Родственные заявки
По настоящей заявке испрашивается приоритет временной патентной заявки США с серийным номером № 61/445455, поданной 22 февраля 2011 года, временной патентной заявки США с серийным номером № 61/443103, поданной 15 февраля 2011 года и временной патентной заявки США с серийным номером № 61/355978, поданной 17 июня 2010 года, содержание которых включено в настоящее описание в качестве ссылки в полном объеме.
Ссылка на список последовательностей, предоставленный через EFS-WEB
Полное содержание следующего предоставленного в электронном виде списка последовательностей через сервер USPTO EFS-WEB, как утверждено и указано в MPEP §1730 II.B.2(a)(C), включено в настоящее описание в качестве ссылки в полном объеме для всех целей. Список последовательностей указан в следующем поданном в электронном виде текстовом файле:
Область техники
Изобретение относится к области пассивной иммунизации против гриппа. Более конкретно, изобретение относится к антителам, которые связываются вблизи консенсусной последовательности участка расщепления при созревании гемагглютинина A, HA0, вируса гриппа, в том числе к антителам, секретируемым клетками человека.
Уровень техники
Белок гемагглютинин вируса гриппа имеет глобулярный головной домен, который является высоко гетерогенным среди штаммов flu, и стеблевую область, содержащую участок слияния, который требуется для вхождения в клетки. Белок гемагглютинин (HA0) активируется, позволяя участку слияния обеспечивать вирулентность путем расщепления на участки HA1 и HA2, которые остаются связанными дисульфидными связями, но претерпевают конформационное изменение. Этот участок расщепления содержит консенсусную последовательность, которая является общей для вируса гриппа A и вируса гриппа B, и для различных штаммов вируса гриппа A и B.
В статье Bianchi, E., et al., J. Virol. (2005) 79:7380-7388 описана “универсальная” вакцина против вируса гриппа B на основе консенсусной последовательности этого участка расщепления, которая была способна индуцировать антитела у мышей, когда она была конъюгирована с белковым комплексом наружной мембраны Neisseria meningitidis. Также описаны моноклональные антитела, которые, по-видимому, связываются с консенсусной последовательностью. Кроме того, был показан успешный пассивный перенос антисыворотки у мышей. Предшествующие вакцины, такие как вакцины, описанные в WO 2004/080403, содержащие пептиды, происходящие из белков M2 и/или HA вируса гриппа, индуцируют антитела, которые либо обладают слабой эффективностью, либо не являются эффективными среди штаммов.
Описание изобретения
Изобретение относится к моноклональным антителам, которые связывают эпитоп, являющийся общим для множества штаммов вируса гриппа, и, более конкретно, которые связывают представителей любой или обеих из группы 1 и группы 2 вируса гриппа A. Такие антитела способны обеспечить пассивный иммунитет в случае пандемии, вызванной, например, ранее неидентифицированным штаммом вируса гриппа или штаммом, против которого не обеспечивается защита доступными в настоящее время сезонными вакцинами. Поскольку антитела связываются со многими штаммами, что указывает на нацеливание на необходимый участок и, таким образом, вероятно включенный даже в ранее не встречавшиеся штаммы, такая вакцина может быть эффективной в этих обстоятельствах. Такие антитела также подходят для смягчения или предупреждения инфекции у индивидуумов, у которых вакцинация не вызвала полный защитный ответ или которые имеют высокий риск вследствие слабой иммунной системы (например, очень молодые, пожилые пациенты, пациенты с трансплантатами, пациенты, подвергаемые лечению химиотерапией против злокачественной опухоли или ВИЧ).
Таким образом, в одном из аспектов изобретение относится к моноклональным антителам или их иммунореактивным фрагментам, которые широко перекрестно реагируют с вирусом гриппа A группы 1, включая H1, H2, H5, H6, H8, H9, H11, H13, H16, или группы 2, включая H3 и H7, в качестве типовых образцов, или которые демонстрируют перекрестную реактивность между группами. Антитела специфично связываются с эпитопом, содержащимся в белке HA0 вируса гриппа, и распознают нативную тримерную форму HA. Как хорошо известно в данной области, белки не на основе иммуноглобулинов могут иметь сходные с антителом свойства распознавания эпитопов и также могут обеспечивать подходящие варианты осуществления, включая связывающие агенты на основе фибронектина, трансферрина, липокалина, или аптамеры на основе нуклеиновых кислот.
В других аспектах изобретение относится к способам применения антител и фрагментов по изобретению для пассивного ингибирования вирусной инфекции у индивидуумов. Изобретение также относится к рекомбинантным материалам и способам продуцирования этих антител или фрагментов.
Краткое описание чертежей
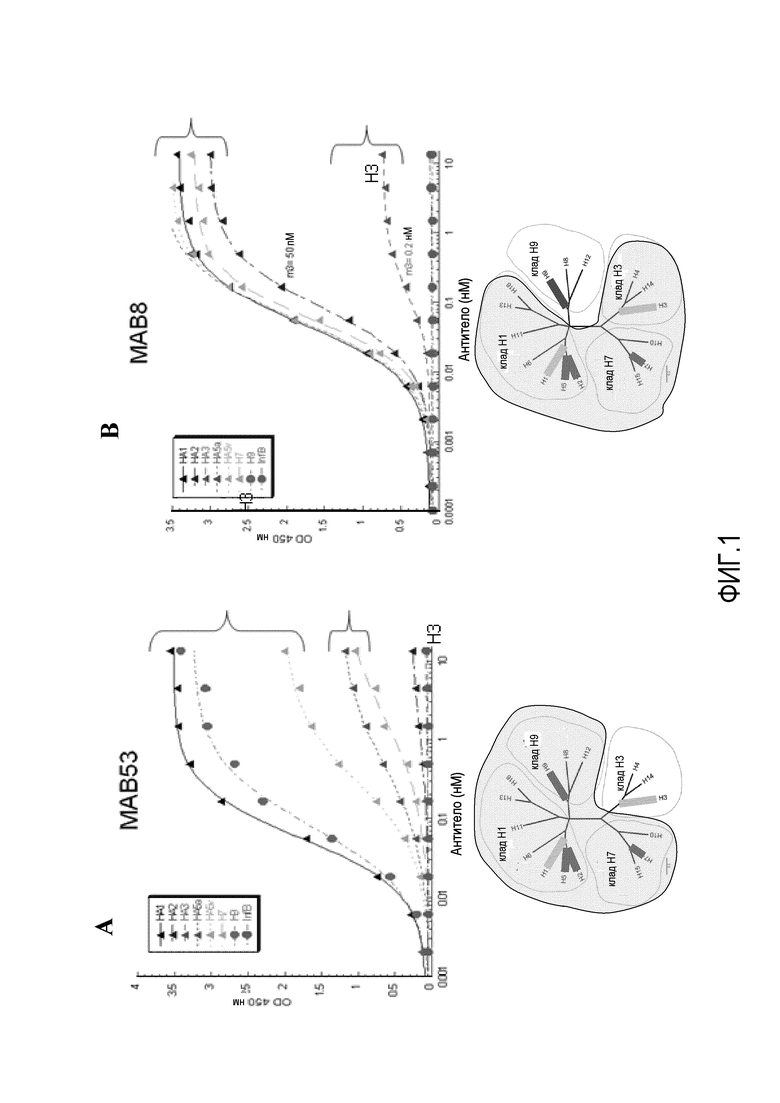
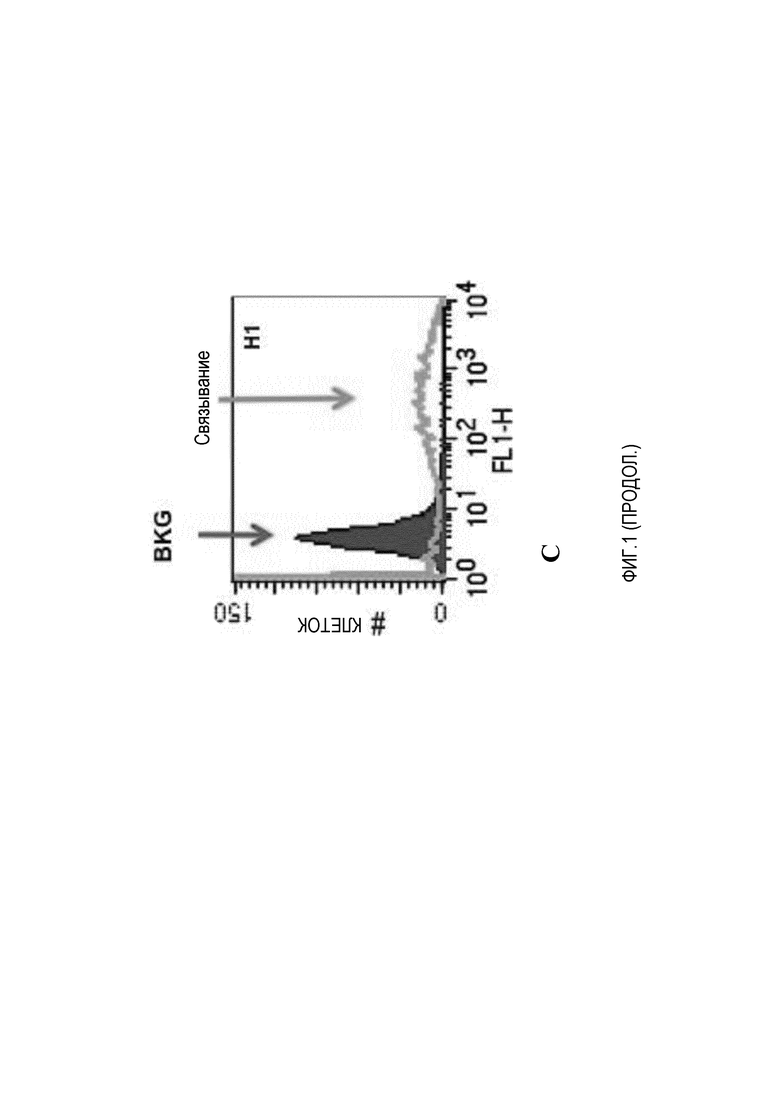
На фиг.1A и 1B представлены результаты связывания MAB53 и MAB8 с белком HA0 из различных кладов вируса гриппа, полученные с помощью ELISA. На фиг.1C показано, что MAB53 связывается с нативным тримером, экспрессируемым в клетках HEK293.
