Родственная заявка
По настоящей заявке испрашивается приоритет патентной заявки США номер 61/567046, поданной 5 декабря 2011, которая включена в настоящее описание в качестве ссылки в полном объеме.
Область техники
Изобретение относится к области пассивной иммунизации против гриппа. В частности, описаны специфические антитела, которые связываются рядом с консенсусной последовательностью сайта расщепления HA0 при посттрансляционном изменении гемагглютинина A вируса гриппа, включая антитела, секретируемые клетками человека.
Уровень техники
Белок гемагглютинин (HA) вируса гриппа содержит домен в виде глобулярной головки, который существенно различается среди штаммов гриппа, и область “стебля”, содержащую сайт, обеспечивающий слияние, который необходим для вхождения вируса внутрь клеток. HA представлен на вирусной оболочке в виде тримера. Нерасщепленная форма белка гемагглютинина (HA0) активируется при расщеплении трипсином с образованием частей HA1 и HA2, что придает эффект вирулентности сайту, обеспечивающему слияние. Две разделяемые части остаются связанными дисульфидными связями, но подвергаются конформационному изменению в среде с низким pH эндосомального компартмента клетки-хозяина, что приводит к слиянию мембран вируса и клетки-хозяина.
Участок расщепления содержит консенсусную последовательность, которая является общей для вируса гриппа А и вируса гриппа B и для различных штаммов вируса гриппа А и вируса гриппа B. Тример нерасщепленного белка гемагглютинина (HA0) обозначается как неактивная форма, тогда как белок гемагглютинин, расщепленный на части HA1 и HA2, обозначается как белок в активной форме.
Bianchi, E., et al., J. Virol. (2005) 79:7380-7388 описывают "универсальную" вакцину против гриппа В на основе консенсусной последовательности указанного сайта расщепления, в которой пептид, содержащий этот сайт, способен вызвать образование антител у мышей, когда он конъюгирован с белковым комплексом внешней мембраны Neisseria meningitidis. Также описаны моноклональные антитела, которые, по-видимому, связываются с консенсусной последовательностью. Кроме того, успешный пассивный перенос антисыворотки наблюдали у мышей. Другие вакцины предшествующего уровня техники, такие как вакцины, описанные в WO 2004/080403, содержащие пептиды, полученные из белков вируса гриппа М2 и/или HA, индуцировали образование антител, которые либо обладают слабой эффективностью, либо не являются эффективными среди штаммов.
Антитела, описанные в данной области, которые связываются с областью “стебля” HA, включают антитела, созданные Crucell, CR6261 и CR8020, описанные в Throsby, M., et al., PLoS One (2008) 3:e3942, Ekiert, D.C, et al., Science (2011) 333:843-850, и Sui, J., et al., Nat. Struct. Mol. Biol. (2009) 16:265-273. MAB также были созданы против консервативного антигена M2E, как описано у Grandea, A.G., et al., PNAS USA (2010) 107: 12658-12663. M2E находится на поверхности инфицированных клеток, а также является мишенью для амантадина и римантадина. К указанным антибиотикам существует устойчивость микроорганизмов, что означает, что указанная мишень не выполняет необходимую функцию.
Группа Lanzavecchia: Corti, D., et al., Science (2011) 333:850-856 описала дополнительное антитело, которое связывает и нейтрализует штаммы вируса гриппа A как группы 1, так и группы 2, но его активность ниже, чем активность антител, описанных далее, как показано в приведенных ниже примерах. Кроме того, MAB, которое иммунореактивно против гриппа A и B, как описано в Dreyfus, C, et al., Science (2012) 337: 1343-1348, обладает меньшей активностью, чем антитела, описанные ниже.
В публикации заявки РСТ WO 2011/160083, которая включена в настоящее описание в качестве ссылки, описаны моноклональные антитела, которые получены из клеток человека и могут использоваться для пассивных вакцин. Антитела характеризуются высокими показателями аффинности связывания с кладой H1 вируса гриппа, который относится к группе 1, и некоторые из антител также характеризуются высокими показателями аффинности к H9, также относящейся к группе 1 и/или к H7 группы 2, и/или к H2 группы 1. Некоторые из описанных антител связываются только с неактивной формой тримера, предположительно, с консенсусным сайтом расщепления, тогда как другие антитела способны связывать активный белок гемагглютинин, который образуется после расщепления.
Сохраняется потребность в антителах, которые связываются с дополнительными кладами и характеризуются повышенной аффинностью к ним.
Описание изобретения
Настоящее изобретение относится к моноклональным антителам, которые с повышенной аффинностью связываются с тримерами, характерными для одной группы или для обеих групп 1 и 2 гриппа А. Такие антитела способны обеспечивать пассивный иммунитет в случае пандемии, вызванной, например, неустановленным ранее штаммом гриппа или штаммом, защиту против которого в настоящее время не вызывают сезонные вакцины.
Учитывая, что по меньшей мере некоторые из антител связываются со многими штаммами, что свидетельствует об их нацеленности на важную область белка, они, вероятно, могут связываться даже с необнаруженными ранее штаммами. Такие антитела также можно использовать для ослабления или профилактики инфекции у пациентов, у которых вакцинация не обеспечивает полностью защитный ответ, или которые подвергаются высокому риску из-за слабой иммунной системы (например, маленькие дети, пожилые люди, пациенты, перенесшие трансплантацию, пациенты, получавшие химиотерапию при раке или ВИЧ).
Таким образом, в одном из аспектов изобретение относится к связывающим реагентам, в частности, к моноклональным антителам или их иммунореактивным фрагментам, которые в целом перекрестно реагируют с вирусом гриппа А группы 1, включая H1, H2, H5, H6, H8, H9, H11, H13, H16, или группы 2, включая H3 и H7, в качестве типичных образцов, или которые характеризуются перекрестной межгрупповой реактивностью. Некоторые из антител, показанных ниже, специфически связываются с эпитопом, содержащимся в белке HA0, и распознают нативную тримерную форму HA, а также активную форму.
Особенно важное значение имеют биспецифичные антитела и их фрагменты, которые способны повышать диапазон специфически связываемых вирусных клад.
Как хорошо известно в данной области, неиммуноглобулиновые белки могут обладать сходными с антителами свойствами распознавания эпитопа, а также могут обеспечивать подходящие варианты осуществления, включающие связывающие реагенты на основе фибронектина, трансферрина или липокалина. Молекулы на основе нуклеиновых кислот, такие как аптомеры, также обладают указанными связывающими свойствами.
В других аспектах настоящее изобретение относится к способам применения связывающих молекул изобретения для пассивного ингибирования вирусной инфекции у пациентов, которые уже подверглись воздействию вируса или которые уже инфицированы. Изобретение также относится к рекомбинантным материалам и к способам получения антител или их фрагментов.
Краткое описание фигур
На фиг. 1 показана известная в данной области классификация вируса гриппа на группы значимых кладов.
На фиг. 2 показаны результаты связывания MAB579 in vitro с различными штаммами H7 и H3, оба представляющие группу 2.
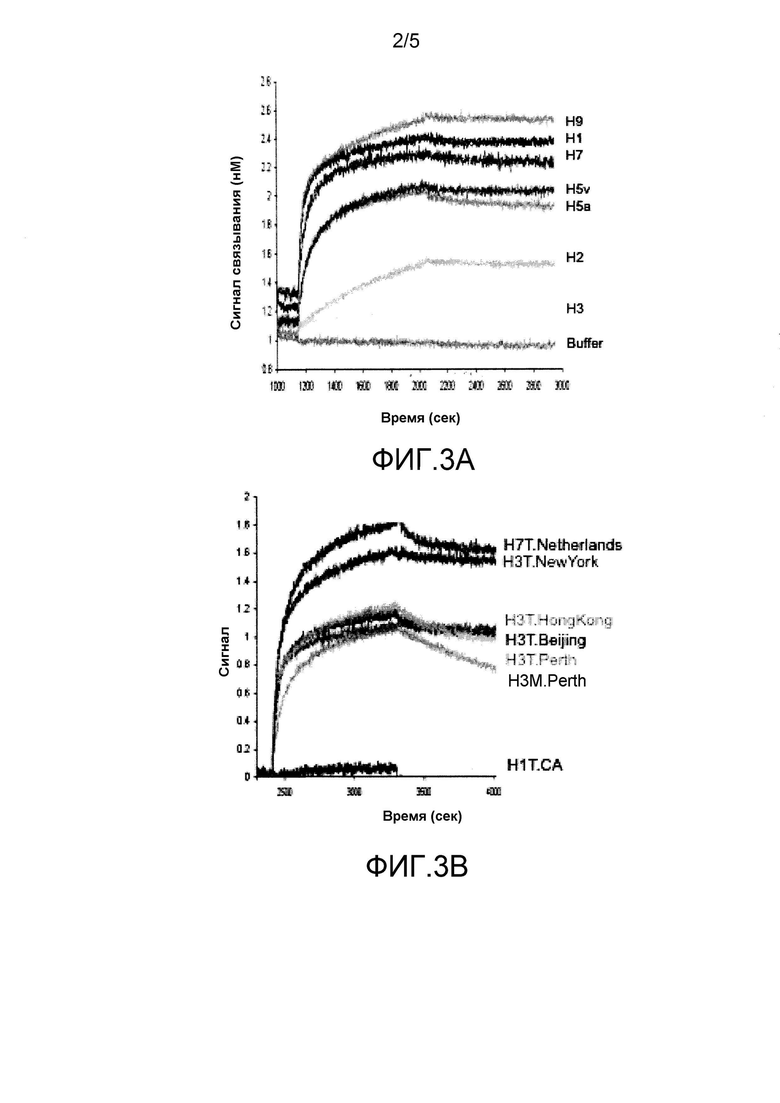
На фиг. 3A и 3B показаны два ведущих mAb, MAB53 (группа 1) и MAB579 (группа 2), обладающие аффинностью менее нМ в кладах, охватывающих соответствующие группы.
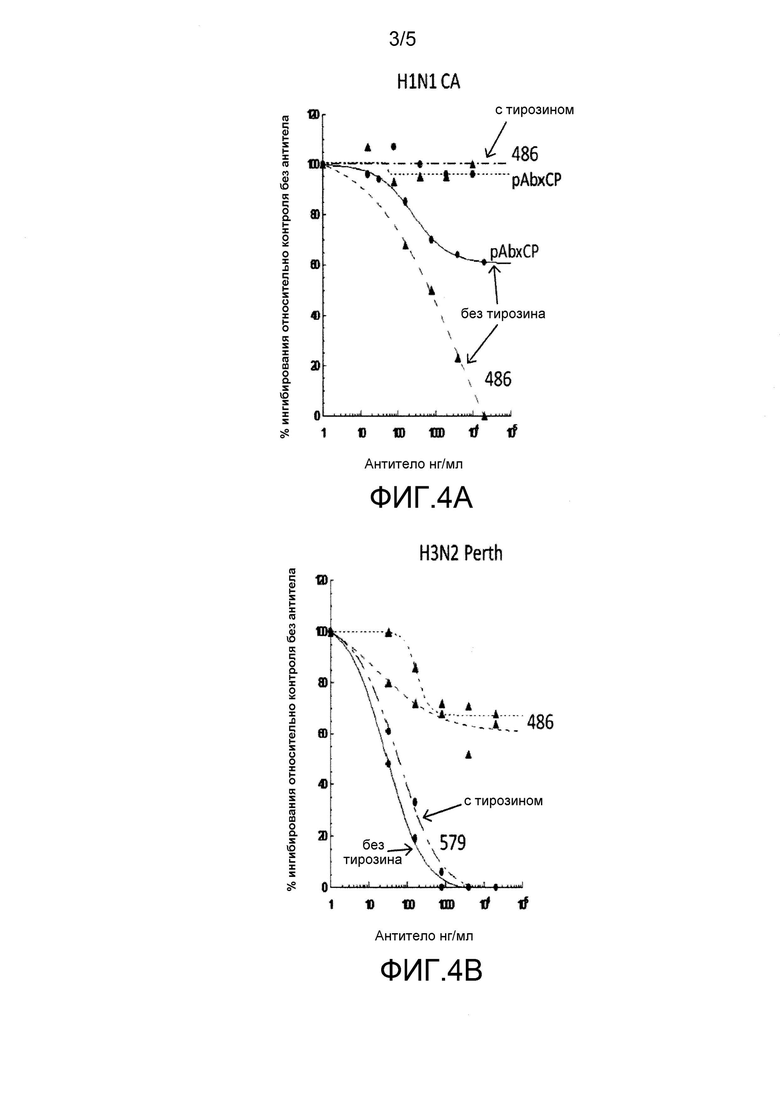
На фиг. 4A и 4B представлено сравнение нейтрализующей активности MAB486 и MAB579. Как показано на фиг. 4A, MAB486, а также поликлональный препарат, полученный в результате иммунизации кролика, эффективно нейтрализуют H1N1 только в отсутствие трипсина. В противоположность этому, на фиг. 4В показано, что MAB579 эффективен как в присутствии, так и в отсутствие трипсина.
