.C.T. соединение (Miles. Inc.) и изготовление срезов проб замороженных тканей с использованием криостата (аппарата для приготовления замороженных срезов). В способе изготовления срезов в парафине фиксированные пробы ткани погружают в заливочный агент, который затем фиксируют для придания посредством этого однородной и подходящей твердости этим срезам. В качестве заливочного агента предпочтительно может быть использован парафин. Фиксированные пробы ткани дегидратируют с использованием этанола. Конкретно, пробы ткани дегидратируют последовательным погружением пробы ткани в 70% этанол, 80% этанол и 100% этанол. Время и количество погружений, требуемое для погружения, может быть выбрано подходящим образом в диапазонах от 1 часа до нескольких дней и один-три раза, соответственно. Кроме того, погружение может выполняться при комнатной температуре или при 4°C. Для погружения (иммерсии) при 4°C предпочтительным является более продолжительное время погружения, например, в течение ночи. Затем эту жидкую фазу заменяют ксилолом, и затем эти пробы ткани заливают в парафин. Время, требуемое для замены ксилолом жидкой фазы, может быть выбрано подходящим образом в диапазоне от одного часа до нескольких часов. В этой процедуре замена может выполняться при комнатной температуре или при 4°C. Для замены при 4°C предпочтительным является более продолжительное время, например, в течение ночи. Время и количество погружений, требуемое для заливки в парафин, может быть выбрано подходящим образом в диапазонах от 1 часа до нескольких часов и один-четыре раза, соответственно. В этой процедуре погружение может выполняться при комнатной температуре или при 4°C. Для погружения при 4°C, предпочтительным является более продолжительное время погружения, например, в течение ночи. Кроме того, пробы ткани могут быть залиты парафином с использованием заливающего в парафин аппарата (например, EG1160, Leica), который автоматически процессирует реакцию заливки в парафин.
Залитые в парафин пробы тканей, как описано выше, присоединяют к основанию для получения ʺблокаʺ, который затем нарезают с использованием микротома до желаемой толщины, выбранной из толщин 1-20 мкм. Тонко нарезанные срезы тканей оставляют стоять на предметном стекле, используемом в качестве прозрачной подложки для связывания. В этом случае, предметные стекла, покрытые 0,01% поли-L-лизином (Sigma) и высушенные, могут быть также использованы для предотвращения отшелушивания срезов ткани. Эти фиксированные срезы ткани сушат на воздухе в течение подходящего периода времени, выбранного от нескольких минут до часа.
Исправление антигена
В способе по данному изобретению реактивность антигена, реактивность антитела которого уменьшалась вследствие фиксации формалином, исправляется. В данном изобретении способ индуцируемого протеазой исправления эпитопа (PIER-способ) применяют к одной из двух проб ткани, тогда как способ индуцируемого нагреванием исправления эпитопа (HIER-способ) применяют к другой пробе. Затем различие в степени окрашивания между ними после реакции антитела оцифровывают.
Способ индуцированного нагреванием исправления эпитопа подходящим образом использует способ нагревания микроволнами (СВЧ-излучением), способ нагревания в автоклаве, способ нагревания обработкой кипячением или т.п. При выполнении обработки кипячением при выходе 780 Вт для поддержания температуры раствора при приблизительно 98°C, время, требуемое для исправления, включающего в себя эту обработку, подходящим образом выбирают между пятью и 60 минутами, и оно составляет, например, десять минут. Обработка исправления антигена может выполняться в 10 мМ натрий-цитратном буфере, а также коммерчески доступном растворе для исправления мишени Target Retrieval Solution (DakoCytomation) или подобном растворе. В Примерах, описанных ниже, используется Target Retrieval Solution. Любой буфер или водный раствор предпочтительно используют, пока эпитоп в этом антигене, узнаваемый антителом Эпирегулина, требует связывания с этим антителом в результате обработки исправления, так что может быть детектирован комплекс антиген-антитело, описанный ниже.
Тип или происхождение протеазы, используемой в способе индуцированного протеазой исправления эпитопа, особо не ограничивается, и обычно доступная протеаза может быть подходящим способом выбрана и использована. Предпочтительные примеры протеазы, подлежащей использованию, включают в себя 0,05% пепсин в 0,01 н хлористоводородной кислоте, 0,1% трипсин, дополнительно содержащий CaCl2 в Трис-буфере при рН 7,6 и 1-50 мг/мл протеазы K в 10 мМ Трис-HCl-буфере при рН 7,8, содержащем 10 мМ ЭДТА и 0,5% ДСН. Кроме того, при использовании протеазы К, рН этого реакционного раствора подходящим образом выбрана между 6,5 и 9,5, и SH-реагент, ингибитор трипсина или ингибитор химотрипсина может быть подходящим образом использован. Протеаза, присоединенная к набору Histofine HER2 (MONO) (Nichrei Biocscience), описанная здесь в Примерах, также включена в такие специфические примеры предпочтительной протеазы. Протеаза-индуцируемое исправление эпитопа обычно выполняют при 37°C. Однако температура реакции может быть изменена подходящим образом в пределах диапазона 25°C-50°C. При выполнении протеаза-индуцированного исправления эпитопа при 37°C, время реакции подходящим образом выбирают, например, из диапазона одна минута-пять часов, и оно равно 15 минутам, 30 минутам, 45 минутам, 1 часу, 2 часам, 3 часам или 4 часам. После завершения обработки исправления, эти обработанные пробы ткани промывают промывочным буфером. PBC (ЗФР, забуференный фосфатом солевой раствор) предпочтительно используют в качестве промывочного буфера. Кроме того, предпочтительно может быть использован Трис-HCl-буфер. Условия промывания обычно заимствуют способ, включающий выполнение 5-минутных промывок при комнатной температуре три раза. Однако время промывания и температура могут быть изменены подходящим образом.
Вещества для детектирования, используемого в данном изобретении, обычно включают в себя антитела, нуклеиновые кислоты и т.п. Например, можно (1) детектировать белок EREG иммунологическим способом с использованием антител EREG, которые могут детектировать белок EREG, и (2) детектировать EREG-кодирующую мРНК с использованием молекулы нуклеиновой кислоты (например, ДНК, РНК или мРНК), комплементарной EREG-кодирующей мРНК, которая может детектировать эту мРНК, или подходящую комплементарную молекулу нуклеиновой кислоты, где это детектирование использует гибридизацию между участвующими молекулами нуклеиновых кислот (например, способ FISH). В данном изобретении предпочтительно используют иммунологические способы, в частности, иммуногистологическое окрашивание.
Реакция проб ткани с антителом Эпирегулина
Проба ткани, подлежащая обработке исправления антигена на основе способа индуцированного нагреванием исправления эпитопа, и проба ткани, подлежащая обработке исправления антигена на основе способа индуцированного протеазой исправления эпитопа, реагируют с антителом Эпирегулина в качестве первичного антитела. Эту реакцию проводят при условиях, подходящих для узнавания эпитопа в этом антигене антителом Эпирегулина и образования комплекса антиген-антитело. Эту реакцию обычно выполняют в течение ночи при 4°C или при 37°C в течение одного часа. Однако условия реакции могут быть изменены подходящим образом в диапазоне, подходящем для узнавания эпитопа в антигене этим антителом и образования комплекса антиген-антитело. Например, температура реакции может быть изменена в диапазоне 4°C-50°C, и время реакции может быть изменено между одной минутой и семью днями. При выполнении этой реакции при низкой температуре, более продолжительное время является предпочтительным. После завершения первичной реакции, пробы ткани промывают в промывочном буфере. PBS (ЗФР) (забуференный фосфатом солевой раствор) используют предпочтительно в качестве промывочного буфера. Кроме того, Трис-HCl-буфер может быть также предпочтительно используемым. Условия промывания обычно заимствуют способ, включающий 5-минутные промывки при комнатной температуре три раза. Однако время промывания и температура могут быть изменены подходящим образом.
Затем пробы ткани, подвергнутые реакции первичного антитела, реагируют с вторичным антителом, которое узнает это первичное антитело. Обычно используют вторичное антитело, н заранее метящим веществом, для визуализации этого вторичного антитела. Предпочтительное метящее вещество включает в себя: флуоресцентные красители, такие как флуоресцеинизотиоцианат (FITC), Cy2 ((Amersham) и Alexa488 (Molecular Probes); ферменты, такие как пероксидаза и щелочная фосфатаза; и коллоидное золото.
Реакцию с вторичным антителом проводят при условиях, подходящих для образования комплекса антиген-антитело антителом Эпирегулина и вторичным антителом, которое узнает это антитело Эпирегулина. Эту реакцию обычно выполняют при комнатной температуре или 37°C в течение от 30 минут до одного часа. Однако условия реакции могут быть изменены подходящим образом в диапазоне, подходящем для образования комплекса антиген-антитело антителом Эпирегулина и вторичным антителом. Например, температура реакции может быть изменена в пределах диапазона 4°C-50°C, и время реакции может быть изменено между одной минутой и семью днями. При проведении этой реакции при низкой температуре предпочтительным является более продолжительное время реакции. После завершения реакции вторичного антитела, пробы ткани промывают промывочным буфером. PBS (ЗФР) (забуференный фосфатом солевой раствор) используют предпочтительно в качестве промывочного буфера. Кроме того, Трис-HCl-буфер может быть также предпочтительно используемым. Условия промывания обычно заимствуют способ, включающий 50-минутные промывки при комнатной температуре три раза. Однако время промывания и температура могут быть изменены подходящим образом.
Затем пробы ткани, подвергнутые реакции вторичного антитела, реагируют с веществом для визуализации метящего вещества. При использовании пероксидазы в качестве метящего вещества для вторичного антитела, пробы ткани инкубируют с реакционным раствором, полученным смешиванием, непосредственно перед инкубированием, равных количеств 0,02% водного раствора пероксида водорода и раствора DAB (диаминобензидина), доведенного до концентрации 0,1% 0,1 М Трис-буфером (рН 7,2). Кроме DAB могут быть выбраны подходящим образом хромогенные субстраты, такие как DAB-Ni и AEC+ (все DAKО). Во время прохождения инкубации степень развития окраски наблюдают под микроскопом во времени. В точке, когда подходящее развитие окраски подтверждается, реакцию визуализации заканчивают погружением пробы ткани в PBS (ЗФР).
При использовании щелочной фосфатазы в качестве метящего вещества для вторичного антитела, пробы ткани инкубируют с BCIP (5-бром-4-хлор-3-индолилфосфат)/NBT (нитро синий тетразолий) (Zymed), раствор субстрата (NBТ при концентрации 0,4 мМ, и BCIP при концентрации 0,38 мМ) растворяют в 50 мМ натрий-карбонатном буфере (рН 9,8), содержащем 10 мМ MgCl2 и 28 мМ NaCl). Кроме того, наряду с BCIP и NBT, Permanent Red (Перманентный красный), Fast Red (Быстрый красный) или Fuchsin+ (Фуксин) (все DAKO) могут быть использованы подходящим образом. Перед этой инкубацией пробы ткани могут быть прединкубированы при комнатной температуре в течение от одной минуты до нескольких часов с 0,1 М Трис-HCl-буфером (рН 9,5), содержащим 0,1 М хлорид натрия, 50 мМ хлорид марганца и хлорид левамизола, который является ингибитором эндогенной щелочной фосфатазы (Nacalai Tesque) в концентрации 1 мМ. Во время прохождения инкубации пробы ткани наблюдают под микроскопом во времени. В точке, где наблюдают остатки пурпурного формазана, конечного продукта реакции, реакцию останавливают промыванием проб ткани водой или добавлением TBS, содержащего 2% поливиниловый спирт. Затем эти пробы ткани промывают TBST (TBS, содержащим 0,1% Твин 20). При использовании золота в качестве метки для вторичного антитела, коллоидное золото визуализируют присоединением металлического серебра к частицам золота усилением серебром. Этот способ усиления серебром известен квалифицированным в данной области специалистам.
Когда любой из флуоресцентных красителей, таких как FITC (флуоресцеинизотиоцианат), Cy2 (Amersham) и Alexa488 (Molecular Probes), используют в качестве вещества для вторичного антитела, стадия реакции для визуализации этого вещества является необязательной. Свет, испускаемый излучением, со светом при длине волны возбуждения этого флуоресцентного вещества, может быть детектируемым подходящим образом с использованием флуоресцентного микроскопа.
В данном изобретении белок Эпирегулин, содержащийся в пробах ткани, выделенных из тест-субъектов, детектируют, как описано выше. Является ли детектируемый белок Эпирегулин, содержащийся в пробах ткани, высоко экспрессируемым в тканях, содержащих раковые клетки, может быть определено оценками в баллах интенсивности окрашивания, которые оцифровывают картины окрашивания, полученные вышеупомянутым иммуногистологическим способом. Следующие критерии могут быть иллюстрированы в качестве неограничивающего варианта преобразования в цифровую форму (оцифровки) картин окрашивания:
- оценка интенсивности окрашивания 0: ни одна или менее 10% опухолевых клеток в тестируемой ткани являются Эпирегулин-положительными клетками;
- оценка интенсивности окрашивания 1: Эпирегулин-положительные клетки составляют 10% или более опухолевых клеток в тестируемой ткани, но имеют слабую интенсивность окрашивания, локализованную в части клеточной мембраны опухоли;
- оценка интенсивности окрашивания 2: Эпирегулин-положительные клетки составляют 30% или более опухолевых клеток в тестируемой ткани, или Эпирегулин-положительные клетки составляют 10% или более опухолевых клеток в тестируемой ткани и имеют умеренную интенсивность окрашивания, локализованную в клеточной мембране опухоли; и
- оценка интенсивности окрашивания 3: Эпирегулин-положительные клетки составляют 60% или более опухолевых клеток в тестируемой ткани, или Эпирегулин-положительные клетки составляют 10% или более опухолевых клеток в тестируемой ткани, и имеют сильную интенсивность окрашивания, локализованную в клеточной мембране опухоли.
Кроме того, в данном изобретении, при определении оценок уровня экспрессии Эпирегулин в пробах ткани, выделенных из тест-субъектов на основе оценок интенсивности окрашивания, которые оцифровывают картины окрашивания, полученные иммуногистологическим способом окрашивания, описанным выше, можно сравнить с изображениями картин окрашивания для предварительно определенных уровней экспрессии белка Эпирегулина, полученных для коррекции детектирования. Для определения этого уровня экспрессии белка Эпирегулина может быть подходящим образом использован известный способ, который использует флуоресцентно меченные гранулы, используемые для получения калибровочной кривой для количественного определения антигена (например, BD Quantibrite PE™).
Является или не является Эпирегулин, содержащийся в пробе ткани, выделенной из тест-субъекта, высокоэкспрессируемым, может быть определено вышеупомянутым способом. Примеры ʺвысокоэкспрессируемыхʺ могут быть выбраны подходящим образом из диапазона 1×104, 2×104, 3×104, 4×104, 5×104, 6×104, 7×104, 8×104, 9×104, 1×105, 2×105, 3×105, 4×105, 5×105, 6×105, 7×105, 8×105, 9×105, 1×106, 2×106, 3×106, 4×106, 5×106, 6×106, 7×106, 8×106, 9×106, 1×107, 2×107, 3×107, 4×107, 5×107, 6×107, 7×107, 8×107, 9×107, в виде количества молекул Эпирегулина, экспрессируемых на клетку.
Все ссылки предыдущего уровня техники, цитируемые в данном описании, включены здесь посредством ссылки.
Примеры
Здесь ниже данное изобретение будет более конкретно описано при помощи Примеров, но оно не ограничивается ими.
Пример 1. Гуманизация химерного антитела ЕР27
1-1. Выбор каркасных последовательностей для гуманизации
Гуманизировали химерное антитело ЕР27 (описанное в WO 2008/047723), содержащее вариабельные районы мыши и константные районы человека. CDR и FR определяли в соответствии с определением Кабата (нумерацией Кабата).
Сначала последовательности вариабельного района антитела человека и последовательности вариабельного района мыши ЕР27 в базе данных сравнивали для выбора FR последовательностей человека ниже, которые могут быть использованы в качестве матрицы гуманизации. В качестве баз данных использовали IMGT Datebase (http://www.imgt.org/) и NCBI GenBank (http:/www.ncbi.nlm.nih.gov.genbank/). Отобранные последовательности FR показаны в Таблице 2.