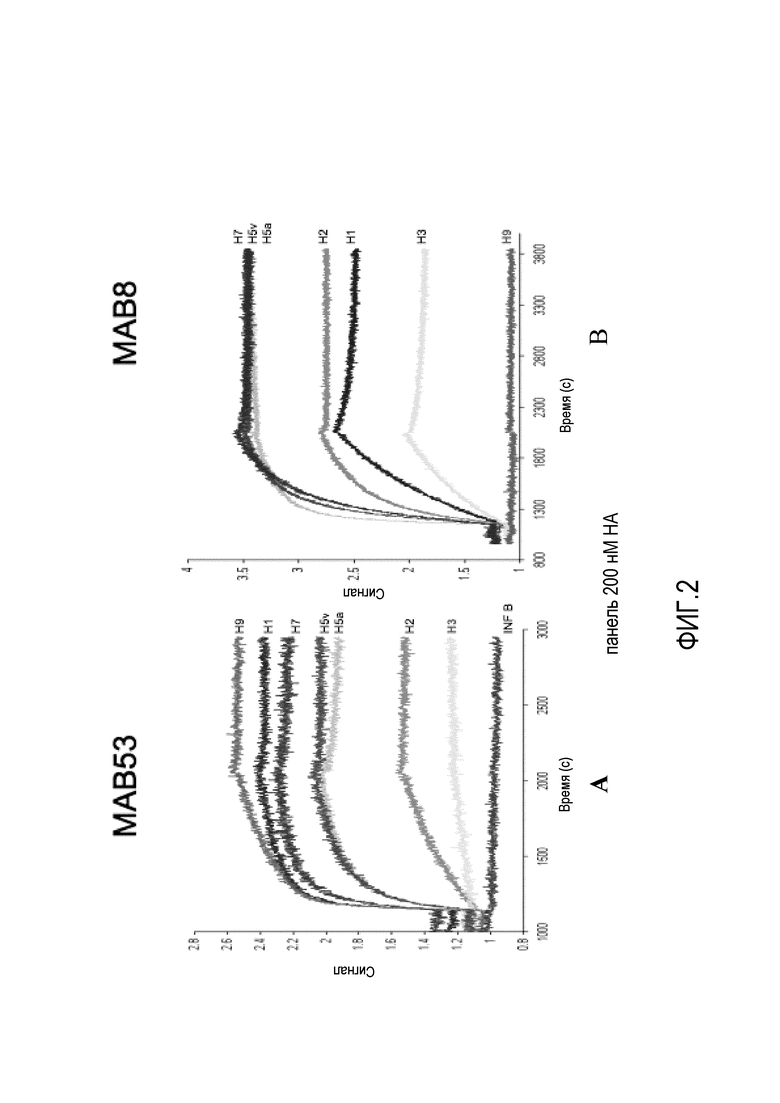
На фиг.2A и 2B представлены результаты связывания MAB53 и MAB8 с белком HA0 из различных кладов, полученные с помощью биосенсора ForteBio®.
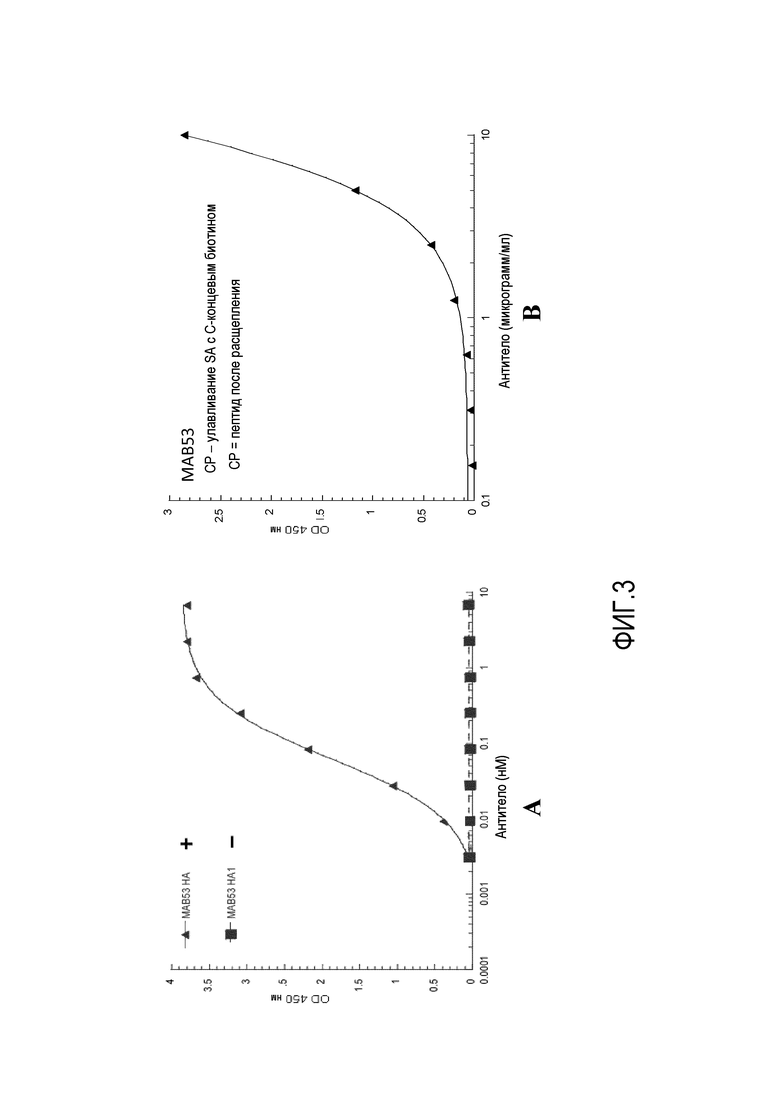
На фиг.3A представлена степень связывания, исследованная с помощью ELISA для MAB53, в отношении HA0 в качестве целого белка и фрагмента HA1 после расщепления. На фиг.3B представлена степень связывания MAB53 с пептидом, обозначенным CP, из HA2.
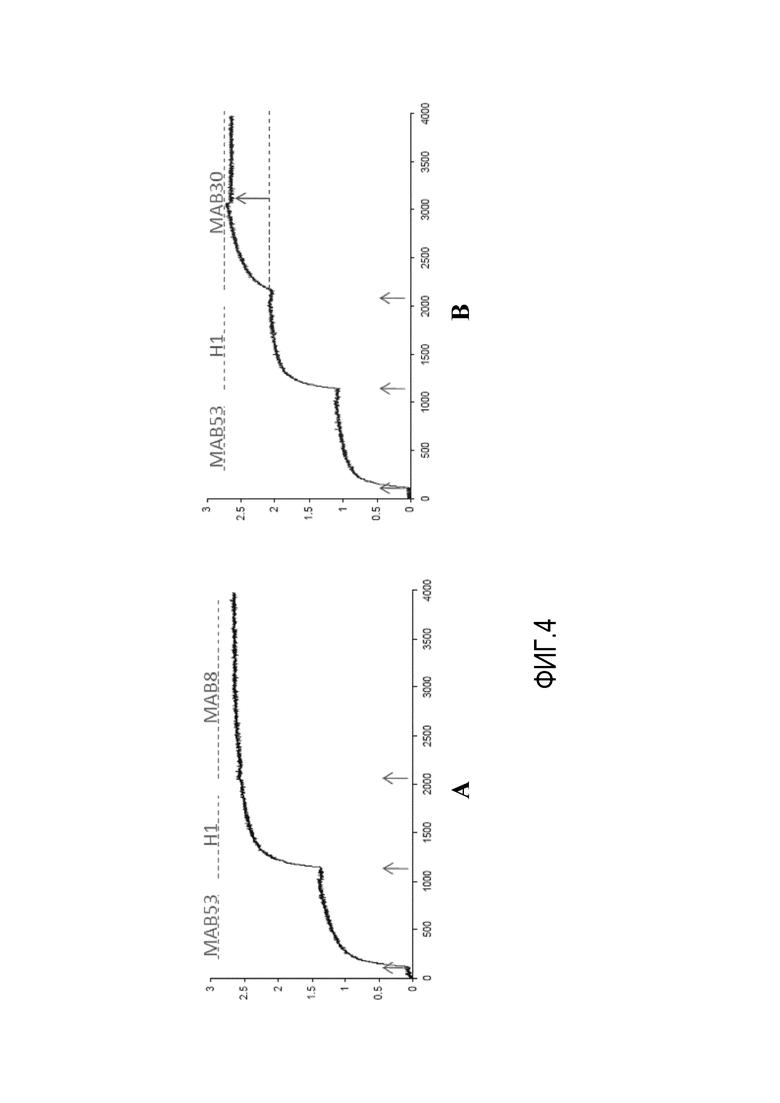
На фиг.4A и 4B представлены результаты анализа ForteBio®, демонстрирующие, что MAB53 конкурирует с MAB8, но не с MAB30.
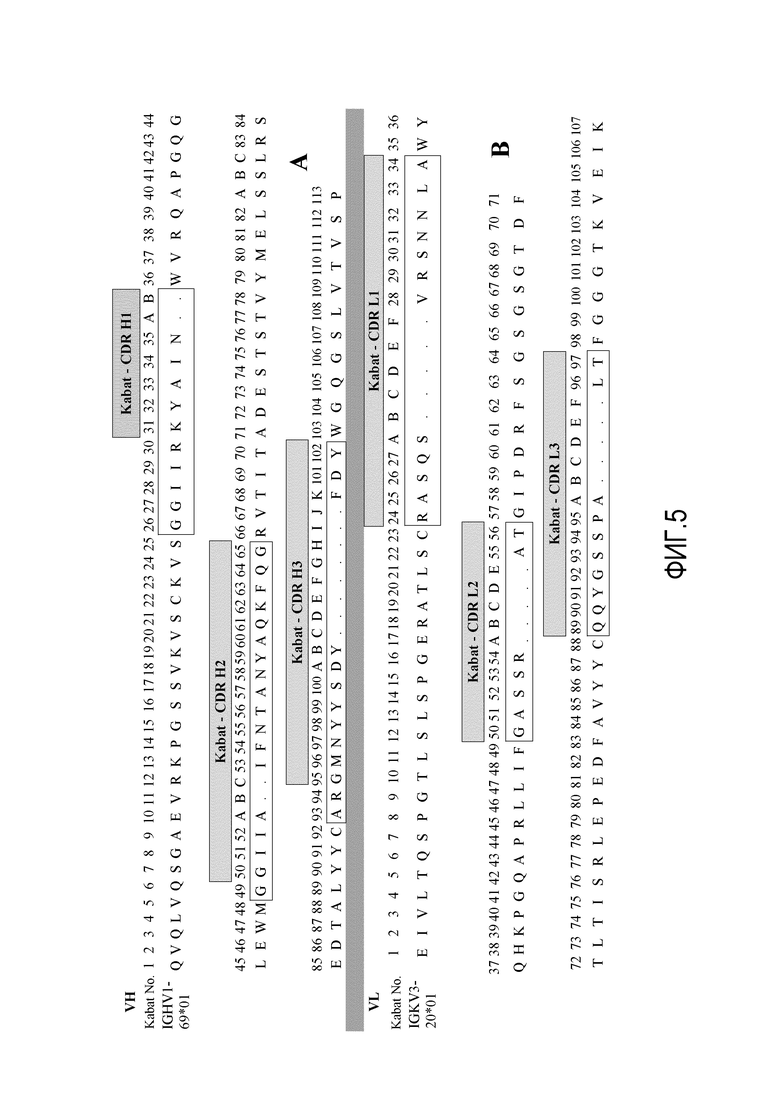
На фиг.5A и 5B представлено картирование CDR согласно номеру по Kabat вариабельных областей тяжелой и легкой цепей MAB53. IGHV1-69*01 представляет собой SEQ ID NO:83 и IGKV3-20*01 представляет собой SEQ ID NO:84.
На фиг.6 представлена нейтрализация H1N1 различными количествами MAB53 при измерении с помощью анализа бляшкообразования in vitro.