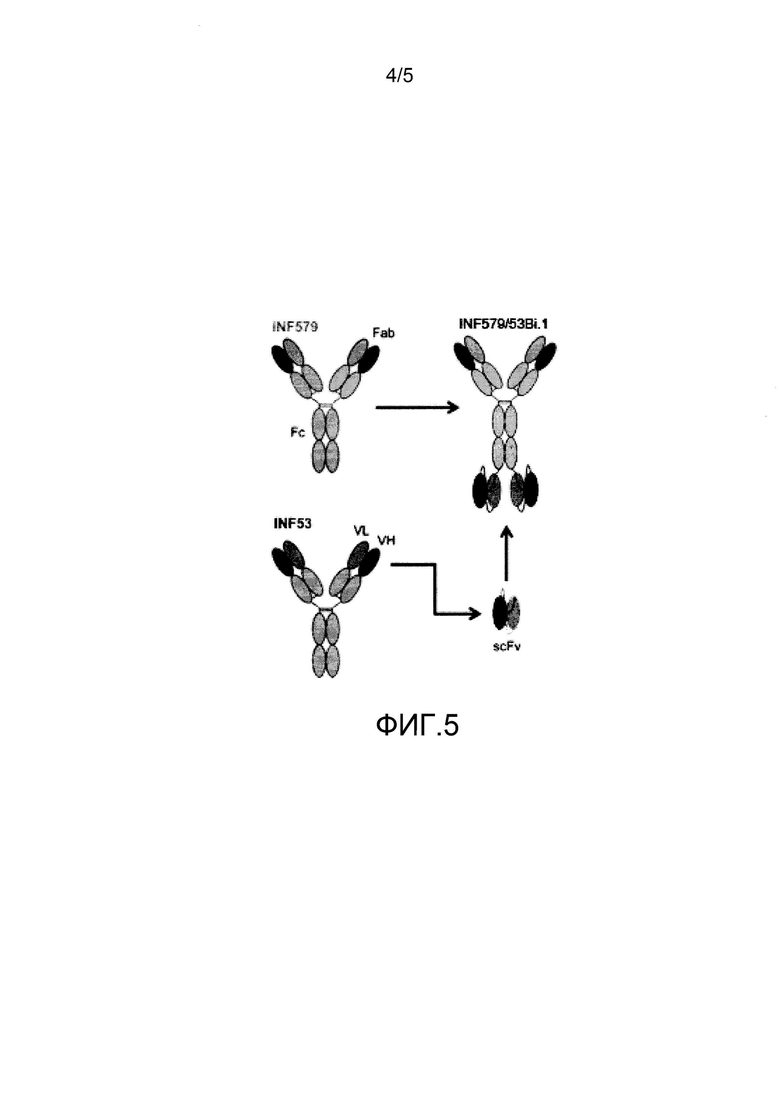
На фиг. 5 представлена схема биспецифического антитела, которое содержит вариабельные участки MAB579 и MAB53.
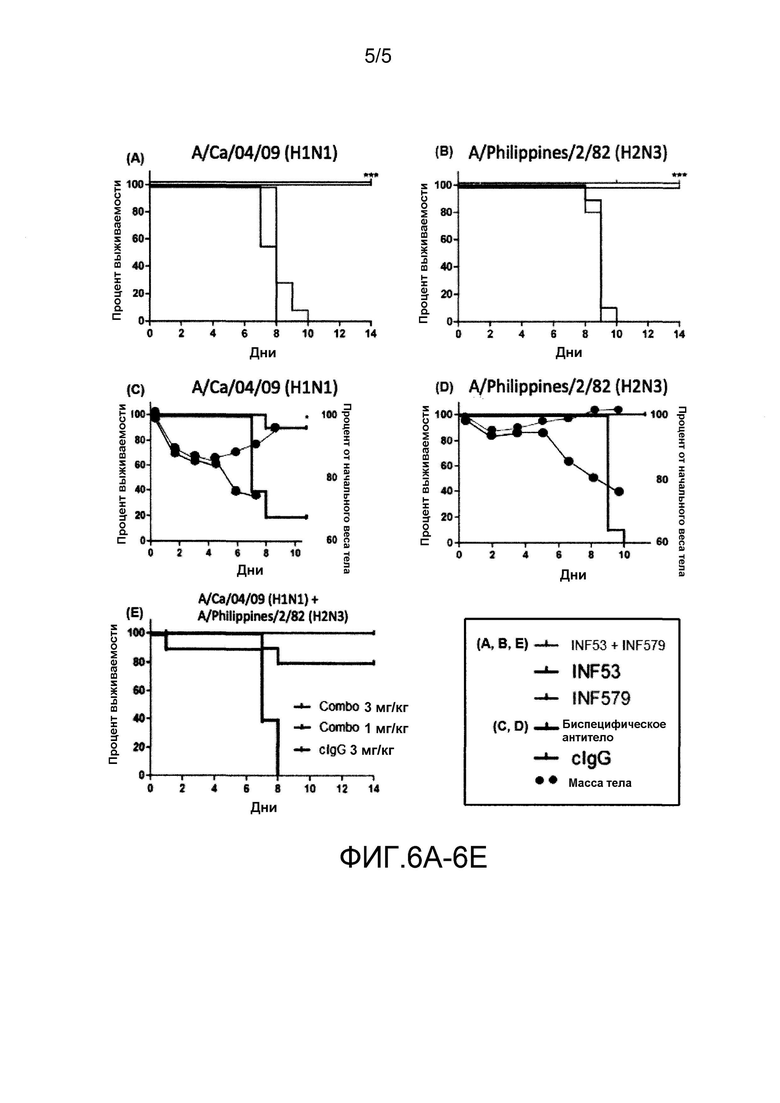
На фиг. 6A-6E показана эффективность in vivo MAB53, MAB579, их смесей и биспецифического антитела, представленного на фиг. 5.
Варианты осуществления изобретения
Настоящее изобретение обеспечивает эффективные связывающие молекулы, включая антитела и их фрагменты, а также эффективные способы идентификации клеток, которые секретируют такие антитела так, что соответствующие кодирующие последовательности могут быть получены и сохранены для последующего легкого рекомбинантного получения таких антител.
Антитела или аналогичные связывающие молекулы изобретения используются как для профилактики, так и для терапии. Таким образом, они могут быть использованы для защиты индивида против заражения вирусом, а также для лечения индивидов, которые уже подверглись воздействию вируса или инфицированы вирусом гриппа. Исследуемые объекты, вызывающие наибольший интерес, представляют собой человека и объекты, предназначенные для использования у человека, человеческие формы или гуманизированные формы связывающих молекул, среди которых предпочтительными являются традиционные природные антитела или их иммунореактивные фрагменты. Однако антитела, обладающие соответствующими характеристиками связывания, которые обусловлены областями CDR, при использовании в исследованиях на лабораторных животных, могут сохранять характеристики нечеловеческих антител. Антитела, используемые в исследованиях приведенных ниже примеров, хотя исследования проводились на мышах, тем не менее содержат как вариабельные, так и константные участки, которые являются человеческими.
Индивиды, для которых связывающие молекулы, включая антитела изобретения, можно использовать для терапии и профилактики, включают, помимо человека, любого индивида, который подвержен заражению гриппом. Таким образом, различные млекопитающие, такие как крупный рогатый скот, свиньи, овцы и другие млекопитающие, включая лошадей и домашних животных, получат пользу от профилактического и терапевтического применения указанных связывающих молекул. Кроме того, как известно, гриппом заражаются различные виды птиц, которым также будут полезны композиции, содержащие антитела изобретения.
Способы применения для профилактики и терапии являются общепринятыми и обычно хорошо известны. Антитела или другие связывающие молекулы, как правило, предоставляются с помощью инъекции, но следует понимать, что пероральные вакцины также являются эффективными. Дозировки и время введения легко оптимизировать и они являются частью профессиональных навыков в данной области.
Клетки человека, которые секретируют подходящие антитела, можно идентифицировать, в частности, способом CellSpot™, описанным в патенте США 7413868, содержание которого включено в настоящее описание в качестве ссылки.
Коротко, способ позволяет осуществить скрининг отдельных клеток, полученных у человека (или других индивидов) в анализах с высокой пропускной способностью, используя маркировки частиц определенными метками и микроскопические исследования.
В одном из характерных вариантов осуществления можно осуществить анализ даже одной клетки в отношении антител, которые она секретирует, путем адсорбции или соединения секретируемых антител с поверхностью и последующей обработки поверхности желаемыми антигенами, каждый из которых соединен с отличительной меткой в виде частицы. Таким образом, «отпечаток» клетки можно идентифицировать с помощью микроскопа.
С использованием такой методики можно провести скрининг миллионов клеток в отношении секреции желаемых антител и можно получить даже редкие антитела, например, антитела, подходящие для пассивной иммунизации против многих штаммов гриппа.
Поскольку люди уже имеют антитела по меньшей мере к некоторым штаммам гриппа, и поскольку антитела, полученные по способу изобретения, связываются с консервативной последовательностью, указанные антитела предназначены для новых штаммов, а также для штаммов, с которыми популяции людей уже встречались.
Способы получения подходящих антител не ограничивается методом CellSpot™ и не ограничиваются людьми. Клетки, которые продуцируют подходящие антитела, можно идентифицировать различными способами, и клетки могут представлять собой клетки лабораторных животных, таких как мыши или другие грызуны.
Могут быть выделены последовательности нуклеиновых кислот, кодирующие указанные антитела, и могут быть получены различные формы продуцируемых антител, включая химерные и гуманизированные формы антител, производимых нечеловеческими клетками. Кроме того, полученные рекомбинантным путем антитела или фрагменты включают одноцепочечные антитела или их области Fab или FAb2. Антитела человека также могут быть получены с использованием хозяев, например, XenoMouse® с гуманизированной иммунной системой. Средства получения антител при использовании скрининга в отношении подходящих связывающих характеристик, хорошо известны в данной области
Аналогичным образом, в данной области также известны средства создания аптамеров с заданными типами связывания.
Как отмечалось выше, антитела или другие связывающие молекулы могут связываться с активной формой, неактивной формой или обеими формами белка гемагглютинина. В ряде случаев полезно, когда эпитоп находится в сайте расщепления указанного белка, поскольку он является относительно консервативным в штаммах, но предпочтительно связывающая молекула связывается как с тримером, так и с активной формой.
Известен сайт расщепления для различных штаммов вируса гриппа А и гриппа В. Например, в приведенной выше статье Bianchi, et al., в таблице 1 показана последовательность вокруг сайта расщепления для нескольких таких штаммов.
(SEQ ID NO:51)
(SEQ ID NO:52)
(SEQ ID NO:53)
(SEQ ID NO:54)
(SEQ ID NO:55)
(SEQ ID NO:56)
b Консенсус является одинаковым для линий Victoria и Yamagata.
Как указано, строгий консенсус наблюдается, начиная с остатка аргинина выше сайта расщепления и, таким образом, предпочтительные консенсусные последовательности, включенные в тестируемые пептиды изобретения, имеют последовательность RGI/L/F FGAIAGFLE (SEQ ID NO:57). Возможно использование только части указанной последовательности в тестируемых пептидах.