Затем последовательности вариабельного района гуманизированного антитела конструировали связыванием CDR-последовательностей (Таблица 3, SEQ ID NO: 9-14) H- и L-цепей вариабельного района ЕР27 с FR человека, выбранных выше. Их обозначали как гуманизированная последовательность вариабельного района H-цепи НА (SEQ ID NO: 15) и гуманизированная последовательность вариабельного района L-цепи LA (SEQ ID NO: 16).
1-2. Конструирование гуманизированного вариабельного района HJ ЕР27
Сообщалось, что в последовательности FR1 Н-цепи, выбранной в Таблице 2, аминокислотные остатки в положениях 2, 4, 24, 27 и 29, показанные нумерацией Кабата, способствуют стабилизации структуры антитела образованием верхнего кора (Ewert et al., Methods, 2004 Oct; 34(2): 184-99). Согласно определению Хотиа, аминокислотные остатки в положениях 26-30, показанных нумерацией Кабата, включены в CDR1 (Honegger et al., J. Mol. Biol. (2001) 309, 657-670). На основе вышеуказанной информации, для приготовления гуманизированного антитела, имеющего антигенсвязывающую активность, эквивалентную антигенсвязывающей активности химерного антитела ЕР27, вышеупомянутые аминокислотные остатки восстанавливали с остатками, присутствующими в последовательности вариабельного района мыши ЕР27. Таким образом, гуманизированный FR1 Н-цепи (Таблица 4, SEQ ID NO: 17) был заново сконструирован восстановлением аминокислотных остатков в положениях 2, 24, 27, 28, 29 и 30, показанных нумерацией Кабата, в FR1 H-цепи человека (SEQ ID NO: 1), выбранной в Таблице 2, с аминокислотными остатками, присутствующими в последовательности вариабельного района мыши ЕР27.
Аминокислотный остаток в положении 93, указанный нумерацией Кабата, в последовательности FR3 Н-цепи, выбранный в Таблице 2, является Ala, тогда как этот остаток в последовательности вариабельного района мыши ЕР27 является Val. На основе базы данных IMGT (http:/www.imgt.org/) последовательностей зародышевой линии мыши и человека, было подтверждено, что только малое количество последовательностей содержат Val в соответствующем положении. Считается, что этот Val в положении 93, указанный нумерацией Кабата, в последовательности вариабельного района мыши ЕР27, участвует в связывании антигена. Таким образом, гуманизированный FR3 Н-цепи (Таблица 4, SEQ ID NO: 18) был заново сконструирован заменой аминокислотного остатка в положении 93, указанным нумерацией Кабата, в FR3 Н-цепи человека (SEQ ID NO: 3), выбранной в Таблице 2, с этим остатком, присутствующим в последовательности вариабельного района цепи мыши ЕР27.
Из гуманизированной последовательности HA вариабельного района Н-цепи (SEQ ID NO: 15) была заново сконструирована гуманизированная последовательность HJ вариабельного района Н-цепи (SEQ ID NO: 19), содержащая FR1 (Таблица 4, SEQ ID NO: 17) и FR3 (Таблица 4, SEQ ID NO: 18).
1-3. Конструирование гуманизированных вариабельных районов LB и L18 ЕР27
Аминокислотный остаток в положении 49, указанный нумерацией Кабата, в последовательности FR2 L-цепи, выбранный в Таблице 2, является Tyr, тогда как этот остаток в последовательности вариабельного района мыши ЕР27 является Gln. На основе базы данных IMGT (http:/www.imgt.org/) последовательностей зародышевой линии мыши и человека, было подтверждено, что только малое количество последовательностей содержат Gln в соответствующем положении. Это предполагает возможность того, что Gln в положении 49, содержащийся в последовательности вариабельного района мыши ЕР27, участвует в связывании антигена. На основе этого, этот гуманизированный FR2 L-цепи (Таблица 5, SEQ ID NO: 20) был заново сконструирован заменой аминокислотного остатка в положении 49, указанного нумерацией Кабата, в FR2 L-цепи человека (SEQ ID NO: 6), выбранной в Таблице 2, аминокислотным остатком, присутствующим в последовательности вариабельного района мыши ЕР27.
Сообщалось, что в последовательности FR3 L-цепи, выбранной в Таблице 2, аминокислотный остаток в положении 71, указанном нумерацией Кабата, способствует стабилизации структуры этого антитела образованием верхнего кора ((Ewert et al., Methods, 2004 Oct; 34(2): 184-99). Для получения гуманизированного антитела, имеющего антигенсвязывающую активность, эквивалентную антигенсвязывающей активности химерного антитела ЕР27, для замены вышеуказанного аминокислотного остатка аминокислотным остатком последовательности вариабельного района мыши ЕР27, этот гуманизированный FR3 L-цепи (Таблица 5, SEQ ID NO: 21) был заново сконструирован заменой аминокислотного остатка в положении 71, указанного нумерацией Кабата, в FR3 L-цепи человека (SEQ ID NO: 7), выбранной в Таблице 2, аминокислотным остатком в последовательности вариабельного района мыши ЕР27.
Из гуманизированной последовательности вариабельного района LA (SEQ ID NO: 16) были заново сконструированы гуманизированная последовательность L-цепи вариабельного района LB (SEQ ID NO: 22), содержащая заново сконструированные FR2 и FR3 (Таблица 5, SEQ ID NO: 20 и 21).
Также, из гуманизированной последовательности вариабельного района LA (SEQ ID NO: 16) были заново сконструированы гуманизированная последовательность L-цепи вариабельного района L18 (SEQ ID NO: 24), содержащая заново сконструированный FR2 (Таблица 5, SEQ ID NO: 20), и последовательность L-цепи антитела человека FR3 (Таблица 5, SEQ ID NO: 23), показанная в Accession No. AB06434 (NCBI GenBank).
1-4 Получение последовательности гуманизированного антитела ЕР27
Сначала, для получения кДНК гуманизированного вариабельного района ЕР27, конструировали синтетические олиго-ДНК для Н (HJ)- и L (LB)-цепей. Затем эти синтетические олиго-ДНК смешивали для амплификации фрагмента гена, кодирующего вариабельный район гуманизированного антитела ЕР27, способом, известным квалифицированным в данной области специалистам, таким как ПЦР сборки. Наконец, этот амплифицированный фрагмент клонировали в экспрессирующем векторе подходящего животного и лигировали с геном константного района человека IgG1. Нуклеотидную последовательность полученного экспрессирующего вектора определяли способом, известным квалифицированным в данной области специалистам.
Что касается гетерогенности, образуемой из С-концевой последовательности (G1, SEQ ID NO: 25) Н-цепи IgG-антитела человека, сообщалась делеция остатка лизина на С-концевой аминокислоте и амидирование С-концевой группы делецией двух С-концевых аминокислот, глицина и лизина (Anal. Biochem. 2007 Jan1; 369(1): 75-83). В качестве способа уменьшения гетерогенности, известен способ, который делетирует две аминокислоты на С-конце Н-цепи, т.е. глицин в положении 446 и лизин в положении 447, показанные EU-нумерацией (WO 2009/041613). Желательно, чтобы гетерогенность, образуемая из С-концевой последовательности Н-цепи, также отсутствовала в гуманизированном антителе ЕР27. Таким образом, IgG1-последовательность (G1d, SEQ ID NO: 26), лишенная глицина в положении 446 и лизина в положении 447, показанная EU-нумерацией, в IgG1 человека может быть использована в качестве последовательности константного района. С другой стороны, нативная цепь κ человека (k, SEQ ID NO: 27) может быть использована в качестве последовательности константного района L-цепи. HJ-G1d (SEQ ID NO: 28) и LB-k (SEQ ID NO: 29) получали соответственно в качестве Н-цепи и L-цепи для последовательности гуманизированного антитела ЕР27, полученного, как описано выше. Для константного района IgG1 сообщались аллотипы (Jefferis et al., mAb 1:4, 1-7; July/August 2009. Кроме вышеупомянутого константного района G1m17,1-типа (SEQ ID NO: 26) IgG1, G1m17 (SEQ ID NO: 30) и G1m3 (SEQ ID NO: 31) могут быть использованы для получения гуманизированного антитела ЕР27.
1-5. Оценивание гуманизированного антитела ЕР27
Для оценивания сконструированной гуманизированной последовательности антитела ЕP27, Н-цепь (HJ-G1d, SEQ ID NO: 28) и L-цепь (LB-k, SEQ ID NO: 29) коэкспрессировали для получения гуманизированного антитела ER27 HJ-G1d/LB-k). Конкретно, Н- и L-цепи экспрессирующих векторов, приготовленных, как упоминалось выше, транзиторно трансфицировали в штамм HEK293H, полученный из клеток карциномы эмбриональной почки человека (Invitrogen) или в клетки FreeStyle293 (Invitrogen) для экспрессии антител. Полученные из культурального супернатанта антитела очищали способом, известным квалифицированным в данной области специалистам, с использованием rProtein A Sepharose™ Fast Flow (GE Healthcare) или т.п. Оптическую плотность антитело-содержащего раствора измеряли при длине волны 280 нм с использованием спектрофотометра, и концентрацию антитела рассчитывали с использованием измеренной оптической плотности и коэффициент оптической плотности определяли способом PACE (Protein Science 1995; 4: 2411-2423). Между тем, cH-G1/cL-k, который является химерным антителом ЕР27, получали в качестве контроля тем же способом с использованием цепей H (cH-G1, SEQ ID NO: 32) и L (cL-k, SEQ ID NO: 33).
Способность гуманизированного антитела ЕР27 связывать эпирегулин человека (SEQ ID NO: 167) оценивали анализом оценивания ингибирования связывания эпирегулина (Ссылочный пример 5) в отношении химерного антитела ЕР27. При установлении концентрации IC50 этого химерного антитела ЕР27 (cH-G1/cL-k) на 100, величина этого гуманизированного антитела ЕР27 HJ-G1d/LB-k была равна 24.
1-6. Приготовление гуманизированных антител ЕР27 с активностью, эквивалентной этому химерному антителу
Последовательности были сконструированы с заменой некоторых аминокислотных остатков FR-последовательности в последовательности Н-цепи HJ-G1d гуманизированного антитела ЕР27 аминокислотными остатками последовательности вариабельного района ЕР27 мыши. Сначала последовательность (SEQ ID NO: 35) конструировали заменой Thr-Asp, которые являются аминокислотными остатками в положениях 75 и 76, показанными нумерацией Кабата, последовательности FR3 человека, остатками Ser-Asn, которые являются аминокислотными остатками, содержащимися в последовательностях вариабельного района мыши ЕР27; и последовательность (SEQ ID NO: 36) конструировали заменой только Asp в положении 76 остатком Asn, который является аминокислотным остатком последовательности вариабельного района мыши ЕР27 (Таблица 6). Затем HS-G1d (SEQ ID NO: 37), которая является гуманизированной последовательностью вариабельного района Н-цепи, имеющей последовательность FR3 (SEQ ID NO: 35), показанной в Таблице 6, может быть получена введением мутации в экспрессирующий вектор, содержащий гуманизированную последовательность вариабельного района Н-цепи HJ-G1d (SEQ ID NO: 28). Замену аминокислотного остатка вводили способом, известным квалифицированным в данной области специалистам, с использованием ПЦР или т.п. Подобным образом, получали HY-G1d (SEQ ID NO: 38), который является гуманизированной последовательностью вариабельного района Н-цепи, содержащей FR3 последовательность (SEQ ID NO: 36), показанную в Таблице 6. Гуманизированные антитела ЕР27, HS-G1d/LB-k, HY-G1d/LB-k и HY-G1d/L18-k, получали таким же способом, который описан в Примере 1-4. Способность полученных антител связываться с эпирегулином человека оценивали ингибированием связывания эпирегулина с химерным антителом ЕР27 (Ссылочный Пример 5). В результате, при установлении концентрации IC50 этого химерного антитела ЕР27 (cH-G1/cL-k) на 100, эти концентрации IC50 гуманизированных антител ЕР27, HS-G1d/LB-k, HY-G1d/LB-k и HY-G1d/L18-k были равны 142, 146 и 100, соответственно.
Таким образом, были обнаружены, как результат гуманизации, последовательности гуманизированного антитела ЕР27, имеющие антигенсвязывающую способность, эквивалентную антигенсвязывающей способности химерного антитела ЕР27, а также уменьшенные риски иммуногенности в сравнении с химерным антителом ЕР27.
Пример 2. Введение мутаций, которые ослабляют реакцию деамидирования, изомеризации и гидролиза
Имеется гетерогенность в антителах, используемых для фармацевтических препаратов, хотя они являются моноклональными антителами, полученными из клонов, произведенных из единственной антитело-продуцирующий клетки. Известно, что такая гетерогенность антител встречается как результат модификаций, таких как окисление, деамидирование, изомеризация и гидролиз, и увеличивается, когда белки, включающие в себя антитела, хранятся в течение продолжительного времени или подвергаются условиям стресса, таким как тепловой стресс и связанный с освещением стресс (Heterogeneity of Monoclonal Antibodies: Journal of Pharmaceutical Sciences, vol. 97, No. 7, 2426-2447). Однако физические свойства белков, в частности, гомогенность и стабильность, являются очень важными для развития антител в качестве фармацевтических препаратов. Желательно, чтобы гетерогенность представляющего интерес вещества была уменьшена для получения единого материала в максимальной степени.
Реакция деамидирования осуществляется неферментативно в боковых цепях аспарагина и глутамина, и эта реакция является реакцией, которая изменяет амид, присутствующий в боковых цепях аспарагина и глутамина, в карбоновую кислоту. Изомеризация обусловлена образованием нестабильного циклического имидного промежуточного продукта вследствие деамидирования аспарагина или дегидратации аспарагиновой кислоты, как результата атаки электронной пары атома азота в С-концевом остатке на карбонильных группах в боковых цепях аспарагина и аспарагиновой кислоты. Этот промежуточный продукт обычно изменяется в изоаспарагиновую кислоту расщеплением, тогда как остаток изменяется в аспарагиновую кислоту. Желательно, чтобы реакции деамидирования и изомеризации, которые встречаются во время хранения белков, таких как антитела, ослаблялись как можно больше, так как они вызывают вышеописанную гетерогенность. Сообщалось, что реакция деамидирования имеет тенденцию осуществляться в сайте, где глицин и аспарагин являются смежными друг с другом (Asn-Pro) (Geiger et al., J. Biol. Chem. 1987; 262: 785-794). Кроме того, сообщается, что пептидная цепь аспарагиновой кислоты расщепляется реакцией гидролиза; и, в частности, вероятно, что последовательность (Asp-Pro), где пролин присутствует на С-концевом конце, деградируется при кислотных условиях (Segalas et al., FEBS Letters 1995; 371: 171-175).
В CDR-последовательности гуманизированной последовательности HY вариабельного района Н-цепи, аспарагин и аспарагиновая кислота присутствуют в положениях 31 (Asp), 52 (Asp), 54 (Asn), 56 (Asn) и 101 (Asp), указанных нумерацией Кабата. В CDR-последовательности гуманизированной последовательности LB вариабельного района L-цепи, они присутствуют в положениях 28 (Asp), 92 (Asp) и 93 (Asn), указанных нумерацией Кабата. Исследовали, могут ли варианты, которые ослабляют деамидирование, изомеризацию и гидролиз, быть получены как результат замены аминокислотных остатков в этих сайтах.