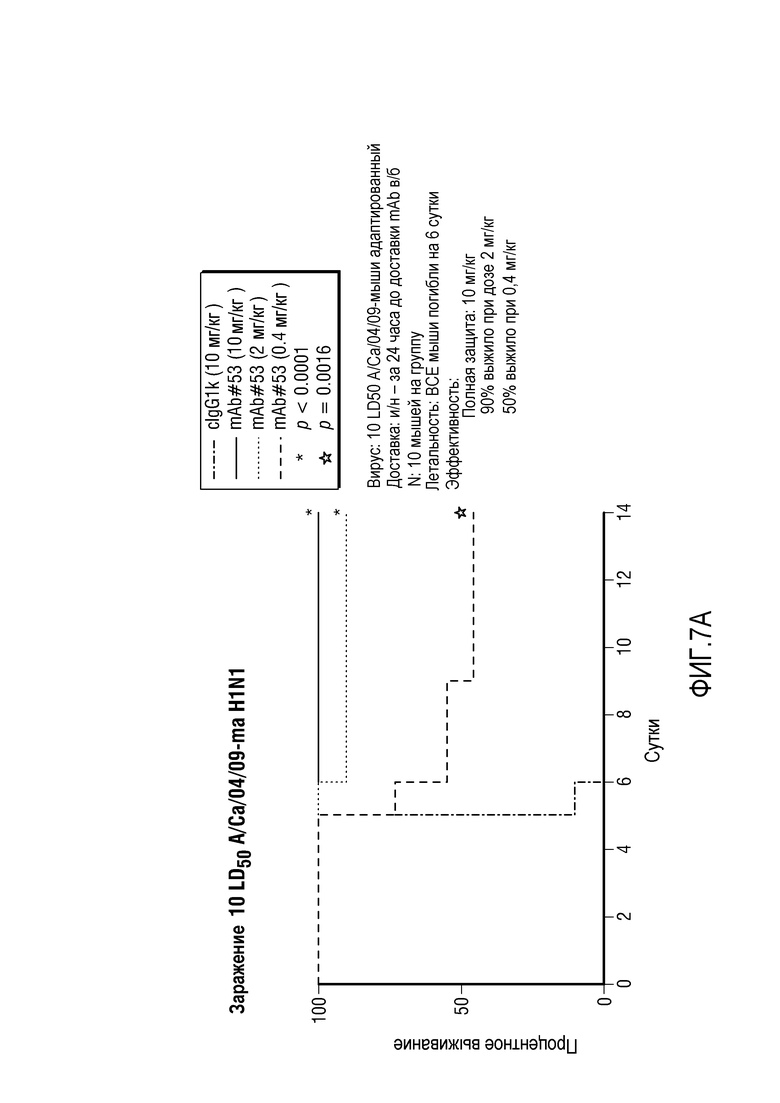
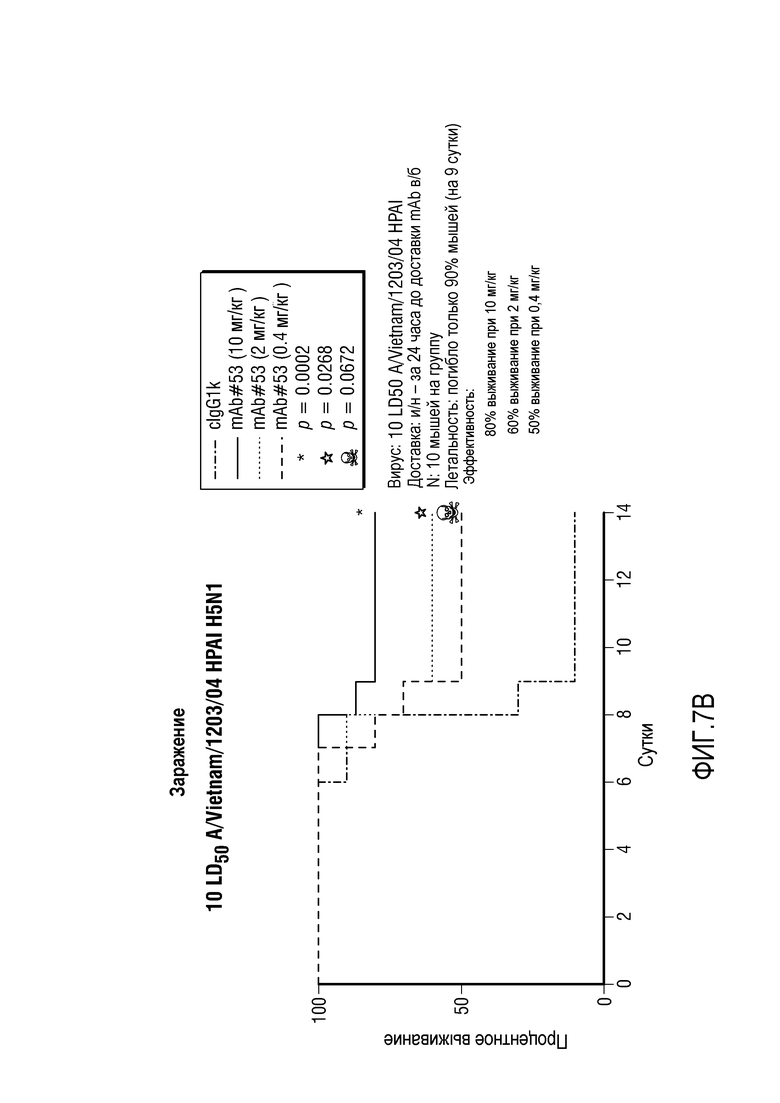
На фиг.7A и 7B представлено время выживания для мышей, которых заразили H1N1 (панель A) или H5N1 (панель B), в качестве функции введения различных количеств MAB53.
На фиг.8 представлен эффект введения MAB53 после инфицирования H5N1.
Способы осуществления изобретения
Настоящее изобретение относится к эффективным антителам, в том числе к получению эффективных средств для идентификации клеток, которые секретируют такие антитела, так чтобы соответствующие кодирующие последовательности можно было извлечь и сохранить для последующего и легкого рекомбинантного продуцирования таких антител. Способ включает модель процедуры скрининга на основе бинарной логики.
Такую процедуру можно легко использовать в отношении клеток человека с использованием, в частности, способа CellSpot™, описанного в патенте США 7413868, содержание которого включено в настоящее описание в качестве ссылки. В кратком изложении, способ способен осуществлять скрининг индивидуальных клеток, полученных от человека (или других индивидуумов) в высокопроизводительных анализах, использующих мечение метками в виде частиц и наблюдение под микроскопом. В одном из иллюстративных вариантов осуществления даже одну клетку можно анализировать в отношении антител, которые она секретирует, путем обеспечения адсорбирования или связывания секретированных антител с поверхностью, а затем обработки поверхности желаемыми антигенами, каждый из которых связан с отдельной меткой в виде частиц. Таким образом, с помощью микроскопа можно идентифицировать отпечаток клетки. С использованием этого способа миллионы клеток можно подвергать скринингу в отношении секреции желаемых антител и даже можно выделять редкие антитела, такие как антитела, желаемые в рамках изобретения для пассивной иммунизации против вируса гриппа между штаммами. Поскольку люди имеют существующие антитела по меньшей мере к некоторым штаммам вируса гриппа и поскольку антитела, получаемые способом по изобретению, связывают консервативную последовательность, эти антитела служат для нацеливания на новые штаммы, а также на штаммы, которые встречались в человеческой популяции.
Изобретение относится к способу идентификации моноклонального антитела, которое связывается с областью вблизи консенсусной последовательности участка расщепления гемагглютинина (HA0). Способ включает приведение в контакт моноклональных антител-кандидатов или фрагментов-кандидатов с: i) пептидом, по существу состоящим из аминокислотной последовательности выше или ниже указанной консенсусной последовательности, но лишенным указанной консенсусной последовательности; ii) пептидом, по существу состоящим из аминокислотной последовательности выше указанной консенсусной последовательности и включающим указанную консенсусную последовательность; и iii) пептидом, по существу состоящим из аминокислотной последовательности ниже указанной консенсусной последовательности и включающим указанную консенсусную последовательность; где моноклональное антитело, которое связывается с пептидом согласно ii) и iii), но не с пептидом согласно i) идентифицируют в качестве пептида, который специфично связывается с консенсусной последовательностью участка расщепления HA0. Также можно использовать другие комбинации, как будет очевидно квалифицированному специалисту, при условии соблюдения бинарной логики. Например, i) может представлять собой пептид, по существу состоящий из аминокислоты выше консенсусной последовательности первого штамма и лишенный консенсусной последовательности, где ii) представляет собой целую последовательность HA0 из первого штамма и iii) представляет собой целую последовательность HA0 из второго штамма. Также можно использовать более короткие участки. Для дальнейшего подтверждения также можно использовать выделенный пептид из консервативной области, хотя полагают, что информация, полученная от более крупных белковых доменов, является более информативной в отношении распознавания интактного антигена.
Этот способ не ограничивается использованием технологии CellSpot™, также как он не ограничивается антителами человека. Бинарную логику этого способа можно использовать в любом альтернативном способе скрининга. Аналогично, ее можно использовать к другим библиотекам разнообразия, помимо природных иммуноглобулинов.
Способ по изобретению основан на бинарной логике, где пептиды, которые содержат желаемую консенсусную последовательность и дополнительные вышележащие и/или нижележащие участки, используют в качестве исследуемых пептидов и оценивают их способность образовать комплекс с антителами по сравнению с областями, лишенными консенсусной последовательности. Таким образом, получают профили, с помощью которых можно сразу идентифицировать клетки, секретирующие соответствующие антитела.
В одном иллюстративном варианте осуществления для оценки популяции секретируемых антител используют три антигена. Первый пептид представляет собой всю или по существу всю аминокислотную последовательность выше консенсусной последовательности, содержащейся в HA0, и связан с меткой в виде частиц, например, красного цвета. Второй исследуемый антиген содержит эти вышележащие последовательности, но также содержит консенсусную последовательность и является меченным частицами другого цвета, например, синего. Третий исследуемый пептид содержит консенсусную последовательность и все или по существу все из нижерасположенных областей белка HA0, и является меченным частицами третьего цвета, например, зеленого. (Под вышележащей частью подразумевают участок в направлении N-конца от консенсусной последовательности и под нижележащей частью подразумевают продолжение аминокислотной последовательности от консенсусной последовательности до C-конца. Под “по существу всеми” подразумевают отсутствие только одной или нескольких не являющихся необходимыми аминокислот.) Антитела, которые связываются с консенсусной последовательностью, связываются с пептидами, меченными как зелеными, так и синими частицами, но они не связываются с меченной красным цветом вышележащей последовательностью, лишенной консенсусной последовательности. Если желательно, специфичность можно подтверждать путем добавления четвертого пептида, представляющего собой только нижележащую часть без консенсусной последовательности, связанного, например, с желтой меткой в виде частиц, где желтая метка в виде частиц не связывается с антителом. Безусловно, неважно, вышележащая или нижележащая область будет выбрана в качестве отрицательного контроля.
Участок расщепления для различных штаммов вируса гриппа A и вируса гриппа B известен. Например, в цитированной выше статье Bianchi, et al., в таблице 1 показана последовательность в области участка расщепления нескольких таких штаммов:
Консенсусная последовательность экспонированной для растворителя области участков связанного с созреванием расщепления вируса гриппа A и B
подтип
(SEQ ID NO:1)
(SEQ ID NO:3)
(SEQ ID NO:5)
b Консенсусная последовательность является одинаковой как для линии Victoria, так и для линии Yamagata.
Как указано, точная консенсусная последовательность начинается с остатка аргинина выше участка расщепления и, таким образом, предпочтительные консенсусные последовательности, включенные в исследуемые пептиды по изобретению, имеют последовательность RGI/L/F FGAIAGFLE (SEQ ID NO:7). В исследуемых пептидах может быть использована только часть этой последовательности.
После идентификации клеток, которые секретируют желаемые антитела, нетрудно извлечь нуклеотидные последовательности, кодирующие их, и продуцировать желаемые антитела рекомбинантными способами в большом масштабе. Это также дает возможность манипулировать с антителами, так чтобы их можно было продуцировать, например, в качестве одноцепочечных антител, или только с точки зрения их вариабельных областей.