Как отмечалось выше, после идентификации клеток, которые секретируют желаемые антитела, легко получить кодирующие их нуклеотидные последовательности и произвести желаемые антитела рекомбинантным путем в большом количестве. Это также позволяет манипулировать антителами, так что они могут быть получены, например, в виде одноцепочечных антител или с точки зрения только их вариабельных областей.
При этом также возможно проведение манипуляций с антителами, например, для их получения в виде одноцепочечных антител или в виде только вариабельных участков. Выделенные нуклеиновые кислоты могут быть физически сохранены и использованы для последующего рекомбинантного производства и/или информация, содержащаяся в последовательности, касающаяся кодирующей последовательности антитела, может быть получена и сохранена для обеспечения последующего синтеза соответствующих нуклеиновых кислот. Наличие информации, содержащейся в кодирующих последовательностях, и способов быстрого синтеза и клонирования наряду с известными способами рекомбинантного получения обеспечивают быстрое производство необходимых антител в случае пандемии или другой чрезвычайной ситуации.
Для сведения, последовательности константных участков обеих тяжелой и легкой цепей антитела человека описаны и представлены в данном описании в виде SEQ ID NO:1-3. В вышеуказанном WO 2011/160083 получены различные моноклональные антитела с вариабельными участками с установленной аминокислотной последовательностью и нуклеотидными кодирующими последовательностями, которые с разной степенью аффинности связываются с белком HA различных штаммов гриппа.
Структуры вариабельных участков как легкой, так и тяжелой цепей, представляющие особый интерес, приведены в данном описании для удобства в виде SEQ ID NO:22-25. Указанные антитела включают MAB8 и MAB53. MAB53 и MAB8 связываются с определенной аффинностью с H1; кроме того, MAB53 связывается с высокой аффинностью с H5, H7 и H9. MAB8 также связывается с H7 и H2. Ни одно из указанных антител не связывается прочно с H3, но MAB579 связывается с H3, описанным в данном описании. H7 и H3 являются особенно привлекательными мишенями.
Более подробно, каждое из указанных MAB связывается по меньшей мере с тремя различными кладами с достаточной или высокой аффинностью. MAB53 связывается с HA0 из клад H1, H9 и H7, и MAB8 связывается с HA0 из клад H1, H7 и менее сильно с H3, о чем свидетельствуют результаты теста ELISA для белка HA0. Показатели аффинности находятся в наномолярном диапазоне. Реакционную способность в отношении нативного тримера HA из всех кладов группы 1 проверяли с использованием HA, экспрессированного в клетках HEK293, при этом связывание антител измеряли с помощью проточной цитометрии.
Указанные результаты подтверждали с помощью альтернативной системы анализа, интерферометрии на биологическом уровне, основанной на количественном анализе связывания, называемой биосенсор ForteBio®. При измерении с помощью указанного более точного метода анализа получают следующие показатели аффинности:
MAB53/H1=60 пM, H5=6 нМ, H7=70 пМ, H9=30 пМ;
MAB8/H1=9 нМ, H3=16 нМ, H5=0,2 нМ.
Дополнительные специфические антитела, идентифицированные в настоящем описании, MAB383, MAB486, MAB579, MAB699, MAB700, MAB708, MAB710, MAB711 и MAB723 представлены SEQ ID NO:4-21 в виде их аминокислотных последовательностей вариабельных участков тяжелой цепи и легкой цепи. Указанные антитела связываются с повышенной аффинностью с дополнительными кладами штаммов вируса гриппа. Например, MAB579 связывается с высокой аффинностью с H3 и H7. Таким образом, указанные антитела дополняют репертуар антител, полезных для профилактики и лечения гриппа.
В настоящее время существует несколько методик создания отдельной молекулы, подобной антителу, которая содержит антиген-специфические домены двух отдельных антител (биспецифическое антитело). Таким образом, можно сконструировать одно антитело, обладающее реакционной способностью в отношении широкого спектра штаммов, с использованием Fab-доменов отдельных антител с широким спектром реактивности в отношении группы 1 и группы 2, соответственно. Подходящие методики описаны Macrogenics (Rockville, MD), Micromet (Bethesda, MD) и Merrimac (Cambridge, MA), (см., например, Orcutt KD, Ackerman ME, Cieslewicz M, Quiroz E, Slusarczyk AL, Frangioni JV, Wittrup KD. A modular IgG-scFv bispecific antibody topology, Protein Eng Des Sel. (2010) 23:221-228; Fitzgerald J, Lugovskoy A. Rational engineering of antibody therapeutics targeting multiple oncogene pathways. MAbs. (2011) 1:3(3); Baeuerle PA, Reinhardt C. Bispecific T-cell engaging antibodies for cancer therapy. Cancer Res. (2009) 69:4941-4944.)
Таким образом, особенно эффективно предоставление антител или других связывающих молекул, которые связываются с различными типами белка гемагглютинина, путем создания биспецифических антител. Особенно эффективными являются комбинации, которые сочетают в себе специфичность связывания с MAB53 (H1, H5, H9) и с MAB579 (H3, H7).
Все антитела настоящего изобретения обладают по меньшей мере специфичностью связывания в отношении одного из впервые описанных антител, описанных выше. Они могут быть объединены с другими различными антителами, включая антитела, которые описаны в вышеуказанном WO 2011/160083, а также с другими членами новой группы антител, описанной в настоящем описании. Все возможные комбинации таких специфичностей связывания входят в объем настоящего изобретения.
В то время как антитело MAB53 связывается с высокой аффинностью с HA0, оно не связывается с HA1, что позволяет предположить его связывание с комплементарным фрагментом HA2, которое было подтверждено. Поскольку MAB53 не связывается с HA0 при тестировании методом вестерн-блоттинга, предполагают, что доминантный эпитоп, по меньшей мере частично, является конформационным. Обнаружено, что MAB8 и MAB53 связываются с одними и теми же или соседними эпитопами, о чем свидетельствует их способность конкурировать друг с другом за связывание с белком HA0 клады H1.
Все антитела, описанные в настоящем описании, включая антитела, раскрытые ранее в вышеуказанном WO 2011/160083, связываются с нативным тримером HA, экспрессированным на поверхности трансфицированных клеток HA. Указанную способность проверяли с помощью HA-кодирующей плазмиды, предоставляемой S. Galloway и D. Steinhauer из Университета Эмори. То есть тример представленный на клеточной поверхности кладов, распознаваемых различными MAB изобретения, распознается указанными MAB.
Показано, что MAB53 и MAB8 отличаются тем, что MAB8 освобождается от белка HA0, когда рН снижается до 6, тогда как MAB53 не освобождается. Это отличие является существенным, поскольку оно может быть прогностическим в отношении нейтрализующей способности антитела. В тестах определения способности нейтрализации вирусной инфекцию H1N1 в реакции нейтрализации бляшкообразования с клетками-мишенями MDCK, MAB53 в низких дозах, 1-5 мкг/мл, нейтрализует инфекцию, вызываемую H1N1, H7N3, H5N1 и H9N2. Однако MAB8 не нейтрализует инфекцию, вызываемую указанными штаммами. Таким образом, можно провести предпочтительный отбор нейтрализующихся штаммов путем отмывки связанного MAB или его фрагмента при pH 6 во время первичного скрининга, при этом снимая с HA0 MAB, которые с низкой вероятностью останутся связанными при поступлении комплекса антитело-вирус в клетку через эндосомальный компартмент и при этом предполагается, что они обладают пониженной способностью к нейтрализации вируса. Например, в методе CellSpot HA0 может быть связан с твердым носителем (флуоресцентные шарики) и может захватываться MAB или смесью MAB с последующей отмывкой при pH 6.
Кроме того, показано, что мыши, предварительно получившие MAB53 в возрастающих дозах, выживают при введении вирусов H1N1 и H5N1, в количествах, вызывающих летальный исход в других случаях, благодаря 100% защите против инфицирования H1N1. Эффективность антитела сравнима с эффективностью антитела известного уровня техники, описанным Crucell, которое не обладает активностью против штаммов группы 2. Throsby, M., (supra) 3:e3942. Антитела Crucell представляют собой гетеросубтипические нейтрализующие моноклональные антитела, обеспечивающие перекрестную защиту против H5N1 и H1N1, полученные из IgM+B-клеток памяти человека. Также антитела MAB53 обеспечивали полную защиту при использовании в дозе 10 мг/кг; 90% выживало при использовании в дозе 2 мг/кг и 50% выживало при использовании в дозе 0,4 мг/кг. Когда при контрольном заражении вместо H5N1 использовали H1N1, применение 10 мг/кг MAB53 обеспечивало 80% выживаемость; 2 мг/кг обеспечивало 60% выживаемость и 0,4 мг/кг обеспечивало 50% выживаемость.
MAB53 и антитела, которые связываются с тем же эпитопом в тех же условиях, то есть, в дальнейшем остаются связанными, когда рН снижается до 6, являются эффективными в качестве пассивных вакцин, подходящих для защиты населения от эпидемии и пандемии, и для профилактического или терапевтического использования против сезонного гриппа у пациентов с ослабленной иммунной системой. Комбинации области связывания эпитопа MAB53 и областей связывания эпитопов, с которыми связываются с высокой аффинностью антитела настоящего изобретения, особенно полезны при создании биспецифических антител. При этом обеспечивается, например, эффективное связывание H7, H3 и H1 одним и тем же антителом, если в антитело включены связывающие области MAB579. В таблице 2 приведены значения IC50, полученные для белка гемагглютинина различных штаммов гриппа при использовании MAB579.
Указанные значения получены в тесте микронейтрализации на монослое MDCK. Аффинность связывания MAB579 для различных штаммов также представлена в графическом виде на фиг. 2. Как показано, тогда как H3 и H7 связываются с высокой аффинностью, в случае H1 обнаружена незначительная аффинность связывания. Таким образом, особенно полезным является комбинирование связывающей области MAB579 и связывающей области MAB с высокой аффинностью связывания к H1. В таком случае получается, что представлены области связывания для вирусов группы 1 и группы 2. Один вариант осуществления настоящего изобретения включает биоспецифическое антитело, которое связывается с эпитопом, с которым связывается MAB53, и с эпитопом, с которым связывается MAB579.
Кроме биспецифических антител, изобретение относится к применению только тяжелой цепи в конструкциях, предназначенных для нейтрализации вирусной инфекции; такие антитела могут быть также биспецифическими. В данной области известно, что специфичность обеспечивается, главным образом, вариабельными участками тяжелых цепей, и при некоторых состояниях тяжелые цепи сами по себе успешно использовались в качестве активных ингредиентов в вакцинах. Альтернативно, тяжелая цепь соответствующей специфичности может быть соединена с различными формами легкой цепи для повышения аффинности или способности нейтрализовать вирус.
Следует особо отметить, что область CDR3 тяжелых цепей антител, описанных в данном описании, увеличена и содержит несколько остатков тирозина. Следует понимать, что такие остатки тирозина могут сульфироваться в качестве посттрансляционного события. Таким образом, частью изобретения также являются вакцины, которые содержат области CDR3 тяжелых цепей MAB579, MAB699, MAB700, MAB708, MAB710, MAB711 или MAB723, где один или более остатков тирозина необязательно сульфирован. Указанные области с сульфированием или без сульфирования также могут быть использованы отдельно в виде пассивных вакцин. Сульфирование области CDR3 соответствует критериям сульфирования, описанным Monigatti, F., et al., Bioinformatics (2002) 18:769-770. Другие примеры успешного использования только областей CDR3 тяжелых цепей для нейтрализации вирусной инфекции описаны Pejchal, R., et al., PNAS (2010) 107: 11483-11488 и Liu, L., et al., J. Virol. (2011) 85:8467-8476.
В контексте настоящего описания термин "антитело" включает иммунореактивные фрагменты обычных антител даже если, в том или ином случае, упоминание "фрагментов" является излишним. Антитела, таким образом, включают Fab-фрагменты, Fv-одноцепочечные антитела, которые по существу содержат только вариабельные участки, биспецифические антитела и их различные фрагментированные формы, которые сохраняют иммуноспецифичность, и в целом белки, которые имитируют активность "природных" антител в результате включения в состав аминокислотных последовательностей или модифицированных аминокислотных последовательностей (т.е. псевдопептиды), которые приблизительно соответствуют активности вариабельных участков более часто встречающихся в природе обычных антител.