Для ослабления вышеуказанных реакций деградации заменой аминокислотных остатков, эти аминокислотные остатки заменяли на различные остатки. Конкретно, Н71-G1d (SEQ ID NO: 49), H57-G1d: 50), H61-G1d (SEQ ID NO: 51), H-65-G1d (SEQ ID NO: 52), H66-G1d (SEQ ID NO: 53), H67-G1d (SEQ ID NO: 54), H23-G1d (SEQ ID NO: 55) и H40-G1d (SEQ ID NO: 56) готовили в виде генов Н-цепи, содержащих CDR-последовательности (SEQ ID NO: 39-46), показанные в Таблице 7. Подобным образом, L30-K (SEQ ID NO: 57) и L32-k; (SEQ ID NO:58) конструировали в виде генов L-цепи, содержащих CDR-последовательности (SEQ ID NO: 47 и 48), показанные в Таблице 7. С использованием способа Примера 1, вектор гена Н-цепи, введенный с замененным остатком, коэкспрессировали с вектором LB-k с получением гуманизированных антител EP27, Н71-G1d/LB-k, H57-G1d/LB-k, H61-G1d/LB-k, H65-G1d/LB-k, H66-G1d/LB-k, H67-G1d/LB-k, Н23-G1d/LB-k и H40-G1d/LB-k. Подобным образом, вектор гена L-цепи, введенный с замененным остатком, коэкспрессировали с вектором HY-G1d с получением гуманизированного антитела ЕР27 HY-G1d/L32-k. Способность полученных антител связываться с эпирегулином человека оценивали как ингибирование связывания эпирегулина с химерным антителом ЕР27 (Ссылочный Пример 5), и этот результат показан в Таблице 8. Это демонстрирует, что антитела, содержащие последовательности, показанные в Таблице 7, имеют связывающую активность, эквивалентную связывающей активности химерного антитела ЕР27. Таким образом, были обнаружены гуманизированные последовательности антитела ЕР27 с ослабленной химической деградацией, такой как деамидирование и изомеризация.
Пример 3. Введение мутаций, которые изменяют изоэлектрическую точку
В качестве способа контролирования полужизни антитела в плазме, известен способ, который модифицирует аминокислотные остатки, экспонированные на поверхности молекулы антитела для контроля заряда поверхности этой молекулы (WO 2007/114319 и WO 2009/041543). Конкретно, известно, что время полужизни в плазме антитела может быть удлинено понижением величины изоэлектрической точки (pI) антитела. С другой стороны, известно, что время полужизни в плазме антитела может быть укорочено увеличением изоэлектрической точки антитела для улучшения тканевой транзиции антитела (Vaisitti et al., J. Biol. Regul. Homeost. Agents. (2005) 19(3-4), 105-112; Partridge et al., J. Pharmacol. Exp. Ther. (1998) 286(1), 548-554).
Из вышеописанного, ожидается, что гуманизированное антитело ЕР27, изоэлектрическая точка которого была изменена, имеет сильную противоопухолевую активность, вследствие его удлиненной полужизни в плазме и улучшенной тканевой транзиции. Таким образом, авторы данного изобретения имеют целью идентификацию аминокислотных остатков, которые позволяют контролировать фармакокинетику антитела коррекцией поверхностного заряда молекулы антитела без воздействия на антигенсвязывающую активность и конформацию ЕР27-гуманизированного антитела. Конкретно, проводили поиск на сайты мутации, которые могут понижать изоэлектрическую точку гуманизированного антитела ЕР27 HY-G1d/LB-k (H-цепь, HY-G1d, SEQ ID NO: 38; и L-цепь, LB-k, SEQ ID NO: 29) без значительного уменьшения его антигенсвязывающей ингибиторной активности.
Использовали трехмерную модель гуманизированного антитела ЕР27 HY-G1d/LB-k для скрининга на остатки, которые могут изменять изоэлектрическую точку вариабельного района без значительного уменьшения связывания с эпирегулином. Конкретно, H3-G1d (SEQ ID NO: 72), H5-G1d (SEQ ID NO: 73), H6-G1d (SEQ ID NO: 74), H7-G1d (SEQ ID NO: 75), H8-G1d (SEQ ID NO: 76), H9-G1d (SEQ ID NO: 77), H10-G1d (SEQ ID NO: 78) и H31-G1d (SEQ ID NO: 79) конструировали в виде генов Н-цепи, содержащих одну или несколько CDR-последовательностей (SEQ ID NO: 59-71, показанных в Таблице 9. Подобным образом, L1-k (SEQ ID NO: 80), L2-k (SEQ ID NO: 81), L12-k (SEQ ID NO: 82), L20-k (SEQ ID NO: 83), L21-k (SEQ ID NO: 84) и L23-k (SEQ ID NO: 85) конструировали в виде генов L-цепи, содержащих CDR-последовательности, показанные в Таблице 9. С использованием способа Примера 1, вектор гена Н-цепи, введенный с замененными остатками, коэкспрессировали с вектором LB-k для получения гуманизированных антител ЕР27, H3-G1d/LB-k, H5-G1d/LB-k, H6-G1d/LB-k, H7-G1d/LB-k, H8-G1d/LB-k, H9-G1d/LB-k, H10-G1d/LB-k и H31-G1d/LB-k. Подобным образом, вектор гена L-цепи, введенный с замененными остатками, коэкспрессировали с вектором HY-G1d для получения гуманизированных антител ЕР27, HY-G1d/L1-k, HY-G1d/L2-k, HY-G1d/L12-k и HY-G1d/L63-k. Способность полученных антител связываться с эпирегулином человека оценивали как ингибирование связывания эпирегулина с химерным антителом ЕР27 (Ссылочный Пример 5). В результате, как показано в Таблице 10, антигенсвязывающие способности гуманизированных антител ЕР27 были эквивалентными антигенсвязывающим способностям химерного антитела ЕР27. Из приведенного выше исследования, были обнаружены гуманизированные последовательности антитела ЕР27, изоэлектрическая точка которых была изменена.
Пример 4. Введение мутаций, которые уменьшают количество агрегата
Контролирование количества агрегата в белковых фармацевтических препаратах является важным в рассмотрении контроля качества и влияний на эффективность и иммуногенность (Curr. Opin. Biotechnol. (2009) 20(6), 708-714). Обычно, на агрегацию влияет как коллоидная стабильность вследствие среды раствора белка, так и конформационная стабильность вследствие структуры белка (J. Pharm. Sci. (2010) 100(4), 1306-1315). Согласно исследованиям на композициях препаратов антител, можно получать желаемые условия, которые являются эффективными для коллоидной стабильности, скринингом концентрации антитела, рН, типа буфера, ионной силы, добавки и т.п. С другой стороны, конформационная стабильность частично зависит от аминокислотных последовательностей; и, следовательно, в случае антител считается важным поддержание характерных структур, таких как каноническая структура CDR, консенсусная структура FR и граница раздела VН/VL (Jung et al., J. Mol. Biol. (2001) 309 (3), 701-716; Xiang et al., J. Mol. Biol. (1995) 253 (3), 385-390; Ewert et al., Methods. (2004) 34(2), 184-199; Vargas-Madrazo et al., J. Mol. Recognit. (2003) 16(3), 113-120; Morea et al., J. Mol. Biol. (1998) 275, 269-294; Vargas-Madrazo et al., J. Mol. Recognit. (2003) 16(3), 113-120).
Из приведенных выше точек зрения, в данном изобретении проводили конструирование в попытке изготовления антител, которые являются структурно более стабильными. В результате, было обнаружено гуманизированное антитело ЕР27 HY-G1d/LB-k. Для дополнительного уменьшения количества агрегатов, содержащихся в HY-G1d/LB-k, гидрофобные остатки, содержащиеся в CDR HY-G1d/LB-k, заменяли гидрофильными остатками, и исследовали ингибиторное действие на агрегацию аттенюированием гидрофобных взаимодействий между молекулами.
Конкретно, H25-G1d (SEQ ID NO: 92), H41-G1d (SEQ ID NO: 93), H42-G1d (SEQ ID NO: 94), H43-G1d (SEQ ID NO: 95), H44-G1d (SEQ ID NO: 96) и H45-G1d (SEQ ID NO: 97) конструировали в виде генов Н-цепи, в которых любая из этих CDR-последовательностей (SEQ ID NO: 86-91), показанных в Таблице 11, вводили в гуманизированное антитело ЕР27 HY-G1d/LB-k (H-цепь HY-G1d/SEQ ID NO: 38; L-цепь LB-k/SEQ ID NO: 29). С использованием способа Примера 1, вектор гена Н-цепи с замененными остатками коэкспрессировали с LB-k-вектором с получением гуманизированных антител ЕР27, H25-G1d/LB-k, H41-G1d/LB-k, H42-G1d/LB-k, H43-G1d/LB-k, H44-G1d/LB-k и H55-G1d/LB-k. Количество агрегатов, содержащееся в гуманизированных антителах ЕР27, определяли гель-фильтрационной хроматографией (Ссылочный Пример 9), и этот результат показан в Таблице 12. Посредством этого были обнаружены гуманизированные последовательности антитела ЕР27 со значимо уменьшенным количество агрегатов в сравнении с химерным антителом ЕР27 (cH-G1d/cL-k).
Способность полученных антител связываться с эпирегулином человека оценивали в виде ингибирования связывания эпирегулина (Ссылочный Пример 5) с химерным антителом ЕР27. Как показано в Таблице 13, антигенсвязывающие способности гуманизированных антител ЕР27 были эквивалентными антигенсвязывающим способностям химерного антитела ЕР27. Из приведенного выше исследования были обнаружены последовательности гуманизированного антитела ЕР27 с уменьшенным количеством агрегатов и антигенсвязывающей способностью, эквивалентной антигенсвязывающей способности химерного антитела Ер27.
Кроме того, следующие антитела, которые были сконструированы в качестве гуманизированных антител ЕР27 произвольным комбинированием последовательностей, содержащих замены аминокислотных остатков, обнаруженные в Примерах 1-4, получали с использованием способа Примера 1:
H87-G1d/LB-k (H-цепь: SEQ ID NO: 98; L-цепь: SEQ ID NO: 29),
H87-G1d/L21-k (H-цепь: SEQ ID NO: 98; L-цепь: SEQ ID NO: 84),
H87-G1d/L37-k (H-цепь: SEQ ID NO: 98; L-цепь: SEQ ID NO: 99).
Их способность связываться с эпирегулином человека оценивали в виде ингибирования связывания эпирегулина (Ссылочный Пример 5) с химерным антителом ЕР27 (Ссылочный пример 5). В качестве результата, как показано в Таблице 14, все эти антитела имели связывающую активность, эквивалентную связывающей способности химерного антитела ЕР27 (cH-G1d/cL-k). Кроме того, количество агрегатов, содержащихся в гуманизированных антителах ЕР27, измеряли гель-фильтрационной хроматографией (Ссылочный Пример 9), температуру тепловой денатурации (Tm) промежуточного продукта Fab измеряли дифференциальным сканирующим калориметром (Ссылочный пример 10), и изоэлектрическую точку (pI) измеряли изоэлектрическим фокусированием (Ссылочный Пример 11). Как показано в Таблице 14, было подтверждено, что количества агрегатов значимо уменьшались и величины Tm увеличивались в сравнении с химерным антителом ЕР27 (cH-G1d/cL-k). Таким образом, были обнаружены гуманизированные последовательности антитела ЕР27, которые достигают рисков уменьшенной иммуногенности, ослабленной химической деградации, пониженной изоэлектрической точки и уменьшенного количества агрегатов посредством гуманизации, как показано в Примерах 1-4.
Пример 5. Введение мутаций, которые улучшают способность связывания с антигеном обезьяны и уменьшают иммуногенность
Во время разработки лекарственного средства, обычно оценивания токсичности и безопасности в клинических исследованиях считаются важными для определения дозы и оцениваний различных рисков. В случае фармацевтических препаратов антитела, их антигенсвязывающая специфичность могла бы ограничивать выбор видов животных, используемых для преклинических исследований. Во многих случаях выбирают приматов, которые являются генетически близкими к человеку. Однако оценивание с использованием приматов иногда не оценивается как подходящее, если представляющее интерес антитело не связывается с антигеном примата, или если способность связывания значительно отличается от их способности связываться с антигеном человека. Чтобы справиться с такими случаями, делались попытки оценивания токсичности и безопасности в преклинических исследованиях с использованием суррогатного антитела, которое связывается с антигеном вида животного, подлежащего использованию, или с использованием трансгенных животных с антигеном человека (Chapman et al., Nat. Rev. Drug Discov. (2007) 6, 120-126).
Считается, что замены аминокислотных остатков способами, известными квалифицированным в данной области специалистам, например, созревания аффинности с использованием библиотеки фагов или дисплея рибосом, являются эффективными для изменения антигенсвязывающей способности представляющего интерес антитела. Кроме того, на основе трехмерных структур антигенов и связывающих сайтов вариабельного района антитела, могут быть использованы способы вычислительной науки для улучшения перекрестной межвидовой реактивности (Farady et al., Bioorganic & Medicinal Chemistry Letters 19 (2009) 3744-3747. Однако не публиковались сообщения об изменении антигенсвязывающей способности антитела против эпирегулина человека без широкомасштабного скрининга и способов вычислительной науки. Кроме того, не были выявлены структурные различия между эпирегулином обезьяны и эпирегулином человека, и, следовательно, не было показано никакого руководства в отношении того, какие аминокислотные остатки антитела против эпирегулина человека должны быть модифицированы для усиления его связывающей активности в отношении эпирегулина обезьяны. В данном изобретении множественные последовательности антител были сконструированы заменой аминокислотных остатков в произвольных сайтах CDR. С использованием способа Примера 1 получали гены антител с этими замененными остатками, и эти антитела экспрессировали. Конкретно, конструировали последовательности, в которых сделана замена остатком Arg в CDR2 и CDR3 H-цепи и CDR3 L-цепи, показанные в Таблице 15, и получали антитела, показанные в Таблице 16. Аффинности в отношении эпирегулина человека и обезьяны оценивали с использованием устройства (BIACORE) с использованием рассеяния поверхностных плазмонов (Ссылочный Пример 6).
В результате, в сравнении с химерным антителом ЕР27, были обнаружены гуманизированные последовательности антитела ЕР27 с неуменьшенной аффинностью в отношении эпирегулина человека, увеличенной аффинностью в отношении эпирегулина обезьяны и низким отношением аффинности (= величина KD в отношении эпирегулина обезьяны/величина KD в отношении эпирегулина человека), которое является индикатором, показывающим перекрестную межвидовую реактивность в отношении эпирегулина обезьяны и в отношении эпирегулина человека (Фиг. 1 и 2).
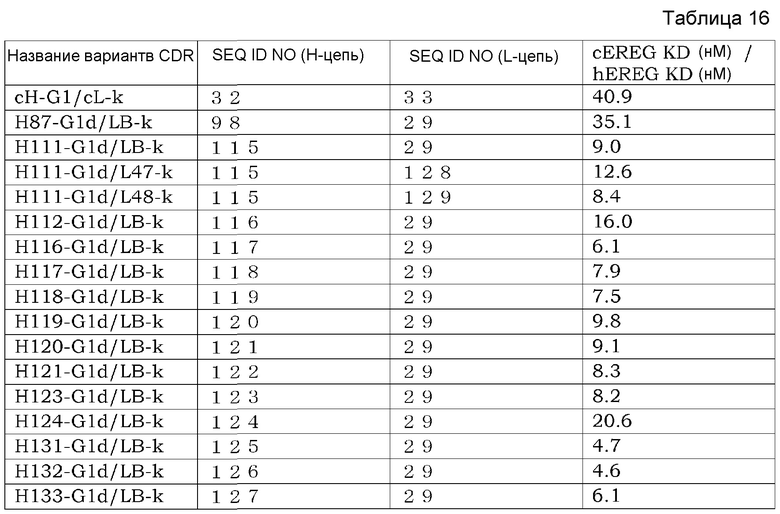
В разработке медицинских фармацевтических препаратов, появление антител против лекарственного средства после его введения создает проблемы в отношении эффективности и безопасности. Таким образом, важно уменьшать иммуногенность представляющего интерес лекарственного средства. Для предсказания иммуногенности биопрепаратов доступны способы in vivo, in vitro и in silico, и были опубликованы сообщения по корреляции со сферой действия антител против лекарственных средств в клинической практике (Baker et al., Curr. Drug. Saf. (2010) 5(4), 308-313; Bryson et al., BioDrugs, (2010) 24(1), 1-8; Groot et al., Curr. Opin. Pharmacol. (2008) 8(5), 620-626). В данном изобретении последовательности гуманизированного антитела EP27, которые уменьшают риски иммуногенности без повреждения антигенсвязывающей способности представляющего интерес антитела, могут быть предсказаны предсказанием Т-клеточных эпитопов с использованием способа, описанного в Ссылочном Примере 13.