Выделенные нуклеиновые кислоты можно физически хранить и выделять для последующего рекомбинантного продуцирования и/или может быть извлечена и сохранена информация о последовательности, касающаяся кодирующей последовательности, для обеспечения последующего синтеза соответствующих нуклеиновых кислот. Доступность информации, содержащейся в кодирующих последовательностях, и способы быстрого синтеза и клонирования вместе с известными способами рекомбинантного продуцирования обеспечивают быстрое продуцирование необходимых антител в случае пандемии или в других неотложных случаях.
Заявители выделили множество моноклональных антител, которые являются иммунореактивными к белку HA0 вируса гриппа из различных кладов (SEQ ID NO:9-23, 26-40, 42-56 и 59-73). Другие последовательности включают аминокислотную последовательность для константной области тяжелой цепи IgG1 человека (SEQ ID NO:8), аминокислотную последовательность для константной области каппа легкой цепи человека (SEQ ID NO:24), аминокислотную последовательность для константной области лямбда легкой цепи человека (SEQ ID NO:25), нуклеотидную последовательность для константной области тяжелой цепи человека (SEQ ID NO:41), нуклеотидную последовательность для константной области каппа легкой цепи человека (SEQ ID NO:57) и нуклеотидную последовательность для константной области лямбда легкой цепи человека (SEQ ID NO:58).
Два из этих mAb, MAB53 и MAB8, имеют значительную перекрестную реактивность среди важных, состоящих в дальнем родстве кладов вируса гриппа. Как показано на фиг.1A и B, каждый из них связывается с тремя различными кладами с умеренной или высокой аффинностью. MAB53 связывается с HA0 из кладов H1, H9 и H7, и MAB8 связывается с белком HA0 из кладов H1, H7 и H3. Результаты, представленные на фиг.1 были получены в анализе ELISA против белка HA0, и подразумевают, что аффинность находится в наномолярном диапазоне. Реактивность к нативному триммеру HA из всех кладов группы 1 подтверждали с использованием HA, экспрессированного в клетках HEK293, и связывание антител измеряли проточной цитометрией.
Эти результаты были подтверждены с использованием альтернативной системы анализа, системы анализа связывания на основе интерферометрии биологических уровней, обозначаемой как биосенсер ForteBio®, как показано на фиг.2A и 2B. При измерении с помощью этого более точного анализа аффинность является следующей:
MAB53 / H1 = 60 пМ, H5 = 6 нМ, H7 = 70 пМ, H9 = 30 пМ;
MAB8 / H1 = 9 нМ, H3 = 16 нМ, H5 = 0,2 нМ.
Как MAB53, так и MAB8, представляют собой полностью человеческие антитела, однако также изобретение относится к сходным антителам, характерным для других видов. В контексте изобретения “антитела” и их фрагменты включают те части молекулы, которые имеют значение для связывания; таким образом, фрагменты могут включать только вариабельные области и “антитела” в качестве общего термина также можно рассматривать как включающие такие фрагменты. Таким образом, включаются Fab-фрагменты, F(ab′)2 и Fv-фрагменты, а также продуцированные рекомбинантными способами одноцепочечные антитела, и слитые молекулы таких конструкций для получения биспецифических агентов. Химерные, гуманизированные антитела и антитела человека находятся в объеме настоящего изобретения, также как и миметики антител на других белковых каркасах, таких как фибронектин, трансферрин или липокалин. Аналогично, в настоящее время существует множество технологий для получения молекулы, подобной единому антителу, которая включает домены антигенной специфичности из двух отдельных антител (биспецифическое антитело). Таким образом, может быть сконструировано единое антитело с очень широкой реактивностью к штаммам с использованием Fab-доменов отдельных антител с широкой реактивностью к группе 1 и группе 2, соответственно. Подходящие технологии описаны Macrogenics (Rockville, MD), Micromet (Bethesda, MD) и Merrimac (Cambridge, MA). (См., например, Orcutt KD, Ackerman ME, Cieslewicz M, Quiroz E, Slusarczyk AL, Frangioni JV, Wittrup KD. A modular IgG-scFv bispecific antibody topology, Protein Eng Des Sel. (2010) 23:221-228; Fitzgerald J, Lugovskoy A. Rational engineering of antibody therapeutics targeting multiple oncogene pathways. MAbs. (2011) 1:3(3); Baeuerle PA, Reinhardt C. Bispecific T-cell engaging antibodies for cancer therapy. Cancer Res. (2009) 69:4941-4944.)
Для идентификации эпитопа, с которым связывается MAB53, проводили анализ ELISA в отношении нерасщепленного белка HA0, фрагмента HA1 и фрагмента HA2. Как показано на фиг.3A и B, хотя MAB53 связывается с HA0 с высокой аффиностью, оно не связывается с HA1, что подразумевает связывание с комплементарным фрагментом HA2. Для подтверждения этой гипотезы пептид, происходящий из HA2, иммобилизовывали на планшете, покрытом стрептавидином, с использованием C-концевого биотина. В частности, исследуемая последовательность представляла собой RGLFGAIAGFIENGW (SEQ ID NO:74). Также использовали незначимые фланкирующие участки. Было подтверждено, что MAB53 способно связываться с этим пептидом. Поскольку MAB53 не связывается с HA0 при исследовании с помощью вестерн-блоттинга, было предположено, что основной эпитоп по меньшей мере частично является конформационным по своей природе.
Также было обнаружено, что MAB8 и MAB53 связываются с одинаковыми или близко расположенными эпитопами, как продемонстрировано по их способности конкурировать друг с другом за связывание белка HA0 клада H1. Это было показано с использованием анализа ForteBio® с использованием 2 мкг/мл антитела и 50 нМ HA0 из H1. Как показано на фиг.4A, сигнал, полученный от MAB53, связанного с поверхностью ForteBio®, усиливается при добавлении 50 нМ раствора HA0. Однако, когда затем добавляют MAB8, не возникает дальнейшего сигнала. Таким образом, MAB53 блокирует эпитоп, связываемый MAB8. Как показано на фиг.4B, однако, другое антитело, которое является иммунореактивным к HA0, MAB30, связывается, по-видимому, с другим эпитопом, поскольку сигнал усиливается, когда его добавляют к связанному MAB53-HA0.
Важно, что MAB53 и MAB8 отличаются тем, что MAB8 освобождается от белка HA0, когда pH снижается до 6, в то время как MAB53 не освобождается. Это отличие является значительным, поскольку это оказалось предсказывающим нейтрализующую способность. В исследованиях на способность MAB8 нейтрализовать вирусную инфекцию H1N1 в бляшкообразующем анализе в клетках-мишенях MDCK, низкие дозы MAB53, составлявшие 1-5 мкг/мл, нейтрализовывали инфекцию H1N1, H7N3, H5N1 и H9N2. Однако, MAB8 не нейтрализует инфекцию этими штаммами. Таким образом, нейтрализующие штаммы можно, предпочтительно, выбирать путем смывания связанного MAB или фрагмента при pH 6 в процессе первичного скрининга, таким образом, удаляя от HA0 MAB, которые с малой вероятностью останутся связанными при вхождении комплекса антитело-вирус в клетку через эндосомальный компартмент и, таким образом, от которых можно ожидать, что они имеют сниженную способность нейтрализовать вирус.
Например, в способе CellSpot HA0 можно связывать с твердой подложкой (флуоресцентные гранулы) и иммобилизовывать с помощью MAB или смеси MAB, а затем промывать при pH 6.
MAB53 было получено рекомбинантными способами и было отсеквенировано. Полноразмерные последовательности тяжелой цепи и легкой цепи являются следующими:

Последовательности, выделенные полужирным шрифтом, представляют собой вариабельные домены, и последовательности, не выделенные полужирным шрифтом, соответствуют константной цепи для тяжелой цепи IgG1 и константной цепи каппа для легкой цепи.
Кроме того, эти вариабельные области анализировали согласно оценке CDR по Kabat на основе соответствия каркасных областей. Как показано на фиг.5A, CDR1, CDR2 и CDR3 тяжелой цепи IGHV1-69*01 (SEQ ID NO:83) представляют собой GGIIRKYAIN (SEQ ID NO:77), GGIIAIFNTANYAQKFQG (SEQ ID NO:78) и ARGMNYYSDYFDY (SEQ ID NO:79), соответственно. Как показано на фиг.5B, CDR1, CDR2 и CDR3 легкой цепи IGKV3-20*01 (SEQ ID NO:84) представляют собой RASQSVRSNNLA (SEQ ID NO:80), GASSRAT (SEQ ID NO:81) и QQYGSSPALT (SEQ ID NO:82), соответственно.
Как показано на фиг.6, MAB53 нейтрализует H1N1 in vitro в анализе бляшкообразования.
Также было показано, что мыши, которым предварительно вводили возрастающие дозы MAB53, выживают после заражения в ином случае летальными титрами вирусов H1N1 и H5N1 со 100% защитой после заражения H1N1, как показано на фиг.7. Эта эффективность сравнима с антителом из уровня техники, описанным Crucell, которое не демонстрирует активности против штаммов группы 2. Throsby M., et al., PLoS One. (2008) 3:e3942. Epub 2008 Dec 16. Они представляют собой гетеросубтипические нейтрализующие моноклональные антитела, обладающие перекрестной защитой против H5N1 и H1N1, выделенные из IgM+ B-клеток памяти человека.
Как показано на фиг.7A, MAB53 обеспечивало полную защиту при 10 мг/кг; 90% выживали при 2 мг/кг и 50% выживали при 0,4 мг/кг. Для сравнения, антитело из уровня техники от Crucell обеспечило полную защиту при 2 мг/кг, но только 20% выжили при введении 0,7 мг/кг. Это произошло, несмотря на тот факт, что летальность вирусной дозы была меньшей, чем летальность в эксперименте, представленном на фиг.7A; только 90% мышей погибли после инфицирования, в то время как в эксперименте, представленном на фиг.7A, все мыши погибли на 6 сутки. Это демонстрирует, что MAB53 является высокоэффективным.
Когда в место заражения H5N1 осуществляли заражение H1N1, для MAB53, представленного на фиг.7B, 10 мг/кг обеспечили 80% выживание; 2 мг/кг обеспечили 60% выживание и 0,4 мг/кг обеспечили 50% выживание. Для сравнения, для антитела уровня техники, 100% выживание было достигнуто при 5 мг/кг и 60% выживание было достигнуто при 1,7 мг/кг. Таким образом, уровни выживания при 1,7 мг/кг и 2 мг/кг были сравнимыми. В этом случае, сама по себе вирусная доза была немного менее эффективной у мышей, исследованных с помощью MAB53.
Как показано на фиг.8, MAB53 (10 мг/кг) вводили в качестве лечения после инфицирования на сутки +3 против высоко патологического штамма H5N1. Контрольное антитело совпадало по изотипу, но не распознавало никакой антиген вируса гриппа. Протокол инфицирования и лечения является таким же, как для фигуры 7A, однако введение проводили на сутки +3 вместо суток -1.
Проводили анализ Pepscan, установивший, что MAB53 и CR6261 связываются со сходными областями HA, но с различными эпитопами (данные не представлены). Это согласуется с отличающейся активностью этих двух антител.
Таким образом, MAB53 и антитела, которые связываются с тем же эпитопом в тех же условиях, являются эффективными в качестве пассивных вакцин, подходящих для защиты популяций против эпидемий и пандемий, и для профилактического или терапевтического применения против сезонного гриппа для пациентов с ослабленной иммунной системой.