Структура антител
Структура антител представлена в следующем порядке:
1. аминокислотные последовательности константного участка тяжелой цепи IgG1 человека, константного участка каппа-цепи человека и константного участка лямбда-цепи человека;
2. аминокислотные последовательности тяжелых и легких цепей из числа вариабельных участков тяжелых и легких цепей MAB 383, 486, 579, 699, 700, 708, 710, 711 и 723 (области CDR подчеркнуты в MAB 579, 699, 700, 708, 710, 711 и 723.);
3. аминокислотные последовательности вариабельных участков тяжелых и легких цепей MAB8 и MAB53, описанные в WO2011/160083 (последовательности LC, представленные в '083, также включали константный участок, и он был удален);
4. нуклеотидные последовательности, кодирующие константный участок тяжелой цепи IgG1 человека, константный участок каппа-цепи человека и константный участок лямбда-цепи человека;
5. нуклеотидные последовательности, кодирующие аминокислотные последовательности тяжелых и легких цепей, входящих в состав вариабельных участков тяжелых и легких цепей MAB 383, 486, 579, 699, 700,708, 710, 711 и 723;
6. нуклеотидные последовательности, кодирующие аминокислотные последовательности вариабельных участков тяжелых и легких цепей MAB 8 и MAB53.
Что касается указанных областей CDR, следует отметить, что существует более чем одна номенклатура для определения CDR. Наиболее часто используется номенклатура Кабата, впервые предложенная Wu, T.T., et al., J. Exp. Med. (1970) 132:211-250. Номенклатура Кабата является широко распространенной системой, которая позволяет идентифицировать определенные положения, которые ассоциированы с CDR. При использовании дополнительной системы, номенклатуры Чотиа, получают несколько другие результаты. Номенклатура Чотиа описана в Al-Lazikani, B., et al., J. Molec. Biol. (1997) 273:927-948. В зависимости от используемой номенклатуры получают несколько отличающиеся результаты для CDR. Например, для MAB53 CDR тяжелой цепи согласно номенклатуре Кабата представляет собой KYAIN, тогда как по номенклатуре Чотиа указанная CDR обозначается GGIIRKYAIN. Область CDR2 тяжелой цепи содержит дополнительный G на N-конце, и CDR3 содержит дополнительные AR на N-конце. Для легкой цепи обозначения CDR являются идентичными в обеих номенклатурах.
Некоторые критические замечания разных исследователей уравнивают обе номенклатуры; таким образом, понятно, что области CDR, обозначенные в описании и в формуле изобретения, могут несколько различаться. При условии, что полученные вариабельные участки сохраняют свою связывающую способность, точное местоположение областей CDR не является существенным, и области, указанные в формуле изобретения, представляют собой области, которые как полагают, включают CDR, идентифицированные согласно любой принятой номенклатуре.
Аминокислотная последовательность константного участка HC IgG1 человека (SEQ ID NO:1)
ASTKGPSVFPLVPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
Аминокислотная последовательность константного участка каппа-цепи LC человека (SEQ ID NO:2)
RTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Аминокислотная последовательность константного участка лямбда-цепи LC человека SEQ (ID NO:3)
GQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVVPAECS
Аминокислотная последовательность вариабельного домена HC MAB383 (SEQ ID NO:4)
QVQLVQSGAEVKRPGASVKVSCRASGYTFTSFGFSWVRQAPGQGLEWMGWISAYNGDTKSPQKLQGRVTMTTDTSTNTAYMELRSLISDDTAVYYCARAPPLYYSSWSSDYWGQGTLLTVSS
Аминокислотная последовательность вариабельного домена LC MAB383 (SEQ ID NO:5)
DIQMTQSPGTLSLSPGERATLSCRASQSVSSNYLAWYQQKHGQAPRPLIYGASRRATDVPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQYGSSPRTFGQGTKLEIK
Аминокислотная последовательность вариабельного домена HC MAB486 (SEQ ID NO:6)
QVQLVESGGGMVQPGGSRRLSCAASGFSFSTYGMHWVRQAPGKGLEWVAVISYDGEKQYYLDSVKGRFTISRDNSKDTLYLQMNSLTAEDTAVYYCVKESARRLLRYFEWLLSSPFDNWGQGALVTVSS
Аминокислотная последовательность вариабельного домена LC MAB486 (SEQ ID NO:7)
DIVMTQSPDSLAVSLGERATINCKSSQTVLYTSNKKNYLAWYQQKPGQPPKLLIYWASTRESGVPDRFSGSGSGTDFTLTISSLQAEDVAVYYCQQYYTSPYTFGQGTKLEIK
Аминокислотная последовательность вариабельного домена HC MAB579 (SEQ ID NO:8)
QVQLVQSGAEVKKPGASVKVSCKTSGYTFTAYTIHWVRQAPGQRLEWMGWINAGNGHTKYSQRFKGRVTITRDTSARTTYMELRSLTSEDTALYFCARGPETYYYDKTNWLNSHPDEYFQHWGHGTQVTVSS
Аминокислотная последовательность вариабельного домена LC MAB579 (SEQ ID NO:9)
DIQMTQSPSTLSASVGDRVTITCRASQTINNYLAWYQQKPGKAPKLLIYKASSLESGVPSRFSGSGSGTEFTLTISSLQPDDFATYYCQEYNNDSPLTFGGGTKVEIK
Аминокислотная последовательность вариабельного домена HC MAB699 (SEQ ID NO:10)
QLQLVQSGAEVKKPGASVKLSCKASGYTFTSYTLHWVRQAPGQTLEWMGWINAGNGKTKYPPKFRGRVTITRDTSATTVDMHLSSLTSEDTAVYFCARGPESYYYDRSDWLNSHPDEYFQYWGQGTLVIVSS
Аминокислотная последовательность вариабельного домена LC MAB699 (SEQ ID NO:11)
DIQMTQSPSTLSASVGDRVTIACRASQSISSWLAWYQQKPGKAPKLLIYKASQLESGVPSRFSGSGSGTEFTLTINSLQPDDFATYYCQLYNVYSPLTFGGGTRVDIK
Аминокислотная последовательность вариабельного домена HC MAB700 (SEQ ID NO:12)
QVQLVESGADVKKPGASVTVSCKASGYTFRSF TMHWVRQVPGQRLEWMGWINAGNGKTKYSQKFQGRVIVTRDTSASTAYMELSSLTSEDTAVYYCARGPETYYYDSSNWLNSHPDEYLQYWGQGTPVTVSS
Аминокислотная последовательность вариабельного домена LC MAB700 (SEQ ID NO:13)
DIVLTQSPSTLSASVGDRVTITCRASQSISSWLAWYQQKPGKAPKLLIYKASTLESGVPSRFSGSGSGTEFTLTISSLQPDDFATYYCQEYNNNSPLTFGGGTKVEIK
Аминокислотная последовательность вариабельного домена HC MAB708 (SEQ ID NO:14)
QVQLVQSGADVKRPGASVTVSCKASGYTFRSF TMHWVRQVPGQRLEWMGWINAGNGKTKYSQKFQGRVIVTRDTSANTAYMELSSLTSEDTAVYYCARGPETYYYDSSNWLNSHPDEYFQHWGQGTPVTVSS
Аминокислотная последовательность вариабельного домена LC MAB708 (SEQ ID NO:15)
DIQMTQSPSTLPASVGDRVTITCRASQSISSWLAWYQQKPGKAPKLLIYKASSLESGVPSRFSGSGSGTEFTLTISSLQPDDFATYYCQEYNNNSPLTFGGGTKVEIK
Аминокислотная последовательность вариабельного домена HC MAB710 (SEQ ID NO:16)
QVQLQESGAEVKKPGASVQVSCKASGYTFTSYSVHWVRQAPGQRPEWMGWINAGNGKTKYPQKFKGRVTITRDTLARTVNIHLSSLTSEDTAVYFCARGPDSYYYDRNDWLNSHPDEYFQHWGQGTVVIVSS
Аминокислотная последовательность вариабельного домена LC MAB710 (SEQ ID NO:17)
DIVMTQSPSTLSASVGDRVTISCRASQSIDSWLAWYQQKPGKAPKLLIYKASNLESGVPSRFSGSGSGTEFTLTISSLQPDDFATYYCQLYNVHLITFGGGTRVDIK
Аминокислотная последовательность вариабельного домена HC MAB711 (SEQ ID NO:18)
QVQLVESGAEVKKPGASVKITCEASGYTFNTYTIHWLRQAPGQRLEWMGWINAANGHTKYSRKLRSRVTIKRDTSARTSYMELSSLGSEDTAVYYCARGPETYYFDKTNWLNSHPDEYFQHWGQGTLVTVSS
Аминокислотная последовательность вариабельного домена LC MAB711 (SEQ ID NO:19)
DIVMTQSPSTLSASVGDRVTITCRASQSISTWLAWYQQKPGKAPKLLIYKASNLESGVPARFSGSGSGTEFTLTISSLQPDDFATYYCQEYNNDSPLILGGGTTVEIK
Аминокислотная последовательность вариабельного домена HC MAB723 (SEQ ID NO:20)
QVQLVQSGAAVNKPGASVKVSCKASGYSFTSYTLHWVRQAPGQRPEWIGWINAGNGKVKYPRKLQGRITITRDVSATTVHMELRSLTSEDTGLYYCARGPESYFFDTSNHLNSHPDEYFQFWGQGTLVTVSS
Аминокислотная последовательность вариабельного домена LC MAB723 (SEQ ID NO:21)
DIQMTQSPSTLSASVGDRVTITCRASQSISSYLAWYQQKPGKAPKLLIYKASNLESGVPSRFSGSGSGTEFTLTISSLQPDDFATYYCQEYNNNSPLTFGAGTKVEIK
Аминокислотная последовательность вариабельного домена HC MAB8 (SEQ ID NO:22)
EVQLVESGGGLVKPGGSLRLSCAASGFTFSTY TMSWVRQAPGQGLEWVSSITRTSSNIYYADSVEGRFTISRDNAKNSLYLQMHSLRVEDTAVYYCARISGVVGPVPFDYWGQGTLITVSS
Аминокислотная последовательность вариабельного домена LC MAB8 (SEQ ID NO:23)
DIQMTQSPSSLSASVGDRVTITCRASQTISKYLNWYQQKPGRAPKLLIYSASSLQSGVPSRFTGSGSGTDFTLTITSLQPEDFATYYCQQSYRPSQITFGPGTKVDIK
Аминокислотная последовательность вариабельного домена HC MAB53 (SEQ ID NO:24)
QVQLVQSGAEVRKPGSSVKVSCKVSGGIIRKYAINWVRQAPGQGLEWMGGIIAIFNTANYAQKFQGRVTITADESTSTVYMELSSLRSEDTALYYCARGMNYYSDYFDYWGQGSLVTVSP
Аминокислотная последовательность LC MAB53 (SEQ ID NO:25)
EIVLTQSPGTLSLSPGERATLSCRASQSVRSNNLAWYQHKPGQAPRLLIFGASSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQYGSSPALTFGGGTKVEIK
Нуклеотидная последовательность, кодирующая HC константного участка IgG1 человека (интроны выделены подчеркиванием) (SEQ ID NO:26)
GCCTCCACCAAGGGCCCATCAGTCTTCCCCCTGGCACCCTCTACCAAGAGCACCTCTGGGGGCACAACGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCGTGGAACTCAGGCGCCCTGACCAGCGGCGTGCACACCTTCCCGGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACCGTGCCCTCCAGCAGCTTGGGCACCCAGACCTACATCTGCAACGTGAATCACAAGCCCAGCAACACCAAGGTGGACAAGAGAGTTGGTGAGAGGCCAGCACAGGGAGGGAGGGTGTCTGCTGGAAGCCAGGCTCAGCGCTCCTGCCTGGACGCATCCCGGCTATGCAGTCCCAGTCCAGGGCAGCAAGGCAGGCCCCGTCTGCCTCTTCACCCGGAGGCCTCTGCCCGCCCCACTCATGCTCAGGGAGAGGGTCTTCTGGCTTTTTCCCCAGGCTCTGGGCAGGCACAGGCTAGGTGCCCCTAACCCAGGCCCTGCACACAAAGGGGCAGGTGCTGGGCTCAGACCTGCCAAGAGCCATATCCGGGAGGACCCTGCCCCTGACCTAAGCCCACCCCAAAGGCCAAACTCTCCACTCCCTCAGCTCGGACACCTTCTCTCCTCCCAGATTCCAGTAACTCCCAATCTTCTCTCTGCAGAGCCCAAATCTTGTGACAAAACTCACACATGCCCACCGTGCCCAGGTAAGCCAGCCCAGGCCTCGCCCTCCAGCTCAAGGCGGGACAGGTGCCCTAGAGTAGCCTGCATCCAGGGACAGGCCCCAGCCGGGTGCTGACACGTCCACCTCCATCTCTTCCTCAGCACCTGAACTCCTGGGGGGACCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGTGGGACCCGTGGGGTGCGAGGGCCACATGGACAGAGGCCGGCTCGGCCCACCCTCTGCCCTGAGAGTGACCGCTGTACCAACCTCTGTCCCTACAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGAGGAGATGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTATAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCCCCGGGTAAATGA
Нуклеотидная последовательность, кодирующая константный участок каппа-цепи LC человека (SEQ ID NO:27)
CGAACTGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCTAGCGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAG
Нуклеотидная последовательность, кодирующая константный участок лямбда-цепи LC человека (SEQ ID NO:28)
GGTCAGCCCAAGGCTGCCCCCTCTGTCACTCTGTTCCCGCCCTCTAGCGAGGAGCTTCAAGCCAACAAGGCCACACTGGTGTGTCTCATAAGTGACTTCTACCCGGGAGCCGTGACAGTGGCCTGGAAGGCAGATAGCAGCCCCGTCAAGGCGGGAGTGGAGACCACCACACCCTCCAAACAAAGCAACAACAAGTACGCGGCCAGCAGCTATCTGAGCCTGACGCCTGAGCAGTGGAAGTCCCACAGAAGCTACAGCTGCCAGGTCACGCATGAAGGGAGCACCGTGGAGAAGACAGTGGTCCCTGCAGAATGCTCT
Нуклеотидная последовательность, кодирующая вариабельный домен MAB383 HC (SEQ ID NO:29)
CAGGTGCAGCTGGTGCAGTCTGGAGCTGAGGTGAAGAGGCCTGGGGCCTCAGTGAAGGTCTCCTGCAGGGCTTCTGGTTACACCTTTACTAGCTTCGGTTTCAGCTGGGTGCGACAGGCCCCAGGACAAGGGCTTGAGTGGATGGGGTGGATCAGCGCTTACAATGGTGACACAAAGTCTCCACAGAAGCTCCAGGGCAGAGTCACCATGACTACAGACACATCCACGAACACAGCCTACATGGAGCTGAGGAGCCTCATATCTGACGACACGGCCGTGTATTATTGTGCGAGAGCCCCCCCCCTGTATTACAGTAGCTGGTCCTCAGACTACTGGGGCCAGGGAACCCTGCTCACCGTCTCCTCA
Нуклеотидная последовательность, кодирующая вариабельный домен MAB383 LC (SEQ ID NO:30)
GATATCCAGATGACGCAGTCTCCAGGCACCCTGTCTTTGTCTCCAGGGGAAAGAGCCACCCTCTCCTGCAGGGCCAGTCAGAGTGTCAGTAGCAACTACTTAGCCTGGTACCAGCAGAAACATGGCCAGGCTCCCAGGCCCCTCATCTACGGTGCATCCAGAAGGGCCACTGACGTCCCAGACAGGTTCAGTGGCAGTGGGTCTGGGACAGACTTCACTCTCACCATCAGCAGACTGGAACCTGAAGATTTTGCAGTGTATTATTGTCAGCAGTATGGTAGTTCACCTCGAACTTTTGGCCAGGGGACCAAACTGGAAATCAAAC
Нуклеотидная последовательность, кодирующая вариабельный домен MAB486 HC (SEQ ID NO:31)
CAGGTGCAGCTGGTGGAGTCTGGGGGAGGCATGGTCCAGCCGGGGGGGTCCCGGAGACTCTCCTGTGCAGCCTCTGGATTCAGCTTCAGTACCTATGGCATGCACTGGGTCCGCCAGGCTCCAGGCAAGGGGCTGGAGTGGGTGGCAGTTATTTCATATGATGGAGAAAAGCAATATTATCTAGACTCCGTGAAGGGACGATTCACCATCTCCAGAGACAATTCCAAGGACACCCTCTATCTGCAAATGAACAGTCTGACAGCTGAGGACACGGCTGTGTATTACTGTGTGAAGGAATCAGCGCGTCGATTATTACGATATTTTGAGTGGTTATTAAGTTCGCCTTTTGACAACTGGGGCCAGGGAGCCCTAGTCACCGTCTCCTCA
Нуклеотидная последовательность, кодирующая вариабельный домен MAB486 LC (SEP ID NO:32)
GATATCGTGATGACCCAGTCTCCAGACTCCCTGGCTGTGTCTTTGGGCGAGAGGGCCACCATCAACTGCAAGTCCAGCCAGACTGTTTTATACACCTCCAACAAGAAAAATTACTTAGCCTGGTACCAACAGAAGCCAGGGCAGCCTCCTAAACTGCTCATTTACTGGGCATCTACCCGGGAATCCGGGGTCCCTGACCGATTCAGTGGCAGCGGGTCTGGGACAGATTTCACTCTCACCATCAGCAGCCTGCAGGCTGAGGATGTGGCAGTTTATTACTGTCAGCAATATTATACGTCTCCCTACACATTTGGCCAGGGGACCAAGCTGGAGATCAAA
Нуклеотидная последовательность, кодирующая вариабельный домен MAB579 HC (SEP ID NO:33)
CAGGTGCAGCTGGTGCAGTCTGGGGCTGAGGTGAAGAAGCCTGGGGCCTCAGTGAAGGTTTCCTGCAAGACTTCTGGATACACCTTCACAGCCTATACTATACACTGGGTGCGCCAGGCCCCCGGACAAAGGCTTGAGTGGATGGGATGGATCAACGCTGGCAATGGTCACACGAAATATTCACAGAGGTTCAAGGGCAGAGTCACCATTACCAGGGACACATCCGCGAGGACAACCTACATGGAGCTGCGCAGTCTGACATCTGAGGACACGGCTCTATATTTCTGTGCGAGAGGGCCCGAGACATATTATTATGATAAAACCAATTGGCTGAACTCCCATCCAGATGAATACTTCCAGCACTGGGGCCACGGCACCCAGGTCACCGTCTCCTCA
Нуклеотидная последовательность, кодирующая вариабельный домен MAB579 LC (SEP ID NP:34)
GATATCCAGATGACCCAGTCTCCTTCCACCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCCAGTCAGACTATTAATAACTACTTGGCCTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAACTCCTGATCTATAAGGCGTCTAGTTTAGAAAGTGGGGTCCCATCAAGATTCAGTGGCAGTGGGTCTGGGACAGAATTCACTCTCACCATCAGCAGCCTGCAGCCTGATGATTTTGCAACTTATTACTGCCAAGAATATAATAATGATTCTCCCCTAACTTTCGGCGGAGGGACCAAAGTGGAGATCAAA
Нуклеотидная последовательность, кодирующая вариабельный домен MAB699 HC (SEP ID NP:35)
CAGGTGCAGCTGGTGCAGTCCGGGGCTGAGGTGAAGAAGCCTGGGGCCTCAGTGAAGCTTTCCTGCAAGGCTTCTGGGTACACCTTCACTTCCTATACTCTACATTGGGTGCGCCAGGCCCCCGGACAGACACTTGAGTGGATGGGATGGATCAACGCTGGCAACGGTAAAACAAAATATCCACCGAAGTTCAGGGGCAGAGTCACCATTACCAGGGACACGTCCGCGACCACAGTCGACATGCATCTAAGCAGCCTGACATCTGAAGACACGGCTGTGTATTTCTGTGCGAGAGGGCCCGAAAGTTATTACTATGATAGAAGTGATTGGCTGAACTCCCATCCAGATGAATACTTCCAGTACTGGGGCCAGGGCACCCTGGTCATCGTCTCCTCA
Нуклеотидная последовательность, кодирующая вариабельный домен MAB699 LC (SEP ID NP:36)
GATATCGTGCTGACGCAGTCTCCTTCCACCCTGTCTGCATCTGTAGGGGACAGAGTCACCATCGCTTGCCGGGCCAGTCAGAGTATTAGCAGCTGGCTGGCCTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAACTCCTGATCTACAAGGCGTCTCAGTTAGAAAGTGGGGTCCCATCAAGATTCAGCGGCAGCGGATCTGGGACAGAGTTCACTCTCACCATCAACAGCCTGCAGCCTGATGATTTTGCAACTTATTACTGCCAACTTTATAATGTTTATTCTCCGCTCACTTTCGGCGGGGGGACCAGGGTGGACATCAAA
Нуклеотидная последовательность, кодирующая вариабельный домен MAB700 HC (SEP ID NO:37)
CAGGTGCAGCTGGTGGAGTCTGGGGCTGACGTGAAGAAGCCTGGGGCCTCAGTGACGGTTTCCTGCAAGGCCTCAGGATACACCTTCAGGAGTTTTACTATGCATTGGGTGCGCCAGGTCCCCGGACAAAGGCTTGAGTGGATGGGATGGATCAACGCTGGCAATGGTAAAACAAAGTATTCTCAGAAGTTCCAGGGCAGAGTCATCGTTACCAGGGACACATCCGCGAGCACAGCCTACATGGAGCTGAGCAGCCTAACATCTGAAGACACGGCTGTTTATTACTGTGCGAGAGGGCCCGAAACATATTACTATGATAGTAGTAATTGGCTGAATTCCCATCCAGATGAATATCTCCAGTACTGGGGCCAGGGCACCCCGGTCACCGTCTCCTCA
Нуклеотидная последовательность, кодирующая вариабельный домен MAB700 LC (SEP ID NO:38)
GATATCCAGATGACCCAGTCTCCTTCCACCCTGTCTGCGTCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCCAGTCAGAGTATTAGTAGCTGGTTGGCCTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAACTCCTGATCTATAAGGCGTCTACTTTAGAAAGTGGGGTCCCATCCAGGTTCAGCGGCAGTGGATCTGGGACAGAATTCACTCTCACCATCAGCAGCCTGCAGCCTGATGATTTTGCAACTTATTACTGCCAAGAGTATAATAATAATTCTCCGCTCACTTTCGGCGGAGGGACCAAGGTGGAGATCAAA
Нуклеотидная последовательность, кодирующая вариабельный домен MAB708 HC (SEP ID NP:39)
CAGGTGCAGCTGGTGCAGTCTGGGGCTGACGTGAAGAGGCCTGGGGCCTCAGTGACGGTTTCCTGCAAGGCTTCAGGATACACCTTCAGGAGCTTTACTATGCATTGGGTGCGCCAGGTCCCCGGACAAAGGCTGGAGTGGATGGGATGGATCAACGCTGGCAATGGTAAAACAAAATATTCCCAGAAGTTTCAGGGCAGAGTCATCGTTACCAGGGACACATCCGCGAACACGGCCTACATGGAGCTGAGCAGCCTGACATCTGAAGACACGGCTGTTTATTACTGTGCGAGAGGGCCCGAAACATATTATTATGATAGTAGTAATTGGCTGAACTCCCATCCAGATGAATATTTCCAGCACTGG
Нуклеотидная последовательность, кодирующая вариабельный домен MAB708 LC (SEP ID NP:40)
GATATCCAGATGACCCAGTCTCCTTCCACCCTGCCTGCGTCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCCAGTCAGAGTATTAGTAGCTGGTTGGCCTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAACTTCTGATCTATAAGGCGTCTAGTTTAGAAAGTGGGGTCCCATCCAGGTTCAGCGGCAGTGGATCTGGGACAGAATTCACTCTCACCATCAGCAGCCTGCAGCCTGATGATTTTGCAACTTATTACTGCCAGGAGTATAATAATAATTCTCCGCTCACTTTCGGCGGAGGGACCAAGGTGGAGATCAAA
Нуклеотидная последовательность, кодирующая вариабельный домен MAB710 HC (SEP ID NP:41)