Для последовательностей, содержащих изменения, обнаруженные в Примерах 1-4, были сконструированы последовательности с произвольной комбинацией мутаций, которые улучшают аффинность в отношении эпирегулина обезьяны и/или уменьшают иммуногенность. Эти гуманизированные антитела EP27, сконструированные таким образом и перечисленные в таблице 17 (SEQ ID NO: 115-150), готовят с использованием способа Примера 1. Мутированные CDR-последовательности гуманизированных антител ЕР27, перечисленные в Таблице 17, показаны в Таблице 18. Аффинности в отношении эпирегулина человека и обезьяны оценивали с использованием устройства (BIOCORE) c применением рассеяния поверхностных плазмонов (Ссылочный Пример 6). Как показано в Фиг. 3, аффинность каждого из гуманизированных антител ЕР27 в отношении эпирегулина человека была эквивалентной аффинности химерного антитела ЕР27 (cH-G1/cL-k). Отношения аффинности (= величина KD в отношении эпирегулина обезьяны/величина KD в отношени эпирегулина человека) сравнивали в качестве индикатора различия в аффинностях для антигенов обезьяны и человека. Было подтверждено, что это различие в аффинностях каждого гуманизированного антитела ER27 в отношении этих двух антигенов значимо уменьшалось в сравнении с различием химерного антитела (Фиг. 4). Таким образом, были обнаружены последовательности гуманизированного антитела ЕР27, которые улучшали аффинность в отношении эпирегулина обезьяны без уменьшения аффинности в отношении эпирегулина человека.
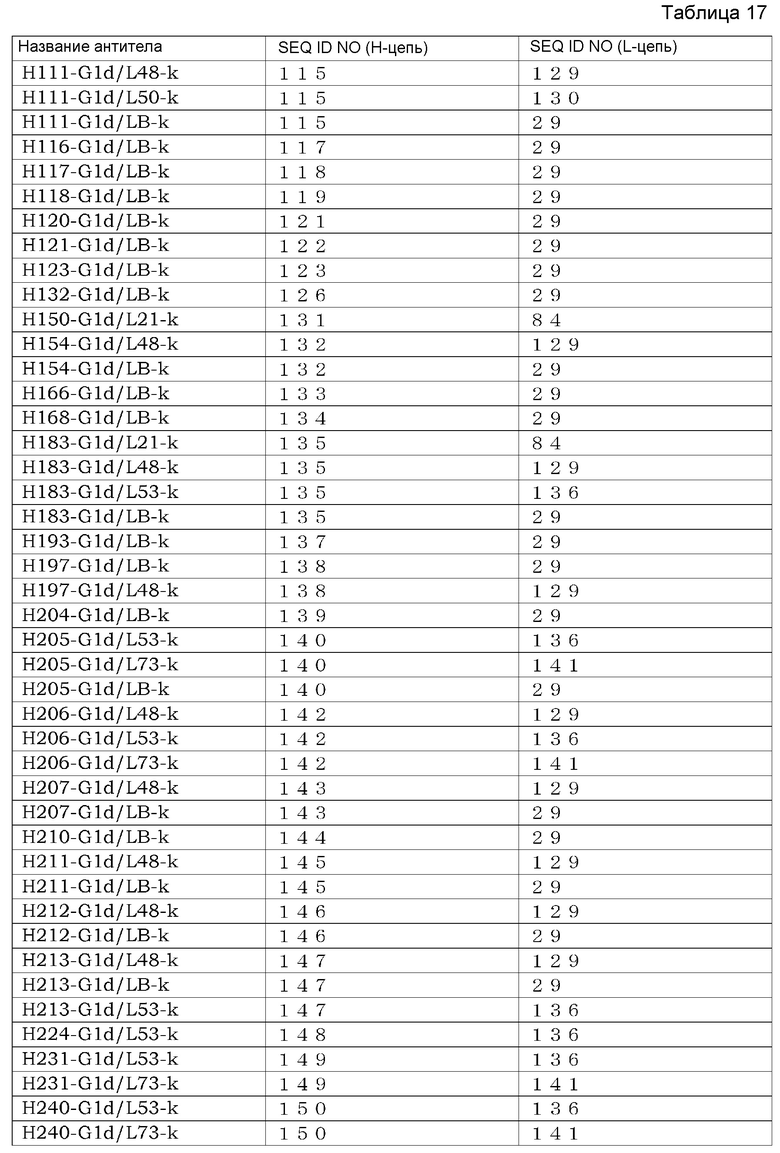
Кроме того, количество агрегата, содержащегося в гуманизированных антителах ЕР27, измеряли гель-фильтрационной хроматографией (Ссылочный Пример 9), и температуру тепловой денатурации (Tm) промежуточного продукта Fab, полученного из гуманизированных антител ЕР27, измеряли дифференциальным сканирующим калориметром (Ссылочный пример 10). Как показано в Таблице 19, было подтверждено, что количества агрегатов значимо уменьшались и величины Tm увеличивались в сравнении с химерным антителом ЕР27 (cH-G1d/cL-k). Таким образом, как показано в примерах 1-5, были обнаружены последовательности гуманизированного антитела ЕР27, содержащие уменьшенные риски иммуногенности, ослабленную химическую деградацию, пониженные изоэлектрические точки, уменьшенные количества агрегатов и улучшенные связывающие способности в отношении антигена обезьян, как результат гуманизации.

Пример 7. Активность ADCC каждого тест-антитела с использованием мононуклеарных клеток периферической крови человека в качестве эффекторных клеток
Мононуклеарные клетки периферической крови человека (далее здесь называемые как PBMC человека) использовали в качестве эффекторных клеток для измерения активности ADCC каждого тест-антитела, как описано здесь ниже. Фракцию мононуклеарных клеток, собранную из периферической крови человека, использовали в отношении эффекторной клетки, происходящей из человека. В результате, было обнаружено, что все гуманизированные антитела ЕР27, используемые в этом тесте, индуцируют ADCC против клеток MIA PaCa-2 (Ссылочный Пример 7) (Фиг. 5).
Пример 8. Измерение активности моноклональных антител против Эпирегулина для нейтрализации стимуляции роста клеток Эпирегулином человека или обезьяны
Последовательности гуманизированного антитела EP27 с увеличенной аффинностью в отношении Эпирегулина обезьяны были обнаружены в Примере 5. Для оценивания их активностей в клетках, гуманизированные антитела ЕР27 измеряли на их активности в отношении нейтрализации стимуляции роста клеток Эпирегулином обезьяны и Эпирегулином человека. Для этих клеток использовали BAF_EGFR. Все из гуманизированных антител EP27, оцениваемых в этот раз, испытывали на их нейтрализующие активности, которые ингибируют зависимый от Эпирегулина человека и обезьяны рост клеток EGFR_BAF (Ссылочный Пример 8), и все из этих гуманизированных антител ЕР27 обнаруживали увеличенные нейтрализующие активности против Эпирегулина обезьяны в сравнении химерным антителом (Фиг. 6 и 7).
Пример 9. Тест на эффективность для гуманизированных антител ЕР27 с использованием модели in vivo.
Противоопухолевую активность in vivo гуманизированных антител ЕР27, активность которых в клетках in vitro была подтверждена в Примерах 7 и 8, оценивали с использованием модели мыши, трансплантированной опухолевыми клетками человека (Ссылочный Пример 14). Противоопухолевые эффекты каждого тест-антитела оценивали в трансплантированных раковыми клетками человека моделях мыши MIA PaCa и DID-12 измерением объема опухоли в день 7 после последнего дня введения пробы. В результате, как показано на Фиг. 8, когда антитело cH-G1/cL-k, антитело H206-G1d/L73-k и Н240/L73-k индивидуально вводили при 0,4 мг/кг и 2 мг/кг, наблюдали превосходную лекарственную эффективность с использованием этих измененных гуманизированных антител EP27 также и в моделях животных.
Пример 10. Иммуногистохимическое окрашивание с использованием анти-Эпирегулин-антитела
Способ иммуногистохимического окрашивания, который отражает уровень экспрессии антигена, устанавливали с использованием антитела Е27. Иммуногистохимическое окрашивание проводили с использованием залитых в парафин блоков ткани, приготовленных с тканями SCID мышей, трансплантированных клеточными линиями, которые заставляют экспрессировать EREG при различных уровнях экспрессии, которые являются SKE-18 (определенное количество антигена было 7,7×103), SKE-23 (определенное количество антигена было 6,1×104) и SKE-4B2 (определенное количество антигена было 2,9×105), и клетка-хозяин SK-HEP-1 (количество антигена было 8,4×102). Экспрессию EREG в этих тонко нарезанных срезах ткани, приготовленных из этих парафиновых блоков, визуализировали инкубированием с использованием антитела ER27 в качестве первичного антитела с последующей реакцией кроличьего антитела против IgG мыши (Jackson Immunoresearch Laboratories) в качестве вторичного антитела и связанного с полимером НRP (DAKO cytomation) козьего антитела против кроличьего IgG-антитела в качестве третичного антитела, и с использованием диаминобензидина в качестве субстрата. Как показано на Фиг. 10, градацию окрашивания наблюдали в окрашенных клетках/ткани в зависимости от количества экспрессии EREG. Кроме того, было подтверждено, что in vitro ADCC-активность антитела H240-G1d/L73-k зависит от уровня экспрессии в этих клетках (Фиг. 11). В вышеупомянутом исследовании низкофукозное антитело использовали для антитела H240-G1d/L73-k. Низкофукозное антитело готовили способом, описанным в WO2004/0654540. Здесь, ниже, это низкофукозное антитело называют также гуманизированным антителом EP27 Glycomab. Активность ADCC in vitro антитела H240-G1d/L73-k (гуманизированного антитела EP27 Clycomab) подтверждали в SKE-15 и SKE-10, которые имеют оцененный уровень антигена 2,8×104, в дополнение к вышеупомянутым четырем клеточным линиям принудительной экспрессии EREG. Подобным Фиг. 11 образом была подтверждена активность ADCC in vitro антитела H240-G1d/L73-k в соответствии с уровнем экспрессии EREG в этих клетках (Фиг. 20). Кроме того, была подтверждена активность in vivo ингибирования роста опухоли антитела H240-G1d/L73-k (гуманизированного антитела EP27 Clycomab) в соответствии с уровнем экспрессии в этих клетках (фиг. 12). То есть оценивание уровня экспрессии EREG иммуногистохимическим окрашиванием подтвердило корреляцию между уровнем экспрессии и лекарственной эффективностью.

Пример 11. Экспрессия EREG в клинических случаях слабо дифференцированного рака ободочной кишки
Для испытания экспрессии EREG в девяти клинических случаях слабо дифференцированного рака ободочной кишки, тонко нарезанные, залитые в парафин образцы некоторых случаев окрашивали способом окрашивания, описанным в Примере 10. В результате, положительные изображения подтверждались в 7/9 случаях (Фиг. 13).
Пример 12. Лекарственная эффективность гуманизированного антитела EP27 Glycomab против онкогенеза стволовых клеток ободочной кишки
Стволовые клетки рака ободочной кишки (1×106 клеток), выделенные из модели опухоли рака ободочной кишки человека PLR123 (WO02021/046797), трансплантировли в паховую область мышей SCID. От следующего дня введения этих стволовых клеток мышам SCID, гуманизированное антитело EP27 Glycomab вводили один раз в неделю при 10 мг/кг. В результате, знаковый критерий рангов Вилкоксона (SAS-система 8.02 TS-уровень 02М0 и SAS преклинический пакет Version 5.00.010720) подтвердил, что в день 29 после трансплантации онкогенность была значимо ослаблена в группе, которой вводили антитело, в сравнении с контрольной группой, как показано на Фиг. 14.
Пример 13. Лекарственная эффективность гуманизированного антитела EP27 Glycomab против метастазирования стволовых клеток рака ободочной кишки
Стволовые клетки рака ободочной кишки (2×106 клеток), выделенные из модели опухоли рака ободочной кишки человека PLR123, трансплантировали в мышей SCID, с врожденным отсутствием естественных клеток-киллеров, из хвостовой вены. Спустя три дня после введения стволовых клеток мышам, гуманизированное антитело EP27 Glycomab вводили один раз в неделю при 10 мг/кг. В результате, было подтверждено, что в день 52 после трансплантации количество опухолевых узелков в легких мыши (количество метастатических повреждений) и размер опухолевого узелка (диаметр метастатического повреждения) были значимо ослаблены в группе, которой вводили антитело, в сравнении с контрольной группой, как показано на Фиг. 15.
Пример 14. Лекарственная эффективность гуманизированного антитела EP27 Glycomab против слабо дифференцированного рака ободочной кишки
Срез опухолевой ткани слабо дифференцированного рака ободочной кишки человека COL-53-JCK (Центрального Института для Экспериментальных животных) трансплантировали в мышь SCID. От дня 14 после трансплантации среза ткани опухоли гуманизированное антитело ЕР27 Glycomab вводили один раз в неделю при 10 мг/кг. В результате, оценивание объема опухоли в день 14 после начала введения антитела подтверждало, что рост опухоли значимо ослаблялся в группе, которой вводили антитело, в сравнении с контрольной группой (Фиг. 16).
Пример 15. Лекарственная эффективность гуманизированного антитела EP27 Glycomab против умеренно дифференцированного рака ободочной кишки
Опухоли в умеренно дифференцированной опухолевой модели рака ободочной кишки человека PLR379 (PCT/JP2012/072852) имели признак морфологически наблюдаемого почкования опухоли, но экспрессию эпирегулина детектировали, невзирая на участок почкования опухоли. Срез опухолевой ткани этого PLR379 трансплантировали в мышь SCID, и от дня 21 после трансплантации этого среза опухолевой ткани, гуманизированное антитело ЕР27 Glycomab вводили один раз в неделю при 10 мг/кг. Оценивание объема опухоли в день 18 после начала ведения антитела подтверждало, что рост опухоли значимо ослаблялся в группе, которой вводили антитела, в сравнении с контрольной группой (Фиг. 17).
Пример 16. Экспрессия EREG в клинической аденокарциноме легких
Для испытания экспрессии EREG в семи клинических случаях аденокарциномы легких, тонко нарезанные залитые в парафин образцы этих случаев окрашивали с использованием способа окрашивания, описанного в Прмере 10. В результате, ясные положительные изображения подтверждались в четырех из семи случаев (Фиг. 18).
Пример 17. Активность ADCC гуманизированного антитела EP27 Glycomab с использованием мононуклеарных клеток периферической крови человека в качестве эффекторных клеток
Активность ADCC оценивали для линии Calu-3 аденокарциномы легких человека с использованием PBMC человека в качестве эффекторных клеток. В результате, было подтверждено, что гуманизированное антитело EP27 Glycomab индуцирует ADCC против клеток Calu-3 (Фиг. 19).
Пример 18. Лекарственная эффективность гуманизированного антитела EP27 Glycomab против аденокарциномы легких
Гуманизированное антитело EP27 Glycomab вводили мыши SCID с имплантированным фрагментом опухолевой ткани модели Calu-3 аденокарциномы легкого человека один раз в неделю 10 мг/кг от дня 13 после трансплантации фрагмента опухолевой ткани. Объем опухоли измеряли в день 28 после начала введения антитела. Было подтверждено, что рост опухоли значимо ослаблялся в группе, которой вводили антитело, в сравнении с контрольной группой (Фиг. 20).