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ:
NVPEKQTR (SEQ ID NO:1)
GIFGAIAGFIE (SEQ ID NO:2)
NIPSIQSR (SEQ ID NO:3)
GLFGAIAGFIE (SEQ ID NO:4)
PAKLLKER (SEQ ID NO:5)
GFFGAIAGFLE (SEQ ID NO:6)
RGI/L/F FGAIAGFLE (SEQ ID NO:7).
Аминокислотная последовательность константной области HC IgG1 человека (SEQ ID NO:8)
ASTKGPSVFPLVPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
Аминокислотная последовательность вариабельного домена HC MAB1 (SEQ ID NO:9)
QVQLQESGPGLVKPSETLSLICRVSGGSISSHYWSWIRQPPGKGLEWIGYISYRGRSNHNPSLGRRVSMSIDTSENQFSLNLSSVIAADTAVYYCARDATGIREINALDIWGQGTTVTVSS
Аминокислотная последовательность вариабельного домена HC MAB8 (SEQ ID NO:10)
EVQLVESGGGLVKPGGSLRLSCAASGFTFSTYTMSWVRQAPGQGLEWVSSITRTSSNIYYADSVEGRFTISRDNAKNSLYLQMHSLRVEDTAVYYCARISGVVGPVPFDYWGQGTLITVSS
Аминокислотная последовательность вариабельного домена HC MAB30 (SEQ ID NO:11)
EVQLVESGGGLVQPGGSLRLSCAASGFTFSDHYMDWVRQAPGKGLEWVGRIRNKAAIYTTEYAASVKGRFTISRDDLKSSVYLQMNSLKTDDTAIYYCARSYGYFDYWGQGTLVTVSS
Аминокислотная последовательность вариабельного домена HC MAB42 (SEQ ID NO:12)
QVQLVQSGAEVKKPGASVKVSCKASGYSFNGYYMHWVRQAPGQGLEWMGWINLSSGGTDYAQKFQGWVTLTRDTSITTAYMELSSLRSNDTAVYYCARIRPRTGGLDSWGQGTLVIVSS
Аминокислотная последовательность вариабельного домена HC MAB48 (SEQ ID NO:13)
QVQLVQSGAEVKKPGSSVKVSCKASGVTFTAYAISWVRQAPGRGLEWMGGISPLFGIVNFGQNFQGRVTITADKSTGAAYMELSSLSSEDTAMYYCARGPYYYDRSHLDYWGQGTLVTVSS
Аминокислотная последовательность вариабельного домена HC MAB49 (SEQ ID NO:14)
QVQLVQSGAEVKRPGSSVKVSCKASGGTFSSYAISWVRQAPGQGLEWMGGIIGMFGTTNYAQKFQGRVTITADEFTSTAYMELTSLRSDDTAMYYCARDRNYYASGTYDHWGQGTLVTVSS
Аминокислотная последовательность вариабельного домена HC MAB52 (SEQ ID NO:15)
QVLLVQSGAEVKKPGSSVNISCKASGGTFSNYAISWVRQAPGQGLDWMGRIIPIFGTANYAQKFQGRLTITADESTSTAYMELSSLRSEDTAVFYCAITKPGSVYALDVWGQGTTVTVSS
Аминокислотная последовательность вариабельного домена HC MAB53 (SEQ ID NO:16)
QVQLVQSGAEVRKPGSSVKVSCKVSGGIIRKYAINWVRQAPGQGLEWMGGIIAIFNTANYAQKFQGRVTITADESTSTVYMELSSLRSEDTALYYCARGMNYYSDYFDYWGQGSLVTVSP
Аминокислотная последовательность вариабельного домена HC MAB285 (SEQ ID NO:17)
QVQLVQSGAEVKKPGASVKVSCRASGYTFTGYYMQWVRQAPGQGLEWMGFINANTGVTNFAQKFQGRVTLTRDTSISTAYMELRRLTSADTAVYYCARAPQWLSYSFDIWGQGTMVTVSS
Аминокислотная последовательность вариабельного домена HC MAB321 (SEQ ID NO:18)
EVQLVESGAEVRSPGASVKLSCKASAYTFINYYLHWVRQAPGQRLEWMGWINPDSGVTEYAQTFQGRVTMTRDTSINTAYLDLERLTSDDTAVYYCARGFIPWGGKYFYLDYWGQGTLVTVSS
Аминокислотная последовательность вариабельного домена HC MAB322 (SEQ ID NO:19)
QVQLQQSGPGLVKPSQTLSLTCSVSGSFIRSGDYNWSWIRQPPGKGLEWIGYIDNSGSTHYNPSLKSRVSISVDTSKNHLSLKLSFVTDADTGVYYCAGEQASDSRGNYYYYAMDVWGQGTPVTVSS
Аминокислотная последовательность вариабельного домена HC MAB375 (SEQ ID NO:20)
QVQLQQSGPGLMKPSETLSLSCTVSGDSVSSFYWSWIRQSPGKGLEWIGYLLYSGNTKYNPSLKSRATISRDTSKNQLSLELTSLTAADTAVYYCARVVRWRHGGDLDVWGQGTMVTVSS
Аминокислотная последовательность вариабельного домена HC MAB376 (SEQ ID NO:21)
QVQLVQSGGDLVQPGGSLRLSCAVSGFIFRKYIMSWVRQAPGKGPEWVAVISSSGDRTFYADSVEGRFIVSRDNSKDTLFLQMNSLRTEDTAMYYCAKDLLGFCSGGDCLKVFDLWGRGTMVTVSS
Аминокислотная последовательность вариабельного домена HC MAB377 (SEQ ID NO:22)
QVQLLQSGPGLIKASETLSLSCSVSNDSVSNYYWSWIRQSPEKGLEWIGYLLYSGNTKYNPSLKSRAIISRDMSKNQLSLRVTSVTAADTAIYYCARVVRWRFGGDMDVWGQGTAVTVST
Аминокислотная последовательность вариабельного домена HC MAB378 (SEQ ID NO:23)
QVQLQQSGPGLIKPSETLSLSCSVSGDSVNNYYWSWIRQPPEKGLEWIGYLQYSGSTKYNPSLKSRVTISRDTSKNQLSLKLTSVTAADTAIYYCARVVRWRHGGDMDVWGQGTAVTVSS
Аминокислотная последовательность константной области каппа LC человека (SEQ ID NO:24)
RTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Аминокислотная последовательность константной области лямбда LC человека (SEQ ID NO:25)
GQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVVPAECS
Аминокислотная последовательность LC MAB1 (SEQ ID NO:26)
DIQMTQSPSSLSASGGDRVTITCRASQSVSTYLNWYQQKPGKAPNLLVYAVSNLQRGVPSRFSGSGSGTHFTLTISSLQPEDFATYYCQQSYSDPLTFGGGTKVEIKR TVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Аминокислотная последовательность LC MAB8 (SEQ ID NO:27)
DIQMTQSPSSLSASVGDRVTITCRASQTISKYLNWYQQKPGRAPKLLIYSASSLQSGVPSRFTGSGSGTDFTLTITSLQPEDFATYYCQQSYRPSQITFGPGTKVDIKR TVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Аминокислотная последовательность LC MAB30 (SEQ ID NO:28)
DIQMTQSPSTLSASVGDRVTITCRASQSISSWLAWYQQKPGNAPNLLIYKASSLESGVPSRFSGSGSGTEFTLTISSLQPDDFATYYCQQYDTYSPTFGQGTKVEIKR TVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Аминокислотная последовательность LC MAB42 (SEQ ID NO:29)
QSALTQPASVSGSAGQSITISCTGTSSDVGAYNFVSWYQHHPGKAPKLMIYDVDNRPSGVSNRFSGSKSGDTASLTISGLQAEDEADYYCSSYRRNGPWVFGGGTKLTVLGQPKAAPTVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVVPAECS
Аминокислотная последовательность LC MAB48 (SEQ ID NO:30)
EIVLTQSPGTLSLSPGERATLSCRASQSVGSSDLAWYQQKPGQAPRLLIYGASSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQYVSSPLTFGGGTKVEIKR TVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Аминокислотная последовательность LC MAB49 (SEQ ID NO:31)
DIQMTQSPSSLSASVGDRVTITCRASQSISRYLNWYQQKPGKAPKLLIYSASSLQSGVPSRFGGSGSGTDFTLTISSLQPEDFALYYCQQTYSIPITFGQGTRLDFKR TVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Аминокислотная последовательность LC MAB52 (SEQ ID NO:32)
DIQMTQSPSSLSASVGDRVTITCRASQTISTYLNWYQQKPGKAPNLLIYTASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQSYDAPTWTFGPGTKVEIKR TVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Аминокислотная последовательность LC MAB53 (SEQ ID NO:33)
EIVLTQSPGTLSLSPGERATLSCRASQSVRSNNLAWYQHKPGQAPRLLIFGASSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQYGSSPALTFGGGTKVEIKR TVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Аминокислотная последовательность LC MAB285 (SEQ ID NO:34)
QSVLTQPPSASGTPGQRVTISCSGSSSNIGSNPVNWYQQLPGTAPRLLIYSNNQRPSGVPDRFSGSKSGTSASLAISGLRSEDEADYYCTSWDDSLNAWVFGGGTRLTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVVPAECS
Аминокислотная последовательность LC MAB321 (SEQ ID NO:35)
DIVLTQSPPSLSASVGDRVTITCRASQSINNYLNWYQQKPGNAPRILIYGASSLVSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQSYRPLYTFGPGTQLDVKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Аминокислотная последовательность LC MAB322 (SEQ ID NO:36)
DIVMTQSPSSLSASVGDRVTITCRASESISAYLNWYQHTPGRAPKLLIYAASSLETGVPSRFSGSGSGTEFTLTISGLQPEDFVTYYCQQTYNTPRTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Аминокислотная последовательность LC MAB375 (SEQ ID NO:37)
DIQMTQSPSFLSASVGDRVTFTCRASQGIASSLAWYQQKAGKAPKLLIYAASTLEDGVPSRFSGSGFGTEFTLTITSLQPEDFATYYCHQVNSYPRTFGPGTTVDINR TVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Аминокислотная последовательность LC MAB376 (SEQ ID NO:38)
DIQMTQSPSTLSASVGDTVTITCRASQSISTWLAWFQQKPGRAPKLLIYQASSLEGGVPSRFSGSGSGTDFNLTISGLQPDDFATYYCLQYNTYSKSFGQGTKVEIKR TVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Аминокислотная последовательность LC MAB377 (SEQ ID NO:39)
DIQMTQSPSFLSASVGDRVTITCRASQGIATSLAWYQQKPGKAPRLLIYAASTLESGVPSRFSGGGSGTDFTLTISSLQPEDFAVYYCQQVNSYPRTFGPGTKLDVKR TVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Аминокислотная последовательность LC MAB378 (SEQ ID NO:40)
DIQMTQSPSFLSASVGDRVTMTCRASQGISSYLAWYQQKPGKAPKLLIYAASTLESGVPSRFSGSGSGTEFTLTISSLQPEDFAIYYCQQVNGYPRTFGPGTKVDIKR TVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Нуклеотидная последовательность константной области HC IgG1 человека (интроны подчеркнуты) (SEQ ID NO:41)

Нуклеотидная последовательность вариабельного домена HC MAB1 (SEQ ID NO:42)