CAAGTGCAGCTGCAGGAGTCGGGGGCTGAGGTGAAGAAGCCTGGGGCCTCAGTGCAGGTTTCCTGCAAGGCTTCTGGGTACACCTTCACGTCCTATAGCGTACATTGGGTGCGCCAGGCCCCCGGACAAAGGCCTGAGTGGATGGGATGGATCAACGCTGGCAACGGAAAGACAAAATATCCACAGAAGTTCAAGGGCAGAGTCACCATAACCAGAGACACATTAGCGCGCACTGTCAACATACATCTAAGCAGCCTGACATCCGAAGACACGGCTGTGTATTTCTGTGCGAGAGGGCCCGATAGTTATTACTATGATAGAAATGATTGGCTGAACTCCCATCCAGATGAATACTTCCAGCACTGGGGCCAGGGCACCGTGGTCATCGTCTCCTCA
Нуклеотидная последовательность, кодирующая вариабельный домен MAB710 LC (SEP ID NO:42)
GATATCGTGATGACCCAGTCTCCTTCCACCCTGTCTGCATCTGTGGGAGACAGAGTCACCATCTCTTGCCGGGCCAGTCAGAGTATTGACAGTTGGTTGGCCTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAACTCCTGATCTATAAGGCGTCTAATTTAGAAAGTGGGGTCCCATCAAGATTCAGCGGCAGCGGATCTGGGACAGAATTCACTCTCACCATCAGCAGCCTGCAGCCTGATGATTTTGCGACTTATTACTGCCAACTCTATAATGTTCATTTGATCACTTTCGGCGGAGGGACCAGGGTGGACATCAAA
Нуклеотидная последовательность, кодирующая вариабельный домен MAB711 HC (SEP ID NO:43)
CAGGTGCAGCTGGTGGAGTCTGGGGCTGAGGTGAAGAAGCCTGGGGCCTCAGTGAAGATCACCTGCGAGGCTTCTGGATACACTTTCAATACCTATACTATACATTGGCTGCGCCAGGCCCCCGGACAAAGACTTGAGTGGATGGGGTGGATCAACGCTGCCAATGGTCATACAAAATATTCACGGAAGCTCAGGTCCAGAGTCACCATTAAGAGGGACACATCCGCGAGGACAAGTTACATGGAGCTGAGCAGCCTGGGATCTGAAGACACGGCTGTCTATTACTGTGCGAGAGGGCCCGAAACATATTACTTTGATAAGACGAATTGGCTGAACTCCCATCCAGATGAATACTTCCAGCACTGGGGCCAGGGCACCCTGGTCACCGTCTCCTCA
Нуклеотидная последовательность, кодирующая вариабельный домен MAB711 LC (SEP ID NP:44)
GATATCGTGATGACGCAGTCTCCTTCCACCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCCAGTCAGAGTATTTCTACCTGGTTGGCCTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATAAGGCGTCCAATTTAGAAAGTGGGGTCCCAGCAAGATTCAGCGGCAGTGGATCTGGGACAGAGTTCACTCTCACCATCAGCAGCCTGCAGCCTGATGATTTTGCAACTTATTACTGCCAAGAATATAATAATGATTCTCCGCTGATTTTAGGCGGAGGGACCACGGTGGAGATCAAA
Нуклеотидная последовательность, кодирующая вариабельный домен MAB723 HC (SEP ID NP:45)
CAGGTGCAGCTGGTGCAGTCTGGGGCTGCGGTGAACAAGCCTGGGGCCTCAGTGAAGGTTTCCTGCAAGGCTTCTGGATACAGCTTCACTAGTTACACTTTGCATTGGGTGCGCCAGGCCCCCGGACAAAGGCCTGAGTGGATAGGGTGGATCAACGCTGGCAATGGTAAAGTAAAATATCCACGGAAGTTGCAGGGCAGAATCACCATAACCAGGGACGTATCCGCTACGACAGTTCACATGGAACTGAGGAGCCTGACATCTGAGGACACGGGTCTATATTACTGTGCGAGAGGGCCCGAAAGTTACTTCTTTGATACTTCTAATCATCTGAACTCCCATCCAGATGAATACTTCCAGTTCTGGGGCCAGGGCACCCTGGTCACCGTCTCCTCA
Нуклеотидная последовательность, кодирующая вариабельный домен MAB723 LC (SEP ID NP:46)
GATATCGTGCTGACGCAGTCTCCTTCCACCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCCAGTCAGAGTATTAGTAGTTACTTGGCCTGGTATCAACAGAAACCAGGGAAAGCCCCTAAACTCCTGATCTATAAGGCGTCTAATTTAGAAAGTGGGGTCCCATCAAGATTCAGCGGCAGTGGATCTGGGACAGAATTCACTCTCACCATCAGCAGCCTGCAGCCTGATGATTTTGCAACTTATTATTGCCAAGAATATAATAATAACTCTCCGCTCACTTTCGGCGCAGGGACCAAGGTGGAGATCAAA
Нуклеотидная последовательность, кодирующая вариабельный домен MAB8 HC (SEQ ID NO: 47)
GAGGTGCAGCTGGTGGAGTCTGGGGGAGGCCTGGTCAAGCCTGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGTTTCACTTTCAGTACCTATACTATGAGTTGGGTCCGCCAGGCTCCAGGGCAGGGGCTAGAGTGGGTCTCGTCCATTACTAGGACTAGTAGTAATATATACTACGCAGACTCAGTGGAGGGCCGATTCACCATCTCCAGAGACAACGCCAAGAACTCACTGTATCTGCAGATGCATAGCCTGAGAGTCGAAGACACGGCTGTGTATTACTGTGCGAGAATCAGCGGGGTAGTGGGACCTGTCCCCTTTGACTACTGGGGCCAGGGAACCCTGATCACCGTCTCCTCT
Нуклеотидная последовательность, кодирующая вариабельный домен MAB8 LC (SEQ ID NO:48)
GACATCCAGATGACCCAGTCTCCATCTTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGACCATTAGCAAGTATTTAAATTGGTATCAGCAGAAGCCAGGGAGAGCCCCTAAACTCCTGATCTACTCTGCGTCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCACTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCACCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAACAGAGTTACAGACCCTCCCAGATCACTTTCGGCCCTGGGACCAAAGTGGATATCAAA
Нуклеотидная последовательность, кодирующая вариабельный домен MAB53 HC (SEQ ID NO:49)
CAGGTGCAGCTGGTGCAGTCTGGGGCTGAGGTGAGGAAGCCGGGGTCCTCGGTGAAGGTCTCCTGCAAGGTTTCTGGAGGCATCATTAGGAAATATGCTATCAACTGGGTGCGACAGGCCCCCGGACAAGGGCTTGAGTGGATGGGAGGGATCATCGCTATCTTTAATACAGCAAACTATGCACAGAAATTCCAGGGCAGAGTCACGATTACCGCGGACGAGTCCACGAGCACAGTCTACATGGAGCTGAGCAGCCTGAGATCTGAAGACACGGCCCTTTATTACTGTGCGAGAGGAATGAATTACTACAGTGACTACTTTGACTACTGGGGCCAGGGAAGCCTTGTCACCGTCTCCCCA
Нуклеотидная последовательность, кодирующая вариабельный домен MAB53 LC (SEQ ID NO:50)
GAAATTGTGTTGACACAGTCTCCAGGCACCCTGTCTTTGTCTCCAGGGGAAAGAGCCACCCTCTCCTGCAGGGCCAGTCAGAGTGTTAGAAGCAACAACTTAGCCTGGTACCAGCACAAACCTGGCCAGGCTCCCAGGCTCCTCATCTTTGGTGCATCCAGCAGGGCCACTGGCATCCCAGACAGGTTCAGTGGCAGTGGGTCTGGGACAGACTTCACTCTCACCATCAGCAGACTGGAGCCTGAAGATTTTGCAGTATATTACTGTCAGCAGTATGGTAGCTCACCTGCGCTCACTTTCGGCGGAGGGACCAAGGTGGAGATCAAA
Следующие примеры представлены с иллюстративной целью, но не для ограничения изобретения.
Пример 1. Аффинность MAB53 и MAB579
Аффинность MAB53 описана в приведенной выше публикации PCT. Указанное антитело связывается с высокой аффинностью с HA из клад H5, H7, H1 и H9, при этом не обладает аффинностью в отношении H2 и H3. MAB579 обладает высокой аффинностью в отношении HA из H7 и H3. На фиг. 3A и 3B представлены конкретные результаты, полученные с помощью стандартного теста ForteBio™ для каждого антитела.
Пример 2. Нейтрализация инфекции с помощью MAB486 и MAB579
MAB 486 и 579 тестировали в реакции ингибирования инфекции, вызываемой H1N1 и H3N2 (A/Perth/16/2009) и в реакции нейтрализации бляшкообразования в монослое клеток MDCK в присутствии трипсина при начальной фазе инфекции или без него. MAB486 и pAb ×CP (поликлональные антитела кролика, полученные против консенсусной последовательности сайта расщепления) нейтрализуют H1N1 (A/California/04/2009) только в отсутствии трипсина, как показано на фиг. 4A, и не способны ингибировать инфекцию и образование бляшек, если вирус сначала активирован трипсином. Данные показывают, что антитела, против участка, обеспечивающего слияние, в котором расположены эпитопы, полученные на основании интактного пептида слияния (т.е. чувствительные к протеазам), не эффективны для контролирования вирусной инфекции. Как показано на фиг. 4B, MAB579 ингибирует инфекцию, как в присутствии трипсина, так и без него.
Способность MAB53 нейтрализовать инфекцию описана ранее, но сравнение показателей аффинности и EC50 с соответствующими показателями для моноклональных антител Crucell CR6261 в реакции нейтрализации in vitro представлено в таблице 3 ниже.
KD (нМ)
EC50 (мкг/мл)
EC50 (мкг/мл)
KD (нМ)
EC50 (мкг/мл)
EC50 (мкг/мл)
Значения EC50 получали, как описано выше.
Пример 3. Определение эпитопов
Использовали технологию Pepscan CLIPS™ для картирования сайтов связывания MAB53 и MAB579. Для H1 и H3 синтезировали приблизительно 6000 уникальных пептидов различной длины и с различными по длине соединительными звеньями, связывающими концы каждого пептида, которые воспроизводят нативную структуру. Связывание MAB53 и MAB579 с областью “стебля” подтверждали с использованием антител кролика к глобулярной головке или к “стеблю” в реакции конкурентного связывания и с помощью прямого связывания с пептидами, полученными из области “стебля”. Как отмечалось выше, MAB486 связывается с вирусами группы 1 и группы 2, но только в предварительно активированном состоянии перед протеазным расщеплением HA0 на HA1 и HA2, связанные дисульфидными связями. Пришли к заключению, что эпитоп для перекрестного связывания кладов представляет собой прерывистый эпитоп, охватывающий два мономера нативного тримерного HA0.
Пример 4. Эффективность in vivo (MAB53) и фармакокинетика (MAB53 и MAB579)
В экспериментах использовали следующие штаммы:
H1N1: A/CA/04/09;
H5N1: A/Vietnam/1203/04/HPAI;
H3N2: A/Perth/16/09;
H7N3: A/Red Knot/NJ/1523470/06.
Для проверки профилактического действия MAB53 вводили мышам внутрибрюшинно в виде однократной дозы 10 мг/кг в день -1, после чего день 0 интраназально вводили вирус 10 раз в дозе LD50. Эффективность MAB53 определяли по обнаружению EC50 при дозе 0,4 мг/кг в сравнении с антителом Crucell CR6261, которое согласно сообщениям характеризуется значением EC50 1-1,5 мг/кг (Koudstaal, W., et al., J. Infect. Dis. (2009) 200: 1870-1873).