Пример 19. Клеточная интернализация гуманизированного антитела EP27
Клеточную интернализацию гуманизированного антитела ЕР27 оценивали следующим способом. Когда первичное антитело становится интернализованным, сапорин, который является рибосома-инактивирующим белком, несомым вторичным антителом, транспортируют в клетку через связывание вторичного антитела с первичным антителом. После его интернализации, сапорин диссоциируется из конъюгата IgG, ингибирует синтез белка и затем вызывает смерть клеток. EREG-экспрессирующую линию клеток DLD-1 или контрольную клеточную линию SK-HEP1 высевали при плотности клеток 1×103 в каждую лунку 96-луночных планшетов, и на следующий день это гуманизированное антитело ЕР27 Glycomab добавляли в качестве первичного антитела, и козий IgG, распознающий моноклональное антитело человека, связанное с рибосома-инактивирующим белком Hum-ZAP (Advanced Targeting Systems), добавляли в качестве вторичного антитела. Спустя четыре дня после добавления, жизнеспособность клеток оценивали с использованием WST8 (Donjindo). Как показано на Фиг. 21, гуманизированное антитело ЕР27 Glycomab индуцировало смерть клеток только против EREG-экспрессирующей клеточной линии DLD-1. С другой стороны, оно не индуцировало смерти клеток против клеточной линии SK-HEP1, которую использовали в качестве контроля.
Ссылочный Пример 1. Выделение генов Эпирегулина человека и собакоподобной обезьяны
Полноразмерную кДНК EREG человека (CR541887, SEQ ID NO: 169) выделяли стандартным способом, и плазмида, продуцируемая клонированием этого фрагмента гена в вектор для экспрессии в клетках млекопитающих (pMCN), была названа hEREG/pМCN. pМCN может индуцировать экспрессию чужеродного гена при промоторе CMV мыши (ACCESSION No. U 68299) и является вектором, инсертированным с геном устойчивости к неомицину. Полноразмерную кДНК EREG собакоподобной обезьяны выделяли стандартным способом из кДНК-библиотеки собакоподобной обезьяны на основе информации ДНК последовательности макака-резус (XM_001102069), и плазмида, продуцируемая клонированием этого фрагмента гена (фрагмента гена, кодирующего последовательность SEQ ID NO: 165) в вектор для экспрессии в клетках млекопитающих (pМCN), была названа cyEREG/pМCN. hEREG/pМCN и cyEREG/pМCN вводили в штамм CHO DG44 (Invitrogen) электропорацией, и селекция с 500 мкг/мл генетицина установила клетки СНО, которые стабильно экспрессируют полноразмерный EREG человека, и СНО-клетки, которые экспрессируют полноразмерный EREG собакоподобной обезьяны, которые называют hEREG_DG и cyEREG_DG, соответственно.
Ссылочный Пример 2. Установление способов для экспрессии зрелых форм Эпирегулинов человека и собакоподобной обезьяны
ПЦР-способ использовали для амплификации кДНК для экспрессии последовательности шести-гистидиновых повторов на С-концах внеклеточных районов зрелых EREG генов Эпирегулина человека и собакоподобной обезьяны (полипептидов, в которых последовательность SEQ ID NO: 171 была слита с N-концами полипептидов SEQ ID NO: 170 и 34, соответственно). Экспрессирующие векторы для использования в клетках млекопитающих, в которые эти кДНК были индивидуально инсертированы, линеаризовали рестрикционными ферментами; и затем введением их в клетки Freestyle 293 с использованием 293 фектина, зрелые Эпирегулины человека и собакоподобной обезьяны транзиторно экспрессировались.
Ссылочный Пример 3. Приготовление Эпирегулина
Зрелый ген Эпирегулина человека и зрелый ген эпирегулина собакоподобной обезьяны инсертировали индивидуально в экспрессирующий вектор для клеток млекопитающих. Зрелые слитые белки EREG-6HIS человека и EREG-6HIS собакоподобной обезьяны, выделенные из культуральных растворов клеток животных, приготовленных для экспрессии каждого из этих генов, очищали.
Внеклеточный район зрелого EREG человека (полипептида, в котором последовательность SEQ ID NO: 171 слита с N-концом SEQ ID NO: 34), слитый в рамке считывания с шести-гистидиновым районом, инсертировали в экспрессирующий вектор для млекопитающих-животных для конструирования экспрессирующего вектора hsEFEG-6His (здесь ниже, этот экспрессированный слитый пептид называют hsEREG-His). Внеклеточный район зрелого EREG собакоподобной обезьяны (полипептида, в котором последовательность SEQ ID NO: 171 слита с N-концом SEQ ID NO: 170), слитый в рамке считывания с шести-гистидиновым районом, инсертировали в экспрессирующий вектор для млекопитающих-животных для конструирования экспрессирующего вектора сsEREG-6His (здесь ниже, этот экспрессированный слитый пептид называют cysEREG-His). Эти hsEREG-6His-экспрессирующий вектор и csEREG-6His-экспрессирующий вектор вводили в клетки FreeStyle 293 (Invitrogen) с использованием 293-фектина (Invitrogen), и трансдуцированные клеточные линии культивировали при селекции Zeocin (500 мкг/мл) в течение шести дней при 37°С в 8% CO2-термостате.
Затем hsEREG-His и cysEREG-His очищали из культурального супернатанта. 4 М имидазол добавляли в культуральный супернатант при конечной концентрации 10 мМ имидазол, и эту жидкую смесь смешивали с Ni-смолой в системе очистки His MicroSpin (Amersham). Эту смолу промывали 20 мМ имидазолом и 50 мМ имидазолом, и затем hsEREG-His или cysEREG-His элюировали с использованием 200 мМ имидазола (фракция 1 элюции), и затем hsEREG-His или cysEREG-His элюировали 400 мМ имидазолом (фракция 2 элюции). Затем буфер, содержащий элюированные hsEREG-His или cysEREG-His, заменяли посредством диализа на PBS (Зфр) с использованием Bio-tech dialysis cup MWCO8000. Очищенный белок определяли количественно с использованием способа PAСE при длине волны 280 нм, и результат преобразовывали в содержание белка.
Электрофоретические картины очищенных белков показаны на Фиг. 9.
Ссылочный Пример 4. Установление ELISA-системы Эпирегулина
Растворимый фрагмент EREG человека, имеющий структуру зрелого EGF-домена (полипептида SEQ ID NO: 34, которая соответствует 63Val-108Leu), покупали из R&D Systems (cat. No. 1195-EP/CF). Этот растворимый фрагмент EREG человека использовали для покрытия nunc-иммунопланшетов, и после блокирования BCA-содержащим раствором анализировали связывающую реактивность очищенных анти-Эпирегулин-антител. Добавление очищенных анти-Эпирегулин-антител сопровождалось одним часом инкубирования, планшеты промывали, и затем в каждую лунку промытых планшетов добавляли меченное щелочной фосфатазой анти-IgG человека-антитело (Zymed) и затем давали реагировать. Каждую лунку промывали, и затем количество связанного антитела определяли добавлением тест-реагента, таблеток Sigma п-нитрофенилфосфата.
Ссылочный Пример 5. Установление системы для ингибирования связывания Эпирегулина с использованием конкурентного ELISA
Растворимый фрагмент EREG человека, имеющий зрелую структуру домена EREG (полипептида SEQ ID NO: 34, которая соответствует 63Val-108Leu), покупали из R&D Systems (cat. No. 1195-EP/CF). Этот растворимый фрагмент EREG человека использовали для покрытия nunc-иммунопланшетов, и после блокирования BCA-содержащим раствором анализировали конкурентную связывающую реактивность между полученным из гибридомы мыши ЕР27 и очищенным гуманизированным анти-EREG-антителом. Полученные из гибридомы мыши Е21 и очищенное гуманизированное анти-Эпирегулин-антитело добавляли в эти планшеты, и после двух часов ингибирования эти планшеты промывали; и меченное щелочной фосфатазой анти-IgG человека-антитело (Zymed) добавляли в каждую лунку этих планшетов и затем позволяли реагировать. Каждую лунку промывали, и затем количество связанного антитела (величина детектирования A405/655) определяли добавлением тест-реагента, таблеток Sigma п-нитрофенилфосфата.
Ссылочный Пример 6. Установление антигенсвязывающей активности (способа для измерения аффинности)
Аффинность и константу скорости ассоциации анти-EREG-антитела в отношении антигена измеряли с использованием анализа резонанса поверхностных плазмонов с кинетикой одного цикла с использованием Biacore™-T100 (GE Healthcare Japan). HBS-EP+ (GE Healthcare Japan) использовали для прохождения буфера, и амин-связывающий набор (GE Healthcare Japan) использовали для ковалентного связывания Протеина А с CM5-чипом (покрытым карбоксиметилдекстраном чипом). HBS-EP+ (Ge Healthcare Japan) использовали для прохождения буфера и амин-связывающий набор (Ge Healthcare Japan) использовали для ковалентного связывания Протеина А с CM5-чипом (покрытым карбоксиметилдекстраном чипом). Каждое анти-EREG антитело готовили таким образом, что приблизительно 350 RU будет захватываться Протеином А. EREG человека или EREG собакоподобной обезьяны, используемых в качестве аналита, готовили при 0, 0,7, 1,4, 2,8, 5,6 и 11,2 нМ с использованием HBS-EP+. Измерение проводили сначала позволением захвата Протеином А раствора антитела, и затем при скорости потока 30 мкл/мин последовательной инжекцией каждого из 0, 0,7, 1,4, 2,8, 5,6 и 11,2 нМ EREG человека или растворов EREG собакоподобной обезьяны в течение трех минут для позволения прохождения реакции. Затем этот раствор переключали на HBS-EP+, и фазу диссоциации измеряли в течение 15 минут. После завершения измерения фазы диссоциации сенсорный чип промывали 25 мМ NaOH и регенерировали. Измерение концентрации при нуле проводили сходным образом позволением захвата Протеином А раствора антитела и проведением трехминутных HBS-EP+-инжекций последовательно в течение 5 раз для позволения прохождения реакции, и затем переключением на HBS-EP+ для измерения фазы диссоциации в течение 15 минут. После завершения измерения фазы диссоциации сенсорный чип промывали 25 мМ NaOH и регенерировали. Программу для анализа данных исключительно для Biacore-T100, Biocore T100 Evaluation Software Version 2.0.1, использовали для выполнения кинетических анализов для расчета константы скорости ассоциации (ka), константы скорости диссоциации (kd) и отношения констант скорости из полученных сенсограмм. Эти результаты показаны в Таблице 21. Для коррекции относительно различий со дня на день в величинах измерений, Фиг. 1-4 и Таблицы 16 и 19 показывают отношения на основе взятия величины контрольной пробы (cH-G1/cL-k), измеренной в тот же самый день, за 1.
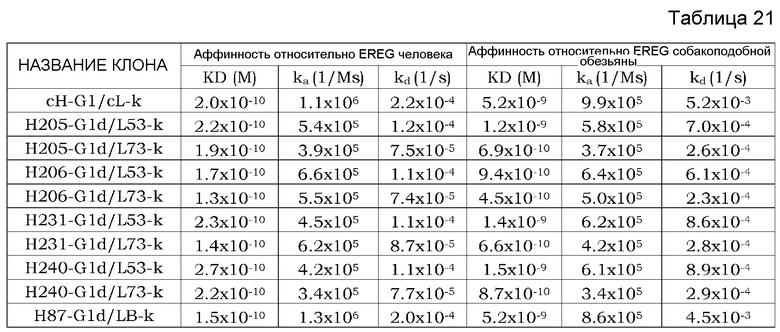
Ссылочный Пример 7. Установление способа для измерения ADCC-активности
Фракцию мононуклеарных клеток, собранную из периферической крови человека, использовали для полученных из человека эффекторных клеток. Из здорового волонтера (взрослого мужчины) собирали периферическую кровь с использованием шприца, предварительно загруженного 200 мкл раствора гепарина 1000 единиц/мл (Novo-Heparin Injection 5000 units, Novo Nordisk). Эту периферическую кровь разбавляли в 2 раза PBS(-), и Ficoll-Paque PLUS инъецировали заранее для проведения центрифугирования. Ее добавляли в пробирку Leucosep для отделения лимфоцитов (Greiner Bio-one) и центрифугировали (при 2150 об/мин в течение десяти минут при комнатной температуре), с последующим сбором слоя фракции мононуклеарных клеток. Эти клетки промывали один раз 10% ФТС/D-MEM и суспендировали в 10% ФТС/D-MEM при плотности клеток 5×106/мл для приготовления раствора суспензии эффекторных клеток. Эта клеточная суспензия служила в качестве раствора PBMC человека в следующем эксперименте.
Этот суспензионный раствор клеток-мишеней готовили для тестирования во время применения. Линию клеток (ATCC) панкреатического рака человека MIA PaCa-2 поддерживали субкультивированием в модифицированной по способу Дульбекко среде Игла (Invitrogen), содержащей 10% ФТС, 2,5% лошадиную сыворотку, 4 ммоль/л L-глутамина (Invitrogen), 4,5 г/л глюкозы (Invitrogen) и 1,5 г/л бикарбоната натрия) (далее называемой здесь субкультуральной средой). Для мечения реагента, 228 мкл 10% ФТС/D-MEM добавляли в пробирку, содержащую 12 мкл исходного раствора Calcein-AM/DMSO (nacalai), приготовленного при 4 мг/мл, для получения раствора Calcein-AM осторожным суспендированием. К 1×106 клеткам клеточной линии MIA PaCa-2, подвергнутой центрифугированию, (при 1200 об/мин в течение пяти минут при 4°C) раствор Calcein-AM, приготовленный, как описано выше, добавляли при 200 мкл на осадок клеток 1×106 для приготовления суспензии клеток. Все количество этого суспензионного раствора переносили в пластиковую пробирку для сбора крови (Niton Pharmaceutical), и эту суспензию инкубировали в СО2-термостате при 37°C в течение двух часов. Эти клетки промывали три раза 10% ФТС/D-MEM, и суспензионный раствор клеток-мишеней готовили суспендированием промытых клеток в 10% ФТС/D-MEM с получением 20×104 клеток/мл (1×104/50 мкл).
Для суспензионного раствора клеток-мишеней, 100 мкл этой среды добавляли в 96-луночный плоскодонный планшет. Затем моноклональные анти-Эпирегулин-антитела (H206-G1d/L73-k (SEQ ID NO: 142/141), H240-G1d/L73-k (SEQ ID NO: 150-141) и химерный EP27 (SEQ ID NO: 32/33)) разводили в среде и затем добавляли к этому планшету при 50 мкл на лунку. Эти антитела добавляли при конечных концентрациях 0,0001 мкг/мл до 10 мкг/мл. Затем раствор PBMC (1×107 клеток/мл) добавляли при 50 мкл на лунку, планшет оставляли стоять в 5% СО2-газ содержащем термостате при 37°С в течение четырех часов, и определяли специфическую скорость высвобождения Calcein-AM. Планшет подвергали центрифугированию (при 1200 об/мин в течение пяти минут при комнатной температуре), и интенсивность флуоресценции (λex = 490 нм и λem = 515 нм) 100 мкл супернатанта, собранного из каждой лунки этого планшета, измеряли с использованием флуорофотометра. Конкретную скорость высвобождения кальцеина определяли из приведенной ниже формулы (Формула 3).
Формула 3
Конкретная скорость высвобождения кальцеина (%) = (А-С)×100/(В-С).
А представляет интенсивность флуоресценции в каждой лунке; В представляет среднюю интенсивность флуоресценции лунок, в которые добавляли 50 мкл 10% ФТС/D-MEM, 50 мкл суспензионного раствора клеток-мишеней и 100 мкл раствора NP-40; и С представляет среднюю интенсивность флуоресценции лунок, в которые добавляли 50 мкл 10% ФТС/D-MEM, 50 мкл суспензионного раствора клеток-мишеней и 100 мкл 10% ФТС/D-MEM. Этот тест выполняли при N=3, и конкретную скорость высвобождения кальцеина для каждой концентрации антитела определяли с использованием Microsoft Office Excel 2007.
Ссылочный Пример 8. Способ для измерения нейтрализующей активности
(1) Установление клеточной линии BAF/F3, которая экспрессирует химерный рецептор EGFR человека (EGFR_ BAF)
С использованием стандартных способов выделяли рецептор EGF человека, имеющий последовательность, показанную в SEQ ID NO: 166 (GenBank Acc. No. NM_005228) (здесь далее называемый как ʺhEGFRʺ), и затем готовили вектор, который может экспрессировать рецептор EGF человека (pCXZD1/EGFR#3).