CAGGTGCAGCTGCAGGAGTCGGGCCCAGGACTGGTGAAGCCTTCGGAGACCCTGTCCCTCATCTGCAGAGTCTCTGGTGGCTCGATCAGTAGTCATTACTGGAGCTGGATCCGGCAGCCCCCAGGGAAGGGACTGGAGTGGATTGGATATATTTCTTATAGGGGGAGAAGCAACCACAATCCTTCCCTTGGGAGACGAGTCTCTATGTCAATAGACACGTCGGAGAACCAGTTCTCCCTGAACCTGAGCTCTGTGATCGCTGCGGACACGGCCGTATATTACTGTGCGAGAGATGCTACTGGGATCAGAGAAATCAATGCTCTTGATATCTGGGGCCAAGGGACAACGGTCACCGTCTCTTCA
Нуклеотидная последовательность вариабельного домена HC MAB8 (SEQ ID NO:43)
GAGGTGCAGCTGGTGGAGTCTGGGGGAGGCCTGGTCAAGCCTGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGTTTCACTTTCAGTACCTATACTATGAGTTGGGTCCGCCAGGCTCCAGGGCAGGGGCTAGAGTGGGTCTCGTCCATTACTAGGACTAGTAGTAATATATACTACGCAGACTCAGTGGAGGGCCGATTCACCATCTCCAGAGACAACGCCAAGAACTCACTGTATCTGCAGATGCATAGCCTGAGAGTCGAAGACACGGCTGTGTATTACTGTGCGAGAATCAGCGGGGTAGTGGGACCTGTCCCCTTTGACTACTGGGGCCAGGGAACCCTGATCACCGTCTCCTCT
Нуклеотидная последовательность вариабельного домена HC MAB30 (SEQ ID NO:44)
GAGGTGCAGCTGGTGGAGTCTGGGGGAGGCTTGGTCCAGCCTGGAGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTCAGTGACCACTACATGGACTGGGTCCGCCAGGCTCCAGGGAAGGGGCTGGAGTGGGTTGGCCGTATTAGAAATAAAGCTGCCATTTACACCACAGAATACGCCGCGTCTGTGAAAGGCAGATTCACCATCTCAAGAGATGATTTAAAGAGCTCAGTGTATCTGCAAATGAACAGTCTGAAAACCGACGACACGGCCATATATTACTGTGCTAGGAGCTATGGATACTTTGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCA
Нуклеотидная последовательность вариабельного домена HC MAB42 (SEQ ID NO:45)
CAGGTGCAGCTGGTACAGTCTGGGGCTGAGGTGAAGAAGCCTGGGGCCTCAGTGAAGGTCTCCTGCAAGGCTTCTGGATATTCCTTCAACGGCTACTATATGCACTGGGTGCGACAGGCCCCTGGACAAGGGCTTGAGTGGATGGGTTGGATCAACCTGAGCAGTGGTGGCACAGATTATGCACAGAAATTTCAGGGGTGGGTCACTTTGACCAGGGACACGTCCATCACCACAGCCTACATGGAGTTGAGCAGCCTGAGATCGAACGACACGGCCGTGTATTACTGTGCGAGAATTAGACCTCGCACTGGTGGACTTGACTCCTGGGGCCAGGGAACCCTGGTCATCGTCTCCTCA
Нуклеотидная последовательность вариабельного домена HC MAB48 (SEQ ID NO:46)
CAGGTGCAGCTGGTGCAGTCTGGGGCTGAGGTGAAGAAGCCTGGGTCCTCGGTGAAAGTCTCCTGCAAGGCTTCTGGAGTCACCTTCACCGCCTATGCTATCAGTTGGGTGCGACAGGCCCCTGGACGAGGGCTTGAGTGGATGGGAGGGATCAGCCCTTTGTTTGGAATAGTAAATTTCGGACAGAACTTCCAGGGCAGAGTCACGATTACCGCGGACAAATCCACGGGCGCAGCCTACATGGAGCTGAGCAGCCTGAGCTCTGAGGACACGGCCATGTATTACTGTGCGAGAGGACCCTATTATTACGATAGAAGTCACCTAGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCA
Нуклеотидная последовательность вариабельного домена HC MAB49 (SEQ ID NO:47)
CAGGTGCAGCTGGTGCAGTCTGGGGCTGAGGTGAAGAGGCCTGGGTCCTCGGTGAAGGTCTCCTGCAAGGCTTCTGGAGGCACCTTCAGCAGTTATGCTATTAGCTGGGTGCGACAGGCCCCTGGACAAGGGCTTGAGTGGATGGGAGGGATCATCGGTATGTTTGGAACAACAAACTACGCACAGAAGTTCCAGGGCAGAGTCACGATTACCGCGGACGAATTCACGAGCACAGCCTACATGGAGCTGACCAGCCTGAGATCTGACGACACGGCCATGTATTACTGTGCGAGAGACCGAAATTACTATGCTTCGGGGACTTATGACCACTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCA
Нуклеотидная последовательность вариабельного домена HC MAB52 (SEQ ID NO:48)
CAAGTGCTGCTGGTGCAGTCTGGGGCTGAAGTGAAGAAGCCTGGGTCCTCGGTGAATATCTCTTGCAAGGCTTCTGGAGGCACTTTCAGCAACTATGCTATCTCCTGGGTGCGACAGGCCCCTGGACAAGGTCTTGACTGGATGGGAAGGATCATCCCTATCTTTGGAACAGCAAACTACGCACAGAAATTCCAGGGCAGACTCACCATTACCGCGGACGAATCCACGAGCACAGCCTACATGGAACTGAGCAGCCTGAGATCTGAAGACACGGCCGTGTTTTACTGTGCGATTACTAAACCGGGGTCTGTCTACGCTTTGGACGTCTGGGGCCAAGGGACCACGGTCACCGTCTCCTCA
Нуклеотидная последовательность вариабельного домена HC MAB53 (SEQ ID NO:49)
CAGGTGCAGCTGGTGCAGTCTGGGGCTGAGGTGAGGAAGCCGGGGTCCTCGGTGAAGGTCTCCTGCAAGGTTTCTGGAGGCATCATTAGGAAATATGCTATCAACTGGGTGCGACAGGCCCCCGGACAAGGGCTTGAGTGGATGGGAGGGATCATCGCTATCTTTAATACAGCAAACTATGCACAGAAATTCCAGGGCAGAGTCACGATTACCGCGGACGAGTCCACGAGCACAGTCTACATGGAGCTGAGCAGCCTGAGATCTGAAGACACGGCCCTTTATTACTGTGCGAGAGGAATGAATTACTACAGTGACTACTTTGACTACTGGGGCCAGGGAAGCCTTGTCACCGTCTCCCCA
Нуклеотидная последовательность вариабельного домена HC MAB285 (SEQ ID NO:50)
CAGGTGCAGCTGGTGCAGTCTGGGGCTGAGGTGAAGAAGCCTGGGGCCTCAGTGAAGGTCTCCTGCCGGGCTTCTGGATACACCTTCACCGGCTACTATATGCAGTGGGTGCGGCAGGCCCCTGGCCAAGGGCTTGAGTGGATGGGATTCATCAATGCTAACACTGGTGTCACAAACTTTGCTCAGAAGTTTCAGGGCAGGGTCACCTTGACCAGGGACACGTCCATCAGCACAGCCTACATGGAGCTGAGGAGGCTGACATCTGCCGACACGGCCGTGTATTACTGTGCGAGAGCGCCCCAGTGGTTATCGTATTCTTTTGATATCTGGGGCCAAGGGACAATGGTCACCGTCTCCTCA
Нуклеотидная последовательность вариабельного домена HC MAB321 (SEQ ID NO:51)
GAGGTGCAGCTGGTGGAGTCTGGGGCTGAGGTGAGGAGCCCTGGGGCCTCAGTGAAGCTCTCCTGCAAGGCTTCTGCATACACCTTCATCAACTACTATCTGCACTGGGTGCGACAGGCCCCTGGACAAAGGCTTGAGTGGATGGGATGGATCAACCCTGACAGTGGTGTCACAGAATATGCACAGACATTTCAGGGCAGGGTCACCATGACCAGGGACACGTCCATCAATACAGCCTACCTGGACCTGGAGAGACTGACATCTGACGACACGGCCGTATATTACTGTGCGAGAGGTTTTATTCCTTGGGGTGGGAAGTACTTCTACCTTGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCA
Нуклеотидная последовательность вариабельного домена HC MAB322 (SEQ ID NO:52)
CAGGTACAGCTGCAGCAGTCAGGGCCAGGACTGGTGAAGCCTTCACAGACCCTGTCCCTCACCTGCAGTGTATCTGGTAGTTTCATCAGAAGTGGAGATTATAATTGGAGTTGGATCCGCCAGCCCCCAGGGAAGGGCCTGGAGTGGATTGGGTACATCGATAATAGCGGGAGCACCCACTACAACCCGTCCCTCAAGAGTCGAGTTAGCATATCAGTGGACACGTCCAAGAACCACTTGTCCCTGAAGCTGAGTTTTGTGACTGACGCAGACACGGGCGTGTATTACTGTGCCGGAGAACAAGCGTCTGATAGTCGTGGTAATTACTACTACTACGCTATGGACGTCTGGGGCCAAGGGACCCCGGTCACCGTCTCCTCA
Нуклеотидная последовательность вариабельного домена HC MAB375 (SEQ ID NO:53)
CAGGTGCAGCTGCAGCAGTCGGGCCCCGGACTGATGAAGCCTTCGGAGACCCTGTCCCTCAGCTGCACTGTCTCTGGTGACTCCGTCAGTAGTTTTTATTGGAGTTGGATTCGGCAGTCTCCAGGAAAGGGACTGGAGTGGATTGGGTATTTGCTTTACAGTGGGAATACCAAGTATAATCCGTCCCTCAAGAGTCGAGCCACCATATCAAGAGACACGTCCAAGAACCAGTTGTCCCTGGAGTTGACCTCTCTGACCGCTGCGGACACGGCCGTCTACTATTGTGCGAGAGTGGTGAGATGGCGACATGGTGGCGATTTGGACGTCTGGGGCCAAGGGACCACGGTCACCGTCTCCTCA
Нуклеотидная последовательность вариабельного домена HC MAB376 (SEQ ID NO:54)
CAGGTGCAGCTGGTGCAGTCCGGGGGGGACTTGGTCCAGCCGGGGGGGTCCCTGAGACTGTCATGTGCAGTCTCTGGATTCATCTTTAGAAAATATATCATGAGTTGGGTCCGGCAGGCTCCAGGGAAGGGGCCGGAGTGGGTCGCAGTTATTAGTTCTAGTGGTGACCGGACATTCTACGCCGACTCCGTGGAGGGCCGCTTCATCGTCTCCAGAGACAATTCCAAGGACACACTGTTTCTGCAAATGAACAGCCTGAGAACCGAGGACACGGCCATGTATTACTGTGCGAAAGACCTTTTGGGATTTTGTAGTGGTGGTGATTGCCTGAAGGTCTTCGATCTCTGGGGCCGAGGCACCATGGTCACTGTCTCCTCA
Нуклеотидная последовательность вариабельного домена HC MAB377 (SEQ ID NO:55)
CAGGTGCAGCTGCTGCAGTCGGGCCCAGGACTGATAAAGGCTTCGGAGACCCTGTCTCTCAGCTGCAGTGTCTCTAATGACTCCGTCAGTAATTATTATTGGAGTTGGATCCGGCAGTCCCCAGAGAAGGGACTGGAGTGGATTGGGTATTTGCTTTATAGTGGGAATACCAAGTACAATCCCTCCCTCAAGAGTCGAGCCATCATATCAAGAGACATGTCCAAAAATCAGTTGTCCCTCAGAGTGACTTCTGTGACCGCTGCGGACACGGCCATATATTATTGTGCGCGAGTGGTGAGATGGCGATTTGGTGGTGATATGGACGTCTGGGGTCAAGGGACCGCGGTCACCGTCTCCACA
Нуклеотидная последовательность вариабельного домена HC MAB378 (SEQ ID NO:56)
CAGGTGCAGCTGCAGCAGTCGGGCCCAGGACTGATAAAGCCTTCGGAGACCCTGTCTCTCAGCTGCTCTGTCTCTGGTGACTCCGTCAATAATTATTATTGGAGTTGGATCCGGCAGCCCCCAGAGAAGGGACTGGAGTGGATTGGGTATCTGCAGTATAGTGGGAGTACAAAGTACAACCCCTCCCTCAAGAGTCGAGTCACCATATCAAGAGACACGTCCAAAAACCAGTTGTCCCTGAAGCTGACCTCTGTGACCGCTGCGGACACGGCCATATATTATTGTGCGAGAGTGGTGAGATGGCGACATGGTGGGGATATGGACGTCTGGGGCCAAGGGACCGCGGTCACCGTCTCCTCT
Нуклеотидная последовательность константной области каппа LC человека (SEQ ID NO:57)
CGAACTGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCTAGCGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAG
Нуклеотидная последовательность константной области лямбда LC человека (SEQ ID NO:58)
GGTCAGCCCAAGGCTGCCCCCTCTGTCACTCTGTTCCCGCCCTCTAGCGAGGAGCTTCAAGCCAACAAGGCCACACTGGTGTGTCTCATAAGTGACTTCTACCCGGGAGCCGTGACAGTGGCCTGGAAGGCAGATAGCAGCCCCGTCAAGGCGGGAGTGGAGACCACCACACCCTCCAAACAAAGCAACAACAAGTACGCGGCCAGCAGCTATCTGAGCCTGACGCCTGAGCAGTGGAAGTCCCACAGAAGCTACAGCTGCCAGGTCACGCATGAAGGGAGCACCGTGGAGAAGACAGTGGTCCCTGCAGAATGCTCT