Для проверки терапевтической эффективности MAB вводили внутрибрюшинно в виде однократной дозы 10 мг/кг в день +3 для большинства штаммов или в день +1 для H7N3. MAB53 характеризовалось полной эффективностью в отношении H1N1 и H5N1, тогда как по существу все мыши контрольной группы погибали к 10-му дню. MAB579 характеризовалось полной эффективностью против H3N2 и H7N3, в то время как все мыши контрольной группы погибали до 10-го дня.
Также измеряли потерю массы, и показатели уменьшения массы составляли не более 20% у мышей, получавших лечение.
При проведении сравнительного исследования с лечением Tamiflu® (осельтамивира фосфат), мышей (10 животных в группе) после проведения анестезии инфицировали интраназально 10 раз вирусом в дозе LD50 (H1N1 Influenza A/Ca/04/09). MAB53 (или контрольный изотипически сходный IgG человека) вводили i.p. в день +1 после заражения. Tamiflu® вводили с помощью желудочного зонда два раза в день в течение 4 дней, начиная со дня +1 после инфицирования. Показатели смертности и заболеваемости (оцениваемой по потери массы) были значительно выше в группе Tamiflu®, по сравнению с группой MAB53.
Все животные контрольных групп погибли в течение восьми дней после заражения. Все животные, получавшие лечение Tamiflu®, за исключением двух мышей, погибли в течение восьми дней после инфицирования; две мыши прожили по меньшей мере до 14-го дня. В группе, получавшей лечение MAB53, восемь из десяти мышей прожили 8-4 дней после заражения.
В отношении потери массы, контрольная группа теряла в массе до 70% от начальный значений спустя восемь дней. Уменьшение массы прекращалось на 4-й день у мышей, получавших MAB53, и у 14-му дню показатели массы превышали исходные значения. У мышей, получавших Tamiflu®, потеря массы прекращалась к 6 дню, но только 92% исходной массы восстанавливалось к 14 дню.
Также у мышей исследовали фармакокинетические параметры MAB53 и MAB579. Показано, что период полувыведения у мышей, составляющий 7-14 дней, соответствует периоду полувыведения у человека продолжительностью 3-4 недели. Указанный результат соответствует результату, характерному для MAB IgG1κ. Для биспецифического антитела MAB579/53Bi (см. пример 5) получают сходное значение периода полувыведения.
Пример 5. Конструкция MAB579/53Bi
Конструкция MAB579/53Bi предоставляет scFv-часть MAB53, соединенную с константным участком MAB579, как показано на фиг. 5. Конструкция таких биспецифических антител хорошо известна в данной области (Marvin, J. S., Acta Pharmacologica Sinica (2005) 26:649-658). Таким образом, MAB579/53Bi обеспечивает бивалентное связывание по обоим концам молекулы, а также предоставляет интактную Fc-область. Из таблицы 4 видно, что биспецифическое антитело сохраняет аффинность отдельных антител, которую измеряли с помощью ForteBio™ в нМ. Биспецифическое антитело также сохраняет нейтрализующую активность отдельных антител, из которых оно состоит, и характеризуется значением EC50 3,5 мкг/мл в отношении H1N1; 6,0 мкг/мл в отношении H5N1, и 2,2 мкг/мл в отношении H3N2.
Пример 6. Эффективность MAB579/53Bi in vivo
Эффективность in vivo измеряли, как описано в целом в примере 4, полученные результаты представлены на фиг. 6A-6E.
Как показано на фиг. 6A и 6C, мышей инфицировали A/Ca/04/09 (H1N1) (представляет собой вирус гриппа группы 1) в день 0, и вводили с помощью IP инъекции отдельно 10 мг/мл MAB53, отдельно 10 мг/мл MAB579 или смесь MAB53 и MAB579 (фиг. 6A) или биспецифическое антитело (фиг. 6C) в день 2, (на фиг. 6C также представлены кривые потери массы). В качестве контролей вводили IgG в дозе 20 мг/кг. Смесь MAB вводили в дозе 10 мг/кг каждый, и биспецифическое антитело вводили в дозе 10 мг/кг. Как показано на фиг. 6A, смесь MAB53 и MAB579, а также MAB53 в чистом виде оказывали протективное действие, тогда как животные, получавшие MAB579 и контроль, погибли к 10 дню. Как показано на фиг. 6C, биспецифическое антитело обладало такой же эффективностью, как и смесь.
Сходные результаты получали в аналогичном протоколе для мышей, инфицированных вирусом гриппа группы 2 Philippines 2/82 (H2N3), как показано на фиг. 6B и 6D. (на фиг. 6D также представлены кривые потери массы). Как показано на фиг. 6B, смесь антител обладала эффективностью против вируса, как MAB579, но применение MAB53 в чистом виде и контроля приводило к смерти животных через 10 дней. На фиг. 6D показано, что биспецифическое антитело обладает такой же эффективностью как и смесь антител.
На фиг. 6E представлены результаты лечения инфекции с использованием аналогичного протокола, в котором контрольное заражение осуществляли вирусами как группы 1, так и группы 2, H1N1 и H2N3, которыми инфицировали одну и ту же мышь. Комбинация антител MAB579 и MAB53 при использовании в дозе 3 мг/кг каждое обеспечивала полную защиту и при использовании в дозе 1 мг/кг каждое обеспечивала 80% выживаемость мышей. Данный результат является существенным, поскольку в природе происходит коинфекция, приводящая к появлению рекомбинированного вируса, который может вызвать пандемию.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА, ПОДХОДЯЩИЕ ДЛЯ ПАССИВНОЙ ИММУНИЗАЦИИ ПРОТИВ ГРИППА | 2011 |
|
RU2635999C2 |
| АНТИТЕЛА, ПОДХОДЯЩИЕ ДЛЯ ПАССИВНОЙ ИММУНИЗАЦИИ ПРОТИВ ГРИППА, И ИХ КОМПОЗИЦИИ, КОМБИНАЦИИ И СПОСОБЫ ПРИМЕНЕНИЯ | 2015 |
|
RU2720282C1 |
| ВЫСОКОАФФИННЫЕ АНТИТЕЛА ЧЕЛОВЕКА К БЕЛКУ GB ЦИТОМЕГАЛОВИРУСУ (CMV) ЧЕЛОВЕКА | 2011 |
|
RU2613421C2 |
| НЕЙТРАЛИЗУЮЩИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, НАПРАВЛЕННЫЕ ПРОТИВ ВИРУСА ГРИППА, И ПУТИ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2721706C2 |
| КОНСТРУКЦИИ, НАПРАВЛЕННЫЕ НА КОМПЛЕКСЫ ПЕПТИДА AFP/МНС, И ВИДЫ ИХ ИСПОЛЬЗОВАНИЯ | 2016 |
|
RU2754041C2 |
| ЛЕЧЕНИЕ РАКА С ПОМОЩЬЮ ХИМЕРНОГО АНТИГЕННОГО РЕЦЕПТОРА К CD33 | 2015 |
|
RU2747384C2 |
| ДВУХВАЛЕНТНЫЕ БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА | 2008 |
|
RU2547615C2 |
| АНТИТЕЛА, СПЕЦИФИЧНЫЕ В ОТНОШЕНИИ НЕКТИНА-2 ЧЕЛОВЕКА | 2020 |
|
RU2820275C2 |
| ДВУХВАЛЕНТНЫЕ БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА | 2008 |
|
RU2575066C2 |
| АНТИТЕЛА, СПЕЦИФИЧНЫЕ К НЕКТИНУ-4 ЧЕЛОВЕКА | 2019 |
|
RU2825839C2 |





























Настоящее изобретение относится к иммунологии. Предложены варианты моноклональных антител и их фрагментов, которые связываются с вирусом гриппа A и нейтрализуют инфекцию, вызванную кладой H3 и по меньшей мере несколькими вариантами клады H7 вируса гриппа A. Также описана фармацевтическая композиция, содержащая такие антитела или их фрагменты, и применение антитела или его фрагмента в способе лечения или профилактики инфекции гриппа. Указанные антитела и их фрагменты пригодны для контролирования эпидемии и пандемии гриппа, а также для обеспечения профилактической или терапевтической защиты против сезонного гриппа. 3 н. и 6 з.п. ф-лы, 6 ил., 4 табл., 6 пр.
1. Моноклональное антитело, включая необязательно биспецифическое антитело или его фрагмент, которое(-ый) связывается с вирусом гриппа A, и которое(-ый) нейтрализует инфекцию, вызванную кладой H3 и по меньшей мере несколькими вариантами клады H7 вируса гриппа A, и которое(-ый) является человеческим или гуманизированным, причем антитело или его фрагмент содержит:
a) вариабельный участок тяжелой цепи, содержащий CDR1 с последовательностью AYTIH и CDR2 с последовательностью WINAGNGHTKYSQRFKGR, и CDR3 с последовательностью GPETYYYDKTNWLNSHPDEYFQH, и вариабельный участок легкой цепи, содержащий CDR1 с последовательность RASQTINNYLA и CDR2 с последовательностью KASSLES, и CDR3 с последовательностью QEYNNDSPLT;
b) вариабельный участок тяжелой цепи, содержащий CDR1 с последовательностью SYTLH и CDR2 с последовательностью WINAGNGKTKYPPKFRGR, и CDR3 с последовательностью GPESYYYDRSDWLNSHPDEYFQY, и вариабельный участок легкой цепи, содержащий DR1 с последовательностью RASQSISSWLA и CDR2 с последовательностью KASQLES, и CDR3 с последовательностью QLYNVYSPLT;
c) вариабельный участок тяжелой цепи, содержащий CDR1 с последовательностью SFTMH и CDR2 с последовательностью WINAGNGKTKYSQKFQGR, и CDR3 с последовательностью GPETYYYDSSNWLNSHPDEYLQY, и вариабельный участок легкой цепи, содержащий CDR1 с последовательностью RASQSISSWLA и CDR2 с последовательностью KASTLES, и CDR3 с последовательностью QEYNNNSPLT;
d) вариабельный участок тяжелой цепи, содержащий CDR1 с последовательностью SFTMH и CDR2 с последовательностью WINAGNGKTKYSQKFQGR, и CDR3 с последовательностью GPETYYYDSSNWLNSHPDEYFQH, и вариабельный участок легкой цепи, содержащий CDR1 с последовательностью RASQSISSWLA и CDR2 с последовательностью KASSLES, и CDR3 с последовательностью QEYNNNSPLT;
e) вариабельный участок тяжелой цепи, содержащий CDR1 с последовательностью SYSVH и CDR2 с последовательностью WINAGNGKTKYPQKFKGR, и CDR3 с последовательностью GPDSYYYDRNDWLNSHPDEYFQH, и вариабельный участок легкой цепи, содержащий CDR1 с последовательностью RASQSIDSWLA и CDR2 с последовательностью KASNLES, и CDR3 с последовательностью QLYNVHLI;
f) вариабельный участок тяжелой цепи, содержащий CDR1 с последовательностью TYTIH и CDR2 с последовательностью WINAANGHTKYSRKLRSR, и CDR3 с последовательностью GPETYYFDKTNWLNSHPDEYFQH, и вариабельный участок легкой цепи, содержащий CDR1 с последовательностью RASQSISTWLA и CDR2 с последовательностью KASNLES, и CDR3 с последовательностью QEYNNDSPLI;
g) вариабельный участок тяжелой цепи, содержащий CDR1 с последовательностью SYTLH и CDR2 с последовательностью WINAGNGKVKYPRKLQGR, и CDR3 с последовательностью GPESYFFDTSNHLNSHPDEYFQF, и вариабельный участок легкой цепи, содержащий CDR1 с последовательностью RASQSISSYLA и CDR2 с последовательностью KASNLES, и CDR3 с последовательностью QEYNNNSPLT;
h) тяжелую цепь, содержащую последовательность QVQLVQSGAEVKRPGASVKVSCRASGYTFTSFGFSWVRQAPGQGLEWMGWISAYNGDTKSPQKLQGRVTMTTDTSTNTAYMELRSLISDDTAVYYCARAPPLYYSSWSSDYWGQGTLLTVSS, и легкую цепь, содержащую последовательность DIQMTQSPGTLSLSPGERATLSCRASQSVSSNYLAWYQQKHGQAPRPLIYGASRRATDVPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQYGSSPRTFGQGTKLEIK; или
i) тяжелую цепь, содержащую последовательность QVQLVESGGGMVQPGGSRRLSCAASGFSFSTYGMHWVRQAPGKGLEWVAVISYDGEKQYYLDSVKGRFTISRDNSKDTLYLQMNSLTAEDTAVYYCVKESARRLLRYFEWLLSSPFDNWGQGALVTVSS, и легкую цепь, содержащую последовательность DIVMTQSPDSLAVSLGERATINCKSSQTVLYTSNKKNYLAWYQQKPGQPPKLLIYWASTRESGVPDRFSGSGSGTDFTLTISSLQAEDVAVYYCQQYYTSPYTFGQGTKLEIK.