Пятнадцать микрограммов линеаризованного экспрессирующего вектора hEGFR (pCXZD1/EGFR#3), полученного расщеплением PvuI, трансфицировали в клетки Ba/F3 электропорацией (Gene Pulser; BioRad) при условиях 0,33 кВ и 950 мкFD. Трансфицированные клетки отбирали в среде RPMI1640, содержащей 10% ФТС, 300 мкг/мл Зеоцина и рекомбинантный Эпирегулин человека (R&AD Systems, Cat: 1195-EP/CF, 200 нг/мл); и выделяли клеточную линию EGFR_BAF.
(2) Активность анти-Эпирегулин-антител для нейтрализации зависимой от Эпирегулина человека или Эпирегулина собакоподобной обезьяны пролиферации клеток клеточной линии EGFR_BAF
Эксперименты для измерения активности анти-Эпирегулин-антител для нейтрализации зависимой от Эпирегулина человека или Эпирегулина собакоподобной обезьяны пролиферации клеток выполняли с использованием клеточной линии EGFR_BAF, выделенной способом, описанным в (1). Клетки промывали для удаления Эпирегулина человека, присутствующего во время культивирования. Затем эти клетки ресуспендировали в среде RPMI1640, содержащей 10% ФТС, а также hsEREG-His или cysEREG-His (конечная концентрация 2,5 нг/мл), и эти клетки высевали в 96-луночный планшет при плотности 2×104 клеток/100 мкл на лунку. Анти-Эпирегулин-антитело, разведенное этой средой, добавляли к этим клеткам при различных концентрациях (0,014 мкг/мл-30 мкг/мл), и затем эти клетки культивировали в 5% СО2-термостате в течение трех дней при 37°С. После культивирования добавляли реагент измерения из набора Cell Counting Kit (Dojindo), и развитие окраски проводили в течение двух часов. Затем оптическую плотность реакционного раствора (450/655 нм) измеряли с использованием Benchmark Plus (BioRad). Величину для концентрации антител 0 мкг/мл использовали в качестве контрольной величины для расчета скорости подавления роста клеток (величина OD при концентрации каждого антитела/величина OD контроля × 100(%)).
Ссылочный Пример 9. Измерение количества агрегата гель-фильтрационной хроматографией
Используя G3000SWxL (размер частиц 5 мкм, 7,9 мм I.D. x 30 см, изготовляемые TOSOH) для колонки и используя 50 мМ натрий-фосфатный буфер (рН7,0), содержащий 300 мМ NaCl в качестве подвижной фазы, величину агрегата измеряли гель-фильтрационной хроматографией. Колонку, связанную с системой Alliance (изготовляемой Waters), уравновешивали при скорости потока 0,5 мл/мин, и затем 5-10 мкг раствора антитела инжектировали в эту колонку. Элюцию антител детектировали с использованием детектора абсорбции ультрафиолета (215 или 280 нм). Из полученной хроматограммы рассчитывали долю площади пика агрегата в общей площади пика.
Ссылочный Пример 10. Измерение средней температуры тепловой денатурации (Tm) с использованием дифференциального сканирующего калориметра
Готовили 20 ммоль/л натрий-ацетатный буфер (рН 6,0), содержащий 150 ммоль/л хлорид натрия, в качестве наружного раствора для диализа, и диализ проводили в течение всего дня пропитыванием в этом наружном растворе диализной мембраны, отгораживающей раствор антитела, эквивалентным количеством 50-100 мкг антитела. Раствор антитела, приготовленный при концентрации 50 мкг/мл-100 мкг/мл антитела, использующий этот наружный раствор для диализа, использовали в качестве раствора пробы для измерения величины Tm.
Для этого эксперимента может быть использовано подходящее оборудование DSC, например, DSC-II (изготовляемое Calorimetry Sciences Corporation) или MicroCal VP-DSC (изготовляемое GE healthcare). Достаточно дегазированный раствор пробы и ссылочный раствор (наружный раствор для диализа) индивидуально заключали в ячейки калориметра и подвергали достаточному тепловому уравновешиванию при 40°С. Затем DSC-скан проходил из 40°С в 100°С со скоростью сканирования приблизительно 1К-2,5 К/мин. Результаты этого измерения даются как верхняя часть пика денатурации в качестве функции температуры. Температуру средней точки тепловой денатурации пробы рассчитывали определением этого пика Fab-домена в соответствии с Непатентным документом (Rodolfo et al., Immunology Letters (1999), p.47-52).
Ссылочный Пример 11. Измерения pI изоэлектрическим фокусированием
С использованием Phastsystem Cassette (Amersham Bioscience), Phast-Gel Dry IFF ((Amersham Bioscience) гель подвергали набуханию в течение 30 минут в растворе для набухания, имеющем состав, описанный ниже.
Этот набухший гель использовали для выполнения электрофореза с использованием PhastSystem (Amersham Bioscience), контролируемой программой, описанной ниже. Пробу добавляли в этот гель на стадии 2. Калибровочный Набор для pI (Amersham Bioscience) использовали для маркеров pI.
После электрофореза этот гель фиксировали 20% TCA и затем проводили окрашивание серебром с использованием Набора для окрашивания серебром, Protein (Amersham Bioscience) в соответствии с инструкциями, обеспеченными этим набором. После окрашивания изоэлектрическую точку рассчитывали на основе известных изоэлектрических точек маркеров pI.