Нуклеотидная последовательность вариабельного домена LC MAB1 (SEQ ID NO:59)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGGAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGTGTTAGTACGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAACCTCCTGGTCTATGCTGTATCCAATTTACAACGTGGCGTGCCATCAAGGTTCAGTGGCAGTGGATCTGGGACACATTTCACTCTCACAATCAGCAGTCTGCAACCTGAGGATTTCGCAACTTACTACTGTCAACAGAGTTACAGTGACCCTCTCACTTTCGGCGGAGGGACCAAGGTGGAGATCAAA
Нуклеотидная последовательность вариабельного домена LC MAB8 (SEQ ID NO:60)
GACATCCAGATGACCCAGTCTCCATCTTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGACCATTAGCAAGTATTTAAATTGGTATCAGCAGAAGCCAGGGAGAGCCCCTAAACTCCTGATCTACTCTGCGTCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCACTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCACCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAACAGAGTTACAGACCCTCCCAGATCACTTTCGGCCCTGGGACCAAAGTGGATATCAAA
Нуклеотидная последовательность вариабельного домена LC MAB30 (SEQ ID NO:61)
GACATCCAGATGACCCAGTCTCCTTCCACCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCCAGTCAGAGTATTAGTAGTTGGTTGGCCTGGTATCAGCAGAAACCAGGGAACGCCCCTAACCTCCTGATCTATAAGGCGTCTAGTTTAGAAAGTGGGGTCCCATCAAGGTTCAGCGGCAGTGGATCTGGGACAGAATTCACTCTCACCATCAGCAGCCTGCAGCCTGATGATTTTGCAACTTATTACTGCCAACAGTATGATACTTATTCTCCGACGTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Нуклеотидная последовательность вариабельного домена LC MAB42 (SEQ ID NO:62)
CAGTCTGCCCTGACTCAGCCTGCCTCCGGGTCTGGGTCTGCTGGACAGGCGATCACCATCTCCTGCACTGGAACCGGCACTGACGTCTGTGCTTATAACTTTGTCTCCTGGTACCAACACCACCCCGGCGAAGCCCCCAAACTCATGATTTATGATGTCGATAATCGGCCCTCATGGGTTTCTAATCGCTTCTCTGGCTCCAAGTCTGGTAACACGGCCTCCCTGACCATCTCTGGGCTCCAGGCTGAGGACGAGGCTGATTACTACTGCAGCTCATATAGAAGGAACGGCCCTTGCTTGTTCGGCGGAGGGACCAAGCTGACCGTCCTG
Нуклеотидная последовательность вариабельного домена LC MAB48 (SEQ ID NO:63)
GAAATTGTGTTGACGCAGTCTCCAGGCACCCTGTCTTTGTCTCCAGGGGAAAGAGCCACCCTCTCCTGCAGGGCCAGTCAGAGTGTTGGCAGCAGCGACTTAGCCTGGTACCAGCAGAAACCTGGCCAGGCTCCCAGGCTCCTCATATATGGTGCATCCAGCCGGGCCACTGGCATCCCAGACAGGTTCAGTGGCAGTGGGTCTGGGACAGACTTCACTCTCACCATCAGCAGACTGGAGCCTGAAGATTTTGCAGTGTATTACTGTCAGCAGTATGTCAGTTCACCCCTCACTTTCGGCGGAGGGACCAAGGTGGAGATCAAG
Нуклеотидная последовательность вариабельного домена LC MAB49 (SEQ ID NO:64)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTAGCAGGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAACTCCTGATCTATTCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCGGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCACTTTACTACTGTCAACAGACTTACAGTATCCCGATCACCTTCGGCCAAGGGACACGACTGGACTTTAAA
Нуклеотидная последовательность вариабельного домена LC MAB52 (SEQ ID NO:65)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACTATCACTTGCCGGGCAAGTCAGACCATTAGCACCTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAACCTCCTGATCTATACTGCATCCAGTTTGCAAAGCGGGGTCCCATCAAGATTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTATTACTGTCAACAGAGTTACGATGCCCCCACGTGGACCTTCGGCCCAGGGACCAAGGTGGAAATCAAA
Нуклеотидная последовательность вариабельного домена LC MAB53 (SEQ ID NO:66)
GAAATTGTGTTGACACAGTCTCCAGGCACCCTGTCTTTGTCTCCAGGGGAAAGAGCCACCCTCTCCTGCAGGGCCAGTCAGAGTGTTAGAAGCAACAACTTAGCCTGGTACCAGCACAAACCTGGCCAGGCTCCCAGGCTCCTCATCTTTGGTGCATCCAGCAGGGCCACTGGCATCCCAGACAGGTTCAGTGGCAGTGGGTCTGGGACAGACTTCACTCTCACCATCAGCAGACTGGAGCCTGAAGATTTTGCAGTATATTACTGTCAGCAGTATGGTAGCTCACCTGCGCTCACTTTCGGCGGAGGGACCAAGGTGGAGATCAAA
Нуклеотидная последовательность вариабельного домена LC MAB285 (SEQ ID NO:67)
CAGTCTGTGCTGACTCAGCCACCCTCAGCGTCTGGGACCCCCGGGCAGAGGGTCACCATCTCTTGTTCTGGAAGCAGCTCCAACATCGGAAGTAATCCTGTAAACTGGTACCAGCAGCTCCCAGGAACGGCCCCCAGACTTCTCATCTATAGTAATAATCAGCGGCCCTCAGGGGTCCCTGACCGATTCTCTGGCTCCAAGTCTGGCACCTCAGCCTCCCTGGCCATCAGTGGGCTCCGGTCCGAGGATGAGGCTGATTACTACTGTACATCATGGGATGACAGCCTGAATGCTTGGGTGTTCGGCGGGGGGACCAGGCTGACCGTCCTA
Нуклеотидная последовательность вариабельного домена LC MAB321 (SEQ ID NO:68)
GATATCGTGTTGACTCAGTCTCCACCCTCCCTGTCTGCATCTGTGGGGGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTAATAACTACTTAAATTGGTATCAACAGAAACCAGGGAACGCCCCAAGAATACTAATCTATGGTGCATCCAGTTTGGTAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACCCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAACAGAGTTACCGGCCCCTGTACACTTTTGGCCCGGGGACCCAGCTGGATGTCAAA
Нуклеотидная последовательность вариабельного домена LC MAB322 (SEQ ID NO:69)
GATATCGTGATGACCCAGTCTCCATCTTCCCTGTCTGCATCTGTGGGAGACAGAGTCACCATCACTTGCCGGGCAAGTGAGAGCATTAGCGCTTATTTAAATTGGTATCAGCACACACCAGGGAGAGCCCCTAAGCTCCTGATCTATGCTGCCTCCAGTTTGGAAACTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGCACAGAATTCACTCTCACCATCAGCGGTCTGCAACCTGAAGATTTTGTCACTTACTACTGTCAACAGACTTACAATACCCCTCGGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Нуклеотидная последовательность вариабельного домена LC MAB375 (SEQ ID NO:70)
GATATCCAGATGACCCAGTCTCCATCCTTCTTGTCTGCATCTGTGGGAGACAGAGTCACCTTCACTTGCCGGGCCAGTCAGGGCATTGCCAGTTCTTTAGCCTGGTATCAGCAAAAAGCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCTTCTACTTTGGAAGATGGGGTCCCATCAAGGTTCAGCGGCAGTGGATTTGGGACAGAATTCACTCTCACAATCACCAGCCTGCAGCCTGAAGATTTTGCAACCTATTACTGTCATCAGGTGAATAGTTACCCTCGGACTTTCGGCCCTGGGACCACAGTGGATATCAAC
Нуклеотидная последовательность вариабельного домена LC MAB376 (SEQ ID NO:71)
GATATCCAGATGACCCAGTCTCCTTCCACCCTGTCTGCATCTGTGGGAGACACAGTCACCATCACTTGCCGGGCCAGTCAGAGTATTAGTACTTGGTTGGCCTGGTTTCAGCAGAAACCAGGGAGAGCCCCTAAACTCCTGATCTATCAGGCGTCTAGTTTGGAAGGTGGGGTCCCATCAAGGTTCAGCGGCAGTGGGTCTGGGACAGACTTCAACCTCACCATCAGCGGCCTGCAGCCTGATGATTTTGCAACTTATTACTGCCTACAATATAACACTTATTCGAAGTCATTCGGCCAAGGGACCAAGGTGGAAATCAAAC
Нуклеотидная последовательность вариабельного домена LC MAB377 (SEQ ID NO:72)
GATATCCAGATGACCCAGTCTCCATCCTTCTTGTCTGCATCTGTCGGAGACAGAGTCACCATCACCTGCCGGGCCAGTCAGGGCATTGCCACTTCTTTAGCCTGGTATCAGCAAAAACCTGGGAAAGCCCCGAGGCTCCTGATCTATGCTGCATCCACTTTGGAAAGTGGGGTCCCATCAAGGTTCAGCGGCGGTGGATCTGGGACAGACTTCACTCTCACAATCAGCAGTCTGCAGCCCGAAGATTTTGCTGTTTATTACTGTCAACAGGTTAACTCCTATCCTCGGACTTTCGGCCCTGGGACCAAACTGGATGTCAAAC
Нуклеотидная последовательность вариабельного домена LC MAB378 (SEQ ID NO:73)
GATATCCAGATGACCCAGTCTCCATCCTTCTTGTCTGCATCTGTAGGAGACAGAGTCACCATGACCTGCCGGGCCAGTCAGGGCATTAGCAGTTATTTAGCCTGGTATCAGCAAAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCGACTTTGGAAAGTGGGGTCCCATCAAGGTTCAGCGGCAGTGGATCTGGGACAGAATTCACTCTCACAATCAGCAGCCTGCAGCCCGAAGATTTTGCAATTTATTACTGTCAACAGGTTAATGGTTACCCTCGGACTTTCGGCCCTGGGACCAAAGTGGATATCAAAC
RGLFGAIAGFIENGW (SEQ ID NO:74)
Тяжелая цепь MAB53 (SEQ ID NO:75)
QVQLVQSGAEVRKPGSSVKVSCKVSGGIIRKYAINWVRQAPGQGLEWMGGIIAIFNTANYAQKFQGRVTITADESTSTVYMELSSLRSEDTALYYCARGMNYYSDYFDYWGQGSLVTVSPASTKGPSVFPLVPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
Легкая цепь MAB53 (SEQ ID NO:76)
EIVLTQSPGTLSLSPGERATLSCRASQSVRSNNLAWYQHKPGQAPRLLIFGASSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQYGSSPALTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
GGIIRKYAIN (SEQ ID NO:77)
GGIIAIFNTANYAQKFQG (SEQ ID NO:78)
ARGMNYYSDYFDY (SEQ ID NO:79)
RASQSVRSNNLA (SEQ ID NO:80)
GASSRAT (SEQ ID NO:81)
QQYGSSPALT (SEQ ID NO:82)
IGHV1-69*01 (SEQ ID NO:83)
QVQLVQSGAEVRK PGSSVKVSCKVSGGIIRKYAINWVRQAPGQG LEWMGGIIAIFNTANYAQKFQGRVTITADESTSTVYMELSSLRSEDTALYYCARGMNYYSDYFDYWGQGSLVTTVS
IGKV3-20*01 (SEQ ID NO:84)
EIVLTQSPGTLSLSPGERATLSCRASQSVRSNNLAWYQHKPGQAPRLLIFGASSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQYGSSPALTFGGGTKVEIK
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА, ИСПОЛЬЗУЕМЫЕ ДЛЯ ПАССИВНОЙ ВАКЦИНАЦИИ ПРОТИВ ГРИППА | 2012 |
|
RU2668802C2 |