2. Антитело или фрагмент по п.1, которое(-ый) представляет собой биспецифическое антитело или его фрагмент и дополнительно содержит второй вариабельный участок тяжелой цепи, который содержит
CDR1 с последовательностью KYAIN и
CDR2 с последовательностью GIIAIFNTANYAQKFQG, и
CDR3 с последовательностью GMNYYSDYFDY (MAB53), и
вариабельный участок легкой цепи, который содержит
CDR1 с последовательностью RASQSVRSNNLA и
CDR2 с последовательностью GASSRAT, и
CDR3 с последовательностью QQYGSSPALT (MAB53).
3. Антитело или фрагмент по п.2, которое(-ый) представляет собой биспецифическое антитело или его фрагмент, которое(-ый) содержит вариабельный участок тяжелой цепи, который содержит
CDR1 с последовательностью AYTIH и
CDR2 с последовательностью WINAGNGHTKYSQRFKGR, и
CDR3 с последовательностью GPETYYYDKTNWLNSHPDEYFQH (MAB579), и
вариабельный участок легкой цепи, который содержит
CDR1 с последовательностью RASQTINNYLA и
CDR2 с последовательностью KASSLES, и
CDR3 с последовательностью QEYNNDSPLT (MAB579).
4. Антитело или фрагмент по п.1, которое(-ый) содержит тяжелую цепь, содержащую последовательность
a) QVQLVQSGAEVKKPGASVKVSCKTSGYTFTAYTIHWVRQAPGQRLEWMGWINAGNGHTKYSQRFKGRVTITRDTSARTTYMELRSLTSEDTALYFCARGPETYYYDKTNWLNSHPDEYFQHWGHGTQVTVSS (MAB579) или
b) QVQLVQSGAEVKRPGASVKVSCRASGYTFTSFGFSWVRQAPGQGLEWMGWISAYNGDTKSPQKLQGRVTMTTDTSTNTAYMELRSLISDDTAVYYCARAPPLYYSSWSSDYWGQGTLLTVSS (MAB383), или
c) QVQLVESGGGMVQPGGSRRLSCAASGFSFSTYGMHWVRQAPGKGLEWVAVISYDGEKQYYLDSVKGRFTISRDNSKDTLYLQMNSLTAEDTAVYYCVKESARRLLRYFEWLLSSPFDNWGQGALVTVSS (MAB486), или
d) QLQLVQSGAEVKKPGASVKLSCKASGYTFTSYTLHWVRQAPGQTLEWMGWINAGNGKTKYPPKFRGRVTITRDTSATTVDMHLSSLTSEDTAVYFCARGPESYYYDRSDWLNSHPDEYFQYWGQGTLVIVSS (MAB699), или
e) QVQLVESGADVKKPGASVTVSCKASGYTFRSFTMHWVRQVPGQRLEWMGWINAGNGKTKYSQKFQGRVIVTRDTSASTAYMELSSLTSEDTAVYYCARGPETYYYDSSNWLNSHPDEYLQYWGQGTPVTVSS (MAB700), или
f) QVQLVQSGADVKRPGASVTVSCKASGYTFRSFTMHWVRQVPGQRLEWMGWINAGNGKTKYSQKFQGRVIVTRDTSANTAYMELSSLTSEDTAVYYCARGPETYYYDSSNWLNSHPDEYFQHWGQGTPVTVSS (MAB708), или
g) QVQLQESGAEVKKPGASVQVSCKASGYTFTSYSVHWVRQAPGQRPEWMGWINAGNGKTKYPQKFKGRVTITRDTLARTVNIHLSSLTSEDTAVYFCARGPDSYYYDRNDWLNSHPDEYFQHWGQGTVVIVSS (MAB710), или
h) QVQLVESGAEVKKPGASVKITCEASGYTFNTYTIHWLRQAPGQRLEWMGWINAANGHTKYSRKLRSRVTIKRDTSARTSYMELSSLGSEDTAVYYCARGPETYYFDKTNWLNSHPDEYFQHWGQGTLVTVSS (MAB711), или
i) QVQLVQSGAAVNKPGASVKVSCKASGYSFTSYTLHWVRQAPGQRPEWIGWINAGNGKVKYPRKLQGRITITRDVSATTVHMELRSLTSEDTGLYYCARGPESYFFDTSNHLNSHPDEYFQFWGQGTLVTVSS (MAB723).
5. Антитело или фрагмент по п.4, которое(-ый)
если содержит тяжелую цепь а), также содержит легкую цепь, содержащую последовательность DIQMTQSPSTLSASVGDRVTITCRASQTINNYLAWYQQKPGKAPKLLIYKASSLESGVPSRFSGSGSGTEFTLTISSLQPDDFATYYCQEYNNDSPLTFGGGTKVEIK (MAB579), или
если содержит тяжелую цепь b), также содержит легкую цепь, содержащую последовательность DIQMTQSPGTLSLSPGERATLSCRASQSVSSNYLAWYQQKHGQAPRPLIYGASRRATDVPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQYGSSPRTFGQGTKLEIK (MAB383), или
если содержит тяжелую цепь c), также содержит легкую цепь, содержащую последовательность DIVMTQSPDSLAVSLGERATINCKSSQTVLYTSNKKNYLAWYQQKPGQPPKLLIYWASTRESGVPDRFSGSGSGTDFTLTISSLQAEDVAVYYCQQYYTSPYTFGQGTKLEIK (MAB486), или
если содержит тяжелую цепь d), также содержит легкую цепь, содержащую последовательность DIQMTQSPSTLSASVGDRVTIACRASQSISSWLAWYQQKPGKAPKLLIYKASQLESGVPSRFSGSGSGTEFTLTINSLQPDDFATYYCQLYNVYSPLTFGGGTRVDIK (MAB699), или
если содержит тяжелую цепь e), также содержит легкую цепь, содержащую последовательность DIVLTQSPSTLSASVGDRVTITCRASQSISSWLAWYQQKPGKAPKLLIYKASTLESGVPSRFSGSGSGTEFTLTISSLQPDDFATYYCQEYNNNSPLTFGGGTKVEIK (MAB700), или
если содержит тяжелую цепь f), также содержит легкую цепь, содержащую последовательность DIQMTQSPSTLPASVGDRVTITCRASQSISSWLAWYQQKPGKAPKLLIYKASSLESGVPSRFSGSGSGTEFTLTISSLQPDDFATYYCQEYNNNSPLTFGGGTKVEIK (MAB708), или
если содержит тяжелую цепь g), также содержит легкую цепь, содержащую последовательность DIVMTQSPSTLSASVGDRVTISCRASQSIDSWLAWYQQKPGKAPKLLIYKASNLESGVPSRFSGSGSGTEFTLTISSLQPDDFATYYCQLYNVHLITFGGGTRVDIK (MAB710), или
если содержит тяжелую цепь h), также содержит легкую цепь, содержащую последовательность DIVMTQSPSTLSASVGDRVTITCRASQSISTWLAWYQQKPGKAPKLLIYKASNLESGVPARFSGSGSGTEFTLTISSLQPDDFATYYCQEYNNDSPLILGGGTTVEIK (MAB711), или
если содержит тяжелую цепь i), также содержит легкую цепь, содержащую последовательность DIQMTQSPSTLSASVGDRVTITCRASQSISSYLAWYQQKPGKAPKLLIYKASNLESGVPSRFSGSGSGTEFTLTISSLQPDDFATYYCQEYNNNSPLTFGAGTKVEIK (MAB723).
6. Антитело или фрагмент по п.4 или 5, которое(-ый) представляет собой биспецифическое антитело или фрагмент, которое(-ый) также содержит вторую тяжелую цепь, содержащую
a) QVQLVQSGAEVRKPGSSVKVSCKVSGGIIRKYAINWVRQAPGQGLEWMGGIIAIFNTANYAQKFQGRVTITADESTSTVYMELSSLRSEDTALYYCARGMNYYSDYFDYWGQGSLVTVSP (MAB53) или
b) EVQLVESGGGLVKPGGSLRLSCAASGFTFSTYTMSWVRQAPGQGLEWVSSITRTSSNIYYADSVEGRFTISRDNAKNSLYLQMHSLRVEDTAVYYCARISGVVGPVPFDYWGQGTLITVSS (MAB8), и
если биспецифическое антитело или фрагмент содержит тяжелую цепь a), также содержит легкую цепь, содержащую последовательность EIVLTQSPGTLSLSPGERATLSCRASQSVRSNNLAWYQHKPGQAPRLLIFGASSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQYGSSPALTFGGGTKVEIK (MAB53) и
если биспецифическое антитело или фрагмент содержит тяжелую цепь b), также содержит легкую цепь, содержащую последовательность DIQMTQSPSSLSASVGDRVTITCRASQTISKYLNWYQQKPGRAPKLLIYSASSLQSGVPSRFTGSGSGTDFTLTITSLQPEDFATYYCQQSYRPSQITFGPGTKVDIK (MAB8).
7. Антитело или фрагмент по п.1, которое(-ый) представляет собой биспецифическое антитело или фрагмент, которое(-ый) содержит вариабельный участок тяжелой цепи, содержащий
QVQLVQSGAEVRKPGSSVKVSCKVSGGIIRKYAINWVRQAPGQGLEWMGGIIAIFNTANYAQKFQGRVTITADESTSTVYMELSSLRSEDTALYYCARGMNYYSDYFDYWGQGSLVTVSP (MAB53) и
легкую цепь, содержащую, последовательность EIVLTQSPGTLSLSPGERATLSCRASQSVRSNNLAWYQHKPGQAPRLLIFGASSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQYGSSPALTFGGGTKVEIK (MAB53), и
которое(-ый) содержит вариабельный участок тяжелой цепи, содержащий
QVQLVQSGAEVKKPGASVKVSCKTSGYTFTAYTIHWVRQAPGQRLEWMGWINAGNGHTKYSQRFKGRVTITRDTSARTTYMELRSLTSEDTALYFCARGPETYYYDKTNWLNSHPDEYFQHWGHGTQVTVSS (MAB579), и
легкую цепь, содержащую последовательность DIQMTQSPSTLSASVGDRVTITCRASQTINNYLAWYQQKPGKAPKLLIYKASSLESGVPSRFSGSGSGTEFTLTISSLQPDDFATYYCQEYNNDSPLTFGGGTKVEIK (MAB579).
8. Фармацевтическая композиция для использования в способе лечения или профилактики инфекции, вызванной гриппом, содержащая антитело или его фрагмент по любому из пп.1-7 в эффективной дозе против клады H3 и по меньшей мере нескольких вариантов клады H7 вируса гриппа A.
9. Применение антитела или его фрагмента по любому из пп.1-7 в способе лечения или профилактики инфекции гриппа у индивида, где указанное антитело или его фрагмент направлено против клады H3 и по меньшей мере нескольких вариантов клады H7 вируса гриппа A.
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| WANG TAIA T | |||
| et al | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО К ВИРУСУ ТИПА А ГРИППА И УСТРОЙСТВО ДЛЯ ИММУННОГО АНАЛИЗА С ИСПОЛЬЗОВАНИЕМ АНТИТЕЛА | 2004 |
|
RU2366662C2 |