Ссылочный Пример 12. Конструирование штамма, экспрессирующего афукозильные антитела
В клетке, где экспрессия обоих генов транспортера фукозы на гомологичных хромосомах искусственно ингибирована, функция транспортера фукозы ингибируется. С использованием такой клетки, может быть получено фукоза-недостаточное антитело (WO 2006/067913 и т.п.). Кроме того, фукоза-недостаточные антитела могут быть также получены, когда антитела продуцируются в клетках с принудительной экспрессией бета-1,4-N-ацетилглюкозаминилтрансферазы III и альфа-маннозидазы II Гольджи (Ferrara et al., Biotechnol. Bioeng. (2006) 93(5), 851-861). Афукозилированные антитела EREG, приготовленные способами, которые известны квалифицированным в данной области специалистам, использовали для этого исследования.
Ссылочный Пример 13. Оценивание риска иммуногенности с использованием инструмента предсказания иммуногенности in silico, Epibase
Клиническая полезность и эффективность фармацевтических препаратов антител ограничены антителами против лекарственного средства (ADA). ADA влияют на эффективность лекарственного средства и кинетику фармацевтических препаратов антител и иногда вызывают серьезные побочные эффекты. Сообщались многие влияющие на иммуногенность факторы, и, в частности, считается важным, что Т-клеточные эпитопы содержатся в антигенах. In silico инструменты, доступные для предсказания таких Т-клеточных эпитопов, включают в себя Epibase (Lonza), iTope/TCED (Antitope) и EpiMatrix (EpiVax). Сообщалось, что последовательности, содержащие Т-клеточные эпитопы, присутствующие в представляющих интерес белках, могут быть предсказаны с использованием вышеописанных инструментов (Expert Opin Biol Ther. 2007 Mar; 7(3): 405-18).
Epibase Light (Lonza) является in silico инструментом для расчета связывающей способности между 9-мерным пептидом и основным аллелем DRB1 с использованием алгоритма FASTER (Expert Opin Biol Ther. 2007 Mar; 7(3): 405-18). Этот инструмент делает возможной идентификации Т-клеточных эпитопов, которые сильно или умеренно связываются с MHС класса II.
Оценка иммуногенности in silico может быть определена для каждого модифицированного антитела в соответствии со следующей формулой (Формула 4) в системе Epibase Light (Lonza).
Формула 4
Оценка иммуногенности = Сумма (каждая величина частоты X критических эпитопов DRB1 аллотипической популяции).
Этот расчет отражает коэффициент копийности аллотипов DRB1. Для этой цели можно использовать следующий коэффициент копийности в Индо-Европейской группе населения.
DRB1*1501(24,5%), DRB1*0301(23,7%), DRB1*0701(23,3%), DRB1*0101(15,0%), DRB1*1101(11,6%), DRB1*1302(8,2%), DRB1*1401/1454(4,9%), DRB1*0901(2,3%), DRB1*1502(0,5%), DRB1*1202(0,1%).
Все эпитопы, содержащиеся в каждой модифицированной последовательности антитела, которые проявляют сильное или умеренное связывание, идентифицируют алгоритмом FASTER, и затем эпитопы после исключения последовательностей зародышевой линии человека и последовательностей соединения между вариабельным районом и константным районом, используют в качестве критических эпитопов в расчете оценки иммуногенности. Когда оценка является меньшей, это означает, что последовательность имеет более низкий риск иммуногенности.
Ссылочная последовательность 14. Тесты лекарственной эффективности на анти-Эпирегулин-антителах с использованием моделей in vivo.
(1) Поддержание клеточных линий, используемых для трансплантации в модели in vivo
Клетки MIA PаCа-2 (АТСС) и DLD-1 (АТСС) использовали для моделей in vivo. Клетки MIA PаCа-2 поддерживали субкультивированием в модифицированной по способу Дульбекко среде Игла (Invitrogen), содержащей 10% ФТС, 2,5% лошадиную сыворотку, 4 ммоль/л L-глутамин (Invitrogen), 4,5 г/л глюкозы (Invitrogen) и 1,5 г/л бикарбоната натрия (Invitrogen) (здесь далее называемой субкультуральной средой). Клетки DLD-1 поддерживали субкультивированием в среде RPMI1640 (Sigma), содержащей 10% ФТС, 10 ммоль/л HEPES (Invitrogen), 4,5 г/л глюкозы (Invitrogen) 1 ммоль/л пируват натрия (Invitrogen) (здесь далее называемой субкультуральной средой).
(2) Получение моделей мышей, трансплантированных клетками MIA PаCа-2 и DLD-1
С использованием раствора, содержащего субкультуральную среду и MATRIGEL Matrics (BD Bioscience) при соотношении 1:1, суспензии MIA PаCа-2 и DLD-1 готовили при 5×107 клеток/мл. 100 мкл этой клеточной суспензии (5×106 клеток на мышь) трансплантировали подкожно в брюшной области мышей SCID (самок, 5-недельного возраста, CLEA Japan, Inc.). Объем опухоли рассчитывали с использованием Формулы 5, и когда средний объем опухоли достигал 130-330 мм3, эту модель мыши считали установленной.
Формула 5