| АНТИТЕЛА, ПОДХОДЯЩИЕ ДЛЯ ПАССИВНОЙ ИММУНИЗАЦИИ ПРОТИВ ГРИППА, И ИХ КОМПОЗИЦИИ, КОМБИНАЦИИ И СПОСОБЫ ПРИМЕНЕНИЯ | 2015 |
|
RU2720282C1 |
| ОПТИМИЗИРОВАННЫЕ С ПОМОЩЬЮ ВЫЧИСЛИТЕЛЬНЫХ СРЕДСТВ АНТИГЕНЫ С ШИРОКИМ СПЕКТРОМ РЕАКТИВНОСТИ ДЛЯ ВИРУСОВ ГРИППА H5N1 И H1N1 | 2013 |
|
RU2639551C2 |
| НЕЙТРАЛИЗУЮЩИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, НАПРАВЛЕННЫЕ ПРОТИВ ВИРУСА ГРИППА, И ПУТИ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2721706C2 |
| ВЫСОКОАФФИННЫЕ АНТИТЕЛА ЧЕЛОВЕКА К БЕЛКУ GB ЦИТОМЕГАЛОВИРУСУ (CMV) ЧЕЛОВЕКА | 2011 |
|
RU2613421C2 |
| ЭНХАНСЕРНЫЕ ЭЛЕМЕНТЫ CPMV | 2015 |
|
RU2699982C2 |
| ХИМЕРНЫЕ ВАКЦИНЫ ПРОТИВ ГРИППА | 2021 |
|
RU2834969C1 |
| НЕЙТРАЛИЗУЮЩИЕ АНТИТЕЛА К ВИРУСУ ГРИППА А И ПУТИ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2683498C2 |
| НЕЙТРАЛИЗУЮЩИЕ АНТИТЕЛА К ВИРУСУ ГРИППА А И ПУТИ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2803850C2 |
| ИММУНОГЕННЫЙ ЭПИТОП ВИРУСА ГРИППА | 2009 |
|
RU2546872C2 |





































Изобретение относится к области биотехнологии и иммунологии. Описаны моноклональные антитела и их фрагменты, которые являются перекрестно реактивными с множеством кладов вируса гриппа, включая представителей как группы 1, так и группы 2. Эти антитела пригодны для контроля эпидемий и пандемий гриппа, а также для предоставления профилактической или терапевтической защиты от сезонного гриппа. Предложенная группа изобретений может быть использована в медицине. 8 н. и 8 з.п. ф-лы, 8 ил., 1 табл.
1. Моноклональное антитело, которое не обязательно может быть биспецифическим антителом или его иммунореактивным фрагментом, причем (i) связывается со стержневой областью белка НА0 из кладов вируса гриппа, которые включают по меньшей мере одну кладу из представителей как группы 1, так и группы 2 вируса гриппа A, (ii) остается связанным с белком НА0 вируса гриппа при рН 6 in vitro, или захватывается через эндосомальный каскад, и (iii) содержит вариабельную область тяжелой цепи, содержащую CDR1 с последовательностью GGIIRKYAIN (SEQ ID NO: 77), CDR2 с последовательностью GGIIAIFNTANYAQKFQG (SEQ ID NO: 78) и CDR3 с последовательностью ARGMNYYSDYFDY (SEQ ID NO: 79), и вариабельную область легкой цепи, содержащую CDR1 с последовательностью RASQSVRSNNLA (SEQ ID NO: 80), CDR2 с последовательностью GASSRAT (SEQ ID NO: 81) и CDR3 с последовательностью QQYGSSPALT (SEQ ID NO: 82).
2. Моноклональное антитело, которое необязательно может быть биспецифическим антителом или его иммунореактивным фрагментом, которое (i) связывается с стержневой областью белка НА0 и НА2 из кладов вируса гриппа, которые включают по меньшей мере одну кладу из представителей как группы 1, так и группы 2 вируса гриппа А, и (ii) содержит вариабельную область тяжелой цепи, содержащую CDR1 с последовательностью GGIIRKYAIN (SEQ ID NO: 77), CDR2 с последовательностью GGIIAIFNTANYAQKFQG (SEQ ID NO: 78) и CDR3 с последовательностью ARGMNYYSDYFDY (SEQ ID NO: 79), и вариабельную область легкой цепи, содержащую CDR1 с последовательностью RASQSVRSNNLA (SEQ ID NO: 80), CDR2 с последовательностью GASSRAT (SEQ ID NO: 81) и CDR3 с последовательностью QQYGSSPALT (SEQ ID NO: 82).
3. Моноклональное антитело, которое необязательно может быть биспецифическим антителом или его иммунореактивным фрагментом, которое содержит вариабельную область тяжелой цепи, содержащую CDR1 с последовательностью GGIIRKYAIN (SEQ ID NO: 77), CDR2 с последовательностью GGIIAIFNTANYAQKFQG (SEQ ID NO: 78) и CDR3 с последовательностью ARGMNYYSDYFDY (SEQ ID NO: 79 вариабельную область легкой цепи, содержащую CDR1 с последовательностью RASQSVRSNNLA (SEQ ID NO: 80), CDR2 с последовательностью GASSRAT (SEQ ID NO: 81) и CDR3 с последовательностью QQYGSSPALT (SEQ ID NO: 82).
4. Моноклональное антитело, которое не обязательно может быть биспецифическим антителом или его иммунореактивным фрагментом, которое (i) связывается со стержневой областью белка НА0 из кладов вируса гриппа, которые включают по меньшей мере одну кладу из представителей как группы 1, так и группы 2 вируса гриппа A, (ii) при этом остается связанным с белком НА0 вируса гриппа при рН 6 in vitro, или захватывается через эндосомальный каскад, и (iii) имеет IC50, меньше или равное 0,5 мкг/мл.
5. Моноклональное антитело по п. 4, которое имеет дополнительное свойство конкурировать за связывание с МАВ53 в конкурентном анализе.
6. Моноклональное антитело по любому из пп. 1-3, которое перекрестно реагирует со стержневой областью белка НА0 из кладов вируса гриппа H1, Н7 и Н9 или с кладами вируса гриппа H1, Н7 и Н3.
7. Моноклональное антитело по любому из пп. 1-3, которое представляет собой антитело или его фрагмент, которые являются человеческими, или гуманизированными, или химеризованными.
8. Моноклональное антитело по любому из пп. 1-3, которое нейтрализует инфекцию вирусом H1N1, H7N3 или H5N1 в клетках MDCK.
9. Моноклональное антитело по любому из пп. 1-4, которое является защитным у мышей против заражения в ином случае летальными титрами H1N1 или H5N1 в однократной дозе 1-10 мг/кг.
10. Моноклональное антитело по любому из пп. 1-3, которое содержит тяжелую цепь, содержащую последовательность QVQLVQSGAEVRKPGSSVKVSCKVSGGIIRKYAINWVRQAPGQGLEWMGGIIAIFNTANYAQKFQGRVTITADESTSTVYMELSSLRSEDTALYYCARGMNYYSDYFDYWGQGSLVTVSP (аминокислоты 1-120 SEQ ID NO: 75).
11. Моноклональное антитело по п. 10, которое содержит легкую цепь, содержащую последовательность EIVLTQSPGTLSLSPGERATLSCRASQSVRSNNLAWYQHKPGQAPRLLIFGASSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQYGSSPALTFGGGTKVEIK (аминокислоты 1-109 SEQ ID NO: 76).
12. Фармацевтическая композиция, содержащая моноклональное антитело по любому из пп. 1-4, 6-11 в количестве, эффективном для выработки пассивного иммунитета против представителя группы А и вируса гриппа группы В.
13. Моноклональное антитело по любому из пп. 1-4, 6-11 или композиция по п. 12 для применения в способе лечения или профилактики инфекции вирусом гриппа у индивидуума.
14. Рекомбинантная экспрессирующая система, которая содержит нуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи или легкой цепи антитела или фрагмента по любому из пп. 1-4, 10 и 11, функционально связанную с последовательностями контроля экспрессии.
15. Рекомбинантная клетка-хозяин, модифицированная так, чтобы она содержала экспрессирующую систему по п. 14, где клетка способна продуцировать антитело или его фрагмент по любому из пп. 1-4.
16. Способ продукции моноклонального антитела или фрагмента, иммунореактивного к вирусу гриппа, который включает культивирование клеток по п. 15 в условиях, в которых экспрессируется указанная нуклеотидная последовательность, и выделение указанного моноклонального антитела или его фрагмента.
| WO 2010010466 A2, 28.01.2010 | |||
| WO 2010010467 A2, 28.01.2010 | |||
| PRABHU N | |||
| et al., "Monoclonal antibodies against the fusion peptide of hemagglutinin protect mice from lethal influenza A virus H5N1 infection", J Virol | |||
| Колосоуборка | 1923 |
|
SU2009A1 |
| ZIEGLER T | |||
| et al., "Type- and subtype-specific detection of influenza viruses in clinical specimens by rapid culture assay", J Clin Microbiol | |||
| Топка с качающимися колосниковыми элементами | 1921 |
|
SU1995A1 |
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО К ВИРУСУ ТИПА А ГРИППА И УСТРОЙСТВО ДЛЯ ИММУННОГО АНАЛИЗА С ИСПОЛЬЗОВАНИЕМ АНТИТЕЛА | 2004 |
|
RU2366662C2 |