Объем опухоли = длинный диаметр × короткий диаметр × короткий диаметр/2.
(3) Получение вводимых проб, содержащих каждое тест-антитело
Вводимые пробы, каждая из которых содержит антитело cH-G1/cL-k, антитело H206-G1d/L7-3k или антитело H240-G1d/L73-k при 0,04 мг/мл (0,4 мг/кг группы введения) или 0,2 мг/мл (2 мг/кг группы введения), готовили с физиологическим солевым раствором в день введения.
(4) Введение антителосодержащих вводимых проб
Вводимые пробы, приготовленные в (3), вводили через хвостовую вену моделям мышей, полученных в (2), один раз в неделю в течение 14 дней, начиная от дня 14 после трансплантации клеток MIA PаCа-2, и один раз в неделю в течение 14 дней, начиная от дня 12 после трансплантации клеток DLD. В качестве отрицательного контроля, физиологический солевой раствор вводили сходным образом в дозе 10 мл/кг через хвостовую вену один раз в неделю в течение соответствующих периодов времени. Все группы имели пять животных в каждой группе, и каждой группе вводили соответствующую антителосодержащую пробу введения.
(5) Оценивание противоопухолевого эффекта каждого тест-антитела
Противоопухолевый эффект каждого тест-антитела оценивали в модели мыши, трансплантированной раковыми клетками человека. Объем опухоли измеряли в день 3 или день 7 после последнего дня введения пробы.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА АНТИ-TNFRSF25 | 2017 |
|
RU2746314C2 |
| АНТИ-PD-L1 АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2783685C2 |
| АНТИ-SIRPα АНТИТЕЛА | 2018 |
|
RU2771174C2 |
| АНТИ-PD-L1 АНТИТЕЛА | 2015 |
|
RU2722212C2 |
| АНТИТЕЛО К PD-1 И ЕГО ПРИМЕНЕНИЕ | 2014 |
|
RU2663795C2 |
| Анти-LAG-3 антитела | 2016 |
|
RU2760582C2 |
| АНТИ-CEACAM1 АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2018 |
|
RU2754876C2 |
| АНТИТЕЛА К ICOS | 2016 |
|
RU2742241C2 |
| АНТИ-C5 АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2774716C2 |
| АНТИ-ГАЛЕКТИН-9 АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2840501C1 |























































































































































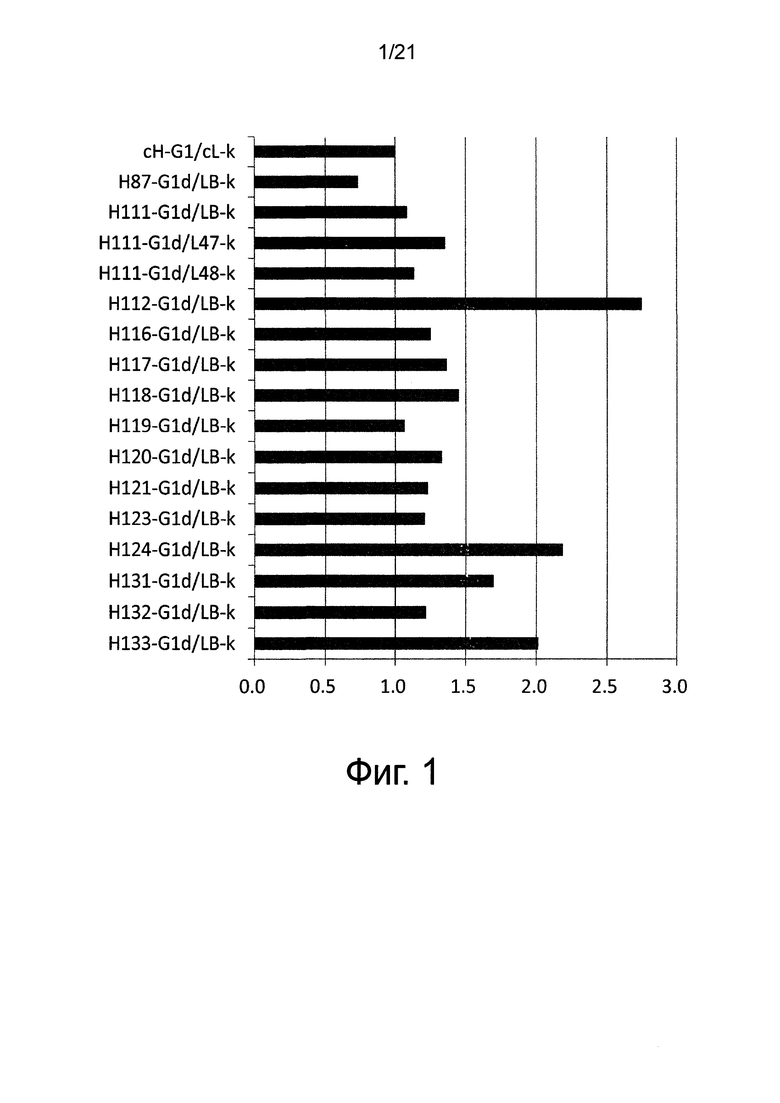
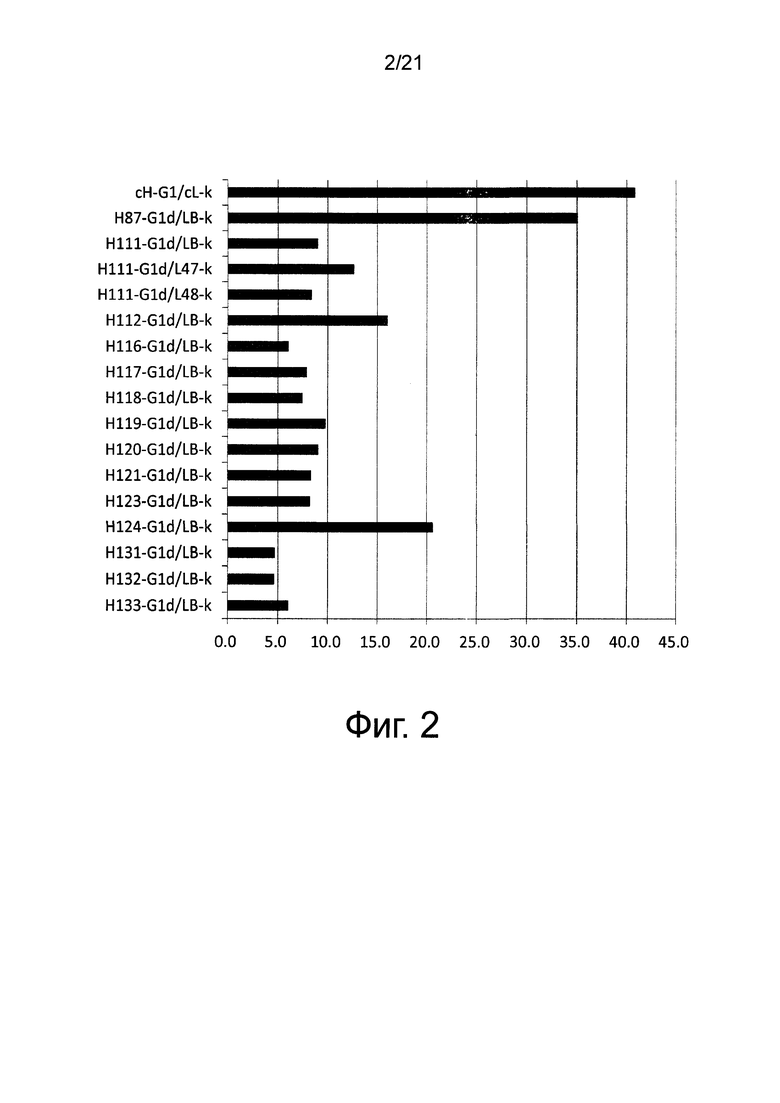

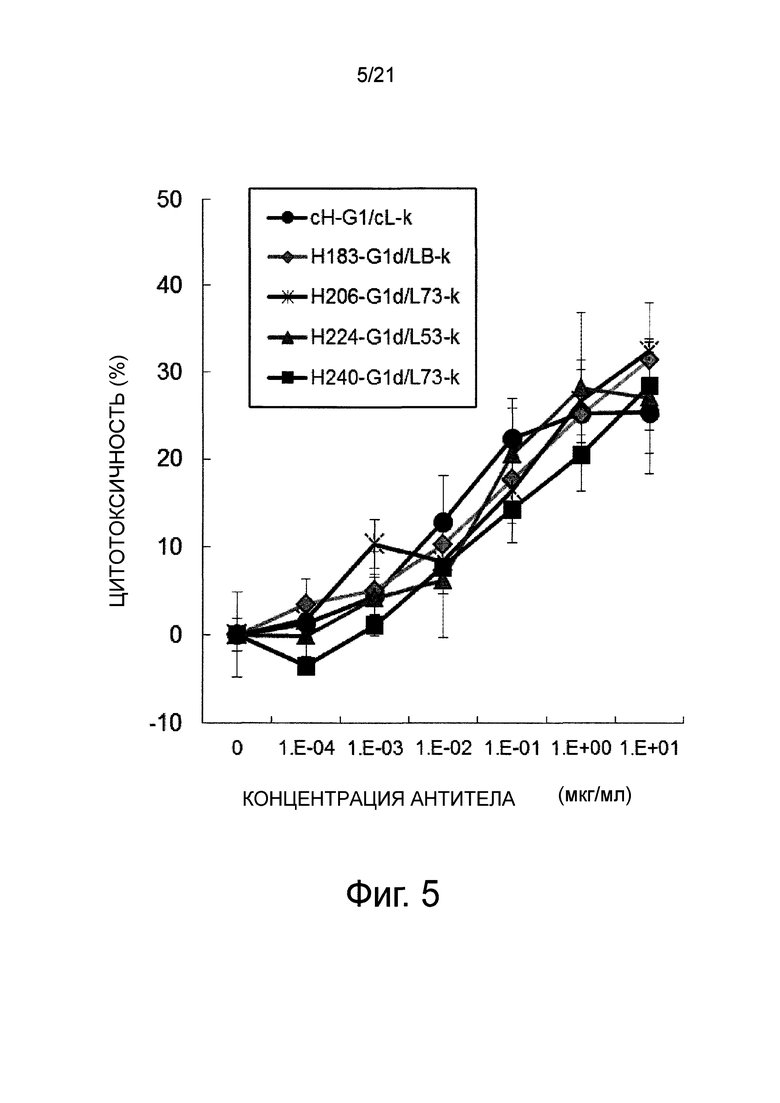
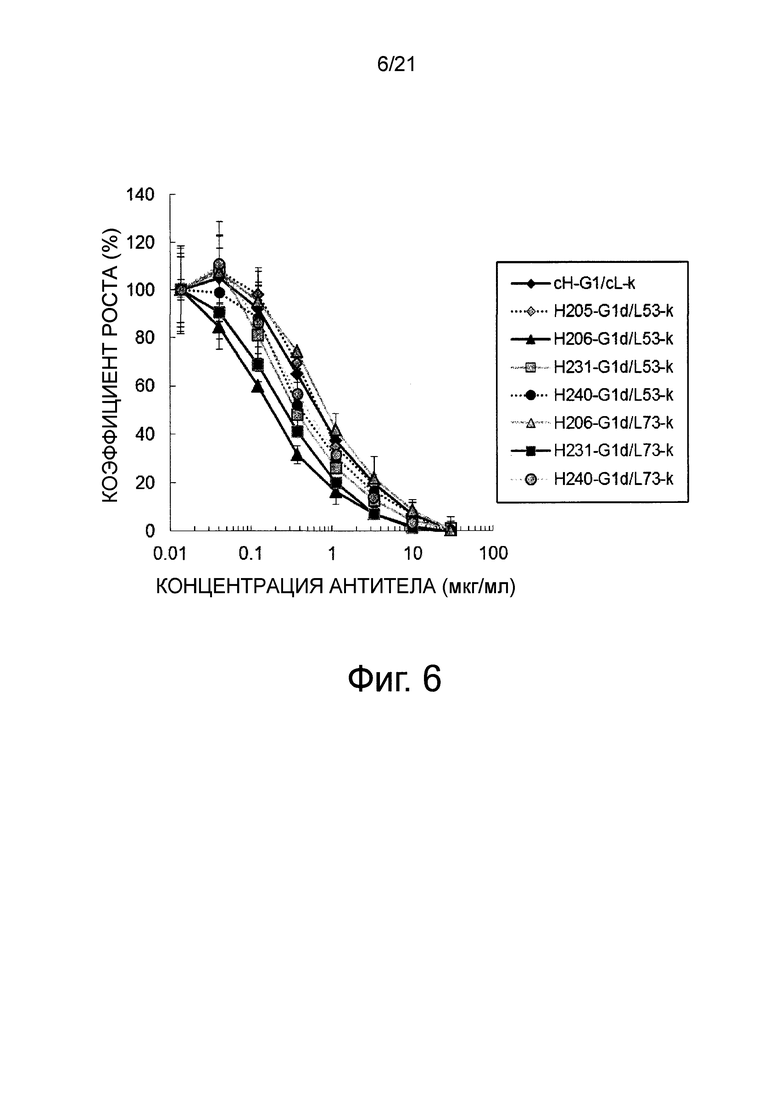

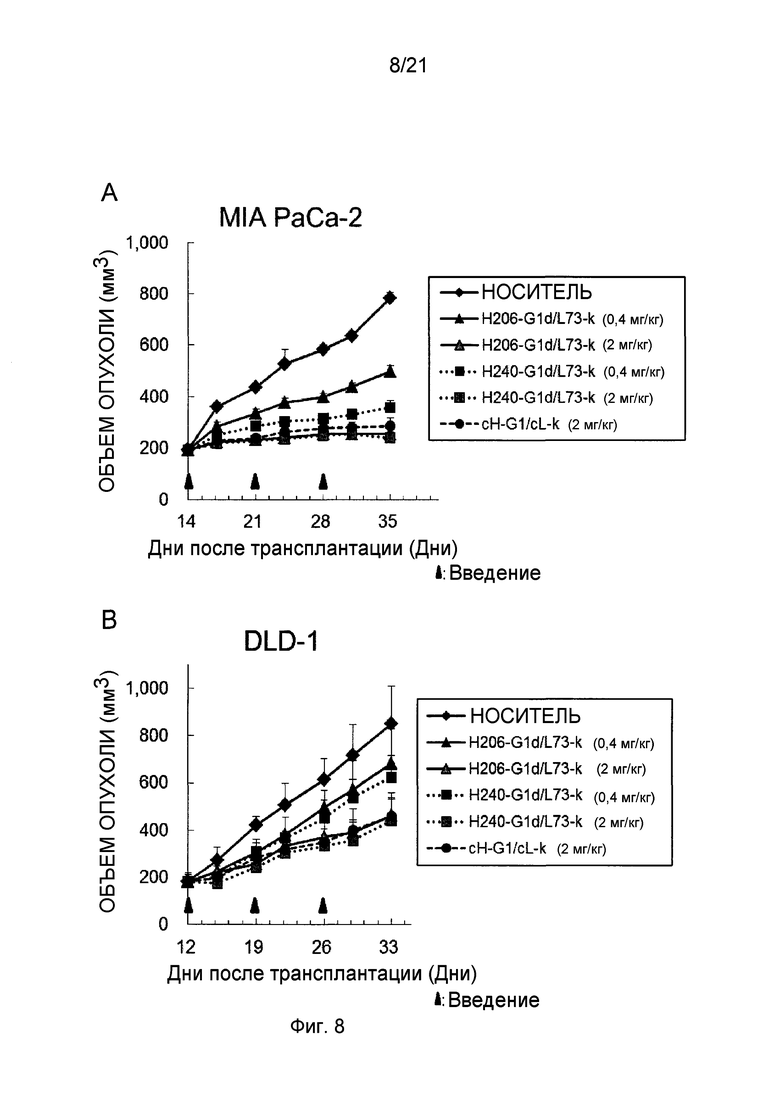

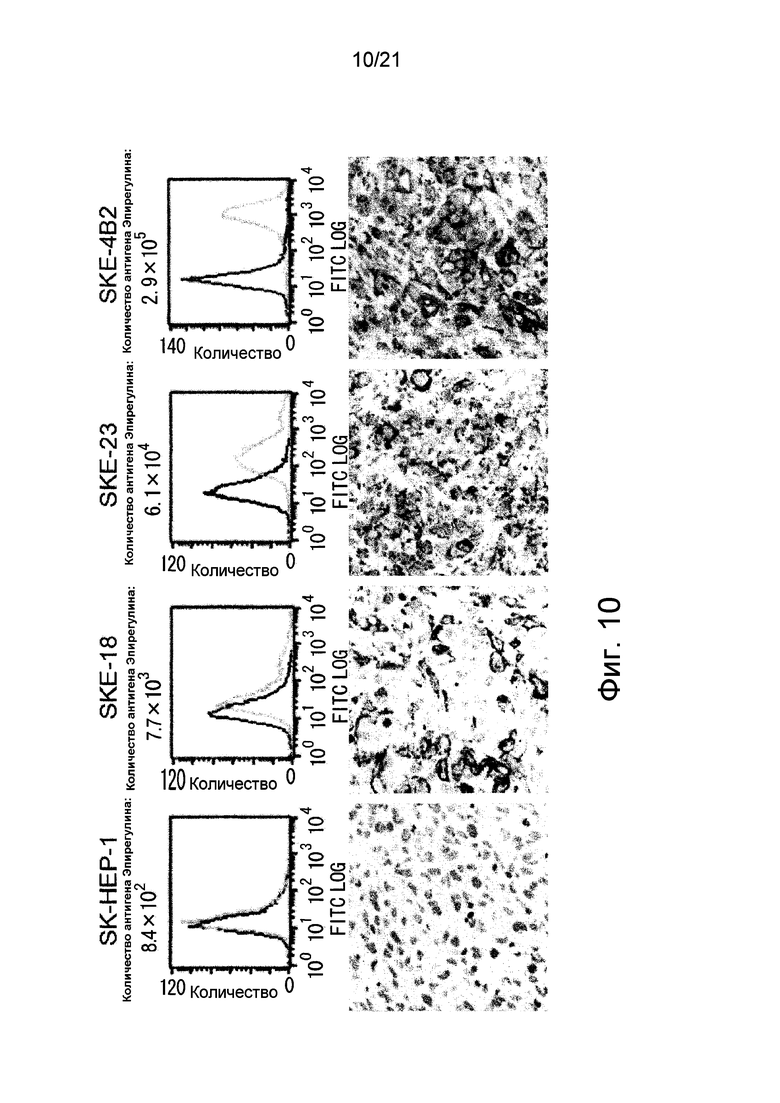
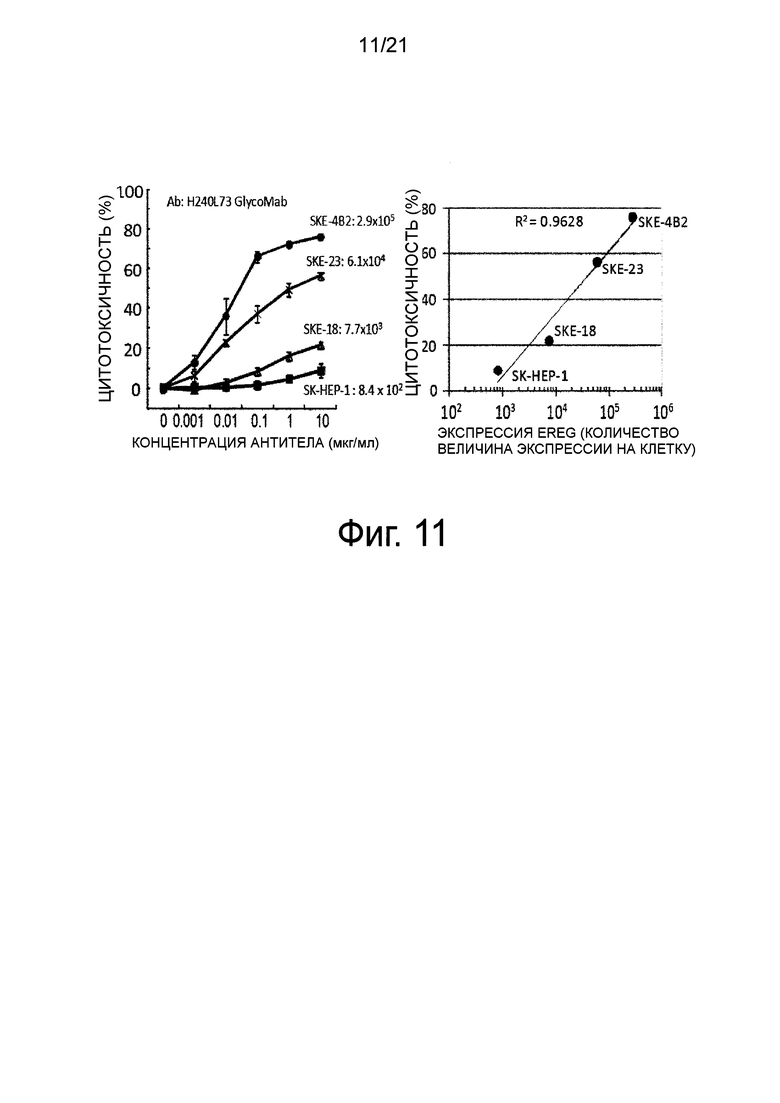
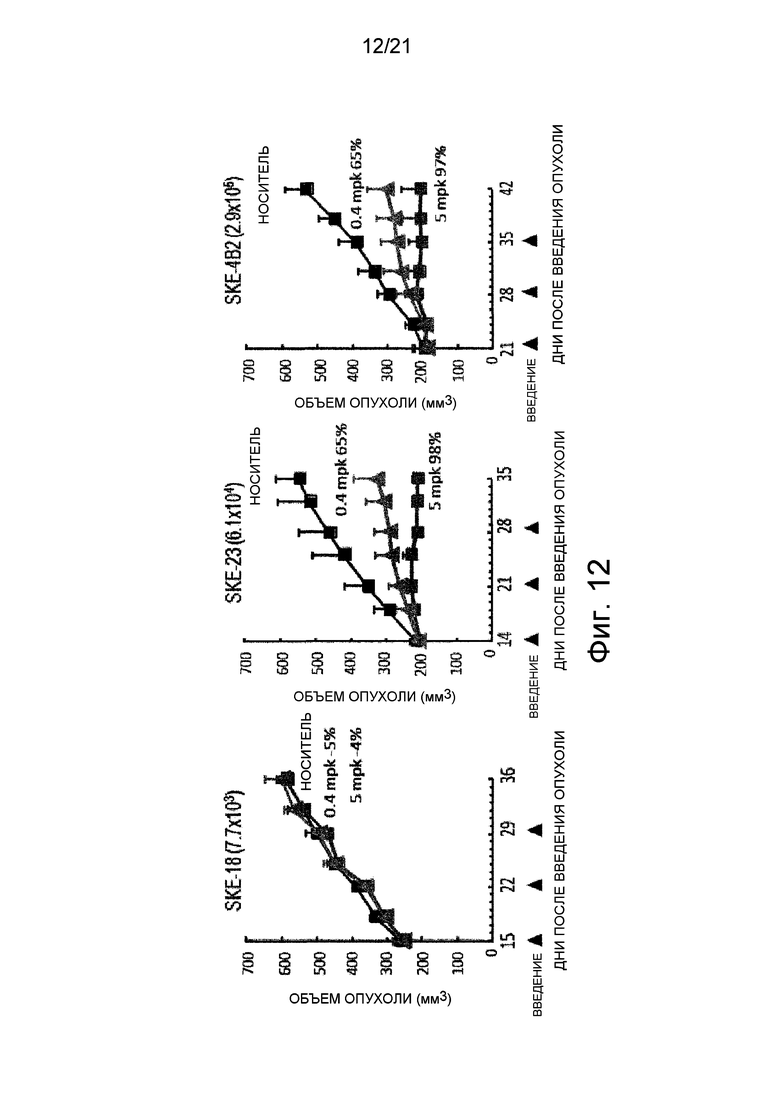

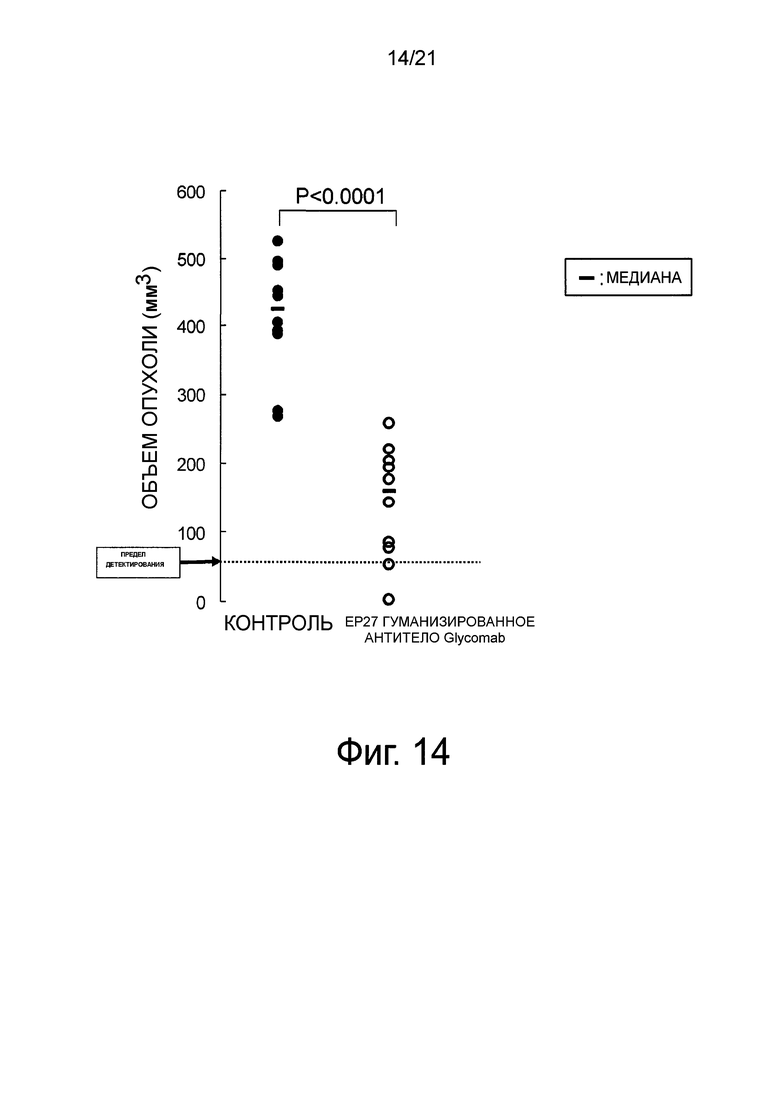
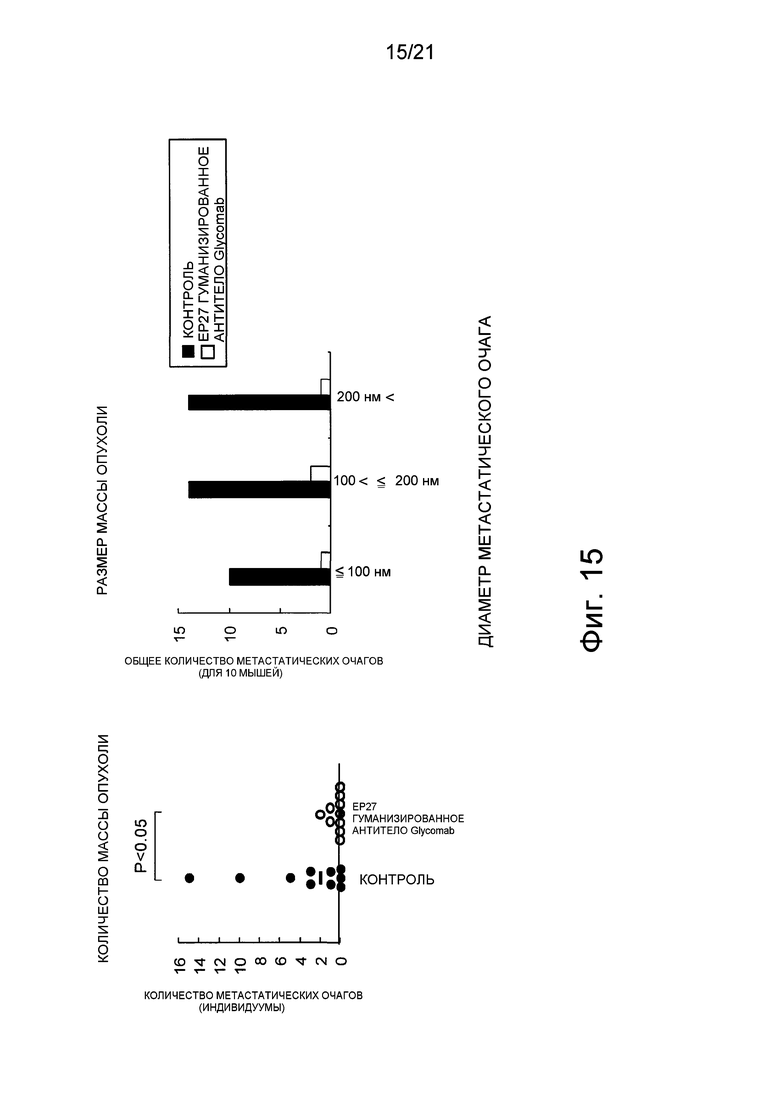



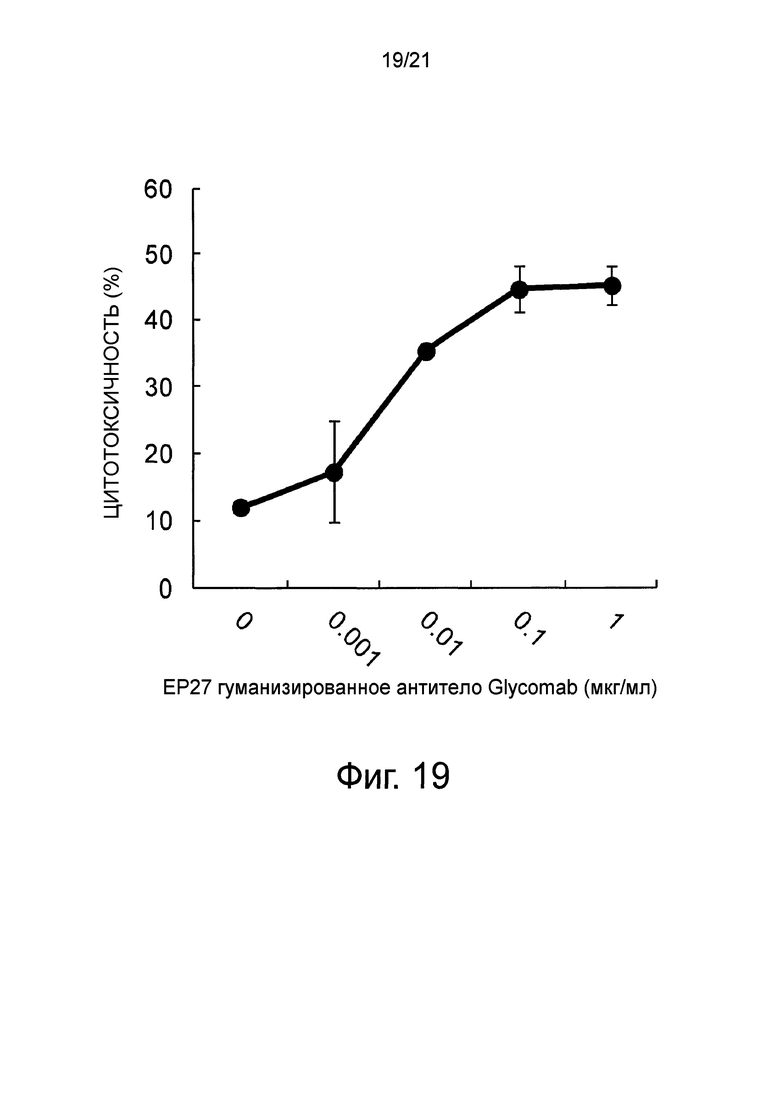

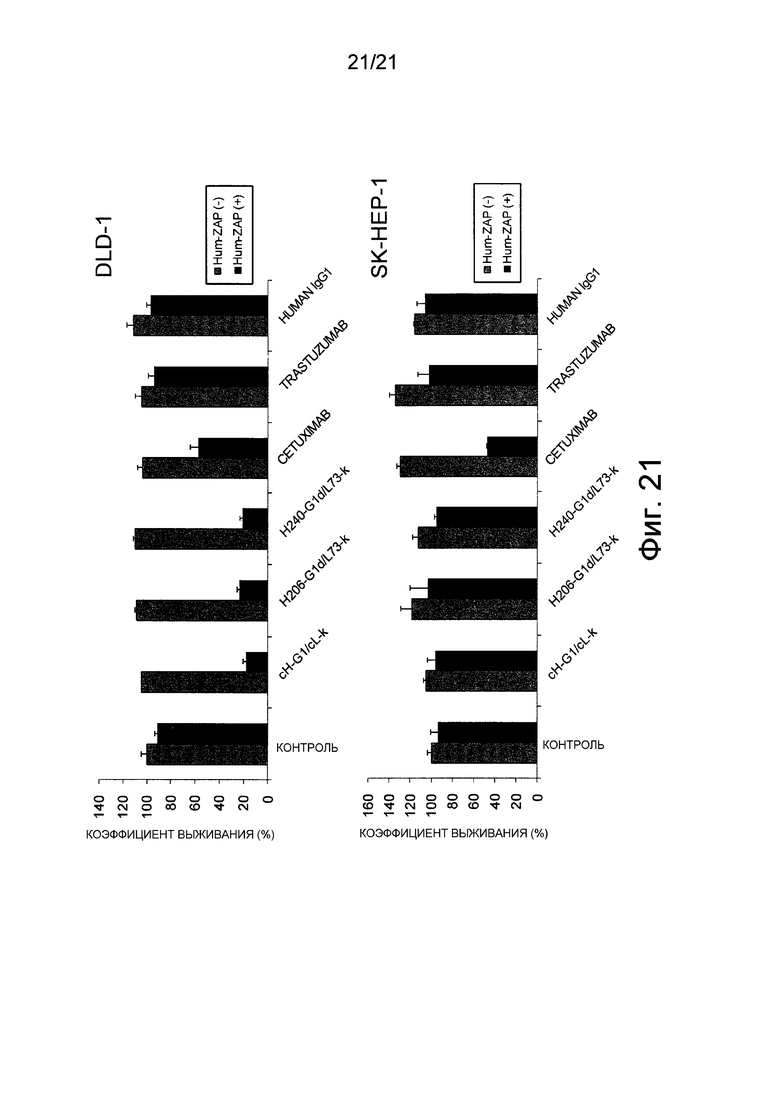
Изобретение относится к медицине. Изобретение представляет собой антитело к Эпирегулину, которое содержит вариабельную область тяжелой цепи, включающую CDR1 тяжелой цепи последовательности SEQ ID NO: 9, CDR2 тяжелой цепи SEQ ID NO: 160 и CDR3 тяжелой цепи SEQ ID NO: 158; и вариабельную область легкой цепи, включающую CDR1 легкой цепи SEQ ID NO: 163, CDR2 легкой цепи SEQ ID NO: 13 и CDR3 легкой цепи SEQ ID NO: 164, которое содержит вариабельную область тяжелой цепи, соответствующую вариабельной области тяжелой цепи в SEQ ID NO: 150, и вариабельную область легкой цепи, соответствующую вариабельной области в SEQ ID NO: 141. константную область тяжелой цепи SEQ ID NO: 30, которое содержит константную область легкой цепи SEQ ID NO: 27, обладает нейтрализующей активностью, цитотоксичностью, где цитотоксичность представляет собой CDC и/или ADCC. Изобретение ингибирует рост раковых клеток проявлением цитотоксической активности и нейтрализующей активности против Эпирегулин-экспрессирующих раковых клеток. 6 з.п. ф-лы, 23 табл., 21 ил., 19 пр.
1. Антитело к Эпирегулину, которое содержит вариабельную область тяжелой цепи, включающую CDR1 тяжелой цепи последовательности SEQ ID NO: 9, CDR2 тяжелой цепи SEQ ID NO: 160 и CDR3 тяжелой цепи SEQ ID NO: 158, и вариабельную область легкой цепи, включающую CDR1 легкой цепи SEQ ID NO: 163, CDR2 легкой цепи SEQ ID NO: 13 и CDR3 легкой цепи SEQ ID NO: 164.
2. Антитело к Эпирегулину по п. 1, которое содержит вариабельную область тяжелой цепи, соответствующую вариабельной области тяжелой цепи в SEQ ID NO: 150, и вариабельную область легкой цепи, соответствующую вариабельной области в SEQ ID NO: 141.
3. Антитело по п. 1, которое содержит константную область тяжелой цепи SEQ ID NO: 30.
4. Антитело по п. 1, которое содержит константную область легкой цепи SEQ ID NO: 27.
5. Антитело по п. 1, которое обладает нейтрализующей активностью.
6. Антитело по п. 1, которое обладает цитотоксичностью.
7. Антитело по п. 6, где цитотоксичность представляет собой CDC и/или ADCC.
| РОТАЦИОННЫЙ ВИСКОЗИМЕТР | 0 |
|
SU231209A1 |
| WO 2008047723 A1, 24.04.2008 | |||
| WO 2010142952 A2, 16.12.2010. | |||

